
皮膚T細胞リンパ腫(CTCL)の新規な診断及び治療法
【課題】Tリンパ腫の新規な診断及び治療法を提供する。
【解決手段】下記の段階を含むことを特徴とする、患者の特定の体内区分内の、悪性CD4+CTCL細胞の存在比率の数値を求める方法。p140分子を発現するCD4+細胞の比率の値を、前記患者から得られた生物学的試料を基に測定し、そして、悪性CD4+CTCL細胞の、現実に前記体内区分に存在する比率の数値は、前記測定値の約±10%の範囲内に入る数値と考える。
【解決手段】下記の段階を含むことを特徴とする、患者の特定の体内区分内の、悪性CD4+CTCL細胞の存在比率の数値を求める方法。p140分子を発現するCD4+細胞の比率の値を、前記患者から得られた生物学的試料を基に測定し、そして、悪性CD4+CTCL細胞の、現実に前記体内区分に存在する比率の数値は、前記測定値の約±10%の範囲内に入る数値と考える。
【発明の詳細な説明】
【技術分野】
【0001】
この出願はTリンパ腫の新規な診断及び治療法、更に詳しくは、皮膚T細胞リンパ腫(CTCLと略称する)の新規な診断及び治療法に関する。本発明は現実に、CTCLに共通する腫瘍マーカーを提供し、そのバイオテクノロジー的及び医学的用途をも含む。
【背景技術】
【0002】
CTCLは本来皮膚に関するTリンパ腫のグループである。CTCLグループは即ち、形質転換菌状息肉症(形質転換MFと略称する)、セザリー症候群(SSと略称する)、リンパ腫丘疹(LPと略称する)及びCD30+リンパ腫からなる。形質転換MFは表現タイプが成長済みTヘルパー細胞に類似するクローン誘導型悪性Tリンパ球の皮膚侵襲に因るものである。LP及びCD30+リンパ腫もまた、皮膚で進展する。より進行性な型のCTCLは、悪性細胞が非表皮性になったとき発達し、特殊な皮膚関連性を示すようになる。SSはCD4+/CD45+RO+T細胞のクローン増大及びこれらの悪性T細胞の血中への出現による、より進行性なCTCLのタイプである。大部分のCTCLはCD4+CTCLであり、稀にCD8+CTCLの場合がある。
細胞表面の特殊なマーカーが欠如しているために、悪性細胞の同定が困難であるから、CTCL病の生物学は、十分には、理解されていない状態が継続している。皮膚の病変において、活動性、浸潤性のリンパ球(非腫瘍性の)の中から、CTCL細胞を識別するのは困難である。
CTCLのようなTリンパ腫の診断は、現時点では、疑わしい体内(脳の細胞核を示す細胞の検知によって、皮膚生検及び/または血液中のSSの存在の外観の組織病理学的観察)から採取された試料中に腫瘍細胞が存在するか、しないかを、細胞学的且つ組織病理学的観察によって行なうことに基づいている。このような診断法は、皮膚リンパ球の悪性リンパ球への形質転換の初期では、特に、十分な信頼性に欠ける。このような診断法は病気を現実のものとして捉えることができない。技術的には時間も浪費する。
今日では、CTCLの治療は、非特異的化学療法による腫瘍細胞アポトーシスの誘導により試験的に行なわれている。
だから、CTCLのような悪性T細胞の増殖に関連する病気の診断及び治療の問題点のより正確で好適な解決策が、まだ、必要とされている。
【0003】
このような従来技術の状況に鑑み、発明者等はCTCLの診断及び治療の問題点を解決する二つの分子を提供する。一つは発明者によってSC5と命名され、他方はp140分子(p140は、また、KIR3DL2である)である。
本発明におけるSC5分子は生化学的、機能的に新規な蛋白質であり、還元状態で、見かけの分子量は96kDである。これは発明者等によって産生されたモノクローナル抗体の抗原として分離された。このモノクローナル抗体を産生するハイブリドーマは2000年10月30日、C.N.C.M.(C.N.C.M.パスツール研究所;25、rue du Docteur Roux、F-75724 Paris Cedex15、France)にブダペスト条約(C.N.C.M.寄託番号I-2575)に従がって寄託された。細胞表面で発現され、集合されたとき、SC5分子は細胞活動及び増殖の阻害受容体として行動する。本発明の第二のマーカー、即ち、p140は既に阻害受容体として知られているが、以前に、NK細胞及び健康なヒトのCD4+CD8+細胞のサブグループであると述べられていた。発明者等は、ここで、p140がCTCL細胞のような腫瘍T細胞の表面で発現されることを示す。
発明者等は、更に、CD4+CTCL細胞のような悪性のCD4+T細胞の表面で、p140が発現するのに対し、NK細胞(p58.1、p58.2、p70KIRs、CD94/NKG2A)の表面で通常観察される他の受容体は、悪性のCD4+T細胞の表面で観察されないことを示す。勿論、p140は皮膚炎(例えば、皮膚結核、苔癬)または毒物による皮膚壊死のような非腫瘍性の皮膚系疾病に罹っている患者から採取されたCD4+T細胞の表面では、発明者によって観察されていないことに留意すべきである。
腫瘍性CD4+T細胞の表面で、p140の二つの対立型が発明者によって同定された。即ち、対立型KIR3Dクローン24(SEQ ID No.1)及び対立型p140クローン1.1(SEQ ID No.3)である。対立型KIR3Dクローン24(SEQ ID No.1)は新規蛋白質(SEQ ID No.2)をコード化する新規なポリヌクレオチドである。KIR3Dクローン24(SEQ ID No.1)は、前記p140クローン1.1DNA配列(SEQ ID No.3)に比較して、5つの相違点が明らかであり、前記p140クローン1.1成熟蛋白(SEQ ID No.4)に比較して、成熟蛋白(SEQ ID No.2)において、4個のアミノ酸置換になっている。
【0004】
発明者等は、SC5及びp140の両者が、以下の同じ技術的特徴を共に有することを示す。
SC5及びp140は共に、悪性T細胞、とりわけ、悪性CD4+T細胞の特徴である、細胞膜分化抗原である。
CD4+CTCLの何れの型が関与しているにしても、SC5またはp140をそれらの表面(通常、SC5及びp140の両者が発現される)に発現する、悪性のCD4+T細胞がある。SC5及びp140のいずれも、CD4+CTCL、特に、セザリー症候群(略語でSS)、形質転換菌状息肉症(略語で形質転換MF)、リンパ腫症丘疹(略語でLP)及びCD30+リンパ腫の全範囲を実際にカバーする。
SC5またはp140のCD4+T細胞の表面での存在と、CD4+CTCLの存在の間には、CD4+CTCL診断が疑わしい体内部分(例えば、形質転換MFが疑われる際の紅皮症の皮膚試料または、SSのような進行性のCTCL型が疑われる際の末梢血液試料)から採取したCD4+細胞表面のSC5またはp140の存在の解析に基づき、この信頼性は90%以上、より好ましくは95%以上、最も好ましくは100%である(これまで試験に供された患者については信頼性100%である)という関連がある。事実、本発明によれば、CD4+T細胞は、CD4+T細胞表面でのp140分子の存在が検知されるか、または、SC5+CD4+T細胞の比率の値が、平均標準レベルより高く測定されれば(平均標準レベル1〜15%、一般には5〜10%の範囲内にある)、即、腫瘍であると決定される。
また、SC5またはp140のCD4+T細胞の表面での存在と、CD4+CTCLの存在の間には、CD4+SC5+T細胞及びCD4+p140+T細胞の両者の存在比率、−これはセザリー症候群(SS)が疑われる患者から採取された試料から測定されるが、−は本質的に、この患者の末梢血液に現実に存在する悪性のSSの存在比率に相当する(SC5+CD4+細胞は±10%の範囲内、p140+CD4+細胞は±5%の範囲内)。だから、本発明におけるSC5及びp140マーカーはSSの進行状況を評価できる共通の特別な特徴を共に有する。
【0005】
発明者の知る限りでは、SC5及びp140分子は共通の技術的特徴を有する最初の分子である。本発明は、かくして、新規なCTCLに共通のマーカーを提供する。これより以前には、このようなCTCLに共通のマーカーは知られていない。これらに最も近似するこの種のものは、CD30であって、このものの、悪性のCD4+T細胞の表面での存在は、患者がCD30+リンパ腫に関連するCD4+CTCLの特殊型を有していると結論付ける。それ故、CD30はCTCLのマーカーであるが、その信頼性はCTCLの特殊型(CD30+リンパ腫)に限られる。そして、CD30はCD4+CTCLの全ての型をカバーしない。即ち、SS、形質転換型MFまたはLPのようなCD4+CTCLでは、表面でCD30を発現する悪性CD4+T細胞は必ずしも存在しない。CD30を使用したCTCLの診断及び治療は、このように、CTCLの特殊型に限られる。それに対して、本発明のマーカーはCD4+CTCLの全範囲を好適にカバーする。
【0006】
p140と共有する技術的特徴に加えて、SC5は、更に、非増殖性のT細胞の増殖及び/または機能的活性(例えば、サイトカインプロフィール)の肯定的指標を有する。T細胞表面のSC5の存在は、事実、正常(CD4+及びCD8+)T細胞またはウイルス感染CD4+T細胞のような非腫瘍性T細胞の活性状態に肯定的に関連する。SC5は、かくして、また、HIV感染症、及び自己免疫性(例えば、脊椎関節炎のようなリューマチ性関節炎または乾癬、湿疹、アトピー性皮膚炎のような免疫関連性皮膚病)のようなT細胞ウイルス性感染症の診断及び治療のための新規手段として有効である。SC5は、また、グラフト宿主反応を調節するための有効な標的である。即ち、SC5の活性化は、エフェクター宿主細胞をグラフトに対して進展させる反応を阻害するために使用できる阻害性シグナルを伝達する。
【0007】
本発明は、また、SC5新分子に結合できる産物を提供し、特に、SC5に対するモノクローナル抗体(抗SC5mAbs)を提供する。また、T細胞、特に悪性CTCL細胞のような腫瘍性T細胞の増殖及び/または機能的活性の調節ができる抗SC5モノクローナル抗体を提供する。特に、ハイブリドーマI−2575で産性されたモノクローナル抗体は、T細胞の増殖及び/または機能的活性の阻害が可能で、特に有効な特徴によれば、悪性CTCL細胞(下記の例参照)のような悪性T細胞の増殖及び/または機能的活性の阻害が可能である。SC5に結合するが、これらを集合させない化合物は、SC5の阻害性シグナルの伝達を妨げるか、または、化合物がSC5+細胞(標的マスキング)に到着するのを妨げる。
【0008】
本発明によれば、一般に、SC5またはp140に結合する化合物は、補体漸増剤、ADCC刺激剤、または、CTCLのようなTリンパ腫を予防し、一時的緩和し、治療するための治療剤用ベクターとして使用される。
【0009】
本発明はそれ故に、抗SC5及び/または抗p140モノクローナル抗体及び直接それらから製造された産物(例えば、ヒト化モノクローナル抗体またはCD4−p140/SC5に対する二重特異性を有するモノクローナル抗体)の診断及び治療への使用に関する。
【発明を実施するための最良の形態】
【0010】
2000年10月30日、寄託番号I−2575として、C.N.C.M.(C.N.C.M.、国立微生物保存機関、パスツール研究所;25、rue du Docteur Roux、F-75724 Paris Cedex15、France)に、ブダペスト条約に従がって、寄託されたハイブリドーマを提供し、ハイブリドーマ細胞の好適な代謝条件(例えば、37℃で3日間、5%炭酸ガス含有湿潤雰囲気下、RPMI1640剤:Gibco BRL Cat.N021875-034;10%牛胎児血清;ぺニシリン100単位/ミリリットル;ストレプトマイシン100μg/ミリリットルからなる好適な培養液に2.106個の細胞を含有したものを使用。)下、ハイブリドーマI−2575の培養を内容とする、抗SC5モノクローナル抗体の調製方法を提供する。
本発明は、それ故、ハイブリドーマI−2575によって産生されるか、またはこの寄託されたハイブリドーマの培養上澄み液から得られるモノクローナル抗体(mAb)を包含する。このmAbは5種類の抗SC5IgMである。これは不適当または好ましくないT細胞の増殖及び/または機能的活性(サイトカインプロフィール)、不適当または好ましくないIL−2産生の診断及び治療のための有効な手段である。これは正常T細胞、ウイルス感染T細胞(例えば、HIV感染CD4+T細胞)及び特に、悪性CTCL細胞のような悪性T細胞の増殖及び/または機能的活性を阻害する。
【0011】
ハイブリドーマI−2575によって産生されるモノクローナル抗体は、発明者によってSC5と命名された蛋白質を分離できる。SC5蛋白質は生化学的、機能的に新規な分子で、見かけ分子量が還元条件下で96kDである。これは多くのPBL細胞中及び細胞上に存在する。即ち、正常細胞(CD4+及びCD8+部分集合)及び特にCD3+T細胞、CD45RO+T細胞、正常NK細胞、B細胞、顆粒球及びマクロファージ(大食細胞)である。更に、SC5はNIH/3T3(ATCC CRL−1658)及びCHO−KI(ATCC CRL−9618)のような他の系統及び他の種の細胞の細胞質中にも存在する。
休眠中の正常PBL細胞では、SC5は主として細胞内に存在する、しかし、CD3刺激はSC5の細胞膜への移行を誘発する。殊に目立つのは、発明者によって、SC5が、悪性CTCL細胞のような悪性T細胞、特に悪性CD4+CTCL細胞で、主として、細胞表面に観られることである(幾ばくかの生体外のCD3刺激なしで)。
【0012】
更に著しい特徴はSC5の表面発現が、現在まで試験された全てのCTCL患者に観られることである(下記例参照)。
本発明の他の更に著しい特徴は、SC5を共発現するCD4+T細胞の比率が、健常者よりも、腫瘍性CD4+T細胞をもつ患者の方が、統計的に高いことである。CD4+CTCLの患者におけるSC5+CD4+の比率は、健常者において観察される比率(SC5+CD4細胞の平均標準比率は1〜15%の範囲内であり、一般には5〜10%の範囲内である)より有意に高い。
特にセザリー症候群(SS)のCD4+CTCLの場合、SC5を共発現するCD4+細胞の比率は、もっと、統計的に、CTCL患者のPBLにおける悪性CTCL細胞の比率に密接に関連する。即ち、SC5+CD4+T細胞の比率は本質的に悪性CTCL細胞の比率と同等である(この比率の±10%以内)。SC5蛋白質は、それ故、細胞表面での経膜的な発現が、非腫瘍性T細胞の活性化及び/または増殖状態に関係があり(例えば、HIV感染細胞及び正常CD4+またはCD8+T細胞のようなウイルス感染細胞)、非活性化T細胞の悪性化状態に関係がある新規分子である。著しい特徴は、CD4+CTCLが陽性との診断を90%以上の信頼性で可能にすることである(実際、最新の試験を行ったヒトでは100%)。
【0013】
本発明の更に顕著な特徴は、SC5新分子が、細胞表面で発現されたとき、細胞増殖及び/または機能的活性のための阻害受容体として活動できることである。即ち、SC5の集合は、それを発現する細胞の増殖及び活性の減少を誘発する。例えば、ハイブリドーマI−2575により産生される、抗SC5モノクローナル抗体[5種類のIg(免疫グロブリンの略語)]は、ヒトまたは動物から採取した、正常CD3+T細胞、または正常CD3+T細胞クローンのようなSC5+細胞の抗CD3誘発増殖を阻害し、また、抗CD3が誘発した、CTCL患者から採取したCTCL細胞の試験管内増殖及び動物モデルでのヒトCTCL細胞の生体内自然増殖を阻害する。このmAbは、また、HIV感染CD4+細胞と結合し、それ故に、感染細胞を殺し、HIVの増殖を抑制させることを目的とする抗HIV薬の実際的原理として有用である。
【0014】
幾つかのSC5蛋白質の分離可能な方法は、本発明のモノクローナル抗体を製造できる専門家にとって使用できるのものである。標準的な方法は、細胞溶解物からの還元下、親和性クロマトグラフまたは電気泳動による回収からなる。ハイブリドーマI−2575により産生するモノクローナル抗体が抗原と認識する全てのヒトまたは動物の細胞、あるいは該モノクローナル抗体と細胞溶解物が免疫沈殿反応する全てのヒトまたは動物の細胞は、SC5分子を分離するための細胞供給源として適当である。そのような細胞には、特に、T細胞、CD3+T細胞、CD45RO+T細胞、B細胞、NK細胞、マクロファージ、顆粒球等の正常細胞(ヒトまたは動物または細胞クローン)を含む。また、悪性T細胞、例えば、CTCL患者から採取されたCD4+CTCL細胞のような腫瘍性細胞、またはCTCLクローンHUT78(ATCC TIB−161)のような腫瘍性クローンも好適な細胞である。好適な細胞源が広範囲なので、粗血球全体またはPBLが何らかの精製を経ずに、直接使用されてもかまわないことが理解される。
【0015】
正常細胞はSC5の細胞内型及び経膜受容体型の両者を分離できる。SC5の受容体型またはその部分の分離が望まれるときは、SC5+細胞源(例えば、それらの表面でSC5を発現する細胞)を使うのが好ましい。そのような細胞源(ソース)としては、前記されたものであって、細胞表面でSC5発現が増加するように、CD3刺激されたものが含まれる(下記CD3刺激法の例参照)。全血球またはPBLがSC5+細胞の供給源として使用されるとき、全血球またはPBLは、T細胞及び単核細胞を含有するから、CD3活性化はPHA(Wellcome社から入手可能)のような、ポリクローナル活性体の約1μg/ミリリットルと培養することにより誘発される。より均一な正常細胞集団がSC5+細胞(例えば、SC5+のみとNK細胞)の供給源に使用されるときは、CD3活性化は、好ましくは固定化抗CD3抗体との培養、または、可溶性抗CD3抗体及びPHAのようなポリクローナル活性体との培養によって為される。培養段階は、選択的に、CD3活性化の前及び/または後に、細胞量を増加させるために行なってよい(培養条件は、例えば、RPMI中、10%牛胎児血清、湿度95%及び5%炭酸ガス雰囲気)。
SC5を細胞内型で分離したいときには、CD3刺激は省かなければならない。
CTCL細胞のような腫瘍性細胞、特に、悪性形質転換MFまたはSS細胞では、生体外CD3活性化を必要とせずに、受容体型のSC5分子を分離することができる。即ち、CTCL細胞は既に経膜区域でSC5を発現する。
【0016】
故に、本発明は下記の段階によって得られる分離蛋白質を包含する。
(i)T細胞、CD3+T細胞、CD45RO+T細胞、B細胞、NK細胞、マクロファージ、顆粒球からなる群から選択される細胞を採取する。採取した細胞を固定化抗CD3抗体、または、可溶性抗CD3抗体及びPHAの1μg/ミリリットルで刺激するか、または、悪性CD+CTCL細胞のような悪性CTCL細胞を採取するか、または、全血球細胞及び末梢血液リンパ球(PBL)からなる群から選択される細胞を採取し、採取した細胞をPHAの約1μg/ミリリットルで刺激する。
(ii) 例えば、TritonX-100の1%含有の強力な溶解バッファー中での培養によって、多種類のポリペプチド複合体を分離できる条件下に上記細胞を溶解する。
(iii)ハイブリドーマI−2575によって産生されるモノクローナル抗体が、このモノクローナル抗体が抗原抗体型(例えば、この抗体によって抗原と認められる化合物)の反応を行なうことができる条件下に、結合する蛋白質化合物を溶解物から回収する。
【0017】
回収された蛋白質化合物のアミノ酸配列は、質量分析のような、なんらかの標準技術によって得ることができる。
上記で示されたように、段階(i)の細胞は活性化細胞であり、それ故に、経膜受容体のときに、SC5の分離ができる。段階(i)で使用されたCS5のソースが休眠中のT細胞(生体外刺激を除く)からなっていれば、このようにして分離された蛋白質化合物はその脂肪質型SC5であろう。
段階(i)で選択される細胞が活性化された、または、悪性の細胞のときは、段階(ii)の溶解は、それ故に、細胞全体で行なわれるか、または、これらの細胞膜の小部分で行なわれる。
溶解段階自体はその休眠中の多数のポリペプチド複合体の分離技術によって行なわれ得る。例えば、細胞またはその膜小部分はTritonX-100のような強力な界面活性剤1%含有溶解バッファー中で、1時間、4℃で培養される。これによって、休眠中の細胞成分からの、SC5蛋白質の分離が行なわれる。もし、ジギトニンの1%またはブリッヂ58の1%のようなマイルドな界面活性剤が使用されれば、SC5は、それが自然に結合している、これらの化合物から分離されないであろう。上記段階(ii)でのマイルドな界面活性剤の使用は、だから、SC5の分離を可能にし、更に、SC5の形質導入、実施体の分離を可能にする。
故に、本発明は、前記プロセスで得られる化合物を包含する。
次いで、段階(ii)で得られた溶解物は、この抗体が、細胞溶解物含有(例えば、ハイブリドーマI−2575によって産生された2μgのモノクローナル抗体は、107個の溶解物に相当する)マイクロウエルに抗体をコートするような、抗原抗体型反応を行なうのに好適な条件下、ハイブリドーマI−2575によって産生されたモノクローナル抗体とともに培養され、次いで、モノクローナル抗体は細胞溶解物とともに2時間、4℃で培養される。
【0018】
ハイブリドーマI−2575によって産生されるモノクローナル抗体が結合する化合物の回収段階(段階iii)はこの分野のエキスパートにとって熟知の技術で、行ない得る。非還元条件下の回収は、本来の形態のSC5の分離が可能で、SC5の本来の生物学的特性の維持が望まれるときに行なうのが好ましい。
SC5蛋白質の非還元条件下での回収の好適な技術は、即ち、親和分離クロマトグラフィ(アフィニティークロマトグラフィー)を含む。好ましくは、濾過前に、濾過を容易にするために、溶解物中の化合物の構造を本質的、部分的に変えて、溶解物を生理学的pH、例えば、pH6〜8の間の、pH7のようなバッファーと混合して、溶解物をハイブリドーマI−2575によって産生されたモノクローナル抗体(mAb)が固定されているカラムにより濾過する(例えば、このmAbを寒天粒に結合させ、この粒を前記カラム内に堆積させる)。カラムを前記生理学的pHのバッファーで洗い、例えば、本来の化合物の構造を、本質的に変えることなく、mAbへの化合物の結合をバッファーで攪乱して、溶出させ、固定されたmAbに結合している化合物を回収する。この溶出バッファーは、通常、弱酸性のpH(通常、pH2〜5の、例えば、pH3)である。抗体分子の通常の方向性を有し、それにより、抗原を高い効率で結合させるマトリックスの調製のため、一段階の精製方法のため、膜蛋白質の分離のために、例えば、Schneider et al.1982(J.Biol Chem.257:10766〜10769)記載の内容が参考になる。このようにして回収された蛋白質化合物はその本来の構造のSC5蛋白質である。
【0019】
一方、段階(iii)では還元条件下、段階(ii)で得られた溶解物とハイブリドーマI−2575によって産生されたモノクローナル抗体とで免疫沈降反応を行なうことができる。このようにして得られた免疫沈降は二十日鼠のIgMと結合できる抗体からなるカラムを通して濾過される。ハイブリドーマI−2575によって産生されたモノクローナル抗体に結合する蛋白質化合物は前記と同じ溶出法で回収される。このようにして得られた蛋白質化合物も、また、本来の構造の分離SC5蛋白質である。休眠中の標準的なSC5蛋白質回収方法[段階(iii)]は、還元条件下の電気泳動法による処理である。これは、特に、四つのステップを包含する。
ハイブリドーマI−2575によって産生されたモノクローナル抗体と段階(ii)で得られた溶解物を、このmAbを抗原抗体型反応させることができる条件下に培養する。かくして、生成した免疫沈降物を採取し、免疫沈降物に含有する化合物を還元条件下に分離し、見掛け分子量96kDの蛋白質化合物を回収する。
一方、所望のSC5蛋白質を電気泳動ゲルまたは膜に局在させる方法(続いて、それをゲルまたは膜から分離する)に続いて、溶解に先立ち、細胞の表面ポリペプチド成分をラベルする(例えば、スルホ−NHS−LC−biotinによる、125ヨードまたは蛍光ラベリング)、ラベリングステップが行なわれる。そして、免疫沈降物からラベルを帯びたこれら化合物を回収する。もし、ラベリングステップが溶解の後に行なわれれば、SC5の細胞内型及び経膜受容体型の両者が分離されることに注目しなければならぬ(この場合、前記のように、非活性化T細胞がSC5源として使用され得る。)。変性条件が使用されるから、このようにして得られた蛋白質化合物は、本来の構造ではない。即ち、まだ、本来のアミノ酸配列ではあるが、本来の生物学的特性が失われている。しかし、本来の構造ではないSC5蛋白質が観察され(調べられ)、試験される生物学的試料が変性条件下(例えば、皮膚サンプルのパラフィン薄片)に処理されたときに、本来の構造ではないSC5蛋白質が観察される(調べられる)。本来の構造ではないSC5蛋白質は、それ故、モノクローナル抗体の産生に直接使用され、このような、モノクローナル抗体は変性サンプルのSC5検出に有用である。もし、本来の構造のSC5蛋白質が所望されれば、変性蛋白質の少なくとも10アミノ酸区分が配列され(例えば、質量分析で)、上記配列区分から理論的に導かれたオリゴヌクレオチッドが合成され、試験的に、SC5+細胞から得られるcDNAライブラリー(DNAコレクション)からのスクリーニングに使用される。このようにして選択されたcDNAが、産生及び、好ましくは、クローン培養によるcDNAコード化産物の排出が可能な条件下、産生され、クローン化される[例えば、ヴァキュロウイルスベクターヘのSC5コード化cDNAの挿入、昆虫細胞、例えば、Sf9細胞の形質転換により、または、ワクチンウイルス及びEBV細胞により、Rindis Bacher 1995、J.biol. Chem. 270(23):14220〜14228]。
【0020】
故に、本発明はその配列が以下のようにして得られる分離蛋白質を包含する。
全血液細胞及び末梢血リンパ球(PBL)からなる群から選択される細胞を採取する。採取した細胞をPHAの1μg/ミリリットルで刺激する。刺激された細胞をビオチンのようなポリペプチド特異性ラベルでラベリングする。ラベルされた細胞をTritonX-100の1%含有溶解バッファー中で、培養して溶解する。溶解物をハイブリドーマI−2575によって産生されたモノクローナル抗体で免疫沈降させる。免疫沈降物から、ラベリングされた蛋白質化合物を回収する。このようにして得られた蛋白質配列をコード化するcDNAを合成する。昆虫細胞にこのcDNAを、導入させる(transfection:transformationとinfectionの合成語、細胞へ外来DNA断片を導入すること)。導入された細胞、例えば、ハイブリドーマI−2575によって産生されたモノクローナル抗体で、産生された蛋白質を回収する。
または、以下のようにして得られる。
全血中細胞及び末梢血中リンパ球(PBL)からなる群から選択される細胞を採取する。採取した細胞をPHAの1μg/ミリリットルで刺激する。ラベリングされた細胞をTritonX-100の1%含有溶解バッファー中で、培養して溶解する。溶解物をハイブリドーマI−2575によって産生されたモノクローナル抗体で免疫沈降させる。免疫沈降物から、還元条件下、見掛け分子量96kDの蛋白質化合物を回収する。このようにして得られた蛋白質配列をコード化するcDNAを合成する。昆虫細胞にこのcDNAを導入させる。導入された細胞、例えば、ハイブリドーマI−2575によって産生されたモノクローナル抗体で、産生された蛋白質を回収する。
【0021】
本発明は前記プロセスによって得られる分離蛋白質化合物、同様に、その分離蛋白質化合物のアミノ酸配列、及び分離蛋白質の配列を包含する。
【0022】
本発明の蛋白質化合物または蛋白質が配置される固体サポートが本発明の範囲内に含まれる。そのような固体サポートは特にセファロース粒を含む。
【0023】
本発明は、また、その配列が本発明の分離蛋白質をコード化する分離DNA(またはcDNA)を包含する。これは、また、DNAを導入することにより作り変えられた細胞、本発明の分離蛋白質を排出する、作り変えられた細胞及び前記作り変えられた細胞の培養液から分離された分離蛋白質に関する。
【0024】
本発明は特に、以下の段階から得られるcDNAを包含する。
ハイブリドーマI−2575によって産生されたモノクローナル抗体が結合する細胞からなる細胞集団を採取し(例えば、それに対しては、このmAbが抗原と認められる)、または、その溶解物がこのモノクローナル抗体と免疫沈降反応するような細胞で、例えば、全血中細胞及びPBLからなる群から選択される細胞を採取する。
もし、SC5の単独の経膜型が所望されるときは、前記細胞集団が主として、非活性化細胞から構成されていれば(例えば、休眠中の非腫瘍性細胞)、SC5発現が細胞表面で増加するように、例えば、採取された細胞集団をCD3活性化体(例えば、PHAのようなポリクローナル活性化体の1μg/ミリリットルを、採取された細胞集団がPBLまたは全血液または固定化、抗CD3抗体またはPHAのようなポリクロナール活性化体及び抗CD3抗体の両者であり、採取された細胞集団が、CD3+T細胞またはCD45RO+細胞またはNK細胞またはマクロファージまたは顆粒球から、主として、構成されている細胞集団のようにより均一であるときは、)と培養することによって、CD3経路を刺激する。
もし、採取された細胞集団が主として、活性化細胞(CD3活性化非腫瘍細胞)から構成されていれば(例えば、休眠中の非腫瘍性細胞、またはCD4+CTCL細胞のような腫瘍細胞)、そのときは、CD3刺激は、SD5の存在が、これら活性化細胞において、既に、主として、経膜的になっているため不要である。
もし、SC5の細胞内型及び経膜型の両者が所望されるときは、非活性化細胞が好まれ、CD3活性化は省略される。
前記細胞集団からのトータルmRNA集団の抽出及び精製(mRNAの抽出及び精製に使用できる市販キットの例:polydT column)。
各相補的cDNAの合成(逆転写酵素で、例えば、Seed B及びArrufo A 1987、Proc.Natl.Acad.Sci.USA 84:3365〜3369、に参考文献が収録されている。)。
このクローンのcDNAの発現が可能になるように、好適なクローン培養条件で各cDNAのクローン化を行い、各クローンを培養する。
ハイブリドーマI−2575によって産生されたモノクローナル抗体が抗原抗体型反応を行なうのに好適な条件下に、ハイブリドーマI−2575によって産生されたモノクローナル抗体が結合する化合物を発現するクローンを選択し、及び/または溶解物がハイブリドーマI−2575によって産生されたモノクローナル抗体と免疫沈降反応を行なう化合物を発現するクローンが選択される。
このようにして選択されたクローンを選択的に増幅し、選択されたクローンから挿入されたcDNAを回収する。
【0025】
このようにして回収されたcDNAは本発明におけるSC5cDNAであり、標準的なDNA配列技術により配列される。
このクローンのcDNAの発現が、好適なクローン培養条件で、各クローン培養によって、可能になるように、各cDNAのクローン化が、専門家にとって実施可能な、通常のクローン化法によって行なわれる。クローン化法の1例は、宿主細胞のcDNAの発現のために好適なプロモーターの制御下に、トランスフェクション(遺伝子導入)ベクターに各cDNAを挿入し、宿主細胞にトランスフェクション(遺伝子導入)ベクターを導入し、cDNAを導入された宿主細胞を、それらの代謝に好適な条件で培養する。好適なトランスフェクションベクター及び宿主細胞はBaculoウイルスのようなウイルス及び昆虫細胞(Sf9細胞)のような宿主細胞またはベクターとして牛痘ウイルス、宿主細胞としてEVB細胞(Rindis Bacher reference supra 参照)。
このようなcDNAは細胞への導入及びこのようにして得られたクローンからのSC5の産生に使用される。これはまた、本来のSC5mRNAの同定及び分離の試験に使用される。
【0026】
故に、本発明は、また、本発明におけるcDNAに相補的の、前記mRNA集団から選択可能な分離mRNAを包含する(例えば、前記cDNAに下記の文献記載の厳密な条件下、ハイブリダイズするmRNAの選択によって、Freemann G.J.etal.1992 J.Immunol.149:3745)。前記細胞集団の全mRNA集団からの回収する試みとして、本発明における分離cDNAを使用して、得られる分離mRNAを包含する。このmRNAは抽出されるcDNAに相補的である。一方、通常の技術の一つが、選択されたクローンから回収されるcDNAを直接配列化し、この配列からmRNAの配列を導き出す。
【0027】
本発明は、また、本発明における分離されたmRNAをコード化するDNAを包含する。特に、ゲノムバンク(www.ucsc.edu.のような)の上記SC5cDNAにマッチするゲノムDNAの調査によって、得られるゲノムDNAを包含する。
【0028】
本発明は、また、本発明における、cDNA、mRNAまたはDNAが導入された工学的細胞を包含する。
【0029】
本発明は、また、SC5特異性試料及びプライマーを包含する。そのような試料及びプライマーは専門家にとって、使用可能な技術によって得られる(例えば、候補配列として、SC5配列の特異な部分を選択し、これらDNA/RNAバンクの候補配列をSC5以外の細胞受容体の配列と対比し、前記対比後、SC5配列が特異であると見なされる、及び/または、前記対比後、SC5配列の鎖長が特異であると見なされる候補配列を選択する)。
【0030】
本発明の範囲内の発明には、cDNA、mRNAまたはゲノムDNAを担持するいかなる固体支持体も含まれる。このような固体は特にDNAチップまたはDNA微小球を含有する。
【0031】
本発明は、また、本発明に記載される分離cDNA、mRNAまたはDNAによってコード化された分離蛋白質を含有する。
【0032】
