
目的のポリヌクレオチドの発現を増強するためのポリヌクレオチド

本発明は、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドに関する。さらに、本発明は前記ポリヌクレオチドを含む宿主細胞、非ヒトトランスジェニック生物、ベクターおよびキットに関する。さらに、本発明は目的のポリヌクレオチドを発現する方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドに関する。さらに本発明は、前記ポリヌクレオチドを含む宿主細胞、非ヒトトランスジェニック生物、ベクターおよびキットに関する。さらに、本発明は、目的のポリヌクレオチドを発現する方法に関する。
【背景技術】
【0002】
基礎研究ならびに商業的応用の両方において、目的の産物(例えば、ポリペプチドまたは機能性RNA)をコードするポリヌクレオチドを発現するために真核細胞および原核細胞が通常用いられる。目的の産物を宿主細胞で合成するために、前記産物をコードする遺伝子(しばしばトランスジーンと呼ばれる)が宿主細胞中に、例えば、トランスフェクションまたは形質転換により導入される。次いで細胞を目的の産物を発現するのに好適な条件下で増殖する。この培養ステップは選択可能なマーカー遺伝子を使用することが多く、該遺伝子を宿主細胞中に目的の産物をコードするポリヌクレオチドと一緒に導入し、前記ポリヌクレオチドを保持する細胞を選択する。培養後、産物、例えば組換えタンパク質を適当な精製ステップにより細胞から精製することができる。
【0003】
一般に、真核細胞と原核細胞の両方を組換えタンパク質の発現に利用することができる。原核生物発現系は、大量のタンパク質を容易に生産するのに有利である。
【0004】
しかし、かかる系で生産される組換えタンパク質は、翻訳後に改変されることなく、不溶性の封入体に含まれうる。機能性になるために徹底した翻訳後改変および/またはフォールディングを経る必要があるタンパク質については、真核生物発現系を使用することが好ましい。
【0005】
目的のポリヌクレオチドによりコードされた所望の産物の合成にとって、前記ポリヌクレオチドの強くかつ信頼しうる安定な発現は、強くかつ安定な発現が費用および時間の点でより効果的であるので、非常に重要である。ポリヌクレオチドの強い発現は、前記ポリヌクレオチドの高度に転写されるクロマチン領域中への組込みおよび/または多重組込みイベントに関係することが多い。しかし、とりわけ、複雑な、いわゆる「逆方向反復(inverted repeat)」パターンと組合わせた多重組込みイベントは、トランスジーンサイレンシング、すなわち、ポリヌクレオチドの不安定なかつ低下した発現を生じがちである。
【0006】
最近の数十年間、多大な努力が、細胞系におけるトランスジーン発現の増強に向けられてきた。例えば、発現の増強は、遺伝子発現を駆動する強いプロモーターを利用することにより達成することができる。さらに、トランスジーンをある特定のクロマチンエレメントと組合わせて宿主細胞に導入すると、トランスジーンの発現を増強することができる。
【0007】
クロマチンは、真核細胞の核内部においてDNA、RNAおよびタンパク質から成る複合体である。一般に、クロマチンにはループとドメインから構成され、色々な凝縮状態で存在する。凝縮の程度が転写、複写またはDNA修復などのDNAにアクセスを必要とする過程の進行に影響を与えると思われる。一般に、比較的凝縮した構造は転写的にサイレントであると考えられ、他方、比較的脱凝縮した(オープンな)構造は効率的な遺伝子発現に欠かせないと思われる。おそらく、共トランスフェクトしたトランスジーン、すなわち目的のポリヌクレオチドの発現を増強できるクロマチンドメイン/エレメントは、クロマチン構造を脱凝縮することができる因子に対する結合部位を有する。これらのドメインの近隣の配列エレメントは、転写関係因子が容易にアクセスしうるので、より高い発現を示すと思われる。
【0008】
WO 02/24930は、哺乳動物細胞におけるトランスジーン発現を増強できる偏在性クロマチン開放性エレメント(UCOE;ubiquitous chromatin opening elements)を開示している。
【0009】
Zahn-Zabalら(J Biotechnol.、2001、87(1):29-42)は、高生産クローンの比率の増加をもたらす共トランスフェクトしたレポーター遺伝子の安定な発現を有意に増加する、ニワトリリゾチームマトリックス付着領域(MAR)を開示している。
【0010】
WO2005/040377は、タンパク質生産を増加する活性を有するDNA配列およびこれらのDNAの、真核細胞におけるタンパク質生産活性を増加するための使用を開示している。
【0011】
KalosおよびFournier(Mol Cell Biol.、1995、15(1):198-207)は、マトリックス付着領域に隣接するアポリポタンパク質B(apoB)クロマチンドメインのエレメントがトランスジーン発現に介在することを開示している。
【0012】
Klehrら(Biochemistry、1991、30(5):1264-70)は、ヒトインターフェロンβドメイン由来のヒトスカフォールド付着領域は遺伝子の発現を増強できることを示唆してる。さらに、スカフォールド付着領域(SAR)は一方向にかつ部分的に距離と無関係に、全般的なプロモーター機能を増強することを示した。
【0013】
Dangら(J Virol.、2000、74(6):2671-8)は、ヒトβインターフェロンスカフォールド付着領域(IFN-SAR)の2kb断片を挿入するとこのベクターでの遺伝子の安定な発現を引き起こすことを開示している。
【0014】
EP 1,676,916は、共トランスフェクトしたトランスジーンの発現を増強するための伸長因子1αの、5'領域の非コーディング12kb核酸配列および3'領域の4kb核酸配列を開示している。
【0015】
生産性を増加するために、増幅可能な遺伝子をトランスフェクトするベクターに対する選択可能なマーカーとして用いる遺伝子増幅細胞エンジニアリング技法を応用することができる。第1のステップで、トランスジーンと増幅可能なマーカー遺伝子を含有するベクターを宿主細胞中に導入する。選択可能でかつ増幅可能な遺伝子を増幅すると、遺伝的に連結されたトランスジーンを含有する配列は共増幅される。増幅は、選択剤の濃度を段階的に増加する、時間のかかる多段階選択プロセスにより誘導する。サブクローニングした耐性細胞を引き続いてスクリーニングすることによって、生産性の高い細胞株を同定することができる。哺乳動物細胞における遺伝子増幅のための増幅可能かつ選択可能なマーカーは、T. Omasha, Journal of Bioscience and Bioengineering (2002) 94、600-605に総括されている。
【0016】
さらに、トランスジーンの発現は、形質転換された細胞を選択するための改変された低活性の選択可能なマーカーを用いることにより増加することができる。WO/2004/050884は細胞集団中の高生産体の比率を増強する発現系を開示している。形質転換されたCHO細胞を選択するマーカーとして、改変されたネオマイシンホスホトランスフェラーゼを使用すると、高生産細胞数-対-全細胞数の比を増強した。
【先行技術文献】
【特許文献】
【0017】
【特許文献1】WO 02/24930
【特許文献2】WO2005/040377
【特許文献3】EP 1,676,916
【非特許文献】
【0018】
【非特許文献1】Zahn-Zabalら、J Biotechnol.、2001、87(1):29-42
【非特許文献2】KalosおよびFournier、Mol Cell Biol.、1995、15(1):198-207
【非特許文献3】Klehrら、Biochemistry、1991、30(5):1264-70
【非特許文献4】Dangら(J Virol.、2000、74(6):2671-8
【非特許文献5】T. Omasha, Journal of Bioscience and Bioengineering (2002) 94, 600-605
【発明の概要】
【発明が解決しようとする課題】
【0019】
しかし、宿主細胞中の目的のポリヌクレオチドの発現のレベルは、記載した技法により増加できるが、高発現はしばしば、発現のサイレンシング、すなわち、低くかつ不安定な発現をもたらす。遺伝子サイレンシング効果によって、培養初期に目的のポリヌクレオチドを強く発現する細胞クローンは、培養の過程で発現の有意な低下を示しうる。さらに、もし細胞集団における発現の不安定な細胞の比率が非常に高ければ、十分な量の所望の産物を得るために、単一の高生産クローンを、時間と費用のかかるスクリーニング手順により単離しなければならないかも知れない。
【0020】
従って、以上の欠点なしに、目的のポリヌクレオチドの発現の改善、すなわち、強くかつ安定な発現を可能にする発現系が大いに所望されている。
【0021】
従って、本発明の根底に横たわる技術的問題は、宿主細胞における遺伝子発現を改善して、高効率でかつ比較的安定な発現を達成する手段と方法を提供することにあると考えることができる。
【課題を解決するための手段】
【0022】
よって、本発明は、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドに関する。好ましいことに、かかるポリヌクレオチドは、宿主細胞中に前記ポリヌクレオチドと組合わせて導入した目的のポリヌクレオチドの発現の増強を可能にする。
【発明を実施するための形態】
【0023】
本明細書に使用した用語「ポリヌクレオチド」は核酸配列に関する。核酸配列はDNAであってもまたはRNA配列であってもよく、好ましくは核酸配列はDNA配列である。
【0024】
本発明のポリヌクレオチドは上述の核酸配列から成るかまたは上述の核酸配列を含むことが必須である。従って、これらはさらなる核酸配列も含有することができる。
【0025】
本発明のポリヌクレオチドは、好ましくは、単離された(すなわち、自然環境から単離された)ポリヌクレオチドとしてまたは遺伝的に改変された形態において提供されよう。本明細書で意味する単離されたポリヌクレオチドはまた、その自然細胞環境以外の細胞環境で存在するポリヌクレオチド、すなわち、異種ポリヌクレオチドを包含する。用語「ポリヌクレオチド」は、1本鎖ならびに2本鎖のポリヌクレオチドを包含する。さらに、グリコシル化またはメチル化ポリヌクレオチドなどの天然の改変されたポリヌクレオチドを含む化学的に改変されたポリヌクレオチド、またはビオチン化ポリヌクレオチドなどの人工の改変されたものも含まれる。
【0026】
本明細書に使用される用語「発現」は、宿主細胞内で起こる転写だけまたは転写と翻訳を意味する。用語「発現」は、好ましくは、宿主細胞中に本発明の少なくとも1つのポリペプチドと一緒に共導入された少なくとも1つの目的のポリヌクレオチドの転写または転写と翻訳に関すると解釈されるべきである。本明細書に使用される用語「共導入された」は、様々なポリヌクレオチドの宿主細胞中への、例えばトランスフェクションまたは形質転換技法による導入が、好ましくは、同じ時点で、すなわち同時に行われることを意味する。それにより、好ましくは、近傍クロマチン領域における、共導入された配列の組込みが可能になろう。
【0027】
本明細書に使用される用語「目的のポリヌクレオチド」は核酸配列に関する。前記核酸配列はRNAまたはDNA配列であってもよく、好ましくは、DNA配列である。本発明の方法によって、目的のポリヌクレオチドは目的の産物をコードすることができる。目的の産物は、目的のポリペプチド、例えば、タンパク質、または目的のRNA、例えば、機能性のRNA、例えば、2本鎖RNA、マイクロRNA、またはsiRNAであってもよい。機能性RNAはしばしば対応する標的遺伝子をサイレントにするために使用される。好ましくは、目的のポリヌクレオチドは、当技術分野で周知でありよく記載されていて、目的のポリヌクレオチドの転写に有効である好適な調節配列(例えば、プロモーター)と機能しうる形で連結されている。
【0028】
宿主細胞における所望の産物の発現のレベルは、細胞中に存在する対応するmRNAの量または目的のポリヌクレオチドによりコードされる所望の産物の量に基づいて決定することができる。例えば、選択した配列から転写されるmRNAは、PCRによりまたはノーザンハイブリダイゼーションにより数値化することができる(Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press (1989)を参照)。ポリペプチドは様々な方法により、例えば、ポリペプチドの生物学的活性をアッセイすることより(例えば、酵素アッセイにより)、または、かかる活性に依らないアッセイ、例えば、タンパク質を認識して結合する抗体を用いるウェスタンブロット、ELISA、またはラジオイムノアッセイを利用することにより、数値化することができる。
【0029】
本明細書で使用される用語「発現を増強できるクロマチンエレメントに対する核酸配列」は、ポリヌクレオチドの発現を増強できるクロマチンエレメントまたはドメインに対する核酸配列に関する。好ましくは、この用語は、宿主細胞中に前記クロマチンエレメントとともに共導入された目的のポリヌクレオチドの発現を増強できるクロマチンエレメントに対する核酸配列に関する。目的のポリヌクレオチドの発現を増強できるクロマチンエレメント/ドメイン(例えば、MARまたは偏在性クロマチンオープニングエレメント)は当技術分野で周知であり、例えば、EP1676916、WO2005/040377、WO0224930、Dangら、Klehrら、Kalosら、Zahn-Zabalら(全て、上記引用文中に)に記載されている。さらに、クロマチンの構造に影響を与えることにより遺伝子活性を調節できる非ランダムなゲノムの分散された遺伝エレメントが、Hodgetts in Current Opinion in Gemtetics & Development (2004) 14 680-685により記載されている。WO2005/040377は、活性を増加するタンパク質生産を有する、例えば、DNA配列、および真核細胞中のタンパク質生産活性を増加するための、これらのDNA配列の使用を記載している。好ましくは、発現を増強できるクロマチンエレメントは、クロマチン中への(すなわち、染色体DNA中への)その組込みが、より脱凝縮した(「オープンな」)クロマチン構造、好ましくは、組込み部位に隣接するクロマチン領域のより脱凝縮した構造を生じさせることができる核酸エレメントである。
