
癌胎児抗原をコードする合成遺伝子およびその使用
ヒト癌胎児抗原(CEA)をコードし、ヒト細胞環境において発現するためにコドンが最適化されている合成ポリヌクレオチドを提供する。CEAをコードする遺伝子は通常、ヒト癌の発症に関連する。本発明は、異常なCEA発現が癌またはその発症に関連するので、CEA腫瘍関連抗原によって発現する蛋白質産物に対する免疫を惹起し、または増強する組成物および方法を提供する。本発明は、具体的には、コドンを最適化したヒトCEAを有するアデノウイルスベクターおよびプラスミド構築物を提供し、癌を予防および治療するためのワクチンおよび医薬品組成物におけるそれらの使用を開示する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は一般的に癌の治療に関する。より具体的には、本発明は、本明細書でhCEAoptと称するヒト腫瘍関連癌胎児抗原をコードする合成ポリヌクレオチドに関し、該ポリペプチドはヒト細胞環境で発現するためにコドンが最適化されている。本発明はまた、組換えベクターおよび前記合成ポリヌクレオチドを含む宿主を提供する。本発明はまた、hCEAoptを有するアデノウイルスベクターおよびプラスミド構築物並びに癌の予防および治療のためのワクチンにおけるそれらの使用並びに医薬品組成物に関する。
【背景技術】
【0002】
免疫グロブリンスーパーファミリー(IgSF)は、様々な機能を備えた蛋白質をコードする数多くの遺伝子から構成され、その機能の1つが細胞間接着である。IgSF蛋白質は、適切な分子間結合相互作用を維持するために重要なIg関連ドメインを少なくとも1つ含有する。このような相互作用は、IgSF構成要素の様々な生物学的機能に不可欠なので、多くのIgSF接着分子の破壊または異常発現は、多くのヒト疾患と相関してきた。
【0003】
癌胎児抗原(CEA)は、細胞表面糖蛋白質を構成するIgスーパーファミリーに属する。CEAサブファミリーの構成要素は、CEA関連細胞接着分子(CEACAM)として知られている。最近の科学文献では、CEA遺伝子はCEACAM5と改名されたが、蛋白質の名称はCEAのままである。機能的には、CEACAMはホモタイプおよびヘテロタイプ両方の細胞間接着分子として作用することが示された(Benchimol他、Cell 57:327〜334(1989))。細胞接着に加えて、CEAは細胞外マトリックスからの細胞脱離によって生じる細胞死を阻害し、Bcl2およびC−Mycなどのある種のプロトオンコジーンに関連した細胞形質転換の一因となり得る(Berinstein、J.Oncol.20(8):2197〜2207参照)。
【0004】
CEAの正常な発現は、胎児の発育中および成人結腸粘膜で検出された。CEA過剰発現は、30年以上前にヒト結腸癌において最初に検出され(Gold and Freedman、J.Exp.Med.121:439〜462(1965))、その後ほとんど全ての結腸直腸癌で見いだされてきた。さらに、CEA過剰発現は、膵臓、胸および肺の腺癌の高い割合で検出可能である。これらの種類の腫瘍ではCEA発現が一般的なので、CEAはこれらの癌の管理および予後において広く臨床的に使用されている。
【0005】
ヒトCEAをコードする配列は、クローニングされ、特徴付けられている(米国特許第5274087号、米国特許第5571710号および米国特許第5843761号。Beauchemin他、Mol.Cell.Biol.7:3221〜3230(1987)、Zimmerman他、Proc.Natl.Acad.Sci.USA 84:920〜924(1987)。Thompson他、Proc.Natl.Acad.Sci.USA 84(9):2965〜69(1987))。
【0006】
CEA発現と転移増殖との間の相関関係によって、結腸直腸癌治療のための分子的および免疫学的介入の標的としての存在価値が引き出された。CEAを標的とする1つの治療的取り組みは、抗CEA抗体の使用であり(Chester他、Cancer Chemother.Pharmacol.46(Suppl):S8〜S12(2000)参照)、もう1つはCEAをベースとしたワクチンを使用してCEA発現腫瘍を攻撃する免疫系を活性化することである(概説としてはBerinstein、前述参照)。
【発明の開示】
【発明が解決しようとする課題】
【0007】
多くのワクチンの開発および商品化は、宿主生物をうまく形質転換する外来遺伝子を高い発現濃度で獲得するのに伴う困難さのために、妨げられている。したがって、前述のCEA蛋白質をコード化する野生型ヌクレオチド配列が同定されたにも関わらず、効率的で、自己寛容によって妨害されない癌ワクチンの開発が可能になる、企図する宿主細胞で発現するために最適化されたCEAをコードするヌクレオチド配列を利用するヒトCEA蛋白質の容易に再生可能な材料を開発することが強く望まれている。
【課題を解決するための手段】
【0008】
本発明は、直腸結腸癌を含めた数多くの腺癌に関連してきたCEA遺伝子によって発現した蛋白質生成物に対する免疫を惹起するか、または増強する組成物および方法に関する。具体的に、本発明は、ヒトCEA蛋白質をコードするポリヌクレオチドを提供する。前記ポリヌクレオチドはヒト細胞において高レベルで発現するようにコドンが最適化されている。本発明はさらに、合成ポリヌクレオチドを含むアデノウイルスベクターおよびプラスミドをベースとしたベクターを提供し、CEAに関連した癌を予防し、かつ/または治療するための免疫原性組成物およびワクチンにおける前記ベクターの使用を開示する。
【0009】
本発明はまた、配列番号2に記載されているヒト癌胎児抗原(以後hCEA)をコードするヌクレオチドの配列を含む合成核酸分子(ポリヌクレオチド)に関し、前記合成核酸分子は、ヒト細胞において高レベルで発現するようにコドンが最適化されている(以後、hCEAopt)。本明細書で開示された核酸分子は、選択した宿主細胞にトランスフェクトすることができ、該組換え宿主細胞は、十分な濃度で機能的hCEA蛋白質(配列番号2)を発現させるための原料となる。
【0010】
本発明はさらに、ヒトCEA蛋白質を発現するmRNAをコードする合成核酸分子に関し、このDNA分子は本明細書で配列番号1として開示したヌクレオチド配列を含む。本発明のこの部分の好ましい態様は、hCEA蛋白質(配列番号2または配列番号16)をコードするDNA分子(配列番号1)を示す図1に開示される。本発明の好ましい核酸分子は、ヒト細胞において高レベルで発現するようにコドンが最適化されている。
【0011】
本発明の他の好ましいDNA分子は、ヒト完全長CEA(配列番号2)のアミノ酸約679からアミノ酸約702に位置するC末端アンカードメイン(AD)が欠失したヒトCEAをコードするヌクレオチドの配列を含み、ヌクレオチドの前記配列はヒト細胞において高レベルで発現するようにコドンが最適化されている。アンカードメインを切断されたCEA変種をコードするDNA分子の例を図15に示す(図10Aを参照)。hCEA−ΔADの対応するアミノ酸配列は、配列番号16に記載する(図10Bを参照)。
【0012】
本発明はまた、この明細書を通じて開示した核酸分子を含有する組換えベクター、並びに原核細胞および真核細胞両方の組換え宿主細胞に関する。
【0013】
本発明はさらに、(a)配列番号1または配列番号15で表した核酸分子を含むベクターを適切な宿主細胞に導入すること、および(b)前記コドン最適化ヒト蛋白質の発現を可能にする条件下で該宿主細胞を培養することを含む、組換え宿主細胞におけるコドン最適化ヒトCEA蛋白質の発現方法に関する。
【0014】
本発明の他の態様は、合成核酸分子を含むワクチンベクターを哺乳類に投与することを含む癌の予防または治療方法であり、該合成核酸分子は配列番号2または配列番号16で表したヒト癌胎児抗原(hCEA)蛋白質をコードするヌクレオチドの配列を含み、該合成核酸分子はヒト細胞において高濃度に発現するようにコドンが最適化されている。
【0015】
本発明はさらに、E1領域に欠失を有し、及びE1領域に挿入部分を有するアデノウイルスゲノムを含むアデノウイルスワクチンベクターに関し、前記挿入部分は、(a)ヒトCEA蛋白質をコードするコドン最適化ポリヌクレオチドと、(b)ポリヌクレオチドに操作可能に結合されたプロモーターとを含む発現カセットを含む。
【0016】
本発明はまた、プラスミド部分と発現カセット部分とを含むワクチンプラスミドに関し、該発現カセット部分は、(a)ヒト細胞において最適に発現させるためにコドンを最適化した、ヒトCEA蛋白質をコードする合成ポリヌクレオチドと、(b)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む。
【0017】
本発明の他の態様は、(a)(i)ヒトCEA蛋白質をコードするコドン最適化ポリヌクレオチドおよび(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターを含む第1のベクターを哺乳類に導入すること、(b)予め決定された時間経過させること、および(c)(i)ヒトCEA蛋白質をコードするコドン最適化ポリヌクレオチドおよび(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターを含む第2のベクターを該哺乳類に導入することを含む、前記哺乳類を癌から予防もしくは癌を治療するか、またはCEAに関連した癌に罹患した前記哺乳類を治療する方法である。
【0018】
明細書および添付の特許請求の範囲を通じて使用したように、単数形「a」、「an」および「the」は、明白に別段の指定がない限り、複数を指すことを含む。
【0019】
明細書および添付の特許請求の範囲を通じて使用したように、以下の定義および略語を適用する。
【0020】
「プロモーター」という用語は、RNAポリメラーゼが結合するDNA鎖の認識部位のことである。該プロモーターは、RNAポリメラーゼとの開始複合体を形成し、転写活動を開始し、推進する。該複合体は、「エンハンサー」と称する活性化配列または「サイレンサー」と称する阻害配列によって変更することができる。
【0021】
「カセット」という用語は、発現させるべき核酸配列を含有する本発明の配列のことである。該カセットは、カセットテープの概念と同じで、各カセットは独自の配列を有する。したがって、カセットを交換することによって、ベクターは異なる配列を発現する。5′および3′末端に制限部位があるために、該カセットは容易に挿入したり、除去したり、他のカセットで置換したりすることができる。
【0022】
「ベクター」という用語は、DNA断片を宿主生物または宿主組織に導入することができるいくつかの手段を意味する。ベクターには、プラスミド、(アデノウイルスを含む)ウイルス、バクテリオファージおよびコスミドを含む様々な種類がある。
【0023】
アデノウイルスベクターに関して使用したような「第1世代」という用語は、複製に欠陥のある前記アデノウイルスベクターを意味する。一般的に、第1世代アデノウイルスベクターは、E1遺伝子領域が欠失しているか、または不活性化しており、好ましくはE3遺伝子領域が欠失しているか、または不活性化している。
【0024】
「pV1J/hCEAopt」という呼称は、イントロンAを有するヒトCMV前初期(IE)プロモーター、完全長コドン最適化ヒトCEA遺伝子、ウシ成長ホルモン由来ポリアデニル化配列および転写終止配列並びに最小pUC主鎖を含む、本明細書で開示したプラスミド構築物である(実施例2参照)。前述のように、「pV1J/hCEA」という呼称は、構築物がコドン最適化ヒトCEA遺伝子の代わりに野生型ヒトCEA遺伝子を含むこと以外は前述のような構築物を意味する。
【0025】
「MRKAd5/hCEAopt」および「MRKAd5/hCEA」という呼称は、E1およびE3領域に欠失があるAd5アデノウイルスゲノムを含む、本明細書で開示した2種類の構築物を意味する。「MRKAd5/hCEAopt」構築物においては、E1領域は、イントロンAを含まないヒトCMVプロモーターの制御下でE1と平行したコドン最適化ヒトCEA遺伝子およびそれに続くウシ成長ホルモンポリアデニル化シグナルによって置換されている。「MRKAd5/hCEA」構築物は、Ad5ゲノムのE1領域が野生型ヒトCEA配列と置換していること以外は、本質的に前述の通りである(実施例2参照)。
【0026】
「有効量」という用語は、免疫応答を生じるように、十分なワクチン組成物が適切な濃度のポリペプチドの産生を誘導することを意味する。当業者は、この濃度は変化し得ることを承知している。
【0027】
「保存的アミノ酸置換」とは、1個のアミノ酸残基がもう1個の化学的に類似したアミノ酸残基によって置換されることを意味する。このような保存的置換の例は、1疎水性残基(イソロイシン、ロイシン、バリンまたはメチオニン)と他の残基との置換、1極性残基と電荷が同じ他の1極性残基との置換(たとえば、アルギニンとリジン、グルタミン酸とアスパラギン酸)である。
【0028】
「hCEA」は、ヒト癌胎児抗原を意味し、「hCEAopt」は、コドン最適化ヒト癌胎児抗原を意味する。
【0029】
「hCEA−ΔAD」という用語は、ヒト完全長CEA(配列番号2)のアミノ酸約679からアミノ酸約702に位置するC末端アンカードメイン(AD)を欠失したヒトCEAの変種を意味する。本発明のhCEA−ΔADをコードするヌクレオチド配列は、ヒト細胞環境において高レベルで発現するようにコドンが最適化されている(本明細書では、hCEAopt−ΔAD”と称する)。アンカードメインを切断されたCEA変種をコードするDNA分子の例を図15に示す(図10Aを参照)。hCEA−ΔADの対応するアミノ酸配列は配列番号16に記載する(図10Bを参照)。hCEA−ΔADをコードするヌクレオチドは、癌の治療および/または予防のための癌ワクチンの開発に有用である。
【0030】
「哺乳類」という用語は、人類を含めた任意の哺乳類を意味する。
【0031】
略語「Ag」は、抗原のことである。
【0032】
略語「Ab」は抗体を意味し、「mAb」はモノクローナル抗体を意味する。
【0033】
略語「ORF]は、遺伝子のオープンリーディングフレームを意味する。
【発明を実施するための最良の形態】
【0034】
癌胎児抗原(CEA)は通常、腺癌の発症に関連する。本発明は、異常なCEA発現が癌または癌の発症に関連するCEA腫瘍関連抗原によって発現した蛋白質生成物に対する免疫を惹起するか、または高める組成物および方法に関する。異常なCEA発現は腫瘍開始時に存在し、腫瘍進行後まで検出されなくてもよく、その逆であってもよいので、異常なCEA発現と癌との関連には、CEA蛋白質が腫瘍組織において、その発症の全ての時点において発現されていることは必要ではない。
【0035】
このために、ヒトCEA蛋白質をコードする合成DNA分子を提供する。好ましい実施形態ではヒト細胞である計画した宿主細胞に好まれるコドンを使用するように、合成分子のコドンを設計する。抗体の中和および細胞性免疫によって、CEA関連癌に対する効果的な免疫予防を提供する組換えアデノウイルスワクチンまたはプラスミドをベースとしたワクチンを開発するために、該合成分子を使用することができる。該合成分子は、免疫原組成物として使用することができる。本発明は、霊長類およびヒトなどの哺乳類を含めた脊椎動物に直接インビボで導入すると、該動物でコード化された蛋白質の発現を誘導するポリペプチドを提供する。
【0036】
野生型ヒトCEAヌクレオチド配列は報告されている(たとえば、米国特許第5274087号、米国特許第5571710号および米国特許第5843761号参照)。本発明は、ヒトCEA蛋白質をコードする合成DNA分子を提供する。本発明の合成分子は、いくつかはヒト細胞によって好まれるコドンを使用するように変更されており、したがってヒト宿主細胞においてCEAを高濃度で発現することが可能であるヌクレオチドの配列を含む。該合成分子は、抗体の中和および細胞性免疫によってCEA関連癌に対する効果的な免疫予防を提供する癌ワクチンで使用することができる、CEA蛋白質の材料として使用することができる。
【0037】
4種類の可能なヌクレオチド塩基の「3連」コドンは、60個を上回る変種の形で存在することが可能である。これらのコドンは、異なるアミノ酸20個(および転写開始および終止)のみの情報を提供するので、いくつかのアミノ酸は、複数のコドンによってコードされることが可能で、コドン重複として公知の現象である。完全には理解されていない理由のために、他のコドンは異なる種類の細胞の内在DNAに均一に存在することはない。実際に、ある種の細胞においてある種のコドンには様々な天然の階層または「選択性」が存在するようである。1例として、アミノ酸ロイシンはCTA、CTC、CTG、CTT、TTAおよびTTGを含めた6種類のDNAコドンのいずれかによって特定される。微生物のゲノムコドン出現頻度の徹底的な分析によって、E.コリの内在性DNAは極めて一般的に、CTGロイシン指定コドンを含有し、一方酵母および粘菌のDNAは極めて一般的にTTAロイシン指定コドンを含むことが明らかになった。この階層的観点から、E.コリ宿主によってロイシンリッチなポリペプチドの高濃度発現が得られる可能性は、使用したコドンの頻度にある程度左右されることが一般に信じられている。たとえば、TTAコドンに富んだ遺伝子はE.コリでは十分に発現せず、一方CTGに富んだ遺伝子はおそらくこの宿主で高く発現するようである。同様に、酵母宿主細胞でロイシンに富んだポリペプチドの発現に好まれるコドンはTTAである。
【0038】
組換えDNA技術に対するコドン選択現象の意味は一目瞭然で、該現象によって、形質転換が成功した宿主生物において外来遺伝子の高濃度発現を実現できなかったかつての多くの失敗を説明することができ、あまり「好ましくない」コドンを挿入遺伝子内に反復して存在させて、宿主細胞の発現機構をあまり効率よく作動できないようにすることが可能である。この現象は、計画した宿主細胞に好まれるコドンを含むように設計された合成遺伝子は、組換えDNA技術を実施するために最適な形態の外来遺伝子材料となることを示唆する。したがって、本発明の1態様は、ヒト細胞で発現するためにコドンを最適化したヒトCEA遺伝子である。本発明の好ましい実施形態では、同じ蛋白質配列をコードする他のコドンを使用することによって、ヒト細胞における外来CEA蛋白質発現の制約を取り除くことができることが見いだされた。
【0039】
本発明によれば、本明細書に参照により組み込まれた、Latheの「アミノ酸配列データから推測されたオリゴヌクレオチドプローブ:理論および実際の考察(Synthetic Oligonucleotide Probes Deduced from Amino Acid Sequence Data:Theoretical and Practical Considerations」J.Molec.Biol.183:1〜12(1985)に記載されたように、ヒトCEA遺伝子配列は、同じ翻訳配列を有するが他のコドンを使用したポリヌクレオチドに変換された。この方法は一般的に、高発現のヒト遺伝子に通常関連しない野生型配列のコドンを同定すること、およびこのコドンとヒト細胞において高く発現するために最適なコドンとを置換することから構成される。次に、新規配列について、これらのコドン置換によって生じた望ましくない配列を調べる(たとえば、「ATTTA」配列、イントロンスプライシング認識部位の偶発的な発生、望ましくない制限酵素部位など)。所望しない配列は、同一アミノ酸をコードする異なるコドンで既存のコドンを置換することによって除去される。次に、合成遺伝子部分の発現が改善されているかを調べる。
【0040】
前述の方法を使用してヒトCEAの合成遺伝子配列を作製して、高い発現濃度に最適化したコドンを含む遺伝子が生じた。前記の方法は、癌ワクチンで使用するコドン最適化遺伝子を設計するための方法を要約したものであるが、該方法を少し変更するか、または配列を少し変化させることによって、同様のワクチンの有効性または遺伝子発現の増加を実現することは、当業者に理解されている。当業者はまた、DNA分子のほんの一部のコドンをコドン最適化して、ヒト細胞において高濃度のCEA発現をもたらす他のDNA分子を構築できることを理解するであろう。
【0041】