更に、休眠中の正常な細胞との対比で、CTCL細胞に固有である部分である限り、本発明は、本発明の蛋白質のあらゆる分離部分を含む。特に、C.N.C.M.に寄託番号I−2575として寄託されているハイブリドーマによって産生されるモノクローナル抗体によって確認されている部分を包含する。また、前記部分が前記細胞によって発現され、ハイブリドーマI−2575によって産生されたmAbによって接触されたとき、この部分が、正常細胞のCD3誘発増殖の調節(阻害または刺激)を誘発でき、及び/または、CTCL細胞のような悪性T細胞の生体内増殖の調節を誘発でき、及び/または、正常T細胞または悪性T細胞のIL−2(インターロイキン−2)産生の調節を誘発できる、ような形態の発明の蛋白質化合物の分離された部分を包含する。特に、この部分が、経膜部分で、悪性T細胞によって発現され、休眠中の非腫瘍性細胞によって、細胞質小球内で、発現される分子の一部であるような形態の発明の蛋白質化合物の分離された部分を包含する。本発明は、特に、本発明におけるSC5蛋白質の酵素的開裂によって得られるポリペプチド断片のバンクに関与する。好ましい酵素は、Tyr、Phe、Trpレベルを開裂するセリンエンドペプチダーゼ(例えば、α−キモトリプシン)、グルタミン酸及びアスパラギン酸のC末端での開裂を行なう酵素のような、蛋白質分解酵素から構成される。例えば、Shesberadaran and Payne 1988、 Proc.Natl.Acad.USA 85:1〜5参照、この内容はここでも、参考として組込まれている。SC5ポリペプチド断片のバンクは、質量分析のような特別な分析に使用される。このような分析は、ポリペプチドバンクを特徴によって分類するのに使用され、SC5の特徴であり、SC5の検出に使用されるポリペプチドのプロフィールを示す。本発明はこのような用法及びプロフィールを包含する。
本発明は、特に、本発明のSC5蛋白質化合物の細胞外、経膜部及び内部細胞質部分を包含する。受容体分子の細胞外、経膜部及び内部細胞質部分の決定は好適な技術に従がって、専門家によって為され得る。これらは分子全体において、グリコシルサイト(細胞外部分)を含む領域、疎水性(経膜部分)領域及び親水性領域、(内部細胞質領域)の同定によって、同定され得る。そして、それらは開裂または合成によって分離され得る。
本発明の蛋白質化合物の細胞外部分を分離する一つの方法は、前記のように、蛋白質化合物全体を処理するもので、これに、分子全体から細胞外部分を開裂させるために酵素処理の段階を付加する。この開裂のための好適な酵素としては、前記のように、V8プロテアーゼまたはα−キモトリプシンが含まれる。
【0033】
このように、本発明に包含されるポリペプチド化合物は以下のようにして得られる。
前記した、例えば、PBLまたは全血液細胞のようなSC5細胞源を採取する。
もし、ポリペプチド化合物が細胞表面で自然に発現されるときの形態で、分離されることを望むときは、または、採取されたSC5細胞源が、主として休眠中の正常細胞を構成するときは、例えば、適当なCD3活性化体と共に培養して(例えば、SC5源が全血液細胞またはPBLであるとき、PHA1μg/ミリリットルによる培養)、CD3経路を刺激する。そして、例えば、ビオチン(全細胞表面のラベリング)のようなポリペプチド特異性ラベルで該細胞をラベリングする。
そして、マルチポリペプチド複合体を分離するために、SC5細胞源(例えば、PBLまたは全血液細胞)を、例えば、Triton X-100のような強力界面活性剤の1%からなる溶解バッファー中での細胞培養により、溶解する(それによって、自然に結合したトランスデューサーとエフェクターからSC5を分離回収する)。
該溶解物を酵素による消化してSC5を部分に解裂させる。例えば、V8プロテアーゼまたはアルファ−キモトリプシンからなる群から選択される酵素である蛋白質分解酵素で該溶解物を培養する。そして、該溶解物を、ハイブリドーマI−2575によって産生されたモノクローナル抗体と免疫沈降反応を行わせ、該免疫沈降物からラベルを帯びたポリペプチド化合物を回収する。
回収された化合物は、完全なSC5細胞外部分または、完全なSC5細胞外部分の一部であるSC5部分であるか、または、そのようなSC5細胞外部分を構成するものである。このようにして得られるポリペプチド化合物のセットがSC5細胞外部分バンクを構成する。そのようなバンク及びその医療的使途、例えば、T関連疾病(例えば、質量分析による)の診断のための使用は本発明の範囲内にある。
この分野の通常の専門家は、その溶解及び酵素的消化を一段階で達成できることを知っている。
細胞全体のラベリングは、細胞外部分、または、細胞外部分の一部、または、その部分を構成するSC5部分を同定するために、細胞溶解に先だって為される。
【0034】
本発明は、また、本発明の蛋白質の部分を構成する分離された化合物を包含する。
【0035】
本発明の抗体(mAb)は、この抗体が結合するSC5エピトープの同定が可能である。通常の強制された突然変異誘発のような技術が好適である(例えば、Chang H.C.et al. 1989、J.Exp.Med.169:2073〜2083、参照、この内容は本発明の明細書に参考として組み入れてある。)。
【0036】
本発明の蛋白質及び本発明によるその部分、及び、特にその細胞外部分は、それらに対するモノクローナル抗体の産生を直接できるようにする。そのようなモノクローナル抗体の例はハイブリドーマI−2575で産生される。本発明のモノクローナル抗体はSC5の検出、CTCLのようなTリンパ腫の診断に有効であり、且つ、正常CD4+及びCD8+T細胞;HIV感染T細胞、CTCLのようなTリンパ腫、の如き、ウイルス感染T細胞;のようなT細胞の好ましくなく、または不適当な増殖、及び/または活性、の調節体として有効である。
故に、本発明は以下の手段により得られるモノクローナル抗体を包含する。
(i)本発明の蛋白質または上記で既定したその部分に対して或動物に免疫性を与える。
(ii)この動物の脾臓細胞からハイブリドーマを産生し、これを培養してその培養上澄み液中にモノクローナル抗体を産生させる。
(iii)段階(i)の免疫源として使用される蛋白質または蛋白質部分に結合し得、且つ、下記のような特性からなる群から選択される少なくとも一種の特性を有する抗体の上澄み液中での存在を調べる(評価する)。
主として細胞質分画中で休眠中の非腫瘍性T細胞に結合し、且つ、主として細胞表面で悪性CD4+CTCL細胞に結合する。
T細胞のCD3活性化経路を調節する。
T細胞からのIL−2産生を調節する。
T細胞のCD3誘発増殖を調節する。
CD3誘発の、CD4+CTCL細胞のような悪性T細胞の試験管内増殖を調節する。
動物中、好ましくはヒト中の、CD4+CTCL細胞のような、悪性T細胞増殖を調節する。
ハイブリドーマI−2575によって産生されたモノクローナル抗体のような本発明のモノクローナル抗体との、本発明の蛋白質への結合または上記で規定したその部分ヘの結合についての競合、及び、特に、該蛋白質の細胞外部分との競合。
(iv)所望の抗体を産生するハイブリドーマの選択及びクローン化。
(v)上記クローンの上澄み液かの抗体の回収。
【0037】
ハイブリドーマの産生は通常の技術で行なわれる。これは、前記動物からの脾臓の除去、脾臓細胞の懸濁液の調製、脾臓細胞を骨髄腫細胞と共に溶融プロモーターの存在下で溶融する過程、溶融細胞を別の容器で、非溶融細胞を生育させない培養液で希釈、培養する過程からなる。本発明は前記免疫化動物から採取される腹水をも包含する。
段階(iii)でT細胞及び悪性T細胞は標準法でヒトから採取される。一方のクローンはCD4+CTCLHUT28細胞系(ATCC TIB−161)などが使用され得る。CD4+腫瘍の場合は、試料はCD4+腫瘍細胞を表示する体の部分から採取される(SSにあたるPBL、形質転換MFにあたる皮膚紅皮症、等)。前記諸特性は、この分野の通常の知識を使用して、専門家によって容易に評価され得る。段階(iii)で挙げられた諸特性の評価のための好適な実験条件の例は、以下の例で見出すことができる。この分野の通常の知識を有する者が、それらを特異な細胞または使用濃度に是正する。細胞増殖の調節は増殖の刺激及び阻害と異なる。CTCL細胞が使用されたとき、増殖阻害は治療用の本発明のmAbは、診断または治療用であるか、またはその両方であるところの全ての抗SC5mAbsである。例えば、蛋白質精製用担体、または、これらができる調節特性の行使用担体のような担体に、これらのmAbの一種または複数種を固定することが所望される用途がある。本出願は本発明のmAbが固定される担体を包含する。そのような担体には、特に、親和クロマトグラフのカラム用の好適なものが包含される(例えば、寒天粒)。
【0038】
本出願は、また、H鎖、L鎖、VH、VL、Fab、F(ab')2、CD1、CDR2、CDR3から選択される本発明のmAbの断片(抗SC5断片)を含む。本出願は、該断片を含むあらゆる化合物、すなわち、あらゆる抗体を含む。本出願は、特に、少なくとも、上記断片を含むヒト化抗体に関する。ヒト化抗体の産生技術は先行技術で広く述べられている(Farah et al.1998、Crit.Rev.Eucaryote Gene Exp.Vol.8 pp 321〜345、参照、この内容は参考として本明細書に組込まれている)。
それは、特に、化合物、及び特に、少なくとも、一つの、ここで規定された抗SC5断片及び、また、H鎖、L鎖、VH、VL、Fab、F(ab')2、CD1、CDR2、CDR3各断片からなる群から選択された少なくとも、一つの、抗CD4断片からなる、抗体及びヒト化(人体に適応させた)された抗体に関する(例えば、WO94/13804、多価、多特異性結合蛋白質及びその製造方法、発明者:Holliger et al.出願人:ケンブリッジ抗体技術有限会社及び医療調査協議会、参照;また、Merchant et al.ヒトの二特異性IgG、Nat.Biotechnol.1998 vol.16 pp 677〜681、参照)。
【0039】
先に示したように、本発明は通常の一般的発明構想で結びつく二つの分子、即ち、SC5及びp140を提供する。SC5は、新規分子で、既に、十分に述べた。p140分子は従来技術で、NK細胞のサブグループ及び健常者(非癌性)からのCD3+CD8+細胞によって発現される阻害受容体として述べられている。本発明は、p140が悪性T細胞、特に、CTCL細胞によって発現され、新規なp140対立遺伝子型(下記例3のクローン24;SEQ ID No.1)を提供し、新規蛋白質(SEQ ID No.2)をコード化する。この新規な対立遺伝子型のcDNA及びアミノ酸配列を以下に示す。
【0040】
SEQID No.1
【0041】

【0042】
SEQID No.2
【0043】
【0044】
SEQ ID No.3
前記のp140対立遺伝子型(クローン1.1)のDNA及びアミノ酸配列を下記に示す。
【0045】
【0046】
SEQ ID No.4
【0047】
【0048】
SEQ ID No.1はSEQ ID No.3と5つの部分で異なる。
得られたSEQ ID No.2は先行技術で開示されたp140(SEQ ID No.4)と四個のアミノ酸置換の種類が異なる(その位置を太いアンダー系で目印する)。即ち、20位のQの代わりにRが、92位のLの代わりにVが、102位のTの代わりにPが、401位のPの代わりにLが示されている(図11参照)。
この出願は、配列がSEQ ID No.1及び/またはSEQ ID No.3であるか、またはSEQ ID No.1及び/またはSEQ ID No.3から構成される化合物を包含する。また、SEQ ID No.1に相補的であるmRNA及びSEQ ID No.2をコード化するゲノムDNAを包含する。
【0049】
好適なp140蛋白質産生技術はこの分野の専門家が使用し得るものである。この技術は培養液中でp140蛋白質が産生されるように加工された細胞の産生を特に含む。そのような技術の一例は、以下の過程よりなる。SEQ ID No.1またはSEQ ID No.3をヴァキュロウイルスベクターに挿入し、このベクターで、Sf9細胞系のような昆虫細胞を移入し、培養液中の産生されたp140蛋白質を回収する(培養液を、例えば、セファデックスカラムのようなカラムで濾過、蛋白質化合物がその分子量の函数として、次いで、140kD溶出物の分離)。
生理学的条件下(生体内または生体内を偽装する試験管内の条件下)で、p140に結合可能な化合物については既に記載した。そして、好適な、この分野の専門家により、使用可能な技術により得ることができる。経膜受容体部分(例えば、前記細胞の二層リピッドの上層にあるSEQ ID No.2及び/またはSEQ ID No.4部分の認識)で、細胞により発現されるとき、p140に結合する化合物、ここで言う、前記生理学的条件下、p140分子を抗原(SEQ ID No.2及び/またはSEQ ID No.4)として認め得る化合物、によって、為される。
そのようなp140に結合する化合物は、特に、HLA−A11及びHLA−A3分子(天然HLA−A11リガンド)及び特に、Fc−HLA−A11のような溶融蛋白質及びFc−HLA−A3のような溶融蛋白質から構成される。
p140に結合する化合物は、また、p140抗血清から構成される(ウサギのような動物にp140に対する免疫性を与え、このようにして産生された抗血清を回収する。付加的な免疫精製段階は、抗血清中のp140の濃度を増加させるためのもので、例えば、p140カラムによる濾過による)。休眠中の、p140に結合する化合物は、例えば、AZ158、Q66、Q241のようなPende et al.1996 (J.Exp.Med.184:505〜518)に述べられている、p140SEQ ID No.4に対するモノクローナル抗体で構成される。そのような抗SEQ ID No.4mAbsとしては、また、新規なSEQ ID No.2が抗原として認められている。
mAb産生に使用可能な技術は、抗p140mAbs産生にも好適である。そのような技術の一例は以下の段階からなる。
(i)或動物にp140に対する免疫性を与える。
(ii)この動物の脾臓細胞からハイブリドーマを産生し、これらの培養液上澄み中にモノクローナル抗体を産生するために、これらを培養する。
(iii)段階(i)で免疫源として使用されるp140分子に結合でき、休眠中の良性T細胞には、主として、その細胞質小球に結合でき、悪性CD4+CTCL細胞には、主として、その表面に結合する抗体の、上澄み液中における存在を評価する(測定する)。
(iv)所望の抗体を産生するハイブリドーマを選択し、クローン化する。
(v)前記クローン上の上澄み液から抗体を回収する。
好ましいp140結合化合物は抗CD4実在物(例えば、二特異性mAb)を構成する。
p140結合化合物はCTCL細胞、特に、CD4+CTCLのような悪性T細胞の検出に有効である。かくして、これらはCD4+癌の検出に有効である。これらは、また、以下で述べるポリペプチド系ベクターとしても有効である。
【0050】
休眠中の観点によれば、本発明では、抗SC5mAbs及び本発明の抗SC5mAb断片からなる群から選択される少なくとも一つの要素からなるポリペプチド系ベクターが提供される。このようなポリペプチド系ベクターは活性化されたT細胞に到達するか、またはその付近の活性、理論的、分子に有効である(例えば、炎症性疾病またはウイルス感染T細胞または悪性CTCL細胞及び特に、悪性CD4+CTCL細胞などにおいて、観られる活性化されたT細胞)。
また、それは、SEQ ID No.2及び/またはSEQ ID No.4の細胞外部分に対するmAbのようなp140結合化合物からなるポリペプチド系ベクターに関する。このようなポリペプチド系ベクターは分子送達、または悪性T細胞、特に、CTCL細胞及び、特に、悪性CD4+CTCL細胞付近に有効である。
分子は、特に、放射性元素または毒素のような細胞死またはアポトーシスを誘発することができる分子からなるCTCL細胞のような悪性T細胞に送られて有効な働きをする。そのような分子は、また、カルボキシペプチダーゼのように、抗有糸分裂性のプロドラッグから活性な医薬へと形質転換させることが可能な酵素からなっている。そのような酵素を運ぶポリペプチド系ベクターは、その際、抗有糸分裂性のプロドラッグの組み合わせによって有効に使用される。本発明はそれ故、そのようなベクター及び抗有糸分裂性のプロドラッグからなる医薬品キットを包含する。
これら隣接する細胞から前記悪性T細胞への抗腫瘍効果を誘発する作用力によって、悪性T細胞の付近にある細胞の免疫機能を刺激することができる分子、特に、構成するCTCL細胞のような悪性T細胞の近傍に送られて有効に働く分子がある。そのような分子の例は、特に、γインターフェロンのようなサイトカイン及びIL−2(例えば、Xu et al.2000,Cancer Research 60:4475〜4484参照)のようなインターロイキンである。
本発明のポリペプチド系ベクターが、CD4+CTCL細胞のようなCD4+細胞を標的にしようとするときは、更に、抗CD4体を含むことが好ましい。
【0051】
本発明は、また、前記ポリペプチド系ベクター、特に、前記ポリペプチド系ベクター含有薬剤で構成される医薬品組成物を包含する。そのような薬剤は不適当または好ましくないT細胞の増殖及び/または機能的活性(好ましくないIL−2の産生を含む)の制御に有効である。それらは、特に、不適当な免疫活性、の予防、一時的緩和、軽減、または治療;及び、特に、CTCLのようなTリンパ腫、及び、特に、CD4+CTCLの妨害及び阻害に有効である。不適当なT細胞の増殖及び/または機能的活性の予防、一時的緩和、軽減、または治療を目的とする、薬剤製造のためのこのようなベクターの使用、及び、CTCLの増殖の妨害及び阻害は、本出願の一つの目的である。
【0052】
SC5に生理学型条件下結合する化合物はSC5の活性を刺激するためのSC5リガンド作用物質として使用可能であり、このSC5分子は細胞阻害受容体として関与する。そのような抗SC5化合物は、即ち、抗SC5抗血清、本発明の抗SC5mAb、本発明のFab、F(ab')2断片、本発明の人体に適応するmAbsからなる。CD4+細胞が特別に関与するときは、前記抗SC5化合物は、更に、幸便に、抗CD4体を含有する。
抗SC5化合物は、特に、SC5+細胞の不適当または好ましくないT細胞の増殖及び/または機能的活性の調節に有効である。これらは、T細胞、CD45RO+細胞、CD3+細胞、CD4+T細胞、CD8+T細胞またはウイルス感染T細胞(例えば、HIV感染CD4+T細胞)またはCTCL細胞及び特にCD4+CTCL細胞のような悪性CTCL細胞の増殖の調節に有効である。これらはT細胞からのIL−2の産生にも有効である。C.N.C.M.に寄託番号I−2575で寄託されたハイブリドーマによって産生されたmAbが使用されたとき(5種類のIgM)、この調節は阻害の方に傾く。故に、CTCLのようなTリンパ腫の阻害に特に好適である。この出願は活性成分として少なくとも一つの抗SC5化合物からなる薬剤を包含する。このような薬剤はTリンパ腫、CTCL、ウイルス感染(例えば、HIV)のようなT感染、自己免疫型の諸症(例えば、リュウマチ性関節炎、脊椎関節症、または、乾癬、湿疹、アトピー性皮膚炎のような皮膚免疫薬物療法性疾病)のような炎症性疾病の予防、一時的緩和、治療が可能である。このような薬剤は移植改善(移植拒絶反応の抑制)も意図され得る。
【0053】
本発明は、補体漸増剤またはADCC活性化剤または刺激剤としての抗SC5化合物の使用を包含する。
前記で規定したp140結合化合物は、CTCL及び特にCD4+CTCLのようなTリンパ腫の予防、一時的緩和、軽減、または治療において、補体漸増剤またはADCC刺激剤または活性化剤として使用され得る。故に、本発明は抗Tリンパ腫薬剤、抗CTCL薬剤、抗CD4+CTCL薬剤の製造におけるp140結合化合物の使用及びその薬剤自体をも包含する。
【0054】
本発明は開発段階でのCTCL(段階化)の評価方法を提供する。即ち、悪性CD4+CTCL細胞の患者の体の或一定区分内における比率(例えば、%)が測定できる。この方法によれば、本発明のSC5蛋白質、本発明のSC5部分(特に、細胞外部分)、及びp140分子からなる群から選択される一要素をその表面で発現するCD4+細胞の比率は、前記体内区分から採取された生物学的試料によって測定される。前記体内区分内の現実のCD4+CTCL細胞の比率は、上記で測定された比率と本質的に同等とみなすことができる。それは、通常、この測定値の概略±10%以内である。
【0055】
本発明はCTCLの診断方法をも提供する。そこでは、本発明のSC5蛋白質、本発明のSC5部分(特に、細胞外部分)、及びp140分子からなる群から選択される一要素をその表面で発現するT細胞の比率は、患者から採取された生物学的試料によって測定され、ヒトの非CTCL(健常者が好ましい)の平均百分率と比較され、前記測定百分率が前記平均百分率より有意に高いとき、CTCL陽性と決定される。
このCTCLの診断方法は、特に、CD4+であるCTCLに適用される。この場合、測定されたT細胞の百分率は、好ましくは、CD4+の百分率である。SC5を発現するCD4+T細胞の平均標準比率は、一般に、1〜15%であり、ヒトの場合で、通常、5〜10%である。SC5+CD4+T細胞の測定百分率が前記平均百分率より高いとき、CD4+CTCLが陽性との診断が結論される。
SC5蛋白質またはその部分が関与する限り、本発明の方法は一般に、前記体内区分内にあるCD4+活性化T細胞の比率が測定できる。これは移植が拒絶反応を誘発するかどうか、及び自己免疫型の諸症(例えば、脊椎間接症のようなリュウマチ性関節炎、乾癬、湿疹、アトピー性皮膚炎のような皮膚免疫薬物療法性疾病)のような炎症性疾病、の診断及び追跡調査の評価に有効である。前記生物学的試料は内部滑液状でもよい。これは、また、非CTCLTリンパ腫の診断及び監視にも有効である(例えば、CD4+ALLまたはCD4+T−LL)。
【0056】
本発明のCTCLの評価/診断方法は全ての種類のCD4+CTCL、特に、形質転換MF、SS、LP及びCD30+リンパ腫に著しく有効である。前記生物学的試料は、例えば、紅皮症(例えば,形質転換MFの疑いある)患者から採取した皮膚試料及び/またはPBLまたは総合血液試料(形質転換MFがより進行性のSSのような形に進展したかどうかを評価するために)であってもよい。
CD4+細胞表面で検出された要素がSC5化合物のときは、細胞学的観察(腫瘍状細胞学的調査)の(前駆的または将来的)段階は、90%以上の確度で、現実のCD4+CTCLであるとの決定が為されることが好ましい。p140がCD4+細胞表面で検出されたときは、追加の段階は不要であり、CD4+T細胞の表面でのp140の検出という単純な事実が、直ちに、CD4+CTCLの陽性診断になる。
【0057】
本発明は好適な要素、特に下記の各要素からなる群から選択される一要素によって実行され得る。即ち、本発明の抗SC5モノクローナル抗体、本発明のFab、F(ab')の2断片、本発明から導かれる人体適応mAb、本発明のSC5細胞外バンク(前記で規定した)、SC5cDNA、mRNA、本発明のゲノムDNA、本発明で規定されるp140結合化合物、p140DNA、SEQ IDNo.1またはSEQ IDNo.3またはコード化SEQ IDNo.2またはSEQ IDNo.4からなるDNA。
このような要素はSC5またはp140細胞表面の発現の検出手段として使用される。前記検出手段はSC5/p140+CD4+細胞を直接一段階で行なうための、抗CD4含有のものである。
cDNA/mRNA物質が検出手段として使用され、このとき、CD4+細胞は、前記細胞に現実に存在するSC5またはp140をハイブリッド化するmRNAを検出するために、溶解されねばならぬ。このようなcDNA/mRNA検出手段は、DNAチップまたはDNA微小球のような固体担体上またはその中に施されるのが好ましい。
mAb物質またはmAbから導かれる物質は検出手段として使用され、これによって、SC5またはp140の表面発現が検出される。好適な抗体抗原技術は好適なものである。その例は特に流動細胞分析である。
【0058】
本発明は、また、緩和、予防、軽減、特に、CTCL細胞、及び特に、CD4+CTCL細胞のような、特に、悪性T細胞の増殖のような、好ましくないか、または、不適当なT細胞の活動の治療に有効な化合物の同定方法を包含する。この方法は、前記規定の、SC5蛋白質、SCポリペプチド化合物、または、p140分子(SEQ IDNo.2及び/またはSEQ IDNo.4のような)に結合できる化合物の検出からなることを特徴とする。このような化合物は、前記のように、ポリペプチド系ベクターとして使用される。付加的にSC5化合物に結合するこれら化合物は、細胞表面にこれらSC5分子を集合することができ、SC5の阻害機能を刺激できる。この同定法を実行する容易な方法は、蛋白質AセファロースCL−4Bのような固体担体上に、SC5及び/またはp140分子を固定化処理する方法である。この方法を続行するための化合物の候補は、特に、SC5またはp140に対して、免疫性のある血清からなる。
【0059】
本発明の例を図により説明するが、下記の特別な具体例が、本発明を限定するものではない。この分野の通常の知識を有する者が、二者択一的、試験的に、具体例を実施し、このような、この分野の通常の知識から導かれた具体例は本発明の範囲内にある。通常の知識を有する者は、各種の標準的免疫技術、各種の標準的分子生物学的技術が彼にとって、有効なものであることを評価する(例えば、抗体:実験手引、Ed Harlow、David lane、Cold Spring Harbor Laboratory 1988;Maniatis 1982 Molecular cloning:a laboratory manual、Cold Spring Harbor、New York、 Cold Spring Harbor Laboratory;Current protocols in Immunology protocols on CD−ROM、John Wiley、West Sussex、England;Immunological techniques made easy、edited by Olivier Cochet et al.、John Wiley、West Sussex、England;and the publication specifically referred to in the present application)。これら公知文献の内容は参考としてこの明細書に組み入れている。ここに、完全な明細が記載されている、実験における一つの特別な型のCD4+CTCLは、この分野の専門家によって、休眠中の何らかのCD4+CTCL型に置き換えることができる。
【0060】
これら実施例では、下記の図が参考のために掲げられている。
図1A、1B及び1Cは健常者のPBLsのSC5分子の細胞膜発現を図解している。健常者のPBMCを抗SC5mAb(腹水1/200)で汚染させ、次いで、FITC接合アイソタイプ特殊ヤギ抗マウスIgM2次試薬及び、抗CD3(図1A)、抗CD56(図1C)及び抗CD45RO(図1B)のようなPE接合mAbsの一種で汚染する。二重汚染されたリンパ球通過規制の細胞の比率は下部右象限に示される(図1A:3.3;図1B:5.1;図1C:0.6)。この実験は研究10ドナーの代表である。
【0061】
図2はPBLsの活性化の間、SC5分子の細胞膜発現を図解する。PBMCは1μg/ミリリットルPHAで刺激され、SC5及びCD69の発現のカイネチックスはCD3+細胞と並列に研究された。
図3はPBLのSC5分子の細胞内所在部を図解する。抗SC5、抗CD35、mAb鎖及び抗BY55アイソタイプ適合mAbが、健常ドナーの透過性及び非透過性の通過規制リンパ球の汚染に使用された。陰を施されたヒストグラムは不適切なコントロールmAbに対する図示mAbsから得られたラベリングを表す。
図4ではSC5分子の生化学的分析結果を図解する。DS6T細胞クローンはビオチン及び1%Triton X-100で、表面をラベルされ、溶解物は、抗SC5mAbを使用して免疫沈降される。試料はSDS10%PAGEで還元条件下分析される。抗SC5mAbは二種類の濃度、1/200(レーン2)及び1/500(レーン3)で使用される。ネガティブコントロールサンプルはヤギ抗マウスIg単独(レーン1)及びアイソタイプ適合mAb(レーン4)で免疫沈降させられる。分子量マーカー(kD)左に示されている。
【0062】
図5A、5B、5C、5Dは、抗CD3誘発増殖及び正常Tリンパ球のサイトカイン分泌を、それらの細胞毒活性の影響なしに、抗SC5mAbが調節する様子を図解する。即ち、
図5A:PBLまたはT細胞クローンLSO及びGDS.3が固定化抗CD3mAb(コートされたmAb1μg/源)またはIL−2(50IU/ミリリットル)で、抗SC5mAb(腹水の、1:200最終希釈)またはアイソタイプ適合、不適切mAbの存在で、刺激される。結果は、少なくとも、3通りの実験の代表及び3通りの平均cpm・SDとして表している。
図5B:GDS.3CD4+T細胞クローンは抗SC5mAbの異なる濃度の存在下、図示された濃度のプレコート抗CD3mAbで刺激される。
図5C:抗SC5またはコントロールmAbの存在下、抗CD3刺激PBLの上澄み中のIL−2の分泌は、下記に挙げた物質の存在下、IL−2依存性T細胞クローンの増殖の測定によって評価される。上記物質は、(a)剤単独、(b)PBL/抗CD3+コントロールmAb、(c)PBL/抗CD3+抗SC5mAb の上澄み。データは3つの代表的実験からの3通りの平均値・SDを表す。
図5D:細胞毒CD8+細胞クローン(JF1)が、抗SC5mAb(腹水の、1:200最終希釈)またはアイソタイプ適合コントロールmAbの存在で、E/T比が5/1での、FcyR+ミューリン腫瘍細胞P815との共培養の前に、培養される。標的の細胞は、表示された抗CD3の最終濃度で前培養される。結果は3通りの源泉の平均値で示される。
【0063】
図6A、6BはCTCL患者のPBLのSC5分子の細胞膜発現を図解する。
8人のセザリー症候群患者、3人の菌状息肉症患者及び8人の健常ドナーの血球の細胞が、CD4+リンパ球によるSC5分子の発現のための2色フルオレスッセンスで分析される。
図6A:セザリー症候群患者のPBLのSC5及びCD4分子の共発現を健常者と比較している。
図6B:ヒストグラムは示された3群におけるSC5発現の平均値を示す。SS患者と休眠中の2群の間には統計的に有意に差があることが解る(Mann-Witny U test、p<0.001)。
図7A、7B、7Cはセザリー症候群患者の悪性細胞の膜表面発現を図解する。
図7A:循環TCRVβ22+悪性T細胞クローンの存在で、SS患者のPBMCの二色蛍光分析。細胞は抗SC5mAbで汚染させ、次いで、PE接合アイソトープ特殊ヤギ抗マウスIgM及びFITC接合抗TCRVβ22mAbによる。
図7B:弱くラベルした、抗SC5mAb(陰を付したヒストグラム)TCRV竈22陽性悪性細胞。
図7C:同じ患者の細胞を、PE接合抗CD69mAb及びFITC接合抗TCRVβ22mAbによって汚染される。
【0064】
図8は抗SC5mAbによるCTCL悪性細胞系の抗CD3誘発増殖の調節を図解する。Pno細胞系は、IL−7(10ng/ミリリットル)、PMA単独(2ng/ミリリットル)、固定化抗CD3mAbまたはPMA/抗CD3mAbの組み合わせによって刺激される。SC5mAbまたはアイソタイプ不適当mAb(腹水の、1:200最終希釈)が培養の開始時に添加される。結果は4種の実験の代表及び3通りの源泉の平均cpm・SDとして示される。
図9A、9Bは、CTCL細胞系(図9A)のp140/KIR分子及び新鮮なリンパ球腫瘍(図9B)の検出を図解する。ダブル免疫汚染流動細胞分析は、Bagot et al.1998(Blood 91:4331〜4341)及びPoszepczynska et al.2000(Blood 96:1056〜1063)の記載によって行なった。
図10はp140分子の生化学的分析を図解する。即ち、健常者ドナーから導かれたNKクローンAM61(p140+/p70−)及びPno細胞系及びCou-Lがビオチンで表面をラベルされ、抗p140及び抗CD8mAbsで免疫沈降される。試料はN−グリコシダーゼFで処理(+)または非処理(−)され、還元条件下、8%(左パネル)または11%(右パネル)SDS−PAGEで分析された。セファロース蛋白−A単独はネガティブコントロールである。分子量マーカー(kD)は右に示される。
図11は蛋白質でコードされたKIR3D cl.24(SEQ ID No.2)及びcl.1.1(SEQ ID No.4)のアミノ酸配列を示す。信号ペプチドに一致するアミノ酸は小文字で示し、一方、経膜通過領域は一致する配列にアンダー系を施した。一致配列と同じアミノ酸はドットで示した。
【実施例1】
【0065】
SC5の分離、SC5の生化学的機能的特徴、抗SC5mAbsの産生。
Tリンパ球免疫応答は抗原特異性クローンを拡大させるポジティブ信号及び抗原自体への過剰な刺激または応答を抑制するネガティブ信号を発する機能的な細胞表面分子によって制御される。休眠中のTリンパ球の活性化は二つの同一の信号を必要とする。T細胞受容体(TCR)の関与によって発される信号は、通常のT細胞で応答を発させるために、2番目のポジティブ信号を同伴しなければならぬ。CD28はTリンパ球活性化及びIL−2合成を誘発し、細胞死を阻害するメジャー共刺激性受容体と見なされる。最近の研究はCD3/TCR刺激が、TCR/CD3が集合し、且つ、DIG結合性の信号導入分子となる、界面活性剤溶解性グリコリピッド強化膜フラクション(DIG)またはラフトの再配布に導くことを明らかにしている。CD28が、TCR関与サイトヘのラフトの再配布を惹起することが発表されている。更に、一連の休眠中の分子は異なる時点で活動し、特別な部分集合に影響し、または明確な実施機能を振興するT細胞に同時刺激を与えることができる。Tリンパ球の応答のネガティブな規制はCD152誘発信号またはTNF上科受容体メンバーを通しての細胞死に影響され得る。最近、NK部分集合及びCD8+リンパ球についての新しいタイプのネガティブな規制が報告された(Lan998、Annu.Rev.Immunol.16:359〜393)。免疫グロブリン様(KIRs)またはC型レクチン構造はこれらの信号に応じて、CTL活性の強化またはサイトカイン産生のような好ましくない実施体機能を阻害する。
