
癌の治療のためのSMYD3トランスフェラーゼを調整する化合物の同定
本発明は、ポリペプチドのメチルトランスフェラーゼ活性を検出する方法およびメチルトランスフェラーゼ活性のモジュレーター、より具体的には、SMYD3による網膜芽細胞腫のメチル化のモジュレーターをスクリーニングする方法を特徴とする。さらに、本発明は、このように同定されたモジュレーターを用いて、結腸直腸癌、肝細胞癌、膀胱癌および/または乳癌を予防または治療するための方法または薬学的組成物を提供する。N末端短縮型SMYD3(aliasZNFN3A1)は、より高いメチル化活性を有する。Lys824は、SMYD3のRB1タンパク質上の好ましいメチル化部位である。
【発明の詳細な説明】
【技術分野】
【0001】
技術分野
本発明は、転写調節、より具体的には、メチルトランスフェラーゼ活性を調整する作用因子、例えば、SMYD3(「ZNFN3A1」としても知られる)による網膜芽細胞腫のメチル化を調整する作用因子の同定に関する。SMYD3は多くの癌種において上方制御されるので、前記のように同定されたSMYD3モジュレーターは、例えば、結腸直腸癌、肝細胞癌、乳癌、および膀胱癌を含む癌の治療において有用なことが判明する可能性がある。
【0002】
本願は、2005年7月1日に提出された米国特許仮出願第60/695,957号の恩典を主張し、その内容はその全体が参照として本明細書に組み入れられる。
【背景技術】
【0003】
背景技術
最近の分子的研究から、細胞周期制御の抑止は広範囲のヒト腫瘍の基礎をなすことが明らかになっている(Sherr, C. J., Science 274, 1672-7 (1996)(非特許文献1))。p53遺伝子、RB1遺伝子、またはp16遺伝子の遺伝子変化がヒト癌の大多数に関与しており、細胞周期進行の調節が解除されることによって無秩序な細胞増殖が起こる(Hanahan, D.& Weinberg, R. A. Cell 100, 57-70 (2000)(非特許文献2); Sherr, C. J.& McCormick, F.Cancer Cell 2, 103-12 (2002)(非特許文献3))。細胞周期の中で、細胞周期が停止し、ゲノムの完全性が調べられ、かつDNA損傷が修復されるG1/S境界は、正常な細胞特性およびゲノム特性の維持に重要である。2つの重要なシグナル伝達経路、すなわち、p53およびRB1は、多くの下流遺伝子を制御することによってG1/S境界の調節に関与している。損傷を受けたDNAを含む細胞は、蓄積された野生型p53タンパク質のトランス活性化を介してp21Cip1を誘導することによって、この境界で停止している(Sherr, C. J. & Roberts, J. M. Genes Dev 13, 1501-12 (1999)(非特許文献4))。RB1は、家族性網膜芽細胞腫の原因遺伝子として単離され(Friend, S. H. et al. Nature 323, 643-6 (1986)(非特許文献5); Fung, Y. K. et al. Science 236, 1657-61 (1987)(非特許文献6); Lee, W. H. et al. Science 235, 1394-9 (1987)(非特許文献7))、細胞周期進行を制御することによって腫瘍サプレッサーとして機能する。細胞周期移行のG1期からS期にかけて、RB1はリン酸化されることで不活性化される。このリン酸化はサイクリン依存キナーゼ(CDK)によって触媒される。不十分にリン酸化されたRB1は、DNA複製および細胞周期進行に必要とされる遺伝子の発現を調整する転写因子であるアクチベーターE2Fの活性化ドメインと直接相互作用し、HDACと複合体化するクロマチン構造を変え、かつリプレッサー複合体を応答遺伝子のプロモーター領域にあるE2F結合部位に動員することによって(Weintraub, S. J., et al., Nature 358, 259-61 (1992)(非特許文献8); Sellers, W. R., et al., Proc Natl Acad Sci USA 92, 11544-8 (1995)(非特許文献9))、E2Fを阻害する(Dannenberg, J. H., et al., Genes Dev 14, 3051-64 (2000)(非特許文献10); Sage, J. et al. Genes Dev 14, 3037-50 (2000)(非特許文献11))。RB1は、CDK4/サイクリンDなどのCDK/サイクリン複合体によってリン酸化されるとE2Fを解離し、次いで、サイクリンE、c-Myb、CDK2、およびBCL2を含む下流遺伝子をトランス活性化する。
【0004】
本発明者らは、以前に、SMYD3が過剰発現するとNIH3T3細胞の増殖が促進され、いくつかの癌細胞において内因性SMYD3発現をノックダウンすると癌細胞の増殖阻害およびアポトーシスが誘導されたことから、SMYD3がヒストンH3のリジン4(H3-K4)に対してジメチルトランスフェラーゼ活性およびトリメチルトランスフェラーゼ活性を有し、SMYD3高発現が結腸直腸癌(CRC)細胞および肝細胞癌(HCC)細胞の増殖において重要な役割を果たすことを報告した(Hamamoto, R. et al., Nat Cell Biol 6, 731-40 (2004)(非特許文献12))。しかしながら、SMYD3の過剰発現によって増殖が促進される正確な機構は未解決のままである。アセチル化、リン酸化、および/またはメチル化によるヒストン修飾は、様々な分子を動員することによって標的遺伝子の転写を活性化または不活性化するクロマチン構造を調節する。ヒストンリジンのメチル化に関して、H3-K4、H3-K36、およびH3-K79の修飾は、コンホメーションをヘテロクロマチン構造からユークロマチン構造に変えることによって転写活性化と関連するのに対して(Im, H. et al., J Biol Chem 278, 18346-52 (2003)(非特許文献13); Bannister, A. J. et al., J Biol Chem 280, 17732-6 (2005)(非特許文献14); Schneider, R. et al., Nat Cell Biol 6, 73-7 (2004)(非特許文献15))、H3-K9、H3-K27、およびH4-K20のメチル化はヘテロクロマチン構造によって転写抑制をもたらす(Schotta, G. et al., Genes Dev 18, 1251-62 (2004)(非特許文献16); Nakayama, J. et al., Science 292, 110-3 (2001)(非特許文献17); Kirmizis, A. et al. Genes Dev 18, 1592-605 (2004)(非特許文献18))。
【0005】
【非特許文献1】Sherr, C. J., Science 274, 1672-7 (1996)
【非特許文献2】Hanahan, D.& Weinberg, R. A. Cell 100, 57-70 (2000)
【非特許文献3】Sherr, C. J.& McCormick, F.Cancer Cell 2, 103-12 (2002)
【非特許文献4】Sherr, C. J. & Roberts, J. M. Genes Dev 13, 1501-12 (1999)
【非特許文献5】Friend, S. H. et al. Nature 323, 643-6 (1986)
【非特許文献6】Fung, Y. K. et al. Science 236, 1657-61 (1987)
【非特許文献7】Lee, W. H. et al. Science 235, 1394-9 (1987)
【非特許文献8】Weintraub, S. J., et al., Nature 358, 259-61 (1992)
【非特許文献9】Sellers, W. R., et al., Proc Natl Acad Sci USA 92, 11544-8 (1995)
【非特許文献10】Dannenberg, J. H., et al., Genes Dev 14, 3051-64 (2000)
【非特許文献11】Sage, J. et al. Genes Dev 14, 3037-50 (2000)
【非特許文献12】Hamamoto, R. et al., Nat Cell Biol 6, 731-40 (2004)
【非特許文献13】Im, H. et al., J Biol Chem 278, 18346-52 (2003)
【非特許文献14】Bannister, A. J. et al., J Biol Chem 280, 17732-6 (2005)
【非特許文献15】Schneider, R. et al., Nat Cell Biol 6, 73-7 (2004)
【非特許文献16】Schotta, G. et al., Genes Dev 18, 1251-62 (2004)
【非特許文献17】Nakayama, J. et al., Science 292, 110-3 (2001)
【非特許文献18】Kirmizis, A. et al. Genes Dev 18, 1592-605 (2004)
【発明の開示】
【0006】
発明の概要
本発明は、少なくとも部分的に、SMYD3によるリジン824のメチル化によってRB1を調節する新規の機構の発見に基づく。SMYD3は、遺伝子名「ZNFN3A1」としても知られ、結腸直腸癌および肝細胞癌の大多数(例えば、WO2003/027413を参照されたい)において、ならびに膀胱癌および乳癌において上方制御されるヒストンH3メチルトランスフェラーゼである。
【0007】
RB1のC末端領域はSMYD3のSETドメインと相互作用する。さらに、HEK293細胞におけるSMYD3発現は、CDK2/サイクリンE複合体またはCDK6/サイクリンD3複合体によるRB1の821/826および807/811のリン酸化をインビトロおよびインビボで増強し、ひいては、その結果として、E2Fの転写活性を増大させた。このデータは、癌細胞におけるSMYD3発現の増強が、RB1を修飾し、その後に、E2F転写を活性化することによって細胞周期進行を促進することを意味している。本知見は、RB1調節の基礎となる新規の機構を示唆している。さらに、本知見は、発癌、より具体的には、結腸直腸、肝細胞、膀胱、および乳房での発癌をより深く理解することに寄与し、従って、これらの腫瘍の新たな治療方針を開発することに寄与する。
【0008】
従って、本発明の目的は、SMYD3による網膜芽細胞腫のメチル化を調整する作用因子を同定するための方法を提供することであり、この方法は、以下の工程を含む:
(a)メチルトランスフェラーゼ活性を有するSMYD3ポリペプチドと、メチル化される網膜芽細胞腫ペプチドおよび補助因子を、網膜芽細胞腫ペプチドのメチル化に適した条件下、試験作用因子の存在下で接触させる工程;
(b)網膜芽細胞腫ペプチドのメチル化レベルを検出する工程;ならびに
(c)工程(b)において検出されたメチル化レベルと、作用因子の非存在下で検出された対照レベルを比較する工程であって、対照レベルと比較したメチル化レベルの増加または減少は、作用因子がSMYD3による網膜芽細胞腫のメチル化を調整することを示す、工程。
【0009】
本発明のさらなる目的は、網膜芽細胞腫のメチル化を調節する試験化合物の能力を検出するためのキットを提供することであり、このようなキットは、(a)メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド、(b)SMYD3ポリペプチドによってメチル化することができる網膜芽細胞腫ペプチド、および(c)網膜芽細胞腫ペプチドのメチル化のための補助因子を含む。さらなる態様において、キットは、任意に、S-アデノシルホモシステイン加水分解酵素(SAHH)を含んでもよい。
【0010】
さらに、本発明は、結腸直腸癌、肝細胞癌、膀胱癌、および乳癌などの癌を治療するための化合物をスクリーニングする方法を提供し、このような方法は、(a)前記方法に従ってメチル化を調整する試験化合物を同定する工程、および(b)試験化合物の非存在下で検出された対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを下げる試験化合物を選択する工程を含む。
【0011】
さらに、本発明は、結腸直腸癌、肝細胞癌、膀胱癌、および乳癌などの癌の症状を軽減するための組成物を提供し、このような組成物は、前記方法によって同定された薬学的に有効な量の化合物および薬学的に許容される担体により構成される。
【0012】
本発明のさらなる目的は、結腸直腸癌、肝細胞癌、膀胱癌、および乳癌などの癌の症状を軽減するための方法を提供することであり、この方法は、癌細胞と、前記方法によって同定された薬学的に有効な量の化合物を接触させる工程を含む。
【0013】
本発明のこれらのおよび他の目的、特徴、および利点は、以下の詳細な説明を、添付の図面および実施例ならびに本明細書に付した特許請求の範囲と共に読めば十分に明らかになるだろう。
【0014】
特別に定義されない限り、本明細書において使用される全ての技術用語および科学用語は、本発明が属する当業者に一般的に理解されるものと同じ意味を有する。さらに、本明細書で使用する単語「a」、「an」、および「the」は、特別に他に指すものがない限り「少なくとも1つ」を意味する。
【0015】
本発明の実施または試行においては、本明細書に記載されたものに類似するかまたは等価の方法および材料が使用され得るが、適当な方法および材料を以下の明細書に記載する。
【0016】
本明細書において言及される出版物、特許出願、特許、およびその他の参考文献は、その全体が参照により組み入れられる。矛盾する場合、定義を含む本明細書によって統制される。さらに、材料、方法、および実施例は、例示的なものに過ぎず、本発明を制限するためのものではない。
【0017】
発明の詳細な説明
SMYD3 cDNAは、配列番号:1に示したように、1284のヌクレオチドのオープンリーディングフレームを含む1622のヌクレオチドからなる。オープンリーディングフレームは、配列番号:2に示したように、ジンクフィンガーモチーフおよびSETドメインを有する428アミノ酸のタンパク質をコードする。ジンクフィンガードメイン(MYND)はアミノ酸49からアミノ酸87にわたり、SET(Su3-9、Enhancer-of-zeste、Trihorrax)ドメインはアミノ酸117からアミノ酸246にわたる。
【0018】
SMYD3タンパク質の細胞内局在は、細胞周期の進行中に、および培養細胞の密度によって変化する。SMYD3タンパク質は、細胞がS期中期から後期にある時、または低密度な条件で培養されている時には核に蓄積する。しかしながら、SMYD3タンパク質は、細胞が細胞周期の他の期にある時、または高密度な条件で培養されている時には細胞質ならびに核に局在する。
【0019】
従って、本発明は、SMYD3メチルトランスフェラーゼ活性を調整する作用因子をスクリーニングする方法を提供する。この方法は、SMYD3ポリペプチドまたはメチルトランスフェラーゼ活性を有するその機能的等価物と網膜芽細胞腫タンパク質とを接触させ、接触されたSMYD3またはその機能的等価物のメチルトランスフェラーゼ活性をアッセイすることによって行われる。これによって、SMYD3または機能的等価物のメチルトランスフェラーゼ活性を調整する作用因子が同定される。
【0020】
本発明において、「機能的に等価な」という用語は、本タンパク質またはポリペプチドが、SMYD3と同じメチルトランスフェラーゼ活性または実質的に同じメチルトランスフェラーゼ活性を有することを意味する。特に、このタンパク質は、網膜芽細胞腫タンパク質またはリジン824を含む網膜芽細胞腫タンパク質断片のメチル化を触媒する。本タンパク質が標的活性を有するか否かは、本発明によって日常的に決定することができる。すなわち、メチルトランスフェラーゼ活性は、(a)ポリペプチドと基質(例えば、網膜芽細胞腫タンパク質またはリジン824を含む断片)および補助因子(例えば、S-アデノシル-L-メチオニン)を、基質のメチル化に適した条件下で接触させ、(b)基質のメチル化レベルを検出することによって決定することができる。
【0021】
本明細書で使用する「網膜芽細胞腫ペプチド」という用語は、完全長の網膜芽細胞腫タンパク質(例えば、配列番号:4)ならびにその変異体および断片を意味する。機能的断片の例には、C末端断片、例えば、配列番号:4のアミノ酸769〜921により構成される断片が含まれるが、これに限定されない。好ましい断片は、位置824にあるリジン残基を含む。機能的変異体の例には、完全長網膜芽細胞腫タンパク質のメチル化能を保持している以下のRB1変異体:K889A、K896A、K791A、K814A、K791A/K824A、およびK814A/K824Aが含まれるが、これに限定されない。
【0022】
所定のタンパク質と機能的に等価であるタンパク質を調製する方法は当業者に周知であり、これにはタンパク質に変異を導入する従来の方法が含まれる。例えば、当業者は、例えば部位特異的突然変異を用いてヒトSMYD3タンパク質のアミノ酸配列内に適切な変異を導入することにより、ヒトSMYD3タンパク質と機能的に等価のタンパク質を調製することができる(Hashimoto-Gotoh, T. et al. (1995), Gene 152, 271〜275;Zoller, MJ, and Smith, M. (1983), Methods Enzymol. 100, 468〜500;Kramer, W. et al. (1984), Nucleic Acids Res. 12, 9441〜9456;Kramer W, and Fritz HJ. (1987) Methods. Enzymol. 154, 350〜367;Kunkel, TA (1985), Proc. Natl. Acad. Sci. USA. 82,488〜492)。アミノ酸変異は自然下でも起こり得る。本発明の状況において有用なSMYD3ポリペプチドは、結果的に生じた変異タンパク質がヒトSMYD3タンパク質と機能的に等価であり、より具体的にはヒトSMYD3タンパク質のメチルトランスフェラーゼ活性を保持するという前提で、1つまたは複数のアミノ酸が変異したヒトSMYD3タンパク質のアミノ酸配列を有するタンパク質を含む。このような変異体において変異させるアミノ酸の数は一般に20アミノ酸またはそれ未満、典型的には10アミノ酸またはそれ未満、好ましくは6アミノ酸またはそれ未満、およびより好ましくは3アミノ酸またはそれ未満である。メチルトランスフェラーゼ活性を維持するために、SETドメイン「NHSCXXN」および[GEELXXXY」が、変異タンパク質のアミノ酸配列中に好ましくは保存されている(「X」は任意のアミノ酸を表す)。
【0023】
変異または改変されたタンパク質、すなわち、特定のアミノ酸配列の一つまたは複数のアミノ酸残基を欠失、付加、および/または置換することによって改変されたアミノ酸配列を有するタンパク質は、当初のタンパク質の生物学的活性を保持することが公知である(Mark, D.F. et al.、Proc. Natl. Acad. Sci. USA(1984)81:5662〜5666;Zoller, M.J.およびSmith, M.、Nucleic Acids Research(1982)10:6487〜6500;Wang, A. et al.、Science 224:1431〜1433;Dalbadie-McFarland, G. et al.、Proc. Natl. Acad. Sci. USA(1982)79:6409〜6413)。
【0024】
変異されるアミノ酸残基は、好ましくは、アミノ酸側鎖の特性が保存され得る異なるアミノ酸に変異される(保存的アミノ酸置換として公知の過程)。アミノ酸側鎖の特性の例は、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、および以下の官能基または共通の特徴を有する側鎖:脂肪族側鎖(G、A、V、L、I、P);ヒドロキシル基含有側鎖(S、T、Y);硫黄原子含有側鎖(C、M);カルボン酸およびアミド含有側鎖(D、N、E、Q);塩基含有側鎖(R、K、H);ならびに芳香族含有側鎖(H、F、Y、W)を含む。括弧内の文字はアミノ酸の一文字コードを示すことに注意されたい。
【0025】
一つまたは複数のアミノ酸残基においてヒトSMYD3タンパク質のアミノ酸配列(配列番号:2)に付加されるタンパク質の例は、ヒトSMYD3タンパク質を含む融合タンパク質である。融合タンパク質は、ヒトSMYD3タンパク質と他のペプチドまたはタンパク質との融合体を含み、これらも本発明に用いられる。融合タンパク質は、当業者に周知の技術、例えば本発明のヒトSMYD3タンパク質をコードするDNAを、他のペプチドまたはタンパク質をコードするDNAに、フレームが一致するように連結させ、融合DNAを発現ベクターに挿入し、宿主でそれを発現させることによって作製することができる。本発明のタンパク質に融合させるペプチドまたはタンパク質に関して制限はない。
【0026】
SMYD3タンパク質に融合されるペプチドとして用いることができる公知のペプチドには、例えば、FLAG(Hopp, T.P.et al.,Biotechnology(1986)6:1204〜1210)、6個のHis(ヒスチジン)残基を含む6×His、10×His、インフルエンザ凝集素(HA)、ヒトc-myc断片、VSP-GP断片、p18HIV断片、T7-タグ、HSV-タグ、E-タグ、SV40T抗原断片、lckタグ、α-チューブリン断片、B-タグ、プロテインC断片等が含まれる。本発明のタンパク質に融合し得るタンパク質の例には、GST(グルタチオン-S-トランスフェラーゼ)、インフルエンザ凝集素(HA)、免疫グロブリン定常領域、β-ガラクトシダーゼ、MBP(マルトース結合タンパク質)等が含まれる。
【0027】
融合タンパク質は、上記の融合ペプチドまたはタンパク質をコードする市販のDNAを、本発明のタンパク質をコードするDNAと融合させて、調製された融合DNAを発現させることによって調製することができる。
【0028】
機能的に等価なタンパク質を単離するための当技術分野で公知のもう一つの方法は、相同的な配列を同定するためのハイブリダイゼーション技術(Sambrook, J.et al.、Molecular Cloning、第2版、9.47〜9.58、コールドスプリングハーバー研究所出版、1989)を用いる方法である。当業者は、ヒトSMYD3タンパク質をコードするSMYD3 DNA配列(例えば、配列番号:1)の全て、または一部と高い相同性を有するDNAを容易に単離することができ、単離されたDNAからヒトSMYD3タンパク質と機能的に等価なタンパク質を単離することができる。本発明のタンパク質には、ヒトSMYD3タンパク質をコードするDNA配列の全てまたは一部とハイブリダイズするDNAによってコードされるタンパク質、およびヒトSMYD3タンパク質と機能的に等価であるタンパク質が含まれる。これらのタンパク質には、ヒトまたはマウスに由来するタンパク質に対応する哺乳類相同体が含まれる(例えば、サル、ラット、ウサギ、およびウシ遺伝子によってコードされるタンパク質)。ヒトSMYD3タンパク質をコードするDNAと高い相同性を示すcDNAを動物から単離する場合、骨格筋、精巣、HCC、または結腸直腸腫瘍由来の組織を用いることが特に好ましい。
【0029】
ヒトSMYD3タンパク質と機能的に等価であるタンパク質をコードするDNAを単離するためのハイブリダイゼーション条件は、当業者によって日常的に選択することができる。例えば、ハイブリダイゼーションは、「Rapid-hyb緩衝液」(Amersham LIFE SCIENCE)を用いて、68℃で30分またはそれ以上プレハイブリダイゼーションを行い、標識プローブを加えて、68℃で1時間またはそれ以上加温することによって行ってもよい。次に続く洗浄段階は、例えば低ストリンジェント条件で行うことができる。低ストリンジェンシー条件は、例えば、42℃、2×SSC、0.1%SDS、または好ましくは50℃、2×SSC、0.1%SDSである。より好ましくは高ストリンジェント条件を用いる。本発明の状況において、高ストリンジェント条件は、例えば、2×SSC、0.01%SDS中で室温20分間の洗浄を3回行い、次いで1×SSC、0.1%SDS中で37℃20分間の洗浄を3回行い、かつ1×SSC、0.1%SDS中で50℃20分間の洗浄を2回行うことを含む。しかしながら、温度および塩濃度のようないくつかの要因がハイブリダイゼーションのストリンジェンシーに影響を及ぼす可能性があり、当業者は必須のストリンジェンシーを達成するために要因を適切に選択することができる。
【0030】
ハイブリダイゼーションの代わりに、遺伝子増幅法、例えば、ヒトSMYD3タンパク質(配列番号:2)をコードするDNA(配列番号:1)の配列情報に基づいて合成されたプライマーを用い、ポリメラーゼ連鎖反応(PCR)法を用いて、ヒトSMYD3タンパク質と機能的に等価なタンパク質をコードするDNAを単離することができる。
【0031】
上記のハイブリダイゼーション技術または遺伝子増幅技術によって単離されたDNAによってコードされる、ヒトSMYD3タンパク質と機能的に等価であるタンパク質は、通常、ヒトSMYD3タンパク質のアミノ酸配列に対して高い相同性を有する。「高い相同性」(「高い同一性」とも呼ばれる)とは典型的に、2つの最適に整列化された配列(ポリペプチド配列またはポリヌクレオチド配列)間の同一性の程度を指す。典型的には、高い相同性または同一性とは、40%またはそれ以上、好ましくは60%またはそれ以上、より好ましくは80%またはそれ以上、さらにより好ましくは85%、90%、95%、98%、99%またはそれ以上の相同性を指す。2つのポリペプチド配列またはポリヌクレオチド配列の間の相同性または同一性の程度は、「Wilbur, W. J. and Lipman, D. J. Proc.Natl. Acad.Sci. USA (1983) 80, 726〜730」におけるアルゴリズムに従って決定することができる。
【0032】
本発明の状況において有用なタンパク質は、その産生のために用いられる細胞もしくは宿主または用いる精製方法に依存して、アミノ酸配列、分子量、等電点、糖鎖の有無または形態に関して差異がある可能性がある。しかしながら、それがヒトSMYD3タンパク質(配列番号:2)の機能と等価である限り、それは本発明において有用である。
【0033】
本発明の状況において有用なタンパク質は、当業者に周知の方法によって、組換え型タンパク質または天然のタンパク質として調製することができる。組換え型タンパク質は、本発明のタンパク質をコードするDNA(例えば、配列番号:1のヌクレオチド配列を含むDNA)を、適当な発現ベクターに挿入し、ベクターを適当な宿主細胞に導入し、抽出物を得、かつ抽出物にクロマトグラフィー、例えばイオン交換クロマトグラフィー、逆相クロマトグラフィー、ゲル濾過、または本発明のタンパク質に対する抗体を固定したカラムを利用するアフィニティクロマトグラフィーを行うか、または上記のカラムの複数を組み合わせることにより、タンパク質を精製することによって調製され得る。
【0034】
さらに、本発明の状況において有用なタンパク質が宿主細胞(例えば、動物細胞および大腸菌(E.coli))内において、グルタチオン-S-トランスフェラーゼタンパク質との融合タンパク質として、または多数のヒスチジンを加えた組換え型タンパク質として発現する場合、発現された組換え型タンパク質は、グルタチオンカラムまたはニッケルカラムを用いて精製することができる。
【0035】
融合タンパク質を精製した後、必要に応じてトロンビンまたは第Xa因子による切断によって、目的とするタンパク質以外の領域を除去することも可能である。
【0036】
天然のタンパク質は、当業者に公知の方法、例えば下記のSMYD3タンパク質に結合する抗体を結合させたアフィニティカラムを、本発明のタンパク質を発現する組織または細胞の抽出物に接触させることによって単離することができる。抗体はポリクローナル抗体またはモノクローナル抗体であり得る。
【0037】
本発明において、SMYD3ポリペプチドのメチルトランスフェラーゼ活性は、当技術分野で公知の方法によって決定することができる。例えば、SMYD3ポリペプチド基質および網膜芽細胞腫ペプチド基質は、標識されたメチル供与体と共に、適したアッセイ条件下でインキュベートされ得る。好ましいメチル供与体の例は、S-アデノシル-[メチル-14C]-L-メチオニンおよびS-アデノシル-[メチル-3H]-L-メチオニンを好ましくは含むが、これらに限定されない。放射性標識の網膜芽細胞腫ペプチドへの移行は、例えば、SDS-PAGE電気泳動およびフルオログラフィーによって検出することができる。または、反応に続き、網膜芽細胞腫ペプチドを濾過によってメチル供与体から分離し、かつフィルター上に保持された放射標識の量をシンチレーション計数法によって定量することができる。メチル供与体と結合し得る他の標識、例えば発色性標識および蛍光標識、ならびにこれらの標識の網膜芽細胞腫ペプチドへの移行を検出する方法は当技術分野で公知である。
【0038】
または、SMYD3のメチルトランスフェラーゼ活性を、非標識メチル供与体(例えば、S-アデノシル-L-メチオニン)、およびメチル化網膜芽細胞腫ペプチドを選択的に認識する試薬を用いて決定することができる。例えば、SMYD3、メチル化しようとする基質、およびメチル供与体を、基質のメチル化に好適な条件下でインキュベートした後、メチル化基質を従来の免疫学的方法を用いて検出することができる。メチル化基質を認識する抗体を用いる任意の免疫学的技術を検出のために用いることができる。
【0039】
さらに、Lys824がメチル化されたRB1において、RB1のSer807およびSer811のリン酸化が増強されたことが確認された。従って、別の態様において、RB1のメチル化レベルはRB1のリン酸化を介して評価されてもよい。CDK2またはCDK6などのキナーゼもRB1のリン酸化に必要なことがある。RB1のリン酸化は、放射標識リン酸供与体を用いて検出されてもよい。あるいは、RB1のリン酸化部位を認識する抗体がRB1のリン酸化レベルの評価に用いられてもよい。
【0040】
さまざまなロースループットおよびハイスループット酵素アッセイ形式が当技術分野で公知であり、かつSMYD3のメチルトランスフェラーゼ活性の検出または測定のために容易に適合させることができる。ハイスループットアッセイ法のために、網膜芽細胞腫ペプチド基質は、マルチウェルプレート、スライドガラス、またはチップなどの固体支持体上に、都合良く固定化され得る。反応に続き、メチル化産物を固体支持体上で上記の方法によって検出することができる。または、メチルトランスフェラーゼ反応は溶液中で行われ得、その後網膜芽細胞腫ペプチドが固体支持体上に固定化され得、およびメチル化産物が検出される。そのようなアッセイ法を容易にするため、固体支持体はストレプトアビジンでコーティングされ得、かつビオチン標識網膜芽細胞腫、または固体支持体は抗網膜芽細胞腫抗体でコーティングされ得る。当業者は、スクリーニングの所望のスループット能力に依存して、適したアッセイ形式を決定することができる。
【0041】
本発明はまた、本発明のタンパク質の部分的ペプチドの使用方法も含む。部分的ペプチドは、SMYD3のタンパク質に特異的なアミノ酸配列を有し、かつ約400アミノ酸未満、通常は約200アミノ酸未満、およびしばしば約100アミノ酸未満であり、ならびに少なくとも約7アミノ酸、好ましくは約8アミノ酸またはそれ以上、およびより好ましくは約9アミノ酸またはそれ以上からなる。部分的ペプチドは、例えば、SMYD3のタンパク質と結合する作用因子または化合物のスクリーニングにおいて、および、SMYD3とその補助因子、例えば、SAMなど、との間の結合の阻害物質のスクリーニングために用いられ得る。そのようなスクリーニングのために、好ましくはSETドメインを含む部分的ペプチドが用いられる。
【0042】
本発明の状況において有用な部分的ペプチドは、遺伝子操作により、ペプチド合成の公知の方法により、または本発明のタンパク質を適切なペプチダーゼで消化することにより作製され得る。ペプチド合成のために、例えば、固相合成または液相合成が用いられ得る。
【0043】
SETドメインの変異を有するSMYD3変異体は、細胞増殖に対する阻害作用を示す。したがって、SMYD3の部分的ペプチドは、好ましくは、SETドメイン「NHSCXXN」および/または「GEELXXXY」を含む。
【0044】