したがって、本発明は、ヒトCEA蛋白質(配列番号2)、またはhCEA−ΔAD(配列番号16)を含むがそれだけに限定されないヒトCEA蛋白質の生物学的に活性のある断片または変異型をコードするヌクレオチドの配列を含む合成ポリヌクレオチドに関し、該ポリヌクレオチド配列はヒト宿主で発現するために最適化されたコドンを含む。CEA蛋白質の前記変異型は、本明細書で集合的に「変種」として称される保存的アミノ酸置換、アミノ末端切断、カルボキシ末端切断、欠失または添加を含むが、それだけに限定されない。このような生物学的活性のある断片および/または変異体はいずれも、配列番号2で示したようなCEA蛋白質の免疫学的特性を少なくとも実質的に模倣した蛋白質または蛋白質断片をコードする。本発明の合成ポリヌクレオチドは、治療用または予防用癌ワクチンを開発するのに有用であるように、機能的なヒトCEA蛋白質を発現するmRNA分子をコードする。
【0042】
前述したように、本発明は、ヒトCEA蛋白質(配列番号2)をコードするヌクレオチドまたはそれらの生物学的に活性のある断片または変異型に関する。このために、本発明は、C末端アンカー配列が欠失したヒトCEA蛋白質を含むhCEA−ΔAD(配列番号16、図10B)をコードするヌクレオチドを提供する。hCEA−ΔADをコードする本発明の核酸分子は、ヒト細胞での発現を高めるためにコドンが最適化されている。hCEA−ΔADをコードする核酸配列の例は、配列番号15で示したヌクレオチドの配列を含む(図10A)。
【0043】
本発明は、配列番号2に記載されている新規hCEA蛋白質を発現するmRNAをコードするヌクレオチドの配列を含む合成核酸分子(ポリヌクレオチド)に関し、前記合成核酸分子は、ヒト宿主細胞で高レベルで発現するようにコドンが最適化されている。本発明の核酸分子は、実質的に他の核酸を含まない。
【0044】
本発明はまた、この明細書を通じて開示した核酸分子を含有する組換えベクター並びに原核細胞および真核細胞の両方の組換え宿主細胞に関する。該合成DNA分子、関連ベクターおよび本発明の宿主は、癌ワクチンの開発に有用である。
【0045】
本発明の好ましいDNA分子は、配列番号2として表した、図2に示したヒトCEA蛋白質をコードする、本明細書で配列番号1として開示した、図1に示したヌクレオチド配列を含む。
【0046】
本発明のさらに好ましいDNA分子は、配列番号16として表した、図10Bに示したC末端アンカー配列を欠失したヒトCEA変種をコードする、本明細書で配列番号15として開示した、図10Aに示したヌクレオチド配列を含む。
【0047】
本発明はまた、ヒトCEA蛋白質を発現するmRNAをコードする配列番号1の生物学的に活性のある断片または変異体を含む。このような生物学的活性のある断片および/または変異体は、配列番号2で表したhCEA蛋白質を含むが、それだけに限定されないhCEA蛋白質の薬理学的特性を少なくとも実質的に模倣した蛋白質または蛋白質断片のいずれかをコードする。このようなポリヌクレオチドはいずれも、ヌクレオチド置換、欠失、添加、アミノ末端切断およびカルボキシ末端切断を含むが、必ずしもそれだけに限定されない。本発明の変異体は、癌ワクチンの開発に有用であるように真核細胞において機能的hCEA蛋白質を発現するmRNA分子をコードする。
【0048】
本発明はまた、合成DNAのヌクレオチド配列が配列番号1のヌクレオチド配列とは著しく異なるが、まだ配列番号2で表したhCEA蛋白質をコードする合成コドン最適化DNA分子に関する。このような合成DNAは、本発明の範囲内にあるものとする。したがって、本発明は、同一の蛋白質を発現する数多くのDNA分子をもたらすことができるコドン重複を開示する。本発明の範囲内にはまた、発現した蛋白質の基本的な物理特性を実質的に変更しないDNA配列の変異が含まれる。たとえば、ロイシンをバリンに、リジンをアルギニンに、またはグルタミンをアスパラギンに置換しても、ポリペプチドの機能に変化は生じる可能性はない。
【0049】
ペプチドをコードするDNA配列は、天然に生じるペプチドと異なる特性を有するペプチドをコードするために改変することができることが知られている。DNA配列の改変方法は、部位特異的変異誘発を含むが、それだけに限定されない。改変された特性の例は、基質に対する酵素またはリガンドに対する受容体の親和性の変化を含むが、それだけに限定されない。
【0050】
本発明はまた、GFP(緑の蛍光蛋白質)、MYCエピトープ、GSTおよびFcを含むが、それだけに限定されない様々なマーカーに結合させたヒトCEA蛋白質の一部を発現する融合構築物を含むが、それだけに限定されないhCEAopt融合構築物に関する。このような融合構築物はいずれも、関心のある細胞系で発現することが可能で、本明細書で開示したヒトCEA蛋白質の修飾因子のスクリーニングに使用することが可能である。DOMおよびhsp70並びにLTBを含むが、それだけに限定されない、ヒトCEAに対する免疫応答を高めるために構築された融合蛋白質もまた、企図する。
【0051】
本発明はさらに、この明細書を通じて開示した合成核酸分子を含む組換えベクターに関する。これらのベクターは、DNAまたはRNAから構成されることが可能である。ほとんどのクローニングのためには、DNAベクターが好まれる。一般的なベクターは、プラスミド、改変ウイルス、バキュロウイルス、バクテリオファージ、コスミド、酵母の人工的染色体、およびhCEA蛋白質をコードすることができるその他の形態のエピソームDNAまたは統合DNAを含む。特定の遺伝子輸送またはその他の用途に適切なベクターを決定することは、十分当業者の範囲内である。
【0052】
hCEA蛋白質をコードするコドン最適化DNAを含有する発現ベクターは、組換え宿主細胞においてhCEAを高濃度に発現するために使用することができる。発現ベクターは、クローニングベクター、改変クローニングベクター、特異的に設計されたプラスミドまたはウイルスを含むが、それだけに限定されない。また、所望するならば、様々な細菌発現ベクターを使用して、細菌細胞で組換えhCEAを発現することができる。さらに、様々な真菌細胞発現ベクターを使用して、真菌細胞で組換えhCEAを発現することができる。さらに、様々な昆虫細胞発現ベクターを使用して、昆虫細胞で組換え蛋白質を発現することができる。
【0053】
本発明はまた、本発明の核酸分子を含むベクターで形質転換またはトランスフェクトした宿主細胞に関する。組換え宿主細胞は、E.コリなどの細菌、酵母などの真菌細胞を含めるがそれだけに限定されない原核細胞または真核細胞、ウシ、ブタ、サルおよび齧歯類由来の細胞系を含むがそれだけに限定されない哺乳類細胞、並びにショウジョウバエおよびカイコ由来の細胞系を含むがそれだけに限定されない昆虫細胞であってよい。このような組換え宿主細胞は、hCEAまたは生物学的に等価な形態を生成するために適した条件下で培養することができる。本発明の好ましい実施形態では、該宿主細胞はヒトである。本明細書で定義したように、「宿主細胞」という用語は、遺伝子を組み換えたヒトの体、ヒト胎児またはヒト胚における宿主細胞を含むものではない。
【0054】
前述のように、hCEA蛋白質をコードするDNAを含有する発現ベクターは、組換え宿主細胞においてhCEAを発現するために使用することができる。したがって、本発明の他の態様は、(a)配列番号1または配列番号15で表した核酸を含むベクターを適切なヒト宿主細胞に導入すること、および(b)前記ヒトCEA蛋白質またはCEA蛋白質変種の発現を可能にする条件下で該宿主細胞を培養することを含む、組換え宿主細胞におけるヒトCEA蛋白質または蛋白質変種の発現方法である。
【0055】
宿主細胞でhCEAが発現した後、hCEA蛋白質を回収して、活性型hCEA蛋白質を提供することができる。いくつかのhCEA蛋白質精製方法が有用で、使用に適している。組換えhCEA蛋白質は、塩分画、イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ハイドロキシアパタイト吸着クロマトグラフィーおよび疎水性相互作用クロマトグラフィーの様々な組み合わせ、または個々の適用によって細胞溶解物および抽出物から精製することができる。さらに、組換えhCEA蛋白質は、完全長hCEA蛋白質またはhCEAのポリペプチド断片に特異的なモノクローナル抗体またはポリクローナル抗体で作製した免疫親和性カラムを使用することによって、その他の細胞蛋白質から分離することができる。
【0056】
本発明の核酸は、ヒト細胞において蛋白質の効果的な発現をもたらすように設計された配列を含む発現カセットに組み込むことができる。
【0057】
該カセットは、コドンを最適化した完全長hCEA遺伝子と共に、プロモーターなど、それに操作可能に結合された転写および翻訳制御配列並びに終止配列を含有することが好ましい。好ましい実施形態では、該プロモーターはイントロンA配列(CMV)を含まないサイトメガロウイルスプロモーターであるが、当業者であれば、強力なイムノグロブリンなどの多数のその他の公知のプロモーター、またはその他の真核細胞遺伝子プロモーターを使用できることを認識するであろう。好ましい転写終結区は、ウシ成長ホルモン終結区であるが、その他の公知の転写終結区もまた使用することができる。CMV−BGH終結区の組み合わせは特に好ましい。
【0058】
本発明によれば、hCEAopt発現カセットは、ベクターに挿入される。該ベクターは、好ましくはアデノウイルスベクターであるが、プロモーターに結合した線状DNAまたはその他のベクター、たとえば、アデノ関連ウイルスまたは改変ワクチンウイルス、レトロウイルスまたはレンチウイルスベクターもまた使用することができる。
【0059】
選択したベクターがアデノウイルスの場合、ベクターがいわゆる第1世代アデノウイルスベクターであることが好ましい。これらのアデノウイルスベクターは、非機能性E1遺伝子領域を有することが特徴で、好ましくはアデノウイルスE1遺伝子領域が欠失していることが好ましい。いくつかの実施形態では、該発現カセットは、E1遺伝子が通常位置する位置に挿入される。さらに、これらのベクターは、場合によって非機能性E3領域を有するか、E3領域を欠失している。使用したアデノウイルスゲノムは、E1およびE3領域の両方が欠失していること(ΔE1ΔE3)が好ましい。該アデノウイルスは、ウイルスE1遺伝子を発現する公知の細胞系、たとえば、293細胞、またはPERC.6細胞、または一時的にもしくは安定して形質転換して外来蛋白質を発現する293またはPERC.6から得られた細胞系で増殖することができる。たとえば、テトラサイクリン制御プロモーター系などの遺伝子発現が制御された構築物を使用するとき、該細胞系は、制御系に関連した成分を発現することができる。このような細胞系の1例は、T−Rex−293であり、その他は当業者に公知である。
【0060】
アデノウイルスベクターの操作に便利なように、該アデノウイルスはシャトルプラスミド型であることが可能である。本発明はまた、プラスミド部分およびアデノウイルス部分を含むシャトルプラスミドベクターを対象としており、該アデノウイルス部分はE1が欠失し、場合によってE3も欠失したアデノウイルスゲノムを含み、コドン最適化ヒトCEAを含む発現カセットが挿入されている。好ましい実施形態では、アデノウイルスベクターが容易に除去され得るようにプラスミドのアデノウイルス部分に隣接して制限部位がある。該シャトルプラスミドは、原核細胞または真核細胞で複製することができる。
【0061】
本発明の好ましい実施形態では、発現カセットはpMRKAd5−HV0アデノウイルスプラスミド(本明細書に参照により組み込まれた、Emini他、WO02/22080参照)に挿入する。このプラスミドは、E1およびE3領域が欠失したAd5アデノウイルスゲノムを含む。pMRKAd5−HV0プラスミドの設計は、5’シス作用パッキング領域をE1遺伝子にさらに伸長し、ウイルスパッケージングを最適化するために重要であることが発見された要素をとりこみ、ウイルス増幅の増強をもたらすことによって、以前のアデノウイルスベクターより改良された。有利なことに、この増強されたアデノウイルスは、何度か継代増殖させた後でも遺伝子安定性を維持することができる。
【0062】
DNA構築物を調製および精製する分子生物学の標準的技術によって、本発明のアデノウイルス、シャトルプラスミドおよびDNA免疫原を調製することが可能である。
【0063】
本発明によって、ヒト細胞において高レベルで発現するようにコドンが最適化された本明細書で説明した合成cDNA分子(配列番号1)は、対応する野生型配列よりも効率的に発現することが測定された。驚くべきことに、コドンを最適化したhCEAのcDNAは、野生型配列よりも効率的にhCEAに対する寛容を打破する。さらに、本明細書では、hCEAoptはhCEAよりも免疫原性が高く、細胞性および体液性両方の免疫応答をより効果的に惹起することが示された。
【0064】
したがって、前述のベクターは、異常なCEA発現に関連した腺癌の発症の予防および/または既存の癌の治療のための免疫原性組成物およびワクチンに使用することができる。本発明のベクターは、うまく形質転換した宿主生物において外来性CEAを高濃度に発現させることに伴う問題を排除し、ワクチン開発および商品化を可能にする。このために、本発明の1態様は、配列番号2に記載されているヒトCEA蛋白質をコードするヌクレオチドの配列を含むコドン最適化合成核酸分子を含むワクチンベクターを哺乳類に投与することを含む癌の予防および治療方法である。
【0065】
前述の方法によれば、該ワクチンベクターはいかなる哺乳類の癌の治療または予防のためにも投与することができる。本発明の好ましい実施形態では、哺乳類はヒトである。
【0066】
さらに、当業者は説明した治療方法および予防方法で使用するための任意の形態のベクターを選択することができる。好ましくは、該ベクターはアデノウイルスベクターまたはプラスミドベクターである。本発明の好ましい実施形態では、該ベクターはアデノウイルスE1領域中に欠失を有し、及びアデノウイルスE1領域中に挿入部分を有するアデノウイルスゲノムを含むアデノウイルスベクターであり、前記挿入は、(a)ヒトCEA蛋白質をコードするコドン最適化合成ポリヌクレオチドと、(b)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む発現カセットを含む。
【0067】
本発明はさらに、E1領域中に欠失を有し、及びE1領域中に挿入部分を有するアデノウイルスゲノムを含むアデノウイルスワクチンベクターに関し、前記挿入は、(a)ヒトCEA蛋白質をコードするコドン最適化合成ポリヌクレオチドと、(b)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む発現カセットを含む。
【0068】
本発明の好ましい実施形態では、該アデノウイルスベクターはAd5ベクターである。
【0069】
本発明の他の好ましい実施形態では、該アデノウイルスベクターはAd6ベクターである。
【0070】
本発明のさらに他の好ましい実施形態では、該アデノウイルスベクターはAd24ベクターである。
【0071】
他の態様では、本発明は、プラスミド部分と発現カセット部分とを含むワクチンプラスミドに関し、該発現カセット部分は、(a)ヒトCEA蛋白質またはその変種をコードするコドン最適化合成ポリヌクレオチドと、(b)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む。
【0072】
本発明のある実施形態では、本明細書で開示した組換えアデノウイルスワクチンは、増強された免疫応答を誘導するため、プラスミドをベースとしたポリヌクレオチドワクチンによる様々な初期/追加免疫の組み合わせで使用される。この場合、この2種類のベクターが「初期および追加免疫」計画で投与される。たとえば、第1の種類のベクターを投与し、次に所与の時間を経た後、たとえば、2週間後、1ヶ月後、2ヶ月後、6ヶ月後またはその他の適切な間隔を空けて、第2の種類のベクターを投与する。好ましくは、該ベクターは同一のポリヌクレオチドまたはポリヌクレオチドの組み合わせをコードする発現カセットを有する。プラスミドDNAもまた使用する実施形態では、該ベクターは哺乳類細胞または昆虫細胞によって認識される1種または複数のプロモーターを含有することが好ましい。好ましい実施形態では、該プラスミドは強力なプロモーターを含有し、たとえば、CMVプロモーターであるが、それだけに限定されない。該合成ヒトCEA遺伝子または発現すべきその他の遺伝子は、このようなプロモーターに結合される。このようなプラスミドの例は、前述のような哺乳類発現プラスミドV1Jnsである(本明細書に参照により組み込まれた、J.Shiver他、DNA Vacchines、M.Liu他著、N.Y.Acad.Sci.,N.Y.,772:198〜208(1996))。
【0073】
前述のように、アデノウイルスベクターワクチンおよびプラスミドワクチンは、免疫応答を惹起する単一の治療計画の一部として脊椎動物に投与することができる。このために、本発明は、(a)(i)ヒトCEA蛋白質またはヒトCEA蛋白質変種をコードするコドン最適化合成ポリヌクレオチドと、(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む第1のベクターを哺乳類に導入すること、(b)予め決定された時間経過させること、および(c)(i)ヒトCEA蛋白質またはヒトCEA蛋白質変種をコードするコドン最適化合成ポリヌクレオチドと、(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む第2のベクターを該哺乳類に導入することを含む、前記哺乳類を癌から防御する方法に関する。
【0074】
前述の防御方法の1実施形態において、第1のベクターはプラスミドであり、第2のベクターはアデノウイルスベクターである。他の実施形態では、第1のベクターはアデノウイルスベクターであり、第2のベクターはプラスミドである。
【0075】
本発明はさらに、(a)(i)ヒトCEA蛋白質またはヒトCEA蛋白質変種をコードするコドン最適化合成ポリヌクレオチドと、(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む第1のベクターを哺乳類に導入すること、(b)予め決定された時間経過させること、および(c)(i)ヒトCEA蛋白質またはヒトCEA蛋白質変種をコードするコドン最適化合成ポリヌクレオチドと、(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む第2のベクターを該哺乳類に導入することを含む、腺癌に罹患した哺乳類を治療する方法に関する。
【0076】
前述の治療方法の1実施形態において、第1のベクターはプラスミドであり、第2のベクターはアデノウイルスベクターである。他の実施形態では、第1のベクターはアデノウイルスベクターであり、第2のベクターはプラスミドである。
【0077】
ワクチン受容体に導入すべき発現可能なDNAまたは転写されたRNAの量は、使用したプロモーターの強度および発現遺伝子産物の免疫原性に部分的に左右される。一般的に、免疫学的または予防的に有効な用量、約1ngから100mg、好ましくは約10μgから300μgのプラスミドワクチンベクターを直接筋肉組織に投与する。組換えアデノウイルスの有効量は、粒子約106個から1012個で、好ましくは約107個から1011個である。皮下注射、皮内注射、皮膚からの圧入およびその他の投与様式、たとえば、腹腔内、静脈内または吸入送達もまた企図される。追加免疫ワクチン接種を行うこともまた、企図される。本発明のワクチンの非経口的導入と同時に、またはその後で、アジュバント、たとえば、インターロイキン12蛋白質を非経口的に、たとえば、静脈内、筋肉内、皮下またはその他の投与手段で投与することもまた、有利である。
【0078】
