
癌細胞にアポトーシスを誘導するための切断型EIF−5A1ポリヌクレオチドの使用

本発明は、ホスト内での内在性遺伝子の発現をノックアウト又はノックダウンするための、内在性遺伝子を標的とするsiRNA、及び、ポリヌクレオチドによりコードされるタンパク質をホスト内で発現させるための、送達媒体/発現ベクター内の遺伝子をコードするポリヌクレオチドのホストへの送達を組み合わせた使用に関する。本発明はまた、細胞、特に哺乳類の癌細胞のアポトーシス及び死滅の誘導に有用なeIF−5A1の切断型にも関する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[関連出願]
本出願は、2008年9月3日に提出された米国仮出願第61/093,749号、及び2009年3月19日に提出された米国特許出願第12/400,742号に対する優先権を主張するものであり、これらはその全体が参照により本明細書に取り込まれる。
【0002】
[背景技術]
比較的最近、二重鎖RNA(「dsRNA」)がタンパク質発現の阻害に使用できることが研究者らにより観察された。遺伝子の発現を停止するこの能力にはヒトの疾病の治療に対する広範な可能性があり、現在多くの研究者及び営利団体がこの技術に基づく治療法の開発にかなりの資金を投じている。
【0003】
二重鎖RNAにより誘導される遺伝子サイレンシングは、少なくとも異なる3段階:(i)RNAにより導かれるDNA又はヒストンのメチル化を指す、転写の不活性化;(ii)siRNAにより誘導されるmRNAの分解;及び(iii)mRNAにより誘導される転写の減衰において起こり得る。
【0004】
一般に、哺乳類細胞においてRNAにより誘導されるサイレンシング(RNA干渉、又はRNAi)の主要な機構は、mRNAの分解であると考えられている。哺乳類細胞におけるRNAi使用の最初の試みは、長鎖のdsRNAの使用に集中していた。しかしながらRNAi誘導のためのこれらの試みは、標的特異的とは対照的に、一つには一般的なタンパク質合成の阻害をもたらすインターフェロン反応の誘導のためにあまり成功しなかった。従って、哺乳類系におけるRNAiに対して長鎖dsRNAは実行可能な選択肢ではない。
【0005】
さらに最近、短い(18〜30bp)RNA二本鎖を培養哺乳類細胞に導入した際、インターフェロン反応を誘導することなく標的mRNAの配列特異的な阻害が実現できることが示された。阻害的低分子RNA(「siRNA」)と称されるこれらの短いdsRNAのあるものは、細胞内で95%を超える標的mRNAを切断するために、モル濃度未満の濃度で触媒的に作用し得る。siRNAの作用機構、同様に幾つかのその応用についての記述は、Provost et al. (2002) Ribonuclease Activity and RNA Binding of Recombinant Human Dicer, EMBO J. 21(21): 5864-5874; Tabara et al. (2002) The dsRNA Binding Protein RDE-4 Interacts with RDE-1, DCR-1 and a DexH-box Helicase to Direct RNAi in C. elegans, Cell 109(7): 861-71; Ketting et al. (2002) Dicer Functions in RNA Interference and in Synthesis of Small RNA Involved in Developmental Timing in C. elegans; Martinez et al., Single-Stranded Antisense siRNAs Guide Target RNA Cleavage in RNAi, Cell 110(5):563; Hutvagner & Zamore (2002) A microRNA in a multiple-turnover RNAi enzyme complex, Science 297:2056に記載される。
【0006】
機構的な視点から、長い二重鎖RNAの植物及び無脊椎動物の細胞内への導入は、ダイサーとして知られるタイプIIIエンドヌクレアーゼによってsiRNAへ分解される。Sharp, RNA interference--2001, Genes Dev. 2001, 15:485. Dicer, a ribonuclease-III-like enzyme, processes the dsRNA into 19-23 base pair short interfering RNAs with characteristic two base 3' overhangs. Bernstein, Caudy, Hammond, & Hannon (2001) Role for a bidentate ribonuclease in the initiation step of RNA interference, Nature 409:363。siRNAは次に、RNA誘導型サイレンシング複合体(RNA-induced silencing complex)(RISC)に取り込まれ、ここで1以上のヘリカーゼがsiRNA二本鎖を巻き戻し、標的の認識を導くための相補的アンチセンス鎖が有効となる。Nykanen, Haley, & Zamore (2001), ATP requirements and small interfering RNA structure in the RNA interference pathway, Cell 107:309。適切な標的mRNAへの結合により、RISC内の1以上のエンドヌクレアーゼが標的を切断してサイレンシングを誘導する。Elbashir, Lendeckel, & Tuschl (2001) RNA interference is mediated by 21- and 22-nucleotide RNAs, Genes Dev. 15:188、図1。
【0007】
干渉効果は長く持続し得り、多数の細胞分裂後にも検出され得る。さらに、RNAiは配列特異性を示す。Kisielow, M. et al. (2002), Isoform-specific knockdown and expression of adaptor protein ShcA using small interfering RNA, J. Biochem. 363:1-5。従ってRNAi機構により、密接に関連するmRNAに影響を及ぼすことなく、一つの型の転写を特異的にノックダウンできる。これらの特性によってsiRNAは、遺伝子発現の阻害、及び遺伝子機能の研究、及び薬物標的の検証に対して価値ある手段となる可能性がある。さらにsiRNAは、(1)遺伝子の過剰発現又は異所性発現により引き起こされる疾病;及び(2)変異を含む遺伝子の発現によりもたらされる疾病に対する治療薬として有用な可能性がある。
【0008】
成功したsiRNA依存性の遺伝子サイレンシングは幾つかの因子に依存する。RNAiにおける最も議論のある問題の一つは、siRNA設計の必要性、すなわち使用するsiRNAの配列を考慮することについての論点である。C. elegans及び植物における初期の研究は、長いdsRNAの導入によって設計の問題を回避した(例えば、Fire, A. et al. (1998) Nature 391 :806-811を参照)。この原始的な生命体において、長いdsRNA分子はダイサーによりsiRNAに切断され、こうして転写の全体を網羅する可能性のある二本鎖の多様な集団が生成される。これらの分子のうち幾つかの分画は非機能的(すなわちサイレンシングをわずかに誘導するか又は誘導しない)であるが、1以上は高度に機能的となる可能性があり、これによって興味ある遺伝子のサイレンシングが起こり、かつsiRNA設計の必要は少なくなる。残念なことに、哺乳類系ではインターフェロン反応のためにこの同じアプローチは使用できない。この効果はダイサーによる切断工程を介さずに、siRNAを直接導入することにより回避できるが、この方策は選択したsiRNA配列が非機能的又は半機能的であるという危険性を持ち越す。
【0009】
多くの研究者が、siRNAの設計はRNAiの決定的な要素ではないという見解を示してきた。一方で当該分野における他の研究者達は、siRNAの設計に注目することにより、RNAiがさらに効率的となる可能性を探求し始めた。
【0010】
様々な疾病又は疾患を治療するためには特定のタンパク質の上昇が望ましいが、それが必要な全てではない可能性もある。例えば、内在性タンパク質又は異なるタンパク質発現をノックダウン又はノックアウトするために、siRNAを組み合わせた使用が必要となり得る。本発明はこの必要性を満たし、癌、特に多発性骨髄腫の治療法を提供する。
【0011】
多発性骨髄腫を含む癌は、アポトーシス誘導能から利益を得られる疾病である。多発性骨髄腫の従来の治療法は、化学療法、幹細胞移植、幹細胞移植を併用する高用量化学療法、及びサルベージ療法を含む。化学療法は、Thalomid(登録商標)(サリドマイド)、ボルテゾミブ、Aredia(登録商標)(パミドロネート)、ステロイド、及びZometa(登録商標)(ゾレドロン酸)による治療を含む。しかしながら多くの化学療法薬は、骨髄、胃及び腸の内面、並びに、毛包のような活発に分裂する非癌細胞に有毒である。従って化学療法は、血液細胞数の減少、悪心、嘔吐、下痢、及び脱毛をもたらし得る。
【0012】
従来の化学療法又は標準用量の化学療法は、典型的には多発性骨髄腫の患者の第一の、又は最初の治療である。患者はまた、高用量化学療法及び幹細胞移植のために用意された化学療法も受け得る。移植前に腫瘍の負荷量を減少させるため、導入療法(幹細胞移植前の従来の化学療法)が使用できる。特定の化学療法薬は骨髄細胞への毒性が少なく、かつ骨髄からの幹細胞のより高い収率をもたらすため、他のものよりも導入療法に適している。導入療法に適した化学療法薬の例は、デキサメタゾン、サリドマイド/デキサメタゾン、VAD(ビンクリスチン、Adriamycin(登録商標)(ドキソルビシン)、及びデキサメタゾンの組み合わせ)、及びDVd(ペグ化リポソームドキソルビシン(Doxil(登録商標)、Caelyx(登録商標))、ビンクリスチン、及び投与頻度縮小型デキサメタゾンの組み合わせ)を含む。
【0013】
多発性骨髄腫の標準的治療は、50%の反応率を達成するメルファランとプレドニゾン(副腎皮質ステロイド薬)の組み合わせである。残念なことにメルファランはアルキル化剤であり、導入療法にはあまり適さない。副腎皮質ステロイド(特にデキサメタゾン)は時々単独で多発性骨髄腫の治療法に用いられ、特に高齢患者及び化学療法を許容できない患者に対して用いられる。デキサメタゾンはまた、単独又はその他の薬剤と組み合わせて導入療法にも使用される。VADは最も一般に用いられる導入療法であるが、最近DVdが導入療法に有効であることが示されている。多発性骨髄腫の治療のために最近ボルテゾミブが承認されたが、これは非常に有毒である。しかしながら、現存の治療法のいずれも治癒の著しい可能性を提供していない。従って、癌及び多発性骨髄腫に適した治療を見出す必要が依然としてある。本発明はこの必要性を満たす。
【0014】
[発明の概要]
本発明は、eIF−5A1の切断型をコードする単離されたポリヌクレオチド、同様に切断型eIF−A1ポリペプチドを提供する。切断型eIF−5A1ポリヌクレオチドは、被験体における癌細胞のアポトーシス及び死滅の誘導に有用である。切断型ポリヌクレオチドは発現ベクター内に使用され得り、これは次に被験体へ投与される。切断型eIF−5Aは哺乳類内に発現し、癌細胞を死滅させる。切断型eIF−5A1タンパク質は、約17kDaである完全長eIF−5A1タンパク質に対し、約16kDAである。切断型eIF5A1タンパク質をコードするポリヌクレオチドは、被験体の癌細胞又は腫瘍にアポトーシスを誘導するために用いられる薬剤の製造に使用されてよい。切断型eIF5A1タンパク質をコードするポリヌクレオチドは、本明細書に記載されるその他の実施形態と協同して(例えば、完全長eIF5A1をコードするポリヌクレオチド、完全長変異eIF5A1をコードするポリヌクレオチド、及び/又はeIF5Aの3’UTRを標的とするsiRNAと組み合わせて)使用されてよい。
【0015】
本発明はまた、ホスト内での内在性遺伝子の発現をノックアウト又はノックダウンするための、内在性遺伝子を標的とするsiRNA、及び、ポリヌクレオチドによりコードされるタンパク質をホスト内で発現させるための、送達媒体/発現ベクター内の遺伝子をコードするポリヌクレオチドのホストへの送達を組み合わせた使用にも関する。正常タンパク質がその必要な機能を実施できるように、正常(欠陥のない)タンパク質をコードするポリヌクレオチド(又はタンパク質それ自体)がホストに投与され、発現する(ポリヌクレオチドの場合)。siRNAは、欠陥のあるタンパク質の内在性の発現をノックダウン又はノックアウトするが、同時に投与された正常タンパク質をコードするポリヌクレオチドの外来性の発現には影響しないような遺伝子領域を標的として好ましく設計される。
【0016】
本発明は、eIF5A1の3’末端を標的とするeIF5A1 siRNA、変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターの複合体を含んでなる組成物を提供し、ここで該変異eIF5A1はヒプシン化できず、かつ該siRNA及び該発現ベクターは、複合体形成のためにポリエチレンイミンと複合される。
【0017】
本発明は、被験体における標的遺伝子の内在性発現を抑制するための、標的遺伝子を標的とするsiRNA、及び被験体内で発現できる標的タンパク質をコードするポリヌクレオチドを含んでなる組成物を提供する。特定の実施形態によれば、ポリヌクレオチドはRNAi耐性プラスミド内(siRNAにより抑制され得ない)にある。siRNA及びプラスミドは、複合体形成のためにポリエチレンイミンと好ましく複合される。
【0018】
特定の実施形態によれば、siRNAは図25に示す配列を有し、ここで変異eIF5A1をコードするポリヌクレオチドはeIF5A1K50Rである。発現ベクターは、変異eIF5A1をコードするポリヌクレオチド、及び被験体内でポリヌクレオチドを発現させるために機能的に連結されたプロモーターを含んでなる。プロモーターは、好ましくは組織特異的又は全身的のいずれかである。例えば組成物が癌の治療に使用される場合、プロモーターは、好ましくは癌が属する組織に対して組織特異的である。例えば多発性骨髄腫を治療するためには、B29のようなB細胞特異的プロモーターの使用が好ましい。特定の実施形態によれば、発現ベクターはpCpGプラスミドを含んでなる。
【0019】
特定の実施形態によれば、eIF5A1 siRNA、及び変異eIF5A1ポリヌクレオチドを含んでなる発現ベクターは、独立してin vivo JetPEI(商標)のようなポリエチレンイミンと複合される。別の実施形態によれば、eIF5A1 siRNA、及び変異eIF5A1ポリヌクレオチドを含んでなる発現ベクターは、一緒にポリエチレンイミンと複合される。
【0020】
本発明はさらに、eIF5A1の3’末端を標的とするeIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターを含んでなる組成物を提供し、ここで該変異eIF5A1はヒプシン化できず、かつ該siRNA及び該発現ベクターは、癌治療のため被験体に送達される。癌は多発性骨髄腫を含むいずれの癌であってもよい。
【0021】
本発明はさらに、本発明の組成物を被験体(哺乳類及びヒトを含むがこれらに限定されない)に投与することを含んでなる癌の治療法を提供する。
【0022】
組成物は、限定はされないが静脈内、腹腔内、皮下、又は腫瘍内のような許容されるいずれの経路で投与されてよい。siRNA及び発現ベクターは、異なる時間に異なる経路を通して投与されてよいか、又は同時に同じ経路を通して一緒に投与されてよい。例えば限定はされないが、これらのsiRNAはネイキッドで、又はin vivo jetPEIのような担体と複合してIVを通して送達されてよく、かつ発現ベクターは腫瘍内に投与されてよいか、又はsiRNA及び発現ベクターの両方がIV若しくは腫瘍内などに投与されてよい。
【0023】
本発明は、癌細胞の生育を阻害及び/又は癌細胞を死滅させる方法を提供する。本発明はまた、癌細胞の転移能を阻害又は遅くさせる方法も提供する。癌生育の阻害は、腫瘍サイズの縮小、腫瘍生育の低下を含み、かつ腫瘍の完全な寛解もまた包含し得る。癌は、大腸癌、結腸直腸腺癌、膀胱癌、子宮頸部腺癌、及び肺癌を含むがこれらに限定されないいずれの癌又は腫瘍であってよい。好ましくは、癌は多発性骨髄腫である。
【0024】
好ましい実施形態によれば、eIF−5Aはヒプシン化できない変異体である。典型的な変異体は本明細書に記載される。
【0025】
eIF−5A、又はeIF−5Aをコードするポリヌクレオチドの被験体への供給(eIF−5Aを発現させるため)に加え、eIF−5Aの内在性発現をノックアウト又はノックダウンするためにsiRNAが供給される。
【0026】
本発明はまた、多発性骨髄腫を有する被験体内の多発性骨髄腫細胞を死滅させ、癌を治療する薬剤の製造のために、eIF5A、eIF5A1をコードするポリヌクレオチド、及びeIF5A1に対するsiRNAの使用も提供する。好ましくは、変異eIF−5Aをコードするポリヌクレオチドはヒプシン化できない。
【0027】
本発明はまた、鎌状赤血球貧血の治療法も提供する。健常ヘモグロビン遺伝子(例えばHBB)をコードするポリヌクレオチドを、鎌状赤血球貧血を患う患者へ投与する。欠陥のあるタンパク質の発現をノックダウン又はノックアウトするため、患者には欠陥のあるヘモグロビン遺伝子をコードする遺伝子(例えば変異HbSをコードする遺伝子)を標的とするsiRNAもまた併用して投与される。治療は、その他の公知の医薬の投与、又は鎌状赤血球貧血を治療するために一般に用いられる治療をさらに含んでなってよい。
【図面の簡単な説明】
【0028】
【図1】ヒトeIF−5A1のアミノ酸配列及び様々な重要部位を示した図である。
【図2】K50及びK67におけるeIF−5A1の変異が、トランスフェクトしたタンパク質の蓄積を増大させることを示した図である。実施例1を参照のこと。
【図3】K47、K50、及びK67におけるeIF5A1の変異が、トランスフェクトしたタンパク質の蓄積を増大させることを示した図である。実施例2を参照のこと。
【図4】K50及びK67におけるeIF5A1の変異が、KAS細胞にトランスフェクトした際にアポトーシスの誘導をもたらすことを示した図である。実施例3を参照のこと。
【図5】K50及びK67におけるeIF5A1の変異が、KAS細胞にトランスフェクトした際にアポトーシスの誘導をもたらすことを示した図である。実施例4を参照のこと。
【図6】K50及びK67におけるeIF5A1の変異が、KAS細胞にトランスフェクトした際にアポトーシスの誘導をもたらすことを示した図である。実施例5を参照のこと。
【図7A】siRNAのトランスフェクション及びeIF−5A1を発現するように改変されたアデノウィルスによる処理が、KAS細胞にアポトーシスをもたらすことを示した図である。実施例6Aを参照のこと。
【図7B】eIF5A1 siRNA(標的#1(配列番号1)に対する)(siRNAコンストラクトの配列を図25に示す)による前処理は内在性eIF5A1の発現を減少させるが、アデノウィルスにより発現されたRNAi耐性のeIF5A1k50Aは蓄積させることを示した図である。実施例6Bを参照のこと。
【図7C】アデノウィルス感染前の、標的#1に対するeIF5A1 siRNAによる前処理が、ヒト多発性骨髄腫細胞におけるリン酸化NF−κBの発現を減少させることを示した図である。実施例6Cを参照のこと。
【図7D】アデノウィルス感染前の、標的#1に対するeIF5A1 siRNAによる前処理が、ヒト多発性骨髄腫細胞におけるリン酸化NF−κB及びICAM−1の発現を減少させることを示した図である。実施例6Dを参照のこと。
【図7E】ヒト多発性骨髄腫細胞における、siRNAにより仲介されるeIF5Aの抑制が、LPSにより仲介されるNFκBのDNA結合活性の誘導を阻害することを示した図である。NF−κBは多くの生存促進性及び抗アポトーシス経路を調節することから、eIF5A siRNAによるNF−κB活性の阻害は、eIF5AK50Rの過剰発現と組み合わせたときに、そのアポトーシス誘導を増大する能力を説明できる。
【図7F】siRNAによるKAS細胞の前処理が、IL−6の存在下でeIF5A1k50R遺伝子送達によるアポトーシスを増大させることを示した図である。実施例6Eを参照のこと。
【図8】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。示されるデータは、各群における全てのマウスの腫瘍容積である。実施例7を参照のこと。
【図9】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。示されるデータは、群あたりの平均腫瘍容積+/−標準誤差である。実施例7を参照のこと。
【図10】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の重量を減少させることを示した図である。実施例7を参照のこと。
【図11】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させ、かつ腫瘍の縮小をもたらすことを示した図である。実施例8を参照のこと。
【図12】eIF5A1 siRNAの静脈内(i.v.)投与、及びPEI/eIF5AlK50Rプラスミド複合体の腫瘍内(i.t.)投与が、多発性骨髄腫皮下腫瘍の縮小をもたらすことを示した図である。実施例9を参照のこと。
【図13A】eIF5A1プラスミド及びeIF5A1 siRNAによる処置が、多発性骨髄腫皮下腫瘍の生育を遅延させ、かつ腫瘍の縮小をもたらすことを示した図である。
【図13B】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が腫瘍の縮小をもたらすことを示した図である。
【図13C】eIF5A1 siRNAの静脈内(i.v.)投与、及びPEI/eIF5AlK50Rプラスミド複合体の腫瘍内(i.t.)投与が、多発性骨髄腫皮下腫瘍の縮小をもたらすことを示した図である。
【図14】eIF5A1プラスミド及びeIF5A1 siRNAの静脈内同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。実施例10を参照のこと。
【図15】eIF5A1 siRNAの静脈内(i.v.)投与、及びPEI/eIF5AlK50Rプラスミド複合体の静脈内(i.v.)又は腹腔内(i.p.)投与が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。実施例11を参照のこと。
【図16】eIF5A1プラスミド及びeIF5A1 siRNAによる処置が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。
【図17】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させ、かつ腫瘍の縮小をもたらすことを示した図である。実施例12を参照のこと。
【図18】eIF5A1 siRNAの静脈内(i.v.)投与、及びPEI/eIF5AlK50Rプラスミド複合体の腫瘍内(i.t.)投与が、多発性骨髄腫皮下腫瘍の縮小をもたらすことを示した図である。実施例13を参照のこと。
【図19】EF1又はB29プロモーターのいずれかにより駆動されるeIF5A1K50Rプラスミド、及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させ、かつ腫瘍の縮小をもたらすことを示した図である(KAS−SQ−5)。実施例14を参照のこと。
【図20】eIF5A1 siRNAの同時投与が、EF1又はB29プロモーターのいずれかにより駆動されるeIF5A1K50Rプラスミドの多発性骨髄腫皮下腫瘍における抗腫瘍効果を増大させ、腫瘍の負荷量を減少させることを示した図である(KAS−SQ−5)。実施例15を参照のこと。
【図21】eIF5A1 siRNAが、肺腺癌細胞におけるAd−eIF5Aの感染によりもたらされたアポトーシスを相乗的に増大させることを示した図である。実施例16を参照のこと。
【図22】その構築について実施例17に記載される、pExp5Aのマップを示した図である。
【図23】pExp5A(3371bp)の予測される配列を示した図である。実施例17を参照のこと。
【図24】様々な細胞株におけるeIF5A1K50Rの再発現を示した図である。実施例18を参照のこと。
【図25】標的配列及び好ましいeIF5A1 siRNAの配列を示した図である。
【図26】多発性骨髄腫における有効性試験の結果を示した図である。実施例21を参照のこと。
【図27】多発性骨髄腫における有効性試験の結果を示した図である。実施例21を参照のこと。
【図28】eIF5A1k50RcDNAの配列を示した図である。
【図29】ヒトeIF5A1k50Rに対するヒトeIF−5A1のアラインメントを示した図である。
【図30】HA−eIF5AK50Rの発現におけるDNA:siRNA比の効果を示した図である。実施例23を参照のこと。
【図31】ナノ粒子のトランスフェクションにより誘導されるアポトーシスにおけるDNA:siRNA比の効果を示した図である。実施例24を参照のこと。
【図32】eIF5A1K50Rプラスミド及びeIF5A1 siRNA(siSTABLE又は非siSTABLE)を含むPEI複合体(N/P=6又は8)の投与が、多発性骨髄腫皮下腫瘍の生育を阻害し、かつ腫瘍の縮小をもたらすことを示した図である。実施例25を参照のこと。
【図33】JET PEI(商標)ナノ粒子が腫瘍組織によって効果的に取り込まれ、該ナノ粒子が同細胞にプラスミド及びsiRNAを送達することを示した図である。実施例26を参照のこと。
【図34】KAS細胞がアクチノマイシンDに反応してアポトーシスを起こすことを示した図である。KAS細胞は無処理か、又は0.5μg/mlのアクチノマイシンDにより処理された。処理開始後24時間目に細胞を回収、洗浄し、アポトーシス細胞をアネキシン/PI(BD Bioscience)を用いて標識してフローサイトメトリーにより解析した。
【図35】eIF−5A1の切断型が、アクチノマイシンD処理に反応して蓄積することを示した図である。KAS細胞を、0.5μg/mlのアクチノマイシンDによって0時間〜30時間の範囲の時間枠で処理した。細胞ライセートを回収し、eIF5A1に対する抗体を用いるウェスタンブロットによって分析した。eIF5A1の、より小さな分子量の(切断された)形の蓄積はアクチノマイシンD処理後おおよそ8時間後から観察され始め、残りの処理の間を通して存在した。同じ膜でのウェスタンブロットによる等しい負荷を、アクチンに対する抗体を用いて示した。
【図36】eIF−5A1の切断型が、完全長eIF−5A1よりも高い等電点を持つことを示した図である。KAS細胞は無処理か、又は0.5μg/mlのアクチノマイシンDにより処理された。処理開始後17時間目に細胞ライセートを回収し、2−Dゲル電気泳動によって分離した後、eIF5A1に対する抗体を用いてウェスタンブロット分析を行った。無処理サンプル中に大量の完全長eIF5A1が観察できるが、アクチノマイシンD処理サンプル中には高めのpIを有するeIF5A1の切断型の蓄積が観察できる。
【図37】切断型eIF−5A1の2−Dゲルの写真を示した図である。KAS細胞は0.5μg/mlのアクチノマイシンDにより処理された。処理開始後17時間目に細胞ライセートを回収し、2−Dゲル電気泳動によって分離した後、クマシーブルーによる染色を行った。マススペクトロメトリー分析のために収集したeIF5A1の切断型の位置を示す。
【図38】切断型eIF−5A1のマススペクトロメトリーによる配列決定結果を示した図である。(A)eIF5A1の完全長アミノ酸配列を黒で示す。eIF5A1の完全長配列と共に整列させた、マススペクトロメトリーにより同定された配列決定ペプチド(B)を下線で示す(A)。配列決定ペプチドから、eIF5A1の最初の6アミノ酸が欠失していることに留意されたい。
【図39】カスパーゼ3、8、及び9の阻害剤がeIF−5A1の切断型の形成を阻害することを示した図である。ヒト多発性骨髄腫KAS細胞を異なるカスパーゼ阻害剤と共に8時間インキュベートし、さらに16時間、0.5μg/mlのアクチノマイシンD(ActD)と共に同時処理した。細胞ライセートを回収した。タンパク質を2D−PAGEによって分離し、eIF5Aに対する抗体を用いるウェスタン分析に供した。使用したカスパーゼ阻害剤はZ−VAD−FMK(一般的なカスパーゼ阻害剤)、Z−LEHD−FMK(カスパーゼ9阻害剤)、Z−DEVD−FMK(カスパーゼ3/7阻害剤)、Z−IETD−FMK(カスパーゼ6/8阻害剤)、及びZ−YVAD−FMK(カスパーゼ1阻害剤)である。データにより、一般的なカスパーゼ阻害剤、並びに、カスパーゼ3、8、及び9の阻害剤は全て、ActDによりKAS細胞に誘導されるアポトーシスの間に蓄積されるeIF5Aの切断型の形成を強力に阻止することが示された。カスパーゼ1の阻害剤もまた、完全な遮断ではないが、切断産物の形成を減少させる。これらの結果は、eIF5A1が遺伝毒性のストレス(アクチノマイシンDにより誘導される)後にカスパーゼにより切断され;eIF5A1の切断はその活性を変化させる、すなわちアポトーシス促進活性を増大させ得ることを示した。
【図40】脱アセチル化酵素阻害剤(ニコチンアミド)が、eIF−5A1の切断型の形成を促進することを示した図である。リジン47がアセチル化され得るeIF5Aは、Sir2関連脱アセチル化酵素であるHst2の基質である(Shirai et al., 2008)。ヒト子宮頸癌Hela S3細胞を、0.5μg/mlアクチノマイシンD(ActD)及び/又はSir2の強力な阻害剤である20μMニコチンアミドにより、示される時間の長さで処理した。細胞ライセートを回収した。タンパク質を2D−PAGEによって分離し、eIF5Aに対する抗体を用いるウェスタン分析に供した。ActD又はニコチンアミド単独によるHela S3細胞の処理は、eIF5Aの切断型の蓄積を誘導しない。しかしながら、アクチノマイシンD及びニコチンアミドの両方により処理されたHeLa細胞では、eIF5Aの切断型はActDへの暴露後少なくとも4時間目に上昇することから、アポトーシスの間にアセチル化がeIF5Aの切断を促進し得ることが示唆される。
【図41】切断型ヒトeIF5A1のDNA配列を示した図である。
【図42】完全長eIF5A1、切断型eIF5A、及び切断部位のタンパク質配列を示した図である。
【図43】アデノウィルスコンストラクトによるHela細胞の感染が、eIF5A導入遺伝子の蓄積をもたらすことを示した図である。Hela S3細胞を、異なるアデノウィルスコンストラクトにより、細胞あたり500感染単位にて感染させた。感染後48及び72時間目に細胞ライセートを回収し、SDS−PAGE、ならびにeIF5A及びアクチンに対する抗体を用いるウェスタンブロット分析に供した。切断型eIF5Aの蓄積が観察され得る。
【図44】アクチノマイシンD、ニトロプルシドナトリウム、又はIL−6の離脱による処理を通したアポトーシスの誘導が、ヒト骨髄腫細胞にeIF5A1の切断型(eIF5A1Δ1−6)の蓄積をもたらすことを示した図である。KAS−6/1細胞を示されるように処理し、次に2D−PAGE、及びeIF5Aに対する抗体を用いるウェスタンブロット分析に供した。アクチノマイシンD、NOドナー、ニトロプルシドナトリウム、又はIL−6の欠乏を通したKAS−6/1細胞におけるアポトーシスの誘導は、全て切断型eIF5A1の蓄積を誘導した。Ad−A1Δ(2−6)又はAd−A1Δ(2−6)/K50Rのいずれかを用いる切断型eIF5A1の過剰発現は、アポトーシスの間に産生される切断型eIF5A1(eIF5A1Δ1−6)よりも低い等電点を有するeIF5A1タンパク質の蓄積を誘導したことから、eIF5A1Δ1−6にはその他の翻訳後修飾が存在することが示される。
【図45】eIF5A1D6E及びeIF5A1D6E/K50Rは、アポトーシスの間にアクチノマイシンDにより誘導される切断に対して耐性であることを示した図である。ヒト多発性骨髄腫KAS−6/1細胞を示されるように処理し、次に2D−PAGE、及びeIF5Aに対する抗体を用いるウェスタンブロット分析に供した。過剰発現されたeIF5A1(Ad−A1)をアクチノマイシンDによる処理の間に切断に供した。しかしながら、eIF5A1D6E又はeIF5A1D6E/K50Rは、アクチノマイシンDによる処理後の切断に耐性である。これらの結果は、eIF5A1のアミノ酸3〜6がカスパーゼ切断部位を構成するという仮説に一致する。
【図46】eIF5A1が、in vitroで組み替えカスパーゼにより切断されることを示した図である。ヒトeIF5A1(A1)及び変異eIF5A1D6E(D6E)及びeIF5A1D6E/K50R(D6E/K50R)を、HA−タグ化タンパク質の発現をもたらすpHM6ベクター(Roche)にサブクローン化した。pHM6−HA−A1、pHM6−HA−A1(D6E)、及びpHM6−HA−A1(D6E/K50R)をin vitroで転写及び翻訳し(TNT T7 Quick Coupled Transcription/Translation Systems; Promega)、次にTranscend Non-Radioactive Translation Detection Systems(Promega)を用い、製造者の指示に従って標識した。各3マイクロリットルのin vitro翻訳産物を、1マイクロリットルの組み替えカスパーゼ(カスパーゼ1、2、3、6、7、8、9、又は10;Calbiochem)と共にカスパーゼ反応緩衝液(50mM Hepes、0.1%CHAPS、10%スクロース、100mM NaCl、10mM DTT、1mM EDTA)中で37℃にて2時間インキュベートした。最終産物をSDS−PAGEに供し、ビオチン化タンパク質をストレプトアビジン−HRPとのインキュベーション、それに続く化学発光の検出により可視化した。野生型eIF5A1は、カスパーゼ2、3、6、7、8、9、及び10により切断されるが、カスパーゼ1によっては切断されない。HA−タグ化eIF5A1D6E及びeIF5A1D6E/K50Rは、カスパーゼ3、7、及び8によってin vitroで効果的に切断されるが、カスパーゼ9及び10によっては効率的に切断されない。しかしながらこの切断は、in vitro系における高度に豊富なカスパーゼ活性、同様にグルタミン酸とアスパラギン酸の間の高い構造類似性による可能性があり、これによって低い効率ではあるが(特にeIF5A1D6E及びeIF5A1D6E/K50Rはin vivoアッセイでの切断には耐性であったことから(図45))、eIF5A1D6Eの切断が許容される。
【図47】Ad−eIF5A1Δ(2−6)又はAd−eIF5A1K50RΔ(2−6)によるHela細胞の感染は、野生型Ad−eIF5A1又はAd−eIF5A1K50Rによる感染に比較してアポトーシスの増大をもたらすことを示した図である。ヒト子宮頸癌Hela S3細胞を、異なるアデノウィルスコンストラクトにより、細胞あたり500感染単位にて感染させた。感染後48又は72時間目に細胞を回収し、アネキシンV/PI染色に供した。次に細胞をフローサイトメトリーにより選別した。アポトーシスの初期段階の細胞(アネキシンV+/PI−)、同様にアポトーシスの後期段階(アネキシンV+/PI+)の細胞のパーセンテージを示す。アミノ酸2〜6の切断によってeIF5A1又はeIF5A1K50Rのアポトーシス活性は増大する。逆に、予測される切断部位の変異によってeIF5A1又はeIF5A1K50Rのアポトーシス活性は低下するが、無にはならない。これらの結果は、カスパーゼにより仲介されるeIF5A1の切断が、eIF5A1の切断型のアポトーシス活性を増大させることを示す。
【図48】アポトーシスにおける、カスパーゼにより仲介されるeIF5A1の切断の寄与についてのモデルを示した図である。
【図49】KASヒト骨髄腫細胞におけるアクチノマイシンD処理後に、切断型eIF5Aが核に蓄積することを示した図である。
【図50】KASヒト骨髄腫細胞におけるアクチノマイシンD処理後に、切断型eIF5Aが核に蓄積することを示した図である。略語一覧:UN=無処理;actD=アクチノマイシンDによる処理;C=細胞質分画;N=核分画、α−チューブリン及びα−PCNAはコントロールである。この図はまた、完全長eIF5Aは細胞質と核の間に均等に分布するが、切断型は主に核内に蓄積することも示す。
【図51】eIF5Aの切断型が複数の細胞株及び細胞型において異なるアポトーシス刺激に反応して蓄積することにより、カスパーゼにより仲介されるeIF5Aの切断はアポトーシスの間の一般的な現象であることが示されることを示した図である。この図はまた、完全長eIF5Aは細胞質と核の間に均等に分布するが、切断型は主に核内に蓄積することも示す。
【図52】切断型eIF5Aが、異なる細胞株において異なるアポトーシス刺激[KASヒト多発性骨髄腫細胞におけるIL−6の欠乏、又はIL−6及びFBSの欠乏による処理後、並びにUACC1598(ヒト卵巣癌細胞株)におけるアクチノマイシンDによる処理後]に反応して産生されるという研究結果を示した図である。
【図53】ヒトeIF5A1及びeIF5A2のヌクレオチド配列を示した図である。
【0029】
[発明の詳細な説明]
本発明は、eIF−5A1の切断型をコードする単離されたポリヌクレオチド、同様に切断型eIF−A1ポリペプチドを提供する。切断型eIF−5A1ポリヌクレオチドは、癌細胞のアポトーシス及び死滅の誘導に有用である。切断型ポリヌクレオチドは発現ベクター内に使用され得り、これは次に哺乳類へ投与される。切断型eIF−5Aは哺乳類内に発現し、癌細胞を死滅させる。約17kDaである完全長eIF−5A1タンパク質に対し、切断型eIF−5A1タンパク質は約16kDAである。
【0030】
アポトーシスを起こす細胞内には、eIF5Aのより小さな分子量型(〜16kDa)の形成が観察されている。このことは、サイトカインカクテルにより処理されたベータ膵島細胞、及び細胞傷害性薬物のアクチノマイシンDにより処理されたヒト骨髄腫細胞(KAS細胞)において観察されている。図35を参照されたい。eIF5Aのより小さな分子量型はまた、Ad−eIF5A1の感染後にアポトーシスを起こしているHeLa子宮頸癌細胞に蓄積することも見出されており、このことからeIF5Aのより小さな分子量型は、選択的スプライシングの結果というよりはプロテアーゼによる切断によるものであると示唆される。eIF5Aのより小さな分子量型の蓄積は完全長ヒプシン化eIF5Aの量の著しい減少を伴うという観察は、eIF5Aのより小さな分子量型が選択的スプライシングというよりは切断によるものであることをさらに支持する。図36を参照されたい。
【0031】
本発明は、切断型eIF5A1タンパク質をコードするeIF5A1ポリヌクレオチドを含んでなる組成物を提供する。この組成物は、被験体の癌細胞又は腫瘍にアポトーシスを誘導するために用いられる薬剤の製造に有用である。eIF5A1ポリヌクレオチドは、好ましくは図38に示される配列番号37に記載のアミノ酸配列を含んでなる、切断型eIF5A1タンパク質をコードする。特定の実施形態によれば、eIF5A1ポリヌクレオチドは約16kDAの切断型eIF5A1タンパク質をコードする。
【0032】
組成物及び又は薬剤は、ヒトを含む哺乳類に投与できる。本明細書で用いられる「被験体」は哺乳類及びヒトを含む。
【0033】
アポトーシスの誘導は、癌細胞又は腫瘍において以下の効果を持ち得る:癌細胞若しくは腫瘍の生育を遅くする、癌細胞若しくは腫瘍の生育を停止する、又は癌細胞を死滅させるか若しくは腫瘍サイズを縮小させる、及び上記のいずれの組み合わせ。いずれの癌も治療でき、特定の実施形態によれば、癌は多発性骨髄腫である。
【0034】
特定の実施形態によれば、eIF5A1ポリヌクレオチドは配列番号38(図41に示される)に記載の配列を含んでなる。特定の実施形態によれば、eIF5A1ポリヌクレオチドはプラスミド又は発現ベクター内に構成される。プラスミド又は発現ベクターについては、以下にさらに詳しく記載される。特定の実施形態によれば、発現ベクターはアデノウィルス発現ベクター又はpHM6である。特定の実施形態によれば、組成物又は薬剤が多発性骨髄腫の治療に使用される場合、発現ベクターはB細胞特異的プロモーター(すなわちB29)のような組織特異的プロモーターを含んでなる。発現ベクターはpCpGプラスミドを含んでなってよい。以下にさらに詳しく考察されるように、発現ベクターはポリエチレンイミンと複合されてよい。
【0035】
組成物又は薬剤は、好ましくは腫瘍内、静脈内、又は皮下投与される。
【0036】
本発明はまた、ポリヌクレオチドが配列番号38(図41に示される)に記載の配列を含んでなる、切断型eIF5A1タンパク質をコードする単離されたポリヌクレオチドも提供する。
【0037】
本発明はまた、切断型タンパク質が配列番号37に記載のアミノ酸配列を含んでなる、切断型eIF5A1タンパク質をコードする単離されたポリヌクレオチドも提供する。
【0038】
以下に考察されるように、eIF5A1はカスパーゼにより仲介される切断によって形成される。従って本発明はさらに、カスパーゼにより仲介されるeIF5A1の切断によって形成される、単離された切断型eIF5A1ポリペプチドを提供する。
【0039】
本発明はさらに、組成物、又は切断型eIF5A1タンパク質をコードするeIF5A1ポリヌクレオチドを、完全長eIF5A1ポリヌクレオチドと組み合わせて含んでなる薬剤の製造のための該組成物の使用を提供する。この組成物は、被験体の癌細胞又は腫瘍にアポトーシスを誘導する薬剤の製造に有用である。切断型eIF5Aタンパク質をコードするeIF5A1ポリヌクレオチドについては上述の通りである。完全長eIF5A1ポリヌクレオチドは、配列番号35に記載のアミノ酸配列を含んでなるタンパク質をコードする。完全長eIF5A1ポリヌクレオチドについては、以下にさらに詳しく記載される。特定の実施形態によれば、eIF5A1ポリヌクレオチドは配列番号38に記載の配列を含んでなり、完全長eIF5A1ポリヌクレオチドは配列番号43(図53に示される)に記載の配列を含んでなる。
【0040】
本明細書に記載されるように、完全長及び切断型ポリヌクレオチドは発現ベクター内に存在してよい。加えてベクターは、本明細書に記載されるプロモーター、同様にその他のいずれの有用なプロモーターを含んでなってよい。
【0041】
特定の実施形態によれば、完全長eIF5Aポリヌクレオチドは変異eIF5A1をコードし、ここで該変異はデオキシヒプシンシンターゼによるヒプシン化を阻止若しくは阻害し、並びに/又は、該変異はユビキチン化部位及び/若しくはアセチル化部位に存在する。変異については、以下にさらに詳しく記載される。特定の実施形態によれば、変異はK50A、K50R、K67A、K47R、K67R、K50A/K67A、K50A/K47R、K50A/K67R、K50R/K67A、K50R/K47R、K50R/K67R、及びK47A/K67Aからなる群より選択される。
【0042】
本発明はまた、切断型eIF5Aをコードするポリヌクレオチド、変異eIF5Aをコードするヌクレオチド、及びeIF5Aの3’UTRを標的とするsiRNAを含んでなる組成物を提供する。変異eIF5Aをコードするヌクレオチド、及びeIF5Aの3’UTRを標的とするsiRNAの組み合わせ/二元的使用については、以下にさらに完全に記載される。該組成物は、癌細胞及び腫瘍にアポトーシスを誘導するため、被験体に投与される薬剤の製造に有用である。
【0043】
特定の好ましい実施形態によれば、siRNAはeIF5A1の配列である5’−GCT GGA CTC CTC CTA CAC A−3’を標的とする。siRNAはdsRNAであり、dsRNAの一本の鎖は5’−GCU GGA CUC CUC CUA CAC A−3’の配列を含んでなる。特定の実施形態によれば、siRNAは血清中での分解を阻止するために安定化される。
【0044】
完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチドは、発現ベクター内に存在してよい。特定の実施形態によれば、完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチド及び/又はsiRNAは、ポリエチレンイミンと複合される。完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチド及び/又はsiRNAは、独立してポリエチレンイミンと複合される。
【0045】
本発明はまた、本明細書に記載される、例えば切断型eIF5A1をコードするヌクレオチドを含んでなり、任意に完全長eIF5Aをコードするヌクレオチド又は完全長変異eIF5Aをコードするヌクレオチドを含んでなり、かつ任意にeIF5Aの3’UTRを標的とするsiRNAを含んでなる組成物又は薬剤を哺乳類に提供することによる、哺乳類の癌細胞又は哺乳類の腫瘍にアポトーシスを誘導する方法も提供する。特定の実施形態によれば、癌は多発性骨髄腫であり、組成物/薬剤は静脈内、腹腔内、又は腫瘍内投与される。
【0046】
本発明はまた、被験体内での内在性遺伝子の発現をノックアウト又はノックダウンするための、内在性遺伝子を標的とするsiRNA、及び、ポリヌクレオチドによりコードされるタンパク質をホスト内で発現させるために被験体へ提供される、送達媒体/発現ベクター内の遺伝子をコードするポリヌクレオチドを組み合わせた使用にも関する。
【0047】
この組み合わせは、欠陥又は変異のあるタンパク質の存在、すなわち被験体内で産生されたタンパク質が、その欠陥のある構造のために、必要な機能を実行できないか、又は代わりに代謝経路若しくは生体分子の相互作用を不調にすることにより起こる、疾病又は状態を有する被験体の治療に有用である。siRNAは欠陥のあるタンパク質をコードする遺伝子を標的として設計され、その欠陥のあるタンパク質の発現をノックダウン又はノックアウトする。正常な(欠陥のない)タンパク質をコードするポリヌクレオチドが被験体に投与され、正常なタンパク質がその必要な機能を実行できるように被験体内で発現する。
【0048】
別の実施形態によれば、所望のタンパク質をコードするポリヌクレオチドの投与の代わりに、被験体へタンパク質が投与される。本明細書では、タンパク質、ペプチド、及びポリペプチドの語は互換的に用いられる。
【0049】
siRNAは、欠陥のあるタンパク質の内在性の発現をノックダウン又はノックアウトするが、同時に投与された正常タンパク質をコードするポリヌクレオチドの外来性の発現に影響しないような遺伝子の特定の領域を標的として好ましく設計される。例えばsiRNAは、投与されたセンスコンストラクト(タンパク質をコードするポリヌクレオチド)の外来性の発現に影響しないように、3’UTRを標的とし得る。欠陥のあるタンパク質の内在性の発現をノックダウン又はノックアウトすることにより、外来性のポリペプチドから発現する正常タンパク質と競合する欠陥のあるタンパク質は、減少するか又は無になるであろう。
【0050】
この応用が有用となり得る疾病状態の一例は、鎌状赤血球貧血に関する。鎌状赤血球貧血は、赤血球(red blood cell:RBC)に見出され、身体全体への酸素の運搬を補助するタンパク質であるヘモグロビンに影響を及ぼす血液疾患である。
【0051】
鎌状赤血球貧血は、ヒトが変異ヘモグロビン(Hbs)の発現をもたらす2つの異常な遺伝子(各両親から1つずつ)を受け継いだ際に発生する。変異ヘモグロビンは、RBCの形を変える原因となる。正常ヘモグロビン(ヘモグロビンA、又はHbA)を有する赤血球は血流を通して容易に移動し、身体の全ての細胞に酸素を送達する。これらは、非常に微小な血管であっても容易に「圧搾」して通り抜けることができる。鎌状赤血球貧血は、ヘモグロビンの異常型(HbS)が、共に凝集する傾向があり、赤血球を粘着性で、堅く、さらに脆弱にさせること、かつそれらを曲がった鎌状にさせることから起こる。
【0052】
数百のHBB遺伝子の変異形が知られているが、鎌状赤血球貧血は、最も一般的にはヘモグロビン変異形HbSにより引き起こされる。この変異形では、疎水性アミノ酸のバリンが、HBBポリペプチド鎖の6番目のアミノ酸の位置における親水性のグルタミン酸に取って代わる。この置換によりタンパク質構造の外側に疎水性の点が作り出され、これは隣接するヘモグロビン分子のベータ鎖の疎水性領域に固着する。HbS分子がこのように共に凝集(重合)することによって強固な線維となり、赤血球を「鎌状」にする。
【0053】
重合は、赤血球が身体全体の様々な組織に運搬する酸素分子を放出した後のみに起こる。ヘモグロビンが酸素を結合できる場である肺に赤血球がいったん帰還すると、HbSの長い線維は脱重合するか、又は単一の分子に分裂する。重合と脱重合の間の循環により、赤血球の膜は強固になる。これらの赤血球の剛性、及び酸素を運搬しないときのそれらの歪んだ形は、微小血管の封鎖をもたらし得る。この封鎖は疼痛の発現の原因となり得り、かつ臓器を損傷し得る。
【0054】
鎌状赤血球貧血は、常染色体劣性遺伝疾患である。疾病が発現するためには、ヒトは2コピーのHbS変異形、又は1コピーのHbS及び1コピーの別の変異形のいずれかを受け継ぐ必要がある。1コピーの正常HBB遺伝子(HbA)及び1コピーのHbSを有する保因者は、鎌状細胞の形質を有すると説明され、疾病の症状を表さない。
【0055】
従って本発明の一実施形態は、鎌状赤血球貧血を有する被験体を治療する方法を提供する。患者には、HBB遺伝子を標的とするsiRNAが投与される。siRNAは、ヘモグロビンのHbS変異形の発現をノックダウン、好ましくはノックアウトするように設計される。被験体が正常ヘモグロビンを発現するように、被験体へ正常ヘモグロビンをコードするポリヌクレオチドが提供される。siRNAはまた、正常ヘモグロビンをコードする外来性ポリヌクレオチドの発現に干渉しないようにも設計される。従って、被験体はもはや変異ヘモグロビンを作らず(又は実質的に少なく作り)、代わりに正常健常ヘモグロビンを作ることによって、正常に機能するさらに正常な赤血球がもたらされる。
【0056】
本発明はまた、タンパク質に、疾病状態を引き起こすか又は導く翻訳後修飾が起こる状況においても有用である。siRNAは内在性タンパク質の発現をノックダウンするために用いられ、これにより翻訳後修飾されるものは無くなるか又は少なくなる。次に外来性発現のために、タンパク質をコードするポリヌクレオチドが患者へ提供される。タンパク質は、翻訳後修飾できないように改変される。次にこのタンパク質はその適切な使用のために身体に利用可能であるが、翻訳後修飾できないために疾病状態を導き得ない。当業者は異なる翻訳後修飾について理解するであろう。例えば翻訳後、アミノ酸の翻訳後修飾は、タンパク質に酢酸塩、リン酸塩、様々な脂質及び炭水化物のようなその他の生化学官能基を付着させることにより、アミノ酸の化学的性質を変化させることにより(例えばシトルリン化)、又はジスルフィド架橋の形成のように構造を変化させることにより、タンパク質の機能の範囲を拡張する。また、酵素はタンパク質のアミノ末端からアミノ酸を除くか、又は中間でペプチド鎖を切断し得る。例えばペプチドホルモンのインスリンは、ジスルフィド結合の形成後に2度切断され、鎖の中間からプロペプチドが除かれ;得られたタンパク質はジスルフィド結合によって連結する2つのポリペプチド鎖から成る。また、最も新生のポリペプチドはアミノ酸のメチオニンから始まるが、これはmRNAの「開始」コドンもまた、このアミノ酸をコードするためである。このアミノ酸は通常、翻訳後修飾の間に取り除かれる。リン酸化のようなその他の修飾は、タンパク質の作用を制御するための共通機構、例えば酵素の活性化又は不活性化の一部である。別の翻訳後修飾は、デオキシヒプシンシンターゼ(DHS)による真核生物翻訳開始因子5A(eIF5A)のヒプシン化を含む。
【0057】
従って本発明は、タンパク質をコードするポリヌクレオチドが患者へ提供され、かつ患者に発現される、被験体内の遺伝子の発現を変化させる方法を提供する。タンパク質は正常/野生型タンパク質、又は変異したタンパク質であってよい。対応する内在性遺伝子の発現は、被験体へ投与されるsiRNAによって抑制される。
【0058】
該方法はさらに、標的タンパク質をコードするポリヌクレオチドを含んでなるコンストラクトを提供することを含んでなり、該ポリヌクレオチドは標的タンパク質を産生するために被験体内に発現される。内在遺伝子が欠陥のあるタンパク質を発現する特定の実施形態によれば、ポリヌクレオチドは正常/健常タンパク質のいずれかをコードするように設計される。siRNAは欠陥のある内在性タンパク質の発現を抑制するために投与される。内在遺伝子が正常健常タンパク質を発現する特定の実施形態によれば、ポリヌクレオチドは、正常/健常タンパク質又は非変異タンパク質に起こるような翻訳後修飾のできない変異タンパク質をコードするように設計される。siRNAは内在性タンパク質の発現を抑制するために投与され、翻訳後修飾に利用できるこのタンパク質は減少する。
【0059】
特定の実施形態によれば、siRNAは外来性ポリヌクレオチドの発現に影響しないような遺伝子の領域を標的として選択又は設計される。例えばsiRNAは3’UTR又は3’末端を標的としてよい。
【0060】
siRNAは、ネイキッドsiRNA又は血清に対して安定化されたネイキッドsiRNAのいずれかとして患者へ送達され得る。siRNAは全身性に、すなわちIP又はIVのいずれかにより注射され得る。あるいは、siRNAは身体の望まれる領域へ局所的に注射又は送達され得る。特定の実施形態によれば、siRNAは、限定はされないがデンドリマー、リポソーム、又はポリマーのような送達ビヒクルにおいて投与され得る。
【0061】
所望のタンパク質をコードするポリヌクレオチドは、ヌクレオチドの発現を提供するか又は許容するいずれの送達手段を通して投与され得る。本明細書で、ポリヌクレオチド及びヌクレオチドの語は互換的に用いられる。送達は、限定はされないがプラスミド、発現ベクター、ウィルス性コンストラクト、アデノウィルスコンストラクト、デンドリマー、リポソーム、又はポリマーのような、いずれのウィルス性又は非ウィルス性機構を通してよい。
【0062】
特定の実施形態によれば、ポリヌクレオチドの発現のために、還元型CpGジヌクレオチドを有する発現プラスミドが用いられる。ポリヌクレオチドの発現を促進できるいずれのプロモーターも使用されてよく、これは治療法に望まれる応用に基づいて選択され得る。例えば、多発性骨髄腫を死滅させるためには、ヒトB29プロモーター/エンハンサーのようなB細胞に特異的なプロモーターが望ましいであろう。別の実施形態によれば、プロモーターは別の組織特異的プロモーターであってよいか、又はシステムプロモーターであってよい。標的タンパク質をコードするポリヌクレオチドは、IV若しくは皮下注射、又はその他のいずれの生物学的に適した送達機構を通して送達され得る。あるいはポリヌクレオチドは、リポソーム、又は標的腫瘍若しくは癌細胞へのDNA(又はプラスミド若しくは発現ベクター)送達のために提供される、その他のいずれの適切な「担体」又は「ビヒクル」において送達され得る。合成DNA送達系についての総説は、例えばLuo, Dan, et al, Nature Biotechnology, Vol. 18, January 2000, pp. 33-37を参照されたい。従って、ヌクレオチド/プラスミド/発現ベクターは、リポソーム、デンドリマー、又は同様の無毒性ナノ粒子のような、ナノメートルサイズのビヒクルを通して送達されることが好ましい可能性がある。ビヒクルは、好ましくは、有効量のヌクレオチド/プラスミド/発現ベクターを被験体、腫瘍、又は癌細胞へ送達する間、ヌクレオチド/プラスミド/発現ベクターを早期クリアランスから、又は免疫反応の原因から保護する。典型的なビヒクルは、ヌクレオチド/プラスミド/発現ベクターに付随する単純なナノ粒子から、特異的な細胞受容体を標的とするためにその表面に付着するリガンドを有するペグ化リポソームのような、さらに複雑なペグ化ビヒクルまでに及び得る。
【0063】
リポソーム及びペグ化リポソームは当該技術分野において公知である。従来のリポソームでは、送達される分子(すなわち小分子薬、タンパク質、ヌクレオチド、又はプラスミド)はリポソームの中央の腔内に含まれる。当業者は、分子送達に有用な「ステルス」標的化、及び陽イオンリポソームの存在もまた認識するであろう。例えば、Hortobagyi, Gabriel N., et al, J. Clinical Oncology, Vol. 19, Issue 14 (July) 2001 :3422-3433 及び Yu, Wei, et al., Nucleic Acids Research. 2004, 32(5);e48を参照されたい。リポソームは静脈内に注射でき、かつそれらの表面がさらに親水性になるように修飾でき(二重層へのポリエチレングリコールの添加(「ペグ化」)により)、血流でのそれらの循環時間は増大する。これらは「ステルス」リポソームとして知られ、ドキソルビシン及びミトキサントロンのような親水性(水溶性)の抗癌剤に対する担体として特に有用である。腫瘍細胞のような標的細胞へ薬物を運搬するリポソームの特異的な結合特性を促進するために、リポソームの表面に抗体、タンパク質、ペプチドなどのような特異的分子が付着され得る。例えば、癌細胞の表面にある受容体に対する抗体が、リポソームを癌細胞に向けるために使用されてよい。多発性骨髄腫を標的にする場合、リポソームを多発性骨髄腫細胞に向けるために、例えば葉酸塩、Il−6、又はトランスフェリンが使用されてよい。
【0064】
デンドリマーもまた当該技術分野において公知であり、好ましい送達ビヒクルを提供する。デンドリマーについての考察は、例えば、Marjoros, Istvan, J., et al, "PAMAM Dendrimer-Based Multifunctional Conjugate for Cancer Therapy: Synthesis, Characterization, and Functionality," Biomacromolecules, Vol. 7, No. 2, 2006; 572-579, 及び Majoros, Istvan J., et al., J. Med. Chem, 2005. 48, 5892-5899を参照されたい。
【0065】
好ましい実施形態によれば、送達ビヒクルはポリエチレンイミンナノ粒子を含んでなる。典型的なポリエチレンイミンナノ粒子は、現在Polyplus Transfection, Incにより製造されるin vivo-jetPEI(商標)である。In vivo-jetPEI(商標)は、DNA及びsiRNAの送達薬剤として有用な陽イオンポリマートランスフェクション剤である。Polyplus Transfectionから市販されているIn vivo-jetPEI(商標)は、動物において信頼性のある核酸送達を提供する直鎖ポリエチレンイミン試薬である。これは遺伝子治療(Ohana et al, 2004. Gene Ther Mol Bio 8:181-192; Vernejoul et al, 2002. Cancer Research 62:6124-31)、RNA干渉(Urbain-Klein et al., 2004. Gene therapy 23:1-6; Grezelinski et al., 2006. Human Gene Therapy 17:751-66)、及び遺伝子ワクチン接種(Garzon et al., 2005. Vaccine 23:1384-92)に用いられる。In vivo JET-PEIは現在、癌の遺伝子治療のための送達ベクターとしてヒトの治験に使用されている(Lemkine et al., 2002. Mol. Cell. Neurosci. 19:165-174)。
【0066】
In vivo-jetPEI(商標)は核酸をおおよそ50nmのナノ粒子に凝縮し、これは数時間安定である。この独特な保護機構の結果として、注射後の血液細胞の凝集は、組織内での制限された拡散、赤血球の凝集、及び微小塞栓を防止するその他の試薬に比較して減少する。これらのナノ粒子は、組織内へ拡散し、エンドサイトーシスによって細胞へ入るためには十分に小さい。In vivo-jetPEI(商標)は、エンドソームからの核酸の放出及び核膜を横断しての移動を支持する。
【0067】
好ましい実施形態によれば、siRNA、及びポリヌクレオチドを含んでなるベクター/プラスミドの両方が、in vivo-jetPEI(商標)複合体を通して被験体に投与される。siRNA、及びポリヌクレオチドを含んでなるベクター/プラスミドは、ポリエチレンイミン又はin vivo-jetPEI(商標)複合体のようなポリマー複合体を通して共に複合されてよいか、又は別々にポリマーと複合されてよい。例えば、siRNA、及びポリヌクレオチドを含んでなるベクター/プラスミドが別々に被験体へ投与される場合(時間及び/又は送達部位が別々であるという意味で)には、異なる担体と複合されたsiRNA及びポリヌクレオチドを用意することが好ましい。同一部位において同時に投与され得る場合には、siRNA及びポリヌクレオチドを共に複合することが好ましい。
【0068】
別の実施形態によれば、被験体内で発現され得るポリヌクレオチドを送達するために投与されるプラスミド又はベクターの代わりに、タンパク質それ自体が被験体へ送達される。タンパク質は単離されてよいか、又は合成されてよい。
【0069】
本発明の一実施形態は、哺乳類及びヒトを含む被験体の癌を治療する方法を提供する。癌の治療は、癌細胞におけるアポトーシスの誘導、癌細胞の死滅、癌細胞の数の減少、及び腫瘍容積/重量の減少を含むがこれらに限定されない。該方法は、eIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドを含んでなる組成物を投与することを含んでなる。組成物並びにeIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドについては以下で考察する。
【0070】
全ての細胞は真核生物翻訳開始因子5A(「eIF−5A」)(又は、本明細書では「因子5A」とも称される)を産生する。哺乳類細胞は、eIF−5A1の2つのアイソフォーム(eIF−5A1及びeIF−5A2)を産生する。eIF−5A1はアポトーシスを起こしている細胞内で上昇していることから、アポトーシス特異的eIF−5Aと称されている。ヒトeIF−5A1は受入番号NM001970を有し、図1に示される。eIF−5A1は、アポトーシスに必要なタンパク質をコードするmRNAの核サブセットの核外輸送の原因であると信じられている。eIF−5A2は、細胞増殖に必要なタンパク質をコードするmRNAの核サブセットの核外輸送の原因であると信じられていることから、増殖eIF−5Aと称されている。Liu & Tartakoff (1997) Supplement to Molecular Biology of the Cell, 8, 426a. Abstract No. 2476, 37th American Society for Cell Biology Annual Meeting, 及び Rosorius et al. (1999) J. Cell Science, 112, 2369-2380を参照されたい。
【0071】
両方の因子5Aは、デオキシヒプシンシンターゼ(「DHS」)により翻訳後修飾される。DHSはeIF−5Aをヒプシン化する。独特なアミノ酸であるヒプシンは調べられた全ての真核生物及び古細菌に見出されるが、真正細菌には見出されず、eIF−5Aは知られている唯一のヒプシン含有タンパク質である。Park (1988) J. Biol. Chem., 263, 7447-7449; Schumann & Klink (1989) System. Appl. Microbiol, 11, 103-107; Bartig et al. (1990) System. Appl. Microbiol, 13, 112-116; Gordon et al (1987a) J. Biol. Chem., 262, 16585-16589。ヒプシン化eIF−5Aは2つの翻訳後工程によって形成され:第1工程は、デオキシヒプシンシンターゼにより触媒される、スペルミジンの4−アミノブチル成分の、前駆体eIF−5Aの特定のリジンのα−アミノ基への転移による、デオキシヒプシン残基の形成である。第2工程は、ヒプシンを形成するための、デオキシヒプシンヒドロキシラーゼによるこの4−アミノブチル成分のヒドロキシル化に関する。