任意の試験作用因子が用いられ得る。例として、細胞抽出物、細胞培養上清、発酵微生物の産物、海洋生物からの抽出物、植物抽出物、精製タンパク質または粗タンパク質、ペプチド、非ペプチド化合物、合成マイクロ分子化合物および天然化合物を含むが、これらに限定されない。
【0045】
本明細書に記載したアッセイ法において有用な試験作用因子または試験化合物は、SMYD3と特異的に結合する抗体、またはメチルトランスフェラーゼ活性を持たない部分的SMYD3ペプチドという形をとり得る。例えば、抗体(例えば、モノクローナル抗体)は、SMYD3とその網膜芽細胞腫基質との間の結合をブロックする能力に関して試験され得る。
【0046】
本発明のスクリーニング方法によって単離された作用因子または化合物は、SMYD3のメチルトランスフェラーゼ活性を阻害する薬剤の候補であり、それゆえ、肝細胞癌、結腸直腸癌、乳癌、および/または膀胱癌の治療または予防に応用することができる。
【0047】
さらに、SMYD3のメチルトランスフェラーゼ活性を阻害する作用因子または化合物の構造の一部が、付加、欠失および/または置換によって変換された作用因子または化合物も、本発明のスクリーニング方法によって得られる作用因子および化合物に含まれる。
【0048】
上に注記したように、SMYD3のメチルトランスフェラーゼ活性を阻害する作用因子または化合物は、SMYD3のメチルトランスフェラーゼ活性を欠く部分的ペプチドのいずれかであり得、またはSMYD3に対する抗体であり得る。本明細書で用いる場合、「抗体」という用語は、抗体を合成するために用いた抗原またはそれと密接に関連した抗原のみと相互作用する(すなわち、結合する)、特異的な構造を有する免疫グロブリン分子のことを指す。さらに、抗体は、SMYD3遺伝子によってコードされるタンパク質に結合する限り、抗体または修飾された抗体の断片であってもよい。例えば、抗体断片は、Fab、F(ab')2、Fv、またはH鎖およびL鎖からのFv断片が適当なリンカーによって連結されている一本鎖Fv(scFv)であってもよい(Huston, J.S.et al.、Proc. Natl. Acad. Sci. U.S.A. 85:5879〜5883(1988))。より具体的には、抗体断片は、パパインまたはペプシンのような酵素によって抗体を処理することによって生成してもよい。または、抗体断片をコードする遺伝子を構築して、発現ベクターに挿入し、適当な宿主細胞において発現してもよい(例えば、Co, M.S.et al.、J. Immunol. 152:2968〜2976(1994);Better, M.およびHorwitz, A.H.、Methods Enzymol. 178:476〜496(1989);Pluckthun, A.およびSkerra, A.、Methods Enzymol. 178:497〜515(1989);Lamoyi, E.、Methods Enzymol. 121:652〜663(1986);Rousseaux, J.et al.、Methods Enzymol. 121:663〜669(1986);Bird, R.E.およびWalker, B.W.、Trends Biotechnol. 9:132〜137(1991)を参照されたい)。
【0049】
抗体を、ポリエチレングリコール(PEG)のなどの多様な分子との結合によって修飾してもよい。本発明は、そのような修飾された抗体を提供する。修飾された抗体は、抗体を化学修飾することによって得ることができる。そのような修飾法は当技術分野において一般的である。または、抗体を、非ヒト抗体に由来する可変領域およびヒト抗体に由来する定常領域を有するキメラ抗体、または、非ヒト抗体に由来する相補性決定領域(CDR)、ヒト抗体に由来するフレームワーク領域(FR)、および定常領域を含むヒト化抗体を含み得る。そのような抗体は、公知の技術を用いて調製することができる。ヒト化は、ヒト抗体の対応する配列の代わりに、齧歯類のCDRまたはCDR配列を用いることによって実施され得る(例えば、Verhoeyen et al.、 Science 239:1534〜1536 (1988)を参照されたい)。したがって、そのようなヒト化抗体は、実際には、ヒト可変ドメインの全域より狭い領域が、非ヒト種由来の対応する配列によって置き換えられたキメラ抗体である。
【0050】
ヒトフレームワーク領域および定常領域に加えて、ヒト可変領域を含む完全ヒト抗体もまた、用いることができる。そのような抗体は、当技術分野で公知のさまざまな技術を用いて作製することができる。例えば、バクテリオファージ上に提示されたヒト抗体断片の組換えライブラリーの使用を含むインビトロの方法を用いてもよい(例えば、Hoogenboom & Winter, J. Mol. Biol. 227:381 (1991))。同様に、ヒト免疫グロブリン座位を、内因性免疫グロブリン遺伝子が部分的または完全に不活性化されたトランスジェニック動物、例えばマウスに導入することによってヒト抗体を作製することができる。このアプローチは例えば、米国特許第6,150,584号、第5,545,807号;第5,545,806号;第5,569,825号;第5,625,126号;第5,633,425号;第5,661,016号に記載されている。
【0051】
本発明の方法によって単離された作用因子または化合物を、ヒトおよび他の哺乳動物、例えばマウス、ラット、モルモット、ウサギ、ネコ、イヌ、ヒツジ、ブタ、ウシ、サル、ヒヒ、およびチンパンジーに対して製剤として投与する場合、単離された作用因子または化合物を直接投与してもよく、または公知の薬学的調製法を用いて剤形に製剤化してもよい。例えば、必要に応じて、薬剤を糖衣錠剤、カプセル剤、エリキシル剤、およびマイクロカプセルとして経口投与することもでき、または水もしくは他の任意の薬学的に許容される液体との滅菌溶液もしくは懸濁液である注射剤の形態として非経口的に投与することもできる。例えば、作用因子または化合物を、一般に認められる薬剤の実現のために必要な単位用量の形で、薬学的に許容される担体または媒体、具体的には滅菌水、生理食塩水、植物油、乳化剤、懸濁剤、界面活性剤、安定剤、香味剤、賦形剤、媒質、保存料、結合剤などと混合することができる。これらの製剤における有効成分の量により、指定された範囲内にある適した投与量が得られる。
【0052】
錠剤およびカプセル剤に混合し得る添加剤の例には、ゼラチン、コーンスターチ、トラガカントゴムおよびアラビアゴムなどの結合剤;結晶セルロースなどの媒質;コーンスターチ、ゼラチンおよびアルギン酸などの膨潤剤;ステアリン酸マグネシウムなどの潤滑剤;スクロース、ラクトースまたはサッカリンなどの甘味料;ならびにペパーミント、冬緑油、およびチェリーなどの香味剤がある。単位用量剤形がカプセル剤である場合、油などの液体担体も上記の成分にさらに含めることができる。注射用の滅菌混合物は、通常の薬剤の実現に倣って、注射用の蒸留水などの媒体を用いて製剤化することができる。
【0053】
生理食塩水、グルコース、ならびにD-ソルビトール、D-マンノース、D-マンニトールおよび塩化ナトリウムなどのアジュバントを含むその他の等張液を、注射用の水溶液として用いることができる。これらは適した可溶化剤、例えばアルコール、具体的にはエタノール、プロピレングリコールおよびポリエチレングリコールなどの多価アルコール、Polysorbate 80(商標)およびHCO-50などの非イオン性界面活性剤などと組み合わせて用いることができる。
【0054】
ゴマ油またはダイズ油は油脂性液体として用いることができ、これらを可溶化剤としての安息香酸ベンジルまたはベンジルアルコールと組み合わせて用いることもでき、さらにこれらをリン酸緩衝液および酢酸ナトリウム緩衝液などの緩衝液;塩酸プロカインなどの鎮痛薬;ベンジルアルコールおよびフェノールなどの安定剤;ならびに抗酸化剤と共に製剤化することもできる。調製された注射剤は適したアンプルに充填することができる。
【0055】
本発明の薬学的組成物を、例えば動脈内注射、静脈内注射、または皮下注射として、および同じく鼻腔内投与、筋肉内投与または経口投与として患者に投与するために、当業者に周知の方法を用いることができる。投与量および投与方法は患者の体重および年齢ならびに投与方法に応じて異なる。しかしながら、当業者は日常的に適切な投与方法を選択することができる。さらに、関心対象の前記作用因子または前記化合物がDNAによってコードされ得る場合、DNAを遺伝子治療用のベクターに挿入し、治療を行うためにそのベクターを患者に投与することができる。投与量および投与方法は患者の体重、年齢および症状に応じて異なるが、当業者はそれらを適切に選択することができる。
【0056】
例えば、SMYD3と結合してその活性を調節する作用因子または化合物の典型的な投与量は、その症状に依存するとはいえ、標準的な成人(体重60 kg)に対して経口投与する場合、約0.1 mg〜約100 mg/日、好ましくは約1.0 mg〜約50 mg/日、およびより好ましくは約1.0 mg〜約20 mg/日の範囲である。
【0057】
標準的な成人(体重60 kg)に対して注射剤の形態として非経口的に投与する場合、患者、標的臓器、症状および投与方法に応じて若干の違いはあるものの、約0.01 mg〜約30 mg/日、好ましくは約0.1〜約20 mg/日、およびより好ましくは約0.1〜約10 mg/日の用量を静脈注射することが好都合である。同じく、他の動物の場合にも、体重60 kgに換算した量を投与することが可能である。
【0058】
本発明はさらに、対象における、肝細胞癌、結腸直腸癌、膀胱癌、および乳癌などの、癌を治療するための方法を提供する。投与は、SMYD3の異常なメチルトランスフェラーゼ活性と関連する障害に対するリスクのある(もしくは感受性のある)またはそのような障害を有する対象に対して、予防的または治療的に行うことができる。本方法は、適当な癌細胞におけるSMYD3の機能を低下させる段階を含む。機能は、本発明のスクリーニング方法によって得られた作用因子または化合物の投与によって阻害することができる。
【0059】
もう一つの局面において、本発明は、本明細書に記載の1つまたは複数の治療的作用因子または治療的化合物を含む、薬学的または治療的な組成物を含む。または、本発明は、癌、より具体的には、肝細胞癌、結腸直腸癌、膀胱癌、および乳癌の治療および/もしくは予防のための薬学的または治療的な組成物を製造するための、本明細書に記載の1つまたは複数の治療的作用因子または治療的化合物の使用方法を提供する。薬学的製剤には、経口、直腸、鼻、局所(頬および舌下を含む)、膣、もしくは非経口(筋肉内、皮下、および静脈内を含む)の投与に適したもの、または吸入もしくは通気による投与に適したものが含まれ得る。製剤は、適宜、個別の投与量単位で都合良く提示でき、薬学分野における周知の方法のいずれかによって調製できる。そのような薬学の方法は、全て、必要に応じて、液状担体もしくは微細に粉砕された固形担体またはその両方と活性化合物とを会合させる段階を含み、かつ必要であれば、次いで、その生成物を所望の製剤へと成形する段階を含む。
【0060】
経口投与に適した薬学的製剤は、所定の量の活性成分を各々含有しているカプセル剤、カシェ剤、もしくは錠剤のような個別の単位として;散剤もしくは顆粒剤として;または液剤、懸濁剤、もしくは乳濁剤として、都合良く提示できる。活性成分は、ボーラス舐剤またはペースト剤として提示してもよく、そして純粋な(即ち、担体を含まない)形態であってもよい。経口投与用の錠剤およびカプセル剤は、結合剤、増量剤、滑沢剤、崩壊剤、または湿潤剤のような従来の賦形剤を含有し得る。錠剤は、任意で、1個以上の製剤化成分を用いて、圧縮または成型により作成してもよい。圧縮錠剤は、結合剤、滑沢剤、不活性希釈剤、滑沢、界面活性、または分散剤と任意に混合された散剤または顆粒剤のような流動性の高い形態の活性成分を適当な機械で圧縮することにより調製できる。成型錠剤は、不活性液状希釈剤で湿らされた粉末化合物の混合物を適当な機械で成型することにより作成できる。錠剤は、当技術分野における周知の方法に従い、剤皮を施してもよい。経口液状調製物は、例えば、水性もしくは油性の懸濁剤、液剤、乳濁剤、シロップ剤、もしくはエリキシル剤の形態であってもよいし、または乾燥生成物として提示し、使用前に水もしくはその他の適当な媒体で調製してもよい。そのような液状調製物は、懸濁化剤、乳化剤、非水性媒体(食用油を含み得る)、または保存剤のような従来の添加剤を含有し得る。錠剤は、任意で、含まれる活性成分を徐放するようにまたは放出が制御されるように製剤化してもよい。
【0061】
非経口投与用の製剤には、抗酸化剤、緩衝剤、静菌剤、および対象となるレシピエントの血液と等張にする溶質を含有し得る水性および非水性の無菌の注射液剤;ならびに懸濁化剤および濃化剤を含み得る水性および非水性の無菌の懸濁剤が含まれる。製剤は、単位用量容器または多回用量容器、例えば、密封されたアンプルおよびバイアルで提示でき、使用直前に無菌の液状担体、例えば、生理食塩水、注射用水の添加のみを必要とする、フリーズドライ(凍結乾燥)状態で保管してもよい。または、製剤は連続注入用に提示してもよい。即席の注射用の液剤および懸濁剤は、前記の種類の無菌の散剤、顆粒剤、および錠剤から調製できる。
【0062】
直腸投与用の製剤は、カカオバターまたはポリエチレングリコールのような通常の担体を含む坐剤として提示できる。口内、例えば、頬または舌下への局所投与用の製剤には、ショ糖およびアラビアゴムもしくはトラガカントゴムのような風味付きの基剤中に活性成分が含まれたロゼンジ剤、ならびにゼラチンおよびグリセリンもしくはショ糖およびアラビアゴムのような基剤中に活性成分が含まれた香錠が挙げられる。鼻腔内投与の場合、本発明の化合物は、液状スプレーもしくは分散可能な散剤として、または滴剤の形態で使用できる。滴剤は、1つ以上の分散剤、可溶化剤、または懸濁化剤も含む水性または非水性の基剤を用いて製剤化できる。液状スプレーは、加圧パックから都合良く送達される。
【0063】
吸入による投与の場合、化合物は、注入器、ネブライザー、加圧パック、またはエアロゾルスプレーを送達するその他の便利な手段から都合良く送達される。加圧パックは、ジクロロジフルオロメタン、トリクロロフルオロメタン、ジクロロテトラフルオロエタン、二酸化炭素、またはその他の適当なガス等の適当な噴霧剤を含み得る。加圧エアロゾル剤の場合、投与量単位はバルブによって決定され、計量された量が送達される。
【0064】
または、吸入または通気による投与の場合、化合物は、乾燥粉末組成物、例えば、化合物と乳糖またはデンプンのような適当な粉末基剤との粉末混合物の形態をとり得る。粉末組成物は、単位剤形、例えば、カプセル、カートリッジ、ゼラチン、またはブリスター包装で提示でき、それらから、吸入器または注入器を使用して粉末を投与できる。
【0065】
所望の場合、活性成分を徐放するよう適合させた上記の製剤が利用されてもよい。薬学的組成物は、抗微生物剤、免疫抑制剤、または保存剤のようなその他の活性成分を含有していてもよい。
【0066】
特に上で言及した成分に加え、本発明の製剤には、問題となる製剤の型を考慮した上で、当技術分野において一般的なその他の薬剤が含まれていてもよいこと、例えば、経口投与に適した薬剤には着香料が含まれていてもよいことが理解される。
【0067】
好ましい単位投与量製剤は、下記のような有効量またはその活性成分の適切な画分を含有しているものとする。
【0068】
前記の各条件のため、組成物は、1日当たり約0.1〜約250 mg/kgの用量で経口的にまたは注射により投与され得る。成人のヒトに対する用量範囲は、一般に、約5 mg〜約17.5 g/日、好ましくは約5 mg〜約10 g/日、最も好ましくは約100 mg〜約3 g/日である。個別の単位で提供される錠剤またはその他の単位剤形には、そのような投与量またはその多回数の投与量で有効量となる量、例えば、約5 mg〜約500 mg、通常約100 mg〜約500 mgを含有する単位が都合良く含まれる。
【0069】
薬学的組成物は、好ましくは、経口的にまたは注射(静脈内もしくは皮下)により投与され、対象へ投与される正確な量は、担当医師の責任であろう。しかしながら、用いられる用量は、対象の年齢および性別、治療が施される正確な障害およびその重傷度を含む多数の要因によるものと考えられる。また、投与経路も、状態およびその重症度によって変動し得る。
【0070】
以下の実施例は単なる例示であり、本発明の範囲を限定することを目的としない。本発明の局面を以下の実施例において説明するが、当業者であれば、本明細書に記載のものと類似のまたは等価な他の方法および材料を、本発明の実施または試験において使用できることを理解するだろう。
【0071】
実施例
材料および方法:
試薬:
抗RB(IF8)抗体、抗リン酸化RB(Ser807/811,sc-16670)抗体、および抗リン酸化RB(Thr821/826)抗体はSanta Cruz Biotechnologyから購入し、抗Flag抗体はSIGMAから購入し、抗パンメチルリジン抗体(ab7315)は Abcam Ltd.から購入した。組換えSMYD3タンパク質、またはジメチル化リジン824もしくはトリメチル化リジン824を含有する合成RB1ペプチド(残基820〜828)をウサギ(SIGMA-ALDRICH、St.Louis、MO)に接種し、免疫したウサギの血清からポリクローナル抗体を精製した。組換えC末端GST-RB1および完全長GST-p53タンパク質はSanta Cruz Biotechnologyから購入し、His結合C末端RB1、CDK2/サイクリンE、およびCDK6/サイクリンD3タンパク質はUpstate Biotechnologyから購入し、完全長組換えRBタンパク質(3108)はQED Bioscienceから購入した。S-(5'-アデノシル)-L-ホモシステイン加水分解酵素(SAHH)はSIGMAから入手した。
【0072】
メチルトランスフェラーゼおよびキナーゼのインビトロアッセイ法:
293T細胞を、Flagタグ化野生型SMYD3を発現するプラスミド(p3XFLAG-CMV-SMYD3)、Flagタグ化変異体SMYD3を発現するプラスミド(p3XFLAG-CMV-SMYD3ΔEEL)でトランスフェクトし、抗Flag抗体を用いた免疫沈降によってタグ化SMYD3タンパク質を精製した。バキュロウイルス系(Clontech)を用いてSf9細胞において、組換えSMYD3タンパク質を調製した。インビトロHMTアーゼアッセイ法は、他の記載に(Strahl, B. D., et al. Proc Natl Acad Sci U S A 96,14967-72 (1999))、軽微な変更を加えて実施した。簡単に述べると、メチルトランスフェラーゼ緩衝液(50mM Tris-HCl pH8.5、100mM NaCl、10mM DTT)の緩衝液中に、免疫沈降したSMYD3タンパク質または組換えSMYD3タンパク質を、メチル供与体として2μCiの[メチル-3H]標識S-アデノシル-L-メチオニン(SAM, Amersham Biosciences)の存在下で、1μgの組換えヒストンH3、RB1、またはp53タンパク質と混合した。反応混合物を30℃で1時間インキュベートした。タンパク質をSDS-PAGEで分離し、標識されたタンパク質をフルオログラフィーにより検出した。CDK2/サイクリンEおよびCDK6/サイクリンD3のインビトロキナーゼアッセイ法は、製造業者(Upstate Biotechnology)のプロトコールに従って実施した。非メチル化RB1およびメチル化RB1(#12-439、Upstate Biotechnology)の両方を反応基質として使用した。
【0073】
インビボメチル化アッセイ法:
メチル化RB1をインビボで測定するために、LiuおよびDreyfuss (Liu, Q. & Dreyfuss, G. Mol Cell Biol 15, 2800-8 (1995))により記載の方法に従い、軽微な変更を加えて、RB1のインビボ標識を[メチル-3H]標識S-アデノシル-L-メチオニンを用いて培養細胞において実施した。HEK293細胞を、100μg/mlのシクロヘキシミドおよび40μg/mlのクロラムフェニコールと共に37℃で30分間インキュベートした。次いで、培地を、10μCi/mlのL-[メチル-3H]メチオニンおよびタンパク質合成阻害剤を含有し非標識メチオニンを含有しない培地と交換し、さらに3時間維持した。全細胞溶解産物を、抗RB抗体(IF8; Santa Cruz Biotechnology)を用いた免疫沈降に供した。免疫沈降したRB1タンパク質をSDS-PAGEで分離し、その後に、ニトロセルロース膜に転写した。ニトロセルロース膜を、BASイメージングシステム(BAS-TR2040、FUJI)またはイムノブロット分析で分析した。
【0074】
免疫細胞化学染色:
チャンバースライド上の培養細胞を、4%パラホルムアルデヒド含有PBSで15分間固定し、次いで、0.1%Triton X-100含有PBSを用いて室温で2.5分間、透過性にした。非特異的ハイブリダイゼーションをブロックするために、細胞を、PBSに溶解した2%BSAを用いて4℃で24時間覆い、次いで、一次抗体として抗SMYD3抗体、抗RB[IF8]抗体、および抗リン酸化RB(Thr 821/826)抗体と共にインキュベートした。二次抗体として、蛍光基質結合抗ウサギまたは抗マウスIgG(Molecular Probes)を使用した。核は、4',6-ジアミジノ-2-フェニルインドール二塩酸塩(DAPI)を用いて対比染色した。TCS-SP2共焦点顕微鏡(Leica)を用いて、蛍光画像を得た。
【0075】
ルシフェラーゼアッセイ法:
ルシフェラーゼアッセイ法は、Dual-Luciferase Reporter Assay Systemを製造業者の説明書(Promega)に従って使用して実施した。
【0076】
細胞株および組織標本:
ヒト胚腎臓293(HEK293)細胞、HEK293T細胞、およびHEK293F細胞はIWAKIから購入した。ヒト肝癌細胞株HepG2、ならびにHCT116およびSW480ヒト結腸癌株は、American Type Culture Collection (ATCC)から入手した。ヒトHCC細胞株SNU423は、韓国細胞株バンクからの贈与物であった。T47DおよびMCF7乳癌細胞株は、財団法人癌研究会癌研究所から厚意により提供された。全ての細胞株を適切な培地において単層で増殖させた。原発性乳癌組織は、患者からインフォームドコンセントを得て入手した(Hamamoto, R. et al. Cancer Sci 97, 113-118 (2006))。
【0077】
プラスミドの調製:
C末端FLAGタグ化SMYD3の調製は以前に説明されている(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004))。さらに、本発明者らは、野生型SMYD3 cDNAかまたは欠失型SMYD3 cDNAかを含む様々なPCR産物をpCMV-HA(Clontech)ベクターまたはp3XFLAG-CMV 14(Sigma)ベクターの適切な部位にクローニングすることによって、N末端HAタグ化SMYD3またはN末端3xFLAGタグ化SMYD3を発現するプラスミドを調製した。野生型プラスミドに使用したプライマーは、5’-AAGCTTGCGGCCGCGATGGAGCCGCTGAAGGTGGAAAAG-3’(配列番号: 5)および5’-GGTACCTCTAGATTAGGATGCTCTGATGTTGGCGTC-3’(配列番号: 6)であり、変異体(FLAG-SMYD3-ΔN44、-ΔN99、-ΔN244、および-Δ34)に使用したプライマーは、それぞれ、5’-GGGGTACCTTAGGATGCTCTGATGTTGGCGTC-3’(配列番号: 7)および5’-CGGAATTCTGGCGCGATGGAGCCGCTGAAGGTGGAAAAG-3’(配列番号: 8)、 5’-CGGAATTCTGACTCCGTTCGACTTCTTGGCAG-3’(配列番号: 9)、5’- CGGAATTCTCGGAAGCAGCTGAGGGACCAGTACTGC-3’(配列番号: 10)、または5’-CGGAATTCACCCTTGGCGTACACGGTGTGCAAGG-3’(配列番号: 11)であった。グリシン15、17、または27での置換を発現する変異体プラスミドは、QuikChange II XL部位特異的変異誘発キットを使用して供給業者のプロトコール(Stratagene、California、USA)に従って調製した。
【0078】
ウエスタンブロット分析:
SMYD3に対するポリクローナル抗体は、他に記載されているように、大腸菌において産生された組換えHisタグ化SMYD3タンパク質で免疫したウサギの血清から精製した。タンパク質を10%SDS-PAGEにより分離し、抗SMYD3抗体、抗HA抗体(Sigma)、抗FLAG抗体(Sigma)、抗GST抗体(Pharmingen)、または抗β-アクチン抗体(Sigma)を用いてイムノブロットした。HRP結合抗ウサギIgG、抗マウスIgG(Amersham Biosciences)、または抗ヤギIgG抗体(Santa Cruz)は、ECL Detection System (Amersham)用の二次抗体とした。
【0079】
切断部位の決定:
C末端FLAGタグ化SMYD3を293F細胞において外因的に発現させた。抗FLAG抗体を用いて細胞から免疫沈降させたSMYD3タンパク質を2枚のSDS-PAGEゲルで分離し、ニトロセルロース膜およびシークエンスグレードPVDF膜に転写した。2種類のSMYD3タンパク質を検出するために、ニトロセルロース膜を、抗FLAG抗体を用いたイムノブロット分析に使用した。本発明者らは、PVDF膜を、酢酸を含まないCBB溶液(0.025%CBBを40%メタノールに溶解したもの)で染色した後に、短いSMYD3に対応するバンドを切り出し、アミノ酸シークエンスに供した。タンパク質のアミノ酸配列は、エドマンアミノ酸シークエンス法(Edman amino acid sequence method)(Shimadzu Biotechnologies、Tokyo、Japan)によって決定した。
【0080】
インビトロヒストンメチルトランスフェラーゼ(HMTアーゼ)アッセイ:
FLAGタグ化SMYD3は、抗FLAG抗体を用いた免疫沈降によって、野生型(p3XFLAG-CMV-SMYD3)を発現する293T細胞または変異体SMYD3(p3XFLAG-ΔN34、-ΔN44、-SETNm1、-SETNm2、および-SETNm3)を発現する293T細胞から精製した。GST融合SMYD3タンパク質は、野生型(GST-SMYD3-wt)を発現する細菌細胞または変異体SMYD3構築物(GST-SMYD3-ΔN9、-ΔN19、-ΔN29、-ΔN44、-ΔN74)を発現する細菌細胞から精製した。インビトロHMTアーゼアッセイ法は、他に記載されているように(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004))実施した。3H-放射能は、液体シンチレーションカウンターで測定した。
【0081】
実施例1:SMYD3の基質としてのRB1
最近の2つの報告から、ヒストンH3-K4メチルトランスフェラーゼSET7/9が基質としてTAF10およびp53を触媒することが示されたため(Chuikov, S. et al., Nature 432, 353-60 (2004))、本発明者らは、ヒストンH3以外のSMYD3(GenBankアクセッション番号AB057595;配列番号;1、2)のさらなる基質を探索した。p53およびRB1は細胞周期進行の周知の制御因子であるので、最初に、これらを候補基質として試験した(GenBankアクセッション番号NM_000321;配列番号;3、4)。研究中に、組換えヒストンH3、野生型p53、およびRB1のC末端領域(コドン769-921)を、メチル供与体である3H標識SAMの存在下で、293T細胞から免疫沈降したSMYD3タンパク質と共にインキュベートした。それに続くPAGEおよびオートラジオグラフィーから、メチル化ヒストンH3に対応するバンドが示された。これは、SMYD3はヒストンH3をメチル化するという知見と一致する。興味深いことに、メチル化RB1に対応するバンドも検出された。しかしながら、メチル化p53に対応するバンドは検出されなかった(図1a)。ヒストンH3およびC末端RB1に対するメチルトランスフェラーゼ(MTアーゼ)活性を、組換えSMYD3タンパク質を用いてさらに測定した。結果から、両基質に対するMTアーゼ活性は用量依存的に増加することが明らかになった(図1b)。特に、C末端RB1に対するMTアーゼ活性は、ヒストンH3と比較してより高かった。さらに、SET7/9も、RB1に対するメチルトランスフェラーゼ活性を有することが発見された(データ示さず)。さらに、SMYD3は完全長RB1をメチル化した(図1c)。このことは、RB1が、インビトロで、2種類のヒストンH3-K4メチルトランスフェラーゼであるSMYD3ならびにSET7/9によってメチル化されることを示唆している。
【0082】
実施例2:RB1タンパク質に対するSMYD3のメチルトランスフェラーゼ活性
SMYD3とRB1タンパク質との間の可能性のある会合を調べるために、HepG2細胞またはHCT116細胞から抽出したタンパク質を抗SMYD3抗体で免疫沈降した。予想通り、抗RB1抗体を用いたイムノブロット分析によって、RB1タンパク質に対応するバンドが観察された(図2a)。会合を担うRB1領域を決定するために、Flagタグ化野生型RB1タンパク質またはFlagタグ化変異体RB1タンパク質を、HEK293細胞においてHAタグ化SMYD3と共に発現させ、抗Flag抗体を用いて免疫沈降を行った。C末端RB1タンパク質のメチル化と一致して、C末端基質ドメイン(コドン772〜928)はSMYD3と相互作用した(図2b)。RB1との結合を担うSMYD3領域を決定するために、野生型SMYD3および様々な型の変異体SMYD3を発現するプラスミドを使用した。野生型SMYD3、ならびにΔ1型(コドン45-428)およびΔ2型(コドン1〜250)の変異体SMYD3はFlagタグ化RB1と相互作用したが、SETドメインを欠くΔ3型(コドン1〜100)はRB1と相互作用しなかった。このことは、SETドメインが会合に必須であることを示唆している(図2c)。先の報告から、ヒストンH3-K9メチルトランスフェラーゼSUV39H1はRBおよびHP1と会合し、その複合体はサイクリンEの転写抑制において役割を果たすことが示された(Nielsen, S. J. et al. Nature 412, 561-565 (2001))。さらに、最近の研究から、ヒストンH4-K20メチルトランスフェラーゼであるSuv4-20h1およびSuv4-20h2の活性は、RB1との相互作用を通じて著しく増強されることが明らかになった(Gonzalo, S. et al. Nat Cell Biol 7, 420-428 (2005))。従って、本発明者らは、RB1がSMYD3のH3-K4メチルトランスフェラーゼ活性を増強するか否かを試験した。結果として、SMYD3を介したヒストンH3のメチル化はRB1による影響を受けなかった(図2d)。特に、SMYD3は、H3-K910またはH4-K20に対してメチルトランスフェラーゼ活性を示さなかった(図2e)。このデータは、SMYD3のH3-K4特異的HMT(ヒストンメチルトランスフェラーゼ)活性を強化し、かつRB1がHMTに依存的な様式でヒストン修飾の役割を果たすことを示唆した。
【0083】
実施例3:RB1のメチル化基質ドメインの同定
RB1の基質ドメインのメチル化を担う残基を決定するために、SET7/9メチルトランスフェラーゼの基質にある保存アミノ酸配列を比較した。メチル化されるリジンの前にはセリンまたはトレオニンがあったので、本発明者らは、候補としてリジン824、リジン889、およびリジン896に焦点を当てた。野生型RB1基質ドメインおよび3種類の変異体RB1基質ドメインの組換えタンパク質を調製した(図3a)。野生型タンパク質と比較して、K889AおよびK896A変異体はSMYD3によってほぼ同じレベルでメチル化された(図3b、c)。しかしながら、K824Aのメチル化は著しく低下した(図3b、c)。さらに、K824A置換でもRB1タンパク質のメチル化は完全に減少しなかったので、チロシンの後にあるリジン791およびリジン814のメチル化を調べた。2つの変異体RB1タンパク質K791AおよびK814Aは、野生型RB1とほぼ同じレベルのメチル化を示した(図3d、e)。さらに、2種類の二重変異体RB1であるK791A/K824AおよびK814A/K824Aは、K824Aタンパク質と同等レベルのメチル化を示した。従って、本発明者らは、このリジン824がメチル化の主要な標的残基であると結論付けた。リジン824のメチル化を確認するために、ジメチル化リジン824またはトリメチル化リジン824を認識するメチル化RB1特異的抗体を調製した。野生型RB1タンパク質のメチル化に従って、この抗体は、SMYD3がヒストンH3リジン4のジメチル化およびトリメチル化を発揮するのと同様に(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004))、イムノブロット分析においてジメチル化およびトリメチル化RB1タンパク質を検出した(図3f)。H3-K4、TAF10、およびp53を含むSET7/9の基質にあるメチル化リジンの前には2つの保存的ペプチド、リジンの位置-2にR/Kおよび位置-1にS/Tがあったが、リジン824の前には-2にPおよび-1にTがあった。RB1はSMYD3ならびにSET7/9によってメチル化されるので、-2のR/Kは必須ではないかもしれないが、-1のS/TはSMYD3またはSET7/9によるメチル化に欠くことができない。
