
病原性の低いPRRS生ウイルスワクチン、ならびにその製造方法
【課題】毒性が低く、投与した豚に対してPRRSに対する有効な免疫を付与する豚用PRRS生ワクチンあるいは変性生ワクチン。
【解決手段】好適なワクチンは、平均プラーク径が約2mm未満で、病原性の低いウイルス分離株を含有している。好適なワクチンの一例は、プラークが小径の系統であるATCC受託番号VR2509を含有している。本発明のワクチンは、繁殖用雌豚あるいは雄豚、あるいは離乳子豚に投与することができ、豚に、この疾患の呼吸器型ならびに繁殖傷害型の双方に対する免疫を有効なかたちで付与することができる。
【解決手段】好適なワクチンは、平均プラーク径が約2mm未満で、病原性の低いウイルス分離株を含有している。好適なワクチンの一例は、プラークが小径の系統であるATCC受託番号VR2509を含有している。本発明のワクチンは、繁殖用雌豚あるいは雄豚、あるいは離乳子豚に投与することができ、豚に、この疾患の呼吸器型ならびに繁殖傷害型の双方に対する免疫を有効なかたちで付与することができる。
【発明の詳細な説明】
【技術分野】
【0001】
病原性の低いPRRS生ウイルスワクチン、ならびにその製造方法
発明の背景
<発明の分野>
本発明は、豚繁殖・呼吸傷害症候群(PRRS)ウイルスによる感染に対する有効な免疫を豚に与える目的で豚に投与する病原性の低い生ウイルスワクチンに関するものである。より詳細には、本発明は、この種の生ワクチン、PRRSウイルスに対する免疫を豚に付与する方法、ならびにこの種のワクチンの製造方法を提供するものである。新規で、実質的に分離、精製されている低病原性のPRRSウイルス(ATCC受託番号VR2509)も、本発明の一部を構成するものである。
【背景技術】
【0002】
<先行技術の記載>
PRRSは、この数年間に、重要な豚ウイルス性疾患として出現した疾患である。
PRRSは、妊娠豚に重篤な生殖障害を起こし、この生殖障害は、早産、死産数の増加、ミイラ化した胎仔や虚弱仔、分娩率の低下、発情期への復帰の遅れとなって顕われる。感染農家でのPRRSの急性生殖障害の徴候は、通常2−4カ月にわたって続くが、この疾患の呼吸器傷害は、何年にもわたって存続し、有意な生産減をもたらす。妊娠後期(妊娠77−95日)の豚でのPRRSウイルス感染の病原性について調べた研究がいくつか報告されている。こうした研究のいずれでも、PRRSウイルスが経胎盤性感染を生じ、胎児に対する病原性を有していることが明らかとなった。しかし、妊娠中期の豚では、胎児への病原性ははっきりせず、子宮内で感染した妊娠中期の胎仔は、総体としては正常なままであった。
野外条件下では、急性の繁殖傷害も、この疾患の慢性呼吸器障害病態も見られないのに、PRRSに関して血清学的に陽性であるような農家もある。こうした場合に、臨床疾患徴候が見られない理由ははっきりとはわかっていない。病原性の低いPRRSウイルスの系統が存在する可能性、そして、PRRSの臨床徴候を示さない豚個体群の感染原因がこうしたウイルスである可能性が示唆されている。さまざまな研究者によって数多くのPRRSウイルス分離株が報告されており、そのいずれも、RNAならびに脂質含有エンベロープを有する一方、各種動物種の赤血球に対して赤血球凝集能を有さないことが示されている。
【発明の開示】
【発明が解決しようとする課題】
【0003】
<発明の開示>
本発明は、以上に概略を示したような問題を解決するものであり、豚投与用の優れたPRRS生ワクチン、あるいは変性生ワクチンを提供するものである。本発明のワクチンは、毒性の野生型PRRSによる感染に対して有効な免疫を豚に付与するうえで十分な量の生あるいは変性生ウイルスを含有している。本発明で使用する場合、「有効な免疫」とは、ワクチンが、実質的な臨床徴候を示すような豚のPRRS感染症を防止しうることをいう。すなわち、免疫された豚は、PRRSに関して血清学的に陽性であろうとなかろうと、実質的な臨床徴候は何ら示さない。
好適な態様では、本発明のワクチンは、MN-Hsと称される新規な生きたウイルスを含んでおり、このウイルスは、米国、20852、メリーランド州ロックビル、パークローン・ドライブ、12301(12301 Parklawn Drive,Rockville,MD, 20852)のアメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)に、1995年7月26日付けで寄託済みで、ATCC受託番号はVR2509であった。このウイルスは、実質的に無毒で、しかも有効な免疫を付与するものであることが示された。本発明のワクチンは、繁殖用未成熟雌豚、成熟雌豚、成熟雄豚、離乳子豚に投与することができ、こうした投与は、筋内注射や経口−鼻腔投与などの任意の好都合な手段によって行うことができる。一般に、1回量のワクチンは、約104−約108PFU(プラーク形成単位)のウイルスを含んでいる。
さらに一般的には、本発明は、PRRS豚ワクチンを製造するにあたって、プラーククローニング法によって、MARK-145細胞のコンフルエントな層で平均直径約2mm未満プラークを形成するようなPRRSウイルスの系統、すなわち分離株を得、そして、この系統から生ワクチンを製造する方法に関するものである。MARK-145株化細胞は、ATCCに寄託されており、ATCC受託番号CRL−12219が付与されている。本発明のワクチンは、実質的に精製されたかたちで得られ、上述のようにプラークが小径のウイルスのみを本質的に含むものが好適である。好適なウイルスであるATCC受託番号VR2509のウイルスは、上述のとおりプラークが小径で、また、上述したように、免疫は付与するにもかかわらず、ほぼ完全に無毒である。
【0004】
<好適実施態様の詳細な説明>
以下の実施例は、病原性の低いPRRSウイルスの系統を分離、同定、クローニングするにあたっての好適な方法、ならびにこの系統からワクチンを製造する際の好適な方法を説明するものであり、以下の情報は例示の目的で示しただけで、こうした情報によって本発明全体としての範囲が限定されるものではないと理解されたい。記載中で言及する文献については、本明細書に引用文献として組み込むものである。
【0005】
(実施例1)
<概略>
この実施例では、豚繁殖・呼吸傷害症候群(PRRS)ウイルスの小型プラーク変異株(MN-Hs)の病原性を、妊娠豚で調べた。MN-Hs系統は、当初、小型プラークと大型プラークのまざったウイルス(MN-H)であるMN-Hウイルスからクローニングされた。胎児に対する病原性を比較するための第1の実験では、MN-Hs、MN-HL、野外ウイルス分離株(MN-W)、細胞培養液(対照)を、それぞれ2匹の妊娠86日の妊娠豚の鼻腔に接種した。対照の豚以外は、いずれの豚も予定日に分娩させた。感染豚は、接種後(PI)7日にはウイルス血症を起こしており、間接蛍光抗体(IFA)試験で調べたところ、接種後(PI)14日には血清が陽性となった。MN-Hsウイルスに感染させた2匹の豚は、14匹の生仔と5匹の死仔を分娩し、一方、MN-HLウイルスに感染させた2匹の豚は、0匹の生仔と25匹の死仔を分娩した。MN-Wウイルスを接種した2匹の豚は、10匹の生仔と20匹の死仔を分娩した。2匹の対照の豚は、妊娠107日に屠殺したところ、16匹の正常な胎仔を有していた。6匹の感染豚の生仔24匹のうち16匹(66.7%)、死仔14匹のうちの9匹(64.3%)、25匹のミイラ化仔のうちの3匹(12.0%)からウイルスが分離された。IFA法で調べたところ、4匹のMN-HL あるいはMN-W感染豚の死仔から得た13の血清のうち6は、1:16−1:1,024の範囲のPRRSウイルス抗体価を有していた。MN-Hsウイルスについての結果を追試するための次の実験では、MN-Hsを鼻腔に、MN-Hsを筋内に、別の野外ウイルス(OVL-173)分離株を鼻腔に投与して、それぞれ2匹の妊娠86日の妊娠豚に対して接種を行い、いずれの豚も予定日に分娩させた。MN-Hsウイルスを鼻腔あるいは筋内に投与して感染させた2匹の豚は、それぞれ、15匹の生仔と6匹の死仔、ならびに25匹の生仔と5匹の死仔を分娩し、一方、OVL-173 に感染させた2匹の豚は、6匹の生仔と24匹の死仔を分娩した。こうした結果からは、PRRSウイルスの豚胎仔に対する病原性がウイルス分離株によって異なっており、PRRSウイルスのMN-Hs系統は、病原性の穏やかなウイルスであることが示唆された。死仔の血清中のPRRSウイルス抗体の検出は、胎児感染を診断するうえで有用な方法であることがわかった。
