
生存可能な無毒性グラム陰性細菌
本発明は、無毒性グラム陰性細菌を提供する。特に、本発明は、外膜内にリポ多糖(LPS、内毒素)を実質的に欠く生存可能なグラム陰性細菌(例えば、大腸菌)を提供する。本発明はさらに、生存可能な無毒性グラム陰性細菌を作製する方法、およびその使用を提供する。本発明はまた、免疫応答を誘導するための、および治療剤を研究かつ開発するための組成物ならびに方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、全体として本明細書に参照により組み入れられた、2006年1月19日に出願された米国特許仮出願第60/760,314号の優先権を主張する。本発明は、National Institutes of Healthによって授与された認可番号GM53609による政府支援でなされた。
【0002】
発明の分野
本発明は、無毒性グラム陰性細菌を提供する。特に、本発明は、外膜内にリポ多糖(LPS、内毒素)を実質的に欠く、生存可能なグラム陰性細菌(例えば、大腸菌(E. coli))を提供する。本発明はさらに、生存可能な無毒性グラム陰性細菌を作製する方法およびその使用を提供する。本発明はまた、免疫応答を誘導するための、および治療剤を研究かつ開発するための組成物ならびに方法を提供する。
【背景技術】
【0003】
発明の背景
リポ多糖(LPS、内毒素)は、グラム陰性細菌の主要抗原である。LPSは、N,O-アシルβ-1,6-D-グルコサミン1,4'-二リン酸として構造的に定義される保存された疎水性領域であるリピドAに共有結合した抗原性の可変サイズの糖鎖からなる糖リン脂質である。LPSの毒性は、リピドAにより哺乳動物免疫系のB細胞およびマクロファージとの相互作用を通して発現され、それは、宿主にとって致命的結果を生じうる、炎症性サイトカイン、主にTNF、の分泌に導く過程である。リピドAはまた、「インビボ」でマウスCD4+およびCD8+ T細胞だけでなく、「インビトロ」でヒトTリンパ球(Th-1)も活性化し、それは宿主の免疫系がLPSの可変サイズの糖鎖に対して特定の既往性IgG抗体応答を開始するのを可能にする性質である。これらに基づいて、LPSは最近、「インビボ」でのT細胞依存性抗原として認識されている。
【0004】
毒性を完全に発現するために、LPSは、リピドAを形成する糖リン脂質単量体の数個の単位の会合を通してのその超分子構造を保持しなければならない。この分子の立体構造的再編成はまた、免疫原性特性の完全な発現のための基礎となる。
【0005】
敗血症および敗血性ショックは、細菌により、および上記病態の原因である細菌によって産生される内毒素であるLPSにより、引き起こされるはっきりした臨床症状である。
【0006】
敗血症および敗血性ショックの臨床的徴候は、存在する内毒素の量、および疾患過程に経過した時間に依存して異なる。感染の最も早い臨床的徴候は、熱、軽度の鬱病、および食欲不振でありうる。疾患過程をさらに進むと、患者は、心拍数の増加、弱い脈圧、脱水症、歯肉の黒ずみ、冷たい足および耳、正常より低い体温、呼吸数の増加、または下痢を含む、より明らかなショックの徴候を示す。いったん患者が内毒素性ショックの徴候を示したならば、それは緊急事態とみなされるべきであり、すぐに医師に連絡すべきである。
【0007】
抗生物質および他の治療手段の慎重な使用にもかかわらず、内毒素関連障害による死亡率は、深刻な問題のままである。細菌の抗生物質抵抗性、基礎疾患過程の重症度、および支持療法の不適切な実施が、部分的に従来の処置の失敗の原因となっている。必要とされることは、内毒素関連障害を引き起こすグラム陰性細菌の理解の向上である。さらに、内毒素関連障害についての処置の改善が必要とされる。
【発明の開示】
【0008】
発明の概要
本発明は、無毒性グラム陰性細菌を提供する。特に、本発明は、外膜内にリポ多糖(LPS、内毒素)を実質的に欠く、生存可能なグラム陰性細菌(例えば、大腸菌(E. coli))を提供する。本発明はさらに、生存可能な無毒性グラム陰性細菌を作製する方法およびその使用を提供する。本発明はまた、免疫応答を誘導するための、および治療剤を研究かつ開発するための組成物ならびに方法を提供する。
【0009】
本発明の態様は、LPSを欠くグラム陰性細菌(例えば、大腸菌)を用いる広範囲の方法および組成物を提供する。例示的な態様は、下の発明の概要、発明の詳細な説明、および下の実施例セクションに記載されている。本発明はこれらの例示的な態様に限定されない。LPSを欠くグラム陰性細菌は、任意の機構により作製されうる。そのような細菌を作製するための多様な種類の異なる機構は本明細書に記載されている。例えば、いくつかの態様において、KDO合成に関与する遺伝子が突然変異されている(機能性タンパク質の発現を低減させる、または排除するために)。いくつかの態様において、KDOのリピドIVAとの会合に関与する遺伝子が突然変異されている。いくつかの態様において、リピドIVA合成に関与する遺伝子が突然変異されている。いくつかの態様において、LPS産生または提示に関与する他の遺伝子が突然変異されている。本発明は遺伝子突然変異に限定されない。いくつかの態様において、発現は、RNA干渉または他の技術を用いて変化させられる。いくつかの態様において、タンパク質機能は、インヒビター(例えば、合成もしくは天然の競合的または非競合的リガンド、抗体など)を供給することにより変化させられる。いくつかの態様において、改変細菌は、LPS状態に影響するようになされた変化のために、健康、増殖などの維持に有用な栄養物、他の改変、または他の成分をさらに与えられる。本発明の態様は、他に規定がない限り、これらの機構に限定されない。本発明は、LPSを欠く細菌が生存可能であること、様々な経路を通して作製されうること、および様々な設定で使用を見出すことを実証する。
【0010】
LPS層は、グラム陰性細菌の外膜の形および機能の両方にとって必須である。グラム陰性病原性の主要な介在物質であることに加えて、少なくともKDO2-リピドA[2-ケト3-デオキシ-D-マンノ-オクツロソン酸(KDO)]からなるLPS(内毒素)構造は、大腸菌において持続的増殖のために必要な最小構造として長年認識されていた。
【0011】
いくつかの態様において、本発明は、ヒトにおけるLPS誘発性敗血症の既知のアンタゴニストである内毒素的に不活性のLPS前駆体リピドIVAをもっぱら産生するだけにもかかわらず、KDOを欠く生存可能なグラム陰性細菌株を提供する。いくつかの態様において、本発明は、D-アラビノース5-リン酸イソメラーゼ(API)発現を欠く生存可能なグラム陰性細菌を提供する。いくつかの態様において、生存可能なグラム陰性細菌は、その菌株がKDOを実質的に含まないように突然変異を含む。いくつかの態様において、突然変異には、KDO合成または改変に関与する1つまたは複数の遺伝子における1つまたは複数の突然変異が挙げられる。いくつかの態様において、生存可能なグラム陰性細菌は突然変異を含み、その突然変異がLPS生合成経路におけるKDO2とリピドIVAの間の会合を妨げて、リピドIVAのみが外膜へ輸送される。いくつかの態様において、KDOにおける1つもしくは複数の突然変異、またはLPS生合成経路における1つもしくは複数の突然変異には、限定されるわけではないが、遺伝子gutQ、kdsD(yrbH)、kdsA、kdsB、waaA、msbA、およびyhjDにおける突然変異が挙げられる。いくつかの態様において、菌株は、KDOタンパク質の合成を欠く、または実質的に欠く。いくつかの態様において、生存可能なグラム陰性細菌の外膜はリピドIVaを発現する。いくつかの態様において、グラム陰性細菌は大腸菌である。
【0012】
特定の態様において、本発明は、生存可能なグラム陰性細菌からリピドIVaを抽出する段階を含む、リピドIVaを作製する方法を提供する。
【0013】
特定の態様において、本発明は、グラム陰性細菌から単離されたリピドIVaを含む組成物を内毒素関連障害をもつ被験体に投与する段階を含む、内毒素関連障害を処置する方法を提供する。
【0014】
特定の態様において、本発明は、グラム陰性細菌に対して免疫応答を誘導するための外膜ワクチンまたは他の組成物を提供し、この組成物は、本発明の生存可能なグラム陰性細菌の外膜を含む。そのような組成物は、研究、薬物スクリーニング、および治療的設定において免疫応答を誘導するために用いられうる。
【0015】
特定の態様において、本発明は、グラム陰性細菌から単離されたリピドIVaを含むアジュバントを提供する。
【0016】
特定の態様において、本発明は、gutQ、kdsD(yrbH)、kdsA、kdsB、waaA、msbA、および/もしくはyhjDの1つもしくは複数の遺伝子の発現、または外膜LPS提示に関連した任意の他の生合成、プロセシング、もしくは輸送遺伝子の発現を欠く生存可能なグラム陰性細菌を提供する。本発明の細菌、またはその部分(例えば、膜画分)は、研究および治療的適用に使用を見出す。
【0017】
定義
本発明の理解を促進するために、いくつかの用語が下に定義されている。
【0018】
本明細書に用いられる場合、「被験体」および「患者」という用語は、イヌ、ネコ、鳥、家畜、および好ましくはヒトのような哺乳動物などの任意の動物を指す。
【0019】
本明細書に用いられる場合、「LPS関連障害」、「内毒素に関係した状態」、「内毒素関係障害」、「内毒素関連障害」、「敗血症」、「敗血症関連障害」という用語、または同類語は、LPSに関連した任意の状態、例えば、菌血症、または血流への、もしくは胃腸外粘膜表面(例えば、肺)上へのリポ多糖の導入に関係した状態を記載する。そのような障害には、限定されるわけではないが、内毒素関連ショック、内毒素関連播種性血管内凝固、内毒素関連貧血、内毒素関連血小板減少症、内毒素関連成人呼吸窮迫症候群、内毒素関連腎不全、内毒素関連肝臓疾患または肝炎、グラム陰性感染に起因する全身性免疫応答症候群(SIRS)、グラム陰性新生児敗血症、グラム陰性髄膜炎、グラム陰性肺炎、グラム陰性感染に起因する好中球減少症および/または白血球減少症、血行動態ショック、ならびに内毒素関連発熱(pyresis)が挙げられる。
【0020】
「生存可能な無毒性グラム陰性細菌」という用語は、LPSを実質的に含まない外膜を含む生存可能なグラム陰性細菌株を指す。
【0021】
「細胞」および「宿主細胞」および「組換え宿主細胞」という用語は、本明細書で交換可能に用いられるが、ベクター、典型的には発現ベクターで形質転換される能力がある、または形質転換された細胞を指す。本明細書に用いられる宿主細胞は、好ましくは、グラム陰性細菌である。そのような用語が特定の対象細胞だけでなく、そのような細胞の子孫または潜在的子孫も指すことは理解される。特定の改変が突然変異かまたは環境的影響のいずれかにより後世に起こりうるため、そのような子孫は、実際、親細胞と同一でない可能性があるが、本明細書に用いられる場合の用語の範囲内にさらに含まれる。
【0022】
「培地」という用語は、当技術分野において認識されており、一般的には、生細胞の培養に用いられる任意の物質または調製物を指す。
【0023】
例えば、グラム陰性細菌の菌株からリピドIVaを得るという文脈において用いられる場合の「に由来する」という用語は、細菌またはタンパク質から得られうるリピドIVaを指し、タンパク質の断片または部分を含むことが意図される。
【0024】
遺伝子または遺伝子発現に関して、本明細書に用いられる場合の「欠陥のある」という用語は、遺伝子が野生型遺伝子ではないこと、ならびに生物体が野生型の遺伝子型および/または野生型の表現型をもたないことを意味する。欠陥のある遺伝子、遺伝子型、または表現型は、その正常な発現が乱されるもしくは失われるような、その遺伝子における、または遺伝子の発現(例えば、転写または転写後の)を制御する遺伝子の、突然変異の結果である可能性がある。「乱された遺伝子発現」は、完全な阻害、および野生型遺伝子発現より低い遺伝子発現の減少(例えば、漏出突然変異におけるような)の両方を含むことが意図される。
【0025】
「グラム陰性細菌」という用語は、当技術分野において認識されており、一般的には、グラム染色(例えば、クリスタルバイオレットとヨウ素の間の着色複合体の沈着)を保持しない細菌を指す。例示的なグラム染色において、細胞を、まず、加熱によりスライドに固定し、すべての細菌(すなわち、グラム陰性およびグラム陽性の両方)により取り込まれる塩基性色素(例えば、クリスタルバイオレット)で染色する。スライドを、その後、染色を定着させるためにヨウ素-KI混合物で処理し、アセトンまたはアルコールで洗浄し、最後に、異なる色の淡染色性色素(例えば、サフラニン)で対比染色する。グラム陽性生物体は、最初のバイオレット染色を保持するが、グラム陰性生物体は、有機溶媒により脱色され、このゆえに、対比染色を示す。例示的なグラム陰性細菌および細胞系には、限定されるわけではないが、エシェリキア種(Escherichia spp.)、シゲラ種(Shigella spp.)、サルモネラ種(Salmonella spp.)、カンピロバクター種(Campylobacter spp.)、ナイセリア種(Neisseria spp.)、ヘモフィルス種(Haemophilus spp.)、アエロモナス種(Aeromonas spp.)、フランシセラ種(Francisella spp.)、エルシニア種(Yersinia spp.)、クレブシエラ種(Klebsiella spp.)、ボルデテラ種(Bordetella spp.)、レジオネラ種(Legionella spp.)、コリネバクテリア種(Corynebacteria spp.)、シトロバクター種(Citrobacter spp.)、クラミジア種(Chlamydia spp.)、ブルセラ種(Brucella spp.)、シュードモナス種(Pseudomonas spp.)、ヘリコバクター種(Helicobacter spp.)、およびビブリオ種(Vibrio spp.)が挙げられる。
【0026】
本明細書に用いられる場合、「突然変異体グラム陰性細菌」、「LPS突然変異体グラム陰性細菌」、「kdsDおよびgutQ突然変異体グラム陰性細菌」、「API突然変異体グラム陰性細菌」という用語、または同類語は、例えば、gutQ、kdsD、kdsA、kdsB、waaA、msbA、yhjD遺伝子、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の1つまたは複数において1回または複数回、突然変異されており、それにより、LPSタンパク質発現を実質的に欠く外膜を生成する、本発明のグラム陰性細菌を含む。
【0027】
「分子の免疫原性部分」は、被験体においてその分子に対して免疫反応を誘発する能力がある分子の部分を指す。
【0028】
LPSまたはリピドIVa分子に適用される場合の「単離される」という用語は、他の細菌成分から、特に外膜から、単離されている(例えば、部分的または完全な単離)LPSまたはリピドIVaを指す。
【0029】
本明細書に用いられる場合、配列(例えば、タンパク質のアミノ酸配列、遺伝子の核酸配列)に関して用いられる時の「部分」という用語は、参照配列の任意の量(例えば、アミノ酸配列または核酸配列の0.001%、0.1%、1%、10%、30%、50%、75%、80%、85%、90%、95%、98%、99.999%)を表す。
【0030】
本明細書に用いられる場合の「調節」という用語は、上方制御(すなわち、活性化または刺激(例えば、アゴナイズする(agonizing)、または増強することにより))および下方制御(すなわち、阻害または抑制(例えば、アンタゴナイズする(antagonizing)、減少させる、または阻害することにより))の両方を指す。「誘導性」という用語は、特に、恒常的ではなく、刺激(例えば、温度、重金属、または他の培地添加物)に応答して起こる遺伝子発現を指す。
【0031】
「非ヒト動物」という用語は、処置されうる、または本発明を試験するのに用いられうる任意の動物を含み、非ヒト霊長類、齧歯類、ヒツジ、イヌ、ウシ、ブタ、ニワトリなどの哺乳動物、および両生類、爬虫類などが挙げられる。好ましい非ヒト動物は、霊長類ファミリーまたは齧歯類ファミリー(例えば、ラットおよびマウス)から選択される。
【0032】
「核酸」という用語は、デオキシリボ核酸(DNA)、および適切な場合にはリボ核酸(RNA)、などのポリヌクレオチドまたはオリゴヌクレオチドを指す。その用語がまた、等価物として、ヌクレオチド類似体から生成したRNAまたはDNAのいずれかの類似体、ならびに記載されることになっている態様に応じて適切に、一本鎖(センスまたはアンチセンス)および二本鎖ポリヌクレオチドを含むことは理解されているはずである。
【0033】
「薬学的に許容される」という用語は、生物学的にまたはその他の点で望ましくないことはない物質、すなわち、その物質は、任意の望ましくない生物学的効果を引き起こすこと、またはそれが含まれる薬学的組成物の他の化合物のいずれかと有害な様式で相互作用することなしに、選択された化合物と共に個体へ投与されうる。
【0034】
「発熱性の」または「発熱性」は、被験体に投与された場合、発熱または発熱応答を誘導する化合物の能力を指す。そのような発熱応答は、一般的に、宿主炎症性サイトカインIL-1、IL-6、および/またはTNF-αにより媒介され、その分泌は、例えば、LPSにより、誘導される。
【0035】
「低下した発熱性」をもつ物質、または「発熱性の低下した誘導体」は、対応物質より低い発熱活性、例えば、対応物質に対して約80%未満の発熱性、好ましくは、約70%未満の発熱性、より好ましくは約60%未満の発熱性、より好ましくは約50°未満の発熱性、より好ましくは約40%未満の発熱性、およびよりいっそう好ましくは約30%未満の発熱性を有する物質を指す。換言すれば、低下した発熱性をもつ物質は、本明細書に記載された、または当技術分野において公知のアッセイ法のいずれかにより測定される場合、対応する物質より少なくとも約20%、30%、40%、50%、60%、または70%低い発熱性である。
【0036】
「実質的に低下した発熱性」または「実質的に低下した発熱性誘導体」は、それが野生型物質に対して20%未満の発熱性、好ましくは、野生型物質に対して、10%未満の発熱性、好ましくは1%未満の発熱性、好ましくは10-1%未満の発熱性、好ましくは10-2%未満の発熱性、好ましくは10-3%未満の発熱性、好ましくは10-4%未満の発熱性、好ましくは10-5%未満の発熱性、および最も好ましくは10-6%未満の発熱性を有するように変化している物質(例えば、生存可能な無毒性グラム陰性細菌により産生される)を指す。換言すれば、実質的に低下した発熱性をもつ物質は、本明細書に記載された、または当技術分野において公知のアッセイ法のいずれかにより測定される場合、対応する未変化物質に対して、少なくとも約90%低い、99%低い、1/10倍、約1/100倍、約1/1000倍、少なくとも約1/1000倍、少なくとも約1/10000倍、少なくとも約1/1000000倍の発熱性である。
【0037】
本明細書に用いられる場合、「トランスフェクション」という用語は、核酸媒介性遺伝子移入によるレシピエント細胞への核酸の導入(例えば、発現ベクターを介して)を意味する。本明細書に用いられる場合、「形質転換」は、細胞の遺伝子型が外因性DNAまたはRNAの細胞取り込みの結果として変化している過程を指す。例証的な態様において、形質転換細胞は、kdsDおよびgutQ遺伝子の1つまたは複数の突然変異体型を発現するものである。形質転換細胞はまた、gutQ、kdsD、kdsA、kdsB、waaA、msbA、ynjD遺伝子、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の発現を干渉する核酸を発現するものであることができる。
【0038】
本明細書に用いられる場合、「トランスジーン」は、細胞へ導入されている核酸(例えば、突然変異体kdsD、gutQ、kdsA、kdsB、waaA、msbA、ynjD遺伝子、または任意の他の生合成、プロセシング、もしくは輸送遺伝子、またはそれらのアンチセンス転写産物)を意味する。トランスジーンは、それが導入されているトランスジェニック動物もしくは細胞に対して、部分的にもしくは完全に異種性、すなわち、外来性でありうる、またはそれが導入されている生物体もしくは細胞の内因性遺伝子に相同性でありうるが、それが挿入されている細胞のゲノムを変化させるように、動物もしくは細胞のゲノムへ挿入されるように設計される、もしくは挿入される。トランスジーンはまた、エピソームの形をとって細胞に存在しうる。
【0039】
本明細書に用いられる場合、状態または疾患について被験体を「処置する」という用語は、状態または疾患の少なくとも1つの症状を治すこと、および寛解させることを含むよう意図される。
【0040】
「ベクター」という用語は、それが連結している別の核酸を輸送する能力がある核酸分子を指す。機能的に連結している遺伝子の発現を方向づける能力があるベクターは、本明細書では「発現ベクター」と呼ばれる。本明細書に用いられる場合、「発現系」という用語は、mRNAが転写されうる、および/またはmRNAがタンパク質、構造RNA、もしくは他の細胞成分へ翻訳されうる条件下での発現ベクターを指す。発現系は、インビトロ発現系であり、市販されているか、もしくは当技術分野公知の技術に従って容易に作製される、または発現ベクターを含む真核細胞もしくは原核細胞などのインビボ発現系でありうる。一般的に、組換えDNA技術における有用な発現ベクターは、しばしば、「プラスミド」の形をとり、そのプラスミドは、一般的に、それらのベクター型において、染色体に結合していない環状二本鎖DNAループを指す。本明細書において、「プラスミド」および「ベクター」は、プラスミドが最も一般的に用いられるベクターの形であるため、交換可能に用いられる。しかしながら、本発明は、等価の機能を果たし、かつ当技術分野において周知である、またはこの点に関して後で当技術分野において知られるようになる、そのような他の型の発現ベクター(例えば、コスミド、ファージミド、およびバクテリオファージベクター)を含むことを意図される。
【0041】
発明の詳細な説明
本発明は、無毒性グラム陰性細菌を提供する。特に、本発明は、外膜内にリポ多糖(LPS、内毒素)を実質的に欠く、生存可能なグラム陰性細菌(例えば、大腸菌)を提供する。本発明はさらに、生存可能な無毒性グラム陰性細菌を作製する方法およびその使用を提供する。本発明はまた、免疫応答を誘導するための、および治療剤を研究かつ開発するための組成物ならびに方法を提供する。
【0042】
グラム陰性細菌は、ペプチドグリカンを囲む非対称脂質二重層である、外膜(OM)を有する。OM内層は、主として様々なグリセロリン脂質で構成されるが、外層は、主に、固有の両親媒性高分子である、リポ多糖(LPS)を含む。大腸菌および他の密接に関連した腸内細菌において、OM総重量の〜30%を占める、総細胞表面積の75%近くを網羅する細胞あたり〜106個のLPS分子がある(例えば、S.M. Galloway, C.R. Raetz, J. Biol. Chem. 265, 6394 (1990); L. Leive, Ann. N.Y. Acad. Sci. 235, 109 (1974); H. Nikaido, Escherichia coli and Salmonella typhimurium: cellular and molecular biology, F.C. Neidhardt, Ed. (American Society for Microbiology, Washington, D.C., 1987), vol. 1, pp. 29-47参照;それぞれは全体として参照により本明細書に組み入れられている)。細菌細胞と水性環境の間の界面での露出された位置は、主要なOM関連表面抗原としてLPSを提示する。LPSは、宿主免疫応答に関連した、多様な範囲の病理学的および生理学的活性に関与している(例えば、A. Wiese, et al., Biol. Chem. 380, 767 (1999); H. Heine, et al., Mol. Biotechnol. 19, 279 (2001)参照;それぞれは全体として参照により本明細書に組み入れられている)。LPSは、グラム陰性病原性および全身性炎症の介在物質として認識された免疫刺激性/炎症性分子であり、それとして、内毒素という用語は、しばしば、LPSと交換可能に用いられる。LPS層は、グラム陰性細菌のOMの形および機能の両方に必須である。従って、グラム陰性病原性におけるキープレーヤーであることに加えて、LPSはまた、細菌の生存の重要な決定因子である。
【0043】
様々なグラム陰性細菌のLPSは、以下の3つの領域へ概念的に分けられる一般的な構造的構築に従う:OMに埋め込まれたリピドA、オリゴ糖コア、続いて、腸内細菌科におけるn回の繰り返し単位からなるO特異的親水性多糖鎖、または髄膜炎菌(Neisseria meningitis)、淋菌(N. gonorrhoeae)、インフルエンザ菌(Haemophilus influenzae)、百日咳菌(Bordetella pertussis)、およびクラミジア種(Chlamydia spp.)などのヒト粘膜病原体を含む特定の細菌における短い分岐オリゴ糖。リピドAはグラム陰性細菌属の間で最も保存されているLPSドメインであり、かつ宿主内での生物活性の原因である構造的構成要素であることは、LPSの内毒素原理を表している。腸内細菌において、リピドAは、エステル-(3,3')結合またはアミド-(2,2')結合で4つの(R)-3-ヒドロキシ-ミリスチン酸でアシル化されるβ-1,6-結合型D-グルコサミン二糖バックボーンからなる。大腸菌野生型株の成熟リピドA分子は、典型的には、非還元グルコサミンの(R)-3-ヒドロキシミリストイル基に付着した、2つの追加のアシル鎖、主としてラウレートおよびミリステートを含み、リピドAの特徴的なアシルオキシアシル単位を形成する。オリゴ糖コアは、リピドAを超可変多糖鎖に接続して、内側および外側のオリゴ糖コア領域へさらに分けられる。外側コアはあまり保存されておらず、糖組成およびグリコシド結合の両方の点で異なっているが、グラム陰性細菌の大部分は、少なくとも1つの2-ケト3-ヒドロキシ-D-マンノ-オクツロソン酸(KDO)分子を含む内側コアを産生する。
【0044】
KDOは、ほとんど全部のLPS構造に見出される保存された残基である、LPSの必須構成要素である(例えば、O. Holst, Trends Glycosci. Glycotechnol. 14,87 (2002)参照;全体として参照により本明細書に組み入れられている)。大腸菌の増殖に必要とされる最小LPS構造は、リピドAに付着した2つのKDO残基(KDO2-リピドAまたはRe内毒素)であり(例えば、C.R. Raetz, C. Whitfield, Annu. Rev. Biochem. 71, 635 (2002); S. Gronow, H. Brade, J. Endotoxin Res. 7, 3(2001)参照;それぞれは全体として参照により本明細書に組み入れられている)、細菌細胞の完全性および生存能力の維持におけるKDOの重要性を強調している。L-APIは、大腸菌K-12におけるkdsD遺伝子によりコードされる(例えば、Meredith, T.C. & Woodard, R.W. (2003) J Biol Chem 278, 32771-7参照;全体として参照により本明細書に組み入れられている)。
【0045】
LPS構造内のKDOの遍在性は、その生合成の解明を促進させた。経路は、酵素d-アラビノース5-リン酸(A5P)イソメラーゼ(API)により惹起され、その酵素は、ペントース経路中間体D-リブロース5-リン酸をA5Pへ変換する。その後、A5Pは、ホスホエノールピルビン酸と縮合してKdo 8-リン酸(Kdo8P)(KdsA)を形成し、Kdo(KdsC)へ加水分解され、最後にCMP-KdoからアクセプターのリピドIVA(WaaA)へ転移される前に、糖ヌクレオチドCMP-Kdo(KdsB)として活性化される(図11)。後期アシルトランスフェラーゼLpxLおよびLpxMは、次に、脂肪酸、ラウレートおよびミリステートを、それぞれ、Kdo2-リピドIVAへ転移し、ヘキサアシル化Kdo-リピドAの特徴的なアシルオキシアシル単位を形成する。大腸菌K-12において、2つのAPI遺伝子(kdsDおよびgutQ)がある。
【0046】
相同性検索に基づいて他のAPIが大腸菌に存在する可能性があることが推測された。特に、グルシトールオペロンgutQの最後のオープンリーディングは、kdsD(以前は、yrbH)と有意な相同性(45%同一性)を有する。G-APIは、7つの収束的に転写される遺伝子を含むgutAEBDMRQオペロンの最終遺伝子産物である(図10)。表1に示されているように、gutQおよびkdsDは、類似した生化学的性質を共有する。
【0047】
(表1)kdsDおよびgutQの生化学的性質

aYamada, M., Yamada, Y. & Saier, M.H., Jr. (1990) DNA Seq 1, 141-5(全体として参照により本明細書に組み入れられている)からのデータ。b31P NMRにより測定され;Haldane関係(Ru5P/A5P)から計算された。c試験された基質について実験手順を参照。d高分解能誘導結合プラズマ質量分析法により測定された、単量体あたりのZn2+の当量。e5%未満の活性残存。f10μM EDTAにより酵素を単離した場合。gエレクトロスプレーイオン化質量分析法により測定され;タンパク質配列から計算された。hゲル濾過により測定された。
【0048】
グルシトールオペロンは、環境からのD-グルシトールの協調的取り込みおよび異化反応に関与するホスホエノールピルビン酸:糖ホスホトランスフェラーゼ系(PTS)を発現する(例えば、T.C. Meredith, R.W. Woodard, J. Biol. Chem. 278, 32771 (2003)参照;全体として参照により本明細書に組み入れられている)。そのオペロンは、最初にLengeler(例えば、C. Galanos, et al., Eur. J. Biochem. 9, 245 (1969); S. Muller-Loennies, et al., J. Biol. Chem. 278, 34090 (2003)参照;それぞれは全体として参照により本明細書に組み入れられている)により、その後、Saier(例えば、K.A. Brozek, C.R. Raetz, J. Biol. Chem. 265, 15410 (1990); H. Nikaido, Microbiol. Mol. Biol. Rev. 67, 593 (2003)参照;それぞれは全体として参照により本明細書に組み入れられている)により、研究され、7つの収束的に転写される遺伝子、gutAEBDMRQからなることが知られている。EIIGut複合体は、gutA(EIIC1ドメイン)、gutE(EIIBC2ドメイン)、およびgutB(EIIAドメイン)により形成され、D-グルシトールを内膜を渡って、D-グルシトール6-リン酸として細胞へ輸送する。D-グルシトール6-リン酸は、その後さらに、NADH依存性脱水素酵素である、gutDにより、解糖中間体D-フルクトース6-リン酸へ代謝される。gutオペロンの発現は、cAMP-CAP(環状アデノシン一リン酸-カタボライト活性化タンパク質)媒介性制御に加えて、転写リプレッサー(gutR)および転写アクチベーター(gutM)からなる複雑な多成分制御系により厳重に調節されている(例えば、C.J. Belunis, et al., J. Biol. Chem. 270, 27646 (1995)参照;全体として参照により本明細書に組み入れられている)。しかしながら、gutQの機能はまだわかっていない(例えば、R.C. Goldman, W.E. Kohlbrenner, J. Bacteriol. 163, 256 (1985)参照;全体として参照により本明細書に組み入れられている)。
【0049】
本発明の態様の開発中に行われた実験において、外膜LPS発現を実質的に欠く生存可能なグラム陰性細菌は、ヒトにおけるLPS誘発性敗血症の公知のアンタゴニストである、内毒素的に不活性のLPS前駆体リピドIVaをもっぱら産生するにもかかわらず、構築された。本発明は、外膜LPS発現を実質的に欠く生存可能なグラム陰性細菌を構築する特定の方法(例えば、API発現の抑制による;gutQおよび/またはkdsD遺伝子の突然変異による;KDO発現の抑制による;KDOとリピドIVAの間の会合の阻害による;kdsAおよび/もしくはkdsBおよび/もしくはwaaAおよび/もしくはmsbAおよび/もしくはyhjD遺伝子、または他の生合成、プロセシング、もしくは輸送遺伝子の突然変異による;リピドIVA発現の抑制による;lpxM遺伝子、またはリピドIVAについての他の生合成、プロセシング、もしくは輸送遺伝子の突然変異による)に限定されない。
【0050】
本発明は、外膜LPS発現を実質的に欠く生存可能なグラム陰性細菌の構築における任意の型のグラム陰性細菌株の使用を企図する。本発明に有用なグラム陰性細菌の例には、限定されるわけではないが、エシェリキア種、シゲラ種、サルモネラ種、カンピロバクター種、ナイセリア種、ヘモフィルス種、アエロモナス種、フランシセラ種、エルシニア種、クレブシエラ種、ボルデテラ種、レジオネラ種、コリネバクテリア種、シトロバクター種、クラミジア種、ブルセラ種、シュードモナス種、ヘリコバクター種、およびビブリオ種が挙げられる。好ましい態様において、大腸菌が用いられる。用いられうるエシェリキア株の例には、限定されるわけではないが、大腸菌(Escherichia coli)(E. coli)株DH5a、HB 101、HS-4、4608-58、1-184-68、53638-C-17、13-80、および6-81(例えば、Sambrook, etal., (Eds.), 1993, Molecular Cloning, Cold Spring Harbor Press, Cold Spring Harbor, N.Y.); Grant, et al., 1990, Proc. Natl. Acad. Sci., USA, 87:4645; Sansonetti, et al., 1982, Ann. Microbiol. (Inst. Pasteur), 132A:351参照)、腸内毒素原性大腸菌(Evans, et al., 1975, Infect. Immun., 12:656)、腸管病原性大腸菌(Donnenberg, et al., 1994, J. Infect. Dis., 169:831;それぞれは全体として参照により本明細書に組み入れられている)、ならびに腸管出血性大腸菌(例えば、McKee and O'Brien, 1995, Infect. Immun., 63:2070)が挙げられる。
【0051】
本発明は、突然変異体グラム陰性細菌株(例えば、kdsDおよび/もしくはgutQ、kdsA、kdsB、waaA、msbA、ynjD遺伝子、または他の生合成、プロセシング、もしくは輸送遺伝子に突然変異を有するグラム陰性細菌株)の増殖のために特定の培養条件に限定されない。例示を目的として、細菌は、増殖されることになっている細菌株に適切である通常の培養技術を用いて、LB培地(Difco, Detroit Mich.)、Nutrientブロス(Difco)、Tryptic Soyブロス(Difco)、またはM9最少ブロス(Difco)などの、細菌増殖に適した任意の標準液体培地において増殖することができる(Miller, 1991, 前記)。代替として、細菌は、L-寒天(Difco)、Nutrient寒天(Difco)、Tryptic Soy寒天(Difco)、またはM9最少寒天(Difco)などの固体培地上で培養されうる。菌株が突然変異kdsDおよび/またはgutQを含むグラム陰性細菌株について、外因性D-アラビノース5-リン酸源が細菌増殖および生存に用いられる(Meredith et al., 2006, ACS Chem. Biol. 1:33-42;全体として参照により本明細書に組み入れられている)。または、本発明のいくつかの態様の開発中に行われた実験は、kdsDおよび/またはgutQ突然変異を含む菌株におけるmsbA遺伝子の過剰発現が細菌増殖および生存のためのD-アラビノース5-リン酸による補充の代替であることを示している。
【0052】
いくつかの態様において、本発明は、gutQ、kdsD(yrbH)、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子における突然変異を有する、生存可能なグラム陰性細菌を提供する。いくつかの態様において、gutQおよびkdsD遺伝子の突然変異は、細菌株内のAPI発現を阻害し、それがKDO発現を阻害し、それが外膜LPS発現を阻害する。いくつかの態様において、本発明は、kdsA遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供する。いくつかの態様において、本発明は、kdsB遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供する。いくつかの態様において、本発明は、waaA遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供する。本発明の態様の開発中に行われた実験は、kdsA、kdsB、および/またはwaaAの突然変異が、KDOの産生またはKDO2とリピドIVAの間の会合を阻止して、リピドIVAのみが外膜へ輸送されることにより、LPS生合成経路を阻害することを示した。細菌細胞は生き延び、LPSを含まず、かつ無毒性である。いくつかの態様において、本発明は、gutQ、kdsD、kdsA、kdsB、および/またはwaaA遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供し、msbA遺伝子における突然変異をさらに含む。いくつかの態様において、本発明は、gutQ、kdsD、kdsA、kdsB、またはwaaA遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供し、yhjD遺伝子における突然変異をさらに含む。
【0053】
本発明は、グラム陰性細菌内に遺伝子突然変異を導入するための任意の技術の使用を企図する。そのような技術には、限定されるわけではないが、N-メチル-N'-ニトロN-ニトロソグアニジン、アクリジンオレンジ、臭化エチジウムなどの化学物質、または紫外線への非致死的曝露を用いる非特異的突然変異誘発が挙げられる(例えば、Miller (Ed.), 1991, A Short Course in Bacterial Genetics, Cold Spring Harbor Press, Cold Spring Harbor, N.Y.参照;全体として参照により本明細書に組み入れられている)。または、突然変異は、Tn10突然変異誘発、バクテリオファージ媒介性形質導入、λファージ媒介性対立遺伝子交換、もしくは接合伝達、または組換えDNA技術を用いる部位特異的突然変異誘発を用いて導入されうる(例えば、Miller (Ed.), 1991, 前記; Hone, et al., 1987, J. Infect. Dis., 156:167; Noriega, et al., 1994, Infect. Immun., 62:5168; Hone, et al., 1991, Vaccine, 9:810; Chatfield, et al., 1992, Vaccine, 10:53; Pickard, et al., 1994, Infect. Immun., 62:3984; Odegaard, et al., 1997, J. Biol. Chem., 272:19688; Lee, et al., 1995, J. Biol. Chem., 270:27151; Garrett, et al., 1998, J. Biol. Chem., 273:12457参照;それぞれは全体として参照により本明細書に組み入れられている)。突然変異を導入するための任意の方法を用いてもよく、その突然変異を、1つまたは複数の追加の突然変異と共に導入することができる。例えば、いくつかの態様において、本発明は、gutQ、kdsD、kdsA、kdsB、waaA、msbA、yhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子における突然変異などの1つより多い突然変異を有する生存可能なグラム陰性細菌を提供する。
