
癌治療のためのヘキソース化合物
治療を必要とする対象に、治療有効量のヘキソース化合物を投与することによる、膠芽細胞腫及び膵臓癌の治療方法を提供する。本発明は、治療を必要とする対象に、治療有効量のマンノース化合物を投与することを含む、脳腫瘍及び膵臓癌の治療方法を含む。本発明は、治療を必要とする対象に、治療有効量の2−FMを投与することを含む、腫瘍の増殖の治療方法を、さらに含む。

【発明の詳細な説明】
【技術分野】
【0001】
本出願は、2006年2月24日提出の米国仮出願第60/776,793号、2006年4月27日提出の米国仮出願第60/795,621号及び2006年4月28日提出の米国仮出願第60/796,173号の優先権を主張するものである。これらの出願全体を、参照により本明細書に組み込む。
【0002】
本発明は、癌の治療に有用なヘキソース化合物、及びこのような化合物を投与することによる、治療を必要とする対象における、癌が仲介する疾患の治療方法を対象とする。
【背景技術】
【0003】
癌の治療は、多くの場合腫瘍耐性の促進の課題を伴う。プログラムされた細胞死の型であるアポトーシスは、細胞形態学及び細胞死を引き起こす一連の生化学的事象に伴って生じる。アポトーシスの過程は、細胞断片の安全な処分のような方法で達成される。しかし、癌療法を介して細胞内シグナル伝達経路を解明することにより、細胞死の誘導にとって重大な構造及び過程に影響を与えることが可能である。実際、アポトーシスの過程に欠陥があることは、多くの疾患に関係がある。過剰なアポトーシスは、虚血性障害のような細胞消失を引き起こす。他方、アポトーシスの量が不十分であると、癌のように細胞増殖の調節が効かなくなる。
【0004】
悪性神経膠腫の進行に伴い生じる変化は、PI−3K/AKT経路の活性化(通常、PTENの消失によって、又はEGFRのような成長因子の活性を介して)に関連すると思われる。この生存経路は、とりわけ、血管形成の刺激、アポトーシスの阻害剤、及び解糖の活性化を促進する代謝転換を含む、多くの適応変化を優先的に活性化する。同様に、膵臓癌治療の新しい標的は、シグナル伝達経路及び血管形成に関連する分子の標的、具体的には、ras癌遺伝子シグナル経路及びマトリックスメタロプロテアーゼ(MMP)の阻害剤を含む。
【0005】
癌悪性神経膠腫及び膵臓癌などの多くの癌は、従来の療法に対して内在的に耐性であり、重要な治療課題を示す。悪性神経膠腫は、年間発生率が100,000人に6.4人であり(Central Brain Tumor Registry of the United States,2002−2003)、初期脳腫瘍の最も一般的な亜型であり、最も致命的なヒトの癌である。その最も悪性の兆候である多形性膠芽腫(GBM)おいては、最善の治療を尽くしても患者の平均生存期間は9から12か月である。実際に、GBM患者のおよそ1/3では、放射線治療及び化学療法にもかかわらず、彼らの腫瘍は成長を続けるであろう。同様に、診断時の腫瘍の広がり次第では、膵臓癌の予後は一般的に不良であると見なされ、診断後さらに5年生存する罹病者はほとんどなく、完全な回復はまれである。
【0006】
さらに、治療に耐性のある腫瘍の進行に加えて、悪性腫瘍の治療における別の問題は、疾患にかかっていない正常な組織に対する、治療による毒性である。多くの場合化学療法は、細胞が正常か又は悪性であるかどうかに関係なく、分裂の早い細胞を殺すことを標的とする。しかし、広範囲にわたる細胞死及び癌治療に随伴する副作用は、腫瘍の成長調節経路を無効にできれば、腫瘍の抑制に不可欠ではないかもしれない。例えば、1つの研究は療法感作の使用である、すなわち、標準的治療の低用量と併用して、腫瘍細胞において重要な過程を特異的に標的とする薬剤を使用し、他の薬剤の効果を増強する。
【0007】
さらに併用療法は、免疫療法の腫瘍細胞の細胞毒性特異性を用いた、化学療法の腫瘍縮小及び免疫調節要素を併用した、ワクチンに基づいた取り組み方法を含む。しかし、併用療法は通常、単一薬剤のみを必要とする療法より、患者及び医師の両方にとってより困難である。さらに特定の腫瘍は、放射線療法に対する内在性の耐性を有し、多くの化学療法のモダリティは、異なる成長パターンによるものと思われ、成長パターンの型の違いは、個々の腫瘍内の低酸素領域の多様な度合いを表し得る。例えば、神経膠腫は、より急速に成長し、コントラストが増強する腫瘤性病巣に対して、MRIスキャンでコントラストの増強がほとんどないし全くなく、主に浸潤性に成長しうる。同様に、膵臓癌の早期段階は察知できない。相対低酸素部位は多くの場合、低酸素に伴うネクローシス領域を有する、急速に成長する腫瘤の中心、並びに、その上腫瘍の浸潤性要素内のいくつかの相対低酸素領域の両方に見ることができる。したがって、これらの相対低酸素領域のいくつかはより低速で循環する細胞を有すると思われ、それ故化学療法剤に対して耐性であると思われる。
【0008】
近年提唱されたある癌療法は、解糖阻害剤の使用を対象とする。この型の阻害剤は、細胞が好気性代謝から嫌気性代謝に切り替える場合に得られる選択性から利益を得るように設計される。腫瘍の成長のため、癌細胞は血液(酸素供給)から除去される。低酸素下では、腫瘍細胞はグルコース輸送体及び糖分解酵素の両方の発現を上方制御し、順に、好気性環境において正常細胞と比較して、グルコース類似体の取り込み増加を促進する。細胞は、酸素使用して、細胞のミトコンドリア内の脂肪及びタンパク質を燃焼し、エネルギーを産生する(ATPなどのエネルギー貯蔵分子を介して)ことによって生存するので、血液内で、細胞における解糖を遮断しても細胞は殺されない。それに反して、低酸素環境下で細胞内の解糖を遮断する場合、酸素がないため細胞はミトコンドリアを介したエネルギーの産生(脂肪及びタンパク質の酸化)ができず、細胞は死ぬ。それ故、解糖阻害剤は特定の癌の治療に対して期待されるが、低酸素環境下に存在する全ての癌細胞にではない。実際、Otto Warburgによる有名な見解により、十分な量の酸素が存在しても、多くの腫瘍が細胞エネルギーの産生に関して、解糖の優先的利用を選択することが明示されている(酸化的解糖又は「Warburg効果」と呼ばれる)。この腫瘍の適応反応は、悪性神経膠腫にもまた当てはまると思われる。
【0009】
したがって、化学療法に対する耐性を示し、異なる成長パターンの若しくは腫瘍内に多様な度合いの低酸素領域を有する、及び/又は血管形成を刺激する若しくはアポトーシスを阻害する生存経路を有する成長パターンを呈する、癌の治療の必要性がある。
【発明の開示】
【発明が解決しようとする課題】
【0010】
癌を予防、阻害及び調節するヘキソース化合物及びこれらの医薬組成物、並びに癌、特に膠芽細胞腫及び膵臓癌を治療するための該化合物の使用が見出された。本発明は、癌並びに癌が仲介する障害及び病態の治療に有用なヘキソース化合物の使用を開示する。膠芽細胞腫及び膵臓癌の治療方法は、治療を必要とする対象に、治療有効量のヘキソース化合物を投与することを含む。特に興味深いのは、治療を必要とする対象に、治療有効量の2−FMを投与することを含む、腫瘍の増殖の治療方法である。本発明は、治療を必要とする対象に、マンノース化合物を投与することによる癌の治療方法を含む。
【課題を解決するための手段】
【0011】
悪性神経膠腫のための治療の選択肢は依然として限られている。これは一部には、利用可能な多くの化学療法の選択肢に対して、細胞が内在的に耐性であることによる。また一部には、悪性腫瘍の示す成長パターンの差にもよると思われる。すなわち、神経膠腫は、主に浸潤性に成長でき、コントラストを増強すると腫瘤性病巣が急速に成長しているのに対して、コントラストを増強しないとMRIスキャンではほとんど見られない。多くの研究により、これらの異なる型の成長パターンもまた、個々の腫瘍内の低酸素領域の様々な度合いを表すことが示されている。相対低酸素部位は、多くの場合ネクローシス領域を伴う急速に成長する腫瘤の中心、並びに、同様に腫瘍の浸潤性要素内のいくつかの相対低酸素領域の両方に見ることができる。したがって、これらの相対低酸素領域のいくつかはより低速で循環する細胞を有すると思われ、それ故多くの化学療法剤に対してより耐性であると思われる。さらに、十分な量の酸素が存在しても、多くの腫瘍が解糖を経ることを選択すること(酸化的解糖又は「Warburg効果」と呼ばれる)を記述したWarburgによる見解は、悪性神経膠腫にもまた当てはまると思われる。本発明者らは、これらの特徴から、神経膠腫及び他の高度に解糖的に維持された腫瘍、例えば膵臓癌は、解糖の阻害剤に対して感受性であり得、腫瘍の成長に関して有意な影響を有する可能性があることを前提とした。
【0012】
したがって、一般的に脳に、特に神経膠腫に特有の追加の特徴は、CNSに糖を貪欲に取り込むグルコース輸送体の発現の増加である。本発明者らは、これらの特徴から、神経膠腫が解糖阻害剤に著しく感受性であるはずの唯一の病状を表すことを前提とした。この仮説を試験するために、本発明者らは、低酸素状態及び正常な酸素状態の両方の下で、インビトロで、多くの神経膠腫細胞系のパネルに対して公知の解糖阻害剤を使用した。さらに薬剤の効果を、同所性の神経膠腫異種移植片を有する動物において、多くの異なる投与スキームを使用して試験した。
【0013】
酸素の存在下でもエネルギー産生のための第1供給源として解糖を優先的に利用する、高悪性度神経膠腫による代謝の転換である「Warburg効果」、これは、一部はHIF−1a及びPI−3キナーゼ経路の活性化により作動する。解糖の有効な阻害剤である2−デオキシグルコースは、エノラーゼによる2−デオキシグルコース−6−リン酸の変換を遮断し、帯電したリン酸基のため細胞内にこの種を蓄積する。
【0014】
細胞の必要とするエネルギーのために解糖系を優先的に使用する、公知の代謝変化は、高悪性度の神経膠腫を含む新生物において起こる。これらの変化は、解糖系に必要な重要な酵素の産生並びにグルコース輸送体の上方制御を誘導する、HIF−1a及びPI−3キナーゼ活性化を含む生存経路により作動される。この解糖系表現型は大きな特徴であり、これらは通常酸素下でも優勢である。この表現型は認知されており、「Warburg効果」としてすでに述べられている。この表現型の変化により、これらの腫瘍は正常細胞より解糖阻害剤に対して感受性が強いはずである。糖をベースとした解糖系阻害剤のグループ及び他のマンノース化合物は、治療薬としての機能を果たすことができる。原型的な糖をベースとした阻害剤である2−デオキシグルコースが、許容でき、且つ有力な抗神経膠腫効果を有することを、本研究において示した。単独又は細胞毒性化学療法との併用のどちらかであるヘキソース化合物は、癌、特に神経膠腫及び膵臓癌の治療に有効である。さらに、この解糖系表現型は、腫瘍環境内で低酸素状態によりまず動作するので、この型の治療は抗血管形成療法と見なされるべきである。実際、抗血管形成療法を「エスケープ」できる腫瘍は、解糖阻害剤及び/又はヘキソース化合物全般に対して優先的により感受性であると思われる。
【0015】
本発明者らは、糖をベースとしたヘキソース化合物が、高悪性度の神経膠腫腫瘍及び膵臓癌の治療に有効であることを示した。さらに、化合物の他の阻害剤型は、CNSに有利に取り込まれるように設計され、2−DGが現在有している有利な経口生体利用性を維持する。細胞毒性の薬剤及び抗血管形成剤の両方と併用したヘキソース化合物に関する継続中の研究は、未来の臨床的コンビナトリアルな試行のための知的な糸口を楽観的に提供するであろう。
【0016】
ヘキソース化合物は、6個の炭素原子を含む任意の単糖類を意味し、且つ含む。ヘキソースの1つのクラスは、アルドヘキソースファミリーであり、これは例えばグルコース、ガラクトース及びマンノースを含む。アルドヘキソースはさらに、2−デオキシグルコース、フコース、シマロース及びラムノースなどの、様々なデオキシ糖もまた含み得る。ヘキソースの他のクラスは、フルクトース及びソルボースを例とする、ケトヘキソースファミリーである。本発明のヘキソースは、通常は天然発生的なD−配置であるが、ヘキソースはさらにL−エナンチオマーでもあり得る。本発明のヘキソースは、αアノマー、βアノマー及びこれらの混合物を含み得る。本発明のヘキソースはいずれも、場合により置換されていてもよい。このような置換は、ヒドロキシル基を、フッ素、塩素又は臭素などのハロゲンと置き換えることを含む。本発明において、置換は通常ヘキソースのC−2炭素においてであり、六員環いす型立体配座においてヘキソースのアクシアル配位又はエクアトリアル配位のどちらかをとり得る。アクシアルであるC−2における置換は、糖をマンノース誘導体又はマンノ配置の糖と指定する。エクアトリアルであるC−2における置換は、糖をグルコース誘導体又はグルコ配置の糖と指定する。
【0017】
本発明の実践に有用なヘキソース化合物は、参照により本明細書に組み込まれる、米国特許第6,670,330号及び米国特許出願第20030181393号、第20050043250号及び第20060025351号に開示された化合物を含む。本発明のある実施形態において、好ましい化合物は、2−デオキシ−グルコース(2−DG)、2−デオキシ−マンノース(2−DM)、2−フルオロ−グルコース(2−FG)及び2−フルオロ−マンノース(2−FM)などの糖をベースとした腫瘍増殖阻害剤である。
【0018】
「中枢神経系の腫瘍」は、良性であろうと悪性であろうと、脳、脊髄又は他の中枢神経系組織内の異常成長組織のいずれをも意味する。特に、毛様細胞性星状細胞腫、低悪性度星状細胞腫、未分化星状細胞腫及び多形性膠芽腫(GBM又は膠芽細胞腫)などの神経膠腫を含む。「中枢神経系の腫瘍」は、さらに脳幹神経膠腫、上衣腫、神経節細胞腫、若年性毛様細胞性星状細胞腫、混合性神経膠腫、希突起膠腫及び視神経膠腫などの、他の型の良性又は悪性の神経膠腫も含む。「中枢神経系の腫瘍」は、さらに脊索腫、頭蓋咽頭腫、髄芽腫、髄膜腫、松果体腫瘍、下垂体腺腫、未分化神経外胚葉性腫瘍、神経鞘腫、血管の腫瘍及び神経線維腫などの非神経膠腫も含む。最後に、中枢神経系の腫瘍はさらに、悪性細胞が体内の他の部分から中枢神経系に広がった、転移性腫瘍も含む。
【0019】
本発明による「治療すること(treating)」、「治療(treatment)」又は「緩和」は、治療的処置及び予防的(prophylactic)又は防止的(preventative)手段の両方を指し、目的は中枢神経系の腫瘍の予防又は成長遅延、腫瘍の大きさの減少、或いは腫瘍を完全に除去することである。治療を必要とする人々は、特定された中枢神経系の腫瘍を有する対象、中枢神経系の腫瘍を有すると疑われる対象、中枢神経系の腫瘍の進行の危険があると特定された対象を含む。本発明の方法によるヘキソース化合物の治療量を受けた後に、以下の状態の1つ又は複数が観察された場合に、対象は中枢神経系の腫瘍を良好に「治療」されている:腫瘍の大きさの減少又は腫瘍がなくなる;腫瘍の成長の阻害又は中止;腫瘍の転移の阻害又は中止;並びに/或いは罹患率及び死亡率の減少又は生活の質の向上などの、腫瘍に伴う1つ又は複数の症状のある程度の緩和。
【0020】
ヘキソース化合物が、既存の脳腫瘍細胞の成長を防止する及び/又は既存の脳腫瘍細胞を殺す程度により、細胞増殖阻害性及び/又は細胞毒性であると見なすことができる。
【0021】
「同時投与すること(coadministering)」又は「同時投与(coadministration)」という用語は、療法の同時の又は連続した投与を包含することを意図する。例えば同時投与は、解糖系阻害剤及び化学療法剤の両方を1つの組成物において投与することを含み得る。同時投与はまた、複数のこのような組成物を同時に投与することも含み得る。或いは、同時投与は複数のこのような組成物を同じ期間中の異なる時間に投与することも含み得る。
【0022】
本発明によるヘキソース化合物は、限定するものではないが、神経膠腫又は他の脳腫瘍において酸化的解糖を阻害できる化合物である、解糖系阻害剤を含み、2−デオキシグルコース、2−フルオロ−グルコース、2−フルオロ−マンノースなどのヘキソース化合物を含み得る。
【0023】
本明細書中で先に定義した抗増殖治療は、単独の療法として適用でき、又は少なくとも1種の本発明の化合物に加えて、1種又は複数種の他の物質及び/又は治療を含み得る。このような治療は、治療用の個別の成分を同時、連続又は別々に投与する手段で達成できる。本発明の化合物はまた、公知の抗癌剤又は細胞毒性薬剤及び放射線療法などの治療との併用において有用であり得る。固定用量として処方する場合、このような併用製品は、本明細書に記載の用量範囲内の本発明の化合物及び承認された範囲内の他の医薬活性剤を用いる。解糖系阻害剤は、他の抗癌剤又は細胞毒性薬剤もまた伴う化学療法の投与計画の一部として連続的に、及び/又は非化学療法治療、例えば外科手術又は放射線療法と併せて使用できる。
【0024】
化学療法剤は、限定するものではないが、治療薬の3つの大きなカテゴリー:(i)抗血管形成薬、例えばリノミド、インテグリン−α−β3の機能の阻害剤、アンジオスタチン、ラゾキサン、(ii)細胞増殖抑制剤、例えば抗エストロゲン(例えば、タモキシフェン、トレミフェン、ラロキシフェン、ドロロキシフェン、ヨードキシフェン)、プロゲストゲン(例えば、酢酸メゲストロール)、アロマターゼ阻害剤(例えば、アナストロゾール、レトロゾール、ボラゾール、エキセメスタン)、抗ホルモン、抗プロゲストゲン、抗アンドロゲン(例えば、フルタミド、ニルタミド、ビカルタミド、酢酸シプロテロン)、LHRHアゴニスト及びアンタゴニスト(例えば、酢酸ゴセレリン、ロイプロリド)、テストステロン5−α−ジヒドロレダクターゼ阻害剤(例えば、フィナステリド)、ファルネシルトランスフェラーゼ阻害剤、抗侵害剤(例えば、マリマスタットのようなメタロプロテイナーゼ阻害剤、及びウロキナーゼプラスミノーゲン活性化因子受容体機能の阻害剤)及び成長因子機能の阻害剤(このような成長因子は、例えばEGF、FGF、血小板由来成長因子及び肝細胞成長因子を含み、このような阻害剤は、成長因子抗体、成長因子受容体抗体(例えば、アバスチン(ベバシズマブ)及びエルビタックス(セツキシマブ))、チロシンキナーゼ阻害剤及びセリン/スレオニンキナーゼ阻害剤を含む)並びに(iii)内科的腫瘍学で使用される抗増殖/抗新生物薬及びこれらの組み合わせ、例えば代謝拮抗剤(例えば、メトトレキサートのような抗葉酸薬、5−フルオロウラシルのようなフルオロピリミジン、プリン及びアデノシン類似体、シトシンアラビノシド)、挿入抗腫瘍抗生物質(例えば、ドキソルビシン、ダウノマイシン、エピルビシン及びイダルビシンのようなアントラサイクリン、マイトマイシン−C、ダクチノマイシン、ミトラマイシン)、白金誘導体(例えば、シスプラチン、カルボプラチン)、アルキル化剤(例えば、ナイトロジェンマスタード、メルファラン、クロラムブシル、ブスルファン、シクロホスファミド、イフォスファミドニトロソウレア、チオテパ)、抗有糸分裂薬(例えば、ビンクリスチンのようなビンカアルカロイド及びタクソール(パクリタキセル)、タキソテール(ドセタキセル)のようなタキソイド及びポチロン類似体、ディスコデルモライド類似体及びエレウセロビン類似体などの最新の微小管(安定)剤(microbtubule agents))、トポイソメラーゼ阻害剤(例えば、エトポシド及びテニポシドのようなエピポドフィロトキシン、アムサクリン、トポテカン)、細胞周期阻害剤、生物反応修飾物質及びベルケイド(ボルテゾミブ)などのプロテアソーム阻害剤、を含む。
【発明を実施するための最良の形態】
【0025】
当分野で普通の技術の1つにより、本発明に開示する治療方法が、複数の投与経路を介して、様々な量/濃度のヘキソース化合物を用いて達成できることが容易に認識されるであろう。好ましい投与経路は、使用するヘキソース化合物により変更でき、このような経路は、限定するものではないが、経口、口腔、筋肉内(i.m.)、静脈内(i.v.)腹腔内(i.p.)、局所又は他の任意のFDAに承認された投与経路を含む。投与濃度又は治療濃度は、治療する対象及び投与するヘキソース化合物により変わるであろう。ある実施形態において、ヘキソース化合物の濃度は、1mgから50gm/kg体重におよぶ。
【0026】
まず、一連の2−フルオロ、2−ブロモ及び2−クロロ−置換グルコース類似体を調製し、解糖経路におけるグルコースに対する有望な競合的基質として、このような類似体が2−デオキシ−グルコース(2−DG)と類似の手段で解糖系阻害剤として作動できることを分析した。本発明者らは、2−フルオロ−D−マンノースが有効な抗腫瘍剤であることを発見した。なぜならその特性が、2−フルオロ−D−マンノース(本明細書中では「2−FM」とも称する)が、フッ素原子と水素の大きさの類似性を考慮すると2−デオキシ−D−グルコース(2−デオキシ−D−マンノースと同じ)と似ており、又は誘導効果及び水素結合の形成の可能性に関しては水素よりマンノースのヒドロキシ基が似ていることにより、D−マンノースと似ている可能性があるという、どちらかの事実に由来すると思われるためである。その後の状況において、2−フルオロ−D−マンノースは、D−マンノースが関連する生物機能、代謝及び生物過程に作用し得る。また、D−グルコース及びD−マンノースの両方に作用できる効果の組み合わせは、細胞過程に関する。
【0027】
実際、図15から18に提供したデータは、2−フルオロ−D−マンノースが2−DGより強力であり、また膵臓Colo357−FG細胞における2−フルオロ−D−グルコースの活性より良い又は同様の活性を有することを示している。さらに2−フルオロ−D−マンノース(2−FM)を、他の2−デオキシ−D−マンノース類似体、すなわち2−クロロ−D−マンノース(2−CM)及び2−ブロモ−D−マンノース(2−BM)と比較した。驚いたことに、また予期せずに2−フルオロ−マンノースはこの一連の他のものより強力である。具体的には、データは、2−フルオロ−D−マンノース(2−FM)が、ブロモ(2−BM)及びクロロ(2−CM)の両方の類似体より、U251膠芽細胞腫脳腫瘍細胞の成長を阻害することにおいて、明らかに優れていることを示している。2−FMはさらに、低酸素状態下より通常酸素下でU87膠芽細胞腫細胞に対して驚くほど良い活性を示した(図17)。さらに、予測不能な、2−FMの作用の少なくとも1つの様式は、すなわち2−FMが腫瘍脳細胞において自食作用を強く誘導する能力を示すことであり、それ故腫瘍細胞系に対するその作用機序の少なくとも1つの説明を提供する。
【0028】
すぐ下に示したように、2−デオキシグルコース(2−DG)は2個の水素を糖のC−2の位置に有する。糖の六員環いす型配座において、これらの2個の水素はアクシアル及びエクアトリアル位を占める。
【化1】
【0029】
基本的に、2−フルオロマンノース(2−FM)は2−デオキシグルコース(2デオキシマンノースと同じことである)のアクシアル水素をフッ素に置き換える。フッ素は、水素と等比体積であると一般的に見なされる。したがっていくつかの態様において、2−FMの化学的性質は2−DGと似ているかもしれない。実際に、2−FMは、この等比体積の根拠に基づく解糖系阻害活性を示すだろう。しかし、フッ素は実質的に水素より電気的に陰性であり、結果として水素結合のモチーフに関係することができる。この点において、2−FMはマンノースにより近く作用でき、それ故2−FMは高マンノース型オリゴ糖合成においてN結合型糖脂質/タンパク質経路を撹乱できると思われる。
【0030】
要するに、2−FMは驚くほど優れた増殖効果を腫瘍細胞に対して示し、2−デオキシ−D−グルコース(本明細書中では「2−DG」とも称する)及び2−デオキシ−2−フルオロ−D−グルコース(本明細書中では「2−FG」とも称する)より強力であると思われる。以下に論じるように、化合物2−FMを、231−GFP乳癌、U251膠芽腫多形成脳腫瘍(図16)及びCo1o357−FG膵臓ヒト癌細胞系(図15)において特に試験した。U251及びCo1o357−FG細胞において、2−FMを、2−DG、2−FG、2−デオキシ−2−クロロ−D−マンノース(本明細書中では時として「2−CM」とも称する)、2−デオキシ−2−ブロモ−マンノース(本明細書中では「2−BM」とも称する)、2−デオキシ−クロロ−D−グルコース(本明細書中では「2−CG」とも称する)及び2−デオキシ−2−ブロモ−D−グルコース(本明細書中では「2−BG」とも称する)と直接比較した。膠芽細胞腫(図16、17及び18)及び膵臓癌(図15)の両方において、2−FMは比較した薬剤の中で最も強力な薬剤であり、観察された差は2−FMとそのクロロ及びブロモ誘導体の間で特に大きかった。この差は、2−DGと比較した場合にもまた有意であった。したがって、該データは2−FMが、腫瘍細胞増殖の阻害において2−DG及び2−FGと異なった働きをする可能性を示す。該データはさらに、2−FMが癌、特に脳及び膵臓の腫瘍のための非常に有効な抗腫瘍治療であり得ることを示す。
【0031】
より具体的には、図15は、2−デオキシ−グルコース(2−DG)、2−フルオロ−グルコース(2−FG)又は2−フルオロ−マンノース(2−FM)を用いた治療のいずれかに反応したCo1o357細胞系の、MTT試験を介した細胞生存能力の用量反応曲線を明示する。見て分かるように、用量反応曲線の左への変化は、2−FMが2−DG又は2−FGのどちらよりも強力であることを示す。図16は、マンノースの2位におけるハロゲンの性質が活性に作用する重要な因子であることを明示する。神経膠腫細胞系U251MGを、2−クロロ−マンノース(2−CM)、2−ブロモ−マンノース(2−BM)又は2−フルオロ−マンノース(2−FM)のどれかを用いて処理した。再度、細胞生存能力をMTT試験により測定したところ、結果は、他のハロゲンをベースとした類似体と比較した場合に、2−FMの優れた活性を明らかに示す。図17は、低酸素状態(酸素1%未満)又は正常な酸素状態(酸素20%)において、2−フルオロ−マンノース(2−FM)を用いて処理したU87細胞系のMTT試験を明示する。見て分かるように、データは、U87細胞系におけるこの薬剤による異常事態が、低酸素状態において感受性がより強くはないことを表す。これは、2−FMに関する代わりの作用機序が殺細胞効果に関与し得ることを示す可能性がある。
【0032】
図18は、2−フルオロ−マンノース(2−FM)の唯一且つこれまでに特定されていない機序を明示する。U87MG神経膠腫細胞系において、2−FMは自食作用を介した細胞死を誘導する。図は、アクリジンオレンジを用いて染色することによる、酸性小胞オルガネラ(Acidic Vesicular Organelles)(AVO)のフローサイトメトリー分析の結果を表しており(手順を参照されたし)、これは自食作用過程に特有であり、特徴的である。結果は、2−FMの曝露量が増えるに従って、自食作用を起こす細胞のパーセントが増加することを示す。この効果を見るためにはわずか40時間の曝露時間は極めて短いため、この自食作用の誘導程度は強い印象を与える。
【0033】
2−DGは、解糖系阻害剤の添加の範囲を評価するために現在臨床試験において投与されており、この阻害剤は固形腫瘍に見出される最も耐性の細胞集団である緩慢に成長する低酸素腫瘍細胞を殺し、迅速に分裂する通常の酸素状態の細胞を標的とする、標準的化学療法の治療有効性を増加できる。本発明は、一部は、2−DG又は2−FMを用いた場合、酸素の存在下でも特定の腫瘍細胞系が殺されるが、2−デオキシ−2−フルオロ−D−グルコース(2−FG)を投与した場合にはそうはならないという発見から始まった。2−FG及び2−DGは両方とも解糖を阻害するため、この効果に関与する、解糖の遮断以外の機序を推測した。
【0034】
1970年代に実施された研究は、2−DG及び2−FMがウィルスのコートタンパク質のN結合型グリコシル化に干渉し、この干渉はマンノースを添加することにより逆転できるという報告を導いた。マンノースとグルコースとの違いは、2−炭素位の水素の配向にあり、2−DGは2位に2個の水素を有する(水素及びヒドロキシル基の代わりに)ため、マンノース及びグルコースの両方の場合のように、2−DGをマンノース又はグルコース類似体のどちらかとして見なすことができる。したがって、2−DGは解糖及びグリコシル化の両方に作用できる。
【0035】
本発明は、細胞が低酸素環境下又は正常な酸素環境下にかかわらず、ヘキソース誘導体を単独又は、限定するものではないが、正常な酸素状態細胞を標的とする細胞毒性薬剤、抗血管形成剤、放射線療法及び外科手術を含む、他の抗腫瘍治療と併用して使用する腫瘍細胞の増殖を阻害する方法を提供する。本発明はさらに、2−DG、2−CM及び2−FMなどの類似体を、正常な酸素状態の(グリコシル化への干渉を介して)及び全ての低酸素(解糖の遮断を介して)の、特定の腫瘍型の癌細胞集団両方を標的にできる細胞毒性薬剤としての臨床使用のための根拠を提供する。
【0036】
以下の実施例は、本発明の有効性を検証し、2−DG、2−CM及び2−FMが毒性であるが2−FGは毒性でなく、正常な酸素状態で成長する腫瘍細胞型を選択することを確認するデータを提供する。実施例に記載した実験のいくつかは、解糖の阻害が通常酸素効果に関与する機序であるのとは対照的に、グリコシル化に干渉するかどうかを決定するように設計した。理論に縛られるつもりはないが、得られた結果は、これらの化合物がグリコシル化を阻害でき、その結果、それらの細胞が低酸素環境下にあるかどうかとは独立して、特定の癌細胞型を殺すことを裏付けるものである。
【0037】
結果はまた、2−FM、2−DG及び2−CMが脂質結合オリゴ糖鎖の組立てを撹乱し、糖タンパク質合成への干渉の指標であり得る、小胞体ストレス応答(unfolded protein response)(UPR)を誘導するが、2−FGはしないという結論を裏付ける。言い換えると、UPRは、感受性であるが耐性ではない細胞のUPR特異的アポトーシスシグナルの活性化を引き起こす。
【0038】
通常酸素下で2DG、2−FM、及び2−CMに感受性である腫瘍細胞型を特定した。細胞を腫瘍から単離し、エクスビボで試験し、細胞が通常酸素下で2DG、2−CM、及び2−FMに感受性であるかどうかを確定した。以下の実施例は、細胞が感受性かどうかを確定する方法を例示する。他の実施形態において、密接に関連のある2−DGに感受性である分子署名及び耐性細胞対を試験細胞系と比較する。ホスホマンノースイソメラーゼと、グリコシル化に関連する他の酵素及び2−DG蓄積に関連する酵素との、濃度及び/又は活性の違いを記載する。
【0039】
酸素の存在下で(正常な酸素状態)、2−DGは腫瘍細胞系のサブセットに有毒である。これまでの研究により、2−DGを用いて正常な酸素状態で処理した場合、腫瘍細胞及び正常細胞は成長を阻害されるが、殺されないということが実証されたので、この結果は意外であった。この成長阻害のこれまでの見解は、解糖経路の中間体濃度が減少するため成長が減少するように、2−DGが、通常酸素下で細胞内の解糖の遮断に十分な高濃度まで蓄積されることによるものと考えられ、これは細胞増殖に関与する様々な同化作用のために使用される。しかし、ミトコンドリアの機能が正常であるため細胞が死なないならば、その後好気的に処理された細胞は2−DGによる解糖の遮断を生き延びられる。どのように細胞が通常酸素下で2−DGに感受性であり得るかの1つの有望な説明は、したがって、細胞はミトコンドリアに欠陥を有するということである。これに関して、腫瘍細胞は、ミトコンドリア呼吸に欠陥があるため、エネルギー(ATP)産生のために酸化的リン酸化の代わりに嫌気的解糖を介してグルコースを利用することが公知である。しかし、さらなる実験により、オキサマート及び2−FGなどの他の解糖阻害剤はこれらの細胞に対して毒性でなく、ミトコンドリア呼吸の血管がそれらの2−DGに対する感受性が原因であるとは思えないことが実証された。したがって、解糖の遮断以外の機序が、これらの選択された細胞系において通常酸素下で、2−DGの毒性に関与すると仮定した。
【0040】
したがって、他の仮説によりこの正常な酸素状態での細胞毒性の機序を説明できる。1つの有力な機序はグリコシル化の干渉である。この有力な機序の裏付けは、1970年代末から一連の研究論文において特定され得、その中で特定のウィルスにおいてN結合型糖タンパク質合成が、2−DGを含む多くの糖類似体によって阻害されたことが報告された。
【0041】
グルコースは、3つの主要な経路:解糖、ペントースリン酸分路、グリコシル化を介して代謝される。図7は解糖及びグリコシル化代謝経路のスキーム図である。グルコースが細胞質に入った後、ヘキソキナーゼがグルコースの炭素6をリン酸化し、グルコース−6−リン酸(G6P)の合成が起こる。G6Pがホスホグルコースイソメラーゼ(PGI)により、フルクトース−6−リン酸に転換した場合、解糖経路において継続でき、ATP及びピルビン酸を産生する。或いは、G6Pはまた、マンノースを含む様々な糖成分の合成に使用でき、脂質結合オリゴ糖の組立てに必要であり、その合成はERで実施される。2−DGは、3つの代謝経路のうちの2つに干渉することが示されている:2−DGはPGIを阻害することによって解糖を遮断でき、又は2−DGは、グアノシン2リン酸(GDP)ドリコールリン酸結合マンノースのN−アセチルグルコサミン残基への転移に干渉することによってN結合型オリゴ糖前駆体の組立てを撹乱でき、及びマンノースを細胞質からERの内腔に運搬するために必要なドリコール−Pを減少させることができる。
【0042】
2−DGは、炭素2の両方の位置に水素を有し、マンノース類似体に似ているため上記の通りである。対照的に、フルオロ類似体のこの位置におけるフッ素の存在は、新しい鏡像異性体の中心を作り出し、それでフルオロ誘導体はグルコース又はマンノースのどちらかの類似体としか見なすことができない;これらの類似体の表現において、フッ素部分をマンノース類似体の炭水化物環の平面の「上に」又は上側に、及びグルコース類似体の炭化水素間の平面の下側に描く。
【0043】
脂質結合オリゴ糖鎖に加えるマンノースは、まずグアノシン2リン酸(GDP)又はドリコールリン酸への転移によって活性化しなくてはならない。2−DGは、2−DG−GDPへの転換を受け、これは、脂質結合オリゴ糖の組立て時に、マンノースをN−アセチルグルコサミン残基に加えるためのマンノース−GDPと競合する。したがって、科学文献に報告された実験において、2−DG処理の結果産生された異常なオリゴ糖はウィルス糖タンパク質の合成を低減させた。これらの実験において、2−DGの阻害効果は外因性のマンノースを加えることによって逆転するがグルコースを加えた場合はせず、さらに2−DGはいくぶんマンノース類似体のように作用することが確認された。これらの調査員らはまた、別のマンノース類似体の2−フルオロ−マンノース(2−FM)が、マンノースによって逆転する2−DGと同様の効果を有することを示し、これらの類似体のマンノース配置はそれらのグリコシル化への干渉にとって重要であり得ることを示した。
【0044】
