
発現ベクター、異種遺伝子生成物の製造方法およびこのような生成物を高濃度で製造する組換え細胞の選別方法
ハムスターユビキチン/S27aプロモータと機能的な結合状態にある、対象とするタンパク質をコードする遺伝子、および蛍光タンパク質をコードする遺伝子を含む、真核生物細胞の発現ベクターを開示する。好ましくは、この発現ベクターは、また増幅可能な選択性マーカー遺伝子をも含む。該発現ベクターでトランスフェクションされた宿主細胞、好ましくは哺乳動物細胞、および高い生産性を持つ細胞を選別する方法をも開示する。

【発明の詳細な説明】
【発明の詳細な説明】
【0001】
技術分野
本発明は、高い生産性を示す組換え細胞の選別方法、異種遺伝子生成物の製造方法および発現ベクター並びに該ベクターでトランスフェクションされた、これらの方法において使用可能な宿主細胞に関するものである。
【0002】
発明の背景
哺乳動物細胞は、複合生物薬学的タンパク質を製造するための好ましい宿主細胞である。というのは、翻訳後に行われる修飾が、機能的にも薬物動力学的にも、ヒトと相溶性であるからである。工業的には、関連する細胞型は、ハイブリドーマ、ミエローマCHO(チャイニーズハムスター卵巣)細胞およびBHK(ベビーハムスター腎臓)細胞である。この宿主の培養は、次第に血清およびタンパク質を含まない条件の下で行われるようになっている。その理由は、付随するコストの減少、該組換えタンパク質の精製における妨害の低下、病原体(例えば、プリオンおよびウイルス)導入の可能性の低下にある。宿主としてのCHO細胞の使用は、これら細胞が、血清およびタンパク質を含まない培地内での懸濁成長に適したものとなるにつれて、より一層普及するようになり、また規制当局により安全な生産細胞と見做され、かつ許容される。
【0003】
対象とする異種遺伝子を発現する、安定な哺乳動物細胞系を製造するためには、一般に選択性のあるマーカー遺伝子、例えばネオマイシントランスフェラーゼと共に、該異種遺伝子を、該所定の細胞系にトランスフェクションにより挿入する。該異種遺伝子および該選択性マーカー遺伝子は、単一のベクターにより一緒に、あるいは同時トランスフェクションされる別々のベクターによって、発現させることができる。トランスフェクションの2〜3日後、選択性の試薬、例えばネオマイシントランスフェラーゼを用いた場合にはG418を含有する培地に、該細胞を移し、これら選択条件下で数週間培養する。次いで、エマージェント(emergent)抵抗性細胞を単離し、かつ所定の遺伝子生成物の発現につき検討を行うことができる。該宿主細胞ゲノムへの無秩序で、方向性の無い組込みの結果として、一群の細胞が得られ、これら細胞は、全く異なる該異種遺伝子発現率を持つ。これらは、非-発現性の細胞をも含むことができ、ここで該選択性マーカーは発現されるが、対象とする該遺伝子は発現されない。従って、対象とする該遺伝子の極めて高い発現性を呈する細胞クローンを同定するためには、多数のクローンを検査し、かつテストする必要があるが、これは時間を要し、また労力を消耗し、しかも経費がかさむ。
【0004】
遺伝子の増幅は、動物細胞の培養において広く普及している現象であり、組換え生物薬学的なタンパク質の製造のために使用されている。この遺伝子増幅は、多数の哺乳動物細胞系の、元々比較的低い生産性を、大幅に改善する。広く利用されている増幅法の一つは、ジヒドロ葉酸レダクターゼ(DHFR)を基本とする、DHFR-欠乏チャイニーズハムスター卵巣(CHO)細胞において、極めて頻繁に使用される遺伝子増幅系である。このDHFR-欠乏CHO細胞、例えばCHO-DUKX(ATCC CRL-9096)またはCHO-DG44(Urlaub等、1983)を、DHFRおよび該対象とするタンパク質をコードする、適当なベクター系でトランスフェクションする。次いで、該トランスフェクタントを、グリシン、ヒポキサンチンおよびチミジンを含まない培地で選別する。この増幅およびその結果としての高い生産性を持つ細胞系の確立は、メトトレキセート(MTX)、即ちジヒドロ葉酸レダクターゼ阻害剤の次第に増加する濃度での添加によって達成される(Kaufman等、1982, US 4,656,134)。得られる高い生産性を示す細胞の、その後の選別も、偶然の原理を条件としており、また確率に基くものであり、その結果として、この選別段階は多大な労力を要し、かつ時間を浪費するものである。
【0005】
より良好にかつより迅速に、遺伝子形質転換および発現を追跡するために、あらゆる種類の方法が開発されている。これらは、まずリポータ分子、例えばクロラムフェニコール-アセチルトランスフェラーゼ、ルシフェラーゼ、β-ガラクトシダーゼ、またはβ-ガラクトシダーゼもしくはルシフェラーゼのコード領域を含む融合タンパク質の使用を含む。これらリポータ遺伝子アッセイの欠点は、該細胞を固定しまたは溶解する必要があり、また外部から添加した物質および補助因子と共にインキュベートする必要がある。従って、該分析された細胞の更なる培養が、当然必要となる。E.コリ酵素β-ガラクトシダーゼの同時発現に基く極最近の方法は、実際に、FACS装置(Nolan等, 1988)を用いて、溶解した細胞の選別を可能とするが、フルオロゲン性基質と共に該細胞を充填するために、低張性予備処理が必要となる。また、該FACSを基本とする選別の前に、その活性を阻害する必要がある。
【0006】
エクオレアビクトリア(Aequorea victoria)由来の蛍光タンパク質(GFP)およびこれからリポータ分子として発展されたGFP変異体の導入により、異種遺伝子を発現する細胞を同定することが、より一層容易になった。GFPの同時発現は、付随的な基質または補助因子を必要とすることなしに、その蛍光に基いた、インビボでの実時間分析、およびトランスフェクタントの選別を可能とした。遺伝子転位を追跡するためのリポータ分子としてのGFPの使用は、様々な刊行物に記載されている。Chalfie等の米国特許第5,491,084号および同第6,146,826号は、対象とするタンパク質を発現する細胞の選別方法を記載している。この方法は、該対象とするタンパク質に対するコード配列を含むDNAおよび該GFP-遺伝子をコードする第二のDNA分子による、細胞の同時トランスフェクションを含む。次いで、該GFP-発現細胞を選別する。Gubin等(1997)は、選択成長条件を使用しない場合の、CHO細胞内でのGFP発現の安定性を検討した。これらの細胞は、GFPおよびネオマイシンホスホトランスフェラーゼ両者を含むプラスミドによりトランスフェクションした。Mosser等(1997)は、GFPおよびターゲット遺伝子(これも興味深い遺伝子として知られる)をコードするビシストロン発現カセットを含むプラスミドを使用して、誘発可能な生成物を発現する細胞の同定および選別した。このターゲット遺伝子は、調節可能なプロモータの制御下にあった。このGFP発現とターゲット遺伝子発現との結合は、ウイルス由来のIRES(内部リボソームエントリーサイト)を用いて達成され、その結果として、GFPおよび該対象とするタンパク質をコードするビシストロンmRNAが発現された。使用したプラスミドそれ自体は、選択性マーカー遺伝子を何等含んでいなかった。従って、これは同時トランスフェクションにおいて、または後のトランスフェクションにおいて、第二のプラスミドによって導入された。これとは対照的に、Levenson等(1998)は、ビシストロン発現カセットを持つレトロウイルスベクターを使用しており、該カセットにおいて、該対象とする遺伝子は、該IRES配列の前でクローニングすることができる。他方、該IRES配列に続く配列は、選択性マーカー遺伝子をコードし、このマーカーには、G418、プロマイシン、ハイグロマイシン(hygromycin) B、ヒスチジノールDまたはフレオマイシンに対する耐性が付与されており、あるいはこれはGFPであった。
【0007】
ピコルナウイルス群由来のIRES要素を含むベクターも、既に記載されており、このIRES要素は、該生成物遺伝子と、選択性マーカー遺伝子との間に配置されている(Pelletier等, 1988; Jang等, 1989; Davies等, 1992)。
また、GFPも首尾よく耐性マーカー遺伝子と融合されている。例えば、Bennett等(1998)は、GFP/ゼオマイシン融合タンパク質を記載している。この二官能性の選択性マーカーは、トランスフェクションされた哺乳動物細胞を同定し、かつ選別するのに、上首尾で使用された。他方Primig等(1998)は、そのエンハンサーの研究に対して、GFPとネオマイシンホスホトランスフェラーゼとの融合タンパク質を使用した。
Meng等(2000)による刊行物および国際特許出願WO 01/04306では、単一のベクター由来の、増幅可能な選択性マーカー遺伝子DHFRおよびGFP遺伝子と共に、該対象とする遺伝子を発現する発現系を使用して、組換えタンパク質の高い発現性を持つ細胞を選別し、かつ同定した。これら3つの遺伝子は、一つの転写単位内で結合されているか、あるいは2つの単位間で分割されていた。単一の発現ベクター内でのこれら3種の遺伝子全ての空間的並びに転写的な結合は、選択圧力の下で同時増幅の確率を高め、かつその結果として高い生産性を呈するクローンを同定し、かつ選別した。この組合せ選別を利用して、増幅可能なDHFR選択性マーカーおよびGFPを基本とするFACS選別によって単離された、最良のクローンは、1日当たり、1細胞当たり3〜4.5pg程度の量で、該対象とするタンパクを発現した。これらの実験は、接着性の細胞を用いて、血清を含む培地内で、即ちかなり大雑把なものであることが知られており、またより高い基本的な生産性によって特徴付けられる細胞および条件を用いて行った。
【0008】
発明の概要
従って、本発明の目的は、以下のような要件を持たす、高い生産性を持つ組換え細胞の選別系を開発することであった:
(1) 生物薬学的タンパク質を生産する、高い生産性を呈する細胞の開発に要する時間の短縮、および同時にこの開発に係る経費の節減;
(2) 設備に関する低い経費で、高い生産性を呈する細胞の選別における高い処理量;
(3) 例えば、高いメトトレキセート濃度における、成長性に対する影響の小さい、「醗酵に関して強固な」高生産生細胞の使用;
(4) 好ましくは血清を含まない培地内での、懸濁培養に適した細胞の、トランスフェクション、選別および培養性;および
(5) 必要とされる遺伝子増幅段階数の減少。
本発明の、更なる目的は、発現ベクター、およびこれによりトランスフェクションされた、このクローン選別系において使用可能な宿主細胞、並びにこれら宿主細胞を使用して、異種遺伝子生成物を製造する方法を提供することにあった。
【0009】
これら本発明の目的は、本発明の一局面に従えば、発現ベクターによって達成され、この発現ベクターは、ハムスターユビキチン/S27aプロモータと機能可能な状態で結合した、対象とするタンパク質をコードする遺伝子(以下「対象とする遺伝子」とも言う)および蛍光タンパク質をコードする遺伝子を含む。
この発現ベクターは、好ましくは更に増幅可能な、選択性マーカー遺伝子、例えばジヒドロ葉酸レダクターゼ(DHFR)をも含む。好ましい発現ベクターは、また他の調節要素、例えば該プロモータと機能可能な状態で結合したエンハンサーをも含む。また、この発現ベクターは、好ましくは内部リボソームエントリーサイト(IRES)をも含み、このサイトは蛍光タンパク質をコードする遺伝子および対象とする遺伝子の、ビシストロン発現を可能とする。
本発明は、また該対象とする遺伝子の代わりに、このような遺伝子を組込むための多重クローニングサイト、即ち制限エンドヌクレアーゼによる多数の切断サイトを持つ配列領域を持つ、基本ベクターにも関連する。
【0010】
本発明のもう一つの局面は宿主細胞に関連し、該宿主細胞は、上記発現ベクターの一種によってトランスフェクションされている。これら宿主細胞は、真核宿主細胞、好ましくは哺乳動物細胞であり、ハムスター細胞等の齧歯目動物の細胞、特にCHO細胞またはBHK細胞が特に好ましい。
本発明のもう一つの局面は、異種遺伝子生成物の製造方法に係り、この方法では、本発明の発現ベクターでトランスフェクションされた宿主細胞を、該異種遺伝子生成物の発現を可能とする条件の下で培養し、かつ該遺伝子生成物を培養物または培地から単離する。
本発明の一特定の態様では、該宿主細胞は、本発明による発現ベクターで、また一種またはそれ以上の他の対象とするタンパクをコードする遺伝子を含む、一種またはそれ以上のベクターでトランスフェクションし、または好ましくは同時にトランスフェクションする。
【0011】
これに関連して、本発明は、ヘテロダイマー型タンパク質の製法を提供するものであり、この方法では、該へテロダイマー型タンパク質の様々なサブユニットをコードする発現ベクターによって同時にトランスフェクションされている、上記類の宿主細胞を該へテロダイマー型タンパクの発現を可能とする条件の下で培養し、かつ該へテロダイマー型タンパクを培養物または培地から単離する。このような方法の、一特定の応用は、抗体およびそのサブユニットの製造である。
もう一つの局面において、本発明は、対象とするタンパクを発現する宿主細胞を選別する方法に係り、この方法では、本発明による発現ベクターによりトランスフェクションされている、一群の宿主細胞を、該対象とするタンパクおよび該蛍光タンパク質の発現を可能とする条件の下で培養し、かつ該蛍光タンパクの最大の発現率を示す該細胞または細胞群を同定し、および/または選別する。この選別は、好ましくは、蛍光-活性化セルソータ(FACS)を用いて行われる。
驚いたことに、本発明の提供するこの系を用いた場合には、極めて短時間の間に、1日当たりかつ1細胞当たり、組換えタンパク質15pg(単鎖タンパク質)または10pg(ヒト化抗体)を越える平均の比生産率(specific productivities)で発現する細胞のプールを、遺伝子増幅段階を経ることなしに、単離することが可能となることを見出した。該比生産率は、一回のDHFRに基く遺伝子増幅段階によって、1日当たりかつ1細胞当たり30pgを越えるレベルまで高めることができた。これら細胞のプールにおいて達成されるこの生産率は、結果的に、これまでに公開された最良の細胞クローンにより達成された最大の生産率よりも、8〜10倍だけ高い。
【0012】
驚いたことに、該対象とするタンパク質および該蛍光タンパク質の発現同士の間には、極めて良好な相関関係がある。このことは、発現された抗体と関連して、2つの免疫グロブリン鎖が、各々それ自身のベクターによって発現される場合の、同時トランスフェクションにおいてさえ正しく、またFACS選別において、選択は該蛍光タンパク質との転写的な結合を考慮して、一方の鎖の発現に関してのみ行うことができる。該蛍光タンパク質の高い発現率は、該細胞の成長および生存率について何の影響も示さない。更に、高い生産性を示す細胞を選別するための、技術開発時間を、従来の段階的な遺伝子増幅戦略と比較して、少なくとも半分にまで減じることができ、結果的に開発設備および経費の大幅な節減を達成できる。
【0013】
発明の詳細な説明および好ましい態様
本発明による発現ベクターは、対象とするタンパク質(対象とする遺伝子)をコードし、機能可能な状態でハムスターユビキチン/S27aプロモータと結合した遺伝子および蛍光性タンパク質をコードする遺伝子を含む。好ましくは、該発現ベクターは、また増幅可能な選択性マーカー遺伝子を含む。
ハムスターユビキチン/S27aプロモータ
このハムスターのユビキチン/S27aプロモータは、WO 97/15664に記載されている強力な相同的プロモータである。このようなプロモータは、好ましくは、以下に列挙する特徴の内の少なくとも一つを持つ:GCに富む配列領域、Sp1結合サイト、ポリピリミジン要素を持ち、TATAボックスを持たない。同様に好ましいものは、構造的に活性化されており、かつ特に血清を含む、低濃度の血清を含むおよび血清を含まない細胞培養条件にて、等しく活性であるプロモータである。もう一つの態様では、これは、誘導性のプロモータ、特に血清の除去により活性化されるプロモータである。
特に有利な態様は、WO 97/15664の図5に含まれているような、ヌクレオチド配列を持つプロモータである。特に好ましいものは、図5の-161〜-45なる範囲の位置の配列を含むプロモータ配列である。
【0014】
本特許出願の実施例において使用するプロモータ各々は、添付した配列表のSEQ ID NO:1の位置1923〜2404に対応する配列を持つDNA分子を含む。この配列は、WO 97/15664の図5のフラグメント:-372〜+111に相当し、また好ましいプロモータを表し、従って好ましいプロモータにはこの配列領域を組込むべきである。もう一つの適当なプロモータフラグメントは、位置:2134〜2406の配列(WO 97/15664の図5における位置:-161〜+111に相当)を含む。位置:2251〜2406の配列のみを含むプロモータは、最早機能性ではない(WO 97/15664の図5における位置:-45〜+111に相当)。このプロモータ配列を、位置:2134から開始して、5'方向に伸ばすことができる。
また、完全ハムスターユビキチン/S27aプロモータ配列の機能性のサブフラグメント並びに例えば置換、挿入または削除によって変性されている、そのサブフラグメントの該完全配列の、機能性の突然変異体/変異型を使用することも可能である。対応するサブフラグメント、突然変異体または変異型は、以下において「変性プロモータ」とも呼ばれる。
【0015】
場合により他の調節要素と結合した、変性プロモータは、好ましくはSEQ ID NO:1に与えられたヌクレオチド配列の位置:1923〜2406のプロモータフラグメントの活性に相当する、転写活性を持つ(WO 97/15664の図5における位置:-372〜+111)。変性プロモータは、これが転写活性を示し、その活性が、比較リポータ遺伝子のフラグメント:1923〜2406(-372〜+111フラグメント)の持つ活性の少なくとも50%、好ましくは少なくとも80%、より好ましくは少なくとも90%および最も好ましくは少なくとも100%である場合には、本発明の目的にとって有用であることが分かる。特に好ましい変性プロモータは、少なくとも80%、好ましくは少なくとも85%、好ましくは少なくとも90%、より好ましくは少なくとも95%および最も好ましくは少なくとも97%なる、上記ハムスターユビキチン/S27aプロモータの、野生型配列:SEQ ID NO:1に対する最低配列相同性を持ち、かつ比較リポータ遺伝子アッセイにおいて対応するプロモータ活性を持つものである。
【0016】
対応する比較リポータ遺伝子アッセイにおいて、該基準配列を含む、テストすべきプロモータフラグメントは、例えばルシフェラーゼをコードする、プロモータを含まないリポータ遺伝子の前にクローニングされ、アルカリホスファターゼまたはグリーン蛍光タンパク質(GFP)を分泌する。これらの構築物(プロモータ配列+リポータ遺伝子)は、後にこのテスト細胞、例えばCHO-DG44に、トランスフェクションによって導入され、また問題とする該プロモータフラグメントによる、該リポータ遺伝子発現の誘導は、該リポータ遺伝子のタンパク含有率を測定することにより決定される。対応するテストは、例えばAusubel等の、分子生物における今日のプロトコール(Current Protocols in Molecular Biology), 1994, 最新版に見出すことができる。
【0017】
ハムスターユビキチン/S27aプロモータのプロモータ配列および例えば5'未翻訳領域または選択されたそのフラグメントをも含んでいても良い、変性プロモータ、およびユビキチン/S27a遺伝子のコード配列またはその選択されたフラグメントは、WO 97/15664に記載されている配列に関する知見を持つ熟練者によって、例えばSambrook等, 1989; Ausubel等, 1994に記載されている、様々な標準的方法を利用して得ることができる。WO 97/15664に記載されている配列から出発し、例えば適当な配列を選択し、この部分の配列を含むオリゴヌクレオチドプローブを化学的に合成することができる。例えば、該ユビキチン/S27a遺伝子またはその5'未翻訳領域またはその他のフラグメントを、例えばハムスターゲノムのライブラリーからハイブリッド化することにより、クローニングする目的で、この種のプローブを使用することができる。上記のリポータ遺伝子アッセイを利用して、当業者は、多大な努力を払うことなしに、プロモータ-活性フラグメントを同定し、かつこれらを本発明の目的において使用することができる。該5'未翻訳領域またはその特定のフラグメントは、ゲノムDNAまたはゲノムライブラリーからの対応するプライマーを用いたPCR増幅により、容易に得ることができる。該5'未翻訳領域のフラグメントも、より大きなDNAフラグメントから、制限エキソヌクレアーゼIIIにより得ることができる。このようなDNA分子も、化学的に合成することができ、あるいは結合によって、化学的に合成されたフラグメントから製造することができる。
【0018】
欠失、挿入および置換突然変異体は、「サイト-特異的突然変異誘発」および/または「PCRに基く突然変異誘発技術」によって製造することができる。対応する方法は、例えば他の参考文献をも含む、LottspeichおよびZorbas, 1998,第36.1章に述べられている。
ハムスターユビキチン/S27a遺伝子の5'未翻訳領域またはハムスターユビキチン/S27a遺伝子のS27a部分由来のプローブを用いた、クロス-ハイブリッド形成により、他の、好ましくは哺乳動物種の、対応する相同遺伝子由来の適当なプロモータ配列を、同定しかつ単離することができる。適当な技術は、一例として、LottspeichおよびZorbas, 1998,第23章に記載されている。遺伝子は、そのヌクレオチド配列が、相同であるとされる該遺伝子のヌクレオチド配列に対して、少なくとも70%、好ましくは少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%および最も好ましくは少なくとも97%の適合性を示す場合には、本発明の目的にとって「相同」である。
【0019】
対象とする遺伝子
本発明の発現ベクターに含まれる該対象とする遺伝子は、対象とするタンパク質をコードする、任意の長さのヌクレオチド配列を含む。この遺伝子生成物または「対象とする生成物」は、一般にタンパク質、ポリペプチド、ペプチドまたはこれらのフラグメントまたは誘導体である。しかし、これはRNAまたはアンチセンスRNAであっても良い。該対象とする遺伝子は、完全な長さで、短縮された形状で、融合遺伝子としてまたは標識遺伝子として存在することができる。これはゲノムDNAまたは好ましくはcDNAまたは融合物の対応するフラグメントであり得る。該対象とする遺伝子は、本来の遺伝子配列であり得、あるいは突然変異または変性されていてもよい。このような変性は、特定の宿主に対する適合化またはヒト化するための、コドン最適化を含む。該対象とする遺伝子は、例えば分泌された、細胞質、核内に位置する、膜と結合した、または細胞表面に結合したポリペプチドをコードすることができる。
用語「ヌクレオチド配列」または「核酸配列」とは、オリゴヌクレオチド、ヌクレオチド、ポリヌクレオチドおよびこれらのフラグメント、並びに一本鎖または二本鎖であり得る、ゲノムまたは合成起源のDNAまたはRNAを意味し、また遺伝子のコード性または非-コード性のストランドを表すこともできる。核酸配列は、サイト-特異的突然変異形成またはPCR-媒介突然変異形成(例えば、Sambrook等, 1989; Ausubel等, 1994に記載されている)等の標準的な技術を用いて変性することができる。
【0020】
用語「コードする」とは、核酸、例えば染色体中の遺伝子またはmRNA内の特定のヌクレオチド配列が、他のポリマーおよび巨大分子、例えばrRNA、tRNA、mRNA、その他のRNA分子、cDNAまたは生物学的な過程におけるポリペプチドを合成するためのマトリックスとして機能する性質または能力を意味する。従って、所定のタンパク質が、転写およびそれに続くmRNAの翻訳によって、細胞または別の生物学的な系内で製造される場合には、遺伝子はタンパク質をコードする。ヌクレオチド配列がそのmRNAの配列と等価であり、通常は配列データバンク、例えばEMBLまたはGenBankにも与えられている、コード用のストランドおよび転写に対するマトリックスとして作用する、遺伝子またはcDNAの非-コード用のストランド両者は、生成物またはタンパク質をコードするものと見做される。タンパク質をコードする核酸は、また縮重遺伝子コードを基準として、異なる順序のヌクレオチド配列を含むが、該タンパク質と同一のアミノ酸配列をもたらす。タンパク質をコードする核酸配列は、またイントロンをも含む。
【0021】
cDNAなる用語は、逆転写およびmRNAまたは遺伝子から作られた他のRNA由来の第二のDNAストランドの合成によって製造される。該cDNAが二本鎖DNA分子として存在する場合、これはコード用ストランドおよび非-コード用ストランド両者を含む。
用語:イントロンとは、任意の長さの非-コード用ヌクレオチド配列を意味する。これらは、自然において数多くの真核生物遺伝子内に見られ、予め転写されるmRNAプリカーサから、スプライシングとして知られる過程によって排除される。これは、5'および3'末端における該イントロンの正確な切除および得られるmRNA末端の正確な結合を必要とし、結果的にタンパク質合成を首尾よく行うための正確な読み取り枠を持つ、成熟プロセスmRNAが作られる。このスプライシング過程に関与する、多くのスプライスドナーおよびスプライスアクセプタサイト、即ち該エクソン-イントロンあるいはイントロン-エクソン界面に直接位置する配列が、特徴付けされている。これに関する概説に関しては、Ohshima等, 1987を参照のこと。
【0022】
対象とするタンパク質
生物薬学的に重要なタンパク質/ポリペプチドは、例えば抗体、酵素、サイトカイン、リンホカイン、接着分子、レセプタおよびこれらの誘導体またはフラグメントを含むが、これらに限定されない。一般に、アゴニストまたはアンタゴニストとして作用するおよび/または治療または診断における用途を持つ、全てのポリペプチドが、価値あるものである。
用語「ポリペプチド」は、アミノ酸配列またはタンパク質について用いられ、また任意の長さを持つアミノ酸のポリマーを意味する。この用語は、またグリコシル化、ホスホリル化、アセチル化またはタンパク質のプロセシング等の反応により、翻訳後に変性されたタンパク質を包含する。このポリペプチドの構造は、該ポリペプチドの生物学的な活性を維持しつつ、例えばアミノ酸の置換、欠失または挿入により、および他のタンパク質と融合することにより変性することができる。
【0023】
治療用のタンパク質の例は、インシュリン、インシュリン-様の成長因子、ヒト成長ホルモン(hGH)および他の成長因子、組織プラスミノーゲン活性化因子(tPA)、エリスロポエチン(EPO)、サイトカイン、例えばIL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-8、IL-9、IL-10、IL-11、IL-12、IL-13、IL-14、IL-15、IL-16、IL-17、IL-18等のインターロイキン(IL)、インターフェロン(IFN)-α、-β、-γ、-ω、または-τ、腫瘍壊死因子(TNF)、例えばTHF-α、-βまたは-γ、TRAIL、G-CSF、GM-CSF、M-CSF、MCP-1およびVEGFである。その他の例は、モノクローナル、ポリクローナル、多重特異的および一本鎖抗体およびそのフラグメント、例えばFab、Fab'、F(ab')2、FcおよびFc'フラグメント、免疫グロブリン軽鎖(L)および重鎖(H)およびその定常、可変および超可変領域並びにFvおよびFdフラグメント(Chamov等, 1999)である。これらの抗体は、ヒト起源または非-ヒト起源のものであり得る。ヒト化またはキメラ抗体も可能である。
Fabフラグメント(抗原結合フラグメント=Fab)は、隣接する定常領域によって、一緒に保持されている2つの鎖の可変領域からなる。これらは、例えば公知の抗体を、パパイン等のプロテアーゼで処理することにより、あるいはDNAクローニングによって製造できる。他の抗体フラグメントは、ペプシンによるタンパク分解消化により製造できる、F(ab')2フラグメントである。
【0024】
