
癌、特に悪性乳癌のための分析標識
【発明の詳細な説明】
本発明は腫瘍関連酵素標識(マーカー)に関する。
本発明は、ストロメリシン−3(Stromelysin−3)をコードするDNAおよびストロメリシン−3に結合し得る抗体を使用することにより、癌、具体的には悪性乳癌の診断法を提供するものである。
世界中の癌による死亡数は毎年主たる懸念材料であり続けており、特定の型の癌について数種の治療法が利用できるだけであって、またこれらが成功するという絶対的な保証もない。ほとんどの治療は迅速に成長する細胞を死滅させる一般的な“ショットガン”法に頼っており、この方法は迅速に成長する癌細胞がこの治療によって死滅するか、もしくは少なくとも癌細胞数が減少することによってその身体の系による残りの癌細胞の排除が可能になるであろうことを期待するものである。
治療法の探索は、異なる形態の癌が異なる治療を必要とするという発見によって妨げられてきた。身体のほとんど全ての部分が癌に冒され得るとすれば、その課題は膨大なものになる。
しかしながら、それらの相違にもかかわらず癌はいくつかの類似点をも有している。その中で最も重要なことは非分化組織の成長である。しかしこれは、ある種の癌細胞がある程度の分化を示すという点で100%正確な訳ではない。これは乳癌や精巣癌などの性癌で示されており、この場合腫瘍はホルモン受容体に関して陽性の場合もあり、陰性の場合もある。これらの腫瘍の治療はホルモン状態に依存し、タモキシフェンTMなどの関連ホルモン拮抗薬を投与する程度に単純な場合もある。
ほとんどの癌が共有しているもう1つの要素は、致死的であるためには癌が転移しなければならないということである。転移が起こる時までは、腫瘍はそれが悪性であっても身体の一領域に限定される。これは不快および/または痛みをもたらしたり、あるいはさらに重度な兆候を導くことさえあるが、その位置が特定できるのであれば、それを外科的に除去することができ、また十分な注意を払えば、さらなる問題を引き起こさない。
しかしいったん転移が始まれば、外科的切除によって原腫瘍を切除することはできるであろうが、癌細胞はその身体に侵入し終わっており、成功の見込みがあるのは化学療法か、もしくは特定の形態の標的療法のいくつかだけである。
したがって、局部的に侵入する可能性と一次腫瘍から離れた器官に転移する(腫瘍進行)可能性はほとんどの癌の過程において致命的な事象である。一次腫瘍を取り巻く細胞外マトリックス(ECM)の変性/分解および腫瘍細胞接着特性の変化は、転移性細胞の一次腫瘍細胞からの解離にとって極めて重要であることが知られている(Liotta,Cancer Res.46:1−7(1986);Hartら、Biochim.Biophys.Acta 989:65−84(1989))。
腫瘍脈管形成は一次腫瘍の拡大と転移性腫瘍の広がりの両方にとって必須であり、脈管形成自体はECM分解を必要とする(Bloodら,Biochem.Biophys.Acta 1032:89−118(1990))。したがって悪性は、新生物細胞とその環境がその病理学的過程において決定的な役割を果す全身性疾患である(Fidler,I.J.,Cancer Metastasis Rev.5:29−49(1986))。
悪性腫瘍に伴う遺伝子発現の変化の同定は、腫瘍の進行に関与するものも含めて、明らかに、癌を完全に理解するためだけでなく、癌に対する新しい合理的療法を開発するためにも欠くことのできない前提条件である。細胞遺伝子の2つの群(癌原遺伝子群および腫瘍抑圧遺伝子群)の発現の変異および/または異常制御が、多段階の過程で正常な成長制御の欠失を導き、転換した細胞表現型の獲得を導くことが示されている(Weinberg,R.A.,Cancer Res.49:3713−3721(1989))。しかし腫瘍の進行を導く分子機構についてははるかに不明瞭である(Nowell,P.C.,Cancer Res.46:2203−2207(1986);Fidler,I.J.,Cytometry 10:673−680(1989))。
したがって、癌細胞に特有の遺伝子が異常に発現した宿主遺伝子であることが極めて多いという点で新たな問題が生じる。極めて多くの場合、ある与えられた癌に関して特定のタンパク質標識がその癌中で過剰発現するが、この標識はその身体中の別の場所でも低レベルであるとはいえ発現する。
癌に関連するタンパク質のいくつかは、細胞群を互いに適切な関係に保つために重要な細胞外マトリックスを破壊する酵素である。このような酵素の1種は金属プロテイナーゼ(MMP)類(Matrisian,L.M.,Trends Genet.6:121−125(1990))であり、これらは亜鉛を結合するのでこのように呼ばれている。しかし、いくつかの存在が暗示され得るとはいうものの、癌あるいは特定の腫瘍の症状を示すことがわかっているものはなかった。
MMP類は、ECM改造および細胞移転が関係するいくつかの生理学的および病理学的過程(例えば形態発生および胚発育、リウマトイド関節炎、および腫瘍の浸潤および転移など)に関与する。MMP阻害剤は、腫瘍の進行にとって極めて重要な腫瘍浸潤および脈管形成を実験モデル中で遮断し得ることが知られている。
マトリックス金属プロテイナーゼファミリー(族)の構成要素はすべてECMの少なくとも一成分を分解するプロテイナーゼであり、潜在型で分泌され、活性になるためには(例えばプラスミンによる)タンパク加水分解などの活性化を必要とする。間質コラーゲナーゼ類は結合組織コラーゲン類(IからIIIまで)を特異的に攻撃し、一方、IV型コラーゲナーゼ類(72kDおよび92kD)はフィブロネクチンおよび基底膜中に存在するコラーゲンを分解する。ストロメリシン類(トランシン(transin)類)−1および−2、そしてポンプ−1(pump−1)も、はるかに広い基質特異性を有し、プロテオグリカン類、ラミニン(laminin)、フィブロネクチン、およびコラーゲン類(IIIからVまで)を分解する。
男性の場合悪性腫瘍のほとんどは癌腫であり、非喫煙者のなかでは乳癌が女性の癌による死亡率の主たる原因である(Willett,W.,Nature 338:389−394(1989))。数種の腫瘍遺伝子の発現が悪性乳細胞および腫瘍中で変化することが報告されているが、一貫して乳癌に伴い得る特定の腫瘍遺伝子/抑圧遺伝子発現様式はない(Gullick,W.J.,Prog.Growth.Factor Res.2:1−13(1990))。
しかし乳腫瘍の新生物細胞はしばしば、その増殖の制御と転移能にとって重要でもあり得る脂肪および間葉支質中に埋め込まれている。実際、支質細胞が正常な乳房上皮の成長を正負両方に変調させ得ることが知られており(Salomonら,“Breast Cancer:Cellular and Molecular Biology"(Lippman,M.E.およびDickson,R.B.編),363〜389頁(Kluwer,Boston,(1988)))、また上皮と支質成分との相互作用が乳腺における上皮発癌に影響を与え得ることが知られている(DeOmeら,Cancer Res.38:2103−2111(1978))。
悪性乳腫瘍中に“活性化された”(Tremblay,G.Exp.Mol.Pathol.31:248−260(1979))および/または異常な(Greyら,Proc.Nat.Acad.Sci.USA 86:2438−2442(1989))線維芽細胞が存在することが推測されており、乳癌は支質要求に対する依存性からの部分的逸脱または支質成分に対する異常に強い応答を表すのであろうと提唱されてきた。
癌組織の性質ゆえに、ある与えられた癌の連続的培養または株化細胞を作成(この操作により、ある与えられた治療法の効果の研究が容易になる)することは比較的簡単である。このような系の少なからぬ欠点はその性質そのものにある:試験的処置はそれが細胞に対して直接作用し得るかどうかを明確にするであろうが、その処置が生体内でどのような効果を有するかは決して明確ではなく、またこのような株の生化学的分析は必然的に、通常生体内の腫瘍を取り巻いている組織の非存在下で行うことになる。
本発明の目的は、乳癌腫中で発現が増大し、それによって乳癌腫がその周辺支質と相互作用する悪性上皮細胞とみなされる遺伝子を同定することにある。
本発明者らは過去に特徴づけられたことのないあるタンパク質が、ある種の浸潤性癌、具体的には乳癌腫、頭部および首部偏平上皮細胞癌腫および皮膚(偏平および基底細胞型)癌腫の症状を示すことを新たに発見した。このタンパク質は見かけ上金属プロテイナーゼ群に属し、ここではこれをストロメリシン−3と呼ぶ。
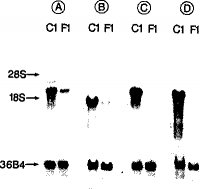
図面の説明図1:C1乳癌腫および線維腺腫から得た全RNAのノーザンブロット分析 実施例1に記述するように、C1乳癌腫および線維腺腫細胞から得たRNAボレート(bolate)を別個に単離した4種のcDNAプローブを用いてプローブした。
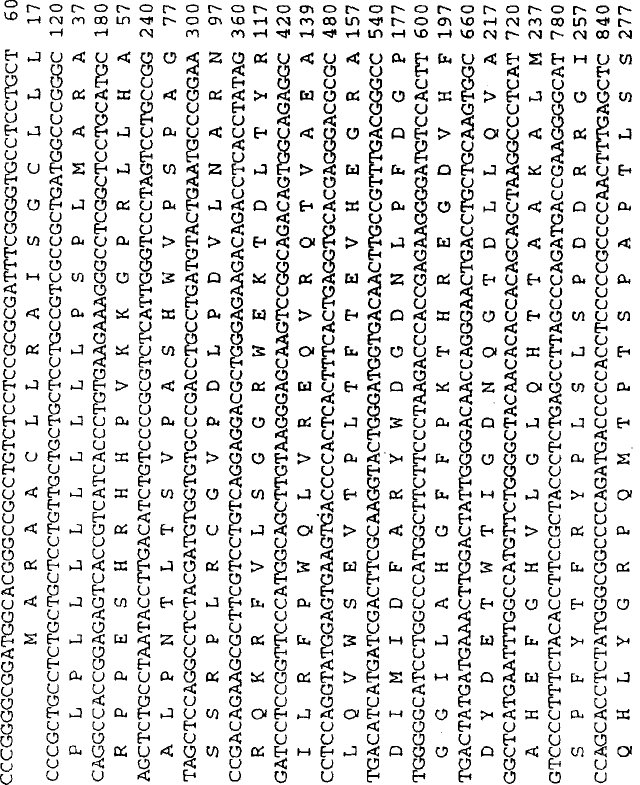
図2:ストロメリシン−3 cDNAのヌクレオチド配列 ストロメリシン−3のcDNAのヌクレオチド配列および推定アミノ酸配列を表す。下線を引いたヌクレオチド配列は5'末端から順に:推定シグナルペプチド;プロ金属プロテイナーゼに特有のPRCGBPD配列、亜鉛結合ドメインの保存されたヒスチジン残基およびポリ(A+)シグナル配列に対応する。
図3:金属プロテイナーゼ配列の比較 実施例3に記述するように、すべて推定上の金属プロテイナーゼであるストロメリシン−3、ストロメリシン−2、ストロメリシン−1およびコラーゲナーゼ−1を並列し、比較する。
図4:ヒト金属プロテイナーゼのノーザンブロット分析 4種のエストロゲン受容体陰性乳癌腫(C1,グレードII;C2,C3およびC4,グレードIII)、6種のエストロゲン受容体陽性乳癌腫(C5,C8およびC9,グレードII;C6およびC7,グレードIII;Cio,グレードI)および4種の乳線維腺腫(F2〜F5)から全RNAを調製した。
そのRNAを(a)ストロメリシン−3 RNA、(b)1型コラーゲナーゼRNA(CO I)、(c)92kd 4型コラーゲナーゼRNA(CO IV 92k)、(d)72kd 4型コラーゲナーゼRNA(CO IV 72k)、(e)ストロメリシン−1および2 RNA(ST1/2)およびポンプ−1 RNA(PU I)で実施例4に記述するようにプローブした。
図5:種々の株化細胞および組織から得たストロメリシン−3 RNAのノーザンブロット分析 (a)乳癌患者から得た5種の転移性腋窩リンパ節および3種の正常腋窩リンパ節;(b)4種のエストロゲン受容体陰性乳癌腫株(BT−20,MDA−231,SK−BR−3,HBL−100)および4種のエストロゲン受容体陽性乳癌腫株(T−47D,BT−474,ZR−75−1,MCF−7);(c)10種の正常ヒト組織;および(d)血清非含培地(1および2)中tPA非存在下(1)またはtPA存在下(2)で培養した、20mg/m■インシュリンを補足した血清非含培地(3〜6)中、PDGF非存在下(3)またはPDGF存在下(4)、EGF存在下(5)、あるいはbFGF存在下(6)で培養した、HFL−1ヒト胎児デプロイド線維芽細胞(ATCC CCL 153)、をストロメリシン−3配列で実施例5に記述するようにプローブした。
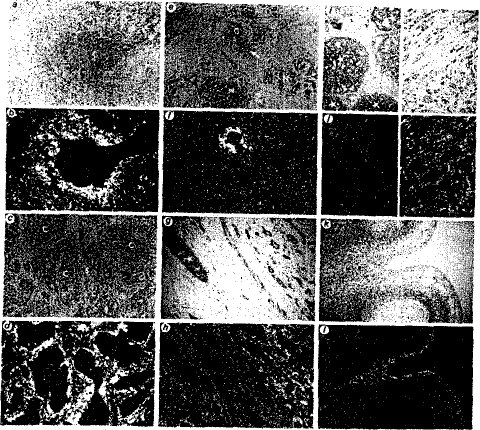
図6:乳癌腫および胎児肢芽の切片中のストロメリシン−3 RNA転写物の局在化 ヘマトキシリンで染色した組織切片の明視野顕微鏡写真(x100)(A,C,E,G,IおよびK);ならびに、実施例6に記述するように原位置でアンチセンス・ストロメリシン−3 cRNAとハイブリッド形成させた後の同切片(やはりヘマトキシリンで染色したもの)の暗視野画像(B,D,F,H,JおよびL)。
図7:マウスST3 cDNAのcDNA配列 マウスST3遺伝子のcDNA配列およびヒトST3 cDNA配列との比較。
本発明は第1の側面として、ストロメリシン−3またはストロメリシン−3をコードするヌクレオチド配列の検出からなる浸潤性癌、具体的には胸部、頭部および首部、ならびに皮膚の癌腫の診断法を提供する。
本発明はもう1つの側面として、浸潤性癌、具体的には胸部、頭部および首部、ならびに皮膚の癌腫の治療または予防における、ストロメリシン−3の合成または活性を妨害する薬剤の使用を提供する。
転移性腫瘍は浸潤性であるが、浸潤性腫瘍が必ずしも転移性ではない(例えば基底細胞皮膚癌腫)ことは理解されるであろう。
ストロメリシン−3遺伝子の発現はECM分解の領域に特異的であり、見かけ上金属プロテイナーゼをコードしているので、そのECM分解活性は腫瘍の転移への進行にとって極めて重要であると考えられる。支質細胞によるストロメリシン−3の発現はECMの重要な部分を破壊し、それによって癌細胞が原腫瘍から転移することを可能にすると思われる。
したがってストロメリシン−3の活性に影響を与え得る薬剤は転移に対する効果を有するであろう。そのような薬剤は、該タンパク質の合成を阻害するか、該タンパク質の成熟を阻害するか、あるいは該酵素の活性を遮断もしくは変化させることによってその活性を変化させるものが適しているであろう。
ストロメリシン−3遺伝子の発現が転移相にある乳癌の症状を示すことが初めてわかった。実際、この結果は切除された種々の腫瘍中のmRNAを検出することによって達成された。乳癌は非喫煙女性集団において癌による最も高い死亡率の原因であるので、乳癌を選択した。
ストロメリシン−3はほぼ確実にMMPファミリーに属する新規タンパク質であり、ホルモン状態にかかわらず浸潤性乳癌腫に関与している。
MMPファミリーの構成要素は活性になるために活性化過程を必要とし、この過程にはプレ−およびプロ−配列の除去が関与し得る。プロ−および成熟−ストロメリシン−3のアミノ酸配列は過去に特徴づけられたMMP類のものとは著しく異なっており、成熟化、活性化およびECM成分に対する特異性に関して異なる特性を示し得る。
ストロメリシン−3遺伝子はすべての一次浸潤性乳癌腫によって発現され、またその転移部のいくつかによっても発現され、そのような発現について分析された広範なECM改造が起こることが知られている組織(子宮、胎盤、および肢芽)中でも発現されるが、乳線維腺腫および正常成人組織中では発現されない。このことはストロメリシン−3遺伝子産物が乳癌の進行において重要な役割を果すことを示唆している。またこの概念と一致して、通常前浸潤性病巣と見なされしばしば微小浸潤に関連するコメド型の原位置癌腫を除いて、ストロメリシン−3遺伝子はほとんどの原位置乳癌腫中で発現されない。したがって、その身体中の子宮または胎盤以外の別の場所に認められるような低濃度ではないストロメリシン−3 RNA転写物の存在は、転移性癌または浸潤性になる危険の高い癌の症状を示す。
ストロメリシン−3は浸潤性癌成長に伴うと思われる溶解過程に関与し得る。あるいは、ほとんどの浸潤性乳癌病巣に伴い、悪性細胞のさらなる拡大を防止するための宿主反応を表す結合織線維増生の形成において、ストロメリシン−3がある役割を果すことも考えられる(Ahamed,A.,Pathol.Annu.25(Pt2):237−286(1990))。このような場合には、ストロメリシン−3活性の増大が有利であろう。
さらに、新生物細胞群(島)のすぐ周囲の支質線維芽細胞におけるストロメリシン−3遺伝子の限定された発現は、いくつかの腫瘍形成性細胞の悪性変換に関連することが知られているもう1つの金属プロテイナーゼであるコーラーゲナーゼIV、および乳癌腫中でその発現が増大するリソソームのアスパルチルプロテアーゼであるカセプシンD(両酵素とも線維芽細胞中では発現されないが、乳癌の新生物上皮細胞中で発現される)とは対照的である(Monteagudoら,Am.J.Phathol.136:585−592(1990);Garciaら,Steroid Biochem.27:439−445(1987))。
新規乳癌標識を同定するために、cDNAライブラリーを構築し、線維腺腫供給源から得たポリ(A+)RNAで抽出した。この操作によって、このcDNAライブラリーは転移性癌に特有の配列に関して濃縮された。
いくつかのクローンを成長させ、転移性腫瘍および線維腺腫からのポリ(A+)RNAから誘導したプローブを用いてスクリーニングした。次いで、転移性癌ポリ(A+)RNAから誘導したプローブに対してかなり強く結合したクローンをさらに成長させた。
この方法で作成したクローンのうちの1つは、悪性乳癌および咽頭癌、頭部、首部、および皮膚(偏平および基底細胞型)癌腫、ならびに子宮および胎盤(これらのすべてにおいてECMの破壊が起こっており、この破壊は癌と関連した場合、癌細胞がその身体中に広がること(転移)を可能にする)中にのみ認められる高い発現率の程度にまで弁別的に発現されることがわかった。
子宮および胎盤の場合にはECMの破壊が自然に起こるが、それ以外の場所で起こる同じ事象は腫瘍成長の特徴であると思われる。
また、ECMの破壊を伴う胎児の肢発芽中の指間分化にストロメリシン−3遺伝子の発現が認められたことに注目することも興味深い。
cDNA配列を特徴づけることによって、読み取り枠の存在が明らかになった。コード化されているタンパク質配列を既知のライブラリーと比較することによって、このタンパク質がECMを破壊することが知られているファミリーに属することが立証された。ストロメリシン−3の配列はそのファミリーの他の構成要素に対して、他の構成要素間に認められるほどの類似性を有さないが、それでもこの酵素の性質の同定に利用できるいくつかの特徴的な領域を呈示している。したがって、このタンパク質はコラーゲナーゼの一種であるかも知れないし、全く異なるECM構成成分を破壊するのかも知れないが、このタンパク質をストロメリシン−3と命名した。
ストロメリシン−3 mRNAの出現を立証するためのヌクレオチドプローブを構築することによって上述の組織分布が明らかになり、また、標識化によってストロメリシン−3遺伝子の発現領域を顕微鏡写真で正確に位置決定することが可能になった。
この方法で作成した顕微鏡写真の分析によって、やや意外なことに、ストロメリシン−3遺伝子が癌細胞そのものでは発現されず、その周辺支質で発現されることが示された。さらに支質は、腫瘍の基底膜がまだ無傷である場合にはストロメリシン−3 mRNAの証拠を全く示さなかった(図6を参照のこと)。ストロメリシン−3遺伝子はすべての一次浸潤性乳癌腫によって発現され、またその転移節のいくつかによっても発現され、そのような発現について分析された広範なECM改造が起こることが知られている組織(子宮、胎盤、および肢芽)中でも発現されるが、乳線維腺腫および正常成人組織中では発現されない。このことはストロメリシン−3遺伝子産物が乳癌の進行において重要な役割を果すことを示唆している。またこの概念と一致して、通常前浸潤性病巣と見なされしばしば微小浸潤に関連するコメド型の原位置癌腫を除いて、ストロメリシン−3遺伝子はほとんどの原位置乳癌腫中で発現されない。ストロメリシン−3は常に転移性癌の支質中に出現し、原位置一次腫瘍(まだ基底膜を有しており、浸潤性でない腫瘍)中には出現しない。したがって、その身体中の子宮または胎盤以外の別の場所に認められるような低濃度ではないストロメリシン−3 RNA転写物の存在は、転移性癌または浸潤性になる危険の高い癌の症状を示す。
さらに、ER−陽性または陰性乳癌株化細胞のいくつかはEGF/TGF−αおよびFGF(ストロメリシン−3遺伝子の発現に関与する因子)に対する受容体を分泌し保持していることが知られているにもかかわらず、ストロメリシン−3遺伝子の発現はいずれのER−陽性または陰性乳癌株化細胞中にも検出されなかった。
したがって、ストロメリシン−3、その前駆体あるいはそれをコードするヌクレオチド配列に標準的な検出技術を適用することにより、転移性癌を診断したり、あるいは一次腫瘍がまだ致命的な転移相に達していないことを確認することができる。
このような技術にはヌクレオチドプローブを例えば上述のように用いる検出法が含まれ、またこのような技術が例えば抗体あるいはその等価物によるストロメリシン−3タンパク質の検出から構成されていてもよい。
ヌクレオチドプローブは、添付の図2に示す配列に対して天然に存在する他の配列に対するより強くハイブリッド形成するものなら何でもよい。プローブの種類にはcDNA、リボプローブ、合成オリゴヌクレオチドおよびゲノムプローブが含まれる。使用するプローブの種類は一般に特定の状況によって決まる(例えば、原位置ハイブリッド形成のためにはリボプローブ、ノーザンブロッティングのためにはcDNA)。最も好ましいプローブは図2のcDNAの(−)鎖に対応するものである。またストロメリシン−3遺伝子中に位置するイントロンを認識するプローブを提供することもできるが、これがRNA転写物の検出と同程度の信頼性を有するとは限らない。
