
発酵微生物からの有機物質の生成を高める方法

本発明は糖化と発酵を同時に行うことにより発酵微生物から有機物質を生成するための方法及び組成物に関する。本発明の1つの側面はセルロース基質、全発酵ブロス、及びエタノール生成微生物を、セルロースをグルコースに加水分解し、且つグルコースをアルコールに転化することができる条件下で、組み合わせることによりアルコールを生成することを提供することである。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は2007年10月12日に出願された米国継続出願No.60/979,720に対して優先権を主張する。該文献を参照により本明細書に援用する。
【0002】
本発明は糖化及び発酵方法及び組成物を用いた発酵微生物からの有機物質の生成に関する。
【背景技術】
【0003】
非再生エネルギーの限界が近づくにつれ、セルラーゼ等の複合ポリサッカライドの再生可能エネルギーとしての可能性が大きくなっている。セルラーゼはセルロースをグルコース等の糖類に転換することができ、工業的なプロセスでバクテリア、酵母、及び糸状菌を含む数多くの微生物によりエネルギー源として用いられている。セルロース分解物質を再生可能な炭素源として用いるには、セルロースを糖へ加水分解することと、分解された糖類をエタノール等の燃料へ転化することの両方の方法について、経済的に利用可能な方法を開発しなければならない。
【発明の概要】
【発明が解決しようとする課題】
【0004】
セルラーゼと呼ばれている酵素によって、セルロースはグルコース、セロビオース、及び他のセロオリゴサッカライド等の生成物に分解することができる。セルラーゼ酵素は相互作用的に働き、セルロールをグルコースに加水分解する。CBHI及びCBHIII等のエキソセロビオヒドロラーゼ(CBH)は通常、セルロースの末端に作用して、セロビオースを生成する。一方エンドグルカナーゼ(EG)はセルロースのランダムな位置に作用する。これらの酵素は一緒にセルロースを加水分解してセロビオース等の小さなセロオリゴサッカライドにする。セロビオースはバクテリアグルコシダーゼにより、グルコースへ加水分解される。多くの微生物がセルロースを加水分解することができるけれども、結晶性セルロースを完全に加水分解する能力のある酵素を大量に生成することができる微生物は僅かである。今日までに、効率的な転換を行って、エタノール等の有機物質を工業的なスケールで生成する株は存在していない。第二ステップは工業的に利用可能な酵母等各種発酵微生物を用いて行い、エタノールを生成する。セルロース系エタノール等の再生可能有機物質の開発は環境的に好ましいけれども、この複合工程は経済的にはオイル及び天然ガス等の非再生炭素源を完全に代替するものではない。従って、源物質からエタノール等を、発酵微生物から有機物質を生成するより効果的なシステムを開発する強い需要が存在する。それゆえ、セルロース系物質の加水分解(糖化)及び発酵の効率的且つ経済的な改善が望まれている。
【課題を解決するための手段】
【0005】
本発明は糖化と醗酵を同時に行って有機物質を生成するための方法及び組成物に関する。本発明の1つの側面は、同時に糖化と醗酵とを行うことによる有機物質の生成を提供することであり、このことは、補助的な窒素源の不存在下で、セルロース基質、全体醗酵ブロス、及び醗酵微生物を結合させる工程、及びセルロースからグルコース及び/又はキシロースの加水分解並びに、グルコース及び/又はキシロースの有機基質への転換の両方を誘発することができる条件で、セルロース系物質、全体醗酵ブロス、及び発酵微生物をインキュベートする工程を含む。本発明の1つの側面は糖化と発酵とを同時に行ってエタノールを生成する方法であり、この方法は、補助的な窒素源の不存在下でセルロース基質、全体醗酵ブロスを組み合わせる工程、及びセルロースからグルコースへの分解及び/又はグルコースからエタノールへの加水分解の両方を行うことができる条件下で、セルロース基質、全体醗酵ブロス、及びエタノール生成微生物をインキュベートする工程を含む。
【0006】
本発明は、セルロース基質、発酵全体ブロス、及び/又は発酵微生物の混合物を含む、発酵微生物から有機基質を生成するための発酵組成物に関する。ここにおいて、反応組成物は補助的な窒素源を実質的に含まない。本発明の他の側面は、エタノール生成のための反応組成物を提供することであり、この組成物はセルロース基質、発酵全体ブロス、エタノール生成微生物の混合物を含む。該発酵組成物は補助的な窒素源を実質的には含まない。
【図面の簡単な説明】
【0007】
図面は説明の目的のみに用いるものであり、本開示の範囲を決して限定するものではないことを当業者は理解するだろう。
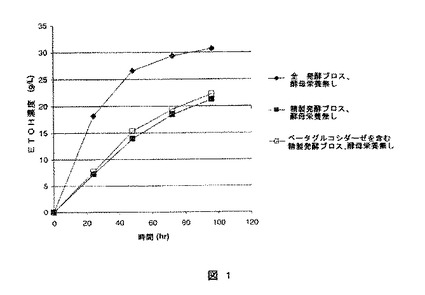
【図1】図1は補助的窒素の不存在下で、酸前処理バッガスの糖化と発酵を同時に行ったエタノール(ETOH)濃度(g/L)の結果を示す。
【図2】図2は全発酵ブロスで前処理バッガスの糖化と発酵とを行ったものに匹敵するエタノール濃度を達成するためには、精製発酵ブロスに追加的な栄養素を補わなければならないことを説明する。
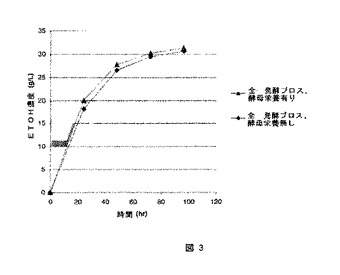
【図3】補助的な栄養素を含まない発酵ブロスを用いた、図3は補助的な栄養素を含む発酵ブロスを用いた同時糖化発酵を同時糖化発酵の比較である。
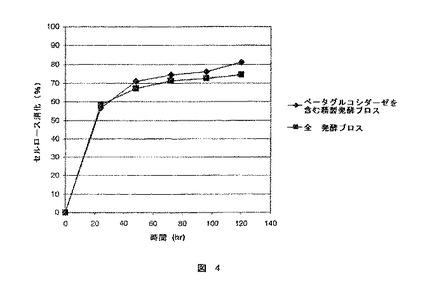
【図4】図4は発酵ブロスを用いて前処理バッガスの糖化の比較及びセルロース分解活性と同じ量のベータグルコシダーゼを補った精製発酵ブロスを提供する。
【発明を実施するための形態】
【0008】
前述の発明の概要及び以下の詳細な説明の両方は、例示及び説明のみであり、本開示の組成物及び方法を限定するものではないことは理解されるであろう。別に定義しない限り、本明細書で用いる図ベースストックテトラキスの技術的及び科学的用語は本発明が属する当業者に通常理解されているのと同じ意味を有する。本出願において、単数形は別に定義しない限り、複数も含む。別に定義しない限り、「又は」、は「及び/又は」を意味する。同様に「含む」及び「含んでいる」(「coprise」、「comprising」、「compreises」、「include」、「including」、及び「includes」)は限定のみを意味するものではない。特許及び刊行物内に開示されている全てのアミノ酸及び核酸配列を含む全ての特許及び刊行物を参照により本明細書に援用する。本明細書で提供されている表題は本発明の各所側面及び態様を限定するものではなく、これらは明細書全体を参照することにより定義されるべきである。
【0009】
本発明は同時糖化発酵を用いた有機物質を生成するための方法及び組成物に関する。
【0010】
本発明は同時糖化発酵により有機基質を精製するための方法に関する。該方法は、(a)補助的な窒素源の不存在下で、セルロース系物質、全発酵ブロス、及び発酵微生物を組み合わせる工程、及び(b)グルコース及び/又はキシロースに分解し、且つグルコース及び/又はキシロースを有機基質に転換することができる条件下で全発酵ブロスと発酵微生物とをインキュベートする工程を含む。
【0011】
本明細書は発酵微生物から有機物質を生成するための反応組成物も提供する。幾つかの態様において、この発酵微生物から有機物質を生成するための発酵組成物は、セルロース系基質、発酵全体ブロス、発酵微生物、及び水の混合物を必ず含む。幾つかの態様において、有機物質の生成のための反応組成物はセルロース系物質、発酵全体ブロス、及び発酵微生物を含む。該反応組成物は補助的な窒素源を実質的に含まない。
【0012】
本明細書で用いる「セルロース系物質」の語はセルロース及び/又はヘミセルロースを含んでいる任意の植物系バイオマスを意味する。セルロース系基質は、セルロース、ヘミセルロース、及びベータグルカンがお互いに及びリグニンと結合している、リグノセルロース系物質でもよい。そのようなセルロース系物質は、ペクチン、タンパク質、デンプン、及び脂質のような他の物質を含むが、好ましくはセルロース、ヘミセルロース、及び/又はベータグルカンを主成分として含む。
【0013】
好適な非限定的な、セルロース系基質の例としては、バイオマス、草類、農業廃棄物、林業廃棄物、都市ゴミ、廃棄紙類、パルプ及び製紙廃棄物を含むがこれらに限定されない。本発明に用いるセルロース系物質の共通の形態は、木、木材及び草、小麦、麦わら、トウキビバッガス、コーン、とうもろこしの皮、穀粒、生成物及び(湿式及び乾式を含む)コーン等の穀物の粉砕からの副生成物を含むコーンカーネル、並びに都市ゴミ、廃棄紙類、及び伐採草木を含むがこれらに限定されない。これらのセルロース系物質は「バージンバイオマス」(木、ブッシュ、草、フルーツ、花、作物、硬質及び軟質木材等)、「非バージンバイオマス」(農業副生成物、商業的有機廃棄物、構築及び解体瓦礫、都市ゴミ、及び伐採草木)、又はバージン及び非バージンバイオマスの混合物である、ブレンドバイオマスから得られる。
【0014】
幾つかの態様において、セルロース系物質は木材、木材パルプ、製紙スラッジ、製紙パルプの廃棄流れ、パーチクルボード、コーンストーバー、コーン繊維、ライス、製紙及びパルプ処理廃棄物、木又は草植物、フルーツパルプ、ベジタブルパルプ、パミス、醸造かす、草、籾殻、トウキビバッガス、綿、ジュート、ヘンプ、麻、バンブー、サイザル、アバカ、ストロー、コーンコブ、葉、小麦わら、ココナツ繊維、藻類、及びこれらの混合物を含む。
【0015】
セルロース系基質は当業者に知られている従来の方法を用いて前処理してもよい。そのような前処理は、化学的、物理的、及び生物学的前処理含む。例えば、物理的前処理技術は、とくに限定がないが、粉砕、クラッシュ、スチーミング/蒸気爆発、放射、及び水熱分解を含む。化学的前処理技術は特に限定がないが、希酸、アルカリ、有機溶剤、アンモニア、二酸化硫黄、二酸化炭素、及びpH制御水熱分解を含む。生物学的前処理技術はリグニン可溶化微生物を含む。
【0016】
本開示において、全発酵ブロスは、セルロース系基質の分解に有用な任意の糸状菌から調製することができる。「糸状菌」という語は、当業者により糸状菌と認識される任意の及び全ての糸状菌を含み、自然発生糸状菌、自然発生又は誘発変異を有する糸状菌、及び遺伝的に修飾された糸状菌を含む。通常、糸状菌は真核微生物であり、ユーマイコチナ(Eumycotina)及びオーミコータ(Oomycota)の再分類の全ての糸状菌形態を含む。これらの糸状菌はキチン、ベータグルカン、及び他の複合ポリサッカライドの複合細胞壁に栄養菌糸により特徴付けられる。幾つかの態様において、本開示の糸状菌は形態学的、物理学的、及び遺伝的に酵母とは異なる。
【0017】
幾つかの態様において、発酵全体ブロスは、アクレモニウム(Acremonium)、アスペルギルス(Aspergillus)、エメリセラ(Emericella)、フサリウム(Fusarium)、フミコーラ(Humicola)、ムコール(Mucor)、 ミセリオフィソーラ(Myceliophthora)、ニューロスポラ(Neurospora)、スキタリジウム(Scytalidium)、シエラビア(Thielavia)、トリポクラジウム(Tolypocladium)、又はトリコデルマ(Trichoderma)から調製される。
【0018】