【0006】
<材料ならびに方法>
<ウイルスならびに細胞の培養>
本研究では、3種の互いに異なったPRRSウイルス分離株を使用した。MN-H分離株は、PRRSの不顕性徴候が見られた農家で飼育されていた健康な豚の血清から得たものである。MN-Hウイルスは、当初、プラークサイズの異なる各種のウイルス集団が混ざったものであった。このMN-Hウイルスから、プラークが小型(MN-Hs)ならびに大型(MN-HL)のウイルスを別々にクローニングし、各ウイルスを、最初の実験については4回、次の実験についてはさらに6回、Himら、Am.J.Vet.Res.,52:1649-1652(1991)にしたがって、プラーククローニング法によってプラーク精製した。プラーク継代の各代では、コンフルエントなMARC-145(アフリカミドリザルの腎株化細胞(MA-104)由来の許容クローン)の単層細胞を60mm×15mmのペトリ皿で生育させ(細胞は、3%ウシ胎仔血清(FCS)、0.15%の炭酸ナトリウム、抗生物質を加えたイーグル最少必須培地(MEM)で維持した(Kimら、Arch.Virol.,133: 477-483(1993))、この培養に、それぞれのウイルスを接種した。培養を37°Cにて60分間インキュベートしてから、イノキュラムを除去し、培養をMEMで1回洗浄した。その後、等容量の2xMEMと1.6%煮沸ノーブル寒天(Disco Laboratories)にジメチルアミノエチル(DEAE)−デキストラン50μg/mlを加えた平板培養のそれぞれに、この培養液の5mlを加えた。さらに、この平板培養を、7°Cで5日間、CO2インキュベーター中でインキュベートし、インキュベーション期間の最後に、1%のニュートラルレッド加PBS3mlを加えることによって、プラーク培養を可視化した。選んだプラークを滅菌パスツールピペットで釣りあげてクローニングし、非感染MARC-145単層細胞に接種することによって継代した。永続的染色をおこなうために、寒天を注意深く除去し、20%エタノールへのクリスタルバイオレットの1%溶液2mlを使用して単層細胞を染色した。この平板培養を水道水で洗浄して、プラークを調べやすいようにした。
PRRSウイルスMN-Wは、典型的な急性PRRSの徴候を示す農家の罹病雌豚から分離したもので、OVL-173は、米国ミネソタ州ワーシントン(Worthington)のオックスフォード獣医学研究施設(Oxford Veterinary Laboratories)から入手したものである。
【0007】
<動物ならびに実験のデザイン>
妊娠約80日の妊娠した雌豚を、これまでに臨床的、血清学的にPRRSウイルスによる感染が確認された経緯のない農家から購入した。この農家では、豚は、年2回、豚パルボウイルス(PPV)ならびにレプトスピラの種に対するワクチンの接種を受けている。各雌豚の正確な繁殖日についての情報も入手した。購入後、各豚は、ミネソタ大学の隔離室内に別々に収容した。妊娠86日に、MN-Hs、MN-HL、MN-Wウイルス(2ml、105.0-5.5TCID50/ml)を、それぞれ2匹の雌豚の鼻腔に接種した。残りの2匹の豚には細胞培養液を接種して対照とした。各豚からは、適当な間隔をおいて血清サンプルを採取して、ウイルスの分離ならびに血清学的性質について調べた。6匹の感染豚は、自然分娩させ、2匹の対照豚は、妊娠107日に屠殺して胎児を調べた。分娩時には、生仔ならびに死仔から血液ならびに肺のサンプルを採取し、ミイラ化胎仔から胸液を採取して、ウイルスの分離と血清学的性質を調べた。第2の実験では、MN-Hsウイルスをさらに6回プラーク精製してから、接種を行った。MN-Hs を鼻腔に、MN-Hsを筋内に、別の野外分離ウイルス(OVL-173)を鼻腔に投与して、それぞれ2匹の妊娠86日の豚に対して接種を行い、いずれの豚も予定日に分娩させた。分娩時には、ミイラ化胎仔ならびに死仔の頭臀長を測定して、死亡時を推定した(Marrableら,J.Agric.Sci.,69:443-447(1967))。血液ならびに肺のサンプルは、第1の実験に記載したようにして採取、分析した。
【0008】
<ウイルスの分離ならびに血清学的性質の検討>
ウイルスの分離にあたっては、各血清サンプルあるいは肺ホモジネートの上清を24穴プレートのウェルに入れ、3%FCS 加MEMに懸濁させたMARK-145細胞(1−2×105個/ml)を加えた。培養は、PRRSウイルスに特有の細胞傷害作用(CPE)について、5−7日にわたって観察した。プレートを、2回にわたって凍結、解凍し、上清を、新鮮なMARK-145細胞懸濁液とともに96穴マイクロプレートのウェルに接種し、3−4日間インキュベートした。CPE と特異的蛍光の有無(PRRSウイルスの参照株に対して陽性の豚血清を使用)を観察することによって、ウイルス感染の事実を調べた。
親豚ならびに子豚から採取した血清は、間接蛍光抗体(IFA)法(Yoonら,J.Vet.Diag.Invest.,4:144-147(1992))によって、抗体に関して調べた。MARK-145株化細胞の細胞を使用して、96穴の試験プレートを調製した。血清のいくつかについては、すでに記載されている方法(Jooら,Aust.Vet.J.,52:422-424(1976))にしたがって、赤血球凝集阻害(HL)試験によって抗PPV抗体について調べた。
【0009】
<結果>
PRRSウイルスの各分離株は、分離の当初から、それぞれに異なったプラークサイズを示していた。MN-H分離株は、プラークのサイズに応じて、MN-HsとMN-HLの2つの異なった集団にクローニングした。クローニング後のプラークのサイズは、MN-HsとMN-HLが、それぞれ常時直径<2mmならびに3−5mmの範囲であったのに対し、MN-Wは、2−3mmであった(図面参照)。
PRRSウイルス分離株による感染後も、112番と153番の雌豚が接種後(PI)5日目に軽い食欲不振を示した以外、雌豚が主だった臨床徴候を示すことはなかった。接種後(PI)7日目には、6匹中6匹の血清サンプルでウイルスが分離され、接種後(PI)14日目には、6匹中1匹の血清サンプルでウイルスが分離された。表1に示すように、接種を行ったいずれの豚でも、接種後(PI)14日には高い抗体価が検出された。
【0010】
【表1】

【0011】
2回にわたって凍結、解凍し、上清を、新鮮なMARK-145細胞懸濁液とともに96穴マイクロプレートのウェルに接種し、3−4日間インキュベートした。CPEと特異的蛍光の有無(PRRSウイルスの参照株に対して陽性の豚血清を使用)を観察することによって、ウイルス感染の事実を調べた。
親豚ならびに子豚から採取した血清は、間接蛍光抗体(IFA)法(Yoonら,J.Vet.Diag.Invest.,4: 144-147(1992))によって、抗体に関して調べた。MARK-145株化細胞の細胞を使用して、96穴の試験プレートを調製した。血清のいくつかについては、すでに記載されている方法(Joo ら,Aust.Vet.J.,52: 422-424(1976))にしたがって、赤血球凝集阻害(HL)試験によって抗PPV抗体について調べた。
【0012】
<結果>
PRRSウイルスの各分離株は、分離の当初から、それぞれに異なったプラークサイズを示していた。MN-H分離株は、プラークのサイズに応じて、MN-HsとMN-HLの2つの異なった集団にクローニングした。クローニング後のプラークのサイズは、MN-HsとMN-HLが、それぞれ常時直径<2mmならびに3−5mmの範囲であったのに対し、MN-Wは、2−3mmであった(図面参照)。
PRRSウイルス分離株による感染後も、112番と153番の雌豚が接種後(PI)5日目に軽い食欲不振を示した以外、雌豚が主だった臨床徴候を示すことはなかった。接種後(PI)7日目には、6匹中6匹の血清サンプルでウイルスが分離され、接種後(PI)14日目には、6匹中1匹の血清サンプルでウイルスが分離された。表1に示すように、接種を行ったいずれの豚でも、接種後(PI)14日には高い抗体価が検出された。
【0013】