【0072】
eIF−5Aのアミノ酸配列は種の間でよく保存されており、eIF−5Aのヒプシン残基周囲のアミノ酸配列は厳密に保存されていることから、この修飾は生存のために重要であるかもしれないことが示唆される。Park et al. (1993) Biofactors, 4, 95-104。この仮定は、これまでに酵母に見出されたeIF−5Aの両方のアイソフォームの不活性化、又はその活性化によって第1工程を触媒するDHS遺伝子の不活性化が細胞分裂を遮断するという観察により、さらに支持される。Schnier et al (1991) Mol Cell. Biol, 11, 3105-3114; Sasaki et al (1996) FEBS Lett., 384, 151-154; Park et al (1998) J. Biol. Chem., 273, 1677-1683。しかしながら、酵母におけるeIF−5Aタンパク質の枯渇がもたらしたものは総タンパク質合成のわずかな減少のみであったことから、eIF−5Aはタンパク質の全体的な合成よりも、mRNAの特定のサブセットの翻訳のために必要とされ得ることが示唆される。Kang et al. (1993), "Effect of initiation factor eIF-5A depletion on cell proliferation and protein synthesis," in Tuite, M. (ed.), Protein Synthesis and Targeting in Yeast, NATO Series H。eIF−5Aを結合するリガンドがより高く保存されたモチーフを共有するという最近の知見もまた、eIF−5Aの重要性を支持する。Xu & Chen (2001) J. Biol. Chem., 276, 2555-2561。加えて、修飾されたeIF−5Aのヒプシン残基が配列特異的なRNAの結合に必須であることが見出され、かつ結合によってリボヌクレアーゼからは保護されなかった。
【0073】
本発明者らは、eIF−5Aをコードするポリヌクレオチドを細胞に投与した際、それらの細胞でアポトーシスが増大することを示した。投与後に癌細胞内で発現するeIF−5A1ポリヌクレオチドを投与することによって、癌細胞をアポトーシスへ押しやることが可能であることが示された。全てそれらの全体が参照により取り込まれる、同時係属出願の10/200,148号;11/287,460号;11/293,391号、及び11/637,835号を参照されたい。
【0074】
本発明者らは、細胞に因子5Aのヒプシン化型が蓄積される際に細胞は生存状態に入り、通常は時間と共に起こるアポトーシスが起こらないことをさらに特定した。特に癌細胞には著しい量の因子5Aのヒプシン化型が存在することから、細胞はアポトーシスを起こさない(死滅しない)。従って、癌細胞を死滅させること(癌細胞をアポトーシス経路へ押しやること)によって癌を治療するために、eIF−5A1をコードするポリヌクレオチドが被験体へ、又は癌細胞若しくは腫瘍へ投与され、癌細胞にアポトーシス、最終的には細胞死及び腫瘍の縮小を順に引き起こすeIF−5A1の発現が増大される。しかしながらeIF−5A1の遺伝子発現の上昇のためにeIF−5A1タンパク質をコードするポリヌクレオチドのみが供給され、eIF−5A1の内在性発現をノックダウンするためのsiRNAが使用されない場合には綱引き:細胞をアポトーシス経路へ向けるeIF−5A1の発現と、細胞を細胞生存経路へ向けるヒプシン化因子5Aの存在が競合する状態となる。本発明はこの綱引きを排除し、eIF−5A1の発現の増大のみによる改良を示す。被験体又は細胞へ投与されたポリヌクレオチドは、得られる発現タンパク質がヒプシン化され得ないように変異導入される。加えて、因子5Aの内在性発現はeIF−5Aを標的とするsiRNAによってノックアウト/ダウンされ、ヒプシン化される内在性eIF−5A1は周囲に存在しなく/少なくなる。従って、細胞内にヒプシン化eIF−5Aが存在しない(又は実質的に少ない)ため、細胞は生存状態に押しやられることはない。
【0075】
変異eIF−5A1をコードするポリヌクレオチドは、ヒプシン化されず、それ故細胞を生存状態にするために利用され得ないように好ましく変異導入される。例えば一実施形態によれば、eIF−5Aをコードするポリヌクレオチドは、通常はDHSによりヒプシン化される位置50のリジン(K)がアラニン(A)(ヒプシン化され得ない)に変わるように変異導入される。この変異体はK50Aと表示される。
【0076】
別の実施形態によれば、位置67のリジンがアルギニン(R)に変更される。この変異体は(K67R)と表示される。別の実施形態によれば、位置67のリジン(K)がアラニン(A)に変更され、これは(K67A)と表示される。別の実施形態によれば、位置50のリジン(K)がアルギニンに変更され(K50R)、別の実施形態によれば、位置47のリジン(K)がアルギニンに変更される(K47R)変異体が提供される。
【0077】
別の実施形態によれば、二重変異体が使用される。一つの二重変異体では位置50のリジン(K)がアルギニン(R)に変更され、かつ位置67のリジン(K)がアルギニン(R)に変更される。この二重変異体はK50R/K67Rと称される。この二重変異体も同様にヒプシン化不可能であるが、このアミノ酸の変更は、eIF−5A1の3−D構造を単一変異体(K50A)ほどには変化させない。従って二重変異体は野生型に非常に類似した3−Dの形及びフォールディングのタンパク質を提供し、それ故に単一変異体よりもさらに安定である。さらに安定になることで、これは体内により長く存在して、治療上の利点をより長く提供する。従って、身体が正常細胞機能に必要とされるがヒプシン化され得ない因子5Aを有し得ることから、細胞は細胞生存状態にならず、かつアポトーシスを回避しない。
【0078】
別の二重変異体では位置47のリジン(K)がアルギニン(R)に変更され、かつ位置50のリジンがアルギニン(R)に変更される。この二重変異体は(K47R/K50R)と表示される。本発明は、位置50のリジン(L)がアラニン(A)に変更され、かつ位置67のリジンがアラニン(A)に変更される、別の二重変異体を提供する。この二重変異体は(K50A/K67A)と表示される。
【0079】
身体は正常細胞の生存及び健常細胞の増殖のために因子5Aを必要とすることから、siRNAが全身性に送達される場合、siRNAによって被験体内の発現を完全に遮断しないことが好ましい。eIF−5A発現の制御は、発現の遮断に良好ではない(すなわち、発現を停止又は減少させるが、発現を完全には遮断しない)siRNAを使用すること、又は代わりに、健常細胞の正常な生育及び機能を許容しかつ癌性細胞をアポトーシスへ押しやるために、発現レベルのバランスを取る投薬及び/又は治療計画を利用することのいずれかによって達成できる。
【0080】
あるいは、siRNAの局所送達が利用されてよい。siRNAが癌細胞又は腫瘍へ局所的に送達された場合、発現は好ましくノックアウトされる。発現をノックアウトすることにより、ヒプシン化可能な因子5Aは周囲に存在しなくなり、従って細胞を生存状態にするヒプシン化eIF−5Aは存在しなくなる。siRNAは癌又は腫瘍へ局所的に送達されることから、通常の細胞生育に利用可能なeIF−5Aを有する必要はない。
【0081】
特定の実施形態によれば、内在性遺伝子はeIF5A1である。内在性eIF−5A1の発現を抑制するため、被験体へeIF5A1を標的とするsiRNAが投与される。特定の実施形態によれば、siRNAは配列番号1又は配列番号2を含んでなるか、又は内在性eIF−5A1の発現を抑制し得る、eIF5A1を標的とするいずれのsiRNAである。特定の実施形態によればeIF5A1はヒトeIF−5A1(図1に示す)であり、被験体はヒトである。ヒトeIF−5A1を標的とするその他のsiRNAは公知であり、同時係属出願の11/134,445号;11/287,460号;11/184,982号;11/293,391号;11/725,520号;11/725,470号;11/637,835号に開示される。別の実施形態によれば被験体は哺乳類であり、eIF5A1は哺乳類に対して特異的である。例えば被験体はイヌであり、eIF5A1はイヌeIF5A1である。特定の実施形態によれば、siRNAは図25に示されるsiRNAコンストラクトから本質的になる。例えば、siRNAはeIF5A1を標的とする核酸を含むが、U又はT核酸のような突出もまた含むか、又はhisタグ(HAタグとも呼ばれ、しばしばin vitroでの研究に用いられる)のようなタグもまた含む。siRNAコンストラクトが標的遺伝子の発現を減少可能である限り、5’又は3’末端のいずれか(又は例えば図25に示される核酸の連続鎖の内側であっても)に付着する分子又はさらなる核酸が含まれてよく、「本質的になる」の範囲内となる。好ましくはsiRNAは、外来性ポリヌクレオチドの発現に影響しないようなeIF5A1遺伝子の領域を標的とする。例えば、eIF5A1 siRNAは3’UTR又は3’末端を標的とする。図25に示すsiRNAは、典型的なeIF5A1 siRNAである。
【0082】
ポリヌクレオチドはeIF5A1をコードし、ここで該ポリヌクレオチドはeIF5A1変異形をコードするように変異導入される。変異eIF5A1は、変異形eIF5A1が翻訳後修飾され得ないように(ヒプシン化され得ないように)設計される。典型的な変異体については上に考察される。
【0083】
固形癌に関与する癌の場合、siRNAを直接腫瘍へ送達することが望ましい可能性がある。siRNAは、時間、同様に送達部位に関してポリヌクレオチドとは別に投与されてよいか、又は、同時及び/若しくは同じ送達部位において共に投与されてよい。当業者はsiRNAの投与のタイミングについて、内在性タンパク質が翻訳される時に投与され、それが既に行われた後ではないことが必要な可能性があることを理解するであろう。
【0084】
本発明者らは以前にeIF5A1が正常組織に対して無毒であることを示したが(その全体が参照により本明細書に取り込まれる、2005年11月28日に提出した係属出願11/293,391号を参照)、送達複合体が(eIF5Aポリヌクレオチド/プラスミド/発現ベクターの直接投与に比較して)好ましい可能性がある。好ましい送達系は有効量のeIF5A1を被験体、腫瘍、又は癌細胞群へ提供し、同様に腫瘍又は癌細胞群への標的送達を好ましく提供する。従って特定の実施形態によれば、eIF5A1ヌクレオチド/プラスミド/発現ベクターを、リポソーム、デンドリマー、又は類似の無毒性ナノ粒子、例えばポリエチレンイミンポリマー(例えばin vivo JetPEI(商標)複合体)のようなナノメートルサイズのビヒクルを通して送達することが好ましい。
【0085】
eIF5A1タンパク質もまた、腫瘍部位へ直接送達されてよい。当業者はeIF5A1タンパク質送達の用量及び治療計画期間を決定できるであろう。
【0086】
eIF5A1によるアポトーシス誘導の分子的基盤について以下に考察する。
【0087】
デスレセプターのシグナル伝達
Ad−eIF5A1(野生型eIF5A1を有するアデノウィルス)又はAd−eIF5A1(K50A)による癌細胞の処理により、デスレセプター−リガンド結合により開始されるカスパーゼ8、及び実行カスパーゼであるカスパーゼ3の活性化が誘導される。これらはeIF5A1の間接的な効果であると考えられ、カスパーゼ8及びカスパーゼ3はヒプシン化され得ないeIF5A1(K50A)による処理後にも活性化されるという事実から、この効果はリジン50eIF5A1に起因し得ることが示される。Ad−eIF5A1による処理はまた、TNFR1の上昇によって以前に示されたように、デスレセプターの上昇ももたらすと思われる。
【0088】
ミトコンドリア経路
アポトーシスへと向かうミトコンドリア経路におけるリジン50eIF5A1の直接又は間接の関与は、eIF5A1又はeIF5A1(K50A)のいずれかによる癌細胞の処理によってカスパーゼ9が活性化されるという知見を含む幾つかの観察によって支持される。同様に、ミトコンドリアのアポトーシス経路の活性化に役割を果たすp53もeIF5A1により制御されると思われる。例えば、癌細胞のアクチノマイシンDによる処理はp53の発現を上昇させ、このp53の上昇はeIF5A1 siRNAによって阻害される。これと一致して、Ad−eIF5A1による癌細胞の処理はp53mRNAを上昇させる。癌細胞のeIF5A1による処理はまた、Baxのサイトゾルからミトコンドリアへの移動も誘導し、これによってミトコンドリア膜電位の減少及びミトコンドリア内腔からサイトゾルへのチトクロムCの放出が保証される。加えて、この処理によって切断型Bcl2、Bim、及びスプライス型Bimの上昇がもたらされ、これらの全てはアポトーシス促進性である。
【0089】
MAPKシグナル伝達
加えて本発明者らは、アポトーシスに関係するMAPKシグナル伝達におけるeIF5A1の関与についての証明を得た。例えばAd−eIF5A1による癌細胞の処理によって、抗アポトーシス性のBcl2を阻害するP−JNKが上昇した。加えて、Ad−eIF5A1及びAd−eIF5A1(K50A)の両方は、TNFR1及びTNF;FAS及びFASL;カスパーゼ8;Bid;チトクロムC、ならびにカスパーゼ3を含む様々なアポトーシス促進因子に影響することによってアポトーシスを開始できる、P−p38の形成を誘導する。
【0090】
NF−κBシグナル伝達
NF−κBシグナル伝達が骨髄腫の生育を支持することの証明が存在する。例えば骨髄ストローマ細胞への骨髄腫細胞の接着により、多発性骨髄腫における生育及び抗アポトーシス因子の両方であるIL−6の、NF−κB依存性の転写上昇が誘導される[Chauhan et al. (1996) Blood 87, 1104]。加えて、骨髄腫細胞により分泌されるTNF−αは骨髄ストローマ細胞内でNF−κBを活性化し、これによってIL−6の転写及び分泌が上昇する。TNF−αはまた骨髄腫細胞内でもNF−κBを活性化し、これにより骨髄腫細胞及び骨髄ストローマ細胞の両方に細胞内接着分子−1(intracellular adhesion molecule-1:ICAM−1;CD45)及び血管細胞接着分子(vascular cell adhesion molecule:VCAM−1;CD106)の上昇がもたらされる[Hideshima et al. (2001) Oncogene 20,4519]。これは次に骨髄腫細胞と骨髄ストローマ細胞との結合を増強する[Hideshima et al. (2001) Oncogene 20,4519]。反対に、これらの効果はTNF−αにより誘導されたNF−κBの活性化を遮断することによって阻害される[Hideshima et al. (2001) Oncogene 20,4519]。実際にNF−κBは、骨髄腫細胞においてTNF−αにより誘導されたアポトーシスに対する防御を仲介するように思われる[Hideshima et al. (2002) JBC 277, 16639]。これら及びその他の観察により、NF−κBシグナル伝達が多発性骨髄腫の治療法の魅力的な標的であり得るという見解が促進された。
【0091】
本発明者らは、eIF5A1 siRNAが、ヒト骨髄腫細胞におけるNF−κBの活性化及びICAM−1形成の両方を阻害することを示した。これらの観察により、eIF5A1がNF−κBの活性化における役割を果たすことが示され、かつNF−κBの活性化に続く効果が事実上生存促進性である限りにおいて、本発明者らはこの活性化がヒプシン化eIF5A1によって直接的又は間接的に仲介されると予想する。
【0092】
IL−1
炎症誘発性サイトカインであるIL−1の骨髄腫細胞による過剰産生は、骨組織の劣化を導く多発性骨髄腫の特徴的な性質である。eIF5A1 siRNAは、マウスへのLPS投与により誘導されるIL−1の過剰産生を劇的に減少させることが示されている。
【0093】
本発明の一実施形態は、多発性骨髄腫を治療する方法を提供する。多発性骨髄腫(「MM」)は、骨髄内の悪性形質細胞の増殖及び溶骨性の病変部の存在によって特徴付けられる、進行性かつ致死性の疾病である。多発性骨髄腫は難病であるが、治療可能な形質細胞の癌である。形質細胞は免疫系の重要な一部であり、感染及び疾病との戦いを援助する免疫グロブリン(抗体)を産生する。多発性骨髄腫は、骨髄内の過剰数の異常な形質細胞、及び完全なままのモノクローナル免疫グロブリン(IgG、IgA、IgD、又はIgE;「M−タンパク質」)又はベンス・ジョーンズタンパク質(遊離のモノクローナル軽鎖)の過剰産生により特徴付けられる。低カルシウム血症、貧血、腎障害、細菌感染への感受性の増大、及び正常免疫グロブリンの産生障害は、多発性骨髄腫の一般的な臨床症状である。多発性骨髄腫はしばしば、通常は骨盤、脊椎、肋骨、及び頭蓋内の広範性骨粗鬆症によっても特徴付けられる。
【0094】
本発明は、多発性骨髄腫において見出される刺激のフィードバックループのために、多発性骨髄腫の治療によく適していると思われる。例えば、多発性骨髄腫は骨髄内で低濃度のIl−1を産生する。Il−1は次にストローマ細胞を刺激してIL−6を産生させ、これが次に多発性骨髄腫の生育を刺激する。本発明者らは以前に、eIF−5A1に向けられたsiRNAが、Il−1;TNF−α、及びIl−8のような炎症誘発性サイトカインの発現を阻害できることを示した(係属出願の11/725,539号及び11/184,982号を参照)。従って、siRNAはヒプシン化のために利用可能なものが少なくなるようにeIF−5Aの発現をノックダウンするのみではなく、Il−1/Il−6のフィードバックループもまた、カットオフ又は減少させ得る。
【0095】
ヒトeIF5Aを標的とするsiRNAを、腫瘍内の内在性ヒプシン化eIF5Aのレベルを抑制するために使用し、一方でヒプシン化不可能なeIF5Aの変異体(eIF5AK50R)を発現するRNAi耐性プラスミドを、in vivoでの非修飾eIF5Aのレベルを上昇させるために使用した。eIF5A siRNAを含むPEIナノ複合体の腫瘍内注射によって、MM腫瘍の生育がコントロールsiRNAを含む複合体に対して80%(***p=0.0003)を超えるまで阻害され、ヒプシン化eIF5Aのレベルの抑制が抗腫瘍効果を有することが示された。eIF5AK50R発現プラスミドを含むPEI複合体は同様の効果を有し、腫瘍の生育はコントロールプラスミドを含む複合体に対して70%(**p0.001)を超えるまで阻害された。従ってMM腫瘍の生育は、生育促進性のヒプシン化eIF5Aの抑制によって、又はアポトーシス促進性である非ヒプシン化型eIF5Aのレベル上昇のいずれかによって阻害できる。eIF5A siRNA及びRNAi耐性eIF5AK50Rプラスミドの両方を含む複合体の腫瘍内送達は、腫瘍の生育に対して相乗的な効果を有し、腫瘍の生育を94%(***p=0.0002)まで阻害する著しい腫瘍の収縮をもたらした。eIF5A siRNA/eIF5AK50RPEI複合体の静脈内送達もまた、腫瘍の生育を95%(**p=0.002)まで効率的に減少させ、治療薬の全身性送達が実行可能であることが示された。
【0096】
eIF5A siRNA/eIF5AK50RpDNA PEI複合体の局所及び全身性送達の両方が、多発性骨髄腫において著しい抗腫瘍反応をもたらした。
【0097】
本発明はさらに、多発性骨髄腫を含む癌の治療に有用な組成物を提供する。好ましい実施形態によれば組成物は、ヒプシン化され得ない点変異eIF5A1をコードするプラスミドDNAと、内在性ヒトeIF5A1を選択的に抑制するが、該プラスミドによってコードされる点変異eIF5A1には影響しないeIF5A1 siRNAとの複合体である。eIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドについては上で考察されている。プラスミドDNA及びsiRNAの両方は、PEI(ポリエチレンイミン)ナノ粒子と好ましく複合される。これらは別々に複合されて別々若しくは共に投与されてよいか、又はこれらは共に複合されてよい。DNA及びRNAはPEI上の陽性に荷電したアミノ基に結合し、ナノ粒子が細胞内に取り込まれる際に放出される。PEI−核酸複合体は、分裂及び非分裂細胞の両方へ効果的に取り込まれることが示されている。
【0098】
プラスミドDNAはeIF5A1(K50R)を好ましくコードし、これはeIF5A1(K50A)のようにヒプシン化され得ないことから強力にアポトーシス原性である。eIF5A1(K50R)の発現は、B細胞特異的プロモーターによって好ましく調節される。
【0099】
eIF5A1 siRNAは、好ましくは内在性ヒトeIF5A1の3’末端に特異的であり、トランスeIF5A1(K50R)の発現に対する効果をもたない。典型的な好ましいeIF5A1 siRNAは、図25に示されるsiRNAを含んでなるか、該siRNAから本質的になるか、又は該siRNAからなる。eIF5A1 siRNAを含むことの理論的根拠は:(1)ほとんど全てがヒプシン化されていることから、生存促進型である内在性eIF5A1を除去するため;(2)NF−κBの活性化を阻害して、IL−6の産生及び細胞内接着分子の形成を減少させるため;及び(3)IL−1の形成を阻害するためである。このeIF5A1 siRNAはeIF5A1(K50R)と相乗的に作用して、骨髄腫細胞にアポトーシスを誘導する。上記の(2)及び(3)が生存促進性の現象である限りにおいて、これらはヒプシン化eIF5A1により仲介されると思われることから、ヒプシン化され得ないeIF5A1(K50R)によっては影響されない。このアプローチにより、多発性骨髄腫細胞を含む癌細胞にアポトーシスを導くが健常細胞にはほとんど効果を示さない、非ヒプシン化eIF5A1のより大きなプールがもたらされる。
【0100】
本明細書での好ましい組成物はSNS01と称される。SNS01は、安全性増大のためにB細胞起源(骨髄腫細胞を含む)の細胞に発現を限定するプロモーターにより駆動されるeIF5A1K50Rをコードする、RNAi耐性プラスミドDNA、及びヌクレアーゼ耐性増大のためにdTdT3’突出を有するヒトeIF5Aを標的とするsiRNAの両方を含む複合体であり、このsiRNA及びプラスミドはin vivo JetPEI(商標)と複合される。
【実施例】
【0101】
[実施例1]
eIF5A1の野生型及び変異形によるHeLaS3細胞へのトランスフェクション
HeLaS3細胞に、野生型eIF5A1(WT)、eIF5A1K50R(K50R)、eIF5A1K67R(K67R)、eIF5A1K67A(K67A)、eIF5A1K47R/K50R(K4750R)、eIF5A1K50R/K67R(K5067R)、又はeIF5A1K50A/K67A(K5067A)を含むHA−タグ化eIF5A1変異形を発現するプラスミドを、Lipofectamine 2000を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後24及び48時間目(A)又は28及び52時間目(B)に、細胞ライセートを回収し、SDS−PAGEによって分画した。トランスフェクトしたeIF5A1の発現レベルを、HAに対する抗体を用いて検出した。結果:eIF5A1の、推定されるユビキチン化部位のリジンにおける変異(K67R)は、野生型(A)を超えてeIF5A1トランス遺伝子の蓄積を増大させた。eIF5A1の、ヒプシン化に必要とされるリジンにおける変異(K50R)もまた、野生型eIF5A1(B)を超えてeIF5A1トランス遺伝子の蓄積を増大させた。eIF5A1(K50A/K67A)の二重変異体型は、非変異の野生型eIF5A1トランス遺伝子(A+B)に比較して特に良好に発現された。図2を参照されたい。
【0102】
[実施例2]
eIF5A1の野生型及び変異形によるKAS細胞へのトランスフェクション
KAS細胞に、野生型eIF5A1(5A1)、eIF5A1K67A(K67A)、eIF5A1K50A/K67A(K50A K67A)、eIF5A1K50R(K50R)、eIF5A1K47R(K47R)、eIF5A1K67R(K67R)、eIF5A1K47R/K50R(K47R K50R)、又はeIF5A1K50R/K67R(K50R K67R)を含むHA−タグ化eIF5A1変異形を発現するプラスミドを、PAMAMデンドリマー(FMD44)を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後48時間目に細胞ライセートを回収し、SDS−PAGEによって分画した。トランスフェクトしたeIF5A1の発現レベルを、HAに対する抗体を用いて検出した。アクチンに対する抗体を用いて等しい負荷を確認した。結果:eIF5A1の、推定されるユビキチン化部位のリジンにおける変異(K67A又はK67R)は、野生型を超えてeIF5A1トランス遺伝子の蓄積を増大させた。eIF5A1の、ヒプシン化に必要とされるリジンにおける変異(K50R)、又はアセチル化部位における変異(K47R)もまた、野生型eIF5Aを超えてeIF5A1トランス遺伝子の蓄積を増大させた。eIF5A1(K50A/K67A)の二重変異体型もまた、非変異の野生型eIF5A1トランス遺伝子に比較して高いレベルで発現された。図3を参照されたい。
【0103】
[実施例3]
PAMAMデンドリマーを用いたKAS細胞へのトランスフェクション
KAS細胞に、eIF5A1K50R(K50R)、eIF5A1K50A/K67A(K50A/K67A)、又はeIF5A1K50R/K67R(K50R K67R)を含むHA−タグ化eIF5A1変異形を発現するプラスミドを、PAMAMデンドリマー(FMD45−2)を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後72時間目に細胞をアネキシン/PIで染色し、FACSにより解析した。アネキシンVによって陽性に染まり、PI(ヨウ化プロピジウム)によって陰性であった細胞はアポトーシスの初期段階(Ann+/PI−)にあるとみなされ、アネキシンV及びPIの両方によって陽性に染まった細胞はアポトーシスの後期段階(Ann+/PI+)にあるとみなされた。結果:eIF5A1の、ヒプシン化部位のリジンにおける変異(K50R)又は推定されるユビキチン化部位のリジンにおける変異(K67R)、同様に二重変異体(K50A/K67A)は、LacZコントロールのレベルを著しく超えてKAS細胞のアポトーシスをもたらした。図4を参照されたい。
【0104】
[実施例4]
eIF−5A1及びeIF−5A1の変異形を発現するプラスミドによるKAS細胞へのトランスフェクション
KAS細胞に、eIF5A1K50A(K50A)、eIF5A1K50R(K50R)、eIF5A1K67R(K67R)、eIF5A1K50A/K67A(K50A/K67A)、又はeIF5A1K50R/K67R(K50R K67R)を含むHA−タグ化eIF5A1変異形を発現するプラスミドを、Lipofectamine 2000を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後72時間目に細胞をアネキシン/PIで染色し、FACSにより解析した。アネキシンVによって陽性に染まり、PI(ヨウ化プロピジウム)によって陰性であった細胞はアポトーシスの初期段階(Ann+/PI−)にあるとみなされ、アネキシンV及びPIの両方によって陽性に染まった細胞はアポトーシスの後期段階(Ann+/PI+)にあるとみなされた。結果:eIF5A1の、ヒプシン化部位のリジンにおける変異(K50R)又は推定されるユビキチン化部位のリジンにおける変異(K67R)、同様に二重変異体(K50A/K67A)は、LacZコントロールのレベルを著しく超えてKAS細胞のアポトーシスをもたらした。図5を参照されたい。
【0105】
[実施例5]
KAS細胞にアポトーシスをもたらす処理のための、変異eIF−5A1の使用
KAS細胞に、HA−タグ化eIF5A1変異形であるeIF5A1K50R(K50R)又はeIF5A1K50A/K67A(K50A/K67A)を発現するプラスミドを、Lipofectamine 2000を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後72時間目に細胞をアネキシン/PIで染色し、FACSにより解析した。アネキシンVによって陽性に染まり、PI(ヨウ化プロピジウム)によって陰性であった細胞はアポトーシスの初期段階(Ann+/PI−)にあるとみなされ、アネキシンV及びPIの両方によって陽性に染まった細胞はアポトーシスの後期段階(Ann+/PI+)にあるとみなされた。結果:eIF5A1のヒプシン化部位のリジンにおける変異(K50R)、又はeIF5A1のヒプシン化部位及び推定されるユビキチン化部位の両方における変異(K50A/K67A)は、LacZコントロールのレベルを著しく超えてKAS細胞のアポトーシスをもたらした。図6を参照されたい。
【0106】
[実施例6A]
siRNA/アデノウィルスにより仲介される多発性骨髄腫細胞の死滅
KAS(ヒト多発性骨髄腫)細胞は、S10培地[4ng/ml IL−6、10%ウシ胎仔血清(FBS)、及びペニシリン/ストレプトマイシン(P/S)を含むRPMI 1640]中で維持した。Lipofectamine 2000(Invitrogen)を用いて、KAS細胞に58.7ピコモルのsiRNAをトランスフェクトした。モックトランスフェクト細胞は、siRNA非存在下でLipofectamine 2000により処理した。トランスフェクションは抗生物質を含まないS10培地中で行った。
a)ヒトeIF5A1を標的とするsiRNA:
eIF5A1 siRNA標的#1(siRNAはヒトeIF5A1のこの領域を標的とする:5’−AAGCTGGACTCCTCCTACACA−3’(配列番号:_))。siRNA配列は図25に示され、これは本明細書ではしばしばh5A1と称される。
eIF5A1 siRNA標的#2(このsiRNAはヒトeIF5A1のこの領域を標的とする:5’−AAAGGAATGACTTCCAGCTGA−3’(配列番号:_))。(siRNA配列は、本明細書ではしばしばash5A1−ALTと称される)
b)コントロールsiRNA:コントロールsiRNAは次の配列を有した:センス鎖、5’−ACACAUCCUCCUCAGGUCGdTdT−3’;及びアンチセンス鎖、3’−dTdTUGUGUAGGAGGAGUCCAGC−5’。
【0107】
使用されたその他のコントロールには、Dharmaconから入手した標的をもたないことが確認された非標的指向化siRNAが含まれるが、その理由は、これらは望まれないオフターゲット効果を制限するためにマイクロアレイによって試験されていることである。例えば、NFκBのin vitro試験のために用いられたコントロールはDharmaconの非標的指向化siRNA(配列D−001700−01)であり、in vivo研究のために用いられたコントロールはDharmaconのものであった(配列D−001810−01)。
【0108】
トランスフェクション後4時間目に細胞をペレットにして、1mlのS10培地に再懸濁した。最初のsiRNAトランスフェクション後72時間目に、トランスフェクトされたKAS細胞を計数し、24ウェルプレート内に300,000細胞/ウェルにてまき、同じsiRNAによる2度目のトランスフェクションを行った。
【0109】
トランスフェクション後4時間目に細胞をペレットにして、3000ifuのAd−LacZ(−ガラクトシダーゼを発現するアデノウィルス)又はAd−5A1M(ヒトeIF5A1K50Aを発現するアデノウィルス)のいずれかを含む1mlのS10培地(IL−6を含まない)に再懸濁した。
【0110】
72時間後に細胞を回収し、アネキシンV−FITC及びPI(BD Bioscience)を用いる染色とそれに続くFACS解析により、アポトーシスについて分析した。
a)初期のアポトーシスは、アネキシン−FITCによって陽性に染まり、PI染色によって陰性であった細胞(Ann+/PI−)として定義した。
b)総アポトーシスは、初期アポトーシス(Ann+/PI−)又は後期アポトーシス/ネクローシス(Ann+/PI+)のいずれかである細胞の総数として定義した。
【0111】
#1を標的とする5A1 siRNAは、ヒトeIF5A1の3’UTRを標的とすることから、アデノウィルス由来のeIF5A1の発現には影響し得ない。#2を標的とする5A1 siRNAは、ヒトeIF5A1の翻訳領域内を標的とすることから、アデノウィルス由来のeIF5A1の発現に潜在的に干渉し得る。
【0112】
結果:siRNAによって処理し、かつeIF5A1 K50A変異形を発現するアデノウィルスを感染させた細胞は、無処理の細胞及びsiRNAのみによって処理された細胞よりも、多数がアポトーシスを起こす。図7を参照されたい。
【0113】
[実施例6B]
eIF5A1標的#1(図25に示す)に対するeIF5A1 siRNAを用いる前処理により、内在性eIF5A1の発現は減少したが、アデノウィルスによって発現されるRNAi耐性eIF5A1k50Aは蓄積される。
KAS細胞に、コントロールsiRNA(C)又はeIF5A1を標的とする2つのsiRNA(#1及び#2)のうちの1つのいずれかを、Lipofectamine 2000を用いてトランスフェクトした。eIF5A1 siRNA#1は、eIF5A1の3’UTRを標的とすることから、eIF5A1の翻訳領域のみを含むアデノウィルス由来のeIF5A1の発現には干渉しない。siRNAの配列を図25に示す。eIF5A1#2 siRNAはeIF5A1の翻訳領域を標的とすることから、内在性及び外来性に発現されるeIF5A1の両方の発現に影響し得る。最初のトランスフェクション時間から72時間目に、細胞へ同じsiRNAによる2度目のトランスフェクションを行った。4時間後にトランスフェクション複合体を細胞から除き、Ad−LacZ(L)又はAd−eIF5A1K50A(5M)のいずれかを含む増殖培地(−)IL6と置き換えた。72時間後に細胞ライセートを回収し、eIF5A及びアクチンに対する抗体を用いるウェスタンブロットによって分析した。図7Bを参照されたい。ウィルスによって発現されたeIF5A1の蓄積が観察でき(レーン1対レーン2)、#1及び#2を標的とするeIF5A1 siRNAによるeIF5A発現の減少が明瞭に観察できる(レーン5及び7対レーン3)。予想されたように、eIF5A1 siRNA#1はウィルスによって発現されたeIF5A1K50Aの蓄積に影響しないが(レーン6対レーン4)、eIF5A1 siRNA#2はウィルスによって発現されたトランス遺伝子の発現に中程度にのみ影響する(レーン8対レーン4)。
【0114】
[実施例6C]
標的#1に対するeIF5A1 siRNAを用いるアデノウィルス感染前の前処理により、ヒト多発性骨髄腫細胞におけるリン酸化NFκBの発現は減少する。
KAS細胞に、コントロールsiRNA(hC)又はeIF5A1を標的とするsiRNA(#1)のいずれかを、Lipofectamine 2000を用いてトランスフェクトした。eIF5A1 siRNA#1は、eIF5A1の3’UTRを標的とすることから、eIF5A1の翻訳領域のみを含むアデノウィルス由来のeIF5A1の発現には干渉し得ない。最初のトランスフェクション時間から72時間目に、細胞へ同じsiRNAによる2度目のトランスフェクションを行った。4時間後にトランスフェクション複合体を細胞から除き、Ad−LacZ(L)又はAd−eIF5A1K50A(5M)のいずれかを含む増殖培地(+)IL6と置き換えた。24時間後に細胞ライセートを回収し、ホスホNF−κB p65(Ser536)及びeIF5Aに対する抗体を用いるウェスタンブロットによって分析した。予想されたように、eIF5A1 siRNA#1はウィルスによって発現されたeIF5A1K50Aの蓄積に影響しない。NF−κB p65のSer536におけるリン酸化は、活性化、核局在化、タンパク質−タンパク質相互作用、及び転写活性を調節する。図7Cを参照されたい。
【0115】
[実施例6D]
eIF5A1 siRNA#1を用いるアデノウィルス感染前の前処理により、ヒト多発性骨髄腫細胞におけるリン酸化NF−kB及びICAM−1の発現は減少する。
KAS細胞に、コントロールsiRNA(C)又はeIF5A1を標的とする2つのsiRNA(#1及び#2)のうちの1つのいずれかを、Lipofectamine 2000を用いてトランスフェクトした。最初のトランスフェクション時間から72時間目に、細胞へ同じsiRNAによる2度目のトランスフェクションを行った。4時間後にトランスフェクション複合体を細胞から除き、増殖培地(+)IL6と置き換えた。2度目のトランスフェクションから24時間後、細胞を40ng/mlのTNF−αで刺激して0、4、又は24時間目に細胞ライセートを回収し、ホスホNF−kB p65(Ser536)、ICAM−1、及びアクチンに対する抗体を用いるウェスタンブロットによって分析した。両方のeIF5A1特異的siRNAのトランスフェクション後、TNF−αにより誘導されたNF−kB p65のリン酸化(Ser536)及びICAM−1の発現の減少が観察された。NF−kB p65のSer536におけるリン酸化は、活性化、核局在化、タンパク質−タンパク質相互作用、及び転写活性を調節する。ICAM−1は、多発性骨髄腫の病因に関与すると信じられている細胞間接着表面糖タンパク質である。図7Dを参照されたい。
【0116】
[実施例6E]
siRNAを用いるKAS細胞の前処理により、IL−6存在下でのeIF5A1k50R遺伝子送達によるアポトーシスは増大する。
KAS細胞に、コントロールsiRNA(hcon)又はヒトeIF5A1 siRNA(h5A1)のいずれかをLipofectamine 2000を用いてトランスフェクトした。72時間後、細胞にsiRNAを再トランスフェクトした。siRNAトランスフェクション培地の除去後4時間目に、空のベクター(mcs)又はeIF5A1k50R(K50R)プラスミドのPEI複合体を細胞に加えた。この研究の間を通して使用した増殖培地にはIL−6が含まれていた。72時間後に、アネキシン/PIを用いる細胞の染色及びFACS解析によってアポトーシスを測定した。図7Fを参照されたい。
【0117】
[実施例7]
eIF−5A1プラスミド及びeIF−5A1 siRNAの同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延する(図8〜10)。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。3〜5週齢のSCID/CB17マウス6匹の右側腹部に、200μLのPBS中のKAS−6/1骨髄腫細胞107個を注射して、腫瘍の最小サイズが4mm3に達した時点で処置を開始した。
【0118】
コントロールマウスには、pCpG−mcs(空のベクター)及びコントロールsiRNAを含むPEI複合体を週2回腫瘍内注射した(コントロール群は3マウス:C−1、C−2、及びC−3によって構成された)。処置マウスには、RNAi耐性プラスミドpCpG−eIF5A1k50R及びeIF5A1 siRNAを含むPEI複合体を週2回腫瘍内注射した(処置群は3マウス:5A−1、5A−2、及び5A−3によって構成された)。注射は逆流を防ぐために腫瘍内の複数個所に行い、取り込みを増大させるために緩速で注射した。図8のデータに、群内のマウス全ての腫瘍容積を示す。図9に示されるデータは、群あたりの平均腫瘍容積+/−標準誤差である。アステリスクは統計的有意性を示す(*=p<0.025;n=3)。
【0119】
図10に、eIF−5A1プラスミド及びeIF−5A1 siRNAの同時投与によって多発性骨髄腫皮下腫瘍の重量が減少することを示す。示されるデータは、群あたりの平均腫瘍重量+/−標準誤差である。アステリスクは統計的有意性を示す(*=p<0.05;n=3)。
【0120】
in vivo試験には2x0.1mlのJET-PEI(PoIyPlus)を使用した。N/P比は8であった。PEI/DNA/siRNA複合体を、5%グルコース中において全量0.1mlで形成した。複合体形成のプロトコルは以下の通りである。
1.成分を室温に置く。無菌状態を保つ。
2.20μgのプラスミドDNA(2mg/mlを〜10μl)及び10μgのsiRNA(1mg/mlを10μl)を全量25μlに希釈する。差分を補うために滅菌水を使用する。
3.25μlの10%グルコースを加え、DNA溶液の量を50μlの5%グルコースに調整する。穏やかにボルテックスした後、短時間遠心する。
4.全量25μlの10%グルコース中へ4.8μlのin vivo JETPEIを希釈する。最終濃度が5%グルコースになるように、滅菌水で量を50μlに調整する。穏やかにボルテックスした後、短時間遠心する。
5.50μlの希釈PEIを50μlの希釈DNAへ直ちに加える(順序を逆にしないこと)。穏やかにボルテックスした後、短時間遠心沈殿する。
6.注射前に15分間インキュベートする。複合体は6時間安定である。
【0121】
CpGフリーのクローニングベクター及びpCpGプラスミドはInvivoGenから得た。これらのプラスミドはpCpGと命名され、CpGジヌクレオチドを完全に欠いている。これらのプラスミドはin vitro及びin vivoの両方で高レベルのトランス遺伝子発現をもたらし、かつCMVに基づくプラスミドとは対照的に、in vivoにおける持続発現を可能にする。pCpGプラスミドは、全てのCpGを除くように改変された、必然的にCpGジヌクレオチドを欠如するエレメント、又は選択性マーカー若しくはレポーターをコードする遺伝子のような、完全に合成されたエレメントのいずれかを含む。遺伝コードを形成する16のジヌクレオチドのうち、CGは唯一必須ではなく置換可能なジヌクレオチドであるという事実によってこれらの新しい対立遺伝子の合成が可能となった。8のコドンが、異なる5アミノ酸をコードするCGを含む。野生型と同一のままであることから、それらの野生型の対応物と同じように活性があるアミノ酸配列を有するタンパク質をコードする新しい対立遺伝子を作り出すため、8コドンの全ては、同じアミノ酸をコードする少なくとも2コドンの選択によって置換できる。これらの新しい対立遺伝子は、それらを容易に切り出すことのできるpMODというプラスミドから個別に入手できる。
【0122】
pCpGプラスミドはin vivoにおける持続性の発現を可能にし、かつTLR9を発現する細胞株を用いたin vivo及びin vitroでの遺伝子発現におけるCpGの効果、同様に自然及び獲得免疫系におけるそれらの効果を研究するために貴重な手段の典型である。空のベクターであるpCpG−mcs(Invivogen)は、発現遺伝子産物を含まないマルチクローニング部位のみのベクターであり、コントロールベクターとして用いた。処置用ベクターpCpG−eIF5A1(K50R)を作るため、LacZ遺伝子を除去したpCpG−LacZベクター(Invivogen)のNcoI及びNheI部位へ、HA−タグ化eIF5A1k50RcDNAをサブクローン化した。DNAはEndo-Free Qiagenキットを用いて調製した。測定したエンドトキシン濃度は、水中で2mg/mlであるはずのDNAについて、<0.03EU/μgDNAであった。
【0123】
実験に用いたコントロールsiRNAは、Dharmacon(D-001810-01)から入手した、マイクロアレイによって確認済みの非標的指向化コントロールsiRNAであった。血清中での分解を防ぐため、改変されたsiRNA(siSTABLE)を得た。
【0124】
実験に用いたeIF5A1 siRNAは、ヒトeIF5A1の3’UTRに対して設計した。該eIF5A1 siRNAとマウスeIF5A1の間に類似性は無いことから、該siRNAはヒト(マウスではない)eIF5A1のみを抑制するはずである。該siRNAはまた、eIF5A2(ヒト又はマウスのいずれか)とも類似性は無い。該siRNAは、血清中での分解を防ぐために改変された(siSTABLE)。該eIF5A1 siRNAは以下の標的配列:5’GCU GGA CUC CUC CUA CAC A(UU)3を有する。
【0125】
siSTABLE siRNAは、1mg/mlにて水に溶解した(一定分量で−20℃において保管した)。
【0126】
長さ(l)及び幅(w)に関する腫瘍の寸法は、デジタルキャリパーを用いて週あたり2〜3回測定した。腫瘍容積は以下の方程式に従って算出した:
l=長さ;最小寸法
w=幅;最大寸法
腫瘍容積(mm3)=l2*w*0.5
【0127】
統計的分析
統計的分析にはスチューデントt−検定を用いた。有意性は95%(p<0.05)を超える信頼水準であるとみなされた。
【0128】
[実施例8]
eIF5A1プラスミド及びeIF5A1 siRNAの同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延し、かつ腫瘍の収縮がもたらされる。
別の研究において、再びKAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。コントロールマウスには、pCpG−mcs(空のベクター)及びコントロールsiRNAを含むPEI複合体を週2回腫瘍内注射した(コントロール群、G−1、G−2、及びG−3)。処置マウスには、RNAi耐性プラスミドpCpG−eIF5A1k50R(20μgのプラスミドDNA)及びeIF5A1 siRNA(10μgのsiRNA)を含むPEI複合体を週2回腫瘍内注射した(処置群、G−4、G−5、及びG−6)。図11に示されるデータは、群あたりの平均腫瘍容積+/−標準誤差である。アステリスクは統計的有意性を示す(*=p<0.025;n=3)。21日間にわたり、6回の注射を行った(赤の矢印)。
【0129】
[実施例9]
eIF5A1 siRNAの静脈内(i.v.)投与及びPEI/eIF5A1K50Rプラスミド複合体の腫瘍内(i.t.)投与により、多発性骨髄腫皮下腫瘍の腫瘍収縮がもたらされる(群2B)。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初にコントロールsiRNA(コントロール群)又はヒトeIF5A1 siRNA(処置群)のいずれか50μgを尾注射した。コントロールマウスは続いて、pCpG−mcs(空のベクター;コントロール群;G−1、G−2、及びG−3)を含むPEI複合体の、週2回の腫瘍内注射によって処置された。処置マウスは続いて、RNAi耐性プラスミドpCpG−eIF5A1k50R(20μgのプラスミドDNA)(処置群;G−4、G−5、及びG−6)を含むPEI複合体の、週2回の腫瘍内注射によって処置された。コントロールマウスは週1回のi.v.注射により、コントロールsiRNAを投与され続けた(コントロール群、R−1、R−2、及びR−3)。処置マウスは週1回のi.v.注射により、ヒトeIF5A1 siRNA(20μg)を投与され続けた(処置群、R−4、R−5、及びR−6)。図12に示されるデータは、各群における全てのマウスの腫瘍容積である。21日間にわたり、PEI/DNAの壁内注射を6回(赤の矢印)、及びsiRNAのi.v.注射を4回(青の矢印)行った。
【0130】
図13に、実施例8及び9に由来するオーバーレイを示す。KAS細胞をSCIDマウスに皮下注射した。図13に示されるデータは、各群におけるマウスの平均腫瘍容積+/−標準誤差である。アステリスクは処置群とコントロール群の間の統計的有意性を示す(**=p<0.01;***=p<0.001;n=3)。
【0131】
PEI複合体形成のためのプロトコル:
1.成分を室温に置く。無菌状態を保つ。
2.プラスミドDNA又はプラスミドDNA+siRNAを全量25μlに希釈する。量を調整するために滅菌水を使用する。
a)プラスミドDNAのみの複合体は:
20μgのプラスミドDNA(2mg/mlを10μl)を全量25μlに希釈する。差分を補うために滅菌水を使用する。
b)プラスミドDNA+siRNAの複合体は:
20μgのプラスミドDNA(2mg/mlを〜10μl)及び10μgのsiRNA(1mg/mlを10μl)を全量25μlに希釈する。差分を補うために滅菌水を使用する。
3.25μlの10%グルコース(PEIキットと共に提供される)を加え、DNA溶液の量を50μlの5%グルコースに調整する。穏やかにボルテックスした後、短時間遠心する。
4.全量25μlの10%グルコース中へin vivo JETPEIを希釈する。
a)プラスミドDNAのみの複合体は:
全量25μlの10%グルコース中へ3.2μlのin vivo JETPEIを希釈する。最終濃度が5%グルコースになるように、滅菌水で量を50μlに調整する。穏やかにボルテックスした後、短時間遠心する。
b)プラスミドDNA+siRNAの複合体は:
全量25μlの10%グルコース中へ4.8μlのin vivo JETPEIを希釈する。最終濃度が5%グルコースになるように、滅菌水で量を50μlに調整する。穏やかにボルテックスした後、短時間遠心する。
5.50μlの希釈PEIを50μlの希釈DNAへ直ちに加える(順序を逆にしないこと!)。穏やかにボルテックスした後、短時間遠心沈殿する。
6.注射前に15分間インキュベートする。複合体は6時間安定である。
【0132】
siRNAの尾静脈注射に関して、最初のsiRNA注射は50マイクログラムであった。siRNAはPBS中で0.4mg/mlに希釈した。マウスあたり125μl(50μg)を尾静脈内へ注射した。続く血清安定化siRNAの注射は、マウスあたり20μgで週2回行った。siRNAはPBS中で0.4mg/mlに希釈した。マウスあたり50μl(20μg)を尾静脈内へ注射した。
【0133】
図13Bに、eIF5A1プラスミド及びeIF5A1 siRNAの同時投与により腫瘍の収縮がもたらされることを示す。KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。マウスには、RNAi耐性プラスミドpCpG−eIF5A1k50R及びeIF5A1 siRNAを含むPEI複合体を週2回腫瘍内注射した(処置群;G−4、G−5、及びG−6)。21日間にわたり、6回の注射を行った。処置開始後42日目にマウスを屠殺し、腫瘍部位下の皮膚を開いて腫瘍生育の形跡について調べた。群2Aの処置マウスのいずれにも腫瘍の生育は観察されなかった。
【0134】
図13Cに、eIF5A1 siRNAの静脈内(i.v.)投与及びPEI/eIF5A1K50Rプラスミド複合体の腫瘍内(i.t.)投与により、多発性骨髄腫皮下腫瘍の腫瘍収縮がもたらされることを示す。KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初に50μgのヒトeIF5A1 siRNAを注射した(処置群)。マウスは続いて、RNAi耐性プラスミドpCpG−eIF5A1k50Rを含むPEI複合体の、週2回の腫瘍内注射によって処置された(処置群;R−4、R−5、及びR−6)。マウスは週1回のi.v.注射により、ヒトeIF5A1 siRNAを投与され続けた。処置開始後21日目に処置を終了した。処置開始後42日目にマウスを屠殺し、腫瘍部位下の皮膚を開いて腫瘍生育の形跡について調べた。処置群(群2B)の1マウスに、腫瘍生育の形跡がみられなかった。
【0135】
[実施例10]
eIF5A1プラスミド及びeIF5A1 siRNAの静脈内同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延する。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初にコントロールsiRNA(コントロール群)又はヒトeIF5A1 siRNA(処置群)のいずれか50μgを注射した。マウスは続いて、pCpG−mcs(空のベクター;コントロール群;A1、A2、及びA3)を含むPEI複合体、又はRNAi耐性プラスミドpCpG−eIF5A1k50R(処置群;A4、A5、及びA6)を含むPEI複合体のいずれかによる、週あたり〜2回の静脈内(赤い矢印)又は腹腔内(緑の矢印)注射によって処置された。マウスは週1回のi.v.注射(青い矢印)により、コントロールsiRNA(コントロール群、A1、A2、及びA3)又はヒトeIF5A1 siRNA(処置群、A4、A5、及びA6)のいずれかを投与され続けた。示されるデータは、各群における全てのマウスの腫瘍容積である。図14に示されるデータは、各群における全てのマウスの腫瘍容積である。
【0136】
[実施例11]
eIF5A1 siRNAの静脈内(i.v.)投与及びPEI/eIF5A1K50Rプラスミド複合体の静脈内(i.v.)投与又は腹腔内(i.p.)投与により、多発性骨髄腫皮下腫瘍の生育は遅延する。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初に50μgのコントロールsiRNA(コントロール群)又はヒトeIF5A1 siRNA(処置群)のいずれかを注射した。コントロールマウスは続いて、pCpG−mcs(空のベクター;コントロール群は3マウスであった;B1、B2、及びB3)を含むPEI複合体の、週あたり〜1回の静脈内又は腹腔内注射によって処置された。処置マウスは続いて、RNAi耐性プラスミドpCpG−eIF5A1K50R(処置群;B4、B5、及びB6)を含むPEI複合体の、週あたり〜1回の静脈内又は腹腔内注射によって処置された。マウスは週1回のi.v.注射により、コントロールsiRNA(コントロール群、B1、B2、及びB3)又はヒトeIF5A1 siRNA(処置群は3マウスであった;B4、B5、及びB6)のいずれかを投与され続けた。実験は50μgのsiRNAの最初の注射によって開始された(図15のグラフの2日目)。続く注射では、毎週1回20マイクログラムのsiRNAを用いた。siRNAはネイキッド、すなわち送達ビヒクルなしで投与された。PEI複合体は20μgのプラスミドDNAを含んだ。最初のPEI注射はi.p.で行われ、続く注射はi.v.で行われた。図15に示されるデータは、各群における全てのマウスの腫瘍容積である。
【0137】
図16に、実施例10及び11のオーバーレイを示す。KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。1セットのマウスは、コントロールsiRNA(コントロール;群A)又はeIF5A1 siRNA(処置;群A)のいずれかの週1回のi.v.注射、及びpCpG−mcsを含むPEI複合体(コントロール;群A)若しくはRNAi耐性プラスミドpCpG−eIF5A1K50Rを含むPEI複合体(処置;群A)のいずれかのi.v.又はi.p.のいずれかを受けた。第2セットのマウスは、pCpG−mcs(空のベクター)及びコントロールsiRNAを含むPEI複合体(コントロール;群B)又はRNAi耐性プラスミドpCpG−eIF5A1k50R及びeIF5A1 siRNAを含むPEI複合体(処置;群B)のいずれかによるi.v.又はi.p.注射を、週あたり〜2回受けた。示されるデータは、各群におけるマウスの平均腫瘍容積+/−標準誤差である。アステリスクは処置群とコントロール群の間の統計的有意性を示す(*=p<0.05;***=p<0.001;n=3)。
【0138】
PEI複合体及びsiRNAを調製するためのプロトコルは、以前の実施例に記載した通りである。
【0139】
[実施例12]
eIF5A1プラスミド及びeIF5A1 siRNAの同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延し、かつ腫瘍の収縮がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。コントロールマウスには、pCpG−mcs(空のベクター)及びコントロールsiRNAを含むPEI複合体を週2回腫瘍内注射した(コントロール群は3マウスを有した;コントロール1、コントロール2、及びコントロール3)。処置マウスには、RNAi耐性プラスミドpCpG−eIF5A1k50R及びeIF5A1 siRNAを含むPEI複合体を週2回腫瘍内注射した(処置群には4マウスが含まれた;5A−1、5A−2、5A−3、及び5A−4)。PEI複合体の腫瘍内拒絶には、20μgのプラスミドDNA及び10μgのsiRNAの両方が含まれた。図17に示されるデータは、各群における全てのマウスの腫瘍容積である。
【0140】
[実施例13]
eIF5A1 siRNAの静脈内(i.v.)投与及びPEI/eIF5A1K50Rプラスミド複合体の腫瘍内(i.t.)投与により、多発性骨髄腫皮下腫瘍の腫瘍収縮がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初にコントロールsiRNA(コントロール群は3マウスを有した;コントロール1、コントロール2、及びコントロール3)又はヒトeIF5A1 siRNA(処置群は3マウスを有した;5A−1、5A−2、5A−3)のいずれか50μgを注射した。コントロールマウスは続いて、pCpG−mcs(20μg)を含むPEI複合体の、週2回の腫瘍内注射によって処置された(コントロール群1〜3)。処置マウスは続いて、RNAi耐性プラスミドpCpG−eIF5A1K50R(20μg)を含むPEI複合体の、週2回の腫瘍内注射によって処置された(5A−1、5A−2、5A−3)。コントロールマウスは週2回の尾静脈へのi.v.注射により、コントロールsiRNA(20μg)を投与され続けた。処置マウスは週2回の尾静脈へのi.v.注射により、ヒトeIF5A1 siRNA(20μg)を投与され続けた。注射は、腫瘍内注射前の48時間に行われた。siRNAはネイキッドsiRNAとして、すなわち送達ビヒクルなしで投与された。図18に示されるデータは、各群における全てのマウスの腫瘍容積である。
【0141】
[実施例14]
EF1又はB29プロモーターのいずれかにより駆動されるeIF5A1K50Rプラスミド及びeIF5A1 siRNAの同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延し、かつ腫瘍の収縮がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。マウスには、コントロールベクター(G1及びG2)又はB29プロモーター(G3及びG4)若しくはEF1プロモーター(G5及びG6)のいずれかにより駆動されるeIF5A1プラスミドのいずれか、並びに、コントロールsiRNA(G1、G3、G5)又はh5A1 siRNA(G2、G4、G6)のいずれかを含むPEI複合体を週2回腫瘍内注射した。示されるデータは、各群の平均腫瘍容積+/−標準誤差である。注記:B29プロモーターはB細胞特異的プロモーターとして意図された。しかしながら、この研究で用いたB29プロモーター/mCMVエンハンサーは、in vitroでKAS細胞にHA−eIF5A1K50Rの高い発現を駆動することが見出されたが、B細胞特異的ではないと思われる(おそらくCMVエンハンサーのため)。図19を参照されたい。
【0142】
[実施例15]
eIF5A1 siRNAの同時投与により、EF1又はB29プロモーターのいずれかにより駆動されるeIF5A1K50Rプラスミドの多発性骨髄腫皮下腫瘍における抗腫瘍効果は増大され、かつ腫瘍の負荷量の減少がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。マウスには、コントロールベクター(G1及びG2)又はB29プロモーター(G3及びG4)若しくはEF1プロモーター(G5及びG6)のいずれかにより駆動されるeIF5A1プラスミドのいずれか、並びに、コントロールsiRNA(G1、G3、G5)又はh5A1 siRNA(G2、G4、G6)のいずれかを含むPEI複合体を週2回腫瘍内注射した。処置開始後24日目にマウスを屠殺し、皮下腫瘍を除去して秤量した。示されるデータは、全ての群の平均腫瘍重量+/−標準誤差である。図20を参照されたい。
【0143】
[実施例16]
eIF5A1 siRNAは、肺腺癌細胞におけるAd−eIF5Aの感染によりもたらされたアポトーシスの誘導を相乗的に増大させる。
A549細胞に、Ad−LacZ又はAd−eIF5Aのいずれかを感染させた。ウィルスの添加後直ちに細胞へトランスフェクション培地を添加することにより、細胞にコントロールsiRNA又はヒトeIF5A1(h5A1)を標的とするsiRNAのいずれかをトランスフェクトした。siRNAによるトランスフェクション及びウィルスによる感染後4時間目に培地を新鮮な培地に置き換え、細胞を72時間インキュベートした後に、アポトーシス性の細胞を検出するためアネキシン/PIにより標識した。注記:この細胞株におけるeIF5Aの過剰発現は、DHS及びDOHHの量が限られていることから非ヒプシン化eIF5Aの蓄積をもたらし、そのためにeIF5AK50Rの過剰発現と同じアポトーシス促進効果をもたらす。これらのデータにより、ヒプシン化eIF5Aの同時抑制及び非ヒプシン化eIF5Aの過剰発現によって引き起こされるアポトーシスにおける相乗的効果が、骨髄腫以外の腫瘍型にも同様に観察されることが示される。図21を参照されたい。
【0144】
[実施例17]
プラスミドpExp5Aの構築
pExp5Aは、eIF5A1K50Rの発現が主にヒトB細胞系列の細胞内で駆動されるように設計された、CpGジヌクレオチドの少ない発現プラスミドである。このベクターは、CpGジヌクレオチドを完全に欠いたプラスミドであるpCpG−LacZ(Invivogen)に由来する。E.Coliの複製及び選択に必要な全てのエレメントはCpGジヌクレオチドを含まない。CpG−LacZベクター由来のオリジナルのCMVエンハンサー/プロモーター及びLacZ遺伝子は、eIF5A1K50RのB細胞特異的な発現を駆動するため、それぞれヒト最小B細胞特異的プロモーター(B29/CD79b;Invivogen)及びヒトeIF5A1K50Rによって置き換えられた。B29プロモーターの活性を増大させ、非B細胞における発現を減少させるために、プラスミドのeIF5A1発現カセットの下流にB29 DHS4.4 3’エンハンサーが導入された(Malone et al. 2006. B29 gene silencing in pituitary cells is regulated by its 3' enhancer. J. Mol. Biol. 362: 173-183)。B29最小プロモーター、eIF5A1K50R、及びB29 DHS4.4 3’エンハンサーの取り込みにより、ベクター内に32のCpGジヌクレオチドが導入された。
【0145】
E.Coliにおける発現のためのエレメント
複製開始点:E.Coli R6Kガンマori
*R6Kガンマ複製開始点の存在により、pCpGプラスミドはpir変異遺伝子を発現する変異株のみにおいて増幅できる。これらは標準のE.Coli株においては複製し得ない。従ってpCpGプラスミドは、Demメチル化も欠くpir変異体であるE.Coli GT115株(Invivogen)と共に供給される。
細菌性プロモーター:細菌性EM7プロモーターのCpGを含まないバージョンであるEM2K。
選択性マーカー:Zeocin(商標)耐性遺伝子;CpGを含まない合成対立遺伝子。
【0146】
哺乳類細胞における発現のためのエレメント
哺乳類プロモーター:B細胞における組織特異的発現のための、ヒト−167bp最小B29(CD79b)プロモーター。5’UTRに合成イントロン(I140)も存在する。
ポリアデニル化シグナル:後期SV40ポリアデニル化シグナルのCpGジヌクレオチドを含まないバージョン。
3’エンハンサー:ヒトB29 DHS4.4 3’エンハンサー。
MAR:細菌性及び哺乳類性の転写ユニットの間に、CpGを含まないマトリックス結合領域(Matrix attached regions:MAR)が2つ存在する。MARの1つはヒトIFN−β遺伝子の5’領域由来であり、1つはβ−グロビン遺伝子の5’領域由来である。
【0147】
pExp5Aの予測される配列(3371bp)を図23に示す。
【0148】
eIF5A1K50Rのアミノ酸配列
*K50R変異を下線で示す。
【0149】
【化1】

