
生物製剤の製造および精製を改善するための薬理学的シャペロンの利用
本発明は、組換えタンパク質のための薬理学的シャペロンの使用によって、組換えタンパク質の産生を改善するための方法を提供する。本発明によって例示されるように、細胞により発現される組換えタンパク質への薬理学的シャペロンの結合はタンパク質を安定化し、細胞の小胞体からのタンパク質の排出を増大させ、細胞によるタンパク質の分泌を増大させることができる。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本出願は、参照によってその全体が本明細書に援用される2009年5月26日に出願された米国仮特許出願第61/181,255号の利益を主張する。
【0002】
本発明は、組換えタンパク質のための薬理学的シャペロンの使用によって、組換えタンパク質の産生を改善するための方法を提供する。細胞により発現される組換えタンパク質への薬理学的シャペロンの結合はタンパク質を安定化し、細胞の小胞体からのタンパク質の排出を増大させ、細胞によるタンパク質の分泌を増大させることができる。
【背景技術】
【0003】
種々の組換えヒトタンパク質は、これらの生物製剤を培地中に過剰発現および分泌する哺乳類細胞培養系を用いて産生され、これは次に、クロマトグラフ法によって精製される。首尾よく産生される組換えタンパク質には、分泌タンパク質(例えば、血液凝固因子、免疫グロブリン、エリスロポエチンおよび他のホルモン、エラスターゼ阻害薬など)、およびリソソーム酵素(例えば、β−グルコセレブロシダーゼ、α−ガラクトシダーゼA、酸性α−グルコシダーゼなど)が含まれる。いくつかの重要な因子はこのような製造方法の効率および収率に影響を及ぼす(細胞株の発現のレベル、分泌される組換えタンパク質が精製の前に細胞培地中でその生物活性を保持するかどうか、そして生物製剤の回収のためのタンパク質精製スキーム)。
【0004】
上記の例は、小胞体(ER)において全てが共通生合成経路を共有するタンパク質である(非特許文献1)。これらのタンパク質(全ての膜タンパク質、分泌タンパク質、ペルオキシソームおよびリソソームタンパク質を含む)は、新しく合成されるタンパク質を付加的な翻訳後修飾のためにゴルジ装置に輸送するERからの共通排出経路も共有し、異なる種類のタンパク質を分類して、これらがその意図される細胞および細胞外の目的地に到達できるようにする(非特許文献2)。これらのタンパク質がその最終目的地に到達するためには、これらはまず、この区画を出る前にERの品質管理(QC)システムを十分に通過する安定した構造に折り畳まれなければならない(非特許文献3)。変異タンパク質は安定に折り畳まれないことが多く、ERのQCシステムによって認識されて保持される(非特許文献3)。これらの変異タンパク質が複数回の試みの後に安定な立体配座に至らなければ、これらは最終的にER関連分解(ERAD)システムによって除去される。安定性の低い変異タンパク質の異常なER保持および過剰なERADは、嚢胞性線維症、2型糖尿病、および種々のリソソーム蓄積症を含む多数の疾患の主要な原因であることが示されている(非特許文献4、非特許文献5、非特許文献6)。
【0005】
野生型タンパク質のかなりの割合が割り当ての時間枠内に安定した立体配座に到達できず、最終的にERADによって除去されるような正常なタンパク質の早期の分解も観察される。最も多く引用される例は、野生型嚢胞性線維症の膜コンダクタンス制御因子(CFTR)塩素イオンチャネルの50〜70%の除去である。大きい複合タンパク質(例えば、CFTR、受容体、凝固因子など)はより小さくより簡単な対応物よりも低い効率で折り畳まれる傾向があり、従って早期に分解しやすいと考えられる。さらに、Randall Kaufmanおよび同僚らにより、組換えヒト第VIII因子の蓄積およびERの異常な膨張、この生物製剤の産生中の特定のキナーゼおよび種々の細胞経路の急性の活性化が記載された。これらの深刻な細胞効果は、現在、複合タンパク質および他の問題のあるタンパク質(例えば、第VIII因子およびZ型α−1アンチトリプシン)の発現に関連するERストレスとして知られている。また、タンパク質の蓄積、過剰分解およびERストレスは組換えタンパク質の産生に悪影響を与え、低いタンパク質収率をもたらすとも考えられる。従って、組換えタンパク質の製造方法を改善することが必要とされている。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Blobelら,1979年
【非特許文献2】Kornfeld,1987年
【非特許文献3】Ellgaard & Helenius,2003年
【非特許文献4】Schmitzら,2005年
【非特許文献5】Fanら,1999年
【非特許文献6】Tropakら,2004年
【発明の概要】
【課題を解決するための手段】
【0007】
本発明は、組換えタンパク質のための薬理学的シャペロン(活性部位特異的シャペロン、ASSCとしても知られている)の使用によって、組換えタンパク質の産生を改善するための方法を提供する。本発明によると、組換えタンパク質(例えば、酸性α−グルコシダーゼ、酸性α−ガラクトシダーゼA、または酸性β−グルコシダーゼ)の産生は、例えば、宿主細胞において発現される組換えタンパク質への、薬理学的シャペロン(例えば、1−デオキシノジリマイシン、1−デオキシガラクトノジリマイシン、またはイソファゴミン)の結合によって改善され得る。
【0008】
1つの非限定的な実施形態では、組換えタンパク質は、細胞株によってインビトロで発現される。別の非限定的な実施形態では、宿主細胞は哺乳類細胞である。別の非限定的な実施形態では、宿主細胞はCHO細胞、HeLa細胞、HEK−293細胞、293T細胞、COS細胞、COS−7細胞、マウス初代筋芽細胞、またはNIH3T3細胞であり得る。
【0009】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、細胞の小胞体からのタンパク質の排出を増大させる。
【0010】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、細胞からのンパク質の分泌を増大させる。
【0011】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、細胞によるタンパク質の分泌後に、細胞の外側でタンパク質を安定化する。
【0012】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、組換えタンパク質の発現に関連する小胞体ストレスを低下させる。
【0013】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、5.0よりも高いpHにおけるタンパク質の安定性を向上させる。
【0014】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、細胞による分泌後の細胞培地からのタンパク質の精製の間、組換えタンパク質を安定化する。さらなる非限定的な実施形態では、細胞による分泌後の細胞培地からのタンパク質の精製の間の薬理学的シャペロンの組換えタンパク質への結合は、精製タンパク質の収率を増大させる。
【0015】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、貯蔵の間、タンパク質を安定化することができる。さらなる非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、貯蔵の間、タンパク質消化および/またはタンパク質に対する化学損傷を低下させる。
【0016】
上記の実施形態のいずれか1つにおいて使用される場合、組換えタンパク質および薬理学的シャペロンという用語は無制限に使用される。しかしながら、1つの非限定的な実施形態では、タンパク質は酵素、例えばリソソーム酵素である。別の非限定的な実施形態では、タンパク質は酸性α−グルコシダーゼ(GAA)であり、薬理学的シャペロンは1−デオキシノジリマイシン(DNJ)である。別の非限定的な実施形態では、タンパク質は酸性α−ガラクトシダーゼA(α−Gal A)であり、薬理学的シャペロンは1−デオキシガラクトノジリマイシン(DGJ)である。別の非限定的な実施形態では、タンパク質は酸性β−グルコシダーゼ(グルコセレブロシダーゼ、Gba、GCase)であり、薬理学的シャペロンはイソファゴミン(IFG)である。特定のタンパク質および薬理学的シャペロンは、本明細書に記載される製造方法のいずれか1つにおいて使用することができる。
【図面の簡単な説明】
【0017】
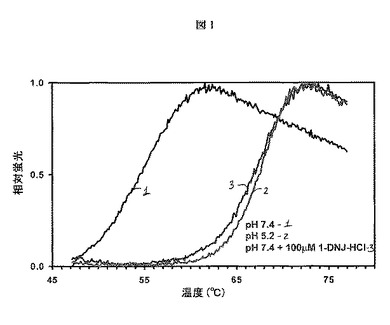
【図1】熱安定性アッセイで決定されるように、100μMの1−デオキシノジリマイシン塩酸塩(1−DNJ−HCl)の存在または不在下で、中性pH(7.4)または酸性pH(5.2)における組換えヒトGAA(Myozyme(登録商標)、Genzyme Corp.)の安定性を示す。熱安定性アッセイは熱を利用してタンパク質の変性を誘発し、これを、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターする。変性のためにより多くの熱を必要とするタンパク質構造は、定義によって、より安定である。Myozymeは、通常、中性pH(7.4)よりもリソソームpH(5.2)においてはるかに安定である。しかしながら、100μMのデオキシノジリマイシンを添加すると、pH7.4における酵素の安定性は、Myozymeだけの場合と比較して著しく向上される。
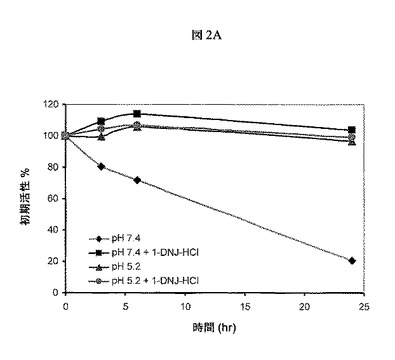
【図2A】37℃で中性pH(7.4)またはリソソームpH(5.2)において、組換えヒトGAA(Myozyme(登録商標)、Genzyme Corp.)活性に対する1−DNJ−HClの効果を示す。GAA活性を評価して、ASSCがrhGAAの活性を時間と共に延長する能力を査定した。Myozyme(45nM)は、50μMの1−DNJを含むまたは含まないpH7.4またはpH5.2の緩衝液中、37℃で24時間にわたってインキュベートした。0、3、6および24時間で、4−MU−α−グルコースを用いて、サンプルをGAA酵素活性についてアッセイし、残留GAA活性を初期活性の%として表した。これらの結果は、1−DNJが中性pH(7.4)におけるGAA酵素活性の損失を改良することを示す。
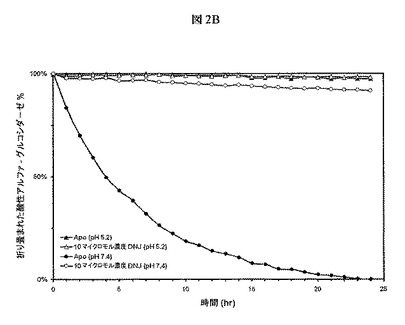
【図2B】図2Aに示される酵素活性の素質、特に中性pH(7.4)におけるMyozyme活性の損失が、タンパク質のアンフォールディングおよび変性と相関するかどうかを決定するために、パラレルなSYPRO Orange熱安定性実験を示す。10μMの1−DNJ−HClを含むまたは含まないpH7.4またはpH5.2の緩衝液中、Myozyme(0.9μM)を37℃でインキュベートし、タンパク質の折り畳みを1時間ごとに24時間にわたってモニターした。図2Aおよび2Bは、GAAの変性が酵素活性の損失と相関することを示す。より重要なことには、これらの結果は、1−DNJが中性pHにおけるGAAの変性および酵素活性の損失を防止できることを示す。
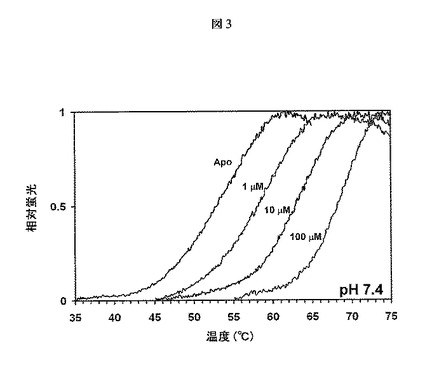
【図3】熱を利用してタンパク質の変性を誘発し、これを、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターする熱安定性アッセイの結果を示す。GAAの融解温度の用量依存的な上昇によって明らかであるように、1−DNJ−HClはGAAの熱安定性を向上させる。実験はpH7.4で行った。
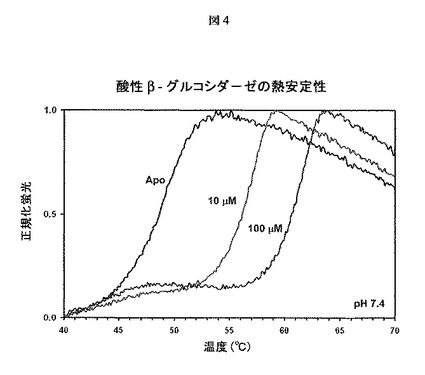
【図4】熱を利用してタンパク質の変性を誘発し、これを、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターする熱安定性アッセイの結果を示す。融解温度の用量依存的な上昇によって明らかであるように、イソファゴミンは酸性β−グルコシダーゼの熱安定性を向上させる。実験はpH7.4で行った。
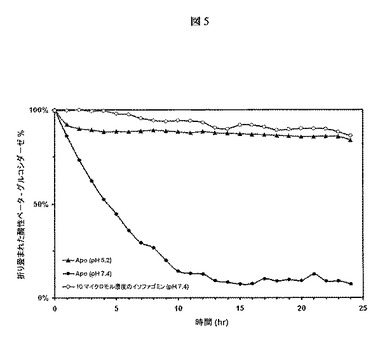
【図5】酸性β−グルコシダーゼ(GCase、Cerezyme(登録商標))のアンフォールディングをモニターするためのSYPRO Orange熱安定性実験を示す。10μMのIFGを含むまたは含まないpH7.4またはpH5.2の緩衝液中、GCase(2μM)を37℃でインキュベートし、タンパク質のアンフォールディングを1時間ごとに24時間にわたってモニターした。これらの結果は、IFGが中性pHにおけるGCaseの変性を防止できることを示す。
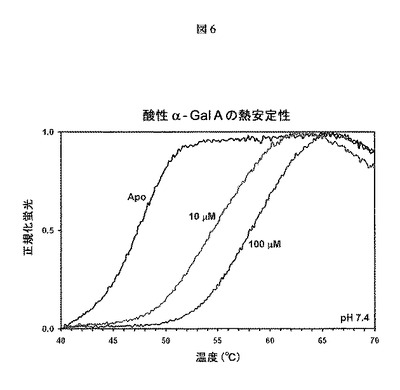
【図6】熱を利用してタンパク質の変性を誘発し、これを、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターする熱安定性アッセイの結果を示す。α−Gal Aの融解温度の上昇によって明らかであるように、1−デオキシガラクトノジリマイシンは、α−Gal A(Fabrazyme(登録商標))の安定性を用量依存的に向上させる。実験はpH7.4で行った。
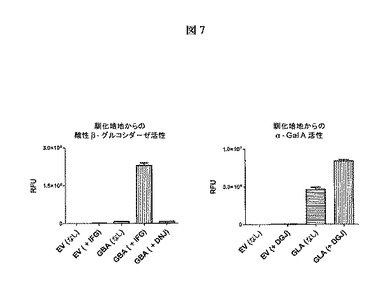
【図7】一過性発現およびそれぞれ100μMのIFGまたはDGJによるインキュベーション中のCOS−7細胞の馴化培地からの酸性β−グルコシダーゼまたはα−Gal Aの活性の増大を示す。これらの結果は、標的タンパク質と結合して安定化する既知の薬理学的シャペロンによるインキュベーションが酵素活性の増大を引き起こすことを実証する。酵素活性レベルの増大は、標的タンパク質のタンパク質分泌の増大および/または馴化培地からの分泌タンパク質の分解および不活性化の防止の結果である。空ベクター(EV)による対照の一過性トランスフェクションと比較して、構造的に類似した薬理学的シャペロン(DNJ)は酸性β−グルコシダーゼ活性の変化を引き起こさなかったので、酸性β−グルコシダーゼの活性の増大は、その薬理学的シャペロン(IFG)に対して特異的である。
【発明を実施するための形態】
【0018】
本発明は、組換えタンパク質のための薬理学的シャペロンの使用によって、組換えタンパク質の産生を改善するための方法を提供する。本発明は、一部分は、薬理学的シャペロンが、リソソーム酵素を発現する細胞の小胞体中で分解されない立体配座においてリソソーム酵素を安定化することができるという発見に基づく。また本発明は、一部分は、薬理学的シャペロンのリソソーム酵素への結合が、温度およびpHストレスに対する酵素の安定性を向上させるという発見にも基づく。
【0019】
明確にするため、そして限定を目的とせずに、この詳細な説明は以下の下位部分に分けられる:
(i) 定義、
(ii) タンパク質欠乏性障害、
(iii) 組換えタンパク質の産生、および
(iv) インビトロ安定性。
【0020】
5.1 定義
本明細書において使用される用語は、一般に、本発明の文脈内および各用語が使用される特定の文脈内で、当該技術分野におけるその通常の意味を有する。特定の用語は以下においてまたは本明細書の他の箇所で論じられており、本発明の組成物および方法、ならびにこれらの製造および使用方法の記載において、実務者にさらなるガイダンスを提供する。
【0021】
「酵素補充療法」または「ERT」という用語は、ネイティブでない精製酵素を、このような酵素が欠乏している個体に導入することを指す。投与される酵素は、天然源から、または組換え発現によって得ることができる。またこの用語は、他の方法で精製酵素の投与を必要としているかその利益を受けている個体、例えば、タンパク質の不足に苦しんでいる個体における精製酵素の導入も指す。導入される酵素は、インビトロで産生された精製組換え酵素であっても、あるいは単離された組織もしくは流体(例えば、胎盤または動物の乳など)から、または植物から精製された酵素であってもよい。
【0022】
「適切な立体配座を安定化する」という用語は、野生型または変異タンパク質の構造がその天然または適切な形態で保持され得るような形で、化合物またはペプチドまたは他の分子が野生型タンパク質と結合する、あるいはインビトロおよびインビボでその野生型機能を果たすことができる変異タンパク質と結合する能力を指す。この効果は、実際には、(i)タンパク質の貯蔵寿命の向上、(ii)タンパク質の単位/量当たりの活性がより高いこと、または(iii)より大きいインビボ効力のうちの1つまたは複数によって現れ得る。これは、発現の間のERからの収率の増大と、温度上昇(例えば、熱安定性アッセイにおいて決定されるように)またはカオトロピック剤の存在、そして同様の手段によるアンフォールディングに対するより優れた耐性とによって、実験的に観察することができる。
【0023】
本明細書で使用される場合、「活性部位」という用語は、いくらかの特異的な生物活性を有するタンパク質の領域を指す。例えば、基質または他の結合パートナーと結合して、化学結合の作製および破壊に直接関与するアミノ酸残基に寄与する部位であり得る。本発明における活性部位は、酵素の触媒部位、抗体の抗原結合部位、受容体のリガンド結合ドメイン、制御因子の結合ドメイン、または分泌タンパク質の受容体結合ドメインを包含することができる。また活性部位は、トランス活性化、タンパク質−タンパク質相互作用、または転写因子および制御因子のDNA結合ドメインも包含することができる。
【0024】
本明細書で使用される場合、「薬理学的シャペロン」または「活性部位特異的シャペロン」という用語は、タンパク質の活性部位と特異的に可逆的相互作用を起こして安定な分子立体配座の形成を高める、タンパク質、ペプチド、核酸、炭水化物などを含む任意の分子を指す。本明細書で使用される場合、「活性部位特異的シャペロン」は、Bip、カルネキシンもしくはカルレティキュリンなどの細胞のER中に存在する内在性の一般的なシャペロン、または重水、DMSO、もしくはTMAOなどの一般的な非特異的化学シャペロンを含まない。
【0025】