【0030】
以上説明したように、好ましいクロマチンエレメントはマトリックス付着活性を有するDNAエレメントを含む。このDNAエレメントは共トランスフェクトされる目的のポリヌクレオチドの発現を増強できることは理解されよう。好ましくは、クロマチンエレメントは発現を増強できるMARを含む。マトリックス付着領域はクロマチンループのベースに位置する特定のDNA配列であって、DNAを核マトリックスの繊維に固着する。マトリックス付着領域はSAR(スカフォールド付着領域)に類似し、従って、SARまたはスカフォールド/マトリックス付着領域(S/MAR)とも呼ばれる。MARを含有する配列はプロモーター活性を抑制できることは公知である(Lauberら J Biol Chem. 1997 Sep 26;272(39):24657-65.)。また、特定のMARエレメントの存在は、トランスジーン発現の平均レベルの位置誘導性変動の低下をもたらさない(Van Leeuwen ら Plant Mol Biol. 2001 Nov;47(4):543-54)。興味深いことに、マウスパーフォリン遺伝子のMAR配列は、グローバルなサイレンシング効果を果たす(Mol Cells. 2002 Feb 28;13(1):61-8)。また、MARβ、T細胞受容体βエンハンサー(Eβ)の上流に位置するマトリックス関連領域は、Eβ介在性TCR活性をサイレント化する重大な役割を果たす(Kaul-Ghanekarら Nucleic Acids Res. 2004 Sep 15;32(16):4862-75)。特定のMAR配列は、遺伝子、例えば、共導入されたトランスジーンの発現を増強することができ、複製起点として機能できることは公知である。MAR全体について明確なコンセンサス配列を規定できないが、いくつかのDNA配列モチーフが、典型的には、MAR様複製起点、TGリッチ配列、湾曲DNAモチーフ、よじれDNAモチーフ、トポイソメラーゼ部位、ATリッチ配列に見出される。これらのモチーフは当技術分野で周知であり、例えば、Singhら(Nucleic Acids Research、1997、25(7): 1419-1425)に掲げられている。本文献は開示したポリヌクレオチド配列に関する参照により本明細書にその全てが組み入れられる。さらに、S/MAR予測プログラムはゲノム配列からMARまたはSARの予測を可能にする(例えば、 Fioriniら, Biochemistry (Moscow), 2006, 71(5): 481-488。
【0031】
本明細書で言及する、発現を増強できるクロマチンエレメントは、好ましくは、配列番号28に示された核酸配列(ヒト白血球由来のゲノムDNAから増幅された;実施例1を参照)を含む。より好ましくは、クロマチンエレメントは、ヒトトポイソメラーゼIのイントロン13のATリッチMII領域を含む配列番号27に示された核酸配列(HeLa細胞由来のゲノムDNAから増幅された)を含む(GenBank L23999.1; GI: 398042; Romigら, 1994、Eur J Biochem. 221:411-419)。
【0032】
ニワトリリゾチーム5'マトリックス付着領域のMARエレメントもトランスジーンの発現を増強できることは公知である(Girodら, Biotechnol Bioeng. 2005 Jul 5;91(1):1-11)。
【0033】
本明細書で使用する「発現を増強する」は、宿主細胞中に本発明のポリヌクレオチドとともに共導入しておいた目的の産物をコードするポリヌクレオチドの転写、または転写および翻訳を含む発現を、改善または増加することを意味する。好ましくは、目的の産物の量が増加する。この増加は、より安定な発現によるかもしくはより強い発現によるかまたはそれらの両方によるものでありうる。増強された発現は、本発明によるポリヌクレオチドと一緒に宿主細胞中に共導入しておいた目的のポリヌクレオチドの発現を、参照宿主細胞中の発現と比較することにより示すことができる。好ましくは、参照宿主細胞は、本発明によるポリヌクレオチドのさらなる導入なしに、同じ目的のポリヌクレオチドを導入することにより作製した宿主細胞である。ポリペプチドの量(例えば、ウェスタンブロットにより、Elisaによりまたは酵素アッセイにより、前掲)およびRNA分子(例えば、PCRまたはノーザンブロットにより、前掲)を決定するためのならびに前記量を比較するための手段と方法は当技術分野において周知である。好ましくは、所望の産物の量は、参照量と比較して統計的に有意に増加し、さらに好ましくは、所望の産物の量は、2倍、3倍、4倍、5倍、10倍、20倍、30倍、50倍、100倍、200倍または1000倍増加する。
【0034】
本明細書で使用する用語「湾曲起点モチーフ」は、少なくとも1つの複製起点シグナル(また「複製起点シグナル」、「起点シグナル」、または「oriシグナル」とも呼ばれる)と少なくとも1つの湾曲DNAシグナルとの組合わせと解釈することとする。好ましくは、湾曲起点モチーフは、1、2、3、4、5、7、10、15、20、21、25、30、35、40、50、55、60、70、80、90、100、200、500、1000以上の複製起点シグナルおよび1、2、3、4、5、7、10、15、20、21、25、30、35、40、50、55、60、70、80、90、100、200、500、1000以上の湾曲DNAシグナルを含む。本発明による湾曲起点モチーフは異なるoriシグナルを含んでもよい。さらに、本発明による湾曲起点モチーフは異なる湾曲DNAシグナルを含んでもよい。本明細書に参照によりその全てが組み入れられるSinghら(Nucleic Acids Research, 1997, 25(7), 1419 - 1425)は、oriシグナルならびに湾曲DNAシグナルを開示している。本明細書で意味する湾曲DNAシグナルは、これらの配列を含有するDNA分子中に湾曲(内因性の)を導入できることで特徴付けられる核酸配列である。当業者にとってDNA分子中に湾曲を導入する核酸配列エレメントは公知である。例えば、湾曲は、例えば、アデニンの短いホモポリマー列(Aトラック)(4〜6個の塩基対)が、一致してらせん型スクリューで繰り返されると起こりうる。1つのらせんスクリューはおよそ10.4bpを含む(Haranら, 1994, Sequences elements responsible for DNA curvature. J Mol Biol, 244: 135-143を参照)。DNA断片の湾曲は、例えば、DNA断片の電気泳動移動度により決定するかまたは好適なコンピューターソフトウエア(例えば、Shpigelmanら, 1993, Comput Appl Biosci.;9(4):435-40に記載)を用いることにより予測することができる。湾曲DNAは、例えば、いくつかのマトリックス付着部位またはその近傍にて同定されている。湾曲DNAはまた、組換え、複製、および転写などのDNAおよびタンパク質の相互作用に関わる核プロセスにおいて重要な役割を果たしうる。DNA配列エレメント中への湾曲は、AAAANNNNNNNAAANNNNNNNAAAA(配列番号18に示した)、またはAAAANNNNNNNAAAANNNNNNNAAAA(配列番号19に示した)、またはTTTTNNNNNNNTTTNNNNNNNTTTT(配列番号20に示した)、またはTTTTNNNNNNNTTTTNNNNNNNTTTT(配列番号21に示した)[ここでNは任意のヌクレオチド、従ってA、T、CまたはGまたは前記ヌクレオチドのいずれかの化学的に改変された誘導体を表し、好ましくはNはGまたはCを表す]の核酸配列を有する湾曲DNAシグナルを含むDNAモチーフにより導入することができる。特に、湾曲はこれらのシグナルの反復配列/多量体により導入される。
【0035】
さらに、DNA配列エレメント中への湾曲は、湾曲DNAシグナル、好ましくは、湾曲DNAシグナルの多量体/反復配列、より好ましくは、おおよそ、らせんスクリューの相にある湾曲DNAシグナルの多量体/反復配列を含むDNAモチーフにより導入することができ、ここで、湾曲DNAシグナルはAAAANNNNNN(配列番号37に示した)、またはAAAANNNNNNN(配列番号38に示した)、またはAAAAANNNNN(配列番号39に示した)、またはAAAAANNNNNN(配列番号40に示した)、AAAAAANNNN(配列番号41に示した)、またはAAAAAANNNNN(配列番号42に示した)、またはTTTTNNNNNN(配列番号43に示した)、またはTTTTNNNNNNN(配列番号44に示した)、またはTTTTTNNNNN(配列番号45に示した)、またはTTTTTNNNNNN(配列番号46に示した)、またはTTTTTTNNNN(配列番号47に示した)、またはTTTTTTNNNNN(配列番号48に示した)[ここでNは任意のヌクレオチド、従ってA、T、CまたはGまたは前記ヌクレオチドのいずれかの化学的に改変された誘導体を表し、好ましくはNはGまたはCを表す]の核酸配列を有する。さらに、DNA配列中への湾曲は、湾曲DNAシグナルTTTAAA(配列番号22に示した)、およびAAATTT(配列番号23に示した)、(例えば、Singhら、前掲)、とりわけこれらのシグナルの多量体/反復配列により導入される。
【0036】
oriシグナルはoriのエレメント、好ましくはATリッチである。当業者にとってori(複製の起点)は公知である。oriはDNA複製を開始することができるDNA配列である。oriのDNA配列は、種毎に、特に原核生物と真核生物では変化しうるが、oriモチーフは一般にいくつかの共通の特性を共有する。本明細書で使用するoriシグナルは、好ましくは、真核生物の複製起点由来の配列モチーフであるが、原核生物の複製起点由来のシグナル/モチーフであってもよい。真核生物のoriはよく記載されていて、例えば、出芽酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)については起点活性(自律複製配列またはARSエレメントと呼ばれる)を与える具体的な配列が単離されている。DNA複製が核マトリックスと関係していることは公知である。また、核マトリックス付着部位、ホメオティックなタンパク質認識および結合部位、ならびに複製起点は、次のoriシグナル:ATTA(配列番号24に示した)、ATTTA(配列番号25に示した)、およびATTTTA(配列番号26に示した)、(例えば、Singhら、上記引用文中)を共有することが実証されている。
【0037】
湾曲起点モチーフの湾曲DNAシグナルと湾曲起点モチーフのoriシグナルは離れていても、すなわち、さらなる核酸配列により分離されていてもよい。さらに、湾曲DNAシグナルとoriシグナルの核酸配列は重複していてもよい、すなわち、oriシグナルのエレメント/部分であるヌクレオチドが湾曲DNAシグナルのエレメントであってもよい。さらに、湾曲DNAシグナルが1個以上のoriシグナルを含んでもよく、その逆もありうる。湾曲DNAシグナルとoriシグナルの重複する例は、例えば、配列番号1、配列番号2、配列番号29、または配列番号31に示した湾曲起点モチーフである(下記参照)。本発明による湾曲起点モチーフが、それを含有するDNA中に湾曲を導入することができるのは理解されよう。湾曲は、上記の手段と方法により決定することができる。さらに、湾曲起点モチーフはまた、様々な湾曲起点モチーフの組合わせを含みうることも理解されよう。
【0038】
本発明のある好ましい実施形態において、湾曲起点モチーフは、配列番号1に示した核酸配列ATTTTANNNNNATTTTANNNNNATTTTA、または少なくとも2つの配列番号29に示した核酸配列ATTTTANNNNNATTTTANNNNNATTTTANNNNNの反復配列(従って、前記核酸配列の少なくとも二量体をもたらす、少なくとも2倍の配列番号29に示した核酸)、配列番号2に示した核酸配列TAAAATNNNNNTAAAATNNNNNTAAAATまたは少なくとも2つの配列番号30に示した核酸配列TAAAATNNNNNTAAAATNNNNNTAAAATNNNNNの反復配列[ここで、Nは任意のヌクレオチド、すなわち、A、T、CまたはG、好ましくはCまたはG、または前記ヌクレオチドのいずれかの化学的に改変された誘導体を表す]を含む。より好ましくは、湾曲起点モチーフは、配列番号31に示した核酸配列ATTTTAGCTCGATTTTAGCTCGATTTTA、または少なくとも2つの配列番号32に示した核酸配列ATTTTAGCTCGATTTTAGCTCGATTTTAGCTCGの反復配列、または配列番号33に示した核酸配列TAAAATCGAGCTAAAATCGAGCTAAAAT、配列番号34に示したTAAAATCGAGCTAAAATCGAGCTAAAATCGAGC、または少なくとも2つの配列番号75に示した核酸配列AAAATCGAGCTAAAATCGAGCTAAAATCGAGCTの反復配列を含む。本明細書で使用する用語「少なくとも2つの反復配列」は、好ましくは、2以上の反復配列(および従って2以上のユニットまたはモノマー)、好ましくは、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7または少なくとも10、少なくとも20、少なくとも30、少なくとも40、少なくとも50、少なくとも100、少なくとも200の反復配列/ユニット/モノマーを意味する。従って、湾曲起点モチーフは、好ましくは、2倍以上、または3倍以上などの、本明細書で先に言及したそれぞれの核酸を含み、従って、好ましくは前記核酸の二量体(2つの反復配列を含む場合)、三量体(3つの反復配列を含む場合)、オリゴマー、多量体などである。本発明のクロマチンエレメント中に含まれる湾曲起点モチーフの、特に好ましい数は7である。本発明のクロマチンエレメント中に含まれる湾曲起点モチーフの、他の特に好ましい数は22である。好ましくは、前記少なくとも2つの反復配列は、正方向の反復配列(頭部から尾部への反復配列、好ましくは、反復配列の間にさらなるヌクレオチドの無いもの)である。