幾つかの態様において、全発酵ブロスの発酵から調製される。他の側面において、発酵ブロスはアスペルギルスアキュレアタス(Aspergillus aculeatus)、アスペルギルスアワモリ(Aspergillus awamori)、アスペルギルスフォエチダス(Aspergillus foetidus)、アスペルギルスジャポニクス(Aspergillus japonicus)、アスペルギルスニヂュランス(Aspergillus nidulans)、アスペルギルスニガー(Aspergillus niger)、又はアスペルギルスオリザエ(Aspergillus oryzae)から調製される。他の側面において、全発酵ブロスはフサリウムセレアリス(Fusarium cerealis)、フサリウムクロウェレンス(Fusarium crookwellense)、フサリウムクルモラム(Fusarium culmorum)、フサリウムグラミネアラム(Fusarium graminearum)、フサリウムグラミウム(Fusarium graminum)、フサリウムヘテロスポラム(Fusarium heterosporum)、フサリウムネグンジ(Fusarium negundi)、フサリウムオキシスポラム(Fusarium oxysporum)、フサリウムレチキョラタム(Fusarium reticulatum)、フサリウムロゼウム(Fusarium roseum)、フサリウムサムブキヌム(Fusarium sambucinum)、フサリウムザルコクロラム(Fusarium sarcochroum)、フサリウムスポロトリコキオイデス(Fusarium sporotrichioides)、フサリウムサルフレウム(Fusarium sulphureum)、フサリウムトルローサム(Fusarium torulosum)、フサリウムトリコセシオイデス(Fusarium trichothecioides)、フサリウムベネナタム(Fusarium venenatum)の発酵から調製される。他の側面において、全発酵ブロスはフミコーラインソレンス(Humicola insolens)、フミコーラランギノーサ(Humicola lanuginosa)、ムコールミエヘイ(Mucor miehei)、ミセリオフトラサーモフィラ(Myceliophthora thermophila)、ニューロスポラクラッサ(Neurospora crassa)、シタリディウムサーモフィルム(Scytalidium thermophilum)、又はチエラビアテルレストリス(Thielavia terrestris)から選択される。他の側面において、全発酵ブロスはトリコデルマハルジアナム(Trichoderma harzianum)、トリコデルマコニンギ(Trichoderma koningii)、トリコデルマロンギブラキアタム(Trichoderma longibrachiatum)、トリコデルマレーシ(Trichoderma reesei)、例えば、RL−P37(Sheir−Neissら、Appl.Microbiol. Biotechnology、20(1984)pp.46−53;Montenecourt B.S.,Can.,1−20,1987)、QM9414(ATCC No.26921)、NRRL15709、ATCC13631、56764、56466、56767、又はトリコデルマビリデ(Trichoderma viride)、例えば、ATCC32098及び32086の発酵から調製される。
【0019】
幾つかの態様において、全発酵ブロスはアスペルギルス(Aspergillus)、アクレモニウム(Acremonium)、オーレオバシジウム(Aureobasidium)、ビューベリア(Beauveria)、セファロスポリウム(Cephalosporium)、セリオリオプシス(Ceriporiopsis)、カエトミウムペシロミセス(Chaetomium paecilomyces)、クリソスポリウム(Chrysosporium)、クラビセプス(Claviceps)、コチオボラス(Cochiobolus)、クリプトコッカス(Cryptococcus)、キュアトス(Cyathus)、エンドシア(Endothia)エンドシアムコール(Endothia mucor)、フサリウム(Fusarium)、ギロクラジウム(Gilocladium)、フミコーラ(Humicola)、マグナポルテ(Magnaporthe)、ミセリオフィソーラ(Myceliophthora)ミロセシウム(Myrothecium)、ムコール(Mucor)、ニューロスポラ(Neurospora)、ファネロカエテ(Phanerochaete)、ポドスポラ(Podospora)、パエシロマイセス(Paecilomyces)、ピリキュラリア(Pyricularia)、リゾムコール(Rhizomucor)リゾプス(Rhizopus)、スキゾフィラム(Schizophylum)、スタゴノスポラ(Stagonospora)、タラロマイセス(Talaromyces)、トリコデルマ(Trichoderma)、セルモマイセス(Thermomyces)、セルモアスクス(Thermoascus)、シエラビア(Thielavia)、トリコクラジウム(Tolypocladium)、トリコフィタム(Trichophyton)、及びトラメテスプレウロタス(Trametes pleurotus)を含むがこれらに限定されない糸状菌の発酵から調製される。幾つかの態様において、全発酵ブロスはアスペルギルスニヂュランス(A.nidulans)、アスペルギルスニガー(A.niger)、アスペルギルスアワモリ(A.awomari)、アスペルギルスアキュレタス(A.aculeatus)、アスペルギルスカワチ(A.kawachi)例えば、NRRL3112、ATCC22342(NRRL3112)、ATCC44733、ATCC14331、及びUVK143f株、アスペルギルスオリザエ(A.oryzae)例えば、ATCC11490、ニューロスポラクラッサ(N.crassa)、トリコデルマレーシ(Trichoderma reesei)、例えば、NRRL15709、ATCC13631、56764、56765、56466、56767、並びにトリコデルマビリデ(Trichoderma viride)例えば、ATCC32098及び32086を含むがこれらに限定されない糸状菌から調製される。好適な実施形態において、全発酵ブロスはトリコデルマ種の発酵から調製される。本発明に用いるのに特に有用な種及び株はトリコデルマレーシRutC30全体セルラーゼである。これはアメリカンタイプカルチャーコレクションより、トリコデルマレーシATCC56765として入手することができる。
【0020】
前述のように、全発酵ブロスは非組換え及び/又は組換え糸状菌の発酵から調製することができる。幾つかの態様において、この糸状菌は1つ以上の相同又は異種遺伝子を含む組み換え糸状菌である。幾つかの態様において、この糸状菌は1つ以上の相同又は異種遺伝子を含む組換え糸状菌であり、この1つ以上の遺伝子はセルロース系物質を分解することができる酵素をコードしている。セルロース系物質を分解する酵素をコードしている遺伝子は当業者に良く知られている。セルロース系基質を分解する酵素をコードする遺伝子の非限定的な例としては、エンドグルカナーゼ、セロビオヒドロラーゼ、グルコヒドロラーゼ、ベータグルコシダーゼ、キシログルカナーゼ、キシラナーゼ、キシロシダーゼ、アルファアラビノフラノシダーゼ、アルファグルクロニダーゼ、アセチルキシランエラスターゼ、マンナナーゼ、マンノシダーゼ、アルファガラクトシダーゼ、マンナンアセチルエステラーゼ、ガラクタナーゼ、アラビナンナーゼ、ペクチントリアーゼ、ペクチンリアーゼ、ポリガラクチュロナーゼ、ペクチンアセチルエステラーゼ、ペクチンメチルエステラーゼ、アルファアラビノフラノシダーゼ、ベータガラクトシダーゼ、ガラクタナーゼ、アラビナナーゼ、アルファアラビノフラノシダーゼ、ラムノガラクチュロナーゼ、ラムノガラクチュロナンリアーゼ、ラムノガラクチュロナンアセチルエステラーゼ、キシロガラクツロノシダーゼ、キシロガラクツロナーゼ、ラムノガラクツロナンリアーゼ、リグニンペルオキシダーゼ、マンガンアーゼ依存ペルオキシダーゼ及びラッカーゼを含む。
【0021】
幾つかの態様において、全発酵ブロスは、内性的には発現されない、または糸状菌によっては非常に低いレベルで発現する1つ以上の酵素活性を補って、セルロース系物質の分解、例えば、グルコース又はキシロース等の発酵可能糖への分解を促進する。補う酵素は全発酵ブロスに補助的に添加することができ、酵素は別の全発酵ブロスとして添加することもでき、精製し、又はわずかに回収して精製してもよい。好適な補助的酵素の非限定的な例は、セロビオヒドロラーゼ、エンドグルカナーゼ、ベータグルコシダーゼ、エンドベータ1,3,(4)グルカナーゼ、グルコヒドロラーゼ、キシログルカナーゼ、キシラナーゼ、キシロシダーゼ、アラビノフラノシダーゼ、アルファグルクロニダーゼ、アセチルキシランエステラーゼ、マンナーゼ、マンノシダーゼ、アルファガラクトシダーゼ、マンナンアセチルエステラーゼ、ガラクタナーゼ、アラビナナーゼ、ペクチンリアーゼ、ペクチナーゼリアーゼ、ポリガラクチュロナーゼ、ペクチンアセチルエステラーゼ、ペクチンメチルエステラーゼ、ベータガラクトシダーゼ、ガラクタナーゼ、アラビナナーゼ、アルファアラビノフラノシダーゼ、ラムノガラクチュロナーゼ、フェルイック酸エステラーゼ、ラムノガラクチュロナンリアーゼ、ラムノガラクチュロナンリアーゼ、ラムノガラクチュロナンアセチルエステラーゼ、キシロガラクチュロノシダーゼ、ラムノガラクチュロナンリアーゼ、リグニンペルオキシダーゼ、マンガナーゼ依存ペルオキシダーゼ、ハイブリッドペルオキシダーゼ、リグニンペルオキシダーゼの組合せペプチド、及びマンガナーゼ依存ペルオキシド、グルコアミラーゼ、アミラーゼ、プロテアーゼ、及びラッカーゼを含む。
【0022】
本発明の幾つかの態様において、全発酵ブロスは、酵素を過剰発現している組換え糸状菌の発酵の全発酵ブロスを含み、セルローススブチリスの分解を改善する。代替的に、全発酵ブロスは非組換え糸状菌の発酵の全発酵ブロスと酵素を過剰発現している糸状菌全発酵ブロスとを含み、セルロース基質の分解を改善する。
【0023】
本発明の幾つかの態様において、全発酵ブロスはベータグルコシダーゼを過剰発現している糸状菌の発酵糸状菌の全発酵ブロスを含む。代替的に、本発明の方法及び反応組成物に用いるための全発酵ブロス非組換え糸状菌の発酵の発酵全体ブロス及びβグルコシダーゼを過剰発現している組換え糸状菌の発酵の発酵全体ブロスを含む。
【0024】
「ベータグルコシダーゼ」の語はEC3.2.1.21として分類されるベータDグルコシダーゼとして本明細書において定義され、及び/又はセロビオースの加水分解を促進してDグルコシダーゼをリリースするGHファミリー1,3,7,9、又は48を含むがこれらに限定されない特定のGHファミリーに分類されるものを含むがこれらに限定されない。これらはセロビオースの加水分解を促進してベータDグルコースを放出する。ベータグルコシダーゼの過剰発現は宿主糸状菌のものと同じかまたは異なる種を形成することができる。特に、過剰発現されたベータグルコシダーゼは糸状菌ベータグルコシダーゼである必要はない。
【0025】
幾つかの態様において、ベータグルコシダーゼはベータグルコシダーゼをコードしている遺伝子の発現により生産することができる。例えば、ベータグルコシダーゼは、例えば、グラム陽性有機体(バチルス又はアクチノマイセテス等)又は原核宿主細胞(例えば、トリコデルマ、アスペルギルス、サッカロマイセス、及びピチア)による、細胞外スペースへ分泌される。幾つかの態様において、ベータグルコシダーゼ組換え微生物内で天然のレベルに対して過剰に発現させることができることは理解されている。幾つかの態様において、もし宿主細胞がベータグルコシダーゼの発現に用いられたなら、この細胞は宿主細胞の内性的な1つ以上のタンパク質の発現を減らすために修飾される。1つの態様において、この細胞は1つ以上のネガティブ遺伝子、特に欠失又は不活性化された1つ以上のプロテアーゼをコードしている遺伝子を含む。例えば、1つ以上のプロテアーゼコード遺伝子(例えば、アスパチルプロテアーゼコード遺伝子; Breakら、Gene 1990 86:153−162及び米国特許No.6,509,171参照)又はセルラーゼコード遺伝子は欠失又は不活性化されている。1つの態様において、トリコデルマ種宿主細胞は、WO05/001036に記載のように、cbh1、cbh2、及びegl1、egl2遺伝子を欠失している。ベータグルコシダーゼをコードしている核酸はトリコデルマ種宿主細胞の核酸ゲノムに存在するか、又は例えば、トリコデルマ宿主細胞で複製するプラスミド中に存在する。
【0026】
用いることができるベータグルコシダーゼの好適な例は、アスペルギルスアキュレアタス(Aspergillus aculeatus)(Kawaguchiら、1996, Gene 173:287−288)、アスペルギルスカワチ(Aspergillus kawachi)(Iwashitaら、1999,Appl.Environ.Microbiol.65:5546−5553)、アスペルギルスオリザエ(Aspergillus oryzae)(WO2002/095014)、セルロモナスビアゾテア(Cellulomonas biazotea)(Wongら、1998,Gene 207:79−86)、サッカロマイセスフィブリゲラ(Saccharomycopsis fibuligera)(Machidaら、1988,Appl.Environ.Microbiol.54:3147−3155)、スキゾサッカロマイセスポムブ(schizosaccharomyces pombe)(Woodら、2002,Nature 415871−880)由来ベータグルコシダーゼ, 並びにトリコデルマレーシベータグルコシダーゼ1(米国特許No.6,022,725)、トリコデルマレーシベータグルコシダーゼ3(米国特許No.6,982,159)トリコデルマレーシベータグルコシダーゼ4(米国特許No.7,045,332)、トリコデルマレーシベータグルコシダーゼ5(米国特許No.7,005,289)、トリコデルマレーシベータグルコシダーゼ6(USPN 20060258554)、トリコデルマレーシベータグルコシダーゼ7(USPN 20040102619)を含む。
【0027】