さらに、遺伝学的研究は、グリコシル化の撹乱が意味深い生物効果を有し得ることを示した。糖タンパク質糖鎖不全症候群1b型に苦しむ患者に、酵素のホスホマンノースイソメラーゼ(PMI)は存在しない。この酵素の不在は血清糖タンパク質のグリコシル化の減少をもたらし、蛋白漏出性腸症を特徴とする血栓症及び胃腸障害を引き起こす。外因性マンノースをこれらの患者の食事に加えた場合、彼らの症状が消え、彼らの血清糖タンパク質は正常に戻り、彼らは病気から回復した。この観察は、実験データが、外因性マンノースが、2−DGを用いて正常な酸素濃度状態で処理した場合に殺される選択された腫瘍細胞を救うことができたことを示すように、本発明において有用な化合物に関する作用機序と一致する。これらの特定の腫瘍は、PMIが下方制御されている、又はこの酵素に欠陥を有するかの、どちらかである可能性がある。他方、N結合型グリコシル化に必要なマンノース中間体を産生する酵素は、これらの細胞において上方制御され得、マンノース−GDPに対して2−DG−GDPをより高い比率でもたらし、その結果正常な酸素濃度状態において2−DGに対するこの異常な感受性の原因となる。
【0045】
機序にかかわらず、本発明は、2−DG並びに他のグルコース及びマンノース類似体を、正常な酸素濃度状態においても腫瘍を治療する単剤として投与することによって癌を治療する方法を提供する。該化合物は、ヒト乳癌(SKBR3)、非小細胞肺(NSCLC)、神経膠腫、膵臓及び骨肉腫の癌細胞系を含む、多くの腫瘍細胞系に対して有効であり、これらの全てが2−DGの相対的低用量を用いて処理した場合に、細胞死を起こすことが実証されている。
【0046】
図3Bは、2−DG、2−FM及び他の薬剤を用いて、通常酸素下で表示の用量で、72時間処理したSKBR3細胞の反応を示したチャートである。細胞毒性を、トリパンブルー排除法により測定した。結果は、2−DG及びマンノース類似体の2−FMが毒性であり、一方グルコース類似体の2−FGは毒性ではないことを示す。さらに、乳酸デヒドロゲナーゼ段階で解糖を遮断するピルビン酸塩類似体であるオキサマートもまた、通常酸素下で成長するこれらの細胞に対して毒性ではない。対照的に、マンノース類似体の2−FMはまた、これらの細胞において毒性であることが証明され、再度マンノース骨格がこの活性を有する化合物にとって重要であったことを示した。
【0047】
2−DGの阻害効果は、外因性マンノースの添加により逆転されたが、グルコースを加えた場合には逆転せず、さらに2−DGがマンノース類似体として作用することが確認された。他の試験により、2−DGもまた、通常酸素下で成長するNSCLCに対して毒性であり、1mMのマンノースの添加がこの毒性を逆転することが示された。
【0048】
このデータは、2−DG及び2−FMは、グリコシル化に干渉するため、通常酸素下で成長する選択腫瘍細胞に対して毒性であることをさらに裏付ける。これらのマンノース類似体がこの機序を介して働き、解糖の遮断を介してではないことの付加的証拠は、ミスフォールド及び又はミスグリコシル化のタンパク質を示す小胞体ストレス応答タンパク質のGRP78及び94が、2−DG及び2−FMによって用量依存性様式で上方制御されるが、2−FGによってはされないことであり、この効果はマンノースの添加によって同様に逆転する。
【0049】
したがって、マンノース類似体の2−DG及び2−FMは、通常酸素下で成長する選択腫瘍細胞型に対して毒性であり、マンノースの添加によりこの毒性は逆転するが、グルコース類似体の2−FGではそうではない。2−FGは、2−DGより解糖をよく阻害するので、グリコシル化への干渉及び解糖を阻害しないことは、この効果に関与すると考えられる機序である。上記のように、2−DGが、ウィルスコートタンパク質のN結合型グリコシル化に干渉すること、及び外因的に加えられたマンノースがこの効果を逆転することは、報告されている。2−DGのSKBR3、NSCLC及び2種の他のヒト腫瘍細胞系に対する通常酸素下での毒性効果は、したがってグリコシル化への干渉によるものと思われる。この機序的理論が正しければ、その後マンノースの添加により、これらの細胞系において2−DGの毒性は逆転されるはずである。実際、被験細胞系の1つ(NSCLC)において、1mMのマンノースは6mMの2−DGの毒性効果を逆転する。
【0050】
マンノースの血中濃度は50〜60μg/mlとして知られているので、2−DGの毒性を逆転するために必要な最小マンノース用量を確定するための、用量反応性実験が実施できる。例えば、FBSは普通、残留量のマンノースを含んでいるため、成長培地に透析されたウシ胎児血清(FBS)を添加した実験によって達成できる。さらに、2−DGの毒性を逆転するためには、マンノースの添加が必要であって、他の糖は必要でないことを確認するために、糖タンパク質合成に関与することが公知の糖、すなわちグルコース、フコース、ガラクトースなどを2−DG毒性の逆転能力に関して試験できる。これらの糖のいずれかが同様に毒性を逆転できる場合、その後それらの活性を、UPRの誘導及びその結果、オリゴ糖鎖の伸長への干渉並びにコンカナバリンAの結合についての、2−DGの効果の逆転に関する以下に記載した実験において、マンノースの活性と比較できる。全体的にこれらの実験は、生理的濃度のマンノースの存在下で抗腫瘍活性をもたらす、インビボで使用できる2−DG又は2−FMの用量を、インビトロで評価することを可能にするものである。しかし、本発明の方法に使用する、経口投与の2−DG、2−FM及び2−CMの治療有効量は、通常は5〜500mg/患者の体重1kg、例えば50〜250mg/kgの範囲であろう。一実施形態において、用量は、約100mg/患者の体重1kgである。
【0051】
本発明はまた、腫瘍又は他の癌が現在の治療方法に感受性の細胞を含むかどうかを確定するために臨床医が使用できる、多くの診断方法を提供する。一実施形態において、腫瘍由来の細胞を、それらが2−DG,2−FM,又は2−CMにより殺されるかどうかを確定するために、通常酸素下で試験する。別の実施形態において、この試験を実施し、その後マンノースを加え、マンノースが細胞毒性効果を逆転させるかどうかを確定する。
【0052】
別の実施形態において、感受性に関する試験を、N結合型グリコシル化を指標として使用して実施する。上記のように、2−DG及び2−FMは、脂質結合オリゴ糖鎖の組立てを撹乱する、(2)正常な糖タンパク質合成への干渉の指標であり得る、小胞体ストレス応答(UPR)の誘導する、及び(3)2−DGに感受性であるが耐性ではない細胞においてUPR特異的アポトーシスシグナルを活性化する、が2−FGはしない。さらに、マンノースはこれらの効果を逆転する。したがって、これらの同じ試験を対象となる腫瘍又は癌細胞に実施でき、その細胞が本方法を用いた治療に感受性であるかどうかを確定できる。
【0053】
上記のように、ウィルスに感染した細胞のERの細胞質表面上で、マンノースの脂質結合オリゴ糖鎖への取り込みが起こり、この取り込みを2−DG又は2−FMのGDP誘導体、すなわちGDP−2DG及びGDP−2−FMにより阻害できる。普通は、5回目のマンノースを加えた後で、脂質結合オリゴ糖鎖は、ERの内腔に向かってフリップする。成長する鎖にマンノースを加え続けるために、ドリコールリン酸(Dol−P)を、細胞質からERのマトリックスにマンノースを輸送するための担体として使用する。2−DG−GDPはドリコールに結合することに関してマンノース−GDPと競合し、その結果N結合型グリコシル化にさらに干渉する。さらにドリコール結合2−DGもまた、ER内のオリゴ糖鎖上へのマンノースの輸送に競合する。したがって、実験を、脂質結合オリゴ糖鎖前駆体及びマンノース誘導体、すなわちマンノース−6−リン酸、マンノース−1−リン酸、GDP−マンノース及びDol−P−マンノースの形成についての2−DG及び2−FMの効果を実証するために、2−DGに感受性及び耐性の両方の細胞系において実施できる。これは同様に2−DG及び2−FMにより阻害される、オリゴ糖組立てにおける1つ又は複数のステップを実証する。これは同様に、2−DG、2−FM、及び/又は2−CMに曝露することで、オリゴ糖を産生する(及び産生しない)ことに基づいて、感受性又は耐性として他の細胞型の特徴付けを可能にするものである。
【0054】
これまでに確立されたクロマトグラフィー法を、SKBR3及びNSCLC細胞内のマンノース誘導体及び脂質結合オリゴ糖前駆体の回収及び量の測定に使用できる。簡潔に言えば、細胞は[2−H3]マンノースを用いて標識でき、細胞溶解物を、クロロホルム/メタノール(3:2)及びクロロホルム/メタノール/水(10:10:3)を、それぞれDol−P−Man及び脂質結合オリゴ糖を回収するために用いて抽出した。
【0055】
Dol−P−Manを含むアリコートを薄層クロマトグラフィーに供与し、一方脂質結合オリゴ糖は、HPLCによって分離できる。溶出分画は、液体シンチレーション計測により分析できる。マンノースリン酸及びGDP−マンノースは、下降ペーパークロマトグラフィーによって、各分画から放出された[2−3H]マンノースは弱酸加水分解によって分離でき、測定できる。2−DG又は2−FMを用いて処理した細胞に由来する値を、未処理の対照と比較し、N結合型オリゴ糖前駆体及びマンノース誘導体に関するこれらの薬剤の効果を実証できる。外因性マンノースは、2−DGの毒性を逆転するので、マンノースがまた観察された、2−DGによるグリコシル化の動揺を逆転するかどうかを試験できる。
【0056】
2−DG及び2−FMに加えて、N結合型グリコシル化の特定のステップを阻害できる、2種の他のグリコシル化阻害剤、ツニカマイシン及びデオキシマンノジリマイシン(DMJ)を陽性対照として使用できる。ツニカマイシンは、第1のN−アセチルグルコサミン残基のドリコールリン酸への付加に干渉し、DMJは、N結合型オリゴ糖鎖の末端の3マンノース残基を切り取る、マンノシダーゼIの特異的阻害剤である。したがって、外因性マンノースは、毒性又はこれらの薬剤のどちらかのグリコシル化に関する効果のうち、いずれかを逆転できないはずである。さらに、グルコース類似体の2−FGは、通常酸素下でSKBR3及びNSCLC細胞を殺さないが、解糖の遮断及び低酸素細胞を殺すことにおいて2−DGより強力であるため、グリコシル化に作用することなく解糖に干渉でき、そのためこのような試験にツールとしても使用できる。
【0057】
小胞体(ER)内のN結合グリコシル化の過程への干渉は、糖タンパク質の不適切なフォールディンクを引き起こし、これは、小胞体ストレス応答(UPR)と呼ばれるERのストレス応答を誘発する。DNAの損傷に対するP53応答によく似て、ERはストレスに対して(1)内在シャペロン(GRP78及びGRP94)の誘導を介したフォールディング能力の増加、(2)タンパク質合成の停止による自身の生合成負荷の減少、及び(3)フォールディングしていないタンパク質の分解の増加、によるほとんど同じ方法で応答する。ストレスが緩和できない場合、アポトーシス経路が開始され、続いて細胞が死ぬ。したがってグリコシル化への干渉の1つの指標は、UPRの上方制御である。
【0058】
SKBR3細胞を2−DGを用いて処理した場合、これらのERストレス応答タンパク質である、GRP78及び94の両方が用量増加の機能として増加するが、マンノースがこの誘導を逆転する。2−FGは、このタンパク質を誘導しない。したがって、本発明の別の実施形態において、この応答を腫瘍又は癌細胞が本方法に従った治療に感受性であるかどうかを確定するために使用する。通常酸素下で2−DGに感受性でない細胞系も、これらのタンパク質の上方制御の不在が、それらの2−DGに対する耐性に相関する、陰性対照として同様に使用できる。
【0059】
ERストレスを克服できない場合に、アポトーシスシグナルが開始される。ERストレスは、CHOP/GADD153を介したミトコンドリア依存性アポトーシス経路、BCL−2を下方制御する核転写因子及びヒトのカスパーゼ4及び5、並びにマウス細胞系のカスパーゼ12によるミトコンドリア依存性経路を誘導する。したがって、実験を実施し、ERストレスに特有のアポトーシスシグナルが、2−DGに感受性であるが耐性ではない細胞において活性化されるかどうかを確定できる。CHOP/GADD153の上方制御及びカスパーゼ4及び5の活性化は、ウェスタンブロットにより検査できる。先の試験と同様に、この上方制御が2−DG感受性系に特有である場合、その後試験癌細胞において観察された上方制御は、細胞が由来した癌が本発明に従った治療に感受性であることの指標として機能を果たす。
【0060】
SKBR3は糖タンパク質ErbB2を豊富に発現するので、2−DGがこのタンパク質のN結合型グリコシル化に作用し、ミスフォールディング及び分解を引き起こすであろうことが期待される。2−DGを用いて処理したSKBR3細胞由来のErbB2のウェスタンブロットを未処理の細胞のものと比較して、このタンパク質の全濃度を測定できる。さらに、2−DG処理後のErbB2のマンノース含有量を、このタンパク質を免疫沈降し、コンカナバリンA、高マンノース型N結合オリゴ糖を認識するレクチンを用いてブロットすることによって分析できる。マンノース類似体はErbB2だけでなく、全N結合型糖タンパク質に含まれるマンノースを阻害できるのと思われるので、これらの細胞から得た全細胞溶解物をこのレクチンを用いてさらに解析できる。全タンパク質に結合するポンソー染色を、陰性対照として使用でき、2−DG及び2−FMが糖タンパク質に特異的に作用し、再度この方法論又は類似の方法論が、癌又は腫瘍の細胞が本発明に従った治療に感受性であるかどうかを確定するために使用できることを検証できる。
【0061】
N結合型グリコシル化への干渉を示すERストレスが、2−DG及び2−FMによって起こることが実際に確認されたとしても、ERとは対照的に細胞質において起こるO−グリコシル化への干渉もまた評価できる。科学文献により、2−DGは、O−グルコシル化転写因子であるSp1からのN−アセチルグルコサミン残基の切り取りを阻害でき、その個別のプロモーターへの結合を阻害することが報告されている。Sp1は、多くの癌遺伝子の活性化に重要な転写因子であり、2−DGをSp1に作用させれば、なぜ通常酸素下で成長するSKBR3細胞が2−DGに感受性なのかを、少なくとも部分的に説明できる。したがって、2−DG及び2−FMを用いた処理の後のSp1のグリコシル化のパターンを、免疫沈降及びWGA、O−グリコシル化タンパク質に特異的に結合するレクチンを用いて解析することによって調査できる。2−DGがSp1及びO−結合型グリコシル化に作用するという点で、グリコシル化のこの変化を測定し、腫瘍又は他の癌細胞系が2−DG仲介殺細胞に感受性であることの指標として使用できる。
【0062】
細胞死は、小胞体ストレス応答によって誘発され、ミスフォールドタンパク質に応答して、各細胞の小胞体において起こり、追加の薬剤であるベルシペロスタチンを投与することによって促進できる。したがって、一実施形態において、2−DG、2−FM及び/又は2−CMを、癌治療を必要とする患者に投与し、ベルシペロスタチンを前記患者に同時投与する。
【0063】
同様に、タンパク質のミスフォールドに応答して起こる細胞死は、ミスフォールド糖タンパク質のタンパク質分解を、プロテオソーム阻害剤を用いて遮断することによって促進できる。したがって、別の実施形態において、本発明は、2−DG、2−FM及び/又は2−CMと併用して、プロテオソーム阻害剤を投与することによる、癌の治療方法を提供する。一実施形態において、プロテオソーム阻害剤はベルケイド(Velcade)である。
【0064】
癌の特定の型は、本方法による治療に他の癌より感受性である可能性がある。このような型を特定するために、本発明の方法に従って様々な細胞型の種類を試験できる。例えば、ATCCから様々な癌細胞系を得ることができ、酸素の存在下で2DG及び2FMなどのマンノース類似体に非常に敏感な他の細胞型を特定するためにそれらを上記のようにスクリーニングできる。5mM以下の2−DG又は2−FM濃度で殺される細胞を、感受性であるとして特定する。これらの感受性腫瘍細胞系は、さらに2−FG及びオキサマートに対する感受性に関して、それぞれ用量20mM及び30mMまで試験できる。グリコシル化への干渉が、2−DG及び2−FMの毒性の様式である場合、それならばこれらの細胞系は、ミトコンドリアの酸化的リン酸化に欠陥を有さない限り、他の解糖系阻害剤である2−FG及びオキサマートに対して耐性であるはずである。これらの細胞のミトコンドリアの機能性を確認するために、呼吸を、例えばクラーク式電極装置を使用して測定できる。2−DG及び2−FMの毒性が、これらの細胞系におけるグリコシル化への干渉によるものであることを確認するために、マンノースによる細胞死の回復を上記のように検査できる。
【0065】
1つの細胞が本方法に耐性であり、別の細胞は耐性でないことに関する分子基礎は、グルコースからGDPマンノースの合成に関与する遺伝子、すなわちグルコース−6−リン酸をマンノース−6−リン酸に転換する、ホスホグルコースイソメラーゼ(PMI)の発現における違いによると思われる(図7)。上で述べたようにPMIの欠失が、グリコシル化症候群1bの原因であり、これは血清糖タンパク質のグリコシル化の減少をもたらし、この欠陥を有すると特定された患者に血栓症及び胃腸障害を引き起こすことを示した。マンノースを食事に加えることにより、患者の症状を軽減し、患者の糖タンパク質を正常化することを示した。したがって、この酵素の欠如又は下方制御により、今までに試験された感受性細胞系における2−DG及び2−FMの毒性及び外因性マンノースによる逆転を説明できる。
【0066】
PMIの下方制御又は欠失が、感受性細胞系において2−DGの毒性を引き起こし得る理由は、この酵素が存在しない状態では、細胞がN結合オリゴ糖前駆体の合成を外因性マンノース(血清中に存在)に依存するからである。哺乳動物の血清中のマンノース濃度(50〜60μg/ml)又はインビトロ研究に使用した培地中のマンノース濃度は、グルコース濃度よりも有意に低いことは公知である。したがって、PMIが欠失した又は下方制御された細胞において、低用量の2−DG及び2−FMは血清中に存在する低量のマンノースと都合よく競合し、この糖のオリゴ糖鎖上への付加を完全に遮断する。他方、正常なPMIを有する細胞はグルコースからGDP−マンノースを産生でき、したがってオリゴ糖の組立てを完全に撹乱するためにはより多くの用量の2−DG又は2−FMが必要である。これにより、大部分の被験細胞が通常酸素下で2−DGに対して耐性であることの理由が説明できる。この酵素の活性の直接測定は本発明に従って使用でき、PMIの欠如又は低濃度のPMIが、通常酸素下で成長する選択細胞において、2−DG及び2−FMに対する感受性に関与するかどうか、またもしそうならば、その時は本方法に従った治療に感受性の腫瘍及び癌細胞の特定に使用できるかどうかを確定できる。あまり見込みはないが、この異常な感受性を説明する別の可能性は、正常な酸素圧の下で成長する場合、これらの選択細胞におけるPMIが2−DG及び2−FMによって大部分の正常な細胞系及びこれらの薬剤に作用されない腫瘍細胞系より阻害されるということである。このことを直接試験するために、細胞抽出液をSKBRの耐性及び感受性の細胞対から単離でき、グルコース−6−Pからマンノース−6−Pへの転換能力を、2−DG及び2−FMの存在する又は存在しない状態で確定できる。
【0067】
PMI活性の減少が、SKBR3の感受性細胞において2−DGによる毒性に関与していない場合、その時にこれを説明する代わりの機序は、オリゴ糖の組立てに使用するマンノース誘導体の産生に関与する酵素、すなわち、ホスホマンノムターゼ(PMM)及びGDP−Manシンターゼをコードする遺伝子の上方制御である(図7)。2−DGに感受性の細胞のグリコシル化が増加し、それ故、これらの酵素のどちらか1種、又は両方が上方制御される可能性がある。このような細胞は2−DG−GDPがさらに蓄積され、それ故グリコシル化が低速又は低い能力で起こった耐性細胞よりも、グリコシル化及びそれに続く細胞死に、より大幅な干渉を引き起こすであろう。
【0068】
グリコシル化の上方制御が機序であり、それにより細胞が2−DGに感受性になるかどうかにかかわらず、細胞内に蓄積された又は取り込まれた2−DGの全量はまた、細胞の感受性の増加の原因となる。したがって、[3H]で標識した2−DGを使用した、取り込み及び蓄積の研究は、グルコース輸送体濃度がより高い細胞が、本方法に従った治療に、より強い感受性を示すかどうかを確定するために実施できる。
【0069】
感受性細胞を2−DGの漸増用量を用いて処理し、生存細胞を選択することによって、感受性細胞から2−DG耐性変異体を得ることができる。耐性変異体及びそれらの親感受性対応物を、記載の方法に使用できる。このような研究は、細胞が2−DGに耐性になる機序を理解する方法を提供するはずであり、それ故この薬剤の臨床的に優れた使用に適用できると思われる。前述の議論は、分子署名を、どの腫瘍細胞型が酸素の存在下で2−DG及び2−FMに感受性であろうか予測するために使用できることを反映している。
【0070】
細胞死の実行は、アポトーシスとネクローシスとの範囲に及ぶ顕著な柔軟性を示す。DNAの切断の型、細胞膜の組成の変化、完全性及び状態を調査することによって細胞死の様式を比較する、確立された方法を使用することにより、グリコシル化への干渉によって及び解糖の阻害によって誘導される細胞死の機序を確定できる。解糖及び酸化的リン酸化の両方の阻害は、深刻なATPの減少をもたらし、その結果アポトーシスからネクローシスへの転換を引き起こす。ATPはカスパーゼの活性化に必要なので、ATPが大幅に減少した場合、アポトーシスは遮断され、最終的にエネルギーがなくなり、細胞はネクローシスを介して死に至る。解糖系阻害剤を用いて処理した好気性細胞は、エネルギー源としてアミノ酸及び/又は脂肪のどちらかに支えられて、酸化的リン酸化を介してATPを産生できる。したがって、2−DGがUPR応答を誘導し、通常酸素下で細胞死を引き起こす場合、細胞はアポトーシスに陥るであろうと考えられる。逆に言えば、低酸素細胞モデルにおいて、2−DGの用量が解糖を遮断するために十分多い場合これらの細胞はATPが減少し、ネクローシスを介して死ぬはずであることが期待される。
【0071】
したがって、アポトーシス及びネクローシスに関する検査の確立された方法を使用でき、2−DGがアポトーシス、ネクローシス及び/又は両方の混合を介して細胞を殺しているかどうかを確定できる。いくつかのアポトーシスパラメーターを、フローサイトメトリー分析の使用によって、ネクローシスとアポトーシスを識別するために検査できる。2−DGによる処理の後で、細胞をアネキシンV及びヨウ化プロピジウムを用いて二重染色し、細胞表面上のホスファチジルセリンの露出及び細胞膜の完全性の喪失をそれぞれ検出できる。アネキシンV単独又はアネキシンV及びヨウ化プロピジウムの両方、のどちらかを用いた染色により、アポトーシスが示され、一方、ヨウ化プロピジウム単独を用いた染色によりネクローシスが示される。さらに、アポトーシスの最終結末の2つである、核DNAの断片化及び一本鎖DNAの形成もまた測定できる。これら2つの後者のパラメーターは、アポトーシス細胞死に独特であることが報告されており、アポトーシスとネクローシスを差別化するために、様々な研究者に使用されている。ATP濃度はまた、それらが検出された死の様式と相関するかどうかを確定するために検査できる。
【0072】
さらに、2−DGが低酸素細胞においてアポトーシス及びネクローシスの両方を誘導する場合、その時低酸素状態の下で2−FGにより誘導された細胞死の様式を確定できる。上に述べたように、2−FGはグリコシル化に干渉せず、2−DGより強力な解糖阻害剤である。したがって、2−FGにより誘導される細胞死は、ネクローシスを介して単独で起こるであろうことが期待される。
【0073】
通常酸素下で、インビトロで2−DG及び/又は2−FM及び/又は2−CMに感受性であると判明した、ヌードマウスにおいて容易に成長する細胞系を、2−DG(及び2−FM及び2−CM)がインビボで与えられた場合、該細胞系に対して単剤として有効であることを実証するために使用できる。腫瘍が特定の大きさに達した後に、2−DGを用いた治療が腹腔内注射を介して適用されるであろう。これらの動物においてあらかじめ確立された最小致死量に従った2−DGの用量及び治療計画を、腫瘍の退縮及び細胞毒性を実証するために使用できる。
【実施例】
【0074】
(実施例1)
材料及び方法
耐性変異体の単離。2−DG感受性SKBR3及びNSCLC細胞を、2−DGの漸増用量に曝露し、耐性コロニーを単離し、適切な用量の2−DGでクローン化する。クローン化した2−DG耐性細胞を、その後分析し、野生型の感受性対応物と、この独自の感受性に関与できる特定の遺伝子の発現に関して比較する。
【0075】
薬剤及び抗体。Rho123、オリゴマイシン、スタウロポリン及び2−DG、2−FG、2−FM、ツニカマイシン、デオキシマンノジリマイシンは、Sigma Chemical Co.から入手する。以下の一次Absを使用できる:モノクロナール抗体を、HIF−la及びLDH−a(BD Biosciences);erbB2(Calbiochem,USA);Grps78&94,(StressGen,USA);カスパーゼ4及び5(StressGen,USA);並びにアクチン(Sigma Chemical Co.)に;ポリクロナール抗体を、GLUT−1(USA Biological)及びGADD153/CHOP(Santa Cruz,USA)に。二次抗体はホースラディッシュペルオキシダーゼ標識ウサギ抗マウス及びヤギ抗ウサギ(Promega,Co.)である。
【0076】
細胞毒性検査及び高速DNA含有量分析。細胞を37℃で24時間、5%CO2においてインキュベートし、その時に薬剤処理を開始し72時間続ける。この時に、吸着細胞をトリプシン処理し、それらの個別の培養培地と混合し、その後400gで5分間遠心分離機にかける。細胞を含むペレットを、細胞毒性検査のために1.5mlの培地/トリパンブルー混合物、又は核DNA含有量及びCoulter XLフローサイトメーターによる細胞周期の測定のためにヨウ化プロピジウム/低張クエン酸塩染色溶液のどちらかに再懸濁する。DNA分布ヒストグラムを作成するために最低10000細胞を分析する。
【0077】
乳酸検査。乳酸を、処理済み培養液又は未処理培養液から0.025mlの脱タンパク質化培地を、0.1mlの乳酸デヒドロゲナーゼ、(1000単位/ml)、2mlのグリシンバッファー(グリシン0.6モル/L及びヒドラジン、pH9.2)及び1.66mg/mlのNADを含む、反応混合物に加えることによって測定する。脱タンパク質化を、試験培養物からの0.5mlの培地を1mlの8%w/v過塩素酸を用いて処理し、30秒ボルテックスにかけ、その後この混合物を4℃で5分間インキュベートし、1500gで10分間遠心分離機にかけることにより起こす。上清をさらに3回遠心分離機にかけ、0.025mlの最終的に不要物を除いた上清を乳酸の測定に使用する。NADHの形成を、Beckman DU r 520 UV/vis分光光度計を用いて340nmで測定し、これは乳酸塩標準曲線により確定された乳酸濃度に直接一致する。
【0078】
2−DGの取り込み。細胞をペトリ皿に播種し、37℃で24時間、5%CO2においてインキュベートする。培地をその後除去し、プレートをグルコース及び血清を含まない培地で洗浄する。2mlの、3H標識2−DGを含み、血清を含まない培地を皿に加え(1γCi/プレート)、プレートを適切な時間インキュベートする。培地をその後除去し、プレートを4℃で、100μMの標識のない2−DGを含み、血清を含まない培地を用いて3回洗浄し、0.5mlの1NのNaOHを加える。37℃で3時間(又は一晩)インキュベートした後で、細胞をこそげ取り超音波により均一化した(10秒)。溶液を、3Hの定量化のためにチューブに回収した(タンパク質検査のために一部は保存)。100μLの蟻酸、250μLの試料及び7mlのシンチレーションカクテルを、3H計測バイアルに混ぜ合わせ、シンチレーションカウンターを用いて読み取る。輸送速度(nモル/mgタンパク質/時間)を全CPM/比放射能/全タンパク質により算出する。
【0079】
ATP定量検査。ATPライトキット(lite kit)(Perkin Elmer社)を、ATP濃度の定量化に使用できる。約50μLの細胞溶解液を、96ウェルプレート(ボトム白)において100μLの細胞懸濁液に加える。プレートを室温で、シェーカーにのせて(700rpm)5分間インキュベートする。50μLの基質溶液を、続けてウェルに加え、さらに5分間室温で振とう(700rpm)する。プレートをその後10分間暗順応させ、発光を測定する。
【0080】
代謝標識並びにDol−P Man及び脂質結合オリゴ糖(LLO)の抽出。Lehleにより記載された手順に従って、細胞を[2−3H]マンノースを用いて30分間標識し、2mlの氷冷メタノールにこそげ取り、超音波処理により溶解する。4mlのクロロホルムを加えた後、材料を超音波で分解し、その後4℃において5000rpmで10分間遠心分離機にかける。上清を回収し、ペレットをクロロホルム/メタノース(3:2)(C/M)を用いて2回抽出する。Dol−P Man及び小型の脂質結合オリゴ糖を含む、混ぜ合わせた上清を、N2下で乾燥させ、3mlのC/Mに溶解し、洗浄し、シリカゲル60アルミニウムシート上でC/M/H2O(65:25:4)を含むランニングバッファーを使用した薄層クロマトグラフィーによって分析する。大型のLLOを含む残りのペレットを、洗浄し、C/M/H2O(10:10:3)を用いて抽出する。C/M及びC/M/H2O抽出物の対応するアリコートを混ぜ合わせ、N2下で乾燥させ、35γl 1−プロパノールに再懸濁する。弱酸加水分解によってオリゴ糖を放出するために、500γl、0.02NのHClを加え、その後100℃で30分間インキュベートする。
【0081】
加水分解した材料をN2下で乾燥させ、その後超音波処理により200γlの水に再懸濁し、遠心分離にかけ、不要物を取り除く。放出されたオリゴ糖を含む上清をHPLC分析に使用する。
【0082】
HPLCによるオリゴ糖のサイズ分画。LLOの分離は、LC−NH2(2cmx4.6mm)プリカラムを含むSupelcosil LC−NH2カラム(25cmx4.6mm;5γm;Supelco社)で実施できる。アセトニトリルが水に対して70%から50%の直線勾配を、流速1ml/分で適用する。溶出分画を液体シンチレーション計測によって分析する。
【0083】
マンノース6−リン酸、マンノース1−リン酸、GDP−マンノースの調製。[2−3H]マンノースを用いて標識した後で、細胞を収穫し、遊離のマンノースをヌクレオチド結合及びリン酸化マンノース誘導体から、Kornerらにより記載されたペーパークロマトグラフィーによって分離する。溶出分画を液体シンチレーション計測によって分析する。
【0084】
ウェスタンブロット分析。細胞を、104細胞/cm2で播種し、薬剤処理下で表示の時間成長させる。処理期間の終りに、細胞を回収し、プロテイナーゼ阻害剤カクテルを添加したRIPAバッファー(150mM NaCl、1%Np−40、0.5%DOC、0.1%SDS、50mM Tris−HCl、ph8.0)を用いて溶解する。DNAを、溶液を21Gの針に10回通過させることにより断片化する。タンパク質濃度をSuper Protein Assay キット(Cytoskeleton社,USA)により測定する。試料を、2×Laemmliサンプルバッファー(Bio−Rad社,USA)と混合し、SDS−ポリアクリルアミドゲルにかける。ゲルをニトロセルロース膜(Amersham社,USA)に転写し、特異的抗体を用いて解析する。解析の後で、膜を洗浄し、HRP標識二次抗体と共にインキュベートする。化学発光をフィルムに感光させることにより検出する。
【0085】
指示がある場合には、膜をストリッピングバッファー(Pierce社,USA)を用いてストリップし、抗アクチン一次抗体を用いて再度解析する。
【0086】
ErbB2の免疫沈降。細胞を24時間処理した後で、細胞をRIPA(15mM NaCl、1%Np−40、0.1%SDS)により溶解し、超音波で分解する。細胞溶解物をモノクロナールErbB2抗体(Calbiochem,USA)に結合したCnBr活性化セファロースビーズ(Amersham社,USA)と共に400gで5分間回転させて、インキュベートする。
【0087】
免疫沈降させたErbB2をSDS−PAGEゲルに添加し、糖タンパク質のマンノース残基に特異的に結合する、コンカナバリンAを用いてブロットする。
【0088】
アポトーシス検査。アポトーシスELISA検査を記載のように使用し、該検査はホルムアミドによるアポトーシス細胞の濃縮クロマチンにおける選択的なDNAの変性及びアポトーシス細胞の一本鎖DNA(ssDNA)と、ssDNAに高度に特異的なモノクロナール抗体との反応性に基づく。これらの抗体は、アポトーシス細胞を特異的に検出し、ネクローシス細胞とは反応しない。
【0089】
フローサイトメトリーによる細胞死の機序の調査。アポトーシスを、An−nexin−V−Fluos Staining キット(Roche社,USA)によりネクローシスと識別する。表示の処理の後で、106細胞を、FITC標識アネキシンV及びヨウ化プロピオジウムを含むインキュベーションバッファーに再懸濁し、ホスファチジルセリン及び細胞膜の完全性をそれぞれ検出する。インキュベーションの後で、細胞を、488nmの励起光及びフルオレセインの検出のために515nmのバンドパスフィルター及びPIの検出のために600nmを超えるフィルターを使用したフローサイトメーターにより分析する。
【0090】
遺伝子発現のプロファイリング。Gene−array kitをSuper Array Inc.より購入できる。選択された細胞系由来の全RNAをdCTP[α−32P](3000Ci/mmol)を用いて、逆転写反応を介して解析する。解析した標識cDNAを、その後予備ハイブリッド形成アレイ膜(pre−hybridized array membrane)に添加し、ハイブリダイゼーションオーブンにおいて一晩インキュベートする。複数回洗浄した後で、遊離のプローブを除去し、膜をX線フィルムに感光し、画像を記録する。
【0091】
インビボ腫瘍実験。Cancer Res.2004(by Lampidis et al)に報告された2DG+Doxに関して記載されたプロトコルを、2−FGを2−DGに置き換えて再現できる。ヌードマウス、CD1系、5〜6週齢、体重30gに、100γlのヒト骨肉腫細胞系143bの107細胞/mlを移植した(S.C.)。腫瘍が50mm3の大きさになった時(9〜10日後)、動物を4つの群(8匹のマウス/群)に、以下のようにペア合わせをする:生理食塩水処理の対照:2−FG単独;Dox単独及びDox+2−FG。0日目、2−FG単独及びDox+2−FG群に、75mg/ml(500mg/kg)の2−FG、0.2mlをi.p.で与え、これを実験期間中、週に3回繰り返した。1日目、Dox及びDox+2−DG群に、0.6mg/ml(6mg/kg)のDox、0.3mlをi.v.で与え、これを週に1度、合わせて3回繰り返した(18mg/kg)。週に3回、マウスの体重を量り、腫瘍の測定をキャリパーにより行う。