【0039】
さらに、湾曲起点モチーフは、核酸配列TAAAATNNNN(配列番号49に示した)、またはTAAAATNNNNN(配列番号50に示した)、またはTAAAAATNNN(配列番号51に示した)、またはTAAAAATNNNN(配列番号52に示した)、TAAAAAATNN(配列番号53に示した)、またはTAAAAAATNNN(配列番号54に示した)、またはATTTTANNNN(配列番号55に示した)、またはATTTTANNNNN(配列番号56に示した)、またはATTTTTANNN(配列番号57に示した)、またはATTTTTANNNN(配列番号58に示した)、またはATTTTTTANN(配列番号59に示した)、またはATTTTTTANNN(配列番号60に示した)の少なくとも1つの反復配列(従って、少なくとも1回の核酸配列)を含むことができる[ここでここでNは任意のヌクレオチド、従ってA、T、CまたはGまたは前記ヌクレオチドのいずれかの化学的に改変された誘導体を表し、好ましくはNはGまたはCを表す]。本明細書で使用する用語「少なくとも1つの反復配列」は、1以上の反復配列/ユニット/モノマー、好ましくは、少なくとも2、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7または少なくとも10、少なくとも20、少なくとも22、少なくとも30、少なくとも40、少なくとも50、少なくとも75、少なくとも100、少なくとも200の反復配列/ユニット/モノマーを意味する。先に記載の通り、本発明のクロマチンエレメント中に含まれるべき湾曲起点モチーフの、特に好ましい数は22である。好ましくは、前記反復配列は正方向の反復配列である(前記参照)。
【0040】
他のさらに好ましい本発明の実施形態において、湾曲起点モチーフは配列番号7または配列番号8、または配列番号9、または配列番号10に示した核酸配列を含む。
【0041】
本発明によれば、湾曲起点モチーフ用の核酸配列はクロマチンエレメントに対する核酸配列と共有結合により連結されている。当業者にとって、共有結合によって2以上の核酸配列を連結するのに使用できる技法、例えば、ライゲーションまたはクローニングなどの組換えDNA技法として知られる技法は公知である。湾曲起点モチーフが、発現を増強できるクロマチンエレメント中にさらに組み込まれることは理解されよう。それにより、「合成」クロマチンエレメントが作製される。湾曲起点モチーフ用核酸配列はクロマチンエレメント用核酸配列のいずれの位置にあってもよいことは理解されよう。好ましくは、湾曲起点モチーフは、配列番号27のヌクレオチドの1900と2200の間、より好ましくは、2000と2100の間、そして最も好ましくは、位置2031の位置に導入される。
【0042】
さらに、共トランスフェクトしたポリヌクレオチドの発現を増強できるクロマチンエレメントは、湾曲起点モチーフ用の核酸配列を、1つ以上の、例えば、2、3、4、5、10、20、30、40、50の位置にまたは100の位置に含んでもよく、好ましくは、前記ポリヌクレオチドは前記核酸配列を1つの位置に含むことが理解されよう。
【0043】
他の最も好ましい本発明の実施形態において、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドは、配列番号3にまたは配列番号4にまたは配列番号5にまたは配列番号6にまたは前記配列番号のいずれか1つの変異体に示した核酸配列を有する。かかる変異体は、好ましくは、配列番号3、4、5または6のいずれか1つと少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%または少なくとも99%同一である核酸配列を有する。2以上の核酸配列の間の同一性の程度(パーセント、%)は、好ましくは、NeedlemanおよびWunschまたはSmithおよびWatermanのアルゴリズムにより決定される。配列アラインメントを行うために、プログラムPileUp(J. Mol. Evolution., 25, 351-360, 1987, Higgins 1989, CABIOS, 5: 151-153)またはプログラムGapおよびBestFit(Needleman 1970, J. Mol. Biol. 48; 443-453 および Smith 1981, Adv. Appl. Math. 2; 482-489)が利用され、これらはGCGソフトウエアパケット(Genetics Computer Group, 575 Science Drive, Madison, Wisconsin, USA 53711, vers. 1991)の一部分である。パーセント(%)で以上に記載される配列同一性値は、好ましくは、プログラムGapを用いて、全配列領域にわたり、次の設定値、ギャップ重み:50、長さ重み:3、平均マッチ:10.000および平均ミスマッチ:0.000を用いて決定し、これは、特に断らない限り、常に配列アラインメント用の標準設定として用いることとする。さらに、前記変異体核酸配列は、好ましくは、先に規定した少なくとも1つの湾曲起点モチーフを含む。好ましくは、かかる変異体核酸配列を含む、ポリヌクレオチドは共トランスフェクトしたポリヌクレオチドの発現を増強することができる。この増強は、添付した実施例に記載のアッセイにより試験することができる。
【0044】
本発明はまた、先に規定した湾曲起点モチーフにも関する。好ましくは、前記モチーフは、共トランスフェクトしたポリヌクレオチドの発現を増強するために使われる。
【0045】
有利なことに、本発明によって、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドと目的のポリヌクレオチドとの宿主細胞中への共導入は、目的のポリヌクレオチドの発現を増強することが見出されている。例えば、ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域を含むポリヌクレオチド(Romig, 1994 Eur J Biochem. 221:411-419)と湾曲起点モチーフを、選択可能なマーカーとしてのレポーター遺伝子EGFP(Enhanced Green Fluorescent Protein;緑色蛍光が増強されたタンパク質)および未改変ネオマイシンホスホトランスフェラーゼ遺伝子を保持するClontechから入手したpEGFP-N1ベクターと一緒に、CHO細胞(Chinese Hamster Ovary;チャイニーズハムスター卵巣)中に同時トランスフェクトした。対照としてpEGFP-N1ベクターを、単独で、またはヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域に対するDNAと湾曲起点モチーフなしで組合わせて、CHO細胞中に導入した。トランスフェクション後、G418を含有する培地(0.6mg/mlG418)で細胞を培養して選択した。10日後、単一のG418耐性クローンがコロニーを形成した。全体の耐性細胞集団を用いて、FACS分析(Fluorescent Activated Cell Sorting;蛍光活性化細胞ソーティング)によりレポーター遺伝子発現の数値化を行った。対照と比較して、高発現細胞の比率、従って発現が、レポーター遺伝子と湾曲起点モチーフを含む前記ATリッチ領域を共導入することにより作製したCHO細胞において、有意に増加することを見出した。従って、共トランスフェクトした目的のポリペプチドの発現をそれ自体増強できるクロマチンエレメント中への、湾曲起点モチーフの挿入は、前記クロマチンエレメントの発現増強効果をさらに増強する。湾曲モチーフを単独で(湾曲起点モチーフの代わりに)前記クロマチンエレメントに組み込んだ場合、前記クロマチンエレメント単独(湾曲モチーフを挿入しないで)と比較すると、発現の増強が驚くほど検出されなかった。この驚く
べき発現増強効果は、例えば、配列番号3、配列番号4および配列番号5に示した核酸配列を有するポリヌクレオチドについて示された。従って、本発明による上述のポリヌクレオチドの使用は、高生産クローンを単離するための時間と費用のかかるスクリーニングステップを軽減するであろう。
【0046】
さらに、本発明は、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするか、または配列番号14、配列番号15および配列番号16のいずれか1つに示した核酸配列を含む、ポリヌクレオチドに関する。
【0047】
前記ポリヌクレオチドは、配列番号35に示したアミノ酸配列を有する未改変の(野生型)ネオマイシンホスホトランスフェラーゼと比較して、アミノ酸配列に1または2個のアミノ酸変化を有する改変されたネオマイシンホスホトランスフェラーゼII(NPT II)をコードする。具体的には、本発明の改変されたNPT IIは、それぞれRのAへの、およびRのVへのアミノ酸置換を、それぞれ配列番号11および配列番号12に記載のNPT IIの226位に対応する位置に有するか、またはそれぞれAのGへの、およびTのSへのアミノ酸置換を、それぞれ配列番号13に記載のNPT IIの224位および225位に対応する位置に有することを想定している。ネオマイシン耐性遺伝子とも呼ばれるNPT II遺伝子は、最初に大腸菌株K12のトランスポゾンTn5から単離された。NPT IIタンパク質はアミノグリコシド-アミノシクリトール抗生物質を位置特異的リン酸化により不活性化する酵素のクラスに属する。これはカナマイシンおよびG418などの様々なアミノグリコシド抗生物質に対する耐性を与える。この遺伝子は、一般に選択可能なマーカーとして、細菌、植物、動物および酵母のような様々な種類の生物を形質転換するために利用される。当技術分野では、NPT IIアミノ酸配列の改変、すなわち、アミノ酸変化がNPT IIの酵素活性を低下させうることは公知である。さらに、酵素活性の低下した突然変異NPT II変異体の形質転換への使用は、未改変のNPT II遺伝子を保持する生物よりもネオマイシンに対する耐性が低いトランスジェニック生物をもたらすことは公知である。さらに、これらの変異体を高生産細胞の選択に利用することができる(例えば、WO2004/050884を参照)。
【0048】
改変されたネオマイシンホスホトランスフェラーゼをコードする本発明のポリヌクレオチドは、融合タンパク質の1つのパートナーが先に挙げた核酸配列によりコードされているポリペプチドである融合タンパク質をコードしうることは理解されよう。かかる融合タンパク質は、さらなる部分として脂肪酸または脂質生合成経路の他の酵素、発現をモニターするためのポリペプチド(例えば、緑色、黄色、青色または赤色蛍光タンパク質、アルカリホスファターゼなど)または検出可能なマーカーとしてまたは精製のための補助的基準として役割を果たしうるいわゆる「タグ」を含んでもよい。色々な目的のためのタグが当技術分野では周知であり、それには、FLAG-タグ、6-ヒスチジン-タグ、MYC-タグなどが含まれる。
【0049】
さらに本発明は、配列番号11、配列番号12および配列番号13のいずれか1つに示されるアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16のいずれか1つに示される核酸配列を含むものである、ポリヌクレオチドによりコードされるポリペプチドに関する。
【0050】
有利なことに、本発明によれば、配列番号14、配列番号15および配列番号16のいずれか1つに示した核酸配列をCHO細胞のトランスフェクションに対する選択可能なマーカー遺伝子として含むポリヌクレオチドの使用は、特に、共導入した目的のポリヌクレオチドの発現を増強することが示されている。例えば、改変されたNPT II遺伝子およびレポーター遺伝子EGPFを含む様々なベクターをCHO細胞中に導入した。トランスフェクション後、前記細胞をG418含有培地(0.6mg/ml)で培養することにより選択した。選択可能なマーカーとして、改変されたNPT IIの代わりに未改変のネオマイシンホスホトランスフェラーゼをコードするポリヌクレオチドを用いる比較実験を実施した。高発現細胞、従ってレポーター遺伝子発現の比率は、改変されたネオマイシンホスホトランスフェラーゼを選択可能なマーカーとして用いることにより作製したトランスジェニック宿主細胞において有意に増強されることを見出した。従って、記載した本発明によるポリヌクレオチドの使用は、高生産クローンを単離するための時間と費用のかかるスクリーニングステップを軽減するであろう。
【0051】
本発明はまた、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列、または、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするポリヌクレオチドまたは配列番号14、配列番号15および配列番号16に示した核酸配列のいずれか1つに示した核酸配列、またはその組合わせを含むものである、以上説明したポリヌクレオチドを含む宿主細胞に関する。
【0052】