各豚の分娩結果を表2にまとめて示す。感染させたウイルス変異株が異なると、分娩結果の顕著な差が観察された。MN-Hsウイルスに感染させた2匹の雌豚からは、生仔14匹、死仔1匹、ミイラした豚4匹が、一方、MN-HLウイルスに感染させた2匹の雌豚からは、生仔0匹、死仔6匹、ミイラした豚19匹が得られた。MN-Wウイルスに感染させた2匹の雌豚からは、生仔10匹、死仔7匹、ミイラした豚13匹が得られた。2匹の対照豚は、妊娠107日に屠殺したところ、26匹の正常な胎児を有していた。実験IIでは、鼻腔ならびに筋内経由でMN-Hsに感染させた2匹の雌豚が、それぞれ、15匹の生仔と6匹の死仔、ならびに、25匹の生仔と5匹の死仔を分娩したのに対し、OVL-173に感染させた2匹の雌豚は、8匹の生仔と24匹の死仔を分娩した(表2)。実験Iと実験IIの分娩結果で比較を行うと、MN-Hsに感染させた6匹、ならびにMN-HL、MN-W、あるいはOVL-173に感染させた豚は、それぞれ、54匹の生仔(77.1%)と16匹の死仔、ならびに、16匹の生仔(18.8%)と69匹の死仔を分娩した。ミイラ化胎児と死亡胎児の頂尾長は、雌豚147と176の3匹のミイラ化胎児以外はいずれも≧24cmで(表2)、推定死亡時期が≧妊娠95日であることが示された。
【0014】
【表2】
【0015】
a 分娩あるいは屠殺の時点での妊娠日数
b ミイラ化胎仔あるいは死産胎仔の頭臀長(cm)
c ウイルスが分離されたサンプルの数/検査サンプル数
d 豚は妊娠107日に屠殺して、胎仔を調べた。
LB:生産、SB:死産、M:ミイラ化、IN:鼻腔投与、IM:筋内注射、ND:測定せず
【0016】
ウイルスは、感染豚の全群で、1以上の胎仔から分離された。実験1の6匹の感染豚の各個体についてのウイルス分離の結果では、調べたうちの約半分の胎仔(63匹中28匹)が、ウイルス分離に関して陽性であった。ウイルスの存在について調べた豚については、24匹の生仔のうち16匹(66.7%)、14匹の死仔のうち9匹(64.3%)、25匹のミイラ化豚のうち3匹(12.0%)がウイルスに関して陽性であった。血清サンプルがウイルスについて陽性であった28匹の豚のうち10匹が、肺のサンプルをウイルスの分離について調べた場合に、ウイルスについて陰性であった。
雌豚153、147、68、112の死仔から採取した血清を、IFAによってPRRSウイルス抗体について、HLによってPPVについて調べた。結果を表3に示す。死仔から採取した血清13サンプルのうち6サンプル、ミイラ化豚から採取した胸液25サンプルのうち2サンプルが、PRRSウイルス抗体を有していた。IFAでの力価は、1:16から1:1,024の範囲であり、PPVに対する抗体を有していた血清はなかった。雌豚17ならびに53の生仔14匹のうち13匹が、PRRSウイルス(IFAの力価、1:16−1:1,024)ならびにPPV(HLの力価、1:512−1:16,384)の双方に対して抗体を有していた。
【0017】
【表3】
【0018】
a 頂尾長(cm)
b IFAあるいはHL力価の逆数
c LB:生産、SB:死産、M:ミイラ化
【0019】
<考察>
本研究によって、PRRSウイルスの各種の分離株が、経胎盤性感染を生じ、分限後期の豚の胎児に対する病原性を有していることが確認された。しかし、PRRSウイルスの各種の分離株の間には、明らかな病原性の差が見られた。複数の多産系の雌豚の鼻腔にPRRSウイルスATTC-VR2332を妊娠93日に接種したところ(Christianson,W.T.ら,Can J.Vet.Res.,57: 262-268(1993))、同腹仔あたり、平均で、5.8匹の生仔と6.0匹の死亡胎仔を分娩した。本研究では、MN-HLあるいは2種の野外ウイルスに感染させた6匹の雌豚が、平均で、2.7匹の生仔と11.5匹の死亡胎仔を分娩しており、したがって、これらのウイルスは毒性が高いと考えられる。ATTC-VR2332と本研究で使用した毒性のウイルスとでは、病原性が異なっていたが、これは、ATTC-VR2332の方が感染させた時期が7日間遅いという感染時の妊娠日数のちがいによる可能性もある。一方、MN-Hs に感染させた6匹の雌豚は、同腹仔あたりの平均で、9.0匹の生仔と2.7匹の死亡胎仔を分娩した。この結果は毒性のウイルスについての結果とは著しく異なっており、MN-Hsウイルスが病原性の穏やかな系統であることが示唆された。
同一起源のMN-Hs ウイルスとMN-HLウイルスに感染させた豚の分娩結果に明らかな差が見られたことは、興味深い。MN-HsウイルスとMN-HLウイルスの場合、同一条件下で、それぞれ、5匹ならびに25匹の豚が死産で生まれた(p<0.005)。本研究の結果からすると、MN-HsとMN-HLの病原性は有意に異なっており、一方が病原性が低い系統で、他方が病原性の高い系統であると結論づけることができる。
PRRSウイルスに感染した同腹仔からのウイルスの分離は比較的容易で、ウイルスは、生産ならびに死産の子豚とも、ほぼ同様の率で分離された。感染同腹仔の全子豚からウイルスが分離されたわけではなかったことからすると、診断目的で行うウイルスの分離の場合には、一同腹仔あたり、少なくとも2匹以上の子豚からウイルス分離を試みる必要がある。また、ウイルスは、肺組織のサンプルからよりは、血清から分離した方が好都合であることもわかった。
毒性ウイルス感染豚の同腹仔では、いずれも、1匹以上の死仔で、PRRSウイルスに対して特異的な抗体が検出可能であった。こうした結果からは、非正常な同腹仔でのPRRSウイルスの感染を診断するうえで、死産あるいは分娩前に吸引された胎仔からの抗体の検出が有用な方法となりうることが示唆される。この方法は、ウイルス分離の技術や設備のない検査施設でも有用な方法となるはずである。
本研究では、死産の子豚13匹のうち6匹が、PRRSウイルスに対して陽性の抗体力価を示し、なおかつPPVに対しては陽性ではなかったので、検出された抗体が胎児由来のものであり、PRRSウイルスに起因するものであることが確認された。
PRRSウイルス感染群であっても、群によっては、臨床徴候を発現しない場合もあることについては、理由はわかっていない。PRRSウイルス感染以前の健康状態が良好であった群は、健康状況がそれより悪かった群より臨床応答が穏やかであることが報告されている。臨床所見が一見して異なっていることに関しては、健康状態一般と、本研究で立証されたような系統の差とがあいまって生じた結果として説明できる可能性がある。また、小型プラークのウイルスと大型プラークのウイルスの相互作用が宿主動物中で生じることによって病原性が弱まったと考えることも可能である。MN-Hウイルスの分離された農家では、病原性の高いPRRSウイルスであるMN-HLがその農家に存在していたにもかかわらず、繁殖傷害が生じていなかったことを勘案すると、この推定が正しい可能性もある。
【0020】
(実施例2)
本発明の好適なワクチンは、繁殖用未成熟雌豚、成熟雌豚、成熟雄豚、離乳子豚に投与することができる。投与は、筋内注射あるいは経口−鼻腔投与によって行うことができ、任意の時点で行うことができる。しかし、ワクチンの接種は、繁殖雌豚の場合、交尾前に行って、全妊娠期間にわたって保護を行い、若年豚の場合、離乳のすぐ後に行って、その後の肥育後期、成長期、仕上げ段階を保護するのが好ましい。
本発明のワクチンは、一般に2mlの用量を投与し、約104−約106プラーク形成単位(PFU)、さらに好ましくは106PFUを含むものとする。免疫は、繁殖用雌豚の場合は、少なくとも妊娠期間1回分は存続し、一方、若齢豚の離乳後にワクチン接種を行った場合には、仕上げ段階に至るまでの保護免疫が付与される。本発明の好適なワクチンは、広範な交差免疫を付与することができる。
本発明のワクチンは、生きたウイルスあるいは変性(弱毒化)ウイルス、ならびに通常の担体、安定剤および/またはアジュバントに帰納することができる。
【0021】
(実施例3)
<はじめに>