1つの非限定的な実施形態では、活性部位特異的シャペロンはタンパク質または酵素の「競合的阻害薬」でよく、ここで、競合的阻害薬は、酵素基質の化学構造および分子構造と構造的に類似しており、基質とほぼ同じ位置で酵素に結合する化合物を指すことができる。従って、阻害薬は基質分子と同じ活性部位を競合するので、Kmが増大する。阻害薬を置換するのに十分な基質分子が利用可能であれば競合的阻害は通常可逆的であり、すなわち、競合的阻害薬は可逆的に結合することができる。従って、酵素阻害の量は、阻害薬の濃度、基質の濃度、ならびに活性部位に対する阻害薬および基質の相対的親和性に依存する。
【0026】
「宿主細胞」という用語は、細胞による物質の産生のため、例えば、遺伝子、DNAまたはRNA配列、タンパク質または酵素の細胞による発現のために、選択され、修飾され、形質転換され、増殖され、もしくは使用され、または何らか形で操作される任意の生物体の任意の細胞を意味する。一実施形態では、宿主細胞は、タンパク質補充療法、例えば酵素補充療法のために使用することができるタンパク質をコードするベクターがトランスフェクトされる。
【0027】
別の非限定的な実施形態では、宿主細胞は、CHO細胞、HeLa細胞、HEK−293細胞、293T細胞、COS細胞、COS−7細胞、マウス初代筋芽細胞、またはNIH3T3細胞であり得る。
【0028】
「精製」という用語は、本明細書で使用される場合、無関係の材料(すなわち、材料が得られたネイティブな材料を含む汚染物質)の存在を低減または除去する条件下で単離された材料を指す。例えば、精製タンパク質は、好ましくは、細胞中で関連のある他のタンパク質または核酸を実質的に含まず、精製核酸分子は、好ましくは、細胞内で一緒に見出すことができるタンパク質または他の無関係の核酸分子を実質的に含まない。本明細書で使用される場合、「実質的に含まない」という用語は、材料の分析試験との関連において、操作上で使用される。好ましくは、汚染物質を実質的に含まない精製材料は、少なくとも95%純粋であり、より好ましくは少なくとも97%純粋であり、さらにより好ましくは少なくとも99%純粋である。純度は、クロマトグラフィ、ゲル電気泳動、イムノアッセイ、組成分析、生物学的アッセイ、および当該技術分野において知られている他の方法によって評価することができる。特定の実施形態では、精製は、汚染物質のレベルがヒトまたは非ヒト動物への安全投与のために規制当局に容認されるレベルよりも低いことを意味する。
【0029】
本明細書で使用される場合、「変異体」および「変異」という用語は、遺伝子材料(例えば、DNA)の検出可能な任意の変化、またはこのような変化の任意のプロセス、メカニズムもしくは結果を意味する。これには、遺伝子の構造(例えば、DNA配列)が変更された遺伝子変異、任意の変異プロセスから生じた任意の遺伝子またはDNA、および改変された遺伝子またはDNA配列によって発現された任意の発現産物(例えば、RNA、タンパク質または酵素)が含まれる。
【0030】
本明細書で使用される場合、「変異タンパク質」という用語は、変更されたタンパク質配列をもたらす遺伝子変異を含有する遺伝子から翻訳されたタンパク質を指す。特定の実施形態では、このような変異の結果、ER中に通常存在する条件下で、タンパク質はその天然の立体配座を達成することができなくなる。この立体配座を達成できないと、その結果、これらのタンパク質は、タンパク質輸送系内のその正常な経路を通って細胞内のその適切な位置へ輸送されるのではなく分解される。他の変異は、活性の低下またはより急速なターンオーバーをもたらし得る。
【0031】
本明細書で使用される場合、「野生型遺伝子」という用語は、インビボで正常な生物学的機能活性を有することができるタンパク質をコードする核酸配列を指す。野生型核酸配列は、その変化が生物活性に少ししかまたは全く影響を及ぼさないアミノ酸置換をもたらす限りは、既知の公表された配列とは異なるヌクレオチド変化を含有し得る。野生型という用語は、内在性または天然タンパク質に対して増大または増強された活性が可能なタンパク質をコードするように操作された核酸配列も含むことができる。
【0032】
本明細書で使用される場合、「野生型タンパク質」という用語は、インビボで発現または導入された場合に機能性生物活性を有することができる野生型遺伝子によってコードされる任意のタンパク質を指す。「正常な野生型活性」という用語は、細胞中のタンパク質の正常な生理機能を指す。このような機能性は、タンパク質の機能性を確立することが知られている任意の手段によって試験することができる。
【0033】
「遺伝子改変」という用語は、コード配列の発現を調節する制御要素と共に、遺伝子産物をコードするコード配列を含む核酸の導入後に特定の遺伝子産物を発現する細胞を指す。核酸の導入は、遺伝子ターゲティングおよび相同組換えを含む当該技術分野において既知の任意の方法によって達成することができる。本明細書で使用される場合、この用語は、例えば遺伝子活性化技術によって、このような細胞によって通常は発現されない内在性遺伝子または遺伝子産物を発現または過剰発現するように操作された細胞も含む。
【0034】
「薬学的に許容可能な」という語句は、本発明の医薬組成物と関連して使用される場合でもそうでなくても、生理学的に容認でき、ヒトに投与された場合に通常有害な反応を生じない分子実体および組成物を指す。好ましくは、本明細書で使用される場合、「薬学的に許容可能な」という用語は、動物、およびより詳細にはヒトでの使用のために、連邦または州政府の規制機関により承認されているか、あるいは米国薬局方または他の一般に認識された薬局方に記載されていることを意味する。「キャリア」という用語は、化合物と一緒に投与される希釈剤、補助剤、賦形剤、または媒体を指す。このような薬学的キャリアは、水および油などの無菌液体であり得る。水または水溶液、生理食塩水溶液、ならびにデキストロースおよびグリセロール水溶液は、好ましくはキャリアとして、特に注射溶液のためのキャリアとして使用される。適切な薬学的キャリアは、E.W.Martinによる「Remington’s Pharmaceutical Sciences」第18版に記載されている。
【0035】
「治療的に有効な用量」および「有効量」という用語は、治療的応答をもたらすのに十分な化合物の量を指す。ASSCおよび酵素が複合体で投与される実施形態では、「治療的に有効な用量」および「有効量」という用語は、治療的応答をもたらすのに十分な複合体の量を指すことができる。治療的応答は、使用者(例えば、臨床医)が治療に対する有効な応答であると認識し得る任意の応答であり得る。従って、治療的応答は、一般に、疾患または障害の1つまたは複数の症状または徴候の寛解であるだろう。
【0036】
「約」および「およそ」という用語は、一般に、測定の性質または精度を考慮して、測定された量に対して許容できる誤差の程度を意味するものとする。通常、例示的な誤差の程度は、所与の値または値の範囲の20パーセント(%)以内、好ましくは10%以内、より好ましくは5%以内である。あるいは、そして特に生物系では、「約」および「およそ」という用語は、一桁分以内である値、好ましくは所与の値の10倍または5倍以内、より好ましくは2倍以内である値を意味してもよい。本明細書において与えられる数量は、他に記載されない限りはおよそのものであり、明確に記載されていない場合でも「約」または「およそ」という用語が推察され得ることを意味する。
【0037】
精製される治療用タンパク質のインビトロ産生、輸送、または貯蔵の間、阻害性であるASSCの濃度は、インビボでの投与時のASSCの希釈(およびその結果として、平衡の変化による結合のシフト)、生物学的利用能および代謝のために、本発明の目的では依然として「有効量」を構成し得ることに注意すべきである。
【0038】
5.2 タンパク質欠乏性障害
タンパク質または酵素の欠乏または特定の組織における機能の損失を特徴とする障害は、理論的にはタンパク質補充療法によって治療可能であり得る。このような障害では、個体の特定の細胞または細胞の全ては十分な機能性タンパク質が欠けており、タンパク質の不活性型を含有するか、あるいは不十分な生物学的機能レベルを含有している。
【0039】
タンパク質または酵素欠乏性障害は、例えば、機能性でないか、あるいは機能が低下または変更されたタンパク質または酵素の発現をもたらすタンパク質または酵素をコードする遺伝子の変異によって生じ得る。また欠乏症は、タンパク質または酵素の発現を少ししかまたは全くもたらさないタンパク質または酵素遺伝子の変異(例えば、ヌル変異)によっても生じ得る。
【0040】
さらに、タンパク質または酵素欠乏症は、タンパク質の折り畳みを変更する変異およびER中の変異タンパク質の遅延によって生じる立体配座の障害(タンパク質の欠乏をもたらす)によるものでもあり得る。このような疾患には、例えば、嚢胞性線維症、α1−アンチトリプシン欠乏症、家族性高コレステロール血症、ファブリー病、アルツハイマー病(Selkoe,Annu.Rev.Neurosci.1994年,17:489−517)、骨形成不全症(Chesslerら,J.Biol.Chem.1993年,268:18226−18233)、糖鎖不全糖タンパク質症候群(Marquardtら,Eur.J.Cell.Biol.1995年,66:268−273)、マロトー・ラミー症候群(Bradfordら,Biochem.J.1999年,341:193−201)、遺伝性盲目(Kaushalら,Biochemistry 1994年,33:6121−8)、グランツマン血小板無力症(Katoら,Blood 1992年,79:3212−8)、遺伝性第VII因子欠乏症(Arbiniら,Blood 1996年,87:5085−94)、眼皮膚白皮症(Halabanら,Proc.Natl.Acad.Sci.USA.2000年,97:5889−94)およびプロテインC欠乏症(Katsumiら,Blood 1996年,87:4164−75)が含まれるが、これらに限定されない。近年、X連鎖疾患の副腎白質ジストロフィー(ALD)における1つの変異が、病的細胞の低温培養によって救済され得る欠損ペルオキシソームトランスポーターの誤った折り畳みをもたらした(Walterら,Am J Hum Genet 2001年,69:35−48)。変異が遺伝子の配列全体にわたって均一に起こることは一般に認められている。従って、欠乏性タンパク質の誤った折り畳みから生じる表現型は、多くの他の遺伝性障害に存在することが予測可能である。
【0041】
遺伝性のタンパク質欠乏性障害の多くは酵素欠乏症である。遺伝性酵素障害の大きな群はリソソーム酵素の変異を伴い、リソソーム蓄積症(LSD)と称される。リソソーム蓄積症は、スフィンゴ糖脂質、グリコーゲン、およびムコ多糖の蓄積によって引き起こされる疾患群である。リソソーム障害の例には、ゴーシェ病(Beutlerら,The Metabolic and Molecular Bases of Inherited Disease,第8版 2001年 Scriverら編 3635−3668頁,McGraw−Hill,New York)、GM1−ガングリオシドーシス(同上 3775−3810頁)、フコシドーシス(The Metabolic and Molecular Bases of Inherited Disease,1995年.Scriver,C.R.,Beaudet,A.L.,Sly,W.S.およびValle,D.編 2529−2561頁,McGraw−Hill,New York)、ムコ多糖症(同上 3421−3452頁)、ポンペ病(同上 3389−3420頁)、ハーラー・シャイエ病(Weismannら,Science 1970年,169,72−74)、AおよびB型ニーマン・ピック病(The Metabolic and Molecular Bases of Inherited Disease 第8版 2001年.Scriverら編,3589−3610頁,McGraw−Hill,New York)、およびファブリー病(同上 3733−3774頁)が含まれるが、これらに限定されない。
【0042】
ファブリー病
「ファブリー病」という用語は、リソソームα−ガラクトシダーゼA(α−Gal A)活性の欠乏によるスフィンゴ糖脂質異化のX連鎖の先天異常を指す。この欠損は、心臓、腎臓、皮膚、およびその他の組織の血管内皮リソソームにおいてグロボトリアオシルセラミド(セラミドトリヘキソシド)および関連のスフィンゴ糖脂質の蓄積を引き起こす。
【0043】
1つの非限定的な実施形態では、α−ガラクトシダーゼAは、GenBank受入番号NM_000169で記載される核酸配列を含むヒトGla遺伝子を指す。あるいは、α−ガラクトシダーゼAは、α−ガラクトシダーゼA遺伝子に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%および100%までの相同性(標準的なソフトウェア、例えばBLASTまたはFASTAによって決定される)を示す任意の核酸分子、ならびに標準条件下でこれらの配列とハイブリッド形成する任意の配列によってコードされ得る。
【0044】
別の非限定的な実施形態では、ヒトα−ガラクトシダーゼA(α−GAL A)は、ヒトGla遺伝子またはこれに対して少なくとも90%相同である任意の他のアミノ酸配列によってコードされる酵素を指す。ヒトα−GAL酵素は429個のアミノ酸からなり、GenBank受入番号U78027およびNP_000160にある。
【0045】
「非定型ファブリー病」という用語は、主にα−GAL欠乏症の心臓症状、すなわち、心臓(特に、左心室)の著しい拡張につながる心筋細胞における進行性のグロボトリアオシルセラミド(GL−3)の蓄積を有する患者を指す。
【0046】
「キャリア」は、欠損α−GAL遺伝子を持つ1つのX染色体と、正常な遺伝子を持つ1つのX染色体とを有し、正常な対立遺伝子のX染色体の不活性化が1つまたは複数の細胞型において存在する女性である。キャリアはファブリー病であると診断されることが多い。
【0047】
1つの非限定的な実施形態では、α−ガラクトシダーゼAのための薬理学的シャペロンは、1−デオキシガラクトノジリマイシン(DGJ)であり得る。ここで、DGJは、以下の構造:
【化1】

を有する化合物である。
【0048】
この用語には、遊離塩基および任意の塩形態、ならびにこれらの任意のプロドラッグが含まれる。
【0049】
α−GAL Aのためのさらにその他の薬理学的シャペロンは、米国特許第6,274,597号明細書、同第6,774,135号明細書、および同第6,599,919号明細(Fanら)に記載されており、α−allo−ホモノジリマイシン、β−1−C−ブチル−デオキシガラクトノジリマイシン、およびα−ガラクト−ホモノジリマイシン、カリステギンA3、カリステギンB2、カリステギンB3、N−メチル−カリステギンA3、N−メチル−カリステギンB2およびN−メチル−カリステギンB3が含まれる。
【0050】
ポンペ病
ポンペ病は、リソソームのグリコーゲン代謝を損なう酸性アルファグルコシダーゼ(GAA、α−グルコシダーゼ)活性の欠乏を特徴とする常染色体劣性LSDである。酵素欠乏症はリソソームのグリコーゲン蓄積をもたらし、その結果、疾患の末期において、進行性の骨格筋の弱化、心臓機能の低下、呼吸不全、および/またはCNS機能障害が起こる。GAA遺伝子の遺伝子変異は発現の低下を引き起こすか、あるいは安定性および/または生物活性が変更された酵素の変異形態を産生し、最終的には疾患に至る(概して、Hirschhorn R,1995年,Glycogen Storage Disease Type II:Acid α−Glucosidase(Acid Maltase)Deficiency,The Metabolic and Molecular Bases of Inherited Disease,Scriverら編,McGraw−Hill,New York,第7版,2443−2464頁を参照)。ポンペ病の3つの認識された臨床形態(小児型、若年型および成人型)は、残留α−グルコシダーゼ活性のレベルと相関している(Reuser A Jら,1995年,Glycogenosis Type II(Acid Maltase Deficiency),Muscle & Nerve Supplement 3、S61−S69)。ASSC(他では「薬理学的シャペロン」とも呼ばれる)は、リソソーム蓄積症(例えば、ポンペ病)などの遺伝性疾患の治療のための有望な新しい治療的アプローチを表す。
【0051】
1つの非限定的な実施形態では、酸性アルファグルコシダーゼ(GAA)は、GenBank受入番号NM_000152、NM_001079803、またはNM_001079804で記載される核酸配列を含むヒトグルコシダーゼ、アルファ、酸性(GAA)遺伝子を指す。あるいは、酸性アルファグルコシダーゼは、酸性α−グルコシダーゼ遺伝子に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%および100%までの相同性(標準的なソフトウェア、例えばBLASTまたはFASTAによって決定される)を示す任意の核酸分子、ならびに標準条件下でこれらの配列とハイブリッド形成する任意の配列によってコードされ得る。
【0052】
別の非限定的な実施形態では、ヒト酸性α−グルコシダーゼは、グリコーゲン、マルトース、およびイソマルトース中に存在するアルファ−1,4−およびアルファ−1,6−結合−D−グルコースポリマーを加水分解する酵素を指す。代替的な名称は以下の通りである:グルコアミラーゼ、1,4−α−D−グルカングルコヒドロラーゼ、アミログルコシダーゼ、ガンマ−アミラーゼ、およびエキソ−1,4−α−グルコシダーゼ。ヒトGAA遺伝子は染色体17q25.2−25.3に位置付けられており、GenBank受入番号Y00839、NP_000143、NP_001073271、またはNP_001073272で記載されるアミノ酸配列、またはこれに対して少なくとも90%相同である任意の他のアミノ酸配列を有する。
【0053】
