
生化学反応ネットワークの進化的設計方法
【課題】生化学反応ネットワークの最適な機能を達成するための方法の提供。
【解決手段】細胞の生化学反応ネットワークの再構築を、反復最適化手順を用いてインシリコで実施する。さらに、インシリコ法を用いて為された決定を確認し、および事に依っては拡張し、並びに最適な機能を備えた培養細胞または細胞集団を産生するための、実験室での培養段階を含む。コンピュータ・システムおよびインシリコ段階を実施するためのコンピュータ可読プログラム・コードを含むコンピュータ製品が含まれる。
【解決手段】細胞の生化学反応ネットワークの再構築を、反復最適化手順を用いてインシリコで実施する。さらに、インシリコ法を用いて為された決定を確認し、および事に依っては拡張し、並びに最適な機能を備えた培養細胞または細胞集団を産生するための、実験室での培養段階を含む。コンピュータ・システムおよびインシリコ段階を実施するためのコンピュータ可読プログラム・コードを含むコンピュータ製品が含まれる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明の分野
本発明は広くは生化学反応ネットワークおよびより具体的には最適な所望の全細胞特性を得るための生物体中の代謝ネットワークの再構築に関する。
【背景技術】
【0002】
背景情報
ゲノム配列解析およびアノテーション技術により細胞を構成する分子成分の詳細なリストが得られ、ハイスループット技術によりこれらの成分がどのように使用されるかに関する情報が得られている。従って、我々は生物学的設計がゲノム規模で可能となる段階に近づいている。ある生物体由来の1遺伝子を別の生物体に「挿入(splice)」することおよび予測可能な結果を起こすのは困難なことが証明されてきた。主な理由としては、長い進化過程を通じて生細胞中のすべての成分が細胞全体の機能に最適に「適合」するように磨かれてきたことが挙げられる。単純に外来遺伝子を導入しても、または存在する遺伝子を欠損させても、予測可能な結果、そしてまた最適な結果に至ることはない。単一のまたは複数の遺伝子欠損または付加が全細胞機能から成るその機能に与える結果を演繹的に予測する方法および予め決定した方法で残りの成分を機能させる方法が必要とされている。これまでに細胞規模の代謝機能に関して若干の進歩が遂げられたが、かかる方法は欠けている。
【0003】
暫くの間、医学および産業目的のため代謝流束に再び関心が向けられた。この関心の結果、代謝工学の分野が生まれた。代謝工学の主な目標は生細胞中で望ましい代謝反応を実行することである。代謝工学に大事なのはコンピュータ技術、遺伝学、およびシステム科学を含む幾つかの科学分野の進歩および適用である。
【0004】
分析および設計のための従来の工学手法では、数学的またはコンピュータモデルを利用する。物質代謝に関しては、基本的な物理化学法則および原理を基礎としたコンピュータモデルが必要となると思われる。代謝技術者は次に、かかる方法が新規且つ改善された生細胞を体系的に「設計する」ために使用できることを望む。そこで組換えDNA技術の方法が、所望の細胞設計を達成するために適用されるはずである。
【0005】
代謝解析の25年から30年間の歴史により、細胞物質代謝の体系的特徴を定量する必要性が実証されてきた(例えば、Fell D.,「物質代謝制御の理解(Understanding the control of cellular metabolism)」(London, Portland Press) (1996)(非特許文献1); Heinrich R.ら,「代謝調節および数学モデル(Metabolic regulation and mathematical models)」Progress in Biophysics and Molecular Biology, 32:1-82, (1977)(非特許文献2); Heinrich R.およびSchuster S.,「細胞システムの調節(The regulation of cellular systems)」(New York, Chapman & Hall), xix, p.372 (1996)(非特許文献3); Savageau M. A.,「生化学系解析. I. 酵素反応構成要素に関する反応速度則のある数学的特性(Biochemical systems analysis. I. Some mathematical properties of the rate law for the ecomponent enzymatic reactions)」J. Theor. Biol. 25(3):365-69 (1969)(非特許文献4)を参照されたい)。代謝力学を研究する有意な動機がある。物質代謝の量的記述および代謝変化の創生能は、特定の治療目標を達成するのに重要であるばかりでなく、細胞生物学を理解するのに広く重要である。重要な用途には、治療学およびその他の生化学を提供するための菌株設計、遺伝的欠陥の代謝影響の評価、感染症と戦うための体系的方法の組み立てなどが含まれる。物質代謝の量的および体系的解析は従って、根本的に重要である。しかし、同分野における批評は「代謝工学における近年の急激な関心の高まりにもかかわらず、利用可能な分子生物学的技術力と生化学ネットワークの合理的な解析能との間には、依然として大きな格差が存在している」(Stephanopoulos G.,「代謝工学(Metabolic engineering)」Current Opinions in Biotechnology, 5:196-200 (1994)(非特許文献5))と結論付けている。この意見は数年前のものであるが、基本的には依然として真実である。我々は、与えられた環境での生物体の最良適合を達成する、何百万年もの自然進化と競争しているので、この結論は驚きではない。
【0006】
少数反応を含む部分的遺伝子調節ネットワークが設計された(Hastyら、「遺伝子調節ネットワークのコンピュータ研究(Computational studies of gene regulatory networks)」: In numero molecular biology, Nature, 2: 268-79 (2001)(非特許文献6)に総説されている)が、性能特性が定義されおよびそれらを引き続き構築した代謝ネットワークのような、生化学調節ネットワークの演繹的設計は実践されなかった。主な理由には、主としてその数値を決定する必要のある反応速度パラメータが非常に多く存在し且つ詳細な反応速度方程式が全般的に未知であって、全代謝ネットワークに対して信頼性のある詳細な反応速度モデルを構築できないことが挙げられる。従って、予測的反応速度モデルを達成できないので、物質代謝のような、最適な生化学反応ネットワークの演繹的設計は可能でない。実際、突然変異および進化過程により速度定数値は時間とともに変化する。
【0007】
従来、進化過程の終点を予測することは、それらが偶発的事象から選択された結果であると予測されるため、不可能であった。本発明は定義された環境における代謝ネットワークの進化終点の演繹的計算を可能とする方法を開示する。生物学的体系における最適化原則に基づく他の数学的モデリング法があるが;即ち、人工頭脳学モデリング法(Varner J.およびRamkrishna D.,「代謝経路の数学的モデル(Mathematical models of metabolic pathways)」Curr. Opin. Biotechnol., 10(2):146-50, (1999)(非特許文献7))、それらは要求されるパラメータ数のため生物学ネットワークの設計に従順ではない。従って、かかる設計を創出し構築するための進化過程の利用に対する基礎が提供される。
【非特許文献1】Fell D.,「物質代謝制御の理解(Understanding the control of cellular metabolism)」(London, Portland Press) (1996)
【非特許文献2】Heinrich R.ら,「代謝調節および数学モデル(Metabolic regulation and mathematical models)」Progress in Biophysics and Molecular Biology, 32:1-82, (1977)
【非特許文献3】Heinrich R.およびSchuster S.,「細胞システムの調節(The regulation of cellular systems)」(New York, Chapman & Hall), xix, p.372 (1996)
【非特許文献4】Savageau M. A.,「生化学系解析. I. 酵素反応構成要素に関する反応速度則のある数学的特性(Biochemical systems analysis. I. Some mathematical properties of the rate law for the ecomponent enzymatic reactions)」J. Theor. Biol. 25(3):365-69 (1969)
【非特許文献5】Stephanopoulos G.,「代謝工学(Metabolic engineering)」Current Opinions in Biotechnology, 5:196-200 (1994)
【非特許文献6】Hastyら、「遺伝子調節ネットワークのコンピュータ研究(Computational studies of gene regulatory networks)」: In numero molecular biology, Nature, 2: 268-79 (2001)
【非特許文献7】Varner J.およびRamkrishna D.,「代謝経路の数学的モデル(Mathematical models of metabolic pathways)」Curr. Opin. Biotechnol., 10(2):146-50, (1999)
【発明の開示】
【0008】
概要
本発明は生細胞中の生化学反応ネットワークの最適な機能を達成する方法に関する。生化学反応ネットワークは包括的な生化学反応ネットワーク、実質的に全ての生化学反応ネットワーク、または全ての生化学反応ネットワークで有り得る。方法は細胞の生化学反応ネットワークの再構築を用いて、インシリコで実施できる。方法はさらに、実験室におけるインシリコ段階を用いてなされた決定を確認するおよび事に依っては拡張する培養段階、並びに最適な機能を備えた培養細胞、または細胞集団を産生する培養段階を含むことができる。
【0009】
方法はネットワーク中の生化学反応をリストにしてコンピュータに表示することにより、例えばネットワーク中の生化学反応データベースを提供する;最適化方法を使用してネットワークの最適特性を計算する;ネットワーク中の反応リストを変更し、かつ最適特性を再計算する;および所望の性能に達するまで前述の工程を繰り返し行うことにより、実施できる。方法にはさらに最適化手順の結果生じる生化学反応が含まれるように、細胞の遺伝子構成を構築する段階;その条件の下で構築した細胞を、特定環境下の培養状態に置く段階;および細胞を、十分な期間、定めた所望の性能に細胞を進化させる条件下で培養する段階を含んでもよい。
【0010】
生化学反応ネットワークは代謝ネットワーク、例えば調節ネットワークで有り得る。さらに、遺伝子構成が構築される細胞は原核細胞または真核細胞;例えば大腸菌、出芽酵母、チャイニーズハムスター卵母細胞などで有り得る。さらに、細胞の遺伝子構成は、細胞中の1つまたはそれ以上の遺伝子を改変することにより、例えば付加もしくは欠失により、またはその調節成分(例えば、プロモーター、転写因子結合部位など)を介して遺伝子調節を改変することにより構築できる。別の局面において、本発明は前述の方法により産生される濃縮細胞集団を提供する。
【0011】
別の局面において、本発明は、ネットワーク中の生化学反応を含むデータベースを提供すること;最適化方法を使用してネットワークの最適特性を計算すること;ネットワーク中の反応を変更するための利用者による選択を受信し、かつ最適特性を再計算すること;所望の特性基準が満たされるまで最適化を繰り返し行うこと; 最適化情報の結果として生化学反応が含まれるように、細胞の遺伝子構成を構築するための最適化結果を表示すること;構築した細胞を、特定環境条件の下で培養すること;および細胞が所望の性能に進化するような十分な期間、細胞を培養することにより、細胞における包括的な生化学反応ネットワークの最適な機能を達成するための方法を提供する。
【0012】
最適化方法は本発明により提供されるコンピュータ・システムを用いて行ってもよい。コンピュータ・システムには一般的に、少なくとも1つの生物体の1つまたはそれ以上の生化学反応ネットワークに関する情報を提供するデータベース;最適化および/または比較のための1つもしくはそれ以上の生化学反応ネットワークの選択を受信でき、および所望の性能の選択を受信できるユーザ・インターフェース;並びに最適化方法の計算および再計算を実行するための機能が含まれる。本発明のコンピュータ・システムは生化学反応ネットワークを再構築する機能を含むことができる。データベースは内部データベースまたは外部データベースで有り得る。
【0013】
別の局面において、本発明はコンピュータ可読プログラム・コードが媒体上で統合された、コンピュータ利用可能媒体を含むコンピュータ・プログラム製品を提供する。プログラム・コードはデータベースと情報のやりとりを行うことができ、コンピューティング・システム内で以下の段階を達成する:ネットワークの所望の性能の選択を受信するためのインターフェースを提供する段階;生化学反応ネットワークの最適特性を再計算する前に、所望の最適特性を決定し、決定の結果を表示し、かつ生化学反応ネットワークを変更し、所望の最適な機能に達するまでその工程を繰り返し行う段階。生化学反応ネットワークの変更は、利用者により手動で入力された変更に基づいて実施されてもよく、またはプログラム・コードにより自動的に実施されてもよい。コンピュータ・プログラムはさらに、再構築された生化学ネットワークの一部であるデータベース入力を識別できるか、または生化学反応ネットワークの再構築を実行できる。
【0014】
発明の詳細な説明
本発明のある局面では、広大な生化学反応ネットワークの特性を設計する方法が提供される。本発明のこの局面の方法を使用することにより、生化学反応ネットワークを特定環境において予め決定された性能に設計できる。本発明のある局面には以下の段階が含まれる:
1) 生化学反応ネットワークの反応構造についてのコンピュータ再構築を使用する段階、
2) 最適化方法を使用して反応ネットワークの最適な機能性を決定する段階、
3) 単一または複数の遺伝子を除去するまたは付加することにより、ネットワークを表示するコンピュータにおいて反応構造を変更し、かつ最適特性を再計算する段階、
4) コンピュータ・シミュレーションにより最適特性が決定した反応ネットワークの構造に、遺伝子操作を使用して生物体中の遺伝子補完を対応させる段階、および
5) 確定した淘汰圧の下での、拡張培養を使用して、コンピュータ・シミュレーションにより予測された最適解の方へ実際の反応ネットワークの機能を進化させる段階。適応性進化過程それ自体により、最適設計を達成する反応速度パラメータの最良セットが決定されると思われる。パラメータ値の2つ以上の同様なセットが進化過程を介して決定できる。
【0015】
本明細書に開示される方法および手順を用いることで、生化学反応ネットワークをコンピュータで演繹的に設計できる。反応ネットワークの設計に続き、進化過程は、遺伝的に改変した生物体またはコンピュータ・シミュレーションに使用したネットワークに対応する野生型菌株に関して、最適な条件の下で、実験室にて実施される。生物体は非独特なやり方で最適な挙動を達成するかもしれない-即ち、そこに等価な最適解が存在するかもしれない。従って、本発明には生細胞の環境内で生化学反応ネットワークの最適性能を達成するコンピュータ設計方法、遺伝子改変、および進化過程の自明でない且つ現存しない組合せが含まれる。
【0016】
別の局面では、本発明は生細胞中の包括的な生化学反応ネットワークの最適な機能を決定するための方法に関する。方法は生細胞の所望の性能を達成するために使用される。方法はネットワーク中の生化学反応をリストにしてコンピュータに表示する段階;ネットワークの最適特性を計算するため最適化方法を使用する段階;ネットワークの反応リストを変更し、かつ最適特性を再計算する段階;および所望の機能が満たされるまで変更段階を繰り返し行う段階により、実施できる。
【0017】
インシリコで実施される段階に加えて、方法はさらに1個の生細胞、または細胞集団の培養を含む段階を包含できる。これらの段階には、所望の性能が満たされるまで変更段階を繰り返し行った結果生じる生化学反応が含まれるように細胞の遺伝子構成を構築する段階;その条件で構築した細胞を特定環境下の培養状態に置く段階;並びに細胞を、十分な期間、および定めた所望の性能にその細胞を進化させる条件の下で培養する段階が含まれる。
【0018】
生化学反応ネットワークは、1つの生化学経路または連関した生化学経路の一部の、相互に関連する一連の生化学反応である。代謝反応ネットワーク、異化反応ネットワーク、ポリペプチドおよび核酸合成反応ネットワーク、アミノ酸合成ネットワーク、エネルギー代謝等のような多数の生化学反応ネットワークが同定されている。生化学反応ネットワークの他の種類には、細胞シグナル伝達ネットワーク、細胞周期ネットワーク、オペロン調節ネットワークのような遺伝子発現の調節に関連する遺伝的ネットワーク、および細胞骨格の一部を産生するアクチン重合ネットワークを含む調節ネットワークが包含される。主要な細胞機能の大部分は、相互に作用する生化学反応のネットワークに依存する。
【0019】
本発明を実践するため、生化学的に設計される生物体中の包括的な、好ましくは実質的に全ての、または最も好ましくは全ての生化学反応ネットワークの反応構造をコンピュータ・シミュレーションために再構築する必要がある。全ての生化学反応ネットワークには、ある生化学機能と関連する細胞の生化学反応の全てが含まれる。例えば、全ての代謝反応ネットワークには、細胞の物質代謝を与える生化学反応の全てが本質的に含まれる。代謝反応ネットワークは、全ての生細胞において幾つかの形態で見られる全生化学反応ネットワークの良い例となる。
【0020】
包括的な生化学反応ネットワークとは、検出可能な特性をもたらす、および生細胞に照らして、検出可能な特性を改する効果に関して予測可能な方法で改変できる、相互に関連した生化学反応群である。例えば、細胞により産生される生体分子の産生をもたらす中心的反応、その中心的反応には生体分子の産生に関連する全ての生化学反応ネットワークにおける一部の反応しか含まれていないが、包括的な生化学反応ネットワークには、生細胞における生体分子の産生に及ぼす中心的生化学反応の変化の効果を予測するためにコンピュータを利用した方法が使用できるような条件で、その中心的反応が含まれうる。
【0021】
実質的に全ての生化学反応ネットワークとは、生細胞の検出可能な特性を担う相互に関連した生化学反応群である。実質的に全ての生化学反応ネットワークには中心的反応のほか、効果は比較的小さい可能性があるが、検出可能な特性に効果を及ぼす二次反応が含まれる。実質的に全ての生化学反応ネットワークにおける変化は、コンピュータを利用した方法を用いて予測できる。本発明はまた、包括的な、実質的に全ての、または全ての生化学反応ネットワークではなく、全ての生化学反応ネットワークにおける大部分の反応を利用することもできる。
【0022】
本発明の方法を用いて決定される最適特性(本明細書ではまた最適な機能とも呼んでいる)には、例えば、グリセロール取込速度、酸素取込速度、増殖速度、胞子形成発生および/または速度、栄養不足条件下での希元素の精錬速度、バイオマス、並びにタンパク質、炭水化物、抗生物質、ビタミン、アミノ酸、発酵生産物(例えば乳酸塩産生物)のような生体分子の産生量が含まれる。最適特性にはまた、例えば、キラル化合物および他の低分子量化合物の産生量も含まれる。最適特性にはまた、例えば、エネルギー運搬ATPまたは酸化還元運搬NADPHおよびNADHのような重要な補因子の最大内部産生量も含まれる。最適特性にはまた、生化学反応ネットワーク中の重要な反応を介した代謝流速度(flux rates)のような特性が含まれることが、細胞技術者により定義され得る。本発明により、これらの特性の1つまたはそれ以上と関連した最適性能が達成される。例えば方法により、特に望ましい増殖速度または特に望ましい産生量の達成が可能となる。
【0023】
典型的には、本発明の方法に関して、再構築された生化学反応ネットワークの生化学反応をコンピュータに表示する。この表示には再構築された生化学反応ネットワークの生化学反応のリスト作成を含むことができる。リスト作成は、ネットワーク・シミュレーションおよび最適特性の計算に相当するコンピュータアルゴリズムと連動できるように、コンピュータデータベース中に、例えば関係型データベースの一連の表として示すことができる。
【0024】
本発明に関して生化学ネットワーク再構築は高品質でなければならない。高品質生化学反応ネットワークの工程、具体的には代謝反応ネットワークでは、再構築はM. W. Covert, C. H. Schilling, I. Famili, J. S. Edwards, I. I. Goryanin, E. Selkov,およびB. O. Palsson,「微生物菌株のインシリコ代謝モデリング(Metabolic modeling of microbial stains in silico)」Trends in Biochemical Sciences, 26:179-186(2001); Edwards J.,およびPalsson, B, 「代謝流束均衡解析および大腸菌K-12遺伝子欠損のインシリコ解析(Metabolic flux balance analysis and the in silico analysis of Escherichia coli K-12 gene deletions)」BMC Structural Biology, 1(2)(2000a); Edwards J. S.,およびPalsson, B, O.,「インフルエンザ菌Rd代謝遺伝子型の体系的特性(Systemic properties of the Haemophilus influenzae Rd metabolic genotype)」Journal of Biological Chemistry, 274(25):17410-16,(1999); Karp P. D.ら,「HinCyc:インフルエンザ菌の完全ゲノムおよび代謝経路の知識基盤(HinCyc: A knowledge base of the complete genome and metabolic pathways of H. influenzae)」ISMB 4:116-24,(1996); Karp P. D.ら,「EcoCycおよびMetaCycデータベース(The EcoCyc and MetaCyc databases)」Nucleic. Acids Res. 28(1):56-59(2000); Ogataら,「KEGG京都大学遺伝子・ゲノム辞典(KEGG: Kyoto encyclopedia of genes and genomes)」Nucleic Acids Res. 27(1):29-34 (1999); Schilling C. H.およびPalsson B. O.,「ゲノム規模の経路解析を介したインフルエンザ菌Rdの代謝能の評価(Assessment of the metabolic capabilities of the Haemophilus influenzae Rd through a genome-scale pathway analysis)」J. Theor. Biol., 203(3):249-83(2000); Selkov E. Jr.ら, 「代謝経路データベース(MPW: the metabolic pathways database)」Nucleic Acids Res., 26(1):43-45(1998); Selkov E.ら,「メタン生成菌の物質代謝の配列データからの再構築(A reconstruction of the metabolism of Methanococcus jannaschii from sequence data)」Gene 197(1-2):GC11-26(1997)により確立された。本工程にはアノテートされたゲノム配列、並びに生化学的および生理学的データの利用が含まれる。これらのアノテートされたゲノム配列並びに生化学的および生理学的データは、以下の本発明のコンピュータ・システムの考察の項で詳細に記載されるような、1つまたはそれ以上の内部または外部データベースで見つけられる。使用される全データソースを照合するには再構築されたネットワークの入念な解析が必要とされる。他の生化学反応ネットワークの再構築に関して、同様の方法が使用できる。