さて、我々は、健常者の末梢血球の小集団を詳細に描写する新しい96kD経膜受容体について述べる。抗SC5mAbは主として、CD8及びCD4小集団並びにNK系統の細胞由来のCD45RO+リンパ球を汚染する。我々は、SC5が、主として、休眠中のTリンパ球の細胞内小球に局在し、細胞活性化後に、その表面膜発現が急速に増加することを発見した。SC5分子の関与が、抗CD3mAbに誘発される、休眠中のTリンパ球またはT細胞クローンの増殖を阻害した。それはこれら細胞のサイトカイン誘発増殖になんの効果も与えない。同時に、CD8+T細胞クローンに左右されるエファクター細胞毒活性は抗SC5mAbによって変更されない。更に、我々は、SC5+CD4+循環リンパ球の比率が、セザリー症候群(SS)患者においては、健常者の血中リンパ球に比較して、著しく増加していることを発見した。これは大切なことだが、我々は、セザリー細胞がSC5分子を発現することを明確に示すことができる。更に、我々は、皮膚T細胞リンパ腫(CTCL)の細胞系におけるSC5分子のライゲーションが、抗CD3mAbにより誘発される悪性細胞増殖の強力な阻害をすることを発見した。このように、SC5分子の発現はSS患者の循環、悪性CD4+細胞の存在の検知に役立つことができる。更に、ライゲーションにネガティブ信号を送る細胞表面分子が一致することは病理学的T細胞の機構の解明の新手段となり、その阻害に使用される。
【0066】
原料及び方法
抗SC5mAの産生
SC5mAは出版されているプロトコル(David et al.1990;J.Immunol.144:1〜6)を使用して、NK細胞系統Ytindiで6wk old Balb/c miceを免疫化して得られる。ハイブリドーマ上澄み液は、免疫化された細胞系及びPnoと称するCTCL細胞系(Poszcepczynska et al.2000;Blood 96:1056〜1063)とともに、間接的免疫蛍光及び流動血球計で、選択的再活性のために濾過された。Pno細胞系が患者のCTCL細胞から由来し、CTCL細胞はCD3+Vβ22+CD4+CD8αα+CD25フェノタイプを有し、IL−70及び抗CD3mAbヘの応答として増殖する。CTCL細胞系はこのスクリーンニングに好適である。休眠中のCTCL細胞系の例は、HUT78細胞系(ATCC TIB−161)を含む。反応性上澄み液は、更に、末梢血中リンパ球(PBL)との再活性を試験される。ハイブリドーマ上澄み液は腫瘍性細胞系及びPBL小集団の両者との、その再活性について選別される。ハイブリドーマは二回クローン化され、クローン化ハイブリドーマはプリスタン投与Balb/c マウス(複数)腹水を産生させるために継代接種された。抗体アイソタイプはIgMと決定され、抗SC5mAbと命名された。腹水はPBSで透析され、限外濾過で滅菌され、更に、1/200で最終希釈された。このハイブリドーマは2000年10月30日、C.N.C.M.(Collection Nationale de Microorganismes パスツール研究所;25、rue du Docteur Roux、F-75724 Paris Cedex15、France)にブダペスト条約(C.N.C.M.寄託番号I-2575)に従がって寄託され、C.N.C.M.寄託番号はI−2575である。この実施例の抗SC5mAbはハイブリドーマI−2575によって産生されたmAbに起因する。
【0067】
患者
インフォームド・コンセントの後、倫理委員会(CCPPRB、Hopital、Henri、Mondor、Creteil、France)の承認を受け、我々は11人のCTCL患者から血液試料を得た。8人の患者は、セザリー症候群に罹っており、10〜80%の循環CD3+、CD4+、CD8−セザリー細胞を有し、3人の患者は形質転換菌状息肉症(MF)で、CD3+、CD4+、CD8−フェノタイプを有する伝播性皮膚腫瘍と診断されていた。患者全員がこれまで化学療法を受けていなかった。
【0068】
細胞及び細胞系統
末梢血中モノクロナール細胞(PBMC)はFicoll-Isopaque の技法(Pharmacia、fine、Chemicals、Piscataway、NJ)で、密度勾配遠心分離で、分離される。ヒトT細胞クローンGDS.3(CD3+TCRαβ+CD4+CD8−)、DS6(CD3+TCRαβ+CD4−CD8−)、LSO(CD3+TCR聿+CD4−CD8−)及びJF1(CD3+TCRαβ+CD4−CD8+)がBensussan等、1984(J.Exp.Med.159:947〜952)及びYssel et al.1984(J.Immunol.Methods 72:219)に記載されているが、これらを8日毎に、同種の放射線照射PBMCと共に、下記からなる培養液に供給した。
RPMI1640(GIBCO、Paisley、UK)、2mmol/l L−グルタミン、ペニシリン(100U/ミリリットル)、ストレプトマイシン(100mg/ミリリットル)、10%熱非活性化ヒト血清、50IU/ミリリットル組替えインターロイキン−2(γIL-2、Sanofi、Synthelabo、Labege、France)、及び1μg/ミリリットル植物性赤血球凝集素(PHA)(Wellcome、Beckenham、UK)。機能的研究は供給7日後に行なわれた。標準的なヒト白血病(患者の)細胞系はマイコプラズマ・フリーで、完全なRPMI剤に10%子牛血清(FCS)及び抗体を補強した培養液中で対数函数的な生育カーブを保持した。ヒトCTCL細胞クローンPnoは、セザリー患者の末梢血液から得て、これを前記のように培養、保持した(Poszcepczynska et al.2000;supra参照)。Pno細胞系はCD3+Vβ22+CD4+CD8αα+CD25−フェノタイプを有し、IL−7及び抗CD3mAb刺激に応答して増殖する。休眠中のCTCL細胞系はHUT78細胞である(ATCC TIB−161)。
【0069】
モノクローナル抗体及び流動細胞計数の研究
間接的免疫蛍光物汚染ハイブリドーマ上澄み液または腹水液とともに、カルタグ研究所(カリフォルニア州、サンフランシスコ)調製のFITC接合ヤギ抗マウスIgMを使用して行なわれた。2色分析には、免疫蛍光汚染が、抗SC5mAbと3×105個の細胞が、30分、4℃の培養によって行なわれた。次いで、細胞は洗浄され、FITC接合ヤギ抗マウスIgM(カルタグ研究所)とともに培養、次いで、IgGアイソタイプのPE−、ECD−、TRI−接合、特殊mAbで培養された。汚染細胞の分析はアルゴン単独流動細胞分析計(Epics XL、Bekman-Coulter、Miami、FL)を使用して、(Schiavon V et al.1999;Tissue Antigens 53:23〜32)に記載のようにして行なった。細胞内ラベリングには、細胞をPBS4%含有パラフォルムアルデヒド中で4℃、20分固定した。次いで洗浄、それから、0.1%サポニン含有汚染緩衝液とともに透過させた。接合抗CD3、抗CD4、抗CD8、抗CD45RO、抗CD69及び抗TCRVβ22mAbsはインミュノテク(Immunotech、Marseille、France)から購入した。
休眠中のmAbsはBeckman Coulterから入手可能である。即ち、下記のカタログ番号参照抗。
CD3(PE)は、細胞分析2000カタログのIM1282、精製抗CD3は細胞分析2000カタログのIM0178、抗CD4は細胞分析2000カタログのIM0449、抗CD8は細胞分析2000カタログのIM0452、抗CD45ROは細胞分析2000カタログのIM1307、抗CD69は細胞分析2000カタログのIM1943、抗CD94は細胞分析2000カタログのIM2276、抗CD158aは細胞分析2000カタログのIM2277、抗CD158bは細胞分析2000カタログのIM2278、抗NKG2aは細胞分析2000カタログのIM3291、抗BY55は細胞分析2000カタログのIM2745。
【0070】
ビオチンラベル表面分子の免疫沈降
2×107個の細胞が、PBS中で2回洗浄され、Sulfo-NHS-LCビオチン(Pier Europe、Interchim、Montlucon、France)(PBS中2mg/ミリリットル)で、20分、4℃で、表面をラベルされた。RPMI1640で20分間、急冷した後、細胞はPBSで2回洗浄、下記の組成の溶解緩衝液中に1時間、4℃で、再懸濁された。
溶解緩衝液組成:(20mM Tris-HCl pH 7.5、150 mM 塩化ナトリウム、1% TritonX-100、1mMナトリウムヴァナデイト、10mM NaF、1mM PMSF、1μg/ミリリットルアプロチニン及び1μg/ミリリットルロイペプチン)。それから、ポスト細胞核上澄みが、2時間、4℃で、96-wellプレート(Maxisorp Nunc immuno-plate)で、ヤギ抗マウスIgG+IgM(カルタグ研究所)で予備コートされ、次いで、抗SC5mAbまたは抗TCRβ(抗TCRβC305アイソタイプに適合するmAb、細胞分析2000カタログのIM1466参照;抗TCRVβ22、細胞分析2000カタログのIM2051参照;抗TCRゼータ、細胞分析2000カタログのIM3169参照;全てはBeckman Coulterから入手可能。)で予備コートされた。免疫沈降物は洗浄用緩衝液(20mM Tris-HCl pH 7.5、150 mM 塩化ナトリウム、0.1% TritonX-100、1mMナトリウムヴァナデイト、10mM NaF、1mM PMSF)で4回洗浄し、沈降蛋白質はSDS−PAGEを使用する。ウエスタンブロット分析はストレプトアビジン−パーオキシダーゼ(インミュノテク)とECL検出システムをメーカーの推薦(Amersham Pharmacia、Orsay、France)に従がって使用して行なった。
【0071】
増殖アッセイ
リンパ球活性化のために、PBMCを、1μg/ミリリットルPHA(Wellcome、Beckenham、UK)添加の、15%ヒト血清含有培養媒体で培養した。増殖アッセイには、5×104個の細胞は3通りの96源泉丸底培養皿(Greiner、Nuertingen、Germany)で、最終容量0.2ミリリットルの培養媒体で行なった。必要なら、培養皿を前記濃度(Poszcepczynska et al.2000;supra参照)の抗CD3mAbで予備コートする。
CTCL細胞系Pnoの刺激のため、予備コートされた抗CD3mAbが2ng/ミリリットルホルボール12β−ミリステート13α−アセテート(PMA、シグマバイオケミカルス)との組合わせで使用される。細胞は4日間培養し、培養の終期の8〜16時間の間に3H[TdR]の1μCiを適用した。3H[TdR]の適用量は液状シンチレーションカウンター(Topcount;Packard Instrument Co,Meriden、CT)で計測した。IL−2産生量の定量では、105PBLが固定化抗CD3mAbで、抗SC5またはコントロールmAbの存在下刺激される。培養の32時間後、IL−2依存T細胞クローンから100μリットルの上澄み液が105個の細胞に加えられた。
【0072】
CD3誘発・転送・細胞毒アッセイ
転送・細胞毒アッセイは、Le Cleach et al.2000、Clin.Exp.Immunol.119:225〜230に記載されたように実施した。標的の二十日鼠肥満腫瘍のP815腫瘍細胞に100μlの51Cr(25mCi/ミリリットル)を37℃、90分付加し、洗浄、次いで抗CD3mAbで、15分間室温で培養した。エフェクター細胞は抗SC5mAb(腹水の最終希釈1:200)で前培養し、次いで、標的細胞に5:1の比で、添加し、最終容積150μlで、96源泉V字型底マイクロ滴定皿で、4時間培養した。遠心分離後、100μl部分標本を51Crの放出量を定量するために、γカウンターで計測した。自発的放出は常に最高値の20%減であった(1%NP40で標的細胞)。特異的51Crの放出量比率はDavid et al.1987、J.Immunol.138:2831〜2836に記載のようにして計算した。
【0073】
統計的分析
この結果の統計的分析は、スタティスティカソフトウエア5.0(StatSoft、Los Angeles、CA)によって行なった。患者とコントロールグループのSC5発現の間の大きな相違は、Mann-Whitney Uテストで評価され、SC5+CD4+細胞の比率及び患者の末梢血中の悪性細胞の比率の間の関係は、Spearman Rank Order Correlations testによって評価された。
【0074】
成果
ここに提出する成果はCTCL細胞系Pno及び患者から採取されたCTCL細胞によって得られたものである。例えば、HUT78(ATCC TIB−161)のような、どのCTCL細胞でも同等の実験が行なわれ、同等の成果が得られた。
【0075】
正常で、休眠中且つ、活性化末梢血リンパ球内のSC5分子の発現。
機能的腫瘍細胞系Pno Ytindiに対して、高められたモノクローナル抗体は、CTCL細胞系Pno(Poszcepczynska et al.2000、supra参照)及び末梢血中のT細胞について、同時反応性が分析された。IgMアイソタイプのSC5特異性mAbは免疫化とCTCL細胞系の両者を汚染した。末梢血リンパ球中、抗SC5mAbは、小集団(図1A、1B、1C参照)の輪郭を描いた。抗SC5mAb反応性分子は、20%を超えない(平均10.5%、SD±5.6)CD3+T細胞の可変比率によって発現される。CD4+及びCD8+リンパ球の両者は、抗SC5mAbで汚染される。個人間の差異(平均19.4%、SD±11.4;表1参照)がより重要ではあるが、末梢血中CD56+NK細胞の小集団も、また、抗SC5mAbで反応する。
【0076】
末梢血リンパ球小集団におけるSC5の発現。
【0077】
【表1】
【0078】
正常ドナー(提供者)のPBLのゲイトリンパ球における、SC5の、CD3またはCD56との、共発現の代表例が図1、1Aに示される。大部分のSC5が図1、1Bに示されるように、活性化/記憶−プールに属することに注意すべきである。SC5陽性細胞の70%以上が共発現CD45ROである。
試験された全IL−2依存性Tリンパ球クローンでSC5分子の発現が検出されたので、我々はTリンパ球活性化の間に、表面発現が誘発されたか、どうかを、立証した。我々は、PBLのPHAによる刺激がCD3+リンパ球におけるSC5の発現を増加させたことを見出した。SC%+T細胞の比率及びSC5発現のレベル、の両者が、刺激の後に急速に増加する(図2参照)。PHAの刺激後4時間で、SC5+CD3+細胞の比率が2倍から3倍に増加し、24〜48時間後に最高値に達した。5〜7日の試験管内の刺激の過程で、SC5発現は基本レベルに戻った。これと並行して、我々はCD69抗原の活性化の最初の発現を研究し、CD3+リンパ球におけるCD69及びSC5の類似した速動性的プロフィールを観察した。CD69分子と異なり、低レベルSC5が既に、刺激前に循環PBLに存在することに注目すべきである(図2)。
活性化誘導抗原発現はデ・ノボ蛋白質合成または細胞内先在分子の細胞表面排出の何れかの帰結をとる。それ故に、我々は、非活性化透過性PBLにおけるSC5発現を研究した。図3に示すように、抗SC5mAbは、非透過性細胞の16%に対して、透過性リンパ球の90%以上を汚染する。抗SC5mAb汚染の特異性が、循環リンパ球の小集団によって発現される、GPI固定細胞表面構造を検出する、アイソタイプ適合mAbBY55を使用して、確認される(Agrawal et al.1999、J.Immunol.162:1223〜1226参照)。BY55+細胞の比率は、透過性及び非透過性リンパ球において見出される。細胞内抗原決定基の検出のための陽性コントロールとして、我々は透過性Tリンパ球のみをラベルする抗CD3ζ鎖mAbを使用した。このようにして、SC5の表面発現は、細胞の活性化状態によってダイナミックに規制され、且つ、T細胞活性化の間の、その誘発は細胞内プールから表面膜への分子の輸送を振興することになる。
【0079】
SC5抗原の生化学的特徴
抗SC5mAb、IL−2依存性TCRγδ+クローン、DS6と呼ばれるものによって同定される構造の分子量を特徴付けるために、表面がビオチンによってラベルされ、細胞溶解物が抗SC5mAbまたはアイソタイプ適合抗TCRβmAb(陰性コントロールとして)の何れかで、免疫沈降される。抗SC5mAb還元条件下、見掛け分子量96kDで単一分子を確認する。腫瘍細胞系(免疫化Ytindi細胞)が、たびたび、炭水化物抗原決定基(20、22)の異常型発現によって、特徴付けられるので、我々は、ニュウラミニダーゼまたは過沃素酸ナトリウムを、公知のシアル化抗原決定基を分解するような濃度で、処理した後の、抗SC5mAbの、SC5+NK細胞系NK3.3及びCTCL細胞系HUT78(ATCC TIB−161)との反応性を研究した。SC5の発現は、ニュウラミニダーゼまたは過沃素酸ナトリウムの処理によっては影響されない。このようにして、我々は、抗原決定基の炭水化物の可能性を除外した。
【0080】
抗CD3mA誘発Tリンパ球の抗SC5mAbによる増殖の阻害。
正常細胞のSC5分子の可能な機能を調べるために、我々は、抗SC5mAb単独の効果を、または、固定化、抗CD3mAbで誘発された、増殖応答の期間にこれを研究した。我々は抗SC5mAb単独またはPMAとの組み合わせが、末梢血中Tリンパ球の増殖を誘発しないことを見出した。興味あることに、溶解性の抗SC5mAbが固定化抗CD3mAbと共存しているときは、リンパ球の増殖の著しい阻害が観察された。阻害は、ドナーによっては、アイソタイプ適合の不適切なコントロールmAbの効果に比較して、33〜66%の間で変化した(図5A上部参照)。SC5分子が単球で発現されるので、阻害効果が単球依存性であるかどうかを決めるのに重要であった。我々は、二つのT細胞クローン、即ち、抗SC5mAbまたはアイソタイプコントロールmAbの存在で、抗CD3mAbによって、刺激される、GDS3及びLSOの増殖を研究した。二つのT細胞クローンから得られた結果は、SC5分子の結合は、抗CD3mAbヘの増殖を阻害する結果となることを示した(図5A下部参照)。抗SC5mAbのT細胞増殖の阻害効果はその濃度及び拮抗的、抗CD3mAbの濃度に依存したことに注目すべきである。抗SC5mAbで得られた阻害は、T細胞が普通に希釈された抗CD3mAbで刺激されたときにのみ、著しいものだった(図5B参照)。このような阻害挙動は抗KIRs mAbsについても報告されている(Cambiaggi et al.1999、Blood 94:2396〜2402)。
【0081】
SC5の結合に続くT細胞増殖の低下は、細胞死、IL−2R発現の摂動またはサイトカイン合成の阻害に起因し得る。我々は、抗SC5mAbがPBLまたはT細胞クローンのIL−2依存性増殖の阻害をしないことを見出した(図5A)。更に、抗SC5mAb(希釈腹水1/200)と抗CD3mAbとの組み合わせによって48時間前培養した、T細胞へのγIL−2の添加はこれらの増殖を復興させる。特異性のmAbがIL−2R発現の阻害をしたり、細胞死の原因になったりすることはあり得ない。終わりに、SC5がサイトカイン産生の阻害の原因になることを解決するために、我々は、IL−2依存性T細胞系を使用して、抗SC5mAbの存在下または不存在下で、CD3活性化PBLにより排出されるIL−2の量を比較した。抗SC5mAbの存在下で産生されたIL−2の品質は、アイソタイプ適合抗体で得られるコントロール値の40〜65%で表される(図5C参照)。このようにして、SC5及びCD3分子の同時結合はサイトカイン産生を減少させる。
次に我々は、FcgR+二十日鼠細胞系P815に対するキラーアッセイに書き換えられた、抗CD3mAbを使用して、T細胞エフェクター細胞毒活性への抗SC5mAbの効果について研究した。図5DはE/T比5/1で、細胞毒のT細胞クローンによって行なった代表的実験を示す。大量のSC5分子を発現する、CD8+T細胞クローンJF1は抗CD3mAb刺激に続いて、二十日鼠標的細胞を殺すために誘発された。その特異的mAbによる、SC5の同時標的化は、E/T比または抗CD3mAbを刺激する濃度に関係なく、JF1細胞毒活性に影響しない。
【0082】
セザリー症候群患者から採取した悪性リンパ球におけるSC5分子の発現。
我々は、最初に、CTCL細胞系との反応性について、抗SC5mAbのスクリーニングを行なったので、更に、8人のセザリー症候群患者から採取した、末梢血中リンパ球のSC5分子の発現を研究した。二色細胞流量計分析が、SC5発現がCD4+小集団で非常に増加したことを示した(図6Aの一患者から得られた代表的結果参照)。平均で、SS患者の血液中のCD4+リンパ球の35.5%SC5を発現し、健常なコントロール(p>0.001)の9.4%に比較される(図6B)。8人のSS患者についての研究で、CD4+細胞のSC5発現が細胞生態学(r=0.91、p=0.0012)によって検出される循環、悪性細胞の比率と関連があることが解った。これらの患者の、トータルCD4+細胞の比率(平均88%、SD±8.8)はセザリー患者の細胞の比率(平均26%、SD±12.8)より高いことに注目すべきである。
我々は、3人の、血液に関連しない、MF患者の末梢血液試料における、SC5分子の発現をも研究した。末梢血中CD4+細胞におけるSC5発現のレベルでは、MF患者と健常ドナーの間に大きな差異はなかった(図6B参照)。増加するSC5+CD4+の比率は、我々が試験できた、これら二人の患者の皮膚試料から検出された(それぞれ、26%、19%)。
一人のSS患者では、我々は、予め、循環悪性、細胞が、CD4+TCRVβ22+フェノタイプ(Poszcepczynska et al.2000 supra参照)のクロナールリンパ球であることを示した。ここで我々は、SC5が、例えば、悪性細胞集団によって、TCRVβ22+細胞で共発現されることを示した(図7A)。TCRVβ22+細胞に結合するSC5mAbのプロフィールが、全ての、悪性、クローン集団における弱い均一な発現に該当することは注目すべきである(図7)。興味あるのは、悪性TCRVβ22+細胞は前記のように(Poszcepczynska et al.2000 supra参照)、共通の活性化マーカーとしてのCD69(図7C)、CD25またはCD30を共発現しないことである。このように、増加した、SSPBLにおけるSC5のレベルは殆ど、循環、悪性、T細胞における抗原の発現に起因する。
【0083】
抗SC5mAbによる抗CD3mAb誘発CTCL細胞系の増殖の阻害
次に、我々はSC5分子が悪性CTCL細胞におけるネガティブ信号を提供できるか否かを調べた。我々は、IL−7の存在下、著しく増殖する長期間培養Pno CTCL腫瘍T細胞系を使用した。先に報告したように、固定化、抗CD3mAbまたは溶解性抗CD3mAbs及びPMAの組み合わせはこれらの増殖を著しく誘発するから、(Poszcepczynska et al.2000 supra参照)、Pno細胞系は機能的T細胞受容体を発現する。我々はPno細胞の増殖でのSC5分子の結合効果を調べた。図8は、抗SC5mAbが、CTCL細胞系の抗CD3mAb誘発増殖を強力に阻害することを示す。対照的に、PMA単独(2ng/ミリリットル)では、抗SC5によっては、それほど影響されない低レベルの増殖を為す。抗SC5mAbの存在下、IL−7で培養したPno細胞はアイソタイプコントロールmAbの存在下と同じ強さで、増殖したので、抗SC5mAbはIL−7依存性増殖に何ら影響を与えない。
かくして、我々は、SC5抗原がCTCL悪性細胞で、機能的であり、且つ、特異的、予見的に、TCRCD3信号経路によって惹起されるこれらの増殖を阻害するとの結論を出した。
次いで、CTCL腫瘍が、機能的、活性なNK細胞及びTリンパ球の両者を、本質的に有しない(欠落または欠損)遺伝子導入マウス(SCIDマウス)に接種された[(Charley
et al.(1990)、CB-17 SCID、 ヒト皮膚T細胞リンパ腫のマウスへの移植、J.Invest.Dermatol.94:381〜384に記載のように]、(またUS5,530,179
Cornelius P.Terhorst and Baoping Wang for obtention of SCID mice参照)。抗SC5mAbについては、マウスの半数に、静脈または皮下に、複数回、マウス体重1gあたり、複数種の濃度の精製mAbが投与された。PBL及び皮膚細胞が、浸潤性,大T細胞の標準細胞観察基準に則って採取され、CTCL細胞の比率が、各供試マウスについて計算された。この方法は専門家が、C.N.C.M.に寄託番号I−2575として寄託されているハイブリドーマによって産生されるmAbのような、抗SC5化合物が、ヒトCTCLの生体内増殖を阻害できること、をチェックできる方法の一例である。
【0084】
討論
多くのダイナミックな表面発現を有する受容体は他の免疫細胞またはサイトカインとの相互作用に影響を与え、その結果としてT細胞免疫応答を制御する。この研究で、我々は、その表面発現が試験管内T細胞活性化の間に急速に誘発されるところの、PBLの矮小集団により発現される、新規な96kD機能性分子を同定した。SC5は類似の分子量及び/またはT細胞制御機能を有する移植分子とは明瞭に異なる。第一に、SC5は、4時間後に検知される、その表面発現の重要な増加によって、非常に早期にT細胞活性化上で誘発される。同様の速動性が、刺激の24時間後に細胞表面で検知される、非常に早期の活性化抗原CD69について観察される。4II−B、OX−40 LIGHTまたはCD−152(CTLA−4)のような、大部分が移植されたT細胞特異的、誘発可能な、受容体が、刺激に続いて、10〜24時間の間に、T細胞表面に現われる。更に、SC5発現はT細胞で限定されていない。抗原はNK小集団及びBリンパ球、単球及び顆粒球上で検知される。このような広い分布形態は、NK細胞、T及びBリンパ球、脊髄性及び他の抗原含有細胞などの上に見出される、最近発表されたAIRM1及びIRp60のような、ある種のキラー細胞Ig様受容体(KIRs)の典型である。免疫グロブリンファミリーは、転写(ILT)阻害受容体のように、全白血球タイプの発現を有し、個々の受容体は異なるNK、T、B及び脊髄性小集団上で検出される。その上、ILT4はSC5に極く近い分子量を有する。しかし、AIRM1及びIRp60、ILTRsはともに細胞の活性化状態に依存しない。
【0085】
我々が細胞透過の後に、大部分の非活性化Tリンパ球で、SC5を迅速に検出したので、SC5分子の活性化T細胞表面での誘発に、次いで、細胞内分子の細胞膜及び副ヴァーサ(versa)への通行に起因する制御緩和が続いた。そのような制御の機構は他のT細胞受容体、CD152、について報告されている。更に、膜蛋白(LAMP)に結合するリゾソームの不均一ファミリーが存在し、包含する分子はエンドサイトーシス、リゾソーム通行及び細胞内顆粒からの排出に関与し、単球、ニュウトロフィルNK、BまたはT細胞の表面で活性化へと、誘発される。細胞内の局在性及びSC5の移出の付加的研究結果は、T細胞活性化及び機能におけるその役割について正確である。抗CD3mAb誘発T細胞増殖を阻害するその特異的mAbによるSC5の惹起を明らかにした。この阻害効果は、PBLにおける最大SC5表面発現を誘発するのに必要な、通常の抗CD3mAb刺激を必要とする(図5B参照)。A:抗SC5腹水の1:200の最終希釈物は、抗CD3mAbの2μg/wellで誘発される増殖を著しく阻害し、この阻害効果は抗CD3mAbの低濃度(1μg/well)で増加する。即ち、0.5μg/well−抗SC5腹水の1:400の最終希釈物は抗CD3mAbの2μg/wellで誘発される増殖を十分には阻害しないが、抗CD3mAbの低濃度(1μg/well;0.5μg/well)では阻害は著しい。この分野の専門家は、ここで述べた特別な結果または彼自身の試みから得られる標準的書き加えによる修正を行なうことができる。
【0086】
同時に、SC5結合が提起した阻害閾値は過度の抗CD3mAb刺激により、迂回される。SC5分子の連結で得られた阻害機能的効果はAPC非依存性機構で観察されることに注目すべきである。更なる実験は、SC5刺激がAPCT細胞または同種被刺激T細胞の増殖に異なる効果を与えるか否かを決定するのに必要である。近年、免疫応答の陰性制御の研究が、特に、Tリンパ球及びNK細胞について、徹底的に行なわれた。最もよく研究されたT細胞特異的阻害受容体、即ち、CD152(CTLA−4)は、CD−28共刺激受容体に機能的に結合する、典型的な活性化誘発抗原である。実に、CD152は、低い抗原濃度で、CD−28に打ち勝ち、免疫応答を阻害するである。CD−28で誘発されるT細胞増殖をIL−2R発現及びIL−2合成に影響することによって、制御緩和及び終結させる。CD152と同様に、SC5は表面発現が細胞の活性化状態によって制御される細胞内分子である。両受容体とも、IL−2合成に影響することによって、それらの効果を左右する。SC5特異的リガンド及び信号経路の同定は、これらの陰性T細胞制御体間の時間的、空間的、関係を要約するだろう。この広い細胞分布を考慮すると、SC5機能効果は、多分、T細胞を制限しないだろう。
数種の観点で、SC5はNK細胞阻害受容体に、T細胞及び他のリンパ球小集団によって発現される点が同じことから、比肩し得る。KIRs及びCD94/NKG2受容体の両者は、抗原特異性及び、細胞毒、増殖及びサイトカイン排出を含むTCRαβ及びTCRγδT細胞のスーパー抗原駆動活性化を阻害することが示されている。T細胞の活性化状態及びT細胞刺激の効力に依存するKIR誘発阻害は、特異的生体内条件下で、迂回され得る。しかし、我々が、抗CD3mAb刺激T細胞の増殖を選択的に阻害したことを、これらのエフェクター細胞毒機能なしで、示したので、SC5は、KIRsとは明らかに別のものであることになる。
【0087】
SC5分子の重要な特徴は、SS患者の末梢血液リンパ球における発現が、健常者のそれに比較して非常に増加することである。SSは成熟したフェノタイプの、初期類表皮悪性T細胞クローンという概念で特徴付けられる。事実、移植されたT細胞機能性受容体は末梢血液SS腫瘍細胞とは同一でないとの結論が出ている。MF/SSは、特殊フェノタイプT細胞小集団、通常のTメモリー小集団に相当する、CD+CD7−が増大したものと考えられている。しかし、近年の研究は、CD+CD7−集団が、SS患者の優勢なT細胞クローンを常には、代表するものではないことを示した(Dummer et al.1999、Arch.Dermatol.Res.291:307〜311)。SSはその刺激性は知られていないが、特異的活性化T細胞の増殖であるとみなされる(Fargnoli et al 1997、Leukemia 11:1338〜1346)。この理由から、SS及び通常のT細胞での、活性化誘発抗原の明瞭な発現が特に興味ある点である。CD69、CD25、CD30のような移植された活性化抗原は、我々自身の研究で確認したように、セザリー細胞に常に検出されるものではない。CD25の発現はむしろ、病気の進行と各種少数ケースの大細胞へのMF/SS細胞の形質転換に関係する。HLA−DR、CD25またはSS中のCD71のようなT細胞活性化分子の発現の増加率は反応性腫瘍浸潤T細胞に優先的に関与する。78kDの活性化が非常に遅い抗原BE2は、正常T細胞では検出されないのに対して、特に、SS末梢血液で増加すると報告されている。事実、BE2は、報告された悪性細胞の比率に関して、末梢血中細胞の低比率のものは、BE2陽性であったのに比較して、研究された全てのSS患者において、常に検出されるわけではない。そして、BE2は、血液関与の臨床的証明なしに、患者において検出される。それと対照的に、SCは研究された8/8SSケースで検出された。悪性細胞の比率は細胞生態学で決められ、患者によってかなり差が出るが(10〜80%)、CD4+細胞のSC5発現はこの比率に関与する。終わりに、CD4+細胞のアップしないSC5発現は、血液の関与なしに、MF患者で検出される。更に、我々はセザリー細胞はクローンタイプTCRVβ鎖共発現SC5分子の発現によって同定されることを示し、SS患者の末梢血液におけるSC5の増加レベルは、悪性T細胞クローンにおけるその発現に原因があることを確認した。
【0088】
非常に重要な観察は、セザリー細胞のSC5トリガーがCD3に影響される腫瘍細胞の増殖の制御緩和を誘発することであり、これは正常T細胞において観察される阻害に類似する。悪性CTCLリンパ球が、サイトカインの異常型産生ではあるが、基礎的な信号経路によって活性化されるような、ある機能的性質を保護することは既に示された。我々は阻害経路がCTCL腫瘍細胞中で保護され、これらの増殖を平衡状態に保持することを示した。特に、阻害効果は抗SC5mAbが抗CD3mAbに結合した後に観察され、CTCLPno細胞で刺激されたCTCLPno細胞は正常T細胞におけるよりも重要である。それがrIL誘発増殖を制御しないので、抗SC5mAbは正常細胞またはセザリーT細胞の何れの細胞死にも、直接関与するようには見えない。更に、抗SC5mAbの存在下で培養されたT細胞はrILの添加によって再刺激され得る。同時に、我々は、少なくとも部分的にそれらの増殖阻害の原因になる、抗SC5mAb処理T細胞によるIL−2産生の減少を観察した。
【0089】
悪性細胞増殖の機能的受容体の阻害効果は、既に観察された。近年、同族NK細胞唾液粘性受容体AIRMI及びCD33骨髄特異性抗原受容体によって影響される正常細胞及び白血病骨髄細胞の増殖阻害が報告された(Vitale et al.1999、Proc.Natl.Acad.Sci.USA 96:15091〜15096)。SC5に類似せず、これらの受容体は、生育因子の存在で、最も可能性ある細胞死の誘発によって阻害される。興味あることは、他のT細胞活性化抗原、正常CD45RO+T細胞の小集団によって発現されるCD30は、MF/SSより優勢なCTCLの退行を左右する。