小児型ポンペ病(IまたはA型)は最も一般的かつ最も重症であり、生後2年以内の成長障害、全身性低血圧症、心臓肥大、および心肺不全を特徴とする。若年型ポンペ病(IIまたはB型)は重症度が中間であり、心肥大のない筋肉症状の優勢を特徴とする。若年型ポンペ病の個体は、通常、呼吸不全のために20歳に達する前に死亡する。成人型ポンペ病(IIIまたはC型)は、多くの場合、10代または遅くても60代に緩徐進行性ミオパシーとして出現する(Felice K Jら,1995年,Clinical Variability in Adult−Onset Acid Maltase Deficiency:Report of Affected Sibs and Review of the Literature、Medicine 74,131−135)。
【0054】
ポンペ病では、α−グルコシダーゼは、グリコシル化、リン酸化、およびタンパク質分解処理によって翻訳後に広範に修飾されることが示されている。最適なグリコーゲン触媒作用のために、リソソームにおけるタンパク質分解による110キロダルトン(kDa)の前駆体から76および70kDaの成熟型への転換が必要とされる。
【0055】
本明細書で使用される場合、「ポンペ病」という用語は、全てのタイプのポンペ病を指す。本出願において開示される製剤および投薬計画は、例えばI型、II型またはIII型ポンペ病の治療のために使用することができる。
【0056】
1つの特定の非限定的な実施形態では、酸性α−グルコシダーゼのための薬理学的シャペロンは、以下の式:
【化2】
によって表される1−デオキシノジリマイシン(1−DNJ)、またはその薬学的に許容可能な塩、そのエステルまたはそのプロドラッグであり得る。一実施形態では、塩は塩酸塩である(すなわち、1−デオキシノジリマイシンHCl)。
【0057】
酸性α−グルコシダーゼのためのさらにその他の薬理学的シャペロンは、米国特許第6,599,919号明細書(Fanら)および米国特許出願公開第2006/0264467号明細書(Mugrageら)に記載されており、α−ホモノジリマイシンおよびカスタノスペルミンが含まれる。
【0058】
ゴーシェ病
本明細書で使用される場合、「ゴーシェ病」という用語は、脂肪グルコセレブロシドを分解するリソソーム酵素グルコセレブロシダーゼの欠乏を指す。そして脂肪は、主に肝臓、脾臓および骨髄に蓄積する。ゴーシェ病は、痛み、疲労、黄疸、骨の損傷、貧血、そしてさらには死をもたらし得る。ゴーシェ病の臨床表現型は3つある。I型の患者は年少期または青年期のいずれかに発症し、容易に傷がつき、貧血、少ない血小板、肝臓および脾臓の拡張、骨格の弱化のために疲労を経験し、場合によっては、肺障害および腎障害を有する。脳障害の徴候は存在しない。II型では、早発性の肝臓および脾臓の拡張が生後3か月までに発生し、広範な脳障害が存在する。2歳までの死亡率は高い。III型は、肝臓および脾臓の拡張ならびに脳の発作を特徴とする。β−グルコセレブロシダーゼ遺伝子はヒト1q21染色体上に位置する。そのタンパク質前駆体は536個のアミノ酸を含有し、その変異タンパク質は497個のアミノ酸の長さである。
【0059】
1つの非限定的な実施形態では、グルコセレブロシダーゼは、GenBank受入番号NM_001005741、NM_001005741、NM_001005749、NM_001005750、またはNM_000157で記載される核酸配列を含むヒトグルコシダーゼ、ベータ(Gba)遺伝子を指す。あるいは、グルコセレブロシダーゼは、グルコセレブロシダーゼ遺伝子に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%および100%までの相同性(標準的なソフトウェア、例えばBLASTまたはFASTAによって決定される)を示す任意の核酸分子、ならびに標準条件下でこれらの配列とハイブリッド形成する任意の配列によってコードされ得る。
【0060】
別の非限定的な実施形態では、グルコセレブロシダーゼは、ヒトグルコシダーゼ、ベータ(Gba)遺伝子(GenBank受入番号NP_001005741、NP_001005742、NP_001005749、NP_001005750、またはNP_000148)、またはこれに対して少なくとも90%相同である任意の他のアミノ酸配列によってコードされる酵素を指す。
【0061】
1つの非限定的な実施形態では、グルコセレブロシダーゼのための薬理学的シャペロンは、以下の構造:
【化3】
を有するイソファゴミン(IFG、(3R,4R,5R)−5−(ヒドロキシメチル)−3,4−ピペリジンジオール)であり得る。
【0062】
酒石酸イソファゴミンは、共同所有される米国特許第7,501,439号明細書(Mugrageら)に最近記載されており、CAS番号919364−56−0が割り当てられている。またイソファゴミンは、様々な有機および無機酸と作られる他の酸付加塩の形態で調製され得る。このような塩には、塩化水素、臭化水素、メタンスルホン酸、硫酸、酢酸、トリフルオロ酢酸、シュウ酸、マレイン酸、ベンゼンスルホン酸、トルエンスルホン酸および種々の他の酸(例えば、硝酸、リン酸、ホウ酸、クエン酸、安息香酸、アスコルビン酸、サリチル酸など)と形成されるものが含まれる。このような塩は、当業者に知られているように形成することができる。
【0063】
グルコセレブロシダーゼのためのさらにその他の薬理学的シャペロンは、米国特許第6,599,919号明細書(Fanら)、同第6,046,214号明細書(Kristiansenら)、同第5,844,102号明細書(Sierksら)、および米国特許出願公開第2008/0009516号明細書(Wustman)に記載されており、C−ベンジルイソファゴミンおよび誘導体、N−アルキル(C9〜12)−DNJ、グルコイミダゾール(および誘導体)、C−アルキル−IFG(および誘導体)、N−アルキル−β−バリエナミン、フルフェノジン(fluphenozine)、N−ドデシル−DNJ、ならびにカリステギンA3、B1、B2およびC1が含まれる。
【0064】
遺伝性障害に加えて、その他の酵素欠乏症は、原発性または続発性の障害に起因する組織または器官への損傷から生じる。例えば、損傷した膵臓組織、または膵炎は、アルコール依存症によって引き起こされ、消化に必要な膵臓酵素の欠乏をもたらす。膵炎は、現在、酵素補充療法を用いて治療されている。
【0065】
タンパク質欠乏の障害は、生物学的過程を増強または刺激する補充タンパク質の投与によって治療することができる。例えば、貧血のある個体は、赤血球の産生を刺激して組織への酸素輸送を増大させるために組換えエリスロポエチン(EPOGEN(登録商標)、PROCRIT(登録商標)、EPOIETIN(登録商標))が投与される。さらに、インターフェロンアルファ2b(INTRON A(登録商標)、PEG−INTRON(登録商標)、またはREBETOL(登録商標))、およびインターフェロンベータ1a(AVONEX(登録商標)、BETASERON(登録商標))などの組換えインターフェロンは、それぞれB型肝炎および多発性硬化症を治療するために投与される。投与されるさらにその他のタンパク質は、組換えヒトデオキシリボヌクレアーゼI(rhDNase−PULMOZYME(登録商標))と、嚢胞性線維症患者の肺機能を改善するために使用されるDNAを選択的に切断する酵素と、ほぼ全部または全部の甲状腺摘出術を受けたことがあり、従って、甲状腺ホルモンを摂取しなければならない甲状腺癌患者において使用するために開発された組換え甲状腺刺激ホルモン(THYROGEN(登録商標))と、化学療法からの好中球減少を治療するための組換えG−CSF(NEUPOGEN(登録商標))と、そして膵炎のある個体における消化酵素である。タンパク質治療の別の重要な領域は、非常に特異的で明確に定義された活性部位を有する抗体による感染症および癌の治療である。抗体治療製品には、呼吸器多核体ウィルスのためのRESPIRGRAM(登録商標)、乳癌のためのHERCEPTIN(登録商標)、関節炎および炎症性疾患のためのREMICAID(登録商標)およびHUMIRA(登録商標)などが含まれる。抗体のためのASSCはよく知られており、標的抗原または構造的に関連のある類似体(例えば、活性標的の修飾型または模倣薬)のいずれかを使用することができる。
【0066】
「患者」は、特定の疾患であると診断されたか、あるいは特定の疾患である疑いのある被験者を指す。患者はヒトであっても動物であってもよい。例えば、「ファブリー病患者」は、ファブリー病であると診断されたか、あるいはファブリー病である疑いのある個体を指し、変異α−GALを有する。ファブリー病の特徴的なマーカーは男性の半接合体および女性キャリアにおいて同じ有病率で発生し得るが、通常、女性は重症度が低い。
【0067】
5.3 組換えタンパク質の産生
タンパク質欠乏症患者の治療(例えば、酵素補充療法による)のために有用な補充タンパク質は、当該技術分野の技能の範囲内の通常の分子生物学、微生物学、および組換えDNA技術を用いて単離および精製することができる。例えば、補充タンパク質をコードする核酸は、文献に記載されるような組換えDNA発現を用いて単離することができる。例えば、Sambrook,Fritsch & Maniatis,Molecular Cloning:A Laboratory Manual,第2版(1989年)Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.(本明細書では「Sambrookら,1989年」)、DNA Cloning:A Practical Approach,IおよびII巻(D.N.Glover編1985年)、Oligonucleotide Synthesis(M.J.Gait編1984年)、Nucleic Acid Hybridization(B.D.Hames & S.J.Higgins編(1985年))、Transcription And Translation(B.D.Hames & S.J.Higgins編(1984年))、Animal Cell Culture(R.I.Freshney編(1986年))、Immobilized Cells And Enzymes(IRL Press,(1986年))、B.E Perbal,A Practical Guide To Molecular Cloning(1984年)、F.M.Ausubelら(編),Current Protocols in Molecular Biology,John Wiley & Sons,Inc.(1994年)を参照されたい。タンパク質をコードする核酸は、遺伝子が生物学的に活性なタンパク質をコードする限りは、全長でも切断されていてもよい。例えば、ファブリー病に関連する欠損酵素α−Gal Aの生物学的に活性な切断型は、米国特許第6,210,666号明細書(Miyamuraら)に記載されている。
【0068】
標的タンパク質をコードする同定および単離された遺伝子は、次に、適切なクローニングベクターに挿入され得る。当該技術分野において知られている多数のベクター−宿主系を使用することができる。可能性のあるベクターにはプラスミドまたは変性ウィルスが含まれる(これらに限定されない)が、ベクター系は、使用される宿主細胞と適合性でなければならない。ベクターの例としては、大腸菌(E.coli)、ラムダ誘導体などのバクテリオファージ、またはpBR322誘導体もしくはpUCプラスミド誘導体、例えばpGEXベクター、pmal−c、pFLAGなどのプラスミドが挙げられるが、これらに限定されない。クローニングベクターへの挿入は、例えば、相補的な付着末端を有するクローニングベクターへDNA断片を連結することによって達成することができる。しかしながら、DNAを断片化するために使用される相補的な制限部位がクローニングベクターに存在しなければ、DNA分子の末端は酵素的に修飾され得る。あるいは、所望される部位はどれもDNA末端へのヌクレオチド配列(リンカー)の連結によって生成することができ、これらの連結されたリンカーは、制限エンドヌクレアーゼの認識配列をコードする特異的な化学合成オリゴヌクレオチドを含むことができる。組換えタンパク質の産生は、分泌を容易にするN末端のシグナルペプチドの包含、またはポリアデニル化部位を含有する3’非翻訳配列の包含などの遺伝子操作によって最大にすることができる。
【0069】
1つの非限定的な実施形態では、宿主細胞を形質導入するために使用される構築物は、アデノウィルス、アデノ随伴ウィルス、ヘルペスウィルス、ムンプスウィルス、ポリオウィルス、レトロウイルス、シンドビスウィルスおよびワクシニアウィルスを含むがこれらに限定されないウィルス由来のベクターである。
【0070】
組換え分子は、遺伝子配列の多数のコピーが作成されるように、形質転換、トランスフェクション、感染、エレクトロポレーションなどによって宿主細胞中に導入され得る。好ましくは、クローン化遺伝子はシャトルベクタープラスミド上に含有され、クローニング細胞、例えば大腸菌中での発現、およびその後の適切な発現細胞株への挿入のための容易な精製(それが所望される場合)を提供する。
【0071】
可能性のある宿主−ベクター系には、ウィルス(例えば、ワクシニアウィルス、アデノウィルスなど)に感染した哺乳類細胞系、ウィルス(例えば、バキュロウイルス)に感染した昆虫細胞系、酵母ベクターを含有する酵母などの微生物、またはバクテリオファージ、DNA、プラスミドDNA、もしくはコスミドDNAで形質転換された細菌が含まれるが、これらに限定されない。ベクターの発現要素は、その強度および特異性が異なる。利用される宿主−ベクター系に応じて、多数の適切な転写および翻訳要素のどれか1つが使用され得る。異なる宿主細胞は、タンパク質の翻訳および翻訳後処理および修飾(例えば、グリコシル化、切断[例えば、シグナル配列の])のための特徴的で特異的なメカニズムを有する。適切な細胞株または宿主系は、グリコシル化、シアリル化(sialyation)およびリン酸化などの、発現される外来性タンパク質の所望の修飾および処理を保証するように選択することができる。例えば、細菌系における発現は、非グリコシル化コアタンパク質産物を産生するために使用することができる。しかしながら、細菌において発現されるタンパク質は適切に折り畳まれないことがある。酵母における発現は、グリコシル化産物を産生することができる。真核細胞における発現は、異種タンパク質の「ネイティブな」グリコシル化および折り畳みの可能性を増大させることができる。さらに、哺乳類細胞における発現は、タンパク質を再構成または構成するための手段を提供することができる。
【0072】
組換えで発現されたタンパク質の精製は、硫酸アンモニウム沈殿、疎水性相互作用樹脂、カチオン交換樹脂、アニオン交換樹脂、およびクロマトフォーカシング樹脂を含有するカラムクロマトグラフィなどの当該技術分野において既知の方法を用いて達成することができる。あるいは、タンパク質、または組換えタンパク質に融合されたタグに特異的に結合する適切なポリクローナルまたはモノクローナル抗体を用いて組換えタンパク質を精製するために、イムノアフィニティクロマトグラフィを使用することもできる。好ましい実施形態では、本発明の方法のために使用される組換えタンパク質の純度は、少なくとも95%、好ましくは少なくとも97%、最も好ましくは98%よりも高いであろう。
【0073】
他の非限定的な実施形態では、タンパク質欠乏症患者の治療(例えば、酵素補充療法による)のために有用な補充タンパク質は、組換え細胞発現系から精製してもよい(例えば、哺乳類細胞または昆虫細胞−概して、米国特許第5,580,757号明細書(Desnickら)、米国特許第6,395,884号明細書および同第6,458,574号明細書(Seldenら)、米国特許第6,461,609号明細書(Calhounら)、米国特許第6,210,666号明細書(Miyamuraら)、米国特許第6,083,725号明細書(Seldenら)、米国特許第6,451,600号明細書(Rasmussenら)、米国特許第5,236,838号明細書(Rasmussenら)、ならびに米国特許第5,879,680号明細書(Ginnsら)を参照、ヒト胎盤、または動物乳(米国特許第6,188,045号明細書(Reuserら)を参照))。
【0074】
薬学的使用に適した補充タンパク質を得るためのその他の合成技術は、例えば、米国特許第7,423,135号明細書、同第6,534,300号明細書、および同第6,537,785号明細書、国際公開第2005/077093号パンフレット、ならびに米国特許出願公開第2007/0280925号明細書および同第2004/0029779号明細書において見出し得る。これらの参考文献は全ての目的で参照によってその全体が本明細書に援用される。
【0075】
さらに、異なるベクター/宿主発現系は、タンパク質切断などの処理反応に異なる程度で影響を与え得る。発現効率は、米国特許第6,274,597号明細書に記載されるような特異的なシャペロンおよび関連のファミリーメンバーの使用によって増大させることができる。
【0076】
上記のように、本発明の1つの態様は、組換えタンパク質の産生を改善するための方法を提供し、本方法は、組換えタンパク質を発現する宿主細胞と、組換えタンパク質に特異的な薬理学的シャペロンとを接触させることを含む。
【0077】
1つの非限定的な実施形態では、薬理学的シャペロンは、タンパク質を発現する細胞のER中で標的の組換えタンパク質を安定化し、タンパク質のERADおよび早期分解を防止し得る。そうすることで、薬理学的シャペロンは、組換えタンパク質の発現に関連するERストレスを低下させ得、これは次に産生細胞株がタンパク質の高い生存率および発現を保持できるようにし得る。
【0078】
他の非限定的な実施形態では、薬理学的シャペロンは、細胞により発現される組換えタンパク質を安定化し、細胞のERからのタンパク質の排出を増大させ、細胞からのタンパク質の分泌を増大させ得る。
【0079】
他の非限定的な実施形態では、薬理学的シャペロンは、タンパク質を発現する細胞の外側で(すなわち、タンパク質が細胞培地中に分泌された後)、標的の組換えタンパク質を安定化させ得る。組換えタンパク質を安定化することによって、薬理学的シャペロンは、安定性を保つために普通はより低いpHで貯蔵されなければならず、そして通常中性pHで処方されるほとんどの細胞培地中で普通は安定ではないタンパク質に利益を与え得る。酵素活性の著しい損失が起こり得るが、これらのタンパク質は収穫および精製の前は細胞培地中に保持される。薬理学的シャペロンは、5.0よりも高いpH、例えば中性pHを有する培地中で、これらのタンパク質を安定化させ、不可逆的な変性およびタンパク質活性の不活性化を防止することができる。