本発明のワクチンは、裸であること、すなわち、いかなる蛋白質、アジュバントまたは受容対象の免疫系に影響を与えるその他の薬剤を伴わないことが可能である。この場合、ワクチンベクターは生理学的に許容される溶液、たとえば、滅菌生理食塩水または滅菌緩衝生理食塩水に入れることが望ましい。あるいは、本発明のワクチンまたは免疫原組成物と共に免疫賦活剤、たとえば、アジュバント、サイトカイン、蛋白質、またはその他の担体を投与することが有利であり得る。したがって、本発明は、本発明の組成物および方法と組み合わせてこのような免疫賦活剤を使用することを含む。本明細書で使用した免疫賦活剤とは、外来抗原に対する免疫応答(抗体および/または細胞性)を増強または助長する本質的に任意の物質である。前記免疫賦活剤は、DNAまたは蛋白質の形態で投与することができる。GM−CSF、IFNα、破傷風毒素、IL12、B7.1、LFA−3およびICAM−1を含むが、それだけに限定されない様々な免疫賦活剤はいずれも、本発明のワクチンおよび免疫組成物と組み合わせて使用することができる。前記免疫賦活剤は、当業界で周知である。
【0079】
DNAの細胞による摂取を助ける薬剤、たとえば、それだけに限定されないがカルシウムイオンはまた、使用することができる。これらの薬剤は一般的に、トランスフェクションを容易にする試薬および薬剤として許容される担体のことである。当業者であれば、特定の免疫賦活剤または薬剤として許容される担体並びに適切な時間および投与様式を決定することができる。
【0080】
本明細書で記載した刊行物は全て、本発明と関連づけて使用することが可能な方法および材料を記載し、開示するために参照により組み込まれる。本発明が先行技術の効力によってその開示が先行する資格を有していないことを許容したと解釈すべきでない。
【0081】
添付した図面を参考にして、本発明の好ましい実施形態を説明したが、本発明はそれらの厳密な実施形態に限定されるのではなく、添付の特許請求の範囲で定義した本発明の範囲または精神を逸脱することなく、当業者が様々な変更および修正を実行できることを理解されたい。
【0082】
以下の実施例は本発明を例示するものであって、限定するものではない。
【実施例1】
【0083】
コドン最適化ヒトCEA配列
完全なhCEAoptコーディング配列を合成して、BIONEXUS(Oakland、CA)によって構築した。5’末端に最適化したKozak配列を有するhCEAoptcDNAは、PCRによって構築したオリゴヌクレオチドを使用して構築した。構築したcDNAは、pCR−Bluntベクター(Invitrogen、Carlsbad、CA)に挿入して、pCR−hCEAoptを得た。hCEAoptcDNAの強度は、両鎖を配列決定することによって測定した。
【実施例2】
【0084】
プラスミド構築物およびアデノウイルスベクター
pVIJ/hCEAopt:プラスミドpCR−hCEAoptは、EcoRIで37℃で1時間消化した。得られた2156bp挿入物を精製して、プラスミドpV1JnsBのEcoRI部位にクローニングした(Montgomery他、DNA Cell Biol.,12(9):777〜83(1993))。
【0085】
pVlJ/hCEA:プラスミドpCI/hCEA(Song他、DNAをベースとしたワクチン接種および非ウイルス性サイトカイン遺伝子移入の組み合わせによるTヘルパー1対Tヘルパー2活性の制御および腫瘍免疫の増強(Regulation of T−helper−1 versus T−helper−2 activity and enhancement of tumour immunity by combined DNA−based vaccination and nonviral cytokine gene transfer.)Gene Therapy 7:481〜492(2000))は、EcoRIで消化した。得られた2109bp挿入物は、プラスミドpV1JnsA(Montgomery他、前述)のEcoRI部位にクローニングした。
【0086】
Ad5/hCEAopt:プラスミドpCR−hCEAoptは、EcoRIで消化した。得られた2156bp挿入物は、精製してpolyMRK−Ad5シャトルプラスミドのEcoRIにクローニングした(本明細書に参照により組み込まれた、Emini他、WO02/22080参照)。
【0087】
Ad5/CEA:Ad5ベクターを生成するためのシャトルプラスミドpMRK−hCEAは、プラスミドpDelta sp1B/hCEAをSspIおよびEcoRVで消化することによって得られた。次に、該9.52kb断片は、プラスミドポリMRKから得られた1272bpのBglII−BamHIで制限した、クレノウ処理生成物と連結した。hCEAの発現カセットおよびE1隣接Ad5領域を含有するpMRK−hCEAおよびpMRK−hCEAoptのPacI/StuI断片は、BJ5183 E.コリ細胞においてClaI直線化プラスミドpAd5に結合し直した。得られたプラスミドはそれぞれ、pAd5−hCEAおよびpAd5−hCEAoptであった。両プラスミドをPacIで切断して、Ad逆方向末端反復(ITR)を遊離し、PerC−6細胞にトランスフェクトした。Ad5ベクター増幅は、一連の手順によって実施した。MRKAd5/hCEAおよびMRKAd5/hCEAoptは、標準的なCsCl勾配精製法によって精製し、A105緩衝液(5mM Tris−Cl pH8.0、MgCl2 1mM、NaCl 75mM、スクロース 5%、0.005 Tween 20)によって集中的に透析した。
【実施例3】
【0088】
CEAの発現および検出
プラスミドおよびAdベクターによるhCEAの発現は、ウエスタンブロット分析によってモニターした。プラスミドは、リポフェクタミン2000(Life Technologies、Carlsbad、CA)によってHela細胞またはPerC.6細胞にトランスフェクトした。PerC.6のアデノウイルス感染は、血清を含まない培地中で、37℃で30分間実施し、その後新鮮な培地を添加した。48時間インキュベートした後、全細胞溶解物および培養上清を収集した。細胞溶解物中に存在するCEA蛋白質は、ウサギポリクローナル抗血清を使用したウエスタンブロット分析によって検出した。該蛋白質は、180kDaから200kDaのバンドとして検出された。分泌型CEAは、Direct Elisa CEAキット(DBC−Diagnostics Biochem Canada Inc.,Ontario、Canada)を使用して、細胞上清中および注射したマウス(注射後3日目)の末梢血中に検出された。
【実施例4】
【0089】
マウスの免疫
雌のC57BL/6マウス(H−2b)は、Charles River(Lecco、Italy)から購入した。CEA.tgマウス(H−2b)は、J.Primus(Vanderblit University)から恵与され、標準的条件で維持した。既に記載されたように(Rizzuto他、Proc.Natl.Acad.Sci.U.S.A.96(11):6417〜22(1999))、プラスミドDNA 50マイクログラムをマウスの大腿4頭筋に50μlの用量で電気注射した。Ad注射は、50μlの用量でマウスの大腿4頭筋に実施した。体液性および細胞性免疫応答は、指定した時点で分析した。
【実施例5】
【0090】
hCEAのコドン最適化cDNAはhCEA発現を有意に増加させた。
【0091】
ヒトCEA(hCEAopt)の合成遺伝子は、各アミノ酸(以後aa)残基についてヒトに好ましい(ヒト化)コドンを組み込むために設計した。コドン最適化cDNAは、元々のクローンに対して76.8%のヌクレオチド同一性を維持するように改変された(図1参照)。該コドン最適化cDNAは、pV1Jベクター(Montgomery他、前述)のKozak最適化配列(5’−GCCGCCACC−3’、配列番号13)の前、ヒトサイトメガロウイルス(CMV)/イントロンAプロモーターおよびウシ成長ホルモン(BGH)終止シグナルの制御下に配置してクローニングした。該構築物は、pV1J/hCEAopt(実施例2参照)と命名した。さらに、アデノウイルス5型ベクターは、CMV/イントロンAプロモーターおよびBGH終止シグナルの隣接したhCEAopt配列を有するように構築した(Ad5/hCEAopt)。比較のために、同等のプラスミドおよびV5ベクターは、pV1J/hCEAおよびAd5/hCEAを生じる野生型hCEA配列を有するように構築した。コドン最適化cDNAを含有するものと同様に、これらのベクターは、CMV/int Aプロモーターおよびウシ成長ホルモン(BGH)終止シグナルの制御下に野生型遺伝子を有する。
【0092】
プラスミドpV1J/hCEAoptでトランスフェクトしたHela細胞のウエスタンブロット分析では、pV1J/hCEA構築物でトランスフェクトした細胞で検出された大きさと区別できない大分子量(180kDaから200lDa)の蛋白質が生じた。同様に、Ad5/hCEAまたはAd5H7h/hCEAoptで感染させたPerC−6細胞溶解物で検出される蛋白質の大きさには、明らかな差は検出できなかった(データは示さなかった)。
【0093】
hCEA発現に対するhCEAopt発現の効率を比較するために、C57BL/6マウス10匹の群の大腿4頭筋に1x107から1x104pfuの範囲の様々な用量のAd5/hCEAoptを注射した。注射して3日後に、CEA蛋白質濃度を測定して、同用量のAd5/hCEAを注射した対照群の濃度と比較した。Ad5/hCEAを注射したマウスと比較してAd/hCEAopt 1x107pfu(48.2μg/l)の注射では、hCEA濃度の相乗平均値の6倍の増加が認められたが、一方、同ウイルスを1x106pfu(10.9μg/l)で注射すると蛋白質濃度の10倍の増加が認められた(図2A)。対照的に、低用量のAd5/hCEAoptを注射しても、Ad5/hCEAと比較して循環CEA濃度の実質的な増加は生じなかった。pV1J/hCEAと比較して、プラスミドpV1J/hCEAop、25μgまたは50μgを電気注射すると、たとえ低い範囲であっても、CEA蛋白質濃度の増強がまた、示された(図2B)。したがって、これらの結果から、利用した遺伝子輸送媒体とは関係なく、コドン最適化cDNAは対応する野生型配列よりも効果的に発現することが示された。
【実施例6】
【0094】
IFN−γELISPOT測定法
滅菌PBSで2.5μg/mlに希釈した精製ラット抗マウスIFN−γ(IgG1、R4−6A2クローン、Pharmingen、San Diego、CA)100μl/ウェルで、96ウェルMAIPプレート(Millipore、Bedford、MA)をコーティングした。PBSで洗浄後、プレートのブロックはR10培地200μl/ウェルで37℃で2時間実施した。
【0095】
脾細胞は、安楽死させたマウスから滅菌的に脾臓を取り出すことによって得た。脾臓破砕は、金属格子上で切断した脾臓をすりつぶすことによって実施した。細胞ペレットに0.1XPBS 1mlを添加し15秒以上激しく撹拌して、浸透圧溶解することによって赤血球細胞を除去した。次に、2XPBS 1mlを添加し、1XPBSで体積を4mlにした。室温で1200rpmで10分間遠心することによって細胞をペレットにし、該ペレットをR10培地1mlに再懸濁した。生細胞をチュルク染色を使用して計数した。
【0096】
脾細胞は、5x105および2x105細胞/ウェルで2連で播種し、各ペプチドの懸濁液1μg/mlと共に37℃で20分間インキュベートした。各マウスの陽性内部対照としてコンカナバリンA(ConA)を5μg/mlで使用した。PBS、0.05%Tween20で洗浄した後、測定用緩衝液で1:2500に希釈したビオチン結合ラット抗マウスIFNγ(RatIgG1、XMG1.2クローン、PharMingen)50μg/ウェルをプレートに入れて、4℃でO/Nインキュベートした。集中的に洗浄した後、プレートにNBT/B−CIP(Pierce Biotechnoogy Inc.,Rockford、IL)を添加して、スポットがはっきりと視認できるほどになるまで発色させた。蒸留水で徹底的にプレートを洗浄することによって反応を停止させた。プレートを空気乾燥して、次にスポットは自動化ELISPOT読み取り機を使用して計数した。
【実施例7】
【0097】
細胞内サイトカイン染色
RPMI 10%FCS 1mlに溶かした100万個から200万個のマウス脾細胞またはPBMCを、ペプチド収集物(各ペプチドの最終濃度5μgから6μg/ml)およびブレフェルジンA(1μg/ml、BD Pharmingen カタログ番号55028/2300kk)と共に、37℃で5%CO2で12時間から16時間インキュベートした。次に、細胞をFACS緩衝液(PBS 1% FBS、NaN3 0.01%)で洗浄して、精製抗マウスCD16/CD32Fcブロック(BD Pharmingen カタログ番号553142)と共に4℃で15分間インキュベートした。次に、細胞を洗浄して、表面抗体、CD4−PE結合抗マウス(BD Pharmingen、カタログ番号553049)、PercP CD8結合抗マウス(BD Pharmingen カタログ番号553036)およびAPC−結合抗マウスCD3e(BD Pharmingen カタログ番号553066)によって暗所で室温で30分間染色した。洗浄後、細胞を固定し、Cytofix−Cytoperm溶液(BD Pharmingen カタログ番号55028/2300kk)で暗所において4℃で20分間透過させた。PermWash溶液(BD Pharmingen カタログ番号55028/2300kk)で洗浄した後、細胞をIFNγ−FITC抗体(BD Pharmingen)と共にインキュベートした。次に、細胞を洗浄して、ホルムアルデヒドの1%PBS溶液で固定して、FACS−Caliburフローサイトメトリーで、CellQuestソフトウェア(Becton Dickinson、San Jose、CA)を使用して分析した。
【実施例8】
【0098】
CEA特異的T細胞を直接列挙するためのエピトープ含有ペプチドの同定および特徴付け
マウスにおいて、CEAに対する遺伝子ワクチンによって惹起された免疫応答をよりよく特徴付けるために、ELISPOT分析をC57BL/6マウスおよびBALB/cマウスについて実施し、CD4+およびCD8+CEA特異的エピトープを同定した。このために、完全な蛋白質を網羅する個々のペプチドに対する応答の同定に利用できる強く免疫されたマウスの作製について、異なる免疫様式を比較した。プラスミドDNA初期免疫−Ad追加免疫の様式を利用することによって、高濃度の細胞性免疫をウイルス抗原および細菌抗原に対して誘導することができることを示した最近の報告を考慮して、同様の免疫方法を本研究に使用した。異なる計画、i)Ad/hCEAの1x109vpを2回投与(Ad/Ad)、ii)プラスミドp1J/hCEAを2回投与(DNA/DNA)およびiii)プラスミドDNA1回投与した後Ad/hCEAを投与(DNA/Ad)でマウスを筋肉内で免疫した。免疫間隔は、2週間とした。
【0099】
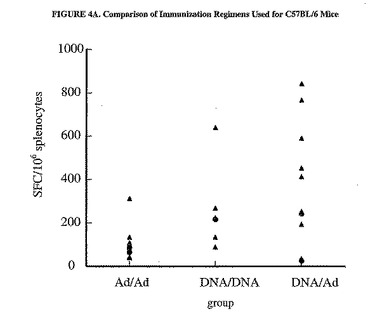
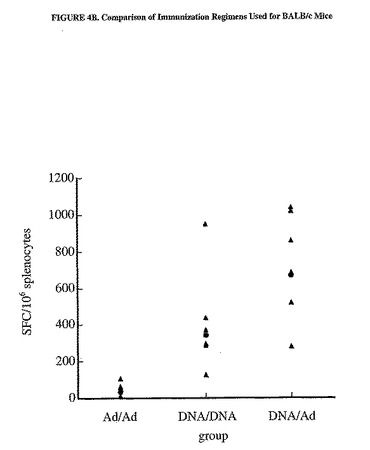
異なる免疫計画によって惹起された細胞性免疫は、追加免疫2週間後にELISPOTによって測定した。異なるワクチン接種計画の免疫原効果を比較するために、11個のaaが重複し、aa497から703を網羅した15マーペプチドの収集物(プールD)を使用して、脾細胞の抗原特異的サイトカイン分泌を刺激した。SFCの高い相乗平均値によって示される最強の応答は、DNA/Adを注射した群のC57BL/6およびBALB/cマウスで認められた(図4)。したがって、この計画を利用して免疫応答をさらに分析した。
【0100】
該免疫応答が、完全なCEA蛋白質に対して等しく分布するかどうかを測定するために、免疫したC57BL/6およびBALB/cマウスの脾細胞を、完全な蛋白質配列を集合的に包含する1個から4個の15マーペプチドの収集物によってインビトロで刺激した。各収集物は、11個の残基によって重複するアミノ酸長15個のペプチドから構成された。凍結乾燥したhCEAペプチドは、Bio−Synthesis(Lewisville、TX)から購入し、DMSOに40mg/mlで再懸濁した。プールDに加えて、プールA(aa1から147)、B(aa137から237)およびC(aa317から507)を本研究で使用した。最終濃度は以下の通りである。プールA=1.2mg/ml、プールB=0.89mg/ml、プールC=0.89mg/mlmプールD=0.8mg/ml。ペプチドは−80℃で保存した。
【0101】
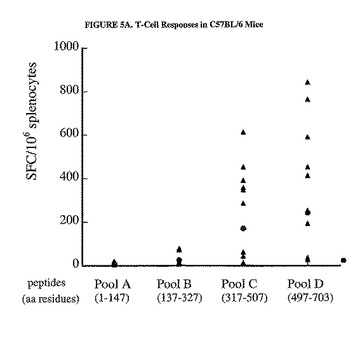
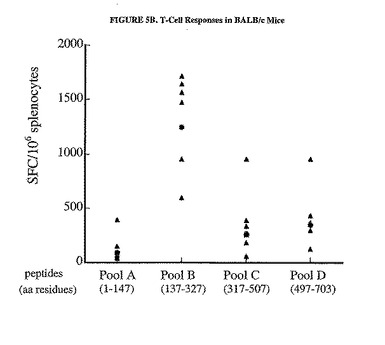
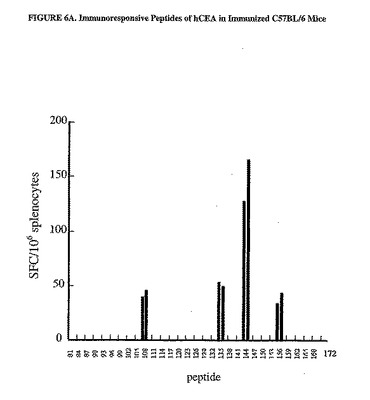
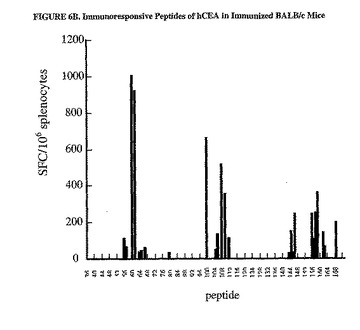
C57BL/6マウスにおいてDNA/Adワクチン接種計画によって惹起された免疫応答は、主に蛋白質のC末端領域に偏っていた(図5A参照)。有意なSFC値はペプチドプールCおよびDで得られ(それぞれの相乗平均値、170および244SFC/106脾細胞)、一方、プールAおよびBではかなり低い値が得られた(それぞれ、10および27SFC/106脾細胞)。対照的に、BALB/cマウスの免疫応答は、プールBで最も高いが(相乗平均値、1236SFC/106脾細胞)、プールA、CおよびDでは有意に高いSFC値が示された(それぞれ、93、364および344)。関連のないペプチドのプールに対する応答は、いずれの群のマウスでも示されなかった(データは示さなかった)。
【0102】
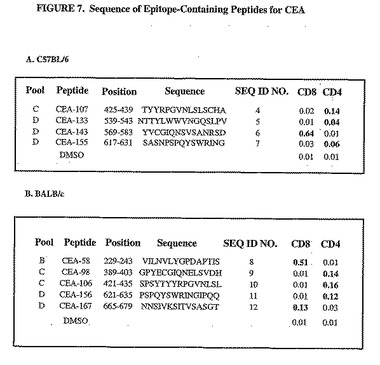
応答を惹起するペプチド収集物に存在する個々のペプチドを同定するために、DNA/Adワクチン接種計画によって免疫した4種類のマウスの脾細胞を、有意な免疫応答が認められたプールを含む個々のペプチドそれぞれに対するIFNγ−ELISPOT測定法で分析した。C57BL/6マウスの脾細胞は、プールCおよびDに含まれるペプチド80から173に対して調べた。BALB/cマウスの脾細胞は、プールB、CおよびDを含むペプチド35から173に対して調べた。C57BL/6マウスにおけるCEA特異的応答は、重複した配列を有する15マーペプチドの4種類の組み合わせに位置づけた(aa431から435および425から439;529から543および533から547;565から579および569から593;613から627および617から631)(図6A)。BALB/cマウスにおけるCEAに対する免疫応答は、22個のペプチドに位置づけられ、そのウチの17個は配列が重複している(aa213から227および213および227;229から243および233から247;409から423および413から427;421から435および425から439;565から579および569から583;573から587;613から627および617から631並びに621から635および625から639;637から651および641から655)(図6B)。
【0103】
選択したペプチド内に含有されるエピトープのT細胞特異性を確認するために、注射したマウスの脾細胞についてIFNγ細胞内染色測定法を実施した。得られた結果を図7に示す。該データは、Tリンパ球の循環濃度を定量するために使用することができるC57BL/6およびBALB/cマウス両方について、CD8+およびCD4+特異的エピトープが同定されたことを示す。
【実施例9】
【0104】
コドンを最適化したhCEA cDNAは、hCEAトランスジェニックマウスにおける寛容を打破する
コドンを最適化したhCEAのcDNAの増強された免疫原性がヒトCEAに対する寛容を効果的に打破するかどうかを測定するために、hCEAトランスジェニックマウスを野生型またはコドン最適化hCEA配列のいずれかを有するベクターで免疫した。これらのトランスジェニックマウスは、完全なヒトCEA遺伝子および隣接配列を有し、盲腸および結腸においてhCEA蛋白質を発現する。したがって、このマウス系は、この腫瘍の自己抗原を対象とする免疫治療計画の安全性および効率を研究するための有用なモデルである(Clarke他、「免疫治療のモデルとしてのヒトCEA遺伝子組換えマウス(Mice transgenic for human CEA as a model for immunotherapy)」Cancer Res.1469〜77(1998))。
【0105】