【0150】
pExp5Aの構築−−構築の概略
工程1:B29 DHS4.4 3’エンハンサーのクローニング及びpGEM T easy(Promega)へのサブクローニング−−pGEM−4.4enh#8の作製。
工程2:最小B29プロモーターのpCpG−LacZ(Invivogen)へのサブクローニング−−B29−5#3の作製。
工程3:HA−eIF5A1K50RのB29−5#3ベクターへのサブクローニング−−B29−5#3−eIF5A1K50Rの作製。
工程4:pGpG−mcs(Invivogen)内の新しいマルチクローニング部位の作製−−pCpG−リンカー4の作製。
工程5:B29 DHS4.4 3’エンハンサーのpCpG−リンカー4へのサブクローニング−−pCpG−DHS4.4の作製。
工程6:B29プロモーター+HA−eIF5A1K50R+SV40 pA発現カセットのpCpG−DHS4.4へのサブクローニングによるpExp−5の作製。
工程7:pExp−5内のHA−eIF5A1K50Rを非HA eIF5A1K50Rに置き換えることによる、最終ベクターpExp5Aの作製。
【0151】
構築の詳細
工程1:B29 DHS4.4 3’エンハンサーのクローニング及びpGEM T easy(Promega)へのサブクローニング−−pGEM−4.4enh#8の作製。
B29 DHS4.4 3’エンハンサーを、以下のプライマー:順方向5’−CAGCAAGGGAGCACCTATG−3’及び逆方向5’−GTTGCAGTGAGCGGAGATG−3’を用いて、KAS細胞(ヒト多発性骨髄腫細胞株)より分離したゲノムDNAからPCRによってクローン化した。プライマーは、ヒトCD79B/GH−1遺伝子間領域(受入番号AB062674)の配列を用いて設計した。得られた608bpのPCR断片を、pGEM(登録商標)T easyクローニングベクター(Promega)にサブクローン化して配列決定した。Komatsu et al. 2002. Novel regulatory regions found downstream of the rat B29/Ig-b gene. Eur. J. Biochem. 269 : 1227-1236。
【0152】
pGEM−4.4enh#8内の、B29 DHS4.4 3’エンハンサーPCR断片(297bp)の配列
【0153】
【化2】
【0154】
○ +4.4の領域に、幾つかの転写因子結合部位が含まれる。
【0155】
【化3】
【0156】
pGEM−4.4enh#8内の、B29 DHS4.4 3’エンハンサーPCR断片(297bp)及びヒトCD79B/GH−1遺伝子間領域(受入番号AB062674)の配列のアラインメント。
【0157】
【化4】
【0158】
【化5】
【0159】
工程2:最小B29プロモーターのpCpG−LacZ(Invivogen)へのサブクローニング−−B29−5#3の作製。
最小−167ヒトB29プロモーターを、以下のプライマー:順方向5’−CCAACTAGTGCGACCGCCAAACCTTAGC−3’;逆方向5’−CAAAAGCTTGACAACGTCCGAGGCTCCTTGG−3’を用いて、完全長ヒトB29プロモーターを有する市販のプラスミド(pDrive-hB29;Invivogen)から増幅した。得られたPCR断片をSpeI及びHindIIIを用いて消化し、pCpG−LacZベクター(Invivogen)のSpeI及びHindIII部位へライゲートして、B29−5#3を作製した。
【0160】
B29−5#3内の、B29最小プロモーターPCR断片(188bp)の配列
【0161】
【化6】
【0162】
B29−5#3内のB29最小プロモーターPCR断片(B29_min)とpDrive-hB29由来の完全長ヒトB29プロモーターとのアラインメント。
【0163】
【化7】
【0164】
【化8】
【0165】
【化9】
【0166】
工程3:HA−eIF5A1K50RのB29−5#3ベクターへのサブクローニング−−pB29−eIF5A1K50R_7の作製。
HA−eIF5A1K50Rを、DNAテンプレートとしてのpHM6−eIF5A1K50R、ならびに以下のプライマー:順方向5’−CGCCATGGACATGTACCCTTACGACGTCCCAGACTACGCTGCAGATGATTTGGACTTCGAG−3’及び逆方向5’−CGCGCTAGCCAGTTATTTTGCCATCGCC−3’を用いて、PCRにより増幅した。得られたPCR断片をNcoI及びNheIを用いて消化し、B29−5#3のNcoI及びNheI部位へサブクローン化して、LacZを置き換えた。
【0167】
pB29−eIF5A1K50R_7内の、HA−eIF5A1K50RPCR断片(497bp)の配列
【0168】
【化10】
【0169】
pB29−eIF5A1K50R_7内の、HA−eIF5A1K50RPCR断片の翻訳
HAエピトープ
eIF5A1K50R
K50R
【0170】
【化11】
【0171】
pB29−eIF5A1K50R_7内の、HA−eIF5A1K50RPCR断片とヒトeIF5A1(受入#NP_001961)とのアラインメント
【0172】
【化12】
【0173】
工程4:pGpG−mcs(Invivogen)内の新しいマルチクローニング部位の作製−−pCpG−リンカー4の作製。
pGpGクローニングベクターであるpGpG−mcs G2(Invivogen)をEcoRIで消化して、mCMVエンハンサー、hEF1プロモーター、合成イントロン、マルチクローニング部位、及びSV40ポリアデニル化シグナルを含む哺乳類発現カセットを除いた。次にEcoRIで消化したpGpG−mcs G2ベクターをEcoRI粘着末端を有する合成リンカーへライゲートして、新しいマルチクローニング部位を有する、プロモーターの無いベクター(pCpG−リンカー4)を作製した。
【0174】
【化13】
【0175】
pCpG−リンカー4内の新しいマルチクローニング部位周囲の領域の配列
【0176】
【化14】
【0177】
上の配列の番号付けは、以下の特徴:βGlo MAR(ヌクレオチド1−380);EcoRI認識配列(ヌクレオチド396−401);XhoI認識配列(ヌクレオチド401−406);ClaI認識配列(ヌクレオチド409−414);NotI認識配列(ヌクレオチド417−424);MluI認識配列(ヌクレオチド427−432);IFNβ S/MAR(ヌクレオチド438−1,030)への参照を容易にするためである。
【0178】
工程5:B29 DS4.4 3’エンハンサーのpCpG−リンカー4へのサブクローニング−−pCpG−DHS4.4の作製。
B29 DHS4.4 3’エンハンサーを、テンプレートとしてのpGEM−4.4enh#8、ならびに以下のプライマー:順方向5’−GAAGCGGCCGCACCACCCTGGGCCAGGCTGG−3’;及び逆方向5’−CCACGCGTAGAGGTGTTAAAAAGTCTTTAGGTAAAG−3’を用いて、PCRにより増幅した。得られたPCR断片をNotI及びMluIを用いて消化し、pCpG−リンカー4の新しいマルチクローニング部位内のNotI及びMluI部位へライゲートして、pCpG−DHS4.4を作製した。
【0179】
>pCpG−DHS4.4完全長配列(2,282bp)
【0180】
【化15】
【0181】
【化16】
【0182】
上の配列の番号付けは、以下の特徴:βGlo MAR(ヌクレオチド1−400);EcoRI認識配列(ヌクレオチド416−421);XhoI認識配列(ヌクレオチド421−426);ClaI認識配列(ヌクレオチド429−434);NotI認識配列(ヌクレオチド437−444);DHS4.4(ヌクレオチド445−741);MluI認識配列(ヌクレオチド742−747);IFNβ S/MAR(ヌクレオチド753−1,569)への参照を容易にするためである。
【0183】
工程6:B29プロモーター+HA−eIF5A1K50R+SV40 pA発現カセットのpCpG−DHS4.4へのサブクローニングによるpExp−5の作製。
最小B29プロモーター、合成イントロン、HA−eIF5A1K50R、及びSV40pAを含むB29−eIF5A1発現カセットを、pB29−eIF5A1K50R_7(工程3)からPCRにより、以下のプライマー:順方向5’−GTTATCGATACTAGTGCGACCGCCAAACC−3’;及び逆方向5’−CAAGCGGCCGCCATACCACATTTGTAGAGGTTTTAC−3’を用いて増幅した。得られたPCR断片をClaI及びNotIを用いて消化し、pCpG−DHS4.4のマルチクローニング部位内のClaI及びNotI部位へサブクローン化して、pExp−5を作製した。
【0184】
工程7:pExp−5内のHA−eIF5A1K50Rを非HA eIF5A1K50Rに置き換えることによる、最終ベクターpExp5Aの作製。
pExp−5プラスミドをNcoI及びNheIにより消化してHA−eIF5A1K50Rを除いた。非HA−タグ化HA−eIF5A1K50RPCR断片を、pHM6−eIF5A1K50RからPCRにより、以下のプライマー:順方向5’−CACCATGGCAGATGATTTGGACTTC−3’;及び逆方向5’−CGCGCTAGCCAGTTATTTTGCCATCGCC−3’を用いて増幅した。得られたPCR産物をNcoI及びNheIを用いて消化し、B29−5#3のNcoI及びNheI部位へライゲートしてB29−K50Rを作製した。B29−K50RをNcoI及びNheIを用いて消化し、470bpのeIF5A1K50R断片をゲル精製した後、NcoI/NheIで消化したpExp−5へライゲートして最終発現ベクターのpExp5Aを作製した。
【0185】
[実施例18]
pExp5Aの試験
様々な細胞株にLipofectamine 2000を用いてプラスミドをトランスフェクトし、トランスフェクション後24時間目に抗HA抗体(Roche)を用いるウェスタンブロットによってHA−eIF5A1K50Rの発現を判定した。試験された異なる細胞株は、P3X63Ag8.653(マウスBリンパ芽球−骨髄腫)、KAS(ヒト骨髄腫)、HepG2(ヒト肝臓肝細胞癌)、T24(ヒト膀胱癌);HT−29(ヒト結腸直腸腺癌)、HEK−293(ヒト胚性腎細胞)、PC3(ヒト前立腺腺癌);HeLa(ヒト子宮頸部腺癌)、及びA549(肺癌)であった。図24を参照されたい。
【0186】
pExp−5プラスミドは、HA−eIF5A1K50Rをヒト及びマウス両方の骨髄腫細胞株に、恒常的なEF1プロモーター(CpG−eIF5A1K50R)を用いるプラスミドに匹敵するレベルで発現する。しかしながら、恒常的なプロモーターによる発現と比較して、pExp−5により駆動されるHA−eIF5A1K50Rの発現は非B細胞に限定される。一例外は、pExp−5のトランスフェクション後にHA−eIF5A1K50Rの発現が高いレベルで観察された、ヒト胚性腎細胞のHEK−293であった−これは細胞株の胚性の性質による可能性があり;現時点で本発明者らには、pExp−5が成体の腎細胞に発現するか否かは明らかでない。毒性試験及び治験に使用する最終プラスミドは、HA−eIF5A1K50Rが非HAタグ化eIF5A1K50Rによって置き換えられたpExp−5のバージョン(pExp−5A)となり得る。pExp−5は、最小ヒトB29プロモーター/エンハンサーの制御下にHA−タグ化eIF5A1K50Rを含み;HA−eIF5A1K50Rの発現は、恒常的に発現するプラスミドにより駆動される発現に対して、同様に完全長B29プロモーターを含むプラスミドに対して比較された。
【0187】
[実施例19]
in vivo JETPEI(商標)ナノ粒子の形成
この実施例は、20gのマウスへ1.5mg/kg(0.1mL)の用量−−1.5mg/kg=1.0mg pExp5A/kg+0.5mg h5A1/kg−−DNA:siRNA比=2:1により注射するための、in vivo JETPEI(商標)ナノ粒子複合体形成のために行われる。
【0188】
プラスミドDNA及びsiRNAを全量25mlに希釈する。量を調整するために滅菌水を使用する。*20mgのプラスミドDNA(2mg/mlを10ml)及び10mgのsiRNA(1mg/mlを10ml)を全量25mlに希釈する。差分を補うために滅菌水を使用する。25mlの10%グルコース(PEIキットと共に提供される)を加え、DNA溶液の量を50mlの5%グルコースに調整する。穏やかにボルテックスした後、短時間遠心する。全量25mlの水中へin vivo JETPEI(商標)を希釈する。*3.6mlのin vivo JETPEI(商標)を水で全量25mlに希釈する。最終濃度が5%グルコースになるように、10%グルコースで最終量を50mlに調整する。穏やかにボルテックスした後、短時間遠心する。50mlの希釈PEIを50mlの希釈DNAへ直ちに加える(順序を逆にしないこと!)。穏やかにボルテックスした後、短時間遠心沈殿する。
【0189】
形成後、複合体は8〜10時間安定である。複合体のN/P比は6でなければならない。N/P比は、DNA及びsiRNAの陰性に荷電したリン酸残基の数に対する、in vivo-jetPEIの陽性に荷電した窒素残基の数の比である。DNA及びsiRNAは、グラムあたり同数のリン酸基を含む。従ってN/P比は複合体内のイオン平衡の尺度である。複合体のN/P比の増大により、複合体の毒性が増大され得る。in vivo JET-PEIは150mM溶液(窒素残基で表される)として供給されるが、DNAは1mg中に3ナノモルの陰イオン性リン酸を含む。最終量中のDNAの最終濃度は0.5mg/mlを超えてはならない。DNAは高い品質で、水中で調製する必要がある。in vivo-jetPEI及び10%グルコースは、使用前に室温に置く必要がある。
【0190】
[実施例20]
マウスにおける静脈内SNS01及びSNS−EF1/UUによる用量設定及び反復投与試験。
SNS01は本発明の一実施形態である−これは多発性骨髄腫の治療を目標とする、癌治療の生物工学製品である。SNS01は3つの成分:eIF5Aのアポトーシス促進性変異体を発現するDNAベクター(図22を参照);癌細胞の生育/抗アポトーシスを促進する野生型のeIF5Aを標的とするsiRNA(図25の配列を参照);及び送達ビヒクルとして作用する、ポリエチレンイミンと称される合成ポリマー(In vivo-jetPEI;Polyplus Transfection Inc.)により構成される。
【0191】
試験の目的は、マウスへの静脈内SNS01の最大許容用量及び治療用量の長期投与の実行可能性を決定することである。別々の2試験を行った。最大許容用量試験(試験ID:MTD)は、マウスに増加性の量のSNS01(2.2mg/kg〜3.7mg/kg)の静脈内用量を2回投与し、臨床徴候、体重、臓器重量、及び肝酵素をモニターすることによって毒性を評価する8日間の試験である。9週間の反復投与試験(試験ID:EX6)は、SNS−EF1/UUの治療用量(1.5mg/kg)及び同様にその様々な成分の、週2回の長期投与後の毒性を評価するために計画された試験である。SNS−EF1/UUはSNS01の前臨床版であり、主に、eIF5AK50Rの発現がSNS01複合体のようにB細胞特異的プロモーターではなく、恒常的なプロモーター(全ての組織にいつでも発現するもの)により駆動されることで異なっている。SNS01におけるB細胞特異的B29プロモーターの使用は、アポトーシス促進性のeIF5A変異体の発現を、骨髄腫細胞を含むB細胞起源の細胞に限定することによって治療上の安全性を高めるために計画された。EX6試験にはマウス特異的なeIF5A siRNAを投与されたマウスの群もまた含まれたが、これはマウス組織においてeIF5Aを抑制することによるいずれの毒性効果があるか否かを判定するためである。反復投与試験における毒性は、臨床徴候、体重、血液、肝酵素、同様に組織病理をモニターすることによって評価した。
【0192】
【表1】
【0193】
【表2】
【0194】
前臨床試験により、SNS01は0.75mg/kg〜1.5mg/kgの用量において治療的であることが示された(試験EX9)。8日間の用量設定試験(試験ID:MTD)において、用量範囲の上限を決定するため、治療用範囲よりも著しく高い用量が試験された。試験品の週2回の静脈内用量は2.2mg/kg及び2.9mg/kgの低めの用量レベルで十分に許容されたが、1マウスは2.9mg/kgにおいて病的状態に至り、安楽死させた。3.3mg/kg以上の用量は、おおよそ20〜25%の生存率をもたらした。従って最大許容用量は2.2mg/kgと2.9mg/kgの間であり、これは0.75mg/kg〜1.5mg/kgの治療用範囲を十分に超えている。
【0195】
9週間の反復投与試験(試験ID:EX6)において、マウスは治療用量(1.5mg/kg)のSNS−EF1/UUを週2回尾静脈注射によって投与され、試験期間にわたって試験品に関連した毒性効果は観察されなかった。この試験においてはDNA及びsiRNAもまた別々に試験され、両方共マウスにより十分に許容された。ヒトeIF5A siRNAはマウスにおいて活性を示さないことから、マウスeIF5Aに特異的なsiRNAもまた試験に含めた。9週間の期間にわたり、マウスeIF5A siRNAの長期的な投与に関連した毒性効果は観察されなかった。これらの結果により、SNS01及びSNS−EF1/UUの治療用量は長期間にわたって投与されたとしてもマウスには無毒であることが示された。
【0196】
【表3】
【0197】
試験系及び試験計画
この試験の全ての態様は、カナダ動物管理協会(Canadian Council on Animal Care)及びオンタリオ州研究動物条例(Province of Ontario Animals for Research Act)により確立された、ウォータールー大学動物管理委員会(University of Waterloo Animal Care Committee)(ウォータールー、オンタリオ州、カナダ)により設定された指針に従って実行された。
【0198】
CD−1及びBALB/CマウスはCharles River Laboratories(ケベック州、カナダ)から入手した。両試験のマウスは試験品を週2回、尾静脈を通した静脈内注射によって投与された。0.2mlを超える量の送達には緩慢な注射(〜2−3分)を用いた。
【0199】
8日間の試験に用いたマウスは、試験開始時におおよそ6〜9週齢であった。9週間の反復投与試験に用いたマウスは、試験開始時におおよそ5〜6週齢であった。
【0200】
【表4】
【0201】
【表5】
【0202】
8日間の最大許容用量試験(MTD)
2用量による8日間の試験は、SNS01の最大許容用量を決定するために計画された用量設定試験である。用量範囲は2.2mg/kg〜3.7mg/kgであり、これは0.75mg/kg〜1.5mg/kgの治療用量範囲を十分に超えている。SNS01の最小用量(2.2mg/kg)において、1マウスがわずかに毛を逆立たせ、活動を低下させたが1時間以内に回復したことを除けば、毒性の臨床徴候は観察されなかった。2.2mg/kgのSNS01を2回目に注射した後には毒性の臨床徴候は観察されなかった。全てのマウスは試験の間中、その体重を維持した。臓器に肉眼的な変化は観察されなかった。体重に対する臓器重量の比は、肝臓重量:体重の比における中程度の増大を除いては、コントロール群からの変化は無かった。しかしながら、この比の増大はより高い用量レベル群のいずれにも観察されなかったことから、試験品に関連したものであるとは考えにくい。
【0203】
5マウスのうち4が、毒性の臨床徴候を示すことなく2.9mg/kgのSNS01を許容した。しかしながら、1マウスが注射後1時間以内に痙攣及び軽度の呼吸促迫症を経験したため人道的に安楽死させる必要があった。残りのマウスには、SNS01の2回目の注射後に毒性の臨床徴候は観察されなかった。マウスは試験の間中、その体重を維持し、臓器の肉眼的な変化、又は体重に対する臓器重量の比における変化は観察されなかった。2用量の2.9mg/kgのSNS01の後に、ALTの血清濃度がわずかに上昇した。
【0204】
予想されたように、3.3mg/kg以上の用量のSNS01は十分に許容されず、両群において1匹を除く全てのマウスを人道的に安楽死させる必要があった。病的状態のためマウスを人道的に安楽死させる必要があった全ての事例において臨床徴候は注射後1時間以内に現れ、これは高用量のPEIを用いたその他の報告された試験と一致していた。3.3mg/kg及び3.7mg/kgにおいて生存したマウスは2回目の用量は投与されなかったが、注射後4時間以内に完全に回復し、試験の間中その体重を維持した。従ってSNS01の最大許容用量は2.2mg/kgと2.9mg/kgの間であると思われる。
【0205】
9週間の反復投与試験(EX6)
9週間の反復投与試験の目的は、SNS01開発の間の前臨床試験のために使用された複合体であるSNS−EF1/UUの、治療用量(1.5mg/kg)の長期的な投与の安全性を評価することである。SNS−EF1/UUはSNS01と大きくは異ならないが、主要な相違は材料が研究グレードであること、及びeIF5AK50Rの発現が全ての細胞型において活性のある恒常的なヒトEF1プロモーターにより駆動されていることである。SNS01はeIF5AK50Rの発現を駆動するためにB細胞特異的なプロモーターを使用しているが、この安全性試験における恒常的なプロモーターの使用により、非B細胞における変異eIF5AK50Rタンパク質の蓄積がもたらす毒性の評価が可能となる。9週間の反復投与試験の別の態様は、SNS−EF1/UUの個別成分の安全性を試験するための群を包含することであった。DNA群(Ex6−G3)にはeIF5Aプラスミド及び非標的指向化コントロールsiRNAを含む複合体が投与され、一方でsiRNA群(Ex6−G4)にはヒトeIF5A(h5A1)siRNA及び非発現プラスミドを含む複合体が投与された。試験品のSNS−EF1/UUには、内在性のマウスeIF5Aの発現に影響し得ないヒトeIF5A siRNAが含まれることから、この試験の別の特色は、非発現プラスミド及びマウスeIF5Aを効率的に標的とするsiRNAを含むPEI複合体を投与された群(Ex6−G6)を包含することであった。この群によって、活性のあるeIF5A siRNAの長期的な投与の安全性についての評価が可能となった。
【0206】
全ての動物は、予定された屠殺日まで生存した。9週間の試験過程にわたり、いずれの群にも毒性の臨床徴候は観察されず、全ての群におけるマウスの体重は試験期間の間に増加し続けた。赤血球及び白血球の数は処置開始後3及び6週間目に測定され、全ての投与群において正常であった。肝酵素の血清濃度は屠殺後に測定され、全てのマウスについて正常範囲内であった。いずれの群にも、臓器の肉眼的外観に変化は観察されなかった。独立した病理学者により、主要な臓器の組織病理学的分析が行われた(試験品に起因する毒性は示されなかった。
【0207】
SNS−EF1/UUの治療用量の長期的な投与はマウスにより十分に許容され、有害作用は観察されなかった。加えて、マウス特異的なeIF5A siRNAの長期的な投与が毒性効果を示さなかったことから、ヒトeIF5A siRNAを含むPEI複合体の投与はヒトに対して安全なはずであることが示された。
【0208】
[実施例21]
多発性骨髄腫を有するマウスにおける、静脈内SNS−B29/UU及びSNS01の治療効果試験
SNS01は上記の通りである。試験品SNS−B29/UUはSNS01の前臨床版である。SNS−B29/UUのSNS01との違いはわずかであり、主要な違いは成分がGLPグレードではなく研究グレードであることである。ここに報告される試験の目的は、SNS−B29/UUの最小有効用量の決定であり、かつSNS01を含んでなるGLPグレードの材料が、前臨床試験で用いられた研究グレードの材料と同様に作用することの確認であった。反復投与腫瘍試験(repeated dose tumor study)(試験ID:EX9)は、SNS01の至適な治療用量を決定する目的で、マウスの皮下腫瘍の生育を阻害する、SNS−B29/UUの次第に縮小する能力を持つ週2回の用量を評価する5週間の試験であった。処置された動物はまた、臨床徴候、体重、及び臓器重量をモニターすることによって毒性の徴候を評価された。
【0209】
【表6】
【0210】
【表7】
【0211】
SNS−B29/UUの治療用範囲を、ヒト多発性骨髄腫皮下腫瘍を有するSCIDマウスにおいて決定した。0.15mg/kgと1.5mg/kgの間のSNS−B29/UUの用量について試験した。試験品の抗腫瘍効果を、週2回の腫瘍容積の測定ならびに屠殺後の腫瘍組織の切除及び秤量によって決定した。0.75mg/kg及び1.5mg/kgのSNS−B29/UUの用量により腫瘍の著しい収縮がもたらされたことから、SNS−B29/UUの治療用範囲は0.75mg/kgと1.5mg/kgの間にあることが示される。腫瘍の収縮は観察されなかったが、皮下腫瘍の生育の効果的な阻害は0.38mg/kgのSNS−B29/UUでもまた観察された。0.15mg/kgという低用量のSNS−B29/UUでさえも腫瘍生育の幾らかの阻害が観察されたことから、治療用範囲は広いことが示される。図26及び27を参照されたい。
【0212】
GLPグレード成分を用いて作られたSNS01の有効性をSNS−B29/UUと比較し、腫瘍生育の阻害において同程度の有効性を持つことが見出された。腫瘍を有するSCIDマウスのSNS01及びSNS−B29/UUによる処置は十分に許容され、マウスの体重は試験期間の間に増加し続けた。
【0213】
【表8】
【0214】
試験系及び試験計画
雌のC.B.17/IcrHsd−Prkdc(SCID)マウスはHarlan(インディアナポリス、インディアナ州、米国)から入手した。皮下腫瘍は、10x106のKAS−6/1(ヒト多発性骨髄腫)生細胞を5〜6週齢のマウスの右側腹部に注射することにより確立した。SNS−B29/UUによる処置は、腫瘍がおおよそ20〜40mm3のサイズに達した時点(腫瘍細胞注射後おおよそ4週間目)に開始した。SNS01による処置は、腫瘍がおおよそ130mm3のサイズに達した時点(腫瘍細胞注射後おおよそ6週間目)に開始した。マウスは試験品を週2回、尾静脈の静脈内注射を通して投与された。
【0215】
【表9】
【0216】
反復投与腫瘍試験(repeated dose tumor study)
反復投与腫瘍試験(repeated dose tumor study)は、SNS−B29/UUの最小有効治療用量を決定し、かつGLPグレードのSNS01試験品が、研究グレードの試験品であるSNS−B29/UUによって示された腫瘍の阻害活性を保持していることを確認するために計画された。第2の目的は、処置されたマウスの臨床徴候、体重、及び臓器重量をモニターすることによって、処置によるいずれの毒性効果を評価することであった。試験品の治療上の抗腫瘍活性は、デジタルキャリパーを用いて腫瘍容積を週2回測定することによりモニターした。屠殺後に腫瘍を切除して秤量した。
【0217】
全てのマウスは予定された屠殺日まで生存した。非発現プラスミド及び非標的指向化siRNAを含むPEIナノ複合体によって処置されたコントロールマウスは、屠殺時に284mm3の平均腫瘍容積を有したが、1.5mg/kgのSNS−B29/UUによって処置されたマウスは、わずかに13mm3の平均腫瘍容積、腫瘍生育における95%(*p=0.026)の減少を有した。しかしながら、1.5mg/kgのSNS−B29/UUによって処置されたマウスからの腫瘍の切除を試みた際、いずれのマウスにも腫瘍の形跡は見出されなかった。SNS−B29/UUの用量を半分の0.75mg/kgまで減少させてもなお、腫瘍容積及び重量それぞれにおいて91%(*p=0.03)及び87%(*p=0.04)の減少がもたらされ、1マウスでは腫瘍が完全に消失していた。従って、SNS−B29/UUの週2回の注射の至適な治療用量は、0.75mg/kgと1.5mg/kgの間であると思われる。SNS−B29/UUの週2回の用量が0.15mg/kgと低くてもなお、最終的な腫瘍容積の60%の減少がもたらされたことから、SNS−B29/UUは広い用量範囲にわたって強力な抗腫瘍活性を有することが示される。
【0218】
腫瘍生育の阻害に加え、0.75mg/kg及び1.5mg/kgのSNS−B29/UU及びSNS01による処置は腫瘍容積の著しい減少をもたらしたことから、この処置が、おそらくは腫瘍へのアポトーシスの誘発を通して腫瘍の退縮を誘導できることが示される。0.75mg/kg及び1.5mg/kgの用量レベルであるSNS−B29/UUによって処置された、腫瘍を有するマウスにおける腫瘍容積のパーセント変化は、それぞれ−244%及び−245%であった。コントロールマウスの腫瘍サイズは、同期間の間に2000%を超えて増大した。SNS01の週2回の注射もまた、多発性骨髄腫を著しく収縮させた。1.5mg/kgのSNS01によって処置されたマウスの腫瘍容積のパーセント変化は−349%であったことから、SNS01はSNS−B29/UUと同様に有効であることが示される。SNS01による処置は、わずかに25日間の処置後に腫瘍容積の349%の減少を達成したが、SNS−B29/UUは35日間の処置後に腫瘍容積の245%の減少を達成したことから、GLPグレード材料の使用によって生物活性が事実上増大された可能性がある。加えて、SNS01によって処置された腫瘍は極めて大きかった(〜130mm3)ことから、SNS01による処置は十分に確立された腫瘍に対して有効であることが示される。
【0219】
処置は全てのマウスにより十分に許容され、毒性の徴候は観察されなかった。全ての群におけるマウスの体重は試験の間を通して増加し続けた。剖検では臓器の肉眼的外観に変化は観察されず、体重に対する臓器重量の比における著しい変化は起こらなかった。
【0220】
従ってSNS01(及びその前臨床版であるSNS−B29/UU)はSCIDマウスによって十分に許容され、週あたり2回の静脈内注射によって送達される際、皮下のヒト多発性骨髄腫の治療に非常に有効である。試験された全てのSNS−B29/UUの用量は腫瘍生育の阻害に有効であったが、最高用量である1.5mg/kgにより、処置された全てのマウスの腫瘍は成功裏に除去された。
【0221】
[実施例22]
プラスミドDNA及びsiRNAポリエチレンイミン(JetPEI)複合体の生体内分布
PEI複合体により送達されたプラスミドDNAの局在を判定するため、緑色蛍光タンパク質(「GFP」)GFP発現コンストラクトを用いた。GFP発現を駆動するため、2つのプロモーター:EF1:遍在性プロモーター(EF1::GFP)又はB29:B細胞特異的プロモーター(B29::GFP)が用いられた。20マイクログラムのGFPプラスミドDNA及び10マイクログラムの蛍光標識(DY547)h5A1 siRNAを含むPEI複合体を、6のN/P比において調製した。BalB/Cマウスに、5%グルコース又はPEI複合体のいずれかを2日間連続して静脈内注射した。最初の注射後72時間目にマウスを安楽死させ、それらの臓器を回収して、GFPの発現及びDY547−siRNAを共焦点顕微鏡法によって解析した。
骨髄:ほとんどの事例でDY547−siRNAの形跡が存在したが、GFP発現は見られなかった。臓器回収の時期はGFP発現のピークと一致しない可能性があり;GFPシグナルは消失したか、又はGFPは発現していない可能性がある。しかしながら幾つかの例で、GFP及びDY547が同じ骨髄細胞に共存することが観察された。従ってこのことにより、静脈内注射によって投与する際、PEIナノ粒子を生きた動物の骨髄細胞にトランスフェクトできることが証明された。
肺:ほとんどの事例でDY547−siRNAの形跡が存在したが、GFP発現は見られなかった。臓器回収の時期はGFP発現のピークと一致しない可能性があるか、又はGFPシグナルは消失したか若しくはGFPは発現していない可能性がある。
脾臓:DY547−siRNAの存在もまた陽性である細胞内で、GFP発現(EF1プロモーターにより駆動される場合)が共存する形跡が見られた。B29プロモーターにより駆動した際、脾細胞内でのGFPの発現はかなり低かった。このことにより、PEIナノ粒子は脾臓の細胞にトランスフェクトされるようであることが示される。
腎臓:GFP又はDY547の発現は観察されなかったことから、ナノ粒子は腎臓には進入し得ないことが示される。
肝臓:ほとんどの事例でDY547−siRNAの形跡が存在したが、GFP発現は見られなかった。このことにより、PEIナノ粒子は肝臓の細胞にトランスフェクトされることが証明される。
心臓:心臓組織におけるEF1::GFPとDY547−siRNAとの共存が観察されたことから、PEIナノ粒子はこの臓器にトランスフェクトし得ることが示される。B29プロモーターを用いた際にはGFPは観察されなかった。
【0222】
[実施例23]
HA−eIF5AK50Rの発現における、DNA:siRNA比の効果
KAS細胞に、B29−HA−eIF5AK50R(B細胞特異的プロモーターにより駆動されるプラスミド)及びh5A1 siRNAを含むナノ粒子をトランスフェクトした。pExp5A及びh5A1 siRNAを異なる比において含むJET PEI(商標)ナノ粒子を作製し、KAS細胞への添加前に室温で4時間インキュベートした。トランスフェクション後4時間目に、ナノ粒子を含む培地を新鮮な培地に交換した。24時間後に細胞ライセートを回収し、HAに対する抗体を用いるウェスタンブロット分析に使用した。DNA:siRNAの比は、標準的な比である2:1から異なっていた。HA−eIF5AK50Rの蓄積は、1:0、3:1、及び2:1の比においてピークであった。図30を参照されたい。
【0223】
[実施例24]
ナノ粒子のトランスフェクションにより誘導されたアポトーシスにおけるDNA:siRNA比の効果
pExp5A及びh5A1 siRNAを異なる比において含むナノ粒子を作製し、KAS細胞への添加前に室温で4時間インキュベートした。トランスフェクション後4時間目に、ナノ粒子を含む培地を新鮮な培地に交換した。48時間後に細胞を回収し、アネキシンV/PIにより標識してFACSによって解析した。アポトーシスの誘導は、標準的なDNA:siRNA比である2:1によるナノ粒子をトランスフェクトした細胞において最も高かった。図31を参照されたい。
【0224】
[実施例25]
eIF5A1K50Rプラスミド及びeIF5A1 siRNA(siSTABLE又は非siSTABLE)を含むPEI複合体(N/P=6又は8)の投与により、多発性骨髄腫皮下腫瘍の生育は阻害され、かつ腫瘍の収縮がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。マウスに:(G1)20mgのpCpG−mcs(空のベクター)及び10mgのコントロールsiRNAをN/P=8において含むPEI複合体(中程度用量);(G5)20mgのRNAi耐性プラスミドであるpCpG−eIF5A1k50R及び10mgのsiSTABLE h5A1 siRNAをN/P=8において含むPEI複合体(中程度用量、siSTABLE);(G8)20mgのRNAi耐性プラスミドであるpCpG−eIF5A1k50R及び10mgのh5A1 siRNAをN/P=6において含むPEI複合体(中程度用量、N/P=6)のいずれかを週あたり2回静脈内注射した。示されるデータは、各群のマウスについての個々の腫瘍容積である。最終の注射は、処置開始後40日目に行った。図32を参照されたい。
【0225】
[実施例26]
JET PEI(商標)ナノ粒子は腫瘍組織によって効果的に取り込まれ、このナノ粒子は同細胞へプラスミド及びsiRNAを送達する。
pExp−GFP(B細胞特異的プロモーターによる制御下のGFP)及びDY547−siRNA(蛍光標識されたsiRNA)を含むナノ粒子の注射後48時間目の腫瘍切片。共焦点顕微鏡法に従って腫瘍切片内にGFP及びDY547の共存した発現が観察されることから、ナノ粒子は腫瘍組織によって効果的に取り込まれ、このナノ粒子は同細胞へプラスミド及びsiRNAを送達することが示される。図33を参照されたい。
【0226】
[実施例27]
切断試験のために作製されたアデノウィルスコンストラクト。
アデノウィルス(血清型5、E3欠失)
1)Ad−eIF5A1Δ(2−6)[Δ(2−6)]−これはアミノ酸2〜6を欠損したヒトeIF5A1である。
2)Ad−eIF5A1K50RΔ(2−6)[Δ(2−6)/K50R]−これはアミノ酸2〜6を欠損したヒトeIF5A1である。加えて、「K50R」は位置50のリジン(K)がアルギニン(R)に変異しており、このためにDHSによりヒプシン化不可能であると信じられていることを示す。
3)Ad−eIF5A1D6E[D6E]−これは予想される切断部位が変異している(D6がEへ)ヒトeIF5A1である。
4)Ad−eIF5A1D6E/K50R[D6E/K50R]−これは予想される切断部位が変異している(D6がEへ)ヒトeIF5A1K50Rである。加えて、「K50R」は位置50のリジン(K)がアルギニン(R)に変異しており、このためにDHSによりヒプシン化不可能であると信じられていることを示す。
【0227】
[実施例28]
カスパーゼにより仲介されるeIF5Aの切断
切断部位を同定するため、アクチノマイシンDにより処理されたKAS細胞から分離したタンパク質ライセートを2−Dゲル電気泳動により分離し(図37)、分子量の小さなeIF5Aの切断産物に相当する点をゲルから切り出して質量分析法により配列決定した(図38B)。切断産物の完全長配列は得られなかったが、切断はタンパク質のN−末端から6番目のアミノ酸の後に起こったことが特定された(図38A)。考えられる切断部位のすぐ前の配列は「DDLD」であり、これはカスパーゼ切断部位として同定された配列である(Chay et al., 2002)。
【0228】
切断断片の産物がカスパーゼ依存性であるか否かを決定するため、ヒト骨髄腫細胞においてアクチノマイシンD誘導性のアポトーシスの間に産生される切断断片の産生を遮断する、カスパーゼ阻害剤の能力を試験した(図41)。一般的なカスパーゼ阻害剤ならびにカスパーゼ3、8、及び9の特異的な阻害剤は全て、KAS細胞においてActD誘導性のアポトーシスの間に蓄積するeIF5Aの切断型の形成を強力に阻止する。カスパーゼ1の阻害剤もまた、切断産物の形成を完全には遮断しなかったものの、減少させた。カスパーゼ阻害剤とのインキュベーションは、切断産物の蓄積を阻止しただけではなく、eIF5Aのヒプシン化型の減少もまた阻害したことから(図41)、アクチノマイシンD誘導性のアポトーシスの間のヒプシン化eIF5Aの減少は切断の結果であることが示される。
【0229】
アポトーシスの間のカスパーゼによるタンパク質の切断(Fisher et al., 2003による総説)は無数の目的:1)生存促進性又は抗アポトーシス性タンパク質の不活性化−例えば:mRNAの5’キャップ構造に結合して、キャップ形成されたmRNAの40Sリボソームサブユニットへの結合を促進する真核生物翻訳開始因子4G1が、カスパーゼによる切断によって不活性化されることで翻訳を阻害する;2)タンパク質のドミナントネガティブ型の作製(例えば、NF−カッパB p65はアポトーシスの間にカスパーゼによって切断され、DNAに結合はするが転写活性化の活性を持たないためにNF−kBのドミナントネガティブ阻害物質として作用する、ドミナントネガティブ断片を作り出す);3)阻害性又は調節性ドメインの除去によって新しい活性又は増大した活性を持つ断片の形成がもたらされることによる、アポトーシス促進性タンパク質の活性化をもたらす機能獲得型の切断(例えば、細胞周期の停止及びアポトーシスを仲介する乳癌抑制タンパク質であるBRCA−1は、アポトーシス促進性切断断片の放出しうるカスパーゼによる切断によって活性化される。eIF4GファミリーのメンバーであるDAP5はカスパーゼによる切断後に活性化され;切断産物はアポトーシス関連タンパク質の翻訳を刺激する);4)抗アポトーシスタンパク質のアポトーシス促進性タンパク質への変換(例えばBcl−xL及びc−IAP1のようなアポトーシス阻害物質は、カスパーゼによる切断を通してアポトーシス促進性タンパク質に変化し得る);及び5)細胞内の再分布(例えばカスパーゼ8による切断に従い、Bidのアポトーシス促進性p15断片では切断によって曝露されたグリシン残基がミリストイル化され、続いてこれによりBidがミトコンドリアへ移動してチトクロムcの放出を増大させることが可能になる)を有し得る。
【0230】
アクチノマイシンD誘導性アポトーシスの間の、カスパーゼにより仲介されるeIF5Aの切断が一般的な現象であるか又は骨髄腫細胞に特異的であるかを決定するために、切断されたeIF5Aの存在下において、アクチノマイシンDによって処理したHeLa細胞を調べた(図42)。アクチノマイシンDによって処理したHeLa細胞内に、eIF5Aの切断断片の蓄積は観察されなかった。eIF5Aの翻訳後修飾(例えばリン酸化又はアセチル化)がカスパーゼによる切断の必要条件であり得るか否かを決定する試みにおいて、脱アセチル化酵素阻害剤であるニコチンアミドの、切断断片の形成における効果について調べた。本発明者らの研究室及びその他(Kim et al., 2006)によるeIF5Aの質量分析によって、eIF5Aのアセチル化が観察された。さらに、酵母eIF5AがSir2関連の脱アセチル化酵素であるHst2の基質として同定され(Shirai et al., 2008)、著者らによりeIF5Aのアセチル化がHst2を欠損した酵母か又はeIF5Aを過剰発現する酵母においてのみ見られることが観察されたことから、eIF5Aは一般には非アセチル化型で存在することが示される。アクチノマイシンDによって処理したHeLa細胞とSir2脱アセチル化酵素阻害剤であるニコチンアミドとのインキュベーションによりeIF5A切断産物の蓄積がもたらされたことから、カスパーゼにより仲介される切断が起こるためにはeIF5Aのアセチル化が必要とされることが示される。これらのデータにより、eIF5Aは通常、アポトーシスシグナルがeIF5Aのアセチル化を引き起こしてカスパーゼにより仲介される切断が可能となるまで、カスパーゼ活性から保護され得ることが示唆される。
【0231】
参考文献
Chay KO, Park SS, Mushinski JF (2002). Linkage of caspase-mediated degradation of paxillin to apoptosis in Ba/F3 murine pro-B lymphocytes. J Biol Chem. 277, 14521-14529.
Fischer U, Janicke RU, Schulze-Osthoff K (2003). Many cuts to ruin: a comprehensive update of caspase substrates. Cell Death and Differentiation 10, 76-100.
Kim, S. C, Sprung, R., Chen, Y., Xu, Y., Ball, H., Pei, J., Cheng, T., Kho, Y., Xiao, H., Xiao, L., Grishin, N. V., White, M., Yang, X. J., and Zhao, Y. (2006) Substrate and Functional Diversity of Lysine Acetylation Revealed by a Proteomics Survey. Mol Cell 23, 607-618.
Shirai et al. (2008). Global analysis of gel mobility of proteins and its use in target identification J. Biol. Chem. 283, 10745-10752.
【発明の詳細な説明】
【0001】
[関連出願]
本出願は、2008年9月3日に提出された米国仮出願第61/093,749号、及び2009年3月19日に提出された米国特許出願第12/400,742号に対する優先権を主張するものであり、これらはその全体が参照により本明細書に取り込まれる。
【0002】
[背景技術]
比較的最近、二重鎖RNA(「dsRNA」)がタンパク質発現の阻害に使用できることが研究者らにより観察された。遺伝子の発現を停止するこの能力にはヒトの疾病の治療に対する広範な可能性があり、現在多くの研究者及び営利団体がこの技術に基づく治療法の開発にかなりの資金を投じている。
【0003】
二重鎖RNAにより誘導される遺伝子サイレンシングは、少なくとも異なる3段階:(i)RNAにより導かれるDNA又はヒストンのメチル化を指す、転写の不活性化;(ii)siRNAにより誘導されるmRNAの分解;及び(iii)mRNAにより誘導される転写の減衰において起こり得る。
【0004】
一般に、哺乳類細胞においてRNAにより誘導されるサイレンシング(RNA干渉、又はRNAi)の主要な機構は、mRNAの分解であると考えられている。哺乳類細胞におけるRNAi使用の最初の試みは、長鎖のdsRNAの使用に集中していた。しかしながらRNAi誘導のためのこれらの試みは、標的特異的とは対照的に、一つには一般的なタンパク質合成の阻害をもたらすインターフェロン反応の誘導のためにあまり成功しなかった。従って、哺乳類系におけるRNAiに対して長鎖dsRNAは実行可能な選択肢ではない。
【0005】
さらに最近、短い(18〜30bp)RNA二本鎖を培養哺乳類細胞に導入した際、インターフェロン反応を誘導することなく標的mRNAの配列特異的な阻害が実現できることが示された。阻害的低分子RNA(「siRNA」)と称されるこれらの短いdsRNAのあるものは、細胞内で95%を超える標的mRNAを切断するために、モル濃度未満の濃度で触媒的に作用し得る。siRNAの作用機構、同様に幾つかのその応用についての記述は、Provost et al. (2002) Ribonuclease Activity and RNA Binding of Recombinant Human Dicer, EMBO J. 21(21): 5864-5874; Tabara et al. (2002) The dsRNA Binding Protein RDE-4 Interacts with RDE-1, DCR-1 and a DexH-box Helicase to Direct RNAi in C. elegans, Cell 109(7): 861-71; Ketting et al. (2002) Dicer Functions in RNA Interference and in Synthesis of Small RNA Involved in Developmental Timing in C. elegans; Martinez et al., Single-Stranded Antisense siRNAs Guide Target RNA Cleavage in RNAi, Cell 110(5):563; Hutvagner & Zamore (2002) A microRNA in a multiple-turnover RNAi enzyme complex, Science 297:2056に記載される。
【0006】
機構的な視点から、長い二重鎖RNAの植物及び無脊椎動物の細胞内への導入は、ダイサーとして知られるタイプIIIエンドヌクレアーゼによってsiRNAへ分解される。Sharp, RNA interference--2001, Genes Dev. 2001, 15:485. Dicer, a ribonuclease-III-like enzyme, processes the dsRNA into 19-23 base pair short interfering RNAs with characteristic two base 3' overhangs. Bernstein, Caudy, Hammond, & Hannon (2001) Role for a bidentate ribonuclease in the initiation step of RNA interference, Nature 409:363。siRNAは次に、RNA誘導型サイレンシング複合体(RNA-induced silencing complex)(RISC)に取り込まれ、ここで1以上のヘリカーゼがsiRNA二本鎖を巻き戻し、標的の認識を導くための相補的アンチセンス鎖が有効となる。Nykanen, Haley, & Zamore (2001), ATP requirements and small interfering RNA structure in the RNA interference pathway, Cell 107:309。適切な標的mRNAへの結合により、RISC内の1以上のエンドヌクレアーゼが標的を切断してサイレンシングを誘導する。Elbashir, Lendeckel, & Tuschl (2001) RNA interference is mediated by 21- and 22-nucleotide RNAs, Genes Dev. 15:188、図1。
【0007】
干渉効果は長く持続し得り、多数の細胞分裂後にも検出され得る。さらに、RNAiは配列特異性を示す。Kisielow, M. et al. (2002), Isoform-specific knockdown and expression of adaptor protein ShcA using small interfering RNA, J. Biochem. 363:1-5。従ってRNAi機構により、密接に関連するmRNAに影響を及ぼすことなく、一つの型の転写を特異的にノックダウンできる。これらの特性によってsiRNAは、遺伝子発現の阻害、及び遺伝子機能の研究、及び薬物標的の検証に対して価値ある手段となる可能性がある。さらにsiRNAは、(1)遺伝子の過剰発現又は異所性発現により引き起こされる疾病;及び(2)変異を含む遺伝子の発現によりもたらされる疾病に対する治療薬として有用な可能性がある。
【0008】
成功したsiRNA依存性の遺伝子サイレンシングは幾つかの因子に依存する。RNAiにおける最も議論のある問題の一つは、siRNA設計の必要性、すなわち使用するsiRNAの配列を考慮することについての論点である。C. elegans及び植物における初期の研究は、長いdsRNAの導入によって設計の問題を回避した(例えば、Fire, A. et al. (1998) Nature 391 :806-811を参照)。この原始的な生命体において、長いdsRNA分子はダイサーによりsiRNAに切断され、こうして転写の全体を網羅する可能性のある二本鎖の多様な集団が生成される。これらの分子のうち幾つかの分画は非機能的(すなわちサイレンシングをわずかに誘導するか又は誘導しない)であるが、1以上は高度に機能的となる可能性があり、これによって興味ある遺伝子のサイレンシングが起こり、かつsiRNA設計の必要は少なくなる。残念なことに、哺乳類系ではインターフェロン反応のためにこの同じアプローチは使用できない。この効果はダイサーによる切断工程を介さずに、siRNAを直接導入することにより回避できるが、この方策は選択したsiRNA配列が非機能的又は半機能的であるという危険性を持ち越す。
【0009】
多くの研究者が、siRNAの設計はRNAiの決定的な要素ではないという見解を示してきた。一方で当該分野における他の研究者達は、siRNAの設計に注目することにより、RNAiがさらに効率的となる可能性を探求し始めた。
【0010】
様々な疾病又は疾患を治療するためには特定のタンパク質の上昇が望ましいが、それが必要な全てではない可能性もある。例えば、内在性タンパク質又は異なるタンパク質発現をノックダウン又はノックアウトするために、siRNAを組み合わせた使用が必要となり得る。本発明はこの必要性を満たし、癌、特に多発性骨髄腫の治療法を提供する。
【0011】
多発性骨髄腫を含む癌は、アポトーシス誘導能から利益を得られる疾病である。多発性骨髄腫の従来の治療法は、化学療法、幹細胞移植、幹細胞移植を併用する高用量化学療法、及びサルベージ療法を含む。化学療法は、Thalomid(登録商標)(サリドマイド)、ボルテゾミブ、Aredia(登録商標)(パミドロネート)、ステロイド、及びZometa(登録商標)(ゾレドロン酸)による治療を含む。しかしながら多くの化学療法薬は、骨髄、胃及び腸の内面、並びに、毛包のような活発に分裂する非癌細胞に有毒である。従って化学療法は、血液細胞数の減少、悪心、嘔吐、下痢、及び脱毛をもたらし得る。
【0012】
従来の化学療法又は標準用量の化学療法は、典型的には多発性骨髄腫の患者の第一の、又は最初の治療である。患者はまた、高用量化学療法及び幹細胞移植のために用意された化学療法も受け得る。移植前に腫瘍の負荷量を減少させるため、導入療法(幹細胞移植前の従来の化学療法)が使用できる。特定の化学療法薬は骨髄細胞への毒性が少なく、かつ骨髄からの幹細胞のより高い収率をもたらすため、他のものよりも導入療法に適している。導入療法に適した化学療法薬の例は、デキサメタゾン、サリドマイド/デキサメタゾン、VAD(ビンクリスチン、Adriamycin(登録商標)(ドキソルビシン)、及びデキサメタゾンの組み合わせ)、及びDVd(ペグ化リポソームドキソルビシン(Doxil(登録商標)、Caelyx(登録商標))、ビンクリスチン、及び投与頻度縮小型デキサメタゾンの組み合わせ)を含む。
【0013】
多発性骨髄腫の標準的治療は、50%の反応率を達成するメルファランとプレドニゾン(副腎皮質ステロイド薬)の組み合わせである。残念なことにメルファランはアルキル化剤であり、導入療法にはあまり適さない。副腎皮質ステロイド(特にデキサメタゾン)は時々単独で多発性骨髄腫の治療法に用いられ、特に高齢患者及び化学療法を許容できない患者に対して用いられる。デキサメタゾンはまた、単独又はその他の薬剤と組み合わせて導入療法にも使用される。VADは最も一般に用いられる導入療法であるが、最近DVdが導入療法に有効であることが示されている。多発性骨髄腫の治療のために最近ボルテゾミブが承認されたが、これは非常に有毒である。しかしながら、現存の治療法のいずれも治癒の著しい可能性を提供していない。従って、癌及び多発性骨髄腫に適した治療を見出す必要が依然としてある。本発明はこの必要性を満たす。
【0014】
[発明の概要]
本発明は、eIF−5A1の切断型をコードする単離されたポリヌクレオチド、同様に切断型eIF−A1ポリペプチドを提供する。切断型eIF−5A1ポリヌクレオチドは、被験体における癌細胞のアポトーシス及び死滅の誘導に有用である。切断型ポリヌクレオチドは発現ベクター内に使用され得り、これは次に被験体へ投与される。切断型eIF−5Aは哺乳類内に発現し、癌細胞を死滅させる。切断型eIF−5A1タンパク質は、約17kDaである完全長eIF−5A1タンパク質に対し、約16kDAである。切断型eIF5A1タンパク質をコードするポリヌクレオチドは、被験体の癌細胞又は腫瘍にアポトーシスを誘導するために用いられる薬剤の製造に使用されてよい。切断型eIF5A1タンパク質をコードするポリヌクレオチドは、本明細書に記載されるその他の実施形態と協同して(例えば、完全長eIF5A1をコードするポリヌクレオチド、完全長変異eIF5A1をコードするポリヌクレオチド、及び/又はeIF5Aの3’UTRを標的とするsiRNAと組み合わせて)使用されてよい。
【0015】
本発明はまた、ホスト内での内在性遺伝子の発現をノックアウト又はノックダウンするための、内在性遺伝子を標的とするsiRNA、及び、ポリヌクレオチドによりコードされるタンパク質をホスト内で発現させるための、送達媒体/発現ベクター内の遺伝子をコードするポリヌクレオチドのホストへの送達を組み合わせた使用にも関する。正常タンパク質がその必要な機能を実施できるように、正常(欠陥のない)タンパク質をコードするポリヌクレオチド(又はタンパク質それ自体)がホストに投与され、発現する(ポリヌクレオチドの場合)。siRNAは、欠陥のあるタンパク質の内在性の発現をノックダウン又はノックアウトするが、同時に投与された正常タンパク質をコードするポリヌクレオチドの外来性の発現には影響しないような遺伝子領域を標的として好ましく設計される。
【0016】
本発明は、eIF5A1の3’末端を標的とするeIF5A1 siRNA、変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターの複合体を含んでなる組成物を提供し、ここで該変異eIF5A1はヒプシン化できず、かつ該siRNA及び該発現ベクターは、複合体形成のためにポリエチレンイミンと複合される。
【0017】
本発明は、被験体における標的遺伝子の内在性発現を抑制するための、標的遺伝子を標的とするsiRNA、及び被験体内で発現できる標的タンパク質をコードするポリヌクレオチドを含んでなる組成物を提供する。特定の実施形態によれば、ポリヌクレオチドはRNAi耐性プラスミド内(siRNAにより抑制され得ない)にある。siRNA及びプラスミドは、複合体形成のためにポリエチレンイミンと好ましく複合される。
【0018】
特定の実施形態によれば、siRNAは図25に示す配列を有し、ここで変異eIF5A1をコードするポリヌクレオチドはeIF5A1K50Rである。発現ベクターは、変異eIF5A1をコードするポリヌクレオチド、及び被験体内でポリヌクレオチドを発現させるために機能的に連結されたプロモーターを含んでなる。プロモーターは、好ましくは組織特異的又は全身的のいずれかである。例えば組成物が癌の治療に使用される場合、プロモーターは、好ましくは癌が属する組織に対して組織特異的である。例えば多発性骨髄腫を治療するためには、B29のようなB細胞特異的プロモーターの使用が好ましい。特定の実施形態によれば、発現ベクターはpCpGプラスミドを含んでなる。
【0019】
特定の実施形態によれば、eIF5A1 siRNA、及び変異eIF5A1ポリヌクレオチドを含んでなる発現ベクターは、独立してin vivo JetPEI(商標)のようなポリエチレンイミンと複合される。別の実施形態によれば、eIF5A1 siRNA、及び変異eIF5A1ポリヌクレオチドを含んでなる発現ベクターは、一緒にポリエチレンイミンと複合される。
【0020】
本発明はさらに、eIF5A1の3’末端を標的とするeIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターを含んでなる組成物を提供し、ここで該変異eIF5A1はヒプシン化できず、かつ該siRNA及び該発現ベクターは、癌治療のため被験体に送達される。癌は多発性骨髄腫を含むいずれの癌であってもよい。