遺伝子クローニングによって、重鎖(VH)および軽鎖(VL)の可変領域のみからなる、短い抗体フラグメントを製造することも可能である。これらは、Fvフラグメント(可変フラグメント=可変部分のフラグメント)として知られている。該定常鎖のシステイン基を介する共有結合は、これらのFvフラグメントでは不可能であることから、これらはしばしば他の幾つかの方法によって安定化される。この目的のために、該重鎖および軽鎖の可変領域は、しばしば約10〜30個のアミノ酸、好ましくは15個のアミノ酸を含む短いペプチドフラグメントで、一緒に結合される。これは、単一のポリペプチド鎖を製造し、そこではVHおよびVLがペプチドリンカーによって一緒に結合されている。このような抗体フラグメントは、また一本鎖Fvフラグメント(scFv)とも呼ばれる。scFv抗体の例は公知であり、例えばHuston等, 1988に記載されている。
過去において、様々な方法が、マルチマーscFv誘導体を製造するために開発されている。その意図は、改良された薬物動力学的特性および高い結合活性を持つ、組換え抗体を製造することにある。該scFvフラグメントのマルチマー化を達成するためには、これらを、マルチマー化ドメインを持つ融合タンパクとして製造する必要がある。該マルチマー化ドメインは、例えばIgGのCH3領域またはロイシンジッパードメイン等のヘリックス構造(螺旋状コイル構造)であり得る。他の方法では、該scFvフラグメントのVHおよびVL領域間の相互作用を、マルチマー化(例えば、二重特異性-、三重特異性-および五重特異性抗体)のために利用する。
【0025】
用語:二重特異性抗体とは、当分野において、二価のホモダイマー型scFv誘導体を表すのに用いられる。該scFv分子内のペプチドリンカーの長さを、5〜10アミノ酸に短縮することは、VH/VL鎖の重ね合わせにより、ホモダイマーの生成をもたらす。これら二重特異性抗体は、更にジスルフィド架橋によって安定化することができる。二重特異性抗体の例は、Perisic等, 1994などの文献に見出すことができる。
用語:ミニ抗体(minibody)とは、当分野において、二価のホモダイマー型scFv誘導体を表すのに用いられる。これは融合タンパク質からなり、該タンパクは免疫グロブリン、好ましくはIgG、最も好ましくはIgG1のCH3領域をダイマー化領域として含む。これは、ヒンジ領域、またIgGおよびリンカー領域によって、該scFvフラグメントと接続している。このようなミニ抗体の例は、Hu等, 1996によって記載されている。
該三重特異性抗体とは、当分野において、三価のホモダイマー型scFv誘導体(Kortt等, 1997)を表すのに用いられる。リンカー配列の使用なしの、VH/VL鎖の直接的な融合は、トリマーの生成をもたらす。
当分野において、二価、三価、または四価の構造を持つミニ抗体として知られるフラグメントは、またscFvフラグメント誘導体でもある。このマルチマー化は、ダイマー、トリマーまたはテトラマー型の螺旋状コイル構造によって達成される(Pack等, 1993およびLovejoy等, 1993)。
【0026】
蛍光タンパク質をコードする遺伝子
本発明による発現ベクターは、対象とする遺伝子と機能可能な状態で結合し、またハムスターユビキチン/S27aプロモータ、変性ハムスターユビキチン/S27aプロモータまたはその同族体の制御下にある、蛍光タンパク質をコードする遺伝子を含有する。
該蛍光タンパク質は、例えば緑、青味を帯びた緑、青、黄色またはその他の色を持つ蛍光タンパク質であり得る。一特定例は、エクオレアビクトリア(Aequorea victoria)またはレニラレニホルミス(Renilla reniformis)およびこれらから発展された突然変異体由来のグリーン蛍光タンパク質(GFP)である(例えば、Bennet等, 1998;Chalfie等, 1994;WO 01/04306およびこれらに引用されている文献)。
他の蛍光タンパク質およびこれらをコードする遺伝子は、WO 00/34318、WO 00/34326、WO 00/34526、WO 01/27150に記載されている。これら文献を、本発明の参考とする。これらの蛍光タンパク質は、アントゾア(Anthozoa)種、例えばアネモニアマジャノ(Anemonia majano)、クラブラリア(Clavularia) sp、ゾアンサス(Zoanthus) sp. I、ゾアンサスsp. II、ディスコゾーマストリアータ(Discosoma striata)、ディスコゾーマsp.「レッド」、ディスコゾーマsp.「グリーン」、ディスコゾーマsp.「マゼンタ」、アネモニアスルカタ(sulcata)等の非-生物発光生物の発蛍光団である。
【0027】
本発明で使用する該蛍光タンパク質は、野生型のタンパク質に加えて、例えば他のタンパク質またはペプチドと融合されている、天然のまたは遺伝子操作した突然変異体および変異種、フラグメント、その誘導体または変異種を含む。使用する突然変異は、例えば励起または発光スペクトル、発色団の形成、該タンパク質の吸光係数または安定性を変更する可能性がある。その上、哺乳動物細胞またはその他の種における発現は、コドン最適化により改善することができる。本発明によれば、該蛍光タンパク質は、また選択性マーカー、好ましくは増幅可能な、選択性マーカー、例えばジヒドロ葉酸レダクターゼ(DHFR)との融合においても使用できる。
該蛍光タンパク質により発せられる蛍光は、例えば蛍光-活性化細胞選別機(FACS)または蛍光顕微鏡を用いた、スルーフロー(throughflow)サイトメトリーによって、該タンパク質の検出を可能とする。
他の調節要素
ハムスター-ユビキチン/S27aプロモータは、他の調節配列と機能可能な状態で結合して、発現カセットにおける転写活性を増大/調節することができる。
例えば、このプロモータは、機能可能な状態で、エンハンサ配列と結合して、該転写活性を高めることができる。このために、一種またはそれ以上のエンハンサおよび/または一つのエンハンサの幾つかのコピー、例えばCMVまたはSV40エンハンサを使用することができる。
【0028】
エンハンサなる用語は、シス-位において、プロモータの活性に作用し、結果的にこのプロモータと機能可能な状態で接続された遺伝子の転写を刺激する、ポリヌクレオチド配列を表す。プロモータとは異なり、エンハンサの効果は、位置および配向とは無関係であり、従ってこれらはイントロン内でまたは該コード領域においてさえ、転写単位の前または背後に位置することができる。該エンハンサは、該転写単位の極近傍および該プロモータからかなり遠く離れた位置の何れにも位置することができる。該プロモータと物理的および機能的な重なりを有していても良い。当業者は、様々な起源由来の多数のエンハンサ(およびGenBank等のデータバンクに寄託されているもの、例えばSV40エンハンサ、CMVエンハンサ、ポリオーマエンハンサ、アデノウイルスエンハンサ)を認識しており、これらは独立した要素として、あるいはポリヌクレオチド配列内でクローニングされた要素(例えば、ATCCに寄託された、または市場のまたは個人的な源からの要素)として入手できる。また、多くのプロモータ配列、例えば頻繁に使用されるCMVプロモータ等が、エンハンサ配列をも含む。ヒトCMVエンハンサは、これまでに同定された中で最強のエンハンサである。誘導エンハンサの一例は、メタロチオネインエンハンサであり、これはグルココルチコイドまたは重金属によって刺激することができる。
【0029】
他の可能な変性は、例えば多数のSp1結合サイトの導入である。また、これらプロモータ配列は、調節配列と結合することができ、この調節配列は、転写活性の制御/調節を可能とする。従って、該プロモータを抑制/誘導することができる。これは、例えばアップ-またはダウン-レギュレーション転写因子に対する結合サイトである、配列と結合することにより実施できる。例えば、上記転写因子Sp1は、該転写活性に対して正の効果を持つ。他の例は、活性化因子タンパク質AP1に対する結合サイトであり、これは転写に対して正および負何れにも作用する。AP1の活性は、あらゆる類の因子、例えば成長因子、サイトカインおよび血清によって制御できる(Faisst等, 1992およびこれに含まれる参考文献)。また1、2、3またはそれ以上の塩基の突然変異(置換、挿入または欠失)により該プロモータ配列を変更し、次いでリポータ遺伝子アッセイにおいて、これが高いプロモータ活性を持つか否かを決定することによって、転写効率を高めることができる。
基本的には、これらの付随的な調節要素は、ハムスター-ユビキチン/S27aプロモータ以外のプロモータ、エンハンサ、終結およびポリアデニル化シグナル並びに他の発現制御要素を含む。誘導および構造的調節要素両者は、様々な細胞型について公知である。「転写-調節要素」は、一般に発現すべき遺伝子配列の上流のプロモータ、転写開始および終止サイトおよびポリアデニル化シグナルを含む。
【0030】
プロモータなる用語は、遺伝子またはこれと機能的に接続した配列の転写を可能とし、かつこれを制御する、ポリヌクレオチド配列を意味する。プロモータは、RNAポリメラーゼを結合するための認識配列および転写用の開始サイト(転写開始サイト)を含む。ある細胞型または宿主細胞において、所定の配列を発現するためには、適当な機能的プロモータを選択する必要がある。当業者は、構造的、誘導性および抑制プロモータを含む、様々な起源由来の種々のプロモータについて十分な馴染みを持つであろう。これらは、例えばGenBank等のデータバンクに寄託されており、またこれらは市場または個人的な源から、別々の要素として、あるいはポリヌクレオチド配列内でクローニングされた要素として得ることができる。誘導プロモータにおいて、該プロモータの活性は、シグナルに応じて低下または増大させることが可能である。誘導プロモータの一例は、テトラサイクリン(tet)プロモータである。これは、テトラサイクリンオペレータ配列(tetO)を含み、この配列はテトラサイクリン-調節トランス活性化因子タンパク質(tTA)により誘導できる。テトラサイクリンの存在下において、tTAとtetOとの結合は阻害される。他の誘導プロモータの例は、jun、fos、メタロチオネインおよび熱ショックプロモータである(同様に、Sambrook等, 1989; Gossen等, 1994をも参照のこと)。真核生物における高い発現性を得るのに、特に適したプロモータとしては、例えばSV40初期プロモータ、アデノウイルス主後期プロモータ、マウスメタロチオネイン-Iプロモータ、ラウス肉腫ウイルスの反復配列領域およびヒトサイトメガロウイルスの初期プロモータが挙げられる。他の哺乳動物プロモータの例は、アクチン、免疫グロブリンまたは熱ショックプロモータである。
【0031】
用語「転写開始サイト」とは、一次転写産物、即ちmRNAプリカーサに組込まれる、第一の核酸に対応する構築物内の、核酸を意味する。該転写開始サイトは、該プロモータ配列と重複していてもよい。
用語「転写終止サイト」とは、通常は、対象とする遺伝子または転写すべき該遺伝子部分の3'末端にあり、かつRNAポリメラーゼによる転写の終結をもたらす、核酸配列を意味する。
「ポリアデニル化シグナル」とは、真核生物mRNAの3'末端における特定のサイトにおいて開裂を生じ、かつ該開裂された3'末端において、約100-200アデニンヌクレオチド(ポリAテール)を持つ配列の、転写後の組み込みを引起すシグナル配列である。このポリアデニル化シグナルは、該開裂サイトの上流約10-30ヌクレオチドに配列AATAAAおよび下流側に位置するある配列を含む。様々なポリアデニル化要素が知られており、その例はtkポリA、SV40後期および初期ポリA、またはBGHポリA等である(例えば、米国特許第5,122,458号に記載されている)。
本発明の好ましい態様では、各転写単位は、プロモータまたはプロモータ/エンハンサ要素、対象とする遺伝子および/またはマーカー遺伝子並びに転写終止要素を持つ。もう一つの好ましい態様では、該転写単位は、更に二つの翻訳調節単位を含む。
【0032】
「翻訳調節要素」は、翻訳開始サイト(AUG)、終止コドン、および発現すべき各ポリペプチドに対するポリAシグナルを含む。最適の発現を得るためには、発現すべき該核酸配列の5'-および/または3'-未翻訳領域を除去、付加または変更して、あらゆる可能な不適当と思われる付随的な翻訳開始コドン、あるいは転写または翻訳レベルにおいて、発現に影響を与える可能性のある他の配列を排除することを推奨できる。発現を促進するために、リボソームコンセンサス結合サイトは、該開始コドンの直ぐ上流位置に、交互に挿入することができる。分泌性のポリペプチドを製造するために、該対象とする遺伝子は、通常ER膜におよびこれを通して、合成されたポリペプチドを搬送する、シグナルプリカーサペプチドをコードする、シグナル配列を含む。
該シグナル配列は、常にではないが、しばしば分泌されたタンパク質のアミノ酸末端に位置しており、また該タンパク質が該ER膜を介して挿入された後に、シグナルペプチダーゼによって開裂される。この遺伝子配列は、必要ではないが、通常それ自身のシグナル配列を含んでいる。この本来のシグナル配列が、存在しない場合には、異種シグナル配列を公知の方法で導入することができる。この種の多数のシグナル配列が、当業者には公知であり、また配列データバンク、例えばGenBankおよびEMBLに寄託されている。
【0033】
本発明による、一つの重要な調節要素は、内部リボソームエントリーサイト(IRES)である。このIRES要素はある配列を含み、これは該遺伝子上流の、5'-末端メチルグアノシニウムキャップ(methylguanosinium cap) (CAP構造)とは無関係に、翻訳開始を機能的に活性化し、かつ動物細胞内で、単一の転写物から、2つのシストロン(オープンリーディングフレーム)の翻訳を可能とする。このIRES要素は、直ぐ下流に位置する、該オープンリーディングフレーム翻訳用の、独立したリボソームエントリーサイトを与える。マルチシストロン性であり得る、即ちこのmRNAによって順次翻訳される多くの異なるポリペプチドまたは製品をコードできるバクテリアmRNAとは対照的に、動物細胞由来の大多数のmRNAはモノシストロン性であり、唯一つのタンパク質または製品をコードするに過ぎない。真核生物細胞におけるマルチシストロン性の転写物の場合には、その翻訳は、最も近接した上流側にある、翻訳開始サイトから開始され、また最初の終止コドンによって停止され、その後該転写物は該リボソームから放出されるであろう。このように、該mRNAによりコードされた、該第一のポリペプチドまたは生成物のみが、翻訳中に生成されるであろう。これと比較して、IRES要素を含むマルチシストロン性の転写物中の第二のまたはこれに続くオープンリーディングフレームと、機能可能な状態で結合している該転写物は、引続きその下流側に位置するオープンリーディングフレームの翻訳を可能とし、結果としてこの同一の転写物によりコードされる、2またはそれ以上のポリペプチドまたは生成物が、該真核生物細胞内で生産される。
【0034】
該IRES要素は、様々な長さおよび様々な起源のものであり得、また例えば脳心筋炎ウイルス(EMCV)または他のピコルナウイルス由来のものであり得る。様々なIRES配列およびベクターの構築におけるその利用は、例えばPelletier等, 1988; Jang等, 1989; Davies等, 1992; Adam等, 1991; Morgan等, 1992; Sugimoto等, 1994; Ramesh等, 1996; Mosser等, 1997等の文献に記載されている。
下流側に位置する遺伝子配列は、該IRES要素の3'-末端と機能可能な状態で結合しており、即ちスペーシングは、該遺伝子の発現が影響されず、あるいはほんの僅かに影響されるに過ぎず、あるいは意図した目的に対する十分な発現を示すように選択すべきである。十分な発現のために、該IRES要素と、その下流側に位置する遺伝子の開始コドンとの間の、最適な許容できる間隔は、リポータ遺伝子アッセイを利用して、該スペーシングを変更し、該スペーシングの関数として、その発現率を測定することからなる、簡単な実験によって決定できる。
記載した手段によって、最適の発現カセットを得ることが可能となり、このカセットは、異種遺伝子生成物の発現にとって極めて価値の高いものである。
【0035】
増幅可能な選択性マーカー遺伝子
本発明の好ましいベクターは、付随的に増幅可能なマーカー遺伝子の増幅および好ましくは上記ハムスター-ユビキチン/S27a遺伝子、該対象とする遺伝子および該蛍光タンパク質に対する遺伝子を含む、転写単位の同時増幅を可能とする、増幅可能な選択性マーカー遺伝子を含む。このために、対応する発現ベクターによりトランスフェクションされた宿主細胞は、適当な選別試薬の存在下で培養され、結果的に少なくとも該増幅可能な選択性マーカー遺伝子の幾つかの遺伝子コピーを持つ、これら宿主細胞のみが、複製できる(複製され得る)。好ましくは、これは、徐々に増大する量の選別試薬の存在下で、これら細胞を段階的に培養することにより達成される。
該増幅可能な選択性マーカー遺伝子は、通常幾つかの培養条件化で、真核生物細胞を生育するのに必要な酵素をコードする。例えば、該増幅可能な選択性マーカー遺伝子は、ジヒドロ葉酸レダクターゼ(DHFR)をコードすることができる。この場合、これによってトランスフェクションされた宿主細胞を、選別試薬であるメトトレキセート(MTX)の存在下で培養する場合に、該遺伝子は増幅される。
以下の表1に、その他の増幅可能な選択性マーカー遺伝子および本発明により使用できる、これに関連する選別試薬の例を与えるが、これらは、Kaufmanの概説:Method in Enzymology, 1990, 185:537-566に記載されている。
【0036】
表1:増幅可能な選択性マーカー遺伝子
【0037】
本発明によれば、使用する該増幅可能な選択性マーカー遺伝子は、好ましくはDHFRの機能を持つポリペプチド、例えばDHFRまたは蛍光タンパク質とDHFRとの融合タンパク質をコードする遺伝子である。DHFRは、プリンの生合成にとって必要である。このDHFR遺伝子を各細胞は、プリン欠乏培地内では成育できない。このDHFR遺伝子は、従ってプリンを含まない培地内で培養した細胞における遺伝子を、選別し、かつ増幅するための、有用な選択性マーカーである。このDHFR遺伝子と組み合わせて使用する選択媒体は、メトトレキセート(MTX)である。本発明は、従って、高い生産性を示す組換え宿主細胞の製造方法を包含し、この方法は以下の諸工程を含む:(i) 少なくとも対象とするタンパク質、蛍光タンパク質およびDHFRをコードする遺伝子で、宿主細胞をトランスフェクションし;(ii) 該細胞を、これら様々な遺伝子の発現を可能とする条件下で培養し;および(iii) 少なくとも該増幅可能な選択性マーカー遺伝子の増幅を可能とする選別試薬、例えばメトトレキセートの存在下で、該細胞を培養することによって、これら同時に組込まれた遺伝子を増幅する工程。好ましくは、これらトランスフェクションされた細胞を、血清の不在下で、徐々に増大する濃度のMTXを添加した、ヒポキサンチン/チミジンを含まない培地内で培養する。好ましくは、第一の増幅段階におけるMTXの濃度は、少なくとも200nMであり、およびより好ましい態様では、これは少なくとも500nMであり、また1μMまで段階的に増やすことができる。各場合において、1μMを超える濃度を使用することができる。
【0038】
哺乳動物細胞、好ましくはマウスミエローマおよびハムスター細胞が、増幅可能な選択性マーカー遺伝子として、DHFRを使用する際に好ましい宿主細胞である。細胞系CHO-DUKX(ATCC CRL-9096)およびCHO-GD44(Urlaub等, 1983)が、特に好ましい。というのは、これらは、突然変異の結果として、それ自体のDHFR活性を持たないからである。自身の内因性DHFR活性を持つ、その他の細胞型においても、このDHFR-誘導増幅を可能とするためには、該トランスフェクション過程において突然変異処理したDHFR遺伝子を用いることができ、該遺伝子は、メトトレキセートに対して低感度でタンパク質をコードする(Simonson等, 1983; Wigler等, 1980; Haber等, 1982)。
【0039】
本発明による発現ベクターの製造
本発明による発現ベクターは、理論的には、当分野において公知の従来法、例えばSambrook等(1989)に記載されているような方法により製造できる。Sambrookは、またベクターの機能的な成分、例えば適当なプロモータ(該ハムスター-ユビキチン/S27aプロモータに加えて)、エンハンサ、停止およびポリアデニル化シグナル、抗生物質耐性遺伝子選択性マーカー、複製開始点およびスプライシングシグナルをも記載している。従来のクローニングベクター、例えばプラスミド、バクテリオファージ、ファージミド、コスミドまたはウイルスベクター、例えばバキュロウイルス、レトロウイルス、アデノウイルス、アデノ-関連ウイルスおよび単純ヘルペスウイルス並びに染色体/ミニ染色体を使用して、これらを製造することができる。真核生物発現ベクターは、また典型的に原核生物の配列、例えばバクテリア内での複製および選別を可能とする、複製起点および抗生物質耐性遺伝子を含む。ポリヌクレオチド配列を導入するための、多重クローニングサイトを含む、幾つかの原核生物の発現ベクターは公知であり、その幾つかは、様々な製造元、例えばUSA、CA、ラジョラのストラタジーヌ(Stratagene)社、USA、CA、カールスバッドのインビトロゲン(Invitrogen)社、USA、WI、マディソンのプロメガ(Promega)社、またはUSA、CA、パロアルトのBDバイオサイエンスズクロンテック(Biosciences Clontech)社から市販品として得ることができる。
【0040】
該ハムスター-ユビキチン/S27aプロモータ、該対象とする遺伝子、蛍光タンパク質をコードする遺伝子および好ましくはまた該増幅可能な選択性マーカー遺伝子、例えばジヒドロ葉酸レダクターゼ、および場合により追加の調節要素、例えば該内部リボソームエントリーサイト(IRES)エンハンサまたはポリアデニル化シグナルを、当業者には馴染み深い方法で、該発現ベクター内に導入する。本発明による発現ベクターは、最低でも、ユビキチン/S27aプロモータ、該対象とする遺伝子および蛍光タンパクをコードする遺伝子を含む。好ましくは、該発現ベクターは、また増幅可能な選択性マーカー遺伝子をも含む。本発明によれば、変性ユビキチン/S27aプロモータ、例えば本出願に記載されている変性ユビキチン/S27aプロモータをも使用する。特に好ましいものは、該ユビキチンプロモータ、該対象とする遺伝子および蛍光タンパク質をコードする遺伝子が、一緒に機能可能に結合し、あるいは機能可能に結合している、発現ベクターである。
【0041】
本発明の範囲内において、「機能的結合」または「機能可能な状態で結合した」とは、2またはそれ以上の核酸配列または部分配列が、その意図した機能を達成できるように配置されている、これらの核酸配列または部分配列を意味する。例えば、プロモータ/エンハンサが、シス位置にある、結合遺伝子配列の転写を制御または変調できる場合には、該プロモータ/エンハンサは、コード遺伝子配列と機能可能な状態で結合している。必要ではないが、一般に機能可能に結合したDNA配列は、相互に近接しており、また2つのコード遺伝子配列が結合している場合、または分泌シグナル配列の場合には、同一の読み取り枠内にある。機能可能に結合したプロモータは、一般に該コード遺伝子配列の上流側に位置するが、これと近接している必要はない。エンハンサは何れかに近接している必要はないが、これらは該遺伝子配列の転写を補助する。この目的のために、これらは、該遺伝子配列の上流および下流側両者に、可能ならばこれと幾分距離を保って位置することができる。ポリアデニル化サイトは、これがある遺伝子配列の3'末端に位置する場合には、転写が該ポリアデニル化シグナルに対するコード配列を介して進行するように、該遺伝子配列と機能可能に結合する。結合は、PCR技術、適当な制限切断サイトにおける結合、またはスプライシング等の従来の組換え法に従って行うことができる。適当な制限切断サイトが利用できない場合には、合成オリゴヌクレオチドリンカーまたはアダプター自体を公知の方法で使用することができる。本発明によれば、この機能的結合は、好ましくはイントロン配列を介して起こることは無い。
【0042】
記載された一態様においては、該ユビキチン/S27aプロモータまたはその変性型、該対象とする遺伝子および該蛍光タンパク質をコードする遺伝子は、一緒に機能的に結合している。このことは、該同一のユビキチン/S27aプロモータまたはその変性型から、例えば該対象とする遺伝子および該蛍光タンパク質をコードする遺伝子両者が、発現されることを意味する。特に好ましい態様において、該機能的結合は、IRES要素を介して起こり、従ってビシストロン性mRNAは、両遺伝子から合成される。本発明による発現ベクターは、更にエンハンサ要素を含み、これは1種またはそれ以上のプロモータに対して機能的に作用する。特に好ましいものは、該ユビキチン/S27aプロモータまたはその変性型がエンハンサ要素、例えばSV40エンハンサまたはCMVエンハンサ要素と結合している、発現ベクターである。
基本的には、発現ベクター内でのこれら遺伝子の発現は、一またはそれ以上の転写単位から開始して起こる。用語:転写単位とは、転写すべき一またはそれ以上の遺伝子を含む領域として定義される。転写単位内のこれら遺伝子は、このような単位内の該遺伝子全てが、同一のプロモータまたはプロモータ/エンハンサによる転写上の制御下にあるように、相互に機能的に結合している。これら遺伝子の該転写結合の結果として、1を越えるタンパク質または生成物が、転写単位から転写され、かくして発現され得る。各転写単位は、調節要素を含み、該要素は、その中に含まれる該遺伝子配列の転写および翻訳にとって必要である。各転写単位は、同一のまたは異なる調節要素を含むことができる。IRES要素またはイントロンは、転写単位内の該遺伝子の機能的な結合のために利用できる。
【0043】
該発現ベクターは、該対象とする遺伝子、該蛍光タンパク質をコードする遺伝子および該増幅可能な選択性マーカーを発現するための、単一の転写単位を含むことができる。あるいはまた、これらの遺伝子を、2またはそれ以上の転写単位内に配置することも可能である。転写単位内での、これら遺伝子の様々な組合せが、可能である。本発明のもう一つの態様では、1、2またはそれ以上の転写単位からなる、1を越える発現ベクターが、同時トランスフェクションまたは任意の所定の順序で連続的にトランスフェクションすることにより、宿主細胞内に挿入することができる。調節要素および遺伝子の、各ベクター上でのあらゆる組合せを選択することができるが、該転写単位の十分な発現が確保できることを条件とする。必要ならば、他の調節要素および遺伝子、例えば追加の対象とする遺伝子または選択性マーカーを、該発現ベクター上に配置できる。
【0044】
従って、本発明による発現ベクターは、該蛍光タンパク質をコードする遺伝子および該増幅可能な選択性マーカー遺伝子を、一つの、または2つの別々の転写単位内に含むことができる。各転写単位は、一またはそれ以上の遺伝子生成物を転写し、発現することができる。両遺伝子が一つの転写単位に含まれている場合、これらの遺伝子は、同一のプロモータまたはプロモータ/エンハンサの制御下にあり、一方で好ましくはIRES要素は、これら全ての成分の機能的な結合を補償するために、使用することができる。該蛍光タンパク質をコードする遺伝子および該増幅可能な選択性マーカー遺伝子が、2つの別々の転写単位内に含まれる場合、これらは、同一のまたは異なるプロモータ/エンハンサの制御下にあり得る。しかし、好ましくは、その天然のまたはより弱い異種プロモータ、例えばSV40初期プロモータを、該選択性マーカー遺伝子に対して使用し、また好ましくはエンハンサを使用しない。2つの別々の転写単位を持つ発現ベクターが、本発明の範囲内において好ましい。一つの(ビシストロン性)転写単位は、該対象とする遺伝子および該蛍光タンパク質をコードする遺伝子を含み、一方で他の転写単位は、該増幅可能な選択性マーカー遺伝子を含む。好ましくは、各転写単位は、ポリAシグナル、好ましくはtkポリA、BGHポリAまたはSV40ポリAをコードする配列によって、3'末端において制限されている。
【0045】
同様に本発明において好ましいものは、上記に対して、該対象とする遺伝子が、多重クローニングサイトのみを持つベクターであり、これらは制限エンドヌクレアーゼに対する認識配列を介して、該対象とする遺伝子のクローニングを可能とする。あらゆる種類の制限エンドヌクレアーゼ並びに関連する制限エンドヌクレアーゼに対する多くの認識配列が、従来技術から公知である。好ましくは、少なくとも6個のヌクレオチドからなる配列を、認識配列として使用する。適当な認識配列のリストは、例えばSambrook等, 1989に見出すことができる。