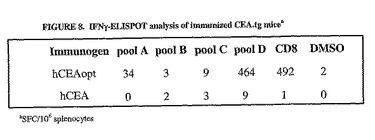
第1の試験として、トランスジェニックマウス10匹のうち5匹にプラスミドDNA50μgを4回電気注射し、次いで最終的にアデノウイルス1x1010ppを注射した。hCEAに対する免疫応答は、注射したマウス4匹から収集した脾細胞についてIFNγ−ELISPOT測定を行うことによって分析した。hCEAに対する免疫応答は、hCEAoptcDNAで免疫したマウスの脾細胞のみで検出された(図8参照)。該免疫応答は、ペプチド143およびプールDで検出されたので、免疫によってC末端エピトープに対する有意なCD8+応答が惹起されたことが示唆された。
【0106】
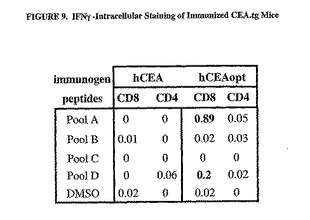
hCEAのコドン最適化cDNAの増強された免疫原性はまた、アデノウイルスベクター1x1010ppを2週間間隔で2回注射したトランスジェニックマウスで試験した。CEA特異的免疫応答は、4種類の免疫マウスから収集したPBMCをIFNγ細胞内染色することによって測定した。hCEAに対する免疫応答は、Ad/hCEAoptで免疫したマウスのみに検出された(図9)。DNAおよびAdコホートで認められたように、CD8+T細胞の誘導はペプチドプールDで検出されたが、有意なCD8+応答はまた、ペプチドプールAで示された。したがって、これらの結果は、hCEAのコドン最適化cDNAは、野生型配列よりも免疫原性が高く、hCEAに対する寛容を効率的に打破することを示す。
【実施例10】
【0107】
抗体検出および力価測定
抗体力価測定用の血清は、眼窩後方出血によって得られた。ELISAプレート(Nunc maxisorpTM)は、CEA蛋白質100ng/ウェルでコーティングして(高純度CEA、Fitzgerald Industires International Inc.,Concord MA)でコーティングして、コーティング緩衝液(50mM NaHCO3、pH9.4)で希釈して、4℃でO/Nインキュベートした。次に、プレートを5%BSAを含有するPBSで37℃で1時間ブロックした。マウス血清は、PBS5%BSA(血清転換率の評価には1/50の希釈度、力価測定には1:10から1:312150までの希釈度)で希釈した。予備免疫血清は、背景として使用した。希釈した血清は、4℃でO/Nインキュベートした。洗浄は、PBS1%BSA、0.05%Tween20で実施した。2次抗体(ヤギ抗マウス、IgGペルオキシダーゼ、Sigma)は、PBS、5%BSAで1/2000に希釈して、室温で2時間から3時間、振盪機上でインキュベートした。洗浄後、プレートをTMB基質100μl/ウェル(Pierce Biotechnology、Inc.,Rockford、IL)で発色させた。該反応は、1M H2SO4溶液25μl/ウェルで停止し、プレートは450nm/620nmで読み取った。抗CEA血清力価は、同じ希釈度の自己由来予備免疫血清の吸収より少なくとも3倍大きな吸収を生じる血清の限界希釈の逆数として算出した。
【実施例11】
【0108】
hCEAoptの高い免疫原性
野生型およびコドン最適化CEA発現ベクターによって誘導されるインビボでの免疫応答を調べるために、C57BL/6マウスを1x105から1x103pfuの範囲の様々な用量のAd5/hCEAoptを筋肉内注射して免疫した。マウスに3週間間隔で2回注射を行った。2回目の免疫を行ってから2週間後に脾細胞を各マウスから単離した。アデノウイルスによる免疫によって生じるIFNγ分泌型CEA特異的CD8T細胞前駆体発生頻度を定量するために、H−2b制限T細胞エピトープCGIQNSVSA(配列番号14、以下参照)についてELISPOT測定法を使用した。1x104pfuで免疫すると、測定可能な免疫応答を惹起し、CGIONSVSAエピトープ(配列番号14)に特異的な53個のIFNγスポット形成細胞(SFC、相乗平均値)が生じたが、一方1x103pfuの注射ではごくわずかなSFC値しか惹起しなかった(図3A)。Ad/hCEAopt 1x105pfuで免疫した群において、SFCは302個まで増加した。対照的に、1x106pfuの用量で免疫したマウスの群において、SFCを168まで増加させる有意なCD8T−細胞前駆体頻度を惹起するために、Ad5/hCEAは少なくとも1x105pfu必要であった。ペプチド特異的IFNγSFCは、Ad5免疫マウスでは検出されなかった(データは示さなかった)。
【0109】
それぞれのhCEAアデノウイルスベクター1x105pfuで免疫したマウスの血清は、基質として精製ヒトCEA蛋白質を使用してELISAで試験した。Ad5/hCEAoptで免疫したマウスのCEA特異的抗体の力価は、免疫したマウス全てにおいて検出され、Ad力価の相乗平均値は46474であった。対照的に、Ad5/hCEA免疫群は、CEA特異的抗体の約100倍低い相乗平均力価を示した(454)。したがって、これらの結果は、CEAのコドン最適化cDNAは細胞性および体液性免疫応答をより効率的に惹起することを示す。
【実施例12】
【0110】
統計解析
指示した場合は、結果はスチューデントt検定によって解析した。p値<0.05は、有意とみなされた。
【図面の簡単な説明】
【0111】
【図1】野生型ヒトCEAcDNA(配列番号3)およびコドン最適化クローン(hCEAopt、配列番号1)のヌクレオチド配列を示す図である。推定アミノ酸配列を上部に示す(配列番号2)。合成コドン最適化cDNAの置換ヌクレオチドをhCEAcDNA配列の下に示す。実施例2参照。
【図1A】野生型ヒトCEAcDNA(配列番号3)およびコドン最適化クローン(hCEAopt、配列番号1)のヌクレオチド配列を示す図である。推定アミノ酸配列を上部に示す(配列番号2)。合成コドン最適化cDNAの置換ヌクレオチドをhCEAcDNA配列の下に示す。実施例2参照。
【図1B】野生型ヒトCEAcDNA(配列番号3)およびコドン最適化クローン(hCEAopt、配列番号1)のヌクレオチド配列を示す図である。推定アミノ酸配列を上部に示す(配列番号2)。合成コドン最適化cDNAの置換ヌクレオチドをhCEAcDNA配列の下に示す。実施例2参照。
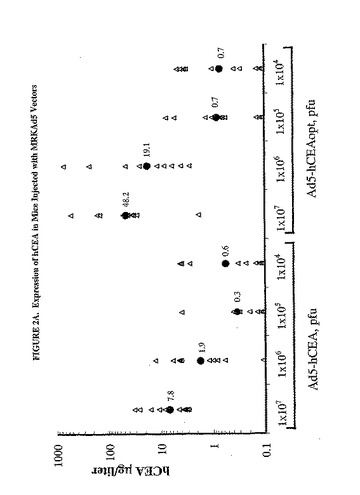
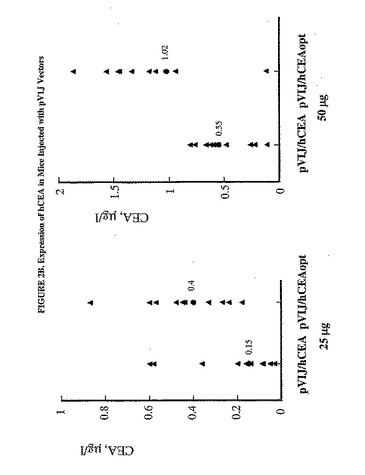
【図2A】注射したマウスにおけるhCEAの発現を示した図である。C57BL/6マウス10匹の群において、様々な用量のMRKAd5/hCEAおよびMRKAd5/hCEAopt(パネルA)または25マイクログラムもしくは50マイクログラムのプラスミドpV1J/hCEAおよびpV1J/hCEAopt(パネルB)のいずれかを大腿4頭筋に注射した。注射して3日後に血液試料を収集し、CEA濃度を測定した。塗りつぶされた三角形は個々のマウスのCEA測定値を表す。相乗平均値もまた示す(塗りつぶされた丸)。
【図2B】注射したマウスにおけるhCEAの発現を示した図である。C57BL/6マウス10匹の群において、様々な用量のMRKAd5/hCEAおよびMRKAd5/hCEAopt(パネルA)または25マイクログラムもしくは50マイクログラムのプラスミドpV1J/hCEAおよびpV1J/hCEAopt(パネルB)のいずれかを大腿4頭筋に注射した。注射して3日後に血液試料を収集し、CEA濃度を測定した。塗りつぶされた三角形は個々のマウスのCEA測定値を表す。相乗平均値もまた示す(塗りつぶされた丸)。
【図3A】コドン最適化がヒトCEAに対する免疫応答を増強することを示した図である。C57BL/6マウス8匹の群において、様々な用量のMRKAd5−hCEAおよびMRKAd5−hCEAoptのいずれかを大腿4頭筋に注射した。ウイルス注射は、0日目と21日目に実施した。パネルA。追加免疫注射の2週間後に、aa569から583を含みCD8+エピトープを含むペプチド143を使用したELISPOTアッセイで、個々のマウスの脾細胞でhCEAに特異的なCD8+IFNγ分泌型T細胞の数を測定した(塗りつぶされた三角形)。2種類の異なる量の脾細胞(2.5x105および5x105)で、脾細胞のそれぞれの試験料について2回反復した。平均値は、ペプチド無しで測定した背景濃度を差し引くことによって算出し(一般的に10SFC/106全脾細胞)、結果はSFCの数/106全脾細胞として表した。個々のマウスの値(塗りつぶされた三角形)および相乗平均値(塗りつぶされた丸)を示した。パネルB。個々のマウスの血清中の抗CEA抗体力価(塗りつぶされた三角形)は、追加免疫10日後の血清試料を使用して測定した。相乗平均力価(GMT)(塗りつぶされた丸)もまた示す。Ad/hCEAoptはAd/hCEAと有意に異なる。
【図3B】コドン最適化がヒトCEAに対する免疫応答を増強することを示した図である。C57BL/6マウス8匹の群において、様々な用量のMRKAd5−hCEAおよびMRKAd5−hCEAoptのいずれかを大腿4頭筋に注射した。ウイルス注射は、0日目と21日目に実施した。パネルA。追加免疫注射の2週間後に、aa569から583を含みCD8+エピトープを含むペプチド143を使用したELISPOTアッセイで、個々のマウスの脾細胞でhCEAに特異的なCD8+IFNγ分泌型T細胞の数を測定した(塗りつぶされた三角形)。2種類の異なる量の脾細胞(2.5x105および5x105)で、脾細胞のそれぞれの試験料について2回反復した。平均値は、ペプチド無しで測定した背景濃度を差し引くことによって算出し(一般的に10SFC/106全脾細胞)、結果はSFCの数/106全脾細胞として表した。個々のマウスの値(塗りつぶされた三角形)および相乗平均値(塗りつぶされた丸)を示した。パネルB。個々のマウスの血清中の抗CEA抗体力価(塗りつぶされた三角形)は、追加免疫10日後の血清試料を使用して測定した。相乗平均力価(GMT)(塗りつぶされた丸)もまた示す。Ad/hCEAoptはAd/hCEAと有意に異なる。
【図4A】異なる免疫計画の比較を示した図である。プラスミドpV1J/hCEA(50μg/大腿4頭筋に電気注射した用量)およびMRKAd5/hCEA(1x109pp/用量)の様々な組み合わせで、C57BL/6(A)またはBALB/c(B)マウスの群を免疫した。個々のマウスの脾細胞におけるIFNγ分泌型T細胞の数は、材料および方法および図3の凡例で説明したようにaa497から703(プールD)に亘るペプチドのプールを使用して測定した。相乗平均値もまた示す(塗りつぶされた丸)。D/DおよびD/Aは、C57BL/6マウスのAd/Ad群と有意に異なる。3群は全て、BALB/cマウスにおいて有意に異なる。
【図4B】異なる免疫計画の比較を示した図である。プラスミドpV1J/hCEA(50μg/大腿4頭筋に電気注射した用量)およびMRKAd5/hCEA(1x109pp/用量)の様々な組み合わせで、C57BL/6(A)またはBALB/c(B)マウスの群を免疫した。個々のマウスの脾細胞におけるIFNγ分泌型T細胞の数は、材料および方法および図3の凡例で説明したようにaa497から703(プールD)に亘るペプチドのプールを使用して測定した。相乗平均値もまた示す(塗りつぶされた丸)。D/DおよびD/Aは、C57BL/6マウスのAd/Ad群と有意に異なる。3群は全て、BALB/cマウスにおいて有意に異なる。
【図5A】hCEA蛋白質の選択領域に対するT細胞応答のマッピングの結果を示した図である。C57BL/6(パネルA)またはBALB/c(パネルB)マウスの群は、プラスミドpV1J/hCEAで免疫し、3週間後にAd/hCEA 1x109で追加免疫した。個々のマウスの脾細胞におけるIFNγ分泌型T細胞の数は、材料および方法および図3の凡例で説明したように、完全な蛋白質を網羅したペプチドのプールを使用して追加免疫の2週間後に測定した。相乗平均値もまた示す(塗りつぶされた丸)。
【図5B】hCEA蛋白質の選択領域に対するT細胞応答のマッピングの結果を示した図である。C57BL/6(パネルA)またはBALB/c(パネルB)マウスの群は、プラスミドpV1J/hCEAで免疫し、3週間後にAd/hCEA 1x109で追加免疫した。個々のマウスの脾細胞におけるIFNγ分泌型T細胞の数は、材料および方法および図3の凡例で説明したように、完全な蛋白質を網羅したペプチドのプールを使用して追加免疫の2週間後に測定した。相乗平均値もまた示す(塗りつぶされた丸)。
【図6A】hCEAの免疫応答ペプチドの同定を示した図である。4匹の免疫したC57/BL/6(パネルA)またはBALB/c(パネルB)マウスから収集した脾細胞の指定したペプチドそれぞれに対するIFNγ分泌について、ELISPOT測定法を使用して測定した(実施例8参照)。
【図6B】hCEAの免疫応答ペプチドの同定を示した図である。4匹の免疫したC57/BL/6(パネルA)またはBALB/c(パネルB)マウスから収集した脾細胞の指定したペプチドそれぞれに対するIFNγ分泌について、ELISPOT測定法を使用して測定した(実施例8参照)。
【図7】C57BL/6マウス(パネルA)およびBALB/cマウス(パネルB)におけるCEAのペプチドを含有するエピトープの配列を示した図である(実施例xx参照)。右に挙げたのは、IFNγ産生CD8+(CD4+)CD3+細胞の割合である。
【図8】実施例9で説明したような免疫したCEAトランスジェニックマウスのIFNγ−ELISPOT測定法から得られた結果を示した図である。マウスは、1週間間隔でプラスミドDNAを4回電気注射し、さらにアデノウイルス注射を1回行って免疫した。各免疫原について、3回注射したマウスから収集した脾細胞でデータを採った。CD8特異的応答は、ペプチド143を使用して測定した。
【図9】免疫したCEA.tgマウスのIFNγ細胞内染色を示した図である。マウスは、2週間間隔でアデノウイルス1x1010vpを2回注射して免疫した。3回注射したマウスから収集した脾細胞で得られたデータを示す。右に挙げたのは、CD8+またはCD4+細胞の割合である。
【図10A】パネルAは、配列番号15で表したアンカードメインを切断したCEA変種をコードするコドン最適化DNA分子の例を示した図である。hCEAopt−ΔADの対応するアミノ酸配列をパネルBに示す(配列番号16)。
【図10B】パネルAは、配列番号15で表したアンカードメインを切断したCEA変種をコードするコドン最適化DNA分子の例を示した図である。hCEAopt−ΔADの対応するアミノ酸配列をパネルBに示す(配列番号16)。
【技術分野】
【0001】
本発明は一般的に癌の治療に関する。より具体的には、本発明は、本明細書でhCEAoptと称するヒト腫瘍関連癌胎児抗原をコードする合成ポリヌクレオチドに関し、該ポリペプチドはヒト細胞環境で発現するためにコドンが最適化されている。本発明はまた、組換えベクターおよび前記合成ポリヌクレオチドを含む宿主を提供する。本発明はまた、hCEAoptを有するアデノウイルスベクターおよびプラスミド構築物並びに癌の予防および治療のためのワクチンにおけるそれらの使用並びに医薬品組成物に関する。
【背景技術】
【0002】
免疫グロブリンスーパーファミリー(IgSF)は、様々な機能を備えた蛋白質をコードする数多くの遺伝子から構成され、その機能の1つが細胞間接着である。IgSF蛋白質は、適切な分子間結合相互作用を維持するために重要なIg関連ドメインを少なくとも1つ含有する。このような相互作用は、IgSF構成要素の様々な生物学的機能に不可欠なので、多くのIgSF接着分子の破壊または異常発現は、多くのヒト疾患と相関してきた。
【0003】
癌胎児抗原(CEA)は、細胞表面糖蛋白質を構成するIgスーパーファミリーに属する。CEAサブファミリーの構成要素は、CEA関連細胞接着分子(CEACAM)として知られている。最近の科学文献では、CEA遺伝子はCEACAM5と改名されたが、蛋白質の名称はCEAのままである。機能的には、CEACAMはホモタイプおよびヘテロタイプ両方の細胞間接着分子として作用することが示された(Benchimol他、Cell 57:327〜334(1989))。細胞接着に加えて、CEAは細胞外マトリックスからの細胞脱離によって生じる細胞死を阻害し、Bcl2およびC−Mycなどのある種のプロトオンコジーンに関連した細胞形質転換の一因となり得る(Berinstein、J.Oncol.20(8):2197〜2207参照)。
【0004】
CEAの正常な発現は、胎児の発育中および成人結腸粘膜で検出された。CEA過剰発現は、30年以上前にヒト結腸癌において最初に検出され(Gold and Freedman、J.Exp.Med.121:439〜462(1965))、その後ほとんど全ての結腸直腸癌で見いだされてきた。さらに、CEA過剰発現は、膵臓、胸および肺の腺癌の高い割合で検出可能である。これらの種類の腫瘍ではCEA発現が一般的なので、CEAはこれらの癌の管理および予後において広く臨床的に使用されている。
【0005】
ヒトCEAをコードする配列は、クローニングされ、特徴付けられている(米国特許第5274087号、米国特許第5571710号および米国特許第5843761号。Beauchemin他、Mol.Cell.Biol.7:3221〜3230(1987)、Zimmerman他、Proc.Natl.Acad.Sci.USA 84:920〜924(1987)。Thompson他、Proc.Natl.Acad.Sci.USA 84(9):2965〜69(1987))。
【0006】
CEA発現と転移増殖との間の相関関係によって、結腸直腸癌治療のための分子的および免疫学的介入の標的としての存在価値が引き出された。CEAを標的とする1つの治療的取り組みは、抗CEA抗体の使用であり(Chester他、Cancer Chemother.Pharmacol.46(Suppl):S8〜S12(2000)参照)、もう1つはCEAをベースとしたワクチンを使用してCEA発現腫瘍を攻撃する免疫系を活性化することである(概説としてはBerinstein、前述参照)。
【発明の開示】
【発明が解決しようとする課題】
【0007】
多くのワクチンの開発および商品化は、宿主生物をうまく形質転換する外来遺伝子を高い発現濃度で獲得するのに伴う困難さのために、妨げられている。したがって、前述のCEA蛋白質をコード化する野生型ヌクレオチド配列が同定されたにも関わらず、効率的で、自己寛容によって妨害されない癌ワクチンの開発が可能になる、企図する宿主細胞で発現するために最適化されたCEAをコードするヌクレオチド配列を利用するヒトCEA蛋白質の容易に再生可能な材料を開発することが強く望まれている。
【課題を解決するための手段】
【0008】
本発明は、直腸結腸癌を含めた数多くの腺癌に関連してきたCEA遺伝子によって発現した蛋白質生成物に対する免疫を惹起するか、または増強する組成物および方法に関する。具体的に、本発明は、ヒトCEA蛋白質をコードするポリヌクレオチドを提供する。前記ポリヌクレオチドはヒト細胞において高レベルで発現するようにコドンが最適化されている。本発明はさらに、合成ポリヌクレオチドを含むアデノウイルスベクターおよびプラスミドをベースとしたベクターを提供し、CEAに関連した癌を予防し、かつ/または治療するための免疫原性組成物およびワクチンにおける前記ベクターの使用を開示する。
【0009】
本発明はまた、配列番号2に記載されているヒト癌胎児抗原(以後hCEA)をコードするヌクレオチドの配列を含む合成核酸分子(ポリヌクレオチド)に関し、前記合成核酸分子は、ヒト細胞において高レベルで発現するようにコドンが最適化されている(以後、hCEAopt)。本明細書で開示された核酸分子は、選択した宿主細胞にトランスフェクトすることができ、該組換え宿主細胞は、十分な濃度で機能的hCEA蛋白質(配列番号2)を発現させるための原料となる。
【0010】
本発明はさらに、ヒトCEA蛋白質を発現するmRNAをコードする合成核酸分子に関し、このDNA分子は本明細書で配列番号1として開示したヌクレオチド配列を含む。本発明のこの部分の好ましい態様は、hCEA蛋白質(配列番号2または配列番号16)をコードするDNA分子(配列番号1)を示す図1に開示される。本発明の好ましい核酸分子は、ヒト細胞において高レベルで発現するようにコドンが最適化されている。
【0011】