【0021】
本発明はさらに、本発明の組成物を被験体(哺乳類及びヒトを含むがこれらに限定されない)に投与することを含んでなる癌の治療法を提供する。
【0022】
組成物は、限定はされないが静脈内、腹腔内、皮下、又は腫瘍内のような許容されるいずれの経路で投与されてよい。siRNA及び発現ベクターは、異なる時間に異なる経路を通して投与されてよいか、又は同時に同じ経路を通して一緒に投与されてよい。例えば限定はされないが、これらのsiRNAはネイキッドで、又はin vivo jetPEIのような担体と複合してIVを通して送達されてよく、かつ発現ベクターは腫瘍内に投与されてよいか、又はsiRNA及び発現ベクターの両方がIV若しくは腫瘍内などに投与されてよい。
【0023】
本発明は、癌細胞の生育を阻害及び/又は癌細胞を死滅させる方法を提供する。本発明はまた、癌細胞の転移能を阻害又は遅くさせる方法も提供する。癌生育の阻害は、腫瘍サイズの縮小、腫瘍生育の低下を含み、かつ腫瘍の完全な寛解もまた包含し得る。癌は、大腸癌、結腸直腸腺癌、膀胱癌、子宮頸部腺癌、及び肺癌を含むがこれらに限定されないいずれの癌又は腫瘍であってよい。好ましくは、癌は多発性骨髄腫である。
【0024】
好ましい実施形態によれば、eIF−5Aはヒプシン化できない変異体である。典型的な変異体は本明細書に記載される。
【0025】
eIF−5A、又はeIF−5Aをコードするポリヌクレオチドの被験体への供給(eIF−5Aを発現させるため)に加え、eIF−5Aの内在性発現をノックアウト又はノックダウンするためにsiRNAが供給される。
【0026】
本発明はまた、多発性骨髄腫を有する被験体内の多発性骨髄腫細胞を死滅させ、癌を治療する薬剤の製造のために、eIF5A、eIF5A1をコードするポリヌクレオチド、及びeIF5A1に対するsiRNAの使用も提供する。好ましくは、変異eIF−5Aをコードするポリヌクレオチドはヒプシン化できない。
【0027】
本発明はまた、鎌状赤血球貧血の治療法も提供する。健常ヘモグロビン遺伝子(例えばHBB)をコードするポリヌクレオチドを、鎌状赤血球貧血を患う患者へ投与する。欠陥のあるタンパク質の発現をノックダウン又はノックアウトするため、患者には欠陥のあるヘモグロビン遺伝子をコードする遺伝子(例えば変異HbSをコードする遺伝子)を標的とするsiRNAもまた併用して投与される。治療は、その他の公知の医薬の投与、又は鎌状赤血球貧血を治療するために一般に用いられる治療をさらに含んでなってよい。
【図面の簡単な説明】
【0028】
【図1】ヒトeIF−5A1のアミノ酸配列及び様々な重要部位を示した図である。
【図2】K50及びK67におけるeIF−5A1の変異が、トランスフェクトしたタンパク質の蓄積を増大させることを示した図である。実施例1を参照のこと。
【図3】K47、K50、及びK67におけるeIF5A1の変異が、トランスフェクトしたタンパク質の蓄積を増大させることを示した図である。実施例2を参照のこと。
【図4】K50及びK67におけるeIF5A1の変異が、KAS細胞にトランスフェクトした際にアポトーシスの誘導をもたらすことを示した図である。実施例3を参照のこと。
【図5】K50及びK67におけるeIF5A1の変異が、KAS細胞にトランスフェクトした際にアポトーシスの誘導をもたらすことを示した図である。実施例4を参照のこと。
【図6】K50及びK67におけるeIF5A1の変異が、KAS細胞にトランスフェクトした際にアポトーシスの誘導をもたらすことを示した図である。実施例5を参照のこと。
【図7A】siRNAのトランスフェクション及びeIF−5A1を発現するように改変されたアデノウィルスによる処理が、KAS細胞にアポトーシスをもたらすことを示した図である。実施例6Aを参照のこと。
【図7B】eIF5A1 siRNA(標的#1(配列番号1)に対する)(siRNAコンストラクトの配列を図25に示す)による前処理は内在性eIF5A1の発現を減少させるが、アデノウィルスにより発現されたRNAi耐性のeIF5A1k50Aは蓄積させることを示した図である。実施例6Bを参照のこと。
【図7C】アデノウィルス感染前の、標的#1に対するeIF5A1 siRNAによる前処理が、ヒト多発性骨髄腫細胞におけるリン酸化NF−κBの発現を減少させることを示した図である。実施例6Cを参照のこと。
【図7D】アデノウィルス感染前の、標的#1に対するeIF5A1 siRNAによる前処理が、ヒト多発性骨髄腫細胞におけるリン酸化NF−κB及びICAM−1の発現を減少させることを示した図である。実施例6Dを参照のこと。
【図7E】ヒト多発性骨髄腫細胞における、siRNAにより仲介されるeIF5Aの抑制が、LPSにより仲介されるNFκBのDNA結合活性の誘導を阻害することを示した図である。NF−κBは多くの生存促進性及び抗アポトーシス経路を調節することから、eIF5A siRNAによるNF−κB活性の阻害は、eIF5AK50Rの過剰発現と組み合わせたときに、そのアポトーシス誘導を増大する能力を説明できる。
【図7F】siRNAによるKAS細胞の前処理が、IL−6の存在下でeIF5A1k50R遺伝子送達によるアポトーシスを増大させることを示した図である。実施例6Eを参照のこと。
【図8】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。示されるデータは、各群における全てのマウスの腫瘍容積である。実施例7を参照のこと。
【図9】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。示されるデータは、群あたりの平均腫瘍容積+/−標準誤差である。実施例7を参照のこと。
【図10】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の重量を減少させることを示した図である。実施例7を参照のこと。
【図11】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させ、かつ腫瘍の縮小をもたらすことを示した図である。実施例8を参照のこと。
【図12】eIF5A1 siRNAの静脈内(i.v.)投与、及びPEI/eIF5AlK50Rプラスミド複合体の腫瘍内(i.t.)投与が、多発性骨髄腫皮下腫瘍の縮小をもたらすことを示した図である。実施例9を参照のこと。
【図13A】eIF5A1プラスミド及びeIF5A1 siRNAによる処置が、多発性骨髄腫皮下腫瘍の生育を遅延させ、かつ腫瘍の縮小をもたらすことを示した図である。
【図13B】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が腫瘍の縮小をもたらすことを示した図である。
【図13C】eIF5A1 siRNAの静脈内(i.v.)投与、及びPEI/eIF5AlK50Rプラスミド複合体の腫瘍内(i.t.)投与が、多発性骨髄腫皮下腫瘍の縮小をもたらすことを示した図である。
【図14】eIF5A1プラスミド及びeIF5A1 siRNAの静脈内同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。実施例10を参照のこと。
【図15】eIF5A1 siRNAの静脈内(i.v.)投与、及びPEI/eIF5AlK50Rプラスミド複合体の静脈内(i.v.)又は腹腔内(i.p.)投与が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。実施例11を参照のこと。
【図16】eIF5A1プラスミド及びeIF5A1 siRNAによる処置が、多発性骨髄腫皮下腫瘍の生育を遅延させることを示した図である。
【図17】eIF5A1プラスミド及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させ、かつ腫瘍の縮小をもたらすことを示した図である。実施例12を参照のこと。
【図18】eIF5A1 siRNAの静脈内(i.v.)投与、及びPEI/eIF5AlK50Rプラスミド複合体の腫瘍内(i.t.)投与が、多発性骨髄腫皮下腫瘍の縮小をもたらすことを示した図である。実施例13を参照のこと。
【図19】EF1又はB29プロモーターのいずれかにより駆動されるeIF5A1K50Rプラスミド、及びeIF5A1 siRNAの同時投与が、多発性骨髄腫皮下腫瘍の生育を遅延させ、かつ腫瘍の縮小をもたらすことを示した図である(KAS−SQ−5)。実施例14を参照のこと。
【図20】eIF5A1 siRNAの同時投与が、EF1又はB29プロモーターのいずれかにより駆動されるeIF5A1K50Rプラスミドの多発性骨髄腫皮下腫瘍における抗腫瘍効果を増大させ、腫瘍の負荷量を減少させることを示した図である(KAS−SQ−5)。実施例15を参照のこと。
【図21】eIF5A1 siRNAが、肺腺癌細胞におけるAd−eIF5Aの感染によりもたらされたアポトーシスを相乗的に増大させることを示した図である。実施例16を参照のこと。
【図22】その構築について実施例17に記載される、pExp5Aのマップを示した図である。
【図23】pExp5A(3371bp)の予測される配列を示した図である。実施例17を参照のこと。
【図24】様々な細胞株におけるeIF5A1K50Rの再発現を示した図である。実施例18を参照のこと。
【図25】標的配列及び好ましいeIF5A1 siRNAの配列を示した図である。
【図26】多発性骨髄腫における有効性試験の結果を示した図である。実施例21を参照のこと。
【図27】多発性骨髄腫における有効性試験の結果を示した図である。実施例21を参照のこと。
【図28】eIF5A1k50RcDNAの配列を示した図である。
【図29】ヒトeIF5A1k50Rに対するヒトeIF−5A1のアラインメントを示した図である。
【図30】HA−eIF5AK50Rの発現におけるDNA:siRNA比の効果を示した図である。実施例23を参照のこと。
【図31】ナノ粒子のトランスフェクションにより誘導されるアポトーシスにおけるDNA:siRNA比の効果を示した図である。実施例24を参照のこと。
【図32】eIF5A1K50Rプラスミド及びeIF5A1 siRNA(siSTABLE又は非siSTABLE)を含むPEI複合体(N/P=6又は8)の投与が、多発性骨髄腫皮下腫瘍の生育を阻害し、かつ腫瘍の縮小をもたらすことを示した図である。実施例25を参照のこと。
【図33】JET PEI(商標)ナノ粒子が腫瘍組織によって効果的に取り込まれ、該ナノ粒子が同細胞にプラスミド及びsiRNAを送達することを示した図である。実施例26を参照のこと。
【図34】KAS細胞がアクチノマイシンDに反応してアポトーシスを起こすことを示した図である。KAS細胞は無処理か、又は0.5μg/mlのアクチノマイシンDにより処理された。処理開始後24時間目に細胞を回収、洗浄し、アポトーシス細胞をアネキシン/PI(BD Bioscience)を用いて標識してフローサイトメトリーにより解析した。
【図35】eIF−5A1の切断型が、アクチノマイシンD処理に反応して蓄積することを示した図である。KAS細胞を、0.5μg/mlのアクチノマイシンDによって0時間〜30時間の範囲の時間枠で処理した。細胞ライセートを回収し、eIF5A1に対する抗体を用いるウェスタンブロットによって分析した。eIF5A1の、より小さな分子量の(切断された)形の蓄積はアクチノマイシンD処理後おおよそ8時間後から観察され始め、残りの処理の間を通して存在した。同じ膜でのウェスタンブロットによる等しい負荷を、アクチンに対する抗体を用いて示した。
【図36】eIF−5A1の切断型が、完全長eIF−5A1よりも高い等電点を持つことを示した図である。KAS細胞は無処理か、又は0.5μg/mlのアクチノマイシンDにより処理された。処理開始後17時間目に細胞ライセートを回収し、2−Dゲル電気泳動によって分離した後、eIF5A1に対する抗体を用いてウェスタンブロット分析を行った。無処理サンプル中に大量の完全長eIF5A1が観察できるが、アクチノマイシンD処理サンプル中には高めのpIを有するeIF5A1の切断型の蓄積が観察できる。
【図37】切断型eIF−5A1の2−Dゲルの写真を示した図である。KAS細胞は0.5μg/mlのアクチノマイシンDにより処理された。処理開始後17時間目に細胞ライセートを回収し、2−Dゲル電気泳動によって分離した後、クマシーブルーによる染色を行った。マススペクトロメトリー分析のために収集したeIF5A1の切断型の位置を示す。
【図38】切断型eIF−5A1のマススペクトロメトリーによる配列決定結果を示した図である。(A)eIF5A1の完全長アミノ酸配列を黒で示す。eIF5A1の完全長配列と共に整列させた、マススペクトロメトリーにより同定された配列決定ペプチド(B)を下線で示す(A)。配列決定ペプチドから、eIF5A1の最初の6アミノ酸が欠失していることに留意されたい。
【図39】カスパーゼ3、8、及び9の阻害剤がeIF−5A1の切断型の形成を阻害することを示した図である。ヒト多発性骨髄腫KAS細胞を異なるカスパーゼ阻害剤と共に8時間インキュベートし、さらに16時間、0.5μg/mlのアクチノマイシンD(ActD)と共に同時処理した。細胞ライセートを回収した。タンパク質を2D−PAGEによって分離し、eIF5Aに対する抗体を用いるウェスタン分析に供した。使用したカスパーゼ阻害剤はZ−VAD−FMK(一般的なカスパーゼ阻害剤)、Z−LEHD−FMK(カスパーゼ9阻害剤)、Z−DEVD−FMK(カスパーゼ3/7阻害剤)、Z−IETD−FMK(カスパーゼ6/8阻害剤)、及びZ−YVAD−FMK(カスパーゼ1阻害剤)である。データにより、一般的なカスパーゼ阻害剤、並びに、カスパーゼ3、8、及び9の阻害剤は全て、ActDによりKAS細胞に誘導されるアポトーシスの間に蓄積されるeIF5Aの切断型の形成を強力に阻止することが示された。カスパーゼ1の阻害剤もまた、完全な遮断ではないが、切断産物の形成を減少させる。これらの結果は、eIF5A1が遺伝毒性のストレス(アクチノマイシンDにより誘導される)後にカスパーゼにより切断され;eIF5A1の切断はその活性を変化させる、すなわちアポトーシス促進活性を増大させ得ることを示した。
【図40】脱アセチル化酵素阻害剤(ニコチンアミド)が、eIF−5A1の切断型の形成を促進することを示した図である。リジン47がアセチル化され得るeIF5Aは、Sir2関連脱アセチル化酵素であるHst2の基質である(Shirai et al., 2008)。ヒト子宮頸癌Hela S3細胞を、0.5μg/mlアクチノマイシンD(ActD)及び/又はSir2の強力な阻害剤である20μMニコチンアミドにより、示される時間の長さで処理した。細胞ライセートを回収した。タンパク質を2D−PAGEによって分離し、eIF5Aに対する抗体を用いるウェスタン分析に供した。ActD又はニコチンアミド単独によるHela S3細胞の処理は、eIF5Aの切断型の蓄積を誘導しない。しかしながら、アクチノマイシンD及びニコチンアミドの両方により処理されたHeLa細胞では、eIF5Aの切断型はActDへの暴露後少なくとも4時間目に上昇することから、アポトーシスの間にアセチル化がeIF5Aの切断を促進し得ることが示唆される。
【図41】切断型ヒトeIF5A1のDNA配列を示した図である。
【図42】完全長eIF5A1、切断型eIF5A、及び切断部位のタンパク質配列を示した図である。
【図43】アデノウィルスコンストラクトによるHela細胞の感染が、eIF5A導入遺伝子の蓄積をもたらすことを示した図である。Hela S3細胞を、異なるアデノウィルスコンストラクトにより、細胞あたり500感染単位にて感染させた。感染後48及び72時間目に細胞ライセートを回収し、SDS−PAGE、ならびにeIF5A及びアクチンに対する抗体を用いるウェスタンブロット分析に供した。切断型eIF5Aの蓄積が観察され得る。
【図44】アクチノマイシンD、ニトロプルシドナトリウム、又はIL−6の離脱による処理を通したアポトーシスの誘導が、ヒト骨髄腫細胞にeIF5A1の切断型(eIF5A1Δ1−6)の蓄積をもたらすことを示した図である。KAS−6/1細胞を示されるように処理し、次に2D−PAGE、及びeIF5Aに対する抗体を用いるウェスタンブロット分析に供した。アクチノマイシンD、NOドナー、ニトロプルシドナトリウム、又はIL−6の欠乏を通したKAS−6/1細胞におけるアポトーシスの誘導は、全て切断型eIF5A1の蓄積を誘導した。Ad−A1Δ(2−6)又はAd−A1Δ(2−6)/K50Rのいずれかを用いる切断型eIF5A1の過剰発現は、アポトーシスの間に産生される切断型eIF5A1(eIF5A1Δ1−6)よりも低い等電点を有するeIF5A1タンパク質の蓄積を誘導したことから、eIF5A1Δ1−6にはその他の翻訳後修飾が存在することが示される。
【図45】eIF5A1D6E及びeIF5A1D6E/K50Rは、アポトーシスの間にアクチノマイシンDにより誘導される切断に対して耐性であることを示した図である。ヒト多発性骨髄腫KAS−6/1細胞を示されるように処理し、次に2D−PAGE、及びeIF5Aに対する抗体を用いるウェスタンブロット分析に供した。過剰発現されたeIF5A1(Ad−A1)をアクチノマイシンDによる処理の間に切断に供した。しかしながら、eIF5A1D6E又はeIF5A1D6E/K50Rは、アクチノマイシンDによる処理後の切断に耐性である。これらの結果は、eIF5A1のアミノ酸3〜6がカスパーゼ切断部位を構成するという仮説に一致する。
【図46】eIF5A1が、in vitroで組み替えカスパーゼにより切断されることを示した図である。ヒトeIF5A1(A1)及び変異eIF5A1D6E(D6E)及びeIF5A1D6E/K50R(D6E/K50R)を、HA−タグ化タンパク質の発現をもたらすpHM6ベクター(Roche)にサブクローン化した。pHM6−HA−A1、pHM6−HA−A1(D6E)、及びpHM6−HA−A1(D6E/K50R)をin vitroで転写及び翻訳し(TNT T7 Quick Coupled Transcription/Translation Systems; Promega)、次にTranscend Non-Radioactive Translation Detection Systems(Promega)を用い、製造者の指示に従って標識した。各3マイクロリットルのin vitro翻訳産物を、1マイクロリットルの組み替えカスパーゼ(カスパーゼ1、2、3、6、7、8、9、又は10;Calbiochem)と共にカスパーゼ反応緩衝液(50mM Hepes、0.1%CHAPS、10%スクロース、100mM NaCl、10mM DTT、1mM EDTA)中で37℃にて2時間インキュベートした。最終産物をSDS−PAGEに供し、ビオチン化タンパク質をストレプトアビジン−HRPとのインキュベーション、それに続く化学発光の検出により可視化した。野生型eIF5A1は、カスパーゼ2、3、6、7、8、9、及び10により切断されるが、カスパーゼ1によっては切断されない。HA−タグ化eIF5A1D6E及びeIF5A1D6E/K50Rは、カスパーゼ3、7、及び8によってin vitroで効果的に切断されるが、カスパーゼ9及び10によっては効率的に切断されない。しかしながらこの切断は、in vitro系における高度に豊富なカスパーゼ活性、同様にグルタミン酸とアスパラギン酸の間の高い構造類似性による可能性があり、これによって低い効率ではあるが(特にeIF5A1D6E及びeIF5A1D6E/K50Rはin vivoアッセイでの切断には耐性であったことから(図45))、eIF5A1D6Eの切断が許容される。
【図47】Ad−eIF5A1Δ(2−6)又はAd−eIF5A1K50RΔ(2−6)によるHela細胞の感染は、野生型Ad−eIF5A1又はAd−eIF5A1K50Rによる感染に比較してアポトーシスの増大をもたらすことを示した図である。ヒト子宮頸癌Hela S3細胞を、異なるアデノウィルスコンストラクトにより、細胞あたり500感染単位にて感染させた。感染後48又は72時間目に細胞を回収し、アネキシンV/PI染色に供した。次に細胞をフローサイトメトリーにより選別した。アポトーシスの初期段階の細胞(アネキシンV+/PI−)、同様にアポトーシスの後期段階(アネキシンV+/PI+)の細胞のパーセンテージを示す。アミノ酸2〜6の切断によってeIF5A1又はeIF5A1K50Rのアポトーシス活性は増大する。逆に、予測される切断部位の変異によってeIF5A1又はeIF5A1K50Rのアポトーシス活性は低下するが、無にはならない。これらの結果は、カスパーゼにより仲介されるeIF5A1の切断が、eIF5A1の切断型のアポトーシス活性を増大させることを示す。
【図48】アポトーシスにおける、カスパーゼにより仲介されるeIF5A1の切断の寄与についてのモデルを示した図である。
【図49】KASヒト骨髄腫細胞におけるアクチノマイシンD処理後に、切断型eIF5Aが核に蓄積することを示した図である。
【図50】KASヒト骨髄腫細胞におけるアクチノマイシンD処理後に、切断型eIF5Aが核に蓄積することを示した図である。略語一覧:UN=無処理;actD=アクチノマイシンDによる処理;C=細胞質分画;N=核分画、α−チューブリン及びα−PCNAはコントロールである。この図はまた、完全長eIF5Aは細胞質と核の間に均等に分布するが、切断型は主に核内に蓄積することも示す。
【図51】eIF5Aの切断型が複数の細胞株及び細胞型において異なるアポトーシス刺激に反応して蓄積することにより、カスパーゼにより仲介されるeIF5Aの切断はアポトーシスの間の一般的な現象であることが示されることを示した図である。この図はまた、完全長eIF5Aは細胞質と核の間に均等に分布するが、切断型は主に核内に蓄積することも示す。
【図52】切断型eIF5Aが、異なる細胞株において異なるアポトーシス刺激[KASヒト多発性骨髄腫細胞におけるIL−6の欠乏、又はIL−6及びFBSの欠乏による処理後、並びにUACC1598(ヒト卵巣癌細胞株)におけるアクチノマイシンDによる処理後]に反応して産生されるという研究結果を示した図である。
【図53】ヒトeIF5A1及びeIF5A2のヌクレオチド配列を示した図である。
【0029】
[発明の詳細な説明]
本発明は、eIF−5A1の切断型をコードする単離されたポリヌクレオチド、同様に切断型eIF−A1ポリペプチドを提供する。切断型eIF−5A1ポリヌクレオチドは、癌細胞のアポトーシス及び死滅の誘導に有用である。切断型ポリヌクレオチドは発現ベクター内に使用され得り、これは次に哺乳類へ投与される。切断型eIF−5Aは哺乳類内に発現し、癌細胞を死滅させる。約17kDaである完全長eIF−5A1タンパク質に対し、切断型eIF−5A1タンパク質は約16kDAである。
【0030】
アポトーシスを起こす細胞内には、eIF5Aのより小さな分子量型(〜16kDa)の形成が観察されている。このことは、サイトカインカクテルにより処理されたベータ膵島細胞、及び細胞傷害性薬物のアクチノマイシンDにより処理されたヒト骨髄腫細胞(KAS細胞)において観察されている。図35を参照されたい。eIF5Aのより小さな分子量型はまた、Ad−eIF5A1の感染後にアポトーシスを起こしているHeLa子宮頸癌細胞に蓄積することも見出されており、このことからeIF5Aのより小さな分子量型は、選択的スプライシングの結果というよりはプロテアーゼによる切断によるものであると示唆される。eIF5Aのより小さな分子量型の蓄積は完全長ヒプシン化eIF5Aの量の著しい減少を伴うという観察は、eIF5Aのより小さな分子量型が選択的スプライシングというよりは切断によるものであることをさらに支持する。図36を参照されたい。
【0031】
本発明は、切断型eIF5A1タンパク質をコードするeIF5A1ポリヌクレオチドを含んでなる組成物を提供する。この組成物は、被験体の癌細胞又は腫瘍にアポトーシスを誘導するために用いられる薬剤の製造に有用である。eIF5A1ポリヌクレオチドは、好ましくは図38に示される配列番号37に記載のアミノ酸配列を含んでなる、切断型eIF5A1タンパク質をコードする。特定の実施形態によれば、eIF5A1ポリヌクレオチドは約16kDAの切断型eIF5A1タンパク質をコードする。
【0032】
組成物及び又は薬剤は、ヒトを含む哺乳類に投与できる。本明細書で用いられる「被験体」は哺乳類及びヒトを含む。
【0033】
アポトーシスの誘導は、癌細胞又は腫瘍において以下の効果を持ち得る:癌細胞若しくは腫瘍の生育を遅くする、癌細胞若しくは腫瘍の生育を停止する、又は癌細胞を死滅させるか若しくは腫瘍サイズを縮小させる、及び上記のいずれの組み合わせ。いずれの癌も治療でき、特定の実施形態によれば、癌は多発性骨髄腫である。
【0034】
特定の実施形態によれば、eIF5A1ポリヌクレオチドは配列番号38(図41に示される)に記載の配列を含んでなる。特定の実施形態によれば、eIF5A1ポリヌクレオチドはプラスミド又は発現ベクター内に構成される。プラスミド又は発現ベクターについては、以下にさらに詳しく記載される。特定の実施形態によれば、発現ベクターはアデノウィルス発現ベクター又はpHM6である。特定の実施形態によれば、組成物又は薬剤が多発性骨髄腫の治療に使用される場合、発現ベクターはB細胞特異的プロモーター(すなわちB29)のような組織特異的プロモーターを含んでなる。発現ベクターはpCpGプラスミドを含んでなってよい。以下にさらに詳しく考察されるように、発現ベクターはポリエチレンイミンと複合されてよい。
【0035】
組成物又は薬剤は、好ましくは腫瘍内、静脈内、又は皮下投与される。
【0036】
本発明はまた、ポリヌクレオチドが配列番号38(図41に示される)に記載の配列を含んでなる、切断型eIF5A1タンパク質をコードする単離されたポリヌクレオチドも提供する。
【0037】
本発明はまた、切断型タンパク質が配列番号37に記載のアミノ酸配列を含んでなる、切断型eIF5A1タンパク質をコードする単離されたポリヌクレオチドも提供する。
【0038】
以下に考察されるように、eIF5A1はカスパーゼにより仲介される切断によって形成される。従って本発明はさらに、カスパーゼにより仲介されるeIF5A1の切断によって形成される、単離された切断型eIF5A1ポリペプチドを提供する。
【0039】
本発明はさらに、組成物、又は切断型eIF5A1タンパク質をコードするeIF5A1ポリヌクレオチドを、完全長eIF5A1ポリヌクレオチドと組み合わせて含んでなる薬剤の製造のための該組成物の使用を提供する。この組成物は、被験体の癌細胞又は腫瘍にアポトーシスを誘導する薬剤の製造に有用である。切断型eIF5Aタンパク質をコードするeIF5A1ポリヌクレオチドについては上述の通りである。完全長eIF5A1ポリヌクレオチドは、配列番号35に記載のアミノ酸配列を含んでなるタンパク質をコードする。完全長eIF5A1ポリヌクレオチドについては、以下にさらに詳しく記載される。特定の実施形態によれば、eIF5A1ポリヌクレオチドは配列番号38に記載の配列を含んでなり、完全長eIF5A1ポリヌクレオチドは配列番号43(図53に示される)に記載の配列を含んでなる。
【0040】
本明細書に記載されるように、完全長及び切断型ポリヌクレオチドは発現ベクター内に存在してよい。加えてベクターは、本明細書に記載されるプロモーター、同様にその他のいずれの有用なプロモーターを含んでなってよい。
【0041】
特定の実施形態によれば、完全長eIF5Aポリヌクレオチドは変異eIF5A1をコードし、ここで該変異はデオキシヒプシンシンターゼによるヒプシン化を阻止若しくは阻害し、並びに/又は、該変異はユビキチン化部位及び/若しくはアセチル化部位に存在する。変異については、以下にさらに詳しく記載される。特定の実施形態によれば、変異はK50A、K50R、K67A、K47R、K67R、K50A/K67A、K50A/K47R、K50A/K67R、K50R/K67A、K50R/K47R、K50R/K67R、及びK47A/K67Aからなる群より選択される。
【0042】
本発明はまた、切断型eIF5Aをコードするポリヌクレオチド、変異eIF5Aをコードするヌクレオチド、及びeIF5Aの3’UTRを標的とするsiRNAを含んでなる組成物を提供する。変異eIF5Aをコードするヌクレオチド、及びeIF5Aの3’UTRを標的とするsiRNAの組み合わせ/二元的使用については、以下にさらに完全に記載される。該組成物は、癌細胞及び腫瘍にアポトーシスを誘導するため、被験体に投与される薬剤の製造に有用である。
【0043】
特定の好ましい実施形態によれば、siRNAはeIF5A1の配列である5’−GCT GGA CTC CTC CTA CAC A−3’を標的とする。siRNAはdsRNAであり、dsRNAの一本の鎖は5’−GCU GGA CUC CUC CUA CAC A−3’の配列を含んでなる。特定の実施形態によれば、siRNAは血清中での分解を阻止するために安定化される。
【0044】
完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチドは、発現ベクター内に存在してよい。特定の実施形態によれば、完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチド及び/又はsiRNAは、ポリエチレンイミンと複合される。完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチド及び/又はsiRNAは、独立してポリエチレンイミンと複合される。
【0045】
本発明はまた、本明細書に記載される、例えば切断型eIF5A1をコードするヌクレオチドを含んでなり、任意に完全長eIF5Aをコードするヌクレオチド又は完全長変異eIF5Aをコードするヌクレオチドを含んでなり、かつ任意にeIF5Aの3’UTRを標的とするsiRNAを含んでなる組成物又は薬剤を哺乳類に提供することによる、哺乳類の癌細胞又は哺乳類の腫瘍にアポトーシスを誘導する方法も提供する。特定の実施形態によれば、癌は多発性骨髄腫であり、組成物/薬剤は静脈内、腹腔内、又は腫瘍内投与される。
【0046】
本発明はまた、被験体内での内在性遺伝子の発現をノックアウト又はノックダウンするための、内在性遺伝子を標的とするsiRNA、及び、ポリヌクレオチドによりコードされるタンパク質をホスト内で発現させるために被験体へ提供される、送達媒体/発現ベクター内の遺伝子をコードするポリヌクレオチドを組み合わせた使用にも関する。
【0047】
この組み合わせは、欠陥又は変異のあるタンパク質の存在、すなわち被験体内で産生されたタンパク質が、その欠陥のある構造のために、必要な機能を実行できないか、又は代わりに代謝経路若しくは生体分子の相互作用を不調にすることにより起こる、疾病又は状態を有する被験体の治療に有用である。siRNAは欠陥のあるタンパク質をコードする遺伝子を標的として設計され、その欠陥のあるタンパク質の発現をノックダウン又はノックアウトする。正常な(欠陥のない)タンパク質をコードするポリヌクレオチドが被験体に投与され、正常なタンパク質がその必要な機能を実行できるように被験体内で発現する。
【0048】
別の実施形態によれば、所望のタンパク質をコードするポリヌクレオチドの投与の代わりに、被験体へタンパク質が投与される。本明細書では、タンパク質、ペプチド、及びポリペプチドの語は互換的に用いられる。
【0049】
siRNAは、欠陥のあるタンパク質の内在性の発現をノックダウン又はノックアウトするが、同時に投与された正常タンパク質をコードするポリヌクレオチドの外来性の発現に影響しないような遺伝子の特定の領域を標的として好ましく設計される。例えばsiRNAは、投与されたセンスコンストラクト(タンパク質をコードするポリヌクレオチド)の外来性の発現に影響しないように、3’UTRを標的とし得る。欠陥のあるタンパク質の内在性の発現をノックダウン又はノックアウトすることにより、外来性のポリペプチドから発現する正常タンパク質と競合する欠陥のあるタンパク質は、減少するか又は無になるであろう。
【0050】
この応用が有用となり得る疾病状態の一例は、鎌状赤血球貧血に関する。鎌状赤血球貧血は、赤血球(red blood cell:RBC)に見出され、身体全体への酸素の運搬を補助するタンパク質であるヘモグロビンに影響を及ぼす血液疾患である。
【0051】
鎌状赤血球貧血は、ヒトが変異ヘモグロビン(Hbs)の発現をもたらす2つの異常な遺伝子(各両親から1つずつ)を受け継いだ際に発生する。変異ヘモグロビンは、RBCの形を変える原因となる。正常ヘモグロビン(ヘモグロビンA、又はHbA)を有する赤血球は血流を通して容易に移動し、身体の全ての細胞に酸素を送達する。これらは、非常に微小な血管であっても容易に「圧搾」して通り抜けることができる。鎌状赤血球貧血は、ヘモグロビンの異常型(HbS)が、共に凝集する傾向があり、赤血球を粘着性で、堅く、さらに脆弱にさせること、かつそれらを曲がった鎌状にさせることから起こる。
【0052】
数百のHBB遺伝子の変異形が知られているが、鎌状赤血球貧血は、最も一般的にはヘモグロビン変異形HbSにより引き起こされる。この変異形では、疎水性アミノ酸のバリンが、HBBポリペプチド鎖の6番目のアミノ酸の位置における親水性のグルタミン酸に取って代わる。この置換によりタンパク質構造の外側に疎水性の点が作り出され、これは隣接するヘモグロビン分子のベータ鎖の疎水性領域に固着する。HbS分子がこのように共に凝集(重合)することによって強固な線維となり、赤血球を「鎌状」にする。
【0053】
重合は、赤血球が身体全体の様々な組織に運搬する酸素分子を放出した後のみに起こる。ヘモグロビンが酸素を結合できる場である肺に赤血球がいったん帰還すると、HbSの長い線維は脱重合するか、又は単一の分子に分裂する。重合と脱重合の間の循環により、赤血球の膜は強固になる。これらの赤血球の剛性、及び酸素を運搬しないときのそれらの歪んだ形は、微小血管の封鎖をもたらし得る。この封鎖は疼痛の発現の原因となり得り、かつ臓器を損傷し得る。
【0054】
鎌状赤血球貧血は、常染色体劣性遺伝疾患である。疾病が発現するためには、ヒトは2コピーのHbS変異形、又は1コピーのHbS及び1コピーの別の変異形のいずれかを受け継ぐ必要がある。1コピーの正常HBB遺伝子(HbA)及び1コピーのHbSを有する保因者は、鎌状細胞の形質を有すると説明され、疾病の症状を表さない。
【0055】
従って本発明の一実施形態は、鎌状赤血球貧血を有する被験体を治療する方法を提供する。患者には、HBB遺伝子を標的とするsiRNAが投与される。siRNAは、ヘモグロビンのHbS変異形の発現をノックダウン、好ましくはノックアウトするように設計される。被験体が正常ヘモグロビンを発現するように、被験体へ正常ヘモグロビンをコードするポリヌクレオチドが提供される。siRNAはまた、正常ヘモグロビンをコードする外来性ポリヌクレオチドの発現に干渉しないようにも設計される。従って、被験体はもはや変異ヘモグロビンを作らず(又は実質的に少なく作り)、代わりに正常健常ヘモグロビンを作ることによって、正常に機能するさらに正常な赤血球がもたらされる。
【0056】
本発明はまた、タンパク質に、疾病状態を引き起こすか又は導く翻訳後修飾が起こる状況においても有用である。siRNAは内在性タンパク質の発現をノックダウンするために用いられ、これにより翻訳後修飾されるものは無くなるか又は少なくなる。次に外来性発現のために、タンパク質をコードするポリヌクレオチドが患者へ提供される。タンパク質は、翻訳後修飾できないように改変される。次にこのタンパク質はその適切な使用のために身体に利用可能であるが、翻訳後修飾できないために疾病状態を導き得ない。当業者は異なる翻訳後修飾について理解するであろう。例えば翻訳後、アミノ酸の翻訳後修飾は、タンパク質に酢酸塩、リン酸塩、様々な脂質及び炭水化物のようなその他の生化学官能基を付着させることにより、アミノ酸の化学的性質を変化させることにより(例えばシトルリン化)、又はジスルフィド架橋の形成のように構造を変化させることにより、タンパク質の機能の範囲を拡張する。また、酵素はタンパク質のアミノ末端からアミノ酸を除くか、又は中間でペプチド鎖を切断し得る。例えばペプチドホルモンのインスリンは、ジスルフィド結合の形成後に2度切断され、鎖の中間からプロペプチドが除かれ;得られたタンパク質はジスルフィド結合によって連結する2つのポリペプチド鎖から成る。また、最も新生のポリペプチドはアミノ酸のメチオニンから始まるが、これはmRNAの「開始」コドンもまた、このアミノ酸をコードするためである。このアミノ酸は通常、翻訳後修飾の間に取り除かれる。リン酸化のようなその他の修飾は、タンパク質の作用を制御するための共通機構、例えば酵素の活性化又は不活性化の一部である。別の翻訳後修飾は、デオキシヒプシンシンターゼ(DHS)による真核生物翻訳開始因子5A(eIF5A)のヒプシン化を含む。
【0057】
従って本発明は、タンパク質をコードするポリヌクレオチドが患者へ提供され、かつ患者に発現される、被験体内の遺伝子の発現を変化させる方法を提供する。タンパク質は正常/野生型タンパク質、又は変異したタンパク質であってよい。対応する内在性遺伝子の発現は、被験体へ投与されるsiRNAによって抑制される。
【0058】
該方法はさらに、標的タンパク質をコードするポリヌクレオチドを含んでなるコンストラクトを提供することを含んでなり、該ポリヌクレオチドは標的タンパク質を産生するために被験体内に発現される。内在遺伝子が欠陥のあるタンパク質を発現する特定の実施形態によれば、ポリヌクレオチドは正常/健常タンパク質のいずれかをコードするように設計される。siRNAは欠陥のある内在性タンパク質の発現を抑制するために投与される。内在遺伝子が正常健常タンパク質を発現する特定の実施形態によれば、ポリヌクレオチドは、正常/健常タンパク質又は非変異タンパク質に起こるような翻訳後修飾のできない変異タンパク質をコードするように設計される。siRNAは内在性タンパク質の発現を抑制するために投与され、翻訳後修飾に利用できるこのタンパク質は減少する。
【0059】
特定の実施形態によれば、siRNAは外来性ポリヌクレオチドの発現に影響しないような遺伝子の領域を標的として選択又は設計される。例えばsiRNAは3’UTR又は3’末端を標的としてよい。
【0060】
siRNAは、ネイキッドsiRNA又は血清に対して安定化されたネイキッドsiRNAのいずれかとして患者へ送達され得る。siRNAは全身性に、すなわちIP又はIVのいずれかにより注射され得る。あるいは、siRNAは身体の望まれる領域へ局所的に注射又は送達され得る。特定の実施形態によれば、siRNAは、限定はされないがデンドリマー、リポソーム、又はポリマーのような送達ビヒクルにおいて投与され得る。
【0061】
所望のタンパク質をコードするポリヌクレオチドは、ヌクレオチドの発現を提供するか又は許容するいずれの送達手段を通して投与され得る。本明細書で、ポリヌクレオチド及びヌクレオチドの語は互換的に用いられる。送達は、限定はされないがプラスミド、発現ベクター、ウィルス性コンストラクト、アデノウィルスコンストラクト、デンドリマー、リポソーム、又はポリマーのような、いずれのウィルス性又は非ウィルス性機構を通してよい。
【0062】
特定の実施形態によれば、ポリヌクレオチドの発現のために、還元型CpGジヌクレオチドを有する発現プラスミドが用いられる。ポリヌクレオチドの発現を促進できるいずれのプロモーターも使用されてよく、これは治療法に望まれる応用に基づいて選択され得る。例えば、多発性骨髄腫を死滅させるためには、ヒトB29プロモーター/エンハンサーのようなB細胞に特異的なプロモーターが望ましいであろう。別の実施形態によれば、プロモーターは別の組織特異的プロモーターであってよいか、又はシステムプロモーターであってよい。標的タンパク質をコードするポリヌクレオチドは、IV若しくは皮下注射、又はその他のいずれの生物学的に適した送達機構を通して送達され得る。あるいはポリヌクレオチドは、リポソーム、又は標的腫瘍若しくは癌細胞へのDNA(又はプラスミド若しくは発現ベクター)送達のために提供される、その他のいずれの適切な「担体」又は「ビヒクル」において送達され得る。合成DNA送達系についての総説は、例えばLuo, Dan, et al, Nature Biotechnology, Vol. 18, January 2000, pp. 33-37を参照されたい。従って、ヌクレオチド/プラスミド/発現ベクターは、リポソーム、デンドリマー、又は同様の無毒性ナノ粒子のような、ナノメートルサイズのビヒクルを通して送達されることが好ましい可能性がある。ビヒクルは、好ましくは、有効量のヌクレオチド/プラスミド/発現ベクターを被験体、腫瘍、又は癌細胞へ送達する間、ヌクレオチド/プラスミド/発現ベクターを早期クリアランスから、又は免疫反応の原因から保護する。典型的なビヒクルは、ヌクレオチド/プラスミド/発現ベクターに付随する単純なナノ粒子から、特異的な細胞受容体を標的とするためにその表面に付着するリガンドを有するペグ化リポソームのような、さらに複雑なペグ化ビヒクルまでに及び得る。
【0063】
リポソーム及びペグ化リポソームは当該技術分野において公知である。従来のリポソームでは、送達される分子(すなわち小分子薬、タンパク質、ヌクレオチド、又はプラスミド)はリポソームの中央の腔内に含まれる。当業者は、分子送達に有用な「ステルス」標的化、及び陽イオンリポソームの存在もまた認識するであろう。例えば、Hortobagyi, Gabriel N., et al, J. Clinical Oncology, Vol. 19, Issue 14 (July) 2001 :3422-3433 及び Yu, Wei, et al., Nucleic Acids Research. 2004, 32(5);e48を参照されたい。リポソームは静脈内に注射でき、かつそれらの表面がさらに親水性になるように修飾でき(二重層へのポリエチレングリコールの添加(「ペグ化」)により)、血流でのそれらの循環時間は増大する。これらは「ステルス」リポソームとして知られ、ドキソルビシン及びミトキサントロンのような親水性(水溶性)の抗癌剤に対する担体として特に有用である。腫瘍細胞のような標的細胞へ薬物を運搬するリポソームの特異的な結合特性を促進するために、リポソームの表面に抗体、タンパク質、ペプチドなどのような特異的分子が付着され得る。例えば、癌細胞の表面にある受容体に対する抗体が、リポソームを癌細胞に向けるために使用されてよい。多発性骨髄腫を標的にする場合、リポソームを多発性骨髄腫細胞に向けるために、例えば葉酸塩、Il−6、又はトランスフェリンが使用されてよい。
【0064】
デンドリマーもまた当該技術分野において公知であり、好ましい送達ビヒクルを提供する。デンドリマーについての考察は、例えば、Marjoros, Istvan, J., et al, "PAMAM Dendrimer-Based Multifunctional Conjugate for Cancer Therapy: Synthesis, Characterization, and Functionality," Biomacromolecules, Vol. 7, No. 2, 2006; 572-579, 及び Majoros, Istvan J., et al., J. Med. Chem, 2005. 48, 5892-5899を参照されたい。
【0065】
好ましい実施形態によれば、送達ビヒクルはポリエチレンイミンナノ粒子を含んでなる。典型的なポリエチレンイミンナノ粒子は、現在Polyplus Transfection, Incにより製造されるin vivo-jetPEI(商標)である。In vivo-jetPEI(商標)は、DNA及びsiRNAの送達薬剤として有用な陽イオンポリマートランスフェクション剤である。Polyplus Transfectionから市販されているIn vivo-jetPEI(商標)は、動物において信頼性のある核酸送達を提供する直鎖ポリエチレンイミン試薬である。これは遺伝子治療(Ohana et al, 2004. Gene Ther Mol Bio 8:181-192; Vernejoul et al, 2002. Cancer Research 62:6124-31)、RNA干渉(Urbain-Klein et al., 2004. Gene therapy 23:1-6; Grezelinski et al., 2006. Human Gene Therapy 17:751-66)、及び遺伝子ワクチン接種(Garzon et al., 2005. Vaccine 23:1384-92)に用いられる。In vivo JET-PEIは現在、癌の遺伝子治療のための送達ベクターとしてヒトの治験に使用されている(Lemkine et al., 2002. Mol. Cell. Neurosci. 19:165-174)。
【0066】
In vivo-jetPEI(商標)は核酸をおおよそ50nmのナノ粒子に凝縮し、これは数時間安定である。この独特な保護機構の結果として、注射後の血液細胞の凝集は、組織内での制限された拡散、赤血球の凝集、及び微小塞栓を防止するその他の試薬に比較して減少する。これらのナノ粒子は、組織内へ拡散し、エンドサイトーシスによって細胞へ入るためには十分に小さい。In vivo-jetPEI(商標)は、エンドソームからの核酸の放出及び核膜を横断しての移動を支持する。
【0067】
好ましい実施形態によれば、siRNA、及びポリヌクレオチドを含んでなるベクター/プラスミドの両方が、in vivo-jetPEI(商標)複合体を通して被験体に投与される。siRNA、及びポリヌクレオチドを含んでなるベクター/プラスミドは、ポリエチレンイミン又はin vivo-jetPEI(商標)複合体のようなポリマー複合体を通して共に複合されてよいか、又は別々にポリマーと複合されてよい。例えば、siRNA、及びポリヌクレオチドを含んでなるベクター/プラスミドが別々に被験体へ投与される場合(時間及び/又は送達部位が別々であるという意味で)には、異なる担体と複合されたsiRNA及びポリヌクレオチドを用意することが好ましい。同一部位において同時に投与され得る場合には、siRNA及びポリヌクレオチドを共に複合することが好ましい。
【0068】
別の実施形態によれば、被験体内で発現され得るポリヌクレオチドを送達するために投与されるプラスミド又はベクターの代わりに、タンパク質それ自体が被験体へ送達される。タンパク質は単離されてよいか、又は合成されてよい。
【0069】
本発明の一実施形態は、哺乳類及びヒトを含む被験体の癌を治療する方法を提供する。癌の治療は、癌細胞におけるアポトーシスの誘導、癌細胞の死滅、癌細胞の数の減少、及び腫瘍容積/重量の減少を含むがこれらに限定されない。該方法は、eIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドを含んでなる組成物を投与することを含んでなる。組成物並びにeIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドについては以下で考察する。
【0070】
全ての細胞は真核生物翻訳開始因子5A(「eIF−5A」)(又は、本明細書では「因子5A」とも称される)を産生する。哺乳類細胞は、eIF−5A1の2つのアイソフォーム(eIF−5A1及びeIF−5A2)を産生する。eIF−5A1はアポトーシスを起こしている細胞内で上昇していることから、アポトーシス特異的eIF−5Aと称されている。ヒトeIF−5A1は受入番号NM001970を有し、図1に示される。eIF−5A1は、アポトーシスに必要なタンパク質をコードするmRNAの核サブセットの核外輸送の原因であると信じられている。eIF−5A2は、細胞増殖に必要なタンパク質をコードするmRNAの核サブセットの核外輸送の原因であると信じられていることから、増殖eIF−5Aと称されている。Liu & Tartakoff (1997) Supplement to Molecular Biology of the Cell, 8, 426a. Abstract No. 2476, 37th American Society for Cell Biology Annual Meeting, 及び Rosorius et al. (1999) J. Cell Science, 112, 2369-2380を参照されたい。
【0071】
両方の因子5Aは、デオキシヒプシンシンターゼ(「DHS」)により翻訳後修飾される。DHSはeIF−5Aをヒプシン化する。独特なアミノ酸であるヒプシンは調べられた全ての真核生物及び古細菌に見出されるが、真正細菌には見出されず、eIF−5Aは知られている唯一のヒプシン含有タンパク質である。Park (1988) J. Biol. Chem., 263, 7447-7449; Schumann & Klink (1989) System. Appl. Microbiol, 11, 103-107; Bartig et al. (1990) System. Appl. Microbiol, 13, 112-116; Gordon et al (1987a) J. Biol. Chem., 262, 16585-16589。ヒプシン化eIF−5Aは2つの翻訳後工程によって形成され:第1工程は、デオキシヒプシンシンターゼにより触媒される、スペルミジンの4−アミノブチル成分の、前駆体eIF−5Aの特定のリジンのα−アミノ基への転移による、デオキシヒプシン残基の形成である。第2工程は、ヒプシンを形成するための、デオキシヒプシンヒドロキシラーゼによるこの4−アミノブチル成分のヒドロキシル化に関する。
【0072】
eIF−5Aのアミノ酸配列は種の間でよく保存されており、eIF−5Aのヒプシン残基周囲のアミノ酸配列は厳密に保存されていることから、この修飾は生存のために重要であるかもしれないことが示唆される。Park et al. (1993) Biofactors, 4, 95-104。この仮定は、これまでに酵母に見出されたeIF−5Aの両方のアイソフォームの不活性化、又はその活性化によって第1工程を触媒するDHS遺伝子の不活性化が細胞分裂を遮断するという観察により、さらに支持される。Schnier et al (1991) Mol Cell. Biol, 11, 3105-3114; Sasaki et al (1996) FEBS Lett., 384, 151-154; Park et al (1998) J. Biol. Chem., 273, 1677-1683。しかしながら、酵母におけるeIF−5Aタンパク質の枯渇がもたらしたものは総タンパク質合成のわずかな減少のみであったことから、eIF−5Aはタンパク質の全体的な合成よりも、mRNAの特定のサブセットの翻訳のために必要とされ得ることが示唆される。Kang et al. (1993), "Effect of initiation factor eIF-5A depletion on cell proliferation and protein synthesis," in Tuite, M. (ed.), Protein Synthesis and Targeting in Yeast, NATO Series H。eIF−5Aを結合するリガンドがより高く保存されたモチーフを共有するという最近の知見もまた、eIF−5Aの重要性を支持する。Xu & Chen (2001) J. Biol. Chem., 276, 2555-2561。加えて、修飾されたeIF−5Aのヒプシン残基が配列特異的なRNAの結合に必須であることが見出され、かつ結合によってリボヌクレアーゼからは保護されなかった。
【0073】
本発明者らは、eIF−5Aをコードするポリヌクレオチドを細胞に投与した際、それらの細胞でアポトーシスが増大することを示した。投与後に癌細胞内で発現するeIF−5A1ポリヌクレオチドを投与することによって、癌細胞をアポトーシスへ押しやることが可能であることが示された。全てそれらの全体が参照により取り込まれる、同時係属出願の10/200,148号;11/287,460号;11/293,391号、及び11/637,835号を参照されたい。
【0074】
本発明者らは、細胞に因子5Aのヒプシン化型が蓄積される際に細胞は生存状態に入り、通常は時間と共に起こるアポトーシスが起こらないことをさらに特定した。特に癌細胞には著しい量の因子5Aのヒプシン化型が存在することから、細胞はアポトーシスを起こさない(死滅しない)。従って、癌細胞を死滅させること(癌細胞をアポトーシス経路へ押しやること)によって癌を治療するために、eIF−5A1をコードするポリヌクレオチドが被験体へ、又は癌細胞若しくは腫瘍へ投与され、癌細胞にアポトーシス、最終的には細胞死及び腫瘍の縮小を順に引き起こすeIF−5A1の発現が増大される。しかしながらeIF−5A1の遺伝子発現の上昇のためにeIF−5A1タンパク質をコードするポリヌクレオチドのみが供給され、eIF−5A1の内在性発現をノックダウンするためのsiRNAが使用されない場合には綱引き:細胞をアポトーシス経路へ向けるeIF−5A1の発現と、細胞を細胞生存経路へ向けるヒプシン化因子5Aの存在が競合する状態となる。本発明はこの綱引きを排除し、eIF−5A1の発現の増大のみによる改良を示す。被験体又は細胞へ投与されたポリヌクレオチドは、得られる発現タンパク質がヒプシン化され得ないように変異導入される。加えて、因子5Aの内在性発現はeIF−5Aを標的とするsiRNAによってノックアウト/ダウンされ、ヒプシン化される内在性eIF−5A1は周囲に存在しなく/少なくなる。従って、細胞内にヒプシン化eIF−5Aが存在しない(又は実質的に少ない)ため、細胞は生存状態に押しやられることはない。
【0075】
変異eIF−5A1をコードするポリヌクレオチドは、ヒプシン化されず、それ故細胞を生存状態にするために利用され得ないように好ましく変異導入される。例えば一実施形態によれば、eIF−5Aをコードするポリヌクレオチドは、通常はDHSによりヒプシン化される位置50のリジン(K)がアラニン(A)(ヒプシン化され得ない)に変わるように変異導入される。この変異体はK50Aと表示される。
【0076】
別の実施形態によれば、位置67のリジンがアルギニン(R)に変更される。この変異体は(K67R)と表示される。別の実施形態によれば、位置67のリジン(K)がアラニン(A)に変更され、これは(K67A)と表示される。別の実施形態によれば、位置50のリジン(K)がアルギニンに変更され(K50R)、別の実施形態によれば、位置47のリジン(K)がアルギニンに変更される(K47R)変異体が提供される。
【0077】
別の実施形態によれば、二重変異体が使用される。一つの二重変異体では位置50のリジン(K)がアルギニン(R)に変更され、かつ位置67のリジン(K)がアルギニン(R)に変更される。この二重変異体はK50R/K67Rと称される。この二重変異体も同様にヒプシン化不可能であるが、このアミノ酸の変更は、eIF−5A1の3−D構造を単一変異体(K50A)ほどには変化させない。従って二重変異体は野生型に非常に類似した3−Dの形及びフォールディングのタンパク質を提供し、それ故に単一変異体よりもさらに安定である。さらに安定になることで、これは体内により長く存在して、治療上の利点をより長く提供する。従って、身体が正常細胞機能に必要とされるがヒプシン化され得ない因子5Aを有し得ることから、細胞は細胞生存状態にならず、かつアポトーシスを回避しない。
【0078】
別の二重変異体では位置47のリジン(K)がアルギニン(R)に変更され、かつ位置50のリジンがアルギニン(R)に変更される。この二重変異体は(K47R/K50R)と表示される。本発明は、位置50のリジン(L)がアラニン(A)に変更され、かつ位置67のリジンがアラニン(A)に変更される、別の二重変異体を提供する。この二重変異体は(K50A/K67A)と表示される。
【0079】
身体は正常細胞の生存及び健常細胞の増殖のために因子5Aを必要とすることから、siRNAが全身性に送達される場合、siRNAによって被験体内の発現を完全に遮断しないことが好ましい。eIF−5A発現の制御は、発現の遮断に良好ではない(すなわち、発現を停止又は減少させるが、発現を完全には遮断しない)siRNAを使用すること、又は代わりに、健常細胞の正常な生育及び機能を許容しかつ癌性細胞をアポトーシスへ押しやるために、発現レベルのバランスを取る投薬及び/又は治療計画を利用することのいずれかによって達成できる。
【0080】
あるいは、siRNAの局所送達が利用されてよい。siRNAが癌細胞又は腫瘍へ局所的に送達された場合、発現は好ましくノックアウトされる。発現をノックアウトすることにより、ヒプシン化可能な因子5Aは周囲に存在しなくなり、従って細胞を生存状態にするヒプシン化eIF−5Aは存在しなくなる。siRNAは癌又は腫瘍へ局所的に送達されることから、通常の細胞生育に利用可能なeIF−5Aを有する必要はない。
【0081】
特定の実施形態によれば、内在性遺伝子はeIF5A1である。内在性eIF−5A1の発現を抑制するため、被験体へeIF5A1を標的とするsiRNAが投与される。特定の実施形態によれば、siRNAは配列番号1又は配列番号2を含んでなるか、又は内在性eIF−5A1の発現を抑制し得る、eIF5A1を標的とするいずれのsiRNAである。特定の実施形態によればeIF5A1はヒトeIF−5A1(図1に示す)であり、被験体はヒトである。ヒトeIF−5A1を標的とするその他のsiRNAは公知であり、同時係属出願の11/134,445号;11/287,460号;11/184,982号;11/293,391号;11/725,520号;11/725,470号;11/637,835号に開示される。別の実施形態によれば被験体は哺乳類であり、eIF5A1は哺乳類に対して特異的である。例えば被験体はイヌであり、eIF5A1はイヌeIF5A1である。特定の実施形態によれば、siRNAは図25に示されるsiRNAコンストラクトから本質的になる。例えば、siRNAはeIF5A1を標的とする核酸を含むが、U又はT核酸のような突出もまた含むか、又はhisタグ(HAタグとも呼ばれ、しばしばin vitroでの研究に用いられる)のようなタグもまた含む。siRNAコンストラクトが標的遺伝子の発現を減少可能である限り、5’又は3’末端のいずれか(又は例えば図25に示される核酸の連続鎖の内側であっても)に付着する分子又はさらなる核酸が含まれてよく、「本質的になる」の範囲内となる。好ましくはsiRNAは、外来性ポリヌクレオチドの発現に影響しないようなeIF5A1遺伝子の領域を標的とする。例えば、eIF5A1 siRNAは3’UTR又は3’末端を標的とする。図25に示すsiRNAは、典型的なeIF5A1 siRNAである。