本明細書で使用する「宿主細胞」には、異種DNAの導入により改変することができる任意の培養可能な細胞が含まれる。好ましくは、宿主細胞は、その細胞において目的のポリヌクレオチドを安定して発現することができ、前記ポリペプチドがコードするポリペプチドを翻訳後に改変して適当なサブ細胞コンパートメントに局在化できる細胞である。適当な宿主細胞の選択はまた、検出シグナルの選択によっても影響を受けうる。本発明の宿主細胞には原核細胞および真核細胞が含まれる。原核生物にはグラム陰性またはグラム陽性生物、例えば、大腸菌または桿菌が含まれる。好ましくは、本発明によるポリヌクレオチド、特に本発明のポリヌクレオチドを含むベクター(下記参照)の増殖には原核細胞が使われることは理解されよう。形質転換のための好適な原核生物の宿主細胞には、例えば、大腸菌、枯草菌、ネズミチフス菌、ならびにシュードモナス菌、ストレプトマイセス菌、およびブドウ球菌属内の様々な他の種が含まれる。真核細胞には、限定されるものでないが、動物細胞、脊椎動物細胞、酵母細胞、植物細胞、真菌細胞、昆虫細胞(例えば、バキュロウイルスにより感染させることができるSF9細胞)、哺乳動物細胞、爬虫類動物細胞、藻類細胞および寄生生物、例えば、トリパノソーマ類の細胞が含まれる。本明細書で使用する、酵母には、厳密に分類学的な意味の酵母、すなわち、単細胞生物だけでなく、糸状真菌の酵母様多細胞真菌が含まれる。種の例には、クライベラ・ラクチス(Kluyvera lactis)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)、およびウスチラゴ・マイディス(Ustilago maydis)が含まれ、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)が好ましい。本発明の実施に使用できる他の酵母は、アカパンカビ(Neurospora crassa)、クロコウジカビ(Aspergillus niger)、アスペルギルス・ニデュランス(Aspergillus nidulans)、ピチア・パストリス(Pichia pastoris)、カンジダ・トロピカリス(Candida tropicalis)、および ハンセヌラ・ポリモルファ(Hansenula polymorpha)である。哺乳動物の宿主細胞培養系には、
COS細胞、L細胞、3T3細胞、HeLa細胞、ベビーハムスター腎(BHK)細胞、チャイニーズハムスター卵巣(CHO)細胞、胚幹細胞などの確立された細胞が含まれる。
【0053】
本発明のある好ましい実施形態において、宿主細胞は哺乳動物細胞である。さらに好ましい実施形態において、宿主細胞はCHO細胞である。CHO細胞培養系には、血清を含有するおよび/または血清を含有しない培地で培養できる全てのCHO細胞、例えば、CHO-K1、CHO-S、CHO DG44、CHO DXB11、CHO(タンパク質を含有しない)ECACC No. 00102307、CHO(dhfr-ACを含有しない)ECACC No. 05011002 細胞が含まれる。
【0054】
さらに、本発明は、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列、または、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするポリヌクレオチドまたは配列番号14、配列番号15および配列番号16に示した核酸配列のいずれか1つに示した核酸配列、またはその組合わせを含む、以上説明したポリヌクレオチドを含む、非ヒトトランスジェニック生物に関する。
【0055】
本発明によれば、非ヒトトランスジェニック生物は非ヒト動物または植物であってもよい。本明細書で意味するトランスジェニック非ヒト動物は、出生前に、例えば、胚段階に非ヒト動物または前記非ヒト動物の祖先中に導入されたトランスジーンを含有する細胞を有するものとする。トランスジーンは、細胞のゲノム中に組み込まれているDNAであり、これからトランスジェニック非ヒト動物が発生し、これが成熟非ヒト動物のゲノム中に残り、それによりトランスジェニック非ヒト動物の1以上の細胞型または組織中のコードされた遺伝子産物の発現を駆動する。好ましい非ヒトトランスジェニック動物は、マウス、ヤギ、ヒツジ、ブタ、ウシ、ウマ、魚類、および非ヒト霊長類からなる群より選択される。非ヒト農場動物は、タンパク質の大規模生産のために有用である(いわゆる「遺伝子農場(gene farm)」)。トランスジェニック非ヒト動物は、例えば、トランスジーンを受精卵母細胞の雄前核中に、例えば、マイクロインジェクションによって導入し、次いで卵母細胞を擬妊娠雌里親動物中で発生させることにより作製することができる。
【0056】
トランスジェニック動物、特にマウスなどの動物を作製する方法は当技術分野で一般的になっていて、例えば、米国特許第4,736,866号および第4,870,009号およびHogan 1986, A Laboratory Manual, Cold Spring Harbor, New York, Cold Spring Harbor Laboratoryに記載されている。トランスジェニック里親動物を用いてトランスジーンを保持するさらなる動物を育種することができる。本発明によるトランスジーンを保持するトランスジーン動物を、他のトランスジーンを保持する他の動物、例えば、テトラサイクリン依存性転写レギュレーターをコードするポリヌクレオチドを発現するトランスジェニックを育種することができる(以下にさらに詳細に考察される)。本発明はまた、本発明のベクターまたは発現制御配列を、好ましくは、先に言及したトランスジーンの形態で含む相同的組換え非ヒト動物も提供する。かかる非ヒト相同的組換え動物においては、前記核酸はゲノムの特定部位中に導入されている、すなわち、核酸分子はゲノムの内因性遺伝子または他の部分と相同的に組換えられている。前記事例において、動物は好ましくはマウスである。かかる相同的組換え動物を作製するために、好ましくは、相同的組換えが起こるゲノムの遺伝子または部分のさらなる核酸にその5'および3'末端で隣接した、導入すべき核酸をコードするDNAを含有するベクターを調製する。融合タンパク質をコードする隣接したさらなる核酸は、真核生物の遺伝子との相同的組換えが成功するのに十分な長さである。典型的には、数キロ塩基の(5'および3'末端の両方で)隣接するDNAがベクター中に含まれる(例えば、相同的組換えベクターの説明については、Thomas, 1987, Cell 51:503を参照)。ベクターを胚幹細胞株中に導入し(例えば、エレクトロポレーションにより)、導入したDNAが内因性DNAと相同的に組換えられた細胞を選択する(例えば、Li 1992, Cell 69:915を参照)。選択した細胞を次いで動物(例えば、マウス)の胚盤胞中に注射して凝縮キメラを形成する(例えば、Bradley, A., Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E.J. Robertson, 編 (IRL, Oxford, 1987) pp.113-152を参照)。キメラ胚を次いで好適な偽妊娠雌里親動物中に移植し、胚を出産させる。相同的に組換えたDNAをその生殖細胞中に保持する子孫を用いて、その全細胞が相同的に組換えられたDNAを含有する動物を育種する。これらの「生殖系列導入」動物をさらにテトラサイクリン依存性転写レギュレーターをコードする遺伝子を保持する動物と交配することができる。以上記載した相同的組換え手法だけでなく、酵素の助けをかりる部位特的組込み系が当技術分野では公知であり、本発明の調節システムの成分に適用して第2の標的DNA分子中の予め定めた位置にDNAを組み込むことができる。かかる酵素の助けをかりる組込み系の例としては、Creリコンビナーゼ-lox標的系(例えば、Baubonis 1993, Nucl. Acids Res. 21:2025-2029;およびFukushige 1992, Proc. Natl. Acad. Sci. USA 89:7905-7909に記載)およびFLPリコンビナーゼ-FRT標的系(例えば、Dang 1992, Dev. Genet. 13:367-375;およびFiering 1993、Proc. Natl. Acad. Sci. USA 90:8469-8473に記載)が挙げられる。
【0057】
本明細書で使用する用語「植物」は植物と藻類を包含する。好ましくは、この用語は多細胞陸生植物に関する。より好ましくは、多細胞陸生植物は作物植物、などの高等植物であり、トウモロコシ、キャノーラ、ダイズ、イネ、マリーゴールド、アブラナ、コムギ、または グリシン(glycine)が含まれる。原理的に、トランスジェニック植物は、Becker 1992, Plant Mol. Biol. 20:1195-1197、Bevan 1984, Nucleic Acids Res. 12:8711-8721、および“Vectors for Gene Transfer in Higher Plants” in: Transgenic Plants, Vol. 1, Engineering and Utilization, 編: Kung、およびR. Wu, Academic Press, 1993, p. 15-38に記載のように得ることができる。好ましくは、植物細胞の形質転換、従って、トランスジェニック植物の作製は、アグロバクテリウム介在性形質転換によりまたは物理的な力(例えば、「遺伝子銃」)により達成しうる。
【0058】
本発明はまた、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列、または、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16に示した核酸配列のいずれか1つに示した核酸配列、またはその組合わせを含むポリヌクレオチドを含むものである、以上に規定したポリヌクレオチドを含むベクターに関する。
【0059】
用語「ベクター」は、好ましくは、ファージ、プラスミド、ウイルスまたはレトロウイルスベクター、ならびに細菌または酵母人工染色体などの人工染色体を包含する。さらに、本用語はまた、標的化構築物のゲノムDNA中への無作為または部位指定組込みを可能にする標的化構築物に関する。かかる標的構築物は、好ましくは、以下に詳細に記載する相同的なまたは異種組換えに十分な長さのDNAを含む。本発明のポリヌクレオチドを包含するベクターは、好ましくは、さらに宿主における繁殖および/または選択のための選択可能なマーカーを含む。ベクターは、宿主細胞中に当技術分野で周知の様々な技法により組み込むことができる。例えば、プラスミドベクターはリン酸カルシウム沈降物または塩化ルビジウム沈降物などの沈降物で、または荷電脂質との複合体で、またはフラーレンなどの炭素に基づくクラスターで導入することができる。あるいは、プラスミドベクターは熱ショックまたはエレクトロポレーション技法により導入することができる。ベクターがウイルスであれば、適当なパッケージング細胞株を用いてin vitroでパックした後、宿主細胞に適用してもよい。レトロウイルスベクターは複製コンピテントまたは複製欠陥性であってもよい。後者の場合、ウイルス繁殖は一般に補完性宿主細胞/細胞でのみ起こりうる。
【0060】
より好ましくは、本発明のベクターにおいて、本発明によるポリヌクレオチドは、原核生物または真核細胞、特に真核細胞またはそれらの単離された画分における発現または繁殖を可能にする発現制御配列と機能しうる形で連結されている。前記ポリヌクレオチドの発現は、ポリヌクレオチドの、好ましくは、翻訳可能なmRNA中への転写を含む。真核細胞、好ましくは哺乳動物の細胞における発現を保証する調節エレメントは当技術分野で周知である。これらは、好ましくは、転写の開始を保証する調節配列および、任意に、転写の終結および転写物の安定化を保証するポリ-Aシグナルを含む。さらなる調節エレメントには、転写ならびに翻訳エンハンサーが含まれてもよい。原核生物宿主細胞における発現を可能にすると思われる調節エレメントとしては、例えば、大腸菌のlac、trpまたはtacプロモーターが含まれ、真核生物宿主細胞における発現を可能にする調節エレメントの例は、酵母のAOX1またはGAL1プロモーターまたは哺乳動物および他の動物細胞のCMV-、SV40-、RSV-プロモーター(ラウス肉腫ウイルス)、CMV-エンハンサー、SV40-エンハンサーまたはグロビン-イントロンである。好適な発現制御配列は当技術分野で周知である。転写の開始に関わるエレメントに加えて、かかる調節エレメントはポリヌクレオチドの下流にSV40-ポリA部位またはtk-ポリA部位などの転写終結シグナルを含んでもよい。この関係において、好適な発現ベクターとしては、Okayama-Berg cDNA発現ベクターpcDV1(Pharmacia)、pBluescript(Stratagene)、pCDM8、pRc/CMV、pcDNA1、pcDNA3(Invitrogene)またはpSPORT1(GIBCO BRL)などが当技術分野において公知である。好ましくは、前記ベクターは発現ベクターおよび遺伝子導入または標的化ベクターである。レトロウイルス、ワクシニアウイルス、アデノ随伴ウイルス、ヘルペスウイルス、またはウシパピローマウイルスなどのウイルスから誘導される発現ベクターを、本発明のポリヌクレオチドまたはベクターを標的化細胞集団中への送達するのに利用することができる当業者に周知の方法を、構築物組換えウイルスベクターを構築するために利用することができる;例えば、Sambrook, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (1989) N.Y. および Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience、N.Y. (1994)に記載の技法を参照されたい。
【0061】
本発明はまた、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列、または、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするポリヌクレオチドまたは配列番号14、配列番号15および配列番号16に示した核酸配列のいずれか1つに示した核酸配列、またはその組合わせを含むものである、以上説明したポリヌクレオチドを含む、キットに関する。