本発明の他の好ましいDNA分子は、ヒト完全長CEA(配列番号2)のアミノ酸約679からアミノ酸約702に位置するC末端アンカードメイン(AD)が欠失したヒトCEAをコードするヌクレオチドの配列を含み、ヌクレオチドの前記配列はヒト細胞において高レベルで発現するようにコドンが最適化されている。アンカードメインを切断されたCEA変種をコードするDNA分子の例を図15に示す(図10Aを参照)。hCEA−ΔADの対応するアミノ酸配列は、配列番号16に記載する(図10Bを参照)。
【0012】
本発明はまた、この明細書を通じて開示した核酸分子を含有する組換えベクター、並びに原核細胞および真核細胞両方の組換え宿主細胞に関する。
【0013】
本発明はさらに、(a)配列番号1または配列番号15で表した核酸分子を含むベクターを適切な宿主細胞に導入すること、および(b)前記コドン最適化ヒト蛋白質の発現を可能にする条件下で該宿主細胞を培養することを含む、組換え宿主細胞におけるコドン最適化ヒトCEA蛋白質の発現方法に関する。
【0014】
本発明の他の態様は、合成核酸分子を含むワクチンベクターを哺乳類に投与することを含む癌の予防または治療方法であり、該合成核酸分子は配列番号2または配列番号16で表したヒト癌胎児抗原(hCEA)蛋白質をコードするヌクレオチドの配列を含み、該合成核酸分子はヒト細胞において高濃度に発現するようにコドンが最適化されている。
【0015】
本発明はさらに、E1領域に欠失を有し、及びE1領域に挿入部分を有するアデノウイルスゲノムを含むアデノウイルスワクチンベクターに関し、前記挿入部分は、(a)ヒトCEA蛋白質をコードするコドン最適化ポリヌクレオチドと、(b)ポリヌクレオチドに操作可能に結合されたプロモーターとを含む発現カセットを含む。
【0016】
本発明はまた、プラスミド部分と発現カセット部分とを含むワクチンプラスミドに関し、該発現カセット部分は、(a)ヒト細胞において最適に発現させるためにコドンを最適化した、ヒトCEA蛋白質をコードする合成ポリヌクレオチドと、(b)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む。
【0017】
本発明の他の態様は、(a)(i)ヒトCEA蛋白質をコードするコドン最適化ポリヌクレオチドおよび(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターを含む第1のベクターを哺乳類に導入すること、(b)予め決定された時間経過させること、および(c)(i)ヒトCEA蛋白質をコードするコドン最適化ポリヌクレオチドおよび(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターを含む第2のベクターを該哺乳類に導入することを含む、前記哺乳類を癌から予防もしくは癌を治療するか、またはCEAに関連した癌に罹患した前記哺乳類を治療する方法である。
【0018】
明細書および添付の特許請求の範囲を通じて使用したように、単数形「a」、「an」および「the」は、明白に別段の指定がない限り、複数を指すことを含む。
【0019】
明細書および添付の特許請求の範囲を通じて使用したように、以下の定義および略語を適用する。
【0020】
「プロモーター」という用語は、RNAポリメラーゼが結合するDNA鎖の認識部位のことである。該プロモーターは、RNAポリメラーゼとの開始複合体を形成し、転写活動を開始し、推進する。該複合体は、「エンハンサー」と称する活性化配列または「サイレンサー」と称する阻害配列によって変更することができる。
【0021】
「カセット」という用語は、発現させるべき核酸配列を含有する本発明の配列のことである。該カセットは、カセットテープの概念と同じで、各カセットは独自の配列を有する。したがって、カセットを交換することによって、ベクターは異なる配列を発現する。5′および3′末端に制限部位があるために、該カセットは容易に挿入したり、除去したり、他のカセットで置換したりすることができる。
【0022】
「ベクター」という用語は、DNA断片を宿主生物または宿主組織に導入することができるいくつかの手段を意味する。ベクターには、プラスミド、(アデノウイルスを含む)ウイルス、バクテリオファージおよびコスミドを含む様々な種類がある。
【0023】
アデノウイルスベクターに関して使用したような「第1世代」という用語は、複製に欠陥のある前記アデノウイルスベクターを意味する。一般的に、第1世代アデノウイルスベクターは、E1遺伝子領域が欠失しているか、または不活性化しており、好ましくはE3遺伝子領域が欠失しているか、または不活性化している。
【0024】
「pV1J/hCEAopt」という呼称は、イントロンAを有するヒトCMV前初期(IE)プロモーター、完全長コドン最適化ヒトCEA遺伝子、ウシ成長ホルモン由来ポリアデニル化配列および転写終止配列並びに最小pUC主鎖を含む、本明細書で開示したプラスミド構築物である(実施例2参照)。前述のように、「pV1J/hCEA」という呼称は、構築物がコドン最適化ヒトCEA遺伝子の代わりに野生型ヒトCEA遺伝子を含むこと以外は前述のような構築物を意味する。
【0025】
「MRKAd5/hCEAopt」および「MRKAd5/hCEA」という呼称は、E1およびE3領域に欠失があるAd5アデノウイルスゲノムを含む、本明細書で開示した2種類の構築物を意味する。「MRKAd5/hCEAopt」構築物においては、E1領域は、イントロンAを含まないヒトCMVプロモーターの制御下でE1と平行したコドン最適化ヒトCEA遺伝子およびそれに続くウシ成長ホルモンポリアデニル化シグナルによって置換されている。「MRKAd5/hCEA」構築物は、Ad5ゲノムのE1領域が野生型ヒトCEA配列と置換していること以外は、本質的に前述の通りである(実施例2参照)。
【0026】
「有効量」という用語は、免疫応答を生じるように、十分なワクチン組成物が適切な濃度のポリペプチドの産生を誘導することを意味する。当業者は、この濃度は変化し得ることを承知している。
【0027】
「保存的アミノ酸置換」とは、1個のアミノ酸残基がもう1個の化学的に類似したアミノ酸残基によって置換されることを意味する。このような保存的置換の例は、1疎水性残基(イソロイシン、ロイシン、バリンまたはメチオニン)と他の残基との置換、1極性残基と電荷が同じ他の1極性残基との置換(たとえば、アルギニンとリジン、グルタミン酸とアスパラギン酸)である。
【0028】
「hCEA」は、ヒト癌胎児抗原を意味し、「hCEAopt」は、コドン最適化ヒト癌胎児抗原を意味する。
【0029】
「hCEA−ΔAD」という用語は、ヒト完全長CEA(配列番号2)のアミノ酸約679からアミノ酸約702に位置するC末端アンカードメイン(AD)を欠失したヒトCEAの変種を意味する。本発明のhCEA−ΔADをコードするヌクレオチド配列は、ヒト細胞環境において高レベルで発現するようにコドンが最適化されている(本明細書では、hCEAopt−ΔAD”と称する)。アンカードメインを切断されたCEA変種をコードするDNA分子の例を図15に示す(図10Aを参照)。hCEA−ΔADの対応するアミノ酸配列は配列番号16に記載する(図10Bを参照)。hCEA−ΔADをコードするヌクレオチドは、癌の治療および/または予防のための癌ワクチンの開発に有用である。
【0030】
「哺乳類」という用語は、人類を含めた任意の哺乳類を意味する。
【0031】
略語「Ag」は、抗原のことである。
【0032】
略語「Ab」は抗体を意味し、「mAb」はモノクローナル抗体を意味する。
【0033】
略語「ORF]は、遺伝子のオープンリーディングフレームを意味する。
【発明を実施するための最良の形態】
【0034】
癌胎児抗原(CEA)は通常、腺癌の発症に関連する。本発明は、異常なCEA発現が癌または癌の発症に関連するCEA腫瘍関連抗原によって発現した蛋白質生成物に対する免疫を惹起するか、または高める組成物および方法に関する。異常なCEA発現は腫瘍開始時に存在し、腫瘍進行後まで検出されなくてもよく、その逆であってもよいので、異常なCEA発現と癌との関連には、CEA蛋白質が腫瘍組織において、その発症の全ての時点において発現されていることは必要ではない。
【0035】
このために、ヒトCEA蛋白質をコードする合成DNA分子を提供する。好ましい実施形態ではヒト細胞である計画した宿主細胞に好まれるコドンを使用するように、合成分子のコドンを設計する。抗体の中和および細胞性免疫によって、CEA関連癌に対する効果的な免疫予防を提供する組換えアデノウイルスワクチンまたはプラスミドをベースとしたワクチンを開発するために、該合成分子を使用することができる。該合成分子は、免疫原組成物として使用することができる。本発明は、霊長類およびヒトなどの哺乳類を含めた脊椎動物に直接インビボで導入すると、該動物でコード化された蛋白質の発現を誘導するポリペプチドを提供する。
【0036】
野生型ヒトCEAヌクレオチド配列は報告されている(たとえば、米国特許第5274087号、米国特許第5571710号および米国特許第5843761号参照)。本発明は、ヒトCEA蛋白質をコードする合成DNA分子を提供する。本発明の合成分子は、いくつかはヒト細胞によって好まれるコドンを使用するように変更されており、したがってヒト宿主細胞においてCEAを高濃度で発現することが可能であるヌクレオチドの配列を含む。該合成分子は、抗体の中和および細胞性免疫によってCEA関連癌に対する効果的な免疫予防を提供する癌ワクチンで使用することができる、CEA蛋白質の材料として使用することができる。
【0037】
4種類の可能なヌクレオチド塩基の「3連」コドンは、60個を上回る変種の形で存在することが可能である。これらのコドンは、異なるアミノ酸20個(および転写開始および終止)のみの情報を提供するので、いくつかのアミノ酸は、複数のコドンによってコードされることが可能で、コドン重複として公知の現象である。完全には理解されていない理由のために、他のコドンは異なる種類の細胞の内在DNAに均一に存在することはない。実際に、ある種の細胞においてある種のコドンには様々な天然の階層または「選択性」が存在するようである。1例として、アミノ酸ロイシンはCTA、CTC、CTG、CTT、TTAおよびTTGを含めた6種類のDNAコドンのいずれかによって特定される。微生物のゲノムコドン出現頻度の徹底的な分析によって、E.コリの内在性DNAは極めて一般的に、CTGロイシン指定コドンを含有し、一方酵母および粘菌のDNAは極めて一般的にTTAロイシン指定コドンを含むことが明らかになった。この階層的観点から、E.コリ宿主によってロイシンリッチなポリペプチドの高濃度発現が得られる可能性は、使用したコドンの頻度にある程度左右されることが一般に信じられている。たとえば、TTAコドンに富んだ遺伝子はE.コリでは十分に発現せず、一方CTGに富んだ遺伝子はおそらくこの宿主で高く発現するようである。同様に、酵母宿主細胞でロイシンに富んだポリペプチドの発現に好まれるコドンはTTAである。
【0038】
組換えDNA技術に対するコドン選択現象の意味は一目瞭然で、該現象によって、形質転換が成功した宿主生物において外来遺伝子の高濃度発現を実現できなかったかつての多くの失敗を説明することができ、あまり「好ましくない」コドンを挿入遺伝子内に反復して存在させて、宿主細胞の発現機構をあまり効率よく作動できないようにすることが可能である。この現象は、計画した宿主細胞に好まれるコドンを含むように設計された合成遺伝子は、組換えDNA技術を実施するために最適な形態の外来遺伝子材料となることを示唆する。したがって、本発明の1態様は、ヒト細胞で発現するためにコドンを最適化したヒトCEA遺伝子である。本発明の好ましい実施形態では、同じ蛋白質配列をコードする他のコドンを使用することによって、ヒト細胞における外来CEA蛋白質発現の制約を取り除くことができることが見いだされた。
【0039】
本発明によれば、本明細書に参照により組み込まれた、Latheの「アミノ酸配列データから推測されたオリゴヌクレオチドプローブ:理論および実際の考察(Synthetic Oligonucleotide Probes Deduced from Amino Acid Sequence Data:Theoretical and Practical Considerations」J.Molec.Biol.183:1〜12(1985)に記載されたように、ヒトCEA遺伝子配列は、同じ翻訳配列を有するが他のコドンを使用したポリヌクレオチドに変換された。この方法は一般的に、高発現のヒト遺伝子に通常関連しない野生型配列のコドンを同定すること、およびこのコドンとヒト細胞において高く発現するために最適なコドンとを置換することから構成される。次に、新規配列について、これらのコドン置換によって生じた望ましくない配列を調べる(たとえば、「ATTTA」配列、イントロンスプライシング認識部位の偶発的な発生、望ましくない制限酵素部位など)。所望しない配列は、同一アミノ酸をコードする異なるコドンで既存のコドンを置換することによって除去される。次に、合成遺伝子部分の発現が改善されているかを調べる。
【0040】
前述の方法を使用してヒトCEAの合成遺伝子配列を作製して、高い発現濃度に最適化したコドンを含む遺伝子が生じた。前記の方法は、癌ワクチンで使用するコドン最適化遺伝子を設計するための方法を要約したものであるが、該方法を少し変更するか、または配列を少し変化させることによって、同様のワクチンの有効性または遺伝子発現の増加を実現することは、当業者に理解されている。当業者はまた、DNA分子のほんの一部のコドンをコドン最適化して、ヒト細胞において高濃度のCEA発現をもたらす他のDNA分子を構築できることを理解するであろう。
【0041】
したがって、本発明は、ヒトCEA蛋白質(配列番号2)、またはhCEA−ΔAD(配列番号16)を含むがそれだけに限定されないヒトCEA蛋白質の生物学的に活性のある断片または変異型をコードするヌクレオチドの配列を含む合成ポリヌクレオチドに関し、該ポリヌクレオチド配列はヒト宿主で発現するために最適化されたコドンを含む。CEA蛋白質の前記変異型は、本明細書で集合的に「変種」として称される保存的アミノ酸置換、アミノ末端切断、カルボキシ末端切断、欠失または添加を含むが、それだけに限定されない。このような生物学的活性のある断片および/または変異体はいずれも、配列番号2で示したようなCEA蛋白質の免疫学的特性を少なくとも実質的に模倣した蛋白質または蛋白質断片をコードする。本発明の合成ポリヌクレオチドは、治療用または予防用癌ワクチンを開発するのに有用であるように、機能的なヒトCEA蛋白質を発現するmRNA分子をコードする。
【0042】
前述したように、本発明は、ヒトCEA蛋白質(配列番号2)をコードするヌクレオチドまたはそれらの生物学的に活性のある断片または変異型に関する。このために、本発明は、C末端アンカー配列が欠失したヒトCEA蛋白質を含むhCEA−ΔAD(配列番号16、図10B)をコードするヌクレオチドを提供する。hCEA−ΔADをコードする本発明の核酸分子は、ヒト細胞での発現を高めるためにコドンが最適化されている。hCEA−ΔADをコードする核酸配列の例は、配列番号15で示したヌクレオチドの配列を含む(図10A)。
【0043】
本発明は、配列番号2に記載されている新規hCEA蛋白質を発現するmRNAをコードするヌクレオチドの配列を含む合成核酸分子(ポリヌクレオチド)に関し、前記合成核酸分子は、ヒト宿主細胞で高レベルで発現するようにコドンが最適化されている。本発明の核酸分子は、実質的に他の核酸を含まない。
【0044】
本発明はまた、この明細書を通じて開示した核酸分子を含有する組換えベクター並びに原核細胞および真核細胞の両方の組換え宿主細胞に関する。該合成DNA分子、関連ベクターおよび本発明の宿主は、癌ワクチンの開発に有用である。
【0045】
本発明の好ましいDNA分子は、配列番号2として表した、図2に示したヒトCEA蛋白質をコードする、本明細書で配列番号1として開示した、図1に示したヌクレオチド配列を含む。
【0046】
本発明のさらに好ましいDNA分子は、配列番号16として表した、図10Bに示したC末端アンカー配列を欠失したヒトCEA変種をコードする、本明細書で配列番号15として開示した、図10Aに示したヌクレオチド配列を含む。
【0047】
本発明はまた、ヒトCEA蛋白質を発現するmRNAをコードする配列番号1の生物学的に活性のある断片または変異体を含む。このような生物学的活性のある断片および/または変異体は、配列番号2で表したhCEA蛋白質を含むが、それだけに限定されないhCEA蛋白質の薬理学的特性を少なくとも実質的に模倣した蛋白質または蛋白質断片のいずれかをコードする。このようなポリヌクレオチドはいずれも、ヌクレオチド置換、欠失、添加、アミノ末端切断およびカルボキシ末端切断を含むが、必ずしもそれだけに限定されない。本発明の変異体は、癌ワクチンの開発に有用であるように真核細胞において機能的hCEA蛋白質を発現するmRNA分子をコードする。
【0048】
本発明はまた、合成DNAのヌクレオチド配列が配列番号1のヌクレオチド配列とは著しく異なるが、まだ配列番号2で表したhCEA蛋白質をコードする合成コドン最適化DNA分子に関する。このような合成DNAは、本発明の範囲内にあるものとする。したがって、本発明は、同一の蛋白質を発現する数多くのDNA分子をもたらすことができるコドン重複を開示する。本発明の範囲内にはまた、発現した蛋白質の基本的な物理特性を実質的に変更しないDNA配列の変異が含まれる。たとえば、ロイシンをバリンに、リジンをアルギニンに、またはグルタミンをアスパラギンに置換しても、ポリペプチドの機能に変化は生じる可能性はない。
【0049】
ペプチドをコードするDNA配列は、天然に生じるペプチドと異なる特性を有するペプチドをコードするために改変することができることが知られている。DNA配列の改変方法は、部位特異的変異誘発を含むが、それだけに限定されない。改変された特性の例は、基質に対する酵素またはリガンドに対する受容体の親和性の変化を含むが、それだけに限定されない。
【0050】
本発明はまた、GFP(緑の蛍光蛋白質)、MYCエピトープ、GSTおよびFcを含むが、それだけに限定されない様々なマーカーに結合させたヒトCEA蛋白質の一部を発現する融合構築物を含むが、それだけに限定されないhCEAopt融合構築物に関する。このような融合構築物はいずれも、関心のある細胞系で発現することが可能で、本明細書で開示したヒトCEA蛋白質の修飾因子のスクリーニングに使用することが可能である。DOMおよびhsp70並びにLTBを含むが、それだけに限定されない、ヒトCEAに対する免疫応答を高めるために構築された融合蛋白質もまた、企図する。
【0051】
本発明はさらに、この明細書を通じて開示した合成核酸分子を含む組換えベクターに関する。これらのベクターは、DNAまたはRNAから構成されることが可能である。ほとんどのクローニングのためには、DNAベクターが好まれる。一般的なベクターは、プラスミド、改変ウイルス、バキュロウイルス、バクテリオファージ、コスミド、酵母の人工的染色体、およびhCEA蛋白質をコードすることができるその他の形態のエピソームDNAまたは統合DNAを含む。特定の遺伝子輸送またはその他の用途に適切なベクターを決定することは、十分当業者の範囲内である。
【0052】
hCEA蛋白質をコードするコドン最適化DNAを含有する発現ベクターは、組換え宿主細胞においてhCEAを高濃度に発現するために使用することができる。発現ベクターは、クローニングベクター、改変クローニングベクター、特異的に設計されたプラスミドまたはウイルスを含むが、それだけに限定されない。また、所望するならば、様々な細菌発現ベクターを使用して、細菌細胞で組換えhCEAを発現することができる。さらに、様々な真菌細胞発現ベクターを使用して、真菌細胞で組換えhCEAを発現することができる。さらに、様々な昆虫細胞発現ベクターを使用して、昆虫細胞で組換え蛋白質を発現することができる。
【0053】