【0092】
SKBR3細胞を、2−DG又は2−FMを用い、ドキソルビシン(Dox)を用いない上記のモデルに移植し、試験した。
【0093】
(実施例2)
マンノース誘導体に対する特定の腫瘍細胞の正常な酸素状態での感受性。
低酸素状態での細胞の成長は、エネルギー産生のための解糖を介したグルコースの代謝にもっぱら依存する。その結果として、この経路が、2−デオキシ−D−グルコース(2−DG)により遮断された場合に、低酸素細胞は死ぬ。対照的に、解糖が、通常酸素下で遮断された場合に、脂肪及びタンパク質をエネルギー源として置換でき、ミトコンドリアの酸化的リン酸化に燃料を供給するため、大部分の細胞は生存する。本発明は、正常な酸素圧の下で、選択されたいくつかの腫瘍細胞系が、比較的低用量の2−DG(4mM)で死ぬという発見に一部基づいている。2−DGが、ウィルスコート糖タンパク質合成において、N結合型グリコシル化の過程に干渉し、これは外因性マンノースの添加により逆転できることが先に示されている。本明細書に記載された通常酸素下での2−DGの細胞毒性は、低用量のマンノース(2mM)によって完全に逆転できるため、解糖ではなく、グリコシル化がこれらの結果に関与する機序であると考えられる。さらに、解糖の遮断及び低酸素細胞を殺すことにおいて2−DGより強力である、2−フルオロ−デオキシ−D−グルコース(2−FDG)は、通常酸素下で2−DGに感受性である、いかなる細胞型にも細胞毒性は示さない。
【0094】
糖タンパク質合成に関する2−DGの効果を調査するために、コンカナバリンA(糖タンパク質のマンノース部分に特異的に結合する)を、2−DGは結合を減少させるが2−FDGはせず、これは外因性マンノースの添加により逆転可能であったことを示す研究に使用した。同様に、N結合型グリコシル化が変化した場合に誘導されることが公知である、小胞体ストレス応答(UPR)タンパク質、grp98及び78が2−DGによって上方制御されるが、2−FDGによってはされず、再度この効果がマンノースによって逆転できることが見出された。さらに、2−DGが、UPR特異的転写因子(GADD154/CHOP)の上方制御を介して細胞死を誘導し、これはアポトーシスを仲介する。したがって、特定の腫瘍細胞型において、2−DGが、固形腫瘍の好気的細胞(グリコシル化への干渉を介して)並びに低酸素細胞(解糖の阻害を介して)の両方を選択的に殺すための単剤として、臨床的に使用できる。
【0095】
血管形成にもかかわらず、急速な腫瘍成長の代謝要求は、多くの場合酸素供給より勝っており、これが大部分の固形腫瘍内における低酸素領域の形成の一因となる。腫瘍の成長につれて起こる酸素濃度の減少は、低酸素部分における細胞の複製速度を遅くし、通常急速に増殖する細胞を標的とする、大部分の化学療法剤に対する耐性をもたらす。Brown,J.M,et al.,「癌治療における腫瘍内低酸素状態の活用(Exploiting Tumor Hypoxia In Cancer Treatment)」,Nat Rev Cancer 2004;4:437−47。低酸素細胞は、緩慢な成長及び活性酸素種を発生するために必要な酸素の不在により、放射線治療にもまた耐性である。Semenza,G.L.,「腫瘍内低低酸素状態、放射線耐性及びHIF−1(Intratumoral Hypoxia,Radiation Resistance,And HIF−1)」,Cancer Cell 2004;5:405−406。癌治療に関するこれらの不都合に加えて、低酸素状態は、腫瘍細胞をエネルギー産生のために解糖に依存させ、生存する。低酸素状態の下で、ATP産生の最も効率の良い方法である酸化的リン酸化が阻害され、ATPの産生のための唯一の方法として解糖が残される。したがって、低酸素腫瘍細胞において解糖を遮断することが細胞死を引き起こすはずである。実際、インビトロの3種の異なる症状の模擬低酸素状態において、腫瘍細胞は解糖阻害剤によって殺すことができることが示されている。Maher,J.C.,et al.,「低酸素対好気的状態で処理された腫瘍細胞における、2−デオキシ−D−グルコースによって誘導された大幅な細胞周期の阻害及び細胞毒性(Greater Cell Cycle Inhibition And Cytotoxicity Induced By 2−Deoxy−D−Glucose In Tumor Cells Treated Under Hypoxic vs Aerobic Conditions)」,Cancer Chemother Pharmacol 2004;53:116−122。さらに、正常な酸素処理細胞において解糖を阻害することは、代わりの炭素源、すなわちアミノ酸及び脂肪がミトコンドリアの酸化的リン酸化の作動に利用できるので、それらのエネルギー産生に有意には作用しない。したがって、解糖系阻害剤は低酸素腫瘍細胞を選択的に標的とするために使用でき、正常細胞又は好気的に成長する腫瘍細胞に対してはあまり毒性を示さない。Boros,L.G.,et al.,「ソマトスタチンによる、酸化的及び非酸化的ペントースリン酸経路の阻害:抗腫瘍作用の有望な機序(Inhibition Of Oxidative And Nonoxidative Pentose Phosphate Pathways By Somatostatin:A Possible Mechanism Of Antitumor Action)」,Med Hypotheses 1998;50:501;LaManna,J.C.,「栄養消費及び代謝摂動(Nutrient Consumption And Metabolic Perturbation)」,Neurosurg Clin N Am I997;8:145−163。
【0096】
実際、インビボ実験により、2−DG(緩慢に成長する低酸素腫瘍細胞を標的にする)が標準的化学療法剤(好気性細胞の急速な増殖を対象とする)の効果を、異なるヒト腫瘍異種移植片において促進することが示された。Maschek,G.,et al.,「2−デオキシ−D−グルコースは、ヒト骨肉腫及び非小細胞肺癌においてアドリアマイシン及びパクリタキセルの効果を、インビボで促進する(2−Deoxy−D−Glucose Increases The Efficacy Of Adriamycin And Paclitaxel In Human Osteosarcoma And Non−Small Cell Lung Cancers In Vivo)」,Cancer Res 2004;64:31−4。これらの研究結果、並びに低酸素状態のインビトロモデルからのデータは、「進行性固形悪性腫瘍を有する対象における、2−デオキシ−D−グルコース単独、及びドセタキセルとの併用についての第1相用量漸増試験」と題した第1相臨床試験の形で、ヒトにおける化学療法プロトコルを改善するためにこの戦略の試験を引き起こし、これは現在継続中である。Maher,J.C.,et al.,「低酸素対好気的状態で処理された腫瘍細胞における、2−デオキシ−D−グルコースによって誘導された大幅な細胞周期の阻害及び細胞毒性(Greater Cell Cycle Inhibition And Cytotoxicity Induced By 2−Deoxy−D−Glucose In Tumor Cells Treated Under Hypoxic vs Aerobic Conditions)」,Cancer Chemother Pharmacol 2004;53:116−122。動物実験からのデータ、並びに第1相臨床試験からの予備試験結果は、2−DGが正常細胞に対して十分耐性であり、比較的非毒性であることを示している。
【0097】
理論上は、ミトコンドリアが酸化的リン酸化を受けられる腫瘍細胞は、解糖系阻害剤の2−DGにより殺されないはずであるが、選択されたいくつかの癌細胞系は酸素の存在下で、低用量のこの糖類似体により死ぬ。毒性の機序は、これらの細胞系が正常なミトコンドリアの呼吸をし、他の解糖系阻害剤に耐性であるため、解糖の遮断を介したものではない。同様の機序は、ウィルスの糖タンパク質合成において示されており、2−DGがN結合型グリコシル化を、脂質結合オリゴ糖の組立てに干渉することにより遮断する。Datema,R.,et al.,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」Biochem J 1979;184:113−123;Datema R.,et al.,「脂質結合オリゴ糖を含む2−デオキシグルコースの形成(Formation Of 2−Deoxyglucose−Containing Lipid−Linked Oligosaccharides)」,Eur J Biochem 1978;90:505−516。正常な酸素状態で成長する選択腫瘍細胞系における2−DGの毒性は、同様の機序によると思われる。
【0098】
本発明によれば、2−DGは、通常酸素下で2−DGに感受性の細胞を含む固形腫瘍を有する特定の患者に、単剤として使用できる。したがって、これらの患者において2−DGは二重の効果(1)グリコシル化への干渉を介して好気性腫瘍細胞集団と標的とする、及び(2)腫瘍の低酸素部分において解糖を阻害する、を有するはずであり、両方の機序は細胞死を引き起こす。
【0099】
材料及び方法
細胞型
p0細胞を、骨肉腫細胞系143B(wt)を、前記のように臭化エチジウムを用いて長期処理することによって単離した。King,M.P.,et al.,「Mtdnaを欠いたヒト細胞:相補性による外因性ミトコンドリアを用いた再生(Hunan Cells Lacking Mtdna:Repopulation With Exogenous Mitochondria By Complementation.)」Science 1989;246:500−503。p0細胞は、ウリジン及びピルビン酸塩要求性体なので、それらを、10%ウシ胎児血清、50μg/mlのウリジン及び100mMのピルビン酸ナトリウムを添加したDMEM(GIBCO社,USA)で成長させる。SKBR3細胞系を、マイアミ大学のJoseph Rosenblatt博士の研究室より入手した。膵臓癌細胞系1420及び1469、卵巣癌細胞系SKOV3、子宮頸部癌細胞系HELA及び骨肉腫細胞系143Bを、ATCCより購入した。非小細胞肺癌及び小細胞肺癌細胞系はマイアミ大学のNiramol Savaraj博士により、患者に由来した。SKBR3及びSKOV3細胞をMcCoy’s 5A培地で成長させ、1420、1469及び143BをDMEM(GIBCO社、USA)で成長させ、HELAをMEM(GIBCO社、USA)で成長させた。培地には10%ウシ胎児血清を添加した。全ての細胞を5%CO2及び37℃で成長させた。
【0100】
薬剤及び化学物質
2−DG、オリゴマイシン及びツニカマイシンは、Sigmaより購入した。2−FDG及び2−FDMは、Priebe博士(MD Anderson Cancer Center,TX)のご厚意により寄贈された。
【0101】
低酸素状態
低酸素状態(モデルC)の研究のために、細胞を播種し、直接細胞毒性検査のために以下に記載のように37℃で24時間、5%CO2でインキュベートする。24時間インキュベートした後で、細胞を薬剤処理し、モデル110酸素コントローラーに取り付けたPro−Oxインビトロチャンバー(Reming Bioinstruments Co.Redfield,NY)に置き、チャンバーを所望のO2濃度(0.1%)にするため、内部には95%窒素及び5%CO2の混合物を使用し、かん流させる。
【0102】
細胞毒性検査
細胞を37℃で24時間、5%CO2においてインキュベートし、その時に薬剤処理を開始し72時間続ける。この時に、吸着細胞をトリプシン処理し、それらの個別の培養培地と混合し、その後400gで5分間遠心分離機にかける。ペレットを1mlのHanks溶液に再懸濁し、Vi−Cell(Beckman Coulter社、USA)細胞生存分析器により分析した。
【0103】
乳酸検査
乳酸を、処理済み培養液又は未処理培養液から0.025mlの脱タンパク質化培地を、0.1mlの乳酸デヒドロゲナーゼ、(1000単位/ml)、2mlのグリシンバッファー(グリシン0.6モル/L及びヒドラジン、pH9.2)及び1.66mg/mlのNADを含む、反応混合物に加えることによって測定する。脱タンパク質化を、試験培養物からの0.5mlの培地を1mlの8%w/v過塩素酸を用いて処理し、30秒ボルテックスにかけ、その後この混合物を4℃で5分間曝し、1500gで10分間遠心分離機にかけることにより起こす。上清をさらに3回遠心分離機にかけ、0.025mlの最終的に不要物を除いた上清を上記のように乳酸の測定に使用する。NADHの形成を、Beckman DU r 520 UV/vis分光光度計を用いて340nmで測定し、これは乳酸塩標準曲線により確定された乳酸濃度に直接一致する。
【0104】
ATP定量化検査。
ATPライトキット(lite kit)(Perkin Elmer社)を、ATP濃度の定量化に使用できる。約50mlの細胞溶解液を、96ウェルプレート(ボトム白)において100mlの細胞懸濁液に加える。プレートを室温で、シェーカーにのせて(700rpm)5分間インキュベートする。約50mlの基質溶液を、続けてウェルに加え、さらに5分間室温で振とう(700rpm)する。プレートをその後10分間暗順応させ、発光を測定する。
【0105】
ウェスタンブロット分析
細胞を、104細胞/cm2で播種し、薬剤処理下で表示の時間成長させる。処理期間の終りに、細胞を回収し、プロテイナーゼ阻害剤カクテルを添加した1%SDSの80mM Tris−HCL(ph7.4)バッファーを用いて溶解する。DNAを超音波で分解することにより断片化し、タンパク質濃度をミクロBCA タンパク質検査キット(Pierce社、USA)により測定する。試料を、2×Laemmliサンプルバッファー(Bio−Rad社,USA)と混合し、SDS−ポリアクリルアミドゲルにかける。ゲルをニトロセルロース膜(Amersham社,USA)に転写し、抗−KDEL(Stressgen社、Canada)(Grp78及びGrp94に関して);ポリクロナール抗−CHOP/GADD154(Santa Cruz社、USA),ポリクロナール抗erbB2(DAKO,USA).を用いて解析する。解析の後で、膜を洗浄し、HRP標識二次抗体と共にインキュベートする。化学発光をフィルムに感光させることにより検出する。指示がある場合には、膜をストリッピングバッファー(Pierce社,USA)を用いてストリップし、抗アクチン(Sigma社,USA)一次抗体を用いて再度解析する。コンコナバリンA(ConA)の結合を分析するため、膜を0.2μg/mlのHRP標識ConAと共にインキュベートし、記載のように化学発光を検出した。
【0106】
結果
2−DC及び2−フルオロ−D−マンノースは通常酸素下で成長するSKBR3細胞を殺すが、2−FDGは殺さない
多くの腫瘍細胞系を、正常な酸素状態対低酸素状態の下での解糖系阻害剤に対するそれらの感受性の差に関する調査において、通常酸素下で成長した場合、ヒト乳癌細胞系SKBR3は2−DGに関して感受性であることが見出された。図1A及びBは、SKBR3を3mMの2−DGで72時間処理した場合、成長は50%阻害され(ID50)、一方12mMでは60%の細胞が殺されることを実証する。これまでの研究により、ミトコンドリアの呼吸が不完全又は化学的に処断される場合、同様の用量の2−DGを用いて処理した腫瘍細胞は死ぬことが示された。したがって、これらの細胞がミトコンドリアの呼吸において不完全であったかどうかを確定するために、それらの酸素消費量を測定した。以下の表1に明示したように、SKBR3細胞と、通常酸素下で成長した場合に2−DGによる処理に対して耐性である他の2種の細胞系の平均酸素消費量の間に有意な差はなかった。他方、ミトコンドリア不全細胞系のp0は、酸素消費量の激減を示し、SKBR3が正常に呼吸していたことが確認された。さらに、通常酸素下で2−DGに対して感受性である、他の2種の細胞系、1420及びHELAは、同様に、又は耐性細胞系よりも盛んに呼吸した(表1)。したがって、通常酸素下でのこれらの細胞における2−DGの毒性は、解糖の遮断以外の機序によるものである。このことを確かめるために、SKBR3細胞を他の2種の解糖系阻害剤、すなわち2−デオキシ−2−フルオロ−グルコース(2−FDG)及びオキサマートを用いて処理した。図1A及びBにおいて、通常酸素下で成長した場合に、これらの薬剤はどちらもSKBR3細胞に対して毒性がなかったことが見られる。
【表1】
【0107】
さらに、2−フルオロ−D−マンノース(2−FDM)は、効果は低いが、SKBR3細胞において細胞毒性を引き起こす点で2−DGと同様であった(図1を参照されたし)。2−DG及び2−FDMは両方ともマンノースの構造と似ているが、2−FDGは似ておらず、その結果マンノースの代謝に干渉できる。このデータは、2−DG及び2−FDMによるマンノースの代謝への干渉を示し、これは主として多くのタンパク質のN結合型グリコシル化に関与し、SKBR3細胞における細胞死並びに成長阻害をもたらす。
【0108】
2−FDGは、2−DGより優れた解糖阻害剤であり、SKBR3細胞においてATPを十分減少させる
これまでの報告において、通常酸素下で成長するSKBR3細胞における2−DGの毒性が、解糖及びATP産生の阻害を介して仲介されることを示唆していた。Aft,R.L.,et al.,「化学療法剤としての2−デオキシ−D−グルコースの評価:細胞死の機序(Evaluation Of 2−Deoxy−D−glucose As A Chemotherapeutic Agent:Mechanism Of Cell Death)」,Br J Cancer 2002;87:805−812。しかし、上に述べたように、別の解糖系阻害剤、2−FDGはこれらの細胞において非毒性である。さらに2−FDG類似体は、2−DGより解糖の阻害及び殺低酸素細胞において優れている。Lampidis,T.J.,et al.,「解糖の遮断及び殺「低酸素腫瘍細胞」における2−ハロゲン置換D−グルコース類似体の有効性(Efficacy of 2−Halogen Substituted D−Glucose Analogs in Blocking Glycolysis and Killing“Hypoxic Tumor Cells,”)」Cancer Chemother Pharmacol(印刷中)。実際に、SKBR3細胞を2−FDG対2−DGを用いて処理した場合、前者は後者より乳酸塩濃度(解糖の基準)を阻害する(図2Aを参照されたし)。さらにATPの減少は、2−FDG処理によりさらに顕著になり、さらにこの糖類似体がこれらの細胞において解糖及びATP産生の優れた阻害剤であることが確認された(図2Bを参照されたし)。さらに、SKBR3細胞を低酸素状態で成長させた場合、2−FDGは2−DGより毒性が強いことが発見され、さらにSKBR3細胞における解糖の優れた阻害剤であることが確認された(データ未掲載)。したがって、これまでの報告とは反対に、通常酸素下で2−DGにより誘導された毒性は、解糖を阻害し、ATPのプールを減少させる能力から独立しているように思われる。
【0109】
通常酸素下でのSKBR3細胞における2−DGの毒性は、外因性マンノースにより逆転できる
ウィルス性タンパク質において、2−DGは、N結合型オリゴ糖の組立てを阻害し、この阻害は外因性マンノースによって逆転できることが示されている。Datema,R.,et al.,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123。図3A及び3Bは、マンノースの添加により、通常酸素下での2−DGへの曝露による細胞死を逆転できるが、他の糖、すなわちグルコース、フルクトース及びフコースの添加では逆転できないことを例示し、細胞死は、同様の機序を介してグリコシル化への干渉により仲介されることを示唆している。Datema,R.,et al.,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123。陰性対照として、マンノースは、同じ状態下でSKBR3細胞におけるツニカマイシン誘導性の毒性を逆転しないことが発見された。これは、オリゴ糖鎖にマンノースを加えるよりも前のステップにおいてツニカマイシンがグリコシル化に干渉し、その結果マンノースの代謝から独立しているという事実により説明できる(データ未掲載)。
【0110】
「低酸素」の3種のモデルにおける2−DGの毒性は、外因性マンノースによっては逆転できない
先に述べたように、低酸素状態下で成長する細胞は、エネルギー産生のためにグリコシル化を唯一解糖系に依存する。したがって、解糖系阻害剤によるこの代謝経路の阻害は、これまでに実証されているように、細胞死を引き起こすはずである。Maher,J.C.,et al,「低酸素対好気的状態で処理された腫瘍細胞における、2−デオキシ−D−グルコースによって誘導された大幅な細胞周期の阻害及び細胞毒性(Greater Cell Cycle Inhibition And Cytotoxicity Induced By 2−Deoxy−D−Glucose In Tumor Cells Treated Under Hypoxic vs Aerobic Conditions)」,Cancer Chemother Pharmacol 2004;53:116−122。2−DGが低酸素下で成長するSKBR3細胞に対して毒性であることの機序を識別するために、マンノースを3種の異なる「低酸素」状態下で成長する細胞に添加した。図3C及びDに示すように、通常の成長培地又は2mMのマンノースを添加した同じ培地のどちらにおいても、成長阻害及び細胞死に有意な差は見出されなかった。これらの結果は、外因性マンノースによる通常酸素下で成長するSKBR3細胞における2−DGの毒性の逆転は解糖とは関連がなく、通常酸素下で成長するこれらの細胞における細胞死の様式として、グリコシル化の干渉に関連している証拠を提供する。
【0111】
2−DG及び2−FDGは、通常酸素状態下で成長する、選択されたいくつかの腫瘍細胞系のみに対して毒性である
通常酸素状態下での2−DGの毒性が、特定の型の癌細胞に限定されたかどうかを調査するために、いくつかの細胞系を試験した。表2に示したこの試験の結果は、6mMの2−DG又は2−FDMのどちらかを用いて処理した場合には、通常酸素圧で成長する、選択されたいくつかの腫瘍細胞系(15種のうちの6種)のみが有意に細胞死に至り、6mMの2−FDGを用いた場合には至らないことを示す。2−DGに対して感受性が見出された細胞系は、SKBR3(乳癌細胞系)、1420(膵臓癌細胞系)、患者に直接由来する2種の非小細胞肺癌細胞系、RT 8226(多発性骨髄腫細胞系)、HELA(子宮頸癌)及びTG98(膠芽腫細胞系)であった。しかし、類似の組織に由来する癌細胞系が、通常酸素圧下で2−DG及び2−FDMの両方に耐性があることが見出され、これらの糖類似体の毒性が必ずしも組織型に特異的とは限らないことが示唆される。
【表2】
【0112】
2−DG及び2−FDMは、SKBR3細胞においてコンカナバリンA(ConA)の結合及び糖タンパク質の分子量を減少させる
ConAは、糖タンパク質のマンノースに特異的に結合するレクチンであり、高マンノース型糖タンパク質の検出に使用されている。「タンパク質精製法:実践的取り組み(Protein Purification Methods:A Practical Approach)」,In:Harris ELV,Angal S,editors.New York IRLPress at Oxford University Press;1994.p270。この技術は、2−DG及び2−FDMの両方、並びにツニカマイシンがいくつかの糖タンパク質においてConAの結合を減少させることを示すために使用された(図4Aを参照されたし)。さらに、外因性マンノースは2−DG及び2−FDMを用いて処理した細胞において対照のConA結合濃度を回復させるがツニカマイシン処理細胞においては回復せず、一方2−FDG処理細胞はConA結合の減少を示さない。さらに、2−DG処理の後でSKBR3細胞においてチロシン−キナーゼ受容体を発現する、公知の糖タンパク質であるerbB2の大きさの変化を、ウェスタンブロットにより分析した。図4Bは、2−DG及び2−FDMの両方がerbB2の分子量を減少させ、一方2−FDGは効果がなかったことを例示する。ConAのデータとの相関関係において、外因性マンノースはタンパク質の大きさを元の重さに復元した。これらのデータはさらに、2−DG及び2−FDMが選択された腫瘍細胞に対してN結合型グリコシル化への干渉を介して毒性であるが、2−FDGは毒性ではなく、この干渉はマンノースによって逆転できるという結論を裏付ける。
【0113】
2−DG又は2−FDMのどちらかによる処理が、通常酸素下でSKBR3細胞における小細胞ストレス応答を引き起こす
タンパク質のグリコシル化の通常過程が作用した場合、小胞体(ER)に蓄積されたミスフォールドタンパク質が、小胞体ストレス応答(UPR)として公知のシグナル伝達カスケードを引き起こす。グリコシル化に干渉する薬剤はUPRの誘導を示しており、シャペロン、すなわちGrp7S/Bip又はGrp94の上方制御を介して、ERのタンパク質のフォールディング能力を増加させる。図5Aに示すように、SKBR3細胞を2−DG、2−FDM又はツニカマイシン(周知のグリコシル化阻害剤)を用いて、通常酸素下で処理する場合、Grp78及びGrp94の両方が上方制御される。さらに2mMのマンノースの添加により、2−DG及び2−FDMのシャペロンの上方制御を逆転するが、ツニカマイシンは逆転しない。2−DGが誘導したUPRのマンノースによる逆転は、2−DGの毒性が外因性マンノースの添加により逆転されることを実証する、図3Dのデータに相関し、同様の結果が2−FDM処理細胞においても見出された(データ未掲載)。期待通り、2−FDGは、2−DG又は2−FDMほどにはこれらのシャペロン濃度を増加せず、通常酸素状態下で処理した場合、SKBR3細胞において細胞死は起こらないことを例示する毒性のデータ(図1B)に相関する。対照的に、2−DG又は2−FDMを、低酸素の3種の異なる実験条件の下で成長させた細胞に適用した場合、両方のシャペロンが上方制御されたモデルCと比較して、モデルA及びBにおいてUPRの有意な上方制御は観察されない。さらに、陽性対照としてのツニカマイシンは、3種のモデル全てにおいてこれらのシャペロンの合成の誘導を示した(図5B)。これらの結果は、細胞を2−DG又は2−FDMを用いて処理した場合、細胞死の機序が「低酸素」(解糖の遮断)対通常酸素(グリコシル化への干渉)状態とで異なることを示している。
【0114】
2−DG及び2−FDMの毒性は、SKBR3細胞におけるUPR特異的アポトーシス経路の誘導に相関する。
細胞がERストレスを克服できない場合、GADD154/CHOPの誘導を介してUPRは特異的アポトーシス経路を誘導することが報告されている。Xu,C.,et al.,「小胞体ストレス:細胞の寿命及び死の決定(Endoplasmic Reticulum Stress:Cell Life And Death Decisions)」,J Clin Invest 2005;115:2656−2664、Obeng,E.A.,et al.,「カスパーゼ12及びカスパーゼ4は、カスパーゼ依存性小胞体ストレス誘導アポトーシスに必要ではない(Caspase−12 And Caspase−4 Are Not Required For Caspase−Dependent Endoplasmic Reticulum Stress−Induced Apoptosis)」,J Biol Chem 2005;280:29578−29587。したがって、2−DG及び2−FDMが通常酸素下でERストレスによりSKBR3細胞を殺すかどうかを確定するために、このUPR特異的アポトーシスタンパク質を、ウェスタンブロット分析を使用して検査した。図6に見られるように、2−DG、2−FDM及びツニカマイシン処理の後でGADD154/CHOPが誘導されるが、2−FDG処理では誘導されない。このアポトーシス経路が2−DG又は2−FDMのどちらかにより誘導された場合、マンノースを用いた同時処理により逆転できる。しかし、ツニカマイシンにより誘導されたGADD154/CHOPはこの糖の添加により逆転できない。図2Bに示されるように、これらのデータは、マンノースによる毒性の逆転に相関する。
【0115】
実施例1及び2の考察
固形腫瘍は、不十分な血管形成、腫瘍の急速な成長及び、腫瘍血管の酸素運搬能力の減少による低酸素並びに通常酸素領域を含む。Gillies,R.J.,et al.,「腫瘍の微小環境のMRI(MRI Of The Tumor Microenviroment)」,J Magn Reson Imaging 2002;16:430−450;Maxwell,P.H.,et al,「低酸素誘導性Facoro−1は、固形腫瘍において遺伝子発現を調節し、血管形成及び腫瘍成長の両方に干渉する(Hypoxia Inducible Facoro−1 Modulates Gene Expression In Solid Tumors And Influences Both Angiogenesis And Tumor Growth)」,PNAS 1997;94:8104−8109;Semenza,G.L.,「癌治療用標的化HIF−1(Targeting HIF−1 For Cancer Therapy)」,Nature Rev 2003;3:721−732。低酸素細胞における唯一のエネルギー産生経路は解糖なので、解糖阻害剤の2−DGがこれらの細胞に対して選択的に毒性であるが、好気性細胞には非毒性であり、成長を阻害するだけであることが示されている。Maher,J.C.,et al.,「低酸素対好気的状態で処理された腫瘍細胞における、2−デオキシ−D−グルコースによって誘導された大幅な細胞周期の阻害及び細胞毒性(Greater Cell Cycle Inhibition And Cytotoxicity Induced By 2−Deoxy−D−Glucose In Tumor Cells Treated Under Hypoxic vs Aerobic Conditions)」,Cancer Chemother Phannacol 2004;53:116−122、Maschek,G.,et al.,「2−デオキシ−D−グルコースは、ヒト骨肉腫及び非小細胞肺癌においてアドリアマイシン及びパクリタキセルの効果を、インビボで促進する(2−Deoxy−D−Glucose Increases The Efficacy Of Adriamycin And Paclitaxel In Human Osteosarcoma And Non−Small Cell Lung Cancers In Vivo)」,Cancer Res 2004;64:31−4、Liu,H.,et al.,「解糖系阻害剤に対する腫瘍細胞の過感作(Hypersensitization Of Tumor Cells To Glycolytic Inhibitors)」,Biochemistry 2001;40:5542−5547;Liu,H.,et al.,「低酸素による解糖系阻害剤に対する腫瘍細胞の感受性の増加:固形腫瘍療法のための戦略(Hypoxia Increases Tumor Cell Sensitivity To Glycolytic Inhibitors:A Strategy For Solid Tumor Therapy)」,Model C.Biochem Pharmacol 2002;64:1745−1751。しかし、選択されたいくつかの腫瘍細胞系は酸素の存在下で2−DGに殺される。これらの感受性細胞型にはヒト乳癌細胞系SKBR3がある。損傷したミトコンドリアを有する細胞における解糖の遮断はATPの濃度を下げ、ネクローシス細胞死を引き起こすので、ミトコンドリア呼吸の不全によりこれらの細胞の2−DGに対する感受性は説明できる。Gramaglia,D.,et al,「アポトソーム形成の下流におけるアポトーシスからネクローシスへの転換は、BCL−XL及びPKB/AKT独立性の様式で解糖及び酸化的リン酸化の両方を阻害する必要がある(Apoptosis To Necrosis Switching Downstream Of Apoptosome Formation Requires Inhibition Of Both Glycolysis And Oxidative Phosphorylation In A BCL−XL And PKB/AKT−Independent Fashion)」,Cell Death Differentiation 2004;11:342−353。しかしこの可能性は、SKBR3細胞が、通常酸素下で2−DG耐性であることが見出された他の2種の細胞系と同様に呼吸することを示した酸素消費量実験に反した(表1)。さらに、通常酸素下で2−DGに感受性である細胞系1420の呼吸率が、2−DG耐性の細胞系よりも高いことが発見された。したがって、通常酸素下のSKBR3における2−DGの毒性は、ミトコンドリア機能の不全では説明できず、細胞死の機序が、解糖の遮断についてのこの糖の効果とは関連がないことが示唆される。
【0116】
これまでに、SKBR3細胞が通常酸素下で、解糖の阻害により2−DGに感受性であり、グルコース輸送体−1の発現増加をもたらし、2−DGを大幅に取り込むATPプールの減少を引き起こすことが報告された。Aft,R.L.,et al.,「化学療法剤としての2−デオキシ−D−グルコースの評価:細胞死の機序(Evaluation Of 2−Deoxy−D−Glucose As A Chemotherapeutic Agent:Mechanism Of Cell Death)」,Br J Cancer 2002;87:805−812。しかし、2−FDGは2−DGより強力な解糖阻害剤であるが(11、図2)、通常酸素下で成長するSKBR3細胞に対しては非毒性であり、さらに2−DGがこれらの細胞を、解糖の遮断及びATP産生の阻害による以外の機序を介して殺すという結論を支持する。
【0117】
SKBR3細胞がさらにマンノース類似体の2−FDMに感受性であることを示すデータは、通常酸素下で成長する選択された腫瘍細胞において、糖類似体のマンノ配置がそれらの毒性活性に重要であることを示す。2−DGの第2炭素において酸素原子が欠失すると、この化合物がグルコース類似体及びマンノースの類似体の両方になるのに反して、2−FDGにおいてフルオロ基が欠失してもグルコース類似体にしかならない。マンノ配置がこれらの糖類似体の毒性に関係があるという結論は、Schwartzが率いるグループによる1970年代終りに発表された仕事により裏付けられている。