【0054】
いくつかの態様において、グラム陰性細菌内の突然変異(例えば、gutQ、kdsD(yrbH)、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の突然変異)における突然変異は、恒常的に発現しているか、または、例えば、プロモーターの温度感受性熱ショックファミリーもしくは嫌気的に誘導されるnirBプロモーター(例えば、Harborne, et al., 1992, Mol. Micro., 6:2805参照;全体として参照により本明細書に組み入れられている)などの誘導性プロモーターまたはuapA(例えば、Gorfinkiel, et al., 1993, J. Biol. Chem., 268:23376参照;全体として参照により本明細書に組み入れられている)もしくはgcv(例えば、Stauffer, et al., 1994, J. Bact, 176:6159参照;全体として参照により本明細書に組み入れられている)などの抑制性プロモーターの制御下にあるかのいずれかである。適切なプロモーターの選択は、宿主細菌株に依存し、当業者にとって明らかであると思われる。
【0055】
いくつかの態様において、本発明は、API発現を欠く生存可能なグラム陰性細菌(例えば、大腸菌)を提供する。本発明は、API発現を阻害する特定の方法に限定されない。いくつかの態様において、API発現は、KDOタンパク質発現の抑制によって阻害される。本発明は、KDOタンパク質発現を抑制する特定の方法に限定されない。いくつかの態様において、KDOタンパク質発現は、例えば、gutQ遺伝子、kdsD遺伝子、kdsA遺伝子、もしくはkdsB遺伝子の突然変異、または任意の他のKDO生合成遺伝子における突然変異によって抑制される。
【0056】
いくつかの態様において、本発明は、生存可能な無毒性(例えば、内毒素を含まない)グラム陰性細菌(例えば、大腸菌)を提供する。本発明は、生存可能な無毒性グラム陰性細菌を提供する特定の方法に限定されない。いくつかの態様において、生存可能な無毒性グラム陰性細菌は、外膜におけるLPS発現の抑制によって提供される。本発明は、外膜におけるLPS発現の抑制についての特定の方法に限定されない。いくつかの態様において、LPS発現は、APIタンパク質発現の抑制によって抑制される。本発明は、API発現を抑制する特定の方法に限定されない。いくつかの態様において、API発現は、KDOタンパク質発現の抑制によって抑制される。本発明は、KDOタンパク質発現を抑制する特定の方法に限定されない。いくつかの態様において、KDOタンパク質発現は、例えば、gutQ遺伝子およびkdsD遺伝子の突然変異によって抑制される。いくつかの態様において、KDOタンパク質がリピドIVAと会合しないために、外膜におけるKDOタンパク質発現が生じず、リピドIVAのみが外膜へ輸送される。例えば、gutQ、kdsD、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の突然変異は、KDO2-リピドIVA複合体の形成または膜提示を排除し、結果として、例えば、リピドIVA分子のみが外膜へ輸送されることを生じ、その後のLPS形成は生じない。
【0057】
いくつかの態様において、生存可能な無毒性グラム陰性細菌は、限定されるわけではないが、他の細菌生物体由来のLPS、固有の脂質誘導体、ヒトタンパク質またはペプチド産生、非ヒトタンパク質またはペプチド産生、ワクチン作製などの非天然タンパク質およびペプチドを発現する、産生する、およびディスプレイするための当業者に公知のクローニング方法(Sambrook et al., Molecular Cloning; A Laboratory Manual, Cold Spring Harbor Laboratory Press参照;全体として参照により本明細書に組み入れられている)により遺伝子操作されうる。そのような産生された産物は、限定されるわけではないが、臨床的治療および基礎研究活動を含む様々な適用に有用性を見出す。
【0058】
いくつかの態様において、本発明は、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌(例えば、大腸菌)を提供する。本発明は、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌を提供する特定の方法に限定されない。いくつかの態様において、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌は、APIタンパク質発現の抑制によって達成される。本発明は、APIタンパク質発現を抑制する特定の方法に限定されない。本発明は、API発現を抑制する特定の方法に限定されない。いくつかの態様において、API発現は、KDOタンパク質発現の抑制によって抑制される。本発明は、KDOタンパク質発現を抑制する特定の方法に限定されない。いくつかの態様において、KDOタンパク質発現は、例えば、gutQ、kdsD、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子の突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の突然変異によって抑制される。いくつかの態様において、リピドIVAを発現する外膜を含む、LPSを含まない生存可能なグラム陰性細菌は、リピドIVAのみが外膜へ輸送される(例えば、KDOなしで)ように、KDOとリピドIVAの間の会合を阻害することにより達成される。本発明は、KDOとリピドIVAの間の会合を阻害する特定の方法に限定されない。いくつかの態様において、KDOとリピドIVAの会合は、例えば、gutQ、kdsD、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の任意の突然変異により阻害される。いくつかの態様において、本発明は、生存可能な無毒性グラム陰性細菌(例えば、大腸菌)から単離されたリピドIVaを提供する。リピドIVaは、例えば、哺乳動物の敗血性ショックシグナル伝達経路を研究することにおいて、およびLPS型分子の合成におけるビルディングブロックとして用いられる。リピドIVaを単離するための現行の方法は、従来の全有機合成、成熟LPSの分解、または所望のリピドIVaの画分を含む異種性LPS層を産生する条件的突然変異体からの精製を含む。そのような方法の欠点には、低いリピドIVa収量および高い労働量が挙げられる。本発明の生存可能な無毒性グラム陰性細菌からのリピドIVaの単離は、リピドIVaの外膜存在によりそのような方法を超える有意な改善を示している。
【0059】
いくつかの態様において、本発明は、生存可能な無毒性グラム陰性細菌(例えば、大腸菌)から単離された外膜小胞を提供する。リピドIVaは、敗血性ショックシグナル伝達経路のアンタゴニストであり、急性敗血症をもつ患者を処置するための実行可能なアプローチは、LPSを含むシグナル伝達経路を遮断することである。いくつかの態様において、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌からの単離された外膜小胞が、敗血症関連障害を処置するために、または予防的に阻止するために用いられる。本発明の生存可能な無毒性グラム陰性細菌(例えば、ΔAPI菌株)から調製された外膜小胞は、LPSアンタゴニストとしてリピドIVaを含む。
【0060】
いくつかの態様において、生存可能なグラム陰性細菌(例えば、大腸菌)から単離された外膜小胞が、外膜小胞に基づいたワクチンの改善を目的として用いられる。OMVに基づいたワクチンは、しばしば、きつい化学処理によりLPSを剥ぎ取ることにより「解毒」される。しかしながら、剥ぎ取り方法は、抗体をターゲットする良い候補でありうる、特に他のグラム陰性病原体由来のクローニングされた外膜タンパク質の、OMVワクチンのタンパク質成分に、有害な影響を及ぼす。解毒は、宿主としてのΔAPI突然変異体株に関しては必須ではなく、追加的な安全レベルのを与える。
【0061】
いくつかの態様において、本発明は、リピドIVaおよびLPSの両方の発現をもつ外膜を含むグラム陰性細菌を提供する。LPSの毒性を免疫刺激性から離すことは、LPSに基づいたアジュバントまたはLPSに基づいたワクチンを開発するための大きな課題である。ΔAPI菌株における遮断はLPS経路の初期であるため、他の細菌由来の酵素(リン酸基、エタノールアミン、L-4-デオキシアラビノース、異なるアシル鎖長でLPSを修飾する)および変化した活性を有する突然変異型酵素が、細胞内で固有の生物活性を有するLPS分子のアレイを作製するために用いられる。そのような遺伝子操作についての多くの方法は、すでに大腸菌において存在している。さらに、成熟LPS合成は、増殖培地におけるD-アラビノース5-リン酸の含有により回復され、LPS誘導体の成熟LPSに対する量および比率を調節かつ最適化することを可能にすることができる。そのようなLPS「ブレンド」は、受け入れられる低レベルの潜在的毒性を保持している間、免疫刺激活性の望ましいバランスを達しうる。
【0062】
いくつかの態様において、生存可能な無毒性グラム陰性細菌は、内毒素を含まない治療用分子の産生のための宿主として用いられる。本発明は、特定の治療用分子に限定されない。従来より、グラム陰性細菌における治療用分子の産生は、それが、ワクチンのためのOM小胞か、アジュバントとして用いられうるLPS型分子(モノホスホリルリピドA(MPLA)など)か、組換え薬学的タンパク質、高分子か、または哺乳動物細胞トランスフェクション遺伝子治療のためのDNAかに関わらず、細菌宿主由来の内毒素の存在に悩まされる。LPSの免疫原性能力は十分立証されているが、治療用分子の内毒素の混入は懸念材料である。内毒素混入を軽減するための現行の産生ストラテジーには、内毒素を含まないDNAプラスミド精製、続いて、内毒素レベルを測定するためのアッセイ法のための販売されているキットなどの様々な精製技術が挙げられる。ΔAPI菌株は内毒素を産生しないので、そのような精製段階が必要とされない。そういうものとして、本発明の生存可能な無毒性グラム陰性細菌株(例えば、ΔAPI菌株)は、内毒素を含まない治療用分子(例えば、リピドIVa)を単離する改善された方法を提供する。例えば、ΔAPI菌株は、よく研究されたグラム陰性細菌を用いて内毒素を含まない環境における商業的に重要な治療用分子の産生のための宿主であることが企図される。さらに、gutQ、kdsD、kdsA、kdsB、waaA、msbA、yhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送細菌遺伝子における突然変異を含む菌株は、グラム陰性細菌を用いて内毒素を含まない環境における商業的に重要な治療用分子の産生のための宿主であることが企図される。
【0063】
いくつかの態様において、生存可能な無毒性グラム陰性細菌は、免疫応答を刺激するワクチンまたは他の組成物の産生のために用いられうる。例えば、腸チフスに対するより毒性の低いワクチンは、本明細書に記載されているようなグラム陰性細菌を用いて作製される。腸チフスについての現行のワクチンは、ワクチン調製物に存在する内毒素のせいで副作用を引き起こす。LPS(例えば、内毒素)が外膜上に提示されない、本明細書に記載されているような生存可能な無毒性グラム陰性細菌またはその部分を利用することが、内毒素が混入されたワクチン調製物により引き起こされる副作用を回避することが企図される。本発明は、内毒素混入が典型的に見出される任意のワクチン調製物または他の組成物において有用性を見出す。本発明の生存可能な無毒性グラム陰性細菌はまた、それらのLPS欠損表現型による弱毒生ワクチンとしての有用性を見出すことが企図される。
【0064】
そういうものとして、本発明は、免疫化および研究を目的として被験体に投与されうる内毒素混入のない免疫応答を誘導するためのOMワクチンおよび他の組成物の開発に使用を見出す。例えば、弱毒またはOMワクチンは、全体として参照により本明細書に組み入れられている、米国特許出願第2005/0013831号または米国特許第6,558,677号に記載されているような、手順を用いて調製されうる。例えば、そのようなワクチンは、外科患者などの敗血性ショック(例えば、大腸菌からの)に罹るリスクがある被験体を免疫するのに有用性を見出す。さらに、内毒素を含まない弱毒またはOMワクチンは、例えば、百日咳(例えば、ボルデテラ種(Bordetella sp.))、ブルセラ症または内毒素性ショック(例えば、ブルセラ種(Brucella sp.))、肺および呼吸器感染(例えば、シュードモナス種(Pseudomonas sp.)、ヘモフィルス種(Haemophilus sp.)、モラクセラ種(Moraxella sp.))、コレラ(例えば、ビブリオ種(Vibrio sp.))、肺炎(例えば、クレブシエラ種(Klebsiella sp.)、ヘモフィルス種)、胃潰瘍(例えば、ヘリコバクター種(Helicobacter sp.))、髄膜炎(例えば、ナイセリア種(Neisseria sp.)、ヘモフィルス種)、中耳炎(例えば、ヘモフィルス種、モラクセラ種)、赤痢および下痢(例えば、シゲラ種(Shigella sp.)、大腸菌、ビブリオ種、カンピロバクター種(Campylobacter sp.)、エルシニア種(Yersenia sp.))、腸熱(例えば、サルモネラ種(Salmonella sp.))、トラコーマおよび性感染症(例えば、クラミジア種(Chlamydia sp.))、ツラレミア(例えば、フランシセラ種(Franciscella sp.))、ならびにペスト(例えば、エルシニア種)に対する免疫化のために開発されうる。
【0065】
いくつかの態様において、本明細書に記載されているような無毒性の生存可能なグラム陰性細菌は、治療的および研究的適用として、治療用抗体の作製に有用性を見出す。例えば、いくつかの態様において、被験体は、無毒性グラム陰性細菌またはその部分(例えば、膜調製物)を用いて能動的に免疫され、ヒト過免疫血清から調製された抗体は、その後、細菌感染および敗血症から被験体を受動的に保護するために用いられる。しかしながら、治療用抗体の作製は、限定されるわけではないが、霊長類、ウサギ、イヌ、モルモット、マウス、ラット、ヒツジ、ヤギなどの宿主動物においてより従来的な方法によって達成される。例えば、治療用抗体は、ヒト被験体への投与について宿主動物において抗体を産生するために自ら免疫原として無毒性の生存可能なグラム陰性細菌を用いて産生される。本明細書に記載されているような無毒性の生存可能なグラム陰性細菌は、さらに、宿主動物において治療用抗体を産生するために用いられる外来抗原(例えば、免疫原性ペプチドまたはタンパク質)を提示するための宿主として有用性を見出す。例えば、無毒性の生存可能なグラム陰性細菌は、LPSが実質的に欠損していることの他に、抗体産生のための免疫原として使用を見出す非天然タンパク質およびペプチドを発現するように遺伝子操作されうる(例えば、当業者に公知の確立されたクローニング方法によって)。そのような免疫原には、限定されるわけではないが、癌細胞および他の病原性細胞へ抗体をターゲットするためのペプチド、ウイルス細胞ターゲティングのためのウイルスのコートタンパク質などが挙げられる。
【0066】
いくつかの態様において、本発明は、治療用抗体産生のために免疫原性タンパク質を提示するのに有用な無毒性の生存可能なグラム陰性細菌を提供する。免疫原性タンパク質に対する抗体は、それが抗原性タンパク質を認識できる限り、任意のモノクローナル抗体またはポリクローナル抗体でありうる。抗体は、当業者に公知の通常の抗体または抗血清調製方法に従って産生されうる。
【0067】
いくつかの態様において、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌(例えば、大腸菌)が、薬学的スクリーニング(例えば、抗発熱性作用物質についてのスクリーニング)のために用いられる。ΔAPI突然変異体菌株は、非常に低い透過性のバリアを有し、特に、通常にはOMを透過できない大きな疎水性薬物分子に対して感受性を高くさせている。化合物ライブラリーのホールセルバイオアッセイ法は、通常では、OMの透過を促進することによりヒットを同定するのを助けるために、トルエン、EDTA、陽イオン性ペプチドなどの透過処理剤を用いる。いったん親のリードヒットが生じたならば、溶解性、分配、サイズなどを改善するために医薬品化学を用いて、抗生物質を作製することができる。これらのスクリーニングからの多くの可能性のあるリードは、その化合物がOMの内側のそのタンパク質標的にアクセスできないため、見逃されている。そのようなスクリーニングにおいて、例えば、ΔAPI突然変異体菌株を用いることは、追加の試薬を供給する必要なしに透過性バリアを低下させることによりOM透過性問題を多少とも解決する。同様に、ΔAPIのそのような低いOM透過性は、DNAライブラリーの作製中に、例えばΔAPI突然変異体菌株をDNAプラスミドで形質転換する場合に、有利である。高形質転換効率細胞は、すべての組換えDNAテクノロジーに必須であり、ΔAPI菌株はそのような適用のための有用な宿主である。
【0068】
実施例
実施例I.
本実施例は、ΔAPI突然変異体TCM15およびKPM22を記載する。G-APIおよびL-APIの両方が欠失した栄養要求性ΔAPI突然変異体、TCM15を構築し、それは、大腸菌についての確立されたKDO2-リピドA定説に従って増殖のための外因性A5Pに依存するようになった。TCM15は、A5Pを含まなければ、増殖培地、インキュベーション温度、または時間にかかわらず、固体培地上にコロニーを形成することができなかった。単一炭素源として0.2%グリセロールを含む液体MOPS最少培地において培養される場合、細胞分裂が、A5Pの欠乏にもかかわらず、32〜48時間遅れて、いつものように再開した。
【0069】
大腸菌KPM22菌株は、細胞抽出物に測定可能なAPI活性は残存していなかったが、37℃で最初の遅延無しに富栄養培地において持続的増殖の能力がある非条件的ΔAPI突然変異体であることが示された。表2に示されているように、倍加時間は、LB培地において親の野生型株のそれより2倍近くまで増加した。
【0070】
(表2)様々な温度におけるLB培地での世代時間
a. 2〜3世代後、増殖速度は非指数関数的であった。
【0071】
非許容的温度(42℃)への移行後、指数関数的増殖速度は、2〜3世代後維持されなかった。増殖は、上昇した温度においてkdsDをコードするプラスミド(KPM25)によりKPM22に回復され、KDO合成における遮断による細胞外被の欠陥を示唆した。
【0072】
KPM22をさらに研究するために、LPS試料を、フェノール-クロロホルム-石油エーテル(PCP)抽出方法を用いて細胞から抽出した(例えば、C. Galanos, et al., Eur. J. Biochem. 9, 245 (1969)参照;全体として参照により本明細書に組み入れられている)。LPS抽出物の糖組成は、内側コア糖成分[KDOおよびL-グリセロ-D-マンノ-ヘプトース(ヘプトース)]およびリピドA[D-グルコサミン(GlcN)]について測定された(図1A参照)。野生型BW30270[1 GlcN:0.9 KDO:2.2 ヘプトース]およびKPM25[1.0 GlcN:1.0 KDO:2.5 ヘプトース]の両方についての比は、大腸菌K-12[1.0 GlcN:1.0 KDO:2.0 ヘプトース]により産生された主なLPS種(グリコフォームI)についての比と一致した(例えば、S. Muller-Loennies, et al., J. Biol. Chem. 278, 34090 (2003)参照;全体として参照により本明細書に組み入れられている)。ほんの微量のKDOまたはヘプトースが、KPM22についての比較において検出されたが、GlcNはなお存在しており、リピドAバックボーンが無傷であることを示唆した。
【0073】
プロテイナーゼK処理された細胞全体可溶化物から調製されたLPS試料の銀染色SDS-PAGE分析により、KPM22についてバンドは検出されなかった(図1B参照;上部パネル)。リピドAの非グリコシル化1,4'-二リン酸化β-1,6-結合GlcN二糖バックボーンを認識するmAb A6抗体で探索される前に、糖コアを切断するために、ブロッティングされた膜を酸で処理した。Re内毒素標準より速いが、合成リピドIVaと同じレベルで移動したKPM22由来の単一バンドは、その抗体により認識された(図1B、中央パネル、それぞれ、レーン6および16)。酸加水分解段階を省いた場合、合成リピドIVaと共にKPM22から調製されたLPS試料のみがmAb A6により認識され、天然のリピドA構造がグリコシル化されていないことを確認した(図1B、下部パネル)。
【0074】
KPM22におけるLPS前駆体のケモタイプは、精製されたLPS試料を用いて負イオン様式でのエレクトロスプレーイオン化フーリエ変換イオンサイクロトロン(ESI FT-ICR)質量分析法により測定された(図2、表3参照)。
【0075】
(表3)ESI FT-ICR MSピークリスト
a. 示された質量数は、電荷デコンヴォルーション後、LPS画分の負イオンESI FT-ICR質量スペクトルから推定された中性分子のモノアイソトピック質量を指す。b. 太字のピークは、図4上でテキストでラベルされている。c. 略語:PE - ホスファチジルエタノールアミン;GlcN - D-グルコサミン;P - リン酸;P-EtN - ホスホエタノールアミン;Gal - D-ガラクトース;Glc - D-グルコース;Hep - L-グリセロ-D-マンノ-ヘプトース;KDO - 2-ケト 3-デオキシ-D-マンノ-オクツロソン酸;Rha - ラムノース;GlcNAc - N-アセチルD-グルコオサミン;LAtri、LAtetra、LApenta、LAhexa - リピドAのアシル化状態。
【0076】
野生型BW30270およびKPM25の両方のスペクトルは、成熟大腸菌K-12 LPSコアの異なるグリコフォームの特徴的な質量範囲[〜3900uから〜4300uまで]内で類似したピークパターンおよび不均一性を示した(図2A、C参照)。KPM22における1つのLPSに関連したピークは、1,4'-二リン酸化テトラアシル化リピドA(リピドIVa、計算された質量1404.854u)の構造と一致する1404.86uの分子量を有した(図2B参照)。
【0077】
リピドIVaは、KDO2-リピドIVaを形成するための2つのKDO残基の逐次付加のアクセプターとしての役割を果たす、LPS経路における中間体であり、後期アシルトランスフェラーゼLpxLおよびLpxMは、次に、脂肪酸ラウレートおよびミリステート、それぞれをKDO2-リピドIVaへ転移させ、ヘキサアシル化KDO2-リピドAを形成する。Raetzおよび共同研究者らは、大腸菌由来の両方の酵素が、活性について脂質基質においてKDOの絶対的基質要求性を示すことを示しており(例えば、K. A. Brozek, C.R. Raetz, J. Biol. Chem. 265, 15410 (1990)参照;全体として参照により本明細書に組み入れられている)、KPM22由来のリピドAにおける二次アシル鎖の欠如を説明している。
【0078】
リピドIVAの細胞内局在性に取り組み、かつそれがKPM22のOMへ輸送されるかどうかを決定するために、不連続性ショ糖勾配遠心分離を用いて、OMを内膜(IM)から分離させた(図3参照)。両方の膜はよく分離されていたが、KPM22についてのOMは、野生型OMほどには移動せず、浮遊密度の減少を示唆した。OMにおける蓄積を犠牲にしてIMに局在したままであるOMポーリン(OMP)タンパク質(〜35kDa)の量の増加は別にして、SDS-PAGEにより分析された、全体的な総タンパク質含有量および構成は類似していた。多くのOMタンパク質は、それらの折り畳みおよび機能の両方についてLPSの分子シャペロン性質に依存することが示されている(例えば、全体として参照により本明細書に組み入れられる、H. de Cock, J. Tommassen, Embo J. 15, 5567(1996);P. V. Bulieris, et al., J. Biol. Chem. 278, 9092(2003);K. Sen, H. Nikaido, J. Bacteriol. 173, 926(1991)参照)ため、OMPの減少は、KPM22のOMへのタンパク質輸送速度および/または挿入効率の減少を反映している可能性がある。単離されたOM画分を、特徴的なLPS/リピドIVA脂肪酸マーカーである、3-ヒドロキシミリスチン酸(3-OH C14:0)の存在についてアッセイした。野生型およびKPM22のOMは、1mgの乾燥膜あたり、それぞれ11.7μgおよび31.1μgの3-OH C14:0を含み、野生型におけるLPSの量と少なくとも等しいリピドIVAの相当量が、実際、KPM22のOMに存在することを示唆した。さらに、ESI FT-ICR質量分析法により、KPM22のOMおよびIMの両方におけるリピドIVAについてのピークが明らかにされたが、リピドIVAに起因しうるピークは、野生型由来のいずれの膜画分にも検出されなかった。ひとまとめにすると、これは、リピドIVAがKPM22のOMへ輸送されるが、リピドIVA輸送の速度がその合成速度と切り離されていることを示す。
【0079】
二次アシル鎖は、機能に必要とされる条件であるアシル鎖の数を増加させることによりOM内での低度の流動性を維持することに関与している(例えば、H. Nikaido, Microbiol. Mol. Biol. Rev. 67, 593 (2003)参照;全体として参照により本明細書に組み入れられている)。飽和アシル鎖の密充填は、それがファンデルワールス力を通してOM外層の完全性を維持する、疎水性相互作用のネットワークを誘導する。4つのみのアシル鎖を含み、かつ内側糖コアを含まないにもかかわらず、リピドIVaは、KPM22において、輸送され、その後、OM生合成を援助する能力がある。KPM22のOMにおけるリピドIVa層の前例のない性質は、腸内細菌科の生存能力のための不可欠なLPS構造を再定義する。
【0080】
KDOは、今まで、大腸菌におけるKDO生合成酵素の条件的突然変異体のみが構築されていたので、機能的LPS層の必須成分と、通常、みなされている(例えば、C.J. Belunis, et al., J. Biol. Chem. 270, 27646 (1995); R.C. Goldman, W.E. Kohlbrenner, J. Bacteriol. 163, 256 (1985); P.D. Rick, M.J. Osborn, Proc. Natl. Acad. Sci. U.S.A. 69, 3756 (1972)参照;全体として参照により本明細書に組み入れられている)。これは、脂質二重層内で低度の流動性を維持することにおけるKDOの役割に(およびおそらく部分的にKDOより遠位に付着した他の糖に)帰するとされている(例えば、H. Nikaido, Microbiol. Mol. Biol. Rev. 67, 593 (2003)参照;全体として参照により本明細書に組み入れられている)。二価陽イオン、すなわち、Mg2+およびCa2+は、リン酸化リピドAバックボーンおよびKDOのカルボキシレートの両方によって与えられる負電荷によりイオン架橋を生じ、静電反発力を最小にし、かつ強い側方の相互作用を作り出すと考えられている。さらに、OMの表面でのKDOの配置は、OMタンパク質に極めて接近してKDOを置き、OMタンパク質の多くは、それらの折り畳みおよび機能の両方についてコア含有LPSの分子シャペロン性質に依存する(例えば、H. de Cock, J. Tommassen, Embo J. 15, 5567 (1996); P.V. Bulieris, et al., J. Biol. Chem. 278, 9092 (2003); K. Sen, H. Nikaido, J. Bacteriol. 173, 926 (1991)参照;全体として参照により本明細書に組み入れられている)。
【0081】
KPM22におけるKDOの非必須性質を検証するために、KDO生合成における最初の関係した段階(kdsA)および最終段階(waaA)をコードする遺伝子を破壊した。KPM22/KPM25(図1B、レーン8、9)と対照的に、外因性A5Pもプラスミドが有するAPIも、KPM31/KPM40(レーン12、15)またはKPM34/KPM42(レーン11、14)のいずれにおいても、それぞれ、成熟LPS合成を回復させず、完全なKDO経路無しに生存するKPM22の能力と一致した。
【0082】
KPM22の細胞形態を、透過電子顕微鏡(TEM)によって調べた。全体として、KPM22の構造は、親の菌株と全く類似していた(図4)。明らかな分裂欠陥はTEMによって観察されず、細胞は正常な桿状形を維持していた。2つの明らかに異なる膜が、KPM22について識別され(図4D)、加えて、周辺質であるその2つの膜の間の領域も識別された。周辺質体積は、野生型と比較して均一に圧縮されていた。OM不安定性は、KPM22の表面に現れる小さな膜小胞により示唆された(図4C)。外膜小胞(OMV)形成は、糖コアの相互作用の安定化、膜湾曲の増加、および結果として生じる細菌表面からの小胞放出によって代償されない、隣接するリピドIVa分子の1,4'-GlcNリン酸の間の静電反発力により引き起こされてた可能性がある。ESI-MS分析により、4-アミノ4-デオキシ-L-アラビノース修飾は検出されず、最小限のホスホエタノールアミン修飾のみが検出された(両方とも正味負電荷の量を低下させる働きをする)ことを考えると、電荷反発力はKPM22について特に関連性があった(例えば、C.R. Raetz, C. Whitfield, Annu. Rev. Biochem. 71, 635 (2002)参照;全体として参照により本明細書に組み入れられている)。
【0083】
極度のLPS切り詰めによるOM完全性の喪失に抵抗するKPM22における代償的な機構は、他のOM結合糖脂質に関する安定化を引き起こした。髄膜炎菌(N. meningitidis)由来のLPS欠陥突然変異体において、莢膜多糖合成が生存能力に絶対的に必要になったと報告された(例えば、P. van der Ley, L. Steeghs, J. Endotoxin. Res. 9, 124 (2003)参照)。大腸菌K-12は莢膜多糖を合成しなかったが、LPSに加えて2つの他の細胞表面多糖、すなわち、ストレス誘導性粘液エキソポリサッカライドコラン酸(M抗原)(例えば、A. Markovitz Surface carbohydrates of the prokaryotic cell I. W. Sutherland, Ed. (Academic Press, Inc., New York, N.Y., 1977), vol. I. pp. 415-462)参照;全体として参照により本明細書に組み入れられている)およびホスホグリセリド結合腸内細菌一般抗原(ECA)(例えば、H.M. Kuhn, et al., FEMS Microbiol. Rev. 4, 195 (1988)参照:全体として参照により本明細書に組み入れられている)、が存在する。コラン酸の構成糖質マーカーである、非透析性メチルペントースのレベルにおける差はなかった(図5A)(例えば、S. Gottesman, et al., J. Bacteriol. 162, 1111 (1985)参照;全体として参照により本明細書に組み入れられている)。細胞溶解物の免疫ブロット分析により、グリセロホスファチジル結合ECAの量は実際、KPM22において減少していることが明らかにされ(図5B)、フェノール抽出物のKPM22スペクトルからの4つの三糖繰り返し単位(2429.89u)を含む環状ECAの消失(図6)と一致した。従って、リピドIVaに加えて、KPM22のOMは、微量レベルのECA、および同等に低い野生型レベルのコラン酸を含んだ。一括して、KPM22外被は、生存能力を維持することができる、大腸菌で報告された最も最小のOM糖脂質含有量である。
【0084】
LPS層の主要な機能は、大きな疎水性分子とデフェンシン(ポリカチオンのペプチド)の両方の細胞への拡散に対する透過性バリアとして働くこと、および周辺質区画の内容物を保持することである。OM内での隣接LPS分子間の強い側方相互作用が、宿主応答に対する非特異的防御の手段を与えることに加えて、LPS層をそのような機能に特に良く適したものにさせている。疎水性小分子、栄養分、および抗生物質の選択的透過は、外膜ポーリン(OMP)タンパク質チャネルを通して達成される。抗生物質および界面活性剤のパネルは、透過性バリアとしてのリピドIVaの有効性を評価するためにKPM22に対してスクリーニングされた(表4参照)。
【0085】
(表4)KPM22の透過性バリア性質
最小阻止濃度(μg/mL)
a. 74,000ユニット/g。b. コール酸ナトリウムおよびデオキシコール酸ナトリウムの混合物。c. コリスチン;20,261ユニット/mg。
【0086】
KPM22は、典型的にはグラム陽性細菌対してのみ妥当な効力を生じるいくつかの大きな疎水性抗生物質に対して超高感受性であった。通常ではそれらの作用部位へのアクセスをOMにより拒否されるが、これらの化合物はKPM22において細胞内標的にアクセスした。膜表面へのアクセスは、糖コアにより妨げられず、さらに、分配、およびその後の、損なわれた脂質二重層を通しての透過を促進した。しかしながら、主にOMPを通してOMの通過を得る小さな(<600Da)比較的疎水性の化合物の最小阻止濃度(MIC)は、せいぜいやや減少するだけであった。注目すべき例外は、正電荷をもつアミノグリコシドカナマイシンであった。アミノグリコシドは、OMPと無関係に、LPSとの最初の電荷対形成相互作用を含む、主に自己促進性取り込み機構を通しての侵入を得ることを示唆している(例えば、R.E. Hancock, et al. Antimicrob. Agents. Chemother. 35, 1309 (1991);全体として参照により本明細書に組み入れられている)。KPM22は、特に界面活性剤に対して感受性が高く、ドデシル硫酸ナトリウムについてMICにおいて4000分の1未満の減少であった。ヒト腸管における胆汁塩(コレステロール代謝物)の濃度は4mMから16mMまで(〜1650μg/mLから6650μg/mLまで)の範囲であるため(例えば、全体として本明細書に参照により組み入れられるB. Borgstrom, Acta Med. Scand. 196, 1(1974)参照)、KPM22の欠陥があるOMはもはや、その宿主環境から細胞を保護するのに適していない。驚くべきことに、界面活性剤様作用機構をもつ陽イオンペプチドである、ポリミキシンE(コリスチン)のMICは、KPM22においてたった〜4分の1に抑制されただけであった。臨界凝集濃度までの膜表面におけるポリミキシンの蓄積は、OMを通しての自己促進性輸送のチャネルとして実質的に働く層状二重層内にミセル損傷を形成することに関係がある(例えば、A. Wiese et al., J. Membr. Biol. 162, 127 (1998)参照;全体として参照により本明細書に組み入れられている)。リピドIVaは、LPSと比較して表面積に対する電荷比が減少しており、ポリミキシン結合における負荷電内側コア残基の役割を目立たせている。選択された抗生物質は様々な作用機構をもつため、疎水性化合物の間でのMICにおける変化は、一般的な適応度または薬物流出機構の反映とは対照的に、透過性における変化の結果である可能性が高い。KPM22の透過特性は、OMバリアの内因性抵抗を低下させることにより、すでに存在する抗生物質の活性のスペクトルの幅を広げるための手段としてのKDO生合成阻害の可能性を示す。
【0087】
細菌内毒素は、ただ微量存在するのみの場合でさえ、ヒトにおいて先天性免疫応答を誘発する強力な炎症性分子である(例えば、E.S. Van Amersfoort, et al., Clin. Microbiol. Rev. 16, 379 (2003)参照;全体として参照により本明細書に組み入れられている)。グラム陰性細菌誘発性敗血性ショックは、不均衡な調節不全の免疫応答に起因する。部分的にこの病態生理学的カスケードは、LPSによるマクロファージの活性化により引き起こされ、マクロファージが次に、一連の炎症性介在物質を分泌する。マクロファージにより放出された最初のサイトカインの一つは、多面的なサイトカイン、TNF-α(腫瘍壊死因子)である。LPS調製物の内毒素性潜在能力を、刺激されたヒト単核細胞からのhTNF-α分泌についてELISAに基づいたアッセイ法を用いて測定した(図7)。KPM22からの調製物は、1μg/mLまでの濃度で内毒素的に不活性であり、クロマトグラフィー精製されたリピドIVaを用いた以前の研究と一致した(例えば、D.T. Golenbock, et al., J. Biol. Chem. 266, 19490 (1991)参照;全体として参照により本明細書に組み入れられている)。大腸菌および関連細菌において、KDO阻害は、細菌の宿主応答および抗生物質の両方に対する感受性を増加させるだけでなく、内毒素負荷量を低下させることにより敗血症のリスクを減少させる可能性も有する。
【0088】
LPSを細胞質からIMの周辺質表面へフリップ反転する(flip)内膜ABC(ATP結合カセット)輸送体は、インビトロでヘキサアシル化LPS/脂質A基質に対して高選択性である(例えば、Zhou Zhou, Z., et al., J. Biol. Chem. 273, 12466-12475 (1998); Doerrler, W.T., et al., J. Biol. Chem. 277, 36697-36705 (2002)参照;それぞれ、全体として参照により本明細書に組み入れられている)。MsbAは、最初に、LpxL(HtrB)温度感受性表現型の多コピー抑制因子として同定された(例えば、Polissi, A., and Georgopoulos, C. Mol. Microbiol. 20, 1221-1233 (1998)参照;全体として参照により本明細書に組み入れられている)。栄養要求性TCM15菌株のKPM22ゲノムDNAのコスミドライブラリーでの補完により、MsbAがΔKdo表現型の多コピー抑制因子であることが明らかにされた。msbA座を含む17個の別々のコスミドクローンを単離した。野生型と同一の無傷の野生型msbA配列のみを有する3.5kb挿入断片を含むコスミドサブクローン(pMMW52)は、A5P栄養要求性の喪失および固体寒天上でのコロニー形成能力の回復により示されているように、推定抑制因子突然変異を発生させる必要なしにTCM15を直接、救出することができた(表5)。TCM15(pMMW52)の増殖速度は、KPM22と類似している(表2および5)。これらの結果は、リピドIVAがインビトロでは弱い基質であるが(Doerrler, W.T., and Raetz, C.R., J. Biol. Chem. 277, 36697-36705 (2002)参照;全体として参照により本明細書に組み入れられている)、リピドIVaは、単純な質量的作用により高濃度で存在する場合、インビボでMsbAの基質になる。
【0089】
(表5)MsbAによるTCM15栄養要求性の多コピー抑制
aCfu値は、直接プレーティング(TCM15)かまたは電気形質転換後のいずれかに対応する。b測定可能の場合、37℃における世代時間(分)が括弧に入れて記載されている。c15μM A5P、10μM G6P。dAmp(100μg mL-1)がプラスミド保有菌株について含まれた。eクローニングベクター。fmsbAを含むサブクローン。
【0090】
実施例II.