ストロメリシン−3をコードする遺伝子そのものの検出は一般的には診断目的には有効でないであろうが、転写物および他の発現産物を検出するための他の形態の検定法は一般に有用であろう。プローブはストロメリシン−3 mRNA転写物を弁別的に認識するために必要な程度に短くてもよく、例えば15塩基長程度でもよい。
プローブの標識化(ラベル化)の形態は適切なものであれば何でもよく、例えば32Pおよび35Sなどの放射性同位体を使用することができる。放射性同位体による標識化は、そのプローブが化学的に合成されたものであれ、生物学的に合成されたものあれ、適切に標識された塩基を用いることによって達成できる。標識化の他の形態には、ELISAの特徴であるように酵素あるいは抗体標識化が含まれ得るが、標識されたプローブによるmRNA転写物の検出には一般にX−ラジオグラフィーが用いられるであろう。
RNA転写物の検出はノーザンブロッティングによって達成できる。この方法では、例えばRNAの調製物を変性化アガロースゲルに流し、適切な支持体(例:活性化セルロース、ニトロセルロースまたは硝子またはナイロン膜)に移し、次いで、放射性標識cDNAまたはRNAをこの調製物にハイブリダイズさせ、洗浄し、オートラジオグラフィーで分析する。
原位置ハイブリッド形成可視化法を用いることもできる(実施例6)。この方法では、[35S]−標識アンチセンスcRNAプローブを生検試料の薄片とハイブリダイズさせ、洗浄し、RNaseで切断し、オートラジオグラフィー用感受性乳剤にさらす。この試料をヘマトキシリンで染色することにより、試料の組織学的組成を明らかにすることができ、適切な光フィルターによる暗視野画像化によって顕色乳剤が明示される。
免疫組織化学を用いて生検試料中のストロメリシン−3の発現を検出することができる。例えば適切な抗体を細胞の薄層と接触させ、洗浄し、次いで第2の標識抗体と接触させる。標識化はペルオキシダーゼ、アビジンなどの酵素、あるいは放射性標識によって行うことができる。色素原標識は顕微鏡下で検出できるので、一般に好ましい。
より一般的に好ましい検出法は、極めて迅速に実行できる免疫検定法(例えば、ELISAまたはRIA)でタンパク質を検出することである。したがって抗体または抗体等価物を用いてストロメリシン−3を検出することが一般に好ましいが、適切に標識されたストロメリシン−3基質の使用も有利であり得る。
酵素過程の生成物がそれ自体の特性として検出可能であり特徴的である場合(例えば過酸化水素など)には、基質を標識する必要はないであろう。しかし基質の標識化が必要な場合には、基質の標識化は酵素標識化、放射性同位体による標識化、抗体標識化、蛍光標識標識化、あるいは当業者にとって容易に理解されるであろう他の適切な形態のいずれによってもよい。
最も好ましいストロメリシン−3発現の検出法は、抗体を使用する方法である。抗体は以下に記述するようにして調製することができ、ストロメリシン−3発現の検出に適したあらゆる方法で使用できる。
抗体に基づく技術にはELISA(酵素結合イムノソルベント検定法)およびRIA(放射線免疫検定法)が含まれる。このような免疫検定法には従来の操作法のいずれを使用してもよい。この操作を以下のように実行するのも適切であり得る:ストロメリシン−3標準を放射性同位体(例:125Iまたは35S)または検定可能な酵素(例:西洋ワサビペルオキシダーゼまたはアルカリ性ホスファターゼ)で標識し、これを非標識試料と共に対応する抗体と接触させ、そこに第2の抗体を用いて第1の抗体を結合させ、放射活性または固定化酵素を検定する(競争的検定法)か、もしくは別法として、試料中のストロメリシン−3を対応する固定化抗体と反応させ、放射性同位体または酵素で標識された抗ストロメリシン−3抗体をこの系と反応させ、放射活性または酵素を検定する(ELISA−サンドイッチ検定法)。他の従来法も使用に適し得る。
上記の技術は基本的に“一段階”または“二段階”検定法として実行できる。“一段階”検定法は、抗原と固定化抗体を接触させ、洗浄することなくこの混合物を標識抗体と接触させることを伴う。“二段階”検定法は、上記の混合物を標識抗体と接触させる前に洗浄することを伴う。他の従来法も使用に適し得る。
ストロメリシン−3および/または抗体の酵素または放射線による標識化は従来の手段によって達成できる。一般的にそのような手段には、問題の抗原または抗体に対する酵素の(例えばグルタルアルデヒドによる)共有結合的連結が含まれる。具体的にはこの連結が酵素の活性に不利な影響を及ぼさないようにする。これは酵素が連結後でもその基質と相互作用できなければならないことを意味するが、検定を達成するに足る十分な活性が残っているのであれば、酵素のすべてが活性である必要はない。実際、酵素を結合させるいくつかの技術は非特異的であり(例えばホルムアルデヒドを用いる方法)、活性酵素の一部を与えるに過ぎないであろう。
普通は、検定系の一成分を支持体上に固定化し、それによってその成分とその系の他の成分との接触を可能にし、困難で時間のかかる労力を伴わずに容易に除去できるようにすることが望ましい。第2層を固定化して第1層から分離することができるが、通常は1層で十分である。
酵素自体を支持体に固定化することができるが、固相酵素が必要な場合には、抗体に結合させ、抗体を支持体に固定することによってこれを達成することが一般に最善の方法であり、そのモデルおよび系は当該技術分野でよく知られている。単純なポリエチレンが適切な支持体を提供し得る。
標識化に使用できる酵素に特に制限はないが、例えばオキシダーゼ群の構成要素から選択することができる。これらの酵素はその基質との反応によって過酸化水素の生成を触媒し、グルコースオキシダーゼはその良好な安定性、利用の容易さおよび安価であること、ならびにその基質(グルコース)が容易に利用できることから頻繁に使用されている。オキシダーゼの活性は、この酵素で標識した抗体を当該技術分野でよく知られている制御された条件下でその基質と反応させた後に生成する過酸化水素の濃度を測定することによって検定できる。
好みによって他の技術をストロメリシン−3の検出に使用することもできる。これらの1つはウエスタンブロッティング(Towbinら,Proc.Nat.Acad.Sci.76:4350(1979))であり、この方法では適切に処理した試料をSDS PAGEゲルに流した後、ニトロセルロースフィルターなどの固体支持体に移す。次に、抗ストロメリシン−3抗体(非標識)をこの支持体と接触させ、標識したプロテインAや抗免疫グロブリン(適切な標識には125I、西洋ワサビペルオキシダーゼおよびアルカリ性ホスファターゼが含まれる)などの第2免疫学的試薬で検定する。
診断用の試料は関連部位のいずれからでも得ることができる。腫瘍から直接得た支質や細胞質などの試料が理想的であろうが、例えば血液から得た試料もまた適切であろう。しかし試料を血液から得た場合には、ストロメリシン−3の量が血流中に希釈されているであろうから、高感度の検定法が必要であろう。このような診断は、腫瘍を切除するための外科手術後などの患者の経過を監視する際に特に重要であり得る。参照値を手術後に読み取り、一定の間隔でもう1つの値を読み取れば、上昇は再発あるいは場合により転移を表し得る。このような値の読み取りは、例えば子宮中の活性を考慮に入れる必要があろう。
抗ストロメリシン−3抗体は画像化のためにも使用できる。酵素に加えて他の適切な標識には、放射性同位体:ヨウ素(125I、121I)、炭素(14C)、硫黄(35S)、トリチウム(3H)、インジウム(112In)、およびテクネチウム(99mTc);フルオレセインおよびローダミンなどの蛍光標識、およびビオチンが含まれる。
しかしインビボ画像化のためにはその位置がより限定的になり、抗体はそのままでは身体の外側から検出できないので、抗体を標識するか、もしくは他の方法で修飾することによって検出できるようにしなければならない。
この目的のための標識は抗体の結合を実質的に妨害せず、外部からの検出を可能にするものあれば何でもよい。適切な標識には、X−ラジオグラフィー、NMRまたはESRで検出できるものが含まれ得る。X−ラジオグラフィー技術に適した標識には、検出可能な放射線を放射するが患者にとって明白に有害でない放射性同位体(例えばバリウムまたはセシウムなど)が含まれる。NMRおよびESRに適した標識には一般に、重水素のように検出可能な特徴的スピンを伴い、例えば関連するハイブリドーマ用の栄養素を適切に標識することによって抗体中に導入できるものが含まれる。
インビボ画像化法の場合には、適切な検出可能画像化部分(放射性同位体(例:131I、112I、99mTc)、放射線不透過性物質、あるいは核磁気共鳴で検出できる物質など)で標識されている抗体または抗体断片を試験すべき対象(ヒトなど)中に(例えば非経口的、皮下的、あるいは腹腔内に)導入する。
対象のサイズおよび使用する画像化系が、診断的画像を作成するのに必要な画像化部分の量を決定するであろう。放射性同位体部分の場合、ヒト対象については、注射される放射活性の量は通常約5〜20ミリキュリーの範囲のテクネチウム−99mであってもよい。
次いで、標識した抗体または抗体断片がストロメリシン−3を含有する細胞の位置に優先的に蓄積するであろう。次いで、この標識抗体または抗体断片を既知の技術を用いて検出することができる。
この技術分野に関する一般的議論については、S.W.Burchielら,“Immunopharmacokinetics of Radiolabelled Antibodies and Their Fragments"(“Tumor Imaging:The Radiochemical Detection of Cancer",S.W.BurchielおよびB.A.Rhodes編,Masson Publishing Inc(1982))の第13章)を参照のこと。
抗体はストロメリシン−3のペプチドに対しても、またその全分子に対しても生じさせることができる。このようなペプチドはアルブミンなどの担体タンパク質と共に動物系に提供するか、あるいはそれが十分長い(例えば25アミノ酸残基)場合には担体なしで動物系に提供できる。ストロメリシン−3は自己タンパク質に相当であろうから、ヒト抗体がストロメリシン−3を認識できるとは思われない。
本明細書において用語“ペプチド”とは、2以上のアミノ酸がペプチド結合を通して連結してなるあらゆる分子を意味する。したがってこの用語はオリゴペプチド、ポリペプチドおよびタンパク質を包含する。
上記の技術で作成したポリクローナル抗体を直接使用してもよく、また適切な抗体産生細胞をその動物から単離し、これを用いて既知の手段(KohlerおよびMilstein,Nature 256:795および以下参照(1975))でハイブリドーマを作成することもできる。適切なハイブリドーマの選択も当業者には明白であろうし、得られる抗体はストロメリシン−3を同定するのに適した検定法で使用できる。
抗体またはその等価物を本発明に従って転移性癌の治療または予防に用いることもできる。抗体の適切な投与量の投与はストロメリシン−3の生産を遮断するか、もしくはストロメリシン−3の有効活性を遮断するよう作用することができ、これは悪性成長を治療するための極めて重要な時間を提供するであろう。
どのような場合にも何が実際に転移を導くのかはわかっていないので、予防はこの疾患のまさに初期段階でも適切であり得る。したがってストロメリシン−3活性を妨害する抗体、その等価物または因子類(例えばTIMP類(MMP類を調節する天然化合物−−金属プロテイナーゼの組織阻害因子))の投与を癌が診断されると同時に実施することができ、必要なかぎり長期間、好ましくはこの疾患のおそれが排除されるまで治療を続けることができる。
治療の好ましい形態はいわゆるマジック・ビュレット技術であり、この方法では腫瘍の領域に誘導する抗体に適切な毒素を結合させる。このような毒素は当該技術分野でよく知られており、1細胞あたりたった1または2分子程度で作用し得るリシンなどの天然分子に限らず、毒性放射性同位体、重金属、酵素および補体活性化因子からなってもよい。このような技術をホルモン拮抗薬あるいは例えば癌の治療に使用できる生理学的に活性な他の適切な化合物の局在化した投与量を送達するために使用することもできる。
本発明に従って使用するための抗体が診断的応用であれ、治療的応用であれ、モノクローナルでもポリクローナルでも適切であり得ることは理解されるであろう。これらの抗体等価物は例えば抗体のFab'断片(Fab、Fab'、F(ab')2およびFvなど);イディオタイプ;あるいはアロトープ移植の産物(患者中での免疫応答を避けるために、動物抗体の認識領域がヒト抗体の適切な領域に移植されているもの)などからなり得る。他の適切な修飾物および/または薬剤は当業者に明白であろう。
ストロメリシン−3を阻害または除去するための抗体の使用に加えて、他の形態の阻害剤を用いることもできるであろう。そのような阻害剤は(例えばECM分解酵素について)汎用であってもよいし、ストロメリシン−3に対して特異的であってもよい。金属プロテイナーゼ(TIMP)類の組織阻害因子はその存在が知られており、ストロメリシン−3に対して特異的なTIMPが存在する可能性は極めて高い。このようなTIMPは標準的な技術によって容易に同定できる。
ストロメリシン−3の合成阻害剤も製造することができ、これらは一般に酵素活性の作用を受ける基質の領域に対応するであろう。このような阻害剤が基質と切断生成物との間の凍結中間体に相当することが一般に好ましいが、その結合部位の立体障害型か、あるいはそれ自体がストロメリシン−3に不可逆的に結合するであろう型の結合部位を提供することもできる。他の適切な阻害剤は当業者に明白であろう。
ストロメリシン−3活性を遮断するために他の方法を用いることもできる。これらは例えば変性剤からなってもよいが、これらは非特異的である傾向が強く、例えば特異的抗体の使用などによってそれらを誘導できる場合にのみ適当に使用できる。ストロメリシン−3遮断活性の他の形態は、プレ−プロタンパク質からタンパク質への進行を遮断することによって達成され得る。この過程は数個の標的段階を提供し、唯一必要なことは他の必須酵素に影響を与えないように独立に遮断し得る段階、あるいは再度標的にされ得る段階を同定することだけである。
ストロメリシン−3の3次構造を選択的に認識し、それによってその酵素活性を遮断するために、ペプチドまたは他の小分子を用いることもできるであろう。このような活性遮断物質が必ずしも活性部位に結合する必要はなく、それでもストロメリシン−3の3次構造を改変または凍結してその活性を破壊、一時停止、または改変するよう作用することができる。またこの遮断物質が必ずしもそれ自体によって作用する必要はなく、この目的のためのもう1つの物質に結合させることができるし、あるいは適切な不活化剤のための認識部位として機能することもできる。
I型コラーゲナーゼおよび92kD IV型コラーゲナーゼのmRNAの出現はもっぱら悪性腫瘍に伴うが、その逆は常に成り立つわけではない(即ち、腫瘍は常にこのタンパク質を伴うわけではない)ことが、本発明者らの研究によって立証された。
浸潤性乳癌腫中でのストロメリシン−3遺伝子の発現とテナシン(tenascin)遺伝子の発現との間には明らかに類似点がある。ECM糖タンパク質テナシン(Chiquet−Ehrismannら,Cell 47:131−139(1986))は上皮間葉細胞相互作用、および正常発育中の細胞移動(器官形成中の乳腺のものを含む)に必須の役割を果していると思われる。
テナシンは悪性乳腫瘍の線維性支質中で過剰発現されることが一貫して認められてきており、ストロメリシン−3に類似した様式で誘導されると思われる。フィブロネクチンと比較した場合、テナシンは乳房腫瘍上皮細胞の付着に関する基質として劣っており、このことはそれらが浸潤性になることを可能にし得ることを示唆している。
したがってストロメリシン−3は乳癌の浸潤相中にテナシンと共同して作用し得る。またストロメリシン−3およびテナシンは胚形成中に、上皮−間葉相互作用が重要な役割を果すことが知られている領域、および細胞移動が起こる領域で同時発現し得る。
したがって本発明は、上記のタンパク質のいずれか、もしくはそれらをコードするヌクレオチド配列の検出をも含む、上に定義した転移性癌の診断法をも提供する。
また本発明は、上記のタンパク質のいずれかを結合する薬剤の使用をも含む転移性癌の治療または予防における使用をも提供する。
さらに、本発明はストロメリシン−3の一部または全体をコードするヌクレオチド配列をも提供する。ストロメリシン−3の配列は添付の図2に示すものが好ましく、またそのヌクレオチド配列も図2に示すものが好ましい。しかし、遺伝子コードの縮重ゆえに、その配列がストロメリシン−3の少なくとも一部をコードしている限りにおいて、ヌクレオチド配列がこの図に示したものとは実質的に異なり得ることは理解されるであろう。
必要な配列はそれを利用する方法によってさらに変化することさえあり得る。生物学的試料中のRNA転写物を検出するための使用を意図する場合には、普通は図2に記載した配列にかなり密接に対応することが好ましいであろう。しかしそれでも、ハイブリッド形成が選択した厳密性条件下で可能である場合には、その配列はさらに変化し得る。
当業者は図2のペプチド配列から逆にプローブを作成することができる。しかし遺伝コードの縮重性ゆえに、逆に作成したある配列が同じペプチドから逆に作成した相補的配列と必ずしも充分にハイブリッド形成するとは限らず、あるいは全くハイブリッド形成しないこともあることは理解されるであろうから、このようなプローブの使用は制限されるであろう。これは当業者の計画には共通の要素であり、いかなる配列の縮重性もしばしば非常に広いためにどのような1配列のためのプローブの数も極めて多数になる。
ヌクレオチド配列がストロメリシン−3ペプチドまたは該酵素全体の発現のために必要な場合には、上述のように遺伝子コードの点、ならびにストロメリシン−3の構造または機能に有意な影響を与えることなくこの酵素のいくつかのアミノ酸配列を変化させ得るという点で、かなり大きな自由度が存在し得る。
このような配列の相違を意図する場合には、活性を決定する極めて重要な領域がこの分子上に存在するであろうことに注意すべきである。通常このような領域は結合部位を形成する残基群か、もしくは結合部位に影響を及ぼす3次構造を形成する残基群からなるであろう。一般論として、類似の機能を発揮する残基を使用する限りにおいて、3次構造を形成する残基群を置換することができる。他の例では、残基の種類は全く重要でないこともある。
したがって本発明は、ストロメリシン−3の例えば抗体の作成に使用するための特徴的領域を示すか、もしくはまだ実際にストロメリシン−3活性を示す、その配列上の変種および変異体をも包含する。このような変種および変異体には欠失体、付加体、挿入体、反転体、反復体および型置換体(例えば、ある親水性残基で別の親水性残基を置換するが、一般に高度に親水性の残基で高度に疎水性の残基を置換しない)が含まれる。小さい変化は一般に、それらがその分子の必須部分でない限り活性にほとんど影響を与えないであろう。また小さい変化は遺伝子操作の副産物でもあり得るし、例えば追加の制限部位を作成する場合などには小さい変化が望まれるべきである。修飾には1以上の残基を他の適切な残基で置換することも含まれ、このような置換は1:1でもよいし、あるいは1以上または1以下の他の適切な比でもよい。
遺伝子操作/発現を異なった方法で補助するため、あるいはストロメリシン−3分子を増大させるか、もしくは他の方法で都合よく修飾するために、コード配列中に点変異および他の変化を導入することによって、例えば制限部位を付加または欠失させることができる。
ストロメリシン−3等価物が他の動物(具体的には哺乳類)中でも発見されるであろうこと、またそのような供給源から得られる配列情報がストロメリシン−3分子の保存された領域を解明するために特に重要であり得ることも理解されるであろう。例えば、マウスの対応する配列ではストロメリシン−3に特有のプロドメイン中の10アミノ酸配列などを含む約80%が保存されている。ヒトストロメリシン−3配列に対応する動物配列が当該技術分野で既知の方法および上述の方法によって容易に検出できるであろうこと、およびそのような配列およびそれらのペプチドがその変異体および変種と共に本発明の一部を構成していることは理解されるであろう。
所望により、制限部位を提供するように本発明の配列を加工することもできる。この加工はコードされているストロメリシン−3のペプチド配列を干渉しないように行うことが可能であり、また最終生成物が望ましい特性を有するという条件下で、望ましい程度あるいは必要な程度に干渉することもできる。
