
病的免疫応答を抑制するサイトカインおよびマイトジェンの使用
【課題】抗体仲介および細胞仲介の両方の異常を含む自己免疫異常を処置する方法の提供。
【解決手段】生体外の末梢血単核細胞(PBMC)中の免疫応答を阻害する方法であって、該方法は、患者由来の末梢血単核細胞(PBMC)を患者から取り出すこと、およびこの細胞をTGF−およびIL−2を含む調節組成物で、免疫細胞による組織損傷を抑制するのに十分な時間処理した後、該処理済細胞を患者に再導入することで、これにより自己免疫の症状が改善する。
【解決手段】生体外の末梢血単核細胞(PBMC)中の免疫応答を阻害する方法であって、該方法は、患者由来の末梢血単核細胞(PBMC)を患者から取り出すこと、およびこの細胞をTGF−およびIL−2を含む調節組成物で、免疫細胞による組織損傷を抑制するのに十分な時間処理した後、該処理済細胞を患者に再導入することで、これにより自己免疫の症状が改善する。
【発明の詳細な説明】
【技術分野】
【0001】
(技術分野)
本発明は一般的に、抗体仲介および細胞仲介の両方の異常を含む自己免疫異常を処置する方法に関する。
【背景技術】
【0002】
(背景技術)
自己免疫疾患は、自己か自己でないかを識別する免疫システムの機能不全によっておこる。これらの疾患で、免疫系は自己組織に対して作用し、その応答は最終的に炎症や組織損傷をもたらす。自己免疫疾患は2つの基本的部類に分類される:全身性エリテマトーデス(SLE)、尋常性天疱瘡、重症筋無力症、溶血性貧血、血小板減少性紫斑病、グレーブス病、シェーグレン症および皮膚筋炎などの抗体仲介疾患と、橋本病、多発性筋炎、炎症性腸疾患、多発性硬化症、真性糖尿病、慢性関節リウマチおよび硬皮症などの細胞仲介疾患である。
【0003】
多くの自己免疫疾患において、組織損傷は元の組織に対する抗体の産生によって引き起こされる。この抗体は自己抗体と言われている。哺乳動物によって産生され、哺乳動物自体の組織に対する結合部位を持つためである。この疾患のうちいくつかは、循環する抗体量の特有な漸増と漸減を有し、やがて徴候に変化をもたらす。
【0004】
抗体仲介自己免疫異常の種々のタイプの中でもSLEは充分研究され考証されている異常である。SLEは一般化した自己免疫の異常であり、核、細胞質および細胞表面の抗原に対する様々な自己抗体でのB細胞の活動過多を特徴とする。この自己免疫疾患は、遺伝的および環境的な促進因子による多因性病原を有する(参照:Hahn, B.H., Dobois' Lupus Erythematosus, 5th Ed. (1997), pp. 69−76 (D.J. Wallace et al. ads., Williams and Wilkins, Baltimore))。SLEで説明されている多数のリンパ球欠失は、調節性T細胞の機能不全であり、B細胞の機能を阻害する(Horwitz, D.A., Dubois' Lupus Erythematosus, 5th Ed. (1997), pp. 155−194 (D.J. Wallace et al. eds., Williams and Wilkins, Baltimore))。インビトロでポリクローナルIgGおよび自己抗体を持続的に産生するには、T細胞の助力が必要である(Shivakumar, S. et al. (1989), J Immunel 143:103−112)。
【0005】
調節性T細胞は、胞溶解性またはサイトカイン仲介メカニズムによる抗体合成を下方制御する。後者はトランスフォーミング成長因子−β(TGF−β)と他の抑制的サイトカインを必要とする(Wahl, S.M. (1994), J Exp Med 180:1587−190)。 循環Bリンパ球自発的分泌抗体は、活動性SLEの患者で増加する(Klinman, D.M. et al. (1991), Arthritis Rheum 34:1404−1410)。
【0006】
SLEの臨床症状は、発疹(特に、顔面の蝶型紅斑)、糸球体腎炎、肋膜炎、心膜炎および中枢神経系関与がある。患者の大部分は女性であり、比較的若い(診断時の平均年齢は29才である)。
【0007】
SLEに対する処置は臨床的症状によって変わる。穏やかな臨床症状の患者は、非ステロイド性抗炎症剤等の簡単な処置で効き目がでる。しかし、さらに重い症状に対しては、通常、プレドニゾンなどの強い抗炎症および免疫抑制の作用をもつステロイドが必要となる。他の強力な免疫抑制剤として、アザチオプリンとシクロホスファミドを使用することができる。ステロイドおよび他の免疫抑制剤では、哺乳動物の免疫システムが全体的に低下するために副作用が生じる。現在、SLEに対する理想的な処置はなく、疾患を治すことができない。
【0008】
現在、SLEに対する感受性または抵抗性を高める遺伝子の同定、疾患を引き起こす抗原性決定子の同定、生存かアポトーシスかを決定づけるT細胞活性化の分子メカニズム、T細胞の機能を決定するサイトカイン、および自己抗体を形成するB細胞の特性が、かなり注目されている。SLEのT細胞調節不全について多くの例証が叙述されている(参照:Horwitz, D.A. et al., Duobois’Lupus Erythematosus, 5th Ed. (1997), pp. 83−96 (D.J. Wallace et al. ads., Wiliams and Wilkins, Baltimore)。あるリンパ球の一次的役割が免疫応答の下方調節であることは充分認められているが、これらの細胞の産生で必要とされる同一性およびメカニズムの解明は進んでいない。
【0009】
インターロイキン−2(IL−2)は、これまで抗原非特異的T抑制細胞の産生で重要な役割を果たすと考えられている。マウスに与えられる抗IL−2抗体(移植片対宿主病の誘導物と一致する)はSLEの特色をもたらす(Via, C.S. et al. (1993), Intemational Immunol. 5:565−572)。IL−2が抑制の産生において直接的または間接的に重要であるかどうかが議論されている(Fast, L.D. (1992), J. Immunol. 149:1510−1515; Hirohata, S. et al. (1989), J. Immunol. 142.3104−3112; Baylor, C.E. (1992), Advances Exp. Med. Biol. 319:125−135)。最近、IL−2が、CD8+細胞を誘発して、CD4+T細胞中のHIV複製を非分解メカニズムにより抑制することがわかった。この効果はサイトカイン仲介性であるが、具体的なサイトカインは同定されていない(Kinter, A.L. et a1. Proc. Nalt Acad. Sci. USA 92:10985−10989; Barker, T.D. et al. (1996), J. Immunol. 156.4478−4483)。IL−2のT細胞産生はSLEで減少する(Horwitz, D.A. et al. (1997), Dubois' Lupus Erythematosus, 5th Ed. (1997), pp. 83−96, D.J. Wallace et al. eds., Williams and Wilkins, Baltimore)。
【0010】
SLE患者由来のCD8+T細胞は、ポリクローナルIgG産生を抑制するよりむしろ維持する(Linker−Israeli, M. et al. (1990), Arthritis Rheum. 33:1216−1225)。健康なドナーのCD8+T細胞を刺激して抗体産生を高めることができる(Takahashi, T. et al. (1991), Clin. Immunol. Immunopath. 58:352−365)。しかしIL−2またはCD4+T細胞のどちらも、それ自体では強力な抑制活性を発生させるCD8+T細胞を産出しないことがわかった。NK細胞を培地に含めると、強力な抑制活性が現われた(Gray, J.D. et al. (1994) J. Exp. Med. 180:1937−1942)。培地中のNK細胞の働きは、トランスフォーミング成長因子ベータ(TGF−β)を活性形態で産出するものと考えられる。非免疫抑制(2−10pg/ml)濃度のこのサイトカインは、IgGおよびIgMの産生に対する強力な抑制作用をつくるための補助因子として働くことが見出された(Gray, J.D. et al (1994) J. Exp. Med 180:1937−1942)。加えて、NK細胞は非刺激リンパ球でTGF−βの主要源であると考えられる(Gray, J.D. etaL (1998), J. Immunol. 160:2248−2254)。
【0011】
TGF−βは、組織修復、炎症および免疫調節における重要なサイトカインの多機能性系統群である(Massague, J. (198O), Ann. Rev. Cell Biol. 6:597)。TGF−βは、他の大部分のサイトカインと異なり、その放出タンパク質が生物学的に不活性であり、特定の受容体と結合することができない(Sporn, M.B. et al. (1987) J. Cell Biol. 105:1039−1045)。潜在の複合体が細胞外で切断され、以下に記すように活性サイトカインを放出する。TGF−βの応答は単核細胞に偏在する2つの表面受容体(TGF−β−R1とTGF−β−R2)の相互作用を必要とする(Massague, J. (1992), Cell 69:1067−1070)。従って、潜在型の活性TGF−βへの変化は、サイトカインの生物学的効果を決定する重大な段階である。
【0012】
SLE患者ではNK細胞によるTGF−βの産生が減少することが分かっている。NK細胞により産生される構成TGF−βの欠失および誘導されたTGF−βの欠失は、38人のSLE患者についての研究で報告されている(Ohtsuka, K. et al. (1998), J. Immunol. 160.2539−2545)。組換え型IL−2やTNF−αまたはIL−10拮抗エフェクターの何れを加えても、SLEでのTGF−β欠失を正常し得なかった。SLEでのTGF−β産生の減少は、疾病の活性と相関しておらず、従って、初期の欠失かも知れない。
【0013】
SLE患者をTGF−β、IL−2またはその両方の組合せで処置する全身的な投与は、深刻な副作用を引き起こす。これらのサイトカインは、種々の体組織に対して非常に多くの影響を与えるので、患者に全身的に送達するのはあまり安全ではない。従って、本発明の対象は、自己抗体調節制御に関与する哺乳類細胞を処置して、自己抗体産生を下方制御する細胞集団を増加させる方法およびキットを提供することである。
【発明の開示】
【発明が解決しようとする課題】
【0014】
(本発明の要旨)
ここに概略する目的のように、本発明は、生体外の末梢血単核細胞 (PBMC)のサンプル中の免疫応答を阻害する方法であって、細胞集団に調節組成物を添加することを含む方法を提供する。
【課題を解決するための手段】
【0015】
更なる態様では、本発明は、患者の自己免疫異常を処置する方法を提供する。その方法は、患者由来の末梢血単核細胞(PBMC)を患者から取り出すこと、およびこの細胞を調節組成物で、免疫細胞による組織損傷を抑制するのに十分な時間、処理することを含む。特に、本発明の方法は、抗体産生を抑制し、または細胞を誘発して、抗体産生を下方調節し、そして抗体仲介自己免疫疾患の患者における細胞仲介免疫応答を高める。次いで、細胞を患者に再導入すると、自己免疫の症状が改善する。調節組成物は、好ましくは、TGF−βおよびT細胞のTGF−βヘの応答を可能にする物質を含む。
【0016】
更なる態様では、本発明は、細胞仲介自己免疫異常を処置する方法を提供する。この方法は、該患者から末梢血単球細胞(PBMC)を取り出し、該細胞を調節組成物で、異常の免疫応答を抑制するのに十分な時間処置することを含む。処置細胞を患者に再導入すると、自己免疫症状の改善が得られる。調節組成物は、好ましくは、TGF−βおよびT細胞のTGF−βへの応答を可能にする物質を含む。
【0017】
更なる態様では、本発明は、患者の自己免疫異常の処置用キットを提供する。このキットは、抗体仲介自己免疫異常または細胞仲介異常の患者の細胞を受容するのに適当な細胞処置容器および少なくとも1用量の調節組成物を含む。
【0018】
(図面の説明)
図1は、SLE患者のPBMCをIL−2およびTGF−βと共にインキュベーションすることにより、自発的免疫グロブリン産生が減少することを示す。PBMC(2×105/well)を、AIM−V血清を含まない培地であって、IL−2(10U/ml)およびTGF−β(10pg/ml)を含むか、または含まない培地中で培養した。3日後、ウェルを3回洗浄し、新たなAIM−V培地を加えた。更に7日後、上清をウェルから回収し、IgG成分をELISAで測定した。
【0019】
図2は、IL−2およびTGF−βの両方により、自発的IgG産生が有意に減少することを示す。その値は、図1の説明に記載の通りに培養した12人のSLE患者のPBMCにより産生した平均±SEMのIgG(μg/ml)を示す。ただし、IL−2(10U/ml)のみ、またはTGF−β(10pg/ml)のみで培養した細胞もある。
【0020】
図3Aおよび3Bは、抗TGF−βがIL−2の効果を逆転することを示す。SLE患者のPBMCを、3日間、IL−2(10U/ml)の存在下(黒色バー)、または非存在下(灰色バー)で培養した。これらの培養液中に含まれていたのは、培地、抗TGF−β(10μg/ml)または対照マウスIgG1(10μg/ml)であった。3日後、ウェルを洗浄し、新たなAIM−V培地を加えた。更に7日後、上清を回収し、ELISAでIgG(図3A)または抗核タンパク質(NP)(図3B)成分を検定した。
【0021】
図4A、4Bおよび4Cは、抗体産生に対するDC8+T細胞の調節効果を示す。(A)健康人のIgG産生の抑制におけるNK細胞とCD8+細胞の相乗作用。CD4+細胞およびB細胞を抗CD2で刺激し、CD8+細胞およびNK細胞の作用を調べた。NKおよびCD8+細胞の組み合わせは、我々が以前報告した(Gray, J.D. et al., (1998), J Immunol 160: 2248−2254; Gray, J.D. et al., (1994), J Exp Med 180; 1937−1942)抗CD2誘発性IgG産生を顕著に阻害した。(B)NK細胞およびCD8+細胞はSLEにおけるIgG合成を促進する。活動性SLEの患者由来のCD4+細胞および正常人の休止B細胞を、抗CD2で刺激した。SLE CD8+細胞によるIgG産生の促進は、NK細胞の添加により顕著に増加した。(C)SLEにおけるCD8+T細胞機能のサイトカインによる正常化。図4Bに示す試験と平行して、この患者のCD4+T細胞を、CD8+T細胞の存在下または非存在下、抗CD2で刺激した。表示のようにIL−2(10U/ml)および/またはTGF−β(2pg/ml)を加えた。これらサイトカインにより、CD8+細胞のヘルパー作用が喪失し、その細胞がIgG産生の75%を阻害した。
【0022】
図5Aおよび5Bは、非刺激細胞および抗CD2刺激細胞によるTGF−β1のリンパ球産生を示す。健常者ドナーならびにSLE患者およびRA患者のPBLを、マイクロタイタープレートに1×105/ウェルで加えた。幾つかのウェルは、抗CD2mAbsGT2(1:40)およびT11(1:80)であった。37℃で2日後、上清を回収し、活性TGF−β1および全TGF−β1を検定した。有意なp値を示す。
【0023】
図6は、TNF−αおよびIL−10のT細胞産生に対するTGF−βの作用を示す。血清なしのAIMV培地中の精製T細胞(1x105細胞/ウエル)を、平底ミクロウエルに加え、低量(0.5μg/ml)または高量(5μg/ml)のConAで、IL−2と共にまたはなしに、TGF−β(1ng/ml)の存在または不存在下に刺激した。上清を2日および5日に採取し、TNF−αおよびIL−10についてELISAで調べた。最大産生がTGF−βは2日に、IL−10は10日にあった。TGF−βはIL−10の産生をなくし、TNF−αの産生を上方調節した。
【0024】
図7は、TGF−βによる調節性T細胞の生成にTNF−αが必須の中間体であることを示す。精製CD8+細胞を、ConA(2.5μg/ml)、IL−2(10U)およびTGF−β(10pg/ml)で一夜インキュベートした。洗浄後これらの細胞をCD4+B細胞に加え、抗CD2で刺激した。いくつかのウエルに抗TNF−α抗体(10μg/ml)またはイソ型対照抗体(10μg/ml)を入れた。7日後に上清をIgG含量についてELISAで調べた。抗TNF−αは条件CD8+細胞の調節活性を逆転した。
【0025】
図8は、TGF−β起動T細胞によるTh1サイトカインの産生の増加がTNF−αに依存性であることを示す。精製ナイーブT細胞をConA(5μg/ml)およびIL−2(10U/ml)とともにTGF−β(1ng/ml)の存在下に培養した。いくつかのウエルには中和抗TNF−α抗体(10μg/ml)またはイソ型対照抗体(10μg/ml)も加えた。5日培養後に細胞を水洗し、1x105細胞/ウエルで新鮮な培地に入れた。翌日にConAおよびIL−2で6時間再刺激し、ブレフェルジンA(10μg/ml)の存在下にCD8およびサイトカインについて着色した。TNF−α、IL−2、IFN−γを発現するCD8+およびCD8−細胞の%を示す。一次培地におけるTNFの中和はTh1サイトカイン産生に対するTGF−β作用の増加を無くする。
【0026】
図9は、細胞毒性T細胞活性の抑制物質の産生におけるTGF−βの作用を示す。Eロゼット法によりつくられたドナーA由来のT細胞を2つに分けた。1つを同種異系混合リンパ球反応(allo−MLR)についての応答物として用いた。他の1つを用いて、細胞を適当なモノクロナール抗体で染め、その細胞を免疫磁性ビーズで除いた後、陰性選択で表示されるT細胞サブセットをつくった。応答T細胞をドナーB由来の刺激細胞と混合し(照射T細胞は末梢血単核細胞を枯渇した)、5日培養してキラー細胞を作った。対照は刺激細胞の有無で5日培養したT細胞サブセットからなる。その後、細胞を洗い、計数し、アロ細胞毒性T細胞活性の検定に使用した。ドナーA由来の応答細胞をドナーB由来のクロミウム標識リンパ芽球とエフェクター質中で混合し、細胞比率を標的にして、クロミウム放出を標準的4時間検定法で測定した(白四角)。刺激物とともに培養したT細胞サブセットを、4応答細胞につき1調節細胞の比率で加えた(白丸)。刺激物と培養のT細胞サブセットへのTGF−βの添加を黒丸で示す。すべての試験で、TGF−βの最大作用はナイーブCD4CD45RA+CD45RO−細胞に対して見られた。
【0027】
図10は、TGF−βにより起動されたCD4細胞のアロ細胞毒性Tリンパ球(CTL)活性に対する作用を示す。刺激物質なしで5日間培養されたCD4CD45RAの付加はCTL活性に対する作用を有しない(結果を示していない)。これらのT細胞を刺激物質と培養すると、低度から中程度の抑制活性となる。すべての試験において、これらのT細胞のTGF−β1ng/mlとの培養は、アロCTL活性を顕著に抑制するか無くする。
【0028】
図11は、調節性T細胞がCTL活性を抑制するために細胞接触を必要とすることを示す。調節性CD4細胞は、上記のように、TGF−βと培養したCD4CD45RAからつくった。これらの細胞のいくつかを応答物質およびクロミウム標識の標的細胞と混合し、他の細胞を膜によりキラー細胞から分離した。細胞毒性Tリンパ球活性(CTL)の抑制を、T細胞がキラー細胞と直接接触したときにのみ認めた。
【0029】
図12は、TGF−βで誘発された調節性CD4+T細胞によるリンパ球増殖の抑制を示す。ドナーA由来のナイーブCD4+T細胞を上記のように刺激細胞と混合し、表示の比率で新鮮な応答物質および刺激物質に加えた。バーは7日間培養後のトリチウム標識チミジンの取り込み±SEMを示す。薄色のバー(Nil)は、添加CD4+細胞なしの応答T細胞の増殖性応答を示す。濃色バーは、刺激細胞と培養した対照CD4+細胞のTGF−βなしでの作用を示す。黒バーは、刺激細胞と培養した対照CD4+細胞のTGF−βあり(1ng/ml)での作用を示す。照射刺激細胞に加えた新鮮な応答細胞の増殖性応答に対するこれらのCD4+細胞の作用を培養7日間後に示す。バーはトリチウム標識チミジンの平均取り込みを示す。
【0030】
図13は、CD25+CD4T細胞の調節活性を示す。CD4+細胞を照射同種異系非T細胞±TGF−β(1ng/ml)で5日間刺激した。水洗後、CD4+細胞をDIIで染め、新鮮な応答T細胞をカルボキシフルオレセイン(CFSE)で染めた。対照またはTGF−β駆動CD+細胞を応答T細胞およびアロ刺激細胞に1:4の比率で加えた。5日後に細胞を採取し、フローサイトメトリーにより分析した。CD8+におけるCFSEの強さをDII陰性細胞でもって決定した。TGF−β駆動CD+細胞の応答T細胞への付加はCD8+細胞による細胞分裂を顕著に低下した。
【0031】
図14は、調節CD+4細胞がその表面でCD25+(IL−2)受容体を発現することを示す。対照またはTGF−β誘発CD4+調節性T細胞を上記のようにつくった。アロ刺激細胞およびTGF−βで条件を整えた後、CD4+細胞を細胞選別によりCD25+とCD25−に分けて、新鮮な応答T細胞および照射刺激細胞に加えた。これらの応答細胞のキラー刺激Tリンパ芽細胞に対する能力を、標準的4時間クロミウム放出検定で示す。
【0032】
図14Aにおいて、白四角は添付のCD4+細胞のないCTL活性を表示する。対照またはTGF−β誘発調節性T細胞を応答細胞とともに1:4の比率で加えた。白丸からすると対照CD4+細胞がCTL活性を変えなかった。黒丸からするとTGF−βが誘発CD4+がCTL活性をほとんど完全に抑制した。黒菱形からするとCD25+サブセットのみが抑制活性を有していた。CD25サブセット(黒四角)は抑制活性を有していなかった。
【0033】
図14Bは、MLRに加えたCD4+調節細胞の数を減少さす作用を示す。わずか3%への減少は抑制作用の減少における最小の作用である。
【0034】
図15は、少量のブドウ球菌性エンテロトキシンB(SEB)でのT細胞の反復刺激がT細胞を誘発してTGF−βの免疫抑制レベルをつくることを示す。CD4+T細胞を、SEB(0.01ng/ml)およびスパー抗原提示細胞として照射B細胞で、TGF−βの存在または不存在で、矢印で表示の時に刺激した。活性TGF−βを2日または5日後に測定した。
【0035】
図16は、低量のSEBでCD4+T細胞の反復刺激によってT細胞がTGF−βの免疫抑制レベルをつくることを示す。CD4+T細胞をSEB(0.01ng/ml)およびスパー抗原提示細胞として照射B細胞で、TGF−βの存在または不存在で、矢印で表示の時に刺激した。活性TGF−βを2日または5日後に測定した。
【0036】
図17は、ナイーブ(CD45RA+CD45RO−)CD4+およびCD8+T細胞に対するSEBの作用を示す。細胞をSEBで5日毎に計3回刺激した。各T細胞サブセットおよびCD25IL−2受容体活性マーカーを発現する細胞の%を各刺激後に測定した。パネルAおよびCは、TGF−β1ng/mlが最初の刺激に含まれていると、CD4+T細胞が反復刺激後に培養基中で優勢になることを示す。パネルBおよびCは、SEB刺激細胞によるCD25発現が対照培養基中で第3刺激で減少することを示す。しかしCD25発現は、T細胞がTGF−βで駆動されていると、高く留まっている。
【0037】
(詳細な説明)
本発明は、全身性エリマトーデス(SLE)などの細胞仲介および抗体仲介の両方の異常を含む自己免疫異常を処置する方法に関する。この方法は、患者から細胞を取り出し、この両方の1つに作用できる組成物でもって細胞を処置する。1つの実施態様において、抗体仲介自己免疫異常の症状を本発明の組成物によって改善する。この組成物は、自己抗体などの抗体の産生を阻害して、B細胞の過剰活性を下方調節する。さらに、この組成物は、SLEや他のある種の抗体仲介自己免疫異常の患者においてしばしば欠失している細胞仲介免疫応答を高める。すなわち、抗体仲介自己免疫異常の患者を処置して、その欠失細胞仲介症状を改善できる。
【0038】
あるいは、この組成物を使用して細胞仲介自己免疫疾患を処置する。この実施態様において、組成物は免疫細胞を誘発し、抑制T細胞をつくる。この抑制T細胞は他のT細胞が細胞毒性になって個体の細胞および組織を攻撃するのを防ぐ。このように、組成物は細胞毒性を低下することにより細胞仲介自己免疫異常の症状を改善する。
【0039】
この方策は、現在のほとんどすべての処置が抗炎症剤や免疫抑制剤を使用すると異なっている。普通よく用いられるコルチコステロイドは、サイトカインの産生を抑制し、組織の損傷を起こす最終事象を阻止するが、基にある自己免疫応答を一般に変えることはない。細胞障害剤またはモノクローナル抗体などの実験的に遺伝子工学的に処理された生物剤も、特殊なリンパ球集団をなくするか、その機能を妨害する。これらの薬剤は、一般に緩和な効果をもたらすのみであって、また重大な有害副作用を起こす。ある種のサイトカインは、患者に全身的に与えられるが、重大な有害副作用に関連する広範な作用を有している。