本実施例は、KPM22およびTCM15を含む研究に用いられる細菌株、プラスミド、およびプライマーを記載する。KPM22およびTCM15を含む研究に用いられる細菌株、プラスミド、およびプライマーは表6に列挙されている。
【0091】
(表6)細菌株、プラスミド、およびプライマー
a. 相同領域は下線が引かれている。
【0092】
全菌株は、単一炭素源として0.2%グリセロールを含む標準Luria-Bertani培地(10gトリプトン、5g酵母抽出物、10g NaCl)またはMOPS最少培地(例えば、F.C. Neidhardt, P.L. Bloch, D.F. Smith, J. Bacteriol. 119, 736 (1974);全体として参照により本明細書に組み入れられている)において増殖した。大腸菌株KPM22が、DatsenkoおよびWanner(例えば、K.A. Datsenko, B.L. Wanner, Proc. Natl. Acad. Sci. U.S.A. 97, 6640 (2000)参照;全体として参照により本明細書に組み入れられている)の手順によるファージλレッド(Red)リコンビナーゼ系を用いる染色体kdsAおよびwaaA遺伝子破壊のための宿主として用いられた。カナマイシンおよびアンピシリンが、それぞれ、15μg/mLおよび100μg/mLで用いられた。鋳型としてのpKD13(kan)を含むP1/P2またはpKD4(kan)を含むプライマー対P3/P4が、それぞれ、KPM31およびKPM40についての挿入カセットを構築するために用いられた。抗生物質抵抗性マーカーは、全プラスミドを37℃で回復させることを除き記載されているように(例えば、K.A. Datsenko, B.L. Wanner, Proc. Natl. Acad. Sci. U.S.A. 97, 6640 (2000)参照;全体として参照により本明細書に組み入れられている)、FLPリコンビナーゼ系を用いて切除された。
【0093】
実施例III.
本実施例はKPM22の増殖を記載する。KPM22の増殖は、37℃における10μM D-グルコース6-リン酸および15μM D-アラビノース5-リン酸を追加したMOPS最少培地中のTCM15の指数関数的に分裂する培養物を、糖リン酸追加を欠く同じ培地へ希釈(1:200v/v)することを含んだ。24〜32時間続いた最初の遅れの後、増殖は再開し、培養物は、LB寒天プレート上でコロニー精製された。
【0094】
実施例IV.
本実施例は、KPM22が関わる実験についての増殖速度測定を記載する。一晩培養物は、30℃で増殖し、新鮮なあらかじめ温められたLB培地(30℃、37℃、または42℃)へ0.05〜0.1に等しいOD600nmまで接種するのに用いられた。増殖をOD600nmにおける変化を測定することによりモニターし、培養物を、指数関数的増殖を維持するために、ODが〜0.7にほぼ等しいように希釈した。倍加時間は表7に列挙されている。
【0095】
(表7)様々な温度におけるLB培地での世代時間
a. 2〜3世代後、増殖速度は非指数関数的であった。
【0096】
実施例V.
本実施例は、KPM22およびTCM15が関わる実験についてのLPS精製を記載する。試料を、500mLの各菌株をLB培地において250rpmの一定の通気、37℃で増殖させることにより、日常的に調製した。静止期培養物由来の細胞を、遠心分離(10min、8000xg、4℃)により収集し、蒸留水で洗浄し、再遠心分離した。そのバイオマスを、以前に記載されているように(例えば、U. Zahringer et al., J. Biol. Chem. 279, 21046 (2004)参照;全体として参照により本明細書に組み入れられている)、エタノール(95%)、アセトン、およびジエチルエーテルでの処理により脱水した。LPSの単離を、フェノール-クロロホルム-石油エーテル手順により乾燥細胞の抽出により行った(例えば、C. Galanos, O. Luderitz, O. Westphal, Eu.r J. Biochem. 9, 245 (1969)参照;全体として参照により本明細書に組み入れられている)。糖組成について分析されるべきフェノール粗抽出物のアリコート(図1A)を、大規模に蒸留水に対して透析し(MWCO=1000Da)、凍結乾燥により収集した。質量分光分析およびヒトTNFαサイトカイン放出の測定のためのLPS試料を、水の滴下式添加による沈殿により粗フェノール相から精製した。綿状沈殿は、BW30270およびKPM25についてのみ形成し、遠心分離により収集し、引き続いて、80%フェノールおよびその後、アセトンで洗浄した。沈殿を水に溶解し、それらの個々のフェノール相母液から別々に透析した。凍結乾燥後、試料を緩衝液(20mM Tris-HCl、pH=7.5、10mM NaCl、10mM MgCl2)に再懸濁し、DNアーゼI(20μg/mL)およびRNアーゼA(20μg/mL)で37℃で8時間、続いて、プロテイナーゼK(100μg/mL)で16時間、処理した。LPS試料を、超遠心分離(SW 41 Tiスウィングバケットローター、200,000xg、2時間、15℃)により収集し、蒸留水で3回、洗浄し、凍結乾燥の前に水に対して大規模に透析した。代表的なLPS精製収量は表8に列挙されている。
【0097】
(表8)LPS精製概要
a. Ppt. - 沈殿。b. DNアーゼI/RNアーゼA/プロテイナーゼK処理後。c. 乾燥細胞質量に基づいて。
【0098】
実施例VI.
本実施例は、KPM22およびTCM15に関わる実験についての糖組成分析を記載する。フェノール粗抽出物からのLPS試料のD-グルコサミン(GlcN)、2-ケト3-デオキシ-D-マンノ-オクツロソン酸(KDO)、およびL-グリセロ-D-マンノ-ヘプトース(ヘプトース)含有量を、比色定量化学的アッセイ法を用いて測定した。GlcN含有量は、LPS試料(〜1mg)の、500μLの4 M HCl中100℃で18時間の加水分解により測定された。遊離GlcNは、アセチルアミノ糖アッセイ法を用いて定量された(例えば、J.L. Strominger, J.T. Park, R.E. Thompson, J. Biol. Chem. 234, 3263 (1959)参照;全体として参照により本明細書に組み入れられている)。KDO含有量はLPS適応化チオバルビツール酸アッセイ法を用いて測定されたが(例えば、Y.D. Karkhanis, J.Y. Zeltner, J.J. Jackson, D.J. Carlo, Anal. Biochem. 85, 595 (1978)参照;全体として参照により本明細書に組み入れられている)、ヘプトースの量は改変システイン-硫酸アッセイ法を用いて推定された(例えば、M.J. Osborn, Proc. Natl. Acad. Sci. U.S.A. 50, 499 (1963)参照;全体として参照により本明細書に組み入れられている)。
【0099】
実施例VII.
本実施例は、KPM22およびTCM15に関わる実験についてのSDS-PAGE電気泳動およびリピドA/ECA免疫ブロットを記載する。細胞全体の可溶化物のLPSプロファイルは、HitchcockおよびBrown(例えば、P.J. Hitchcock, T.M. Brown, J. Bacteriol. 154, 269 (1983)参照;全体として参照により本明細書に組み入れられている)の方法によるSDS-PAGEにより分析された。簡潔には、各試料のコロニーを、LB寒天プレートから掻爬し、ダルベッコ(Dulbecco)のリン酸緩衝食塩水に等しい濁度へ懸濁した。洗浄された細胞ペレットを遠心分離により収集し、溶解緩衝液(50μl 62.5mM Tris-HCl、pH6.8、2% SDS、5% 2-メルカプトエタノール、10%グリセロール、0.002%ブロムフェノールブルー)を加え、試料を沸騰水浴中で10分間、加熱した。プロテイナーゼK(25μg、2.5mg/mlの10μl)を各細胞全体可溶化物に加え、56℃で1時間、インキュベートした。同一の容量を、13%SDS-PAGEゲルへローディングし、その後、一定電流(15mA)で流した。ゲルをLPS分析のために銀染色した(例えば、P.J. Hitchcock, T.M. Brown, J. Bacteriol. 154, 269 (1983)参照;全体として参照により本明細書に組み入れられている)、または記載されているように(例えば、H. Towbin, T. Staehelin, J. Gordon, Proc. Natl. Acad. Sci. U.S.A. 76, 4350 (1979)参照;全体として参照により本明細書に組み入れられている)、Tris-グリシン緩衝液(20mM Tris、150mMグリシン、pH8.3、20%メタノール)を用いてゲルからポリ二フッ化ビニリデン膜へ一定電圧(26V)で電気的転写した。リピドAの非グリコシル化1,4'-二リン酸化β1,6-結合GlcN二糖バックボーンを認識するmAb A6とのブロットのインキュベーションの前に(例えば、L. Brade, O. Holst, H. Brade, Infect. Immun. 61, 4514 (1993)参照;全体として参照により本明細書に組み入れられている)、膜を1%酢酸中で1時間、煮沸して、通常の免疫手順により発色される前にα2,6-KDO-GlcN結合を切断した(例えば、R. Pantophlet, L. Brade, H. Brade, J. Endotoxin Res. 4, 89(1997)参照;全体として参照により本明細書に組み入れられている)。真正の合成リピドIVa(化合物406)を標準として用いた(例えば、M. Imoto et al., Bull. Chem. Soc. Japan 60, 2197 (1987)参照;全体として参照により本明細書に組み入れられている)。腸内細菌一般抗原(ECA)免疫ブロットを、mAb 898を用いて探索した(例えば、H. Peters et al., Infect. Immun. 50, 459 (1985)参照;全体として参照により本明細書に組み入れられている)。免疫ブロットを、アルカリホスファターゼ結合ヤギ抗マウスIgG(H+L)とインキュベートし、ニトロブルーテトラゾリウムおよび5-ブロモ-4-クロロ-3-インドリルリン酸基質の存在下で発色させた。
【0100】
実施例VIII.
本実施例は、本発明の過程において行われた実験に用いられたエレクトロスプレーイオン化フーリエ変換イオンサイクロトロン質量分析(ESI FT-ICR MS)を記載する。ESI FT-ICR MSは、7テスラのアクティブシールド型磁石およびアポロ(Apollo)イオン源を備えたAPEX II装置(Bruker Daltonics, Billerica, USA)を用いて負イオン様式で行われた。質量スペクトルは、製造会社により提供された標準実験配列を用いて得られた。試料を、2-プロパノール、水、およびトリエチルアミンの50:50:0.001(v/v/v)混合物に〜10ng/μlの濃度で溶解し、2μl/minの流速でスプレーした。キャピラリー入口電圧を3.8kVに、および乾性ガス温度を150℃に設定した。スペクトルは電荷デコンヴォルーションされ、示された質量数は中性モノアイソトピック質量を指す。ピーク指定は、大腸菌K-12株W3100由来のLPSの以前に発表された詳細な構造分析(例えば、S. Muller-Loennies, B. Lindner, H. Brade, J. Biol. Chem. 278, 34090 (2003)参照;全体として参照により本明細書に組み入れられている)に基づいて解釈された。明白に同定することができない重複アイソトピックピークを有するいくつかの分子種があったため、最も豊富なイオンのみが、表9に要約されている。
【0101】
(表9)ESI FT-ICR MSピークリスト
a. 示された質量数は、電荷デコンヴォルーション後、LPS画分の負イオンESI FT-ICR質量スペクトルから推定された中性分子のモノアイソトピック質量を指す。b. 太字ピークは、図4上でテキストで標識されている。c. 略語:PE - ホスファチジルエタノールアミン;GlcN - D-グルコサミン;P - リン酸;P-EtN - ホスホエタノールアミン;Gal - D-ガラクトース;Glc - D-グルコース;Hep - L-グリセロ-D-マンノ-ヘプトース;KDO - 2-ケト3-デオキシ-D-マンノ-オクツロソン酸;Rha - ラムノース;GlcNAc - N-アセチルD-グルコオサミン;LAtri、LAtetra、LApenta、LAhexa - リピドAのアシル化状態。
【0102】
実施例IX.
本実施例は、KPM22およびTCM15に関わる実験におけるコラン酸の定量化を記載する。コラン酸は、KangおよびMarkovitz(例えば、S. Kang, A. Markovitz, J. Bacteriol. 93, 584 (1967)参照;全体として参照により本明細書に組み入れられている)により報告された方法の改変により推定された。LB寒天プレートからのコロニーを掻爬し、同一の濁度(OD600nm)まで10mLの蒸留水に再懸濁し、細胞外多糖を遊離させるために沸騰水浴に15分間、浸し、遠心分離(10min、8000xg)により清澄にした。上清を、コラン酸の成分であるメチルペントース(L-フコース)について、標準として真正L-フコースを用いる特定の比色定量反応によりアッセイした(例えば、Z. Dische, L.B. Shettles, J. Biol. Chem. 175, 595 (1948)参照;全体として参照により本明細書に組み入れられている)。BW30270のムコイド単離物が陽性対照として含まれた。
【0103】
実施例X.
本実施例は、本発明の過程において行われた実験に用いられた透過電子顕微鏡法(TEM)を記載する。37℃、LB培地において対数期初期で増殖中の細胞の培養物を、2%四酸化オスミウムに室温で90分間、固定した。細胞を、室温で1時間、2%酢酸ウラニルとインキュベートする前に、蒸留水で3回、洗浄した。細胞を、もう一度、蒸留水で3回、洗浄し、その後、一連の増加性エタノール洗浄(30%、50%、70%、90%、および無水エタノール、室温でそれぞれ15分間)により脱水した。脱水した細胞を、2回、室温で各15分間、酸化プロピレンに浸し、続いて、4℃での一晩インキュベーションにより酸化プロピレン/エポン混合物(1:1、v/v)に含浸させた。その後、重合を60℃で一晩、行った。その塊を超薄切片(80〜100nm)へスライスし、グリッド上に置き、クエン酸鉛溶液において対比した。画像は、自動コンピュステージ(compustage)およびKodak 1.6 Megaplus高解像度デジタルカメラを備えたPhilips CM-100透過電子顕微鏡上で得られた。
【0104】
実施例XI.
本実施例は、本発明の過程において行われた実験に用いられた最小阻止濃度(MIC)測定を記載する。用いられた抗生物質は、MicroSource Discovery Systemsから入手されたセファロリジンを除いて、Sigmaからであった。抗生物質は、それらの作用および細胞への侵入の様々な様式に基づいて選択された。研究されたすべての抗生物質および薬物のMICは、記載されているように(例えば、R. Vuorio, M. Vaara, Antimicrob. Agents Chemother. 36, 826 (1992)参照;全体として参照により本明細書に組み入れられている)、標準段階微量希釈方法を用いてLB培地で測定された。LB寒天プレートからのコロニーを掻爬し、様々な濃度の抗生物質を含む培地(1mLあたり〜104細胞)に懸濁した。培養物を、振盪(〜200rpm)しながら37℃で18時間、インキュベートし、その時点で増殖を目視検査により点数化した。報告されたMIC値を、完全に増殖を阻止した薬物の最低濃度として解釈した。
【0105】
実施例XII.
本実施例は、本発明の過程において行われた実験に用いられたヒトTNFαサイトカインアッセイ法を記載する。上記のように単離されたLPS調製物のヒト単核細胞(MNC)への腫瘍壊死因子(TNF)αサイトカイン誘導性能力を、酵素結合免疫吸着アッセイ法(ELISA)を用いて測定した。LPS試料を、激しいボルテックスによりハンクス平衝塩溶液(Hanks' Balanced Salt Solution)に再懸濁し、使用直前の超音波処理/ボルテックスに供する前に、4℃で一晩、寝かした。健康なドナーから採取されたヘパリン処理された血液を、等容量のハンクス平衝塩溶液と直接、混合し、Greiner Bio-OneからのLymphoprep培地でのLeucosepシステムを用いて製造会社の使用説明書に従い、示差的勾配遠心分離により単離した。MNCをRPMI 1640(3mM L-グルタミン、100ユニット/mL ペニシリン、100μg/mL ストレプトマイシン)で2回、洗浄し、96ウェル培養プレートへ移した(7.5x105細胞/ウェル)。MNCの刺激は、以前に記載されているように(例えば、M. Mueller et al., J. Biol. Chem. 279, 26307 (2004)参照;全体として参照により本明細書に組み入れられている)、行われ、上清は、4℃で一晩、保存された。hTNFα産生は、Copelandら(例えば、S. Copeland, H.S. Warren, S.F. Lowry, S.E. Calvano, D. Remick, Clin. Diagn. Lab. Immunol. 12, 60 (2005)参照;全体として参照により本明細書に組み入れられている)により記載されているように、ELISAにより測定された。データは、3つの別々の実験において二回通り収集され、代表的なデータセットが図7に報告されている。
【0106】
実施例XIII.
本実施例は、KPM22コスミドライブラリーの構築を記載する。コスミドライブラリーは、製造会社(Stratagene)により記載されているように、Sau3Aでの部分的消化、SuperCos1へのライゲーションによりKPM22ゲノムDNAから構築され、Gigapack III XLパッケージング抽出物を用いてパッケージングされた。0.2%(w/v)マルトースおよび10mM MgSO4を含み、加えてさらに、A5PおよびG6Pを追加した、LB培地における増殖により、TCM15をファージ感染のために調製した。形質転換体を、コスミドベクター抗生物質抵抗性マーカー(100μg mL-1 Amp)と共の、追加の糖リン酸を欠くLBプレート上での増殖について選択した。コスミドを、部分的Sau3A消化、続いて中位のコピー数pMBL19クローニングベクターのBamHI部位へのライゲーションによりサブクローニングした(例えば、Nakano, Y., et al., Gene 162, 157-158 (1995)参照;全体として参照により本明細書に組み入れられている)。
【0107】
実施例XIV.
本実施例は、gutQ遺伝子に関わる実験に用いられた材料を記載する。プライマーはInvitrogenにより合成された。ゲノム大腸菌K-12 MG1655 DNAは、American Type Culture Collection(ATCC 700926D)から購入された。Promega Wizard DNA精製キットは、プラスミド精製に利用された。化学的コンピテントな大腸菌XL1-Blue(Stratagene)および大腸菌BL21(DE3)(Novagen)は、それぞれ、宿主プラスミドおよびタンパク質発現に向いていた。大腸菌K-12 MG1655の誘導体である菌株BW30270(rph+、fnr+)は、大腸菌Genetic Stock Center(CGSC#7925)から入手された。糖および糖リン酸は、Sigma-Aldrichから購入されたが、例外として、D-グルシトール6-リン酸は、D-グルコース6-リン酸の水素化四ホウ酸ナトリウム還元により調製され(例えば、Bigham, E.C., et al., (1984) J Med Chem 27, 717-26参照;全体として参照により本明細書に組み入れられている)、陰イオン交換クロマトグラフィー(AG MP-1、Bio-Rad)により精製され、ゲル濾過(Bio-Gel P-2、Bio-Rad)により脱塩された。タンパク質濃度は、標準としてBSAを用いるBio-Rad Protein Assay Reagentを使用して、測定された。
【0108】
実施例XV.
本実施例は、gutQ遺伝子のクローニング、過剰発現、および精製を記載する。gutQ遺伝子を、F-Rプライマー対(表10)での標準PCR方法を用いて増幅し、NdeIおよびBamHIで制限酵素処理し、仔ウシアルカリホスファターゼで処理されている、同様に制限酵素処理された直線化pT7-7発現ベクターへ直接、ライゲーションした。
【0109】
(表10)プライマーのヌクレオチド配列
aNdeI部位は下線が引かれている。bBamHI部位は下線が引かれている。
【0110】
ライゲーション混合物は、化学的コンピテントな大腸菌XL1-Blue細胞を形質転換するために用いられ、pT7-gutQプラスミドを有する形質転換体は、制限酵素分析およびDNAシーケンシングにより同定された。大腸菌BL21(DE3)細胞は、プラスミドで形質転換され、制限酵素分析により再チェックされ、-80℃で保存された。大腸菌BL21(DE3)/pT7-gutQ細胞を、アンピシリン(100mg/L)を含む2xYT培地において37℃で振盪(250rpm)しながら、増殖させた。いったん培養が対数増殖期の中頃(OD600〜0.7-0.9)に達したならば、培養物を、0.4mMの最終濃度でのイソプロピル-β-D-チオガラクトシドで誘導される前に、18℃まで冷却させた。18℃での16時間の増殖後、細胞を遠心分離(6,500xg、15min、4℃)により収集した。細胞ペレットを、20mLの緩衝液A(20mM Tris-HCl;1mM DL-ジチオスレイトール(DTT);pH=8.0)に懸濁し、その後、氷上で超音波処理した(5x30秒間;パルスの間に2分間の一時的停止)。細胞片を遠心分離(29,000xg、40min、4℃)により除去し、上清を0.22μM Millex(登録商標)フィルターを通して濾過した。溶液を、緩衝液Aとあらかじめ平衡に達しているHi Load(商標)(16/10)Q Sepharose高速流カラムへローディングした。タンパク質を、緩衝液AにおけるNaClの0〜900mM勾配を用いて120分間に渡り溶出した。SDS-PAGEにより測定された場合主に組換えタンパク質(〜33kDa)を含む画分を、プールした。硫酸アンモニウムの飽和溶液を、15%飽和に達するまで室温で撹拌しながらゆっくり加えた。溶液を遠心分離(29,000xg、30min、22℃)により清澄化し、上清を30%飽和に至らせた。タンパク質ペレットを遠心分離(29,000xg、30min、22℃)により収集し、緩衝液Aに再懸濁し、2Lの緩衝液Aに対して4℃で一晩、透析した。調製物は、SDS-PAGEにより判断される場合、〜95%より高い均一性であり、180mg gutQ/1L細胞培養物の収率であった。
【0111】
実施例XVI.
本実施例は、本発明の過程において行われた実験に用いられたゲル電気泳動方法を記載する。SDS-PAGEは、12%ポリアクリルアミド上で還元条件下、タンパク質試料(〜5-10μg)に行われ、0.25%クーマシーブリリアントブルーR250溶液で染色した。LPS試料を、トリシン-SDS PAGE(スタッキング4%T、3%C;分離16.5%T、6%C)により分析し(例えば、Lesse, A.J., et al., (1990) J Immunol Methods 126, 109-17参照;全体として参照により本明細書に組み入れられている)、銀染色により可視化した(例えば、Hitchcock, P.J. & Brown, T.M. (1983) J Bacteriol 154, 269-77参照;全体として参照により本明細書に組み入れられている)。
【0112】
実施例XVII.
本実施例は、本発明の過程において行われた実験に用いられた酵素アッセイ法を記載する。API活性は、以前に記載されているように(例えば、Meredith, T.C. & Woodard, R.W. (2003) J Biol Chem 278, 32771-7参照;全体として参照により本明細書に組み入れられている)、96ウェルのマイクロプレートに適応した非連続的システイン-カルバゾール比色定量アッセイ法(例えば、Dische, Z., Borenfreund, E. (1951) J Biol Chem 192, 583-587参照;全体として参照により本明細書に組み入れられている)により測定された。全プレートは、内部Ru5P標準および適切なA5P対照を3回通り含んだ。酵素活性の1ユニットは、37℃で1分間あたり1μmolの糖リン酸の変換として定義される。
【0113】
シロイヌナズナ(Arabidopsis thaliana)由来の3-デオキシ-D-マンノ-オクツロソン酸8-リン酸シンターゼ(kdsA)を利用する、第二のより感度の高い結合アッセイ法が、細胞粗抽出物においてAPI活性を測定するために開発された。この酵素は、A5PおよびPEPの不可逆性立体特異的縮合を触媒して、3-デオキシ-D-マンノ-オクツロソン酸8-リン酸(KDO8P)および無機リン酸を形成する。40uLの100mM Tris-HCl(pH=8.25)中に精製kdsA溶液(3mg/mL;10U/mg)、10mM Ru5P、6mM PEP、および1mM EDTAの5μLを含むの反応混合物を、37℃で3分間、インキュベートした。反応を、10μLの細胞抽出物の添加により惹起した。5分後、50μLの10%(w/v)トリクロロ酢酸の添加により反応を停止した。生成されたKDO8Pを、Aminoff過ヨウ素酸-チオバルビツール酸アッセイ法により測定した(例えば、Sheflyan, G.Y., et al., (1998) Journal of the American Chemical Society 120, 11027-11032参照;全体として参照により本明細書に組み入れられている)。これらの条件下で、kdsAは、KDO8Pの形成において律速ではなかった。
【0114】
D-グルシトール6-リン酸デヒドロゲナーゼ(gutD)活性は、340nmでNADHの形成をモニターすることによる連続的な分光光度アッセイ法を用いて測定された。反応を20mMの最終濃度でのD-グルシトール6-リン酸の添加により惹起する前に、酵素溶液(100mM Tris-HCl、pH=8.7、5mM NAD+)を25℃で2分間、プレインキュベートした。
【0115】
実施例XVIII.