上述のようにハイブリッド形成は信頼性の低い配列相同性の指標であり得るが、好ましい配列は一般に図2の配列に対して50%以上、好ましくは70%以上、より好ましくは80%以上の相同性を示すものであろう。
他の金属プロテイナーゼ類の場合と同様に、ストロメリシン−3は最初はプレ−プロ酵素として発現される。したがって生体内では2つの切断段階が観測される。切断はインビトロ発現にとって必ずしも必要条件ではなく、例えば大腸菌は成熟タンパク質を発現することができるであろう。
ストロメリシン−3またはその特徴的なペプチドを発現させることを望む場合、適切な糸はすべて使用することができる。適切なベクター、発現ベクターおよびその構築法の一般的性質は当業者には理解できるであろう。
“特徴的な”という表現はストロメリシン−3に特有の配列を有するあらゆるペプチドを意味する。このような配列はストロメリシン−3活性にとって重要であってもよいし、あるいはただ単に他のペプチドには認められない配列であるだけでもよい。しかし一般にストロメリシン−3活性にとって重要な配列が好ましい。というのはこれらは母集団内で保存されている可能性がより高いからである。
適切な発現ベクターはファージまたはプラスミドに基づいていてもよく、これらは両者とも一般に宿主特異的であるが、これらを他の宿主のために加工することもしばしば可能である。他の適切なベクターにはコスミドおよびレトロウイルスならびに他のあらゆる媒体が含まれ、これらは与えられた系に関して特異的であってもよく、また特異的でなくてもよい。また認識、プロモーター、オペレーター、誘導因子、終止因子、および発現の制御に不可欠および/または有用な他の配列などの制御配列は当業者には容易に理解できるであろう。またこれらの制御配列は天然のストロメリシン−3配列に伴っているものでもよいし、あるいは使用するベクターに伴っているものでもよく、あるいは他の供給源から適切に誘導することができる。これらのベクターはあらゆる適切な方法で修飾または加工することができる。
ヌクレオチド配列の正しい調製は、例えばSangerらの方法(Proc.Natl.Acad.Sci.USA 74:5463−7(1977))によって確認することができる。
本発明のストロメリシン−3をコードしているcDNA断片は適切なベクター中に容易に挿入できる。受容ベクターが容易な挿入に適した制限部位を有することが理想的であるが、例えば平滑末端連結も使用できる。ただしこれは読み枠および挿入の方向に関する不確定性を導くであろう。このような場合には発現に関して形質転換体を試験することは当然のことであり、これらの形質転換体は6個中1個の割合で正しい読み枠を有するはずである。当然のことながら、当業者は望ましい発現系に応じて適切なベクターを選択することができる。
適切な生物(好ましくはHeLaなどの真核細胞株)を得られたプラスミドで形質転換し、アンピシリンまたは必要ならば他の適切な手段によって形質転換体を選択し、トリプトファンまたは必要ならば他の適切なプロモーター−誘導物質(インドールアクリル酸など)を添加することによって、目的のストロメリシン−3を発現させることができる。発現の程度をSDSポリアクリルアミドゲル電気泳動:SDS−PAGE(Lemelli,Nature 227:680−685(1970))で分析することができる。
培養などの生育および形質転換に適した方法は例えばManiatis(“Molecular Cloning,A Laboratory Notebook",Maniatisら編,Cold Spring Harbor Labs,NY(1989))に実用的に明示されている。
ストロメリシン−3またはそのペプチドの生産に有用な培養はあらゆる生存細胞の培養が適切であり得、原核生物発現系から真核生物発現系までに及び得る。好ましい原核生物系の1つは、その操作の容易さゆえに大腸菌の系である。しかし、より高等な系(哺乳類細胞株など)も真核タンパク質の発現に使用できる。現在のところ仮発現とって好ましい細胞株はHeLaおよびCos細胞株である。他の発現系にはチャイニーズハムスター卵巣(CHO)細胞株が含まれる。
価値ある系の1つはバクロウイルス系であり、この系では蝶細胞をストロメリシン−3または適切なペプチドをコードしているDNAベクターおよびバクロウイルスDNAで同時トランスフェクションする。組換えがその細胞内で起こり、適切なバクロウイルス組換体を標準的な技術で選択することができる。その後、その組換体を用いて望ましい細胞株を感染させることができ、感染によってストロメリシン−3またはペプチドが発現される。この系に特有の利点は生産されるタンパク質の量であり、その量は約1〜約500mg/■の範囲に及び得る。
このような系は大腸菌系ほど使用しやすくない傾向にあるが、その利点は一次合成後のタンパク質のプロセシングにある。例えば大腸菌はプレ−プロタンパク質のプロセシングに哺乳類細胞と同じ系を使用しない。
使用し得る他の発現系には、例えばストレプトミセス科の放線菌、およびサッカロミセス種、具体的にはサッカロミセス・セレビシェなどの酵素が含まれる。一般に操作者が何を要求するかに依存して、所望によりどのような系も使用できる。適切な系を用いて遺伝物質を増幅することもできるが、そのDNAの増殖だけが必要なこの目的には一般に大腸菌の使用が便利である。
成熟酵素の配列はプロ−およびプレプロ−配列にも共通であるので、抗体を生じさせるためには成熟酵素だけを生産することが有利であろう。しかし、プロおよびプレプロ部分の切断がこの分子の3次構造を変化させるかも知れず、したがって成熟酵素に対して生じた抗体が例えばプロ酵素を検出できないこともあり得ることは理解されるであろう。初期段階のいずれかにある酵素および/または切断されるプレ−またはプロ−ペプチドに対して生じた抗体も有用となるであろう。
ペプチドまたはヌクレオチド配列は、それを使用する目的が考慮されたストロメリシン−3に特徴的なものであれば何でもよい。配列はストロメリシン−3に完全に特徴的であることが理想的であろうが、そのような配列の長さはストロメリシン−3分子のその領域に応じて様々であり得る。最も好ましい領域は高度に保存されている領域、および他のタンパク質に共通しない領域であるが、その配列がMMP類、もしくはより具体的には浸潤性腫瘍に関連するMMP類に特徴的な配列である場合にはそれが有利であり得る。
本発明は上記のペプチドおよびヌクレオチド配列の等価物を包含し、これに関連する。ここで用語“等価物”は前述の意味で使用されており、換言すればC−またはN−末端あるいは他のいずれかの位置に置換を有する配列という意味の等価物である。
本発明は本配列の変異体をも包含する。ここで用語“変異体”は、上述の制限を前提とする配列中のアミノ酸残基または塩基の欠失体、付加体、挿入体、反転体および置換体に関して使用されている。
さらに本発明は配列の変種をも包含し、この用語は、図2に示す配列と基本的には同じ配列を共有するが、大きい母集団内で予期される程度に変化している、ときおり発見されるであろう他の天然に存在するストロメリシン−3に関連して使用されている。この定義内にはアレル変種および類似型の活性を示し関連する配列を有する他の種由来のペプチドが含まれる。またあまり好ましくはないが、動物配列も含まれる。
本発明者らはストロメリシン−3の発現が例えば成長因子および腫瘍プロモーターによって刺激され得ることをも発見した。このような因子の典型例にはEGF FGFおよびPDGFおよびTPAが含まれる。したがって上述の方法と共に、腫瘍試料中のこれらの因子のいずれかの検出は癌の転移状態を診断するためにも役立ち得る。
したがって本発明は、ストロメリシン−3遺伝子の発現を変化させることによる転移性癌の治療をも提供する。これはストロメリシン−3の生産を刺激するために必要な因子を、例えばこの因子に対する特異的抗体を使用するなどして妨害することによって達成することができ、目的の結果を達成するためにこの抗体をさらに修飾してもよい。この因子の受容体を遮断すること(これは例えば抗体または合成ペプチドであり得る必要な結合剤の局在化によってより容易に達成され得る)も可能であろう。
ストロメリシン−3遺伝子の発現に影響を与えることは、例えばゲノムDNA上のプロモーターなどの部位の遮断などによってより直接的にも達成され得る。
例えば抗体を患者に投与するために本発明を用いる場合には、これは適切な経路のいずれによってもよい。腫瘍がまだ局在化していると思われるか、もしくはそのように診断される場合には、適切な投与法はその部位への直接的な注射による方法であり得る。標的が乳癌である場合には乳房への注射で十分であり得、また埋め込み剤を用いてもよい。例えばTIMP類を投与すべき場合には、長期間持続する投与のために皮膚パッチを使用することもできるであろう。
咽頭癌の場合には、さらなる選択枝として例えばうがい剤を用いる経口投与であってもよい。
いずれかの例において、投与を別法として、あるいは追加的に、皮下、筋肉内、静脈内および皮内注射を含む注射によって行ってもよい。
製剤はその投与経路に適したものであれば何でもよく、当業者には自明であろう。本製剤は食塩水などの適切な担体を含有してもよく、またバルキング剤、他の医薬調製物、佐剤および他のあらゆる適切な医薬成分を含有してもよい。
適切な調製物にはストロメリシン−3またはその特徴的なペプチドを含有するワクチンも含まれ得る。このようなワクチンは能動型であっても受動型であってもよいが、ストロメリシン−3の発現が子宮内で起こり、抗ストロメリシン−3抗体への不明確な暴露が望ましくない効果を有するかも知れないので、一般的には受動型が好ましい。しかし能動予防接種が有利なこともあり、具体的には患者が既に子宮切開術を受けている場合にはどの組織もストロメリシン−3を正常には発現しないであろうから、能動予防接種が有利であり得る。他の適切なワクチンにはストロメリシン−3またはその特徴的なペプチドをコードしているヌクレオチド配列を含有する組換えウイルスが含まれる。このようなウイルスの適切なものの1つはワクシニアウイルスである。
以下の実施例は本発明を明示するよう機能するものであり、いかなる形でも本発明を制限することを意図するものではない。
実施例1乳癌特異的cDNAのクローニング 一次乳癌の外科的切除試料(腫瘍C1と呼ぶ)から得たポリ(A+)RNAを用いて乳癌cDNAライブラリーをλgt10ベクター中に構築した。それぞれC1−ポリ(A+)RNAおよび乳線維腺腫(F1と呼ぶ)から得たポリ(A+)RNAから逆転写したcDNAに対応する(+)および(−)プローブを用いて50000プラークを個別にスクリーニングした。
図1にC1−乳癌腫およびF1−線維腺腫から得た全RNAの、線維腺腫中より癌腫中でより高い発現レベルを示す4腫の遺伝子のcDNAプローブ(A〜D)を用いたノーザンブロット分析を示す。各レーンは8μgの全RNAを含有する。偏在的に発現する遺伝子に対応する36B4プローブを用いてこれらのフィルターを再プローブした(Rioら,Proc.Nat.Acad.Sci.USA 84:9243−9247(1987))。
具体的には、全RNAを液化窒素中に保存した外科標本から調製し(Chirgwinら,Biochemistry 18:5294−5299(1979))、オリゴ(dT)−セルロース・クロマトグラフィーによってポリ(A+)RNAを選択した。全細胞集団の約50%が支質細胞であるエストロゲン受容体−陰性(グレードII)導管癌腫(C1と呼ぶ)から調製したcDNAを用いて、乳癌−濃縮cDNAライブラリーを構築した。
クローニングに先立って、一本鎖cDNAを過剰量の乳線維腺腫(F1と呼ぶ)から得たポリ(A+)RNAを用いて減じ、その一本鎖濃縮物質をヒドロキシアパタイトクロマトグラフィーで精製した(Davisら,Proc.Nat.Acad.Sci.USA 81:2194−2198(1984);Rhynerら,Neuroscience Res.16:167−181(1986))。
この乳癌−濃縮cDNAを二本鎖にし、λgt10ベクターのEcoR I部位中にクローン化した。3百万の組換えファージが得られ、約5000cDNAクローンを含有するプレートから得た複製ナイロンフィルター(Biodyne A,Pall Corporation)を用いて約50000を個別にスクリーニングした。
C1−乳癌cDNAおよびF1−乳線維腺腫cDNAを用いて、それぞれ(+)および(−)プローブを作成した。両プローブとも過剰量の全ヒト肝臓RNAで減じ(Davisら,Proc.Nat.Acad.Sci.USA 81:2194−2198(1984);Rhynerら,Neuroscience Res.16:167−181(1986))、その後ランダムプライミング合成を用いて[32P]−標識した。
ハイブリッド形成を厳密条件下(50%ホルムアミド、42℃)で2日間行い、2xSSC、0.1%SDS中22℃で洗浄した後、0.1xSSC、0.1%SDS中55℃で洗浄した。第2スクリーニングのために、弁別的に標識されたプラーク130個を選択した。
無作為に取り上げた5個の異なるプラークのcDNA挿入物をPCR増幅法で精製し、[32P]−標識し、関連クローンを同定するためにすべての異なるプラークにハイブリダイズさせた。この操作を無作為に取り上げた異なるプラークで数回繰り返し、最終的にF1−線維腺腫中よりもC1−癌腫中でより高レベルな発現を示したA〜Dと呼ぶ4種の遺伝子を得た。ホルムアルデヒドを含有する1%アガロース中での電気泳動によって分離した全RNA(8μg)を用いて、C1−乳癌およびF1−乳線維腺腫に関するノーザンブロットを調製し、Hybond−Nフィルター(Amersham)に移した。
このブロットをメチレンブルーで染色した後、転移させたRNAの完全性と量を調べるためにプレハイブリッド形成させた。ハイブリッド形成(18時間)および洗浄を、A〜D遺伝子に対応する[32P]−標識cDNA挿入物を用いて、上述のように標準的条件下で行った。
正常結腸でも発現された(示していない)遺伝子AおよびBはこれ以上調べなかった。
C遺伝子は結腸で発現した(示していない)が、その高レベルの弁別的発現(図1)ゆえにこの遺伝子を部分的に特徴づけた。これは種々の形質転換上皮細胞株および正常ヒト皮膚中でも発現された(示していない)。1つのCクローンのcDNAの配列決定は、対応する遺伝子がケラチン遺伝子スーパーファミリーに属することを示した(データは示していない)。
最後に、D遺伝子(ここではストロメリシン−3遺伝子とも呼ぶ)の発現はC1−癌腫およびF1−線維腺腫間で著しく異なり(図1)、またこの遺伝子は正常ヒト結腸および他のいくつかのヒト組織中では発現されなかった(後述)ので、D遺伝子をさらに調べた。
実施例2ストロメリシン−3遺伝子およびコードされているタンパク質の配列決定 D cDNA挿入物をプローブとして用いて数個の別個のクローンを非削減C1−乳癌λgt10cDNAライブラリーから単離し、配列を決定した。図2に全長D cDNAのヌクレオチド配列および対応するタンパク質配列を示す。
488アミノ酸長のタンパク質をコードしているcDNA読み取り枠の後ろには、そのRNAの3'−末端から14塩基上流に位置するポリ(A)付加シグナルを含む714塩基の3'−非翻訳領域が続いている。推定される開始メチオニンはヌクレオチド位置10〜12に位置する。対応するAUGはKozak共通モチーフに一致する配列を伴っておらず、その中にも位置していないが、この配列は予期される性質である(後述)疎水性先導ペプチドの配列のすぐ下流に対応しているので、翻訳はおそらくこのAUGから開始されるのであろう。
ストロメリシン−3 cDNAのヌクレオチド配列および推定アミノ酸配列を示す図2において、ヌクレオチド残基は5'から3'に向かって番号が付与されており、読み取り枠中の推定アミノ酸は、その1文字コードで指定されている。下線を付したヌクレオチド配列は5'−末端から順に:推定シグナルペプチド(2つの潜在的切断部位を矢印で示している);プロ金属プロテイナーゼ類に特有のPRCGVPD配列;亜鉛結合ドメインの保存されたヒスチジン残基(Matrisian,L.M.,Trends Genet.6:21−125(1990));およびポリ(A)付加シグナル配列に対応している。
具体的には、D cDNAの3'−部分に対応するcDNA挿入物[19bpのポリ(AT)領域を含む250bp]をランダムプライミング合成によって[32P]−標識し、これを用いて、C1−乳腫瘍ポリ(A+)RNAから作成した非削減λgt10 cDNAライブラリーをGublerおよびHoffmannの方法(Gene 25:262−269(1983))でスクリーニングした。数個の別個のクローンを同定し、M13シークエンシングベクター中にサブクローニングした。US Biochemical製のデアザ−dGTP試薬キットおよびシークエナーゼを用いるジデオキシ法でDNA配列を決定した。この配列をPC/GENEソフトウェアパッケージを用いて分析した。
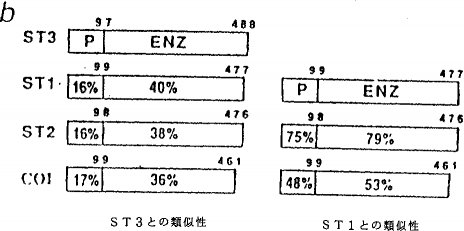
実施例3推定上の金属プロテイナーゼ:ストロメリシン−3 図3にヒト・ストロメリシン類とヒトI型コラーゲナーゼの予想アミノ酸配列の比較を示す。
(a)多数整列(multialignment)プログラム(Higginsら,Gene 73:237−244(1988))を用いてアミノ酸配列を並べた。4つの配列のすべてで一致するアミノ酸残基に星印を付けてある。矢印はストロメリシン−3の推定上のシグナルペプチド切断部位を表す。矢じりはI型プロコラーゲナーゼおよびプロストロメリシン類の活性化に際して起こる切断を示す。この切断部位のレベルでストロメリシン−3に特異的な10アミノ酸残基を囲んである。PRCGVPD配列および推定上の亜鉛結合ドメインの保存残基に下線を付してある。
(b)左:ストロメリシン−3、ストロメリシン−1(ST1,Whithamら,Biochem.J.240:913−916(1986))、ストロメリシン−2(ST2,Mullerら,Biochem.J.253:187−192(1988))およびI型コラーゲナーゼ(CO I,Whithamら,Biochem.J.240:913−916(1986))間の類似性(アミノ酸一致率(%))領域;
(b)右:ST1、ST2およびCO I間の類似性領域;Pはシグナルペプチドおよびプロドメインを示す;ENZは成熟活性酵素に対応するドメインを示す。
このように推定タンパク質配列とSwissprotデータライブラリー(14版)との比較は、この新しいタンパク質が分泌されるマトリックス金属プロテイナーゼ(MMP)類に属することを示した(図3a)。したがってこの新しいタンパク質は疎水性N−末端先導配列候補(図2の下線部)を有しており、MMP類のプロドメインの特徴である高度に保存された配列PRCGVPD(アミノ酸残基78〜84)を示し、またMMP類の亜鉛結合部位(アミノ酸残基212〜225:図3a)を有する(Matrisian,L.M.,Trends Genet.6:121−125(1990))。
このファミリーの他の構成要素との類似性から、成熟タンパク質のN−末端アミノ酸はそのプレ−プロタンパク質のフェニルアラニン98に対応すると思われる(Whithamら,Biochem.J.240:913−916(1986))(図3a)。最適に並べた後、推定成熟タンパク質との類似性はストロメリシン−1(Whithamら,Biochem.J.240:913−916(1986))と40%、ストロメリシン−2(Mullerら,Biochem.J.253:187−197(1988))と38%、I型コラーゲナーゼ(Goldbergら,J.Biol.Chem.261:6600−6605(1986))と36%である(図3b)。
この新タンパク質の基質特異性はわかっていない。この新タンパク質のストロメリシン−1との類似性(40%)は明らかにストロメリシン−1とストロメリシン−2との間に存在する類似性(79%)よりはるかに低く、またI型コラーゲナーゼとストロメリシン−1の間に存在する類似性(53%)よりさらに低いが、本明細書ではこれをストロメリシン−3と呼ぶことにする。したがってこのタンパク質はMMPの一種であって、“ストロメリシン”という名称は必ずしも厳密に正確ではないが、この名称が便利である。
さらに、PRCGVPD配列の上流域では、ストロメリシン−3と既に比較した他のMMP類との間に有意な類似性はない(図3)。しかしストロメリシン−3は、I型コラーゲナーゼおよびストロメリシン類のプロタンパク質切断部位(Whithamら,上述)と本質的に正確に対応する位置に独特の短い配列(アミノ酸残基88〜97)を有する。さらにストロメリシン−3はI型コラーゲナーゼおよび他のストロメリシン類の場合と同様に、IV型コラーゲナーゼ類に特有のフィブロネクチン様ドメイン(Wilhelmら,J.Biol.Chem.264:17213−17221(1989))を示さない。
