
癌抗原NYESO−1由来の新規MHCクラスII拘束T細胞エピトープ

【課題】癌抗原NY−ESO−1由来の新規MHCクラスIIエピトープの同定及び単離方法の提供。
【解決手段】NY−EsO−1由来の新規癌ペプチドであるMHCクラスIIエピトープは、HLAクラスII拘束様式で、特にHLA−DR又はHLA−DP拘束様式でCD4+Tリンパ球によって認識される。少なくとも1つの単離されたNY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドをコードする核酸配列遺伝子産物は、癌患者の予防、治療及び診断のための免疫治療的戦略の有望な候補である。
【解決手段】NY−EsO−1由来の新規癌ペプチドであるMHCクラスIIエピトープは、HLAクラスII拘束様式で、特にHLA−DR又はHLA−DP拘束様式でCD4+Tリンパ球によって認識される。少なくとも1つの単離されたNY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドをコードする核酸配列遺伝子産物は、癌患者の予防、治療及び診断のための免疫治療的戦略の有望な候補である。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、癌診断及び癌ワクチンを含む癌治療の領域に関する。より詳細には、本発明は、癌ペプチドNY−ESO−1由来の新規なヒトMHCクラスII拘束T細胞エピトープ及びそのアナログ並びにMHCIIクラスII拘束T細胞エピトープ又はその部分をコードするDNA配列の単離と精製に関する。特に、本発明は、NY−ESO−1由来のHLA−DR及びHLA−DP拘束T細胞エピトープに関する。本発明は更に、個体における癌及び前癌の検出、診断及び治療の方法に関する。
【背景技術】
【0002】
発明の背景
T細胞は、腫瘍増殖を制御し、腫瘍退行を媒介するのに重要な役割を果たす。T細胞媒介抗腫瘍免疫の分子的基盤を理解するために、CD8+T細胞によって認識される多くの腫瘍抗原が、メラノーマや他の種類の癌で同定されてきた(1〜3)。これらの研究は、分子的に明確な腫瘍抗原由来のペプチドを用いる幾つかの臨床試験に至った(4〜7)。gp100由来の修飾ペプチドを用いる臨床試験は、転移性メラノーマの患者の治療に対し治療的効力の幾分かの証拠をもたらしたが(4)、これらの研究は主にCD8+T細胞の使用に焦点をあてていた。ヒト及び動物研究の両方からの証拠の増大は、最適の癌ワクチンがCD4+及びCD8+T細胞の両方の参加を必要とすることを示した(8,9)。更に、腫瘍特異的CD4+T細胞は、MHCクラスII陰性腫瘍細胞に対する防御免疫を産生するのに必要とされる(10,11)。それ故、このような抗原の同定は、癌ワクチンの開発及びCD4+T細胞が宿主免疫応答を制御する機構を我々が理解するために重要である。
【0003】
今までに、ほんの限られた数のMHCクラスII拘束腫瘍抗原が同定された。チロシナーゼ、gp100及びMAGE−3などの幾つかの公知のMHCクラスI拘束腫瘍抗原は、CD4+T細胞によって認識されるMHCクラスII拘束エピトープを含むことが示された(12〜15)。腫瘍特異的CD4+T細胞を用いて、未知のMHCクラスII拘束腫瘍抗原を同定するために、最近、遺伝的アプローチが開発された。これは、CDC27、TPI及びLDFPを含む幾つかの変異腫瘍抗原の同定に至った(16,17)。それらのうち、TPIは、生化学的アプローチによって独立に同定された変異抗原である(18)。
【0004】
NY−ESO−1遺伝子は、抗体スクリーニングによって以前に同定され(19)、最近、MHCクラスI拘束腫瘍抗原としても同定された(20,21)。NY−ESO−1に対する高力価の抗体がまた、癌患者から検出された(22)。NY−ESO−1 cDNAは、2つの重複するオープンリーディングフレーム由来の2つの遺伝子産物をコードしていた(20)。正常精巣での発現を除くその厳密な腫瘍特異的発現パターン、並びにメラノーマ、乳癌、前立腺癌、肺癌及び他の癌を含む多くの腫瘍での高頻度の発現(18,20,23)の故に、NY−ESO−1は、種々の癌タイプに対する免疫治療の開発のための重要な免疫標的の可能性がある(24)。
【発明の概要】
【発明が解決しようとする課題】
【0005】
NY−ESO−1に対するCTL及び抗体免疫応答の両方が癌患者で示されたが、NY−ESO−1タンパク質におけるMHCクラスII拘束T細胞エピトープは報告されなかった。
【課題を解決するための手段】
【0006】
発明の概要
本発明の1つの目的は、CD4+Tリンパ球によってMHCクラスII拘束T細胞エピトープとして認識される新規ペプチド及びその部分を提供することである。本発明の抗原性癌ペプチドは、NY−ESO−1(本明細書ではCAG−3と交換可能に使用される用語)遺伝子(配列番号1)(Genbank登録番号 AF038567; 8:9)内もしくは部分によって、又はその変異体もしくはホモログ(例えば、LAGE遺伝子)(Genbank登録番号 AJ223040, AJ223041及びAJ223093)内もしくは部分によってコードされる。
【0007】
本発明の1局面は、癌の進展及び転移からレシピエントを防御するCD4+Tリンパ球を誘発できる癌ワクチンとして有用である、NY−ESO−1遺伝子によってコードされるMHCクラスII拘束T細胞エピトープ又はその変異体である。本発明はまた、癌の阻害又は予防のために、あるいはNY−ESO−1遺伝子産物を発現している細胞の増殖を阻害するために、有効量の癌ワクチンを投与する方法に関する。
【0008】
本発明の1局面は、NY−ESO−1遺伝子によってコードされるHLA−DR及びHLA−DP拘束T細胞エピトープ、又はその変異体及びホモログである。これらは、CD4+Tリンパ球及び抗NY−ESO−1抗体応答を誘発でき、次いで、癌の進展からレシピエントに防御及び/又は治療的利益を提供し、かつ転移からの防御を提供する免疫原及び癌ワクチンとして有用である。本発明はまた、癌を阻害又は予防し、NY−ESO−1遺伝子産物を発現している細胞の増殖を阻害し、かつ転移を阻害するために、有効量の癌ワクチンを投与する方法に関する。
【0009】
本発明の別の局面は、NY−ESO−1由来のMHCクラスII拘束T細胞エピトープ又はその変異体を単独で、又は1つ以上の免疫刺激分子と組み合わせて含む医薬組成物である。医薬組成物は、腫瘍及び癌に対する免疫応答を誘発するために、少なくとも1つのNY−ESO−1 MHCクラスII拘束T細胞エピトープを含むか、又はNY−ESO−1抗原特異的CD4+T細胞を刺激するエピトープの組み合わせを含む。医薬組成物は、CD8+Tリンパ球の産生のために、NY−ESO−1由来の1つ以上のMHCクラスI拘束T細胞エピトープを更に含んでもよい。NY−ESO−1MHCクラスII拘束T細胞エピトープとNY−ESO−1MHCクラスI拘束T細胞エピトープは各々、別のエピトープとして提供されてもよいし、又は一緒に結合されて、マルチマーの形態で提供されてもよい。癌エピトープ又はその変異体は、癌の予防又は治療のために、免疫原又はワクチンとして提供されてもよい。医薬組成物は、哺乳動物の癌を治療又は予防する方法に有用である。治療方法において、医薬組成物は、哺乳動物の癌を予防又は阻害する有効な量において哺乳動物に投与される。
【0010】
本発明の別の局面は、NY−ESO−1由来のHLA−DR及び/又はHLA−DP拘束T細胞エピトープ又はその変異体を単独で、又は1つ以上の免疫刺激分子と組み合わせて含む医薬組成物である。医薬組成物は、腫瘍及び癌に対する免疫原反応を誘発するために、少なくとも1つのNY−ESO−1 HLA−DR拘束T細胞エピトープ又は少なくとも1つのNY−ESO−1 HLA−DP拘束T細胞エピトープ、又はNY−ESO−1抗原特異的CD4+T細胞を刺激するMHCクラスII拘束T細胞エピトープの組み合わせを含む。医薬組成物は、CD8+Tリンパ球の産生のために、NY−ESO−1由来の1つ以上のMHCクラスI拘束T細胞エピトープを更に含んでもよい。NY−ESO−1 HLA−DR拘束T細胞エピトープ又はNY−ESO−1 HLA−DP拘束T細胞エピトープ、及びNY−ESO−1 MHCクラスI拘束T細胞エピトープは各々、別個のエピトープとして提供されてもよいし、一緒に結合されてもよい。癌エピトープ又はその変異体は、癌の予防又は治療のために、免疫原又はワクチンとして提供されてもよい。医薬組成物は、哺乳動物における癌の治療又は予防の方法において有用である。治療方法において、医薬組成物は、哺乳動物の癌の予防又は阻害のためにCD4+T細胞及び/又は抗NY−ESO−1抗体応答を誘発するのに有効な量において哺乳動物に投与される。
【0011】
本発明の別の目的は、NY−ESO−1遺伝子、その部分又はホモログに由来するNY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体をコードしているDNA配列の翻訳によって、同上を製造する方法である。
【0012】
本発明の別の目的は、NY−ESO−1遺伝子、その部分又はホモログに由来するNY−ESO−1のHLA−DR拘束T細胞エピトープもしくはHLA−DP拘束T細胞エピトープ又はそれらの変異体又は誘導体をコードしているDNA配列の翻訳によって、同上を製造する方法である。
【0013】
本発明の更なる局面は、NY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその変異体をコードする単離されたDNA又はRNA配列、及びそれらの相補配列、並びにワクチンとしての該DNA又はRNA配列の使用、及びNY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体の製造方法における該DNA又はRNA配列の使用である。本発明は更に、プローブ、プライマー又はアンチセンスとしての使用のために、該DNA又はRNA配列のオリゴヌクレオチドを提供する。
【0014】
本発明の更なる局面は、NY−ESO−1のHLA−DR拘束T細胞エピトープ、HLA−DP拘束T細胞エピトープ、又はそれらの変異体及びそれらの組み合わせをコードする単離されたDNA又はRNA配列、並びにNY−ESO−1のHLA−DR拘束T細胞エピトープ、HLA−DP拘束T細胞エピトープ、又はそれらの変異体及びそれらの組み合わせの製造方法における該DNA又はRNA配列の使用である。本発明は更に、プローブ、プライマー又はアンチセンスとしての使用のために、該DNA又はRNA配列のオリゴヌクレオチドを提供する。
【0015】
本発明は更に、NY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその変異体をコードする核酸配列のみを含むか、又は少なくとも1つの免疫刺激分子をコードする第2のDNA配列を組み合わせて含むベクターを提供する。
【0016】
本発明は更に、NY−ESO−1の少なくとも1つのHLA−DR拘束細胞エピトープ又は少なくとも1つのHLA−DP拘束T細胞エピトープ又はそれらの変異体若しくは組み合わせをコードする核酸配列のみを含むか、又は少なくとも1つの免疫刺激分子をコードする第2のDNA配列を組み合わせて含むベクターを提供する。
【0017】
本発明はまた、NY−ESO−1の少なくとも1つのMHCクラスII拘束細胞エピトープ又はその変異体をコードするDNA配列のみを含むか、又は少なくとも1つの免疫刺激分子をコードする第2のDNA配列を組み合わせて含むベクターによってトランスフェクトされた、又は形質導入された宿主細胞を提供する。ベクター及び宿主細胞は、ワクチンとして役立ちうる。ワクチンにおいて、MHCクラスII拘束T細胞エピトープの発現により、ワクチンで免疫化した哺乳動物において腫瘍抗原特異的CD4+Tリンパ球の刺激が生ずる。
【0018】
本発明はまた、NY−ESO−1の少なくとも1つのHLA−DR拘束細胞エピトープ又は少なくとも1つのHLA−DP拘束T細胞エピトープ又はそれらの変異体若しくは組み合わせをコードするDNA配列のみを含むか、又は少なくとも1つの免疫刺激分子をコードする第2のDNA配列を組み合わせて含むベクターによってトランスフェクトされた、又は形質導入された宿主細胞を提供する。ベクター及び宿主細胞は、ワクチンとして役立ちうる。ワクチンにおいて、HLA−DR拘束T細胞エピトープ及び/又はHLA−DP拘束T細胞エピトープの発現により、ワクチンで免疫化した哺乳動物において腫瘍抗原特異的CD4+Tリンパ球の刺激が生ずる。
【0019】
本発明は、NY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体の検出により、哺乳動物における癌又は前癌の診断方法を提供する。
【0020】
本発明は、NY−ESO−1のHLA−DR拘束T細胞エピトープ及び/又はHLA−DP拘束T細胞エピトープ又はそれらの変異体の検出により、哺乳動物における癌又は前癌の診断方法を提供する。
【0021】
ヒト前腫瘍性及び腫瘍性細胞及び組織の診断方法を提供することは、本発明の更に別の目的である。本発明によれば、この方法は、ヒトから細胞、組織、又はその抽出物を単離し、そしてNY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体をコードするDNA配列、RNA配列、又はそれらの部分を検出するか、又はDNA配列もしくはRNA配列によって発現されたエピトープ若しくはその変異体を検出することを含む。ここで、DNA配列、RNA配列又は発現産物の増大の検出/増大は、前腫瘍及び腫瘍の指標となる。
【0022】
ヒト前腫瘍性及び腫瘍性細胞及び組織の診断方法を提供することは、本発明の更に別の目的である。本発明によれば、この方法は、ヒトから細胞、組織、又はその抽出物を単離し、そしてNY−ESO−1のHLA−DR拘束T細胞エピトープ又はHLA−DP拘束T細胞エピトープ又はそれらの変異体をコードするDNA配列、RNA配列、又はその部分を検出するか、又はそれらのDNA配列もしくはRNA配列によって発現されたエピトープ若しくはその変異体若しくは組み合わせを検出することを含む。ここで、DNA配列、RNA配列又は発現産物の増大の検出/増大は、前腫瘍及び腫瘍の指標となる。
【0023】
本発明の別の目的は、NY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその変異体をコードするDNA配列の1つ以上のコピーをそのゲノムに組み込んだトランスジェニック動物を提供することである。DNA配列の組み込みにより、エピトープの発現又は過剰発現が生じる。このようなトランスジェニック動物は、癌の治療に有用な治療薬のスクリーニングのために有用である。
【0024】
本発明の更に別の目的は、NY−ESO−1の少なくとも1つのHLA−DR拘束T細胞エピトープ又は少なくとも1つのHLA−DR拘束T細胞エピトープ又はそれらの変異体又は組み合わせをコードするDNA配列の1つ以上のコピーをそのゲノムに組み込んだトランスジェニック動物を提供することである。DNA配列の組み込みにより、エピトープの発現又は過剰発現が生じる。このようなトランスジェニック動物は、癌の治療に有用な治療薬のスクリーニングのために有用である。
【0025】
本発明の更に別の局面は、治療薬としての使用に関する、並びに診断及び検出アッセイにおける使用に関する、NY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体と反応性のあるモノクローナル、ポリクローナル及び組換え抗体である。モノクローナル及びポリクローナル抗体は、単独でキットの形態で、又は診断及び検出アッセイで通常使用される他の試薬と一緒にキットの形態で提供されてもよい。
【0026】
本発明の更に別の局面は、治療薬としての使用に関する、並びに診断及び検出アッセイにおける使用に関する、NY−ESO−1のHLA−DR拘束T細胞エピトープ又はHLA−DP拘束T細胞エピトープと反応性のあるか、あるいはHLA−DP分子との組み合わせにおいてHLA−DPエピトープと、又はHLA−DR分子との組み合わせにおいてHLA−DRエピトープと反応性のあるか、あるいはそれらの変異体と反応性のあるモノクローナル、ポリクローナル及び組換え抗体である。モノクローナル及びポリクローナル抗体は、単独でキットの形態で、又は診断及び検出アッセイで通常使用される他の試薬と一緒にキットの形態で提供されてもよい。
【発明の効果】
【0027】
本発明は、CD4+T細胞によって認識されるNY−ESO−1由来の新規なMHCクラスII拘束T細胞エピトープの同定と単離である。本発明の癌エピトープは、哺乳動物において癌を阻害又は予防する免疫原及びワクチンとして有用であり、癌又は前癌を検出する診断薬として有用である。
【図面の簡単な説明】
【0028】
【図1−A】NY−ESO−1のヌクレオチド配列及びアミノ酸配列。NY−ESO−1のヌクレオチド配列の番号付けは、5’非翻訳領域の最初のヌクレオチドから始まる。
【図1−B】NY−ESO−1のヌクレオチド配列及びアミノ酸配列。NY−ESO−1のヌクレオチド配列の番号付けは、5’非翻訳領域の最初のヌクレオチドから始まる。
【図2−A】(2A)Ni2+クロマトグラフィーカラムを用いる完全長NY−ESO−1タンパク質の精製。SDSポリアクリルアミドゲルは、pET28ベクター(レーン1)、pNY−ESO−1(レーン2)を担うE.coli株BL21(DE3)からの粗抽出物、精製NY−ESO−1タンパク質(レーン3)、トランケートNY−ESO−1をコードする細菌抽出物(レーン4)、及び精製されたトランケートNY−ESO−1タンパク質であるESO1−74(レーン5)を示した。
【図2−B】(2B)NY−ESO−1に対する抗体の特異性を確認するためのウエスタンブロット。2人の代表的な患者からの1/2000希釈の血清(1つは、ELISAによって検出できるNY−ESO−1抗体をもつもの(レーン1、2及び3)及び1つは、該抗体無しのもの(レーン4、5及び6))を、ベクターだけをコードする細菌抽出物(レーン1、4)、NY−ESO−1をコードする細菌抽出物(レーン2、5)及び精製NY−ESO−1タンパク質(レーン3、6)に対して用いた。
【図2−C】(2C)患者TEは、NY−ESO−1タンパク質に対して抗体を有するメラノーマ患者の1人であった。NY−ESO−1(コントロールタンパク質としてBSA)に対する抗体の存在について88人の患者からの血清を用いてELISAを行った。血清希釈の1:25、1:250、1:2500でのO.D.450の値をプロットした。正常ドナーからの血清をコントロールとして用いた。それらの平均OD値もプロットした(ND)。
【図3】NY−ESO−1タンパク質で免疫化したHLA−DR4−Tgマウスを用いる推定NY−ESO−1エピトープの試験。HLA−DR4に対する予測された結合アフィニティに基づく8つのペプチドを、免疫化マウスからのリンパ球のインビトロ感作のために用いた。マウスリンパ球を、培地だけ、1359EBV B(HLA DR4+)細胞だけ、又はインビトロ刺激のために使用されたペプチドでパルスした1359EBV B細胞に対しIFN産生について試験した。
【図4−A】TE4−1 CD4+T細胞株の特徴付け。(4A)TE4−1は、ESOp116−135ペプチド又は精製NY−ESO−1タンパク質でパルスした1088EBV B細胞(HLA−DR+)を特異的に認識したが、推定エピトープを欠くESO1−74タンパク質を認識しなかった。
【図4−B】TE4−1 CD4+T細胞株の特徴付け。(4B)HLA DR拘束は、TE4−1によるNY−ESO−1の認識のために必要であった。293IMDR細胞にパルスしたとき、2つの重複するペプチドESOp111−130とp116−135は、認識された。GM−CSF分泌を測定する前に、1×105標的細胞を、4×104のTE4−1細胞と一晩共培養した。
【図4−C】TE4−1 CD4+T細胞株の特徴付け。(4C)ESOp116−135ペプチドでパルスした293IMDRの認識は、抗HLA−DR抗体(HB55)によって特異的に阻害されたが、抗クラスI抗体(HB95)によっては阻害されなかった。抗体の非存在下でTE4−1によって分泌されたGM−CSFの量を参照として用い、この参照に対し、抗体存在下のGM−CSF放出のパーセントを計算した。コントロール(マウスIgG2a)と抗MHC−クラスI抗体(HB95)による阻害は殆ど効果を有しなかった。
【図4−D】TE4−1 CD4+T細胞株の特徴付け。CTLC3G1(C.Macalliの好意)は、624.38melを認識するgp100特異的CD8+T細胞株であり、HB95の活性についてのコントロールとして使用した(図4D)。
【図4−E】TE4−1 CD4+T細胞株の特徴付け。T3−80は、1362melを認識するCD4+T細胞株であり、HB55の活性についてのコントロールとして使用した(図4E)。
【図5】CD4+T細胞株TE4−1による腫瘍細胞の認識。TE4−1の標的として使用した全てのメラノーマ株は、FACSとRT−PCRによってそれぞれ、HLA−DR4とNY−ESO−1の発現について分析された。TE4−1は、HLA−DR4分子を構成的に発現しているNY−ESO−1+腫瘍株(1359mel及びF049mel)を認識できた。DR1及びNY−ESO−1を発現しているF050melもT細胞によって認識された。コントロール標的526mel(DR4陽性及びNY−ESO−1陰性)、397mel、624.38mel(DR陰性及びNY−ESO−1陽性)、又は1300mel(DR1陽性及びNY−ESO−1弱陽性)のいずれに対しても反応性は無かった。
【図6−A】TE4−1によって認識されるNY−ESO−1ペプチドエピトープの特徴付け。(6A)HLA DR拘束NY−ESO−1エピトープのアンカー位置の決定。1088EBV B細胞を記載されたペプチド20Mでパルスした。GM−CSFを測定する前に、TE4−1細胞を一晩、標的細胞と共培養した。
【図6−B】TE4−1によって認識されるNY−ESO−1ペプチドエピトープの特徴付け。(6B)ESOp119−130を用いるペプチド力価測定実験。ESOp119−130は、図6Aに示すように、その認識に基づいて選択された。記載された濃度で希釈したESOp119−130を、1088EBV B細胞上にパルスした。この1088EBV B細胞は、TE4−1による認識のための標的として用いられた。コントロールペプチドESOp91−110の認識は、33Mの最高濃度でのみ測定された。
【図7−A】(7A)APCとしてDR1+EBVBを用いる、TE4−1によるNY−ESO−1タンパク質とESOp116−135の認識。NY−ESO−1タンパク質(5μg/ml)とペプチド(33μM)を、586EBV B(DR1+)上にパルスし、2度洗浄した。次いで、GM−CSFをアッセイする前に、TE4−1 T細胞を加え、一晩共培養した。
【図7−B】(7B)ESOp116−135(33μM)でパルスした586EBV Bを用いて、ブロッキング抗体の存在下及び非存在下でTE−4を刺激した。IID95(抗クラスI抗体)、IIB55(抗DR抗体)及びアイソタイプコントロール抗体を用いた。
【図8−A】合成ペプチドによるインビトロ刺激後のPBMCからのCD4+T細胞の産生。(図8A)3回のインビトロ刺激後、特異的ペプチド反応性をマルチプルウェルで検出した。96ウェルプレートで各々2.5×105PBMCを含む合計24ウェルを3週間弱く刺激した。24ウェルの15ウェルは、顕著な増殖を示し、そして比活性について試験した。各ウェルからのT細胞を、1088EBVB細胞、及びESOp161−180ペプチドでパルスした1088EBVB細胞とそれぞれインキュベートした。GM−CSF放出を上清から測定した。
【図8−B】合成ペプチドによるインビトロ刺激後のPBMCからのCD4+T細胞の産生。(図8B)TE4−2 T細胞は、NY−ESO−1ペプチド及びタンパク質と特異的に反応した。重複するペプチドESOp161−180とESOp156−175を、20μg/mlで90分間、1088(DR4+)と586EBVB(DR1+)細胞にパルスした。ESOp91−110をパルスについての無関係ペプチドとして用いた。精製NY−ESO−1とESO1−75タンパク質を、同じモル比を維持するために、それぞれ5μg/mlと2μg/mlで一晩パルスした。3度の洗浄後、TE4−2 T細胞を加え、一晩インキュベートした。GM−CSF放出を測定した。
【図8−C】合成ペプチドによるインビトロ刺激後のPBMCからのCD4+T細胞の産生。(図8C)ESOp161−180でパルスしたEBVB細胞のパネルを、TE4−2 CD4+T細胞の標的として用いた。これらのEBVB株は、異なるHLA DR及びDQ対立遺伝子を発現することが知られていた。それらのHLA DP対立遺伝子を、本研究では分子的にタイプ分けした(表2)。
【図9−A】抗DP抗体によるNY−ESO−1エピトープのT細胞認識のブロッキング。20μg/ml ESOp161−180ペプチドでパルスした1088EBVB細胞を、異なるブロッキング抗体存在下、標的細胞として使用した。使用した抗体の特異性は次のようであった:抗MHCクラスI(HLA A,B,C)抗体(W6/32)、抗MHCクラスII(HLA DP,DQ,DR)抗体(IVA12)、抗HLA DP抗体(B4/21)、抗HLA DR抗体(L243)及び抗HLA DQ抗体(Genox3.53(抗DQw1)及びIVD12(抗DQw3)の混合物)。全ての抗体を、各々、最終濃度20μg/mlで用いた。(図9A)CK3H6 T細胞は、HLA A2の状況においてgp100p209−218ペプチドを特異的に認識し、そして抗MHCクラスI抗体のための特異性コントロールとして用いた(図9B)。gp100p209−218ペプチドでパルスした1088EBVB(A2+)を標的として用いた。CD4+T細胞株(T3−80)は、HLA−DR拘束様式で1362melを認識し、そして抗MHCクラスII及び抗DR抗体のための特異性コントロールとして用いた(図9C)。
【図9−B】抗DP抗体によるNY−ESO−1エピトープのT細胞認識のブロッキング。20μg/ml ESOp161−180ペプチドでパルスした1088EBVB細胞を、異なるブロッキング抗体存在下、標的細胞として使用した。使用した抗体の特異性は次のようであった:抗MHCクラスI(HLA A,B,C)抗体(W6/32)、抗MHCクラスII(HLA DP,DQ,DR)抗体(IVA12)、抗HLA DP抗体(B4/21)、抗HLA DR抗体(L243)及び抗HLA DQ抗体(Genox3.53(抗DQw1)及びIVD12(抗DQw3)の混合物)。全ての抗体を、各々、最終濃度20μg/mlで用いた。(図9A)CK3H6 T細胞は、HLA A2の状況においてgp100p209−218ペプチドを特異的に認識し、そして抗MHCクラスI抗体のための特異性コントロールとして用いた(図9B)。gp100p209−218ペプチドでパルスした1088EBVB(A2+)を標的として用いた。CD4+T細胞株(T3−80)は、HLA−DR拘束様式で1362melを認識し、そして抗MHCクラスII及び抗DR抗体のための特異性コントロールとして用いた(図9C)。
【図9−C】抗DP抗体によるNY−ESO−1エピトープのT細胞認識のブロッキング。20μg/ml ESOp161−180ペプチドでパルスした1088EBVB細胞を、異なるブロッキング抗体存在下、標的細胞として使用した。使用した抗体の特異性は次のようであった:抗MHCクラスI(HLA A,B,C)抗体(W6/32)、抗MHCクラスII(HLA DP,DQ,DR)抗体(IVA12)、抗HLA DP抗体(B4/21)、抗HLA DR抗体(L243)及び抗HLA DQ抗体(Genox3.53(抗DQw1)及びIVD12(抗DQw3)の混合物)。全ての抗体を、各々、最終濃度20μg/mlで用いた。(図9A)CK3H6 T細胞は、HLA A2の状況においてgp100p209−218ペプチドを特異的に認識し、そして抗MHCクラスI抗体のための特異性コントロールとして用いた(図9B)。gp100p209−218ペプチドでパルスした1088EBVB(A2+)を標的として用いた。CD4+T細胞株(T3−80)は、HLA−DR拘束様式で1362melを認識し、そして抗MHCクラスII及び抗DR抗体のための特異性コントロールとして用いた(図9C)。
【図10−A】TE4−2 CD4+T細胞による腫瘍細胞と293CIITA/NY−ESO−1の認識。(図10A)TE4−2は、NY−ESO−1とDP4の両方を発現しているメラノーマ株を認識した。公知のNY−ESO−1発現(RT−PCRによって)とHLA DPタイプ(RT−PCRと配列決定によって決定)をもつメラノーマ株を、標的として使用した。この実験で、MHCクラスII発現をアップレギュレートするために、一晩のIFN−γ処理(500ユニット/ml)を、F026mel、526mel、及び397melについて行った。サイトカイン放出を測定する前に、TE4−2 T細胞を腫瘍細胞と一晩共培養した。
【図10−B】TE4−2 CD4+T細胞による腫瘍細胞と293CIITA/NY−ESO−1の認識。(図10B)TE4−2 CD4+T細胞株は、インバリアント鎖(Ii)ターゲティング配列の存在下又は非存在下においてNY−ESO−1でトランスフェクトされた293CIITAを認識した。親の293細胞と293CIITA細胞を、NY−ESO−1(pESO)、Ii−NY−ESO−1(pIi−ESO)、又はGFP(pGFP)をそれぞれコードするプラスミドでトランスフェクトした。サイトカイン放出をアッセイする前に、TE4−2 T細胞を、トランスフェクタントと共に一晩、共培養した。
【図11−A】TE4−2によって認識されるT細胞エピトープの特徴付け。(図11A)T細胞認識のためのNY−ESO−1エピトープのアンカー残基と最小長さの決定。N又はC末端にアミノ酸欠失を有する合成ペプチドを用いて、40μg/mlで1088EBVB細胞をパルスした。次いで、EBVB細胞を徹底的に洗浄し、TE4−2 T細胞を刺激するために、標的細胞として用いた。2つの別々の実験を行い、この図の上のパネルと下のパネルとして示した。
【図11−B】TE4−2によって認識されるT細胞エピトープの特徴付け。(図11B)T細胞認識に必要な最小ペプチド濃度の決定。ESOp157−170ペプチドを用いて、種々の濃度で1088EBVB細胞をパルスした。次いで、ペプチドをパルスされた細胞を洗浄し、TE4−2株を刺激するために、標的として用いた。コントロールペプチドESOp91−110を、最高濃度33μMでのみ用いた。
【図11−C】TE4−2によって認識されるT細胞エピトープの特徴付け。(図11C)NY−ESO−1特異的CD8+T細胞によるDPB1*0401−拘束CD4+T細胞エピトープの認識。TE8−1細胞株を、ESOp157−167ペプチドによるインビトロ刺激によって、親TEのPBMCから作製した。L023EBVB細胞(HLA−A2+,DP4-)を、無血清培地において、DPB1*0401エピトープ領域をカバーするペプチドでパルスし、洗浄し、TE8−1 T細胞を刺激するために用いた。
【図12−A】TE4−2 CD4+T細胞とTE8−1 CD8+T細胞による認識についてのペプチドの力価測定(図12A)586EBVB細胞(A2−,DP4+)を抗原提示細胞として用い、種々の濃度で、記載されたペプチドでパルスした。細胞を洗浄し、次いで、サイトカイン放出をアッセイする前に、TE4−2 CD4+T細胞とインキュベートした。
【図12−B】TE4−2 CD4+T細胞とTE8−1 CD8+T細胞による認識についてのペプチドの力価測定(図12B)L023EBVB細胞(A2+,DP4−)を抗原提示細胞として用い、種々の濃度で、記載されたペプチドでパルスした。細胞を洗浄し、次いで、サイトカイン放出をアッセイする前に、TE4−2 CD4+T細胞とインキュベートした。
【発明を実施するための形態】
【0029】
発明の詳細な説明
本発明は、免疫系のMHCクラスII拘束CD4+Tリンパ球によって免疫学的に認識されるNY−ESO−1の癌エピトープ、その部分、誘導体又は変異体を包含する。本発明の癌エピトープは、免疫系のCD4+T細胞との相互作用によって、体液性媒介免疫応答を特異的に引き起こす。抗原性癌エピトープとCD4+T細胞とのこの相互作用は、ヒトを含む哺乳動物における癌の予防、除去、又は減少において、CD4+T細胞が抗原性癌エピトープに反応し、免疫系の他の細胞をリクルートすることを引き起こす。
【0030】
本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープは、原発性もしくは転移性メラノーマ、胸腺腫、リンパ腫、肉腫、肺癌、肝癌、非ホジキンリンパ腫、ホジキンリンパ腫、白血病、子宮癌、子宮頚癌、膀胱癌、腎癌、頭部及び頚部癌、神経芽腫及び腺癌(例えば、乳癌、前立腺癌、卵巣癌、膵癌、甲状腺癌など)を含むが、それらに限定されない癌の一部を形成するか、又はそれらの癌に由来する。
【0031】
メラノーマという用語は、メラノーマ、転移性メラノーマ、メラノサイト又はメラノサイト関連母斑細胞由来のメラノーマ、悪性メラノーマ、黒色上皮腫、黒色肉腫、インサイチュメラノーマ(melanoma in situ)、表在拡大型メラノーマ、結節型(nodular)メラノーマ、悪性ほくろ性メラノーマ、末端性ほくろ性メラノーマ、浸潤性メラノーマ又は家族性非定型奇胎及びメラノーマ(familial atypical mole and melanoma)(FAM−M)症候群を含むが、それらに限定されない。
【0032】
癌、特にメラノーマの患者の自己CD4+Tリンパ球によって認識される癌エピトープ又はその誘導体は特に興味深い。MHC(又はHLA)クラスII拘束CD4+Tリンパ球、特にHLA−DR拘束Tリンパ球及び/又はHLA−DP拘束Tリンパ球によって認識される癌エピトープ又はその誘導体は更に興味深い。一実施態様では、NY−ESO癌エピトープは、HLA−DR分子の状況においてCD4+Tリンパ球によって認識される。別の実施態様では、NY−ESO癌エピトープは、HLA−DP分子の状況においてCD4+Tリンパ球によって認識される。
【0033】
本発明の「癌エピトープ」は、HLA−DR拘束及びHLA−DP拘束Tリンパ球などのMHCクラスII拘束Tリンパ球を誘発するNY−ESO−1の部分又は変異体部分を包含する。このようなリンパ球は、腫瘍細胞に由来する完全長NY−ESO−1タンパク質、MHCクラスII拘束T細胞エピトープ、及び天然でプロセスされた抗原と特異的に反応しうる。
【0034】
本発明のNY−ESO−1のMHCクラスII拘束T細胞エピトープは、サイズにおいて約9アミノ酸乃至約30アミノ酸、好ましくは、長さにおいて約10乃至約15アミノ酸に変わりうる。
【0035】
好適な実施態様では、本発明のNY−ESO−1のMHCクラスII拘束T細胞エピトープは、長さにおいて約9乃至10アミノ酸である。
【0036】
一実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:Xaa1KEFTVSXaa2(配列番号4)及びその変異体又は誘導体を含む、長さにおいて少なくとも約10アミノ酸のペプチドである。Xaa1及びXaa2のアミノ酸、及びN末端及びC末端でのこれらの位置におけるアミノ酸数は、エピトープが、CD4+Tリンパ球に結合し、刺激する能力を保持するかぎり、変わりうる。エピトープは、約10乃至約30アミノ酸、好ましくは20アミノ酸未満、より好ましくは約10乃至約15アミノ酸を含みうる。
【0037】
本発明の別の実施態様では、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、式:Xaa1VLLKEFTVSGXaa2(配列番号5)(式中、Xaa1はアミノ酸ではないか、又は1乃至約10個の天然のアミノ酸であり、好ましくは1乃至約5個のアミノ酸である;Xaa2はアミノ酸ではないか、又は長さにおいて1乃至約7個のアミノ酸である)によって表されることができる。
【0038】
本発明の別の実施態様では、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:QDAPPLPVPGVLLKEFTVSGNILTIRL(配列番号6)又はそのフラグメント若しくは誘導体を含む。
【0039】
また、本発明の範囲には、配列番号4、5又は6と部分配列相同性を有する癌ペプチド又はその部分が包含される。部分アミノ酸配列相同性とは、配列番号4、5又は6と少なくとも85%の配列相同性、好ましくは、少なくとも95%の配列相同性又はそれ以上を有し、かつNY−ESO−1 MHCクラスII拘束特異的CD4+Tリンパ球を刺激するという生物学的機能を有するペプチドを意味する。哺乳動物ホモログは、霊長類及びマウスホモログを含むがそれらに限定されない本発明の範囲に含まれる。
【0040】
一実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:
APPLPVPGVLLKEFTVSGNILTIRL(配列番号7);
APPLPVPGVLLKEFTVS(配列番号8);
APPLPVPGVLLKEFTV(配列番号9);
LPVPGVLLKEFTVSG(配列番号10);
PVPGVLLKEFTVSG(配列番号11);
VPGVLLKEFTVSG(配列番号12);
PGVLLKEFTVSG(配列番号13);
GVLLKEFTVSG(配列番号14);
LLKEFTVSGNILTIR(配列番号15);
LKEFTVSGNILTIRL(配列番号16);
KEFTVSGNILTIRL(配列番号17);
LPVPGVLLKEFTVSGNILTI(配列番号18);
又はそれらの変異体若しくは誘導体を含む。
【0041】
好適な実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープはアミノ酸配列:
【0042】
【数1】

【0043】
及びその変異体若しくは誘導体を含む。
【0044】
上記配列内に置換を有するエピトープは、本発明に包含される。置換アミノ酸により、配列番号19と比較して等価な又は増大した結合が生じるならば、天然に存在するアミノ酸の置換は、1個以上のアンカー位置でありうる。配列番号19のアンカー位置は、位置1−Leu、位置4−Glu、位置6−Thr、位置7−Valであると予測される。一実施態様では、アラニンは、位置1でロイシンの代わりに置換される。
【0045】
本発明の別の実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:DHRQLQLSIS SCLQQLSLLM(配列番号20)、又はその部分、変異体及び誘導体を含む。一実施態様において、配列番号20の部分はアミノ酸配列:
DHRQLQLSIS SCLQQLS(配列番号29);
DHRQLQLSIS SCLQ(配列番号30);
QLQLSIS SCLQQL(配列番号31)
又はそれらの変異体及び誘導体を含む。
【0046】
本発明の別の実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:
WITQCFLPVF LAQPPSGQRR(配列番号21)
その部分、変異体及び誘導体を含む。一実施態様では、配列番号21の部分は、アミノ酸配列:
QCFLPVF LAQPPSGQRR(配列番号32);
LPVF LAQPPSGQRR(配列番号33);
CFLPVF LAQPPSGQ(配列番号34);
又はそれらの変異体及び誘導体を含む。
【0047】
NY−ESO−1癌エピトープ及びその誘導体は、MHCクラスII拘束CD4+Tリンパ球によって認識される。NY−ESO−1エピトープと組み合わせて認識されるクラスII分子としては、HLA−DR1、HLA−DR3、HLA−DR4及びクラスIIペプチド結合の縮重のために機能する他のクラスII分子などの少なくとも1つのHLA−DRが挙げられるが、それに限定されない。一実施態様では、癌ペプチドによって認識されるHLAサブタイプは、HLA−DR4サブタイプである。別の実施態様では、NY−ESO−1癌エピトープは、HLA−DR1及びHLA−DR4と結合する。
【0048】
別の実施態様では、エピトープは、以下:
Xaa1ITQXaa2FXaa3PXaa4(配列番号51)
(式中、Xaa1は、少なくとも1つの天然に存在するアミノ酸、好ましくは、Trp、Phe、Tyr、Met、Ile、Val、Ala及びそれらの組み合わせからなる群から選択されるアミノ酸であり;
Xaa2は、少なくとも1つの天然に存在するアミノ酸、好ましくは、Cys、Ser、Val、Ala、Thr、及びそれらの組み合わせからなる群から選択されるアミノ酸であり;
Xaa3は、少なくとも1つの天然に存在するアミノ酸、好ましくは、Leu、Phe、Tyr、Met、Ile、Val、Ala、及びそれらの組み合わせからなる群から選択されるアミノ酸であり;
Xaa4は、少なくとも1つの天然に存在するアミノ酸、好ましくは、Val、Tyr、Ile、Ala、Leu、Pro、及びそれらの組み合わせからなる群から選択されるアミノ酸である)の一般的なアミノ酸モチーフを含む、NY−ESO−1のHLA−DP拘束T細胞エピトープである。
【0049】
また、配列番号51と部分配列相同性を共有する癌ペプチド又はその部分は、本発明の範囲に包含される。部分アミノ酸配列相同性とは、配列番号51と少なくとも85%の配列相同性、好ましくは、少なくとも95%の配列相同性又はそれ以上を有し、かつNY−ESO−1 MHCクラスII拘束特異的CD4+Tリンパ球、好ましくはHLA−DP拘束Tリンパ球を刺激するという生物学的機能を有するペプチドを意味する。哺乳動物ホモログは、霊長類及びマウスホモログを含むがそれらに限定されない本発明の範囲に含まれる。ホモログはまた、LAGE遺伝子などのNY−ESO−1遺伝子以外の遺伝子によってコードされうる配列番号51と配列相同性を有するペプチドを含む。
【0050】
上記配列内に置換を有するエピトープは、本発明に包含される。置換アミノ酸により、配列番号51と比較して等価な又は増大した結合が生じるならば、天然に存在するアミノ酸の置換は、1個以上のアンカー位置でありうる。配列番号51のアンカー位置は、位置1、7、及び9、即ち、それぞれW、L、及びV残基であると推定される。これらのアンカー残基の置換は、「W」残基の代わりにF、Y、M、I、V及びA;「L」残基の代わりにF、Y、M、V、I及びA;「V」残基の代わりにY、I、A、L及びPでありうるが、それらに限定されない。別の置換は、配列番号1の5位としてC残基を含みうるが、それは、残基S、V、A及びTでありうるが、それらに限定されない。
【0051】
一実施態様では、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:Xaa1WITQCFLPVFXaa2(配列番号52)及びその変異体又は誘導体を含む、長さで少なくとも約10アミノ酸のペプチドである。Xaa1及びXaa2は、アミノ酸ではないか、又は1つ以上の同じ、又は種々の天然に存在するアミノ酸でありうる。エピトープがCD4+Tリンパ球、特にHLA−DP拘束CD4+Tリンパ球と結合/及びそれらを刺激する能力を保持するかぎりは、N末端及びC末端でのこれらの位置でのアミノ酸の数は変りうる。エピトープは、約10乃至約30アミノ酸、好ましくは20アミノ酸未満、より好ましくは約10乃至約15アミノ酸を含みうる。
【0052】
一実施態様では、NY−ESO−1のHLA−DP拘束T細胞エピトープは、アミノ酸配列:
151-SCLQQLSLLMWITQCFLPVFLAQPPSG-177(配列番号53);
SLLMWITQCFLPVF(配列番号54);
LLMWITQCFLPVFL(配列番号55);