本実施例は、gutQの特徴付けを記載する。gutQの特徴付けは、ksdDについて報告された方法に従って同様に行われた(例えば、Meredith, T.C. & Woodard, R.W. (2003) J Biol Chem 278, 32771-7参照;全体として参照により本明細書に組み入れられている)。簡潔には、基質特異性について、酵素試料を、100mM Tizma-HCl緩衝液(pH=8.25)に希釈し、反応を基質(15nM gutQ、10mM糖、1mM EDTA)で惹起することによりアッセイした。37℃での10分後、可能性のある代替基質、D-アラビノース、D-リボース5-リン酸、D-グルコース6-リン酸(G6P)、D-グルコース1-リン酸、D-グルコサミン6-リン酸、またはD-マンノース6-リン酸を含む反応を停止し、ケトースの存在を測定した。D/L-グリセルアルデヒド3-リン酸、D-エリトロース4-リン酸、およびD-フルクトース6-リン酸についての生成物出現を、31P NMRによりアッセイした。速度定数は、非連続的マイクロプレートアッセイ法を用いて37℃で測定され、基質の添加により惹起された。濃度は、典型的には、0.2Kmから10Kmまでの範囲であった。2分後、反応(pH=8.25における50mM Tris-HCl、5nM gutQ、1mM EDTA)を停止し、その時点で、基質の約10%未満が消費されていた。初速度(v0)を、3回通り測定し、非線形最小二乗回帰を用いて標準ミカエリス・メンテン式にフィットさせて、Ru5Pの形成および消失の両方についてのKmおよびkcat値を決定した。平衡定数(Keq)は、kdsDについて記載されているように、31P NMRを用いて決定した(例えば、Meredith, T.C. & Woodard, R.W. (2003) J Biol Chem 278, 32771-7参照;全体として参照により本明細書に組み入れられている)。gutQのpH最適値は、様々なpH値(37℃で調整されたpH=6.25〜10)のBTP緩衝溶液に酵素を希釈することにより測定された。活性は、3分間の反応時間で3回通り、上に概要を説明されているように、測定された(100mM BTP、15nM gutQ、10mM A5P、1mM EDTA)。単離されたgutQの酵素試料を、100mM Trizma-HCl緩衝液(pH=8.25)に希釈し、様々な二価金属またはEDTAと4℃で30分間、インキュベートした。その後、残存活性を、3分間の反応時間、3回通り、飽和基質条件下、37℃でアッセイした(15nM gutQ、10mM A5P、10μM金属またはEDTA)。
【0116】
実施例XIX.
本実施例は、gutQに関わる実験についての大腸菌株構築および増殖条件を記載する。大腸菌株BW30270が、DatsenkoおよびWanner(例えば、Datsenko, K.A. & Wanner, B.L. (2000) Proc Natl Acad Sci USA 97, 6640-5参照;全体として参照により本明細書に組み入れられている)の手順によるファージλレッドリコンビナーゼ系を用いる染色体gutQおよびkdsD遺伝子破壊のための宿主として用いられた。カナマイシンおよびクロラムフェニコールは50μg/mLで用いられた。鋳型としてのpKD13(kan)かまたはpKD3(cat)のいずれかに関するプライマー対GQF-GQRおよびKDF-KDRが、それぞれ、BW30270(ΔgutQ::kan)およびBW30270(ΔkdsD::cat)を構築するために用いられ、表10に列挙されている。抵抗性マーカーを、その後、記載されているように(例えば、Datsenko, K.A. & Wanner, B.L. (2000) Proc Natl Acad Sci USA 97, 6640-5参照;全体として参照により本明細書に組み入れられている)、FLPリコンビナーゼ系を用いて切除した。BW30270(ΔgutQ ΔkdsD)は、電気形質転換後に行われる次の操作のために培地およびプレートがG6P(10μM)およびA5P(15μM)を常に補充されている点を除いて、GQF-GQR PCR産物挿入断片を用いてBW30270(ΔkdsD)から同様に構築された。用いられた全菌株はコロニー精製され、すべての抗生物質抵抗性の喪失について試験され、関連座位は、予想される欠失部位を確認するためにシーケンシングされた。
【0117】
培養物を、チアミン(1μg/mL)および示された炭素源を追加したM9最少培地(26)かまたはMOPS最少培地(例えば、Neidhardt, F.C., Bloch, P.L. & Smith, D.F. (1974) J Bacteriol 119, 736-47参照;全体として参照により本明細書に組み入れられている)のいずれかで37℃で振盪(250rpm)しながら、増殖させた。BW30270(ΔgutQ ΔkdsD)培養物は、G6P(10μM)およびA5P(5〜50μM)をさらに追加された。アンピシリン(100μg/mL)が、pT7-7(AmpR)プラスミドを有する菌株に加えられた。
【0118】
実施例XX.
本実施例は、酵素アッセイ法、LPS分析、およびRT-PCRのための細胞抽出物の調製を記載する。一晩培養物は、単一炭素源としてのグリセロール(0.2%)および示されたサプリメントを含む最少培地で増殖した。培養物を新鮮な最少培地へ希釈し(1:20 v/v)、37℃で2時間、振盪して、細菌を指数関数的増殖へ戻らせた。BW30270(ΔgutQ ΔkdsD)の培養物を、A5P(5μM)およびG6P(10μM)を加えることにより、ヘキソースリン酸輸送系(uhp)を上方制御するためにこの期間中、事前誘導した。微量のG6Pを除去するために細胞を遠心分離(6,500xg、5min、22℃)によりペレット化し、その後、新鮮培地へ接種した。示されている場合、10mM D-グルシトールを培養物へ加え、増殖をさらに4〜6時間、継続して、gutオペロンの上方制御を可能にし、その時点で、すべての培養物は、対数増殖の初期〜中期にあった。細胞を遠心分離(6,500xg、5min、4℃)により収集した。APIおよびgutD活性についてアッセイされるべき画分を、冷却した1%NaCl溶液で2回、洗浄し、その後、緩衝液(20mM Tris-HCl、1mM DTT、pH=8.0)に再懸濁した。細胞を、超音波処理により破壊し、遠心分離(29,000xg、20min、4℃)により清澄化し、凍結した。LPS分析のための試料を、ダルベッコのリン酸緩衝食塩水で2回、洗浄し、ペレットを溶解緩衝液(200mM Tris(pH=6.8)、2%SDS、4%2-メルカプトエタノール、10%グリセロール)に再懸濁した。OD600nmに基づいた等しい数の細胞を、HitchcockおよびBrown(例えば、Hitchcock, P.J. & Brown, T.M. (1983) J Bacteriol 154, 269-77参照;全体として参照により本明細書に組み入れられている)の方法により処理した。RNAについて分析されるべき細胞ペレットを、Max Bacterial Enhancement試薬に迅速に再懸濁し、TRIzol(Invitrogen)を用いて製造会社のプロトコールにより抽出した。RNA試料をさらに、RNアーゼを含まないDNアーゼでの消化により精製し、RNeasyミニキット(Qiagen)を用いて単離した。RNAの質は、アガロース電気泳動により検査され、260nmにおけるUV吸光度により定量化された。定性RT-PCRを、Superscript II One-Step RT-PCRシステム(Invitrogen)を用いて、説明書に従って、鋳型としての1pgの精製された全RNA、およびgutD遺伝子の最初の342塩基対を増幅するためのGDF-GDRプライマー(0.2μM)と共に行った。
【0119】
実施例XXI.
本実施例は、gutQの精製および特徴付けを記載する。均一性への精製は、Q-Sepharose陰イオン交換クロマトグラフィー、続いて硫酸アンモニウム沈殿を用いる2段階で達成された。タンパク質は、SDS-PAGEによる単一の鋭い高分子量バンド(〜33kDa)として現れ、比活性度は329U/mgであった。gutQの生化学的性質は、kdsDのそれらと類似していることが測定された。速度パラメーター、pH最適値、補因子要求性の喪失、および四次構造がすべて類似していた。A5Pと共通の機能性を共有する単糖を、gutQについての可能性のある代替基質として試験した。システイン-カラバゾール比色定量アッセイ法において、2-ケトヘキソースおよび2-ケトペントースは、540nmでの光を吸収するパープル-レッド発色団を形成する(例えば、Dische, Z., Borenfreund, E. (1951) J Biol Chem 192, 583-587参照;全体として参照により本明細書に組み入れられている)。アルドースのケトースへの変換は、対照に対する試料のA540nmでの吸光度の比率における増加を測定することにより観察されうる。試験された糖のいずれも、それらのそれぞれのケトース型へ変換されなかった。短鎖リン酸化アルドースD/L-グリセルアルデヒド3-リン酸およびD-エリトロース4-リン酸、加えてD-フラクトース6-リン酸が、31P-NMRにより測定された場合、代替基質としての役割を果たした。検出の範囲内で、gutQは、A5PおよびRu5Pについての特異的リン酸糖アルドール-ケトールイソメラーゼであることが示された。
【0120】
実施例XXII.
本実施例は、gutQがリポ多糖生合成を持続させる能力があることを実証する。gutQのインビボでAPIとして機能する能力を評価するために、BW30270(ΔgutQ)およびBW30270(ΔkdsD)を、λレッド(γ、β、exo)相同組換え系を用いて構築した(例えば、Datsenko, K.A. & Wanner, B.L. (2000) Proc Natl Acad Sci USA 97, 6640-5参照;全体として参照により本明細書に組み入れられている)。どちらの突然変異も致死性ではなく、必須のLPS生合成に必要とされる十分な量のA5Pを供給することができる他のAPIコード遺伝子の存在を示している。LPSゲルは、gutオペロンが誘導されたかどうかに関わらず、野生型K-12LPSコアのほとんど等しい量を示し(図8A参照)、A5P合成がこれらの増殖条件下で菌株のいずれにおいても律速ではなかったことを示唆している。BW30270(ΔkdsD)におけるgutQの基底レベルは、生存能力を維持し、かつ機能性LPS層を産生するのに十分なA5Pを供給するのに適切であり、gutQが細胞内でAPIとして機能することを強く示唆している。
【0121】
実施例XXIII.
本実施例は、ΔAPI菌株BW30270(ΔgutQ ΔkdsD)におけるLPS生合成を記載する。BW3270におけるgutQおよびkdsD遺伝子の両方は、外因性A5Pを供給するためにG6P誘導性ヘキソースリン酸輸送体(uhp)を利用することにより、破壊された。A5Pは、ヘキソースリン酸輸送系(uhp)の、非誘導性にもかかわらず、高親和性の基質である(例えば、Kadner, R.J., Murphy, G.P. & Stephens, C.M. (1992) J Gen Microbiol 138(Pt 10), 2007-14; Rick, P.D. & Osborn, M.J. (1972) Proc Natl Acad Sci USA 69, 3756-60; Eidels, L., Rick, P.D., Stimler, N.P. & Osborn, M.J. (1974) J Bacteriol 119, 138-43参照;全体として参照により本明細書に組み入れられている)。低濃度の無機リン酸(1.3mM)を有するMOPS最少培地が、無機リン酸によるuhp媒介性輸送の阻害を防ぐために用いられた(例えば、Shattuck-Eidens, D.M. & Kadner, R.J. (1981) J Bacteriol 148, 203-9参照;全体として参照により本明細書に組み入れられている)。uhp輸送体G6Pの天然基質が、A5Pの効率的な誘導および細胞への輸送に必要とされた。A5PおよびG6Pの両方が培地に含まれない限り、研究の時間経過において検出可能な増殖はなかったため、A5PまたはG6P単独は、増殖を回復させることができなかった(図9A参照)。従って、gutQおよびkdsDは、KDO合成のためのA5Pの唯一の細胞内源であった。培養は、リポ多糖生合成を可能にするために培地にA5Pを補充された。接種材料としてA5Pが培地から消耗されている一晩培養物および延長されたインキュベーション時間を用いることにより、BW30270(ΔgutQ ΔkdsD)において合成されることになっている成熟LPSの量は、培地に含まれるA5Pの量に依存した(図9B)。
【0122】
実施例XXIV.
本実施例は、gutオペロンの発現を記載する。BW30270、BW30270(ΔgutQ)、およびBW30270(pT7-gutQ)を、二重の炭素源、D-グルコースおよびD-グルシトールを含むM9最少培地において増殖させた。すべての3つの菌株は、ほとんど同一の速度で増殖し、D-グルコースが培地から消耗された後の約40分の特徴的な通常の長いディオーキシー遅延時間を示した(例えば、Lengeler, J. & Lin, E.C. (1972) J Bacteriol 112, 840-8参照;全体として参照により本明細書に組み入れられている)。これらの条件下で、誘導は、gutQにより影響されなかった。菌株BW30270、BW30270(ΔgutQ)、およびBW30270(ΔkdsD)を、炭素源としてグリセロールを含むM9最少培地で増殖させた。グリセロールは、クラスB炭素源であり、有意な異化代謝産物抑制を引き起こさず(例えば、Lengeler, J.W. (1986) Methods Enzymol 125, 473-85参照;全体として参照により本明細書に組み入れられている)、cAMPレベルの上昇によってD-グルシトールによるgutオペロンの誘導を促進した。総API(kdsDおよび/またはgutQ)およびgutD比活性度は、すべての3つの菌株において測定された(表11)。
【0123】
(表11)細胞抽出物におけるgutDおよびAPIの比活性度
a菌株は炭素源として0.2%グリセロールを含むM9最少培地で増殖した。bD-グルシトールは、収集の4時間前に(+)で示された培地へ10mMで加えられた。cnモル/min/mgで報告された比活性度。d値はkdsDおよび/またはgutQ活性を含む。
【0124】
BW30270(ΔgutQ)およびBW30270(ΔkdsD)のgutオペロンは依然として誘導可能のままであり、親のBW30270菌株と比較した時、gutD活性により推定された場合、誘導の程度においてたった2倍の差であった。総API活性レベルは、D-グルシトールが培地へ添加された場合、BW30270およびBW30270(ΔkdsD)の両方において増加し、gutQがgutDと共に上方制御されることを示している。BW30270(ΔgutQ)においてD-グルシトールの添加により、その菌株が依然としてgutDを上方制御する能力があるにもかかわらず、観察されたAPIレベルにおける変化がなかった。API活性の大部分は、D-グルシトールを欠損する培地においてkdsDに起因したが、kdsDの恒常的に発現したLPS生合成酵素としての同定を確認した。BW30270(pT7-gutQ)は、上昇したAPIレベルのgutオペロンへの効果を調べるために用いられた(表10)。D-グルシトールが培地に供給されない限りオペロンが抑制されたままであるため、gutDレベルに感知できるほどの差が観察されなかったにもかかわらず、APIレベルの基底レベルはBW30270(pT7-gutQ)において〜250倍、増加した。
【0125】
実施例XXV.
本実施例は、A5Pがgutオペロンの上方制御に重要であることを示す。単一のAPI遺伝子が破壊された場合、制御に差が観察されなかったため、表現型を直接観察できなかったことは、APIの第二のコピーによる抑制のせいであった可能性がある。gutオペロンの誘導能は、BW30270(ΔgutQ ΔkdsD)において調べられた。 一晩培養物はMOPS最少培地(0.2%グリセロール、15μM A5P、10μM G6P)で増殖し、細胞を指数関数的増殖へ戻すために新鮮培地(0.2%グリセロール、5μM A5P、10μM G6P)へ希釈された。2時間の振盪後、細胞を収集し、グリセロールおよびA5Pのみを含む培地へ接種するのに用いた。細胞をuhp輸送体遺伝子について事前誘導したため、G6Pを加えなかった。LPSおよび増殖速度のレベルにおける差が研究の時間経過において最小であるように、2つの濃度のA5P(5μMおよび50μM)を選択した。50μMにおいて、A5P、gutDは、ほとんど野生型レベルまで誘導可能のままであった(表12)。
【0126】
(表12)ΔAPI細胞抽出物におけるgutDおよびgutQ321の比活性度
a菌株は0.2%グリセロールを含むMOPS最少培地で増殖し、10μM G6P/5μM A5Pで事前誘導された。
bD-グルシトールは、収集の4時間前に(+)で示された培地へ10mMで加えられた。cnモル/min/mgで報告された比活性度。dN.D.=検出された活性無し。
【0127】
gutQタンパク質産物自体は、発現に必要ではなかった。A5P濃度が5μMまで減少した場合、D-グルシトール増殖細胞においてgutD活性の顕著かつ再現性のある減少があった。しかしながら、LPSのレベルは、相対的にほんのわずかのみ低下した(図9C)。これは、A5PレベルとgutDの量の間の直接的相関、および差が枯渇したLPS層から生じる多面発現効果の結果によるのではないことを示した。gutD遺伝子の発現レベルの分析は、gutDの測定された比活性度の減少がmRNAの量と相関することを示した(図9D)。gutオペロンは、gutQをコードするプラスミドにより補完された場合、同じ増殖条件下で誘導可能のままであった。
【0128】
実施例XXVI.
本実施例は、過剰発現した場合、遺伝子msbAが、ΔKDO大腸菌細胞をD-アラビノース5-リン酸培地補充なしの寒天上で増殖するのを可能にすることを示す。
【0129】
MsbAは、最初、LpxL(HtrB)温度感受性表現型の多コピー抑制因子として同定された(全体として参照により本明細書に組み入れられている、Polissi et al., 1996, Mol. Microbiol. 20:1221-1233)。栄養要求性TCM15(大腸菌)株のKPM22ゲノムDNAのコスミドライブラリーでの補完により、msbAがΔKdo表現型の多コピー抑制因子であることが明らかにされた。msbA座を含む17個の別々のコスミドクローンを単離した。野生型と同一の無傷の野生型msbA配列のみを有する3.5kb挿入断片を含むコスミドサブクローン(pMMW52)は、A5P栄養要求性の喪失および固体寒天上でのコロニー形成能力の回復により判断されるように、TCM15を直接、救出することができた。TCM15(pMMW52)の増殖速度は、KPM22大腸菌株と著しく類似している(全体として参照により本明細書に組み入れられている、Meredith et al., 2006, ACS Chem. Biol. 1:33-42)。
【0130】
上記明細書に挙げられたすべての刊行物および特許は、参照により本明細書組み入れられている。本発明は特定の好ましい態様に関して記載されているが、主張される本発明がそのような特定の態様に不当に限定されるべきではないことは理解されているはずである。実際、当業者に明らかである、本発明を行うための記載された様式の様々な改変が、特許請求の範囲の範囲内にあることを意図される。
【図面の簡単な説明】
【0131】
【図1】KPM22から抽出されたLPS試料の特徴付けを示す。図1A:透析後のフェノール抽出物由来のLPSの内側コア-リピドA糖組成。GlcN - D-グルコサミン;KDO - 2-ケト3-デオキシ-D-マンノ-オクツロソン酸;L-グリセロ-D-マンノ-ヘプトース - ヘプトース。図1B:プロテイナーゼK処理された細胞全体の可溶化物由来のLPSのSDS-PAGE分析。上部パネルは銀染色されたが、中央および下部パネルは、リピドAの非グリコシル化1,4'-二リン酸化β-1,6-結合GlcN二糖バックボーンに対して方向づけられたmAB A6を用いて発色した免疫ブロットである。中央パネル膜は、免疫学的反応の前に、リピドAを遊離させるために1%酢酸で処理された。レーン1〜5は、異なるLPSケモタイプのサルモネラ・エンテリカ(Salmonella enterica)血液型亜型である[1. 3749 (Ra); 2. 3750 (Rb2); 3. 3748 (Rb3); 4. 3769 (Rd1); 5. 1102 (Re)]、6. 野生型BW30270、7. KPM22、8. KPM25、9. 増殖培地にA5Pを含むKPM22、10. KPM31、11. KPM34、12. 増殖培地にA5Pを含むKPM31、13. KPM40、14. KPM42、15. 増殖培地にA5Pを含むKPM40、16. 200ngの化学合成されたリピドIVa(化合物406)。
【図2】KPM22におけるLPS前駆体の特徴付けを示す。精製されたLPS試料の負イオン様式での電荷デコンヴォルーションされたエレクトロスプレーイオン化フーリエ変換イオンサイクロトロン(ESI FT-ICR)質量スペクトル。示された質量数は、中性分子のモノアイソトピック質量を指す。図2A:BW30270(挿入図、グリコフォームI;3915.71 uのアイソトピック分布)。図2B:KPM22(挿入図、リピドIVa;1404.86 uの構造)。図2C:KPM25(挿入図、描かれたKDO2-リピドA(Re内毒素)の化学構造を有する野生型LPS、および矢印で示されたヘプトース付着点。赤色、青色、および緑色のピーク標識は、それぞれ、グリコフォームI、IV、およびIIのピークファミリーに対応する(例えば、S. Muller-Loennies, B. Lindner, H. Brade, J. Biol. Chem. 278, 34090 (2003)参照;全体として参照により本明細書に組み入れられている)。個々の構造ピーク割当は、表9に列挙されている。PE - ホスファチジルエタノールアミン;P - リン酸;P-EtN - ホスホエタノールアミン;LAtri、LAtetra、LApenta、LAhexa - リピドAのアシル化状態。
【図3】野生型BW30270(a)およびKPM22(b)の内膜および外膜のスクロース勾配分離を示す。画分は、総タンパク質含量(X)、外膜ホスホリパーゼA(OMPLA)(O)、および内膜NADHオキシダーゼ(▲)についてアッセイされた。タンパク質試料のSDS-PAGEゲル(12%)は、還元条件下で流された。分子量タンパク質マーカー(kDa)は、各ゲルの左側に記載されている。矢印は、OMPタンパク質(〜35kDa)の位置を示す[Q2]。
【図4】KPM22の特徴付けを示す。野生型BW30270(パネルAおよびB)およびKPM22(パネルCおよびD)の透過電子顕微鏡(TEM)像。矢印は、KPM22(パネルC)のOM表面における外膜小胞(OMV)を示す。IM - 内膜、OM - 外膜、PG - ペプチドグリカン。スケールバー=50nm。
【図5】KPM22の特徴付けを示す。図5A:細胞浮遊液の培養物のODあたり1mLあたりのメチルペントース(L-フコース)のμgとして推定されたコラン酸産生。図5B:mAb 898抗体を用いての腸内細菌一般抗原(ECA)の免疫ブロット。レーン1(BW30270)、レーン2(KPM22)、レーン3(KPM25)。
【図6】フェノール相抽出物のESI FT-ICR質量スペクトルを示す。BW30270(A)、KPM22(B)、およびKPM25(C)からのフェノール相の電荷デコンヴォルーションされた負イオンESI FT-ICR質量スペクトル。LPSを、水の滴下式添加によりフェノール粗抽出物から沈殿させた。遠心分離による清澄化後、フェノール上清を上記のように透析および処理した。リピドIVaはこの手順中に水によりフェノール相から沈殿しなかったことに留意されたい(B)。ECAcyc - 環状腸内細菌一般抗原(ECA)。
【図7】LPS調製物のhTNFαサイトカイン誘導能力を示す。ヒト単核細胞(MNC)を、上記のように単離された様々な濃度のLPS調製物に曝露した。hTNFα放出はELISAに基づいたアッセイ法を用いて定量化された。データ点は二回通り収集された(暗色の棒 - BW30270、空白の棒 - KPM22、あや目引きの棒 - KPM25)。
【図8】gutQのD-グルシトール利用およびLPS生合成への効果を示す。(A)BW30270(□)、BW30270(ΔgutQ)(○)、およびBW30270(pT7-gutQ)(△)についてのディオーキシー増殖曲線。1μg/mLチアミンおよび10mM D-グルコースを追加したM9最少培地で増殖した一晩培養物を、二重炭素源として2mM D-グルコースおよび2mM D-グルシトールを含む新鮮培地へ希釈した。細胞増殖は、600nmにおける濁度を測定することによりモニターされた。(B)BW30270(WT)、BW30270(ΔgutQ)、およびBW30270(ΔkdsD)由来のプロテイナーゼK処理された細胞全体の可溶化物の銀染色されたトリシンSDS-PAGE LPSゲル。D-グルシトール(10mM)を含む(+)または含まない(-)最少培地(0.2%グリセロール)において増殖した等量の細菌細胞を、対数期初期に収集し、実験手順に記載されているように、処理した。
【図9】ΔAPI菌株BW30270(ΔgutQ ΔkdsD)における増殖およびLPS合成を示す。(A)チアミン(1ug/mL)および単一炭素源としてのグリセロール(0.1%)を含むMOPS最少培地における大腸菌BW30270(ΔgutQ ΔyrbH)の増殖曲線。糖リン酸は、10μM G6P(△)、15μM A5P(□)のいずれかを、または両方(○)を培地に追加された。(B)A5PでのLPSの滴定。分裂を中止しているMOPS最少培地(0.2%グリセロール、5μM A5P、10μM G6P)で増殖した静止期培養物を、G6P(10μM)および様々な濃度のA5P(0.1、1、10、50、および100μM)を含む新鮮培地へ希釈し、6時間、振盪した。LPS試料は、ODに基づいて同数の細胞から調製され、トリシンSDS-PAGEおよび銀染色により分析された。野生型BW30270(レーン1および2)およびBW30270(ΔgutQ ΔkdsD)(レーン3および4)から調製された試料のgutD分析の(C)LPSトリシンSDS-PAGEおよび(D)定性RT-PCR。BW30270(ΔgutQ ΔkdsD)を、10μM G6Pおよび5μM A5Pで事前誘導し、ペレット化し、A5Pのみおよび示されているように10mM D-グルシトールを含む新鮮なMOPS最少培地(0.2%グリセロール)に再懸濁し、分析のための収集前にさらに4時間、振盪した。S - 0.1〜1kb DNA分子量マーカー;Con - 鋳型としてのゲノムDNA。
【図10】大腸菌K-12MG1655のgutオペロンを示す。転写開始部位は、YamadaおよびSaier(例えば、Yamada, M. & Saier, M.H., Jr. (1988) J Mol Biol 203, 569-83参照;全体として本明細書に組み入れられている)による逆転写酵素マッピングを用いて決定され、矢印で示されている。
【図11】Kdoの生合成およびLPSへの組み入れを示す。関わる酵素は、(1)D-アラビノース5-リン酸イソメラーゼ(KdsD/GutQ)、(2)Kdo8Pシンターゼ(KdsA)、(3)Kdo8Pホスファターゼ(KdsC)、および(4)CMP-Kdoシンテターゼ(KdsB)である。大腸菌において、活性化Kdoの2つの分子は、その後、二次アシル鎖(6)ラウレート(LpxL)および(7)ミリステート(LpxM)の段階的付加の前に、(5)Kdoトランスフェラーゼ(WaaA)によりリピドIVAへ連続的に転移する。
【技術分野】
【0001】
本出願は、全体として本明細書に参照により組み入れられた、2006年1月19日に出願された米国特許仮出願第60/760,314号の優先権を主張する。本発明は、National Institutes of Healthによって授与された認可番号GM53609による政府支援でなされた。
【0002】
発明の分野
本発明は、無毒性グラム陰性細菌を提供する。特に、本発明は、外膜内にリポ多糖(LPS、内毒素)を実質的に欠く、生存可能なグラム陰性細菌(例えば、大腸菌(E. coli))を提供する。本発明はさらに、生存可能な無毒性グラム陰性細菌を作製する方法およびその使用を提供する。本発明はまた、免疫応答を誘導するための、および治療剤を研究かつ開発するための組成物ならびに方法を提供する。
【背景技術】
【0003】
発明の背景
リポ多糖(LPS、内毒素)は、グラム陰性細菌の主要抗原である。LPSは、N,O-アシルβ-1,6-D-グルコサミン1,4'-二リン酸として構造的に定義される保存された疎水性領域であるリピドAに共有結合した抗原性の可変サイズの糖鎖からなる糖リン脂質である。LPSの毒性は、リピドAにより哺乳動物免疫系のB細胞およびマクロファージとの相互作用を通して発現され、それは、宿主にとって致命的結果を生じうる、炎症性サイトカイン、主にTNF、の分泌に導く過程である。リピドAはまた、「インビボ」でマウスCD4+およびCD8+ T細胞だけでなく、「インビトロ」でヒトTリンパ球(Th-1)も活性化し、それは宿主の免疫系がLPSの可変サイズの糖鎖に対して特定の既往性IgG抗体応答を開始するのを可能にする性質である。これらに基づいて、LPSは最近、「インビボ」でのT細胞依存性抗原として認識されている。
【0004】
毒性を完全に発現するために、LPSは、リピドAを形成する糖リン脂質単量体の数個の単位の会合を通してのその超分子構造を保持しなければならない。この分子の立体構造的再編成はまた、免疫原性特性の完全な発現のための基礎となる。
【0005】
敗血症および敗血性ショックは、細菌により、および上記病態の原因である細菌によって産生される内毒素であるLPSにより、引き起こされるはっきりした臨床症状である。
【0006】
敗血症および敗血性ショックの臨床的徴候は、存在する内毒素の量、および疾患過程に経過した時間に依存して異なる。感染の最も早い臨床的徴候は、熱、軽度の鬱病、および食欲不振でありうる。疾患過程をさらに進むと、患者は、心拍数の増加、弱い脈圧、脱水症、歯肉の黒ずみ、冷たい足および耳、正常より低い体温、呼吸数の増加、または下痢を含む、より明らかなショックの徴候を示す。いったん患者が内毒素性ショックの徴候を示したならば、それは緊急事態とみなされるべきであり、すぐに医師に連絡すべきである。
【0007】
抗生物質および他の治療手段の慎重な使用にもかかわらず、内毒素関連障害による死亡率は、深刻な問題のままである。細菌の抗生物質抵抗性、基礎疾患過程の重症度、および支持療法の不適切な実施が、部分的に従来の処置の失敗の原因となっている。必要とされることは、内毒素関連障害を引き起こすグラム陰性細菌の理解の向上である。さらに、内毒素関連障害についての処置の改善が必要とされる。
【発明の開示】
【0008】
発明の概要
本発明は、無毒性グラム陰性細菌を提供する。特に、本発明は、外膜内にリポ多糖(LPS、内毒素)を実質的に欠く、生存可能なグラム陰性細菌(例えば、大腸菌(E. coli))を提供する。本発明はさらに、生存可能な無毒性グラム陰性細菌を作製する方法およびその使用を提供する。本発明はまた、免疫応答を誘導するための、および治療剤を研究かつ開発するための組成物ならびに方法を提供する。
【0009】
本発明の態様は、LPSを欠くグラム陰性細菌(例えば、大腸菌)を用いる広範囲の方法および組成物を提供する。例示的な態様は、下の発明の概要、発明の詳細な説明、および下の実施例セクションに記載されている。本発明はこれらの例示的な態様に限定されない。LPSを欠くグラム陰性細菌は、任意の機構により作製されうる。そのような細菌を作製するための多様な種類の異なる機構は本明細書に記載されている。例えば、いくつかの態様において、KDO合成に関与する遺伝子が突然変異されている(機能性タンパク質の発現を低減させる、または排除するために)。いくつかの態様において、KDOのリピドIVAとの会合に関与する遺伝子が突然変異されている。いくつかの態様において、リピドIVA合成に関与する遺伝子が突然変異されている。いくつかの態様において、LPS産生または提示に関与する他の遺伝子が突然変異されている。本発明は遺伝子突然変異に限定されない。いくつかの態様において、発現は、RNA干渉または他の技術を用いて変化させられる。いくつかの態様において、タンパク質機能は、インヒビター(例えば、合成もしくは天然の競合的または非競合的リガンド、抗体など)を供給することにより変化させられる。いくつかの態様において、改変細菌は、LPS状態に影響するようになされた変化のために、健康、増殖などの維持に有用な栄養物、他の改変、または他の成分をさらに与えられる。本発明の態様は、他に規定がない限り、これらの機構に限定されない。本発明は、LPSを欠く細菌が生存可能であること、様々な経路を通して作製されうること、および様々な設定で使用を見出すことを実証する。
【0010】
LPS層は、グラム陰性細菌の外膜の形および機能の両方にとって必須である。グラム陰性病原性の主要な介在物質であることに加えて、少なくともKDO2-リピドA[2-ケト3-デオキシ-D-マンノ-オクツロソン酸(KDO)]からなるLPS(内毒素)構造は、大腸菌において持続的増殖のために必要な最小構造として長年認識されていた。
【0011】
いくつかの態様において、本発明は、ヒトにおけるLPS誘発性敗血症の既知のアンタゴニストである内毒素的に不活性のLPS前駆体リピドIVAをもっぱら産生するだけにもかかわらず、KDOを欠く生存可能なグラム陰性細菌株を提供する。いくつかの態様において、本発明は、D-アラビノース5-リン酸イソメラーゼ(API)発現を欠く生存可能なグラム陰性細菌を提供する。いくつかの態様において、生存可能なグラム陰性細菌は、その菌株がKDOを実質的に含まないように突然変異を含む。いくつかの態様において、突然変異には、KDO合成または改変に関与する1つまたは複数の遺伝子における1つまたは複数の突然変異が挙げられる。いくつかの態様において、生存可能なグラム陰性細菌は突然変異を含み、その突然変異がLPS生合成経路におけるKDO2とリピドIVAの間の会合を妨げて、リピドIVAのみが外膜へ輸送される。いくつかの態様において、KDOにおける1つもしくは複数の突然変異、またはLPS生合成経路における1つもしくは複数の突然変異には、限定されるわけではないが、遺伝子gutQ、kdsD(yrbH)、kdsA、kdsB、waaA、msbA、およびyhjDにおける突然変異が挙げられる。いくつかの態様において、菌株は、KDOタンパク質の合成を欠く、または実質的に欠く。いくつかの態様において、生存可能なグラム陰性細菌の外膜はリピドIVaを発現する。いくつかの態様において、グラム陰性細菌は大腸菌である。
【0012】
特定の態様において、本発明は、生存可能なグラム陰性細菌からリピドIVaを抽出する段階を含む、リピドIVaを作製する方法を提供する。
【0013】
特定の態様において、本発明は、グラム陰性細菌から単離されたリピドIVaを含む組成物を内毒素関連障害をもつ被験体に投与する段階を含む、内毒素関連障害を処置する方法を提供する。
【0014】
特定の態様において、本発明は、グラム陰性細菌に対して免疫応答を誘導するための外膜ワクチンまたは他の組成物を提供し、この組成物は、本発明の生存可能なグラム陰性細菌の外膜を含む。そのような組成物は、研究、薬物スクリーニング、および治療的設定において免疫応答を誘導するために用いられうる。
【0015】
特定の態様において、本発明は、グラム陰性細菌から単離されたリピドIVaを含むアジュバントを提供する。
【0016】