【0025】
本発明のこの局面の方法は次に、指定のおよび様々な環境条件の下で、包括的な、実質的に全ての、または全ての生化学反応ネットワークの最適特性を決定するため、その再構築された包括的な、実質的に全ての、または全ての生化学反応ネットワークを利用する。この決定により、既定の環境で所望の性能を達成する生化学反応ネットワークの設計が可能となる。これにはまた、所望の性能を達成する、組換え型細胞または細胞集団を増殖させる方法を提供するため、下記に記載の、細胞の遺伝子構成を構築するおよびその細胞を培養する段階と組み合わせることができる。
【0026】
一連の特定環境下の包括的な、実質的に全ての、または全ての生化学反応ネットワークの最適特性は、最適化方法として知られるコンピュータによる方法を用いて決定できる。最適化方法は当技術分野において周知である(例えば、EdwardsおよびPalsson(1999)を参照されたい)。本発明の方法で使用される最適化方法では、例えば、限定することを意図するものではないが、下記にてさらに詳細に記載される、流束均衡解析(FBA)、相空間解析(PhPP)を組合せて、および最適線(LO)の決定を利用することができる。
【0027】
再構築された代謝ネットワークは次に、確立された方法(Bonariusら,「過小決定代謝ネットワークの流束解析:失われた制約の探索(Flux analysis of underdetermined metabolic networks: The guest for the missing constraints)」Trends in Biotechnology, 15(8):308-14(1997); Edwards J.S.ら,「代謝流束均衡解析(Metabolic flux Balance Analysis)」In: (Lee S.Y., Papoutsakis E.T.編), Metabolic Engineering: Marcel Deker. P13-57(1999); Varma A.およびPalsson B.O,「代謝流束均衡:基本概念, 科学的および実践的使用(Metabolic flux balancing: Basic concepts, Scientific and practical use)」Bio/Technology 12:994-98(1994a))を用い、定常状態における代謝流分布の定量的シミュレーションを実施するために使用できる。代謝ネットワークのコンピュータ・シミュレーションはどんな条件の下でも実施できる。さらに、どの反応リストでもその環境およびその反応リストの中身を記述するパラメータを変更することによりコンピュータで模擬できる。
【0028】
再構築された代謝ネットワークの代謝能は、流束均衡解析(FBA)(Bonariusら,(1997);Edwardsら,(1999);VarmaおよびPalsson(1994a))の確立された方法を用いて評価できる。FBAは、ネットワーク中の個々の反応の定常状態および能力制約(その反応を介した最大代謝流)にある代謝ネットワークの質量保存に基づいている。さらに、実験的に決定した菌株特異的なパラメータ、バイオマス組成(Pramanik J.およびKeasling J.D.,「大腸菌物質代謝の化学量論的モデル:増殖速度依存的なバイオマス組成および力学的エネルギー量の組込み(Stochiometric model of Escherichia coli metabolism: Incorporation of growth-rate dependent biomass composition and mechanistic energy requirements)」Biotechnology and Bioengineering, 56(4):398-421(1997))および維持エネルギー量(the maintenance requirements)(Varma A.およびPalsson B.O.,「化学量論的代謝流均衡モデルにより野生型大腸菌W3110において増殖および代謝副生成物の分泌が定量的に予測される(Stochiometric flux balance models quantitatively predict growth and metabolic by-product secretion in wild-type Escherichia coli W3110)」Applied and Environmental Microbiology, 60(10):3724-31(1994b))もまた必要とされる。これらの因子は次に、再構築された代謝ネットワークを介して流束分布を計算するために使用される。
【0029】
より具体的には、これらの因子の定義により、全ての実施可能な解が存する方程式に対する閉じた解空間(closed solution space)に数学的に至る。従って、その課題に対して多数の可能な解(流束分布)がある。全許容解のセットの中で「最良」または最適解は次いで、最適化手順および規定の目的を用いて決定できる。使用される最適化手順は線形計画法であり、その目的は全てのバイオマス成分を同時に産生するための生化学反応ネットワークの最適使用である。これらの最適化手順が確立され、公表されている(VarmaおよびPalsson(1994a); Bonarious(1997); およびEdwardsら,(1999))。最適増殖を目的として計算された挙動の実験データとの比較は、大部分の場合において好適である(Varma(1994b); Edwards J.S., Ibarra R.U., およびPalsson B.O.(参照として本明細書に組み入れられる),「大腸菌代謝能のインシリコ予測は実験データと一致する(In silico predictions of Escherichia coli metabolic capabilities are consistent with experimental data)」Nat Biotechnol., 19(2):125-30(2001a); 並びにEdwards, Ramakrishna,およびPalsson,「表現型可変性の特徴付け:表現型相空間解析(Characterizing phenotypic plasticity: A phenotype phase plane analysis)」Biotech Bioeng, 印刷中, (2001b))。換言すると、これらの解の制約および最適化手順により、細胞増殖を補助するおよび進化前の菌株に対し実際の生化学機能を望ましく推定するための、生化学反応ネットワークの最適使用の予測に繋がる。
【0030】
胞子形成、および栄養不足条件下での希元素の探知のような、別の生存目的の使用については、記述されていない。これらの条件の下での競争および進化はまた、最適なネットワーク機能を限定し且つ創生するために使用できる。
【0031】
これらの手順により、単一増殖条件下における最適な機能の計算に繋がる。これは非常に限定的であり、多数の増殖条件を分析するための手段が必要とされる。
【0032】
上記のように、全ての定常状態代謝流束分布は、規定の再構築された代謝ネットワークに対して定義される解空間(その解空間における各解はそのネットワークを介した特定の代謝流分布または特定の代謝表現型に相当する)に数学的に制約される(EdwardsおよびPalsson(1999))。単一の指定増殖条件の下、コーン(錐状)のその最適代謝流束分布は線形計画法(LP)またはかかる課題の最適解を計算する他の関連手法を用いて決定できる。その制約が変化すれば、コーンの形は変化し、最適流束ベクトルは質的に変化するかもしれない。表現型相空間(PhPP)解析は2つまたはそれ以上の制約環境変数における全ての変動可能性を考慮する。本方法をここに開示する。
【0033】
2栄養素の取込速度(炭素基質および酸素のような)は(x,y)空間上の2軸として定義でき、前述の手順を用い、その最適流束分布はこの空間中の全点に対して計算できる。生物学的に現実的な、再構築された代謝ネットワークに対してこの手順を実施する場合、かかる空間に存在する、有限数の質的に異なる最適代謝流束地図、または代謝表現型があることが判る。その相空間の境界は線形最適化の限界価値(shadow price)(Chvatal V.,「線形計画法(linear Programming)」(New York: W. H. Freeman and Co.)(1983))を用いて定義できる。記載の方法により、代謝ネットワークの最適使用が質的に異なる(x,y)空間中の異なる領域、または「相」の定義に繋がる。各相はPnx,y,として指定でき、ここでPは特定の表現型または流束分布を表し、nはこの表現型に対する区画領域に任意に割り当てられた数であり、およびその2つの取込速度がその空間軸を形成する。
【0034】
相空間は2-パラメータ空間の全体にわたって限界価値を計算することにより構築でき、一定の限界価値の領域を区別するために直線を描く。限界価値により、目的関数に対する各代謝産物の実態価値が定義される(Chvatal(1983))。xおよびy軸に相当する取込速度の特定数値により表されるように、限界価値は既定の環境条件下での最適増殖に対する代謝産物の価値に依存して、マイナス、ゼロまたはプラスのどちらかとなる。限界価値がゼロとなる場合、取込速度値が変更されるように最適代謝地図の質的転換が起こる。この基準によりPhPPにおける直線が定義される。
【0035】
最適線(LO)はPhPPの軸に相当する2つの取込流束間の最適な相関を表す直線と定義される。好気性に関して、この直線は最大バイオマス産生を支えるための基質の完全酸化に対する最適な酸素取り込みと解釈できる。
【0036】
代謝再構築および表現型相空間解析手順を、次いで、望ましい代謝挙動、例えば最大取込速度が最適となると思われる条件を予測するために使用する。換言すれば、最適性能を決定するために代謝再構築およびPhPPを使用する。最大取込速度により、相空間における有限の長方形領域の定義に繋がる。この長方形内の最適増殖条件は次いで、規定の制約内での進化過程の予測結果になると思われる。
【0037】
最適化手順を使用しても、その対応する実際の生化学反応ネットワークの特性は、最適でないまたは実用上の見地から望まれるものと同一でないかもしれない。生物体において模擬した再構築ネットワークおよびその合成は、所望の最適解(本明細書では同様に所望の最適性能または所望の最適な機能とも呼んでいる)を示さないかもしれない。最適性欠如は以下の事実による可能性がある:
1. 完全なネットワークを備えた天然生物体は、関心のある環境条件を経験していない並びにこの環境における増殖競争および淘汰を経験していない、または
2. ネットワークからの/ネットワークへの新規反応の除去/追加を介した、遺伝子操作により人工ネットワークはその進化的に決定された最適状態から混乱させられる。
【0038】
本発明のインシリコ方法は、所望の性能が達成されるまでネットワーク中の反応を変更することにより、最適性欠如のこの第2番目の原因を解決するように設計される。次に、培養方法、これは以下に詳細に記載されているように本発明の方法に含めることができるが、増殖競争および淘汰に関連する最適性欠如の第1番目の原因を解決するために使用できる。
【0039】
上記のように、最適特性の計算後、所望の性能を達成するため、代謝技術者はネットワーク中の反応リストを変更できる、または反応リスト中の1つまたはそれ以上の反応を自動的に変更するアルゴリズムが開発され得る。生化学リストの変更後、既定の環境条件下のこのネットワークの最適特性を計算できる。これは、所望の性能に達するまで反応リストの多数の別版(バージョン)が必要とされる可能性がある、会話型設計手段である。所望の性能は、質的特徴または最適化手段を用いて計算された特性に関する定量値である。所望の性能が達成され得る多くの特性は、同分野において周知である。例えば、所望の性能は、所望の増殖速度または酵素もしくは抗生物質のような生体分子の所望の産生量で有り得る。
【0040】
最適化方法は本発明により提供されるコンピュータ・システムを用いて実施できる。コンピュータ・システムには典型的に、少なくとも1生物体の1つまたはそれ以上の生化学反応ネットワークに関する情報を提供するデータベース;最適化および/または比較のため1つまたはそれ以上の生化学反応ネットワークの選択を受信できる、および好ましい性能の選択を受信できるユーザ・インターフェース;並びに最適化方法の計算および再計算を実行するための機能が含まれる。さらに、本発明のコンピュータ・システムは、上述の生化学反応ネットワークの再構築を実行するための機能を含んでいてもよい。
【0041】
コンピュータ・システムはスタンド・アロンのコンピュータまたはクライアント/サーバー環境および1つまたはそれ以上のデータベース・サーバを含む従来型のネットワーク・システムで有り得る。ローカルエリア・ネットワーク(LAN)または広域ネットワーク(WAN)を含む、多くの従来型のネットワーク・システムが同分野において周知である。さらに、クライアント/サーバー環境、データベース・サーバ、およびネットワークは技術、営業用および特許文献中に十分に記録されている。例えば、データベース・サーバはリレーショナル・データベース管理システム、ワールドワイド・ウェブ・アプリケーション、およびワールドワイド・ウェブサーバを作動させる、UNIX(登録商標)のようなオペレーティング・システム上で作動できる。
【0042】
典型的に、本発明のコンピュータ・システムのデータベースには、包括的な生化学反応ネットワーク、実質的に全ての生化学反応ネットワーク、または全ての生化学反応ネットワーク中の生化学反応に関する情報が含まれる。この情報は少なくともネットワークの生化学反応の一部と関連する、生体分子反応物質、産物、補因子、酵素、補酵素の反応速度等の同定を含むことができる。この情報は特定の生化学反応に関与する化合物の分子数を示す化学量論係数を含むことができる。この情報は生物体中に見つけられる任意のおよび全てのイソ酵素を含むことができる。この情報は単一酵素により触媒され得る全ての関連する生化学反応を含むことができる。この情報は、生化学反応を遂行できる複合体の形成のため多くの異なるタンパク質分子が非共有的に会合する必要がある場合、そのオリゴマー酵素複合体の構造を含むことができる。この情報は反応を遂行する酵素の場所(即ち、膜中に、細胞質中に、またはミトコンドリアのような細胞内小器官内部に存在する場合)を含むことができる。この情報はまた、種々の条件下での実験由来のまたは計算による反応速度、バイオマス組成、および維持エネルギー量(maintenance requirements)を含むことができる。この情報は非力学的増殖(non-mechanistic growth)および非増殖関連維持エネルギー量(non-growth associated maintenance requirements)を含むことができる。この情報はタンパク質合成における非効率性のような力学的維持情報(mechanistic maintenance information)を含むことができる。この情報はゲノム規模のmRNAまたはタンパク質発現プロファイル由来のデータを含むことができる。この情報は特定遺伝子の発現と関連したオペロンまたは調節構造に関するデータを含むことができる。この情報は酵素特性の変化を反映する配列変異を含むことができる。この情報は例えば、遺伝子が関心のある条件下で発現していない場合に応じて、条件依存的な生化学反応の組込みを含むことができる。生化学反応ネットワークが代謝反応ネットワークである場合、例えばその情報には既知消費速度、副生成物産生速度、および取込速度を含むことができる。
【0043】
データベースの情報は生化学反応ネットワークの生化学反応に関連する生体分子に関する生体分子配列情報、例えばゲノム配列のような多数の生体分子配列に関する情報を含んでもよい。少なくともゲノム配列の一部は、1つまたはそれ以上の生物体の各ゲノム上にて1つまたはそれ以上の連続配列に沿って位置する読み枠に相当しうる。生化学反応ネットワークに関する情報は、ある生体分子配列が関与するそれらの生化学反応ネットワークおよびその生体分子を含む生化学反応ネットワーク中の特定反応を同定する情報を含むことができる。
【0044】
データベースは生化学反応に関するどんな種類の生物学的配列情報も含むことができる。例えば、データベースはESTを含む核酸配列データベースおよび/またはより好ましくは完全長配列、またはアミノ酸配列データベースで有り得る。好ましくは、データベースは包括的な、実質的に全ての、または全ての生化学反応ネットワークに関する情報を提供する。例えば、データベースは全ての代謝反応ネットワークに関する情報を提供できる。データベースは生物体の完全ゲノムからなる核酸および/またはアミノ酸配列を提供できる。
【0045】
データベースはいかなる生物由来の生化学情報および配列情報も含むことができ、二部に分割できる。一方は配列を記憶するための部分および他方は配列に関する情報を記憶するための部分である。例えば、データベースは動物(例えば、ヒト、霊長類、齧歯類、両生類、昆虫等)、植物、または微生物に関する生化学反応情報および配列情報を提供できる。データベースは例えば、データベースの生体分子の機能、特に生化学機能に関する情報がアノテートされていることが好ましい。アノテーションはそのデータベースの生体分子の生化学、例えばある生体分子が関連する特定反応において、その生体分子は酵素であるかまたは酵素をコードするかどうか、その配列は野生型配列かどうか等、を研究している公開報告から得られる情報を含むことができる。
【0046】
データベースのアノテーションおよび配列は、同定する生物体での選択した生化学的遺伝子型(biochemical genotype)に関する十分な情報を提供する。生化学的遺伝子型は生物体での選択した生化学過程中の全ての核酸またはアミノ酸配列の集団である。例えば、代謝遺伝子型は物質代謝に関連するタンパク質の全ての核酸および/またはアミノ酸配列の集団である。代謝遺伝子型の同定方法は文献に記載されている(例えば、EdwardsおよびPalsson 1999を参照されたい)。
【0047】
データベースはカード型データベースまたはリレーショナル・データベースで有り得る。データベースは内部データベース、または利用者がアクセスできる外部データベース、例えばGenbankまたはGenpeptのような、公共の生物学的配列データベースで有り得る。内部データベースは個人データベースとして保守される、典型的には企業により、ファイアウォール後方で保守されるデータベースである。外部データベースは内部データベースの外側に位置し、典型的には内部データベースとは異なる実在者により保守される。公共の生物学的配列外部データベースの多くが利用可能であり、本発明で使用できる。例えば、米国立バイオテクノロジー情報センター(NCBI)、米国立医学図書館の一部から入手可能な生物学的配列データベースの多くが本発明で使用できる。外部データベースの他の実施例にはシアトルのフレッドハッチンソン・ガン研究センターにより保守されているBlocksデータベース、およびジュネーブ大学により保守されているSwiss-Protサイトが含まれる。さらに、外部データベースは生化学反応について記載し且つ分析している公開済み参考文献のデータベースを含め、生化学反応に関する情報を提供するデータベースを含むことができる。本発明のコンピュータ・システムに含まれるデータベースが特定の生化学反応ネットワークに関連する情報を同定していない公共のコンピュータ・データベースである場合、コンピュータ・システムには生化学反応ネットワークの再構築を実行する機能が含まれるか、または特定の生化学反応ネットワークに関するデータベース登録の確認が含まれる。さらに、生化学経路情報を備えた幾つかのデータベースがあり、これらのデータベースには、例えばこれに限定されないが、EcoCyc、KEGG、WITおよびEMPが含まれる。これらのデータベースは代謝モデルを再構築するための情報を提供するために使用できる。
【0048】
上記のデータベースに加え、本発明のコンピュータ・システムには最適化および/または比較のため1つもしくはそれ以上の生化学反応ネットワークの選択を受信できる、並びに最適性能の選択を受信できるユーザ・インターフェースが含まれる。例えば、一連のメニュー、ダイアログ・ボックス、および/または選択可能ボタンを用いて選択が為される場合、インターフェースはグラフィカル・ユーザ・インターフェースで有り得る。インターフェースは典型的に、主画面から始まる一連の画面を介し、利用者を案内する。ユーザ・インターフェースは利用者が生化学反応ネットワークに関連する追加情報にアクセスするために選択できるリンクを含むことができる。
【0049】
最適化方法を実行する本発明のコンピュータ・システムの機能には典型的に、コンピュータ・プログラム製品(それ自体、本発明の別の局面を表しているが、コンピュータ利用可能媒体上で統合され、かつ処理装置に接続する記憶装置に渡されたコンピュータ可読プログラム・コードが含まれる)を実行する処理装置が含まれる。記憶装置は例えば、ディスク・ドライブ、ランダム・アクセス・メモリー、リード・オンリー・メモリー、またはフラッシュ・メモリーで有り得る。
【0050】
本発明のコンピュータ・システムの処理装置により読み込まれ、かつ実行されるコンピュータ・プログラム製品には、コンピュータが利用可能な媒体上で統合されたコンピュータ可読プログラム・コードが含まれる。プログラム・コードは、データベースと情報のやりとりを行うことができ、かつコンピューティング・システム内で以下の段階を達成する:ネットワークの望ましい性能の選択を受信するためのインターフェースを提供する段階;生化学反応ネットワークの最適特性を再計算して所望の最適な機能に達するまでその工程を繰り返し行う前に、所望の最適特性を決定し、その決定の結果を表示し、かつ生化学反応ネットワークを変更する段階。生化学反応ネットワークの変更は利用者により認識される変更に基づき実施してもよく、またはプログラム・コードにより自動的に実行してもよい。コンピュータ・プログラムはさらに、再構築された生化学ネットワークの一部にあたるデータベース登録を確認してもよく、または生化学反応ネットワークの再構築を実行してもよい。さらに、本発明のコンピュータ・プログラムは、成分および特性の相違を同定するため生化学反応ネットワークを比較する機能を提供できる。