【0084】
実施例4:インビボメチル化アッセイ法
インビボでのSMYD3によるRB1メチル化をさらに調べるために、SMYD3を発現しないHEK293細胞を使用して、インビボメチル化アッセイ法を行った(Liu, Q. & Dreyfuss, G. Mol Cell Biol 15, 2800-8 (1995))。SMYD3を発現するHEK293細胞株(HEK-SMYD3-1および-2)(図4a)を樹立し、細胞を、タンパク質合成阻害剤の存在下でL-[メチル-3H]メチオニンとインキュベートした。次いで、細胞からの抽出物を抗RB1モノクローナル抗体で免疫精製し(immunopurified)、免疫沈降したタンパク質をSDS-PAGEで、その後にオートラジオグラフィーで分析した。モックをトランスフェクトしたHEK293細胞(HEK-モック-1および-2)と比較して、HEK-SMYD3(HEK-SMYD3-1および-2)細胞からの抽出物は、メチル化RB1に対応する著しく強いバンドを示した。免疫沈降したRB1の量は細胞株間で変化しなかった(図4b)。抗パンメチル-リジン抗体、抗ジメチル化RB1-リジン824抗体、または抗トリメチル化RB1-リジン824抗体を使用したウエスタンブロット分析によって、HEK-モック細胞と比較してHEK-SMYD3細胞におけるメチル化RB1の増加が一貫して観察された(図4c)。HEK-SMYD3細胞の免疫細胞化学染色から、多量のSMYD3を発現する細胞は、抗ジメチル化または抗トリメチル化RB1-リジン824抗体で、より少量のSMYD3を発現する細胞よりも強く染色された(それぞれ、図4d〜g、4h〜k)ことが示された。このデータは、インビボでのSMYD3によるRB1-リジン824のメチル化を裏付けている。
【0085】
実施例5:インビボリン酸化アッセイ法
RB1のリジン824は、トレオニン821stとトレオニン826thの間に位置する。トレオニン821st残基およびトレオニン826th残基はCDK/サイクリン複合体によってリン酸化され、中央のポケットドメインのコンホメーションを変えることによってRB1とE2Fとの間の相互作用を調節する。これらの周囲のトレオニンリン酸化に及ぼすRB1メチル化の影響を調べるために、メチル化または非メチル化RB1タンパク質を用いて、インビボリン酸化アッセイ法を行った。組換えC末端RB1を、SMYD3の存在下または非存在下で3H標識SAMとインキュベートし、次いで、32P-γATPを組換えCDK2/サイクリンEまたはCDK6/サイクリンD3と組み合わせて混合した。組換えRB1のメチル化およびリン酸化を、液体シンチレーションカウンターによって同時に測定した。C末端RB1タンパク質は、SMYD3存在下では、SMYD3非存在下の4〜6倍の量の3H標識メチル供与体を組み込んだ(データ示さず)。重要なことに、SMYD3は、CDK2/サイクリンE複合体によるRB1のリン酸化を用量依存的な様式で増強したのに対して、SMYD3のみではリン酸化を高めなかった(図5a)。さらに、RB1のリン酸化は、SMYD3存在下では、SMYD3非存在下と比較してCDK6/サイクリンD3によって増大することが発見された(図5b)。しかしながら、CDK2/サイクリンEまたはCDK6/サイクリンD3によるK824A変異体RB1のリン酸化は、野生型RB1と比較して有意に抑制された(それぞれ、図5c、d)。このデータは、RB1リン酸化は、SMYD3によるリジン824のメチル化によって増強されることを示唆している。抗リン酸化RB1抗体を用いたさらなるイムノブロット分析から、トレオニン821/826のリン酸化はSMYD3によって誘導されることが明らかになった。興味深いことに、セリン807/811のリン酸化もSMYD3によって増強された(図5e)。従って、リジン824のメチル化はセリン807/811のリン酸化を増加させる。または、さらなるメチル化残基がリン酸化を増強する可能性がある。
【0086】
インビボでの増強されたRB1のリン酸化を調べるために、抗リン酸化RB1抗体で、HEK-SMYD3細胞およびHEK-モック細胞からの抽出物を用いて、ウエスタンブロット分析を行った。インビトロでRB1タンパク質のリン酸化が増強されたことと一致して、HEK-SMYD細胞ではセリン807/811およびトレオニン821/826の両方のリン酸化が対照細胞と比較して高いことが検出された(図5f)。抗リン酸化トレオニン821/826抗体および抗SMYD3抗体を用いた免疫細胞化学染色から、SMYD3を発現する細胞は、抗リン酸化トレオニン821/826抗体によって、SMYD3を発現しない細胞より強く染色されることが明らかになった(図5g〜j)。さらに、SNU475細胞における野生型SMYD3の外因性発現は、メチルトランスフェラーゼ活性を欠く変異体SMYD3(SMYD3-ΔEELまたはSMYD3-ΔNHSC)の外因性発現と比較して、RB1リジン824のジメチル化およびトリメチル化を増大させた(Hamamoto, R. et al. Nat Cell Biol 6, 731-40 (2004))。RB1リジン824のメチル化と相関づけて、本発明者らは、この細胞において、トレオニン821/826でのリン酸化の著しい増加およびセリン807/811でのリン酸化の中程度の増加をそれぞれ観察した(図6a)。重要なことに、ウエスタンブロット分析から、対応する非癌性乳房組織と比較して、SMYD3を多く発現する乳癌組織では、セリン807/811およびトレオニン821/826のリン酸化が増加すると共に、RB1リジン824のメチル化が増強することが示された(図6b)。このデータは、インビボでのSMYD3によるセリン807/811およびトレオニン821/826のリン酸化が増強されたことを要約したものであった。RB1のリン酸化は、RB1からのE2Fの解離につながるポケットドメインを調整するので、MercuryTM細胞周期プロファイリングシステムを用いて、E2Fを介した転写のレポーター活性をHEK-SMYD3細胞において比較した。HEK-モック細胞と比較して、HEK-SMYD3細胞は高いE2F転写活性を示した(図7)。このデータは、SMYD3は、リジン824をメチル化することによってRB1のリン酸化を増強し、E2F転写活性を高めることを示す。
【0087】
実施例6:ヒト癌細胞における切断型SMYD3タンパク質
本発明者らは、本発明者らの最近の研究において、SMYD3タンパク質の発現レベルがヒト肝細胞癌(HCC)、結腸直腸癌(CRC)、および乳癌において高いことを示した(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004), Hamamoto, R. et al. Cancer Sci 97, 113-118 (2006))。興味深いことに、抗SMYD3抗体を用いたウエスタンブロット分析から、調べた全ての乳癌組織には45kDaおよび42kDaの2つのバンドが示されたが、正常乳腺には2つのバンドがどちらも無いことが検出された。45kDaおよび42kDaのバンドは2つともHCC、CRC、および乳癌細胞株(図8a)、ならびに正常精巣(データ示さず)において観察された。SMYD3の推定分子量は45kDaであり、本発明者らは、RT-PCR分析においてSMYD3転写物が変化した形を発見しなかった。従って、本発明者らは、42kDaのバンドが完全長SMYD3タンパク質の切断に起因する可能性があると仮説を立てた。SMYD3の切断を調べるために、本発明者らは、N末端HAタグ化SMYD3またはC末端FLAGタグ化SMYD3を発現するプラスミドを調製した。HAタグ化SMYD3タンパク質を発現するまたはFLAGタグ化SMYD3タンパク質を発現する、HEK293細胞の抽出物を、それぞれ抗HA抗体または抗FLAG抗体をを用いたイムノブロット分析に使用した。結果として、本発明者らは、抗HA抗体を用いてN末端HAタグ化タンパク質に対応する46kDaバンドを入手した。一方で、本発明者らは、抗FLAG抗体を用いて、46kDaおよび43kDaタンパク質の2つのバンドを発見した(図8b)。この結果は、完全長タンパク質がN末端領域で切断されることを示唆した。切断部位を調べるために、本発明者らは、内因性SMYD3を発現しないHEK293細胞において、N末端3XFLAG-タグを含む、野生型SMYD3および欠失変異体SMYD3を外因的に発現させた(図8c)。図8bのデータと一致して、抗FLAG抗体を用いたウエスタンブロット分析からは、野生型SMYD3を発現する細胞では48kDa FLAGタグ化完全長タンパク質に対応するバンドしか認められなかった。しかしながら、同じ抽出物を使用した、抗SMYD3抗体を用いた分析によって、48kDa FLAGタグ化SMYD3および42kDaタンパク質に対応する2つのバンドが検出された。抗SMYD3抗体を用い、N末端欠失型SMYD3(FLAG-SMYD3-ΔN44、-ΔN99、および-ΔN244)を発現する細胞からの抽出物を使用したウエスタンブロット分析からは、1本のバンドが認められた。これらのデータは、SMYD3の切断部位がコドン1と45との間に位置することを示唆した。
【0088】
実施例7:SMYD3タンパク質の切断部位の決定
SMYD3の正確な切断部位を決定しようと、本発明者らは、免疫沈降したFLAGタグ化SMYD3タンパク質を転写したPVDF膜から42kDaタンパク質を精製し(図8b)、そのアミノ酸配列を決定した。結果として、本発明者らは、N末端34アミノ酸を欠く欠失型SMYD3タンパク質を同定した。このことから、コドン34(アスパラギン酸)とコドン35(プロリン)との間に切断部位があることが明らかになった(図9a)。SMYD3は、コドン5と27との間にSET-N領域と呼ばれるアミノ酸配列を含む。SET-N領域はSETドメインタンパク質において保存されている(Marmorstein, R. Trends in Biochem. Sci., Vol.8 no.2, (2003); Kouzarides, T. Curr. Opin. Genet. Dev. 12, 198-209 (2002); Lachner, M and Jenuwein, T. Curr. Opin. Cell biol. 14, 286-298 (2002))。SET-N領域のアミノ酸配列のアラインメントは、SMYD3および他のメチルトランスフェラーゼにある領域の高い類似性を示した(図9b)。このことは、この領域が重要なことを意味している。
【0089】
実施例8:切断されたSMYD3のHMTアーゼ活性は野生型タンパク質と比較して高い
切断SMYD3タンパク質のメチルトランスフェラーゼ活性を調べるために、本発明者らは、HEK293細胞において、3xFLAGタグ化野生型SMYD3または34アミノ酸欠失型SMYD3もしくは44アミノ酸欠失型SMYD3を外因的に発現させ、これらのタンパク質を免疫沈降した(図10a)。本発明者らは、これらのタンパク質を酵素源として使用して、ヒストンメチルトランスフェラーゼ(HMTアーゼ)アッセイ法を行い、野生型SMYD3のHMTアーゼ活性は用量依存的な様式で増加することを示した(図10b)。メチル供与体であるS-アデノシルメチオニン(SAM)の存在下でのメチル化反応は、メチルトランスフェラーゼ反応を競合的に阻害する可能性のあるS-アデノシルホモシステイン(SAH)の産生を伴う。従って、本発明者らは、反応混合物に、SAHをホモシステインおよびアデノシンに加水分解するS-アデノシルホモシステイン加水分解酵素(SAHH)を添加した。予想したとおり、本発明者らは、SAHH存在下では、非存在下と比較してHMT活性が著しく増加することを観察した(図10b)。この知見は、SMYD3のメチルトランスフェラーゼ阻害剤のスクリーニングに有用である。驚くべきことに、切断されたSMYD3タンパク質のHMTアーゼ活性は、完全長タンパク質と比較して有意に高かった(図10b)。この結果から、翻訳後の切断はヒト細胞におけるSMYD3 HMTアーゼ活性の調節に関与し、N末端SET-N領域はHMTアーゼ活性に対して抑制的な役割を有し得ることが分かる。
【0090】
実施例9:SET-N領域におけるグリシン15および17はHMT活性に重要である
酵素活性の抑制に関するSET-N領域にある保存アミノ酸配列の重要性を確かめるために、本発明者らは、野生型N末端FLAGタグ化SMYD3タンパク質を発現するプラスミド、または変異体N末端FLAGタグ化SMYD3タンパク質、SMYD3-SETNm1、-SETNm2、もしくは-SETNm3を発現するプラスミドを調製した。SMYD3-SETNm1、-SETNm2、または-SETNm3は、それぞれ、Gly15AlaおよびGly17Alaの両方、Gly15Ala、またはGly27Alaの置換を含む(図11a)。これらの変異体を発現するHEK293細胞からの溶解産物のウエスタンブロットによって、置換はSMYD3タンパク質の切断に影響を及ぼさないことが示された(図11b、上パネル)。本発明者らは、免疫沈降したSMYD3タンパク質を用いてHMTアーゼアッセイ法を行った。結果として、Gly15AlaまたはGly27Alaを含む変異体タンパク質(SMYD3-SETNm2または-SETNm3)のHMTアーゼ活性は野生型タンパク質とほぼ同じであった (図11c)。これに対して、Gly15AlaおよびGly17Alaの2つの置換を含む変異体タンパク質(SMYD3-SETNm1)の活性は野生型タンパク質と比較して有意に増強された(図11c)。これらのデータは、SMYD3のHMTアーゼ活性の調節にグリシン15および17が重要な役割を果たしている可能性があることを示唆している。
【0091】
実施例10:N末端10アミノ酸の欠失はHMTアーゼ活性の増強に必須である
N末端領域が酵素活性を増強したことから、本発明者らは、2つの可能性のある機構について仮説を立てた。すなわち、N末端領域が酵素活性に関する未決定の負の調節因子と会合し得るか、または欠失によって、増強された酵素活性につながるタンパク質のコンホメーション変化が起こり得るという仮説を立てた。さらなる負の調節因子が酵素活性において役割を果たしている可能性があるか否かを確かめるために、本発明者らは、野生型SMYD3およびN末端欠失SMYD3の組換えタンパク質を調製し、そのHMTアーゼ活性をインビトロで調べた。図12に示したように、全ての欠失変異体(SMYD3-ΔN9、-ΔN19、-ΔN29、-ΔN44、-ΔN74)は、野生型タンパク質と比較して4〜5倍に増強されたメチルトランスフェラーゼ活性を示した(図12)。この結果は、さらなる因子が、切断されたSMYD3の高活性に関与している可能性は低く、N末端10アミノ酸がメチルトランスフェラーゼ活性の抑制に非常に重要な役割を果たしている可能性があることを示唆している。
【0092】
考察
SMYD3には、インビトロおよびインビボでRB1のリジン824に対するメチルトランスフェラーゼ活性があり、メチル化RB1は、CDK/サイクリン複合体によるリン酸化に対して非メチル化RB1より感受性であるという知見が本明細書において開示される。結果として、SMYD3を発現するHEK293-SMYD3細胞は、HEK293-モック細胞と比較して高いE2F転写活性を示した。このことは、SMYD3の増殖促進作用とよく一致している。なぜなら、E2F-1の過剰発現は、細胞増殖に必須の産物の一連の遺伝子を調節することによって、細胞周期のG1期からS期への移行を促進できるからである。Harbourらは、G1からSへ進行する間にRB1がリン酸化されるモデルを示した。このモデルでは、RB1のリン酸化は、C末端領域とポケットドメインとの間の連続した分子内相互作用を開始する(Harbour, J. W. et al., D. C. Cell 98, 859-69 (1999))。G1期の間に、CDK4/6-サイクリンDによってRB1のC末端領域がリン酸化されると、中央のポケットドメインとの分子内相互作用が誘発される。これによって、HDAC結合が阻害され、活発な転写抑制がブロックされる。この相互作用は、CDK2/サイクリンEがRB1のセリン567に接近するのを促し、その結果として、A/B境界面が壊れ、RB1とE2Fとの相互作用が阻止される。このモデルでは、E2Fが解離するために、RB1がCDK4/6-サイクリンD複合体およびCDK2/サイクリンE複合体の両方によって連続してリン酸化されることが必要とされる(Lundberg, A. S. & Weinberg, R. A. Mol Cell Biol 18, 753-61 (1998))。報告によると、RB1にあるThr821およびThr826がリン酸化されると、A/Bポケットドメインと、E2FおよびHDACを含むLXCXEモチーフ含有タンパク質との相互作用が不活性化されるのに対して、Ser807およびSer811がリン酸化されるとC末端ドメインが不活性化される。このデータは、SMYD3によるRB1のメチル化によって、CDK2/サイクリンE複合体またはCDK6/サイクリンD複合体によるリン酸化が増強され、Thr821/826のリン酸化が高まるので、SMYD3を発現する細胞が高いE2F転写活性を示すという本発見と一致する。あるいは、リジン824のメチル化は、C末端領域のコンホメーションを直接変え、それによって、ポケットドメインとE2Fとの会合を阻害するかもしれない。なぜなら、ヒストンおよびp53のリジンの両方がメチル化されるとコンホメーションが変化するからである(Tsuge, M. et al. Nat Genet 37, 1104-7 (2005))。SMYD3はE2F転写活性を増加させるので、多量のSMYD3は正のフィードバック機構としてE2F1活性を高めるのかもしれない。従って、SMYD3を介したRB1不活性化は、ヒトの発癌において非常に重要な役割を果たしている可能性が高い。
【0093】
RB1が、いくつかの機構を介して転写抑制において役割を果たすことは注目すべきことである。RB1は転写因子と相互作用し、かつ転写因子の活性を直接抑制する。RB1がプロモーター領域に動員されると、開始前複合体の集合がブロックされる。RB1はクラスI HDAC(HDAC-1、-2、および-3)とも会合し、かつヒストンの脱アセチル化を誘導して、ヘテロクロマチン状態へのコンホメーション変化をもたらす。RB1は、標的遺伝子のプロモーター領域におけるDNAメチル化につながるDNMT1と複合体を形成する(Harbour, J. W. & Dean, D. C. Genes Dev 14, 2393-409 (2000); Robertson, K. D. et al. Nat Genet 25, 338-42 (2000))。これらの機構に加えて、ヒストンメチル化に関する最近の研究から、RB1は、SUV39HおよびSuv4-20h1またはSuv4-20h1を含むヒストンメチルトランスフェラーゼとも会合することが開示された。SUV39HおよびSuv4-20h1またはSuv4-20h1は、それぞれ、H3-K9およびH4-K20のメチル化に関与する(Gonzalo, S. et al. Nat Cell Biol 7, 420-8 (2005); Nielsen, S. J. et al. Nature 412, 561-5 (2001))。これらのメチルトランスフェラーゼに結合すると、RB1は、HP1またはCBXを複合体に動員することによってヘテロクロマチン形成を安定化する。本知見は、ヒストンH3-K4メチルトランスフェラーゼの転写活性化の調節への新規の洞察をもたらす。リジン824がメチル化したRB1は、RB1リン酸化を増強し、およびその後に、恐らくE2F1を中央のポケットドメインから放出させることによってE2F1のトランス活性化を増強する。さらに、メチル化されたRB1は、コンホメーションを変え、それによって、HDAC、ヘテロクロマチンタンパク質1(HP1)、および/またはクロモボックスタンパク質(CBX)を、SUV39Hおよび/またはSuv4-20h1メチルトランスフェラーゼの複合体から解離し、H3-K9およびH4-K20のメチル化を低下させるかもしれない。さらなる研究が必要だが、本明細書のデータは、E2Fを担う遺伝子の調節におけるRB1メチル化の重要性を強調する。RB1は様々なメチルトランスフェラーゼに結合するので、RB1におけるメチル化の位置および程度はメチルトランスフェラーゼによって異なる可能性がある。RB1が様々な残基でリン酸化されることもあわせると、本明細書のデータは、RB1における複数の修飾の組み合わせがRB1の生物学的特性を規定する可能性があることを示唆している。このことは、ヒストンおよびp53の修飾を連想させる。
【0094】
RB1の変異は、散発性および家族性の網膜芽細胞腫症例に関与するだけでなく(Weinberg, R. A. Science 254, 1138-46 (1991))、他のヒト癌にも関与している(Classon, M. & Harlow, E. Nat Rev Cancer 2, 910-7 (2002))。いくつかの腫瘍ウイルスタンパク質、例えば、アデノウイルスE1A、HPV-E7、およびシミアンウイルス40(SV40)ラージT抗原は一部の癌のタイプにおいてRB1と会合し、RB1とE2Fとの間の相互作用を阻害して、E2Fを解離させる(Chellappan, S. P., et al. Cell 65, 1053-61 (1991); Bagchi, S., et al. Cell 65, 1063-72 (1991))。サイクリン依存性キナーゼ4阻害剤であるp16は、p16プロモーターのメチル化によって頻繁に不活性化される。これによって、癌細胞ではCDK/サイクリン複合体によるRB1のリン酸化が増強される(Nuovo, G. J., et al. Proc Natl Acad Sci U S A 96, 12754-9 (1999))。これらの欠陥は、結腸直腸癌および肝細胞癌の一部に関与していると報告されており(Chaubert, P. et al. Hepatology 25, 1376-81 (1997); Toyota, M. et al. Proc Natl Acad Sci U S A 96, 8681-6 (1999))、これらの腫瘍タイプにおける全症例の原因ではないかもしれない。本明細書において、新規のRB1不活性化機構、すなわち、RB1不活性化はRB1のメチル化およびその後の増強されたリン酸化によって引き起こされることが開示されている。SMYD3の発現は結腸直腸癌および肝細胞癌の大部分において増強されているので、SMYD3は、RB1の腫瘍抑制因子機能を抑止してE2Fをトランス活性化することにより癌細胞増殖において非常に重要な役割を果たしている可能性がある。興味深いことに、本発明者らは、最近、SMYD3の発現が、E2F-1とSMYD3プロモーター領域内のE2F-1結合エレメントの相互作用を介してE2F-1によって調節され、およびこのエレメントがE2F-1結合モチーフの2タンデムリピートまたは3タンデムリピートより構成されることを発見した。日本人の結腸直腸癌患者(n=350)、肝臓癌患者(n=360)、および乳癌患者(n=334)における3リピートの対立遺伝子頻度は、一般的な日本人集団からの健常対照(n=730)より有意に高かった。このデータは、一旦SMYD3が活性化されると、RB1の修飾を介してE2F転写活性を増強し、結果として、正のフィードバックによりSMYD3を上方制御することを示唆している。従って、E2F-1結合エレメントの3リピートを含む人は、2リピートを含む人よりSMYD3によるRB1不活性化を受けやすい。さらに、SMYD3の阻害は、正のフィードバックループをブロックし、それによって、RB1のリン酸化によるE2F-1を介した分裂促進活性を効率的に抑制するので、結腸直腸癌および肝臓癌ならびに膀胱癌および乳癌の有望な治療方針であるように思われる。
【0095】
本明細書において、SMYD3によるRB1のメチル化は、CDK/サイクリン複合体によるRB1のリン酸化の増強を介してG1期からS期への細胞周期の進行を速める可能性があることが明らかになった。このデータから、リジンのメチル化はヒストンだけでなく、p53およびRB1などの他の非ヒストンタンパク質にとっても重要なことが分かる。さらに、本発明者らの知見は、ヒト発癌に関与するRB1調節の新規機構の解明に役立った。
【0096】
エピジェネティックな調節の混乱がヒト発癌と関連することが示されている。細胞周期、DNA修復、および細胞接着を調節する遺伝子のプロモーター領域における異常なDNAメチル化に加えて、最近の研究から、ヒストンのメチル化もヒト発癌において抑止されることが開示された。ヒストンのメチル化は、クロマチン構造の変化を介して遺伝子発現の調節において非常に重要な役割を果たしている。本発明者らは、ヒストンH3-リジン4-特異的メチルトランスフェラーゼであるSMYD3が、HCC、CRC、および乳癌を含む、いくつかのヒト癌において過剰発現していることを報告した(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004): Hamamoto, R. et al. Cancer Sci 97, 113-118 (2006))。本発明者らの以前の論文において、本発明者らは、様々なヒト癌において頻繁に増強される転写因子であるE2F1の転写活性化によって、SMYD3の発現が上昇することを示した。
【0097】
タンパク質の機能は、転写後レベルでだけでなく、タンパク質の切断ならびにアセチル化、リン酸化、メチル化、グリコシル化、およびユビキチン化などの他の広く知られている修飾を含む翻訳後修飾によっても調節される。これらの修飾は、タンパク質を活性化または不活性化する、タンパク質安定性、タンパク質のコンホメーション、および/またはタンパク質-タンパク質相互作用と関連する。本発明者らは、SMYD3の切断がSMYD3のHMTアーゼ活性を高めることを発見した。このことは、ペプシン、インシュリン、カスパーゼ、PARP、およびMMPなどの重要な酵素の調節を思い出させる。なぜなら、これらのタンパク質は切断されると酵素活性が増加するからである。さらに、この知見は、未決定のSMYD3切断機構がHMTアーゼ活性の調整において役割を果たしている可能性があることを示唆している。従って、切断を担うプロテアーゼの同定、および調節機構の解明が、SMYD3活性を抑制する新規の治療アプローチの開発に寄与するだろう。さらに、切断型SMYD3は、完全長タンパク質と比較してSMYD3阻害剤のスクリーニングに有用な可能性がある。
【0098】
本発明者らは、本研究において、SMYD3 N末端領域が欠失すると、インビトロにおいてSMYD3の酵素活性が増強されることを発見した。このことは、欠失は、増強された酵素活性につながるSMYD3のコンホメーション変化を付与し得ることを示唆している。興味深いことに、HSP90はSMYD3のN末端領域に結合して、SMYD3のHMTアーゼ活性を増加させる。このデータは、コンホメーション変化がHMTアーゼ活性に関与するという考えとよく一致している。なぜなら、HSP90は、正常なタンパク質構造の安定化に寄与するシャペロン様機能を発揮するからである。本発明者らの知見はまた、HMTアーゼ活性の調節のための保存されたSET-N領域の重要性も強調する。この保存領域は、他のSETドメイン含有タンパク質においてHMTアーゼの負の制御因子としても働く可能性がある。さらなる研究によって、SETドメイン含有タンパク質におけるHMTアーゼ活性の調節機構が見出されるだろう。
【0099】
本発明者らは、本明細書において、癌細胞においてN末端切断型SMYD3タンパク質が発現され、かつ切断されたタンパク質のHMTアーゼ活性が完全長タンパク質より著しく高いことを示した。これらのデータは、翻訳後調節システムが、可能性のあるタンパク質コンホメーション変化を介してHMTアーゼ活性を調節することを意味した。さらに、本発明者らは、SAHHが添加されるとSMYD3のメチルトランスフェラーゼ活性が増加することを発見した。本発明者らの知見は、SMYD3活性調節機構のより深い理解の助けとなり、かつHMTアーゼ活性を阻害する新規の治療方針の同定に寄与する可能性がある。
【0100】
産業上の利用可能性:
本明細書に記載の方法は、結腸直腸癌、肝細胞癌、乳癌、および膀胱癌を含む様々な癌を予防、診断、および治療するためのさらなる分子標的の同定に有用である。さらに、本明細書において報告されたデータは、癌の包括的な理解を深め、新規の診断法の開発を促し、かつ治療薬剤および予防剤の分子標的を同定する手がかりを与える。このような情報は腫瘍形成のさらに深い理解に寄与し、ならびに新規の癌診断法、癌治療法、および最終的には癌予防法を開発する指標を与える。本発明は詳細に、かつ本発明の同定の態様に関して説明されたが、前述の記載は本質的に例示および説明であり、本発明および本発明の好ましい態様を例証することを目的とすることが理解されるはずである。当業者であれば、日常的な実験によって、本発明の精神および範囲から逸脱することなく、本明細書に様々な変化および変更を加えることができることを容易に理解するだろう。従って、本発明は、前述の記載では定義されず、特許請求の範囲およびその均等物によって定義されることが意図される。
【図面の簡単な説明】
【0101】
【図1】組換えRB1タンパク質に対するSMYD3のMTアーゼ活性を示す。パートaは、基質として組換えヒストンH3、p53、またはRB1のC末端領域を用いたインビトロMTアーゼアッセイ法の結果を示す。等量の基質を、免疫沈降したFlagタグ化SMYD3およびメチル供与体である3H標識SAMとインキュベートした。タンパク質をSDS-PAGEで分離し、メチル化された基質をフルオログラムによって検出した。基質の総量は、特異的抗体を用いたイムノブロット分析によって調べた。パートbは、ヒストンH3およびC末端RB1タンパク質に対する組換えSMYD3の用量依存的MTアーゼ活性を示す。パートcは、C末端RB1および完全長RB1(それぞれ、レーン2および4)に対するSMYD3のMTアーゼ活性を示す。保存アミノ酸の欠失を含む変異体SMYD3(SMYD3ΔEEL)はMTアーゼ活性を著しく低下させた(レーン3)。
【図2】インビボでのSMYD3とRB1との間の会合を示す。パートaはイムノアッセイの結果を示す。具体的には、抗SMYD3抗体を用いたHepG2細胞またはHCT116細胞の溶解産物からの免疫沈降物を、抗RB1抗体でイムノブロットした。パートbは、HEK293細胞における野生型RB1ならびに欠失型RB1(RB1Δ1およびRB1Δ2)とSMYD3との間の相互作用を示す(下)。RB1の保存領域および発現構築物を上パネルに示す。パートcは、RB1との相互作用を担うSMYD3の領域を示す。SMYD3の保存領域および発現構築物を上パネルに示す。パートdは、組換えRB1の存在下/非存在下でのヒストンH3に対するSMYD3のインビトロメチルトランスフェラーゼ活性を示す。ヒストンH3のメチル化はRB1による影響を受けなかった(上パネル)。等量の組換えヒトヒストンH3タンパク質を基質として使用した(下パネル)。パートeは、ヒストンH4-K20メチル化のインビトロ分析を示す。免疫沈降したSMYD3タンパク質または組換えSMYD3タンパク質を、基質として組換えヒトヒストンH4とインキュベートした。免疫沈降したSuv4-20h2タンパク質は正の対照とした。抗トリメチルH4-K20抗体を用いて、メチル化されたH4-K20が検出された。