【0062】
本発明のポリヌクレオチドは、本発明の方法を行うのに適応したキットとして、宿主細胞における宿主細胞の核酸の発現に必要な他の成分と一緒に提供することができる。キットの他の成分は、好ましくは、発現のために利用する宿主細胞および/またはポリヌクレオチドの細胞中への導入のために用いる物質である。
【0063】
本明細書で使用する用語「キット」は、上述した本発明の化合物、手段または試薬のコレクションを意味する。キットの成分は、別のバイアルに含まれていてもよいし(すなわち、別の部品のキットとして)または単一バイアルで提供されてもよい。さらに、本発明のキットは本明細書で以上言及した方法を実施するために使われることは理解されよう。好ましくは、以上言及した方法を実施するために直ぐ使用できる方式で提供されると想定している。さらに、本キットは好ましくは、前記方法を実行するための取扱説明書を含む。取扱説明書は、紙または電子形式のユーザーマニュアルで提供することができる。例えば、マニュアルは、本発明のキットを用いて上述の方法を行ったときに得られる結果の解釈に対する指示書を含むものである。
【0064】
以上記載した用語の定義と説明は、以下の実施形態に対して必要な変更を加えて適用される。
【0065】
本発明はまた、少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)宿主細胞中に
(i)発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチド、および
(ii)少なくとも1つの目的のポリヌクレオチド
を導入するステップ、ならびに
(b)前記宿主細胞を培養するステップ
を含んでなる前記方法に関する。
【0066】
本発明による方法においては、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドを用いて少なくとも1つの目的のポリヌクレオチドを発現する。好ましくは、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドを用いて少なくとも1つの目的のポリヌクレオチドの発現を増強する。本明細書で使用する用語「少なくとも1つの目的のポリヌクレオチド」は、好ましくは、1以上の、例えば、少なくとも2、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7または少なくとも10、または少なくとも20の目的のポリヌクレオチドを意味し、好ましくは用語「少なくとも1つの目的のポリヌクレオチド」は1つの目的のポリヌクレオチドを意味する。好ましくは、本発明の方法のポリヌクレオチドおよび目的のポリヌクレオチドは宿主細胞中に共導入(同時に導入)される。本発明のポリヌクレオチドは、宿主細胞中にポリヌクレオチドを導入するのに好適ないずれの技法により宿主細胞中に導入してもよく、かかる技法は当技術分野で周知でありかつよく記載されている。例えば、ポリヌクレオチドはエレクトロポレーションにより、または、マイクロインジェクションにより、リン酸カルシウム沈降物または塩化ルビジウム沈降物などの沈降物で、または脂質に基づくトランスフェクションにより、または荷電脂質との複合体で、またはフラーレンなどの炭素に基づくクラスターで導入することができる。さらに、ポリペプチドは、レトロウイルスなどのウイルスを用いることにより組み込むことができる。本発明のポリヌクレオチドは。いずれの宿主細胞中に導入してもよく、好ましくは、本出願の文脈で記述したいずれの宿主細胞中に導入してもよい。より好ましくは、ポリヌクレオチドはCHO細胞中に脂質に基づくトランスフェクションにより導入される。宿主細胞中に導入されるべき目的のポリヌクレオチドおよび本発明の方法によるポリヌクレオチドは同じ核酸分子、例えば、DNAまたはRNAベクター上に、または異なる核酸分子上に位置してもよい。好ましくは、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドと目的のポリヌクレオチドは異なる核酸分子上に存在する。さらに、宿主細胞中に導入すべきポリヌクレオチドは、直鎖または環状型で存在してもよい。核酸分子、特に、DNA分子に関する用語「直鎖」または用語「環状」は当業者により理解されている。好ましくは、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドは、直鎖または環状型で存在し、目的のポリヌクレオチドまたはいずれかの他のポリヌクレオチド(例えば、マーカーをコードするポリヌクレオチド)は環状型、例えば、プラスミドで存在する。より好ましくは、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドは直鎖型で存在し、目的のポリヌクレオチドまたはいずれかの他のポリヌクレオチド(例えば、マーカーをコードするポリヌクレオチド)は直鎖型、例えば、直鎖プラスミドで存在する。
【0067】
本発明の方法によると、好ましくは、参照宿主細胞における目的のポリヌクレオチドの発現と比較して、目的のポリヌクレオチドの発現が増強されるであろうことは理解されよう。かかる参照細胞は、本発明のポリヌクレオチドを導入せずに目的のポリヌクレオチドを導入すること(例えば、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドなしに、目的のヌクレオチドをトランスフェクトすること)により作製することができる。発現の増強は本明細書に先に記載した手段と方法により示すことができる。
【0068】
本明細書で使用する用語「前記宿主細胞を培養する」は、好適な条件下で細胞を培養する方法および手段に関する。細胞を培養するための好適な条件は当技術分野で周知であり、よく記載されている。好ましくは、培養は、細胞が増殖しまたは成長し、目的のポリヌクレオチドを発現することを可能にする。例えば、培養に用いる培地は最小限の成長速度を可能にする培地(最小培地)であってもまたはさらに高い成長速度を可能にする培地であってもよい。さらに、培地の組成は生物/宿主 細胞の亜集団を選択することを可能にしてもよい。好ましくは、選択によって、導入された選択可能なマーカー遺伝子を保持しかつ十分な量の選択可能なマーカー遺伝子を発現する宿主細胞だけが、増殖することができる。選択は、選択剤、例えば、ある特定の抗生物質を、培養に用いる培地に加えることにより、または物質、例えば、ある特定の栄養分を培地に加えないことにより達成することができる。本発明の方法によって用いることができる選択剤は、酵素ネオマイシンホスホトランスフェラーゼにより不活性化されるアミノグリコシド抗生物質、好ましくはジェネティシン(G418)である。従って、上述の方法の関係で使用することができる選択可能なマーカー遺伝子は、ネオマイシンホスホトランスフェラーゼ、ハイグロマイシンホスホトランスフェラーゼ、ピューロマイシン-N-アセチル-トランスフェラーゼ、ジヒドロ葉酸レダクターゼ、グルタミンシンテターゼ、アスパラギン酸トランスカルバミラーゼ、メタロチオネイン、アデノシンデアミナーゼ、アデニル酸デアミナーゼ、UMPシンテターゼ、P-糖タンパク質(p-gp、MDR)、アスパラギンシンテターゼまたはオルニチンデカルボキシラーゼである。本明細書で以下に記載するように、マーカー遺伝子は、選択能力の低い選択可能なマーカータンパク質をコードする遺伝子であってもよい。先に記載の通り、宿主はCHO細胞であることが好ましい。好ましくは、CHO細胞を、L-グルタミン(例えば、PAA Laboratories GmbHより供給される)と10%ウシ胎児血清(FBS)を加えたHamのF-12培地において37℃にて、5%CO2雰囲気中で増殖する。サブコンフルエントな培養物(70〜80%)を1:4〜1:10に分割して、すなわち、0.25%トリプシン/EDTA(PAA Laboratories GmbH)を用いて1〜2x10,000細胞/cm2にて接種した。細胞の培養には、単離するステップおよび引き続いての個々の細胞、特に高発現する細胞を繁殖するステップなどのさらなるステップが含まれることは理解されよう。
【0069】
有利なことに、本発明の上述の方法によって、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドと目的のポリペプチドとの宿主細胞中への共導入は、目的のポリヌクレオチドの発現を増強することが見出されている。例えば、ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域(Romig,1994, Eur J Biochem. 221:411-419)およびさらなる湾曲起点モチーフを含むポリヌクレオチドを、レポーター遺伝子EGFP(Enhanced Green Fluorescent Protein;増強緑色蛍光タンパク質)およびNPT耐性遺伝子を含むpEGFP-N1ベクター(Clontech)と一緒に、CHO細胞(Chinese Hamster Ovary)中に共トランスフェクト(すなわち共導入)した。対照として、(a)ベクターpEGFP-N1単独、または(b)ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域のDNAと組合わせた、さらなる湾曲起点モチーフを伴わないベクターをCHO細胞中に導入した。G418(0.6mg/ml)による選択を含む培養ステップの後に、個々の細胞のレポーター遺伝子発現をFACS分析(Fluorescent Activated Cell Sorting)により数値化した。対照と比較して、高発現細胞の比率、従ってその発現は、レポーター遺伝子およびさらなる湾曲起点モチーフを含む前記ATリッチ領域を共導入することにより作製したCHO細胞において、有意に増加することが見出された。従って、目的のポリヌクレオチド(ここではレポーター遺伝子)および湾曲起点モチーフを含む、発現を増強できる前記ATリッチ領域をコードするDNAの共導入は、目的のポリヌクレオチドの高くかつ安定な発現を可能にする。従って、共トランスフェクトした目的のポリペプチドの発現をそれ自体で増強できるクロマチンエレメント中への、湾曲起点モチーフの挿入は、さらに前記クロマチンエレメントの発現増強効果を増強する。
【0070】
有意に増強しかつ比較的安定な発現の驚くべき効果は、例えば、配列番号3および配列番号4に示した核酸配列を有するポリヌクレオチドについて、レポーター遺伝子による共トランスフェクションと耐性細胞の選択の後に示された。従って、本発明の方法は単一の高生産クローンに対する時間と費用のかかるスクリーニングステップを軽減しうる。
【0071】
本発明はまた、少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)宿主細胞中に
(i)配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16のいずれか1つに示した核酸配列を含む、少なくとも1つのポリヌクレオチド、および
(ii)少なくとも1つの目的のポリヌクレオチド
を導入するステップ、ならびに
(b)前記宿主細胞を培養するステップ
を含んでなる前記方法に関する。
【0072】
上述の方法は、少なくとも1つの目的のポリヌクレオチドおよび少なくとも1つの改変されたネオマイシンホスホトランスフェラーゼ遺伝子をコードする本発明によるポリヌクレオチドの宿主細胞中への共導入を含んでなる。用語「宿主細胞中に導入する」、「宿主細胞」、「少なくとも1つの目的のポリヌクレオチド」は既に本明細書において先に記載されている。上述の方法の文脈において前記宿主細胞を培養するとは、宿主細胞を好適な培養条件下でインキュベートすることを意味する(前記参照)。上述の方法の場合、選択可能なマーカー遺伝子はネオマイシンホスホトランスフェラーゼであり、選択剤は、酵素ネオマイシンホスホトランスフェラーゼにより不活性化されるアミノグリコシド抗生物質であってもよい。好ましくは、選択剤はG418である。
【0073】
上述の方法によれば、宿主細胞における目的のポリヌクレオチドの発現は参照宿主細胞と比較して増強されるであろうことは理解されよう。好ましくは、上述の方法の文脈において、参照宿主細胞は、未改変のNPT II遺伝子と共導入された対応する目的のポリヌクレオチドを発現する宿主細胞である。
【0074】
さらに、上述の方法の文脈において、NPT IIをコードするポリヌクレオチドおよび目的のポリヌクレオチドは、宿主細胞中に導入される前に、同じ核酸分子上にまたは異なる核酸分子上に位置してもよいことは理解されよう。好ましくは、選択可能なマーカーをコードするポリヌクレオチドと目的のポリヌクレオチドは1分子上に存在する。
【0075】
有利なことに、本発明によれば、本明細書の随所に記載されるNPT II突然変異体ポリペプチドをコードするポリヌクレオチド(すなわち、配列番号11、配列番号12および配列番号13のいずれかの1つに示したアミノ酸配列を含むポリペプチドをコードするか、または配列番号14、配列番号15および配列番号16のいずれかの1つに示した核酸配列を含むものであるポリヌクレオチド)の、CHO細胞を形質転換するための選択可能なマーカー遺伝子としての使用は、共導入したレポーター遺伝子の発現を増強することが見出されている。上述の核酸配列を含むDNA配列をEGPFレポーター遺伝子と共にCHO細胞中に共導入した。トランスフェクション後、細胞をG418を含有する培地で増殖した。比較実験を行い、ここでは改変されたネオマイシンホスホトランスフェラーゼをコードする本発明によるポリヌクレオチドの代わりに、未改変のNPT II遺伝子(配列番号17に示した核酸配列)を選択可能なマーカー遺伝子として用いた。未改変のマーカーでトランスフェクトした宿主細胞の大部分と比較して、改変されたNPT IIでトランスフェクトした宿主細胞の大部分においてレポーター遺伝子発現が有意に増強されることが見出された。従って、本発明による上述の方法は、単一の高生産クローンに対する時間と費用のかかるスクリーニングステップを軽減しうる。