【0118】
このグループは、2−DG、2−FDG及び2−FDMが、家禽ペストウィルスにより感染する、鶏胚線維芽細胞においてN結合型グリコシル化に干渉でき、糖タンパク質合成及びウィルスの復元を減少させることを示した。Datema,R,et al,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123、Datema,R.,et al,「脂質結合オリゴ糖を含む2−デオキシグルコースの形成(Formation Of 2−Deoxyglucose−Containing Lipid−Linked Oligosaccharides)」,Eur J Biochem 1978;90:505−516、Datema,R.,et al,「フルオロ−グルコースによる、インビボにおけるタンパク質のグルコシル化の阻害(Fluoro−Glucose Inhibition Of Protein Glycosylation In Vivo)」,Eur J Biochem 1980;109:331−341、Schmidt,M.F.G.,et al,「動物細胞における2−デオキシ−D−グルコースのヌクレオシド2リン酸誘導体(Nucleoside−diphosphate Derivatives Of 2−Deoxy−D−Glucose In Animal Cells)」,Eur J Biochem 1974;49:237−247、Schmidt,M.F.G.,et al.,「酵母及び鶏胚における2−デオキシ−2−フルオロ−D−[3H]グルコース及び2−デオキシ−2−フルオロ−D−[3H]マンノースの代謝(Metabolism Of 2−Deoxy−2 Fluoro−D−[3H]Glucose And 2−Deoxy−2−Fluoro−D−[3H]Mannose In Yeast And Chick−Embryo Cells)」,Eur J Biochem 1978;87:55−68、McDowell,W.,et al.,「抗ウィルス性糖類似体である2−デオキシ−2−フルオロ−D−マンノースによるタンパク質のグリコシル化の阻害の機序:グアノシン2リン酸エステルによるMan(Gicnac)2 PP−Dolの合成の阻害(Mechanism Of Inhibition Of Protein Glycosylation By The Antiviral Sugar Analogue 2−Deoxy 2−Fluoro−D−Mannose:Inhibition Of Synthesis Of Man(Gicnac)2 PP−Dol By The Guanosine Diphosphate Ester)」,Biochemistry 1985;24:8145−8152。それらの報告は、2−DGが、細胞の小胞体内のタンパク質に転写されるべき脂質結合オリゴ糖の組立てを阻害できると結んでいる。2−DG、GDP−MGの代謝産物は、オリゴ糖の組立ての中途終了を起こすことができ、タンパク質への転写には不適切な短い脂質結合オリゴ糖をもたらし得ることが実証された。Datema,R.,et al.,「脂質結合オリゴ糖を含む2−デオキシグルコースの形成(Formation Of 2−Deoxyglucose−Containing Lipid−Linked Oligosaccharides)」,Eur J Biochem 1978;90:505−516。全般的に、これらの結果は、これら類似体のウィルス性糖タンパク質合成阻害の能力が、2−DG>2−FDM>2−FDGの順であったことを示し、これらは、通常酸素下で成長するSKBR3細胞においてこれら類似体の毒性が似ている。Datema,R.,et al,「フルオロ−グルコースによる、インビボにおけるタンパク質のグルコシル化の阻害(Fluoro−Glucose Inhibition Of Protein Glycosylation In Vivo)」,Eur J Biochem 1980;109:331−341。このグループはさらに、これらの類似体の阻害効果が低用量の外因性マンノースの添加により逆転できることを報告した。Datema,R.,et al,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123。同様に、2mMのマンノースは、SKBR3細胞における2−DG及び2−FDMの毒性をを完全に逆転し、両方のマンノース類似体がこれらの細胞をN結合型グリコシル化への干渉を介して殺すことを示す。Datema,R.,et al,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123、Datema,R.,et al.,「脂質結合オリゴ糖を含む2−デオキシグルコースの形成(Formation Of 2−Deoxyglucose−Containing Lipid−Linked Oligosaccharides)」,Eur J Biochem 1978;90:505−516、Datema,R.,et al,「フルオロ−グルコースによる、インビボにおけるタンパク質のグルコシル化の阻害(Fluoro−Glucose Inhibition Of Protein Glycosylation In Vivo)」,Eur J Biochem 1980;109:331−341、Schmidt,M.F.G.,et al,「動物細胞における2−デオキシ−D−グルコースのヌクレオシド2リン酸誘導体(Nucleoside−diphosphate Derivatives Of 2−Deoxy−D−Glucose In Animal Cells)」,Eur J Biochem 1974;49:237−247、Schmidt,M.F.G.,et al.,「酵母及び鶏胚における2−デオキシ−2−フルオロ−D−[3H]グルコース及び2−デオキシ−2−フルオロ−D−[3H]マンノースの代謝(Metabolism Of 2−Deoxy−2−Fluoro−D−[3H]Glucose And 2−Deoxy−2−Fluoro−D−[3H]Mannose In Yeast And Chick−Embryo Cells)」,Eur J Biochem 1978;87:55−68、McDowell,W.,et al.,「抗ウィルス性糖類似体である2−デオキシ−2−フルオロ−D−マンノースによるタンパク質のグリコシル化の阻害の機序:グアノシン2リン酸エステルによるMan(Gicnac)2 PP−Dolの合成の阻害(Mechanism Of Inhibition Of Protein Glycosylation By The Antiviral Sugar Analogue 2−Deoxy 2−Fluoro−D−Mannose:Inhibition Of Synthesis Of Man(Gicnac)2 PP−Dol By The Guanosine Diphosphate Ester)」,Biochemistry 1985;24:8145−8152。
【0119】
マンノースは、N結合型グリコシル化タンパク質における核となる糖であるが、また、マンノースはホスホマンノイソメラーゼによりフルクトース−6−リン酸に転換できるため、解糖経路にも関与している。したがって、マンノースがSKBR3細胞において、2−DGが阻害する解糖系段階を避けることによって2−DGの毒性を逆転できるという可能性は残っている(図7)。しかし、2mMのマンノースが「低酸素」モデルA及びBにおいて2−DGにより誘導された成長阻害及び細胞死を逆転せず(図3C及び3Dを参照されたし)、一方低酸素状態で細胞が実際に成長したモデルCにおいて、わずかに回復効果があったため、この可能性はありそうもない。このわずかな回復は、(1)2−DG及び2−FDMが低酸素状態下でもなおグリコシル化に干渉した、及び/又は(2)これらの細胞が0.5%の低酸素状態下でもなお、少ないながらも酸化的リン酸化を行うので、マンノースはモデルCにおいて解糖の阻害を逆転した、と説明できた。全般的に、通常酸素下でこれらの糖類似体に対して感受性の細胞において、マンノースにより2−DG及び2−FGの毒性が逆転し、ミトコンドリアが停止した細胞(モデルA及びB)において逆転しないことにより、グリコシル化に干渉し、解糖の阻害がないことが、通常酸素下の低酸素状態に関与することが裏付けられる。
【0120】
N結合型グリコシル化が阻害された場合、タンパク質は適切にフォールドできず、小胞体内に残される。Ellgaard,L.,et al.,「小胞体における品質管理(Quality Control In The Endoplasmic Reticulum)」,Nat Rev Mol Cell Biol 2003;4:181−191;Parodi,A.J.,「タンパク質のグリコシル化及びタンパク質のフォールディングにおけるその役割(Protein Glycosylation And Its Role In Protein Folding)」,Annu Rev Biochem 2000;69:69−93。フォールドしていないタンパク質が蓄積すると、オルガネラの弛緩並びにタンパク質翻訳の混乱が起こる。このような事象において、細胞は、複合を開始するが、小胞体ストレス応答(UPR)として公知のシグナル伝達カスケードがなお保存されており、ERに恒常性を回復する。3種のER膜貫通タンパク質:イノシトール要求性酵素1(IRE1)、二本鎖RNA活性化タンパク質キナーゼ(PERK)及び活性化転写因子6(ATF6)が、フォールドしていないタンパク質のシグナルを核に伝達する。Schroder,M.,et al 「ERストレス及びフォールドしていないタンパク質の応答(ER Stress And Unfolded Protein Response)」,Mutat Res 2005;569:29−63。フォールドしていないタンパク質がERに蓄積した場合、分子シャペロンのグルコース制御タンパク質78(Grp78/Bip)は、これらの3種のER膜貫通タンパク質から解離し、その結果それらを活性化する。Pahl,H.L.,「小胞体から細胞核へのシグナル伝達(Signal Transduction From The Endoplasmic Reticulum To The Cell Nucleus)」,Physiol Rev 1999;79:683−701。これは、糖輸送体の上方制御、リン脂質の合成の増加、アミノ酸輸送及び分子シャペロンGrp78/Bip及びGrp94の発現を含む、いくつかの代謝及び分子の変化をもたらす。Ma,Y.,et al,「小胞体ストレス応答が明らかになる話(The Unfolding Tale Of The Unfolded Protein Response)」,Cell 2001;107:827−830;Doerrler.W.T.,et al,「小胞体ストレス応答によるヒト線維芽細胞におけるドリコール経路の制御(Regulation Of Dolichol Pathway In Human Fibroblasts By The Endoplasmic Reticulum Unfolded Protein Response)」,PNAS 1999;96:13050−13055;Breckenridge,D.G.,et al.,「小胞体経路によるアポトーシスの制御(Regulation Of Apoptosis By Endoplasmic Reticulum Pathways)」,Oncogene 2003;22:8608−8618。
【0121】
2−DG及び2−FDMは、通常酸素状態下で成長するSKBR3細胞においてGrp78及びGrp94の両方の発現を上方制御し、これは外因性マンノースの添加により逆転でき、これらの糖類似体がN結合型グリコシル化に干渉し、フォールドしていないタンパク質をもたらし、その結果UPRを開始することを強く裏付ける。さらに、2−DG又は2−FDMのどちらよりも優れた解糖阻害剤である2−FDGは、UPR応答の誘導に有効ではない。これらの類似体に対するUPR応答の規模は、グリコシル化への干渉の程度を反映しているように思われ、これはウィルスコートタンパク質における脂質結合オリゴ糖の組立ての遮断において2−DG>2−FDM>2−FDGであることを実証する報告に一致する。Datema,R.,et al,「フルオロ−グルコースによる、インビボにおけるタンパク質のグルコシル化の阻害(Fluoro−Glucose Inhibition Of Protein Glycosylation In Vivo)」,Eur J Biochem 1980;109:331−341、Schmidt,M.F.G.,et al,「動物細胞における2−デオキシ−D−グルコースのヌクレオシド2リン酸誘導体(Nucleoside−diphosphate Derivatives Of 2−Deoxy−D−Glucose In Animal Cells)」,Eur J Biochem 1974;49:237−247、Schmidt,M.F.G.,et al.,「酵母及び鶏胚における2−デオキシ−2 フルオロ−D−[3H]グルコース及び2−デオキシ−2−フルオロ−D−[3H]マンノースの代謝(Metabolism Of 2−Deoxy−2−Fluoro−D−[3H]Glucose And 2−Deoxy−2−Fluoro−D−[3H]Mannose In Yeast And Chick−Embryo Cells)」,Eur J Biochem 1978;87:55−68、McDowell,W.,et al.,「抗ウィルス性糖類似体である2−デオキシ−2−フルオロ−D−マンノースによるタンパク質のグリコシル化の阻害の機序:グアノシン2リン酸エステルによるMan(Gicnac)2 PP−Dolの合成の阻害(Mechanism Of Inhibition Of Protein Glycosylation By The Antiviral Sugar Analogue 2−Deoxy−2−Fluoro−D−Mannose:Inhibition Of Synthesis Of Man(Gicnac)2PP−Dol By The Guanosine Diphosphate Ester)」,Biochemistry 1985;24:8145−8152。さらにこのUPRのデータは、細胞毒性の結果と相関し、これは通常酸素下でSKBR3細胞の成長阻害及び殺害において2−DG>2−FDM>>>2−FDGであることを同様に示す。
【0122】
一方、「低酸素」モデルA及びBにおいて、Grp78及びGrp94は2−DGによって上方制御されず、これらの細胞は解糖の阻害を介して死亡し、グリコシル化への干渉を介してではないことを示している。UPRがこれらのモデルにおいて誘導されない理由を説明する有望な機序は、Grp78/Bipのフォールドしていないタンパク質への結合及びその結果のUPRの活性化に必要であることが公知のATPの濃度に関する。モデルA及びBとは対照的に、UPRはモデルCにおいて誘導され(図5B)、モデルCにおいてはATP濃度は2−DGにより減少する。さらに、ATP濃度に有意な影響を与えないことが公知のツニカマイシンは、「低酸素」モデルにおいてシャペロンを上方制御し、これらの細胞における機能的UPR経路を実証する。
【0123】
UPRはp53に酷似しており、P53はDNAの損傷シグナルが細胞周期を停止させ、DNA修復酵素を活性化し、これらの過程の結末であるアポトーシスに依存する。したがって、UPRが小胞内の恒常性の確立に失敗した場合、ERストレス特異的アポトーシス経路が活性化される。Breckenridge,D.G.,et al.,「小胞体経路によるアポトーシスの制御(Regulation Of Apoptosis By Endoplasmic Reticulum Pathways)」,Oncogene 2003;22:8608−8618。カスパーゼ4、カスパーゼ12及びCHOP/GADD154を含むアポトーシス経路のメディエーターの中で、後者の活性化の増加は、他と比較してER誘導性哺乳動物アポトーシス経路の優れた指標であることが示されている。Obeng,E.A.,et al.,「カスパーゼ12及びカスパーゼ4は、カスパーゼ依存性小胞体ストレス誘導アポトーシスに必要ではない(Caspase−12 And Caspase−4 Are Not Required For Caspase−Dependent Endaplasmic Reticulum Stress−Induced Apoptosis)」,J Biol Chem 2005;280:29578−29587。したがって、CHOP/GADD154の発現が、通常酸素下で成長するSKBR3細胞における2−DG及び2−FDMの細胞毒性と相関することを示す図6は、これらの糖類似体がグリコシル化への干渉を介して毒性であり、ERのストレスを引き起こすことを裏付ける。さらに、マンノースの添加によりCHOP/GADD154の誘導が逆転し、グルコースによっては逆転しないことは、2−DG及び2−FDMがこの機序を介して毒性であることをさらに裏付ける。
【0124】
根本的な疑問は、O2の存在下で2−DGを用いて処理した場合、大部分の腫瘍並びに正常細胞は死なないのになぜ特定の腫瘍細胞型は死ぬのかである。この疑問に対する答えは、遺伝学研究によってもたらされ、酵素であるホスホマンノースイソメラーゼが、糖タンパク質糖鎖不全症候群1b型(Carbohydrate−Deficient Glycoprotein Syndrome Type 1b) Niehues,R.,et al.,糖タンパク質糖鎖不全症候群1b型(Carbohydrate−Deficient Glycoprotein Syndrome Type 1b),J Clin Invest 1998;101:1414−1420;Freeze,H.H.,「Nグリコシル化及び動物モデルにおけるヒトの障害(Human Disorders in N−glycosylation and Animal Models)」,Biochim Biophys Acta 2002;1573:388−93に記載のように苦しんでいる患者において欠失していることが示されている。この酵素の欠失は、血清糖タンパク質のグリコシル化の減少をもたらし、蛋白漏出性腸症を特徴とする血栓症及び胃腸障害を引き起こす。外因性マンノースをこれらの患者の食事に加えた場合、彼らの血清糖タンパク質は正常に戻り、彼らの症状は消えた。Freeze,H.H.,「甘味溶液:糖による救済(Sweet Solution:Sugars to the Rescue)」,J Cell Biol 2002;158:615−616;Paneerselvam,K.,et al,「マンノースが、糖タンパク質糖鎖不全症候群の神経芽細胞におけるNグリコシル化の変化を修正する(Mannose Corrects Altered N−glycosylation in Carbohydrate−Deficient Glycoprotein Syndrome Fibroblasts)」J Clin Invest 1996;97:1478−1487。これは、外因性マンノースが、通常酸素下で2−DGを用いて処理した場合に殺される、選択された腫瘍細胞を救済することを示す当該データに相関する。これらの型の腫瘍細胞は、ホスホマンノイソメラーゼが下方制御される又は不完全であるかのどちらかであり、或いは通常酸素下で2−DGによる処理に耐性があることが示された大部分の他の細胞よりこれらの腫瘍細胞において、2−DGはこの酵素に大きく作用することが考えられる。しかし、図7に示すように、2−DG及び2−FDMがN結合型グリコシル化に関与するマンノースの代謝を阻害できる多くの他のステップがある。
【0125】
2−DG、2−CM及び2−FDM(2−FM)は、グリコシル化に干渉し、ERストレス及びアポトーシスを引き起こすことを介して、特定の腫瘍型を殺す。2−FDGがこれらの細胞を殺さないという発見は、2−DG及び2−FDMの毒性が解糖の阻害及びATPの減少によるものである可能性を排除する。これらの薬剤は、選択された固形腫瘍の治療において、単剤療法として使用できる(図7を参照されたし)。
【0126】
(実施例3)
図8及び9に示すように、複数のMTT検査により、選択された高悪性度の神経膠腫細胞系の感受性が実証され、様々な糖をベースとした解糖系阻害剤を図で示す。通常酸素状態及び低酸素状態のどちらかに曝すことを含む、様々な状態、及びこれらの化合物に対する感受性についてのその作用を使用する。結果は、糖をベースとした様々な解糖系阻害剤の感受性が比較的均一であることを示す(いくらか微妙な差がある)。低酸素状態における感受性の作用に関して、いくつかの細胞系に明らかな差がある。全般的に、低酸素状態で成長した場合、予測されたように、大部分の細胞系は解糖系阻害剤に対してより感受性である。しかし、U87MGなどのいくつかの細胞系は、乳酸濃度(解糖の代理マーカー)が通常酸素状態において最大であり、低酸素状態においては増加しない(以下の乳酸塩のデータを参照されたし)という好気性解糖表現型(完全な「Warburg効果」)に完全に傾倒している。この状況下で、感受性の差は、低酸素状態で成長した細胞は成長が遅く、したがって細胞の要求するエネルギーがおそらく少ないという経験的観測により説明される。
【0127】
図8Aは、低酸素下(<1% 酸素)又は通常酸素下(20% 酸素)で2−FGを用いて処理したU87ヒト脳腫瘍細胞系のMTT検査を示す。図8B及び8Cの両方は、同様の実験を表すが、糖をベースとした解糖阻害剤が異なる。パネルBの場合2−DGを使用し、パネルCでは2−FMを用いる。見て分かるようにU87は、その代謝要求に関して一貫して解糖を利用し、したがってこの細胞系は、低酸素下でこれらの薬剤に対して感受性の増加を示さない、独特の表現型を表す。
【0128】
図9は、2−FG又は2−FMのどちらかの存在下における、6日にわたる成長曲線を示す。このパネルは、U87細胞系の有意な成長阻害を実証し、2−FGは2−FMよりわずかに効果的と思われる。パネルB及びパネルCは、低酸素状態及び通常酸素状態の両方において成長した細胞系D−54に関する成長曲線の同様の阻害を実証する。この場合、細胞が低酸素状態で成長すると、明らかに増大された効果があり、これは、低酸素下のこの特定の細胞系に関するさらなる解糖系代謝を刺激する能力に関連がある。
【0129】
(実施例4)
図10及び11は、2−DGに曝した、通常酸素状態及び低酸素状態におけるヒトU87MG膠芽細胞腫星状細胞腫細胞系(U87)対D−54ヒト神経膠腫細胞系の感受性の差を示す。U87MG細胞は、低酸素状態又は好気性状態のどちらかにおいて高い率の解糖(酸化的解糖又は「Warburg効果」)を示す、したがって、それらが低酸素状態で成長した場合、2−DGに対するU87MG細胞の感受性は変化しない。他方、D54細胞は好気性成長状態において解糖系代謝を一部転換し、それ故この細胞系を低酸素状態で成長させた場合2−DGに対する感受性は高くなる。
【0130】
図10は、これらの2種の細胞系の有意な差、及びはるかに大きい、U87の相対無感覚を示す。
【0131】
図11は、U87とD54とのこの表現型の差の背後にある理由を示す。このパネルは、D54による大幅な解糖の誘導を実証するのに反して、U87はすでに最大限の乳酸塩濃度を産生している。
【0132】
図10及び11に示した結果は、細胞系を解糖系阻害剤を用いて処理した場合の、低酸素の効果が異なることを実証する。高度に解糖系に依存した細胞系(U87MGなど)は、すでに最大限に解糖系阻害剤に対して感作されており、感受性を示すために無酸素環境におく必要はない。これは、U87MGなどの細胞系による、高く、変化のない乳酸塩の産生濃度により実証されるが、一方D54は、その感受性及び低酸素に反応した乳酸塩濃度の両方を増加する。
【0133】
特筆すべきことに、神経膠腫細胞系は対酸素状態に完全に耐性である。図12を見て分かるように、通常酸素状態又は完全な低酸素状態(<1%)のどちらかにおいて成長した細胞系は、細胞の要求するエネルギーを提供するために、解糖を非常に頼りにして適度に成長を続けることができる。
【0134】
(実施例5)
2−DG類似体が2−フルオロ18−グルコース(2−F18G)の腫瘍により取り込まれることの実証。図13は、所定のPETスキャン研究の間に2−F18Gの神経膠腫内への取り込みが増強することを実証する。多形性膠芽細胞腫を有する患者のPETスキャンにより、この腫瘍内に2−FGが有意に取り込まれることが実証される。パネルは、患者に17mCiの2−F18Gを与えた後の、CT非増強(A)、増強したCT(B)及びCT一体型PETスキャンを示す。薬力学的事象は、これらの腫瘍が糖をベースとした解糖阻害剤に特に適しているという劇的な証明を提供する。
【0135】
(実施例6)
マウスにおけるヒト神経膠腫の治療。ヒト神経膠腫のマウス同所性異種移植片を、2−DG単独、又はテモゾロマイド(テモダール)と共に用いて、のどちらかで治療した。これらの動物は、高悪性度の神経膠腫の同所性の異種移植片モデルを表す。これらの実験を3回繰り返し、図14に示すように、同様の結果であった。動物にU87MG細胞を頭蓋内に移植し、5日後に、陰性対照(PBS)、陽性対照(テモダール)、実験的単剤(2−DG)又は実験的併用(2−DG+テモダール)のいずれかを用いて治療した。
【0136】
図12に示した結果は、同所性の腫瘍モデルに対する2−DGの単剤有効性を初めて実証した。これらの結果を繰り返し、各群に全18匹の動物を用いた連続した3回の動物実験に一致した(データ未掲載)。新規の治験薬により、この特定の動物モデルが非常に厳しく、生存者がわずかに増えたにすぎないことが明らかになった。見て分かるように、2−DGは現在脳腫瘍に利用できる、最も良い薬剤のテモゾロマイドと同等の効果である(陽性対照として見られる)。
【0137】
驚いたことに、2−DGはテモドールと同等に有効であり、併用は単剤療法より優れていた。2−DGを経口的に与え、十分に耐性であった。テモドールは脳腫瘍の治療に関して現在「gold standard(最も基準となる(信頼できる)もの)」であるので、2−DG及びテモドールの機能的同義性は顕著であった。最終的に、このクラスの薬剤に関して、単剤の有効性が実証され、これはこの疾患に固有である。
【0138】
これらの結果は、使用した陽性対照のテモゾロマイドと同様に、解糖の阻害が動物の生存を増加させたことを実証した。この療法に動物は十分耐性であり、毒性の証拠は示されなかった。最終的に、本発明者らは、最有力候補選択に関して、関連する糖をベースとした解糖系阻害剤の群から化合物を選択しており、これはインビトロ及びインビボの有効性、薬物速度論的特性、化学的安定性及び化学合成のコストに基づくであろう。最有力選択について、さらなる先端研究並びに正式な動物毒物試験が開始されるであろう。
【0139】
本発明の具体的な実施形態が、本発明の原理の適用を例示するために示され、詳細に記載されているが、このような原理を逸脱することがなければ本発明を具体化できると理解されるであろう。
【図面の簡単な説明】
【0140】
【図1A】SKBR3細胞における、2−DG、2−FDG、2−FDM及びオキサマートによる、24時間にわたる、腫瘍成長阻害試験の結果を表した図である。各値は、3連の試料の平均±SDである。
【図1B】SKBR3細胞における、2−DG、2−FDG、2−FDM又はオキサマートによる、24時間の細胞毒性試験の結果を表した図である。各値は、3連の試料の平均±SDである。
【図2A】2mMの2−DG又は2−FDGが存在しない又はいずれかが存在する状態で24時間SKBR3細胞を成長させ、培地中の乳酸塩濃度の結果を表した図である。
【図2B】図2Aで使用したのと同じ濃度の2−DG又は2−FDGのいずれかが存在する状態で6時間SKBR3細胞を成長させ、その後全細胞溶解物中のATPを定量化した結果を表した図である。
【図3A】様々な糖の存在下で、2−DGを用いて処理した後のSKBR3細胞における成長阻害試験の結果を表した図である。各値は、3連の試料の平均±SDである。
【図3B】様々な糖の存在下で、2−DGを用いて処理した後のSKBR3細胞における細胞毒性試験の結果を表した図である。各値は、3連の試料の平均±SDである。
【図3C】2mMのマンノースが存在する又は存在しない状態で、2−DGを用いて処理した後の、3種の異なる「低酸素」モデルにおける成長阻害試験の結果を表した図である。
【技術分野】
【0001】
本出願は、2006年2月24日提出の米国仮出願第60/776,793号、2006年4月27日提出の米国仮出願第60/795,621号及び2006年4月28日提出の米国仮出願第60/796,173号の優先権を主張するものである。これらの出願全体を、参照により本明細書に組み込む。
【0002】
本発明は、癌の治療に有用なヘキソース化合物、及びこのような化合物を投与することによる、治療を必要とする対象における、癌が仲介する疾患の治療方法を対象とする。
【背景技術】
【0003】
癌の治療は、多くの場合腫瘍耐性の促進の課題を伴う。プログラムされた細胞死の型であるアポトーシスは、細胞形態学及び細胞死を引き起こす一連の生化学的事象に伴って生じる。アポトーシスの過程は、細胞断片の安全な処分のような方法で達成される。しかし、癌療法を介して細胞内シグナル伝達経路を解明することにより、細胞死の誘導にとって重大な構造及び過程に影響を与えることが可能である。実際、アポトーシスの過程に欠陥があることは、多くの疾患に関係がある。過剰なアポトーシスは、虚血性障害のような細胞消失を引き起こす。他方、アポトーシスの量が不十分であると、癌のように細胞増殖の調節が効かなくなる。
【0004】
悪性神経膠腫の進行に伴い生じる変化は、PI−3K/AKT経路の活性化(通常、PTENの消失によって、又はEGFRのような成長因子の活性を介して)に関連すると思われる。この生存経路は、とりわけ、血管形成の刺激、アポトーシスの阻害剤、及び解糖の活性化を促進する代謝転換を含む、多くの適応変化を優先的に活性化する。同様に、膵臓癌治療の新しい標的は、シグナル伝達経路及び血管形成に関連する分子の標的、具体的には、ras癌遺伝子シグナル経路及びマトリックスメタロプロテアーゼ(MMP)の阻害剤を含む。
【0005】
癌悪性神経膠腫及び膵臓癌などの多くの癌は、従来の療法に対して内在的に耐性であり、重要な治療課題を示す。悪性神経膠腫は、年間発生率が100,000人に6.4人であり(Central Brain Tumor Registry of the United States,2002−2003)、初期脳腫瘍の最も一般的な亜型であり、最も致命的なヒトの癌である。その最も悪性の兆候である多形性膠芽腫(GBM)おいては、最善の治療を尽くしても患者の平均生存期間は9から12か月である。実際に、GBM患者のおよそ1/3では、放射線治療及び化学療法にもかかわらず、彼らの腫瘍は成長を続けるであろう。同様に、診断時の腫瘍の広がり次第では、膵臓癌の予後は一般的に不良であると見なされ、診断後さらに5年生存する罹病者はほとんどなく、完全な回復はまれである。
【0006】
さらに、治療に耐性のある腫瘍の進行に加えて、悪性腫瘍の治療における別の問題は、疾患にかかっていない正常な組織に対する、治療による毒性である。多くの場合化学療法は、細胞が正常か又は悪性であるかどうかに関係なく、分裂の早い細胞を殺すことを標的とする。しかし、広範囲にわたる細胞死及び癌治療に随伴する副作用は、腫瘍の成長調節経路を無効にできれば、腫瘍の抑制に不可欠ではないかもしれない。例えば、1つの研究は療法感作の使用である、すなわち、標準的治療の低用量と併用して、腫瘍細胞において重要な過程を特異的に標的とする薬剤を使用し、他の薬剤の効果を増強する。
【0007】
さらに併用療法は、免疫療法の腫瘍細胞の細胞毒性特異性を用いた、化学療法の腫瘍縮小及び免疫調節要素を併用した、ワクチンに基づいた取り組み方法を含む。しかし、併用療法は通常、単一薬剤のみを必要とする療法より、患者及び医師の両方にとってより困難である。さらに特定の腫瘍は、放射線療法に対する内在性の耐性を有し、多くの化学療法のモダリティは、異なる成長パターンによるものと思われ、成長パターンの型の違いは、個々の腫瘍内の低酸素領域の多様な度合いを表し得る。例えば、神経膠腫は、より急速に成長し、コントラストが増強する腫瘤性病巣に対して、MRIスキャンでコントラストの増強がほとんどないし全くなく、主に浸潤性に成長しうる。同様に、膵臓癌の早期段階は察知できない。相対低酸素部位は多くの場合、低酸素に伴うネクローシス領域を有する、急速に成長する腫瘤の中心、並びに、その上腫瘍の浸潤性要素内のいくつかの相対低酸素領域の両方に見ることができる。したがって、これらの相対低酸素領域のいくつかはより低速で循環する細胞を有すると思われ、それ故化学療法剤に対して耐性であると思われる。
【0008】
近年提唱されたある癌療法は、解糖阻害剤の使用を対象とする。この型の阻害剤は、細胞が好気性代謝から嫌気性代謝に切り替える場合に得られる選択性から利益を得るように設計される。腫瘍の成長のため、癌細胞は血液(酸素供給)から除去される。低酸素下では、腫瘍細胞はグルコース輸送体及び糖分解酵素の両方の発現を上方制御し、順に、好気性環境において正常細胞と比較して、グルコース類似体の取り込み増加を促進する。細胞は、酸素使用して、細胞のミトコンドリア内の脂肪及びタンパク質を燃焼し、エネルギーを産生する(ATPなどのエネルギー貯蔵分子を介して)ことによって生存するので、血液内で、細胞における解糖を遮断しても細胞は殺されない。それに反して、低酸素環境下で細胞内の解糖を遮断する場合、酸素がないため細胞はミトコンドリアを介したエネルギーの産生(脂肪及びタンパク質の酸化)ができず、細胞は死ぬ。それ故、解糖阻害剤は特定の癌の治療に対して期待されるが、低酸素環境下に存在する全ての癌細胞にではない。実際、Otto Warburgによる有名な見解により、十分な量の酸素が存在しても、多くの腫瘍が細胞エネルギーの産生に関して、解糖の優先的利用を選択することが明示されている(酸化的解糖又は「Warburg効果」と呼ばれる)。この腫瘍の適応反応は、悪性神経膠腫にもまた当てはまると思われる。
【0009】
したがって、化学療法に対する耐性を示し、異なる成長パターンの若しくは腫瘍内に多様な度合いの低酸素領域を有する、及び/又は血管形成を刺激する若しくはアポトーシスを阻害する生存経路を有する成長パターンを呈する、癌の治療の必要性がある。
【発明の開示】
【発明が解決しようとする課題】
【0010】