宿主細胞
本発明の発現ベクターでトランスフェクションするために、真核生物の宿主細胞、好ましくは哺乳動物細胞、およびより好ましくは齧歯目動物の細胞、例えばマウス、ラットおよびハムスターの細胞系を使用する。対応する細胞の、本発明の発現ベクターによる首尾良いトランスフェクションは、形質転換された、一般には変性された、組換えまたはトランスジェニック細胞をもたらし、これらも本発明の課題である。
【0046】
本発明の目的にとって、好ましい宿主細胞は、ハムスター細胞、例えばBHK21、BHK TK-、CHO、CHO-K1、CHO-DUKX、CHO-DUKX B1およびCHO-DG44細胞またはこれら細胞系の誘導体/子孫である。特に好ましいものは、CHO-DG44、CHO-DUKX、CHO-K1およびBHK21細胞であり、特にCHO-DG44およびCHO-DUKX細胞が好ましい。同様に適当な細胞は、マウス由来のミエローマ細胞、好ましくはNS0およびSp2/0細胞およびこれら細胞系の誘導体/子孫である。
本発明において使用することのできるハムスターおよびマウス細胞の例を、以下の表2に与える。しかし、これら細胞、他の哺乳動物の細胞、例えばヒト、マウス、ラット、サル、齧歯目動物の細胞系(これらに限定されない)、または真核生物細胞、例えば酵母、昆虫、および植物細胞(これらに限定されない)の誘導体および子孫も、生物薬学的タンパク質を製造するための、宿主細胞として使用することができる。
【0047】
表2:ハムスターおよびマウスの産生細胞系
【0048】
ポリヌクレオチドまたは本発明の発現ベクターの一つを含む、真核宿主細胞のトランスフェクションは、従来の方法に従って行われる(Sambrook等, 1989; Ausubel等, 1994)。適当なトランスフェクション法は、例えばリポソーム媒介トランスフェクション、リン酸カルシウム共沈、エレクトロポレーション、ポリカチオン-(例えば、DEAEデキストラン)-媒介トランスフェクション、プロトプラストヒュージョン、マイクロインジェクションおよびウイルス感染法を包含する。本発明によれば、好ましくは安定なトランスフェクションが行われ、そこでは該構築物が、該宿主細胞のゲノムあるいは人工的な染色体/ミニ染色体内に組込まれるか、もしくは安定な方法で該宿主細胞内にエピソーム的に含められる。好ましいトランスフェクション法は、問題とする該宿主細胞内で、該異種遺伝子の最適なトランスフェクション頻度および発現を与えるものである。定義によれば、宿主細胞内に挿入された各配列または各遺伝子は、該宿主細胞との関連で、「異種配列」または「異種遺伝子」と呼ばれる。これは、導入すべき該配列または導入すべき該遺伝子が、該宿主細胞の内在性配列または内在性遺伝子と等価である場合にも成り立つ。例えば、ハムスター宿主細胞に導入されたハムスターアクチン遺伝子は、定義によれば異種遺伝子である。
【0049】
ヘテロダイマータンパク、例えばモノクローナル抗体(mAb)の組換え生産において、適当な宿主細胞のトランスフェクションは、理論的には2つの異なる方法により実施できる。この種のmAbは、幾つかのサブユニット、即ち重鎖および軽鎖で構成されている。これらのサブユニットをコードする遺伝子は、単一のプラスミド上の、独立したまたはマルチシストロン性の転写単位内に収容されており、次いで該プラスミドによって、該宿主細胞をトランスフェクションする。これは、該宿主細胞のゲノム内への組み込み後に、該遺伝子の化学量論的な発現量を確保するためである。しかし、独立した転写単位の場合、異なるタンパク質をコードするmRNAが、結果として同一の安定性および転写並びに翻訳効率を示すことを保証する必要がある。第二の場合において、該遺伝子の発現は、単一のプロモータによって、マルチシストロン性転写単位内で起こり、またただ一つの転写物が形成される。IRES要素を用いることにより、該遺伝子の内部翻訳開始の高い効率が、第二のおよびその後のシストロンにおいて得られる。しかし、これらシストロンに関する発現率は、第一のシストロンの発現率よりも低く、その翻訳開始は、所謂「キャップ」-依存性プレイニシェーション(pre-initiation)錯体によって、実質的にIRES-依存性の翻訳開始よりも、一層効率的になる。これらシストロンの真の等モル量発現を達成するためには、追加のシストロン間要素を導入することができ、これらは、例えば該IRES要素との組合せで、均一な発現率を確保する(WO 94/05785)。
【0050】
幾つかの異種タンパク質を同時に製造するための、本発明にとって好ましいもう一つの可能な方法は、同時トランスフェクション法であり、ここでは、遺伝子は、異なる発現ベクター内に別々に組込まれている。この方法は、ある比率の遺伝子および遺伝子生成物を、相互に調節することができ、結果的に該mRNAにおけるおよび転写および翻訳の効率におけるあらゆる差異を釣り合わせることを可能とする。更に、該発現ベクターは、そのサイズが小さいことから、より安定であり、またクローニングおよびトランスフェクション両者において、取り扱いが容易である。
従って、本発明の一特定の態様では、該宿主細胞を、1またはそれ以上の他の対象とするタンパク質をコードする遺伝子を持つ、1またはそれ以上のベクターで付随的にトランスフェクション、好ましくは同時トランスフェクションする。該同時トランスフェクションにおいて使用する該他のベクター(1または複数)は、例えば同一のプロモータ/エンハンサの組合せによる制御下において、対象とする他のタンパク質(1または複数)を、および少なくとも1種の他の選択性マーカー、例えばネオマイシンホスホトランスフェラーゼをコードする。
【0051】
本発明によれば、該宿主細胞は、血清を含まない条件化で、場合によっては動物タンパク質/ペプチドを含まない培地内で樹立され、適合されかつ培養される。市販品として入手できる媒体の例は、Ham's F12 (DE、ダイセンホフェン(Deisenhofen)のシグマ(Sigma)社)、RPMI-1640(シグマ社)、ドゥルベコ変性イーグル培地(DMEM;シグマ)、最少必須培地(MEM;シグマ)、イスコフス(Iscove's)変性ドゥルベコ培地(IMDM;シグマ)、CD-CHO(USA、CA、カールスバッドのインビトロゲン(Invitrogen))、CHO-S-SFMII(インビトロゲン)、血清を含まないCHO-培地(シグマ)およびタンパク質を含まないCHO-培地(シグマ)を包含する。これら培地各々は、場合により様々な化合物、例えばホルモンおよび/または他の成長因子(例えば、インシュリン、トランスフェリン、上皮増殖因子、インシュリン-様の成長因子)、塩(例えば、塩化ナトリウム、カルシウム、マグネシウム、リン酸塩)、バッファー(例えば、HEPES)、ヌクレオシド(例えば、アデノシン、チミジン)、グルタミン、グルコース、または他の等価な栄養物、抗生物質および/または痕跡元素を補充することもできる。血清を含まない培地が、本発明において好ましいが、該宿主細胞は、適当量の血清と混合した培地を用いて培養することも可能である。1またはそれ以上の選択性のマーカー遺伝子を発現する、遺伝的に変性された細胞を選別するために、1またはそれ以上の選別剤を、該培地に添加する。
【0052】
該用語「選別剤」とは、問題とする該選択性マーカー遺伝子に欠ける、宿主細胞の成育または生存に影響を及ぼす物質を意味する。例えば、発現された抗生物質耐性遺伝子、例えばネオマイシンホスホトランスフェラーゼの存在について選別するためには、抗生物質ジェネティシン(geneticin; G418)を、該培地添加物として使用することが好ましい。該選別剤は、また使用した該選択性マーカー遺伝子が、増幅可能な選択性マーカー遺伝子である場合には、該マーカー遺伝子の増幅を誘発する物質であっても良い(表1参照)。例えば、メトトレキセートは、該DHFR遺伝子を増幅するのに適した選択培地である。増幅を誘発するその他の選別剤の例は、表1に列挙してある。
【0053】
選択性マーカー遺伝子は、培地に対応する選別剤を添加することによって、この遺伝子を含む細胞の特異的な選別を可能とする遺伝子である。一例として、抗生物質耐性遺伝子は、正の選択性マーカーとして使用できる。この遺伝子で形質転換されている細胞のみが、該対応する抗生物質の存在下で生育でき、その結果該細胞が選別される。他方において、未形質転換細胞は、これらの選択条件下では生育または生存することができない。正、負および二官能性の選択性マーカーが存在する。正の選択性マーカーは、該選別剤に対する耐性を付与することにより、あるいは該宿主細胞における代謝上のまたは異化欠陥を補償することにより、形質転換細胞の選別およびその結果としての該細胞の富化を可能とする。これとは対照的に、この選択性マーカー用の遺伝子を受け取った細胞は、負の選択性マーカーによって、選択的に排除され得る。その一例は、単純疱疹ウイルスのチミジンキナーゼ遺伝子であり、同時に付加されたアシクロビルまたはガンシクロビルをも含む細胞内でのその発現は、その排除に導く。該増幅可能な選択性マーカーをも含む、本発明において使用するこれらの選択性マーカーは、一般に変性された突然変異体および変異型、フラグメント、機能的な等価物、誘導体、類似体および他のタンパク質またはペプチドとの融合体を包含するが、該選択性マーカーが、その選択特性を保持することを条件とする。このような誘導体は、選択性であると思われる領域またはドメインにおけるアミノ酸配列について、かなりの相同性を示す。文献には、多数の選択性マーカー遺伝子、例えば二官能性(正/負)マーカー(例えば、WO 92/08796およびWO 94/28143を参照のこと)が記載されている。真核細胞において通常使用される選択性マーカーの例は、アミノグリコシドホスホトランスフェラーゼ(APH)、ハイグロマイシンホスホトランスフェラーゼ(HYG)、ジヒドロ葉酸レダクターゼ(DHFR)、チミジンキナーゼ(TK)、グルタミンシンセターゼ、アスパラギンシンセターゼに関する遺伝子、およびネオマイシン(G418)、プロマイシン、ヒスチジノールD、ベロマイシン(belomycin)、フレオマイシン(phleomycin)およびゼオシン(zeocin)に対する耐性を付与する遺伝子を包含する。
【0054】
蛍光-活性化細胞選別(FACS)によって、形質転換した細胞を選別することも可能である。そのために、細菌性のβ-ガラクトシダーゼ、細胞表面マーカーまたは蛍光タンパク質(例えば、エクオレアビクトリア(Aequorea victoria)またはレニラレニホルミス(Renilla reniformis)または他の種を由来とする、グリーン蛍光タンパク質(GFP)およびその変異型、非-生体発光性生物、例えばディスコゾーマ(Discosoma) sp、アネモニア(Anemonia) sp、クラブラリア(Clavularia) sp、ゾアンサス(Zoanthus) sp.)を、形質転換細胞の選別のために使用することができる。
【0055】
本発明においては、該DHFR遺伝子の使用が、遺伝的に変性された(組換え)宿主細胞を選別するための、該増幅可能な選択性マーカー遺伝子として好ましい。このマーカーは、DHFR-負の基本的な細胞、例えばCHO-DG44またはCHO-DUKXを使用する場合に、選別並びにその後の増幅のために特に適している。というのは、これらの細胞が内在性のDHFRを発現せず、その結果プリンを含まない培地中で生育しないからである。結果的に、該DHFR遺伝子は、ここでは支配的な選択性マーカーとして使用することができ、また該形質転換された細胞は、ハイポキサンチン/チミジンを含まない培地内で選別される。DHFR-媒介遺伝子および増幅を行うために、メトトレキセート(MTX)を使用する。この生育特性は、MTXの添加によって臨界的に影響を受ける。通常、該MTXの濃度および増幅段階の数が増えるに従って、該細胞の醗酵強度における実質的な低下が観測される。しかし、驚いたことに、本発明によるクローン選別系を用いた場合、高濃度のMTXの存在下で、より一層強固な挙動を示す、組換え宿主細胞が豊富となる(図7を参照のこと)。このように、蛍光-活性化セルソータ(FACS)を用いて同定し、かつ選別した宿主細胞が、500nM、好ましくは1μMのMTXの存在下で培養し、かつ増幅でき、大幅に高められた生産性を与えた。このように、高い生産性を示す宿主細胞の選別法は、とりわけ本発明による成果であると考えられ、本発明の発現ベクターにより形質転換され、かつ少なくとも対象とする遺伝子、該蛍光タンパク質およびDHFR遺伝子を発現する宿主細胞が、FACS選別によって選別され、また少なくとも500nM、好ましくは1μMのMTXの存在下で、遺伝子増幅段階に付される場合には、高い生産性を示す宿主細胞の選別法は、とりわけ本発明による成果であると考えられる。
【0056】
「醗酵強度」なる用語は、上記細胞の生育特性、例えばある成長速度の維持、「規模拡大」(バイオリアクタのより大きなサイズ)に対する強靭さ、および規模拡大の際の工業的生産率を満たすための、ストックの保持における高い細胞計測数および生存能力の達成を意味する。
発現
発現なる用語は、宿主細胞内における、異種遺伝子配列の転写および/または翻訳に関連する。発現率は、一般に該宿主細胞内に存在する、対応するmRNAの量、または該対象とする遺伝子によりコードされ、製造される遺伝子生成物の量に基いて決定できる。選択されたヌクレオチド配列の転写により製造されるmRNAの量は、例えばノーザンブロットハイブリッド化、リボヌクレアーゼ-RNA-プロテクション、細胞性RNAのその場でのハイブリッド化、あるいはPCR法によって決定できる(Sambrook等, 1989; Ausubel等, 1994)。選択されたヌクレオチド配列によってコードされるタンパク質も、様々な方法、例えばELISA、ウエスタンブロット、ラジオイムノアッセイ、免疫沈降、該タンパク質の生物学的な活性の検出または該タンパク質の免疫染色およびこれに続くFACS分析により決定できる(Sambrook等, 1989; Ausubel等, 1994)。
【0057】
用語「高い発現レベル(または率)、高い発現、高められた発現または高(い)生産性」とは、長期間持続する、かつ十分に高い発現、または宿主細胞に導入された異種配列、例えば治療用のタンパク質をコードする遺伝子の合成を意味する。高められたまたは高い発現または高い発現レベルまたは発現率もしくは高い生産性は、本発明による細胞が、ここに記載する本発明による方法の一つで培養し、またこの細胞が、1日当たり少なくとも約5pg(5pg/日/細胞)を越える所定の遺伝子生成物を製造する場合に存在する。高められたまたは高い発現または高い発現レベルまたは発現率もしくは高い生産性は、本発明による細胞が、1日当たり少なくとも約10pg(10pg/日/細胞)を越える所定の遺伝子生成物を製造する場合にも存在する。高められたまたは高い発現または高い発現レベルまたは発現率もしくは高い生産性は、特に、本発明による細胞が、1日当たり少なくとも約20pg(20pg/日/細胞)を越える所定の遺伝子生成物を製造する場合にも存在する。特別に高められたまたは高い発現、または特別に高い発現レベルまたは発現率もしくは特別に高い生産性は、本発明による細胞が、1日当たり少なくとも約30pg(30pg/日/細胞)を越える所定の遺伝子生成物を製造する場合に存在する。
【0058】
本発明による、高いまたは高められた発現、高い生産性または高い発現レベルまたは発現率は、様々な方法で達成することができる。例えば、増幅可能な選択性マーカーに対する遺伝子を含む、対象とする該遺伝子の同時発現によって、高度に該異種遺伝子を発現する細胞を選別し、かつ同定することが可能である。該増幅可能な選択性マーカーは、安定にトランスフェクションされた宿主細胞の選択を可能とするばかりでなく、対象とする該異種遺伝子の、遺伝子増幅をも可能とする。該核酸の付随的なコピーは、該宿主細胞のゲノムに、付随的な人工的/ミニ染色体にあるいはエピソームとして位置するポリヌクレオチドに組込むことができる。この手順は、付随的な選択性マーカーとして、1またはそれ以上の蛍光タンパク質(例えば、GFP)または細胞表面マーカーを含む組換え宿主細胞の、FACS-アシスト選別と組み合わせることができる。高められた発現を達成するための他の方法(および異なる方法の組合せも利用可能である)は、例えば(人工的な)転写因子の利用、より調節性の良い内在性または異種遺伝子発現用の、天然または合成薬物による該細胞の処理、mRNAまたは該タンパク質の安定性(半減期)の改善、mRNA翻訳開始の改善、エピソームプラスミドの使用による、遺伝子用量の増大(複製起源としてのウイルス配列、例えばSV40、ポリオーマ、アデノウイルス、EBVまたはBPVの使用に基く)、増幅-促進配列の利用(Hemann等, 1994)またはDNAコンカテマーに基くインビトロ増幅系(Monaco等, 1996)。
【0059】
本発明によれば、該対象とする遺伝子と該蛍光タンパク質をコードする遺伝子との、組み合わせた転写を行う。得られるビシストロン性のmRNAは、該対象とするタンパク質および該蛍光タンパク質両者を発現する。該対象とするタンパク質および該蛍光タンパク質の発現に関するこの組合せに基いて、本発明によれば、発現された蛍光性タンパク質によって、例えば蛍光-活性化セルソータ(FACS)を用いた選別によって、高い生産性を示す組換え宿主細胞を選択し、かつ単離することが、容易に可能となる。
高い生存能力および該所定の遺伝子生成物の、高い発現率を示す、組換え宿主細胞の選別は、多段階法である。本発明による発現ベクターでトランスフェクションされ、あるいは場合により、他のベクターにより同時トランスフェクションされている宿主細胞は、例えば蛍光タンパク質をコードし、該対象とする遺伝子と結合した遺伝子の発現について、少なくとも検討されるが、これは蛍光タンパク質の最大の発現率を示す細胞/細胞集団を同定し、かつ選別するためである。好ましくは、蛍光タンパク質の最大の発現率を示す細胞の10%に属する細胞のみが、選別され、かつ更に培養される。事実、このことは、該蛍光性細胞の最も明るい10%が、選別され、かつ更に培養される。従って、細胞混合物の該蛍光性細胞の最も明るい5%、好ましくは最も明るい3%またはさらには最も明るい1%を、選別し、かつ複製することも可能である。特に好ましい態様では、該蛍光性細胞の最も明るい0.5%または最も明るい0.1%のみを、選別し、かつ複製する。
【0060】
この目的のために、本発明の発現ベクターで前もって形質転換されている細胞を、選択培地で培養する。この選択培地は、場合により該増幅可能な選択性マーカーに対して特異的な選別剤をも含む。段階的に高められる選別剤の濃度を使用して、徐々に高くなる選択圧力を及ぼすことができる。
この選別段階は、細胞プールについて、あるいは予め選別した細胞プール/細胞クローンについて行うこともできる。1またはそれ以上、好ましくは2またはそれ以上および特に3またはそれ以上の選別段階を実施し、一方で各選別段階間で、所定の期間、例えばプールの場合にはほぼ2週間に渡り、該細胞を培養し、かつ複製する。
少なくとも対象とする遺伝子および該増幅可能な選択性マーカー遺伝子のコピー数を増大させるために、所定の宿主細胞を、1またはそれ以上の遺伝子増幅段階に付すことができる。メトトレキセートを用いた、段階的な遺伝子増幅方法は、例えば米国特許第5,179,017号に記載されている。この発明によれば、達成することのできる高い生産性は、遺伝子コピー数の多さとは結び付けられない。これは寧ろ、該高性能クローンの高められた安定性および醗酵強度の現れである。従って、これは、必要とされる遺伝子増幅段階数を減じ、また例えばただ一回の遺伝子増幅段階の実施を可能とする。
【0061】
従って、本発明は、細胞を選別する方法に関連し、この方法は以下に列挙する諸工程を含む:
(i) 少なくとも、本発明によるベクターの一つを含む、適当な宿主細胞の形質転換工程、ここで該発現ベクターのDNAは、好ましくは該宿主細胞のゲノム内に、あるいは人工的な染色体/ミニ染色体内に安定に組込まれている;
(ii)該対象とする遺伝子および該蛍光タンパク質の発現を可能とする条件下で、該形質転換した細胞を培養する工程;
(iii) 該細胞を、少なくとも1種の選別剤の存在下で培養し、該選別剤の存在下で成育できる細胞のみを複製する工程;
(iv) 蛍光タンパク質に対して最大の発現率を示す細胞を、細胞混合物から選別する工程、ここで該細胞は、蛍光-活性化セルソータ(FACS)によって、検出されかつ選別される;および
(v) 該蛍光タンパク質に対して最高の発現率を示す、該選別された細胞を培養する工程。
場合により、上記工程ii)〜v)は、上記工程v)に従って得た細胞について、1またはそれ以上の回数に渡り繰返すことができる。更に、該形質転換細胞は、場合により該増幅可能な選択性マーカー遺伝子の増幅へと導く、選別剤の存在下で培養することにより、1またはそれ以上の遺伝子増幅段階に付すことも可能である。この段階は、まだ選別されていない細胞および既に1またはそれ以上の回数に渡り予備的に選別されている細胞両者について実施できる。
【0062】
本発明は、また同様に選別された細胞を、複製し、かつ対象とするコード性の該遺伝子生成物を製造する方法にも関連する。好ましくは、該選択された高生産性の細胞は、血清を含まない培地内で培養し、および好ましくは該対象とする遺伝子の発現を可能とする条件の下で、懸濁培養により培養する。該対象とするタンパク質は、好ましくは分泌された遺伝子生成物として、該細胞培養培地から得られる。しかし、このタンパク質が、分泌シグナル無しに発現される場合には、該遺伝子生成物は、また細胞溶解物から単離することもできる。他の組換えタンパク質および宿主細胞タンパク質を実質的に含まない、純粋な均質な製品を得るために、公知の精製段階を実施する。まず、通常は、細胞および細胞デブリを、該培地または溶解物から除去する。次いで、所定の遺伝子生成物を、例えばイムノアフィニティーおよびイオン交換カラム上での分別、エタノール沈殿、逆相HPLCまたはセファデックス、シリカまたはDEAE等のカチオン交換樹脂上でのクロマトグラフィーによって、混入した溶解性のタンパク質、ポリペプチドおよび核酸から分離することができる。組換え細胞により発現される異種タンパク質の精製法は、当業者には公知であり、文献、例えばHarris等(1995)およびScopes (1988)に記載されている。
【0063】
以下、本発明を、幾つかの非-限定的な例示的態様を参照して、より一層詳細に説明する。
実施例
略号:
AP:アルカリホスファターゼ;bp:塩基対;CHO:チャイニーズハムスター卵巣;DHFR:ジヒドロ葉酸レダクターゼ;ELISA:酵素結合イムノソルベント検定法;FACS:蛍光-活性化セルソータ;FAP:繊維芽細胞-活性化タンパク質;GFP:グリーン蛍光タンパク質;HBSS:ハンクスの平衡塩類溶液;HT:ハイポキサンチン/チミジン;HRPO:西洋ワサビパーオキシダーゼ;IRES:内部リボソームエントリーサイト;kb:キロベース;mAb:モノクローナル抗体;MTX:メトトレキセート;PCR:ポリメラーゼ連鎖反応;slCAM:可溶性細胞内接着分子
【0064】
方法
1. 細胞培養およびトランスフェクション
細胞CHO-DG44/dhfr-I-(Ulraub等, 1983)を、細胞培養フラスコ内で、湿潤大気および5%CO2中で37℃にて、ハイポキサンチンおよびチミジンを補充した、血清を含まないCHO-S-SFMII培地(DE、カールスルーエ(Karlsruhe)のインビトロゲンGmbH)内で、懸濁細胞として、永続的に培養した。生細胞数および生存率を、CASY1細胞計数器(DE、シェルフェシステム(Schaerfe System))を用いて、あるいはトリパン青染色により決定し、また次に該細胞を1-3×105/mLなる濃度で播種し、2-3日毎に操作(alle 2-3 Tage passagiert)した。
【0065】
リポフェクタミンプラス試薬(Lipofectamine Plus Reagent; インビトロゲンGmbH)を、CHO-DG44のトランスフェクションのために使用した。各トランスフェクション混合物に対して、全体で1μgのプラスミド-DNA、4μLのリポフェクタミンおよび6μLのプラス試薬を、製造業者の指示に従って一緒に混合し、200μLなる容量で、0.8mLのHT-補充CHO-S-SFMII培地内の、6×105個の指数増殖するCHO-DG44細胞に添加した。細胞インキュベータ内で37℃にて3時間インキュベータした後、2mLのHT-補充CHO-S-SFMII培地を添加した。安定にトランスフェクションされたCHO-DG44細胞の、上記DHFRに基く選別のために、トランスフェクションの2日後に、該細胞を、ハイポキサンチンおよびチミジンが全く添加されていない、CHO-S-SFMII培地に移し、3〜4日ごとに培地を交換した。一つの発現ベクターが、DHFRおよびネオマイシンホスホトランスフェラーゼ選択性マーカーを含む他の発現ベクターを含む、同時トランスフェクションの場合の、DHFRに基くおよびネオマイシンホスホトランスフェラーゼに基く選別においては、G418(インビトロゲン)をも、400μg/mLなる濃度にて、該培地に添加した。
該組込み異種遺伝子のDHFRに基く遺伝子増幅は、HTを含まないCHO-S-SFMII培地に、5-2000nMなる濃度で、選別剤MTX(DE、ダイセンホーフェン(Deisenhofen)のシグマ社)を添加することにより達成した。
【0066】
2. 発現ベクター
該発現を解析するために、pAD-CMVベクター(Werner等, 1998)を基本とし、CMVエンハンサ/ハムスターユビキチン/S27aプロモータ(WO 97/15664)の組合せにより、異種遺伝子の構成性発現を媒介する、真核細胞発現ベクターを使用した。その基本ベクターpBIDは、該ベクターpBIN内で、増幅可能な選択性マーカーとして機能する(例えば、EP 0 393 438)DHFRミニ遺伝子を含むが、該DHFRミニ遺伝子は、ネオマイシン耐性遺伝子で置換えられている(図2)。この目的のために、SV40初期プロモータおよびTK-ポリアデニル化シグナルを含む、該選択性マーカーネオマイシンホスホトランスフェラーゼを、1640bpのBsu36Iフラグメントとして、市販のプラスミドpBK-CMV(USA、CA、ラジョラのストラタジーヌ)から単離した。該フラグメントの末端を、クレノウ-DNA-ポリメラーゼで満たすための反応の後に、該フラグメントを、同様にクレノウ-DNA-ポリメラーゼで処理した、該ベクターpBIDの3750bpのBsu36I・StuIフラグメントを用いて結合した。
【0067】
このビシストロン性基本ベクターpBIDG(図2)において、そのIRES-GFP遺伝子領域は、ベクターpIRES2-EGFP(USA、CA、パロアルトのクロンテック社)から単離し、また該ベクターpBID内のCMVエンハンサ/プロモータの制御下に置き、結果的に該プロモータ領域とIRES-要素との間の、多重クローニングサイトを保持した。以下の手順を使用した。該プラスミドpIRES2-EGFPが、鋳型として機能するPCR突然変異誘発においては、一方で、該IRES配列内のHindIII切断サイトAAGCTTが、突然変異原性のプライマーを用いることにより配列ATGCTTに転化され、かつ脱離された。他方、Xbal切断サイトは、該IRES配列の5'末端に対して相補性を持つプライマーによって挿入され、あるいはSpel切断サイトが、該GFP配列の3'末端に対して相補性を持つプライマーによって導入された。完全なIRESおよびGFP配列を含む、得られるPCRフラグメントを、XbalおよびSpelで消化し、該ベクターpBIDの多重クローニングサイトの3'末端における、単一のXbal切断サイト内にクローニングした。
ヒトslCAM遺伝子を、pAD-slCAM(Werner等, 1998)由来のHindIII/Sallフラグメントとして単離し、該ベクターpBIDGの対応する切断サイトにクローニングして、ベクターpBIDG-slCAM(図3)を得た。
【0068】
モノクローナルヒト化F19抗体を発現するために、その重鎖を、プラスミドpG1D105F19HC(NAGENESEQ: AAZ32786)から、1.5kbのNael/HindIIIフラグメントとして単離し、またEcoRI(クレノウ-DNA-ポリメラーゼで満たされている)およびHindIIIで消化した、該ベクターpBIDG内にクローニングして、ベクターpBIDG-F19HC(図3)を得た。他方で、その軽鎖は、プラスミドpKN100F19LC(NAGENESEQ: AAZ32784)から、1.3kbのHindIII/EcoRIフラグメントとして単離し、該ベクターpBINの対応する切断サイト内にクローニングして、ベクターpBIN-F19LC(図3)を得た。