この実施例では、北米型PRRSウイルスのMN-Hs系統から構成されるワクチンが、3週齢の子豚で、北米型PRRSウイルスの毒性系統であるMN-HL系統の感染によって生じるウイルス血症を防止する能力を調べた。北米型PRRSウイルスの感染に伴う呼吸器疾患は、基本的には、養豚農家に存在している二次病原体によって生じるものである。しかし、こうした二次病原体は実験条件下では不在なので、北米型PRRSウイルスに感染した豚での呼吸器系の臨床徴候が定常的に再現されることはない。したがって、ウイルス血症が、北米型PRRSによる感染の最良の指標となる。MN-HL系統の投与後にウイルス血症が認められなければ、MN-Hs 系統をワクチンとして接種した子豚がMN-HL系統による感染に対して免疫を有していることが示唆されたことになる。
【0022】
<材料ならびに方法>
3週齢の子豚20匹を、PRRSのない農家から購入した。そのうち12匹の子豚の鼻腔に、MN-Hs系統(2ml、104.5 TCID50/ml)をワクチン接種し、残りの8匹の子豚を対照とした。ワクチンを接種した子豚と対照の子豚は、隔離ユニットの別々の部屋に収容した。ワクチン接種子豚6匹(豚番号81−86)と対照子豚4匹(豚番号93−96)には、ワクチン接種の2週間後に、MN-HL系統(2ml、104.5TCID50/ml)を鼻腔から投与した(第I群)。残りのワクチン接種子豚6匹(豚番号87−92)と対照子豚4匹(豚番号97−100)には、ワクチン接種の6週間後に、同様にしてMN-HL系統のウイルスを投与した(第II群)。子豚はすべて、臨床徴候を毎日観察した。さらに、血液サンプルも各子豚から毎週採取して、(1)MARK-145細胞培養を使用したウイルスの単離(Yoonら,J.Vet.Diag.Invest.,6: 289-282(1994))、(2)血清中和(SN)抗体の力価の測定(Parkら,Am.J.Vet.Res.,57:320-323(1996))を行った。ワクチン接種子豚でSN抗体が検出されることは、毒性の北米型PRRSウイルスに対する保護免疫が成立していることを示唆するものである。
【0023】
<結果>
ワクチンの接種後、第I群と第II群の子豚からは、ワクチンウイルスが、ワクチンの接種後3週間にわたって分離された。MN-HLウイルスの投与を行ったところ、第I群ならびに第II群の対照子豚ならびにワクチン接種子豚のいずれでも、臨床徴候は観察されなかった。
第I群:MN-HLウイルス投与の時点では、SN抗体は、どの子豚からも検出されなかった。MN-HLウイルスの投与後は、ワクチン接種子豚と対照の子豚の双方から投与ウイルスが検出された。さらに、ワクチン接種子豚と対照の子豚の双方が、ウイルス血症を発症した(表1)。
第II群:ワクチン接種子豚は、ワクチン接種後3週間から中和抗体を産生し、MN-HLウイルス投与の時点では、力価は、1:2から1:8の範囲であった。MN-HLウイルスの投与後は、ワクチン接種子豚からは、投与ウイルスが分離されなかったのに対し、対照の子豚はウイルス血症を発症した(表4)。こうした結果から、ワクチンを接種した子豚が、毒性の北米型PRRSウイルスに対する保護免疫を獲得していることが例証された。
【0024】
【表4】
【0025】
V=ワクチン接種、C=対照
a=MN-HLウイルスを投与
b=ウイルスの分離の有無/SN抗体の力価
第I群ならびに第II群の子豚は、それぞれ、ワクチン接種後4週ならびに8週が経過した時点で屠殺した。
【図面の簡単な説明】
【0026】
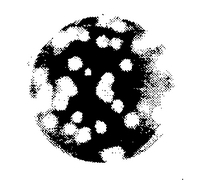
【図1】MARK-145株化細胞で培養したPRRSウイルスのMN-Hs分離株のプラークの形態ならびにサイズ(<2mm)を示す写真である。
【図2】MARK-145株化細胞で培養したPRRSウイルスのMN-HL 分離株のプラークの形態ならびにサイズ(3−5mm)を示す写真である。
【図3】MARK-145株化細胞で培養したPRRSウイルスのMN-W分離株のプラークの形態ならびにサイズ(2−3mm)を示す写真である。
【技術分野】
【0001】
病原性の低いPRRS生ウイルスワクチン、ならびにその製造方法
発明の背景
<発明の分野>
本発明は、豚繁殖・呼吸傷害症候群(PRRS)ウイルスによる感染に対する有効な免疫を豚に与える目的で豚に投与する病原性の低い生ウイルスワクチンに関するものである。より詳細には、本発明は、この種の生ワクチン、PRRSウイルスに対する免疫を豚に付与する方法、ならびにこの種のワクチンの製造方法を提供するものである。新規で、実質的に分離、精製されている低病原性のPRRSウイルス(ATCC受託番号VR2509)も、本発明の一部を構成するものである。
【背景技術】
【0002】
<先行技術の記載>
PRRSは、この数年間に、重要な豚ウイルス性疾患として出現した疾患である。
PRRSは、妊娠豚に重篤な生殖障害を起こし、この生殖障害は、早産、死産数の増加、ミイラ化した胎仔や虚弱仔、分娩率の低下、発情期への復帰の遅れとなって顕われる。感染農家でのPRRSの急性生殖障害の徴候は、通常2−4カ月にわたって続くが、この疾患の呼吸器傷害は、何年にもわたって存続し、有意な生産減をもたらす。妊娠後期(妊娠77−95日)の豚でのPRRSウイルス感染の病原性について調べた研究がいくつか報告されている。こうした研究のいずれでも、PRRSウイルスが経胎盤性感染を生じ、胎児に対する病原性を有していることが明らかとなった。しかし、妊娠中期の豚では、胎児への病原性ははっきりせず、子宮内で感染した妊娠中期の胎仔は、総体としては正常なままであった。
野外条件下では、急性の繁殖傷害も、この疾患の慢性呼吸器障害病態も見られないのに、PRRSに関して血清学的に陽性であるような農家もある。こうした場合に、臨床疾患徴候が見られない理由ははっきりとはわかっていない。病原性の低いPRRSウイルスの系統が存在する可能性、そして、PRRSの臨床徴候を示さない豚個体群の感染原因がこうしたウイルスである可能性が示唆されている。さまざまな研究者によって数多くのPRRSウイルス分離株が報告されており、そのいずれも、RNAならびに脂質含有エンベロープを有する一方、各種動物種の赤血球に対して赤血球凝集能を有さないことが示されている。
【発明の開示】
【発明が解決しようとする課題】
【0003】
<発明の開示>
本発明は、以上に概略を示したような問題を解決するものであり、豚投与用の優れたPRRS生ワクチン、あるいは変性生ワクチンを提供するものである。本発明のワクチンは、毒性の野生型PRRSによる感染に対して有効な免疫を豚に付与するうえで十分な量の生あるいは変性生ウイルスを含有している。本発明で使用する場合、「有効な免疫」とは、ワクチンが、実質的な臨床徴候を示すような豚のPRRS感染症を防止しうることをいう。すなわち、免疫された豚は、PRRSに関して血清学的に陽性であろうとなかろうと、実質的な臨床徴候は何ら示さない。
好適な態様では、本発明のワクチンは、MN-Hsと称される新規な生きたウイルスを含んでおり、このウイルスは、米国、20852、メリーランド州ロックビル、パークローン・ドライブ、12301(12301 Parklawn Drive,Rockville,MD, 20852)のアメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)に、1995年7月26日付けで寄託済みで、ATCC受託番号はVR2509であった。このウイルスは、実質的に無毒で、しかも有効な免疫を付与するものであることが示された。本発明のワクチンは、繁殖用未成熟雌豚、成熟雌豚、成熟雄豚、離乳子豚に投与することができ、こうした投与は、筋内注射や経口−鼻腔投与などの任意の好都合な手段によって行うことができる。一般に、1回量のワクチンは、約104−約108PFU(プラーク形成単位)のウイルスを含んでいる。
さらに一般的には、本発明は、PRRS豚ワクチンを製造するにあたって、プラーククローニング法によって、MARK-145細胞のコンフルエントな層で平均直径約2mm未満プラークを形成するようなPRRSウイルスの系統、すなわち分離株を得、そして、この系統から生ワクチンを製造する方法に関するものである。MARK-145株化細胞は、ATCCに寄託されており、ATCC受託番号CRL−12219が付与されている。本発明のワクチンは、実質的に精製されたかたちで得られ、上述のようにプラークが小径のウイルスのみを本質的に含むものが好適である。好適なウイルスであるATCC受託番号VR2509のウイルスは、上述のとおりプラークが小径で、また、上述したように、免疫は付与するにもかかわらず、ほぼ完全に無毒である。