好適な態様において、全発酵ブロスは少なくとも400pNPGU/gのベータグルコシダーゼ活性を有している。活性の1pNPG単位は50℃、pH4.8において10分間にパラニトロフェニル−B−Dグルコプラノシドから遊離した1μモルのニトロフェノールを示す。ベータグルコシダーゼのこの活性及び全体セルラーゼ調製物の活性は当分野において既知の方法で測定することができる。測定においては、以下の条件を用いることができる。ベータグルコシダーゼ活性は、Chen.H.、Hayn.M.、Esterbauer.H、“Purification and Characterization of two extracellular β−glucosidases from Trichoderma reesei”,Biochimica et Biophysica Acta,1992,1121,54−60の様な、本分野で知られている任意の手段を用いて測定することができる。1pNPGは50℃、pH4.8において、10分間にパラニトロフェニルβ−Dグルコピラノシドから遊離するニトロフェノール1モルを意味する。全体セルラーゼ調製物のセルラーゼ活性はカルボキシメチルセルロース(CMC)を基質として用いて決定することができる。全体セルラーゼ活性の決定はCMC活性の観点から測定される。この方法はCMCに作用する酵素混合物により生成される還元糖の生成を測定する。1単位は1分間に1モルの生成物を遊離する酵素の量である(Ghose,T.K.,Measurement of Cellulse Activities,Pure&Appl.Chem.59,pp.257−268,1987)。
【0028】
幾つかの態様において、全発酵ブロスは少なくとも2500CMCU/gエンドグルカナーゼ活性を有する。活性の1CMC単位は50℃、pH4.8において1分間に、還元糖1モルを遊離することを示す。全体セルラーゼ活性は本発明に重要であるが、ベータグルコシダーゼは、他のセルラーゼの活性別のやり方で負の影響を与える最終生成物を加水分解するので、ベータグルコシダーゼ活性に対する全体セルラーゼ活性の割合は重要である。幾つかの態様において、全体ブロスは約0.5乃至25pNPG/MCM単位の範囲の酵素活性比を含む。幾つかの態様において、全発酵ブロスは約2乃至20pNPG/CMC単位、又は約1.5pNPG/CMC単位、又は約2乃至10pNPG/CMC単位、又は約2。5乃至8pNPG/CMC単位、又は約3乃至7pNPG/CMC単位、又は約3.5乃至6.5pNPG/CMC単位、又は約4乃至6pNPG/CMC単位、又は約4.5乃至5.5pNPG/CMC単位、又は約5乃至6pNPG/CMC単位の酵素化成比を含む。特に、態様において、この比は約5.5pNPG/CMC単位である。
【0029】
好適な投与レベル及び操作条件は特に本発明の明細書の発明な詳細を参照することで、当業者に明らかになる。全発酵ブロスの好適な投与レベルはセルロース基質及び用いる前処理技術により変化する。pH、温度、及び反応時間等の操作条件は、エタノール生成の速度に影響を与える。好ましくは、反応組成物はセルロース1グラム当たり、0.006乃至6mlの全発酵ブロスを含み、最も好ましくは0.03乃至0.6mlの全発酵ブロスを含む。代替的に、全発酵ブロスの量はシステム内のバイオマスの総量に基づいて決定される。そのような場合、反応組成物は好ましくは、0.003乃至3mlの全発酵ブロスをバイオマス1グラム当たりに含み、より好ましくは0.075乃至0.75mlの全発酵ブロス、及びより好ましくは0.015乃至0.3mlの全発酵ブロス、をバイオマス1グラム当たりに含む。他の態様において、全発酵ブロスはバイオマス固形分基質に対して約0.3乃至300.0重量%、より好ましくは約0.75%乃至75%、及び最も好ましくは1.5%乃至30重量%、の効果的な量で固形バイオマス基質に添加される。代替的に、全発酵ブロスの量は、システムに提供される全発酵ブロス由来の細胞の総量に基づいて決定される。そのような場合、反応組成物は好ましくはバイオマス基質1グラム当たり0.0001乃至0.1グラムの細胞量由来の全発酵ブロス、より好ましくは0.00025乃至0.025グラムの細胞量由来の全発酵ブロス、より好ましくは0.0005乃至0.01グラムの細胞量由来の全発酵ブロスを含む。
【0030】
本明細書において、全発酵ブロスはセルラーゼ基質を加水分解することができる酵素を発現する既知の糸状菌培地から得ることができる。発酵は振とうフラスコ培養、連続、バッチ、フェドバッチ等のスモール又はラージスケール発酵、セルラーゼを発現することができる好適な培地又は条件下で行われる実験室的又は工業的発酵に及びメル固形状態発酵を含む。通常、全発酵ブロスは、(i)エンドグルカナーゼ(EG)又は1,4−d−グルカン−4−グルカノヒドロラーゼ(EC3.2.1.4)、(ii)1,4−d−グルカングルカノヒドロラーゼ(セロデキストリナーゼとしても知られている)(EC3.2.1.74)及び1,4−d−グルカンセロビオヒドロラーゼ(エキソ−セロビオヒドロラーゼ、CBH)(EC3.2.1.91)、並びに(iii)ベータグルコシダーゼ(BG)又はベータグルコシダーゼグルコヒドロラーゼ(EC3.2.1.21)を含むがこれらに限定されない。
【0031】
通常、糸状菌はセルロース基質を加水分解することができる酵素の生成に好適な細胞培地中で培養される。培養は炭素及び窒素の源、並びに無機塩を含む好適な栄養培地で、当業者に既知の方法を用いて行われる。育成及びセルラーゼ生成に好適な培地、温度範囲、並びに他の条件は当業者に知られている。
【0032】
好ましくは、糸状菌の発酵は、炭素含有培地を制限因子として制御できるように行われ、それ故、転化される基質の相当量を用いて、細胞へ炭素含有基質の好適な転化を提供し、細胞の汚染を避けることができる。任意の残渣を容易に水洗することができるので、水溶性基質を用いる場合はコンタミは重要な問題とはならない。しかしながら、非水溶性基質の場合は問題であり、好適な洗浄工程等の追加的な生成処理工程を必要とする。
【0033】
全発酵ブロスは糸状菌を静止期まで育成し、1つ以上のセルラーゼ又はベータグルコシダーゼを発現するのに充分な期間制限された炭素条件下で糸状菌を維持することにより行われる。セルラーゼ等の酵素が一旦糸状菌により発酵培地に分泌されると、全発酵ブロスを用いることができる。
【0034】
本発明の全発酵ブロスは、糸状菌を含む。幾つかの態様において、全発酵ブロスは発酵の終わりに発酵物質由来の分画されていない内容物を含む。通常、全発酵ブロスは、糸状菌が静止期まで育成され、炭素制限条件下でインキュベートされてタンパク質合成(特にセルラーゼ及び/又はグルコシダーゼ)をした後に、使用済みの培養培地及び細胞の残骸を含む。幾つかの態様において、全発酵ブロスは使用済みの培養培地、細胞外酵素、及び/又は糸状菌を含む。幾つかの態様において、全発酵ブロス中に存在している糸状菌は既知の方法で、溶解、透過処理、又は殺して、細胞が死んでいる全発酵ブロスを生成することができる。
【0035】
幾つかの態様において、全発酵ブロスである糸状菌細胞を含む全発酵ブロス中の細胞は溶解又は殺されている。幾つかの態様において、この細胞は化学的及び/又はpH制御により糸状菌を溶解して、殺すことにより、糸状菌の発酵の細胞を殺した全体ブロスを生成する。幾つかの態様において、この細胞は化学的及び/又はpH制御により糸状菌を溶解して殺し、並びに殺された細胞の発酵混合物のpHを約4乃至6の範囲内に調整して、糸状菌の発酵の細胞を殺した全体ブロスを生成する。
【0036】
ソルビトール、塩化ナトリウム、ソルビン酸カリウム、及び当業者の知る他のものを含むがこれらに限定されない、追加的な保存料及び/又は静菌剤を全発酵ブロス又は細胞を殺した全発酵ブロスに添加することができる。
【0037】
理論に拘束されることは意図しないけれども、精製されていない全発酵ブロスはエタノール生成微生物に残りの栄養素を提供すると信じられている。このことはエタノール発酵を導き、エタノール生産を改善する。糖化されたセルロースに加えてエタノール生成微生物に、栄養ブロスを提供する必要性を排除又は補助的な栄養素の量を少なくできる可能性はエタノール発酵工程の原料のコストを低くする結果となるだろう。
【0038】
本明細書で説明する方法法及び組成物は発酵微生物に対しての補助的な栄養素及び/又は栄養素源が無いか実質的に存在しない。幾つかの態様において、この方法及び/又は組成物は酵素抽出物、ペプトン、及び/又は尿素が無いか、これらを実質的に含まない。当業者は本発明の方法及び組成物が補助的な窒素源が無いか、これを実質的に含まないけれども、窒素及び/又は栄養源を不純物として存在するか、又は糸状菌に対して全発酵ブロスの栄養価が実質的に増えない程度に添加されてもよいことを理解するだろう。
【0039】
本明細書の方法及び組成物は糸状菌に対する補助的な窒素源の量及び/又はタイプを減らすことができる。幾つかの態様において、本発明の方法及び組成物は糸状菌に用いる酵素抽出物、ペプトン、及び/又は尿素を減らすことができる。
【0040】
本発明の方法及び組成物は糸状菌に対する補助的な窒素源の量及び/又はタイプを減らすことができることは、当業者に理解されるだろう。
【0041】
発酵培地の栄養価は糸状菌の発酵の間に減少又は消費されることから、全発酵ブロスはエタノール生成微生物に対して栄養価を有しているという発見は従来の予想とは異なる。幾つかの研究はろ過されたブロスと比較してろ過していないセルラーゼブロスが高いエタノール生成を示す事を報告している。しかしながら、高められたエタノール生成はろ過工程の間に除去された糸状菌の細胞壁に付着している追加的なベータグルコシダーゼに起因するものであると予測されていた。Schellら、“Whole Broth Cellulase Production for Use in Simultaneous Saccharification and Fermentation of Cellulase to Ethanol,” Appl.Biochem. Biotech. 24/25:287−298 (1990)。驚くべきことに、発明者らは全発酵ブロスが、酵母等のエタノール生成微生物に対する窒素源として通常必要とされていた補助的な窒素源(通常パプトン、トリプトン、酵素抽出物、又は尿素の形態)を代替できることを発見した。精製発酵ブロスにベータグルコシダーゼのみを補っても全発酵ブロスの栄養的な効果(利点)は達成できない(図1及び実施例)。従って、本発明の1つの側面は、補助的な窒素源を用いずに、セルロース系基質、全発酵ブロス、及びエタノール生成微生物を組合せ、インキュベーションする方法である。他の態様において、このセルロース系物質、全発酵ブロス、及びエタノール生成微生物は補助的窒素源を用いずに組み合わせられ、インキュベートされる。
【0042】
本発明の1つの側面は、糖化及び発酵を同時に行うことによりエタノールを生成する方法を提供する。この方法は(a)補助的な窒素源の不存在下で、セルロース系基質、全発酵ブロスを組み合わせる工程及び(b)セルロース系基質、全発酵ブロス、及び/又はエタノール生成微生物をセルロースを加水分解し、グルコースにして、グルココースをエタノールに転化する条件下で、インキュベートする工程を含む。
【0043】
本発明の1つの側面は、糖化及び発酵を同時に行うことによって、エタノールを生成する方法を提供する。この方法は(a)補助的な窒素源の不存在下で、セルロース系物質、全発酵ブロス、及びエタノール生成微生物を組み合わせる工程、及び(b)セルロース基質、全発酵ブロス、及びエタノール生成微生物をセルロース加水分解して、グルコース及び/又はキシロースに転換し、グルコース及び/又はキシロースをエタノールに転化する条件下でインキュベートする工程を含む。
【0044】