【0040】
一方、本発明の方法による治癒効果では、正常な調節性細胞機能が回復し、免疫系が「再置」される。本方法の他の重要な利点は、重大な有害副作用があまり起きないことである。サイトカインなどの調節組成物のごく少量を患者に戻すにすぎないので、毒性が最小となる。
【0041】
IgGを自発的に分泌する循環Bリンパ球が活動性SLEの患者で増加する(Blaese, R.M., et al.(1980), Am J. Med. 69: 345−350; Klinman, D.M. et al.(1991) Arthritis Rheum 34: 1404−1410)。インビトロでのポリクローナルIgGおよび自己抗体の持続的な産生には、T細胞の助けを必要とする(Shivakumar, S. et al.(1989), J Immunol 143: 103−112)。自発的IgG産生のT細胞による調節についての以前の研究によると、健常人ではCD8+T細胞が抗体産生を阻害するのに対し、SLEではこの細胞が代りにB細胞機能を支持する(Linker−Israeli, M. et al.(1990), Arthritis Rheum 33: 1216−1225)。他の自己免疫疾患、例えば、慢性関節リウマチや多発性硬化症において、抗体よりもむしろT細胞が組織損傷の原因であり炎症をもたらす(Panayl GS et al. Anthritis Rheum (1992) 35: 725−775, Allegretta M et al. Science (1990) 247: 718−722)。
【0042】
従って、本発明は、抗体−およびT細胞−仲介自己免疫疾患を処置する方法に関し、自己免疫異常の患者から末梢血単核細胞(PBMC)を取りだし、その細胞を調節組成物で処置する。
【0043】
理論に拘束されるわけでないが、本発明方法が作動するには、いくつかの経路があるようである。その第1は、調節組成物で細胞を処置すると、処置細胞における抗体産生が直接抑制され、自己免疫症状の改善につながり得る。あるいは、または加えて、細胞の処置は調節性細胞を誘導して、他の細胞における抗体産生を下方調節する。本明細書における抗体はすべての形態の抗体、例えば、IgA、IgM、IgG、IgEなどを含む。明らかな結果は系における抗体量の低下である。
【0044】
さらに、細胞の処置は、抗体仲介自己免疫症状の患者において細胞仲介免疫応答を高める。理論に縛られるものでないが、細胞の処置がIL−10とTNF−αとの均衡を修復して、Th1サイトカインの産生を高め、細胞仲介免疫を正常化するようである。
【0045】
さらに、TGF−βを含む調節組成物で免疫細胞を刺激すると、細胞仲介免疫応答が抑制される。理論に縛られるものでないが、CD4+T細胞を刺激すると、活性TGF−βの免疫抑制レベルをつくり、細胞仲介免疫応答を抑制するようである。あるいは、CD4+T細胞を刺激すると、他のT細胞の活性化および/またはエフェクター機能を作用の接触依存性メカニズムによって抑制する。これらの作用には、TGF−βの存在で活性化されるCD4+細胞を必要とする。
【0046】
このように、本発明は、異常な免疫応答を阻害する。抗体仲介自己免疫異常の患者において、本発明は、調節組成物で末梢血T細胞を生体外で処置することにより、T細胞の能力を修復し、抗体産生を下方調節し、細胞仲介応答を修復する。細胞仲介異常の患者において本発明は、他のT細胞において細胞毒性T細胞活性を抑制する調節性T細胞をつくる。
【0047】
「免疫応答」とは、外部または自己の抗原に対する宿主の応答を意味する。「異常免疫応答」とは、自己と非自己とを識別する免疫系のないこと、または外部抗原に応答しないことを意味する。換言すると、異常免疫応答とは、患者の症状をもたらしている不適当に調節された免疫応答である。「不適当に調節された」とは、不適当に誘発されること、不適当に抑制されること、および/または応答性のないことを意味する。異常な免疫応答は、限定でないが、生体自身の組織に対する抗体の産生により生じる組織損傷や炎症、IL−2、TNF−α、IFN−γの産生の障害、作用の細胞毒性または非毒性メカニズムにより生じる組織障害を含む。
【0048】
従って、好ましい実施態様において、本発明は患者の抗体仲介自己免疫異常を処置する方法を提供する。「抗体仲介自己免疫疾患」とは、個体が自己の細胞または組織の構成成分に対する抗体を発生する疾患を意味する。抗体仲介自己免疫疾患には、限定するのではないが、全身性エリマトーデス(SLE)、天疱瘡、重症筋無力症、溶血性貧血、血小板減少性紫斑症、グレーブス症、皮膚真菌症、シェーグレン症などがある。本発明方法による処置に好適な疾患はSLEである。
【0049】
さらに、抗体仲介異常の患者は細胞仲介免疫応答において欠失をしばしば有する。「細胞仲介免疫応答の欠失」とは、感染に対する宿主防御の障害を意味する。感染に対する宿主防御の障害は、限定でないが、遅い過剰反応の障害、T細胞の細胞毒性の障害、TGF−β産生の障害を含む。他の欠失は、限定でないが、IL−10産生の増加、およびIL−2、TNF−α、IFN−γの産生の減少を含む。本発明の方法を用いて、精製T細胞を刺激して、IL−2、TNF−α、IFN−γの産生を増加させ、IL−10の産生を低下させる。この方法で刺激し得るT細胞は、限定でないが、CD4+およびCD8+を含む。
【0050】
ある実施態様においては、抗体仲介異常を処置しない。
好ましい実施態様において、本発明は、患者における細胞仲介自己免疫異常を処置する方法を提供する。「細胞仲介自己免疫疾患」とは、個体の細胞が活性化または刺激されて、細胞毒性となり、自己の細胞または組織を攻撃する疾患を意味する。あるいは、個体の自己免疫細胞が他の細胞を刺激して作用の細胞毒性または非細胞毒性メカニズムによって組織障害を起こす。細胞仲介自己免疫疾患は、限定でないが、橋本病、多発性筋炎、炎症性腸疾患、多発性硬化症、真性糖尿病、慢性関節リウマチおよび硬皮症などを含む。
【0051】
自己免疫異常の「処置」とは、自己免疫異常の少なくとも1つの症状が、本明細書に記載の方法で改善されることである。その評価を種々の方法でなす。この評価には、患者側の主観的または客観的な因子が含まれる。例えば、疾患の免疫原性徴候を調べる。例えば、自発的抗体および自己抗体産生のレベル、SLEの場合は特にIgG産生のレベルを低下する。全Ig抗体または自己抗体のレベルを測定する。限定するものでないが、抗2本鎖DNA(dsDNA)抗体、抗核タンパク質抗体、抗Sm、抗Rhoおよび抗Laである。身体症状が変化する。例えば、SLEの皮疹の消失または減少である。腎機能検査を行って変化を調べる。炎症に関する組織障害について臨床検査を行う。循環免疫複合体レベルおよび血清補体レベルの低下も改善の証拠である。SLEの場合、貧血の減少も見られる。免疫抑制剤などの薬剤に対する患者の必要性が低下することも処置効果の証である。処置効果について他の判定方法は、自己免疫疾患分野の当業者にとって明らかであろう。
【0052】
「患者」とは、処置の対象となる哺乳動物を意味するが、ヒトが好ましい。ある場合において、本発明方法は、実験動物、獣医学分野、疾患モデル動物にも使用され、これらの動物には、限定ではないが、マウスラット、ハムスターなどのゲッシ類および霊長類がある。
【0053】
本発明では、患者から血液細胞を取り出す。一般に、標準的な方法で患者から末梢血単核細胞(PBMC)を取り出す。「末梢血単核細胞」すなわち「PBMC」とは、リンパ球(T細胞、B細胞、NK細胞など)および単球を意味する。詳しく下記するように、調節組成物の主要な作用は、CD8+T細胞がIG産生を抑制することである。好ましくは、PBMCのみを採取する。赤血球または多形核白血球を患者に残すようにするか、これらを患者にもどす。これは既知の方法、例えば、白血球泳動(leukophoresis)で行う。一般に5から7リットルの白血球泳動工程を行う。患者からPBMCを基本的に採取し、他の血液成分を戻す。細胞サンプルの採取は、ヘパリンなど抗凝固剤の存在下で、既知方法により行う。
【0054】
いくつかの実施態様においては、白血球泳動工程を必要としない。
一般に、PBMC含有のサンプルを種々の方法であらかじめ処理する。まず細胞を採取し、次いで採取に際して自動的に濃縮されていなければ、濃縮し、さらに精製および/または濃縮する。細胞を水洗し、計数し、緩衝液中に入れる。
【0055】
一般的にPBMCを標準的方法で処置のために濃縮する。好ましい態様において、白血球泳動採取工程によってPBMCの濃縮サンプルを無菌の血球パックに得、ある種の試薬および/または調節組成物を含有せしめる。詳しくは下記する。一般に、追加の濃縮/精製工程を行なう。既知のFicoll−Hypaque密度勾配遠心法などである。
【0056】
好ましい態様において、PBMCを洗い、血清タンパク質および溶性血液成分、例えば自己抗体、阻害剤などを既知方法で取り出す。一般に、生理媒質または緩衝液を加え、遠心法を行なう。必要に応じて繰り返す。PBMCを生理培養液、好ましくはAIM−V血液なしの培地(Technologie)に再懸濁するが(血清がかなりの量の阻害剤を含有するので)、Hanksバランス塩溶液(HBBS)や生理塩緩衝液(PBS)も使用する。
【0057】
一般に、次いで細胞数を数える。一般に1×109から2×109の白血球を5−7リットルの白血球泳動工程から採取する。これらの細胞を大略200mlの緩衝液または培地に移す。
【0058】
好ましい態様において、PBMCを1以上の細胞型について富ます。例えば、PBMSをCD8+T細胞またはCD4+T細胞ついて富ます。このことは既知である(Gray et al.(1998),J.lmmunol.160:2248、出典明示により本明細書の一部とする。)。一般に、これは市販の免疫吸収カラムを用いるか、研究的方法で行なう(PBMCをナイロンウールカラムに加え、溶出の非接着細胞をCD4、CD16、CD116、CD74に対する抗体で処理し、免疫磁気ビーズで処理して、CD8+T細胞に富んだ集団を取る)。
【0059】
好ましい実施態様において、PBMCを自動化閉鎖系、例えば、Nexell Isolex 300i Magnet Cell Selection System において分離する。一般的に、これは、細胞の分離、活性化、抑制細胞機能の開発のために使用する方法の無菌性の確保および標準化の確保のために行う。
【0060】
細胞に必要な前処理を一旦ほどこしてから、細胞を調節組成物で処置する。「処置」とは、細胞を調節組成物と共に十分な時間でインキュベートし、抗体および自己抗体産生を阻害する能力が、特に患者にもどしたときに、発生するようにすることである。インキュベーションは一般に生理的温度で行なわれる。上記したように、阻害が起きるのは、処置細胞による抗体産生の直接的抑制の結果か、患者リンパ臓器における抗体産生を下方調節する調節細胞の誘導の結果である。
【0061】
「調節組成物」または「抗体産生阻害組成物」または「液素性阻害組成物」「非特異的免疫細胞阻害物質」または「特異的T細胞阻害物質」または「阻害組成物」または「抑制組成物」は、免疫応答の抑制を起こし得る組成物を意味する。この抑制は、T細胞活性化の阻害、自発的抗体および自己抗体の産生または細胞毒性の阻害を含む。一般に、これらの組成物はサイトカインである。適切な調節組成物は、限定でないが、抗CD2などのT細胞アクチベーター、例えば抗CD2抗体およびCD2リガンド、LFA−3、T細胞アクチベーターの混合物や組合せ物、例えば、コンカナバリン Concanavalin A(ConA)、ブドウ球菌エンテロトキシンB(SEB)、抗CD3、抗CD28、およびサイトカイン、例えば、IL−2、IL−4、TGF−β、TNF−αを含む。抗体抑制に好ましい調節組成物は、T細胞アクチベーター、IL−2、TGF−βの混合物である。細胞毒性の抑制に好ましい組成物はTGF−βである。
【0062】
調節組成物の濃度は、組成物の性質によって変ってくる。好ましい態様においては、TFG−βが調節組成物の成分である。「トランスフォーミング成長因子−β」すなわち「TGF−β」は、TGF−βファミリーのいずれか1つを意味する。3種のイソ型、TGF−β1、TGF−β2、TGF−β3がある。参照、Massague,J.(1980),J.Ann.Rev.Cell Biol 6:597。リンパ球および単球は、このサイトカインのβ1のイソ型である(Kehrl,J.H.et al.(1991),lnt J Cell Cloning 9:438−450)。TGF−βは、処置される哺乳動物に対して活性であるTFG−βのすべての型であり得る。ヒトでは、組換えTFG−βが現在のところ好ましい。好ましいヒトTGF−βは、Genzyme Pharmaceuticals,Farmington,MA.から購入できる。一般に、TGF−βの濃度は、細胞懸濁液の約2picogram/mlから約5nanogramであって、約10pgから約4ngが好ましく、約100pgから約2ngが特に好ましく、1ng/mlが理想的である。
【0063】
好ましい態様において、IL−2を調節組成物で用いる。IL−2は、処置される哺乳動物に対して活性であるIL−2のすべての型であり得る。ヒトでは組換えIL−2が現在のところ好ましい。組換えIL−2は Cetus,Emerville、CA.から購入できる。一般に、使用されるIL−2の濃度は、細胞懸濁液の約1Unit/mlから約100U/mlであり、約5U/mlから約25U/mlが好ましく、10U/mlが特に好ましい。好ましい態様においてIL−2は単独で用いない。
【0064】
好ましい態様において、抗CD2抗体またはCD2リガンドLFA−3などのCDアクチベーターを、調節組成物として用いる。CD2はTリンパ球により発現される細胞表面グリコタンパク質である。「CD2アクチベーター」とは、CD2情報伝達経路を開始する化合物を意味する。好ましいCD2アクチベーターには、抗CD抗体(OKT11,American Type Culture Collection,Rockville MD)が含まれる。一般に、CD2アクチベーターの濃度は、TGF−βの産生を誘導するのに十分なものである。抗CD2抗体の濃度は、約1ng/mlから約10μg/mlの範囲にあり、約10ng/mlから約100ng/mlが特に好ましい。
【0065】
いくつかの態様においては、細胞を活性化するためにマイトジェンの使用が望ましい。すなわち、多くの休止相細胞は多量のサイトカイン受容体を含有していない。ConcanavalinAなどのマイトジェンを使用すると細胞が刺激されて、サイトカイン受容体が産生し、本発明方法をさらに効果的にする。マイトジェンを用いるとき、既知方法で知られているように、その濃度範囲は1μg/mlから約10μg/mlである。さらに、既知方法のように、CoAを離す成分、例えばα−メチルマンノシッドと共に細胞を洗うのが望ましい。
【0066】
好ましい実施態様において、T細胞を、抗CD2、抗CD3、抗CD28などのマイトジェンあるいはモノクロナール抗体の組み合せ、すなわち自己抗原と補刺激物質としてのCD28やIL-2でもって刺激する。ConAもT細胞の刺激に使用する。第2培養におけるTGF−βのありなしでのT細胞の反復刺激は最大抑制活性を得るのに必要なようである。
【0067】
好ましい実施態様において、本発明は、T細胞をTGF−βで調整することを含む。このT細胞には、限定でないが、CD8+TやCD4+TおよびCD8−CD4−などの他のマイナーT細胞サブセット、NK細胞などが含まれる。これらのT細胞は、他のT細胞が細胞毒性エフェクター細胞になるのを防ぐ。
【0068】
好ましい実施態様において、本発明は、CD8+TまたはCD4+T細胞をTGF−βで調整して、免疫抑制レベルのTGF−βをつくる方法を含む。
好ましい実施態様において、本発明は、CD8+TまたはCD4+T細胞をTGF−βで調整して、接触依存性メカニズムにより抑制されるT細胞をつくる方法を含む。
【0069】
好ましい実施態様において、本発明は、ナイーブCD4+T細胞を刺激物質で処置して、該CD4+細胞が免疫抑制レベルの活性TGF−βをつくるようにすることを含む方法を提供する。
【0070】
好ましい実施態様において、本発明は、ナイーブCD4+T細胞をTGF−βの存在下に刺激してCD4+細胞数を増やすことを含む方法を提供する。
好ましい実施態様において、本発明は、IL−10の産生を減少し、併せてTNF−αの産生を増加する方法を提供する。
【0071】
調節組成物を細胞と共に、効果を発揮するに十分な時間インキュベートする。好ましい態様において、細胞は、調節組成物での処置に続いて、患者にもどす。好ましい態様において、細胞と調節組成物とのインキュベーションは、約12から約7日行なう。この時間は望む抑制活性により変る。抗体産生の抑制については、48時間が特に好ましい。細胞毒性の抑制については、5日が特に好ましい。
【0072】
ある態様において、細胞を一定の時間インキュベートし、水洗して調節組成物を除去し、再びインキュベートする。患者に導入する前に、ここに概記したように細胞を好ましくは水洗し、調節組成物を除去する。試験または検査のために、さらなるインキュベーションも行うことができ、数時間から数日間である。患者への導入前に抗体産生の検査をしょうとするときは、細胞を数日間インキュベートすると、抗体産生(またはその非産生)が明らかになる。
【0073】
細胞の処置後すぐに、患者に自動的に再移行する前に細胞を検査することができる。例えば、細胞を採取して行う。無菌検査、グラム染色、微生物学的検査、LAL試験、マイコフラズマ検査、細胞型同定のためのフローサイトメトリー、機能検査などである。同様に、これらの検査などの試験を処置の前や後に行うことができる。
【0074】
好ましい態様において、抗体産生の定量や定性(すなわちタイプについて)をなし得る。例えば、抗体の全レベルあるいは抗体の具体的なタイプを調べる。例えば、IgA、IgG、IgM、抗DNA自己抗体、抗核タンパク質(NP)抗体などの具体的なタイプについてである。調節性T細胞も、T細胞活性を抑制する活性、インビトロで特異的な標的細胞に対するT細胞の細胞毒性を防ぐ活性について、検査できる。
【0075】
好ましい態様において、抗体、特にIgGのレベルの検査をELISAアッセイなどの既知の方法(Abo et al.(1987),Clin Exp.lmmunol.67:544 and Linker−Israeli et al.(1990),Arthritis Rheum 33:1216, 出典明示により本明細書の一部とする。)で行なう。これらの技術は自己抗体などの特異的な抗体のレベルを検出するのにも用い得る。
【0076】
好ましい態様において、処置によって産生IgGおよび自己抗体の量の有意な低下が起きる。低下は、少なくとも10%が好ましく、少なくとも25%が特に好ましく、少なくとも50%が特別に好ましい。多くの態様において75%以上の低下がみられる。
【0077】
好ましい態様において、移行前に全または活性のTGF−βの量も検査できる。記載のように、TGF−βを移行後に活性化される潜在的前駆物質としてつくる。
【0078】
処置後、細胞を患者に移行すなわち再導入する。一般に既知のよう行ない、通常は静脈投与によって処置細胞を患者に注入すなわち導入する。例えば、無菌シリンジなどの移行手段を用いて注射により50mlのFenwall注入バッグに細胞を入れる。次いで細胞をすぐに静脈投与によって一定の時間、例えば、15分間で患者の自由に流れている静脈に注入する。ある態様では、緩衝剤や塩などの追加の試薬も加え得る。
【0079】
細胞を患者に再導入後に、処置の効果を所望に応じて、上記の方法で検査できる。すなわち、疾患の免疫原性徴候を検査する。例えば、全抗体または特異的な免疫グロブリンの力価、腎機能、組織障害などを検査する。T細胞の機能についての試験、例えば、T細胞数、表現型、活性状態、抗体および/またはマイトジェンに応答する能力などについても検査し得る。
【0080】
処置は必要に応じて繰り返えす。例えば、数週の期間で週1回、または時間の期間で数回、例えば、2週間に3−5回行う。一般に、自己免疫疾患の症状の改善はなんらかの期間、好ましくは少なくとも数カ月続く。その間に患者が症状の再起を感じると、その時点で処置を繰り返えす。
【0081】
好ましい態様において、本発明方法を実行ため、すなわち細胞を調節組成物と共にインキュベートするためのキットを提供する。キットはいくつかの成分を含有する。キットは、抗体仲介自己免疫疾患の患者からの細胞を受容するのに用いられる細胞処置容器を含む。この容器は無菌とする。ある態様において、細胞処置容器を細胞採取に用いる。例えば、入口を介して白血球泳動器と共に用いる。他の態様では、分離した細胞採取容器を用い得る。
【0082】
好ましい実施態様において、キットは、細胞仲介異常の患者からの細胞を受けるように調整された細胞処置容器を含む。キットはまた、特異的T細胞サブセットを精製し、それを患者に戻すように増やすために、自動化閉鎖系での使用に調整できる。
【0083】
細胞処置容器の形および材質は様々であり、当業者によく知られている。一般に、容器はいくつかの異なる形態を取る。例えば、IVバッグのような柔かいバッグであり、または細胞培養器のような硬い容器である。攪拌できるようになっている。一般に、容器の材質は何でもよいが、生物学的に非活性の物質であり、例えばガラス、ポリプロピレンやポリエチレンなどを含むプラスチックである。細胞処置容器は1以上の入口または出口を有し得る。細胞、試薬、調節組成物などの導入または取り出しのためのものである。例えば、容器は、患者に再導入する前の分析のために細胞のフラクションを採取するためのサンプル採取口を有する。同様に、容器は、患者に細胞を導入するための出口を有する。例えば、容器はIV装置につけるためのアダプターを含み得る。
【0084】
キットは調節組成物の少なくとも1つの用量を含む。本明細書での「用量」とは、サイトカインなどの調節組成物の1つの量で、作用を発揮するのに十分な量を意味する。ある場合には、複数の用量物を含む。ある態様において、用量物の細胞処置容器への添加は入口を介して行なう。あるいは好ましい態様において、用量物は細胞処置容器にすでに存在している。好ましい態様において、用量物は安定性のために乳濁形態にあり、細胞培養液や他の試薬を用いて再生する。
【0085】
ある態様において、キットは少なくとも1種の試薬を追加的に含む。それには緩衝液、塩、培養液、タンパク質、薬剤などがある。例えば、マイトジエン、モノクローナル抗体、細胞分離のための処置磁気ビードを含み得る。
ある態様において、キットはその使用に際しての説明書を追加的に含む。
【0086】
下記の実施例は、上記の本発明をより完全に記述したものであり、また、本発明の種々の態様を実施するために考えられる最上の方法を記述した。これらの実施例は本発明の範囲を限定するものでなく、説明の目的のために提示される。出典明示されたすべての引用文献は全体を本明細書の一部とする。
【0087】
実施例
実施例1
IL-2およびTFG-βの混合物によるPBMCの処置
実施例1によって示されるのは、SLE患者のPBMCをIL-2およびTFG-βで比較的簡単に処置することにより、自発的ポリクローナルIgGおよび自己抗体産生が顕著に阻害されることである。下記で考察するように、12人の活動性SLE患者のPBMCを、TGF-βの存在または非存在で3日間IL-2に曝露し、洗浄し、そして7日以上培養した。IgG分泌の平均減少は79%であった。最も強力な阻害効果は、最も顕著なB細胞の活動過剰の場合に観察された。抗核タンパク質(NP)抗体の自発的産生が観察されたのは4例あり、PBMCのサイトカイン処置により自己抗体産生は、50から96%減少した。IL-2は、個々の患者におけるTGF-β依存性または非依存性の何れかの機構による抗体産生を阻害した。活動性SLE患者における抗CD2刺激性IgG産生の試験では、IL-2およびTGF-βがIgG産生におけるCD8+T細胞の促進効果を逆転し、代わりに抑制活性を誘発し得る。
【0088】
方法
自発的抗体合成についての被験者
12人の被験者をSLE診断で選択した。それらはSLEを分類するARA基準(Arnett, F.C. et al., (1998), Arthritis Rheum 31: 315-324)を満たしていた。これらの患者はすべて女性であり、スペイン人8人、アフリカ系アメリカ人2人、そしてアジア人2人であった。それぞれの患者の年齢および病歴を表1に示す。5人が入院患者で、7人は外来患者であった。コルチコイド(コルチコステロイド)および抗マラリア薬を投与されている患者も表示した。8人の患者が未処置であった。疾患活動性を、SLAM(Liang, M.H. et al., (1989), Arthritis Rheum 32:1107-1148)およびSLEDAI(Bombardier, C. et al., (1992), Arthritis Rheum 35:630-640)で評価すると、それぞれ平均値16.5および13.4であった。