【0051】
コンピュータ可読プログラム・コードは、本発明の方法を実行するソフトウェアを書き込むために使用されるプログラミング言語と互換性のあるいずれか周知のコンパイラを用いて作成できる。本発明の方法を実行するソフトウェアを書き込むために使用できる多くのプログラミング言語は周知である。
【0052】
上記のように、本発明のこの局面の方法にはさらに、望ましい性能を達成するための培養菌株の適応性進化を含む段階を包含できる。実質的には、例えば原核細胞、または真菌細胞もしくは動物株化細胞の細胞を含む動物細胞のような真核細胞を含むどの細胞も、本発明の方法で使用できる。しかし、細胞または近縁生物体の細胞の生化学反応ネットワークは、コンピュータで包括的な、実質的に全ての、および/または全ての生化学反応ネットワークの高品質な再構築を可能とするため十分に特徴付ける必要がある。本質的に生物体の全ゲノムが配列解読されていることおよび生化学ネットワークに関連する生体分子、典型的にはタンパク質をコードする遺伝子が同定されていることが好ましい。
【0053】
細胞の遺伝子構成は所望の性能を満たす潜在性のある細胞を産生するため所望の性能に合う生化学反応を含むように構築できる。これは細胞の固有反応リストを用いて、および上述のインシリコで実行される段階により特定される所望の性能基準を達成できる反応リストに達するよう遺伝子操作を用いて、このリストに反応を追加するおよびこのリストから反応を差引くことにより達成できる。例えば、反応に関連する1つ以上の生体分子をコードする1つ以上の遺伝子の全部もしくは一部を付加する、改変する、または除去することにより、例えば、1つ以上の遺伝子のタンパク質コード領域を付加する、改変する、もしくは除去することにより、または1つ以上の遺伝子の調節領域を付加する、改変する、もしくは除去することにより、リストに反応を追加するまたはリストから反応を差引くことができる。さらに、例えば、リストの1つ以上の反応に関連する1つ以上の生体分子の発現を変化させる調節成分(例えば、転写因子)の発現を改変することにより、リストに反応を追加するまたはリストから反応を差引くことができる。その結果として作成された人工細胞は、上述の反復最適化手順を用いるインシリコ法により予め計算した最適特性を示してもまたは示さなくてもよい。
【0054】
細胞の遺伝子操作について説明する多くの方法が当技術分野に存在する(例えば、Datsenko K. A.およびWanner B. L.,「PCR産物を用いた大腸菌K-12の染色体遺伝子の一段階不活性化法(One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products)」Proc. Natl. Acad. Sci. U.S.A., 97(12):6640-45 (2000); Linkら,「野生型大腸菌のゲノムにおける正確な除去および挿入をもたらす方法:読み枠の特徴付けへの適用(Methods for generating precise deletions and insertions in the genome of wild-type Escherichia coli: Application to open reading frame characterization)」J. of Bacteriology, 179:6228-37 (1997)を参照されたい)。既存のどの遺伝子操作手順も開示される本発明を実践するために使用できる。
【0055】
所望の性能を満たす潜在性を有するように細胞を構築した後、特定環境の下での培養状態に置く。特定環境は最適化手順を用いて決定される。即ち、前述のように、最適化手順により種々の環境下でのネットワーク特性を計算し、所望性能が達成される特定環境を明らかにする。
【0056】
細胞を十分な期間、および所望の性能にその細胞を進化させる条件の下で培養する。即ち、天然または人工菌株の適応性進化は、一般的な最適化方法または手順による誘導に従い実行される。特定の環境を経験していない天然菌株または遺伝的に改変された菌株は、上記に開示されたネットワーク再構築および最適化手順により解析できる。これらの菌株を次いで、その生物体の所望の機能と整合性が取れた淘汰圧の下に置き、予め決定した性能特性へ進化させることができる。その細胞はさらなる適応性進化をせずに、所望の性能を達成できる。即ち、細胞を培養するための十分な期間とは、さらなる適応性進化を必要とせず本発明の方法を用いて、その細胞の遺伝子構成が構築される直後までであってもよい。
【0057】
換言すれば、非最適または非進化株の拡張培養は、定義した環境条件の下で達成できる最適解へ、代謝ネットワークを最適化するまたは進化させるために実施できる。この進化過程の実践には、進化過程の開始時に代謝ネットワークが最適条件からどの程度離れているか、並びに無作為変異および遺伝子発現調節の改変により必要な変更を達成することがどの程度困難であるかに依存して、代謝ネットワークを最適化するのにおよそ数週間から数年程度が必要とされる。この過程は化学的突然変異誘発要因および/または放射線の使用により促進できる。さらに過程は、上述のインシリコ法により決定される、所望の性能を達成する生化学反応物質を含むように生細胞を遺伝学的に改変することにより、促進される。
【0058】
特定環境条件の下で細胞を培養する方法は、同技術分野において周知である。例えば、細胞が大腸菌であれば、所望の性能は所望の増殖速度であり、下記の手順が使用できる。この手順は他の細菌細胞および/または当技術分野において周知の他の性能基準での使用に容易に適応できる。さらに、手順は動物細胞のような他の細胞種での使用に容易に発展できる。例えば、方法は細胞が培養容器に接着する大規模系のような、他の培養系での使用に容易に適応できる。培養方法は当技術分野において周知の、ハイスループット解析に適応されうる。
【0059】
所望の性能を達成するよう、菌株を一方向に進化させる必要がある場合、選択した宿主菌株における代謝反応ネットワークの構築に続き、細胞を典型的には30%グリセロールと共に-80℃で凍結保存する。各適応性進化過程に対し、凍結ストックをLB寒天培地に播き、37℃で一晩増殖させる。プレートからは、単一細胞より生じた単一コロニーを同定できる。前培養として知られているが、単一コロニーは液体培地に植菌するために使用できる。各菌株の単一コロニーから植菌された前培養液を、その後の進化過程のための規定培地中で一晩増殖させる。適応性進化が起こる環境を規定する培養条件に菌株を植菌するため、前培養試料を翌日、典型的には増殖の中間対数期で(対数増殖の中間で)取り出す。次いで、バッチ処理用生物反応器または他の適した培養容器で開始する。これは典型的には、温度制御恒温槽内、電磁攪拌プレート(マグネティック・スターラー・プレート)上に置いたマイクロキャリア・スピナー・フラスコ中、250mLの容量で、十分な曝気を確実とする適当な-典型的には高い-速度および使用菌株の最適増殖温度(野生型大腸菌では37℃)の設定で、行われると思われる。当技術分野において周知の、他のよく利用される培養方法もまた使用できる。
【0060】
適当な時間後、大腸菌では典型的には翌日(培養が静止期に達する前)、中間対数期にある培養液の一部を新鮮な培地を含む新たなスピナー・フラスコに連続的に移入する。もし培養が増殖速度に関して最適化されているところであれば、選択基準が増殖速度であることを確実とするため、静止期は回避しなければならない。次いで、中間対数期の固定時間間隔(増殖速度にもよるが典型的には24時間毎)で連続的に移入し、それから新たな培養容器への植菌容量は増殖速度の増加に基づいて調整する。
【0061】
従って、次に連続的に移入する際に使用する植菌の適容量を決定するため、増殖速度を頻繁に、典型的には毎日、監視する。この連続培養過程を、培地組成による既定条件の下で、細胞をその最適達成可能増殖へ進化させるのに十分なだけ頻繁に繰り返す。
【0062】
時間とともにその集団がどのように進化しているかを決定するため、適応性進化の間、その増殖および代謝挙動を監視する。固定時間間隔、典型的には数日毎に、酸素取込速度、基質取込速度および増殖速度を測定することにより、代謝および増殖挙動に関して培養を試験する。その結果を次いで、表現型相空間にデータ点としてプロットする。最適線に対するそのようにして決定されるデータ点の動きにより、最適増殖挙動への進化が示されると思われる。データ点がその最適条件に位置するようなその代謝ネットワークを、細胞が稼動していることが観察されるまで、増殖速度に加え、膜輸送流束のこれらの測定を繰り返す。その後、その進化過程は最適性能、例えば増殖速度のさらなる増加がなくなるまで、継続できる。さらなる変化が観察されない場合、既定条件での最大増殖速度が達成されたことになる。
【0063】
副産物分泌はHPLCまたは最適増殖挙動への進化に関与する物質代謝の変化を評価する他の適切な分析化学法により監視できる。これらの研究のため、その進化菌株に対する乾燥重量対光学濃度の相関関係を決定することは、これが野生型とは異なっていると思われるので、同様にして必須である。その増殖速度および定常状態の増殖を監視することに加え、何らかの汚染が発生した場合の予備として冷蔵しておいた日毎の培養液に対し、汚染の可能性のあらゆる兆候または突然変異による部分母集団(mutant subpopulation-aliquots)に伴う共進化に関してその培養液を検査する、並びにその培養試料を播き、かつあらゆるコロニー形態の相違または種々の突然変異体について検査することにより、その培養の表現型を確かめる。毎日、その培養の光学濃度、植菌時間、植菌容量、増殖速度、および汚染のあらゆる兆候を記録する。さらなる使用の可能性のため、日毎の培養試料もまた、30%グリセロール中、-80℃にて凍結する。
【0064】
本発明の方法を用いて産生された細胞が最適性能を達成するように進化した後、それらの細胞をさらに特徴付けてもよい。例えば、細胞の生化学反応ネットワークから特徴付けをすることができる。この特徴付けは、生細胞の生化学反応ネットワークの、成分を含むその特性を、インシリコ法を用いて予測されたその特性と比較するために使用できる。
【0065】
下記の実施例は本発明を例証するが、本発明を限定しないことを意図するものである。
【0066】
実施例1
大腸菌の最適な機能および進化の決定方法の詳細
本実施例は、菌株の最適性能を決定し、および適応性進化過程を実行するために使用できる培養手順を提供する。
【0067】
大腸菌の増殖挙動は下記の標準手順により決定される。増殖は炭素源を添加したM9最小培地(Maniatisら, Molecular Cloning: A Laboratory Manual, (Cold Springs Harbor, N.Y., Cold Spring Harbor Laboratory, 545(1982))(表1)で行う。細胞増殖速度は環境条件を変更することにより、即ち約0.05〜4g/Lの範囲で炭素源濃度を変更し、温度(27.5℃〜37℃)、および酸素(空気に対し0〜100%飽和)を変更することにより変えられる。バッチ培養は通気を備えた500mLフラスコ中、250mL容量にて生物反応器中に設定する。これらの培養に際し、排気分析器を用いて酸素に対する物質移動係数(k1a)を測定するか、または当技術分野において周知の方法を用いて呼吸計試験槽中の酸素分圧を監視するかにより、酸素取込速度(OUR)をオンラインで監視する。温度は循環水槽(Haake, Berlin, Germany)を用いて制御する。全ての測定およびデータ解析は増殖の対数期に制限する。バイオマス並びに培地中の基質および代謝副産物の濃度は、当技術分野において周知の方法を用いて実験の間中、監視する。細胞増殖は、600nmおよび420nmでの光学密度(OD)の測定並びに細胞カウントにより監視する。細胞乾燥重量に対するODの相関関係は、2つの異なる方法により決定する;(1)培養試料50mLを遠沈させて75℃で一定重量まで乾燥させる、および(2)試料25mL(培養の間中、採取する)を0.45umフィルタを通してろ過し、一定重量まで乾燥させる。培地中の代謝産物濃度はHPLCで決定する。Aminex社製HPX-87Hイオン交換炭水化物-有機酸カラム(66℃にて)は、移動相として脱気した5mM H2SO4およびUV検出とともに使用する。酵素アッセイ法はまた、基質取込速度および副産物分泌速度を決定するために使用する。培養液中に溶解した酸素は、ポーラログラフ溶存酸素センサを用いて監視する。酸素消費量は3つの異なる方法により測定する;(1)排ガスを1440C Servomex社製酸素分析器(Servomex Co., Inc. Norwood, MA)に通す、(2)溶解酸素測定値およびK1a測定値から計算する、および(3)個別の50mLフラスコ中の呼吸計試験槽中で測定する。酸素取込速度の測定に使用される3方法ともに、同様のおよび再現可能な結果をもたらす。
【0068】
本実施例で提供される培養手順は、菌株の最適性能を決定するおよび適応性進化を実行するために使用できる。
【0069】
【表1】

【0070】
実施例2
最適特性および表現型相空間の計算
本実施例は、再構築されたネットワークの最適特性の計算方法およびその結果の表現型相空間への表現方法を示す。
【0071】
代謝ネットワーク能は流束均衡解析(FBA)(Bonariusら,(1997); Edwardsら,(1999); Varmaら,(1994a); Varmaら,(1994b))を用いて評価できる。FBAは主に代謝ネットワークの質量保存に基づいている。その保存必要条件は化学量論均衡方程式により実行される;従って、FBAは代謝ネットワークの化学量論特性に依存する。
【0072】
流束均衡方程式はS・v = bvであり(ここでSは化学量論行列であり、ベクトルvは代謝流束を定義し、およびbvは通常はゼロである)従って、一組の質量均衡を通じて、同時に起こる質量、エネルギーおよび酸化還元均衡の制約が強要される。ゼロからの bvベクトル変数は限界価値解析で使用した(下記に述べる)。大腸菌の代謝ネットワークでは、代謝流束数は質量均衡数より大きかったが、それ故に、行列Sのヌル空間に位置する実行可能な流束分布が多数になった。代謝ネットワークの各流束の実施可能値はまた、さらに制約された
。これらの制約を内部反応の可逆性を定義するためおよび輸送反応に対する取込速度を確定するために利用した。無機リン酸塩、アンモニア、二酸化炭素、硫酸塩、カリウムおよびナトリウムの輸送は抑制されなかった;ところが条件として指定した炭素源および酸素の取り込みは抑制された。細胞外へ輸送されることが知られている全ての代謝副産物(即ち、酢酸塩、エタノール、ギ酸塩、ピルビン酸塩、コハク酸塩、乳酸塩など)は常に、その代謝系から離脱させた。本解析において、全ての不可逆流束に対して内部流束を表すαiはゼロに設定され、並びに全ての可逆流束は順方向および逆方向で制限されなかった(各反応の可逆性は補助情報のウェブサイト上に定義されている)。ヌル空間および線形不等式により定義される領域の交点が代謝ネットワーク能を定義し、流束コーンと呼ばれている(Schillingら, 1999)。
【0073】
流束コーン内に位置する最適代謝流束分布の決定は、特定の代謝目的を最小にする解が同定される、線形計画(LP)問題として策定した(Bonarius, H.P.J.ら, 「質量均衡を用いた、異なる培地中のハイブリドーマ細胞の代謝流束解析(Metabolic flux analysis of hybridoma cells in different culture media using mass balances)」Biotechnology and Bioengineering 50:299-318(1996); Edwardsら,(1999); Pramanikら,(1997); Varmaら,(1994a); Varmaら,(1994b))。増殖流束を目的として定義した。増殖流束は最適比率で生合成前駆物質、Xmを利用する代謝流束と定義され、
、ここでdmは代謝生成物Xmのバイオマス分画である。バイオマス組成は文献(Neidhardt, F C, Ed., 大腸菌およびサルモネラ: 細胞分子生物学(ASM Press, Washington, D.C., 1996); Neidhardt, F C, Umbarger, H E, 大腸菌およびサルモネラ中: 細胞分子生物学F. C. Neidhardt, Ed. (ASM Press, Washington, D.C., 1996), vol. 1, pp. 13-16 (1996)) を基に定義した。
【0074】
全ての定常状態代謝流束分布は、規定の代謝地図に対して定義される流束コーン(流束コーン中の各解は特定の内部流束分布または特定の代謝表現型に相当する)に数学的に限定される(Varmaら,(1994a); Varmaら, (1994b))。指定の増殖条件の下で、コーン中の最適表現型は線形計画法(LP)を用いて決定できる。その制約が変化すれば、コーンの形状が変化し且つその最適流束ベクトルは質的に変化するかもしれない;例えば、不活性流束が活性化されるかもしれず、その逆もまた同様である。相空間解析は2つの制約環境変数における全可能性変動を考慮するために発展させられる。
【0075】
表現型相空間(PhPPs)の定義:2栄養素(炭素基質および酸素のような)の取込速度を(x,y)空間の2軸に定義し、この空間中の全点についてその最適流束分布を計算した。かかる空間に存在する、有限数の質的に異なる最適代謝流束地図、または代謝表現型がある。その相空間の境界は限界価値解析(Varma, A, Boesch, BW, Palsson, BO.,「種々の酸化速度の下での大腸菌グルコース異化作用の化学量論的解釈(Stoichiometric interpretation of Escherichia coli glucose catabolism under various oxygenation rates)」Applied and Environmental Microbiology 59, 2465-73(1993); Varma, A, Palsson, BO.,「大腸菌代謝能: II. 最適増殖様式(Metabolic capabilities of Escherichia coli: II. Optimal growth patterns)」Journal of Theoretical Biology 165, 503-522(1993))により定義した。この手順により、代謝経路の最適利用が質的に異なった、空間中の別個の領域または「相」を定義するに至った。各相はPnx,yとして記述し、ここでPは表現型を示し、nはこの表現型に対する境界領域数(対応する図1に示されるように)、およびx,yはその空間軸の2つの取込速度である。
【0076】
相空間の計算:相空間は2-パラメータ空間の全体にわたって限界価値を計算することにより構築し、一定の限界価値の領域を区別するために直線を描いた。限界価値により、目的関数に対する各代謝産物の実態価値が定義された。限界価値の変化を代謝挙動の解釈のために利用した。
【0077】
数学的には、限界価値は以下のように定義され、
ネットワーク中の各代謝産物と関連する。限界価値により、各代謝産物の利用可能性の変化に対する目的関数(Z)の感度係数が定義された(bνiは質量均衡制約の破れを定義し且つ取込反応に等価である)。限界価値はその代謝産物の価値に依存して、マイナス、ゼロまたはプラスのどちらかとなった。相空間の各領域中の限界価値ベクトルの方向および大きさは、異なっていた(相空間の定義により);従って、代謝系の状態は各領域で異なっていた。
【0078】
アイソクライン:代謝表現型を解釈するためにまた、アイソクラインを定義した。目的関数の同一値を規定する二次元空間内の点の軌跡を示すために、アイソクラインを定義した。各領域内のアイソクラインの傾きは、限界価値より計算した;従って、定義によるアイソクラインの傾きは、PhPPの各領域で異なっていた。アイソクラインの傾き(ρ)を定義するために、限界価値の比率を用いた:
【0079】
方程式2のマイナス符号はその解釈を予想して導入した。ρと定義される、条件依存的な基質の相対値は、その代謝ネットワークの制約要因を解釈するために用いた。ρがマイナスとなった領域では、基質が二元的に制限された。異なる条件の下では、アイソクラインはまた一元的な基質制限領域を表す、ある相空間領域において水平または垂直の何れかであり、これらの領域におけるρ値はそれぞれゼロまたは無限大であった。また、PhPPのある領域はρがプラスとなった;これらの領域は「意味のない」領域と名付けられ、基質のうち1つの増加取り込みにより、これらの領域の目的関数にマイナスの影響が及ぼされた。最後に、化学量論的制限により、PhPPに実行不可能な定常状態領域が存在した。
【0080】
最適線:最適線(LO)はPhPP軸に相当する2つの代謝流束間の最適相関を示す直線と定義した。本明細書に提示した結果に関して、この直線は最大バイオマス産生を支えるための基質の完全酸化に対する最適な酸素取り込みと解釈できる。
【0081】
これらの手順を(EdwardsおよびPalsson, 大腸菌MG1655のインシリコ代謝遺伝子型: その定義、特性、および能力, Proc. Natl. Acad. Sci. U.S.A., 97(10):5528-33(2000b))により規定した大腸菌K-12に関する反応リストに適応した。それにより、以下の実施例で使用される二次元および三次元相空間が作成される。
【0082】
実施例3
規定条件の下での大腸菌の最適挙動
本実施例は、使用菌株が最初の基質として酢酸塩およびコハク酸塩を用い、適応性進化なしに最適な好気性増殖を示したことを明らかにする。
【0083】
大腸菌K-12 M1655で起こる代謝反応リストが構築された(EdwardsおよびPalsson(2000b))。このリストを基に、化学量論行列を定式化した。酸素(y軸上)および炭素基質(x軸上)に対する最大取込速度を用い、上述の手順を利用して表現型相空間を計算した。具体的には、2つの炭素源、酢酸塩およびコハク酸塩を用いた。次いで、最適増殖条件を決定するために計算した相空間を使用し、それから一連の増殖実験を実施した。計算(即ち、インシリコ)および実験結果を次に、比較した。
【0084】
酢酸塩
酢酸塩に関する最適増殖性能をインシリコで調べ、それから作成したその予測を実験データと比較した。インシリコ研究は大腸菌物質代謝能を示す流束コーンの二次元投射の軸として定義した、酢酸塩および酸素取込速度に関する表現型相空間(PhPP)解析から開始した(図1)。流束コーンは全ての許容可能な定常状態代謝流束分布の領域である(流束コーンの十分な解説としては、参考文献(Schillingら,「代謝経路解析: ポストゲノム時代における基本概念および科学的応用(Metabolic pathway analysis: basic concepts and scientific applications in the post-genomic era)」Biotechnol. Prog., 15(3):296 (1999))を参照されたい)。さらに、第三次元として定義した増殖速度に関する流束コーンの三次元投射を利用した(図1)。酢酸塩-酸素PhPPのインシリコ解析は前記実施例2で説明した。酢酸塩および酸素取込速度間の最適関係(細胞増殖に関して)であり、およびこの直線は最適線(LO)と呼ばれる。