LMWITWCFLPVFLA(配列番号56);
MWITQCFLPVFLAQ(配列番号57);
WITQCFLPVFLAQP(配列番号58);
ITQCFLPVFLAQPP(配列番号59);
QQLSLLMWITQCFL(配列番号60);
QLSLLMWITQCFLP(配列番号61);
LSLLMWITQCFLPV(配列番号62);及び
それらの誘導体を含む。
【0053】
NY−ESO−1癌エピトープ及びその誘導体は、HLA−DP拘束CD4+Tリンパ球、及びその誘導体、好ましくはHLA−DP4拘束CD4+Tリンパ球によって認識される。NY−ESO−1エピトープと組み合わせて認識されるクラスII分子としては、HLA−DP及びクラスIIペプチド結合の縮重のために機能するクラスII分子を含む。癌ペプチドによって認識される好適なHLAサブタイプは、HLA−DPB1*0401−0402対立遺伝子及び機能縮重のために、該ペプチドに結合しうる他の対立遺伝子である。
【0054】
本発明の別の実施態様は、MHCクラスII拘束CD4+Tリンパ球を誘発するように有効に作用するために、NY−ESO−1のMHCクラスII拘束T細胞エピトープに十分な相同性を有する該エピトープの誘導体及び変異体を包含する。このようなペプチドは、1つ以上の位置、特にアンカー位置で保存的アミノ酸変化を有しうる。保存的アミノ酸変化とは、特定の位置での、元々存在した同じタイプのアミノ酸でのアミノ酸の変化、即ち、疎水性アミノ酸の代わりに交換された疎水性アミノ酸、塩基性アミノ酸の代わりに塩基性アミノ酸などを意味する。このようなアミノ酸変化は、ペプチドの全体の電荷やコンフィギュレーションを有意には変えず、それ故、このような変異体は、癌ペプチドの抗癌活性を維持するか、又は増大させる。このような保存的変化の例は、当業者に周知であり、本発明の範囲内である。
【0055】
本発明はまた、NY−ESO−1 MHCクラスII拘束T細胞エピトープの機能的に等価な変異体に関する。「機能的に等価な変異体」としては、部分配列相同性を有するペプチド、1つ以上の特異的な保存及び/又は非保存的アミノ酸変化を有するペプチド、ペプチド結合体、キメラタンパク質、融合タンパク質及びペプチドが挙げられる。
【0056】
本発明の別の局面は、HLA−DP拘束T細胞及びHLAクラスI拘束CD8+Tリンパ球の両方、特にHLA−A拘束Tリンパ球、好ましくはHLA−A2拘束Tリンパ球のためのエピトープとして機能するNY−ESO−1ペプチドである。一実施態様では、NY−ESO−1のHLA−DP拘束及びHLAクラスI拘束エピトープとしては、アミノ酸配列:SLLMWITQCFLPVF(配列番号54)並びにその変異体及びホモログが挙げられる。変異体としては、RSLLMWITQCFLPV(配列番号63)を含むESOp156R−169及びSLLMWITQCFLPVR(配列番号64)を含むESOp157−170Rを含むがそれらに限定されない、SLLMWITQCFLPVF(配列番号54)における1つ以上の置換を有するペプチドが挙げられるがそれらに限定されない。このような置換は、ネイティブ配列の免疫学的機能活性を保持しつつ、ペプチドを水溶液により可溶性にする。高度に水溶性のペプチドは90%超の純度まで容易に精製されうる。
【0057】
NY−ESO−1 MHCクラスII拘束T細胞エピトープは、初代(primary)臨床単離体、細胞株などからのような天然源から精製単離できる。NY−ESO−1 MHCクラスII拘束T細胞エピトープは、少なくとも90%純粋、好ましくは少なくとも95%純粋及びほぼ100%純粋である。エピトープは、当業界公知の化学合成又は組換えDNA技術によっても得ることができる。化学合成の技術は、J.M.Steward and J.D.Young, “Solid Phase Peptide Synthesis”, W.H.Freeman & Co., San Francisco, 1969; M.Bodansky et al “Peptide Synthesis”, John Wiley & Sons, Second Edition, 1976,及びJ.Meienhofer, “Hormonal Proteins and Peptides”, Vol.2, p.46, Academic Press, New York, 1983 及びE.Schroder and K.Kubke, “The Peptides", Vol.1, Academic Press, New York, 1965に記載されている。
【0058】
NY−ESO−1 クラスII拘束T細胞エピトープは、当業界公知の方法によって、医薬として許容できる担体と共に医薬組成物へと製剤化できる。本組成物は、NY−ESO−1特異的CD4+Tリンパ球を誘発するために免疫原として有用であり、そして抗NY−ESO−1抗体を誘発するのに有用でありうる。本組成物はまた、癌を予防又は治療するためのワクチンとして有用である。本組成物は更に、少なくとも1つの免疫刺激分子を含みうる。MHCクラスII特異的T細胞応答を刺激するために、癌エピトープ又はその部分と一緒に使用される免疫刺激分子としては、1つ以上の主要組織適合性複合体(MHC)クラスII分子、又はMHCクラスII分子を発現している細胞が挙げられるがそれらに限定されない。本組成物は、免疫増強のために、B7.1、B7.2、ICAM−1、ICAM−2、LFA−1、LFA−3、CD72など、並びにIL−1〜IL−15、TNFα、IFNγ、RANTES、G−CSF、M−CSF、IFNα、CTAPIII、ENA−78、GRO、I−309、PF−4、IP−10、LD−78、MGSA、MIP−1α、MIP−1β、又はそれらの組み合わせを含むがそれらに限定されないサイトカインなどを含む他の刺激分子を更に含んでもよい。
【0059】
刺激分子は、物理的に分離した実体として提供され得るか、又はB細胞、マクロファージ若しくは樹状細胞などの抗原提示細胞の膜中に提供され得るか、リポソームの膜中に提供され得るか、又は形質導入若しくはトランスフェクトされた細胞の表面上に発現され得る。MHCクラスII免疫刺激分子のDNA配列は、GenBankなどから入手できる。HLA−DP免疫刺激分子のDNA配列は、GenBank/EMBL/DNA Data Bank of Japan(DDBJ) at GenBank, National Center for Biotechnology Information, 8600 Rockville Pike, Bethesda, MD 20894 USA 又はそのウエブサイトから入手できる。
【0060】
本発明の医薬組成物は、NY−ESO−1 HLA−DR又はそのHLA−DP拘束T細胞癌ペプチドに加えて、NY−ESO−1からの幾つかの異なるMHCクラスII拘束T細胞エピトープを含んでもよい。これらには、LPVPGVLLKEFTVSG(配列番号10)、VLLKEFTVSGNILTIRLT(配列番号65)、AADHRQLQLSISSCLQQL(配列番号66)、及びそれらの組み合わせなどのHLA−DR拘束エピトープ及びその変異体が挙げられるがそれらに限定されない。
【0061】
本発明の医薬組成物は、MHCクラスII拘束CD4+Tリンパ球を誘発することに加えて、MHCクラスI拘束細胞傷害性Tリンパ球を誘発するために、MHCクラスI拘束NY−ESO−1癌ペプチドを必要に応じて含んでもよい。MHCクラスI拘束NY−ESO−1癌ペプチドとしては、式:
Xaa1Xaa2Xaa3GPGGGAPXaa4(配列番号22)
(式中、Xaa1は、アミノ酸ではないか、又は1乃至約20個の天然のアミノ酸、好ましくは1乃至5個のアミノ酸であり;
Xaa2は、Ala、Thr、Val、Leu、又はArgであり;
Xaa3は、Ser、又はAla、Val、Ile、Leu、Thrなどの保存的置換であり;
Xaa4は、Arg、Lys、好ましくはArg)並びにそれらのフラグメント及び誘導体によって表される癌ペプチドが挙げられるがそれらに限定されない。一実施態様では、医薬組成物で使用されるMHCクラスI拘束NY−ESO−1癌ペプチドとしては、アミノ酸配列:ASGPGGGAPR(配列番号23)が挙げられる。
【0062】
NY−ESO−1 MHCクラスII拘束T細胞エピトープ及びNY−ESO−1 MHCクラスI拘束T細胞エピトープは各々、別個のエピトープとして提供されてもよいし、単一ペプチドとして一緒に結合されてもよい。エピトープは、当業界公知の方法によって、一緒に結合されてもよいし、化学的に合成されてもよい。化学リンカー、ペプチドリンカー、ペプチド結合などは、エピトープを結合するために使用できる。一実施態様では、MHCクラスIIエピトープのC末端は、ペプチド結合により、MHCクラスIエピトープのN末端に直接結合される。
【0063】
NY−ESO−1 MHCクラスII拘束T細胞エピトープは、癌の予防又は治療方法において有用であり、そしてヒトを含む哺乳動物における癌又は前癌を検出するための診断アッセイにおいて有用である。診断アッセイにおいて、本発明のNY−ESO−1 HLAクラスII拘束T細胞エピトープペプチド、その変異体及び誘導体は、腫瘍抗原NY−ESO−1に対するヘルパー免疫応答を検出するのに有用である。NY−ESO−1は腫瘍細胞(免疫が特別免除された部位である正常精巣を除く)に専ら発現されるので、該タンパク質に対する免疫応答は、患者において早期の癌の検出のための指標として使用できる。ヘルパーT細胞応答の発展は、該タンパク質に対して検出できる抗体の発展よりも早い出来事でありうるので、NY−ESO−1 HLAクラスII拘束T細胞エピトープペプチドに対するヘルパーT細胞応答の検出は、早期の癌の検出に有用である。NY−ESO−1に対するヘルパーT細胞応答の検出方法において、NY−ESO−1 HLAクラスII拘束T細胞エピトープペプチドは、基板又は固体支持体に適用される。患者からのリンパ球は、インフルエンザウイルス由来のペプチドなどのコントロールペプチドと並行して、NY−ESO−1 MHCクラスII拘束T細胞エピトープペプチドの存在下で生育される。次いで、ELISPOT及びELISAなどの技術を用いて、特異的なサイトカイン放出を測定する。陰性コントロールと比較した、ヘルパーT細胞免疫応答の増大の検出は、患者における前癌又は早期の癌の指標となる。
【0064】
本発明のNY−ESO−1 MHCクラスII拘束結合ペプチドは、任意の一定の標的抗原及び/又はハプテンに対する抗体及び/又はCD8+T細胞応答の産生を増大させるために使用できる。詳細には、これらのペプチドは、抗体及び/又はCD8+T細胞応答が意図される標的抗原ペプチド、タンパク質又は任意の他のハプテンに対して結合又は共有結合することができる。任意の標的ハプテン又はタンパク質とのNY−ESO−1 MHCクラスII拘束エピトープペプチドの結合は、破傷風トキソイド、アルブミンなどの通常のT細胞癌タンパク質と同様に、任意の標的抗原、ハプテン、又はタンパク質の免疫原性の増大において、免疫学的T細胞キャリアペプチドとして作用するはずである。増大は、標的ハプテン又はタンパク質に対するより高力価の抗体、IgMからIgG又はIgA抗体へのイムノグロブリンクラススイッチング、及び/又はCD8+T細胞応答の誘発において明白でありうる。このような標的抗原、ハプテン又はタンパク質の例としては、TRP2、GP100、TRP1、gp120及び他のHIV抗原、マラリア抗原、それらのエピトープなどが挙げられるがそれらに限定されない。同様に、NY−ESO−1 MHCクラスII拘束エピトープをコードする核酸配列は、標的抗原又はそのエピトープへの免疫原性を増大させるために、標的抗原又はそのエピトープをコードする核酸配列と共に、操作されたワクチン構築物に組み込まれることができる。この局面に関する例としては、クラスIIエピトープの核酸配列を、標的遺伝子とフレームを合わせて、裸のDNAもしくはRNA、ワクシニアウイルス、アデノウイルス、鶏痘ウイルスなどの形態の任意のワクチン構築物に組み込むことが挙げられる。例えば、プラスミドワクチンの形態のTRP2抗原の免疫原性を増大させるために、NY−ESO−1 クラスIIエピトープの核酸配列は、TRP2遺伝子と同じオープンリーディングフレームにおいて融合されることができる。次いで、TRP2のみをコードするプラスミドを用いる代わりに、患者を免疫化するためにハイブリッドプラスミドが使用される。
【0065】
癌エピトープ又はその変異体は、放射性標識、ビオチン、フルオレセインなどの他の成分が結合している誘導体の形態でありうる。特異的な器官、腫瘍、又は細胞型への特異的ターゲティングを可能とするターゲティング剤をエピトープに結合させることもできる。このようなターゲティング剤は、ホルモン、サイトカイン、細胞レセプターなどでありうる。エピトープは、単独で、又は他の試薬と組み合わせて、キットの形態で製造できる。
【0066】
本発明の別の局面は、本発明のNY−ESO−1 MHCクラスII拘束エピトープを用いて、癌に対する腫瘍特異的体液性媒介免疫を誘導するのに有用な免疫原又はワクチンである。免疫原及びワクチンは、NY−ESO−1特異的CD4+Tリンパ球及び抗NY−ESO−1抗体を誘発する。必要に応じて、免疫原及びワクチンは、CD8+Tリンパ球を誘発するために、NY−ESO−1 MHCクラスI拘束エピトープを含んでもよい。
【0067】
癌免疫治療へのアプローチは、能動又は受動カテゴリーに分けることができる。能動免疫治療は、腫瘍に対する免疫応答を増大させる試みにおいて、癌患者に対する癌抗原による直接的な免疫化を含む。受動免疫治療は、直接的に抗腫瘍応答を媒介するという目標をもって、抗腫瘍反応性を有する免疫細胞又は抗体などの免疫試薬の投与を指す。
【0068】
能動免疫治療における大部分の従来の試みは、免疫応答を刺激できる量と形態で腫瘍抗原を含むという期待をもって、完全な癌細胞又は癌細胞抽出物のいずれかを用いた。しかし、癌抗原とエピトープの分子的同定は、ヒト癌の治療に関する免疫治療を開発するための新しい可能性を有する。これらのアプローチの幾つかの要約を表1に示す。
【0069】
【表1】
【0070】
少なくとも1つのNY−ESO−1 MHCクラスII特異的T細胞エピトープをコードする遺伝子の、E.coli、酵母又はバキュロウイルスなどの高効率発現系への挿入は、免疫化において使用するための精製腫瘍エピトープを大量に得る機会を提供する。あるいは、イムノドミナントエピトープは、インビトロで容易に合成でき、そして免疫化のために、単独で、又はアジュバントとの組み合わせ、脂質/リポソームもしくはヘルパーペプチドとの結合などの免疫原性を増大させるために意図された形態で大量に精製でき、あるいは、抗原提示細胞にパルスすることもできる。MHCクラスII抗原への結合効率を増大させるための、イムノドミナントペプチドの個々のアミノ酸の修飾は、ネイティブタンパク質と比較して、免疫原性を増大させる可能性を有しうる。
【0071】
筋肉内又は皮膚内に直接注射される「裸の」DNAを用いる最近の技術は、コードされた抗原に対し細胞免疫応答と体液性免疫応答の両方を生じさせることが示されている(Cooney,E.L., A.C.Collier, P.D.Greenberg, R.W.Coombs, J.Zarling, D.E.Arditti, M.C.Hoffman, S.L.Hu, 及び L.Correy, 1991, Lancet 337:567; Wolff,J.A., R.W.Malone, P.Williams, W.Chong, G.Acsadi, A.Jani, 及び P.L.Felgner, Science 247:1465; Davis,H.L., R.G.Whalen, 及び B.A.Demeniex, 1993, Hum.Gene.Ther. 4:151;Yang,N.S., J.Burkholder, B.Roberts, B.Martinelli, 及びD.McCabe, 1990, Proc.Natl.Acad.Sci. USA 87:9568;Williams,R.S., S.A.Johnston, M.Riedy, M.J.DeVit, S.G.McElligott, 及び J.C.Sanford, 1991,Proc.Natl.Acad.Sci. USA 88:2726; Fynan,E.R., Webster, D.H.Fuller, J.R.Haynes, J.C.Santoro, 及び H.L.Robinson, 1995, Proc.Natl.Acad.Sci. USA 90:11478; Eisenbraum,M.D., D.H.Fuller, 及び J.R.Haynes, 1993, DNA and Cell Bio. 12:791; Fuller,D.H. 及び J.R.Haynes, 1994, AIDS Res.Hum.Retrovir. 10(11):1433; Acsadi,G., G.Dickson, D.R.Love, A.Jani, F.S.Walsh, A.Gurusinghe, J.A.Wolff, 及び K.E.Davies, 1991, Nature 352:815)。非生存DNAベクターを用いる技術は、製造の容易さと投与の安全性という利点を有する。本発明の核酸配列は、癌に対する免疫原として、及びDNAワクチンとして有用である。NY-ESO-1 MHCクラスII特異的T細胞エピトープの本発明の核酸配列、又は1つ以上の変異体NY-ESO-1 MHCクラスII拘束T細胞エピトープを有する完全長NY-ESO-1タンパク質をコードする核酸配列は、癌細胞に対する体液性反応を誘発させる量で、遺伝子銃を用いて投与できる。ナノグラム量がこのような目的のために有用である。
【0072】
免疫化の有効な形態は、免疫原性分子をコードする遺伝子の、BCG、サルモネラ又はリステリアなどの組換え細菌への、又はワクシニア、鶏痘、又はアデノウイルスなどの組換えウイルスへの組み込みを含む。NY-ESO-1 MHCクラスII特異的T細胞エピトープをコードする遺伝子は、単独で、又は免疫刺激分子をコードする遺伝子もしくは感染後に免疫応答を増大させることができる他の遺伝子と組み合わせて発現させることができる。構築物は、更なるNY-ESO-1 MHCクラスII拘束T細胞エピトープ及び/又は少なくとも1つのNY−ESO−1 MHCクラスI特異的T細胞エピトープをコードする遺伝子を更に含んでもよい。
【0073】
マウスモデルでのモデル腫瘍抗原についての研究は、インターロイキン−2(IL−2)又はB.7.1の遺伝子の組み込みが、モデル腫瘍エピトープの免疫原性を増大でき、これらのエピトープを担う樹立された肺転移の退行を媒介さえできることを示した。能動免疫治療、次いでIL−2、IL−6、IL−10、IL−12又はIL−15などの免疫刺激サイトカインの外来性投与はまた、免疫応答を改善するために使用できる。
【0074】
ヒト腫瘍抗原を認識できる遺伝子的に修飾された免疫細胞による受動免疫治療(通常、養子免疫治療と言われる)は、転移性メラノーマの選択された患者における癌の退行を媒介するのに有効である。抗原提示細胞上に存在する腫瘍抗原イムノドミナントエピトープに対しヒトリンパ球をインビトロで感作するインビトロ技術が開発された。反復性のインビトロ刺激によって、ヒト腫瘍抗原をコードする遺伝子をクローンするために使用されたTILよりも、これらの抗原を認識するずっと大きな能力をもった細胞が得られる。このように、癌ペプチドによる反復性のインビトロ感作によって、TILの50乃至100倍の能力をもったリンパ球を得ることができる。これらの細胞の養子移転(adoptive transfer)は、従来の増殖したTILよりも、インビボで腫瘍退行を媒介するのに有効でありうる。
【0075】
一実施態様において、末梢血単核球(PBMC)は、DR4−IEトランスジェニックマウスの免疫化後、同定された幾つかの候補DRB1*0401ペプチドで刺激された。NY−ESO−1特異的CD4+T細胞は、合成ペプチドESOp161−180によるインビトロ感作で産生された。このCD4+T細胞株は、HLA DP4(コーカソイドの約43〜70%に存在する優勢なMHCクラスII対立遺伝子(52))によって提示されるNY−ESO−1ペプチドを認識した。更に、HLA DP4ハプロタイプは、NY−ESO−1に対し高力価のAbを産生するメラノーマ患者の91%(11人のうち10人)が有していたが、NY−ESO−1陽性腫瘍を有し、検出可能なAbをプロセシングしない3人の患者では発現されなかった。インビトロ刺激の結果は、HLA DP−4拘束T細胞が、NY−ESO−1 Abを有する6人の患者のうち5人の患者から産生できることを示した。これらの結果は、DP4の状況において、CD4+T細胞によるNY−ESO−1の認識は、この抗原に対し抗体応答を開始するこれらの患者の能力と関連しうることを示唆した。
【0076】
癌の予防又は阻害の方法において、NY−ESO−1 MHCクラスII拘束T細胞エピトープは、静脈内、筋肉内、皮下、皮膚内、腹腔内、クモ膜下、胸膜腔内、子宮内、直腸、膣、局所、腫瘍内などを含むがそれらに限定されない幾つかの経路の1つを介し投与できる。
【0077】
投与は、経粘膜又は経皮膚手段によってされてもよい。経粘膜又は経皮膚投与のために、透過すべきバリアに適切な浸透剤が製剤において使用される。このような浸透剤は一般的に当業界公知であり、例えば、経粘膜のための胆汁塩及びフシジン酸誘導体が挙げられる。更に、界面活性剤が透過を容易にするために使用されうる。経粘膜投与は、例えば、鼻腔スプレー、又は坐剤によってされ得る。経口投与の場合、癌ペプチド、腫瘍抗原、その部分又は変異体が、カプセル、錠剤及びトニック(tonic)などの通常の経口投与形態に製剤化される。
【0078】
一般的に、レシピエントに、レシピエントの体重1kg当たり少なくとも約1ng、好ましくは体重1kg当たり少なくとも約1mg、より好ましくは体重1kg当たり少なくとも約10mg又はそれ以上のNY−ESO−1 MHCクラスII拘束T細胞エピトープの投与量を提供することが望ましい。体重1kg当たり約1mg乃至約100mgの範囲が好適であるが、より低い又はより高い投与量も投与できる。投与量は、NY−ESO−1 MHCクラスII特異的CD4+Tリンパ球のクローナル増殖(clonal expansion)をプライムし、刺激し、及び/又は引き起こすのに有効であり、次いで、それは、レシピエントの癌の予防又は阻害をすることができる。
【0079】
投与は、少なくとも1回行われ、またボーラス又は連続的投与として与えられることができる。数週間乃至数ヶ月の期間での複数回の投与は好ましくありうる。その後の投与は、示されたように投与されうる。
【0080】
治療方法において、NY−ESO−1クラスII拘束T細胞エピトープを含むワクチンは、哺乳動物において癌を予防するのに有効な量、又は局所的な癌を有する哺乳動物において転移を予防するのに有効な量で哺乳動物に投与される。必要に応じて、ワクチンは、細胞傷害性Tリンパ球を刺激するために、複数の異なるNY−ESO−1 MHCクラスII拘束T細胞エピトープ及び/又はNY−ESO−1 クラスI拘束癌ペプチド又はエピトープを含みうる。
【0081】
腫瘍を有する動物における腫瘍負荷を減少させる方法において、本方法は、腫瘍負荷の部位にNY−ESO−1 MHCクラスII拘束T細胞エピトープの有効量を投与することを含み、該量は、該部位で腫瘍のサイズを減少させるのに有効であり、かつ腫瘍部位からの転移を阻害しうる。
【0082】
治療方法の別の実施態様において、NY−ESO−1 HLA−DP拘束T細胞エピトープを含む免疫原は、NY−ESO−1 HLA−DP拘束CD4+Tリンパ球と抗NY−ESO−1抗体を誘発するのに有効な量で哺乳動物に投与される。免疫原は、単独で、又はアジュバント、免疫調節剤などと組み合わせて提供されうる。
【0083】
治療の別の方法において、自己リンパ球又は腫瘍浸潤リンパ球が、癌患者から得られうる。リンパ球は、培養で増殖され、そして癌エピトープ特異的CD4+リンパ球は、NY−ESO−1 MHCクラスII拘束T細胞エピトープ単独の存在下で、又はサイトカインの少なくとも1つの免疫刺激分子と組み合わせての存在下で、培養することによって増殖される。次いで、エピトープ特異的CD4+リンパ球は、患者の癌を減少又は除去するのに有効な量において、単独で、又はエピトープと組み合わされて、注入によって患者に戻される。
【0084】
免疫化後、ワクチンの効力は、抗体力価、特異的な溶解活性、特異的サイトカイン産生、腫瘍退行、又はこれらの組み合わせによって評価されるように、NY−ESO−1 MHCクラスII T細胞エピトープを認識する免疫細胞の産生によって評価されうる。免疫化される哺乳動物が既に癌又は転移性癌に罹患しているならば、ワクチンは、免疫調節剤(例えばIL−2、IL−6、IL−10、IL−12、IL−15、インターフェロン、腫瘍壊死因子など)、シスプラチナムなどの化学療法剤、ガンシクロビルなどの抗ウイルス剤、アンホテリシンB、抗生物質などのような他の治療と共に投与されうる。
【0085】
本発明の別の局面は、NY−ESO−1のMHCクラスII拘束T細胞エピトープをコードするNY−ESO−1遺伝子のDNA配列である。
【0086】
一実施態様では、DNA配列は、CD4+Tリンパ球によって認識されるMHCクラスII拘束T細胞エピトープをコードする配列番号1又は2の部分及びその機能的に等価な配列変異体を含む。また、MHCクラスII拘束T細胞エピトープをコードする配列番号1又は2の部分と相補的な核酸配列及び反相補的な核酸配列も本発明に包含される。
【0087】
一実施態様では、DNA配列は、配列番号4〜21又は29〜34の少なくとも1つを含むMHCクラスII拘束T細胞エピトープをコードする。
【0088】
別の実施態様では、MHCクラスII拘束T細胞エピトープをコードするDNA配列は、以下:
CAG GAT GCC CCA CCG CTT CCC GTG
CCA GGG GTG CTT CTG AAG GAG TTC
ACT GTG TCC GGC AAC ATA CTG ACT
ATC CGA CTC(配列番号24)又はその機能性部分又は変異体
を含む。
【0089】
別の実施態様では、MHCクラスII拘束T細胞エピトープをコードするDNA配列は、以下:
AGA CCA CCG CCA ACT GCA GCT
CTC CAT CAG CTC CTG TCT CCA GCA
GCT TTC CCT GTT GAT(配列番号25)又はその機能性部分又は変異体を含む。
【0090】
別の実施態様では、DNA配列は、以下:
TGG ATC ACG CAG TGC TTT CTG CCC
GTG TTT TTG GCT CAG CCT CCC
TCA GGG CAG AGG CGC(配列番号26)又はその機能性部分又は変異体を含む。
【0091】
本発明の別の局面は、NY−ESO−1のHLA−DP拘束CD4+T細胞エピトープをコードするNY−ESO−1遺伝子のDNA配列である。
【0092】
一実施態様では、DNA配列は、CD4+Tリンパ球によって認識されるHLA−DP拘束T細胞エピトープをコードする1つ以上の配列番号51〜64及びその機能性等価配列変異体をコードする核酸配列を含む。HLA−DP拘束T細胞エピトープをコードする核酸配列と相補的及び反相補的な核酸配列も本発明に包含される。
【0093】
包括的な暗号(generic code)の縮重に起因して、DNA配列の変異により、等価なNY−ESO−1エピトープの翻訳がなされよう。結果として、置換によって、NY−ESO−1癌抗原HLA−クラスII拘束CD4+T細胞によって認識されるNY−ESO−1エピトープの発現が生じる限り、置換は本発明の範囲に含まれる。
【0094】
NY−ESO−1遺伝子に由来するオープンリーディングフレームDNA配列の全て又は部分は、他の哺乳動物種におけるNY−ESO−1 MHCクラスII拘束T細胞エピトープのホモログを同定、単離するために、プローブとして使用できる。一実施態様では、ヒトcDNA配列は、マウスホモログ核酸配列のために哺乳動物cDNAライブラリーをスクリーニングするために使用される。陽性クローンを選択し、配列決定する。cDNAライブラリーが合成できる組織源の例としては、真皮、表皮、固形腫瘍、メラノーマ、メラノサイトなどが挙げられるがそれらに限定されない。当業者は、ホモログを検出するのに使用される適切なハイブリダイゼーション条件を理解しよう。ライブラリーの核酸ハイブリダイゼーション構築のための通常の方法及びクローニング技術は、Sambrook et al, (eds)(1989)“Molecular Cloning. A Laboratory Manual” Cold Spring Harbor Press, Plainview, New York; Ausubel al(eds) “Current Protocols in Molecular Biology”(1987), John Wiley and Sons, New York, New Yorkに記載されている。
【0095】
本発明の別の局面は、癌を有する哺乳動物から単離された生物学的サンプルにおける本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープを転写するRNAの検出と定量のための核酸プローブである。コントロールRNAサンプルに対するRNAレベルの変化は、哺乳動物における該疾病の診断や予後に有用である。
【0096】
一実施態様では、mRNAは、癌又は前癌を有すると疑われる患者の組織から得られ、そして健康なコントロール患者由来のmRNAと比較される。コントロールと比較した場合、患者における本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードするmRNAの定量的及び/又は定性的増大は、患者における癌又は前癌の指標となる。mRNAは、mRNAとハイブリダイズできるオリゴヌクレオチドプローブを用いて検出できる。
【0097】
本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードする配列に基づくオリゴヌクレオチド対の組み合わせは、選択されたRNA配列を増大させる逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)方法を用いて、生物学的サンプルにおけるmRNAを検出するためのPCRプライマーとして使用できる。本発明はまた、NY−ESO−1 MHCクラスII拘束T細胞エピトープをコードするDNAとRNAの検出のためのインサイチュPCRとインサイチュRT−PCRを包含する。標的核酸のコピー数が非常に小さいとき、又は核酸の異なる形態を区別しなければならないとき、該技術は好適である。該方法は、正常細胞から前癌細胞及び癌細胞を検出し、区別するのに特に有用である。
【0098】
本発明はまた、mRNA上のある相補(「センス」)領域と結合し、NY−ESO−1の合成阻害に至るアンチセンスオリゴヌクレオチドを包含する。このようなアンチセンスオリゴヌクレオチドは、約12乃至約25モノヌクレオチドの1本鎖核酸であり、本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードする配列に対しアンチセンスである。このようなアンチセンスオリゴヌクレオチドは、Molecular Biology and Biotechnology R.A.Meyers(編 VCH Publishers,Inc., New York, NY, 1995, pp.38-44のUhlmann,E.ら, Antisense oligonucleotides, structure and function に記載のように、当業界公知の方法によって製造できる。
【0099】
本発明はまた、少なくとも1つ以上のNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードするDNA配列を含むベクターを包含する。ベクターは、1つ以上の変異体NY−ESO−1 MHCクラスII拘束T細胞エピトープを有する完全長NY−ESO−1タンパク質をコードするDNA配列を含みうる。必要に応じて、ベクターはまた、少なくとも1つの免疫刺激分子をコードするDNA配列を含んでもよい。ベクターはまた、少なくとも1つ以上のNY−ESO−1 MHCクラスI拘束T細胞エピトープをコードするDNA配列を含んでもよい。ベクターはまた、細胞や組織におけるNY−ESO−1 MHCクラスII拘束T細胞エピトープの局在を検出するのに使用される緑色蛍光タンパク質をコードする遺伝子を含んでもよい。
【0100】
真核発現ベクターとしては、レトロウイルスベクター、ワクシニアウイルスベクター、アデノウイルスベクター、ヘルペスウイルスベクター、鶏痘ウイルスベクター、バキュロウイルスベクター、ヒトパピローマウイルスベクター、ウマ脳炎ベクター、インフルエンザウイルスベクターなどが挙げられるがそれらに限定されない。
【0101】
本発明は、遺伝子、そのフラグメント又は変異体のオープンリーディングフレーム核酸配列によってコードされた少なくとも1つのNY−ESO−1 MHCクラスII拘束T細胞エピトープを発現する新規組換えウイルスを包含する。組換えウイルスはまた、少なくとも1つの免疫刺激分子を発現してもよい。組換えウイルスは、哺乳動物、特にヒトにおける癌の予防又は治療の目的のために、哺乳動物において体液性免疫応答を誘発又はアップレギュレートできる。
【0102】
組換えウイルスに感染した宿主細胞は、1つ以上のNY−ESO−1 MHCクラスII拘束T細胞エピトープを単独で、又は少なくとも1つの免疫刺激分子と組み合わせて発現する。宿主細胞はまた、HLAクラスII分子を発現する組換えウイルスに感染されてもよい。
【0103】
組換えワクシニアウイルスベクターから外因性遺伝子産物を構築、発現する方法は、Perkusら Science 229:981-984, 1985; Kaufmanら Int.J.Cancer 48:900-907,1991; Moss Science 252:1662,1991; Smith及びMoss Bio Techniques Nov/Dec, p.306-312,1984;並びに米国特許第4,738,846号に開示されている。Sutter及びMoss(Proc.Nat'l Acad Sci. U.S.A. 89:10847-10851,1992)及びSutterら(Virology 1994)は、本発明でウイルスベクターとして使用できる非複製組換えアンカラウイルス(MVA,修飾ワクシニアアンカラ)のベクターとしての構築と使用を開示する。Baxby及びPaoletti(Vaccine 10:8-9, 1992)は、本発明のウイルスベクターとして使用のための、カナリア痘ウイルス、鶏痘ウイルス及び他のトリ種を含む非複製ポックスウイルスのベクターとしての構築と使用を開示する。
【0104】
本発明のベクターは、NY−ESO−1 MHCクラスII拘束T細胞エピトープの発現のために適切な宿主細胞中に置かれうる。真核宿主細胞株としては、COS細胞、CHO細胞、Hela細胞、NIH/3T3細胞、昆虫細胞、樹状細胞などの抗原提示細胞などが挙げられるがそれらに限定されない。必要に応じて、宿主細胞はまた、刺激分子を発現してもよい。宿主細胞が、少なくとも1つのMHC(又はHLA)クラスII分子と組み合わせてNY−ESO−1 MHCクラスII拘束T細胞エピトープの両方を発現する場合、真核発現系は、適切なグリコシル化を可能にするように使用されるのが好ましい。宿主細胞による癌エピトープと免疫刺激分子の両方の発現は、抗原認識、及び抗原特異的T細胞の増殖又はクローナル増殖において助けとなるように、特異的T細胞に必要なMHCクラスII拘束ペプチドを提供し、T細胞に適切なシグナルを提供する。全体としての結果は、免疫系のアップレギュレーションである。免疫応答のアップレギュレーションは、癌又は前癌細胞の増殖の阻害のための、癌抗原特異的CD4+リンパ球の増加及び体液性免疫の他のエフェクター細胞の増加によって明白となる。
【0105】
DNAは、当業界公知の方法によるトランスフェクション、形質導入、リポソームなどによって宿主細胞に挿入できる(Sambrook et al, 1989, “Molecular Cloning A Laboratory Manual”, Cold Spring Harbor press, Plainview, New York)。リポソームの場合、Nabel,E.G.et al, 1992, Hum.Gene.Ther. 3:367-275; Nabel,G.J. et al, 1992, Hum.Gene.Ther. 3:649-656; Stewart,M.J. et al 1992, Hum.Gene.Ther. 3:399-410; Nabel,G.J. et al 1993 Proc.Natl.Acad.Sci.USA90:11307-11311;及びHarrison,G.S. et al 1995 Bio Techniques
19:816-823に開示のように、カチオン性脂質が好ましく、例えば、中性リン脂質ジオレオイルホスファチジルエタノールアミン(DOPE)と複合した、ポリカチオン性脂質であるジミリスチルオキシプロピル−3−ジメチル−ヒドロキシエチルアンモニウム(DMRIE)が好ましい。
【0106】
宿主細胞によって発現される組換えNY−ESO−1 MHCクラスII拘束T細胞エピトープは、当業界公知の標準的タンパク質精製方法によって細胞溶解液又は細胞上清から精製できる。これらとしては、分子ふるいクロマトグラフィー、イオン交換クロマトグラフィー、等電点フォーカシング、ゲル電気泳動、アフィニティクロマトグラフィー、HPLC、逆相HPLCなどが挙げられるがそれらに限定されない(Ausubel et al, 1987, Current Protocols in Molecular Biology, John Wiley and Sons, New York, NY)。イムノアフィニティクロマトグラフィーも、イムノアフィニティ剤として、本明細書記載の抗癌タンパク質抗体又はその抗原結合フラグメントを用いて精製のために使用できる。
【0107】
組換えウイルスはまた、治療薬又はワクチンとして使用できる。このような使用において、より低い、又はより高い投与量も投与できるが、レシピエントに、哺乳動物1mg当たり約105乃至約1010プラーク形成ユニットの範囲における組換えウイルスの投与量を提供することが望ましい。
【0108】
組換えウイルスベクターは、メラノーマなどの癌の如何なる証拠の前に、又はメラノーマなどの癌に罹患した哺乳動物における該疾病の退行を媒介するために、哺乳動物に導入できる。哺乳動物にウイルスベクターを投与する方法の例としては、エキソビボで細胞を組換えウイルスに曝すこと、又は冒された組織内に組換えウイルスを注入すること、又はウイルスの静脈内、皮下、皮内、筋肉内などへの投与が挙げられるがそれらに限定されない。あるいは、組換えウイルスベクター又は組換えウイルスベクターの組み合わせは、癌病巣中への直接的注入、又は適切な医薬として許容できる担体中での局所適用によって、局所投与されることができる。問題の核酸配列を担持する組換えウイルスベクターの量は、ウイルス粒子の力価に基づく。免疫化のための好適な範囲は、哺乳動物、好ましくはヒト当たり約105乃至1010ウイルス粒子である。
【0109】
本発明は、少なくとも1つのNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードするDNA配列の1つ以上のコピーをゲノム中に組み込まれたトランスジェニック動物を提供する。トランスジェニック動物作出の一般的方法は、Krimpenfortら,米国特許第5,175,384号、Leder ら,米国特許第5,175,383号、Wagnerら,米国特許第5,175,385号、Evansら,米国特許第4,870,009号、及びBernsら,米国特許第5,174,986号に記載されている。遺伝子の組み込みにより、NY−ESO−1 MHCクラスII拘束T細胞エピトープの過剰発現、発現の変化、又はNY−ESO−1 MHCクラスII拘束T細胞エピトープのマルチプル形態もしくは変異体の発現が生じる。得られるトランスジェニック動物は、癌の進行又は本発明の腫瘍抗原の研究において有用である。動物モデルは、癌治療のためのワクチンや化学療法剤をスクリーニングするのに有用である。トランスジェニック動物はまた、癌の進行の研究において有用である。
【0110】
本発明は更に、本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープによる免疫化によって誘発される抗体又は抗原結合部分を含む。NY−ESO−1 MHCクラスII拘束T細胞エピトープがほんの数個のアミノ酸からなる場合、エピトープは、抗体応答を誘発するために、キャリアタンパク質に結合されてもよい。KLH、破傷風トキソイド、アルブミンなどのキャリアタンパク質及び結合方法は、当業界公知である。抗体は、インタクトなNY−ESO−1タンパク質及びNY−ESO−1タンパク質の天然のプロセスされた形態と同様に、本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープに対する特異性を有し、それらと反応もしくは結合する。
【0111】
典型的な抗体分子は、インタクトなイムノグロブリン分子、実質的にインタクトなイムノグロブリン分子、又は抗原結合部位を含むイムノグロブリン分子のこれらの部分(F(ab)、F(ab’)、F(ab’)2、ヒト化キメラ抗体、及びF(v)のような当業界公知のイムノグロブリン分子の部分を含む)である。ポリクローナル又はモノクローナル抗体は、当業界公知の方法によって製造できる(Kohler and Milstein (1975) Nature 256,495-497; Burdonら(編)(1985) “Laboratory Techniques in Biochemistry and Molecular Biology”, Vol.13, Elsevier Science Publishers, アムステルダム中のCambell “Monoclonal Antibody Technology, the Production and Characterization of Rodent and Human Hybridomas”)。抗体又は抗原結合フラグメントはまた、遺伝子工学によって製造できる。重鎖及び軽鎖遺伝子両方の発現のための技術は、PCT特許出願公開番号WO 901443、WO 9014424、Huse et al(1989)Science 246:1275-1281及び米国特許第4,946,778号の主題である。1つ以上の相補的決定領域を有するヒト化イムノグロブリン及び該抗体の製造方法は、米国特許第5,585,089号及び第5,530,101号に開示されている。
【0112】