表1
SLE患者の概略
【表1】

【0089】
試薬
組換え体TGF-βおよびモノクローナル抗TGF-β(1D11.16)抗体、ネズミIgG1は、Dr. Bruce Pratt(Genzyme Pharmaceuticals, Farmington, MA)から好意的に提供された。組換え体IL-10およびモノクローナル抗IL-10(JES3-19F1)抗体および対照ラットIgG2aは、Dr. Satwant Narula(Schering Plough Pharmaceuticals, Kenilworth, NJ)から好意的に提供された。対照マウスIgG1骨髄腫タンパク質は、Calbiochem, San Diego, CAから購入した。組換え体ヒトIL-2は、Chiron, Emmeryville, CAより購入した。OKT11を用いた抗CD2分泌ハイブリドーマ抗体は、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(ATCC)Rockville、MDから得、GT2は、A. Bernard, Nice, Franceから好意的に提供された。他の抗体には、J. Unkeless, New York, NYから好意的に提供された抗CD4(OKT4、ATCC)、抗CD8(OKT8、ATCC;CD8、Dako, Carpentaria, CA)、抗CD11b(OKM1、ATCC)、抗CD16(3G8);抗CD20(Leu 16、Becton Dickinson, San Jose, CA)および抗CD74(L243、ATCC)が含まれていた。
【0090】
血中単核細胞の単離
末梢血単核細胞(PBMC)を、Ficoll-Hypaque(Pharmacia, Piscataway, NJ)密度勾配遠心分離によりヘパリン処理静脈血から調製した。単核細胞を、5mM EDTAを含むPBS(Life Technologies, Grand Island, NY)で洗浄し、血小板を取り除いた。それはTGF-βを多く含んでいる。
【0091】
細胞培養の手順
細胞培養の手順は以前述べられている通りである(Wahl, S.M. (1994), J Exp Med 180: 1587-1590; Gray, J.D. et al., (1998), J Immunol 160: 2248-2254)。簡単に言うと、2x105のPBMCを、血清を含まないAIM-V培養培地(Life Technologies)中、記載のサイトカインを含むか、または含まない96-ウェルの底の平らなマイクロタイタープレートにおいて培養した。培養3日後、PBMCを3回洗浄し、次いで、新たに血清を含まない培地を添加した。更に37℃で7日後、上清を回収し、以前に述べられている(Linker-Israeli, M. et al., (1990), Arthritis Rheum 33: 1216-1225)通り固相酵素結合免疫収着検定(ELISA)によりウシ胸腺核タンパク質(NP)と反応する全IgGおよび自己抗体を検定する。光学的密度(OD)の読み値を、陽性および陰性標準を用いる標準曲線より、units/ml(U/ml)に変換した。SLE患者(高力価の抗NP抗体を有する)および健康な被験者のPBMC培養の上清を対照として用いた。
【0092】
統計学的分析
データをGraph Pad, Prism software(San Diego, CA)を用い分析した。データの対数変換および非パラメーター性マン‐ホイットニー試験後、分散(ANOVA)分析を使用した。
【0093】
抗CD2誘発IgG合成
抗CD2刺激CD4+T細胞およびB細胞に対する、NK細胞と共にまたはNK細胞を伴わずに培養したCD8+T細胞の効果を、SLE患者および正常対照者で試験した。CD4+およびCD8+細胞を、以前述べられている通りに、免疫磁性ビーズを用いる陰性選択により、ナイロン非付着性リンパ球から調製した(Gray, J.D. et al., (1998), J Immunol 160:2248-2254)。CD4+細胞の場合、ナイロン非付着性細胞をCD8、CD16、CD11bおよびCD74に対する抗体で染色した。同じ抗体を用い、CD8+細胞が得られた。ただし、CD4をCD8と置き換える。CD4+細胞の純度は95%であり、CD8+細胞は89%であった。NK細胞を得るため、PBMCをナイロンウールカラムに加え、そして溶出した非付着性細胞をAET処置ヒツジ赤血球細胞で即座にロゼットした。次いで、非ロゼット画分を抗CD3および抗CD74(抗HLA-DR)抗体で染色し、免疫磁性ビーズ(Dynal)を用い反応細胞をなくした。得られた群にはCD56+が98%およびCD3が0.5%未満およびCD20+リンパ球が0.5%未満含まれていた。SLE B細胞が大量のIgGを自発的に分泌するため、そして、これらの試験に多数のB細胞を調製するのに多量の血液を必要とするため、本試験では、健康な被験者ドナーの休止B細胞を患者のB細胞の代用とした。B細胞を得るため、ナイロンウール付着性細胞を、直ぐにSRBCでロゼットし、任意のT細胞を除去し、単球および機能性NK細胞を完全に除去するため5mM L-ロイシンメチルエステルで処理した。得た集団ではCD20+が92%を超え、CD3+が0.5%未満であった。
【0094】
結果
12人の試験患者において、自発的IgGは0.4から13.7μg/mlの範囲であった(図1)。PBMCをIL-2±TGF-βに72時間曝露することにより、IgG合成が、試験した12例のうち8例で、少なくとも50%減少した(平均減少79%、p=0.008、Mann Whitney)。最も劇的な減少は、最も顕著なB細胞の活動過剰の場合に観察された。分泌するIgG量とIL-2およびTGF-βによる阻害%との相関関係は、r=0.647、p=0.02であった。
【0095】
IL-2およびTGF-β単独の効果を、IL-2およびTGF-βの組み合わせと比較した。図2によって示されるのは、これらサイトカインのそれぞれがまたIL-2産生を阻害することである。しかし、対数変換によるデータの正規分布の達成および複数比較のためのBonnferoni補正適用後では、分散分析から、IL-2およびTGF-βの組合せによってのみ有意な阻害(p=0.05)が生ずることがわかる。
【0096】
IL-10産生はSLE中で増加しており(Llorente, L. et al., (1993), Eur Cytokine Network 4:421-427)、このサイトカインはIL-2およびTGF-βの両方の産生を阻害し得る。9例において、我々はまた、抗IL-10の効果を調べたが、IgG合成の幾分の低下が数例で観察されたのみであり、この違いは統計的に有意でなかった。同様に、TNF−α産生も一部のSLE患者で減少する(Jacob, C.O. et al., (1990), Proc Natl Acad Sci 87:1233-1237)。また、このサイトカインは活性TGF-βの産生を増加するが(Ohtsuka, K. et al., (198), J Immunol 160:2539-2545)、TNF−αを培養に加えることにより最小の効果を生じた(結果は示していない)。
【0097】
抗核タンパク質(NP)自己抗体の自発的産生につきSLE PBMCを試験し、有意な力価が4例みられた。PBMCをIL-2またはIL-2とTGF-βのどちらかに曝露した場合の全例において、少なくとも50%の抗NP産生が阻害された。TGF-β自身は効果がなかった(表2)。これらの場合、IL-2自身の効果は、IL-2とTGF-βとの組み合わせの場合と同等であった。
【0098】
【表2】
【0099】
SLE患者のPBMCをIL-2(10u/ml)およびTGF-β(10pg/ml)に72時間さらした。その細胞を洗浄し、更に7日間培養した。上清中に放出された抗NPをELISAで測定した。
【0100】
以前、我々が報告したのは、IL-2により生物学的に活性なTGF-β(Ohtsuka, K. et al., (1998), J Immunol 160: 2539-2545)の産生が増加することである。そのため、自発的抗体合成におけるIL-2の少なくとも幾つかの効果がTGF-βで仲介される可能性があった。この可能性について、IL-2の効果が抗TGF-β中性化抗体により逆転され得るかどうかを測定することにより調べた。図3Aに示した例では、抗TGF-βの添加は自発的IgG合成に影響を与えなかった。しかし、TGF-βの拮抗物はIgG合成におけるIL-2の阻害効果をなくした。この患者(表2のケースC)のPBMCもまた、抗NP抗体を自発的に産生した。ここで、抗TGF-βも、抗NP産生におけるIL-2の阻害効果をなくした(図3B)。即ち、この被験者おいて、自発的IgGおよび自己抗体合成に対するIL-2の阻害効果はTGF-βを介していた。抗TGF-βのこの効果は、試験した8例のうち4例で記録された。故に、IL-2の阻害効果は、TGF-β依存性または非依存性であり得る。各効果の例を表3に示す。
【表3】
【0101】
ステロイド治療開始後28日のSLE患者において試験を繰り返す機会を得た(表4)。処置前には、自発的IgG合成はIgG2μg/mlよりも多かった。PBMCをIL-2に曝露すると、これがIgG産生を顕著に阻害し、TGF-βは適度な効果を有した。コルチコステロイド治療の後、自発的IgG産生は75%減少した。上記の通り、PBMCをIL-2±TGF-βに曝露すると、IgG産生は50%減少した。しかし、この阻害は抗TGF-βにより逆転した。ここでさらに、IL-2のこの効果は、内在性活性TGF-βの上方制御によるものと説明できよう。
【0102】
【表4】
【0103】
健康な被験者についての我々の先の試験において、IL−2およびTGF−βが活性化CD+T細胞を誘導し、Ig産生を下方制御することから、本試験でSLE患者からのCD8+T細胞の単離および処置を試みた。先の試験が不成功であったのは、自発的抗体合成に顕著な可変性があること、および細胞分離法のために活動性SLE患者からの大量の血液を必要とすることであった。しかし、我々は、活動性SLEの患者の1人から十分な血液を得ることができ、抗CD2誘導IgG合成のCD8+T細胞調節に対するIL−2およびTGF−βの効果を調べた。抗CD3とは異なり、抗CD2モノクローナル抗体のマイトジェン性組合わせが、PBLを誘導しないで、IgGを産生することを、我々は最近報告した(Gray, J. D. et al. (1988), J Immunol 160:2248-2254)。例を図4Aに示す。これは、抗CD2がNK細胞を刺激してTGF−βを産生し、それが順番にCD8+T細胞を誘導し、抗体産生を下方制御するためであった(Gray, J. D. et al. (1988), J Immunol 160:2248-2254)。この患者において、我々が以前に報告したように(Gray J. D. et al. (1994), J Exp Med 180:1937-1972)、CD8+T細胞がIgG合成を増加し、NK細胞とCD8+T細胞の組合わせにより増加が著しく強化された(図4A)。一方、IL−2およびTGF−βは、SLE CD8+T細胞のヘルパー効果を無くし、これらの細胞がIgG産生を抑制することを可能にした。IL−2およびTGF−βのこの阻害効果はCD8+T細胞の存在に依存した(図4B)。このように、IL−2およびTGF−β効果がCD8+T細胞により仲介され得るという証拠が得られた。
【0104】
これらの試験は、PBMCのIL−2およびTGF−βへの短い暴露により、SLE、特に重症な疾病の患者および著しいB細胞活動亢進の患者において、続く自発的ポリクローナルIgGおよび自己抗体産生を大きく減少できることを証明した。この試験は、IL−2が抗体産生を阻害できることを示す先の報告(Hirohata, S. et al. (1989), J Immunol 142:3104-3112およびFast, L. D. (1992), J Immunol 149:1510-1515)を確認し、ピコモル濃度のTGF−βがこの下方制御の一因となり得ることを明らかにする。試験した12名の患者のグループにおいて、IL−2およびTGF−βのポリクローナルIgG合成に対する阻害効果は、IL−2単独より大きかった。しかし、IL−2の阻害活性は異なっていた。試験した8名のうち4名において、阻害はTGF−β依存的であり、中和化抗TGF−β mAbが作用を無くした。残りの例において、IL−2の下方制御効果はTGF−β非依存的であった。同様に、自発的抗NP自己抗体産生におけるTGF−β依存的および非依存的阻害の両方が証明された。また、我々がIL−10の拮抗およびTNF−α添加についての効果を調べたのは、SLEにおけるこれらサイトカイン産生の異常について、先に記載されていたためである(Llorente L. et al. (1993), Eur Cytokine Network 4:421-427; Jacob, C.O. et al. (1990), Proc Natl Acad Sci 87:1233-1237)。しかし、これらの方法は、リンパ球が先に活性化されているとき、自発的抗体合成に対する最小の効果を示した。
【0105】
SLEにおけるB細胞活動亢進の程度が疾病活動性と相関するという他の報告がある(Blaese, R. M. et al. (1980), Am J Med 69:345-350; Klinman, D. M. et al. (1991), Arthritis Rheum 34:1404-1410)。このことが本試験で起きなかったのは、同時薬剤治療によるためであろう。概して、著しい自発的抗体合成の患者は処置せず、かわりにB細胞活性が低い患者にはプレドニゾンを普通に投与した。我々はコルチコステロイド治療開始後に著しく自発的IgG合成が減少した一つの例を提示する。この患者のB細胞は、処置前に抗NP抗体も分泌したが、この自己抗体の産生はステロイド治療後に検出されなくなった(データは示していない)。
【0106】
TGF−βは、組織修復、炎症および免疫調節において重要なサイトカインの多機能性ファミリーから成る(Massague, J. (1990), Annu Rev Cell Biol 6597-641)。TGF−βは、不活性前駆体分子として分泌され、細胞外で生物学的活性形に変換する点で、多くの他のサイトカインと異なる(Massague, J. (1990), Annu Rev Cell Biol 6597-641; Flaumenhaft, R. et al. (1993), Adv Pharmacol 24:51-76)。実験的自己免疫脳炎(Weiner, H. L. et al. (1994), Annu Rev Immunol 12:809-837)および大腸炎(Neurath, M. F. et al. (1996), J Exp Med 183:2605-2516)のような種々の実験的自己免疫モデルは、このサイトカインを産生する。TGF−βは、ナノモル濃度で免疫抑制性であり、TおよびB細胞増殖、NK細胞の細胞毒性活性およびT細胞の細胞毒性発生を阻害できる(Letterio, J. J. et al. (1998), Ann Rev Immunol 16:137-162)。対照的に、TGF−βはマウスCD4+細胞およびCD8+細胞の増殖を促進する(Kehrl, J. H. et al. (1986), J Exp Med 163:1037-1050; Lee, H. M. et al. (1993), J Immunol 151:668-677)、およびB細胞分化を促進できる(Van Vlasselaer, P. et al. (1992), J Immunol 148:2062-2067)と報告されている。
【0107】
健康な被験者から得たリンパ球で調節性T細胞を発生させるという先の試験において、使用したTGF−βの濃度は、TまたはB細胞機能の阻害に必要とされるピコモル濃度より低かった(Gray, J. D.ら(1998),J Immunol 160:2248-2254;Gray,J. D.ら(1994),J Exp Med 180:1937-1942)。同様の濃度をSLE患者での本発明の試験において使用すると、TGF−β自体が抗体合成に対して僅かな阻害効果を有していた。前記の通り、TGF−βとIL−2の組み合わせが最も強力な阻害をもたらした。先の試験において、この効果はCD8+ T細胞により仲介された。
【0108】
IL−2について、T抑制細胞活性の誘発に対する効果が十分確立されている(Hirohata,S. ら(1989),J Immunol 142:3104−3112;Fast,L. D.,J Immunol 149:1510-1515)が、これらの効果が直接的なものであるか、または間接的なものであるかどうかは明らかではない。マウスにおいて、IL−2遺伝子の欠失は、大量のリンパ球増殖および自己免疫疾患を生ずる(Sadlack,B.ら(1995),Eur J Immunol 25:3053-3059)。SLEにおいて、IL−2レベルとB細胞機能亢進との負の相関関係が報告されている(Huang,Y. P.ら(1988),J Immunol 141:827-833)。以前、SLEでIL−2によって自発的Ig産生を阻害しようと試みたが、しかしながら、その結果は極めて可変的であった。強い阻害を幾つかの場合において観察したが、一方で他のIL−2では抗体産生が著しく増大した。我々は、時期およびサイトカイン環境により、この試験で見られる阻害よりもさらに一定した阻害が説明されると考える。ここで、IL−2およびTGF−βは、全培養期間というよりはむしろ、最初の72時間の培養の間にしか存在していなかった。後者の事例での抗体合成の増加は、B細胞分化に対するIL−2の正の効果によって説明し得る(Coffman,R. L.ら(1988),Immunol Rev 102:5-28)。IL−2は、抗体産生を幾つかの機構により下方制御することができる。該報告に記載されているTGF−β回路に加えて、IL−2が誘発する阻害は、IFN−γの上方制御(Noble,A.ら(1998),J Immunol 160:566-571)により、または細胞溶解機構(Stohl,W.ら(1998),J Immunol 160:5231-5238;Esser,M. T.ら(1997),J Immunol 158:5612-5618)により起こり得る。
【0109】
以前、我々は、抗体合成に対するNK細胞の調節効果を調べ、NK細胞の直接的効果がIgG合成を上方制御することである(Kinter,A.ら(1995),Proc Natl Acad Sci USA 92:10985-10989)が、これらのリンパ球が、正常な被験者において、CD8+ T細胞との培養の場合、正反対の効果を有すると報告した(Gray,J. D.ら(1994),J Exp Med 180:1937-1942)。しかしながら、SLE患者において、CD8+ T細胞とNK細胞との組み合わせがIgG産生を高める(Linker-Israeli,M.ら(1990),Arthritis Rheum 33:1216-1225)。これは、本発明の報告において再び観察された。正常な被験者では、NK細胞をCD8+ T細胞に添加すると、抗CD2刺激IgG合成を著しく阻害したが、正反対のことがSLEにおいて観察された。標準的試験から、NK細胞から得たTGF−βがCD8+T細胞の同時刺激を誘発して、IgGおよびIgM産生を下方制御することがわかった(Gray,J. D.ら(1998),J Immunol 160:2248-2254)。この試験において、IL−2およびTGF−βは、CD8+ T細胞による中程度の抑制活性を誘発した。従って、SLEにおいて、IL−2およびTGF−βがB細胞活性を阻害する少なくとも1つの方法は、調節性T細胞を発生させることによるものであるらしい。加えて、これらまたは他のサイトカインで処理した他のリンパ球群もまた、SLEにおけるB細胞活性を下方制御し得る。
【0110】
実施例2
疾患の活性および重症度とTGF−β産生との相関関係
全体および活性型のTGF−βのリンパ球産生が減少することが分かったので、次に、これらの欠失が疾患の活性および/または重症度と関連があるかどうかを調べた。17人の検査したSLE患者から得た血液リンパ球によるTGF−β1産生を、10人の慢性関節リウマチ(RA)患者および23人の適応する健康な対照と比較した。RAにおける活性TGF−β1のレベルは、対照より低かったが、SLEにおける程度までは減少しなかった。ピコモル量で検出される構造性TGF−β1および抗CD2刺激された活性TGF−β1のレベルは、最近発症の非常に活性で重症なSLEのために入院した6人の未処置患者において著しく減少し、処置され活性の低い疾患の11人の患者においても同様に減少した。このように、活性TGF−β1産生の減少は、疾患の活性とは関連がなかった。これとは対照的に、全TGF−β1産生の減少は、疾患の活性と逆の相関関係があった。このように、TGF−β1の潜在性前駆体のリンパ球分泌の欠陥は、疾患活性の結果として生じ得るが、前駆体分子をその活性型に転換する能力は、内因性細胞欠失となり得るようである。活性化される時点でのピコモル濃度量のTGF−β1へのT細胞の不十分な暴露は、抗体合成の下方制御の欠陥を生じ得る。このように、SLEにおける活性TGF−β1のリンパ球産生の減少は、この疾患に特徴的なB細胞機能亢進に起因するのであろう。
【0111】
方法
試験の被験者
SLEの分類に関するアメリカ大学のリウマチ病学基準(Tan,E. M.ら(1982)、Arthritis Rheum 25:1271-1277)を満たすSLE診断を受けた17人の被験者、RAの分類に関するACR 1987 改定基準 (Arnett,F. C.ら(1988),Arthritis Rheum 31:315-324)を満たすRA診断を受けた10人の被験者、および23人の健康なドナーを試験した。SLEグループは、女性15人および男性2人(スペイン人15人、アフリカ系アメリカ人1人、アジア人1人)からなっていた。平均年齢は、34.5歳(範囲:20−75歳)であった。6人の患者を入院させて、11人を外来診療所に通わせた。入院患者は皆、入院するまでは未処置であり、試験した後、彼らに第一回用量のコルチコステロイドを投与した。外来患者には、プレドニゾンを20mg未満を投与し、誰にも細胞毒性薬物を投与しなかった。疾患の活性を、平均値が各々6.6および7.6であるSLAM(Liang,M. H.ら(1989)、Arthritis Rheum 32:1107-1118)およびSLEDAI(Bombardier,C.ら(1992)、Arthritis Rheum 35:630-640)指数で評価した。RAグループは、女性9人および男性1人(スペイン人9人、アジア人1人)からなっていた。平均年齢は、50.9歳(範囲:39−67歳)であった。患者は皆、外来診療所に通わせ、軽度〜中程度に疾患活性を有していた。疾患の平均期間は9.5年であった。1人の患者にマイオクリシン(myochrysine)を投与し、3人の患者にプレドニゾン(1、1および20mg)を投与し、3人の患者にメトトキサレートを投与し、1人の患者にスルファサラジンを投与した。健康なドナーを対照として、年齢、性別、および民族グループに関して可能な限り厳密に適応させた。
【0112】
表5
SLE患者の2つのグループの臨床的特徴
【表5】
【0113】
試薬
使用した抗体は、抗CD2を分泌するハイブリドーマ(OKT11、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(ATCC),Rockville,MD、およびGT2は、Alain Bernard博士,Nice,Franceにより入手可能となった)の上清であった。TGF−βイソ型 1、2、および3(1D11)を認識するモノクローナル抗体、TGF−βイソ型 2および3(3C7)に対する抗体、並びにrTGF−β2は、Bruce Pratt博士(Genzyme Pharmaceuticals,Farmington,MA)により快く提供された。
【0114】
血液リンパ球の単離
先に記載した方法(Ohtsuka,K.ら(1998),J Immunol 160:2539-2545)を使用して、Ficoll−Hypaque(Pharmacia, Piscataway, NJ)密度勾配遠心分離により、末梢血単核細胞(PBMC)をヘパリン処置した静脈血から調製した。単核細胞を、5mM EDTA(Life Technologies, Grand Island,NY)を含むPBSで洗浄して、TGF−βの豊富な原料である血小板を取り除いた。連続Percoll(Pharmacia)密度勾配によって、末梢血リンパ球(PBL)を遠心分離によりPBMCから分離した。高密度のリンパ球に富む画分に残留する単球のパーセンテージは、SLEにおいて幾分高かった(8.5% 対 4.3%)。
【0115】
細胞培養方法