【0004】
<好適実施態様の詳細な説明>
以下の実施例は、病原性の低いPRRSウイルスの系統を分離、同定、クローニングするにあたっての好適な方法、ならびにこの系統からワクチンを製造する際の好適な方法を説明するものであり、以下の情報は例示の目的で示しただけで、こうした情報によって本発明全体としての範囲が限定されるものではないと理解されたい。記載中で言及する文献については、本明細書に引用文献として組み込むものである。
【0005】
(実施例1)
<概略>
この実施例では、豚繁殖・呼吸傷害症候群(PRRS)ウイルスの小型プラーク変異株(MN-Hs)の病原性を、妊娠豚で調べた。MN-Hs系統は、当初、小型プラークと大型プラークのまざったウイルス(MN-H)であるMN-Hウイルスからクローニングされた。胎児に対する病原性を比較するための第1の実験では、MN-Hs、MN-HL、野外ウイルス分離株(MN-W)、細胞培養液(対照)を、それぞれ2匹の妊娠86日の妊娠豚の鼻腔に接種した。対照の豚以外は、いずれの豚も予定日に分娩させた。感染豚は、接種後(PI)7日にはウイルス血症を起こしており、間接蛍光抗体(IFA)試験で調べたところ、接種後(PI)14日には血清が陽性となった。MN-Hsウイルスに感染させた2匹の豚は、14匹の生仔と5匹の死仔を分娩し、一方、MN-HLウイルスに感染させた2匹の豚は、0匹の生仔と25匹の死仔を分娩した。MN-Wウイルスを接種した2匹の豚は、10匹の生仔と20匹の死仔を分娩した。2匹の対照の豚は、妊娠107日に屠殺したところ、16匹の正常な胎仔を有していた。6匹の感染豚の生仔24匹のうち16匹(66.7%)、死仔14匹のうちの9匹(64.3%)、25匹のミイラ化仔のうちの3匹(12.0%)からウイルスが分離された。IFA法で調べたところ、4匹のMN-HL あるいはMN-W感染豚の死仔から得た13の血清のうち6は、1:16−1:1,024の範囲のPRRSウイルス抗体価を有していた。MN-Hsウイルスについての結果を追試するための次の実験では、MN-Hsを鼻腔に、MN-Hsを筋内に、別の野外ウイルス(OVL-173)分離株を鼻腔に投与して、それぞれ2匹の妊娠86日の妊娠豚に対して接種を行い、いずれの豚も予定日に分娩させた。MN-Hsウイルスを鼻腔あるいは筋内に投与して感染させた2匹の豚は、それぞれ、15匹の生仔と6匹の死仔、ならびに25匹の生仔と5匹の死仔を分娩し、一方、OVL-173 に感染させた2匹の豚は、6匹の生仔と24匹の死仔を分娩した。こうした結果からは、PRRSウイルスの豚胎仔に対する病原性がウイルス分離株によって異なっており、PRRSウイルスのMN-Hs系統は、病原性の穏やかなウイルスであることが示唆された。死仔の血清中のPRRSウイルス抗体の検出は、胎児感染を診断するうえで有用な方法であることがわかった。
【0006】
<材料ならびに方法>
<ウイルスならびに細胞の培養>
本研究では、3種の互いに異なったPRRSウイルス分離株を使用した。MN-H分離株は、PRRSの不顕性徴候が見られた農家で飼育されていた健康な豚の血清から得たものである。MN-Hウイルスは、当初、プラークサイズの異なる各種のウイルス集団が混ざったものであった。このMN-Hウイルスから、プラークが小型(MN-Hs)ならびに大型(MN-HL)のウイルスを別々にクローニングし、各ウイルスを、最初の実験については4回、次の実験についてはさらに6回、Himら、Am.J.Vet.Res.,52:1649-1652(1991)にしたがって、プラーククローニング法によってプラーク精製した。プラーク継代の各代では、コンフルエントなMARC-145(アフリカミドリザルの腎株化細胞(MA-104)由来の許容クローン)の単層細胞を60mm×15mmのペトリ皿で生育させ(細胞は、3%ウシ胎仔血清(FCS)、0.15%の炭酸ナトリウム、抗生物質を加えたイーグル最少必須培地(MEM)で維持した(Kimら、Arch.Virol.,133: 477-483(1993))、この培養に、それぞれのウイルスを接種した。培養を37°Cにて60分間インキュベートしてから、イノキュラムを除去し、培養をMEMで1回洗浄した。その後、等容量の2xMEMと1.6%煮沸ノーブル寒天(Disco Laboratories)にジメチルアミノエチル(DEAE)−デキストラン50μg/mlを加えた平板培養のそれぞれに、この培養液の5mlを加えた。さらに、この平板培養を、7°Cで5日間、CO2インキュベーター中でインキュベートし、インキュベーション期間の最後に、1%のニュートラルレッド加PBS3mlを加えることによって、プラーク培養を可視化した。選んだプラークを滅菌パスツールピペットで釣りあげてクローニングし、非感染MARC-145単層細胞に接種することによって継代した。永続的染色をおこなうために、寒天を注意深く除去し、20%エタノールへのクリスタルバイオレットの1%溶液2mlを使用して単層細胞を染色した。この平板培養を水道水で洗浄して、プラークを調べやすいようにした。
PRRSウイルスMN-Wは、典型的な急性PRRSの徴候を示す農家の罹病雌豚から分離したもので、OVL-173は、米国ミネソタ州ワーシントン(Worthington)のオックスフォード獣医学研究施設(Oxford Veterinary Laboratories)から入手したものである。
【0007】
<動物ならびに実験のデザイン>
妊娠約80日の妊娠した雌豚を、これまでに臨床的、血清学的にPRRSウイルスによる感染が確認された経緯のない農家から購入した。この農家では、豚は、年2回、豚パルボウイルス(PPV)ならびにレプトスピラの種に対するワクチンの接種を受けている。各雌豚の正確な繁殖日についての情報も入手した。購入後、各豚は、ミネソタ大学の隔離室内に別々に収容した。妊娠86日に、MN-Hs、MN-HL、MN-Wウイルス(2ml、105.0-5.5TCID50/ml)を、それぞれ2匹の雌豚の鼻腔に接種した。残りの2匹の豚には細胞培養液を接種して対照とした。各豚からは、適当な間隔をおいて血清サンプルを採取して、ウイルスの分離ならびに血清学的性質について調べた。6匹の感染豚は、自然分娩させ、2匹の対照豚は、妊娠107日に屠殺して胎児を調べた。分娩時には、生仔ならびに死仔から血液ならびに肺のサンプルを採取し、ミイラ化胎仔から胸液を採取して、ウイルスの分離と血清学的性質を調べた。第2の実験では、MN-Hsウイルスをさらに6回プラーク精製してから、接種を行った。MN-Hs を鼻腔に、MN-Hsを筋内に、別の野外分離ウイルス(OVL-173)を鼻腔に投与して、それぞれ2匹の妊娠86日の豚に対して接種を行い、いずれの豚も予定日に分娩させた。分娩時には、ミイラ化胎仔ならびに死仔の頭臀長を測定して、死亡時を推定した(Marrableら,J.Agric.Sci.,69:443-447(1967))。血液ならびに肺のサンプルは、第1の実験に記載したようにして採取、分析した。
【0008】
<ウイルスの分離ならびに血清学的性質の検討>
ウイルスの分離にあたっては、各血清サンプルあるいは肺ホモジネートの上清を24穴プレートのウェルに入れ、3%FCS 加MEMに懸濁させたMARK-145細胞(1−2×105個/ml)を加えた。培養は、PRRSウイルスに特有の細胞傷害作用(CPE)について、5−7日にわたって観察した。プレートを、2回にわたって凍結、解凍し、上清を、新鮮なMARK-145細胞懸濁液とともに96穴マイクロプレートのウェルに接種し、3−4日間インキュベートした。CPE と特異的蛍光の有無(PRRSウイルスの参照株に対して陽性の豚血清を使用)を観察することによって、ウイルス感染の事実を調べた。
親豚ならびに子豚から採取した血清は、間接蛍光抗体(IFA)法(Yoonら,J.Vet.Diag.Invest.,4:144-147(1992))によって、抗体に関して調べた。MARK-145株化細胞の細胞を使用して、96穴の試験プレートを調製した。血清のいくつかについては、すでに記載されている方法(Jooら,Aust.Vet.J.,52:422-424(1976))にしたがって、赤血球凝集阻害(HL)試験によって抗PPV抗体について調べた。
【0009】
<結果>