この効果はCD30(CD30L)のナチュラルリガンドに関与する。これは悪性細胞の細胞質内に共存し、細胞死をFas/FasLに無関係に誘発する(Mori et al.1999、Blood 94:3077〜3083)。この観点で、SC5リガンドの同定はその阻害行動のメカニズムを解明するであろう。
【0090】
結論として、我々は早期のフェノタイプ的生化学及び機能的基準が移植されたTリンパ球受容体と異なる活性化抗原を同定した。アンダーラインしたように、SC5分子はポスト−活性化/記憶末梢血液T細胞小集団によって発現される。同様に、KIRsの継続的発現が腫瘍T細胞クローン上に発現され、この発現は特異的抗原によるTCRの占有に依存するように見える。近年、そのようなクローンは自己反応性で、且つ、KIRs信号によって、耐性が保存されることが報告されている。我々は、セザリー症候群腫瘍細胞はそのようなクローンが起源となる可能性があることを示唆し、SC5発現も自己反応性T細胞上で保護されるものと推定した。CTCL悪性T細胞上でのSC5発現が、それらの起源、及び通常のT細胞が制御不能な増殖をするように駆り立てる信号の性質をよりよく規定するのに役立つことが重要である。更に、腫瘍T細胞上の抗SC5mAbの阻害効果は、進行性CTCL患者に対する新規な治療法を構成する。
更に、SC5発現も、CD8+形質転換MFのようなCD8+CTCLの表面で観察される。
【実施例2】
【0091】
P140はCD4+CTCL細胞によって発現されること及び新規P140対立遺伝子型の同定。
皮膚T細胞リンパ種(CTCL)は主として、皮膚に関与するリンパ腫の外生群である。菌状息肉症(MF)は本来のT細胞ヘルパー細胞に類似する、フェノタイプの、クローン由来の悪性CD4+Tリンパ球の皮膚侵襲が特徴である。よりアグレッシブなCTCL型は悪性細胞が類皮的になり、皮膚異常に関与する。セザリー症候群(SS)は、CD4+/CD45RO+T細胞のクローン拡大及びこれら悪性細胞の血液中への出現が特徴であるところの、CTCLのより進行性のタイプである。特異的細胞表面のマーカーがないために、悪性細胞を同定するのが困難なので、病気の生物学はよく理解されない状態が続いている。皮膚の病変では、CD4+CTCL細胞を反応性、浸潤性CD4+Tリンパ球から区別することは困難である。バゴット(Bagot)等は、以前に、CTCL病変由来のユニークなCD4+T細胞系について述べている(Bagot et al.1998、Blood 91:4331〜4341)。この細胞系及び生体内腫瘍細胞は、TCR−Vβ転写の相補的に決定する領域3と同一サイズで発現した。極く最近、Poszepczynska等は(Blood 2000、96:1056〜1063)他の皮膚紅皮症CTCL患者の血液から採取された、IL−7依存性CD4+CD8αα+腫瘍T細胞系を機能的に特徴付けた。TCR−Vβ22の発現及びTCRβ−VDJ配列の同一性によって示されるので、この腫瘍T細胞系は大循環性T細胞集団と同一だった。ここで、我々はこれら二つの異なるCTCL系が、HLA−A対立遺伝子のためのP140/KIR阻害受容体を発現することを示す。重要なのは、同じ患者由来の新しい分離腫瘍細胞で検出されることである。更にその上、P140/KIRは他の7人のSS患者のCD4+リンパ球のメジャーな集団及び進行した2人の追加MF患者の皮膚CD4+腫瘍の両者によって共発現される。P140/KIRは健常者のマイナーNK細胞小集団及び稀な末梢血中CD3+CD8+細胞で検出される。その上、皮膚炎及び毒物による表皮融解のような皮膚病患者の皮膚から得られたT細胞は、この受容体を発現しない(Le Cleach et al.2000、Clin.Exp.Immunol.119:225〜230)。このようにして、我々の研究成果は、P140/KIRがCTCLの同定のための、CD4+細胞上の好適なマーカーを表すことを示した。
【0092】
材料及び方法
患者
インフォームド・コンセントの後、倫理委員会の承認を受け、我々は11人のCTCL患者から皮膚及び血液試料を得た。8人の患者は、セザリー症候群に罹っており、10〜45%の循環性セザリー細胞を血液中に有していた。7例で、腫瘍細胞のフェノタイプがCD3+、CD4+、CD8−であった。1例がCD3+、CD4+、CD8αα+(患者Pno)であった。3人の患者は形質転換菌状息肉症(Lez、Cou、Bic)で、CD3+、CD4+、CD8−フェノタイプを有する伝播性皮膚腫瘍の症状を呈していた。患者全員がこれまで化学療法を受けていなかった。
【0093】
腫瘍リンパ球の分離
新鮮な腫瘍細胞は腫瘍断片を機械的に単細胞懸濁液に分散させたものから得た(Bagot et al.1998、Blood 91:4331〜4341)。次で、単核細胞は洗浄され、後の使用のために、ヒト血清プラス10%ジメチルスルフォキシド中で凍結された。
セザリー症候群患者の場合は、単核血中細胞がフィコール・アイソパクウ技法(Pharmacia fine Chemicals、Piscataway、NJ)で分離された。
【0094】
腫瘍細胞系の長時間培養
我々は前記の(Bagot et al.1998、Blood 91:4331〜4341)患者の末梢血液からの試験管内Pno細胞系(TCR−Vβ22+、CD3+、CD4+、CD8αα+、MHCクラスI、MHCクラスII)の長時間培養を実施した。我々は患者の血中を循環する悪性クローン及び培養されたT細胞系の両者がそれらの細胞表面フェノタイプ及びそれらのサイズ及びTCRβVDJ領域(Poszepczynska et al.2000、cf.supra)に関して同一であることを示した。SS細胞系の他の供給源はATCC(例えば、HUT 78 ATCC TIB-161)で見つけることができる。
Cou−L細胞系(TCRVβ13+)は試験管内でγIL−2と共に、3年以上培養された。それは前記のCD4+Cou−LSCTCL系のサブクローンに相当する(Bagot et al.1998、cf.supra)。Cou−L細胞系、本来のTCRVβ13+CD4+Cou−LSCTCL及び皮膚から新たに分離された腫瘍細胞は、同じサイズで、且つTCRβVDJ領域の配列を分担する(Bagot et al.1998、cf.supra)。
【0095】
モノクローナル抗体(mAbs)及び流動細胞計数の研究
1色または2色の免疫蛍光分析が前記のように(Bagot et al.1998、cf.supra)行なわれた。抗CD3(細胞分析2000カタログ番号6604629)、抗CD4(細胞分析2000カタログ番号6602138)、抗MNCクラスI(細胞分析2000カタログ番号IM0107)及びII(細胞分析2000カタログ番号IM0108)はBeckman
Coulter(細胞分析2000カタログ番号IM2014)から入手可能である。
抗TCRVβ13はBioadvance(Emerainville、France、catalogue番号TCR1738)、TCR−Vβ22+mAb はBeckman Coulterから入手された(Marseille、France、細胞分析2000カタログ番号IM1484/IM1365)。抗CD8αβ2ST8.5H7mAbはBeckman Coulter(細胞分析2000カタログ番号Im2014)から入手可能である。抗P140mAbsとして、Q66(IgM、抗P140)及びAZ158(IgG2a、p70/NKB1及びP140の両者を確認)(これらは、Pande et al.1996により報告された。J.Exp.Med.184:505〜518)が使用された。しかし、抗P140結合化合物が好適である(例えば、抗P140抗血清、Fc-HLA-A11 Fc-HLA-A3溶融蛋白)。Z27(IgG1、抗p70/NKB1)、EB6(IgG1、抗p58.1/p50.1)、GL183(IgG1、抗p58.2/p50.2)、XA185(IgG1、抗CD94)、Z199及びZ270(IgG2b、及びIgG2aの各々、抗NKG2A)及びB9.4(IgG2b、抗CD8)mAbsは全て、Beckman Coulter(細胞分析2000カタログ番号IM2748、IM2277、IM2278、IM1610、IM2750、IM0102)から入手可能である。
ヤギ抗マウスIgMFITC及びヤギ抗マウスIgMPEもまた、Beckman Coulterから入手され得る(細胞分析カタログ番号IM08及びIM0555参照)。
【0096】
生化学的特徴
20×106個の細胞が、培養され(15分、20℃)250μgのEZ−LINK SULFO−NHS−LC−BIOTIN(Pierce、Rockford、IL)含有1ミリリットルPBSpH8中で培養され(15分、20℃)、洗浄用バッファー(10mM TRIS pH8、0.14M塩化ナトリウム)で3回洗浄された。次いで、細胞は1%NP40に溶解され、次いで、セファロースPA(Pharmacia Biotech Inc.Piscataway、NJ)結合AZ158(IgG2a抗p70/、140)またはB9.4(IgG2b抗CD8)mAbで免疫沈降された。試料は不連続SDS−PAGEを使用し、N−グリコシダーゼ(ベーリンガーマンハイム、GmbH、Germany、製)による消化または非消化で、分析された。次いで、Immobilon P(ミリポアCorp.Bedform MAの商品名)ヘ移した。Neutravidin(Pierceの商品名)で汚染後、検出にはルネッサンス・ケミルミネッセンス・キット(NeN、Boston、MAの商品名)が使用された。NK細胞クローンはペンデ(Pende、1996)等が(J.Exp.Med.184:505〜518)報告したように、限定希釈で得られた。
【0097】
RT−PCR分析
トータルRNAはCTCL細胞系Pno及びCou-LCD8ααからRNAクリーンシステム(AGS GmbH、ハイデルベルグ、ドイツ)を使用して抽出した。cDNA合成はオリゴットTプライミング(oligod T priming)を使用して行なった。KIRディスプレー3Ig様ドナーの完全なORF(open reading frame)cDNA増幅のために使用された。使用されたプライマーを以下に示す。5'CATGT(CT)GCTCA(CT)GGTCGTC(Ig3 UP;SEQ ID No.5)及び5'GGTTTTGAGACAGGGCTG(Ig3 DOWN;SEQ ID No.6)。増幅は30秒サイクル(30秒94℃、30秒55℃、30秒72℃)、次いで、Ampli TAQ(パーキンエルマー−Applied Biosystems、Foster City、CA)を使用して7分、72℃の培養を行なった。PCR産物はpcDNA3.1/V5-HisTOPOベクター(Invitrogen、Carlsbad、CA製)のサブクローン化物であった。DNA配列はd-ローダミン(生物体汚染用顔料)ターミネーターサイクル配列キット及び377Applied Biosystems自動配列機(パーキンエルマー−Applied Biosystems)を使用して行なった。
【0098】
過渡的移入
COS−7細胞がpcDNA3.1TOPO−KIR3Dcl.24またはpCR3cl.1.1(Pende et al.1996、J.Exp.Med.184:505〜518)でFugene-6 (Roche、Monza Italy)を使用して移入された。細胞を5×105/プレートの量でシードした。24時間後、これらを6μgプラスミド及び10μlのFugene-6試薬と共に、DMEM/10%FCS中で培養した。48時間から72時間後、移入細胞は血球弗素計量分析にかけられた。移入細胞はQ66及びAZ158mAbsで汚染され、次いで、フィコエリトリン被接合ヤギ抗体接合マウスのIgG2aまたはIgMで汚染され、血球流量計により、FACソート(Becton Dickinson、San、Jose、CA)を使用して分析した。
【0099】
結果
CTCL細胞系がp140/KIRヘの抗体で汚染される。
二つの長期CTCL腫瘍系Pno(抗TCR−Vβ22mAbでラベル)(Poszepczynska et al.2000、cf.supra)及びCou-L(抗CD3mAbでラベル)(Bagot et al.1998、cf.supra)は反応的に、異なる抗KIRmAbsで分析された。
我々は両細胞系がmAbsQ66(図9A参照)及びAZ158と反応性であり、両者はp140/KIRと結合することを見出した。対照的に、これらの細胞系は他のp58.1、p58.2、p70KIRsを含有するHLAクラスI分子に特異的な阻害受容体及びCD94/NKG2Aレクチン様受容体(Poszepczynska et al.2000、cf.supra)を発現しない。
【0100】
CTCL患者から新たに採取された腫瘍Tリンパ球の抗p140/KIRmAbsによる汚染。
p140/KIRが新たに採取された腫瘍細胞により発現されるか否かを決定すべく、我々は、Q66mAbの、SS患者Pno及びMF患者Couの腫瘍性皮膚断片から採取した非培養腫瘍細胞との反応性を試験した。我々は腫瘍細胞の大部分がこのmAbによって汚染されることを確認した(図9B参照)。特に、我々は、患者Pnoの血液から採取された大部分のTCRVβ22+腫瘍リンパ球がQ66抗体と反応性があり、患者Couの皮膚から採取された大部分のTCRVβ13+腫瘍リンパ球が同じ抗体と反応性があることを観察した(図9B参照)。次に、我々は、その循環性CD4+リンパ球の10〜45%が悪性細胞である、追加の7人のセザリー症候群患者から採取した腫瘍Tリンパ球及び他の2人のMF患者の皮膚腫瘍から採取した腫瘍Tリンパ球、の両者のフェノタイプを研究した。供試全患者が共発現CD4及びp140/KIRの大集団を顕著に示していた(下記表2参照)。
【0101】
形質転換菌状息肉症患者の皮膚、及びセザリー症候群患者の血液、から採取されたCD4+リンパ球汚染、抗Q66mAb。
【0102】
【表2】
【0103】
全てのQ66+細胞がCD4+細胞集団に含まれていることに注目すべきである。かくして、健常者のp140/KIR発現は、皮膚及び血液中にあるCTCL腫瘍CD4+Tリンパ球の特徴を表すNK及びCD8+集団から得られる、リンパ球小集団に限定される。Tリンパ球発現の多種類のKIRの比率が低いところの、他の皮膚病、毒物による皮膚溶解(Le Cleach et al.2000、Clin.Exp.Immunol.119:225〜230)、由来の皮膚T細胞はp140/KIRの発現制御ができない。
【0104】
CTCLによって発現されるp140受容体の分子的特徴
Pno及びCou-L細胞系はビオチンでその表面をラベルされており、細胞溶解物は抗p140(AZ158mAb)で免疫沈降する。図10に示すように、このmAbはその分子量が還元条件下で約70kDである、NKクローン及びPno及びCou-L細胞系から免疫沈降される。N−グリコシダーゼによる処理によって、NKクローンに比較してPno及びCou-Lの移動性がやや高く、分子量が約50kDである蛋白質主鎖を明らかにした。これらのデータは、これらCTCL腫瘍細胞系によって発現されるp140阻害受容体が、前記正常NK細胞で検出されたものと殆ど同じであることを示唆した(Pende et al.1996、J.Exp.Med.184:505〜518)。
【0105】
次に、我々はPno及びCou-L細胞系上のQ66及びAZ158mAbが前記p140/KIR(SEQ ID No.3)をコード化するcDNAの一つに相当するかどうかを決定した。この末尾で、RT−PCRは、三個のIg様ドメインを有するKIRをコード化する全てのcDNAを増幅できるプライマーのセットを使用して、これらの細胞から由来するRNA上で行なわれた。Pno細胞系から、KIR3D cl.24と名付けられた、フル鎖長のcDNAを採取した(SEQ ID No.1)。そのヌクレオチッド配列と、三つの細胞外Ig様ドメインが特徴である、全てのKIRをコード化するDNA配列との比較は、KIR3D cl.24pが、140受容体の新規な対立遺伝子型を表すことを明らかにした。特に、そのヌクレオチド配列は前記の成熟蛋白質(図11参照、SEQ ID No.2:クローン24蛋白質;SEQ ID No.4:クローン1.1蛋白質)における4個のアミノ酸置換となる1.1 cDNAに比較される5個の異なるものを表示する(Pende et al.1996、J.Exp.Med.184:505〜518)。4個の置換のうち3個は細胞外Ig様ドメイン(成熟蛋白質のうちのpos.20、92及び111)であり、他方はサイトプラズマ領域に局在する(pos.401)。Cou-LCTCL細胞系で行なわれるRT−PCRはp140/KIRの二つの異なる対立遺伝子型を明らかにした。一つはcl.1.1cDNAに相当し、他方は、KIR3D cl.24(Pno細胞系から分離)と同一である。CTCL細胞系由来のcDNAはCOS−7細胞へ移入される。期待したように、全ての移入された細胞は、Q66及びAZ158mAbによって明瞭に汚染される。
一方、これらはネガティブコントロールとして使用されるp70/KIR特異性Z27mAbと反応しない。終わりに、RT−PCRが三人のSS患者(患者のPno含有)由来のPBLから抽出及び一人の追加のMF患者の皮膚由来のT細胞から抽出されたRNA上で行なわれる。また、これらの試料で、上記のp140/KIRの一つまたは他の対立遺伝子型が同定される。
同じ結果が患者から採取された悪性細胞上で得られた。
【0106】
討議
CTCLの皮膚病変は、殆どCD4+で、非新形成の腫瘍浸潤性Tリンパ球である悪性T細胞からなる異成分のリンパ球の浸潤物を含む。我々は、以前に、CD4+細胞毒腫瘍浸潤性Tリンパ球は特異的に、自家組織の悪性CTCLCD4+細胞系を支配することを報告した。しかし、腫瘍細胞で発現されるクローン型のTCRのみならず、非腫瘍とされている細胞表面構造がCTCL上で同定された。非悪性、反応性CD4+リンパ球と悪性のものとを区別する標準法を使用することは困難である。
我々は、この研究で、最初に、MF及びSS患者から採取した腫瘍細胞が、p140/KIRを発現すると報告した。この受容体は長期培養CTCL系と同様に、CTCL患者からの皮膚及び血液中の腫瘍細胞と同定された。二色蛍光分析はp140/KIR発現は、以前に腫瘍細胞で同定され、与えられた、TCRβ−VDJにより、特徴付けられているT細胞として限定される。このようにして、p140/KIRは、Tリンパ球CD4+集団中の腫瘍反応性細胞からの腫瘍細胞の同定を迅速化させる。これは、特に、その腫瘍T細胞クローンが末梢血中CD4+リンパ球集団内で、常には、容易に同定されないようなSS患者に対しては有効である。更に、病気の間または処置後に、CD4+集団の増加が腫瘍細胞集団の増加が原因なのか、反応性Tリンパ球の増加が原因なのかを評価するのは困難である。抗p140/KIRmAbが、既に解析された全CTCL患者の腫瘍細胞を認知するために出現するので、それらは、CTCLの直接的同定のためには公的な手段と考えられる。その上、CD4+CTCL細胞上での、p140/KIR共発現の使用は悪性細胞と正常細胞を区別するユニークな手段を提供する。これは個々の患者にとって独特であるTCR決定法とは対照的である。その上、p140/KIRは、CTCLの新規な特異的免疫療法の発展のための可能なターゲットである。
【0107】
これまでの研究は、この受容体がHLA.A3及びHLA.A11対立遺伝子の認知への阻害信号を送ることができることを示した。しかし、解析された各種の患者におけるp140/KIR発現それら自体のHLAクラスIハロタイプには明らかに依存しないと見なされることに留意すべきである。それにも拘らずp140/KIRのCTCLの異常病態生理学における現実の役割は、それ自体への耐性での受容体の潜在的役割を考慮することで、研究が続けられるという重要な特徴である。結論として、この研究は最初に、CD4+CTCL細胞におけるp140/KIRの発現及び腫瘍細胞におけるこの受容体の新規な対立遺伝子型の病変を示している。この発見はCTCL患者の異常病態生理学及び臨床管理の両者についての重要な論点である。更に、p140発現がCD8+形質転換MFのようなCD8+CTCLの表面に観察される。
【実施例3】
【0108】
抗CTCL薬の製造
当該分野の専門家は、下記の図の意図による概要に従がって抗CTCL薬を製造できる。
CTCL細胞近傍の細胞の免疫システムを刺激するために、抗SC5または抗p140mAbをサイトカインと共に使用する。
抗CTCL成分をCTCL細胞に向けて送り出すためのベクターとして、これらの抗SC5または抗p140mAbを使用する。抗CTCL成分の例は、放射性元素、トキシンなどのキミオセラピーで使用される標準化合物がある。
放射性元素用mAbベクターは、本発明の抗SC5mAbに使用された標準技法により産生できる[Press OW et al.置換B細胞リンパ腫への自家幹細胞移植を伴うフェイス2試行沃素131B1(抗CD20)抗体療法、Lancet 1995 vol.346 pp336〜340参照、この内容は,この明細書に参考のために記載されている。]。
トキシン用mAbベクターは、本発明の抗SC5mAbに使用された標準技法により産生できる[Ghetie U et al.ヒト組換えCD4及び脱グリコシルリシンA鎖構成免疫接合物の大スケール調製、J.Immunol.Methods 1990、vol.126 pp 135〜141、この内容は,この明細書に参考のために記載されている。]。
抗有糸分裂性のプロドラッグ及び抗SC5mAb及び/または抗p140mAbベクターの両者の結合されたものを使用すると、抗有糸分裂性のプロドラッグを活性な医薬型(機能型の抗有糸分裂性製剤)へと形質転換することができる酵素を送り届ける。
抗有糸分裂性のプロドラッグを活性な医薬型へ形質転換することができる酵素を送り届ける抗SC5mAb及び抗p140mAbベクターは、本発明の抗SC5mAb及び抗p140mAbに適用される次の標準的技術で産生され得る(ADEPTtechnique、for Antibody Directed Enzyme Prodrug Therapy、described in Plakey TC et al. Anti-tumor effects of an antibodycarboxypeptidase G2 conjuguate in combination with phenol mustard prodrugs、British J. Cancer 1995 vol.72 pp 1083〜1088、of which content is herein incorporated by reference、参照)。
リガンド作用物質(SC5刺激またはp140刺激)としての抗SC5または抗p140/抗CD4の使用。
補体漸増剤としての抗SC5または抗p140mAbの使用。
上記用途のために抗SC5/抗CD4または抗p140/抗CD4特異性の共用のmAbが好ましい。
ADCC(Antibody Dependent Cell Cytotoxicity)の活性化体としての、抗SC5または抗p140(CD4特異性は不要)の使用:多核細胞、マクロファージ、Fc受容体含有NK細胞抗はSC5及び抗p140mAbのFc含量によって活性化され、それ故、これらmAbはCTCLの近傍でADCC活性化ができる。
【産業上の利用可能性】
【0109】
本発明はCTCLの診断及び治療の問題点を解決する二つの分子を提供する。
これは、特に、悪性CTCL細胞のような悪性T細胞の増殖及び/または機能的活性を阻害する。また、T細胞、特に悪性CTCL細胞のような腫瘍性T細胞の増殖及び/または機能的活性の調節ができる、ハイブリドーマI−2575で産性された抗SC5モノクローナル抗体を提供する。
【図面の簡単な説明】
【0110】
【図1】図1A、1B及び1Cは健常者のPBLsのSC5分子の細胞膜発現を図解している。 図1A、1B及び1Cはそれぞれ、健常者のPBMCを抗SC5mAb(腹水1/200)で汚染させ、次いで、FITC接合アイソタイプ特殊ヤギ抗マウスIgM2次試薬及び、抗CD3(図1A)、抗CD56(図1C)及び抗CD45RO(図1B)のようなPE接合mAbsの一種で汚染した場合である。
【図2】図2はPBLsの活性化の間、SC5分子の細胞膜発現を図解している。
【図3】図3はPBLのSC5分子の細胞内所在部を図解している。
【図4】図4はSC5分子の生化学的分析結果を図解している。
【図5】図5A、5B、5C、5Dは、抗CD3誘発増殖及び正常Tリンパ球のサイトカイン分泌を、それらの細胞毒活性の影響なしに、抗SC5mAbが調節する様子を図解している。図5A:PBLまたはT細胞クローンLSO及びGDS.3が固定化抗CD3mAb(コートされたmAb1μg/源)またはIL−2(50IU/ミリリットル)で、抗SC5mAb(腹水の、1:200最終希釈)またはアイソタイプ適合、不適切mAbの存在で、刺激した場合。図5B:GDS.3CD4+T細胞クローンは抗SC5mAbの異なる濃度の存在下、図示された濃度のプレコート抗CD3mAbで刺激した場合。 図5C:抗SC5またはコントロールmAbの存在下、抗CD3刺激の場合。図5D:細胞毒CD8+細胞クローン(JF1)が、抗SC5mAb(腹水の、1:200最終希釈)またはアイソタイプ適合コントロールmAbの存在で、E/T比が5/1での、FcyR+ミューリン腫瘍細胞P815との共培養の前に、培養した場合。
【図6】図6A、6BはCTCL患者のPBLのSC5分子の細胞膜発現を図解する。図6A:セザリー症候群患者のPBLのSC5及びCD4分子の共発現を健常者と比較した場合。図6B:ヒストグラムは示された3群におけるSC5発現の平均値を示す
【図7】図7A、7B、7Cはセザリー症候群患者の悪性細胞の膜表面発現を図解する。 図7A:循環TCRVβ22+悪性T細胞クローンの存在で、SS患者のPBMCの二色蛍光分析。図7B:弱くラベルした、抗SC5mAb(陰を付したヒストグラム)TCRVβ22陽性悪性細胞。図7C:同じ患者の細胞を、PE接合抗CD69mAb及びFITC接合抗TCRVβ22mAbによって汚染させた場合。
【図8】図8は抗SC5mAbによるCTCL悪性細胞系の抗CD3誘発増殖の調節を図解している。
【図9】図9A、9Bは、CTCL細胞系(図9A)のp140/KIR分子及び新鮮なリンパ球腫瘍(図9B)の検出を図解いている。
【図10】図10はp140分子の生化学的分析を図解している。
【図11】図11は蛋白質でコードされたKIR3D cl.24(SEQ ID No.2)及びcl.1.1(SEQ ID No.4)のアミノ酸配列を示す。
【0111】
略語
APC:抗原提示細胞、CTCL:皮膚T細胞リンパ腫、DIG:界面活性剤不溶グリコリピッド富化分画、ILT:免疫グロブリン様転写、KIR:キラー細胞免疫グロブリン受容体、mAb:モノクローナル抗体、MF:菌状息肉症、PBL:末梢血中リンパ球、PBMC:末梢血中単核細胞、PHA:フィトヘマグルチニン、PMA:ホルボール12β−ミリステート13α−アセテート、SS:セザリー症候群、TCR:T細胞受容体。
【技術分野】
【0001】
この出願はTリンパ腫の新規な診断及び治療法、更に詳しくは、皮膚T細胞リンパ腫(CTCLと略称する)の新規な診断及び治療法に関する。本発明は現実に、CTCLに共通する腫瘍マーカーを提供し、そのバイオテクノロジー的及び医学的用途をも含む。
【背景技術】
【0002】
CTCLは本来皮膚に関するTリンパ腫のグループである。CTCLグループは即ち、形質転換菌状息肉症(形質転換MFと略称する)、セザリー症候群(SSと略称する)、リンパ腫丘疹(LPと略称する)及びCD30+リンパ腫からなる。形質転換MFは表現タイプが成長済みTヘルパー細胞に類似するクローン誘導型悪性Tリンパ球の皮膚侵襲に因るものである。LP及びCD30+リンパ腫もまた、皮膚で進展する。より進行性な型のCTCLは、悪性細胞が非表皮性になったとき発達し、特殊な皮膚関連性を示すようになる。SSはCD4+/CD45+RO+T細胞のクローン増大及びこれらの悪性T細胞の血中への出現による、より進行性なCTCLのタイプである。大部分のCTCLはCD4+CTCLであり、稀にCD8+CTCLの場合がある。
細胞表面の特殊なマーカーが欠如しているために、悪性細胞の同定が困難であるから、CTCL病の生物学は、十分には、理解されていない状態が継続している。皮膚の病変において、活動性、浸潤性のリンパ球(非腫瘍性の)の中から、CTCL細胞を識別するのは困難である。
CTCLのようなTリンパ腫の診断は、現時点では、疑わしい体内(脳の細胞核を示す細胞の検知によって、皮膚生検及び/または血液中のSSの存在の外観の組織病理学的観察)から採取された試料中に腫瘍細胞が存在するか、しないかを、細胞学的且つ組織病理学的観察によって行なうことに基づいている。このような診断法は、皮膚リンパ球の悪性リンパ球への形質転換の初期では、特に、十分な信頼性に欠ける。このような診断法は病気を現実のものとして捉えることができない。技術的には時間も浪費する。
今日では、CTCLの治療は、非特異的化学療法による腫瘍細胞アポトーシスの誘導により試験的に行なわれている。
だから、CTCLのような悪性T細胞の増殖に関連する病気の診断及び治療の問題点のより正確で好適な解決策が、まだ、必要とされている。
【0003】
このような従来技術の状況に鑑み、発明者等はCTCLの診断及び治療の問題点を解決する二つの分子を提供する。一つは発明者によってSC5と命名され、他方はp140分子(p140は、また、KIR3DL2である)である。
本発明におけるSC5分子は生化学的、機能的に新規な蛋白質であり、還元状態で、見かけの分子量は96kDである。これは発明者等によって産生されたモノクローナル抗体の抗原として分離された。このモノクローナル抗体を産生するハイブリドーマは2000年10月30日、C.N.C.M.(C.N.C.M.パスツール研究所;25、rue du Docteur Roux、F-75724 Paris Cedex15、France)にブダペスト条約(C.N.C.M.寄託番号I-2575)に従がって寄託された。細胞表面で発現され、集合されたとき、SC5分子は細胞活動及び増殖の阻害受容体として行動する。本発明の第二のマーカー、即ち、p140は既に阻害受容体として知られているが、以前に、NK細胞及び健康なヒトのCD4+CD8+細胞のサブグループであると述べられていた。発明者等は、ここで、p140がCTCL細胞のような腫瘍T細胞の表面で発現されることを示す。
発明者等は、更に、CD4+CTCL細胞のような悪性のCD4+T細胞の表面で、p140が発現するのに対し、NK細胞(p58.1、p58.2、p70KIRs、CD94/NKG2A)の表面で通常観察される他の受容体は、悪性のCD4+T細胞の表面で観察されないことを示す。勿論、p140は皮膚炎(例えば、皮膚結核、苔癬)または毒物による皮膚壊死のような非腫瘍性の皮膚系疾病に罹っている患者から採取されたCD4+T細胞の表面では、発明者によって観察されていないことに留意すべきである。
腫瘍性CD4+T細胞の表面で、p140の二つの対立型が発明者によって同定された。即ち、対立型KIR3Dクローン24(SEQ ID No.1)及び対立型p140クローン1.1(SEQ ID No.3)である。対立型KIR3Dクローン24(SEQ ID No.1)は新規蛋白質(SEQ ID No.2)をコード化する新規なポリヌクレオチドである。KIR3Dクローン24(SEQ ID No.1)は、前記p140クローン1.1DNA配列(SEQ ID No.3)に比較して、5つの相違点が明らかであり、前記p140クローン1.1成熟蛋白(SEQ ID No.4)に比較して、成熟蛋白(SEQ ID No.2)において、4個のアミノ酸置換になっている。
【0004】
発明者等は、SC5及びp140の両者が、以下の同じ技術的特徴を共に有することを示す。
SC5及びp140は共に、悪性T細胞、とりわけ、悪性CD4+T細胞の特徴である、細胞膜分化抗原である。
CD4+CTCLの何れの型が関与しているにしても、SC5またはp140をそれらの表面(通常、SC5及びp140の両者が発現される)に発現する、悪性のCD4+T細胞がある。SC5及びp140のいずれも、CD4+CTCL、特に、セザリー症候群(略語でSS)、形質転換菌状息肉症(略語で形質転換MF)、リンパ腫症丘疹(略語でLP)及びCD30+リンパ腫の全範囲を実際にカバーする。
SC5またはp140のCD4+T細胞の表面での存在と、CD4+CTCLの存在の間には、CD4+CTCL診断が疑わしい体内部分(例えば、形質転換MFが疑われる際の紅皮症の皮膚試料または、SSのような進行性のCTCL型が疑われる際の末梢血液試料)から採取したCD4+細胞表面のSC5またはp140の存在の解析に基づき、この信頼性は90%以上、より好ましくは95%以上、最も好ましくは100%である(これまで試験に供された患者については信頼性100%である)という関連がある。事実、本発明によれば、CD4+T細胞は、CD4+T細胞表面でのp140分子の存在が検知されるか、または、SC5+CD4+T細胞の比率の値が、平均標準レベルより高く測定されれば(平均標準レベル1〜15%、一般には5〜10%の範囲内にある)、即、腫瘍であると決定される。
また、SC5またはp140のCD4+T細胞の表面での存在と、CD4+CTCLの存在の間には、CD4+SC5+T細胞及びCD4+p140+T細胞の両者の存在比率、−これはセザリー症候群(SS)が疑われる患者から採取された試料から測定されるが、−は本質的に、この患者の末梢血液に現実に存在する悪性のSSの存在比率に相当する(SC5+CD4+細胞は±10%の範囲内、p140+CD4+細胞は±5%の範囲内)。だから、本発明におけるSC5及びp140マーカーはSSの進行状況を評価できる共通の特別な特徴を共に有する。
【0005】