癌を予防、阻害及び調節するヘキソース化合物及びこれらの医薬組成物、並びに癌、特に膠芽細胞腫及び膵臓癌を治療するための該化合物の使用が見出された。本発明は、癌並びに癌が仲介する障害及び病態の治療に有用なヘキソース化合物の使用を開示する。膠芽細胞腫及び膵臓癌の治療方法は、治療を必要とする対象に、治療有効量のヘキソース化合物を投与することを含む。特に興味深いのは、治療を必要とする対象に、治療有効量の2−FMを投与することを含む、腫瘍の増殖の治療方法である。本発明は、治療を必要とする対象に、マンノース化合物を投与することによる癌の治療方法を含む。
【課題を解決するための手段】
【0011】
悪性神経膠腫のための治療の選択肢は依然として限られている。これは一部には、利用可能な多くの化学療法の選択肢に対して、細胞が内在的に耐性であることによる。また一部には、悪性腫瘍の示す成長パターンの差にもよると思われる。すなわち、神経膠腫は、主に浸潤性に成長でき、コントラストを増強すると腫瘤性病巣が急速に成長しているのに対して、コントラストを増強しないとMRIスキャンではほとんど見られない。多くの研究により、これらの異なる型の成長パターンもまた、個々の腫瘍内の低酸素領域の様々な度合いを表すことが示されている。相対低酸素部位は、多くの場合ネクローシス領域を伴う急速に成長する腫瘤の中心、並びに、同様に腫瘍の浸潤性要素内のいくつかの相対低酸素領域の両方に見ることができる。したがって、これらの相対低酸素領域のいくつかはより低速で循環する細胞を有すると思われ、それ故多くの化学療法剤に対してより耐性であると思われる。さらに、十分な量の酸素が存在しても、多くの腫瘍が解糖を経ることを選択すること(酸化的解糖又は「Warburg効果」と呼ばれる)を記述したWarburgによる見解は、悪性神経膠腫にもまた当てはまると思われる。本発明者らは、これらの特徴から、神経膠腫及び他の高度に解糖的に維持された腫瘍、例えば膵臓癌は、解糖の阻害剤に対して感受性であり得、腫瘍の成長に関して有意な影響を有する可能性があることを前提とした。
【0012】
したがって、一般的に脳に、特に神経膠腫に特有の追加の特徴は、CNSに糖を貪欲に取り込むグルコース輸送体の発現の増加である。本発明者らは、これらの特徴から、神経膠腫が解糖阻害剤に著しく感受性であるはずの唯一の病状を表すことを前提とした。この仮説を試験するために、本発明者らは、低酸素状態及び正常な酸素状態の両方の下で、インビトロで、多くの神経膠腫細胞系のパネルに対して公知の解糖阻害剤を使用した。さらに薬剤の効果を、同所性の神経膠腫異種移植片を有する動物において、多くの異なる投与スキームを使用して試験した。
【0013】
酸素の存在下でもエネルギー産生のための第1供給源として解糖を優先的に利用する、高悪性度神経膠腫による代謝の転換である「Warburg効果」、これは、一部はHIF−1a及びPI−3キナーゼ経路の活性化により作動する。解糖の有効な阻害剤である2−デオキシグルコースは、エノラーゼによる2−デオキシグルコース−6−リン酸の変換を遮断し、帯電したリン酸基のため細胞内にこの種を蓄積する。
【0014】
細胞の必要とするエネルギーのために解糖系を優先的に使用する、公知の代謝変化は、高悪性度の神経膠腫を含む新生物において起こる。これらの変化は、解糖系に必要な重要な酵素の産生並びにグルコース輸送体の上方制御を誘導する、HIF−1a及びPI−3キナーゼ活性化を含む生存経路により作動される。この解糖系表現型は大きな特徴であり、これらは通常酸素下でも優勢である。この表現型は認知されており、「Warburg効果」としてすでに述べられている。この表現型の変化により、これらの腫瘍は正常細胞より解糖阻害剤に対して感受性が強いはずである。糖をベースとした解糖系阻害剤のグループ及び他のマンノース化合物は、治療薬としての機能を果たすことができる。原型的な糖をベースとした阻害剤である2−デオキシグルコースが、許容でき、且つ有力な抗神経膠腫効果を有することを、本研究において示した。単独又は細胞毒性化学療法との併用のどちらかであるヘキソース化合物は、癌、特に神経膠腫及び膵臓癌の治療に有効である。さらに、この解糖系表現型は、腫瘍環境内で低酸素状態によりまず動作するので、この型の治療は抗血管形成療法と見なされるべきである。実際、抗血管形成療法を「エスケープ」できる腫瘍は、解糖阻害剤及び/又はヘキソース化合物全般に対して優先的により感受性であると思われる。
【0015】
本発明者らは、糖をベースとしたヘキソース化合物が、高悪性度の神経膠腫腫瘍及び膵臓癌の治療に有効であることを示した。さらに、化合物の他の阻害剤型は、CNSに有利に取り込まれるように設計され、2−DGが現在有している有利な経口生体利用性を維持する。細胞毒性の薬剤及び抗血管形成剤の両方と併用したヘキソース化合物に関する継続中の研究は、未来の臨床的コンビナトリアルな試行のための知的な糸口を楽観的に提供するであろう。
【0016】
ヘキソース化合物は、6個の炭素原子を含む任意の単糖類を意味し、且つ含む。ヘキソースの1つのクラスは、アルドヘキソースファミリーであり、これは例えばグルコース、ガラクトース及びマンノースを含む。アルドヘキソースはさらに、2−デオキシグルコース、フコース、シマロース及びラムノースなどの、様々なデオキシ糖もまた含み得る。ヘキソースの他のクラスは、フルクトース及びソルボースを例とする、ケトヘキソースファミリーである。本発明のヘキソースは、通常は天然発生的なD−配置であるが、ヘキソースはさらにL−エナンチオマーでもあり得る。本発明のヘキソースは、αアノマー、βアノマー及びこれらの混合物を含み得る。本発明のヘキソースはいずれも、場合により置換されていてもよい。このような置換は、ヒドロキシル基を、フッ素、塩素又は臭素などのハロゲンと置き換えることを含む。本発明において、置換は通常ヘキソースのC−2炭素においてであり、六員環いす型立体配座においてヘキソースのアクシアル配位又はエクアトリアル配位のどちらかをとり得る。アクシアルであるC−2における置換は、糖をマンノース誘導体又はマンノ配置の糖と指定する。エクアトリアルであるC−2における置換は、糖をグルコース誘導体又はグルコ配置の糖と指定する。
【0017】
本発明の実践に有用なヘキソース化合物は、参照により本明細書に組み込まれる、米国特許第6,670,330号及び米国特許出願第20030181393号、第20050043250号及び第20060025351号に開示された化合物を含む。本発明のある実施形態において、好ましい化合物は、2−デオキシ−グルコース(2−DG)、2−デオキシ−マンノース(2−DM)、2−フルオロ−グルコース(2−FG)及び2−フルオロ−マンノース(2−FM)などの糖をベースとした腫瘍増殖阻害剤である。
【0018】
「中枢神経系の腫瘍」は、良性であろうと悪性であろうと、脳、脊髄又は他の中枢神経系組織内の異常成長組織のいずれをも意味する。特に、毛様細胞性星状細胞腫、低悪性度星状細胞腫、未分化星状細胞腫及び多形性膠芽腫(GBM又は膠芽細胞腫)などの神経膠腫を含む。「中枢神経系の腫瘍」は、さらに脳幹神経膠腫、上衣腫、神経節細胞腫、若年性毛様細胞性星状細胞腫、混合性神経膠腫、希突起膠腫及び視神経膠腫などの、他の型の良性又は悪性の神経膠腫も含む。「中枢神経系の腫瘍」は、さらに脊索腫、頭蓋咽頭腫、髄芽腫、髄膜腫、松果体腫瘍、下垂体腺腫、未分化神経外胚葉性腫瘍、神経鞘腫、血管の腫瘍及び神経線維腫などの非神経膠腫も含む。最後に、中枢神経系の腫瘍はさらに、悪性細胞が体内の他の部分から中枢神経系に広がった、転移性腫瘍も含む。
【0019】
本発明による「治療すること(treating)」、「治療(treatment)」又は「緩和」は、治療的処置及び予防的(prophylactic)又は防止的(preventative)手段の両方を指し、目的は中枢神経系の腫瘍の予防又は成長遅延、腫瘍の大きさの減少、或いは腫瘍を完全に除去することである。治療を必要とする人々は、特定された中枢神経系の腫瘍を有する対象、中枢神経系の腫瘍を有すると疑われる対象、中枢神経系の腫瘍の進行の危険があると特定された対象を含む。本発明の方法によるヘキソース化合物の治療量を受けた後に、以下の状態の1つ又は複数が観察された場合に、対象は中枢神経系の腫瘍を良好に「治療」されている:腫瘍の大きさの減少又は腫瘍がなくなる;腫瘍の成長の阻害又は中止;腫瘍の転移の阻害又は中止;並びに/或いは罹患率及び死亡率の減少又は生活の質の向上などの、腫瘍に伴う1つ又は複数の症状のある程度の緩和。
【0020】
ヘキソース化合物が、既存の脳腫瘍細胞の成長を防止する及び/又は既存の脳腫瘍細胞を殺す程度により、細胞増殖阻害性及び/又は細胞毒性であると見なすことができる。
【0021】
「同時投与すること(coadministering)」又は「同時投与(coadministration)」という用語は、療法の同時の又は連続した投与を包含することを意図する。例えば同時投与は、解糖系阻害剤及び化学療法剤の両方を1つの組成物において投与することを含み得る。同時投与はまた、複数のこのような組成物を同時に投与することも含み得る。或いは、同時投与は複数のこのような組成物を同じ期間中の異なる時間に投与することも含み得る。
【0022】
本発明によるヘキソース化合物は、限定するものではないが、神経膠腫又は他の脳腫瘍において酸化的解糖を阻害できる化合物である、解糖系阻害剤を含み、2−デオキシグルコース、2−フルオロ−グルコース、2−フルオロ−マンノースなどのヘキソース化合物を含み得る。
【0023】
本明細書中で先に定義した抗増殖治療は、単独の療法として適用でき、又は少なくとも1種の本発明の化合物に加えて、1種又は複数種の他の物質及び/又は治療を含み得る。このような治療は、治療用の個別の成分を同時、連続又は別々に投与する手段で達成できる。本発明の化合物はまた、公知の抗癌剤又は細胞毒性薬剤及び放射線療法などの治療との併用において有用であり得る。固定用量として処方する場合、このような併用製品は、本明細書に記載の用量範囲内の本発明の化合物及び承認された範囲内の他の医薬活性剤を用いる。解糖系阻害剤は、他の抗癌剤又は細胞毒性薬剤もまた伴う化学療法の投与計画の一部として連続的に、及び/又は非化学療法治療、例えば外科手術又は放射線療法と併せて使用できる。
【0024】
化学療法剤は、限定するものではないが、治療薬の3つの大きなカテゴリー:(i)抗血管形成薬、例えばリノミド、インテグリン−α−β3の機能の阻害剤、アンジオスタチン、ラゾキサン、(ii)細胞増殖抑制剤、例えば抗エストロゲン(例えば、タモキシフェン、トレミフェン、ラロキシフェン、ドロロキシフェン、ヨードキシフェン)、プロゲストゲン(例えば、酢酸メゲストロール)、アロマターゼ阻害剤(例えば、アナストロゾール、レトロゾール、ボラゾール、エキセメスタン)、抗ホルモン、抗プロゲストゲン、抗アンドロゲン(例えば、フルタミド、ニルタミド、ビカルタミド、酢酸シプロテロン)、LHRHアゴニスト及びアンタゴニスト(例えば、酢酸ゴセレリン、ロイプロリド)、テストステロン5−α−ジヒドロレダクターゼ阻害剤(例えば、フィナステリド)、ファルネシルトランスフェラーゼ阻害剤、抗侵害剤(例えば、マリマスタットのようなメタロプロテイナーゼ阻害剤、及びウロキナーゼプラスミノーゲン活性化因子受容体機能の阻害剤)及び成長因子機能の阻害剤(このような成長因子は、例えばEGF、FGF、血小板由来成長因子及び肝細胞成長因子を含み、このような阻害剤は、成長因子抗体、成長因子受容体抗体(例えば、アバスチン(ベバシズマブ)及びエルビタックス(セツキシマブ))、チロシンキナーゼ阻害剤及びセリン/スレオニンキナーゼ阻害剤を含む)並びに(iii)内科的腫瘍学で使用される抗増殖/抗新生物薬及びこれらの組み合わせ、例えば代謝拮抗剤(例えば、メトトレキサートのような抗葉酸薬、5−フルオロウラシルのようなフルオロピリミジン、プリン及びアデノシン類似体、シトシンアラビノシド)、挿入抗腫瘍抗生物質(例えば、ドキソルビシン、ダウノマイシン、エピルビシン及びイダルビシンのようなアントラサイクリン、マイトマイシン−C、ダクチノマイシン、ミトラマイシン)、白金誘導体(例えば、シスプラチン、カルボプラチン)、アルキル化剤(例えば、ナイトロジェンマスタード、メルファラン、クロラムブシル、ブスルファン、シクロホスファミド、イフォスファミドニトロソウレア、チオテパ)、抗有糸分裂薬(例えば、ビンクリスチンのようなビンカアルカロイド及びタクソール(パクリタキセル)、タキソテール(ドセタキセル)のようなタキソイド及びポチロン類似体、ディスコデルモライド類似体及びエレウセロビン類似体などの最新の微小管(安定)剤(microbtubule agents))、トポイソメラーゼ阻害剤(例えば、エトポシド及びテニポシドのようなエピポドフィロトキシン、アムサクリン、トポテカン)、細胞周期阻害剤、生物反応修飾物質及びベルケイド(ボルテゾミブ)などのプロテアソーム阻害剤、を含む。
【発明を実施するための最良の形態】
【0025】
当分野で普通の技術の1つにより、本発明に開示する治療方法が、複数の投与経路を介して、様々な量/濃度のヘキソース化合物を用いて達成できることが容易に認識されるであろう。好ましい投与経路は、使用するヘキソース化合物により変更でき、このような経路は、限定するものではないが、経口、口腔、筋肉内(i.m.)、静脈内(i.v.)腹腔内(i.p.)、局所又は他の任意のFDAに承認された投与経路を含む。投与濃度又は治療濃度は、治療する対象及び投与するヘキソース化合物により変わるであろう。ある実施形態において、ヘキソース化合物の濃度は、1mgから50gm/kg体重におよぶ。
【0026】
まず、一連の2−フルオロ、2−ブロモ及び2−クロロ−置換グルコース類似体を調製し、解糖経路におけるグルコースに対する有望な競合的基質として、このような類似体が2−デオキシ−グルコース(2−DG)と類似の手段で解糖系阻害剤として作動できることを分析した。本発明者らは、2−フルオロ−D−マンノースが有効な抗腫瘍剤であることを発見した。なぜならその特性が、2−フルオロ−D−マンノース(本明細書中では「2−FM」とも称する)が、フッ素原子と水素の大きさの類似性を考慮すると2−デオキシ−D−グルコース(2−デオキシ−D−マンノースと同じ)と似ており、又は誘導効果及び水素結合の形成の可能性に関しては水素よりマンノースのヒドロキシ基が似ていることにより、D−マンノースと似ている可能性があるという、どちらかの事実に由来すると思われるためである。その後の状況において、2−フルオロ−D−マンノースは、D−マンノースが関連する生物機能、代謝及び生物過程に作用し得る。また、D−グルコース及びD−マンノースの両方に作用できる効果の組み合わせは、細胞過程に関する。
【0027】
実際、図15から18に提供したデータは、2−フルオロ−D−マンノースが2−DGより強力であり、また膵臓Colo357−FG細胞における2−フルオロ−D−グルコースの活性より良い又は同様の活性を有することを示している。さらに2−フルオロ−D−マンノース(2−FM)を、他の2−デオキシ−D−マンノース類似体、すなわち2−クロロ−D−マンノース(2−CM)及び2−ブロモ−D−マンノース(2−BM)と比較した。驚いたことに、また予期せずに2−フルオロ−マンノースはこの一連の他のものより強力である。具体的には、データは、2−フルオロ−D−マンノース(2−FM)が、ブロモ(2−BM)及びクロロ(2−CM)の両方の類似体より、U251膠芽細胞腫脳腫瘍細胞の成長を阻害することにおいて、明らかに優れていることを示している。2−FMはさらに、低酸素状態下より通常酸素下でU87膠芽細胞腫細胞に対して驚くほど良い活性を示した(図17)。さらに、予測不能な、2−FMの作用の少なくとも1つの様式は、すなわち2−FMが腫瘍脳細胞において自食作用を強く誘導する能力を示すことであり、それ故腫瘍細胞系に対するその作用機序の少なくとも1つの説明を提供する。
【0028】
すぐ下に示したように、2−デオキシグルコース(2−DG)は2個の水素を糖のC−2の位置に有する。糖の六員環いす型配座において、これらの2個の水素はアクシアル及びエクアトリアル位を占める。
【化1】
【0029】
基本的に、2−フルオロマンノース(2−FM)は2−デオキシグルコース(2デオキシマンノースと同じことである)のアクシアル水素をフッ素に置き換える。フッ素は、水素と等比体積であると一般的に見なされる。したがっていくつかの態様において、2−FMの化学的性質は2−DGと似ているかもしれない。実際に、2−FMは、この等比体積の根拠に基づく解糖系阻害活性を示すだろう。しかし、フッ素は実質的に水素より電気的に陰性であり、結果として水素結合のモチーフに関係することができる。この点において、2−FMはマンノースにより近く作用でき、それ故2−FMは高マンノース型オリゴ糖合成においてN結合型糖脂質/タンパク質経路を撹乱できると思われる。
【0030】
要するに、2−FMは驚くほど優れた増殖効果を腫瘍細胞に対して示し、2−デオキシ−D−グルコース(本明細書中では「2−DG」とも称する)及び2−デオキシ−2−フルオロ−D−グルコース(本明細書中では「2−FG」とも称する)より強力であると思われる。以下に論じるように、化合物2−FMを、231−GFP乳癌、U251膠芽腫多形成脳腫瘍(図16)及びCo1o357−FG膵臓ヒト癌細胞系(図15)において特に試験した。U251及びCo1o357−FG細胞において、2−FMを、2−DG、2−FG、2−デオキシ−2−クロロ−D−マンノース(本明細書中では時として「2−CM」とも称する)、2−デオキシ−2−ブロモ−マンノース(本明細書中では「2−BM」とも称する)、2−デオキシ−クロロ−D−グルコース(本明細書中では「2−CG」とも称する)及び2−デオキシ−2−ブロモ−D−グルコース(本明細書中では「2−BG」とも称する)と直接比較した。膠芽細胞腫(図16、17及び18)及び膵臓癌(図15)の両方において、2−FMは比較した薬剤の中で最も強力な薬剤であり、観察された差は2−FMとそのクロロ及びブロモ誘導体の間で特に大きかった。この差は、2−DGと比較した場合にもまた有意であった。したがって、該データは2−FMが、腫瘍細胞増殖の阻害において2−DG及び2−FGと異なった働きをする可能性を示す。該データはさらに、2−FMが癌、特に脳及び膵臓の腫瘍のための非常に有効な抗腫瘍治療であり得ることを示す。
【0031】
より具体的には、図15は、2−デオキシ−グルコース(2−DG)、2−フルオロ−グルコース(2−FG)又は2−フルオロ−マンノース(2−FM)を用いた治療のいずれかに反応したCo1o357細胞系の、MTT試験を介した細胞生存能力の用量反応曲線を明示する。見て分かるように、用量反応曲線の左への変化は、2−FMが2−DG又は2−FGのどちらよりも強力であることを示す。図16は、マンノースの2位におけるハロゲンの性質が活性に作用する重要な因子であることを明示する。神経膠腫細胞系U251MGを、2−クロロ−マンノース(2−CM)、2−ブロモ−マンノース(2−BM)又は2−フルオロ−マンノース(2−FM)のどれかを用いて処理した。再度、細胞生存能力をMTT試験により測定したところ、結果は、他のハロゲンをベースとした類似体と比較した場合に、2−FMの優れた活性を明らかに示す。図17は、低酸素状態(酸素1%未満)又は正常な酸素状態(酸素20%)において、2−フルオロ−マンノース(2−FM)を用いて処理したU87細胞系のMTT試験を明示する。見て分かるように、データは、U87細胞系におけるこの薬剤による異常事態が、低酸素状態において感受性がより強くはないことを表す。これは、2−FMに関する代わりの作用機序が殺細胞効果に関与し得ることを示す可能性がある。
【0032】
図18は、2−フルオロ−マンノース(2−FM)の唯一且つこれまでに特定されていない機序を明示する。U87MG神経膠腫細胞系において、2−FMは自食作用を介した細胞死を誘導する。図は、アクリジンオレンジを用いて染色することによる、酸性小胞オルガネラ(Acidic Vesicular Organelles)(AVO)のフローサイトメトリー分析の結果を表しており(手順を参照されたし)、これは自食作用過程に特有であり、特徴的である。結果は、2−FMの曝露量が増えるに従って、自食作用を起こす細胞のパーセントが増加することを示す。この効果を見るためにはわずか40時間の曝露時間は極めて短いため、この自食作用の誘導程度は強い印象を与える。
【0033】
2−DGは、解糖系阻害剤の添加の範囲を評価するために現在臨床試験において投与されており、この阻害剤は固形腫瘍に見出される最も耐性の細胞集団である緩慢に成長する低酸素腫瘍細胞を殺し、迅速に分裂する通常の酸素状態の細胞を標的とする、標準的化学療法の治療有効性を増加できる。本発明は、一部は、2−DG又は2−FMを用いた場合、酸素の存在下でも特定の腫瘍細胞系が殺されるが、2−デオキシ−2−フルオロ−D−グルコース(2−FG)を投与した場合にはそうはならないという発見から始まった。2−FG及び2−DGは両方とも解糖を阻害するため、この効果に関与する、解糖の遮断以外の機序を推測した。
【0034】
1970年代に実施された研究は、2−DG及び2−FMがウィルスのコートタンパク質のN結合型グリコシル化に干渉し、この干渉はマンノースを添加することにより逆転できるという報告を導いた。マンノースとグルコースとの違いは、2−炭素位の水素の配向にあり、2−DGは2位に2個の水素を有する(水素及びヒドロキシル基の代わりに)ため、マンノース及びグルコースの両方の場合のように、2−DGをマンノース又はグルコース類似体のどちらかとして見なすことができる。したがって、2−DGは解糖及びグリコシル化の両方に作用できる。
【0035】
本発明は、細胞が低酸素環境下又は正常な酸素環境下にかかわらず、ヘキソース誘導体を単独又は、限定するものではないが、正常な酸素状態細胞を標的とする細胞毒性薬剤、抗血管形成剤、放射線療法及び外科手術を含む、他の抗腫瘍治療と併用して使用する腫瘍細胞の増殖を阻害する方法を提供する。本発明はさらに、2−DG、2−CM及び2−FMなどの類似体を、正常な酸素状態の(グリコシル化への干渉を介して)及び全ての低酸素(解糖の遮断を介して)の、特定の腫瘍型の癌細胞集団両方を標的にできる細胞毒性薬剤としての臨床使用のための根拠を提供する。
【0036】
以下の実施例は、本発明の有効性を検証し、2−DG、2−CM及び2−FMが毒性であるが2−FGは毒性でなく、正常な酸素状態で成長する腫瘍細胞型を選択することを確認するデータを提供する。実施例に記載した実験のいくつかは、解糖の阻害が通常酸素効果に関与する機序であるのとは対照的に、グリコシル化に干渉するかどうかを決定するように設計した。理論に縛られるつもりはないが、得られた結果は、これらの化合物がグリコシル化を阻害でき、その結果、それらの細胞が低酸素環境下にあるかどうかとは独立して、特定の癌細胞型を殺すことを裏付けるものである。
【0037】
結果はまた、2−FM、2−DG及び2−CMが脂質結合オリゴ糖鎖の組立てを撹乱し、糖タンパク質合成への干渉の指標であり得る、小胞体ストレス応答(unfolded protein response)(UPR)を誘導するが、2−FGはしないという結論を裏付ける。言い換えると、UPRは、感受性であるが耐性ではない細胞のUPR特異的アポトーシスシグナルの活性化を引き起こす。
【0038】
通常酸素下で2DG、2−FM、及び2−CMに感受性である腫瘍細胞型を特定した。細胞を腫瘍から単離し、エクスビボで試験し、細胞が通常酸素下で2DG、2−CM、及び2−FMに感受性であるかどうかを確定した。以下の実施例は、細胞が感受性かどうかを確定する方法を例示する。他の実施形態において、密接に関連のある2−DGに感受性である分子署名及び耐性細胞対を試験細胞系と比較する。ホスホマンノースイソメラーゼと、グリコシル化に関連する他の酵素及び2−DG蓄積に関連する酵素との、濃度及び/又は活性の違いを記載する。
【0039】
酸素の存在下で(正常な酸素状態)、2−DGは腫瘍細胞系のサブセットに有毒である。これまでの研究により、2−DGを用いて正常な酸素状態で処理した場合、腫瘍細胞及び正常細胞は成長を阻害されるが、殺されないということが実証されたので、この結果は意外であった。この成長阻害のこれまでの見解は、解糖経路の中間体濃度が減少するため成長が減少するように、2−DGが、通常酸素下で細胞内の解糖の遮断に十分な高濃度まで蓄積されることによるものと考えられ、これは細胞増殖に関与する様々な同化作用のために使用される。しかし、ミトコンドリアの機能が正常であるため細胞が死なないならば、その後好気的に処理された細胞は2−DGによる解糖の遮断を生き延びられる。どのように細胞が通常酸素下で2−DGに感受性であり得るかの1つの有望な説明は、したがって、細胞はミトコンドリアに欠陥を有するということである。これに関して、腫瘍細胞は、ミトコンドリア呼吸に欠陥があるため、エネルギー(ATP)産生のために酸化的リン酸化の代わりに嫌気的解糖を介してグルコースを利用することが公知である。しかし、さらなる実験により、オキサマート及び2−FGなどの他の解糖阻害剤はこれらの細胞に対して毒性でなく、ミトコンドリア呼吸の血管がそれらの2−DGに対する感受性が原因であるとは思えないことが実証された。したがって、解糖の遮断以外の機序が、これらの選択された細胞系において通常酸素下で、2−DGの毒性に関与すると仮定した。
【0040】
したがって、他の仮説によりこの正常な酸素状態での細胞毒性の機序を説明できる。1つの有力な機序はグリコシル化の干渉である。この有力な機序の裏付けは、1970年代末から一連の研究論文において特定され得、その中で特定のウィルスにおいてN結合型糖タンパク質合成が、2−DGを含む多くの糖類似体によって阻害されたことが報告された。
【0041】
グルコースは、3つの主要な経路:解糖、ペントースリン酸分路、グリコシル化を介して代謝される。図7は解糖及びグリコシル化代謝経路のスキーム図である。グルコースが細胞質に入った後、ヘキソキナーゼがグルコースの炭素6をリン酸化し、グルコース−6−リン酸(G6P)の合成が起こる。G6Pがホスホグルコースイソメラーゼ(PGI)により、フルクトース−6−リン酸に転換した場合、解糖経路において継続でき、ATP及びピルビン酸を産生する。或いは、G6Pはまた、マンノースを含む様々な糖成分の合成に使用でき、脂質結合オリゴ糖の組立てに必要であり、その合成はERで実施される。2−DGは、3つの代謝経路のうちの2つに干渉することが示されている:2−DGはPGIを阻害することによって解糖を遮断でき、又は2−DGは、グアノシン2リン酸(GDP)ドリコールリン酸結合マンノースのN−アセチルグルコサミン残基への転移に干渉することによってN結合型オリゴ糖前駆体の組立てを撹乱でき、及びマンノースを細胞質からERの内腔に運搬するために必要なドリコール−Pを減少させることができる。
【0042】
2−DGは、炭素2の両方の位置に水素を有し、マンノース類似体に似ているため上記の通りである。対照的に、フルオロ類似体のこの位置におけるフッ素の存在は、新しい鏡像異性体の中心を作り出し、それでフルオロ誘導体はグルコース又はマンノースのどちらかの類似体としか見なすことができない;これらの類似体の表現において、フッ素部分をマンノース類似体の炭水化物環の平面の「上に」又は上側に、及びグルコース類似体の炭化水素間の平面の下側に描く。
【0043】
脂質結合オリゴ糖鎖に加えるマンノースは、まずグアノシン2リン酸(GDP)又はドリコールリン酸への転移によって活性化しなくてはならない。2−DGは、2−DG−GDPへの転換を受け、これは、脂質結合オリゴ糖の組立て時に、マンノースをN−アセチルグルコサミン残基に加えるためのマンノース−GDPと競合する。したがって、科学文献に報告された実験において、2−DG処理の結果産生された異常なオリゴ糖はウィルス糖タンパク質の合成を低減させた。これらの実験において、2−DGの阻害効果は外因性のマンノースを加えることによって逆転するがグルコースを加えた場合はせず、さらに2−DGはいくぶんマンノース類似体のように作用することが確認された。これらの調査員らはまた、別のマンノース類似体の2−フルオロ−マンノース(2−FM)が、マンノースによって逆転する2−DGと同様の効果を有することを示し、これらの類似体のマンノース配置はそれらのグリコシル化への干渉にとって重要であり得ることを示した。
【0044】
さらに、遺伝学的研究は、グリコシル化の撹乱が意味深い生物効果を有し得ることを示した。糖タンパク質糖鎖不全症候群1b型に苦しむ患者に、酵素のホスホマンノースイソメラーゼ(PMI)は存在しない。この酵素の不在は血清糖タンパク質のグリコシル化の減少をもたらし、蛋白漏出性腸症を特徴とする血栓症及び胃腸障害を引き起こす。外因性マンノースをこれらの患者の食事に加えた場合、彼らの症状が消え、彼らの血清糖タンパク質は正常に戻り、彼らは病気から回復した。この観察は、実験データが、外因性マンノースが、2−DGを用いて正常な酸素濃度状態で処理した場合に殺される選択された腫瘍細胞を救うことができたことを示すように、本発明において有用な化合物に関する作用機序と一致する。これらの特定の腫瘍は、PMIが下方制御されている、又はこの酵素に欠陥を有するかの、どちらかである可能性がある。他方、N結合型グリコシル化に必要なマンノース中間体を産生する酵素は、これらの細胞において上方制御され得、マンノース−GDPに対して2−DG−GDPをより高い比率でもたらし、その結果正常な酸素濃度状態において2−DGに対するこの異常な感受性の原因となる。
【0045】
機序にかかわらず、本発明は、2−DG並びに他のグルコース及びマンノース類似体を、正常な酸素濃度状態においても腫瘍を治療する単剤として投与することによって癌を治療する方法を提供する。該化合物は、ヒト乳癌(SKBR3)、非小細胞肺(NSCLC)、神経膠腫、膵臓及び骨肉腫の癌細胞系を含む、多くの腫瘍細胞系に対して有効であり、これらの全てが2−DGの相対的低用量を用いて処理した場合に、細胞死を起こすことが実証されている。
【0046】
図3Bは、2−DG、2−FM及び他の薬剤を用いて、通常酸素下で表示の用量で、72時間処理したSKBR3細胞の反応を示したチャートである。細胞毒性を、トリパンブルー排除法により測定した。結果は、2−DG及びマンノース類似体の2−FMが毒性であり、一方グルコース類似体の2−FGは毒性ではないことを示す。さらに、乳酸デヒドロゲナーゼ段階で解糖を遮断するピルビン酸塩類似体であるオキサマートもまた、通常酸素下で成長するこれらの細胞に対して毒性ではない。対照的に、マンノース類似体の2−FMはまた、これらの細胞において毒性であることが証明され、再度マンノース骨格がこの活性を有する化合物にとって重要であったことを示した。
【0047】
2−DGの阻害効果は、外因性マンノースの添加により逆転されたが、グルコースを加えた場合には逆転せず、さらに2−DGがマンノース類似体として作用することが確認された。他の試験により、2−DGもまた、通常酸素下で成長するNSCLCに対して毒性であり、1mMのマンノースの添加がこの毒性を逆転することが示された。
【0048】
このデータは、2−DG及び2−FMは、グリコシル化に干渉するため、通常酸素下で成長する選択腫瘍細胞に対して毒性であることをさらに裏付ける。これらのマンノース類似体がこの機序を介して働き、解糖の遮断を介してではないことの付加的証拠は、ミスフォールド及び又はミスグリコシル化のタンパク質を示す小胞体ストレス応答タンパク質のGRP78及び94が、2−DG及び2−FMによって用量依存性様式で上方制御されるが、2−FGによってはされないことであり、この効果はマンノースの添加によって同様に逆転する。
【0049】
したがって、マンノース類似体の2−DG及び2−FMは、通常酸素下で成長する選択腫瘍細胞型に対して毒性であり、マンノースの添加によりこの毒性は逆転するが、グルコース類似体の2−FGではそうではない。2−FGは、2−DGより解糖をよく阻害するので、グリコシル化への干渉及び解糖を阻害しないことは、この効果に関与すると考えられる機序である。上記のように、2−DGが、ウィルスコートタンパク質のN結合型グリコシル化に干渉すること、及び外因的に加えられたマンノースがこの効果を逆転することは、報告されている。2−DGのSKBR3、NSCLC及び2種の他のヒト腫瘍細胞系に対する通常酸素下での毒性効果は、したがってグリコシル化への干渉によるものと思われる。この機序的理論が正しければ、その後マンノースの添加により、これらの細胞系において2−DGの毒性は逆転されるはずである。実際、被験細胞系の1つ(NSCLC)において、1mMのマンノースは6mMの2−DGの毒性効果を逆転する。
【0050】
マンノースの血中濃度は50〜60μg/mlとして知られているので、2−DGの毒性を逆転するために必要な最小マンノース用量を確定するための、用量反応性実験が実施できる。例えば、FBSは普通、残留量のマンノースを含んでいるため、成長培地に透析されたウシ胎児血清(FBS)を添加した実験によって達成できる。さらに、2−DGの毒性を逆転するためには、マンノースの添加が必要であって、他の糖は必要でないことを確認するために、糖タンパク質合成に関与することが公知の糖、すなわちグルコース、フコース、ガラクトースなどを2−DG毒性の逆転能力に関して試験できる。これらの糖のいずれかが同様に毒性を逆転できる場合、その後それらの活性を、UPRの誘導及びその結果、オリゴ糖鎖の伸長への干渉並びにコンカナバリンAの結合についての、2−DGの効果の逆転に関する以下に記載した実験において、マンノースの活性と比較できる。全体的にこれらの実験は、生理的濃度のマンノースの存在下で抗腫瘍活性をもたらす、インビボで使用できる2−DG又は2−FMの用量を、インビトロで評価することを可能にするものである。しかし、本発明の方法に使用する、経口投与の2−DG、2−FM及び2−CMの治療有効量は、通常は5〜500mg/患者の体重1kg、例えば50〜250mg/kgの範囲であろう。一実施形態において、用量は、約100mg/患者の体重1kgである。
【0051】
本発明はまた、腫瘍又は他の癌が現在の治療方法に感受性の細胞を含むかどうかを確定するために臨床医が使用できる、多くの診断方法を提供する。一実施形態において、腫瘍由来の細胞を、それらが2−DG,2−FM,又は2−CMにより殺されるかどうかを確定するために、通常酸素下で試験する。別の実施形態において、この試験を実施し、その後マンノースを加え、マンノースが細胞毒性効果を逆転させるかどうかを確定する。
【0052】
別の実施形態において、感受性に関する試験を、N結合型グリコシル化を指標として使用して実施する。上記のように、2−DG及び2−FMは、脂質結合オリゴ糖鎖の組立てを撹乱する、(2)正常な糖タンパク質合成への干渉の指標であり得る、小胞体ストレス応答(UPR)の誘導する、及び(3)2−DGに感受性であるが耐性ではない細胞においてUPR特異的アポトーシスシグナルを活性化する、が2−FGはしない。さらに、マンノースはこれらの効果を逆転する。したがって、これらの同じ試験を対象となる腫瘍又は癌細胞に実施でき、その細胞が本方法を用いた治療に感受性であるかどうかを確定できる。
【0053】