【図3】RB1のC末端領域にあるK824のメチル化を示す。パートaは、RB1の保存ドメイン、ならびに野生型および変異型のC末端RB1タンパク質(K824A、K889A、およびK896A)の模式図である。パートbは、オートラジオグラフィーによる、SDS-PAGEで分離されたメチル化C末端RB1の検出を示す。パートcは、液体シンチレーションカウンターによって測定されたMTアーゼ活性を示す。パートdは、組換え野生型RB1タンパク質、ならびにK791A、K814A、K824A、K791/K824A、およびK814/K824Aを含む組換え変異型RB1タンパク質のインビトロメチル化を示す。RB1を、3H標識SAMの存在下で組換えSMYD3タンパク質とインキュベートした。メチル化RB1をSDS-PAGEで分離し、フルオログラムによって検出した。パートeは、液体シンチレーションカウンターによって測定されたメチル化RB1を示す。パートfは、SMYD3によるRB1リジン824のジメチル化およびトリメチル化を示す。SMYD3の存在下または非存在下でのメチル化野生型RB1タンパク質を3H-BASイメージングシステムによって検出した(上パネル)。抗ジメチル化リジン824(2番目のパネル)抗体または抗トリメチル化リジン824(3番目のパネル)抗体を用いたRB1タンパク質のウエスタンブロット分析。RB1の総量は、抗RB1抗体を用いて定量した(4番目のパネル)。
【図4】インビボでのSMYD3によるRB1のメチル化を示す。パートaは、HEK-SMYD3(HEK-SMYD3-1および-2)細胞ならびにHEK-モック(HEK-モック-1および-2)細胞におけるSMYD3の発現を示す(上パネル)。パートbは、抗RB1抗体によるHEK-SMYD3細胞およびHEK-モック細胞からの免疫沈降物を用いた、ラジオグラムによるインビボでのメチル化RB1の検出を示す(上パネル)。免疫沈降したRB1の量は変化しなかった。細胞を、タンパク質合成阻害剤の存在下で3H標識SAMで処理した。免疫沈降したRB1の量は変化しなかった(下パネル)。パートcは、免疫沈降したRB1のメチル化を、抗パンメチルリジン抗体、抗ジメチルリジン824抗体、および抗トリメチルリジン824抗体を用いて分析したウエスタンブロットの結果を示す。パートd〜kは、抗ジメチルリジン824(d)抗体または抗トリメチルリジン824(h)抗体を用いた、HEK293-SMYD3細胞の免疫細胞化学染色の結果を示す。パートeおよびiは、抗SMYD3抗体を用いて調べたSMYD3の発現を示す。パートfおよびjは、DAPIによる核染色の結果を示す。パートgおよびkは、d〜f(g)またはh〜j(k)の合成画像を構成する。SMYD3を豊富に発現している細胞は、インビボでRB1 Lys824のジメチル化およびトリメチル化が増強されていることを示した。
【図5】SMYD3によりRB1のリン酸化が増強されることを示す。パートaは、SMYD3の存在下または非存在下でのCDK2/サイクリンEによるC末端RB1のインビトロリン酸化を示す。SMYD3のみではリン酸化を増加しなかった。パートbは、SMYD3の存在下または非存在下でのCDK6/サイクリンD3によるC末端RB1のインビトロリン酸化を示す。SMYD3によるRB1リン酸化の増強は、K824A置換RB1を用いて抑えられた。パートcは、CDK2/サイクリンEによるC末端RB1のインビトロリン酸化を示す。基質として、野生型(Wt)およびK824A変異体(K824A)を比較した。パートdは、CDK6/サイクリンD3によるC末端RB1のインビトロリン酸化を示す。基質として、WtおよびK824Aを比較した。パートeは、SMYD3の存在下での、CDK2/サイクリンE複合体またはCDK6/サイクリンD3複合体によるSer807/811およびThr821/826リン酸化の増加を示す。パートfは、HEK-モック細胞と比較して、HEK-SMYD3細胞におけるSer807/811リン酸化およびThr821/826リン酸化が高いことを示す。外因性SMYD3を発現するHEK293細胞の免疫細胞化学染色。パートgは、抗リン酸化RB1(Thr821/826)抗体を用いて、細胞内のリン酸化RB1を染色した結果を示す。パートhは、細胞におけるSMYD3の発現を示す。パートiは、DAPIによる核染色の結果を示す。パートjはg〜iの合成画像を構成する。SMYD3を発現する細胞は、インビボでのThr821/826リン酸化が増強されていることを示した。
【図6】SMYD3によるRB1のメチル化および増強されたリン酸化を示す。パートaでは、野生型プラスミド(p3xFlag-SMYD3)または変異体SMYD3プラスミド(p3xFlag-SMYD3ΔEELおよびp3xFlag-SMYD3ΔNHSC)でトランスフェクトしたSNU475細胞から、RB1タンパク質を免疫沈降した。ウエスタンブロット分析は、沈降物を使用し、抗ジメチル化リジン824(最上部パネル)抗体、抗トリメチル化リジン824(2番目のパネル)抗体、抗リン酸化セリン807/811(3番目のパネル)抗体、または抗リン酸化トレオニン821/824(4番目のパネル)抗体を用いて行った。抗RB1抗体を用いたイムノブロット分析は量の対照として役立った(最下部パネル)。パートbは、2つの乳癌組織におけるリジン824のジメチル化およびトリメチル化ならびにSer807/811およびThr821/826のリン酸化を示す。ウエスタンブロット分析は、抗ジメチル化リジン824抗体、抗トリメチル化リジン824抗体、抗リン酸化RB(Ser807/811)抗体、または抗リン酸化RB(Thr821/824)抗体を用いて行った。
【図7】HEK-SMYD3細胞においてE2F転写活性が増大していることを示す。HEK-SMYD3細胞およびHEK-モック細胞においてE2F-ルシフェラーゼベクターでトランスフェクトした24時間後に、ルシフェラーゼ活性を測定した。外因性SMYD3を発現するHEK293細胞の免疫細胞化学染色。細胞内のリン酸化RB1を、抗リン酸化RB1(Thr821/826)抗体で染色した。パートhは、細胞におけるSMYD3の発現を示す。パートiは、DAPIによる核染色の結果を示す。パートjは、a〜cの合成画像を構成する。SMYD3を発現する細胞は、インビボでのThr821/826リン酸化が増強されていることを示した。
【図8】SMYD3タンパク質の発現パターン。パートaは、ヒト癌細胞株および組織におけるSMYD3タンパク質の発現を示す。ウエスタンブロット分析は、抗SMYD3抗体を用いて行った。β-アクチンの発現は量の対照とした。パートbは、HAタグ化SMYD3(左パネル)およびFLAGタグ化(右パネル)のイムノブロット分析を示す。ウエスタンブロット分析は、抗HA抗体およびN末端領域にHAタグ化SMYD3を発現する細胞からの抽出物、または抗FLAG抗体およびC末端領域にFlagタグ化SMYD3を発現する細胞からの抽出物を、それぞれ用いて行った。パートcは、欠失型SMYD3の模式図を示す。N末端領域に一連のFLAGタグ化SMYD3を発現するプラスミドを、内因性SMYD3を発現しないHEK293細胞にトランスフェクトした。パートdは、抗SMYD3抗体(上パネル)または抗FLAG抗体(下パネル)を用いて行った、細胞からの抽出物のウエスタンブロット分析を示す。矢印は完全長SMYD3タンパク質を示し、星印は切断型SMYD3に対応する。
【図9】SET含有タンパク質におけるSMYD3切断部位およびSET-N領域の保存アミノ酸配列の決定。パートaは、エドマンアミノ酸シークエンスによって決定された、N末端領域の34アミノ酸が欠失したSMYD3タンパク質を示す。パートbは、ヒストンメチルトランスフェラーゼにおけるSET-Nアミノ酸配列のアラインメントを示す。高度に保存されたアミノ酸を黒色の囲みで示し、中程度に保存されたアミノ酸を影付きの囲みで示した。
【図10】野生型タンパク質と比較して増加している切断型SMYD3のHMTアーゼ活性。パートaは、抗FLAG抗体(上パネル)および抗SMYD3抗体(中パネル)を用いた、野生型SMYD3タンパク質または欠失型(ΔN34およびΔN44)SMYD3タンパク質のウエスタンブロット分析を示す。FLAGタグ化SMYD3タンパク質を発現する細胞から、タンパク質を抽出した。免疫沈降したSMYD3タンパク質をHMTアーゼアッセイ法に使用した。パートbは、完全長SMYD3タンパク質および切断されたSMYD3タンパク質のHMTアーゼ活性の用量反応増加を示す。SAHH(S-アデノシルホモシステイン加水分解酵素)を添加すると活性が増加した。3H-放射能を液体シンチレーションカウンターによって測定した。
【図11】SET-N領域の中の抑制されたHMTアーゼ活性の原因となる領域の決定。パートaは、SET-N領域の保存アミノ酸に置換を含む変異体SMYD3構築物の模式図を示す。パートbは、抗SMYD3(上パネル)抗体または抗FLAG(中パネル)抗体を用いた、FLAGタグ化野生型SMYD3タンパク質または変異体(ΔN34、SETNm1、SETNm2、およびSETNm3)SMYD3タンパク質のイムノブロット分析を示す。FLAGタグ化SMYD3を発現するHEK293F細胞から抗Flag抗体を用いて免疫沈降したタンパク質を、HMTアーゼアッセイ法の酵素源として使用した。パートcは、野生型SMYD3、欠失型SMYD3のHMTアーゼ活性を示す。3H-放射能を液体シンチレーションカウンターによって測定した。
【図12】SMYD3におけるN末端領域の欠失によるHMTアーゼ活性の増強。パートaは、N末端領域が欠失した型のSMYD3の模式図を示す。一連のGST融合SMYD3タンパク質を発現するプラスミドを調製した。パートbは、抗GST抗体を用いた組換えSMYD3タンパク質のイムノブロット分析を示す。GSTと融合した、野生型および変異体組換えSMYD3タンパク質を細菌細胞において発現させ、その細胞から精製した。パートcは、タンパク質のインビトロHMTアーゼ活性を示す。3H-放射能を液体シンチレーションカウンターによって測定した。
【技術分野】
【0001】
技術分野
本発明は、転写調節、より具体的には、メチルトランスフェラーゼ活性を調整する作用因子、例えば、SMYD3(「ZNFN3A1」としても知られる)による網膜芽細胞腫のメチル化を調整する作用因子の同定に関する。SMYD3は多くの癌種において上方制御されるので、前記のように同定されたSMYD3モジュレーターは、例えば、結腸直腸癌、肝細胞癌、乳癌、および膀胱癌を含む癌の治療において有用なことが判明する可能性がある。
【0002】
本願は、2005年7月1日に提出された米国特許仮出願第60/695,957号の恩典を主張し、その内容はその全体が参照として本明細書に組み入れられる。
【背景技術】
【0003】
背景技術
最近の分子的研究から、細胞周期制御の抑止は広範囲のヒト腫瘍の基礎をなすことが明らかになっている(Sherr, C. J., Science 274, 1672-7 (1996)(非特許文献1))。p53遺伝子、RB1遺伝子、またはp16遺伝子の遺伝子変化がヒト癌の大多数に関与しており、細胞周期進行の調節が解除されることによって無秩序な細胞増殖が起こる(Hanahan, D.& Weinberg, R. A. Cell 100, 57-70 (2000)(非特許文献2); Sherr, C. J.& McCormick, F.Cancer Cell 2, 103-12 (2002)(非特許文献3))。細胞周期の中で、細胞周期が停止し、ゲノムの完全性が調べられ、かつDNA損傷が修復されるG1/S境界は、正常な細胞特性およびゲノム特性の維持に重要である。2つの重要なシグナル伝達経路、すなわち、p53およびRB1は、多くの下流遺伝子を制御することによってG1/S境界の調節に関与している。損傷を受けたDNAを含む細胞は、蓄積された野生型p53タンパク質のトランス活性化を介してp21Cip1を誘導することによって、この境界で停止している(Sherr, C. J. & Roberts, J. M. Genes Dev 13, 1501-12 (1999)(非特許文献4))。RB1は、家族性網膜芽細胞腫の原因遺伝子として単離され(Friend, S. H. et al. Nature 323, 643-6 (1986)(非特許文献5); Fung, Y. K. et al. Science 236, 1657-61 (1987)(非特許文献6); Lee, W. H. et al. Science 235, 1394-9 (1987)(非特許文献7))、細胞周期進行を制御することによって腫瘍サプレッサーとして機能する。細胞周期移行のG1期からS期にかけて、RB1はリン酸化されることで不活性化される。このリン酸化はサイクリン依存キナーゼ(CDK)によって触媒される。不十分にリン酸化されたRB1は、DNA複製および細胞周期進行に必要とされる遺伝子の発現を調整する転写因子であるアクチベーターE2Fの活性化ドメインと直接相互作用し、HDACと複合体化するクロマチン構造を変え、かつリプレッサー複合体を応答遺伝子のプロモーター領域にあるE2F結合部位に動員することによって(Weintraub, S. J., et al., Nature 358, 259-61 (1992)(非特許文献8); Sellers, W. R., et al., Proc Natl Acad Sci USA 92, 11544-8 (1995)(非特許文献9))、E2Fを阻害する(Dannenberg, J. H., et al., Genes Dev 14, 3051-64 (2000)(非特許文献10); Sage, J. et al. Genes Dev 14, 3037-50 (2000)(非特許文献11))。RB1は、CDK4/サイクリンDなどのCDK/サイクリン複合体によってリン酸化されるとE2Fを解離し、次いで、サイクリンE、c-Myb、CDK2、およびBCL2を含む下流遺伝子をトランス活性化する。
【0004】
本発明者らは、以前に、SMYD3が過剰発現するとNIH3T3細胞の増殖が促進され、いくつかの癌細胞において内因性SMYD3発現をノックダウンすると癌細胞の増殖阻害およびアポトーシスが誘導されたことから、SMYD3がヒストンH3のリジン4(H3-K4)に対してジメチルトランスフェラーゼ活性およびトリメチルトランスフェラーゼ活性を有し、SMYD3高発現が結腸直腸癌(CRC)細胞および肝細胞癌(HCC)細胞の増殖において重要な役割を果たすことを報告した(Hamamoto, R. et al., Nat Cell Biol 6, 731-40 (2004)(非特許文献12))。しかしながら、SMYD3の過剰発現によって増殖が促進される正確な機構は未解決のままである。アセチル化、リン酸化、および/またはメチル化によるヒストン修飾は、様々な分子を動員することによって標的遺伝子の転写を活性化または不活性化するクロマチン構造を調節する。ヒストンリジンのメチル化に関して、H3-K4、H3-K36、およびH3-K79の修飾は、コンホメーションをヘテロクロマチン構造からユークロマチン構造に変えることによって転写活性化と関連するのに対して(Im, H. et al., J Biol Chem 278, 18346-52 (2003)(非特許文献13); Bannister, A. J. et al., J Biol Chem 280, 17732-6 (2005)(非特許文献14); Schneider, R. et al., Nat Cell Biol 6, 73-7 (2004)(非特許文献15))、H3-K9、H3-K27、およびH4-K20のメチル化はヘテロクロマチン構造によって転写抑制をもたらす(Schotta, G. et al., Genes Dev 18, 1251-62 (2004)(非特許文献16); Nakayama, J. et al., Science 292, 110-3 (2001)(非特許文献17); Kirmizis, A. et al. Genes Dev 18, 1592-605 (2004)(非特許文献18))。
【0005】
【非特許文献1】Sherr, C. J., Science 274, 1672-7 (1996)
【非特許文献2】Hanahan, D.& Weinberg, R. A. Cell 100, 57-70 (2000)
【非特許文献3】Sherr, C. J.& McCormick, F.Cancer Cell 2, 103-12 (2002)
【非特許文献4】Sherr, C. J. & Roberts, J. M. Genes Dev 13, 1501-12 (1999)
【非特許文献5】Friend, S. H. et al. Nature 323, 643-6 (1986)
【非特許文献6】Fung, Y. K. et al. Science 236, 1657-61 (1987)
【非特許文献7】Lee, W. H. et al. Science 235, 1394-9 (1987)
【非特許文献8】Weintraub, S. J., et al., Nature 358, 259-61 (1992)
【非特許文献9】Sellers, W. R., et al., Proc Natl Acad Sci USA 92, 11544-8 (1995)
【非特許文献10】Dannenberg, J. H., et al., Genes Dev 14, 3051-64 (2000)
【非特許文献11】Sage, J. et al. Genes Dev 14, 3037-50 (2000)
【非特許文献12】Hamamoto, R. et al., Nat Cell Biol 6, 731-40 (2004)
【非特許文献13】Im, H. et al., J Biol Chem 278, 18346-52 (2003)
【非特許文献14】Bannister, A. J. et al., J Biol Chem 280, 17732-6 (2005)
【非特許文献15】Schneider, R. et al., Nat Cell Biol 6, 73-7 (2004)
【非特許文献16】Schotta, G. et al., Genes Dev 18, 1251-62 (2004)
【非特許文献17】Nakayama, J. et al., Science 292, 110-3 (2001)
【非特許文献18】Kirmizis, A. et al. Genes Dev 18, 1592-605 (2004)
【発明の開示】
【0006】
発明の概要
本発明は、少なくとも部分的に、SMYD3によるリジン824のメチル化によってRB1を調節する新規の機構の発見に基づく。SMYD3は、遺伝子名「ZNFN3A1」としても知られ、結腸直腸癌および肝細胞癌の大多数(例えば、WO2003/027413を参照されたい)において、ならびに膀胱癌および乳癌において上方制御されるヒストンH3メチルトランスフェラーゼである。
【0007】
RB1のC末端領域はSMYD3のSETドメインと相互作用する。さらに、HEK293細胞におけるSMYD3発現は、CDK2/サイクリンE複合体またはCDK6/サイクリンD3複合体によるRB1の821/826および807/811のリン酸化をインビトロおよびインビボで増強し、ひいては、その結果として、E2Fの転写活性を増大させた。このデータは、癌細胞におけるSMYD3発現の増強が、RB1を修飾し、その後に、E2F転写を活性化することによって細胞周期進行を促進することを意味している。本知見は、RB1調節の基礎となる新規の機構を示唆している。さらに、本知見は、発癌、より具体的には、結腸直腸、肝細胞、膀胱、および乳房での発癌をより深く理解することに寄与し、従って、これらの腫瘍の新たな治療方針を開発することに寄与する。
【0008】
従って、本発明の目的は、SMYD3による網膜芽細胞腫のメチル化を調整する作用因子を同定するための方法を提供することであり、この方法は、以下の工程を含む:
(a)メチルトランスフェラーゼ活性を有するSMYD3ポリペプチドと、メチル化される網膜芽細胞腫ペプチドおよび補助因子を、網膜芽細胞腫ペプチドのメチル化に適した条件下、試験作用因子の存在下で接触させる工程;
(b)網膜芽細胞腫ペプチドのメチル化レベルを検出する工程;ならびに
(c)工程(b)において検出されたメチル化レベルと、作用因子の非存在下で検出された対照レベルを比較する工程であって、対照レベルと比較したメチル化レベルの増加または減少は、作用因子がSMYD3による網膜芽細胞腫のメチル化を調整することを示す、工程。
【0009】
本発明のさらなる目的は、網膜芽細胞腫のメチル化を調節する試験化合物の能力を検出するためのキットを提供することであり、このようなキットは、(a)メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド、(b)SMYD3ポリペプチドによってメチル化することができる網膜芽細胞腫ペプチド、および(c)網膜芽細胞腫ペプチドのメチル化のための補助因子を含む。さらなる態様において、キットは、任意に、S-アデノシルホモシステイン加水分解酵素(SAHH)を含んでもよい。
【0010】
さらに、本発明は、結腸直腸癌、肝細胞癌、膀胱癌、および乳癌などの癌を治療するための化合物をスクリーニングする方法を提供し、このような方法は、(a)前記方法に従ってメチル化を調整する試験化合物を同定する工程、および(b)試験化合物の非存在下で検出された対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを下げる試験化合物を選択する工程を含む。
【0011】
さらに、本発明は、結腸直腸癌、肝細胞癌、膀胱癌、および乳癌などの癌の症状を軽減するための組成物を提供し、このような組成物は、前記方法によって同定された薬学的に有効な量の化合物および薬学的に許容される担体により構成される。
【0012】
本発明のさらなる目的は、結腸直腸癌、肝細胞癌、膀胱癌、および乳癌などの癌の症状を軽減するための方法を提供することであり、この方法は、癌細胞と、前記方法によって同定された薬学的に有効な量の化合物を接触させる工程を含む。
【0013】
本発明のこれらのおよび他の目的、特徴、および利点は、以下の詳細な説明を、添付の図面および実施例ならびに本明細書に付した特許請求の範囲と共に読めば十分に明らかになるだろう。
【0014】
特別に定義されない限り、本明細書において使用される全ての技術用語および科学用語は、本発明が属する当業者に一般的に理解されるものと同じ意味を有する。さらに、本明細書で使用する単語「a」、「an」、および「the」は、特別に他に指すものがない限り「少なくとも1つ」を意味する。
【0015】
本発明の実施または試行においては、本明細書に記載されたものに類似するかまたは等価の方法および材料が使用され得るが、適当な方法および材料を以下の明細書に記載する。
【0016】
本明細書において言及される出版物、特許出願、特許、およびその他の参考文献は、その全体が参照により組み入れられる。矛盾する場合、定義を含む本明細書によって統制される。さらに、材料、方法、および実施例は、例示的なものに過ぎず、本発明を制限するためのものではない。
【0017】
発明の詳細な説明
SMYD3 cDNAは、配列番号:1に示したように、1284のヌクレオチドのオープンリーディングフレームを含む1622のヌクレオチドからなる。オープンリーディングフレームは、配列番号:2に示したように、ジンクフィンガーモチーフおよびSETドメインを有する428アミノ酸のタンパク質をコードする。ジンクフィンガードメイン(MYND)はアミノ酸49からアミノ酸87にわたり、SET(Su3-9、Enhancer-of-zeste、Trihorrax)ドメインはアミノ酸117からアミノ酸246にわたる。
【0018】
SMYD3タンパク質の細胞内局在は、細胞周期の進行中に、および培養細胞の密度によって変化する。SMYD3タンパク質は、細胞がS期中期から後期にある時、または低密度な条件で培養されている時には核に蓄積する。しかしながら、SMYD3タンパク質は、細胞が細胞周期の他の期にある時、または高密度な条件で培養されている時には細胞質ならびに核に局在する。
【0019】
従って、本発明は、SMYD3メチルトランスフェラーゼ活性を調整する作用因子をスクリーニングする方法を提供する。この方法は、SMYD3ポリペプチドまたはメチルトランスフェラーゼ活性を有するその機能的等価物と網膜芽細胞腫タンパク質とを接触させ、接触されたSMYD3またはその機能的等価物のメチルトランスフェラーゼ活性をアッセイすることによって行われる。これによって、SMYD3または機能的等価物のメチルトランスフェラーゼ活性を調整する作用因子が同定される。
【0020】
本発明において、「機能的に等価な」という用語は、本タンパク質またはポリペプチドが、SMYD3と同じメチルトランスフェラーゼ活性または実質的に同じメチルトランスフェラーゼ活性を有することを意味する。特に、このタンパク質は、網膜芽細胞腫タンパク質またはリジン824を含む網膜芽細胞腫タンパク質断片のメチル化を触媒する。本タンパク質が標的活性を有するか否かは、本発明によって日常的に決定することができる。すなわち、メチルトランスフェラーゼ活性は、(a)ポリペプチドと基質(例えば、網膜芽細胞腫タンパク質またはリジン824を含む断片)および補助因子(例えば、S-アデノシル-L-メチオニン)を、基質のメチル化に適した条件下で接触させ、(b)基質のメチル化レベルを検出することによって決定することができる。
【0021】
本明細書で使用する「網膜芽細胞腫ペプチド」という用語は、完全長の網膜芽細胞腫タンパク質(例えば、配列番号:4)ならびにその変異体および断片を意味する。機能的断片の例には、C末端断片、例えば、配列番号:4のアミノ酸769〜921により構成される断片が含まれるが、これに限定されない。好ましい断片は、位置824にあるリジン残基を含む。機能的変異体の例には、完全長網膜芽細胞腫タンパク質のメチル化能を保持している以下のRB1変異体:K889A、K896A、K791A、K814A、K791A/K824A、およびK814A/K824Aが含まれるが、これに限定されない。
【0022】
所定のタンパク質と機能的に等価であるタンパク質を調製する方法は当業者に周知であり、これにはタンパク質に変異を導入する従来の方法が含まれる。例えば、当業者は、例えば部位特異的突然変異を用いてヒトSMYD3タンパク質のアミノ酸配列内に適切な変異を導入することにより、ヒトSMYD3タンパク質と機能的に等価のタンパク質を調製することができる(Hashimoto-Gotoh, T. et al. (1995), Gene 152, 271〜275;Zoller, MJ, and Smith, M. (1983), Methods Enzymol. 100, 468〜500;Kramer, W. et al. (1984), Nucleic Acids Res. 12, 9441〜9456;Kramer W, and Fritz HJ. (1987) Methods. Enzymol. 154, 350〜367;Kunkel, TA (1985), Proc. Natl. Acad. Sci. USA. 82,488〜492)。アミノ酸変異は自然下でも起こり得る。本発明の状況において有用なSMYD3ポリペプチドは、結果的に生じた変異タンパク質がヒトSMYD3タンパク質と機能的に等価であり、より具体的にはヒトSMYD3タンパク質のメチルトランスフェラーゼ活性を保持するという前提で、1つまたは複数のアミノ酸が変異したヒトSMYD3タンパク質のアミノ酸配列を有するタンパク質を含む。このような変異体において変異させるアミノ酸の数は一般に20アミノ酸またはそれ未満、典型的には10アミノ酸またはそれ未満、好ましくは6アミノ酸またはそれ未満、およびより好ましくは3アミノ酸またはそれ未満である。メチルトランスフェラーゼ活性を維持するために、SETドメイン「NHSCXXN」および[GEELXXXY」が、変異タンパク質のアミノ酸配列中に好ましくは保存されている(「X」は任意のアミノ酸を表す)。
【0023】
変異または改変されたタンパク質、すなわち、特定のアミノ酸配列の一つまたは複数のアミノ酸残基を欠失、付加、および/または置換することによって改変されたアミノ酸配列を有するタンパク質は、当初のタンパク質の生物学的活性を保持することが公知である(Mark, D.F. et al.、Proc. Natl. Acad. Sci. USA(1984)81:5662〜5666;Zoller, M.J.およびSmith, M.、Nucleic Acids Research(1982)10:6487〜6500;Wang, A. et al.、Science 224:1431〜1433;Dalbadie-McFarland, G. et al.、Proc. Natl. Acad. Sci. USA(1982)79:6409〜6413)。
【0024】
変異されるアミノ酸残基は、好ましくは、アミノ酸側鎖の特性が保存され得る異なるアミノ酸に変異される(保存的アミノ酸置換として公知の過程)。アミノ酸側鎖の特性の例は、疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)、および以下の官能基または共通の特徴を有する側鎖:脂肪族側鎖(G、A、V、L、I、P);ヒドロキシル基含有側鎖(S、T、Y);硫黄原子含有側鎖(C、M);カルボン酸およびアミド含有側鎖(D、N、E、Q);塩基含有側鎖(R、K、H);ならびに芳香族含有側鎖(H、F、Y、W)を含む。括弧内の文字はアミノ酸の一文字コードを示すことに注意されたい。
【0025】
一つまたは複数のアミノ酸残基においてヒトSMYD3タンパク質のアミノ酸配列(配列番号:2)に付加されるタンパク質の例は、ヒトSMYD3タンパク質を含む融合タンパク質である。融合タンパク質は、ヒトSMYD3タンパク質と他のペプチドまたはタンパク質との融合体を含み、これらも本発明に用いられる。融合タンパク質は、当業者に周知の技術、例えば本発明のヒトSMYD3タンパク質をコードするDNAを、他のペプチドまたはタンパク質をコードするDNAに、フレームが一致するように連結させ、融合DNAを発現ベクターに挿入し、宿主でそれを発現させることによって作製することができる。本発明のタンパク質に融合させるペプチドまたはタンパク質に関して制限はない。
【0026】
SMYD3タンパク質に融合されるペプチドとして用いることができる公知のペプチドには、例えば、FLAG(Hopp, T.P.et al.,Biotechnology(1986)6:1204〜1210)、6個のHis(ヒスチジン)残基を含む6×His、10×His、インフルエンザ凝集素(HA)、ヒトc-myc断片、VSP-GP断片、p18HIV断片、T7-タグ、HSV-タグ、E-タグ、SV40T抗原断片、lckタグ、α-チューブリン断片、B-タグ、プロテインC断片等が含まれる。本発明のタンパク質に融合し得るタンパク質の例には、GST(グルタチオン-S-トランスフェラーゼ)、インフルエンザ凝集素(HA)、免疫グロブリン定常領域、β-ガラクトシダーゼ、MBP(マルトース結合タンパク質)等が含まれる。
【0027】
融合タンパク質は、上記の融合ペプチドまたはタンパク質をコードする市販のDNAを、本発明のタンパク質をコードするDNAと融合させて、調製された融合DNAを発現させることによって調製することができる。
【0028】
機能的に等価なタンパク質を単離するための当技術分野で公知のもう一つの方法は、相同的な配列を同定するためのハイブリダイゼーション技術(Sambrook, J.et al.、Molecular Cloning、第2版、9.47〜9.58、コールドスプリングハーバー研究所出版、1989)を用いる方法である。当業者は、ヒトSMYD3タンパク質をコードするSMYD3 DNA配列(例えば、配列番号:1)の全て、または一部と高い相同性を有するDNAを容易に単離することができ、単離されたDNAからヒトSMYD3タンパク質と機能的に等価なタンパク質を単離することができる。本発明のタンパク質には、ヒトSMYD3タンパク質をコードするDNA配列の全てまたは一部とハイブリダイズするDNAによってコードされるタンパク質、およびヒトSMYD3タンパク質と機能的に等価であるタンパク質が含まれる。これらのタンパク質には、ヒトまたはマウスに由来するタンパク質に対応する哺乳類相同体が含まれる(例えば、サル、ラット、ウサギ、およびウシ遺伝子によってコードされるタンパク質)。ヒトSMYD3タンパク質をコードするDNAと高い相同性を示すcDNAを動物から単離する場合、骨格筋、精巣、HCC、または結腸直腸腫瘍由来の組織を用いることが特に好ましい。
【0029】