発明者の知る限りでは、SC5及びp140分子は共通の技術的特徴を有する最初の分子である。本発明は、かくして、新規なCTCLに共通のマーカーを提供する。これより以前には、このようなCTCLに共通のマーカーは知られていない。これらに最も近似するこの種のものは、CD30であって、このものの、悪性のCD4+T細胞の表面での存在は、患者がCD30+リンパ腫に関連するCD4+CTCLの特殊型を有していると結論付ける。それ故、CD30はCTCLのマーカーであるが、その信頼性はCTCLの特殊型(CD30+リンパ腫)に限られる。そして、CD30はCD4+CTCLの全ての型をカバーしない。即ち、SS、形質転換型MFまたはLPのようなCD4+CTCLでは、表面でCD30を発現する悪性CD4+T細胞は必ずしも存在しない。CD30を使用したCTCLの診断及び治療は、このように、CTCLの特殊型に限られる。それに対して、本発明のマーカーはCD4+CTCLの全範囲を好適にカバーする。
【0006】
p140と共有する技術的特徴に加えて、SC5は、更に、非増殖性のT細胞の増殖及び/または機能的活性(例えば、サイトカインプロフィール)の肯定的指標を有する。T細胞表面のSC5の存在は、事実、正常(CD4+及びCD8+)T細胞またはウイルス感染CD4+T細胞のような非腫瘍性T細胞の活性状態に肯定的に関連する。SC5は、かくして、また、HIV感染症、及び自己免疫性(例えば、脊椎関節炎のようなリューマチ性関節炎または乾癬、湿疹、アトピー性皮膚炎のような免疫関連性皮膚病)のようなT細胞ウイルス性感染症の診断及び治療のための新規手段として有効である。SC5は、また、グラフト宿主反応を調節するための有効な標的である。即ち、SC5の活性化は、エフェクター宿主細胞をグラフトに対して進展させる反応を阻害するために使用できる阻害性シグナルを伝達する。
【0007】
本発明は、また、SC5新分子に結合できる産物を提供し、特に、SC5に対するモノクローナル抗体(抗SC5mAbs)を提供する。また、T細胞、特に悪性CTCL細胞のような腫瘍性T細胞の増殖及び/または機能的活性の調節ができる抗SC5モノクローナル抗体を提供する。特に、ハイブリドーマI−2575で産性されたモノクローナル抗体は、T細胞の増殖及び/または機能的活性の阻害が可能で、特に有効な特徴によれば、悪性CTCL細胞(下記の例参照)のような悪性T細胞の増殖及び/または機能的活性の阻害が可能である。SC5に結合するが、これらを集合させない化合物は、SC5の阻害性シグナルの伝達を妨げるか、または、化合物がSC5+細胞(標的マスキング)に到着するのを妨げる。
【0008】
本発明によれば、一般に、SC5またはp140に結合する化合物は、補体漸増剤、ADCC刺激剤、または、CTCLのようなTリンパ腫を予防し、一時的緩和し、治療するための治療剤用ベクターとして使用される。
【0009】
本発明はそれ故に、抗SC5及び/または抗p140モノクローナル抗体及び直接それらから製造された産物(例えば、ヒト化モノクローナル抗体またはCD4−p140/SC5に対する二重特異性を有するモノクローナル抗体)の診断及び治療への使用に関する。
【発明を実施するための最良の形態】
【0010】
2000年10月30日、寄託番号I−2575として、C.N.C.M.(C.N.C.M.、国立微生物保存機関、パスツール研究所;25、rue du Docteur Roux、F-75724 Paris Cedex15、France)に、ブダペスト条約に従がって、寄託されたハイブリドーマを提供し、ハイブリドーマ細胞の好適な代謝条件(例えば、37℃で3日間、5%炭酸ガス含有湿潤雰囲気下、RPMI1640剤:Gibco BRL Cat.N021875-034;10%牛胎児血清;ぺニシリン100単位/ミリリットル;ストレプトマイシン100μg/ミリリットルからなる好適な培養液に2.106個の細胞を含有したものを使用。)下、ハイブリドーマI−2575の培養を内容とする、抗SC5モノクローナル抗体の調製方法を提供する。
本発明は、それ故、ハイブリドーマI−2575によって産生されるか、またはこの寄託されたハイブリドーマの培養上澄み液から得られるモノクローナル抗体(mAb)を包含する。このmAbは5種類の抗SC5IgMである。これは不適当または好ましくないT細胞の増殖及び/または機能的活性(サイトカインプロフィール)、不適当または好ましくないIL−2産生の診断及び治療のための有効な手段である。これは正常T細胞、ウイルス感染T細胞(例えば、HIV感染CD4+T細胞)及び特に、悪性CTCL細胞のような悪性T細胞の増殖及び/または機能的活性を阻害する。
【0011】
ハイブリドーマI−2575によって産生されるモノクローナル抗体は、発明者によってSC5と命名された蛋白質を分離できる。SC5蛋白質は生化学的、機能的に新規な分子で、見かけ分子量が還元条件下で96kDである。これは多くのPBL細胞中及び細胞上に存在する。即ち、正常細胞(CD4+及びCD8+部分集合)及び特にCD3+T細胞、CD45RO+T細胞、正常NK細胞、B細胞、顆粒球及びマクロファージ(大食細胞)である。更に、SC5はNIH/3T3(ATCC CRL−1658)及びCHO−KI(ATCC CRL−9618)のような他の系統及び他の種の細胞の細胞質中にも存在する。
休眠中の正常PBL細胞では、SC5は主として細胞内に存在する、しかし、CD3刺激はSC5の細胞膜への移行を誘発する。殊に目立つのは、発明者によって、SC5が、悪性CTCL細胞のような悪性T細胞、特に悪性CD4+CTCL細胞で、主として、細胞表面に観られることである(幾ばくかの生体外のCD3刺激なしで)。
【0012】
更に著しい特徴はSC5の表面発現が、現在まで試験された全てのCTCL患者に観られることである(下記例参照)。
本発明の他の更に著しい特徴は、SC5を共発現するCD4+T細胞の比率が、健常者よりも、腫瘍性CD4+T細胞をもつ患者の方が、統計的に高いことである。CD4+CTCLの患者におけるSC5+CD4+の比率は、健常者において観察される比率(SC5+CD4細胞の平均標準比率は1〜15%の範囲内であり、一般には5〜10%の範囲内である)より有意に高い。
特にセザリー症候群(SS)のCD4+CTCLの場合、SC5を共発現するCD4+細胞の比率は、もっと、統計的に、CTCL患者のPBLにおける悪性CTCL細胞の比率に密接に関連する。即ち、SC5+CD4+T細胞の比率は本質的に悪性CTCL細胞の比率と同等である(この比率の±10%以内)。SC5蛋白質は、それ故、細胞表面での経膜的な発現が、非腫瘍性T細胞の活性化及び/または増殖状態に関係があり(例えば、HIV感染細胞及び正常CD4+またはCD8+T細胞のようなウイルス感染細胞)、非活性化T細胞の悪性化状態に関係がある新規分子である。著しい特徴は、CD4+CTCLが陽性との診断を90%以上の信頼性で可能にすることである(実際、最新の試験を行ったヒトでは100%)。
【0013】
本発明の更に顕著な特徴は、SC5新分子が、細胞表面で発現されたとき、細胞増殖及び/または機能的活性のための阻害受容体として活動できることである。即ち、SC5の集合は、それを発現する細胞の増殖及び活性の減少を誘発する。例えば、ハイブリドーマI−2575により産生される、抗SC5モノクローナル抗体[5種類のIg(免疫グロブリンの略語)]は、ヒトまたは動物から採取した、正常CD3+T細胞、または正常CD3+T細胞クローンのようなSC5+細胞の抗CD3誘発増殖を阻害し、また、抗CD3が誘発した、CTCL患者から採取したCTCL細胞の試験管内増殖及び動物モデルでのヒトCTCL細胞の生体内自然増殖を阻害する。このmAbは、また、HIV感染CD4+細胞と結合し、それ故に、感染細胞を殺し、HIVの増殖を抑制させることを目的とする抗HIV薬の実際的原理として有用である。
【0014】
幾つかのSC5蛋白質の分離可能な方法は、本発明のモノクローナル抗体を製造できる専門家にとって使用できるのものである。標準的な方法は、細胞溶解物からの還元下、親和性クロマトグラフまたは電気泳動による回収からなる。ハイブリドーマI−2575により産生するモノクローナル抗体が抗原と認識する全てのヒトまたは動物の細胞、あるいは該モノクローナル抗体と細胞溶解物が免疫沈殿反応する全てのヒトまたは動物の細胞は、SC5分子を分離するための細胞供給源として適当である。そのような細胞には、特に、T細胞、CD3+T細胞、CD45RO+T細胞、B細胞、NK細胞、マクロファージ、顆粒球等の正常細胞(ヒトまたは動物または細胞クローン)を含む。また、悪性T細胞、例えば、CTCL患者から採取されたCD4+CTCL細胞のような腫瘍性細胞、またはCTCLクローンHUT78(ATCC TIB−161)のような腫瘍性クローンも好適な細胞である。好適な細胞源が広範囲なので、粗血球全体またはPBLが何らかの精製を経ずに、直接使用されてもかまわないことが理解される。
【0015】
正常細胞はSC5の細胞内型及び経膜受容体型の両者を分離できる。SC5の受容体型またはその部分の分離が望まれるときは、SC5+細胞源(例えば、それらの表面でSC5を発現する細胞)を使うのが好ましい。そのような細胞源(ソース)としては、前記されたものであって、細胞表面でSC5発現が増加するように、CD3刺激されたものが含まれる(下記CD3刺激法の例参照)。全血球またはPBLがSC5+細胞の供給源として使用されるとき、全血球またはPBLは、T細胞及び単核細胞を含有するから、CD3活性化はPHA(Wellcome社から入手可能)のような、ポリクローナル活性体の約1μg/ミリリットルと培養することにより誘発される。より均一な正常細胞集団がSC5+細胞(例えば、SC5+のみとNK細胞)の供給源に使用されるときは、CD3活性化は、好ましくは固定化抗CD3抗体との培養、または、可溶性抗CD3抗体及びPHAのようなポリクローナル活性体との培養によって為される。培養段階は、選択的に、CD3活性化の前及び/または後に、細胞量を増加させるために行なってよい(培養条件は、例えば、RPMI中、10%牛胎児血清、湿度95%及び5%炭酸ガス雰囲気)。
SC5を細胞内型で分離したいときには、CD3刺激は省かなければならない。
CTCL細胞のような腫瘍性細胞、特に、悪性形質転換MFまたはSS細胞では、生体外CD3活性化を必要とせずに、受容体型のSC5分子を分離することができる。即ち、CTCL細胞は既に経膜区域でSC5を発現する。
【0016】
故に、本発明は下記の段階によって得られる分離蛋白質を包含する。
(i)T細胞、CD3+T細胞、CD45RO+T細胞、B細胞、NK細胞、マクロファージ、顆粒球からなる群から選択される細胞を採取する。採取した細胞を固定化抗CD3抗体、または、可溶性抗CD3抗体及びPHAの1μg/ミリリットルで刺激するか、または、悪性CD+CTCL細胞のような悪性CTCL細胞を採取するか、または、全血球細胞及び末梢血液リンパ球(PBL)からなる群から選択される細胞を採取し、採取した細胞をPHAの約1μg/ミリリットルで刺激する。
(ii) 例えば、TritonX-100の1%含有の強力な溶解バッファー中での培養によって、多種類のポリペプチド複合体を分離できる条件下に上記細胞を溶解する。
(iii)ハイブリドーマI−2575によって産生されるモノクローナル抗体が、このモノクローナル抗体が抗原抗体型(例えば、この抗体によって抗原と認められる化合物)の反応を行なうことができる条件下に、結合する蛋白質化合物を溶解物から回収する。
【0017】
回収された蛋白質化合物のアミノ酸配列は、質量分析のような、なんらかの標準技術によって得ることができる。
上記で示されたように、段階(i)の細胞は活性化細胞であり、それ故に、経膜受容体のときに、SC5の分離ができる。段階(i)で使用されたCS5のソースが休眠中のT細胞(生体外刺激を除く)からなっていれば、このようにして分離された蛋白質化合物はその脂肪質型SC5であろう。
段階(i)で選択される細胞が活性化された、または、悪性の細胞のときは、段階(ii)の溶解は、それ故に、細胞全体で行なわれるか、または、これらの細胞膜の小部分で行なわれる。
溶解段階自体はその休眠中の多数のポリペプチド複合体の分離技術によって行なわれ得る。例えば、細胞またはその膜小部分はTritonX-100のような強力な界面活性剤1%含有溶解バッファー中で、1時間、4℃で培養される。これによって、休眠中の細胞成分からの、SC5蛋白質の分離が行なわれる。もし、ジギトニンの1%またはブリッヂ58の1%のようなマイルドな界面活性剤が使用されれば、SC5は、それが自然に結合している、これらの化合物から分離されないであろう。上記段階(ii)でのマイルドな界面活性剤の使用は、だから、SC5の分離を可能にし、更に、SC5の形質導入、実施体の分離を可能にする。
故に、本発明は、前記プロセスで得られる化合物を包含する。
次いで、段階(ii)で得られた溶解物は、この抗体が、細胞溶解物含有(例えば、ハイブリドーマI−2575によって産生された2μgのモノクローナル抗体は、107個の溶解物に相当する)マイクロウエルに抗体をコートするような、抗原抗体型反応を行なうのに好適な条件下、ハイブリドーマI−2575によって産生されたモノクローナル抗体とともに培養され、次いで、モノクローナル抗体は細胞溶解物とともに2時間、4℃で培養される。
【0018】
ハイブリドーマI−2575によって産生されるモノクローナル抗体が結合する化合物の回収段階(段階iii)はこの分野のエキスパートにとって熟知の技術で、行ない得る。非還元条件下の回収は、本来の形態のSC5の分離が可能で、SC5の本来の生物学的特性の維持が望まれるときに行なうのが好ましい。
SC5蛋白質の非還元条件下での回収の好適な技術は、即ち、親和分離クロマトグラフィ(アフィニティークロマトグラフィー)を含む。好ましくは、濾過前に、濾過を容易にするために、溶解物中の化合物の構造を本質的、部分的に変えて、溶解物を生理学的pH、例えば、pH6〜8の間の、pH7のようなバッファーと混合して、溶解物をハイブリドーマI−2575によって産生されたモノクローナル抗体(mAb)が固定されているカラムにより濾過する(例えば、このmAbを寒天粒に結合させ、この粒を前記カラム内に堆積させる)。カラムを前記生理学的pHのバッファーで洗い、例えば、本来の化合物の構造を、本質的に変えることなく、mAbへの化合物の結合をバッファーで攪乱して、溶出させ、固定されたmAbに結合している化合物を回収する。この溶出バッファーは、通常、弱酸性のpH(通常、pH2〜5の、例えば、pH3)である。抗体分子の通常の方向性を有し、それにより、抗原を高い効率で結合させるマトリックスの調製のため、一段階の精製方法のため、膜蛋白質の分離のために、例えば、Schneider et al.1982(J.Biol Chem.257:10766〜10769)記載の内容が参考になる。このようにして回収された蛋白質化合物はその本来の構造のSC5蛋白質である。
【0019】
一方、段階(iii)では還元条件下、段階(ii)で得られた溶解物とハイブリドーマI−2575によって産生されたモノクローナル抗体とで免疫沈降反応を行なうことができる。このようにして得られた免疫沈降は二十日鼠のIgMと結合できる抗体からなるカラムを通して濾過される。ハイブリドーマI−2575によって産生されたモノクローナル抗体に結合する蛋白質化合物は前記と同じ溶出法で回収される。このようにして得られた蛋白質化合物も、また、本来の構造の分離SC5蛋白質である。休眠中の標準的なSC5蛋白質回収方法[段階(iii)]は、還元条件下の電気泳動法による処理である。これは、特に、四つのステップを包含する。
ハイブリドーマI−2575によって産生されたモノクローナル抗体と段階(ii)で得られた溶解物を、このmAbを抗原抗体型反応させることができる条件下に培養する。かくして、生成した免疫沈降物を採取し、免疫沈降物に含有する化合物を還元条件下に分離し、見掛け分子量96kDの蛋白質化合物を回収する。
一方、所望のSC5蛋白質を電気泳動ゲルまたは膜に局在させる方法(続いて、それをゲルまたは膜から分離する)に続いて、溶解に先立ち、細胞の表面ポリペプチド成分をラベルする(例えば、スルホ−NHS−LC−biotinによる、125ヨードまたは蛍光ラベリング)、ラベリングステップが行なわれる。そして、免疫沈降物からラベルを帯びたこれら化合物を回収する。もし、ラベリングステップが溶解の後に行なわれれば、SC5の細胞内型及び経膜受容体型の両者が分離されることに注目しなければならぬ(この場合、前記のように、非活性化T細胞がSC5源として使用され得る。)。変性条件が使用されるから、このようにして得られた蛋白質化合物は、本来の構造ではない。即ち、まだ、本来のアミノ酸配列ではあるが、本来の生物学的特性が失われている。しかし、本来の構造ではないSC5蛋白質が観察され(調べられ)、試験される生物学的試料が変性条件下(例えば、皮膚サンプルのパラフィン薄片)に処理されたときに、本来の構造ではないSC5蛋白質が観察される(調べられる)。本来の構造ではないSC5蛋白質は、それ故、モノクローナル抗体の産生に直接使用され、このような、モノクローナル抗体は変性サンプルのSC5検出に有用である。もし、本来の構造のSC5蛋白質が所望されれば、変性蛋白質の少なくとも10アミノ酸区分が配列され(例えば、質量分析で)、上記配列区分から理論的に導かれたオリゴヌクレオチッドが合成され、試験的に、SC5+細胞から得られるcDNAライブラリー(DNAコレクション)からのスクリーニングに使用される。このようにして選択されたcDNAが、産生及び、好ましくは、クローン培養によるcDNAコード化産物の排出が可能な条件下、産生され、クローン化される[例えば、ヴァキュロウイルスベクターヘのSC5コード化cDNAの挿入、昆虫細胞、例えば、Sf9細胞の形質転換により、または、ワクチンウイルス及びEBV細胞により、Rindis Bacher 1995、J.biol. Chem. 270(23):14220〜14228]。
【0020】
故に、本発明はその配列が以下のようにして得られる分離蛋白質を包含する。
全血液細胞及び末梢血リンパ球(PBL)からなる群から選択される細胞を採取する。採取した細胞をPHAの1μg/ミリリットルで刺激する。刺激された細胞をビオチンのようなポリペプチド特異性ラベルでラベリングする。ラベルされた細胞をTritonX-100の1%含有溶解バッファー中で、培養して溶解する。溶解物をハイブリドーマI−2575によって産生されたモノクローナル抗体で免疫沈降させる。免疫沈降物から、ラベリングされた蛋白質化合物を回収する。このようにして得られた蛋白質配列をコード化するcDNAを合成する。昆虫細胞にこのcDNAを、導入させる(transfection:transformationとinfectionの合成語、細胞へ外来DNA断片を導入すること)。導入された細胞、例えば、ハイブリドーマI−2575によって産生されたモノクローナル抗体で、産生された蛋白質を回収する。
または、以下のようにして得られる。
全血中細胞及び末梢血中リンパ球(PBL)からなる群から選択される細胞を採取する。採取した細胞をPHAの1μg/ミリリットルで刺激する。ラベリングされた細胞をTritonX-100の1%含有溶解バッファー中で、培養して溶解する。溶解物をハイブリドーマI−2575によって産生されたモノクローナル抗体で免疫沈降させる。免疫沈降物から、還元条件下、見掛け分子量96kDの蛋白質化合物を回収する。このようにして得られた蛋白質配列をコード化するcDNAを合成する。昆虫細胞にこのcDNAを導入させる。導入された細胞、例えば、ハイブリドーマI−2575によって産生されたモノクローナル抗体で、産生された蛋白質を回収する。
【0021】
本発明は前記プロセスによって得られる分離蛋白質化合物、同様に、その分離蛋白質化合物のアミノ酸配列、及び分離蛋白質の配列を包含する。
【0022】
本発明の蛋白質化合物または蛋白質が配置される固体サポートが本発明の範囲内に含まれる。そのような固体サポートは特にセファロース粒を含む。
【0023】
本発明は、また、その配列が本発明の分離蛋白質をコード化する分離DNA(またはcDNA)を包含する。これは、また、DNAを導入することにより作り変えられた細胞、本発明の分離蛋白質を排出する、作り変えられた細胞及び前記作り変えられた細胞の培養液から分離された分離蛋白質に関する。
【0024】
本発明は特に、以下の段階から得られるcDNAを包含する。
ハイブリドーマI−2575によって産生されたモノクローナル抗体が結合する細胞からなる細胞集団を採取し(例えば、それに対しては、このmAbが抗原と認められる)、または、その溶解物がこのモノクローナル抗体と免疫沈降反応するような細胞で、例えば、全血中細胞及びPBLからなる群から選択される細胞を採取する。
もし、SC5の単独の経膜型が所望されるときは、前記細胞集団が主として、非活性化細胞から構成されていれば(例えば、休眠中の非腫瘍性細胞)、SC5発現が細胞表面で増加するように、例えば、採取された細胞集団をCD3活性化体(例えば、PHAのようなポリクローナル活性化体の1μg/ミリリットルを、採取された細胞集団がPBLまたは全血液または固定化、抗CD3抗体またはPHAのようなポリクロナール活性化体及び抗CD3抗体の両者であり、採取された細胞集団が、CD3+T細胞またはCD45RO+細胞またはNK細胞またはマクロファージまたは顆粒球から、主として、構成されている細胞集団のようにより均一であるときは、)と培養することによって、CD3経路を刺激する。
もし、採取された細胞集団が主として、活性化細胞(CD3活性化非腫瘍細胞)から構成されていれば(例えば、休眠中の非腫瘍性細胞、またはCD4+CTCL細胞のような腫瘍細胞)、そのときは、CD3刺激は、SD5の存在が、これら活性化細胞において、既に、主として、経膜的になっているため不要である。
もし、SC5の細胞内型及び経膜型の両者が所望されるときは、非活性化細胞が好まれ、CD3活性化は省略される。
前記細胞集団からのトータルmRNA集団の抽出及び精製(mRNAの抽出及び精製に使用できる市販キットの例:polydT column)。
各相補的cDNAの合成(逆転写酵素で、例えば、Seed B及びArrufo A 1987、Proc.Natl.Acad.Sci.USA 84:3365〜3369、に参考文献が収録されている。)。
このクローンのcDNAの発現が可能になるように、好適なクローン培養条件で各cDNAのクローン化を行い、各クローンを培養する。
ハイブリドーマI−2575によって産生されたモノクローナル抗体が抗原抗体型反応を行なうのに好適な条件下に、ハイブリドーマI−2575によって産生されたモノクローナル抗体が結合する化合物を発現するクローンを選択し、及び/または溶解物がハイブリドーマI−2575によって産生されたモノクローナル抗体と免疫沈降反応を行なう化合物を発現するクローンが選択される。
このようにして選択されたクローンを選択的に増幅し、選択されたクローンから挿入されたcDNAを回収する。
【0025】
このようにして回収されたcDNAは本発明におけるSC5cDNAであり、標準的なDNA配列技術により配列される。
このクローンのcDNAの発現が、好適なクローン培養条件で、各クローン培養によって、可能になるように、各cDNAのクローン化が、専門家にとって実施可能な、通常のクローン化法によって行なわれる。クローン化法の1例は、宿主細胞のcDNAの発現のために好適なプロモーターの制御下に、トランスフェクション(遺伝子導入)ベクターに各cDNAを挿入し、宿主細胞にトランスフェクション(遺伝子導入)ベクターを導入し、cDNAを導入された宿主細胞を、それらの代謝に好適な条件で培養する。好適なトランスフェクションベクター及び宿主細胞はBaculoウイルスのようなウイルス及び昆虫細胞(Sf9細胞)のような宿主細胞またはベクターとして牛痘ウイルス、宿主細胞としてEVB細胞(Rindis Bacher reference supra 参照)。
このようなcDNAは細胞への導入及びこのようにして得られたクローンからのSC5の産生に使用される。これはまた、本来のSC5mRNAの同定及び分離の試験に使用される。
【0026】
故に、本発明は、また、本発明におけるcDNAに相補的の、前記mRNA集団から選択可能な分離mRNAを包含する(例えば、前記cDNAに下記の文献記載の厳密な条件下、ハイブリダイズするmRNAの選択によって、Freemann G.J.etal.1992 J.Immunol.149:3745)。前記細胞集団の全mRNA集団からの回収する試みとして、本発明における分離cDNAを使用して、得られる分離mRNAを包含する。このmRNAは抽出されるcDNAに相補的である。一方、通常の技術の一つが、選択されたクローンから回収されるcDNAを直接配列化し、この配列からmRNAの配列を導き出す。
【0027】
本発明は、また、本発明における分離されたmRNAをコード化するDNAを包含する。特に、ゲノムバンク(www.ucsc.edu.のような)の上記SC5cDNAにマッチするゲノムDNAの調査によって、得られるゲノムDNAを包含する。
【0028】
本発明は、また、本発明における、cDNA、mRNAまたはDNAが導入された工学的細胞を包含する。
【0029】
本発明は、また、SC5特異性試料及びプライマーを包含する。そのような試料及びプライマーは専門家にとって、使用可能な技術によって得られる(例えば、候補配列として、SC5配列の特異な部分を選択し、これらDNA/RNAバンクの候補配列をSC5以外の細胞受容体の配列と対比し、前記対比後、SC5配列が特異であると見なされる、及び/または、前記対比後、SC5配列の鎖長が特異であると見なされる候補配列を選択する)。
【0030】
本発明の範囲内の発明には、cDNA、mRNAまたはゲノムDNAを担持するいかなる固体支持体も含まれる。このような固体は特にDNAチップまたはDNA微小球を含有する。
【0031】
本発明は、また、本発明に記載される分離cDNA、mRNAまたはDNAによってコード化された分離蛋白質を含有する。
【0032】
更に、休眠中の正常な細胞との対比で、CTCL細胞に固有である部分である限り、本発明は、本発明の蛋白質のあらゆる分離部分を含む。特に、C.N.C.M.に寄託番号I−2575として寄託されているハイブリドーマによって産生されるモノクローナル抗体によって確認されている部分を包含する。また、前記部分が前記細胞によって発現され、ハイブリドーマI−2575によって産生されたmAbによって接触されたとき、この部分が、正常細胞のCD3誘発増殖の調節(阻害または刺激)を誘発でき、及び/または、CTCL細胞のような悪性T細胞の生体内増殖の調節を誘発でき、及び/または、正常T細胞または悪性T細胞のIL−2(インターロイキン−2)産生の調節を誘発できる、ような形態の発明の蛋白質化合物の分離された部分を包含する。特に、この部分が、経膜部分で、悪性T細胞によって発現され、休眠中の非腫瘍性細胞によって、細胞質小球内で、発現される分子の一部であるような形態の発明の蛋白質化合物の分離された部分を包含する。本発明は、特に、本発明におけるSC5蛋白質の酵素的開裂によって得られるポリペプチド断片のバンクに関与する。好ましい酵素は、Tyr、Phe、Trpレベルを開裂するセリンエンドペプチダーゼ(例えば、α−キモトリプシン)、グルタミン酸及びアスパラギン酸のC末端での開裂を行なう酵素のような、蛋白質分解酵素から構成される。例えば、Shesberadaran and Payne 1988、 Proc.Natl.Acad.USA 85:1〜5参照、この内容はここでも、参考として組込まれている。SC5ポリペプチド断片のバンクは、質量分析のような特別な分析に使用される。このような分析は、ポリペプチドバンクを特徴によって分類するのに使用され、SC5の特徴であり、SC5の検出に使用されるポリペプチドのプロフィールを示す。本発明はこのような用法及びプロフィールを包含する。
本発明は、特に、本発明のSC5蛋白質化合物の細胞外、経膜部及び内部細胞質部分を包含する。受容体分子の細胞外、経膜部及び内部細胞質部分の決定は好適な技術に従がって、専門家によって為され得る。これらは分子全体において、グリコシルサイト(細胞外部分)を含む領域、疎水性(経膜部分)領域及び親水性領域、(内部細胞質領域)の同定によって、同定され得る。そして、それらは開裂または合成によって分離され得る。
本発明の蛋白質化合物の細胞外部分を分離する一つの方法は、前記のように、蛋白質化合物全体を処理するもので、これに、分子全体から細胞外部分を開裂させるために酵素処理の段階を付加する。この開裂のための好適な酵素としては、前記のように、V8プロテアーゼまたはα−キモトリプシンが含まれる。
【0033】
このように、本発明に包含されるポリペプチド化合物は以下のようにして得られる。
前記した、例えば、PBLまたは全血液細胞のようなSC5細胞源を採取する。
もし、ポリペプチド化合物が細胞表面で自然に発現されるときの形態で、分離されることを望むときは、または、採取されたSC5細胞源が、主として休眠中の正常細胞を構成するときは、例えば、適当なCD3活性化体と共に培養して(例えば、SC5源が全血液細胞またはPBLであるとき、PHA1μg/ミリリットルによる培養)、CD3経路を刺激する。そして、例えば、ビオチン(全細胞表面のラベリング)のようなポリペプチド特異性ラベルで該細胞をラベリングする。
そして、マルチポリペプチド複合体を分離するために、SC5細胞源(例えば、PBLまたは全血液細胞)を、例えば、Triton X-100のような強力界面活性剤の1%からなる溶解バッファー中での細胞培養により、溶解する(それによって、自然に結合したトランスデューサーとエフェクターからSC5を分離回収する)。
該溶解物を酵素による消化してSC5を部分に解裂させる。例えば、V8プロテアーゼまたはアルファ−キモトリプシンからなる群から選択される酵素である蛋白質分解酵素で該溶解物を培養する。そして、該溶解物を、ハイブリドーマI−2575によって産生されたモノクローナル抗体と免疫沈降反応を行わせ、該免疫沈降物からラベルを帯びたポリペプチド化合物を回収する。
回収された化合物は、完全なSC5細胞外部分または、完全なSC5細胞外部分の一部であるSC5部分であるか、または、そのようなSC5細胞外部分を構成するものである。このようにして得られるポリペプチド化合物のセットがSC5細胞外部分バンクを構成する。そのようなバンク及びその医療的使途、例えば、T関連疾病(例えば、質量分析による)の診断のための使用は本発明の範囲内にある。
この分野の通常の専門家は、その溶解及び酵素的消化を一段階で達成できることを知っている。
細胞全体のラベリングは、細胞外部分、または、細胞外部分の一部、または、その部分を構成するSC5部分を同定するために、細胞溶解に先だって為される。
【0034】
本発明は、また、本発明の蛋白質の部分を構成する分離された化合物を包含する。
【0035】
本発明の抗体(mAb)は、この抗体が結合するSC5エピトープの同定が可能である。通常の強制された突然変異誘発のような技術が好適である(例えば、Chang H.C.et al. 1989、J.Exp.Med.169:2073〜2083、参照、この内容は本発明の明細書に参考として組み入れてある。)。
【0036】
本発明の蛋白質及び本発明によるその部分、及び、特にその細胞外部分は、それらに対するモノクローナル抗体の産生を直接できるようにする。そのようなモノクローナル抗体の例はハイブリドーマI−2575で産生される。本発明のモノクローナル抗体はSC5の検出、CTCLのようなTリンパ腫の診断に有効であり、且つ、正常CD4+及びCD8+T細胞;HIV感染T細胞、CTCLのようなTリンパ腫、の如き、ウイルス感染T細胞;のようなT細胞の好ましくなく、または不適当な増殖、及び/または活性、の調節体として有効である。
故に、本発明は以下の手段により得られるモノクローナル抗体を包含する。
(i)本発明の蛋白質または上記で既定したその部分に対して或動物に免疫性を与える。
(ii)この動物の脾臓細胞からハイブリドーマを産生し、これを培養してその培養上澄み液中にモノクローナル抗体を産生させる。
(iii)段階(i)の免疫源として使用される蛋白質または蛋白質部分に結合し得、且つ、下記のような特性からなる群から選択される少なくとも一種の特性を有する抗体の上澄み液中での存在を調べる(評価する)。
主として細胞質分画中で休眠中の非腫瘍性T細胞に結合し、且つ、主として細胞表面で悪性CD4+CTCL細胞に結合する。
T細胞のCD3活性化経路を調節する。
T細胞からのIL−2産生を調節する。
T細胞のCD3誘発増殖を調節する。
CD3誘発の、CD4+CTCL細胞のような悪性T細胞の試験管内増殖を調節する。
動物中、好ましくはヒト中の、CD4+CTCL細胞のような、悪性T細胞増殖を調節する。
ハイブリドーマI−2575によって産生されたモノクローナル抗体のような本発明のモノクローナル抗体との、本発明の蛋白質への結合または上記で規定したその部分ヘの結合についての競合、及び、特に、該蛋白質の細胞外部分との競合。
(iv)所望の抗体を産生するハイブリドーマの選択及びクローン化。
(v)上記クローンの上澄み液かの抗体の回収。
【0037】
ハイブリドーマの産生は通常の技術で行なわれる。これは、前記動物からの脾臓の除去、脾臓細胞の懸濁液の調製、脾臓細胞を骨髄腫細胞と共に溶融プロモーターの存在下で溶融する過程、溶融細胞を別の容器で、非溶融細胞を生育させない培養液で希釈、培養する過程からなる。本発明は前記免疫化動物から採取される腹水をも包含する。
段階(iii)でT細胞及び悪性T細胞は標準法でヒトから採取される。一方のクローンはCD4+CTCLHUT28細胞系(ATCC TIB−161)などが使用され得る。CD4+腫瘍の場合は、試料はCD4+腫瘍細胞を表示する体の部分から採取される(SSにあたるPBL、形質転換MFにあたる皮膚紅皮症、等)。前記諸特性は、この分野の通常の知識を使用して、専門家によって容易に評価され得る。段階(iii)で挙げられた諸特性の評価のための好適な実験条件の例は、以下の例で見出すことができる。この分野の通常の知識を有する者が、それらを特異な細胞または使用濃度に是正する。細胞増殖の調節は増殖の刺激及び阻害と異なる。CTCL細胞が使用されたとき、増殖阻害は治療用の本発明のmAbは、診断または治療用であるか、またはその両方であるところの全ての抗SC5mAbsである。例えば、蛋白質精製用担体、または、これらができる調節特性の行使用担体のような担体に、これらのmAbの一種または複数種を固定することが所望される用途がある。本出願は本発明のmAbが固定される担体を包含する。そのような担体には、特に、親和クロマトグラフのカラム用の好適なものが包含される(例えば、寒天粒)。