一実施態様において、本発明の抗体は、生物学的サンプルにおけるMHCクラスII拘束T細胞エピトープを含むNY-ESO-1ペプチド又は部分を検出するイムノアッセイで使用される。抗体又はその抗原結合フラグメントは、癌に冒された哺乳動物からの組織生検サンプル中で、癌ペプチドを検出するために使用されることができる。病気の組織内でのNY−ESO−1 MHCクラスII拘束T細胞エピトープの評価は、哺乳動物での病気の悪化を予測するために使用できるか、又は治療の効力を診断しうる。イムノアッセイは、ラジオイムノアッセイ、ウエスタンブロットアッセイ、免疫蛍光アッセイ、酵素免疫アッセイ、化学発光アッセイ、免疫組織化学アッセイなどであり得、そしてインビトロ、インビボ、又はインサイチュで行うことができる。ELISAについて当業界公知の標準的技術は、“Methods in Immunodiagnosis”(第2版), Rose及びBigazzi(編), John Wiley & Sons, 1980; Campbellら,“Methods and Immunology”, W.A.Benjamin, Inc., 1964;及びOellerich,M. 1984, J.Clin.Chem.Clin.Biochem. 22:895-904に記載されている。免疫組織化学の通常の方法は、Harlow及びLane(編)(1988) “Antibodies A Laboratory Manual”, Cold Spring Harbor Press, Cold Spring Harbor, New York; Ausbelら(編)(1987) Current Protocols In Molecular Biology, John Wiley and Sons (New York, NY)に記載されている。このような検出アッセイのために適切な生物学的サンプルとしては、細胞、組織生検材料、全血、血漿、血清、痰、脳脊髄液、胸膜液、尿などが挙げられるがそれらに限定されない。
【0113】
本発明の抗体又は抗原結合フラグメントはまた、免疫治療でも使用できる。抗体又はその抗原結合フラグメントは、癌の重篤度、程度又は持続時間を防止、軽減又は減衰させるために十分な量で哺乳動物に提供される。
【0114】
引用された全ての論文及び特許は、引用により本明細書に含まれるものとする。
【0115】
本発明を、ある特定の実施態様に関し上に記載するが、多数のバリエーションが可能で、代わりの材料や試薬を、本発明を逸脱すること無しに使用できることが理解されよう。幾つかの場合に、このようなバリエーション及び置換は何らかの実験を必要とするかも知れないが、慣用的な試験のみを含むであろう。
【0116】
特定の実施態様の上の記載は、本発明の一般的な性質を十分に示し、他の者は、現在の知識を適用することによって、種々の適用のために、一般的概念を逸脱すること無しに、このような特定の実施態様を容易に改変し、及び/又は採用できる。それ故、このような改変や修飾は、開示された実施態様の等価物の意味内と範囲内に包含されるように意図される。
【実施例】
【0117】
実施例1
材料および方法
組換えNY−ESO−1タンパク質の生成および解析:完全長NY−ESO−1遺伝子をコードする細菌発現ベクターを構築するために、本発明者らは一対のプライマー、NdeI部位を含むESO−5p(5’GCTCCGGACATATGCAGGCCG AAGGCCGGGG)(配列番号35)およびXhoI部位を含むESO−3p(5’AAGGGGCTCGAGGCT GGGCTTAGCGCCTCT)(配列番号36)を用いて、PCRフラグメントを作製した。PCR産物の制限酵素による消化およびゲル精製の後、NY−ESO−1をコードするDNAフラグメントをポリヒスチジンペプチドをコードするDNAとpET−28(+)(Novagen,マジソン,WI)内でインフレームに融合させた。同様のストラテジーを、最初の74のアミノ酸残基のみを含むトランケートNY−ESO−1であるESO1−74の発現ベクターを構築するのにも使用した。正確なプラスミド構築物を保持するE.coli株BL21(DE3)を37℃で対数期まで増殖させ、次いで、イソプロピル−d−チオガラクトシド(IPTG)を終濃度0.5mMまで添加し3時間振盪することによってタンパク質産生を誘導した。細菌抽出物の可溶性画分を得た。またNY−ESO−1をNi2+アフィニティークロマトグラフィーにより精製した。精製タンパク質のSDS−PAGE解析を先の報告(25)のように行った。精製タンパク質のN末端配列を自動Edman分解により決定した。
【0118】
血清およびPBMC:転移性メラノーマの患者由来の血清を−80℃で保存した。正常ドナーの血清をNIHのクリニカルセンターの血液バンクから得た。転移性メラノーマの患者TEのMHCクラスII遺伝子型はHLA−DR1*0401,1*1501であった。この患者をgp100:209−217(210M)ペプチド+高用量のIL−2で処置し、目的である腫瘍退行の実験に付した。
【0119】
NY−ESO−1タンパク質に対する抗体の検出:50 lのPBST(0.1%Tween20を含むリン酸緩衝化生理食塩水)に希釈した約50ngの精製NY−ESO−1タンパク質を96ウェルMaxiSorpプレート(Nunc,デンマーク)の各ウェルに室温で終夜吸着させた。コントロールプレートは1ウェルあたり150ngのBSAでコートした。プレートをPBST中5%ドライミルクで少なくとも2時間ブロックし、洗浄し、そして100 lの希釈血清サンプルをロードした。全ての血清サンプルはPBST中3%ドライミルクを用いて1:25、1:250、および1:2500に希釈した。この3つの異なる希釈の各サンプルをNY−ESO−1コートしたプレートおよびBSAコートしたプレート上にロードした。室温で1時間インキュベーションした後、プレートを洗浄し、PBST中1%ドライミルクで希釈した二次抗体(西洋ワサビペルオキシダーゼを結合したヤギ抗ヒトIgG,Sigma Co.,セントルイス,MO)をロードした。プレートを0.5時間インキュベーションした後発色させ、450nmでの吸光度をELISAリーダー(Dynatech,シャンティイー,VA)を用いて読み取った。陽性反応を、1:25と1:250の両方の血清希釈で、正常ドナーの平均O.D.値+標準偏差の3倍を超えるNY−ESO−1に対するO.D.値として定義した。ウエスタンブロットを(24)の記載のように行い、いくつかの代表的血清サンプルにおける抗体の特異性を確認した。
【0120】
細胞株および抗体:メラノーマ株F049およびF050は、NCIの外科のAdam Rikerから提供された細針状アスパレート(asparate)サンプルの初期培養物であった。他の全てのメラノーマ株およびEBV B株は10%ウシ胎仔血清(Biofluid,Inc.,ゲーサーズバーグ,MD)を補充したRPMI1640(Life Technologies,ロックヴィル,MD)で作製し維持した。293IMDR1および293IMDR4をヒトインバリアント鎖であるDMA、DMBおよびDR分子を発現するよう遺伝子組換えし、10%ウシ胎仔血清を補充したRPMI1640で培養した(15)。マウスリンパ球の培養培地は、Hyclone Inc.(ローガン,UT)より供給される、0.05mMメルカプトエタノール、5CU/ml IL−2および10%ウシ胎仔血清を含むRPMI1640だった。ヒトT細胞培養に用いた培地は、Sigma Co.(セントルイス,MO)により供給される、0.05mMメルカプトエタノール、50CU/ml IL−2および10%ヒトAB血清を含むRPMI1640だった。抗体ブロッキング実験を先の記載(15)のように行った。ハイブリドーマHB55およびHB95はAmerican Type of Cell Culture(ATCC,マナッサス,VA)から得た。コントロール抗体はPharmingen Int.(サンディエゴ,CA)から購入した。
【0121】
トランスジェニック動物および免疫手順:HLA−DR4トランスジェニック(DR4−Tg)マウスはマウスクラスII欠損であり、HLA−DR−IE−およびHLA−DR1*0401−IE−キメラ分子を発現した(26)。初代マウスはCase Western Reserve UniversityのPaul Lehmannを通じて得た。マウスは近交系であり、Biocon Inc.(ロックビル,MD)で維持された。6〜10週齢の雌性マウスを完全長組換えNY−ESO−1タンパク質で免疫した。精製タンパク質(約50μg)を完全フロインドアジュバント(CFA)中に懸濁し、均等に分け、後肢足蹠および尾の付け根に皮下注射して各マウスに投与した。注射11日後、マウスを屠殺し、そして両方の後肢膝窩および鼠径部リンパ節を採取した。単一の細胞懸濁液を2匹の免疫動物のリンパ節から得、インビトロ刺激を行った。
【0122】
ペプチド合成:本研究で使用した合成ペプチドはNCIの外科のペプチド合成機(Gilson Co. Inc.,ワージントン,OH)で固相法を用いて作製した。各ペプチドの純度は質量分析(Bio−synthesis, Inc.,ルイスヴィル,TX)により評価した。
【0123】
インビトロ感作(IVS)手順およびサイトカイン放出アッセイ:サイトカイン放出アッセイを行う前1週間、終濃度10Mのペプチドを2.5×105マウスリンパ球と混合した。ヒトPBMCのIVSのために、2.5×105細胞を10M濃度のペプチドでパルスし、平底96ウェルプレートの各ウェル中でインキュベートした。2回のインビトロ刺激の後、細胞を様々な標的に対して試験し、上清をサイトカイン放出アッセイのために収集した。ヒトT細胞の急速な増殖およびクローニングは記載(20)のように行った。
【0124】
終濃度10Mのペプチドまたは終濃度5g/mlのタンパク質を標的細胞にパルスした。4時間のインキュベーション後、細胞を無血清RPMI培地中で洗浄し、およそ3×104の標的細胞を同数のTE4−1細胞とともに終夜インキュベーションし、サイトカイン放出をヒトについてはGM−CSF ELISAキット(R&D Systems,ミネアポリス,MN)、またはマウスについてはIFNキット(Endogen, Inc. ウォバーン,MA)を用いて測定した。ヒトIFN−,IL−10,TNF−,およびIL−4等の他のサイトカインは、製造業者の説明書に従って、Endogen Inc.またはR&D SystemsのELISAキットを用いて測定した。
【0125】
実施例2
組換えNY−ESO−1タンパク質およびNY−ESO−1反応性抗体の検出
NY−ESO−1反応性抗体およびCTLは、癌の患者において報告されている(19〜22)。従って、NY−ESO−1特異的CD4+T細胞がNY−ESO−1抗原に対する抗体の発生およびCTLの調整に関与しているかもしれないようである。しかし、NY−ESO−1由来のCD4+T細胞エピトープはこれまで報告されていない。MHCクラスII拘束CD4+T細胞エピトープを同定するために、本発明者らは、細菌発現系からNY−ESO−1タンパク質を出発物質として精製することから始めた。NY−ESO−1発現およびタンパク質精製を促進するため、NY−ESO−1をコードするcDNAフラグメントを、pET28発現ベクターのN末端に位置するポリヒスチジンタグにインフレームで融合し、組換えタンパク質の高レベル産生を得た。数ミリグラムのNY−ESO−1タンパク質をNi2+荷電アフィニティークロマトグラフィーカラムを使用して精製した。精製タンパク質はSDSポリアクリルアミドゲル上およそ26kDaの見かけの分子量を示した(図2A)。精製タンパク質の同定を確認するために、タンパク質のN末端マイクロシークエンシングを自動Edman分解により行った。Edman分解により得られた25アミノ酸残基すべては予測したアミノ酸配列と一致した(データ示さず)。最初の74アミノ酸残基を含む短い形のNY−ESO−1(ESO1−74)もまた同じアプローチにより精製した(図2A)。
【0126】
メラノーマ患者がNY−ESO−1タンパク質に対する抗体を生じたかどうかを決定するために、NCIの外科において癌ワクチン治療プロトコルに登録された88人の転移性メラノーマ患者由来の血清をスクリーニングした。8人の正常ドナー由来の血清をスクリーニングのコントロールとして使用した。88人の患者のうち11人(13%)が、高力価のNY−ESO−1に対する抗体を持っていることが分かった(図2C)。このデータは他の群により得られた結果と一致した(22)。患者の血清が精製NY−ESO−1タンパク質に存在するわずかな夾雑物と反応する可能性を除去するため、代表的な血清サンプルを用いてウエスタンブロットを行った。図2Bは、患者由来のNY−ESO−1反応性の血清が、NY−ESO−1発現細菌の細胞溶解物および精製NY−ESO−1タンパク質だけと反応し、コントロールベクターを含む細菌由来の抽出物とは反応しなかったことを示した。非反応性の血清サンプルもまた試験した(図2B、レーン4,5,6)。
【0127】
実施例3
HLA−DR4−トランスジェニックマウス由来の推定MHCクラスII拘束エピトープの同定
CD4+T細胞エピトープを同定するために、DR4トランスジェニックマウスをCFA中およそ50gの完全長NY−ESO−1タンパク質で、尾の付け根および後肢足蹠に免疫した。注射後11日目に、2匹の免疫したマウスの両後肢膝窩および鼠径部リンパ節から得た単一の細胞懸濁液を調製し、HLA−DR4分子の推定ペプチド結合能に基づくNY−ESO−1タンパク質由来合成ペプチドでのインビトロ感作に使用した(27)。
【0128】
HLA−DR4に結合することが予測されるアミノ酸配列セグメントを含む8つの高結合ペプチドをインビトロ感作実験に使用した。最初のインビトロ感作後6日目に、マウスリンパ球を、ヒトHLA−DR4陽性1359EBV B細胞のみおよび刺激に使用した対応するペプチドでパルスした1359EBV Bに対するサイトカイン放出について試験した。3つのペプチドは、T細胞からのサイトカイン分泌に基づきマウスT細胞により認識されたが、他の5つのペプチドは認識を示さなかった(図3)。ESO p116−135は陽性ペプチドのうちで最も強い活性を示した。これは、このペプチドが、T細胞認識のためにHLA−DR4分子により提示されるエピトープを含んでいるかもしれないことを示唆している。従って、このペプチドをさらなる解析のために選択した。
【0129】
実施例4
NY−ESO−1特異的なヒトCD4+T細胞の作製
NY−ESO−1に対する高力価の抗体を持っていた患者TE(図2C)由来のPBMCを、ESO p116−135ペプチドを用いたインビトロ刺激に使用した。インビトロ刺激の1週間後、患者TE由来のPBMCは著しい増殖を示した。IL−2を刺激の2週目に添加した。このように樹立した細胞株をTE4−1と命名し、これを20 CU/mlのIL−2の存在下、2週間超、増殖させ続けた。TE4−1 T細胞は、FACS解析に基づくと、90%CD4+T細胞であった。TE4−1はTh1型CD4+T細胞を含んでいた。というのは、それらがGM−CSF、IFN−およびTNFを分泌し、IL−10またはIL−4を分泌しなかったからである(データ示さず)。CD8+T細胞を数%減少させた後、CD4+T細胞の精製集団は依然その反応性を保持していた。TE4−1細胞株由来のT細胞クローンにもESO p116−135ペプチドを認識することを示すものもあった(データ示さず)。
【0130】
TE4−1は、HLA−DR4との関連で完全長NY−ESO−1タンパク質およびESO p116−135ペプチドでパルスしたEBV B細胞は認識したが、最初の74アミノ酸を含むトランケートNY−ESO−1タンパク質でパルスしたEBV B細胞を認識しなかった(図4A)。TE4−1細胞株もまたNY−ESO−1をコードするアデノウイルスに感染させたDR4陽性樹状細胞とは特異的に反応性であったが、緑色蛍光タンパク質をコードするアデノウイルスに感染させたDR4陽性樹状細胞とは特異的に反応性ではなかった(データ示さず)。
【0131】
TE4−1により認識されるT細胞がHLA−DR4により拘束されるかどうかを試験するため、2つの重複するペプチド(ESO p116−135およびESO p111−130)ならびにコントロールペプチド(ESO p91−110)を無血清培地中293IMDR1および293IMDR4細胞にパルスした。細胞を洗浄し続いてTE4−1細胞と終夜インキュベートした。図4Bに示されるように、ペプチド116−135およびペプチド111−130の両方がHLA−DR4との関連でTE4−1により認識された。興味深いことに、ペプチド116−135もまた、293IMDR1細胞にパルスしたとき、T細胞からのサイトカイン分泌を刺激することができた。コントロールESO p91−110ペプチドでパルスした293IMDR4では活性は検出されなかった(図4B)。ESO−p116−135パルス293IMDR4の認識は抗HLA−DR抗体(HB55)により完全に阻害されたが、コントロールおよび抗HLAクラスI抗体(HB95)では阻害されなかった(図4C)。gp100特異的CD8+T細胞株(CTL−C3G1)およびHLA−DR1拘束CD4+T細胞株(T3−80)を抗体ブロッキングの特異性コントロールとして使用した。
【0132】
実施例5
TE4−1による腫瘍細胞の認識
ペプチド特異的CD4+およびCD8+T細胞活性は推定腫瘍抗原に対してしばしば生じ得たが、多くの場合には、腫瘍反応性は、T細胞の低親和性または腫瘍細胞表面上で天然にプロセシングされるペプチドの提示の失敗のいずれかに起因して示され得なかった(3)。TE4−1が腫瘍細胞により天然にプロセシングされ提示されるNY−ESO−1エピトープを認識することができたかどうかを試験するために、いくつかのメラノーマ株を標的として使用した。各株におけるNY−ESO−1の発現はRT−PCRにより決定し、HLA−DR対立遺伝子の発現はFACS解析により決定した(データ示さず)。図5に示されるように、TE4−1はNY−ESO−1/HLA−DR4陽性腫瘍(1359melおよびF049mel)を認識することができたが、腫瘍細胞株397melおよび624.38mel(NY−ESO−1+/HLA−DR-)、ならびに526mel(NY−ESO−1-/HLA−DR4+)は認識することができなかった。興味深いことに、TE4−1もまたF050mel(DR1+/NY−ESO−1+)を認識したが、DR1および低レベルのNY−ESO−1を発現する1300melは認識しなかった。1つの可能な説明は、CD4+T細胞が異なるDR分子により提示される同一のペプチドを認識するかもしれないということである。F049melの認識は、抗HLA−DR抗体の存在下特異的にブロックされ得るが、抗MHCクラスI抗体の存在下にはされない(データ示さず)。これらの研究は、TE4−1細胞株が腫瘍細胞表面で天然にプロセシングされたペプチドを認識することを示唆した。
【0133】
実施例6
TE4−1により認識されるNY−ESO−1エピトープの特徴付け
2つの反応性ペプチドが15アミノ酸(LPVPGVLLKEFTVSG)(配列番号10)を共有するので、ペプチドの最小長を一連のN−およびC−末端トランケートペプチドを試験することによって決定した。ペプチドをDR4+1088EBV B細胞にパルスし、TE4−1細胞を刺激するその能力を試験した。128位のバリン残基がT細胞認識に重要であることが見出された(図6A)。123位のロイシン残基までのN末端欠失を有するペプチドは、T細胞認識に影響を与えなかったが、さらに欠失を有するペプチドはT細胞を刺激するその能力を一部失った。P1、P4、P6およびP7残基がMHCクラスII分子へのペプチド結合に寄与したので、123位のロイシン残基はP1アンカー残基であり得る。さらなる欠失が、MHCクラスII分子に結合するための重要な残基を決定するために必要である。
【0134】
欠失実験に基づいて本発明者らは、短い形のペプチドESO p119−130を用いて、TE4−1により認識されるペプチドの結合親和性を決定した。異なるペプチド濃度で標的として1088EBV B細胞(HLA−DR4+)にペプチドをパルスした。図5Bに示されるように、33nM以下のESO p119−130ペプチド濃度では全くまたはほとんどT細胞活性は観察されず、0.33Mのペプチド濃度で高い活性が検出され、T細胞活性は33Mのペプチド濃度でプラトーに達しなかった。コントロールペプチドは33Mのペプチド濃度でさえTE4−1により認識されなかった。
【0135】
実施例7
本発明者らは、T4−1 CD4+T細胞株がDR4およびおそらくDR1にもまた関連してESO p116−135を認識することを示した。ここで、TE4−1がHLA−DR1に関連してペプチドだけでなくタンパク質も認識できることが示される。認識は抗DR抗体によりブロックされる(図7Aおよび7B)。この結果は、ESO p116−135が雑多なペプチドであり得、DR4およびDR1に結合できることを示す。従って、このペプチドワクチンの適用可能な集団は非常に大きい。
【0136】
実施例8
HLA−DP研究の材料および方法
研究に使用した細胞株、組織培養試薬、および抗体
293CIITAは、MHCクラスIIトランスアクチベーターをコードするレトロウイルスを有する293細胞の形質導入により作製した細胞株である(53)(このレトロウイルスプラスミドはthe University of South Florida, タンパ, FLのDr. George Blancの好意である)。すべてのメラノーマ株およびEBVB株を、10%ウシ胎仔血清(Biofluid, Inc.,ゲーサーズバーグ,MD)を補充したRPMI1640(Life Technologies,ロックビル,MD)中で作製し維持した。リンパ球の培養培地は、0.05mM β−メルカプトエタノール、50CU/ml IL−2+10%ヒト男性AB血清(Valley Biochemicals Inc.(ウィンチェスター,VA)により供給された)を含むRPMI1640であった。ブロッキングアッセイに使用した抗体は以下の供給源から得た:W6/32(HLAクラスI)およびL243(HLA DR)はLoftstrand Labs Lt.(ゲーサーズバーグ,MD)により精製されたハイブリドーマ上清であった;抗体クローンIVA12(HLAクラスII)、B7/21(HLA DP)、Genox3.53およびIVD12(両方ともHLA DQ)はBeckton Dickinson Immunocytometry Systems(サンノゼ,CA)から購入した。
【0137】
プラスミドの構築
pESOプラスミドは、先の記載(45)のようにCMVプロモーターにより駆動されるNY−ESO−1cDNAを含む発現ベクターであった。pIi−ESOプラスミドを、全NY−ESO−1cDNAのNheIおよびNotI消化PCR産物を同じ酵素で消化したpTi80ベクターに挿入することで構築した(54)。NY−ESO−1cDNAを、インバリアント鎖(Ii)リーダー配列の最初の80アミノ酸残基とそのN末端でインフレームに融合した。NY−ESO−1の増幅に使用したPCRプライマーは以下の通りであった:NheI部位を含むフォワードプライマー5’cattgctagcATG CAG GCC GAA GGC CGG GGC A3’(配列番号73)およびNotI部位を含むリバースプライマー5’aaggctacattGC GGC CGC TTA GCG CCT CTG CCC TGA G3’(配列番号74)。
【0138】
ペプチドおよびCD4+T細胞の産生
本研究に使用した合成ペプチドは、National Cancer Institute,ベセズダ,MDの外科のペプチド合成機(Gilson Co. Inc.,ワージントン,OH)において、固相法を用いて作製した。脱保護後、各ペプチドの純度を、質量分析(Bio−synthesis, Inc.,ルイスヴィル,TX)により評価した。合成ペプチドを凍結乾燥し、20mg/mlでDMSO中再構築し、そして示した濃度まで希釈した。
【0139】
インビトロ感作手順を先の記載(50)のように行った。簡潔にいうと、およそ2.5×105PBMCを20μg/mlのペプチド存在下96ウェル平底プレートにプレートした。7日目と14日目に、1×105非照射PBMCを20μg/mlのペプチドでパルスし、2回洗浄し、各ウェルに加え、そして20CU/mlのIL−2を8日目、11日目、15日目、18日目に加えた。21日目、細胞を回収し、標的細胞と終夜インキュベートし、その後、上清をサイトカイン放出アッセイのために採取した。
【0140】
特異的活性を有するウェル由来のT細胞をプールし、OKT−3急速増殖法(OKT−3 rap1d expansion method)を用いて増殖させた(55)。増殖後、CD8+T細胞を磁気ビーズ選択(Dynal Inc,レイクサクセス,NY)により培養物から枯渇させ、引き続いて細胞株をCD4+およびCD8+発現に関してフローサイトメトリーにより解析した。
【0141】
サイトカイン放出アッセイ
タンパク質またはペプチドをパルスした標的を調製するために、ペプチドは終濃度20μg/mlで使用し、タンパク質は終濃度10μmg/mlで使用した。細胞を無血清RPMI培地で洗浄し、4時間血清の不在下37Cでパルスし、2回洗浄した。明記しない限り、およそ3×104標的細胞を同数のT細胞と少なくとも16時間インキュベートし、その後、サイトカイン放出アッセイを行った。サイトカイン分泌はGM−CSF ELISAキット(R&D Systems,ミネアポリス,MN)を用いて測定した。
【0142】
ヒトIL−4、TNF−αおよびTGF−βレベルの定量をR&D Systemsから得たサイトカインキットを使用して行い、IFN−γELISAキットをEndogen Inc(ウォバーン,MA)から購入した。アッセイは製造業者の説明書に従って行った。
【0143】
HLA DP分子の分子タイピング
トータルRNAをEBVB細胞、CD40リガンド刺激B細胞、CD4+T細胞、またはMHCクラスII陽性メラノーマ株からタイピングのために得た。トータルRNAはRNeasyキット(Qiagen,ドイツ)を使用して精製し、100ng〜1μgのRNAをオリゴdTプライム第一鎖cDNA合成に使用した。cDNA産物の1/10を使用し、Clontech(パロアルト,CA)のアドバンテージPCR系を用いてPCR増幅を行った。以下のプライマー対をHLA DP−AおよびDP BPCRに使用した:DPAフォワードプライマー5’ATG CGC CCT GAA GAC AGA ATG T 3’(配列番号75)、DPAリバースプライマー5’TCA CAG GGT CCC CTG GGC CCG GGG GA3’(配列番号76)、DPBフォワードプライマー5’ATG ATG GTT CTG CAG GTT TCT G3’(配列番号77)、およびDPBリバースプライマー5’TTA TGC AGA TCC TCG TTG AAC TTT C3’(配列番号78)。引き続いて、PCR産物を精製し、PCRを行うために使用した同一プライマーを用いてシークエンシングした。単一の配列がPCR産物から得られたので、多数の患者が非常に一般的なHLA DPB1*0401遺伝子産物にホモ接合性であると思われた。ヘテロ接合性の患者の場合、PCR産物をまずpCR4ベクター(Invitrogen,カールズバッド,CA)にクローニングし、ベクター配列に相補的な5’および3’プライマーを用いてシークエンシングした。最終配列をIMGT−HLAデータベースでサーチしてHLA DPであることを確認した(http://www3.ebi.ac.uk/Services/imgt/hla)。
【0144】
実施例9
NY−ESO−1に対するCD4+T細胞株TE4−2の作製
最初の研究をHLA DR4対立遺伝子により拘束されたNY−ESO−1エピトープを同定するために行った。予測される9マーのDR4結合モチーフを含む8つの20マーのペプチドを、NY−ESO−1組換えタンパク質で免疫しインビトロで刺激したHLA−DR4−IEトランスジェニックマウス由来のリンパ球による認識について試験した(50)。3つの20マーのペプチドがこれらの実験に陽性であることが分かった。これらのうちの一つはDRB1*0401とDRB1*0101との夾雑エピトープとして特徴付けされた(50)。他の2つのペプチド(ESO p161−180およびESO p141−160)をさらに特徴付けするために、本発明者らはそれらを使用して、高力価の抗体を有するDRB1*0401患者(TE)由来のPBMCならびにNY−ESO−1に対するCD4+およびCD8+T細胞を刺激した(50)。24ミクロ培養ウェル全体を各ペプチドについて使用した。弱い刺激を3回した後、24ウェルのうち15ウェルがESO p161−180で刺激されたPBMC由来の顕著な増殖を示した。試験した15ウェルの増殖が陽性であるウェルのうち9ウェルが、ペプチドパルスDRB1*0401発現1088EBVB細胞に対する特異的サイトカイン放出を示した(図8A)。特異的CD4+T細胞もまた、ESO p141−160で刺激したPBMCから作製したが、本研究では述べない(データ示さず)。
【0145】
次いで、ESO p161−180ペプチド刺激に特異的に応答した培養物由来のT細胞を合わせ、先の記載(55)のプロトコルを使用して増殖させた。CD8+T細胞の枯渇後、この培養物(TE4−2と命名する)は、FACS解析により評価したところ、95%を超えるCD4+T細胞を含んだ(データ示さず)。
【0146】
TE4−2のサイトカイン分泌プロファイルの分析によりこのT細胞株がペプチドパルス標的に応答してIFN−γ、TNF−α、IL−4およびGM−CSFを分泌しTGF−βを分泌しなかったことが示された(データ示さず)。従って、CD4+T細胞のTh1およびTh2型の両方がこの細胞株に存在するかもしれない。あるいは、Th0表現型の細胞がこの培養物に存在するかもしれない。
【0147】
実施例10
HLA DPB1*0401−0402に関連するTE4−2によるNY−ESO−1の認識
ESO p161−180、重複ペプチドであるESO p156−175、および完全長NY−ESO−1タンパク質それぞれでパルスしたDR4発現標的細胞に対する応答について、TE4−2 T細胞を試験した。DR1を発現するがDR4を発現しない586EBVB細胞もまたAPCとして使用した。無関係なペプチド、ESO p91−110、および精製トランケート組換えタンパク質(ESO1−74;アミノ酸1〜74を含む)(50)をコントロールとして使用した。TE4−2 T細胞は、完全長NY−ESO−1タンパク質でパルスした場合DR4+1088EBVB株を特異的に認識したが、トランケートESO1−74タンパク質でパルスした場合認識しなかった(図8B)。ESO p161−180およびp156−175の両方がTE4−2により認識され、これは最小ペプチドエピトープがアミノ酸161と175との間にあることを示している。対照的に、最初予測したDR4結合モチーフはアミノ酸167と175との間にある。予測しなかったことに、TE4−2 T細胞株は、586EBVB細胞にパルスしたペプチドおよびタンパク質(これらはDRB1*0101を発現するがDRB1*0401を発現しない)に同じくらい良好に応答するようであった。この結果は、類似のペプチドが複数のMHCクラスII拘束エレメントによって提示されたか、またはTE4−2 T細胞に対してペプチドを提示するMHCクラスII拘束エレメントを共有する1088および586EBVB細胞株によって提示されたかのいずれかであったことを示唆していた。これらの可能性を試験するために、公知のHLA DRおよびDQ型を含む多数の他のEBVB細胞もまた、TE4−2 T細胞により利用される拘束エレメントを同定するためにAPCとして使用した。試験したEBVB細胞株のうち一つを除く全てがTE4−2に対してESO p161−180ペプチドを提示することができた(図8C)。
【0148】
次いで、ペプチドのT細胞認識を、異なるMHC拘束エレメントによって拘束されたペプチドの認識をブロックする特異的抗体の存在下で行った。図9A〜9Cの結果は、すべてのMHCクラスII対立遺伝子(IVA12)をブロックする抗体および全てのHLA DP対立遺伝子(B4/21)をブロックする特異性を有する抗体が、TE4−2 T細胞がESO p161−180を認識する能力を無効としたことを示した。HLA−A、BおよびC対立遺伝子(W6/32)に対する抗体ならびにMHCクラスIIDR(L243)およびDQ(Genox3.53およびIVD12のミックス)対立遺伝子に対する抗体は、TE4−2 T細胞の刺激に対してほとんどまたは全く影響しない。従って、これらの結果は、TE4−2 T細胞が本研究で使用したEBVB細胞株により共有される非常に一般的なHLA DP対立遺伝子に関してESO p161−180を認識したことを示唆していた。
【0149】
次いで、HLA−DP対立遺伝子を分子的にクローニングし、図8Cで使用した細胞株に対してシークエンシングした。これらの研究は、1088および586EBVB株がともに、HLA DPB1*0401遺伝子産物、患者TEが発現したDPB1*0401ならびに未知のDP対立遺伝子にホモ接合性であったことを示していた(表2)。L023EBVB細胞株(これはTE4−2に対してESO p161−180ペプチを提示しなかった)はHLA DP対立遺伝子(これはDPB1*0401および0402とは異なる)にホモ接合性として型を決められた。
【0150】
1363、1088、836およびL007 EBVB細胞株はすべてDPB1*0401を発現したが、L041EBVB細胞株はDPB1*0402(84位と85位の2つのアミノ酸残基がDPB1*0401分子と異なる)を発現した。従って、DPB1*0401とDPB1*0402の両方がTE4−2 CD4+T細胞に対するESO p161−180エピトープを提示することができるようであった。
【0151】
【表2】
【0152】
特異的DPA鎖がTE4−2 T細胞に対するエピトープを提示するのに必要であるかどうかを決定するために、DPB1*0401−0402を発現するEBVB細胞中のHLA DPA分子もまた解析した。図8Cで使用したDPB1*0401−0402を発現するEBVB細胞は1タイプ超のHLA DPA分子(データ示さず)を持っていたが、すべてがTE4−2 T細胞にNY−ESO−1エピトープを同じように良好に提示することが可能であった。
【0153】
実施例11
腫瘍細胞上の天然にプロセシングされたNY−ESO−1エピトープのTE4−2による認識
TE4−2により認識されるT細胞エピトープが腫瘍細胞の表面で天然にプロセシングされ提示されるかどうかを調べるために、NY−ESO−1およびDPB1*0401を発現した腫瘍株を標的として用いた。TE4−2 T細胞は、NY−ESO−1とDPB1*0401の両方を発現する複数の腫瘍株を認識したが、NY−ESO−1を発現するがHLA DPB1*0401および0402対立遺伝子のいずれも発現しない腫瘍株(1362mel)は認識しなかった(図10A)。さらに、TE4−2 T細胞はDPB1*0401陰性およびNY−ESO−1陰性の腫瘍(526mel)を認識しなかった。HLA DPB1*0401を発現するがNY−ESO−1を発現しない1つのメラノーマ株(1102mel)はまた、TE4−2 T細胞により認識された。RT−PCR解析の結果は、1102melがLAGE−1遺伝子(NY ESO−1に対しておよそ90%のアミノ酸類似性を有する癌/精巣抗原)を発現したことを示していた(57)。ESO p161−175と同一の配列はまた、LAGE−1タンパク質にも存在した。これらの結果は、TE4−2により認識されるエピトープが腫瘍細胞表面に存在し、NY−ESO−1と、密接に関連した腫瘍抗原LAGE−1との間で共有されていることを示唆した。
【0154】
NY−ESO−1発現メラノーマ株に加えて、TE4−2 T細胞もまた、NY−ESO−1トランスフェクト293CIITA細胞の認識について試験した。293細胞にMHCクラスIIトランスアクチベーター遺伝子(CIITA)を発現するレトロウイルスを用いて形質導入し、293CIITA細胞株を作製した(53)。RT−PCRにより決定されたように、293CIITA細胞はホモ接合性のHLA DPB1*0401分子を発現したが、親の293細胞は発現しなかった(データ示さず)。TE4−2 T細胞は、NY−ESO−1トランスフェクト293CIITA細胞と特異的に反応した(図10B)。対照的に、TE4−2 T細胞はpGFPプラスミドをトランスフェクトした293CIITA細胞またはIi−NY−ESO−1をトランスフェクトした親の293細胞のいずれも認識しなかった。Iiターゲッテイング配列はNY−ESO−1のプロセシングおよび認識に必要ではなかったが、T細胞認識をわずかに増強した(図10B)。これらの結果はTE4−2 T細胞が天然にプロセシングされたNY−ESO−1エピトープを認識したことをさらに示していた。
【0155】
実施例12
HLA−A2拘束エピトープと重複するHLA DP4拘束エピトープ
2つの重複するペプチド(ESO p161−180およびp156−175)でパルスした標的細胞は、TE4−2 T細胞により同じように良好に認識された。これは、最小T細胞エピトープがアミノ酸161から175までの範囲の領域に位置していることを示していた(図8B)。
【0156】
アミノ酸161と175との間にあるアンカー残基を同定するために、重複13マーペプチドの1シリーズを1088EBVB細胞をパルスするのに使用し、TE4−2 T細胞を刺激するその能力を試験した。図11Aに示すように、161位のW残基を除去したときに活性の部分的損失を観察し、162位のI残基を除去したときに活性の完全な損失を観察した。167位のC末端L残基の欠失もまた、TE4−2 T細胞によるペプチド認識を取り除いた。さらに、169位の残基Vも、この残基の欠失がペプチド刺激活性を2倍減少させたので、重要であると思われた。これらの結果は、161位のW残基がP1アンカーであり得、167位のL残基がP7アンカーの代表であることを示した。169位のV残基もまた、ペプチドエピトープの刺激性能に寄与しているようであり、それがP9アンカー残基を示し得ることを示している。これらの推定アンカー残基は先に記載されたコンセンサスHLA DPB1*0401結合モチーフ(57)に非常に一致している。
【0157】
3つすべてのアンカー残基を含むESO p157−170ペプチドを力価測定実験に使用して、ペプチドに対する最小刺激濃度を決定した。その結果は、ESO p157−170が、3nMと33nMとの間の最小濃度でTE4−2 T細胞からの著しいサイトカイン放出を刺激することができることを示していた(図11B)。これらの結果は、TE4−2 T細胞が高い親和性でESO p157−170を認識することを示していた。この見かけの親和性は非変異ペプチド(例えば、gp100(58)、チロシナーゼ(59)、およびCDC−27(54)由来のもの)由来の最もよく知られた公知のMHCクラスII結合エピトープよりも優れている。同じ領域にわたる他のペプチド(例えば、ESO p161−180およびp156−175)もまた、TE4−2 T細胞に関して同様な最小刺激濃度を有した(データ示さず)。
【0158】
興味深いことに、先に同定されたHLA−A2エピトープ、ESO p157−167(47)は、DPB1*0401−0402エピトープ、ESO p157−170内に含まれた。HLA−DPエピトープがHLA−A2により提示されCD8+T細胞と交差反応し得るかどうかを判断するために、ESO p157−170をTE8−1による認識について試験した(CD8+T細胞株はHLA−A2エピトープESO p157−167を特異的に認識する)。L023 EBVB細胞(これは、HLA−A2を発現するがDPB1*0401−0402対立遺伝子を発現しない)にパルスした時、ESO p157−170はTE8−1 T細胞からの顕著なサイトカイン放出を刺激することができた(図11C)。この実験は、ESO p157−170エピトープがMHCクラスIおよびクラスII2つの特異性を持っており、NY−ESO−1タンパク質を認識するCD4+およびCD8+T細胞両方を刺激することができることを示していた。従って、ESO p157−170は、腫瘍細胞を特異的に認識するCD4+およびCD8+T細胞両方を誘発することを目的とする癌ワクチンの興味深い候補物であり得た。
【0159】
実施例13
NY−ESO−1抗体産生とHLA DPB1*0401−0402との関連性
HLA DPB1*0401−0402は、白人の大部分(別の人種を含めた集団調査では43%〜70%)に存在する主要なMHCクラスII対立遺伝子である(52)。先の研究(48、50)は、正常ドナーおよびNY−ESO−1発現腫瘍を有さない癌患者がNY−ESO−1に対する抗体を生じないことを示している。対照的に、NY ESO−1発現腫瘍を有する患者の50%は、NY−ESO−1特異的Abを生じた。血清サンプルを試験した88人のメラノーマ患者のパネルのうち、11人の患者が高力価のNY−ESO−1抗体を有することが見出された(50)。多くの患者がDR4対立遺伝子を全く発現しなかったことから、先に同定したDR4拘束CD4+T細胞ペプチドは、NY−ESO−1特異的Abの産生を説明できなかった(表2)。NY−ESO−1特異的DP4拘束CD4+T細胞がこれらのメラノーマ患者におけるNY−ESO−1特異的Abの産生と関連しているかどうかをさらに調査するため、本発明者らはまずこれらのHLA DPサブタイプを解析した。NY−ESO−1抗体を有する11人の患者のうち10人がDPB1*0401および/または0402を発現したが、主要なDQまたはDR拘束エレメントはこの患者のグループでは同定できなかった(表2)。公知のNY−ESO−1発現腫瘍を有しているが検出可能な抗体を有さないこのパネルの3人の患者はDPB1*0401−0402対立遺伝子を発現しなかった。残りの74人の患者由来の腫瘍細胞株はこれらの患者由来のNY ESO−1発現を評価するためには入手できなかったので、そのHLA DP型を同定するためのさらなる研究は行わなかった。0.011のp値をフィッシャーの直接確率検定により得、これは抗体応答とHLA−DPB1*0401−0402発現との関係の重要さを示していた。NY−ESO−1は25〜30%の腫瘍細胞株で発現され、DP4は集団の43〜70%で発現されるので、NY−ESO−1とDP4の両方を発現し、かつ抗体応答を生じる可能性のある患者の割合は、10〜21%の範囲である。この仮説的予想はNY−ESO−1抗体を有する患者の観察された10〜13%の頻度と非常に近い。
【0160】
NY−ESO−1抗体応答とDPB1*0401−0402発現との関連に関する更なる証拠を手に入れるため、NY−ESO−1抗体を有する11人の患者のうち6人からのPBMCを、ESO p161−180ペプチドによるインビトロ刺激に用いた。インビトロ感作もまた検出可能なNY−ESO−1抗体を有さない2人のDPBI*0401+患者からのPBMCを用いて行った。2または3回のインビトロ刺激の後、ESO p161−180ペプチドでパルスした293CIITA細胞に対するその応答について、T細胞を試験した。NY−ESO−1抗体を有する6人の患者のうち5人(TEを含む)由来のT細胞は、293CIITA細胞により提示されるESO p161−180エピトープ(DP4+およびHLA−A2-)の特異的認識を示した(表3)。患者CTおよびBL由来の複数のウェルは、ペプチドをパルスした標的と反応するようであった。これらの2人の患者由来の感作したPBMCもまた、CD4+T細胞のさらなる濃縮なしにDPBI*0401+およびNY−ESO−l+メラノーマ株の顕著な腫瘍認識を示した(データ示さず)。対照的に、NY−ESO−1反応性T細胞は、3回の刺激後検出可能なNY−ESO−1抗体を有さない2人の患者(WCおよびEW)由来のPBMCを用いて作製されなかった。これらの結果は、抗NY−ESO−1抗体を生じた患者もまた、比較的高い前駆体頻度のDPB1*0401拘束エピトープと反応性のT細胞を含んでいたことを示唆していた。これらのNY−ESO−1特異的CD4+T細胞はNY−ESO−1癌/精巣抗原に対する抗体応答の発生に寄与していたかもしれない。
【0161】
【表3】
【0162】
実施例14
野生型HLA DP4ペプチドの1つに改変を施した。この改変は、ペプチドをより可溶性にし90%超の均質性(これはペプチド臨床試験に関してFDAにより求められている)まで精製し得るようにするためのものであった。野生型および改変ペプチドは以下の通りである:
【0163】
野生型ESOp157−170;SLLMWITQCFLPVF(配列番号54);