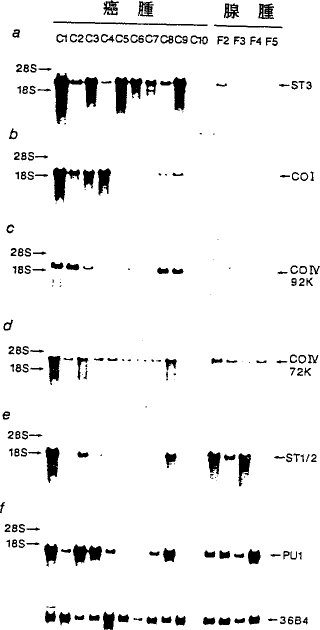
実施例4乳癌腫中での過剰発現 ストロメリシン−3 RNA転写物の出現を30種の乳癌腫および5腫の乳線維腺腫の切除試料で調べた。
図4に乳腫瘍中のヒト金属プロテイナーゼRNA類のノーザンブロット分析を示す:(a)ストロメリシン−3 RNA;(b)I型コラーゲナーゼRNA(CO I);
(c)92kD IV型コラーゲナーゼRNA(CO IV 92K);
(d)72kD IV型コラーゲナーゼRNA(CO IV 72K);
(e)ストロメリシン−1および−2 RNA(ST1/2);
(f)ポンプ−1 RNA(PU I)。
全RNAを4種のエストロゲン受容体−陰性乳癌腫(C1,グレードII;C2,C3およびC4,グレードIII)、6種のエストロゲン受容体−陽性乳癌腫(C5,C8およびC9,グレードII;C6およびC7,グレードIII;CO I,グレードI)および4種の乳線維腺腫(F2〜F5)から調製した。各レーンは8μgのRNAを含有する。36B4シグナルは対照遺伝子のRNAに対応する(図1)。
具体的には、図1の場合と同様に、数個のノーザンブロットを同一のRNA試料で同時に調製し、次のcDNAプローブのいずれかとハイブリダイズさせた:(a)ストロメリシン−3 cDNAの3'−部分を覆う1.6kb挿入物、(b)CO I cDNA、(e)ST2 cDNA(ST1 RNAと交差ハイブリッド形成するもの)、(f)PU1 cDNA(CO I、ST2およびPU1プローブはR.Breathnach氏の好意により供与された,Mullerら,Biochem.J.253:187−192(1988))、または(c)CO IV 92K(ヌクレオチド2144〜2223,Wilhelmら,J.Biol.Chem.2641:7213−17221(1989))および(d)CO IV 72K(ヌクレオチド1937〜2016,Collierら,J.Biol.Chem.263:6579−6587(1988))に対応する80マーのアンチセンスオリゴヌクレオチドプローブ(複数)。
ランダムプライミング合成を用いてこれらのcDNAプローブを(32P)−標識し(約5x105cpm/μg)し、5'−末端キナーゼ処理を用いてこれらのオリゴヌクレオチドを標識した(約108cpm/μg)。ハイブリッド形成を厳密条件下(42℃、50%ホルムアミド)で約106cpm/mlを用いて実行した。次にフィルターを2xSSC、0.1%SDS中22℃で洗浄した後、0.1xSSC、0.1%SDS中55℃で洗浄した。オートラジオグラフィーを(a)18時間、(b)20時間、(c)、(d)および(e)4日間、(f)2日間、−80℃で増感板を用いて行った。
ストロメリシン−3 mRNAはすべての乳癌腫中で、その癌腫がエストラジオール受容体(ER)陽性(C5〜C10)であるか陰性(C1〜C4)であるかにかかわらず認められた(図4a)が、発現レベルが乳癌腫中で観測された最低レベルと同等であった1つの例外(F2)を除いて、線維腺腫試料中では認められなかった。
MMP遺伝子ファミリーの他の構成要素のRNA転写物の出現をも同じ試料中で調べた(図4b〜f)。MMPファミリーのこれらの他の構成要素はそのヒト乳腫瘍中での発現様式に従って明確に2つの種類に分類することができる。第1の種類には72kD IV型コラーゲナーゼ(CO IV 72K、図4d)、ストロメリシン−1および−2(ST1/2、図4e)およびポンプ−1(PU1、図4f)が含まれ、これらの遺伝子はすべて悪性および良性腫瘍の両方で発現された。対照的に、ストロメリシン−3(図4a)、I型コラーゲナーゼ(CO I、図4b)および92kD IV型コラーゲナーゼ(CO IV 92K、図4c)遺伝子を包含する第2の種類は、乳癌腫中でのみ過剰発現を示す。ただし乳癌腫に一貫して伴うのはストロメリシン−3だけである。
発現様式は第2の種類に属する3つの遺伝子間で同一ではなかった。I型コラーゲナーゼRNA転写物はC5、C6、C7およびCIO癌腫中では検出されず、92kD IV型コラーゲナーゼRNA転写物はC7およびCIO試料中には認められなかったが、ストロメリシン−3 RNA転写物はすべての腫瘍中で明瞭に検出された。
したがってストロメリシン−3は浸潤性乳癌腫の症状を示すと思われ、一方I型コラーゲナーゼおよび92kD IV型コラーゲナーゼもまた場合によって乳癌の進行に特異的に関与するのであろう。
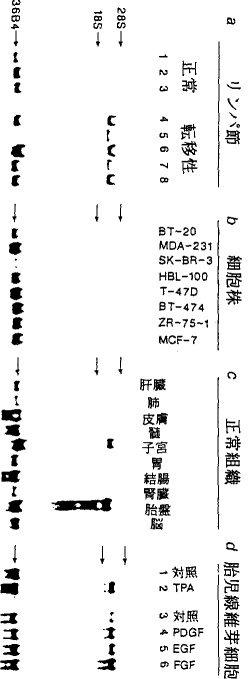
実施例5種々の供給源から得た細胞中での発現 図5に種々の細胞株および組織中のストロメリシン−3 RNAのノーザンブロット分析を示す。
(a)乳癌の患者から得た5種の転移性腋窩リンパ節および3種の正常な腋窩リンパ節;
(b)4種のエストロゲン受容体−陰性(BT−20、MDA−231、SK−BR−3、HBL−100)および4種のエストロゲン−陽性(T−47D、BT−474、ZR−75−1、MCF−7)乳癌株化細胞;
(c)10種の正常ヒト組織;
(d)血清非含培地(1および2)中TPA(10ng/ml)の非存在下(1)または存在下(2)で培養した、あるいは20μg/mlインシュリンを補足した血清非含培地(3〜6)中PDGF(20ng/ml、British Biotechnology)の非存在下(3)または存在下(4)、またはEGF(20ng/ml、Collaborative Research)の存在下(5)、またはbFGF(10ng/ml、Pettmann氏の好意により供与された(FEBS Lett.189:102−108(1985)))の存在下(6)で培養した、HFL−1ヒト胎児二倍体線維芽細胞(ATCC CCL 153)。
(a)では、レーン5(2μg)およびレーン6(20μg)を除いて各レーンは10μgの全RNAを含有する。(b)および(c)では、各レーンとも8μgの全RNAを含有し、(d)では各レーンは5μgの細胞質RNAを含有する。
具体的には、(a)、(b)および(c)については図4でストロメリシン−3について示したようにブロットを調製し、加工した。(d)では、全面成長HFL−1線維芽細胞を血清非含DMEM培養培地中に維持した。24時間後、新鮮な培地を加え、TPAまたは成長因子を上記のように補足するか、もしくは補足しなかった。24時間の培養後、細胞を回収し、細胞質RNAを調製した(Gough,N.M.,Analyt.Biochem.173:93−95(1988))。
次に図4でストロメリシン−3について示したようにブロットを調製し、加工したが、オートラジオグラフィーを3日間とした。
92kDコラーゲナーゼIV RNA転写物は3種の正常および5種の乳癌転移性リンパ節中に認められたが、ストロメリシン−3 RNA転写物は転移性節中でのみ検出された(図5aおよび開示していないデータ)。
一次悪性乳腫瘍および転移性リンパ節で得た結果とは対照的に、8種のヒト乳癌細胞株中ではそれらのER状態にかかわらず、類似の条件下でストロメリシン−3 RNA転写物を検出できなかった(図5b)。同様にストロメリシン−3 RNA転写物は、子宮および胎盤という顕著な2つの例外を除いて、いくつかの正常ヒト成人組織中でも検出できなかった。
ストロメリシン−3はすべての癌に明らかに伴うわけではなく、結腸、卵巣、腎臓および肺癌から得たRNA試料中では低レベルのストロメリシン−3 RNA転写物が認められるに過ぎない。しかし、乳癌中に認められるレベルと同等に高いレベルの発現が喉頭癌RNA試料中に観測された(非開示データ)。
実施例6浸潤性腫瘍の支質細胞中での特異的発現 原始乳癌腫中でストロメリシン−3遺伝子が発現され、いくつかの確立された乳癌株化細胞中では発現されないことは、この遺伝子が新生物細胞そのものではなく、むしろその腫瘍を取り巻く支質細胞中で発現されたことを示唆している。
[35S]−標識ストロメリシン−3アンチセンスリボプローブを用いる原位置ハイブリッド形成実験を6種の癌腫(図4に関して命名した腫瘍C1、C3、C5、C9、C10および図4には示さなかったER−陽性癌腫である腫瘍C11)から得た切片を用いて行った。
具体的には、原位置ハイブリッド形成をCoxらが記述したように行った(Dev.Biol.101:485−502(1984))。脱パラフィン化酸処理切片(6μm厚)をプロテイナーゼKで処理し、Bluescript II(Stratagene)中にサブクローニングしたストロメリシン−3 cDNA挿入物(ヌクレオチド1128から1594までの467bp)から得た[35S]−標識アンチセンス転写物を用いて終夜ハイブリッド形成させた。ハイブリッド形成の後、RNase処理(20μg/ml、30分間、37℃)を行い、厳密に洗浄(2xSSC、50%ホルムアミド、60℃、2時間)した後、NTB2乳剤(Kodak)を用いてオートラジオグラフィーにかけた。オートラジオグラフィーを15日間行った。センスリボプローブを用いる類似の条件下でバックグラウンド以上の有意な標識化は観測されなかった(示していない)。
図6に乳癌腫および胚肢芽の切片中のストロメリシン−3 RNA転写物の存在を示す。a、c、e、g、iおよびk:ヘマトキシリンで染色した組織切片の明視野(x100);b、d、f、h、jおよびl:アンチセンス・ストロメリシン−3 cRNAプローブとの原位置ハイブリッド形成および暗視野画像化後の同切片(やはりヘマトキシリンで染色したもの)。
aおよびb、グレードII導管乳癌腫(腫瘍C1、図4参照のこと):浸潤性癌細胞(C)が紡錘状細胞(S)の豊富な支質に取り巻かれている;ストロメリシン−3 RNA転写物は新生物上皮細胞のすぐ周囲の支質細胞中にもっとも豊富である。cおよびd、グレードIII導管乳癌腫(腫瘍C3、図4参照のこと):浸潤性乳癌細胞(C)の複数の島が支質細胞によって取り巻かれている;ストロメリシン−3遺伝子の発現はほとんどの支質小柱(S)の中心部(即ち、新生物細胞から最も遠い領域)で、より弱い。eおよびf:導管癌腫(腫瘍C3、図4参照のこと)および2つの正常小葉(N);ストロメリシン−3 RNA転写物は、リンパ球(矢印)が豊富な小領域を例外として、浸潤性癌細胞(C)を並置させている支質中で独占的に検出された。gおよびh、導管癌腫(腫瘍C10、図4参照のこと):ストロメリシン−3 RNA転写物は浸潤性乳癌細胞を取り巻く支質細胞(右上角)中でバックグラウンド以上に検出できたが、原位置(星印)乳癌細胞では検出できなかった。iおよびj、導管癌腫(腫瘍C11、ER−陽性、グレードII、癌腫);左:原位置の癌腫(星印)、支質細胞中でストロメリシン−3 RNA転写物を検出できなかった;右:ストロメリシン−3遺伝子を発現している支質細胞によって取り巻かれている浸潤性新生物。kおよびl、8週齢ヒト胚肢芽の指間領域:ストロメリシン−3 RNA転写物が原始表皮の基礎をなす中胚葉中で検出され、指間領域(M)中で最も顕著である;原始表皮(矢印)、形成中の軟骨(PC)、および周辺中胚葉は標識されないことに注目のこと。
すべての場合においてストロメリシン−3 RNA転写物は、腫瘍の浸潤性成分を形成している悪性上皮細胞の島を取り巻く支質細胞中でのみ検出された(図6:パネルaおよびb(腫瘍C1について);パネルc、d、eおよびf(腫瘍C3について);パネルgおよびh(腫瘍C10について);パネルiおよびj、右側(腫瘍C11について);腫瘍C5およびC9についてのデータは示していない)。
転移性リンパ節(C5と同じ供給源)でも、ストロメリシン−3遺伝子の発現がその転移性上皮細胞を取り巻く支質細胞に限定されていた(データは示していない)。
すべての場合において、悪性上皮細胞そのものは標識されなかったこと、ならびに最も高レベルの発現が悪性細胞に並列している支質細胞中で観測されたことが特に注目に値する。著しく対照的に、まだ基底膜によって取り巻かれている原位置癌腫病巣を取り巻く支質細胞中では有意な発現を検出できず(パネルgおよびh(腫瘍C10について;パネルiおよびj、左側(腫瘍C11について))、一方同じ腫瘍の浸潤性成分中では支質細胞の標識化を明確に観測することができた(パネルgおよびh(腫瘍C1Oについて);パネルiおよびj、右側(腫瘍C11について))。
また、癌細胞から離れて位置する支質細胞および正常導管および小管を取り巻く支質細胞中では有意な発現を検出できなかった(例えばパネルeおよびf)。ストロメリシン−3転写物の分離した焦点は、ノーザンブロットでストロメリシン−3 RNAに関して弱く陽性であるF2線維腺腫の切片中で検出されなかった(図4aおよび非開示データ)。
線維芽細胞と筋線維芽細胞が共に浸潤性乳癌腫の支質中に存在することが知られている(Ahmed,A.Pathol.Annu.25(Pt2):237−286(1990))。本発明者らの原位置ハイブリッド形成技術では、これらの細胞型のうち1つだけがストロメリシン−3遺伝子を発現したのか、それとも両方の細胞型がストロメリシン−3遺伝子を発現したのかを決定することは不可能であった。
実施例7成長因子による刺激 上記の結果は、支質細胞中でのストロメリシン−3遺伝子の発現が新生物細胞が分泌する拡散性因子によって誘導されているらしいことを示している。EGF、FGFおよびPDGFなどの成長因子およびいくつかのサイトカイン類(IL−1α、β、およびTNF−α)、および腫瘍プロモーター類(例:TPA)がMMP遺伝子類の転写を活性化することが知られている(Kerrら,Science 242:1424−1427(1988))。数種の供給源から得られた腫瘍細胞がヒト線維芽細胞によるコラーゲナーゼIの生産を刺激する因子(単数もしくは複数)を生産することも報告されている(Lippmanら,Recent Prog.Hormone Res.45:383−440(1990))。乳癌細胞がインビトロでPDGF、FGFおよびTGF−α活性を分泌することが知られている(Salomonら,“Breast cancer:cellular and molecular biology"(Lippman,M.E.およびDickson,R.B.編),363〜389頁(Kluwer,Boston,(1988)))。
ストロメリシン−3遺伝子の発現を外因性の刺激物質で変調させ得るかどうかを調べるために、ヒト胎児二倍体線維芽細胞をPDGF、bFGFおよびEGFならびに腫瘍プロモーターTPAの存在下または非存在下で成長させた。これらの成長因子のいずれか、もしくはTPAの添加が線維芽細胞におけるストロメリシン−3 RNA転写物の増大をもたらし、最も強い刺激がbFGFで観測された(図5d)。
実施例8胚におけるストロメリシン−3の発現 ストロメリシン−3遺伝子が成長因子の刺激に応答して胎児線維芽細胞中で発現されたので、本発明者らはこの遺伝子が胚発育中に通常に発現されるかどうかを調べた。
ストロメリシン−3転写物が8週齢ヒト胚の数個の別個の領域で検出され、胚形成のこの段階で予定された細胞の死を伴うことが知られている領域(Milaire,J.,“Organogenesis"(De Haan,R.L.およびUrsprung,H.編),283〜300頁(Holt,Rinehart and Winston,New York,1965))である胚芽の指間領域で顕著であった(図6、パネルkおよびl、ならびに非開示データ)
標識化は、標識されずに残った原始表皮の基礎をなす胚の中胚葉で観測された。エピブラストから離れて位置する間葉細胞もmRNA−陰性であったことは注目に値する。
このように正常な胚生育中に組織改造が十分証明されている領域でストロメリシン−3遺伝子が発現するという発見は、乳腫瘍におけるストロメリシン−3の発現が癌の進行に関連するECM改造過程においてある役割を果すことを示唆している。
実施例9ST3をコードするマウスcDNAのクローニング ST3をコードするヒトcDNAを含有するプローブを用いて、マウス胎盤cDNAライブラリーを標準的方法(Sambrookら,“Molecular Cloning:A Laboratory Manual"(Cold Spring Harbor Press(1989)))でスクリーニングした。このスクリーニングの結果、ST3をコードする全長マウスcDNAが得られた(図7)。
これら2つの配列の分析によって成熟型ST3のアミノ酸配列には89%、プレおよびプロドメインには55%の相同性が存在することが明らかになった(図7)。
実施例4〜8に記述した方法を用いて、種々のマウス細胞中での発現様式を決定した。
マウスにおけるST3の発現様式は組織特異性に関してヒト組織に認められた様式と同一であることがわかった。最も高い発現レベルは胎盤および子宮組織に認められた。
本発明は腫瘍関連酵素標識(マーカー)に関する。
本発明は、ストロメリシン−3(Stromelysin−3)をコードするDNAおよびストロメリシン−3に結合し得る抗体を使用することにより、癌、具体的には悪性乳癌の診断法を提供するものである。
世界中の癌による死亡数は毎年主たる懸念材料であり続けており、特定の型の癌について数種の治療法が利用できるだけであって、またこれらが成功するという絶対的な保証もない。ほとんどの治療は迅速に成長する細胞を死滅させる一般的な“ショットガン”法に頼っており、この方法は迅速に成長する癌細胞がこの治療によって死滅するか、もしくは少なくとも癌細胞数が減少することによってその身体の系による残りの癌細胞の排除が可能になるであろうことを期待するものである。
治療法の探索は、異なる形態の癌が異なる治療を必要とするという発見によって妨げられてきた。身体のほとんど全ての部分が癌に冒され得るとすれば、その課題は膨大なものになる。
しかしながら、それらの相違にもかかわらず癌はいくつかの類似点をも有している。その中で最も重要なことは非分化組織の成長である。しかしこれは、ある種の癌細胞がある程度の分化を示すという点で100%正確な訳ではない。これは乳癌や精巣癌などの性癌で示されており、この場合腫瘍はホルモン受容体に関して陽性の場合もあり、陰性の場合もある。これらの腫瘍の治療はホルモン状態に依存し、タモキシフェンTMなどの関連ホルモン拮抗薬を投与する程度に単純な場合もある。
ほとんどの癌が共有しているもう1つの要素は、致死的であるためには癌が転移しなければならないということである。転移が起こる時までは、腫瘍はそれが悪性であっても身体の一領域に限定される。これは不快および/または痛みをもたらしたり、あるいはさらに重度な兆候を導くことさえあるが、その位置が特定できるのであれば、それを外科的に除去することができ、また十分な注意を払えば、さらなる問題を引き起こさない。
しかしいったん転移が始まれば、外科的切除によって原腫瘍を切除することはできるであろうが、癌細胞はその身体に侵入し終わっており、成功の見込みがあるのは化学療法か、もしくは特定の形態の標的療法のいくつかだけである。
したがって、局部的に侵入する可能性と一次腫瘍から離れた器官に転移する(腫瘍進行)可能性はほとんどの癌の過程において致命的な事象である。一次腫瘍を取り巻く細胞外マトリックス(ECM)の変性/分解および腫瘍細胞接着特性の変化は、転移性細胞の一次腫瘍細胞からの解離にとって極めて重要であることが知られている(Liotta,Cancer Res.46:1−7(1986);Hartら、Biochim.Biophys.Acta 989:65−84(1989))。
腫瘍脈管形成は一次腫瘍の拡大と転移性腫瘍の広がりの両方にとって必須であり、脈管形成自体はECM分解を必要とする(Bloodら,Biochem.Biophys.Acta 1032:89−118(1990))。したがって悪性は、新生物細胞とその環境がその病理学的過程において決定的な役割を果す全身性疾患である(Fidler,I.J.,Cancer Metastasis Rev.5:29−49(1986))。
悪性腫瘍に伴う遺伝子発現の変化の同定は、腫瘍の進行に関与するものも含めて、明らかに、癌を完全に理解するためだけでなく、癌に対する新しい合理的療法を開発するためにも欠くことのできない前提条件である。細胞遺伝子の2つの群(癌原遺伝子群および腫瘍抑圧遺伝子群)の発現の変異および/または異常制御が、多段階の過程で正常な成長制御の欠失を導き、転換した細胞表現型の獲得を導くことが示されている(Weinberg,R.A.,Cancer Res.49:3713−3721(1989))。しかし腫瘍の進行を導く分子機構についてははるかに不明瞭である(Nowell,P.C.,Cancer Res.46:2203−2207(1986);Fidler,I.J.,Cytometry 10:673−680(1989))。