特定の態様において、本発明は、gutQ、kdsD(yrbH)、kdsA、kdsB、waaA、msbA、および/もしくはyhjDの1つもしくは複数の遺伝子の発現、または外膜LPS提示に関連した任意の他の生合成、プロセシング、もしくは輸送遺伝子の発現を欠く生存可能なグラム陰性細菌を提供する。本発明の細菌、またはその部分(例えば、膜画分)は、研究および治療的適用に使用を見出す。
【0017】
定義
本発明の理解を促進するために、いくつかの用語が下に定義されている。
【0018】
本明細書に用いられる場合、「被験体」および「患者」という用語は、イヌ、ネコ、鳥、家畜、および好ましくはヒトのような哺乳動物などの任意の動物を指す。
【0019】
本明細書に用いられる場合、「LPS関連障害」、「内毒素に関係した状態」、「内毒素関係障害」、「内毒素関連障害」、「敗血症」、「敗血症関連障害」という用語、または同類語は、LPSに関連した任意の状態、例えば、菌血症、または血流への、もしくは胃腸外粘膜表面(例えば、肺)上へのリポ多糖の導入に関係した状態を記載する。そのような障害には、限定されるわけではないが、内毒素関連ショック、内毒素関連播種性血管内凝固、内毒素関連貧血、内毒素関連血小板減少症、内毒素関連成人呼吸窮迫症候群、内毒素関連腎不全、内毒素関連肝臓疾患または肝炎、グラム陰性感染に起因する全身性免疫応答症候群(SIRS)、グラム陰性新生児敗血症、グラム陰性髄膜炎、グラム陰性肺炎、グラム陰性感染に起因する好中球減少症および/または白血球減少症、血行動態ショック、ならびに内毒素関連発熱(pyresis)が挙げられる。
【0020】
「生存可能な無毒性グラム陰性細菌」という用語は、LPSを実質的に含まない外膜を含む生存可能なグラム陰性細菌株を指す。
【0021】
「細胞」および「宿主細胞」および「組換え宿主細胞」という用語は、本明細書で交換可能に用いられるが、ベクター、典型的には発現ベクターで形質転換される能力がある、または形質転換された細胞を指す。本明細書に用いられる宿主細胞は、好ましくは、グラム陰性細菌である。そのような用語が特定の対象細胞だけでなく、そのような細胞の子孫または潜在的子孫も指すことは理解される。特定の改変が突然変異かまたは環境的影響のいずれかにより後世に起こりうるため、そのような子孫は、実際、親細胞と同一でない可能性があるが、本明細書に用いられる場合の用語の範囲内にさらに含まれる。
【0022】
「培地」という用語は、当技術分野において認識されており、一般的には、生細胞の培養に用いられる任意の物質または調製物を指す。
【0023】
例えば、グラム陰性細菌の菌株からリピドIVaを得るという文脈において用いられる場合の「に由来する」という用語は、細菌またはタンパク質から得られうるリピドIVaを指し、タンパク質の断片または部分を含むことが意図される。
【0024】
遺伝子または遺伝子発現に関して、本明細書に用いられる場合の「欠陥のある」という用語は、遺伝子が野生型遺伝子ではないこと、ならびに生物体が野生型の遺伝子型および/または野生型の表現型をもたないことを意味する。欠陥のある遺伝子、遺伝子型、または表現型は、その正常な発現が乱されるもしくは失われるような、その遺伝子における、または遺伝子の発現(例えば、転写または転写後の)を制御する遺伝子の、突然変異の結果である可能性がある。「乱された遺伝子発現」は、完全な阻害、および野生型遺伝子発現より低い遺伝子発現の減少(例えば、漏出突然変異におけるような)の両方を含むことが意図される。
【0025】
「グラム陰性細菌」という用語は、当技術分野において認識されており、一般的には、グラム染色(例えば、クリスタルバイオレットとヨウ素の間の着色複合体の沈着)を保持しない細菌を指す。例示的なグラム染色において、細胞を、まず、加熱によりスライドに固定し、すべての細菌(すなわち、グラム陰性およびグラム陽性の両方)により取り込まれる塩基性色素(例えば、クリスタルバイオレット)で染色する。スライドを、その後、染色を定着させるためにヨウ素-KI混合物で処理し、アセトンまたはアルコールで洗浄し、最後に、異なる色の淡染色性色素(例えば、サフラニン)で対比染色する。グラム陽性生物体は、最初のバイオレット染色を保持するが、グラム陰性生物体は、有機溶媒により脱色され、このゆえに、対比染色を示す。例示的なグラム陰性細菌および細胞系には、限定されるわけではないが、エシェリキア種(Escherichia spp.)、シゲラ種(Shigella spp.)、サルモネラ種(Salmonella spp.)、カンピロバクター種(Campylobacter spp.)、ナイセリア種(Neisseria spp.)、ヘモフィルス種(Haemophilus spp.)、アエロモナス種(Aeromonas spp.)、フランシセラ種(Francisella spp.)、エルシニア種(Yersinia spp.)、クレブシエラ種(Klebsiella spp.)、ボルデテラ種(Bordetella spp.)、レジオネラ種(Legionella spp.)、コリネバクテリア種(Corynebacteria spp.)、シトロバクター種(Citrobacter spp.)、クラミジア種(Chlamydia spp.)、ブルセラ種(Brucella spp.)、シュードモナス種(Pseudomonas spp.)、ヘリコバクター種(Helicobacter spp.)、およびビブリオ種(Vibrio spp.)が挙げられる。
【0026】
本明細書に用いられる場合、「突然変異体グラム陰性細菌」、「LPS突然変異体グラム陰性細菌」、「kdsDおよびgutQ突然変異体グラム陰性細菌」、「API突然変異体グラム陰性細菌」という用語、または同類語は、例えば、gutQ、kdsD、kdsA、kdsB、waaA、msbA、yhjD遺伝子、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の1つまたは複数において1回または複数回、突然変異されており、それにより、LPSタンパク質発現を実質的に欠く外膜を生成する、本発明のグラム陰性細菌を含む。
【0027】
「分子の免疫原性部分」は、被験体においてその分子に対して免疫反応を誘発する能力がある分子の部分を指す。
【0028】
LPSまたはリピドIVa分子に適用される場合の「単離される」という用語は、他の細菌成分から、特に外膜から、単離されている(例えば、部分的または完全な単離)LPSまたはリピドIVaを指す。
【0029】
本明細書に用いられる場合、配列(例えば、タンパク質のアミノ酸配列、遺伝子の核酸配列)に関して用いられる時の「部分」という用語は、参照配列の任意の量(例えば、アミノ酸配列または核酸配列の0.001%、0.1%、1%、10%、30%、50%、75%、80%、85%、90%、95%、98%、99.999%)を表す。
【0030】
本明細書に用いられる場合の「調節」という用語は、上方制御(すなわち、活性化または刺激(例えば、アゴナイズする(agonizing)、または増強することにより))および下方制御(すなわち、阻害または抑制(例えば、アンタゴナイズする(antagonizing)、減少させる、または阻害することにより))の両方を指す。「誘導性」という用語は、特に、恒常的ではなく、刺激(例えば、温度、重金属、または他の培地添加物)に応答して起こる遺伝子発現を指す。
【0031】
「非ヒト動物」という用語は、処置されうる、または本発明を試験するのに用いられうる任意の動物を含み、非ヒト霊長類、齧歯類、ヒツジ、イヌ、ウシ、ブタ、ニワトリなどの哺乳動物、および両生類、爬虫類などが挙げられる。好ましい非ヒト動物は、霊長類ファミリーまたは齧歯類ファミリー(例えば、ラットおよびマウス)から選択される。
【0032】
「核酸」という用語は、デオキシリボ核酸(DNA)、および適切な場合にはリボ核酸(RNA)、などのポリヌクレオチドまたはオリゴヌクレオチドを指す。その用語がまた、等価物として、ヌクレオチド類似体から生成したRNAまたはDNAのいずれかの類似体、ならびに記載されることになっている態様に応じて適切に、一本鎖(センスまたはアンチセンス)および二本鎖ポリヌクレオチドを含むことは理解されているはずである。
【0033】
「薬学的に許容される」という用語は、生物学的にまたはその他の点で望ましくないことはない物質、すなわち、その物質は、任意の望ましくない生物学的効果を引き起こすこと、またはそれが含まれる薬学的組成物の他の化合物のいずれかと有害な様式で相互作用することなしに、選択された化合物と共に個体へ投与されうる。
【0034】
「発熱性の」または「発熱性」は、被験体に投与された場合、発熱または発熱応答を誘導する化合物の能力を指す。そのような発熱応答は、一般的に、宿主炎症性サイトカインIL-1、IL-6、および/またはTNF-αにより媒介され、その分泌は、例えば、LPSにより、誘導される。
【0035】
「低下した発熱性」をもつ物質、または「発熱性の低下した誘導体」は、対応物質より低い発熱活性、例えば、対応物質に対して約80%未満の発熱性、好ましくは、約70%未満の発熱性、より好ましくは約60%未満の発熱性、より好ましくは約50°未満の発熱性、より好ましくは約40%未満の発熱性、およびよりいっそう好ましくは約30%未満の発熱性を有する物質を指す。換言すれば、低下した発熱性をもつ物質は、本明細書に記載された、または当技術分野において公知のアッセイ法のいずれかにより測定される場合、対応する物質より少なくとも約20%、30%、40%、50%、60%、または70%低い発熱性である。
【0036】
「実質的に低下した発熱性」または「実質的に低下した発熱性誘導体」は、それが野生型物質に対して20%未満の発熱性、好ましくは、野生型物質に対して、10%未満の発熱性、好ましくは1%未満の発熱性、好ましくは10-1%未満の発熱性、好ましくは10-2%未満の発熱性、好ましくは10-3%未満の発熱性、好ましくは10-4%未満の発熱性、好ましくは10-5%未満の発熱性、および最も好ましくは10-6%未満の発熱性を有するように変化している物質(例えば、生存可能な無毒性グラム陰性細菌により産生される)を指す。換言すれば、実質的に低下した発熱性をもつ物質は、本明細書に記載された、または当技術分野において公知のアッセイ法のいずれかにより測定される場合、対応する未変化物質に対して、少なくとも約90%低い、99%低い、1/10倍、約1/100倍、約1/1000倍、少なくとも約1/1000倍、少なくとも約1/10000倍、少なくとも約1/1000000倍の発熱性である。
【0037】
本明細書に用いられる場合、「トランスフェクション」という用語は、核酸媒介性遺伝子移入によるレシピエント細胞への核酸の導入(例えば、発現ベクターを介して)を意味する。本明細書に用いられる場合、「形質転換」は、細胞の遺伝子型が外因性DNAまたはRNAの細胞取り込みの結果として変化している過程を指す。例証的な態様において、形質転換細胞は、kdsDおよびgutQ遺伝子の1つまたは複数の突然変異体型を発現するものである。形質転換細胞はまた、gutQ、kdsD、kdsA、kdsB、waaA、msbA、ynjD遺伝子、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の発現を干渉する核酸を発現するものであることができる。
【0038】
本明細書に用いられる場合、「トランスジーン」は、細胞へ導入されている核酸(例えば、突然変異体kdsD、gutQ、kdsA、kdsB、waaA、msbA、ynjD遺伝子、または任意の他の生合成、プロセシング、もしくは輸送遺伝子、またはそれらのアンチセンス転写産物)を意味する。トランスジーンは、それが導入されているトランスジェニック動物もしくは細胞に対して、部分的にもしくは完全に異種性、すなわち、外来性でありうる、またはそれが導入されている生物体もしくは細胞の内因性遺伝子に相同性でありうるが、それが挿入されている細胞のゲノムを変化させるように、動物もしくは細胞のゲノムへ挿入されるように設計される、もしくは挿入される。トランスジーンはまた、エピソームの形をとって細胞に存在しうる。
【0039】
本明細書に用いられる場合、状態または疾患について被験体を「処置する」という用語は、状態または疾患の少なくとも1つの症状を治すこと、および寛解させることを含むよう意図される。
【0040】
「ベクター」という用語は、それが連結している別の核酸を輸送する能力がある核酸分子を指す。機能的に連結している遺伝子の発現を方向づける能力があるベクターは、本明細書では「発現ベクター」と呼ばれる。本明細書に用いられる場合、「発現系」という用語は、mRNAが転写されうる、および/またはmRNAがタンパク質、構造RNA、もしくは他の細胞成分へ翻訳されうる条件下での発現ベクターを指す。発現系は、インビトロ発現系であり、市販されているか、もしくは当技術分野公知の技術に従って容易に作製される、または発現ベクターを含む真核細胞もしくは原核細胞などのインビボ発現系でありうる。一般的に、組換えDNA技術における有用な発現ベクターは、しばしば、「プラスミド」の形をとり、そのプラスミドは、一般的に、それらのベクター型において、染色体に結合していない環状二本鎖DNAループを指す。本明細書において、「プラスミド」および「ベクター」は、プラスミドが最も一般的に用いられるベクターの形であるため、交換可能に用いられる。しかしながら、本発明は、等価の機能を果たし、かつ当技術分野において周知である、またはこの点に関して後で当技術分野において知られるようになる、そのような他の型の発現ベクター(例えば、コスミド、ファージミド、およびバクテリオファージベクター)を含むことを意図される。
【0041】
発明の詳細な説明
本発明は、無毒性グラム陰性細菌を提供する。特に、本発明は、外膜内にリポ多糖(LPS、内毒素)を実質的に欠く、生存可能なグラム陰性細菌(例えば、大腸菌)を提供する。本発明はさらに、生存可能な無毒性グラム陰性細菌を作製する方法およびその使用を提供する。本発明はまた、免疫応答を誘導するための、および治療剤を研究かつ開発するための組成物ならびに方法を提供する。
【0042】
グラム陰性細菌は、ペプチドグリカンを囲む非対称脂質二重層である、外膜(OM)を有する。OM内層は、主として様々なグリセロリン脂質で構成されるが、外層は、主に、固有の両親媒性高分子である、リポ多糖(LPS)を含む。大腸菌および他の密接に関連した腸内細菌において、OM総重量の〜30%を占める、総細胞表面積の75%近くを網羅する細胞あたり〜106個のLPS分子がある(例えば、S.M. Galloway, C.R. Raetz, J. Biol. Chem. 265, 6394 (1990); L. Leive, Ann. N.Y. Acad. Sci. 235, 109 (1974); H. Nikaido, Escherichia coli and Salmonella typhimurium: cellular and molecular biology, F.C. Neidhardt, Ed. (American Society for Microbiology, Washington, D.C., 1987), vol. 1, pp. 29-47参照;それぞれは全体として参照により本明細書に組み入れられている)。細菌細胞と水性環境の間の界面での露出された位置は、主要なOM関連表面抗原としてLPSを提示する。LPSは、宿主免疫応答に関連した、多様な範囲の病理学的および生理学的活性に関与している(例えば、A. Wiese, et al., Biol. Chem. 380, 767 (1999); H. Heine, et al., Mol. Biotechnol. 19, 279 (2001)参照;それぞれは全体として参照により本明細書に組み入れられている)。LPSは、グラム陰性病原性および全身性炎症の介在物質として認識された免疫刺激性/炎症性分子であり、それとして、内毒素という用語は、しばしば、LPSと交換可能に用いられる。LPS層は、グラム陰性細菌のOMの形および機能の両方に必須である。従って、グラム陰性病原性におけるキープレーヤーであることに加えて、LPSはまた、細菌の生存の重要な決定因子である。
【0043】
様々なグラム陰性細菌のLPSは、以下の3つの領域へ概念的に分けられる一般的な構造的構築に従う:OMに埋め込まれたリピドA、オリゴ糖コア、続いて、腸内細菌科におけるn回の繰り返し単位からなるO特異的親水性多糖鎖、または髄膜炎菌(Neisseria meningitis)、淋菌(N. gonorrhoeae)、インフルエンザ菌(Haemophilus influenzae)、百日咳菌(Bordetella pertussis)、およびクラミジア種(Chlamydia spp.)などのヒト粘膜病原体を含む特定の細菌における短い分岐オリゴ糖。リピドAはグラム陰性細菌属の間で最も保存されているLPSドメインであり、かつ宿主内での生物活性の原因である構造的構成要素であることは、LPSの内毒素原理を表している。腸内細菌において、リピドAは、エステル-(3,3')結合またはアミド-(2,2')結合で4つの(R)-3-ヒドロキシ-ミリスチン酸でアシル化されるβ-1,6-結合型D-グルコサミン二糖バックボーンからなる。大腸菌野生型株の成熟リピドA分子は、典型的には、非還元グルコサミンの(R)-3-ヒドロキシミリストイル基に付着した、2つの追加のアシル鎖、主としてラウレートおよびミリステートを含み、リピドAの特徴的なアシルオキシアシル単位を形成する。オリゴ糖コアは、リピドAを超可変多糖鎖に接続して、内側および外側のオリゴ糖コア領域へさらに分けられる。外側コアはあまり保存されておらず、糖組成およびグリコシド結合の両方の点で異なっているが、グラム陰性細菌の大部分は、少なくとも1つの2-ケト3-ヒドロキシ-D-マンノ-オクツロソン酸(KDO)分子を含む内側コアを産生する。
【0044】
KDOは、ほとんど全部のLPS構造に見出される保存された残基である、LPSの必須構成要素である(例えば、O. Holst, Trends Glycosci. Glycotechnol. 14,87 (2002)参照;全体として参照により本明細書に組み入れられている)。大腸菌の増殖に必要とされる最小LPS構造は、リピドAに付着した2つのKDO残基(KDO2-リピドAまたはRe内毒素)であり(例えば、C.R. Raetz, C. Whitfield, Annu. Rev. Biochem. 71, 635 (2002); S. Gronow, H. Brade, J. Endotoxin Res. 7, 3(2001)参照;それぞれは全体として参照により本明細書に組み入れられている)、細菌細胞の完全性および生存能力の維持におけるKDOの重要性を強調している。L-APIは、大腸菌K-12におけるkdsD遺伝子によりコードされる(例えば、Meredith, T.C. & Woodard, R.W. (2003) J Biol Chem 278, 32771-7参照;全体として参照により本明細書に組み入れられている)。
【0045】
LPS構造内のKDOの遍在性は、その生合成の解明を促進させた。経路は、酵素d-アラビノース5-リン酸(A5P)イソメラーゼ(API)により惹起され、その酵素は、ペントース経路中間体D-リブロース5-リン酸をA5Pへ変換する。その後、A5Pは、ホスホエノールピルビン酸と縮合してKdo 8-リン酸(Kdo8P)(KdsA)を形成し、Kdo(KdsC)へ加水分解され、最後にCMP-KdoからアクセプターのリピドIVA(WaaA)へ転移される前に、糖ヌクレオチドCMP-Kdo(KdsB)として活性化される(図11)。後期アシルトランスフェラーゼLpxLおよびLpxMは、次に、脂肪酸、ラウレートおよびミリステートを、それぞれ、Kdo2-リピドIVAへ転移し、ヘキサアシル化Kdo-リピドAの特徴的なアシルオキシアシル単位を形成する。大腸菌K-12において、2つのAPI遺伝子(kdsDおよびgutQ)がある。
【0046】
相同性検索に基づいて他のAPIが大腸菌に存在する可能性があることが推測された。特に、グルシトールオペロンgutQの最後のオープンリーディングは、kdsD(以前は、yrbH)と有意な相同性(45%同一性)を有する。G-APIは、7つの収束的に転写される遺伝子を含むgutAEBDMRQオペロンの最終遺伝子産物である(図10)。表1に示されているように、gutQおよびkdsDは、類似した生化学的性質を共有する。
【0047】
(表1)kdsDおよびgutQの生化学的性質
aYamada, M., Yamada, Y. & Saier, M.H., Jr. (1990) DNA Seq 1, 141-5(全体として参照により本明細書に組み入れられている)からのデータ。b31P NMRにより測定され;Haldane関係(Ru5P/A5P)から計算された。c試験された基質について実験手順を参照。d高分解能誘導結合プラズマ質量分析法により測定された、単量体あたりのZn2+の当量。e5%未満の活性残存。f10μM EDTAにより酵素を単離した場合。gエレクトロスプレーイオン化質量分析法により測定され;タンパク質配列から計算された。hゲル濾過により測定された。
【0048】
グルシトールオペロンは、環境からのD-グルシトールの協調的取り込みおよび異化反応に関与するホスホエノールピルビン酸:糖ホスホトランスフェラーゼ系(PTS)を発現する(例えば、T.C. Meredith, R.W. Woodard, J. Biol. Chem. 278, 32771 (2003)参照;全体として参照により本明細書に組み入れられている)。そのオペロンは、最初にLengeler(例えば、C. Galanos, et al., Eur. J. Biochem. 9, 245 (1969); S. Muller-Loennies, et al., J. Biol. Chem. 278, 34090 (2003)参照;それぞれは全体として参照により本明細書に組み入れられている)により、その後、Saier(例えば、K.A. Brozek, C.R. Raetz, J. Biol. Chem. 265, 15410 (1990); H. Nikaido, Microbiol. Mol. Biol. Rev. 67, 593 (2003)参照;それぞれは全体として参照により本明細書に組み入れられている)により、研究され、7つの収束的に転写される遺伝子、gutAEBDMRQからなることが知られている。EIIGut複合体は、gutA(EIIC1ドメイン)、gutE(EIIBC2ドメイン)、およびgutB(EIIAドメイン)により形成され、D-グルシトールを内膜を渡って、D-グルシトール6-リン酸として細胞へ輸送する。D-グルシトール6-リン酸は、その後さらに、NADH依存性脱水素酵素である、gutDにより、解糖中間体D-フルクトース6-リン酸へ代謝される。gutオペロンの発現は、cAMP-CAP(環状アデノシン一リン酸-カタボライト活性化タンパク質)媒介性制御に加えて、転写リプレッサー(gutR)および転写アクチベーター(gutM)からなる複雑な多成分制御系により厳重に調節されている(例えば、C.J. Belunis, et al., J. Biol. Chem. 270, 27646 (1995)参照;全体として参照により本明細書に組み入れられている)。しかしながら、gutQの機能はまだわかっていない(例えば、R.C. Goldman, W.E. Kohlbrenner, J. Bacteriol. 163, 256 (1985)参照;全体として参照により本明細書に組み入れられている)。
【0049】
本発明の態様の開発中に行われた実験において、外膜LPS発現を実質的に欠く生存可能なグラム陰性細菌は、ヒトにおけるLPS誘発性敗血症の公知のアンタゴニストである、内毒素的に不活性のLPS前駆体リピドIVaをもっぱら産生するにもかかわらず、構築された。本発明は、外膜LPS発現を実質的に欠く生存可能なグラム陰性細菌を構築する特定の方法(例えば、API発現の抑制による;gutQおよび/またはkdsD遺伝子の突然変異による;KDO発現の抑制による;KDOとリピドIVAの間の会合の阻害による;kdsAおよび/もしくはkdsBおよび/もしくはwaaAおよび/もしくはmsbAおよび/もしくはyhjD遺伝子、または他の生合成、プロセシング、もしくは輸送遺伝子の突然変異による;リピドIVA発現の抑制による;lpxM遺伝子、またはリピドIVAについての他の生合成、プロセシング、もしくは輸送遺伝子の突然変異による)に限定されない。
【0050】
本発明は、外膜LPS発現を実質的に欠く生存可能なグラム陰性細菌の構築における任意の型のグラム陰性細菌株の使用を企図する。本発明に有用なグラム陰性細菌の例には、限定されるわけではないが、エシェリキア種、シゲラ種、サルモネラ種、カンピロバクター種、ナイセリア種、ヘモフィルス種、アエロモナス種、フランシセラ種、エルシニア種、クレブシエラ種、ボルデテラ種、レジオネラ種、コリネバクテリア種、シトロバクター種、クラミジア種、ブルセラ種、シュードモナス種、ヘリコバクター種、およびビブリオ種が挙げられる。好ましい態様において、大腸菌が用いられる。用いられうるエシェリキア株の例には、限定されるわけではないが、大腸菌(Escherichia coli)(E. coli)株DH5a、HB 101、HS-4、4608-58、1-184-68、53638-C-17、13-80、および6-81(例えば、Sambrook, etal., (Eds.), 1993, Molecular Cloning, Cold Spring Harbor Press, Cold Spring Harbor, N.Y.); Grant, et al., 1990, Proc. Natl. Acad. Sci., USA, 87:4645; Sansonetti, et al., 1982, Ann. Microbiol. (Inst. Pasteur), 132A:351参照)、腸内毒素原性大腸菌(Evans, et al., 1975, Infect. Immun., 12:656)、腸管病原性大腸菌(Donnenberg, et al., 1994, J. Infect. Dis., 169:831;それぞれは全体として参照により本明細書に組み入れられている)、ならびに腸管出血性大腸菌(例えば、McKee and O'Brien, 1995, Infect. Immun., 63:2070)が挙げられる。
【0051】
本発明は、突然変異体グラム陰性細菌株(例えば、kdsDおよび/もしくはgutQ、kdsA、kdsB、waaA、msbA、ynjD遺伝子、または他の生合成、プロセシング、もしくは輸送遺伝子に突然変異を有するグラム陰性細菌株)の増殖のために特定の培養条件に限定されない。例示を目的として、細菌は、増殖されることになっている細菌株に適切である通常の培養技術を用いて、LB培地(Difco, Detroit Mich.)、Nutrientブロス(Difco)、Tryptic Soyブロス(Difco)、またはM9最少ブロス(Difco)などの、細菌増殖に適した任意の標準液体培地において増殖することができる(Miller, 1991, 前記)。代替として、細菌は、L-寒天(Difco)、Nutrient寒天(Difco)、Tryptic Soy寒天(Difco)、またはM9最少寒天(Difco)などの固体培地上で培養されうる。菌株が突然変異kdsDおよび/またはgutQを含むグラム陰性細菌株について、外因性D-アラビノース5-リン酸源が細菌増殖および生存に用いられる(Meredith et al., 2006, ACS Chem. Biol. 1:33-42;全体として参照により本明細書に組み入れられている)。または、本発明のいくつかの態様の開発中に行われた実験は、kdsDおよび/またはgutQ突然変異を含む菌株におけるmsbA遺伝子の過剰発現が細菌増殖および生存のためのD-アラビノース5-リン酸による補充の代替であることを示している。
【0052】
いくつかの態様において、本発明は、gutQ、kdsD(yrbH)、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子における突然変異を有する、生存可能なグラム陰性細菌を提供する。いくつかの態様において、gutQおよびkdsD遺伝子の突然変異は、細菌株内のAPI発現を阻害し、それがKDO発現を阻害し、それが外膜LPS発現を阻害する。いくつかの態様において、本発明は、kdsA遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供する。いくつかの態様において、本発明は、kdsB遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供する。いくつかの態様において、本発明は、waaA遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供する。本発明の態様の開発中に行われた実験は、kdsA、kdsB、および/またはwaaAの突然変異が、KDOの産生またはKDO2とリピドIVAの間の会合を阻止して、リピドIVAのみが外膜へ輸送されることにより、LPS生合成経路を阻害することを示した。細菌細胞は生き延び、LPSを含まず、かつ無毒性である。いくつかの態様において、本発明は、gutQ、kdsD、kdsA、kdsB、および/またはwaaA遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供し、msbA遺伝子における突然変異をさらに含む。いくつかの態様において、本発明は、gutQ、kdsD、kdsA、kdsB、またはwaaA遺伝子における突然変異を有する生存可能なグラム陰性細菌を提供し、yhjD遺伝子における突然変異をさらに含む。
【0053】
本発明は、グラム陰性細菌内に遺伝子突然変異を導入するための任意の技術の使用を企図する。そのような技術には、限定されるわけではないが、N-メチル-N'-ニトロN-ニトロソグアニジン、アクリジンオレンジ、臭化エチジウムなどの化学物質、または紫外線への非致死的曝露を用いる非特異的突然変異誘発が挙げられる(例えば、Miller (Ed.), 1991, A Short Course in Bacterial Genetics, Cold Spring Harbor Press, Cold Spring Harbor, N.Y.参照;全体として参照により本明細書に組み入れられている)。または、突然変異は、Tn10突然変異誘発、バクテリオファージ媒介性形質導入、λファージ媒介性対立遺伝子交換、もしくは接合伝達、または組換えDNA技術を用いる部位特異的突然変異誘発を用いて導入されうる(例えば、Miller (Ed.), 1991, 前記; Hone, et al., 1987, J. Infect. Dis., 156:167; Noriega, et al., 1994, Infect. Immun., 62:5168; Hone, et al., 1991, Vaccine, 9:810; Chatfield, et al., 1992, Vaccine, 10:53; Pickard, et al., 1994, Infect. Immun., 62:3984; Odegaard, et al., 1997, J. Biol. Chem., 272:19688; Lee, et al., 1995, J. Biol. Chem., 270:27151; Garrett, et al., 1998, J. Biol. Chem., 273:12457参照;それぞれは全体として参照により本明細書に組み入れられている)。突然変異を導入するための任意の方法を用いてもよく、その突然変異を、1つまたは複数の追加の突然変異と共に導入することができる。例えば、いくつかの態様において、本発明は、gutQ、kdsD、kdsA、kdsB、waaA、msbA、yhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子における突然変異などの1つより多い突然変異を有する生存可能なグラム陰性細菌を提供する。
【0054】
いくつかの態様において、グラム陰性細菌内の突然変異(例えば、gutQ、kdsD(yrbH)、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の突然変異)における突然変異は、恒常的に発現しているか、または、例えば、プロモーターの温度感受性熱ショックファミリーもしくは嫌気的に誘導されるnirBプロモーター(例えば、Harborne, et al., 1992, Mol. Micro., 6:2805参照;全体として参照により本明細書に組み入れられている)などの誘導性プロモーターまたはuapA(例えば、Gorfinkiel, et al., 1993, J. Biol. Chem., 268:23376参照;全体として参照により本明細書に組み入れられている)もしくはgcv(例えば、Stauffer, et al., 1994, J. Bact, 176:6159参照;全体として参照により本明細書に組み入れられている)などの抑制性プロモーターの制御下にあるかのいずれかである。適切なプロモーターの選択は、宿主細菌株に依存し、当業者にとって明らかであると思われる。
【0055】
いくつかの態様において、本発明は、API発現を欠く生存可能なグラム陰性細菌(例えば、大腸菌)を提供する。本発明は、API発現を阻害する特定の方法に限定されない。いくつかの態様において、API発現は、KDOタンパク質発現の抑制によって阻害される。本発明は、KDOタンパク質発現を抑制する特定の方法に限定されない。いくつかの態様において、KDOタンパク質発現は、例えば、gutQ遺伝子、kdsD遺伝子、kdsA遺伝子、もしくはkdsB遺伝子の突然変異、または任意の他のKDO生合成遺伝子における突然変異によって抑制される。
【0056】
いくつかの態様において、本発明は、生存可能な無毒性(例えば、内毒素を含まない)グラム陰性細菌(例えば、大腸菌)を提供する。本発明は、生存可能な無毒性グラム陰性細菌を提供する特定の方法に限定されない。いくつかの態様において、生存可能な無毒性グラム陰性細菌は、外膜におけるLPS発現の抑制によって提供される。本発明は、外膜におけるLPS発現の抑制についての特定の方法に限定されない。いくつかの態様において、LPS発現は、APIタンパク質発現の抑制によって抑制される。本発明は、API発現を抑制する特定の方法に限定されない。いくつかの態様において、API発現は、KDOタンパク質発現の抑制によって抑制される。本発明は、KDOタンパク質発現を抑制する特定の方法に限定されない。いくつかの態様において、KDOタンパク質発現は、例えば、gutQ遺伝子およびkdsD遺伝子の突然変異によって抑制される。いくつかの態様において、KDOタンパク質がリピドIVAと会合しないために、外膜におけるKDOタンパク質発現が生じず、リピドIVAのみが外膜へ輸送される。例えば、gutQ、kdsD、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の突然変異は、KDO2-リピドIVA複合体の形成または膜提示を排除し、結果として、例えば、リピドIVA分子のみが外膜へ輸送されることを生じ、その後のLPS形成は生じない。
【0057】
いくつかの態様において、生存可能な無毒性グラム陰性細菌は、限定されるわけではないが、他の細菌生物体由来のLPS、固有の脂質誘導体、ヒトタンパク質またはペプチド産生、非ヒトタンパク質またはペプチド産生、ワクチン作製などの非天然タンパク質およびペプチドを発現する、産生する、およびディスプレイするための当業者に公知のクローニング方法(Sambrook et al., Molecular Cloning; A Laboratory Manual, Cold Spring Harbor Laboratory Press参照;全体として参照により本明細書に組み入れられている)により遺伝子操作されうる。そのような産生された産物は、限定されるわけではないが、臨床的治療および基礎研究活動を含む様々な適用に有用性を見出す。