【0085】
PhPPは代謝ネットワークの働きを解析および解釈するために利用した。例えば、酸素制限の下での代謝ネットワークの特性は、酢酸塩取込速度が酸素取込速度との最適関係を上回る、PhPPの領域2(図1および2)により定義されてもよい。図1から、代謝ネットワークが領域2内で働いている場合、酢酸塩取込速度をLO上のある点まで減少させることにより、増殖を補助する最適能は増大すると理解される。領域1内の点に関して、酸素および酢酸塩の役割を換えることで、同様の解釈が為される。故に、領域1または領域2中の点を規定する代謝流束ベクトルは、利用可能資源の非効率利用を示していると思われる。従って、インシリコPhPP解析により、唯一の炭素源として酢酸塩を用い増殖補助のために最適に作動するよう、大腸菌代謝ネットワーク調節が進化した場合、酢酸塩および酸素取込速度と増殖速度間との関係は、LOにより定義される筈である(図1および2)という結論に至った。
【0086】
大腸菌MG1655を酢酸塩最小培地で培養することにより、酢酸塩および酸素の取込速度と増殖速度間との関係を実験的に調べた。最小培地中の酢酸塩濃度を変化させることで酢酸塩取込速度を実験的に制御した。酢酸塩および酸素取込速度並びに増殖速度を測定し、それから実験データ点をPhPPにプロットした(図1および2)。次に、酢酸塩および酸素取込速度間の計算による最適関係を実験データと比較した(図1)。実験データのインシリコ予測との比較により、酢酸塩最小培地での好気性増殖に関して、実験データに対する回帰直線の傾き(0.91)およびインシリコ定義によるLOの傾き(1.04)間で14%の相違が示された。
【0087】
測定および計算した増殖速度を酢酸塩-酸素PhPP上の三次元としてプロットした(図2)。色分けされた表面は流束コーンの三次元投射を示す。換言すれば、色分けされた表面は解空間を定義し、および全ての実行可能な定常状態代謝流束分布はその表面内に限定される。まさしく可能である。二次元相空間のLO(図1)は、三次元表面の縁のx,y空間(酢酸塩取込速度、酸素取込速度)への投射である。実験データを三次元空間にプロットした(図2)。三次元におけるデータ点のLOへの接近性を量的に視覚化するため、インシリコ予測値および実験データをその基底ベクトルにより形成される各空間上へ投射した。
【0088】
三次元LOおよびその実験データ点の(x,y)、(x,z)、および(y,z)(x軸:酢酸塩取込速度;y軸:酸素取込速度;z軸:増殖速度)空間への投射をそれぞれ、図3および4(回帰直線の適合度を相関係数により示し、且つそのデータをインシリコ予測値と比較している)に示した。各点に対して予測したおよび観測した代謝流束(基質および酸素取込速度並びに増殖速度)を直接比較し、そしてそのインシリコ予測では全体の平均誤差が5.8%であった。この点に関して、代謝ネットワークを構築するために使用した情報は、本実験とは独立して得られたことに注目すべきである(EdwardsおよびPalsson,(2000b))。計算したPhPPは本研究で得られたデータの先験的な解釈および予測を示している。
【0089】
コハク酸塩
コハク酸塩-酸素PhPP(図5)は酢酸塩-酸素PhPPよりもっと複雑であった。コハク酸塩-酸素PhPP(図5)には、質的に異なる最適代謝ネットワーク利用からなる4つの別領域があった。コハク酸塩-酸素PhPPの領域1および4は、酢酸塩-酸素PhPPの領域1および2と類似であった。例えば、図5から、コハク酸取り込みを領域3、4の境界により定義される点に減少させた場合、領域4の流束ベクトルに関する最大増殖流束は増大され得ると理解できる。さらに、PhPP解析から、領域3は1基質制限領域と定義される。1基質制限領域では、コハク酸塩取込速度は領域3の最大増殖流束にほとんど影響しないが、酸素取込速度は増殖速度にプラスに影響することが示される。
【0090】
領域2においてコハク酸塩取込速度が増加した場合、代謝ネットワークは増殖の増加を補助できるので、領域2は2基質制限領域と定義される。インシリコ解析により代謝ネットワークを領域2のLOの外へ操作すること、部分的に好気性の物質代謝および代謝副産物の分泌を行うことにより、細胞増殖速度を増加できることが示される。最適代謝副産物は酢酸塩であると計算された。領域2の代謝副産物の減少産生はしかし、全体のバイオマス産生を減少させる。故に、PhPP解析を基に、代謝ネットワーク調節が唯一の炭素源としてコハク酸塩を伴う最適増殖の方向へ進化した場合、その代謝ネットワークはLOに沿った流束ベクトルで動作することが推測されうる。しかし、増殖速度は流束ベクトルを領域2中に移動させることにより増加でき、従って、酸素が制限され且つコハク酸塩が十分である場合、先に述べたの仮説が真実であるならば、そのネットワークは領域2中でのみ作動する筈であると予想される。
【0091】
インシリコ解析における上記の仮説を批判的に調べるため、コハク酸塩最小M9培地での大腸菌増殖実験を実施した。コハク酸塩取込速度の範囲を測るため、多くのバッチ培養物を種々のコハク酸塩濃度および温度で増殖させた。全培養物で一貫した最大酸素普及率を維持するため、曝気および攪拌を一定に保った。コハク酸塩および酸素取込速度並びに増殖速度を、それぞれ独立した増殖実験に対して別々に測定した。その実験データを、次いで、インシリコ予測と直接比較した(図5)。
【0092】
実験データ点は先述の仮説と一致していた:臨界値以下の酸素取込速度(〜18.8ミリモル/g-DW/時間)に対しては、流束ベクトルは常にLOに沿った点を示した。さらに、LO沿いの点を示した培養物は、代謝副産物として酢酸塩をほとんどまたは全く産生しなかった(インシリコ解析により予測されたように-図5を参照されたい)。仮説の通り、酸素は制限されているがコハク酸塩は十分ある実験系に対し、実験データは流束ベクトルが領域2内へ水平移動することを示している。実験データの区切り点は、最大酸素取込速度18.8±0.5ミリモル/g-DW/時間に一致すると決定された。領域3または4内の流束ベクトルは観測されなかった。領域2に同定された培養物に対し、酢酸塩産生を測定した、さらに図6で、その酢酸塩産生をそのインシリコ予測と定量的に比較する。
【0093】
最適増殖速度表面をコハク酸塩-酸素PhPP上方に構築した、すると測定流束ベクトルは、LOに相当するポリトープの縁近くに移動した(図7)。また、流束ベクトルにより、領域2における相表面上の点の軌跡が、系の最大酸素取込限界と同等の一定酸素取込速度であることが示された。インシリコ解析の予測能およびそのインシリコ由来の仮説を定量的に調べるために、本発明者らは、本発明者らの仮説および実験的に観測した流束ベクトルを記述する区分線形モデルを使用した。区分線形モデルは以下のように定義される:本発明者らはコハク酸塩取込速度および酸素取込限界の範囲に対する流束ベクトルにより定義される点の軌跡を同定した。酸素取込限界よりも下では点の軌跡はLO沿いに位置し、および酸素取込限界よりも上では点の軌跡は一定酸素取込速度(酸素取込限界)の相表面沿いに位置する。区分線形モデルに基づき、コハク酸取込速度は酸素取込速度および増殖速度を予測するために使用され、並びにその他2つの順列もまた考慮した。この解析から、インシリコ予測および実験データ間の全体の平均誤差は10.7%となった。
【0094】
本実施例は、使用した菌株が、主な基質として酢酸塩およびコハク酸塩を用いて最適な好気性増殖を示したことを明らかにしている。この最適性能を達成するために適応性進化は必要とされなかった。
【0095】
実施例4
準最適性大腸菌菌株の最適性への進化
本実施例は、大腸菌が準最適増殖状態からインシリコ決定の最適状態まで幾つかの表現型適応を受けられることを実証する。
【0096】
グルコース
グルコース-酸素PhPPには異なる6領域が含まれる(図8)。コハク酸塩-酸素PhPPと同様、領域1は無益回路および準最適な増殖性能を示すが、領域2は酢酸塩過剰な物質代謝により特徴付けられる。その2領域はLOにより分けられる。
【0097】
すでに述べたように、細胞増殖速度、OUR、およびグルコース接触速度(GUR)を各種のグルコース濃度および温度にわたって実験的に決定した。GUR、OURの実験決定値のほとんどは、PhPPのLO上または僅かに領域2(ここでは予測した酢酸塩分泌が実験的に観測された)の点に相当した(図8)。
【0098】
三次元において、測定した増殖速度はLOに相当する縁近傍の解空間の表面上に位置しているが、そこに密集はしていない(図9)。本発明者らはこのように一定の増殖条件の下で代謝表現型が進化するかどうか決定するため、連続移入を用いて40日間に渡り(約750世代)その菌株を対数増殖(16)状態に維持した(図10および11)。LOと平行した実験点の移動によって示されるように、適合性は確かに増加したが、表現型に質的な変化はなかった。
【0099】
実施例5
準最適性大腸菌菌株の最適性への進化
本実施例は大腸菌が準最適増殖状態からインシリコ決定の最適状態への有効な表現型適応を受けられることを実証する。
【0100】
グリセロール
グリセロール-酸素PhPPは、酢酸塩、コハク酸塩およびグルコースに対するPhPPで見られるのと似た特徴を伴う5つの領域からなる。特に、無益回路を伴う領域(相1)はLOにより酢酸塩過剰領域(相2)と分けられる。
【0101】
各種グリセロール濃度にわたる増殖性能を、すでに述べたように、実験的に決定した。しかし、リンゴ酸塩またはグルコースにおける増殖とはかなり異なって、増殖の実験データはLO(図12)および最適性表面(図13)からかけ離れた、相1の全体にわたり分散していた。調べた他の基質と異なり、グリセロールは従って、大腸菌K-12の準最適増殖のみを補助する。
【0102】
すでに述べたように、本発明者らは従って、今度は唯一の炭素源としてグリセロールを用い、長期の適応性増殖実験を実施した。温度30℃、グリセロール濃度2g/L、および十分な酸素供給を維持しながら、連続移入(17)により、原菌株を再度、40日間の長期の対数増殖状態に保持した。増殖速度、グリセロール取込速度(GlUR)およびOURを10日毎に決定した。
【0103】
40日間の進化経路(E1)は相1中に描かれ、最終的にはLOに収束した(図14)。この間、増殖速度は0.23/時間から0.55/時間へと倍以上に増えた(図15)。結果として生じた進化菌株(凍結保存されていた)をさらに調べたところ、比増殖速度およびバイオマス産生が親菌株よりも高いことが明らかとなった。得られた全データはLO上または近傍に移動し、進化菌株が長期培養の最終日にそのデータを示した(図16および17)ことより、その進化菌株がインシリコ予測と一致する、グリセロールにおける最適増殖性能に到達していたことを示している。2回目の別個の適応性実験は似ているが、同一でない進化軌道(E2)を示し、同じ終点近くに収束した。大腸菌は従って、準最適増殖状態からインシリコ決定の最適状態への有効な表現型適応を受けられる。
【0104】
このように細菌の最適性増殖の実行により、インシリコ解析による予測が確認されると思われる。酢酸塩およびコハク酸塩のようないくつかの基質においては、細胞は最適増殖を示すかもしれないが、グルコースおよびグリセロールのような他の基質においては、そうでないかもしれない。後者においては、増殖は相空間予測の最適増殖の方向へ進化する。
【0105】
本明細書に引用した参考文献の全てが参照として組み入れられる。上記実施例に関連して本発明を説明したが、本発明の精神および範囲内での変更および変形が包含されると理解される。従って、本発明は特許請求の範囲によってのみ制限される。
【図面の簡単な説明】
【0106】
【図1】酢酸塩取込速度(AUR 単位;ミリモル/g-DW/時間、g-DWはグラム乾燥重量である)対酸素取込速度(OUR 単位;ミリモル/g-DW/時間)表現型相空間。インシリコ定義の最適線(LO)が図中に示してある。同直線の傾きもまた図中に示してある。実験データ点は図上に表示してある。誤差指示線は単一の標準偏差を表し、誤差指示線は酢酸塩および酸素取込速度測定値の両方に対して表示してある。実験的に再構築された最適線を定義するため、データ点について直線回帰を行った。曲線に適合する相関係数R2値は0.92である。領域1および2は別個の非最適代謝表現型を表している。
【図2】酢酸塩における大腸菌の増殖に対する相表面の三次元図。xおよびy軸は図1中と同じ変量を示している。第三次元(z-次元)は細胞増殖速度に相当する。z軸値は対応する凡例に定量的に表示された最適増殖速度値を伴う灰色の尺度である。三次元中の最適線(LO)が表示してある。三次元中のLOの助変数方程式が本明細書中に示されている。黒線は流束コーンの三次元投射における代謝能表面を定義し、酢酸塩取込速度または酸素取込速度の定常値を示している。酢酸塩取込速度を増加させること(酸素取込速度の比例的増加なく)による細胞増殖の可能性への定量効果が視覚化できる。データ点もまた三次元図にプロットしてあり、誤差指示線は省略されている。
【図3】酢酸塩取込速度および増殖速度により形成される空間上に投射された、酢酸塩における増殖の最適線。データ点もまた投射してあり、実験的に最適線を定義するため直線回帰は二次元空間中で行った。最適線は灰線として示し、回帰直線は黒線として示してある。
【図4】酸素取込速度および増殖速度により形成される空間上に投射された、酢酸塩における増殖の最適線。データ点もまた投射してあり、実験的に最適線を定義するため直線回帰は二次元空間中で行った。最適線は灰線として示し、回帰直線は黒線として示してある。
【図5】表現型相空間で表されたコハク酸塩取込速度(ミリモル/g-DW/時間)対酸素取込速度(ミリモル/g-DW/時間) 。ラベルの付いた直線はインシリコ定義の最適線(LO)である。実験データ点は図上に示してある。誤差指示線はコハク酸塩および酸素取込速度測定値の両方に対して表示してあり、単一の標準偏差を表している。酢酸塩が0.3ミリモル/gDW/時間の閾値以上に産生された培養は白丸で示してあり、黒丸は酢酸塩産生なしまたは産生が閾値以下のどちらかを表している。黒色の破線は酢酸塩産生なしのデータ点の回帰直線を示している。
【図6】酢酸塩産生の実測値対図5にグラフ化された各点に対するインシリコ予測値。データ点はコハク酸塩取込速度の大きさにより順序付けた階級である。
【図7】コハク酸塩における大腸菌増殖の三次元表現型相空間。xおよびy軸は図6中と同じ変量を示している。第三次元(z軸)は細胞増殖速度に相当する。z軸値は図中の対応する凡例の灰色の尺度である。着色領域を分ける境界線は一定の酸素および酢酸塩取込速度を示し、最適線から離れる定量効果が視覚化され得る。誤差指示線を除き、データ点がこの三次元図にプロットしてある。
【図8】グルコースにおける大腸菌K-12の増殖の計算および実験値。グルコース取込速度、GUR(ミリモル/gDW/時間)および酸素取込速度、OUR(ミリモル/gDW/時間)を表現型相空間に示した。LOが表示してある。データ点はLOまたは酢酸塩過剰領域(ここでは酢酸塩分泌がインシリコ予測され、且つ実験的に観測される)に限定されている。
【図9】グルコースにおける増殖の相空間上方にグラフ化された増殖速度の三次元図。xおよびy軸は図8中と同じ変量を示している。z軸は色分けされた数値および凡例に表示されたその最適増殖速度と共に、細胞増殖速度を示す。
【図10】適応性進化実験に関する実験値を用いてOURに対してプロットしたGUR。データ点はLO近傍および酢酸塩過剰が予測される領域2に位置している。
【図11】グルコース表現型相空間上方の進化後の3D増殖表面の三次元図。全データ点はLO上または近傍に密集している。
【図12】グリセロールにおける増殖の計算および実験値。グリセロール取込速度、GlUR(ミリモル/gDW/時間)、および酸素取込速度、OUR(ミリモル/gDW/時間)を表現型相空間に示した。LOが表示してある。実験データ点は、無益回路および準最適増殖速度を特徴とする、領域1に限定されている。
【図13】グリセロール表現型相空間上方にグラフ化された増殖速度の三次元図。xおよびy軸は図12中と同じ変量を示している。z軸は細胞増殖速度、および凡例に表示された最適増殖速度に相当する。LO近傍にデータ点はない。
【図14】適応性進化実験に関する実験値を用いて酸素取込速度(OUR)に対してプロットしたグリセロール取込速度(GlUR)。進化の出発点が示してある(0日)。1回目の進化軌道(E1)の実験値は青色で表示してあり、2回目の進化軌道(E2)の実験値は緑色で表示してある。両実験とも、初期菌株は最適増殖速度を示す、LO上の同一終点に収束している。
【図15】グリセロールにおける適応性進化実験の時間による、単位または1時間当たりの増殖速度の変化。両実験とも適合性の増大および増殖速度の倍加を伴う同様の適応性の様相を明らかとしている。
【図16】適応性進化実験後のグリセロールにおける増殖の実験値によるグリセロール-酸素表現型相空間。全数値はLO上または近傍に密集している。データは炭素源滴定およびLO沿いに移動する量的効果を表している。
【図17】進化後の相表面の三次元図。全データ点はLO上または近傍に密集している。
【技術分野】
【0001】
本発明の分野
本発明は広くは生化学反応ネットワークおよびより具体的には最適な所望の全細胞特性を得るための生物体中の代謝ネットワークの再構築に関する。
【背景技術】
【0002】
背景情報
ゲノム配列解析およびアノテーション技術により細胞を構成する分子成分の詳細なリストが得られ、ハイスループット技術によりこれらの成分がどのように使用されるかに関する情報が得られている。従って、我々は生物学的設計がゲノム規模で可能となる段階に近づいている。ある生物体由来の1遺伝子を別の生物体に「挿入(splice)」することおよび予測可能な結果を起こすのは困難なことが証明されてきた。主な理由としては、長い進化過程を通じて生細胞中のすべての成分が細胞全体の機能に最適に「適合」するように磨かれてきたことが挙げられる。単純に外来遺伝子を導入しても、または存在する遺伝子を欠損させても、予測可能な結果、そしてまた最適な結果に至ることはない。単一のまたは複数の遺伝子欠損または付加が全細胞機能から成るその機能に与える結果を演繹的に予測する方法および予め決定した方法で残りの成分を機能させる方法が必要とされている。これまでに細胞規模の代謝機能に関して若干の進歩が遂げられたが、かかる方法は欠けている。
【0003】
暫くの間、医学および産業目的のため代謝流束に再び関心が向けられた。この関心の結果、代謝工学の分野が生まれた。代謝工学の主な目標は生細胞中で望ましい代謝反応を実行することである。代謝工学に大事なのはコンピュータ技術、遺伝学、およびシステム科学を含む幾つかの科学分野の進歩および適用である。
【0004】
分析および設計のための従来の工学手法では、数学的またはコンピュータモデルを利用する。物質代謝に関しては、基本的な物理化学法則および原理を基礎としたコンピュータモデルが必要となると思われる。代謝技術者は次に、かかる方法が新規且つ改善された生細胞を体系的に「設計する」ために使用できることを望む。そこで組換えDNA技術の方法が、所望の細胞設計を達成するために適用されるはずである。
【0005】
代謝解析の25年から30年間の歴史により、細胞物質代謝の体系的特徴を定量する必要性が実証されてきた(例えば、Fell D.,「物質代謝制御の理解(Understanding the control of cellular metabolism)」(London, Portland Press) (1996)(非特許文献1); Heinrich R.ら,「代謝調節および数学モデル(Metabolic regulation and mathematical models)」Progress in Biophysics and Molecular Biology, 32:1-82, (1977)(非特許文献2); Heinrich R.およびSchuster S.,「細胞システムの調節(The regulation of cellular systems)」(New York, Chapman & Hall), xix, p.372 (1996)(非特許文献3); Savageau M. A.,「生化学系解析. I. 酵素反応構成要素に関する反応速度則のある数学的特性(Biochemical systems analysis. I. Some mathematical properties of the rate law for the ecomponent enzymatic reactions)」J. Theor. Biol. 25(3):365-69 (1969)(非特許文献4)を参照されたい)。代謝力学を研究する有意な動機がある。物質代謝の量的記述および代謝変化の創生能は、特定の治療目標を達成するのに重要であるばかりでなく、細胞生物学を理解するのに広く重要である。重要な用途には、治療学およびその他の生化学を提供するための菌株設計、遺伝的欠陥の代謝影響の評価、感染症と戦うための体系的方法の組み立てなどが含まれる。物質代謝の量的および体系的解析は従って、根本的に重要である。しかし、同分野における批評は「代謝工学における近年の急激な関心の高まりにもかかわらず、利用可能な分子生物学的技術力と生化学ネットワークの合理的な解析能との間には、依然として大きな格差が存在している」(Stephanopoulos G.,「代謝工学(Metabolic engineering)」Current Opinions in Biotechnology, 5:196-200 (1994)(非特許文献5))と結論付けている。この意見は数年前のものであるが、基本的には依然として真実である。我々は、与えられた環境での生物体の最良適合を達成する、何百万年もの自然進化と競争しているので、この結論は驚きではない。
【0006】
少数反応を含む部分的遺伝子調節ネットワークが設計された(Hastyら、「遺伝子調節ネットワークのコンピュータ研究(Computational studies of gene regulatory networks)」: In numero molecular biology, Nature, 2: 268-79 (2001)(非特許文献6)に総説されている)が、性能特性が定義されおよびそれらを引き続き構築した代謝ネットワークのような、生化学調節ネットワークの演繹的設計は実践されなかった。主な理由には、主としてその数値を決定する必要のある反応速度パラメータが非常に多く存在し且つ詳細な反応速度方程式が全般的に未知であって、全代謝ネットワークに対して信頼性のある詳細な反応速度モデルを構築できないことが挙げられる。従って、予測的反応速度モデルを達成できないので、物質代謝のような、最適な生化学反応ネットワークの演繹的設計は可能でない。実際、突然変異および進化過程により速度定数値は時間とともに変化する。
【0007】
従来、進化過程の終点を予測することは、それらが偶発的事象から選択された結果であると予測されるため、不可能であった。本発明は定義された環境における代謝ネットワークの進化終点の演繹的計算を可能とする方法を開示する。生物学的体系における最適化原則に基づく他の数学的モデリング法があるが;即ち、人工頭脳学モデリング法(Varner J.およびRamkrishna D.,「代謝経路の数学的モデル(Mathematical models of metabolic pathways)」Curr. Opin. Biotechnol., 10(2):146-50, (1999)(非特許文献7))、それらは要求されるパラメータ数のため生物学ネットワークの設計に従順ではない。従って、かかる設計を創出し構築するための進化過程の利用に対する基礎が提供される。