野生型ESOp157−167;SLLMWITQCFL (配列番号79);ESOp156R−169;RSLLMWITQCFLPV(配列番号63);および
ESOp157−170R;SLLMWITQCFLPVR(配列番号64)。
【0164】
これらの改変ペプチドがT細胞により同じように良好に認識されるかどうかを試験するために実験を行った。ESO p157−170がHLA−A2およびHLA−DP4 2つの結合特異性を示したので、TE4−2 CD4+T細胞およびTE8−1 CD8+T細胞それぞれによるDP4(図12A)およびA2(図12B)両方の拘束様式における認識を測定した。
【0165】
結果は、これらの改変ペプチドがCD4+T細胞およびCD8+T細胞により野生型として同じように良好に認識されたことを示していた。
【0166】
実施例15
NY−ESO−1エピトープ特異的CD4+Tリンパ球免疫療法
メラノーマ抗原に対して前感作したTリンパ球は、メラノーマに罹患した哺乳動物を治療処置するのに有効であり得る。Tリンパ球を末梢血またはメラノーマの腫瘍懸濁液から単離し、インビトロで培養する(Kawakami,Y.ら、1988,J.Exp.Med.168:2183−2191)。
【0167】
Tリンパ球をエピトープVLLKEFTVSG(配列番号19)またはエピトープWITQCFLPVF(配列番号80)に濃度1μg/mlのみまたはIL−2の存在下で暴露し、前感作し、そして培養液中で増殖させる。エピトープに暴露したCD4+Tリンパ球を1哺乳動物あたり約109〜1012リンパ球で哺乳動物に投与する。このリンパ球を、静脈内、腹腔内、または病巣内のいずれかで投与する。この処置は、サイトカイン、メラノーマ病変の外科的切除および化学療法剤等の他の治療処置と同時に投与してもよい。NY−ESO−1特異的CD8+Tリンパ球はCD4+Tリンパ球と同時に投与してもよい。
【0168】
実施例16
転移性メラノーマを有する患者の処置
このプロトコルでは、進行メラノーマの患者を抗原性癌エピトープで免疫する。
【0169】
試験にふさわしい患者は、標準的な有効治療が失敗した測定可能または評価可能な転移性メラノーマの徴候を持っていなければならない。患者は、腫瘍細胞RNAのPCRまたはノザンブロット解析により立証されるようなNY−ESO−1抗原を発現する腫瘍を有さなければならない。
【0170】
患者に、体重1kgあたり1ng、1μg、1mgまたは500mgいずれかのMHCクラスII拘束T細胞エピトープを、0日、7日、14日に単独で、またはIL2および/または免疫刺激分子とともに、静脈内に投与する。患者を、毒性、免疫効果および治療有効性について評価する。患者に、NY−ESO−1クラスI拘束T細胞エピトープをさらに投与してもよい。
【0171】
処置患者から採取したリンパ球を、NY−ESO−1癌抗原またはMHCクラスII拘束T細胞エピトープに対する特異的応答について試験する。
【0172】
完全な応答を、少なくとも4週間続く疾患の全臨床的徴候の消失と定義する。部分的応答は、新たな病変またはいずれの病変における増加の出現なしに、少なくとも4週間すべての測定可能な病変の垂直直径の積の合計における50%以上の減少である。軽微な応答は、新たな病変の出現およびいずれの病変における増加なしに、すべての測定可能な病変の垂直直径の積の合計における25〜49%の減少として定義する。部分応答未満のいずれの患者をも非応答者と考える。新たな病変の出現あるいは部分的または完全な応答後の先の病変の垂直直径の積における25%超の増加を再発と考える。
【0173】
考察
NY−ESO−1は、体液性および細胞性両方の免疫応答を引き起こすので、重要な免疫標的である(19〜21)。その発現パターンはMAGE遺伝子ファミリーの抗原と類似しているが、NY−ESO−1はMAGEファミリーのいずれのものよりも乳癌、前立腺癌、肺癌で頻繁に発現される(19、20、23)。さらに興味深いことに、非常に低い割合の患者がMAGE抗原または分化抗原(例えば、チロシナーゼ、gp100、TRP−1およびTRP−2)に対する高力価の抗体を生じた(データ示さず、(22))にもかかわらず、高力価のNY−ESO−1反応性抗体は、癌の患者においてしばしば検出された(図2Bおよび2C)。これらの研究は、NY−ESO−1反応性CD4+T細胞が抗体産生およびCTL増殖に関わっているかもしれないことを示唆する。本研究で、本発明者らは、HLA−DR4トランスジェニックマウスおよび候補ペプチドを用いたヒトPBMCのインビトロ刺激を使用して、NY−ESO−1由来のHLA−DR4拘束T細胞エピトープを同定した。我々の知る限りでは、これは、NY−ESO−1由来T細胞エピトープがCD4+T細胞に対してMHCクラスII分子により提示されることを示した最初の証明である。NY−ESO−1特異的抗体およびCTLは異なるHLA遺伝子型を持つ患者において検出されたので、HLA−DR4以外のHLAクラスII分子により提示される他のCD4+T細胞エピトープは本発明において同定された。
【0174】
最近、2つのグループが、公知のMHCクラスI拘束腫瘍抗原であるMAGE−3からのMHCクラスII拘束T細胞エピトープの同定を報告した。精製MAGE−3タンパク質でパルスされたDCで刺激されたPBMCから作製したCD4+T細胞クローンは、HLA−DR13適合EBV B細胞にパルスされたペプチドまたはタンパク質を認識したが、MAGE−3+/DR13+腫瘍細胞にパルスされたものを認識しなかった(14)。しかし、別の研究では、コンピューターを使ったアルゴリズムによって予測されるペプチドで刺激したPMBCから作製したCD4+T細胞は、EBV B細胞およびMAGE−3+/DR11+腫瘍細胞両方にパルスしたペプチドを認識することができた(15)。NY−ESO−1の場合、本発明者らはここに、CD4+T細胞がDR4マッチEBV B細胞にパルスしたNY−ESO−1タンパク質またはペプチドおよびNY−ESO−1を発現する腫瘍細胞を認識できることを示している(図4および5)。HLA−DRトランスジェニックマウスの利用は、免疫したトランスジェニックマウスが特異的反応性T細胞の高い前駆体頻度をおそらく有しているので、推定ペプチドの同定において利点を有し得る。候補ペプチドを一旦同定すれば、CD4+T細胞を合成候補ペプチドで刺激したPBMCから作製できる。従って、全タンパク質で免疫し、コンピューターを使用したアルゴリズムにより予測したペプチドで刺激したトランスジェニックマウスの組み合わせ使用は、多数のペプチドおよび複数回のインビトロ刺激でヒトPBMCを刺激する必要性を回避し得る。さらに、免疫したトランスジェニックマウスにより同定される候補ペプチドは、天然にプロセシングされ細胞表面上に提示されるペプチドであるようである。これは、ペプチド特異的CD4+T細胞が腫瘍細胞も同様に認識することができる可能性を上昇させ得る。最後に、NY−ESO−1に対する高力価の抗体および高前駆体頻度のCTLを生じた患者(TE)由来のPBMCの使用は、抗体産生およびCTLの両方がCD4+T細胞の助けを必要とするので、腫瘍特異的CD4+T細胞を作製するのをより容易にし得る。このアプローチは、自己免疫疾患に関与する公知の自己抗原から多数のMHCクラスII拘束T細胞エピトープを同定するのに使用されている(28)。従って、本研究で使用するストラテジーは多くの他の公知のMHCクラスI拘束腫瘍抗原に適用可能であり得つつ、直接遺伝子クローニングアプローチ等他のストラテジーが未知のMHCクラスII拘束腫瘍抗原の同定を容易にし得る。
【0175】
gp100等組織特異的分化抗原由来のペプチドを使用する臨床試験は、メラノーマの患者の処置において治療有効性のいくらかの証拠を示した(4)。顕著な毒性の副作用は改変gp100ペプチドで処置した患者において観察されていないが、白斑または色素脱失は、治療に反応を示した患者においてしばしば見られた(29)。このことは、自己抗原による免疫によって誘導された抗腫瘍免疫が自己免疫を生じ得ることを示唆している。TRP−1を免疫標的として使用した動物研究では、類似の結果(抗腫瘍免疫および毛の色素脱失)も得られた(30〜32)。興味深いことに、gp75/TRP−1に媒介される抗腫瘍免疫および自己免疫はCD4+T細胞および抗体を伴っているようであった(33)。mTRP−2(35)ではなくhTRP−2(34)でのマウスの免疫は、自己抗原ならびにCD4+およびCD8+T細胞の両方の関与を必要とする抗腫瘍免疫に対する耐性を壊した(33)。これらの研究は、抗腫瘍免疫が抗体またはCD8+T細胞のいずれかにより媒介され得るが、両方ともCD4+T細胞の決定的な助けを必要とし得ることを示唆していた(24、33)。
【0176】
本研究で同定したMHCクラスII拘束NY−ESO−1ペプチドは、NY−ESO−1に対するCTLおよび抗体が癌の患者において検出されたので、臨床用途に有用であり得る。MHCクラスIおよびII拘束ペプチドまたは精製NY−ESO−1タンパク質での免疫は、NY−ESO−1特異的CD4+、CD8+T細胞および抗体を誘導し得る。あるいは、患者は、クラスIおよびIIの両ペプチドをロードするかまたはNY−ESO−1遺伝子をコードする組換えウイルスを感染させた樹状細胞で免疫され得る。精巣生殖細胞はMHCクラスIおよびIIの分子を発現しないので(36)、NY−ESO−1に対する免疫応答は腫瘍細胞に特異的であるはずであり、従って、ほとんどまたは全く自己免疫応答を生じない。MAGE−3のMHCクラスI拘束ペプチドまたは樹状細胞にパルスしたペプチドを使用した同様の研究は、抗腫瘍免疫(CTL応答)および緩慢な腫瘍の退行が示されたにもかかわらず、色素脱失/白斑または他の顕著な副作用が観察されなかったことを示した(6、7)。抗腫瘍免疫は、腫瘍特異的CD4+T細胞の助けを提供することによって向上され得る。
【0177】
【表4−1】
【表4−2】
【表4−3】
【表4−4】
【表4−5】
【表4−6】
【表4−7】
【0178】
本発明のこれらの及び他の目的、性質及び付随の利点の多くは、以上の詳細な説明を読む際に、添付された図面と関連づけて考えると、より良く理解されよう。
【技術分野】
【0001】
発明の分野
本発明は、癌診断及び癌ワクチンを含む癌治療の領域に関する。より詳細には、本発明は、癌ペプチドNY−ESO−1由来の新規なヒトMHCクラスII拘束T細胞エピトープ及びそのアナログ並びにMHCIIクラスII拘束T細胞エピトープ又はその部分をコードするDNA配列の単離と精製に関する。特に、本発明は、NY−ESO−1由来のHLA−DR及びHLA−DP拘束T細胞エピトープに関する。本発明は更に、個体における癌及び前癌の検出、診断及び治療の方法に関する。
【背景技術】
【0002】
発明の背景
T細胞は、腫瘍増殖を制御し、腫瘍退行を媒介するのに重要な役割を果たす。T細胞媒介抗腫瘍免疫の分子的基盤を理解するために、CD8+T細胞によって認識される多くの腫瘍抗原が、メラノーマや他の種類の癌で同定されてきた(1〜3)。これらの研究は、分子的に明確な腫瘍抗原由来のペプチドを用いる幾つかの臨床試験に至った(4〜7)。gp100由来の修飾ペプチドを用いる臨床試験は、転移性メラノーマの患者の治療に対し治療的効力の幾分かの証拠をもたらしたが(4)、これらの研究は主にCD8+T細胞の使用に焦点をあてていた。ヒト及び動物研究の両方からの証拠の増大は、最適の癌ワクチンがCD4+及びCD8+T細胞の両方の参加を必要とすることを示した(8,9)。更に、腫瘍特異的CD4+T細胞は、MHCクラスII陰性腫瘍細胞に対する防御免疫を産生するのに必要とされる(10,11)。それ故、このような抗原の同定は、癌ワクチンの開発及びCD4+T細胞が宿主免疫応答を制御する機構を我々が理解するために重要である。
【0003】
今までに、ほんの限られた数のMHCクラスII拘束腫瘍抗原が同定された。チロシナーゼ、gp100及びMAGE−3などの幾つかの公知のMHCクラスI拘束腫瘍抗原は、CD4+T細胞によって認識されるMHCクラスII拘束エピトープを含むことが示された(12〜15)。腫瘍特異的CD4+T細胞を用いて、未知のMHCクラスII拘束腫瘍抗原を同定するために、最近、遺伝的アプローチが開発された。これは、CDC27、TPI及びLDFPを含む幾つかの変異腫瘍抗原の同定に至った(16,17)。それらのうち、TPIは、生化学的アプローチによって独立に同定された変異抗原である(18)。
【0004】
NY−ESO−1遺伝子は、抗体スクリーニングによって以前に同定され(19)、最近、MHCクラスI拘束腫瘍抗原としても同定された(20,21)。NY−ESO−1に対する高力価の抗体がまた、癌患者から検出された(22)。NY−ESO−1 cDNAは、2つの重複するオープンリーディングフレーム由来の2つの遺伝子産物をコードしていた(20)。正常精巣での発現を除くその厳密な腫瘍特異的発現パターン、並びにメラノーマ、乳癌、前立腺癌、肺癌及び他の癌を含む多くの腫瘍での高頻度の発現(18,20,23)の故に、NY−ESO−1は、種々の癌タイプに対する免疫治療の開発のための重要な免疫標的の可能性がある(24)。
【発明の概要】
【発明が解決しようとする課題】
【0005】
NY−ESO−1に対するCTL及び抗体免疫応答の両方が癌患者で示されたが、NY−ESO−1タンパク質におけるMHCクラスII拘束T細胞エピトープは報告されなかった。
【課題を解決するための手段】
【0006】
発明の概要
本発明の1つの目的は、CD4+Tリンパ球によってMHCクラスII拘束T細胞エピトープとして認識される新規ペプチド及びその部分を提供することである。本発明の抗原性癌ペプチドは、NY−ESO−1(本明細書ではCAG−3と交換可能に使用される用語)遺伝子(配列番号1)(Genbank登録番号 AF038567; 8:9)内もしくは部分によって、又はその変異体もしくはホモログ(例えば、LAGE遺伝子)(Genbank登録番号 AJ223040, AJ223041及びAJ223093)内もしくは部分によってコードされる。
【0007】
本発明の1局面は、癌の進展及び転移からレシピエントを防御するCD4+Tリンパ球を誘発できる癌ワクチンとして有用である、NY−ESO−1遺伝子によってコードされるMHCクラスII拘束T細胞エピトープ又はその変異体である。本発明はまた、癌の阻害又は予防のために、あるいはNY−ESO−1遺伝子産物を発現している細胞の増殖を阻害するために、有効量の癌ワクチンを投与する方法に関する。
【0008】
本発明の1局面は、NY−ESO−1遺伝子によってコードされるHLA−DR及びHLA−DP拘束T細胞エピトープ、又はその変異体及びホモログである。これらは、CD4+Tリンパ球及び抗NY−ESO−1抗体応答を誘発でき、次いで、癌の進展からレシピエントに防御及び/又は治療的利益を提供し、かつ転移からの防御を提供する免疫原及び癌ワクチンとして有用である。本発明はまた、癌を阻害又は予防し、NY−ESO−1遺伝子産物を発現している細胞の増殖を阻害し、かつ転移を阻害するために、有効量の癌ワクチンを投与する方法に関する。
【0009】
本発明の別の局面は、NY−ESO−1由来のMHCクラスII拘束T細胞エピトープ又はその変異体を単独で、又は1つ以上の免疫刺激分子と組み合わせて含む医薬組成物である。医薬組成物は、腫瘍及び癌に対する免疫応答を誘発するために、少なくとも1つのNY−ESO−1 MHCクラスII拘束T細胞エピトープを含むか、又はNY−ESO−1抗原特異的CD4+T細胞を刺激するエピトープの組み合わせを含む。医薬組成物は、CD8+Tリンパ球の産生のために、NY−ESO−1由来の1つ以上のMHCクラスI拘束T細胞エピトープを更に含んでもよい。NY−ESO−1MHCクラスII拘束T細胞エピトープとNY−ESO−1MHCクラスI拘束T細胞エピトープは各々、別のエピトープとして提供されてもよいし、又は一緒に結合されて、マルチマーの形態で提供されてもよい。癌エピトープ又はその変異体は、癌の予防又は治療のために、免疫原又はワクチンとして提供されてもよい。医薬組成物は、哺乳動物の癌を治療又は予防する方法に有用である。治療方法において、医薬組成物は、哺乳動物の癌を予防又は阻害する有効な量において哺乳動物に投与される。
【0010】
本発明の別の局面は、NY−ESO−1由来のHLA−DR及び/又はHLA−DP拘束T細胞エピトープ又はその変異体を単独で、又は1つ以上の免疫刺激分子と組み合わせて含む医薬組成物である。医薬組成物は、腫瘍及び癌に対する免疫原反応を誘発するために、少なくとも1つのNY−ESO−1 HLA−DR拘束T細胞エピトープ又は少なくとも1つのNY−ESO−1 HLA−DP拘束T細胞エピトープ、又はNY−ESO−1抗原特異的CD4+T細胞を刺激するMHCクラスII拘束T細胞エピトープの組み合わせを含む。医薬組成物は、CD8+Tリンパ球の産生のために、NY−ESO−1由来の1つ以上のMHCクラスI拘束T細胞エピトープを更に含んでもよい。NY−ESO−1 HLA−DR拘束T細胞エピトープ又はNY−ESO−1 HLA−DP拘束T細胞エピトープ、及びNY−ESO−1 MHCクラスI拘束T細胞エピトープは各々、別個のエピトープとして提供されてもよいし、一緒に結合されてもよい。癌エピトープ又はその変異体は、癌の予防又は治療のために、免疫原又はワクチンとして提供されてもよい。医薬組成物は、哺乳動物における癌の治療又は予防の方法において有用である。治療方法において、医薬組成物は、哺乳動物の癌の予防又は阻害のためにCD4+T細胞及び/又は抗NY−ESO−1抗体応答を誘発するのに有効な量において哺乳動物に投与される。
【0011】
本発明の別の目的は、NY−ESO−1遺伝子、その部分又はホモログに由来するNY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体をコードしているDNA配列の翻訳によって、同上を製造する方法である。
【0012】
本発明の別の目的は、NY−ESO−1遺伝子、その部分又はホモログに由来するNY−ESO−1のHLA−DR拘束T細胞エピトープもしくはHLA−DP拘束T細胞エピトープ又はそれらの変異体又は誘導体をコードしているDNA配列の翻訳によって、同上を製造する方法である。
【0013】
本発明の更なる局面は、NY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその変異体をコードする単離されたDNA又はRNA配列、及びそれらの相補配列、並びにワクチンとしての該DNA又はRNA配列の使用、及びNY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体の製造方法における該DNA又はRNA配列の使用である。本発明は更に、プローブ、プライマー又はアンチセンスとしての使用のために、該DNA又はRNA配列のオリゴヌクレオチドを提供する。
【0014】
本発明の更なる局面は、NY−ESO−1のHLA−DR拘束T細胞エピトープ、HLA−DP拘束T細胞エピトープ、又はそれらの変異体及びそれらの組み合わせをコードする単離されたDNA又はRNA配列、並びにNY−ESO−1のHLA−DR拘束T細胞エピトープ、HLA−DP拘束T細胞エピトープ、又はそれらの変異体及びそれらの組み合わせの製造方法における該DNA又はRNA配列の使用である。本発明は更に、プローブ、プライマー又はアンチセンスとしての使用のために、該DNA又はRNA配列のオリゴヌクレオチドを提供する。
【0015】
本発明は更に、NY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその変異体をコードする核酸配列のみを含むか、又は少なくとも1つの免疫刺激分子をコードする第2のDNA配列を組み合わせて含むベクターを提供する。
【0016】
本発明は更に、NY−ESO−1の少なくとも1つのHLA−DR拘束細胞エピトープ又は少なくとも1つのHLA−DP拘束T細胞エピトープ又はそれらの変異体若しくは組み合わせをコードする核酸配列のみを含むか、又は少なくとも1つの免疫刺激分子をコードする第2のDNA配列を組み合わせて含むベクターを提供する。
【0017】
本発明はまた、NY−ESO−1の少なくとも1つのMHCクラスII拘束細胞エピトープ又はその変異体をコードするDNA配列のみを含むか、又は少なくとも1つの免疫刺激分子をコードする第2のDNA配列を組み合わせて含むベクターによってトランスフェクトされた、又は形質導入された宿主細胞を提供する。ベクター及び宿主細胞は、ワクチンとして役立ちうる。ワクチンにおいて、MHCクラスII拘束T細胞エピトープの発現により、ワクチンで免疫化した哺乳動物において腫瘍抗原特異的CD4+Tリンパ球の刺激が生ずる。
【0018】
本発明はまた、NY−ESO−1の少なくとも1つのHLA−DR拘束細胞エピトープ又は少なくとも1つのHLA−DP拘束T細胞エピトープ又はそれらの変異体若しくは組み合わせをコードするDNA配列のみを含むか、又は少なくとも1つの免疫刺激分子をコードする第2のDNA配列を組み合わせて含むベクターによってトランスフェクトされた、又は形質導入された宿主細胞を提供する。ベクター及び宿主細胞は、ワクチンとして役立ちうる。ワクチンにおいて、HLA−DR拘束T細胞エピトープ及び/又はHLA−DP拘束T細胞エピトープの発現により、ワクチンで免疫化した哺乳動物において腫瘍抗原特異的CD4+Tリンパ球の刺激が生ずる。
【0019】
本発明は、NY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体の検出により、哺乳動物における癌又は前癌の診断方法を提供する。
【0020】
本発明は、NY−ESO−1のHLA−DR拘束T細胞エピトープ及び/又はHLA−DP拘束T細胞エピトープ又はそれらの変異体の検出により、哺乳動物における癌又は前癌の診断方法を提供する。
【0021】
ヒト前腫瘍性及び腫瘍性細胞及び組織の診断方法を提供することは、本発明の更に別の目的である。本発明によれば、この方法は、ヒトから細胞、組織、又はその抽出物を単離し、そしてNY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体をコードするDNA配列、RNA配列、又はそれらの部分を検出するか、又はDNA配列もしくはRNA配列によって発現されたエピトープ若しくはその変異体を検出することを含む。ここで、DNA配列、RNA配列又は発現産物の増大の検出/増大は、前腫瘍及び腫瘍の指標となる。
【0022】
ヒト前腫瘍性及び腫瘍性細胞及び組織の診断方法を提供することは、本発明の更に別の目的である。本発明によれば、この方法は、ヒトから細胞、組織、又はその抽出物を単離し、そしてNY−ESO−1のHLA−DR拘束T細胞エピトープ又はHLA−DP拘束T細胞エピトープ又はそれらの変異体をコードするDNA配列、RNA配列、又はその部分を検出するか、又はそれらのDNA配列もしくはRNA配列によって発現されたエピトープ若しくはその変異体若しくは組み合わせを検出することを含む。ここで、DNA配列、RNA配列又は発現産物の増大の検出/増大は、前腫瘍及び腫瘍の指標となる。
【0023】
本発明の別の目的は、NY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその変異体をコードするDNA配列の1つ以上のコピーをそのゲノムに組み込んだトランスジェニック動物を提供することである。DNA配列の組み込みにより、エピトープの発現又は過剰発現が生じる。このようなトランスジェニック動物は、癌の治療に有用な治療薬のスクリーニングのために有用である。
【0024】
本発明の更に別の目的は、NY−ESO−1の少なくとも1つのHLA−DR拘束T細胞エピトープ又は少なくとも1つのHLA−DR拘束T細胞エピトープ又はそれらの変異体又は組み合わせをコードするDNA配列の1つ以上のコピーをそのゲノムに組み込んだトランスジェニック動物を提供することである。DNA配列の組み込みにより、エピトープの発現又は過剰発現が生じる。このようなトランスジェニック動物は、癌の治療に有用な治療薬のスクリーニングのために有用である。
【0025】
本発明の更に別の局面は、治療薬としての使用に関する、並びに診断及び検出アッセイにおける使用に関する、NY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその変異体と反応性のあるモノクローナル、ポリクローナル及び組換え抗体である。モノクローナル及びポリクローナル抗体は、単独でキットの形態で、又は診断及び検出アッセイで通常使用される他の試薬と一緒にキットの形態で提供されてもよい。
【0026】
本発明の更に別の局面は、治療薬としての使用に関する、並びに診断及び検出アッセイにおける使用に関する、NY−ESO−1のHLA−DR拘束T細胞エピトープ又はHLA−DP拘束T細胞エピトープと反応性のあるか、あるいはHLA−DP分子との組み合わせにおいてHLA−DPエピトープと、又はHLA−DR分子との組み合わせにおいてHLA−DRエピトープと反応性のあるか、あるいはそれらの変異体と反応性のあるモノクローナル、ポリクローナル及び組換え抗体である。モノクローナル及びポリクローナル抗体は、単独でキットの形態で、又は診断及び検出アッセイで通常使用される他の試薬と一緒にキットの形態で提供されてもよい。
【発明の効果】
【0027】
本発明は、CD4+T細胞によって認識されるNY−ESO−1由来の新規なMHCクラスII拘束T細胞エピトープの同定と単離である。本発明の癌エピトープは、哺乳動物において癌を阻害又は予防する免疫原及びワクチンとして有用であり、癌又は前癌を検出する診断薬として有用である。
【図面の簡単な説明】
【0028】
【図1−A】NY−ESO−1のヌクレオチド配列及びアミノ酸配列。NY−ESO−1のヌクレオチド配列の番号付けは、5’非翻訳領域の最初のヌクレオチドから始まる。
【図1−B】NY−ESO−1のヌクレオチド配列及びアミノ酸配列。NY−ESO−1のヌクレオチド配列の番号付けは、5’非翻訳領域の最初のヌクレオチドから始まる。
【図2−A】(2A)Ni2+クロマトグラフィーカラムを用いる完全長NY−ESO−1タンパク質の精製。SDSポリアクリルアミドゲルは、pET28ベクター(レーン1)、pNY−ESO−1(レーン2)を担うE.coli株BL21(DE3)からの粗抽出物、精製NY−ESO−1タンパク質(レーン3)、トランケートNY−ESO−1をコードする細菌抽出物(レーン4)、及び精製されたトランケートNY−ESO−1タンパク質であるESO1−74(レーン5)を示した。
【図2−B】(2B)NY−ESO−1に対する抗体の特異性を確認するためのウエスタンブロット。2人の代表的な患者からの1/2000希釈の血清(1つは、ELISAによって検出できるNY−ESO−1抗体をもつもの(レーン1、2及び3)及び1つは、該抗体無しのもの(レーン4、5及び6))を、ベクターだけをコードする細菌抽出物(レーン1、4)、NY−ESO−1をコードする細菌抽出物(レーン2、5)及び精製NY−ESO−1タンパク質(レーン3、6)に対して用いた。
【図2−C】(2C)患者TEは、NY−ESO−1タンパク質に対して抗体を有するメラノーマ患者の1人であった。NY−ESO−1(コントロールタンパク質としてBSA)に対する抗体の存在について88人の患者からの血清を用いてELISAを行った。血清希釈の1:25、1:250、1:2500でのO.D.450の値をプロットした。正常ドナーからの血清をコントロールとして用いた。それらの平均OD値もプロットした(ND)。
【図3】NY−ESO−1タンパク質で免疫化したHLA−DR4−Tgマウスを用いる推定NY−ESO−1エピトープの試験。HLA−DR4に対する予測された結合アフィニティに基づく8つのペプチドを、免疫化マウスからのリンパ球のインビトロ感作のために用いた。マウスリンパ球を、培地だけ、1359EBV B(HLA DR4+)細胞だけ、又はインビトロ刺激のために使用されたペプチドでパルスした1359EBV B細胞に対しIFN産生について試験した。
【図4−A】TE4−1 CD4+T細胞株の特徴付け。(4A)TE4−1は、ESOp116−135ペプチド又は精製NY−ESO−1タンパク質でパルスした1088EBV B細胞(HLA−DR+)を特異的に認識したが、推定エピトープを欠くESO1−74タンパク質を認識しなかった。
【図4−B】TE4−1 CD4+T細胞株の特徴付け。(4B)HLA DR拘束は、TE4−1によるNY−ESO−1の認識のために必要であった。293IMDR細胞にパルスしたとき、2つの重複するペプチドESOp111−130とp116−135は、認識された。GM−CSF分泌を測定する前に、1×105標的細胞を、4×104のTE4−1細胞と一晩共培養した。
【図4−C】TE4−1 CD4+T細胞株の特徴付け。(4C)ESOp116−135ペプチドでパルスした293IMDRの認識は、抗HLA−DR抗体(HB55)によって特異的に阻害されたが、抗クラスI抗体(HB95)によっては阻害されなかった。抗体の非存在下でTE4−1によって分泌されたGM−CSFの量を参照として用い、この参照に対し、抗体存在下のGM−CSF放出のパーセントを計算した。コントロール(マウスIgG2a)と抗MHC−クラスI抗体(HB95)による阻害は殆ど効果を有しなかった。
【図4−D】TE4−1 CD4+T細胞株の特徴付け。CTLC3G1(C.Macalliの好意)は、624.38melを認識するgp100特異的CD8+T細胞株であり、HB95の活性についてのコントロールとして使用した(図4D)。
【図4−E】TE4−1 CD4+T細胞株の特徴付け。T3−80は、1362melを認識するCD4+T細胞株であり、HB55の活性についてのコントロールとして使用した(図4E)。
【図5】CD4+T細胞株TE4−1による腫瘍細胞の認識。TE4−1の標的として使用した全てのメラノーマ株は、FACSとRT−PCRによってそれぞれ、HLA−DR4とNY−ESO−1の発現について分析された。TE4−1は、HLA−DR4分子を構成的に発現しているNY−ESO−1+腫瘍株(1359mel及びF049mel)を認識できた。DR1及びNY−ESO−1を発現しているF050melもT細胞によって認識された。コントロール標的526mel(DR4陽性及びNY−ESO−1陰性)、397mel、624.38mel(DR陰性及びNY−ESO−1陽性)、又は1300mel(DR1陽性及びNY−ESO−1弱陽性)のいずれに対しても反応性は無かった。
【図6−A】TE4−1によって認識されるNY−ESO−1ペプチドエピトープの特徴付け。(6A)HLA DR拘束NY−ESO−1エピトープのアンカー位置の決定。1088EBV B細胞を記載されたペプチド20Mでパルスした。GM−CSFを測定する前に、TE4−1細胞を一晩、標的細胞と共培養した。
【図6−B】TE4−1によって認識されるNY−ESO−1ペプチドエピトープの特徴付け。(6B)ESOp119−130を用いるペプチド力価測定実験。ESOp119−130は、図6Aに示すように、その認識に基づいて選択された。記載された濃度で希釈したESOp119−130を、1088EBV B細胞上にパルスした。この1088EBV B細胞は、TE4−1による認識のための標的として用いられた。コントロールペプチドESOp91−110の認識は、33Mの最高濃度でのみ測定された。
【図7−A】(7A)APCとしてDR1+EBVBを用いる、TE4−1によるNY−ESO−1タンパク質とESOp116−135の認識。NY−ESO−1タンパク質(5μg/ml)とペプチド(33μM)を、586EBV B(DR1+)上にパルスし、2度洗浄した。次いで、GM−CSFをアッセイする前に、TE4−1 T細胞を加え、一晩共培養した。
【図7−B】(7B)ESOp116−135(33μM)でパルスした586EBV Bを用いて、ブロッキング抗体の存在下及び非存在下でTE−4を刺激した。IID95(抗クラスI抗体)、IIB55(抗DR抗体)及びアイソタイプコントロール抗体を用いた。
【図8−A】合成ペプチドによるインビトロ刺激後のPBMCからのCD4+T細胞の産生。(図8A)3回のインビトロ刺激後、特異的ペプチド反応性をマルチプルウェルで検出した。96ウェルプレートで各々2.5×105PBMCを含む合計24ウェルを3週間弱く刺激した。24ウェルの15ウェルは、顕著な増殖を示し、そして比活性について試験した。各ウェルからのT細胞を、1088EBVB細胞、及びESOp161−180ペプチドでパルスした1088EBVB細胞とそれぞれインキュベートした。GM−CSF放出を上清から測定した。
【図8−B】合成ペプチドによるインビトロ刺激後のPBMCからのCD4+T細胞の産生。(図8B)TE4−2 T細胞は、NY−ESO−1ペプチド及びタンパク質と特異的に反応した。重複するペプチドESOp161−180とESOp156−175を、20μg/mlで90分間、1088(DR4+)と586EBVB(DR1+)細胞にパルスした。ESOp91−110をパルスについての無関係ペプチドとして用いた。精製NY−ESO−1とESO1−75タンパク質を、同じモル比を維持するために、それぞれ5μg/mlと2μg/mlで一晩パルスした。3度の洗浄後、TE4−2 T細胞を加え、一晩インキュベートした。GM−CSF放出を測定した。
【図8−C】合成ペプチドによるインビトロ刺激後のPBMCからのCD4+T細胞の産生。(図8C)ESOp161−180でパルスしたEBVB細胞のパネルを、TE4−2 CD4+T細胞の標的として用いた。これらのEBVB株は、異なるHLA DR及びDQ対立遺伝子を発現することが知られていた。それらのHLA DP対立遺伝子を、本研究では分子的にタイプ分けした(表2)。
【図9−A】抗DP抗体によるNY−ESO−1エピトープのT細胞認識のブロッキング。20μg/ml ESOp161−180ペプチドでパルスした1088EBVB細胞を、異なるブロッキング抗体存在下、標的細胞として使用した。使用した抗体の特異性は次のようであった:抗MHCクラスI(HLA A,B,C)抗体(W6/32)、抗MHCクラスII(HLA DP,DQ,DR)抗体(IVA12)、抗HLA DP抗体(B4/21)、抗HLA DR抗体(L243)及び抗HLA DQ抗体(Genox3.53(抗DQw1)及びIVD12(抗DQw3)の混合物)。全ての抗体を、各々、最終濃度20μg/mlで用いた。(図9A)CK3H6 T細胞は、HLA A2の状況においてgp100p209−218ペプチドを特異的に認識し、そして抗MHCクラスI抗体のための特異性コントロールとして用いた(図9B)。gp100p209−218ペプチドでパルスした1088EBVB(A2+)を標的として用いた。CD4+T細胞株(T3−80)は、HLA−DR拘束様式で1362melを認識し、そして抗MHCクラスII及び抗DR抗体のための特異性コントロールとして用いた(図9C)。
【図9−B】抗DP抗体によるNY−ESO−1エピトープのT細胞認識のブロッキング。20μg/ml ESOp161−180ペプチドでパルスした1088EBVB細胞を、異なるブロッキング抗体存在下、標的細胞として使用した。使用した抗体の特異性は次のようであった:抗MHCクラスI(HLA A,B,C)抗体(W6/32)、抗MHCクラスII(HLA DP,DQ,DR)抗体(IVA12)、抗HLA DP抗体(B4/21)、抗HLA DR抗体(L243)及び抗HLA DQ抗体(Genox3.53(抗DQw1)及びIVD12(抗DQw3)の混合物)。全ての抗体を、各々、最終濃度20μg/mlで用いた。(図9A)CK3H6 T細胞は、HLA A2の状況においてgp100p209−218ペプチドを特異的に認識し、そして抗MHCクラスI抗体のための特異性コントロールとして用いた(図9B)。gp100p209−218ペプチドでパルスした1088EBVB(A2+)を標的として用いた。CD4+T細胞株(T3−80)は、HLA−DR拘束様式で1362melを認識し、そして抗MHCクラスII及び抗DR抗体のための特異性コントロールとして用いた(図9C)。
【図9−C】抗DP抗体によるNY−ESO−1エピトープのT細胞認識のブロッキング。20μg/ml ESOp161−180ペプチドでパルスした1088EBVB細胞を、異なるブロッキング抗体存在下、標的細胞として使用した。使用した抗体の特異性は次のようであった:抗MHCクラスI(HLA A,B,C)抗体(W6/32)、抗MHCクラスII(HLA DP,DQ,DR)抗体(IVA12)、抗HLA DP抗体(B4/21)、抗HLA DR抗体(L243)及び抗HLA DQ抗体(Genox3.53(抗DQw1)及びIVD12(抗DQw3)の混合物)。全ての抗体を、各々、最終濃度20μg/mlで用いた。(図9A)CK3H6 T細胞は、HLA A2の状況においてgp100p209−218ペプチドを特異的に認識し、そして抗MHCクラスI抗体のための特異性コントロールとして用いた(図9B)。gp100p209−218ペプチドでパルスした1088EBVB(A2+)を標的として用いた。CD4+T細胞株(T3−80)は、HLA−DR拘束様式で1362melを認識し、そして抗MHCクラスII及び抗DR抗体のための特異性コントロールとして用いた(図9C)。
【図10−A】TE4−2 CD4+T細胞による腫瘍細胞と293CIITA/NY−ESO−1の認識。(図10A)TE4−2は、NY−ESO−1とDP4の両方を発現しているメラノーマ株を認識した。公知のNY−ESO−1発現(RT−PCRによって)とHLA DPタイプ(RT−PCRと配列決定によって決定)をもつメラノーマ株を、標的として使用した。この実験で、MHCクラスII発現をアップレギュレートするために、一晩のIFN−γ処理(500ユニット/ml)を、F026mel、526mel、及び397melについて行った。サイトカイン放出を測定する前に、TE4−2 T細胞を腫瘍細胞と一晩共培養した。
【図10−B】TE4−2 CD4+T細胞による腫瘍細胞と293CIITA/NY−ESO−1の認識。(図10B)TE4−2 CD4+T細胞株は、インバリアント鎖(Ii)ターゲティング配列の存在下又は非存在下においてNY−ESO−1でトランスフェクトされた293CIITAを認識した。親の293細胞と293CIITA細胞を、NY−ESO−1(pESO)、Ii−NY−ESO−1(pIi−ESO)、又はGFP(pGFP)をそれぞれコードするプラスミドでトランスフェクトした。サイトカイン放出をアッセイする前に、TE4−2 T細胞を、トランスフェクタントと共に一晩、共培養した。
【図11−A】TE4−2によって認識されるT細胞エピトープの特徴付け。(図11A)T細胞認識のためのNY−ESO−1エピトープのアンカー残基と最小長さの決定。N又はC末端にアミノ酸欠失を有する合成ペプチドを用いて、40μg/mlで1088EBVB細胞をパルスした。次いで、EBVB細胞を徹底的に洗浄し、TE4−2 T細胞を刺激するために、標的細胞として用いた。2つの別々の実験を行い、この図の上のパネルと下のパネルとして示した。
【図11−B】TE4−2によって認識されるT細胞エピトープの特徴付け。(図11B)T細胞認識に必要な最小ペプチド濃度の決定。ESOp157−170ペプチドを用いて、種々の濃度で1088EBVB細胞をパルスした。次いで、ペプチドをパルスされた細胞を洗浄し、TE4−2株を刺激するために、標的として用いた。コントロールペプチドESOp91−110を、最高濃度33μMでのみ用いた。
【図11−C】TE4−2によって認識されるT細胞エピトープの特徴付け。(図11C)NY−ESO−1特異的CD8+T細胞によるDPB1*0401−拘束CD4+T細胞エピトープの認識。TE8−1細胞株を、ESOp157−167ペプチドによるインビトロ刺激によって、親TEのPBMCから作製した。L023EBVB細胞(HLA−A2+,DP4-)を、無血清培地において、DPB1*0401エピトープ領域をカバーするペプチドでパルスし、洗浄し、TE8−1 T細胞を刺激するために用いた。
【図12−A】TE4−2 CD4+T細胞とTE8−1 CD8+T細胞による認識についてのペプチドの力価測定(図12A)586EBVB細胞(A2−,DP4+)を抗原提示細胞として用い、種々の濃度で、記載されたペプチドでパルスした。細胞を洗浄し、次いで、サイトカイン放出をアッセイする前に、TE4−2 CD4+T細胞とインキュベートした。
【図12−B】TE4−2 CD4+T細胞とTE8−1 CD8+T細胞による認識についてのペプチドの力価測定(図12B)L023EBVB細胞(A2+,DP4−)を抗原提示細胞として用い、種々の濃度で、記載されたペプチドでパルスした。細胞を洗浄し、次いで、サイトカイン放出をアッセイする前に、TE4−2 CD4+T細胞とインキュベートした。
【発明を実施するための形態】
【0029】
発明の詳細な説明
本発明は、免疫系のMHCクラスII拘束CD4+Tリンパ球によって免疫学的に認識されるNY−ESO−1の癌エピトープ、その部分、誘導体又は変異体を包含する。本発明の癌エピトープは、免疫系のCD4+T細胞との相互作用によって、体液性媒介免疫応答を特異的に引き起こす。抗原性癌エピトープとCD4+T細胞とのこの相互作用は、ヒトを含む哺乳動物における癌の予防、除去、又は減少において、CD4+T細胞が抗原性癌エピトープに反応し、免疫系の他の細胞をリクルートすることを引き起こす。
【0030】
本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープは、原発性もしくは転移性メラノーマ、胸腺腫、リンパ腫、肉腫、肺癌、肝癌、非ホジキンリンパ腫、ホジキンリンパ腫、白血病、子宮癌、子宮頚癌、膀胱癌、腎癌、頭部及び頚部癌、神経芽腫及び腺癌(例えば、乳癌、前立腺癌、卵巣癌、膵癌、甲状腺癌など)を含むが、それらに限定されない癌の一部を形成するか、又はそれらの癌に由来する。
【0031】
メラノーマという用語は、メラノーマ、転移性メラノーマ、メラノサイト又はメラノサイト関連母斑細胞由来のメラノーマ、悪性メラノーマ、黒色上皮腫、黒色肉腫、インサイチュメラノーマ(melanoma in situ)、表在拡大型メラノーマ、結節型(nodular)メラノーマ、悪性ほくろ性メラノーマ、末端性ほくろ性メラノーマ、浸潤性メラノーマ又は家族性非定型奇胎及びメラノーマ(familial atypical mole and melanoma)(FAM−M)症候群を含むが、それらに限定されない。
【0032】
癌、特にメラノーマの患者の自己CD4+Tリンパ球によって認識される癌エピトープ又はその誘導体は特に興味深い。MHC(又はHLA)クラスII拘束CD4+Tリンパ球、特にHLA−DR拘束Tリンパ球及び/又はHLA−DP拘束Tリンパ球によって認識される癌エピトープ又はその誘導体は更に興味深い。一実施態様では、NY−ESO癌エピトープは、HLA−DR分子の状況においてCD4+Tリンパ球によって認識される。別の実施態様では、NY−ESO癌エピトープは、HLA−DP分子の状況においてCD4+Tリンパ球によって認識される。
【0033】
本発明の「癌エピトープ」は、HLA−DR拘束及びHLA−DP拘束Tリンパ球などのMHCクラスII拘束Tリンパ球を誘発するNY−ESO−1の部分又は変異体部分を包含する。このようなリンパ球は、腫瘍細胞に由来する完全長NY−ESO−1タンパク質、MHCクラスII拘束T細胞エピトープ、及び天然でプロセスされた抗原と特異的に反応しうる。
【0034】
本発明のNY−ESO−1のMHCクラスII拘束T細胞エピトープは、サイズにおいて約9アミノ酸乃至約30アミノ酸、好ましくは、長さにおいて約10乃至約15アミノ酸に変わりうる。
【0035】
好適な実施態様では、本発明のNY−ESO−1のMHCクラスII拘束T細胞エピトープは、長さにおいて約9乃至10アミノ酸である。