細胞培養方法は、以前に記載されている(Ohtsuka,K.ら(1998),J Immunol 160:2539-2545)。簡単に言えば、1×105個のリンパ球を96ウェルの平底マイクロタイタープレート(Greiner Rocky Mountain Scientific,Salt Lake City UT)のウェルに加えた。血清が著しい量の潜在性TGF−βを含むことから、培養をAIM−V血清不含有培地(Life Technologies)で行った。抗CD2を最適濃度で使用して、TGF−β産生(GT2 1:40およびT11 1:80)ハイブリドーマ培養上清を誘導した。先の試験は、抗CD2がPBLを強く刺激して、TGF−βを産生することを明らかにしている(Gray,J. D.ら(1998),J Immunol 160:2248-2254)。
【0116】
TGF−βアッセイ
ルシフェラーゼ・リポーター遺伝子に融合したプラスミノーゲン活性化因子阻害剤(PAI−1)プロモーターを含む発現構築物をトランスフェクトしたイタチ肺上皮細胞(MLEC)は、D. B. Rifkin博士,New York,NYにより快く提供された。2×104/ウェルのMLECを上清200μlと共に37℃で18時間インキュベートした。ルシフェラーゼ活性についてアッセイするために、MLECを細胞溶解試薬(Analytical Luminescence,Ann Arbor,MI)により溶解した。次いで、細胞溶解物をアッセイ緩衝液およびルシフェリン溶液(両方とも、Analytical Luminescenceから得た)と反応させた後、直ちに、照度計(Lumat,Berthold Analytical Instruments Inc.,Nashua,NH)で測定した。全TGF−β活性を測定するために、試料を80℃で3分間加熱して、潜在性複合体から活性サイトカインを放出させた。上清を加熱することなく、活性TGF−βの活性を測定した。全てのアッセイおいて、幾つかの濃度のrTGF−βが標準曲線を作成するために含まれていた。同型培養の可変性は10%未満であった(Ohtsuka,K.ら(1998),J Immunol 160:2539-2545)。
【0117】
統計解析
GBSTATソフトウェア(Professional Statistics and Graphics Computer Program,Dynamic Microsystems Inc.,Silver Spring,MD)を使用して行うマン‐ホイットニー(Mann−Whitney)試験およびSpearman順位相関関係を使用して、結果の有意性を解析した。
【0118】
結果
SLEまたはRAの患者由来のPBLにより産生される構造的および刺激されたTGF−β1を測定し、正常対照の値と比較した。培養上清中で検出されるサイトカインは、イソ形1、2および3を認識するmAbで中和されたが、イソ形2および3に対するmABではされず、結果としてTGF−β1の産生を確認した。正常対照と比較して、活性TGF−β1の構造的産生はSLEで有意に減少した(14±4対56±21pg/ml、p=0.02、図5)。抗CD2刺激活性TGF−β1も減少した(87±22対399±103pg/ml、p=0.003)。RAにおいて、構造的TGF−β1の平均値は、SLE(19±5pg/mg)と同様であり、抗CD2により刺激された後、正常とSLEの中間値であった(197±54pg/ml)。
【0119】
リンパ球により産生された構造的全TGF−β1も、正常グループと比較してSLEで減少した(286±82対631±185pg/ml、p=0.05)。RAでの値は、正常とSLEの中間値であった(435±161pg/ml)。抗CD2の添加に続き、全TGF−β1はSLEで正常対照より幾分増加したが、差異は統計学的に有意でなかった。RAグループでの値も正常とSLEグループの中間値であった。
【0120】
TGF−β1の減少したレベルと疾病活動性の相関関係の可能性を調べるために、SLEの入院患者と外来患者とを比較した。これらの2つのグループの臨床的特徴は、表5に要約する。入院患者の方が若かった;6名中5名が、3ヶ月以内の症状を示し;そして、著しく活動的な疾病に罹患していた;そして殆どが、腎炎および/または溶血性貧血を伴う重症SLEであった。比較して、外来患者は、処置により活動性が低くなった慢性疾病であった。疾病異種性、期間、活動性および重症度の著しい差異にもかかわらず、構造的および刺激TGF−β1産生は、正常対照と比較して両グループで有意に減少した(表6)。
【0121】
【表6】
【0122】
活性および全TGF−β1のレベルと疾病活動性との相関関係をみると、全TGF−β1の抗CD2刺激産生は、SLEDAI指数と有意な負の相関関係があるが(r=0.55、p=0.03)、SLAM指数とは相関がなかった(−0.43、p=11)。SLEDAI指数は、中枢神経系関与および腎臓疾患に重きを置く。このように、TGF−β1の前駆体形を分泌するリンパ球の能力の減少は、重症な疾病と関連するように見える。活性TGF−β1のレベルは、疾病活動性と相関しなかった。
【0123】
本例での主な発見は、SLEでの活性TGF−β1の産生減少が、疾病活動性または重症度と相関しないことである。構造的および刺激活性のTGF−β1量の減少は、最近発症した疾病および確立した疾病の両方の患者で見られた。更に、この値は、SLAMおよびSLEDAI指数で測定して活動性と、または生体臓器関与により評価した重症度と相関しなかった。しかし、全TGF−β1産生がSLEで減少もするが、この欠失は疾病活動性と相関するように見えた。これは、主として入院SLE患者で見られた。全TGF−β1産生が主要な臓器系関与に重きを置くSLEDAI指数と最も相関するというこの発見はまた、疾病重症度との関係を示す。
【0124】
この試験はまた、確立された疾病のSLE患者と疾病活動性が同等であるRA患者の対照グループを含む。RAグループでのTGF−β1値は、構造的活性TGF−β1以外、正常対照より幾分低いが、欠失の強度はSLE程著しくなく、統計的に有意ではなかった。
【0125】
以前に、我々は、NK細胞がTGF−βの主なリンパ球源であり、このサイトカインを活性形で構造的に産生する唯一のリンパ球集団であることを証明した(Gray, J. D. et al., (1988), J. Immunol. 169:2248-2254)。従って、NK細胞由来TGF−βの構造的産生はSLEで減少しているとの発見は、興味深かった。我々はまたIL−2およびTNF−αの両方が活性TGF−βの産生を促進できることを知った。これら両サイトカイン産生はSLEで減少する(Gray J. D. et al., (1994), J Exp Med 180:1937-1942)。しかし、殆どの患者において、外来性IL−2およびTNF−αはTGF−βを正常に回復できなかった(実施例2)。IL−10産生はSLEで増加し(Llorente, L. et al. (1993), Eur Cytokine Network 4:421))、増加したレベルと疾病活動性の相関関係が報告されている(Housslau, F. A. et al. (1995), Lupus 4:393-395; Haglwara, E. et al. (1996), Arthritis Rheum 39:379)。IL−10はIL−2、TNF−αおよびTGF−β産生を阻害できる(実施例2およびMoore, K. W. et al. (1993), Ann Rev Immunol 11:165-190)。活性TGF−βの産生が軽いおよび活動性の疾病の患者で減少したこと、およびIL−10の拮抗によって産生欠失を部分的にしか回復できなかったことは(実施例2)、増加したIL−10産生自体がSLEにおけるTGF−β1のリンパ球産生の減少について説明となり得ないことを示す。数個の機構が恐らく関与するのであろう。潜伏性前駆体の成熟活動性形への細胞外変換における1個またはそれ以上の欠失がこの異常性を説明し得る。
【0126】
TGF−βは、リンパ球増殖およびエフェクター細胞機能に対する阻害特性が詳細に説明されているが(Letterio, J. J. et al. (1998), Ann Rev. Immunol 16:137-162)、刺激特性も報告されている(Lee, H. M. et al. (1991), J Immunol 151:668-677)。TGF−βは、刺激T細胞およびその産生の上方制御によりサイトカイン産生を調節する。マウスにおいて、TGF−β1は、CD8+T細胞を選択的に活性化して増殖させ(Lee, H. M. et al. (1991), J Immunol 151:668-667)、ナイーブ細胞からメモリーT細胞への成熟を増加する(Lee, M. H. et al. (1991), J. Immunol 147:1127-1133)。ヒトにおいて、TGF−β1はエフェクターT細胞の強い誘発物である(Cerwenka, A. et al. (1994), J Immunol 153:4367-4377)。免疫抑制作用には大量(ナノグラム/ml)が必要であるが、我々が示したように、抗体産生の下方制御効果に関してCD8+T細胞を共刺激するのには、少量(ピコグラム/ml)でよい(Gray, J. D. et al. (1998), J Immunol 160:2248-2254)。
【0127】
そのため、これらの試験によると、TGF−β1の潜伏性前駆体のリンパ球分泌の欠失が疾病活動性の結果として起こり得るが、SLEにおける活性TGF−β1産生の減少は、さらに複雑であり、数個の異なる機構からもたらされる。ナイーブT細胞が抗体産生を下方制御するように計画するには、T細胞が活性化される時点でpg/ml量の活性TGF−βの存在を必要とすると我々は考え、またこの考えを支持する証拠がある(Gray, J. D. et al. (1998), J Immunol 160:2248-2254)。従って、決定的な時点での局所環境におけるピコモル量の活性TGF−βを欠くことは、SLEにおけるBリンパ球活性を制御するT細胞調節機能の無効によるのであろう。
【0128】
実施例3
マイトジェンでのSLEの処置
本実施例において、IgG産生は、調節組成物で細胞を処理することにより下方制御される。この調節組成物はマイトジェン抗CD2モノクローナル抗体組合せ物のようにマイトジェンを含む。これら抗体は、可溶化形態で添加するか、ビーズ上に固定化して、T細胞およびNK細胞上の受容体と橋かけ結合せしめる。これらの細胞は、上記の実施例で説明したように調製し、次いで、マイトジェンとインキュベートし、増幅した。抗体産生が下方制御された細胞数が増加する。ConAはシグマから入手できる(St. Louis, MO)。
【0129】
抗CD2がどのように機能するかは知られていないが、これら抗体がPBMC調製物中においてNK細胞を誘導し、活性TGF−βを分泌し(Ohtsuka, K. et al.(198), J. Imunol 160: 2539-2545)、次いでTGF−βが働いて、T細胞が抗体抑制細胞となると考えられている。
次いで、細胞を必要であれば洗浄し、患者に移植し戻す。
【0130】
実施例4
サイトカインとマイトジェンの混合物での細胞の処理
細胞を上記のように調製して、抗体産生を下方制御する細胞集団を誘導するために、マイトジェンとサイトカインの混合物を含む調節組成物で細胞をインキュベートした。この手法の例は、図4Cにおいて示す。この実施例において、CD4+細胞およびCD8+細胞をConA、IL−2およびTGF−βで処理して、抑制に関する最大誘導を得た。
【0131】
患者に再移送される調節性T細胞の調製のために、抗CD2および/または抗CD3モノクローナル抗体をConAの代わりに使用して、T細胞を活性化する。調節組成物は、IL−2を伴いまたは伴なわずに、TGF−βを含有する。この細胞を、例えば、Nexell 300i. Magnetic Cell Selection System のような閉鎖系中で標準インキュベーション技術を用いて4〜7時間、組成物と共にインキュベーションした。
【0132】
インキュベートの後、細胞をHBBSと洗浄して、溶液中のサイトカインおよびマイトジェンを除去する。所望により、ビーズ上に固定した抗CD3±抗CD28と培養して、細胞数を増加さす。次いで、細胞を200−500mlのHBBS中に懸濁し、患者に再挿入する。
【0133】
実施例5
細胞の細胞仲介免疫を正常化する処置
SLEの自己抗体産生に対する要因は、IL−10とTNF−αとのアンバランスにある。IL−10のレベルは過多で、TNF−αレベルは低い[Llorente et al. 1995. J. Exp. Med. 181:839-44,Houssiau, F.A. et al., 1995. Lupus 4:393-5.(Ishida, H. et al. 1994. J. Exp. Med. 179:305-10)(Jacob, C. O. and McDevitt, H. O.,1988. Nature 331: 356-358)]。我々は、このアンバランスが、TGF−βの存在下でT細胞を強く活性化することによって補正されるという証拠を得て、近年この効果の作用機構を解明した。
【0134】
精製されたT細胞を、上記概要に沿って調製し、TGF−βの存在または不存在で、ConAとIL−2でインキュベーションした。図6では、TGF−βの非存在下でのT細胞の刺激によって、IL−10の産生が増加したことを示す。しかし、TGF−αを刺激されたT細胞に加えると、IL−10の産生が遮断され、TNF−αの産生が増加した。さらに、TNFR2の発現が有意に増加した。理論にしばられるわけではないが、TGF−βによって誘導されるTNFR2を介するTGF−α情報伝達の促進が、抗体産生を抑制する調節性T細胞を生じると考える。我々の結果は、この考察を支持するものであった。
【0135】
TGF−βによるTNF−αの上方制御が調節性T細胞の誘導に重要であることを説明してきた。図7は、TGF−βを活性化CD8+T細胞に添加することで、IgG産生の顕著な抑制が起こることを示す。この抑制活性は、主な中間体としてTNF−αに依存する。これらの各試験において、抗TNF−α抗体を中和することによって、CD8+調節性T細胞(CD8reg)抑制効果を完全になくした。
【0136】
SLE患者は、IL−2、TNF−αおよびIFN−γの産生減少を伴なう細胞仲介免疫に著しい欠失を有する(Horwitz, D.A. et al.,(1997), Dubois' Lupus Erythematosus, 5th Ed. (1997), pp.83-96, D.J. Wallace et al. eds., Williams and Wilkins, Baltimore)。理論にしばられるわけではないが、TGF−βのリンパ球産生における欠失は、IL−2、TNF−αおよびIFN−γの産生減少に対する部分的な要因である。TGF−βの存在下でのT細胞の刺激によって、これらの細胞を再び刺激すると、IL−2、TNF−αおよびIFN−γの産生が顕著に増加することを見出した。さらに、この結果は、TGF−βによるTNF−αの上方制御にも依存する(図8を参照)。
【0137】
TGF−βの産生がSLEで低下し、その欠失がIL−10とTNF−αとのアンバランスによるものであるということを示す。理論にしばられるわけではないが、SLEにおけるIL−10の高いレベルが、自己抗体産生を保ち、TGF−α、IL−2、IFN−γ産生の原因であると考える。これらサイトカインの産生低下がSLEにおける欠失性細胞免疫の原因である。明記した条件下で、TGF−βはIL−10を下方制御し、TNF−αの産生を強めることを示す。TGF−βによるIL−10の下方制御とTNF−β産生の増強は、SLEにおける調節性T細胞の正常化、細胞仲介免疫の修復、疾病の軽減に関して重大な役割をになう。
【0138】
実施例6
細胞仲介自己免疫を抑制する調節性T細胞の生成
前記実施例では、調節組成物を使用して、抗体介在性自己抗免疫疾患を処置した。類似組成物を使用して、CD4+に加えてCD+8T細胞を誘導し、細胞仲介自己免疫疾患を抑制した。TGF−βによって調節されたCD8+またはCD4+細胞が、T細胞の細胞毒性の生成を抑制することを示す。
【0139】
調節性T細胞を誘導するためにマイトジェンを使用する代わりに、同種異系性混合リンパ球反応を、この目的のために使用した。この反応では、1個体からのT細胞が、他の個体のPBMCによって提示された外来の組織適合性抗原を認識し、応答する。これらの応答T細胞は、増加して、これらの標的細胞を殺傷する能力を発展させる。
【0140】
抑制T細胞を創生するために、一個体(ドナーA)からの様々なCD4+およびCD8+T細胞のサブセットを、別の個体(ドナーB)からの照射されたT細胞なしの単球性細胞と共に培養した。その細胞を、懸濁液中のTGF−β(1ng/ml)の存在または不存在で、5日間培養した。これ以降は、TGF−βを除き、該細胞を、新たなドナーAからのT細胞とドナーB細胞からの非T細胞に添加した。図9は、TGF−βが、CD4+およびCD8+T細胞サブセットの両方を誘導し、細胞仲介細胞毒性を抑制する能力を開発することを示す。図10は、TGF−βによって誘導されたCD4+調節性T細胞を用いる2つの追加試験を示す。
【0141】
さらなる試験によると、この方法でつくった調節CD4+T細胞が、該作用に関する独特の作用を示す。前につくったCD8+およびCD4+両方のT細胞が阻害性サイトカインの分泌により抑制をおこすのと異なり、これらアロ特異的調節性CD4+T細胞は、接触依存性の作用機構を示す(図11)。理論にしばられるわけではないが、これらの調節性T細胞は、別のT細胞が活性化されるのを抑制すると考える。これらのT細胞の応答T細胞およびアロ刺激細胞への添加は、増殖を抑制し(図12)、応答CD8+キラー前駆体細胞が活性化される能力を低下させた(図13)。
【0142】
次のことも分かった。これら調節性CD4+細胞は、IL−2受容体(CD25)をそれら細胞表面で発現し、非常に強力である(図14)。調節CD4+細胞の応答T細胞との割合が、1:4(20%)から1:32(3%)に低下すると、これら細胞の阻害効果がごく僅か低下した。
【0143】
これら細胞の少量のみを強い下方制御効果に必要とするので、十分な数を患者に移送し、自己免疫性もしくは他の所望の免疫抑制効果、例えば移植拒絶反応を抑制することができる。
【0144】
実施例7
CD4+T細胞を刺激してTGF−βの免疫抑制レベルを生成する
TGF−βの免疫抑制レベルを生成するCD4+T細胞はTh3細胞と称されているが、その発生に関する機構は殆ど理解されていない。われわれが得た証拠によると、スーパー抗原、スタフィロコッカス・エンテロトキシン(SEB)によるCD4+細胞の強い刺激、または低濃度のSEBよるCD4+細胞の反復刺激が、これらの細胞を誘導し、活性TGF−βの免疫抑制レベルを生成した。
【0145】
図15は、活性および全TGF−βの産生増加が、SEB濃度の増加によって刺激されたCD4+T細胞によりなされたことを示す。図16は、SEBの低用量によってCD4+T細胞の反復刺激に関する効果を示す。T細胞をSEBで3回刺激すると、顕著な量のTGF−βの活性型が生成した。
【0146】
図17は、ナイーブ(CD45RA+CD45RO−)CD4+およびCD8+T細胞に対するSEBの効果を示す。この細胞をSEBで5日ごとに合計3回刺激した。各T細胞のサブセットおよびCD25IL−2受容体活性マーカーを発現する細胞の%を各々の刺激後に測定した。パネルAおよびCは、最初の刺激にTGF−β 1ng/mlが含まれていると、CD4+T細胞が反復刺激の培養物中で優勢なサブセットとなったことを示す。パネルBおよびDは、SEB刺激細胞によるCD25の発現が、対照培養物中で3回目の刺激によって低下したことを示す。しかし、CD25の発現は、T細胞がTGF−βで駆動されると、非常に高いままであった。即ち、TGF−βは、これらのT細胞が反復刺激されると、CD4+細胞に対する優先効果を有するようであり、これらの細胞の殆ど全てが20日間の培養後にCD25+であった。
【0147】
要約すると、T細胞の刺激の結果として、TGF−βの優勢な調節効果が、CD8+細胞に与えられる。反復刺激によって、このサイトカインはCD4+細胞を誘導して、調節性細胞とする。この細胞は、抑制活性に関してCD8+細胞よりも強力である。
【図面の簡単な説明】
【0148】
【図1】図1は、SLE患者のPBMCをIL−2およびTGF−βと共にインキュベーションすることにより、自発的免疫グロブリン産生が減少することを示す。
【図2】図2は、IL−2およびTGF−βの両方により、自発的IgG産生が有意に減少することを示す。
【図3】図3Aおよび3Bは、抗TGF−βがIL−2の効果を逆転することを示す。
【図4】図4A、4Bおよび4Cは、抗体産生に対するDC8+T細胞の調節効果を示す。
【図5】図5Aおよび5Bは、非刺激細胞および抗CD2刺激細胞によるTGF−β1のリンパ球産生を示す。
【図6】図6は、TNF−αおよびIL−10のT細胞産生に対するTGF−βの作用を示す。
【図7】図7は、TGF−βによる調節性T細胞の生成にTNF−αが必須の中間体であることを示す。
【図8】図8は、TGF−β起動T細胞によるTh1サイトカインの産生の増加がTNF−αに依存性であることを示す。
【図9】図9は、細胞毒性T細胞活性の抑制物質の産生におけるTGF−βの作用を示す。
【図10】図10は、TGF−βにより起動されたCD4細胞のアロ細胞毒性Tリンパ球(CTL)活性に対する作用を示す。
【図11】図11は、調節性T細胞がCTL活性を抑制するために細胞接触を必要とすることを示す。
【図12】図12は、TGF−βで誘発された調節性CD4+T細胞によるリンパ球増殖の抑制を示す。
【図13】図13は、CD25+CD4T細胞の調節活性を示す。
【図14】図14は、調節CD+4細胞がその表面でCD25+(IL−2)受容体を発現することを示す。
【図15】図15は、少量のブドウ球菌性エンテロトキシンB(SEB)でのT細胞の反復刺激がT細胞を誘発してTGF−βの免疫抑制レベルをつくることを示す。
【図16】図16は、低量のSEBでCD4+T細胞の反復刺激によってT細胞がTGF−βの免疫抑制レベルをつくることを示す。
【図17】図17は、ナイーブ(CD45RA+CD45RO−)CD4+およびCD8+T細胞に対するSEBの作用を示す。
【技術分野】
【0001】
(技術分野)
本発明は一般的に、抗体仲介および細胞仲介の両方の異常を含む自己免疫異常を処置する方法に関する。
【背景技術】
【0002】
(背景技術)
自己免疫疾患は、自己か自己でないかを識別する免疫システムの機能不全によっておこる。これらの疾患で、免疫系は自己組織に対して作用し、その応答は最終的に炎症や組織損傷をもたらす。自己免疫疾患は2つの基本的部類に分類される:全身性エリテマトーデス(SLE)、尋常性天疱瘡、重症筋無力症、溶血性貧血、血小板減少性紫斑病、グレーブス病、シェーグレン症および皮膚筋炎などの抗体仲介疾患と、橋本病、多発性筋炎、炎症性腸疾患、多発性硬化症、真性糖尿病、慢性関節リウマチおよび硬皮症などの細胞仲介疾患である。
【0003】
多くの自己免疫疾患において、組織損傷は元の組織に対する抗体の産生によって引き起こされる。この抗体は自己抗体と言われている。哺乳動物によって産生され、哺乳動物自体の組織に対する結合部位を持つためである。この疾患のうちいくつかは、循環する抗体量の特有な漸増と漸減を有し、やがて徴候に変化をもたらす。
【0004】
抗体仲介自己免疫異常の種々のタイプの中でもSLEは充分研究され考証されている異常である。SLEは一般化した自己免疫の異常であり、核、細胞質および細胞表面の抗原に対する様々な自己抗体でのB細胞の活動過多を特徴とする。この自己免疫疾患は、遺伝的および環境的な促進因子による多因性病原を有する(参照:Hahn, B.H., Dobois' Lupus Erythematosus, 5th Ed. (1997), pp. 69−76 (D.J. Wallace et al. ads., Williams and Wilkins, Baltimore))。SLEで説明されている多数のリンパ球欠失は、調節性T細胞の機能不全であり、B細胞の機能を阻害する(Horwitz, D.A., Dubois' Lupus Erythematosus, 5th Ed. (1997), pp. 155−194 (D.J. Wallace et al. eds., Williams and Wilkins, Baltimore))。インビトロでポリクローナルIgGおよび自己抗体を持続的に産生するには、T細胞の助力が必要である(Shivakumar, S. et al. (1989), J Immunel 143:103−112)。
【0005】