PRRSウイルスの各分離株は、分離の当初から、それぞれに異なったプラークサイズを示していた。MN-H分離株は、プラークのサイズに応じて、MN-HsとMN-HLの2つの異なった集団にクローニングした。クローニング後のプラークのサイズは、MN-HsとMN-HLが、それぞれ常時直径<2mmならびに3−5mmの範囲であったのに対し、MN-Wは、2−3mmであった(図面参照)。
PRRSウイルス分離株による感染後も、112番と153番の雌豚が接種後(PI)5日目に軽い食欲不振を示した以外、雌豚が主だった臨床徴候を示すことはなかった。接種後(PI)7日目には、6匹中6匹の血清サンプルでウイルスが分離され、接種後(PI)14日目には、6匹中1匹の血清サンプルでウイルスが分離された。表1に示すように、接種を行ったいずれの豚でも、接種後(PI)14日には高い抗体価が検出された。
【0010】
【表1】
【0011】
2回にわたって凍結、解凍し、上清を、新鮮なMARK-145細胞懸濁液とともに96穴マイクロプレートのウェルに接種し、3−4日間インキュベートした。CPEと特異的蛍光の有無(PRRSウイルスの参照株に対して陽性の豚血清を使用)を観察することによって、ウイルス感染の事実を調べた。
親豚ならびに子豚から採取した血清は、間接蛍光抗体(IFA)法(Yoonら,J.Vet.Diag.Invest.,4: 144-147(1992))によって、抗体に関して調べた。MARK-145株化細胞の細胞を使用して、96穴の試験プレートを調製した。血清のいくつかについては、すでに記載されている方法(Joo ら,Aust.Vet.J.,52: 422-424(1976))にしたがって、赤血球凝集阻害(HL)試験によって抗PPV抗体について調べた。
【0012】
<結果>
PRRSウイルスの各分離株は、分離の当初から、それぞれに異なったプラークサイズを示していた。MN-H分離株は、プラークのサイズに応じて、MN-HsとMN-HLの2つの異なった集団にクローニングした。クローニング後のプラークのサイズは、MN-HsとMN-HLが、それぞれ常時直径<2mmならびに3−5mmの範囲であったのに対し、MN-Wは、2−3mmであった(図面参照)。
PRRSウイルス分離株による感染後も、112番と153番の雌豚が接種後(PI)5日目に軽い食欲不振を示した以外、雌豚が主だった臨床徴候を示すことはなかった。接種後(PI)7日目には、6匹中6匹の血清サンプルでウイルスが分離され、接種後(PI)14日目には、6匹中1匹の血清サンプルでウイルスが分離された。表1に示すように、接種を行ったいずれの豚でも、接種後(PI)14日には高い抗体価が検出された。
【0013】
各豚の分娩結果を表2にまとめて示す。感染させたウイルス変異株が異なると、分娩結果の顕著な差が観察された。MN-Hsウイルスに感染させた2匹の雌豚からは、生仔14匹、死仔1匹、ミイラした豚4匹が、一方、MN-HLウイルスに感染させた2匹の雌豚からは、生仔0匹、死仔6匹、ミイラした豚19匹が得られた。MN-Wウイルスに感染させた2匹の雌豚からは、生仔10匹、死仔7匹、ミイラした豚13匹が得られた。2匹の対照豚は、妊娠107日に屠殺したところ、26匹の正常な胎児を有していた。実験IIでは、鼻腔ならびに筋内経由でMN-Hsに感染させた2匹の雌豚が、それぞれ、15匹の生仔と6匹の死仔、ならびに、25匹の生仔と5匹の死仔を分娩したのに対し、OVL-173に感染させた2匹の雌豚は、8匹の生仔と24匹の死仔を分娩した(表2)。実験Iと実験IIの分娩結果で比較を行うと、MN-Hsに感染させた6匹、ならびにMN-HL、MN-W、あるいはOVL-173に感染させた豚は、それぞれ、54匹の生仔(77.1%)と16匹の死仔、ならびに、16匹の生仔(18.8%)と69匹の死仔を分娩した。ミイラ化胎児と死亡胎児の頂尾長は、雌豚147と176の3匹のミイラ化胎児以外はいずれも≧24cmで(表2)、推定死亡時期が≧妊娠95日であることが示された。
【0014】
【表2】
【0015】
a 分娩あるいは屠殺の時点での妊娠日数
b ミイラ化胎仔あるいは死産胎仔の頭臀長(cm)
c ウイルスが分離されたサンプルの数/検査サンプル数
d 豚は妊娠107日に屠殺して、胎仔を調べた。
LB:生産、SB:死産、M:ミイラ化、IN:鼻腔投与、IM:筋内注射、ND:測定せず
【0016】
ウイルスは、感染豚の全群で、1以上の胎仔から分離された。実験1の6匹の感染豚の各個体についてのウイルス分離の結果では、調べたうちの約半分の胎仔(63匹中28匹)が、ウイルス分離に関して陽性であった。ウイルスの存在について調べた豚については、24匹の生仔のうち16匹(66.7%)、14匹の死仔のうち9匹(64.3%)、25匹のミイラ化豚のうち3匹(12.0%)がウイルスに関して陽性であった。血清サンプルがウイルスについて陽性であった28匹の豚のうち10匹が、肺のサンプルをウイルスの分離について調べた場合に、ウイルスについて陰性であった。
雌豚153、147、68、112の死仔から採取した血清を、IFAによってPRRSウイルス抗体について、HLによってPPVについて調べた。結果を表3に示す。死仔から採取した血清13サンプルのうち6サンプル、ミイラ化豚から採取した胸液25サンプルのうち2サンプルが、PRRSウイルス抗体を有していた。IFAでの力価は、1:16から1:1,024の範囲であり、PPVに対する抗体を有していた血清はなかった。雌豚17ならびに53の生仔14匹のうち13匹が、PRRSウイルス(IFAの力価、1:16−1:1,024)ならびにPPV(HLの力価、1:512−1:16,384)の双方に対して抗体を有していた。
【0017】
【表3】
【0018】
a 頂尾長(cm)
b IFAあるいはHL力価の逆数
c LB:生産、SB:死産、M:ミイラ化
【0019】
<考察>
本研究によって、PRRSウイルスの各種の分離株が、経胎盤性感染を生じ、分限後期の豚の胎児に対する病原性を有していることが確認された。しかし、PRRSウイルスの各種の分離株の間には、明らかな病原性の差が見られた。複数の多産系の雌豚の鼻腔にPRRSウイルスATTC-VR2332を妊娠93日に接種したところ(Christianson,W.T.ら,Can J.Vet.Res.,57: 262-268(1993))、同腹仔あたり、平均で、5.8匹の生仔と6.0匹の死亡胎仔を分娩した。本研究では、MN-HLあるいは2種の野外ウイルスに感染させた6匹の雌豚が、平均で、2.7匹の生仔と11.5匹の死亡胎仔を分娩しており、したがって、これらのウイルスは毒性が高いと考えられる。ATTC-VR2332と本研究で使用した毒性のウイルスとでは、病原性が異なっていたが、これは、ATTC-VR2332の方が感染させた時期が7日間遅いという感染時の妊娠日数のちがいによる可能性もある。一方、MN-Hs に感染させた6匹の雌豚は、同腹仔あたりの平均で、9.0匹の生仔と2.7匹の死亡胎仔を分娩した。この結果は毒性のウイルスについての結果とは著しく異なっており、MN-Hsウイルスが病原性の穏やかな系統であることが示唆された。
同一起源のMN-Hs ウイルスとMN-HLウイルスに感染させた豚の分娩結果に明らかな差が見られたことは、興味深い。MN-HsウイルスとMN-HLウイルスの場合、同一条件下で、それぞれ、5匹ならびに25匹の豚が死産で生まれた(p<0.005)。本研究の結果からすると、MN-HsとMN-HLの病原性は有意に異なっており、一方が病原性が低い系統で、他方が病原性の高い系統であると結論づけることができる。
PRRSウイルスに感染した同腹仔からのウイルスの分離は比較的容易で、ウイルスは、生産ならびに死産の子豚とも、ほぼ同様の率で分離された。感染同腹仔の全子豚からウイルスが分離されたわけではなかったことからすると、診断目的で行うウイルスの分離の場合には、一同腹仔あたり、少なくとも2匹以上の子豚からウイルス分離を試みる必要がある。また、ウイルスは、肺組織のサンプルからよりは、血清から分離した方が好都合であることもわかった。
毒性ウイルス感染豚の同腹仔では、いずれも、1匹以上の死仔で、PRRSウイルスに対して特異的な抗体が検出可能であった。こうした結果からは、非正常な同腹仔でのPRRSウイルスの感染を診断するうえで、死産あるいは分娩前に吸引された胎仔からの抗体の検出が有用な方法となりうることが示唆される。この方法は、ウイルス分離の技術や設備のない検査施設でも有用な方法となるはずである。