本明細書において、発酵微生物は有機物質の生成に好適な発酵プロセスに用いる任意の好適な微生物を意味する。好適な非限定的な糸状菌微生物はグルコース、キシロース、ガラクトース、アラビノース、マンノース、又はオリゴサッカライド等の糖類を所望の発酵生成物に発酵又は転化することができるものである。好適な非限定的な糸状菌微生物の例としては、酵母及びバクテリアを含む。好適な態様において、発酵微生物はエタノール生成微生物である。「エタノール生成」の語は炭化水素から、第一発酵生成物としてエタノールを生成する能力のある微生物を含む。しかしながら、本明細書でのエタノール生成微生物はたの有機物質も生成するのに用いられることを当業者は理解するだろう。この語は自然発生エタノール生成微生物、自然発生又は誘発変異体のエタノール生成微生物、及び/又は遺伝的に修飾されたエタノール生成微生物を含む。セルラーゼ物質をグルコース及び他の小糖類に加水分解する工程は重要な工程であるけれども、同時糖化及び発酵(SSF)は、これらの糖をエタノールに転化する、エタノール生成微生物が生存している培養工程に依存している。幾つかの態様において、このエタノール生成微生物は効果的にグルコースをエタノールに発酵ことができる、サッカロナイセスセレビジアエ(Saccharomyces cerevisiae)、サッカロマイセスウバルム(S.uvarum)、クリベロマイセスファギリス(Kluyveromyces fagilis)、カンジダプセウドトロピカリス(Candida pseudotropicalis)、及びパキソレンタンノフィラス(Pachysolen tannophilus)等の酵母細胞である。好適な株はサッカロマイセスデレビシアエ(S.cerevisiae)D5A (ATCC200062)、サッカロマイセスデレビシアエY567(ATCC24858)、ACA174(ATCC60868)、MY91(ATCC201301)、MY138(ATCC201302)、C5(ATCC201298)、ET7(ATCC201299)、LA6(ATCC201300)、OSB21(ATCC201303)、F23(サッカロマイセスグロボサス(S.globosus)ATCC90920)、ACA174(ATCC60868)、A54(ATCC90921)、NRCC202036(ATCC46534)、ATCC24858、ATCC24858、G3706(ATCC42594)、NRRL、Y−265(ATCC60593)、Sa28(ATCC26603)、ATCC24845−ATCC24860を含むがこれらに限定されない。本発明に用いるための他のセレビシエではない酵母株はピチアパストリス(Pichia pastoris)(tozonyID4922)、サッカロマイセスパストリアナス(S.pastorianusSA23(サッカロマイセスカールスベンシス(S.carlsbergensis)ATCC26602)、サッカロマイセスパストリアナス(S.pastorianus(サッカロマイセスカールスベルゲンシス(S.carlsbergensisATCC2345)、カンジダアシドデルモフィリウム(Candida acidothermophilum(イサキタニアオリエンタス(Issatchenkia orientalis、ATCC20381)を含む。幾つかの態様において、エタノール生成微生物は組換え酵母株である。好適な組換え酵母株はキシロースリダクターゼ、キシリトールデヒドロゲナーゼ、及び/又はキシルロキナーゼをコードしている遺伝子を含む(例えば、米国特許No.5,789,210参照)。
【0045】
本発明の幾つかの態様において、エタノール生成微生物はバクテリア細胞、好ましくはグラム陰性、通性嫌気性、及び腸内細菌科のファミリーである。他の関連する態様において、このエタノール生成微生物はエシェリキア(Escherichia)又はクレブシエラ(Klebsiella属)であり、好ましくは、エシェリキア又はクレブシエラ、好ましくはE.coliB、E.coliDH5a、E.coliKO4(ATCC55123)、E.coliKO11(ATCC55124)、E.coliKO12(ATCC55125)、E.coliLY01、クレブシエラオキシトーカ(K.oxytoca)M5A1又はクレブシエラオキシトーカP2(ATCC55307)である。幾つかの態様において、エタノール生成微生物はザイモモナス(Zymomonas)種又はザイモモナスモビリス(Zymomonas mobilis)(ATCC31821)由来である。幾つかの態様において、組換えザイモモナス(Zymomonas)株は、例としてキシロースイソメラーゼ、キシルクキナーゼ、トランスアルドラーゼ、及びトランスケトラーゼをコードする遺伝子を含む。
【0046】
発酵微生物は通常加水分解物を転化して、発酵は12乃至96時間、例えば、30乃至80時間行われる。温度は通常26℃乃至40℃である。特にpH3乃至6で約32℃で行われる。
【0047】
発酵に続いて、所望の有機物質を当業者に既知の任意の手段を用いて回収する。そのような方法としては、蒸留、抽出、クロマトグラフィー、電気泳動、溶解差を含むがこれらに限定されない。例えば、エタノール発酵において、アルコールは従来の蒸留方法により、分離及び精製される。本発明の方法によりえられたエタノールは燃料エタノール、飲料エタノール、又は工業エタノールとして用いられる。
【0048】
本発明の側面は更に、以下の実施例を参照することで更に理解されるが、これらは本発明の範囲を限定するものではない。本開示を逸脱せずに、物質及び方法の両方において多くの変更が可能であることを当業者は理解しているだろう。
【実施例】
【0049】
実施例1:全発酵ブロスの調製
グルコース/ソホロースの調製
60%(W/W)グルコース溶液を121℃、30分で滅菌した。この温度を65℃に下げ、1リットル当たり10gの総タンパク質(全体セルラーゼはT.reeseiにより生成された)を添加した。この混合物をゆっくり撹拌し、65℃で3日間維持した。ソホロース含量はこの60%グルコース溶液中、12g/Lであると測定された。
【0050】
0.8Lの培地を1.5mlのトリコデルマレーシ(Trichoderma reesei)RLP−37凍結胞子で接種して、種フラスコとした。このフラスコを2つの0.4L部分にわけ、48時間後に、2つの別々の14LのBiolafite fermentor中の2×7Lの発酵培地へ写した。この育成培地は以下の組成を有していた:

【0051】
微量元素*:5g/L FeSO4−7H2O、1.6g/L MnSO4−H2O、1.4g/L ZnSO4−7H2O
【0052】
発酵は25℃、750RPM、及び1分間当たり8標準リットル(SLM)の空気流れで行った。
【0053】
バッチグルコースは約20時間で消費され、この時点で細胞の成長が停止し、炭素制限供給が始まった。40%グルコース/ソホロース供給を0.25g/分で添加した。セルラーゼ生成に直接関与する総タンパク質(セルラーゼ活性対総細胞外タンパク質の我々の比較に基づく)をバットフェーズの直後に導入した。この細胞は化学的及びpH処理を組合せ、溶解させて殺した。必要であれば、殺された全発酵ブロスを、より中性、例えば、約pH4乃至6の間に調整する。
【0054】
実施例2:精製発酵ブロスの調製
糸状菌を実施例1で説明したように育成した。細胞を溶解するよりはむしろ、全発酵ブロスの内容物をろ過して、細胞及び大きな細胞の残骸をろ過して精製発酵ブロスを調製した。所望のセルラーゼ及びグルコシダーゼはトリコデルマ細胞から分泌されているので、セルラーゼ分解活性は精製された発酵ブロスの中に残っている。精製発酵ブロスの中に含まれている酵素は10kDaカットオフメンブランでウルトラろ過により濃縮された。
【0055】
実施例3:同時糖化発酵
同時糖化発酵(SSF)は250mlフラスコにおいて、標準的な酵母発酵条件(例えば、セルモサック(Thermosacc)酵母、pH5.0、38℃)で、2回行った。 酸で前処理されたバッガスを20mMのクエン酸ナトリム緩衝液(pH5.0)で7%のセルロース含量になるように調整した。酵母の栄養は1.0g/L酵母抽出物、1.0g/Lペプトン、及び1.0g/L尿素の最終濃度になるように添加した。全発酵ブロス又は精製発酵ブロスを(0.4CMC U/g酸前処理シュガーコーンバッガスの濃度)に添加して、同時に酵母を用いて発酵を行った。懸濁したときに、精製した発酵ブロスは、最終的に0.063pNPG/g酸処理バッガスと同様のベータブルコシダーゼ活性を有していた。このフラスコを150rpmで撹拌した。サンプルを異なる時間で採取し、エタノール、グリセロール、酢酸、酪酸、及び残りの糖類をHPLC法で解析した。
【0056】
図1は、酵母栄養がない場合、SSFと全発酵ブロスは精製発酵ブロス(ベータグルコシダーゼ添加又は無添加)を用いたものよりも充分に高いエタノール生成を提供した。例えば、96時間において、ベータグルコシダーゼを含む精製発酵ブロスを用いた場合のエタノール濃度は22.2g/Lであるのに対して、全発酵ブロスは30.7g/Lであった。
【0057】
図2は全発酵ブロスを用いたSSFパフォーマンスと精製発酵ブロスを用いたSSFパフォーマンスの比較である。全てのケースにおいて、酵母栄養を同じレベルで添加した。全発酵ブロスは発酵速度が速くなり、エタノール生成が僅かに高くなったことを観察した。
【0058】
図3は酵母栄養を添加した、又は添加してない全発酵ブロス精製物を用いたSSFパフォーマンスの比較である。SSFパフォーマンスは両者とも同レベルであることが確認された。96時間において、酵母栄養を含む全発酵ブロスのエタノール濃度は31.7g/Lであるのに対して、酵母栄養を含まない全発酵ブロスのエタノール濃度は30.7g/Lであった。この結果はエタノールへの発酵について、全発酵ブロスからのの窒素源を酵母が利用することができることを示している。
【0059】
実施例4:糖化
酸前処理バッガスの糖化を250mlスラスコで、2回行った。酸前処理バッガスは20mMクエン酸ナトリウム緩衝液(pH5.0)を用いて7%セルロースの付加になるように調整した。ベータグルコシダーゼを補充した、全発酵ブロス又は精製発酵ブロスを添加して酵素加水分解を開始した。同じ量のセルロース活性を用いた。即ち、それぞれ、0.4CMC U/g酸処理バッガス及び0.063pNPG/g酸処理バッガスを用いた。このフラスコを150rpmで撹拌した。サンプルを異なる時間間隔で採取し、グルコース、セロビオース、及びキシロースについて、HPLC方法で解析した。
【0060】
図4は、酸処理したバッガスの同程度の糖化が観察されたことにより、全発酵ブロス及びベータグルコシダーゼを補充した精製発酵ブロスのセルロース分解能力は同一であることを示している。それゆえ、Schellら、”Whole Broth Cellulase Production for Use in Simultaneous Saccharification And Fermentation of Cellulase to Ethanol,” Appl. Biochem. Biotech.24/25:287−298(1990)で提案されていたのとは異なり、高められたエタノール生成は全発酵ブロスによるベータグルコシダーゼに依存するものではないことが証明された。
【技術分野】
【0001】
本出願は2007年10月12日に出願された米国継続出願No.60/979,720に対して優先権を主張する。該文献を参照により本明細書に援用する。
【0002】
本発明は糖化及び発酵方法及び組成物を用いた発酵微生物からの有機物質の生成に関する。
【背景技術】
【0003】
非再生エネルギーの限界が近づくにつれ、セルラーゼ等の複合ポリサッカライドの再生可能エネルギーとしての可能性が大きくなっている。セルラーゼはセルロースをグルコース等の糖類に転換することができ、工業的なプロセスでバクテリア、酵母、及び糸状菌を含む数多くの微生物によりエネルギー源として用いられている。セルロース分解物質を再生可能な炭素源として用いるには、セルロースを糖へ加水分解することと、分解された糖類をエタノール等の燃料へ転化することの両方の方法について、経済的に利用可能な方法を開発しなければならない。
【発明の概要】
【発明が解決しようとする課題】
【0004】
セルラーゼと呼ばれている酵素によって、セルロースはグルコース、セロビオース、及び他のセロオリゴサッカライド等の生成物に分解することができる。セルラーゼ酵素は相互作用的に働き、セルロールをグルコースに加水分解する。CBHI及びCBHIII等のエキソセロビオヒドロラーゼ(CBH)は通常、セルロースの末端に作用して、セロビオースを生成する。一方エンドグルカナーゼ(EG)はセルロースのランダムな位置に作用する。これらの酵素は一緒にセルロースを加水分解してセロビオース等の小さなセロオリゴサッカライドにする。セロビオースはバクテリアグルコシダーゼにより、グルコースへ加水分解される。多くの微生物がセルロースを加水分解することができるけれども、結晶性セルロースを完全に加水分解する能力のある酵素を大量に生成することができる微生物は僅かである。今日までに、効率的な転換を行って、エタノール等の有機物質を工業的なスケールで生成する株は存在していない。第二ステップは工業的に利用可能な酵母等各種発酵微生物を用いて行い、エタノールを生成する。セルロース系エタノール等の再生可能有機物質の開発は環境的に好ましいけれども、この複合工程は経済的にはオイル及び天然ガス等の非再生炭素源を完全に代替するものではない。従って、源物質からエタノール等を、発酵微生物から有機物質を生成するより効果的なシステムを開発する強い需要が存在する。それゆえ、セルロース系物質の加水分解(糖化)及び発酵の効率的且つ経済的な改善が望まれている。
【課題を解決するための手段】
【0005】