上記のように、ウィルスに感染した細胞のERの細胞質表面上で、マンノースの脂質結合オリゴ糖鎖への取り込みが起こり、この取り込みを2−DG又は2−FMのGDP誘導体、すなわちGDP−2DG及びGDP−2−FMにより阻害できる。普通は、5回目のマンノースを加えた後で、脂質結合オリゴ糖鎖は、ERの内腔に向かってフリップする。成長する鎖にマンノースを加え続けるために、ドリコールリン酸(Dol−P)を、細胞質からERのマトリックスにマンノースを輸送するための担体として使用する。2−DG−GDPはドリコールに結合することに関してマンノース−GDPと競合し、その結果N結合型グリコシル化にさらに干渉する。さらにドリコール結合2−DGもまた、ER内のオリゴ糖鎖上へのマンノースの輸送に競合する。したがって、実験を、脂質結合オリゴ糖鎖前駆体及びマンノース誘導体、すなわちマンノース−6−リン酸、マンノース−1−リン酸、GDP−マンノース及びDol−P−マンノースの形成についての2−DG及び2−FMの効果を実証するために、2−DGに感受性及び耐性の両方の細胞系において実施できる。これは同様に2−DG及び2−FMにより阻害される、オリゴ糖組立てにおける1つ又は複数のステップを実証する。これは同様に、2−DG、2−FM、及び/又は2−CMに曝露することで、オリゴ糖を産生する(及び産生しない)ことに基づいて、感受性又は耐性として他の細胞型の特徴付けを可能にするものである。
【0054】
これまでに確立されたクロマトグラフィー法を、SKBR3及びNSCLC細胞内のマンノース誘導体及び脂質結合オリゴ糖前駆体の回収及び量の測定に使用できる。簡潔に言えば、細胞は[2−H3]マンノースを用いて標識でき、細胞溶解物を、クロロホルム/メタノール(3:2)及びクロロホルム/メタノール/水(10:10:3)を、それぞれDol−P−Man及び脂質結合オリゴ糖を回収するために用いて抽出した。
【0055】
Dol−P−Manを含むアリコートを薄層クロマトグラフィーに供与し、一方脂質結合オリゴ糖は、HPLCによって分離できる。溶出分画は、液体シンチレーション計測により分析できる。マンノースリン酸及びGDP−マンノースは、下降ペーパークロマトグラフィーによって、各分画から放出された[2−3H]マンノースは弱酸加水分解によって分離でき、測定できる。2−DG又は2−FMを用いて処理した細胞に由来する値を、未処理の対照と比較し、N結合型オリゴ糖前駆体及びマンノース誘導体に関するこれらの薬剤の効果を実証できる。外因性マンノースは、2−DGの毒性を逆転するので、マンノースがまた観察された、2−DGによるグリコシル化の動揺を逆転するかどうかを試験できる。
【0056】
2−DG及び2−FMに加えて、N結合型グリコシル化の特定のステップを阻害できる、2種の他のグリコシル化阻害剤、ツニカマイシン及びデオキシマンノジリマイシン(DMJ)を陽性対照として使用できる。ツニカマイシンは、第1のN−アセチルグルコサミン残基のドリコールリン酸への付加に干渉し、DMJは、N結合型オリゴ糖鎖の末端の3マンノース残基を切り取る、マンノシダーゼIの特異的阻害剤である。したがって、外因性マンノースは、毒性又はこれらの薬剤のどちらかのグリコシル化に関する効果のうち、いずれかを逆転できないはずである。さらに、グルコース類似体の2−FGは、通常酸素下でSKBR3及びNSCLC細胞を殺さないが、解糖の遮断及び低酸素細胞を殺すことにおいて2−DGより強力であるため、グリコシル化に作用することなく解糖に干渉でき、そのためこのような試験にツールとしても使用できる。
【0057】
小胞体(ER)内のN結合グリコシル化の過程への干渉は、糖タンパク質の不適切なフォールディンクを引き起こし、これは、小胞体ストレス応答(UPR)と呼ばれるERのストレス応答を誘発する。DNAの損傷に対するP53応答によく似て、ERはストレスに対して(1)内在シャペロン(GRP78及びGRP94)の誘導を介したフォールディング能力の増加、(2)タンパク質合成の停止による自身の生合成負荷の減少、及び(3)フォールディングしていないタンパク質の分解の増加、によるほとんど同じ方法で応答する。ストレスが緩和できない場合、アポトーシス経路が開始され、続いて細胞が死ぬ。したがってグリコシル化への干渉の1つの指標は、UPRの上方制御である。
【0058】
SKBR3細胞を2−DGを用いて処理した場合、これらのERストレス応答タンパク質である、GRP78及び94の両方が用量増加の機能として増加するが、マンノースがこの誘導を逆転する。2−FGは、このタンパク質を誘導しない。したがって、本発明の別の実施形態において、この応答を腫瘍又は癌細胞が本方法に従った治療に感受性であるかどうかを確定するために使用する。通常酸素下で2−DGに感受性でない細胞系も、これらのタンパク質の上方制御の不在が、それらの2−DGに対する耐性に相関する、陰性対照として同様に使用できる。
【0059】
ERストレスを克服できない場合に、アポトーシスシグナルが開始される。ERストレスは、CHOP/GADD153を介したミトコンドリア依存性アポトーシス経路、BCL−2を下方制御する核転写因子及びヒトのカスパーゼ4及び5、並びにマウス細胞系のカスパーゼ12によるミトコンドリア依存性経路を誘導する。したがって、実験を実施し、ERストレスに特有のアポトーシスシグナルが、2−DGに感受性であるが耐性ではない細胞において活性化されるかどうかを確定できる。CHOP/GADD153の上方制御及びカスパーゼ4及び5の活性化は、ウェスタンブロットにより検査できる。先の試験と同様に、この上方制御が2−DG感受性系に特有である場合、その後試験癌細胞において観察された上方制御は、細胞が由来した癌が本発明に従った治療に感受性であることの指標として機能を果たす。
【0060】
SKBR3は糖タンパク質ErbB2を豊富に発現するので、2−DGがこのタンパク質のN結合型グリコシル化に作用し、ミスフォールディング及び分解を引き起こすであろうことが期待される。2−DGを用いて処理したSKBR3細胞由来のErbB2のウェスタンブロットを未処理の細胞のものと比較して、このタンパク質の全濃度を測定できる。さらに、2−DG処理後のErbB2のマンノース含有量を、このタンパク質を免疫沈降し、コンカナバリンA、高マンノース型N結合オリゴ糖を認識するレクチンを用いてブロットすることによって分析できる。マンノース類似体はErbB2だけでなく、全N結合型糖タンパク質に含まれるマンノースを阻害できるのと思われるので、これらの細胞から得た全細胞溶解物をこのレクチンを用いてさらに解析できる。全タンパク質に結合するポンソー染色を、陰性対照として使用でき、2−DG及び2−FMが糖タンパク質に特異的に作用し、再度この方法論又は類似の方法論が、癌又は腫瘍の細胞が本発明に従った治療に感受性であるかどうかを確定するために使用できることを検証できる。
【0061】
N結合型グリコシル化への干渉を示すERストレスが、2−DG及び2−FMによって起こることが実際に確認されたとしても、ERとは対照的に細胞質において起こるO−グリコシル化への干渉もまた評価できる。科学文献により、2−DGは、O−グルコシル化転写因子であるSp1からのN−アセチルグルコサミン残基の切り取りを阻害でき、その個別のプロモーターへの結合を阻害することが報告されている。Sp1は、多くの癌遺伝子の活性化に重要な転写因子であり、2−DGをSp1に作用させれば、なぜ通常酸素下で成長するSKBR3細胞が2−DGに感受性なのかを、少なくとも部分的に説明できる。したがって、2−DG及び2−FMを用いた処理の後のSp1のグリコシル化のパターンを、免疫沈降及びWGA、O−グリコシル化タンパク質に特異的に結合するレクチンを用いて解析することによって調査できる。2−DGがSp1及びO−結合型グリコシル化に作用するという点で、グリコシル化のこの変化を測定し、腫瘍又は他の癌細胞系が2−DG仲介殺細胞に感受性であることの指標として使用できる。
【0062】
細胞死は、小胞体ストレス応答によって誘発され、ミスフォールドタンパク質に応答して、各細胞の小胞体において起こり、追加の薬剤であるベルシペロスタチンを投与することによって促進できる。したがって、一実施形態において、2−DG、2−FM及び/又は2−CMを、癌治療を必要とする患者に投与し、ベルシペロスタチンを前記患者に同時投与する。
【0063】
同様に、タンパク質のミスフォールドに応答して起こる細胞死は、ミスフォールド糖タンパク質のタンパク質分解を、プロテオソーム阻害剤を用いて遮断することによって促進できる。したがって、別の実施形態において、本発明は、2−DG、2−FM及び/又は2−CMと併用して、プロテオソーム阻害剤を投与することによる、癌の治療方法を提供する。一実施形態において、プロテオソーム阻害剤はベルケイド(Velcade)である。
【0064】
癌の特定の型は、本方法による治療に他の癌より感受性である可能性がある。このような型を特定するために、本発明の方法に従って様々な細胞型の種類を試験できる。例えば、ATCCから様々な癌細胞系を得ることができ、酸素の存在下で2DG及び2FMなどのマンノース類似体に非常に敏感な他の細胞型を特定するためにそれらを上記のようにスクリーニングできる。5mM以下の2−DG又は2−FM濃度で殺される細胞を、感受性であるとして特定する。これらの感受性腫瘍細胞系は、さらに2−FG及びオキサマートに対する感受性に関して、それぞれ用量20mM及び30mMまで試験できる。グリコシル化への干渉が、2−DG及び2−FMの毒性の様式である場合、それならばこれらの細胞系は、ミトコンドリアの酸化的リン酸化に欠陥を有さない限り、他の解糖系阻害剤である2−FG及びオキサマートに対して耐性であるはずである。これらの細胞のミトコンドリアの機能性を確認するために、呼吸を、例えばクラーク式電極装置を使用して測定できる。2−DG及び2−FMの毒性が、これらの細胞系におけるグリコシル化への干渉によるものであることを確認するために、マンノースによる細胞死の回復を上記のように検査できる。
【0065】
1つの細胞が本方法に耐性であり、別の細胞は耐性でないことに関する分子基礎は、グルコースからGDPマンノースの合成に関与する遺伝子、すなわちグルコース−6−リン酸をマンノース−6−リン酸に転換する、ホスホグルコースイソメラーゼ(PMI)の発現における違いによると思われる(図7)。上で述べたようにPMIの欠失が、グリコシル化症候群1bの原因であり、これは血清糖タンパク質のグリコシル化の減少をもたらし、この欠陥を有すると特定された患者に血栓症及び胃腸障害を引き起こすことを示した。マンノースを食事に加えることにより、患者の症状を軽減し、患者の糖タンパク質を正常化することを示した。したがって、この酵素の欠如又は下方制御により、今までに試験された感受性細胞系における2−DG及び2−FMの毒性及び外因性マンノースによる逆転を説明できる。
【0066】
PMIの下方制御又は欠失が、感受性細胞系において2−DGの毒性を引き起こし得る理由は、この酵素が存在しない状態では、細胞がN結合オリゴ糖前駆体の合成を外因性マンノース(血清中に存在)に依存するからである。哺乳動物の血清中のマンノース濃度(50〜60μg/ml)又はインビトロ研究に使用した培地中のマンノース濃度は、グルコース濃度よりも有意に低いことは公知である。したがって、PMIが欠失した又は下方制御された細胞において、低用量の2−DG及び2−FMは血清中に存在する低量のマンノースと都合よく競合し、この糖のオリゴ糖鎖上への付加を完全に遮断する。他方、正常なPMIを有する細胞はグルコースからGDP−マンノースを産生でき、したがってオリゴ糖の組立てを完全に撹乱するためにはより多くの用量の2−DG又は2−FMが必要である。これにより、大部分の被験細胞が通常酸素下で2−DGに対して耐性であることの理由が説明できる。この酵素の活性の直接測定は本発明に従って使用でき、PMIの欠如又は低濃度のPMIが、通常酸素下で成長する選択細胞において、2−DG及び2−FMに対する感受性に関与するかどうか、またもしそうならば、その時は本方法に従った治療に感受性の腫瘍及び癌細胞の特定に使用できるかどうかを確定できる。あまり見込みはないが、この異常な感受性を説明する別の可能性は、正常な酸素圧の下で成長する場合、これらの選択細胞におけるPMIが2−DG及び2−FMによって大部分の正常な細胞系及びこれらの薬剤に作用されない腫瘍細胞系より阻害されるということである。このことを直接試験するために、細胞抽出液をSKBRの耐性及び感受性の細胞対から単離でき、グルコース−6−Pからマンノース−6−Pへの転換能力を、2−DG及び2−FMの存在する又は存在しない状態で確定できる。
【0067】
PMI活性の減少が、SKBR3の感受性細胞において2−DGによる毒性に関与していない場合、その時にこれを説明する代わりの機序は、オリゴ糖の組立てに使用するマンノース誘導体の産生に関与する酵素、すなわち、ホスホマンノムターゼ(PMM)及びGDP−Manシンターゼをコードする遺伝子の上方制御である(図7)。2−DGに感受性の細胞のグリコシル化が増加し、それ故、これらの酵素のどちらか1種、又は両方が上方制御される可能性がある。このような細胞は2−DG−GDPがさらに蓄積され、それ故グリコシル化が低速又は低い能力で起こった耐性細胞よりも、グリコシル化及びそれに続く細胞死に、より大幅な干渉を引き起こすであろう。
【0068】
グリコシル化の上方制御が機序であり、それにより細胞が2−DGに感受性になるかどうかにかかわらず、細胞内に蓄積された又は取り込まれた2−DGの全量はまた、細胞の感受性の増加の原因となる。したがって、[3H]で標識した2−DGを使用した、取り込み及び蓄積の研究は、グルコース輸送体濃度がより高い細胞が、本方法に従った治療に、より強い感受性を示すかどうかを確定するために実施できる。
【0069】
感受性細胞を2−DGの漸増用量を用いて処理し、生存細胞を選択することによって、感受性細胞から2−DG耐性変異体を得ることができる。耐性変異体及びそれらの親感受性対応物を、記載の方法に使用できる。このような研究は、細胞が2−DGに耐性になる機序を理解する方法を提供するはずであり、それ故この薬剤の臨床的に優れた使用に適用できると思われる。前述の議論は、分子署名を、どの腫瘍細胞型が酸素の存在下で2−DG及び2−FMに感受性であろうか予測するために使用できることを反映している。
【0070】
細胞死の実行は、アポトーシスとネクローシスとの範囲に及ぶ顕著な柔軟性を示す。DNAの切断の型、細胞膜の組成の変化、完全性及び状態を調査することによって細胞死の様式を比較する、確立された方法を使用することにより、グリコシル化への干渉によって及び解糖の阻害によって誘導される細胞死の機序を確定できる。解糖及び酸化的リン酸化の両方の阻害は、深刻なATPの減少をもたらし、その結果アポトーシスからネクローシスへの転換を引き起こす。ATPはカスパーゼの活性化に必要なので、ATPが大幅に減少した場合、アポトーシスは遮断され、最終的にエネルギーがなくなり、細胞はネクローシスを介して死に至る。解糖系阻害剤を用いて処理した好気性細胞は、エネルギー源としてアミノ酸及び/又は脂肪のどちらかに支えられて、酸化的リン酸化を介してATPを産生できる。したがって、2−DGがUPR応答を誘導し、通常酸素下で細胞死を引き起こす場合、細胞はアポトーシスに陥るであろうと考えられる。逆に言えば、低酸素細胞モデルにおいて、2−DGの用量が解糖を遮断するために十分多い場合これらの細胞はATPが減少し、ネクローシスを介して死ぬはずであることが期待される。
【0071】
したがって、アポトーシス及びネクローシスに関する検査の確立された方法を使用でき、2−DGがアポトーシス、ネクローシス及び/又は両方の混合を介して細胞を殺しているかどうかを確定できる。いくつかのアポトーシスパラメーターを、フローサイトメトリー分析の使用によって、ネクローシスとアポトーシスを識別するために検査できる。2−DGによる処理の後で、細胞をアネキシンV及びヨウ化プロピジウムを用いて二重染色し、細胞表面上のホスファチジルセリンの露出及び細胞膜の完全性の喪失をそれぞれ検出できる。アネキシンV単独又はアネキシンV及びヨウ化プロピジウムの両方、のどちらかを用いた染色により、アポトーシスが示され、一方、ヨウ化プロピジウム単独を用いた染色によりネクローシスが示される。さらに、アポトーシスの最終結末の2つである、核DNAの断片化及び一本鎖DNAの形成もまた測定できる。これら2つの後者のパラメーターは、アポトーシス細胞死に独特であることが報告されており、アポトーシスとネクローシスを差別化するために、様々な研究者に使用されている。ATP濃度はまた、それらが検出された死の様式と相関するかどうかを確定するために検査できる。
【0072】
さらに、2−DGが低酸素細胞においてアポトーシス及びネクローシスの両方を誘導する場合、その時低酸素状態の下で2−FGにより誘導された細胞死の様式を確定できる。上に述べたように、2−FGはグリコシル化に干渉せず、2−DGより強力な解糖阻害剤である。したがって、2−FGにより誘導される細胞死は、ネクローシスを介して単独で起こるであろうことが期待される。
【0073】
通常酸素下で、インビトロで2−DG及び/又は2−FM及び/又は2−CMに感受性であると判明した、ヌードマウスにおいて容易に成長する細胞系を、2−DG(及び2−FM及び2−CM)がインビボで与えられた場合、該細胞系に対して単剤として有効であることを実証するために使用できる。腫瘍が特定の大きさに達した後に、2−DGを用いた治療が腹腔内注射を介して適用されるであろう。これらの動物においてあらかじめ確立された最小致死量に従った2−DGの用量及び治療計画を、腫瘍の退縮及び細胞毒性を実証するために使用できる。
【実施例】
【0074】
(実施例1)
材料及び方法
耐性変異体の単離。2−DG感受性SKBR3及びNSCLC細胞を、2−DGの漸増用量に曝露し、耐性コロニーを単離し、適切な用量の2−DGでクローン化する。クローン化した2−DG耐性細胞を、その後分析し、野生型の感受性対応物と、この独自の感受性に関与できる特定の遺伝子の発現に関して比較する。
【0075】
薬剤及び抗体。Rho123、オリゴマイシン、スタウロポリン及び2−DG、2−FG、2−FM、ツニカマイシン、デオキシマンノジリマイシンは、Sigma Chemical Co.から入手する。以下の一次Absを使用できる:モノクロナール抗体を、HIF−la及びLDH−a(BD Biosciences);erbB2(Calbiochem,USA);Grps78&94,(StressGen,USA);カスパーゼ4及び5(StressGen,USA);並びにアクチン(Sigma Chemical Co.)に;ポリクロナール抗体を、GLUT−1(USA Biological)及びGADD153/CHOP(Santa Cruz,USA)に。二次抗体はホースラディッシュペルオキシダーゼ標識ウサギ抗マウス及びヤギ抗ウサギ(Promega,Co.)である。
【0076】
細胞毒性検査及び高速DNA含有量分析。細胞を37℃で24時間、5%CO2においてインキュベートし、その時に薬剤処理を開始し72時間続ける。この時に、吸着細胞をトリプシン処理し、それらの個別の培養培地と混合し、その後400gで5分間遠心分離機にかける。細胞を含むペレットを、細胞毒性検査のために1.5mlの培地/トリパンブルー混合物、又は核DNA含有量及びCoulter XLフローサイトメーターによる細胞周期の測定のためにヨウ化プロピジウム/低張クエン酸塩染色溶液のどちらかに再懸濁する。DNA分布ヒストグラムを作成するために最低10000細胞を分析する。
【0077】
乳酸検査。乳酸を、処理済み培養液又は未処理培養液から0.025mlの脱タンパク質化培地を、0.1mlの乳酸デヒドロゲナーゼ、(1000単位/ml)、2mlのグリシンバッファー(グリシン0.6モル/L及びヒドラジン、pH9.2)及び1.66mg/mlのNADを含む、反応混合物に加えることによって測定する。脱タンパク質化を、試験培養物からの0.5mlの培地を1mlの8%w/v過塩素酸を用いて処理し、30秒ボルテックスにかけ、その後この混合物を4℃で5分間インキュベートし、1500gで10分間遠心分離機にかけることにより起こす。上清をさらに3回遠心分離機にかけ、0.025mlの最終的に不要物を除いた上清を乳酸の測定に使用する。NADHの形成を、Beckman DU r 520 UV/vis分光光度計を用いて340nmで測定し、これは乳酸塩標準曲線により確定された乳酸濃度に直接一致する。
【0078】
2−DGの取り込み。細胞をペトリ皿に播種し、37℃で24時間、5%CO2においてインキュベートする。培地をその後除去し、プレートをグルコース及び血清を含まない培地で洗浄する。2mlの、3H標識2−DGを含み、血清を含まない培地を皿に加え(1γCi/プレート)、プレートを適切な時間インキュベートする。培地をその後除去し、プレートを4℃で、100μMの標識のない2−DGを含み、血清を含まない培地を用いて3回洗浄し、0.5mlの1NのNaOHを加える。37℃で3時間(又は一晩)インキュベートした後で、細胞をこそげ取り超音波により均一化した(10秒)。溶液を、3Hの定量化のためにチューブに回収した(タンパク質検査のために一部は保存)。100μLの蟻酸、250μLの試料及び7mlのシンチレーションカクテルを、3H計測バイアルに混ぜ合わせ、シンチレーションカウンターを用いて読み取る。輸送速度(nモル/mgタンパク質/時間)を全CPM/比放射能/全タンパク質により算出する。
【0079】
ATP定量検査。ATPライトキット(lite kit)(Perkin Elmer社)を、ATP濃度の定量化に使用できる。約50μLの細胞溶解液を、96ウェルプレート(ボトム白)において100μLの細胞懸濁液に加える。プレートを室温で、シェーカーにのせて(700rpm)5分間インキュベートする。50μLの基質溶液を、続けてウェルに加え、さらに5分間室温で振とう(700rpm)する。プレートをその後10分間暗順応させ、発光を測定する。
【0080】
代謝標識並びにDol−P Man及び脂質結合オリゴ糖(LLO)の抽出。Lehleにより記載された手順に従って、細胞を[2−3H]マンノースを用いて30分間標識し、2mlの氷冷メタノールにこそげ取り、超音波処理により溶解する。4mlのクロロホルムを加えた後、材料を超音波で分解し、その後4℃において5000rpmで10分間遠心分離機にかける。上清を回収し、ペレットをクロロホルム/メタノース(3:2)(C/M)を用いて2回抽出する。Dol−P Man及び小型の脂質結合オリゴ糖を含む、混ぜ合わせた上清を、N2下で乾燥させ、3mlのC/Mに溶解し、洗浄し、シリカゲル60アルミニウムシート上でC/M/H2O(65:25:4)を含むランニングバッファーを使用した薄層クロマトグラフィーによって分析する。大型のLLOを含む残りのペレットを、洗浄し、C/M/H2O(10:10:3)を用いて抽出する。C/M及びC/M/H2O抽出物の対応するアリコートを混ぜ合わせ、N2下で乾燥させ、35γl 1−プロパノールに再懸濁する。弱酸加水分解によってオリゴ糖を放出するために、500γl、0.02NのHClを加え、その後100℃で30分間インキュベートする。
【0081】
加水分解した材料をN2下で乾燥させ、その後超音波処理により200γlの水に再懸濁し、遠心分離にかけ、不要物を取り除く。放出されたオリゴ糖を含む上清をHPLC分析に使用する。
【0082】
HPLCによるオリゴ糖のサイズ分画。LLOの分離は、LC−NH2(2cmx4.6mm)プリカラムを含むSupelcosil LC−NH2カラム(25cmx4.6mm;5γm;Supelco社)で実施できる。アセトニトリルが水に対して70%から50%の直線勾配を、流速1ml/分で適用する。溶出分画を液体シンチレーション計測によって分析する。
【0083】
マンノース6−リン酸、マンノース1−リン酸、GDP−マンノースの調製。[2−3H]マンノースを用いて標識した後で、細胞を収穫し、遊離のマンノースをヌクレオチド結合及びリン酸化マンノース誘導体から、Kornerらにより記載されたペーパークロマトグラフィーによって分離する。溶出分画を液体シンチレーション計測によって分析する。
【0084】
ウェスタンブロット分析。細胞を、104細胞/cm2で播種し、薬剤処理下で表示の時間成長させる。処理期間の終りに、細胞を回収し、プロテイナーゼ阻害剤カクテルを添加したRIPAバッファー(150mM NaCl、1%Np−40、0.5%DOC、0.1%SDS、50mM Tris−HCl、ph8.0)を用いて溶解する。DNAを、溶液を21Gの針に10回通過させることにより断片化する。タンパク質濃度をSuper Protein Assay キット(Cytoskeleton社,USA)により測定する。試料を、2×Laemmliサンプルバッファー(Bio−Rad社,USA)と混合し、SDS−ポリアクリルアミドゲルにかける。ゲルをニトロセルロース膜(Amersham社,USA)に転写し、特異的抗体を用いて解析する。解析の後で、膜を洗浄し、HRP標識二次抗体と共にインキュベートする。化学発光をフィルムに感光させることにより検出する。
【0085】
指示がある場合には、膜をストリッピングバッファー(Pierce社,USA)を用いてストリップし、抗アクチン一次抗体を用いて再度解析する。
【0086】
ErbB2の免疫沈降。細胞を24時間処理した後で、細胞をRIPA(15mM NaCl、1%Np−40、0.1%SDS)により溶解し、超音波で分解する。細胞溶解物をモノクロナールErbB2抗体(Calbiochem,USA)に結合したCnBr活性化セファロースビーズ(Amersham社,USA)と共に400gで5分間回転させて、インキュベートする。
【0087】
免疫沈降させたErbB2をSDS−PAGEゲルに添加し、糖タンパク質のマンノース残基に特異的に結合する、コンカナバリンAを用いてブロットする。
【0088】
アポトーシス検査。アポトーシスELISA検査を記載のように使用し、該検査はホルムアミドによるアポトーシス細胞の濃縮クロマチンにおける選択的なDNAの変性及びアポトーシス細胞の一本鎖DNA(ssDNA)と、ssDNAに高度に特異的なモノクロナール抗体との反応性に基づく。これらの抗体は、アポトーシス細胞を特異的に検出し、ネクローシス細胞とは反応しない。
【0089】
フローサイトメトリーによる細胞死の機序の調査。アポトーシスを、An−nexin−V−Fluos Staining キット(Roche社,USA)によりネクローシスと識別する。表示の処理の後で、106細胞を、FITC標識アネキシンV及びヨウ化プロピオジウムを含むインキュベーションバッファーに再懸濁し、ホスファチジルセリン及び細胞膜の完全性をそれぞれ検出する。インキュベーションの後で、細胞を、488nmの励起光及びフルオレセインの検出のために515nmのバンドパスフィルター及びPIの検出のために600nmを超えるフィルターを使用したフローサイトメーターにより分析する。
【0090】
遺伝子発現のプロファイリング。Gene−array kitをSuper Array Inc.より購入できる。選択された細胞系由来の全RNAをdCTP[α−32P](3000Ci/mmol)を用いて、逆転写反応を介して解析する。解析した標識cDNAを、その後予備ハイブリッド形成アレイ膜(pre−hybridized array membrane)に添加し、ハイブリダイゼーションオーブンにおいて一晩インキュベートする。複数回洗浄した後で、遊離のプローブを除去し、膜をX線フィルムに感光し、画像を記録する。
【0091】
インビボ腫瘍実験。Cancer Res.2004(by Lampidis et al)に報告された2DG+Doxに関して記載されたプロトコルを、2−FGを2−DGに置き換えて再現できる。ヌードマウス、CD1系、5〜6週齢、体重30gに、100γlのヒト骨肉腫細胞系143bの107細胞/mlを移植した(S.C.)。腫瘍が50mm3の大きさになった時(9〜10日後)、動物を4つの群(8匹のマウス/群)に、以下のようにペア合わせをする:生理食塩水処理の対照:2−FG単独;Dox単独及びDox+2−FG。0日目、2−FG単独及びDox+2−FG群に、75mg/ml(500mg/kg)の2−FG、0.2mlをi.p.で与え、これを実験期間中、週に3回繰り返した。1日目、Dox及びDox+2−DG群に、0.6mg/ml(6mg/kg)のDox、0.3mlをi.v.で与え、これを週に1度、合わせて3回繰り返した(18mg/kg)。週に3回、マウスの体重を量り、腫瘍の測定をキャリパーにより行う。
【0092】
SKBR3細胞を、2−DG又は2−FMを用い、ドキソルビシン(Dox)を用いない上記のモデルに移植し、試験した。
【0093】
(実施例2)
マンノース誘導体に対する特定の腫瘍細胞の正常な酸素状態での感受性。
低酸素状態での細胞の成長は、エネルギー産生のための解糖を介したグルコースの代謝にもっぱら依存する。その結果として、この経路が、2−デオキシ−D−グルコース(2−DG)により遮断された場合に、低酸素細胞は死ぬ。対照的に、解糖が、通常酸素下で遮断された場合に、脂肪及びタンパク質をエネルギー源として置換でき、ミトコンドリアの酸化的リン酸化に燃料を供給するため、大部分の細胞は生存する。本発明は、正常な酸素圧の下で、選択されたいくつかの腫瘍細胞系が、比較的低用量の2−DG(4mM)で死ぬという発見に一部基づいている。2−DGが、ウィルスコート糖タンパク質合成において、N結合型グリコシル化の過程に干渉し、これは外因性マンノースの添加により逆転できることが先に示されている。本明細書に記載された通常酸素下での2−DGの細胞毒性は、低用量のマンノース(2mM)によって完全に逆転できるため、解糖ではなく、グリコシル化がこれらの結果に関与する機序であると考えられる。さらに、解糖の遮断及び低酸素細胞を殺すことにおいて2−DGより強力である、2−フルオロ−デオキシ−D−グルコース(2−FDG)は、通常酸素下で2−DGに感受性である、いかなる細胞型にも細胞毒性は示さない。
【0094】
糖タンパク質合成に関する2−DGの効果を調査するために、コンカナバリンA(糖タンパク質のマンノース部分に特異的に結合する)を、2−DGは結合を減少させるが2−FDGはせず、これは外因性マンノースの添加により逆転可能であったことを示す研究に使用した。同様に、N結合型グリコシル化が変化した場合に誘導されることが公知である、小胞体ストレス応答(UPR)タンパク質、grp98及び78が2−DGによって上方制御されるが、2−FDGによってはされず、再度この効果がマンノースによって逆転できることが見出された。さらに、2−DGが、UPR特異的転写因子(GADD154/CHOP)の上方制御を介して細胞死を誘導し、これはアポトーシスを仲介する。したがって、特定の腫瘍細胞型において、2−DGが、固形腫瘍の好気的細胞(グリコシル化への干渉を介して)並びに低酸素細胞(解糖の阻害を介して)の両方を選択的に殺すための単剤として、臨床的に使用できる。
【0095】
血管形成にもかかわらず、急速な腫瘍成長の代謝要求は、多くの場合酸素供給より勝っており、これが大部分の固形腫瘍内における低酸素領域の形成の一因となる。腫瘍の成長につれて起こる酸素濃度の減少は、低酸素部分における細胞の複製速度を遅くし、通常急速に増殖する細胞を標的とする、大部分の化学療法剤に対する耐性をもたらす。Brown,J.M,et al.,「癌治療における腫瘍内低酸素状態の活用(Exploiting Tumor Hypoxia In Cancer Treatment)」,Nat Rev Cancer 2004;4:437−47。低酸素細胞は、緩慢な成長及び活性酸素種を発生するために必要な酸素の不在により、放射線治療にもまた耐性である。Semenza,G.L.,「腫瘍内低低酸素状態、放射線耐性及びHIF−1(Intratumoral Hypoxia,Radiation Resistance,And HIF−1)」,Cancer Cell 2004;5:405−406。癌治療に関するこれらの不都合に加えて、低酸素状態は、腫瘍細胞をエネルギー産生のために解糖に依存させ、生存する。低酸素状態の下で、ATP産生の最も効率の良い方法である酸化的リン酸化が阻害され、ATPの産生のための唯一の方法として解糖が残される。したがって、低酸素腫瘍細胞において解糖を遮断することが細胞死を引き起こすはずである。実際、インビトロの3種の異なる症状の模擬低酸素状態において、腫瘍細胞は解糖阻害剤によって殺すことができることが示されている。Maher,J.C.,et al.,「低酸素対好気的状態で処理された腫瘍細胞における、2−デオキシ−D−グルコースによって誘導された大幅な細胞周期の阻害及び細胞毒性(Greater Cell Cycle Inhibition And Cytotoxicity Induced By 2−Deoxy−D−Glucose In Tumor Cells Treated Under Hypoxic vs Aerobic Conditions)」,Cancer Chemother Pharmacol 2004;53:116−122。さらに、正常な酸素処理細胞において解糖を阻害することは、代わりの炭素源、すなわちアミノ酸及び脂肪がミトコンドリアの酸化的リン酸化の作動に利用できるので、それらのエネルギー産生に有意には作用しない。したがって、解糖系阻害剤は低酸素腫瘍細胞を選択的に標的とするために使用でき、正常細胞又は好気的に成長する腫瘍細胞に対してはあまり毒性を示さない。Boros,L.G.,et al.,「ソマトスタチンによる、酸化的及び非酸化的ペントースリン酸経路の阻害:抗腫瘍作用の有望な機序(Inhibition Of Oxidative And Nonoxidative Pentose Phosphate Pathways By Somatostatin:A Possible Mechanism Of Antitumor Action)」,Med Hypotheses 1998;50:501;LaManna,J.C.,「栄養消費及び代謝摂動(Nutrient Consumption And Metabolic Perturbation)」,Neurosurg Clin N Am I997;8:145−163。
【0096】