本発明はまた、本発明の核酸分子を含むベクターで形質転換またはトランスフェクトした宿主細胞に関する。組換え宿主細胞は、E.コリなどの細菌、酵母などの真菌細胞を含めるがそれだけに限定されない原核細胞または真核細胞、ウシ、ブタ、サルおよび齧歯類由来の細胞系を含むがそれだけに限定されない哺乳類細胞、並びにショウジョウバエおよびカイコ由来の細胞系を含むがそれだけに限定されない昆虫細胞であってよい。このような組換え宿主細胞は、hCEAまたは生物学的に等価な形態を生成するために適した条件下で培養することができる。本発明の好ましい実施形態では、該宿主細胞はヒトである。本明細書で定義したように、「宿主細胞」という用語は、遺伝子を組み換えたヒトの体、ヒト胎児またはヒト胚における宿主細胞を含むものではない。
【0054】
前述のように、hCEA蛋白質をコードするDNAを含有する発現ベクターは、組換え宿主細胞においてhCEAを発現するために使用することができる。したがって、本発明の他の態様は、(a)配列番号1または配列番号15で表した核酸を含むベクターを適切なヒト宿主細胞に導入すること、および(b)前記ヒトCEA蛋白質またはCEA蛋白質変種の発現を可能にする条件下で該宿主細胞を培養することを含む、組換え宿主細胞におけるヒトCEA蛋白質または蛋白質変種の発現方法である。
【0055】
宿主細胞でhCEAが発現した後、hCEA蛋白質を回収して、活性型hCEA蛋白質を提供することができる。いくつかのhCEA蛋白質精製方法が有用で、使用に適している。組換えhCEA蛋白質は、塩分画、イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ハイドロキシアパタイト吸着クロマトグラフィーおよび疎水性相互作用クロマトグラフィーの様々な組み合わせ、または個々の適用によって細胞溶解物および抽出物から精製することができる。さらに、組換えhCEA蛋白質は、完全長hCEA蛋白質またはhCEAのポリペプチド断片に特異的なモノクローナル抗体またはポリクローナル抗体で作製した免疫親和性カラムを使用することによって、その他の細胞蛋白質から分離することができる。
【0056】
本発明の核酸は、ヒト細胞において蛋白質の効果的な発現をもたらすように設計された配列を含む発現カセットに組み込むことができる。
【0057】
該カセットは、コドンを最適化した完全長hCEA遺伝子と共に、プロモーターなど、それに操作可能に結合された転写および翻訳制御配列並びに終止配列を含有することが好ましい。好ましい実施形態では、該プロモーターはイントロンA配列(CMV)を含まないサイトメガロウイルスプロモーターであるが、当業者であれば、強力なイムノグロブリンなどの多数のその他の公知のプロモーター、またはその他の真核細胞遺伝子プロモーターを使用できることを認識するであろう。好ましい転写終結区は、ウシ成長ホルモン終結区であるが、その他の公知の転写終結区もまた使用することができる。CMV−BGH終結区の組み合わせは特に好ましい。
【0058】
本発明によれば、hCEAopt発現カセットは、ベクターに挿入される。該ベクターは、好ましくはアデノウイルスベクターであるが、プロモーターに結合した線状DNAまたはその他のベクター、たとえば、アデノ関連ウイルスまたは改変ワクチンウイルス、レトロウイルスまたはレンチウイルスベクターもまた使用することができる。
【0059】
選択したベクターがアデノウイルスの場合、ベクターがいわゆる第1世代アデノウイルスベクターであることが好ましい。これらのアデノウイルスベクターは、非機能性E1遺伝子領域を有することが特徴で、好ましくはアデノウイルスE1遺伝子領域が欠失していることが好ましい。いくつかの実施形態では、該発現カセットは、E1遺伝子が通常位置する位置に挿入される。さらに、これらのベクターは、場合によって非機能性E3領域を有するか、E3領域を欠失している。使用したアデノウイルスゲノムは、E1およびE3領域の両方が欠失していること(ΔE1ΔE3)が好ましい。該アデノウイルスは、ウイルスE1遺伝子を発現する公知の細胞系、たとえば、293細胞、またはPERC.6細胞、または一時的にもしくは安定して形質転換して外来蛋白質を発現する293またはPERC.6から得られた細胞系で増殖することができる。たとえば、テトラサイクリン制御プロモーター系などの遺伝子発現が制御された構築物を使用するとき、該細胞系は、制御系に関連した成分を発現することができる。このような細胞系の1例は、T−Rex−293であり、その他は当業者に公知である。
【0060】
アデノウイルスベクターの操作に便利なように、該アデノウイルスはシャトルプラスミド型であることが可能である。本発明はまた、プラスミド部分およびアデノウイルス部分を含むシャトルプラスミドベクターを対象としており、該アデノウイルス部分はE1が欠失し、場合によってE3も欠失したアデノウイルスゲノムを含み、コドン最適化ヒトCEAを含む発現カセットが挿入されている。好ましい実施形態では、アデノウイルスベクターが容易に除去され得るようにプラスミドのアデノウイルス部分に隣接して制限部位がある。該シャトルプラスミドは、原核細胞または真核細胞で複製することができる。
【0061】
本発明の好ましい実施形態では、発現カセットはpMRKAd5−HV0アデノウイルスプラスミド(本明細書に参照により組み込まれた、Emini他、WO02/22080参照)に挿入する。このプラスミドは、E1およびE3領域が欠失したAd5アデノウイルスゲノムを含む。pMRKAd5−HV0プラスミドの設計は、5’シス作用パッキング領域をE1遺伝子にさらに伸長し、ウイルスパッケージングを最適化するために重要であることが発見された要素をとりこみ、ウイルス増幅の増強をもたらすことによって、以前のアデノウイルスベクターより改良された。有利なことに、この増強されたアデノウイルスは、何度か継代増殖させた後でも遺伝子安定性を維持することができる。
【0062】
DNA構築物を調製および精製する分子生物学の標準的技術によって、本発明のアデノウイルス、シャトルプラスミドおよびDNA免疫原を調製することが可能である。
【0063】
本発明によって、ヒト細胞において高レベルで発現するようにコドンが最適化された本明細書で説明した合成cDNA分子(配列番号1)は、対応する野生型配列よりも効率的に発現することが測定された。驚くべきことに、コドンを最適化したhCEAのcDNAは、野生型配列よりも効率的にhCEAに対する寛容を打破する。さらに、本明細書では、hCEAoptはhCEAよりも免疫原性が高く、細胞性および体液性両方の免疫応答をより効果的に惹起することが示された。
【0064】
したがって、前述のベクターは、異常なCEA発現に関連した腺癌の発症の予防および/または既存の癌の治療のための免疫原性組成物およびワクチンに使用することができる。本発明のベクターは、うまく形質転換した宿主生物において外来性CEAを高濃度に発現させることに伴う問題を排除し、ワクチン開発および商品化を可能にする。このために、本発明の1態様は、配列番号2に記載されているヒトCEA蛋白質をコードするヌクレオチドの配列を含むコドン最適化合成核酸分子を含むワクチンベクターを哺乳類に投与することを含む癌の予防および治療方法である。
【0065】
前述の方法によれば、該ワクチンベクターはいかなる哺乳類の癌の治療または予防のためにも投与することができる。本発明の好ましい実施形態では、哺乳類はヒトである。
【0066】
さらに、当業者は説明した治療方法および予防方法で使用するための任意の形態のベクターを選択することができる。好ましくは、該ベクターはアデノウイルスベクターまたはプラスミドベクターである。本発明の好ましい実施形態では、該ベクターはアデノウイルスE1領域中に欠失を有し、及びアデノウイルスE1領域中に挿入部分を有するアデノウイルスゲノムを含むアデノウイルスベクターであり、前記挿入は、(a)ヒトCEA蛋白質をコードするコドン最適化合成ポリヌクレオチドと、(b)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む発現カセットを含む。
【0067】
本発明はさらに、E1領域中に欠失を有し、及びE1領域中に挿入部分を有するアデノウイルスゲノムを含むアデノウイルスワクチンベクターに関し、前記挿入は、(a)ヒトCEA蛋白質をコードするコドン最適化合成ポリヌクレオチドと、(b)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む発現カセットを含む。
【0068】
本発明の好ましい実施形態では、該アデノウイルスベクターはAd5ベクターである。
【0069】
本発明の他の好ましい実施形態では、該アデノウイルスベクターはAd6ベクターである。
【0070】
本発明のさらに他の好ましい実施形態では、該アデノウイルスベクターはAd24ベクターである。
【0071】
他の態様では、本発明は、プラスミド部分と発現カセット部分とを含むワクチンプラスミドに関し、該発現カセット部分は、(a)ヒトCEA蛋白質またはその変種をコードするコドン最適化合成ポリヌクレオチドと、(b)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む。
【0072】
本発明のある実施形態では、本明細書で開示した組換えアデノウイルスワクチンは、増強された免疫応答を誘導するため、プラスミドをベースとしたポリヌクレオチドワクチンによる様々な初期/追加免疫の組み合わせで使用される。この場合、この2種類のベクターが「初期および追加免疫」計画で投与される。たとえば、第1の種類のベクターを投与し、次に所与の時間を経た後、たとえば、2週間後、1ヶ月後、2ヶ月後、6ヶ月後またはその他の適切な間隔を空けて、第2の種類のベクターを投与する。好ましくは、該ベクターは同一のポリヌクレオチドまたはポリヌクレオチドの組み合わせをコードする発現カセットを有する。プラスミドDNAもまた使用する実施形態では、該ベクターは哺乳類細胞または昆虫細胞によって認識される1種または複数のプロモーターを含有することが好ましい。好ましい実施形態では、該プラスミドは強力なプロモーターを含有し、たとえば、CMVプロモーターであるが、それだけに限定されない。該合成ヒトCEA遺伝子または発現すべきその他の遺伝子は、このようなプロモーターに結合される。このようなプラスミドの例は、前述のような哺乳類発現プラスミドV1Jnsである(本明細書に参照により組み込まれた、J.Shiver他、DNA Vacchines、M.Liu他著、N.Y.Acad.Sci.,N.Y.,772:198〜208(1996))。
【0073】
前述のように、アデノウイルスベクターワクチンおよびプラスミドワクチンは、免疫応答を惹起する単一の治療計画の一部として脊椎動物に投与することができる。このために、本発明は、(a)(i)ヒトCEA蛋白質またはヒトCEA蛋白質変種をコードするコドン最適化合成ポリヌクレオチドと、(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む第1のベクターを哺乳類に導入すること、(b)予め決定された時間経過させること、および(c)(i)ヒトCEA蛋白質またはヒトCEA蛋白質変種をコードするコドン最適化合成ポリヌクレオチドと、(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む第2のベクターを該哺乳類に導入することを含む、前記哺乳類を癌から防御する方法に関する。
【0074】
前述の防御方法の1実施形態において、第1のベクターはプラスミドであり、第2のベクターはアデノウイルスベクターである。他の実施形態では、第1のベクターはアデノウイルスベクターであり、第2のベクターはプラスミドである。
【0075】
本発明はさらに、(a)(i)ヒトCEA蛋白質またはヒトCEA蛋白質変種をコードするコドン最適化合成ポリヌクレオチドと、(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む第1のベクターを哺乳類に導入すること、(b)予め決定された時間経過させること、および(c)(i)ヒトCEA蛋白質またはヒトCEA蛋白質変種をコードするコドン最適化合成ポリヌクレオチドと、(ii)該ポリヌクレオチドに操作可能に結合されたプロモーターとを含む第2のベクターを該哺乳類に導入することを含む、腺癌に罹患した哺乳類を治療する方法に関する。
【0076】
前述の治療方法の1実施形態において、第1のベクターはプラスミドであり、第2のベクターはアデノウイルスベクターである。他の実施形態では、第1のベクターはアデノウイルスベクターであり、第2のベクターはプラスミドである。
【0077】
ワクチン受容体に導入すべき発現可能なDNAまたは転写されたRNAの量は、使用したプロモーターの強度および発現遺伝子産物の免疫原性に部分的に左右される。一般的に、免疫学的または予防的に有効な用量、約1ngから100mg、好ましくは約10μgから300μgのプラスミドワクチンベクターを直接筋肉組織に投与する。組換えアデノウイルスの有効量は、粒子約106個から1012個で、好ましくは約107個から1011個である。皮下注射、皮内注射、皮膚からの圧入およびその他の投与様式、たとえば、腹腔内、静脈内または吸入送達もまた企図される。追加免疫ワクチン接種を行うこともまた、企図される。本発明のワクチンの非経口的導入と同時に、またはその後で、アジュバント、たとえば、インターロイキン12蛋白質を非経口的に、たとえば、静脈内、筋肉内、皮下またはその他の投与手段で投与することもまた、有利である。
【0078】
本発明のワクチンは、裸であること、すなわち、いかなる蛋白質、アジュバントまたは受容対象の免疫系に影響を与えるその他の薬剤を伴わないことが可能である。この場合、ワクチンベクターは生理学的に許容される溶液、たとえば、滅菌生理食塩水または滅菌緩衝生理食塩水に入れることが望ましい。あるいは、本発明のワクチンまたは免疫原組成物と共に免疫賦活剤、たとえば、アジュバント、サイトカイン、蛋白質、またはその他の担体を投与することが有利であり得る。したがって、本発明は、本発明の組成物および方法と組み合わせてこのような免疫賦活剤を使用することを含む。本明細書で使用した免疫賦活剤とは、外来抗原に対する免疫応答(抗体および/または細胞性)を増強または助長する本質的に任意の物質である。前記免疫賦活剤は、DNAまたは蛋白質の形態で投与することができる。GM−CSF、IFNα、破傷風毒素、IL12、B7.1、LFA−3およびICAM−1を含むが、それだけに限定されない様々な免疫賦活剤はいずれも、本発明のワクチンおよび免疫組成物と組み合わせて使用することができる。前記免疫賦活剤は、当業界で周知である。
【0079】
DNAの細胞による摂取を助ける薬剤、たとえば、それだけに限定されないがカルシウムイオンはまた、使用することができる。これらの薬剤は一般的に、トランスフェクションを容易にする試薬および薬剤として許容される担体のことである。当業者であれば、特定の免疫賦活剤または薬剤として許容される担体並びに適切な時間および投与様式を決定することができる。
【0080】
本明細書で記載した刊行物は全て、本発明と関連づけて使用することが可能な方法および材料を記載し、開示するために参照により組み込まれる。本発明が先行技術の効力によってその開示が先行する資格を有していないことを許容したと解釈すべきでない。
【0081】
添付した図面を参考にして、本発明の好ましい実施形態を説明したが、本発明はそれらの厳密な実施形態に限定されるのではなく、添付の特許請求の範囲で定義した本発明の範囲または精神を逸脱することなく、当業者が様々な変更および修正を実行できることを理解されたい。
【0082】
以下の実施例は本発明を例示するものであって、限定するものではない。
【実施例1】
【0083】
コドン最適化ヒトCEA配列
完全なhCEAoptコーディング配列を合成して、BIONEXUS(Oakland、CA)によって構築した。5’末端に最適化したKozak配列を有するhCEAoptcDNAは、PCRによって構築したオリゴヌクレオチドを使用して構築した。構築したcDNAは、pCR−Bluntベクター(Invitrogen、Carlsbad、CA)に挿入して、pCR−hCEAoptを得た。hCEAoptcDNAの強度は、両鎖を配列決定することによって測定した。
【実施例2】
【0084】
プラスミド構築物およびアデノウイルスベクター
pVIJ/hCEAopt:プラスミドpCR−hCEAoptは、EcoRIで37℃で1時間消化した。得られた2156bp挿入物を精製して、プラスミドpV1JnsBのEcoRI部位にクローニングした(Montgomery他、DNA Cell Biol.,12(9):777〜83(1993))。
【0085】
pVlJ/hCEA:プラスミドpCI/hCEA(Song他、DNAをベースとしたワクチン接種および非ウイルス性サイトカイン遺伝子移入の組み合わせによるTヘルパー1対Tヘルパー2活性の制御および腫瘍免疫の増強(Regulation of T−helper−1 versus T−helper−2 activity and enhancement of tumour immunity by combined DNA−based vaccination and nonviral cytokine gene transfer.)Gene Therapy 7:481〜492(2000))は、EcoRIで消化した。得られた2109bp挿入物は、プラスミドpV1JnsA(Montgomery他、前述)のEcoRI部位にクローニングした。
【0086】
Ad5/hCEAopt:プラスミドpCR−hCEAoptは、EcoRIで消化した。得られた2156bp挿入物は、精製してpolyMRK−Ad5シャトルプラスミドのEcoRIにクローニングした(本明細書に参照により組み込まれた、Emini他、WO02/22080参照)。
【0087】
Ad5/CEA:Ad5ベクターを生成するためのシャトルプラスミドpMRK−hCEAは、プラスミドpDelta sp1B/hCEAをSspIおよびEcoRVで消化することによって得られた。次に、該9.52kb断片は、プラスミドポリMRKから得られた1272bpのBglII−BamHIで制限した、クレノウ処理生成物と連結した。hCEAの発現カセットおよびE1隣接Ad5領域を含有するpMRK−hCEAおよびpMRK−hCEAoptのPacI/StuI断片は、BJ5183 E.コリ細胞においてClaI直線化プラスミドpAd5に結合し直した。得られたプラスミドはそれぞれ、pAd5−hCEAおよびpAd5−hCEAoptであった。両プラスミドをPacIで切断して、Ad逆方向末端反復(ITR)を遊離し、PerC−6細胞にトランスフェクトした。Ad5ベクター増幅は、一連の手順によって実施した。MRKAd5/hCEAおよびMRKAd5/hCEAoptは、標準的なCsCl勾配精製法によって精製し、A105緩衝液(5mM Tris−Cl pH8.0、MgCl2 1mM、NaCl 75mM、スクロース 5%、0.005 Tween 20)によって集中的に透析した。
【実施例3】
【0088】
CEAの発現および検出
プラスミドおよびAdベクターによるhCEAの発現は、ウエスタンブロット分析によってモニターした。プラスミドは、リポフェクタミン2000(Life Technologies、Carlsbad、CA)によってHela細胞またはPerC.6細胞にトランスフェクトした。PerC.6のアデノウイルス感染は、血清を含まない培地中で、37℃で30分間実施し、その後新鮮な培地を添加した。48時間インキュベートした後、全細胞溶解物および培養上清を収集した。細胞溶解物中に存在するCEA蛋白質は、ウサギポリクローナル抗血清を使用したウエスタンブロット分析によって検出した。該蛋白質は、180kDaから200kDaのバンドとして検出された。分泌型CEAは、Direct Elisa CEAキット(DBC−Diagnostics Biochem Canada Inc.,Ontario、Canada)を使用して、細胞上清中および注射したマウス(注射後3日目)の末梢血中に検出された。
【実施例4】
【0089】
マウスの免疫
雌のC57BL/6マウス(H−2b)は、Charles River(Lecco、Italy)から購入した。CEA.tgマウス(H−2b)は、J.Primus(Vanderblit University)から恵与され、標準的条件で維持した。既に記載されたように(Rizzuto他、Proc.Natl.Acad.Sci.U.S.A.96(11):6417〜22(1999))、プラスミドDNA 50マイクログラムをマウスの大腿4頭筋に50μlの用量で電気注射した。Ad注射は、50μlの用量でマウスの大腿4頭筋に実施した。体液性および細胞性免疫応答は、指定した時点で分析した。