【0076】
本発明はまた、少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)宿主細胞中に
(i)少なくとも1つの、低下した選択能力を有する選択可能なマーカーをコードするポリヌクレオチド、および
(ii)少なくとも1つの、遺伝子発現を増強することができる核酸配列を含む、ポリヌクレオチド、および
(iii)少なくとも1つの目的のポリヌクレオチドを導入するステップ
を導入するステップ、ならびに
(b)前記宿主細胞を培養するステップ
を含んでなる前記方法に関する。
【0077】
用語「遺伝子発現を増強できる核酸配列」は、共トランスフェクトした目的のポリヌクレオチドの発現を増強できるエンハンサー、MAR、SARまたはUCOE配列などの共トランスフェクトした目的のポリヌクレオチドの発現を増強できるいずれかの核酸配列を含み、好ましくは、遺伝子発現を増強できる核酸配列を含むポリヌクレオチドは発現を増強できるクロマチンエレメントに対する核酸配列を含むポリヌクレオチドであり、より好ましくは、遺伝子発現を増強できる核酸配列を含むポリヌクレオチドは本明細書に先に記載した発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものである先に説明したポリヌクレオチドであり、最も好ましくは、遺伝子発現を増強できる核酸配列を含むポリヌクレオチドは配列番号3に、配列番号4に、配列番号5に、または配列番号6に示した核酸配列を含むものであるポリヌクレオチドである。上述の方法の文脈において、遺伝子発現を増強できる核酸配列を含む、ポリヌクレオチドはさらなる湾曲起点モチーフを含んでもまたは含まなくてもよいことは理解されよう。
【0078】
本明細書で使用する用語「低下した選択能力を有する選択可能なマーカー」は、その選択能力がそれぞれの未改変のマーカー(例えば、野生型)と比較して低下した、遺伝的に改変された選択可能なマーカーに関する。WO 2004/050884は、共トランスフェクトした目的のポリヌクレオチドの発現を増強する、低下した選択能力を有するマーカーを開示している。低下した選択能力のもとでは、マーカータンパク質の酵素活性が低下していることは理解されよう。酵素活性は、マーカー遺伝子タンパク質のアミノ酸配列を改変することにより、例えば、前記マーカータンパク質をコードするポリヌクレオチド配列を変異させることにより低下させることができる。酵素活性は、当技術分野で周知でありかつよく記載された酵素アッセイにより決定することができる。全ての酵素アッセイは、好適な基質の消費または産物の生産を経時的に測定する。基質および産物の濃度を測定する多数の色々な方法が存在し、多くの酵素をいくつかの異なる方法でアッセイすることができる。本明細書で意味する酵素アッセイは、次の手法により行うことができる:初期速度実験、進行曲線実験、過渡的動力学実験、緩和実験。基質の消費または産物の生産は、分光学的手段、蛍光測定的手段、熱量測定的手段または化学発光的手段、放射線測定的手段およびクロマトグラフィ的手段により決定することができる。アッセイは連続的に(生産と消費を連続的に測定する)または非連続的に(サンプルを間隔をおいて採取する)行うことができる。酵素活性を比較するために、第1の酵素(例えば、改変されたマーカータンパク質)の活性を第2の酵素(例えば、未改変のマーカータンパク質)の酵素活性と、同じ条件下で比較する。酵素は精製されていてもまたは細胞抽出物の成分であってもよい。好ましくは、上述の方法の文脈において低下した選択能力を有する選択可能なマーカーの酵素活性は、未改変の選択可能なマーカーの酵素活性と比較すると、1〜80%低下、より好ましくは1〜50%低下、最も好ましくは1〜20%低下している。選択能力の低下は、野生型マーカー遺伝子の核酸配列を改変して、対応するアミノ酸配列がそれぞれの野生型マーカーのアミノ酸配列と比較して少なくとも1つのアミノ酸変化があるようにすることにより達成することができる。本明細書で使用する用語「少なくとも1つのアミノ酸変化」は、好ましくは、1以上、例えば、少なくとも2、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7または少なくとも10、少なくとも20、少なくとも30、少なくとも40、少なくとも50、または少なくとも100のアミノ酸変化を意味する。本発明の文脈において、選択能力の低下はまた、他の発現制御配列と比較して選択可能なマーカー遺伝子の発現を低下させる発現制御配列、例えば、プロモーターを用いることにより引き起こしてもよい。本発明の上述の方法によると、選択可能なマーカー遺伝子は、ネオマイシンホスホトランスフェラーゼ、ハイグロマイシンホスホトランスフェラーゼ、ピューロマイシン-N-アセチル-トランスフェラーゼ、ジヒドロ葉酸レダクターゼ、グルタミンシンテターゼ、アスパラギン酸トランスカルバミラーゼ、メタロチオネイン、アデノシンデアミナーゼ、アデニル酸デアミナーゼ、UMPシンテターゼ、P-糖タンパク質(p-gp、MDR)、アスパラギンシンテターゼまたはオルニチンデカルボキシラーゼであってもよい。
【0079】
上述の方法のある好ましい実施形態において、選択可能なマーカーをコードするポリヌクレオチドは、改変されたネオマイシンホスホトランスフェラーゼをコードするポリヌクレオチドである。上述の方法で用いることができる改変されたネオマイシンホスホトランスフェラーゼ遺伝子は、例えば、WO 2004/050884に記載されていて、これは本明細書に参照によりその全てが組み入れられる。
【0080】
上述の方法の他のさらに好ましい実施形態において、低下した選択能力を有する選択可能なマーカーをコードするポリヌクレオチドは、配列番号11、配列番号12および配列番号13のいずれかの1つに示したアミノ酸配列を含むポリペプチドをコードするか、または配列番号14、配列番号15および配列番号16のいずれかの1つに示した核酸配列を含むものであるポリヌクレオチドである。最も好ましくは、前記ポリヌクレオチドは配列番号13に示したアミノ酸配列を含むポリペプチドをコードするポリヌクレオチド、または配列番号16に示した核酸配列を含むものであるポリヌクレオチドである。また、本発明の方法のための低下した選択能力を有する選択可能なマーカーをコードするポリヌクレオチドと考えられるのは、野生型NPT II(配列番号35を参照)と比較して、野生型NPT IIポリペプチドのアミノ酸182位にてEがDへ、190位にてDがGへ、208位にてDがGへ、227位にてDがGへ、182位にてEがGへ、91位にてWがAへ、198位にてVがGへ、227位にてDがAへ、227位にてDがVへ、261位にてDがGへ、261位にてDがNへ、または240位にてFがIへのアミノ酸置換を有する、改変されたNPT IIポリヌクレオチドをコードするポリヌクレオチドである(これらは全て、Sauter, BIOTECHNOLOGY AND BIOENGINEERING, VOL. 89, NO. 5, 2005により開示されている)。さらに好ましいのは、野生型NPT II(配列番号35を参照)と比較して、位置にてVがMへ、182位にてEがDへ、205位にてGがEへ、210位にてGがAへ、211位においてRがPへ、211位においてRがKへ、211位においてRがHへのアミノ酸置換を有する、改変されたNPT IIポリペプチドをコードするポリヌクレオチドである。
【0081】
有利なことに、本発明によって、改変されたNPT II選択可能なマーカー遺伝子をコードするポリヌクレオチド、および遺伝子発現を増強できる核酸配列を含むポリヌクレオチド、および目的のポリヌクレオチドの宿主細胞中への共導入は、目的のポリヌクレオチドの発現を有意に増強することが見出されている。例えば、配列番号13に示したアミノ酸配列を含む改変されたNPT IIマーカーをコードするポリヌクレオチド、ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域(Romig、1994、Eur J Biochem. 221:411-419)およびさらなる湾曲起点モチーフを含むポリヌクレオチド(例えば、配列番号3に示した核酸配列)およびレポーター遺伝子EGFPをCHO(チャイニーズハムスター卵巣)細胞中に共導入した。対照として、a)EGFP遺伝子および未改変のNPT II遺伝子を保持するベクターpEGFP-N1、またはb)湾曲起点モチーフを含むヒトトポイソメラーゼIのイントロン13のMII領域の対応するATリッチ領域に対するDNA(配列番号3)と組合わせたpEGFP-N1ベクターを、CHO細胞中に導入した。さらに、SautterおよびEnekel(変異体D227V、Sautter および Enekel, Biotechnology and Bioengineering, 89, (2005) 530-538)により記載されている改変されたNPT II遺伝子を、さらなる湾曲起点モチーフを伴うヒトトポイソメラーゼIのイントロン13のMII領域の対応するATリッチ領域に対するDNA(配列番号3)と組合わせて、レポーター遺伝子とCHO細胞に共導入した。トランスフェクション後、耐性細胞を、G418を含有する培地における培養中に選択した。個々のG418耐性宿主細胞のレポーター遺伝子発現をFACS分析(Fluorescent Activated Cell Sorting)により数値化した。対照と比較して、高い発現細胞の比率、従って発現が、EGFP遺伝子、前記ATリッチ領域および湾曲起点モチーフを含むポリヌクレオチド、ならびに改変されたNPT II選択可能なマーカー遺伝子をコードするポリヌクレオチドを共導入することにより作製したCHO細胞において、有意に増加したことを見出した。さらに、改変されたNPT II D227V突然変異体の使用は、目的のポリヌクレオチドの発現を未改変のNPT遺伝子の使用と比較して増強する、しかしながら、改変されたNPT II D227V遺伝子の使用は、例えば配列番号13に示したアミノ酸配列を含む改変されたNPT IIマーカーより有意に低い遺伝子発現レベルをもたらした。従って、改変されたNPT IIをコードするポリヌクレオチドのような低下した選択能力を有する選択可能なマーカーをコードする少なくとも1つのポリヌクレオチド、発現を増強できるクロマチンエレメントに対する第1の核酸配列と少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含む先に規定したポリヌクレオチドのような遺伝子発現を増強できる核酸配列を含む少なくとも1つのポリヌクレオチド、および目的のポリヌクレオチドを宿主細胞中へ共導入すること、ならびに、前記細胞を好適な選択剤を用いて培養することは、目的のポリヌクレオチドの発現を有意に増強する。さらに、この効果はCHO細胞に対してだけでなく、爬虫類細胞(毒ヘビ細胞株VH2)に対しても示された。本発明のポリヌクレオチドの宿主細胞中への組合わせた導入の発現に与える増強効果は、個々の成分単独での効果、または個々の効果の合計よりもより大きく、従って、前記組合わせた導入は予想外の相乗効果を示すものである。例えば、配列番号13に示したアミノ酸配列を有する改変されたNPT IIマーカーを用いると、G418での選択後に、大部分が高生産クローンである耐性クローンをもたらすが、低生産クローンの大部分は選択過程で失われた。
【0082】
従って、上述の方法は、高生産細胞を単離するための効果的かつ迅速な手順を可能にし、費用と時間のかかる遺伝子増幅およびスクリーニングステップの必要性を軽減する。
【0083】
さらに、前記のように作製した細胞は、目的のポリヌクレオチドの驚くべき安定な発現を示した。単一クローンを、前記の通り[改変されたNPT IIマーカー(例えば、配列番号13に示したアミノ酸配列を有する)をコードするポリヌクレオチド、ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域および湾曲起点モチーフを含む、例えば、配列番号3に示した核酸配列を有するポリヌクレオチドを共トランスフェクトし、選択圧力下で培養することによって]作製した細胞から成る耐性コロニーから単離し、そして8週間選択圧力なしに連続培養した。驚くべきことに、選択圧力なしの8週間培養後のFACS分析は、試験したクローンの50%が変わらないレポーター遺伝子発現を示す一方、クローンの他の50%は、8週間培養の開始時のレポーター発現と比較してわずかに低下したレポーター遺伝子発現を示した(開始時の発現の50%〜90%)。従って、上述の方法は、有利なことに、選択可能なマーカー剤の不在のもとですら強くかつ安定な発現をするクローンの時間効率的な生産を可能にする。
【0084】
本発明の方法のある好ましい実施形態においては、宿主細胞を無血清培地または血清含有培地にて培養することができる。より好ましくは、宿主細胞を無血清培地で培養する。宿主細胞を無血清培地で培養することは、血清含有培地での培養から生じる感染のリスクを軽減できるので有利である。さらに、無血清培地の使用は、特に大規模な手法にとって、さらに費用効果が高い。用語「血清」の意味は当業者によく理解されている。本明細書で使用する用語「無血清培地」は、いずれの型の血清も、例えば、ウシ胎児血清または血清から分離されるタンパク質などのいずれの血清成分も含まない、任意の宿主細胞を培養するための増殖培地を意味する。対照的に、本明細書で意味する血清含有培地は、血清または血清から分離されるタンパク質などの血清成分を含有する培地に関する。
【0085】
本発明の方法の他の実施形態において、目的のポリヌクレオチドは、ヒト化抗体などの免疫グロブリン、治療タンパク質、膜タンパク質および酵素、ホルモン、イオンチャネルタンパク質およびプロテアーゼからなる群より選択されるポリペプチドをコードするものである。
【0086】
さらに、本発明は、目的のポリペプチドを製造する方法であって、上述の方法のいずれかのステップ、および目的のポリヌクレオチドがコードするポリペプチドを得るさらなるステップを含んでなる前記方法に関する。