【0082】
ポリヌクレオチドはeIF5A1をコードし、ここで該ポリヌクレオチドはeIF5A1変異形をコードするように変異導入される。変異eIF5A1は、変異形eIF5A1が翻訳後修飾され得ないように(ヒプシン化され得ないように)設計される。典型的な変異体については上に考察される。
【0083】
固形癌に関与する癌の場合、siRNAを直接腫瘍へ送達することが望ましい可能性がある。siRNAは、時間、同様に送達部位に関してポリヌクレオチドとは別に投与されてよいか、又は、同時及び/若しくは同じ送達部位において共に投与されてよい。当業者はsiRNAの投与のタイミングについて、内在性タンパク質が翻訳される時に投与され、それが既に行われた後ではないことが必要な可能性があることを理解するであろう。
【0084】
本発明者らは以前にeIF5A1が正常組織に対して無毒であることを示したが(その全体が参照により本明細書に取り込まれる、2005年11月28日に提出した係属出願11/293,391号を参照)、送達複合体が(eIF5Aポリヌクレオチド/プラスミド/発現ベクターの直接投与に比較して)好ましい可能性がある。好ましい送達系は有効量のeIF5A1を被験体、腫瘍、又は癌細胞群へ提供し、同様に腫瘍又は癌細胞群への標的送達を好ましく提供する。従って特定の実施形態によれば、eIF5A1ヌクレオチド/プラスミド/発現ベクターを、リポソーム、デンドリマー、又は類似の無毒性ナノ粒子、例えばポリエチレンイミンポリマー(例えばin vivo JetPEI(商標)複合体)のようなナノメートルサイズのビヒクルを通して送達することが好ましい。
【0085】
eIF5A1タンパク質もまた、腫瘍部位へ直接送達されてよい。当業者はeIF5A1タンパク質送達の用量及び治療計画期間を決定できるであろう。
【0086】
eIF5A1によるアポトーシス誘導の分子的基盤について以下に考察する。
【0087】
デスレセプターのシグナル伝達
Ad−eIF5A1(野生型eIF5A1を有するアデノウィルス)又はAd−eIF5A1(K50A)による癌細胞の処理により、デスレセプター−リガンド結合により開始されるカスパーゼ8、及び実行カスパーゼであるカスパーゼ3の活性化が誘導される。これらはeIF5A1の間接的な効果であると考えられ、カスパーゼ8及びカスパーゼ3はヒプシン化され得ないeIF5A1(K50A)による処理後にも活性化されるという事実から、この効果はリジン50eIF5A1に起因し得ることが示される。Ad−eIF5A1による処理はまた、TNFR1の上昇によって以前に示されたように、デスレセプターの上昇ももたらすと思われる。
【0088】
ミトコンドリア経路
アポトーシスへと向かうミトコンドリア経路におけるリジン50eIF5A1の直接又は間接の関与は、eIF5A1又はeIF5A1(K50A)のいずれかによる癌細胞の処理によってカスパーゼ9が活性化されるという知見を含む幾つかの観察によって支持される。同様に、ミトコンドリアのアポトーシス経路の活性化に役割を果たすp53もeIF5A1により制御されると思われる。例えば、癌細胞のアクチノマイシンDによる処理はp53の発現を上昇させ、このp53の上昇はeIF5A1 siRNAによって阻害される。これと一致して、Ad−eIF5A1による癌細胞の処理はp53mRNAを上昇させる。癌細胞のeIF5A1による処理はまた、Baxのサイトゾルからミトコンドリアへの移動も誘導し、これによってミトコンドリア膜電位の減少及びミトコンドリア内腔からサイトゾルへのチトクロムCの放出が保証される。加えて、この処理によって切断型Bcl2、Bim、及びスプライス型Bimの上昇がもたらされ、これらの全てはアポトーシス促進性である。
【0089】
MAPKシグナル伝達
加えて本発明者らは、アポトーシスに関係するMAPKシグナル伝達におけるeIF5A1の関与についての証明を得た。例えばAd−eIF5A1による癌細胞の処理によって、抗アポトーシス性のBcl2を阻害するP−JNKが上昇した。加えて、Ad−eIF5A1及びAd−eIF5A1(K50A)の両方は、TNFR1及びTNF;FAS及びFASL;カスパーゼ8;Bid;チトクロムC、ならびにカスパーゼ3を含む様々なアポトーシス促進因子に影響することによってアポトーシスを開始できる、P−p38の形成を誘導する。
【0090】
NF−κBシグナル伝達
NF−κBシグナル伝達が骨髄腫の生育を支持することの証明が存在する。例えば骨髄ストローマ細胞への骨髄腫細胞の接着により、多発性骨髄腫における生育及び抗アポトーシス因子の両方であるIL−6の、NF−κB依存性の転写上昇が誘導される[Chauhan et al. (1996) Blood 87, 1104]。加えて、骨髄腫細胞により分泌されるTNF−αは骨髄ストローマ細胞内でNF−κBを活性化し、これによってIL−6の転写及び分泌が上昇する。TNF−αはまた骨髄腫細胞内でもNF−κBを活性化し、これにより骨髄腫細胞及び骨髄ストローマ細胞の両方に細胞内接着分子−1(intracellular adhesion molecule-1:ICAM−1;CD45)及び血管細胞接着分子(vascular cell adhesion molecule:VCAM−1;CD106)の上昇がもたらされる[Hideshima et al. (2001) Oncogene 20,4519]。これは次に骨髄腫細胞と骨髄ストローマ細胞との結合を増強する[Hideshima et al. (2001) Oncogene 20,4519]。反対に、これらの効果はTNF−αにより誘導されたNF−κBの活性化を遮断することによって阻害される[Hideshima et al. (2001) Oncogene 20,4519]。実際にNF−κBは、骨髄腫細胞においてTNF−αにより誘導されたアポトーシスに対する防御を仲介するように思われる[Hideshima et al. (2002) JBC 277, 16639]。これら及びその他の観察により、NF−κBシグナル伝達が多発性骨髄腫の治療法の魅力的な標的であり得るという見解が促進された。
【0091】
本発明者らは、eIF5A1 siRNAが、ヒト骨髄腫細胞におけるNF−κBの活性化及びICAM−1形成の両方を阻害することを示した。これらの観察により、eIF5A1がNF−κBの活性化における役割を果たすことが示され、かつNF−κBの活性化に続く効果が事実上生存促進性である限りにおいて、本発明者らはこの活性化がヒプシン化eIF5A1によって直接的又は間接的に仲介されると予想する。
【0092】
IL−1
炎症誘発性サイトカインであるIL−1の骨髄腫細胞による過剰産生は、骨組織の劣化を導く多発性骨髄腫の特徴的な性質である。eIF5A1 siRNAは、マウスへのLPS投与により誘導されるIL−1の過剰産生を劇的に減少させることが示されている。
【0093】
本発明の一実施形態は、多発性骨髄腫を治療する方法を提供する。多発性骨髄腫(「MM」)は、骨髄内の悪性形質細胞の増殖及び溶骨性の病変部の存在によって特徴付けられる、進行性かつ致死性の疾病である。多発性骨髄腫は難病であるが、治療可能な形質細胞の癌である。形質細胞は免疫系の重要な一部であり、感染及び疾病との戦いを援助する免疫グロブリン(抗体)を産生する。多発性骨髄腫は、骨髄内の過剰数の異常な形質細胞、及び完全なままのモノクローナル免疫グロブリン(IgG、IgA、IgD、又はIgE;「M−タンパク質」)又はベンス・ジョーンズタンパク質(遊離のモノクローナル軽鎖)の過剰産生により特徴付けられる。低カルシウム血症、貧血、腎障害、細菌感染への感受性の増大、及び正常免疫グロブリンの産生障害は、多発性骨髄腫の一般的な臨床症状である。多発性骨髄腫はしばしば、通常は骨盤、脊椎、肋骨、及び頭蓋内の広範性骨粗鬆症によっても特徴付けられる。
【0094】
本発明は、多発性骨髄腫において見出される刺激のフィードバックループのために、多発性骨髄腫の治療によく適していると思われる。例えば、多発性骨髄腫は骨髄内で低濃度のIl−1を産生する。Il−1は次にストローマ細胞を刺激してIL−6を産生させ、これが次に多発性骨髄腫の生育を刺激する。本発明者らは以前に、eIF−5A1に向けられたsiRNAが、Il−1;TNF−α、及びIl−8のような炎症誘発性サイトカインの発現を阻害できることを示した(係属出願の11/725,539号及び11/184,982号を参照)。従って、siRNAはヒプシン化のために利用可能なものが少なくなるようにeIF−5Aの発現をノックダウンするのみではなく、Il−1/Il−6のフィードバックループもまた、カットオフ又は減少させ得る。
【0095】
ヒトeIF5Aを標的とするsiRNAを、腫瘍内の内在性ヒプシン化eIF5Aのレベルを抑制するために使用し、一方でヒプシン化不可能なeIF5Aの変異体(eIF5AK50R)を発現するRNAi耐性プラスミドを、in vivoでの非修飾eIF5Aのレベルを上昇させるために使用した。eIF5A siRNAを含むPEIナノ複合体の腫瘍内注射によって、MM腫瘍の生育がコントロールsiRNAを含む複合体に対して80%(***p=0.0003)を超えるまで阻害され、ヒプシン化eIF5Aのレベルの抑制が抗腫瘍効果を有することが示された。eIF5AK50R発現プラスミドを含むPEI複合体は同様の効果を有し、腫瘍の生育はコントロールプラスミドを含む複合体に対して70%(**p0.001)を超えるまで阻害された。従ってMM腫瘍の生育は、生育促進性のヒプシン化eIF5Aの抑制によって、又はアポトーシス促進性である非ヒプシン化型eIF5Aのレベル上昇のいずれかによって阻害できる。eIF5A siRNA及びRNAi耐性eIF5AK50Rプラスミドの両方を含む複合体の腫瘍内送達は、腫瘍の生育に対して相乗的な効果を有し、腫瘍の生育を94%(***p=0.0002)まで阻害する著しい腫瘍の収縮をもたらした。eIF5A siRNA/eIF5AK50RPEI複合体の静脈内送達もまた、腫瘍の生育を95%(**p=0.002)まで効率的に減少させ、治療薬の全身性送達が実行可能であることが示された。
【0096】
eIF5A siRNA/eIF5AK50RpDNA PEI複合体の局所及び全身性送達の両方が、多発性骨髄腫において著しい抗腫瘍反応をもたらした。
【0097】
本発明はさらに、多発性骨髄腫を含む癌の治療に有用な組成物を提供する。好ましい実施形態によれば組成物は、ヒプシン化され得ない点変異eIF5A1をコードするプラスミドDNAと、内在性ヒトeIF5A1を選択的に抑制するが、該プラスミドによってコードされる点変異eIF5A1には影響しないeIF5A1 siRNAとの複合体である。eIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドについては上で考察されている。プラスミドDNA及びsiRNAの両方は、PEI(ポリエチレンイミン)ナノ粒子と好ましく複合される。これらは別々に複合されて別々若しくは共に投与されてよいか、又はこれらは共に複合されてよい。DNA及びRNAはPEI上の陽性に荷電したアミノ基に結合し、ナノ粒子が細胞内に取り込まれる際に放出される。PEI−核酸複合体は、分裂及び非分裂細胞の両方へ効果的に取り込まれることが示されている。
【0098】
プラスミドDNAはeIF5A1(K50R)を好ましくコードし、これはeIF5A1(K50A)のようにヒプシン化され得ないことから強力にアポトーシス原性である。eIF5A1(K50R)の発現は、B細胞特異的プロモーターによって好ましく調節される。
【0099】
eIF5A1 siRNAは、好ましくは内在性ヒトeIF5A1の3’末端に特異的であり、トランスeIF5A1(K50R)の発現に対する効果をもたない。典型的な好ましいeIF5A1 siRNAは、図25に示されるsiRNAを含んでなるか、該siRNAから本質的になるか、又は該siRNAからなる。eIF5A1 siRNAを含むことの理論的根拠は:(1)ほとんど全てがヒプシン化されていることから、生存促進型である内在性eIF5A1を除去するため;(2)NF−κBの活性化を阻害して、IL−6の産生及び細胞内接着分子の形成を減少させるため;及び(3)IL−1の形成を阻害するためである。このeIF5A1 siRNAはeIF5A1(K50R)と相乗的に作用して、骨髄腫細胞にアポトーシスを誘導する。上記の(2)及び(3)が生存促進性の現象である限りにおいて、これらはヒプシン化eIF5A1により仲介されると思われることから、ヒプシン化され得ないeIF5A1(K50R)によっては影響されない。このアプローチにより、多発性骨髄腫細胞を含む癌細胞にアポトーシスを導くが健常細胞にはほとんど効果を示さない、非ヒプシン化eIF5A1のより大きなプールがもたらされる。
【0100】
本明細書での好ましい組成物はSNS01と称される。SNS01は、安全性増大のためにB細胞起源(骨髄腫細胞を含む)の細胞に発現を限定するプロモーターにより駆動されるeIF5A1K50Rをコードする、RNAi耐性プラスミドDNA、及びヌクレアーゼ耐性増大のためにdTdT3’突出を有するヒトeIF5Aを標的とするsiRNAの両方を含む複合体であり、このsiRNA及びプラスミドはin vivo JetPEI(商標)と複合される。
【実施例】
【0101】
[実施例1]
eIF5A1の野生型及び変異形によるHeLaS3細胞へのトランスフェクション
HeLaS3細胞に、野生型eIF5A1(WT)、eIF5A1K50R(K50R)、eIF5A1K67R(K67R)、eIF5A1K67A(K67A)、eIF5A1K47R/K50R(K4750R)、eIF5A1K50R/K67R(K5067R)、又はeIF5A1K50A/K67A(K5067A)を含むHA−タグ化eIF5A1変異形を発現するプラスミドを、Lipofectamine 2000を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後24及び48時間目(A)又は28及び52時間目(B)に、細胞ライセートを回収し、SDS−PAGEによって分画した。トランスフェクトしたeIF5A1の発現レベルを、HAに対する抗体を用いて検出した。結果:eIF5A1の、推定されるユビキチン化部位のリジンにおける変異(K67R)は、野生型(A)を超えてeIF5A1トランス遺伝子の蓄積を増大させた。eIF5A1の、ヒプシン化に必要とされるリジンにおける変異(K50R)もまた、野生型eIF5A1(B)を超えてeIF5A1トランス遺伝子の蓄積を増大させた。eIF5A1(K50A/K67A)の二重変異体型は、非変異の野生型eIF5A1トランス遺伝子(A+B)に比較して特に良好に発現された。図2を参照されたい。
【0102】
[実施例2]
eIF5A1の野生型及び変異形によるKAS細胞へのトランスフェクション
KAS細胞に、野生型eIF5A1(5A1)、eIF5A1K67A(K67A)、eIF5A1K50A/K67A(K50A K67A)、eIF5A1K50R(K50R)、eIF5A1K47R(K47R)、eIF5A1K67R(K67R)、eIF5A1K47R/K50R(K47R K50R)、又はeIF5A1K50R/K67R(K50R K67R)を含むHA−タグ化eIF5A1変異形を発現するプラスミドを、PAMAMデンドリマー(FMD44)を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後48時間目に細胞ライセートを回収し、SDS−PAGEによって分画した。トランスフェクトしたeIF5A1の発現レベルを、HAに対する抗体を用いて検出した。アクチンに対する抗体を用いて等しい負荷を確認した。結果:eIF5A1の、推定されるユビキチン化部位のリジンにおける変異(K67A又はK67R)は、野生型を超えてeIF5A1トランス遺伝子の蓄積を増大させた。eIF5A1の、ヒプシン化に必要とされるリジンにおける変異(K50R)、又はアセチル化部位における変異(K47R)もまた、野生型eIF5Aを超えてeIF5A1トランス遺伝子の蓄積を増大させた。eIF5A1(K50A/K67A)の二重変異体型もまた、非変異の野生型eIF5A1トランス遺伝子に比較して高いレベルで発現された。図3を参照されたい。
【0103】
[実施例3]
PAMAMデンドリマーを用いたKAS細胞へのトランスフェクション
KAS細胞に、eIF5A1K50R(K50R)、eIF5A1K50A/K67A(K50A/K67A)、又はeIF5A1K50R/K67R(K50R K67R)を含むHA−タグ化eIF5A1変異形を発現するプラスミドを、PAMAMデンドリマー(FMD45−2)を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後72時間目に細胞をアネキシン/PIで染色し、FACSにより解析した。アネキシンVによって陽性に染まり、PI(ヨウ化プロピジウム)によって陰性であった細胞はアポトーシスの初期段階(Ann+/PI−)にあるとみなされ、アネキシンV及びPIの両方によって陽性に染まった細胞はアポトーシスの後期段階(Ann+/PI+)にあるとみなされた。結果:eIF5A1の、ヒプシン化部位のリジンにおける変異(K50R)又は推定されるユビキチン化部位のリジンにおける変異(K67R)、同様に二重変異体(K50A/K67A)は、LacZコントロールのレベルを著しく超えてKAS細胞のアポトーシスをもたらした。図4を参照されたい。
【0104】
[実施例4]
eIF−5A1及びeIF−5A1の変異形を発現するプラスミドによるKAS細胞へのトランスフェクション
KAS細胞に、eIF5A1K50A(K50A)、eIF5A1K50R(K50R)、eIF5A1K67R(K67R)、eIF5A1K50A/K67A(K50A/K67A)、又はeIF5A1K50R/K67R(K50R K67R)を含むHA−タグ化eIF5A1変異形を発現するプラスミドを、Lipofectamine 2000を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後72時間目に細胞をアネキシン/PIで染色し、FACSにより解析した。アネキシンVによって陽性に染まり、PI(ヨウ化プロピジウム)によって陰性であった細胞はアポトーシスの初期段階(Ann+/PI−)にあるとみなされ、アネキシンV及びPIの両方によって陽性に染まった細胞はアポトーシスの後期段階(Ann+/PI+)にあるとみなされた。結果:eIF5A1の、ヒプシン化部位のリジンにおける変異(K50R)又は推定されるユビキチン化部位のリジンにおける変異(K67R)、同様に二重変異体(K50A/K67A)は、LacZコントロールのレベルを著しく超えてKAS細胞のアポトーシスをもたらした。図5を参照されたい。
【0105】
[実施例5]
KAS細胞にアポトーシスをもたらす処理のための、変異eIF−5A1の使用
KAS細胞に、HA−タグ化eIF5A1変異形であるeIF5A1K50R(K50R)又はeIF5A1K50A/K67A(K50A/K67A)を発現するプラスミドを、Lipofectamine 2000を用いてトランスフェクトした。LacZを発現するプラスミドをコントロールとして使用した。トランスフェクション後72時間目に細胞をアネキシン/PIで染色し、FACSにより解析した。アネキシンVによって陽性に染まり、PI(ヨウ化プロピジウム)によって陰性であった細胞はアポトーシスの初期段階(Ann+/PI−)にあるとみなされ、アネキシンV及びPIの両方によって陽性に染まった細胞はアポトーシスの後期段階(Ann+/PI+)にあるとみなされた。結果:eIF5A1のヒプシン化部位のリジンにおける変異(K50R)、又はeIF5A1のヒプシン化部位及び推定されるユビキチン化部位の両方における変異(K50A/K67A)は、LacZコントロールのレベルを著しく超えてKAS細胞のアポトーシスをもたらした。図6を参照されたい。
【0106】
[実施例6A]
siRNA/アデノウィルスにより仲介される多発性骨髄腫細胞の死滅
KAS(ヒト多発性骨髄腫)細胞は、S10培地[4ng/ml IL−6、10%ウシ胎仔血清(FBS)、及びペニシリン/ストレプトマイシン(P/S)を含むRPMI 1640]中で維持した。Lipofectamine 2000(Invitrogen)を用いて、KAS細胞に58.7ピコモルのsiRNAをトランスフェクトした。モックトランスフェクト細胞は、siRNA非存在下でLipofectamine 2000により処理した。トランスフェクションは抗生物質を含まないS10培地中で行った。
a)ヒトeIF5A1を標的とするsiRNA:
eIF5A1 siRNA標的#1(siRNAはヒトeIF5A1のこの領域を標的とする:5’−AAGCTGGACTCCTCCTACACA−3’(配列番号:_))。siRNA配列は図25に示され、これは本明細書ではしばしばh5A1と称される。
eIF5A1 siRNA標的#2(このsiRNAはヒトeIF5A1のこの領域を標的とする:5’−AAAGGAATGACTTCCAGCTGA−3’(配列番号:_))。(siRNA配列は、本明細書ではしばしばash5A1−ALTと称される)
b)コントロールsiRNA:コントロールsiRNAは次の配列を有した:センス鎖、5’−ACACAUCCUCCUCAGGUCGdTdT−3’;及びアンチセンス鎖、3’−dTdTUGUGUAGGAGGAGUCCAGC−5’。
【0107】
使用されたその他のコントロールには、Dharmaconから入手した標的をもたないことが確認された非標的指向化siRNAが含まれるが、その理由は、これらは望まれないオフターゲット効果を制限するためにマイクロアレイによって試験されていることである。例えば、NFκBのin vitro試験のために用いられたコントロールはDharmaconの非標的指向化siRNA(配列D−001700−01)であり、in vivo研究のために用いられたコントロールはDharmaconのものであった(配列D−001810−01)。
【0108】
トランスフェクション後4時間目に細胞をペレットにして、1mlのS10培地に再懸濁した。最初のsiRNAトランスフェクション後72時間目に、トランスフェクトされたKAS細胞を計数し、24ウェルプレート内に300,000細胞/ウェルにてまき、同じsiRNAによる2度目のトランスフェクションを行った。
【0109】
トランスフェクション後4時間目に細胞をペレットにして、3000ifuのAd−LacZ(−ガラクトシダーゼを発現するアデノウィルス)又はAd−5A1M(ヒトeIF5A1K50Aを発現するアデノウィルス)のいずれかを含む1mlのS10培地(IL−6を含まない)に再懸濁した。
【0110】
72時間後に細胞を回収し、アネキシンV−FITC及びPI(BD Bioscience)を用いる染色とそれに続くFACS解析により、アポトーシスについて分析した。
a)初期のアポトーシスは、アネキシン−FITCによって陽性に染まり、PI染色によって陰性であった細胞(Ann+/PI−)として定義した。
b)総アポトーシスは、初期アポトーシス(Ann+/PI−)又は後期アポトーシス/ネクローシス(Ann+/PI+)のいずれかである細胞の総数として定義した。
【0111】
#1を標的とする5A1 siRNAは、ヒトeIF5A1の3’UTRを標的とすることから、アデノウィルス由来のeIF5A1の発現には影響し得ない。#2を標的とする5A1 siRNAは、ヒトeIF5A1の翻訳領域内を標的とすることから、アデノウィルス由来のeIF5A1の発現に潜在的に干渉し得る。
【0112】
結果:siRNAによって処理し、かつeIF5A1 K50A変異形を発現するアデノウィルスを感染させた細胞は、無処理の細胞及びsiRNAのみによって処理された細胞よりも、多数がアポトーシスを起こす。図7を参照されたい。
【0113】
[実施例6B]
eIF5A1標的#1(図25に示す)に対するeIF5A1 siRNAを用いる前処理により、内在性eIF5A1の発現は減少したが、アデノウィルスによって発現されるRNAi耐性eIF5A1k50Aは蓄積される。
KAS細胞に、コントロールsiRNA(C)又はeIF5A1を標的とする2つのsiRNA(#1及び#2)のうちの1つのいずれかを、Lipofectamine 2000を用いてトランスフェクトした。eIF5A1 siRNA#1は、eIF5A1の3’UTRを標的とすることから、eIF5A1の翻訳領域のみを含むアデノウィルス由来のeIF5A1の発現には干渉しない。siRNAの配列を図25に示す。eIF5A1#2 siRNAはeIF5A1の翻訳領域を標的とすることから、内在性及び外来性に発現されるeIF5A1の両方の発現に影響し得る。最初のトランスフェクション時間から72時間目に、細胞へ同じsiRNAによる2度目のトランスフェクションを行った。4時間後にトランスフェクション複合体を細胞から除き、Ad−LacZ(L)又はAd−eIF5A1K50A(5M)のいずれかを含む増殖培地(−)IL6と置き換えた。72時間後に細胞ライセートを回収し、eIF5A及びアクチンに対する抗体を用いるウェスタンブロットによって分析した。図7Bを参照されたい。ウィルスによって発現されたeIF5A1の蓄積が観察でき(レーン1対レーン2)、#1及び#2を標的とするeIF5A1 siRNAによるeIF5A発現の減少が明瞭に観察できる(レーン5及び7対レーン3)。予想されたように、eIF5A1 siRNA#1はウィルスによって発現されたeIF5A1K50Aの蓄積に影響しないが(レーン6対レーン4)、eIF5A1 siRNA#2はウィルスによって発現されたトランス遺伝子の発現に中程度にのみ影響する(レーン8対レーン4)。
【0114】
[実施例6C]
標的#1に対するeIF5A1 siRNAを用いるアデノウィルス感染前の前処理により、ヒト多発性骨髄腫細胞におけるリン酸化NFκBの発現は減少する。
KAS細胞に、コントロールsiRNA(hC)又はeIF5A1を標的とするsiRNA(#1)のいずれかを、Lipofectamine 2000を用いてトランスフェクトした。eIF5A1 siRNA#1は、eIF5A1の3’UTRを標的とすることから、eIF5A1の翻訳領域のみを含むアデノウィルス由来のeIF5A1の発現には干渉し得ない。最初のトランスフェクション時間から72時間目に、細胞へ同じsiRNAによる2度目のトランスフェクションを行った。4時間後にトランスフェクション複合体を細胞から除き、Ad−LacZ(L)又はAd−eIF5A1K50A(5M)のいずれかを含む増殖培地(+)IL6と置き換えた。24時間後に細胞ライセートを回収し、ホスホNF−κB p65(Ser536)及びeIF5Aに対する抗体を用いるウェスタンブロットによって分析した。予想されたように、eIF5A1 siRNA#1はウィルスによって発現されたeIF5A1K50Aの蓄積に影響しない。NF−κB p65のSer536におけるリン酸化は、活性化、核局在化、タンパク質−タンパク質相互作用、及び転写活性を調節する。図7Cを参照されたい。
【0115】
[実施例6D]
eIF5A1 siRNA#1を用いるアデノウィルス感染前の前処理により、ヒト多発性骨髄腫細胞におけるリン酸化NF−kB及びICAM−1の発現は減少する。
KAS細胞に、コントロールsiRNA(C)又はeIF5A1を標的とする2つのsiRNA(#1及び#2)のうちの1つのいずれかを、Lipofectamine 2000を用いてトランスフェクトした。最初のトランスフェクション時間から72時間目に、細胞へ同じsiRNAによる2度目のトランスフェクションを行った。4時間後にトランスフェクション複合体を細胞から除き、増殖培地(+)IL6と置き換えた。2度目のトランスフェクションから24時間後、細胞を40ng/mlのTNF−αで刺激して0、4、又は24時間目に細胞ライセートを回収し、ホスホNF−kB p65(Ser536)、ICAM−1、及びアクチンに対する抗体を用いるウェスタンブロットによって分析した。両方のeIF5A1特異的siRNAのトランスフェクション後、TNF−αにより誘導されたNF−kB p65のリン酸化(Ser536)及びICAM−1の発現の減少が観察された。NF−kB p65のSer536におけるリン酸化は、活性化、核局在化、タンパク質−タンパク質相互作用、及び転写活性を調節する。ICAM−1は、多発性骨髄腫の病因に関与すると信じられている細胞間接着表面糖タンパク質である。図7Dを参照されたい。
【0116】
[実施例6E]
siRNAを用いるKAS細胞の前処理により、IL−6存在下でのeIF5A1k50R遺伝子送達によるアポトーシスは増大する。
KAS細胞に、コントロールsiRNA(hcon)又はヒトeIF5A1 siRNA(h5A1)のいずれかをLipofectamine 2000を用いてトランスフェクトした。72時間後、細胞にsiRNAを再トランスフェクトした。siRNAトランスフェクション培地の除去後4時間目に、空のベクター(mcs)又はeIF5A1k50R(K50R)プラスミドのPEI複合体を細胞に加えた。この研究の間を通して使用した増殖培地にはIL−6が含まれていた。72時間後に、アネキシン/PIを用いる細胞の染色及びFACS解析によってアポトーシスを測定した。図7Fを参照されたい。
【0117】
[実施例7]
eIF−5A1プラスミド及びeIF−5A1 siRNAの同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延する(図8〜10)。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。3〜5週齢のSCID/CB17マウス6匹の右側腹部に、200μLのPBS中のKAS−6/1骨髄腫細胞107個を注射して、腫瘍の最小サイズが4mm3に達した時点で処置を開始した。
【0118】
コントロールマウスには、pCpG−mcs(空のベクター)及びコントロールsiRNAを含むPEI複合体を週2回腫瘍内注射した(コントロール群は3マウス:C−1、C−2、及びC−3によって構成された)。処置マウスには、RNAi耐性プラスミドpCpG−eIF5A1k50R及びeIF5A1 siRNAを含むPEI複合体を週2回腫瘍内注射した(処置群は3マウス:5A−1、5A−2、及び5A−3によって構成された)。注射は逆流を防ぐために腫瘍内の複数個所に行い、取り込みを増大させるために緩速で注射した。図8のデータに、群内のマウス全ての腫瘍容積を示す。図9に示されるデータは、群あたりの平均腫瘍容積+/−標準誤差である。アステリスクは統計的有意性を示す(*=p<0.025;n=3)。
【0119】
図10に、eIF−5A1プラスミド及びeIF−5A1 siRNAの同時投与によって多発性骨髄腫皮下腫瘍の重量が減少することを示す。示されるデータは、群あたりの平均腫瘍重量+/−標準誤差である。アステリスクは統計的有意性を示す(*=p<0.05;n=3)。
【0120】
in vivo試験には2x0.1mlのJET-PEI(PoIyPlus)を使用した。N/P比は8であった。PEI/DNA/siRNA複合体を、5%グルコース中において全量0.1mlで形成した。複合体形成のプロトコルは以下の通りである。
1.成分を室温に置く。無菌状態を保つ。
2.20μgのプラスミドDNA(2mg/mlを〜10μl)及び10μgのsiRNA(1mg/mlを10μl)を全量25μlに希釈する。差分を補うために滅菌水を使用する。
3.25μlの10%グルコースを加え、DNA溶液の量を50μlの5%グルコースに調整する。穏やかにボルテックスした後、短時間遠心する。
4.全量25μlの10%グルコース中へ4.8μlのin vivo JETPEIを希釈する。最終濃度が5%グルコースになるように、滅菌水で量を50μlに調整する。穏やかにボルテックスした後、短時間遠心する。
5.50μlの希釈PEIを50μlの希釈DNAへ直ちに加える(順序を逆にしないこと)。穏やかにボルテックスした後、短時間遠心沈殿する。
6.注射前に15分間インキュベートする。複合体は6時間安定である。
【0121】
CpGフリーのクローニングベクター及びpCpGプラスミドはInvivoGenから得た。これらのプラスミドはpCpGと命名され、CpGジヌクレオチドを完全に欠いている。これらのプラスミドはin vitro及びin vivoの両方で高レベルのトランス遺伝子発現をもたらし、かつCMVに基づくプラスミドとは対照的に、in vivoにおける持続発現を可能にする。pCpGプラスミドは、全てのCpGを除くように改変された、必然的にCpGジヌクレオチドを欠如するエレメント、又は選択性マーカー若しくはレポーターをコードする遺伝子のような、完全に合成されたエレメントのいずれかを含む。遺伝コードを形成する16のジヌクレオチドのうち、CGは唯一必須ではなく置換可能なジヌクレオチドであるという事実によってこれらの新しい対立遺伝子の合成が可能となった。8のコドンが、異なる5アミノ酸をコードするCGを含む。野生型と同一のままであることから、それらの野生型の対応物と同じように活性があるアミノ酸配列を有するタンパク質をコードする新しい対立遺伝子を作り出すため、8コドンの全ては、同じアミノ酸をコードする少なくとも2コドンの選択によって置換できる。これらの新しい対立遺伝子は、それらを容易に切り出すことのできるpMODというプラスミドから個別に入手できる。
【0122】
pCpGプラスミドはin vivoにおける持続性の発現を可能にし、かつTLR9を発現する細胞株を用いたin vivo及びin vitroでの遺伝子発現におけるCpGの効果、同様に自然及び獲得免疫系におけるそれらの効果を研究するために貴重な手段の典型である。空のベクターであるpCpG−mcs(Invivogen)は、発現遺伝子産物を含まないマルチクローニング部位のみのベクターであり、コントロールベクターとして用いた。処置用ベクターpCpG−eIF5A1(K50R)を作るため、LacZ遺伝子を除去したpCpG−LacZベクター(Invivogen)のNcoI及びNheI部位へ、HA−タグ化eIF5A1k50RcDNAをサブクローン化した。DNAはEndo-Free Qiagenキットを用いて調製した。測定したエンドトキシン濃度は、水中で2mg/mlであるはずのDNAについて、<0.03EU/μgDNAであった。
【0123】
実験に用いたコントロールsiRNAは、Dharmacon(D-001810-01)から入手した、マイクロアレイによって確認済みの非標的指向化コントロールsiRNAであった。血清中での分解を防ぐため、改変されたsiRNA(siSTABLE)を得た。
【0124】
実験に用いたeIF5A1 siRNAは、ヒトeIF5A1の3’UTRに対して設計した。該eIF5A1 siRNAとマウスeIF5A1の間に類似性は無いことから、該siRNAはヒト(マウスではない)eIF5A1のみを抑制するはずである。該siRNAはまた、eIF5A2(ヒト又はマウスのいずれか)とも類似性は無い。該siRNAは、血清中での分解を防ぐために改変された(siSTABLE)。該eIF5A1 siRNAは以下の標的配列:5’GCU GGA CUC CUC CUA CAC A(UU)3を有する。
【0125】
siSTABLE siRNAは、1mg/mlにて水に溶解した(一定分量で−20℃において保管した)。
【0126】
長さ(l)及び幅(w)に関する腫瘍の寸法は、デジタルキャリパーを用いて週あたり2〜3回測定した。腫瘍容積は以下の方程式に従って算出した:
l=長さ;最小寸法
w=幅;最大寸法
腫瘍容積(mm3)=l2*w*0.5
【0127】
統計的分析
統計的分析にはスチューデントt−検定を用いた。有意性は95%(p<0.05)を超える信頼水準であるとみなされた。
【0128】
[実施例8]
eIF5A1プラスミド及びeIF5A1 siRNAの同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延し、かつ腫瘍の収縮がもたらされる。
別の研究において、再びKAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。コントロールマウスには、pCpG−mcs(空のベクター)及びコントロールsiRNAを含むPEI複合体を週2回腫瘍内注射した(コントロール群、G−1、G−2、及びG−3)。処置マウスには、RNAi耐性プラスミドpCpG−eIF5A1k50R(20μgのプラスミドDNA)及びeIF5A1 siRNA(10μgのsiRNA)を含むPEI複合体を週2回腫瘍内注射した(処置群、G−4、G−5、及びG−6)。図11に示されるデータは、群あたりの平均腫瘍容積+/−標準誤差である。アステリスクは統計的有意性を示す(*=p<0.025;n=3)。21日間にわたり、6回の注射を行った(赤の矢印)。
【0129】
[実施例9]
eIF5A1 siRNAの静脈内(i.v.)投与及びPEI/eIF5A1K50Rプラスミド複合体の腫瘍内(i.t.)投与により、多発性骨髄腫皮下腫瘍の腫瘍収縮がもたらされる(群2B)。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初にコントロールsiRNA(コントロール群)又はヒトeIF5A1 siRNA(処置群)のいずれか50μgを尾注射した。コントロールマウスは続いて、pCpG−mcs(空のベクター;コントロール群;G−1、G−2、及びG−3)を含むPEI複合体の、週2回の腫瘍内注射によって処置された。処置マウスは続いて、RNAi耐性プラスミドpCpG−eIF5A1k50R(20μgのプラスミドDNA)(処置群;G−4、G−5、及びG−6)を含むPEI複合体の、週2回の腫瘍内注射によって処置された。コントロールマウスは週1回のi.v.注射により、コントロールsiRNAを投与され続けた(コントロール群、R−1、R−2、及びR−3)。処置マウスは週1回のi.v.注射により、ヒトeIF5A1 siRNA(20μg)を投与され続けた(処置群、R−4、R−5、及びR−6)。図12に示されるデータは、各群における全てのマウスの腫瘍容積である。21日間にわたり、PEI/DNAの壁内注射を6回(赤の矢印)、及びsiRNAのi.v.注射を4回(青の矢印)行った。
【0130】
図13に、実施例8及び9に由来するオーバーレイを示す。KAS細胞をSCIDマウスに皮下注射した。図13に示されるデータは、各群におけるマウスの平均腫瘍容積+/−標準誤差である。アステリスクは処置群とコントロール群の間の統計的有意性を示す(**=p<0.01;***=p<0.001;n=3)。
【0131】
PEI複合体形成のためのプロトコル:
1.成分を室温に置く。無菌状態を保つ。
2.プラスミドDNA又はプラスミドDNA+siRNAを全量25μlに希釈する。量を調整するために滅菌水を使用する。
a)プラスミドDNAのみの複合体は:
20μgのプラスミドDNA(2mg/mlを10μl)を全量25μlに希釈する。差分を補うために滅菌水を使用する。
b)プラスミドDNA+siRNAの複合体は:
20μgのプラスミドDNA(2mg/mlを〜10μl)及び10μgのsiRNA(1mg/mlを10μl)を全量25μlに希釈する。差分を補うために滅菌水を使用する。
3.25μlの10%グルコース(PEIキットと共に提供される)を加え、DNA溶液の量を50μlの5%グルコースに調整する。穏やかにボルテックスした後、短時間遠心する。
4.全量25μlの10%グルコース中へin vivo JETPEIを希釈する。
a)プラスミドDNAのみの複合体は:
全量25μlの10%グルコース中へ3.2μlのin vivo JETPEIを希釈する。最終濃度が5%グルコースになるように、滅菌水で量を50μlに調整する。穏やかにボルテックスした後、短時間遠心する。
b)プラスミドDNA+siRNAの複合体は:
全量25μlの10%グルコース中へ4.8μlのin vivo JETPEIを希釈する。最終濃度が5%グルコースになるように、滅菌水で量を50μlに調整する。穏やかにボルテックスした後、短時間遠心する。
5.50μlの希釈PEIを50μlの希釈DNAへ直ちに加える(順序を逆にしないこと!)。穏やかにボルテックスした後、短時間遠心沈殿する。
6.注射前に15分間インキュベートする。複合体は6時間安定である。
【0132】
siRNAの尾静脈注射に関して、最初のsiRNA注射は50マイクログラムであった。siRNAはPBS中で0.4mg/mlに希釈した。マウスあたり125μl(50μg)を尾静脈内へ注射した。続く血清安定化siRNAの注射は、マウスあたり20μgで週2回行った。siRNAはPBS中で0.4mg/mlに希釈した。マウスあたり50μl(20μg)を尾静脈内へ注射した。
【0133】
図13Bに、eIF5A1プラスミド及びeIF5A1 siRNAの同時投与により腫瘍の収縮がもたらされることを示す。KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。マウスには、RNAi耐性プラスミドpCpG−eIF5A1k50R及びeIF5A1 siRNAを含むPEI複合体を週2回腫瘍内注射した(処置群;G−4、G−5、及びG−6)。21日間にわたり、6回の注射を行った。処置開始後42日目にマウスを屠殺し、腫瘍部位下の皮膚を開いて腫瘍生育の形跡について調べた。群2Aの処置マウスのいずれにも腫瘍の生育は観察されなかった。
【0134】
図13Cに、eIF5A1 siRNAの静脈内(i.v.)投与及びPEI/eIF5A1K50Rプラスミド複合体の腫瘍内(i.t.)投与により、多発性骨髄腫皮下腫瘍の腫瘍収縮がもたらされることを示す。KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初に50μgのヒトeIF5A1 siRNAを注射した(処置群)。マウスは続いて、RNAi耐性プラスミドpCpG−eIF5A1k50Rを含むPEI複合体の、週2回の腫瘍内注射によって処置された(処置群;R−4、R−5、及びR−6)。マウスは週1回のi.v.注射により、ヒトeIF5A1 siRNAを投与され続けた。処置開始後21日目に処置を終了した。処置開始後42日目にマウスを屠殺し、腫瘍部位下の皮膚を開いて腫瘍生育の形跡について調べた。処置群(群2B)の1マウスに、腫瘍生育の形跡がみられなかった。
【0135】
[実施例10]
eIF5A1プラスミド及びeIF5A1 siRNAの静脈内同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延する。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初にコントロールsiRNA(コントロール群)又はヒトeIF5A1 siRNA(処置群)のいずれか50μgを注射した。マウスは続いて、pCpG−mcs(空のベクター;コントロール群;A1、A2、及びA3)を含むPEI複合体、又はRNAi耐性プラスミドpCpG−eIF5A1k50R(処置群;A4、A5、及びA6)を含むPEI複合体のいずれかによる、週あたり〜2回の静脈内(赤い矢印)又は腹腔内(緑の矢印)注射によって処置された。マウスは週1回のi.v.注射(青い矢印)により、コントロールsiRNA(コントロール群、A1、A2、及びA3)又はヒトeIF5A1 siRNA(処置群、A4、A5、及びA6)のいずれかを投与され続けた。示されるデータは、各群における全てのマウスの腫瘍容積である。図14に示されるデータは、各群における全てのマウスの腫瘍容積である。
【0136】
[実施例11]
eIF5A1 siRNAの静脈内(i.v.)投与及びPEI/eIF5A1K50Rプラスミド複合体の静脈内(i.v.)投与又は腹腔内(i.p.)投与により、多発性骨髄腫皮下腫瘍の生育は遅延する。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初に50μgのコントロールsiRNA(コントロール群)又はヒトeIF5A1 siRNA(処置群)のいずれかを注射した。コントロールマウスは続いて、pCpG−mcs(空のベクター;コントロール群は3マウスであった;B1、B2、及びB3)を含むPEI複合体の、週あたり〜1回の静脈内又は腹腔内注射によって処置された。処置マウスは続いて、RNAi耐性プラスミドpCpG−eIF5A1K50R(処置群;B4、B5、及びB6)を含むPEI複合体の、週あたり〜1回の静脈内又は腹腔内注射によって処置された。マウスは週1回のi.v.注射により、コントロールsiRNA(コントロール群、B1、B2、及びB3)又はヒトeIF5A1 siRNA(処置群は3マウスであった;B4、B5、及びB6)のいずれかを投与され続けた。実験は50μgのsiRNAの最初の注射によって開始された(図15のグラフの2日目)。続く注射では、毎週1回20マイクログラムのsiRNAを用いた。siRNAはネイキッド、すなわち送達ビヒクルなしで投与された。PEI複合体は20μgのプラスミドDNAを含んだ。最初のPEI注射はi.p.で行われ、続く注射はi.v.で行われた。図15に示されるデータは、各群における全てのマウスの腫瘍容積である。
【0137】
図16に、実施例10及び11のオーバーレイを示す。KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。1セットのマウスは、コントロールsiRNA(コントロール;群A)又はeIF5A1 siRNA(処置;群A)のいずれかの週1回のi.v.注射、及びpCpG−mcsを含むPEI複合体(コントロール;群A)若しくはRNAi耐性プラスミドpCpG−eIF5A1K50Rを含むPEI複合体(処置;群A)のいずれかのi.v.又はi.p.のいずれかを受けた。第2セットのマウスは、pCpG−mcs(空のベクター)及びコントロールsiRNAを含むPEI複合体(コントロール;群B)又はRNAi耐性プラスミドpCpG−eIF5A1k50R及びeIF5A1 siRNAを含むPEI複合体(処置;群B)のいずれかによるi.v.又はi.p.注射を、週あたり〜2回受けた。示されるデータは、各群におけるマウスの平均腫瘍容積+/−標準誤差である。アステリスクは処置群とコントロール群の間の統計的有意性を示す(*=p<0.05;***=p<0.001;n=3)。
【0138】
PEI複合体及びsiRNAを調製するためのプロトコルは、以前の実施例に記載した通りである。
【0139】
[実施例12]
eIF5A1プラスミド及びeIF5A1 siRNAの同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延し、かつ腫瘍の収縮がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。コントロールマウスには、pCpG−mcs(空のベクター)及びコントロールsiRNAを含むPEI複合体を週2回腫瘍内注射した(コントロール群は3マウスを有した;コントロール1、コントロール2、及びコントロール3)。処置マウスには、RNAi耐性プラスミドpCpG−eIF5A1k50R及びeIF5A1 siRNAを含むPEI複合体を週2回腫瘍内注射した(処置群には4マウスが含まれた;5A−1、5A−2、5A−3、及び5A−4)。PEI複合体の腫瘍内拒絶には、20μgのプラスミドDNA及び10μgのsiRNAの両方が含まれた。図17に示されるデータは、各群における全てのマウスの腫瘍容積である。
【0140】
[実施例13]
eIF5A1 siRNAの静脈内(i.v.)投与及びPEI/eIF5A1K50Rプラスミド複合体の腫瘍内(i.t.)投与により、多発性骨髄腫皮下腫瘍の腫瘍収縮がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始し、最初にコントロールsiRNA(コントロール群は3マウスを有した;コントロール1、コントロール2、及びコントロール3)又はヒトeIF5A1 siRNA(処置群は3マウスを有した;5A−1、5A−2、5A−3)のいずれか50μgを注射した。コントロールマウスは続いて、pCpG−mcs(20μg)を含むPEI複合体の、週2回の腫瘍内注射によって処置された(コントロール群1〜3)。処置マウスは続いて、RNAi耐性プラスミドpCpG−eIF5A1K50R(20μg)を含むPEI複合体の、週2回の腫瘍内注射によって処置された(5A−1、5A−2、5A−3)。コントロールマウスは週2回の尾静脈へのi.v.注射により、コントロールsiRNA(20μg)を投与され続けた。処置マウスは週2回の尾静脈へのi.v.注射により、ヒトeIF5A1 siRNA(20μg)を投与され続けた。注射は、腫瘍内注射前の48時間に行われた。siRNAはネイキッドsiRNAとして、すなわち送達ビヒクルなしで投与された。図18に示されるデータは、各群における全てのマウスの腫瘍容積である。
【0141】
[実施例14]
EF1又はB29プロモーターのいずれかにより駆動されるeIF5A1K50Rプラスミド及びeIF5A1 siRNAの同時投与により、多発性骨髄腫皮下腫瘍の生育は遅延し、かつ腫瘍の収縮がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。マウスには、コントロールベクター(G1及びG2)又はB29プロモーター(G3及びG4)若しくはEF1プロモーター(G5及びG6)のいずれかにより駆動されるeIF5A1プラスミドのいずれか、並びに、コントロールsiRNA(G1、G3、G5)又はh5A1 siRNA(G2、G4、G6)のいずれかを含むPEI複合体を週2回腫瘍内注射した。示されるデータは、各群の平均腫瘍容積+/−標準誤差である。注記:B29プロモーターはB細胞特異的プロモーターとして意図された。しかしながら、この研究で用いたB29プロモーター/mCMVエンハンサーは、in vitroでKAS細胞にHA−eIF5A1K50Rの高い発現を駆動することが見出されたが、B細胞特異的ではないと思われる(おそらくCMVエンハンサーのため)。図19を参照されたい。
【0142】
[実施例15]
eIF5A1 siRNAの同時投与により、EF1又はB29プロモーターのいずれかにより駆動されるeIF5A1K50Rプラスミドの多発性骨髄腫皮下腫瘍における抗腫瘍効果は増大され、かつ腫瘍の負荷量の減少がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。マウスには、コントロールベクター(G1及びG2)又はB29プロモーター(G3及びG4)若しくはEF1プロモーター(G5及びG6)のいずれかにより駆動されるeIF5A1プラスミドのいずれか、並びに、コントロールsiRNA(G1、G3、G5)又はh5A1 siRNA(G2、G4、G6)のいずれかを含むPEI複合体を週2回腫瘍内注射した。処置開始後24日目にマウスを屠殺し、皮下腫瘍を除去して秤量した。示されるデータは、全ての群の平均腫瘍重量+/−標準誤差である。図20を参照されたい。
【0143】
[実施例16]
eIF5A1 siRNAは、肺腺癌細胞におけるAd−eIF5Aの感染によりもたらされたアポトーシスの誘導を相乗的に増大させる。
A549細胞に、Ad−LacZ又はAd−eIF5Aのいずれかを感染させた。ウィルスの添加後直ちに細胞へトランスフェクション培地を添加することにより、細胞にコントロールsiRNA又はヒトeIF5A1(h5A1)を標的とするsiRNAのいずれかをトランスフェクトした。siRNAによるトランスフェクション及びウィルスによる感染後4時間目に培地を新鮮な培地に置き換え、細胞を72時間インキュベートした後に、アポトーシス性の細胞を検出するためアネキシン/PIにより標識した。注記:この細胞株におけるeIF5Aの過剰発現は、DHS及びDOHHの量が限られていることから非ヒプシン化eIF5Aの蓄積をもたらし、そのためにeIF5AK50Rの過剰発現と同じアポトーシス促進効果をもたらす。これらのデータにより、ヒプシン化eIF5Aの同時抑制及び非ヒプシン化eIF5Aの過剰発現によって引き起こされるアポトーシスにおける相乗的効果が、骨髄腫以外の腫瘍型にも同様に観察されることが示される。図21を参照されたい。
【0144】
[実施例17]
プラスミドpExp5Aの構築
pExp5Aは、eIF5A1K50Rの発現が主にヒトB細胞系列の細胞内で駆動されるように設計された、CpGジヌクレオチドの少ない発現プラスミドである。このベクターは、CpGジヌクレオチドを完全に欠いたプラスミドであるpCpG−LacZ(Invivogen)に由来する。E.Coliの複製及び選択に必要な全てのエレメントはCpGジヌクレオチドを含まない。CpG−LacZベクター由来のオリジナルのCMVエンハンサー/プロモーター及びLacZ遺伝子は、eIF5A1K50RのB細胞特異的な発現を駆動するため、それぞれヒト最小B細胞特異的プロモーター(B29/CD79b;Invivogen)及びヒトeIF5A1K50Rによって置き換えられた。B29プロモーターの活性を増大させ、非B細胞における発現を減少させるために、プラスミドのeIF5A1発現カセットの下流にB29 DHS4.4 3’エンハンサーが導入された(Malone et al. 2006. B29 gene silencing in pituitary cells is regulated by its 3' enhancer. J. Mol. Biol. 362: 173-183)。B29最小プロモーター、eIF5A1K50R、及びB29 DHS4.4 3’エンハンサーの取り込みにより、ベクター内に32のCpGジヌクレオチドが導入された。
【0145】
E.Coliにおける発現のためのエレメント
複製開始点:E.Coli R6Kガンマori
*R6Kガンマ複製開始点の存在により、pCpGプラスミドはpir変異遺伝子を発現する変異株のみにおいて増幅できる。これらは標準のE.Coli株においては複製し得ない。従ってpCpGプラスミドは、Demメチル化も欠くpir変異体であるE.Coli GT115株(Invivogen)と共に供給される。
細菌性プロモーター:細菌性EM7プロモーターのCpGを含まないバージョンであるEM2K。
選択性マーカー:Zeocin(商標)耐性遺伝子;CpGを含まない合成対立遺伝子。
【0146】
哺乳類細胞における発現のためのエレメント
哺乳類プロモーター:B細胞における組織特異的発現のための、ヒト−167bp最小B29(CD79b)プロモーター。5’UTRに合成イントロン(I140)も存在する。
ポリアデニル化シグナル:後期SV40ポリアデニル化シグナルのCpGジヌクレオチドを含まないバージョン。
3’エンハンサー:ヒトB29 DHS4.4 3’エンハンサー。
MAR:細菌性及び哺乳類性の転写ユニットの間に、CpGを含まないマトリックス結合領域(Matrix attached regions:MAR)が2つ存在する。MARの1つはヒトIFN−β遺伝子の5’領域由来であり、1つはβ−グロビン遺伝子の5’領域由来である。
【0147】
pExp5Aの予測される配列(3371bp)を図23に示す。
【0148】
eIF5A1K50Rのアミノ酸配列
*K50R変異を下線で示す。
【0149】
【化1】
【0150】
pExp5Aの構築−−構築の概略
工程1:B29 DHS4.4 3’エンハンサーのクローニング及びpGEM T easy(Promega)へのサブクローニング−−pGEM−4.4enh#8の作製。
工程2:最小B29プロモーターのpCpG−LacZ(Invivogen)へのサブクローニング−−B29−5#3の作製。
工程3:HA−eIF5A1K50RのB29−5#3ベクターへのサブクローニング−−B29−5#3−eIF5A1K50Rの作製。
工程4:pGpG−mcs(Invivogen)内の新しいマルチクローニング部位の作製−−pCpG−リンカー4の作製。
工程5:B29 DHS4.4 3’エンハンサーのpCpG−リンカー4へのサブクローニング−−pCpG−DHS4.4の作製。
工程6:B29プロモーター+HA−eIF5A1K50R+SV40 pA発現カセットのpCpG−DHS4.4へのサブクローニングによるpExp−5の作製。
工程7:pExp−5内のHA−eIF5A1K50Rを非HA eIF5A1K50Rに置き換えることによる、最終ベクターpExp5Aの作製。
【0151】
構築の詳細
工程1:B29 DHS4.4 3’エンハンサーのクローニング及びpGEM T easy(Promega)へのサブクローニング−−pGEM−4.4enh#8の作製。
B29 DHS4.4 3’エンハンサーを、以下のプライマー:順方向5’−CAGCAAGGGAGCACCTATG−3’及び逆方向5’−GTTGCAGTGAGCGGAGATG−3’を用いて、KAS細胞(ヒト多発性骨髄腫細胞株)より分離したゲノムDNAからPCRによってクローン化した。プライマーは、ヒトCD79B/GH−1遺伝子間領域(受入番号AB062674)の配列を用いて設計した。得られた608bpのPCR断片を、pGEM(登録商標)T easyクローニングベクター(Promega)にサブクローン化して配列決定した。Komatsu et al. 2002. Novel regulatory regions found downstream of the rat B29/Ig-b gene. Eur. J. Biochem. 269 : 1227-1236。
【0152】
pGEM−4.4enh#8内の、B29 DHS4.4 3’エンハンサーPCR断片(297bp)の配列
【0153】
【化2】
【0154】
○ +4.4の領域に、幾つかの転写因子結合部位が含まれる。
【0155】
【化3】
【0156】
pGEM−4.4enh#8内の、B29 DHS4.4 3’エンハンサーPCR断片(297bp)及びヒトCD79B/GH−1遺伝子間領域(受入番号AB062674)の配列のアラインメント。
【0157】
【化4】
【0158】
【化5】
【0159】
工程2:最小B29プロモーターのpCpG−LacZ(Invivogen)へのサブクローニング−−B29−5#3の作製。
最小−167ヒトB29プロモーターを、以下のプライマー:順方向5’−CCAACTAGTGCGACCGCCAAACCTTAGC−3’;逆方向5’−CAAAAGCTTGACAACGTCCGAGGCTCCTTGG−3’を用いて、完全長ヒトB29プロモーターを有する市販のプラスミド(pDrive-hB29;Invivogen)から増幅した。得られたPCR断片をSpeI及びHindIIIを用いて消化し、pCpG−LacZベクター(Invivogen)のSpeI及びHindIII部位へライゲートして、B29−5#3を作製した。
【0160】
B29−5#3内の、B29最小プロモーターPCR断片(188bp)の配列
【0161】
【化6】
【0162】