ヒトSMYD3タンパク質と機能的に等価であるタンパク質をコードするDNAを単離するためのハイブリダイゼーション条件は、当業者によって日常的に選択することができる。例えば、ハイブリダイゼーションは、「Rapid-hyb緩衝液」(Amersham LIFE SCIENCE)を用いて、68℃で30分またはそれ以上プレハイブリダイゼーションを行い、標識プローブを加えて、68℃で1時間またはそれ以上加温することによって行ってもよい。次に続く洗浄段階は、例えば低ストリンジェント条件で行うことができる。低ストリンジェンシー条件は、例えば、42℃、2×SSC、0.1%SDS、または好ましくは50℃、2×SSC、0.1%SDSである。より好ましくは高ストリンジェント条件を用いる。本発明の状況において、高ストリンジェント条件は、例えば、2×SSC、0.01%SDS中で室温20分間の洗浄を3回行い、次いで1×SSC、0.1%SDS中で37℃20分間の洗浄を3回行い、かつ1×SSC、0.1%SDS中で50℃20分間の洗浄を2回行うことを含む。しかしながら、温度および塩濃度のようないくつかの要因がハイブリダイゼーションのストリンジェンシーに影響を及ぼす可能性があり、当業者は必須のストリンジェンシーを達成するために要因を適切に選択することができる。
【0030】
ハイブリダイゼーションの代わりに、遺伝子増幅法、例えば、ヒトSMYD3タンパク質(配列番号:2)をコードするDNA(配列番号:1)の配列情報に基づいて合成されたプライマーを用い、ポリメラーゼ連鎖反応(PCR)法を用いて、ヒトSMYD3タンパク質と機能的に等価なタンパク質をコードするDNAを単離することができる。
【0031】
上記のハイブリダイゼーション技術または遺伝子増幅技術によって単離されたDNAによってコードされる、ヒトSMYD3タンパク質と機能的に等価であるタンパク質は、通常、ヒトSMYD3タンパク質のアミノ酸配列に対して高い相同性を有する。「高い相同性」(「高い同一性」とも呼ばれる)とは典型的に、2つの最適に整列化された配列(ポリペプチド配列またはポリヌクレオチド配列)間の同一性の程度を指す。典型的には、高い相同性または同一性とは、40%またはそれ以上、好ましくは60%またはそれ以上、より好ましくは80%またはそれ以上、さらにより好ましくは85%、90%、95%、98%、99%またはそれ以上の相同性を指す。2つのポリペプチド配列またはポリヌクレオチド配列の間の相同性または同一性の程度は、「Wilbur, W. J. and Lipman, D. J. Proc.Natl. Acad.Sci. USA (1983) 80, 726〜730」におけるアルゴリズムに従って決定することができる。
【0032】
本発明の状況において有用なタンパク質は、その産生のために用いられる細胞もしくは宿主または用いる精製方法に依存して、アミノ酸配列、分子量、等電点、糖鎖の有無または形態に関して差異がある可能性がある。しかしながら、それがヒトSMYD3タンパク質(配列番号:2)の機能と等価である限り、それは本発明において有用である。
【0033】
本発明の状況において有用なタンパク質は、当業者に周知の方法によって、組換え型タンパク質または天然のタンパク質として調製することができる。組換え型タンパク質は、本発明のタンパク質をコードするDNA(例えば、配列番号:1のヌクレオチド配列を含むDNA)を、適当な発現ベクターに挿入し、ベクターを適当な宿主細胞に導入し、抽出物を得、かつ抽出物にクロマトグラフィー、例えばイオン交換クロマトグラフィー、逆相クロマトグラフィー、ゲル濾過、または本発明のタンパク質に対する抗体を固定したカラムを利用するアフィニティクロマトグラフィーを行うか、または上記のカラムの複数を組み合わせることにより、タンパク質を精製することによって調製され得る。
【0034】
さらに、本発明の状況において有用なタンパク質が宿主細胞(例えば、動物細胞および大腸菌(E.coli))内において、グルタチオン-S-トランスフェラーゼタンパク質との融合タンパク質として、または多数のヒスチジンを加えた組換え型タンパク質として発現する場合、発現された組換え型タンパク質は、グルタチオンカラムまたはニッケルカラムを用いて精製することができる。
【0035】
融合タンパク質を精製した後、必要に応じてトロンビンまたは第Xa因子による切断によって、目的とするタンパク質以外の領域を除去することも可能である。
【0036】
天然のタンパク質は、当業者に公知の方法、例えば下記のSMYD3タンパク質に結合する抗体を結合させたアフィニティカラムを、本発明のタンパク質を発現する組織または細胞の抽出物に接触させることによって単離することができる。抗体はポリクローナル抗体またはモノクローナル抗体であり得る。
【0037】
本発明において、SMYD3ポリペプチドのメチルトランスフェラーゼ活性は、当技術分野で公知の方法によって決定することができる。例えば、SMYD3ポリペプチド基質および網膜芽細胞腫ペプチド基質は、標識されたメチル供与体と共に、適したアッセイ条件下でインキュベートされ得る。好ましいメチル供与体の例は、S-アデノシル-[メチル-14C]-L-メチオニンおよびS-アデノシル-[メチル-3H]-L-メチオニンを好ましくは含むが、これらに限定されない。放射性標識の網膜芽細胞腫ペプチドへの移行は、例えば、SDS-PAGE電気泳動およびフルオログラフィーによって検出することができる。または、反応に続き、網膜芽細胞腫ペプチドを濾過によってメチル供与体から分離し、かつフィルター上に保持された放射標識の量をシンチレーション計数法によって定量することができる。メチル供与体と結合し得る他の標識、例えば発色性標識および蛍光標識、ならびにこれらの標識の網膜芽細胞腫ペプチドへの移行を検出する方法は当技術分野で公知である。
【0038】
または、SMYD3のメチルトランスフェラーゼ活性を、非標識メチル供与体(例えば、S-アデノシル-L-メチオニン)、およびメチル化網膜芽細胞腫ペプチドを選択的に認識する試薬を用いて決定することができる。例えば、SMYD3、メチル化しようとする基質、およびメチル供与体を、基質のメチル化に好適な条件下でインキュベートした後、メチル化基質を従来の免疫学的方法を用いて検出することができる。メチル化基質を認識する抗体を用いる任意の免疫学的技術を検出のために用いることができる。
【0039】
さらに、Lys824がメチル化されたRB1において、RB1のSer807およびSer811のリン酸化が増強されたことが確認された。従って、別の態様において、RB1のメチル化レベルはRB1のリン酸化を介して評価されてもよい。CDK2またはCDK6などのキナーゼもRB1のリン酸化に必要なことがある。RB1のリン酸化は、放射標識リン酸供与体を用いて検出されてもよい。あるいは、RB1のリン酸化部位を認識する抗体がRB1のリン酸化レベルの評価に用いられてもよい。
【0040】
さまざまなロースループットおよびハイスループット酵素アッセイ形式が当技術分野で公知であり、かつSMYD3のメチルトランスフェラーゼ活性の検出または測定のために容易に適合させることができる。ハイスループットアッセイ法のために、網膜芽細胞腫ペプチド基質は、マルチウェルプレート、スライドガラス、またはチップなどの固体支持体上に、都合良く固定化され得る。反応に続き、メチル化産物を固体支持体上で上記の方法によって検出することができる。または、メチルトランスフェラーゼ反応は溶液中で行われ得、その後網膜芽細胞腫ペプチドが固体支持体上に固定化され得、およびメチル化産物が検出される。そのようなアッセイ法を容易にするため、固体支持体はストレプトアビジンでコーティングされ得、かつビオチン標識網膜芽細胞腫、または固体支持体は抗網膜芽細胞腫抗体でコーティングされ得る。当業者は、スクリーニングの所望のスループット能力に依存して、適したアッセイ形式を決定することができる。
【0041】
本発明はまた、本発明のタンパク質の部分的ペプチドの使用方法も含む。部分的ペプチドは、SMYD3のタンパク質に特異的なアミノ酸配列を有し、かつ約400アミノ酸未満、通常は約200アミノ酸未満、およびしばしば約100アミノ酸未満であり、ならびに少なくとも約7アミノ酸、好ましくは約8アミノ酸またはそれ以上、およびより好ましくは約9アミノ酸またはそれ以上からなる。部分的ペプチドは、例えば、SMYD3のタンパク質と結合する作用因子または化合物のスクリーニングにおいて、および、SMYD3とその補助因子、例えば、SAMなど、との間の結合の阻害物質のスクリーニングために用いられ得る。そのようなスクリーニングのために、好ましくはSETドメインを含む部分的ペプチドが用いられる。
【0042】
本発明の状況において有用な部分的ペプチドは、遺伝子操作により、ペプチド合成の公知の方法により、または本発明のタンパク質を適切なペプチダーゼで消化することにより作製され得る。ペプチド合成のために、例えば、固相合成または液相合成が用いられ得る。
【0043】
SETドメインの変異を有するSMYD3変異体は、細胞増殖に対する阻害作用を示す。したがって、SMYD3の部分的ペプチドは、好ましくは、SETドメイン「NHSCXXN」および/または「GEELXXXY」を含む。
【0044】
任意の試験作用因子が用いられ得る。例として、細胞抽出物、細胞培養上清、発酵微生物の産物、海洋生物からの抽出物、植物抽出物、精製タンパク質または粗タンパク質、ペプチド、非ペプチド化合物、合成マイクロ分子化合物および天然化合物を含むが、これらに限定されない。
【0045】
本明細書に記載したアッセイ法において有用な試験作用因子または試験化合物は、SMYD3と特異的に結合する抗体、またはメチルトランスフェラーゼ活性を持たない部分的SMYD3ペプチドという形をとり得る。例えば、抗体(例えば、モノクローナル抗体)は、SMYD3とその網膜芽細胞腫基質との間の結合をブロックする能力に関して試験され得る。
【0046】
本発明のスクリーニング方法によって単離された作用因子または化合物は、SMYD3のメチルトランスフェラーゼ活性を阻害する薬剤の候補であり、それゆえ、肝細胞癌、結腸直腸癌、乳癌、および/または膀胱癌の治療または予防に応用することができる。
【0047】
さらに、SMYD3のメチルトランスフェラーゼ活性を阻害する作用因子または化合物の構造の一部が、付加、欠失および/または置換によって変換された作用因子または化合物も、本発明のスクリーニング方法によって得られる作用因子および化合物に含まれる。
【0048】
上に注記したように、SMYD3のメチルトランスフェラーゼ活性を阻害する作用因子または化合物は、SMYD3のメチルトランスフェラーゼ活性を欠く部分的ペプチドのいずれかであり得、またはSMYD3に対する抗体であり得る。本明細書で用いる場合、「抗体」という用語は、抗体を合成するために用いた抗原またはそれと密接に関連した抗原のみと相互作用する(すなわち、結合する)、特異的な構造を有する免疫グロブリン分子のことを指す。さらに、抗体は、SMYD3遺伝子によってコードされるタンパク質に結合する限り、抗体または修飾された抗体の断片であってもよい。例えば、抗体断片は、Fab、F(ab')2、Fv、またはH鎖およびL鎖からのFv断片が適当なリンカーによって連結されている一本鎖Fv(scFv)であってもよい(Huston, J.S.et al.、Proc. Natl. Acad. Sci. U.S.A. 85:5879〜5883(1988))。より具体的には、抗体断片は、パパインまたはペプシンのような酵素によって抗体を処理することによって生成してもよい。または、抗体断片をコードする遺伝子を構築して、発現ベクターに挿入し、適当な宿主細胞において発現してもよい(例えば、Co, M.S.et al.、J. Immunol. 152:2968〜2976(1994);Better, M.およびHorwitz, A.H.、Methods Enzymol. 178:476〜496(1989);Pluckthun, A.およびSkerra, A.、Methods Enzymol. 178:497〜515(1989);Lamoyi, E.、Methods Enzymol. 121:652〜663(1986);Rousseaux, J.et al.、Methods Enzymol. 121:663〜669(1986);Bird, R.E.およびWalker, B.W.、Trends Biotechnol. 9:132〜137(1991)を参照されたい)。
【0049】
抗体を、ポリエチレングリコール(PEG)のなどの多様な分子との結合によって修飾してもよい。本発明は、そのような修飾された抗体を提供する。修飾された抗体は、抗体を化学修飾することによって得ることができる。そのような修飾法は当技術分野において一般的である。または、抗体を、非ヒト抗体に由来する可変領域およびヒト抗体に由来する定常領域を有するキメラ抗体、または、非ヒト抗体に由来する相補性決定領域(CDR)、ヒト抗体に由来するフレームワーク領域(FR)、および定常領域を含むヒト化抗体を含み得る。そのような抗体は、公知の技術を用いて調製することができる。ヒト化は、ヒト抗体の対応する配列の代わりに、齧歯類のCDRまたはCDR配列を用いることによって実施され得る(例えば、Verhoeyen et al.、 Science 239:1534〜1536 (1988)を参照されたい)。したがって、そのようなヒト化抗体は、実際には、ヒト可変ドメインの全域より狭い領域が、非ヒト種由来の対応する配列によって置き換えられたキメラ抗体である。
【0050】
ヒトフレームワーク領域および定常領域に加えて、ヒト可変領域を含む完全ヒト抗体もまた、用いることができる。そのような抗体は、当技術分野で公知のさまざまな技術を用いて作製することができる。例えば、バクテリオファージ上に提示されたヒト抗体断片の組換えライブラリーの使用を含むインビトロの方法を用いてもよい(例えば、Hoogenboom & Winter, J. Mol. Biol. 227:381 (1991))。同様に、ヒト免疫グロブリン座位を、内因性免疫グロブリン遺伝子が部分的または完全に不活性化されたトランスジェニック動物、例えばマウスに導入することによってヒト抗体を作製することができる。このアプローチは例えば、米国特許第6,150,584号、第5,545,807号;第5,545,806号;第5,569,825号;第5,625,126号;第5,633,425号;第5,661,016号に記載されている。
【0051】
本発明の方法によって単離された作用因子または化合物を、ヒトおよび他の哺乳動物、例えばマウス、ラット、モルモット、ウサギ、ネコ、イヌ、ヒツジ、ブタ、ウシ、サル、ヒヒ、およびチンパンジーに対して製剤として投与する場合、単離された作用因子または化合物を直接投与してもよく、または公知の薬学的調製法を用いて剤形に製剤化してもよい。例えば、必要に応じて、薬剤を糖衣錠剤、カプセル剤、エリキシル剤、およびマイクロカプセルとして経口投与することもでき、または水もしくは他の任意の薬学的に許容される液体との滅菌溶液もしくは懸濁液である注射剤の形態として非経口的に投与することもできる。例えば、作用因子または化合物を、一般に認められる薬剤の実現のために必要な単位用量の形で、薬学的に許容される担体または媒体、具体的には滅菌水、生理食塩水、植物油、乳化剤、懸濁剤、界面活性剤、安定剤、香味剤、賦形剤、媒質、保存料、結合剤などと混合することができる。これらの製剤における有効成分の量により、指定された範囲内にある適した投与量が得られる。
【0052】
錠剤およびカプセル剤に混合し得る添加剤の例には、ゼラチン、コーンスターチ、トラガカントゴムおよびアラビアゴムなどの結合剤;結晶セルロースなどの媒質;コーンスターチ、ゼラチンおよびアルギン酸などの膨潤剤;ステアリン酸マグネシウムなどの潤滑剤;スクロース、ラクトースまたはサッカリンなどの甘味料;ならびにペパーミント、冬緑油、およびチェリーなどの香味剤がある。単位用量剤形がカプセル剤である場合、油などの液体担体も上記の成分にさらに含めることができる。注射用の滅菌混合物は、通常の薬剤の実現に倣って、注射用の蒸留水などの媒体を用いて製剤化することができる。
【0053】
生理食塩水、グルコース、ならびにD-ソルビトール、D-マンノース、D-マンニトールおよび塩化ナトリウムなどのアジュバントを含むその他の等張液を、注射用の水溶液として用いることができる。これらは適した可溶化剤、例えばアルコール、具体的にはエタノール、プロピレングリコールおよびポリエチレングリコールなどの多価アルコール、Polysorbate 80(商標)およびHCO-50などの非イオン性界面活性剤などと組み合わせて用いることができる。
【0054】
ゴマ油またはダイズ油は油脂性液体として用いることができ、これらを可溶化剤としての安息香酸ベンジルまたはベンジルアルコールと組み合わせて用いることもでき、さらにこれらをリン酸緩衝液および酢酸ナトリウム緩衝液などの緩衝液;塩酸プロカインなどの鎮痛薬;ベンジルアルコールおよびフェノールなどの安定剤;ならびに抗酸化剤と共に製剤化することもできる。調製された注射剤は適したアンプルに充填することができる。
【0055】
本発明の薬学的組成物を、例えば動脈内注射、静脈内注射、または皮下注射として、および同じく鼻腔内投与、筋肉内投与または経口投与として患者に投与するために、当業者に周知の方法を用いることができる。投与量および投与方法は患者の体重および年齢ならびに投与方法に応じて異なる。しかしながら、当業者は日常的に適切な投与方法を選択することができる。さらに、関心対象の前記作用因子または前記化合物がDNAによってコードされ得る場合、DNAを遺伝子治療用のベクターに挿入し、治療を行うためにそのベクターを患者に投与することができる。投与量および投与方法は患者の体重、年齢および症状に応じて異なるが、当業者はそれらを適切に選択することができる。
【0056】
例えば、SMYD3と結合してその活性を調節する作用因子または化合物の典型的な投与量は、その症状に依存するとはいえ、標準的な成人(体重60 kg)に対して経口投与する場合、約0.1 mg〜約100 mg/日、好ましくは約1.0 mg〜約50 mg/日、およびより好ましくは約1.0 mg〜約20 mg/日の範囲である。
【0057】
標準的な成人(体重60 kg)に対して注射剤の形態として非経口的に投与する場合、患者、標的臓器、症状および投与方法に応じて若干の違いはあるものの、約0.01 mg〜約30 mg/日、好ましくは約0.1〜約20 mg/日、およびより好ましくは約0.1〜約10 mg/日の用量を静脈注射することが好都合である。同じく、他の動物の場合にも、体重60 kgに換算した量を投与することが可能である。
【0058】
本発明はさらに、対象における、肝細胞癌、結腸直腸癌、膀胱癌、および乳癌などの、癌を治療するための方法を提供する。投与は、SMYD3の異常なメチルトランスフェラーゼ活性と関連する障害に対するリスクのある(もしくは感受性のある)またはそのような障害を有する対象に対して、予防的または治療的に行うことができる。本方法は、適当な癌細胞におけるSMYD3の機能を低下させる段階を含む。機能は、本発明のスクリーニング方法によって得られた作用因子または化合物の投与によって阻害することができる。
【0059】
もう一つの局面において、本発明は、本明細書に記載の1つまたは複数の治療的作用因子または治療的化合物を含む、薬学的または治療的な組成物を含む。または、本発明は、癌、より具体的には、肝細胞癌、結腸直腸癌、膀胱癌、および乳癌の治療および/もしくは予防のための薬学的または治療的な組成物を製造するための、本明細書に記載の1つまたは複数の治療的作用因子または治療的化合物の使用方法を提供する。薬学的製剤には、経口、直腸、鼻、局所(頬および舌下を含む)、膣、もしくは非経口(筋肉内、皮下、および静脈内を含む)の投与に適したもの、または吸入もしくは通気による投与に適したものが含まれ得る。製剤は、適宜、個別の投与量単位で都合良く提示でき、薬学分野における周知の方法のいずれかによって調製できる。そのような薬学の方法は、全て、必要に応じて、液状担体もしくは微細に粉砕された固形担体またはその両方と活性化合物とを会合させる段階を含み、かつ必要であれば、次いで、その生成物を所望の製剤へと成形する段階を含む。
【0060】
経口投与に適した薬学的製剤は、所定の量の活性成分を各々含有しているカプセル剤、カシェ剤、もしくは錠剤のような個別の単位として;散剤もしくは顆粒剤として;または液剤、懸濁剤、もしくは乳濁剤として、都合良く提示できる。活性成分は、ボーラス舐剤またはペースト剤として提示してもよく、そして純粋な(即ち、担体を含まない)形態であってもよい。経口投与用の錠剤およびカプセル剤は、結合剤、増量剤、滑沢剤、崩壊剤、または湿潤剤のような従来の賦形剤を含有し得る。錠剤は、任意で、1個以上の製剤化成分を用いて、圧縮または成型により作成してもよい。圧縮錠剤は、結合剤、滑沢剤、不活性希釈剤、滑沢、界面活性、または分散剤と任意に混合された散剤または顆粒剤のような流動性の高い形態の活性成分を適当な機械で圧縮することにより調製できる。成型錠剤は、不活性液状希釈剤で湿らされた粉末化合物の混合物を適当な機械で成型することにより作成できる。錠剤は、当技術分野における周知の方法に従い、剤皮を施してもよい。経口液状調製物は、例えば、水性もしくは油性の懸濁剤、液剤、乳濁剤、シロップ剤、もしくはエリキシル剤の形態であってもよいし、または乾燥生成物として提示し、使用前に水もしくはその他の適当な媒体で調製してもよい。そのような液状調製物は、懸濁化剤、乳化剤、非水性媒体(食用油を含み得る)、または保存剤のような従来の添加剤を含有し得る。錠剤は、任意で、含まれる活性成分を徐放するようにまたは放出が制御されるように製剤化してもよい。
【0061】
非経口投与用の製剤には、抗酸化剤、緩衝剤、静菌剤、および対象となるレシピエントの血液と等張にする溶質を含有し得る水性および非水性の無菌の注射液剤;ならびに懸濁化剤および濃化剤を含み得る水性および非水性の無菌の懸濁剤が含まれる。製剤は、単位用量容器または多回用量容器、例えば、密封されたアンプルおよびバイアルで提示でき、使用直前に無菌の液状担体、例えば、生理食塩水、注射用水の添加のみを必要とする、フリーズドライ(凍結乾燥)状態で保管してもよい。または、製剤は連続注入用に提示してもよい。即席の注射用の液剤および懸濁剤は、前記の種類の無菌の散剤、顆粒剤、および錠剤から調製できる。
【0062】
直腸投与用の製剤は、カカオバターまたはポリエチレングリコールのような通常の担体を含む坐剤として提示できる。口内、例えば、頬または舌下への局所投与用の製剤には、ショ糖およびアラビアゴムもしくはトラガカントゴムのような風味付きの基剤中に活性成分が含まれたロゼンジ剤、ならびにゼラチンおよびグリセリンもしくはショ糖およびアラビアゴムのような基剤中に活性成分が含まれた香錠が挙げられる。鼻腔内投与の場合、本発明の化合物は、液状スプレーもしくは分散可能な散剤として、または滴剤の形態で使用できる。滴剤は、1つ以上の分散剤、可溶化剤、または懸濁化剤も含む水性または非水性の基剤を用いて製剤化できる。液状スプレーは、加圧パックから都合良く送達される。
【0063】
吸入による投与の場合、化合物は、注入器、ネブライザー、加圧パック、またはエアロゾルスプレーを送達するその他の便利な手段から都合良く送達される。加圧パックは、ジクロロジフルオロメタン、トリクロロフルオロメタン、ジクロロテトラフルオロエタン、二酸化炭素、またはその他の適当なガス等の適当な噴霧剤を含み得る。加圧エアロゾル剤の場合、投与量単位はバルブによって決定され、計量された量が送達される。
【0064】
または、吸入または通気による投与の場合、化合物は、乾燥粉末組成物、例えば、化合物と乳糖またはデンプンのような適当な粉末基剤との粉末混合物の形態をとり得る。粉末組成物は、単位剤形、例えば、カプセル、カートリッジ、ゼラチン、またはブリスター包装で提示でき、それらから、吸入器または注入器を使用して粉末を投与できる。
【0065】
所望の場合、活性成分を徐放するよう適合させた上記の製剤が利用されてもよい。薬学的組成物は、抗微生物剤、免疫抑制剤、または保存剤のようなその他の活性成分を含有していてもよい。
【0066】
特に上で言及した成分に加え、本発明の製剤には、問題となる製剤の型を考慮した上で、当技術分野において一般的なその他の薬剤が含まれていてもよいこと、例えば、経口投与に適した薬剤には着香料が含まれていてもよいことが理解される。
【0067】
好ましい単位投与量製剤は、下記のような有効量またはその活性成分の適切な画分を含有しているものとする。
【0068】
前記の各条件のため、組成物は、1日当たり約0.1〜約250 mg/kgの用量で経口的にまたは注射により投与され得る。成人のヒトに対する用量範囲は、一般に、約5 mg〜約17.5 g/日、好ましくは約5 mg〜約10 g/日、最も好ましくは約100 mg〜約3 g/日である。個別の単位で提供される錠剤またはその他の単位剤形には、そのような投与量またはその多回数の投与量で有効量となる量、例えば、約5 mg〜約500 mg、通常約100 mg〜約500 mgを含有する単位が都合良く含まれる。
【0069】
薬学的組成物は、好ましくは、経口的にまたは注射(静脈内もしくは皮下)により投与され、対象へ投与される正確な量は、担当医師の責任であろう。しかしながら、用いられる用量は、対象の年齢および性別、治療が施される正確な障害およびその重傷度を含む多数の要因によるものと考えられる。また、投与経路も、状態およびその重症度によって変動し得る。
【0070】
以下の実施例は単なる例示であり、本発明の範囲を限定することを目的としない。本発明の局面を以下の実施例において説明するが、当業者であれば、本明細書に記載のものと類似のまたは等価な他の方法および材料を、本発明の実施または試験において使用できることを理解するだろう。
【0071】
実施例
材料および方法:
試薬:
抗RB(IF8)抗体、抗リン酸化RB(Ser807/811,sc-16670)抗体、および抗リン酸化RB(Thr821/826)抗体はSanta Cruz Biotechnologyから購入し、抗Flag抗体はSIGMAから購入し、抗パンメチルリジン抗体(ab7315)は Abcam Ltd.から購入した。組換えSMYD3タンパク質、またはジメチル化リジン824もしくはトリメチル化リジン824を含有する合成RB1ペプチド(残基820〜828)をウサギ(SIGMA-ALDRICH、St.Louis、MO)に接種し、免疫したウサギの血清からポリクローナル抗体を精製した。組換えC末端GST-RB1および完全長GST-p53タンパク質はSanta Cruz Biotechnologyから購入し、His結合C末端RB1、CDK2/サイクリンE、およびCDK6/サイクリンD3タンパク質はUpstate Biotechnologyから購入し、完全長組換えRBタンパク質(3108)はQED Bioscienceから購入した。S-(5'-アデノシル)-L-ホモシステイン加水分解酵素(SAHH)はSIGMAから入手した。
【0072】
メチルトランスフェラーゼおよびキナーゼのインビトロアッセイ法:
293T細胞を、Flagタグ化野生型SMYD3を発現するプラスミド(p3XFLAG-CMV-SMYD3)、Flagタグ化変異体SMYD3を発現するプラスミド(p3XFLAG-CMV-SMYD3ΔEEL)でトランスフェクトし、抗Flag抗体を用いた免疫沈降によってタグ化SMYD3タンパク質を精製した。バキュロウイルス系(Clontech)を用いてSf9細胞において、組換えSMYD3タンパク質を調製した。インビトロHMTアーゼアッセイ法は、他の記載に(Strahl, B. D., et al. Proc Natl Acad Sci U S A 96,14967-72 (1999))、軽微な変更を加えて実施した。簡単に述べると、メチルトランスフェラーゼ緩衝液(50mM Tris-HCl pH8.5、100mM NaCl、10mM DTT)の緩衝液中に、免疫沈降したSMYD3タンパク質または組換えSMYD3タンパク質を、メチル供与体として2μCiの[メチル-3H]標識S-アデノシル-L-メチオニン(SAM, Amersham Biosciences)の存在下で、1μgの組換えヒストンH3、RB1、またはp53タンパク質と混合した。反応混合物を30℃で1時間インキュベートした。タンパク質をSDS-PAGEで分離し、標識されたタンパク質をフルオログラフィーにより検出した。CDK2/サイクリンEおよびCDK6/サイクリンD3のインビトロキナーゼアッセイ法は、製造業者(Upstate Biotechnology)のプロトコールに従って実施した。非メチル化RB1およびメチル化RB1(#12-439、Upstate Biotechnology)の両方を反応基質として使用した。
【0073】
インビボメチル化アッセイ法:
メチル化RB1をインビボで測定するために、LiuおよびDreyfuss (Liu, Q. & Dreyfuss, G. Mol Cell Biol 15, 2800-8 (1995))により記載の方法に従い、軽微な変更を加えて、RB1のインビボ標識を[メチル-3H]標識S-アデノシル-L-メチオニンを用いて培養細胞において実施した。HEK293細胞を、100μg/mlのシクロヘキシミドおよび40μg/mlのクロラムフェニコールと共に37℃で30分間インキュベートした。次いで、培地を、10μCi/mlのL-[メチル-3H]メチオニンおよびタンパク質合成阻害剤を含有し非標識メチオニンを含有しない培地と交換し、さらに3時間維持した。全細胞溶解産物を、抗RB抗体(IF8; Santa Cruz Biotechnology)を用いた免疫沈降に供した。免疫沈降したRB1タンパク質をSDS-PAGEで分離し、その後に、ニトロセルロース膜に転写した。ニトロセルロース膜を、BASイメージングシステム(BAS-TR2040、FUJI)またはイムノブロット分析で分析した。
【0074】
免疫細胞化学染色:
チャンバースライド上の培養細胞を、4%パラホルムアルデヒド含有PBSで15分間固定し、次いで、0.1%Triton X-100含有PBSを用いて室温で2.5分間、透過性にした。非特異的ハイブリダイゼーションをブロックするために、細胞を、PBSに溶解した2%BSAを用いて4℃で24時間覆い、次いで、一次抗体として抗SMYD3抗体、抗RB[IF8]抗体、および抗リン酸化RB(Thr 821/826)抗体と共にインキュベートした。二次抗体として、蛍光基質結合抗ウサギまたは抗マウスIgG(Molecular Probes)を使用した。核は、4',6-ジアミジノ-2-フェニルインドール二塩酸塩(DAPI)を用いて対比染色した。TCS-SP2共焦点顕微鏡(Leica)を用いて、蛍光画像を得た。
【0075】
ルシフェラーゼアッセイ法:
ルシフェラーゼアッセイ法は、Dual-Luciferase Reporter Assay Systemを製造業者の説明書(Promega)に従って使用して実施した。
【0076】
細胞株および組織標本:
ヒト胚腎臓293(HEK293)細胞、HEK293T細胞、およびHEK293F細胞はIWAKIから購入した。ヒト肝癌細胞株HepG2、ならびにHCT116およびSW480ヒト結腸癌株は、American Type Culture Collection (ATCC)から入手した。ヒトHCC細胞株SNU423は、韓国細胞株バンクからの贈与物であった。T47DおよびMCF7乳癌細胞株は、財団法人癌研究会癌研究所から厚意により提供された。全ての細胞株を適切な培地において単層で増殖させた。原発性乳癌組織は、患者からインフォームドコンセントを得て入手した(Hamamoto, R. et al. Cancer Sci 97, 113-118 (2006))。
【0077】
プラスミドの調製:
C末端FLAGタグ化SMYD3の調製は以前に説明されている(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004))。さらに、本発明者らは、野生型SMYD3 cDNAかまたは欠失型SMYD3 cDNAかを含む様々なPCR産物をpCMV-HA(Clontech)ベクターまたはp3XFLAG-CMV 14(Sigma)ベクターの適切な部位にクローニングすることによって、N末端HAタグ化SMYD3またはN末端3xFLAGタグ化SMYD3を発現するプラスミドを調製した。野生型プラスミドに使用したプライマーは、5’-AAGCTTGCGGCCGCGATGGAGCCGCTGAAGGTGGAAAAG-3’(配列番号: 5)および5’-GGTACCTCTAGATTAGGATGCTCTGATGTTGGCGTC-3’(配列番号: 6)であり、変異体(FLAG-SMYD3-ΔN44、-ΔN99、-ΔN244、および-Δ34)に使用したプライマーは、それぞれ、5’-GGGGTACCTTAGGATGCTCTGATGTTGGCGTC-3’(配列番号: 7)および5’-CGGAATTCTGGCGCGATGGAGCCGCTGAAGGTGGAAAAG-3’(配列番号: 8)、 5’-CGGAATTCTGACTCCGTTCGACTTCTTGGCAG-3’(配列番号: 9)、5’- CGGAATTCTCGGAAGCAGCTGAGGGACCAGTACTGC-3’(配列番号: 10)、または5’-CGGAATTCACCCTTGGCGTACACGGTGTGCAAGG-3’(配列番号: 11)であった。グリシン15、17、または27での置換を発現する変異体プラスミドは、QuikChange II XL部位特異的変異誘発キットを使用して供給業者のプロトコール(Stratagene、California、USA)に従って調製した。