したがって、癌細胞に特有の遺伝子が異常に発現した宿主遺伝子であることが極めて多いという点で新たな問題が生じる。極めて多くの場合、ある与えられた癌に関して特定のタンパク質標識がその癌中で過剰発現するが、この標識はその身体中の別の場所でも低レベルであるとはいえ発現する。
癌に関連するタンパク質のいくつかは、細胞群を互いに適切な関係に保つために重要な細胞外マトリックスを破壊する酵素である。このような酵素の1種は金属プロテイナーゼ(MMP)類(Matrisian,L.M.,Trends Genet.6:121−125(1990))であり、これらは亜鉛を結合するのでこのように呼ばれている。しかし、いくつかの存在が暗示され得るとはいうものの、癌あるいは特定の腫瘍の症状を示すことがわかっているものはなかった。
MMP類は、ECM改造および細胞移転が関係するいくつかの生理学的および病理学的過程(例えば形態発生および胚発育、リウマトイド関節炎、および腫瘍の浸潤および転移など)に関与する。MMP阻害剤は、腫瘍の進行にとって極めて重要な腫瘍浸潤および脈管形成を実験モデル中で遮断し得ることが知られている。
マトリックス金属プロテイナーゼファミリー(族)の構成要素はすべてECMの少なくとも一成分を分解するプロテイナーゼであり、潜在型で分泌され、活性になるためには(例えばプラスミンによる)タンパク加水分解などの活性化を必要とする。間質コラーゲナーゼ類は結合組織コラーゲン類(IからIIIまで)を特異的に攻撃し、一方、IV型コラーゲナーゼ類(72kDおよび92kD)はフィブロネクチンおよび基底膜中に存在するコラーゲンを分解する。ストロメリシン類(トランシン(transin)類)−1および−2、そしてポンプ−1(pump−1)も、はるかに広い基質特異性を有し、プロテオグリカン類、ラミニン(laminin)、フィブロネクチン、およびコラーゲン類(IIIからVまで)を分解する。
男性の場合悪性腫瘍のほとんどは癌腫であり、非喫煙者のなかでは乳癌が女性の癌による死亡率の主たる原因である(Willett,W.,Nature 338:389−394(1989))。数種の腫瘍遺伝子の発現が悪性乳細胞および腫瘍中で変化することが報告されているが、一貫して乳癌に伴い得る特定の腫瘍遺伝子/抑圧遺伝子発現様式はない(Gullick,W.J.,Prog.Growth.Factor Res.2:1−13(1990))。
しかし乳腫瘍の新生物細胞はしばしば、その増殖の制御と転移能にとって重要でもあり得る脂肪および間葉支質中に埋め込まれている。実際、支質細胞が正常な乳房上皮の成長を正負両方に変調させ得ることが知られており(Salomonら,“Breast Cancer:Cellular and Molecular Biology"(Lippman,M.E.およびDickson,R.B.編),363〜389頁(Kluwer,Boston,(1988)))、また上皮と支質成分との相互作用が乳腺における上皮発癌に影響を与え得ることが知られている(DeOmeら,Cancer Res.38:2103−2111(1978))。
悪性乳腫瘍中に“活性化された”(Tremblay,G.Exp.Mol.Pathol.31:248−260(1979))および/または異常な(Greyら,Proc.Nat.Acad.Sci.USA 86:2438−2442(1989))線維芽細胞が存在することが推測されており、乳癌は支質要求に対する依存性からの部分的逸脱または支質成分に対する異常に強い応答を表すのであろうと提唱されてきた。
癌組織の性質ゆえに、ある与えられた癌の連続的培養または株化細胞を作成(この操作により、ある与えられた治療法の効果の研究が容易になる)することは比較的簡単である。このような系の少なからぬ欠点はその性質そのものにある:試験的処置はそれが細胞に対して直接作用し得るかどうかを明確にするであろうが、その処置が生体内でどのような効果を有するかは決して明確ではなく、またこのような株の生化学的分析は必然的に、通常生体内の腫瘍を取り巻いている組織の非存在下で行うことになる。
本発明の目的は、乳癌腫中で発現が増大し、それによって乳癌腫がその周辺支質と相互作用する悪性上皮細胞とみなされる遺伝子を同定することにある。
本発明者らは過去に特徴づけられたことのないあるタンパク質が、ある種の浸潤性癌、具体的には乳癌腫、頭部および首部偏平上皮細胞癌腫および皮膚(偏平および基底細胞型)癌腫の症状を示すことを新たに発見した。このタンパク質は見かけ上金属プロテイナーゼ群に属し、ここではこれをストロメリシン−3と呼ぶ。
図面の説明図1:C1乳癌腫および線維腺腫から得た全RNAのノーザンブロット分析 実施例1に記述するように、C1乳癌腫および線維腺腫細胞から得たRNAボレート(bolate)を別個に単離した4種のcDNAプローブを用いてプローブした。
図2:ストロメリシン−3 cDNAのヌクレオチド配列 ストロメリシン−3のcDNAのヌクレオチド配列および推定アミノ酸配列を表す。下線を引いたヌクレオチド配列は5'末端から順に:推定シグナルペプチド;プロ金属プロテイナーゼに特有のPRCGBPD配列、亜鉛結合ドメインの保存されたヒスチジン残基およびポリ(A+)シグナル配列に対応する。
図3:金属プロテイナーゼ配列の比較 実施例3に記述するように、すべて推定上の金属プロテイナーゼであるストロメリシン−3、ストロメリシン−2、ストロメリシン−1およびコラーゲナーゼ−1を並列し、比較する。
図4:ヒト金属プロテイナーゼのノーザンブロット分析 4種のエストロゲン受容体陰性乳癌腫(C1,グレードII;C2,C3およびC4,グレードIII)、6種のエストロゲン受容体陽性乳癌腫(C5,C8およびC9,グレードII;C6およびC7,グレードIII;Cio,グレードI)および4種の乳線維腺腫(F2〜F5)から全RNAを調製した。
そのRNAを(a)ストロメリシン−3 RNA、(b)1型コラーゲナーゼRNA(CO I)、(c)92kd 4型コラーゲナーゼRNA(CO IV 92k)、(d)72kd 4型コラーゲナーゼRNA(CO IV 72k)、(e)ストロメリシン−1および2 RNA(ST1/2)およびポンプ−1 RNA(PU I)で実施例4に記述するようにプローブした。
図5:種々の株化細胞および組織から得たストロメリシン−3 RNAのノーザンブロット分析 (a)乳癌患者から得た5種の転移性腋窩リンパ節および3種の正常腋窩リンパ節;(b)4種のエストロゲン受容体陰性乳癌腫株(BT−20,MDA−231,SK−BR−3,HBL−100)および4種のエストロゲン受容体陽性乳癌腫株(T−47D,BT−474,ZR−75−1,MCF−7);(c)10種の正常ヒト組織;および(d)血清非含培地(1および2)中tPA非存在下(1)またはtPA存在下(2)で培養した、20mg/m
図6:乳癌腫および胎児肢芽の切片中のストロメリシン−3 RNA転写物の局在化 ヘマトキシリンで染色した組織切片の明視野顕微鏡写真(x100)(A,C,E,G,IおよびK);ならびに、実施例6に記述するように原位置でアンチセンス・ストロメリシン−3 cRNAとハイブリッド形成させた後の同切片(やはりヘマトキシリンで染色したもの)の暗視野画像(B,D,F,H,JおよびL)。
図7:マウスST3 cDNAのcDNA配列 マウスST3遺伝子のcDNA配列およびヒトST3 cDNA配列との比較。
本発明は第1の側面として、ストロメリシン−3またはストロメリシン−3をコードするヌクレオチド配列の検出からなる浸潤性癌、具体的には胸部、頭部および首部、ならびに皮膚の癌腫の診断法を提供する。
本発明はもう1つの側面として、浸潤性癌、具体的には胸部、頭部および首部、ならびに皮膚の癌腫の治療または予防における、ストロメリシン−3の合成または活性を妨害する薬剤の使用を提供する。
転移性腫瘍は浸潤性であるが、浸潤性腫瘍が必ずしも転移性ではない(例えば基底細胞皮膚癌腫)ことは理解されるであろう。
ストロメリシン−3遺伝子の発現はECM分解の領域に特異的であり、見かけ上金属プロテイナーゼをコードしているので、そのECM分解活性は腫瘍の転移への進行にとって極めて重要であると考えられる。支質細胞によるストロメリシン−3の発現はECMの重要な部分を破壊し、それによって癌細胞が原腫瘍から転移することを可能にすると思われる。
したがってストロメリシン−3の活性に影響を与え得る薬剤は転移に対する効果を有するであろう。そのような薬剤は、該タンパク質の合成を阻害するか、該タンパク質の成熟を阻害するか、あるいは該酵素の活性を遮断もしくは変化させることによってその活性を変化させるものが適しているであろう。
ストロメリシン−3遺伝子の発現が転移相にある乳癌の症状を示すことが初めてわかった。実際、この結果は切除された種々の腫瘍中のmRNAを検出することによって達成された。乳癌は非喫煙女性集団において癌による最も高い死亡率の原因であるので、乳癌を選択した。
ストロメリシン−3はほぼ確実にMMPファミリーに属する新規タンパク質であり、ホルモン状態にかかわらず浸潤性乳癌腫に関与している。
MMPファミリーの構成要素は活性になるために活性化過程を必要とし、この過程にはプレ−およびプロ−配列の除去が関与し得る。プロ−および成熟−ストロメリシン−3のアミノ酸配列は過去に特徴づけられたMMP類のものとは著しく異なっており、成熟化、活性化およびECM成分に対する特異性に関して異なる特性を示し得る。
ストロメリシン−3遺伝子はすべての一次浸潤性乳癌腫によって発現され、またその転移部のいくつかによっても発現され、そのような発現について分析された広範なECM改造が起こることが知られている組織(子宮、胎盤、および肢芽)中でも発現されるが、乳線維腺腫および正常成人組織中では発現されない。このことはストロメリシン−3遺伝子産物が乳癌の進行において重要な役割を果すことを示唆している。またこの概念と一致して、通常前浸潤性病巣と見なされしばしば微小浸潤に関連するコメド型の原位置癌腫を除いて、ストロメリシン−3遺伝子はほとんどの原位置乳癌腫中で発現されない。したがって、その身体中の子宮または胎盤以外の別の場所に認められるような低濃度ではないストロメリシン−3 RNA転写物の存在は、転移性癌または浸潤性になる危険の高い癌の症状を示す。
ストロメリシン−3は浸潤性癌成長に伴うと思われる溶解過程に関与し得る。あるいは、ほとんどの浸潤性乳癌病巣に伴い、悪性細胞のさらなる拡大を防止するための宿主反応を表す結合織線維増生の形成において、ストロメリシン−3がある役割を果すことも考えられる(Ahamed,A.,Pathol.Annu.25(Pt2):237−286(1990))。このような場合には、ストロメリシン−3活性の増大が有利であろう。
さらに、新生物細胞群(島)のすぐ周囲の支質線維芽細胞におけるストロメリシン−3遺伝子の限定された発現は、いくつかの腫瘍形成性細胞の悪性変換に関連することが知られているもう1つの金属プロテイナーゼであるコーラーゲナーゼIV、および乳癌腫中でその発現が増大するリソソームのアスパルチルプロテアーゼであるカセプシンD(両酵素とも線維芽細胞中では発現されないが、乳癌の新生物上皮細胞中で発現される)とは対照的である(Monteagudoら,Am.J.Phathol.136:585−592(1990);Garciaら,Steroid Biochem.27:439−445(1987))。
新規乳癌標識を同定するために、cDNAライブラリーを構築し、線維腺腫供給源から得たポリ(A+)RNAで抽出した。この操作によって、このcDNAライブラリーは転移性癌に特有の配列に関して濃縮された。
いくつかのクローンを成長させ、転移性腫瘍および線維腺腫からのポリ(A+)RNAから誘導したプローブを用いてスクリーニングした。次いで、転移性癌ポリ(A+)RNAから誘導したプローブに対してかなり強く結合したクローンをさらに成長させた。
この方法で作成したクローンのうちの1つは、悪性乳癌および咽頭癌、頭部、首部、および皮膚(偏平および基底細胞型)癌腫、ならびに子宮および胎盤(これらのすべてにおいてECMの破壊が起こっており、この破壊は癌と関連した場合、癌細胞がその身体中に広がること(転移)を可能にする)中にのみ認められる高い発現率の程度にまで弁別的に発現されることがわかった。
子宮および胎盤の場合にはECMの破壊が自然に起こるが、それ以外の場所で起こる同じ事象は腫瘍成長の特徴であると思われる。
また、ECMの破壊を伴う胎児の肢発芽中の指間分化にストロメリシン−3遺伝子の発現が認められたことに注目することも興味深い。
cDNA配列を特徴づけることによって、読み取り枠の存在が明らかになった。コード化されているタンパク質配列を既知のライブラリーと比較することによって、このタンパク質がECMを破壊することが知られているファミリーに属することが立証された。ストロメリシン−3の配列はそのファミリーの他の構成要素に対して、他の構成要素間に認められるほどの類似性を有さないが、それでもこの酵素の性質の同定に利用できるいくつかの特徴的な領域を呈示している。したがって、このタンパク質はコラーゲナーゼの一種であるかも知れないし、全く異なるECM構成成分を破壊するのかも知れないが、このタンパク質をストロメリシン−3と命名した。
ストロメリシン−3 mRNAの出現を立証するためのヌクレオチドプローブを構築することによって上述の組織分布が明らかになり、また、標識化によってストロメリシン−3遺伝子の発現領域を顕微鏡写真で正確に位置決定することが可能になった。
この方法で作成した顕微鏡写真の分析によって、やや意外なことに、ストロメリシン−3遺伝子が癌細胞そのものでは発現されず、その周辺支質で発現されることが示された。さらに支質は、腫瘍の基底膜がまだ無傷である場合にはストロメリシン−3 mRNAの証拠を全く示さなかった(図6を参照のこと)。ストロメリシン−3遺伝子はすべての一次浸潤性乳癌腫によって発現され、またその転移節のいくつかによっても発現され、そのような発現について分析された広範なECM改造が起こることが知られている組織(子宮、胎盤、および肢芽)中でも発現されるが、乳線維腺腫および正常成人組織中では発現されない。このことはストロメリシン−3遺伝子産物が乳癌の進行において重要な役割を果すことを示唆している。またこの概念と一致して、通常前浸潤性病巣と見なされしばしば微小浸潤に関連するコメド型の原位置癌腫を除いて、ストロメリシン−3遺伝子はほとんどの原位置乳癌腫中で発現されない。ストロメリシン−3は常に転移性癌の支質中に出現し、原位置一次腫瘍(まだ基底膜を有しており、浸潤性でない腫瘍)中には出現しない。したがって、その身体中の子宮または胎盤以外の別の場所に認められるような低濃度ではないストロメリシン−3 RNA転写物の存在は、転移性癌または浸潤性になる危険の高い癌の症状を示す。
さらに、ER−陽性または陰性乳癌株化細胞のいくつかはEGF/TGF−αおよびFGF(ストロメリシン−3遺伝子の発現に関与する因子)に対する受容体を分泌し保持していることが知られているにもかかわらず、ストロメリシン−3遺伝子の発現はいずれのER−陽性または陰性乳癌株化細胞中にも検出されなかった。
したがって、ストロメリシン−3、その前駆体あるいはそれをコードするヌクレオチド配列に標準的な検出技術を適用することにより、転移性癌を診断したり、あるいは一次腫瘍がまだ致命的な転移相に達していないことを確認することができる。
このような技術にはヌクレオチドプローブを例えば上述のように用いる検出法が含まれ、またこのような技術が例えば抗体あるいはその等価物によるストロメリシン−3タンパク質の検出から構成されていてもよい。
ヌクレオチドプローブは、添付の図2に示す配列に対して天然に存在する他の配列に対するより強くハイブリッド形成するものなら何でもよい。プローブの種類にはcDNA、リボプローブ、合成オリゴヌクレオチドおよびゲノムプローブが含まれる。使用するプローブの種類は一般に特定の状況によって決まる(例えば、原位置ハイブリッド形成のためにはリボプローブ、ノーザンブロッティングのためにはcDNA)。最も好ましいプローブは図2のcDNAの(−)鎖に対応するものである。またストロメリシン−3遺伝子中に位置するイントロンを認識するプローブを提供することもできるが、これがRNA転写物の検出と同程度の信頼性を有するとは限らない。
ストロメリシン−3をコードする遺伝子そのものの検出は一般的には診断目的には有効でないであろうが、転写物および他の発現産物を検出するための他の形態の検定法は一般に有用であろう。プローブはストロメリシン−3 mRNA転写物を弁別的に認識するために必要な程度に短くてもよく、例えば15塩基長程度でもよい。
プローブの標識化(ラベル化)の形態は適切なものであれば何でもよく、例えば32Pおよび35Sなどの放射性同位体を使用することができる。放射性同位体による標識化は、そのプローブが化学的に合成されたものであれ、生物学的に合成されたものあれ、適切に標識された塩基を用いることによって達成できる。標識化の他の形態には、ELISAの特徴であるように酵素あるいは抗体標識化が含まれ得るが、標識されたプローブによるmRNA転写物の検出には一般にX−ラジオグラフィーが用いられるであろう。
RNA転写物の検出はノーザンブロッティングによって達成できる。この方法では、例えばRNAの調製物を変性化アガロースゲルに流し、適切な支持体(例:活性化セルロース、ニトロセルロースまたは硝子またはナイロン膜)に移し、次いで、放射性標識cDNAまたはRNAをこの調製物にハイブリダイズさせ、洗浄し、オートラジオグラフィーで分析する。
原位置ハイブリッド形成可視化法を用いることもできる(実施例6)。この方法では、[35S]−標識アンチセンスcRNAプローブを生検試料の薄片とハイブリダイズさせ、洗浄し、RNaseで切断し、オートラジオグラフィー用感受性乳剤にさらす。この試料をヘマトキシリンで染色することにより、試料の組織学的組成を明らかにすることができ、適切な光フィルターによる暗視野画像化によって顕色乳剤が明示される。
免疫組織化学を用いて生検試料中のストロメリシン−3の発現を検出することができる。例えば適切な抗体を細胞の薄層と接触させ、洗浄し、次いで第2の標識抗体と接触させる。標識化はペルオキシダーゼ、アビジンなどの酵素、あるいは放射性標識によって行うことができる。色素原標識は顕微鏡下で検出できるので、一般に好ましい。
より一般的に好ましい検出法は、極めて迅速に実行できる免疫検定法(例えば、ELISAまたはRIA)でタンパク質を検出することである。したがって抗体または抗体等価物を用いてストロメリシン−3を検出することが一般に好ましいが、適切に標識されたストロメリシン−3基質の使用も有利であり得る。
酵素過程の生成物がそれ自体の特性として検出可能であり特徴的である場合(例えば過酸化水素など)には、基質を標識する必要はないであろう。しかし基質の標識化が必要な場合には、基質の標識化は酵素標識化、放射性同位体による標識化、抗体標識化、蛍光標識標識化、あるいは当業者にとって容易に理解されるであろう他の適切な形態のいずれによってもよい。
最も好ましいストロメリシン−3発現の検出法は、抗体を使用する方法である。抗体は以下に記述するようにして調製することができ、ストロメリシン−3発現の検出に適したあらゆる方法で使用できる。
抗体に基づく技術にはELISA(酵素結合イムノソルベント検定法)およびRIA(放射線免疫検定法)が含まれる。このような免疫検定法には従来の操作法のいずれを使用してもよい。この操作を以下のように実行するのも適切であり得る:ストロメリシン−3標準を放射性同位体(例:125Iまたは35S)または検定可能な酵素(例:西洋ワサビペルオキシダーゼまたはアルカリ性ホスファターゼ)で標識し、これを非標識試料と共に対応する抗体と接触させ、そこに第2の抗体を用いて第1の抗体を結合させ、放射活性または固定化酵素を検定する(競争的検定法)か、もしくは別法として、試料中のストロメリシン−3を対応する固定化抗体と反応させ、放射性同位体または酵素で標識された抗ストロメリシン−3抗体をこの系と反応させ、放射活性または酵素を検定する(ELISA−サンドイッチ検定法)。他の従来法も使用に適し得る。
上記の技術は基本的に“一段階”または“二段階”検定法として実行できる。“一段階”検定法は、抗原と固定化抗体を接触させ、洗浄することなくこの混合物を標識抗体と接触させることを伴う。“二段階”検定法は、上記の混合物を標識抗体と接触させる前に洗浄することを伴う。他の従来法も使用に適し得る。
ストロメリシン−3および/または抗体の酵素または放射線による標識化は従来の手段によって達成できる。一般的にそのような手段には、問題の抗原または抗体に対する酵素の(例えばグルタルアルデヒドによる)共有結合的連結が含まれる。具体的にはこの連結が酵素の活性に不利な影響を及ぼさないようにする。これは酵素が連結後でもその基質と相互作用できなければならないことを意味するが、検定を達成するに足る十分な活性が残っているのであれば、酵素のすべてが活性である必要はない。実際、酵素を結合させるいくつかの技術は非特異的であり(例えばホルムアルデヒドを用いる方法)、活性酵素の一部を与えるに過ぎないであろう。