3. FACS
フローサイトメトリー分析および選別を、コールターエピックスアルトラ(Coulter Epics Altra)装置を用いて実施した。このFACSは、励起波長488nmを持つヘリウム-アルゴンレーザーを備えている。その蛍光強度は、蛍光タンパク質と適合する波長において吸収され、付随するソフトウエア:コールターエキスポ32(Coulter Expo32)によって処理する。選別は、8000-10000事象/秒なる速度にて実施した。該懸濁された細胞を遠心分離(180xgにて5分間)し、HBSS内での細胞密度を、1-1.5×107/mLに調節した。次いで、これらの細胞を、その蛍光タンパク質シグナルに従って選別した。これらの細胞を、既に培地を含む試験管に採り、次いで遠心分離処理し、選別された細胞の数に応じて、適当な培養容器に播種した。
【0069】
4. ELISA
安定にトランスフェクションされたCHO-DG44細胞の上澄み中のslCAM力価は、一方でヤギ抗-ヒトIgG Fcフラグメント(DE、ハンブルグのディアノバ(Dianova))および他方でAP-複合ヤギ抗-ヒトκ軽鎖抗体(シグマ社)を使用して、標準的な手順に従って、ELISA法により定量した。精製されたF19抗体を、基準として用いた。
生産性(pg/細胞/日)は、式:pg/((Ct-Co)t/ln(Ct-Co))に従って、算出した。ここで、CoおよびCtは、夫々播種した際および収穫した際の生細胞数であり、またtは培養時間である。
【0070】
実施例1:CMVおよびハムスターユビキチン/S27a-プロモータ活性の比較
ハムスターユビキチン/S27a-プロモータの活性と、真核生物の発現ベクターにおいて頻繁に使用されるCMVの活性とを比較するために、CHO-DG44細胞を、様々な組換えベクターでトランスフェクションした。異種遺伝子生成物、即ち一方ではリソソーム酵素および他方ではIgG1-抗体が、該CMV-プロモータの制御下で、あるいは該ハムスターユビキチン/S27a-プロモータの制御下で発現された。これら2種のプロモータは、機能的にCMV-エンハンサと結合していた。BGH ポリAを、該異種遺伝子に対する終止シグナルとして使用した。該CMV-プロモータを含む該発現ベクターは、変性されたpcDNA3(「CMV」、インビトロゲン)またはpBluescriptベクター(「CMV2」、ストラタジーヌ)に基いており、また付随的に該増幅可能な選択性マーカージヒドロ葉酸レダクターゼをコードした。他方で該ハムスタープロモータを含む発現ベクターは、pAD-CVMベクター(Werner等, 1998)に基くものであった。この抗体の重鎖および軽鎖を発現するために、選択性マーカーとして、ネオマイシン耐性遺伝子を含む、第二のベクターを用いて同時トランスフェクションを行った。しかし、該CMVエンハンサは、SV40エンハンサで置換することもできる。
【0071】
96ウエルプレートでの限界希釈によって、細胞クローンを選別し、またHTを含まない培地(同時トランスフェクションの場合には、400μg/mLのG418を添加した)中で単離した。組換えタンパク質の最大の生産率を持つ細胞クローンを、各場合において、稀釈クローニングと組み合わせて、メトトレキセート濃度を段階的に5nMから、50nMを経て、500nM〜2μMまで高めることにより、段階的なDHFRに基く遺伝子増幅にかけた。各増幅段階において、該最大の生産率を持つ約20〜30クローンを選択した。
一般に、該ハムスタープロモータが、最大の性能を持つことが分かった。該リソソーム酵素発現および該抗体の発現両者において、生産性または力価は、異種遺伝子が、CMVプロモータの制御下で発現される細胞において観測される、これらの値の、2〜5倍高い値として得られた。図1は、この特定の増幅段階における、最良の細胞クローンの相対的な力価および相対的な比生産率を、一例として示す。ここで、該CMVプロモータに基く、特定の異種遺伝子に関する発現を1に設定した(該リソソーム酵素に対してCMV1、該抗-体に対してCMV2)。
【0072】
実施例2:GFPに基くFACS選別による、高い発現性を示すslCAM細胞の単離
細胞間接着分子ICAM1、slCAMの可溶性型は、放射性同位元素を含まないものに関する可能な処理である。というのは、これが、ライノウイルスとの結合に関してICAMレセプタと競合し、このようにして該細胞の侵入およびその後の感染に対する必要条件となる、該ICAMレセプタとの相互作用を低下もしくは阻害さえも起こす(Bella等, 1999; Marlin等, 1990)。
【0073】
slCAMを、CHO細胞において、単鎖タンパク質(480アミノ酸)を発現するための一例として選択した。このために、CHO-DG44をpBIDG-slCAMでトランスフェクションした(図3)。pBIDG-slCAMでトランスフェクションした細胞内での、GFPの付随的な発現は、FACSを基本とする選別法の利用を可能にする。これら治療用タンパク質slCAMおよびGFPは、ビシストロン性転写単位により連帯的に発現され、また該DHFRは別の転写単位により発現される。HTを含まないCHO-S-SFMII培地における最初の選別の2〜3週間後に、最大のGFP蛍光を示す細胞の5%を選別した。約2週間の培養後、最大のGFP蛍光を示す5%の細胞を再度単離した。この周期的な選別操作を全体で6回行った。slCAMの生産性とGFP蛍光との間の良好な相関関係が明らかとなった(図4)。このようにして、FACS-アシスト選別のみによって、如何なるMTX増幅段階も使用することなしに、16pg/細胞/日までの高い比生産率を持つ細胞のプールが、極めて短い期間内に単離された(図5)。該GFPに基く選別と、単一回のこれに続くMTX増幅段階とを組合せることによって、生産性を30pg/細胞/日まで高めることさえ可能となった(図6)。
【0074】
これらの生産性は、第4回目の選別後の、細胞プールの500nMのMTXによる増幅、および第6回目の選別後の、細胞プールの2μMのMTXによる増幅両者により達成された。通常、5-20nMのMTXという、極めて低いMTX濃度にて開始される、段階的な増幅とは対照的に、増幅効果を達成するために、最初からより高いMTX濃度を使用する必要があった。従って、5または50nMのMTXを、第4回目の選別から細胞に添加した結果、あるいは500nMのMTXを、第6回目の選別から細胞に添加した結果として、生産性における有意な増加は何等観測されなかった(図6)。明らかに、出発プールにおけるDHFRの濃度は、既に極めて高く、その結果全DHFR阻害は、MTXの高い用量によってのみ達成できた。更に、予備的に選別した細胞プールは、高い初期MTX用量にも拘らず極めて良好に該選別段階を生き永らえた。即ち、高い生存率を示す細胞群が、従来の段階的な遺伝子増幅法により達成されるよりも、短期間内に得られた(図7)。
【0075】
実施例3:GFPを基本とするFACS選別による、mAb F19の高い発現性を持つ細胞の単離
同時トランスフェクションにおいて、CHO-DG44細胞を、プラスミドの組合せ、pBIDG-F19HCおよびpBIN-F19LCによってトランスフェクションした(図3)。この発現されたヒト化抗体F19は、反応性ストローマ繊維芽細胞によって合成される、表面分子FAPに向けられる(EP 0 953 639をも参照)。使用したベクターの構成において、該抗体の2つのタンパク質鎖は、それら自身のベクターによって発現され、該ベクターは、また付随的に別の転写単位内の、ネオマイシンホスホトランスフェラーゼ選択性マーカーまたはDHFRをもコードする。さらに、他の選択性マーカー、GFPは、ベクターpBIDG-F19HC内に含まれている。IRES要素による該GFPおよび重鎖発現の転写による結合によって、ベクターpBIDG-F19HC/pBIN-F19LCによる、CHO-DG44細胞の同時トランスフェクションにおいては、該抗体F19の高い発現性を示す細胞は、周期的なFACS選別を利用して、単に高いGFP含有率を持つ細胞を選別することにより、迅速に単離することができた。
【0076】
このため、400μg/mLのG418を添加した、HTを含まないCHO-S-SFMII培地内で、該トランスフェクションした細胞プールの初めの2〜3週間の選別後に、最大のGFP蛍光を示す細胞の5%を、FACSにより選別した。この選別は、全体として6回に渡り行い、各選別操作間に約2週間の培養期間を置いた。驚いたことに、F19の生産性と、GFP蛍光との間には、良好な相関関係があることが分かった(図8)が、両タンパク質鎖は、それ自身のベクターから発現され、またGFPを基本とするFACS選別においては、GFPとの転写による結合の結果として、重鎖のみの発現について選別できたに過ぎなかった。この生産性は、10pg/細胞/日(図9)まで高めることができ、また更に第5回目の選別による細胞プールから、1000nMのMTXを該選択培地に添加することにより、後のただ一回のMTX増幅段階によって、平均値37pg/細胞/日まで高めることができた。また、比較データは、ハムスタープロモータと、該CMVエンハンサの代わりのSV40エンハンサとを、機能的に結合することにより得ることができた。同時に、増大する量のMTXを用い、一般に4つの増幅段階を含む、従来の段階的な遺伝子増幅法と比較して、高い生産性を示す細胞を選別するための開発時間を、半分だけ約120日まで短縮し、また付随的に、開発設備およびコストにおる大幅な減少をもたらすことができた。
【0077】
文献
Adam, M.A. et al., J Virol 1991, 65, 4985 - 4990
Ausubel, F.M. et al., Current Protocols in molecular biology. New York : Greene Publishing Associates and Wiley-Interscience. 1994 (updated)
Bella, J. et al., J. Struct. Biol. 1999, 128, 69 - 74
Bennett, R.P. et al., BioTechniques 1998, 24, 478 482
Chalfie, M. et al., Science 1994, 263, 802 - 805
Chamov, S.M. et al., Antibody Fusion Proteins, Wiley-Liss Inc., 1999
Davies, M.V. et al., J Virol 1992, 66, 1924 1932
Faisst, S. et al., Nucleic Acids Research 1992, 20, 3 26
Gossen, M. et al., Curr Opi Biotech 1994, 5, 516 - 520
Gubin, A.N. et al., Biochem Biophys Res Commun 1997, 236, 347 350
Haber, D.A. et al., Somatic Cell Genetics 1982, 8, 499 - 508
Harris et al., Protein Purification : A Practical Approach, Pickwood and Hames, eds., IRL Press, 1995
Hemann, C. et al., DNA Cell Biol 1994, 13 (4), 437 - 445
Huston, C. et al., Proc Natl Acad Sci USA 1988, 85 (16), 5879 - 5883
Hu, S. et al., Cancer Res. 1996, 56 (13), 3055 - 3061
Jang, S.K. et al., J Virol 1989, 63, 1651 1660
Kaufman, R.J. et al., J Mol Biol 1982, 159, 601 - 621
Kaufman, R.J., Methods in Enzymology 1990, 185, 537 - 566
Kortt, A.A. et al., Protein Engineering 1997, 10 (4), 423 - 433
Levenson, V.V. et al., Human Gene Therapy 1998, 9, 1233 1236
Lottspeich F. and Zorbas H. eds., Bioanalytic, Spektrum Akad. Verl., 1998
Lovejoy, B. et al., Science 1993, 259, 1288 - 1293
Marlin, S.D. et al., Nature 1990, 344, 70 - 77
Meng, Y.G. et al., Gene 2000, 242, 201 - 207
Monaco, L. et al., Gene 1996, 180, 145 - 150
Morgan, R.A. et al., Nucleic Acids Research 1992, 20, 1293 - 1299
Mosser, D.D. et al., BioTechniques 1997, 22, 150 - 161
Nolan, G.P. et al., Proc Natl Acad Sci USA 1988, 85, 2603 2607
Ohshima, Y. et al., J Mol Biol 1987, 195, 247 - 259
Pack, P. et al., Biotechnology 1993, 11, 1271 1277
Pack, P. et al., J Mol Biol 1995, 246 (11), 28 - 34
Pelletier, J. et al., Nature 1988, 334, 320 325
Perisic, O. et al., Structure 1994, 2, 1217 - 1226
Primig, M. et al., Gene 1998, 215, 181 189
Ramesh, N. et al., Nucleic Acids Research 1996, 24, 2697 2700
Sambrook, J., Fritsch, E.F. & Maniatis, T.,Molecular Cloning: A Laboratory Manual Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1989
Scopes, R., Protein Purification, Springer Verlag, 1988
Simonson, C.C. et al., Proc Natl Acad Sci USA 1983, 80, 2495 - 2499
Sugimoto et al., Biotechnology 1994, 12, 694 - 698
Urlaub, G. et al., Cell 1983, 33, 405 - 412
Werner, R.G. et al., Arzneim.-Forsch./Drug.Res. 1998, 48, 870 - 880
Wigler, M. et al., Proc Natl Acad Sci USA 1980, 77, 3567 3570
【図面の簡単な説明】
【0078】
【図1】異種遺伝子生成物が、CMVプロモータの制御下、またはハムスターユビキチン/S27aプロモータの制御下の何れかで発現される、組換え細胞を用いて得た、発現レベルの比較を示す。これら二つのプロモータは、機能的にCMVエンハンサおよび終結配列と結合しており、BGHポリAは、何れの場合においても同一である。CMV1の場合には、発現ベクターはpcDNA3を基本とし(DE、カルスルーエ(Kalsruhe)のインビトロゲン)、CMV2の場合には、これはpBluescriptを基本とする発現ベクターであり(US、CA、ラジョラのストラタジーヌ)、およびCHOの場合、これは、pAD-CMVを基本とする発現ベクターである(Werner等, 1998)。リソソーム酵素の発現において、全ての発現ベクターは、増幅可能な選択性マーカー、ジヒドロ葉酸レダクターゼ(DHFR)を含み、また該異種遺伝子の発現は、メトトレキセート(MTX)による後の増幅段階により高められた。抗体(Ab)の二つの鎖を発現するために、同時トランスフェクションを、該選択性のマーカーとしてネオマイシン耐性遺伝子を含む第二のベクターを用いて行った。得られる力価または比生産率は、CMVプロモータを基本とする発現との関連で与えられ、後者の発現を1に設定する(酵素に対してCMV1、Abに対してCMV2)。
【図2】CHO-DG44細胞内で、組換えタンパクを発現するのに使用した基本ベクターの概略的な表示である。「P/E」は、CMVエンハンサとハムスターユビキチン/S27aプロモータとの組合せであり、「P」は、単独でプロモータ要素を表し、「T」は、転写に関する終止シグナルであり、転写されたmRNAのポリアデニル化のために必要とされる。各転写単位における転写開始位置および方向は、矢印で示されている。該異種遺伝子をクローニングするために、制限エンドヌクレアーゼに関する多重切断サイト(多重クローニングサイト-mcs)を持つ配列領域を、該プロモータ要素の背後に挿入する。該増幅可能な選択性マーカージヒドロ葉酸レダクターゼを「dhfr」と略記し、また該選択性マーカーネオマイシンホスホトランスフェラーゼを「neo」と略記する。脳心筋炎ウイルス由来の「IRES」要素は、ビシストロン性転写単位内の内部リボソームエントリーサイトとして機能し、またこれに伴うグリーン蛍光タンパク質「GFP」の翻訳を可能とする。
【図3】各々生物薬学的なタンパク質をコードし、かつCHO-DG44細胞をトランスフェクションするために使用された、真菌発現ベクターを、概略的に示した図である。「P/E」は、CMVエンハンサとハムスターユビキチン/S27aプロモータとの組合せであり、「P」は、単独でプロモータ要素を表し、「T」は、転写に関する終止シグナルであり、転写されたmRNAのポリアデニル化のために必要とされる。各転写単位における転写開始位置および方向は、矢印で示されている。該増幅可能な選択性マーカージヒドロ葉酸レダクターゼを「dhfr」と略記し、また該選択性マーカーネオマイシンホスホトランスフェラーゼを「neo」と略記する。脳心筋炎ウイルス由来の「IRES」要素は、ビシストロン性転写単位内の内部リボソームエントリーサイトとして機能し、またこれに伴うグリーン蛍光タンパク質「GFP」の翻訳を可能とする。「slCAM」は、可溶性の細胞内接着分子をコードし(US 5,412,216)、一方「F19HC」および「F19LC」は、ヒト化抗体F19の重鎖および軽鎖を夫々コードする(EP 953 639)。
【図4】細胞プールZB1を例とした、該slCAMの生産性と該GFP蛍光との相関関係を示す図である。この細胞プールは、ベクターpBIDG-slCAMによるトランスフェクションによって得られたものであり、ここでこれらの治療用タンパク質slCAMおよびGFPは、ビシストロン性転写単位によって協同的に発現される。このプールを、周期的なGFPを基本とするFACS選別にかけた。各選別段階(sort)の後に、該プールの細胞培養上澄中のslCAMの濃度を、ELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率を算出した(pg/c*d)。各データ点は、少なくとも3回の培養実験の平均値である。全体で6回の選別を行った。
【図5】細胞プールZB1を例として用いた、高い発現性を示すslCAM細胞の、GFPを基本とするFACS選別による単離を示す図である。この細胞プールは、ベクターpBIDG-slCAMによるトランスフェクションによって得られたものであり、ここでこれらの治療用タンパク質slCAMおよびGFPは、ビシストロン性転写単位によって一緒に発現される。このプールを、周期的なGFPを基本とするFACS選別にかけた。各選別後に、該プールの細胞培養上澄中のslCAMの濃度を、ELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率を算出した(pg/c*d)。各データ点は、少なくとも3回の培養実験の平均値である。全体で6回の選別を行った。
【図6】細胞プールZB1を例として用いた、MTX増幅段階とGFPを基本とする選別との組合せにより達成された、slCAMの生産性における増大を示す図である。ベクターpBIDG-slCAMによるトランスフェクションによって得られたものである、この細胞プールを、周期的なGFPを基本とするFACS選別にかけた。第4回目および第6回目の選別後に、該培地にメトトレキセート(MTX)を添加(5nM、50nM、500nMまたは2μMのMTX)することによって、DHFR-媒介遺伝子増幅を行った。該プールの細胞培養上澄中のslCAMの濃度を、ELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率(pg/c*d)を算出した。各データ点は、少なくとも3回の培養実験の平均値を表す。
【図7】様々な用量のメトトレキセートを該培地に添加した後の、細胞プールの生存率を示す図である。ベクターpBIDG-slCAMによるトランスフェクション(図3)によって得た、該細胞プールZB1を、周期的なGFPを基本とするFACS選別にかけた。第4回目および第6回目の選別後に、該培地にメトトレキセート(MTX)を添加することにより、DHFR-媒介遺伝子増幅を行った。細胞数および生存率は、トリプタンブルーで染色することにより、選別段階中に測定し、また培養中の数日(dic)に渡り追跡した。
【図8】細胞プールZB1を例として用いた、抗体の生産性(mAb F19)と、GFP蛍光性との間の相関関係を示す図である。この細胞プールは、ベクターpBIDG-F19HCとpBIN-F19LCとの組合せによるトランスフェクションによって得た(図3)。このプールを、周期的なGFPを基本とするFACS選別に掛けた。各選別後に、該プールの細胞培養上澄中の抗体F19の濃度を、ELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率(pg/c*d)を算出した。各データ点は、少なくとも3回の培養実験の平均値を表す。全体で6回の選別を行った。
【図9】細胞プールZB1を例として用いた、FACSを用いたGFPを基本とする選別による、高い発現性を持つmAbF19細胞プールの単離を示す図である。ベクターpBIDG-F19HCとpBIN-F19LCとの同時トランスフェクション(図3)によって得た、この細胞プールを、周期的なGFPを基本とするFACS選別にかけた。該プールの細胞培養上澄中の抗体F19の濃度を、各選別後にELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率(pg/c*d)を算出した。各データ点は、少なくとも3回の培養実験の平均値を表す。
【発明の詳細な説明】
【0001】
技術分野
本発明は、高い生産性を示す組換え細胞の選別方法、異種遺伝子生成物の製造方法および発現ベクター並びに該ベクターでトランスフェクションされた、これらの方法において使用可能な宿主細胞に関するものである。
【0002】
発明の背景
哺乳動物細胞は、複合生物薬学的タンパク質を製造するための好ましい宿主細胞である。というのは、翻訳後に行われる修飾が、機能的にも薬物動力学的にも、ヒトと相溶性であるからである。工業的には、関連する細胞型は、ハイブリドーマ、ミエローマCHO(チャイニーズハムスター卵巣)細胞およびBHK(ベビーハムスター腎臓)細胞である。この宿主の培養は、次第に血清およびタンパク質を含まない条件の下で行われるようになっている。その理由は、付随するコストの減少、該組換えタンパク質の精製における妨害の低下、病原体(例えば、プリオンおよびウイルス)導入の可能性の低下にある。宿主としてのCHO細胞の使用は、これら細胞が、血清およびタンパク質を含まない培地内での懸濁成長に適したものとなるにつれて、より一層普及するようになり、また規制当局により安全な生産細胞と見做され、かつ許容される。
【0003】
対象とする異種遺伝子を発現する、安定な哺乳動物細胞系を製造するためには、一般に選択性のあるマーカー遺伝子、例えばネオマイシントランスフェラーゼと共に、該異種遺伝子を、該所定の細胞系にトランスフェクションにより挿入する。該異種遺伝子および該選択性マーカー遺伝子は、単一のベクターにより一緒に、あるいは同時トランスフェクションされる別々のベクターによって、発現させることができる。トランスフェクションの2〜3日後、選択性の試薬、例えばネオマイシントランスフェラーゼを用いた場合にはG418を含有する培地に、該細胞を移し、これら選択条件下で数週間培養する。次いで、エマージェント(emergent)抵抗性細胞を単離し、かつ所定の遺伝子生成物の発現につき検討を行うことができる。該宿主細胞ゲノムへの無秩序で、方向性の無い組込みの結果として、一群の細胞が得られ、これら細胞は、全く異なる該異種遺伝子発現率を持つ。これらは、非-発現性の細胞をも含むことができ、ここで該選択性マーカーは発現されるが、対象とする該遺伝子は発現されない。従って、対象とする該遺伝子の極めて高い発現性を呈する細胞クローンを同定するためには、多数のクローンを検査し、かつテストする必要があるが、これは時間を要し、また労力を消耗し、しかも経費がかさむ。
【0004】
遺伝子の増幅は、動物細胞の培養において広く普及している現象であり、組換え生物薬学的なタンパク質の製造のために使用されている。この遺伝子増幅は、多数の哺乳動物細胞系の、元々比較的低い生産性を、大幅に改善する。広く利用されている増幅法の一つは、ジヒドロ葉酸レダクターゼ(DHFR)を基本とする、DHFR-欠乏チャイニーズハムスター卵巣(CHO)細胞において、極めて頻繁に使用される遺伝子増幅系である。このDHFR-欠乏CHO細胞、例えばCHO-DUKX(ATCC CRL-9096)またはCHO-DG44(Urlaub等、1983)を、DHFRおよび該対象とするタンパク質をコードする、適当なベクター系でトランスフェクションする。次いで、該トランスフェクタントを、グリシン、ヒポキサンチンおよびチミジンを含まない培地で選別する。この増幅およびその結果としての高い生産性を持つ細胞系の確立は、メトトレキセート(MTX)、即ちジヒドロ葉酸レダクターゼ阻害剤の次第に増加する濃度での添加によって達成される(Kaufman等、1982, US 4,656,134)。得られる高い生産性を示す細胞の、その後の選別も、偶然の原理を条件としており、また確率に基くものであり、その結果として、この選別段階は多大な労力を要し、かつ時間を浪費するものである。
【0005】
より良好にかつより迅速に、遺伝子形質転換および発現を追跡するために、あらゆる種類の方法が開発されている。これらは、まずリポータ分子、例えばクロラムフェニコール-アセチルトランスフェラーゼ、ルシフェラーゼ、β-ガラクトシダーゼ、またはβ-ガラクトシダーゼもしくはルシフェラーゼのコード領域を含む融合タンパク質の使用を含む。これらリポータ遺伝子アッセイの欠点は、該細胞を固定しまたは溶解する必要があり、また外部から添加した物質および補助因子と共にインキュベートする必要がある。従って、該分析された細胞の更なる培養が、当然必要となる。E.コリ酵素β-ガラクトシダーゼの同時発現に基く極最近の方法は、実際に、FACS装置(Nolan等, 1988)を用いて、溶解した細胞の選別を可能とするが、フルオロゲン性基質と共に該細胞を充填するために、低張性予備処理が必要となる。また、該FACSを基本とする選別の前に、その活性を阻害する必要がある。
【0006】