【0058】
いくつかの態様において、本発明は、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌(例えば、大腸菌)を提供する。本発明は、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌を提供する特定の方法に限定されない。いくつかの態様において、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌は、APIタンパク質発現の抑制によって達成される。本発明は、APIタンパク質発現を抑制する特定の方法に限定されない。本発明は、API発現を抑制する特定の方法に限定されない。いくつかの態様において、API発現は、KDOタンパク質発現の抑制によって抑制される。本発明は、KDOタンパク質発現を抑制する特定の方法に限定されない。いくつかの態様において、KDOタンパク質発現は、例えば、gutQ、kdsD、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子の突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の突然変異によって抑制される。いくつかの態様において、リピドIVAを発現する外膜を含む、LPSを含まない生存可能なグラム陰性細菌は、リピドIVAのみが外膜へ輸送される(例えば、KDOなしで)ように、KDOとリピドIVAの間の会合を阻害することにより達成される。本発明は、KDOとリピドIVAの間の会合を阻害する特定の方法に限定されない。いくつかの態様において、KDOとリピドIVAの会合は、例えば、gutQ、kdsD、kdsA、kdsB、waaA、msbA、および/もしくはyhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送遺伝子の任意の突然変異により阻害される。いくつかの態様において、本発明は、生存可能な無毒性グラム陰性細菌(例えば、大腸菌)から単離されたリピドIVaを提供する。リピドIVaは、例えば、哺乳動物の敗血性ショックシグナル伝達経路を研究することにおいて、およびLPS型分子の合成におけるビルディングブロックとして用いられる。リピドIVaを単離するための現行の方法は、従来の全有機合成、成熟LPSの分解、または所望のリピドIVaの画分を含む異種性LPS層を産生する条件的突然変異体からの精製を含む。そのような方法の欠点には、低いリピドIVa収量および高い労働量が挙げられる。本発明の生存可能な無毒性グラム陰性細菌からのリピドIVaの単離は、リピドIVaの外膜存在によりそのような方法を超える有意な改善を示している。
【0059】
いくつかの態様において、本発明は、生存可能な無毒性グラム陰性細菌(例えば、大腸菌)から単離された外膜小胞を提供する。リピドIVaは、敗血性ショックシグナル伝達経路のアンタゴニストであり、急性敗血症をもつ患者を処置するための実行可能なアプローチは、LPSを含むシグナル伝達経路を遮断することである。いくつかの態様において、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌からの単離された外膜小胞が、敗血症関連障害を処置するために、または予防的に阻止するために用いられる。本発明の生存可能な無毒性グラム陰性細菌(例えば、ΔAPI菌株)から調製された外膜小胞は、LPSアンタゴニストとしてリピドIVaを含む。
【0060】
いくつかの態様において、生存可能なグラム陰性細菌(例えば、大腸菌)から単離された外膜小胞が、外膜小胞に基づいたワクチンの改善を目的として用いられる。OMVに基づいたワクチンは、しばしば、きつい化学処理によりLPSを剥ぎ取ることにより「解毒」される。しかしながら、剥ぎ取り方法は、抗体をターゲットする良い候補でありうる、特に他のグラム陰性病原体由来のクローニングされた外膜タンパク質の、OMVワクチンのタンパク質成分に、有害な影響を及ぼす。解毒は、宿主としてのΔAPI突然変異体株に関しては必須ではなく、追加的な安全レベルのを与える。
【0061】
いくつかの態様において、本発明は、リピドIVaおよびLPSの両方の発現をもつ外膜を含むグラム陰性細菌を提供する。LPSの毒性を免疫刺激性から離すことは、LPSに基づいたアジュバントまたはLPSに基づいたワクチンを開発するための大きな課題である。ΔAPI菌株における遮断はLPS経路の初期であるため、他の細菌由来の酵素(リン酸基、エタノールアミン、L-4-デオキシアラビノース、異なるアシル鎖長でLPSを修飾する)および変化した活性を有する突然変異型酵素が、細胞内で固有の生物活性を有するLPS分子のアレイを作製するために用いられる。そのような遺伝子操作についての多くの方法は、すでに大腸菌において存在している。さらに、成熟LPS合成は、増殖培地におけるD-アラビノース5-リン酸の含有により回復され、LPS誘導体の成熟LPSに対する量および比率を調節かつ最適化することを可能にすることができる。そのようなLPS「ブレンド」は、受け入れられる低レベルの潜在的毒性を保持している間、免疫刺激活性の望ましいバランスを達しうる。
【0062】
いくつかの態様において、生存可能な無毒性グラム陰性細菌は、内毒素を含まない治療用分子の産生のための宿主として用いられる。本発明は、特定の治療用分子に限定されない。従来より、グラム陰性細菌における治療用分子の産生は、それが、ワクチンのためのOM小胞か、アジュバントとして用いられうるLPS型分子(モノホスホリルリピドA(MPLA)など)か、組換え薬学的タンパク質、高分子か、または哺乳動物細胞トランスフェクション遺伝子治療のためのDNAかに関わらず、細菌宿主由来の内毒素の存在に悩まされる。LPSの免疫原性能力は十分立証されているが、治療用分子の内毒素の混入は懸念材料である。内毒素混入を軽減するための現行の産生ストラテジーには、内毒素を含まないDNAプラスミド精製、続いて、内毒素レベルを測定するためのアッセイ法のための販売されているキットなどの様々な精製技術が挙げられる。ΔAPI菌株は内毒素を産生しないので、そのような精製段階が必要とされない。そういうものとして、本発明の生存可能な無毒性グラム陰性細菌株(例えば、ΔAPI菌株)は、内毒素を含まない治療用分子(例えば、リピドIVa)を単離する改善された方法を提供する。例えば、ΔAPI菌株は、よく研究されたグラム陰性細菌を用いて内毒素を含まない環境における商業的に重要な治療用分子の産生のための宿主であることが企図される。さらに、gutQ、kdsD、kdsA、kdsB、waaA、msbA、yhjD遺伝子における突然変異、または任意の他の生合成、プロセシング、もしくは輸送細菌遺伝子における突然変異を含む菌株は、グラム陰性細菌を用いて内毒素を含まない環境における商業的に重要な治療用分子の産生のための宿主であることが企図される。
【0063】
いくつかの態様において、生存可能な無毒性グラム陰性細菌は、免疫応答を刺激するワクチンまたは他の組成物の産生のために用いられうる。例えば、腸チフスに対するより毒性の低いワクチンは、本明細書に記載されているようなグラム陰性細菌を用いて作製される。腸チフスについての現行のワクチンは、ワクチン調製物に存在する内毒素のせいで副作用を引き起こす。LPS(例えば、内毒素)が外膜上に提示されない、本明細書に記載されているような生存可能な無毒性グラム陰性細菌またはその部分を利用することが、内毒素が混入されたワクチン調製物により引き起こされる副作用を回避することが企図される。本発明は、内毒素混入が典型的に見出される任意のワクチン調製物または他の組成物において有用性を見出す。本発明の生存可能な無毒性グラム陰性細菌はまた、それらのLPS欠損表現型による弱毒生ワクチンとしての有用性を見出すことが企図される。
【0064】
そういうものとして、本発明は、免疫化および研究を目的として被験体に投与されうる内毒素混入のない免疫応答を誘導するためのOMワクチンおよび他の組成物の開発に使用を見出す。例えば、弱毒またはOMワクチンは、全体として参照により本明細書に組み入れられている、米国特許出願第2005/0013831号または米国特許第6,558,677号に記載されているような、手順を用いて調製されうる。例えば、そのようなワクチンは、外科患者などの敗血性ショック(例えば、大腸菌からの)に罹るリスクがある被験体を免疫するのに有用性を見出す。さらに、内毒素を含まない弱毒またはOMワクチンは、例えば、百日咳(例えば、ボルデテラ種(Bordetella sp.))、ブルセラ症または内毒素性ショック(例えば、ブルセラ種(Brucella sp.))、肺および呼吸器感染(例えば、シュードモナス種(Pseudomonas sp.)、ヘモフィルス種(Haemophilus sp.)、モラクセラ種(Moraxella sp.))、コレラ(例えば、ビブリオ種(Vibrio sp.))、肺炎(例えば、クレブシエラ種(Klebsiella sp.)、ヘモフィルス種)、胃潰瘍(例えば、ヘリコバクター種(Helicobacter sp.))、髄膜炎(例えば、ナイセリア種(Neisseria sp.)、ヘモフィルス種)、中耳炎(例えば、ヘモフィルス種、モラクセラ種)、赤痢および下痢(例えば、シゲラ種(Shigella sp.)、大腸菌、ビブリオ種、カンピロバクター種(Campylobacter sp.)、エルシニア種(Yersenia sp.))、腸熱(例えば、サルモネラ種(Salmonella sp.))、トラコーマおよび性感染症(例えば、クラミジア種(Chlamydia sp.))、ツラレミア(例えば、フランシセラ種(Franciscella sp.))、ならびにペスト(例えば、エルシニア種)に対する免疫化のために開発されうる。
【0065】
いくつかの態様において、本明細書に記載されているような無毒性の生存可能なグラム陰性細菌は、治療的および研究的適用として、治療用抗体の作製に有用性を見出す。例えば、いくつかの態様において、被験体は、無毒性グラム陰性細菌またはその部分(例えば、膜調製物)を用いて能動的に免疫され、ヒト過免疫血清から調製された抗体は、その後、細菌感染および敗血症から被験体を受動的に保護するために用いられる。しかしながら、治療用抗体の作製は、限定されるわけではないが、霊長類、ウサギ、イヌ、モルモット、マウス、ラット、ヒツジ、ヤギなどの宿主動物においてより従来的な方法によって達成される。例えば、治療用抗体は、ヒト被験体への投与について宿主動物において抗体を産生するために自ら免疫原として無毒性の生存可能なグラム陰性細菌を用いて産生される。本明細書に記載されているような無毒性の生存可能なグラム陰性細菌は、さらに、宿主動物において治療用抗体を産生するために用いられる外来抗原(例えば、免疫原性ペプチドまたはタンパク質)を提示するための宿主として有用性を見出す。例えば、無毒性の生存可能なグラム陰性細菌は、LPSが実質的に欠損していることの他に、抗体産生のための免疫原として使用を見出す非天然タンパク質およびペプチドを発現するように遺伝子操作されうる(例えば、当業者に公知の確立されたクローニング方法によって)。そのような免疫原には、限定されるわけではないが、癌細胞および他の病原性細胞へ抗体をターゲットするためのペプチド、ウイルス細胞ターゲティングのためのウイルスのコートタンパク質などが挙げられる。
【0066】
いくつかの態様において、本発明は、治療用抗体産生のために免疫原性タンパク質を提示するのに有用な無毒性の生存可能なグラム陰性細菌を提供する。免疫原性タンパク質に対する抗体は、それが抗原性タンパク質を認識できる限り、任意のモノクローナル抗体またはポリクローナル抗体でありうる。抗体は、当業者に公知の通常の抗体または抗血清調製方法に従って産生されうる。
【0067】
いくつかの態様において、リピドIVaを発現する外膜を含む生存可能なグラム陰性細菌(例えば、大腸菌)が、薬学的スクリーニング(例えば、抗発熱性作用物質についてのスクリーニング)のために用いられる。ΔAPI突然変異体菌株は、非常に低い透過性のバリアを有し、特に、通常にはOMを透過できない大きな疎水性薬物分子に対して感受性を高くさせている。化合物ライブラリーのホールセルバイオアッセイ法は、通常では、OMの透過を促進することによりヒットを同定するのを助けるために、トルエン、EDTA、陽イオン性ペプチドなどの透過処理剤を用いる。いったん親のリードヒットが生じたならば、溶解性、分配、サイズなどを改善するために医薬品化学を用いて、抗生物質を作製することができる。これらのスクリーニングからの多くの可能性のあるリードは、その化合物がOMの内側のそのタンパク質標的にアクセスできないため、見逃されている。そのようなスクリーニングにおいて、例えば、ΔAPI突然変異体菌株を用いることは、追加の試薬を供給する必要なしに透過性バリアを低下させることによりOM透過性問題を多少とも解決する。同様に、ΔAPIのそのような低いOM透過性は、DNAライブラリーの作製中に、例えばΔAPI突然変異体菌株をDNAプラスミドで形質転換する場合に、有利である。高形質転換効率細胞は、すべての組換えDNAテクノロジーに必須であり、ΔAPI菌株はそのような適用のための有用な宿主である。
【0068】
実施例
実施例I.
本実施例は、ΔAPI突然変異体TCM15およびKPM22を記載する。G-APIおよびL-APIの両方が欠失した栄養要求性ΔAPI突然変異体、TCM15を構築し、それは、大腸菌についての確立されたKDO2-リピドA定説に従って増殖のための外因性A5Pに依存するようになった。TCM15は、A5Pを含まなければ、増殖培地、インキュベーション温度、または時間にかかわらず、固体培地上にコロニーを形成することができなかった。単一炭素源として0.2%グリセロールを含む液体MOPS最少培地において培養される場合、細胞分裂が、A5Pの欠乏にもかかわらず、32〜48時間遅れて、いつものように再開した。
【0069】
大腸菌KPM22菌株は、細胞抽出物に測定可能なAPI活性は残存していなかったが、37℃で最初の遅延無しに富栄養培地において持続的増殖の能力がある非条件的ΔAPI突然変異体であることが示された。表2に示されているように、倍加時間は、LB培地において親の野生型株のそれより2倍近くまで増加した。
【0070】
(表2)様々な温度におけるLB培地での世代時間
a. 2〜3世代後、増殖速度は非指数関数的であった。
【0071】
非許容的温度(42℃)への移行後、指数関数的増殖速度は、2〜3世代後維持されなかった。増殖は、上昇した温度においてkdsDをコードするプラスミド(KPM25)によりKPM22に回復され、KDO合成における遮断による細胞外被の欠陥を示唆した。
【0072】
KPM22をさらに研究するために、LPS試料を、フェノール-クロロホルム-石油エーテル(PCP)抽出方法を用いて細胞から抽出した(例えば、C. Galanos, et al., Eur. J. Biochem. 9, 245 (1969)参照;全体として参照により本明細書に組み入れられている)。LPS抽出物の糖組成は、内側コア糖成分[KDOおよびL-グリセロ-D-マンノ-ヘプトース(ヘプトース)]およびリピドA[D-グルコサミン(GlcN)]について測定された(図1A参照)。野生型BW30270[1 GlcN:0.9 KDO:2.2 ヘプトース]およびKPM25[1.0 GlcN:1.0 KDO:2.5 ヘプトース]の両方についての比は、大腸菌K-12[1.0 GlcN:1.0 KDO:2.0 ヘプトース]により産生された主なLPS種(グリコフォームI)についての比と一致した(例えば、S. Muller-Loennies, et al., J. Biol. Chem. 278, 34090 (2003)参照;全体として参照により本明細書に組み入れられている)。ほんの微量のKDOまたはヘプトースが、KPM22についての比較において検出されたが、GlcNはなお存在しており、リピドAバックボーンが無傷であることを示唆した。
【0073】
プロテイナーゼK処理された細胞全体可溶化物から調製されたLPS試料の銀染色SDS-PAGE分析により、KPM22についてバンドは検出されなかった(図1B参照;上部パネル)。リピドAの非グリコシル化1,4'-二リン酸化β-1,6-結合GlcN二糖バックボーンを認識するmAb A6抗体で探索される前に、糖コアを切断するために、ブロッティングされた膜を酸で処理した。Re内毒素標準より速いが、合成リピドIVaと同じレベルで移動したKPM22由来の単一バンドは、その抗体により認識された(図1B、中央パネル、それぞれ、レーン6および16)。酸加水分解段階を省いた場合、合成リピドIVaと共にKPM22から調製されたLPS試料のみがmAb A6により認識され、天然のリピドA構造がグリコシル化されていないことを確認した(図1B、下部パネル)。
【0074】
KPM22におけるLPS前駆体のケモタイプは、精製されたLPS試料を用いて負イオン様式でのエレクトロスプレーイオン化フーリエ変換イオンサイクロトロン(ESI FT-ICR)質量分析法により測定された(図2、表3参照)。
【0075】
(表3)ESI FT-ICR MSピークリスト
a. 示された質量数は、電荷デコンヴォルーション後、LPS画分の負イオンESI FT-ICR質量スペクトルから推定された中性分子のモノアイソトピック質量を指す。b. 太字のピークは、図4上でテキストでラベルされている。c. 略語:PE - ホスファチジルエタノールアミン;GlcN - D-グルコサミン;P - リン酸;P-EtN - ホスホエタノールアミン;Gal - D-ガラクトース;Glc - D-グルコース;Hep - L-グリセロ-D-マンノ-ヘプトース;KDO - 2-ケト 3-デオキシ-D-マンノ-オクツロソン酸;Rha - ラムノース;GlcNAc - N-アセチルD-グルコオサミン;LAtri、LAtetra、LApenta、LAhexa - リピドAのアシル化状態。
【0076】
野生型BW30270およびKPM25の両方のスペクトルは、成熟大腸菌K-12 LPSコアの異なるグリコフォームの特徴的な質量範囲[〜3900uから〜4300uまで]内で類似したピークパターンおよび不均一性を示した(図2A、C参照)。KPM22における1つのLPSに関連したピークは、1,4'-二リン酸化テトラアシル化リピドA(リピドIVa、計算された質量1404.854u)の構造と一致する1404.86uの分子量を有した(図2B参照)。
【0077】
リピドIVaは、KDO2-リピドIVaを形成するための2つのKDO残基の逐次付加のアクセプターとしての役割を果たす、LPS経路における中間体であり、後期アシルトランスフェラーゼLpxLおよびLpxMは、次に、脂肪酸ラウレートおよびミリステート、それぞれをKDO2-リピドIVaへ転移させ、ヘキサアシル化KDO2-リピドAを形成する。Raetzおよび共同研究者らは、大腸菌由来の両方の酵素が、活性について脂質基質においてKDOの絶対的基質要求性を示すことを示しており(例えば、K. A. Brozek, C.R. Raetz, J. Biol. Chem. 265, 15410 (1990)参照;全体として参照により本明細書に組み入れられている)、KPM22由来のリピドAにおける二次アシル鎖の欠如を説明している。
【0078】
リピドIVAの細胞内局在性に取り組み、かつそれがKPM22のOMへ輸送されるかどうかを決定するために、不連続性ショ糖勾配遠心分離を用いて、OMを内膜(IM)から分離させた(図3参照)。両方の膜はよく分離されていたが、KPM22についてのOMは、野生型OMほどには移動せず、浮遊密度の減少を示唆した。OMにおける蓄積を犠牲にしてIMに局在したままであるOMポーリン(OMP)タンパク質(〜35kDa)の量の増加は別にして、SDS-PAGEにより分析された、全体的な総タンパク質含有量および構成は類似していた。多くのOMタンパク質は、それらの折り畳みおよび機能の両方についてLPSの分子シャペロン性質に依存することが示されている(例えば、全体として参照により本明細書に組み入れられる、H. de Cock, J. Tommassen, Embo J. 15, 5567(1996);P. V. Bulieris, et al., J. Biol. Chem. 278, 9092(2003);K. Sen, H. Nikaido, J. Bacteriol. 173, 926(1991)参照)ため、OMPの減少は、KPM22のOMへのタンパク質輸送速度および/または挿入効率の減少を反映している可能性がある。単離されたOM画分を、特徴的なLPS/リピドIVA脂肪酸マーカーである、3-ヒドロキシミリスチン酸(3-OH C14:0)の存在についてアッセイした。野生型およびKPM22のOMは、1mgの乾燥膜あたり、それぞれ11.7μgおよび31.1μgの3-OH C14:0を含み、野生型におけるLPSの量と少なくとも等しいリピドIVAの相当量が、実際、KPM22のOMに存在することを示唆した。さらに、ESI FT-ICR質量分析法により、KPM22のOMおよびIMの両方におけるリピドIVAについてのピークが明らかにされたが、リピドIVAに起因しうるピークは、野生型由来のいずれの膜画分にも検出されなかった。ひとまとめにすると、これは、リピドIVAがKPM22のOMへ輸送されるが、リピドIVA輸送の速度がその合成速度と切り離されていることを示す。
【0079】
二次アシル鎖は、機能に必要とされる条件であるアシル鎖の数を増加させることによりOM内での低度の流動性を維持することに関与している(例えば、H. Nikaido, Microbiol. Mol. Biol. Rev. 67, 593 (2003)参照;全体として参照により本明細書に組み入れられている)。飽和アシル鎖の密充填は、それがファンデルワールス力を通してOM外層の完全性を維持する、疎水性相互作用のネットワークを誘導する。4つのみのアシル鎖を含み、かつ内側糖コアを含まないにもかかわらず、リピドIVaは、KPM22において、輸送され、その後、OM生合成を援助する能力がある。KPM22のOMにおけるリピドIVa層の前例のない性質は、腸内細菌科の生存能力のための不可欠なLPS構造を再定義する。
【0080】
KDOは、今まで、大腸菌におけるKDO生合成酵素の条件的突然変異体のみが構築されていたので、機能的LPS層の必須成分と、通常、みなされている(例えば、C.J. Belunis, et al., J. Biol. Chem. 270, 27646 (1995); R.C. Goldman, W.E. Kohlbrenner, J. Bacteriol. 163, 256 (1985); P.D. Rick, M.J. Osborn, Proc. Natl. Acad. Sci. U.S.A. 69, 3756 (1972)参照;全体として参照により本明細書に組み入れられている)。これは、脂質二重層内で低度の流動性を維持することにおけるKDOの役割に(およびおそらく部分的にKDOより遠位に付着した他の糖に)帰するとされている(例えば、H. Nikaido, Microbiol. Mol. Biol. Rev. 67, 593 (2003)参照;全体として参照により本明細書に組み入れられている)。二価陽イオン、すなわち、Mg2+およびCa2+は、リン酸化リピドAバックボーンおよびKDOのカルボキシレートの両方によって与えられる負電荷によりイオン架橋を生じ、静電反発力を最小にし、かつ強い側方の相互作用を作り出すと考えられている。さらに、OMの表面でのKDOの配置は、OMタンパク質に極めて接近してKDOを置き、OMタンパク質の多くは、それらの折り畳みおよび機能の両方についてコア含有LPSの分子シャペロン性質に依存する(例えば、H. de Cock, J. Tommassen, Embo J. 15, 5567 (1996); P.V. Bulieris, et al., J. Biol. Chem. 278, 9092 (2003); K. Sen, H. Nikaido, J. Bacteriol. 173, 926 (1991)参照;全体として参照により本明細書に組み入れられている)。
【0081】
KPM22におけるKDOの非必須性質を検証するために、KDO生合成における最初の関係した段階(kdsA)および最終段階(waaA)をコードする遺伝子を破壊した。KPM22/KPM25(図1B、レーン8、9)と対照的に、外因性A5Pもプラスミドが有するAPIも、KPM31/KPM40(レーン12、15)またはKPM34/KPM42(レーン11、14)のいずれにおいても、それぞれ、成熟LPS合成を回復させず、完全なKDO経路無しに生存するKPM22の能力と一致した。
【0082】
KPM22の細胞形態を、透過電子顕微鏡(TEM)によって調べた。全体として、KPM22の構造は、親の菌株と全く類似していた(図4)。明らかな分裂欠陥はTEMによって観察されず、細胞は正常な桿状形を維持していた。2つの明らかに異なる膜が、KPM22について識別され(図4D)、加えて、周辺質であるその2つの膜の間の領域も識別された。周辺質体積は、野生型と比較して均一に圧縮されていた。OM不安定性は、KPM22の表面に現れる小さな膜小胞により示唆された(図4C)。外膜小胞(OMV)形成は、糖コアの相互作用の安定化、膜湾曲の増加、および結果として生じる細菌表面からの小胞放出によって代償されない、隣接するリピドIVa分子の1,4'-GlcNリン酸の間の静電反発力により引き起こされてた可能性がある。ESI-MS分析により、4-アミノ4-デオキシ-L-アラビノース修飾は検出されず、最小限のホスホエタノールアミン修飾のみが検出された(両方とも正味負電荷の量を低下させる働きをする)ことを考えると、電荷反発力はKPM22について特に関連性があった(例えば、C.R. Raetz, C. Whitfield, Annu. Rev. Biochem. 71, 635 (2002)参照;全体として参照により本明細書に組み入れられている)。
【0083】
極度のLPS切り詰めによるOM完全性の喪失に抵抗するKPM22における代償的な機構は、他のOM結合糖脂質に関する安定化を引き起こした。髄膜炎菌(N. meningitidis)由来のLPS欠陥突然変異体において、莢膜多糖合成が生存能力に絶対的に必要になったと報告された(例えば、P. van der Ley, L. Steeghs, J. Endotoxin. Res. 9, 124 (2003)参照)。大腸菌K-12は莢膜多糖を合成しなかったが、LPSに加えて2つの他の細胞表面多糖、すなわち、ストレス誘導性粘液エキソポリサッカライドコラン酸(M抗原)(例えば、A. Markovitz Surface carbohydrates of the prokaryotic cell I. W. Sutherland, Ed. (Academic Press, Inc., New York, N.Y., 1977), vol. I. pp. 415-462)参照;全体として参照により本明細書に組み入れられている)およびホスホグリセリド結合腸内細菌一般抗原(ECA)(例えば、H.M. Kuhn, et al., FEMS Microbiol. Rev. 4, 195 (1988)参照:全体として参照により本明細書に組み入れられている)、が存在する。コラン酸の構成糖質マーカーである、非透析性メチルペントースのレベルにおける差はなかった(図5A)(例えば、S. Gottesman, et al., J. Bacteriol. 162, 1111 (1985)参照;全体として参照により本明細書に組み入れられている)。細胞溶解物の免疫ブロット分析により、グリセロホスファチジル結合ECAの量は実際、KPM22において減少していることが明らかにされ(図5B)、フェノール抽出物のKPM22スペクトルからの4つの三糖繰り返し単位(2429.89u)を含む環状ECAの消失(図6)と一致した。従って、リピドIVaに加えて、KPM22のOMは、微量レベルのECA、および同等に低い野生型レベルのコラン酸を含んだ。一括して、KPM22外被は、生存能力を維持することができる、大腸菌で報告された最も最小のOM糖脂質含有量である。
【0084】
LPS層の主要な機能は、大きな疎水性分子とデフェンシン(ポリカチオンのペプチド)の両方の細胞への拡散に対する透過性バリアとして働くこと、および周辺質区画の内容物を保持することである。OM内での隣接LPS分子間の強い側方相互作用が、宿主応答に対する非特異的防御の手段を与えることに加えて、LPS層をそのような機能に特に良く適したものにさせている。疎水性小分子、栄養分、および抗生物質の選択的透過は、外膜ポーリン(OMP)タンパク質チャネルを通して達成される。抗生物質および界面活性剤のパネルは、透過性バリアとしてのリピドIVaの有効性を評価するためにKPM22に対してスクリーニングされた(表4参照)。
【0085】
(表4)KPM22の透過性バリア性質
最小阻止濃度(μg/mL)
a. 74,000ユニット/g。b. コール酸ナトリウムおよびデオキシコール酸ナトリウムの混合物。c. コリスチン;20,261ユニット/mg。
【0086】
KPM22は、典型的にはグラム陽性細菌対してのみ妥当な効力を生じるいくつかの大きな疎水性抗生物質に対して超高感受性であった。通常ではそれらの作用部位へのアクセスをOMにより拒否されるが、これらの化合物はKPM22において細胞内標的にアクセスした。膜表面へのアクセスは、糖コアにより妨げられず、さらに、分配、およびその後の、損なわれた脂質二重層を通しての透過を促進した。しかしながら、主にOMPを通してOMの通過を得る小さな(<600Da)比較的疎水性の化合物の最小阻止濃度(MIC)は、せいぜいやや減少するだけであった。注目すべき例外は、正電荷をもつアミノグリコシドカナマイシンであった。アミノグリコシドは、OMPと無関係に、LPSとの最初の電荷対形成相互作用を含む、主に自己促進性取り込み機構を通しての侵入を得ることを示唆している(例えば、R.E. Hancock, et al. Antimicrob. Agents. Chemother. 35, 1309 (1991);全体として参照により本明細書に組み入れられている)。KPM22は、特に界面活性剤に対して感受性が高く、ドデシル硫酸ナトリウムについてMICにおいて4000分の1未満の減少であった。ヒト腸管における胆汁塩(コレステロール代謝物)の濃度は4mMから16mMまで(〜1650μg/mLから6650μg/mLまで)の範囲であるため(例えば、全体として本明細書に参照により組み入れられるB. Borgstrom, Acta Med. Scand. 196, 1(1974)参照)、KPM22の欠陥があるOMはもはや、その宿主環境から細胞を保護するのに適していない。驚くべきことに、界面活性剤様作用機構をもつ陽イオンペプチドである、ポリミキシンE(コリスチン)のMICは、KPM22においてたった〜4分の1に抑制されただけであった。臨界凝集濃度までの膜表面におけるポリミキシンの蓄積は、OMを通しての自己促進性輸送のチャネルとして実質的に働く層状二重層内にミセル損傷を形成することに関係がある(例えば、A. Wiese et al., J. Membr. Biol. 162, 127 (1998)参照;全体として参照により本明細書に組み入れられている)。リピドIVaは、LPSと比較して表面積に対する電荷比が減少しており、ポリミキシン結合における負荷電内側コア残基の役割を目立たせている。選択された抗生物質は様々な作用機構をもつため、疎水性化合物の間でのMICにおける変化は、一般的な適応度または薬物流出機構の反映とは対照的に、透過性における変化の結果である可能性が高い。KPM22の透過特性は、OMバリアの内因性抵抗を低下させることにより、すでに存在する抗生物質の活性のスペクトルの幅を広げるための手段としてのKDO生合成阻害の可能性を示す。
【0087】
細菌内毒素は、ただ微量存在するのみの場合でさえ、ヒトにおいて先天性免疫応答を誘発する強力な炎症性分子である(例えば、E.S. Van Amersfoort, et al., Clin. Microbiol. Rev. 16, 379 (2003)参照;全体として参照により本明細書に組み入れられている)。グラム陰性細菌誘発性敗血性ショックは、不均衡な調節不全の免疫応答に起因する。部分的にこの病態生理学的カスケードは、LPSによるマクロファージの活性化により引き起こされ、マクロファージが次に、一連の炎症性介在物質を分泌する。マクロファージにより放出された最初のサイトカインの一つは、多面的なサイトカイン、TNF-α(腫瘍壊死因子)である。LPS調製物の内毒素性潜在能力を、刺激されたヒト単核細胞からのhTNF-α分泌についてELISAに基づいたアッセイ法を用いて測定した(図7)。KPM22からの調製物は、1μg/mLまでの濃度で内毒素的に不活性であり、クロマトグラフィー精製されたリピドIVaを用いた以前の研究と一致した(例えば、D.T. Golenbock, et al., J. Biol. Chem. 266, 19490 (1991)参照;全体として参照により本明細書に組み入れられている)。大腸菌および関連細菌において、KDO阻害は、細菌の宿主応答および抗生物質の両方に対する感受性を増加させるだけでなく、内毒素負荷量を低下させることにより敗血症のリスクを減少させる可能性も有する。
【0088】
LPSを細胞質からIMの周辺質表面へフリップ反転する(flip)内膜ABC(ATP結合カセット)輸送体は、インビトロでヘキサアシル化LPS/脂質A基質に対して高選択性である(例えば、Zhou Zhou, Z., et al., J. Biol. Chem. 273, 12466-12475 (1998); Doerrler, W.T., et al., J. Biol. Chem. 277, 36697-36705 (2002)参照;それぞれ、全体として参照により本明細書に組み入れられている)。MsbAは、最初に、LpxL(HtrB)温度感受性表現型の多コピー抑制因子として同定された(例えば、Polissi, A., and Georgopoulos, C. Mol. Microbiol. 20, 1221-1233 (1998)参照;全体として参照により本明細書に組み入れられている)。栄養要求性TCM15菌株のKPM22ゲノムDNAのコスミドライブラリーでの補完により、MsbAがΔKdo表現型の多コピー抑制因子であることが明らかにされた。msbA座を含む17個の別々のコスミドクローンを単離した。野生型と同一の無傷の野生型msbA配列のみを有する3.5kb挿入断片を含むコスミドサブクローン(pMMW52)は、A5P栄養要求性の喪失および固体寒天上でのコロニー形成能力の回復により示されているように、推定抑制因子突然変異を発生させる必要なしにTCM15を直接、救出することができた(表5)。TCM15(pMMW52)の増殖速度は、KPM22と類似している(表2および5)。これらの結果は、リピドIVAがインビトロでは弱い基質であるが(Doerrler, W.T., and Raetz, C.R., J. Biol. Chem. 277, 36697-36705 (2002)参照;全体として参照により本明細書に組み入れられている)、リピドIVaは、単純な質量的作用により高濃度で存在する場合、インビボでMsbAの基質になる。
【0089】
(表5)MsbAによるTCM15栄養要求性の多コピー抑制
aCfu値は、直接プレーティング(TCM15)かまたは電気形質転換後のいずれかに対応する。b測定可能の場合、37℃における世代時間(分)が括弧に入れて記載されている。c15μM A5P、10μM G6P。dAmp(100μg mL-1)がプラスミド保有菌株について含まれた。eクローニングベクター。fmsbAを含むサブクローン。
【0090】
実施例II.