【0080】
他の非限定的な実施形態では、薬理学的シャペロンは、細胞培地からのタンパク質の精製の間、標的組換えタンパク質を保護し得る。組換えタンパク質は、培地中で安定でなければ、変性するかもしれず、精製混合物中の汚染プロテアーゼによるタンパク質分解を受けやすい。薬理学的シャペロンは、タンパク質を安定化させ、タンパク質分解部位が暴露されるのを防止することによって、このタンパク質分解を防止し得る。従って、薬理学的シャペロンは、精製プロセスの間のタンパク質の完全性および/または酵素活性を改善し得る。
【0081】
他の非限定的な実施形態では、薬理学的シャペロンは、医薬製剤の貯蔵および/または調製の間、組換えタンパク質を保護し得る。例えば、薬理学的シャペロンは、このようなタンパク質が貯蔵および処方の間にタンパク質消化されることを保証するために凝固因子(例えば、プロテアーゼ)と結合してその酵素活性を阻害することができる。薬理学的シャペロンは、処方の間、他のタイプの不可逆的化学損傷(例えば、酸化、加水分解または脱アミド)、または凝集、沈殿および表面への吸着などの物理的な不安定性も防止し得る。さらに、薬理学的シャペロンはタンパク質を安定化させ、pH、温度、シェアストレス、凍結/解凍ストレスおよびこれらのストレスの組み合わせなどのストレス(そうでなければ、タンパク質の分解に寄与し得る)から保護し得る。
【0082】
1つの非限定的な実施形態では、補充タンパク質は、この遺伝子の観察される9個のハプロタイプの最も優勢なものによってコードされる組換え酸性α−グルコシダーゼ(GAA)であり、チャイニーズハムスター卵巣細胞株において組換えDNA技術により産生される。組換えGAAは、例えば、Kakkisら,2008年,「An improved alpha−glucosidase enzyme for Pompe disease」,Abstract,第58回Annual Meeting of the ASHG、Kishnaniら,2007年,Neurology.68(2):99−109、米国特許第6,118,045号明細書(Reuserら)、同第7,056,712号明細書(Chen)、および同第7,351,410号明細書(van Bree)(これらはそれぞれ、全ての目的で参照によってその全体が援用される)に記載されるような組換えGAAであり得る。
【0083】
他の好ましい非限定的な実施形態では、ASSCは1−デオキシノジリマイシン(1−DNJ)であり、GAAは組換えGAAである。代替の実施形態では、ASSCはα−ホモノジリマイシンであり、GAAは組換えGAAである。別の代替の実施形態では、ASSCはカスタノスペルミンであり、GAAは組換えGAAである。ASSC(例えば、1−デオキシノジリマイシン、α−ホモノジリマイシンおよびカスタノスペルミン)は、本発明に従う潜在的ASSCの源を提供する合成ライブラリから得ることができる(例えば、Needelsら,Proc.Natl.Acad.Sci.USA 1993年,90:10700−4、Ohlmeyerら,Proc.Natl.Acad.Sci.USA 1993年,90:10922−10926、Lamら,PCT公報国際公開第92/00252号パンフレット、Kocisら,PCT公報国際公開第号パンフレット94/28028を参照)。合成化合物ライブラリは、Maybridge Chemical Co.(Trevillet,Cornwall,UK)、Comgenex(Princeton,N.J.)、Brandon Associates(Merrimack,N.H.)、およびMicrosource(New Milford,Conn.)から市販されている。珍しい化学ライブラリは、Aldrich(Milwaukee,Wis.)から入手可能である。あるいは、細菌、真菌、植物および動物抽出物の形態の天然化合物のライブラリは、例えばPan Laboratories(Bothell,Wash.)またはMycoSearch(NC)から入手可能であるか、または容易に産生可能である。さらに、天然および合成的に産生されたライブラリおよび化合物は、Res.1986年,155:119−29により容易に修飾される。
【0084】
5.4 インビトロ安定性
その貯蔵期間中の補充タンパク質製剤の安定性を保証することは大きな課題である。例えば、組換え酵素のバイアルは、多くの場合、1回だけの使用のためのものであり、未使用の製品は廃棄されなければならない。さらに、組換え酵素は、多くの場合、ヘルスケアの専門家によって再構成、希釈、および投与されなければならず、その投与は遅れてはならない。組換え酵素は、多くの場合、低温、例えば2〜8℃で貯蔵されなければならず、製品は限られた時間、例えば24時間までしか安定でない。
【0085】
ASSCおよび補充タンパク質が同じ組成物中に存在する場合、処方された本発明の組成物は、より安定した組成物を提供する。インビボで投与されるタンパク質の安定化に加えて、ASSCはインビトロで補充タンパク質の立体配座に可逆的に結合して安定化し、これにより、凝集および分解が防止され、製剤の貯蔵寿命が長くなる。ASSC/補充タンパク質の相互作用の分析は、例えば、示差走査熱量測定、または円二色性などの当該技術分野においてよく知られている技術を用いて評価してもよい。
【0086】
例えば、組成物の注射可能な水性製剤が、ニードルおよびシリンジを用いて内容物を引き抜くのに適した栓付バイアル中で供給される場合、ASSCの存在は、補充タンパク質の凝集を阻害する。バイアルは、1回の使用のためまたは多重使用のためのいずれかであり得る。また製剤は、予め充填されたシリンジとして供給することもできる。別の実施形態では、製剤は乾燥または凍結乾燥状態であり、標準または供給される生理学的希釈剤により液体状態に再構成することを必要とし得る。この場合、ASSCの存在は、再構成中および再構成後に、凝集を防止するために補充タンパク質を安定化し得る。製剤が静脈内投与のための液体(例えば、静脈内投与ラインまたはカテーテルへ接続するための無菌バック中など)である実施形態では、ASSCの存在は同じ利益を与え得る。
【0087】
投与される補充タンパク質の安定化に加えて、ASSCの存在は、補充タンパク質製剤を約7.0〜7.5の中性pHで貯蔵可能にし得る。これにより、安定性を保つために普通はより低いpHで貯蔵されなければならないタンパク質に利益が与えられる。例えば、GAAなどのリソソーム酵素は、通常、低pH(例えば、5.0以下)において安定した立体配座を保持する。しかしながら、低pHにおける補充酵素の長期の貯蔵は、酵素および/または製剤の分解を促進し得る。
【実施例】
【0088】
本発明は以下に提示される実施例によってさらに説明される。このような実施例の使用は説明のためだけのものであって、本発明または例示された任意の用語の範囲および意味を決して限定しない。同様に、本発明は、本明細書に記載される特定の好ましい実施形態のどれにも限定されない。実際、本明細書を読めば、本発明の多くの修正および変更は当業者には明らかになるであろう。従って、本発明は、特許請求の範囲が権利を与えられる均等物の全範囲と共に、特許請求の範囲の用語によってのみ限定されるべきである。
【0089】
実施例1: 熱的課題における酸性α−グルコシダーゼの熱安定性
100μMのASSCの1−デオキシノジリマイシン塩酸塩(DNJ)を含む場合および含まない場合の組換えヒトGAA(Myozyme(登録商標)、Genzyme Corp.)の安定性は、熱を利用してタンパク質の変性を誘発させる熱安定性アッセイによって決定した。変性は、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターされる。
【0090】
熱安定性アッセイはERのpHに相当するpH7.4において2つの製剤に対して実施した。図1に示されるように、pH7.4で100μMのDNJを含有する製剤は、pH7.4でASSCを含まない製剤と比較して、変性するために著しく多い熱を必要としたので、より安定である。
【0091】
実施例2: 1−デオキシノジリマイシン(DNJ)は37℃で長時間インキュベートしたときに酸性α−グルコシダーゼの活性損失を防止する
残留GAA活性は、4つの製剤について決定した:
(1)pH7.4においてMyzozymeのみ、
(2)pH7.4においてMyzozyme + 50μMのDNJ、
(3)pH5.2においてMyzozymeのみ、
(4)pH5.2においてMyzozyme + 50μMのDNJ。
【0092】
初期活性(t=0)の%に基づいて活性を24時間にわたって測定した。蛍光発生基質4−MU−α−グルコースの加水分解に基づいて、0、3、6および24時間で、サンプルをGAA酵素活性についてアッセイした。GAA活性は、初期活性の%、すなわち残留活性で表した。
【0093】
図2Aに示されるように、上記製剤(1)(ASSなし)は時間と共に活性を失い、投与の24時間後にその初期活性の約20%しかない。対照的に、製剤(2)は、初期活性の全てではないがほとんどを24時間にわたって保持する。ph5.2の両方の製剤(上記製剤(3)および(4))は、その初期活性のほとんどを24時間にわたって保持する。
【0094】
初期酵素活性の損失が適切な立体配座を保持できないことと相関するかどうかを決定するために、実施例1において説明したように、上記のサンプルにおいてSYPRO Orange熱安定性実験を実施した。この熱安定性実験では、製剤(2)および(4)中のDNJの濃度を10μMに低減した。この実験に基づいて、折り畳まれたGAAの%を推定し、図2Bにプロットした。図2Bにおける製剤(1)の折り畳まれたGAAの量の24時間にわたる減少は、この同じ一般的な製剤について図2Aに示される活性の損失と相関する。
【0095】
実施例3: DNJは熱的課題におけるGAA安定性を向上させる。
実施例1に概略的に説明されるような熱安定性実験を4つの組成物おいて実施した:
(1)Myozymeのみの組成物、
(2)Myozyme + 1μMの1−DNJ−HCl、
(3)Myozyme + 10μMの1−DNJ−HCl、
(4)Myozyme + 100μMの1−DNJ−HCl。
【0096】
図3に示されるように、DNJ−HClは、GAAの融解温度の用量依存的な上昇によって明らかであるように、GAAの熱安定性を向上させる。
【0097】
実施例4: イソファゴミン(IFG)は熱的課題における酸性β−グルコシダーゼの安定性を向上させる
実施例1に概略的に説明されるような熱安定性実験をGCase(Cerezyme(登録商標))の3つの組成物において実施した:
(1)GCaseのみの組成物、pH7.4、
(2)GCase + 10μMのIFG、pH7.4、
(3)GCase + 100μMのIFG、pH7.4。
【0098】
図4に示されるように、IFGは、タンパク質の融解温度の上昇によって明らかであるように、GCaseの熱安定性を用量依存的に向上させる。
【0099】
実施例5: IFGの存在下におけるGCaseの熱安定性
折り畳まれていないGCaseのパーセントを3つの製剤について決定した:
(1)pH5.2においてGCaseのみ、
(2)pH7.4においてGCaseのみ、
(3)pH7.4において10μMのIFGを含むGCase。
【0100】
37℃および中性pHにおいてIFGがGCaseのアンフォールディングを防止するかどうかを決定するために、実施例1において概略的に説明したように、上記のサンプルにおいてSYPRO Orange熱安定性実験を実施した。この熱安定性実験では、IFGの濃度は製剤(3)中10μMであった。図5の結果に基づいて、IFGは、特定の条件下でGCaseのアンフォールディングを防止した。
【0101】
実施例6: 1−デオキシガラクトノジリマシシン(DGJ)は、熱的課題におけるα−Gal Aの安定性を向上させる。
実施例1に概略的に説明されるような熱安定性実験を、α−Gal A(Fabrazyme(登録商標))の3つの組成物において実施した:
(1)α−Gal Aのみの組成物、pH7.4、
(2)α−Gal A + 10μMのDGJ、pH7.4、
(3)α−Gal A + 100μMのDGJ、pH7.4、
【0102】
図6に示されるように、DGJは、タンパク質の融解温度の上昇によって明らかであるように、α−Gal Aの熱安定性を用量依存的に向上させる。
【0103】
実施例7: 薬理学的シャペロンは一過性にトランスフェクトされたCOS−7細胞からの組換えタンパク質の活性レベルを向上させる。
COS−7細胞に空のベクター、GBA遺伝子をコードするプラスミド、またはGLA遺伝子をコードするプラスミドを一過性にトランスフェクトした。種々の一過性トランスフェクションを100μMの表示される薬理学的シャペロン(IFG、DGJまたはDNJ)と共にインキュベートした。タンパク質発現の48時間後に、それぞれのトランスフェクションからの馴化培地を収穫し、コンカナバリンA−アガロースビーズによる分泌タンパク質の捕獲後に、酸性β−グルコシダーゼまたはα−Gal Aの活性のレベルを査定した。このコンカナバリンAの捕獲工程は、適切な蛍光発生基質(GCaseに対しては4−MU−β−グルコース、α−Gal Aに対しては4−MU−β−ガラクト―ス)を用いる活性決定の過程において、薬理学的シャペロンによる可能性のある酵素活性の阻害を除去するために必要であった。
【0104】
図7に示されるように、IFGまたはDGJによるインキュベーションは、それぞれ、酸性β−グルコシダーゼまたはα−Gal Aの活性を増大させた。酸性β−グルコシダーゼの一過性発現においてDNJが含まれる場合、酵素活性の増大は見られなかった。この観察は、酵素活性の増大がタンパク質の既知の薬理学的シャペロンおよび/または阻害薬との特異的な相互作用によるものであることを示す。
【0105】
本発明は、本明細書に記載される特定の実施形態によって範囲が限定されてはならない。実際、本明細書に記載されるものに加えて、上記の説明および添付図面から当業者には本発明の種々の修正が明らかになるであろう。このような修正は特許請求の範囲内に入ることが意図される。
【0106】
特許、特許出願、公報、製品説明、GenBank受入番号、およびプロトコルが本出願全体にわたって引用されており、これらの開示は全ての目的で参照によってその全体が本明細書中に援用される。
【技術分野】
【0001】
関連出願の相互参照
本出願は、参照によってその全体が本明細書に援用される2009年5月26日に出願された米国仮特許出願第61/181,255号の利益を主張する。
【0002】
本発明は、組換えタンパク質のための薬理学的シャペロンの使用によって、組換えタンパク質の産生を改善するための方法を提供する。細胞により発現される組換えタンパク質への薬理学的シャペロンの結合はタンパク質を安定化し、細胞の小胞体からのタンパク質の排出を増大させ、細胞によるタンパク質の分泌を増大させることができる。
【背景技術】
【0003】
種々の組換えヒトタンパク質は、これらの生物製剤を培地中に過剰発現および分泌する哺乳類細胞培養系を用いて産生され、これは次に、クロマトグラフ法によって精製される。首尾よく産生される組換えタンパク質には、分泌タンパク質(例えば、血液凝固因子、免疫グロブリン、エリスロポエチンおよび他のホルモン、エラスターゼ阻害薬など)、およびリソソーム酵素(例えば、β−グルコセレブロシダーゼ、α−ガラクトシダーゼA、酸性α−グルコシダーゼなど)が含まれる。いくつかの重要な因子はこのような製造方法の効率および収率に影響を及ぼす(細胞株の発現のレベル、分泌される組換えタンパク質が精製の前に細胞培地中でその生物活性を保持するかどうか、そして生物製剤の回収のためのタンパク質精製スキーム)。
【0004】
上記の例は、小胞体(ER)において全てが共通生合成経路を共有するタンパク質である(非特許文献1)。これらのタンパク質(全ての膜タンパク質、分泌タンパク質、ペルオキシソームおよびリソソームタンパク質を含む)は、新しく合成されるタンパク質を付加的な翻訳後修飾のためにゴルジ装置に輸送するERからの共通排出経路も共有し、異なる種類のタンパク質を分類して、これらがその意図される細胞および細胞外の目的地に到達できるようにする(非特許文献2)。これらのタンパク質がその最終目的地に到達するためには、これらはまず、この区画を出る前にERの品質管理(QC)システムを十分に通過する安定した構造に折り畳まれなければならない(非特許文献3)。変異タンパク質は安定に折り畳まれないことが多く、ERのQCシステムによって認識されて保持される(非特許文献3)。これらの変異タンパク質が複数回の試みの後に安定な立体配座に至らなければ、これらは最終的にER関連分解(ERAD)システムによって除去される。安定性の低い変異タンパク質の異常なER保持および過剰なERADは、嚢胞性線維症、2型糖尿病、および種々のリソソーム蓄積症を含む多数の疾患の主要な原因であることが示されている(非特許文献4、非特許文献5、非特許文献6)。
【0005】
野生型タンパク質のかなりの割合が割り当ての時間枠内に安定した立体配座に到達できず、最終的にERADによって除去されるような正常なタンパク質の早期の分解も観察される。最も多く引用される例は、野生型嚢胞性線維症の膜コンダクタンス制御因子(CFTR)塩素イオンチャネルの50〜70%の除去である。大きい複合タンパク質(例えば、CFTR、受容体、凝固因子など)はより小さくより簡単な対応物よりも低い効率で折り畳まれる傾向があり、従って早期に分解しやすいと考えられる。さらに、Randall Kaufmanおよび同僚らにより、組換えヒト第VIII因子の蓄積およびERの異常な膨張、この生物製剤の産生中の特定のキナーゼおよび種々の細胞経路の急性の活性化が記載された。これらの深刻な細胞効果は、現在、複合タンパク質および他の問題のあるタンパク質(例えば、第VIII因子およびZ型α−1アンチトリプシン)の発現に関連するERストレスとして知られている。また、タンパク質の蓄積、過剰分解およびERストレスは組換えタンパク質の産生に悪影響を与え、低いタンパク質収率をもたらすとも考えられる。従って、組換えタンパク質の製造方法を改善することが必要とされている。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Blobelら,1979年