エクオレアビクトリア(Aequorea victoria)由来の蛍光タンパク質(GFP)およびこれからリポータ分子として発展されたGFP変異体の導入により、異種遺伝子を発現する細胞を同定することが、より一層容易になった。GFPの同時発現は、付随的な基質または補助因子を必要とすることなしに、その蛍光に基いた、インビボでの実時間分析、およびトランスフェクタントの選別を可能とした。遺伝子転位を追跡するためのリポータ分子としてのGFPの使用は、様々な刊行物に記載されている。Chalfie等の米国特許第5,491,084号および同第6,146,826号は、対象とするタンパク質を発現する細胞の選別方法を記載している。この方法は、該対象とするタンパク質に対するコード配列を含むDNAおよび該GFP-遺伝子をコードする第二のDNA分子による、細胞の同時トランスフェクションを含む。次いで、該GFP-発現細胞を選別する。Gubin等(1997)は、選択成長条件を使用しない場合の、CHO細胞内でのGFP発現の安定性を検討した。これらの細胞は、GFPおよびネオマイシンホスホトランスフェラーゼ両者を含むプラスミドによりトランスフェクションした。Mosser等(1997)は、GFPおよびターゲット遺伝子(これも興味深い遺伝子として知られる)をコードするビシストロン発現カセットを含むプラスミドを使用して、誘発可能な生成物を発現する細胞の同定および選別した。このターゲット遺伝子は、調節可能なプロモータの制御下にあった。このGFP発現とターゲット遺伝子発現との結合は、ウイルス由来のIRES(内部リボソームエントリーサイト)を用いて達成され、その結果として、GFPおよび該対象とするタンパク質をコードするビシストロンmRNAが発現された。使用したプラスミドそれ自体は、選択性マーカー遺伝子を何等含んでいなかった。従って、これは同時トランスフェクションにおいて、または後のトランスフェクションにおいて、第二のプラスミドによって導入された。これとは対照的に、Levenson等(1998)は、ビシストロン発現カセットを持つレトロウイルスベクターを使用しており、該カセットにおいて、該対象とする遺伝子は、該IRES配列の前でクローニングすることができる。他方、該IRES配列に続く配列は、選択性マーカー遺伝子をコードし、このマーカーには、G418、プロマイシン、ハイグロマイシン(hygromycin) B、ヒスチジノールDまたはフレオマイシンに対する耐性が付与されており、あるいはこれはGFPであった。
【0007】
ピコルナウイルス群由来のIRES要素を含むベクターも、既に記載されており、このIRES要素は、該生成物遺伝子と、選択性マーカー遺伝子との間に配置されている(Pelletier等, 1988; Jang等, 1989; Davies等, 1992)。
また、GFPも首尾よく耐性マーカー遺伝子と融合されている。例えば、Bennett等(1998)は、GFP/ゼオマイシン融合タンパク質を記載している。この二官能性の選択性マーカーは、トランスフェクションされた哺乳動物細胞を同定し、かつ選別するのに、上首尾で使用された。他方Primig等(1998)は、そのエンハンサーの研究に対して、GFPとネオマイシンホスホトランスフェラーゼとの融合タンパク質を使用した。
Meng等(2000)による刊行物および国際特許出願WO 01/04306では、単一のベクター由来の、増幅可能な選択性マーカー遺伝子DHFRおよびGFP遺伝子と共に、該対象とする遺伝子を発現する発現系を使用して、組換えタンパク質の高い発現性を持つ細胞を選別し、かつ同定した。これら3つの遺伝子は、一つの転写単位内で結合されているか、あるいは2つの単位間で分割されていた。単一の発現ベクター内でのこれら3種の遺伝子全ての空間的並びに転写的な結合は、選択圧力の下で同時増幅の確率を高め、かつその結果として高い生産性を呈するクローンを同定し、かつ選別した。この組合せ選別を利用して、増幅可能なDHFR選択性マーカーおよびGFPを基本とするFACS選別によって単離された、最良のクローンは、1日当たり、1細胞当たり3〜4.5pg程度の量で、該対象とするタンパクを発現した。これらの実験は、接着性の細胞を用いて、血清を含む培地内で、即ちかなり大雑把なものであることが知られており、またより高い基本的な生産性によって特徴付けられる細胞および条件を用いて行った。
【0008】
発明の概要
従って、本発明の目的は、以下のような要件を持たす、高い生産性を持つ組換え細胞の選別系を開発することであった:
(1) 生物薬学的タンパク質を生産する、高い生産性を呈する細胞の開発に要する時間の短縮、および同時にこの開発に係る経費の節減;
(2) 設備に関する低い経費で、高い生産性を呈する細胞の選別における高い処理量;
(3) 例えば、高いメトトレキセート濃度における、成長性に対する影響の小さい、「醗酵に関して強固な」高生産生細胞の使用;
(4) 好ましくは血清を含まない培地内での、懸濁培養に適した細胞の、トランスフェクション、選別および培養性;および
(5) 必要とされる遺伝子増幅段階数の減少。
本発明の、更なる目的は、発現ベクター、およびこれによりトランスフェクションされた、このクローン選別系において使用可能な宿主細胞、並びにこれら宿主細胞を使用して、異種遺伝子生成物を製造する方法を提供することにあった。
【0009】
これら本発明の目的は、本発明の一局面に従えば、発現ベクターによって達成され、この発現ベクターは、ハムスターユビキチン/S27aプロモータと機能可能な状態で結合した、対象とするタンパク質をコードする遺伝子(以下「対象とする遺伝子」とも言う)および蛍光タンパク質をコードする遺伝子を含む。
この発現ベクターは、好ましくは更に増幅可能な、選択性マーカー遺伝子、例えばジヒドロ葉酸レダクターゼ(DHFR)をも含む。好ましい発現ベクターは、また他の調節要素、例えば該プロモータと機能可能な状態で結合したエンハンサーをも含む。また、この発現ベクターは、好ましくは内部リボソームエントリーサイト(IRES)をも含み、このサイトは蛍光タンパク質をコードする遺伝子および対象とする遺伝子の、ビシストロン発現を可能とする。
本発明は、また該対象とする遺伝子の代わりに、このような遺伝子を組込むための多重クローニングサイト、即ち制限エンドヌクレアーゼによる多数の切断サイトを持つ配列領域を持つ、基本ベクターにも関連する。
【0010】
本発明のもう一つの局面は宿主細胞に関連し、該宿主細胞は、上記発現ベクターの一種によってトランスフェクションされている。これら宿主細胞は、真核宿主細胞、好ましくは哺乳動物細胞であり、ハムスター細胞等の齧歯目動物の細胞、特にCHO細胞またはBHK細胞が特に好ましい。
本発明のもう一つの局面は、異種遺伝子生成物の製造方法に係り、この方法では、本発明の発現ベクターでトランスフェクションされた宿主細胞を、該異種遺伝子生成物の発現を可能とする条件の下で培養し、かつ該遺伝子生成物を培養物または培地から単離する。
本発明の一特定の態様では、該宿主細胞は、本発明による発現ベクターで、また一種またはそれ以上の他の対象とするタンパクをコードする遺伝子を含む、一種またはそれ以上のベクターでトランスフェクションし、または好ましくは同時にトランスフェクションする。
【0011】
これに関連して、本発明は、ヘテロダイマー型タンパク質の製法を提供するものであり、この方法では、該へテロダイマー型タンパク質の様々なサブユニットをコードする発現ベクターによって同時にトランスフェクションされている、上記類の宿主細胞を該へテロダイマー型タンパクの発現を可能とする条件の下で培養し、かつ該へテロダイマー型タンパクを培養物または培地から単離する。このような方法の、一特定の応用は、抗体およびそのサブユニットの製造である。
もう一つの局面において、本発明は、対象とするタンパクを発現する宿主細胞を選別する方法に係り、この方法では、本発明による発現ベクターによりトランスフェクションされている、一群の宿主細胞を、該対象とするタンパクおよび該蛍光タンパク質の発現を可能とする条件の下で培養し、かつ該蛍光タンパクの最大の発現率を示す該細胞または細胞群を同定し、および/または選別する。この選別は、好ましくは、蛍光-活性化セルソータ(FACS)を用いて行われる。
驚いたことに、本発明の提供するこの系を用いた場合には、極めて短時間の間に、1日当たりかつ1細胞当たり、組換えタンパク質15pg(単鎖タンパク質)または10pg(ヒト化抗体)を越える平均の比生産率(specific productivities)で発現する細胞のプールを、遺伝子増幅段階を経ることなしに、単離することが可能となることを見出した。該比生産率は、一回のDHFRに基く遺伝子増幅段階によって、1日当たりかつ1細胞当たり30pgを越えるレベルまで高めることができた。これら細胞のプールにおいて達成されるこの生産率は、結果的に、これまでに公開された最良の細胞クローンにより達成された最大の生産率よりも、8〜10倍だけ高い。
【0012】
驚いたことに、該対象とするタンパク質および該蛍光タンパク質の発現同士の間には、極めて良好な相関関係がある。このことは、発現された抗体と関連して、2つの免疫グロブリン鎖が、各々それ自身のベクターによって発現される場合の、同時トランスフェクションにおいてさえ正しく、またFACS選別において、選択は該蛍光タンパク質との転写的な結合を考慮して、一方の鎖の発現に関してのみ行うことができる。該蛍光タンパク質の高い発現率は、該細胞の成長および生存率について何の影響も示さない。更に、高い生産性を示す細胞を選別するための、技術開発時間を、従来の段階的な遺伝子増幅戦略と比較して、少なくとも半分にまで減じることができ、結果的に開発設備および経費の大幅な節減を達成できる。
【0013】
発明の詳細な説明および好ましい態様
本発明による発現ベクターは、対象とするタンパク質(対象とする遺伝子)をコードし、機能可能な状態でハムスターユビキチン/S27aプロモータと結合した遺伝子および蛍光性タンパク質をコードする遺伝子を含む。好ましくは、該発現ベクターは、また増幅可能な選択性マーカー遺伝子を含む。
ハムスターユビキチン/S27aプロモータ
このハムスターのユビキチン/S27aプロモータは、WO 97/15664に記載されている強力な相同的プロモータである。このようなプロモータは、好ましくは、以下に列挙する特徴の内の少なくとも一つを持つ:GCに富む配列領域、Sp1結合サイト、ポリピリミジン要素を持ち、TATAボックスを持たない。同様に好ましいものは、構造的に活性化されており、かつ特に血清を含む、低濃度の血清を含むおよび血清を含まない細胞培養条件にて、等しく活性であるプロモータである。もう一つの態様では、これは、誘導性のプロモータ、特に血清の除去により活性化されるプロモータである。
特に有利な態様は、WO 97/15664の図5に含まれているような、ヌクレオチド配列を持つプロモータである。特に好ましいものは、図5の-161〜-45なる範囲の位置の配列を含むプロモータ配列である。
【0014】
本特許出願の実施例において使用するプロモータ各々は、添付した配列表のSEQ ID NO:1の位置1923〜2404に対応する配列を持つDNA分子を含む。この配列は、WO 97/15664の図5のフラグメント:-372〜+111に相当し、また好ましいプロモータを表し、従って好ましいプロモータにはこの配列領域を組込むべきである。もう一つの適当なプロモータフラグメントは、位置:2134〜2406の配列(WO 97/15664の図5における位置:-161〜+111に相当)を含む。位置:2251〜2406の配列のみを含むプロモータは、最早機能性ではない(WO 97/15664の図5における位置:-45〜+111に相当)。このプロモータ配列を、位置:2134から開始して、5'方向に伸ばすことができる。
また、完全ハムスターユビキチン/S27aプロモータ配列の機能性のサブフラグメント並びに例えば置換、挿入または削除によって変性されている、そのサブフラグメントの該完全配列の、機能性の突然変異体/変異型を使用することも可能である。対応するサブフラグメント、突然変異体または変異型は、以下において「変性プロモータ」とも呼ばれる。
【0015】
場合により他の調節要素と結合した、変性プロモータは、好ましくはSEQ ID NO:1に与えられたヌクレオチド配列の位置:1923〜2406のプロモータフラグメントの活性に相当する、転写活性を持つ(WO 97/15664の図5における位置:-372〜+111)。変性プロモータは、これが転写活性を示し、その活性が、比較リポータ遺伝子のフラグメント:1923〜2406(-372〜+111フラグメント)の持つ活性の少なくとも50%、好ましくは少なくとも80%、より好ましくは少なくとも90%および最も好ましくは少なくとも100%である場合には、本発明の目的にとって有用であることが分かる。特に好ましい変性プロモータは、少なくとも80%、好ましくは少なくとも85%、好ましくは少なくとも90%、より好ましくは少なくとも95%および最も好ましくは少なくとも97%なる、上記ハムスターユビキチン/S27aプロモータの、野生型配列:SEQ ID NO:1に対する最低配列相同性を持ち、かつ比較リポータ遺伝子アッセイにおいて対応するプロモータ活性を持つものである。
【0016】
対応する比較リポータ遺伝子アッセイにおいて、該基準配列を含む、テストすべきプロモータフラグメントは、例えばルシフェラーゼをコードする、プロモータを含まないリポータ遺伝子の前にクローニングされ、アルカリホスファターゼまたはグリーン蛍光タンパク質(GFP)を分泌する。これらの構築物(プロモータ配列+リポータ遺伝子)は、後にこのテスト細胞、例えばCHO-DG44に、トランスフェクションによって導入され、また問題とする該プロモータフラグメントによる、該リポータ遺伝子発現の誘導は、該リポータ遺伝子のタンパク含有率を測定することにより決定される。対応するテストは、例えばAusubel等の、分子生物における今日のプロトコール(Current Protocols in Molecular Biology), 1994, 最新版に見出すことができる。
【0017】
ハムスターユビキチン/S27aプロモータのプロモータ配列および例えば5'未翻訳領域または選択されたそのフラグメントをも含んでいても良い、変性プロモータ、およびユビキチン/S27a遺伝子のコード配列またはその選択されたフラグメントは、WO 97/15664に記載されている配列に関する知見を持つ熟練者によって、例えばSambrook等, 1989; Ausubel等, 1994に記載されている、様々な標準的方法を利用して得ることができる。WO 97/15664に記載されている配列から出発し、例えば適当な配列を選択し、この部分の配列を含むオリゴヌクレオチドプローブを化学的に合成することができる。例えば、該ユビキチン/S27a遺伝子またはその5'未翻訳領域またはその他のフラグメントを、例えばハムスターゲノムのライブラリーからハイブリッド化することにより、クローニングする目的で、この種のプローブを使用することができる。上記のリポータ遺伝子アッセイを利用して、当業者は、多大な努力を払うことなしに、プロモータ-活性フラグメントを同定し、かつこれらを本発明の目的において使用することができる。該5'未翻訳領域またはその特定のフラグメントは、ゲノムDNAまたはゲノムライブラリーからの対応するプライマーを用いたPCR増幅により、容易に得ることができる。該5'未翻訳領域のフラグメントも、より大きなDNAフラグメントから、制限エキソヌクレアーゼIIIにより得ることができる。このようなDNA分子も、化学的に合成することができ、あるいは結合によって、化学的に合成されたフラグメントから製造することができる。
【0018】
欠失、挿入および置換突然変異体は、「サイト-特異的突然変異誘発」および/または「PCRに基く突然変異誘発技術」によって製造することができる。対応する方法は、例えば他の参考文献をも含む、LottspeichおよびZorbas, 1998,第36.1章に述べられている。
ハムスターユビキチン/S27a遺伝子の5'未翻訳領域またはハムスターユビキチン/S27a遺伝子のS27a部分由来のプローブを用いた、クロス-ハイブリッド形成により、他の、好ましくは哺乳動物種の、対応する相同遺伝子由来の適当なプロモータ配列を、同定しかつ単離することができる。適当な技術は、一例として、LottspeichおよびZorbas, 1998,第23章に記載されている。遺伝子は、そのヌクレオチド配列が、相同であるとされる該遺伝子のヌクレオチド配列に対して、少なくとも70%、好ましくは少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%および最も好ましくは少なくとも97%の適合性を示す場合には、本発明の目的にとって「相同」である。
【0019】
対象とする遺伝子
本発明の発現ベクターに含まれる該対象とする遺伝子は、対象とするタンパク質をコードする、任意の長さのヌクレオチド配列を含む。この遺伝子生成物または「対象とする生成物」は、一般にタンパク質、ポリペプチド、ペプチドまたはこれらのフラグメントまたは誘導体である。しかし、これはRNAまたはアンチセンスRNAであっても良い。該対象とする遺伝子は、完全な長さで、短縮された形状で、融合遺伝子としてまたは標識遺伝子として存在することができる。これはゲノムDNAまたは好ましくはcDNAまたは融合物の対応するフラグメントであり得る。該対象とする遺伝子は、本来の遺伝子配列であり得、あるいは突然変異または変性されていてもよい。このような変性は、特定の宿主に対する適合化またはヒト化するための、コドン最適化を含む。該対象とする遺伝子は、例えば分泌された、細胞質、核内に位置する、膜と結合した、または細胞表面に結合したポリペプチドをコードすることができる。
用語「ヌクレオチド配列」または「核酸配列」とは、オリゴヌクレオチド、ヌクレオチド、ポリヌクレオチドおよびこれらのフラグメント、並びに一本鎖または二本鎖であり得る、ゲノムまたは合成起源のDNAまたはRNAを意味し、また遺伝子のコード性または非-コード性のストランドを表すこともできる。核酸配列は、サイト-特異的突然変異形成またはPCR-媒介突然変異形成(例えば、Sambrook等, 1989; Ausubel等, 1994に記載されている)等の標準的な技術を用いて変性することができる。
【0020】
用語「コードする」とは、核酸、例えば染色体中の遺伝子またはmRNA内の特定のヌクレオチド配列が、他のポリマーおよび巨大分子、例えばrRNA、tRNA、mRNA、その他のRNA分子、cDNAまたは生物学的な過程におけるポリペプチドを合成するためのマトリックスとして機能する性質または能力を意味する。従って、所定のタンパク質が、転写およびそれに続くmRNAの翻訳によって、細胞または別の生物学的な系内で製造される場合には、遺伝子はタンパク質をコードする。ヌクレオチド配列がそのmRNAの配列と等価であり、通常は配列データバンク、例えばEMBLまたはGenBankにも与えられている、コード用のストランドおよび転写に対するマトリックスとして作用する、遺伝子またはcDNAの非-コード用のストランド両者は、生成物またはタンパク質をコードするものと見做される。タンパク質をコードする核酸は、また縮重遺伝子コードを基準として、異なる順序のヌクレオチド配列を含むが、該タンパク質と同一のアミノ酸配列をもたらす。タンパク質をコードする核酸配列は、またイントロンをも含む。
【0021】
cDNAなる用語は、逆転写およびmRNAまたは遺伝子から作られた他のRNA由来の第二のDNAストランドの合成によって製造される。該cDNAが二本鎖DNA分子として存在する場合、これはコード用ストランドおよび非-コード用ストランド両者を含む。
用語:イントロンとは、任意の長さの非-コード用ヌクレオチド配列を意味する。これらは、自然において数多くの真核生物遺伝子内に見られ、予め転写されるmRNAプリカーサから、スプライシングとして知られる過程によって排除される。これは、5'および3'末端における該イントロンの正確な切除および得られるmRNA末端の正確な結合を必要とし、結果的にタンパク質合成を首尾よく行うための正確な読み取り枠を持つ、成熟プロセスmRNAが作られる。このスプライシング過程に関与する、多くのスプライスドナーおよびスプライスアクセプタサイト、即ち該エクソン-イントロンあるいはイントロン-エクソン界面に直接位置する配列が、特徴付けされている。これに関する概説に関しては、Ohshima等, 1987を参照のこと。
【0022】
対象とするタンパク質
生物薬学的に重要なタンパク質/ポリペプチドは、例えば抗体、酵素、サイトカイン、リンホカイン、接着分子、レセプタおよびこれらの誘導体またはフラグメントを含むが、これらに限定されない。一般に、アゴニストまたはアンタゴニストとして作用するおよび/または治療または診断における用途を持つ、全てのポリペプチドが、価値あるものである。
用語「ポリペプチド」は、アミノ酸配列またはタンパク質について用いられ、また任意の長さを持つアミノ酸のポリマーを意味する。この用語は、またグリコシル化、ホスホリル化、アセチル化またはタンパク質のプロセシング等の反応により、翻訳後に変性されたタンパク質を包含する。このポリペプチドの構造は、該ポリペプチドの生物学的な活性を維持しつつ、例えばアミノ酸の置換、欠失または挿入により、および他のタンパク質と融合することにより変性することができる。
【0023】
治療用のタンパク質の例は、インシュリン、インシュリン-様の成長因子、ヒト成長ホルモン(hGH)および他の成長因子、組織プラスミノーゲン活性化因子(tPA)、エリスロポエチン(EPO)、サイトカイン、例えばIL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-8、IL-9、IL-10、IL-11、IL-12、IL-13、IL-14、IL-15、IL-16、IL-17、IL-18等のインターロイキン(IL)、インターフェロン(IFN)-α、-β、-γ、-ω、または-τ、腫瘍壊死因子(TNF)、例えばTHF-α、-βまたは-γ、TRAIL、G-CSF、GM-CSF、M-CSF、MCP-1およびVEGFである。その他の例は、モノクローナル、ポリクローナル、多重特異的および一本鎖抗体およびそのフラグメント、例えばFab、Fab'、F(ab')2、FcおよびFc'フラグメント、免疫グロブリン軽鎖(L)および重鎖(H)およびその定常、可変および超可変領域並びにFvおよびFdフラグメント(Chamov等, 1999)である。これらの抗体は、ヒト起源または非-ヒト起源のものであり得る。ヒト化またはキメラ抗体も可能である。
Fabフラグメント(抗原結合フラグメント=Fab)は、隣接する定常領域によって、一緒に保持されている2つの鎖の可変領域からなる。これらは、例えば公知の抗体を、パパイン等のプロテアーゼで処理することにより、あるいはDNAクローニングによって製造できる。他の抗体フラグメントは、ペプシンによるタンパク分解消化により製造できる、F(ab')2フラグメントである。
【0024】
遺伝子クローニングによって、重鎖(VH)および軽鎖(VL)の可変領域のみからなる、短い抗体フラグメントを製造することも可能である。これらは、Fvフラグメント(可変フラグメント=可変部分のフラグメント)として知られている。該定常鎖のシステイン基を介する共有結合は、これらのFvフラグメントでは不可能であることから、これらはしばしば他の幾つかの方法によって安定化される。この目的のために、該重鎖および軽鎖の可変領域は、しばしば約10〜30個のアミノ酸、好ましくは15個のアミノ酸を含む短いペプチドフラグメントで、一緒に結合される。これは、単一のポリペプチド鎖を製造し、そこではVHおよびVLがペプチドリンカーによって一緒に結合されている。このような抗体フラグメントは、また一本鎖Fvフラグメント(scFv)とも呼ばれる。scFv抗体の例は公知であり、例えばHuston等, 1988に記載されている。
過去において、様々な方法が、マルチマーscFv誘導体を製造するために開発されている。その意図は、改良された薬物動力学的特性および高い結合活性を持つ、組換え抗体を製造することにある。該scFvフラグメントのマルチマー化を達成するためには、これらを、マルチマー化ドメインを持つ融合タンパクとして製造する必要がある。該マルチマー化ドメインは、例えばIgGのCH3領域またはロイシンジッパードメイン等のヘリックス構造(螺旋状コイル構造)であり得る。他の方法では、該scFvフラグメントのVHおよびVL領域間の相互作用を、マルチマー化(例えば、二重特異性-、三重特異性-および五重特異性抗体)のために利用する。
【0025】
用語:二重特異性抗体とは、当分野において、二価のホモダイマー型scFv誘導体を表すのに用いられる。該scFv分子内のペプチドリンカーの長さを、5〜10アミノ酸に短縮することは、VH/VL鎖の重ね合わせにより、ホモダイマーの生成をもたらす。これら二重特異性抗体は、更にジスルフィド架橋によって安定化することができる。二重特異性抗体の例は、Perisic等, 1994などの文献に見出すことができる。
用語:ミニ抗体(minibody)とは、当分野において、二価のホモダイマー型scFv誘導体を表すのに用いられる。これは融合タンパク質からなり、該タンパクは免疫グロブリン、好ましくはIgG、最も好ましくはIgG1のCH3領域をダイマー化領域として含む。これは、ヒンジ領域、またIgGおよびリンカー領域によって、該scFvフラグメントと接続している。このようなミニ抗体の例は、Hu等, 1996によって記載されている。
該三重特異性抗体とは、当分野において、三価のホモダイマー型scFv誘導体(Kortt等, 1997)を表すのに用いられる。リンカー配列の使用なしの、VH/VL鎖の直接的な融合は、トリマーの生成をもたらす。
当分野において、二価、三価、または四価の構造を持つミニ抗体として知られるフラグメントは、またscFvフラグメント誘導体でもある。このマルチマー化は、ダイマー、トリマーまたはテトラマー型の螺旋状コイル構造によって達成される(Pack等, 1993およびLovejoy等, 1993)。
【0026】
蛍光タンパク質をコードする遺伝子
本発明による発現ベクターは、対象とする遺伝子と機能可能な状態で結合し、またハムスターユビキチン/S27aプロモータ、変性ハムスターユビキチン/S27aプロモータまたはその同族体の制御下にある、蛍光タンパク質をコードする遺伝子を含有する。
該蛍光タンパク質は、例えば緑、青味を帯びた緑、青、黄色またはその他の色を持つ蛍光タンパク質であり得る。一特定例は、エクオレアビクトリア(Aequorea victoria)またはレニラレニホルミス(Renilla reniformis)およびこれらから発展された突然変異体由来のグリーン蛍光タンパク質(GFP)である(例えば、Bennet等, 1998;Chalfie等, 1994;WO 01/04306およびこれらに引用されている文献)。
他の蛍光タンパク質およびこれらをコードする遺伝子は、WO 00/34318、WO 00/34326、WO 00/34526、WO 01/27150に記載されている。これら文献を、本発明の参考とする。これらの蛍光タンパク質は、アントゾア(Anthozoa)種、例えばアネモニアマジャノ(Anemonia majano)、クラブラリア(Clavularia) sp、ゾアンサス(Zoanthus) sp. I、ゾアンサスsp. II、ディスコゾーマストリアータ(Discosoma striata)、ディスコゾーマsp.「レッド」、ディスコゾーマsp.「グリーン」、ディスコゾーマsp.「マゼンタ」、アネモニアスルカタ(sulcata)等の非-生物発光生物の発蛍光団である。
【0027】
本発明で使用する該蛍光タンパク質は、野生型のタンパク質に加えて、例えば他のタンパク質またはペプチドと融合されている、天然のまたは遺伝子操作した突然変異体および変異種、フラグメント、その誘導体または変異種を含む。使用する突然変異は、例えば励起または発光スペクトル、発色団の形成、該タンパク質の吸光係数または安定性を変更する可能性がある。その上、哺乳動物細胞またはその他の種における発現は、コドン最適化により改善することができる。本発明によれば、該蛍光タンパク質は、また選択性マーカー、好ましくは増幅可能な、選択性マーカー、例えばジヒドロ葉酸レダクターゼ(DHFR)との融合においても使用できる。
該蛍光タンパク質により発せられる蛍光は、例えば蛍光-活性化細胞選別機(FACS)または蛍光顕微鏡を用いた、スルーフロー(throughflow)サイトメトリーによって、該タンパク質の検出を可能とする。
他の調節要素
ハムスター-ユビキチン/S27aプロモータは、他の調節配列と機能可能な状態で結合して、発現カセットにおける転写活性を増大/調節することができる。
例えば、このプロモータは、機能可能な状態で、エンハンサ配列と結合して、該転写活性を高めることができる。このために、一種またはそれ以上のエンハンサおよび/または一つのエンハンサの幾つかのコピー、例えばCMVまたはSV40エンハンサを使用することができる。
【0028】