B29−5#3内のB29最小プロモーターPCR断片(B29_min)とpDrive-hB29由来の完全長ヒトB29プロモーターとのアラインメント。
【0163】
【化7】
【0164】
【化8】
【0165】
【化9】
【0166】
工程3:HA−eIF5A1K50RのB29−5#3ベクターへのサブクローニング−−pB29−eIF5A1K50R_7の作製。
HA−eIF5A1K50Rを、DNAテンプレートとしてのpHM6−eIF5A1K50R、ならびに以下のプライマー:順方向5’−CGCCATGGACATGTACCCTTACGACGTCCCAGACTACGCTGCAGATGATTTGGACTTCGAG−3’及び逆方向5’−CGCGCTAGCCAGTTATTTTGCCATCGCC−3’を用いて、PCRにより増幅した。得られたPCR断片をNcoI及びNheIを用いて消化し、B29−5#3のNcoI及びNheI部位へサブクローン化して、LacZを置き換えた。
【0167】
pB29−eIF5A1K50R_7内の、HA−eIF5A1K50RPCR断片(497bp)の配列
【0168】
【化10】
【0169】
pB29−eIF5A1K50R_7内の、HA−eIF5A1K50RPCR断片の翻訳
HAエピトープ
eIF5A1K50R
K50R
【0170】
【化11】
【0171】
pB29−eIF5A1K50R_7内の、HA−eIF5A1K50RPCR断片とヒトeIF5A1(受入#NP_001961)とのアラインメント
【0172】
【化12】
【0173】
工程4:pGpG−mcs(Invivogen)内の新しいマルチクローニング部位の作製−−pCpG−リンカー4の作製。
pGpGクローニングベクターであるpGpG−mcs G2(Invivogen)をEcoRIで消化して、mCMVエンハンサー、hEF1プロモーター、合成イントロン、マルチクローニング部位、及びSV40ポリアデニル化シグナルを含む哺乳類発現カセットを除いた。次にEcoRIで消化したpGpG−mcs G2ベクターをEcoRI粘着末端を有する合成リンカーへライゲートして、新しいマルチクローニング部位を有する、プロモーターの無いベクター(pCpG−リンカー4)を作製した。
【0174】
【化13】
【0175】
pCpG−リンカー4内の新しいマルチクローニング部位周囲の領域の配列
【0176】
【化14】
【0177】
上の配列の番号付けは、以下の特徴:βGlo MAR(ヌクレオチド1−380);EcoRI認識配列(ヌクレオチド396−401);XhoI認識配列(ヌクレオチド401−406);ClaI認識配列(ヌクレオチド409−414);NotI認識配列(ヌクレオチド417−424);MluI認識配列(ヌクレオチド427−432);IFNβ S/MAR(ヌクレオチド438−1,030)への参照を容易にするためである。
【0178】
工程5:B29 DS4.4 3’エンハンサーのpCpG−リンカー4へのサブクローニング−−pCpG−DHS4.4の作製。
B29 DHS4.4 3’エンハンサーを、テンプレートとしてのpGEM−4.4enh#8、ならびに以下のプライマー:順方向5’−GAAGCGGCCGCACCACCCTGGGCCAGGCTGG−3’;及び逆方向5’−CCACGCGTAGAGGTGTTAAAAAGTCTTTAGGTAAAG−3’を用いて、PCRにより増幅した。得られたPCR断片をNotI及びMluIを用いて消化し、pCpG−リンカー4の新しいマルチクローニング部位内のNotI及びMluI部位へライゲートして、pCpG−DHS4.4を作製した。
【0179】
>pCpG−DHS4.4完全長配列(2,282bp)
【0180】
【化15】
【0181】
【化16】
【0182】
上の配列の番号付けは、以下の特徴:βGlo MAR(ヌクレオチド1−400);EcoRI認識配列(ヌクレオチド416−421);XhoI認識配列(ヌクレオチド421−426);ClaI認識配列(ヌクレオチド429−434);NotI認識配列(ヌクレオチド437−444);DHS4.4(ヌクレオチド445−741);MluI認識配列(ヌクレオチド742−747);IFNβ S/MAR(ヌクレオチド753−1,569)への参照を容易にするためである。
【0183】
工程6:B29プロモーター+HA−eIF5A1K50R+SV40 pA発現カセットのpCpG−DHS4.4へのサブクローニングによるpExp−5の作製。
最小B29プロモーター、合成イントロン、HA−eIF5A1K50R、及びSV40pAを含むB29−eIF5A1発現カセットを、pB29−eIF5A1K50R_7(工程3)からPCRにより、以下のプライマー:順方向5’−GTTATCGATACTAGTGCGACCGCCAAACC−3’;及び逆方向5’−CAAGCGGCCGCCATACCACATTTGTAGAGGTTTTAC−3’を用いて増幅した。得られたPCR断片をClaI及びNotIを用いて消化し、pCpG−DHS4.4のマルチクローニング部位内のClaI及びNotI部位へサブクローン化して、pExp−5を作製した。
【0184】
工程7:pExp−5内のHA−eIF5A1K50Rを非HA eIF5A1K50Rに置き換えることによる、最終ベクターpExp5Aの作製。
pExp−5プラスミドをNcoI及びNheIにより消化してHA−eIF5A1K50Rを除いた。非HA−タグ化HA−eIF5A1K50RPCR断片を、pHM6−eIF5A1K50RからPCRにより、以下のプライマー:順方向5’−CACCATGGCAGATGATTTGGACTTC−3’;及び逆方向5’−CGCGCTAGCCAGTTATTTTGCCATCGCC−3’を用いて増幅した。得られたPCR産物をNcoI及びNheIを用いて消化し、B29−5#3のNcoI及びNheI部位へライゲートしてB29−K50Rを作製した。B29−K50RをNcoI及びNheIを用いて消化し、470bpのeIF5A1K50R断片をゲル精製した後、NcoI/NheIで消化したpExp−5へライゲートして最終発現ベクターのpExp5Aを作製した。
【0185】
[実施例18]
pExp5Aの試験
様々な細胞株にLipofectamine 2000を用いてプラスミドをトランスフェクトし、トランスフェクション後24時間目に抗HA抗体(Roche)を用いるウェスタンブロットによってHA−eIF5A1K50Rの発現を判定した。試験された異なる細胞株は、P3X63Ag8.653(マウスBリンパ芽球−骨髄腫)、KAS(ヒト骨髄腫)、HepG2(ヒト肝臓肝細胞癌)、T24(ヒト膀胱癌);HT−29(ヒト結腸直腸腺癌)、HEK−293(ヒト胚性腎細胞)、PC3(ヒト前立腺腺癌);HeLa(ヒト子宮頸部腺癌)、及びA549(肺癌)であった。図24を参照されたい。
【0186】
pExp−5プラスミドは、HA−eIF5A1K50Rをヒト及びマウス両方の骨髄腫細胞株に、恒常的なEF1プロモーター(CpG−eIF5A1K50R)を用いるプラスミドに匹敵するレベルで発現する。しかしながら、恒常的なプロモーターによる発現と比較して、pExp−5により駆動されるHA−eIF5A1K50Rの発現は非B細胞に限定される。一例外は、pExp−5のトランスフェクション後にHA−eIF5A1K50Rの発現が高いレベルで観察された、ヒト胚性腎細胞のHEK−293であった−これは細胞株の胚性の性質による可能性があり;現時点で本発明者らには、pExp−5が成体の腎細胞に発現するか否かは明らかでない。毒性試験及び治験に使用する最終プラスミドは、HA−eIF5A1K50Rが非HAタグ化eIF5A1K50Rによって置き換えられたpExp−5のバージョン(pExp−5A)となり得る。pExp−5は、最小ヒトB29プロモーター/エンハンサーの制御下にHA−タグ化eIF5A1K50Rを含み;HA−eIF5A1K50Rの発現は、恒常的に発現するプラスミドにより駆動される発現に対して、同様に完全長B29プロモーターを含むプラスミドに対して比較された。
【0187】
[実施例19]
in vivo JETPEI(商標)ナノ粒子の形成
この実施例は、20gのマウスへ1.5mg/kg(0.1mL)の用量−−1.5mg/kg=1.0mg pExp5A/kg+0.5mg h5A1/kg−−DNA:siRNA比=2:1により注射するための、in vivo JETPEI(商標)ナノ粒子複合体形成のために行われる。
【0188】
プラスミドDNA及びsiRNAを全量25mlに希釈する。量を調整するために滅菌水を使用する。*20mgのプラスミドDNA(2mg/mlを10ml)及び10mgのsiRNA(1mg/mlを10ml)を全量25mlに希釈する。差分を補うために滅菌水を使用する。25mlの10%グルコース(PEIキットと共に提供される)を加え、DNA溶液の量を50mlの5%グルコースに調整する。穏やかにボルテックスした後、短時間遠心する。全量25mlの水中へin vivo JETPEI(商標)を希釈する。*3.6mlのin vivo JETPEI(商標)を水で全量25mlに希釈する。最終濃度が5%グルコースになるように、10%グルコースで最終量を50mlに調整する。穏やかにボルテックスした後、短時間遠心する。50mlの希釈PEIを50mlの希釈DNAへ直ちに加える(順序を逆にしないこと!)。穏やかにボルテックスした後、短時間遠心沈殿する。
【0189】
形成後、複合体は8〜10時間安定である。複合体のN/P比は6でなければならない。N/P比は、DNA及びsiRNAの陰性に荷電したリン酸残基の数に対する、in vivo-jetPEIの陽性に荷電した窒素残基の数の比である。DNA及びsiRNAは、グラムあたり同数のリン酸基を含む。従ってN/P比は複合体内のイオン平衡の尺度である。複合体のN/P比の増大により、複合体の毒性が増大され得る。in vivo JET-PEIは150mM溶液(窒素残基で表される)として供給されるが、DNAは1mg中に3ナノモルの陰イオン性リン酸を含む。最終量中のDNAの最終濃度は0.5mg/mlを超えてはならない。DNAは高い品質で、水中で調製する必要がある。in vivo-jetPEI及び10%グルコースは、使用前に室温に置く必要がある。
【0190】
[実施例20]
マウスにおける静脈内SNS01及びSNS−EF1/UUによる用量設定及び反復投与試験。
SNS01は本発明の一実施形態である−これは多発性骨髄腫の治療を目標とする、癌治療の生物工学製品である。SNS01は3つの成分:eIF5Aのアポトーシス促進性変異体を発現するDNAベクター(図22を参照);癌細胞の生育/抗アポトーシスを促進する野生型のeIF5Aを標的とするsiRNA(図25の配列を参照);及び送達ビヒクルとして作用する、ポリエチレンイミンと称される合成ポリマー(In vivo-jetPEI;Polyplus Transfection Inc.)により構成される。
【0191】
試験の目的は、マウスへの静脈内SNS01の最大許容用量及び治療用量の長期投与の実行可能性を決定することである。別々の2試験を行った。最大許容用量試験(試験ID:MTD)は、マウスに増加性の量のSNS01(2.2mg/kg〜3.7mg/kg)の静脈内用量を2回投与し、臨床徴候、体重、臓器重量、及び肝酵素をモニターすることによって毒性を評価する8日間の試験である。9週間の反復投与試験(試験ID:EX6)は、SNS−EF1/UUの治療用量(1.5mg/kg)及び同様にその様々な成分の、週2回の長期投与後の毒性を評価するために計画された試験である。SNS−EF1/UUはSNS01の前臨床版であり、主に、eIF5AK50Rの発現がSNS01複合体のようにB細胞特異的プロモーターではなく、恒常的なプロモーター(全ての組織にいつでも発現するもの)により駆動されることで異なっている。SNS01におけるB細胞特異的B29プロモーターの使用は、アポトーシス促進性のeIF5A変異体の発現を、骨髄腫細胞を含むB細胞起源の細胞に限定することによって治療上の安全性を高めるために計画された。EX6試験にはマウス特異的なeIF5A siRNAを投与されたマウスの群もまた含まれたが、これはマウス組織においてeIF5Aを抑制することによるいずれの毒性効果があるか否かを判定するためである。反復投与試験における毒性は、臨床徴候、体重、血液、肝酵素、同様に組織病理をモニターすることによって評価した。
【0192】
【表1】
【0193】
【表2】
【0194】
前臨床試験により、SNS01は0.75mg/kg〜1.5mg/kgの用量において治療的であることが示された(試験EX9)。8日間の用量設定試験(試験ID:MTD)において、用量範囲の上限を決定するため、治療用範囲よりも著しく高い用量が試験された。試験品の週2回の静脈内用量は2.2mg/kg及び2.9mg/kgの低めの用量レベルで十分に許容されたが、1マウスは2.9mg/kgにおいて病的状態に至り、安楽死させた。3.3mg/kg以上の用量は、おおよそ20〜25%の生存率をもたらした。従って最大許容用量は2.2mg/kgと2.9mg/kgの間であり、これは0.75mg/kg〜1.5mg/kgの治療用範囲を十分に超えている。
【0195】
9週間の反復投与試験(試験ID:EX6)において、マウスは治療用量(1.5mg/kg)のSNS−EF1/UUを週2回尾静脈注射によって投与され、試験期間にわたって試験品に関連した毒性効果は観察されなかった。この試験においてはDNA及びsiRNAもまた別々に試験され、両方共マウスにより十分に許容された。ヒトeIF5A siRNAはマウスにおいて活性を示さないことから、マウスeIF5Aに特異的なsiRNAもまた試験に含めた。9週間の期間にわたり、マウスeIF5A siRNAの長期的な投与に関連した毒性効果は観察されなかった。これらの結果により、SNS01及びSNS−EF1/UUの治療用量は長期間にわたって投与されたとしてもマウスには無毒であることが示された。
【0196】
【表3】
【0197】
試験系及び試験計画
この試験の全ての態様は、カナダ動物管理協会(Canadian Council on Animal Care)及びオンタリオ州研究動物条例(Province of Ontario Animals for Research Act)により確立された、ウォータールー大学動物管理委員会(University of Waterloo Animal Care Committee)(ウォータールー、オンタリオ州、カナダ)により設定された指針に従って実行された。
【0198】
CD−1及びBALB/CマウスはCharles River Laboratories(ケベック州、カナダ)から入手した。両試験のマウスは試験品を週2回、尾静脈を通した静脈内注射によって投与された。0.2mlを超える量の送達には緩慢な注射(〜2−3分)を用いた。
【0199】
8日間の試験に用いたマウスは、試験開始時におおよそ6〜9週齢であった。9週間の反復投与試験に用いたマウスは、試験開始時におおよそ5〜6週齢であった。
【0200】
【表4】
【0201】
【表5】
【0202】
8日間の最大許容用量試験(MTD)
2用量による8日間の試験は、SNS01の最大許容用量を決定するために計画された用量設定試験である。用量範囲は2.2mg/kg〜3.7mg/kgであり、これは0.75mg/kg〜1.5mg/kgの治療用量範囲を十分に超えている。SNS01の最小用量(2.2mg/kg)において、1マウスがわずかに毛を逆立たせ、活動を低下させたが1時間以内に回復したことを除けば、毒性の臨床徴候は観察されなかった。2.2mg/kgのSNS01を2回目に注射した後には毒性の臨床徴候は観察されなかった。全てのマウスは試験の間中、その体重を維持した。臓器に肉眼的な変化は観察されなかった。体重に対する臓器重量の比は、肝臓重量:体重の比における中程度の増大を除いては、コントロール群からの変化は無かった。しかしながら、この比の増大はより高い用量レベル群のいずれにも観察されなかったことから、試験品に関連したものであるとは考えにくい。
【0203】
5マウスのうち4が、毒性の臨床徴候を示すことなく2.9mg/kgのSNS01を許容した。しかしながら、1マウスが注射後1時間以内に痙攣及び軽度の呼吸促迫症を経験したため人道的に安楽死させる必要があった。残りのマウスには、SNS01の2回目の注射後に毒性の臨床徴候は観察されなかった。マウスは試験の間中、その体重を維持し、臓器の肉眼的な変化、又は体重に対する臓器重量の比における変化は観察されなかった。2用量の2.9mg/kgのSNS01の後に、ALTの血清濃度がわずかに上昇した。
【0204】
予想されたように、3.3mg/kg以上の用量のSNS01は十分に許容されず、両群において1匹を除く全てのマウスを人道的に安楽死させる必要があった。病的状態のためマウスを人道的に安楽死させる必要があった全ての事例において臨床徴候は注射後1時間以内に現れ、これは高用量のPEIを用いたその他の報告された試験と一致していた。3.3mg/kg及び3.7mg/kgにおいて生存したマウスは2回目の用量は投与されなかったが、注射後4時間以内に完全に回復し、試験の間中その体重を維持した。従ってSNS01の最大許容用量は2.2mg/kgと2.9mg/kgの間であると思われる。
【0205】
9週間の反復投与試験(EX6)
9週間の反復投与試験の目的は、SNS01開発の間の前臨床試験のために使用された複合体であるSNS−EF1/UUの、治療用量(1.5mg/kg)の長期的な投与の安全性を評価することである。SNS−EF1/UUはSNS01と大きくは異ならないが、主要な相違は材料が研究グレードであること、及びeIF5AK50Rの発現が全ての細胞型において活性のある恒常的なヒトEF1プロモーターにより駆動されていることである。SNS01はeIF5AK50Rの発現を駆動するためにB細胞特異的なプロモーターを使用しているが、この安全性試験における恒常的なプロモーターの使用により、非B細胞における変異eIF5AK50Rタンパク質の蓄積がもたらす毒性の評価が可能となる。9週間の反復投与試験の別の態様は、SNS−EF1/UUの個別成分の安全性を試験するための群を包含することであった。DNA群(Ex6−G3)にはeIF5Aプラスミド及び非標的指向化コントロールsiRNAを含む複合体が投与され、一方でsiRNA群(Ex6−G4)にはヒトeIF5A(h5A1)siRNA及び非発現プラスミドを含む複合体が投与された。試験品のSNS−EF1/UUには、内在性のマウスeIF5Aの発現に影響し得ないヒトeIF5A siRNAが含まれることから、この試験の別の特色は、非発現プラスミド及びマウスeIF5Aを効率的に標的とするsiRNAを含むPEI複合体を投与された群(Ex6−G6)を包含することであった。この群によって、活性のあるeIF5A siRNAの長期的な投与の安全性についての評価が可能となった。
【0206】
全ての動物は、予定された屠殺日まで生存した。9週間の試験過程にわたり、いずれの群にも毒性の臨床徴候は観察されず、全ての群におけるマウスの体重は試験期間の間に増加し続けた。赤血球及び白血球の数は処置開始後3及び6週間目に測定され、全ての投与群において正常であった。肝酵素の血清濃度は屠殺後に測定され、全てのマウスについて正常範囲内であった。いずれの群にも、臓器の肉眼的外観に変化は観察されなかった。独立した病理学者により、主要な臓器の組織病理学的分析が行われた(試験品に起因する毒性は示されなかった。
【0207】
SNS−EF1/UUの治療用量の長期的な投与はマウスにより十分に許容され、有害作用は観察されなかった。加えて、マウス特異的なeIF5A siRNAの長期的な投与が毒性効果を示さなかったことから、ヒトeIF5A siRNAを含むPEI複合体の投与はヒトに対して安全なはずであることが示された。
【0208】
[実施例21]
多発性骨髄腫を有するマウスにおける、静脈内SNS−B29/UU及びSNS01の治療効果試験
SNS01は上記の通りである。試験品SNS−B29/UUはSNS01の前臨床版である。SNS−B29/UUのSNS01との違いはわずかであり、主要な違いは成分がGLPグレードではなく研究グレードであることである。ここに報告される試験の目的は、SNS−B29/UUの最小有効用量の決定であり、かつSNS01を含んでなるGLPグレードの材料が、前臨床試験で用いられた研究グレードの材料と同様に作用することの確認であった。反復投与腫瘍試験(repeated dose tumor study)(試験ID:EX9)は、SNS01の至適な治療用量を決定する目的で、マウスの皮下腫瘍の生育を阻害する、SNS−B29/UUの次第に縮小する能力を持つ週2回の用量を評価する5週間の試験であった。処置された動物はまた、臨床徴候、体重、及び臓器重量をモニターすることによって毒性の徴候を評価された。
【0209】
【表6】
【0210】
【表7】
【0211】
SNS−B29/UUの治療用範囲を、ヒト多発性骨髄腫皮下腫瘍を有するSCIDマウスにおいて決定した。0.15mg/kgと1.5mg/kgの間のSNS−B29/UUの用量について試験した。試験品の抗腫瘍効果を、週2回の腫瘍容積の測定ならびに屠殺後の腫瘍組織の切除及び秤量によって決定した。0.75mg/kg及び1.5mg/kgのSNS−B29/UUの用量により腫瘍の著しい収縮がもたらされたことから、SNS−B29/UUの治療用範囲は0.75mg/kgと1.5mg/kgの間にあることが示される。腫瘍の収縮は観察されなかったが、皮下腫瘍の生育の効果的な阻害は0.38mg/kgのSNS−B29/UUでもまた観察された。0.15mg/kgという低用量のSNS−B29/UUでさえも腫瘍生育の幾らかの阻害が観察されたことから、治療用範囲は広いことが示される。図26及び27を参照されたい。
【0212】
GLPグレード成分を用いて作られたSNS01の有効性をSNS−B29/UUと比較し、腫瘍生育の阻害において同程度の有効性を持つことが見出された。腫瘍を有するSCIDマウスのSNS01及びSNS−B29/UUによる処置は十分に許容され、マウスの体重は試験期間の間に増加し続けた。
【0213】
【表8】
【0214】
試験系及び試験計画
雌のC.B.17/IcrHsd−Prkdc(SCID)マウスはHarlan(インディアナポリス、インディアナ州、米国)から入手した。皮下腫瘍は、10x106のKAS−6/1(ヒト多発性骨髄腫)生細胞を5〜6週齢のマウスの右側腹部に注射することにより確立した。SNS−B29/UUによる処置は、腫瘍がおおよそ20〜40mm3のサイズに達した時点(腫瘍細胞注射後おおよそ4週間目)に開始した。SNS01による処置は、腫瘍がおおよそ130mm3のサイズに達した時点(腫瘍細胞注射後おおよそ6週間目)に開始した。マウスは試験品を週2回、尾静脈の静脈内注射を通して投与された。
【0215】
【表9】
【0216】
反復投与腫瘍試験(repeated dose tumor study)
反復投与腫瘍試験(repeated dose tumor study)は、SNS−B29/UUの最小有効治療用量を決定し、かつGLPグレードのSNS01試験品が、研究グレードの試験品であるSNS−B29/UUによって示された腫瘍の阻害活性を保持していることを確認するために計画された。第2の目的は、処置されたマウスの臨床徴候、体重、及び臓器重量をモニターすることによって、処置によるいずれの毒性効果を評価することであった。試験品の治療上の抗腫瘍活性は、デジタルキャリパーを用いて腫瘍容積を週2回測定することによりモニターした。屠殺後に腫瘍を切除して秤量した。
【0217】
全てのマウスは予定された屠殺日まで生存した。非発現プラスミド及び非標的指向化siRNAを含むPEIナノ複合体によって処置されたコントロールマウスは、屠殺時に284mm3の平均腫瘍容積を有したが、1.5mg/kgのSNS−B29/UUによって処置されたマウスは、わずかに13mm3の平均腫瘍容積、腫瘍生育における95%(*p=0.026)の減少を有した。しかしながら、1.5mg/kgのSNS−B29/UUによって処置されたマウスからの腫瘍の切除を試みた際、いずれのマウスにも腫瘍の形跡は見出されなかった。SNS−B29/UUの用量を半分の0.75mg/kgまで減少させてもなお、腫瘍容積及び重量それぞれにおいて91%(*p=0.03)及び87%(*p=0.04)の減少がもたらされ、1マウスでは腫瘍が完全に消失していた。従って、SNS−B29/UUの週2回の注射の至適な治療用量は、0.75mg/kgと1.5mg/kgの間であると思われる。SNS−B29/UUの週2回の用量が0.15mg/kgと低くてもなお、最終的な腫瘍容積の60%の減少がもたらされたことから、SNS−B29/UUは広い用量範囲にわたって強力な抗腫瘍活性を有することが示される。
【0218】
腫瘍生育の阻害に加え、0.75mg/kg及び1.5mg/kgのSNS−B29/UU及びSNS01による処置は腫瘍容積の著しい減少をもたらしたことから、この処置が、おそらくは腫瘍へのアポトーシスの誘発を通して腫瘍の退縮を誘導できることが示される。0.75mg/kg及び1.5mg/kgの用量レベルであるSNS−B29/UUによって処置された、腫瘍を有するマウスにおける腫瘍容積のパーセント変化は、それぞれ−244%及び−245%であった。コントロールマウスの腫瘍サイズは、同期間の間に2000%を超えて増大した。SNS01の週2回の注射もまた、多発性骨髄腫を著しく収縮させた。1.5mg/kgのSNS01によって処置されたマウスの腫瘍容積のパーセント変化は−349%であったことから、SNS01はSNS−B29/UUと同様に有効であることが示される。SNS01による処置は、わずかに25日間の処置後に腫瘍容積の349%の減少を達成したが、SNS−B29/UUは35日間の処置後に腫瘍容積の245%の減少を達成したことから、GLPグレード材料の使用によって生物活性が事実上増大された可能性がある。加えて、SNS01によって処置された腫瘍は極めて大きかった(〜130mm3)ことから、SNS01による処置は十分に確立された腫瘍に対して有効であることが示される。
【0219】
処置は全てのマウスにより十分に許容され、毒性の徴候は観察されなかった。全ての群におけるマウスの体重は試験の間を通して増加し続けた。剖検では臓器の肉眼的外観に変化は観察されず、体重に対する臓器重量の比における著しい変化は起こらなかった。
【0220】
従ってSNS01(及びその前臨床版であるSNS−B29/UU)はSCIDマウスによって十分に許容され、週あたり2回の静脈内注射によって送達される際、皮下のヒト多発性骨髄腫の治療に非常に有効である。試験された全てのSNS−B29/UUの用量は腫瘍生育の阻害に有効であったが、最高用量である1.5mg/kgにより、処置された全てのマウスの腫瘍は成功裏に除去された。
【0221】
[実施例22]
プラスミドDNA及びsiRNAポリエチレンイミン(JetPEI)複合体の生体内分布
PEI複合体により送達されたプラスミドDNAの局在を判定するため、緑色蛍光タンパク質(「GFP」)GFP発現コンストラクトを用いた。GFP発現を駆動するため、2つのプロモーター:EF1:遍在性プロモーター(EF1::GFP)又はB29:B細胞特異的プロモーター(B29::GFP)が用いられた。20マイクログラムのGFPプラスミドDNA及び10マイクログラムの蛍光標識(DY547)h5A1 siRNAを含むPEI複合体を、6のN/P比において調製した。BalB/Cマウスに、5%グルコース又はPEI複合体のいずれかを2日間連続して静脈内注射した。最初の注射後72時間目にマウスを安楽死させ、それらの臓器を回収して、GFPの発現及びDY547−siRNAを共焦点顕微鏡法によって解析した。
骨髄:ほとんどの事例でDY547−siRNAの形跡が存在したが、GFP発現は見られなかった。臓器回収の時期はGFP発現のピークと一致しない可能性があり;GFPシグナルは消失したか、又はGFPは発現していない可能性がある。しかしながら幾つかの例で、GFP及びDY547が同じ骨髄細胞に共存することが観察された。従ってこのことにより、静脈内注射によって投与する際、PEIナノ粒子を生きた動物の骨髄細胞にトランスフェクトできることが証明された。
肺:ほとんどの事例でDY547−siRNAの形跡が存在したが、GFP発現は見られなかった。臓器回収の時期はGFP発現のピークと一致しない可能性があるか、又はGFPシグナルは消失したか若しくはGFPは発現していない可能性がある。
脾臓:DY547−siRNAの存在もまた陽性である細胞内で、GFP発現(EF1プロモーターにより駆動される場合)が共存する形跡が見られた。B29プロモーターにより駆動した際、脾細胞内でのGFPの発現はかなり低かった。このことにより、PEIナノ粒子は脾臓の細胞にトランスフェクトされるようであることが示される。
腎臓:GFP又はDY547の発現は観察されなかったことから、ナノ粒子は腎臓には進入し得ないことが示される。
肝臓:ほとんどの事例でDY547−siRNAの形跡が存在したが、GFP発現は見られなかった。このことにより、PEIナノ粒子は肝臓の細胞にトランスフェクトされることが証明される。
心臓:心臓組織におけるEF1::GFPとDY547−siRNAとの共存が観察されたことから、PEIナノ粒子はこの臓器にトランスフェクトし得ることが示される。B29プロモーターを用いた際にはGFPは観察されなかった。
【0222】
[実施例23]
HA−eIF5AK50Rの発現における、DNA:siRNA比の効果
KAS細胞に、B29−HA−eIF5AK50R(B細胞特異的プロモーターにより駆動されるプラスミド)及びh5A1 siRNAを含むナノ粒子をトランスフェクトした。pExp5A及びh5A1 siRNAを異なる比において含むJET PEI(商標)ナノ粒子を作製し、KAS細胞への添加前に室温で4時間インキュベートした。トランスフェクション後4時間目に、ナノ粒子を含む培地を新鮮な培地に交換した。24時間後に細胞ライセートを回収し、HAに対する抗体を用いるウェスタンブロット分析に使用した。DNA:siRNAの比は、標準的な比である2:1から異なっていた。HA−eIF5AK50Rの蓄積は、1:0、3:1、及び2:1の比においてピークであった。図30を参照されたい。
【0223】
[実施例24]
ナノ粒子のトランスフェクションにより誘導されたアポトーシスにおけるDNA:siRNA比の効果
pExp5A及びh5A1 siRNAを異なる比において含むナノ粒子を作製し、KAS細胞への添加前に室温で4時間インキュベートした。トランスフェクション後4時間目に、ナノ粒子を含む培地を新鮮な培地に交換した。48時間後に細胞を回収し、アネキシンV/PIにより標識してFACSによって解析した。アポトーシスの誘導は、標準的なDNA:siRNA比である2:1によるナノ粒子をトランスフェクトした細胞において最も高かった。図31を参照されたい。
【0224】
[実施例25]
eIF5A1K50Rプラスミド及びeIF5A1 siRNA(siSTABLE又は非siSTABLE)を含むPEI複合体(N/P=6又は8)の投与により、多発性骨髄腫皮下腫瘍の生育は阻害され、かつ腫瘍の収縮がもたらされる。
KAS細胞をSCIDマウスに皮下注射した。触知可能な腫瘍が観察された時点で処置を開始した。マウスに:(G1)20mgのpCpG−mcs(空のベクター)及び10mgのコントロールsiRNAをN/P=8において含むPEI複合体(中程度用量);(G5)20mgのRNAi耐性プラスミドであるpCpG−eIF5A1k50R及び10mgのsiSTABLE h5A1 siRNAをN/P=8において含むPEI複合体(中程度用量、siSTABLE);(G8)20mgのRNAi耐性プラスミドであるpCpG−eIF5A1k50R及び10mgのh5A1 siRNAをN/P=6において含むPEI複合体(中程度用量、N/P=6)のいずれかを週あたり2回静脈内注射した。示されるデータは、各群のマウスについての個々の腫瘍容積である。最終の注射は、処置開始後40日目に行った。図32を参照されたい。
【0225】
[実施例26]
JET PEI(商標)ナノ粒子は腫瘍組織によって効果的に取り込まれ、このナノ粒子は同細胞へプラスミド及びsiRNAを送達する。
pExp−GFP(B細胞特異的プロモーターによる制御下のGFP)及びDY547−siRNA(蛍光標識されたsiRNA)を含むナノ粒子の注射後48時間目の腫瘍切片。共焦点顕微鏡法に従って腫瘍切片内にGFP及びDY547の共存した発現が観察されることから、ナノ粒子は腫瘍組織によって効果的に取り込まれ、このナノ粒子は同細胞へプラスミド及びsiRNAを送達することが示される。図33を参照されたい。
【0226】
[実施例27]
切断試験のために作製されたアデノウィルスコンストラクト。
アデノウィルス(血清型5、E3欠失)
1)Ad−eIF5A1Δ(2−6)[Δ(2−6)]−これはアミノ酸2〜6を欠損したヒトeIF5A1である。
2)Ad−eIF5A1K50RΔ(2−6)[Δ(2−6)/K50R]−これはアミノ酸2〜6を欠損したヒトeIF5A1である。加えて、「K50R」は位置50のリジン(K)がアルギニン(R)に変異しており、このためにDHSによりヒプシン化不可能であると信じられていることを示す。
3)Ad−eIF5A1D6E[D6E]−これは予想される切断部位が変異している(D6がEへ)ヒトeIF5A1である。
4)Ad−eIF5A1D6E/K50R[D6E/K50R]−これは予想される切断部位が変異している(D6がEへ)ヒトeIF5A1K50Rである。加えて、「K50R」は位置50のリジン(K)がアルギニン(R)に変異しており、このためにDHSによりヒプシン化不可能であると信じられていることを示す。
【0227】
[実施例28]
カスパーゼにより仲介されるeIF5Aの切断
切断部位を同定するため、アクチノマイシンDにより処理されたKAS細胞から分離したタンパク質ライセートを2−Dゲル電気泳動により分離し(図37)、分子量の小さなeIF5Aの切断産物に相当する点をゲルから切り出して質量分析法により配列決定した(図38B)。切断産物の完全長配列は得られなかったが、切断はタンパク質のN−末端から6番目のアミノ酸の後に起こったことが特定された(図38A)。考えられる切断部位のすぐ前の配列は「DDLD」であり、これはカスパーゼ切断部位として同定された配列である(Chay et al., 2002)。
【0228】
切断断片の産物がカスパーゼ依存性であるか否かを決定するため、ヒト骨髄腫細胞においてアクチノマイシンD誘導性のアポトーシスの間に産生される切断断片の産生を遮断する、カスパーゼ阻害剤の能力を試験した(図41)。一般的なカスパーゼ阻害剤ならびにカスパーゼ3、8、及び9の特異的な阻害剤は全て、KAS細胞においてActD誘導性のアポトーシスの間に蓄積するeIF5Aの切断型の形成を強力に阻止する。カスパーゼ1の阻害剤もまた、切断産物の形成を完全には遮断しなかったものの、減少させた。カスパーゼ阻害剤とのインキュベーションは、切断産物の蓄積を阻止しただけではなく、eIF5Aのヒプシン化型の減少もまた阻害したことから(図41)、アクチノマイシンD誘導性のアポトーシスの間のヒプシン化eIF5Aの減少は切断の結果であることが示される。
【0229】
アポトーシスの間のカスパーゼによるタンパク質の切断(Fisher et al., 2003による総説)は無数の目的:1)生存促進性又は抗アポトーシス性タンパク質の不活性化−例えば:mRNAの5’キャップ構造に結合して、キャップ形成されたmRNAの40Sリボソームサブユニットへの結合を促進する真核生物翻訳開始因子4G1が、カスパーゼによる切断によって不活性化されることで翻訳を阻害する;2)タンパク質のドミナントネガティブ型の作製(例えば、NF−カッパB p65はアポトーシスの間にカスパーゼによって切断され、DNAに結合はするが転写活性化の活性を持たないためにNF−kBのドミナントネガティブ阻害物質として作用する、ドミナントネガティブ断片を作り出す);3)阻害性又は調節性ドメインの除去によって新しい活性又は増大した活性を持つ断片の形成がもたらされることによる、アポトーシス促進性タンパク質の活性化をもたらす機能獲得型の切断(例えば、細胞周期の停止及びアポトーシスを仲介する乳癌抑制タンパク質であるBRCA−1は、アポトーシス促進性切断断片の放出しうるカスパーゼによる切断によって活性化される。eIF4GファミリーのメンバーであるDAP5はカスパーゼによる切断後に活性化され;切断産物はアポトーシス関連タンパク質の翻訳を刺激する);4)抗アポトーシスタンパク質のアポトーシス促進性タンパク質への変換(例えばBcl−xL及びc−IAP1のようなアポトーシス阻害物質は、カスパーゼによる切断を通してアポトーシス促進性タンパク質に変化し得る);及び5)細胞内の再分布(例えばカスパーゼ8による切断に従い、Bidのアポトーシス促進性p15断片では切断によって曝露されたグリシン残基がミリストイル化され、続いてこれによりBidがミトコンドリアへ移動してチトクロムcの放出を増大させることが可能になる)を有し得る。
【0230】
アクチノマイシンD誘導性アポトーシスの間の、カスパーゼにより仲介されるeIF5Aの切断が一般的な現象であるか又は骨髄腫細胞に特異的であるかを決定するために、切断されたeIF5Aの存在下において、アクチノマイシンDによって処理したHeLa細胞を調べた(図42)。アクチノマイシンDによって処理したHeLa細胞内に、eIF5Aの切断断片の蓄積は観察されなかった。eIF5Aの翻訳後修飾(例えばリン酸化又はアセチル化)がカスパーゼによる切断の必要条件であり得るか否かを決定する試みにおいて、脱アセチル化酵素阻害剤であるニコチンアミドの、切断断片の形成における効果について調べた。本発明者らの研究室及びその他(Kim et al., 2006)によるeIF5Aの質量分析によって、eIF5Aのアセチル化が観察された。さらに、酵母eIF5AがSir2関連の脱アセチル化酵素であるHst2の基質として同定され(Shirai et al., 2008)、著者らによりeIF5Aのアセチル化がHst2を欠損した酵母か又はeIF5Aを過剰発現する酵母においてのみ見られることが観察されたことから、eIF5Aは一般には非アセチル化型で存在することが示される。アクチノマイシンDによって処理したHeLa細胞とSir2脱アセチル化酵素阻害剤であるニコチンアミドとのインキュベーションによりeIF5A切断産物の蓄積がもたらされたことから、カスパーゼにより仲介される切断が起こるためにはeIF5Aのアセチル化が必要とされることが示される。これらのデータにより、eIF5Aは通常、アポトーシスシグナルがeIF5Aのアセチル化を引き起こしてカスパーゼにより仲介される切断が可能となるまで、カスパーゼ活性から保護され得ることが示唆される。
【0231】
参考文献
Chay KO, Park SS, Mushinski JF (2002). Linkage of caspase-mediated degradation of paxillin to apoptosis in Ba/F3 murine pro-B lymphocytes. J Biol Chem. 277, 14521-14529.
Fischer U, Janicke RU, Schulze-Osthoff K (2003). Many cuts to ruin: a comprehensive update of caspase substrates. Cell Death and Differentiation 10, 76-100.
Kim, S. C, Sprung, R., Chen, Y., Xu, Y., Ball, H., Pei, J., Cheng, T., Kho, Y., Xiao, H., Xiao, L., Grishin, N. V., White, M., Yang, X. J., and Zhao, Y. (2006) Substrate and Functional Diversity of Lysine Acetylation Revealed by a Proteomics Survey. Mol Cell 23, 607-618.
Shirai et al. (2008). Global analysis of gel mobility of proteins and its use in target identification J. Biol. Chem. 283, 10745-10752.
【特許請求の範囲】
【請求項1】
被験体の癌細胞又は腫瘍にアポトーシスを誘導するために用いられる薬剤を製造するための、切断型eIF5A1タンパク質をコードするeIF5A1ポリヌクレオチドの使用。
【請求項2】
eIF5A1ポリヌクレオチドが、図38に示される配列番号37に記載のアミノ酸配列を含んでなる切断型eIF5A1タンパク質をコードする、請求項1に記載の使用。
【請求項3】
eIF5A1ポリヌクレオチドが約16kDAの切断型eIF5A1タンパク質をコードする、請求項1に記載の使用。
【請求項4】
被験体が哺乳類である、請求項1に記載の使用。
【請求項5】
哺乳類がヒトである、請求項4に記載の使用。
【請求項6】
癌細胞又は腫瘍におけるアポトーシスの誘導が、癌細胞又は腫瘍の生育を遅くする、癌細胞又は腫瘍の生育を停止する、又は癌細胞を死滅させる若しくは腫瘍サイズを縮小させる、請求項1に記載の使用。
【請求項7】
癌が多発性骨髄腫である、請求項1に記載の使用。
【請求項8】
eIF5A1ポリヌクレオチドが、配列番号38に記載の(図41に示される)配列を含んでなる、請求項1に記載の使用。
【請求項9】
eIF5A1ポリヌクレオチドがプラスミド又は発現ベクター内に含まれる、請求項1に記載の使用。
【請求項10】
発現ベクターがアデノウィルス発現ベクター又はpHM6である、請求項1に記載の使用。
【請求項11】
発現ベクターが組織特異的プロモーターを含んでなる、請求項10に記載の使用。
【請求項12】
組織特異的プロモーターがB細胞特異的プロモーターである、請求項11に記載の使用。
【請求項13】
B細胞特異的プロモーターがB29である、請求項12に記載の使用。
【請求項14】
発現ベクターがpCpGプラスミドを含んでなる、請求項10に記載の使用。
【請求項15】
発現ベクターがポリエチレンイミンと複合される、請求項10に記載の使用。
【請求項16】
発現ベクターが還元型CpGジヌクレオチドを有する、請求項10に記載の使用。
【請求項17】
薬剤が腫瘍内、静脈内、又は皮下投与される、請求項1に記載の使用。
【請求項18】
切断型eIF5A1タンパク質をコードする単離されたポリヌクレオチドであって、配列番号38に記載の(図41に示される)配列を含んでなる、ポリヌクレオチド。
【請求項19】
切断型eIF5A1タンパク質をコードする単離されたポリヌクレオチドであって、該切断型タンパク質が配列番号37に記載の(図38に示される)アミノ酸配列を含んでなる、ポリヌクレオチド。
【請求項20】
カスパーゼにより仲介されるeIF5A1の切断によって形成される、単離された切断型eIF5A1ポリペプチド。
【請求項21】
被験体の癌細胞又は腫瘍にアポトーシスを誘導するために用いられる薬剤を製造するための、完全長eIF5A1ポリヌクレオチドと組み合わせた、切断型eIF5A1タンパク質をコードするeIF5A1ポリヌクレオチドの使用。
【請求項22】
eIF5A1ポリヌクレオチドが配列番号37に記載のアミノ酸配列を含んでなる切断型eIF5Aタンパク質をコードし、かつ完全長eIF5A1ポリヌクレオチドが配列番号35に記載のアミノ酸配列を含んでなるタンパク質をコードする、請求項21に記載の使用。
【請求項23】
eIF5A1ポリヌクレオチドが約16kDAの切断型eIF5Aタンパク質をコードする、請求項21に記載の使用。
【請求項24】
被験体が哺乳類である、請求項21に記載の使用。
【請求項25】
哺乳類がヒトである、請求項24に記載の使用。
【請求項26】
アポトーシスの誘導が、癌細胞又は腫瘍の生育を遅くする、癌細胞又は腫瘍の生育を停止する、又は癌細胞を死滅させる若しくは腫瘍サイズを縮小させる、請求項21に記載の使用。
【請求項27】
癌が多発性骨髄腫である、請求項21に記載の使用。
【請求項28】
eIF5A1ポリヌクレオチドが配列番号38に記載の配列を含んでなり、かつ完全長eIF5A1ポリヌクレオチドが配列番号43に記載の(図53に示される)配列を含んでなる、請求項21に記載の使用。
【請求項29】
完全長eIF5Aポリヌクレオチドが変異eIF5A1をコードする、請求項23に記載の使用であって、該変異がデオキシヒプシンシンターゼによるヒプシン化を阻止若しくは阻害し、並びに/又は、該変異がユビキチン化部位及び/若しくはアセチル化部位に存在する使用。
【請求項30】
変異がK50A、K50R、K67A、K47R、K67R、K50A/K67A、K50A/K47R、K50A/K67R、K50R/K67A、K50R/K47R、K50R/K67R、及びK47A/K67Aからなる群より選択される、請求項23に記載の使用。
【請求項31】
eIF5Aの3’UTRを標的とするsiRNAをさらに含んでなる、請求項29に記載の使用。
【請求項32】
siRNAがeIF5A1の配列である5’−GCT GGA CTC CTC CTA CAC A−3’を標的とする、請求項31に記載の使用。
【請求項33】
siRNAがdsRNAであり、dsRNAの一本の鎖が5’−GCU GGA CUC CUC CUA CAC A−3’の配列を含んでなる、請求項32に記載の使用。
【請求項34】
siRNAが血清中での分解を阻止するために安定化される、請求項32に記載の使用。
【請求項35】
完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチドが発現ベクター内に存在する、請求項31に記載の使用。
【請求項36】
完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチド及び/又はsiRNAがポリエチレンイミンと複合される、請求項35に記載の使用。
【請求項37】
完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチド及び/又はsiRNAが独立してポリエチレンイミンと複合される、請求項36に記載の使用。
【請求項38】
請求項1〜38のいずれか一項に記載の組成物のいずれか一つを哺乳類に提供することによる、哺乳類の癌細胞又は哺乳類の腫瘍にアポトーシスを誘導する方法。
【請求項39】
組成物が静脈内、腹腔内、又は腫瘍内投与される、請求項38に記載の方法。
【請求項40】
癌が多発性骨髄腫である、請求項38に記載の方法。
【請求項41】
eIF5A1の3’末端を標的とするeIF5A1 siRNA、変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターの複合体を含んでなる組成物であって、該変異eIF5A1がヒプシン化できず、並びに、該siRNA及び該発現ベクターが複合体形成のためにポリエチレンイミンと複合される、組成物。
【請求項42】
被験体における標的遺伝子の内在性発現を抑制するための、標的遺伝子を標的とするsiRNA;及びRNAI耐性プラスミド内の、被験体内で発現できる標的タンパク質をコードするポリヌクレオチドを含んでなる組成物であって、該siRNA及び該プラスミドが、複合体形成のためにポリエチレンイミンと複合される、組成物。
【請求項43】
siRNAが図25に示す配列を有し、かつ変異eIF5A1をコードするポリヌクレオチドがeIF5A1K50Rである、請求項41に記載の組成物。
【請求項44】
組織特異的プロモーターを含んでなる、請求項43に記載の組成物。
【請求項45】
B細胞特異的プロモーターを含んでなる、請求項44に記載の組成物。
【請求項46】
B細胞プロモーターがB29である、請求項45に記載の組成物。
【請求項47】
発現ベクターがpCpGプラスミドを含んでなる、請求項43に記載の組成物。
【請求項48】
eIF5A1 siRNA、及び変異eIF5A1ポリヌクレオチドを含んでなる発現ベクターが、独立してポリエチレンイミンと複合される、請求項41に記載の組成物。
【請求項49】
eIF5A1 siRNA、及び変異eIF5A1ポリヌクレオチドを含んでなる発現ベクターが、一緒にポリエチレンイミンと複合される、請求項41に記載の組成物。
【請求項50】
eIF5A1の3’末端を標的とするeIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターを含んでなる組成物であって、該変異eIF5A1がヒプシン化できず、並びに、該siRNA及び該発現ベクターが癌治療のため被験体に送達される、組成物。
【請求項51】
癌が多発性骨髄腫である、請求項50に記載の組成物。
【請求項52】
請求項50に記載の組成物を被験体に投与することを含んでなる癌の治療法。
【請求項53】
請求項41に記載の組成物を被験体に投与することを含んでなる癌の治療法。
【請求項54】
組成物が静脈内、腹腔内、又は腫瘍内投与される、請求項52に記載の方法。
【請求項55】
eIF5A1の3’末端を標的とするsiRNA及び変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターが異なる経路を通して送達される、請求項53に記載の方法。
【請求項56】
組成物が約0.15mg/kg〜約1.5mg/kgの用量で週2回の注射で供給される、請求項52に記載の方法。
【請求項57】
組成物が約0.75mg/kg〜約1.5mg/kgの用量で週2回の注射で供給される、請求項52に記載の方法。
【請求項1】
被験体の癌細胞又は腫瘍にアポトーシスを誘導するために用いられる薬剤を製造するための、切断型eIF5A1タンパク質をコードするeIF5A1ポリヌクレオチドの使用。
【請求項2】
eIF5A1ポリヌクレオチドが、図38に示される配列番号37に記載のアミノ酸配列を含んでなる切断型eIF5A1タンパク質をコードする、請求項1に記載の使用。
【請求項3】
eIF5A1ポリヌクレオチドが約16kDAの切断型eIF5A1タンパク質をコードする、請求項1に記載の使用。
【請求項4】
被験体が哺乳類である、請求項1に記載の使用。
【請求項5】
哺乳類がヒトである、請求項4に記載の使用。
【請求項6】
癌細胞又は腫瘍におけるアポトーシスの誘導が、癌細胞又は腫瘍の生育を遅くする、癌細胞又は腫瘍の生育を停止する、又は癌細胞を死滅させる若しくは腫瘍サイズを縮小させる、請求項1に記載の使用。
【請求項7】
癌が多発性骨髄腫である、請求項1に記載の使用。
【請求項8】
eIF5A1ポリヌクレオチドが、配列番号38に記載の(図41に示される)配列を含んでなる、請求項1に記載の使用。
【請求項9】
eIF5A1ポリヌクレオチドがプラスミド又は発現ベクター内に含まれる、請求項1に記載の使用。
【請求項10】
発現ベクターがアデノウィルス発現ベクター又はpHM6である、請求項1に記載の使用。
【請求項11】
発現ベクターが組織特異的プロモーターを含んでなる、請求項10に記載の使用。
【請求項12】
組織特異的プロモーターがB細胞特異的プロモーターである、請求項11に記載の使用。
【請求項13】
B細胞特異的プロモーターがB29である、請求項12に記載の使用。
【請求項14】
発現ベクターがpCpGプラスミドを含んでなる、請求項10に記載の使用。
【請求項15】
発現ベクターがポリエチレンイミンと複合される、請求項10に記載の使用。
【請求項16】
発現ベクターが還元型CpGジヌクレオチドを有する、請求項10に記載の使用。
【請求項17】
薬剤が腫瘍内、静脈内、又は皮下投与される、請求項1に記載の使用。
【請求項18】
切断型eIF5A1タンパク質をコードする単離されたポリヌクレオチドであって、配列番号38に記載の(図41に示される)配列を含んでなる、ポリヌクレオチド。
【請求項19】
切断型eIF5A1タンパク質をコードする単離されたポリヌクレオチドであって、該切断型タンパク質が配列番号37に記載の(図38に示される)アミノ酸配列を含んでなる、ポリヌクレオチド。
【請求項20】
カスパーゼにより仲介されるeIF5A1の切断によって形成される、単離された切断型eIF5A1ポリペプチド。
【請求項21】
被験体の癌細胞又は腫瘍にアポトーシスを誘導するために用いられる薬剤を製造するための、完全長eIF5A1ポリヌクレオチドと組み合わせた、切断型eIF5A1タンパク質をコードするeIF5A1ポリヌクレオチドの使用。
【請求項22】
eIF5A1ポリヌクレオチドが配列番号37に記載のアミノ酸配列を含んでなる切断型eIF5Aタンパク質をコードし、かつ完全長eIF5A1ポリヌクレオチドが配列番号35に記載のアミノ酸配列を含んでなるタンパク質をコードする、請求項21に記載の使用。
【請求項23】
eIF5A1ポリヌクレオチドが約16kDAの切断型eIF5Aタンパク質をコードする、請求項21に記載の使用。
【請求項24】
被験体が哺乳類である、請求項21に記載の使用。
【請求項25】
哺乳類がヒトである、請求項24に記載の使用。
【請求項26】
アポトーシスの誘導が、癌細胞又は腫瘍の生育を遅くする、癌細胞又は腫瘍の生育を停止する、又は癌細胞を死滅させる若しくは腫瘍サイズを縮小させる、請求項21に記載の使用。
【請求項27】
癌が多発性骨髄腫である、請求項21に記載の使用。
【請求項28】
eIF5A1ポリヌクレオチドが配列番号38に記載の配列を含んでなり、かつ完全長eIF5A1ポリヌクレオチドが配列番号43に記載の(図53に示される)配列を含んでなる、請求項21に記載の使用。
【請求項29】
完全長eIF5Aポリヌクレオチドが変異eIF5A1をコードする、請求項23に記載の使用であって、該変異がデオキシヒプシンシンターゼによるヒプシン化を阻止若しくは阻害し、並びに/又は、該変異がユビキチン化部位及び/若しくはアセチル化部位に存在する使用。
【請求項30】
変異がK50A、K50R、K67A、K47R、K67R、K50A/K67A、K50A/K47R、K50A/K67R、K50R/K67A、K50R/K47R、K50R/K67R、及びK47A/K67Aからなる群より選択される、請求項23に記載の使用。
【請求項31】
eIF5Aの3’UTRを標的とするsiRNAをさらに含んでなる、請求項29に記載の使用。
【請求項32】
siRNAがeIF5A1の配列である5’−GCT GGA CTC CTC CTA CAC A−3’を標的とする、請求項31に記載の使用。
【請求項33】
siRNAがdsRNAであり、dsRNAの一本の鎖が5’−GCU GGA CUC CUC CUA CAC A−3’の配列を含んでなる、請求項32に記載の使用。
【請求項34】
siRNAが血清中での分解を阻止するために安定化される、請求項32に記載の使用。
【請求項35】
完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチドが発現ベクター内に存在する、請求項31に記載の使用。
【請求項36】
完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチド及び/又はsiRNAがポリエチレンイミンと複合される、請求項35に記載の使用。
【請求項37】
完全長eIF5A1ポリヌクレオチド及び/又は切断型eIF5A1タンパク質をコードするeIF5Aポリヌクレオチド及び/又はsiRNAが独立してポリエチレンイミンと複合される、請求項36に記載の使用。
【請求項38】
請求項1〜38のいずれか一項に記載の組成物のいずれか一つを哺乳類に提供することによる、哺乳類の癌細胞又は哺乳類の腫瘍にアポトーシスを誘導する方法。
【請求項39】
組成物が静脈内、腹腔内、又は腫瘍内投与される、請求項38に記載の方法。
【請求項40】
癌が多発性骨髄腫である、請求項38に記載の方法。
【請求項41】
eIF5A1の3’末端を標的とするeIF5A1 siRNA、変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターの複合体を含んでなる組成物であって、該変異eIF5A1がヒプシン化できず、並びに、該siRNA及び該発現ベクターが複合体形成のためにポリエチレンイミンと複合される、組成物。
【請求項42】
被験体における標的遺伝子の内在性発現を抑制するための、標的遺伝子を標的とするsiRNA;及びRNAI耐性プラスミド内の、被験体内で発現できる標的タンパク質をコードするポリヌクレオチドを含んでなる組成物であって、該siRNA及び該プラスミドが、複合体形成のためにポリエチレンイミンと複合される、組成物。
【請求項43】
siRNAが図25に示す配列を有し、かつ変異eIF5A1をコードするポリヌクレオチドがeIF5A1K50Rである、請求項41に記載の組成物。
【請求項44】
組織特異的プロモーターを含んでなる、請求項43に記載の組成物。
【請求項45】
B細胞特異的プロモーターを含んでなる、請求項44に記載の組成物。
【請求項46】
B細胞プロモーターがB29である、請求項45に記載の組成物。
【請求項47】
発現ベクターがpCpGプラスミドを含んでなる、請求項43に記載の組成物。
【請求項48】
eIF5A1 siRNA、及び変異eIF5A1ポリヌクレオチドを含んでなる発現ベクターが、独立してポリエチレンイミンと複合される、請求項41に記載の組成物。
【請求項49】
eIF5A1 siRNA、及び変異eIF5A1ポリヌクレオチドを含んでなる発現ベクターが、一緒にポリエチレンイミンと複合される、請求項41に記載の組成物。
【請求項50】
eIF5A1の3’末端を標的とするeIF5A1 siRNA及び変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターを含んでなる組成物であって、該変異eIF5A1がヒプシン化できず、並びに、該siRNA及び該発現ベクターが癌治療のため被験体に送達される、組成物。
【請求項51】
癌が多発性骨髄腫である、請求項50に記載の組成物。
【請求項52】
請求項50に記載の組成物を被験体に投与することを含んでなる癌の治療法。
【請求項53】
請求項41に記載の組成物を被験体に投与することを含んでなる癌の治療法。
【請求項54】
組成物が静脈内、腹腔内、又は腫瘍内投与される、請求項52に記載の方法。
【請求項55】
eIF5A1の3’末端を標的とするsiRNA及び変異eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターが異なる経路を通して送達される、請求項53に記載の方法。
【請求項56】
組成物が約0.15mg/kg〜約1.5mg/kgの用量で週2回の注射で供給される、請求項52に記載の方法。
【請求項57】
組成物が約0.75mg/kg〜約1.5mg/kgの用量で週2回の注射で供給される、請求項52に記載の方法。
【図1】