【0078】
ウエスタンブロット分析:
SMYD3に対するポリクローナル抗体は、他に記載されているように、大腸菌において産生された組換えHisタグ化SMYD3タンパク質で免疫したウサギの血清から精製した。タンパク質を10%SDS-PAGEにより分離し、抗SMYD3抗体、抗HA抗体(Sigma)、抗FLAG抗体(Sigma)、抗GST抗体(Pharmingen)、または抗β-アクチン抗体(Sigma)を用いてイムノブロットした。HRP結合抗ウサギIgG、抗マウスIgG(Amersham Biosciences)、または抗ヤギIgG抗体(Santa Cruz)は、ECL Detection System (Amersham)用の二次抗体とした。
【0079】
切断部位の決定:
C末端FLAGタグ化SMYD3を293F細胞において外因的に発現させた。抗FLAG抗体を用いて細胞から免疫沈降させたSMYD3タンパク質を2枚のSDS-PAGEゲルで分離し、ニトロセルロース膜およびシークエンスグレードPVDF膜に転写した。2種類のSMYD3タンパク質を検出するために、ニトロセルロース膜を、抗FLAG抗体を用いたイムノブロット分析に使用した。本発明者らは、PVDF膜を、酢酸を含まないCBB溶液(0.025%CBBを40%メタノールに溶解したもの)で染色した後に、短いSMYD3に対応するバンドを切り出し、アミノ酸シークエンスに供した。タンパク質のアミノ酸配列は、エドマンアミノ酸シークエンス法(Edman amino acid sequence method)(Shimadzu Biotechnologies、Tokyo、Japan)によって決定した。
【0080】
インビトロヒストンメチルトランスフェラーゼ(HMTアーゼ)アッセイ:
FLAGタグ化SMYD3は、抗FLAG抗体を用いた免疫沈降によって、野生型(p3XFLAG-CMV-SMYD3)を発現する293T細胞または変異体SMYD3(p3XFLAG-ΔN34、-ΔN44、-SETNm1、-SETNm2、および-SETNm3)を発現する293T細胞から精製した。GST融合SMYD3タンパク質は、野生型(GST-SMYD3-wt)を発現する細菌細胞または変異体SMYD3構築物(GST-SMYD3-ΔN9、-ΔN19、-ΔN29、-ΔN44、-ΔN74)を発現する細菌細胞から精製した。インビトロHMTアーゼアッセイ法は、他に記載されているように(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004))実施した。3H-放射能は、液体シンチレーションカウンターで測定した。
【0081】
実施例1:SMYD3の基質としてのRB1
最近の2つの報告から、ヒストンH3-K4メチルトランスフェラーゼSET7/9が基質としてTAF10およびp53を触媒することが示されたため(Chuikov, S. et al., Nature 432, 353-60 (2004))、本発明者らは、ヒストンH3以外のSMYD3(GenBankアクセッション番号AB057595;配列番号;1、2)のさらなる基質を探索した。p53およびRB1は細胞周期進行の周知の制御因子であるので、最初に、これらを候補基質として試験した(GenBankアクセッション番号NM_000321;配列番号;3、4)。研究中に、組換えヒストンH3、野生型p53、およびRB1のC末端領域(コドン769-921)を、メチル供与体である3H標識SAMの存在下で、293T細胞から免疫沈降したSMYD3タンパク質と共にインキュベートした。それに続くPAGEおよびオートラジオグラフィーから、メチル化ヒストンH3に対応するバンドが示された。これは、SMYD3はヒストンH3をメチル化するという知見と一致する。興味深いことに、メチル化RB1に対応するバンドも検出された。しかしながら、メチル化p53に対応するバンドは検出されなかった(図1a)。ヒストンH3およびC末端RB1に対するメチルトランスフェラーゼ(MTアーゼ)活性を、組換えSMYD3タンパク質を用いてさらに測定した。結果から、両基質に対するMTアーゼ活性は用量依存的に増加することが明らかになった(図1b)。特に、C末端RB1に対するMTアーゼ活性は、ヒストンH3と比較してより高かった。さらに、SET7/9も、RB1に対するメチルトランスフェラーゼ活性を有することが発見された(データ示さず)。さらに、SMYD3は完全長RB1をメチル化した(図1c)。このことは、RB1が、インビトロで、2種類のヒストンH3-K4メチルトランスフェラーゼであるSMYD3ならびにSET7/9によってメチル化されることを示唆している。
【0082】
実施例2:RB1タンパク質に対するSMYD3のメチルトランスフェラーゼ活性
SMYD3とRB1タンパク質との間の可能性のある会合を調べるために、HepG2細胞またはHCT116細胞から抽出したタンパク質を抗SMYD3抗体で免疫沈降した。予想通り、抗RB1抗体を用いたイムノブロット分析によって、RB1タンパク質に対応するバンドが観察された(図2a)。会合を担うRB1領域を決定するために、Flagタグ化野生型RB1タンパク質またはFlagタグ化変異体RB1タンパク質を、HEK293細胞においてHAタグ化SMYD3と共に発現させ、抗Flag抗体を用いて免疫沈降を行った。C末端RB1タンパク質のメチル化と一致して、C末端基質ドメイン(コドン772〜928)はSMYD3と相互作用した(図2b)。RB1との結合を担うSMYD3領域を決定するために、野生型SMYD3および様々な型の変異体SMYD3を発現するプラスミドを使用した。野生型SMYD3、ならびにΔ1型(コドン45-428)およびΔ2型(コドン1〜250)の変異体SMYD3はFlagタグ化RB1と相互作用したが、SETドメインを欠くΔ3型(コドン1〜100)はRB1と相互作用しなかった。このことは、SETドメインが会合に必須であることを示唆している(図2c)。先の報告から、ヒストンH3-K9メチルトランスフェラーゼSUV39H1はRBおよびHP1と会合し、その複合体はサイクリンEの転写抑制において役割を果たすことが示された(Nielsen, S. J. et al. Nature 412, 561-565 (2001))。さらに、最近の研究から、ヒストンH4-K20メチルトランスフェラーゼであるSuv4-20h1およびSuv4-20h2の活性は、RB1との相互作用を通じて著しく増強されることが明らかになった(Gonzalo, S. et al. Nat Cell Biol 7, 420-428 (2005))。従って、本発明者らは、RB1がSMYD3のH3-K4メチルトランスフェラーゼ活性を増強するか否かを試験した。結果として、SMYD3を介したヒストンH3のメチル化はRB1による影響を受けなかった(図2d)。特に、SMYD3は、H3-K910またはH4-K20に対してメチルトランスフェラーゼ活性を示さなかった(図2e)。このデータは、SMYD3のH3-K4特異的HMT(ヒストンメチルトランスフェラーゼ)活性を強化し、かつRB1がHMTに依存的な様式でヒストン修飾の役割を果たすことを示唆した。
【0083】
実施例3:RB1のメチル化基質ドメインの同定
RB1の基質ドメインのメチル化を担う残基を決定するために、SET7/9メチルトランスフェラーゼの基質にある保存アミノ酸配列を比較した。メチル化されるリジンの前にはセリンまたはトレオニンがあったので、本発明者らは、候補としてリジン824、リジン889、およびリジン896に焦点を当てた。野生型RB1基質ドメインおよび3種類の変異体RB1基質ドメインの組換えタンパク質を調製した(図3a)。野生型タンパク質と比較して、K889AおよびK896A変異体はSMYD3によってほぼ同じレベルでメチル化された(図3b、c)。しかしながら、K824Aのメチル化は著しく低下した(図3b、c)。さらに、K824A置換でもRB1タンパク質のメチル化は完全に減少しなかったので、チロシンの後にあるリジン791およびリジン814のメチル化を調べた。2つの変異体RB1タンパク質K791AおよびK814Aは、野生型RB1とほぼ同じレベルのメチル化を示した(図3d、e)。さらに、2種類の二重変異体RB1であるK791A/K824AおよびK814A/K824Aは、K824Aタンパク質と同等レベルのメチル化を示した。従って、本発明者らは、このリジン824がメチル化の主要な標的残基であると結論付けた。リジン824のメチル化を確認するために、ジメチル化リジン824またはトリメチル化リジン824を認識するメチル化RB1特異的抗体を調製した。野生型RB1タンパク質のメチル化に従って、この抗体は、SMYD3がヒストンH3リジン4のジメチル化およびトリメチル化を発揮するのと同様に(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004))、イムノブロット分析においてジメチル化およびトリメチル化RB1タンパク質を検出した(図3f)。H3-K4、TAF10、およびp53を含むSET7/9の基質にあるメチル化リジンの前には2つの保存的ペプチド、リジンの位置-2にR/Kおよび位置-1にS/Tがあったが、リジン824の前には-2にPおよび-1にTがあった。RB1はSMYD3ならびにSET7/9によってメチル化されるので、-2のR/Kは必須ではないかもしれないが、-1のS/TはSMYD3またはSET7/9によるメチル化に欠くことができない。
【0084】
実施例4:インビボメチル化アッセイ法
インビボでのSMYD3によるRB1メチル化をさらに調べるために、SMYD3を発現しないHEK293細胞を使用して、インビボメチル化アッセイ法を行った(Liu, Q. & Dreyfuss, G. Mol Cell Biol 15, 2800-8 (1995))。SMYD3を発現するHEK293細胞株(HEK-SMYD3-1および-2)(図4a)を樹立し、細胞を、タンパク質合成阻害剤の存在下でL-[メチル-3H]メチオニンとインキュベートした。次いで、細胞からの抽出物を抗RB1モノクローナル抗体で免疫精製し(immunopurified)、免疫沈降したタンパク質をSDS-PAGEで、その後にオートラジオグラフィーで分析した。モックをトランスフェクトしたHEK293細胞(HEK-モック-1および-2)と比較して、HEK-SMYD3(HEK-SMYD3-1および-2)細胞からの抽出物は、メチル化RB1に対応する著しく強いバンドを示した。免疫沈降したRB1の量は細胞株間で変化しなかった(図4b)。抗パンメチル-リジン抗体、抗ジメチル化RB1-リジン824抗体、または抗トリメチル化RB1-リジン824抗体を使用したウエスタンブロット分析によって、HEK-モック細胞と比較してHEK-SMYD3細胞におけるメチル化RB1の増加が一貫して観察された(図4c)。HEK-SMYD3細胞の免疫細胞化学染色から、多量のSMYD3を発現する細胞は、抗ジメチル化または抗トリメチル化RB1-リジン824抗体で、より少量のSMYD3を発現する細胞よりも強く染色された(それぞれ、図4d〜g、4h〜k)ことが示された。このデータは、インビボでのSMYD3によるRB1-リジン824のメチル化を裏付けている。
【0085】
実施例5:インビボリン酸化アッセイ法
RB1のリジン824は、トレオニン821stとトレオニン826thの間に位置する。トレオニン821st残基およびトレオニン826th残基はCDK/サイクリン複合体によってリン酸化され、中央のポケットドメインのコンホメーションを変えることによってRB1とE2Fとの間の相互作用を調節する。これらの周囲のトレオニンリン酸化に及ぼすRB1メチル化の影響を調べるために、メチル化または非メチル化RB1タンパク質を用いて、インビボリン酸化アッセイ法を行った。組換えC末端RB1を、SMYD3の存在下または非存在下で3H標識SAMとインキュベートし、次いで、32P-γATPを組換えCDK2/サイクリンEまたはCDK6/サイクリンD3と組み合わせて混合した。組換えRB1のメチル化およびリン酸化を、液体シンチレーションカウンターによって同時に測定した。C末端RB1タンパク質は、SMYD3存在下では、SMYD3非存在下の4〜6倍の量の3H標識メチル供与体を組み込んだ(データ示さず)。重要なことに、SMYD3は、CDK2/サイクリンE複合体によるRB1のリン酸化を用量依存的な様式で増強したのに対して、SMYD3のみではリン酸化を高めなかった(図5a)。さらに、RB1のリン酸化は、SMYD3存在下では、SMYD3非存在下と比較してCDK6/サイクリンD3によって増大することが発見された(図5b)。しかしながら、CDK2/サイクリンEまたはCDK6/サイクリンD3によるK824A変異体RB1のリン酸化は、野生型RB1と比較して有意に抑制された(それぞれ、図5c、d)。このデータは、RB1リン酸化は、SMYD3によるリジン824のメチル化によって増強されることを示唆している。抗リン酸化RB1抗体を用いたさらなるイムノブロット分析から、トレオニン821/826のリン酸化はSMYD3によって誘導されることが明らかになった。興味深いことに、セリン807/811のリン酸化もSMYD3によって増強された(図5e)。従って、リジン824のメチル化はセリン807/811のリン酸化を増加させる。または、さらなるメチル化残基がリン酸化を増強する可能性がある。
【0086】
インビボでの増強されたRB1のリン酸化を調べるために、抗リン酸化RB1抗体で、HEK-SMYD3細胞およびHEK-モック細胞からの抽出物を用いて、ウエスタンブロット分析を行った。インビトロでRB1タンパク質のリン酸化が増強されたことと一致して、HEK-SMYD細胞ではセリン807/811およびトレオニン821/826の両方のリン酸化が対照細胞と比較して高いことが検出された(図5f)。抗リン酸化トレオニン821/826抗体および抗SMYD3抗体を用いた免疫細胞化学染色から、SMYD3を発現する細胞は、抗リン酸化トレオニン821/826抗体によって、SMYD3を発現しない細胞より強く染色されることが明らかになった(図5g〜j)。さらに、SNU475細胞における野生型SMYD3の外因性発現は、メチルトランスフェラーゼ活性を欠く変異体SMYD3(SMYD3-ΔEELまたはSMYD3-ΔNHSC)の外因性発現と比較して、RB1リジン824のジメチル化およびトリメチル化を増大させた(Hamamoto, R. et al. Nat Cell Biol 6, 731-40 (2004))。RB1リジン824のメチル化と相関づけて、本発明者らは、この細胞において、トレオニン821/826でのリン酸化の著しい増加およびセリン807/811でのリン酸化の中程度の増加をそれぞれ観察した(図6a)。重要なことに、ウエスタンブロット分析から、対応する非癌性乳房組織と比較して、SMYD3を多く発現する乳癌組織では、セリン807/811およびトレオニン821/826のリン酸化が増加すると共に、RB1リジン824のメチル化が増強することが示された(図6b)。このデータは、インビボでのSMYD3によるセリン807/811およびトレオニン821/826のリン酸化が増強されたことを要約したものであった。RB1のリン酸化は、RB1からのE2Fの解離につながるポケットドメインを調整するので、MercuryTM細胞周期プロファイリングシステムを用いて、E2Fを介した転写のレポーター活性をHEK-SMYD3細胞において比較した。HEK-モック細胞と比較して、HEK-SMYD3細胞は高いE2F転写活性を示した(図7)。このデータは、SMYD3は、リジン824をメチル化することによってRB1のリン酸化を増強し、E2F転写活性を高めることを示す。
【0087】
実施例6:ヒト癌細胞における切断型SMYD3タンパク質
本発明者らは、本発明者らの最近の研究において、SMYD3タンパク質の発現レベルがヒト肝細胞癌(HCC)、結腸直腸癌(CRC)、および乳癌において高いことを示した(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004), Hamamoto, R. et al. Cancer Sci 97, 113-118 (2006))。興味深いことに、抗SMYD3抗体を用いたウエスタンブロット分析から、調べた全ての乳癌組織には45kDaおよび42kDaの2つのバンドが示されたが、正常乳腺には2つのバンドがどちらも無いことが検出された。45kDaおよび42kDaのバンドは2つともHCC、CRC、および乳癌細胞株(図8a)、ならびに正常精巣(データ示さず)において観察された。SMYD3の推定分子量は45kDaであり、本発明者らは、RT-PCR分析においてSMYD3転写物が変化した形を発見しなかった。従って、本発明者らは、42kDaのバンドが完全長SMYD3タンパク質の切断に起因する可能性があると仮説を立てた。SMYD3の切断を調べるために、本発明者らは、N末端HAタグ化SMYD3またはC末端FLAGタグ化SMYD3を発現するプラスミドを調製した。HAタグ化SMYD3タンパク質を発現するまたはFLAGタグ化SMYD3タンパク質を発現する、HEK293細胞の抽出物を、それぞれ抗HA抗体または抗FLAG抗体をを用いたイムノブロット分析に使用した。結果として、本発明者らは、抗HA抗体を用いてN末端HAタグ化タンパク質に対応する46kDaバンドを入手した。一方で、本発明者らは、抗FLAG抗体を用いて、46kDaおよび43kDaタンパク質の2つのバンドを発見した(図8b)。この結果は、完全長タンパク質がN末端領域で切断されることを示唆した。切断部位を調べるために、本発明者らは、内因性SMYD3を発現しないHEK293細胞において、N末端3XFLAG-タグを含む、野生型SMYD3および欠失変異体SMYD3を外因的に発現させた(図8c)。図8bのデータと一致して、抗FLAG抗体を用いたウエスタンブロット分析からは、野生型SMYD3を発現する細胞では48kDa FLAGタグ化完全長タンパク質に対応するバンドしか認められなかった。しかしながら、同じ抽出物を使用した、抗SMYD3抗体を用いた分析によって、48kDa FLAGタグ化SMYD3および42kDaタンパク質に対応する2つのバンドが検出された。抗SMYD3抗体を用い、N末端欠失型SMYD3(FLAG-SMYD3-ΔN44、-ΔN99、および-ΔN244)を発現する細胞からの抽出物を使用したウエスタンブロット分析からは、1本のバンドが認められた。これらのデータは、SMYD3の切断部位がコドン1と45との間に位置することを示唆した。
【0088】
実施例7:SMYD3タンパク質の切断部位の決定
SMYD3の正確な切断部位を決定しようと、本発明者らは、免疫沈降したFLAGタグ化SMYD3タンパク質を転写したPVDF膜から42kDaタンパク質を精製し(図8b)、そのアミノ酸配列を決定した。結果として、本発明者らは、N末端34アミノ酸を欠く欠失型SMYD3タンパク質を同定した。このことから、コドン34(アスパラギン酸)とコドン35(プロリン)との間に切断部位があることが明らかになった(図9a)。SMYD3は、コドン5と27との間にSET-N領域と呼ばれるアミノ酸配列を含む。SET-N領域はSETドメインタンパク質において保存されている(Marmorstein, R. Trends in Biochem. Sci., Vol.8 no.2, (2003); Kouzarides, T. Curr. Opin. Genet. Dev. 12, 198-209 (2002); Lachner, M and Jenuwein, T. Curr. Opin. Cell biol. 14, 286-298 (2002))。SET-N領域のアミノ酸配列のアラインメントは、SMYD3および他のメチルトランスフェラーゼにある領域の高い類似性を示した(図9b)。このことは、この領域が重要なことを意味している。
【0089】
実施例8:切断されたSMYD3のHMTアーゼ活性は野生型タンパク質と比較して高い
切断SMYD3タンパク質のメチルトランスフェラーゼ活性を調べるために、本発明者らは、HEK293細胞において、3xFLAGタグ化野生型SMYD3または34アミノ酸欠失型SMYD3もしくは44アミノ酸欠失型SMYD3を外因的に発現させ、これらのタンパク質を免疫沈降した(図10a)。本発明者らは、これらのタンパク質を酵素源として使用して、ヒストンメチルトランスフェラーゼ(HMTアーゼ)アッセイ法を行い、野生型SMYD3のHMTアーゼ活性は用量依存的な様式で増加することを示した(図10b)。メチル供与体であるS-アデノシルメチオニン(SAM)の存在下でのメチル化反応は、メチルトランスフェラーゼ反応を競合的に阻害する可能性のあるS-アデノシルホモシステイン(SAH)の産生を伴う。従って、本発明者らは、反応混合物に、SAHをホモシステインおよびアデノシンに加水分解するS-アデノシルホモシステイン加水分解酵素(SAHH)を添加した。予想したとおり、本発明者らは、SAHH存在下では、非存在下と比較してHMT活性が著しく増加することを観察した(図10b)。この知見は、SMYD3のメチルトランスフェラーゼ阻害剤のスクリーニングに有用である。驚くべきことに、切断されたSMYD3タンパク質のHMTアーゼ活性は、完全長タンパク質と比較して有意に高かった(図10b)。この結果から、翻訳後の切断はヒト細胞におけるSMYD3 HMTアーゼ活性の調節に関与し、N末端SET-N領域はHMTアーゼ活性に対して抑制的な役割を有し得ることが分かる。
【0090】
実施例9:SET-N領域におけるグリシン15および17はHMT活性に重要である
酵素活性の抑制に関するSET-N領域にある保存アミノ酸配列の重要性を確かめるために、本発明者らは、野生型N末端FLAGタグ化SMYD3タンパク質を発現するプラスミド、または変異体N末端FLAGタグ化SMYD3タンパク質、SMYD3-SETNm1、-SETNm2、もしくは-SETNm3を発現するプラスミドを調製した。SMYD3-SETNm1、-SETNm2、または-SETNm3は、それぞれ、Gly15AlaおよびGly17Alaの両方、Gly15Ala、またはGly27Alaの置換を含む(図11a)。これらの変異体を発現するHEK293細胞からの溶解産物のウエスタンブロットによって、置換はSMYD3タンパク質の切断に影響を及ぼさないことが示された(図11b、上パネル)。本発明者らは、免疫沈降したSMYD3タンパク質を用いてHMTアーゼアッセイ法を行った。結果として、Gly15AlaまたはGly27Alaを含む変異体タンパク質(SMYD3-SETNm2または-SETNm3)のHMTアーゼ活性は野生型タンパク質とほぼ同じであった (図11c)。これに対して、Gly15AlaおよびGly17Alaの2つの置換を含む変異体タンパク質(SMYD3-SETNm1)の活性は野生型タンパク質と比較して有意に増強された(図11c)。これらのデータは、SMYD3のHMTアーゼ活性の調節にグリシン15および17が重要な役割を果たしている可能性があることを示唆している。
【0091】
実施例10:N末端10アミノ酸の欠失はHMTアーゼ活性の増強に必須である
N末端領域が酵素活性を増強したことから、本発明者らは、2つの可能性のある機構について仮説を立てた。すなわち、N末端領域が酵素活性に関する未決定の負の調節因子と会合し得るか、または欠失によって、増強された酵素活性につながるタンパク質のコンホメーション変化が起こり得るという仮説を立てた。さらなる負の調節因子が酵素活性において役割を果たしている可能性があるか否かを確かめるために、本発明者らは、野生型SMYD3およびN末端欠失SMYD3の組換えタンパク質を調製し、そのHMTアーゼ活性をインビトロで調べた。図12に示したように、全ての欠失変異体(SMYD3-ΔN9、-ΔN19、-ΔN29、-ΔN44、-ΔN74)は、野生型タンパク質と比較して4〜5倍に増強されたメチルトランスフェラーゼ活性を示した(図12)。この結果は、さらなる因子が、切断されたSMYD3の高活性に関与している可能性は低く、N末端10アミノ酸がメチルトランスフェラーゼ活性の抑制に非常に重要な役割を果たしている可能性があることを示唆している。
【0092】
考察
SMYD3には、インビトロおよびインビボでRB1のリジン824に対するメチルトランスフェラーゼ活性があり、メチル化RB1は、CDK/サイクリン複合体によるリン酸化に対して非メチル化RB1より感受性であるという知見が本明細書において開示される。結果として、SMYD3を発現するHEK293-SMYD3細胞は、HEK293-モック細胞と比較して高いE2F転写活性を示した。このことは、SMYD3の増殖促進作用とよく一致している。なぜなら、E2F-1の過剰発現は、細胞増殖に必須の産物の一連の遺伝子を調節することによって、細胞周期のG1期からS期への移行を促進できるからである。Harbourらは、G1からSへ進行する間にRB1がリン酸化されるモデルを示した。このモデルでは、RB1のリン酸化は、C末端領域とポケットドメインとの間の連続した分子内相互作用を開始する(Harbour, J. W. et al., D. C. Cell 98, 859-69 (1999))。G1期の間に、CDK4/6-サイクリンDによってRB1のC末端領域がリン酸化されると、中央のポケットドメインとの分子内相互作用が誘発される。これによって、HDAC結合が阻害され、活発な転写抑制がブロックされる。この相互作用は、CDK2/サイクリンEがRB1のセリン567に接近するのを促し、その結果として、A/B境界面が壊れ、RB1とE2Fとの相互作用が阻止される。このモデルでは、E2Fが解離するために、RB1がCDK4/6-サイクリンD複合体およびCDK2/サイクリンE複合体の両方によって連続してリン酸化されることが必要とされる(Lundberg, A. S. & Weinberg, R. A. Mol Cell Biol 18, 753-61 (1998))。報告によると、RB1にあるThr821およびThr826がリン酸化されると、A/Bポケットドメインと、E2FおよびHDACを含むLXCXEモチーフ含有タンパク質との相互作用が不活性化されるのに対して、Ser807およびSer811がリン酸化されるとC末端ドメインが不活性化される。このデータは、SMYD3によるRB1のメチル化によって、CDK2/サイクリンE複合体またはCDK6/サイクリンD複合体によるリン酸化が増強され、Thr821/826のリン酸化が高まるので、SMYD3を発現する細胞が高いE2F転写活性を示すという本発見と一致する。あるいは、リジン824のメチル化は、C末端領域のコンホメーションを直接変え、それによって、ポケットドメインとE2Fとの会合を阻害するかもしれない。なぜなら、ヒストンおよびp53のリジンの両方がメチル化されるとコンホメーションが変化するからである(Tsuge, M. et al. Nat Genet 37, 1104-7 (2005))。SMYD3はE2F転写活性を増加させるので、多量のSMYD3は正のフィードバック機構としてE2F1活性を高めるのかもしれない。従って、SMYD3を介したRB1不活性化は、ヒトの発癌において非常に重要な役割を果たしている可能性が高い。
【0093】
RB1が、いくつかの機構を介して転写抑制において役割を果たすことは注目すべきことである。RB1は転写因子と相互作用し、かつ転写因子の活性を直接抑制する。RB1がプロモーター領域に動員されると、開始前複合体の集合がブロックされる。RB1はクラスI HDAC(HDAC-1、-2、および-3)とも会合し、かつヒストンの脱アセチル化を誘導して、ヘテロクロマチン状態へのコンホメーション変化をもたらす。RB1は、標的遺伝子のプロモーター領域におけるDNAメチル化につながるDNMT1と複合体を形成する(Harbour, J. W. & Dean, D. C. Genes Dev 14, 2393-409 (2000); Robertson, K. D. et al. Nat Genet 25, 338-42 (2000))。これらの機構に加えて、ヒストンメチル化に関する最近の研究から、RB1は、SUV39HおよびSuv4-20h1またはSuv4-20h1を含むヒストンメチルトランスフェラーゼとも会合することが開示された。SUV39HおよびSuv4-20h1またはSuv4-20h1は、それぞれ、H3-K9およびH4-K20のメチル化に関与する(Gonzalo, S. et al. Nat Cell Biol 7, 420-8 (2005); Nielsen, S. J. et al. Nature 412, 561-5 (2001))。これらのメチルトランスフェラーゼに結合すると、RB1は、HP1またはCBXを複合体に動員することによってヘテロクロマチン形成を安定化する。本知見は、ヒストンH3-K4メチルトランスフェラーゼの転写活性化の調節への新規の洞察をもたらす。リジン824がメチル化したRB1は、RB1リン酸化を増強し、およびその後に、恐らくE2F1を中央のポケットドメインから放出させることによってE2F1のトランス活性化を増強する。さらに、メチル化されたRB1は、コンホメーションを変え、それによって、HDAC、ヘテロクロマチンタンパク質1(HP1)、および/またはクロモボックスタンパク質(CBX)を、SUV39Hおよび/またはSuv4-20h1メチルトランスフェラーゼの複合体から解離し、H3-K9およびH4-K20のメチル化を低下させるかもしれない。さらなる研究が必要だが、本明細書のデータは、E2Fを担う遺伝子の調節におけるRB1メチル化の重要性を強調する。RB1は様々なメチルトランスフェラーゼに結合するので、RB1におけるメチル化の位置および程度はメチルトランスフェラーゼによって異なる可能性がある。RB1が様々な残基でリン酸化されることもあわせると、本明細書のデータは、RB1における複数の修飾の組み合わせがRB1の生物学的特性を規定する可能性があることを示唆している。このことは、ヒストンおよびp53の修飾を連想させる。
【0094】
RB1の変異は、散発性および家族性の網膜芽細胞腫症例に関与するだけでなく(Weinberg, R. A. Science 254, 1138-46 (1991))、他のヒト癌にも関与している(Classon, M. & Harlow, E. Nat Rev Cancer 2, 910-7 (2002))。いくつかの腫瘍ウイルスタンパク質、例えば、アデノウイルスE1A、HPV-E7、およびシミアンウイルス40(SV40)ラージT抗原は一部の癌のタイプにおいてRB1と会合し、RB1とE2Fとの間の相互作用を阻害して、E2Fを解離させる(Chellappan, S. P., et al. Cell 65, 1053-61 (1991); Bagchi, S., et al. Cell 65, 1063-72 (1991))。サイクリン依存性キナーゼ4阻害剤であるp16は、p16プロモーターのメチル化によって頻繁に不活性化される。これによって、癌細胞ではCDK/サイクリン複合体によるRB1のリン酸化が増強される(Nuovo, G. J., et al. Proc Natl Acad Sci U S A 96, 12754-9 (1999))。これらの欠陥は、結腸直腸癌および肝細胞癌の一部に関与していると報告されており(Chaubert, P. et al. Hepatology 25, 1376-81 (1997); Toyota, M. et al. Proc Natl Acad Sci U S A 96, 8681-6 (1999))、これらの腫瘍タイプにおける全症例の原因ではないかもしれない。本明細書において、新規のRB1不活性化機構、すなわち、RB1不活性化はRB1のメチル化およびその後の増強されたリン酸化によって引き起こされることが開示されている。SMYD3の発現は結腸直腸癌および肝細胞癌の大部分において増強されているので、SMYD3は、RB1の腫瘍抑制因子機能を抑止してE2Fをトランス活性化することにより癌細胞増殖において非常に重要な役割を果たしている可能性がある。興味深いことに、本発明者らは、最近、SMYD3の発現が、E2F-1とSMYD3プロモーター領域内のE2F-1結合エレメントの相互作用を介してE2F-1によって調節され、およびこのエレメントがE2F-1結合モチーフの2タンデムリピートまたは3タンデムリピートより構成されることを発見した。日本人の結腸直腸癌患者(n=350)、肝臓癌患者(n=360)、および乳癌患者(n=334)における3リピートの対立遺伝子頻度は、一般的な日本人集団からの健常対照(n=730)より有意に高かった。このデータは、一旦SMYD3が活性化されると、RB1の修飾を介してE2F転写活性を増強し、結果として、正のフィードバックによりSMYD3を上方制御することを示唆している。従って、E2F-1結合エレメントの3リピートを含む人は、2リピートを含む人よりSMYD3によるRB1不活性化を受けやすい。さらに、SMYD3の阻害は、正のフィードバックループをブロックし、それによって、RB1のリン酸化によるE2F-1を介した分裂促進活性を効率的に抑制するので、結腸直腸癌および肝臓癌ならびに膀胱癌および乳癌の有望な治療方針であるように思われる。