エンハンサなる用語は、シス-位において、プロモータの活性に作用し、結果的にこのプロモータと機能可能な状態で接続された遺伝子の転写を刺激する、ポリヌクレオチド配列を表す。プロモータとは異なり、エンハンサの効果は、位置および配向とは無関係であり、従ってこれらはイントロン内でまたは該コード領域においてさえ、転写単位の前または背後に位置することができる。該エンハンサは、該転写単位の極近傍および該プロモータからかなり遠く離れた位置の何れにも位置することができる。該プロモータと物理的および機能的な重なりを有していても良い。当業者は、様々な起源由来の多数のエンハンサ(およびGenBank等のデータバンクに寄託されているもの、例えばSV40エンハンサ、CMVエンハンサ、ポリオーマエンハンサ、アデノウイルスエンハンサ)を認識しており、これらは独立した要素として、あるいはポリヌクレオチド配列内でクローニングされた要素(例えば、ATCCに寄託された、または市場のまたは個人的な源からの要素)として入手できる。また、多くのプロモータ配列、例えば頻繁に使用されるCMVプロモータ等が、エンハンサ配列をも含む。ヒトCMVエンハンサは、これまでに同定された中で最強のエンハンサである。誘導エンハンサの一例は、メタロチオネインエンハンサであり、これはグルココルチコイドまたは重金属によって刺激することができる。
【0029】
他の可能な変性は、例えば多数のSp1結合サイトの導入である。また、これらプロモータ配列は、調節配列と結合することができ、この調節配列は、転写活性の制御/調節を可能とする。従って、該プロモータを抑制/誘導することができる。これは、例えばアップ-またはダウン-レギュレーション転写因子に対する結合サイトである、配列と結合することにより実施できる。例えば、上記転写因子Sp1は、該転写活性に対して正の効果を持つ。他の例は、活性化因子タンパク質AP1に対する結合サイトであり、これは転写に対して正および負何れにも作用する。AP1の活性は、あらゆる類の因子、例えば成長因子、サイトカインおよび血清によって制御できる(Faisst等, 1992およびこれに含まれる参考文献)。また1、2、3またはそれ以上の塩基の突然変異(置換、挿入または欠失)により該プロモータ配列を変更し、次いでリポータ遺伝子アッセイにおいて、これが高いプロモータ活性を持つか否かを決定することによって、転写効率を高めることができる。
基本的には、これらの付随的な調節要素は、ハムスター-ユビキチン/S27aプロモータ以外のプロモータ、エンハンサ、終結およびポリアデニル化シグナル並びに他の発現制御要素を含む。誘導および構造的調節要素両者は、様々な細胞型について公知である。「転写-調節要素」は、一般に発現すべき遺伝子配列の上流のプロモータ、転写開始および終止サイトおよびポリアデニル化シグナルを含む。
【0030】
プロモータなる用語は、遺伝子またはこれと機能的に接続した配列の転写を可能とし、かつこれを制御する、ポリヌクレオチド配列を意味する。プロモータは、RNAポリメラーゼを結合するための認識配列および転写用の開始サイト(転写開始サイト)を含む。ある細胞型または宿主細胞において、所定の配列を発現するためには、適当な機能的プロモータを選択する必要がある。当業者は、構造的、誘導性および抑制プロモータを含む、様々な起源由来の種々のプロモータについて十分な馴染みを持つであろう。これらは、例えばGenBank等のデータバンクに寄託されており、またこれらは市場または個人的な源から、別々の要素として、あるいはポリヌクレオチド配列内でクローニングされた要素として得ることができる。誘導プロモータにおいて、該プロモータの活性は、シグナルに応じて低下または増大させることが可能である。誘導プロモータの一例は、テトラサイクリン(tet)プロモータである。これは、テトラサイクリンオペレータ配列(tetO)を含み、この配列はテトラサイクリン-調節トランス活性化因子タンパク質(tTA)により誘導できる。テトラサイクリンの存在下において、tTAとtetOとの結合は阻害される。他の誘導プロモータの例は、jun、fos、メタロチオネインおよび熱ショックプロモータである(同様に、Sambrook等, 1989; Gossen等, 1994をも参照のこと)。真核生物における高い発現性を得るのに、特に適したプロモータとしては、例えばSV40初期プロモータ、アデノウイルス主後期プロモータ、マウスメタロチオネイン-Iプロモータ、ラウス肉腫ウイルスの反復配列領域およびヒトサイトメガロウイルスの初期プロモータが挙げられる。他の哺乳動物プロモータの例は、アクチン、免疫グロブリンまたは熱ショックプロモータである。
【0031】
用語「転写開始サイト」とは、一次転写産物、即ちmRNAプリカーサに組込まれる、第一の核酸に対応する構築物内の、核酸を意味する。該転写開始サイトは、該プロモータ配列と重複していてもよい。
用語「転写終止サイト」とは、通常は、対象とする遺伝子または転写すべき該遺伝子部分の3'末端にあり、かつRNAポリメラーゼによる転写の終結をもたらす、核酸配列を意味する。
「ポリアデニル化シグナル」とは、真核生物mRNAの3'末端における特定のサイトにおいて開裂を生じ、かつ該開裂された3'末端において、約100-200アデニンヌクレオチド(ポリAテール)を持つ配列の、転写後の組み込みを引起すシグナル配列である。このポリアデニル化シグナルは、該開裂サイトの上流約10-30ヌクレオチドに配列AATAAAおよび下流側に位置するある配列を含む。様々なポリアデニル化要素が知られており、その例はtkポリA、SV40後期および初期ポリA、またはBGHポリA等である(例えば、米国特許第5,122,458号に記載されている)。
本発明の好ましい態様では、各転写単位は、プロモータまたはプロモータ/エンハンサ要素、対象とする遺伝子および/またはマーカー遺伝子並びに転写終止要素を持つ。もう一つの好ましい態様では、該転写単位は、更に二つの翻訳調節単位を含む。
【0032】
「翻訳調節要素」は、翻訳開始サイト(AUG)、終止コドン、および発現すべき各ポリペプチドに対するポリAシグナルを含む。最適の発現を得るためには、発現すべき該核酸配列の5'-および/または3'-未翻訳領域を除去、付加または変更して、あらゆる可能な不適当と思われる付随的な翻訳開始コドン、あるいは転写または翻訳レベルにおいて、発現に影響を与える可能性のある他の配列を排除することを推奨できる。発現を促進するために、リボソームコンセンサス結合サイトは、該開始コドンの直ぐ上流位置に、交互に挿入することができる。分泌性のポリペプチドを製造するために、該対象とする遺伝子は、通常ER膜におよびこれを通して、合成されたポリペプチドを搬送する、シグナルプリカーサペプチドをコードする、シグナル配列を含む。
該シグナル配列は、常にではないが、しばしば分泌されたタンパク質のアミノ酸末端に位置しており、また該タンパク質が該ER膜を介して挿入された後に、シグナルペプチダーゼによって開裂される。この遺伝子配列は、必要ではないが、通常それ自身のシグナル配列を含んでいる。この本来のシグナル配列が、存在しない場合には、異種シグナル配列を公知の方法で導入することができる。この種の多数のシグナル配列が、当業者には公知であり、また配列データバンク、例えばGenBankおよびEMBLに寄託されている。
【0033】
本発明による、一つの重要な調節要素は、内部リボソームエントリーサイト(IRES)である。このIRES要素はある配列を含み、これは該遺伝子上流の、5'-末端メチルグアノシニウムキャップ(methylguanosinium cap) (CAP構造)とは無関係に、翻訳開始を機能的に活性化し、かつ動物細胞内で、単一の転写物から、2つのシストロン(オープンリーディングフレーム)の翻訳を可能とする。このIRES要素は、直ぐ下流に位置する、該オープンリーディングフレーム翻訳用の、独立したリボソームエントリーサイトを与える。マルチシストロン性であり得る、即ちこのmRNAによって順次翻訳される多くの異なるポリペプチドまたは製品をコードできるバクテリアmRNAとは対照的に、動物細胞由来の大多数のmRNAはモノシストロン性であり、唯一つのタンパク質または製品をコードするに過ぎない。真核生物細胞におけるマルチシストロン性の転写物の場合には、その翻訳は、最も近接した上流側にある、翻訳開始サイトから開始され、また最初の終止コドンによって停止され、その後該転写物は該リボソームから放出されるであろう。このように、該mRNAによりコードされた、該第一のポリペプチドまたは生成物のみが、翻訳中に生成されるであろう。これと比較して、IRES要素を含むマルチシストロン性の転写物中の第二のまたはこれに続くオープンリーディングフレームと、機能可能な状態で結合している該転写物は、引続きその下流側に位置するオープンリーディングフレームの翻訳を可能とし、結果としてこの同一の転写物によりコードされる、2またはそれ以上のポリペプチドまたは生成物が、該真核生物細胞内で生産される。
【0034】
該IRES要素は、様々な長さおよび様々な起源のものであり得、また例えば脳心筋炎ウイルス(EMCV)または他のピコルナウイルス由来のものであり得る。様々なIRES配列およびベクターの構築におけるその利用は、例えばPelletier等, 1988; Jang等, 1989; Davies等, 1992; Adam等, 1991; Morgan等, 1992; Sugimoto等, 1994; Ramesh等, 1996; Mosser等, 1997等の文献に記載されている。
下流側に位置する遺伝子配列は、該IRES要素の3'-末端と機能可能な状態で結合しており、即ちスペーシングは、該遺伝子の発現が影響されず、あるいはほんの僅かに影響されるに過ぎず、あるいは意図した目的に対する十分な発現を示すように選択すべきである。十分な発現のために、該IRES要素と、その下流側に位置する遺伝子の開始コドンとの間の、最適な許容できる間隔は、リポータ遺伝子アッセイを利用して、該スペーシングを変更し、該スペーシングの関数として、その発現率を測定することからなる、簡単な実験によって決定できる。
記載した手段によって、最適の発現カセットを得ることが可能となり、このカセットは、異種遺伝子生成物の発現にとって極めて価値の高いものである。
【0035】
増幅可能な選択性マーカー遺伝子
本発明の好ましいベクターは、付随的に増幅可能なマーカー遺伝子の増幅および好ましくは上記ハムスター-ユビキチン/S27a遺伝子、該対象とする遺伝子および該蛍光タンパク質に対する遺伝子を含む、転写単位の同時増幅を可能とする、増幅可能な選択性マーカー遺伝子を含む。このために、対応する発現ベクターによりトランスフェクションされた宿主細胞は、適当な選別試薬の存在下で培養され、結果的に少なくとも該増幅可能な選択性マーカー遺伝子の幾つかの遺伝子コピーを持つ、これら宿主細胞のみが、複製できる(複製され得る)。好ましくは、これは、徐々に増大する量の選別試薬の存在下で、これら細胞を段階的に培養することにより達成される。
該増幅可能な選択性マーカー遺伝子は、通常幾つかの培養条件化で、真核生物細胞を生育するのに必要な酵素をコードする。例えば、該増幅可能な選択性マーカー遺伝子は、ジヒドロ葉酸レダクターゼ(DHFR)をコードすることができる。この場合、これによってトランスフェクションされた宿主細胞を、選別試薬であるメトトレキセート(MTX)の存在下で培養する場合に、該遺伝子は増幅される。
以下の表1に、その他の増幅可能な選択性マーカー遺伝子および本発明により使用できる、これに関連する選別試薬の例を与えるが、これらは、Kaufmanの概説:Method in Enzymology, 1990, 185:537-566に記載されている。
【0036】
表1:増幅可能な選択性マーカー遺伝子
【0037】
本発明によれば、使用する該増幅可能な選択性マーカー遺伝子は、好ましくはDHFRの機能を持つポリペプチド、例えばDHFRまたは蛍光タンパク質とDHFRとの融合タンパク質をコードする遺伝子である。DHFRは、プリンの生合成にとって必要である。このDHFR遺伝子を各細胞は、プリン欠乏培地内では成育できない。このDHFR遺伝子は、従ってプリンを含まない培地内で培養した細胞における遺伝子を、選別し、かつ増幅するための、有用な選択性マーカーである。このDHFR遺伝子と組み合わせて使用する選択媒体は、メトトレキセート(MTX)である。本発明は、従って、高い生産性を示す組換え宿主細胞の製造方法を包含し、この方法は以下の諸工程を含む:(i) 少なくとも対象とするタンパク質、蛍光タンパク質およびDHFRをコードする遺伝子で、宿主細胞をトランスフェクションし;(ii) 該細胞を、これら様々な遺伝子の発現を可能とする条件下で培養し;および(iii) 少なくとも該増幅可能な選択性マーカー遺伝子の増幅を可能とする選別試薬、例えばメトトレキセートの存在下で、該細胞を培養することによって、これら同時に組込まれた遺伝子を増幅する工程。好ましくは、これらトランスフェクションされた細胞を、血清の不在下で、徐々に増大する濃度のMTXを添加した、ヒポキサンチン/チミジンを含まない培地内で培養する。好ましくは、第一の増幅段階におけるMTXの濃度は、少なくとも200nMであり、およびより好ましい態様では、これは少なくとも500nMであり、また1μMまで段階的に増やすことができる。各場合において、1μMを超える濃度を使用することができる。
【0038】
哺乳動物細胞、好ましくはマウスミエローマおよびハムスター細胞が、増幅可能な選択性マーカー遺伝子として、DHFRを使用する際に好ましい宿主細胞である。細胞系CHO-DUKX(ATCC CRL-9096)およびCHO-GD44(Urlaub等, 1983)が、特に好ましい。というのは、これらは、突然変異の結果として、それ自体のDHFR活性を持たないからである。自身の内因性DHFR活性を持つ、その他の細胞型においても、このDHFR-誘導増幅を可能とするためには、該トランスフェクション過程において突然変異処理したDHFR遺伝子を用いることができ、該遺伝子は、メトトレキセートに対して低感度でタンパク質をコードする(Simonson等, 1983; Wigler等, 1980; Haber等, 1982)。
【0039】
本発明による発現ベクターの製造
本発明による発現ベクターは、理論的には、当分野において公知の従来法、例えばSambrook等(1989)に記載されているような方法により製造できる。Sambrookは、またベクターの機能的な成分、例えば適当なプロモータ(該ハムスター-ユビキチン/S27aプロモータに加えて)、エンハンサ、停止およびポリアデニル化シグナル、抗生物質耐性遺伝子選択性マーカー、複製開始点およびスプライシングシグナルをも記載している。従来のクローニングベクター、例えばプラスミド、バクテリオファージ、ファージミド、コスミドまたはウイルスベクター、例えばバキュロウイルス、レトロウイルス、アデノウイルス、アデノ-関連ウイルスおよび単純ヘルペスウイルス並びに染色体/ミニ染色体を使用して、これらを製造することができる。真核生物発現ベクターは、また典型的に原核生物の配列、例えばバクテリア内での複製および選別を可能とする、複製起点および抗生物質耐性遺伝子を含む。ポリヌクレオチド配列を導入するための、多重クローニングサイトを含む、幾つかの原核生物の発現ベクターは公知であり、その幾つかは、様々な製造元、例えばUSA、CA、ラジョラのストラタジーヌ(Stratagene)社、USA、CA、カールスバッドのインビトロゲン(Invitrogen)社、USA、WI、マディソンのプロメガ(Promega)社、またはUSA、CA、パロアルトのBDバイオサイエンスズクロンテック(Biosciences Clontech)社から市販品として得ることができる。
【0040】
該ハムスター-ユビキチン/S27aプロモータ、該対象とする遺伝子、蛍光タンパク質をコードする遺伝子および好ましくはまた該増幅可能な選択性マーカー遺伝子、例えばジヒドロ葉酸レダクターゼ、および場合により追加の調節要素、例えば該内部リボソームエントリーサイト(IRES)エンハンサまたはポリアデニル化シグナルを、当業者には馴染み深い方法で、該発現ベクター内に導入する。本発明による発現ベクターは、最低でも、ユビキチン/S27aプロモータ、該対象とする遺伝子および蛍光タンパクをコードする遺伝子を含む。好ましくは、該発現ベクターは、また増幅可能な選択性マーカー遺伝子をも含む。本発明によれば、変性ユビキチン/S27aプロモータ、例えば本出願に記載されている変性ユビキチン/S27aプロモータをも使用する。特に好ましいものは、該ユビキチンプロモータ、該対象とする遺伝子および蛍光タンパク質をコードする遺伝子が、一緒に機能可能に結合し、あるいは機能可能に結合している、発現ベクターである。
【0041】
本発明の範囲内において、「機能的結合」または「機能可能な状態で結合した」とは、2またはそれ以上の核酸配列または部分配列が、その意図した機能を達成できるように配置されている、これらの核酸配列または部分配列を意味する。例えば、プロモータ/エンハンサが、シス位置にある、結合遺伝子配列の転写を制御または変調できる場合には、該プロモータ/エンハンサは、コード遺伝子配列と機能可能な状態で結合している。必要ではないが、一般に機能可能に結合したDNA配列は、相互に近接しており、また2つのコード遺伝子配列が結合している場合、または分泌シグナル配列の場合には、同一の読み取り枠内にある。機能可能に結合したプロモータは、一般に該コード遺伝子配列の上流側に位置するが、これと近接している必要はない。エンハンサは何れかに近接している必要はないが、これらは該遺伝子配列の転写を補助する。この目的のために、これらは、該遺伝子配列の上流および下流側両者に、可能ならばこれと幾分距離を保って位置することができる。ポリアデニル化サイトは、これがある遺伝子配列の3'末端に位置する場合には、転写が該ポリアデニル化シグナルに対するコード配列を介して進行するように、該遺伝子配列と機能可能に結合する。結合は、PCR技術、適当な制限切断サイトにおける結合、またはスプライシング等の従来の組換え法に従って行うことができる。適当な制限切断サイトが利用できない場合には、合成オリゴヌクレオチドリンカーまたはアダプター自体を公知の方法で使用することができる。本発明によれば、この機能的結合は、好ましくはイントロン配列を介して起こることは無い。
【0042】
記載された一態様においては、該ユビキチン/S27aプロモータまたはその変性型、該対象とする遺伝子および該蛍光タンパク質をコードする遺伝子は、一緒に機能的に結合している。このことは、該同一のユビキチン/S27aプロモータまたはその変性型から、例えば該対象とする遺伝子および該蛍光タンパク質をコードする遺伝子両者が、発現されることを意味する。特に好ましい態様において、該機能的結合は、IRES要素を介して起こり、従ってビシストロン性mRNAは、両遺伝子から合成される。本発明による発現ベクターは、更にエンハンサ要素を含み、これは1種またはそれ以上のプロモータに対して機能的に作用する。特に好ましいものは、該ユビキチン/S27aプロモータまたはその変性型がエンハンサ要素、例えばSV40エンハンサまたはCMVエンハンサ要素と結合している、発現ベクターである。
基本的には、発現ベクター内でのこれら遺伝子の発現は、一またはそれ以上の転写単位から開始して起こる。用語:転写単位とは、転写すべき一またはそれ以上の遺伝子を含む領域として定義される。転写単位内のこれら遺伝子は、このような単位内の該遺伝子全てが、同一のプロモータまたはプロモータ/エンハンサによる転写上の制御下にあるように、相互に機能的に結合している。これら遺伝子の該転写結合の結果として、1を越えるタンパク質または生成物が、転写単位から転写され、かくして発現され得る。各転写単位は、調節要素を含み、該要素は、その中に含まれる該遺伝子配列の転写および翻訳にとって必要である。各転写単位は、同一のまたは異なる調節要素を含むことができる。IRES要素またはイントロンは、転写単位内の該遺伝子の機能的な結合のために利用できる。
【0043】
該発現ベクターは、該対象とする遺伝子、該蛍光タンパク質をコードする遺伝子および該増幅可能な選択性マーカーを発現するための、単一の転写単位を含むことができる。あるいはまた、これらの遺伝子を、2またはそれ以上の転写単位内に配置することも可能である。転写単位内での、これら遺伝子の様々な組合せが、可能である。本発明のもう一つの態様では、1、2またはそれ以上の転写単位からなる、1を越える発現ベクターが、同時トランスフェクションまたは任意の所定の順序で連続的にトランスフェクションすることにより、宿主細胞内に挿入することができる。調節要素および遺伝子の、各ベクター上でのあらゆる組合せを選択することができるが、該転写単位の十分な発現が確保できることを条件とする。必要ならば、他の調節要素および遺伝子、例えば追加の対象とする遺伝子または選択性マーカーを、該発現ベクター上に配置できる。
【0044】
従って、本発明による発現ベクターは、該蛍光タンパク質をコードする遺伝子および該増幅可能な選択性マーカー遺伝子を、一つの、または2つの別々の転写単位内に含むことができる。各転写単位は、一またはそれ以上の遺伝子生成物を転写し、発現することができる。両遺伝子が一つの転写単位に含まれている場合、これらの遺伝子は、同一のプロモータまたはプロモータ/エンハンサの制御下にあり、一方で好ましくはIRES要素は、これら全ての成分の機能的な結合を補償するために、使用することができる。該蛍光タンパク質をコードする遺伝子および該増幅可能な選択性マーカー遺伝子が、2つの別々の転写単位内に含まれる場合、これらは、同一のまたは異なるプロモータ/エンハンサの制御下にあり得る。しかし、好ましくは、その天然のまたはより弱い異種プロモータ、例えばSV40初期プロモータを、該選択性マーカー遺伝子に対して使用し、また好ましくはエンハンサを使用しない。2つの別々の転写単位を持つ発現ベクターが、本発明の範囲内において好ましい。一つの(ビシストロン性)転写単位は、該対象とする遺伝子および該蛍光タンパク質をコードする遺伝子を含み、一方で他の転写単位は、該増幅可能な選択性マーカー遺伝子を含む。好ましくは、各転写単位は、ポリAシグナル、好ましくはtkポリA、BGHポリAまたはSV40ポリAをコードする配列によって、3'末端において制限されている。
【0045】
同様に本発明において好ましいものは、上記に対して、該対象とする遺伝子が、多重クローニングサイトのみを持つベクターであり、これらは制限エンドヌクレアーゼに対する認識配列を介して、該対象とする遺伝子のクローニングを可能とする。あらゆる種類の制限エンドヌクレアーゼ並びに関連する制限エンドヌクレアーゼに対する多くの認識配列が、従来技術から公知である。好ましくは、少なくとも6個のヌクレオチドからなる配列を、認識配列として使用する。適当な認識配列のリストは、例えばSambrook等, 1989に見出すことができる。
宿主細胞
本発明の発現ベクターでトランスフェクションするために、真核生物の宿主細胞、好ましくは哺乳動物細胞、およびより好ましくは齧歯目動物の細胞、例えばマウス、ラットおよびハムスターの細胞系を使用する。対応する細胞の、本発明の発現ベクターによる首尾良いトランスフェクションは、形質転換された、一般には変性された、組換えまたはトランスジェニック細胞をもたらし、これらも本発明の課題である。
【0046】
本発明の目的にとって、好ましい宿主細胞は、ハムスター細胞、例えばBHK21、BHK TK-、CHO、CHO-K1、CHO-DUKX、CHO-DUKX B1およびCHO-DG44細胞またはこれら細胞系の誘導体/子孫である。特に好ましいものは、CHO-DG44、CHO-DUKX、CHO-K1およびBHK21細胞であり、特にCHO-DG44およびCHO-DUKX細胞が好ましい。同様に適当な細胞は、マウス由来のミエローマ細胞、好ましくはNS0およびSp2/0細胞およびこれら細胞系の誘導体/子孫である。
本発明において使用することのできるハムスターおよびマウス細胞の例を、以下の表2に与える。しかし、これら細胞、他の哺乳動物の細胞、例えばヒト、マウス、ラット、サル、齧歯目動物の細胞系(これらに限定されない)、または真核生物細胞、例えば酵母、昆虫、および植物細胞(これらに限定されない)の誘導体および子孫も、生物薬学的タンパク質を製造するための、宿主細胞として使用することができる。
【0047】
表2:ハムスターおよびマウスの産生細胞系
【0048】
ポリヌクレオチドまたは本発明の発現ベクターの一つを含む、真核宿主細胞のトランスフェクションは、従来の方法に従って行われる(Sambrook等, 1989; Ausubel等, 1994)。適当なトランスフェクション法は、例えばリポソーム媒介トランスフェクション、リン酸カルシウム共沈、エレクトロポレーション、ポリカチオン-(例えば、DEAEデキストラン)-媒介トランスフェクション、プロトプラストヒュージョン、マイクロインジェクションおよびウイルス感染法を包含する。本発明によれば、好ましくは安定なトランスフェクションが行われ、そこでは該構築物が、該宿主細胞のゲノムあるいは人工的な染色体/ミニ染色体内に組込まれるか、もしくは安定な方法で該宿主細胞内にエピソーム的に含められる。好ましいトランスフェクション法は、問題とする該宿主細胞内で、該異種遺伝子の最適なトランスフェクション頻度および発現を与えるものである。定義によれば、宿主細胞内に挿入された各配列または各遺伝子は、該宿主細胞との関連で、「異種配列」または「異種遺伝子」と呼ばれる。これは、導入すべき該配列または導入すべき該遺伝子が、該宿主細胞の内在性配列または内在性遺伝子と等価である場合にも成り立つ。例えば、ハムスター宿主細胞に導入されたハムスターアクチン遺伝子は、定義によれば異種遺伝子である。
【0049】
ヘテロダイマータンパク、例えばモノクローナル抗体(mAb)の組換え生産において、適当な宿主細胞のトランスフェクションは、理論的には2つの異なる方法により実施できる。この種のmAbは、幾つかのサブユニット、即ち重鎖および軽鎖で構成されている。これらのサブユニットをコードする遺伝子は、単一のプラスミド上の、独立したまたはマルチシストロン性の転写単位内に収容されており、次いで該プラスミドによって、該宿主細胞をトランスフェクションする。これは、該宿主細胞のゲノム内への組み込み後に、該遺伝子の化学量論的な発現量を確保するためである。しかし、独立した転写単位の場合、異なるタンパク質をコードするmRNAが、結果として同一の安定性および転写並びに翻訳効率を示すことを保証する必要がある。第二の場合において、該遺伝子の発現は、単一のプロモータによって、マルチシストロン性転写単位内で起こり、またただ一つの転写物が形成される。IRES要素を用いることにより、該遺伝子の内部翻訳開始の高い効率が、第二のおよびその後のシストロンにおいて得られる。しかし、これらシストロンに関する発現率は、第一のシストロンの発現率よりも低く、その翻訳開始は、所謂「キャップ」-依存性プレイニシェーション(pre-initiation)錯体によって、実質的にIRES-依存性の翻訳開始よりも、一層効率的になる。これらシストロンの真の等モル量発現を達成するためには、追加のシストロン間要素を導入することができ、これらは、例えば該IRES要素との組合せで、均一な発現率を確保する(WO 94/05785)。
【0050】
幾つかの異種タンパク質を同時に製造するための、本発明にとって好ましいもう一つの可能な方法は、同時トランスフェクション法であり、ここでは、遺伝子は、異なる発現ベクター内に別々に組込まれている。この方法は、ある比率の遺伝子および遺伝子生成物を、相互に調節することができ、結果的に該mRNAにおけるおよび転写および翻訳の効率におけるあらゆる差異を釣り合わせることを可能とする。更に、該発現ベクターは、そのサイズが小さいことから、より安定であり、またクローニングおよびトランスフェクション両者において、取り扱いが容易である。
従って、本発明の一特定の態様では、該宿主細胞を、1またはそれ以上の他の対象とするタンパク質をコードする遺伝子を持つ、1またはそれ以上のベクターで付随的にトランスフェクション、好ましくは同時トランスフェクションする。該同時トランスフェクションにおいて使用する該他のベクター(1または複数)は、例えば同一のプロモータ/エンハンサの組合せによる制御下において、対象とする他のタンパク質(1または複数)を、および少なくとも1種の他の選択性マーカー、例えばネオマイシンホスホトランスフェラーゼをコードする。
【0051】