【0087】
本発明の方法の他のさらに好ましい実施形態において、宿主細胞は動物細胞、植物細胞、酵母細胞、真菌細胞および藻類細胞である。より好ましくは、前記細胞は哺乳動物細胞または脊椎動物細胞である。最も好ましくは、先に記載した通り、宿主細胞はCHO細胞である。本発明で想定されるのはまた、爬虫類細胞、好ましくは毒ヘビ細胞、好ましくは細胞株VH2(Viper Russells心臓、ECACC No. 90102539)である。
【0088】
さらに、本発明は、発現を増強できるクロマチンエレメントに対する第1の核酸配列と少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列とを含むポリヌクレオチドおよび/または配列番号11、配列番号12および配列番号13のいずれかの1つに示したアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16のいずれかの1つに示した核酸配列を含む、ポリヌクレオチドを含むものである医薬組成物に関する。さらに、医薬組成物は治療効果を有する産物をコードする目的のポリヌクレオチドを含んでもよい。
【0089】
本明細書で使用する用語「医薬組成物」は、本発明の化合物および任意に1種以上の製薬上許容される担体を含む。本発明の化合物は製薬上許容される塩として製剤することができる。許容される塩には、酢酸塩、塩酸塩、硫酸塩、塩化物などが含まれる。医薬組成物は、好ましくは、局所にまたは全身に投与される。薬物投与用に好都合な、好適な投与の経路は、経口、静脈内または非経口投与、ならびに吸入である。しかし、化合物の性質と作用様式に応じて、医薬組成物を他の経路により投与することもできる。例えば、ポリヌクレオチド化合物は、遺伝子治療手法で、ウイルスベクターまたはウイルスまたはリポソームを用いることにより投与することができる。
【0090】
化合物は、好ましくは、薬物を標準医薬品担体と組合わせることにより調製した慣用の投与剤形で、慣用の手順によって投与される。これらの手順は、所望の調製に適当なように、成分を混合、造粒および圧縮または溶解することに関わりうる。製薬上許容される担体または希釈剤の形態と特性は、組み合わせるべき活性成分の量、投与の経路および他の周知の変数により規定されることは理解されるであろう。
【0091】
担体は、製剤の他成分と共存しうるおよびそのレシピエントに有害でないという意味で、許容されなければならない。使用される医薬品担体は、例えば、固体、ゲルまたは液体でありうる。固体担体の例は、ラクトース、白土、スクロース、タルク、ゼラチン、寒天、ペクチン、アカシア、ステアリン酸マグネシウム、ステアリン酸などである。液体担体の例は、リン酸緩衝生理食塩水溶液、シロップ、ピーナッツ油およびオリーブ油などの油、水、乳濁液、様々なタイプの湿潤剤、無菌溶液などである。同様に、担体または希釈剤は、モノステアリン酸グリセリルまたはジステアリン酸グリセリル単独でまたはワックス併用でなどの当技術分野で周知の時間遅延材料を含んでもよい。前記好適な担体には、前記および他の当技術分野で周知の担体が含まれ、例えば、Remington's Pharmaceutical Sciences, Mack Publishing Company, Easton, Pennsylvaniaを参照されたい。
【0092】
希釈剤は、組合わせの生物学的物性に影響を与えないように選択される。かかる希釈剤の例は、蒸留水、生理食塩水、リンゲル液、ブドウ糖液、およびハンク(Hank)液である。さらに、医薬組成物または製剤はまた、他の担体、アジュバント、または無毒性、無治療性、無免疫原性の安定剤などを含んでもよい。
【0093】
治療上有効な用量は、本明細書で言及した疾患に随伴する症候群または症状を予防、改善または治療する本発明の医薬組成物で使われる化合物の量を意味する。かかる化合物の治療効力および毒性は、細胞培養または実験動物における標準医薬品手順、例えば、ED50(集団の50%にとって治療上有効な用量)およびLD50(集団の50%が死に至る用量)により決定することができる。治療と毒性効果の間の用量比が治療指数であり、比LD50/ED50で表される。
【0094】
投薬レジメンは担当医によりおよび他の臨床的因子;好ましくは以上記載した方法のいずれか1つによって決定されるであろう。医療業界では周知のように、いずれかの一患者に対する投薬は、患者サイズ、身体表面積、年齢、投与する特定の化合物、性別、時間および投与の経路、一般的な健康状態、および同時に投与される他の薬物を含む多くの因子に依存する。進行状態は定期的な評価を行うことによりモニターすることができる。
【0095】
本明細書で意味する医薬組成物および製剤は少なくとも1回、本明細書に挙げた疾患または症状を治療または改善または予防するために投与される。しかし、前記医薬組成物を1回以上、例えば毎日1〜4回、日数に限定されることなく、投与することができる。
【0096】
さらに、本発明は、遺伝子治療に用いる医薬組成物を製造するためのポリヌクレオチドの使用であって、該ポリヌクレオチドが、発現を増強できるクロマチンエレメントに対する第1の核酸配列と少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列とを含むポリヌクレオチドならびに/または配列番号11、配列番号12および配列番号13のいずれかの1つに示したアミノ酸配列を含むポリペプチドをコードするかもしくは配列番号14、配列番号15および配列番号16のいずれかの1つに示した核酸配列を含む、ポリヌクレオチドを含むものである前記使用に関する。
【0097】
本明細書で使用する用語「遺伝子治療」は、少なくとも1つの本発明のポリヌクレオチドをin vivoで、または宿主細胞中へのin vitro導入と次いで被験者(例えば、ヒトまたは非ヒト動物)中への再移植により被験者の宿主細胞中に導入する治療の方法として理解されよう。好ましくは、本発明のポリヌクレオチドを遺伝子治療に用いる宿主細胞中に、好ましくは治療効果を有する目的の産物をコードする目的のポリヌクレオチドと一緒に共導入する。本明細書で使用する用語「遺伝子治療」は、宿主細胞遺伝子の機能の欠如を補償するために目的のポリヌクレオチドを細胞中に導入することを意味してもよい。さらに、異常な遺伝子を、相同的組換えを介して機能性遺伝子により置換えることができる。さらに、調節、すなわち、特定の遺伝子の発現を、例えば、調節RNAにより、例えばRNAi手法で改変することができる。好ましくは、導入は、当技術分野で周知の技法により行われる。より好ましくは、導入は、ウイルスベクターにより行われる。この目的のためには、ウイルスベクターを、ウイルス疾患を生じる遺伝子を取り除いて上述した本発明のポリヌクレオチドを挿入することにより、遺伝的に改変することができる。ウイルスベクターには、限定されるものでないが、レトロウイルス、アデノウイルス、アデノ随伴ウイルスが含まれる。上述したポリヌクレオチドは、裸のDNAによるかまたはリポソームの使用によるトランスフェクションなどの非ウイルス的な方法により送達することもできる。遺伝子療法により治療される疾患は、限定されるものでないが、癌、神経疾患、代謝性疾患、免疫不全疾患でありうる。非ウイルス遺伝子療法に対する出現中のベクターおよび標的化方法は、Lavigne および Gorecki(Expert Opinion of Emerging Drugs (2006) 11、541-557)に記載されている。
【0098】
本明細書に引用される全ての参照は、それらの全開示内容および本明細書に具体的に記載した開示内容に関する参照により、本明細書に組み入れられる。
【図面の簡単な説明】
【0099】
【図1】配列番号3に示した、挿入された湾曲起点モチーフを含むヒトトポイソメラーゼのイントロン13のATリッチ領域の核酸配列である。湾曲起点モチーフに下線が引かれている。
【図2】配列番号4に示した、挿入された湾曲起点モチーフを含むヒトトポイソメラーゼのイントロン13のATリッチ領域の核酸配列である。湾曲起点モチーフに下線が引かれている。
【図3】次のトランスフェクション後のCHO細胞におけるレポーター遺伝子EGFPの発現である:環状pEGFP-N1 100ng単独(「o」)によるトランスフェクション、または環状pEGFP-N1ベクター(Clontech)100ngと配列番号27に示した核酸配列を含むヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域を含むものである直鎖DNA 100ngとの共トランスフェクション(「X」)、または環状pEGFP-N1ベクター(Clontech)100ngと配列番号28に示した核酸配列を含むヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域をコードする直鎖DNA 100ngとの共トランスフェクション(黒丸)、または環状pEGFP-N1ベクター(Clontech)100 ngと配列番号3に示した湾曲起点モチーフを含むヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域を含む直鎖DNA 100ngとの共トランスフェクション(黒四角)。ベクターpEGPF-N1は、EGPFレポーター遺伝子および野生型NPT II遺伝子を含む。およそ8週間の選択圧力(0.6 mg/ml G418)下の培養後、個々の細胞のレポーター遺伝子発現をFACS分析(蛍光検出の感度:4 decades、gain 300)により数値化した。レポーター遺伝子発現が比較的強い細胞の比率は、細胞を湾曲起点モチーフを含む発現を増強できるクロマチンエレメントを用いて共トランスフェクトした場合に、pEGPF-N1単独を用いてまたはpEGPF-N1と湾曲起点モチーフなしの発現を増強できるクロマチンエレメントを用いてトランスフェクトした細胞と比較して、有意に増強する。
【図4】次のトランスフェクション後のCHO細胞におけるEGFPの発現である:直鎖pEGFP-N1単独のDNA 90ngを用いたトランスフェクション(「o」)、または直鎖pEGFP-N1 90ngとヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域およびさらなる湾曲起点モチーフ(配列番号3に示した核酸配列)DNA 300ngとの共トランスフェクション(黒四角)、またはEGFPレポーター遺伝子および配列番号16に示した核酸配列を含む改変されたネオマイシンホスホトランスフェラーゼを保持するベクター(pAG)90ngとヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域およびさらなる配列番号3に示した核酸配列をもつ湾曲起点モチーフをコードするDNA 300ngとの共トランスフェクション(黒丸)。およそ8週間の選択圧力(0.6 mg/ml G418)下の培養後、個々の細胞のレポーター遺伝子発現をFACS分析(計器設定:光学検出器のgain=300、増幅モード=4 decade logarithmic)により数値化した。強いレポーター遺伝子発現を示す細胞の比率は、レポーター遺伝子をCHO細胞中に、さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のATリッチ領域をコードするDNAと組合わせて導入した場合、pEGFP-N1を単独でトランスフェクトした場合より有意に高かった。さらに、強いレポーター遺伝子発現をもつ細胞の比率は、湾曲起点モチーフを含むヒトトポイソメラーゼIのイントロン13のATリッチ領域と、改変されたネオマイシンホスホトランスフェラーゼ遺伝子(未改変のネオマイシンホスホトランスフェラーゼの代わりに、pEGFP-N1プラスミド上に局在する)をコードするDNAとの両方をCHO細胞中に導入した場合に、最高であった。配列番号16に示した核酸配列を有する改変されたネオマイシンホスホトランスフェラーゼと湾曲起点モチーフを含むATリッチ領域(配列番号3に示した核酸配列)とを用いてトランスフェクトした細胞における安定な発現は予想外に高く、蛍光検出の範囲は、低発現細胞と高発現細胞を同時に検出するには不十分であった。低発現細胞と高発現細胞の間の蛍光強度の差は104のファクターより大きいので、細胞を低感度で再測定した(図5:計器設定:光学検出器のgain=200、増幅モード=4 decade logarithmic)。
【図5】図4の凡例に示したようにトランスフェクトしかつ選択したCHO細胞の集団である。蛍光検出の感度(4 decades)は200のgainで測定した。pAGおよびATリッチ領域(配列番号3に示した核酸配列)を用いて共トランスフェクトしたCHO細胞(「O」)の大部分は、pEGFP-N1およびATリッチ領域(配列番号3に示した核酸配列)を用いて共トランスフェクトしたCHO細胞(「X」)よりおよそ10倍高い発現を示した。従って、さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のATリッチ領域に対するDNAと、改変されたネオマイシンホスホトランスフェラーゼ遺伝子をコードするDNAとの両方をCHO細胞に導入した場合に、強いレポーター遺伝子発現をもつ細胞の比率が有意に増強された。
【図6】図3の凡例に示したようにレポーター遺伝子EGFPを様々なネオマイシンホスホトランスフェラーゼ耐性遺伝子(NPT II)と組合わせてトランスフェクトし、選択した後のCHO細胞におけるEGFPの発現である。次のネオマイシンホスホトランスフェラーゼ耐性遺伝子の共トランスフェクトしたレポーター遺伝子の発現に与える効果を分析した:それぞれ、配列番号17(X)に示した核酸配列を含む未改変のNPT II遺伝子、ならびに配列番号14(O)、配列番号15(黒丸)、配列番号16(黒四角)に示した核酸配列を含む改変されたNPT II遺伝子に対する結果である。個々の細胞のレポーター遺伝子発現は、十分な量のG418を含有する選択培地で8週間培養後にFACS分析により測定した。レポーター遺伝子の比較的強い発現を示す細胞の比率は、改変されたネオマイシンホスホトランスフェラーゼでトランスフェクトした細胞については、野生型ネオマイシンホスホトランスフェラーゼでトランスフェクトした細胞と比較して有意に高かった。レポーター遺伝子の比較的強い発現を示す細胞の比率は、配列番号16に示した核酸配列を有する改変されたネオマイシンホスホトランスフェラーゼでトランスフェクトした細胞が最高であった。