【実施例5】
【0090】
hCEAのコドン最適化cDNAはhCEA発現を有意に増加させた。
【0091】
ヒトCEA(hCEAopt)の合成遺伝子は、各アミノ酸(以後aa)残基についてヒトに好ましい(ヒト化)コドンを組み込むために設計した。コドン最適化cDNAは、元々のクローンに対して76.8%のヌクレオチド同一性を維持するように改変された(図1参照)。該コドン最適化cDNAは、pV1Jベクター(Montgomery他、前述)のKozak最適化配列(5’−GCCGCCACC−3’、配列番号13)の前、ヒトサイトメガロウイルス(CMV)/イントロンAプロモーターおよびウシ成長ホルモン(BGH)終止シグナルの制御下に配置してクローニングした。該構築物は、pV1J/hCEAopt(実施例2参照)と命名した。さらに、アデノウイルス5型ベクターは、CMV/イントロンAプロモーターおよびBGH終止シグナルの隣接したhCEAopt配列を有するように構築した(Ad5/hCEAopt)。比較のために、同等のプラスミドおよびV5ベクターは、pV1J/hCEAおよびAd5/hCEAを生じる野生型hCEA配列を有するように構築した。コドン最適化cDNAを含有するものと同様に、これらのベクターは、CMV/int Aプロモーターおよびウシ成長ホルモン(BGH)終止シグナルの制御下に野生型遺伝子を有する。
【0092】
プラスミドpV1J/hCEAoptでトランスフェクトしたHela細胞のウエスタンブロット分析では、pV1J/hCEA構築物でトランスフェクトした細胞で検出された大きさと区別できない大分子量(180kDaから200lDa)の蛋白質が生じた。同様に、Ad5/hCEAまたはAd5H7h/hCEAoptで感染させたPerC−6細胞溶解物で検出される蛋白質の大きさには、明らかな差は検出できなかった(データは示さなかった)。
【0093】
hCEA発現に対するhCEAopt発現の効率を比較するために、C57BL/6マウス10匹の群の大腿4頭筋に1x107から1x104pfuの範囲の様々な用量のAd5/hCEAoptを注射した。注射して3日後に、CEA蛋白質濃度を測定して、同用量のAd5/hCEAを注射した対照群の濃度と比較した。Ad5/hCEAを注射したマウスと比較してAd/hCEAopt 1x107pfu(48.2μg/l)の注射では、hCEA濃度の相乗平均値の6倍の増加が認められたが、一方、同ウイルスを1x106pfu(10.9μg/l)で注射すると蛋白質濃度の10倍の増加が認められた(図2A)。対照的に、低用量のAd5/hCEAoptを注射しても、Ad5/hCEAと比較して循環CEA濃度の実質的な増加は生じなかった。pV1J/hCEAと比較して、プラスミドpV1J/hCEAop、25μgまたは50μgを電気注射すると、たとえ低い範囲であっても、CEA蛋白質濃度の増強がまた、示された(図2B)。したがって、これらの結果から、利用した遺伝子輸送媒体とは関係なく、コドン最適化cDNAは対応する野生型配列よりも効果的に発現することが示された。
【実施例6】
【0094】
IFN−γELISPOT測定法
滅菌PBSで2.5μg/mlに希釈した精製ラット抗マウスIFN−γ(IgG1、R4−6A2クローン、Pharmingen、San Diego、CA)100μl/ウェルで、96ウェルMAIPプレート(Millipore、Bedford、MA)をコーティングした。PBSで洗浄後、プレートのブロックはR10培地200μl/ウェルで37℃で2時間実施した。
【0095】
脾細胞は、安楽死させたマウスから滅菌的に脾臓を取り出すことによって得た。脾臓破砕は、金属格子上で切断した脾臓をすりつぶすことによって実施した。細胞ペレットに0.1XPBS 1mlを添加し15秒以上激しく撹拌して、浸透圧溶解することによって赤血球細胞を除去した。次に、2XPBS 1mlを添加し、1XPBSで体積を4mlにした。室温で1200rpmで10分間遠心することによって細胞をペレットにし、該ペレットをR10培地1mlに再懸濁した。生細胞をチュルク染色を使用して計数した。
【0096】
脾細胞は、5x105および2x105細胞/ウェルで2連で播種し、各ペプチドの懸濁液1μg/mlと共に37℃で20分間インキュベートした。各マウスの陽性内部対照としてコンカナバリンA(ConA)を5μg/mlで使用した。PBS、0.05%Tween20で洗浄した後、測定用緩衝液で1:2500に希釈したビオチン結合ラット抗マウスIFNγ(RatIgG1、XMG1.2クローン、PharMingen)50μg/ウェルをプレートに入れて、4℃でO/Nインキュベートした。集中的に洗浄した後、プレートにNBT/B−CIP(Pierce Biotechnoogy Inc.,Rockford、IL)を添加して、スポットがはっきりと視認できるほどになるまで発色させた。蒸留水で徹底的にプレートを洗浄することによって反応を停止させた。プレートを空気乾燥して、次にスポットは自動化ELISPOT読み取り機を使用して計数した。
【実施例7】
【0097】
細胞内サイトカイン染色
RPMI 10%FCS 1mlに溶かした100万個から200万個のマウス脾細胞またはPBMCを、ペプチド収集物(各ペプチドの最終濃度5μgから6μg/ml)およびブレフェルジンA(1μg/ml、BD Pharmingen カタログ番号55028/2300kk)と共に、37℃で5%CO2で12時間から16時間インキュベートした。次に、細胞をFACS緩衝液(PBS 1% FBS、NaN3 0.01%)で洗浄して、精製抗マウスCD16/CD32Fcブロック(BD Pharmingen カタログ番号553142)と共に4℃で15分間インキュベートした。次に、細胞を洗浄して、表面抗体、CD4−PE結合抗マウス(BD Pharmingen、カタログ番号553049)、PercP CD8結合抗マウス(BD Pharmingen カタログ番号553036)およびAPC−結合抗マウスCD3e(BD Pharmingen カタログ番号553066)によって暗所で室温で30分間染色した。洗浄後、細胞を固定し、Cytofix−Cytoperm溶液(BD Pharmingen カタログ番号55028/2300kk)で暗所において4℃で20分間透過させた。PermWash溶液(BD Pharmingen カタログ番号55028/2300kk)で洗浄した後、細胞をIFNγ−FITC抗体(BD Pharmingen)と共にインキュベートした。次に、細胞を洗浄して、ホルムアルデヒドの1%PBS溶液で固定して、FACS−Caliburフローサイトメトリーで、CellQuestソフトウェア(Becton Dickinson、San Jose、CA)を使用して分析した。
【実施例8】
【0098】
CEA特異的T細胞を直接列挙するためのエピトープ含有ペプチドの同定および特徴付け
マウスにおいて、CEAに対する遺伝子ワクチンによって惹起された免疫応答をよりよく特徴付けるために、ELISPOT分析をC57BL/6マウスおよびBALB/cマウスについて実施し、CD4+およびCD8+CEA特異的エピトープを同定した。このために、完全な蛋白質を網羅する個々のペプチドに対する応答の同定に利用できる強く免疫されたマウスの作製について、異なる免疫様式を比較した。プラスミドDNA初期免疫−Ad追加免疫の様式を利用することによって、高濃度の細胞性免疫をウイルス抗原および細菌抗原に対して誘導することができることを示した最近の報告を考慮して、同様の免疫方法を本研究に使用した。異なる計画、i)Ad/hCEAの1x109vpを2回投与(Ad/Ad)、ii)プラスミドp1J/hCEAを2回投与(DNA/DNA)およびiii)プラスミドDNA1回投与した後Ad/hCEAを投与(DNA/Ad)でマウスを筋肉内で免疫した。免疫間隔は、2週間とした。
【0099】
異なる免疫計画によって惹起された細胞性免疫は、追加免疫2週間後にELISPOTによって測定した。異なるワクチン接種計画の免疫原効果を比較するために、11個のaaが重複し、aa497から703を網羅した15マーペプチドの収集物(プールD)を使用して、脾細胞の抗原特異的サイトカイン分泌を刺激した。SFCの高い相乗平均値によって示される最強の応答は、DNA/Adを注射した群のC57BL/6およびBALB/cマウスで認められた(図4)。したがって、この計画を利用して免疫応答をさらに分析した。
【0100】
該免疫応答が、完全なCEA蛋白質に対して等しく分布するかどうかを測定するために、免疫したC57BL/6およびBALB/cマウスの脾細胞を、完全な蛋白質配列を集合的に包含する1個から4個の15マーペプチドの収集物によってインビトロで刺激した。各収集物は、11個の残基によって重複するアミノ酸長15個のペプチドから構成された。凍結乾燥したhCEAペプチドは、Bio−Synthesis(Lewisville、TX)から購入し、DMSOに40mg/mlで再懸濁した。プールDに加えて、プールA(aa1から147)、B(aa137から237)およびC(aa317から507)を本研究で使用した。最終濃度は以下の通りである。プールA=1.2mg/ml、プールB=0.89mg/ml、プールC=0.89mg/mlmプールD=0.8mg/ml。ペプチドは−80℃で保存した。
【0101】
C57BL/6マウスにおいてDNA/Adワクチン接種計画によって惹起された免疫応答は、主に蛋白質のC末端領域に偏っていた(図5A参照)。有意なSFC値はペプチドプールCおよびDで得られ(それぞれの相乗平均値、170および244SFC/106脾細胞)、一方、プールAおよびBではかなり低い値が得られた(それぞれ、10および27SFC/106脾細胞)。対照的に、BALB/cマウスの免疫応答は、プールBで最も高いが(相乗平均値、1236SFC/106脾細胞)、プールA、CおよびDでは有意に高いSFC値が示された(それぞれ、93、364および344)。関連のないペプチドのプールに対する応答は、いずれの群のマウスでも示されなかった(データは示さなかった)。
【0102】
応答を惹起するペプチド収集物に存在する個々のペプチドを同定するために、DNA/Adワクチン接種計画によって免疫した4種類のマウスの脾細胞を、有意な免疫応答が認められたプールを含む個々のペプチドそれぞれに対するIFNγ−ELISPOT測定法で分析した。C57BL/6マウスの脾細胞は、プールCおよびDに含まれるペプチド80から173に対して調べた。BALB/cマウスの脾細胞は、プールB、CおよびDを含むペプチド35から173に対して調べた。C57BL/6マウスにおけるCEA特異的応答は、重複した配列を有する15マーペプチドの4種類の組み合わせに位置づけた(aa431から435および425から439;529から543および533から547;565から579および569から593;613から627および617から631)(図6A)。BALB/cマウスにおけるCEAに対する免疫応答は、22個のペプチドに位置づけられ、そのウチの17個は配列が重複している(aa213から227および213および227;229から243および233から247;409から423および413から427;421から435および425から439;565から579および569から583;573から587;613から627および617から631並びに621から635および625から639;637から651および641から655)(図6B)。
【0103】
選択したペプチド内に含有されるエピトープのT細胞特異性を確認するために、注射したマウスの脾細胞についてIFNγ細胞内染色測定法を実施した。得られた結果を図7に示す。該データは、Tリンパ球の循環濃度を定量するために使用することができるC57BL/6およびBALB/cマウス両方について、CD8+およびCD4+特異的エピトープが同定されたことを示す。
【実施例9】
【0104】
コドンを最適化したhCEA cDNAは、hCEAトランスジェニックマウスにおける寛容を打破する
コドンを最適化したhCEAのcDNAの増強された免疫原性がヒトCEAに対する寛容を効果的に打破するかどうかを測定するために、hCEAトランスジェニックマウスを野生型またはコドン最適化hCEA配列のいずれかを有するベクターで免疫した。これらのトランスジェニックマウスは、完全なヒトCEA遺伝子および隣接配列を有し、盲腸および結腸においてhCEA蛋白質を発現する。したがって、このマウス系は、この腫瘍の自己抗原を対象とする免疫治療計画の安全性および効率を研究するための有用なモデルである(Clarke他、「免疫治療のモデルとしてのヒトCEA遺伝子組換えマウス(Mice transgenic for human CEA as a model for immunotherapy)」Cancer Res.1469〜77(1998))。
【0105】
第1の試験として、トランスジェニックマウス10匹のうち5匹にプラスミドDNA50μgを4回電気注射し、次いで最終的にアデノウイルス1x1010ppを注射した。hCEAに対する免疫応答は、注射したマウス4匹から収集した脾細胞についてIFNγ−ELISPOT測定を行うことによって分析した。hCEAに対する免疫応答は、hCEAoptcDNAで免疫したマウスの脾細胞のみで検出された(図8参照)。該免疫応答は、ペプチド143およびプールDで検出されたので、免疫によってC末端エピトープに対する有意なCD8+応答が惹起されたことが示唆された。
【0106】
hCEAのコドン最適化cDNAの増強された免疫原性はまた、アデノウイルスベクター1x1010ppを2週間間隔で2回注射したトランスジェニックマウスで試験した。CEA特異的免疫応答は、4種類の免疫マウスから収集したPBMCをIFNγ細胞内染色することによって測定した。hCEAに対する免疫応答は、Ad/hCEAoptで免疫したマウスのみに検出された(図9)。DNAおよびAdコホートで認められたように、CD8+T細胞の誘導はペプチドプールDで検出されたが、有意なCD8+応答はまた、ペプチドプールAで示された。したがって、これらの結果は、hCEAのコドン最適化cDNAは、野生型配列よりも免疫原性が高く、hCEAに対する寛容を効率的に打破することを示す。
【実施例10】
【0107】
抗体検出および力価測定
抗体力価測定用の血清は、眼窩後方出血によって得られた。ELISAプレート(Nunc maxisorpTM)は、CEA蛋白質100ng/ウェルでコーティングして(高純度CEA、Fitzgerald Industires International Inc.,Concord MA)でコーティングして、コーティング緩衝液(50mM NaHCO3、pH9.4)で希釈して、4℃でO/Nインキュベートした。次に、プレートを5%BSAを含有するPBSで37℃で1時間ブロックした。マウス血清は、PBS5%BSA(血清転換率の評価には1/50の希釈度、力価測定には1:10から1:312150までの希釈度)で希釈した。予備免疫血清は、背景として使用した。希釈した血清は、4℃でO/Nインキュベートした。洗浄は、PBS1%BSA、0.05%Tween20で実施した。2次抗体(ヤギ抗マウス、IgGペルオキシダーゼ、Sigma)は、PBS、5%BSAで1/2000に希釈して、室温で2時間から3時間、振盪機上でインキュベートした。洗浄後、プレートをTMB基質100μl/ウェル(Pierce Biotechnology、Inc.,Rockford、IL)で発色させた。該反応は、1M H2SO4溶液25μl/ウェルで停止し、プレートは450nm/620nmで読み取った。抗CEA血清力価は、同じ希釈度の自己由来予備免疫血清の吸収より少なくとも3倍大きな吸収を生じる血清の限界希釈の逆数として算出した。
【実施例11】
【0108】
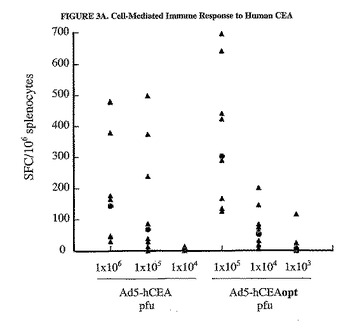
hCEAoptの高い免疫原性
野生型およびコドン最適化CEA発現ベクターによって誘導されるインビボでの免疫応答を調べるために、C57BL/6マウスを1x105から1x103pfuの範囲の様々な用量のAd5/hCEAoptを筋肉内注射して免疫した。マウスに3週間間隔で2回注射を行った。2回目の免疫を行ってから2週間後に脾細胞を各マウスから単離した。アデノウイルスによる免疫によって生じるIFNγ分泌型CEA特異的CD8T細胞前駆体発生頻度を定量するために、H−2b制限T細胞エピトープCGIQNSVSA(配列番号14、以下参照)についてELISPOT測定法を使用した。1x104pfuで免疫すると、測定可能な免疫応答を惹起し、CGIONSVSAエピトープ(配列番号14)に特異的な53個のIFNγスポット形成細胞(SFC、相乗平均値)が生じたが、一方1x103pfuの注射ではごくわずかなSFC値しか惹起しなかった(図3A)。Ad/hCEAopt 1x105pfuで免疫した群において、SFCは302個まで増加した。対照的に、1x106pfuの用量で免疫したマウスの群において、SFCを168まで増加させる有意なCD8T−細胞前駆体頻度を惹起するために、Ad5/hCEAは少なくとも1x105pfu必要であった。ペプチド特異的IFNγSFCは、Ad5免疫マウスでは検出されなかった(データは示さなかった)。
【0109】
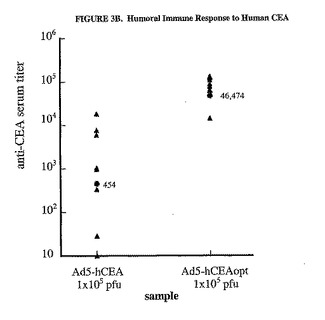
それぞれのhCEAアデノウイルスベクター1x105pfuで免疫したマウスの血清は、基質として精製ヒトCEA蛋白質を使用してELISAで試験した。Ad5/hCEAoptで免疫したマウスのCEA特異的抗体の力価は、免疫したマウス全てにおいて検出され、Ad力価の相乗平均値は46474であった。対照的に、Ad5/hCEA免疫群は、CEA特異的抗体の約100倍低い相乗平均力価を示した(454)。したがって、これらの結果は、CEAのコドン最適化cDNAは細胞性および体液性免疫応答をより効率的に惹起することを示す。
【実施例12】
【0110】
統計解析
指示した場合は、結果はスチューデントt検定によって解析した。p値<0.05は、有意とみなされた。
【図面の簡単な説明】
【0111】
【図1】野生型ヒトCEAcDNA(配列番号3)およびコドン最適化クローン(hCEAopt、配列番号1)のヌクレオチド配列を示す図である。推定アミノ酸配列を上部に示す(配列番号2)。合成コドン最適化cDNAの置換ヌクレオチドをhCEAcDNA配列の下に示す。実施例2参照。
【図1A】野生型ヒトCEAcDNA(配列番号3)およびコドン最適化クローン(hCEAopt、配列番号1)のヌクレオチド配列を示す図である。推定アミノ酸配列を上部に示す(配列番号2)。合成コドン最適化cDNAの置換ヌクレオチドをhCEAcDNA配列の下に示す。実施例2参照。
【図1B】野生型ヒトCEAcDNA(配列番号3)およびコドン最適化クローン(hCEAopt、配列番号1)のヌクレオチド配列を示す図である。推定アミノ酸配列を上部に示す(配列番号2)。合成コドン最適化cDNAの置換ヌクレオチドをhCEAcDNA配列の下に示す。実施例2参照。
【図2A】注射したマウスにおけるhCEAの発現を示した図である。C57BL/6マウス10匹の群において、様々な用量のMRKAd5/hCEAおよびMRKAd5/hCEAopt(パネルA)または25マイクログラムもしくは50マイクログラムのプラスミドpV1J/hCEAおよびpV1J/hCEAopt(パネルB)のいずれかを大腿4頭筋に注射した。注射して3日後に血液試料を収集し、CEA濃度を測定した。塗りつぶされた三角形は個々のマウスのCEA測定値を表す。相乗平均値もまた示す(塗りつぶされた丸)。
【図2B】注射したマウスにおけるhCEAの発現を示した図である。C57BL/6マウス10匹の群において、様々な用量のMRKAd5/hCEAおよびMRKAd5/hCEAopt(パネルA)または25マイクログラムもしくは50マイクログラムのプラスミドpV1J/hCEAおよびpV1J/hCEAopt(パネルB)のいずれかを大腿4頭筋に注射した。注射して3日後に血液試料を収集し、CEA濃度を測定した。塗りつぶされた三角形は個々のマウスのCEA測定値を表す。相乗平均値もまた示す(塗りつぶされた丸)。