【0036】
一実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:Xaa1KEFTVSXaa2(配列番号4)及びその変異体又は誘導体を含む、長さにおいて少なくとも約10アミノ酸のペプチドである。Xaa1及びXaa2のアミノ酸、及びN末端及びC末端でのこれらの位置におけるアミノ酸数は、エピトープが、CD4+Tリンパ球に結合し、刺激する能力を保持するかぎり、変わりうる。エピトープは、約10乃至約30アミノ酸、好ましくは20アミノ酸未満、より好ましくは約10乃至約15アミノ酸を含みうる。
【0037】
本発明の別の実施態様では、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、式:Xaa1VLLKEFTVSGXaa2(配列番号5)(式中、Xaa1はアミノ酸ではないか、又は1乃至約10個の天然のアミノ酸であり、好ましくは1乃至約5個のアミノ酸である;Xaa2はアミノ酸ではないか、又は長さにおいて1乃至約7個のアミノ酸である)によって表されることができる。
【0038】
本発明の別の実施態様では、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:QDAPPLPVPGVLLKEFTVSGNILTIRL(配列番号6)又はそのフラグメント若しくは誘導体を含む。
【0039】
また、本発明の範囲には、配列番号4、5又は6と部分配列相同性を有する癌ペプチド又はその部分が包含される。部分アミノ酸配列相同性とは、配列番号4、5又は6と少なくとも85%の配列相同性、好ましくは、少なくとも95%の配列相同性又はそれ以上を有し、かつNY−ESO−1 MHCクラスII拘束特異的CD4+Tリンパ球を刺激するという生物学的機能を有するペプチドを意味する。哺乳動物ホモログは、霊長類及びマウスホモログを含むがそれらに限定されない本発明の範囲に含まれる。
【0040】
一実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:
APPLPVPGVLLKEFTVSGNILTIRL(配列番号7);
APPLPVPGVLLKEFTVS(配列番号8);
APPLPVPGVLLKEFTV(配列番号9);
LPVPGVLLKEFTVSG(配列番号10);
PVPGVLLKEFTVSG(配列番号11);
VPGVLLKEFTVSG(配列番号12);
PGVLLKEFTVSG(配列番号13);
GVLLKEFTVSG(配列番号14);
LLKEFTVSGNILTIR(配列番号15);
LKEFTVSGNILTIRL(配列番号16);
KEFTVSGNILTIRL(配列番号17);
LPVPGVLLKEFTVSGNILTI(配列番号18);
又はそれらの変異体若しくは誘導体を含む。
【0041】
好適な実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープはアミノ酸配列:
【0042】
【数1】
【0043】
及びその変異体若しくは誘導体を含む。
【0044】
上記配列内に置換を有するエピトープは、本発明に包含される。置換アミノ酸により、配列番号19と比較して等価な又は増大した結合が生じるならば、天然に存在するアミノ酸の置換は、1個以上のアンカー位置でありうる。配列番号19のアンカー位置は、位置1−Leu、位置4−Glu、位置6−Thr、位置7−Valであると予測される。一実施態様では、アラニンは、位置1でロイシンの代わりに置換される。
【0045】
本発明の別の実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:DHRQLQLSIS SCLQQLSLLM(配列番号20)、又はその部分、変異体及び誘導体を含む。一実施態様において、配列番号20の部分はアミノ酸配列:
DHRQLQLSIS SCLQQLS(配列番号29);
DHRQLQLSIS SCLQ(配列番号30);
QLQLSIS SCLQQL(配列番号31)
又はそれらの変異体及び誘導体を含む。
【0046】
本発明の別の実施態様において、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:
WITQCFLPVF LAQPPSGQRR(配列番号21)
その部分、変異体及び誘導体を含む。一実施態様では、配列番号21の部分は、アミノ酸配列:
QCFLPVF LAQPPSGQRR(配列番号32);
LPVF LAQPPSGQRR(配列番号33);
CFLPVF LAQPPSGQ(配列番号34);
又はそれらの変異体及び誘導体を含む。
【0047】
NY−ESO−1癌エピトープ及びその誘導体は、MHCクラスII拘束CD4+Tリンパ球によって認識される。NY−ESO−1エピトープと組み合わせて認識されるクラスII分子としては、HLA−DR1、HLA−DR3、HLA−DR4及びクラスIIペプチド結合の縮重のために機能する他のクラスII分子などの少なくとも1つのHLA−DRが挙げられるが、それに限定されない。一実施態様では、癌ペプチドによって認識されるHLAサブタイプは、HLA−DR4サブタイプである。別の実施態様では、NY−ESO−1癌エピトープは、HLA−DR1及びHLA−DR4と結合する。
【0048】
別の実施態様では、エピトープは、以下:
Xaa1ITQXaa2FXaa3PXaa4(配列番号51)
(式中、Xaa1は、少なくとも1つの天然に存在するアミノ酸、好ましくは、Trp、Phe、Tyr、Met、Ile、Val、Ala及びそれらの組み合わせからなる群から選択されるアミノ酸であり;
Xaa2は、少なくとも1つの天然に存在するアミノ酸、好ましくは、Cys、Ser、Val、Ala、Thr、及びそれらの組み合わせからなる群から選択されるアミノ酸であり;
Xaa3は、少なくとも1つの天然に存在するアミノ酸、好ましくは、Leu、Phe、Tyr、Met、Ile、Val、Ala、及びそれらの組み合わせからなる群から選択されるアミノ酸であり;
Xaa4は、少なくとも1つの天然に存在するアミノ酸、好ましくは、Val、Tyr、Ile、Ala、Leu、Pro、及びそれらの組み合わせからなる群から選択されるアミノ酸である)の一般的なアミノ酸モチーフを含む、NY−ESO−1のHLA−DP拘束T細胞エピトープである。
【0049】
また、配列番号51と部分配列相同性を共有する癌ペプチド又はその部分は、本発明の範囲に包含される。部分アミノ酸配列相同性とは、配列番号51と少なくとも85%の配列相同性、好ましくは、少なくとも95%の配列相同性又はそれ以上を有し、かつNY−ESO−1 MHCクラスII拘束特異的CD4+Tリンパ球、好ましくはHLA−DP拘束Tリンパ球を刺激するという生物学的機能を有するペプチドを意味する。哺乳動物ホモログは、霊長類及びマウスホモログを含むがそれらに限定されない本発明の範囲に含まれる。ホモログはまた、LAGE遺伝子などのNY−ESO−1遺伝子以外の遺伝子によってコードされうる配列番号51と配列相同性を有するペプチドを含む。
【0050】
上記配列内に置換を有するエピトープは、本発明に包含される。置換アミノ酸により、配列番号51と比較して等価な又は増大した結合が生じるならば、天然に存在するアミノ酸の置換は、1個以上のアンカー位置でありうる。配列番号51のアンカー位置は、位置1、7、及び9、即ち、それぞれW、L、及びV残基であると推定される。これらのアンカー残基の置換は、「W」残基の代わりにF、Y、M、I、V及びA;「L」残基の代わりにF、Y、M、V、I及びA;「V」残基の代わりにY、I、A、L及びPでありうるが、それらに限定されない。別の置換は、配列番号1の5位としてC残基を含みうるが、それは、残基S、V、A及びTでありうるが、それらに限定されない。
【0051】
一実施態様では、NY−ESO−1のMHCクラスII拘束T細胞エピトープは、アミノ酸配列:Xaa1WITQCFLPVFXaa2(配列番号52)及びその変異体又は誘導体を含む、長さで少なくとも約10アミノ酸のペプチドである。Xaa1及びXaa2は、アミノ酸ではないか、又は1つ以上の同じ、又は種々の天然に存在するアミノ酸でありうる。エピトープがCD4+Tリンパ球、特にHLA−DP拘束CD4+Tリンパ球と結合/及びそれらを刺激する能力を保持するかぎりは、N末端及びC末端でのこれらの位置でのアミノ酸の数は変りうる。エピトープは、約10乃至約30アミノ酸、好ましくは20アミノ酸未満、より好ましくは約10乃至約15アミノ酸を含みうる。
【0052】
一実施態様では、NY−ESO−1のHLA−DP拘束T細胞エピトープは、アミノ酸配列:
151-SCLQQLSLLMWITQCFLPVFLAQPPSG-177(配列番号53);
SLLMWITQCFLPVF(配列番号54);
LLMWITQCFLPVFL(配列番号55);
LMWITWCFLPVFLA(配列番号56);
MWITQCFLPVFLAQ(配列番号57);
WITQCFLPVFLAQP(配列番号58);
ITQCFLPVFLAQPP(配列番号59);
QQLSLLMWITQCFL(配列番号60);
QLSLLMWITQCFLP(配列番号61);
LSLLMWITQCFLPV(配列番号62);及び
それらの誘導体を含む。
【0053】
NY−ESO−1癌エピトープ及びその誘導体は、HLA−DP拘束CD4+Tリンパ球、及びその誘導体、好ましくはHLA−DP4拘束CD4+Tリンパ球によって認識される。NY−ESO−1エピトープと組み合わせて認識されるクラスII分子としては、HLA−DP及びクラスIIペプチド結合の縮重のために機能するクラスII分子を含む。癌ペプチドによって認識される好適なHLAサブタイプは、HLA−DPB1*0401−0402対立遺伝子及び機能縮重のために、該ペプチドに結合しうる他の対立遺伝子である。
【0054】
本発明の別の実施態様は、MHCクラスII拘束CD4+Tリンパ球を誘発するように有効に作用するために、NY−ESO−1のMHCクラスII拘束T細胞エピトープに十分な相同性を有する該エピトープの誘導体及び変異体を包含する。このようなペプチドは、1つ以上の位置、特にアンカー位置で保存的アミノ酸変化を有しうる。保存的アミノ酸変化とは、特定の位置での、元々存在した同じタイプのアミノ酸でのアミノ酸の変化、即ち、疎水性アミノ酸の代わりに交換された疎水性アミノ酸、塩基性アミノ酸の代わりに塩基性アミノ酸などを意味する。このようなアミノ酸変化は、ペプチドの全体の電荷やコンフィギュレーションを有意には変えず、それ故、このような変異体は、癌ペプチドの抗癌活性を維持するか、又は増大させる。このような保存的変化の例は、当業者に周知であり、本発明の範囲内である。
【0055】
本発明はまた、NY−ESO−1 MHCクラスII拘束T細胞エピトープの機能的に等価な変異体に関する。「機能的に等価な変異体」としては、部分配列相同性を有するペプチド、1つ以上の特異的な保存及び/又は非保存的アミノ酸変化を有するペプチド、ペプチド結合体、キメラタンパク質、融合タンパク質及びペプチドが挙げられる。
【0056】
本発明の別の局面は、HLA−DP拘束T細胞及びHLAクラスI拘束CD8+Tリンパ球の両方、特にHLA−A拘束Tリンパ球、好ましくはHLA−A2拘束Tリンパ球のためのエピトープとして機能するNY−ESO−1ペプチドである。一実施態様では、NY−ESO−1のHLA−DP拘束及びHLAクラスI拘束エピトープとしては、アミノ酸配列:SLLMWITQCFLPVF(配列番号54)並びにその変異体及びホモログが挙げられる。変異体としては、RSLLMWITQCFLPV(配列番号63)を含むESOp156R−169及びSLLMWITQCFLPVR(配列番号64)を含むESOp157−170Rを含むがそれらに限定されない、SLLMWITQCFLPVF(配列番号54)における1つ以上の置換を有するペプチドが挙げられるがそれらに限定されない。このような置換は、ネイティブ配列の免疫学的機能活性を保持しつつ、ペプチドを水溶液により可溶性にする。高度に水溶性のペプチドは90%超の純度まで容易に精製されうる。
【0057】
NY−ESO−1 MHCクラスII拘束T細胞エピトープは、初代(primary)臨床単離体、細胞株などからのような天然源から精製単離できる。NY−ESO−1 MHCクラスII拘束T細胞エピトープは、少なくとも90%純粋、好ましくは少なくとも95%純粋及びほぼ100%純粋である。エピトープは、当業界公知の化学合成又は組換えDNA技術によっても得ることができる。化学合成の技術は、J.M.Steward and J.D.Young, “Solid Phase Peptide Synthesis”, W.H.Freeman & Co., San Francisco, 1969; M.Bodansky et al “Peptide Synthesis”, John Wiley & Sons, Second Edition, 1976,及びJ.Meienhofer, “Hormonal Proteins and Peptides”, Vol.2, p.46, Academic Press, New York, 1983 及びE.Schroder and K.Kubke, “The Peptides", Vol.1, Academic Press, New York, 1965に記載されている。
【0058】
NY−ESO−1 クラスII拘束T細胞エピトープは、当業界公知の方法によって、医薬として許容できる担体と共に医薬組成物へと製剤化できる。本組成物は、NY−ESO−1特異的CD4+Tリンパ球を誘発するために免疫原として有用であり、そして抗NY−ESO−1抗体を誘発するのに有用でありうる。本組成物はまた、癌を予防又は治療するためのワクチンとして有用である。本組成物は更に、少なくとも1つの免疫刺激分子を含みうる。MHCクラスII特異的T細胞応答を刺激するために、癌エピトープ又はその部分と一緒に使用される免疫刺激分子としては、1つ以上の主要組織適合性複合体(MHC)クラスII分子、又はMHCクラスII分子を発現している細胞が挙げられるがそれらに限定されない。本組成物は、免疫増強のために、B7.1、B7.2、ICAM−1、ICAM−2、LFA−1、LFA−3、CD72など、並びにIL−1〜IL−15、TNFα、IFNγ、RANTES、G−CSF、M−CSF、IFNα、CTAPIII、ENA−78、GRO、I−309、PF−4、IP−10、LD−78、MGSA、MIP−1α、MIP−1β、又はそれらの組み合わせを含むがそれらに限定されないサイトカインなどを含む他の刺激分子を更に含んでもよい。
【0059】
刺激分子は、物理的に分離した実体として提供され得るか、又はB細胞、マクロファージ若しくは樹状細胞などの抗原提示細胞の膜中に提供され得るか、リポソームの膜中に提供され得るか、又は形質導入若しくはトランスフェクトされた細胞の表面上に発現され得る。MHCクラスII免疫刺激分子のDNA配列は、GenBankなどから入手できる。HLA−DP免疫刺激分子のDNA配列は、GenBank/EMBL/DNA Data Bank of Japan(DDBJ) at GenBank, National Center for Biotechnology Information, 8600 Rockville Pike, Bethesda, MD 20894 USA 又はそのウエブサイトから入手できる。
【0060】
本発明の医薬組成物は、NY−ESO−1 HLA−DR又はそのHLA−DP拘束T細胞癌ペプチドに加えて、NY−ESO−1からの幾つかの異なるMHCクラスII拘束T細胞エピトープを含んでもよい。これらには、LPVPGVLLKEFTVSG(配列番号10)、VLLKEFTVSGNILTIRLT(配列番号65)、AADHRQLQLSISSCLQQL(配列番号66)、及びそれらの組み合わせなどのHLA−DR拘束エピトープ及びその変異体が挙げられるがそれらに限定されない。
【0061】
本発明の医薬組成物は、MHCクラスII拘束CD4+Tリンパ球を誘発することに加えて、MHCクラスI拘束細胞傷害性Tリンパ球を誘発するために、MHCクラスI拘束NY−ESO−1癌ペプチドを必要に応じて含んでもよい。MHCクラスI拘束NY−ESO−1癌ペプチドとしては、式:
Xaa1Xaa2Xaa3GPGGGAPXaa4(配列番号22)
(式中、Xaa1は、アミノ酸ではないか、又は1乃至約20個の天然のアミノ酸、好ましくは1乃至5個のアミノ酸であり;
Xaa2は、Ala、Thr、Val、Leu、又はArgであり;
Xaa3は、Ser、又はAla、Val、Ile、Leu、Thrなどの保存的置換であり;
Xaa4は、Arg、Lys、好ましくはArg)並びにそれらのフラグメント及び誘導体によって表される癌ペプチドが挙げられるがそれらに限定されない。一実施態様では、医薬組成物で使用されるMHCクラスI拘束NY−ESO−1癌ペプチドとしては、アミノ酸配列:ASGPGGGAPR(配列番号23)が挙げられる。
【0062】
NY−ESO−1 MHCクラスII拘束T細胞エピトープ及びNY−ESO−1 MHCクラスI拘束T細胞エピトープは各々、別個のエピトープとして提供されてもよいし、単一ペプチドとして一緒に結合されてもよい。エピトープは、当業界公知の方法によって、一緒に結合されてもよいし、化学的に合成されてもよい。化学リンカー、ペプチドリンカー、ペプチド結合などは、エピトープを結合するために使用できる。一実施態様では、MHCクラスIIエピトープのC末端は、ペプチド結合により、MHCクラスIエピトープのN末端に直接結合される。
【0063】
NY−ESO−1 MHCクラスII拘束T細胞エピトープは、癌の予防又は治療方法において有用であり、そしてヒトを含む哺乳動物における癌又は前癌を検出するための診断アッセイにおいて有用である。診断アッセイにおいて、本発明のNY−ESO−1 HLAクラスII拘束T細胞エピトープペプチド、その変異体及び誘導体は、腫瘍抗原NY−ESO−1に対するヘルパー免疫応答を検出するのに有用である。NY−ESO−1は腫瘍細胞(免疫が特別免除された部位である正常精巣を除く)に専ら発現されるので、該タンパク質に対する免疫応答は、患者において早期の癌の検出のための指標として使用できる。ヘルパーT細胞応答の発展は、該タンパク質に対して検出できる抗体の発展よりも早い出来事でありうるので、NY−ESO−1 HLAクラスII拘束T細胞エピトープペプチドに対するヘルパーT細胞応答の検出は、早期の癌の検出に有用である。NY−ESO−1に対するヘルパーT細胞応答の検出方法において、NY−ESO−1 HLAクラスII拘束T細胞エピトープペプチドは、基板又は固体支持体に適用される。患者からのリンパ球は、インフルエンザウイルス由来のペプチドなどのコントロールペプチドと並行して、NY−ESO−1 MHCクラスII拘束T細胞エピトープペプチドの存在下で生育される。次いで、ELISPOT及びELISAなどの技術を用いて、特異的なサイトカイン放出を測定する。陰性コントロールと比較した、ヘルパーT細胞免疫応答の増大の検出は、患者における前癌又は早期の癌の指標となる。
【0064】
本発明のNY−ESO−1 MHCクラスII拘束結合ペプチドは、任意の一定の標的抗原及び/又はハプテンに対する抗体及び/又はCD8+T細胞応答の産生を増大させるために使用できる。詳細には、これらのペプチドは、抗体及び/又はCD8+T細胞応答が意図される標的抗原ペプチド、タンパク質又は任意の他のハプテンに対して結合又は共有結合することができる。任意の標的ハプテン又はタンパク質とのNY−ESO−1 MHCクラスII拘束エピトープペプチドの結合は、破傷風トキソイド、アルブミンなどの通常のT細胞癌タンパク質と同様に、任意の標的抗原、ハプテン、又はタンパク質の免疫原性の増大において、免疫学的T細胞キャリアペプチドとして作用するはずである。増大は、標的ハプテン又はタンパク質に対するより高力価の抗体、IgMからIgG又はIgA抗体へのイムノグロブリンクラススイッチング、及び/又はCD8+T細胞応答の誘発において明白でありうる。このような標的抗原、ハプテン又はタンパク質の例としては、TRP2、GP100、TRP1、gp120及び他のHIV抗原、マラリア抗原、それらのエピトープなどが挙げられるがそれらに限定されない。同様に、NY−ESO−1 MHCクラスII拘束エピトープをコードする核酸配列は、標的抗原又はそのエピトープへの免疫原性を増大させるために、標的抗原又はそのエピトープをコードする核酸配列と共に、操作されたワクチン構築物に組み込まれることができる。この局面に関する例としては、クラスIIエピトープの核酸配列を、標的遺伝子とフレームを合わせて、裸のDNAもしくはRNA、ワクシニアウイルス、アデノウイルス、鶏痘ウイルスなどの形態の任意のワクチン構築物に組み込むことが挙げられる。例えば、プラスミドワクチンの形態のTRP2抗原の免疫原性を増大させるために、NY−ESO−1 クラスIIエピトープの核酸配列は、TRP2遺伝子と同じオープンリーディングフレームにおいて融合されることができる。次いで、TRP2のみをコードするプラスミドを用いる代わりに、患者を免疫化するためにハイブリッドプラスミドが使用される。
【0065】
癌エピトープ又はその変異体は、放射性標識、ビオチン、フルオレセインなどの他の成分が結合している誘導体の形態でありうる。特異的な器官、腫瘍、又は細胞型への特異的ターゲティングを可能とするターゲティング剤をエピトープに結合させることもできる。このようなターゲティング剤は、ホルモン、サイトカイン、細胞レセプターなどでありうる。エピトープは、単独で、又は他の試薬と組み合わせて、キットの形態で製造できる。
【0066】
本発明の別の局面は、本発明のNY−ESO−1 MHCクラスII拘束エピトープを用いて、癌に対する腫瘍特異的体液性媒介免疫を誘導するのに有用な免疫原又はワクチンである。免疫原及びワクチンは、NY−ESO−1特異的CD4+Tリンパ球及び抗NY−ESO−1抗体を誘発する。必要に応じて、免疫原及びワクチンは、CD8+Tリンパ球を誘発するために、NY−ESO−1 MHCクラスI拘束エピトープを含んでもよい。
【0067】
癌免疫治療へのアプローチは、能動又は受動カテゴリーに分けることができる。能動免疫治療は、腫瘍に対する免疫応答を増大させる試みにおいて、癌患者に対する癌抗原による直接的な免疫化を含む。受動免疫治療は、直接的に抗腫瘍応答を媒介するという目標をもって、抗腫瘍反応性を有する免疫細胞又は抗体などの免疫試薬の投与を指す。
【0068】
能動免疫治療における大部分の従来の試みは、免疫応答を刺激できる量と形態で腫瘍抗原を含むという期待をもって、完全な癌細胞又は癌細胞抽出物のいずれかを用いた。しかし、癌抗原とエピトープの分子的同定は、ヒト癌の治療に関する免疫治療を開発するための新しい可能性を有する。これらのアプローチの幾つかの要約を表1に示す。
【0069】
【表1】
【0070】
少なくとも1つのNY−ESO−1 MHCクラスII特異的T細胞エピトープをコードする遺伝子の、E.coli、酵母又はバキュロウイルスなどの高効率発現系への挿入は、免疫化において使用するための精製腫瘍エピトープを大量に得る機会を提供する。あるいは、イムノドミナントエピトープは、インビトロで容易に合成でき、そして免疫化のために、単独で、又はアジュバントとの組み合わせ、脂質/リポソームもしくはヘルパーペプチドとの結合などの免疫原性を増大させるために意図された形態で大量に精製でき、あるいは、抗原提示細胞にパルスすることもできる。MHCクラスII抗原への結合効率を増大させるための、イムノドミナントペプチドの個々のアミノ酸の修飾は、ネイティブタンパク質と比較して、免疫原性を増大させる可能性を有しうる。
【0071】
筋肉内又は皮膚内に直接注射される「裸の」DNAを用いる最近の技術は、コードされた抗原に対し細胞免疫応答と体液性免疫応答の両方を生じさせることが示されている(Cooney,E.L., A.C.Collier, P.D.Greenberg, R.W.Coombs, J.Zarling, D.E.Arditti, M.C.Hoffman, S.L.Hu, 及び L.Correy, 1991, Lancet 337:567; Wolff,J.A., R.W.Malone, P.Williams, W.Chong, G.Acsadi, A.Jani, 及び P.L.Felgner, Science 247:1465; Davis,H.L., R.G.Whalen, 及び B.A.Demeniex, 1993, Hum.Gene.Ther. 4:151;Yang,N.S., J.Burkholder, B.Roberts, B.Martinelli, 及びD.McCabe, 1990, Proc.Natl.Acad.Sci. USA 87:9568;Williams,R.S., S.A.Johnston, M.Riedy, M.J.DeVit, S.G.McElligott, 及び J.C.Sanford, 1991,Proc.Natl.Acad.Sci. USA 88:2726; Fynan,E.R., Webster, D.H.Fuller, J.R.Haynes, J.C.Santoro, 及び H.L.Robinson, 1995, Proc.Natl.Acad.Sci. USA 90:11478; Eisenbraum,M.D., D.H.Fuller, 及び J.R.Haynes, 1993, DNA and Cell Bio. 12:791; Fuller,D.H. 及び J.R.Haynes, 1994, AIDS Res.Hum.Retrovir. 10(11):1433; Acsadi,G., G.Dickson, D.R.Love, A.Jani, F.S.Walsh, A.Gurusinghe, J.A.Wolff, 及び K.E.Davies, 1991, Nature 352:815)。非生存DNAベクターを用いる技術は、製造の容易さと投与の安全性という利点を有する。本発明の核酸配列は、癌に対する免疫原として、及びDNAワクチンとして有用である。NY-ESO-1 MHCクラスII特異的T細胞エピトープの本発明の核酸配列、又は1つ以上の変異体NY-ESO-1 MHCクラスII拘束T細胞エピトープを有する完全長NY-ESO-1タンパク質をコードする核酸配列は、癌細胞に対する体液性反応を誘発させる量で、遺伝子銃を用いて投与できる。ナノグラム量がこのような目的のために有用である。
【0072】
免疫化の有効な形態は、免疫原性分子をコードする遺伝子の、BCG、サルモネラ又はリステリアなどの組換え細菌への、又はワクシニア、鶏痘、又はアデノウイルスなどの組換えウイルスへの組み込みを含む。NY-ESO-1 MHCクラスII特異的T細胞エピトープをコードする遺伝子は、単独で、又は免疫刺激分子をコードする遺伝子もしくは感染後に免疫応答を増大させることができる他の遺伝子と組み合わせて発現させることができる。構築物は、更なるNY-ESO-1 MHCクラスII拘束T細胞エピトープ及び/又は少なくとも1つのNY−ESO−1 MHCクラスI特異的T細胞エピトープをコードする遺伝子を更に含んでもよい。
【0073】
マウスモデルでのモデル腫瘍抗原についての研究は、インターロイキン−2(IL−2)又はB.7.1の遺伝子の組み込みが、モデル腫瘍エピトープの免疫原性を増大でき、これらのエピトープを担う樹立された肺転移の退行を媒介さえできることを示した。能動免疫治療、次いでIL−2、IL−6、IL−10、IL−12又はIL−15などの免疫刺激サイトカインの外来性投与はまた、免疫応答を改善するために使用できる。
【0074】
ヒト腫瘍抗原を認識できる遺伝子的に修飾された免疫細胞による受動免疫治療(通常、養子免疫治療と言われる)は、転移性メラノーマの選択された患者における癌の退行を媒介するのに有効である。抗原提示細胞上に存在する腫瘍抗原イムノドミナントエピトープに対しヒトリンパ球をインビトロで感作するインビトロ技術が開発された。反復性のインビトロ刺激によって、ヒト腫瘍抗原をコードする遺伝子をクローンするために使用されたTILよりも、これらの抗原を認識するずっと大きな能力をもった細胞が得られる。このように、癌ペプチドによる反復性のインビトロ感作によって、TILの50乃至100倍の能力をもったリンパ球を得ることができる。これらの細胞の養子移転(adoptive transfer)は、従来の増殖したTILよりも、インビボで腫瘍退行を媒介するのに有効でありうる。
【0075】
一実施態様において、末梢血単核球(PBMC)は、DR4−IEトランスジェニックマウスの免疫化後、同定された幾つかの候補DRB1*0401ペプチドで刺激された。NY−ESO−1特異的CD4+T細胞は、合成ペプチドESOp161−180によるインビトロ感作で産生された。このCD4+T細胞株は、HLA DP4(コーカソイドの約43〜70%に存在する優勢なMHCクラスII対立遺伝子(52))によって提示されるNY−ESO−1ペプチドを認識した。更に、HLA DP4ハプロタイプは、NY−ESO−1に対し高力価のAbを産生するメラノーマ患者の91%(11人のうち10人)が有していたが、NY−ESO−1陽性腫瘍を有し、検出可能なAbをプロセシングしない3人の患者では発現されなかった。インビトロ刺激の結果は、HLA DP−4拘束T細胞が、NY−ESO−1 Abを有する6人の患者のうち5人の患者から産生できることを示した。これらの結果は、DP4の状況において、CD4+T細胞によるNY−ESO−1の認識は、この抗原に対し抗体応答を開始するこれらの患者の能力と関連しうることを示唆した。
【0076】
癌の予防又は阻害の方法において、NY−ESO−1 MHCクラスII拘束T細胞エピトープは、静脈内、筋肉内、皮下、皮膚内、腹腔内、クモ膜下、胸膜腔内、子宮内、直腸、膣、局所、腫瘍内などを含むがそれらに限定されない幾つかの経路の1つを介し投与できる。
【0077】
投与は、経粘膜又は経皮膚手段によってされてもよい。経粘膜又は経皮膚投与のために、透過すべきバリアに適切な浸透剤が製剤において使用される。このような浸透剤は一般的に当業界公知であり、例えば、経粘膜のための胆汁塩及びフシジン酸誘導体が挙げられる。更に、界面活性剤が透過を容易にするために使用されうる。経粘膜投与は、例えば、鼻腔スプレー、又は坐剤によってされ得る。経口投与の場合、癌ペプチド、腫瘍抗原、その部分又は変異体が、カプセル、錠剤及びトニック(tonic)などの通常の経口投与形態に製剤化される。
【0078】
一般的に、レシピエントに、レシピエントの体重1kg当たり少なくとも約1ng、好ましくは体重1kg当たり少なくとも約1mg、より好ましくは体重1kg当たり少なくとも約10mg又はそれ以上のNY−ESO−1 MHCクラスII拘束T細胞エピトープの投与量を提供することが望ましい。体重1kg当たり約1mg乃至約100mgの範囲が好適であるが、より低い又はより高い投与量も投与できる。投与量は、NY−ESO−1 MHCクラスII特異的CD4+Tリンパ球のクローナル増殖(clonal expansion)をプライムし、刺激し、及び/又は引き起こすのに有効であり、次いで、それは、レシピエントの癌の予防又は阻害をすることができる。
【0079】
投与は、少なくとも1回行われ、またボーラス又は連続的投与として与えられることができる。数週間乃至数ヶ月の期間での複数回の投与は好ましくありうる。その後の投与は、示されたように投与されうる。
【0080】
治療方法において、NY−ESO−1クラスII拘束T細胞エピトープを含むワクチンは、哺乳動物において癌を予防するのに有効な量、又は局所的な癌を有する哺乳動物において転移を予防するのに有効な量で哺乳動物に投与される。必要に応じて、ワクチンは、細胞傷害性Tリンパ球を刺激するために、複数の異なるNY−ESO−1 MHCクラスII拘束T細胞エピトープ及び/又はNY−ESO−1 クラスI拘束癌ペプチド又はエピトープを含みうる。
【0081】
腫瘍を有する動物における腫瘍負荷を減少させる方法において、本方法は、腫瘍負荷の部位にNY−ESO−1 MHCクラスII拘束T細胞エピトープの有効量を投与することを含み、該量は、該部位で腫瘍のサイズを減少させるのに有効であり、かつ腫瘍部位からの転移を阻害しうる。
【0082】
治療方法の別の実施態様において、NY−ESO−1 HLA−DP拘束T細胞エピトープを含む免疫原は、NY−ESO−1 HLA−DP拘束CD4+Tリンパ球と抗NY−ESO−1抗体を誘発するのに有効な量で哺乳動物に投与される。免疫原は、単独で、又はアジュバント、免疫調節剤などと組み合わせて提供されうる。
【0083】
治療の別の方法において、自己リンパ球又は腫瘍浸潤リンパ球が、癌患者から得られうる。リンパ球は、培養で増殖され、そして癌エピトープ特異的CD4+リンパ球は、NY−ESO−1 MHCクラスII拘束T細胞エピトープ単独の存在下で、又はサイトカインの少なくとも1つの免疫刺激分子と組み合わせての存在下で、培養することによって増殖される。次いで、エピトープ特異的CD4+リンパ球は、患者の癌を減少又は除去するのに有効な量において、単独で、又はエピトープと組み合わされて、注入によって患者に戻される。
【0084】
免疫化後、ワクチンの効力は、抗体力価、特異的な溶解活性、特異的サイトカイン産生、腫瘍退行、又はこれらの組み合わせによって評価されるように、NY−ESO−1 MHCクラスII T細胞エピトープを認識する免疫細胞の産生によって評価されうる。免疫化される哺乳動物が既に癌又は転移性癌に罹患しているならば、ワクチンは、免疫調節剤(例えばIL−2、IL−6、IL−10、IL−12、IL−15、インターフェロン、腫瘍壊死因子など)、シスプラチナムなどの化学療法剤、ガンシクロビルなどの抗ウイルス剤、アンホテリシンB、抗生物質などのような他の治療と共に投与されうる。
【0085】
本発明の別の局面は、NY−ESO−1のMHCクラスII拘束T細胞エピトープをコードするNY−ESO−1遺伝子のDNA配列である。
【0086】
一実施態様では、DNA配列は、CD4+Tリンパ球によって認識されるMHCクラスII拘束T細胞エピトープをコードする配列番号1又は2の部分及びその機能的に等価な配列変異体を含む。また、MHCクラスII拘束T細胞エピトープをコードする配列番号1又は2の部分と相補的な核酸配列及び反相補的な核酸配列も本発明に包含される。
【0087】
一実施態様では、DNA配列は、配列番号4〜21又は29〜34の少なくとも1つを含むMHCクラスII拘束T細胞エピトープをコードする。
【0088】
別の実施態様では、MHCクラスII拘束T細胞エピトープをコードするDNA配列は、以下:
CAG GAT GCC CCA CCG CTT CCC GTG
CCA GGG GTG CTT CTG AAG GAG TTC
ACT GTG TCC GGC AAC ATA CTG ACT
ATC CGA CTC(配列番号24)又はその機能性部分又は変異体
を含む。
【0089】
別の実施態様では、MHCクラスII拘束T細胞エピトープをコードするDNA配列は、以下:
AGA CCA CCG CCA ACT GCA GCT
CTC CAT CAG CTC CTG TCT CCA GCA
GCT TTC CCT GTT GAT(配列番号25)又はその機能性部分又は変異体を含む。
【0090】
別の実施態様では、DNA配列は、以下:
TGG ATC ACG CAG TGC TTT CTG CCC
GTG TTT TTG GCT CAG CCT CCC
TCA GGG CAG AGG CGC(配列番号26)又はその機能性部分又は変異体を含む。
【0091】
本発明の別の局面は、NY−ESO−1のHLA−DP拘束CD4+T細胞エピトープをコードするNY−ESO−1遺伝子のDNA配列である。
【0092】
一実施態様では、DNA配列は、CD4+Tリンパ球によって認識されるHLA−DP拘束T細胞エピトープをコードする1つ以上の配列番号51〜64及びその機能性等価配列変異体をコードする核酸配列を含む。HLA−DP拘束T細胞エピトープをコードする核酸配列と相補的及び反相補的な核酸配列も本発明に包含される。
【0093】
包括的な暗号(generic code)の縮重に起因して、DNA配列の変異により、等価なNY−ESO−1エピトープの翻訳がなされよう。結果として、置換によって、NY−ESO−1癌抗原HLA−クラスII拘束CD4+T細胞によって認識されるNY−ESO−1エピトープの発現が生じる限り、置換は本発明の範囲に含まれる。
【0094】
NY−ESO−1遺伝子に由来するオープンリーディングフレームDNA配列の全て又は部分は、他の哺乳動物種におけるNY−ESO−1 MHCクラスII拘束T細胞エピトープのホモログを同定、単離するために、プローブとして使用できる。一実施態様では、ヒトcDNA配列は、マウスホモログ核酸配列のために哺乳動物cDNAライブラリーをスクリーニングするために使用される。陽性クローンを選択し、配列決定する。cDNAライブラリーが合成できる組織源の例としては、真皮、表皮、固形腫瘍、メラノーマ、メラノサイトなどが挙げられるがそれらに限定されない。当業者は、ホモログを検出するのに使用される適切なハイブリダイゼーション条件を理解しよう。ライブラリーの核酸ハイブリダイゼーション構築のための通常の方法及びクローニング技術は、Sambrook et al, (eds)(1989)“Molecular Cloning. A Laboratory Manual” Cold Spring Harbor Press, Plainview, New York; Ausubel al(eds) “Current Protocols in Molecular Biology”(1987), John Wiley and Sons, New York, New Yorkに記載されている。
【0095】
本発明の別の局面は、癌を有する哺乳動物から単離された生物学的サンプルにおける本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープを転写するRNAの検出と定量のための核酸プローブである。コントロールRNAサンプルに対するRNAレベルの変化は、哺乳動物における該疾病の診断や予後に有用である。
【0096】
一実施態様では、mRNAは、癌又は前癌を有すると疑われる患者の組織から得られ、そして健康なコントロール患者由来のmRNAと比較される。コントロールと比較した場合、患者における本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードするmRNAの定量的及び/又は定性的増大は、患者における癌又は前癌の指標となる。mRNAは、mRNAとハイブリダイズできるオリゴヌクレオチドプローブを用いて検出できる。
【0097】
本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードする配列に基づくオリゴヌクレオチド対の組み合わせは、選択されたRNA配列を増大させる逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)方法を用いて、生物学的サンプルにおけるmRNAを検出するためのPCRプライマーとして使用できる。本発明はまた、NY−ESO−1 MHCクラスII拘束T細胞エピトープをコードするDNAとRNAの検出のためのインサイチュPCRとインサイチュRT−PCRを包含する。標的核酸のコピー数が非常に小さいとき、又は核酸の異なる形態を区別しなければならないとき、該技術は好適である。該方法は、正常細胞から前癌細胞及び癌細胞を検出し、区別するのに特に有用である。
【0098】
本発明はまた、mRNA上のある相補(「センス」)領域と結合し、NY−ESO−1の合成阻害に至るアンチセンスオリゴヌクレオチドを包含する。このようなアンチセンスオリゴヌクレオチドは、約12乃至約25モノヌクレオチドの1本鎖核酸であり、本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードする配列に対しアンチセンスである。このようなアンチセンスオリゴヌクレオチドは、Molecular Biology and Biotechnology R.A.Meyers(編 VCH Publishers,Inc., New York, NY, 1995, pp.38-44のUhlmann,E.ら, Antisense oligonucleotides, structure and function に記載のように、当業界公知の方法によって製造できる。
【0099】
本発明はまた、少なくとも1つ以上のNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードするDNA配列を含むベクターを包含する。ベクターは、1つ以上の変異体NY−ESO−1 MHCクラスII拘束T細胞エピトープを有する完全長NY−ESO−1タンパク質をコードするDNA配列を含みうる。必要に応じて、ベクターはまた、少なくとも1つの免疫刺激分子をコードするDNA配列を含んでもよい。ベクターはまた、少なくとも1つ以上のNY−ESO−1 MHCクラスI拘束T細胞エピトープをコードするDNA配列を含んでもよい。ベクターはまた、細胞や組織におけるNY−ESO−1 MHCクラスII拘束T細胞エピトープの局在を検出するのに使用される緑色蛍光タンパク質をコードする遺伝子を含んでもよい。
【0100】
真核発現ベクターとしては、レトロウイルスベクター、ワクシニアウイルスベクター、アデノウイルスベクター、ヘルペスウイルスベクター、鶏痘ウイルスベクター、バキュロウイルスベクター、ヒトパピローマウイルスベクター、ウマ脳炎ベクター、インフルエンザウイルスベクターなどが挙げられるがそれらに限定されない。
【0101】
本発明は、遺伝子、そのフラグメント又は変異体のオープンリーディングフレーム核酸配列によってコードされた少なくとも1つのNY−ESO−1 MHCクラスII拘束T細胞エピトープを発現する新規組換えウイルスを包含する。組換えウイルスはまた、少なくとも1つの免疫刺激分子を発現してもよい。組換えウイルスは、哺乳動物、特にヒトにおける癌の予防又は治療の目的のために、哺乳動物において体液性免疫応答を誘発又はアップレギュレートできる。
【0102】
組換えウイルスに感染した宿主細胞は、1つ以上のNY−ESO−1 MHCクラスII拘束T細胞エピトープを単独で、又は少なくとも1つの免疫刺激分子と組み合わせて発現する。宿主細胞はまた、HLAクラスII分子を発現する組換えウイルスに感染されてもよい。
【0103】
組換えワクシニアウイルスベクターから外因性遺伝子産物を構築、発現する方法は、Perkusら Science 229:981-984, 1985; Kaufmanら Int.J.Cancer 48:900-907,1991; Moss Science 252:1662,1991; Smith及びMoss Bio Techniques Nov/Dec, p.306-312,1984;並びに米国特許第4,738,846号に開示されている。Sutter及びMoss(Proc.Nat'l Acad Sci. U.S.A. 89:10847-10851,1992)及びSutterら(Virology 1994)は、本発明でウイルスベクターとして使用できる非複製組換えアンカラウイルス(MVA,修飾ワクシニアアンカラ)のベクターとしての構築と使用を開示する。Baxby及びPaoletti(Vaccine 10:8-9, 1992)は、本発明のウイルスベクターとして使用のための、カナリア痘ウイルス、鶏痘ウイルス及び他のトリ種を含む非複製ポックスウイルスのベクターとしての構築と使用を開示する。
【0104】