実際、インビボ実験により、2−DG(緩慢に成長する低酸素腫瘍細胞を標的にする)が標準的化学療法剤(好気性細胞の急速な増殖を対象とする)の効果を、異なるヒト腫瘍異種移植片において促進することが示された。Maschek,G.,et al.,「2−デオキシ−D−グルコースは、ヒト骨肉腫及び非小細胞肺癌においてアドリアマイシン及びパクリタキセルの効果を、インビボで促進する(2−Deoxy−D−Glucose Increases The Efficacy Of Adriamycin And Paclitaxel In Human Osteosarcoma And Non−Small Cell Lung Cancers In Vivo)」,Cancer Res 2004;64:31−4。これらの研究結果、並びに低酸素状態のインビトロモデルからのデータは、「進行性固形悪性腫瘍を有する対象における、2−デオキシ−D−グルコース単独、及びドセタキセルとの併用についての第1相用量漸増試験」と題した第1相臨床試験の形で、ヒトにおける化学療法プロトコルを改善するためにこの戦略の試験を引き起こし、これは現在継続中である。Maher,J.C.,et al.,「低酸素対好気的状態で処理された腫瘍細胞における、2−デオキシ−D−グルコースによって誘導された大幅な細胞周期の阻害及び細胞毒性(Greater Cell Cycle Inhibition And Cytotoxicity Induced By 2−Deoxy−D−Glucose In Tumor Cells Treated Under Hypoxic vs Aerobic Conditions)」,Cancer Chemother Pharmacol 2004;53:116−122。動物実験からのデータ、並びに第1相臨床試験からの予備試験結果は、2−DGが正常細胞に対して十分耐性であり、比較的非毒性であることを示している。
【0097】
理論上は、ミトコンドリアが酸化的リン酸化を受けられる腫瘍細胞は、解糖系阻害剤の2−DGにより殺されないはずであるが、選択されたいくつかの癌細胞系は酸素の存在下で、低用量のこの糖類似体により死ぬ。毒性の機序は、これらの細胞系が正常なミトコンドリアの呼吸をし、他の解糖系阻害剤に耐性であるため、解糖の遮断を介したものではない。同様の機序は、ウィルスの糖タンパク質合成において示されており、2−DGがN結合型グリコシル化を、脂質結合オリゴ糖の組立てに干渉することにより遮断する。Datema,R.,et al.,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」Biochem J 1979;184:113−123;Datema R.,et al.,「脂質結合オリゴ糖を含む2−デオキシグルコースの形成(Formation Of 2−Deoxyglucose−Containing Lipid−Linked Oligosaccharides)」,Eur J Biochem 1978;90:505−516。正常な酸素状態で成長する選択腫瘍細胞系における2−DGの毒性は、同様の機序によると思われる。
【0098】
本発明によれば、2−DGは、通常酸素下で2−DGに感受性の細胞を含む固形腫瘍を有する特定の患者に、単剤として使用できる。したがって、これらの患者において2−DGは二重の効果(1)グリコシル化への干渉を介して好気性腫瘍細胞集団と標的とする、及び(2)腫瘍の低酸素部分において解糖を阻害する、を有するはずであり、両方の機序は細胞死を引き起こす。
【0099】
材料及び方法
細胞型
p0細胞を、骨肉腫細胞系143B(wt)を、前記のように臭化エチジウムを用いて長期処理することによって単離した。King,M.P.,et al.,「Mtdnaを欠いたヒト細胞:相補性による外因性ミトコンドリアを用いた再生(Hunan Cells Lacking Mtdna:Repopulation With Exogenous Mitochondria By Complementation.)」Science 1989;246:500−503。p0細胞は、ウリジン及びピルビン酸塩要求性体なので、それらを、10%ウシ胎児血清、50μg/mlのウリジン及び100mMのピルビン酸ナトリウムを添加したDMEM(GIBCO社,USA)で成長させる。SKBR3細胞系を、マイアミ大学のJoseph Rosenblatt博士の研究室より入手した。膵臓癌細胞系1420及び1469、卵巣癌細胞系SKOV3、子宮頸部癌細胞系HELA及び骨肉腫細胞系143Bを、ATCCより購入した。非小細胞肺癌及び小細胞肺癌細胞系はマイアミ大学のNiramol Savaraj博士により、患者に由来した。SKBR3及びSKOV3細胞をMcCoy’s 5A培地で成長させ、1420、1469及び143BをDMEM(GIBCO社、USA)で成長させ、HELAをMEM(GIBCO社、USA)で成長させた。培地には10%ウシ胎児血清を添加した。全ての細胞を5%CO2及び37℃で成長させた。
【0100】
薬剤及び化学物質
2−DG、オリゴマイシン及びツニカマイシンは、Sigmaより購入した。2−FDG及び2−FDMは、Priebe博士(MD Anderson Cancer Center,TX)のご厚意により寄贈された。
【0101】
低酸素状態
低酸素状態(モデルC)の研究のために、細胞を播種し、直接細胞毒性検査のために以下に記載のように37℃で24時間、5%CO2でインキュベートする。24時間インキュベートした後で、細胞を薬剤処理し、モデル110酸素コントローラーに取り付けたPro−Oxインビトロチャンバー(Reming Bioinstruments Co.Redfield,NY)に置き、チャンバーを所望のO2濃度(0.1%)にするため、内部には95%窒素及び5%CO2の混合物を使用し、かん流させる。
【0102】
細胞毒性検査
細胞を37℃で24時間、5%CO2においてインキュベートし、その時に薬剤処理を開始し72時間続ける。この時に、吸着細胞をトリプシン処理し、それらの個別の培養培地と混合し、その後400gで5分間遠心分離機にかける。ペレットを1mlのHanks溶液に再懸濁し、Vi−Cell(Beckman Coulter社、USA)細胞生存分析器により分析した。
【0103】
乳酸検査
乳酸を、処理済み培養液又は未処理培養液から0.025mlの脱タンパク質化培地を、0.1mlの乳酸デヒドロゲナーゼ、(1000単位/ml)、2mlのグリシンバッファー(グリシン0.6モル/L及びヒドラジン、pH9.2)及び1.66mg/mlのNADを含む、反応混合物に加えることによって測定する。脱タンパク質化を、試験培養物からの0.5mlの培地を1mlの8%w/v過塩素酸を用いて処理し、30秒ボルテックスにかけ、その後この混合物を4℃で5分間曝し、1500gで10分間遠心分離機にかけることにより起こす。上清をさらに3回遠心分離機にかけ、0.025mlの最終的に不要物を除いた上清を上記のように乳酸の測定に使用する。NADHの形成を、Beckman DU r 520 UV/vis分光光度計を用いて340nmで測定し、これは乳酸塩標準曲線により確定された乳酸濃度に直接一致する。
【0104】
ATP定量化検査。
ATPライトキット(lite kit)(Perkin Elmer社)を、ATP濃度の定量化に使用できる。約50mlの細胞溶解液を、96ウェルプレート(ボトム白)において100mlの細胞懸濁液に加える。プレートを室温で、シェーカーにのせて(700rpm)5分間インキュベートする。約50mlの基質溶液を、続けてウェルに加え、さらに5分間室温で振とう(700rpm)する。プレートをその後10分間暗順応させ、発光を測定する。
【0105】
ウェスタンブロット分析
細胞を、104細胞/cm2で播種し、薬剤処理下で表示の時間成長させる。処理期間の終りに、細胞を回収し、プロテイナーゼ阻害剤カクテルを添加した1%SDSの80mM Tris−HCL(ph7.4)バッファーを用いて溶解する。DNAを超音波で分解することにより断片化し、タンパク質濃度をミクロBCA タンパク質検査キット(Pierce社、USA)により測定する。試料を、2×Laemmliサンプルバッファー(Bio−Rad社,USA)と混合し、SDS−ポリアクリルアミドゲルにかける。ゲルをニトロセルロース膜(Amersham社,USA)に転写し、抗−KDEL(Stressgen社、Canada)(Grp78及びGrp94に関して);ポリクロナール抗−CHOP/GADD154(Santa Cruz社、USA),ポリクロナール抗erbB2(DAKO,USA).を用いて解析する。解析の後で、膜を洗浄し、HRP標識二次抗体と共にインキュベートする。化学発光をフィルムに感光させることにより検出する。指示がある場合には、膜をストリッピングバッファー(Pierce社,USA)を用いてストリップし、抗アクチン(Sigma社,USA)一次抗体を用いて再度解析する。コンコナバリンA(ConA)の結合を分析するため、膜を0.2μg/mlのHRP標識ConAと共にインキュベートし、記載のように化学発光を検出した。
【0106】
結果
2−DC及び2−フルオロ−D−マンノースは通常酸素下で成長するSKBR3細胞を殺すが、2−FDGは殺さない
多くの腫瘍細胞系を、正常な酸素状態対低酸素状態の下での解糖系阻害剤に対するそれらの感受性の差に関する調査において、通常酸素下で成長した場合、ヒト乳癌細胞系SKBR3は2−DGに関して感受性であることが見出された。図1A及びBは、SKBR3を3mMの2−DGで72時間処理した場合、成長は50%阻害され(ID50)、一方12mMでは60%の細胞が殺されることを実証する。これまでの研究により、ミトコンドリアの呼吸が不完全又は化学的に処断される場合、同様の用量の2−DGを用いて処理した腫瘍細胞は死ぬことが示された。したがって、これらの細胞がミトコンドリアの呼吸において不完全であったかどうかを確定するために、それらの酸素消費量を測定した。以下の表1に明示したように、SKBR3細胞と、通常酸素下で成長した場合に2−DGによる処理に対して耐性である他の2種の細胞系の平均酸素消費量の間に有意な差はなかった。他方、ミトコンドリア不全細胞系のp0は、酸素消費量の激減を示し、SKBR3が正常に呼吸していたことが確認された。さらに、通常酸素下で2−DGに対して感受性である、他の2種の細胞系、1420及びHELAは、同様に、又は耐性細胞系よりも盛んに呼吸した(表1)。したがって、通常酸素下でのこれらの細胞における2−DGの毒性は、解糖の遮断以外の機序によるものである。このことを確かめるために、SKBR3細胞を他の2種の解糖系阻害剤、すなわち2−デオキシ−2−フルオロ−グルコース(2−FDG)及びオキサマートを用いて処理した。図1A及びBにおいて、通常酸素下で成長した場合に、これらの薬剤はどちらもSKBR3細胞に対して毒性がなかったことが見られる。
【表1】
【0107】
さらに、2−フルオロ−D−マンノース(2−FDM)は、効果は低いが、SKBR3細胞において細胞毒性を引き起こす点で2−DGと同様であった(図1を参照されたし)。2−DG及び2−FDMは両方ともマンノースの構造と似ているが、2−FDGは似ておらず、その結果マンノースの代謝に干渉できる。このデータは、2−DG及び2−FDMによるマンノースの代謝への干渉を示し、これは主として多くのタンパク質のN結合型グリコシル化に関与し、SKBR3細胞における細胞死並びに成長阻害をもたらす。
【0108】
2−FDGは、2−DGより優れた解糖阻害剤であり、SKBR3細胞においてATPを十分減少させる
これまでの報告において、通常酸素下で成長するSKBR3細胞における2−DGの毒性が、解糖及びATP産生の阻害を介して仲介されることを示唆していた。Aft,R.L.,et al.,「化学療法剤としての2−デオキシ−D−グルコースの評価:細胞死の機序(Evaluation Of 2−Deoxy−D−glucose As A Chemotherapeutic Agent:Mechanism Of Cell Death)」,Br J Cancer 2002;87:805−812。しかし、上に述べたように、別の解糖系阻害剤、2−FDGはこれらの細胞において非毒性である。さらに2−FDG類似体は、2−DGより解糖の阻害及び殺低酸素細胞において優れている。Lampidis,T.J.,et al.,「解糖の遮断及び殺「低酸素腫瘍細胞」における2−ハロゲン置換D−グルコース類似体の有効性(Efficacy of 2−Halogen Substituted D−Glucose Analogs in Blocking Glycolysis and Killing“Hypoxic Tumor Cells,”)」Cancer Chemother Pharmacol(印刷中)。実際に、SKBR3細胞を2−FDG対2−DGを用いて処理した場合、前者は後者より乳酸塩濃度(解糖の基準)を阻害する(図2Aを参照されたし)。さらにATPの減少は、2−FDG処理によりさらに顕著になり、さらにこの糖類似体がこれらの細胞において解糖及びATP産生の優れた阻害剤であることが確認された(図2Bを参照されたし)。さらに、SKBR3細胞を低酸素状態で成長させた場合、2−FDGは2−DGより毒性が強いことが発見され、さらにSKBR3細胞における解糖の優れた阻害剤であることが確認された(データ未掲載)。したがって、これまでの報告とは反対に、通常酸素下で2−DGにより誘導された毒性は、解糖を阻害し、ATPのプールを減少させる能力から独立しているように思われる。
【0109】
通常酸素下でのSKBR3細胞における2−DGの毒性は、外因性マンノースにより逆転できる
ウィルス性タンパク質において、2−DGは、N結合型オリゴ糖の組立てを阻害し、この阻害は外因性マンノースによって逆転できることが示されている。Datema,R.,et al.,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123。図3A及び3Bは、マンノースの添加により、通常酸素下での2−DGへの曝露による細胞死を逆転できるが、他の糖、すなわちグルコース、フルクトース及びフコースの添加では逆転できないことを例示し、細胞死は、同様の機序を介してグリコシル化への干渉により仲介されることを示唆している。Datema,R.,et al.,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123。陰性対照として、マンノースは、同じ状態下でSKBR3細胞におけるツニカマイシン誘導性の毒性を逆転しないことが発見された。これは、オリゴ糖鎖にマンノースを加えるよりも前のステップにおいてツニカマイシンがグリコシル化に干渉し、その結果マンノースの代謝から独立しているという事実により説明できる(データ未掲載)。
【0110】
「低酸素」の3種のモデルにおける2−DGの毒性は、外因性マンノースによっては逆転できない
先に述べたように、低酸素状態下で成長する細胞は、エネルギー産生のためにグリコシル化を唯一解糖系に依存する。したがって、解糖系阻害剤によるこの代謝経路の阻害は、これまでに実証されているように、細胞死を引き起こすはずである。Maher,J.C.,et al,「低酸素対好気的状態で処理された腫瘍細胞における、2−デオキシ−D−グルコースによって誘導された大幅な細胞周期の阻害及び細胞毒性(Greater Cell Cycle Inhibition And Cytotoxicity Induced By 2−Deoxy−D−Glucose In Tumor Cells Treated Under Hypoxic vs Aerobic Conditions)」,Cancer Chemother Pharmacol 2004;53:116−122。2−DGが低酸素下で成長するSKBR3細胞に対して毒性であることの機序を識別するために、マンノースを3種の異なる「低酸素」状態下で成長する細胞に添加した。図3C及びDに示すように、通常の成長培地又は2mMのマンノースを添加した同じ培地のどちらにおいても、成長阻害及び細胞死に有意な差は見出されなかった。これらの結果は、外因性マンノースによる通常酸素下で成長するSKBR3細胞における2−DGの毒性の逆転は解糖とは関連がなく、通常酸素下で成長するこれらの細胞における細胞死の様式として、グリコシル化の干渉に関連している証拠を提供する。
【0111】
2−DG及び2−FDGは、通常酸素状態下で成長する、選択されたいくつかの腫瘍細胞系のみに対して毒性である
通常酸素状態下での2−DGの毒性が、特定の型の癌細胞に限定されたかどうかを調査するために、いくつかの細胞系を試験した。表2に示したこの試験の結果は、6mMの2−DG又は2−FDMのどちらかを用いて処理した場合には、通常酸素圧で成長する、選択されたいくつかの腫瘍細胞系(15種のうちの6種)のみが有意に細胞死に至り、6mMの2−FDGを用いた場合には至らないことを示す。2−DGに対して感受性が見出された細胞系は、SKBR3(乳癌細胞系)、1420(膵臓癌細胞系)、患者に直接由来する2種の非小細胞肺癌細胞系、RT 8226(多発性骨髄腫細胞系)、HELA(子宮頸癌)及びTG98(膠芽腫細胞系)であった。しかし、類似の組織に由来する癌細胞系が、通常酸素圧下で2−DG及び2−FDMの両方に耐性があることが見出され、これらの糖類似体の毒性が必ずしも組織型に特異的とは限らないことが示唆される。
【表2】
【0112】
2−DG及び2−FDMは、SKBR3細胞においてコンカナバリンA(ConA)の結合及び糖タンパク質の分子量を減少させる
ConAは、糖タンパク質のマンノースに特異的に結合するレクチンであり、高マンノース型糖タンパク質の検出に使用されている。「タンパク質精製法:実践的取り組み(Protein Purification Methods:A Practical Approach)」,In:Harris ELV,Angal S,editors.New York IRLPress at Oxford University Press;1994.p270。この技術は、2−DG及び2−FDMの両方、並びにツニカマイシンがいくつかの糖タンパク質においてConAの結合を減少させることを示すために使用された(図4Aを参照されたし)。さらに、外因性マンノースは2−DG及び2−FDMを用いて処理した細胞において対照のConA結合濃度を回復させるがツニカマイシン処理細胞においては回復せず、一方2−FDG処理細胞はConA結合の減少を示さない。さらに、2−DG処理の後でSKBR3細胞においてチロシン−キナーゼ受容体を発現する、公知の糖タンパク質であるerbB2の大きさの変化を、ウェスタンブロットにより分析した。図4Bは、2−DG及び2−FDMの両方がerbB2の分子量を減少させ、一方2−FDGは効果がなかったことを例示する。ConAのデータとの相関関係において、外因性マンノースはタンパク質の大きさを元の重さに復元した。これらのデータはさらに、2−DG及び2−FDMが選択された腫瘍細胞に対してN結合型グリコシル化への干渉を介して毒性であるが、2−FDGは毒性ではなく、この干渉はマンノースによって逆転できるという結論を裏付ける。
【0113】
2−DG又は2−FDMのどちらかによる処理が、通常酸素下でSKBR3細胞における小細胞ストレス応答を引き起こす
タンパク質のグリコシル化の通常過程が作用した場合、小胞体(ER)に蓄積されたミスフォールドタンパク質が、小胞体ストレス応答(UPR)として公知のシグナル伝達カスケードを引き起こす。グリコシル化に干渉する薬剤はUPRの誘導を示しており、シャペロン、すなわちGrp7S/Bip又はGrp94の上方制御を介して、ERのタンパク質のフォールディング能力を増加させる。図5Aに示すように、SKBR3細胞を2−DG、2−FDM又はツニカマイシン(周知のグリコシル化阻害剤)を用いて、通常酸素下で処理する場合、Grp78及びGrp94の両方が上方制御される。さらに2mMのマンノースの添加により、2−DG及び2−FDMのシャペロンの上方制御を逆転するが、ツニカマイシンは逆転しない。2−DGが誘導したUPRのマンノースによる逆転は、2−DGの毒性が外因性マンノースの添加により逆転されることを実証する、図3Dのデータに相関し、同様の結果が2−FDM処理細胞においても見出された(データ未掲載)。期待通り、2−FDGは、2−DG又は2−FDMほどにはこれらのシャペロン濃度を増加せず、通常酸素状態下で処理した場合、SKBR3細胞において細胞死は起こらないことを例示する毒性のデータ(図1B)に相関する。対照的に、2−DG又は2−FDMを、低酸素の3種の異なる実験条件の下で成長させた細胞に適用した場合、両方のシャペロンが上方制御されたモデルCと比較して、モデルA及びBにおいてUPRの有意な上方制御は観察されない。さらに、陽性対照としてのツニカマイシンは、3種のモデル全てにおいてこれらのシャペロンの合成の誘導を示した(図5B)。これらの結果は、細胞を2−DG又は2−FDMを用いて処理した場合、細胞死の機序が「低酸素」(解糖の遮断)対通常酸素(グリコシル化への干渉)状態とで異なることを示している。
【0114】
2−DG及び2−FDMの毒性は、SKBR3細胞におけるUPR特異的アポトーシス経路の誘導に相関する。
細胞がERストレスを克服できない場合、GADD154/CHOPの誘導を介してUPRは特異的アポトーシス経路を誘導することが報告されている。Xu,C.,et al.,「小胞体ストレス:細胞の寿命及び死の決定(Endoplasmic Reticulum Stress:Cell Life And Death Decisions)」,J Clin Invest 2005;115:2656−2664、Obeng,E.A.,et al.,「カスパーゼ12及びカスパーゼ4は、カスパーゼ依存性小胞体ストレス誘導アポトーシスに必要ではない(Caspase−12 And Caspase−4 Are Not Required For Caspase−Dependent Endoplasmic Reticulum Stress−Induced Apoptosis)」,J Biol Chem 2005;280:29578−29587。したがって、2−DG及び2−FDMが通常酸素下でERストレスによりSKBR3細胞を殺すかどうかを確定するために、このUPR特異的アポトーシスタンパク質を、ウェスタンブロット分析を使用して検査した。図6に見られるように、2−DG、2−FDM及びツニカマイシン処理の後でGADD154/CHOPが誘導されるが、2−FDG処理では誘導されない。このアポトーシス経路が2−DG又は2−FDMのどちらかにより誘導された場合、マンノースを用いた同時処理により逆転できる。しかし、ツニカマイシンにより誘導されたGADD154/CHOPはこの糖の添加により逆転できない。図2Bに示されるように、これらのデータは、マンノースによる毒性の逆転に相関する。
【0115】
実施例1及び2の考察
固形腫瘍は、不十分な血管形成、腫瘍の急速な成長及び、腫瘍血管の酸素運搬能力の減少による低酸素並びに通常酸素領域を含む。Gillies,R.J.,et al.,「腫瘍の微小環境のMRI(MRI Of The Tumor Microenviroment)」,J Magn Reson Imaging 2002;16:430−450;Maxwell,P.H.,et al,「低酸素誘導性Facoro−1は、固形腫瘍において遺伝子発現を調節し、血管形成及び腫瘍成長の両方に干渉する(Hypoxia Inducible Facoro−1 Modulates Gene Expression In Solid Tumors And Influences Both Angiogenesis And Tumor Growth)」,PNAS 1997;94:8104−8109;Semenza,G.L.,「癌治療用標的化HIF−1(Targeting HIF−1 For Cancer Therapy)」,Nature Rev 2003;3:721−732。低酸素細胞における唯一のエネルギー産生経路は解糖なので、解糖阻害剤の2−DGがこれらの細胞に対して選択的に毒性であるが、好気性細胞には非毒性であり、成長を阻害するだけであることが示されている。Maher,J.C.,et al.,「低酸素対好気的状態で処理された腫瘍細胞における、2−デオキシ−D−グルコースによって誘導された大幅な細胞周期の阻害及び細胞毒性(Greater Cell Cycle Inhibition And Cytotoxicity Induced By 2−Deoxy−D−Glucose In Tumor Cells Treated Under Hypoxic vs Aerobic Conditions)」,Cancer Chemother Phannacol 2004;53:116−122、Maschek,G.,et al.,「2−デオキシ−D−グルコースは、ヒト骨肉腫及び非小細胞肺癌においてアドリアマイシン及びパクリタキセルの効果を、インビボで促進する(2−Deoxy−D−Glucose Increases The Efficacy Of Adriamycin And Paclitaxel In Human Osteosarcoma And Non−Small Cell Lung Cancers In Vivo)」,Cancer Res 2004;64:31−4、Liu,H.,et al.,「解糖系阻害剤に対する腫瘍細胞の過感作(Hypersensitization Of Tumor Cells To Glycolytic Inhibitors)」,Biochemistry 2001;40:5542−5547;Liu,H.,et al.,「低酸素による解糖系阻害剤に対する腫瘍細胞の感受性の増加:固形腫瘍療法のための戦略(Hypoxia Increases Tumor Cell Sensitivity To Glycolytic Inhibitors:A Strategy For Solid Tumor Therapy)」,Model C.Biochem Pharmacol 2002;64:1745−1751。しかし、選択されたいくつかの腫瘍細胞系は酸素の存在下で2−DGに殺される。これらの感受性細胞型にはヒト乳癌細胞系SKBR3がある。損傷したミトコンドリアを有する細胞における解糖の遮断はATPの濃度を下げ、ネクローシス細胞死を引き起こすので、ミトコンドリア呼吸の不全によりこれらの細胞の2−DGに対する感受性は説明できる。Gramaglia,D.,et al,「アポトソーム形成の下流におけるアポトーシスからネクローシスへの転換は、BCL−XL及びPKB/AKT独立性の様式で解糖及び酸化的リン酸化の両方を阻害する必要がある(Apoptosis To Necrosis Switching Downstream Of Apoptosome Formation Requires Inhibition Of Both Glycolysis And Oxidative Phosphorylation In A BCL−XL And PKB/AKT−Independent Fashion)」,Cell Death Differentiation 2004;11:342−353。しかしこの可能性は、SKBR3細胞が、通常酸素下で2−DG耐性であることが見出された他の2種の細胞系と同様に呼吸することを示した酸素消費量実験に反した(表1)。さらに、通常酸素下で2−DGに感受性である細胞系1420の呼吸率が、2−DG耐性の細胞系よりも高いことが発見された。したがって、通常酸素下のSKBR3における2−DGの毒性は、ミトコンドリア機能の不全では説明できず、細胞死の機序が、解糖の遮断についてのこの糖の効果とは関連がないことが示唆される。
【0116】
これまでに、SKBR3細胞が通常酸素下で、解糖の阻害により2−DGに感受性であり、グルコース輸送体−1の発現増加をもたらし、2−DGを大幅に取り込むATPプールの減少を引き起こすことが報告された。Aft,R.L.,et al.,「化学療法剤としての2−デオキシ−D−グルコースの評価:細胞死の機序(Evaluation Of 2−Deoxy−D−Glucose As A Chemotherapeutic Agent:Mechanism Of Cell Death)」,Br J Cancer 2002;87:805−812。しかし、2−FDGは2−DGより強力な解糖阻害剤であるが(11、図2)、通常酸素下で成長するSKBR3細胞に対しては非毒性であり、さらに2−DGがこれらの細胞を、解糖の遮断及びATP産生の阻害による以外の機序を介して殺すという結論を支持する。
【0117】
SKBR3細胞がさらにマンノース類似体の2−FDMに感受性であることを示すデータは、通常酸素下で成長する選択された腫瘍細胞において、糖類似体のマンノ配置がそれらの毒性活性に重要であることを示す。2−DGの第2炭素において酸素原子が欠失すると、この化合物がグルコース類似体及びマンノースの類似体の両方になるのに反して、2−FDGにおいてフルオロ基が欠失してもグルコース類似体にしかならない。マンノ配置がこれらの糖類似体の毒性に関係があるという結論は、Schwartzが率いるグループによる1970年代終りに発表された仕事により裏付けられている。
【0118】
このグループは、2−DG、2−FDG及び2−FDMが、家禽ペストウィルスにより感染する、鶏胚線維芽細胞においてN結合型グリコシル化に干渉でき、糖タンパク質合成及びウィルスの復元を減少させることを示した。Datema,R,et al,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123、Datema,R.,et al,「脂質結合オリゴ糖を含む2−デオキシグルコースの形成(Formation Of 2−Deoxyglucose−Containing Lipid−Linked Oligosaccharides)」,Eur J Biochem 1978;90:505−516、Datema,R.,et al,「フルオロ−グルコースによる、インビボにおけるタンパク質のグルコシル化の阻害(Fluoro−Glucose Inhibition Of Protein Glycosylation In Vivo)」,Eur J Biochem 1980;109:331−341、Schmidt,M.F.G.,et al,「動物細胞における2−デオキシ−D−グルコースのヌクレオシド2リン酸誘導体(Nucleoside−diphosphate Derivatives Of 2−Deoxy−D−Glucose In Animal Cells)」,Eur J Biochem 1974;49:237−247、Schmidt,M.F.G.,et al.,「酵母及び鶏胚における2−デオキシ−2−フルオロ−D−[3H]グルコース及び2−デオキシ−2−フルオロ−D−[3H]マンノースの代謝(Metabolism Of 2−Deoxy−2 Fluoro−D−[3H]Glucose And 2−Deoxy−2−Fluoro−D−[3H]Mannose In Yeast And Chick−Embryo Cells)」,Eur J Biochem 1978;87:55−68、McDowell,W.,et al.,「抗ウィルス性糖類似体である2−デオキシ−2−フルオロ−D−マンノースによるタンパク質のグリコシル化の阻害の機序:グアノシン2リン酸エステルによるMan(Gicnac)2 PP−Dolの合成の阻害(Mechanism Of Inhibition Of Protein Glycosylation By The Antiviral Sugar Analogue 2−Deoxy 2−Fluoro−D−Mannose:Inhibition Of Synthesis Of Man(Gicnac)2 PP−Dol By The Guanosine Diphosphate Ester)」,Biochemistry 1985;24:8145−8152。それらの報告は、2−DGが、細胞の小胞体内のタンパク質に転写されるべき脂質結合オリゴ糖の組立てを阻害できると結んでいる。2−DG、GDP−MGの代謝産物は、オリゴ糖の組立ての中途終了を起こすことができ、タンパク質への転写には不適切な短い脂質結合オリゴ糖をもたらし得ることが実証された。Datema,R.,et al.,「脂質結合オリゴ糖を含む2−デオキシグルコースの形成(Formation Of 2−Deoxyglucose−Containing Lipid−Linked Oligosaccharides)」,Eur J Biochem 1978;90:505−516。全般的に、これらの結果は、これら類似体のウィルス性糖タンパク質合成阻害の能力が、2−DG>2−FDM>2−FDGの順であったことを示し、これらは、通常酸素下で成長するSKBR3細胞においてこれら類似体の毒性が似ている。Datema,R.,et al,「フルオロ−グルコースによる、インビボにおけるタンパク質のグルコシル化の阻害(Fluoro−Glucose Inhibition Of Protein Glycosylation In Vivo)」,Eur J Biochem 1980;109:331−341。このグループはさらに、これらの類似体の阻害効果が低用量の外因性マンノースの添加により逆転できることを報告した。Datema,R.,et al,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123。同様に、2mMのマンノースは、SKBR3細胞における2−DG及び2−FDMの毒性をを完全に逆転し、両方のマンノース類似体がこれらの細胞をN結合型グリコシル化への干渉を介して殺すことを示す。Datema,R.,et al,「糖タンパク質のグリコシル化への干渉(Interference With Glycosylation Of Glycoproteins)」,Biochem J 1979;184:113−123、Datema,R.,et al.,「脂質結合オリゴ糖を含む2−デオキシグルコースの形成(Formation Of 2−Deoxyglucose−Containing Lipid−Linked Oligosaccharides)」,Eur J Biochem 1978;90:505−516、Datema,R.,et al,「フルオロ−グルコースによる、インビボにおけるタンパク質のグルコシル化の阻害(Fluoro−Glucose Inhibition Of Protein Glycosylation In Vivo)」,Eur J Biochem 1980;109:331−341、Schmidt,M.F.G.,et al,「動物細胞における2−デオキシ−D−グルコースのヌクレオシド2リン酸誘導体(Nucleoside−diphosphate Derivatives Of 2−Deoxy−D−Glucose In Animal Cells)」,Eur J Biochem 1974;49:237−247、Schmidt,M.F.G.,et al.,「酵母及び鶏胚における2−デオキシ−2−フルオロ−D−[3H]グルコース及び2−デオキシ−2−フルオロ−D−[3H]マンノースの代謝(Metabolism Of 2−Deoxy−2−Fluoro−D−[3H]Glucose And 2−Deoxy−2−Fluoro−D−[3H]Mannose In Yeast And Chick−Embryo Cells)」,Eur J Biochem 1978;87:55−68、McDowell,W.,et al.,「抗ウィルス性糖類似体である2−デオキシ−2−フルオロ−D−マンノースによるタンパク質のグリコシル化の阻害の機序:グアノシン2リン酸エステルによるMan(Gicnac)2 PP−Dolの合成の阻害(Mechanism Of Inhibition Of Protein Glycosylation By The Antiviral Sugar Analogue 2−Deoxy 2−Fluoro−D−Mannose:Inhibition Of Synthesis Of Man(Gicnac)2 PP−Dol By The Guanosine Diphosphate Ester)」,Biochemistry 1985;24:8145−8152。