普通は、検定系の一成分を支持体上に固定化し、それによってその成分とその系の他の成分との接触を可能にし、困難で時間のかかる労力を伴わずに容易に除去できるようにすることが望ましい。第2層を固定化して第1層から分離することができるが、通常は1層で十分である。
酵素自体を支持体に固定化することができるが、固相酵素が必要な場合には、抗体に結合させ、抗体を支持体に固定することによってこれを達成することが一般に最善の方法であり、そのモデルおよび系は当該技術分野でよく知られている。単純なポリエチレンが適切な支持体を提供し得る。
標識化に使用できる酵素に特に制限はないが、例えばオキシダーゼ群の構成要素から選択することができる。これらの酵素はその基質との反応によって過酸化水素の生成を触媒し、グルコースオキシダーゼはその良好な安定性、利用の容易さおよび安価であること、ならびにその基質(グルコース)が容易に利用できることから頻繁に使用されている。オキシダーゼの活性は、この酵素で標識した抗体を当該技術分野でよく知られている制御された条件下でその基質と反応させた後に生成する過酸化水素の濃度を測定することによって検定できる。
好みによって他の技術をストロメリシン−3の検出に使用することもできる。これらの1つはウエスタンブロッティング(Towbinら,Proc.Nat.Acad.Sci.76:4350(1979))であり、この方法では適切に処理した試料をSDS PAGEゲルに流した後、ニトロセルロースフィルターなどの固体支持体に移す。次に、抗ストロメリシン−3抗体(非標識)をこの支持体と接触させ、標識したプロテインAや抗免疫グロブリン(適切な標識には125I、西洋ワサビペルオキシダーゼおよびアルカリ性ホスファターゼが含まれる)などの第2免疫学的試薬で検定する。
診断用の試料は関連部位のいずれからでも得ることができる。腫瘍から直接得た支質や細胞質などの試料が理想的であろうが、例えば血液から得た試料もまた適切であろう。しかし試料を血液から得た場合には、ストロメリシン−3の量が血流中に希釈されているであろうから、高感度の検定法が必要であろう。このような診断は、腫瘍を切除するための外科手術後などの患者の経過を監視する際に特に重要であり得る。参照値を手術後に読み取り、一定の間隔でもう1つの値を読み取れば、上昇は再発あるいは場合により転移を表し得る。このような値の読み取りは、例えば子宮中の活性を考慮に入れる必要があろう。
抗ストロメリシン−3抗体は画像化のためにも使用できる。酵素に加えて他の適切な標識には、放射性同位体:ヨウ素(125I、121I)、炭素(14C)、硫黄(35S)、トリチウム(3H)、インジウム(112In)、およびテクネチウム(99mTc);フルオレセインおよびローダミンなどの蛍光標識、およびビオチンが含まれる。
しかしインビボ画像化のためにはその位置がより限定的になり、抗体はそのままでは身体の外側から検出できないので、抗体を標識するか、もしくは他の方法で修飾することによって検出できるようにしなければならない。
この目的のための標識は抗体の結合を実質的に妨害せず、外部からの検出を可能にするものあれば何でもよい。適切な標識には、X−ラジオグラフィー、NMRまたはESRで検出できるものが含まれ得る。X−ラジオグラフィー技術に適した標識には、検出可能な放射線を放射するが患者にとって明白に有害でない放射性同位体(例えばバリウムまたはセシウムなど)が含まれる。NMRおよびESRに適した標識には一般に、重水素のように検出可能な特徴的スピンを伴い、例えば関連するハイブリドーマ用の栄養素を適切に標識することによって抗体中に導入できるものが含まれる。
インビボ画像化法の場合には、適切な検出可能画像化部分(放射性同位体(例:131I、112I、99mTc)、放射線不透過性物質、あるいは核磁気共鳴で検出できる物質など)で標識されている抗体または抗体断片を試験すべき対象(ヒトなど)中に(例えば非経口的、皮下的、あるいは腹腔内に)導入する。
対象のサイズおよび使用する画像化系が、診断的画像を作成するのに必要な画像化部分の量を決定するであろう。放射性同位体部分の場合、ヒト対象については、注射される放射活性の量は通常約5〜20ミリキュリーの範囲のテクネチウム−99mであってもよい。
次いで、標識した抗体または抗体断片がストロメリシン−3を含有する細胞の位置に優先的に蓄積するであろう。次いで、この標識抗体または抗体断片を既知の技術を用いて検出することができる。
この技術分野に関する一般的議論については、S.W.Burchielら,“Immunopharmacokinetics of Radiolabelled Antibodies and Their Fragments"(“Tumor Imaging:The Radiochemical Detection of Cancer",S.W.BurchielおよびB.A.Rhodes編,Masson Publishing Inc(1982))の第13章)を参照のこと。
抗体はストロメリシン−3のペプチドに対しても、またその全分子に対しても生じさせることができる。このようなペプチドはアルブミンなどの担体タンパク質と共に動物系に提供するか、あるいはそれが十分長い(例えば25アミノ酸残基)場合には担体なしで動物系に提供できる。ストロメリシン−3は自己タンパク質に相当であろうから、ヒト抗体がストロメリシン−3を認識できるとは思われない。
本明細書において用語“ペプチド”とは、2以上のアミノ酸がペプチド結合を通して連結してなるあらゆる分子を意味する。したがってこの用語はオリゴペプチド、ポリペプチドおよびタンパク質を包含する。
上記の技術で作成したポリクローナル抗体を直接使用してもよく、また適切な抗体産生細胞をその動物から単離し、これを用いて既知の手段(KohlerおよびMilstein,Nature 256:795および以下参照(1975))でハイブリドーマを作成することもできる。適切なハイブリドーマの選択も当業者には明白であろうし、得られる抗体はストロメリシン−3を同定するのに適した検定法で使用できる。
抗体またはその等価物を本発明に従って転移性癌の治療または予防に用いることもできる。抗体の適切な投与量の投与はストロメリシン−3の生産を遮断するか、もしくはストロメリシン−3の有効活性を遮断するよう作用することができ、これは悪性成長を治療するための極めて重要な時間を提供するであろう。
どのような場合にも何が実際に転移を導くのかはわかっていないので、予防はこの疾患のまさに初期段階でも適切であり得る。したがってストロメリシン−3活性を妨害する抗体、その等価物または因子類(例えばTIMP類(MMP類を調節する天然化合物−−金属プロテイナーゼの組織阻害因子))の投与を癌が診断されると同時に実施することができ、必要なかぎり長期間、好ましくはこの疾患のおそれが排除されるまで治療を続けることができる。
治療の好ましい形態はいわゆるマジック・ビュレット技術であり、この方法では腫瘍の領域に誘導する抗体に適切な毒素を結合させる。このような毒素は当該技術分野でよく知られており、1細胞あたりたった1または2分子程度で作用し得るリシンなどの天然分子に限らず、毒性放射性同位体、重金属、酵素および補体活性化因子からなってもよい。このような技術をホルモン拮抗薬あるいは例えば癌の治療に使用できる生理学的に活性な他の適切な化合物の局在化した投与量を送達するために使用することもできる。
本発明に従って使用するための抗体が診断的応用であれ、治療的応用であれ、モノクローナルでもポリクローナルでも適切であり得ることは理解されるであろう。これらの抗体等価物は例えば抗体のFab'断片(Fab、Fab'、F(ab')2およびFvなど);イディオタイプ;あるいはアロトープ移植の産物(患者中での免疫応答を避けるために、動物抗体の認識領域がヒト抗体の適切な領域に移植されているもの)などからなり得る。他の適切な修飾物および/または薬剤は当業者に明白であろう。
ストロメリシン−3を阻害または除去するための抗体の使用に加えて、他の形態の阻害剤を用いることもできるであろう。そのような阻害剤は(例えばECM分解酵素について)汎用であってもよいし、ストロメリシン−3に対して特異的であってもよい。金属プロテイナーゼ(TIMP)類の組織阻害因子はその存在が知られており、ストロメリシン−3に対して特異的なTIMPが存在する可能性は極めて高い。このようなTIMPは標準的な技術によって容易に同定できる。
ストロメリシン−3の合成阻害剤も製造することができ、これらは一般に酵素活性の作用を受ける基質の領域に対応するであろう。このような阻害剤が基質と切断生成物との間の凍結中間体に相当することが一般に好ましいが、その結合部位の立体障害型か、あるいはそれ自体がストロメリシン−3に不可逆的に結合するであろう型の結合部位を提供することもできる。他の適切な阻害剤は当業者に明白であろう。
ストロメリシン−3活性を遮断するために他の方法を用いることもできる。これらは例えば変性剤からなってもよいが、これらは非特異的である傾向が強く、例えば特異的抗体の使用などによってそれらを誘導できる場合にのみ適当に使用できる。ストロメリシン−3遮断活性の他の形態は、プレ−プロタンパク質からタンパク質への進行を遮断することによって達成され得る。この過程は数個の標的段階を提供し、唯一必要なことは他の必須酵素に影響を与えないように独立に遮断し得る段階、あるいは再度標的にされ得る段階を同定することだけである。
ストロメリシン−3の3次構造を選択的に認識し、それによってその酵素活性を遮断するために、ペプチドまたは他の小分子を用いることもできるであろう。このような活性遮断物質が必ずしも活性部位に結合する必要はなく、それでもストロメリシン−3の3次構造を改変または凍結してその活性を破壊、一時停止、または改変するよう作用することができる。またこの遮断物質が必ずしもそれ自体によって作用する必要はなく、この目的のためのもう1つの物質に結合させることができるし、あるいは適切な不活化剤のための認識部位として機能することもできる。
I型コラーゲナーゼおよび92kD IV型コラーゲナーゼのmRNAの出現はもっぱら悪性腫瘍に伴うが、その逆は常に成り立つわけではない(即ち、腫瘍は常にこのタンパク質を伴うわけではない)ことが、本発明者らの研究によって立証された。
浸潤性乳癌腫中でのストロメリシン−3遺伝子の発現とテナシン(tenascin)遺伝子の発現との間には明らかに類似点がある。ECM糖タンパク質テナシン(Chiquet−Ehrismannら,Cell 47:131−139(1986))は上皮間葉細胞相互作用、および正常発育中の細胞移動(器官形成中の乳腺のものを含む)に必須の役割を果していると思われる。
テナシンは悪性乳腫瘍の線維性支質中で過剰発現されることが一貫して認められてきており、ストロメリシン−3に類似した様式で誘導されると思われる。フィブロネクチンと比較した場合、テナシンは乳房腫瘍上皮細胞の付着に関する基質として劣っており、このことはそれらが浸潤性になることを可能にし得ることを示唆している。
したがってストロメリシン−3は乳癌の浸潤相中にテナシンと共同して作用し得る。またストロメリシン−3およびテナシンは胚形成中に、上皮−間葉相互作用が重要な役割を果すことが知られている領域、および細胞移動が起こる領域で同時発現し得る。
したがって本発明は、上記のタンパク質のいずれか、もしくはそれらをコードするヌクレオチド配列の検出をも含む、上に定義した転移性癌の診断法をも提供する。
また本発明は、上記のタンパク質のいずれかを結合する薬剤の使用をも含む転移性癌の治療または予防における使用をも提供する。
さらに、本発明はストロメリシン−3の一部または全体をコードするヌクレオチド配列をも提供する。ストロメリシン−3の配列は添付の図2に示すものが好ましく、またそのヌクレオチド配列も図2に示すものが好ましい。しかし、遺伝子コードの縮重ゆえに、その配列がストロメリシン−3の少なくとも一部をコードしている限りにおいて、ヌクレオチド配列がこの図に示したものとは実質的に異なり得ることは理解されるであろう。
必要な配列はそれを利用する方法によってさらに変化することさえあり得る。生物学的試料中のRNA転写物を検出するための使用を意図する場合には、普通は図2に記載した配列にかなり密接に対応することが好ましいであろう。しかしそれでも、ハイブリッド形成が選択した厳密性条件下で可能である場合には、その配列はさらに変化し得る。
当業者は図2のペプチド配列から逆にプローブを作成することができる。しかし遺伝コードの縮重性ゆえに、逆に作成したある配列が同じペプチドから逆に作成した相補的配列と必ずしも充分にハイブリッド形成するとは限らず、あるいは全くハイブリッド形成しないこともあることは理解されるであろうから、このようなプローブの使用は制限されるであろう。これは当業者の計画には共通の要素であり、いかなる配列の縮重性もしばしば非常に広いためにどのような1配列のためのプローブの数も極めて多数になる。
ヌクレオチド配列がストロメリシン−3ペプチドまたは該酵素全体の発現のために必要な場合には、上述のように遺伝子コードの点、ならびにストロメリシン−3の構造または機能に有意な影響を与えることなくこの酵素のいくつかのアミノ酸配列を変化させ得るという点で、かなり大きな自由度が存在し得る。
このような配列の相違を意図する場合には、活性を決定する極めて重要な領域がこの分子上に存在するであろうことに注意すべきである。通常このような領域は結合部位を形成する残基群か、もしくは結合部位に影響を及ぼす3次構造を形成する残基群からなるであろう。一般論として、類似の機能を発揮する残基を使用する限りにおいて、3次構造を形成する残基群を置換することができる。他の例では、残基の種類は全く重要でないこともある。
したがって本発明は、ストロメリシン−3の例えば抗体の作成に使用するための特徴的領域を示すか、もしくはまだ実際にストロメリシン−3活性を示す、その配列上の変種および変異体をも包含する。このような変種および変異体には欠失体、付加体、挿入体、反転体、反復体および型置換体(例えば、ある親水性残基で別の親水性残基を置換するが、一般に高度に親水性の残基で高度に疎水性の残基を置換しない)が含まれる。小さい変化は一般に、それらがその分子の必須部分でない限り活性にほとんど影響を与えないであろう。また小さい変化は遺伝子操作の副産物でもあり得るし、例えば追加の制限部位を作成する場合などには小さい変化が望まれるべきである。修飾には1以上の残基を他の適切な残基で置換することも含まれ、このような置換は1:1でもよいし、あるいは1以上または1以下の他の適切な比でもよい。
遺伝子操作/発現を異なった方法で補助するため、あるいはストロメリシン−3分子を増大させるか、もしくは他の方法で都合よく修飾するために、コード配列中に点変異および他の変化を導入することによって、例えば制限部位を付加または欠失させることができる。
ストロメリシン−3等価物が他の動物(具体的には哺乳類)中でも発見されるであろうこと、またそのような供給源から得られる配列情報がストロメリシン−3分子の保存された領域を解明するために特に重要であり得ることも理解されるであろう。例えば、マウスの対応する配列ではストロメリシン−3に特有のプロドメイン中の10アミノ酸配列などを含む約80%が保存されている。ヒトストロメリシン−3配列に対応する動物配列が当該技術分野で既知の方法および上述の方法によって容易に検出できるであろうこと、およびそのような配列およびそれらのペプチドがその変異体および変種と共に本発明の一部を構成していることは理解されるであろう。
所望により、制限部位を提供するように本発明の配列を加工することもできる。この加工はコードされているストロメリシン−3のペプチド配列を干渉しないように行うことが可能であり、また最終生成物が望ましい特性を有するという条件下で、望ましい程度あるいは必要な程度に干渉することもできる。
上述のようにハイブリッド形成は信頼性の低い配列相同性の指標であり得るが、好ましい配列は一般に図2の配列に対して50%以上、好ましくは70%以上、より好ましくは80%以上の相同性を示すものであろう。
他の金属プロテイナーゼ類の場合と同様に、ストロメリシン−3は最初はプレ−プロ酵素として発現される。したがって生体内では2つの切断段階が観測される。切断はインビトロ発現にとって必ずしも必要条件ではなく、例えば大腸菌は成熟タンパク質を発現することができるであろう。
ストロメリシン−3またはその特徴的なペプチドを発現させることを望む場合、適切な糸はすべて使用することができる。適切なベクター、発現ベクターおよびその構築法の一般的性質は当業者には理解できるであろう。
“特徴的な”という表現はストロメリシン−3に特有の配列を有するあらゆるペプチドを意味する。このような配列はストロメリシン−3活性にとって重要であってもよいし、あるいはただ単に他のペプチドには認められない配列であるだけでもよい。しかし一般にストロメリシン−3活性にとって重要な配列が好ましい。というのはこれらは母集団内で保存されている可能性がより高いからである。
適切な発現ベクターはファージまたはプラスミドに基づいていてもよく、これらは両者とも一般に宿主特異的であるが、これらを他の宿主のために加工することもしばしば可能である。他の適切なベクターにはコスミドおよびレトロウイルスならびに他のあらゆる媒体が含まれ、これらは与えられた系に関して特異的であってもよく、また特異的でなくてもよい。また認識、プロモーター、オペレーター、誘導因子、終止因子、および発現の制御に不可欠および/または有用な他の配列などの制御配列は当業者には容易に理解できるであろう。またこれらの制御配列は天然のストロメリシン−3配列に伴っているものでもよいし、あるいは使用するベクターに伴っているものでもよく、あるいは他の供給源から適切に誘導することができる。これらのベクターはあらゆる適切な方法で修飾または加工することができる。
ヌクレオチド配列の正しい調製は、例えばSangerらの方法(Proc.Natl.Acad.Sci.USA 74:5463−7(1977))によって確認することができる。
本発明のストロメリシン−3をコードしているcDNA断片は適切なベクター中に容易に挿入できる。受容ベクターが容易な挿入に適した制限部位を有することが理想的であるが、例えば平滑末端連結も使用できる。ただしこれは読み枠および挿入の方向に関する不確定性を導くであろう。このような場合には発現に関して形質転換体を試験することは当然のことであり、これらの形質転換体は6個中1個の割合で正しい読み枠を有するはずである。当然のことながら、当業者は望ましい発現系に応じて適切なベクターを選択することができる。
適切な生物(好ましくはHeLaなどの真核細胞株)を得られたプラスミドで形質転換し、アンピシリンまたは必要ならば他の適切な手段によって形質転換体を選択し、トリプトファンまたは必要ならば他の適切なプロモーター−誘導物質(インドールアクリル酸など)を添加することによって、目的のストロメリシン−3を発現させることができる。発現の程度をSDSポリアクリルアミドゲル電気泳動:SDS−PAGE(Lemelli,Nature 227:680−685(1970))で分析することができる。
培養などの生育および形質転換に適した方法は例えばManiatis(“Molecular Cloning,A Laboratory Notebook",Maniatisら編,Cold Spring Harbor Labs,NY(1989))に実用的に明示されている。
ストロメリシン−3またはそのペプチドの生産に有用な培養はあらゆる生存細胞の培養が適切であり得、原核生物発現系から真核生物発現系までに及び得る。好ましい原核生物系の1つは、その操作の容易さゆえに大腸菌の系である。しかし、より高等な系(哺乳類細胞株など)も真核タンパク質の発現に使用できる。現在のところ仮発現とって好ましい細胞株はHeLaおよびCos細胞株である。他の発現系にはチャイニーズハムスター卵巣(CHO)細胞株が含まれる。
価値ある系の1つはバクロウイルス系であり、この系では蝶細胞をストロメリシン−3または適切なペプチドをコードしているDNAベクターおよびバクロウイルスDNAで同時トランスフェクションする。組換えがその細胞内で起こり、適切なバクロウイルス組換体を標準的な技術で選択することができる。その後、その組換体を用いて望ましい細胞株を感染させることができ、感染によってストロメリシン−3またはペプチドが発現される。この系に特有の利点は生産されるタンパク質の量であり、その量は約1〜約500mg/