【0038】
本出願は、また、H鎖、L鎖、VH、VL、Fab、F(ab')2、CD1、CDR2、CDR3から選択される本発明のmAbの断片(抗SC5断片)を含む。本出願は、該断片を含むあらゆる化合物、すなわち、あらゆる抗体を含む。本出願は、特に、少なくとも、上記断片を含むヒト化抗体に関する。ヒト化抗体の産生技術は先行技術で広く述べられている(Farah et al.1998、Crit.Rev.Eucaryote Gene Exp.Vol.8 pp 321〜345、参照、この内容は参考として本明細書に組込まれている)。
それは、特に、化合物、及び特に、少なくとも、一つの、ここで規定された抗SC5断片及び、また、H鎖、L鎖、VH、VL、Fab、F(ab')2、CD1、CDR2、CDR3各断片からなる群から選択された少なくとも、一つの、抗CD4断片からなる、抗体及びヒト化(人体に適応させた)された抗体に関する(例えば、WO94/13804、多価、多特異性結合蛋白質及びその製造方法、発明者:Holliger et al.出願人:ケンブリッジ抗体技術有限会社及び医療調査協議会、参照;また、Merchant et al.ヒトの二特異性IgG、Nat.Biotechnol.1998 vol.16 pp 677〜681、参照)。
【0039】
先に示したように、本発明は通常の一般的発明構想で結びつく二つの分子、即ち、SC5及びp140を提供する。SC5は、新規分子で、既に、十分に述べた。p140分子は従来技術で、NK細胞のサブグループ及び健常者(非癌性)からのCD3+CD8+細胞によって発現される阻害受容体として述べられている。本発明は、p140が悪性T細胞、特に、CTCL細胞によって発現され、新規なp140対立遺伝子型(下記例3のクローン24;SEQ ID No.1)を提供し、新規蛋白質(SEQ ID No.2)をコード化する。この新規な対立遺伝子型のcDNA及びアミノ酸配列を以下に示す。
【0040】
SEQID No.1
【0041】
【0042】
SEQID No.2
【0043】
【0044】
SEQ ID No.3
前記のp140対立遺伝子型(クローン1.1)のDNA及びアミノ酸配列を下記に示す。
【0045】
【0046】
SEQ ID No.4
【0047】
【0048】
SEQ ID No.1はSEQ ID No.3と5つの部分で異なる。
得られたSEQ ID No.2は先行技術で開示されたp140(SEQ ID No.4)と四個のアミノ酸置換の種類が異なる(その位置を太いアンダー系で目印する)。即ち、20位のQの代わりにRが、92位のLの代わりにVが、102位のTの代わりにPが、401位のPの代わりにLが示されている(図11参照)。
この出願は、配列がSEQ ID No.1及び/またはSEQ ID No.3であるか、またはSEQ ID No.1及び/またはSEQ ID No.3から構成される化合物を包含する。また、SEQ ID No.1に相補的であるmRNA及びSEQ ID No.2をコード化するゲノムDNAを包含する。
【0049】
好適なp140蛋白質産生技術はこの分野の専門家が使用し得るものである。この技術は培養液中でp140蛋白質が産生されるように加工された細胞の産生を特に含む。そのような技術の一例は、以下の過程よりなる。SEQ ID No.1またはSEQ ID No.3をヴァキュロウイルスベクターに挿入し、このベクターで、Sf9細胞系のような昆虫細胞を移入し、培養液中の産生されたp140蛋白質を回収する(培養液を、例えば、セファデックスカラムのようなカラムで濾過、蛋白質化合物がその分子量の函数として、次いで、140kD溶出物の分離)。
生理学的条件下(生体内または生体内を偽装する試験管内の条件下)で、p140に結合可能な化合物については既に記載した。そして、好適な、この分野の専門家により、使用可能な技術により得ることができる。経膜受容体部分(例えば、前記細胞の二層リピッドの上層にあるSEQ ID No.2及び/またはSEQ ID No.4部分の認識)で、細胞により発現されるとき、p140に結合する化合物、ここで言う、前記生理学的条件下、p140分子を抗原(SEQ ID No.2及び/またはSEQ ID No.4)として認め得る化合物、によって、為される。
そのようなp140に結合する化合物は、特に、HLA−A11及びHLA−A3分子(天然HLA−A11リガンド)及び特に、Fc−HLA−A11のような溶融蛋白質及びFc−HLA−A3のような溶融蛋白質から構成される。
p140に結合する化合物は、また、p140抗血清から構成される(ウサギのような動物にp140に対する免疫性を与え、このようにして産生された抗血清を回収する。付加的な免疫精製段階は、抗血清中のp140の濃度を増加させるためのもので、例えば、p140カラムによる濾過による)。休眠中の、p140に結合する化合物は、例えば、AZ158、Q66、Q241のようなPende et al.1996 (J.Exp.Med.184:505〜518)に述べられている、p140SEQ ID No.4に対するモノクローナル抗体で構成される。そのような抗SEQ ID No.4mAbsとしては、また、新規なSEQ ID No.2が抗原として認められている。
mAb産生に使用可能な技術は、抗p140mAbs産生にも好適である。そのような技術の一例は以下の段階からなる。
(i)或動物にp140に対する免疫性を与える。
(ii)この動物の脾臓細胞からハイブリドーマを産生し、これらの培養液上澄み中にモノクローナル抗体を産生するために、これらを培養する。
(iii)段階(i)で免疫源として使用されるp140分子に結合でき、休眠中の良性T細胞には、主として、その細胞質小球に結合でき、悪性CD4+CTCL細胞には、主として、その表面に結合する抗体の、上澄み液中における存在を評価する(測定する)。
(iv)所望の抗体を産生するハイブリドーマを選択し、クローン化する。
(v)前記クローン上の上澄み液から抗体を回収する。
好ましいp140結合化合物は抗CD4実在物(例えば、二特異性mAb)を構成する。
p140結合化合物はCTCL細胞、特に、CD4+CTCLのような悪性T細胞の検出に有効である。かくして、これらはCD4+癌の検出に有効である。これらは、また、以下で述べるポリペプチド系ベクターとしても有効である。
【0050】
休眠中の観点によれば、本発明では、抗SC5mAbs及び本発明の抗SC5mAb断片からなる群から選択される少なくとも一つの要素からなるポリペプチド系ベクターが提供される。このようなポリペプチド系ベクターは活性化されたT細胞に到達するか、またはその付近の活性、理論的、分子に有効である(例えば、炎症性疾病またはウイルス感染T細胞または悪性CTCL細胞及び特に、悪性CD4+CTCL細胞などにおいて、観られる活性化されたT細胞)。
また、それは、SEQ ID No.2及び/またはSEQ ID No.4の細胞外部分に対するmAbのようなp140結合化合物からなるポリペプチド系ベクターに関する。このようなポリペプチド系ベクターは分子送達、または悪性T細胞、特に、CTCL細胞及び、特に、悪性CD4+CTCL細胞付近に有効である。
分子は、特に、放射性元素または毒素のような細胞死またはアポトーシスを誘発することができる分子からなるCTCL細胞のような悪性T細胞に送られて有効な働きをする。そのような分子は、また、カルボキシペプチダーゼのように、抗有糸分裂性のプロドラッグから活性な医薬へと形質転換させることが可能な酵素からなっている。そのような酵素を運ぶポリペプチド系ベクターは、その際、抗有糸分裂性のプロドラッグの組み合わせによって有効に使用される。本発明はそれ故、そのようなベクター及び抗有糸分裂性のプロドラッグからなる医薬品キットを包含する。
これら隣接する細胞から前記悪性T細胞への抗腫瘍効果を誘発する作用力によって、悪性T細胞の付近にある細胞の免疫機能を刺激することができる分子、特に、構成するCTCL細胞のような悪性T細胞の近傍に送られて有効に働く分子がある。そのような分子の例は、特に、γインターフェロンのようなサイトカイン及びIL−2(例えば、Xu et al.2000,Cancer Research 60:4475〜4484参照)のようなインターロイキンである。
本発明のポリペプチド系ベクターが、CD4+CTCL細胞のようなCD4+細胞を標的にしようとするときは、更に、抗CD4体を含むことが好ましい。
【0051】
本発明は、また、前記ポリペプチド系ベクター、特に、前記ポリペプチド系ベクター含有薬剤で構成される医薬品組成物を包含する。そのような薬剤は不適当または好ましくないT細胞の増殖及び/または機能的活性(好ましくないIL−2の産生を含む)の制御に有効である。それらは、特に、不適当な免疫活性、の予防、一時的緩和、軽減、または治療;及び、特に、CTCLのようなTリンパ腫、及び、特に、CD4+CTCLの妨害及び阻害に有効である。不適当なT細胞の増殖及び/または機能的活性の予防、一時的緩和、軽減、または治療を目的とする、薬剤製造のためのこのようなベクターの使用、及び、CTCLの増殖の妨害及び阻害は、本出願の一つの目的である。
【0052】
SC5に生理学型条件下結合する化合物はSC5の活性を刺激するためのSC5リガンド作用物質として使用可能であり、このSC5分子は細胞阻害受容体として関与する。そのような抗SC5化合物は、即ち、抗SC5抗血清、本発明の抗SC5mAb、本発明のFab、F(ab')2断片、本発明の人体に適応するmAbsからなる。CD4+細胞が特別に関与するときは、前記抗SC5化合物は、更に、幸便に、抗CD4体を含有する。
抗SC5化合物は、特に、SC5+細胞の不適当または好ましくないT細胞の増殖及び/または機能的活性の調節に有効である。これらは、T細胞、CD45RO+細胞、CD3+細胞、CD4+T細胞、CD8+T細胞またはウイルス感染T細胞(例えば、HIV感染CD4+T細胞)またはCTCL細胞及び特にCD4+CTCL細胞のような悪性CTCL細胞の増殖の調節に有効である。これらはT細胞からのIL−2の産生にも有効である。C.N.C.M.に寄託番号I−2575で寄託されたハイブリドーマによって産生されたmAbが使用されたとき(5種類のIgM)、この調節は阻害の方に傾く。故に、CTCLのようなTリンパ腫の阻害に特に好適である。この出願は活性成分として少なくとも一つの抗SC5化合物からなる薬剤を包含する。このような薬剤はTリンパ腫、CTCL、ウイルス感染(例えば、HIV)のようなT感染、自己免疫型の諸症(例えば、リュウマチ性関節炎、脊椎関節症、または、乾癬、湿疹、アトピー性皮膚炎のような皮膚免疫薬物療法性疾病)のような炎症性疾病の予防、一時的緩和、治療が可能である。このような薬剤は移植改善(移植拒絶反応の抑制)も意図され得る。
【0053】
本発明は、補体漸増剤またはADCC活性化剤または刺激剤としての抗SC5化合物の使用を包含する。
前記で規定したp140結合化合物は、CTCL及び特にCD4+CTCLのようなTリンパ腫の予防、一時的緩和、軽減、または治療において、補体漸増剤またはADCC刺激剤または活性化剤として使用され得る。故に、本発明は抗Tリンパ腫薬剤、抗CTCL薬剤、抗CD4+CTCL薬剤の製造におけるp140結合化合物の使用及びその薬剤自体をも包含する。
【0054】
本発明は開発段階でのCTCL(段階化)の評価方法を提供する。即ち、悪性CD4+CTCL細胞の患者の体の或一定区分内における比率(例えば、%)が測定できる。この方法によれば、本発明のSC5蛋白質、本発明のSC5部分(特に、細胞外部分)、及びp140分子からなる群から選択される一要素をその表面で発現するCD4+細胞の比率は、前記体内区分から採取された生物学的試料によって測定される。前記体内区分内の現実のCD4+CTCL細胞の比率は、上記で測定された比率と本質的に同等とみなすことができる。それは、通常、この測定値の概略±10%以内である。
【0055】
本発明はCTCLの診断方法をも提供する。そこでは、本発明のSC5蛋白質、本発明のSC5部分(特に、細胞外部分)、及びp140分子からなる群から選択される一要素をその表面で発現するT細胞の比率は、患者から採取された生物学的試料によって測定され、ヒトの非CTCL(健常者が好ましい)の平均百分率と比較され、前記測定百分率が前記平均百分率より有意に高いとき、CTCL陽性と決定される。
このCTCLの診断方法は、特に、CD4+であるCTCLに適用される。この場合、測定されたT細胞の百分率は、好ましくは、CD4+の百分率である。SC5を発現するCD4+T細胞の平均標準比率は、一般に、1〜15%であり、ヒトの場合で、通常、5〜10%である。SC5+CD4+T細胞の測定百分率が前記平均百分率より高いとき、CD4+CTCLが陽性との診断が結論される。
SC5蛋白質またはその部分が関与する限り、本発明の方法は一般に、前記体内区分内にあるCD4+活性化T細胞の比率が測定できる。これは移植が拒絶反応を誘発するかどうか、及び自己免疫型の諸症(例えば、脊椎間接症のようなリュウマチ性関節炎、乾癬、湿疹、アトピー性皮膚炎のような皮膚免疫薬物療法性疾病)のような炎症性疾病、の診断及び追跡調査の評価に有効である。前記生物学的試料は内部滑液状でもよい。これは、また、非CTCLTリンパ腫の診断及び監視にも有効である(例えば、CD4+ALLまたはCD4+T−LL)。
【0056】
本発明のCTCLの評価/診断方法は全ての種類のCD4+CTCL、特に、形質転換MF、SS、LP及びCD30+リンパ腫に著しく有効である。前記生物学的試料は、例えば、紅皮症(例えば,形質転換MFの疑いある)患者から採取した皮膚試料及び/またはPBLまたは総合血液試料(形質転換MFがより進行性のSSのような形に進展したかどうかを評価するために)であってもよい。
CD4+細胞表面で検出された要素がSC5化合物のときは、細胞学的観察(腫瘍状細胞学的調査)の(前駆的または将来的)段階は、90%以上の確度で、現実のCD4+CTCLであるとの決定が為されることが好ましい。p140がCD4+細胞表面で検出されたときは、追加の段階は不要であり、CD4+T細胞の表面でのp140の検出という単純な事実が、直ちに、CD4+CTCLの陽性診断になる。
【0057】
本発明は好適な要素、特に下記の各要素からなる群から選択される一要素によって実行され得る。即ち、本発明の抗SC5モノクローナル抗体、本発明のFab、F(ab')の2断片、本発明から導かれる人体適応mAb、本発明のSC5細胞外バンク(前記で規定した)、SC5cDNA、mRNA、本発明のゲノムDNA、本発明で規定されるp140結合化合物、p140DNA、SEQ IDNo.1またはSEQ IDNo.3またはコード化SEQ IDNo.2またはSEQ IDNo.4からなるDNA。
このような要素はSC5またはp140細胞表面の発現の検出手段として使用される。前記検出手段はSC5/p140+CD4+細胞を直接一段階で行なうための、抗CD4含有のものである。
cDNA/mRNA物質が検出手段として使用され、このとき、CD4+細胞は、前記細胞に現実に存在するSC5またはp140をハイブリッド化するmRNAを検出するために、溶解されねばならぬ。このようなcDNA/mRNA検出手段は、DNAチップまたはDNA微小球のような固体担体上またはその中に施されるのが好ましい。
mAb物質またはmAbから導かれる物質は検出手段として使用され、これによって、SC5またはp140の表面発現が検出される。好適な抗体抗原技術は好適なものである。その例は特に流動細胞分析である。
【0058】
本発明は、また、緩和、予防、軽減、特に、CTCL細胞、及び特に、CD4+CTCL細胞のような、特に、悪性T細胞の増殖のような、好ましくないか、または、不適当なT細胞の活動の治療に有効な化合物の同定方法を包含する。この方法は、前記規定の、SC5蛋白質、SCポリペプチド化合物、または、p140分子(SEQ IDNo.2及び/またはSEQ IDNo.4のような)に結合できる化合物の検出からなることを特徴とする。このような化合物は、前記のように、ポリペプチド系ベクターとして使用される。付加的にSC5化合物に結合するこれら化合物は、細胞表面にこれらSC5分子を集合することができ、SC5の阻害機能を刺激できる。この同定法を実行する容易な方法は、蛋白質AセファロースCL−4Bのような固体担体上に、SC5及び/またはp140分子を固定化処理する方法である。この方法を続行するための化合物の候補は、特に、SC5またはp140に対して、免疫性のある血清からなる。
【0059】
本発明の例を図により説明するが、下記の特別な具体例が、本発明を限定するものではない。この分野の通常の知識を有する者が、二者択一的、試験的に、具体例を実施し、このような、この分野の通常の知識から導かれた具体例は本発明の範囲内にある。通常の知識を有する者は、各種の標準的免疫技術、各種の標準的分子生物学的技術が彼にとって、有効なものであることを評価する(例えば、抗体:実験手引、Ed Harlow、David lane、Cold Spring Harbor Laboratory 1988;Maniatis 1982 Molecular cloning:a laboratory manual、Cold Spring Harbor、New York、 Cold Spring Harbor Laboratory;Current protocols in Immunology protocols on CD−ROM、John Wiley、West Sussex、England;Immunological techniques made easy、edited by Olivier Cochet et al.、John Wiley、West Sussex、England;and the publication specifically referred to in the present application)。これら公知文献の内容は参考としてこの明細書に組み入れている。ここに、完全な明細が記載されている、実験における一つの特別な型のCD4+CTCLは、この分野の専門家によって、休眠中の何らかのCD4+CTCL型に置き換えることができる。
【0060】
これら実施例では、下記の図が参考のために掲げられている。
図1A、1B及び1Cは健常者のPBLsのSC5分子の細胞膜発現を図解している。健常者のPBMCを抗SC5mAb(腹水1/200)で汚染させ、次いで、FITC接合アイソタイプ特殊ヤギ抗マウスIgM2次試薬及び、抗CD3(図1A)、抗CD56(図1C)及び抗CD45RO(図1B)のようなPE接合mAbsの一種で汚染する。二重汚染されたリンパ球通過規制の細胞の比率は下部右象限に示される(図1A:3.3;図1B:5.1;図1C:0.6)。この実験は研究10ドナーの代表である。
【0061】
図2はPBLsの活性化の間、SC5分子の細胞膜発現を図解する。PBMCは1μg/ミリリットルPHAで刺激され、SC5及びCD69の発現のカイネチックスはCD3+細胞と並列に研究された。
図3はPBLのSC5分子の細胞内所在部を図解する。抗SC5、抗CD35、mAb鎖及び抗BY55アイソタイプ適合mAbが、健常ドナーの透過性及び非透過性の通過規制リンパ球の汚染に使用された。陰を施されたヒストグラムは不適切なコントロールmAbに対する図示mAbsから得られたラベリングを表す。
図4ではSC5分子の生化学的分析結果を図解する。DS6T細胞クローンはビオチン及び1%Triton X-100で、表面をラベルされ、溶解物は、抗SC5mAbを使用して免疫沈降される。試料はSDS10%PAGEで還元条件下分析される。抗SC5mAbは二種類の濃度、1/200(レーン2)及び1/500(レーン3)で使用される。ネガティブコントロールサンプルはヤギ抗マウスIg単独(レーン1)及びアイソタイプ適合mAb(レーン4)で免疫沈降させられる。分子量マーカー(kD)左に示されている。
【0062】
図5A、5B、5C、5Dは、抗CD3誘発増殖及び正常Tリンパ球のサイトカイン分泌を、それらの細胞毒活性の影響なしに、抗SC5mAbが調節する様子を図解する。即ち、
図5A:PBLまたはT細胞クローンLSO及びGDS.3が固定化抗CD3mAb(コートされたmAb1μg/源)またはIL−2(50IU/ミリリットル)で、抗SC5mAb(腹水の、1:200最終希釈)またはアイソタイプ適合、不適切mAbの存在で、刺激される。結果は、少なくとも、3通りの実験の代表及び3通りの平均cpm・SDとして表している。
図5B:GDS.3CD4+T細胞クローンは抗SC5mAbの異なる濃度の存在下、図示された濃度のプレコート抗CD3mAbで刺激される。
図5C:抗SC5またはコントロールmAbの存在下、抗CD3刺激PBLの上澄み中のIL−2の分泌は、下記に挙げた物質の存在下、IL−2依存性T細胞クローンの増殖の測定によって評価される。上記物質は、(a)剤単独、(b)PBL/抗CD3+コントロールmAb、(c)PBL/抗CD3+抗SC5mAb の上澄み。データは3つの代表的実験からの3通りの平均値・SDを表す。
図5D:細胞毒CD8+細胞クローン(JF1)が、抗SC5mAb(腹水の、1:200最終希釈)またはアイソタイプ適合コントロールmAbの存在で、E/T比が5/1での、FcyR+ミューリン腫瘍細胞P815との共培養の前に、培養される。標的の細胞は、表示された抗CD3の最終濃度で前培養される。結果は3通りの源泉の平均値で示される。
【0063】
図6A、6BはCTCL患者のPBLのSC5分子の細胞膜発現を図解する。
8人のセザリー症候群患者、3人の菌状息肉症患者及び8人の健常ドナーの血球の細胞が、CD4+リンパ球によるSC5分子の発現のための2色フルオレスッセンスで分析される。
図6A:セザリー症候群患者のPBLのSC5及びCD4分子の共発現を健常者と比較している。
図6B:ヒストグラムは示された3群におけるSC5発現の平均値を示す。SS患者と休眠中の2群の間には統計的に有意に差があることが解る(Mann-Witny U test、p<0.001)。
図7A、7B、7Cはセザリー症候群患者の悪性細胞の膜表面発現を図解する。
図7A:循環TCRVβ22+悪性T細胞クローンの存在で、SS患者のPBMCの二色蛍光分析。細胞は抗SC5mAbで汚染させ、次いで、PE接合アイソトープ特殊ヤギ抗マウスIgM及びFITC接合抗TCRVβ22mAbによる。
図7B:弱くラベルした、抗SC5mAb(陰を付したヒストグラム)TCRV竈22陽性悪性細胞。
図7C:同じ患者の細胞を、PE接合抗CD69mAb及びFITC接合抗TCRVβ22mAbによって汚染される。
【0064】
図8は抗SC5mAbによるCTCL悪性細胞系の抗CD3誘発増殖の調節を図解する。Pno細胞系は、IL−7(10ng/ミリリットル)、PMA単独(2ng/ミリリットル)、固定化抗CD3mAbまたはPMA/抗CD3mAbの組み合わせによって刺激される。SC5mAbまたはアイソタイプ不適当mAb(腹水の、1:200最終希釈)が培養の開始時に添加される。結果は4種の実験の代表及び3通りの源泉の平均cpm・SDとして示される。
図9A、9Bは、CTCL細胞系(図9A)のp140/KIR分子及び新鮮なリンパ球腫瘍(図9B)の検出を図解する。ダブル免疫汚染流動細胞分析は、Bagot et al.1998(Blood 91:4331〜4341)及びPoszepczynska et al.2000(Blood 96:1056〜1063)の記載によって行なった。
図10はp140分子の生化学的分析を図解する。即ち、健常者ドナーから導かれたNKクローンAM61(p140+/p70−)及びPno細胞系及びCou-Lがビオチンで表面をラベルされ、抗p140及び抗CD8mAbsで免疫沈降される。試料はN−グリコシダーゼFで処理(+)または非処理(−)され、還元条件下、8%(左パネル)または11%(右パネル)SDS−PAGEで分析された。セファロース蛋白−A単独はネガティブコントロールである。分子量マーカー(kD)は右に示される。
図11は蛋白質でコードされたKIR3D cl.24(SEQ ID No.2)及びcl.1.1(SEQ ID No.4)のアミノ酸配列を示す。信号ペプチドに一致するアミノ酸は小文字で示し、一方、経膜通過領域は一致する配列にアンダー系を施した。一致配列と同じアミノ酸はドットで示した。
【実施例1】
【0065】
SC5の分離、SC5の生化学的機能的特徴、抗SC5mAbsの産生。
Tリンパ球免疫応答は抗原特異性クローンを拡大させるポジティブ信号及び抗原自体への過剰な刺激または応答を抑制するネガティブ信号を発する機能的な細胞表面分子によって制御される。休眠中のTリンパ球の活性化は二つの同一の信号を必要とする。T細胞受容体(TCR)の関与によって発される信号は、通常のT細胞で応答を発させるために、2番目のポジティブ信号を同伴しなければならぬ。CD28はTリンパ球活性化及びIL−2合成を誘発し、細胞死を阻害するメジャー共刺激性受容体と見なされる。最近の研究はCD3/TCR刺激が、TCR/CD3が集合し、且つ、DIG結合性の信号導入分子となる、界面活性剤溶解性グリコリピッド強化膜フラクション(DIG)またはラフトの再配布に導くことを明らかにしている。CD28が、TCR関与サイトヘのラフトの再配布を惹起することが発表されている。更に、一連の休眠中の分子は異なる時点で活動し、特別な部分集合に影響し、または明確な実施機能を振興するT細胞に同時刺激を与えることができる。Tリンパ球の応答のネガティブな規制はCD152誘発信号またはTNF上科受容体メンバーを通しての細胞死に影響され得る。最近、NK部分集合及びCD8+リンパ球についての新しいタイプのネガティブな規制が報告された(Lan998、Annu.Rev.Immunol.16:359〜393)。免疫グロブリン様(KIRs)またはC型レクチン構造はこれらの信号に応じて、CTL活性の強化またはサイトカイン産生のような好ましくない実施体機能を阻害する。
さて、我々は、健常者の末梢血球の小集団を詳細に描写する新しい96kD経膜受容体について述べる。抗SC5mAbは主として、CD8及びCD4小集団並びにNK系統の細胞由来のCD45RO+リンパ球を汚染する。我々は、SC5が、主として、休眠中のTリンパ球の細胞内小球に局在し、細胞活性化後に、その表面膜発現が急速に増加することを発見した。SC5分子の関与が、抗CD3mAbに誘発される、休眠中のTリンパ球またはT細胞クローンの増殖を阻害した。それはこれら細胞のサイトカイン誘発増殖になんの効果も与えない。同時に、CD8+T細胞クローンに左右されるエファクター細胞毒活性は抗SC5mAbによって変更されない。更に、我々は、SC5+CD4+循環リンパ球の比率が、セザリー症候群(SS)患者においては、健常者の血中リンパ球に比較して、著しく増加していることを発見した。これは大切なことだが、我々は、セザリー細胞がSC5分子を発現することを明確に示すことができる。更に、我々は、皮膚T細胞リンパ腫(CTCL)の細胞系におけるSC5分子のライゲーションが、抗CD3mAbにより誘発される悪性細胞増殖の強力な阻害をすることを発見した。このように、SC5分子の発現はSS患者の循環、悪性CD4+細胞の存在の検知に役立つことができる。更に、ライゲーションにネガティブ信号を送る細胞表面分子が一致することは病理学的T細胞の機構の解明の新手段となり、その阻害に使用される。
【0066】
原料及び方法
抗SC5mAの産生
SC5mAは出版されているプロトコル(David et al.1990;J.Immunol.144:1〜6)を使用して、NK細胞系統Ytindiで6wk old Balb/c miceを免疫化して得られる。ハイブリドーマ上澄み液は、免疫化された細胞系及びPnoと称するCTCL細胞系(Poszcepczynska et al.2000;Blood 96:1056〜1063)とともに、間接的免疫蛍光及び流動血球計で、選択的再活性のために濾過された。Pno細胞系が患者のCTCL細胞から由来し、CTCL細胞はCD3+Vβ22+CD4+CD8αα+CD25フェノタイプを有し、IL−70及び抗CD3mAbヘの応答として増殖する。CTCL細胞系はこのスクリーンニングに好適である。休眠中のCTCL細胞系の例は、HUT78細胞系(ATCC TIB−161)を含む。反応性上澄み液は、更に、末梢血中リンパ球(PBL)との再活性を試験される。ハイブリドーマ上澄み液は腫瘍性細胞系及びPBL小集団の両者との、その再活性について選別される。ハイブリドーマは二回クローン化され、クローン化ハイブリドーマはプリスタン投与Balb/c マウス(複数)腹水を産生させるために継代接種された。抗体アイソタイプはIgMと決定され、抗SC5mAbと命名された。腹水はPBSで透析され、限外濾過で滅菌され、更に、1/200で最終希釈された。このハイブリドーマは2000年10月30日、C.N.C.M.(Collection Nationale de Microorganismes パスツール研究所;25、rue du Docteur Roux、F-75724 Paris Cedex15、France)にブダペスト条約(C.N.C.M.寄託番号I-2575)に従がって寄託され、C.N.C.M.寄託番号はI−2575である。この実施例の抗SC5mAbはハイブリドーマI−2575によって産生されたmAbに起因する。
【0067】
患者
インフォームド・コンセントの後、倫理委員会(CCPPRB、Hopital、Henri、Mondor、Creteil、France)の承認を受け、我々は11人のCTCL患者から血液試料を得た。8人の患者は、セザリー症候群に罹っており、10〜80%の循環CD3+、CD4+、CD8−セザリー細胞を有し、3人の患者は形質転換菌状息肉症(MF)で、CD3+、CD4+、CD8−フェノタイプを有する伝播性皮膚腫瘍と診断されていた。患者全員がこれまで化学療法を受けていなかった。
【0068】
細胞及び細胞系統
末梢血中モノクロナール細胞(PBMC)はFicoll-Isopaque の技法(Pharmacia、fine、Chemicals、Piscataway、NJ)で、密度勾配遠心分離で、分離される。ヒトT細胞クローンGDS.3(CD3+TCRαβ+CD4+CD8−)、DS6(CD3+TCRαβ+CD4−CD8−)、LSO(CD3+TCR聿+CD4−CD8−)及びJF1(CD3+TCRαβ+CD4−CD8+)がBensussan等、1984(J.Exp.Med.159:947〜952)及びYssel et al.1984(J.Immunol.Methods 72:219)に記載されているが、これらを8日毎に、同種の放射線照射PBMCと共に、下記からなる培養液に供給した。
RPMI1640(GIBCO、Paisley、UK)、2mmol/l L−グルタミン、ペニシリン(100U/ミリリットル)、ストレプトマイシン(100mg/ミリリットル)、10%熱非活性化ヒト血清、50IU/ミリリットル組替えインターロイキン−2(γIL-2、Sanofi、Synthelabo、Labege、France)、及び1μg/ミリリットル植物性赤血球凝集素(PHA)(Wellcome、Beckenham、UK)。機能的研究は供給7日後に行なわれた。標準的なヒト白血病(患者の)細胞系はマイコプラズマ・フリーで、完全なRPMI剤に10%子牛血清(FCS)及び抗体を補強した培養液中で対数函数的な生育カーブを保持した。ヒトCTCL細胞クローンPnoは、セザリー患者の末梢血液から得て、これを前記のように培養、保持した(Poszcepczynska et al.2000;supra参照)。Pno細胞系はCD3+Vβ22+CD4+CD8αα+CD25−フェノタイプを有し、IL−7及び抗CD3mAb刺激に応答して増殖する。休眠中のCTCL細胞系はHUT78細胞である(ATCC TIB−161)。
【0069】
モノクローナル抗体及び流動細胞計数の研究
間接的免疫蛍光物汚染ハイブリドーマ上澄み液または腹水液とともに、カルタグ研究所(カリフォルニア州、サンフランシスコ)調製のFITC接合ヤギ抗マウスIgMを使用して行なわれた。2色分析には、免疫蛍光汚染が、抗SC5mAbと3×105個の細胞が、30分、4℃の培養によって行なわれた。次いで、細胞は洗浄され、FITC接合ヤギ抗マウスIgM(カルタグ研究所)とともに培養、次いで、IgGアイソタイプのPE−、ECD−、TRI−接合、特殊mAbで培養された。汚染細胞の分析はアルゴン単独流動細胞分析計(Epics XL、Bekman-Coulter、Miami、FL)を使用して、(Schiavon V et al.1999;Tissue Antigens 53:23〜32)に記載のようにして行なった。細胞内ラベリングには、細胞をPBS4%含有パラフォルムアルデヒド中で4℃、20分固定した。次いで洗浄、それから、0.1%サポニン含有汚染緩衝液とともに透過させた。接合抗CD3、抗CD4、抗CD8、抗CD45RO、抗CD69及び抗TCRVβ22mAbsはインミュノテク(Immunotech、Marseille、France)から購入した。
休眠中のmAbsはBeckman Coulterから入手可能である。即ち、下記のカタログ番号参照抗。
CD3(PE)は、細胞分析2000カタログのIM1282、精製抗CD3は細胞分析2000カタログのIM0178、抗CD4は細胞分析2000カタログのIM0449、抗CD8は細胞分析2000カタログのIM0452、抗CD45ROは細胞分析2000カタログのIM1307、抗CD69は細胞分析2000カタログのIM1943、抗CD94は細胞分析2000カタログのIM2276、抗CD158aは細胞分析2000カタログのIM2277、抗CD158bは細胞分析2000カタログのIM2278、抗NKG2aは細胞分析2000カタログのIM3291、抗BY55は細胞分析2000カタログのIM2745。