本発明によれば、該宿主細胞は、血清を含まない条件化で、場合によっては動物タンパク質/ペプチドを含まない培地内で樹立され、適合されかつ培養される。市販品として入手できる媒体の例は、Ham's F12 (DE、ダイセンホフェン(Deisenhofen)のシグマ(Sigma)社)、RPMI-1640(シグマ社)、ドゥルベコ変性イーグル培地(DMEM;シグマ)、最少必須培地(MEM;シグマ)、イスコフス(Iscove's)変性ドゥルベコ培地(IMDM;シグマ)、CD-CHO(USA、CA、カールスバッドのインビトロゲン(Invitrogen))、CHO-S-SFMII(インビトロゲン)、血清を含まないCHO-培地(シグマ)およびタンパク質を含まないCHO-培地(シグマ)を包含する。これら培地各々は、場合により様々な化合物、例えばホルモンおよび/または他の成長因子(例えば、インシュリン、トランスフェリン、上皮増殖因子、インシュリン-様の成長因子)、塩(例えば、塩化ナトリウム、カルシウム、マグネシウム、リン酸塩)、バッファー(例えば、HEPES)、ヌクレオシド(例えば、アデノシン、チミジン)、グルタミン、グルコース、または他の等価な栄養物、抗生物質および/または痕跡元素を補充することもできる。血清を含まない培地が、本発明において好ましいが、該宿主細胞は、適当量の血清と混合した培地を用いて培養することも可能である。1またはそれ以上の選択性のマーカー遺伝子を発現する、遺伝的に変性された細胞を選別するために、1またはそれ以上の選別剤を、該培地に添加する。
【0052】
該用語「選別剤」とは、問題とする該選択性マーカー遺伝子に欠ける、宿主細胞の成育または生存に影響を及ぼす物質を意味する。例えば、発現された抗生物質耐性遺伝子、例えばネオマイシンホスホトランスフェラーゼの存在について選別するためには、抗生物質ジェネティシン(geneticin; G418)を、該培地添加物として使用することが好ましい。該選別剤は、また使用した該選択性マーカー遺伝子が、増幅可能な選択性マーカー遺伝子である場合には、該マーカー遺伝子の増幅を誘発する物質であっても良い(表1参照)。例えば、メトトレキセートは、該DHFR遺伝子を増幅するのに適した選択培地である。増幅を誘発するその他の選別剤の例は、表1に列挙してある。
【0053】
選択性マーカー遺伝子は、培地に対応する選別剤を添加することによって、この遺伝子を含む細胞の特異的な選別を可能とする遺伝子である。一例として、抗生物質耐性遺伝子は、正の選択性マーカーとして使用できる。この遺伝子で形質転換されている細胞のみが、該対応する抗生物質の存在下で生育でき、その結果該細胞が選別される。他方において、未形質転換細胞は、これらの選択条件下では生育または生存することができない。正、負および二官能性の選択性マーカーが存在する。正の選択性マーカーは、該選別剤に対する耐性を付与することにより、あるいは該宿主細胞における代謝上のまたは異化欠陥を補償することにより、形質転換細胞の選別およびその結果としての該細胞の富化を可能とする。これとは対照的に、この選択性マーカー用の遺伝子を受け取った細胞は、負の選択性マーカーによって、選択的に排除され得る。その一例は、単純疱疹ウイルスのチミジンキナーゼ遺伝子であり、同時に付加されたアシクロビルまたはガンシクロビルをも含む細胞内でのその発現は、その排除に導く。該増幅可能な選択性マーカーをも含む、本発明において使用するこれらの選択性マーカーは、一般に変性された突然変異体および変異型、フラグメント、機能的な等価物、誘導体、類似体および他のタンパク質またはペプチドとの融合体を包含するが、該選択性マーカーが、その選択特性を保持することを条件とする。このような誘導体は、選択性であると思われる領域またはドメインにおけるアミノ酸配列について、かなりの相同性を示す。文献には、多数の選択性マーカー遺伝子、例えば二官能性(正/負)マーカー(例えば、WO 92/08796およびWO 94/28143を参照のこと)が記載されている。真核細胞において通常使用される選択性マーカーの例は、アミノグリコシドホスホトランスフェラーゼ(APH)、ハイグロマイシンホスホトランスフェラーゼ(HYG)、ジヒドロ葉酸レダクターゼ(DHFR)、チミジンキナーゼ(TK)、グルタミンシンセターゼ、アスパラギンシンセターゼに関する遺伝子、およびネオマイシン(G418)、プロマイシン、ヒスチジノールD、ベロマイシン(belomycin)、フレオマイシン(phleomycin)およびゼオシン(zeocin)に対する耐性を付与する遺伝子を包含する。
【0054】
蛍光-活性化細胞選別(FACS)によって、形質転換した細胞を選別することも可能である。そのために、細菌性のβ-ガラクトシダーゼ、細胞表面マーカーまたは蛍光タンパク質(例えば、エクオレアビクトリア(Aequorea victoria)またはレニラレニホルミス(Renilla reniformis)または他の種を由来とする、グリーン蛍光タンパク質(GFP)およびその変異型、非-生体発光性生物、例えばディスコゾーマ(Discosoma) sp、アネモニア(Anemonia) sp、クラブラリア(Clavularia) sp、ゾアンサス(Zoanthus) sp.)を、形質転換細胞の選別のために使用することができる。
【0055】
本発明においては、該DHFR遺伝子の使用が、遺伝的に変性された(組換え)宿主細胞を選別するための、該増幅可能な選択性マーカー遺伝子として好ましい。このマーカーは、DHFR-負の基本的な細胞、例えばCHO-DG44またはCHO-DUKXを使用する場合に、選別並びにその後の増幅のために特に適している。というのは、これらの細胞が内在性のDHFRを発現せず、その結果プリンを含まない培地中で生育しないからである。結果的に、該DHFR遺伝子は、ここでは支配的な選択性マーカーとして使用することができ、また該形質転換された細胞は、ハイポキサンチン/チミジンを含まない培地内で選別される。DHFR-媒介遺伝子および増幅を行うために、メトトレキセート(MTX)を使用する。この生育特性は、MTXの添加によって臨界的に影響を受ける。通常、該MTXの濃度および増幅段階の数が増えるに従って、該細胞の醗酵強度における実質的な低下が観測される。しかし、驚いたことに、本発明によるクローン選別系を用いた場合、高濃度のMTXの存在下で、より一層強固な挙動を示す、組換え宿主細胞が豊富となる(図7を参照のこと)。このように、蛍光-活性化セルソータ(FACS)を用いて同定し、かつ選別した宿主細胞が、500nM、好ましくは1μMのMTXの存在下で培養し、かつ増幅でき、大幅に高められた生産性を与えた。このように、高い生産性を示す宿主細胞の選別法は、とりわけ本発明による成果であると考えられ、本発明の発現ベクターにより形質転換され、かつ少なくとも対象とする遺伝子、該蛍光タンパク質およびDHFR遺伝子を発現する宿主細胞が、FACS選別によって選別され、また少なくとも500nM、好ましくは1μMのMTXの存在下で、遺伝子増幅段階に付される場合には、高い生産性を示す宿主細胞の選別法は、とりわけ本発明による成果であると考えられる。
【0056】
「醗酵強度」なる用語は、上記細胞の生育特性、例えばある成長速度の維持、「規模拡大」(バイオリアクタのより大きなサイズ)に対する強靭さ、および規模拡大の際の工業的生産率を満たすための、ストックの保持における高い細胞計測数および生存能力の達成を意味する。
発現
発現なる用語は、宿主細胞内における、異種遺伝子配列の転写および/または翻訳に関連する。発現率は、一般に該宿主細胞内に存在する、対応するmRNAの量、または該対象とする遺伝子によりコードされ、製造される遺伝子生成物の量に基いて決定できる。選択されたヌクレオチド配列の転写により製造されるmRNAの量は、例えばノーザンブロットハイブリッド化、リボヌクレアーゼ-RNA-プロテクション、細胞性RNAのその場でのハイブリッド化、あるいはPCR法によって決定できる(Sambrook等, 1989; Ausubel等, 1994)。選択されたヌクレオチド配列によってコードされるタンパク質も、様々な方法、例えばELISA、ウエスタンブロット、ラジオイムノアッセイ、免疫沈降、該タンパク質の生物学的な活性の検出または該タンパク質の免疫染色およびこれに続くFACS分析により決定できる(Sambrook等, 1989; Ausubel等, 1994)。
【0057】
用語「高い発現レベル(または率)、高い発現、高められた発現または高(い)生産性」とは、長期間持続する、かつ十分に高い発現、または宿主細胞に導入された異種配列、例えば治療用のタンパク質をコードする遺伝子の合成を意味する。高められたまたは高い発現または高い発現レベルまたは発現率もしくは高い生産性は、本発明による細胞が、ここに記載する本発明による方法の一つで培養し、またこの細胞が、1日当たり少なくとも約5pg(5pg/日/細胞)を越える所定の遺伝子生成物を製造する場合に存在する。高められたまたは高い発現または高い発現レベルまたは発現率もしくは高い生産性は、本発明による細胞が、1日当たり少なくとも約10pg(10pg/日/細胞)を越える所定の遺伝子生成物を製造する場合にも存在する。高められたまたは高い発現または高い発現レベルまたは発現率もしくは高い生産性は、特に、本発明による細胞が、1日当たり少なくとも約20pg(20pg/日/細胞)を越える所定の遺伝子生成物を製造する場合にも存在する。特別に高められたまたは高い発現、または特別に高い発現レベルまたは発現率もしくは特別に高い生産性は、本発明による細胞が、1日当たり少なくとも約30pg(30pg/日/細胞)を越える所定の遺伝子生成物を製造する場合に存在する。
【0058】
本発明による、高いまたは高められた発現、高い生産性または高い発現レベルまたは発現率は、様々な方法で達成することができる。例えば、増幅可能な選択性マーカーに対する遺伝子を含む、対象とする該遺伝子の同時発現によって、高度に該異種遺伝子を発現する細胞を選別し、かつ同定することが可能である。該増幅可能な選択性マーカーは、安定にトランスフェクションされた宿主細胞の選択を可能とするばかりでなく、対象とする該異種遺伝子の、遺伝子増幅をも可能とする。該核酸の付随的なコピーは、該宿主細胞のゲノムに、付随的な人工的/ミニ染色体にあるいはエピソームとして位置するポリヌクレオチドに組込むことができる。この手順は、付随的な選択性マーカーとして、1またはそれ以上の蛍光タンパク質(例えば、GFP)または細胞表面マーカーを含む組換え宿主細胞の、FACS-アシスト選別と組み合わせることができる。高められた発現を達成するための他の方法(および異なる方法の組合せも利用可能である)は、例えば(人工的な)転写因子の利用、より調節性の良い内在性または異種遺伝子発現用の、天然または合成薬物による該細胞の処理、mRNAまたは該タンパク質の安定性(半減期)の改善、mRNA翻訳開始の改善、エピソームプラスミドの使用による、遺伝子用量の増大(複製起源としてのウイルス配列、例えばSV40、ポリオーマ、アデノウイルス、EBVまたはBPVの使用に基く)、増幅-促進配列の利用(Hemann等, 1994)またはDNAコンカテマーに基くインビトロ増幅系(Monaco等, 1996)。
【0059】
本発明によれば、該対象とする遺伝子と該蛍光タンパク質をコードする遺伝子との、組み合わせた転写を行う。得られるビシストロン性のmRNAは、該対象とするタンパク質および該蛍光タンパク質両者を発現する。該対象とするタンパク質および該蛍光タンパク質の発現に関するこの組合せに基いて、本発明によれば、発現された蛍光性タンパク質によって、例えば蛍光-活性化セルソータ(FACS)を用いた選別によって、高い生産性を示す組換え宿主細胞を選択し、かつ単離することが、容易に可能となる。
高い生存能力および該所定の遺伝子生成物の、高い発現率を示す、組換え宿主細胞の選別は、多段階法である。本発明による発現ベクターでトランスフェクションされ、あるいは場合により、他のベクターにより同時トランスフェクションされている宿主細胞は、例えば蛍光タンパク質をコードし、該対象とする遺伝子と結合した遺伝子の発現について、少なくとも検討されるが、これは蛍光タンパク質の最大の発現率を示す細胞/細胞集団を同定し、かつ選別するためである。好ましくは、蛍光タンパク質の最大の発現率を示す細胞の10%に属する細胞のみが、選別され、かつ更に培養される。事実、このことは、該蛍光性細胞の最も明るい10%が、選別され、かつ更に培養される。従って、細胞混合物の該蛍光性細胞の最も明るい5%、好ましくは最も明るい3%またはさらには最も明るい1%を、選別し、かつ複製することも可能である。特に好ましい態様では、該蛍光性細胞の最も明るい0.5%または最も明るい0.1%のみを、選別し、かつ複製する。
【0060】
この目的のために、本発明の発現ベクターで前もって形質転換されている細胞を、選択培地で培養する。この選択培地は、場合により該増幅可能な選択性マーカーに対して特異的な選別剤をも含む。段階的に高められる選別剤の濃度を使用して、徐々に高くなる選択圧力を及ぼすことができる。
この選別段階は、細胞プールについて、あるいは予め選別した細胞プール/細胞クローンについて行うこともできる。1またはそれ以上、好ましくは2またはそれ以上および特に3またはそれ以上の選別段階を実施し、一方で各選別段階間で、所定の期間、例えばプールの場合にはほぼ2週間に渡り、該細胞を培養し、かつ複製する。
少なくとも対象とする遺伝子および該増幅可能な選択性マーカー遺伝子のコピー数を増大させるために、所定の宿主細胞を、1またはそれ以上の遺伝子増幅段階に付すことができる。メトトレキセートを用いた、段階的な遺伝子増幅方法は、例えば米国特許第5,179,017号に記載されている。この発明によれば、達成することのできる高い生産性は、遺伝子コピー数の多さとは結び付けられない。これは寧ろ、該高性能クローンの高められた安定性および醗酵強度の現れである。従って、これは、必要とされる遺伝子増幅段階数を減じ、また例えばただ一回の遺伝子増幅段階の実施を可能とする。
【0061】
従って、本発明は、細胞を選別する方法に関連し、この方法は以下に列挙する諸工程を含む:
(i) 少なくとも、本発明によるベクターの一つを含む、適当な宿主細胞の形質転換工程、ここで該発現ベクターのDNAは、好ましくは該宿主細胞のゲノム内に、あるいは人工的な染色体/ミニ染色体内に安定に組込まれている;
(ii)該対象とする遺伝子および該蛍光タンパク質の発現を可能とする条件下で、該形質転換した細胞を培養する工程;
(iii) 該細胞を、少なくとも1種の選別剤の存在下で培養し、該選別剤の存在下で成育できる細胞のみを複製する工程;
(iv) 蛍光タンパク質に対して最大の発現率を示す細胞を、細胞混合物から選別する工程、ここで該細胞は、蛍光-活性化セルソータ(FACS)によって、検出されかつ選別される;および
(v) 該蛍光タンパク質に対して最高の発現率を示す、該選別された細胞を培養する工程。
場合により、上記工程ii)〜v)は、上記工程v)に従って得た細胞について、1またはそれ以上の回数に渡り繰返すことができる。更に、該形質転換細胞は、場合により該増幅可能な選択性マーカー遺伝子の増幅へと導く、選別剤の存在下で培養することにより、1またはそれ以上の遺伝子増幅段階に付すことも可能である。この段階は、まだ選別されていない細胞および既に1またはそれ以上の回数に渡り予備的に選別されている細胞両者について実施できる。
【0062】
本発明は、また同様に選別された細胞を、複製し、かつ対象とするコード性の該遺伝子生成物を製造する方法にも関連する。好ましくは、該選択された高生産性の細胞は、血清を含まない培地内で培養し、および好ましくは該対象とする遺伝子の発現を可能とする条件の下で、懸濁培養により培養する。該対象とするタンパク質は、好ましくは分泌された遺伝子生成物として、該細胞培養培地から得られる。しかし、このタンパク質が、分泌シグナル無しに発現される場合には、該遺伝子生成物は、また細胞溶解物から単離することもできる。他の組換えタンパク質および宿主細胞タンパク質を実質的に含まない、純粋な均質な製品を得るために、公知の精製段階を実施する。まず、通常は、細胞および細胞デブリを、該培地または溶解物から除去する。次いで、所定の遺伝子生成物を、例えばイムノアフィニティーおよびイオン交換カラム上での分別、エタノール沈殿、逆相HPLCまたはセファデックス、シリカまたはDEAE等のカチオン交換樹脂上でのクロマトグラフィーによって、混入した溶解性のタンパク質、ポリペプチドおよび核酸から分離することができる。組換え細胞により発現される異種タンパク質の精製法は、当業者には公知であり、文献、例えばHarris等(1995)およびScopes (1988)に記載されている。
【0063】
以下、本発明を、幾つかの非-限定的な例示的態様を参照して、より一層詳細に説明する。
実施例
略号:
AP:アルカリホスファターゼ;bp:塩基対;CHO:チャイニーズハムスター卵巣;DHFR:ジヒドロ葉酸レダクターゼ;ELISA:酵素結合イムノソルベント検定法;FACS:蛍光-活性化セルソータ;FAP:繊維芽細胞-活性化タンパク質;GFP:グリーン蛍光タンパク質;HBSS:ハンクスの平衡塩類溶液;HT:ハイポキサンチン/チミジン;HRPO:西洋ワサビパーオキシダーゼ;IRES:内部リボソームエントリーサイト;kb:キロベース;mAb:モノクローナル抗体;MTX:メトトレキセート;PCR:ポリメラーゼ連鎖反応;slCAM:可溶性細胞内接着分子
【0064】
方法
1. 細胞培養およびトランスフェクション
細胞CHO-DG44/dhfr-I-(Ulraub等, 1983)を、細胞培養フラスコ内で、湿潤大気および5%CO2中で37℃にて、ハイポキサンチンおよびチミジンを補充した、血清を含まないCHO-S-SFMII培地(DE、カールスルーエ(Karlsruhe)のインビトロゲンGmbH)内で、懸濁細胞として、永続的に培養した。生細胞数および生存率を、CASY1細胞計数器(DE、シェルフェシステム(Schaerfe System))を用いて、あるいはトリパン青染色により決定し、また次に該細胞を1-3×105/mLなる濃度で播種し、2-3日毎に操作(alle 2-3 Tage passagiert)した。
【0065】
リポフェクタミンプラス試薬(Lipofectamine Plus Reagent; インビトロゲンGmbH)を、CHO-DG44のトランスフェクションのために使用した。各トランスフェクション混合物に対して、全体で1μgのプラスミド-DNA、4μLのリポフェクタミンおよび6μLのプラス試薬を、製造業者の指示に従って一緒に混合し、200μLなる容量で、0.8mLのHT-補充CHO-S-SFMII培地内の、6×105個の指数増殖するCHO-DG44細胞に添加した。細胞インキュベータ内で37℃にて3時間インキュベータした後、2mLのHT-補充CHO-S-SFMII培地を添加した。安定にトランスフェクションされたCHO-DG44細胞の、上記DHFRに基く選別のために、トランスフェクションの2日後に、該細胞を、ハイポキサンチンおよびチミジンが全く添加されていない、CHO-S-SFMII培地に移し、3〜4日ごとに培地を交換した。一つの発現ベクターが、DHFRおよびネオマイシンホスホトランスフェラーゼ選択性マーカーを含む他の発現ベクターを含む、同時トランスフェクションの場合の、DHFRに基くおよびネオマイシンホスホトランスフェラーゼに基く選別においては、G418(インビトロゲン)をも、400μg/mLなる濃度にて、該培地に添加した。
該組込み異種遺伝子のDHFRに基く遺伝子増幅は、HTを含まないCHO-S-SFMII培地に、5-2000nMなる濃度で、選別剤MTX(DE、ダイセンホーフェン(Deisenhofen)のシグマ社)を添加することにより達成した。
【0066】
2. 発現ベクター
該発現を解析するために、pAD-CMVベクター(Werner等, 1998)を基本とし、CMVエンハンサ/ハムスターユビキチン/S27aプロモータ(WO 97/15664)の組合せにより、異種遺伝子の構成性発現を媒介する、真核細胞発現ベクターを使用した。その基本ベクターpBIDは、該ベクターpBIN内で、増幅可能な選択性マーカーとして機能する(例えば、EP 0 393 438)DHFRミニ遺伝子を含むが、該DHFRミニ遺伝子は、ネオマイシン耐性遺伝子で置換えられている(図2)。この目的のために、SV40初期プロモータおよびTK-ポリアデニル化シグナルを含む、該選択性マーカーネオマイシンホスホトランスフェラーゼを、1640bpのBsu36Iフラグメントとして、市販のプラスミドpBK-CMV(USA、CA、ラジョラのストラタジーヌ)から単離した。該フラグメントの末端を、クレノウ-DNA-ポリメラーゼで満たすための反応の後に、該フラグメントを、同様にクレノウ-DNA-ポリメラーゼで処理した、該ベクターpBIDの3750bpのBsu36I・StuIフラグメントを用いて結合した。
【0067】
このビシストロン性基本ベクターpBIDG(図2)において、そのIRES-GFP遺伝子領域は、ベクターpIRES2-EGFP(USA、CA、パロアルトのクロンテック社)から単離し、また該ベクターpBID内のCMVエンハンサ/プロモータの制御下に置き、結果的に該プロモータ領域とIRES-要素との間の、多重クローニングサイトを保持した。以下の手順を使用した。該プラスミドpIRES2-EGFPが、鋳型として機能するPCR突然変異誘発においては、一方で、該IRES配列内のHindIII切断サイトAAGCTTが、突然変異原性のプライマーを用いることにより配列ATGCTTに転化され、かつ脱離された。他方、Xbal切断サイトは、該IRES配列の5'末端に対して相補性を持つプライマーによって挿入され、あるいはSpel切断サイトが、該GFP配列の3'末端に対して相補性を持つプライマーによって導入された。完全なIRESおよびGFP配列を含む、得られるPCRフラグメントを、XbalおよびSpelで消化し、該ベクターpBIDの多重クローニングサイトの3'末端における、単一のXbal切断サイト内にクローニングした。
ヒトslCAM遺伝子を、pAD-slCAM(Werner等, 1998)由来のHindIII/Sallフラグメントとして単離し、該ベクターpBIDGの対応する切断サイトにクローニングして、ベクターpBIDG-slCAM(図3)を得た。
【0068】
モノクローナルヒト化F19抗体を発現するために、その重鎖を、プラスミドpG1D105F19HC(NAGENESEQ: AAZ32786)から、1.5kbのNael/HindIIIフラグメントとして単離し、またEcoRI(クレノウ-DNA-ポリメラーゼで満たされている)およびHindIIIで消化した、該ベクターpBIDG内にクローニングして、ベクターpBIDG-F19HC(図3)を得た。他方で、その軽鎖は、プラスミドpKN100F19LC(NAGENESEQ: AAZ32784)から、1.3kbのHindIII/EcoRIフラグメントとして単離し、該ベクターpBINの対応する切断サイト内にクローニングして、ベクターpBIN-F19LC(図3)を得た。
3. FACS
フローサイトメトリー分析および選別を、コールターエピックスアルトラ(Coulter Epics Altra)装置を用いて実施した。このFACSは、励起波長488nmを持つヘリウム-アルゴンレーザーを備えている。その蛍光強度は、蛍光タンパク質と適合する波長において吸収され、付随するソフトウエア:コールターエキスポ32(Coulter Expo32)によって処理する。選別は、8000-10000事象/秒なる速度にて実施した。該懸濁された細胞を遠心分離(180xgにて5分間)し、HBSS内での細胞密度を、1-1.5×107/mLに調節した。次いで、これらの細胞を、その蛍光タンパク質シグナルに従って選別した。これらの細胞を、既に培地を含む試験管に採り、次いで遠心分離処理し、選別された細胞の数に応じて、適当な培養容器に播種した。
【0069】
4. ELISA
安定にトランスフェクションされたCHO-DG44細胞の上澄み中のslCAM力価は、一方でヤギ抗-ヒトIgG Fcフラグメント(DE、ハンブルグのディアノバ(Dianova))および他方でAP-複合ヤギ抗-ヒトκ軽鎖抗体(シグマ社)を使用して、標準的な手順に従って、ELISA法により定量した。精製されたF19抗体を、基準として用いた。
生産性(pg/細胞/日)は、式:pg/((Ct-Co)t/ln(Ct-Co))に従って、算出した。ここで、CoおよびCtは、夫々播種した際および収穫した際の生細胞数であり、またtは培養時間である。
【0070】
実施例1:CMVおよびハムスターユビキチン/S27a-プロモータ活性の比較
ハムスターユビキチン/S27a-プロモータの活性と、真核生物の発現ベクターにおいて頻繁に使用されるCMVの活性とを比較するために、CHO-DG44細胞を、様々な組換えベクターでトランスフェクションした。異種遺伝子生成物、即ち一方ではリソソーム酵素および他方ではIgG1-抗体が、該CMV-プロモータの制御下で、あるいは該ハムスターユビキチン/S27a-プロモータの制御下で発現された。これら2種のプロモータは、機能的にCMV-エンハンサと結合していた。BGH ポリAを、該異種遺伝子に対する終止シグナルとして使用した。該CMV-プロモータを含む該発現ベクターは、変性されたpcDNA3(「CMV」、インビトロゲン)またはpBluescriptベクター(「CMV2」、ストラタジーヌ)に基いており、また付随的に該増幅可能な選択性マーカージヒドロ葉酸レダクターゼをコードした。他方で該ハムスタープロモータを含む発現ベクターは、pAD-CVMベクター(Werner等, 1998)に基くものであった。この抗体の重鎖および軽鎖を発現するために、選択性マーカーとして、ネオマイシン耐性遺伝子を含む、第二のベクターを用いて同時トランスフェクションを行った。しかし、該CMVエンハンサは、SV40エンハンサで置換することもできる。
【0071】
96ウエルプレートでの限界希釈によって、細胞クローンを選別し、またHTを含まない培地(同時トランスフェクションの場合には、400μg/mLのG418を添加した)中で単離した。組換えタンパク質の最大の生産率を持つ細胞クローンを、各場合において、稀釈クローニングと組み合わせて、メトトレキセート濃度を段階的に5nMから、50nMを経て、500nM〜2μMまで高めることにより、段階的なDHFRに基く遺伝子増幅にかけた。各増幅段階において、該最大の生産率を持つ約20〜30クローンを選択した。
一般に、該ハムスタープロモータが、最大の性能を持つことが分かった。該リソソーム酵素発現および該抗体の発現両者において、生産性または力価は、異種遺伝子が、CMVプロモータの制御下で発現される細胞において観測される、これらの値の、2〜5倍高い値として得られた。図1は、この特定の増幅段階における、最良の細胞クローンの相対的な力価および相対的な比生産率を、一例として示す。ここで、該CMVプロモータに基く、特定の異種遺伝子に関する発現を1に設定した(該リソソーム酵素に対してCMV1、該抗-体に対してCMV2)。
【0072】
実施例2:GFPに基くFACS選別による、高い発現性を示すslCAM細胞の単離
細胞間接着分子ICAM1、slCAMの可溶性型は、放射性同位元素を含まないものに関する可能な処理である。というのは、これが、ライノウイルスとの結合に関してICAMレセプタと競合し、このようにして該細胞の侵入およびその後の感染に対する必要条件となる、該ICAMレセプタとの相互作用を低下もしくは阻害さえも起こす(Bella等, 1999; Marlin等, 1990)。
【0073】
slCAMを、CHO細胞において、単鎖タンパク質(480アミノ酸)を発現するための一例として選択した。このために、CHO-DG44をpBIDG-slCAMでトランスフェクションした(図3)。pBIDG-slCAMでトランスフェクションした細胞内での、GFPの付随的な発現は、FACSを基本とする選別法の利用を可能にする。これら治療用タンパク質slCAMおよびGFPは、ビシストロン性転写単位により連帯的に発現され、また該DHFRは別の転写単位により発現される。HTを含まないCHO-S-SFMII培地における最初の選別の2〜3週間後に、最大のGFP蛍光を示す細胞の5%を選別した。約2週間の培養後、最大のGFP蛍光を示す5%の細胞を再度単離した。この周期的な選別操作を全体で6回行った。slCAMの生産性とGFP蛍光との間の良好な相関関係が明らかとなった(図4)。このようにして、FACS-アシスト選別のみによって、如何なるMTX増幅段階も使用することなしに、16pg/細胞/日までの高い比生産率を持つ細胞のプールが、極めて短い期間内に単離された(図5)。該GFPに基く選別と、単一回のこれに続くMTX増幅段階とを組合せることによって、生産性を30pg/細胞/日まで高めることさえ可能となった(図6)。