本発明のベクターは、NY−ESO−1 MHCクラスII拘束T細胞エピトープの発現のために適切な宿主細胞中に置かれうる。真核宿主細胞株としては、COS細胞、CHO細胞、Hela細胞、NIH/3T3細胞、昆虫細胞、樹状細胞などの抗原提示細胞などが挙げられるがそれらに限定されない。必要に応じて、宿主細胞はまた、刺激分子を発現してもよい。宿主細胞が、少なくとも1つのMHC(又はHLA)クラスII分子と組み合わせてNY−ESO−1 MHCクラスII拘束T細胞エピトープの両方を発現する場合、真核発現系は、適切なグリコシル化を可能にするように使用されるのが好ましい。宿主細胞による癌エピトープと免疫刺激分子の両方の発現は、抗原認識、及び抗原特異的T細胞の増殖又はクローナル増殖において助けとなるように、特異的T細胞に必要なMHCクラスII拘束ペプチドを提供し、T細胞に適切なシグナルを提供する。全体としての結果は、免疫系のアップレギュレーションである。免疫応答のアップレギュレーションは、癌又は前癌細胞の増殖の阻害のための、癌抗原特異的CD4+リンパ球の増加及び体液性免疫の他のエフェクター細胞の増加によって明白となる。
【0105】
DNAは、当業界公知の方法によるトランスフェクション、形質導入、リポソームなどによって宿主細胞に挿入できる(Sambrook et al, 1989, “Molecular Cloning A Laboratory Manual”, Cold Spring Harbor press, Plainview, New York)。リポソームの場合、Nabel,E.G.et al, 1992, Hum.Gene.Ther. 3:367-275; Nabel,G.J. et al, 1992, Hum.Gene.Ther. 3:649-656; Stewart,M.J. et al 1992, Hum.Gene.Ther. 3:399-410; Nabel,G.J. et al 1993 Proc.Natl.Acad.Sci.USA90:11307-11311;及びHarrison,G.S. et al 1995 Bio Techniques
19:816-823に開示のように、カチオン性脂質が好ましく、例えば、中性リン脂質ジオレオイルホスファチジルエタノールアミン(DOPE)と複合した、ポリカチオン性脂質であるジミリスチルオキシプロピル−3−ジメチル−ヒドロキシエチルアンモニウム(DMRIE)が好ましい。
【0106】
宿主細胞によって発現される組換えNY−ESO−1 MHCクラスII拘束T細胞エピトープは、当業界公知の標準的タンパク質精製方法によって細胞溶解液又は細胞上清から精製できる。これらとしては、分子ふるいクロマトグラフィー、イオン交換クロマトグラフィー、等電点フォーカシング、ゲル電気泳動、アフィニティクロマトグラフィー、HPLC、逆相HPLCなどが挙げられるがそれらに限定されない(Ausubel et al, 1987, Current Protocols in Molecular Biology, John Wiley and Sons, New York, NY)。イムノアフィニティクロマトグラフィーも、イムノアフィニティ剤として、本明細書記載の抗癌タンパク質抗体又はその抗原結合フラグメントを用いて精製のために使用できる。
【0107】
組換えウイルスはまた、治療薬又はワクチンとして使用できる。このような使用において、より低い、又はより高い投与量も投与できるが、レシピエントに、哺乳動物1mg当たり約105乃至約1010プラーク形成ユニットの範囲における組換えウイルスの投与量を提供することが望ましい。
【0108】
組換えウイルスベクターは、メラノーマなどの癌の如何なる証拠の前に、又はメラノーマなどの癌に罹患した哺乳動物における該疾病の退行を媒介するために、哺乳動物に導入できる。哺乳動物にウイルスベクターを投与する方法の例としては、エキソビボで細胞を組換えウイルスに曝すこと、又は冒された組織内に組換えウイルスを注入すること、又はウイルスの静脈内、皮下、皮内、筋肉内などへの投与が挙げられるがそれらに限定されない。あるいは、組換えウイルスベクター又は組換えウイルスベクターの組み合わせは、癌病巣中への直接的注入、又は適切な医薬として許容できる担体中での局所適用によって、局所投与されることができる。問題の核酸配列を担持する組換えウイルスベクターの量は、ウイルス粒子の力価に基づく。免疫化のための好適な範囲は、哺乳動物、好ましくはヒト当たり約105乃至1010ウイルス粒子である。
【0109】
本発明は、少なくとも1つのNY−ESO−1 MHCクラスII拘束T細胞エピトープをコードするDNA配列の1つ以上のコピーをゲノム中に組み込まれたトランスジェニック動物を提供する。トランスジェニック動物作出の一般的方法は、Krimpenfortら,米国特許第5,175,384号、Leder ら,米国特許第5,175,383号、Wagnerら,米国特許第5,175,385号、Evansら,米国特許第4,870,009号、及びBernsら,米国特許第5,174,986号に記載されている。遺伝子の組み込みにより、NY−ESO−1 MHCクラスII拘束T細胞エピトープの過剰発現、発現の変化、又はNY−ESO−1 MHCクラスII拘束T細胞エピトープのマルチプル形態もしくは変異体の発現が生じる。得られるトランスジェニック動物は、癌の進行又は本発明の腫瘍抗原の研究において有用である。動物モデルは、癌治療のためのワクチンや化学療法剤をスクリーニングするのに有用である。トランスジェニック動物はまた、癌の進行の研究において有用である。
【0110】
本発明は更に、本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープによる免疫化によって誘発される抗体又は抗原結合部分を含む。NY−ESO−1 MHCクラスII拘束T細胞エピトープがほんの数個のアミノ酸からなる場合、エピトープは、抗体応答を誘発するために、キャリアタンパク質に結合されてもよい。KLH、破傷風トキソイド、アルブミンなどのキャリアタンパク質及び結合方法は、当業界公知である。抗体は、インタクトなNY−ESO−1タンパク質及びNY−ESO−1タンパク質の天然のプロセスされた形態と同様に、本発明のNY−ESO−1 MHCクラスII拘束T細胞エピトープに対する特異性を有し、それらと反応もしくは結合する。
【0111】
典型的な抗体分子は、インタクトなイムノグロブリン分子、実質的にインタクトなイムノグロブリン分子、又は抗原結合部位を含むイムノグロブリン分子のこれらの部分(F(ab)、F(ab’)、F(ab’)2、ヒト化キメラ抗体、及びF(v)のような当業界公知のイムノグロブリン分子の部分を含む)である。ポリクローナル又はモノクローナル抗体は、当業界公知の方法によって製造できる(Kohler and Milstein (1975) Nature 256,495-497; Burdonら(編)(1985) “Laboratory Techniques in Biochemistry and Molecular Biology”, Vol.13, Elsevier Science Publishers, アムステルダム中のCambell “Monoclonal Antibody Technology, the Production and Characterization of Rodent and Human Hybridomas”)。抗体又は抗原結合フラグメントはまた、遺伝子工学によって製造できる。重鎖及び軽鎖遺伝子両方の発現のための技術は、PCT特許出願公開番号WO 901443、WO 9014424、Huse et al(1989)Science 246:1275-1281及び米国特許第4,946,778号の主題である。1つ以上の相補的決定領域を有するヒト化イムノグロブリン及び該抗体の製造方法は、米国特許第5,585,089号及び第5,530,101号に開示されている。
【0112】
一実施態様において、本発明の抗体は、生物学的サンプルにおけるMHCクラスII拘束T細胞エピトープを含むNY-ESO-1ペプチド又は部分を検出するイムノアッセイで使用される。抗体又はその抗原結合フラグメントは、癌に冒された哺乳動物からの組織生検サンプル中で、癌ペプチドを検出するために使用されることができる。病気の組織内でのNY−ESO−1 MHCクラスII拘束T細胞エピトープの評価は、哺乳動物での病気の悪化を予測するために使用できるか、又は治療の効力を診断しうる。イムノアッセイは、ラジオイムノアッセイ、ウエスタンブロットアッセイ、免疫蛍光アッセイ、酵素免疫アッセイ、化学発光アッセイ、免疫組織化学アッセイなどであり得、そしてインビトロ、インビボ、又はインサイチュで行うことができる。ELISAについて当業界公知の標準的技術は、“Methods in Immunodiagnosis”(第2版), Rose及びBigazzi(編), John Wiley & Sons, 1980; Campbellら,“Methods and Immunology”, W.A.Benjamin, Inc., 1964;及びOellerich,M. 1984, J.Clin.Chem.Clin.Biochem. 22:895-904に記載されている。免疫組織化学の通常の方法は、Harlow及びLane(編)(1988) “Antibodies A Laboratory Manual”, Cold Spring Harbor Press, Cold Spring Harbor, New York; Ausbelら(編)(1987) Current Protocols In Molecular Biology, John Wiley and Sons (New York, NY)に記載されている。このような検出アッセイのために適切な生物学的サンプルとしては、細胞、組織生検材料、全血、血漿、血清、痰、脳脊髄液、胸膜液、尿などが挙げられるがそれらに限定されない。
【0113】
本発明の抗体又は抗原結合フラグメントはまた、免疫治療でも使用できる。抗体又はその抗原結合フラグメントは、癌の重篤度、程度又は持続時間を防止、軽減又は減衰させるために十分な量で哺乳動物に提供される。
【0114】
引用された全ての論文及び特許は、引用により本明細書に含まれるものとする。
【0115】
本発明を、ある特定の実施態様に関し上に記載するが、多数のバリエーションが可能で、代わりの材料や試薬を、本発明を逸脱すること無しに使用できることが理解されよう。幾つかの場合に、このようなバリエーション及び置換は何らかの実験を必要とするかも知れないが、慣用的な試験のみを含むであろう。
【0116】
特定の実施態様の上の記載は、本発明の一般的な性質を十分に示し、他の者は、現在の知識を適用することによって、種々の適用のために、一般的概念を逸脱すること無しに、このような特定の実施態様を容易に改変し、及び/又は採用できる。それ故、このような改変や修飾は、開示された実施態様の等価物の意味内と範囲内に包含されるように意図される。
【実施例】
【0117】
実施例1
材料および方法
組換えNY−ESO−1タンパク質の生成および解析:完全長NY−ESO−1遺伝子をコードする細菌発現ベクターを構築するために、本発明者らは一対のプライマー、NdeI部位を含むESO−5p(5’GCTCCGGACATATGCAGGCCG AAGGCCGGGG)(配列番号35)およびXhoI部位を含むESO−3p(5’AAGGGGCTCGAGGCT GGGCTTAGCGCCTCT)(配列番号36)を用いて、PCRフラグメントを作製した。PCR産物の制限酵素による消化およびゲル精製の後、NY−ESO−1をコードするDNAフラグメントをポリヒスチジンペプチドをコードするDNAとpET−28(+)(Novagen,マジソン,WI)内でインフレームに融合させた。同様のストラテジーを、最初の74のアミノ酸残基のみを含むトランケートNY−ESO−1であるESO1−74の発現ベクターを構築するのにも使用した。正確なプラスミド構築物を保持するE.coli株BL21(DE3)を37℃で対数期まで増殖させ、次いで、イソプロピル−d−チオガラクトシド(IPTG)を終濃度0.5mMまで添加し3時間振盪することによってタンパク質産生を誘導した。細菌抽出物の可溶性画分を得た。またNY−ESO−1をNi2+アフィニティークロマトグラフィーにより精製した。精製タンパク質のSDS−PAGE解析を先の報告(25)のように行った。精製タンパク質のN末端配列を自動Edman分解により決定した。
【0118】
血清およびPBMC:転移性メラノーマの患者由来の血清を−80℃で保存した。正常ドナーの血清をNIHのクリニカルセンターの血液バンクから得た。転移性メラノーマの患者TEのMHCクラスII遺伝子型はHLA−DR1*0401,1*1501であった。この患者をgp100:209−217(210M)ペプチド+高用量のIL−2で処置し、目的である腫瘍退行の実験に付した。
【0119】
NY−ESO−1タンパク質に対する抗体の検出:50 lのPBST(0.1%Tween20を含むリン酸緩衝化生理食塩水)に希釈した約50ngの精製NY−ESO−1タンパク質を96ウェルMaxiSorpプレート(Nunc,デンマーク)の各ウェルに室温で終夜吸着させた。コントロールプレートは1ウェルあたり150ngのBSAでコートした。プレートをPBST中5%ドライミルクで少なくとも2時間ブロックし、洗浄し、そして100 lの希釈血清サンプルをロードした。全ての血清サンプルはPBST中3%ドライミルクを用いて1:25、1:250、および1:2500に希釈した。この3つの異なる希釈の各サンプルをNY−ESO−1コートしたプレートおよびBSAコートしたプレート上にロードした。室温で1時間インキュベーションした後、プレートを洗浄し、PBST中1%ドライミルクで希釈した二次抗体(西洋ワサビペルオキシダーゼを結合したヤギ抗ヒトIgG,Sigma Co.,セントルイス,MO)をロードした。プレートを0.5時間インキュベーションした後発色させ、450nmでの吸光度をELISAリーダー(Dynatech,シャンティイー,VA)を用いて読み取った。陽性反応を、1:25と1:250の両方の血清希釈で、正常ドナーの平均O.D.値+標準偏差の3倍を超えるNY−ESO−1に対するO.D.値として定義した。ウエスタンブロットを(24)の記載のように行い、いくつかの代表的血清サンプルにおける抗体の特異性を確認した。
【0120】
細胞株および抗体:メラノーマ株F049およびF050は、NCIの外科のAdam Rikerから提供された細針状アスパレート(asparate)サンプルの初期培養物であった。他の全てのメラノーマ株およびEBV B株は10%ウシ胎仔血清(Biofluid,Inc.,ゲーサーズバーグ,MD)を補充したRPMI1640(Life Technologies,ロックヴィル,MD)で作製し維持した。293IMDR1および293IMDR4をヒトインバリアント鎖であるDMA、DMBおよびDR分子を発現するよう遺伝子組換えし、10%ウシ胎仔血清を補充したRPMI1640で培養した(15)。マウスリンパ球の培養培地は、Hyclone Inc.(ローガン,UT)より供給される、0.05mMメルカプトエタノール、5CU/ml IL−2および10%ウシ胎仔血清を含むRPMI1640だった。ヒトT細胞培養に用いた培地は、Sigma Co.(セントルイス,MO)により供給される、0.05mMメルカプトエタノール、50CU/ml IL−2および10%ヒトAB血清を含むRPMI1640だった。抗体ブロッキング実験を先の記載(15)のように行った。ハイブリドーマHB55およびHB95はAmerican Type of Cell Culture(ATCC,マナッサス,VA)から得た。コントロール抗体はPharmingen Int.(サンディエゴ,CA)から購入した。
【0121】
トランスジェニック動物および免疫手順:HLA−DR4トランスジェニック(DR4−Tg)マウスはマウスクラスII欠損であり、HLA−DR−IE−およびHLA−DR1*0401−IE−キメラ分子を発現した(26)。初代マウスはCase Western Reserve UniversityのPaul Lehmannを通じて得た。マウスは近交系であり、Biocon Inc.(ロックビル,MD)で維持された。6〜10週齢の雌性マウスを完全長組換えNY−ESO−1タンパク質で免疫した。精製タンパク質(約50μg)を完全フロインドアジュバント(CFA)中に懸濁し、均等に分け、後肢足蹠および尾の付け根に皮下注射して各マウスに投与した。注射11日後、マウスを屠殺し、そして両方の後肢膝窩および鼠径部リンパ節を採取した。単一の細胞懸濁液を2匹の免疫動物のリンパ節から得、インビトロ刺激を行った。
【0122】
ペプチド合成:本研究で使用した合成ペプチドはNCIの外科のペプチド合成機(Gilson Co. Inc.,ワージントン,OH)で固相法を用いて作製した。各ペプチドの純度は質量分析(Bio−synthesis, Inc.,ルイスヴィル,TX)により評価した。
【0123】
インビトロ感作(IVS)手順およびサイトカイン放出アッセイ:サイトカイン放出アッセイを行う前1週間、終濃度10Mのペプチドを2.5×105マウスリンパ球と混合した。ヒトPBMCのIVSのために、2.5×105細胞を10M濃度のペプチドでパルスし、平底96ウェルプレートの各ウェル中でインキュベートした。2回のインビトロ刺激の後、細胞を様々な標的に対して試験し、上清をサイトカイン放出アッセイのために収集した。ヒトT細胞の急速な増殖およびクローニングは記載(20)のように行った。
【0124】
終濃度10Mのペプチドまたは終濃度5g/mlのタンパク質を標的細胞にパルスした。4時間のインキュベーション後、細胞を無血清RPMI培地中で洗浄し、およそ3×104の標的細胞を同数のTE4−1細胞とともに終夜インキュベーションし、サイトカイン放出をヒトについてはGM−CSF ELISAキット(R&D Systems,ミネアポリス,MN)、またはマウスについてはIFNキット(Endogen, Inc. ウォバーン,MA)を用いて測定した。ヒトIFN−,IL−10,TNF−,およびIL−4等の他のサイトカインは、製造業者の説明書に従って、Endogen Inc.またはR&D SystemsのELISAキットを用いて測定した。
【0125】
実施例2
組換えNY−ESO−1タンパク質およびNY−ESO−1反応性抗体の検出
NY−ESO−1反応性抗体およびCTLは、癌の患者において報告されている(19〜22)。従って、NY−ESO−1特異的CD4+T細胞がNY−ESO−1抗原に対する抗体の発生およびCTLの調整に関与しているかもしれないようである。しかし、NY−ESO−1由来のCD4+T細胞エピトープはこれまで報告されていない。MHCクラスII拘束CD4+T細胞エピトープを同定するために、本発明者らは、細菌発現系からNY−ESO−1タンパク質を出発物質として精製することから始めた。NY−ESO−1発現およびタンパク質精製を促進するため、NY−ESO−1をコードするcDNAフラグメントを、pET28発現ベクターのN末端に位置するポリヒスチジンタグにインフレームで融合し、組換えタンパク質の高レベル産生を得た。数ミリグラムのNY−ESO−1タンパク質をNi2+荷電アフィニティークロマトグラフィーカラムを使用して精製した。精製タンパク質はSDSポリアクリルアミドゲル上およそ26kDaの見かけの分子量を示した(図2A)。精製タンパク質の同定を確認するために、タンパク質のN末端マイクロシークエンシングを自動Edman分解により行った。Edman分解により得られた25アミノ酸残基すべては予測したアミノ酸配列と一致した(データ示さず)。最初の74アミノ酸残基を含む短い形のNY−ESO−1(ESO1−74)もまた同じアプローチにより精製した(図2A)。
【0126】
メラノーマ患者がNY−ESO−1タンパク質に対する抗体を生じたかどうかを決定するために、NCIの外科において癌ワクチン治療プロトコルに登録された88人の転移性メラノーマ患者由来の血清をスクリーニングした。8人の正常ドナー由来の血清をスクリーニングのコントロールとして使用した。88人の患者のうち11人(13%)が、高力価のNY−ESO−1に対する抗体を持っていることが分かった(図2C)。このデータは他の群により得られた結果と一致した(22)。患者の血清が精製NY−ESO−1タンパク質に存在するわずかな夾雑物と反応する可能性を除去するため、代表的な血清サンプルを用いてウエスタンブロットを行った。図2Bは、患者由来のNY−ESO−1反応性の血清が、NY−ESO−1発現細菌の細胞溶解物および精製NY−ESO−1タンパク質だけと反応し、コントロールベクターを含む細菌由来の抽出物とは反応しなかったことを示した。非反応性の血清サンプルもまた試験した(図2B、レーン4,5,6)。
【0127】
実施例3
HLA−DR4−トランスジェニックマウス由来の推定MHCクラスII拘束エピトープの同定
CD4+T細胞エピトープを同定するために、DR4トランスジェニックマウスをCFA中およそ50gの完全長NY−ESO−1タンパク質で、尾の付け根および後肢足蹠に免疫した。注射後11日目に、2匹の免疫したマウスの両後肢膝窩および鼠径部リンパ節から得た単一の細胞懸濁液を調製し、HLA−DR4分子の推定ペプチド結合能に基づくNY−ESO−1タンパク質由来合成ペプチドでのインビトロ感作に使用した(27)。
【0128】
HLA−DR4に結合することが予測されるアミノ酸配列セグメントを含む8つの高結合ペプチドをインビトロ感作実験に使用した。最初のインビトロ感作後6日目に、マウスリンパ球を、ヒトHLA−DR4陽性1359EBV B細胞のみおよび刺激に使用した対応するペプチドでパルスした1359EBV Bに対するサイトカイン放出について試験した。3つのペプチドは、T細胞からのサイトカイン分泌に基づきマウスT細胞により認識されたが、他の5つのペプチドは認識を示さなかった(図3)。ESO p116−135は陽性ペプチドのうちで最も強い活性を示した。これは、このペプチドが、T細胞認識のためにHLA−DR4分子により提示されるエピトープを含んでいるかもしれないことを示唆している。従って、このペプチドをさらなる解析のために選択した。
【0129】
実施例4
NY−ESO−1特異的なヒトCD4+T細胞の作製
NY−ESO−1に対する高力価の抗体を持っていた患者TE(図2C)由来のPBMCを、ESO p116−135ペプチドを用いたインビトロ刺激に使用した。インビトロ刺激の1週間後、患者TE由来のPBMCは著しい増殖を示した。IL−2を刺激の2週目に添加した。このように樹立した細胞株をTE4−1と命名し、これを20 CU/mlのIL−2の存在下、2週間超、増殖させ続けた。TE4−1 T細胞は、FACS解析に基づくと、90%CD4+T細胞であった。TE4−1はTh1型CD4+T細胞を含んでいた。というのは、それらがGM−CSF、IFN−およびTNFを分泌し、IL−10またはIL−4を分泌しなかったからである(データ示さず)。CD8+T細胞を数%減少させた後、CD4+T細胞の精製集団は依然その反応性を保持していた。TE4−1細胞株由来のT細胞クローンにもESO p116−135ペプチドを認識することを示すものもあった(データ示さず)。
【0130】
TE4−1は、HLA−DR4との関連で完全長NY−ESO−1タンパク質およびESO p116−135ペプチドでパルスしたEBV B細胞は認識したが、最初の74アミノ酸を含むトランケートNY−ESO−1タンパク質でパルスしたEBV B細胞を認識しなかった(図4A)。TE4−1細胞株もまたNY−ESO−1をコードするアデノウイルスに感染させたDR4陽性樹状細胞とは特異的に反応性であったが、緑色蛍光タンパク質をコードするアデノウイルスに感染させたDR4陽性樹状細胞とは特異的に反応性ではなかった(データ示さず)。
【0131】
TE4−1により認識されるT細胞がHLA−DR4により拘束されるかどうかを試験するため、2つの重複するペプチド(ESO p116−135およびESO p111−130)ならびにコントロールペプチド(ESO p91−110)を無血清培地中293IMDR1および293IMDR4細胞にパルスした。細胞を洗浄し続いてTE4−1細胞と終夜インキュベートした。図4Bに示されるように、ペプチド116−135およびペプチド111−130の両方がHLA−DR4との関連でTE4−1により認識された。興味深いことに、ペプチド116−135もまた、293IMDR1細胞にパルスしたとき、T細胞からのサイトカイン分泌を刺激することができた。コントロールESO p91−110ペプチドでパルスした293IMDR4では活性は検出されなかった(図4B)。ESO−p116−135パルス293IMDR4の認識は抗HLA−DR抗体(HB55)により完全に阻害されたが、コントロールおよび抗HLAクラスI抗体(HB95)では阻害されなかった(図4C)。gp100特異的CD8+T細胞株(CTL−C3G1)およびHLA−DR1拘束CD4+T細胞株(T3−80)を抗体ブロッキングの特異性コントロールとして使用した。
【0132】
実施例5
TE4−1による腫瘍細胞の認識
ペプチド特異的CD4+およびCD8+T細胞活性は推定腫瘍抗原に対してしばしば生じ得たが、多くの場合には、腫瘍反応性は、T細胞の低親和性または腫瘍細胞表面上で天然にプロセシングされるペプチドの提示の失敗のいずれかに起因して示され得なかった(3)。TE4−1が腫瘍細胞により天然にプロセシングされ提示されるNY−ESO−1エピトープを認識することができたかどうかを試験するために、いくつかのメラノーマ株を標的として使用した。各株におけるNY−ESO−1の発現はRT−PCRにより決定し、HLA−DR対立遺伝子の発現はFACS解析により決定した(データ示さず)。図5に示されるように、TE4−1はNY−ESO−1/HLA−DR4陽性腫瘍(1359melおよびF049mel)を認識することができたが、腫瘍細胞株397melおよび624.38mel(NY−ESO−1+/HLA−DR-)、ならびに526mel(NY−ESO−1-/HLA−DR4+)は認識することができなかった。興味深いことに、TE4−1もまたF050mel(DR1+/NY−ESO−1+)を認識したが、DR1および低レベルのNY−ESO−1を発現する1300melは認識しなかった。1つの可能な説明は、CD4+T細胞が異なるDR分子により提示される同一のペプチドを認識するかもしれないということである。F049melの認識は、抗HLA−DR抗体の存在下特異的にブロックされ得るが、抗MHCクラスI抗体の存在下にはされない(データ示さず)。これらの研究は、TE4−1細胞株が腫瘍細胞表面で天然にプロセシングされたペプチドを認識することを示唆した。
【0133】
実施例6
TE4−1により認識されるNY−ESO−1エピトープの特徴付け
2つの反応性ペプチドが15アミノ酸(LPVPGVLLKEFTVSG)(配列番号10)を共有するので、ペプチドの最小長を一連のN−およびC−末端トランケートペプチドを試験することによって決定した。ペプチドをDR4+1088EBV B細胞にパルスし、TE4−1細胞を刺激するその能力を試験した。128位のバリン残基がT細胞認識に重要であることが見出された(図6A)。123位のロイシン残基までのN末端欠失を有するペプチドは、T細胞認識に影響を与えなかったが、さらに欠失を有するペプチドはT細胞を刺激するその能力を一部失った。P1、P4、P6およびP7残基がMHCクラスII分子へのペプチド結合に寄与したので、123位のロイシン残基はP1アンカー残基であり得る。さらなる欠失が、MHCクラスII分子に結合するための重要な残基を決定するために必要である。
【0134】
欠失実験に基づいて本発明者らは、短い形のペプチドESO p119−130を用いて、TE4−1により認識されるペプチドの結合親和性を決定した。異なるペプチド濃度で標的として1088EBV B細胞(HLA−DR4+)にペプチドをパルスした。図5Bに示されるように、33nM以下のESO p119−130ペプチド濃度では全くまたはほとんどT細胞活性は観察されず、0.33Mのペプチド濃度で高い活性が検出され、T細胞活性は33Mのペプチド濃度でプラトーに達しなかった。コントロールペプチドは33Mのペプチド濃度でさえTE4−1により認識されなかった。
【0135】
実施例7
本発明者らは、T4−1 CD4+T細胞株がDR4およびおそらくDR1にもまた関連してESO p116−135を認識することを示した。ここで、TE4−1がHLA−DR1に関連してペプチドだけでなくタンパク質も認識できることが示される。認識は抗DR抗体によりブロックされる(図7Aおよび7B)。この結果は、ESO p116−135が雑多なペプチドであり得、DR4およびDR1に結合できることを示す。従って、このペプチドワクチンの適用可能な集団は非常に大きい。
【0136】
実施例8
HLA−DP研究の材料および方法
研究に使用した細胞株、組織培養試薬、および抗体
293CIITAは、MHCクラスIIトランスアクチベーターをコードするレトロウイルスを有する293細胞の形質導入により作製した細胞株である(53)(このレトロウイルスプラスミドはthe University of South Florida, タンパ, FLのDr. George Blancの好意である)。すべてのメラノーマ株およびEBVB株を、10%ウシ胎仔血清(Biofluid, Inc.,ゲーサーズバーグ,MD)を補充したRPMI1640(Life Technologies,ロックビル,MD)中で作製し維持した。リンパ球の培養培地は、0.05mM β−メルカプトエタノール、50CU/ml IL−2+10%ヒト男性AB血清(Valley Biochemicals Inc.(ウィンチェスター,VA)により供給された)を含むRPMI1640であった。ブロッキングアッセイに使用した抗体は以下の供給源から得た:W6/32(HLAクラスI)およびL243(HLA DR)はLoftstrand Labs Lt.(ゲーサーズバーグ,MD)により精製されたハイブリドーマ上清であった;抗体クローンIVA12(HLAクラスII)、B7/21(HLA DP)、Genox3.53およびIVD12(両方ともHLA DQ)はBeckton Dickinson Immunocytometry Systems(サンノゼ,CA)から購入した。
【0137】
プラスミドの構築
pESOプラスミドは、先の記載(45)のようにCMVプロモーターにより駆動されるNY−ESO−1cDNAを含む発現ベクターであった。pIi−ESOプラスミドを、全NY−ESO−1cDNAのNheIおよびNotI消化PCR産物を同じ酵素で消化したpTi80ベクターに挿入することで構築した(54)。NY−ESO−1cDNAを、インバリアント鎖(Ii)リーダー配列の最初の80アミノ酸残基とそのN末端でインフレームに融合した。NY−ESO−1の増幅に使用したPCRプライマーは以下の通りであった:NheI部位を含むフォワードプライマー5’cattgctagcATG CAG GCC GAA GGC CGG GGC A3’(配列番号73)およびNotI部位を含むリバースプライマー5’aaggctacattGC GGC CGC TTA GCG CCT CTG CCC TGA G3’(配列番号74)。
【0138】
ペプチドおよびCD4+T細胞の産生
本研究に使用した合成ペプチドは、National Cancer Institute,ベセズダ,MDの外科のペプチド合成機(Gilson Co. Inc.,ワージントン,OH)において、固相法を用いて作製した。脱保護後、各ペプチドの純度を、質量分析(Bio−synthesis, Inc.,ルイスヴィル,TX)により評価した。合成ペプチドを凍結乾燥し、20mg/mlでDMSO中再構築し、そして示した濃度まで希釈した。
【0139】
インビトロ感作手順を先の記載(50)のように行った。簡潔にいうと、およそ2.5×105PBMCを20μg/mlのペプチド存在下96ウェル平底プレートにプレートした。7日目と14日目に、1×105非照射PBMCを20μg/mlのペプチドでパルスし、2回洗浄し、各ウェルに加え、そして20CU/mlのIL−2を8日目、11日目、15日目、18日目に加えた。21日目、細胞を回収し、標的細胞と終夜インキュベートし、その後、上清をサイトカイン放出アッセイのために採取した。
【0140】
特異的活性を有するウェル由来のT細胞をプールし、OKT−3急速増殖法(OKT−3 rap1d expansion method)を用いて増殖させた(55)。増殖後、CD8+T細胞を磁気ビーズ選択(Dynal Inc,レイクサクセス,NY)により培養物から枯渇させ、引き続いて細胞株をCD4+およびCD8+発現に関してフローサイトメトリーにより解析した。
【0141】
サイトカイン放出アッセイ
タンパク質またはペプチドをパルスした標的を調製するために、ペプチドは終濃度20μg/mlで使用し、タンパク質は終濃度10μmg/mlで使用した。細胞を無血清RPMI培地で洗浄し、4時間血清の不在下37Cでパルスし、2回洗浄した。明記しない限り、およそ3×104標的細胞を同数のT細胞と少なくとも16時間インキュベートし、その後、サイトカイン放出アッセイを行った。サイトカイン分泌はGM−CSF ELISAキット(R&D Systems,ミネアポリス,MN)を用いて測定した。
【0142】
ヒトIL−4、TNF−αおよびTGF−βレベルの定量をR&D Systemsから得たサイトカインキットを使用して行い、IFN−γELISAキットをEndogen Inc(ウォバーン,MA)から購入した。アッセイは製造業者の説明書に従って行った。
【0143】
HLA DP分子の分子タイピング
トータルRNAをEBVB細胞、CD40リガンド刺激B細胞、CD4+T細胞、またはMHCクラスII陽性メラノーマ株からタイピングのために得た。トータルRNAはRNeasyキット(Qiagen,ドイツ)を使用して精製し、100ng〜1μgのRNAをオリゴdTプライム第一鎖cDNA合成に使用した。cDNA産物の1/10を使用し、Clontech(パロアルト,CA)のアドバンテージPCR系を用いてPCR増幅を行った。以下のプライマー対をHLA DP−AおよびDP BPCRに使用した:DPAフォワードプライマー5’ATG CGC CCT GAA GAC AGA ATG T 3’(配列番号75)、DPAリバースプライマー5’TCA CAG GGT CCC CTG GGC CCG GGG GA3’(配列番号76)、DPBフォワードプライマー5’ATG ATG GTT CTG CAG GTT TCT G3’(配列番号77)、およびDPBリバースプライマー5’TTA TGC AGA TCC TCG TTG AAC TTT C3’(配列番号78)。引き続いて、PCR産物を精製し、PCRを行うために使用した同一プライマーを用いてシークエンシングした。単一の配列がPCR産物から得られたので、多数の患者が非常に一般的なHLA DPB1*0401遺伝子産物にホモ接合性であると思われた。ヘテロ接合性の患者の場合、PCR産物をまずpCR4ベクター(Invitrogen,カールズバッド,CA)にクローニングし、ベクター配列に相補的な5’および3’プライマーを用いてシークエンシングした。最終配列をIMGT−HLAデータベースでサーチしてHLA DPであることを確認した(http://www3.ebi.ac.uk/Services/imgt/hla)。
【0144】
実施例9
NY−ESO−1に対するCD4+T細胞株TE4−2の作製
最初の研究をHLA DR4対立遺伝子により拘束されたNY−ESO−1エピトープを同定するために行った。予測される9マーのDR4結合モチーフを含む8つの20マーのペプチドを、NY−ESO−1組換えタンパク質で免疫しインビトロで刺激したHLA−DR4−IEトランスジェニックマウス由来のリンパ球による認識について試験した(50)。3つの20マーのペプチドがこれらの実験に陽性であることが分かった。これらのうちの一つはDRB1*0401とDRB1*0101との夾雑エピトープとして特徴付けされた(50)。他の2つのペプチド(ESO p161−180およびESO p141−160)をさらに特徴付けするために、本発明者らはそれらを使用して、高力価の抗体を有するDRB1*0401患者(TE)由来のPBMCならびにNY−ESO−1に対するCD4+およびCD8+T細胞を刺激した(50)。24ミクロ培養ウェル全体を各ペプチドについて使用した。弱い刺激を3回した後、24ウェルのうち15ウェルがESO p161−180で刺激されたPBMC由来の顕著な増殖を示した。試験した15ウェルの増殖が陽性であるウェルのうち9ウェルが、ペプチドパルスDRB1*0401発現1088EBVB細胞に対する特異的サイトカイン放出を示した(図8A)。特異的CD4+T細胞もまた、ESO p141−160で刺激したPBMCから作製したが、本研究では述べない(データ示さず)。
【0145】
次いで、ESO p161−180ペプチド刺激に特異的に応答した培養物由来のT細胞を合わせ、先の記載(55)のプロトコルを使用して増殖させた。CD8+T細胞の枯渇後、この培養物(TE4−2と命名する)は、FACS解析により評価したところ、95%を超えるCD4+T細胞を含んだ(データ示さず)。
【0146】
TE4−2のサイトカイン分泌プロファイルの分析によりこのT細胞株がペプチドパルス標的に応答してIFN−γ、TNF−α、IL−4およびGM−CSFを分泌しTGF−βを分泌しなかったことが示された(データ示さず)。従って、CD4+T細胞のTh1およびTh2型の両方がこの細胞株に存在するかもしれない。あるいは、Th0表現型の細胞がこの培養物に存在するかもしれない。
【0147】
実施例10
HLA DPB1*0401−0402に関連するTE4−2によるNY−ESO−1の認識
ESO p161−180、重複ペプチドであるESO p156−175、および完全長NY−ESO−1タンパク質それぞれでパルスしたDR4発現標的細胞に対する応答について、TE4−2 T細胞を試験した。DR1を発現するがDR4を発現しない586EBVB細胞もまたAPCとして使用した。無関係なペプチド、ESO p91−110、および精製トランケート組換えタンパク質(ESO1−74;アミノ酸1〜74を含む)(50)をコントロールとして使用した。TE4−2 T細胞は、完全長NY−ESO−1タンパク質でパルスした場合DR4+1088EBVB株を特異的に認識したが、トランケートESO1−74タンパク質でパルスした場合認識しなかった(図8B)。ESO p161−180およびp156−175の両方がTE4−2により認識され、これは最小ペプチドエピトープがアミノ酸161と175との間にあることを示している。対照的に、最初予測したDR4結合モチーフはアミノ酸167と175との間にある。予測しなかったことに、TE4−2 T細胞株は、586EBVB細胞にパルスしたペプチドおよびタンパク質(これらはDRB1*0101を発現するがDRB1*0401を発現しない)に同じくらい良好に応答するようであった。この結果は、類似のペプチドが複数のMHCクラスII拘束エレメントによって提示されたか、またはTE4−2 T細胞に対してペプチドを提示するMHCクラスII拘束エレメントを共有する1088および586EBVB細胞株によって提示されたかのいずれかであったことを示唆していた。これらの可能性を試験するために、公知のHLA DRおよびDQ型を含む多数の他のEBVB細胞もまた、TE4−2 T細胞により利用される拘束エレメントを同定するためにAPCとして使用した。試験したEBVB細胞株のうち一つを除く全てがTE4−2に対してESO p161−180ペプチドを提示することができた(図8C)。
【0148】
次いで、ペプチドのT細胞認識を、異なるMHC拘束エレメントによって拘束されたペプチドの認識をブロックする特異的抗体の存在下で行った。図9A〜9Cの結果は、すべてのMHCクラスII対立遺伝子(IVA12)をブロックする抗体および全てのHLA DP対立遺伝子(B4/21)をブロックする特異性を有する抗体が、TE4−2 T細胞がESO p161−180を認識する能力を無効としたことを示した。HLA−A、BおよびC対立遺伝子(W6/32)に対する抗体ならびにMHCクラスIIDR(L243)およびDQ(Genox3.53およびIVD12のミックス)対立遺伝子に対する抗体は、TE4−2 T細胞の刺激に対してほとんどまたは全く影響しない。従って、これらの結果は、TE4−2 T細胞が本研究で使用したEBVB細胞株により共有される非常に一般的なHLA DP対立遺伝子に関してESO p161−180を認識したことを示唆していた。
【0149】
次いで、HLA−DP対立遺伝子を分子的にクローニングし、図8Cで使用した細胞株に対してシークエンシングした。これらの研究は、1088および586EBVB株がともに、HLA DPB1*0401遺伝子産物、患者TEが発現したDPB1*0401ならびに未知のDP対立遺伝子にホモ接合性であったことを示していた(表2)。L023EBVB細胞株(これはTE4−2に対してESO p161−180ペプチを提示しなかった)はHLA DP対立遺伝子(これはDPB1*0401および0402とは異なる)にホモ接合性として型を決められた。
【0150】
1363、1088、836およびL007 EBVB細胞株はすべてDPB1*0401を発現したが、L041EBVB細胞株はDPB1*0402(84位と85位の2つのアミノ酸残基がDPB1*0401分子と異なる)を発現した。従って、DPB1*0401とDPB1*0402の両方がTE4−2 CD4+T細胞に対するESO p161−180エピトープを提示することができるようであった。
【0151】
【表2】
【0152】
特異的DPA鎖がTE4−2 T細胞に対するエピトープを提示するのに必要であるかどうかを決定するために、DPB1*0401−0402を発現するEBVB細胞中のHLA DPA分子もまた解析した。図8Cで使用したDPB1*0401−0402を発現するEBVB細胞は1タイプ超のHLA DPA分子(データ示さず)を持っていたが、すべてがTE4−2 T細胞にNY−ESO−1エピトープを同じように良好に提示することが可能であった。
【0153】
実施例11
腫瘍細胞上の天然にプロセシングされたNY−ESO−1エピトープのTE4−2による認識
TE4−2により認識されるT細胞エピトープが腫瘍細胞の表面で天然にプロセシングされ提示されるかどうかを調べるために、NY−ESO−1およびDPB1*0401を発現した腫瘍株を標的として用いた。TE4−2 T細胞は、NY−ESO−1とDPB1*0401の両方を発現する複数の腫瘍株を認識したが、NY−ESO−1を発現するがHLA DPB1*0401および0402対立遺伝子のいずれも発現しない腫瘍株(1362mel)は認識しなかった(図10A)。さらに、TE4−2 T細胞はDPB1*0401陰性およびNY−ESO−1陰性の腫瘍(526mel)を認識しなかった。HLA DPB1*0401を発現するがNY−ESO−1を発現しない1つのメラノーマ株(1102mel)はまた、TE4−2 T細胞により認識された。RT−PCR解析の結果は、1102melがLAGE−1遺伝子(NY ESO−1に対しておよそ90%のアミノ酸類似性を有する癌/精巣抗原)を発現したことを示していた(57)。ESO p161−175と同一の配列はまた、LAGE−1タンパク質にも存在した。これらの結果は、TE4−2により認識されるエピトープが腫瘍細胞表面に存在し、NY−ESO−1と、密接に関連した腫瘍抗原LAGE−1との間で共有されていることを示唆した。
【0154】