【図3A】コドン最適化がヒトCEAに対する免疫応答を増強することを示した図である。C57BL/6マウス8匹の群において、様々な用量のMRKAd5−hCEAおよびMRKAd5−hCEAoptのいずれかを大腿4頭筋に注射した。ウイルス注射は、0日目と21日目に実施した。パネルA。追加免疫注射の2週間後に、aa569から583を含みCD8+エピトープを含むペプチド143を使用したELISPOTアッセイで、個々のマウスの脾細胞でhCEAに特異的なCD8+IFNγ分泌型T細胞の数を測定した(塗りつぶされた三角形)。2種類の異なる量の脾細胞(2.5x105および5x105)で、脾細胞のそれぞれの試験料について2回反復した。平均値は、ペプチド無しで測定した背景濃度を差し引くことによって算出し(一般的に10SFC/106全脾細胞)、結果はSFCの数/106全脾細胞として表した。個々のマウスの値(塗りつぶされた三角形)および相乗平均値(塗りつぶされた丸)を示した。パネルB。個々のマウスの血清中の抗CEA抗体力価(塗りつぶされた三角形)は、追加免疫10日後の血清試料を使用して測定した。相乗平均力価(GMT)(塗りつぶされた丸)もまた示す。Ad/hCEAoptはAd/hCEAと有意に異なる。
【図3B】コドン最適化がヒトCEAに対する免疫応答を増強することを示した図である。C57BL/6マウス8匹の群において、様々な用量のMRKAd5−hCEAおよびMRKAd5−hCEAoptのいずれかを大腿4頭筋に注射した。ウイルス注射は、0日目と21日目に実施した。パネルA。追加免疫注射の2週間後に、aa569から583を含みCD8+エピトープを含むペプチド143を使用したELISPOTアッセイで、個々のマウスの脾細胞でhCEAに特異的なCD8+IFNγ分泌型T細胞の数を測定した(塗りつぶされた三角形)。2種類の異なる量の脾細胞(2.5x105および5x105)で、脾細胞のそれぞれの試験料について2回反復した。平均値は、ペプチド無しで測定した背景濃度を差し引くことによって算出し(一般的に10SFC/106全脾細胞)、結果はSFCの数/106全脾細胞として表した。個々のマウスの値(塗りつぶされた三角形)および相乗平均値(塗りつぶされた丸)を示した。パネルB。個々のマウスの血清中の抗CEA抗体力価(塗りつぶされた三角形)は、追加免疫10日後の血清試料を使用して測定した。相乗平均力価(GMT)(塗りつぶされた丸)もまた示す。Ad/hCEAoptはAd/hCEAと有意に異なる。
【図4A】異なる免疫計画の比較を示した図である。プラスミドpV1J/hCEA(50μg/大腿4頭筋に電気注射した用量)およびMRKAd5/hCEA(1x109pp/用量)の様々な組み合わせで、C57BL/6(A)またはBALB/c(B)マウスの群を免疫した。個々のマウスの脾細胞におけるIFNγ分泌型T細胞の数は、材料および方法および図3の凡例で説明したようにaa497から703(プールD)に亘るペプチドのプールを使用して測定した。相乗平均値もまた示す(塗りつぶされた丸)。D/DおよびD/Aは、C57BL/6マウスのAd/Ad群と有意に異なる。3群は全て、BALB/cマウスにおいて有意に異なる。
【図4B】異なる免疫計画の比較を示した図である。プラスミドpV1J/hCEA(50μg/大腿4頭筋に電気注射した用量)およびMRKAd5/hCEA(1x109pp/用量)の様々な組み合わせで、C57BL/6(A)またはBALB/c(B)マウスの群を免疫した。個々のマウスの脾細胞におけるIFNγ分泌型T細胞の数は、材料および方法および図3の凡例で説明したようにaa497から703(プールD)に亘るペプチドのプールを使用して測定した。相乗平均値もまた示す(塗りつぶされた丸)。D/DおよびD/Aは、C57BL/6マウスのAd/Ad群と有意に異なる。3群は全て、BALB/cマウスにおいて有意に異なる。
【図5A】hCEA蛋白質の選択領域に対するT細胞応答のマッピングの結果を示した図である。C57BL/6(パネルA)またはBALB/c(パネルB)マウスの群は、プラスミドpV1J/hCEAで免疫し、3週間後にAd/hCEA 1x109で追加免疫した。個々のマウスの脾細胞におけるIFNγ分泌型T細胞の数は、材料および方法および図3の凡例で説明したように、完全な蛋白質を網羅したペプチドのプールを使用して追加免疫の2週間後に測定した。相乗平均値もまた示す(塗りつぶされた丸)。
【図5B】hCEA蛋白質の選択領域に対するT細胞応答のマッピングの結果を示した図である。C57BL/6(パネルA)またはBALB/c(パネルB)マウスの群は、プラスミドpV1J/hCEAで免疫し、3週間後にAd/hCEA 1x109で追加免疫した。個々のマウスの脾細胞におけるIFNγ分泌型T細胞の数は、材料および方法および図3の凡例で説明したように、完全な蛋白質を網羅したペプチドのプールを使用して追加免疫の2週間後に測定した。相乗平均値もまた示す(塗りつぶされた丸)。
【図6A】hCEAの免疫応答ペプチドの同定を示した図である。4匹の免疫したC57/BL/6(パネルA)またはBALB/c(パネルB)マウスから収集した脾細胞の指定したペプチドそれぞれに対するIFNγ分泌について、ELISPOT測定法を使用して測定した(実施例8参照)。
【図6B】hCEAの免疫応答ペプチドの同定を示した図である。4匹の免疫したC57/BL/6(パネルA)またはBALB/c(パネルB)マウスから収集した脾細胞の指定したペプチドそれぞれに対するIFNγ分泌について、ELISPOT測定法を使用して測定した(実施例8参照)。
【図7】C57BL/6マウス(パネルA)およびBALB/cマウス(パネルB)におけるCEAのペプチドを含有するエピトープの配列を示した図である(実施例xx参照)。右に挙げたのは、IFNγ産生CD8+(CD4+)CD3+細胞の割合である。
【図8】実施例9で説明したような免疫したCEAトランスジェニックマウスのIFNγ−ELISPOT測定法から得られた結果を示した図である。マウスは、1週間間隔でプラスミドDNAを4回電気注射し、さらにアデノウイルス注射を1回行って免疫した。各免疫原について、3回注射したマウスから収集した脾細胞でデータを採った。CD8特異的応答は、ペプチド143を使用して測定した。
【図9】免疫したCEA.tgマウスのIFNγ細胞内染色を示した図である。マウスは、2週間間隔でアデノウイルス1x1010vpを2回注射して免疫した。3回注射したマウスから収集した脾細胞で得られたデータを示す。右に挙げたのは、CD8+またはCD4+細胞の割合である。
【図10A】パネルAは、配列番号15で表したアンカードメインを切断したCEA変種をコードするコドン最適化DNA分子の例を示した図である。hCEAopt−ΔADの対応するアミノ酸配列をパネルBに示す(配列番号16)。
【図10B】パネルAは、配列番号15で表したアンカードメインを切断したCEA変種をコードするコドン最適化DNA分子の例を示した図である。hCEAopt−ΔADの対応するアミノ酸配列をパネルBに示す(配列番号16)。
【特許請求の範囲】
【請求項1】
配列番号2に記載されているヒト癌胎児抗原(CEA)蛋白質をコードするヌクレオチドの配列を含み、ヒト細胞において高レベルで発現するためにコドンが最適化されている合成核酸分子。
【請求項2】
前記核酸がDNAである、請求項1に記載の合成核酸分子。
【請求項3】
前記核酸がmRNAである、請求項1に記載の合成核酸分子。
【請求項4】
前記核酸がcDNAである、請求項1に記載の合成核酸分子。
【請求項5】
前記ヌクレオチドの配列が配列番号1で表したヌクレオチドの配列を含む、請求項1に記載の合成核酸分子。
【請求項6】
請求項1に記載の核酸分子を含むベクター。
【請求項7】
請求項6に記載のベクターを含む宿主細胞。
【請求項8】
(a)請求項1に記載の核酸を含むベクターを適切な宿主細胞中に導入すること、および
(b)前記ヒトCEA蛋白質の発現を可能にする条件下で前記宿主細胞を培養すること、を含む、組換え宿主細胞中でヒト癌胎児抗原(CEA)蛋白質を発現する方法。
【請求項9】
配列番号2で表したヒト癌胎児抗原(hCEA)蛋白質または配列番号16で表したCEA蛋白質変種をコードするヌクレオチドの配列を含むコドン最適化合成核酸分子を含むワクチンベクターを哺乳類に投与することを含む、癌の予防または治療方法。
【請求項10】
前記哺乳類がヒトである、請求項9に記載の方法。
【請求項11】
前記ベクターがアデノウイルスベクターまたはプラスミドベクターである、請求項9に記載の方法。
【請求項12】
前記ベクターが、E1領域中に欠失を有し、及び前記E1領域中に挿入部分を有するアデノウイルスゲノムを含むアデノウイルスベクターであり、
前記挿入部分が、
(a)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(b)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む発現カセットを含む、請求項9に記載の方法。
【請求項13】
前記ベクターが、
プラスミド部分と、
(a)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチド、および
(b)該ポリヌクレオチドに操作可能に結合されたプロモーターを含む発現カセットと、
を含むプラスミドワクチンベクターである、請求項9に記載の方法。
【請求項14】
E1領域中に欠失を有し、及び前記E1領域中に挿入部分を有するアデノウイルスゲノムを含み、
前記挿入部分が、
(a)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(b)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む発現カセットを含む、アデノウイルスワクチンベクター。
【請求項15】
Ad5ベクターである、請求項14に記載のアデノウイルスベクター。
【請求項16】
Ad6ベクターである、請求項14に記載のアデノウイルスベクター。
【請求項17】
Ad24ベクターである、請求項14に記載のアデノウイルスベクター。
【請求項18】
プラスミド部分と発現カセット部分を含み、
前記発現カセット部分が、
(a)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(b)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む、ワクチンプラスミド。
【請求項19】
(a)(i)ヒト癌胎児抗原(CEA)蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(ii)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む第1のベクターを哺乳類に導入すること、
(b)予め決定された時間経過させること、および
(c)(i)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(ii)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む第2のベクターを前記哺乳類に導入すること、
を含む、前記哺乳類を癌から防御する方法。
【請求項20】
前記第1のベクターがプラスミドであり、前記第2のベクターがアデノウイルスベクターである、請求項19に記載の方法。
【請求項21】
前記第1のベクターがアデノウイルスベクターであり、前記第2のベクターがプラスミドである、請求項19に記載の方法。
【請求項22】
(a)(i)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(ii)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む第1のベクターを哺乳類に導入すること、
(b)予め決定された時間経過させること、および
(c)(i)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(ii)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む第2のベクターを前記哺乳類に導入すること、
を含む、結腸癌に罹患した前記哺乳類の治療方法。
【請求項23】
前記第1のベクターがプラスミドであり、前記第2のベクターがアデノウイルスベクターである、請求項22に記載の方法。
【請求項24】
前記第1のベクターがアデノウイルスベクターであり、前記第2のベクターがプラスミドである、請求項22に記載の方法。
【請求項25】
配列番号16に記載されているヒト癌胎児抗原(CEA)蛋白質変種をコードするヌクレオチドの配列を含み、ヒト細胞において高レベルで発現するためにコドンが最適化されている合成核酸分子。
【請求項26】
前記ヌクレオチドの配列が配列番号15で表したヌクレオチドの配列を含む、請求項25に記載の合成核酸分子。
【請求項27】
請求項25に記載の核酸分子を含むベクター。
【請求項28】
請求項27に記載のベクターを含む宿主細胞。
【請求項1】
配列番号2に記載されているヒト癌胎児抗原(CEA)蛋白質をコードするヌクレオチドの配列を含み、ヒト細胞において高レベルで発現するためにコドンが最適化されている合成核酸分子。
【請求項2】
前記核酸がDNAである、請求項1に記載の合成核酸分子。
【請求項3】
前記核酸がmRNAである、請求項1に記載の合成核酸分子。
【請求項4】
前記核酸がcDNAである、請求項1に記載の合成核酸分子。
【請求項5】
前記ヌクレオチドの配列が配列番号1で表したヌクレオチドの配列を含む、請求項1に記載の合成核酸分子。
【請求項6】
請求項1に記載の核酸分子を含むベクター。
【請求項7】
請求項6に記載のベクターを含む宿主細胞。
【請求項8】
(a)請求項1に記載の核酸を含むベクターを適切な宿主細胞中に導入すること、および
(b)前記ヒトCEA蛋白質の発現を可能にする条件下で前記宿主細胞を培養すること、を含む、組換え宿主細胞中でヒト癌胎児抗原(CEA)蛋白質を発現する方法。
【請求項9】
配列番号2で表したヒト癌胎児抗原(hCEA)蛋白質または配列番号16で表したCEA蛋白質変種をコードするヌクレオチドの配列を含むコドン最適化合成核酸分子を含むワクチンベクターを哺乳類に投与することを含む、癌の予防または治療方法。
【請求項10】
前記哺乳類がヒトである、請求項9に記載の方法。
【請求項11】
前記ベクターがアデノウイルスベクターまたはプラスミドベクターである、請求項9に記載の方法。
【請求項12】
前記ベクターが、E1領域中に欠失を有し、及び前記E1領域中に挿入部分を有するアデノウイルスゲノムを含むアデノウイルスベクターであり、
前記挿入部分が、
(a)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(b)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む発現カセットを含む、請求項9に記載の方法。
【請求項13】
前記ベクターが、
プラスミド部分と、
(a)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチド、および
(b)該ポリヌクレオチドに操作可能に結合されたプロモーターを含む発現カセットと、
を含むプラスミドワクチンベクターである、請求項9に記載の方法。
【請求項14】
E1領域中に欠失を有し、及び前記E1領域中に挿入部分を有するアデノウイルスゲノムを含み、
前記挿入部分が、
(a)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(b)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む発現カセットを含む、アデノウイルスワクチンベクター。
【請求項15】
Ad5ベクターである、請求項14に記載のアデノウイルスベクター。
【請求項16】
Ad6ベクターである、請求項14に記載のアデノウイルスベクター。
【請求項17】
Ad24ベクターである、請求項14に記載のアデノウイルスベクター。
【請求項18】
プラスミド部分と発現カセット部分を含み、
前記発現カセット部分が、
(a)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(b)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む、ワクチンプラスミド。
【請求項19】
(a)(i)ヒト癌胎児抗原(CEA)蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(ii)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む第1のベクターを哺乳類に導入すること、
(b)予め決定された時間経過させること、および
(c)(i)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(ii)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む第2のベクターを前記哺乳類に導入すること、
を含む、前記哺乳類を癌から防御する方法。
【請求項20】
前記第1のベクターがプラスミドであり、前記第2のベクターがアデノウイルスベクターである、請求項19に記載の方法。
【請求項21】
前記第1のベクターがアデノウイルスベクターであり、前記第2のベクターがプラスミドである、請求項19に記載の方法。
【請求項22】
(a)(i)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(ii)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む第1のベクターを哺乳類に導入すること、
(b)予め決定された時間経過させること、および
(c)(i)ヒトCEA蛋白質またはそれらの変種をコードするコドン最適化ポリヌクレオチドと、
(ii)前記ポリヌクレオチドに操作可能に結合されたプロモーターと、
を含む第2のベクターを前記哺乳類に導入すること、
を含む、結腸癌に罹患した前記哺乳類の治療方法。
【請求項23】
前記第1のベクターがプラスミドであり、前記第2のベクターがアデノウイルスベクターである、請求項22に記載の方法。
【請求項24】
前記第1のベクターがアデノウイルスベクターであり、前記第2のベクターがプラスミドである、請求項22に記載の方法。
【請求項25】
配列番号16に記載されているヒト癌胎児抗原(CEA)蛋白質変種をコードするヌクレオチドの配列を含み、ヒト細胞において高レベルで発現するためにコドンが最適化されている合成核酸分子。
【請求項26】
前記ヌクレオチドの配列が配列番号15で表したヌクレオチドの配列を含む、請求項25に記載の合成核酸分子。
【請求項27】
請求項25に記載の核酸分子を含むベクター。
【請求項28】
請求項27に記載のベクターを含む宿主細胞。
【図1】




【図7】
【図8】
【図9】


【図7】
【図8】
【図9】
【公表番号】特表2007−523610(P2007−523610A)
【公表日】平成19年8月23日(2007.8.23)
【国際特許分類】
【出願番号】特願2006−505382(P2006−505382)
【出願日】平成16年5月3日(2004.5.3)
【国際出願番号】PCT/EP2004/004802
【国際公開番号】WO2004/099247
【国際公開日】平成16年11月18日(2004.11.18)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
【公表日】平成19年8月23日(2007.8.23)
【国際特許分類】
【出願日】平成16年5月3日(2004.5.3)
【国際出願番号】PCT/EP2004/004802
【国際公開番号】WO2004/099247
【国際公開日】平成16年11月18日(2004.11.18)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
[ Back to top ]