調節性T細胞は、胞溶解性またはサイトカイン仲介メカニズムによる抗体合成を下方制御する。後者はトランスフォーミング成長因子−β(TGF−β)と他の抑制的サイトカインを必要とする(Wahl, S.M. (1994), J Exp Med 180:1587−190)。 循環Bリンパ球自発的分泌抗体は、活動性SLEの患者で増加する(Klinman, D.M. et al. (1991), Arthritis Rheum 34:1404−1410)。
【0006】
SLEの臨床症状は、発疹(特に、顔面の蝶型紅斑)、糸球体腎炎、肋膜炎、心膜炎および中枢神経系関与がある。患者の大部分は女性であり、比較的若い(診断時の平均年齢は29才である)。
【0007】
SLEに対する処置は臨床的症状によって変わる。穏やかな臨床症状の患者は、非ステロイド性抗炎症剤等の簡単な処置で効き目がでる。しかし、さらに重い症状に対しては、通常、プレドニゾンなどの強い抗炎症および免疫抑制の作用をもつステロイドが必要となる。他の強力な免疫抑制剤として、アザチオプリンとシクロホスファミドを使用することができる。ステロイドおよび他の免疫抑制剤では、哺乳動物の免疫システムが全体的に低下するために副作用が生じる。現在、SLEに対する理想的な処置はなく、疾患を治すことができない。
【0008】
現在、SLEに対する感受性または抵抗性を高める遺伝子の同定、疾患を引き起こす抗原性決定子の同定、生存かアポトーシスかを決定づけるT細胞活性化の分子メカニズム、T細胞の機能を決定するサイトカイン、および自己抗体を形成するB細胞の特性が、かなり注目されている。SLEのT細胞調節不全について多くの例証が叙述されている(参照:Horwitz, D.A. et al., Duobois’Lupus Erythematosus, 5th Ed. (1997), pp. 83−96 (D.J. Wallace et al. ads., Wiliams and Wilkins, Baltimore)。あるリンパ球の一次的役割が免疫応答の下方調節であることは充分認められているが、これらの細胞の産生で必要とされる同一性およびメカニズムの解明は進んでいない。
【0009】
インターロイキン−2(IL−2)は、これまで抗原非特異的T抑制細胞の産生で重要な役割を果たすと考えられている。マウスに与えられる抗IL−2抗体(移植片対宿主病の誘導物と一致する)はSLEの特色をもたらす(Via, C.S. et al. (1993), Intemational Immunol. 5:565−572)。IL−2が抑制の産生において直接的または間接的に重要であるかどうかが議論されている(Fast, L.D. (1992), J. Immunol. 149:1510−1515; Hirohata, S. et al. (1989), J. Immunol. 142.3104−3112; Baylor, C.E. (1992), Advances Exp. Med. Biol. 319:125−135)。最近、IL−2が、CD8+細胞を誘発して、CD4+T細胞中のHIV複製を非分解メカニズムにより抑制することがわかった。この効果はサイトカイン仲介性であるが、具体的なサイトカインは同定されていない(Kinter, A.L. et a1. Proc. Nalt Acad. Sci. USA 92:10985−10989; Barker, T.D. et al. (1996), J. Immunol. 156.4478−4483)。IL−2のT細胞産生はSLEで減少する(Horwitz, D.A. et al. (1997), Dubois' Lupus Erythematosus, 5th Ed. (1997), pp. 83−96, D.J. Wallace et al. eds., Williams and Wilkins, Baltimore)。
【0010】
SLE患者由来のCD8+T細胞は、ポリクローナルIgG産生を抑制するよりむしろ維持する(Linker−Israeli, M. et al. (1990), Arthritis Rheum. 33:1216−1225)。健康なドナーのCD8+T細胞を刺激して抗体産生を高めることができる(Takahashi, T. et al. (1991), Clin. Immunol. Immunopath. 58:352−365)。しかしIL−2またはCD4+T細胞のどちらも、それ自体では強力な抑制活性を発生させるCD8+T細胞を産出しないことがわかった。NK細胞を培地に含めると、強力な抑制活性が現われた(Gray, J.D. et al. (1994) J. Exp. Med. 180:1937−1942)。培地中のNK細胞の働きは、トランスフォーミング成長因子ベータ(TGF−β)を活性形態で産出するものと考えられる。非免疫抑制(2−10pg/ml)濃度のこのサイトカインは、IgGおよびIgMの産生に対する強力な抑制作用をつくるための補助因子として働くことが見出された(Gray, J.D. et al (1994) J. Exp. Med 180:1937−1942)。加えて、NK細胞は非刺激リンパ球でTGF−βの主要源であると考えられる(Gray, J.D. etaL (1998), J. Immunol. 160:2248−2254)。
【0011】
TGF−βは、組織修復、炎症および免疫調節における重要なサイトカインの多機能性系統群である(Massague, J. (198O), Ann. Rev. Cell Biol. 6:597)。TGF−βは、他の大部分のサイトカインと異なり、その放出タンパク質が生物学的に不活性であり、特定の受容体と結合することができない(Sporn, M.B. et al. (1987) J. Cell Biol. 105:1039−1045)。潜在の複合体が細胞外で切断され、以下に記すように活性サイトカインを放出する。TGF−βの応答は単核細胞に偏在する2つの表面受容体(TGF−β−R1とTGF−β−R2)の相互作用を必要とする(Massague, J. (1992), Cell 69:1067−1070)。従って、潜在型の活性TGF−βへの変化は、サイトカインの生物学的効果を決定する重大な段階である。
【0012】
SLE患者ではNK細胞によるTGF−βの産生が減少することが分かっている。NK細胞により産生される構成TGF−βの欠失および誘導されたTGF−βの欠失は、38人のSLE患者についての研究で報告されている(Ohtsuka, K. et al. (1998), J. Immunol. 160.2539−2545)。組換え型IL−2やTNF−αまたはIL−10拮抗エフェクターの何れを加えても、SLEでのTGF−β欠失を正常し得なかった。SLEでのTGF−β産生の減少は、疾病の活性と相関しておらず、従って、初期の欠失かも知れない。
【0013】
SLE患者をTGF−β、IL−2またはその両方の組合せで処置する全身的な投与は、深刻な副作用を引き起こす。これらのサイトカインは、種々の体組織に対して非常に多くの影響を与えるので、患者に全身的に送達するのはあまり安全ではない。従って、本発明の対象は、自己抗体調節制御に関与する哺乳類細胞を処置して、自己抗体産生を下方制御する細胞集団を増加させる方法およびキットを提供することである。
【発明の開示】
【発明が解決しようとする課題】
【0014】
(本発明の要旨)
ここに概略する目的のように、本発明は、生体外の末梢血単核細胞 (PBMC)のサンプル中の免疫応答を阻害する方法であって、細胞集団に調節組成物を添加することを含む方法を提供する。
【課題を解決するための手段】
【0015】
更なる態様では、本発明は、患者の自己免疫異常を処置する方法を提供する。その方法は、患者由来の末梢血単核細胞(PBMC)を患者から取り出すこと、およびこの細胞を調節組成物で、免疫細胞による組織損傷を抑制するのに十分な時間、処理することを含む。特に、本発明の方法は、抗体産生を抑制し、または細胞を誘発して、抗体産生を下方調節し、そして抗体仲介自己免疫疾患の患者における細胞仲介免疫応答を高める。次いで、細胞を患者に再導入すると、自己免疫の症状が改善する。調節組成物は、好ましくは、TGF−βおよびT細胞のTGF−βヘの応答を可能にする物質を含む。
【0016】
更なる態様では、本発明は、細胞仲介自己免疫異常を処置する方法を提供する。この方法は、該患者から末梢血単球細胞(PBMC)を取り出し、該細胞を調節組成物で、異常の免疫応答を抑制するのに十分な時間処置することを含む。処置細胞を患者に再導入すると、自己免疫症状の改善が得られる。調節組成物は、好ましくは、TGF−βおよびT細胞のTGF−βへの応答を可能にする物質を含む。
【0017】
更なる態様では、本発明は、患者の自己免疫異常の処置用キットを提供する。このキットは、抗体仲介自己免疫異常または細胞仲介異常の患者の細胞を受容するのに適当な細胞処置容器および少なくとも1用量の調節組成物を含む。
【0018】
(図面の説明)
図1は、SLE患者のPBMCをIL−2およびTGF−βと共にインキュベーションすることにより、自発的免疫グロブリン産生が減少することを示す。PBMC(2×105/well)を、AIM−V血清を含まない培地であって、IL−2(10U/ml)およびTGF−β(10pg/ml)を含むか、または含まない培地中で培養した。3日後、ウェルを3回洗浄し、新たなAIM−V培地を加えた。更に7日後、上清をウェルから回収し、IgG成分をELISAで測定した。
【0019】
図2は、IL−2およびTGF−βの両方により、自発的IgG産生が有意に減少することを示す。その値は、図1の説明に記載の通りに培養した12人のSLE患者のPBMCにより産生した平均±SEMのIgG(μg/ml)を示す。ただし、IL−2(10U/ml)のみ、またはTGF−β(10pg/ml)のみで培養した細胞もある。
【0020】
図3Aおよび3Bは、抗TGF−βがIL−2の効果を逆転することを示す。SLE患者のPBMCを、3日間、IL−2(10U/ml)の存在下(黒色バー)、または非存在下(灰色バー)で培養した。これらの培養液中に含まれていたのは、培地、抗TGF−β(10μg/ml)または対照マウスIgG1(10μg/ml)であった。3日後、ウェルを洗浄し、新たなAIM−V培地を加えた。更に7日後、上清を回収し、ELISAでIgG(図3A)または抗核タンパク質(NP)(図3B)成分を検定した。
【0021】
図4A、4Bおよび4Cは、抗体産生に対するDC8+T細胞の調節効果を示す。(A)健康人のIgG産生の抑制におけるNK細胞とCD8+細胞の相乗作用。CD4+細胞およびB細胞を抗CD2で刺激し、CD8+細胞およびNK細胞の作用を調べた。NKおよびCD8+細胞の組み合わせは、我々が以前報告した(Gray, J.D. et al., (1998), J Immunol 160: 2248−2254; Gray, J.D. et al., (1994), J Exp Med 180; 1937−1942)抗CD2誘発性IgG産生を顕著に阻害した。(B)NK細胞およびCD8+細胞はSLEにおけるIgG合成を促進する。活動性SLEの患者由来のCD4+細胞および正常人の休止B細胞を、抗CD2で刺激した。SLE CD8+細胞によるIgG産生の促進は、NK細胞の添加により顕著に増加した。(C)SLEにおけるCD8+T細胞機能のサイトカインによる正常化。図4Bに示す試験と平行して、この患者のCD4+T細胞を、CD8+T細胞の存在下または非存在下、抗CD2で刺激した。表示のようにIL−2(10U/ml)および/またはTGF−β(2pg/ml)を加えた。これらサイトカインにより、CD8+細胞のヘルパー作用が喪失し、その細胞がIgG産生の75%を阻害した。
【0022】
図5Aおよび5Bは、非刺激細胞および抗CD2刺激細胞によるTGF−β1のリンパ球産生を示す。健常者ドナーならびにSLE患者およびRA患者のPBLを、マイクロタイタープレートに1×105/ウェルで加えた。幾つかのウェルは、抗CD2mAbsGT2(1:40)およびT11(1:80)であった。37℃で2日後、上清を回収し、活性TGF−β1および全TGF−β1を検定した。有意なp値を示す。
【0023】
図6は、TNF−αおよびIL−10のT細胞産生に対するTGF−βの作用を示す。血清なしのAIMV培地中の精製T細胞(1x105細胞/ウエル)を、平底ミクロウエルに加え、低量(0.5μg/ml)または高量(5μg/ml)のConAで、IL−2と共にまたはなしに、TGF−β(1ng/ml)の存在または不存在下に刺激した。上清を2日および5日に採取し、TNF−αおよびIL−10についてELISAで調べた。最大産生がTGF−βは2日に、IL−10は10日にあった。TGF−βはIL−10の産生をなくし、TNF−αの産生を上方調節した。
【0024】
図7は、TGF−βによる調節性T細胞の生成にTNF−αが必須の中間体であることを示す。精製CD8+細胞を、ConA(2.5μg/ml)、IL−2(10U)およびTGF−β(10pg/ml)で一夜インキュベートした。洗浄後これらの細胞をCD4+B細胞に加え、抗CD2で刺激した。いくつかのウエルに抗TNF−α抗体(10μg/ml)またはイソ型対照抗体(10μg/ml)を入れた。7日後に上清をIgG含量についてELISAで調べた。抗TNF−αは条件CD8+細胞の調節活性を逆転した。
【0025】
図8は、TGF−β起動T細胞によるTh1サイトカインの産生の増加がTNF−αに依存性であることを示す。精製ナイーブT細胞をConA(5μg/ml)およびIL−2(10U/ml)とともにTGF−β(1ng/ml)の存在下に培養した。いくつかのウエルには中和抗TNF−α抗体(10μg/ml)またはイソ型対照抗体(10μg/ml)も加えた。5日培養後に細胞を水洗し、1x105細胞/ウエルで新鮮な培地に入れた。翌日にConAおよびIL−2で6時間再刺激し、ブレフェルジンA(10μg/ml)の存在下にCD8およびサイトカインについて着色した。TNF−α、IL−2、IFN−γを発現するCD8+およびCD8−細胞の%を示す。一次培地におけるTNFの中和はTh1サイトカイン産生に対するTGF−β作用の増加を無くする。
【0026】
図9は、細胞毒性T細胞活性の抑制物質の産生におけるTGF−βの作用を示す。Eロゼット法によりつくられたドナーA由来のT細胞を2つに分けた。1つを同種異系混合リンパ球反応(allo−MLR)についての応答物として用いた。他の1つを用いて、細胞を適当なモノクロナール抗体で染め、その細胞を免疫磁性ビーズで除いた後、陰性選択で表示されるT細胞サブセットをつくった。応答T細胞をドナーB由来の刺激細胞と混合し(照射T細胞は末梢血単核細胞を枯渇した)、5日培養してキラー細胞を作った。対照は刺激細胞の有無で5日培養したT細胞サブセットからなる。その後、細胞を洗い、計数し、アロ細胞毒性T細胞活性の検定に使用した。ドナーA由来の応答細胞をドナーB由来のクロミウム標識リンパ芽球とエフェクター質中で混合し、細胞比率を標的にして、クロミウム放出を標準的4時間検定法で測定した(白四角)。刺激物とともに培養したT細胞サブセットを、4応答細胞につき1調節細胞の比率で加えた(白丸)。刺激物と培養のT細胞サブセットへのTGF−βの添加を黒丸で示す。すべての試験で、TGF−βの最大作用はナイーブCD4CD45RA+CD45RO−細胞に対して見られた。
【0027】
図10は、TGF−βにより起動されたCD4細胞のアロ細胞毒性Tリンパ球(CTL)活性に対する作用を示す。刺激物質なしで5日間培養されたCD4CD45RAの付加はCTL活性に対する作用を有しない(結果を示していない)。これらのT細胞を刺激物質と培養すると、低度から中程度の抑制活性となる。すべての試験において、これらのT細胞のTGF−β1ng/mlとの培養は、アロCTL活性を顕著に抑制するか無くする。
【0028】
図11は、調節性T細胞がCTL活性を抑制するために細胞接触を必要とすることを示す。調節性CD4細胞は、上記のように、TGF−βと培養したCD4CD45RAからつくった。これらの細胞のいくつかを応答物質およびクロミウム標識の標的細胞と混合し、他の細胞を膜によりキラー細胞から分離した。細胞毒性Tリンパ球活性(CTL)の抑制を、T細胞がキラー細胞と直接接触したときにのみ認めた。
【0029】
図12は、TGF−βで誘発された調節性CD4+T細胞によるリンパ球増殖の抑制を示す。ドナーA由来のナイーブCD4+T細胞を上記のように刺激細胞と混合し、表示の比率で新鮮な応答物質および刺激物質に加えた。バーは7日間培養後のトリチウム標識チミジンの取り込み±SEMを示す。薄色のバー(Nil)は、添加CD4+細胞なしの応答T細胞の増殖性応答を示す。濃色バーは、刺激細胞と培養した対照CD4+細胞のTGF−βなしでの作用を示す。黒バーは、刺激細胞と培養した対照CD4+細胞のTGF−βあり(1ng/ml)での作用を示す。照射刺激細胞に加えた新鮮な応答細胞の増殖性応答に対するこれらのCD4+細胞の作用を培養7日間後に示す。バーはトリチウム標識チミジンの平均取り込みを示す。
【0030】
図13は、CD25+CD4T細胞の調節活性を示す。CD4+細胞を照射同種異系非T細胞±TGF−β(1ng/ml)で5日間刺激した。水洗後、CD4+細胞をDIIで染め、新鮮な応答T細胞をカルボキシフルオレセイン(CFSE)で染めた。対照またはTGF−β駆動CD+細胞を応答T細胞およびアロ刺激細胞に1:4の比率で加えた。5日後に細胞を採取し、フローサイトメトリーにより分析した。CD8+におけるCFSEの強さをDII陰性細胞でもって決定した。TGF−β駆動CD+細胞の応答T細胞への付加はCD8+細胞による細胞分裂を顕著に低下した。
【0031】
図14は、調節CD+4細胞がその表面でCD25+(IL−2)受容体を発現することを示す。対照またはTGF−β誘発CD4+調節性T細胞を上記のようにつくった。アロ刺激細胞およびTGF−βで条件を整えた後、CD4+細胞を細胞選別によりCD25+とCD25−に分けて、新鮮な応答T細胞および照射刺激細胞に加えた。これらの応答細胞のキラー刺激Tリンパ芽細胞に対する能力を、標準的4時間クロミウム放出検定で示す。
【0032】
図14Aにおいて、白四角は添付のCD4+細胞のないCTL活性を表示する。対照またはTGF−β誘発調節性T細胞を応答細胞とともに1:4の比率で加えた。白丸からすると対照CD4+細胞がCTL活性を変えなかった。黒丸からするとTGF−βが誘発CD4+がCTL活性をほとんど完全に抑制した。黒菱形からするとCD25+サブセットのみが抑制活性を有していた。CD25サブセット(黒四角)は抑制活性を有していなかった。
【0033】
図14Bは、MLRに加えたCD4+調節細胞の数を減少さす作用を示す。わずか3%への減少は抑制作用の減少における最小の作用である。
【0034】
図15は、少量のブドウ球菌性エンテロトキシンB(SEB)でのT細胞の反復刺激がT細胞を誘発してTGF−βの免疫抑制レベルをつくることを示す。CD4+T細胞を、SEB(0.01ng/ml)およびスパー抗原提示細胞として照射B細胞で、TGF−βの存在または不存在で、矢印で表示の時に刺激した。活性TGF−βを2日または5日後に測定した。
【0035】
図16は、低量のSEBでCD4+T細胞の反復刺激によってT細胞がTGF−βの免疫抑制レベルをつくることを示す。CD4+T細胞をSEB(0.01ng/ml)およびスパー抗原提示細胞として照射B細胞で、TGF−βの存在または不存在で、矢印で表示の時に刺激した。活性TGF−βを2日または5日後に測定した。
【0036】
図17は、ナイーブ(CD45RA+CD45RO−)CD4+およびCD8+T細胞に対するSEBの作用を示す。細胞をSEBで5日毎に計3回刺激した。各T細胞サブセットおよびCD25IL−2受容体活性マーカーを発現する細胞の%を各刺激後に測定した。パネルAおよびCは、TGF−β1ng/mlが最初の刺激に含まれていると、CD4+T細胞が反復刺激後に培養基中で優勢になることを示す。パネルBおよびCは、SEB刺激細胞によるCD25発現が対照培養基中で第3刺激で減少することを示す。しかしCD25発現は、T細胞がTGF−βで駆動されていると、高く留まっている。
【0037】
(詳細な説明)
本発明は、全身性エリマトーデス(SLE)などの細胞仲介および抗体仲介の両方の異常を含む自己免疫異常を処置する方法に関する。この方法は、患者から細胞を取り出し、この両方の1つに作用できる組成物でもって細胞を処置する。1つの実施態様において、抗体仲介自己免疫異常の症状を本発明の組成物によって改善する。この組成物は、自己抗体などの抗体の産生を阻害して、B細胞の過剰活性を下方調節する。さらに、この組成物は、SLEや他のある種の抗体仲介自己免疫異常の患者においてしばしば欠失している細胞仲介免疫応答を高める。すなわち、抗体仲介自己免疫異常の患者を処置して、その欠失細胞仲介症状を改善できる。
【0038】
あるいは、この組成物を使用して細胞仲介自己免疫疾患を処置する。この実施態様において、組成物は免疫細胞を誘発し、抑制T細胞をつくる。この抑制T細胞は他のT細胞が細胞毒性になって個体の細胞および組織を攻撃するのを防ぐ。このように、組成物は細胞毒性を低下することにより細胞仲介自己免疫異常の症状を改善する。
【0039】
この方策は、現在のほとんどすべての処置が抗炎症剤や免疫抑制剤を使用すると異なっている。普通よく用いられるコルチコステロイドは、サイトカインの産生を抑制し、組織の損傷を起こす最終事象を阻止するが、基にある自己免疫応答を一般に変えることはない。細胞障害剤またはモノクローナル抗体などの実験的に遺伝子工学的に処理された生物剤も、特殊なリンパ球集団をなくするか、その機能を妨害する。これらの薬剤は、一般に緩和な効果をもたらすのみであって、また重大な有害副作用を起こす。ある種のサイトカインは、患者に全身的に与えられるが、重大な有害副作用に関連する広範な作用を有している。
【0040】
一方、本発明の方法による治癒効果では、正常な調節性細胞機能が回復し、免疫系が「再置」される。本方法の他の重要な利点は、重大な有害副作用があまり起きないことである。サイトカインなどの調節組成物のごく少量を患者に戻すにすぎないので、毒性が最小となる。
【0041】
IgGを自発的に分泌する循環Bリンパ球が活動性SLEの患者で増加する(Blaese, R.M., et al.(1980), Am J. Med. 69: 345−350; Klinman, D.M. et al.(1991) Arthritis Rheum 34: 1404−1410)。インビトロでのポリクローナルIgGおよび自己抗体の持続的な産生には、T細胞の助けを必要とする(Shivakumar, S. et al.(1989), J Immunol 143: 103−112)。自発的IgG産生のT細胞による調節についての以前の研究によると、健常人ではCD8+T細胞が抗体産生を阻害するのに対し、SLEではこの細胞が代りにB細胞機能を支持する(Linker−Israeli, M. et al.(1990), Arthritis Rheum 33: 1216−1225)。他の自己免疫疾患、例えば、慢性関節リウマチや多発性硬化症において、抗体よりもむしろT細胞が組織損傷の原因であり炎症をもたらす(Panayl GS et al. Anthritis Rheum (1992) 35: 725−775, Allegretta M et al. Science (1990) 247: 718−722)。
【0042】
従って、本発明は、抗体−およびT細胞−仲介自己免疫疾患を処置する方法に関し、自己免疫異常の患者から末梢血単核細胞(PBMC)を取りだし、その細胞を調節組成物で処置する。
【0043】
理論に拘束されるわけでないが、本発明方法が作動するには、いくつかの経路があるようである。その第1は、調節組成物で細胞を処置すると、処置細胞における抗体産生が直接抑制され、自己免疫症状の改善につながり得る。あるいは、または加えて、細胞の処置は調節性細胞を誘導して、他の細胞における抗体産生を下方調節する。本明細書における抗体はすべての形態の抗体、例えば、IgA、IgM、IgG、IgEなどを含む。明らかな結果は系における抗体量の低下である。
【0044】
さらに、細胞の処置は、抗体仲介自己免疫症状の患者において細胞仲介免疫応答を高める。理論に縛られるものでないが、細胞の処置がIL−10とTNF−αとの均衡を修復して、Th1サイトカインの産生を高め、細胞仲介免疫を正常化するようである。
【0045】
さらに、TGF−βを含む調節組成物で免疫細胞を刺激すると、細胞仲介免疫応答が抑制される。理論に縛られるものでないが、CD4+T細胞を刺激すると、活性TGF−βの免疫抑制レベルをつくり、細胞仲介免疫応答を抑制するようである。あるいは、CD4+T細胞を刺激すると、他のT細胞の活性化および/またはエフェクター機能を作用の接触依存性メカニズムによって抑制する。これらの作用には、TGF−βの存在で活性化されるCD4+細胞を必要とする。
【0046】
このように、本発明は、異常な免疫応答を阻害する。抗体仲介自己免疫異常の患者において、本発明は、調節組成物で末梢血T細胞を生体外で処置することにより、T細胞の能力を修復し、抗体産生を下方調節し、細胞仲介応答を修復する。細胞仲介異常の患者において本発明は、他のT細胞において細胞毒性T細胞活性を抑制する調節性T細胞をつくる。