NY−ESO−1発現メラノーマ株に加えて、TE4−2 T細胞もまた、NY−ESO−1トランスフェクト293CIITA細胞の認識について試験した。293細胞にMHCクラスIIトランスアクチベーター遺伝子(CIITA)を発現するレトロウイルスを用いて形質導入し、293CIITA細胞株を作製した(53)。RT−PCRにより決定されたように、293CIITA細胞はホモ接合性のHLA DPB1*0401分子を発現したが、親の293細胞は発現しなかった(データ示さず)。TE4−2 T細胞は、NY−ESO−1トランスフェクト293CIITA細胞と特異的に反応した(図10B)。対照的に、TE4−2 T細胞はpGFPプラスミドをトランスフェクトした293CIITA細胞またはIi−NY−ESO−1をトランスフェクトした親の293細胞のいずれも認識しなかった。Iiターゲッテイング配列はNY−ESO−1のプロセシングおよび認識に必要ではなかったが、T細胞認識をわずかに増強した(図10B)。これらの結果はTE4−2 T細胞が天然にプロセシングされたNY−ESO−1エピトープを認識したことをさらに示していた。
【0155】
実施例12
HLA−A2拘束エピトープと重複するHLA DP4拘束エピトープ
2つの重複するペプチド(ESO p161−180およびp156−175)でパルスした標的細胞は、TE4−2 T細胞により同じように良好に認識された。これは、最小T細胞エピトープがアミノ酸161から175までの範囲の領域に位置していることを示していた(図8B)。
【0156】
アミノ酸161と175との間にあるアンカー残基を同定するために、重複13マーペプチドの1シリーズを1088EBVB細胞をパルスするのに使用し、TE4−2 T細胞を刺激するその能力を試験した。図11Aに示すように、161位のW残基を除去したときに活性の部分的損失を観察し、162位のI残基を除去したときに活性の完全な損失を観察した。167位のC末端L残基の欠失もまた、TE4−2 T細胞によるペプチド認識を取り除いた。さらに、169位の残基Vも、この残基の欠失がペプチド刺激活性を2倍減少させたので、重要であると思われた。これらの結果は、161位のW残基がP1アンカーであり得、167位のL残基がP7アンカーの代表であることを示した。169位のV残基もまた、ペプチドエピトープの刺激性能に寄与しているようであり、それがP9アンカー残基を示し得ることを示している。これらの推定アンカー残基は先に記載されたコンセンサスHLA DPB1*0401結合モチーフ(57)に非常に一致している。
【0157】
3つすべてのアンカー残基を含むESO p157−170ペプチドを力価測定実験に使用して、ペプチドに対する最小刺激濃度を決定した。その結果は、ESO p157−170が、3nMと33nMとの間の最小濃度でTE4−2 T細胞からの著しいサイトカイン放出を刺激することができることを示していた(図11B)。これらの結果は、TE4−2 T細胞が高い親和性でESO p157−170を認識することを示していた。この見かけの親和性は非変異ペプチド(例えば、gp100(58)、チロシナーゼ(59)、およびCDC−27(54)由来のもの)由来の最もよく知られた公知のMHCクラスII結合エピトープよりも優れている。同じ領域にわたる他のペプチド(例えば、ESO p161−180およびp156−175)もまた、TE4−2 T細胞に関して同様な最小刺激濃度を有した(データ示さず)。
【0158】
興味深いことに、先に同定されたHLA−A2エピトープ、ESO p157−167(47)は、DPB1*0401−0402エピトープ、ESO p157−170内に含まれた。HLA−DPエピトープがHLA−A2により提示されCD8+T細胞と交差反応し得るかどうかを判断するために、ESO p157−170をTE8−1による認識について試験した(CD8+T細胞株はHLA−A2エピトープESO p157−167を特異的に認識する)。L023 EBVB細胞(これは、HLA−A2を発現するがDPB1*0401−0402対立遺伝子を発現しない)にパルスした時、ESO p157−170はTE8−1 T細胞からの顕著なサイトカイン放出を刺激することができた(図11C)。この実験は、ESO p157−170エピトープがMHCクラスIおよびクラスII2つの特異性を持っており、NY−ESO−1タンパク質を認識するCD4+およびCD8+T細胞両方を刺激することができることを示していた。従って、ESO p157−170は、腫瘍細胞を特異的に認識するCD4+およびCD8+T細胞両方を誘発することを目的とする癌ワクチンの興味深い候補物であり得た。
【0159】
実施例13
NY−ESO−1抗体産生とHLA DPB1*0401−0402との関連性
HLA DPB1*0401−0402は、白人の大部分(別の人種を含めた集団調査では43%〜70%)に存在する主要なMHCクラスII対立遺伝子である(52)。先の研究(48、50)は、正常ドナーおよびNY−ESO−1発現腫瘍を有さない癌患者がNY−ESO−1に対する抗体を生じないことを示している。対照的に、NY ESO−1発現腫瘍を有する患者の50%は、NY−ESO−1特異的Abを生じた。血清サンプルを試験した88人のメラノーマ患者のパネルのうち、11人の患者が高力価のNY−ESO−1抗体を有することが見出された(50)。多くの患者がDR4対立遺伝子を全く発現しなかったことから、先に同定したDR4拘束CD4+T細胞ペプチドは、NY−ESO−1特異的Abの産生を説明できなかった(表2)。NY−ESO−1特異的DP4拘束CD4+T細胞がこれらのメラノーマ患者におけるNY−ESO−1特異的Abの産生と関連しているかどうかをさらに調査するため、本発明者らはまずこれらのHLA DPサブタイプを解析した。NY−ESO−1抗体を有する11人の患者のうち10人がDPB1*0401および/または0402を発現したが、主要なDQまたはDR拘束エレメントはこの患者のグループでは同定できなかった(表2)。公知のNY−ESO−1発現腫瘍を有しているが検出可能な抗体を有さないこのパネルの3人の患者はDPB1*0401−0402対立遺伝子を発現しなかった。残りの74人の患者由来の腫瘍細胞株はこれらの患者由来のNY ESO−1発現を評価するためには入手できなかったので、そのHLA DP型を同定するためのさらなる研究は行わなかった。0.011のp値をフィッシャーの直接確率検定により得、これは抗体応答とHLA−DPB1*0401−0402発現との関係の重要さを示していた。NY−ESO−1は25〜30%の腫瘍細胞株で発現され、DP4は集団の43〜70%で発現されるので、NY−ESO−1とDP4の両方を発現し、かつ抗体応答を生じる可能性のある患者の割合は、10〜21%の範囲である。この仮説的予想はNY−ESO−1抗体を有する患者の観察された10〜13%の頻度と非常に近い。
【0160】
NY−ESO−1抗体応答とDPB1*0401−0402発現との関連に関する更なる証拠を手に入れるため、NY−ESO−1抗体を有する11人の患者のうち6人からのPBMCを、ESO p161−180ペプチドによるインビトロ刺激に用いた。インビトロ感作もまた検出可能なNY−ESO−1抗体を有さない2人のDPBI*0401+患者からのPBMCを用いて行った。2または3回のインビトロ刺激の後、ESO p161−180ペプチドでパルスした293CIITA細胞に対するその応答について、T細胞を試験した。NY−ESO−1抗体を有する6人の患者のうち5人(TEを含む)由来のT細胞は、293CIITA細胞により提示されるESO p161−180エピトープ(DP4+およびHLA−A2-)の特異的認識を示した(表3)。患者CTおよびBL由来の複数のウェルは、ペプチドをパルスした標的と反応するようであった。これらの2人の患者由来の感作したPBMCもまた、CD4+T細胞のさらなる濃縮なしにDPBI*0401+およびNY−ESO−l+メラノーマ株の顕著な腫瘍認識を示した(データ示さず)。対照的に、NY−ESO−1反応性T細胞は、3回の刺激後検出可能なNY−ESO−1抗体を有さない2人の患者(WCおよびEW)由来のPBMCを用いて作製されなかった。これらの結果は、抗NY−ESO−1抗体を生じた患者もまた、比較的高い前駆体頻度のDPB1*0401拘束エピトープと反応性のT細胞を含んでいたことを示唆していた。これらのNY−ESO−1特異的CD4+T細胞はNY−ESO−1癌/精巣抗原に対する抗体応答の発生に寄与していたかもしれない。
【0161】
【表3】
【0162】
実施例14
野生型HLA DP4ペプチドの1つに改変を施した。この改変は、ペプチドをより可溶性にし90%超の均質性(これはペプチド臨床試験に関してFDAにより求められている)まで精製し得るようにするためのものであった。野生型および改変ペプチドは以下の通りである:
【0163】
野生型ESOp157−170;SLLMWITQCFLPVF(配列番号54);
野生型ESOp157−167;SLLMWITQCFL (配列番号79);ESOp156R−169;RSLLMWITQCFLPV(配列番号63);および
ESOp157−170R;SLLMWITQCFLPVR(配列番号64)。
【0164】
これらの改変ペプチドがT細胞により同じように良好に認識されるかどうかを試験するために実験を行った。ESO p157−170がHLA−A2およびHLA−DP4 2つの結合特異性を示したので、TE4−2 CD4+T細胞およびTE8−1 CD8+T細胞それぞれによるDP4(図12A)およびA2(図12B)両方の拘束様式における認識を測定した。
【0165】
結果は、これらの改変ペプチドがCD4+T細胞およびCD8+T細胞により野生型として同じように良好に認識されたことを示していた。
【0166】
実施例15
NY−ESO−1エピトープ特異的CD4+Tリンパ球免疫療法
メラノーマ抗原に対して前感作したTリンパ球は、メラノーマに罹患した哺乳動物を治療処置するのに有効であり得る。Tリンパ球を末梢血またはメラノーマの腫瘍懸濁液から単離し、インビトロで培養する(Kawakami,Y.ら、1988,J.Exp.Med.168:2183−2191)。
【0167】
Tリンパ球をエピトープVLLKEFTVSG(配列番号19)またはエピトープWITQCFLPVF(配列番号80)に濃度1μg/mlのみまたはIL−2の存在下で暴露し、前感作し、そして培養液中で増殖させる。エピトープに暴露したCD4+Tリンパ球を1哺乳動物あたり約109〜1012リンパ球で哺乳動物に投与する。このリンパ球を、静脈内、腹腔内、または病巣内のいずれかで投与する。この処置は、サイトカイン、メラノーマ病変の外科的切除および化学療法剤等の他の治療処置と同時に投与してもよい。NY−ESO−1特異的CD8+Tリンパ球はCD4+Tリンパ球と同時に投与してもよい。
【0168】
実施例16
転移性メラノーマを有する患者の処置
このプロトコルでは、進行メラノーマの患者を抗原性癌エピトープで免疫する。
【0169】
試験にふさわしい患者は、標準的な有効治療が失敗した測定可能または評価可能な転移性メラノーマの徴候を持っていなければならない。患者は、腫瘍細胞RNAのPCRまたはノザンブロット解析により立証されるようなNY−ESO−1抗原を発現する腫瘍を有さなければならない。
【0170】
患者に、体重1kgあたり1ng、1μg、1mgまたは500mgいずれかのMHCクラスII拘束T細胞エピトープを、0日、7日、14日に単独で、またはIL2および/または免疫刺激分子とともに、静脈内に投与する。患者を、毒性、免疫効果および治療有効性について評価する。患者に、NY−ESO−1クラスI拘束T細胞エピトープをさらに投与してもよい。
【0171】
処置患者から採取したリンパ球を、NY−ESO−1癌抗原またはMHCクラスII拘束T細胞エピトープに対する特異的応答について試験する。
【0172】
完全な応答を、少なくとも4週間続く疾患の全臨床的徴候の消失と定義する。部分的応答は、新たな病変またはいずれの病変における増加の出現なしに、少なくとも4週間すべての測定可能な病変の垂直直径の積の合計における50%以上の減少である。軽微な応答は、新たな病変の出現およびいずれの病変における増加なしに、すべての測定可能な病変の垂直直径の積の合計における25〜49%の減少として定義する。部分応答未満のいずれの患者をも非応答者と考える。新たな病変の出現あるいは部分的または完全な応答後の先の病変の垂直直径の積における25%超の増加を再発と考える。
【0173】
考察
NY−ESO−1は、体液性および細胞性両方の免疫応答を引き起こすので、重要な免疫標的である(19〜21)。その発現パターンはMAGE遺伝子ファミリーの抗原と類似しているが、NY−ESO−1はMAGEファミリーのいずれのものよりも乳癌、前立腺癌、肺癌で頻繁に発現される(19、20、23)。さらに興味深いことに、非常に低い割合の患者がMAGE抗原または分化抗原(例えば、チロシナーゼ、gp100、TRP−1およびTRP−2)に対する高力価の抗体を生じた(データ示さず、(22))にもかかわらず、高力価のNY−ESO−1反応性抗体は、癌の患者においてしばしば検出された(図2Bおよび2C)。これらの研究は、NY−ESO−1反応性CD4+T細胞が抗体産生およびCTL増殖に関わっているかもしれないことを示唆する。本研究で、本発明者らは、HLA−DR4トランスジェニックマウスおよび候補ペプチドを用いたヒトPBMCのインビトロ刺激を使用して、NY−ESO−1由来のHLA−DR4拘束T細胞エピトープを同定した。我々の知る限りでは、これは、NY−ESO−1由来T細胞エピトープがCD4+T細胞に対してMHCクラスII分子により提示されることを示した最初の証明である。NY−ESO−1特異的抗体およびCTLは異なるHLA遺伝子型を持つ患者において検出されたので、HLA−DR4以外のHLAクラスII分子により提示される他のCD4+T細胞エピトープは本発明において同定された。
【0174】
最近、2つのグループが、公知のMHCクラスI拘束腫瘍抗原であるMAGE−3からのMHCクラスII拘束T細胞エピトープの同定を報告した。精製MAGE−3タンパク質でパルスされたDCで刺激されたPBMCから作製したCD4+T細胞クローンは、HLA−DR13適合EBV B細胞にパルスされたペプチドまたはタンパク質を認識したが、MAGE−3+/DR13+腫瘍細胞にパルスされたものを認識しなかった(14)。しかし、別の研究では、コンピューターを使ったアルゴリズムによって予測されるペプチドで刺激したPMBCから作製したCD4+T細胞は、EBV B細胞およびMAGE−3+/DR11+腫瘍細胞両方にパルスしたペプチドを認識することができた(15)。NY−ESO−1の場合、本発明者らはここに、CD4+T細胞がDR4マッチEBV B細胞にパルスしたNY−ESO−1タンパク質またはペプチドおよびNY−ESO−1を発現する腫瘍細胞を認識できることを示している(図4および5)。HLA−DRトランスジェニックマウスの利用は、免疫したトランスジェニックマウスが特異的反応性T細胞の高い前駆体頻度をおそらく有しているので、推定ペプチドの同定において利点を有し得る。候補ペプチドを一旦同定すれば、CD4+T細胞を合成候補ペプチドで刺激したPBMCから作製できる。従って、全タンパク質で免疫し、コンピューターを使用したアルゴリズムにより予測したペプチドで刺激したトランスジェニックマウスの組み合わせ使用は、多数のペプチドおよび複数回のインビトロ刺激でヒトPBMCを刺激する必要性を回避し得る。さらに、免疫したトランスジェニックマウスにより同定される候補ペプチドは、天然にプロセシングされ細胞表面上に提示されるペプチドであるようである。これは、ペプチド特異的CD4+T細胞が腫瘍細胞も同様に認識することができる可能性を上昇させ得る。最後に、NY−ESO−1に対する高力価の抗体および高前駆体頻度のCTLを生じた患者(TE)由来のPBMCの使用は、抗体産生およびCTLの両方がCD4+T細胞の助けを必要とするので、腫瘍特異的CD4+T細胞を作製するのをより容易にし得る。このアプローチは、自己免疫疾患に関与する公知の自己抗原から多数のMHCクラスII拘束T細胞エピトープを同定するのに使用されている(28)。従って、本研究で使用するストラテジーは多くの他の公知のMHCクラスI拘束腫瘍抗原に適用可能であり得つつ、直接遺伝子クローニングアプローチ等他のストラテジーが未知のMHCクラスII拘束腫瘍抗原の同定を容易にし得る。
【0175】
gp100等組織特異的分化抗原由来のペプチドを使用する臨床試験は、メラノーマの患者の処置において治療有効性のいくらかの証拠を示した(4)。顕著な毒性の副作用は改変gp100ペプチドで処置した患者において観察されていないが、白斑または色素脱失は、治療に反応を示した患者においてしばしば見られた(29)。このことは、自己抗原による免疫によって誘導された抗腫瘍免疫が自己免疫を生じ得ることを示唆している。TRP−1を免疫標的として使用した動物研究では、類似の結果(抗腫瘍免疫および毛の色素脱失)も得られた(30〜32)。興味深いことに、gp75/TRP−1に媒介される抗腫瘍免疫および自己免疫はCD4+T細胞および抗体を伴っているようであった(33)。mTRP−2(35)ではなくhTRP−2(34)でのマウスの免疫は、自己抗原ならびにCD4+およびCD8+T細胞の両方の関与を必要とする抗腫瘍免疫に対する耐性を壊した(33)。これらの研究は、抗腫瘍免疫が抗体またはCD8+T細胞のいずれかにより媒介され得るが、両方ともCD4+T細胞の決定的な助けを必要とし得ることを示唆していた(24、33)。
【0176】
本研究で同定したMHCクラスII拘束NY−ESO−1ペプチドは、NY−ESO−1に対するCTLおよび抗体が癌の患者において検出されたので、臨床用途に有用であり得る。MHCクラスIおよびII拘束ペプチドまたは精製NY−ESO−1タンパク質での免疫は、NY−ESO−1特異的CD4+、CD8+T細胞および抗体を誘導し得る。あるいは、患者は、クラスIおよびIIの両ペプチドをロードするかまたはNY−ESO−1遺伝子をコードする組換えウイルスを感染させた樹状細胞で免疫され得る。精巣生殖細胞はMHCクラスIおよびIIの分子を発現しないので(36)、NY−ESO−1に対する免疫応答は腫瘍細胞に特異的であるはずであり、従って、ほとんどまたは全く自己免疫応答を生じない。MAGE−3のMHCクラスI拘束ペプチドまたは樹状細胞にパルスしたペプチドを使用した同様の研究は、抗腫瘍免疫(CTL応答)および緩慢な腫瘍の退行が示されたにもかかわらず、色素脱失/白斑または他の顕著な副作用が観察されなかったことを示した(6、7)。抗腫瘍免疫は、腫瘍特異的CD4+T細胞の助けを提供することによって向上され得る。
【0177】
【表4−1】
【表4−2】
【表4−3】
【表4−4】
【表4−5】
【表4−6】
【表4−7】
【0178】
本発明のこれらの及び他の目的、性質及び付随の利点の多くは、以上の詳細な説明を読む際に、添付された図面と関連づけて考えると、より良く理解されよう。
【特許請求の範囲】
【請求項1】
NY−ESO−1癌ペプチド、即ち配列番号1及びその変異体からなる核酸配列内にコードされるMHCクラスII拘束T細胞エピトープ、その機能性部分、変異体又は誘導体。
【請求項2】
エピトープが、配列番号2、そのホモログ又は機能性部分からなる配列によってコードされる、請求項1に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ、その機能性部分又は誘導体。
【請求項3】
ペプチドが、配列番号27又はその部分からなる配列によってコードされる、請求項1に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ、その機能性部分又は誘導体。
【請求項4】
配列番号4〜21及び29〜34、それらの機能性部分、誘導体又は組み合わせの少なくとも1つを含むNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項5】
配列番号5〜18、それらの機能性部分、誘導体又は組み合わせからなる群から選択される、請求項4に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項6】
VLLKEFTVSG(配列番号19)、その変異体又は誘導体を含む、請求項4に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項7】
HLA−DR拘束Tリンパ球によって免疫学的に認識される、請求項1〜5又は6のいずれかに記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項8】
Tリンパ球がCD4+である、請求項1〜5又は6のいずれかに記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項9】
癌ペプチドが、非ホジキンリンパ腫、白血病、ホジキンリンパ腫、肺癌、肝癌、転移、メラノーマ、頭部癌、頚部癌、神経芽腫、腺癌、胸腺腫、結腸癌、子宮癌、乳癌、前立腺癌、卵巣癌、子宮頚癌、膀胱癌、腎癌、膵癌、及び肉腫からなる群から選択される癌由来である、請求項4に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項10】
AAG GAG TTC ACT GTG TCC(配列番号28)を含む核酸配列によってコードされる、請求項4に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項11】
請求項1〜9又は10のいずれかに記載のNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ及び医薬として許容できる担体を含む医薬組成物。
【請求項12】
更に、免疫刺激分子を含む、請求項11に記載の医薬組成物。
【請求項13】
免疫刺激分子が、HLA−クラスII分子上、又はHLAクラスII分子を発現している細胞上にある、請求項12に記載の医薬組成物。
【請求項14】
更に、B7.1、B7.2、ICAM−1、ICAM−2、LFA−1、LFA−3、サイトカイン又はそれらの組み合わせを含む、請求項11に記載の医薬組成物。
【請求項15】
更に、NY−ESO−1の少なくとも1つのMHCクラスI拘束T細胞エピトープを含む、請求項11に記載の医薬組成物。
【請求項16】
MHCクラスI拘束T細胞エピトープが配列番号22又は配列番号23を含む、請求項15に記載の医薬組成物。
【請求項17】
請求項1〜9又は10のいずれかに記載のNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープを単独で又は組み合わせて含み、かつ必要に応じて、少なくとも1つの免疫刺激分子を含む免疫原。
【請求項18】
免疫刺激分子がHLAクラスII分子である、請求項17に記載の免疫原。
【請求項19】
更に、NY−ESO−1のMHCクラスI拘束T細胞エピトープを含む、請求項17に記載の免疫原。
【請求項20】
MHCクラスII拘束T細胞エピトープとMHCクラスI拘束T細胞エピトープがペプチド結合により一緒に結合している、請求項19に記載の免疫原。
【請求項21】
少なくとも1つのMHCクラスII拘束T細胞エピトープをコードしている、配列番号27、その機能性部分又はホモログを含む単離された核酸配列。
【請求項22】
配列番号24、25、26、又はその部分又は変異体からなる群から選択される、請求項21に記載の単離された核酸配列。
【請求項23】
配列番号4〜21、29〜34、それらの組み合わせ、機能性部分又は誘導体のいずれか1つのアミノ酸配列を含む、NY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープをコードする単離された核酸配列。
【請求項24】
更に、NY−ESO−1の少なくとも1つのMHCクラスI拘束T細胞エピトープをコードする核酸配列を含む、請求項23に記載の単離された核酸配列。
【請求項25】
請求項21、22、23又は24のいずれかに記載の核酸配列を含む組換え発現ベクター。
【請求項26】
請求項25に記載の組換え発現ベクターによって形質転換又はトランスフェクトされた宿主生物。
【請求項27】
抗原提示細胞である、請求項26に記載の宿主生物。
【請求項28】
請求項21、22又は23のいずれかに記載の核酸配列に相補的な核酸配列からなるオリゴヌクレオチド。
【請求項29】
請求項21、22、23又は24のいずれかに記載の核酸配列を含む組換えウイルス。
【請求項30】
レトロウイルス、バキュロウイルス、アンカラウイルス、鶏痘ウイルス、アデノウイルス、ワクシニアウイルス、レンチウイルス、及びレプリカーゼ−ベース−アルファウイルスからなる群から選択される、請求項29に記載の組換えウイルス。
【請求項31】
免疫刺激分子をコードする少なくとも1つの遺伝子を更に含む、請求項29に記載の組換えウイルス。
【請求項32】
免疫刺激分子がHLAクラスII分子である、請求項31に記載の組換えウイルス。
【請求項33】
請求項29〜31又は32のいずれかに記載の組換えウイルスで形質転換又はトランスフェクトされた宿主生物。
【請求項34】
請求項1〜5又は6のいずれかに記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープと結合する、単離された抗体又はその抗原結合部分。
【請求項35】
NY−ESO−1の組換えMHCクラスII拘束T細胞エピトープの製造方法であって、以下:
a.請求項21、22又は23のいずれかに記載のヌクレオチド配列、その機能性部分又は変異体を、発現ベクターに挿入すること;
b.該発現ベクターを宿主細胞に移入すること;
c.エピトープ又はその機能性部分の発現に適切な条件下、宿主細胞を培養すること;及び
d.組換えエピトープ又はその機能性部分を取得すること;
を含む、方法。
【請求項36】
工程(a)において、HLAクラスII分子又はその部分をコードするヌクレオチド配列を発現ベクターに挿入することを更に含む、請求項35に記載の方法。
【請求項37】
哺乳動物における癌又は前癌の存在の検出方法であって、以下:
a.請求項21、22又は23のいずれかに記載の核酸配列、それらの機能性部分又は変異体と、哺乳動物から採取したmRNAの試験用生物学的サンプルとを、該配列と該mRNAとの間で複合体が形成される条件下で接触させること;
b.複合体を検出すること;
c.試験用サンプル中のmRNA量を、既知の正常な生物学的サンプルからのmRNA量と比較することであって、試験用サンプルからのmRNA量の増大が癌又は前癌の指標となること;
を含む、方法。
【請求項38】
癌又は前癌がメラノーマである、請求項37に記載の方法。
【請求項39】
生物学的サンプル中のNY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその部分の検出方法であって、以下:
a.サンプルと、請求項34に記載の該エピトープに特異的な抗体又はその抗原結合部分とを、該抗体と該エピトープとの間で免疫複合体が形成される条件下で接触させること;及び
b.免疫複合体の存在を検出することであって、その存在が生物学的サンプル中のエピトープの指標となること;
を含む、方法。
【請求項40】
哺乳動物における癌の予防又は阻害の方法であって、請求項1〜5又は6のいずれかに記載のNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその機能性部分の有効量を単独で、又はHLAクラスII分子と組み合わせて該哺乳動物に投与することを含み、該量が哺乳動物における癌の予防又は阻害に有効である、方法。
【請求項41】
NY−ESO−1の少なくとも1つのMHCクラスI拘束T細胞エピトープを投与することを更に含む、請求項40に記載の方法。
【請求項42】
哺乳動物におけるメラノーマの阻害方法であって、以下:
a.インビトロでT細胞を、請求項1〜5又は6のいずれかに記載のNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその機能性部分に単独で、又はMHCクラスII分子と組み合わせて、MHCクラスII拘束Tリンパ球を誘発するのに十分な時間、曝すこと;
b.該MHCクラスII拘束Tリンパ球を単独で、又はNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ、その機能性部分又はネイティブNY−ESO−1タンパク質と組み合わせて、メラノーマを阻害するために十分な量で哺乳動物に投与すること;
を含む、方法。
【請求項43】
哺乳動物における癌の予防又は阻害の方法であって、有効量の、請求項29に記載の組換えウイルスを単独で、又は外因性の免疫刺激分子と組み合わせて、哺乳動物に投与することを含み、該量が癌を予防又は阻害するために有効である、方法。
【請求項44】
哺乳動物がHLAクラスIIDR分子を発現している、請求項43に記載の方法。
【請求項45】
NY−ESO−1のMHCクラスI拘束T細胞エピトープをコードする組換えウイルスの投与を更に含む、請求項43に記載の方法。
【請求項46】
請求項29に記載の組換えウイルスを単独で、又は外因性の免疫刺激分子、化学療法剤、抗生物質、抗真菌剤、抗ウイルス剤又はそれらの組み合わせと組み合わせて含み、かつ医薬として許容できる担体を含む医薬組成物。
【請求項47】
少なくとも、配列番号27のフラグメント、又はその機能性部分又は変異体を含むNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープをコードする遺伝子を担持し、かつ発現するトランスジェニック動物。
【請求項48】
請求項1〜5又は6のいずれかに記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープによって誘発される癌抗原特異的ヒトCD4+Tリンパ球。
【請求項49】
HLA−クラスII分子を認識する、請求項48に記載の癌抗原特異的ヒトCD4+Tリンパ球。
【請求項50】
HLA−DR分子を認識する、請求項48に記載の癌抗原特異的ヒトCD4+Tリンパ球。
【請求項51】
ペプチドが、アミノ酸モチーフ:
Xaa1ITQXaa2FXaa3PXaa4(配列番号51)及びそのホモログ(式中、Xaa1、Xaa2、Xaa3、及びXaa4は各々少なくとも1つのアミノ酸である)を含み、該ペプチドは、HLA−DP拘束CD4+Tリンパ球によって免疫学的に認識される、請求項1に記載の癌ペプチド。
【請求項52】
Xaa1が、Trp、Phe、Tyr、Met、Ile、Val、Ala、及びそれらの組み合わせからなる群から選択される、請求項51に記載の癌ペプチド。
【請求項53】
Xaa2が、Cys、Ser、Val、Ala、Thr、及びそれらの組み合わせからなる群から選択される、請求項51に記載の癌ペプチド。
【請求項54】
Xaa3が、Leu、Phe、Tyr、Met、Ile、Val、Ala、及びそれらの組み合わせからなる群から選択される、請求項51に記載の癌ペプチド。
【請求項55】
Xaa4が、Val、Tyr、Ile、Ala、Leu、Pro、及びそれらの組み合わせからなる群から選択される、請求項51に記載の癌ペプチド。
【請求項56】
約9乃至約30個のアミノ酸を含む、請求項51に記載の癌ペプチド。
【請求項57】
約10乃至約20個のアミノ酸を含む、請求項51に記載の癌ペプチド。
【請求項58】
約10乃至約15個のアミノ酸を含む、請求項51に記載の癌ペプチド。
【請求項59】
ホモログがLAGE遺伝子由来である、請求項51に記載の癌ペプチド。
【請求項60】
アミノ酸配列:
Xaa1WITQCFLPVF−Xaa2(配列番号52)及びその変異体
(式中、Xaa1及びXaa2は各々、アミノ酸ではないか、又は1個以上の天然に存在するアミノ酸である)を含む、請求項51に記載の癌ペプチド。
【請求項61】
約10乃至約30個のアミノ酸を含む、請求項60に記載の癌ペプチド。
【請求項62】
約10乃至約20個のアミノ酸を含む、請求項60に記載の癌ペプチド。
【請求項63】
約10乃至約15個のアミノ酸を含む、請求項60に記載の癌ペプチド。
【請求項64】
WITQCFLPVF(配列番号80)からなる、請求項60に記載の癌ペプチド。
【請求項65】
LLMWITQCFLPVFL(配列番号55)、LMWITQCFLPVFLA(配列番号56)、MWITQCFLPVFLAQ(配列番号57)、及びWITQCFLPVFLAQP(配列番号58)を含む、請求項1に記載の癌ペプチド。
【請求項66】
HLA−DPB1*0401−0402拘束CD4+Tリンパ球によって免疫学的に認識される、請求項51に記載のペプチド。
【請求項67】
キャリアタンパク質に結合している、請求項51に記載のペプチド。
【請求項68】
キャリアタンパク質が、KLH、破傷風トキソイド及びアルブミンからなる群から選択される、請求項67に記載のペプチド。
【請求項69】
請求項51、60、64又は65のいずれかに記載のペプチドをコードする単離された核酸配列。
【請求項70】
配列番号53、55〜64のいずれか1つ、又はそれらの組み合わせを含む少なくとも1つのMHCクラスII拘束T細胞エピトープペプチドをコードする単離された核酸配列。
【請求項71】
ストリンジェント条件下、請求項69又は70に記載の核酸配列とハイブリダイズする核酸配列を含む単離されたポリヌクレオチド。
【請求項72】
アミノ酸モチーフSLLMWITQCFLPVF(配列番号54)又はその変異体を有し、HLA−DP拘束CD4+Tリンパ球とMHCクラスI拘束CD8+Tリンパ球の両方によって免疫学的に認識される癌ペプチド。
【請求項73】
変異体が、アルギニン残基の付加をアミノ末端に有する、請求項72に記載の癌ペプチド。
【請求項74】
変異体が、フェニルアラニンの代わりに置換されたアルギニンをカルボキシル末端に有する、請求項72に記載の癌ペプチド。
【請求項75】
HLA−DPB1*0401−0402拘束Tリンパ球及びHLA−A2拘束Tリンパ球によって免疫学的に認識される、請求項72に記載のペプチド。
【請求項76】
請求項72に記載のペプチドをコードする単離された核酸配列。
【請求項77】
配列番号54のペプチドをコードする単離された核酸配列。
【請求項78】
ストリンジェント条件下、請求項76又は77に記載の核酸配列とハイブリダイズする核酸配列を含む単離されたポリヌクレオチド。
【請求項79】
請求項69に記載の核酸配列を含む組換え発現ベクター。
【請求項80】
請求項70に記載の核酸配列を含む組換え発現ベクター。
【請求項81】
請求項76に記載の核酸配列を含む組換え発現ベクター。
【請求項82】
請求項77に記載の核酸配列を含む組換え発現ベクター。
【請求項83】
少なくとも1つのNY−ESO−1 HLA−DR拘束T細胞癌ペプチドをコードする核酸配列を更に含む、請求項79〜82のいずれか1項に記載の組換え発現ベクター。
【請求項84】
NY−ESO−1 HLA−DR拘束T細胞癌ペプチドが、以下:
LPVPGVLLKEFTVSG(配列番号10);
VLLKEFTVSGNILTIRLT(配列番号65);
AADHRQLQLSISSCLQQL(配列番号66);及び
それらの組み合わせからなる群から選択されるアミノ酸配列を含む、請求項83に記載の組換え発現ベクター。
【請求項85】
少なくとも1つの単離されたNY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドをコードする核酸配列を更に含む、請求項83に記載の組換え発現ベクター。
【請求項86】
NY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドが、以下:
Xaa1Xaa2Xaa3GPGGGAPXaa4(配列番号22)
(式中、Xaa1は、アミノ酸ではないか、又は1乃至約20個の天然に存在するアミノ酸であり;
Xaa2は、Ale、Thr、Val、Leu又はArgであり;
Xaa3は、Ser又は保存的置換であり;
Xaa4は、Arg又はLysである)
及びそれらの誘導体を含む、請求項85に記載の組換え発現ベクター。
【請求項87】
核酸配列が、ASGPGGGAPR(配列番号23)をコードする、請求項86に記載の組換え発現ベクター。
【請求項88】
請求項79〜87のいずれか1項に記載のベクターによって形質転換又はトランスフェクトされた宿主細胞又は宿主生物。
【請求項89】
請求項51、60、64、65又は72のいずれか1項に記載の少なくとも1つの癌ペプチドをコードする組換えウイルス。
【請求項90】
請求項89に記載の組換えウイルスによって形質転換された宿主細胞又は宿主生物。
【請求項91】
配列番号51〜64、80のいずれか1つ又はそれらの組み合わせを含む、少なくとも1つのMHCクラスII拘束T細胞エピトープペプチドをコードする単離された核酸配列を含む組換えウイルス。
【請求項92】
ワクシニア、鶏痘、アデノウイルス、レトロウイルス、レンチウイルス、バキュロウイルス、アンカラウイルス、及びレプリカーゼ−ベース−アルファウイルスからなる群から選択される、請求項91に記載の組換えウイルス。
【請求項93】
HLA−DP分子をコードする核酸配列を更に含む、請求項91に記載の組換えウイルス。
【請求項94】
少なくとも1つのNY−ESO−1 HLA−DR拘束T細胞癌ペプチドをコードする核酸配列を更に含む、請求項91に記載の組換えウイルス。
【請求項95】
NY−ESO−1 HLA−DR拘束T細胞癌ペプチドが、以下:
LPVPGVLLKEFTVSG(配列番号10);
VLLKEFTVSGNILTIRLT(配列番号65);
AADHRQLQLSISSCLQQL(配列番号66);
それらの変異体及び組み合わせからなる群から選択されるアミノ酸配列を含む、請求項94に記載の組換えウイルス。
【請求項96】
少なくとも1つの単離されたNY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドをコードする核酸配列を更に含む、請求項91に記載の組換えウイルス。
【請求項97】
NY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドが、以下:
Xaa1Xaa2Xaa3GPGGGAPXaa4(配列番号22)
(式中、Xaa1は、アミノ酸ではないか、又は1乃至約20個の天然に存在するアミノ酸であり;
Xaa2は、Ale、Thr、Val、Leu又はArgであり;
Xaa3は、Ser又は保存的置換であり;
Xaa4は、Arg又はLysである)
及びそれらの誘導体を含む、請求項96に記載の組換えウイルス。
【請求項98】
核酸配列が、アミノ酸配列ASGPGGGAPR(配列番号23)を含むペプチドをコードする、請求項97に記載の組換え発現ベクター。
【請求項99】
請求項91〜98のいずれか1項に記載のウイルスによって形質転換又はトランスフェクトされた宿主細胞又は宿主生物。
【請求項100】
HLA−DP分子をコードする核酸配列を更に含む、請求項99に記載の宿主細胞又は宿主生物。
【請求項101】
請求項51、60、64又は65のいずれか1項に記載のペプチド及び医薬として許容できる担体を含む医薬組成物。
【請求項102】
請求項72に記載のペプチド及び医薬として許容できる担体を含む医薬組成物。
【請求項103】
請求項79〜87のいずれか1項に記載の組換え発現ベクター及び医薬として許容できる担体を含む医薬組成物。
【請求項104】
請求項91〜95のいずれか1項に記載の組換えウイルス及び医薬として許容できる担体を含む医薬組成物。
【請求項105】
免疫刺激分子を更に含む、請求項101に記載の医薬組成物。
【請求項106】
免疫刺激分子が、B7.1、B7.2、ICAM−1、ICAM−2、LFA−1、LFA−3、サイトカイン又はそれらの組み合わせである、請求項105に記載の医薬組成物。
【請求項107】
HLA−クラスII分子又はHLA−クラスII分子を発現している細胞を更に含む、請求項101に記載の医薬組成物。
【請求項108】
HLA−クラスII分子がHLA−DPである、請求項107に記載の医薬組成物。
【請求項109】
HLA−クラスII分子がHLA−DPB1*0401−0402である、請求項108に記載の医薬組成物。
【請求項110】
少なくとも1つのNY−ESO−1 HLA−DR拘束T細胞癌ペプチドを更に含む、請求項101に記載の医薬組成物。
【請求項111】
NY−ESO−1 HLA−DR拘束T細胞癌ペプチドが、以下:
LPVPGVLLKEFTVSG(配列番号10);
VLLKEFTVSGNILTIRLT(配列番号65);
AADHRQLQLSISSCLQQL(配列番号66);及び
それらの組み合わせからなる群から選択されるアミノ酸配列を含む、請求項110に記載の医薬組成物。
【請求項112】
少なくとも1つの単離されたNY−ESO−1 HLAクラスI拘束T細胞癌ペプチドを更に含む、請求項101に記載の医薬組成物。
【請求項113】
NY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドが、以下:
Xaa1Xaa2Xaa3GPGGGAPXaa4(配列番号22)
(式中、Xaa1は、アミノ酸ではないか、又は1乃至約20個の天然に存在するアミノ酸であり;
Xaa2は、Ala、Thr、Val、Leu又はArgであり;
Xaa3は、Ser又は保存的置換であり;
Xaa4は、Arg又はLysである)及びそれらの誘導体を含む、請求項112に記載の医薬組成物。
【請求項114】
NY−ESO−1 HLAクラスI拘束T細胞癌ペプチドは、ASGPGGGAPR(配列番号23)を含む、請求項113に記載の医薬組成物。
【請求項115】
請求項69、70、76又は77のいずれか1項に記載の核酸配列及び医薬として許容できる担体を含む医薬組成物。
【請求項116】
NY−ESO−1 HLA−DP拘束CD4+Tリンパ球を誘発する、請求項51、60、64、65又は72のいずれか1項に記載の癌ペプチドを含む免疫原。
【請求項117】
抗NY−ESO−1抗体を誘発する、請求項116に記載の免疫原。
【請求項118】
アジュバントを更に含む、請求項116に記載の免疫原。
【請求項119】
ペプチドがキャリアタンパク質に結合している、請求項116に記載の免疫原。
【請求項120】
請求項51、60、64、65又は72のいずれか1項に記載の癌ペプチドを含む癌ワクチン。
【請求項121】
請求項51、60、64、65又は72のいずれか1項に記載のNY−ESO−1のHLA−DP拘束T細胞エピトープと結合する単離された抗体又はその抗原結合部分。
【請求項122】
ポリクローナル抗体である、請求項121に記載の抗体。
【請求項123】
モノクローナル抗体である、請求項121に記載の抗体。
【請求項124】
検出可能標識で標識されている、請求項121に記載の抗体。
【請求項125】
検出可能標識が放射性同位体である、請求項121に記載の抗体。
【請求項126】
細胞毒性剤又は細胞増殖抑制剤の付加によって修飾されている、請求項121に記載の抗体。
【請求項127】
請求項121に記載の抗体及び医薬として許容できる担体を含む医薬組成物。
【請求項128】
請求項121に記載の抗体を含むキット。
【請求項129】
イムノアッセイ試薬を更に含む、請求項128に記載のキット。
【請求項130】
抗体が更に、HLA−DP分子と結合する、請求項121に記載の抗体又はその抗原結合部分。
【請求項131】
NY−ESO−1の組換えHLA−DP拘束CD4+T細胞エピトープの製造方法であって、以下:
a.配列番号51〜64、80又はその部分若しくは変異体の少なくとも1つをコードするヌクレオチド配列を、発現ベクターに挿入すること;
b.発現ベクターを宿主細胞に移すこと;
c.エピトープ又はその部分の発現のために適切な条件下で宿主細胞を培養すること;及び