【非特許文献2】Kornfeld,1987年
【非特許文献3】Ellgaard & Helenius,2003年
【非特許文献4】Schmitzら,2005年
【非特許文献5】Fanら,1999年
【非特許文献6】Tropakら,2004年
【発明の概要】
【課題を解決するための手段】
【0007】
本発明は、組換えタンパク質のための薬理学的シャペロン(活性部位特異的シャペロン、ASSCとしても知られている)の使用によって、組換えタンパク質の産生を改善するための方法を提供する。本発明によると、組換えタンパク質(例えば、酸性α−グルコシダーゼ、酸性α−ガラクトシダーゼA、または酸性β−グルコシダーゼ)の産生は、例えば、宿主細胞において発現される組換えタンパク質への、薬理学的シャペロン(例えば、1−デオキシノジリマイシン、1−デオキシガラクトノジリマイシン、またはイソファゴミン)の結合によって改善され得る。
【0008】
1つの非限定的な実施形態では、組換えタンパク質は、細胞株によってインビトロで発現される。別の非限定的な実施形態では、宿主細胞は哺乳類細胞である。別の非限定的な実施形態では、宿主細胞はCHO細胞、HeLa細胞、HEK−293細胞、293T細胞、COS細胞、COS−7細胞、マウス初代筋芽細胞、またはNIH3T3細胞であり得る。
【0009】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、細胞の小胞体からのタンパク質の排出を増大させる。
【0010】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、細胞からのンパク質の分泌を増大させる。
【0011】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、細胞によるタンパク質の分泌後に、細胞の外側でタンパク質を安定化する。
【0012】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、組換えタンパク質の発現に関連する小胞体ストレスを低下させる。
【0013】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、5.0よりも高いpHにおけるタンパク質の安定性を向上させる。
【0014】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、細胞による分泌後の細胞培地からのタンパク質の精製の間、組換えタンパク質を安定化する。さらなる非限定的な実施形態では、細胞による分泌後の細胞培地からのタンパク質の精製の間の薬理学的シャペロンの組換えタンパク質への結合は、精製タンパク質の収率を増大させる。
【0015】
別の非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、貯蔵の間、タンパク質を安定化することができる。さらなる非限定的な実施形態では、薬理学的シャペロンの組換えタンパク質への結合は、貯蔵の間、タンパク質消化および/またはタンパク質に対する化学損傷を低下させる。
【0016】
上記の実施形態のいずれか1つにおいて使用される場合、組換えタンパク質および薬理学的シャペロンという用語は無制限に使用される。しかしながら、1つの非限定的な実施形態では、タンパク質は酵素、例えばリソソーム酵素である。別の非限定的な実施形態では、タンパク質は酸性α−グルコシダーゼ(GAA)であり、薬理学的シャペロンは1−デオキシノジリマイシン(DNJ)である。別の非限定的な実施形態では、タンパク質は酸性α−ガラクトシダーゼA(α−Gal A)であり、薬理学的シャペロンは1−デオキシガラクトノジリマイシン(DGJ)である。別の非限定的な実施形態では、タンパク質は酸性β−グルコシダーゼ(グルコセレブロシダーゼ、Gba、GCase)であり、薬理学的シャペロンはイソファゴミン(IFG)である。特定のタンパク質および薬理学的シャペロンは、本明細書に記載される製造方法のいずれか1つにおいて使用することができる。
【図面の簡単な説明】
【0017】
【図1】熱安定性アッセイで決定されるように、100μMの1−デオキシノジリマイシン塩酸塩(1−DNJ−HCl)の存在または不在下で、中性pH(7.4)または酸性pH(5.2)における組換えヒトGAA(Myozyme(登録商標)、Genzyme Corp.)の安定性を示す。熱安定性アッセイは熱を利用してタンパク質の変性を誘発し、これを、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターする。変性のためにより多くの熱を必要とするタンパク質構造は、定義によって、より安定である。Myozymeは、通常、中性pH(7.4)よりもリソソームpH(5.2)においてはるかに安定である。しかしながら、100μMのデオキシノジリマイシンを添加すると、pH7.4における酵素の安定性は、Myozymeだけの場合と比較して著しく向上される。
【図2A】37℃で中性pH(7.4)またはリソソームpH(5.2)において、組換えヒトGAA(Myozyme(登録商標)、Genzyme Corp.)活性に対する1−DNJ−HClの効果を示す。GAA活性を評価して、ASSCがrhGAAの活性を時間と共に延長する能力を査定した。Myozyme(45nM)は、50μMの1−DNJを含むまたは含まないpH7.4またはpH5.2の緩衝液中、37℃で24時間にわたってインキュベートした。0、3、6および24時間で、4−MU−α−グルコースを用いて、サンプルをGAA酵素活性についてアッセイし、残留GAA活性を初期活性の%として表した。これらの結果は、1−DNJが中性pH(7.4)におけるGAA酵素活性の損失を改良することを示す。
【図2B】図2Aに示される酵素活性の素質、特に中性pH(7.4)におけるMyozyme活性の損失が、タンパク質のアンフォールディングおよび変性と相関するかどうかを決定するために、パラレルなSYPRO Orange熱安定性実験を示す。10μMの1−DNJ−HClを含むまたは含まないpH7.4またはpH5.2の緩衝液中、Myozyme(0.9μM)を37℃でインキュベートし、タンパク質の折り畳みを1時間ごとに24時間にわたってモニターした。図2Aおよび2Bは、GAAの変性が酵素活性の損失と相関することを示す。より重要なことには、これらの結果は、1−DNJが中性pHにおけるGAAの変性および酵素活性の損失を防止できることを示す。
【図3】熱を利用してタンパク質の変性を誘発し、これを、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターする熱安定性アッセイの結果を示す。GAAの融解温度の用量依存的な上昇によって明らかであるように、1−DNJ−HClはGAAの熱安定性を向上させる。実験はpH7.4で行った。
【図4】熱を利用してタンパク質の変性を誘発し、これを、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターする熱安定性アッセイの結果を示す。融解温度の用量依存的な上昇によって明らかであるように、イソファゴミンは酸性β−グルコシダーゼの熱安定性を向上させる。実験はpH7.4で行った。
【図5】酸性β−グルコシダーゼ(GCase、Cerezyme(登録商標))のアンフォールディングをモニターするためのSYPRO Orange熱安定性実験を示す。10μMのIFGを含むまたは含まないpH7.4またはpH5.2の緩衝液中、GCase(2μM)を37℃でインキュベートし、タンパク質のアンフォールディングを1時間ごとに24時間にわたってモニターした。これらの結果は、IFGが中性pHにおけるGCaseの変性を防止できることを示す。
【図6】熱を利用してタンパク質の変性を誘発し、これを、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターする熱安定性アッセイの結果を示す。α−Gal Aの融解温度の上昇によって明らかであるように、1−デオキシガラクトノジリマイシンは、α−Gal A(Fabrazyme(登録商標))の安定性を用量依存的に向上させる。実験はpH7.4で行った。
【図7】一過性発現およびそれぞれ100μMのIFGまたはDGJによるインキュベーション中のCOS−7細胞の馴化培地からの酸性β−グルコシダーゼまたはα−Gal Aの活性の増大を示す。これらの結果は、標的タンパク質と結合して安定化する既知の薬理学的シャペロンによるインキュベーションが酵素活性の増大を引き起こすことを実証する。酵素活性レベルの増大は、標的タンパク質のタンパク質分泌の増大および/または馴化培地からの分泌タンパク質の分解および不活性化の防止の結果である。空ベクター(EV)による対照の一過性トランスフェクションと比較して、構造的に類似した薬理学的シャペロン(DNJ)は酸性β−グルコシダーゼ活性の変化を引き起こさなかったので、酸性β−グルコシダーゼの活性の増大は、その薬理学的シャペロン(IFG)に対して特異的である。
【発明を実施するための形態】
【0018】
本発明は、組換えタンパク質のための薬理学的シャペロンの使用によって、組換えタンパク質の産生を改善するための方法を提供する。本発明は、一部分は、薬理学的シャペロンが、リソソーム酵素を発現する細胞の小胞体中で分解されない立体配座においてリソソーム酵素を安定化することができるという発見に基づく。また本発明は、一部分は、薬理学的シャペロンのリソソーム酵素への結合が、温度およびpHストレスに対する酵素の安定性を向上させるという発見にも基づく。
【0019】
明確にするため、そして限定を目的とせずに、この詳細な説明は以下の下位部分に分けられる:
(i) 定義、
(ii) タンパク質欠乏性障害、
(iii) 組換えタンパク質の産生、および
(iv) インビトロ安定性。
【0020】
5.1 定義
本明細書において使用される用語は、一般に、本発明の文脈内および各用語が使用される特定の文脈内で、当該技術分野におけるその通常の意味を有する。特定の用語は以下においてまたは本明細書の他の箇所で論じられており、本発明の組成物および方法、ならびにこれらの製造および使用方法の記載において、実務者にさらなるガイダンスを提供する。
【0021】
「酵素補充療法」または「ERT」という用語は、ネイティブでない精製酵素を、このような酵素が欠乏している個体に導入することを指す。投与される酵素は、天然源から、または組換え発現によって得ることができる。またこの用語は、他の方法で精製酵素の投与を必要としているかその利益を受けている個体、例えば、タンパク質の不足に苦しんでいる個体における精製酵素の導入も指す。導入される酵素は、インビトロで産生された精製組換え酵素であっても、あるいは単離された組織もしくは流体(例えば、胎盤または動物の乳など)から、または植物から精製された酵素であってもよい。
【0022】
「適切な立体配座を安定化する」という用語は、野生型または変異タンパク質の構造がその天然または適切な形態で保持され得るような形で、化合物またはペプチドまたは他の分子が野生型タンパク質と結合する、あるいはインビトロおよびインビボでその野生型機能を果たすことができる変異タンパク質と結合する能力を指す。この効果は、実際には、(i)タンパク質の貯蔵寿命の向上、(ii)タンパク質の単位/量当たりの活性がより高いこと、または(iii)より大きいインビボ効力のうちの1つまたは複数によって現れ得る。これは、発現の間のERからの収率の増大と、温度上昇(例えば、熱安定性アッセイにおいて決定されるように)またはカオトロピック剤の存在、そして同様の手段によるアンフォールディングに対するより優れた耐性とによって、実験的に観察することができる。
【0023】
本明細書で使用される場合、「活性部位」という用語は、いくらかの特異的な生物活性を有するタンパク質の領域を指す。例えば、基質または他の結合パートナーと結合して、化学結合の作製および破壊に直接関与するアミノ酸残基に寄与する部位であり得る。本発明における活性部位は、酵素の触媒部位、抗体の抗原結合部位、受容体のリガンド結合ドメイン、制御因子の結合ドメイン、または分泌タンパク質の受容体結合ドメインを包含することができる。また活性部位は、トランス活性化、タンパク質−タンパク質相互作用、または転写因子および制御因子のDNA結合ドメインも包含することができる。
【0024】
本明細書で使用される場合、「薬理学的シャペロン」または「活性部位特異的シャペロン」という用語は、タンパク質の活性部位と特異的に可逆的相互作用を起こして安定な分子立体配座の形成を高める、タンパク質、ペプチド、核酸、炭水化物などを含む任意の分子を指す。本明細書で使用される場合、「活性部位特異的シャペロン」は、Bip、カルネキシンもしくはカルレティキュリンなどの細胞のER中に存在する内在性の一般的なシャペロン、または重水、DMSO、もしくはTMAOなどの一般的な非特異的化学シャペロンを含まない。
【0025】
1つの非限定的な実施形態では、活性部位特異的シャペロンはタンパク質または酵素の「競合的阻害薬」でよく、ここで、競合的阻害薬は、酵素基質の化学構造および分子構造と構造的に類似しており、基質とほぼ同じ位置で酵素に結合する化合物を指すことができる。従って、阻害薬は基質分子と同じ活性部位を競合するので、Kmが増大する。阻害薬を置換するのに十分な基質分子が利用可能であれば競合的阻害は通常可逆的であり、すなわち、競合的阻害薬は可逆的に結合することができる。従って、酵素阻害の量は、阻害薬の濃度、基質の濃度、ならびに活性部位に対する阻害薬および基質の相対的親和性に依存する。
【0026】
「宿主細胞」という用語は、細胞による物質の産生のため、例えば、遺伝子、DNAまたはRNA配列、タンパク質または酵素の細胞による発現のために、選択され、修飾され、形質転換され、増殖され、もしくは使用され、または何らか形で操作される任意の生物体の任意の細胞を意味する。一実施形態では、宿主細胞は、タンパク質補充療法、例えば酵素補充療法のために使用することができるタンパク質をコードするベクターがトランスフェクトされる。
【0027】
別の非限定的な実施形態では、宿主細胞は、CHO細胞、HeLa細胞、HEK−293細胞、293T細胞、COS細胞、COS−7細胞、マウス初代筋芽細胞、またはNIH3T3細胞であり得る。
【0028】
「精製」という用語は、本明細書で使用される場合、無関係の材料(すなわち、材料が得られたネイティブな材料を含む汚染物質)の存在を低減または除去する条件下で単離された材料を指す。例えば、精製タンパク質は、好ましくは、細胞中で関連のある他のタンパク質または核酸を実質的に含まず、精製核酸分子は、好ましくは、細胞内で一緒に見出すことができるタンパク質または他の無関係の核酸分子を実質的に含まない。本明細書で使用される場合、「実質的に含まない」という用語は、材料の分析試験との関連において、操作上で使用される。好ましくは、汚染物質を実質的に含まない精製材料は、少なくとも95%純粋であり、より好ましくは少なくとも97%純粋であり、さらにより好ましくは少なくとも99%純粋である。純度は、クロマトグラフィ、ゲル電気泳動、イムノアッセイ、組成分析、生物学的アッセイ、および当該技術分野において知られている他の方法によって評価することができる。特定の実施形態では、精製は、汚染物質のレベルがヒトまたは非ヒト動物への安全投与のために規制当局に容認されるレベルよりも低いことを意味する。
【0029】
本明細書で使用される場合、「変異体」および「変異」という用語は、遺伝子材料(例えば、DNA)の検出可能な任意の変化、またはこのような変化の任意のプロセス、メカニズムもしくは結果を意味する。これには、遺伝子の構造(例えば、DNA配列)が変更された遺伝子変異、任意の変異プロセスから生じた任意の遺伝子またはDNA、および改変された遺伝子またはDNA配列によって発現された任意の発現産物(例えば、RNA、タンパク質または酵素)が含まれる。
【0030】
本明細書で使用される場合、「変異タンパク質」という用語は、変更されたタンパク質配列をもたらす遺伝子変異を含有する遺伝子から翻訳されたタンパク質を指す。特定の実施形態では、このような変異の結果、ER中に通常存在する条件下で、タンパク質はその天然の立体配座を達成することができなくなる。この立体配座を達成できないと、その結果、これらのタンパク質は、タンパク質輸送系内のその正常な経路を通って細胞内のその適切な位置へ輸送されるのではなく分解される。他の変異は、活性の低下またはより急速なターンオーバーをもたらし得る。
【0031】
本明細書で使用される場合、「野生型遺伝子」という用語は、インビボで正常な生物学的機能活性を有することができるタンパク質をコードする核酸配列を指す。野生型核酸配列は、その変化が生物活性に少ししかまたは全く影響を及ぼさないアミノ酸置換をもたらす限りは、既知の公表された配列とは異なるヌクレオチド変化を含有し得る。野生型という用語は、内在性または天然タンパク質に対して増大または増強された活性が可能なタンパク質をコードするように操作された核酸配列も含むことができる。
【0032】
本明細書で使用される場合、「野生型タンパク質」という用語は、インビボで発現または導入された場合に機能性生物活性を有することができる野生型遺伝子によってコードされる任意のタンパク質を指す。「正常な野生型活性」という用語は、細胞中のタンパク質の正常な生理機能を指す。このような機能性は、タンパク質の機能性を確立することが知られている任意の手段によって試験することができる。
【0033】
「遺伝子改変」という用語は、コード配列の発現を調節する制御要素と共に、遺伝子産物をコードするコード配列を含む核酸の導入後に特定の遺伝子産物を発現する細胞を指す。核酸の導入は、遺伝子ターゲティングおよび相同組換えを含む当該技術分野において既知の任意の方法によって達成することができる。本明細書で使用される場合、この用語は、例えば遺伝子活性化技術によって、このような細胞によって通常は発現されない内在性遺伝子または遺伝子産物を発現または過剰発現するように操作された細胞も含む。
【0034】