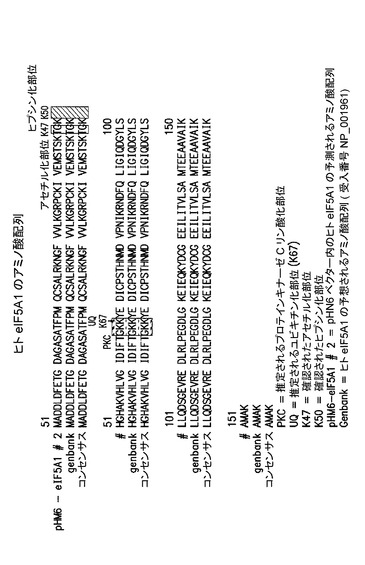
【図2A】
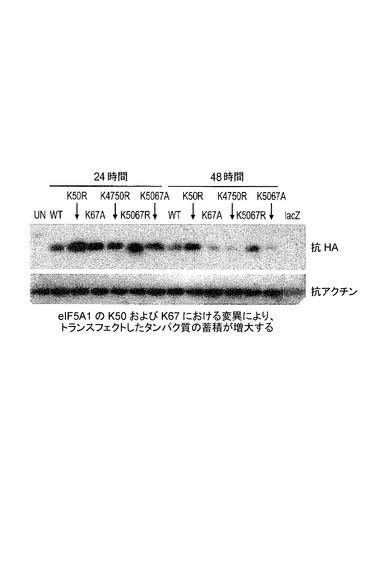
【図2B】
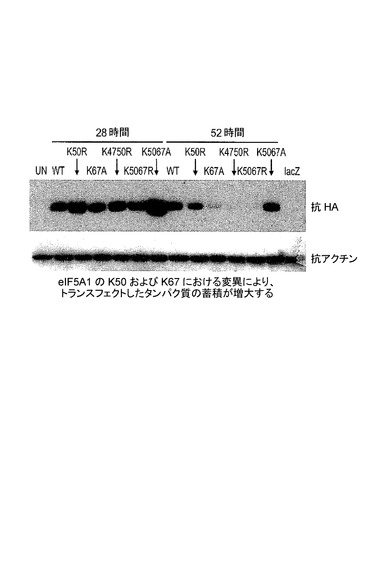
【図3】
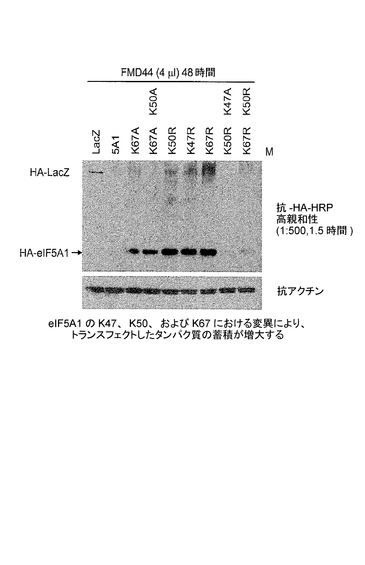
【図4】
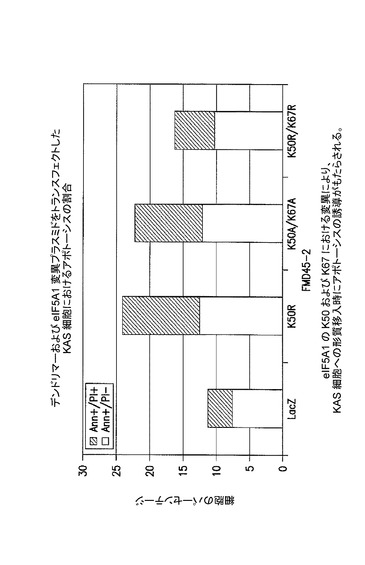
【図5】
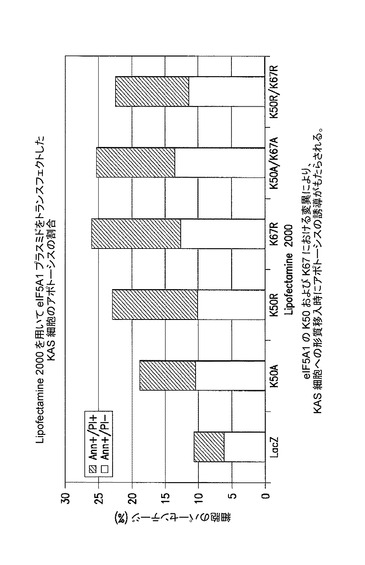
【図6】
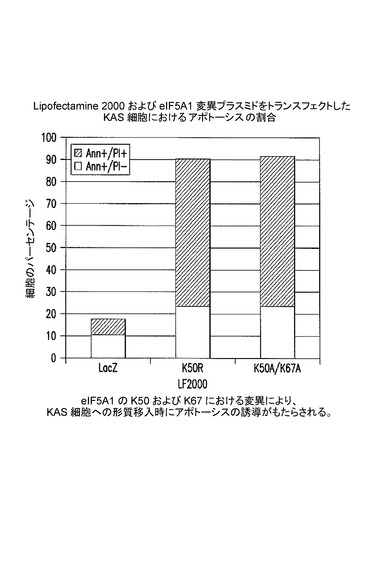
【図7A】
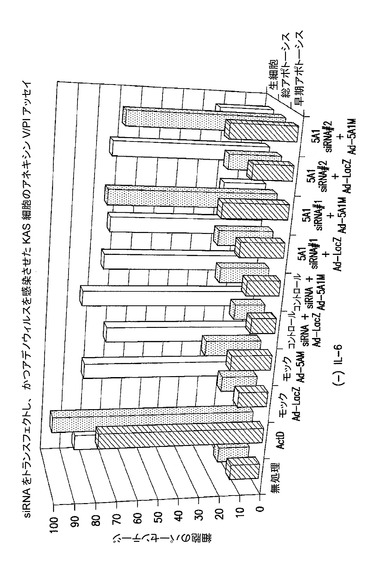
【図7B】