本研究では、死産の子豚13匹のうち6匹が、PRRSウイルスに対して陽性の抗体力価を示し、なおかつPPVに対しては陽性ではなかったので、検出された抗体が胎児由来のものであり、PRRSウイルスに起因するものであることが確認された。
PRRSウイルス感染群であっても、群によっては、臨床徴候を発現しない場合もあることについては、理由はわかっていない。PRRSウイルス感染以前の健康状態が良好であった群は、健康状況がそれより悪かった群より臨床応答が穏やかであることが報告されている。臨床所見が一見して異なっていることに関しては、健康状態一般と、本研究で立証されたような系統の差とがあいまって生じた結果として説明できる可能性がある。また、小型プラークのウイルスと大型プラークのウイルスの相互作用が宿主動物中で生じることによって病原性が弱まったと考えることも可能である。MN-Hウイルスの分離された農家では、病原性の高いPRRSウイルスであるMN-HLがその農家に存在していたにもかかわらず、繁殖傷害が生じていなかったことを勘案すると、この推定が正しい可能性もある。
【0020】
(実施例2)
本発明の好適なワクチンは、繁殖用未成熟雌豚、成熟雌豚、成熟雄豚、離乳子豚に投与することができる。投与は、筋内注射あるいは経口−鼻腔投与によって行うことができ、任意の時点で行うことができる。しかし、ワクチンの接種は、繁殖雌豚の場合、交尾前に行って、全妊娠期間にわたって保護を行い、若年豚の場合、離乳のすぐ後に行って、その後の肥育後期、成長期、仕上げ段階を保護するのが好ましい。
本発明のワクチンは、一般に2mlの用量を投与し、約104−約106プラーク形成単位(PFU)、さらに好ましくは106PFUを含むものとする。免疫は、繁殖用雌豚の場合は、少なくとも妊娠期間1回分は存続し、一方、若齢豚の離乳後にワクチン接種を行った場合には、仕上げ段階に至るまでの保護免疫が付与される。本発明の好適なワクチンは、広範な交差免疫を付与することができる。
本発明のワクチンは、生きたウイルスあるいは変性(弱毒化)ウイルス、ならびに通常の担体、安定剤および/またはアジュバントに帰納することができる。
【0021】
(実施例3)
<はじめに>
この実施例では、北米型PRRSウイルスのMN-Hs系統から構成されるワクチンが、3週齢の子豚で、北米型PRRSウイルスの毒性系統であるMN-HL系統の感染によって生じるウイルス血症を防止する能力を調べた。北米型PRRSウイルスの感染に伴う呼吸器疾患は、基本的には、養豚農家に存在している二次病原体によって生じるものである。しかし、こうした二次病原体は実験条件下では不在なので、北米型PRRSウイルスに感染した豚での呼吸器系の臨床徴候が定常的に再現されることはない。したがって、ウイルス血症が、北米型PRRSによる感染の最良の指標となる。MN-HL系統の投与後にウイルス血症が認められなければ、MN-Hs 系統をワクチンとして接種した子豚がMN-HL系統による感染に対して免疫を有していることが示唆されたことになる。
【0022】
<材料ならびに方法>
3週齢の子豚20匹を、PRRSのない農家から購入した。そのうち12匹の子豚の鼻腔に、MN-Hs系統(2ml、104.5 TCID50/ml)をワクチン接種し、残りの8匹の子豚を対照とした。ワクチンを接種した子豚と対照の子豚は、隔離ユニットの別々の部屋に収容した。ワクチン接種子豚6匹(豚番号81−86)と対照子豚4匹(豚番号93−96)には、ワクチン接種の2週間後に、MN-HL系統(2ml、104.5TCID50/ml)を鼻腔から投与した(第I群)。残りのワクチン接種子豚6匹(豚番号87−92)と対照子豚4匹(豚番号97−100)には、ワクチン接種の6週間後に、同様にしてMN-HL系統のウイルスを投与した(第II群)。子豚はすべて、臨床徴候を毎日観察した。さらに、血液サンプルも各子豚から毎週採取して、(1)MARK-145細胞培養を使用したウイルスの単離(Yoonら,J.Vet.Diag.Invest.,6: 289-282(1994))、(2)血清中和(SN)抗体の力価の測定(Parkら,Am.J.Vet.Res.,57:320-323(1996))を行った。ワクチン接種子豚でSN抗体が検出されることは、毒性の北米型PRRSウイルスに対する保護免疫が成立していることを示唆するものである。
【0023】
<結果>
ワクチンの接種後、第I群と第II群の子豚からは、ワクチンウイルスが、ワクチンの接種後3週間にわたって分離された。MN-HLウイルスの投与を行ったところ、第I群ならびに第II群の対照子豚ならびにワクチン接種子豚のいずれでも、臨床徴候は観察されなかった。
第I群:MN-HLウイルス投与の時点では、SN抗体は、どの子豚からも検出されなかった。MN-HLウイルスの投与後は、ワクチン接種子豚と対照の子豚の双方から投与ウイルスが検出された。さらに、ワクチン接種子豚と対照の子豚の双方が、ウイルス血症を発症した(表1)。
第II群:ワクチン接種子豚は、ワクチン接種後3週間から中和抗体を産生し、MN-HLウイルス投与の時点では、力価は、1:2から1:8の範囲であった。MN-HLウイルスの投与後は、ワクチン接種子豚からは、投与ウイルスが分離されなかったのに対し、対照の子豚はウイルス血症を発症した(表4)。こうした結果から、ワクチンを接種した子豚が、毒性の北米型PRRSウイルスに対する保護免疫を獲得していることが例証された。
【0024】
【表4】
【0025】
V=ワクチン接種、C=対照
a=MN-HLウイルスを投与
b=ウイルスの分離の有無/SN抗体の力価
第I群ならびに第II群の子豚は、それぞれ、ワクチン接種後4週ならびに8週が経過した時点で屠殺した。
【図面の簡単な説明】
【0026】
【図1】MARK-145株化細胞で培養したPRRSウイルスのMN-Hs分離株のプラークの形態ならびにサイズ(<2mm)を示す写真である。
【図2】MARK-145株化細胞で培養したPRRSウイルスのMN-HL 分離株のプラークの形態ならびにサイズ(3−5mm)を示す写真である。
【図3】MARK-145株化細胞で培養したPRRSウイルスのMN-W分離株のプラークの形態ならびにサイズ(2−3mm)を示す写真である。
【特許請求の範囲】
【請求項1】
(A)プラーククローニングによって北米型PRRSウイルスの特定の系統を得、そして
(B)工程(a)で得た系統からワクチンを調製する工程よりなるワクチンの製造方法であって、ここで、上記系統が、
(a)上記系統を一群の細胞に接種し(ここで、この細胞はCRL12219の名称を有し、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液で生育させた細胞のコンフルエントな単層から得たものであり、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液1mlあたり、1−2×105の細胞を含むものであり)、
(b)上記細胞を37°Cで60分インキュベートし、
(c)上記細胞からイノキュラムを取り除き、
(d)上記細胞をイーグル最少必須培養液で洗浄し、
(e)上記細胞を、250μg のジエチルアミノエチル(DEAE)−デキストランを加えた2×イーグル最少必須培養液ならびに1.6%煮沸ノーブル寒天5mlに再懸濁し、
(f)上記細胞を平板培養し、そして
(g)上記細胞を、さらに、37°Cで5日間、CO2インキュベーター中でインキュベートしてプラークを得る工程からなる方法によってプラークを得た場合に、平均直径が2mm未満のプラークを生じる系統であるようなワクチンの製造方法。
【請求項2】
ワクチンが、生きた北米型PRRSウイルスの系統を含むものである請求項1に記載の方法。
【請求項3】
ワクチンの製造方法が、上記系統を精製形態で得る工程を含むものである請求項1に記載の方法。
【請求項4】
上記系統が、ATCC VR2509 の名称を有するものである請求項1に記載の方法。
【請求項5】
北米型PRRSウイルスの特定の系統を含むワクチンであって、この系統が、以下の工程、すなわち、