【0070】
ビオチンラベル表面分子の免疫沈降
2×107個の細胞が、PBS中で2回洗浄され、Sulfo-NHS-LCビオチン(Pier Europe、Interchim、Montlucon、France)(PBS中2mg/ミリリットル)で、20分、4℃で、表面をラベルされた。RPMI1640で20分間、急冷した後、細胞はPBSで2回洗浄、下記の組成の溶解緩衝液中に1時間、4℃で、再懸濁された。
溶解緩衝液組成:(20mM Tris-HCl pH 7.5、150 mM 塩化ナトリウム、1% TritonX-100、1mMナトリウムヴァナデイト、10mM NaF、1mM PMSF、1μg/ミリリットルアプロチニン及び1μg/ミリリットルロイペプチン)。それから、ポスト細胞核上澄みが、2時間、4℃で、96-wellプレート(Maxisorp Nunc immuno-plate)で、ヤギ抗マウスIgG+IgM(カルタグ研究所)で予備コートされ、次いで、抗SC5mAbまたは抗TCRβ(抗TCRβC305アイソタイプに適合するmAb、細胞分析2000カタログのIM1466参照;抗TCRVβ22、細胞分析2000カタログのIM2051参照;抗TCRゼータ、細胞分析2000カタログのIM3169参照;全てはBeckman Coulterから入手可能。)で予備コートされた。免疫沈降物は洗浄用緩衝液(20mM Tris-HCl pH 7.5、150 mM 塩化ナトリウム、0.1% TritonX-100、1mMナトリウムヴァナデイト、10mM NaF、1mM PMSF)で4回洗浄し、沈降蛋白質はSDS−PAGEを使用する。ウエスタンブロット分析はストレプトアビジン−パーオキシダーゼ(インミュノテク)とECL検出システムをメーカーの推薦(Amersham Pharmacia、Orsay、France)に従がって使用して行なった。
【0071】
増殖アッセイ
リンパ球活性化のために、PBMCを、1μg/ミリリットルPHA(Wellcome、Beckenham、UK)添加の、15%ヒト血清含有培養媒体で培養した。増殖アッセイには、5×104個の細胞は3通りの96源泉丸底培養皿(Greiner、Nuertingen、Germany)で、最終容量0.2ミリリットルの培養媒体で行なった。必要なら、培養皿を前記濃度(Poszcepczynska et al.2000;supra参照)の抗CD3mAbで予備コートする。
CTCL細胞系Pnoの刺激のため、予備コートされた抗CD3mAbが2ng/ミリリットルホルボール12β−ミリステート13α−アセテート(PMA、シグマバイオケミカルス)との組合わせで使用される。細胞は4日間培養し、培養の終期の8〜16時間の間に3H[TdR]の1μCiを適用した。3H[TdR]の適用量は液状シンチレーションカウンター(Topcount;Packard Instrument Co,Meriden、CT)で計測した。IL−2産生量の定量では、105PBLが固定化抗CD3mAbで、抗SC5またはコントロールmAbの存在下刺激される。培養の32時間後、IL−2依存T細胞クローンから100μリットルの上澄み液が105個の細胞に加えられた。
【0072】
CD3誘発・転送・細胞毒アッセイ
転送・細胞毒アッセイは、Le Cleach et al.2000、Clin.Exp.Immunol.119:225〜230に記載されたように実施した。標的の二十日鼠肥満腫瘍のP815腫瘍細胞に100μlの51Cr(25mCi/ミリリットル)を37℃、90分付加し、洗浄、次いで抗CD3mAbで、15分間室温で培養した。エフェクター細胞は抗SC5mAb(腹水の最終希釈1:200)で前培養し、次いで、標的細胞に5:1の比で、添加し、最終容積150μlで、96源泉V字型底マイクロ滴定皿で、4時間培養した。遠心分離後、100μl部分標本を51Crの放出量を定量するために、γカウンターで計測した。自発的放出は常に最高値の20%減であった(1%NP40で標的細胞)。特異的51Crの放出量比率はDavid et al.1987、J.Immunol.138:2831〜2836に記載のようにして計算した。
【0073】
統計的分析
この結果の統計的分析は、スタティスティカソフトウエア5.0(StatSoft、Los Angeles、CA)によって行なった。患者とコントロールグループのSC5発現の間の大きな相違は、Mann-Whitney Uテストで評価され、SC5+CD4+細胞の比率及び患者の末梢血中の悪性細胞の比率の間の関係は、Spearman Rank Order Correlations testによって評価された。
【0074】
成果
ここに提出する成果はCTCL細胞系Pno及び患者から採取されたCTCL細胞によって得られたものである。例えば、HUT78(ATCC TIB−161)のような、どのCTCL細胞でも同等の実験が行なわれ、同等の成果が得られた。
【0075】
正常で、休眠中且つ、活性化末梢血リンパ球内のSC5分子の発現。
機能的腫瘍細胞系Pno Ytindiに対して、高められたモノクローナル抗体は、CTCL細胞系Pno(Poszcepczynska et al.2000、supra参照)及び末梢血中のT細胞について、同時反応性が分析された。IgMアイソタイプのSC5特異性mAbは免疫化とCTCL細胞系の両者を汚染した。末梢血リンパ球中、抗SC5mAbは、小集団(図1A、1B、1C参照)の輪郭を描いた。抗SC5mAb反応性分子は、20%を超えない(平均10.5%、SD±5.6)CD3+T細胞の可変比率によって発現される。CD4+及びCD8+リンパ球の両者は、抗SC5mAbで汚染される。個人間の差異(平均19.4%、SD±11.4;表1参照)がより重要ではあるが、末梢血中CD56+NK細胞の小集団も、また、抗SC5mAbで反応する。
【0076】
末梢血リンパ球小集団におけるSC5の発現。
【0077】
【表1】
【0078】
正常ドナー(提供者)のPBLのゲイトリンパ球における、SC5の、CD3またはCD56との、共発現の代表例が図1、1Aに示される。大部分のSC5が図1、1Bに示されるように、活性化/記憶−プールに属することに注意すべきである。SC5陽性細胞の70%以上が共発現CD45ROである。
試験された全IL−2依存性Tリンパ球クローンでSC5分子の発現が検出されたので、我々はTリンパ球活性化の間に、表面発現が誘発されたか、どうかを、立証した。我々は、PBLのPHAによる刺激がCD3+リンパ球におけるSC5の発現を増加させたことを見出した。SC%+T細胞の比率及びSC5発現のレベル、の両者が、刺激の後に急速に増加する(図2参照)。PHAの刺激後4時間で、SC5+CD3+細胞の比率が2倍から3倍に増加し、24〜48時間後に最高値に達した。5〜7日の試験管内の刺激の過程で、SC5発現は基本レベルに戻った。これと並行して、我々はCD69抗原の活性化の最初の発現を研究し、CD3+リンパ球におけるCD69及びSC5の類似した速動性的プロフィールを観察した。CD69分子と異なり、低レベルSC5が既に、刺激前に循環PBLに存在することに注目すべきである(図2)。
活性化誘導抗原発現はデ・ノボ蛋白質合成または細胞内先在分子の細胞表面排出の何れかの帰結をとる。それ故に、我々は、非活性化透過性PBLにおけるSC5発現を研究した。図3に示すように、抗SC5mAbは、非透過性細胞の16%に対して、透過性リンパ球の90%以上を汚染する。抗SC5mAb汚染の特異性が、循環リンパ球の小集団によって発現される、GPI固定細胞表面構造を検出する、アイソタイプ適合mAbBY55を使用して、確認される(Agrawal et al.1999、J.Immunol.162:1223〜1226参照)。BY55+細胞の比率は、透過性及び非透過性リンパ球において見出される。細胞内抗原決定基の検出のための陽性コントロールとして、我々は透過性Tリンパ球のみをラベルする抗CD3ζ鎖mAbを使用した。このようにして、SC5の表面発現は、細胞の活性化状態によってダイナミックに規制され、且つ、T細胞活性化の間の、その誘発は細胞内プールから表面膜への分子の輸送を振興することになる。
【0079】
SC5抗原の生化学的特徴
抗SC5mAb、IL−2依存性TCRγδ+クローン、DS6と呼ばれるものによって同定される構造の分子量を特徴付けるために、表面がビオチンによってラベルされ、細胞溶解物が抗SC5mAbまたはアイソタイプ適合抗TCRβmAb(陰性コントロールとして)の何れかで、免疫沈降される。抗SC5mAb還元条件下、見掛け分子量96kDで単一分子を確認する。腫瘍細胞系(免疫化Ytindi細胞)が、たびたび、炭水化物抗原決定基(20、22)の異常型発現によって、特徴付けられるので、我々は、ニュウラミニダーゼまたは過沃素酸ナトリウムを、公知のシアル化抗原決定基を分解するような濃度で、処理した後の、抗SC5mAbの、SC5+NK細胞系NK3.3及びCTCL細胞系HUT78(ATCC TIB−161)との反応性を研究した。SC5の発現は、ニュウラミニダーゼまたは過沃素酸ナトリウムの処理によっては影響されない。このようにして、我々は、抗原決定基の炭水化物の可能性を除外した。
【0080】
抗CD3mA誘発Tリンパ球の抗SC5mAbによる増殖の阻害。
正常細胞のSC5分子の可能な機能を調べるために、我々は、抗SC5mAb単独の効果を、または、固定化、抗CD3mAbで誘発された、増殖応答の期間にこれを研究した。我々は抗SC5mAb単独またはPMAとの組み合わせが、末梢血中Tリンパ球の増殖を誘発しないことを見出した。興味あることに、溶解性の抗SC5mAbが固定化抗CD3mAbと共存しているときは、リンパ球の増殖の著しい阻害が観察された。阻害は、ドナーによっては、アイソタイプ適合の不適切なコントロールmAbの効果に比較して、33〜66%の間で変化した(図5A上部参照)。SC5分子が単球で発現されるので、阻害効果が単球依存性であるかどうかを決めるのに重要であった。我々は、二つのT細胞クローン、即ち、抗SC5mAbまたはアイソタイプコントロールmAbの存在で、抗CD3mAbによって、刺激される、GDS3及びLSOの増殖を研究した。二つのT細胞クローンから得られた結果は、SC5分子の結合は、抗CD3mAbヘの増殖を阻害する結果となることを示した(図5A下部参照)。抗SC5mAbのT細胞増殖の阻害効果はその濃度及び拮抗的、抗CD3mAbの濃度に依存したことに注目すべきである。抗SC5mAbで得られた阻害は、T細胞が普通に希釈された抗CD3mAbで刺激されたときにのみ、著しいものだった(図5B参照)。このような阻害挙動は抗KIRs mAbsについても報告されている(Cambiaggi et al.1999、Blood 94:2396〜2402)。
【0081】
SC5の結合に続くT細胞増殖の低下は、細胞死、IL−2R発現の摂動またはサイトカイン合成の阻害に起因し得る。我々は、抗SC5mAbがPBLまたはT細胞クローンのIL−2依存性増殖の阻害をしないことを見出した(図5A)。更に、抗SC5mAb(希釈腹水1/200)と抗CD3mAbとの組み合わせによって48時間前培養した、T細胞へのγIL−2の添加はこれらの増殖を復興させる。特異性のmAbがIL−2R発現の阻害をしたり、細胞死の原因になったりすることはあり得ない。終わりに、SC5がサイトカイン産生の阻害の原因になることを解決するために、我々は、IL−2依存性T細胞系を使用して、抗SC5mAbの存在下または不存在下で、CD3活性化PBLにより排出されるIL−2の量を比較した。抗SC5mAbの存在下で産生されたIL−2の品質は、アイソタイプ適合抗体で得られるコントロール値の40〜65%で表される(図5C参照)。このようにして、SC5及びCD3分子の同時結合はサイトカイン産生を減少させる。
次に我々は、FcgR+二十日鼠細胞系P815に対するキラーアッセイに書き換えられた、抗CD3mAbを使用して、T細胞エフェクター細胞毒活性への抗SC5mAbの効果について研究した。図5DはE/T比5/1で、細胞毒のT細胞クローンによって行なった代表的実験を示す。大量のSC5分子を発現する、CD8+T細胞クローンJF1は抗CD3mAb刺激に続いて、二十日鼠標的細胞を殺すために誘発された。その特異的mAbによる、SC5の同時標的化は、E/T比または抗CD3mAbを刺激する濃度に関係なく、JF1細胞毒活性に影響しない。
【0082】
セザリー症候群患者から採取した悪性リンパ球におけるSC5分子の発現。
我々は、最初に、CTCL細胞系との反応性について、抗SC5mAbのスクリーニングを行なったので、更に、8人のセザリー症候群患者から採取した、末梢血中リンパ球のSC5分子の発現を研究した。二色細胞流量計分析が、SC5発現がCD4+小集団で非常に増加したことを示した(図6Aの一患者から得られた代表的結果参照)。平均で、SS患者の血液中のCD4+リンパ球の35.5%SC5を発現し、健常なコントロール(p>0.001)の9.4%に比較される(図6B)。8人のSS患者についての研究で、CD4+細胞のSC5発現が細胞生態学(r=0.91、p=0.0012)によって検出される循環、悪性細胞の比率と関連があることが解った。これらの患者の、トータルCD4+細胞の比率(平均88%、SD±8.8)はセザリー患者の細胞の比率(平均26%、SD±12.8)より高いことに注目すべきである。
我々は、3人の、血液に関連しない、MF患者の末梢血液試料における、SC5分子の発現をも研究した。末梢血中CD4+細胞におけるSC5発現のレベルでは、MF患者と健常ドナーの間に大きな差異はなかった(図6B参照)。増加するSC5+CD4+の比率は、我々が試験できた、これら二人の患者の皮膚試料から検出された(それぞれ、26%、19%)。
一人のSS患者では、我々は、予め、循環悪性、細胞が、CD4+TCRVβ22+フェノタイプ(Poszcepczynska et al.2000 supra参照)のクロナールリンパ球であることを示した。ここで我々は、SC5が、例えば、悪性細胞集団によって、TCRVβ22+細胞で共発現されることを示した(図7A)。TCRVβ22+細胞に結合するSC5mAbのプロフィールが、全ての、悪性、クローン集団における弱い均一な発現に該当することは注目すべきである(図7)。興味あるのは、悪性TCRVβ22+細胞は前記のように(Poszcepczynska et al.2000 supra参照)、共通の活性化マーカーとしてのCD69(図7C)、CD25またはCD30を共発現しないことである。このように、増加した、SSPBLにおけるSC5のレベルは殆ど、循環、悪性、T細胞における抗原の発現に起因する。
【0083】
抗SC5mAbによる抗CD3mAb誘発CTCL細胞系の増殖の阻害
次に、我々はSC5分子が悪性CTCL細胞におけるネガティブ信号を提供できるか否かを調べた。我々は、IL−7の存在下、著しく増殖する長期間培養Pno CTCL腫瘍T細胞系を使用した。先に報告したように、固定化、抗CD3mAbまたは溶解性抗CD3mAbs及びPMAの組み合わせはこれらの増殖を著しく誘発するから、(Poszcepczynska et al.2000 supra参照)、Pno細胞系は機能的T細胞受容体を発現する。我々はPno細胞の増殖でのSC5分子の結合効果を調べた。図8は、抗SC5mAbが、CTCL細胞系の抗CD3mAb誘発増殖を強力に阻害することを示す。対照的に、PMA単独(2ng/ミリリットル)では、抗SC5によっては、それほど影響されない低レベルの増殖を為す。抗SC5mAbの存在下、IL−7で培養したPno細胞はアイソタイプコントロールmAbの存在下と同じ強さで、増殖したので、抗SC5mAbはIL−7依存性増殖に何ら影響を与えない。
かくして、我々は、SC5抗原がCTCL悪性細胞で、機能的であり、且つ、特異的、予見的に、TCRCD3信号経路によって惹起されるこれらの増殖を阻害するとの結論を出した。
次いで、CTCL腫瘍が、機能的、活性なNK細胞及びTリンパ球の両者を、本質的に有しない(欠落または欠損)遺伝子導入マウス(SCIDマウス)に接種された[(Charley
et al.(1990)、CB-17 SCID、 ヒト皮膚T細胞リンパ腫のマウスへの移植、J.Invest.Dermatol.94:381〜384に記載のように]、(またUS5,530,179
Cornelius P.Terhorst and Baoping Wang for obtention of SCID mice参照)。抗SC5mAbについては、マウスの半数に、静脈または皮下に、複数回、マウス体重1gあたり、複数種の濃度の精製mAbが投与された。PBL及び皮膚細胞が、浸潤性,大T細胞の標準細胞観察基準に則って採取され、CTCL細胞の比率が、各供試マウスについて計算された。この方法は専門家が、C.N.C.M.に寄託番号I−2575として寄託されているハイブリドーマによって産生されるmAbのような、抗SC5化合物が、ヒトCTCLの生体内増殖を阻害できること、をチェックできる方法の一例である。
【0084】
討論
多くのダイナミックな表面発現を有する受容体は他の免疫細胞またはサイトカインとの相互作用に影響を与え、その結果としてT細胞免疫応答を制御する。この研究で、我々は、その表面発現が試験管内T細胞活性化の間に急速に誘発されるところの、PBLの矮小集団により発現される、新規な96kD機能性分子を同定した。SC5は類似の分子量及び/またはT細胞制御機能を有する移植分子とは明瞭に異なる。第一に、SC5は、4時間後に検知される、その表面発現の重要な増加によって、非常に早期にT細胞活性化上で誘発される。同様の速動性が、刺激の24時間後に細胞表面で検知される、非常に早期の活性化抗原CD69について観察される。4II−B、OX−40 LIGHTまたはCD−152(CTLA−4)のような、大部分が移植されたT細胞特異的、誘発可能な、受容体が、刺激に続いて、10〜24時間の間に、T細胞表面に現われる。更に、SC5発現はT細胞で限定されていない。抗原はNK小集団及びBリンパ球、単球及び顆粒球上で検知される。このような広い分布形態は、NK細胞、T及びBリンパ球、脊髄性及び他の抗原含有細胞などの上に見出される、最近発表されたAIRM1及びIRp60のような、ある種のキラー細胞Ig様受容体(KIRs)の典型である。免疫グロブリンファミリーは、転写(ILT)阻害受容体のように、全白血球タイプの発現を有し、個々の受容体は異なるNK、T、B及び脊髄性小集団上で検出される。その上、ILT4はSC5に極く近い分子量を有する。しかし、AIRM1及びIRp60、ILTRsはともに細胞の活性化状態に依存しない。
【0085】
我々が細胞透過の後に、大部分の非活性化Tリンパ球で、SC5を迅速に検出したので、SC5分子の活性化T細胞表面での誘発に、次いで、細胞内分子の細胞膜及び副ヴァーサ(versa)への通行に起因する制御緩和が続いた。そのような制御の機構は他のT細胞受容体、CD152、について報告されている。更に、膜蛋白(LAMP)に結合するリゾソームの不均一ファミリーが存在し、包含する分子はエンドサイトーシス、リゾソーム通行及び細胞内顆粒からの排出に関与し、単球、ニュウトロフィルNK、BまたはT細胞の表面で活性化へと、誘発される。細胞内の局在性及びSC5の移出の付加的研究結果は、T細胞活性化及び機能におけるその役割について正確である。抗CD3mAb誘発T細胞増殖を阻害するその特異的mAbによるSC5の惹起を明らかにした。この阻害効果は、PBLにおける最大SC5表面発現を誘発するのに必要な、通常の抗CD3mAb刺激を必要とする(図5B参照)。A:抗SC5腹水の1:200の最終希釈物は、抗CD3mAbの2μg/wellで誘発される増殖を著しく阻害し、この阻害効果は抗CD3mAbの低濃度(1μg/well)で増加する。即ち、0.5μg/well−抗SC5腹水の1:400の最終希釈物は抗CD3mAbの2μg/wellで誘発される増殖を十分には阻害しないが、抗CD3mAbの低濃度(1μg/well;0.5μg/well)では阻害は著しい。この分野の専門家は、ここで述べた特別な結果または彼自身の試みから得られる標準的書き加えによる修正を行なうことができる。
【0086】
同時に、SC5結合が提起した阻害閾値は過度の抗CD3mAb刺激により、迂回される。SC5分子の連結で得られた阻害機能的効果はAPC非依存性機構で観察されることに注目すべきである。更なる実験は、SC5刺激がAPCT細胞または同種被刺激T細胞の増殖に異なる効果を与えるか否かを決定するのに必要である。近年、免疫応答の陰性制御の研究が、特に、Tリンパ球及びNK細胞について、徹底的に行なわれた。最もよく研究されたT細胞特異的阻害受容体、即ち、CD152(CTLA−4)は、CD−28共刺激受容体に機能的に結合する、典型的な活性化誘発抗原である。実に、CD152は、低い抗原濃度で、CD−28に打ち勝ち、免疫応答を阻害するである。CD−28で誘発されるT細胞増殖をIL−2R発現及びIL−2合成に影響することによって、制御緩和及び終結させる。CD152と同様に、SC5は表面発現が細胞の活性化状態によって制御される細胞内分子である。両受容体とも、IL−2合成に影響することによって、それらの効果を左右する。SC5特異的リガンド及び信号経路の同定は、これらの陰性T細胞制御体間の時間的、空間的、関係を要約するだろう。この広い細胞分布を考慮すると、SC5機能効果は、多分、T細胞を制限しないだろう。
数種の観点で、SC5はNK細胞阻害受容体に、T細胞及び他のリンパ球小集団によって発現される点が同じことから、比肩し得る。KIRs及びCD94/NKG2受容体の両者は、抗原特異性及び、細胞毒、増殖及びサイトカイン排出を含むTCRαβ及びTCRγδT細胞のスーパー抗原駆動活性化を阻害することが示されている。T細胞の活性化状態及びT細胞刺激の効力に依存するKIR誘発阻害は、特異的生体内条件下で、迂回され得る。しかし、我々が、抗CD3mAb刺激T細胞の増殖を選択的に阻害したことを、これらのエフェクター細胞毒機能なしで、示したので、SC5は、KIRsとは明らかに別のものであることになる。
【0087】
SC5分子の重要な特徴は、SS患者の末梢血液リンパ球における発現が、健常者のそれに比較して非常に増加することである。SSは成熟したフェノタイプの、初期類表皮悪性T細胞クローンという概念で特徴付けられる。事実、移植されたT細胞機能性受容体は末梢血液SS腫瘍細胞とは同一でないとの結論が出ている。MF/SSは、特殊フェノタイプT細胞小集団、通常のTメモリー小集団に相当する、CD+CD7−が増大したものと考えられている。しかし、近年の研究は、CD+CD7−集団が、SS患者の優勢なT細胞クローンを常には、代表するものではないことを示した(Dummer et al.1999、Arch.Dermatol.Res.291:307〜311)。SSはその刺激性は知られていないが、特異的活性化T細胞の増殖であるとみなされる(Fargnoli et al 1997、Leukemia 11:1338〜1346)。この理由から、SS及び通常のT細胞での、活性化誘発抗原の明瞭な発現が特に興味ある点である。CD69、CD25、CD30のような移植された活性化抗原は、我々自身の研究で確認したように、セザリー細胞に常に検出されるものではない。CD25の発現はむしろ、病気の進行と各種少数ケースの大細胞へのMF/SS細胞の形質転換に関係する。HLA−DR、CD25またはSS中のCD71のようなT細胞活性化分子の発現の増加率は反応性腫瘍浸潤T細胞に優先的に関与する。78kDの活性化が非常に遅い抗原BE2は、正常T細胞では検出されないのに対して、特に、SS末梢血液で増加すると報告されている。事実、BE2は、報告された悪性細胞の比率に関して、末梢血中細胞の低比率のものは、BE2陽性であったのに比較して、研究された全てのSS患者において、常に検出されるわけではない。そして、BE2は、血液関与の臨床的証明なしに、患者において検出される。それと対照的に、SCは研究された8/8SSケースで検出された。悪性細胞の比率は細胞生態学で決められ、患者によってかなり差が出るが(10〜80%)、CD4+細胞のSC5発現はこの比率に関与する。終わりに、CD4+細胞のアップしないSC5発現は、血液の関与なしに、MF患者で検出される。更に、我々はセザリー細胞はクローンタイプTCRVβ鎖共発現SC5分子の発現によって同定されることを示し、SS患者の末梢血液におけるSC5の増加レベルは、悪性T細胞クローンにおけるその発現に原因があることを確認した。
【0088】
非常に重要な観察は、セザリー細胞のSC5トリガーがCD3に影響される腫瘍細胞の増殖の制御緩和を誘発することであり、これは正常T細胞において観察される阻害に類似する。悪性CTCLリンパ球が、サイトカインの異常型産生ではあるが、基礎的な信号経路によって活性化されるような、ある機能的性質を保護することは既に示された。我々は阻害経路がCTCL腫瘍細胞中で保護され、これらの増殖を平衡状態に保持することを示した。特に、阻害効果は抗SC5mAbが抗CD3mAbに結合した後に観察され、CTCLPno細胞で刺激されたCTCLPno細胞は正常T細胞におけるよりも重要である。それがrIL誘発増殖を制御しないので、抗SC5mAbは正常細胞またはセザリーT細胞の何れの細胞死にも、直接関与するようには見えない。更に、抗SC5mAbの存在下で培養されたT細胞はrILの添加によって再刺激され得る。同時に、我々は、少なくとも部分的にそれらの増殖阻害の原因になる、抗SC5mAb処理T細胞によるIL−2産生の減少を観察した。
【0089】
悪性細胞増殖の機能的受容体の阻害効果は、既に観察された。近年、同族NK細胞唾液粘性受容体AIRMI及びCD33骨髄特異性抗原受容体によって影響される正常細胞及び白血病骨髄細胞の増殖阻害が報告された(Vitale et al.1999、Proc.Natl.Acad.Sci.USA 96:15091〜15096)。SC5に類似せず、これらの受容体は、生育因子の存在で、最も可能性ある細胞死の誘発によって阻害される。興味あることは、他のT細胞活性化抗原、正常CD45RO+T細胞の小集団によって発現されるCD30は、MF/SSより優勢なCTCLの退行を左右する。
この効果はCD30(CD30L)のナチュラルリガンドに関与する。これは悪性細胞の細胞質内に共存し、細胞死をFas/FasLに無関係に誘発する(Mori et al.1999、Blood 94:3077〜3083)。この観点で、SC5リガンドの同定はその阻害行動のメカニズムを解明するであろう。
【0090】
結論として、我々は早期のフェノタイプ的生化学及び機能的基準が移植されたTリンパ球受容体と異なる活性化抗原を同定した。アンダーラインしたように、SC5分子はポスト−活性化/記憶末梢血液T細胞小集団によって発現される。同様に、KIRsの継続的発現が腫瘍T細胞クローン上に発現され、この発現は特異的抗原によるTCRの占有に依存するように見える。近年、そのようなクローンは自己反応性で、且つ、KIRs信号によって、耐性が保存されることが報告されている。我々は、セザリー症候群腫瘍細胞はそのようなクローンが起源となる可能性があることを示唆し、SC5発現も自己反応性T細胞上で保護されるものと推定した。CTCL悪性T細胞上でのSC5発現が、それらの起源、及び通常のT細胞が制御不能な増殖をするように駆り立てる信号の性質をよりよく規定するのに役立つことが重要である。更に、腫瘍T細胞上の抗SC5mAbの阻害効果は、進行性CTCL患者に対する新規な治療法を構成する。
更に、SC5発現も、CD8+形質転換MFのようなCD8+CTCLの表面で観察される。
【実施例2】
【0091】
P140はCD4+CTCL細胞によって発現されること及び新規P140対立遺伝子型の同定。
皮膚T細胞リンパ種(CTCL)は主として、皮膚に関与するリンパ腫の外生群である。菌状息肉症(MF)は本来のT細胞ヘルパー細胞に類似する、フェノタイプの、クローン由来の悪性CD4+Tリンパ球の皮膚侵襲が特徴である。よりアグレッシブなCTCL型は悪性細胞が類皮的になり、皮膚異常に関与する。セザリー症候群(SS)は、CD4+/CD45RO+T細胞のクローン拡大及びこれら悪性細胞の血液中への出現が特徴であるところの、CTCLのより進行性のタイプである。特異的細胞表面のマーカーがないために、悪性細胞を同定するのが困難なので、病気の生物学はよく理解されない状態が続いている。皮膚の病変では、CD4+CTCL細胞を反応性、浸潤性CD4+Tリンパ球から区別することは困難である。バゴット(Bagot)等は、以前に、CTCL病変由来のユニークなCD4+T細胞系について述べている(Bagot et al.1998、Blood 91:4331〜4341)。この細胞系及び生体内腫瘍細胞は、TCR−Vβ転写の相補的に決定する領域3と同一サイズで発現した。極く最近、Poszepczynska等は(Blood 2000、96:1056〜1063)他の皮膚紅皮症CTCL患者の血液から採取された、IL−7依存性CD4+CD8αα+腫瘍T細胞系を機能的に特徴付けた。TCR−Vβ22の発現及びTCRβ−VDJ配列の同一性によって示されるので、この腫瘍T細胞系は大循環性T細胞集団と同一だった。ここで、我々はこれら二つの異なるCTCL系が、HLA−A対立遺伝子のためのP140/KIR阻害受容体を発現することを示す。重要なのは、同じ患者由来の新しい分離腫瘍細胞で検出されることである。更にその上、P140/KIRは他の7人のSS患者のCD4+リンパ球のメジャーな集団及び進行した2人の追加MF患者の皮膚CD4+腫瘍の両者によって共発現される。P140/KIRは健常者のマイナーNK細胞小集団及び稀な末梢血中CD3+CD8+細胞で検出される。その上、皮膚炎及び毒物による表皮融解のような皮膚病患者の皮膚から得られたT細胞は、この受容体を発現しない(Le Cleach et al.2000、Clin.Exp.Immunol.119:225〜230)。このようにして、我々の研究成果は、P140/KIRがCTCLの同定のための、CD4+細胞上の好適なマーカーを表すことを示した。
【0092】
材料及び方法
患者
インフォームド・コンセントの後、倫理委員会の承認を受け、我々は11人のCTCL患者から皮膚及び血液試料を得た。8人の患者は、セザリー症候群に罹っており、10〜45%の循環性セザリー細胞を血液中に有していた。7例で、腫瘍細胞のフェノタイプがCD3+、CD4+、CD8−であった。1例がCD3+、CD4+、CD8αα+(患者Pno)であった。3人の患者は形質転換菌状息肉症(Lez、Cou、Bic)で、CD3+、CD4+、CD8−フェノタイプを有する伝播性皮膚腫瘍の症状を呈していた。患者全員がこれまで化学療法を受けていなかった。
【0093】
腫瘍リンパ球の分離
新鮮な腫瘍細胞は腫瘍断片を機械的に単細胞懸濁液に分散させたものから得た(Bagot et al.1998、Blood 91:4331〜4341)。次で、単核細胞は洗浄され、後の使用のために、ヒト血清プラス10%ジメチルスルフォキシド中で凍結された。
セザリー症候群患者の場合は、単核血中細胞がフィコール・アイソパクウ技法(Pharmacia fine Chemicals、Piscataway、NJ)で分離された。
【0094】
腫瘍細胞系の長時間培養
我々は前記の(Bagot et al.1998、Blood 91:4331〜4341)患者の末梢血液からの試験管内Pno細胞系(TCR−Vβ22+、CD3+、CD4+、CD8αα+、MHCクラスI、MHCクラスII)の長時間培養を実施した。我々は患者の血中を循環する悪性クローン及び培養されたT細胞系の両者がそれらの細胞表面フェノタイプ及びそれらのサイズ及びTCRβVDJ領域(Poszepczynska et al.2000、cf.supra)に関して同一であることを示した。SS細胞系の他の供給源はATCC(例えば、HUT 78 ATCC TIB-161)で見つけることができる。
Cou−L細胞系(TCRVβ13+)は試験管内でγIL−2と共に、3年以上培養された。それは前記のCD4+Cou−LSCTCL系のサブクローンに相当する(Bagot et al.1998、cf.supra)。Cou−L細胞系、本来のTCRVβ13+CD4+Cou−LSCTCL及び皮膚から新たに分離された腫瘍細胞は、同じサイズで、且つTCRβVDJ領域の配列を分担する(Bagot et al.1998、cf.supra)。
【0095】
モノクローナル抗体(mAbs)及び流動細胞計数の研究
1色または2色の免疫蛍光分析が前記のように(Bagot et al.1998、cf.supra)行なわれた。抗CD3(細胞分析2000カタログ番号6604629)、抗CD4(細胞分析2000カタログ番号6602138)、抗MNCクラスI(細胞分析2000カタログ番号IM0107)及びII(細胞分析2000カタログ番号IM0108)はBeckman
Coulter(細胞分析2000カタログ番号IM2014)から入手可能である。
抗TCRVβ13はBioadvance(Emerainville、France、catalogue番号TCR1738)、TCR−Vβ22+mAb はBeckman Coulterから入手された(Marseille、France、細胞分析2000カタログ番号IM1484/IM1365)。抗CD8αβ2ST8.5H7mAbはBeckman Coulter(細胞分析2000カタログ番号Im2014)から入手可能である。抗P140mAbsとして、Q66(IgM、抗P140)及びAZ158(IgG2a、p70/NKB1及びP140の両者を確認)(これらは、Pande et al.1996により報告された。J.Exp.Med.184:505〜518)が使用された。しかし、抗P140結合化合物が好適である(例えば、抗P140抗血清、Fc-HLA-A11 Fc-HLA-A3溶融蛋白)。Z27(IgG1、抗p70/NKB1)、EB6(IgG1、抗p58.1/p50.1)、GL183(IgG1、抗p58.2/p50.2)、XA185(IgG1、抗CD94)、Z199及びZ270(IgG2b、及びIgG2aの各々、抗NKG2A)及びB9.4(IgG2b、抗CD8)mAbsは全て、Beckman Coulter(細胞分析2000カタログ番号IM2748、IM2277、IM2278、IM1610、IM2750、IM0102)から入手可能である。