【図7C】
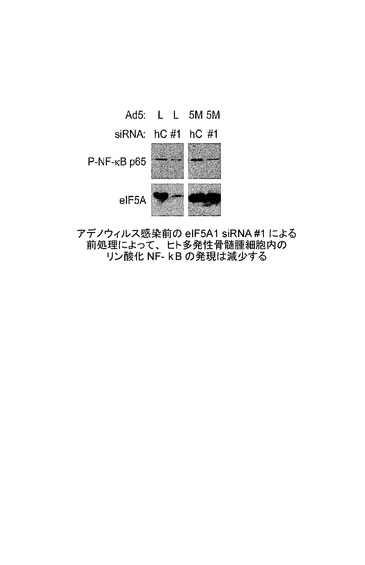
【図7D】

【図7E】
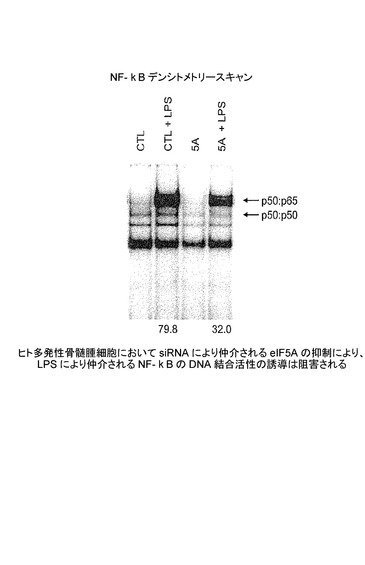
【図7F】
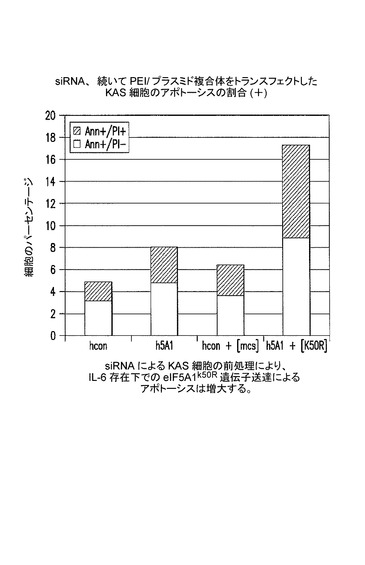
【図8】
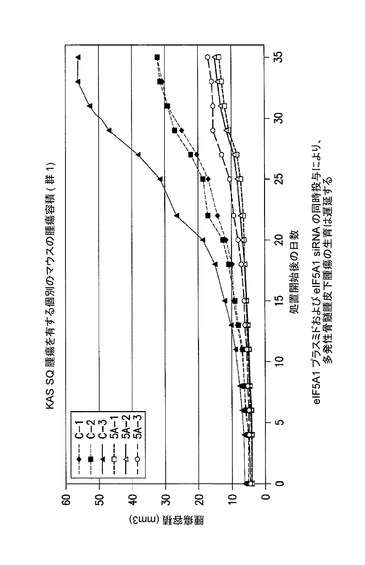
【図9】
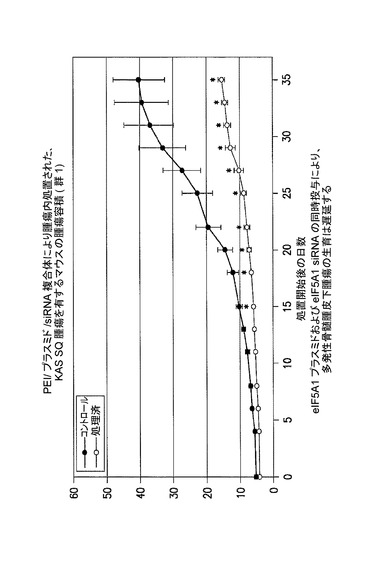
【図10】
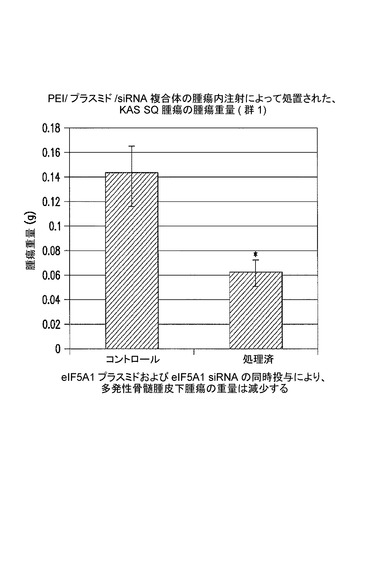
【図11】
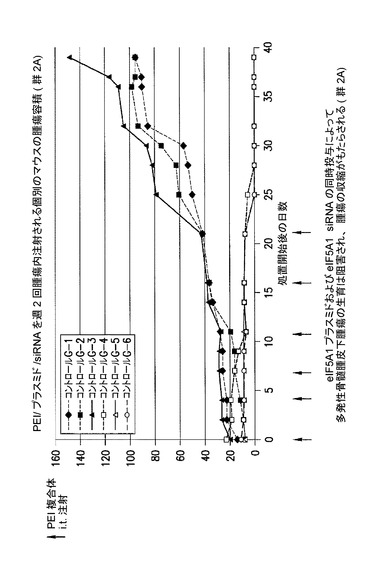
【図12】
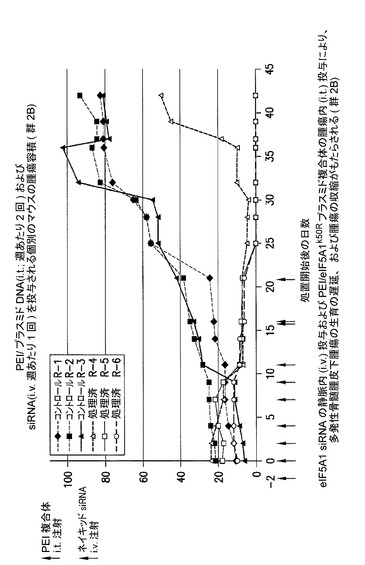
【図13A】
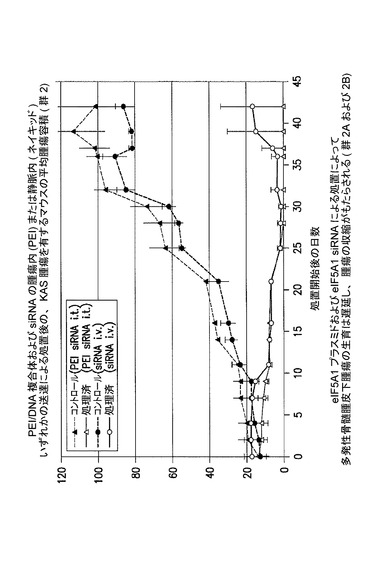
【図13B】
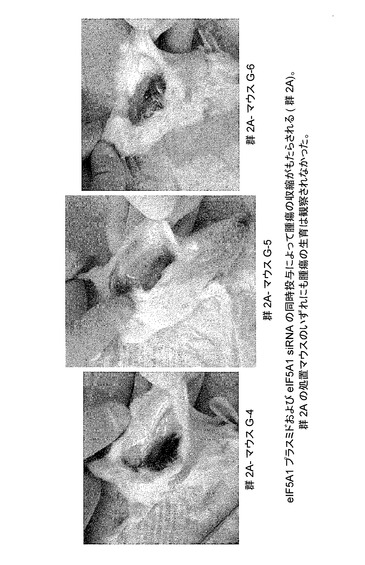
【図13C】
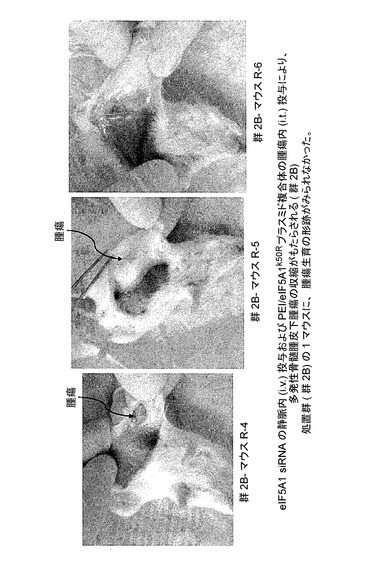
【図14】
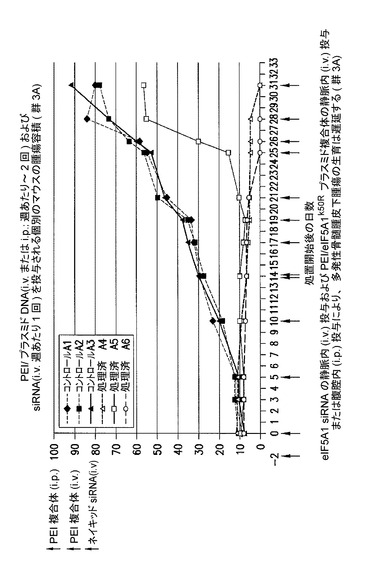
【図15】
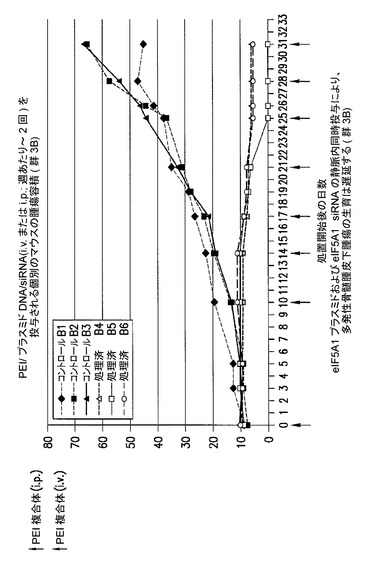
【図16】
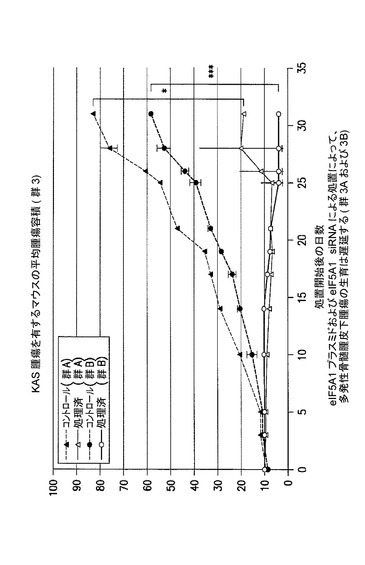
【図17A】
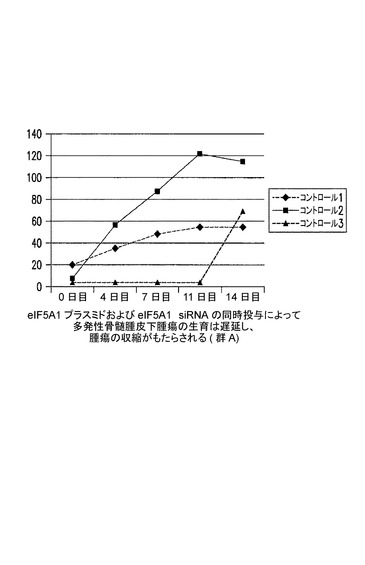
【図17B】
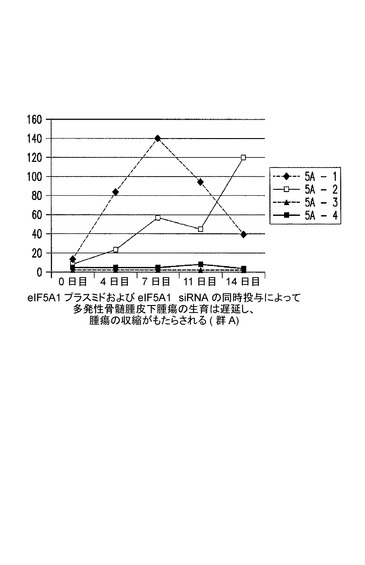
【図18A】
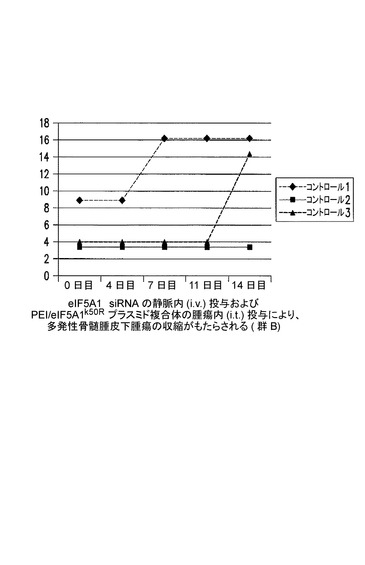
【図18B】
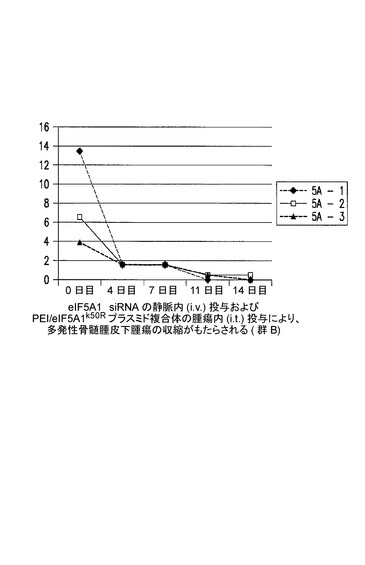
【図19】
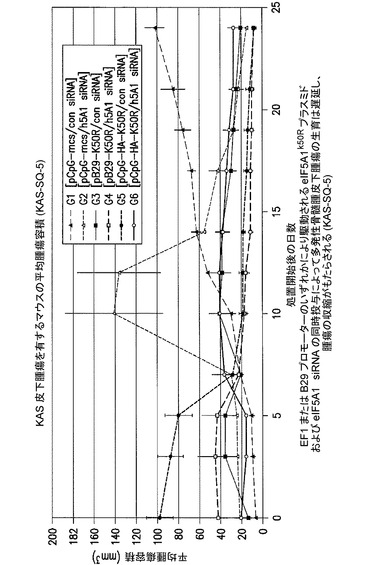
【図20】
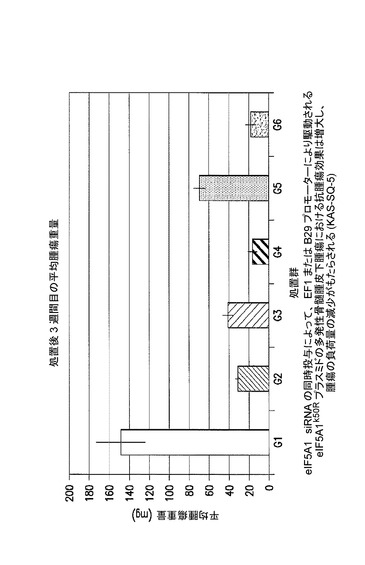
【図21】
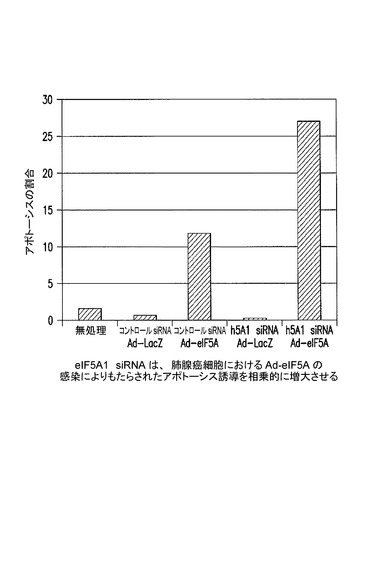
【図22】
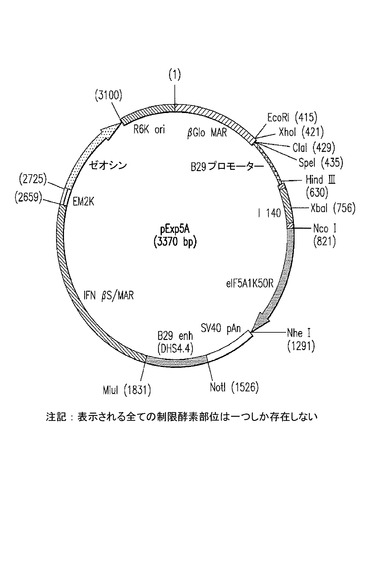
【図23A】
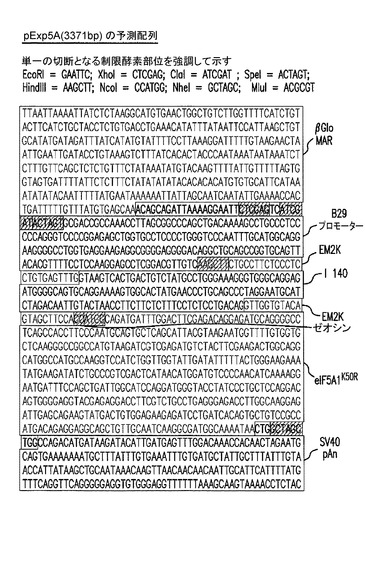
【図23B】
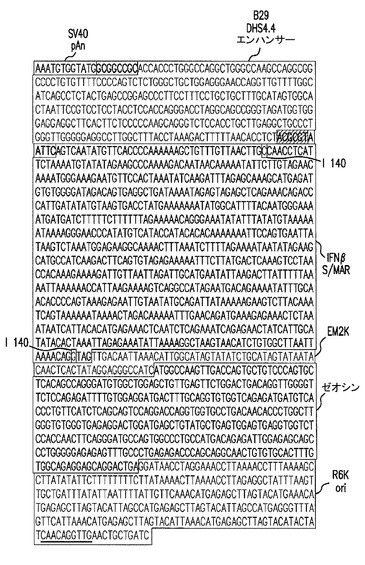
【図24A】
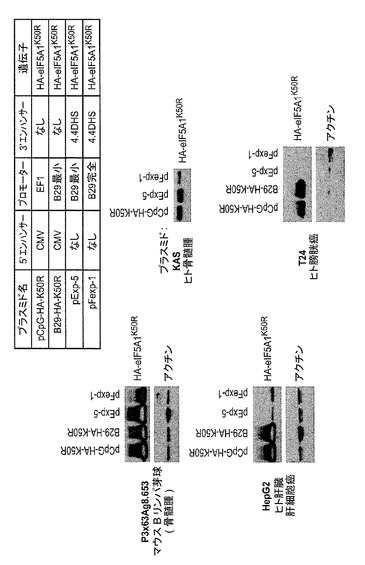
【図24B】
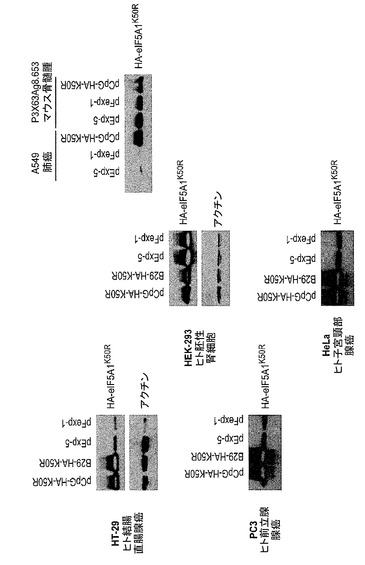
【図25】

【図26】
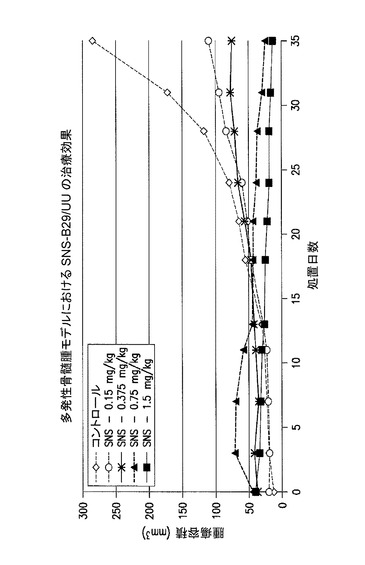
【図27】
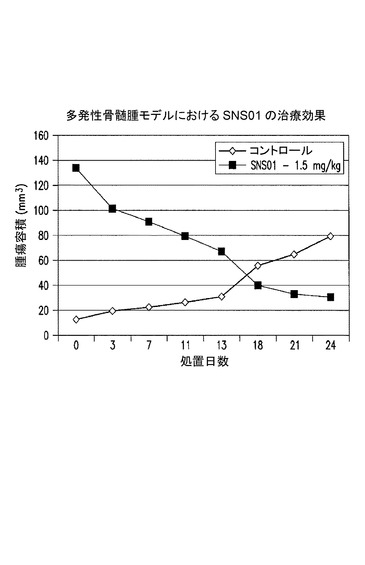
【図28】

【図29】

【図30】

【図31】
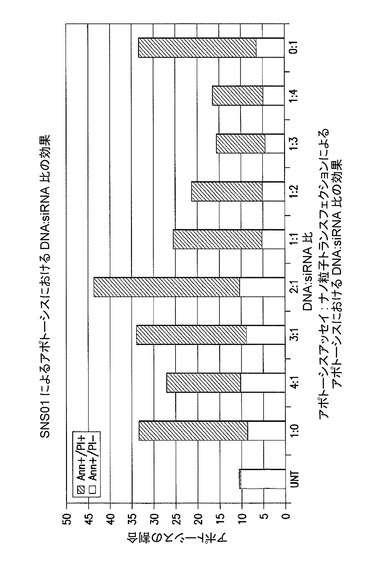
【図32】
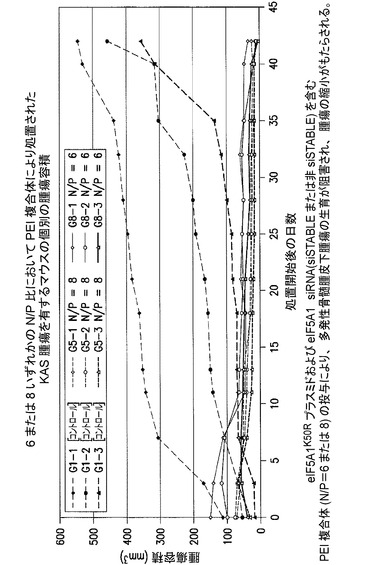
【図33】
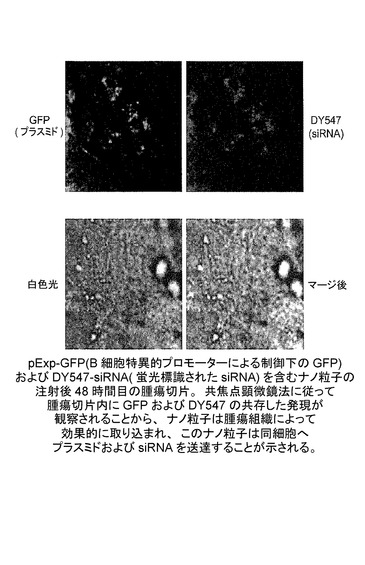
【図34】
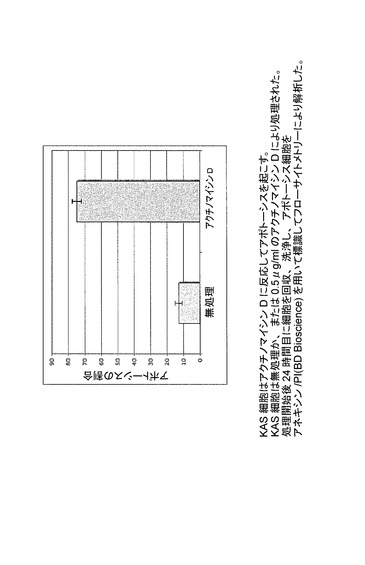
【図35】
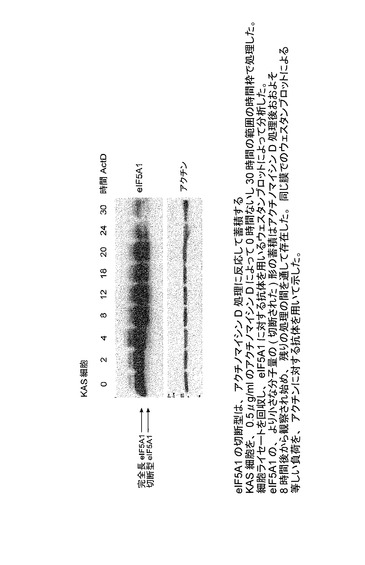
【図36】
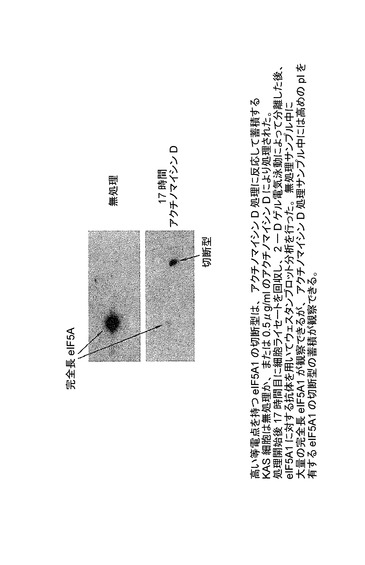
【図37】

【図38】

【図39】
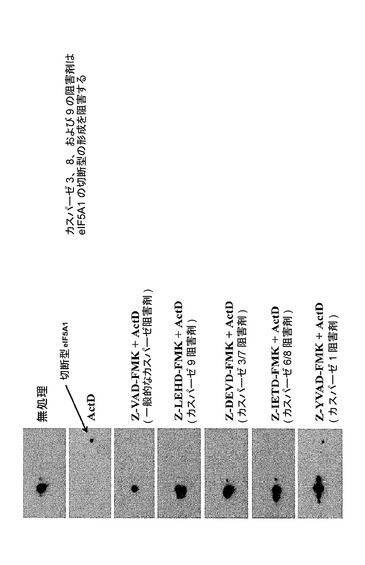
【図40】
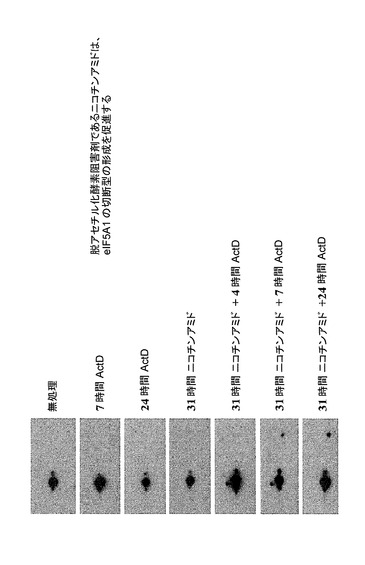
【図41】

【図42】

【図43】
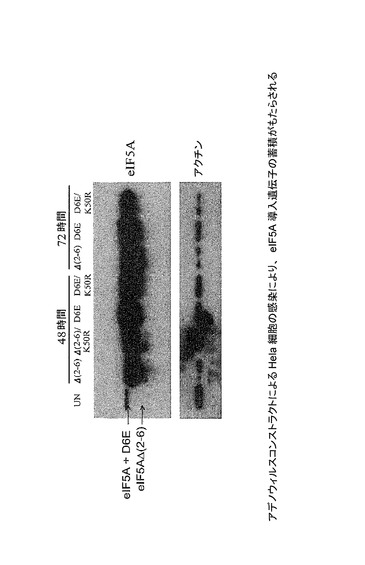
【図44】
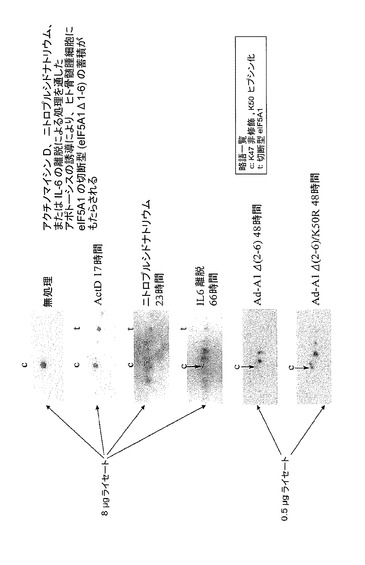
【図45】
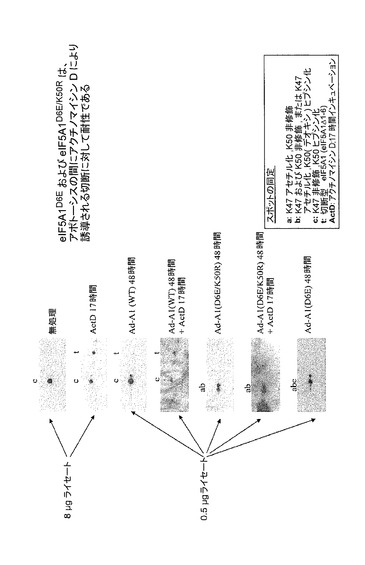
【図46】
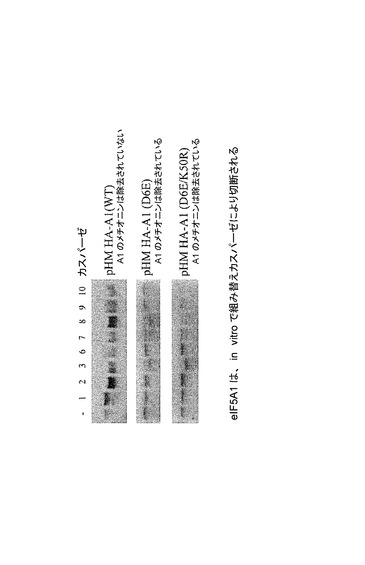
【図47】

【図48】
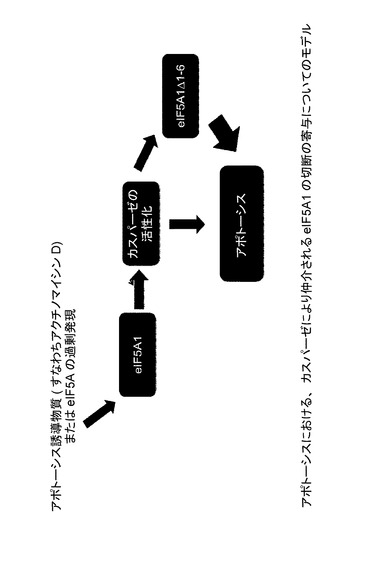
【図49】
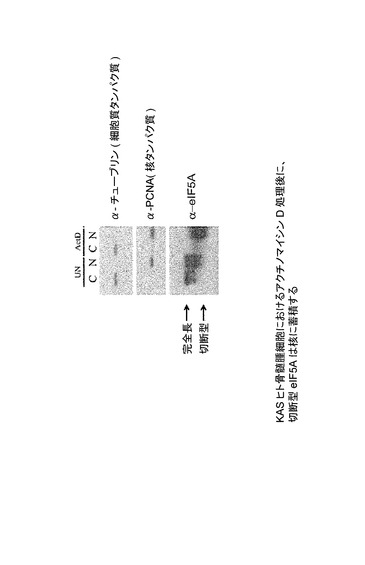
【図50】

【図51】
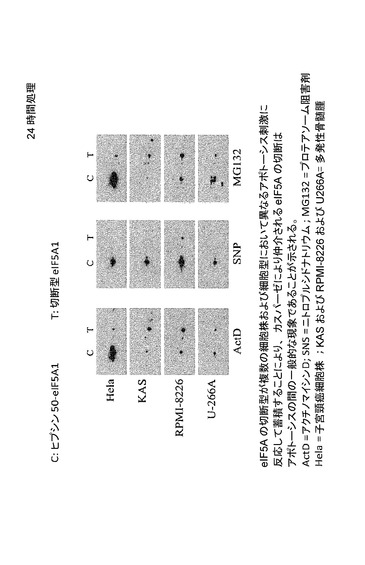
【図52】
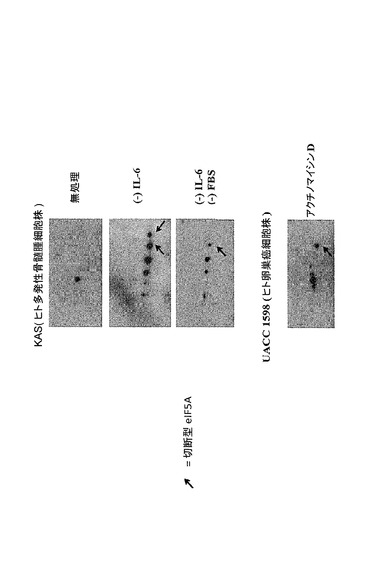
【図53】

【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13A】
【図13B】
【図13C】
【図14】
【図15】
【図16】
【図17A】
【図17B】
【図18A】
【図18B】
【図19】
【図20】
【図21】
【図22】
【図23A】
【図23B】
【図24A】
【図24B】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【図45】
【図46】
【図47】
【図48】
【図49】
【図50】
【図51】
【図52】
【図53】
【公表番号】特表2012−501650(P2012−501650A)
【公表日】平成24年1月26日(2012.1.26)
【国際特許分類】
【出願番号】特願2011−526177(P2011−526177)
【出願日】平成21年9月2日(2009.9.2)
【国際出願番号】PCT/US2009/055791
【国際公開番号】WO2010/028093
【国際公開日】平成22年3月11日(2010.3.11)
【出願人】(502458132)セネスコ テクノロジーズ,インコーポレイティド (16)
【Fターム(参考)】
【公表日】平成24年1月26日(2012.1.26)
【国際特許分類】
【出願日】平成21年9月2日(2009.9.2)
【国際出願番号】PCT/US2009/055791
【国際公開番号】WO2010/028093
【国際公開日】平成22年3月11日(2010.3.11)
【出願人】(502458132)セネスコ テクノロジーズ,インコーポレイティド (16)
【Fターム(参考)】
[ Back to top ]