本発明は糖化と醗酵を同時に行って有機物質を生成するための方法及び組成物に関する。本発明の1つの側面は、同時に糖化と醗酵とを行うことによる有機物質の生成を提供することであり、このことは、補助的な窒素源の不存在下で、セルロース基質、全体醗酵ブロス、及び醗酵微生物を結合させる工程、及びセルロースからグルコース及び/又はキシロースの加水分解並びに、グルコース及び/又はキシロースの有機基質への転換の両方を誘発することができる条件で、セルロース系物質、全体醗酵ブロス、及び発酵微生物をインキュベートする工程を含む。本発明の1つの側面は糖化と発酵とを同時に行ってエタノールを生成する方法であり、この方法は、補助的な窒素源の不存在下でセルロース基質、全体醗酵ブロスを組み合わせる工程、及びセルロースからグルコースへの分解及び/又はグルコースからエタノールへの加水分解の両方を行うことができる条件下で、セルロース基質、全体醗酵ブロス、及びエタノール生成微生物をインキュベートする工程を含む。
【0006】
本発明は、セルロース基質、発酵全体ブロス、及び/又は発酵微生物の混合物を含む、発酵微生物から有機基質を生成するための発酵組成物に関する。ここにおいて、反応組成物は補助的な窒素源を実質的に含まない。本発明の他の側面は、エタノール生成のための反応組成物を提供することであり、この組成物はセルロース基質、発酵全体ブロス、エタノール生成微生物の混合物を含む。該発酵組成物は補助的な窒素源を実質的には含まない。
【図面の簡単な説明】
【0007】
図面は説明の目的のみに用いるものであり、本開示の範囲を決して限定するものではないことを当業者は理解するだろう。
【図1】図1は補助的窒素の不存在下で、酸前処理バッガスの糖化と発酵を同時に行ったエタノール(ETOH)濃度(g/L)の結果を示す。
【図2】図2は全発酵ブロスで前処理バッガスの糖化と発酵とを行ったものに匹敵するエタノール濃度を達成するためには、精製発酵ブロスに追加的な栄養素を補わなければならないことを説明する。
【図3】補助的な栄養素を含まない発酵ブロスを用いた、図3は補助的な栄養素を含む発酵ブロスを用いた同時糖化発酵を同時糖化発酵の比較である。
【図4】図4は発酵ブロスを用いて前処理バッガスの糖化の比較及びセルロース分解活性と同じ量のベータグルコシダーゼを補った精製発酵ブロスを提供する。
【発明を実施するための形態】
【0008】
前述の発明の概要及び以下の詳細な説明の両方は、例示及び説明のみであり、本開示の組成物及び方法を限定するものではないことは理解されるであろう。別に定義しない限り、本明細書で用いる図ベースストックテトラキスの技術的及び科学的用語は本発明が属する当業者に通常理解されているのと同じ意味を有する。本出願において、単数形は別に定義しない限り、複数も含む。別に定義しない限り、「又は」、は「及び/又は」を意味する。同様に「含む」及び「含んでいる」(「coprise」、「comprising」、「compreises」、「include」、「including」、及び「includes」)は限定のみを意味するものではない。特許及び刊行物内に開示されている全てのアミノ酸及び核酸配列を含む全ての特許及び刊行物を参照により本明細書に援用する。本明細書で提供されている表題は本発明の各所側面及び態様を限定するものではなく、これらは明細書全体を参照することにより定義されるべきである。
【0009】
本発明は同時糖化発酵を用いた有機物質を生成するための方法及び組成物に関する。
【0010】
本発明は同時糖化発酵により有機基質を精製するための方法に関する。該方法は、(a)補助的な窒素源の不存在下で、セルロース系物質、全発酵ブロス、及び発酵微生物を組み合わせる工程、及び(b)グルコース及び/又はキシロースに分解し、且つグルコース及び/又はキシロースを有機基質に転換することができる条件下で全発酵ブロスと発酵微生物とをインキュベートする工程を含む。
【0011】
本明細書は発酵微生物から有機物質を生成するための反応組成物も提供する。幾つかの態様において、この発酵微生物から有機物質を生成するための発酵組成物は、セルロース系基質、発酵全体ブロス、発酵微生物、及び水の混合物を必ず含む。幾つかの態様において、有機物質の生成のための反応組成物はセルロース系物質、発酵全体ブロス、及び発酵微生物を含む。該反応組成物は補助的な窒素源を実質的に含まない。
【0012】
本明細書で用いる「セルロース系物質」の語はセルロース及び/又はヘミセルロースを含んでいる任意の植物系バイオマスを意味する。セルロース系基質は、セルロース、ヘミセルロース、及びベータグルカンがお互いに及びリグニンと結合している、リグノセルロース系物質でもよい。そのようなセルロース系物質は、ペクチン、タンパク質、デンプン、及び脂質のような他の物質を含むが、好ましくはセルロース、ヘミセルロース、及び/又はベータグルカンを主成分として含む。
【0013】
好適な非限定的な、セルロース系基質の例としては、バイオマス、草類、農業廃棄物、林業廃棄物、都市ゴミ、廃棄紙類、パルプ及び製紙廃棄物を含むがこれらに限定されない。本発明に用いるセルロース系物質の共通の形態は、木、木材及び草、小麦、麦わら、トウキビバッガス、コーン、とうもろこしの皮、穀粒、生成物及び(湿式及び乾式を含む)コーン等の穀物の粉砕からの副生成物を含むコーンカーネル、並びに都市ゴミ、廃棄紙類、及び伐採草木を含むがこれらに限定されない。これらのセルロース系物質は「バージンバイオマス」(木、ブッシュ、草、フルーツ、花、作物、硬質及び軟質木材等)、「非バージンバイオマス」(農業副生成物、商業的有機廃棄物、構築及び解体瓦礫、都市ゴミ、及び伐採草木)、又はバージン及び非バージンバイオマスの混合物である、ブレンドバイオマスから得られる。
【0014】
幾つかの態様において、セルロース系物質は木材、木材パルプ、製紙スラッジ、製紙パルプの廃棄流れ、パーチクルボード、コーンストーバー、コーン繊維、ライス、製紙及びパルプ処理廃棄物、木又は草植物、フルーツパルプ、ベジタブルパルプ、パミス、醸造かす、草、籾殻、トウキビバッガス、綿、ジュート、ヘンプ、麻、バンブー、サイザル、アバカ、ストロー、コーンコブ、葉、小麦わら、ココナツ繊維、藻類、及びこれらの混合物を含む。
【0015】
セルロース系基質は当業者に知られている従来の方法を用いて前処理してもよい。そのような前処理は、化学的、物理的、及び生物学的前処理含む。例えば、物理的前処理技術は、とくに限定がないが、粉砕、クラッシュ、スチーミング/蒸気爆発、放射、及び水熱分解を含む。化学的前処理技術は特に限定がないが、希酸、アルカリ、有機溶剤、アンモニア、二酸化硫黄、二酸化炭素、及びpH制御水熱分解を含む。生物学的前処理技術はリグニン可溶化微生物を含む。
【0016】
本開示において、全発酵ブロスは、セルロース系基質の分解に有用な任意の糸状菌から調製することができる。「糸状菌」という語は、当業者により糸状菌と認識される任意の及び全ての糸状菌を含み、自然発生糸状菌、自然発生又は誘発変異を有する糸状菌、及び遺伝的に修飾された糸状菌を含む。通常、糸状菌は真核微生物であり、ユーマイコチナ(Eumycotina)及びオーミコータ(Oomycota)の再分類の全ての糸状菌形態を含む。これらの糸状菌はキチン、ベータグルカン、及び他の複合ポリサッカライドの複合細胞壁に栄養菌糸により特徴付けられる。幾つかの態様において、本開示の糸状菌は形態学的、物理学的、及び遺伝的に酵母とは異なる。
【0017】
幾つかの態様において、発酵全体ブロスは、アクレモニウム(Acremonium)、アスペルギルス(Aspergillus)、エメリセラ(Emericella)、フサリウム(Fusarium)、フミコーラ(Humicola)、ムコール(Mucor)、 ミセリオフィソーラ(Myceliophthora)、ニューロスポラ(Neurospora)、スキタリジウム(Scytalidium)、シエラビア(Thielavia)、トリポクラジウム(Tolypocladium)、又はトリコデルマ(Trichoderma)から調製される。
【0018】
幾つかの態様において、全発酵ブロスの発酵から調製される。他の側面において、発酵ブロスはアスペルギルスアキュレアタス(Aspergillus aculeatus)、アスペルギルスアワモリ(Aspergillus awamori)、アスペルギルスフォエチダス(Aspergillus foetidus)、アスペルギルスジャポニクス(Aspergillus japonicus)、アスペルギルスニヂュランス(Aspergillus nidulans)、アスペルギルスニガー(Aspergillus niger)、又はアスペルギルスオリザエ(Aspergillus oryzae)から調製される。他の側面において、全発酵ブロスはフサリウムセレアリス(Fusarium cerealis)、フサリウムクロウェレンス(Fusarium crookwellense)、フサリウムクルモラム(Fusarium culmorum)、フサリウムグラミネアラム(Fusarium graminearum)、フサリウムグラミウム(Fusarium graminum)、フサリウムヘテロスポラム(Fusarium heterosporum)、フサリウムネグンジ(Fusarium negundi)、フサリウムオキシスポラム(Fusarium oxysporum)、フサリウムレチキョラタム(Fusarium reticulatum)、フサリウムロゼウム(Fusarium roseum)、フサリウムサムブキヌム(Fusarium sambucinum)、フサリウムザルコクロラム(Fusarium sarcochroum)、フサリウムスポロトリコキオイデス(Fusarium sporotrichioides)、フサリウムサルフレウム(Fusarium sulphureum)、フサリウムトルローサム(Fusarium torulosum)、フサリウムトリコセシオイデス(Fusarium trichothecioides)、フサリウムベネナタム(Fusarium venenatum)の発酵から調製される。他の側面において、全発酵ブロスはフミコーラインソレンス(Humicola insolens)、フミコーラランギノーサ(Humicola lanuginosa)、ムコールミエヘイ(Mucor miehei)、ミセリオフトラサーモフィラ(Myceliophthora thermophila)、ニューロスポラクラッサ(Neurospora crassa)、シタリディウムサーモフィルム(Scytalidium thermophilum)、又はチエラビアテルレストリス(Thielavia terrestris)から選択される。他の側面において、全発酵ブロスはトリコデルマハルジアナム(Trichoderma harzianum)、トリコデルマコニンギ(Trichoderma koningii)、トリコデルマロンギブラキアタム(Trichoderma longibrachiatum)、トリコデルマレーシ(Trichoderma reesei)、例えば、RL−P37(Sheir−Neissら、Appl.Microbiol. Biotechnology、20(1984)pp.46−53;Montenecourt B.S.,Can.,1−20,1987)、QM9414(ATCC No.26921)、NRRL15709、ATCC13631、56764、56466、56767、又はトリコデルマビリデ(Trichoderma viride)、例えば、ATCC32098及び32086の発酵から調製される。
【0019】
幾つかの態様において、全発酵ブロスはアスペルギルス(Aspergillus)、アクレモニウム(Acremonium)、オーレオバシジウム(Aureobasidium)、ビューベリア(Beauveria)、セファロスポリウム(Cephalosporium)、セリオリオプシス(Ceriporiopsis)、カエトミウムペシロミセス(Chaetomium paecilomyces)、クリソスポリウム(Chrysosporium)、クラビセプス(Claviceps)、コチオボラス(Cochiobolus)、クリプトコッカス(Cryptococcus)、キュアトス(Cyathus)、エンドシア(Endothia)エンドシアムコール(Endothia mucor)、フサリウム(Fusarium)、ギロクラジウム(Gilocladium)、フミコーラ(Humicola)、マグナポルテ(Magnaporthe)、ミセリオフィソーラ(Myceliophthora)ミロセシウム(Myrothecium)、ムコール(Mucor)、ニューロスポラ(Neurospora)、ファネロカエテ(Phanerochaete)、ポドスポラ(Podospora)、パエシロマイセス(Paecilomyces)、ピリキュラリア(Pyricularia)、リゾムコール(Rhizomucor)リゾプス(Rhizopus)、スキゾフィラム(Schizophylum)、スタゴノスポラ(Stagonospora)、タラロマイセス(Talaromyces)、トリコデルマ(Trichoderma)、セルモマイセス(Thermomyces)、セルモアスクス(Thermoascus)、シエラビア(Thielavia)、トリコクラジウム(Tolypocladium)、トリコフィタム(Trichophyton)、及びトラメテスプレウロタス(Trametes pleurotus)を含むがこれらに限定されない糸状菌の発酵から調製される。幾つかの態様において、全発酵ブロスはアスペルギルスニヂュランス(A.nidulans)、アスペルギルスニガー(A.niger)、アスペルギルスアワモリ(A.awomari)、アスペルギルスアキュレタス(A.aculeatus)、アスペルギルスカワチ(A.kawachi)例えば、NRRL3112、ATCC22342(NRRL3112)、ATCC44733、ATCC14331、及びUVK143f株、アスペルギルスオリザエ(A.oryzae)例えば、ATCC11490、ニューロスポラクラッサ(N.crassa)、トリコデルマレーシ(Trichoderma reesei)、例えば、NRRL15709、ATCC13631、56764、56765、56466、56767、並びにトリコデルマビリデ(Trichoderma viride)例えば、ATCC32098及び32086を含むがこれらに限定されない糸状菌から調製される。好適な実施形態において、全発酵ブロスはトリコデルマ種の発酵から調製される。本発明に用いるのに特に有用な種及び株はトリコデルマレーシRutC30全体セルラーゼである。これはアメリカンタイプカルチャーコレクションより、トリコデルマレーシATCC56765として入手することができる。