【非特許文献1】Fell D.,「物質代謝制御の理解(Understanding the control of cellular metabolism)」(London, Portland Press) (1996)
【非特許文献2】Heinrich R.ら,「代謝調節および数学モデル(Metabolic regulation and mathematical models)」Progress in Biophysics and Molecular Biology, 32:1-82, (1977)
【非特許文献3】Heinrich R.およびSchuster S.,「細胞システムの調節(The regulation of cellular systems)」(New York, Chapman & Hall), xix, p.372 (1996)
【非特許文献4】Savageau M. A.,「生化学系解析. I. 酵素反応構成要素に関する反応速度則のある数学的特性(Biochemical systems analysis. I. Some mathematical properties of the rate law for the ecomponent enzymatic reactions)」J. Theor. Biol. 25(3):365-69 (1969)
【非特許文献5】Stephanopoulos G.,「代謝工学(Metabolic engineering)」Current Opinions in Biotechnology, 5:196-200 (1994)
【非特許文献6】Hastyら、「遺伝子調節ネットワークのコンピュータ研究(Computational studies of gene regulatory networks)」: In numero molecular biology, Nature, 2: 268-79 (2001)
【非特許文献7】Varner J.およびRamkrishna D.,「代謝経路の数学的モデル(Mathematical models of metabolic pathways)」Curr. Opin. Biotechnol., 10(2):146-50, (1999)
【発明の開示】
【0008】
概要
本発明は生細胞中の生化学反応ネットワークの最適な機能を達成する方法に関する。生化学反応ネットワークは包括的な生化学反応ネットワーク、実質的に全ての生化学反応ネットワーク、または全ての生化学反応ネットワークで有り得る。方法は細胞の生化学反応ネットワークの再構築を用いて、インシリコで実施できる。方法はさらに、実験室におけるインシリコ段階を用いてなされた決定を確認するおよび事に依っては拡張する培養段階、並びに最適な機能を備えた培養細胞、または細胞集団を産生する培養段階を含むことができる。
【0009】
方法はネットワーク中の生化学反応をリストにしてコンピュータに表示することにより、例えばネットワーク中の生化学反応データベースを提供する;最適化方法を使用してネットワークの最適特性を計算する;ネットワーク中の反応リストを変更し、かつ最適特性を再計算する;および所望の性能に達するまで前述の工程を繰り返し行うことにより、実施できる。方法にはさらに最適化手順の結果生じる生化学反応が含まれるように、細胞の遺伝子構成を構築する段階;その条件の下で構築した細胞を、特定環境下の培養状態に置く段階;および細胞を、十分な期間、定めた所望の性能に細胞を進化させる条件下で培養する段階を含んでもよい。
【0010】
生化学反応ネットワークは代謝ネットワーク、例えば調節ネットワークで有り得る。さらに、遺伝子構成が構築される細胞は原核細胞または真核細胞;例えば大腸菌、出芽酵母、チャイニーズハムスター卵母細胞などで有り得る。さらに、細胞の遺伝子構成は、細胞中の1つまたはそれ以上の遺伝子を改変することにより、例えば付加もしくは欠失により、またはその調節成分(例えば、プロモーター、転写因子結合部位など)を介して遺伝子調節を改変することにより構築できる。別の局面において、本発明は前述の方法により産生される濃縮細胞集団を提供する。
【0011】
別の局面において、本発明は、ネットワーク中の生化学反応を含むデータベースを提供すること;最適化方法を使用してネットワークの最適特性を計算すること;ネットワーク中の反応を変更するための利用者による選択を受信し、かつ最適特性を再計算すること;所望の特性基準が満たされるまで最適化を繰り返し行うこと; 最適化情報の結果として生化学反応が含まれるように、細胞の遺伝子構成を構築するための最適化結果を表示すること;構築した細胞を、特定環境条件の下で培養すること;および細胞が所望の性能に進化するような十分な期間、細胞を培養することにより、細胞における包括的な生化学反応ネットワークの最適な機能を達成するための方法を提供する。
【0012】
最適化方法は本発明により提供されるコンピュータ・システムを用いて行ってもよい。コンピュータ・システムには一般的に、少なくとも1つの生物体の1つまたはそれ以上の生化学反応ネットワークに関する情報を提供するデータベース;最適化および/または比較のための1つもしくはそれ以上の生化学反応ネットワークの選択を受信でき、および所望の性能の選択を受信できるユーザ・インターフェース;並びに最適化方法の計算および再計算を実行するための機能が含まれる。本発明のコンピュータ・システムは生化学反応ネットワークを再構築する機能を含むことができる。データベースは内部データベースまたは外部データベースで有り得る。
【0013】
別の局面において、本発明はコンピュータ可読プログラム・コードが媒体上で統合された、コンピュータ利用可能媒体を含むコンピュータ・プログラム製品を提供する。プログラム・コードはデータベースと情報のやりとりを行うことができ、コンピューティング・システム内で以下の段階を達成する:ネットワークの所望の性能の選択を受信するためのインターフェースを提供する段階;生化学反応ネットワークの最適特性を再計算する前に、所望の最適特性を決定し、決定の結果を表示し、かつ生化学反応ネットワークを変更し、所望の最適な機能に達するまでその工程を繰り返し行う段階。生化学反応ネットワークの変更は、利用者により手動で入力された変更に基づいて実施されてもよく、またはプログラム・コードにより自動的に実施されてもよい。コンピュータ・プログラムはさらに、再構築された生化学ネットワークの一部であるデータベース入力を識別できるか、または生化学反応ネットワークの再構築を実行できる。
【0014】
発明の詳細な説明
本発明のある局面では、広大な生化学反応ネットワークの特性を設計する方法が提供される。本発明のこの局面の方法を使用することにより、生化学反応ネットワークを特定環境において予め決定された性能に設計できる。本発明のある局面には以下の段階が含まれる:
1) 生化学反応ネットワークの反応構造についてのコンピュータ再構築を使用する段階、
2) 最適化方法を使用して反応ネットワークの最適な機能性を決定する段階、
3) 単一または複数の遺伝子を除去するまたは付加することにより、ネットワークを表示するコンピュータにおいて反応構造を変更し、かつ最適特性を再計算する段階、
4) コンピュータ・シミュレーションにより最適特性が決定した反応ネットワークの構造に、遺伝子操作を使用して生物体中の遺伝子補完を対応させる段階、および
5) 確定した淘汰圧の下での、拡張培養を使用して、コンピュータ・シミュレーションにより予測された最適解の方へ実際の反応ネットワークの機能を進化させる段階。適応性進化過程それ自体により、最適設計を達成する反応速度パラメータの最良セットが決定されると思われる。パラメータ値の2つ以上の同様なセットが進化過程を介して決定できる。
【0015】
本明細書に開示される方法および手順を用いることで、生化学反応ネットワークをコンピュータで演繹的に設計できる。反応ネットワークの設計に続き、進化過程は、遺伝的に改変した生物体またはコンピュータ・シミュレーションに使用したネットワークに対応する野生型菌株に関して、最適な条件の下で、実験室にて実施される。生物体は非独特なやり方で最適な挙動を達成するかもしれない-即ち、そこに等価な最適解が存在するかもしれない。従って、本発明には生細胞の環境内で生化学反応ネットワークの最適性能を達成するコンピュータ設計方法、遺伝子改変、および進化過程の自明でない且つ現存しない組合せが含まれる。
【0016】
別の局面では、本発明は生細胞中の包括的な生化学反応ネットワークの最適な機能を決定するための方法に関する。方法は生細胞の所望の性能を達成するために使用される。方法はネットワーク中の生化学反応をリストにしてコンピュータに表示する段階;ネットワークの最適特性を計算するため最適化方法を使用する段階;ネットワークの反応リストを変更し、かつ最適特性を再計算する段階;および所望の機能が満たされるまで変更段階を繰り返し行う段階により、実施できる。
【0017】
インシリコで実施される段階に加えて、方法はさらに1個の生細胞、または細胞集団の培養を含む段階を包含できる。これらの段階には、所望の性能が満たされるまで変更段階を繰り返し行った結果生じる生化学反応が含まれるように細胞の遺伝子構成を構築する段階;その条件で構築した細胞を特定環境下の培養状態に置く段階;並びに細胞を、十分な期間、および定めた所望の性能にその細胞を進化させる条件の下で培養する段階が含まれる。
【0018】
生化学反応ネットワークは、1つの生化学経路または連関した生化学経路の一部の、相互に関連する一連の生化学反応である。代謝反応ネットワーク、異化反応ネットワーク、ポリペプチドおよび核酸合成反応ネットワーク、アミノ酸合成ネットワーク、エネルギー代謝等のような多数の生化学反応ネットワークが同定されている。生化学反応ネットワークの他の種類には、細胞シグナル伝達ネットワーク、細胞周期ネットワーク、オペロン調節ネットワークのような遺伝子発現の調節に関連する遺伝的ネットワーク、および細胞骨格の一部を産生するアクチン重合ネットワークを含む調節ネットワークが包含される。主要な細胞機能の大部分は、相互に作用する生化学反応のネットワークに依存する。
【0019】
本発明を実践するため、生化学的に設計される生物体中の包括的な、好ましくは実質的に全ての、または最も好ましくは全ての生化学反応ネットワークの反応構造をコンピュータ・シミュレーションために再構築する必要がある。全ての生化学反応ネットワークには、ある生化学機能と関連する細胞の生化学反応の全てが含まれる。例えば、全ての代謝反応ネットワークには、細胞の物質代謝を与える生化学反応の全てが本質的に含まれる。代謝反応ネットワークは、全ての生細胞において幾つかの形態で見られる全生化学反応ネットワークの良い例となる。
【0020】
包括的な生化学反応ネットワークとは、検出可能な特性をもたらす、および生細胞に照らして、検出可能な特性を改する効果に関して予測可能な方法で改変できる、相互に関連した生化学反応群である。例えば、細胞により産生される生体分子の産生をもたらす中心的反応、その中心的反応には生体分子の産生に関連する全ての生化学反応ネットワークにおける一部の反応しか含まれていないが、包括的な生化学反応ネットワークには、生細胞における生体分子の産生に及ぼす中心的生化学反応の変化の効果を予測するためにコンピュータを利用した方法が使用できるような条件で、その中心的反応が含まれうる。
【0021】
実質的に全ての生化学反応ネットワークとは、生細胞の検出可能な特性を担う相互に関連した生化学反応群である。実質的に全ての生化学反応ネットワークには中心的反応のほか、効果は比較的小さい可能性があるが、検出可能な特性に効果を及ぼす二次反応が含まれる。実質的に全ての生化学反応ネットワークにおける変化は、コンピュータを利用した方法を用いて予測できる。本発明はまた、包括的な、実質的に全ての、または全ての生化学反応ネットワークではなく、全ての生化学反応ネットワークにおける大部分の反応を利用することもできる。
【0022】
本発明の方法を用いて決定される最適特性(本明細書ではまた最適な機能とも呼んでいる)には、例えば、グリセロール取込速度、酸素取込速度、増殖速度、胞子形成発生および/または速度、栄養不足条件下での希元素の精錬速度、バイオマス、並びにタンパク質、炭水化物、抗生物質、ビタミン、アミノ酸、発酵生産物(例えば乳酸塩産生物)のような生体分子の産生量が含まれる。最適特性にはまた、例えば、キラル化合物および他の低分子量化合物の産生量も含まれる。最適特性にはまた、例えば、エネルギー運搬ATPまたは酸化還元運搬NADPHおよびNADHのような重要な補因子の最大内部産生量も含まれる。最適特性にはまた、生化学反応ネットワーク中の重要な反応を介した代謝流速度(flux rates)のような特性が含まれることが、細胞技術者により定義され得る。本発明により、これらの特性の1つまたはそれ以上と関連した最適性能が達成される。例えば方法により、特に望ましい増殖速度または特に望ましい産生量の達成が可能となる。
【0023】
典型的には、本発明の方法に関して、再構築された生化学反応ネットワークの生化学反応をコンピュータに表示する。この表示には再構築された生化学反応ネットワークの生化学反応のリスト作成を含むことができる。リスト作成は、ネットワーク・シミュレーションおよび最適特性の計算に相当するコンピュータアルゴリズムと連動できるように、コンピュータデータベース中に、例えば関係型データベースの一連の表として示すことができる。
【0024】
本発明に関して生化学ネットワーク再構築は高品質でなければならない。高品質生化学反応ネットワークの工程、具体的には代謝反応ネットワークでは、再構築はM. W. Covert, C. H. Schilling, I. Famili, J. S. Edwards, I. I. Goryanin, E. Selkov,およびB. O. Palsson,「微生物菌株のインシリコ代謝モデリング(Metabolic modeling of microbial stains in silico)」Trends in Biochemical Sciences, 26:179-186(2001); Edwards J.,およびPalsson, B, 「代謝流束均衡解析および大腸菌K-12遺伝子欠損のインシリコ解析(Metabolic flux balance analysis and the in silico analysis of Escherichia coli K-12 gene deletions)」BMC Structural Biology, 1(2)(2000a); Edwards J. S.,およびPalsson, B, O.,「インフルエンザ菌Rd代謝遺伝子型の体系的特性(Systemic properties of the Haemophilus influenzae Rd metabolic genotype)」Journal of Biological Chemistry, 274(25):17410-16,(1999); Karp P. D.ら,「HinCyc:インフルエンザ菌の完全ゲノムおよび代謝経路の知識基盤(HinCyc: A knowledge base of the complete genome and metabolic pathways of H. influenzae)」ISMB 4:116-24,(1996); Karp P. D.ら,「EcoCycおよびMetaCycデータベース(The EcoCyc and MetaCyc databases)」Nucleic. Acids Res. 28(1):56-59(2000); Ogataら,「KEGG京都大学遺伝子・ゲノム辞典(KEGG: Kyoto encyclopedia of genes and genomes)」Nucleic Acids Res. 27(1):29-34 (1999); Schilling C. H.およびPalsson B. O.,「ゲノム規模の経路解析を介したインフルエンザ菌Rdの代謝能の評価(Assessment of the metabolic capabilities of the Haemophilus influenzae Rd through a genome-scale pathway analysis)」J. Theor. Biol., 203(3):249-83(2000); Selkov E. Jr.ら, 「代謝経路データベース(MPW: the metabolic pathways database)」Nucleic Acids Res., 26(1):43-45(1998); Selkov E.ら,「メタン生成菌の物質代謝の配列データからの再構築(A reconstruction of the metabolism of Methanococcus jannaschii from sequence data)」Gene 197(1-2):GC11-26(1997)により確立された。本工程にはアノテートされたゲノム配列、並びに生化学的および生理学的データの利用が含まれる。これらのアノテートされたゲノム配列並びに生化学的および生理学的データは、以下の本発明のコンピュータ・システムの考察の項で詳細に記載されるような、1つまたはそれ以上の内部または外部データベースで見つけられる。使用される全データソースを照合するには再構築されたネットワークの入念な解析が必要とされる。他の生化学反応ネットワークの再構築に関して、同様の方法が使用できる。
【0025】
本発明のこの局面の方法は次に、指定のおよび様々な環境条件の下で、包括的な、実質的に全ての、または全ての生化学反応ネットワークの最適特性を決定するため、その再構築された包括的な、実質的に全ての、または全ての生化学反応ネットワークを利用する。この決定により、既定の環境で所望の性能を達成する生化学反応ネットワークの設計が可能となる。これにはまた、所望の性能を達成する、組換え型細胞または細胞集団を増殖させる方法を提供するため、下記に記載の、細胞の遺伝子構成を構築するおよびその細胞を培養する段階と組み合わせることができる。
【0026】
一連の特定環境下の包括的な、実質的に全ての、または全ての生化学反応ネットワークの最適特性は、最適化方法として知られるコンピュータによる方法を用いて決定できる。最適化方法は当技術分野において周知である(例えば、EdwardsおよびPalsson(1999)を参照されたい)。本発明の方法で使用される最適化方法では、例えば、限定することを意図するものではないが、下記にてさらに詳細に記載される、流束均衡解析(FBA)、相空間解析(PhPP)を組合せて、および最適線(LO)の決定を利用することができる。
【0027】
再構築された代謝ネットワークは次に、確立された方法(Bonariusら,「過小決定代謝ネットワークの流束解析:失われた制約の探索(Flux analysis of underdetermined metabolic networks: The guest for the missing constraints)」Trends in Biotechnology, 15(8):308-14(1997); Edwards J.S.ら,「代謝流束均衡解析(Metabolic flux Balance Analysis)」In: (Lee S.Y., Papoutsakis E.T.編), Metabolic Engineering: Marcel Deker. P13-57(1999); Varma A.およびPalsson B.O,「代謝流束均衡:基本概念, 科学的および実践的使用(Metabolic flux balancing: Basic concepts, Scientific and practical use)」Bio/Technology 12:994-98(1994a))を用い、定常状態における代謝流分布の定量的シミュレーションを実施するために使用できる。代謝ネットワークのコンピュータ・シミュレーションはどんな条件の下でも実施できる。さらに、どの反応リストでもその環境およびその反応リストの中身を記述するパラメータを変更することによりコンピュータで模擬できる。
【0028】
再構築された代謝ネットワークの代謝能は、流束均衡解析(FBA)(Bonariusら,(1997);Edwardsら,(1999);VarmaおよびPalsson(1994a))の確立された方法を用いて評価できる。FBAは、ネットワーク中の個々の反応の定常状態および能力制約(その反応を介した最大代謝流)にある代謝ネットワークの質量保存に基づいている。さらに、実験的に決定した菌株特異的なパラメータ、バイオマス組成(Pramanik J.およびKeasling J.D.,「大腸菌物質代謝の化学量論的モデル:増殖速度依存的なバイオマス組成および力学的エネルギー量の組込み(Stochiometric model of Escherichia coli metabolism: Incorporation of growth-rate dependent biomass composition and mechanistic energy requirements)」Biotechnology and Bioengineering, 56(4):398-421(1997))および維持エネルギー量(the maintenance requirements)(Varma A.およびPalsson B.O.,「化学量論的代謝流均衡モデルにより野生型大腸菌W3110において増殖および代謝副生成物の分泌が定量的に予測される(Stochiometric flux balance models quantitatively predict growth and metabolic by-product secretion in wild-type Escherichia coli W3110)」Applied and Environmental Microbiology, 60(10):3724-31(1994b))もまた必要とされる。これらの因子は次に、再構築された代謝ネットワークを介して流束分布を計算するために使用される。
【0029】
より具体的には、これらの因子の定義により、全ての実施可能な解が存する方程式に対する閉じた解空間(closed solution space)に数学的に至る。従って、その課題に対して多数の可能な解(流束分布)がある。全許容解のセットの中で「最良」または最適解は次いで、最適化手順および規定の目的を用いて決定できる。使用される最適化手順は線形計画法であり、その目的は全てのバイオマス成分を同時に産生するための生化学反応ネットワークの最適使用である。これらの最適化手順が確立され、公表されている(VarmaおよびPalsson(1994a); Bonarious(1997); およびEdwardsら,(1999))。最適増殖を目的として計算された挙動の実験データとの比較は、大部分の場合において好適である(Varma(1994b); Edwards J.S., Ibarra R.U., およびPalsson B.O.(参照として本明細書に組み入れられる),「大腸菌代謝能のインシリコ予測は実験データと一致する(In silico predictions of Escherichia coli metabolic capabilities are consistent with experimental data)」Nat Biotechnol., 19(2):125-30(2001a); 並びにEdwards, Ramakrishna,およびPalsson,「表現型可変性の特徴付け:表現型相空間解析(Characterizing phenotypic plasticity: A phenotype phase plane analysis)」Biotech Bioeng, 印刷中, (2001b))。換言すると、これらの解の制約および最適化手順により、細胞増殖を補助するおよび進化前の菌株に対し実際の生化学機能を望ましく推定するための、生化学反応ネットワークの最適使用の予測に繋がる。