「薬学的に許容可能な」という語句は、本発明の医薬組成物と関連して使用される場合でもそうでなくても、生理学的に容認でき、ヒトに投与された場合に通常有害な反応を生じない分子実体および組成物を指す。好ましくは、本明細書で使用される場合、「薬学的に許容可能な」という用語は、動物、およびより詳細にはヒトでの使用のために、連邦または州政府の規制機関により承認されているか、あるいは米国薬局方または他の一般に認識された薬局方に記載されていることを意味する。「キャリア」という用語は、化合物と一緒に投与される希釈剤、補助剤、賦形剤、または媒体を指す。このような薬学的キャリアは、水および油などの無菌液体であり得る。水または水溶液、生理食塩水溶液、ならびにデキストロースおよびグリセロール水溶液は、好ましくはキャリアとして、特に注射溶液のためのキャリアとして使用される。適切な薬学的キャリアは、E.W.Martinによる「Remington’s Pharmaceutical Sciences」第18版に記載されている。
【0035】
「治療的に有効な用量」および「有効量」という用語は、治療的応答をもたらすのに十分な化合物の量を指す。ASSCおよび酵素が複合体で投与される実施形態では、「治療的に有効な用量」および「有効量」という用語は、治療的応答をもたらすのに十分な複合体の量を指すことができる。治療的応答は、使用者(例えば、臨床医)が治療に対する有効な応答であると認識し得る任意の応答であり得る。従って、治療的応答は、一般に、疾患または障害の1つまたは複数の症状または徴候の寛解であるだろう。
【0036】
「約」および「およそ」という用語は、一般に、測定の性質または精度を考慮して、測定された量に対して許容できる誤差の程度を意味するものとする。通常、例示的な誤差の程度は、所与の値または値の範囲の20パーセント(%)以内、好ましくは10%以内、より好ましくは5%以内である。あるいは、そして特に生物系では、「約」および「およそ」という用語は、一桁分以内である値、好ましくは所与の値の10倍または5倍以内、より好ましくは2倍以内である値を意味してもよい。本明細書において与えられる数量は、他に記載されない限りはおよそのものであり、明確に記載されていない場合でも「約」または「およそ」という用語が推察され得ることを意味する。
【0037】
精製される治療用タンパク質のインビトロ産生、輸送、または貯蔵の間、阻害性であるASSCの濃度は、インビボでの投与時のASSCの希釈(およびその結果として、平衡の変化による結合のシフト)、生物学的利用能および代謝のために、本発明の目的では依然として「有効量」を構成し得ることに注意すべきである。
【0038】
5.2 タンパク質欠乏性障害
タンパク質または酵素の欠乏または特定の組織における機能の損失を特徴とする障害は、理論的にはタンパク質補充療法によって治療可能であり得る。このような障害では、個体の特定の細胞または細胞の全ては十分な機能性タンパク質が欠けており、タンパク質の不活性型を含有するか、あるいは不十分な生物学的機能レベルを含有している。
【0039】
タンパク質または酵素欠乏性障害は、例えば、機能性でないか、あるいは機能が低下または変更されたタンパク質または酵素の発現をもたらすタンパク質または酵素をコードする遺伝子の変異によって生じ得る。また欠乏症は、タンパク質または酵素の発現を少ししかまたは全くもたらさないタンパク質または酵素遺伝子の変異(例えば、ヌル変異)によっても生じ得る。
【0040】
さらに、タンパク質または酵素欠乏症は、タンパク質の折り畳みを変更する変異およびER中の変異タンパク質の遅延によって生じる立体配座の障害(タンパク質の欠乏をもたらす)によるものでもあり得る。このような疾患には、例えば、嚢胞性線維症、α1−アンチトリプシン欠乏症、家族性高コレステロール血症、ファブリー病、アルツハイマー病(Selkoe,Annu.Rev.Neurosci.1994年,17:489−517)、骨形成不全症(Chesslerら,J.Biol.Chem.1993年,268:18226−18233)、糖鎖不全糖タンパク質症候群(Marquardtら,Eur.J.Cell.Biol.1995年,66:268−273)、マロトー・ラミー症候群(Bradfordら,Biochem.J.1999年,341:193−201)、遺伝性盲目(Kaushalら,Biochemistry 1994年,33:6121−8)、グランツマン血小板無力症(Katoら,Blood 1992年,79:3212−8)、遺伝性第VII因子欠乏症(Arbiniら,Blood 1996年,87:5085−94)、眼皮膚白皮症(Halabanら,Proc.Natl.Acad.Sci.USA.2000年,97:5889−94)およびプロテインC欠乏症(Katsumiら,Blood 1996年,87:4164−75)が含まれるが、これらに限定されない。近年、X連鎖疾患の副腎白質ジストロフィー(ALD)における1つの変異が、病的細胞の低温培養によって救済され得る欠損ペルオキシソームトランスポーターの誤った折り畳みをもたらした(Walterら,Am J Hum Genet 2001年,69:35−48)。変異が遺伝子の配列全体にわたって均一に起こることは一般に認められている。従って、欠乏性タンパク質の誤った折り畳みから生じる表現型は、多くの他の遺伝性障害に存在することが予測可能である。
【0041】
遺伝性のタンパク質欠乏性障害の多くは酵素欠乏症である。遺伝性酵素障害の大きな群はリソソーム酵素の変異を伴い、リソソーム蓄積症(LSD)と称される。リソソーム蓄積症は、スフィンゴ糖脂質、グリコーゲン、およびムコ多糖の蓄積によって引き起こされる疾患群である。リソソーム障害の例には、ゴーシェ病(Beutlerら,The Metabolic and Molecular Bases of Inherited Disease,第8版 2001年 Scriverら編 3635−3668頁,McGraw−Hill,New York)、GM1−ガングリオシドーシス(同上 3775−3810頁)、フコシドーシス(The Metabolic and Molecular Bases of Inherited Disease,1995年.Scriver,C.R.,Beaudet,A.L.,Sly,W.S.およびValle,D.編 2529−2561頁,McGraw−Hill,New York)、ムコ多糖症(同上 3421−3452頁)、ポンペ病(同上 3389−3420頁)、ハーラー・シャイエ病(Weismannら,Science 1970年,169,72−74)、AおよびB型ニーマン・ピック病(The Metabolic and Molecular Bases of Inherited Disease 第8版 2001年.Scriverら編,3589−3610頁,McGraw−Hill,New York)、およびファブリー病(同上 3733−3774頁)が含まれるが、これらに限定されない。
【0042】
ファブリー病
「ファブリー病」という用語は、リソソームα−ガラクトシダーゼA(α−Gal A)活性の欠乏によるスフィンゴ糖脂質異化のX連鎖の先天異常を指す。この欠損は、心臓、腎臓、皮膚、およびその他の組織の血管内皮リソソームにおいてグロボトリアオシルセラミド(セラミドトリヘキソシド)および関連のスフィンゴ糖脂質の蓄積を引き起こす。
【0043】
1つの非限定的な実施形態では、α−ガラクトシダーゼAは、GenBank受入番号NM_000169で記載される核酸配列を含むヒトGla遺伝子を指す。あるいは、α−ガラクトシダーゼAは、α−ガラクトシダーゼA遺伝子に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%および100%までの相同性(標準的なソフトウェア、例えばBLASTまたはFASTAによって決定される)を示す任意の核酸分子、ならびに標準条件下でこれらの配列とハイブリッド形成する任意の配列によってコードされ得る。
【0044】
別の非限定的な実施形態では、ヒトα−ガラクトシダーゼA(α−GAL A)は、ヒトGla遺伝子またはこれに対して少なくとも90%相同である任意の他のアミノ酸配列によってコードされる酵素を指す。ヒトα−GAL酵素は429個のアミノ酸からなり、GenBank受入番号U78027およびNP_000160にある。
【0045】
「非定型ファブリー病」という用語は、主にα−GAL欠乏症の心臓症状、すなわち、心臓(特に、左心室)の著しい拡張につながる心筋細胞における進行性のグロボトリアオシルセラミド(GL−3)の蓄積を有する患者を指す。
【0046】
「キャリア」は、欠損α−GAL遺伝子を持つ1つのX染色体と、正常な遺伝子を持つ1つのX染色体とを有し、正常な対立遺伝子のX染色体の不活性化が1つまたは複数の細胞型において存在する女性である。キャリアはファブリー病であると診断されることが多い。
【0047】
1つの非限定的な実施形態では、α−ガラクトシダーゼAのための薬理学的シャペロンは、1−デオキシガラクトノジリマイシン(DGJ)であり得る。ここで、DGJは、以下の構造:
【化1】
を有する化合物である。
【0048】
この用語には、遊離塩基および任意の塩形態、ならびにこれらの任意のプロドラッグが含まれる。
【0049】
α−GAL Aのためのさらにその他の薬理学的シャペロンは、米国特許第6,274,597号明細書、同第6,774,135号明細書、および同第6,599,919号明細(Fanら)に記載されており、α−allo−ホモノジリマイシン、β−1−C−ブチル−デオキシガラクトノジリマイシン、およびα−ガラクト−ホモノジリマイシン、カリステギンA3、カリステギンB2、カリステギンB3、N−メチル−カリステギンA3、N−メチル−カリステギンB2およびN−メチル−カリステギンB3が含まれる。
【0050】
ポンペ病
ポンペ病は、リソソームのグリコーゲン代謝を損なう酸性アルファグルコシダーゼ(GAA、α−グルコシダーゼ)活性の欠乏を特徴とする常染色体劣性LSDである。酵素欠乏症はリソソームのグリコーゲン蓄積をもたらし、その結果、疾患の末期において、進行性の骨格筋の弱化、心臓機能の低下、呼吸不全、および/またはCNS機能障害が起こる。GAA遺伝子の遺伝子変異は発現の低下を引き起こすか、あるいは安定性および/または生物活性が変更された酵素の変異形態を産生し、最終的には疾患に至る(概して、Hirschhorn R,1995年,Glycogen Storage Disease Type II:Acid α−Glucosidase(Acid Maltase)Deficiency,The Metabolic and Molecular Bases of Inherited Disease,Scriverら編,McGraw−Hill,New York,第7版,2443−2464頁を参照)。ポンペ病の3つの認識された臨床形態(小児型、若年型および成人型)は、残留α−グルコシダーゼ活性のレベルと相関している(Reuser A Jら,1995年,Glycogenosis Type II(Acid Maltase Deficiency),Muscle & Nerve Supplement 3、S61−S69)。ASSC(他では「薬理学的シャペロン」とも呼ばれる)は、リソソーム蓄積症(例えば、ポンペ病)などの遺伝性疾患の治療のための有望な新しい治療的アプローチを表す。
【0051】
1つの非限定的な実施形態では、酸性アルファグルコシダーゼ(GAA)は、GenBank受入番号NM_000152、NM_001079803、またはNM_001079804で記載される核酸配列を含むヒトグルコシダーゼ、アルファ、酸性(GAA)遺伝子を指す。あるいは、酸性アルファグルコシダーゼは、酸性α−グルコシダーゼ遺伝子に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%および100%までの相同性(標準的なソフトウェア、例えばBLASTまたはFASTAによって決定される)を示す任意の核酸分子、ならびに標準条件下でこれらの配列とハイブリッド形成する任意の配列によってコードされ得る。
【0052】
別の非限定的な実施形態では、ヒト酸性α−グルコシダーゼは、グリコーゲン、マルトース、およびイソマルトース中に存在するアルファ−1,4−およびアルファ−1,6−結合−D−グルコースポリマーを加水分解する酵素を指す。代替的な名称は以下の通りである:グルコアミラーゼ、1,4−α−D−グルカングルコヒドロラーゼ、アミログルコシダーゼ、ガンマ−アミラーゼ、およびエキソ−1,4−α−グルコシダーゼ。ヒトGAA遺伝子は染色体17q25.2−25.3に位置付けられており、GenBank受入番号Y00839、NP_000143、NP_001073271、またはNP_001073272で記載されるアミノ酸配列、またはこれに対して少なくとも90%相同である任意の他のアミノ酸配列を有する。
【0053】
小児型ポンペ病(IまたはA型)は最も一般的かつ最も重症であり、生後2年以内の成長障害、全身性低血圧症、心臓肥大、および心肺不全を特徴とする。若年型ポンペ病(IIまたはB型)は重症度が中間であり、心肥大のない筋肉症状の優勢を特徴とする。若年型ポンペ病の個体は、通常、呼吸不全のために20歳に達する前に死亡する。成人型ポンペ病(IIIまたはC型)は、多くの場合、10代または遅くても60代に緩徐進行性ミオパシーとして出現する(Felice K Jら,1995年,Clinical Variability in Adult−Onset Acid Maltase Deficiency:Report of Affected Sibs and Review of the Literature、Medicine 74,131−135)。
【0054】
ポンペ病では、α−グルコシダーゼは、グリコシル化、リン酸化、およびタンパク質分解処理によって翻訳後に広範に修飾されることが示されている。最適なグリコーゲン触媒作用のために、リソソームにおけるタンパク質分解による110キロダルトン(kDa)の前駆体から76および70kDaの成熟型への転換が必要とされる。
【0055】
本明細書で使用される場合、「ポンペ病」という用語は、全てのタイプのポンペ病を指す。本出願において開示される製剤および投薬計画は、例えばI型、II型またはIII型ポンペ病の治療のために使用することができる。
【0056】
1つの特定の非限定的な実施形態では、酸性α−グルコシダーゼのための薬理学的シャペロンは、以下の式:
【化2】
によって表される1−デオキシノジリマイシン(1−DNJ)、またはその薬学的に許容可能な塩、そのエステルまたはそのプロドラッグであり得る。一実施形態では、塩は塩酸塩である(すなわち、1−デオキシノジリマイシンHCl)。
【0057】
酸性α−グルコシダーゼのためのさらにその他の薬理学的シャペロンは、米国特許第6,599,919号明細書(Fanら)および米国特許出願公開第2006/0264467号明細書(Mugrageら)に記載されており、α−ホモノジリマイシンおよびカスタノスペルミンが含まれる。
【0058】
ゴーシェ病
本明細書で使用される場合、「ゴーシェ病」という用語は、脂肪グルコセレブロシドを分解するリソソーム酵素グルコセレブロシダーゼの欠乏を指す。そして脂肪は、主に肝臓、脾臓および骨髄に蓄積する。ゴーシェ病は、痛み、疲労、黄疸、骨の損傷、貧血、そしてさらには死をもたらし得る。ゴーシェ病の臨床表現型は3つある。I型の患者は年少期または青年期のいずれかに発症し、容易に傷がつき、貧血、少ない血小板、肝臓および脾臓の拡張、骨格の弱化のために疲労を経験し、場合によっては、肺障害および腎障害を有する。脳障害の徴候は存在しない。II型では、早発性の肝臓および脾臓の拡張が生後3か月までに発生し、広範な脳障害が存在する。2歳までの死亡率は高い。III型は、肝臓および脾臓の拡張ならびに脳の発作を特徴とする。β−グルコセレブロシダーゼ遺伝子はヒト1q21染色体上に位置する。そのタンパク質前駆体は536個のアミノ酸を含有し、その変異タンパク質は497個のアミノ酸の長さである。
【0059】
1つの非限定的な実施形態では、グルコセレブロシダーゼは、GenBank受入番号NM_001005741、NM_001005741、NM_001005749、NM_001005750、またはNM_000157で記載される核酸配列を含むヒトグルコシダーゼ、ベータ(Gba)遺伝子を指す。あるいは、グルコセレブロシダーゼは、グルコセレブロシダーゼ遺伝子に対して少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%および100%までの相同性(標準的なソフトウェア、例えばBLASTまたはFASTAによって決定される)を示す任意の核酸分子、ならびに標準条件下でこれらの配列とハイブリッド形成する任意の配列によってコードされ得る。
【0060】
別の非限定的な実施形態では、グルコセレブロシダーゼは、ヒトグルコシダーゼ、ベータ(Gba)遺伝子(GenBank受入番号NP_001005741、NP_001005742、NP_001005749、NP_001005750、またはNP_000148)、またはこれに対して少なくとも90%相同である任意の他のアミノ酸配列によってコードされる酵素を指す。
【0061】
1つの非限定的な実施形態では、グルコセレブロシダーゼのための薬理学的シャペロンは、以下の構造:
【化3】
を有するイソファゴミン(IFG、(3R,4R,5R)−5−(ヒドロキシメチル)−3,4−ピペリジンジオール)であり得る。
【0062】
酒石酸イソファゴミンは、共同所有される米国特許第7,501,439号明細書(Mugrageら)に最近記載されており、CAS番号919364−56−0が割り当てられている。またイソファゴミンは、様々な有機および無機酸と作られる他の酸付加塩の形態で調製され得る。このような塩には、塩化水素、臭化水素、メタンスルホン酸、硫酸、酢酸、トリフルオロ酢酸、シュウ酸、マレイン酸、ベンゼンスルホン酸、トルエンスルホン酸および種々の他の酸(例えば、硝酸、リン酸、ホウ酸、クエン酸、安息香酸、アスコルビン酸、サリチル酸など)と形成されるものが含まれる。このような塩は、当業者に知られているように形成することができる。
【0063】