【0047】
「免疫応答」とは、外部または自己の抗原に対する宿主の応答を意味する。「異常免疫応答」とは、自己と非自己とを識別する免疫系のないこと、または外部抗原に応答しないことを意味する。換言すると、異常免疫応答とは、患者の症状をもたらしている不適当に調節された免疫応答である。「不適当に調節された」とは、不適当に誘発されること、不適当に抑制されること、および/または応答性のないことを意味する。異常な免疫応答は、限定でないが、生体自身の組織に対する抗体の産生により生じる組織損傷や炎症、IL−2、TNF−α、IFN−γの産生の障害、作用の細胞毒性または非毒性メカニズムにより生じる組織障害を含む。
【0048】
従って、好ましい実施態様において、本発明は患者の抗体仲介自己免疫異常を処置する方法を提供する。「抗体仲介自己免疫疾患」とは、個体が自己の細胞または組織の構成成分に対する抗体を発生する疾患を意味する。抗体仲介自己免疫疾患には、限定するのではないが、全身性エリマトーデス(SLE)、天疱瘡、重症筋無力症、溶血性貧血、血小板減少性紫斑症、グレーブス症、皮膚真菌症、シェーグレン症などがある。本発明方法による処置に好適な疾患はSLEである。
【0049】
さらに、抗体仲介異常の患者は細胞仲介免疫応答において欠失をしばしば有する。「細胞仲介免疫応答の欠失」とは、感染に対する宿主防御の障害を意味する。感染に対する宿主防御の障害は、限定でないが、遅い過剰反応の障害、T細胞の細胞毒性の障害、TGF−β産生の障害を含む。他の欠失は、限定でないが、IL−10産生の増加、およびIL−2、TNF−α、IFN−γの産生の減少を含む。本発明の方法を用いて、精製T細胞を刺激して、IL−2、TNF−α、IFN−γの産生を増加させ、IL−10の産生を低下させる。この方法で刺激し得るT細胞は、限定でないが、CD4+およびCD8+を含む。
【0050】
ある実施態様においては、抗体仲介異常を処置しない。
好ましい実施態様において、本発明は、患者における細胞仲介自己免疫異常を処置する方法を提供する。「細胞仲介自己免疫疾患」とは、個体の細胞が活性化または刺激されて、細胞毒性となり、自己の細胞または組織を攻撃する疾患を意味する。あるいは、個体の自己免疫細胞が他の細胞を刺激して作用の細胞毒性または非細胞毒性メカニズムによって組織障害を起こす。細胞仲介自己免疫疾患は、限定でないが、橋本病、多発性筋炎、炎症性腸疾患、多発性硬化症、真性糖尿病、慢性関節リウマチおよび硬皮症などを含む。
【0051】
自己免疫異常の「処置」とは、自己免疫異常の少なくとも1つの症状が、本明細書に記載の方法で改善されることである。その評価を種々の方法でなす。この評価には、患者側の主観的または客観的な因子が含まれる。例えば、疾患の免疫原性徴候を調べる。例えば、自発的抗体および自己抗体産生のレベル、SLEの場合は特にIgG産生のレベルを低下する。全Ig抗体または自己抗体のレベルを測定する。限定するものでないが、抗2本鎖DNA(dsDNA)抗体、抗核タンパク質抗体、抗Sm、抗Rhoおよび抗Laである。身体症状が変化する。例えば、SLEの皮疹の消失または減少である。腎機能検査を行って変化を調べる。炎症に関する組織障害について臨床検査を行う。循環免疫複合体レベルおよび血清補体レベルの低下も改善の証拠である。SLEの場合、貧血の減少も見られる。免疫抑制剤などの薬剤に対する患者の必要性が低下することも処置効果の証である。処置効果について他の判定方法は、自己免疫疾患分野の当業者にとって明らかであろう。
【0052】
「患者」とは、処置の対象となる哺乳動物を意味するが、ヒトが好ましい。ある場合において、本発明方法は、実験動物、獣医学分野、疾患モデル動物にも使用され、これらの動物には、限定ではないが、マウスラット、ハムスターなどのゲッシ類および霊長類がある。
【0053】
本発明では、患者から血液細胞を取り出す。一般に、標準的な方法で患者から末梢血単核細胞(PBMC)を取り出す。「末梢血単核細胞」すなわち「PBMC」とは、リンパ球(T細胞、B細胞、NK細胞など)および単球を意味する。詳しく下記するように、調節組成物の主要な作用は、CD8+T細胞がIG産生を抑制することである。好ましくは、PBMCのみを採取する。赤血球または多形核白血球を患者に残すようにするか、これらを患者にもどす。これは既知の方法、例えば、白血球泳動(leukophoresis)で行う。一般に5から7リットルの白血球泳動工程を行う。患者からPBMCを基本的に採取し、他の血液成分を戻す。細胞サンプルの採取は、ヘパリンなど抗凝固剤の存在下で、既知方法により行う。
【0054】
いくつかの実施態様においては、白血球泳動工程を必要としない。
一般に、PBMC含有のサンプルを種々の方法であらかじめ処理する。まず細胞を採取し、次いで採取に際して自動的に濃縮されていなければ、濃縮し、さらに精製および/または濃縮する。細胞を水洗し、計数し、緩衝液中に入れる。
【0055】
一般的にPBMCを標準的方法で処置のために濃縮する。好ましい態様において、白血球泳動採取工程によってPBMCの濃縮サンプルを無菌の血球パックに得、ある種の試薬および/または調節組成物を含有せしめる。詳しくは下記する。一般に、追加の濃縮/精製工程を行なう。既知のFicoll−Hypaque密度勾配遠心法などである。
【0056】
好ましい態様において、PBMCを洗い、血清タンパク質および溶性血液成分、例えば自己抗体、阻害剤などを既知方法で取り出す。一般に、生理媒質または緩衝液を加え、遠心法を行なう。必要に応じて繰り返す。PBMCを生理培養液、好ましくはAIM−V血液なしの培地(Technologie)に再懸濁するが(血清がかなりの量の阻害剤を含有するので)、Hanksバランス塩溶液(HBBS)や生理塩緩衝液(PBS)も使用する。
【0057】
一般に、次いで細胞数を数える。一般に1×109から2×109の白血球を5−7リットルの白血球泳動工程から採取する。これらの細胞を大略200mlの緩衝液または培地に移す。
【0058】
好ましい態様において、PBMCを1以上の細胞型について富ます。例えば、PBMSをCD8+T細胞またはCD4+T細胞ついて富ます。このことは既知である(Gray et al.(1998),J.lmmunol.160:2248、出典明示により本明細書の一部とする。)。一般に、これは市販の免疫吸収カラムを用いるか、研究的方法で行なう(PBMCをナイロンウールカラムに加え、溶出の非接着細胞をCD4、CD16、CD116、CD74に対する抗体で処理し、免疫磁気ビーズで処理して、CD8+T細胞に富んだ集団を取る)。
【0059】
好ましい実施態様において、PBMCを自動化閉鎖系、例えば、Nexell Isolex 300i Magnet Cell Selection System において分離する。一般的に、これは、細胞の分離、活性化、抑制細胞機能の開発のために使用する方法の無菌性の確保および標準化の確保のために行う。
【0060】
細胞に必要な前処理を一旦ほどこしてから、細胞を調節組成物で処置する。「処置」とは、細胞を調節組成物と共に十分な時間でインキュベートし、抗体および自己抗体産生を阻害する能力が、特に患者にもどしたときに、発生するようにすることである。インキュベーションは一般に生理的温度で行なわれる。上記したように、阻害が起きるのは、処置細胞による抗体産生の直接的抑制の結果か、患者リンパ臓器における抗体産生を下方調節する調節細胞の誘導の結果である。
【0061】
「調節組成物」または「抗体産生阻害組成物」または「液素性阻害組成物」「非特異的免疫細胞阻害物質」または「特異的T細胞阻害物質」または「阻害組成物」または「抑制組成物」は、免疫応答の抑制を起こし得る組成物を意味する。この抑制は、T細胞活性化の阻害、自発的抗体および自己抗体の産生または細胞毒性の阻害を含む。一般に、これらの組成物はサイトカインである。適切な調節組成物は、限定でないが、抗CD2などのT細胞アクチベーター、例えば抗CD2抗体およびCD2リガンド、LFA−3、T細胞アクチベーターの混合物や組合せ物、例えば、コンカナバリン Concanavalin A(ConA)、ブドウ球菌エンテロトキシンB(SEB)、抗CD3、抗CD28、およびサイトカイン、例えば、IL−2、IL−4、TGF−β、TNF−αを含む。抗体抑制に好ましい調節組成物は、T細胞アクチベーター、IL−2、TGF−βの混合物である。細胞毒性の抑制に好ましい組成物はTGF−βである。
【0062】
調節組成物の濃度は、組成物の性質によって変ってくる。好ましい態様においては、TFG−βが調節組成物の成分である。「トランスフォーミング成長因子−β」すなわち「TGF−β」は、TGF−βファミリーのいずれか1つを意味する。3種のイソ型、TGF−β1、TGF−β2、TGF−β3がある。参照、Massague,J.(1980),J.Ann.Rev.Cell Biol 6:597。リンパ球および単球は、このサイトカインのβ1のイソ型である(Kehrl,J.H.et al.(1991),lnt J Cell Cloning 9:438−450)。TGF−βは、処置される哺乳動物に対して活性であるTFG−βのすべての型であり得る。ヒトでは、組換えTFG−βが現在のところ好ましい。好ましいヒトTGF−βは、Genzyme Pharmaceuticals,Farmington,MA.から購入できる。一般に、TGF−βの濃度は、細胞懸濁液の約2picogram/mlから約5nanogramであって、約10pgから約4ngが好ましく、約100pgから約2ngが特に好ましく、1ng/mlが理想的である。
【0063】
好ましい態様において、IL−2を調節組成物で用いる。IL−2は、処置される哺乳動物に対して活性であるIL−2のすべての型であり得る。ヒトでは組換えIL−2が現在のところ好ましい。組換えIL−2は Cetus,Emerville、CA.から購入できる。一般に、使用されるIL−2の濃度は、細胞懸濁液の約1Unit/mlから約100U/mlであり、約5U/mlから約25U/mlが好ましく、10U/mlが特に好ましい。好ましい態様においてIL−2は単独で用いない。
【0064】
好ましい態様において、抗CD2抗体またはCD2リガンドLFA−3などのCDアクチベーターを、調節組成物として用いる。CD2はTリンパ球により発現される細胞表面グリコタンパク質である。「CD2アクチベーター」とは、CD2情報伝達経路を開始する化合物を意味する。好ましいCD2アクチベーターには、抗CD抗体(OKT11,American Type Culture Collection,Rockville MD)が含まれる。一般に、CD2アクチベーターの濃度は、TGF−βの産生を誘導するのに十分なものである。抗CD2抗体の濃度は、約1ng/mlから約10μg/mlの範囲にあり、約10ng/mlから約100ng/mlが特に好ましい。
【0065】
いくつかの態様においては、細胞を活性化するためにマイトジェンの使用が望ましい。すなわち、多くの休止相細胞は多量のサイトカイン受容体を含有していない。ConcanavalinAなどのマイトジェンを使用すると細胞が刺激されて、サイトカイン受容体が産生し、本発明方法をさらに効果的にする。マイトジェンを用いるとき、既知方法で知られているように、その濃度範囲は1μg/mlから約10μg/mlである。さらに、既知方法のように、CoAを離す成分、例えばα−メチルマンノシッドと共に細胞を洗うのが望ましい。
【0066】
好ましい実施態様において、T細胞を、抗CD2、抗CD3、抗CD28などのマイトジェンあるいはモノクロナール抗体の組み合せ、すなわち自己抗原と補刺激物質としてのCD28やIL-2でもって刺激する。ConAもT細胞の刺激に使用する。第2培養におけるTGF−βのありなしでのT細胞の反復刺激は最大抑制活性を得るのに必要なようである。
【0067】
好ましい実施態様において、本発明は、T細胞をTGF−βで調整することを含む。このT細胞には、限定でないが、CD8+TやCD4+TおよびCD8−CD4−などの他のマイナーT細胞サブセット、NK細胞などが含まれる。これらのT細胞は、他のT細胞が細胞毒性エフェクター細胞になるのを防ぐ。
【0068】
好ましい実施態様において、本発明は、CD8+TまたはCD4+T細胞をTGF−βで調整して、免疫抑制レベルのTGF−βをつくる方法を含む。
好ましい実施態様において、本発明は、CD8+TまたはCD4+T細胞をTGF−βで調整して、接触依存性メカニズムにより抑制されるT細胞をつくる方法を含む。
【0069】
好ましい実施態様において、本発明は、ナイーブCD4+T細胞を刺激物質で処置して、該CD4+細胞が免疫抑制レベルの活性TGF−βをつくるようにすることを含む方法を提供する。
【0070】
好ましい実施態様において、本発明は、ナイーブCD4+T細胞をTGF−βの存在下に刺激してCD4+細胞数を増やすことを含む方法を提供する。
好ましい実施態様において、本発明は、IL−10の産生を減少し、併せてTNF−αの産生を増加する方法を提供する。
【0071】
調節組成物を細胞と共に、効果を発揮するに十分な時間インキュベートする。好ましい態様において、細胞は、調節組成物での処置に続いて、患者にもどす。好ましい態様において、細胞と調節組成物とのインキュベーションは、約12から約7日行なう。この時間は望む抑制活性により変る。抗体産生の抑制については、48時間が特に好ましい。細胞毒性の抑制については、5日が特に好ましい。
【0072】
ある態様において、細胞を一定の時間インキュベートし、水洗して調節組成物を除去し、再びインキュベートする。患者に導入する前に、ここに概記したように細胞を好ましくは水洗し、調節組成物を除去する。試験または検査のために、さらなるインキュベーションも行うことができ、数時間から数日間である。患者への導入前に抗体産生の検査をしょうとするときは、細胞を数日間インキュベートすると、抗体産生(またはその非産生)が明らかになる。
【0073】
細胞の処置後すぐに、患者に自動的に再移行する前に細胞を検査することができる。例えば、細胞を採取して行う。無菌検査、グラム染色、微生物学的検査、LAL試験、マイコフラズマ検査、細胞型同定のためのフローサイトメトリー、機能検査などである。同様に、これらの検査などの試験を処置の前や後に行うことができる。
【0074】
好ましい態様において、抗体産生の定量や定性(すなわちタイプについて)をなし得る。例えば、抗体の全レベルあるいは抗体の具体的なタイプを調べる。例えば、IgA、IgG、IgM、抗DNA自己抗体、抗核タンパク質(NP)抗体などの具体的なタイプについてである。調節性T細胞も、T細胞活性を抑制する活性、インビトロで特異的な標的細胞に対するT細胞の細胞毒性を防ぐ活性について、検査できる。
【0075】
好ましい態様において、抗体、特にIgGのレベルの検査をELISAアッセイなどの既知の方法(Abo et al.(1987),Clin Exp.lmmunol.67:544 and Linker−Israeli et al.(1990),Arthritis Rheum 33:1216, 出典明示により本明細書の一部とする。)で行なう。これらの技術は自己抗体などの特異的な抗体のレベルを検出するのにも用い得る。
【0076】
好ましい態様において、処置によって産生IgGおよび自己抗体の量の有意な低下が起きる。低下は、少なくとも10%が好ましく、少なくとも25%が特に好ましく、少なくとも50%が特別に好ましい。多くの態様において75%以上の低下がみられる。
【0077】
好ましい態様において、移行前に全または活性のTGF−βの量も検査できる。記載のように、TGF−βを移行後に活性化される潜在的前駆物質としてつくる。
【0078】
処置後、細胞を患者に移行すなわち再導入する。一般に既知のよう行ない、通常は静脈投与によって処置細胞を患者に注入すなわち導入する。例えば、無菌シリンジなどの移行手段を用いて注射により50mlのFenwall注入バッグに細胞を入れる。次いで細胞をすぐに静脈投与によって一定の時間、例えば、15分間で患者の自由に流れている静脈に注入する。ある態様では、緩衝剤や塩などの追加の試薬も加え得る。
【0079】
細胞を患者に再導入後に、処置の効果を所望に応じて、上記の方法で検査できる。すなわち、疾患の免疫原性徴候を検査する。例えば、全抗体または特異的な免疫グロブリンの力価、腎機能、組織障害などを検査する。T細胞の機能についての試験、例えば、T細胞数、表現型、活性状態、抗体および/またはマイトジェンに応答する能力などについても検査し得る。
【0080】
処置は必要に応じて繰り返えす。例えば、数週の期間で週1回、または時間の期間で数回、例えば、2週間に3−5回行う。一般に、自己免疫疾患の症状の改善はなんらかの期間、好ましくは少なくとも数カ月続く。その間に患者が症状の再起を感じると、その時点で処置を繰り返えす。
【0081】
好ましい態様において、本発明方法を実行ため、すなわち細胞を調節組成物と共にインキュベートするためのキットを提供する。キットはいくつかの成分を含有する。キットは、抗体仲介自己免疫疾患の患者からの細胞を受容するのに用いられる細胞処置容器を含む。この容器は無菌とする。ある態様において、細胞処置容器を細胞採取に用いる。例えば、入口を介して白血球泳動器と共に用いる。他の態様では、分離した細胞採取容器を用い得る。
【0082】
好ましい実施態様において、キットは、細胞仲介異常の患者からの細胞を受けるように調整された細胞処置容器を含む。キットはまた、特異的T細胞サブセットを精製し、それを患者に戻すように増やすために、自動化閉鎖系での使用に調整できる。
【0083】
細胞処置容器の形および材質は様々であり、当業者によく知られている。一般に、容器はいくつかの異なる形態を取る。例えば、IVバッグのような柔かいバッグであり、または細胞培養器のような硬い容器である。攪拌できるようになっている。一般に、容器の材質は何でもよいが、生物学的に非活性の物質であり、例えばガラス、ポリプロピレンやポリエチレンなどを含むプラスチックである。細胞処置容器は1以上の入口または出口を有し得る。細胞、試薬、調節組成物などの導入または取り出しのためのものである。例えば、容器は、患者に再導入する前の分析のために細胞のフラクションを採取するためのサンプル採取口を有する。同様に、容器は、患者に細胞を導入するための出口を有する。例えば、容器はIV装置につけるためのアダプターを含み得る。
【0084】
キットは調節組成物の少なくとも1つの用量を含む。本明細書での「用量」とは、サイトカインなどの調節組成物の1つの量で、作用を発揮するのに十分な量を意味する。ある場合には、複数の用量物を含む。ある態様において、用量物の細胞処置容器への添加は入口を介して行なう。あるいは好ましい態様において、用量物は細胞処置容器にすでに存在している。好ましい態様において、用量物は安定性のために乳濁形態にあり、細胞培養液や他の試薬を用いて再生する。
【0085】
ある態様において、キットは少なくとも1種の試薬を追加的に含む。それには緩衝液、塩、培養液、タンパク質、薬剤などがある。例えば、マイトジエン、モノクローナル抗体、細胞分離のための処置磁気ビードを含み得る。
ある態様において、キットはその使用に際しての説明書を追加的に含む。
【0086】
下記の実施例は、上記の本発明をより完全に記述したものであり、また、本発明の種々の態様を実施するために考えられる最上の方法を記述した。これらの実施例は本発明の範囲を限定するものでなく、説明の目的のために提示される。出典明示されたすべての引用文献は全体を本明細書の一部とする。
【0087】
実施例
実施例1
IL-2およびTFG-βの混合物によるPBMCの処置
実施例1によって示されるのは、SLE患者のPBMCをIL-2およびTFG-βで比較的簡単に処置することにより、自発的ポリクローナルIgGおよび自己抗体産生が顕著に阻害されることである。下記で考察するように、12人の活動性SLE患者のPBMCを、TGF-βの存在または非存在で3日間IL-2に曝露し、洗浄し、そして7日以上培養した。IgG分泌の平均減少は79%であった。最も強力な阻害効果は、最も顕著なB細胞の活動過剰の場合に観察された。抗核タンパク質(NP)抗体の自発的産生が観察されたのは4例あり、PBMCのサイトカイン処置により自己抗体産生は、50から96%減少した。IL-2は、個々の患者におけるTGF-β依存性または非依存性の何れかの機構による抗体産生を阻害した。活動性SLE患者における抗CD2刺激性IgG産生の試験では、IL-2およびTGF-βがIgG産生におけるCD8+T細胞の促進効果を逆転し、代わりに抑制活性を誘発し得る。
【0088】
方法
自発的抗体合成についての被験者
12人の被験者をSLE診断で選択した。それらはSLEを分類するARA基準(Arnett, F.C. et al., (1998), Arthritis Rheum 31: 315-324)を満たしていた。これらの患者はすべて女性であり、スペイン人8人、アフリカ系アメリカ人2人、そしてアジア人2人であった。それぞれの患者の年齢および病歴を表1に示す。5人が入院患者で、7人は外来患者であった。コルチコイド(コルチコステロイド)および抗マラリア薬を投与されている患者も表示した。8人の患者が未処置であった。疾患活動性を、SLAM(Liang, M.H. et al., (1989), Arthritis Rheum 32:1107-1148)およびSLEDAI(Bombardier, C. et al., (1992), Arthritis Rheum 35:630-640)で評価すると、それぞれ平均値16.5および13.4であった。
表1
SLE患者の概略
【表1】
【0089】
試薬
組換え体TGF-βおよびモノクローナル抗TGF-β(1D11.16)抗体、ネズミIgG1は、Dr. Bruce Pratt(Genzyme Pharmaceuticals, Farmington, MA)から好意的に提供された。組換え体IL-10およびモノクローナル抗IL-10(JES3-19F1)抗体および対照ラットIgG2aは、Dr. Satwant Narula(Schering Plough Pharmaceuticals, Kenilworth, NJ)から好意的に提供された。対照マウスIgG1骨髄腫タンパク質は、Calbiochem, San Diego, CAから購入した。組換え体ヒトIL-2は、Chiron, Emmeryville, CAより購入した。OKT11を用いた抗CD2分泌ハイブリドーマ抗体は、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(ATCC)Rockville、MDから得、GT2は、A. Bernard, Nice, Franceから好意的に提供された。他の抗体には、J. Unkeless, New York, NYから好意的に提供された抗CD4(OKT4、ATCC)、抗CD8(OKT8、ATCC;CD8、Dako, Carpentaria, CA)、抗CD11b(OKM1、ATCC)、抗CD16(3G8);抗CD20(Leu 16、Becton Dickinson, San Jose, CA)および抗CD74(L243、ATCC)が含まれていた。
【0090】
血中単核細胞の単離
末梢血単核細胞(PBMC)を、Ficoll-Hypaque(Pharmacia, Piscataway, NJ)密度勾配遠心分離によりヘパリン処理静脈血から調製した。単核細胞を、5mM EDTAを含むPBS(Life Technologies, Grand Island, NY)で洗浄し、血小板を取り除いた。それはTGF-βを多く含んでいる。
【0091】
細胞培養の手順
細胞培養の手順は以前述べられている通りである(Wahl, S.M. (1994), J Exp Med 180: 1587-1590; Gray, J.D. et al., (1998), J Immunol 160: 2248-2254)。簡単に言うと、2x105のPBMCを、血清を含まないAIM-V培養培地(Life Technologies)中、記載のサイトカインを含むか、または含まない96-ウェルの底の平らなマイクロタイタープレートにおいて培養した。培養3日後、PBMCを3回洗浄し、次いで、新たに血清を含まない培地を添加した。更に37℃で7日後、上清を回収し、以前に述べられている(Linker-Israeli, M. et al., (1990), Arthritis Rheum 33: 1216-1225)通り固相酵素結合免疫収着検定(ELISA)によりウシ胸腺核タンパク質(NP)と反応する全IgGおよび自己抗体を検定する。光学的密度(OD)の読み値を、陽性および陰性標準を用いる標準曲線より、units/ml(U/ml)に変換した。SLE患者(高力価の抗NP抗体を有する)および健康な被験者のPBMC培養の上清を対照として用いた。
【0092】
統計学的分析
データをGraph Pad, Prism software(San Diego, CA)を用い分析した。データの対数変換および非パラメーター性マン‐ホイットニー試験後、分散(ANOVA)分析を使用した。