本実施例は、KPM22およびTCM15を含む研究に用いられる細菌株、プラスミド、およびプライマーを記載する。KPM22およびTCM15を含む研究に用いられる細菌株、プラスミド、およびプライマーは表6に列挙されている。
【0091】
(表6)細菌株、プラスミド、およびプライマー
a. 相同領域は下線が引かれている。
【0092】
全菌株は、単一炭素源として0.2%グリセロールを含む標準Luria-Bertani培地(10gトリプトン、5g酵母抽出物、10g NaCl)またはMOPS最少培地(例えば、F.C. Neidhardt, P.L. Bloch, D.F. Smith, J. Bacteriol. 119, 736 (1974);全体として参照により本明細書に組み入れられている)において増殖した。大腸菌株KPM22が、DatsenkoおよびWanner(例えば、K.A. Datsenko, B.L. Wanner, Proc. Natl. Acad. Sci. U.S.A. 97, 6640 (2000)参照;全体として参照により本明細書に組み入れられている)の手順によるファージλレッド(Red)リコンビナーゼ系を用いる染色体kdsAおよびwaaA遺伝子破壊のための宿主として用いられた。カナマイシンおよびアンピシリンが、それぞれ、15μg/mLおよび100μg/mLで用いられた。鋳型としてのpKD13(kan)を含むP1/P2またはpKD4(kan)を含むプライマー対P3/P4が、それぞれ、KPM31およびKPM40についての挿入カセットを構築するために用いられた。抗生物質抵抗性マーカーは、全プラスミドを37℃で回復させることを除き記載されているように(例えば、K.A. Datsenko, B.L. Wanner, Proc. Natl. Acad. Sci. U.S.A. 97, 6640 (2000)参照;全体として参照により本明細書に組み入れられている)、FLPリコンビナーゼ系を用いて切除された。
【0093】
実施例III.
本実施例はKPM22の増殖を記載する。KPM22の増殖は、37℃における10μM D-グルコース6-リン酸および15μM D-アラビノース5-リン酸を追加したMOPS最少培地中のTCM15の指数関数的に分裂する培養物を、糖リン酸追加を欠く同じ培地へ希釈(1:200v/v)することを含んだ。24〜32時間続いた最初の遅れの後、増殖は再開し、培養物は、LB寒天プレート上でコロニー精製された。
【0094】
実施例IV.
本実施例は、KPM22が関わる実験についての増殖速度測定を記載する。一晩培養物は、30℃で増殖し、新鮮なあらかじめ温められたLB培地(30℃、37℃、または42℃)へ0.05〜0.1に等しいOD600nmまで接種するのに用いられた。増殖をOD600nmにおける変化を測定することによりモニターし、培養物を、指数関数的増殖を維持するために、ODが〜0.7にほぼ等しいように希釈した。倍加時間は表7に列挙されている。
【0095】
(表7)様々な温度におけるLB培地での世代時間
a. 2〜3世代後、増殖速度は非指数関数的であった。
【0096】
実施例V.
本実施例は、KPM22およびTCM15が関わる実験についてのLPS精製を記載する。試料を、500mLの各菌株をLB培地において250rpmの一定の通気、37℃で増殖させることにより、日常的に調製した。静止期培養物由来の細胞を、遠心分離(10min、8000xg、4℃)により収集し、蒸留水で洗浄し、再遠心分離した。そのバイオマスを、以前に記載されているように(例えば、U. Zahringer et al., J. Biol. Chem. 279, 21046 (2004)参照;全体として参照により本明細書に組み入れられている)、エタノール(95%)、アセトン、およびジエチルエーテルでの処理により脱水した。LPSの単離を、フェノール-クロロホルム-石油エーテル手順により乾燥細胞の抽出により行った(例えば、C. Galanos, O. Luderitz, O. Westphal, Eu.r J. Biochem. 9, 245 (1969)参照;全体として参照により本明細書に組み入れられている)。糖組成について分析されるべきフェノール粗抽出物のアリコート(図1A)を、大規模に蒸留水に対して透析し(MWCO=1000Da)、凍結乾燥により収集した。質量分光分析およびヒトTNFαサイトカイン放出の測定のためのLPS試料を、水の滴下式添加による沈殿により粗フェノール相から精製した。綿状沈殿は、BW30270およびKPM25についてのみ形成し、遠心分離により収集し、引き続いて、80%フェノールおよびその後、アセトンで洗浄した。沈殿を水に溶解し、それらの個々のフェノール相母液から別々に透析した。凍結乾燥後、試料を緩衝液(20mM Tris-HCl、pH=7.5、10mM NaCl、10mM MgCl2)に再懸濁し、DNアーゼI(20μg/mL)およびRNアーゼA(20μg/mL)で37℃で8時間、続いて、プロテイナーゼK(100μg/mL)で16時間、処理した。LPS試料を、超遠心分離(SW 41 Tiスウィングバケットローター、200,000xg、2時間、15℃)により収集し、蒸留水で3回、洗浄し、凍結乾燥の前に水に対して大規模に透析した。代表的なLPS精製収量は表8に列挙されている。
【0097】
(表8)LPS精製概要
a. Ppt. - 沈殿。b. DNアーゼI/RNアーゼA/プロテイナーゼK処理後。c. 乾燥細胞質量に基づいて。
【0098】
実施例VI.
本実施例は、KPM22およびTCM15に関わる実験についての糖組成分析を記載する。フェノール粗抽出物からのLPS試料のD-グルコサミン(GlcN)、2-ケト3-デオキシ-D-マンノ-オクツロソン酸(KDO)、およびL-グリセロ-D-マンノ-ヘプトース(ヘプトース)含有量を、比色定量化学的アッセイ法を用いて測定した。GlcN含有量は、LPS試料(〜1mg)の、500μLの4 M HCl中100℃で18時間の加水分解により測定された。遊離GlcNは、アセチルアミノ糖アッセイ法を用いて定量された(例えば、J.L. Strominger, J.T. Park, R.E. Thompson, J. Biol. Chem. 234, 3263 (1959)参照;全体として参照により本明細書に組み入れられている)。KDO含有量はLPS適応化チオバルビツール酸アッセイ法を用いて測定されたが(例えば、Y.D. Karkhanis, J.Y. Zeltner, J.J. Jackson, D.J. Carlo, Anal. Biochem. 85, 595 (1978)参照;全体として参照により本明細書に組み入れられている)、ヘプトースの量は改変システイン-硫酸アッセイ法を用いて推定された(例えば、M.J. Osborn, Proc. Natl. Acad. Sci. U.S.A. 50, 499 (1963)参照;全体として参照により本明細書に組み入れられている)。
【0099】
実施例VII.
本実施例は、KPM22およびTCM15に関わる実験についてのSDS-PAGE電気泳動およびリピドA/ECA免疫ブロットを記載する。細胞全体の可溶化物のLPSプロファイルは、HitchcockおよびBrown(例えば、P.J. Hitchcock, T.M. Brown, J. Bacteriol. 154, 269 (1983)参照;全体として参照により本明細書に組み入れられている)の方法によるSDS-PAGEにより分析された。簡潔には、各試料のコロニーを、LB寒天プレートから掻爬し、ダルベッコ(Dulbecco)のリン酸緩衝食塩水に等しい濁度へ懸濁した。洗浄された細胞ペレットを遠心分離により収集し、溶解緩衝液(50μl 62.5mM Tris-HCl、pH6.8、2% SDS、5% 2-メルカプトエタノール、10%グリセロール、0.002%ブロムフェノールブルー)を加え、試料を沸騰水浴中で10分間、加熱した。プロテイナーゼK(25μg、2.5mg/mlの10μl)を各細胞全体可溶化物に加え、56℃で1時間、インキュベートした。同一の容量を、13%SDS-PAGEゲルへローディングし、その後、一定電流(15mA)で流した。ゲルをLPS分析のために銀染色した(例えば、P.J. Hitchcock, T.M. Brown, J. Bacteriol. 154, 269 (1983)参照;全体として参照により本明細書に組み入れられている)、または記載されているように(例えば、H. Towbin, T. Staehelin, J. Gordon, Proc. Natl. Acad. Sci. U.S.A. 76, 4350 (1979)参照;全体として参照により本明細書に組み入れられている)、Tris-グリシン緩衝液(20mM Tris、150mMグリシン、pH8.3、20%メタノール)を用いてゲルからポリ二フッ化ビニリデン膜へ一定電圧(26V)で電気的転写した。リピドAの非グリコシル化1,4'-二リン酸化β1,6-結合GlcN二糖バックボーンを認識するmAb A6とのブロットのインキュベーションの前に(例えば、L. Brade, O. Holst, H. Brade, Infect. Immun. 61, 4514 (1993)参照;全体として参照により本明細書に組み入れられている)、膜を1%酢酸中で1時間、煮沸して、通常の免疫手順により発色される前にα2,6-KDO-GlcN結合を切断した(例えば、R. Pantophlet, L. Brade, H. Brade, J. Endotoxin Res. 4, 89(1997)参照;全体として参照により本明細書に組み入れられている)。真正の合成リピドIVa(化合物406)を標準として用いた(例えば、M. Imoto et al., Bull. Chem. Soc. Japan 60, 2197 (1987)参照;全体として参照により本明細書に組み入れられている)。腸内細菌一般抗原(ECA)免疫ブロットを、mAb 898を用いて探索した(例えば、H. Peters et al., Infect. Immun. 50, 459 (1985)参照;全体として参照により本明細書に組み入れられている)。免疫ブロットを、アルカリホスファターゼ結合ヤギ抗マウスIgG(H+L)とインキュベートし、ニトロブルーテトラゾリウムおよび5-ブロモ-4-クロロ-3-インドリルリン酸基質の存在下で発色させた。
【0100】
実施例VIII.
本実施例は、本発明の過程において行われた実験に用いられたエレクトロスプレーイオン化フーリエ変換イオンサイクロトロン質量分析(ESI FT-ICR MS)を記載する。ESI FT-ICR MSは、7テスラのアクティブシールド型磁石およびアポロ(Apollo)イオン源を備えたAPEX II装置(Bruker Daltonics, Billerica, USA)を用いて負イオン様式で行われた。質量スペクトルは、製造会社により提供された標準実験配列を用いて得られた。試料を、2-プロパノール、水、およびトリエチルアミンの50:50:0.001(v/v/v)混合物に〜10ng/μlの濃度で溶解し、2μl/minの流速でスプレーした。キャピラリー入口電圧を3.8kVに、および乾性ガス温度を150℃に設定した。スペクトルは電荷デコンヴォルーションされ、示された質量数は中性モノアイソトピック質量を指す。ピーク指定は、大腸菌K-12株W3100由来のLPSの以前に発表された詳細な構造分析(例えば、S. Muller-Loennies, B. Lindner, H. Brade, J. Biol. Chem. 278, 34090 (2003)参照;全体として参照により本明細書に組み入れられている)に基づいて解釈された。明白に同定することができない重複アイソトピックピークを有するいくつかの分子種があったため、最も豊富なイオンのみが、表9に要約されている。
【0101】
(表9)ESI FT-ICR MSピークリスト
a. 示された質量数は、電荷デコンヴォルーション後、LPS画分の負イオンESI FT-ICR質量スペクトルから推定された中性分子のモノアイソトピック質量を指す。b. 太字ピークは、図4上でテキストで標識されている。c. 略語:PE - ホスファチジルエタノールアミン;GlcN - D-グルコサミン;P - リン酸;P-EtN - ホスホエタノールアミン;Gal - D-ガラクトース;Glc - D-グルコース;Hep - L-グリセロ-D-マンノ-ヘプトース;KDO - 2-ケト3-デオキシ-D-マンノ-オクツロソン酸;Rha - ラムノース;GlcNAc - N-アセチルD-グルコオサミン;LAtri、LAtetra、LApenta、LAhexa - リピドAのアシル化状態。
【0102】
実施例IX.
本実施例は、KPM22およびTCM15に関わる実験におけるコラン酸の定量化を記載する。コラン酸は、KangおよびMarkovitz(例えば、S. Kang, A. Markovitz, J. Bacteriol. 93, 584 (1967)参照;全体として参照により本明細書に組み入れられている)により報告された方法の改変により推定された。LB寒天プレートからのコロニーを掻爬し、同一の濁度(OD600nm)まで10mLの蒸留水に再懸濁し、細胞外多糖を遊離させるために沸騰水浴に15分間、浸し、遠心分離(10min、8000xg)により清澄にした。上清を、コラン酸の成分であるメチルペントース(L-フコース)について、標準として真正L-フコースを用いる特定の比色定量反応によりアッセイした(例えば、Z. Dische, L.B. Shettles, J. Biol. Chem. 175, 595 (1948)参照;全体として参照により本明細書に組み入れられている)。BW30270のムコイド単離物が陽性対照として含まれた。
【0103】
実施例X.
本実施例は、本発明の過程において行われた実験に用いられた透過電子顕微鏡法(TEM)を記載する。37℃、LB培地において対数期初期で増殖中の細胞の培養物を、2%四酸化オスミウムに室温で90分間、固定した。細胞を、室温で1時間、2%酢酸ウラニルとインキュベートする前に、蒸留水で3回、洗浄した。細胞を、もう一度、蒸留水で3回、洗浄し、その後、一連の増加性エタノール洗浄(30%、50%、70%、90%、および無水エタノール、室温でそれぞれ15分間)により脱水した。脱水した細胞を、2回、室温で各15分間、酸化プロピレンに浸し、続いて、4℃での一晩インキュベーションにより酸化プロピレン/エポン混合物(1:1、v/v)に含浸させた。その後、重合を60℃で一晩、行った。その塊を超薄切片(80〜100nm)へスライスし、グリッド上に置き、クエン酸鉛溶液において対比した。画像は、自動コンピュステージ(compustage)およびKodak 1.6 Megaplus高解像度デジタルカメラを備えたPhilips CM-100透過電子顕微鏡上で得られた。
【0104】
実施例XI.
本実施例は、本発明の過程において行われた実験に用いられた最小阻止濃度(MIC)測定を記載する。用いられた抗生物質は、MicroSource Discovery Systemsから入手されたセファロリジンを除いて、Sigmaからであった。抗生物質は、それらの作用および細胞への侵入の様々な様式に基づいて選択された。研究されたすべての抗生物質および薬物のMICは、記載されているように(例えば、R. Vuorio, M. Vaara, Antimicrob. Agents Chemother. 36, 826 (1992)参照;全体として参照により本明細書に組み入れられている)、標準段階微量希釈方法を用いてLB培地で測定された。LB寒天プレートからのコロニーを掻爬し、様々な濃度の抗生物質を含む培地(1mLあたり〜104細胞)に懸濁した。培養物を、振盪(〜200rpm)しながら37℃で18時間、インキュベートし、その時点で増殖を目視検査により点数化した。報告されたMIC値を、完全に増殖を阻止した薬物の最低濃度として解釈した。
【0105】
実施例XII.
本実施例は、本発明の過程において行われた実験に用いられたヒトTNFαサイトカインアッセイ法を記載する。上記のように単離されたLPS調製物のヒト単核細胞(MNC)への腫瘍壊死因子(TNF)αサイトカイン誘導性能力を、酵素結合免疫吸着アッセイ法(ELISA)を用いて測定した。LPS試料を、激しいボルテックスによりハンクス平衝塩溶液(Hanks' Balanced Salt Solution)に再懸濁し、使用直前の超音波処理/ボルテックスに供する前に、4℃で一晩、寝かした。健康なドナーから採取されたヘパリン処理された血液を、等容量のハンクス平衝塩溶液と直接、混合し、Greiner Bio-OneからのLymphoprep培地でのLeucosepシステムを用いて製造会社の使用説明書に従い、示差的勾配遠心分離により単離した。MNCをRPMI 1640(3mM L-グルタミン、100ユニット/mL ペニシリン、100μg/mL ストレプトマイシン)で2回、洗浄し、96ウェル培養プレートへ移した(7.5x105細胞/ウェル)。MNCの刺激は、以前に記載されているように(例えば、M. Mueller et al., J. Biol. Chem. 279, 26307 (2004)参照;全体として参照により本明細書に組み入れられている)、行われ、上清は、4℃で一晩、保存された。hTNFα産生は、Copelandら(例えば、S. Copeland, H.S. Warren, S.F. Lowry, S.E. Calvano, D. Remick, Clin. Diagn. Lab. Immunol. 12, 60 (2005)参照;全体として参照により本明細書に組み入れられている)により記載されているように、ELISAにより測定された。データは、3つの別々の実験において二回通り収集され、代表的なデータセットが図7に報告されている。
【0106】
実施例XIII.
本実施例は、KPM22コスミドライブラリーの構築を記載する。コスミドライブラリーは、製造会社(Stratagene)により記載されているように、Sau3Aでの部分的消化、SuperCos1へのライゲーションによりKPM22ゲノムDNAから構築され、Gigapack III XLパッケージング抽出物を用いてパッケージングされた。0.2%(w/v)マルトースおよび10mM MgSO4を含み、加えてさらに、A5PおよびG6Pを追加した、LB培地における増殖により、TCM15をファージ感染のために調製した。形質転換体を、コスミドベクター抗生物質抵抗性マーカー(100μg mL-1 Amp)と共の、追加の糖リン酸を欠くLBプレート上での増殖について選択した。コスミドを、部分的Sau3A消化、続いて中位のコピー数pMBL19クローニングベクターのBamHI部位へのライゲーションによりサブクローニングした(例えば、Nakano, Y., et al., Gene 162, 157-158 (1995)参照;全体として参照により本明細書に組み入れられている)。
【0107】
実施例XIV.
本実施例は、gutQ遺伝子に関わる実験に用いられた材料を記載する。プライマーはInvitrogenにより合成された。ゲノム大腸菌K-12 MG1655 DNAは、American Type Culture Collection(ATCC 700926D)から購入された。Promega Wizard DNA精製キットは、プラスミド精製に利用された。化学的コンピテントな大腸菌XL1-Blue(Stratagene)および大腸菌BL21(DE3)(Novagen)は、それぞれ、宿主プラスミドおよびタンパク質発現に向いていた。大腸菌K-12 MG1655の誘導体である菌株BW30270(rph+、fnr+)は、大腸菌Genetic Stock Center(CGSC#7925)から入手された。糖および糖リン酸は、Sigma-Aldrichから購入されたが、例外として、D-グルシトール6-リン酸は、D-グルコース6-リン酸の水素化四ホウ酸ナトリウム還元により調製され(例えば、Bigham, E.C., et al., (1984) J Med Chem 27, 717-26参照;全体として参照により本明細書に組み入れられている)、陰イオン交換クロマトグラフィー(AG MP-1、Bio-Rad)により精製され、ゲル濾過(Bio-Gel P-2、Bio-Rad)により脱塩された。タンパク質濃度は、標準としてBSAを用いるBio-Rad Protein Assay Reagentを使用して、測定された。
【0108】
実施例XV.
本実施例は、gutQ遺伝子のクローニング、過剰発現、および精製を記載する。gutQ遺伝子を、F-Rプライマー対(表10)での標準PCR方法を用いて増幅し、NdeIおよびBamHIで制限酵素処理し、仔ウシアルカリホスファターゼで処理されている、同様に制限酵素処理された直線化pT7-7発現ベクターへ直接、ライゲーションした。
【0109】
(表10)プライマーのヌクレオチド配列
aNdeI部位は下線が引かれている。bBamHI部位は下線が引かれている。
【0110】
ライゲーション混合物は、化学的コンピテントな大腸菌XL1-Blue細胞を形質転換するために用いられ、pT7-gutQプラスミドを有する形質転換体は、制限酵素分析およびDNAシーケンシングにより同定された。大腸菌BL21(DE3)細胞は、プラスミドで形質転換され、制限酵素分析により再チェックされ、-80℃で保存された。大腸菌BL21(DE3)/pT7-gutQ細胞を、アンピシリン(100mg/L)を含む2xYT培地において37℃で振盪(250rpm)しながら、増殖させた。いったん培養が対数増殖期の中頃(OD600〜0.7-0.9)に達したならば、培養物を、0.4mMの最終濃度でのイソプロピル-β-D-チオガラクトシドで誘導される前に、18℃まで冷却させた。18℃での16時間の増殖後、細胞を遠心分離(6,500xg、15min、4℃)により収集した。細胞ペレットを、20mLの緩衝液A(20mM Tris-HCl;1mM DL-ジチオスレイトール(DTT);pH=8.0)に懸濁し、その後、氷上で超音波処理した(5x30秒間;パルスの間に2分間の一時的停止)。細胞片を遠心分離(29,000xg、40min、4℃)により除去し、上清を0.22μM Millex(登録商標)フィルターを通して濾過した。溶液を、緩衝液Aとあらかじめ平衡に達しているHi Load(商標)(16/10)Q Sepharose高速流カラムへローディングした。タンパク質を、緩衝液AにおけるNaClの0〜900mM勾配を用いて120分間に渡り溶出した。SDS-PAGEにより測定された場合主に組換えタンパク質(〜33kDa)を含む画分を、プールした。硫酸アンモニウムの飽和溶液を、15%飽和に達するまで室温で撹拌しながらゆっくり加えた。溶液を遠心分離(29,000xg、30min、22℃)により清澄化し、上清を30%飽和に至らせた。タンパク質ペレットを遠心分離(29,000xg、30min、22℃)により収集し、緩衝液Aに再懸濁し、2Lの緩衝液Aに対して4℃で一晩、透析した。調製物は、SDS-PAGEにより判断される場合、〜95%より高い均一性であり、180mg gutQ/1L細胞培養物の収率であった。
【0111】
実施例XVI.
本実施例は、本発明の過程において行われた実験に用いられたゲル電気泳動方法を記載する。SDS-PAGEは、12%ポリアクリルアミド上で還元条件下、タンパク質試料(〜5-10μg)に行われ、0.25%クーマシーブリリアントブルーR250溶液で染色した。LPS試料を、トリシン-SDS PAGE(スタッキング4%T、3%C;分離16.5%T、6%C)により分析し(例えば、Lesse, A.J., et al., (1990) J Immunol Methods 126, 109-17参照;全体として参照により本明細書に組み入れられている)、銀染色により可視化した(例えば、Hitchcock, P.J. & Brown, T.M. (1983) J Bacteriol 154, 269-77参照;全体として参照により本明細書に組み入れられている)。
【0112】
実施例XVII.
本実施例は、本発明の過程において行われた実験に用いられた酵素アッセイ法を記載する。API活性は、以前に記載されているように(例えば、Meredith, T.C. & Woodard, R.W. (2003) J Biol Chem 278, 32771-7参照;全体として参照により本明細書に組み入れられている)、96ウェルのマイクロプレートに適応した非連続的システイン-カルバゾール比色定量アッセイ法(例えば、Dische, Z., Borenfreund, E. (1951) J Biol Chem 192, 583-587参照;全体として参照により本明細書に組み入れられている)により測定された。全プレートは、内部Ru5P標準および適切なA5P対照を3回通り含んだ。酵素活性の1ユニットは、37℃で1分間あたり1μmolの糖リン酸の変換として定義される。
【0113】
シロイヌナズナ(Arabidopsis thaliana)由来の3-デオキシ-D-マンノ-オクツロソン酸8-リン酸シンターゼ(kdsA)を利用する、第二のより感度の高い結合アッセイ法が、細胞粗抽出物においてAPI活性を測定するために開発された。この酵素は、A5PおよびPEPの不可逆性立体特異的縮合を触媒して、3-デオキシ-D-マンノ-オクツロソン酸8-リン酸(KDO8P)および無機リン酸を形成する。40uLの100mM Tris-HCl(pH=8.25)中に精製kdsA溶液(3mg/mL;10U/mg)、10mM Ru5P、6mM PEP、および1mM EDTAの5μLを含むの反応混合物を、37℃で3分間、インキュベートした。反応を、10μLの細胞抽出物の添加により惹起した。5分後、50μLの10%(w/v)トリクロロ酢酸の添加により反応を停止した。生成されたKDO8Pを、Aminoff過ヨウ素酸-チオバルビツール酸アッセイ法により測定した(例えば、Sheflyan, G.Y., et al., (1998) Journal of the American Chemical Society 120, 11027-11032参照;全体として参照により本明細書に組み入れられている)。これらの条件下で、kdsAは、KDO8Pの形成において律速ではなかった。
【0114】
D-グルシトール6-リン酸デヒドロゲナーゼ(gutD)活性は、340nmでNADHの形成をモニターすることによる連続的な分光光度アッセイ法を用いて測定された。反応を20mMの最終濃度でのD-グルシトール6-リン酸の添加により惹起する前に、酵素溶液(100mM Tris-HCl、pH=8.7、5mM NAD+)を25℃で2分間、プレインキュベートした。
【0115】
実施例XVIII.
本実施例は、gutQの特徴付けを記載する。gutQの特徴付けは、ksdDについて報告された方法に従って同様に行われた(例えば、Meredith, T.C. & Woodard, R.W. (2003) J Biol Chem 278, 32771-7参照;全体として参照により本明細書に組み入れられている)。簡潔には、基質特異性について、酵素試料を、100mM Tizma-HCl緩衝液(pH=8.25)に希釈し、反応を基質(15nM gutQ、10mM糖、1mM EDTA)で惹起することによりアッセイした。37℃での10分後、可能性のある代替基質、D-アラビノース、D-リボース5-リン酸、D-グルコース6-リン酸(G6P)、D-グルコース1-リン酸、D-グルコサミン6-リン酸、またはD-マンノース6-リン酸を含む反応を停止し、ケトースの存在を測定した。D/L-グリセルアルデヒド3-リン酸、D-エリトロース4-リン酸、およびD-フルクトース6-リン酸についての生成物出現を、31P NMRによりアッセイした。速度定数は、非連続的マイクロプレートアッセイ法を用いて37℃で測定され、基質の添加により惹起された。濃度は、典型的には、0.2Kmから10Kmまでの範囲であった。2分後、反応(pH=8.25における50mM Tris-HCl、5nM gutQ、1mM EDTA)を停止し、その時点で、基質の約10%未満が消費されていた。初速度(v0)を、3回通り測定し、非線形最小二乗回帰を用いて標準ミカエリス・メンテン式にフィットさせて、Ru5Pの形成および消失の両方についてのKmおよびkcat値を決定した。平衡定数(Keq)は、kdsDについて記載されているように、31P NMRを用いて決定した(例えば、Meredith, T.C. & Woodard, R.W. (2003) J Biol Chem 278, 32771-7参照;全体として参照により本明細書に組み入れられている)。gutQのpH最適値は、様々なpH値(37℃で調整されたpH=6.25〜10)のBTP緩衝溶液に酵素を希釈することにより測定された。活性は、3分間の反応時間で3回通り、上に概要を説明されているように、測定された(100mM BTP、15nM gutQ、10mM A5P、1mM EDTA)。単離されたgutQの酵素試料を、100mM Trizma-HCl緩衝液(pH=8.25)に希釈し、様々な二価金属またはEDTAと4℃で30分間、インキュベートした。その後、残存活性を、3分間の反応時間、3回通り、飽和基質条件下、37℃でアッセイした(15nM gutQ、10mM A5P、10μM金属またはEDTA)。
【0116】
実施例XIX.
本実施例は、gutQに関わる実験についての大腸菌株構築および増殖条件を記載する。大腸菌株BW30270が、DatsenkoおよびWanner(例えば、Datsenko, K.A. & Wanner, B.L. (2000) Proc Natl Acad Sci USA 97, 6640-5参照;全体として参照により本明細書に組み入れられている)の手順によるファージλレッドリコンビナーゼ系を用いる染色体gutQおよびkdsD遺伝子破壊のための宿主として用いられた。カナマイシンおよびクロラムフェニコールは50μg/mLで用いられた。鋳型としてのpKD13(kan)かまたはpKD3(cat)のいずれかに関するプライマー対GQF-GQRおよびKDF-KDRが、それぞれ、BW30270(ΔgutQ::kan)およびBW30270(ΔkdsD::cat)を構築するために用いられ、表10に列挙されている。抵抗性マーカーを、その後、記載されているように(例えば、Datsenko, K.A. & Wanner, B.L. (2000) Proc Natl Acad Sci USA 97, 6640-5参照;全体として参照により本明細書に組み入れられている)、FLPリコンビナーゼ系を用いて切除した。BW30270(ΔgutQ ΔkdsD)は、電気形質転換後に行われる次の操作のために培地およびプレートがG6P(10μM)およびA5P(15μM)を常に補充されている点を除いて、GQF-GQR PCR産物挿入断片を用いてBW30270(ΔkdsD)から同様に構築された。用いられた全菌株はコロニー精製され、すべての抗生物質抵抗性の喪失について試験され、関連座位は、予想される欠失部位を確認するためにシーケンシングされた。
【0117】
培養物を、チアミン(1μg/mL)および示された炭素源を追加したM9最少培地(26)かまたはMOPS最少培地(例えば、Neidhardt, F.C., Bloch, P.L. & Smith, D.F. (1974) J Bacteriol 119, 736-47参照;全体として参照により本明細書に組み入れられている)のいずれかで37℃で振盪(250rpm)しながら、増殖させた。BW30270(ΔgutQ ΔkdsD)培養物は、G6P(10μM)およびA5P(5〜50μM)をさらに追加された。アンピシリン(100μg/mL)が、pT7-7(AmpR)プラスミドを有する菌株に加えられた。
【0118】
実施例XX.