【0074】
これらの生産性は、第4回目の選別後の、細胞プールの500nMのMTXによる増幅、および第6回目の選別後の、細胞プールの2μMのMTXによる増幅両者により達成された。通常、5-20nMのMTXという、極めて低いMTX濃度にて開始される、段階的な増幅とは対照的に、増幅効果を達成するために、最初からより高いMTX濃度を使用する必要があった。従って、5または50nMのMTXを、第4回目の選別から細胞に添加した結果、あるいは500nMのMTXを、第6回目の選別から細胞に添加した結果として、生産性における有意な増加は何等観測されなかった(図6)。明らかに、出発プールにおけるDHFRの濃度は、既に極めて高く、その結果全DHFR阻害は、MTXの高い用量によってのみ達成できた。更に、予備的に選別した細胞プールは、高い初期MTX用量にも拘らず極めて良好に該選別段階を生き永らえた。即ち、高い生存率を示す細胞群が、従来の段階的な遺伝子増幅法により達成されるよりも、短期間内に得られた(図7)。
【0075】
実施例3:GFPを基本とするFACS選別による、mAb F19の高い発現性を持つ細胞の単離
同時トランスフェクションにおいて、CHO-DG44細胞を、プラスミドの組合せ、pBIDG-F19HCおよびpBIN-F19LCによってトランスフェクションした(図3)。この発現されたヒト化抗体F19は、反応性ストローマ繊維芽細胞によって合成される、表面分子FAPに向けられる(EP 0 953 639をも参照)。使用したベクターの構成において、該抗体の2つのタンパク質鎖は、それら自身のベクターによって発現され、該ベクターは、また付随的に別の転写単位内の、ネオマイシンホスホトランスフェラーゼ選択性マーカーまたはDHFRをもコードする。さらに、他の選択性マーカー、GFPは、ベクターpBIDG-F19HC内に含まれている。IRES要素による該GFPおよび重鎖発現の転写による結合によって、ベクターpBIDG-F19HC/pBIN-F19LCによる、CHO-DG44細胞の同時トランスフェクションにおいては、該抗体F19の高い発現性を示す細胞は、周期的なFACS選別を利用して、単に高いGFP含有率を持つ細胞を選別することにより、迅速に単離することができた。
【0076】
このため、400μg/mLのG418を添加した、HTを含まないCHO-S-SFMII培地内で、該トランスフェクションした細胞プールの初めの2〜3週間の選別後に、最大のGFP蛍光を示す細胞の5%を、FACSにより選別した。この選別は、全体として6回に渡り行い、各選別操作間に約2週間の培養期間を置いた。驚いたことに、F19の生産性と、GFP蛍光との間には、良好な相関関係があることが分かった(図8)が、両タンパク質鎖は、それ自身のベクターから発現され、またGFPを基本とするFACS選別においては、GFPとの転写による結合の結果として、重鎖のみの発現について選別できたに過ぎなかった。この生産性は、10pg/細胞/日(図9)まで高めることができ、また更に第5回目の選別による細胞プールから、1000nMのMTXを該選択培地に添加することにより、後のただ一回のMTX増幅段階によって、平均値37pg/細胞/日まで高めることができた。また、比較データは、ハムスタープロモータと、該CMVエンハンサの代わりのSV40エンハンサとを、機能的に結合することにより得ることができた。同時に、増大する量のMTXを用い、一般に4つの増幅段階を含む、従来の段階的な遺伝子増幅法と比較して、高い生産性を示す細胞を選別するための開発時間を、半分だけ約120日まで短縮し、また付随的に、開発設備およびコストにおる大幅な減少をもたらすことができた。
【0077】
文献
Adam, M.A. et al., J Virol 1991, 65, 4985 - 4990
Ausubel, F.M. et al., Current Protocols in molecular biology. New York : Greene Publishing Associates and Wiley-Interscience. 1994 (updated)
Bella, J. et al., J. Struct. Biol. 1999, 128, 69 - 74
Bennett, R.P. et al., BioTechniques 1998, 24, 478 482
Chalfie, M. et al., Science 1994, 263, 802 - 805
Chamov, S.M. et al., Antibody Fusion Proteins, Wiley-Liss Inc., 1999
Davies, M.V. et al., J Virol 1992, 66, 1924 1932
Faisst, S. et al., Nucleic Acids Research 1992, 20, 3 26
Gossen, M. et al., Curr Opi Biotech 1994, 5, 516 - 520
Gubin, A.N. et al., Biochem Biophys Res Commun 1997, 236, 347 350
Haber, D.A. et al., Somatic Cell Genetics 1982, 8, 499 - 508
Harris et al., Protein Purification : A Practical Approach, Pickwood and Hames, eds., IRL Press, 1995
Hemann, C. et al., DNA Cell Biol 1994, 13 (4), 437 - 445
Huston, C. et al., Proc Natl Acad Sci USA 1988, 85 (16), 5879 - 5883
Hu, S. et al., Cancer Res. 1996, 56 (13), 3055 - 3061
Jang, S.K. et al., J Virol 1989, 63, 1651 1660
Kaufman, R.J. et al., J Mol Biol 1982, 159, 601 - 621
Kaufman, R.J., Methods in Enzymology 1990, 185, 537 - 566
Kortt, A.A. et al., Protein Engineering 1997, 10 (4), 423 - 433
Levenson, V.V. et al., Human Gene Therapy 1998, 9, 1233 1236
Lottspeich F. and Zorbas H. eds., Bioanalytic, Spektrum Akad. Verl., 1998
Lovejoy, B. et al., Science 1993, 259, 1288 - 1293
Marlin, S.D. et al., Nature 1990, 344, 70 - 77
Meng, Y.G. et al., Gene 2000, 242, 201 - 207
Monaco, L. et al., Gene 1996, 180, 145 - 150
Morgan, R.A. et al., Nucleic Acids Research 1992, 20, 1293 - 1299
Mosser, D.D. et al., BioTechniques 1997, 22, 150 - 161
Nolan, G.P. et al., Proc Natl Acad Sci USA 1988, 85, 2603 2607
Ohshima, Y. et al., J Mol Biol 1987, 195, 247 - 259
Pack, P. et al., Biotechnology 1993, 11, 1271 1277
Pack, P. et al., J Mol Biol 1995, 246 (11), 28 - 34
Pelletier, J. et al., Nature 1988, 334, 320 325
Perisic, O. et al., Structure 1994, 2, 1217 - 1226
Primig, M. et al., Gene 1998, 215, 181 189
Ramesh, N. et al., Nucleic Acids Research 1996, 24, 2697 2700
Sambrook, J., Fritsch, E.F. & Maniatis, T.,Molecular Cloning: A Laboratory Manual Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1989
Scopes, R., Protein Purification, Springer Verlag, 1988
Simonson, C.C. et al., Proc Natl Acad Sci USA 1983, 80, 2495 - 2499
Sugimoto et al., Biotechnology 1994, 12, 694 - 698
Urlaub, G. et al., Cell 1983, 33, 405 - 412
Werner, R.G. et al., Arzneim.-Forsch./Drug.Res. 1998, 48, 870 - 880
Wigler, M. et al., Proc Natl Acad Sci USA 1980, 77, 3567 3570
【図面の簡単な説明】
【0078】
【図1】異種遺伝子生成物が、CMVプロモータの制御下、またはハムスターユビキチン/S27aプロモータの制御下の何れかで発現される、組換え細胞を用いて得た、発現レベルの比較を示す。これら二つのプロモータは、機能的にCMVエンハンサおよび終結配列と結合しており、BGHポリAは、何れの場合においても同一である。CMV1の場合には、発現ベクターはpcDNA3を基本とし(DE、カルスルーエ(Kalsruhe)のインビトロゲン)、CMV2の場合には、これはpBluescriptを基本とする発現ベクターであり(US、CA、ラジョラのストラタジーヌ)、およびCHOの場合、これは、pAD-CMVを基本とする発現ベクターである(Werner等, 1998)。リソソーム酵素の発現において、全ての発現ベクターは、増幅可能な選択性マーカー、ジヒドロ葉酸レダクターゼ(DHFR)を含み、また該異種遺伝子の発現は、メトトレキセート(MTX)による後の増幅段階により高められた。抗体(Ab)の二つの鎖を発現するために、同時トランスフェクションを、該選択性のマーカーとしてネオマイシン耐性遺伝子を含む第二のベクターを用いて行った。得られる力価または比生産率は、CMVプロモータを基本とする発現との関連で与えられ、後者の発現を1に設定する(酵素に対してCMV1、Abに対してCMV2)。
【図2】CHO-DG44細胞内で、組換えタンパクを発現するのに使用した基本ベクターの概略的な表示である。「P/E」は、CMVエンハンサとハムスターユビキチン/S27aプロモータとの組合せであり、「P」は、単独でプロモータ要素を表し、「T」は、転写に関する終止シグナルであり、転写されたmRNAのポリアデニル化のために必要とされる。各転写単位における転写開始位置および方向は、矢印で示されている。該異種遺伝子をクローニングするために、制限エンドヌクレアーゼに関する多重切断サイト(多重クローニングサイト-mcs)を持つ配列領域を、該プロモータ要素の背後に挿入する。該増幅可能な選択性マーカージヒドロ葉酸レダクターゼを「dhfr」と略記し、また該選択性マーカーネオマイシンホスホトランスフェラーゼを「neo」と略記する。脳心筋炎ウイルス由来の「IRES」要素は、ビシストロン性転写単位内の内部リボソームエントリーサイトとして機能し、またこれに伴うグリーン蛍光タンパク質「GFP」の翻訳を可能とする。
【図3】各々生物薬学的なタンパク質をコードし、かつCHO-DG44細胞をトランスフェクションするために使用された、真菌発現ベクターを、概略的に示した図である。「P/E」は、CMVエンハンサとハムスターユビキチン/S27aプロモータとの組合せであり、「P」は、単独でプロモータ要素を表し、「T」は、転写に関する終止シグナルであり、転写されたmRNAのポリアデニル化のために必要とされる。各転写単位における転写開始位置および方向は、矢印で示されている。該増幅可能な選択性マーカージヒドロ葉酸レダクターゼを「dhfr」と略記し、また該選択性マーカーネオマイシンホスホトランスフェラーゼを「neo」と略記する。脳心筋炎ウイルス由来の「IRES」要素は、ビシストロン性転写単位内の内部リボソームエントリーサイトとして機能し、またこれに伴うグリーン蛍光タンパク質「GFP」の翻訳を可能とする。「slCAM」は、可溶性の細胞内接着分子をコードし(US 5,412,216)、一方「F19HC」および「F19LC」は、ヒト化抗体F19の重鎖および軽鎖を夫々コードする(EP 953 639)。
【図4】細胞プールZB1を例とした、該slCAMの生産性と該GFP蛍光との相関関係を示す図である。この細胞プールは、ベクターpBIDG-slCAMによるトランスフェクションによって得られたものであり、ここでこれらの治療用タンパク質slCAMおよびGFPは、ビシストロン性転写単位によって協同的に発現される。このプールを、周期的なGFPを基本とするFACS選別にかけた。各選別段階(sort)の後に、該プールの細胞培養上澄中のslCAMの濃度を、ELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率を算出した(pg/c*d)。各データ点は、少なくとも3回の培養実験の平均値である。全体で6回の選別を行った。
【図5】細胞プールZB1を例として用いた、高い発現性を示すslCAM細胞の、GFPを基本とするFACS選別による単離を示す図である。この細胞プールは、ベクターpBIDG-slCAMによるトランスフェクションによって得られたものであり、ここでこれらの治療用タンパク質slCAMおよびGFPは、ビシストロン性転写単位によって一緒に発現される。このプールを、周期的なGFPを基本とするFACS選別にかけた。各選別後に、該プールの細胞培養上澄中のslCAMの濃度を、ELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率を算出した(pg/c*d)。各データ点は、少なくとも3回の培養実験の平均値である。全体で6回の選別を行った。
【図6】細胞プールZB1を例として用いた、MTX増幅段階とGFPを基本とする選別との組合せにより達成された、slCAMの生産性における増大を示す図である。ベクターpBIDG-slCAMによるトランスフェクションによって得られたものである、この細胞プールを、周期的なGFPを基本とするFACS選別にかけた。第4回目および第6回目の選別後に、該培地にメトトレキセート(MTX)を添加(5nM、50nM、500nMまたは2μMのMTX)することによって、DHFR-媒介遺伝子増幅を行った。該プールの細胞培養上澄中のslCAMの濃度を、ELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率(pg/c*d)を算出した。各データ点は、少なくとも3回の培養実験の平均値を表す。
【図7】様々な用量のメトトレキセートを該培地に添加した後の、細胞プールの生存率を示す図である。ベクターpBIDG-slCAMによるトランスフェクション(図3)によって得た、該細胞プールZB1を、周期的なGFPを基本とするFACS選別にかけた。第4回目および第6回目の選別後に、該培地にメトトレキセート(MTX)を添加することにより、DHFR-媒介遺伝子増幅を行った。細胞数および生存率は、トリプタンブルーで染色することにより、選別段階中に測定し、また培養中の数日(dic)に渡り追跡した。
【図8】細胞プールZB1を例として用いた、抗体の生産性(mAb F19)と、GFP蛍光性との間の相関関係を示す図である。この細胞プールは、ベクターpBIDG-F19HCとpBIN-F19LCとの組合せによるトランスフェクションによって得た(図3)。このプールを、周期的なGFPを基本とするFACS選別に掛けた。各選別後に、該プールの細胞培養上澄中の抗体F19の濃度を、ELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率(pg/c*d)を算出した。各データ点は、少なくとも3回の培養実験の平均値を表す。全体で6回の選別を行った。
【図9】細胞プールZB1を例として用いた、FACSを用いたGFPを基本とする選別による、高い発現性を持つmAbF19細胞プールの単離を示す図である。ベクターpBIDG-F19HCとpBIN-F19LCとの同時トランスフェクション(図3)によって得た、この細胞プールを、周期的なGFPを基本とするFACS選別にかけた。該プールの細胞培養上澄中の抗体F19の濃度を、各選別後にELISAにより測定し、また1細胞当たりのかつ1日当たりの比生産率(pg/c*d)を算出した。各データ点は、少なくとも3回の培養実験の平均値を表す。
【特許請求の範囲】
【請求項1】
ハムスター-ユビキチン/S27a-プロモータと機能可能な状態で結合した、対象とするタンパク質をコードする遺伝子、および蛍光タンパク質をコードする遺伝子を含むことを特徴とする、発現ベクター。
【請求項2】
増幅可能な、選択性マーカー遺伝子を含む、請求項1記載の発現ベクター。
【請求項3】
1または複数の上記プロモータと機能可能な状態で結合した、一種またはそれ以上のエンハンサを含む、請求項1または2記載の発現ベクター。
【請求項4】
更に、蛍光タンパク質をコードする該遺伝子、および対象とするタンパク質/生成物をコードする遺伝子のビシストロン発現を可能とする、内部リボソームエントリーサイト(IRES)をも含む、請求項1〜3の何れか1項に記載の発現ベクター。
【請求項5】
蛍光タンパク質をコードする該遺伝子および該増幅可能な、選択性マーカー遺伝子が、一つのまたは二つの別々の転写単位内に位置している、請求項1〜4の何れか1項に記載の発現ベクター。
【請求項6】
該機能性結合は、イントロン配列を介して起こることがない、請求項1〜5の何れか1項に記載の発現ベクター。
【請求項7】
該増幅可能な、選択性マーカー遺伝子が、ジヒドロ葉酸レダクターゼ(DHFR)または該蛍光タンパク質とDHFRとの融合タンパク質をコードする、請求項1〜6の何れか1項に記載の発現ベクター。
【請求項8】
該エンハンサがCMVまたはSV40エンハンサである、請求項3〜7の何れか1項に記載の発現ベクター。
【請求項9】
更に、少なくとも1種のポリアデニル化シグナルをも含む、請求項1〜8の何れか1項に記載の発現ベクター。
【請求項10】
対象とするタンパク質/生成物をコードする該遺伝子の代わりに、対象とするタンパク質をコードする遺伝子を組込むための、多重クローニングサイトを含む、請求項1〜9の何れか1項に記載の発現ベクター。
【請求項11】
請求項1〜10の何れか1項に記載の発現ベクターを用いてトランスフェクションされていることを特徴とする、真核宿主細胞。
【請求項12】
哺乳動物細胞、好ましくはCHO細胞である、請求項11記載の宿主細胞。
【請求項13】
一またはそれ以上の他の対象とするタンパク質/生成物をコードする遺伝子および少なくとも1種の他の選択性マーカーを含む、一またはそれ以上のベクターによって、更にトランスフェクションされている、請求項11または12記載の宿主細胞。
【請求項14】
異種遺伝子生成物の発現を可能とする条件の下で、請求項11〜13の何れか1項記載の宿主細胞を培養し、該遺伝子生成物を培養物または培地から単離することを特徴とする、異種遺伝子生成物の製造方法。
【請求項15】
ヘテロマータンパク質/生成物の異なるサブユニットをコードする発現ベクターにより同時にトランスフェクションされている、請求項13記載の宿主細胞を、該ヘテロマータンパク質/生成物の発現を可能とする条件の下で培養し、該ヘテロマータンパク質/生成物を、培養物または培地から単離することを特徴とする、ヘテロマータンパク質/生成物の製造方法。
【請求項16】
該ヘテロマータンパク質が抗体である、請求項15記載の方法。
【請求項17】
該宿主細胞が、一またはそれ以上の遺伝子増幅工程に付されている、請求項14〜16の何れか1項に記載の方法。
【請求項18】
該増幅可能な、選択性マーカーが、DHFRであり、かつ該増幅剤がメトトレキセートである、請求項17記載の方法。
【請求項19】
該宿主細胞が、メトトレキセートによる、一段階のみの遺伝子増幅工程に付されている、請求項18記載の方法。
【請求項20】
該遺伝子増幅が、血清を含まない培地内で行われる、請求項14〜19の何れか1項に記載の方法。
【請求項21】
該宿主細胞を懸濁培地内で培養する、請求項14〜20の何れか1項に記載の方法。
【請求項22】
対象とするタンパク質/生成物を発現する、宿主細胞を選択する方法であって、請求項11〜13の何れか1項に記載の、一群の宿主細胞を、該対象とするタンパク質/生成物および該蛍光タンパク質の発現を可能とする条件の下で培養し、該蛍光タンパク質に対して、最大の発現レベルを呈する該細胞を同定しおよび/または選別する工程を含むことを特徴とする、上記方法。
【請求項23】
上記選別を、蛍光-活性化セルソータ(FACS)を用いて実施する、請求項22記載の方法。
【請求項24】
該当する該選別された宿主細胞を、一またはそれ以上の、追加の遺伝子増幅工程に付す、請求項22または23記載の方法。
【請求項25】
該増幅可能な、選択性マーカーがDHFRであり、かつ該増幅剤がメトトレキセートである、請求項24記載の方法。
【請求項1】
ハムスター-ユビキチン/S27a-プロモータと機能可能な状態で結合した、対象とするタンパク質をコードする遺伝子、および蛍光タンパク質をコードする遺伝子を含むことを特徴とする、発現ベクター。
【請求項2】
増幅可能な、選択性マーカー遺伝子を含む、請求項1記載の発現ベクター。
【請求項3】
1または複数の上記プロモータと機能可能な状態で結合した、一種またはそれ以上のエンハンサを含む、請求項1または2記載の発現ベクター。
【請求項4】
更に、蛍光タンパク質をコードする該遺伝子、および対象とするタンパク質/生成物をコードする遺伝子のビシストロン発現を可能とする、内部リボソームエントリーサイト(IRES)をも含む、請求項1〜3の何れか1項に記載の発現ベクター。
【請求項5】
蛍光タンパク質をコードする該遺伝子および該増幅可能な、選択性マーカー遺伝子が、一つのまたは二つの別々の転写単位内に位置している、請求項1〜4の何れか1項に記載の発現ベクター。
【請求項6】
該機能性結合は、イントロン配列を介して起こることがない、請求項1〜5の何れか1項に記載の発現ベクター。
【請求項7】
該増幅可能な、選択性マーカー遺伝子が、ジヒドロ葉酸レダクターゼ(DHFR)または該蛍光タンパク質とDHFRとの融合タンパク質をコードする、請求項1〜6の何れか1項に記載の発現ベクター。
【請求項8】
該エンハンサがCMVまたはSV40エンハンサである、請求項3〜7の何れか1項に記載の発現ベクター。
【請求項9】
更に、少なくとも1種のポリアデニル化シグナルをも含む、請求項1〜8の何れか1項に記載の発現ベクター。
【請求項10】
対象とするタンパク質/生成物をコードする該遺伝子の代わりに、対象とするタンパク質をコードする遺伝子を組込むための、多重クローニングサイトを含む、請求項1〜9の何れか1項に記載の発現ベクター。
【請求項11】
請求項1〜10の何れか1項に記載の発現ベクターを用いてトランスフェクションされていることを特徴とする、真核宿主細胞。
【請求項12】
哺乳動物細胞、好ましくはCHO細胞である、請求項11記載の宿主細胞。
【請求項13】
一またはそれ以上の他の対象とするタンパク質/生成物をコードする遺伝子および少なくとも1種の他の選択性マーカーを含む、一またはそれ以上のベクターによって、更にトランスフェクションされている、請求項11または12記載の宿主細胞。
【請求項14】
異種遺伝子生成物の発現を可能とする条件の下で、請求項11〜13の何れか1項記載の宿主細胞を培養し、該遺伝子生成物を培養物または培地から単離することを特徴とする、異種遺伝子生成物の製造方法。
【請求項15】
ヘテロマータンパク質/生成物の異なるサブユニットをコードする発現ベクターにより同時にトランスフェクションされている、請求項13記載の宿主細胞を、該ヘテロマータンパク質/生成物の発現を可能とする条件の下で培養し、該ヘテロマータンパク質/生成物を、培養物または培地から単離することを特徴とする、ヘテロマータンパク質/生成物の製造方法。
【請求項16】
該ヘテロマータンパク質が抗体である、請求項15記載の方法。
【請求項17】
該宿主細胞が、一またはそれ以上の遺伝子増幅工程に付されている、請求項14〜16の何れか1項に記載の方法。
【請求項18】
該増幅可能な、選択性マーカーが、DHFRであり、かつ該増幅剤がメトトレキセートである、請求項17記載の方法。
【請求項19】
該宿主細胞が、メトトレキセートによる、一段階のみの遺伝子増幅工程に付されている、請求項18記載の方法。
【請求項20】
該遺伝子増幅が、血清を含まない培地内で行われる、請求項14〜19の何れか1項に記載の方法。
【請求項21】
該宿主細胞を懸濁培地内で培養する、請求項14〜20の何れか1項に記載の方法。
【請求項22】
対象とするタンパク質/生成物を発現する、宿主細胞を選択する方法であって、請求項11〜13の何れか1項に記載の、一群の宿主細胞を、該対象とするタンパク質/生成物および該蛍光タンパク質の発現を可能とする条件の下で培養し、該蛍光タンパク質に対して、最大の発現レベルを呈する該細胞を同定しおよび/または選別する工程を含むことを特徴とする、上記方法。
【請求項23】
上記選別を、蛍光-活性化セルソータ(FACS)を用いて実施する、請求項22記載の方法。
【請求項24】
該当する該選別された宿主細胞を、一またはそれ以上の、追加の遺伝子増幅工程に付す、請求項22または23記載の方法。
【請求項25】
該増幅可能な、選択性マーカーがDHFRであり、かつ該増幅剤がメトトレキセートである、請求項24記載の方法。
【図1】

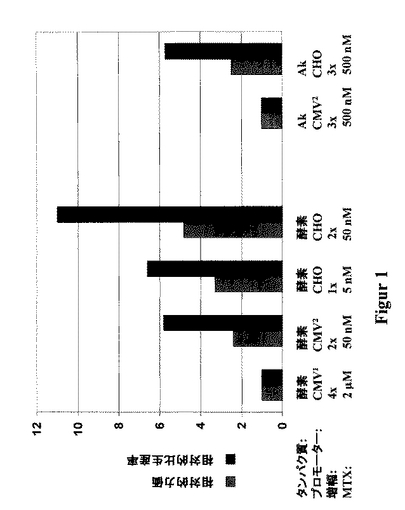
【図2】

【図3】
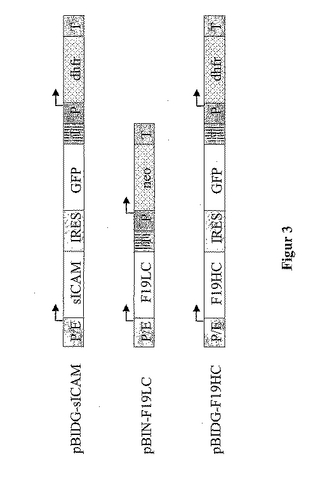
【図4】
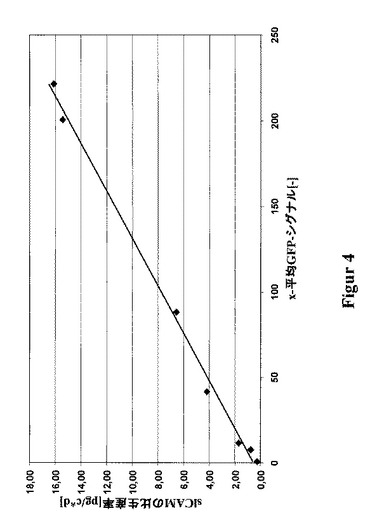
【図5】
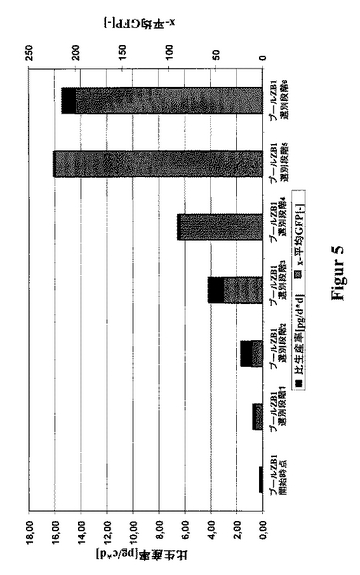
【図6】
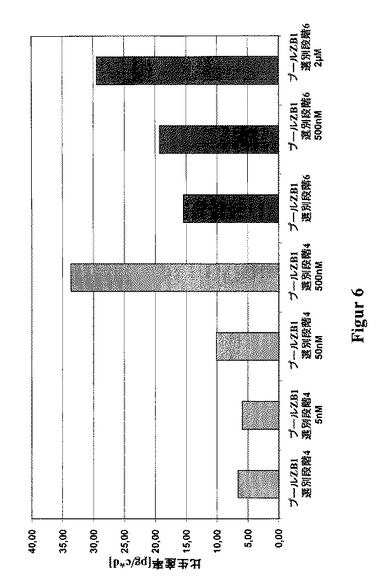
【図7】
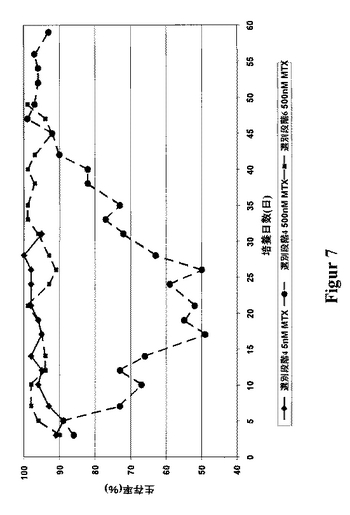
【図8】
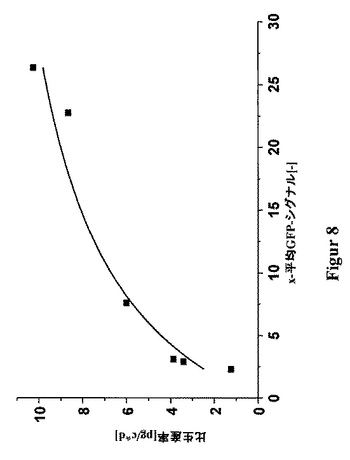
【図9】
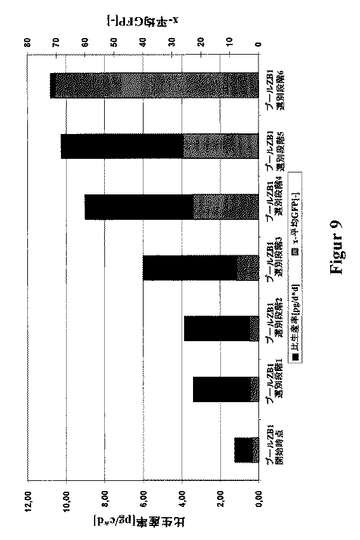
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2006−507829(P2006−507829A)
【公表日】平成18年3月9日(2006.3.9)
【国際特許分類】
【出願番号】特願2004−556182(P2004−556182)
【出願日】平成15年11月25日(2003.11.25)
【国際出願番号】PCT/EP2003/013225
【国際公開番号】WO2004/050879
【国際公開日】平成16年6月17日(2004.6.17)
【出願人】(503137975)ベーリンガー インゲルハイム ファルマ ゲゼルシャフト ミット ベシュレンクテル ハフツング ウント コンパニー コマンディトゲゼルシャフト (129)
【Fターム(参考)】
【公表日】平成18年3月9日(2006.3.9)
【国際特許分類】
【出願日】平成15年11月25日(2003.11.25)
【国際出願番号】PCT/EP2003/013225
【国際公開番号】WO2004/050879
【国際公開日】平成16年6月17日(2004.6.17)
【出願人】(503137975)ベーリンガー インゲルハイム ファルマ ゲゼルシャフト ミット ベシュレンクテル ハフツング ウント コンパニー コマンディトゲゼルシャフト (129)
【Fターム(参考)】
[ Back to top ]