このような系は大腸菌系ほど使用しやすくない傾向にあるが、その利点は一次合成後のタンパク質のプロセシングにある。例えば大腸菌はプレ−プロタンパク質のプロセシングに哺乳類細胞と同じ系を使用しない。
使用し得る他の発現系には、例えばストレプトミセス科の放線菌、およびサッカロミセス種、具体的にはサッカロミセス・セレビシェなどの酵素が含まれる。一般に操作者が何を要求するかに依存して、所望によりどのような系も使用できる。適切な系を用いて遺伝物質を増幅することもできるが、そのDNAの増殖だけが必要なこの目的には一般に大腸菌の使用が便利である。
成熟酵素の配列はプロ−およびプレプロ−配列にも共通であるので、抗体を生じさせるためには成熟酵素だけを生産することが有利であろう。しかし、プロおよびプレプロ部分の切断がこの分子の3次構造を変化させるかも知れず、したがって成熟酵素に対して生じた抗体が例えばプロ酵素を検出できないこともあり得ることは理解されるであろう。初期段階のいずれかにある酵素および/または切断されるプレ−またはプロ−ペプチドに対して生じた抗体も有用となるであろう。
ペプチドまたはヌクレオチド配列は、それを使用する目的が考慮されたストロメリシン−3に特徴的なものであれば何でもよい。配列はストロメリシン−3に完全に特徴的であることが理想的であろうが、そのような配列の長さはストロメリシン−3分子のその領域に応じて様々であり得る。最も好ましい領域は高度に保存されている領域、および他のタンパク質に共通しない領域であるが、その配列がMMP類、もしくはより具体的には浸潤性腫瘍に関連するMMP類に特徴的な配列である場合にはそれが有利であり得る。
本発明は上記のペプチドおよびヌクレオチド配列の等価物を包含し、これに関連する。ここで用語“等価物”は前述の意味で使用されており、換言すればC−またはN−末端あるいは他のいずれかの位置に置換を有する配列という意味の等価物である。
本発明は本配列の変異体をも包含する。ここで用語“変異体”は、上述の制限を前提とする配列中のアミノ酸残基または塩基の欠失体、付加体、挿入体、反転体および置換体に関して使用されている。
さらに本発明は配列の変種をも包含し、この用語は、図2に示す配列と基本的には同じ配列を共有するが、大きい母集団内で予期される程度に変化している、ときおり発見されるであろう他の天然に存在するストロメリシン−3に関連して使用されている。この定義内にはアレル変種および類似型の活性を示し関連する配列を有する他の種由来のペプチドが含まれる。またあまり好ましくはないが、動物配列も含まれる。
本発明者らはストロメリシン−3の発現が例えば成長因子および腫瘍プロモーターによって刺激され得ることをも発見した。このような因子の典型例にはEGF FGFおよびPDGFおよびTPAが含まれる。したがって上述の方法と共に、腫瘍試料中のこれらの因子のいずれかの検出は癌の転移状態を診断するためにも役立ち得る。
したがって本発明は、ストロメリシン−3遺伝子の発現を変化させることによる転移性癌の治療をも提供する。これはストロメリシン−3の生産を刺激するために必要な因子を、例えばこの因子に対する特異的抗体を使用するなどして妨害することによって達成することができ、目的の結果を達成するためにこの抗体をさらに修飾してもよい。この因子の受容体を遮断すること(これは例えば抗体または合成ペプチドであり得る必要な結合剤の局在化によってより容易に達成され得る)も可能であろう。
ストロメリシン−3遺伝子の発現に影響を与えることは、例えばゲノムDNA上のプロモーターなどの部位の遮断などによってより直接的にも達成され得る。
例えば抗体を患者に投与するために本発明を用いる場合には、これは適切な経路のいずれによってもよい。腫瘍がまだ局在化していると思われるか、もしくはそのように診断される場合には、適切な投与法はその部位への直接的な注射による方法であり得る。標的が乳癌である場合には乳房への注射で十分であり得、また埋め込み剤を用いてもよい。例えばTIMP類を投与すべき場合には、長期間持続する投与のために皮膚パッチを使用することもできるであろう。
咽頭癌の場合には、さらなる選択枝として例えばうがい剤を用いる経口投与であってもよい。
いずれかの例において、投与を別法として、あるいは追加的に、皮下、筋肉内、静脈内および皮内注射を含む注射によって行ってもよい。
製剤はその投与経路に適したものであれば何でもよく、当業者には自明であろう。本製剤は食塩水などの適切な担体を含有してもよく、またバルキング剤、他の医薬調製物、佐剤および他のあらゆる適切な医薬成分を含有してもよい。
適切な調製物にはストロメリシン−3またはその特徴的なペプチドを含有するワクチンも含まれ得る。このようなワクチンは能動型であっても受動型であってもよいが、ストロメリシン−3の発現が子宮内で起こり、抗ストロメリシン−3抗体への不明確な暴露が望ましくない効果を有するかも知れないので、一般的には受動型が好ましい。しかし能動予防接種が有利なこともあり、具体的には患者が既に子宮切開術を受けている場合にはどの組織もストロメリシン−3を正常には発現しないであろうから、能動予防接種が有利であり得る。他の適切なワクチンにはストロメリシン−3またはその特徴的なペプチドをコードしているヌクレオチド配列を含有する組換えウイルスが含まれる。このようなウイルスの適切なものの1つはワクシニアウイルスである。
以下の実施例は本発明を明示するよう機能するものであり、いかなる形でも本発明を制限することを意図するものではない。
実施例1乳癌特異的cDNAのクローニング 一次乳癌の外科的切除試料(腫瘍C1と呼ぶ)から得たポリ(A+)RNAを用いて乳癌cDNAライブラリーをλgt10ベクター中に構築した。それぞれC1−ポリ(A+)RNAおよび乳線維腺腫(F1と呼ぶ)から得たポリ(A+)RNAから逆転写したcDNAに対応する(+)および(−)プローブを用いて50000プラークを個別にスクリーニングした。
図1にC1−乳癌腫およびF1−線維腺腫から得た全RNAの、線維腺腫中より癌腫中でより高い発現レベルを示す4腫の遺伝子のcDNAプローブ(A〜D)を用いたノーザンブロット分析を示す。各レーンは8μgの全RNAを含有する。偏在的に発現する遺伝子に対応する36B4プローブを用いてこれらのフィルターを再プローブした(Rioら,Proc.Nat.Acad.Sci.USA 84:9243−9247(1987))。
具体的には、全RNAを液化窒素中に保存した外科標本から調製し(Chirgwinら,Biochemistry 18:5294−5299(1979))、オリゴ(dT)−セルロース・クロマトグラフィーによってポリ(A+)RNAを選択した。全細胞集団の約50%が支質細胞であるエストロゲン受容体−陰性(グレードII)導管癌腫(C1と呼ぶ)から調製したcDNAを用いて、乳癌−濃縮cDNAライブラリーを構築した。
クローニングに先立って、一本鎖cDNAを過剰量の乳線維腺腫(F1と呼ぶ)から得たポリ(A+)RNAを用いて減じ、その一本鎖濃縮物質をヒドロキシアパタイトクロマトグラフィーで精製した(Davisら,Proc.Nat.Acad.Sci.USA 81:2194−2198(1984);Rhynerら,Neuroscience Res.16:167−181(1986))。
この乳癌−濃縮cDNAを二本鎖にし、λgt10ベクターのEcoR I部位中にクローン化した。3百万の組換えファージが得られ、約5000cDNAクローンを含有するプレートから得た複製ナイロンフィルター(Biodyne A,Pall Corporation)を用いて約50000を個別にスクリーニングした。
C1−乳癌cDNAおよびF1−乳線維腺腫cDNAを用いて、それぞれ(+)および(−)プローブを作成した。両プローブとも過剰量の全ヒト肝臓RNAで減じ(Davisら,Proc.Nat.Acad.Sci.USA 81:2194−2198(1984);Rhynerら,Neuroscience Res.16:167−181(1986))、その後ランダムプライミング合成を用いて[32P]−標識した。
ハイブリッド形成を厳密条件下(50%ホルムアミド、42℃)で2日間行い、2xSSC、0.1%SDS中22℃で洗浄した後、0.1xSSC、0.1%SDS中55℃で洗浄した。第2スクリーニングのために、弁別的に標識されたプラーク130個を選択した。
無作為に取り上げた5個の異なるプラークのcDNA挿入物をPCR増幅法で精製し、[32P]−標識し、関連クローンを同定するためにすべての異なるプラークにハイブリダイズさせた。この操作を無作為に取り上げた異なるプラークで数回繰り返し、最終的にF1−線維腺腫中よりもC1−癌腫中でより高レベルな発現を示したA〜Dと呼ぶ4種の遺伝子を得た。ホルムアルデヒドを含有する1%アガロース中での電気泳動によって分離した全RNA(8μg)を用いて、C1−乳癌およびF1−乳線維腺腫に関するノーザンブロットを調製し、Hybond−Nフィルター(Amersham)に移した。
このブロットをメチレンブルーで染色した後、転移させたRNAの完全性と量を調べるためにプレハイブリッド形成させた。ハイブリッド形成(18時間)および洗浄を、A〜D遺伝子に対応する[32P]−標識cDNA挿入物を用いて、上述のように標準的条件下で行った。
正常結腸でも発現された(示していない)遺伝子AおよびBはこれ以上調べなかった。
C遺伝子は結腸で発現した(示していない)が、その高レベルの弁別的発現(図1)ゆえにこの遺伝子を部分的に特徴づけた。これは種々の形質転換上皮細胞株および正常ヒト皮膚中でも発現された(示していない)。1つのCクローンのcDNAの配列決定は、対応する遺伝子がケラチン遺伝子スーパーファミリーに属することを示した(データは示していない)。
最後に、D遺伝子(ここではストロメリシン−3遺伝子とも呼ぶ)の発現はC1−癌腫およびF1−線維腺腫間で著しく異なり(図1)、またこの遺伝子は正常ヒト結腸および他のいくつかのヒト組織中では発現されなかった(後述)ので、D遺伝子をさらに調べた。
実施例2ストロメリシン−3遺伝子およびコードされているタンパク質の配列決定 D cDNA挿入物をプローブとして用いて数個の別個のクローンを非削減C1−乳癌λgt10cDNAライブラリーから単離し、配列を決定した。図2に全長D cDNAのヌクレオチド配列および対応するタンパク質配列を示す。
488アミノ酸長のタンパク質をコードしているcDNA読み取り枠の後ろには、そのRNAの3'−末端から14塩基上流に位置するポリ(A)付加シグナルを含む714塩基の3'−非翻訳領域が続いている。推定される開始メチオニンはヌクレオチド位置10〜12に位置する。対応するAUGはKozak共通モチーフに一致する配列を伴っておらず、その中にも位置していないが、この配列は予期される性質である(後述)疎水性先導ペプチドの配列のすぐ下流に対応しているので、翻訳はおそらくこのAUGから開始されるのであろう。
ストロメリシン−3 cDNAのヌクレオチド配列および推定アミノ酸配列を示す図2において、ヌクレオチド残基は5'から3'に向かって番号が付与されており、読み取り枠中の推定アミノ酸は、その1文字コードで指定されている。下線を付したヌクレオチド配列は5'−末端から順に:推定シグナルペプチド(2つの潜在的切断部位を矢印で示している);プロ金属プロテイナーゼ類に特有のPRCGVPD配列;亜鉛結合ドメインの保存されたヒスチジン残基(Matrisian,L.M.,Trends Genet.6:21−125(1990));およびポリ(A)付加シグナル配列に対応している。
具体的には、D cDNAの3'−部分に対応するcDNA挿入物[19bpのポリ(AT)領域を含む250bp]をランダムプライミング合成によって[32P]−標識し、これを用いて、C1−乳腫瘍ポリ(A+)RNAから作成した非削減λgt10 cDNAライブラリーをGublerおよびHoffmannの方法(Gene 25:262−269(1983))でスクリーニングした。数個の別個のクローンを同定し、M13シークエンシングベクター中にサブクローニングした。US Biochemical製のデアザ−dGTP試薬キットおよびシークエナーゼを用いるジデオキシ法でDNA配列を決定した。この配列をPC/GENEソフトウェアパッケージを用いて分析した。
実施例3推定上の金属プロテイナーゼ:ストロメリシン−3 図3にヒト・ストロメリシン類とヒトI型コラーゲナーゼの予想アミノ酸配列の比較を示す。
(a)多数整列(multialignment)プログラム(Higginsら,Gene 73:237−244(1988))を用いてアミノ酸配列を並べた。4つの配列のすべてで一致するアミノ酸残基に星印を付けてある。矢印はストロメリシン−3の推定上のシグナルペプチド切断部位を表す。矢じりはI型プロコラーゲナーゼおよびプロストロメリシン類の活性化に際して起こる切断を示す。この切断部位のレベルでストロメリシン−3に特異的な10アミノ酸残基を囲んである。PRCGVPD配列および推定上の亜鉛結合ドメインの保存残基に下線を付してある。
(b)左:ストロメリシン−3、ストロメリシン−1(ST1,Whithamら,Biochem.J.240:913−916(1986))、ストロメリシン−2(ST2,Mullerら,Biochem.J.253:187−192(1988))およびI型コラーゲナーゼ(CO I,Whithamら,Biochem.J.240:913−916(1986))間の類似性(アミノ酸一致率(%))領域;
(b)右:ST1、ST2およびCO I間の類似性領域;Pはシグナルペプチドおよびプロドメインを示す;ENZは成熟活性酵素に対応するドメインを示す。
このように推定タンパク質配列とSwissprotデータライブラリー(14版)との比較は、この新しいタンパク質が分泌されるマトリックス金属プロテイナーゼ(MMP)類に属することを示した(図3a)。したがってこの新しいタンパク質は疎水性N−末端先導配列候補(図2の下線部)を有しており、MMP類のプロドメインの特徴である高度に保存された配列PRCGVPD(アミノ酸残基78〜84)を示し、またMMP類の亜鉛結合部位(アミノ酸残基212〜225:図3a)を有する(Matrisian,L.M.,Trends Genet.6:121−125(1990))。
このファミリーの他の構成要素との類似性から、成熟タンパク質のN−末端アミノ酸はそのプレ−プロタンパク質のフェニルアラニン98に対応すると思われる(Whithamら,Biochem.J.240:913−916(1986))(図3a)。最適に並べた後、推定成熟タンパク質との類似性はストロメリシン−1(Whithamら,Biochem.J.240:913−916(1986))と40%、ストロメリシン−2(Mullerら,Biochem.J.253:187−197(1988))と38%、I型コラーゲナーゼ(Goldbergら,J.Biol.Chem.261:6600−6605(1986))と36%である(図3b)。
この新タンパク質の基質特異性はわかっていない。この新タンパク質のストロメリシン−1との類似性(40%)は明らかにストロメリシン−1とストロメリシン−2との間に存在する類似性(79%)よりはるかに低く、またI型コラーゲナーゼとストロメリシン−1の間に存在する類似性(53%)よりさらに低いが、本明細書ではこれをストロメリシン−3と呼ぶことにする。したがってこのタンパク質はMMPの一種であって、“ストロメリシン”という名称は必ずしも厳密に正確ではないが、この名称が便利である。
さらに、PRCGVPD配列の上流域では、ストロメリシン−3と既に比較した他のMMP類との間に有意な類似性はない(図3)。しかしストロメリシン−3は、I型コラーゲナーゼおよびストロメリシン類のプロタンパク質切断部位(Whithamら,上述)と本質的に正確に対応する位置に独特の短い配列(アミノ酸残基88〜97)を有する。さらにストロメリシン−3はI型コラーゲナーゼおよび他のストロメリシン類の場合と同様に、IV型コラーゲナーゼ類に特有のフィブロネクチン様ドメイン(Wilhelmら,J.Biol.Chem.264:17213−17221(1989))を示さない。
実施例4乳癌腫中での過剰発現 ストロメリシン−3 RNA転写物の出現を30種の乳癌腫および5腫の乳線維腺腫の切除試料で調べた。
図4に乳腫瘍中のヒト金属プロテイナーゼRNA類のノーザンブロット分析を示す:(a)ストロメリシン−3 RNA;(b)I型コラーゲナーゼRNA(CO I);
(c)92kD IV型コラーゲナーゼRNA(CO IV 92K);
(d)72kD IV型コラーゲナーゼRNA(CO IV 72K);
(e)ストロメリシン−1および−2 RNA(ST1/2);
(f)ポンプ−1 RNA(PU I)。
全RNAを4種のエストロゲン受容体−陰性乳癌腫(C1,グレードII;C2,C3およびC4,グレードIII)、6種のエストロゲン受容体−陽性乳癌腫(C5,C8およびC9,グレードII;C6およびC7,グレードIII;CO I,グレードI)および4種の乳線維腺腫(F2〜F5)から調製した。各レーンは8μgのRNAを含有する。36B4シグナルは対照遺伝子のRNAに対応する(図1)。
具体的には、図1の場合と同様に、数個のノーザンブロットを同一のRNA試料で同時に調製し、次のcDNAプローブのいずれかとハイブリダイズさせた:(a)ストロメリシン−3 cDNAの3'−部分を覆う1.6kb挿入物、(b)CO I cDNA、(e)ST2 cDNA(ST1 RNAと交差ハイブリッド形成するもの)、(f)PU1 cDNA(CO I、ST2およびPU1プローブはR.Breathnach氏の好意により供与された,Mullerら,Biochem.J.253:187−192(1988))、または(c)CO IV 92K(ヌクレオチド2144〜2223,Wilhelmら,J.Biol.Chem.2641:7213−17221(1989))および(d)CO IV 72K(ヌクレオチド1937〜2016,Collierら,J.Biol.Chem.263:6579−6587(1988))に対応する80マーのアンチセンスオリゴヌクレオチドプローブ(複数)。
ランダムプライミング合成を用いてこれらのcDNAプローブを(32P)−標識し(約5x105cpm/μg)し、5'−末端キナーゼ処理を用いてこれらのオリゴヌクレオチドを標識した(約108cpm/μg)。ハイブリッド形成を厳密条件下(42℃、50%ホルムアミド)で約106cpm/mlを用いて実行した。次にフィルターを2xSSC、0.1%SDS中22℃で洗浄した後、0.1xSSC、0.1%SDS中55℃で洗浄した。オートラジオグラフィーを(a)18時間、(b)20時間、(c)、(d)および(e)4日間、(f)2日間、−80℃で増感板を用いて行った。
ストロメリシン−3 mRNAはすべての乳癌腫中で、その癌腫がエストラジオール受容体(ER)陽性(C5〜C10)であるか陰性(C1〜C4)であるかにかかわらず認められた(図4a)が、発現レベルが乳癌腫中で観測された最低レベルと同等であった1つの例外(F2)を除いて、線維腺腫試料中では認められなかった。
MMP遺伝子ファミリーの他の構成要素のRNA転写物の出現をも同じ試料中で調べた(図4b〜f)。MMPファミリーのこれらの他の構成要素はそのヒト乳腫瘍中での発現様式に従って明確に2つの種類に分類することができる。第1の種類には72kD IV型コラーゲナーゼ(CO IV 72K、図4d)、ストロメリシン−1および−2(ST1/2、図4e)およびポンプ−1(PU1、図4f)が含まれ、これらの遺伝子はすべて悪性および良性腫瘍の両方で発現された。対照的に、ストロメリシン−3(図4a)、I型コラーゲナーゼ(CO I、図4b)および92kD IV型コラーゲナーゼ(CO IV 92K、図4c)遺伝子を包含する第2の種類は、乳癌腫中でのみ過剰発現を示す。ただし乳癌腫に一貫して伴うのはストロメリシン−3だけである。
発現様式は第2の種類に属する3つの遺伝子間で同一ではなかった。I型コラーゲナーゼRNA転写物はC5、C6、C7およびCIO癌腫中では検出されず、92kD IV型コラーゲナーゼRNA転写物はC7およびCIO試料中には認められなかったが、ストロメリシン−3 RNA転写物はすべての腫瘍中で明瞭に検出された。
したがってストロメリシン−3は浸潤性乳癌腫の症状を示すと思われ、一方I型コラーゲナーゼおよび92kD IV型コラーゲナーゼもまた場合によって乳癌の進行に特異的に関与するのであろう。
実施例5種々の供給源から得た細胞中での発現 図5に種々の細胞株および組織中のストロメリシン−3 RNAのノーザンブロット分析を示す。
(a)乳癌の患者から得た5種の転移性腋窩リンパ節および3種の正常な腋窩リンパ節;