【0020】
前述のように、全発酵ブロスは非組換え及び/又は組換え糸状菌の発酵から調製することができる。幾つかの態様において、この糸状菌は1つ以上の相同又は異種遺伝子を含む組み換え糸状菌である。幾つかの態様において、この糸状菌は1つ以上の相同又は異種遺伝子を含む組換え糸状菌であり、この1つ以上の遺伝子はセルロース系物質を分解することができる酵素をコードしている。セルロース系物質を分解する酵素をコードしている遺伝子は当業者に良く知られている。セルロース系基質を分解する酵素をコードする遺伝子の非限定的な例としては、エンドグルカナーゼ、セロビオヒドロラーゼ、グルコヒドロラーゼ、ベータグルコシダーゼ、キシログルカナーゼ、キシラナーゼ、キシロシダーゼ、アルファアラビノフラノシダーゼ、アルファグルクロニダーゼ、アセチルキシランエラスターゼ、マンナナーゼ、マンノシダーゼ、アルファガラクトシダーゼ、マンナンアセチルエステラーゼ、ガラクタナーゼ、アラビナンナーゼ、ペクチントリアーゼ、ペクチンリアーゼ、ポリガラクチュロナーゼ、ペクチンアセチルエステラーゼ、ペクチンメチルエステラーゼ、アルファアラビノフラノシダーゼ、ベータガラクトシダーゼ、ガラクタナーゼ、アラビナナーゼ、アルファアラビノフラノシダーゼ、ラムノガラクチュロナーゼ、ラムノガラクチュロナンリアーゼ、ラムノガラクチュロナンアセチルエステラーゼ、キシロガラクツロノシダーゼ、キシロガラクツロナーゼ、ラムノガラクツロナンリアーゼ、リグニンペルオキシダーゼ、マンガンアーゼ依存ペルオキシダーゼ及びラッカーゼを含む。
【0021】
幾つかの態様において、全発酵ブロスは、内性的には発現されない、または糸状菌によっては非常に低いレベルで発現する1つ以上の酵素活性を補って、セルロース系物質の分解、例えば、グルコース又はキシロース等の発酵可能糖への分解を促進する。補う酵素は全発酵ブロスに補助的に添加することができ、酵素は別の全発酵ブロスとして添加することもでき、精製し、又はわずかに回収して精製してもよい。好適な補助的酵素の非限定的な例は、セロビオヒドロラーゼ、エンドグルカナーゼ、ベータグルコシダーゼ、エンドベータ1,3,(4)グルカナーゼ、グルコヒドロラーゼ、キシログルカナーゼ、キシラナーゼ、キシロシダーゼ、アラビノフラノシダーゼ、アルファグルクロニダーゼ、アセチルキシランエステラーゼ、マンナーゼ、マンノシダーゼ、アルファガラクトシダーゼ、マンナンアセチルエステラーゼ、ガラクタナーゼ、アラビナナーゼ、ペクチンリアーゼ、ペクチナーゼリアーゼ、ポリガラクチュロナーゼ、ペクチンアセチルエステラーゼ、ペクチンメチルエステラーゼ、ベータガラクトシダーゼ、ガラクタナーゼ、アラビナナーゼ、アルファアラビノフラノシダーゼ、ラムノガラクチュロナーゼ、フェルイック酸エステラーゼ、ラムノガラクチュロナンリアーゼ、ラムノガラクチュロナンリアーゼ、ラムノガラクチュロナンアセチルエステラーゼ、キシロガラクチュロノシダーゼ、ラムノガラクチュロナンリアーゼ、リグニンペルオキシダーゼ、マンガナーゼ依存ペルオキシダーゼ、ハイブリッドペルオキシダーゼ、リグニンペルオキシダーゼの組合せペプチド、及びマンガナーゼ依存ペルオキシド、グルコアミラーゼ、アミラーゼ、プロテアーゼ、及びラッカーゼを含む。
【0022】
本発明の幾つかの態様において、全発酵ブロスは、酵素を過剰発現している組換え糸状菌の発酵の全発酵ブロスを含み、セルローススブチリスの分解を改善する。代替的に、全発酵ブロスは非組換え糸状菌の発酵の全発酵ブロスと酵素を過剰発現している糸状菌全発酵ブロスとを含み、セルロース基質の分解を改善する。
【0023】
本発明の幾つかの態様において、全発酵ブロスはベータグルコシダーゼを過剰発現している糸状菌の発酵糸状菌の全発酵ブロスを含む。代替的に、本発明の方法及び反応組成物に用いるための全発酵ブロス非組換え糸状菌の発酵の発酵全体ブロス及びβグルコシダーゼを過剰発現している組換え糸状菌の発酵の発酵全体ブロスを含む。
【0024】
「ベータグルコシダーゼ」の語はEC3.2.1.21として分類されるベータDグルコシダーゼとして本明細書において定義され、及び/又はセロビオースの加水分解を促進してDグルコシダーゼをリリースするGHファミリー1,3,7,9、又は48を含むがこれらに限定されない特定のGHファミリーに分類されるものを含むがこれらに限定されない。これらはセロビオースの加水分解を促進してベータDグルコースを放出する。ベータグルコシダーゼの過剰発現は宿主糸状菌のものと同じかまたは異なる種を形成することができる。特に、過剰発現されたベータグルコシダーゼは糸状菌ベータグルコシダーゼである必要はない。
【0025】
幾つかの態様において、ベータグルコシダーゼはベータグルコシダーゼをコードしている遺伝子の発現により生産することができる。例えば、ベータグルコシダーゼは、例えば、グラム陽性有機体(バチルス又はアクチノマイセテス等)又は原核宿主細胞(例えば、トリコデルマ、アスペルギルス、サッカロマイセス、及びピチア)による、細胞外スペースへ分泌される。幾つかの態様において、ベータグルコシダーゼ組換え微生物内で天然のレベルに対して過剰に発現させることができることは理解されている。幾つかの態様において、もし宿主細胞がベータグルコシダーゼの発現に用いられたなら、この細胞は宿主細胞の内性的な1つ以上のタンパク質の発現を減らすために修飾される。1つの態様において、この細胞は1つ以上のネガティブ遺伝子、特に欠失又は不活性化された1つ以上のプロテアーゼをコードしている遺伝子を含む。例えば、1つ以上のプロテアーゼコード遺伝子(例えば、アスパチルプロテアーゼコード遺伝子; Breakら、Gene 1990 86:153−162及び米国特許No.6,509,171参照)又はセルラーゼコード遺伝子は欠失又は不活性化されている。1つの態様において、トリコデルマ種宿主細胞は、WO05/001036に記載のように、cbh1、cbh2、及びegl1、egl2遺伝子を欠失している。ベータグルコシダーゼをコードしている核酸はトリコデルマ種宿主細胞の核酸ゲノムに存在するか、又は例えば、トリコデルマ宿主細胞で複製するプラスミド中に存在する。
【0026】
用いることができるベータグルコシダーゼの好適な例は、アスペルギルスアキュレアタス(Aspergillus aculeatus)(Kawaguchiら、1996, Gene 173:287−288)、アスペルギルスカワチ(Aspergillus kawachi)(Iwashitaら、1999,Appl.Environ.Microbiol.65:5546−5553)、アスペルギルスオリザエ(Aspergillus oryzae)(WO2002/095014)、セルロモナスビアゾテア(Cellulomonas biazotea)(Wongら、1998,Gene 207:79−86)、サッカロマイセスフィブリゲラ(Saccharomycopsis fibuligera)(Machidaら、1988,Appl.Environ.Microbiol.54:3147−3155)、スキゾサッカロマイセスポムブ(schizosaccharomyces pombe)(Woodら、2002,Nature 415871−880)由来ベータグルコシダーゼ, 並びにトリコデルマレーシベータグルコシダーゼ1(米国特許No.6,022,725)、トリコデルマレーシベータグルコシダーゼ3(米国特許No.6,982,159)トリコデルマレーシベータグルコシダーゼ4(米国特許No.7,045,332)、トリコデルマレーシベータグルコシダーゼ5(米国特許No.7,005,289)、トリコデルマレーシベータグルコシダーゼ6(USPN 20060258554)、トリコデルマレーシベータグルコシダーゼ7(USPN 20040102619)を含む。
【0027】
好適な態様において、全発酵ブロスは少なくとも400pNPGU/gのベータグルコシダーゼ活性を有している。活性の1pNPG単位は50℃、pH4.8において10分間にパラニトロフェニル−B−Dグルコプラノシドから遊離した1μモルのニトロフェノールを示す。ベータグルコシダーゼのこの活性及び全体セルラーゼ調製物の活性は当分野において既知の方法で測定することができる。測定においては、以下の条件を用いることができる。ベータグルコシダーゼ活性は、Chen.H.、Hayn.M.、Esterbauer.H、“Purification and Characterization of two extracellular β−glucosidases from Trichoderma reesei”,Biochimica et Biophysica Acta,1992,1121,54−60の様な、本分野で知られている任意の手段を用いて測定することができる。1pNPGは50℃、pH4.8において、10分間にパラニトロフェニルβ−Dグルコピラノシドから遊離するニトロフェノール1モルを意味する。全体セルラーゼ調製物のセルラーゼ活性はカルボキシメチルセルロース(CMC)を基質として用いて決定することができる。全体セルラーゼ活性の決定はCMC活性の観点から測定される。この方法はCMCに作用する酵素混合物により生成される還元糖の生成を測定する。1単位は1分間に1モルの生成物を遊離する酵素の量である(Ghose,T.K.,Measurement of Cellulse Activities,Pure&Appl.Chem.59,pp.257−268,1987)。
【0028】
幾つかの態様において、全発酵ブロスは少なくとも2500CMCU/gエンドグルカナーゼ活性を有する。活性の1CMC単位は50℃、pH4.8において1分間に、還元糖1モルを遊離することを示す。全体セルラーゼ活性は本発明に重要であるが、ベータグルコシダーゼは、他のセルラーゼの活性別のやり方で負の影響を与える最終生成物を加水分解するので、ベータグルコシダーゼ活性に対する全体セルラーゼ活性の割合は重要である。幾つかの態様において、全体ブロスは約0.5乃至25pNPG/MCM単位の範囲の酵素活性比を含む。幾つかの態様において、全発酵ブロスは約2乃至20pNPG/CMC単位、又は約1.5pNPG/CMC単位、又は約2乃至10pNPG/CMC単位、又は約2。5乃至8pNPG/CMC単位、又は約3乃至7pNPG/CMC単位、又は約3.5乃至6.5pNPG/CMC単位、又は約4乃至6pNPG/CMC単位、又は約4.5乃至5.5pNPG/CMC単位、又は約5乃至6pNPG/CMC単位の酵素化成比を含む。特に、態様において、この比は約5.5pNPG/CMC単位である。
【0029】
好適な投与レベル及び操作条件は特に本発明の明細書の発明な詳細を参照することで、当業者に明らかになる。全発酵ブロスの好適な投与レベルはセルロース基質及び用いる前処理技術により変化する。pH、温度、及び反応時間等の操作条件は、エタノール生成の速度に影響を与える。好ましくは、反応組成物はセルロース1グラム当たり、0.006乃至6mlの全発酵ブロスを含み、最も好ましくは0.03乃至0.6mlの全発酵ブロスを含む。代替的に、全発酵ブロスの量はシステム内のバイオマスの総量に基づいて決定される。そのような場合、反応組成物は好ましくは、0.003乃至3mlの全発酵ブロスをバイオマス1グラム当たりに含み、より好ましくは0.075乃至0.75mlの全発酵ブロス、及びより好ましくは0.015乃至0.3mlの全発酵ブロス、をバイオマス1グラム当たりに含む。他の態様において、全発酵ブロスはバイオマス固形分基質に対して約0.3乃至300.0重量%、より好ましくは約0.75%乃至75%、及び最も好ましくは1.5%乃至30重量%、の効果的な量で固形バイオマス基質に添加される。代替的に、全発酵ブロスの量は、システムに提供される全発酵ブロス由来の細胞の総量に基づいて決定される。そのような場合、反応組成物は好ましくはバイオマス基質1グラム当たり0.0001乃至0.1グラムの細胞量由来の全発酵ブロス、より好ましくは0.00025乃至0.025グラムの細胞量由来の全発酵ブロス、より好ましくは0.0005乃至0.01グラムの細胞量由来の全発酵ブロスを含む。