【0119】
マンノースは、N結合型グリコシル化タンパク質における核となる糖であるが、また、マンノースはホスホマンノイソメラーゼによりフルクトース−6−リン酸に転換できるため、解糖経路にも関与している。したがって、マンノースがSKBR3細胞において、2−DGが阻害する解糖系段階を避けることによって2−DGの毒性を逆転できるという可能性は残っている(図7)。しかし、2mMのマンノースが「低酸素」モデルA及びBにおいて2−DGにより誘導された成長阻害及び細胞死を逆転せず(図3C及び3Dを参照されたし)、一方低酸素状態で細胞が実際に成長したモデルCにおいて、わずかに回復効果があったため、この可能性はありそうもない。このわずかな回復は、(1)2−DG及び2−FDMが低酸素状態下でもなおグリコシル化に干渉した、及び/又は(2)これらの細胞が0.5%の低酸素状態下でもなお、少ないながらも酸化的リン酸化を行うので、マンノースはモデルCにおいて解糖の阻害を逆転した、と説明できた。全般的に、通常酸素下でこれらの糖類似体に対して感受性の細胞において、マンノースにより2−DG及び2−FGの毒性が逆転し、ミトコンドリアが停止した細胞(モデルA及びB)において逆転しないことにより、グリコシル化に干渉し、解糖の阻害がないことが、通常酸素下の低酸素状態に関与することが裏付けられる。
【0120】
N結合型グリコシル化が阻害された場合、タンパク質は適切にフォールドできず、小胞体内に残される。Ellgaard,L.,et al.,「小胞体における品質管理(Quality Control In The Endoplasmic Reticulum)」,Nat Rev Mol Cell Biol 2003;4:181−191;Parodi,A.J.,「タンパク質のグリコシル化及びタンパク質のフォールディングにおけるその役割(Protein Glycosylation And Its Role In Protein Folding)」,Annu Rev Biochem 2000;69:69−93。フォールドしていないタンパク質が蓄積すると、オルガネラの弛緩並びにタンパク質翻訳の混乱が起こる。このような事象において、細胞は、複合を開始するが、小胞体ストレス応答(UPR)として公知のシグナル伝達カスケードがなお保存されており、ERに恒常性を回復する。3種のER膜貫通タンパク質:イノシトール要求性酵素1(IRE1)、二本鎖RNA活性化タンパク質キナーゼ(PERK)及び活性化転写因子6(ATF6)が、フォールドしていないタンパク質のシグナルを核に伝達する。Schroder,M.,et al 「ERストレス及びフォールドしていないタンパク質の応答(ER Stress And Unfolded Protein Response)」,Mutat Res 2005;569:29−63。フォールドしていないタンパク質がERに蓄積した場合、分子シャペロンのグルコース制御タンパク質78(Grp78/Bip)は、これらの3種のER膜貫通タンパク質から解離し、その結果それらを活性化する。Pahl,H.L.,「小胞体から細胞核へのシグナル伝達(Signal Transduction From The Endoplasmic Reticulum To The Cell Nucleus)」,Physiol Rev 1999;79:683−701。これは、糖輸送体の上方制御、リン脂質の合成の増加、アミノ酸輸送及び分子シャペロンGrp78/Bip及びGrp94の発現を含む、いくつかの代謝及び分子の変化をもたらす。Ma,Y.,et al,「小胞体ストレス応答が明らかになる話(The Unfolding Tale Of The Unfolded Protein Response)」,Cell 2001;107:827−830;Doerrler.W.T.,et al,「小胞体ストレス応答によるヒト線維芽細胞におけるドリコール経路の制御(Regulation Of Dolichol Pathway In Human Fibroblasts By The Endoplasmic Reticulum Unfolded Protein Response)」,PNAS 1999;96:13050−13055;Breckenridge,D.G.,et al.,「小胞体経路によるアポトーシスの制御(Regulation Of Apoptosis By Endoplasmic Reticulum Pathways)」,Oncogene 2003;22:8608−8618。
【0121】
2−DG及び2−FDMは、通常酸素状態下で成長するSKBR3細胞においてGrp78及びGrp94の両方の発現を上方制御し、これは外因性マンノースの添加により逆転でき、これらの糖類似体がN結合型グリコシル化に干渉し、フォールドしていないタンパク質をもたらし、その結果UPRを開始することを強く裏付ける。さらに、2−DG又は2−FDMのどちらよりも優れた解糖阻害剤である2−FDGは、UPR応答の誘導に有効ではない。これらの類似体に対するUPR応答の規模は、グリコシル化への干渉の程度を反映しているように思われ、これはウィルスコートタンパク質における脂質結合オリゴ糖の組立ての遮断において2−DG>2−FDM>2−FDGであることを実証する報告に一致する。Datema,R.,et al,「フルオロ−グルコースによる、インビボにおけるタンパク質のグルコシル化の阻害(Fluoro−Glucose Inhibition Of Protein Glycosylation In Vivo)」,Eur J Biochem 1980;109:331−341、Schmidt,M.F.G.,et al,「動物細胞における2−デオキシ−D−グルコースのヌクレオシド2リン酸誘導体(Nucleoside−diphosphate Derivatives Of 2−Deoxy−D−Glucose In Animal Cells)」,Eur J Biochem 1974;49:237−247、Schmidt,M.F.G.,et al.,「酵母及び鶏胚における2−デオキシ−2 フルオロ−D−[3H]グルコース及び2−デオキシ−2−フルオロ−D−[3H]マンノースの代謝(Metabolism Of 2−Deoxy−2−Fluoro−D−[3H]Glucose And 2−Deoxy−2−Fluoro−D−[3H]Mannose In Yeast And Chick−Embryo Cells)」,Eur J Biochem 1978;87:55−68、McDowell,W.,et al.,「抗ウィルス性糖類似体である2−デオキシ−2−フルオロ−D−マンノースによるタンパク質のグリコシル化の阻害の機序:グアノシン2リン酸エステルによるMan(Gicnac)2 PP−Dolの合成の阻害(Mechanism Of Inhibition Of Protein Glycosylation By The Antiviral Sugar Analogue 2−Deoxy−2−Fluoro−D−Mannose:Inhibition Of Synthesis Of Man(Gicnac)2PP−Dol By The Guanosine Diphosphate Ester)」,Biochemistry 1985;24:8145−8152。さらにこのUPRのデータは、細胞毒性の結果と相関し、これは通常酸素下でSKBR3細胞の成長阻害及び殺害において2−DG>2−FDM>>>2−FDGであることを同様に示す。
【0122】
一方、「低酸素」モデルA及びBにおいて、Grp78及びGrp94は2−DGによって上方制御されず、これらの細胞は解糖の阻害を介して死亡し、グリコシル化への干渉を介してではないことを示している。UPRがこれらのモデルにおいて誘導されない理由を説明する有望な機序は、Grp78/Bipのフォールドしていないタンパク質への結合及びその結果のUPRの活性化に必要であることが公知のATPの濃度に関する。モデルA及びBとは対照的に、UPRはモデルCにおいて誘導され(図5B)、モデルCにおいてはATP濃度は2−DGにより減少する。さらに、ATP濃度に有意な影響を与えないことが公知のツニカマイシンは、「低酸素」モデルにおいてシャペロンを上方制御し、これらの細胞における機能的UPR経路を実証する。
【0123】
UPRはp53に酷似しており、P53はDNAの損傷シグナルが細胞周期を停止させ、DNA修復酵素を活性化し、これらの過程の結末であるアポトーシスに依存する。したがって、UPRが小胞内の恒常性の確立に失敗した場合、ERストレス特異的アポトーシス経路が活性化される。Breckenridge,D.G.,et al.,「小胞体経路によるアポトーシスの制御(Regulation Of Apoptosis By Endoplasmic Reticulum Pathways)」,Oncogene 2003;22:8608−8618。カスパーゼ4、カスパーゼ12及びCHOP/GADD154を含むアポトーシス経路のメディエーターの中で、後者の活性化の増加は、他と比較してER誘導性哺乳動物アポトーシス経路の優れた指標であることが示されている。Obeng,E.A.,et al.,「カスパーゼ12及びカスパーゼ4は、カスパーゼ依存性小胞体ストレス誘導アポトーシスに必要ではない(Caspase−12 And Caspase−4 Are Not Required For Caspase−Dependent Endaplasmic Reticulum Stress−Induced Apoptosis)」,J Biol Chem 2005;280:29578−29587。したがって、CHOP/GADD154の発現が、通常酸素下で成長するSKBR3細胞における2−DG及び2−FDMの細胞毒性と相関することを示す図6は、これらの糖類似体がグリコシル化への干渉を介して毒性であり、ERのストレスを引き起こすことを裏付ける。さらに、マンノースの添加によりCHOP/GADD154の誘導が逆転し、グルコースによっては逆転しないことは、2−DG及び2−FDMがこの機序を介して毒性であることをさらに裏付ける。
【0124】
根本的な疑問は、O2の存在下で2−DGを用いて処理した場合、大部分の腫瘍並びに正常細胞は死なないのになぜ特定の腫瘍細胞型は死ぬのかである。この疑問に対する答えは、遺伝学研究によってもたらされ、酵素であるホスホマンノースイソメラーゼが、糖タンパク質糖鎖不全症候群1b型(Carbohydrate−Deficient Glycoprotein Syndrome Type 1b) Niehues,R.,et al.,糖タンパク質糖鎖不全症候群1b型(Carbohydrate−Deficient Glycoprotein Syndrome Type 1b),J Clin Invest 1998;101:1414−1420;Freeze,H.H.,「Nグリコシル化及び動物モデルにおけるヒトの障害(Human Disorders in N−glycosylation and Animal Models)」,Biochim Biophys Acta 2002;1573:388−93に記載のように苦しんでいる患者において欠失していることが示されている。この酵素の欠失は、血清糖タンパク質のグリコシル化の減少をもたらし、蛋白漏出性腸症を特徴とする血栓症及び胃腸障害を引き起こす。外因性マンノースをこれらの患者の食事に加えた場合、彼らの血清糖タンパク質は正常に戻り、彼らの症状は消えた。Freeze,H.H.,「甘味溶液:糖による救済(Sweet Solution:Sugars to the Rescue)」,J Cell Biol 2002;158:615−616;Paneerselvam,K.,et al,「マンノースが、糖タンパク質糖鎖不全症候群の神経芽細胞におけるNグリコシル化の変化を修正する(Mannose Corrects Altered N−glycosylation in Carbohydrate−Deficient Glycoprotein Syndrome Fibroblasts)」J Clin Invest 1996;97:1478−1487。これは、外因性マンノースが、通常酸素下で2−DGを用いて処理した場合に殺される、選択された腫瘍細胞を救済することを示す当該データに相関する。これらの型の腫瘍細胞は、ホスホマンノイソメラーゼが下方制御される又は不完全であるかのどちらかであり、或いは通常酸素下で2−DGによる処理に耐性があることが示された大部分の他の細胞よりこれらの腫瘍細胞において、2−DGはこの酵素に大きく作用することが考えられる。しかし、図7に示すように、2−DG及び2−FDMがN結合型グリコシル化に関与するマンノースの代謝を阻害できる多くの他のステップがある。
【0125】
2−DG、2−CM及び2−FDM(2−FM)は、グリコシル化に干渉し、ERストレス及びアポトーシスを引き起こすことを介して、特定の腫瘍型を殺す。2−FDGがこれらの細胞を殺さないという発見は、2−DG及び2−FDMの毒性が解糖の阻害及びATPの減少によるものである可能性を排除する。これらの薬剤は、選択された固形腫瘍の治療において、単剤療法として使用できる(図7を参照されたし)。
【0126】
(実施例3)
図8及び9に示すように、複数のMTT検査により、選択された高悪性度の神経膠腫細胞系の感受性が実証され、様々な糖をベースとした解糖系阻害剤を図で示す。通常酸素状態及び低酸素状態のどちらかに曝すことを含む、様々な状態、及びこれらの化合物に対する感受性についてのその作用を使用する。結果は、糖をベースとした様々な解糖系阻害剤の感受性が比較的均一であることを示す(いくらか微妙な差がある)。低酸素状態における感受性の作用に関して、いくつかの細胞系に明らかな差がある。全般的に、低酸素状態で成長した場合、予測されたように、大部分の細胞系は解糖系阻害剤に対してより感受性である。しかし、U87MGなどのいくつかの細胞系は、乳酸濃度(解糖の代理マーカー)が通常酸素状態において最大であり、低酸素状態においては増加しない(以下の乳酸塩のデータを参照されたし)という好気性解糖表現型(完全な「Warburg効果」)に完全に傾倒している。この状況下で、感受性の差は、低酸素状態で成長した細胞は成長が遅く、したがって細胞の要求するエネルギーがおそらく少ないという経験的観測により説明される。
【0127】
図8Aは、低酸素下(<1% 酸素)又は通常酸素下(20% 酸素)で2−FGを用いて処理したU87ヒト脳腫瘍細胞系のMTT検査を示す。図8B及び8Cの両方は、同様の実験を表すが、糖をベースとした解糖阻害剤が異なる。パネルBの場合2−DGを使用し、パネルCでは2−FMを用いる。見て分かるようにU87は、その代謝要求に関して一貫して解糖を利用し、したがってこの細胞系は、低酸素下でこれらの薬剤に対して感受性の増加を示さない、独特の表現型を表す。
【0128】
図9は、2−FG又は2−FMのどちらかの存在下における、6日にわたる成長曲線を示す。このパネルは、U87細胞系の有意な成長阻害を実証し、2−FGは2−FMよりわずかに効果的と思われる。パネルB及びパネルCは、低酸素状態及び通常酸素状態の両方において成長した細胞系D−54に関する成長曲線の同様の阻害を実証する。この場合、細胞が低酸素状態で成長すると、明らかに増大された効果があり、これは、低酸素下のこの特定の細胞系に関するさらなる解糖系代謝を刺激する能力に関連がある。
【0129】
(実施例4)
図10及び11は、2−DGに曝した、通常酸素状態及び低酸素状態におけるヒトU87MG膠芽細胞腫星状細胞腫細胞系(U87)対D−54ヒト神経膠腫細胞系の感受性の差を示す。U87MG細胞は、低酸素状態又は好気性状態のどちらかにおいて高い率の解糖(酸化的解糖又は「Warburg効果」)を示す、したがって、それらが低酸素状態で成長した場合、2−DGに対するU87MG細胞の感受性は変化しない。他方、D54細胞は好気性成長状態において解糖系代謝を一部転換し、それ故この細胞系を低酸素状態で成長させた場合2−DGに対する感受性は高くなる。
【0130】
図10は、これらの2種の細胞系の有意な差、及びはるかに大きい、U87の相対無感覚を示す。
【0131】
図11は、U87とD54とのこの表現型の差の背後にある理由を示す。このパネルは、D54による大幅な解糖の誘導を実証するのに反して、U87はすでに最大限の乳酸塩濃度を産生している。
【0132】
図10及び11に示した結果は、細胞系を解糖系阻害剤を用いて処理した場合の、低酸素の効果が異なることを実証する。高度に解糖系に依存した細胞系(U87MGなど)は、すでに最大限に解糖系阻害剤に対して感作されており、感受性を示すために無酸素環境におく必要はない。これは、U87MGなどの細胞系による、高く、変化のない乳酸塩の産生濃度により実証されるが、一方D54は、その感受性及び低酸素に反応した乳酸塩濃度の両方を増加する。
【0133】
特筆すべきことに、神経膠腫細胞系は対酸素状態に完全に耐性である。図12を見て分かるように、通常酸素状態又は完全な低酸素状態(<1%)のどちらかにおいて成長した細胞系は、細胞の要求するエネルギーを提供するために、解糖を非常に頼りにして適度に成長を続けることができる。
【0134】
(実施例5)
2−DG類似体が2−フルオロ18−グルコース(2−F18G)の腫瘍により取り込まれることの実証。図13は、所定のPETスキャン研究の間に2−F18Gの神経膠腫内への取り込みが増強することを実証する。多形性膠芽細胞腫を有する患者のPETスキャンにより、この腫瘍内に2−FGが有意に取り込まれることが実証される。パネルは、患者に17mCiの2−F18Gを与えた後の、CT非増強(A)、増強したCT(B)及びCT一体型PETスキャンを示す。薬力学的事象は、これらの腫瘍が糖をベースとした解糖阻害剤に特に適しているという劇的な証明を提供する。
【0135】
(実施例6)
マウスにおけるヒト神経膠腫の治療。ヒト神経膠腫のマウス同所性異種移植片を、2−DG単独、又はテモゾロマイド(テモダール)と共に用いて、のどちらかで治療した。これらの動物は、高悪性度の神経膠腫の同所性の異種移植片モデルを表す。これらの実験を3回繰り返し、図14に示すように、同様の結果であった。動物にU87MG細胞を頭蓋内に移植し、5日後に、陰性対照(PBS)、陽性対照(テモダール)、実験的単剤(2−DG)又は実験的併用(2−DG+テモダール)のいずれかを用いて治療した。
【0136】
図12に示した結果は、同所性の腫瘍モデルに対する2−DGの単剤有効性を初めて実証した。これらの結果を繰り返し、各群に全18匹の動物を用いた連続した3回の動物実験に一致した(データ未掲載)。新規の治験薬により、この特定の動物モデルが非常に厳しく、生存者がわずかに増えたにすぎないことが明らかになった。見て分かるように、2−DGは現在脳腫瘍に利用できる、最も良い薬剤のテモゾロマイドと同等の効果である(陽性対照として見られる)。
【0137】
驚いたことに、2−DGはテモドールと同等に有効であり、併用は単剤療法より優れていた。2−DGを経口的に与え、十分に耐性であった。テモドールは脳腫瘍の治療に関して現在「gold standard(最も基準となる(信頼できる)もの)」であるので、2−DG及びテモドールの機能的同義性は顕著であった。最終的に、このクラスの薬剤に関して、単剤の有効性が実証され、これはこの疾患に固有である。
【0138】
これらの結果は、使用した陽性対照のテモゾロマイドと同様に、解糖の阻害が動物の生存を増加させたことを実証した。この療法に動物は十分耐性であり、毒性の証拠は示されなかった。最終的に、本発明者らは、最有力候補選択に関して、関連する糖をベースとした解糖系阻害剤の群から化合物を選択しており、これはインビトロ及びインビボの有効性、薬物速度論的特性、化学的安定性及び化学合成のコストに基づくであろう。最有力選択について、さらなる先端研究並びに正式な動物毒物試験が開始されるであろう。
【0139】
本発明の具体的な実施形態が、本発明の原理の適用を例示するために示され、詳細に記載されているが、このような原理を逸脱することがなければ本発明を具体化できると理解されるであろう。
【図面の簡単な説明】
【0140】
【図1A】SKBR3細胞における、2−DG、2−FDG、2−FDM及びオキサマートによる、24時間にわたる、腫瘍成長阻害試験の結果を表した図である。各値は、3連の試料の平均±SDである。
【図1B】SKBR3細胞における、2−DG、2−FDG、2−FDM又はオキサマートによる、24時間の細胞毒性試験の結果を表した図である。各値は、3連の試料の平均±SDである。
【図2A】2mMの2−DG又は2−FDGが存在しない又はいずれかが存在する状態で24時間SKBR3細胞を成長させ、培地中の乳酸塩濃度の結果を表した図である。
【図2B】図2Aで使用したのと同じ濃度の2−DG又は2−FDGのいずれかが存在する状態で6時間SKBR3細胞を成長させ、その後全細胞溶解物中のATPを定量化した結果を表した図である。
【図3A】様々な糖の存在下で、2−DGを用いて処理した後のSKBR3細胞における成長阻害試験の結果を表した図である。各値は、3連の試料の平均±SDである。
【図3B】様々な糖の存在下で、2−DGを用いて処理した後のSKBR3細胞における細胞毒性試験の結果を表した図である。各値は、3連の試料の平均±SDである。
【図3C】2mMのマンノースが存在する又は存在しない状態で、2−DGを用いて処理した後の、3種の異なる「低酸素」モデルにおける成長阻害試験の結果を表した図である。
【特許請求の範囲】
【請求項1】
治療を必要とする対象に、治療有効量のヘキソース化合物を投与することを含む、膠芽細胞腫の治療方法。
【請求項2】
治療を必要とする対象に、治療有効量のヘキソース化合物を投与することを含む、膵臓癌の治療方法。
【請求項3】
治療を必要とする対象に、治療有効量の2−FMを投与することを含む、腫瘍の増殖を治療する方法。
【請求項4】
治療を必要とする対象に、治療有効量のマンノース化合物を投与することを含む、膵臓癌の治療方法。
【請求項5】
治療を必要とする対象に、治療有効量のマンノース化合物を投与することを含む、脳腫瘍の治療方法。
【請求項6】
治療有効量のヘキソース化合物を投与することを含み、効果は膵臓癌及び膠芽細胞腫からなる群から選択される、患者において効果を発揮する方法。
【請求項1】
治療を必要とする対象に、治療有効量のヘキソース化合物を投与することを含む、膠芽細胞腫の治療方法。
【請求項2】
治療を必要とする対象に、治療有効量のヘキソース化合物を投与することを含む、膵臓癌の治療方法。
【請求項3】
治療を必要とする対象に、治療有効量の2−FMを投与することを含む、腫瘍の増殖を治療する方法。
【請求項4】
治療を必要とする対象に、治療有効量のマンノース化合物を投与することを含む、膵臓癌の治療方法。
【請求項5】
治療を必要とする対象に、治療有効量のマンノース化合物を投与することを含む、脳腫瘍の治療方法。
【請求項6】
治療有効量のヘキソース化合物を投与することを含み、効果は膵臓癌及び膠芽細胞腫からなる群から選択される、患者において効果を発揮する方法。
【図3C】2mMのマンノースが存在する又は存在しない状態で、2−DGを用いて処理した後の、3種の異なる「低酸素」モデルにおける細胞毒性試験の結果を表した図である。
【図4A】SKBR3細胞を、各レーンに示した様々な薬剤を用いて48時間処理し、全細胞抽出液を得、HRP標識ConAを用いてブロットした結果を表した図である。等量のタンパク質を各レーンに添加し、βアクチンにより解析した。糖タンパク質(矢印で区切られた)は、8mMの2−DG及び2−FDMにより糖タンパク質とConAとの結合が低下するが、2−FDGによっては低下せず、この低減はマンノースにより逆転できることを示している。
【図4B】erbB2にブロットした図4Aの細胞が、高度に糖タンパク質を発現した結果を表した図である。このタンパク質の分子量の変化は、類似の用量の2−DG及び2−FDMにより引き起こされる。
【図5A】SKBR3細胞を、8mMの2−DG、2−FDG又は2−FDMのいずれかを用いて24時間処理し、全細胞溶解物を2種の分子シャペロンのGrp78及びGrp94に関してブロットした結果を表した図である。1μg/mlのツニカマイシン(TUN)を陽性対照として使用した。タンパク質のローディングをβアクチンにより解析した。
【図5B】「低酸素」モデルA、B及びCの細胞を、類似の用量の糖類似体を用いて処理した場合の、被験タンパク質のウェスタンブロットを示した図である。
【図6】SKBR3細胞を、8mMの2−DG、2−FDG又は2−FDMのいずれかを用いて24時間処理し、全細胞溶解物をCHOP/GADD154に関して解析した結果を表した図である。2−DG及び2−FDMの両方により誘導されたCHOP/GADD154の誘導は、外因性のマンノースを添加することにより逆転し、一方グルコースはこのタンパク質の量に関して何の効果も示さなかった。ツニカマイシンを陽性対照として使用した、タンパク質のローディングをβアクチンに関して解析した。
【図7】2−DG、2−FDM又は2−FDGがホスホグルコイソメラーゼを阻害でき、解糖の遮断をもたらし、低酸素腫瘍細胞において細胞死を確実にすることを説明した、解糖経路及びN結合型グリコシル化経路を示した図である。しかし、好気性状態下の特定の腫瘍細胞型において、2−DG及び2−FDMは、それらの構造がマンノース並びにグルコースに似ているため、オリゴ糖の脂質結合組立てに干渉でき、小胞体ストレス応答(unfolded protein response)及び毒性の誘導を引き起こす(三角形=グルコース、六角形=マンノース、及び四角形=N−アセチル−グルコサミン)。
【図8A】MTT試験に実証よりされた、選択された神経膠腫細胞系と、本発明の特定のヘキソース化合物との感受性を示した図である。
【図8B】MTT試験に実証よりされた、選択された神経膠腫細胞系と、本発明の特定のヘキソース化合物との感受性を示した図である。
【図8C】MTT試験に実証よりされた、選択された神経膠腫細胞系と、本発明の特定のヘキソース化合物との感受性を示した図である。
【図9A】様々なヘキソース化合物を用いて処理した神経膠腫細胞の成長を表した図である。
【図9B】2−DGを用いた処理によるD54細胞の成長の抑制を表した図である。
【図9C】2−FGを用いた処理によるD54細胞の成長の抑制を表した図である。
【図10】2−DGを用いて処理した細胞についての低酸素効果の違いを実証する図である。
【図11】低酸素状態及び正常な酸素状態におけるヒト膠芽細胞腫細胞系の乳酸塩産生を示した図である。
【図12】低酸素状態及び正常な酸素状態における神経膠腫細胞系の成長結果を示した図である。
【図13】神経膠腫細胞における2−FGの取り込みを実証した図である。
【図14】マウスの神経膠腫を2−DGを用いて処理した結果を示した図である。
【図15】Colo357−FG膵臓癌細胞に対する2−FM活性を示した図である。
【図16】U251神経膠腫細胞に対する2−ハロ−D−マンノース活性を示した図である。
【図17】U87細胞の成長が、2−FMにより抑制されることを示した図である。
【図18】2−フルオロ−マンノースを用いて処理した後のU87神経膠腫細胞に自食作用が誘導されるパーセントを表したチャートを提供する図である。
【図1】

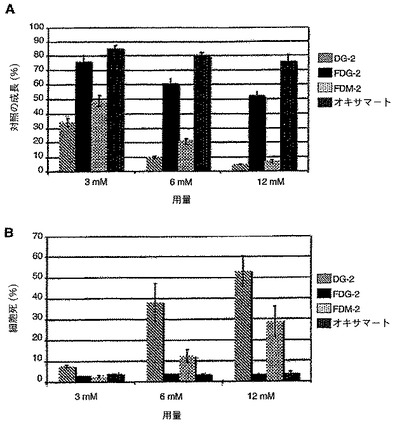
【図2】
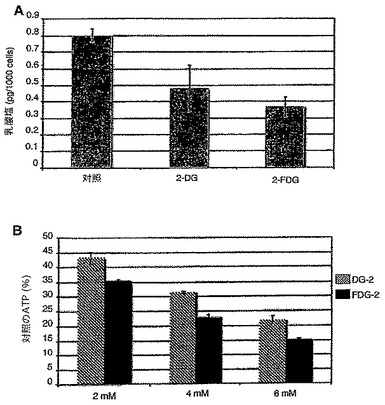
【図3】
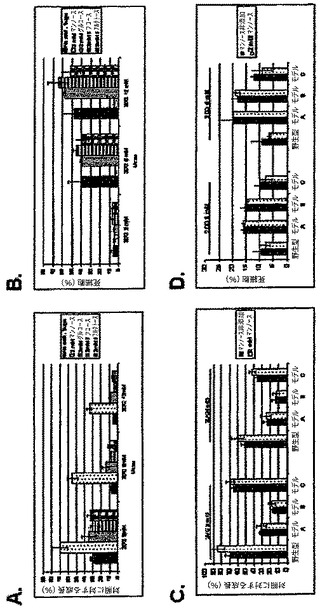
【図4】
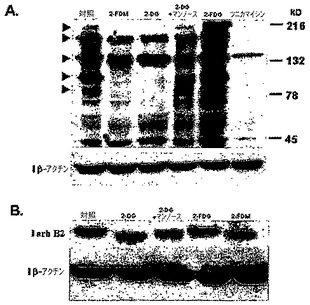
【図5】
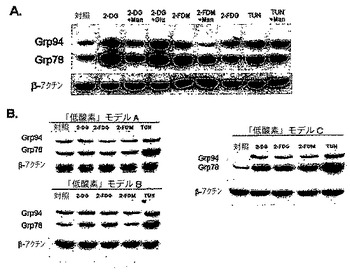
【図6】
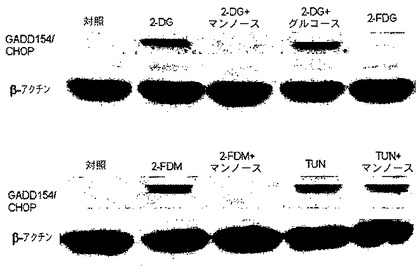
【図7】
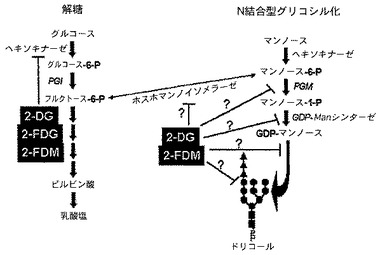
【図8】
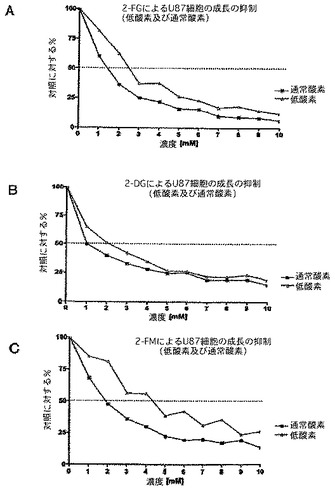
【図9】
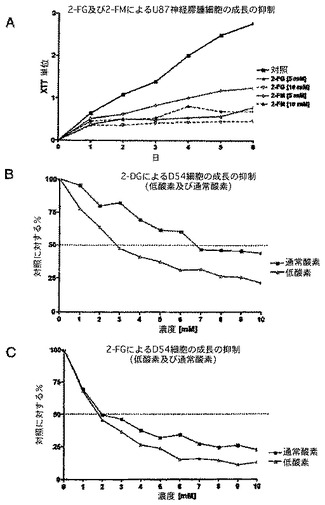
【図10】
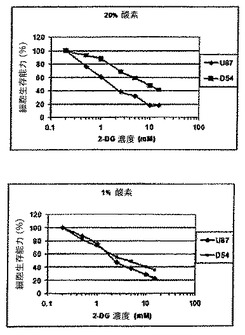
【図11】
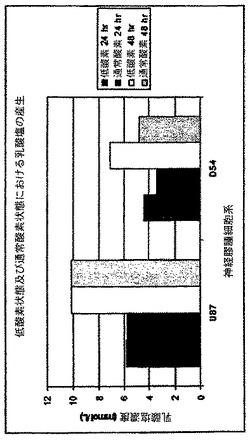
【図12】
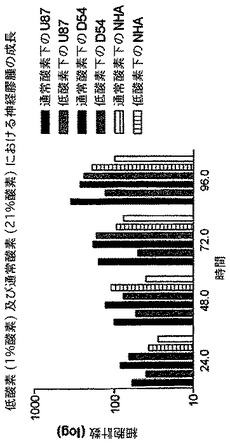
【図13】
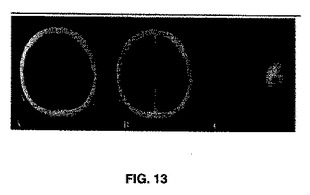
【図14】
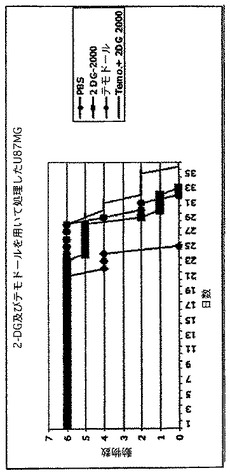
【図15】
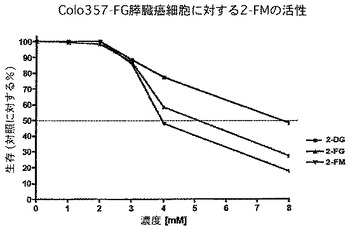
【図16】
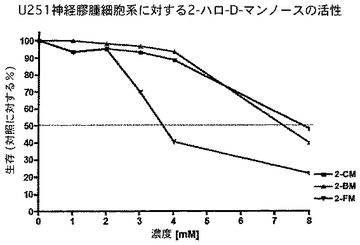
【図17】
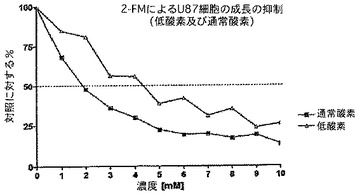
【図18】
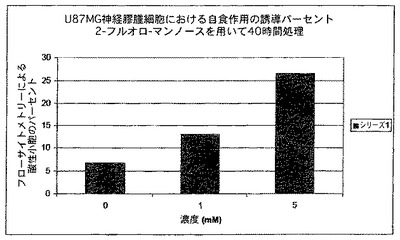
【図4A】SKBR3細胞を、各レーンに示した様々な薬剤を用いて48時間処理し、全細胞抽出液を得、HRP標識ConAを用いてブロットした結果を表した図である。等量のタンパク質を各レーンに添加し、βアクチンにより解析した。糖タンパク質(矢印で区切られた)は、8mMの2−DG及び2−FDMにより糖タンパク質とConAとの結合が低下するが、2−FDGによっては低下せず、この低減はマンノースにより逆転できることを示している。
【図4B】erbB2にブロットした図4Aの細胞が、高度に糖タンパク質を発現した結果を表した図である。このタンパク質の分子量の変化は、類似の用量の2−DG及び2−FDMにより引き起こされる。
【図5A】SKBR3細胞を、8mMの2−DG、2−FDG又は2−FDMのいずれかを用いて24時間処理し、全細胞溶解物を2種の分子シャペロンのGrp78及びGrp94に関してブロットした結果を表した図である。1μg/mlのツニカマイシン(TUN)を陽性対照として使用した。タンパク質のローディングをβアクチンにより解析した。
【図5B】「低酸素」モデルA、B及びCの細胞を、類似の用量の糖類似体を用いて処理した場合の、被験タンパク質のウェスタンブロットを示した図である。
【図6】SKBR3細胞を、8mMの2−DG、2−FDG又は2−FDMのいずれかを用いて24時間処理し、全細胞溶解物をCHOP/GADD154に関して解析した結果を表した図である。2−DG及び2−FDMの両方により誘導されたCHOP/GADD154の誘導は、外因性のマンノースを添加することにより逆転し、一方グルコースはこのタンパク質の量に関して何の効果も示さなかった。ツニカマイシンを陽性対照として使用した、タンパク質のローディングをβアクチンに関して解析した。
【図7】2−DG、2−FDM又は2−FDGがホスホグルコイソメラーゼを阻害でき、解糖の遮断をもたらし、低酸素腫瘍細胞において細胞死を確実にすることを説明した、解糖経路及びN結合型グリコシル化経路を示した図である。しかし、好気性状態下の特定の腫瘍細胞型において、2−DG及び2−FDMは、それらの構造がマンノース並びにグルコースに似ているため、オリゴ糖の脂質結合組立てに干渉でき、小胞体ストレス応答(unfolded protein response)及び毒性の誘導を引き起こす(三角形=グルコース、六角形=マンノース、及び四角形=N−アセチル−グルコサミン)。
【図8A】MTT試験に実証よりされた、選択された神経膠腫細胞系と、本発明の特定のヘキソース化合物との感受性を示した図である。
【図8B】MTT試験に実証よりされた、選択された神経膠腫細胞系と、本発明の特定のヘキソース化合物との感受性を示した図である。
【図8C】MTT試験に実証よりされた、選択された神経膠腫細胞系と、本発明の特定のヘキソース化合物との感受性を示した図である。
【図9A】様々なヘキソース化合物を用いて処理した神経膠腫細胞の成長を表した図である。
【図9B】2−DGを用いた処理によるD54細胞の成長の抑制を表した図である。
【図9C】2−FGを用いた処理によるD54細胞の成長の抑制を表した図である。
【図10】2−DGを用いて処理した細胞についての低酸素効果の違いを実証する図である。
【図11】低酸素状態及び正常な酸素状態におけるヒト膠芽細胞腫細胞系の乳酸塩産生を示した図である。
【図12】低酸素状態及び正常な酸素状態における神経膠腫細胞系の成長結果を示した図である。
【図13】神経膠腫細胞における2−FGの取り込みを実証した図である。
【図14】マウスの神経膠腫を2−DGを用いて処理した結果を示した図である。
【図15】Colo357−FG膵臓癌細胞に対する2−FM活性を示した図である。
【図16】U251神経膠腫細胞に対する2−ハロ−D−マンノース活性を示した図である。
【図17】U87細胞の成長が、2−FMにより抑制されることを示した図である。
【図18】2−フルオロ−マンノースを用いて処理した後のU87神経膠腫細胞に自食作用が誘導されるパーセントを表したチャートを提供する図である。
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公表番号】特表2009−531301(P2009−531301A)
【公表日】平成21年9月3日(2009.9.3)
【国際特許分類】
【出願番号】特願2008−556577(P2008−556577)
【出願日】平成19年2月26日(2007.2.26)
【国際出願番号】PCT/US2007/062789
【国際公開番号】WO2007/101148
【国際公開日】平成19年9月7日(2007.9.7)
【出願人】(501100582)ボード オブ リージェンツ オブ ザ ユニバーシティ オブ テキサス システム (19)
【Fターム(参考)】
【公表日】平成21年9月3日(2009.9.3)
【国際特許分類】
【出願日】平成19年2月26日(2007.2.26)
【国際出願番号】PCT/US2007/062789
【国際公開番号】WO2007/101148
【国際公開日】平成19年9月7日(2007.9.7)
【出願人】(501100582)ボード オブ リージェンツ オブ ザ ユニバーシティ オブ テキサス システム (19)
【Fターム(参考)】
[ Back to top ]