【0093】
抗CD2誘発IgG合成
抗CD2刺激CD4+T細胞およびB細胞に対する、NK細胞と共にまたはNK細胞を伴わずに培養したCD8+T細胞の効果を、SLE患者および正常対照者で試験した。CD4+およびCD8+細胞を、以前述べられている通りに、免疫磁性ビーズを用いる陰性選択により、ナイロン非付着性リンパ球から調製した(Gray, J.D. et al., (1998), J Immunol 160:2248-2254)。CD4+細胞の場合、ナイロン非付着性細胞をCD8、CD16、CD11bおよびCD74に対する抗体で染色した。同じ抗体を用い、CD8+細胞が得られた。ただし、CD4をCD8と置き換える。CD4+細胞の純度は95%であり、CD8+細胞は89%であった。NK細胞を得るため、PBMCをナイロンウールカラムに加え、そして溶出した非付着性細胞をAET処置ヒツジ赤血球細胞で即座にロゼットした。次いで、非ロゼット画分を抗CD3および抗CD74(抗HLA-DR)抗体で染色し、免疫磁性ビーズ(Dynal)を用い反応細胞をなくした。得られた群にはCD56+が98%およびCD3が0.5%未満およびCD20+リンパ球が0.5%未満含まれていた。SLE B細胞が大量のIgGを自発的に分泌するため、そして、これらの試験に多数のB細胞を調製するのに多量の血液を必要とするため、本試験では、健康な被験者ドナーの休止B細胞を患者のB細胞の代用とした。B細胞を得るため、ナイロンウール付着性細胞を、直ぐにSRBCでロゼットし、任意のT細胞を除去し、単球および機能性NK細胞を完全に除去するため5mM L-ロイシンメチルエステルで処理した。得た集団ではCD20+が92%を超え、CD3+が0.5%未満であった。
【0094】
結果
12人の試験患者において、自発的IgGは0.4から13.7μg/mlの範囲であった(図1)。PBMCをIL-2±TGF-βに72時間曝露することにより、IgG合成が、試験した12例のうち8例で、少なくとも50%減少した(平均減少79%、p=0.008、Mann Whitney)。最も劇的な減少は、最も顕著なB細胞の活動過剰の場合に観察された。分泌するIgG量とIL-2およびTGF-βによる阻害%との相関関係は、r=0.647、p=0.02であった。
【0095】
IL-2およびTGF-β単独の効果を、IL-2およびTGF-βの組み合わせと比較した。図2によって示されるのは、これらサイトカインのそれぞれがまたIL-2産生を阻害することである。しかし、対数変換によるデータの正規分布の達成および複数比較のためのBonnferoni補正適用後では、分散分析から、IL-2およびTGF-βの組合せによってのみ有意な阻害(p=0.05)が生ずることがわかる。
【0096】
IL-10産生はSLE中で増加しており(Llorente, L. et al., (1993), Eur Cytokine Network 4:421-427)、このサイトカインはIL-2およびTGF-βの両方の産生を阻害し得る。9例において、我々はまた、抗IL-10の効果を調べたが、IgG合成の幾分の低下が数例で観察されたのみであり、この違いは統計的に有意でなかった。同様に、TNF−α産生も一部のSLE患者で減少する(Jacob, C.O. et al., (1990), Proc Natl Acad Sci 87:1233-1237)。また、このサイトカインは活性TGF-βの産生を増加するが(Ohtsuka, K. et al., (198), J Immunol 160:2539-2545)、TNF−αを培養に加えることにより最小の効果を生じた(結果は示していない)。
【0097】
抗核タンパク質(NP)自己抗体の自発的産生につきSLE PBMCを試験し、有意な力価が4例みられた。PBMCをIL-2またはIL-2とTGF-βのどちらかに曝露した場合の全例において、少なくとも50%の抗NP産生が阻害された。TGF-β自身は効果がなかった(表2)。これらの場合、IL-2自身の効果は、IL-2とTGF-βとの組み合わせの場合と同等であった。
【0098】
【表2】
【0099】
SLE患者のPBMCをIL-2(10u/ml)およびTGF-β(10pg/ml)に72時間さらした。その細胞を洗浄し、更に7日間培養した。上清中に放出された抗NPをELISAで測定した。
【0100】
以前、我々が報告したのは、IL-2により生物学的に活性なTGF-β(Ohtsuka, K. et al., (1998), J Immunol 160: 2539-2545)の産生が増加することである。そのため、自発的抗体合成におけるIL-2の少なくとも幾つかの効果がTGF-βで仲介される可能性があった。この可能性について、IL-2の効果が抗TGF-β中性化抗体により逆転され得るかどうかを測定することにより調べた。図3Aに示した例では、抗TGF-βの添加は自発的IgG合成に影響を与えなかった。しかし、TGF-βの拮抗物はIgG合成におけるIL-2の阻害効果をなくした。この患者(表2のケースC)のPBMCもまた、抗NP抗体を自発的に産生した。ここで、抗TGF-βも、抗NP産生におけるIL-2の阻害効果をなくした(図3B)。即ち、この被験者おいて、自発的IgGおよび自己抗体合成に対するIL-2の阻害効果はTGF-βを介していた。抗TGF-βのこの効果は、試験した8例のうち4例で記録された。故に、IL-2の阻害効果は、TGF-β依存性または非依存性であり得る。各効果の例を表3に示す。
【表3】
【0101】
ステロイド治療開始後28日のSLE患者において試験を繰り返す機会を得た(表4)。処置前には、自発的IgG合成はIgG2μg/mlよりも多かった。PBMCをIL-2に曝露すると、これがIgG産生を顕著に阻害し、TGF-βは適度な効果を有した。コルチコステロイド治療の後、自発的IgG産生は75%減少した。上記の通り、PBMCをIL-2±TGF-βに曝露すると、IgG産生は50%減少した。しかし、この阻害は抗TGF-βにより逆転した。ここでさらに、IL-2のこの効果は、内在性活性TGF-βの上方制御によるものと説明できよう。
【0102】
【表4】
【0103】
健康な被験者についての我々の先の試験において、IL−2およびTGF−βが活性化CD+T細胞を誘導し、Ig産生を下方制御することから、本試験でSLE患者からのCD8+T細胞の単離および処置を試みた。先の試験が不成功であったのは、自発的抗体合成に顕著な可変性があること、および細胞分離法のために活動性SLE患者からの大量の血液を必要とすることであった。しかし、我々は、活動性SLEの患者の1人から十分な血液を得ることができ、抗CD2誘導IgG合成のCD8+T細胞調節に対するIL−2およびTGF−βの効果を調べた。抗CD3とは異なり、抗CD2モノクローナル抗体のマイトジェン性組合わせが、PBLを誘導しないで、IgGを産生することを、我々は最近報告した(Gray, J. D. et al. (1988), J Immunol 160:2248-2254)。例を図4Aに示す。これは、抗CD2がNK細胞を刺激してTGF−βを産生し、それが順番にCD8+T細胞を誘導し、抗体産生を下方制御するためであった(Gray, J. D. et al. (1988), J Immunol 160:2248-2254)。この患者において、我々が以前に報告したように(Gray J. D. et al. (1994), J Exp Med 180:1937-1972)、CD8+T細胞がIgG合成を増加し、NK細胞とCD8+T細胞の組合わせにより増加が著しく強化された(図4A)。一方、IL−2およびTGF−βは、SLE CD8+T細胞のヘルパー効果を無くし、これらの細胞がIgG産生を抑制することを可能にした。IL−2およびTGF−βのこの阻害効果はCD8+T細胞の存在に依存した(図4B)。このように、IL−2およびTGF−β効果がCD8+T細胞により仲介され得るという証拠が得られた。
【0104】
これらの試験は、PBMCのIL−2およびTGF−βへの短い暴露により、SLE、特に重症な疾病の患者および著しいB細胞活動亢進の患者において、続く自発的ポリクローナルIgGおよび自己抗体産生を大きく減少できることを証明した。この試験は、IL−2が抗体産生を阻害できることを示す先の報告(Hirohata, S. et al. (1989), J Immunol 142:3104-3112およびFast, L. D. (1992), J Immunol 149:1510-1515)を確認し、ピコモル濃度のTGF−βがこの下方制御の一因となり得ることを明らかにする。試験した12名の患者のグループにおいて、IL−2およびTGF−βのポリクローナルIgG合成に対する阻害効果は、IL−2単独より大きかった。しかし、IL−2の阻害活性は異なっていた。試験した8名のうち4名において、阻害はTGF−β依存的であり、中和化抗TGF−β mAbが作用を無くした。残りの例において、IL−2の下方制御効果はTGF−β非依存的であった。同様に、自発的抗NP自己抗体産生におけるTGF−β依存的および非依存的阻害の両方が証明された。また、我々がIL−10の拮抗およびTNF−α添加についての効果を調べたのは、SLEにおけるこれらサイトカイン産生の異常について、先に記載されていたためである(Llorente L. et al. (1993), Eur Cytokine Network 4:421-427; Jacob, C.O. et al. (1990), Proc Natl Acad Sci 87:1233-1237)。しかし、これらの方法は、リンパ球が先に活性化されているとき、自発的抗体合成に対する最小の効果を示した。
【0105】
SLEにおけるB細胞活動亢進の程度が疾病活動性と相関するという他の報告がある(Blaese, R. M. et al. (1980), Am J Med 69:345-350; Klinman, D. M. et al. (1991), Arthritis Rheum 34:1404-1410)。このことが本試験で起きなかったのは、同時薬剤治療によるためであろう。概して、著しい自発的抗体合成の患者は処置せず、かわりにB細胞活性が低い患者にはプレドニゾンを普通に投与した。我々はコルチコステロイド治療開始後に著しく自発的IgG合成が減少した一つの例を提示する。この患者のB細胞は、処置前に抗NP抗体も分泌したが、この自己抗体の産生はステロイド治療後に検出されなくなった(データは示していない)。
【0106】
TGF−βは、組織修復、炎症および免疫調節において重要なサイトカインの多機能性ファミリーから成る(Massague, J. (1990), Annu Rev Cell Biol 6597-641)。TGF−βは、不活性前駆体分子として分泌され、細胞外で生物学的活性形に変換する点で、多くの他のサイトカインと異なる(Massague, J. (1990), Annu Rev Cell Biol 6597-641; Flaumenhaft, R. et al. (1993), Adv Pharmacol 24:51-76)。実験的自己免疫脳炎(Weiner, H. L. et al. (1994), Annu Rev Immunol 12:809-837)および大腸炎(Neurath, M. F. et al. (1996), J Exp Med 183:2605-2516)のような種々の実験的自己免疫モデルは、このサイトカインを産生する。TGF−βは、ナノモル濃度で免疫抑制性であり、TおよびB細胞増殖、NK細胞の細胞毒性活性およびT細胞の細胞毒性発生を阻害できる(Letterio, J. J. et al. (1998), Ann Rev Immunol 16:137-162)。対照的に、TGF−βはマウスCD4+細胞およびCD8+細胞の増殖を促進する(Kehrl, J. H. et al. (1986), J Exp Med 163:1037-1050; Lee, H. M. et al. (1993), J Immunol 151:668-677)、およびB細胞分化を促進できる(Van Vlasselaer, P. et al. (1992), J Immunol 148:2062-2067)と報告されている。
【0107】
健康な被験者から得たリンパ球で調節性T細胞を発生させるという先の試験において、使用したTGF−βの濃度は、TまたはB細胞機能の阻害に必要とされるピコモル濃度より低かった(Gray, J. D.ら(1998),J Immunol 160:2248-2254;Gray,J. D.ら(1994),J Exp Med 180:1937-1942)。同様の濃度をSLE患者での本発明の試験において使用すると、TGF−β自体が抗体合成に対して僅かな阻害効果を有していた。前記の通り、TGF−βとIL−2の組み合わせが最も強力な阻害をもたらした。先の試験において、この効果はCD8+ T細胞により仲介された。
【0108】
IL−2について、T抑制細胞活性の誘発に対する効果が十分確立されている(Hirohata,S. ら(1989),J Immunol 142:3104−3112;Fast,L. D.,J Immunol 149:1510-1515)が、これらの効果が直接的なものであるか、または間接的なものであるかどうかは明らかではない。マウスにおいて、IL−2遺伝子の欠失は、大量のリンパ球増殖および自己免疫疾患を生ずる(Sadlack,B.ら(1995),Eur J Immunol 25:3053-3059)。SLEにおいて、IL−2レベルとB細胞機能亢進との負の相関関係が報告されている(Huang,Y. P.ら(1988),J Immunol 141:827-833)。以前、SLEでIL−2によって自発的Ig産生を阻害しようと試みたが、しかしながら、その結果は極めて可変的であった。強い阻害を幾つかの場合において観察したが、一方で他のIL−2では抗体産生が著しく増大した。我々は、時期およびサイトカイン環境により、この試験で見られる阻害よりもさらに一定した阻害が説明されると考える。ここで、IL−2およびTGF−βは、全培養期間というよりはむしろ、最初の72時間の培養の間にしか存在していなかった。後者の事例での抗体合成の増加は、B細胞分化に対するIL−2の正の効果によって説明し得る(Coffman,R. L.ら(1988),Immunol Rev 102:5-28)。IL−2は、抗体産生を幾つかの機構により下方制御することができる。該報告に記載されているTGF−β回路に加えて、IL−2が誘発する阻害は、IFN−γの上方制御(Noble,A.ら(1998),J Immunol 160:566-571)により、または細胞溶解機構(Stohl,W.ら(1998),J Immunol 160:5231-5238;Esser,M. T.ら(1997),J Immunol 158:5612-5618)により起こり得る。
【0109】
以前、我々は、抗体合成に対するNK細胞の調節効果を調べ、NK細胞の直接的効果がIgG合成を上方制御することである(Kinter,A.ら(1995),Proc Natl Acad Sci USA 92:10985-10989)が、これらのリンパ球が、正常な被験者において、CD8+ T細胞との培養の場合、正反対の効果を有すると報告した(Gray,J. D.ら(1994),J Exp Med 180:1937-1942)。しかしながら、SLE患者において、CD8+ T細胞とNK細胞との組み合わせがIgG産生を高める(Linker-Israeli,M.ら(1990),Arthritis Rheum 33:1216-1225)。これは、本発明の報告において再び観察された。正常な被験者では、NK細胞をCD8+ T細胞に添加すると、抗CD2刺激IgG合成を著しく阻害したが、正反対のことがSLEにおいて観察された。標準的試験から、NK細胞から得たTGF−βがCD8+T細胞の同時刺激を誘発して、IgGおよびIgM産生を下方制御することがわかった(Gray,J. D.ら(1998),J Immunol 160:2248-2254)。この試験において、IL−2およびTGF−βは、CD8+ T細胞による中程度の抑制活性を誘発した。従って、SLEにおいて、IL−2およびTGF−βがB細胞活性を阻害する少なくとも1つの方法は、調節性T細胞を発生させることによるものであるらしい。加えて、これらまたは他のサイトカインで処理した他のリンパ球群もまた、SLEにおけるB細胞活性を下方制御し得る。
【0110】
実施例2
疾患の活性および重症度とTGF−β産生との相関関係
全体および活性型のTGF−βのリンパ球産生が減少することが分かったので、次に、これらの欠失が疾患の活性および/または重症度と関連があるかどうかを調べた。17人の検査したSLE患者から得た血液リンパ球によるTGF−β1産生を、10人の慢性関節リウマチ(RA)患者および23人の適応する健康な対照と比較した。RAにおける活性TGF−β1のレベルは、対照より低かったが、SLEにおける程度までは減少しなかった。ピコモル量で検出される構造性TGF−β1および抗CD2刺激された活性TGF−β1のレベルは、最近発症の非常に活性で重症なSLEのために入院した6人の未処置患者において著しく減少し、処置され活性の低い疾患の11人の患者においても同様に減少した。このように、活性TGF−β1産生の減少は、疾患の活性とは関連がなかった。これとは対照的に、全TGF−β1産生の減少は、疾患の活性と逆の相関関係があった。このように、TGF−β1の潜在性前駆体のリンパ球分泌の欠陥は、疾患活性の結果として生じ得るが、前駆体分子をその活性型に転換する能力は、内因性細胞欠失となり得るようである。活性化される時点でのピコモル濃度量のTGF−β1へのT細胞の不十分な暴露は、抗体合成の下方制御の欠陥を生じ得る。このように、SLEにおける活性TGF−β1のリンパ球産生の減少は、この疾患に特徴的なB細胞機能亢進に起因するのであろう。
【0111】
方法
試験の被験者
SLEの分類に関するアメリカ大学のリウマチ病学基準(Tan,E. M.ら(1982)、Arthritis Rheum 25:1271-1277)を満たすSLE診断を受けた17人の被験者、RAの分類に関するACR 1987 改定基準 (Arnett,F. C.ら(1988),Arthritis Rheum 31:315-324)を満たすRA診断を受けた10人の被験者、および23人の健康なドナーを試験した。SLEグループは、女性15人および男性2人(スペイン人15人、アフリカ系アメリカ人1人、アジア人1人)からなっていた。平均年齢は、34.5歳(範囲:20−75歳)であった。6人の患者を入院させて、11人を外来診療所に通わせた。入院患者は皆、入院するまでは未処置であり、試験した後、彼らに第一回用量のコルチコステロイドを投与した。外来患者には、プレドニゾンを20mg未満を投与し、誰にも細胞毒性薬物を投与しなかった。疾患の活性を、平均値が各々6.6および7.6であるSLAM(Liang,M. H.ら(1989)、Arthritis Rheum 32:1107-1118)およびSLEDAI(Bombardier,C.ら(1992)、Arthritis Rheum 35:630-640)指数で評価した。RAグループは、女性9人および男性1人(スペイン人9人、アジア人1人)からなっていた。平均年齢は、50.9歳(範囲:39−67歳)であった。患者は皆、外来診療所に通わせ、軽度〜中程度に疾患活性を有していた。疾患の平均期間は9.5年であった。1人の患者にマイオクリシン(myochrysine)を投与し、3人の患者にプレドニゾン(1、1および20mg)を投与し、3人の患者にメトトキサレートを投与し、1人の患者にスルファサラジンを投与した。健康なドナーを対照として、年齢、性別、および民族グループに関して可能な限り厳密に適応させた。
【0112】
表5
SLE患者の2つのグループの臨床的特徴
【表5】
【0113】
試薬
使用した抗体は、抗CD2を分泌するハイブリドーマ(OKT11、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(ATCC),Rockville,MD、およびGT2は、Alain Bernard博士,Nice,Franceにより入手可能となった)の上清であった。TGF−βイソ型 1、2、および3(1D11)を認識するモノクローナル抗体、TGF−βイソ型 2および3(3C7)に対する抗体、並びにrTGF−β2は、Bruce Pratt博士(Genzyme Pharmaceuticals,Farmington,MA)により快く提供された。
【0114】
血液リンパ球の単離
先に記載した方法(Ohtsuka,K.ら(1998),J Immunol 160:2539-2545)を使用して、Ficoll−Hypaque(Pharmacia, Piscataway, NJ)密度勾配遠心分離により、末梢血単核細胞(PBMC)をヘパリン処置した静脈血から調製した。単核細胞を、5mM EDTA(Life Technologies, Grand Island,NY)を含むPBSで洗浄して、TGF−βの豊富な原料である血小板を取り除いた。連続Percoll(Pharmacia)密度勾配によって、末梢血リンパ球(PBL)を遠心分離によりPBMCから分離した。高密度のリンパ球に富む画分に残留する単球のパーセンテージは、SLEにおいて幾分高かった(8.5% 対 4.3%)。
【0115】
細胞培養方法
細胞培養方法は、以前に記載されている(Ohtsuka,K.ら(1998),J Immunol 160:2539-2545)。簡単に言えば、1×105個のリンパ球を96ウェルの平底マイクロタイタープレート(Greiner Rocky Mountain Scientific,Salt Lake City UT)のウェルに加えた。血清が著しい量の潜在性TGF−βを含むことから、培養をAIM−V血清不含有培地(Life Technologies)で行った。抗CD2を最適濃度で使用して、TGF−β産生(GT2 1:40およびT11 1:80)ハイブリドーマ培養上清を誘導した。先の試験は、抗CD2がPBLを強く刺激して、TGF−βを産生することを明らかにしている(Gray,J. D.ら(1998),J Immunol 160:2248-2254)。
【0116】
TGF−βアッセイ
ルシフェラーゼ・リポーター遺伝子に融合したプラスミノーゲン活性化因子阻害剤(PAI−1)プロモーターを含む発現構築物をトランスフェクトしたイタチ肺上皮細胞(MLEC)は、D. B. Rifkin博士,New York,NYにより快く提供された。2×104/ウェルのMLECを上清200μlと共に37℃で18時間インキュベートした。ルシフェラーゼ活性についてアッセイするために、MLECを細胞溶解試薬(Analytical Luminescence,Ann Arbor,MI)により溶解した。次いで、細胞溶解物をアッセイ緩衝液およびルシフェリン溶液(両方とも、Analytical Luminescenceから得た)と反応させた後、直ちに、照度計(Lumat,Berthold Analytical Instruments Inc.,Nashua,NH)で測定した。全TGF−β活性を測定するために、試料を80℃で3分間加熱して、潜在性複合体から活性サイトカインを放出させた。上清を加熱することなく、活性TGF−βの活性を測定した。全てのアッセイおいて、幾つかの濃度のrTGF−βが標準曲線を作成するために含まれていた。同型培養の可変性は10%未満であった(Ohtsuka,K.ら(1998),J Immunol 160:2539-2545)。
【0117】
統計解析
GBSTATソフトウェア(Professional Statistics and Graphics Computer Program,Dynamic Microsystems Inc.,Silver Spring,MD)を使用して行うマン‐ホイットニー(Mann−Whitney)試験およびSpearman順位相関関係を使用して、結果の有意性を解析した。
【0118】
結果
SLEまたはRAの患者由来のPBLにより産生される構造的および刺激されたTGF−β1を測定し、正常対照の値と比較した。培養上清中で検出されるサイトカインは、イソ形1、2および3を認識するmAbで中和されたが、イソ形2および3に対するmABではされず、結果としてTGF−β1の産生を確認した。正常対照と比較して、活性TGF−β1の構造的産生はSLEで有意に減少した(14±4対56±21pg/ml、p=0.02、図5)。抗CD2刺激活性TGF−β1も減少した(87±22対399±103pg/ml、p=0.003)。RAにおいて、構造的TGF−β1の平均値は、SLE(19±5pg/mg)と同様であり、抗CD2により刺激された後、正常とSLEの中間値であった(197±54pg/ml)。
【0119】