(a)上記系統を一群の細胞に接種し(ここで、この細胞はCRL12219の名称を有し、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液で生育させた細胞のコンフルエントな単層から得たものであり、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液1mlあたり、1−2×105の細胞を含むものであり)、
(b)上記細胞を37°Cで60分インキュベートし、
(c)上記細胞からイノキュラムを取り除き、
(d)上記細胞をイーグル最少必須培養液で洗浄し、
(e)上記細胞を、250μg のジエチルアミノエチル(DEAE)−デキストランを加えた2×イーグル最少必須培養液ならびに1.6%煮沸ノーブル寒天5mlに再懸濁し、
(f)上記細胞を平板培養し、そして
(g)上記細胞を、さらに、37°Cで5日間、CO2インキュベーター中でインキュベートしてプラークを得る工程からなる方法によってプラークを得た場合に、平均直径が2mm未満のプラークを生じる系統であるようなワクチン。
【請求項6】
ワクチンが、生きた北米型PRRSウイルスの系統を含むものである請求項5に記載のワクチン。
【請求項7】
上記系統が、ATCC VR2509 の名称を有するものである請求項5に記載のワクチン。
【請求項8】
分離、精製された北米型PRRSウイルスの系統であって、ATCC VR2509 の名称を有する系統。
【請求項9】
上記系統が生きたウイルスである請求項8に記載の系統。
【請求項10】
分離、精製された北米型PRRSウイルスの特定の系統であって、
(a)上記系統を一群の細胞に接種し(ここで、この細胞はCRL12219の名称を有し、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液で生育させた細胞のコンフルエントな単層から得たものであり、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液1mlあたり、1−2×105の細胞を含むものであり)、
(b)上記細胞を37°Cで60分インキュベートし、
(c)上記細胞からイノキュラムを取り除き、
(d)上記細胞をイーグル最少必須培養液で洗浄し、
(e)上記細胞を、250μgのジエチルアミノエチル(DEAE)−デキストランを加えた2×イーグル最少必須培養液ならびに1.6%煮沸ノーブル寒天5mlに再懸濁し、
(f)上記細胞を平板培養し、そして
(g)上記細胞を、さらに、37°Cで5日間、CO2インキュベーター中でインキュベートしてプラークを得る 工程からなる方法によってプラークを得た場合に、平均直径が2mm未満のプラークを生じるような系統。
【請求項11】
上記系統が生きたウイルスである請求項10に記載の系統。
【請求項1】
(A)プラーククローニングによって北米型PRRSウイルスの特定の系統を得、そして
(B)工程(a)で得た系統からワクチンを調製する工程よりなるワクチンの製造方法であって、ここで、上記系統が、
(a)上記系統を一群の細胞に接種し(ここで、この細胞はCRL12219の名称を有し、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液で生育させた細胞のコンフルエントな単層から得たものであり、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液1mlあたり、1−2×105の細胞を含むものであり)、
(b)上記細胞を37°Cで60分インキュベートし、
(c)上記細胞からイノキュラムを取り除き、
(d)上記細胞をイーグル最少必須培養液で洗浄し、
(e)上記細胞を、250μg のジエチルアミノエチル(DEAE)−デキストランを加えた2×イーグル最少必須培養液ならびに1.6%煮沸ノーブル寒天5mlに再懸濁し、
(f)上記細胞を平板培養し、そして
(g)上記細胞を、さらに、37°Cで5日間、CO2インキュベーター中でインキュベートしてプラークを得る工程からなる方法によってプラークを得た場合に、平均直径が2mm未満のプラークを生じる系統であるようなワクチンの製造方法。
【請求項2】
ワクチンが、生きた北米型PRRSウイルスの系統を含むものである請求項1に記載の方法。
【請求項3】
ワクチンの製造方法が、上記系統を精製形態で得る工程を含むものである請求項1に記載の方法。
【請求項4】
上記系統が、ATCC VR2509 の名称を有するものである請求項1に記載の方法。
【請求項5】
北米型PRRSウイルスの特定の系統を含むワクチンであって、この系統が、以下の工程、すなわち、
(a)上記系統を一群の細胞に接種し(ここで、この細胞はCRL12219の名称を有し、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液で生育させた細胞のコンフルエントな単層から得たものであり、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液1mlあたり、1−2×105の細胞を含むものであり)、
(b)上記細胞を37°Cで60分インキュベートし、
(c)上記細胞からイノキュラムを取り除き、
(d)上記細胞をイーグル最少必須培養液で洗浄し、
(e)上記細胞を、250μg のジエチルアミノエチル(DEAE)−デキストランを加えた2×イーグル最少必須培養液ならびに1.6%煮沸ノーブル寒天5mlに再懸濁し、
(f)上記細胞を平板培養し、そして
(g)上記細胞を、さらに、37°Cで5日間、CO2インキュベーター中でインキュベートしてプラークを得る工程からなる方法によってプラークを得た場合に、平均直径が2mm未満のプラークを生じる系統であるようなワクチン。
【請求項6】
ワクチンが、生きた北米型PRRSウイルスの系統を含むものである請求項5に記載のワクチン。
【請求項7】
上記系統が、ATCC VR2509 の名称を有するものである請求項5に記載のワクチン。
【請求項8】
分離、精製された北米型PRRSウイルスの系統であって、ATCC VR2509 の名称を有する系統。
【請求項9】
上記系統が生きたウイルスである請求項8に記載の系統。
【請求項10】
分離、精製された北米型PRRSウイルスの特定の系統であって、
(a)上記系統を一群の細胞に接種し(ここで、この細胞はCRL12219の名称を有し、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液で生育させた細胞のコンフルエントな単層から得たものであり、この細胞は、3%のウシ胎仔血清ならびに0.15%の炭酸ナトリウムを加えたイーグル最少必須培養液1mlあたり、1−2×105の細胞を含むものであり)、
(b)上記細胞を37°Cで60分インキュベートし、
(c)上記細胞からイノキュラムを取り除き、
(d)上記細胞をイーグル最少必須培養液で洗浄し、
(e)上記細胞を、250μgのジエチルアミノエチル(DEAE)−デキストランを加えた2×イーグル最少必須培養液ならびに1.6%煮沸ノーブル寒天5mlに再懸濁し、
(f)上記細胞を平板培養し、そして
(g)上記細胞を、さらに、37°Cで5日間、CO2インキュベーター中でインキュベートしてプラークを得る 工程からなる方法によってプラークを得た場合に、平均直径が2mm未満のプラークを生じるような系統。
【請求項11】
上記系統が生きたウイルスである請求項10に記載の系統。
【図1】


【図2】

【図3】
【図2】
【図3】
【公開番号】特開2006−306894(P2006−306894A)
【公開日】平成18年11月9日(2006.11.9)
【国際特許分類】
【出願番号】特願2006−211493(P2006−211493)
【出願日】平成18年8月2日(2006.8.2)
【分割の表示】特願平9−503926の分割
【原出願日】平成8年6月19日(1996.6.19)
【出願人】(503411037)リージェンツ オブ ザ ユニバーシティ オブ ミネソタ (2)
【Fターム(参考)】
【公開日】平成18年11月9日(2006.11.9)
【国際特許分類】
【出願日】平成18年8月2日(2006.8.2)
【分割の表示】特願平9−503926の分割
【原出願日】平成8年6月19日(1996.6.19)
【出願人】(503411037)リージェンツ オブ ザ ユニバーシティ オブ ミネソタ (2)
【Fターム(参考)】
[ Back to top ]