【0030】
胞子形成、および栄養不足条件下での希元素の探知のような、別の生存目的の使用については、記述されていない。これらの条件の下での競争および進化はまた、最適なネットワーク機能を限定し且つ創生するために使用できる。
【0031】
これらの手順により、単一増殖条件下における最適な機能の計算に繋がる。これは非常に限定的であり、多数の増殖条件を分析するための手段が必要とされる。
【0032】
上記のように、全ての定常状態代謝流束分布は、規定の再構築された代謝ネットワークに対して定義される解空間(その解空間における各解はそのネットワークを介した特定の代謝流分布または特定の代謝表現型に相当する)に数学的に制約される(EdwardsおよびPalsson(1999))。単一の指定増殖条件の下、コーン(錐状)のその最適代謝流束分布は線形計画法(LP)またはかかる課題の最適解を計算する他の関連手法を用いて決定できる。その制約が変化すれば、コーンの形は変化し、最適流束ベクトルは質的に変化するかもしれない。表現型相空間(PhPP)解析は2つまたはそれ以上の制約環境変数における全ての変動可能性を考慮する。本方法をここに開示する。
【0033】
2栄養素の取込速度(炭素基質および酸素のような)は(x,y)空間上の2軸として定義でき、前述の手順を用い、その最適流束分布はこの空間中の全点に対して計算できる。生物学的に現実的な、再構築された代謝ネットワークに対してこの手順を実施する場合、かかる空間に存在する、有限数の質的に異なる最適代謝流束地図、または代謝表現型があることが判る。その相空間の境界は線形最適化の限界価値(shadow price)(Chvatal V.,「線形計画法(linear Programming)」(New York: W. H. Freeman and Co.)(1983))を用いて定義できる。記載の方法により、代謝ネットワークの最適使用が質的に異なる(x,y)空間中の異なる領域、または「相」の定義に繋がる。各相はPnx,y,として指定でき、ここでPは特定の表現型または流束分布を表し、nはこの表現型に対する区画領域に任意に割り当てられた数であり、およびその2つの取込速度がその空間軸を形成する。
【0034】
相空間は2-パラメータ空間の全体にわたって限界価値を計算することにより構築でき、一定の限界価値の領域を区別するために直線を描く。限界価値により、目的関数に対する各代謝産物の実態価値が定義される(Chvatal(1983))。xおよびy軸に相当する取込速度の特定数値により表されるように、限界価値は既定の環境条件下での最適増殖に対する代謝産物の価値に依存して、マイナス、ゼロまたはプラスのどちらかとなる。限界価値がゼロとなる場合、取込速度値が変更されるように最適代謝地図の質的転換が起こる。この基準によりPhPPにおける直線が定義される。
【0035】
最適線(LO)はPhPPの軸に相当する2つの取込流束間の最適な相関を表す直線と定義される。好気性に関して、この直線は最大バイオマス産生を支えるための基質の完全酸化に対する最適な酸素取り込みと解釈できる。
【0036】
代謝再構築および表現型相空間解析手順を、次いで、望ましい代謝挙動、例えば最大取込速度が最適となると思われる条件を予測するために使用する。換言すれば、最適性能を決定するために代謝再構築およびPhPPを使用する。最大取込速度により、相空間における有限の長方形領域の定義に繋がる。この長方形内の最適増殖条件は次いで、規定の制約内での進化過程の予測結果になると思われる。
【0037】
最適化手順を使用しても、その対応する実際の生化学反応ネットワークの特性は、最適でないまたは実用上の見地から望まれるものと同一でないかもしれない。生物体において模擬した再構築ネットワークおよびその合成は、所望の最適解(本明細書では同様に所望の最適性能または所望の最適な機能とも呼んでいる)を示さないかもしれない。最適性欠如は以下の事実による可能性がある:
1. 完全なネットワークを備えた天然生物体は、関心のある環境条件を経験していない並びにこの環境における増殖競争および淘汰を経験していない、または
2. ネットワークからの/ネットワークへの新規反応の除去/追加を介した、遺伝子操作により人工ネットワークはその進化的に決定された最適状態から混乱させられる。
【0038】
本発明のインシリコ方法は、所望の性能が達成されるまでネットワーク中の反応を変更することにより、最適性欠如のこの第2番目の原因を解決するように設計される。次に、培養方法、これは以下に詳細に記載されているように本発明の方法に含めることができるが、増殖競争および淘汰に関連する最適性欠如の第1番目の原因を解決するために使用できる。
【0039】
上記のように、最適特性の計算後、所望の性能を達成するため、代謝技術者はネットワーク中の反応リストを変更できる、または反応リスト中の1つまたはそれ以上の反応を自動的に変更するアルゴリズムが開発され得る。生化学リストの変更後、既定の環境条件下のこのネットワークの最適特性を計算できる。これは、所望の性能に達するまで反応リストの多数の別版(バージョン)が必要とされる可能性がある、会話型設計手段である。所望の性能は、質的特徴または最適化手段を用いて計算された特性に関する定量値である。所望の性能が達成され得る多くの特性は、同分野において周知である。例えば、所望の性能は、所望の増殖速度または酵素もしくは抗生物質のような生体分子の所望の産生量で有り得る。
【0040】
最適化方法は本発明により提供されるコンピュータ・システムを用いて実施できる。コンピュータ・システムには典型的に、少なくとも1生物体の1つまたはそれ以上の生化学反応ネットワークに関する情報を提供するデータベース;最適化および/または比較のため1つまたはそれ以上の生化学反応ネットワークの選択を受信できる、および好ましい性能の選択を受信できるユーザ・インターフェース;並びに最適化方法の計算および再計算を実行するための機能が含まれる。さらに、本発明のコンピュータ・システムは、上述の生化学反応ネットワークの再構築を実行するための機能を含んでいてもよい。
【0041】
コンピュータ・システムはスタンド・アロンのコンピュータまたはクライアント/サーバー環境および1つまたはそれ以上のデータベース・サーバを含む従来型のネットワーク・システムで有り得る。ローカルエリア・ネットワーク(LAN)または広域ネットワーク(WAN)を含む、多くの従来型のネットワーク・システムが同分野において周知である。さらに、クライアント/サーバー環境、データベース・サーバ、およびネットワークは技術、営業用および特許文献中に十分に記録されている。例えば、データベース・サーバはリレーショナル・データベース管理システム、ワールドワイド・ウェブ・アプリケーション、およびワールドワイド・ウェブサーバを作動させる、UNIX(登録商標)のようなオペレーティング・システム上で作動できる。
【0042】
典型的に、本発明のコンピュータ・システムのデータベースには、包括的な生化学反応ネットワーク、実質的に全ての生化学反応ネットワーク、または全ての生化学反応ネットワーク中の生化学反応に関する情報が含まれる。この情報は少なくともネットワークの生化学反応の一部と関連する、生体分子反応物質、産物、補因子、酵素、補酵素の反応速度等の同定を含むことができる。この情報は特定の生化学反応に関与する化合物の分子数を示す化学量論係数を含むことができる。この情報は生物体中に見つけられる任意のおよび全てのイソ酵素を含むことができる。この情報は単一酵素により触媒され得る全ての関連する生化学反応を含むことができる。この情報は、生化学反応を遂行できる複合体の形成のため多くの異なるタンパク質分子が非共有的に会合する必要がある場合、そのオリゴマー酵素複合体の構造を含むことができる。この情報は反応を遂行する酵素の場所(即ち、膜中に、細胞質中に、またはミトコンドリアのような細胞内小器官内部に存在する場合)を含むことができる。この情報はまた、種々の条件下での実験由来のまたは計算による反応速度、バイオマス組成、および維持エネルギー量(maintenance requirements)を含むことができる。この情報は非力学的増殖(non-mechanistic growth)および非増殖関連維持エネルギー量(non-growth associated maintenance requirements)を含むことができる。この情報はタンパク質合成における非効率性のような力学的維持情報(mechanistic maintenance information)を含むことができる。この情報はゲノム規模のmRNAまたはタンパク質発現プロファイル由来のデータを含むことができる。この情報は特定遺伝子の発現と関連したオペロンまたは調節構造に関するデータを含むことができる。この情報は酵素特性の変化を反映する配列変異を含むことができる。この情報は例えば、遺伝子が関心のある条件下で発現していない場合に応じて、条件依存的な生化学反応の組込みを含むことができる。生化学反応ネットワークが代謝反応ネットワークである場合、例えばその情報には既知消費速度、副生成物産生速度、および取込速度を含むことができる。
【0043】
データベースの情報は生化学反応ネットワークの生化学反応に関連する生体分子に関する生体分子配列情報、例えばゲノム配列のような多数の生体分子配列に関する情報を含んでもよい。少なくともゲノム配列の一部は、1つまたはそれ以上の生物体の各ゲノム上にて1つまたはそれ以上の連続配列に沿って位置する読み枠に相当しうる。生化学反応ネットワークに関する情報は、ある生体分子配列が関与するそれらの生化学反応ネットワークおよびその生体分子を含む生化学反応ネットワーク中の特定反応を同定する情報を含むことができる。
【0044】
データベースは生化学反応に関するどんな種類の生物学的配列情報も含むことができる。例えば、データベースはESTを含む核酸配列データベースおよび/またはより好ましくは完全長配列、またはアミノ酸配列データベースで有り得る。好ましくは、データベースは包括的な、実質的に全ての、または全ての生化学反応ネットワークに関する情報を提供する。例えば、データベースは全ての代謝反応ネットワークに関する情報を提供できる。データベースは生物体の完全ゲノムからなる核酸および/またはアミノ酸配列を提供できる。
【0045】
データベースはいかなる生物由来の生化学情報および配列情報も含むことができ、二部に分割できる。一方は配列を記憶するための部分および他方は配列に関する情報を記憶するための部分である。例えば、データベースは動物(例えば、ヒト、霊長類、齧歯類、両生類、昆虫等)、植物、または微生物に関する生化学反応情報および配列情報を提供できる。データベースは例えば、データベースの生体分子の機能、特に生化学機能に関する情報がアノテートされていることが好ましい。アノテーションはそのデータベースの生体分子の生化学、例えばある生体分子が関連する特定反応において、その生体分子は酵素であるかまたは酵素をコードするかどうか、その配列は野生型配列かどうか等、を研究している公開報告から得られる情報を含むことができる。
【0046】
データベースのアノテーションおよび配列は、同定する生物体での選択した生化学的遺伝子型(biochemical genotype)に関する十分な情報を提供する。生化学的遺伝子型は生物体での選択した生化学過程中の全ての核酸またはアミノ酸配列の集団である。例えば、代謝遺伝子型は物質代謝に関連するタンパク質の全ての核酸および/またはアミノ酸配列の集団である。代謝遺伝子型の同定方法は文献に記載されている(例えば、EdwardsおよびPalsson 1999を参照されたい)。
【0047】
データベースはカード型データベースまたはリレーショナル・データベースで有り得る。データベースは内部データベース、または利用者がアクセスできる外部データベース、例えばGenbankまたはGenpeptのような、公共の生物学的配列データベースで有り得る。内部データベースは個人データベースとして保守される、典型的には企業により、ファイアウォール後方で保守されるデータベースである。外部データベースは内部データベースの外側に位置し、典型的には内部データベースとは異なる実在者により保守される。公共の生物学的配列外部データベースの多くが利用可能であり、本発明で使用できる。例えば、米国立バイオテクノロジー情報センター(NCBI)、米国立医学図書館の一部から入手可能な生物学的配列データベースの多くが本発明で使用できる。外部データベースの他の実施例にはシアトルのフレッドハッチンソン・ガン研究センターにより保守されているBlocksデータベース、およびジュネーブ大学により保守されているSwiss-Protサイトが含まれる。さらに、外部データベースは生化学反応について記載し且つ分析している公開済み参考文献のデータベースを含め、生化学反応に関する情報を提供するデータベースを含むことができる。本発明のコンピュータ・システムに含まれるデータベースが特定の生化学反応ネットワークに関連する情報を同定していない公共のコンピュータ・データベースである場合、コンピュータ・システムには生化学反応ネットワークの再構築を実行する機能が含まれるか、または特定の生化学反応ネットワークに関するデータベース登録の確認が含まれる。さらに、生化学経路情報を備えた幾つかのデータベースがあり、これらのデータベースには、例えばこれに限定されないが、EcoCyc、KEGG、WITおよびEMPが含まれる。これらのデータベースは代謝モデルを再構築するための情報を提供するために使用できる。
【0048】
上記のデータベースに加え、本発明のコンピュータ・システムには最適化および/または比較のため1つもしくはそれ以上の生化学反応ネットワークの選択を受信できる、並びに最適性能の選択を受信できるユーザ・インターフェースが含まれる。例えば、一連のメニュー、ダイアログ・ボックス、および/または選択可能ボタンを用いて選択が為される場合、インターフェースはグラフィカル・ユーザ・インターフェースで有り得る。インターフェースは典型的に、主画面から始まる一連の画面を介し、利用者を案内する。ユーザ・インターフェースは利用者が生化学反応ネットワークに関連する追加情報にアクセスするために選択できるリンクを含むことができる。
【0049】
最適化方法を実行する本発明のコンピュータ・システムの機能には典型的に、コンピュータ・プログラム製品(それ自体、本発明の別の局面を表しているが、コンピュータ利用可能媒体上で統合され、かつ処理装置に接続する記憶装置に渡されたコンピュータ可読プログラム・コードが含まれる)を実行する処理装置が含まれる。記憶装置は例えば、ディスク・ドライブ、ランダム・アクセス・メモリー、リード・オンリー・メモリー、またはフラッシュ・メモリーで有り得る。
【0050】
本発明のコンピュータ・システムの処理装置により読み込まれ、かつ実行されるコンピュータ・プログラム製品には、コンピュータが利用可能な媒体上で統合されたコンピュータ可読プログラム・コードが含まれる。プログラム・コードは、データベースと情報のやりとりを行うことができ、かつコンピューティング・システム内で以下の段階を達成する:ネットワークの望ましい性能の選択を受信するためのインターフェースを提供する段階;生化学反応ネットワークの最適特性を再計算して所望の最適な機能に達するまでその工程を繰り返し行う前に、所望の最適特性を決定し、その決定の結果を表示し、かつ生化学反応ネットワークを変更する段階。生化学反応ネットワークの変更は利用者により認識される変更に基づき実施してもよく、またはプログラム・コードにより自動的に実行してもよい。コンピュータ・プログラムはさらに、再構築された生化学ネットワークの一部にあたるデータベース登録を確認してもよく、または生化学反応ネットワークの再構築を実行してもよい。さらに、本発明のコンピュータ・プログラムは、成分および特性の相違を同定するため生化学反応ネットワークを比較する機能を提供できる。
【0051】
コンピュータ可読プログラム・コードは、本発明の方法を実行するソフトウェアを書き込むために使用されるプログラミング言語と互換性のあるいずれか周知のコンパイラを用いて作成できる。本発明の方法を実行するソフトウェアを書き込むために使用できる多くのプログラミング言語は周知である。
【0052】
上記のように、本発明のこの局面の方法にはさらに、望ましい性能を達成するための培養菌株の適応性進化を含む段階を包含できる。実質的には、例えば原核細胞、または真菌細胞もしくは動物株化細胞の細胞を含む動物細胞のような真核細胞を含むどの細胞も、本発明の方法で使用できる。しかし、細胞または近縁生物体の細胞の生化学反応ネットワークは、コンピュータで包括的な、実質的に全ての、および/または全ての生化学反応ネットワークの高品質な再構築を可能とするため十分に特徴付ける必要がある。本質的に生物体の全ゲノムが配列解読されていることおよび生化学ネットワークに関連する生体分子、典型的にはタンパク質をコードする遺伝子が同定されていることが好ましい。
【0053】
細胞の遺伝子構成は所望の性能を満たす潜在性のある細胞を産生するため所望の性能に合う生化学反応を含むように構築できる。これは細胞の固有反応リストを用いて、および上述のインシリコで実行される段階により特定される所望の性能基準を達成できる反応リストに達するよう遺伝子操作を用いて、このリストに反応を追加するおよびこのリストから反応を差引くことにより達成できる。例えば、反応に関連する1つ以上の生体分子をコードする1つ以上の遺伝子の全部もしくは一部を付加する、改変する、または除去することにより、例えば、1つ以上の遺伝子のタンパク質コード領域を付加する、改変する、もしくは除去することにより、または1つ以上の遺伝子の調節領域を付加する、改変する、もしくは除去することにより、リストに反応を追加するまたはリストから反応を差引くことができる。さらに、例えば、リストの1つ以上の反応に関連する1つ以上の生体分子の発現を変化させる調節成分(例えば、転写因子)の発現を改変することにより、リストに反応を追加するまたはリストから反応を差引くことができる。その結果として作成された人工細胞は、上述の反復最適化手順を用いるインシリコ法により予め計算した最適特性を示してもまたは示さなくてもよい。
【0054】
細胞の遺伝子操作について説明する多くの方法が当技術分野に存在する(例えば、Datsenko K. A.およびWanner B. L.,「PCR産物を用いた大腸菌K-12の染色体遺伝子の一段階不活性化法(One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products)」Proc. Natl. Acad. Sci. U.S.A., 97(12):6640-45 (2000); Linkら,「野生型大腸菌のゲノムにおける正確な除去および挿入をもたらす方法:読み枠の特徴付けへの適用(Methods for generating precise deletions and insertions in the genome of wild-type Escherichia coli: Application to open reading frame characterization)」J. of Bacteriology, 179:6228-37 (1997)を参照されたい)。既存のどの遺伝子操作手順も開示される本発明を実践するために使用できる。
【0055】
所望の性能を満たす潜在性を有するように細胞を構築した後、特定環境の下での培養状態に置く。特定環境は最適化手順を用いて決定される。即ち、前述のように、最適化手順により種々の環境下でのネットワーク特性を計算し、所望性能が達成される特定環境を明らかにする。
【0056】
細胞を十分な期間、および所望の性能にその細胞を進化させる条件の下で培養する。即ち、天然または人工菌株の適応性進化は、一般的な最適化方法または手順による誘導に従い実行される。特定の環境を経験していない天然菌株または遺伝的に改変された菌株は、上記に開示されたネットワーク再構築および最適化手順により解析できる。これらの菌株を次いで、その生物体の所望の機能と整合性が取れた淘汰圧の下に置き、予め決定した性能特性へ進化させることができる。その細胞はさらなる適応性進化をせずに、所望の性能を達成できる。即ち、細胞を培養するための十分な期間とは、さらなる適応性進化を必要とせず本発明の方法を用いて、その細胞の遺伝子構成が構築される直後までであってもよい。
【0057】
換言すれば、非最適または非進化株の拡張培養は、定義した環境条件の下で達成できる最適解へ、代謝ネットワークを最適化するまたは進化させるために実施できる。この進化過程の実践には、進化過程の開始時に代謝ネットワークが最適条件からどの程度離れているか、並びに無作為変異および遺伝子発現調節の改変により必要な変更を達成することがどの程度困難であるかに依存して、代謝ネットワークを最適化するのにおよそ数週間から数年程度が必要とされる。この過程は化学的突然変異誘発要因および/または放射線の使用により促進できる。さらに過程は、上述のインシリコ法により決定される、所望の性能を達成する生化学反応物質を含むように生細胞を遺伝学的に改変することにより、促進される。
【0058】
特定環境条件の下で細胞を培養する方法は、同技術分野において周知である。例えば、細胞が大腸菌であれば、所望の性能は所望の増殖速度であり、下記の手順が使用できる。この手順は他の細菌細胞および/または当技術分野において周知の他の性能基準での使用に容易に適応できる。さらに、手順は動物細胞のような他の細胞種での使用に容易に発展できる。例えば、方法は細胞が培養容器に接着する大規模系のような、他の培養系での使用に容易に適応できる。培養方法は当技術分野において周知の、ハイスループット解析に適応されうる。
【0059】
所望の性能を達成するよう、菌株を一方向に進化させる必要がある場合、選択した宿主菌株における代謝反応ネットワークの構築に続き、細胞を典型的には30%グリセロールと共に-80℃で凍結保存する。各適応性進化過程に対し、凍結ストックをLB寒天培地に播き、37℃で一晩増殖させる。プレートからは、単一細胞より生じた単一コロニーを同定できる。前培養として知られているが、単一コロニーは液体培地に植菌するために使用できる。各菌株の単一コロニーから植菌された前培養液を、その後の進化過程のための規定培地中で一晩増殖させる。適応性進化が起こる環境を規定する培養条件に菌株を植菌するため、前培養試料を翌日、典型的には増殖の中間対数期で(対数増殖の中間で)取り出す。次いで、バッチ処理用生物反応器または他の適した培養容器で開始する。これは典型的には、温度制御恒温槽内、電磁攪拌プレート(マグネティック・スターラー・プレート)上に置いたマイクロキャリア・スピナー・フラスコ中、250mLの容量で、十分な曝気を確実とする適当な-典型的には高い-速度および使用菌株の最適増殖温度(野生型大腸菌では37℃)の設定で、行われると思われる。当技術分野において周知の、他のよく利用される培養方法もまた使用できる。