グルコセレブロシダーゼのためのさらにその他の薬理学的シャペロンは、米国特許第6,599,919号明細書(Fanら)、同第6,046,214号明細書(Kristiansenら)、同第5,844,102号明細書(Sierksら)、および米国特許出願公開第2008/0009516号明細書(Wustman)に記載されており、C−ベンジルイソファゴミンおよび誘導体、N−アルキル(C9〜12)−DNJ、グルコイミダゾール(および誘導体)、C−アルキル−IFG(および誘導体)、N−アルキル−β−バリエナミン、フルフェノジン(fluphenozine)、N−ドデシル−DNJ、ならびにカリステギンA3、B1、B2およびC1が含まれる。
【0064】
遺伝性障害に加えて、その他の酵素欠乏症は、原発性または続発性の障害に起因する組織または器官への損傷から生じる。例えば、損傷した膵臓組織、または膵炎は、アルコール依存症によって引き起こされ、消化に必要な膵臓酵素の欠乏をもたらす。膵炎は、現在、酵素補充療法を用いて治療されている。
【0065】
タンパク質欠乏の障害は、生物学的過程を増強または刺激する補充タンパク質の投与によって治療することができる。例えば、貧血のある個体は、赤血球の産生を刺激して組織への酸素輸送を増大させるために組換えエリスロポエチン(EPOGEN(登録商標)、PROCRIT(登録商標)、EPOIETIN(登録商標))が投与される。さらに、インターフェロンアルファ2b(INTRON A(登録商標)、PEG−INTRON(登録商標)、またはREBETOL(登録商標))、およびインターフェロンベータ1a(AVONEX(登録商標)、BETASERON(登録商標))などの組換えインターフェロンは、それぞれB型肝炎および多発性硬化症を治療するために投与される。投与されるさらにその他のタンパク質は、組換えヒトデオキシリボヌクレアーゼI(rhDNase−PULMOZYME(登録商標))と、嚢胞性線維症患者の肺機能を改善するために使用されるDNAを選択的に切断する酵素と、ほぼ全部または全部の甲状腺摘出術を受けたことがあり、従って、甲状腺ホルモンを摂取しなければならない甲状腺癌患者において使用するために開発された組換え甲状腺刺激ホルモン(THYROGEN(登録商標))と、化学療法からの好中球減少を治療するための組換えG−CSF(NEUPOGEN(登録商標))と、そして膵炎のある個体における消化酵素である。タンパク質治療の別の重要な領域は、非常に特異的で明確に定義された活性部位を有する抗体による感染症および癌の治療である。抗体治療製品には、呼吸器多核体ウィルスのためのRESPIRGRAM(登録商標)、乳癌のためのHERCEPTIN(登録商標)、関節炎および炎症性疾患のためのREMICAID(登録商標)およびHUMIRA(登録商標)などが含まれる。抗体のためのASSCはよく知られており、標的抗原または構造的に関連のある類似体(例えば、活性標的の修飾型または模倣薬)のいずれかを使用することができる。
【0066】
「患者」は、特定の疾患であると診断されたか、あるいは特定の疾患である疑いのある被験者を指す。患者はヒトであっても動物であってもよい。例えば、「ファブリー病患者」は、ファブリー病であると診断されたか、あるいはファブリー病である疑いのある個体を指し、変異α−GALを有する。ファブリー病の特徴的なマーカーは男性の半接合体および女性キャリアにおいて同じ有病率で発生し得るが、通常、女性は重症度が低い。
【0067】
5.3 組換えタンパク質の産生
タンパク質欠乏症患者の治療(例えば、酵素補充療法による)のために有用な補充タンパク質は、当該技術分野の技能の範囲内の通常の分子生物学、微生物学、および組換えDNA技術を用いて単離および精製することができる。例えば、補充タンパク質をコードする核酸は、文献に記載されるような組換えDNA発現を用いて単離することができる。例えば、Sambrook,Fritsch & Maniatis,Molecular Cloning:A Laboratory Manual,第2版(1989年)Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.(本明細書では「Sambrookら,1989年」)、DNA Cloning:A Practical Approach,IおよびII巻(D.N.Glover編1985年)、Oligonucleotide Synthesis(M.J.Gait編1984年)、Nucleic Acid Hybridization(B.D.Hames & S.J.Higgins編(1985年))、Transcription And Translation(B.D.Hames & S.J.Higgins編(1984年))、Animal Cell Culture(R.I.Freshney編(1986年))、Immobilized Cells And Enzymes(IRL Press,(1986年))、B.E Perbal,A Practical Guide To Molecular Cloning(1984年)、F.M.Ausubelら(編),Current Protocols in Molecular Biology,John Wiley & Sons,Inc.(1994年)を参照されたい。タンパク質をコードする核酸は、遺伝子が生物学的に活性なタンパク質をコードする限りは、全長でも切断されていてもよい。例えば、ファブリー病に関連する欠損酵素α−Gal Aの生物学的に活性な切断型は、米国特許第6,210,666号明細書(Miyamuraら)に記載されている。
【0068】
標的タンパク質をコードする同定および単離された遺伝子は、次に、適切なクローニングベクターに挿入され得る。当該技術分野において知られている多数のベクター−宿主系を使用することができる。可能性のあるベクターにはプラスミドまたは変性ウィルスが含まれる(これらに限定されない)が、ベクター系は、使用される宿主細胞と適合性でなければならない。ベクターの例としては、大腸菌(E.coli)、ラムダ誘導体などのバクテリオファージ、またはpBR322誘導体もしくはpUCプラスミド誘導体、例えばpGEXベクター、pmal−c、pFLAGなどのプラスミドが挙げられるが、これらに限定されない。クローニングベクターへの挿入は、例えば、相補的な付着末端を有するクローニングベクターへDNA断片を連結することによって達成することができる。しかしながら、DNAを断片化するために使用される相補的な制限部位がクローニングベクターに存在しなければ、DNA分子の末端は酵素的に修飾され得る。あるいは、所望される部位はどれもDNA末端へのヌクレオチド配列(リンカー)の連結によって生成することができ、これらの連結されたリンカーは、制限エンドヌクレアーゼの認識配列をコードする特異的な化学合成オリゴヌクレオチドを含むことができる。組換えタンパク質の産生は、分泌を容易にするN末端のシグナルペプチドの包含、またはポリアデニル化部位を含有する3’非翻訳配列の包含などの遺伝子操作によって最大にすることができる。
【0069】
1つの非限定的な実施形態では、宿主細胞を形質導入するために使用される構築物は、アデノウィルス、アデノ随伴ウィルス、ヘルペスウィルス、ムンプスウィルス、ポリオウィルス、レトロウイルス、シンドビスウィルスおよびワクシニアウィルスを含むがこれらに限定されないウィルス由来のベクターである。
【0070】
組換え分子は、遺伝子配列の多数のコピーが作成されるように、形質転換、トランスフェクション、感染、エレクトロポレーションなどによって宿主細胞中に導入され得る。好ましくは、クローン化遺伝子はシャトルベクタープラスミド上に含有され、クローニング細胞、例えば大腸菌中での発現、およびその後の適切な発現細胞株への挿入のための容易な精製(それが所望される場合)を提供する。
【0071】
可能性のある宿主−ベクター系には、ウィルス(例えば、ワクシニアウィルス、アデノウィルスなど)に感染した哺乳類細胞系、ウィルス(例えば、バキュロウイルス)に感染した昆虫細胞系、酵母ベクターを含有する酵母などの微生物、またはバクテリオファージ、DNA、プラスミドDNA、もしくはコスミドDNAで形質転換された細菌が含まれるが、これらに限定されない。ベクターの発現要素は、その強度および特異性が異なる。利用される宿主−ベクター系に応じて、多数の適切な転写および翻訳要素のどれか1つが使用され得る。異なる宿主細胞は、タンパク質の翻訳および翻訳後処理および修飾(例えば、グリコシル化、切断[例えば、シグナル配列の])のための特徴的で特異的なメカニズムを有する。適切な細胞株または宿主系は、グリコシル化、シアリル化(sialyation)およびリン酸化などの、発現される外来性タンパク質の所望の修飾および処理を保証するように選択することができる。例えば、細菌系における発現は、非グリコシル化コアタンパク質産物を産生するために使用することができる。しかしながら、細菌において発現されるタンパク質は適切に折り畳まれないことがある。酵母における発現は、グリコシル化産物を産生することができる。真核細胞における発現は、異種タンパク質の「ネイティブな」グリコシル化および折り畳みの可能性を増大させることができる。さらに、哺乳類細胞における発現は、タンパク質を再構成または構成するための手段を提供することができる。
【0072】
組換えで発現されたタンパク質の精製は、硫酸アンモニウム沈殿、疎水性相互作用樹脂、カチオン交換樹脂、アニオン交換樹脂、およびクロマトフォーカシング樹脂を含有するカラムクロマトグラフィなどの当該技術分野において既知の方法を用いて達成することができる。あるいは、タンパク質、または組換えタンパク質に融合されたタグに特異的に結合する適切なポリクローナルまたはモノクローナル抗体を用いて組換えタンパク質を精製するために、イムノアフィニティクロマトグラフィを使用することもできる。好ましい実施形態では、本発明の方法のために使用される組換えタンパク質の純度は、少なくとも95%、好ましくは少なくとも97%、最も好ましくは98%よりも高いであろう。
【0073】
他の非限定的な実施形態では、タンパク質欠乏症患者の治療(例えば、酵素補充療法による)のために有用な補充タンパク質は、組換え細胞発現系から精製してもよい(例えば、哺乳類細胞または昆虫細胞−概して、米国特許第5,580,757号明細書(Desnickら)、米国特許第6,395,884号明細書および同第6,458,574号明細書(Seldenら)、米国特許第6,461,609号明細書(Calhounら)、米国特許第6,210,666号明細書(Miyamuraら)、米国特許第6,083,725号明細書(Seldenら)、米国特許第6,451,600号明細書(Rasmussenら)、米国特許第5,236,838号明細書(Rasmussenら)、ならびに米国特許第5,879,680号明細書(Ginnsら)を参照、ヒト胎盤、または動物乳(米国特許第6,188,045号明細書(Reuserら)を参照))。
【0074】
薬学的使用に適した補充タンパク質を得るためのその他の合成技術は、例えば、米国特許第7,423,135号明細書、同第6,534,300号明細書、および同第6,537,785号明細書、国際公開第2005/077093号パンフレット、ならびに米国特許出願公開第2007/0280925号明細書および同第2004/0029779号明細書において見出し得る。これらの参考文献は全ての目的で参照によってその全体が本明細書に援用される。
【0075】
さらに、異なるベクター/宿主発現系は、タンパク質切断などの処理反応に異なる程度で影響を与え得る。発現効率は、米国特許第6,274,597号明細書に記載されるような特異的なシャペロンおよび関連のファミリーメンバーの使用によって増大させることができる。
【0076】
上記のように、本発明の1つの態様は、組換えタンパク質の産生を改善するための方法を提供し、本方法は、組換えタンパク質を発現する宿主細胞と、組換えタンパク質に特異的な薬理学的シャペロンとを接触させることを含む。
【0077】
1つの非限定的な実施形態では、薬理学的シャペロンは、タンパク質を発現する細胞のER中で標的の組換えタンパク質を安定化し、タンパク質のERADおよび早期分解を防止し得る。そうすることで、薬理学的シャペロンは、組換えタンパク質の発現に関連するERストレスを低下させ得、これは次に産生細胞株がタンパク質の高い生存率および発現を保持できるようにし得る。
【0078】
他の非限定的な実施形態では、薬理学的シャペロンは、細胞により発現される組換えタンパク質を安定化し、細胞のERからのタンパク質の排出を増大させ、細胞からのタンパク質の分泌を増大させ得る。
【0079】
他の非限定的な実施形態では、薬理学的シャペロンは、タンパク質を発現する細胞の外側で(すなわち、タンパク質が細胞培地中に分泌された後)、標的の組換えタンパク質を安定化させ得る。組換えタンパク質を安定化することによって、薬理学的シャペロンは、安定性を保つために普通はより低いpHで貯蔵されなければならず、そして通常中性pHで処方されるほとんどの細胞培地中で普通は安定ではないタンパク質に利益を与え得る。酵素活性の著しい損失が起こり得るが、これらのタンパク質は収穫および精製の前は細胞培地中に保持される。薬理学的シャペロンは、5.0よりも高いpH、例えば中性pHを有する培地中で、これらのタンパク質を安定化させ、不可逆的な変性およびタンパク質活性の不活性化を防止することができる。
【0080】
他の非限定的な実施形態では、薬理学的シャペロンは、細胞培地からのタンパク質の精製の間、標的組換えタンパク質を保護し得る。組換えタンパク質は、培地中で安定でなければ、変性するかもしれず、精製混合物中の汚染プロテアーゼによるタンパク質分解を受けやすい。薬理学的シャペロンは、タンパク質を安定化させ、タンパク質分解部位が暴露されるのを防止することによって、このタンパク質分解を防止し得る。従って、薬理学的シャペロンは、精製プロセスの間のタンパク質の完全性および/または酵素活性を改善し得る。
【0081】
他の非限定的な実施形態では、薬理学的シャペロンは、医薬製剤の貯蔵および/または調製の間、組換えタンパク質を保護し得る。例えば、薬理学的シャペロンは、このようなタンパク質が貯蔵および処方の間にタンパク質消化されることを保証するために凝固因子(例えば、プロテアーゼ)と結合してその酵素活性を阻害することができる。薬理学的シャペロンは、処方の間、他のタイプの不可逆的化学損傷(例えば、酸化、加水分解または脱アミド)、または凝集、沈殿および表面への吸着などの物理的な不安定性も防止し得る。さらに、薬理学的シャペロンはタンパク質を安定化させ、pH、温度、シェアストレス、凍結/解凍ストレスおよびこれらのストレスの組み合わせなどのストレス(そうでなければ、タンパク質の分解に寄与し得る)から保護し得る。
【0082】
1つの非限定的な実施形態では、補充タンパク質は、この遺伝子の観察される9個のハプロタイプの最も優勢なものによってコードされる組換え酸性α−グルコシダーゼ(GAA)であり、チャイニーズハムスター卵巣細胞株において組換えDNA技術により産生される。組換えGAAは、例えば、Kakkisら,2008年,「An improved alpha−glucosidase enzyme for Pompe disease」,Abstract,第58回Annual Meeting of the ASHG、Kishnaniら,2007年,Neurology.68(2):99−109、米国特許第6,118,045号明細書(Reuserら)、同第7,056,712号明細書(Chen)、および同第7,351,410号明細書(van Bree)(これらはそれぞれ、全ての目的で参照によってその全体が援用される)に記載されるような組換えGAAであり得る。
【0083】
他の好ましい非限定的な実施形態では、ASSCは1−デオキシノジリマイシン(1−DNJ)であり、GAAは組換えGAAである。代替の実施形態では、ASSCはα−ホモノジリマイシンであり、GAAは組換えGAAである。別の代替の実施形態では、ASSCはカスタノスペルミンであり、GAAは組換えGAAである。ASSC(例えば、1−デオキシノジリマイシン、α−ホモノジリマイシンおよびカスタノスペルミン)は、本発明に従う潜在的ASSCの源を提供する合成ライブラリから得ることができる(例えば、Needelsら,Proc.Natl.Acad.Sci.USA 1993年,90:10700−4、Ohlmeyerら,Proc.Natl.Acad.Sci.USA 1993年,90:10922−10926、Lamら,PCT公報国際公開第92/00252号パンフレット、Kocisら,PCT公報国際公開第号パンフレット94/28028を参照)。合成化合物ライブラリは、Maybridge Chemical Co.(Trevillet,Cornwall,UK)、Comgenex(Princeton,N.J.)、Brandon Associates(Merrimack,N.H.)、およびMicrosource(New Milford,Conn.)から市販されている。珍しい化学ライブラリは、Aldrich(Milwaukee,Wis.)から入手可能である。あるいは、細菌、真菌、植物および動物抽出物の形態の天然化合物のライブラリは、例えばPan Laboratories(Bothell,Wash.)またはMycoSearch(NC)から入手可能であるか、または容易に産生可能である。さらに、天然および合成的に産生されたライブラリおよび化合物は、Res.1986年,155:119−29により容易に修飾される。
【0084】
5.4 インビトロ安定性
その貯蔵期間中の補充タンパク質製剤の安定性を保証することは大きな課題である。例えば、組換え酵素のバイアルは、多くの場合、1回だけの使用のためのものであり、未使用の製品は廃棄されなければならない。さらに、組換え酵素は、多くの場合、ヘルスケアの専門家によって再構成、希釈、および投与されなければならず、その投与は遅れてはならない。組換え酵素は、多くの場合、低温、例えば2〜8℃で貯蔵されなければならず、製品は限られた時間、例えば24時間までしか安定でない。
【0085】