ヤギ抗マウスIgMFITC及びヤギ抗マウスIgMPEもまた、Beckman Coulterから入手され得る(細胞分析カタログ番号IM08及びIM0555参照)。
【0096】
生化学的特徴
20×106個の細胞が、培養され(15分、20℃)250μgのEZ−LINK SULFO−NHS−LC−BIOTIN(Pierce、Rockford、IL)含有1ミリリットルPBSpH8中で培養され(15分、20℃)、洗浄用バッファー(10mM TRIS pH8、0.14M塩化ナトリウム)で3回洗浄された。次いで、細胞は1%NP40に溶解され、次いで、セファロースPA(Pharmacia Biotech Inc.Piscataway、NJ)結合AZ158(IgG2a抗p70/、140)またはB9.4(IgG2b抗CD8)mAbで免疫沈降された。試料は不連続SDS−PAGEを使用し、N−グリコシダーゼ(ベーリンガーマンハイム、GmbH、Germany、製)による消化または非消化で、分析された。次いで、Immobilon P(ミリポアCorp.Bedform MAの商品名)ヘ移した。Neutravidin(Pierceの商品名)で汚染後、検出にはルネッサンス・ケミルミネッセンス・キット(NeN、Boston、MAの商品名)が使用された。NK細胞クローンはペンデ(Pende、1996)等が(J.Exp.Med.184:505〜518)報告したように、限定希釈で得られた。
【0097】
RT−PCR分析
トータルRNAはCTCL細胞系Pno及びCou-LCD8ααからRNAクリーンシステム(AGS GmbH、ハイデルベルグ、ドイツ)を使用して抽出した。cDNA合成はオリゴットTプライミング(oligod T priming)を使用して行なった。KIRディスプレー3Ig様ドナーの完全なORF(open reading frame)cDNA増幅のために使用された。使用されたプライマーを以下に示す。5'CATGT(CT)GCTCA(CT)GGTCGTC(Ig3 UP;SEQ ID No.5)及び5'GGTTTTGAGACAGGGCTG(Ig3 DOWN;SEQ ID No.6)。増幅は30秒サイクル(30秒94℃、30秒55℃、30秒72℃)、次いで、Ampli TAQ(パーキンエルマー−Applied Biosystems、Foster City、CA)を使用して7分、72℃の培養を行なった。PCR産物はpcDNA3.1/V5-HisTOPOベクター(Invitrogen、Carlsbad、CA製)のサブクローン化物であった。DNA配列はd-ローダミン(生物体汚染用顔料)ターミネーターサイクル配列キット及び377Applied Biosystems自動配列機(パーキンエルマー−Applied Biosystems)を使用して行なった。
【0098】
過渡的移入
COS−7細胞がpcDNA3.1TOPO−KIR3Dcl.24またはpCR3cl.1.1(Pende et al.1996、J.Exp.Med.184:505〜518)でFugene-6 (Roche、Monza Italy)を使用して移入された。細胞を5×105/プレートの量でシードした。24時間後、これらを6μgプラスミド及び10μlのFugene-6試薬と共に、DMEM/10%FCS中で培養した。48時間から72時間後、移入細胞は血球弗素計量分析にかけられた。移入細胞はQ66及びAZ158mAbsで汚染され、次いで、フィコエリトリン被接合ヤギ抗体接合マウスのIgG2aまたはIgMで汚染され、血球流量計により、FACソート(Becton Dickinson、San、Jose、CA)を使用して分析した。
【0099】
結果
CTCL細胞系がp140/KIRヘの抗体で汚染される。
二つの長期CTCL腫瘍系Pno(抗TCR−Vβ22mAbでラベル)(Poszepczynska et al.2000、cf.supra)及びCou-L(抗CD3mAbでラベル)(Bagot et al.1998、cf.supra)は反応的に、異なる抗KIRmAbsで分析された。
我々は両細胞系がmAbsQ66(図9A参照)及びAZ158と反応性であり、両者はp140/KIRと結合することを見出した。対照的に、これらの細胞系は他のp58.1、p58.2、p70KIRsを含有するHLAクラスI分子に特異的な阻害受容体及びCD94/NKG2Aレクチン様受容体(Poszepczynska et al.2000、cf.supra)を発現しない。
【0100】
CTCL患者から新たに採取された腫瘍Tリンパ球の抗p140/KIRmAbsによる汚染。
p140/KIRが新たに採取された腫瘍細胞により発現されるか否かを決定すべく、我々は、Q66mAbの、SS患者Pno及びMF患者Couの腫瘍性皮膚断片から採取した非培養腫瘍細胞との反応性を試験した。我々は腫瘍細胞の大部分がこのmAbによって汚染されることを確認した(図9B参照)。特に、我々は、患者Pnoの血液から採取された大部分のTCRVβ22+腫瘍リンパ球がQ66抗体と反応性があり、患者Couの皮膚から採取された大部分のTCRVβ13+腫瘍リンパ球が同じ抗体と反応性があることを観察した(図9B参照)。次に、我々は、その循環性CD4+リンパ球の10〜45%が悪性細胞である、追加の7人のセザリー症候群患者から採取した腫瘍Tリンパ球及び他の2人のMF患者の皮膚腫瘍から採取した腫瘍Tリンパ球、の両者のフェノタイプを研究した。供試全患者が共発現CD4及びp140/KIRの大集団を顕著に示していた(下記表2参照)。
【0101】
形質転換菌状息肉症患者の皮膚、及びセザリー症候群患者の血液、から採取されたCD4+リンパ球汚染、抗Q66mAb。
【0102】
【表2】
【0103】
全てのQ66+細胞がCD4+細胞集団に含まれていることに注目すべきである。かくして、健常者のp140/KIR発現は、皮膚及び血液中にあるCTCL腫瘍CD4+Tリンパ球の特徴を表すNK及びCD8+集団から得られる、リンパ球小集団に限定される。Tリンパ球発現の多種類のKIRの比率が低いところの、他の皮膚病、毒物による皮膚溶解(Le Cleach et al.2000、Clin.Exp.Immunol.119:225〜230)、由来の皮膚T細胞はp140/KIRの発現制御ができない。
【0104】
CTCLによって発現されるp140受容体の分子的特徴
Pno及びCou-L細胞系はビオチンでその表面をラベルされており、細胞溶解物は抗p140(AZ158mAb)で免疫沈降する。図10に示すように、このmAbはその分子量が還元条件下で約70kDである、NKクローン及びPno及びCou-L細胞系から免疫沈降される。N−グリコシダーゼによる処理によって、NKクローンに比較してPno及びCou-Lの移動性がやや高く、分子量が約50kDである蛋白質主鎖を明らかにした。これらのデータは、これらCTCL腫瘍細胞系によって発現されるp140阻害受容体が、前記正常NK細胞で検出されたものと殆ど同じであることを示唆した(Pende et al.1996、J.Exp.Med.184:505〜518)。
【0105】
次に、我々はPno及びCou-L細胞系上のQ66及びAZ158mAbが前記p140/KIR(SEQ ID No.3)をコード化するcDNAの一つに相当するかどうかを決定した。この末尾で、RT−PCRは、三個のIg様ドメインを有するKIRをコード化する全てのcDNAを増幅できるプライマーのセットを使用して、これらの細胞から由来するRNA上で行なわれた。Pno細胞系から、KIR3D cl.24と名付けられた、フル鎖長のcDNAを採取した(SEQ ID No.1)。そのヌクレオチッド配列と、三つの細胞外Ig様ドメインが特徴である、全てのKIRをコード化するDNA配列との比較は、KIR3D cl.24pが、140受容体の新規な対立遺伝子型を表すことを明らかにした。特に、そのヌクレオチド配列は前記の成熟蛋白質(図11参照、SEQ ID No.2:クローン24蛋白質;SEQ ID No.4:クローン1.1蛋白質)における4個のアミノ酸置換となる1.1 cDNAに比較される5個の異なるものを表示する(Pende et al.1996、J.Exp.Med.184:505〜518)。4個の置換のうち3個は細胞外Ig様ドメイン(成熟蛋白質のうちのpos.20、92及び111)であり、他方はサイトプラズマ領域に局在する(pos.401)。Cou-LCTCL細胞系で行なわれるRT−PCRはp140/KIRの二つの異なる対立遺伝子型を明らかにした。一つはcl.1.1cDNAに相当し、他方は、KIR3D cl.24(Pno細胞系から分離)と同一である。CTCL細胞系由来のcDNAはCOS−7細胞へ移入される。期待したように、全ての移入された細胞は、Q66及びAZ158mAbによって明瞭に汚染される。
一方、これらはネガティブコントロールとして使用されるp70/KIR特異性Z27mAbと反応しない。終わりに、RT−PCRが三人のSS患者(患者のPno含有)由来のPBLから抽出及び一人の追加のMF患者の皮膚由来のT細胞から抽出されたRNA上で行なわれる。また、これらの試料で、上記のp140/KIRの一つまたは他の対立遺伝子型が同定される。
同じ結果が患者から採取された悪性細胞上で得られた。
【0106】
討議
CTCLの皮膚病変は、殆どCD4+で、非新形成の腫瘍浸潤性Tリンパ球である悪性T細胞からなる異成分のリンパ球の浸潤物を含む。我々は、以前に、CD4+細胞毒腫瘍浸潤性Tリンパ球は特異的に、自家組織の悪性CTCLCD4+細胞系を支配することを報告した。しかし、腫瘍細胞で発現されるクローン型のTCRのみならず、非腫瘍とされている細胞表面構造がCTCL上で同定された。非悪性、反応性CD4+リンパ球と悪性のものとを区別する標準法を使用することは困難である。
我々は、この研究で、最初に、MF及びSS患者から採取した腫瘍細胞が、p140/KIRを発現すると報告した。この受容体は長期培養CTCL系と同様に、CTCL患者からの皮膚及び血液中の腫瘍細胞と同定された。二色蛍光分析はp140/KIR発現は、以前に腫瘍細胞で同定され、与えられた、TCRβ−VDJにより、特徴付けられているT細胞として限定される。このようにして、p140/KIRは、Tリンパ球CD4+集団中の腫瘍反応性細胞からの腫瘍細胞の同定を迅速化させる。これは、特に、その腫瘍T細胞クローンが末梢血中CD4+リンパ球集団内で、常には、容易に同定されないようなSS患者に対しては有効である。更に、病気の間または処置後に、CD4+集団の増加が腫瘍細胞集団の増加が原因なのか、反応性Tリンパ球の増加が原因なのかを評価するのは困難である。抗p140/KIRmAbが、既に解析された全CTCL患者の腫瘍細胞を認知するために出現するので、それらは、CTCLの直接的同定のためには公的な手段と考えられる。その上、CD4+CTCL細胞上での、p140/KIR共発現の使用は悪性細胞と正常細胞を区別するユニークな手段を提供する。これは個々の患者にとって独特であるTCR決定法とは対照的である。その上、p140/KIRは、CTCLの新規な特異的免疫療法の発展のための可能なターゲットである。
【0107】
これまでの研究は、この受容体がHLA.A3及びHLA.A11対立遺伝子の認知への阻害信号を送ることができることを示した。しかし、解析された各種の患者におけるp140/KIR発現それら自体のHLAクラスIハロタイプには明らかに依存しないと見なされることに留意すべきである。それにも拘らずp140/KIRのCTCLの異常病態生理学における現実の役割は、それ自体への耐性での受容体の潜在的役割を考慮することで、研究が続けられるという重要な特徴である。結論として、この研究は最初に、CD4+CTCL細胞におけるp140/KIRの発現及び腫瘍細胞におけるこの受容体の新規な対立遺伝子型の病変を示している。この発見はCTCL患者の異常病態生理学及び臨床管理の両者についての重要な論点である。更に、p140発現がCD8+形質転換MFのようなCD8+CTCLの表面に観察される。
【実施例3】
【0108】
抗CTCL薬の製造
当該分野の専門家は、下記の図の意図による概要に従がって抗CTCL薬を製造できる。
CTCL細胞近傍の細胞の免疫システムを刺激するために、抗SC5または抗p140mAbをサイトカインと共に使用する。
抗CTCL成分をCTCL細胞に向けて送り出すためのベクターとして、これらの抗SC5または抗p140mAbを使用する。抗CTCL成分の例は、放射性元素、トキシンなどのキミオセラピーで使用される標準化合物がある。
放射性元素用mAbベクターは、本発明の抗SC5mAbに使用された標準技法により産生できる[Press OW et al.置換B細胞リンパ腫への自家幹細胞移植を伴うフェイス2試行沃素131B1(抗CD20)抗体療法、Lancet 1995 vol.346 pp336〜340参照、この内容は,この明細書に参考のために記載されている。]。
トキシン用mAbベクターは、本発明の抗SC5mAbに使用された標準技法により産生できる[Ghetie U et al.ヒト組換えCD4及び脱グリコシルリシンA鎖構成免疫接合物の大スケール調製、J.Immunol.Methods 1990、vol.126 pp 135〜141、この内容は,この明細書に参考のために記載されている。]。
抗有糸分裂性のプロドラッグ及び抗SC5mAb及び/または抗p140mAbベクターの両者の結合されたものを使用すると、抗有糸分裂性のプロドラッグを活性な医薬型(機能型の抗有糸分裂性製剤)へと形質転換することができる酵素を送り届ける。
抗有糸分裂性のプロドラッグを活性な医薬型へ形質転換することができる酵素を送り届ける抗SC5mAb及び抗p140mAbベクターは、本発明の抗SC5mAb及び抗p140mAbに適用される次の標準的技術で産生され得る(ADEPTtechnique、for Antibody Directed Enzyme Prodrug Therapy、described in Plakey TC et al. Anti-tumor effects of an antibodycarboxypeptidase G2 conjuguate in combination with phenol mustard prodrugs、British J. Cancer 1995 vol.72 pp 1083〜1088、of which content is herein incorporated by reference、参照)。
リガンド作用物質(SC5刺激またはp140刺激)としての抗SC5または抗p140/抗CD4の使用。
補体漸増剤としての抗SC5または抗p140mAbの使用。
上記用途のために抗SC5/抗CD4または抗p140/抗CD4特異性の共用のmAbが好ましい。
ADCC(Antibody Dependent Cell Cytotoxicity)の活性化体としての、抗SC5または抗p140(CD4特異性は不要)の使用:多核細胞、マクロファージ、Fc受容体含有NK細胞抗はSC5及び抗p140mAbのFc含量によって活性化され、それ故、これらmAbはCTCLの近傍でADCC活性化ができる。
【産業上の利用可能性】
【0109】
本発明はCTCLの診断及び治療の問題点を解決する二つの分子を提供する。
これは、特に、悪性CTCL細胞のような悪性T細胞の増殖及び/または機能的活性を阻害する。また、T細胞、特に悪性CTCL細胞のような腫瘍性T細胞の増殖及び/または機能的活性の調節ができる、ハイブリドーマI−2575で産性された抗SC5モノクローナル抗体を提供する。
【図面の簡単な説明】
【0110】
【図1】図1A、1B及び1Cは健常者のPBLsのSC5分子の細胞膜発現を図解している。 図1A、1B及び1Cはそれぞれ、健常者のPBMCを抗SC5mAb(腹水1/200)で汚染させ、次いで、FITC接合アイソタイプ特殊ヤギ抗マウスIgM2次試薬及び、抗CD3(図1A)、抗CD56(図1C)及び抗CD45RO(図1B)のようなPE接合mAbsの一種で汚染した場合である。
【図2】図2はPBLsの活性化の間、SC5分子の細胞膜発現を図解している。
【図3】図3はPBLのSC5分子の細胞内所在部を図解している。
【図4】図4はSC5分子の生化学的分析結果を図解している。
【図5】図5A、5B、5C、5Dは、抗CD3誘発増殖及び正常Tリンパ球のサイトカイン分泌を、それらの細胞毒活性の影響なしに、抗SC5mAbが調節する様子を図解している。図5A:PBLまたはT細胞クローンLSO及びGDS.3が固定化抗CD3mAb(コートされたmAb1μg/源)またはIL−2(50IU/ミリリットル)で、抗SC5mAb(腹水の、1:200最終希釈)またはアイソタイプ適合、不適切mAbの存在で、刺激した場合。図5B:GDS.3CD4+T細胞クローンは抗SC5mAbの異なる濃度の存在下、図示された濃度のプレコート抗CD3mAbで刺激した場合。 図5C:抗SC5またはコントロールmAbの存在下、抗CD3刺激の場合。図5D:細胞毒CD8+細胞クローン(JF1)が、抗SC5mAb(腹水の、1:200最終希釈)またはアイソタイプ適合コントロールmAbの存在で、E/T比が5/1での、FcyR+ミューリン腫瘍細胞P815との共培養の前に、培養した場合。
【図6】図6A、6BはCTCL患者のPBLのSC5分子の細胞膜発現を図解する。図6A:セザリー症候群患者のPBLのSC5及びCD4分子の共発現を健常者と比較した場合。図6B:ヒストグラムは示された3群におけるSC5発現の平均値を示す
【図7】図7A、7B、7Cはセザリー症候群患者の悪性細胞の膜表面発現を図解する。 図7A:循環TCRVβ22+悪性T細胞クローンの存在で、SS患者のPBMCの二色蛍光分析。図7B:弱くラベルした、抗SC5mAb(陰を付したヒストグラム)TCRVβ22陽性悪性細胞。図7C:同じ患者の細胞を、PE接合抗CD69mAb及びFITC接合抗TCRVβ22mAbによって汚染させた場合。
【図8】図8は抗SC5mAbによるCTCL悪性細胞系の抗CD3誘発増殖の調節を図解している。
【図9】図9A、9Bは、CTCL細胞系(図9A)のp140/KIR分子及び新鮮なリンパ球腫瘍(図9B)の検出を図解いている。
【図10】図10はp140分子の生化学的分析を図解している。
【図11】図11は蛋白質でコードされたKIR3D cl.24(SEQ ID No.2)及びcl.1.1(SEQ ID No.4)のアミノ酸配列を示す。
【0111】
略語
APC:抗原提示細胞、CTCL:皮膚T細胞リンパ腫、DIG:界面活性剤不溶グリコリピッド富化分画、ILT:免疫グロブリン様転写、KIR:キラー細胞免疫グロブリン受容体、mAb:モノクローナル抗体、MF:菌状息肉症、PBL:末梢血中リンパ球、PBMC:末梢血中単核細胞、PHA:フィトヘマグルチニン、PMA:ホルボール12β−ミリステート13α−アセテート、SS:セザリー症候群、TCR:T細胞受容体。
【特許請求の範囲】
【請求項1】
下記の段階を含むことを特徴とする、患者の特定の体内区分内の、悪性CD4+CTCL細胞の存在比率の数値を求める方法。
p140分子を発現するCD4+細胞の比率の値を、前記患者から得られた生物学的試料を基に測定し、そして、悪性CD4+CTCL細胞の、現実に前記体内区分に存在する比率の数値は、前記測定値の約±10%の範囲内に入る数値と考える。
【請求項2】
p140分子を発現するT細胞の比率の値を、前記患者から得られた生物学的試料を基に測定し、そして、この値を非CTCLのヒトで観察される平均比率の値と比較し、前記測定値が平均比率の値より有意に高いとき、CTCLが陽性であるという診断を下すことを特徴とするCTCLの診断方法。
【請求項3】
p140を発現するCD4+細胞の存在を、潜在的にCTCL細胞を含む生物学的試料で調べ、その存在が検知されるとき、CTCLの診断を終了することを特徴とするCTCLの診断方法。
【請求項4】
前記比率の測定が、p140結合化合物、p140DNA、及びSEQIDNo.4のタンパク質からなる群から選択される一つの要素を使用することを特徴とする請求項1〜3のいずれか1項に記載の方法。
【請求項5】
p140結合化合物、p140DNA、SEQIDNo.4の蛋白質からなる群から選択される一つの要素を含む、CTCLの進行レベルの評価又はCTCLの診断のためのキット。
【請求項6】
p140分子に結合可能な化合物の検知を含むことを特徴とする、悪性CTCL細胞の増殖に対する一時的緩和、予防、軽減、治療に有効な化合物の同定方法。
【請求項7】
p140分子が、SEQIDNo.2またはSEQIDNo.4の蛋白質である請求項6に記載の方法。
【請求項8】
CTCLの治療薬を調製するためのポリペプチドベクターの使用であって、該ベクターはp140結合化合物を含むポリペプチドベクターの使用。
【請求項9】
抗p140結合化合物が抗体である請求項8に記載の使用。
【請求項10】
p140結合化合物が、SEQIDNo.2の蛋白質に対する抗体である請求項9に記載の使用。
【請求項11】
p140結合化合物が、SEQIDNo.4の蛋白質に対する抗体である請求項9に記載の使用。
【請求項12】
p140結合化合物が、SEQIDNo.2の蛋白質及びSEQIDNo.4の蛋白質に対する抗体である請求項10又は11に記載の使用。
【請求項13】
ベクターがさらに毒性元素及び放射性元素からなる群より選択される元素を含む請求項8〜12のいずれか1項に記載の使用。
【請求項14】
p140結合化合物が、補体の漸増を介在する請求項8〜12のいずれか1項に記載の使用。
【請求項15】
p140結合化合物が、抗体依存性細胞傷害を介在する請求項8〜12のいずれか1項に記載の使用。
【請求項16】
CTCLが、セザリー症候群である請求項8〜12のいずれか1項に記載の使用。
【請求項17】
CTCLが、菌状息肉症である請求項8〜12のいずれか1項に記載の使用。
【請求項18】
CTCLを治療するためのポリペプチドベクターであり、該ポリペプチドベクターはp140結合化合物を含むポリペプチドベクター。
【請求項19】
抗p140結合化合物が抗体である請求項18に記載のポリペプチドベクター。
【請求項20】
p140結合化合物が、SEQIDNo.2の蛋白質に対する抗体である請求項19に記載のポリペプチドベクター。
【請求項21】
p140結合化合物が、SEQIDNo.4の蛋白質に対する抗体である請求項19に記載のポリペプチドベクター。
【請求項22】
p140結合化合物が、SEQIDNo.2の蛋白質及びSEQIDNo.4の蛋白質に対する抗体である請求項20又は21に記載のポリペプチドベクター。
【請求項23】
ベクターがさらに毒性元素及び放射性元素からなる群より選択される元素を含む請求項18〜22のいずれか1項に記載のポリペプチドベクター。
【請求項24】
p140結合化合物が、補体の漸増を介在する請求項18〜22のいずれか1項に記載のポリペプチドベクター。
【請求項25】
p140結合化合物が、抗体依存性細胞傷害を介在する請求項18〜22のいずれか1項に記載のポリペプチドベクター。
【請求項26】
CTCLが、セザリー症候群である請求項18〜22のいずれか1項に記載のポリペプチドベクター。
【請求項27】
CTCLが、菌状息肉症である請求項18〜22のいずれか1項に記載のベクター。
【請求項28】
抗p140化合物を含む抗CTCL薬。
【請求項29】
抗p140結合化合物が抗体である請求項28に記載の抗CTCL薬。
【請求項30】
p140結合化合物が、SEQIDNo.2の蛋白質に対する抗体である請求項29に記載の抗CTCL薬。
【請求項31】
p140結合化合物が、SEQIDNo.4の蛋白質に対する抗体である請求項29に記載の抗CTCL薬。
【請求項32】
p140結合化合物が、SEQIDNo.2の蛋白質及びSEQIDNo.4の蛋白質に対する抗体である請求項28又は29に記載の抗CTCL薬。
【請求項33】
ベクターがさらに毒性元素及び放射性元素からなる群より選択される元素を含む請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項34】
p140結合化合物が、補体の漸増を介在する請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項35】
p140結合化合物が、抗体依存性細胞傷害を介在する請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項36】
CTCLが、セザリー症候群である請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項37】
CTCLが、菌状息肉症である請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項38】
抗p140結合化合物が抗体である請求項5に記載のキット。
【請求項39】
p140結合化合物が、SEQIDNo.2の蛋白質に対する抗体である請求項38に記載のキット。
【請求項40】
p140結合化合物が、SEQIDNo.4の蛋白質に対する抗体である請求項38に記載のキット。
【請求項1】
下記の段階を含むことを特徴とする、患者の特定の体内区分内の、悪性CD4+CTCL細胞の存在比率の数値を求める方法。
p140分子を発現するCD4+細胞の比率の値を、前記患者から得られた生物学的試料を基に測定し、そして、悪性CD4+CTCL細胞の、現実に前記体内区分に存在する比率の数値は、前記測定値の約±10%の範囲内に入る数値と考える。
【請求項2】
p140分子を発現するT細胞の比率の値を、前記患者から得られた生物学的試料を基に測定し、そして、この値を非CTCLのヒトで観察される平均比率の値と比較し、前記測定値が平均比率の値より有意に高いとき、CTCLが陽性であるという診断を下すことを特徴とするCTCLの診断方法。
【請求項3】
p140を発現するCD4+細胞の存在を、潜在的にCTCL細胞を含む生物学的試料で調べ、その存在が検知されるとき、CTCLの診断を終了することを特徴とするCTCLの診断方法。
【請求項4】
前記比率の測定が、p140結合化合物、p140DNA、及びSEQIDNo.4のタンパク質からなる群から選択される一つの要素を使用することを特徴とする請求項1〜3のいずれか1項に記載の方法。
【請求項5】
p140結合化合物、p140DNA、SEQIDNo.4の蛋白質からなる群から選択される一つの要素を含む、CTCLの進行レベルの評価又はCTCLの診断のためのキット。
【請求項6】
p140分子に結合可能な化合物の検知を含むことを特徴とする、悪性CTCL細胞の増殖に対する一時的緩和、予防、軽減、治療に有効な化合物の同定方法。
【請求項7】
p140分子が、SEQIDNo.2またはSEQIDNo.4の蛋白質である請求項6に記載の方法。
【請求項8】
CTCLの治療薬を調製するためのポリペプチドベクターの使用であって、該ベクターはp140結合化合物を含むポリペプチドベクターの使用。
【請求項9】
抗p140結合化合物が抗体である請求項8に記載の使用。
【請求項10】
p140結合化合物が、SEQIDNo.2の蛋白質に対する抗体である請求項9に記載の使用。
【請求項11】
p140結合化合物が、SEQIDNo.4の蛋白質に対する抗体である請求項9に記載の使用。
【請求項12】
p140結合化合物が、SEQIDNo.2の蛋白質及びSEQIDNo.4の蛋白質に対する抗体である請求項10又は11に記載の使用。
【請求項13】
ベクターがさらに毒性元素及び放射性元素からなる群より選択される元素を含む請求項8〜12のいずれか1項に記載の使用。
【請求項14】
p140結合化合物が、補体の漸増を介在する請求項8〜12のいずれか1項に記載の使用。
【請求項15】
p140結合化合物が、抗体依存性細胞傷害を介在する請求項8〜12のいずれか1項に記載の使用。
【請求項16】
CTCLが、セザリー症候群である請求項8〜12のいずれか1項に記載の使用。
【請求項17】
CTCLが、菌状息肉症である請求項8〜12のいずれか1項に記載の使用。
【請求項18】
CTCLを治療するためのポリペプチドベクターであり、該ポリペプチドベクターはp140結合化合物を含むポリペプチドベクター。
【請求項19】
抗p140結合化合物が抗体である請求項18に記載のポリペプチドベクター。
【請求項20】
p140結合化合物が、SEQIDNo.2の蛋白質に対する抗体である請求項19に記載のポリペプチドベクター。
【請求項21】
p140結合化合物が、SEQIDNo.4の蛋白質に対する抗体である請求項19に記載のポリペプチドベクター。
【請求項22】
p140結合化合物が、SEQIDNo.2の蛋白質及びSEQIDNo.4の蛋白質に対する抗体である請求項20又は21に記載のポリペプチドベクター。
【請求項23】
ベクターがさらに毒性元素及び放射性元素からなる群より選択される元素を含む請求項18〜22のいずれか1項に記載のポリペプチドベクター。
【請求項24】
p140結合化合物が、補体の漸増を介在する請求項18〜22のいずれか1項に記載のポリペプチドベクター。
【請求項25】
p140結合化合物が、抗体依存性細胞傷害を介在する請求項18〜22のいずれか1項に記載のポリペプチドベクター。
【請求項26】
CTCLが、セザリー症候群である請求項18〜22のいずれか1項に記載のポリペプチドベクター。
【請求項27】
CTCLが、菌状息肉症である請求項18〜22のいずれか1項に記載のベクター。
【請求項28】
抗p140化合物を含む抗CTCL薬。
【請求項29】
抗p140結合化合物が抗体である請求項28に記載の抗CTCL薬。
【請求項30】
p140結合化合物が、SEQIDNo.2の蛋白質に対する抗体である請求項29に記載の抗CTCL薬。
【請求項31】
p140結合化合物が、SEQIDNo.4の蛋白質に対する抗体である請求項29に記載の抗CTCL薬。
【請求項32】
p140結合化合物が、SEQIDNo.2の蛋白質及びSEQIDNo.4の蛋白質に対する抗体である請求項28又は29に記載の抗CTCL薬。
【請求項33】
ベクターがさらに毒性元素及び放射性元素からなる群より選択される元素を含む請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項34】
p140結合化合物が、補体の漸増を介在する請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項35】
p140結合化合物が、抗体依存性細胞傷害を介在する請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項36】
CTCLが、セザリー症候群である請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項37】
CTCLが、菌状息肉症である請求項28〜32のいずれか1項に記載の抗CTCL薬。
【請求項38】
抗p140結合化合物が抗体である請求項5に記載のキット。
【請求項39】
p140結合化合物が、SEQIDNo.2の蛋白質に対する抗体である請求項38に記載のキット。
【請求項40】
p140結合化合物が、SEQIDNo.4の蛋白質に対する抗体である請求項38に記載のキット。
【図1】

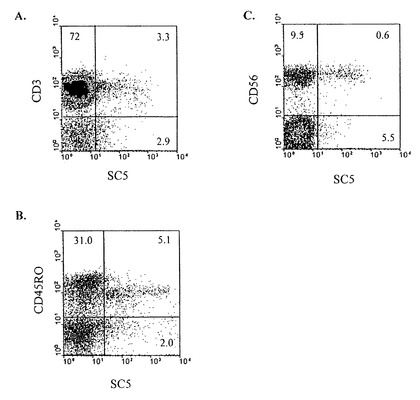
【図2】
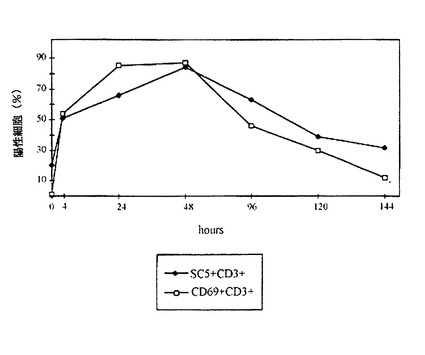
【図3】
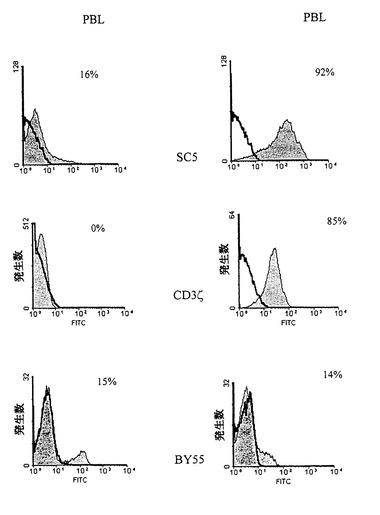
【図4】
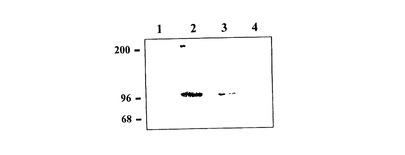
【図5】
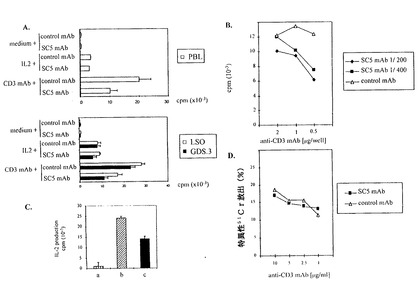
【図6】
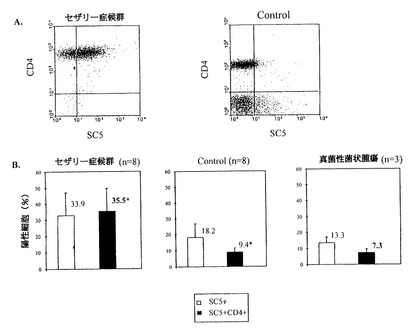
【図7】
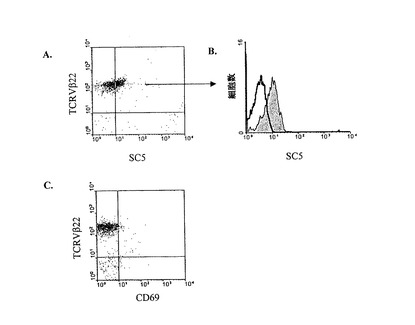
【図8】
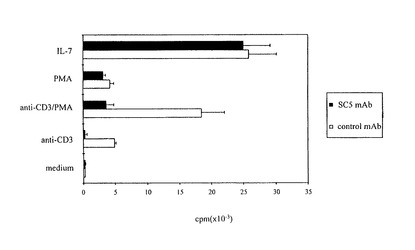
【図9】
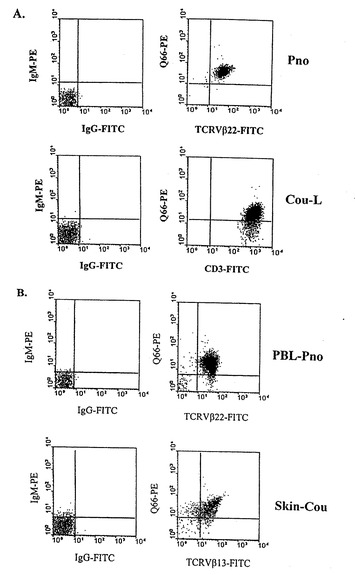
【図10】
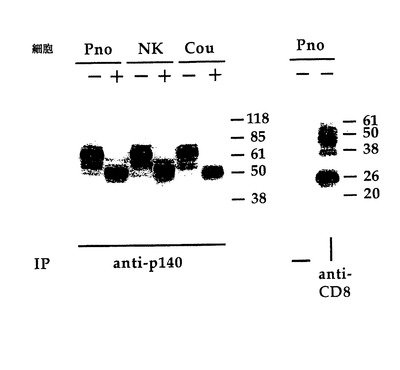
【図11】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2008−261875(P2008−261875A)
【公開日】平成20年10月30日(2008.10.30)
【国際特許分類】
【出願番号】特願2008−149642(P2008−149642)
【出願日】平成20年6月6日(2008.6.6)
【分割の表示】特願2002−552015(P2002−552015)の分割
【原出願日】平成13年12月18日(2001.12.18)
【出願人】(503218632)アーンスティテュ ナシオナール ド ラ サント エ ド ラ ルシェルシェ メディカル(イー.エンヌ.エス.ウー.エール.エンム.) (2)
【Fターム(参考)】
【公開日】平成20年10月30日(2008.10.30)
【国際特許分類】
【出願日】平成20年6月6日(2008.6.6)
【分割の表示】特願2002−552015(P2002−552015)の分割
【原出願日】平成13年12月18日(2001.12.18)
【出願人】(503218632)アーンスティテュ ナシオナール ド ラ サント エ ド ラ ルシェルシェ メディカル(イー.エンヌ.エス.ウー.エール.エンム.) (2)
【Fターム(参考)】
[ Back to top ]