【0095】
本明細書において、SMYD3によるRB1のメチル化は、CDK/サイクリン複合体によるRB1のリン酸化の増強を介してG1期からS期への細胞周期の進行を速める可能性があることが明らかになった。このデータから、リジンのメチル化はヒストンだけでなく、p53およびRB1などの他の非ヒストンタンパク質にとっても重要なことが分かる。さらに、本発明者らの知見は、ヒト発癌に関与するRB1調節の新規機構の解明に役立った。
【0096】
エピジェネティックな調節の混乱がヒト発癌と関連することが示されている。細胞周期、DNA修復、および細胞接着を調節する遺伝子のプロモーター領域における異常なDNAメチル化に加えて、最近の研究から、ヒストンのメチル化もヒト発癌において抑止されることが開示された。ヒストンのメチル化は、クロマチン構造の変化を介して遺伝子発現の調節において非常に重要な役割を果たしている。本発明者らは、ヒストンH3-リジン4-特異的メチルトランスフェラーゼであるSMYD3が、HCC、CRC、および乳癌を含む、いくつかのヒト癌において過剰発現していることを報告した(Hamamoto, R. et al. Nat Cell Biol 6, 731-740 (2004): Hamamoto, R. et al. Cancer Sci 97, 113-118 (2006))。本発明者らの以前の論文において、本発明者らは、様々なヒト癌において頻繁に増強される転写因子であるE2F1の転写活性化によって、SMYD3の発現が上昇することを示した。
【0097】
タンパク質の機能は、転写後レベルでだけでなく、タンパク質の切断ならびにアセチル化、リン酸化、メチル化、グリコシル化、およびユビキチン化などの他の広く知られている修飾を含む翻訳後修飾によっても調節される。これらの修飾は、タンパク質を活性化または不活性化する、タンパク質安定性、タンパク質のコンホメーション、および/またはタンパク質-タンパク質相互作用と関連する。本発明者らは、SMYD3の切断がSMYD3のHMTアーゼ活性を高めることを発見した。このことは、ペプシン、インシュリン、カスパーゼ、PARP、およびMMPなどの重要な酵素の調節を思い出させる。なぜなら、これらのタンパク質は切断されると酵素活性が増加するからである。さらに、この知見は、未決定のSMYD3切断機構がHMTアーゼ活性の調整において役割を果たしている可能性があることを示唆している。従って、切断を担うプロテアーゼの同定、および調節機構の解明が、SMYD3活性を抑制する新規の治療アプローチの開発に寄与するだろう。さらに、切断型SMYD3は、完全長タンパク質と比較してSMYD3阻害剤のスクリーニングに有用な可能性がある。
【0098】
本発明者らは、本研究において、SMYD3 N末端領域が欠失すると、インビトロにおいてSMYD3の酵素活性が増強されることを発見した。このことは、欠失は、増強された酵素活性につながるSMYD3のコンホメーション変化を付与し得ることを示唆している。興味深いことに、HSP90はSMYD3のN末端領域に結合して、SMYD3のHMTアーゼ活性を増加させる。このデータは、コンホメーション変化がHMTアーゼ活性に関与するという考えとよく一致している。なぜなら、HSP90は、正常なタンパク質構造の安定化に寄与するシャペロン様機能を発揮するからである。本発明者らの知見はまた、HMTアーゼ活性の調節のための保存されたSET-N領域の重要性も強調する。この保存領域は、他のSETドメイン含有タンパク質においてHMTアーゼの負の制御因子としても働く可能性がある。さらなる研究によって、SETドメイン含有タンパク質におけるHMTアーゼ活性の調節機構が見出されるだろう。
【0099】
本発明者らは、本明細書において、癌細胞においてN末端切断型SMYD3タンパク質が発現され、かつ切断されたタンパク質のHMTアーゼ活性が完全長タンパク質より著しく高いことを示した。これらのデータは、翻訳後調節システムが、可能性のあるタンパク質コンホメーション変化を介してHMTアーゼ活性を調節することを意味した。さらに、本発明者らは、SAHHが添加されるとSMYD3のメチルトランスフェラーゼ活性が増加することを発見した。本発明者らの知見は、SMYD3活性調節機構のより深い理解の助けとなり、かつHMTアーゼ活性を阻害する新規の治療方針の同定に寄与する可能性がある。
【0100】
産業上の利用可能性:
本明細書に記載の方法は、結腸直腸癌、肝細胞癌、乳癌、および膀胱癌を含む様々な癌を予防、診断、および治療するためのさらなる分子標的の同定に有用である。さらに、本明細書において報告されたデータは、癌の包括的な理解を深め、新規の診断法の開発を促し、かつ治療薬剤および予防剤の分子標的を同定する手がかりを与える。このような情報は腫瘍形成のさらに深い理解に寄与し、ならびに新規の癌診断法、癌治療法、および最終的には癌予防法を開発する指標を与える。本発明は詳細に、かつ本発明の同定の態様に関して説明されたが、前述の記載は本質的に例示および説明であり、本発明および本発明の好ましい態様を例証することを目的とすることが理解されるはずである。当業者であれば、日常的な実験によって、本発明の精神および範囲から逸脱することなく、本明細書に様々な変化および変更を加えることができることを容易に理解するだろう。従って、本発明は、前述の記載では定義されず、特許請求の範囲およびその均等物によって定義されることが意図される。
【図面の簡単な説明】
【0101】
【図1】組換えRB1タンパク質に対するSMYD3のMTアーゼ活性を示す。パートaは、基質として組換えヒストンH3、p53、またはRB1のC末端領域を用いたインビトロMTアーゼアッセイ法の結果を示す。等量の基質を、免疫沈降したFlagタグ化SMYD3およびメチル供与体である3H標識SAMとインキュベートした。タンパク質をSDS-PAGEで分離し、メチル化された基質をフルオログラムによって検出した。基質の総量は、特異的抗体を用いたイムノブロット分析によって調べた。パートbは、ヒストンH3およびC末端RB1タンパク質に対する組換えSMYD3の用量依存的MTアーゼ活性を示す。パートcは、C末端RB1および完全長RB1(それぞれ、レーン2および4)に対するSMYD3のMTアーゼ活性を示す。保存アミノ酸の欠失を含む変異体SMYD3(SMYD3ΔEEL)はMTアーゼ活性を著しく低下させた(レーン3)。
【図2】インビボでのSMYD3とRB1との間の会合を示す。パートaはイムノアッセイの結果を示す。具体的には、抗SMYD3抗体を用いたHepG2細胞またはHCT116細胞の溶解産物からの免疫沈降物を、抗RB1抗体でイムノブロットした。パートbは、HEK293細胞における野生型RB1ならびに欠失型RB1(RB1Δ1およびRB1Δ2)とSMYD3との間の相互作用を示す(下)。RB1の保存領域および発現構築物を上パネルに示す。パートcは、RB1との相互作用を担うSMYD3の領域を示す。SMYD3の保存領域および発現構築物を上パネルに示す。パートdは、組換えRB1の存在下/非存在下でのヒストンH3に対するSMYD3のインビトロメチルトランスフェラーゼ活性を示す。ヒストンH3のメチル化はRB1による影響を受けなかった(上パネル)。等量の組換えヒトヒストンH3タンパク質を基質として使用した(下パネル)。パートeは、ヒストンH4-K20メチル化のインビトロ分析を示す。免疫沈降したSMYD3タンパク質または組換えSMYD3タンパク質を、基質として組換えヒトヒストンH4とインキュベートした。免疫沈降したSuv4-20h2タンパク質は正の対照とした。抗トリメチルH4-K20抗体を用いて、メチル化されたH4-K20が検出された。
【図3】RB1のC末端領域にあるK824のメチル化を示す。パートaは、RB1の保存ドメイン、ならびに野生型および変異型のC末端RB1タンパク質(K824A、K889A、およびK896A)の模式図である。パートbは、オートラジオグラフィーによる、SDS-PAGEで分離されたメチル化C末端RB1の検出を示す。パートcは、液体シンチレーションカウンターによって測定されたMTアーゼ活性を示す。パートdは、組換え野生型RB1タンパク質、ならびにK791A、K814A、K824A、K791/K824A、およびK814/K824Aを含む組換え変異型RB1タンパク質のインビトロメチル化を示す。RB1を、3H標識SAMの存在下で組換えSMYD3タンパク質とインキュベートした。メチル化RB1をSDS-PAGEで分離し、フルオログラムによって検出した。パートeは、液体シンチレーションカウンターによって測定されたメチル化RB1を示す。パートfは、SMYD3によるRB1リジン824のジメチル化およびトリメチル化を示す。SMYD3の存在下または非存在下でのメチル化野生型RB1タンパク質を3H-BASイメージングシステムによって検出した(上パネル)。抗ジメチル化リジン824(2番目のパネル)抗体または抗トリメチル化リジン824(3番目のパネル)抗体を用いたRB1タンパク質のウエスタンブロット分析。RB1の総量は、抗RB1抗体を用いて定量した(4番目のパネル)。
【図4】インビボでのSMYD3によるRB1のメチル化を示す。パートaは、HEK-SMYD3(HEK-SMYD3-1および-2)細胞ならびにHEK-モック(HEK-モック-1および-2)細胞におけるSMYD3の発現を示す(上パネル)。パートbは、抗RB1抗体によるHEK-SMYD3細胞およびHEK-モック細胞からの免疫沈降物を用いた、ラジオグラムによるインビボでのメチル化RB1の検出を示す(上パネル)。免疫沈降したRB1の量は変化しなかった。細胞を、タンパク質合成阻害剤の存在下で3H標識SAMで処理した。免疫沈降したRB1の量は変化しなかった(下パネル)。パートcは、免疫沈降したRB1のメチル化を、抗パンメチルリジン抗体、抗ジメチルリジン824抗体、および抗トリメチルリジン824抗体を用いて分析したウエスタンブロットの結果を示す。パートd〜kは、抗ジメチルリジン824(d)抗体または抗トリメチルリジン824(h)抗体を用いた、HEK293-SMYD3細胞の免疫細胞化学染色の結果を示す。パートeおよびiは、抗SMYD3抗体を用いて調べたSMYD3の発現を示す。パートfおよびjは、DAPIによる核染色の結果を示す。パートgおよびkは、d〜f(g)またはh〜j(k)の合成画像を構成する。SMYD3を豊富に発現している細胞は、インビボでRB1 Lys824のジメチル化およびトリメチル化が増強されていることを示した。
【図5】SMYD3によりRB1のリン酸化が増強されることを示す。パートaは、SMYD3の存在下または非存在下でのCDK2/サイクリンEによるC末端RB1のインビトロリン酸化を示す。SMYD3のみではリン酸化を増加しなかった。パートbは、SMYD3の存在下または非存在下でのCDK6/サイクリンD3によるC末端RB1のインビトロリン酸化を示す。SMYD3によるRB1リン酸化の増強は、K824A置換RB1を用いて抑えられた。パートcは、CDK2/サイクリンEによるC末端RB1のインビトロリン酸化を示す。基質として、野生型(Wt)およびK824A変異体(K824A)を比較した。パートdは、CDK6/サイクリンD3によるC末端RB1のインビトロリン酸化を示す。基質として、WtおよびK824Aを比較した。パートeは、SMYD3の存在下での、CDK2/サイクリンE複合体またはCDK6/サイクリンD3複合体によるSer807/811およびThr821/826リン酸化の増加を示す。パートfは、HEK-モック細胞と比較して、HEK-SMYD3細胞におけるSer807/811リン酸化およびThr821/826リン酸化が高いことを示す。外因性SMYD3を発現するHEK293細胞の免疫細胞化学染色。パートgは、抗リン酸化RB1(Thr821/826)抗体を用いて、細胞内のリン酸化RB1を染色した結果を示す。パートhは、細胞におけるSMYD3の発現を示す。パートiは、DAPIによる核染色の結果を示す。パートjはg〜iの合成画像を構成する。SMYD3を発現する細胞は、インビボでのThr821/826リン酸化が増強されていることを示した。
【図6】SMYD3によるRB1のメチル化および増強されたリン酸化を示す。パートaでは、野生型プラスミド(p3xFlag-SMYD3)または変異体SMYD3プラスミド(p3xFlag-SMYD3ΔEELおよびp3xFlag-SMYD3ΔNHSC)でトランスフェクトしたSNU475細胞から、RB1タンパク質を免疫沈降した。ウエスタンブロット分析は、沈降物を使用し、抗ジメチル化リジン824(最上部パネル)抗体、抗トリメチル化リジン824(2番目のパネル)抗体、抗リン酸化セリン807/811(3番目のパネル)抗体、または抗リン酸化トレオニン821/824(4番目のパネル)抗体を用いて行った。抗RB1抗体を用いたイムノブロット分析は量の対照として役立った(最下部パネル)。パートbは、2つの乳癌組織におけるリジン824のジメチル化およびトリメチル化ならびにSer807/811およびThr821/826のリン酸化を示す。ウエスタンブロット分析は、抗ジメチル化リジン824抗体、抗トリメチル化リジン824抗体、抗リン酸化RB(Ser807/811)抗体、または抗リン酸化RB(Thr821/824)抗体を用いて行った。
【図7】HEK-SMYD3細胞においてE2F転写活性が増大していることを示す。HEK-SMYD3細胞およびHEK-モック細胞においてE2F-ルシフェラーゼベクターでトランスフェクトした24時間後に、ルシフェラーゼ活性を測定した。外因性SMYD3を発現するHEK293細胞の免疫細胞化学染色。細胞内のリン酸化RB1を、抗リン酸化RB1(Thr821/826)抗体で染色した。パートhは、細胞におけるSMYD3の発現を示す。パートiは、DAPIによる核染色の結果を示す。パートjは、a〜cの合成画像を構成する。SMYD3を発現する細胞は、インビボでのThr821/826リン酸化が増強されていることを示した。
【図8】SMYD3タンパク質の発現パターン。パートaは、ヒト癌細胞株および組織におけるSMYD3タンパク質の発現を示す。ウエスタンブロット分析は、抗SMYD3抗体を用いて行った。β-アクチンの発現は量の対照とした。パートbは、HAタグ化SMYD3(左パネル)およびFLAGタグ化(右パネル)のイムノブロット分析を示す。ウエスタンブロット分析は、抗HA抗体およびN末端領域にHAタグ化SMYD3を発現する細胞からの抽出物、または抗FLAG抗体およびC末端領域にFlagタグ化SMYD3を発現する細胞からの抽出物を、それぞれ用いて行った。パートcは、欠失型SMYD3の模式図を示す。N末端領域に一連のFLAGタグ化SMYD3を発現するプラスミドを、内因性SMYD3を発現しないHEK293細胞にトランスフェクトした。パートdは、抗SMYD3抗体(上パネル)または抗FLAG抗体(下パネル)を用いて行った、細胞からの抽出物のウエスタンブロット分析を示す。矢印は完全長SMYD3タンパク質を示し、星印は切断型SMYD3に対応する。
【図9】SET含有タンパク質におけるSMYD3切断部位およびSET-N領域の保存アミノ酸配列の決定。パートaは、エドマンアミノ酸シークエンスによって決定された、N末端領域の34アミノ酸が欠失したSMYD3タンパク質を示す。パートbは、ヒストンメチルトランスフェラーゼにおけるSET-Nアミノ酸配列のアラインメントを示す。高度に保存されたアミノ酸を黒色の囲みで示し、中程度に保存されたアミノ酸を影付きの囲みで示した。
【図10】野生型タンパク質と比較して増加している切断型SMYD3のHMTアーゼ活性。パートaは、抗FLAG抗体(上パネル)および抗SMYD3抗体(中パネル)を用いた、野生型SMYD3タンパク質または欠失型(ΔN34およびΔN44)SMYD3タンパク質のウエスタンブロット分析を示す。FLAGタグ化SMYD3タンパク質を発現する細胞から、タンパク質を抽出した。免疫沈降したSMYD3タンパク質をHMTアーゼアッセイ法に使用した。パートbは、完全長SMYD3タンパク質および切断されたSMYD3タンパク質のHMTアーゼ活性の用量反応増加を示す。SAHH(S-アデノシルホモシステイン加水分解酵素)を添加すると活性が増加した。3H-放射能を液体シンチレーションカウンターによって測定した。
【図11】SET-N領域の中の抑制されたHMTアーゼ活性の原因となる領域の決定。パートaは、SET-N領域の保存アミノ酸に置換を含む変異体SMYD3構築物の模式図を示す。パートbは、抗SMYD3(上パネル)抗体または抗FLAG(中パネル)抗体を用いた、FLAGタグ化野生型SMYD3タンパク質または変異体(ΔN34、SETNm1、SETNm2、およびSETNm3)SMYD3タンパク質のイムノブロット分析を示す。FLAGタグ化SMYD3を発現するHEK293F細胞から抗Flag抗体を用いて免疫沈降したタンパク質を、HMTアーゼアッセイ法の酵素源として使用した。パートcは、野生型SMYD3、欠失型SMYD3のHMTアーゼ活性を示す。3H-放射能を液体シンチレーションカウンターによって測定した。
【図12】SMYD3におけるN末端領域の欠失によるHMTアーゼ活性の増強。パートaは、N末端領域が欠失した型のSMYD3の模式図を示す。一連のGST融合SMYD3タンパク質を発現するプラスミドを調製した。パートbは、抗GST抗体を用いた組換えSMYD3タンパク質のイムノブロット分析を示す。GSTと融合した、野生型および変異体組換えSMYD3タンパク質を細菌細胞において発現させ、その細胞から精製した。パートcは、タンパク質のインビトロHMTアーゼ活性を示す。3H-放射能を液体シンチレーションカウンターによって測定した。
【特許請求の範囲】
【請求項1】
SMYD3による網膜芽細胞腫のメチル化を調整する作用因子を同定するための方法であって、以下の工程を含む方法:
a.以下からなる群より選択される、メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド:
i.配列番号:2のアミノ酸配列を含むポリペプチド;
ii.1つまたは複数のアミノ酸が置換、欠失、または挿入された、配列番号:2のアミノ酸配列を含むポリペプチドであって、さらに、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
iii.配列番号:2と少なくとも約80%の相同性を有するアミノ酸配列を含むポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
vi.ストリンジェントな条件下で配列番号:1のヌクレオチド配列からなるポリヌクレオチドとハイブリダイズするポリヌクレオチドによってコードされるポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;および
v.配列番号:2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
と、メチル化される網膜芽細胞腫ペプチドおよび補助因子を、網膜芽細胞腫ペプチドのメチル化に適した条件下、作用因子の存在下で接触させる工程;
b.網膜芽細胞腫ペプチドのメチル化レベルを検出する工程;ならびに
c.工程(b)のメチル化レベルと、作用因子の非存在下で検出された対照レベルを比較する工程であって、対照レベルと比較したメチル化レベルの増加または減少は、作用因子がSMYD3による網膜芽細胞腫のメチル化を調整することを示す、工程。
【請求項2】
網膜芽細胞腫ペプチドが、配列番号:4のアミノ酸配列を含むポリペプチド、またはその機能的な変異体もしくは断片である、請求項1記載の方法。
【請求項3】
補助因子がS-アデノシルホモシステイン加水分解酵素(SAHH)である、請求項1記載の方法。
【請求項4】
網膜芽細胞腫のメチル化を調節する試験化合物の能力を検出するためのキットであって、以下の構成成分を含むキット:
a.以下からなる群より選択される、メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド:
i.配列番号:2のアミノ酸配列を含むポリペプチド;
ii. 1つまたは複数のアミノ酸が置換、欠失、または挿入された、配列番号:2のアミノ酸配列を含むポリペプチドであって、さらに、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
iii.配列番号:2と少なくとも約80%の相同性を有するアミノ酸配列を含むポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
vi.ストリンジェントな条件下で配列番号:1のヌクレオチド配列からなるポリヌクレオチドとハイブリダイズするポリヌクレオチドによってコードされるポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;および
v.配列番号:2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
b.(a)のポリペプチドによってメチル化することができる網膜芽細胞腫ペプチド;ならびに
c.網膜芽細胞腫ペプチドのメチル化のための補助因子。
【請求項5】
網膜芽細胞腫ペプチドが、配列番号:4のアミノ酸配列を含むポリペプチド、またはその機能的な変異体断片である、請求項4記載のキット。
【請求項6】
a.S-アデノシルホモシステイン加水分解酵素(SAHH)
の要素をさらに含む、請求項4記載のキット。
【請求項7】
結腸直腸癌、肝細胞癌、膀胱癌、および乳癌からなる群より選択される癌を治療するための化合物をスクリーニングする方法であって、以下の工程を含む方法:
a.請求項1記載の方法を用いて、メチル化を調整する試験化合物を同定する工程、および
b.試験化合物の非存在下で検出される対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを下げる試験化合物を選択する工程。
【請求項8】
結腸直腸癌、肝細胞癌、膀胱癌、および乳癌からなる群より選択される癌の症状を軽減するための組成物であって、請求項7記載の方法によって同定された薬学的に有効な量の化合物および薬学的に許容される担体を含む、組成物。
【請求項9】
結腸直腸癌、肝細胞癌、膀胱癌、および乳癌からなる群より選択される癌の症状を軽減するための方法であって、癌細胞と、請求項7記載の方法によって同定された薬学的に有効な量の化合物を接触させる工程を含む、方法。
【請求項10】
パート(iii)において定義されるポリペプチドが、配列番号:2と少なくとも約95%の相同性を有するアミノ酸配列を含む、請求項1記載の方法。
【請求項11】
パート(ii)において定義されるポリペプチドが、20までの保存的アミノ酸置換を含む配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項12】
パート(ii)において定義されるポリペプチドが、配列番号:2のアミノ酸配列の位置1〜250のアミノ酸配列を含む、請求項1記載の方法。
【請求項13】
パート(ii)において定義されるポリペプチドが、配列番号:2のアミノ酸配列の位置45〜428のアミノ酸配列を含む、請求項1記載の方法。
【請求項14】
パート(ii)において定義されるポリペプチドが、位置1〜30のアミノ酸が欠失している配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項15】
パート(ii)において定義されるポリペプチドが、位置1〜44のアミノ酸が欠失している配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項16】
パート(ii)において定義されるポリペプチドが、位置1〜20のアミノ酸が欠失している配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項17】
パート(ii)において定義されるポリペプチドが、位置1〜10のアミノ酸が欠失している配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項18】
パート(iv)において定義されるポリペプチドが、高ストリンジェントな条件下で、配列番号:1のヌクレオチド配列からなるポリヌクレオチドと特異的にハイブリダイズする、請求項1記載の方法。
【請求項19】
機能的な網膜芽細胞腫断片がC末端断片を含む、請求項2記載の方法。
【請求項20】
機能的な網膜芽細胞腫断片が、配列番号:4のアミノ酸配列の位置769〜921のアミノ酸配列からなる、請求項2記載の方法。
【請求項21】
機能的な網膜芽細胞腫変異体が、以下の変異:K889A、K896A、K791A、K814A、K791A/K824A、およびK814A/K824Aの1つまたは複数を含む配列番号:4のアミノ酸配列を含む、請求項2記載の方法。
【請求項1】
SMYD3による網膜芽細胞腫のメチル化を調整する作用因子を同定するための方法であって、以下の工程を含む方法:
a.以下からなる群より選択される、メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド:
i.配列番号:2のアミノ酸配列を含むポリペプチド;
ii.1つまたは複数のアミノ酸が置換、欠失、または挿入された、配列番号:2のアミノ酸配列を含むポリペプチドであって、さらに、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
iii.配列番号:2と少なくとも約80%の相同性を有するアミノ酸配列を含むポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
vi.ストリンジェントな条件下で配列番号:1のヌクレオチド配列からなるポリヌクレオチドとハイブリダイズするポリヌクレオチドによってコードされるポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;および
v.配列番号:2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
と、メチル化される網膜芽細胞腫ペプチドおよび補助因子を、網膜芽細胞腫ペプチドのメチル化に適した条件下、作用因子の存在下で接触させる工程;
b.網膜芽細胞腫ペプチドのメチル化レベルを検出する工程;ならびに
c.工程(b)のメチル化レベルと、作用因子の非存在下で検出された対照レベルを比較する工程であって、対照レベルと比較したメチル化レベルの増加または減少は、作用因子がSMYD3による網膜芽細胞腫のメチル化を調整することを示す、工程。
【請求項2】
網膜芽細胞腫ペプチドが、配列番号:4のアミノ酸配列を含むポリペプチド、またはその機能的な変異体もしくは断片である、請求項1記載の方法。
【請求項3】
補助因子がS-アデノシルホモシステイン加水分解酵素(SAHH)である、請求項1記載の方法。
【請求項4】
網膜芽細胞腫のメチル化を調節する試験化合物の能力を検出するためのキットであって、以下の構成成分を含むキット:
a.以下からなる群より選択される、メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド:
i.配列番号:2のアミノ酸配列を含むポリペプチド;
ii. 1つまたは複数のアミノ酸が置換、欠失、または挿入された、配列番号:2のアミノ酸配列を含むポリペプチドであって、さらに、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
iii.配列番号:2と少なくとも約80%の相同性を有するアミノ酸配列を含むポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
vi.ストリンジェントな条件下で配列番号:1のヌクレオチド配列からなるポリヌクレオチドとハイブリダイズするポリヌクレオチドによってコードされるポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;および
v.配列番号:2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、該ポリペプチドが配列番号:2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
b.(a)のポリペプチドによってメチル化することができる網膜芽細胞腫ペプチド;ならびに
c.網膜芽細胞腫ペプチドのメチル化のための補助因子。
【請求項5】
網膜芽細胞腫ペプチドが、配列番号:4のアミノ酸配列を含むポリペプチド、またはその機能的な変異体断片である、請求項4記載のキット。
【請求項6】
a.S-アデノシルホモシステイン加水分解酵素(SAHH)
の要素をさらに含む、請求項4記載のキット。
【請求項7】
結腸直腸癌、肝細胞癌、膀胱癌、および乳癌からなる群より選択される癌を治療するための化合物をスクリーニングする方法であって、以下の工程を含む方法:
a.請求項1記載の方法を用いて、メチル化を調整する試験化合物を同定する工程、および
b.試験化合物の非存在下で検出される対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを下げる試験化合物を選択する工程。
【請求項8】
結腸直腸癌、肝細胞癌、膀胱癌、および乳癌からなる群より選択される癌の症状を軽減するための組成物であって、請求項7記載の方法によって同定された薬学的に有効な量の化合物および薬学的に許容される担体を含む、組成物。
【請求項9】
結腸直腸癌、肝細胞癌、膀胱癌、および乳癌からなる群より選択される癌の症状を軽減するための方法であって、癌細胞と、請求項7記載の方法によって同定された薬学的に有効な量の化合物を接触させる工程を含む、方法。
【請求項10】
パート(iii)において定義されるポリペプチドが、配列番号:2と少なくとも約95%の相同性を有するアミノ酸配列を含む、請求項1記載の方法。
【請求項11】
パート(ii)において定義されるポリペプチドが、20までの保存的アミノ酸置換を含む配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項12】
パート(ii)において定義されるポリペプチドが、配列番号:2のアミノ酸配列の位置1〜250のアミノ酸配列を含む、請求項1記載の方法。
【請求項13】
パート(ii)において定義されるポリペプチドが、配列番号:2のアミノ酸配列の位置45〜428のアミノ酸配列を含む、請求項1記載の方法。
【請求項14】
パート(ii)において定義されるポリペプチドが、位置1〜30のアミノ酸が欠失している配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項15】
パート(ii)において定義されるポリペプチドが、位置1〜44のアミノ酸が欠失している配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項16】
パート(ii)において定義されるポリペプチドが、位置1〜20のアミノ酸が欠失している配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項17】
パート(ii)において定義されるポリペプチドが、位置1〜10のアミノ酸が欠失している配列番号:2のアミノ酸配列を含む、請求項1記載の方法。
【請求項18】
パート(iv)において定義されるポリペプチドが、高ストリンジェントな条件下で、配列番号:1のヌクレオチド配列からなるポリヌクレオチドと特異的にハイブリダイズする、請求項1記載の方法。
【請求項19】
機能的な網膜芽細胞腫断片がC末端断片を含む、請求項2記載の方法。
【請求項20】
機能的な網膜芽細胞腫断片が、配列番号:4のアミノ酸配列の位置769〜921のアミノ酸配列からなる、請求項2記載の方法。
【請求項21】
機能的な網膜芽細胞腫変異体が、以下の変異:K889A、K896A、K791A、K814A、K791A/K824A、およびK814A/K824Aの1つまたは複数を含む配列番号:4のアミノ酸配列を含む、請求項2記載の方法。
【図1】