ASSCおよび補充タンパク質が同じ組成物中に存在する場合、処方された本発明の組成物は、より安定した組成物を提供する。インビボで投与されるタンパク質の安定化に加えて、ASSCはインビトロで補充タンパク質の立体配座に可逆的に結合して安定化し、これにより、凝集および分解が防止され、製剤の貯蔵寿命が長くなる。ASSC/補充タンパク質の相互作用の分析は、例えば、示差走査熱量測定、または円二色性などの当該技術分野においてよく知られている技術を用いて評価してもよい。
【0086】
例えば、組成物の注射可能な水性製剤が、ニードルおよびシリンジを用いて内容物を引き抜くのに適した栓付バイアル中で供給される場合、ASSCの存在は、補充タンパク質の凝集を阻害する。バイアルは、1回の使用のためまたは多重使用のためのいずれかであり得る。また製剤は、予め充填されたシリンジとして供給することもできる。別の実施形態では、製剤は乾燥または凍結乾燥状態であり、標準または供給される生理学的希釈剤により液体状態に再構成することを必要とし得る。この場合、ASSCの存在は、再構成中および再構成後に、凝集を防止するために補充タンパク質を安定化し得る。製剤が静脈内投与のための液体(例えば、静脈内投与ラインまたはカテーテルへ接続するための無菌バック中など)である実施形態では、ASSCの存在は同じ利益を与え得る。
【0087】
投与される補充タンパク質の安定化に加えて、ASSCの存在は、補充タンパク質製剤を約7.0〜7.5の中性pHで貯蔵可能にし得る。これにより、安定性を保つために普通はより低いpHで貯蔵されなければならないタンパク質に利益が与えられる。例えば、GAAなどのリソソーム酵素は、通常、低pH(例えば、5.0以下)において安定した立体配座を保持する。しかしながら、低pHにおける補充酵素の長期の貯蔵は、酵素および/または製剤の分解を促進し得る。
【実施例】
【0088】
本発明は以下に提示される実施例によってさらに説明される。このような実施例の使用は説明のためだけのものであって、本発明または例示された任意の用語の範囲および意味を決して限定しない。同様に、本発明は、本明細書に記載される特定の好ましい実施形態のどれにも限定されない。実際、本明細書を読めば、本発明の多くの修正および変更は当業者には明らかになるであろう。従って、本発明は、特許請求の範囲が権利を与えられる均等物の全範囲と共に、特許請求の範囲の用語によってのみ限定されるべきである。
【0089】
実施例1: 熱的課題における酸性α−グルコシダーゼの熱安定性
100μMのASSCの1−デオキシノジリマイシン塩酸塩(DNJ)を含む場合および含まない場合の組換えヒトGAA(Myozyme(登録商標)、Genzyme Corp.)の安定性は、熱を利用してタンパク質の変性を誘発させる熱安定性アッセイによって決定した。変性は、疎水性アミノ酸(折り畳みタンパク質では露出されない)に結合すると蛍光を発するSYPRO Orange色素を用いてモニターされる。
【0090】
熱安定性アッセイはERのpHに相当するpH7.4において2つの製剤に対して実施した。図1に示されるように、pH7.4で100μMのDNJを含有する製剤は、pH7.4でASSCを含まない製剤と比較して、変性するために著しく多い熱を必要としたので、より安定である。
【0091】
実施例2: 1−デオキシノジリマイシン(DNJ)は37℃で長時間インキュベートしたときに酸性α−グルコシダーゼの活性損失を防止する
残留GAA活性は、4つの製剤について決定した:
(1)pH7.4においてMyzozymeのみ、
(2)pH7.4においてMyzozyme + 50μMのDNJ、
(3)pH5.2においてMyzozymeのみ、
(4)pH5.2においてMyzozyme + 50μMのDNJ。
【0092】
初期活性(t=0)の%に基づいて活性を24時間にわたって測定した。蛍光発生基質4−MU−α−グルコースの加水分解に基づいて、0、3、6および24時間で、サンプルをGAA酵素活性についてアッセイした。GAA活性は、初期活性の%、すなわち残留活性で表した。
【0093】
図2Aに示されるように、上記製剤(1)(ASSなし)は時間と共に活性を失い、投与の24時間後にその初期活性の約20%しかない。対照的に、製剤(2)は、初期活性の全てではないがほとんどを24時間にわたって保持する。ph5.2の両方の製剤(上記製剤(3)および(4))は、その初期活性のほとんどを24時間にわたって保持する。
【0094】
初期酵素活性の損失が適切な立体配座を保持できないことと相関するかどうかを決定するために、実施例1において説明したように、上記のサンプルにおいてSYPRO Orange熱安定性実験を実施した。この熱安定性実験では、製剤(2)および(4)中のDNJの濃度を10μMに低減した。この実験に基づいて、折り畳まれたGAAの%を推定し、図2Bにプロットした。図2Bにおける製剤(1)の折り畳まれたGAAの量の24時間にわたる減少は、この同じ一般的な製剤について図2Aに示される活性の損失と相関する。
【0095】
実施例3: DNJは熱的課題におけるGAA安定性を向上させる。
実施例1に概略的に説明されるような熱安定性実験を4つの組成物おいて実施した:
(1)Myozymeのみの組成物、
(2)Myozyme + 1μMの1−DNJ−HCl、
(3)Myozyme + 10μMの1−DNJ−HCl、
(4)Myozyme + 100μMの1−DNJ−HCl。
【0096】
図3に示されるように、DNJ−HClは、GAAの融解温度の用量依存的な上昇によって明らかであるように、GAAの熱安定性を向上させる。
【0097】
実施例4: イソファゴミン(IFG)は熱的課題における酸性β−グルコシダーゼの安定性を向上させる
実施例1に概略的に説明されるような熱安定性実験をGCase(Cerezyme(登録商標))の3つの組成物において実施した:
(1)GCaseのみの組成物、pH7.4、
(2)GCase + 10μMのIFG、pH7.4、
(3)GCase + 100μMのIFG、pH7.4。
【0098】
図4に示されるように、IFGは、タンパク質の融解温度の上昇によって明らかであるように、GCaseの熱安定性を用量依存的に向上させる。
【0099】
実施例5: IFGの存在下におけるGCaseの熱安定性
折り畳まれていないGCaseのパーセントを3つの製剤について決定した:
(1)pH5.2においてGCaseのみ、
(2)pH7.4においてGCaseのみ、
(3)pH7.4において10μMのIFGを含むGCase。
【0100】
37℃および中性pHにおいてIFGがGCaseのアンフォールディングを防止するかどうかを決定するために、実施例1において概略的に説明したように、上記のサンプルにおいてSYPRO Orange熱安定性実験を実施した。この熱安定性実験では、IFGの濃度は製剤(3)中10μMであった。図5の結果に基づいて、IFGは、特定の条件下でGCaseのアンフォールディングを防止した。
【0101】
実施例6: 1−デオキシガラクトノジリマシシン(DGJ)は、熱的課題におけるα−Gal Aの安定性を向上させる。
実施例1に概略的に説明されるような熱安定性実験を、α−Gal A(Fabrazyme(登録商標))の3つの組成物において実施した:
(1)α−Gal Aのみの組成物、pH7.4、
(2)α−Gal A + 10μMのDGJ、pH7.4、
(3)α−Gal A + 100μMのDGJ、pH7.4、
【0102】
図6に示されるように、DGJは、タンパク質の融解温度の上昇によって明らかであるように、α−Gal Aの熱安定性を用量依存的に向上させる。
【0103】
実施例7: 薬理学的シャペロンは一過性にトランスフェクトされたCOS−7細胞からの組換えタンパク質の活性レベルを向上させる。
COS−7細胞に空のベクター、GBA遺伝子をコードするプラスミド、またはGLA遺伝子をコードするプラスミドを一過性にトランスフェクトした。種々の一過性トランスフェクションを100μMの表示される薬理学的シャペロン(IFG、DGJまたはDNJ)と共にインキュベートした。タンパク質発現の48時間後に、それぞれのトランスフェクションからの馴化培地を収穫し、コンカナバリンA−アガロースビーズによる分泌タンパク質の捕獲後に、酸性β−グルコシダーゼまたはα−Gal Aの活性のレベルを査定した。このコンカナバリンAの捕獲工程は、適切な蛍光発生基質(GCaseに対しては4−MU−β−グルコース、α−Gal Aに対しては4−MU−β−ガラクト―ス)を用いる活性決定の過程において、薬理学的シャペロンによる可能性のある酵素活性の阻害を除去するために必要であった。
【0104】
図7に示されるように、IFGまたはDGJによるインキュベーションは、それぞれ、酸性β−グルコシダーゼまたはα−Gal Aの活性を増大させた。酸性β−グルコシダーゼの一過性発現においてDNJが含まれる場合、酵素活性の増大は見られなかった。この観察は、酵素活性の増大がタンパク質の既知の薬理学的シャペロンおよび/または阻害薬との特異的な相互作用によるものであることを示す。
【0105】
本発明は、本明細書に記載される特定の実施形態によって範囲が限定されてはならない。実際、本明細書に記載されるものに加えて、上記の説明および添付図面から当業者には本発明の種々の修正が明らかになるであろう。このような修正は特許請求の範囲内に入ることが意図される。
【0106】
特許、特許出願、公報、製品説明、GenBank受入番号、およびプロトコルが本出願全体にわたって引用されており、これらの開示は全ての目的で参照によってその全体が本明細書中に援用される。
【特許請求の範囲】
【請求項1】
組換えタンパク質の産生を改善するための方法であって、前記組換えタンパク質を発現する宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとを接触させることを含む方法。
【請求項2】
前記宿主細胞が哺乳類細胞である、請求項1に記載の方法。
【請求項3】
前記タンパク質が酵素である、請求項1に記載の方法。
【請求項4】
前記酵素がリソソーム酵素である、請求項3に記載の方法。
【請求項5】
前記リソソーム酵素がα−ガラクトシダーゼAである、請求項4に記載の方法。
【請求項6】
前記薬理学的シャペロンが、1−デオキシガラクトノジリマイシン、α−allo−ホモノジリマイシン、β−1−C−ブチル−デオキシガラクトノジリマイシン、およびα−ガラクト−ホモノジリマイシン、カリステギンA3、カリステギンB2、カリステギンB3、N−メチル−カリステギンA3、N−メチル−カリステギンB2およびN−メチル−カリステギンB3からなる群から選択される、請求項5に記載の方法。
【請求項7】
前記薬理学的シャペロンが1−デオキシガラクトノジリマイシンである、請求項6に記載の方法。
【請求項8】
前記リソソーム酵素がβ−グルコシダーゼである、請求項4に記載の方法。
【請求項9】
前記薬理学的シャペロンが、イソファゴミン、C−ベンジルイソファゴミン、N−アルキル(C9〜12)−DNJ、グルコイミダゾール、C−アルキル−IFG、N−アルキル−β−バリエナミン、フルフェノジン、N−ドデシル−DNJ、カリステギンA3、カリステギンB1、カリステギンB2およびカリステギンC1からなる群から選択される、請求項8に記載の方法。
【請求項10】
前記薬理学的シャペロンがイソファゴミンである、請求項9に記載の方法。
【請求項11】
前記リソソーム酵素が酸性α−グルコシダーゼである、請求項4に記載の方法。
【請求項12】
前記薬理学的シャペロンが、1−デオキシノジリマイシン、α−ホモノジリマイシンおよびカスタノスペルミンからなる群から選択される、請求項11に記載の方法。
【請求項13】
前記薬理学的シャペロンが1−デオキシノジリマイシンである、請求項12に記載の方法。
【請求項14】
前記宿主細胞が、CHO細胞、HeLa細胞、HEK−293細胞、293T細胞、COS細胞、COS−7細胞、マウス初代筋芽細胞、およびNIH3T3細胞からなる群から選択される、請求項1に記載の方法。
【請求項15】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、前記細胞の小胞体内で分解されない適切な野生型立体配座において前記タンパク質を安定化する、請求項1に記載の方法。
【請求項16】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、前記組換えタンパク質の過剰発現に関連する小胞体ストレスを低下させる、請求項1に記載の方法。
【請求項17】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、前記細胞の小胞体からの前記タンパク質の排出を増大させる、請求項1に記載の方法。
【請求項18】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、前記細胞からの前記タンパク質の分泌を増大させる、請求項1に記載の方法。
【請求項19】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、5.0よりも高いpHにおける前記タンパク質の安定性を向上させる、請求項1に記載の方法。
【請求項20】
組換え酸性α−グルコシダーゼの産生を改善するための方法であって、前記組換え酸性α−グルコシダーゼを発現するCHO宿主細胞と、1−デオキシノジリマイシンまたはその塩とを接触させることを含む方法。
【請求項1】
組換えタンパク質の産生を改善するための方法であって、前記組換えタンパク質を発現する宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとを接触させることを含む方法。
【請求項2】
前記宿主細胞が哺乳類細胞である、請求項1に記載の方法。
【請求項3】
前記タンパク質が酵素である、請求項1に記載の方法。
【請求項4】
前記酵素がリソソーム酵素である、請求項3に記載の方法。
【請求項5】
前記リソソーム酵素がα−ガラクトシダーゼAである、請求項4に記載の方法。
【請求項6】
前記薬理学的シャペロンが、1−デオキシガラクトノジリマイシン、α−allo−ホモノジリマイシン、β−1−C−ブチル−デオキシガラクトノジリマイシン、およびα−ガラクト−ホモノジリマイシン、カリステギンA3、カリステギンB2、カリステギンB3、N−メチル−カリステギンA3、N−メチル−カリステギンB2およびN−メチル−カリステギンB3からなる群から選択される、請求項5に記載の方法。
【請求項7】
前記薬理学的シャペロンが1−デオキシガラクトノジリマイシンである、請求項6に記載の方法。
【請求項8】
前記リソソーム酵素がβ−グルコシダーゼである、請求項4に記載の方法。
【請求項9】
前記薬理学的シャペロンが、イソファゴミン、C−ベンジルイソファゴミン、N−アルキル(C9〜12)−DNJ、グルコイミダゾール、C−アルキル−IFG、N−アルキル−β−バリエナミン、フルフェノジン、N−ドデシル−DNJ、カリステギンA3、カリステギンB1、カリステギンB2およびカリステギンC1からなる群から選択される、請求項8に記載の方法。
【請求項10】
前記薬理学的シャペロンがイソファゴミンである、請求項9に記載の方法。
【請求項11】
前記リソソーム酵素が酸性α−グルコシダーゼである、請求項4に記載の方法。
【請求項12】
前記薬理学的シャペロンが、1−デオキシノジリマイシン、α−ホモノジリマイシンおよびカスタノスペルミンからなる群から選択される、請求項11に記載の方法。
【請求項13】
前記薬理学的シャペロンが1−デオキシノジリマイシンである、請求項12に記載の方法。
【請求項14】
前記宿主細胞が、CHO細胞、HeLa細胞、HEK−293細胞、293T細胞、COS細胞、COS−7細胞、マウス初代筋芽細胞、およびNIH3T3細胞からなる群から選択される、請求項1に記載の方法。
【請求項15】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、前記細胞の小胞体内で分解されない適切な野生型立体配座において前記タンパク質を安定化する、請求項1に記載の方法。
【請求項16】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、前記組換えタンパク質の過剰発現に関連する小胞体ストレスを低下させる、請求項1に記載の方法。
【請求項17】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、前記細胞の小胞体からの前記タンパク質の排出を増大させる、請求項1に記載の方法。
【請求項18】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、前記細胞からの前記タンパク質の分泌を増大させる、請求項1に記載の方法。
【請求項19】
前記宿主細胞と、前記組換えタンパク質に特異的な薬理学的シャペロンとの接触が、5.0よりも高いpHにおける前記タンパク質の安定性を向上させる、請求項1に記載の方法。
【請求項20】
組換え酸性α−グルコシダーゼの産生を改善するための方法であって、前記組換え酸性α−グルコシダーゼを発現するCHO宿主細胞と、1−デオキシノジリマイシンまたはその塩とを接触させることを含む方法。
【図1】

【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2012−527900(P2012−527900A)
【公表日】平成24年11月12日(2012.11.12)
【国際特許分類】
【出願番号】特願2012−513221(P2012−513221)
【出願日】平成22年5月26日(2010.5.26)
【国際出願番号】PCT/US2010/036225
【国際公開番号】WO2010/138608
【国際公開日】平成22年12月2日(2010.12.2)
【出願人】(507170099)アミカス セラピューティックス インコーポレイテッド (21)
【Fターム(参考)】
【公表日】平成24年11月12日(2012.11.12)
【国際特許分類】
【出願日】平成22年5月26日(2010.5.26)
【国際出願番号】PCT/US2010/036225
【国際公開番号】WO2010/138608
【国際公開日】平成22年12月2日(2010.12.2)
【出願人】(507170099)アミカス セラピューティックス インコーポレイテッド (21)
【Fターム(参考)】
[ Back to top ]