【0030】
本明細書において、全発酵ブロスはセルラーゼ基質を加水分解することができる酵素を発現する既知の糸状菌培地から得ることができる。発酵は振とうフラスコ培養、連続、バッチ、フェドバッチ等のスモール又はラージスケール発酵、セルラーゼを発現することができる好適な培地又は条件下で行われる実験室的又は工業的発酵に及びメル固形状態発酵を含む。通常、全発酵ブロスは、(i)エンドグルカナーゼ(EG)又は1,4−d−グルカン−4−グルカノヒドロラーゼ(EC3.2.1.4)、(ii)1,4−d−グルカングルカノヒドロラーゼ(セロデキストリナーゼとしても知られている)(EC3.2.1.74)及び1,4−d−グルカンセロビオヒドロラーゼ(エキソ−セロビオヒドロラーゼ、CBH)(EC3.2.1.91)、並びに(iii)ベータグルコシダーゼ(BG)又はベータグルコシダーゼグルコヒドロラーゼ(EC3.2.1.21)を含むがこれらに限定されない。
【0031】
通常、糸状菌はセルロース基質を加水分解することができる酵素の生成に好適な細胞培地中で培養される。培養は炭素及び窒素の源、並びに無機塩を含む好適な栄養培地で、当業者に既知の方法を用いて行われる。育成及びセルラーゼ生成に好適な培地、温度範囲、並びに他の条件は当業者に知られている。
【0032】
好ましくは、糸状菌の発酵は、炭素含有培地を制限因子として制御できるように行われ、それ故、転化される基質の相当量を用いて、細胞へ炭素含有基質の好適な転化を提供し、細胞の汚染を避けることができる。任意の残渣を容易に水洗することができるので、水溶性基質を用いる場合はコンタミは重要な問題とはならない。しかしながら、非水溶性基質の場合は問題であり、好適な洗浄工程等の追加的な生成処理工程を必要とする。
【0033】
全発酵ブロスは糸状菌を静止期まで育成し、1つ以上のセルラーゼ又はベータグルコシダーゼを発現するのに充分な期間制限された炭素条件下で糸状菌を維持することにより行われる。セルラーゼ等の酵素が一旦糸状菌により発酵培地に分泌されると、全発酵ブロスを用いることができる。
【0034】
本発明の全発酵ブロスは、糸状菌を含む。幾つかの態様において、全発酵ブロスは発酵の終わりに発酵物質由来の分画されていない内容物を含む。通常、全発酵ブロスは、糸状菌が静止期まで育成され、炭素制限条件下でインキュベートされてタンパク質合成(特にセルラーゼ及び/又はグルコシダーゼ)をした後に、使用済みの培養培地及び細胞の残骸を含む。幾つかの態様において、全発酵ブロスは使用済みの培養培地、細胞外酵素、及び/又は糸状菌を含む。幾つかの態様において、全発酵ブロス中に存在している糸状菌は既知の方法で、溶解、透過処理、又は殺して、細胞が死んでいる全発酵ブロスを生成することができる。
【0035】
幾つかの態様において、全発酵ブロスである糸状菌細胞を含む全発酵ブロス中の細胞は溶解又は殺されている。幾つかの態様において、この細胞は化学的及び/又はpH制御により糸状菌を溶解して、殺すことにより、糸状菌の発酵の細胞を殺した全体ブロスを生成する。幾つかの態様において、この細胞は化学的及び/又はpH制御により糸状菌を溶解して殺し、並びに殺された細胞の発酵混合物のpHを約4乃至6の範囲内に調整して、糸状菌の発酵の細胞を殺した全体ブロスを生成する。
【0036】
ソルビトール、塩化ナトリウム、ソルビン酸カリウム、及び当業者の知る他のものを含むがこれらに限定されない、追加的な保存料及び/又は静菌剤を全発酵ブロス又は細胞を殺した全発酵ブロスに添加することができる。
【0037】
理論に拘束されることは意図しないけれども、精製されていない全発酵ブロスはエタノール生成微生物に残りの栄養素を提供すると信じられている。このことはエタノール発酵を導き、エタノール生産を改善する。糖化されたセルロースに加えてエタノール生成微生物に、栄養ブロスを提供する必要性を排除又は補助的な栄養素の量を少なくできる可能性はエタノール発酵工程の原料のコストを低くする結果となるだろう。
【0038】
本明細書で説明する方法法及び組成物は発酵微生物に対しての補助的な栄養素及び/又は栄養素源が無いか実質的に存在しない。幾つかの態様において、この方法及び/又は組成物は酵素抽出物、ペプトン、及び/又は尿素が無いか、これらを実質的に含まない。当業者は本発明の方法及び組成物が補助的な窒素源が無いか、これを実質的に含まないけれども、窒素及び/又は栄養源を不純物として存在するか、又は糸状菌に対して全発酵ブロスの栄養価が実質的に増えない程度に添加されてもよいことを理解するだろう。
【0039】
本明細書の方法及び組成物は糸状菌に対する補助的な窒素源の量及び/又はタイプを減らすことができる。幾つかの態様において、本発明の方法及び組成物は糸状菌に用いる酵素抽出物、ペプトン、及び/又は尿素を減らすことができる。
【0040】
本発明の方法及び組成物は糸状菌に対する補助的な窒素源の量及び/又はタイプを減らすことができることは、当業者に理解されるだろう。
【0041】
発酵培地の栄養価は糸状菌の発酵の間に減少又は消費されることから、全発酵ブロスはエタノール生成微生物に対して栄養価を有しているという発見は従来の予想とは異なる。幾つかの研究はろ過されたブロスと比較してろ過していないセルラーゼブロスが高いエタノール生成を示す事を報告している。しかしながら、高められたエタノール生成はろ過工程の間に除去された糸状菌の細胞壁に付着している追加的なベータグルコシダーゼに起因するものであると予測されていた。Schellら、“Whole Broth Cellulase Production for Use in Simultaneous Saccharification and Fermentation of Cellulase to Ethanol,” Appl.Biochem. Biotech. 24/25:287−298 (1990)。驚くべきことに、発明者らは全発酵ブロスが、酵母等のエタノール生成微生物に対する窒素源として通常必要とされていた補助的な窒素源(通常パプトン、トリプトン、酵素抽出物、又は尿素の形態)を代替できることを発見した。精製発酵ブロスにベータグルコシダーゼのみを補っても全発酵ブロスの栄養的な効果(利点)は達成できない(図1及び実施例)。従って、本発明の1つの側面は、補助的な窒素源を用いずに、セルロース系基質、全発酵ブロス、及びエタノール生成微生物を組合せ、インキュベーションする方法である。他の態様において、このセルロース系物質、全発酵ブロス、及びエタノール生成微生物は補助的窒素源を用いずに組み合わせられ、インキュベートされる。
【0042】
本発明の1つの側面は、糖化及び発酵を同時に行うことによりエタノールを生成する方法を提供する。この方法は(a)補助的な窒素源の不存在下で、セルロース系基質、全発酵ブロスを組み合わせる工程及び(b)セルロース系基質、全発酵ブロス、及び/又はエタノール生成微生物をセルロースを加水分解し、グルコースにして、グルココースをエタノールに転化する条件下で、インキュベートする工程を含む。
【0043】
本発明の1つの側面は、糖化及び発酵を同時に行うことによって、エタノールを生成する方法を提供する。この方法は(a)補助的な窒素源の不存在下で、セルロース系物質、全発酵ブロス、及びエタノール生成微生物を組み合わせる工程、及び(b)セルロース基質、全発酵ブロス、及びエタノール生成微生物をセルロース加水分解して、グルコース及び/又はキシロースに転換し、グルコース及び/又はキシロースをエタノールに転化する条件下でインキュベートする工程を含む。
【0044】
本明細書において、発酵微生物は有機物質の生成に好適な発酵プロセスに用いる任意の好適な微生物を意味する。好適な非限定的な糸状菌微生物はグルコース、キシロース、ガラクトース、アラビノース、マンノース、又はオリゴサッカライド等の糖類を所望の発酵生成物に発酵又は転化することができるものである。好適な非限定的な糸状菌微生物の例としては、酵母及びバクテリアを含む。好適な態様において、発酵微生物はエタノール生成微生物である。「エタノール生成」の語は炭化水素から、第一発酵生成物としてエタノールを生成する能力のある微生物を含む。しかしながら、本明細書でのエタノール生成微生物はたの有機物質も生成するのに用いられることを当業者は理解するだろう。この語は自然発生エタノール生成微生物、自然発生又は誘発変異体のエタノール生成微生物、及び/又は遺伝的に修飾されたエタノール生成微生物を含む。セルラーゼ物質をグルコース及び他の小糖類に加水分解する工程は重要な工程であるけれども、同時糖化及び発酵(SSF)は、これらの糖をエタノールに転化する、エタノール生成微生物が生存している培養工程に依存している。幾つかの態様において、このエタノール生成微生物は効果的にグルコースをエタノールに発酵ことができる、サッカロナイセスセレビジアエ(Saccharomyces cerevisiae)、サッカロマイセスウバルム(S.uvarum)、クリベロマイセスファギリス(Kluyveromyces fagilis)、カンジダプセウドトロピカリス(Candida pseudotropicalis)、及びパキソレンタンノフィラス(Pachysolen tannophilus)等の酵母細胞である。好適な株はサッカロマイセスデレビシアエ(S.cerevisiae)D5A (ATCC200062)、サッカロマイセスデレビシアエY567(ATCC24858)、ACA174(ATCC60868)、MY91(ATCC201301)、MY138(ATCC201302)、C5(ATCC201298)、ET7(ATCC201299)、LA6(ATCC201300)、OSB21(ATCC201303)、F23(サッカロマイセスグロボサス(S.globosus)ATCC90920)、ACA174(ATCC60868)、A54(ATCC90921)、NRCC202036(ATCC46534)、ATCC24858、ATCC24858、G3706(ATCC42594)、NRRL、Y−265(ATCC60593)、Sa28(ATCC26603)、ATCC24845−ATCC24860を含むがこれらに限定されない。本発明に用いるための他のセレビシエではない酵母株はピチアパストリス(Pichia pastoris)(tozonyID4922)、サッカロマイセスパストリアナス(S.pastorianusSA23(サッカロマイセスカールスベンシス(S.carlsbergensis)ATCC26602)、サッカロマイセスパストリアナス(S.pastorianus(サッカロマイセスカールスベルゲンシス(S.carlsbergensisATCC2345)、カンジダアシドデルモフィリウム(Candida acidothermophilum(イサキタニアオリエンタス(Issatchenkia orientalis、ATCC20381)を含む。幾つかの態様において、エタノール生成微生物は組換え酵母株である。好適な組換え酵母株はキシロースリダクターゼ、キシリトールデヒドロゲナーゼ、及び/又はキシルロキナーゼをコードしている遺伝子を含む(例えば、米国特許No.5,789,210参照)。
【0045】
本発明の幾つかの態様において、エタノール生成微生物はバクテリア細胞、好ましくはグラム陰性、通性嫌気性、及び腸内細菌科のファミリーである。他の関連する態様において、このエタノール生成微生物はエシェリキア(Escherichia)又はクレブシエラ(Klebsiella属)であり、好ましくは、エシェリキア又はクレブシエラ、好ましくはE.coliB、E.coliDH5a、E.coliKO4(ATCC55123)、E.coliKO11(ATCC55124)、E.coliKO12(ATCC55125)、E.coliLY01、クレブシエラオキシトーカ(K.oxytoca)M5A1又はクレブシエラオキシトーカP2(ATCC55307)である。幾つかの態様において、エタノール生成微生物はザイモモナス(Zymomonas)種又はザイモモナスモビリス(Zymomonas mobilis)(ATCC31821)由来である。幾つかの態様において、組換えザイモモナス(Zymomonas)株は、例としてキシロースイソメラーゼ、キシルクキナーゼ、トランスアルドラーゼ、及びトランスケトラーゼをコードする遺伝子を含む。
【0046】
発酵微生物は通常加水分解物を転化して、発酵は12乃至96時間、例えば、30乃至80時間行われる。温度は通常26℃乃至40℃である。特にpH3乃至6で約32℃で行われる。