【0060】
適当な時間後、大腸菌では典型的には翌日(培養が静止期に達する前)、中間対数期にある培養液の一部を新鮮な培地を含む新たなスピナー・フラスコに連続的に移入する。もし培養が増殖速度に関して最適化されているところであれば、選択基準が増殖速度であることを確実とするため、静止期は回避しなければならない。次いで、中間対数期の固定時間間隔(増殖速度にもよるが典型的には24時間毎)で連続的に移入し、それから新たな培養容器への植菌容量は増殖速度の増加に基づいて調整する。
【0061】
従って、次に連続的に移入する際に使用する植菌の適容量を決定するため、増殖速度を頻繁に、典型的には毎日、監視する。この連続培養過程を、培地組成による既定条件の下で、細胞をその最適達成可能増殖へ進化させるのに十分なだけ頻繁に繰り返す。
【0062】
時間とともにその集団がどのように進化しているかを決定するため、適応性進化の間、その増殖および代謝挙動を監視する。固定時間間隔、典型的には数日毎に、酸素取込速度、基質取込速度および増殖速度を測定することにより、代謝および増殖挙動に関して培養を試験する。その結果を次いで、表現型相空間にデータ点としてプロットする。最適線に対するそのようにして決定されるデータ点の動きにより、最適増殖挙動への進化が示されると思われる。データ点がその最適条件に位置するようなその代謝ネットワークを、細胞が稼動していることが観察されるまで、増殖速度に加え、膜輸送流束のこれらの測定を繰り返す。その後、その進化過程は最適性能、例えば増殖速度のさらなる増加がなくなるまで、継続できる。さらなる変化が観察されない場合、既定条件での最大増殖速度が達成されたことになる。
【0063】
副産物分泌はHPLCまたは最適増殖挙動への進化に関与する物質代謝の変化を評価する他の適切な分析化学法により監視できる。これらの研究のため、その進化菌株に対する乾燥重量対光学濃度の相関関係を決定することは、これが野生型とは異なっていると思われるので、同様にして必須である。その増殖速度および定常状態の増殖を監視することに加え、何らかの汚染が発生した場合の予備として冷蔵しておいた日毎の培養液に対し、汚染の可能性のあらゆる兆候または突然変異による部分母集団(mutant subpopulation-aliquots)に伴う共進化に関してその培養液を検査する、並びにその培養試料を播き、かつあらゆるコロニー形態の相違または種々の突然変異体について検査することにより、その培養の表現型を確かめる。毎日、その培養の光学濃度、植菌時間、植菌容量、増殖速度、および汚染のあらゆる兆候を記録する。さらなる使用の可能性のため、日毎の培養試料もまた、30%グリセロール中、-80℃にて凍結する。
【0064】
本発明の方法を用いて産生された細胞が最適性能を達成するように進化した後、それらの細胞をさらに特徴付けてもよい。例えば、細胞の生化学反応ネットワークから特徴付けをすることができる。この特徴付けは、生細胞の生化学反応ネットワークの、成分を含むその特性を、インシリコ法を用いて予測されたその特性と比較するために使用できる。
【0065】
下記の実施例は本発明を例証するが、本発明を限定しないことを意図するものである。
【0066】
実施例1
大腸菌の最適な機能および進化の決定方法の詳細
本実施例は、菌株の最適性能を決定し、および適応性進化過程を実行するために使用できる培養手順を提供する。
【0067】
大腸菌の増殖挙動は下記の標準手順により決定される。増殖は炭素源を添加したM9最小培地(Maniatisら, Molecular Cloning: A Laboratory Manual, (Cold Springs Harbor, N.Y., Cold Spring Harbor Laboratory, 545(1982))(表1)で行う。細胞増殖速度は環境条件を変更することにより、即ち約0.05〜4g/Lの範囲で炭素源濃度を変更し、温度(27.5℃〜37℃)、および酸素(空気に対し0〜100%飽和)を変更することにより変えられる。バッチ培養は通気を備えた500mLフラスコ中、250mL容量にて生物反応器中に設定する。これらの培養に際し、排気分析器を用いて酸素に対する物質移動係数(k1a)を測定するか、または当技術分野において周知の方法を用いて呼吸計試験槽中の酸素分圧を監視するかにより、酸素取込速度(OUR)をオンラインで監視する。温度は循環水槽(Haake, Berlin, Germany)を用いて制御する。全ての測定およびデータ解析は増殖の対数期に制限する。バイオマス並びに培地中の基質および代謝副産物の濃度は、当技術分野において周知の方法を用いて実験の間中、監視する。細胞増殖は、600nmおよび420nmでの光学密度(OD)の測定並びに細胞カウントにより監視する。細胞乾燥重量に対するODの相関関係は、2つの異なる方法により決定する;(1)培養試料50mLを遠沈させて75℃で一定重量まで乾燥させる、および(2)試料25mL(培養の間中、採取する)を0.45umフィルタを通してろ過し、一定重量まで乾燥させる。培地中の代謝産物濃度はHPLCで決定する。Aminex社製HPX-87Hイオン交換炭水化物-有機酸カラム(66℃にて)は、移動相として脱気した5mM H2SO4およびUV検出とともに使用する。酵素アッセイ法はまた、基質取込速度および副産物分泌速度を決定するために使用する。培養液中に溶解した酸素は、ポーラログラフ溶存酸素センサを用いて監視する。酸素消費量は3つの異なる方法により測定する;(1)排ガスを1440C Servomex社製酸素分析器(Servomex Co., Inc. Norwood, MA)に通す、(2)溶解酸素測定値およびK1a測定値から計算する、および(3)個別の50mLフラスコ中の呼吸計試験槽中で測定する。酸素取込速度の測定に使用される3方法ともに、同様のおよび再現可能な結果をもたらす。
【0068】
本実施例で提供される培養手順は、菌株の最適性能を決定するおよび適応性進化を実行するために使用できる。
【0069】
【表1】
【0070】
実施例2
最適特性および表現型相空間の計算
本実施例は、再構築されたネットワークの最適特性の計算方法およびその結果の表現型相空間への表現方法を示す。
【0071】
代謝ネットワーク能は流束均衡解析(FBA)(Bonariusら,(1997); Edwardsら,(1999); Varmaら,(1994a); Varmaら,(1994b))を用いて評価できる。FBAは主に代謝ネットワークの質量保存に基づいている。その保存必要条件は化学量論均衡方程式により実行される;従って、FBAは代謝ネットワークの化学量論特性に依存する。
【0072】
流束均衡方程式はS・v = bvであり(ここでSは化学量論行列であり、ベクトルvは代謝流束を定義し、およびbvは通常はゼロである)従って、一組の質量均衡を通じて、同時に起こる質量、エネルギーおよび酸化還元均衡の制約が強要される。ゼロからの bvベクトル変数は限界価値解析で使用した(下記に述べる)。大腸菌の代謝ネットワークでは、代謝流束数は質量均衡数より大きかったが、それ故に、行列Sのヌル空間に位置する実行可能な流束分布が多数になった。代謝ネットワークの各流束の実施可能値はまた、さらに制約された
。これらの制約を内部反応の可逆性を定義するためおよび輸送反応に対する取込速度を確定するために利用した。無機リン酸塩、アンモニア、二酸化炭素、硫酸塩、カリウムおよびナトリウムの輸送は抑制されなかった;ところが条件として指定した炭素源および酸素の取り込みは抑制された。細胞外へ輸送されることが知られている全ての代謝副産物(即ち、酢酸塩、エタノール、ギ酸塩、ピルビン酸塩、コハク酸塩、乳酸塩など)は常に、その代謝系から離脱させた。本解析において、全ての不可逆流束に対して内部流束を表すαiはゼロに設定され、並びに全ての可逆流束は順方向および逆方向で制限されなかった(各反応の可逆性は補助情報のウェブサイト上に定義されている)。ヌル空間および線形不等式により定義される領域の交点が代謝ネットワーク能を定義し、流束コーンと呼ばれている(Schillingら, 1999)。
【0073】
流束コーン内に位置する最適代謝流束分布の決定は、特定の代謝目的を最小にする解が同定される、線形計画(LP)問題として策定した(Bonarius, H.P.J.ら, 「質量均衡を用いた、異なる培地中のハイブリドーマ細胞の代謝流束解析(Metabolic flux analysis of hybridoma cells in different culture media using mass balances)」Biotechnology and Bioengineering 50:299-318(1996); Edwardsら,(1999); Pramanikら,(1997); Varmaら,(1994a); Varmaら,(1994b))。増殖流束を目的として定義した。増殖流束は最適比率で生合成前駆物質、Xmを利用する代謝流束と定義され、
、ここでdmは代謝生成物Xmのバイオマス分画である。バイオマス組成は文献(Neidhardt, F C, Ed., 大腸菌およびサルモネラ: 細胞分子生物学(ASM Press, Washington, D.C., 1996); Neidhardt, F C, Umbarger, H E, 大腸菌およびサルモネラ中: 細胞分子生物学F. C. Neidhardt, Ed. (ASM Press, Washington, D.C., 1996), vol. 1, pp. 13-16 (1996)) を基に定義した。
【0074】
全ての定常状態代謝流束分布は、規定の代謝地図に対して定義される流束コーン(流束コーン中の各解は特定の内部流束分布または特定の代謝表現型に相当する)に数学的に限定される(Varmaら,(1994a); Varmaら, (1994b))。指定の増殖条件の下で、コーン中の最適表現型は線形計画法(LP)を用いて決定できる。その制約が変化すれば、コーンの形状が変化し且つその最適流束ベクトルは質的に変化するかもしれない;例えば、不活性流束が活性化されるかもしれず、その逆もまた同様である。相空間解析は2つの制約環境変数における全可能性変動を考慮するために発展させられる。
【0075】
表現型相空間(PhPPs)の定義:2栄養素(炭素基質および酸素のような)の取込速度を(x,y)空間の2軸に定義し、この空間中の全点についてその最適流束分布を計算した。かかる空間に存在する、有限数の質的に異なる最適代謝流束地図、または代謝表現型がある。その相空間の境界は限界価値解析(Varma, A, Boesch, BW, Palsson, BO.,「種々の酸化速度の下での大腸菌グルコース異化作用の化学量論的解釈(Stoichiometric interpretation of Escherichia coli glucose catabolism under various oxygenation rates)」Applied and Environmental Microbiology 59, 2465-73(1993); Varma, A, Palsson, BO.,「大腸菌代謝能: II. 最適増殖様式(Metabolic capabilities of Escherichia coli: II. Optimal growth patterns)」Journal of Theoretical Biology 165, 503-522(1993))により定義した。この手順により、代謝経路の最適利用が質的に異なった、空間中の別個の領域または「相」を定義するに至った。各相はPnx,yとして記述し、ここでPは表現型を示し、nはこの表現型に対する境界領域数(対応する図1に示されるように)、およびx,yはその空間軸の2つの取込速度である。
【0076】
相空間の計算:相空間は2-パラメータ空間の全体にわたって限界価値を計算することにより構築し、一定の限界価値の領域を区別するために直線を描いた。限界価値により、目的関数に対する各代謝産物の実態価値が定義された。限界価値の変化を代謝挙動の解釈のために利用した。
【0077】
数学的には、限界価値は以下のように定義され、
ネットワーク中の各代謝産物と関連する。限界価値により、各代謝産物の利用可能性の変化に対する目的関数(Z)の感度係数が定義された(bνiは質量均衡制約の破れを定義し且つ取込反応に等価である)。限界価値はその代謝産物の価値に依存して、マイナス、ゼロまたはプラスのどちらかとなった。相空間の各領域中の限界価値ベクトルの方向および大きさは、異なっていた(相空間の定義により);従って、代謝系の状態は各領域で異なっていた。
【0078】
アイソクライン:代謝表現型を解釈するためにまた、アイソクラインを定義した。目的関数の同一値を規定する二次元空間内の点の軌跡を示すために、アイソクラインを定義した。各領域内のアイソクラインの傾きは、限界価値より計算した;従って、定義によるアイソクラインの傾きは、PhPPの各領域で異なっていた。アイソクラインの傾き(ρ)を定義するために、限界価値の比率を用いた:
【0079】
方程式2のマイナス符号はその解釈を予想して導入した。ρと定義される、条件依存的な基質の相対値は、その代謝ネットワークの制約要因を解釈するために用いた。ρがマイナスとなった領域では、基質が二元的に制限された。異なる条件の下では、アイソクラインはまた一元的な基質制限領域を表す、ある相空間領域において水平または垂直の何れかであり、これらの領域におけるρ値はそれぞれゼロまたは無限大であった。また、PhPPのある領域はρがプラスとなった;これらの領域は「意味のない」領域と名付けられ、基質のうち1つの増加取り込みにより、これらの領域の目的関数にマイナスの影響が及ぼされた。最後に、化学量論的制限により、PhPPに実行不可能な定常状態領域が存在した。
【0080】
最適線:最適線(LO)はPhPP軸に相当する2つの代謝流束間の最適相関を示す直線と定義した。本明細書に提示した結果に関して、この直線は最大バイオマス産生を支えるための基質の完全酸化に対する最適な酸素取り込みと解釈できる。
【0081】
これらの手順を(EdwardsおよびPalsson, 大腸菌MG1655のインシリコ代謝遺伝子型: その定義、特性、および能力, Proc. Natl. Acad. Sci. U.S.A., 97(10):5528-33(2000b))により規定した大腸菌K-12に関する反応リストに適応した。それにより、以下の実施例で使用される二次元および三次元相空間が作成される。
【0082】
実施例3
規定条件の下での大腸菌の最適挙動
本実施例は、使用菌株が最初の基質として酢酸塩およびコハク酸塩を用い、適応性進化なしに最適な好気性増殖を示したことを明らかにする。
【0083】
大腸菌K-12 M1655で起こる代謝反応リストが構築された(EdwardsおよびPalsson(2000b))。このリストを基に、化学量論行列を定式化した。酸素(y軸上)および炭素基質(x軸上)に対する最大取込速度を用い、上述の手順を利用して表現型相空間を計算した。具体的には、2つの炭素源、酢酸塩およびコハク酸塩を用いた。次いで、最適増殖条件を決定するために計算した相空間を使用し、それから一連の増殖実験を実施した。計算(即ち、インシリコ)および実験結果を次に、比較した。
【0084】
酢酸塩
酢酸塩に関する最適増殖性能をインシリコで調べ、それから作成したその予測を実験データと比較した。インシリコ研究は大腸菌物質代謝能を示す流束コーンの二次元投射の軸として定義した、酢酸塩および酸素取込速度に関する表現型相空間(PhPP)解析から開始した(図1)。流束コーンは全ての許容可能な定常状態代謝流束分布の領域である(流束コーンの十分な解説としては、参考文献(Schillingら,「代謝経路解析: ポストゲノム時代における基本概念および科学的応用(Metabolic pathway analysis: basic concepts and scientific applications in the post-genomic era)」Biotechnol. Prog., 15(3):296 (1999))を参照されたい)。さらに、第三次元として定義した増殖速度に関する流束コーンの三次元投射を利用した(図1)。酢酸塩-酸素PhPPのインシリコ解析は前記実施例2で説明した。酢酸塩および酸素取込速度間の最適関係(細胞増殖に関して)であり、およびこの直線は最適線(LO)と呼ばれる。
【0085】
PhPPは代謝ネットワークの働きを解析および解釈するために利用した。例えば、酸素制限の下での代謝ネットワークの特性は、酢酸塩取込速度が酸素取込速度との最適関係を上回る、PhPPの領域2(図1および2)により定義されてもよい。図1から、代謝ネットワークが領域2内で働いている場合、酢酸塩取込速度をLO上のある点まで減少させることにより、増殖を補助する最適能は増大すると理解される。領域1内の点に関して、酸素および酢酸塩の役割を換えることで、同様の解釈が為される。故に、領域1または領域2中の点を規定する代謝流束ベクトルは、利用可能資源の非効率利用を示していると思われる。従って、インシリコPhPP解析により、唯一の炭素源として酢酸塩を用い増殖補助のために最適に作動するよう、大腸菌代謝ネットワーク調節が進化した場合、酢酸塩および酸素取込速度と増殖速度間との関係は、LOにより定義される筈である(図1および2)という結論に至った。
【0086】
大腸菌MG1655を酢酸塩最小培地で培養することにより、酢酸塩および酸素の取込速度と増殖速度間との関係を実験的に調べた。最小培地中の酢酸塩濃度を変化させることで酢酸塩取込速度を実験的に制御した。酢酸塩および酸素取込速度並びに増殖速度を測定し、それから実験データ点をPhPPにプロットした(図1および2)。次に、酢酸塩および酸素取込速度間の計算による最適関係を実験データと比較した(図1)。実験データのインシリコ予測との比較により、酢酸塩最小培地での好気性増殖に関して、実験データに対する回帰直線の傾き(0.91)およびインシリコ定義によるLOの傾き(1.04)間で14%の相違が示された。
【0087】
測定および計算した増殖速度を酢酸塩-酸素PhPP上の三次元としてプロットした(図2)。色分けされた表面は流束コーンの三次元投射を示す。換言すれば、色分けされた表面は解空間を定義し、および全ての実行可能な定常状態代謝流束分布はその表面内に限定される。まさしく可能である。二次元相空間のLO(図1)は、三次元表面の縁のx,y空間(酢酸塩取込速度、酸素取込速度)への投射である。実験データを三次元空間にプロットした(図2)。三次元におけるデータ点のLOへの接近性を量的に視覚化するため、インシリコ予測値および実験データをその基底ベクトルにより形成される各空間上へ投射した。
【0088】
三次元LOおよびその実験データ点の(x,y)、(x,z)、および(y,z)(x軸:酢酸塩取込速度;y軸:酸素取込速度;z軸:増殖速度)空間への投射をそれぞれ、図3および4(回帰直線の適合度を相関係数により示し、且つそのデータをインシリコ予測値と比較している)に示した。各点に対して予測したおよび観測した代謝流束(基質および酸素取込速度並びに増殖速度)を直接比較し、そしてそのインシリコ予測では全体の平均誤差が5.8%であった。この点に関して、代謝ネットワークを構築するために使用した情報は、本実験とは独立して得られたことに注目すべきである(EdwardsおよびPalsson,(2000b))。計算したPhPPは本研究で得られたデータの先験的な解釈および予測を示している。
【0089】
コハク酸塩
コハク酸塩-酸素PhPP(図5)は酢酸塩-酸素PhPPよりもっと複雑であった。コハク酸塩-酸素PhPP(図5)には、質的に異なる最適代謝ネットワーク利用からなる4つの別領域があった。コハク酸塩-酸素PhPPの領域1および4は、酢酸塩-酸素PhPPの領域1および2と類似であった。例えば、図5から、コハク酸取り込みを領域3、4の境界により定義される点に減少させた場合、領域4の流束ベクトルに関する最大増殖流束は増大され得ると理解できる。さらに、PhPP解析から、領域3は1基質制限領域と定義される。1基質制限領域では、コハク酸塩取込速度は領域3の最大増殖流束にほとんど影響しないが、酸素取込速度は増殖速度にプラスに影響することが示される。
【0090】
領域2においてコハク酸塩取込速度が増加した場合、代謝ネットワークは増殖の増加を補助できるので、領域2は2基質制限領域と定義される。インシリコ解析により代謝ネットワークを領域2のLOの外へ操作すること、部分的に好気性の物質代謝および代謝副産物の分泌を行うことにより、細胞増殖速度を増加できることが示される。最適代謝副産物は酢酸塩であると計算された。領域2の代謝副産物の減少産生はしかし、全体のバイオマス産生を減少させる。故に、PhPP解析を基に、代謝ネットワーク調節が唯一の炭素源としてコハク酸塩を伴う最適増殖の方向へ進化した場合、その代謝ネットワークはLOに沿った流束ベクトルで動作することが推測されうる。しかし、増殖速度は流束ベクトルを領域2中に移動させることにより増加でき、従って、酸素が制限され且つコハク酸塩が十分である場合、先に述べたの仮説が真実であるならば、そのネットワークは領域2中でのみ作動する筈であると予想される。
【0091】
インシリコ解析における上記の仮説を批判的に調べるため、コハク酸塩最小M9培地での大腸菌増殖実験を実施した。コハク酸塩取込速度の範囲を測るため、多くのバッチ培養物を種々のコハク酸塩濃度および温度で増殖させた。全培養物で一貫した最大酸素普及率を維持するため、曝気および攪拌を一定に保った。コハク酸塩および酸素取込速度並びに増殖速度を、それぞれ独立した増殖実験に対して別々に測定した。その実験データを、次いで、インシリコ予測と直接比較した(図5)。
【0092】
実験データ点は先述の仮説と一致していた:臨界値以下の酸素取込速度(〜18.8ミリモル/g-DW/時間)に対しては、流束ベクトルは常にLOに沿った点を示した。さらに、LO沿いの点を示した培養物は、代謝副産物として酢酸塩をほとんどまたは全く産生しなかった(インシリコ解析により予測されたように-図5を参照されたい)。仮説の通り、酸素は制限されているがコハク酸塩は十分ある実験系に対し、実験データは流束ベクトルが領域2内へ水平移動することを示している。実験データの区切り点は、最大酸素取込速度18.8±0.5ミリモル/g-DW/時間に一致すると決定された。領域3または4内の流束ベクトルは観測されなかった。領域2に同定された培養物に対し、酢酸塩産生を測定した、さらに図6で、その酢酸塩産生をそのインシリコ予測と定量的に比較する。
【0093】
最適増殖速度表面をコハク酸塩-酸素PhPP上方に構築した、すると測定流束ベクトルは、LOに相当するポリトープの縁近くに移動した(図7)。また、流束ベクトルにより、領域2における相表面上の点の軌跡が、系の最大酸素取込限界と同等の一定酸素取込速度であることが示された。インシリコ解析の予測能およびそのインシリコ由来の仮説を定量的に調べるために、本発明者らは、本発明者らの仮説および実験的に観測した流束ベクトルを記述する区分線形モデルを使用した。区分線形モデルは以下のように定義される:本発明者らはコハク酸塩取込速度および酸素取込限界の範囲に対する流束ベクトルにより定義される点の軌跡を同定した。酸素取込限界よりも下では点の軌跡はLO沿いに位置し、および酸素取込限界よりも上では点の軌跡は一定酸素取込速度(酸素取込限界)の相表面沿いに位置する。区分線形モデルに基づき、コハク酸取込速度は酸素取込速度および増殖速度を予測するために使用され、並びにその他2つの順列もまた考慮した。この解析から、インシリコ予測および実験データ間の全体の平均誤差は10.7%となった。