【図2A】
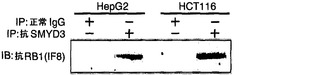
【図2B】

【図2C】

【図2D】
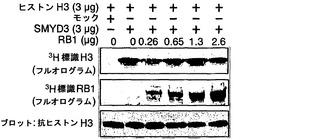
【図2E】
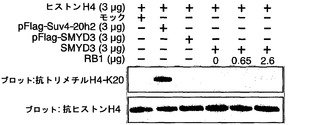
【図3A】

【図3B】
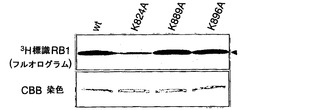
【図3C】
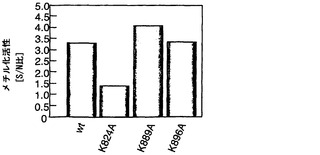
【図3D】

【図3E】

【図3F】
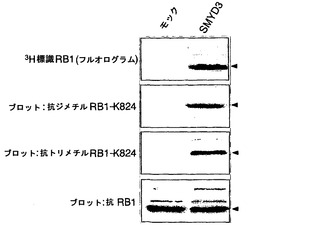
【図4A】
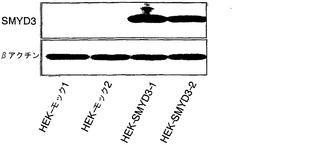
【図4B】
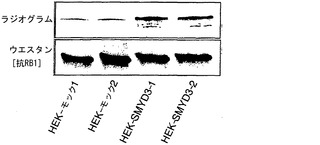
【図4C】
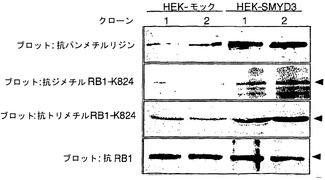
【図4D】

【図4E】

【図4F】

【図4G】

【図4H】

【図4I】

【図4J】

【図4K】

【図5A】
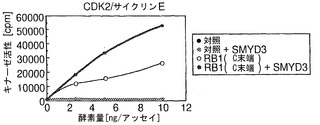
【図5B】
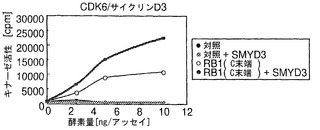
【図5C】
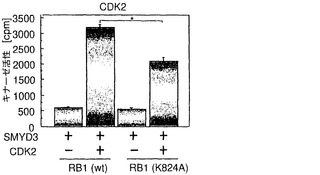
【図5D】

【図5E】

【図5F】
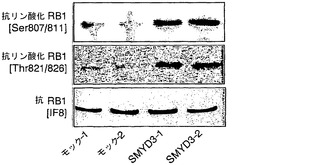
【図5G】

【図5H】
【図5I】

【図5J】

【図6】
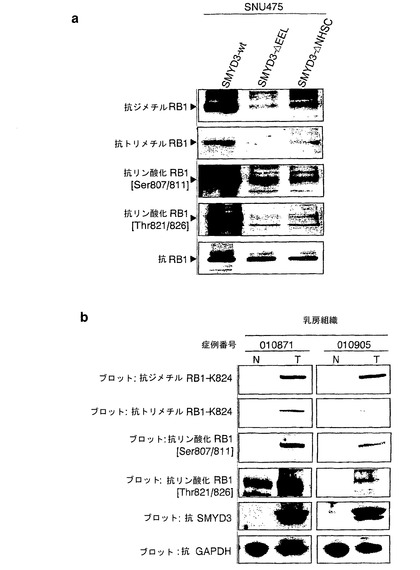
【図7】
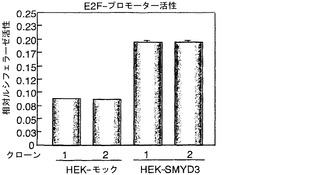
【図8】
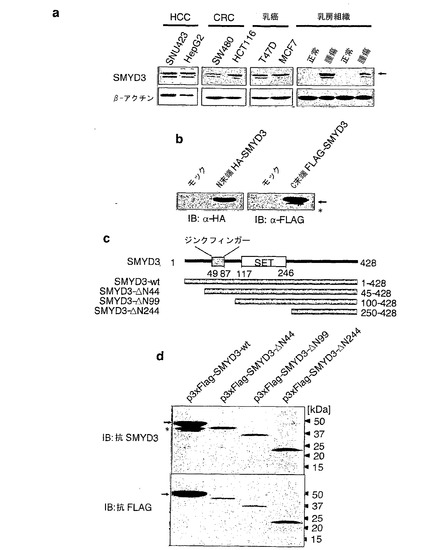
【図9】

【図10】
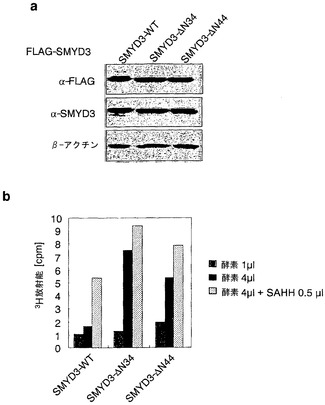
【図11】
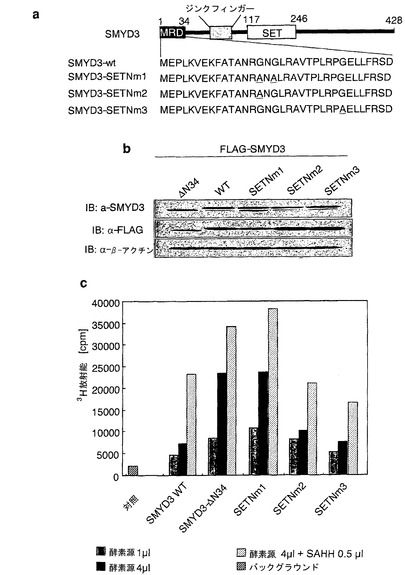
【図12】
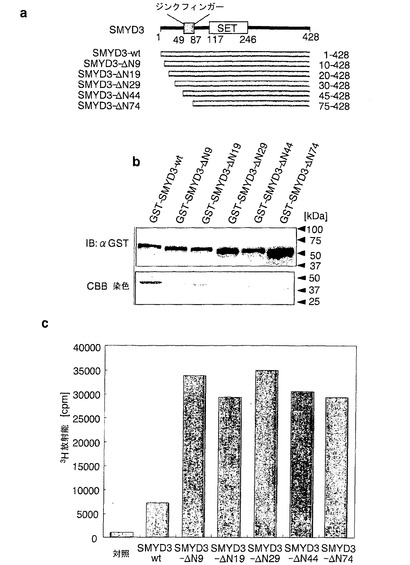
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図3F】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図4G】
【図4H】
【図4I】
【図4J】
【図4K】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図5H】
【図5I】
【図5J】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2009−500002(P2009−500002A)
【公表日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願番号】特願2007−557265(P2007−557265)
【出願日】平成18年6月23日(2006.6.23)
【国際出願番号】PCT/JP2006/313038
【国際公開番号】WO2007/004526
【国際公開日】平成19年1月11日(2007.1.11)
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【Fターム(参考)】
【公表日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願日】平成18年6月23日(2006.6.23)
【国際出願番号】PCT/JP2006/313038
【国際公開番号】WO2007/004526
【国際公開日】平成19年1月11日(2007.1.11)
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【Fターム(参考)】
[ Back to top ]