本実施例は、酵素アッセイ法、LPS分析、およびRT-PCRのための細胞抽出物の調製を記載する。一晩培養物は、単一炭素源としてのグリセロール(0.2%)および示されたサプリメントを含む最少培地で増殖した。培養物を新鮮な最少培地へ希釈し(1:20 v/v)、37℃で2時間、振盪して、細菌を指数関数的増殖へ戻らせた。BW30270(ΔgutQ ΔkdsD)の培養物を、A5P(5μM)およびG6P(10μM)を加えることにより、ヘキソースリン酸輸送系(uhp)を上方制御するためにこの期間中、事前誘導した。微量のG6Pを除去するために細胞を遠心分離(6,500xg、5min、22℃)によりペレット化し、その後、新鮮培地へ接種した。示されている場合、10mM D-グルシトールを培養物へ加え、増殖をさらに4〜6時間、継続して、gutオペロンの上方制御を可能にし、その時点で、すべての培養物は、対数増殖の初期〜中期にあった。細胞を遠心分離(6,500xg、5min、4℃)により収集した。APIおよびgutD活性についてアッセイされるべき画分を、冷却した1%NaCl溶液で2回、洗浄し、その後、緩衝液(20mM Tris-HCl、1mM DTT、pH=8.0)に再懸濁した。細胞を、超音波処理により破壊し、遠心分離(29,000xg、20min、4℃)により清澄化し、凍結した。LPS分析のための試料を、ダルベッコのリン酸緩衝食塩水で2回、洗浄し、ペレットを溶解緩衝液(200mM Tris(pH=6.8)、2%SDS、4%2-メルカプトエタノール、10%グリセロール)に再懸濁した。OD600nmに基づいた等しい数の細胞を、HitchcockおよびBrown(例えば、Hitchcock, P.J. & Brown, T.M. (1983) J Bacteriol 154, 269-77参照;全体として参照により本明細書に組み入れられている)の方法により処理した。RNAについて分析されるべき細胞ペレットを、Max Bacterial Enhancement試薬に迅速に再懸濁し、TRIzol(Invitrogen)を用いて製造会社のプロトコールにより抽出した。RNA試料をさらに、RNアーゼを含まないDNアーゼでの消化により精製し、RNeasyミニキット(Qiagen)を用いて単離した。RNAの質は、アガロース電気泳動により検査され、260nmにおけるUV吸光度により定量化された。定性RT-PCRを、Superscript II One-Step RT-PCRシステム(Invitrogen)を用いて、説明書に従って、鋳型としての1pgの精製された全RNA、およびgutD遺伝子の最初の342塩基対を増幅するためのGDF-GDRプライマー(0.2μM)と共に行った。
【0119】
実施例XXI.
本実施例は、gutQの精製および特徴付けを記載する。均一性への精製は、Q-Sepharose陰イオン交換クロマトグラフィー、続いて硫酸アンモニウム沈殿を用いる2段階で達成された。タンパク質は、SDS-PAGEによる単一の鋭い高分子量バンド(〜33kDa)として現れ、比活性度は329U/mgであった。gutQの生化学的性質は、kdsDのそれらと類似していることが測定された。速度パラメーター、pH最適値、補因子要求性の喪失、および四次構造がすべて類似していた。A5Pと共通の機能性を共有する単糖を、gutQについての可能性のある代替基質として試験した。システイン-カラバゾール比色定量アッセイ法において、2-ケトヘキソースおよび2-ケトペントースは、540nmでの光を吸収するパープル-レッド発色団を形成する(例えば、Dische, Z., Borenfreund, E. (1951) J Biol Chem 192, 583-587参照;全体として参照により本明細書に組み入れられている)。アルドースのケトースへの変換は、対照に対する試料のA540nmでの吸光度の比率における増加を測定することにより観察されうる。試験された糖のいずれも、それらのそれぞれのケトース型へ変換されなかった。短鎖リン酸化アルドースD/L-グリセルアルデヒド3-リン酸およびD-エリトロース4-リン酸、加えてD-フラクトース6-リン酸が、31P-NMRにより測定された場合、代替基質としての役割を果たした。検出の範囲内で、gutQは、A5PおよびRu5Pについての特異的リン酸糖アルドール-ケトールイソメラーゼであることが示された。
【0120】
実施例XXII.
本実施例は、gutQがリポ多糖生合成を持続させる能力があることを実証する。gutQのインビボでAPIとして機能する能力を評価するために、BW30270(ΔgutQ)およびBW30270(ΔkdsD)を、λレッド(γ、β、exo)相同組換え系を用いて構築した(例えば、Datsenko, K.A. & Wanner, B.L. (2000) Proc Natl Acad Sci USA 97, 6640-5参照;全体として参照により本明細書に組み入れられている)。どちらの突然変異も致死性ではなく、必須のLPS生合成に必要とされる十分な量のA5Pを供給することができる他のAPIコード遺伝子の存在を示している。LPSゲルは、gutオペロンが誘導されたかどうかに関わらず、野生型K-12LPSコアのほとんど等しい量を示し(図8A参照)、A5P合成がこれらの増殖条件下で菌株のいずれにおいても律速ではなかったことを示唆している。BW30270(ΔkdsD)におけるgutQの基底レベルは、生存能力を維持し、かつ機能性LPS層を産生するのに十分なA5Pを供給するのに適切であり、gutQが細胞内でAPIとして機能することを強く示唆している。
【0121】
実施例XXIII.
本実施例は、ΔAPI菌株BW30270(ΔgutQ ΔkdsD)におけるLPS生合成を記載する。BW3270におけるgutQおよびkdsD遺伝子の両方は、外因性A5Pを供給するためにG6P誘導性ヘキソースリン酸輸送体(uhp)を利用することにより、破壊された。A5Pは、ヘキソースリン酸輸送系(uhp)の、非誘導性にもかかわらず、高親和性の基質である(例えば、Kadner, R.J., Murphy, G.P. & Stephens, C.M. (1992) J Gen Microbiol 138(Pt 10), 2007-14; Rick, P.D. & Osborn, M.J. (1972) Proc Natl Acad Sci USA 69, 3756-60; Eidels, L., Rick, P.D., Stimler, N.P. & Osborn, M.J. (1974) J Bacteriol 119, 138-43参照;全体として参照により本明細書に組み入れられている)。低濃度の無機リン酸(1.3mM)を有するMOPS最少培地が、無機リン酸によるuhp媒介性輸送の阻害を防ぐために用いられた(例えば、Shattuck-Eidens, D.M. & Kadner, R.J. (1981) J Bacteriol 148, 203-9参照;全体として参照により本明細書に組み入れられている)。uhp輸送体G6Pの天然基質が、A5Pの効率的な誘導および細胞への輸送に必要とされた。A5PおよびG6Pの両方が培地に含まれない限り、研究の時間経過において検出可能な増殖はなかったため、A5PまたはG6P単独は、増殖を回復させることができなかった(図9A参照)。従って、gutQおよびkdsDは、KDO合成のためのA5Pの唯一の細胞内源であった。培養は、リポ多糖生合成を可能にするために培地にA5Pを補充された。接種材料としてA5Pが培地から消耗されている一晩培養物および延長されたインキュベーション時間を用いることにより、BW30270(ΔgutQ ΔkdsD)において合成されることになっている成熟LPSの量は、培地に含まれるA5Pの量に依存した(図9B)。
【0122】
実施例XXIV.
本実施例は、gutオペロンの発現を記載する。BW30270、BW30270(ΔgutQ)、およびBW30270(pT7-gutQ)を、二重の炭素源、D-グルコースおよびD-グルシトールを含むM9最少培地において増殖させた。すべての3つの菌株は、ほとんど同一の速度で増殖し、D-グルコースが培地から消耗された後の約40分の特徴的な通常の長いディオーキシー遅延時間を示した(例えば、Lengeler, J. & Lin, E.C. (1972) J Bacteriol 112, 840-8参照;全体として参照により本明細書に組み入れられている)。これらの条件下で、誘導は、gutQにより影響されなかった。菌株BW30270、BW30270(ΔgutQ)、およびBW30270(ΔkdsD)を、炭素源としてグリセロールを含むM9最少培地で増殖させた。グリセロールは、クラスB炭素源であり、有意な異化代謝産物抑制を引き起こさず(例えば、Lengeler, J.W. (1986) Methods Enzymol 125, 473-85参照;全体として参照により本明細書に組み入れられている)、cAMPレベルの上昇によってD-グルシトールによるgutオペロンの誘導を促進した。総API(kdsDおよび/またはgutQ)およびgutD比活性度は、すべての3つの菌株において測定された(表11)。
【0123】
(表11)細胞抽出物におけるgutDおよびAPIの比活性度
a菌株は炭素源として0.2%グリセロールを含むM9最少培地で増殖した。bD-グルシトールは、収集の4時間前に(+)で示された培地へ10mMで加えられた。cnモル/min/mgで報告された比活性度。d値はkdsDおよび/またはgutQ活性を含む。
【0124】
BW30270(ΔgutQ)およびBW30270(ΔkdsD)のgutオペロンは依然として誘導可能のままであり、親のBW30270菌株と比較した時、gutD活性により推定された場合、誘導の程度においてたった2倍の差であった。総API活性レベルは、D-グルシトールが培地へ添加された場合、BW30270およびBW30270(ΔkdsD)の両方において増加し、gutQがgutDと共に上方制御されることを示している。BW30270(ΔgutQ)においてD-グルシトールの添加により、その菌株が依然としてgutDを上方制御する能力があるにもかかわらず、観察されたAPIレベルにおける変化がなかった。API活性の大部分は、D-グルシトールを欠損する培地においてkdsDに起因したが、kdsDの恒常的に発現したLPS生合成酵素としての同定を確認した。BW30270(pT7-gutQ)は、上昇したAPIレベルのgutオペロンへの効果を調べるために用いられた(表10)。D-グルシトールが培地に供給されない限りオペロンが抑制されたままであるため、gutDレベルに感知できるほどの差が観察されなかったにもかかわらず、APIレベルの基底レベルはBW30270(pT7-gutQ)において〜250倍、増加した。
【0125】
実施例XXV.
本実施例は、A5Pがgutオペロンの上方制御に重要であることを示す。単一のAPI遺伝子が破壊された場合、制御に差が観察されなかったため、表現型を直接観察できなかったことは、APIの第二のコピーによる抑制のせいであった可能性がある。gutオペロンの誘導能は、BW30270(ΔgutQ ΔkdsD)において調べられた。 一晩培養物はMOPS最少培地(0.2%グリセロール、15μM A5P、10μM G6P)で増殖し、細胞を指数関数的増殖へ戻すために新鮮培地(0.2%グリセロール、5μM A5P、10μM G6P)へ希釈された。2時間の振盪後、細胞を収集し、グリセロールおよびA5Pのみを含む培地へ接種するのに用いた。細胞をuhp輸送体遺伝子について事前誘導したため、G6Pを加えなかった。LPSおよび増殖速度のレベルにおける差が研究の時間経過において最小であるように、2つの濃度のA5P(5μMおよび50μM)を選択した。50μMにおいて、A5P、gutDは、ほとんど野生型レベルまで誘導可能のままであった(表12)。
【0126】
(表12)ΔAPI細胞抽出物におけるgutDおよびgutQ321の比活性度
a菌株は0.2%グリセロールを含むMOPS最少培地で増殖し、10μM G6P/5μM A5Pで事前誘導された。
bD-グルシトールは、収集の4時間前に(+)で示された培地へ10mMで加えられた。cnモル/min/mgで報告された比活性度。dN.D.=検出された活性無し。
【0127】
gutQタンパク質産物自体は、発現に必要ではなかった。A5P濃度が5μMまで減少した場合、D-グルシトール増殖細胞においてgutD活性の顕著かつ再現性のある減少があった。しかしながら、LPSのレベルは、相対的にほんのわずかのみ低下した(図9C)。これは、A5PレベルとgutDの量の間の直接的相関、および差が枯渇したLPS層から生じる多面発現効果の結果によるのではないことを示した。gutD遺伝子の発現レベルの分析は、gutDの測定された比活性度の減少がmRNAの量と相関することを示した(図9D)。gutオペロンは、gutQをコードするプラスミドにより補完された場合、同じ増殖条件下で誘導可能のままであった。
【0128】
実施例XXVI.
本実施例は、過剰発現した場合、遺伝子msbAが、ΔKDO大腸菌細胞をD-アラビノース5-リン酸培地補充なしの寒天上で増殖するのを可能にすることを示す。
【0129】
MsbAは、最初、LpxL(HtrB)温度感受性表現型の多コピー抑制因子として同定された(全体として参照により本明細書に組み入れられている、Polissi et al., 1996, Mol. Microbiol. 20:1221-1233)。栄養要求性TCM15(大腸菌)株のKPM22ゲノムDNAのコスミドライブラリーでの補完により、msbAがΔKdo表現型の多コピー抑制因子であることが明らかにされた。msbA座を含む17個の別々のコスミドクローンを単離した。野生型と同一の無傷の野生型msbA配列のみを有する3.5kb挿入断片を含むコスミドサブクローン(pMMW52)は、A5P栄養要求性の喪失および固体寒天上でのコロニー形成能力の回復により判断されるように、TCM15を直接、救出することができた。TCM15(pMMW52)の増殖速度は、KPM22大腸菌株と著しく類似している(全体として参照により本明細書に組み入れられている、Meredith et al., 2006, ACS Chem. Biol. 1:33-42)。
【0130】
上記明細書に挙げられたすべての刊行物および特許は、参照により本明細書組み入れられている。本発明は特定の好ましい態様に関して記載されているが、主張される本発明がそのような特定の態様に不当に限定されるべきではないことは理解されているはずである。実際、当業者に明らかである、本発明を行うための記載された様式の様々な改変が、特許請求の範囲の範囲内にあることを意図される。
【図面の簡単な説明】
【0131】
【図1】KPM22から抽出されたLPS試料の特徴付けを示す。図1A:透析後のフェノール抽出物由来のLPSの内側コア-リピドA糖組成。GlcN - D-グルコサミン;KDO - 2-ケト3-デオキシ-D-マンノ-オクツロソン酸;L-グリセロ-D-マンノ-ヘプトース - ヘプトース。図1B:プロテイナーゼK処理された細胞全体の可溶化物由来のLPSのSDS-PAGE分析。上部パネルは銀染色されたが、中央および下部パネルは、リピドAの非グリコシル化1,4'-二リン酸化β-1,6-結合GlcN二糖バックボーンに対して方向づけられたmAB A6を用いて発色した免疫ブロットである。中央パネル膜は、免疫学的反応の前に、リピドAを遊離させるために1%酢酸で処理された。レーン1〜5は、異なるLPSケモタイプのサルモネラ・エンテリカ(Salmonella enterica)血液型亜型である[1. 3749 (Ra); 2. 3750 (Rb2); 3. 3748 (Rb3); 4. 3769 (Rd1); 5. 1102 (Re)]、6. 野生型BW30270、7. KPM22、8. KPM25、9. 増殖培地にA5Pを含むKPM22、10. KPM31、11. KPM34、12. 増殖培地にA5Pを含むKPM31、13. KPM40、14. KPM42、15. 増殖培地にA5Pを含むKPM40、16. 200ngの化学合成されたリピドIVa(化合物406)。
【図2】KPM22におけるLPS前駆体の特徴付けを示す。精製されたLPS試料の負イオン様式での電荷デコンヴォルーションされたエレクトロスプレーイオン化フーリエ変換イオンサイクロトロン(ESI FT-ICR)質量スペクトル。示された質量数は、中性分子のモノアイソトピック質量を指す。図2A:BW30270(挿入図、グリコフォームI;3915.71 uのアイソトピック分布)。図2B:KPM22(挿入図、リピドIVa;1404.86 uの構造)。図2C:KPM25(挿入図、描かれたKDO2-リピドA(Re内毒素)の化学構造を有する野生型LPS、および矢印で示されたヘプトース付着点。赤色、青色、および緑色のピーク標識は、それぞれ、グリコフォームI、IV、およびIIのピークファミリーに対応する(例えば、S. Muller-Loennies, B. Lindner, H. Brade, J. Biol. Chem. 278, 34090 (2003)参照;全体として参照により本明細書に組み入れられている)。個々の構造ピーク割当は、表9に列挙されている。PE - ホスファチジルエタノールアミン;P - リン酸;P-EtN - ホスホエタノールアミン;LAtri、LAtetra、LApenta、LAhexa - リピドAのアシル化状態。
【図3】野生型BW30270(a)およびKPM22(b)の内膜および外膜のスクロース勾配分離を示す。画分は、総タンパク質含量(X)、外膜ホスホリパーゼA(OMPLA)(O)、および内膜NADHオキシダーゼ(▲)についてアッセイされた。タンパク質試料のSDS-PAGEゲル(12%)は、還元条件下で流された。分子量タンパク質マーカー(kDa)は、各ゲルの左側に記載されている。矢印は、OMPタンパク質(〜35kDa)の位置を示す[Q2]。
【図4】KPM22の特徴付けを示す。野生型BW30270(パネルAおよびB)およびKPM22(パネルCおよびD)の透過電子顕微鏡(TEM)像。矢印は、KPM22(パネルC)のOM表面における外膜小胞(OMV)を示す。IM - 内膜、OM - 外膜、PG - ペプチドグリカン。スケールバー=50nm。
【図5】KPM22の特徴付けを示す。図5A:細胞浮遊液の培養物のODあたり1mLあたりのメチルペントース(L-フコース)のμgとして推定されたコラン酸産生。図5B:mAb 898抗体を用いての腸内細菌一般抗原(ECA)の免疫ブロット。レーン1(BW30270)、レーン2(KPM22)、レーン3(KPM25)。
【図6】フェノール相抽出物のESI FT-ICR質量スペクトルを示す。BW30270(A)、KPM22(B)、およびKPM25(C)からのフェノール相の電荷デコンヴォルーションされた負イオンESI FT-ICR質量スペクトル。LPSを、水の滴下式添加によりフェノール粗抽出物から沈殿させた。遠心分離による清澄化後、フェノール上清を上記のように透析および処理した。リピドIVaはこの手順中に水によりフェノール相から沈殿しなかったことに留意されたい(B)。ECAcyc - 環状腸内細菌一般抗原(ECA)。
【図7】LPS調製物のhTNFαサイトカイン誘導能力を示す。ヒト単核細胞(MNC)を、上記のように単離された様々な濃度のLPS調製物に曝露した。hTNFα放出はELISAに基づいたアッセイ法を用いて定量化された。データ点は二回通り収集された(暗色の棒 - BW30270、空白の棒 - KPM22、あや目引きの棒 - KPM25)。
【図8】gutQのD-グルシトール利用およびLPS生合成への効果を示す。(A)BW30270(□)、BW30270(ΔgutQ)(○)、およびBW30270(pT7-gutQ)(△)についてのディオーキシー増殖曲線。1μg/mLチアミンおよび10mM D-グルコースを追加したM9最少培地で増殖した一晩培養物を、二重炭素源として2mM D-グルコースおよび2mM D-グルシトールを含む新鮮培地へ希釈した。細胞増殖は、600nmにおける濁度を測定することによりモニターされた。(B)BW30270(WT)、BW30270(ΔgutQ)、およびBW30270(ΔkdsD)由来のプロテイナーゼK処理された細胞全体の可溶化物の銀染色されたトリシンSDS-PAGE LPSゲル。D-グルシトール(10mM)を含む(+)または含まない(-)最少培地(0.2%グリセロール)において増殖した等量の細菌細胞を、対数期初期に収集し、実験手順に記載されているように、処理した。
【図9】ΔAPI菌株BW30270(ΔgutQ ΔkdsD)における増殖およびLPS合成を示す。(A)チアミン(1ug/mL)および単一炭素源としてのグリセロール(0.1%)を含むMOPS最少培地における大腸菌BW30270(ΔgutQ ΔyrbH)の増殖曲線。糖リン酸は、10μM G6P(△)、15μM A5P(□)のいずれかを、または両方(○)を培地に追加された。(B)A5PでのLPSの滴定。分裂を中止しているMOPS最少培地(0.2%グリセロール、5μM A5P、10μM G6P)で増殖した静止期培養物を、G6P(10μM)および様々な濃度のA5P(0.1、1、10、50、および100μM)を含む新鮮培地へ希釈し、6時間、振盪した。LPS試料は、ODに基づいて同数の細胞から調製され、トリシンSDS-PAGEおよび銀染色により分析された。野生型BW30270(レーン1および2)およびBW30270(ΔgutQ ΔkdsD)(レーン3および4)から調製された試料のgutD分析の(C)LPSトリシンSDS-PAGEおよび(D)定性RT-PCR。BW30270(ΔgutQ ΔkdsD)を、10μM G6Pおよび5μM A5Pで事前誘導し、ペレット化し、A5Pのみおよび示されているように10mM D-グルシトールを含む新鮮なMOPS最少培地(0.2%グリセロール)に再懸濁し、分析のための収集前にさらに4時間、振盪した。S - 0.1〜1kb DNA分子量マーカー;Con - 鋳型としてのゲノムDNA。
【図10】大腸菌K-12MG1655のgutオペロンを示す。転写開始部位は、YamadaおよびSaier(例えば、Yamada, M. & Saier, M.H., Jr. (1988) J Mol Biol 203, 569-83参照;全体として本明細書に組み入れられている)による逆転写酵素マッピングを用いて決定され、矢印で示されている。
【図11】Kdoの生合成およびLPSへの組み入れを示す。関わる酵素は、(1)D-アラビノース5-リン酸イソメラーゼ(KdsD/GutQ)、(2)Kdo8Pシンターゼ(KdsA)、(3)Kdo8Pホスファターゼ(KdsC)、および(4)CMP-Kdoシンテターゼ(KdsB)である。大腸菌において、活性化Kdoの2つの分子は、その後、二次アシル鎖(6)ラウレート(LpxL)および(7)ミリステート(LpxM)の段階的付加の前に、(5)Kdoトランスフェラーゼ(WaaA)によりリピドIVAへ連続的に転移する。
【特許請求の範囲】
【請求項1】
リポ多糖を実質的に欠く生存可能な大腸菌(Escherichia coli)の外膜を含む、組成物。
【請求項2】
リポ多糖を実質的に欠く生存可能な大腸菌を含む、請求項1記載の組成物。
【請求項3】
KDO2-リピドIVA生合成経路において破壊を有する生存可能なグラム陰性細菌を含む、組成物。
【請求項4】
生存可能なグラム陰性細菌がD-アラビノース5-リン酸イソメラーゼ(API)発現を欠く、請求項3記載の組成物。
【請求項5】
菌株がKDOを実質的に含まないように、生存可能なグラム陰性細菌が突然変異を含む、請求項3記載の組成物。
【請求項6】
突然変異が、突然変異されたgutQ遺伝子および突然変異されたkdsD遺伝子からなる群より選択される、請求項5記載の組成物。
【請求項7】
突然変異がKDO2とリピドIVAの間の会合を排除するように、生存可能なグラム陰性細菌が突然変異を含む、請求項3記載の組成物。
【請求項8】
突然変異が、突然変異されたmsbA遺伝子および突然変異されたkdsA遺伝子を含む、請求項7記載の組成物。
【請求項9】
突然変異が、突然変異されたmsbA遺伝子および突然変異されたkdsB遺伝子を含む、請求項7記載の組成物。
【請求項10】
突然変異が、突然変異されたmsbA遺伝子および突然変異されたwaaA遺伝子を含む、請求項7記載の組成物。
【請求項11】
突然変異が、突然変異されたyhjD遺伝子および突然変異されたkdsA遺伝子を含む、請求項7記載の組成物。
【請求項12】
突然変異が、突然変異されたyhjD遺伝子および突然変異されたkdsB遺伝子を含む、請求項7記載の組成物。
【請求項13】
突然変異が、突然変異されたyhjD遺伝子および突然変異されたwaaA遺伝子を含む、請求項7記載の組成物。
【請求項14】
生存可能なグラム陰性細菌の外膜がリピドIVAを発現する、請求項3記載の組成物。
【請求項15】
請求項14記載の生存可能なグラム陰性細菌からリピドIVAを抽出する段階を含む、リピドIVAを作製する方法。
【請求項16】
請求項1記載の組成物を含む、免疫応答を誘導するための組成物。
【請求項17】
gutQおよびkdsD遺伝子発現を欠く生存可能なグラム陰性細菌を含む、組成物。
【請求項18】
外因性D-アラビノース5-リン酸をさらに含む、請求項17記載の組成物。
【請求項19】
グラム陰性細菌がmsbAをさらに過剰発現する、請求項17記載の組成物。
【請求項20】
リピドIVAを含む外膜を含む生存可能な無毒性グラム陰性細菌を含む、組成物。
【請求項21】
外膜がLPS発現を実質的に含まない、請求項20記載の組成物。
【請求項22】
投与が免疫応答を誘導するように、請求項1記載の組成物を被験体に投与する段階を含む、被験体において免疫応答を誘導するための方法。
【請求項1】
リポ多糖を実質的に欠く生存可能な大腸菌(Escherichia coli)の外膜を含む、組成物。
【請求項2】
リポ多糖を実質的に欠く生存可能な大腸菌を含む、請求項1記載の組成物。
【請求項3】
KDO2-リピドIVA生合成経路において破壊を有する生存可能なグラム陰性細菌を含む、組成物。
【請求項4】
生存可能なグラム陰性細菌がD-アラビノース5-リン酸イソメラーゼ(API)発現を欠く、請求項3記載の組成物。
【請求項5】
菌株がKDOを実質的に含まないように、生存可能なグラム陰性細菌が突然変異を含む、請求項3記載の組成物。
【請求項6】
突然変異が、突然変異されたgutQ遺伝子および突然変異されたkdsD遺伝子からなる群より選択される、請求項5記載の組成物。
【請求項7】
突然変異がKDO2とリピドIVAの間の会合を排除するように、生存可能なグラム陰性細菌が突然変異を含む、請求項3記載の組成物。
【請求項8】
突然変異が、突然変異されたmsbA遺伝子および突然変異されたkdsA遺伝子を含む、請求項7記載の組成物。
【請求項9】
突然変異が、突然変異されたmsbA遺伝子および突然変異されたkdsB遺伝子を含む、請求項7記載の組成物。
【請求項10】
突然変異が、突然変異されたmsbA遺伝子および突然変異されたwaaA遺伝子を含む、請求項7記載の組成物。
【請求項11】
突然変異が、突然変異されたyhjD遺伝子および突然変異されたkdsA遺伝子を含む、請求項7記載の組成物。
【請求項12】
突然変異が、突然変異されたyhjD遺伝子および突然変異されたkdsB遺伝子を含む、請求項7記載の組成物。
【請求項13】
突然変異が、突然変異されたyhjD遺伝子および突然変異されたwaaA遺伝子を含む、請求項7記載の組成物。
【請求項14】
生存可能なグラム陰性細菌の外膜がリピドIVAを発現する、請求項3記載の組成物。
【請求項15】
請求項14記載の生存可能なグラム陰性細菌からリピドIVAを抽出する段階を含む、リピドIVAを作製する方法。
【請求項16】
請求項1記載の組成物を含む、免疫応答を誘導するための組成物。
【請求項17】
gutQおよびkdsD遺伝子発現を欠く生存可能なグラム陰性細菌を含む、組成物。
【請求項18】
外因性D-アラビノース5-リン酸をさらに含む、請求項17記載の組成物。
【請求項19】
グラム陰性細菌がmsbAをさらに過剰発現する、請求項17記載の組成物。
【請求項20】
リピドIVAを含む外膜を含む生存可能な無毒性グラム陰性細菌を含む、組成物。
【請求項21】
外膜がLPS発現を実質的に含まない、請求項20記載の組成物。
【請求項22】
投与が免疫応答を誘導するように、請求項1記載の組成物を被験体に投与する段階を含む、被験体において免疫応答を誘導するための方法。
【図1】

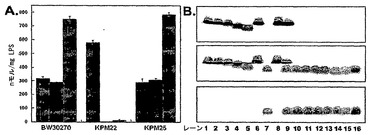
【図2】
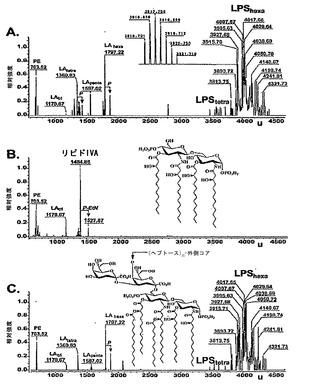
【図3】
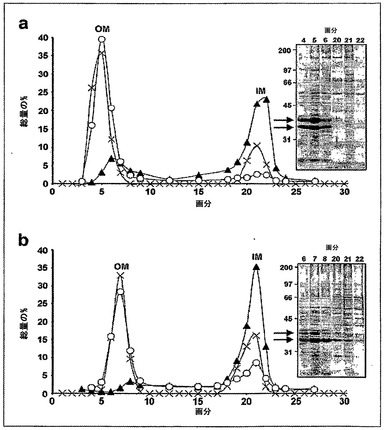
【図4】
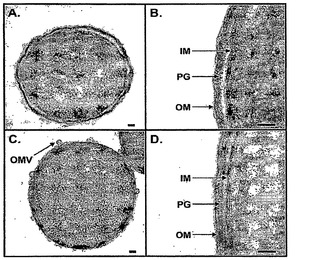
【図5】
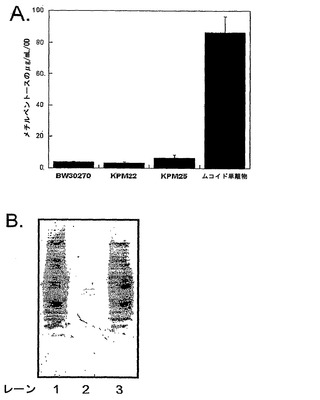
【図6】
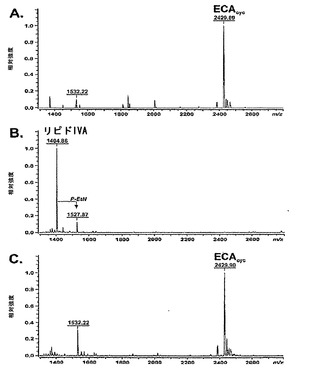
【図7】
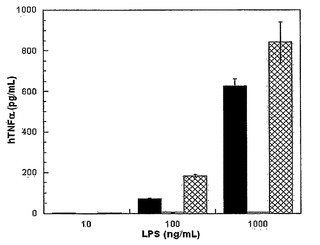
【図8】
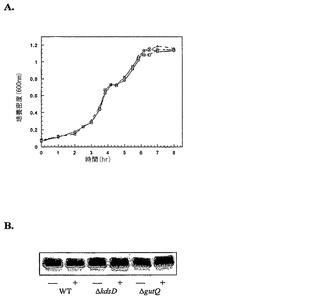
【図9】
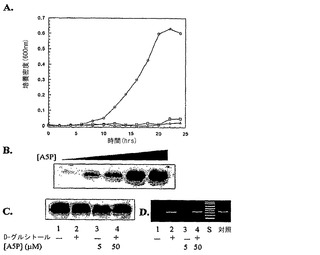
【図10】

【図11】
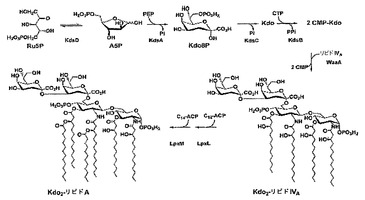
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2009−523810(P2009−523810A)
【公表日】平成21年6月25日(2009.6.25)
【国際特許分類】
【出願番号】特願2008−551397(P2008−551397)
【出願日】平成19年1月19日(2007.1.19)
【国際出願番号】PCT/US2007/001367
【国際公開番号】WO2007/084633
【国際公開日】平成19年7月26日(2007.7.26)
【出願人】(507286909)ザ リージェンツ オブ ザ ユニバーシティ オブ ミシガン (13)
【Fターム(参考)】
【公表日】平成21年6月25日(2009.6.25)
【国際特許分類】
【出願日】平成19年1月19日(2007.1.19)
【国際出願番号】PCT/US2007/001367
【国際公開番号】WO2007/084633
【国際公開日】平成19年7月26日(2007.7.26)
【出願人】(507286909)ザ リージェンツ オブ ザ ユニバーシティ オブ ミシガン (13)
【Fターム(参考)】
[ Back to top ]