リンパ球により産生された構造的全TGF−β1も、正常グループと比較してSLEで減少した(286±82対631±185pg/ml、p=0.05)。RAでの値は、正常とSLEの中間値であった(435±161pg/ml)。抗CD2の添加に続き、全TGF−β1はSLEで正常対照より幾分増加したが、差異は統計学的に有意でなかった。RAグループでの値も正常とSLEグループの中間値であった。
【0120】
TGF−β1の減少したレベルと疾病活動性の相関関係の可能性を調べるために、SLEの入院患者と外来患者とを比較した。これらの2つのグループの臨床的特徴は、表5に要約する。入院患者の方が若かった;6名中5名が、3ヶ月以内の症状を示し;そして、著しく活動的な疾病に罹患していた;そして殆どが、腎炎および/または溶血性貧血を伴う重症SLEであった。比較して、外来患者は、処置により活動性が低くなった慢性疾病であった。疾病異種性、期間、活動性および重症度の著しい差異にもかかわらず、構造的および刺激TGF−β1産生は、正常対照と比較して両グループで有意に減少した(表6)。
【0121】
【表6】
【0122】
活性および全TGF−β1のレベルと疾病活動性との相関関係をみると、全TGF−β1の抗CD2刺激産生は、SLEDAI指数と有意な負の相関関係があるが(r=0.55、p=0.03)、SLAM指数とは相関がなかった(−0.43、p=11)。SLEDAI指数は、中枢神経系関与および腎臓疾患に重きを置く。このように、TGF−β1の前駆体形を分泌するリンパ球の能力の減少は、重症な疾病と関連するように見える。活性TGF−β1のレベルは、疾病活動性と相関しなかった。
【0123】
本例での主な発見は、SLEでの活性TGF−β1の産生減少が、疾病活動性または重症度と相関しないことである。構造的および刺激活性のTGF−β1量の減少は、最近発症した疾病および確立した疾病の両方の患者で見られた。更に、この値は、SLAMおよびSLEDAI指数で測定して活動性と、または生体臓器関与により評価した重症度と相関しなかった。しかし、全TGF−β1産生がSLEで減少もするが、この欠失は疾病活動性と相関するように見えた。これは、主として入院SLE患者で見られた。全TGF−β1産生が主要な臓器系関与に重きを置くSLEDAI指数と最も相関するというこの発見はまた、疾病重症度との関係を示す。
【0124】
この試験はまた、確立された疾病のSLE患者と疾病活動性が同等であるRA患者の対照グループを含む。RAグループでのTGF−β1値は、構造的活性TGF−β1以外、正常対照より幾分低いが、欠失の強度はSLE程著しくなく、統計的に有意ではなかった。
【0125】
以前に、我々は、NK細胞がTGF−βの主なリンパ球源であり、このサイトカインを活性形で構造的に産生する唯一のリンパ球集団であることを証明した(Gray, J. D. et al., (1988), J. Immunol. 169:2248-2254)。従って、NK細胞由来TGF−βの構造的産生はSLEで減少しているとの発見は、興味深かった。我々はまたIL−2およびTNF−αの両方が活性TGF−βの産生を促進できることを知った。これら両サイトカイン産生はSLEで減少する(Gray J. D. et al., (1994), J Exp Med 180:1937-1942)。しかし、殆どの患者において、外来性IL−2およびTNF−αはTGF−βを正常に回復できなかった(実施例2)。IL−10産生はSLEで増加し(Llorente, L. et al. (1993), Eur Cytokine Network 4:421))、増加したレベルと疾病活動性の相関関係が報告されている(Housslau, F. A. et al. (1995), Lupus 4:393-395; Haglwara, E. et al. (1996), Arthritis Rheum 39:379)。IL−10はIL−2、TNF−αおよびTGF−β産生を阻害できる(実施例2およびMoore, K. W. et al. (1993), Ann Rev Immunol 11:165-190)。活性TGF−βの産生が軽いおよび活動性の疾病の患者で減少したこと、およびIL−10の拮抗によって産生欠失を部分的にしか回復できなかったことは(実施例2)、増加したIL−10産生自体がSLEにおけるTGF−β1のリンパ球産生の減少について説明となり得ないことを示す。数個の機構が恐らく関与するのであろう。潜伏性前駆体の成熟活動性形への細胞外変換における1個またはそれ以上の欠失がこの異常性を説明し得る。
【0126】
TGF−βは、リンパ球増殖およびエフェクター細胞機能に対する阻害特性が詳細に説明されているが(Letterio, J. J. et al. (1998), Ann Rev. Immunol 16:137-162)、刺激特性も報告されている(Lee, H. M. et al. (1991), J Immunol 151:668-677)。TGF−βは、刺激T細胞およびその産生の上方制御によりサイトカイン産生を調節する。マウスにおいて、TGF−β1は、CD8+T細胞を選択的に活性化して増殖させ(Lee, H. M. et al. (1991), J Immunol 151:668-667)、ナイーブ細胞からメモリーT細胞への成熟を増加する(Lee, M. H. et al. (1991), J. Immunol 147:1127-1133)。ヒトにおいて、TGF−β1はエフェクターT細胞の強い誘発物である(Cerwenka, A. et al. (1994), J Immunol 153:4367-4377)。免疫抑制作用には大量(ナノグラム/ml)が必要であるが、我々が示したように、抗体産生の下方制御効果に関してCD8+T細胞を共刺激するのには、少量(ピコグラム/ml)でよい(Gray, J. D. et al. (1998), J Immunol 160:2248-2254)。
【0127】
そのため、これらの試験によると、TGF−β1の潜伏性前駆体のリンパ球分泌の欠失が疾病活動性の結果として起こり得るが、SLEにおける活性TGF−β1産生の減少は、さらに複雑であり、数個の異なる機構からもたらされる。ナイーブT細胞が抗体産生を下方制御するように計画するには、T細胞が活性化される時点でpg/ml量の活性TGF−βの存在を必要とすると我々は考え、またこの考えを支持する証拠がある(Gray, J. D. et al. (1998), J Immunol 160:2248-2254)。従って、決定的な時点での局所環境におけるピコモル量の活性TGF−βを欠くことは、SLEにおけるBリンパ球活性を制御するT細胞調節機能の無効によるのであろう。
【0128】
実施例3
マイトジェンでのSLEの処置
本実施例において、IgG産生は、調節組成物で細胞を処理することにより下方制御される。この調節組成物はマイトジェン抗CD2モノクローナル抗体組合せ物のようにマイトジェンを含む。これら抗体は、可溶化形態で添加するか、ビーズ上に固定化して、T細胞およびNK細胞上の受容体と橋かけ結合せしめる。これらの細胞は、上記の実施例で説明したように調製し、次いで、マイトジェンとインキュベートし、増幅した。抗体産生が下方制御された細胞数が増加する。ConAはシグマから入手できる(St. Louis, MO)。
【0129】
抗CD2がどのように機能するかは知られていないが、これら抗体がPBMC調製物中においてNK細胞を誘導し、活性TGF−βを分泌し(Ohtsuka, K. et al.(198), J. Imunol 160: 2539-2545)、次いでTGF−βが働いて、T細胞が抗体抑制細胞となると考えられている。
次いで、細胞を必要であれば洗浄し、患者に移植し戻す。
【0130】
実施例4
サイトカインとマイトジェンの混合物での細胞の処理
細胞を上記のように調製して、抗体産生を下方制御する細胞集団を誘導するために、マイトジェンとサイトカインの混合物を含む調節組成物で細胞をインキュベートした。この手法の例は、図4Cにおいて示す。この実施例において、CD4+細胞およびCD8+細胞をConA、IL−2およびTGF−βで処理して、抑制に関する最大誘導を得た。
【0131】
患者に再移送される調節性T細胞の調製のために、抗CD2および/または抗CD3モノクローナル抗体をConAの代わりに使用して、T細胞を活性化する。調節組成物は、IL−2を伴いまたは伴なわずに、TGF−βを含有する。この細胞を、例えば、Nexell 300i. Magnetic Cell Selection System のような閉鎖系中で標準インキュベーション技術を用いて4〜7時間、組成物と共にインキュベーションした。
【0132】
インキュベートの後、細胞をHBBSと洗浄して、溶液中のサイトカインおよびマイトジェンを除去する。所望により、ビーズ上に固定した抗CD3±抗CD28と培養して、細胞数を増加さす。次いで、細胞を200−500mlのHBBS中に懸濁し、患者に再挿入する。
【0133】
実施例5
細胞の細胞仲介免疫を正常化する処置
SLEの自己抗体産生に対する要因は、IL−10とTNF−αとのアンバランスにある。IL−10のレベルは過多で、TNF−αレベルは低い[Llorente et al. 1995. J. Exp. Med. 181:839-44,Houssiau, F.A. et al., 1995. Lupus 4:393-5.(Ishida, H. et al. 1994. J. Exp. Med. 179:305-10)(Jacob, C. O. and McDevitt, H. O.,1988. Nature 331: 356-358)]。我々は、このアンバランスが、TGF−βの存在下でT細胞を強く活性化することによって補正されるという証拠を得て、近年この効果の作用機構を解明した。
【0134】
精製されたT細胞を、上記概要に沿って調製し、TGF−βの存在または不存在で、ConAとIL−2でインキュベーションした。図6では、TGF−βの非存在下でのT細胞の刺激によって、IL−10の産生が増加したことを示す。しかし、TGF−αを刺激されたT細胞に加えると、IL−10の産生が遮断され、TNF−αの産生が増加した。さらに、TNFR2の発現が有意に増加した。理論にしばられるわけではないが、TGF−βによって誘導されるTNFR2を介するTGF−α情報伝達の促進が、抗体産生を抑制する調節性T細胞を生じると考える。我々の結果は、この考察を支持するものであった。
【0135】
TGF−βによるTNF−αの上方制御が調節性T細胞の誘導に重要であることを説明してきた。図7は、TGF−βを活性化CD8+T細胞に添加することで、IgG産生の顕著な抑制が起こることを示す。この抑制活性は、主な中間体としてTNF−αに依存する。これらの各試験において、抗TNF−α抗体を中和することによって、CD8+調節性T細胞(CD8reg)抑制効果を完全になくした。
【0136】
SLE患者は、IL−2、TNF−αおよびIFN−γの産生減少を伴なう細胞仲介免疫に著しい欠失を有する(Horwitz, D.A. et al.,(1997), Dubois' Lupus Erythematosus, 5th Ed. (1997), pp.83-96, D.J. Wallace et al. eds., Williams and Wilkins, Baltimore)。理論にしばられるわけではないが、TGF−βのリンパ球産生における欠失は、IL−2、TNF−αおよびIFN−γの産生減少に対する部分的な要因である。TGF−βの存在下でのT細胞の刺激によって、これらの細胞を再び刺激すると、IL−2、TNF−αおよびIFN−γの産生が顕著に増加することを見出した。さらに、この結果は、TGF−βによるTNF−αの上方制御にも依存する(図8を参照)。
【0137】
TGF−βの産生がSLEで低下し、その欠失がIL−10とTNF−αとのアンバランスによるものであるということを示す。理論にしばられるわけではないが、SLEにおけるIL−10の高いレベルが、自己抗体産生を保ち、TGF−α、IL−2、IFN−γ産生の原因であると考える。これらサイトカインの産生低下がSLEにおける欠失性細胞免疫の原因である。明記した条件下で、TGF−βはIL−10を下方制御し、TNF−αの産生を強めることを示す。TGF−βによるIL−10の下方制御とTNF−β産生の増強は、SLEにおける調節性T細胞の正常化、細胞仲介免疫の修復、疾病の軽減に関して重大な役割をになう。
【0138】
実施例6
細胞仲介自己免疫を抑制する調節性T細胞の生成
前記実施例では、調節組成物を使用して、抗体介在性自己抗免疫疾患を処置した。類似組成物を使用して、CD4+に加えてCD+8T細胞を誘導し、細胞仲介自己免疫疾患を抑制した。TGF−βによって調節されたCD8+またはCD4+細胞が、T細胞の細胞毒性の生成を抑制することを示す。
【0139】
調節性T細胞を誘導するためにマイトジェンを使用する代わりに、同種異系性混合リンパ球反応を、この目的のために使用した。この反応では、1個体からのT細胞が、他の個体のPBMCによって提示された外来の組織適合性抗原を認識し、応答する。これらの応答T細胞は、増加して、これらの標的細胞を殺傷する能力を発展させる。
【0140】
抑制T細胞を創生するために、一個体(ドナーA)からの様々なCD4+およびCD8+T細胞のサブセットを、別の個体(ドナーB)からの照射されたT細胞なしの単球性細胞と共に培養した。その細胞を、懸濁液中のTGF−β(1ng/ml)の存在または不存在で、5日間培養した。これ以降は、TGF−βを除き、該細胞を、新たなドナーAからのT細胞とドナーB細胞からの非T細胞に添加した。図9は、TGF−βが、CD4+およびCD8+T細胞サブセットの両方を誘導し、細胞仲介細胞毒性を抑制する能力を開発することを示す。図10は、TGF−βによって誘導されたCD4+調節性T細胞を用いる2つの追加試験を示す。
【0141】
さらなる試験によると、この方法でつくった調節CD4+T細胞が、該作用に関する独特の作用を示す。前につくったCD8+およびCD4+両方のT細胞が阻害性サイトカインの分泌により抑制をおこすのと異なり、これらアロ特異的調節性CD4+T細胞は、接触依存性の作用機構を示す(図11)。理論にしばられるわけではないが、これらの調節性T細胞は、別のT細胞が活性化されるのを抑制すると考える。これらのT細胞の応答T細胞およびアロ刺激細胞への添加は、増殖を抑制し(図12)、応答CD8+キラー前駆体細胞が活性化される能力を低下させた(図13)。
【0142】
次のことも分かった。これら調節性CD4+細胞は、IL−2受容体(CD25)をそれら細胞表面で発現し、非常に強力である(図14)。調節CD4+細胞の応答T細胞との割合が、1:4(20%)から1:32(3%)に低下すると、これら細胞の阻害効果がごく僅か低下した。
【0143】
これら細胞の少量のみを強い下方制御効果に必要とするので、十分な数を患者に移送し、自己免疫性もしくは他の所望の免疫抑制効果、例えば移植拒絶反応を抑制することができる。
【0144】
実施例7
CD4+T細胞を刺激してTGF−βの免疫抑制レベルを生成する
TGF−βの免疫抑制レベルを生成するCD4+T細胞はTh3細胞と称されているが、その発生に関する機構は殆ど理解されていない。われわれが得た証拠によると、スーパー抗原、スタフィロコッカス・エンテロトキシン(SEB)によるCD4+細胞の強い刺激、または低濃度のSEBよるCD4+細胞の反復刺激が、これらの細胞を誘導し、活性TGF−βの免疫抑制レベルを生成した。
【0145】
図15は、活性および全TGF−βの産生増加が、SEB濃度の増加によって刺激されたCD4+T細胞によりなされたことを示す。図16は、SEBの低用量によってCD4+T細胞の反復刺激に関する効果を示す。T細胞をSEBで3回刺激すると、顕著な量のTGF−βの活性型が生成した。
【0146】
図17は、ナイーブ(CD45RA+CD45RO−)CD4+およびCD8+T細胞に対するSEBの効果を示す。この細胞をSEBで5日ごとに合計3回刺激した。各T細胞のサブセットおよびCD25IL−2受容体活性マーカーを発現する細胞の%を各々の刺激後に測定した。パネルAおよびCは、最初の刺激にTGF−β 1ng/mlが含まれていると、CD4+T細胞が反復刺激の培養物中で優勢なサブセットとなったことを示す。パネルBおよびDは、SEB刺激細胞によるCD25の発現が、対照培養物中で3回目の刺激によって低下したことを示す。しかし、CD25の発現は、T細胞がTGF−βで駆動されると、非常に高いままであった。即ち、TGF−βは、これらのT細胞が反復刺激されると、CD4+細胞に対する優先効果を有するようであり、これらの細胞の殆ど全てが20日間の培養後にCD25+であった。
【0147】
要約すると、T細胞の刺激の結果として、TGF−βの優勢な調節効果が、CD8+細胞に与えられる。反復刺激によって、このサイトカインはCD4+細胞を誘導して、調節性細胞とする。この細胞は、抑制活性に関してCD8+細胞よりも強力である。
【図面の簡単な説明】
【0148】
【図1】図1は、SLE患者のPBMCをIL−2およびTGF−βと共にインキュベーションすることにより、自発的免疫グロブリン産生が減少することを示す。
【図2】図2は、IL−2およびTGF−βの両方により、自発的IgG産生が有意に減少することを示す。
【図3】図3Aおよび3Bは、抗TGF−βがIL−2の効果を逆転することを示す。
【図4】図4A、4Bおよび4Cは、抗体産生に対するDC8+T細胞の調節効果を示す。
【図5】図5Aおよび5Bは、非刺激細胞および抗CD2刺激細胞によるTGF−β1のリンパ球産生を示す。
【図6】図6は、TNF−αおよびIL−10のT細胞産生に対するTGF−βの作用を示す。
【図7】図7は、TGF−βによる調節性T細胞の生成にTNF−αが必須の中間体であることを示す。
【図8】図8は、TGF−β起動T細胞によるTh1サイトカインの産生の増加がTNF−αに依存性であることを示す。
【図9】図9は、細胞毒性T細胞活性の抑制物質の産生におけるTGF−βの作用を示す。
【図10】図10は、TGF−βにより起動されたCD4細胞のアロ細胞毒性Tリンパ球(CTL)活性に対する作用を示す。
【図11】図11は、調節性T細胞がCTL活性を抑制するために細胞接触を必要とすることを示す。
【図12】図12は、TGF−βで誘発された調節性CD4+T細胞によるリンパ球増殖の抑制を示す。
【図13】図13は、CD25+CD4T細胞の調節活性を示す。
【図14】図14は、調節CD+4細胞がその表面でCD25+(IL−2)受容体を発現することを示す。
【図15】図15は、少量のブドウ球菌性エンテロトキシンB(SEB)でのT細胞の反復刺激がT細胞を誘発してTGF−βの免疫抑制レベルをつくることを示す。
【図16】図16は、低量のSEBでCD4+T細胞の反復刺激によってT細胞がTGF−βの免疫抑制レベルをつくることを示す。
【図17】図17は、ナイーブ(CD45RA+CD45RO−)CD4+およびCD8+T細胞に対するSEBの作用を示す。
【特許請求の範囲】
【請求項1】
TGF-βおよびIL−2を含む組成物を用いて、細胞仲介自己免疫異常患者から得たPBMCを処置することを含む、該患者を処理するための処理済末梢血単核細胞(PBMC)の調製方法。
【請求項2】
処理済PBMCが、細胞仲介自己免疫異常に関連する細胞毒性を減少せしめる作用を有する、請求項1記載の方法。
【請求項3】
細胞仲介自己免疫異常が、橋本病、多発性筋炎、炎症性腸疾患、多発性硬化症、真性糖尿病、慢性関節リウマチおよび硬皮症よりなる群から選ばれる、請求項1の方法。
【請求項4】
PBMCがCD8+T細胞に富む、請求項1−3のいずれか記載の方法。
【請求項5】
PBMCがCD4+細胞に富む、請求項1−3のいずれか記載の方法。
【請求項6】
PBMCがナイーブCD4+細胞に富む、請求項1−3のいずれか記載の方法。
【請求項7】
PBMCがCD8+細胞およびCD4+細胞に富む、請求項1−3のいずれか記載の方法。
【請求項8】
PBMCがNK細胞に富む、請求項1−3のいずれか記載の方法。
【請求項9】
調節用組成物がT細胞アクチベーターを含む、請求項1−8のいずれか記載の方法。
【請求項10】
T細胞アクチベーターが抗CD2抗体である、請求項9の方法。
【請求項11】
T細胞アクチベーターが抗CD3抗体である、請求項9の方法。
【請求項12】
T細胞アクチベーターが抗CD28抗体である、請求項9の方法。
【請求項13】
処理済PBMCが、該患者における他のT細胞が細胞毒性エフェクター細胞になるのを防止する作用を有する、請求項1−12いずれかに記載の方法。
【請求項14】
処置済細胞が、誘発されて、活性TGF-βの免疫抑制レベルをつくる、請求項1−13のいずれかに記載の方法。
【請求項1】
TGF-βおよびIL−2を含む組成物を用いて、細胞仲介自己免疫異常患者から得たPBMCを処置することを含む、該患者を処理するための処理済末梢血単核細胞(PBMC)の調製方法。
【請求項2】
処理済PBMCが、細胞仲介自己免疫異常に関連する細胞毒性を減少せしめる作用を有する、請求項1記載の方法。
【請求項3】
細胞仲介自己免疫異常が、橋本病、多発性筋炎、炎症性腸疾患、多発性硬化症、真性糖尿病、慢性関節リウマチおよび硬皮症よりなる群から選ばれる、請求項1の方法。
【請求項4】
PBMCがCD8+T細胞に富む、請求項1−3のいずれか記載の方法。
【請求項5】
PBMCがCD4+細胞に富む、請求項1−3のいずれか記載の方法。
【請求項6】
PBMCがナイーブCD4+細胞に富む、請求項1−3のいずれか記載の方法。
【請求項7】
PBMCがCD8+細胞およびCD4+細胞に富む、請求項1−3のいずれか記載の方法。
【請求項8】
PBMCがNK細胞に富む、請求項1−3のいずれか記載の方法。
【請求項9】
調節用組成物がT細胞アクチベーターを含む、請求項1−8のいずれか記載の方法。
【請求項10】
T細胞アクチベーターが抗CD2抗体である、請求項9の方法。
【請求項11】
T細胞アクチベーターが抗CD3抗体である、請求項9の方法。
【請求項12】
T細胞アクチベーターが抗CD28抗体である、請求項9の方法。
【請求項13】
処理済PBMCが、該患者における他のT細胞が細胞毒性エフェクター細胞になるのを防止する作用を有する、請求項1−12いずれかに記載の方法。
【請求項14】
処置済細胞が、誘発されて、活性TGF-βの免疫抑制レベルをつくる、請求項1−13のいずれかに記載の方法。
【図1】

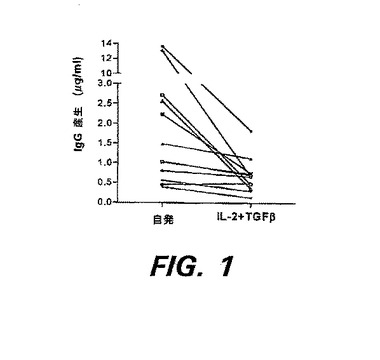
【図2】
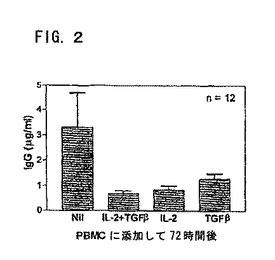
【図3】
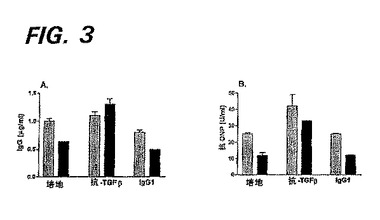
【図4】
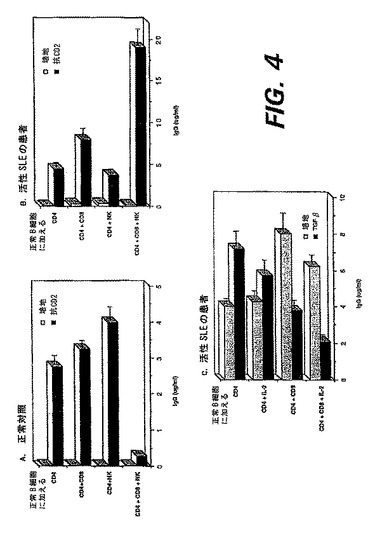
【図5】

【図6】
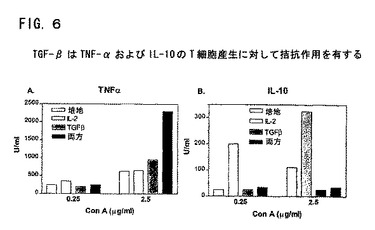
【図7】
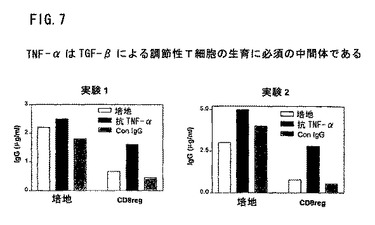
【図8】
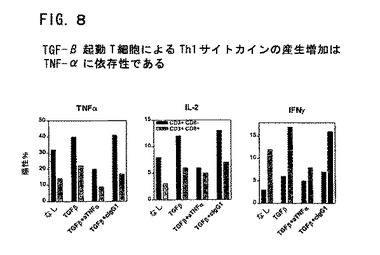
【図9】
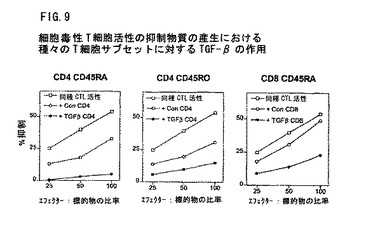
【図10】
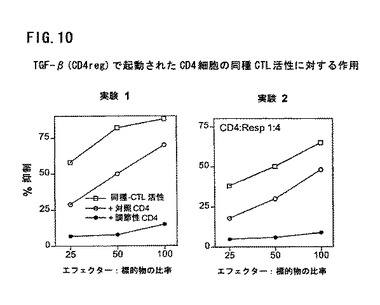
【図11】
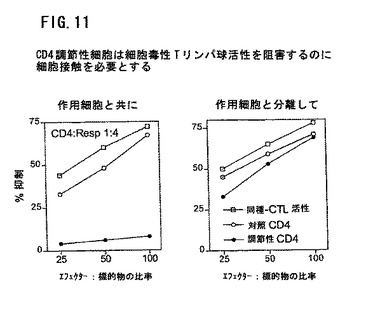
【図12】
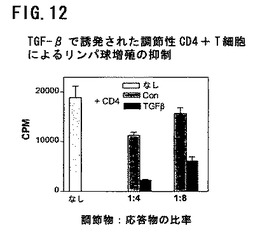
【図13】
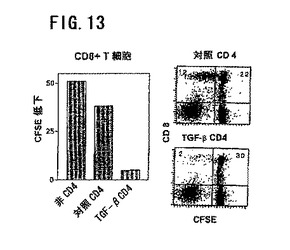
【図14】
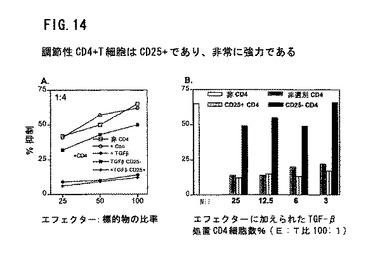
【図15】
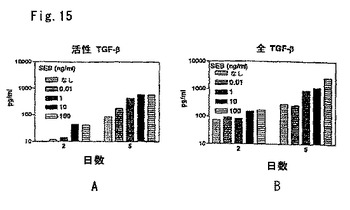
【図16】
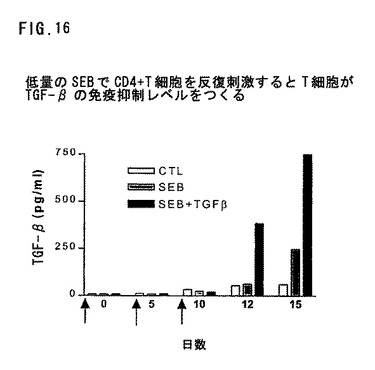
【図17】
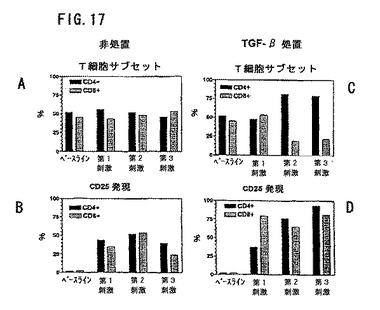
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公開番号】特開2008−174564(P2008−174564A)
【公開日】平成20年7月31日(2008.7.31)
【国際特許分類】
【出願番号】特願2008−33141(P2008−33141)
【出願日】平成20年2月14日(2008.2.14)
【分割の表示】特願2000−615042(P2000−615042)の分割
【原出願日】平成12年5月5日(2000.5.5)
【出願人】(592048844)ユニバーシティ オブ サザン カリフォルニア (26)
【Fターム(参考)】
【公開日】平成20年7月31日(2008.7.31)
【国際特許分類】
【出願日】平成20年2月14日(2008.2.14)
【分割の表示】特願2000−615042(P2000−615042)の分割
【原出願日】平成12年5月5日(2000.5.5)
【出願人】(592048844)ユニバーシティ オブ サザン カリフォルニア (26)
【Fターム(参考)】
[ Back to top ]