(b)4種のエストロゲン受容体−陰性(BT−20、MDA−231、SK−BR−3、HBL−100)および4種のエストロゲン−陽性(T−47D、BT−474、ZR−75−1、MCF−7)乳癌株化細胞;
(c)10種の正常ヒト組織;
(d)血清非含培地(1および2)中TPA(10ng/ml)の非存在下(1)または存在下(2)で培養した、あるいは20μg/mlインシュリンを補足した血清非含培地(3〜6)中PDGF(20ng/ml、British Biotechnology)の非存在下(3)または存在下(4)、またはEGF(20ng/ml、Collaborative Research)の存在下(5)、またはbFGF(10ng/ml、Pettmann氏の好意により供与された(FEBS Lett.189:102−108(1985)))の存在下(6)で培養した、HFL−1ヒト胎児二倍体線維芽細胞(ATCC CCL 153)。
(a)では、レーン5(2μg)およびレーン6(20μg)を除いて各レーンは10μgの全RNAを含有する。(b)および(c)では、各レーンとも8μgの全RNAを含有し、(d)では各レーンは5μgの細胞質RNAを含有する。
具体的には、(a)、(b)および(c)については図4でストロメリシン−3について示したようにブロットを調製し、加工した。(d)では、全面成長HFL−1線維芽細胞を血清非含DMEM培養培地中に維持した。24時間後、新鮮な培地を加え、TPAまたは成長因子を上記のように補足するか、もしくは補足しなかった。24時間の培養後、細胞を回収し、細胞質RNAを調製した(Gough,N.M.,Analyt.Biochem.173:93−95(1988))。
次に図4でストロメリシン−3について示したようにブロットを調製し、加工したが、オートラジオグラフィーを3日間とした。
92kDコラーゲナーゼIV RNA転写物は3種の正常および5種の乳癌転移性リンパ節中に認められたが、ストロメリシン−3 RNA転写物は転移性節中でのみ検出された(図5aおよび開示していないデータ)。
一次悪性乳腫瘍および転移性リンパ節で得た結果とは対照的に、8種のヒト乳癌細胞株中ではそれらのER状態にかかわらず、類似の条件下でストロメリシン−3 RNA転写物を検出できなかった(図5b)。同様にストロメリシン−3 RNA転写物は、子宮および胎盤という顕著な2つの例外を除いて、いくつかの正常ヒト成人組織中でも検出できなかった。
ストロメリシン−3はすべての癌に明らかに伴うわけではなく、結腸、卵巣、腎臓および肺癌から得たRNA試料中では低レベルのストロメリシン−3 RNA転写物が認められるに過ぎない。しかし、乳癌中に認められるレベルと同等に高いレベルの発現が喉頭癌RNA試料中に観測された(非開示データ)。
実施例6浸潤性腫瘍の支質細胞中での特異的発現 原始乳癌腫中でストロメリシン−3遺伝子が発現され、いくつかの確立された乳癌株化細胞中では発現されないことは、この遺伝子が新生物細胞そのものではなく、むしろその腫瘍を取り巻く支質細胞中で発現されたことを示唆している。
[35S]−標識ストロメリシン−3アンチセンスリボプローブを用いる原位置ハイブリッド形成実験を6種の癌腫(図4に関して命名した腫瘍C1、C3、C5、C9、C10および図4には示さなかったER−陽性癌腫である腫瘍C11)から得た切片を用いて行った。
具体的には、原位置ハイブリッド形成をCoxらが記述したように行った(Dev.Biol.101:485−502(1984))。脱パラフィン化酸処理切片(6μm厚)をプロテイナーゼKで処理し、Bluescript II(Stratagene)中にサブクローニングしたストロメリシン−3 cDNA挿入物(ヌクレオチド1128から1594までの467bp)から得た[35S]−標識アンチセンス転写物を用いて終夜ハイブリッド形成させた。ハイブリッド形成の後、RNase処理(20μg/ml、30分間、37℃)を行い、厳密に洗浄(2xSSC、50%ホルムアミド、60℃、2時間)した後、NTB2乳剤(Kodak)を用いてオートラジオグラフィーにかけた。オートラジオグラフィーを15日間行った。センスリボプローブを用いる類似の条件下でバックグラウンド以上の有意な標識化は観測されなかった(示していない)。
図6に乳癌腫および胚肢芽の切片中のストロメリシン−3 RNA転写物の存在を示す。a、c、e、g、iおよびk:ヘマトキシリンで染色した組織切片の明視野(x100);b、d、f、h、jおよびl:アンチセンス・ストロメリシン−3 cRNAプローブとの原位置ハイブリッド形成および暗視野画像化後の同切片(やはりヘマトキシリンで染色したもの)。
aおよびb、グレードII導管乳癌腫(腫瘍C1、図4参照のこと):浸潤性癌細胞(C)が紡錘状細胞(S)の豊富な支質に取り巻かれている;ストロメリシン−3 RNA転写物は新生物上皮細胞のすぐ周囲の支質細胞中にもっとも豊富である。cおよびd、グレードIII導管乳癌腫(腫瘍C3、図4参照のこと):浸潤性乳癌細胞(C)の複数の島が支質細胞によって取り巻かれている;ストロメリシン−3遺伝子の発現はほとんどの支質小柱(S)の中心部(即ち、新生物細胞から最も遠い領域)で、より弱い。eおよびf:導管癌腫(腫瘍C3、図4参照のこと)および2つの正常小葉(N);ストロメリシン−3 RNA転写物は、リンパ球(矢印)が豊富な小領域を例外として、浸潤性癌細胞(C)を並置させている支質中で独占的に検出された。gおよびh、導管癌腫(腫瘍C10、図4参照のこと):ストロメリシン−3 RNA転写物は浸潤性乳癌細胞を取り巻く支質細胞(右上角)中でバックグラウンド以上に検出できたが、原位置(星印)乳癌細胞では検出できなかった。iおよびj、導管癌腫(腫瘍C11、ER−陽性、グレードII、癌腫);左:原位置の癌腫(星印)、支質細胞中でストロメリシン−3 RNA転写物を検出できなかった;右:ストロメリシン−3遺伝子を発現している支質細胞によって取り巻かれている浸潤性新生物。kおよびl、8週齢ヒト胚肢芽の指間領域:ストロメリシン−3 RNA転写物が原始表皮の基礎をなす中胚葉中で検出され、指間領域(M)中で最も顕著である;原始表皮(矢印)、形成中の軟骨(PC)、および周辺中胚葉は標識されないことに注目のこと。
すべての場合においてストロメリシン−3 RNA転写物は、腫瘍の浸潤性成分を形成している悪性上皮細胞の島を取り巻く支質細胞中でのみ検出された(図6:パネルaおよびb(腫瘍C1について);パネルc、d、eおよびf(腫瘍C3について);パネルgおよびh(腫瘍C10について);パネルiおよびj、右側(腫瘍C11について);腫瘍C5およびC9についてのデータは示していない)。
転移性リンパ節(C5と同じ供給源)でも、ストロメリシン−3遺伝子の発現がその転移性上皮細胞を取り巻く支質細胞に限定されていた(データは示していない)。
すべての場合において、悪性上皮細胞そのものは標識されなかったこと、ならびに最も高レベルの発現が悪性細胞に並列している支質細胞中で観測されたことが特に注目に値する。著しく対照的に、まだ基底膜によって取り巻かれている原位置癌腫病巣を取り巻く支質細胞中では有意な発現を検出できず(パネルgおよびh(腫瘍C10について;パネルiおよびj、左側(腫瘍C11について))、一方同じ腫瘍の浸潤性成分中では支質細胞の標識化を明確に観測することができた(パネルgおよびh(腫瘍C1Oについて);パネルiおよびj、右側(腫瘍C11について))。
また、癌細胞から離れて位置する支質細胞および正常導管および小管を取り巻く支質細胞中では有意な発現を検出できなかった(例えばパネルeおよびf)。ストロメリシン−3転写物の分離した焦点は、ノーザンブロットでストロメリシン−3 RNAに関して弱く陽性であるF2線維腺腫の切片中で検出されなかった(図4aおよび非開示データ)。
線維芽細胞と筋線維芽細胞が共に浸潤性乳癌腫の支質中に存在することが知られている(Ahmed,A.Pathol.Annu.25(Pt2):237−286(1990))。本発明者らの原位置ハイブリッド形成技術では、これらの細胞型のうち1つだけがストロメリシン−3遺伝子を発現したのか、それとも両方の細胞型がストロメリシン−3遺伝子を発現したのかを決定することは不可能であった。
実施例7成長因子による刺激 上記の結果は、支質細胞中でのストロメリシン−3遺伝子の発現が新生物細胞が分泌する拡散性因子によって誘導されているらしいことを示している。EGF、FGFおよびPDGFなどの成長因子およびいくつかのサイトカイン類(IL−1α、β、およびTNF−α)、および腫瘍プロモーター類(例:TPA)がMMP遺伝子類の転写を活性化することが知られている(Kerrら,Science 242:1424−1427(1988))。数種の供給源から得られた腫瘍細胞がヒト線維芽細胞によるコラーゲナーゼIの生産を刺激する因子(単数もしくは複数)を生産することも報告されている(Lippmanら,Recent Prog.Hormone Res.45:383−440(1990))。乳癌細胞がインビトロでPDGF、FGFおよびTGF−α活性を分泌することが知られている(Salomonら,“Breast cancer:cellular and molecular biology"(Lippman,M.E.およびDickson,R.B.編),363〜389頁(Kluwer,Boston,(1988)))。
ストロメリシン−3遺伝子の発現を外因性の刺激物質で変調させ得るかどうかを調べるために、ヒト胎児二倍体線維芽細胞をPDGF、bFGFおよびEGFならびに腫瘍プロモーターTPAの存在下または非存在下で成長させた。これらの成長因子のいずれか、もしくはTPAの添加が線維芽細胞におけるストロメリシン−3 RNA転写物の増大をもたらし、最も強い刺激がbFGFで観測された(図5d)。
実施例8胚におけるストロメリシン−3の発現 ストロメリシン−3遺伝子が成長因子の刺激に応答して胎児線維芽細胞中で発現されたので、本発明者らはこの遺伝子が胚発育中に通常に発現されるかどうかを調べた。
ストロメリシン−3転写物が8週齢ヒト胚の数個の別個の領域で検出され、胚形成のこの段階で予定された細胞の死を伴うことが知られている領域(Milaire,J.,“Organogenesis"(De Haan,R.L.およびUrsprung,H.編),283〜300頁(Holt,Rinehart and Winston,New York,1965))である胚芽の指間領域で顕著であった(図6、パネルkおよびl、ならびに非開示データ)
標識化は、標識されずに残った原始表皮の基礎をなす胚の中胚葉で観測された。エピブラストから離れて位置する間葉細胞もmRNA−陰性であったことは注目に値する。
このように正常な胚生育中に組織改造が十分証明されている領域でストロメリシン−3遺伝子が発現するという発見は、乳腫瘍におけるストロメリシン−3の発現が癌の進行に関連するECM改造過程においてある役割を果すことを示唆している。
実施例9ST3をコードするマウスcDNAのクローニング ST3をコードするヒトcDNAを含有するプローブを用いて、マウス胎盤cDNAライブラリーを標準的方法(Sambrookら,“Molecular Cloning:A Laboratory Manual"(Cold Spring Harbor Press(1989)))でスクリーニングした。このスクリーニングの結果、ST3をコードする全長マウスcDNAが得られた(図7)。
これら2つの配列の分析によって成熟型ST3のアミノ酸配列には89%、プレおよびプロドメインには55%の相同性が存在することが明らかになった(図7)。
実施例4〜8に記述した方法を用いて、種々のマウス細胞中での発現様式を決定した。
マウスにおけるST3の発現様式は組織特異性に関してヒト組織に認められた様式と同一であることがわかった。最も高い発現レベルは胎盤および子宮組織に認められた。
【特許請求の範囲】
【請求項1】生物学的試料中の、以下に示されるアミノ酸配列
を有するストロメリシン−3をコードする核酸分子と該核酸分子とストリンジェントな条件下でハイブリダイズするヌクレオチドプローブとのハイブリッド形成を試験管内で検定することを含む、ストロメリシン−3の発現を検出する方法。
【請求項2】前記ヌクレオチドプローブが、ストロメリシン−3の特徴的な部分または全体をコードする核酸分子のヌクレオチドとハイブリッド形成する、請求項1記載の方法。
【請求項3】前記ヌクレオチドプローブが、ストロメリシン−3のプレ−またはプロ−ペプチドをコードしていない核酸分子のヌクレオチドとハイブリッド形成する、請求項1または2記載の方法。
【請求項4】前記ヌクレオチドプローブが、ストロメリシン−3をコードするcDNAの一部または全体を有するか、もしくは遺伝子コードの縮重によってそれに対応するものであるか、もしくは検定条件下でそれとハイブリッド形成するものである、請求項1から3のいずれかに記載の方法。
【請求項5】前記ヌクレオチドプローブが放射能標識されている、請求項1から4のいずれかに記載の方法。
【請求項6】前記ヌクレオチドプローブがリボプローブである、請求項1から5のいずれかに記載の方法。
【請求項7】以下に示されるアミノ酸配列
を有するポリペプチドをコードする単離された核酸分子。
【請求項8】請求項7において定義されるアミノ酸配列の第32〜488番のアミノ酸を含んでなるポリペプチドをコードする単離された核酸分子。
【請求項9】請求項7において定義されるアミノ酸配列の第34〜488番のアミノ酸を含んでなるポリペプチドをコードする単離された核酸分子。
【請求項10】請求項7において定義されるアミノ酸配列の第98〜488番のアミノ酸を含んでなるポリペプチドをコードする単離された核酸分子。
【請求項11】以下に示されるヌクレオチド配列
にストリンジェントな条件下でハイブリッド形成するヌクレオチド配列に相補的なポリヌクレオチドを含んでなる単離された核酸分子。
【請求項12】請求項11において定義されるヌクレオチド配列の第109〜1473番のヌクレオチドを含んでなる単離された核酸分子。
【請求項13】請求項11において定義されるヌクレオチド配列の第103〜1473番のヌクレオチドを含んでなる単離された核酸分子。
【請求項14】請求項11において定義されるヌクレオチド配列の第10〜1473番のヌクレオチドを含んでなる単離された核酸分子。
【請求項15】請求項11において定義されるヌクレオチド配列の第1〜1473番のヌクレオチドを含んでなる単離された核酸分子。
【請求項16】請求項11において定義されるヌクレオチド配列の第1〜1500番のヌクレオチドを含んでなる単離された核酸分子。
【請求項17】請求項7〜16のいずれかに記載の核酸分子に相補的なポリヌクレオチドを含んでなる単離された核酸分子。
【請求項18】前記核酸分子がcDNAである、請求項7〜17のいずれかに記載の核酸分子。
【請求項19】請求項7〜18のいずれかに記載の核酸分子を含むベクター。
【請求項20】適切な宿主細胞および発現ベクターである請求項19のベクターを含んでなる、請求項7〜10のいずれかにおいて定義されるポリペプチドを産出する発現系。
【請求項1】生物学的試料中の、以下に示されるアミノ酸配列
を有するストロメリシン−3をコードする核酸分子と該核酸分子とストリンジェントな条件下でハイブリダイズするヌクレオチドプローブとのハイブリッド形成を試験管内で検定することを含む、ストロメリシン−3の発現を検出する方法。
【請求項2】前記ヌクレオチドプローブが、ストロメリシン−3の特徴的な部分または全体をコードする核酸分子のヌクレオチドとハイブリッド形成する、請求項1記載の方法。
【請求項3】前記ヌクレオチドプローブが、ストロメリシン−3のプレ−またはプロ−ペプチドをコードしていない核酸分子のヌクレオチドとハイブリッド形成する、請求項1または2記載の方法。
【請求項4】前記ヌクレオチドプローブが、ストロメリシン−3をコードするcDNAの一部または全体を有するか、もしくは遺伝子コードの縮重によってそれに対応するものであるか、もしくは検定条件下でそれとハイブリッド形成するものである、請求項1から3のいずれかに記載の方法。
【請求項5】前記ヌクレオチドプローブが放射能標識されている、請求項1から4のいずれかに記載の方法。
【請求項6】前記ヌクレオチドプローブがリボプローブである、請求項1から5のいずれかに記載の方法。
【請求項7】以下に示されるアミノ酸配列
を有するポリペプチドをコードする単離された核酸分子。
【請求項8】請求項7において定義されるアミノ酸配列の第32〜488番のアミノ酸を含んでなるポリペプチドをコードする単離された核酸分子。
【請求項9】請求項7において定義されるアミノ酸配列の第34〜488番のアミノ酸を含んでなるポリペプチドをコードする単離された核酸分子。
【請求項10】請求項7において定義されるアミノ酸配列の第98〜488番のアミノ酸を含んでなるポリペプチドをコードする単離された核酸分子。
【請求項11】以下に示されるヌクレオチド配列
にストリンジェントな条件下でハイブリッド形成するヌクレオチド配列に相補的なポリヌクレオチドを含んでなる単離された核酸分子。
【請求項12】請求項11において定義されるヌクレオチド配列の第109〜1473番のヌクレオチドを含んでなる単離された核酸分子。
【請求項13】請求項11において定義されるヌクレオチド配列の第103〜1473番のヌクレオチドを含んでなる単離された核酸分子。
【請求項14】請求項11において定義されるヌクレオチド配列の第10〜1473番のヌクレオチドを含んでなる単離された核酸分子。
【請求項15】請求項11において定義されるヌクレオチド配列の第1〜1473番のヌクレオチドを含んでなる単離された核酸分子。
【請求項16】請求項11において定義されるヌクレオチド配列の第1〜1500番のヌクレオチドを含んでなる単離された核酸分子。
【請求項17】請求項7〜16のいずれかに記載の核酸分子に相補的なポリヌクレオチドを含んでなる単離された核酸分子。
【請求項18】前記核酸分子がcDNAである、請求項7〜17のいずれかに記載の核酸分子。
【請求項19】請求項7〜18のいずれかに記載の核酸分子を含むベクター。
【請求項20】適切な宿主細胞および発現ベクターである請求項19のベクターを含んでなる、請求項7〜10のいずれかにおいて定義されるポリペプチドを産出する発現系。
【第1図】

【第3b図】
【第4図】
【第2図】



【第3a図】


【第5図】
【第6図】
【第7図】


【第3b図】
【第4図】
【第2図】
【第3a図】
【第5図】
【第6図】
【第7図】
【特許番号】特許第3516448号(P3516448)
【登録日】平成16年1月30日(2004.1.30)
【発行日】平成16年4月5日(2004.4.5)
【国際特許分類】
【出願番号】特願平4−501188
【出願日】平成3年11月21日(1991.11.21)
【公表番号】特表平5−505530
【公表日】平成5年8月19日(1993.8.19)
【国際出願番号】PCT/FR91/00924
【国際公開番号】WO92/009701
【国際公開日】平成4年6月11日(1992.6.11)
【審査請求日】平成10年11月11日(1998.11.11)
【前置審査】 前置審査
【出願人】(592236234)アンスティテュー・ナシオナル・ドゥ・ラ・サンテ・エ・ドゥ・ラ・ルシェルシュ・メディカル・(イ・エヌ・エス・ウ・エール・エム) (12)
【氏名又は名称原語表記】INSTITUT NATIONAL DE LA SANTE ET DE LA RECHERCHE MEDICALE (I.N.S.E.R.M.)
【参考文献】
【文献】Nature(1990 Dec),Vol.348,No.6303,p.699−704
【登録日】平成16年1月30日(2004.1.30)
【発行日】平成16年4月5日(2004.4.5)
【国際特許分類】
【出願日】平成3年11月21日(1991.11.21)
【公表番号】特表平5−505530
【公表日】平成5年8月19日(1993.8.19)
【国際出願番号】PCT/FR91/00924
【国際公開番号】WO92/009701
【国際公開日】平成4年6月11日(1992.6.11)
【審査請求日】平成10年11月11日(1998.11.11)
【前置審査】 前置審査
【出願人】(592236234)アンスティテュー・ナシオナル・ドゥ・ラ・サンテ・エ・ドゥ・ラ・ルシェルシュ・メディカル・(イ・エヌ・エス・ウ・エール・エム) (12)
【氏名又は名称原語表記】INSTITUT NATIONAL DE LA SANTE ET DE LA RECHERCHE MEDICALE (I.N.S.E.R.M.)
【参考文献】
【文献】Nature(1990 Dec),Vol.348,No.6303,p.699−704
[ Back to top ]