d.組換えエピトープ又はその部分を取得すること;
を含む、方法。
【請求項132】
工程(a)において、HLAクラスII分子又はその部分をコードするヌクレオチド配列を発現ベクターに挿入することを更に含む、請求項131に記載の方法。
【請求項133】
HLAクラスII分子がHLA−DPである、請求項132に記載の方法。
【請求項134】
配列番号51〜64又は80の少なくとも1つのアミノ酸配列を含む癌ペプチドを発現するベクターの製造方法であって、プロモーターと機能可能に連結した配列番号51〜64又は80の少なくとも1つをコードする単離された核酸配列を組み込むことを含む、方法。
【請求項135】
哺乳動物における癌又は前癌の存在の検出方法であって、以下:
a.配列番号51〜64若しくは80又はその部分若しくは変異体の少なくとも1つをコードする核酸配列と、哺乳動物から採取したmRNAの試験用生物学的サンプルとを、該配列と該mRNAとの間で複合体が形成される条件下で接触させること;
b.複合体を検出すること;
c.試験用サンプル中のmRNA量を、既知の正常な生物学的サンプルからのmRNA量と比較することであって、試験用サンプルからのmRNA量の増大が癌又は前癌の指標となること;
を含む、方法。
【請求項136】
癌又は前癌がメラノーマである、請求項135に記載の方法。
【請求項137】
哺乳動物における癌又は前癌の存在の検出方法であって、以下:
a)請求項51、60、64、65又は72のいずれかに記載の癌ペプチドを、該哺乳動物から得たリンパ球の試験用生物学的サンプルと接触させること;
b)該癌ペプチドと免疫応答性のCD4+Tリンパ球を検出することであって、陰性コントロールサンプルと比較した、該癌ペプチドと免疫反応性のCD4+Tリンパ球の数の増大が癌又は前癌の指標となること;
を含む方法。
【請求項138】
生物学的サンプルにおけるNY−ESO−1のHLA−DP拘束CD4+T細胞エピトープ又はその部分の検出方法であって、以下:
a.サンプルを、請求項121〜125のいずれかに記載の、該エピトープに特異的な抗体又はその抗原結合部分と、該抗体と該エピトープとの間で免疫複合体が形成される条件下で接触させること;及び
b.免疫複合体の存在を検出することであって、その存在が、生物学的サンプルにおけるエピトープの指標となること;
を含む、方法。
【請求項139】
哺乳動物における癌の予防又は阻害の方法であって、
請求項51、60、64、69又は72のいずれかに記載の、NY−ESO−1のHLA−DP拘束T細胞エピトープ又はその部分の有効量を単独で、又はHLA−DP分子と組み合わせて哺乳動物に投与することを含み、該量が該哺乳動物において癌の予防又は阻害に有効である、方法。
【請求項140】
HLA−DP分子がHLA−DPB*0401−0402である、請求項139に記載の方法。
【請求項141】
NY−ESO−1のHLA−DR拘束T細胞エピトープを単独で、又はHLA−DR分子と組み合わせて投与することを更に含む、請求項139に記載の方法。
【請求項142】
NY−ESO−1のHLA−DR拘束T細胞エピトープが配列番号10、65又は66の少なくとも1つを含む、請求項141に記載の方法。
【請求項143】
NY−ESO−1のHLAクラスI拘束T細胞エピトープを投与することを更に含む、請求項139に記載の方法。
【請求項144】
HLAクラスI拘束T細胞エピトープが配列番号22又は23を含む、請求項143に記載の方法。
【請求項145】
NY−ESO−1を発現している腫瘍の増殖の阻害方法であって、請求項51、60、64、65又は72のいずれか1項に記載の癌ペプチドのある量を投与することを含み、該量がNY−ESO−1を発現している腫瘍の増殖の阻害に有効である、方法。
【請求項146】
哺乳動物におけるメラノーマの阻害方法であって、以下:
a.インビトロでTリンパ球を、請求項51、60、64、65又は72のいずれか1項に記載のNY−ESO−1のHLA−DP拘束T細胞癌ペプチド又はその部分に単独で、又はHLA−DP分子と組み合わせて、NY−ESO−1 HLA−DP拘束Tリンパ球を誘発する十分な時間、曝すこと;
b.メラノーマを阻害するために十分な量で、哺乳動物に該Tリンパ球を投与すること;
を含む、方法。
【請求項147】
哺乳動物に癌ペプチド又はネイティブNY−ESO−1タンパク質を投与することを更に含む、請求項146に記載の方法。
【請求項148】
哺乳動物における癌の予防又は阻害の方法であって、請求項91に記載の組換えウイルスの有効量を単独で、又は外因性の免疫刺激分子と組み合わせて、哺乳動物に投与することを含み、該量が癌の予防又は阻害に有効である、方法。
【請求項149】
哺乳動物はHLA−DP分子を発現している、請求項148に記載の方法。
【請求項150】
NY−ESO−1のMHCクラスI拘束T細胞エピトープをコードする組換えウイルスを投与することを更に含む、請求項148に記載の方法。
【請求項151】
NY−ESO−1を発現している腫瘍の腫瘍増殖の阻害方法であって、請求項69、70、76又は77のいずれか1項に記載の核酸のある量を投与することを含み、該量が腫瘍の増殖の阻害に有効である、方法。
【請求項152】
請求項91に記載の組換えウイルスを単独で、又は外因性の免疫刺激分子、化学療法剤、抗生物質、抗真菌剤、抗ウイルス剤又はそれらの組み合わせと組み合わせて含み、かつ医薬として許容できる担体を含む医薬組成物。
【請求項153】
配列番号51〜64、80、又はその機能性部分の少なくとも1つを含むNY−ESO−1のHLA−DP拘束CD4+T細胞エピトープをコードする遺伝子を担持し、かつ発現するトランスジェニック動物。
【請求項154】
請求項51、60、64、65又は72のいずれかに記載のNY−ESO−1のHLA−DP拘束T細胞エピトープと免疫応答性である癌抗原特異的ヒトCD4+Tリンパ球。
【請求項155】
HLA DPB1*0401−0402分子を認識する、請求項154に記載の癌抗原特異的ヒトCD4+Tリンパ球。
【請求項156】
配列番号51〜64又は80の少なくとも1つを含む癌ペプチドキャリアと結合した標的抗原又はその標的エピトープを含む、標的抗原−癌ペプチドキャリア結合体。
【請求項157】
標的抗原が、TRP2、GP−100、TRP1、HIV抗原、gp120、マラリア抗原及びそれらのエピトープからなる群から選択される、請求項156に記載の標的抗原−癌ペプチドキャリア結合体。
【請求項158】
配列番号51〜64又は80の少なくとも1つを含む癌ペプチドキャリアをコードする核酸配列、標的抗原又はその標的エピトープをコードする少なくとも1つの核酸配列を含む核酸ワクチン構築物。
【請求項159】
標的抗原又はその標的エピトープが、TRP2、GP−100、TRP1、HIV抗原、gp120、マラリア抗原及びそれらのエピトープからなる群から選択される、請求項158に記載の核酸ワクチン構築物。
【請求項160】
NY−ESO−1を発現している細胞の増殖の阻害方法であって、請求項29又は91に記載の組換えウイルスのある量を投与することを含み、該量が細胞の増殖の阻害に有効である、方法。
【請求項161】
NY−ESO−1を発現している細胞の増殖の阻害方法であって、請求項23、69又は77のいずれか1項に記載の核酸配列のある量を投与することを含み、該量が細胞の増殖の阻害に有効である、方法。
【請求項1】
NY−ESO−1癌ペプチド、即ち配列番号1及びその変異体からなる核酸配列内にコードされるMHCクラスII拘束T細胞エピトープ、その機能性部分、変異体又は誘導体。
【請求項2】
エピトープが、配列番号2、そのホモログ又は機能性部分からなる配列によってコードされる、請求項1に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ、その機能性部分又は誘導体。
【請求項3】
ペプチドが、配列番号27又はその部分からなる配列によってコードされる、請求項1に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ、その機能性部分又は誘導体。
【請求項4】
配列番号4〜21及び29〜34、それらの機能性部分、誘導体又は組み合わせの少なくとも1つを含むNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項5】
配列番号5〜18、それらの機能性部分、誘導体又は組み合わせからなる群から選択される、請求項4に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項6】
VLLKEFTVSG(配列番号19)、その変異体又は誘導体を含む、請求項4に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項7】
HLA−DR拘束Tリンパ球によって免疫学的に認識される、請求項1〜5又は6のいずれかに記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項8】
Tリンパ球がCD4+である、請求項1〜5又は6のいずれかに記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項9】
癌ペプチドが、非ホジキンリンパ腫、白血病、ホジキンリンパ腫、肺癌、肝癌、転移、メラノーマ、頭部癌、頚部癌、神経芽腫、腺癌、胸腺腫、結腸癌、子宮癌、乳癌、前立腺癌、卵巣癌、子宮頚癌、膀胱癌、腎癌、膵癌、及び肉腫からなる群から選択される癌由来である、請求項4に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項10】
AAG GAG TTC ACT GTG TCC(配列番号28)を含む核酸配列によってコードされる、請求項4に記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープ。
【請求項11】
請求項1〜9又は10のいずれかに記載のNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ及び医薬として許容できる担体を含む医薬組成物。
【請求項12】
更に、免疫刺激分子を含む、請求項11に記載の医薬組成物。
【請求項13】
免疫刺激分子が、HLA−クラスII分子上、又はHLAクラスII分子を発現している細胞上にある、請求項12に記載の医薬組成物。
【請求項14】
更に、B7.1、B7.2、ICAM−1、ICAM−2、LFA−1、LFA−3、サイトカイン又はそれらの組み合わせを含む、請求項11に記載の医薬組成物。
【請求項15】
更に、NY−ESO−1の少なくとも1つのMHCクラスI拘束T細胞エピトープを含む、請求項11に記載の医薬組成物。
【請求項16】
MHCクラスI拘束T細胞エピトープが配列番号22又は配列番号23を含む、請求項15に記載の医薬組成物。
【請求項17】
請求項1〜9又は10のいずれかに記載のNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープを単独で又は組み合わせて含み、かつ必要に応じて、少なくとも1つの免疫刺激分子を含む免疫原。
【請求項18】
免疫刺激分子がHLAクラスII分子である、請求項17に記載の免疫原。
【請求項19】
更に、NY−ESO−1のMHCクラスI拘束T細胞エピトープを含む、請求項17に記載の免疫原。
【請求項20】
MHCクラスII拘束T細胞エピトープとMHCクラスI拘束T細胞エピトープがペプチド結合により一緒に結合している、請求項19に記載の免疫原。
【請求項21】
少なくとも1つのMHCクラスII拘束T細胞エピトープをコードしている、配列番号27、その機能性部分又はホモログを含む単離された核酸配列。
【請求項22】
配列番号24、25、26、又はその部分又は変異体からなる群から選択される、請求項21に記載の単離された核酸配列。
【請求項23】
配列番号4〜21、29〜34、それらの組み合わせ、機能性部分又は誘導体のいずれか1つのアミノ酸配列を含む、NY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープをコードする単離された核酸配列。
【請求項24】
更に、NY−ESO−1の少なくとも1つのMHCクラスI拘束T細胞エピトープをコードする核酸配列を含む、請求項23に記載の単離された核酸配列。
【請求項25】
請求項21、22、23又は24のいずれかに記載の核酸配列を含む組換え発現ベクター。
【請求項26】
請求項25に記載の組換え発現ベクターによって形質転換又はトランスフェクトされた宿主生物。
【請求項27】
抗原提示細胞である、請求項26に記載の宿主生物。
【請求項28】
請求項21、22又は23のいずれかに記載の核酸配列に相補的な核酸配列からなるオリゴヌクレオチド。
【請求項29】
請求項21、22、23又は24のいずれかに記載の核酸配列を含む組換えウイルス。
【請求項30】
レトロウイルス、バキュロウイルス、アンカラウイルス、鶏痘ウイルス、アデノウイルス、ワクシニアウイルス、レンチウイルス、及びレプリカーゼ−ベース−アルファウイルスからなる群から選択される、請求項29に記載の組換えウイルス。
【請求項31】
免疫刺激分子をコードする少なくとも1つの遺伝子を更に含む、請求項29に記載の組換えウイルス。
【請求項32】
免疫刺激分子がHLAクラスII分子である、請求項31に記載の組換えウイルス。
【請求項33】
請求項29〜31又は32のいずれかに記載の組換えウイルスで形質転換又はトランスフェクトされた宿主生物。
【請求項34】
請求項1〜5又は6のいずれかに記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープと結合する、単離された抗体又はその抗原結合部分。
【請求項35】
NY−ESO−1の組換えMHCクラスII拘束T細胞エピトープの製造方法であって、以下:
a.請求項21、22又は23のいずれかに記載のヌクレオチド配列、その機能性部分又は変異体を、発現ベクターに挿入すること;
b.該発現ベクターを宿主細胞に移入すること;
c.エピトープ又はその機能性部分の発現に適切な条件下、宿主細胞を培養すること;及び
d.組換えエピトープ又はその機能性部分を取得すること;
を含む、方法。
【請求項36】
工程(a)において、HLAクラスII分子又はその部分をコードするヌクレオチド配列を発現ベクターに挿入することを更に含む、請求項35に記載の方法。
【請求項37】
哺乳動物における癌又は前癌の存在の検出方法であって、以下:
a.請求項21、22又は23のいずれかに記載の核酸配列、それらの機能性部分又は変異体と、哺乳動物から採取したmRNAの試験用生物学的サンプルとを、該配列と該mRNAとの間で複合体が形成される条件下で接触させること;
b.複合体を検出すること;
c.試験用サンプル中のmRNA量を、既知の正常な生物学的サンプルからのmRNA量と比較することであって、試験用サンプルからのmRNA量の増大が癌又は前癌の指標となること;
を含む、方法。
【請求項38】
癌又は前癌がメラノーマである、請求項37に記載の方法。
【請求項39】
生物学的サンプル中のNY−ESO−1のMHCクラスII拘束T細胞エピトープ又はその部分の検出方法であって、以下:
a.サンプルと、請求項34に記載の該エピトープに特異的な抗体又はその抗原結合部分とを、該抗体と該エピトープとの間で免疫複合体が形成される条件下で接触させること;及び
b.免疫複合体の存在を検出することであって、その存在が生物学的サンプル中のエピトープの指標となること;
を含む、方法。
【請求項40】
哺乳動物における癌の予防又は阻害の方法であって、請求項1〜5又は6のいずれかに記載のNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその機能性部分の有効量を単独で、又はHLAクラスII分子と組み合わせて該哺乳動物に投与することを含み、該量が哺乳動物における癌の予防又は阻害に有効である、方法。
【請求項41】
NY−ESO−1の少なくとも1つのMHCクラスI拘束T細胞エピトープを投与することを更に含む、請求項40に記載の方法。
【請求項42】
哺乳動物におけるメラノーマの阻害方法であって、以下:
a.インビトロでT細胞を、請求項1〜5又は6のいずれかに記載のNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ又はその機能性部分に単独で、又はMHCクラスII分子と組み合わせて、MHCクラスII拘束Tリンパ球を誘発するのに十分な時間、曝すこと;
b.該MHCクラスII拘束Tリンパ球を単独で、又はNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープ、その機能性部分又はネイティブNY−ESO−1タンパク質と組み合わせて、メラノーマを阻害するために十分な量で哺乳動物に投与すること;
を含む、方法。
【請求項43】
哺乳動物における癌の予防又は阻害の方法であって、有効量の、請求項29に記載の組換えウイルスを単独で、又は外因性の免疫刺激分子と組み合わせて、哺乳動物に投与することを含み、該量が癌を予防又は阻害するために有効である、方法。
【請求項44】
哺乳動物がHLAクラスIIDR分子を発現している、請求項43に記載の方法。
【請求項45】
NY−ESO−1のMHCクラスI拘束T細胞エピトープをコードする組換えウイルスの投与を更に含む、請求項43に記載の方法。
【請求項46】
請求項29に記載の組換えウイルスを単独で、又は外因性の免疫刺激分子、化学療法剤、抗生物質、抗真菌剤、抗ウイルス剤又はそれらの組み合わせと組み合わせて含み、かつ医薬として許容できる担体を含む医薬組成物。
【請求項47】
少なくとも、配列番号27のフラグメント、又はその機能性部分又は変異体を含むNY−ESO−1の少なくとも1つのMHCクラスII拘束T細胞エピトープをコードする遺伝子を担持し、かつ発現するトランスジェニック動物。
【請求項48】
請求項1〜5又は6のいずれかに記載のNY−ESO−1のMHCクラスII拘束T細胞エピトープによって誘発される癌抗原特異的ヒトCD4+Tリンパ球。
【請求項49】
HLA−クラスII分子を認識する、請求項48に記載の癌抗原特異的ヒトCD4+Tリンパ球。
【請求項50】
HLA−DR分子を認識する、請求項48に記載の癌抗原特異的ヒトCD4+Tリンパ球。
【請求項51】
ペプチドが、アミノ酸モチーフ:
Xaa1ITQXaa2FXaa3PXaa4(配列番号51)及びそのホモログ(式中、Xaa1、Xaa2、Xaa3、及びXaa4は各々少なくとも1つのアミノ酸である)を含み、該ペプチドは、HLA−DP拘束CD4+Tリンパ球によって免疫学的に認識される、請求項1に記載の癌ペプチド。
【請求項52】
Xaa1が、Trp、Phe、Tyr、Met、Ile、Val、Ala、及びそれらの組み合わせからなる群から選択される、請求項51に記載の癌ペプチド。
【請求項53】
Xaa2が、Cys、Ser、Val、Ala、Thr、及びそれらの組み合わせからなる群から選択される、請求項51に記載の癌ペプチド。
【請求項54】
Xaa3が、Leu、Phe、Tyr、Met、Ile、Val、Ala、及びそれらの組み合わせからなる群から選択される、請求項51に記載の癌ペプチド。
【請求項55】
Xaa4が、Val、Tyr、Ile、Ala、Leu、Pro、及びそれらの組み合わせからなる群から選択される、請求項51に記載の癌ペプチド。
【請求項56】
約9乃至約30個のアミノ酸を含む、請求項51に記載の癌ペプチド。
【請求項57】
約10乃至約20個のアミノ酸を含む、請求項51に記載の癌ペプチド。
【請求項58】
約10乃至約15個のアミノ酸を含む、請求項51に記載の癌ペプチド。
【請求項59】
ホモログがLAGE遺伝子由来である、請求項51に記載の癌ペプチド。
【請求項60】
アミノ酸配列:
Xaa1WITQCFLPVF−Xaa2(配列番号52)及びその変異体
(式中、Xaa1及びXaa2は各々、アミノ酸ではないか、又は1個以上の天然に存在するアミノ酸である)を含む、請求項51に記載の癌ペプチド。
【請求項61】
約10乃至約30個のアミノ酸を含む、請求項60に記載の癌ペプチド。
【請求項62】
約10乃至約20個のアミノ酸を含む、請求項60に記載の癌ペプチド。
【請求項63】
約10乃至約15個のアミノ酸を含む、請求項60に記載の癌ペプチド。
【請求項64】
WITQCFLPVF(配列番号80)からなる、請求項60に記載の癌ペプチド。
【請求項65】
LLMWITQCFLPVFL(配列番号55)、LMWITQCFLPVFLA(配列番号56)、MWITQCFLPVFLAQ(配列番号57)、及びWITQCFLPVFLAQP(配列番号58)を含む、請求項1に記載の癌ペプチド。
【請求項66】
HLA−DPB1*0401−0402拘束CD4+Tリンパ球によって免疫学的に認識される、請求項51に記載のペプチド。
【請求項67】
キャリアタンパク質に結合している、請求項51に記載のペプチド。
【請求項68】
キャリアタンパク質が、KLH、破傷風トキソイド及びアルブミンからなる群から選択される、請求項67に記載のペプチド。
【請求項69】
請求項51、60、64又は65のいずれかに記載のペプチドをコードする単離された核酸配列。
【請求項70】
配列番号53、55〜64のいずれか1つ、又はそれらの組み合わせを含む少なくとも1つのMHCクラスII拘束T細胞エピトープペプチドをコードする単離された核酸配列。
【請求項71】
ストリンジェント条件下、請求項69又は70に記載の核酸配列とハイブリダイズする核酸配列を含む単離されたポリヌクレオチド。
【請求項72】
アミノ酸モチーフSLLMWITQCFLPVF(配列番号54)又はその変異体を有し、HLA−DP拘束CD4+Tリンパ球とMHCクラスI拘束CD8+Tリンパ球の両方によって免疫学的に認識される癌ペプチド。
【請求項73】
変異体が、アルギニン残基の付加をアミノ末端に有する、請求項72に記載の癌ペプチド。
【請求項74】
変異体が、フェニルアラニンの代わりに置換されたアルギニンをカルボキシル末端に有する、請求項72に記載の癌ペプチド。
【請求項75】
HLA−DPB1*0401−0402拘束Tリンパ球及びHLA−A2拘束Tリンパ球によって免疫学的に認識される、請求項72に記載のペプチド。
【請求項76】
請求項72に記載のペプチドをコードする単離された核酸配列。
【請求項77】
配列番号54のペプチドをコードする単離された核酸配列。
【請求項78】
ストリンジェント条件下、請求項76又は77に記載の核酸配列とハイブリダイズする核酸配列を含む単離されたポリヌクレオチド。
【請求項79】
請求項69に記載の核酸配列を含む組換え発現ベクター。
【請求項80】
請求項70に記載の核酸配列を含む組換え発現ベクター。
【請求項81】
請求項76に記載の核酸配列を含む組換え発現ベクター。
【請求項82】
請求項77に記載の核酸配列を含む組換え発現ベクター。
【請求項83】
少なくとも1つのNY−ESO−1 HLA−DR拘束T細胞癌ペプチドをコードする核酸配列を更に含む、請求項79〜82のいずれか1項に記載の組換え発現ベクター。
【請求項84】
NY−ESO−1 HLA−DR拘束T細胞癌ペプチドが、以下:
LPVPGVLLKEFTVSG(配列番号10);
VLLKEFTVSGNILTIRLT(配列番号65);
AADHRQLQLSISSCLQQL(配列番号66);及び
それらの組み合わせからなる群から選択されるアミノ酸配列を含む、請求項83に記載の組換え発現ベクター。
【請求項85】
少なくとも1つの単離されたNY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドをコードする核酸配列を更に含む、請求項83に記載の組換え発現ベクター。
【請求項86】
NY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドが、以下:
Xaa1Xaa2Xaa3GPGGGAPXaa4(配列番号22)
(式中、Xaa1は、アミノ酸ではないか、又は1乃至約20個の天然に存在するアミノ酸であり;
Xaa2は、Ale、Thr、Val、Leu又はArgであり;
Xaa3は、Ser又は保存的置換であり;
Xaa4は、Arg又はLysである)
及びそれらの誘導体を含む、請求項85に記載の組換え発現ベクター。
【請求項87】
核酸配列が、ASGPGGGAPR(配列番号23)をコードする、請求項86に記載の組換え発現ベクター。
【請求項88】
請求項79〜87のいずれか1項に記載のベクターによって形質転換又はトランスフェクトされた宿主細胞又は宿主生物。
【請求項89】
請求項51、60、64、65又は72のいずれか1項に記載の少なくとも1つの癌ペプチドをコードする組換えウイルス。
【請求項90】
請求項89に記載の組換えウイルスによって形質転換された宿主細胞又は宿主生物。
【請求項91】
配列番号51〜64、80のいずれか1つ又はそれらの組み合わせを含む、少なくとも1つのMHCクラスII拘束T細胞エピトープペプチドをコードする単離された核酸配列を含む組換えウイルス。
【請求項92】
ワクシニア、鶏痘、アデノウイルス、レトロウイルス、レンチウイルス、バキュロウイルス、アンカラウイルス、及びレプリカーゼ−ベース−アルファウイルスからなる群から選択される、請求項91に記載の組換えウイルス。
【請求項93】
HLA−DP分子をコードする核酸配列を更に含む、請求項91に記載の組換えウイルス。
【請求項94】
少なくとも1つのNY−ESO−1 HLA−DR拘束T細胞癌ペプチドをコードする核酸配列を更に含む、請求項91に記載の組換えウイルス。
【請求項95】
NY−ESO−1 HLA−DR拘束T細胞癌ペプチドが、以下:
LPVPGVLLKEFTVSG(配列番号10);
VLLKEFTVSGNILTIRLT(配列番号65);
AADHRQLQLSISSCLQQL(配列番号66);
それらの変異体及び組み合わせからなる群から選択されるアミノ酸配列を含む、請求項94に記載の組換えウイルス。
【請求項96】
少なくとも1つの単離されたNY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドをコードする核酸配列を更に含む、請求項91に記載の組換えウイルス。
【請求項97】
NY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドが、以下:
Xaa1Xaa2Xaa3GPGGGAPXaa4(配列番号22)
(式中、Xaa1は、アミノ酸ではないか、又は1乃至約20個の天然に存在するアミノ酸であり;
Xaa2は、Ale、Thr、Val、Leu又はArgであり;
Xaa3は、Ser又は保存的置換であり;
Xaa4は、Arg又はLysである)
及びそれらの誘導体を含む、請求項96に記載の組換えウイルス。
【請求項98】
核酸配列が、アミノ酸配列ASGPGGGAPR(配列番号23)を含むペプチドをコードする、請求項97に記載の組換え発現ベクター。
【請求項99】
請求項91〜98のいずれか1項に記載のウイルスによって形質転換又はトランスフェクトされた宿主細胞又は宿主生物。
【請求項100】
HLA−DP分子をコードする核酸配列を更に含む、請求項99に記載の宿主細胞又は宿主生物。
【請求項101】
請求項51、60、64又は65のいずれか1項に記載のペプチド及び医薬として許容できる担体を含む医薬組成物。
【請求項102】
請求項72に記載のペプチド及び医薬として許容できる担体を含む医薬組成物。
【請求項103】
請求項79〜87のいずれか1項に記載の組換え発現ベクター及び医薬として許容できる担体を含む医薬組成物。
【請求項104】
請求項91〜95のいずれか1項に記載の組換えウイルス及び医薬として許容できる担体を含む医薬組成物。
【請求項105】
免疫刺激分子を更に含む、請求項101に記載の医薬組成物。
【請求項106】
免疫刺激分子が、B7.1、B7.2、ICAM−1、ICAM−2、LFA−1、LFA−3、サイトカイン又はそれらの組み合わせである、請求項105に記載の医薬組成物。
【請求項107】
HLA−クラスII分子又はHLA−クラスII分子を発現している細胞を更に含む、請求項101に記載の医薬組成物。
【請求項108】
HLA−クラスII分子がHLA−DPである、請求項107に記載の医薬組成物。
【請求項109】
HLA−クラスII分子がHLA−DPB1*0401−0402である、請求項108に記載の医薬組成物。
【請求項110】
少なくとも1つのNY−ESO−1 HLA−DR拘束T細胞癌ペプチドを更に含む、請求項101に記載の医薬組成物。
【請求項111】
NY−ESO−1 HLA−DR拘束T細胞癌ペプチドが、以下:
LPVPGVLLKEFTVSG(配列番号10);
VLLKEFTVSGNILTIRLT(配列番号65);
AADHRQLQLSISSCLQQL(配列番号66);及び
それらの組み合わせからなる群から選択されるアミノ酸配列を含む、請求項110に記載の医薬組成物。
【請求項112】
少なくとも1つの単離されたNY−ESO−1 HLAクラスI拘束T細胞癌ペプチドを更に含む、請求項101に記載の医薬組成物。
【請求項113】
NY−ESO−1 HLA−クラスI拘束T細胞癌ペプチドが、以下:
Xaa1Xaa2Xaa3GPGGGAPXaa4(配列番号22)
(式中、Xaa1は、アミノ酸ではないか、又は1乃至約20個の天然に存在するアミノ酸であり;
Xaa2は、Ala、Thr、Val、Leu又はArgであり;
Xaa3は、Ser又は保存的置換であり;
Xaa4は、Arg又はLysである)及びそれらの誘導体を含む、請求項112に記載の医薬組成物。
【請求項114】
NY−ESO−1 HLAクラスI拘束T細胞癌ペプチドは、ASGPGGGAPR(配列番号23)を含む、請求項113に記載の医薬組成物。
【請求項115】
請求項69、70、76又は77のいずれか1項に記載の核酸配列及び医薬として許容できる担体を含む医薬組成物。
【請求項116】
NY−ESO−1 HLA−DP拘束CD4+Tリンパ球を誘発する、請求項51、60、64、65又は72のいずれか1項に記載の癌ペプチドを含む免疫原。
【請求項117】
抗NY−ESO−1抗体を誘発する、請求項116に記載の免疫原。
【請求項118】
アジュバントを更に含む、請求項116に記載の免疫原。
【請求項119】
ペプチドがキャリアタンパク質に結合している、請求項116に記載の免疫原。
【請求項120】
請求項51、60、64、65又は72のいずれか1項に記載の癌ペプチドを含む癌ワクチン。
【請求項121】
請求項51、60、64、65又は72のいずれか1項に記載のNY−ESO−1のHLA−DP拘束T細胞エピトープと結合する単離された抗体又はその抗原結合部分。
【請求項122】
ポリクローナル抗体である、請求項121に記載の抗体。
【請求項123】
モノクローナル抗体である、請求項121に記載の抗体。
【請求項124】
検出可能標識で標識されている、請求項121に記載の抗体。
【請求項125】
検出可能標識が放射性同位体である、請求項121に記載の抗体。
【請求項126】
細胞毒性剤又は細胞増殖抑制剤の付加によって修飾されている、請求項121に記載の抗体。
【請求項127】
請求項121に記載の抗体及び医薬として許容できる担体を含む医薬組成物。
【請求項128】
請求項121に記載の抗体を含むキット。
【請求項129】
イムノアッセイ試薬を更に含む、請求項128に記載のキット。
【請求項130】
抗体が更に、HLA−DP分子と結合する、請求項121に記載の抗体又はその抗原結合部分。
【請求項131】
NY−ESO−1の組換えHLA−DP拘束CD4+T細胞エピトープの製造方法であって、以下:
a.配列番号51〜64、80又はその部分若しくは変異体の少なくとも1つをコードするヌクレオチド配列を、発現ベクターに挿入すること;
b.発現ベクターを宿主細胞に移すこと;
c.エピトープ又はその部分の発現のために適切な条件下で宿主細胞を培養すること;及び
d.組換えエピトープ又はその部分を取得すること;
を含む、方法。
【請求項132】
工程(a)において、HLAクラスII分子又はその部分をコードするヌクレオチド配列を発現ベクターに挿入することを更に含む、請求項131に記載の方法。
【請求項133】
HLAクラスII分子がHLA−DPである、請求項132に記載の方法。
【請求項134】
配列番号51〜64又は80の少なくとも1つのアミノ酸配列を含む癌ペプチドを発現するベクターの製造方法であって、プロモーターと機能可能に連結した配列番号51〜64又は80の少なくとも1つをコードする単離された核酸配列を組み込むことを含む、方法。
【請求項135】
哺乳動物における癌又は前癌の存在の検出方法であって、以下:
a.配列番号51〜64若しくは80又はその部分若しくは変異体の少なくとも1つをコードする核酸配列と、哺乳動物から採取したmRNAの試験用生物学的サンプルとを、該配列と該mRNAとの間で複合体が形成される条件下で接触させること;
b.複合体を検出すること;
c.試験用サンプル中のmRNA量を、既知の正常な生物学的サンプルからのmRNA量と比較することであって、試験用サンプルからのmRNA量の増大が癌又は前癌の指標となること;
を含む、方法。
【請求項136】
癌又は前癌がメラノーマである、請求項135に記載の方法。
【請求項137】
哺乳動物における癌又は前癌の存在の検出方法であって、以下:
a)請求項51、60、64、65又は72のいずれかに記載の癌ペプチドを、該哺乳動物から得たリンパ球の試験用生物学的サンプルと接触させること;
b)該癌ペプチドと免疫応答性のCD4+Tリンパ球を検出することであって、陰性コントロールサンプルと比較した、該癌ペプチドと免疫反応性のCD4+Tリンパ球の数の増大が癌又は前癌の指標となること;
を含む方法。
【請求項138】
生物学的サンプルにおけるNY−ESO−1のHLA−DP拘束CD4+T細胞エピトープ又はその部分の検出方法であって、以下:
a.サンプルを、請求項121〜125のいずれかに記載の、該エピトープに特異的な抗体又はその抗原結合部分と、該抗体と該エピトープとの間で免疫複合体が形成される条件下で接触させること;及び
b.免疫複合体の存在を検出することであって、その存在が、生物学的サンプルにおけるエピトープの指標となること;
を含む、方法。
【請求項139】
哺乳動物における癌の予防又は阻害の方法であって、
請求項51、60、64、69又は72のいずれかに記載の、NY−ESO−1のHLA−DP拘束T細胞エピトープ又はその部分の有効量を単独で、又はHLA−DP分子と組み合わせて哺乳動物に投与することを含み、該量が該哺乳動物において癌の予防又は阻害に有効である、方法。
【請求項140】
HLA−DP分子がHLA−DPB*0401−0402である、請求項139に記載の方法。
【請求項141】
NY−ESO−1のHLA−DR拘束T細胞エピトープを単独で、又はHLA−DR分子と組み合わせて投与することを更に含む、請求項139に記載の方法。
【請求項142】
NY−ESO−1のHLA−DR拘束T細胞エピトープが配列番号10、65又は66の少なくとも1つを含む、請求項141に記載の方法。
【請求項143】
NY−ESO−1のHLAクラスI拘束T細胞エピトープを投与することを更に含む、請求項139に記載の方法。
【請求項144】
HLAクラスI拘束T細胞エピトープが配列番号22又は23を含む、請求項143に記載の方法。
【請求項145】
NY−ESO−1を発現している腫瘍の増殖の阻害方法であって、請求項51、60、64、65又は72のいずれか1項に記載の癌ペプチドのある量を投与することを含み、該量がNY−ESO−1を発現している腫瘍の増殖の阻害に有効である、方法。
【請求項146】
哺乳動物におけるメラノーマの阻害方法であって、以下:
a.インビトロでTリンパ球を、請求項51、60、64、65又は72のいずれか1項に記載のNY−ESO−1のHLA−DP拘束T細胞癌ペプチド又はその部分に単独で、又はHLA−DP分子と組み合わせて、NY−ESO−1 HLA−DP拘束Tリンパ球を誘発する十分な時間、曝すこと;
b.メラノーマを阻害するために十分な量で、哺乳動物に該Tリンパ球を投与すること;
を含む、方法。
【請求項147】
哺乳動物に癌ペプチド又はネイティブNY−ESO−1タンパク質を投与することを更に含む、請求項146に記載の方法。
【請求項148】
哺乳動物における癌の予防又は阻害の方法であって、請求項91に記載の組換えウイルスの有効量を単独で、又は外因性の免疫刺激分子と組み合わせて、哺乳動物に投与することを含み、該量が癌の予防又は阻害に有効である、方法。
【請求項149】
哺乳動物はHLA−DP分子を発現している、請求項148に記載の方法。
【請求項150】
NY−ESO−1のMHCクラスI拘束T細胞エピトープをコードする組換えウイルスを投与することを更に含む、請求項148に記載の方法。
【請求項151】
NY−ESO−1を発現している腫瘍の腫瘍増殖の阻害方法であって、請求項69、70、76又は77のいずれか1項に記載の核酸のある量を投与することを含み、該量が腫瘍の増殖の阻害に有効である、方法。
【請求項152】
請求項91に記載の組換えウイルスを単独で、又は外因性の免疫刺激分子、化学療法剤、抗生物質、抗真菌剤、抗ウイルス剤又はそれらの組み合わせと組み合わせて含み、かつ医薬として許容できる担体を含む医薬組成物。
【請求項153】
配列番号51〜64、80、又はその機能性部分の少なくとも1つを含むNY−ESO−1のHLA−DP拘束CD4+T細胞エピトープをコードする遺伝子を担持し、かつ発現するトランスジェニック動物。
【請求項154】
請求項51、60、64、65又は72のいずれかに記載のNY−ESO−1のHLA−DP拘束T細胞エピトープと免疫応答性である癌抗原特異的ヒトCD4+Tリンパ球。
【請求項155】
HLA DPB1*0401−0402分子を認識する、請求項154に記載の癌抗原特異的ヒトCD4+Tリンパ球。
【請求項156】
配列番号51〜64又は80の少なくとも1つを含む癌ペプチドキャリアと結合した標的抗原又はその標的エピトープを含む、標的抗原−癌ペプチドキャリア結合体。
【請求項157】
標的抗原が、TRP2、GP−100、TRP1、HIV抗原、gp120、マラリア抗原及びそれらのエピトープからなる群から選択される、請求項156に記載の標的抗原−癌ペプチドキャリア結合体。
【請求項158】
配列番号51〜64又は80の少なくとも1つを含む癌ペプチドキャリアをコードする核酸配列、標的抗原又はその標的エピトープをコードする少なくとも1つの核酸配列を含む核酸ワクチン構築物。
【請求項159】
標的抗原又はその標的エピトープが、TRP2、GP−100、TRP1、HIV抗原、gp120、マラリア抗原及びそれらのエピトープからなる群から選択される、請求項158に記載の核酸ワクチン構築物。
【請求項160】
NY−ESO−1を発現している細胞の増殖の阻害方法であって、請求項29又は91に記載の組換えウイルスのある量を投与することを含み、該量が細胞の増殖の阻害に有効である、方法。
【請求項161】
NY−ESO−1を発現している細胞の増殖の阻害方法であって、請求項23、69又は77のいずれか1項に記載の核酸配列のある量を投与することを含み、該量が細胞の増殖の阻害に有効である、方法。
【図1−A】


【図1−B】

【図2−A】
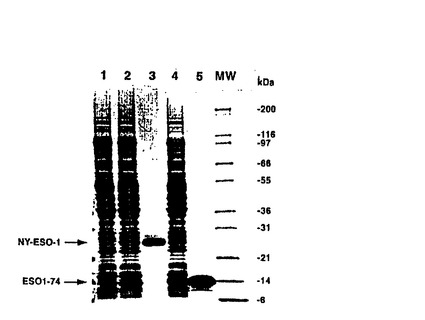
【図2−B】
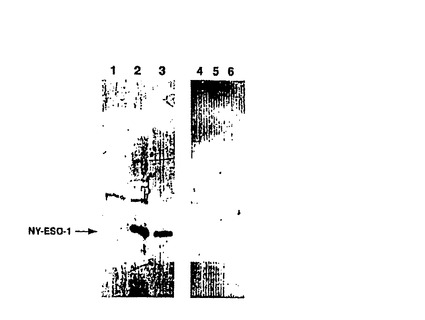
【図2−C】
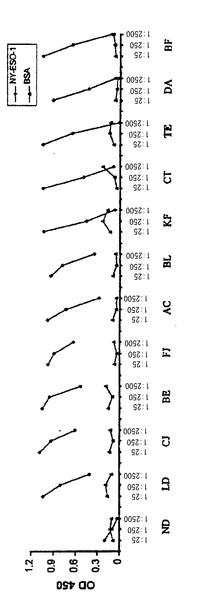
【図3】
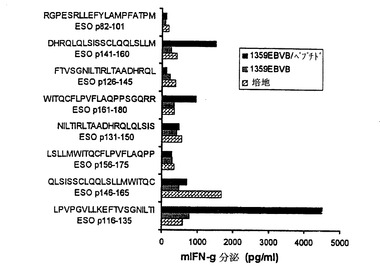
【図4−A】
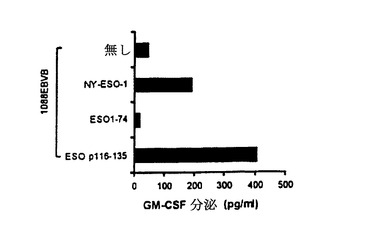
【図4−B】
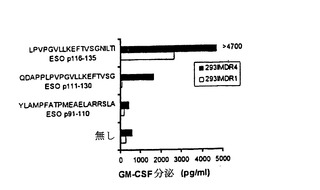
【図4−C】
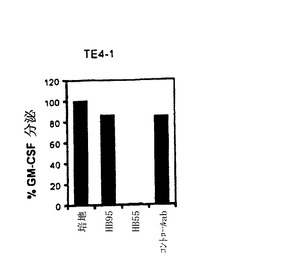
【図4−D】
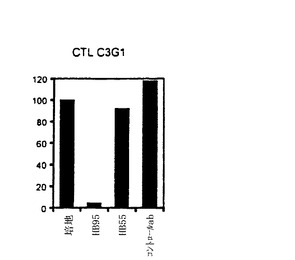
【図4−E】
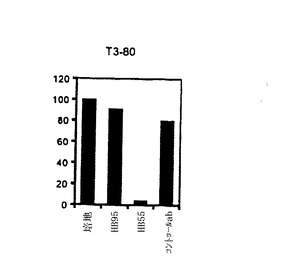
【図5】
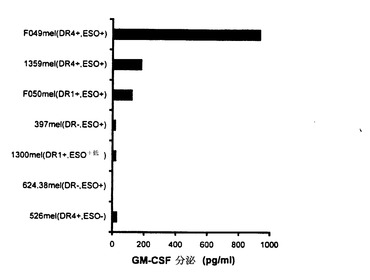
【図6−A】
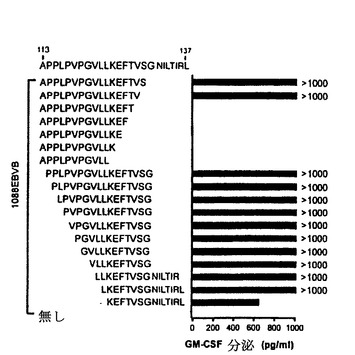
【図6−B】
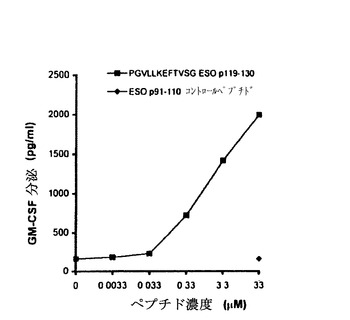
【図7−A】
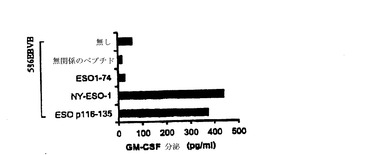
【図7−B】

【図8−A】
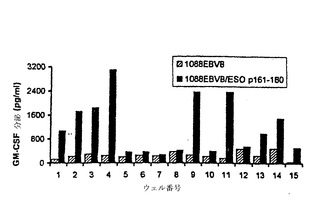
【図8−B】
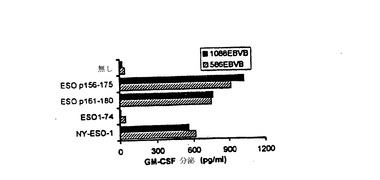
【図8−C】
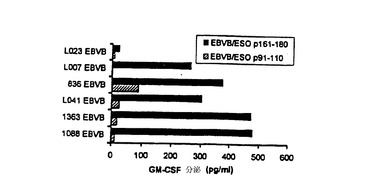
【図9−A】
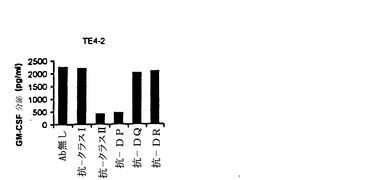
【図9−B】
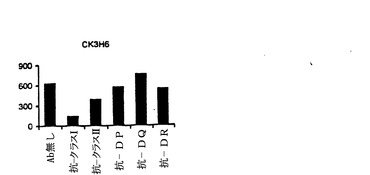
【図9−C】
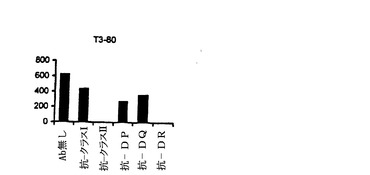
【図10−A】
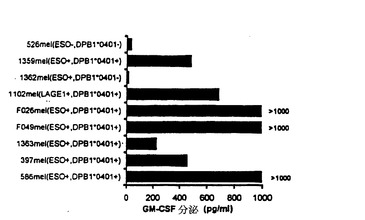
【図10−B】
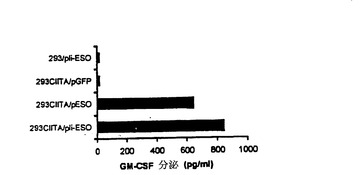
【図11−A】
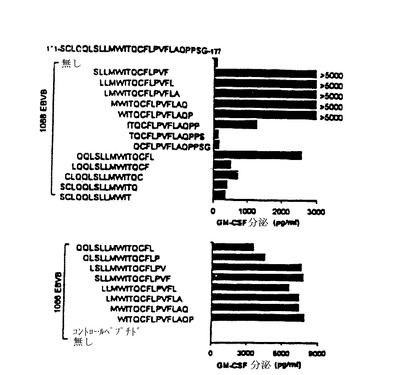
【図11−B】
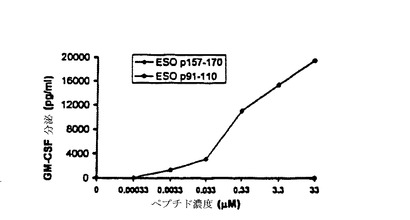
【図11−C】
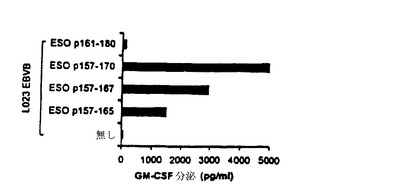
【図12−A】
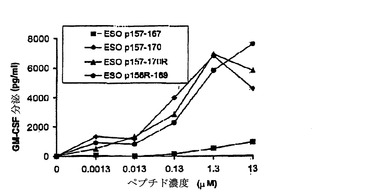
【図12−B】
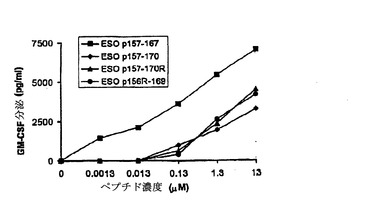
【図1−B】
【図2−A】
【図2−B】
【図2−C】
【図3】
【図4−A】
【図4−B】
【図4−C】
【図4−D】
【図4−E】
【図5】
【図6−A】
【図6−B】
【図7−A】
【図7−B】
【図8−A】
【図8−B】
【図8−C】
【図9−A】
【図9−B】
【図9−C】
【図10−A】
【図10−B】
【図11−A】
【図11−B】
【図11−C】
【図12−A】
【図12−B】
【公開番号】特開2011−135874(P2011−135874A)
【公開日】平成23年7月14日(2011.7.14)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−2419(P2011−2419)
【出願日】平成23年1月7日(2011.1.7)
【分割の表示】特願2001−554422(P2001−554422)の分割
【原出願日】平成13年1月26日(2001.1.26)
【出願人】(398029728)アメリカ合衆国 (3)
【Fターム(参考)】
【公開日】平成23年7月14日(2011.7.14)
【国際特許分類】
【出願番号】特願2011−2419(P2011−2419)
【出願日】平成23年1月7日(2011.1.7)
【分割の表示】特願2001−554422(P2001−554422)の分割
【原出願日】平成13年1月26日(2001.1.26)
【出願人】(398029728)アメリカ合衆国 (3)
【Fターム(参考)】
[ Back to top ]