【0094】
本実施例は、使用した菌株が、主な基質として酢酸塩およびコハク酸塩を用いて最適な好気性増殖を示したことを明らかにしている。この最適性能を達成するために適応性進化は必要とされなかった。
【0095】
実施例4
準最適性大腸菌菌株の最適性への進化
本実施例は、大腸菌が準最適増殖状態からインシリコ決定の最適状態まで幾つかの表現型適応を受けられることを実証する。
【0096】
グルコース
グルコース-酸素PhPPには異なる6領域が含まれる(図8)。コハク酸塩-酸素PhPPと同様、領域1は無益回路および準最適な増殖性能を示すが、領域2は酢酸塩過剰な物質代謝により特徴付けられる。その2領域はLOにより分けられる。
【0097】
すでに述べたように、細胞増殖速度、OUR、およびグルコース接触速度(GUR)を各種のグルコース濃度および温度にわたって実験的に決定した。GUR、OURの実験決定値のほとんどは、PhPPのLO上または僅かに領域2(ここでは予測した酢酸塩分泌が実験的に観測された)の点に相当した(図8)。
【0098】
三次元において、測定した増殖速度はLOに相当する縁近傍の解空間の表面上に位置しているが、そこに密集はしていない(図9)。本発明者らはこのように一定の増殖条件の下で代謝表現型が進化するかどうか決定するため、連続移入を用いて40日間に渡り(約750世代)その菌株を対数増殖(16)状態に維持した(図10および11)。LOと平行した実験点の移動によって示されるように、適合性は確かに増加したが、表現型に質的な変化はなかった。
【0099】
実施例5
準最適性大腸菌菌株の最適性への進化
本実施例は大腸菌が準最適増殖状態からインシリコ決定の最適状態への有効な表現型適応を受けられることを実証する。
【0100】
グリセロール
グリセロール-酸素PhPPは、酢酸塩、コハク酸塩およびグルコースに対するPhPPで見られるのと似た特徴を伴う5つの領域からなる。特に、無益回路を伴う領域(相1)はLOにより酢酸塩過剰領域(相2)と分けられる。
【0101】
各種グリセロール濃度にわたる増殖性能を、すでに述べたように、実験的に決定した。しかし、リンゴ酸塩またはグルコースにおける増殖とはかなり異なって、増殖の実験データはLO(図12)および最適性表面(図13)からかけ離れた、相1の全体にわたり分散していた。調べた他の基質と異なり、グリセロールは従って、大腸菌K-12の準最適増殖のみを補助する。
【0102】
すでに述べたように、本発明者らは従って、今度は唯一の炭素源としてグリセロールを用い、長期の適応性増殖実験を実施した。温度30℃、グリセロール濃度2g/L、および十分な酸素供給を維持しながら、連続移入(17)により、原菌株を再度、40日間の長期の対数増殖状態に保持した。増殖速度、グリセロール取込速度(GlUR)およびOURを10日毎に決定した。
【0103】
40日間の進化経路(E1)は相1中に描かれ、最終的にはLOに収束した(図14)。この間、増殖速度は0.23/時間から0.55/時間へと倍以上に増えた(図15)。結果として生じた進化菌株(凍結保存されていた)をさらに調べたところ、比増殖速度およびバイオマス産生が親菌株よりも高いことが明らかとなった。得られた全データはLO上または近傍に移動し、進化菌株が長期培養の最終日にそのデータを示した(図16および17)ことより、その進化菌株がインシリコ予測と一致する、グリセロールにおける最適増殖性能に到達していたことを示している。2回目の別個の適応性実験は似ているが、同一でない進化軌道(E2)を示し、同じ終点近くに収束した。大腸菌は従って、準最適増殖状態からインシリコ決定の最適状態への有効な表現型適応を受けられる。
【0104】
このように細菌の最適性増殖の実行により、インシリコ解析による予測が確認されると思われる。酢酸塩およびコハク酸塩のようないくつかの基質においては、細胞は最適増殖を示すかもしれないが、グルコースおよびグリセロールのような他の基質においては、そうでないかもしれない。後者においては、増殖は相空間予測の最適増殖の方向へ進化する。
【0105】
本明細書に引用した参考文献の全てが参照として組み入れられる。上記実施例に関連して本発明を説明したが、本発明の精神および範囲内での変更および変形が包含されると理解される。従って、本発明は特許請求の範囲によってのみ制限される。
【図面の簡単な説明】
【0106】
【図1】酢酸塩取込速度(AUR 単位;ミリモル/g-DW/時間、g-DWはグラム乾燥重量である)対酸素取込速度(OUR 単位;ミリモル/g-DW/時間)表現型相空間。インシリコ定義の最適線(LO)が図中に示してある。同直線の傾きもまた図中に示してある。実験データ点は図上に表示してある。誤差指示線は単一の標準偏差を表し、誤差指示線は酢酸塩および酸素取込速度測定値の両方に対して表示してある。実験的に再構築された最適線を定義するため、データ点について直線回帰を行った。曲線に適合する相関係数R2値は0.92である。領域1および2は別個の非最適代謝表現型を表している。
【図2】酢酸塩における大腸菌の増殖に対する相表面の三次元図。xおよびy軸は図1中と同じ変量を示している。第三次元(z-次元)は細胞増殖速度に相当する。z軸値は対応する凡例に定量的に表示された最適増殖速度値を伴う灰色の尺度である。三次元中の最適線(LO)が表示してある。三次元中のLOの助変数方程式が本明細書中に示されている。黒線は流束コーンの三次元投射における代謝能表面を定義し、酢酸塩取込速度または酸素取込速度の定常値を示している。酢酸塩取込速度を増加させること(酸素取込速度の比例的増加なく)による細胞増殖の可能性への定量効果が視覚化できる。データ点もまた三次元図にプロットしてあり、誤差指示線は省略されている。
【図3】酢酸塩取込速度および増殖速度により形成される空間上に投射された、酢酸塩における増殖の最適線。データ点もまた投射してあり、実験的に最適線を定義するため直線回帰は二次元空間中で行った。最適線は灰線として示し、回帰直線は黒線として示してある。
【図4】酸素取込速度および増殖速度により形成される空間上に投射された、酢酸塩における増殖の最適線。データ点もまた投射してあり、実験的に最適線を定義するため直線回帰は二次元空間中で行った。最適線は灰線として示し、回帰直線は黒線として示してある。
【図5】表現型相空間で表されたコハク酸塩取込速度(ミリモル/g-DW/時間)対酸素取込速度(ミリモル/g-DW/時間) 。ラベルの付いた直線はインシリコ定義の最適線(LO)である。実験データ点は図上に示してある。誤差指示線はコハク酸塩および酸素取込速度測定値の両方に対して表示してあり、単一の標準偏差を表している。酢酸塩が0.3ミリモル/gDW/時間の閾値以上に産生された培養は白丸で示してあり、黒丸は酢酸塩産生なしまたは産生が閾値以下のどちらかを表している。黒色の破線は酢酸塩産生なしのデータ点の回帰直線を示している。
【図6】酢酸塩産生の実測値対図5にグラフ化された各点に対するインシリコ予測値。データ点はコハク酸塩取込速度の大きさにより順序付けた階級である。
【図7】コハク酸塩における大腸菌増殖の三次元表現型相空間。xおよびy軸は図6中と同じ変量を示している。第三次元(z軸)は細胞増殖速度に相当する。z軸値は図中の対応する凡例の灰色の尺度である。着色領域を分ける境界線は一定の酸素および酢酸塩取込速度を示し、最適線から離れる定量効果が視覚化され得る。誤差指示線を除き、データ点がこの三次元図にプロットしてある。
【図8】グルコースにおける大腸菌K-12の増殖の計算および実験値。グルコース取込速度、GUR(ミリモル/gDW/時間)および酸素取込速度、OUR(ミリモル/gDW/時間)を表現型相空間に示した。LOが表示してある。データ点はLOまたは酢酸塩過剰領域(ここでは酢酸塩分泌がインシリコ予測され、且つ実験的に観測される)に限定されている。
【図9】グルコースにおける増殖の相空間上方にグラフ化された増殖速度の三次元図。xおよびy軸は図8中と同じ変量を示している。z軸は色分けされた数値および凡例に表示されたその最適増殖速度と共に、細胞増殖速度を示す。
【図10】適応性進化実験に関する実験値を用いてOURに対してプロットしたGUR。データ点はLO近傍および酢酸塩過剰が予測される領域2に位置している。
【図11】グルコース表現型相空間上方の進化後の3D増殖表面の三次元図。全データ点はLO上または近傍に密集している。
【図12】グリセロールにおける増殖の計算および実験値。グリセロール取込速度、GlUR(ミリモル/gDW/時間)、および酸素取込速度、OUR(ミリモル/gDW/時間)を表現型相空間に示した。LOが表示してある。実験データ点は、無益回路および準最適増殖速度を特徴とする、領域1に限定されている。
【図13】グリセロール表現型相空間上方にグラフ化された増殖速度の三次元図。xおよびy軸は図12中と同じ変量を示している。z軸は細胞増殖速度、および凡例に表示された最適増殖速度に相当する。LO近傍にデータ点はない。
【図14】適応性進化実験に関する実験値を用いて酸素取込速度(OUR)に対してプロットしたグリセロール取込速度(GlUR)。進化の出発点が示してある(0日)。1回目の進化軌道(E1)の実験値は青色で表示してあり、2回目の進化軌道(E2)の実験値は緑色で表示してある。両実験とも、初期菌株は最適増殖速度を示す、LO上の同一終点に収束している。
【図15】グリセロールにおける適応性進化実験の時間による、単位または1時間当たりの増殖速度の変化。両実験とも適合性の増大および増殖速度の倍加を伴う同様の適応性の様相を明らかとしている。
【図16】適応性進化実験後のグリセロールにおける増殖の実験値によるグリセロール-酸素表現型相空間。全数値はLO上または近傍に密集している。データは炭素源滴定およびLO沿いに移動する量的効果を表している。
【図17】進化後の相表面の三次元図。全データ点はLO上または近傍に密集している。
【特許請求の範囲】
【請求項1】
細胞中の生化学反応ネットワークの最適な機能を達成する方法であって:
(a) 最適化方法を使用して生化学反応ネットワークの最適特性を計算する段階;
(b) 生化学反応ネットワークの反応のリスト作成を変更し、かつ最適特性を再計算する段階;
(c) 所望の最適な機能に達するまで段階(b)を繰り返し行う段階;
(d) 段階(c)の結果生じる生化学反応が含まれるように、細胞の遺伝子構成を構築する段階;
(e) 段階(d)にて構築した細胞を特定環境下の培養状態に置いて、細胞集団を得る段階;ならびに
(f) 段階(e)の細胞を、十分な期間、および段階(c)で定めた所望の最適な機能にその細胞を進化させる条件下で培養する段階を含み、
生化学反応ネットワークが包括的な生化学反応ネットワークを含む方法。
【請求項2】
生化学ネットワークが代謝ネットワークである、請求項1記載の方法。
【請求項3】
生化学ネットワークが調節ネットワークである、請求項1記載の方法。
【請求項4】
細胞が原核細胞である、請求項1記載の方法。
【請求項5】
細胞が真核細胞である、請求項1記載の方法。
【請求項6】
真核細胞が真菌細胞、動物細胞または細胞株である、請求項5記載の方法。
【請求項7】
段階(d)に、細胞中の1つまたはそれ以上の遺伝子の改変が含まれる、請求項1記載の方法。
【請求項8】
改変に、細胞への1つの遺伝子または複数の遺伝子導入が含まれる、請求項7記載の方法。
【請求項9】
改変に、細胞中の1つの内在性遺伝子または複数の内在性遺伝子の修飾が含まれる、請求項7記載の方法。
【請求項10】
生化学反応ネットワークに、実質的に全ての生化学反応ネットワークが含まれる、請求項1記載の方法。
【請求項11】
請求項1記載の方法により産生される濃縮細胞集団。
【請求項12】
請求項1記載の方法により産生される濃縮生体分子。
【請求項13】
以下の段階を含む、細胞における実質的に全ての生化学反応ネットワークの最適な機能を構築する方法:
(a) ネットワーク中の生化学反応を含むデータベースを提供する段階;
(b) 最適化方法を使用してネットワークの最適特性を計算する段階;
(c) ネットワーク中の反応を変更するための利用者による選択を受信し、かつ最適特性を再計算する段階;
(d) 所望の特性基準が満たされるまで最適化を繰り返し行う段階;
(e) 最適化情報の結果として生化学反応が含まれるように、細胞の遺伝子構成を構築するための最適化結果を表示する段階;
(f) 構築した細胞を、特定環境条件下で培養する段階;および
(g) 細胞が所望の最適性能にまで進化するように、十分な期間細胞を培養する段階。
【請求項14】
以下を含む、コンピュータ・システム:
少なくとも1つの生物体の生化学反応ネットワークを含むデータベース;および
少なくとも1つの生化学反応ネットワークを比較および最適化するための該生化学反応ネットワークについて2つまたはそれ以上の選択を受信できるユーザ・インターフェース。
【請求項15】
データベースが内部データベースである、請求項14記載のシステム。
【請求項16】
データベースが外部データベースである、請求項14記載のシステム。
【請求項17】
遺伝子配列がEST、全長配列、またはその組合せより選択される、請求項14記載のコンピュータ・システム。
【請求項18】
配列にアミノ酸配列が含まれる、請求項14記載のコンピュータ・システム。
【請求項19】
ユーザ・インターフェースに、生化学反応ネットワークに関する追加情報にアクセスするために利用者によって選択されうるリンクがさらに含まれる、請求項14記載のコンピュータ・システム。
【請求項20】
1つまたはそれ以上の生物体の生化学反応ネットワークを含むデータベースに関してコンピュータ可読プログラム・コードが媒体上で統合された、コンピュータ利用可能な媒体を含むコンピュータ・プログラム製品であって、該ネットワークは、多数のゲノム配列によりコードされ、少なくともそのうちの幾つかは、1つまたはそれ以上の生物体の各ゲノム上に1つまたはそれ以上の連続配列に沿って位置する読み枠を意味しており、コンピュータ・プログラム製品は、コンピューティング・システム内で以下の段階を達成するためのコンピュータ可読プログラム・コードを含む、コンピュータ・プログラム製品:
該ネットワークの所望の最適特性の選択を受信するためのインターフェースを提供する段階;
該ネットワークの最適特性を決定する段階;および
該決定の結果を表示する段階。
【請求項21】
以下の段階を含む、生細胞における包括的な生化学反応ネットワークの所望の最適な機能を達成するための方法:
(a) ネットワーク中の生化学反応をリストにしてコンピュータに表示する段階;
(b) 最適化方法を使用してネットワークの最適特性を計算する段階;
(c) ネットワーク中の反応リストを変更し、かつ最適特性を再計算する段階;および
(d) 所望の最適な機能に達するまで段階(c)を繰り返し行う段階。
【請求項22】
さらに以下の段階を含む、請求項21記載の方法:
(e) 段階(d)の結果生じる生化学反応が含まれるように、細胞の遺伝子構成を構築する段階;
(f) 段階(e)にて構築した細胞を特定環境下の培養状態に置く段階;ならびに
(g) 段階(f)の細胞を、十分な期間、および段階(d)にて定めた所望の最適な機能にその細胞を進化させる条件下で培養する段階。
【請求項1】
細胞中の生化学反応ネットワークの最適な機能を達成する方法であって:
(a) 最適化方法を使用して生化学反応ネットワークの最適特性を計算する段階;
(b) 生化学反応ネットワークの反応のリスト作成を変更し、かつ最適特性を再計算する段階;
(c) 所望の最適な機能に達するまで段階(b)を繰り返し行う段階;
(d) 段階(c)の結果生じる生化学反応が含まれるように、細胞の遺伝子構成を構築する段階;
(e) 段階(d)にて構築した細胞を特定環境下の培養状態に置いて、細胞集団を得る段階;ならびに
(f) 段階(e)の細胞を、十分な期間、および段階(c)で定めた所望の最適な機能にその細胞を進化させる条件下で培養する段階を含み、
生化学反応ネットワークが包括的な生化学反応ネットワークを含む方法。
【請求項2】
生化学ネットワークが代謝ネットワークである、請求項1記載の方法。
【請求項3】
生化学ネットワークが調節ネットワークである、請求項1記載の方法。
【請求項4】
細胞が原核細胞である、請求項1記載の方法。
【請求項5】
細胞が真核細胞である、請求項1記載の方法。
【請求項6】
真核細胞が真菌細胞、動物細胞または細胞株である、請求項5記載の方法。
【請求項7】
段階(d)に、細胞中の1つまたはそれ以上の遺伝子の改変が含まれる、請求項1記載の方法。
【請求項8】
改変に、細胞への1つの遺伝子または複数の遺伝子導入が含まれる、請求項7記載の方法。
【請求項9】
改変に、細胞中の1つの内在性遺伝子または複数の内在性遺伝子の修飾が含まれる、請求項7記載の方法。
【請求項10】
生化学反応ネットワークに、実質的に全ての生化学反応ネットワークが含まれる、請求項1記載の方法。
【請求項11】
請求項1記載の方法により産生される濃縮細胞集団。
【請求項12】
請求項1記載の方法により産生される濃縮生体分子。
【請求項13】
以下の段階を含む、細胞における実質的に全ての生化学反応ネットワークの最適な機能を構築する方法:
(a) ネットワーク中の生化学反応を含むデータベースを提供する段階;
(b) 最適化方法を使用してネットワークの最適特性を計算する段階;
(c) ネットワーク中の反応を変更するための利用者による選択を受信し、かつ最適特性を再計算する段階;
(d) 所望の特性基準が満たされるまで最適化を繰り返し行う段階;
(e) 最適化情報の結果として生化学反応が含まれるように、細胞の遺伝子構成を構築するための最適化結果を表示する段階;
(f) 構築した細胞を、特定環境条件下で培養する段階;および
(g) 細胞が所望の最適性能にまで進化するように、十分な期間細胞を培養する段階。
【請求項14】
以下を含む、コンピュータ・システム:
少なくとも1つの生物体の生化学反応ネットワークを含むデータベース;および
少なくとも1つの生化学反応ネットワークを比較および最適化するための該生化学反応ネットワークについて2つまたはそれ以上の選択を受信できるユーザ・インターフェース。
【請求項15】
データベースが内部データベースである、請求項14記載のシステム。
【請求項16】
データベースが外部データベースである、請求項14記載のシステム。
【請求項17】
遺伝子配列がEST、全長配列、またはその組合せより選択される、請求項14記載のコンピュータ・システム。
【請求項18】
配列にアミノ酸配列が含まれる、請求項14記載のコンピュータ・システム。
【請求項19】
ユーザ・インターフェースに、生化学反応ネットワークに関する追加情報にアクセスするために利用者によって選択されうるリンクがさらに含まれる、請求項14記載のコンピュータ・システム。
【請求項20】
1つまたはそれ以上の生物体の生化学反応ネットワークを含むデータベースに関してコンピュータ可読プログラム・コードが媒体上で統合された、コンピュータ利用可能な媒体を含むコンピュータ・プログラム製品であって、該ネットワークは、多数のゲノム配列によりコードされ、少なくともそのうちの幾つかは、1つまたはそれ以上の生物体の各ゲノム上に1つまたはそれ以上の連続配列に沿って位置する読み枠を意味しており、コンピュータ・プログラム製品は、コンピューティング・システム内で以下の段階を達成するためのコンピュータ可読プログラム・コードを含む、コンピュータ・プログラム製品:
該ネットワークの所望の最適特性の選択を受信するためのインターフェースを提供する段階;
該ネットワークの最適特性を決定する段階;および
該決定の結果を表示する段階。
【請求項21】
以下の段階を含む、生細胞における包括的な生化学反応ネットワークの所望の最適な機能を達成するための方法:
(a) ネットワーク中の生化学反応をリストにしてコンピュータに表示する段階;
(b) 最適化方法を使用してネットワークの最適特性を計算する段階;
(c) ネットワーク中の反応リストを変更し、かつ最適特性を再計算する段階;および
(d) 所望の最適な機能に達するまで段階(c)を繰り返し行う段階。
【請求項22】
さらに以下の段階を含む、請求項21記載の方法:
(e) 段階(d)の結果生じる生化学反応が含まれるように、細胞の遺伝子構成を構築する段階;
(f) 段階(e)にて構築した細胞を特定環境下の培養状態に置く段階;ならびに
(g) 段階(f)の細胞を、十分な期間、および段階(d)にて定めた所望の最適な機能にその細胞を進化させる条件下で培養する段階。
【図1】

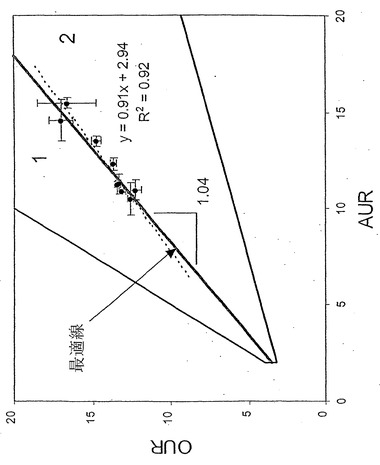
【図2】
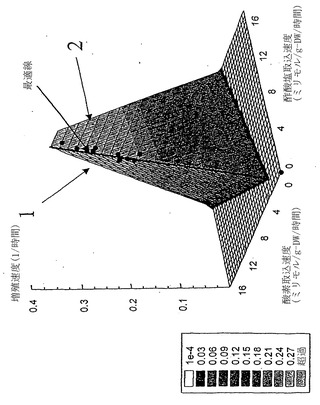
【図3】
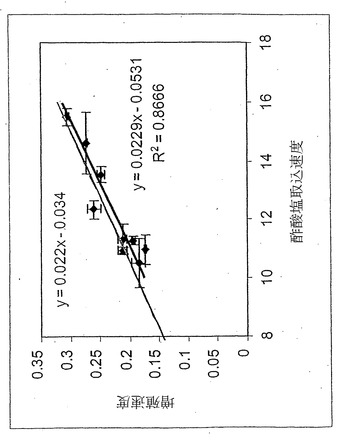
【図4】
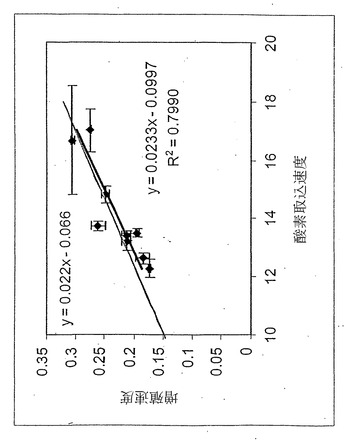
【図5】
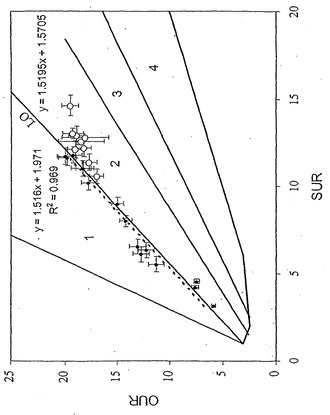
【図6】
【図7】
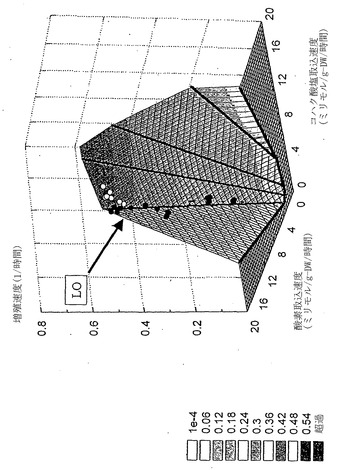
【図8】
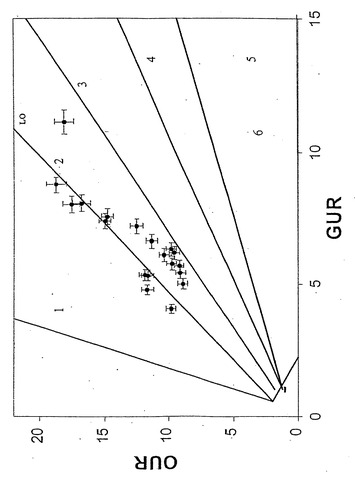
【図9】
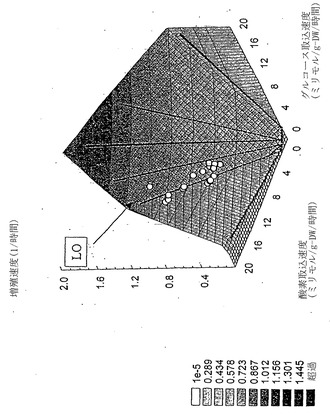
【図10】
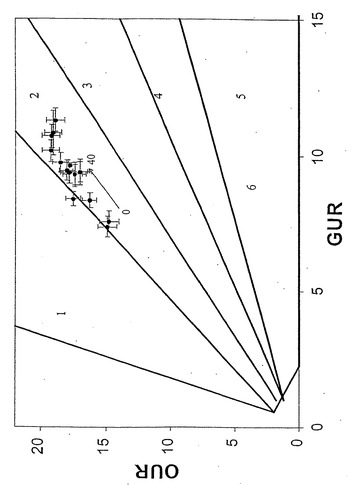
【図11】
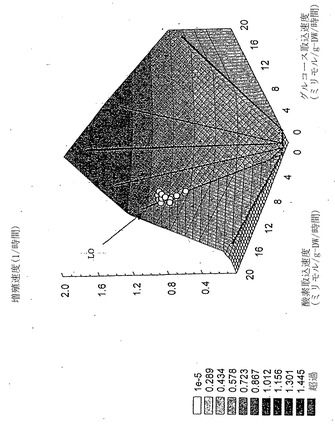
【図12】
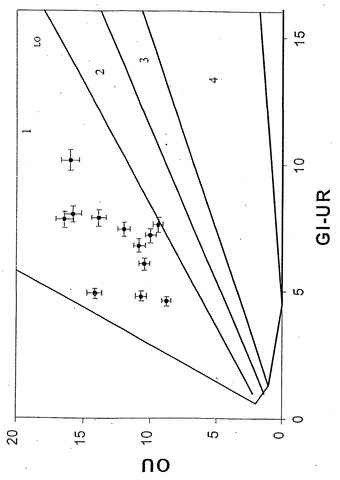
【図13】
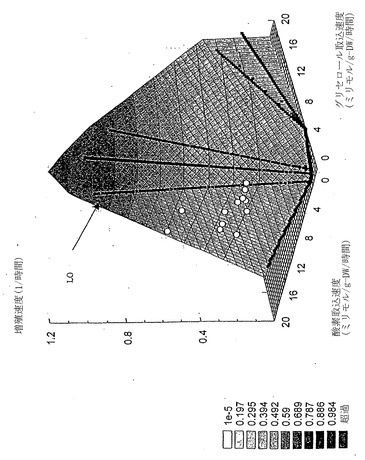
【図14】
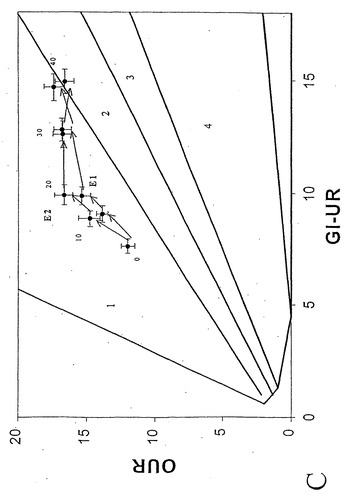
【図15】
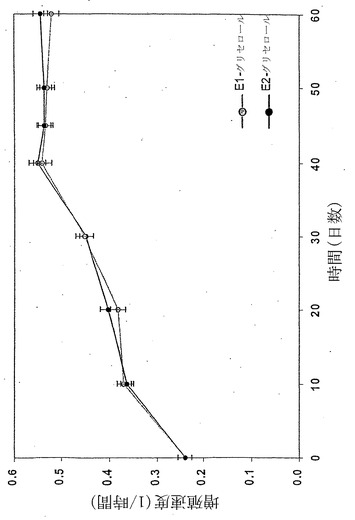
【図16】
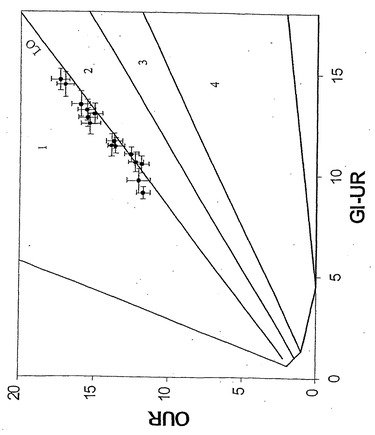
【図17】
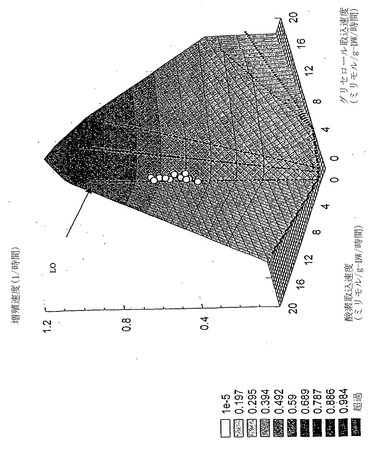
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公開番号】特開2008−178415(P2008−178415A)
【公開日】平成20年8月7日(2008.8.7)
【国際特許分類】
【出願番号】特願2008−41411(P2008−41411)
【出願日】平成20年2月22日(2008.2.22)
【分割の表示】特願2002−561050(P2002−561050)の分割
【原出願日】平成14年1月31日(2002.1.31)
【出願人】(502212660)ザ リージェンツ オブ ザ ユニバーシティー オブ カリフォルニア (1)
【Fターム(参考)】
【公開日】平成20年8月7日(2008.8.7)
【国際特許分類】
【出願日】平成20年2月22日(2008.2.22)
【分割の表示】特願2002−561050(P2002−561050)の分割
【原出願日】平成14年1月31日(2002.1.31)
【出願人】(502212660)ザ リージェンツ オブ ザ ユニバーシティー オブ カリフォルニア (1)
【Fターム(参考)】
[ Back to top ]