【0047】
発酵に続いて、所望の有機物質を当業者に既知の任意の手段を用いて回収する。そのような方法としては、蒸留、抽出、クロマトグラフィー、電気泳動、溶解差を含むがこれらに限定されない。例えば、エタノール発酵において、アルコールは従来の蒸留方法により、分離及び精製される。本発明の方法によりえられたエタノールは燃料エタノール、飲料エタノール、又は工業エタノールとして用いられる。
【0048】
本発明の側面は更に、以下の実施例を参照することで更に理解されるが、これらは本発明の範囲を限定するものではない。本開示を逸脱せずに、物質及び方法の両方において多くの変更が可能であることを当業者は理解しているだろう。
【実施例】
【0049】
実施例1:全発酵ブロスの調製
グルコース/ソホロースの調製
60%(W/W)グルコース溶液を121℃、30分で滅菌した。この温度を65℃に下げ、1リットル当たり10gの総タンパク質(全体セルラーゼはT.reeseiにより生成された)を添加した。この混合物をゆっくり撹拌し、65℃で3日間維持した。ソホロース含量はこの60%グルコース溶液中、12g/Lであると測定された。
【0050】
0.8Lの培地を1.5mlのトリコデルマレーシ(Trichoderma reesei)RLP−37凍結胞子で接種して、種フラスコとした。このフラスコを2つの0.4L部分にわけ、48時間後に、2つの別々の14LのBiolafite fermentor中の2×7Lの発酵培地へ写した。この育成培地は以下の組成を有していた:
【0051】
微量元素*:5g/L FeSO4−7H2O、1.6g/L MnSO4−H2O、1.4g/L ZnSO4−7H2O
【0052】
発酵は25℃、750RPM、及び1分間当たり8標準リットル(SLM)の空気流れで行った。
【0053】
バッチグルコースは約20時間で消費され、この時点で細胞の成長が停止し、炭素制限供給が始まった。40%グルコース/ソホロース供給を0.25g/分で添加した。セルラーゼ生成に直接関与する総タンパク質(セルラーゼ活性対総細胞外タンパク質の我々の比較に基づく)をバットフェーズの直後に導入した。この細胞は化学的及びpH処理を組合せ、溶解させて殺した。必要であれば、殺された全発酵ブロスを、より中性、例えば、約pH4乃至6の間に調整する。
【0054】
実施例2:精製発酵ブロスの調製
糸状菌を実施例1で説明したように育成した。細胞を溶解するよりはむしろ、全発酵ブロスの内容物をろ過して、細胞及び大きな細胞の残骸をろ過して精製発酵ブロスを調製した。所望のセルラーゼ及びグルコシダーゼはトリコデルマ細胞から分泌されているので、セルラーゼ分解活性は精製された発酵ブロスの中に残っている。精製発酵ブロスの中に含まれている酵素は10kDaカットオフメンブランでウルトラろ過により濃縮された。
【0055】
実施例3:同時糖化発酵
同時糖化発酵(SSF)は250mlフラスコにおいて、標準的な酵母発酵条件(例えば、セルモサック(Thermosacc)酵母、pH5.0、38℃)で、2回行った。 酸で前処理されたバッガスを20mMのクエン酸ナトリム緩衝液(pH5.0)で7%のセルロース含量になるように調整した。酵母の栄養は1.0g/L酵母抽出物、1.0g/Lペプトン、及び1.0g/L尿素の最終濃度になるように添加した。全発酵ブロス又は精製発酵ブロスを(0.4CMC U/g酸前処理シュガーコーンバッガスの濃度)に添加して、同時に酵母を用いて発酵を行った。懸濁したときに、精製した発酵ブロスは、最終的に0.063pNPG/g酸処理バッガスと同様のベータブルコシダーゼ活性を有していた。このフラスコを150rpmで撹拌した。サンプルを異なる時間で採取し、エタノール、グリセロール、酢酸、酪酸、及び残りの糖類をHPLC法で解析した。
【0056】
図1は、酵母栄養がない場合、SSFと全発酵ブロスは精製発酵ブロス(ベータグルコシダーゼ添加又は無添加)を用いたものよりも充分に高いエタノール生成を提供した。例えば、96時間において、ベータグルコシダーゼを含む精製発酵ブロスを用いた場合のエタノール濃度は22.2g/Lであるのに対して、全発酵ブロスは30.7g/Lであった。
【0057】
図2は全発酵ブロスを用いたSSFパフォーマンスと精製発酵ブロスを用いたSSFパフォーマンスの比較である。全てのケースにおいて、酵母栄養を同じレベルで添加した。全発酵ブロスは発酵速度が速くなり、エタノール生成が僅かに高くなったことを観察した。
【0058】
図3は酵母栄養を添加した、又は添加してない全発酵ブロス精製物を用いたSSFパフォーマンスの比較である。SSFパフォーマンスは両者とも同レベルであることが確認された。96時間において、酵母栄養を含む全発酵ブロスのエタノール濃度は31.7g/Lであるのに対して、酵母栄養を含まない全発酵ブロスのエタノール濃度は30.7g/Lであった。この結果はエタノールへの発酵について、全発酵ブロスからのの窒素源を酵母が利用することができることを示している。
【0059】
実施例4:糖化
酸前処理バッガスの糖化を250mlスラスコで、2回行った。酸前処理バッガスは20mMクエン酸ナトリウム緩衝液(pH5.0)を用いて7%セルロースの付加になるように調整した。ベータグルコシダーゼを補充した、全発酵ブロス又は精製発酵ブロスを添加して酵素加水分解を開始した。同じ量のセルロース活性を用いた。即ち、それぞれ、0.4CMC U/g酸処理バッガス及び0.063pNPG/g酸処理バッガスを用いた。このフラスコを150rpmで撹拌した。サンプルを異なる時間間隔で採取し、グルコース、セロビオース、及びキシロースについて、HPLC方法で解析した。
【0060】
図4は、酸処理したバッガスの同程度の糖化が観察されたことにより、全発酵ブロス及びベータグルコシダーゼを補充した精製発酵ブロスのセルロース分解能力は同一であることを示している。それゆえ、Schellら、”Whole Broth Cellulase Production for Use in Simultaneous Saccharification And Fermentation of Cellulase to Ethanol,” Appl. Biochem. Biotech.24/25:287−298(1990)で提案されていたのとは異なり、高められたエタノール生成は全発酵ブロスによるベータグルコシダーゼに依存するものではないことが証明された。
【特許請求の範囲】
【請求項1】
発酵及び糖化を同時に行って有機物質を生成する方法であって、
(a)補充される窒素源の不存在下で、セルロース系基質、全発酵ブロス、及び発酵微生物を組み合わせる工程、及び
(b)セルロース基質、全発酵ブロス、及び発酵微生物を、セルロースからグルコース及び/又はキシロースへの加水分解並びにグルコース及び/又はキシロースから有機物質への転換を行う条件下でインキュベートする工程を含む、方法。
【請求項2】
前記セルロース系物質が、木材、木材パルプ、製紙スラッジ、製紙パルプの廃棄流れ、パーチクルボード、コーンストーバー、コーンファイバー、コーンコブ、ライス、製紙及びパルプ処理廃棄物、木又は草植物、フルーツパルプ、ベジタブルパルプ、パミス、醸造かす、草、籾殻、トウキビバッガス、綿、ジュート、ヘンプ、麻、バンブー、サイザル、アバカ、ストロー、コーンコブ、葉、小麦わら、ココナツ繊維、藻類、及びこれらの混合物からなる群より選択される1つ以上のセルロース源を含む、請求項1の方法。
【請求項3】
前記セルロース系物質が機械的又は化学的に前処理されている、請求項1の方法。
【請求項4】
前記全発酵ブロスが糸状菌の発酵から調製される、請求項1の方法。
【請求項5】
前記発酵微生物が酵母又はバクテリア細胞である、請求項1の方法。
【請求項6】
前記発酵微生物がエタノール生成微生物である、請求項1の方法。
【請求項7】
前記有機物質がアルコールである、請求項1の方法。
【請求項8】
前記有機物質がエタノールである、請求項1の方法。
【請求項9】
セルロース系物質、全発酵ブロス、及び発酵微生物の混合物を含む、有機基質を生成するための反応組成物であって、前記反応組成物には実質的に追加的な窒素源が含まれていないことを特徴とする、方法。
【請求項10】
前記セルロース系物質が、木材、木材パルプ、製紙スラッジ、製紙パルプの廃棄流れ、パーチクルボード、コーンストーバー、コーンファイバー、ライス、製紙及びパルプ処理廃棄物、木又は草植物、フルーツパルプ、ベジタブルパルプ、パミス、醸造かす、草、籾殻、トウキビバッガス、綿、ジュート、ヘンプ、麻、バンブー、サイザル、アバカ、ストロー、コーンコブ、葉、小麦わら、ココナツ繊維、藻類、及びこれらの混合物からなる群より選択される1つ以上のセルロース源を含む、請求項9の反応組成物。
【請求項11】
前記セルロース系物質が機械的又は化学的に前処理されている、請求項9の反応組成物。
【請求項12】
前記全発酵ブロスが糸状菌全発酵ブロスである、請求項1の反応組成物。
【請求項13】
前記発酵微生物が酵母又はバクテリア細胞である、請求項9の反応組成物。
【請求項14】
前記発酵微生物がエタノール生成微生物である、請求項9の反応組成物。
【請求項1】
発酵及び糖化を同時に行って有機物質を生成する方法であって、
(a)補充される窒素源の不存在下で、セルロース系基質、全発酵ブロス、及び発酵微生物を組み合わせる工程、及び
(b)セルロース基質、全発酵ブロス、及び発酵微生物を、セルロースからグルコース及び/又はキシロースへの加水分解並びにグルコース及び/又はキシロースから有機物質への転換を行う条件下でインキュベートする工程を含む、方法。
【請求項2】
前記セルロース系物質が、木材、木材パルプ、製紙スラッジ、製紙パルプの廃棄流れ、パーチクルボード、コーンストーバー、コーンファイバー、コーンコブ、ライス、製紙及びパルプ処理廃棄物、木又は草植物、フルーツパルプ、ベジタブルパルプ、パミス、醸造かす、草、籾殻、トウキビバッガス、綿、ジュート、ヘンプ、麻、バンブー、サイザル、アバカ、ストロー、コーンコブ、葉、小麦わら、ココナツ繊維、藻類、及びこれらの混合物からなる群より選択される1つ以上のセルロース源を含む、請求項1の方法。
【請求項3】
前記セルロース系物質が機械的又は化学的に前処理されている、請求項1の方法。
【請求項4】
前記全発酵ブロスが糸状菌の発酵から調製される、請求項1の方法。
【請求項5】
前記発酵微生物が酵母又はバクテリア細胞である、請求項1の方法。
【請求項6】
前記発酵微生物がエタノール生成微生物である、請求項1の方法。
【請求項7】
前記有機物質がアルコールである、請求項1の方法。
【請求項8】
前記有機物質がエタノールである、請求項1の方法。
【請求項9】
セルロース系物質、全発酵ブロス、及び発酵微生物の混合物を含む、有機基質を生成するための反応組成物であって、前記反応組成物には実質的に追加的な窒素源が含まれていないことを特徴とする、方法。
【請求項10】
前記セルロース系物質が、木材、木材パルプ、製紙スラッジ、製紙パルプの廃棄流れ、パーチクルボード、コーンストーバー、コーンファイバー、ライス、製紙及びパルプ処理廃棄物、木又は草植物、フルーツパルプ、ベジタブルパルプ、パミス、醸造かす、草、籾殻、トウキビバッガス、綿、ジュート、ヘンプ、麻、バンブー、サイザル、アバカ、ストロー、コーンコブ、葉、小麦わら、ココナツ繊維、藻類、及びこれらの混合物からなる群より選択される1つ以上のセルロース源を含む、請求項9の反応組成物。
【請求項11】
前記セルロース系物質が機械的又は化学的に前処理されている、請求項9の反応組成物。
【請求項12】
前記全発酵ブロスが糸状菌全発酵ブロスである、請求項1の反応組成物。
【請求項13】
前記発酵微生物が酵母又はバクテリア細胞である、請求項9の反応組成物。
【請求項14】
前記発酵微生物がエタノール生成微生物である、請求項9の反応組成物。
【図1】

【図2】

【図3】
【図4】
【図2】
【図3】
【図4】
【公表番号】特表2011−500029(P2011−500029A)
【公表日】平成23年1月6日(2011.1.6)
【国際特許分類】
【出願番号】特願2010−529047(P2010−529047)
【出願日】平成20年10月9日(2008.10.9)
【国際出願番号】PCT/US2008/079377
【国際公開番号】WO2009/049067
【国際公開日】平成21年4月16日(2009.4.16)
【出願人】(509240479)ダニスコ・ユーエス・インク (81)
【Fターム(参考)】
【公表日】平成23年1月6日(2011.1.6)
【国際特許分類】
【出願日】平成20年10月9日(2008.10.9)
【国際出願番号】PCT/US2008/079377
【国際公開番号】WO2009/049067
【国際公開日】平成21年4月16日(2009.4.16)
【出願人】(509240479)ダニスコ・ユーエス・インク (81)
【Fターム(参考)】
[ Back to top ]