【図7】レポーター遺伝子EGFPを、改変されたネオマイシンホスホトランスフェラーゼ耐性遺伝子D227V(配列番号36)およびさらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域に対するDNA(配列番号3)と組合わせて共トランスフェクトした後のCHO細胞におけるEGFPの発現である(O)。対照として、未改変のネオマイシンホスホトランスフェラーゼ(pEGFP-N1)を、湾曲起点モチーフを伴うヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域に対するDNA(配列番号3)を用いて共トランスフェクトした(X)。実施例3に記載の通り、細胞を培養し、FuGENE6でトランスフェクトし、G418培地で選択した。0.6mg/ml G418を補充した培地における6週間の培養後、個々の細胞の遺伝子発現を、実施例3に記載の通りFACS分析により測定した。(計器設定:光学検出器のgain=200、増幅モード=4decadelogarithmic)(図7)。ATリッチ領域(配列番号3)と共トランスフェクトした記載の改変されたNPT IID227V突然変異体は、ATリッチ領域(配列番号3)と共トランスフェクトした未改変のNPT遺伝子と比較して、目的のポリヌクレオチドの発現を増強するが、その増加は、例えば、配列番号16に示した核酸配列を有する本発明の改変されたネオマイシンホスホトランスフェラーゼの共導入に対してより低かった。
【0100】
以下の実施例は単に本発明を説明するものである。これらは、どういう場合でも、本発明の範囲を限定すると解釈してはならない。
【実施例】
【0101】
(実施例1)
湾曲起点モチーフをもつMAR配列の作製
ヒト白血球由来およびHeLa細胞由来のゲノムDNAを、「血液および細胞培養DNAキット」(QIAGEN GmbH, Hilden, Germany)を用いて、製造業者取扱説明書に従い精製した。ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域(Romig, Eur J Biochem. 221:411-419)を、2回の逐次PCR反応(ネストPCR)によって増幅し、ここで、テンプレートとしてHeLa細胞からまたはヒト血液から抽出したゲノムDNAのおよそ200ngを、ポリメラーゼとしてPfuポリメラーゼを使用し、そして次のプライマーを用いた:
プライマー1(フォワード):5'-CCAGCTAACTGCTATACACAATAGACATTTGTGC(配列番号61)
プライマー2(リバース):5'-CTGCCAACAGTAATTAGCACATGAATTCACTTCAGGTAAC(配列番号62)
第2のPCRについては、
プライマー3(フォワード):5'-GTGCCTGACTTGAACTGCAATGGG(配列番号63)
プライマー4(リバース):5'-CAGTAATTAGCACATGAATTCAC(配列番号64)。
【0102】
下線部は、後のクローニングステップで使用するEcoRI制限部位の配列である。
【0103】
第1のPCR反応は次の通り実施した:30サイクルの94℃における2分間変性、50℃における35秒間のアニーリングおよび72℃における2分間の伸長、その後に72℃における5分間の最後の伸長。
【0104】
第2のPCR反応は第1のPCRの一部分をテンプレートとして用いて、次の通り実施した:30サイクルの94℃における1分間変性、50℃における35秒間のアニーリングおよび72℃における2分間の伸長、その後に72℃における5分間の最後の伸長。
【0105】
以下のステップで、PCR産物をベクターpPCR-Script-Amp-SK(+)(Stratagene、LaJolla、USA)のSrfI部位(SrfIは平滑末端を作製する)中に、PCR-Script(登録商標)Ampクローニングキットを用いることにより製造業者の取扱説明書に従ってライゲートした。ライゲーションの後、ライゲーション産物を、コンピテント大腸菌細胞(XL-GoldKan、PCR-Script(登録商標)Ampクローニングキット)中に、製造業者取扱説明書に従って形質転換した。形質転換の後、細胞を、首尾よく形質転換した細胞を選択できるように、およそ20時間、37℃にて、100μg/mlアンピシリンを補充したLuria-ブロス-培地においてインキュベートした。次いで、得られるPCR断片を保持するベクターの2.9kb BamH1/EcoR1 血液ATリッチ核酸断片および2.9kb BamH1/EcoR1 HeLa ATリッチ核酸断片を、ベクターpUC18のBamH1/EcoR1制限部位中にクローニングした。クローニングしたPCR産物を、「血液ATリッチ核酸」(起源として、血液から抽出したゲノムDNAから増幅した;配列番号28に示した核酸配列)および「HeLaATリッチ核酸」(起源として、HeLa細胞から抽出したゲノムDNAから増幅した;配列番号27に示した核酸配列)と名付けた。両方のATリッチ配列は、高い配列同一性を示すがいくつかの位置で配列差を有する(613位、1079〜1082位、1128位、1160-1163位、1176位、1188位、1197位、1205位、1227位、1236位、1333位、1905位、1919位、1931位、1950位、1959位、1975位、2191位、2234位、2495位、2507位、2634位、2652位、2733位、2762位および2768位)。部位指定変異誘発によって、制限エンドヌクレアーゼAvrIIに対する制限部位を、HeLaおよび血液ATリッチ核酸配列の両方の中に、QuickChange(登録商標)部位特異的変異誘発キット(Stratagene)を用いて製造業者の取扱説明書に従い導入した。部位指定変異誘発に用いたプライマーの核酸配列は次の通りである:
プライマーavr_sense ggtcttgtatctgcctaggagatacaagaggtgctc(配列番号65)
プライマーavr_asense gagcacctcttgtatctcctaggcagatacaagacc(配列番号66)
湾曲起点モチーフを、GeneartAG(Regensburg)により、Nalezkovaaら(2005)Protein Expression and Purification 39、296-306に記載の方法を用いて合成した。
【0106】
湾曲起点モチーフを含むAvrII断片を、両方向に、新しく作製したHeLaおよび血液AT-リッチ配列のAvrII制限部位中にクローニングした。4種の得られる構築物の核酸配列を配列番号3、配列番号4、配列番号5および配列番号6に示す。
【0107】
(実施例2)
改変されたネオマイシンホスホトランスフェラーゼII(NPT II)をコードするポリヌクレオチドの作製
1つまたは2つのアミノ酸変化を有する改変されたNPT IIタンパク質をコードするNPT IIポリヌクレオチドを、部位特異的変異誘発により、未改変のNPT II(野生型)遺伝子をテンプレートとして保持するベクターpEGFP-N1を用いて作製した。次のプライマー対を部位特異的変異誘発に用いて、QuickChange(登録商標)部位特異的変異誘発キット(Stratagene)を製造業者の取扱説明書に従って使用した。
【0108】
NPT II R226A突然変異体(配列番号14に示した核酸配列を得る)について
R226A_sense:catagcgttggctaccgctgatatcgctgaagagcttggc(配列番号67)
R226A_asense:gccaagctcttcagcgatatcagcggtagccaacgctatg(配列番号68)
R226V突然変異体(配列番号15に示した核酸配列を得る)について
R226V_sense:catagcgttggctaccgttgatatcgctgaagagcttggc(配列番号69)
R226V_asense:gccaagctcttcagcgatatcaacggtagccaacgctatg(配列番号70)
NPT II A224G;T225S二重突然変異体(配列番号16に示した核酸配列を得る)について
A224G_sense:ctatcaggacatagcgttgggatcccgtgatattgctgaagagc(配列番号71)
A224G_asense:gctcttcagcaatatcacgggatcccaacgctatgtcctgatag(配列番号72)
FortheNPT IID227V突然変異体(配列番号36に示した核酸配列を得る)
D227V_sense:ggacatagcgttggctacacgcgttattgctgaagagcttgg(配列番号73)
D227V_asense:ccaagctcttcagcaataacgcgtgtagccaacgctatgtcc(配列番号74)
得られるベクターはEGFP遺伝子と改変されたネオマイシンホスホトランスフェラーゼII遺伝子の両方を含む。
【0109】
(実施例3)
さらなる湾曲起点モチーフを伴うヒトトポイソメラーゼIのイントロン13のATリッチ領域とレポーター遺伝子とのCHO細胞中への共導入は、高いレポーター遺伝子活性を示す細胞の比率を有意に増強する
ヒトトポイソメラーゼIのイントロン13のATリッチ領域と湾曲起点モチーフを含むポリヌクレオチドの、共トランスフェクトしたレポーター遺伝子の発現に与える効果を分析した。この理由で、レポーター遺伝子5EGFPおよび未改変ネオマイシンホスホトランスフェラーゼ遺伝子を保持するpEGFP-N1ベクター100ngをCHO細胞中に次のMAR構築物(各100ng)と一緒にトランスフェクトした。
【0110】
配列番号28に示したATリッチ核酸(血液ATリッチ配列)
配列番号27に示したATリッチ核酸(HeLaATリッチ配列)
配列番号3に示したATリッチ核酸(挿入された湾曲起点モチーフを伴うATリッチ配列)
対照として、pEGFP-N1ベクターを単独でトランスフェクトした。
【0111】
CHO-K1細胞株を、10%FBS(PAALaboratoriesGmbH)を補充したL-グルタミン(PAALaboratoriesGmbH)とHamF-12培地で培養した。EGFPを発現する安定なCHO細胞のプールを、FuGENE6(Roche)でのトランスフェクションにより、製造業者取扱説明書に従いかつ1:4のDNA:FuGENE比(μgDNA:μlFuGENE6)で得た。細胞を24ウエルプレートで100〜200000細胞/ウエルにて接種し、一晩付着させた。100ngのpEGFP-N1を100ngの直鎖ATリッチ配列と共トランスフェクトした。直鎖ATリッチ配列(配列番号3、配列番号27、配列番号28)は制限酵素消化およびゲル溶出によるゲル電気泳動後に単離した。48時間後、培地を0.6mg/ml G418(G418硫酸塩、Calbiochem)を補充した培地と変更した。引き続いて、培地を0.6mg/ml G418(G418硫酸塩、Calbiochem)を補充した培地と変更し、クローンのプールを8週間の選択後に試験した。レポーター遺伝子(EGFP)発現、すなわち、個々の細胞の蛍光を、FACS分析によりPARTECフローサイトメーター(PartecAnalysingSystemPAS、PartecGmbH、Muenster)を用いて数値化した。蛍光イベントは、緑色蛍光タンパク質シグナルに対する530/15帯域通過フィルターを用いて得た。(計器設定:光学検出器のgain=300、増幅モード=4decadelogarithmic)。
【0112】
レポーター遺伝子シグナルの強度は発現を示し、すなわち、強い蛍光シグナルは高いレポーター遺伝子発現を示す。特定の蛍光を有する個々の細胞の数(イベントとも呼ぶ)を蛍光のレベルに対してプロットした。様々なトランスフェクション実験の結果を図3に示す。GPF遺伝子を保持するベクターをCHO細胞中に単独でトランスフェクトした場合、大多数の細胞は比較的低いレポーター遺伝子発現を示した。そのベクターに加えてトポイソメラーゼ遺伝子のATリッチ領域(配列番号28に示した「血液ATリッチ」構築物、または配列番号27に示した「HeLaATリッチ」構築物)をコードするDNAを細胞中にトランスフェクトした場合、比較的高いレポーター遺伝子発現を示す細胞の比率はベクター単独のトランスフェクションと比較して有意に増強された。予想外なこととして、ATリッチ配列を共トランスフェクトしたほとんど全ての細胞がEGFP発現を示した。おそらく、遺伝子サイレンシングを示したのは非常に少量の細胞だけであった。対照的に、EGFP-N1ベクター単独でトランスフェクトしたG418耐性細胞プールのほぼ全ての細胞がEGFP発現を示さなかった。従って、遺伝子サイレンシングを示さなかったのは、EGFP-N1ベクター単独でトランスフェクトした細胞のうちの非常に少量だけであり、この細胞集団はFACS分析で101〜102の蛍光強度で検出された。pEGFP-N1ベクターに加えて、さらなる核酸配列番号3に示した湾曲起点モチーフをもつヒトトポイソメラーゼのATリッチ領域をCHO細胞中にトランスフェクトした場合、レポーター遺伝子発現は最高であった。発現は、配列番号28で共トランスフェクトした細胞と比較して、およそ10〜20のファクターで増強された(共トランスフェクトした配列番号3)。対応する結果はまた、湾曲起点が反対方向にあるATリッチ構築物についても観察された(配列番号4に示した核酸配列)。従って、クロマチンエレメント(単独で発現を増強できる)とさらなる湾曲起点モチーフとのトランスフェクションは、レポーター遺伝子およびさらなる湾曲起点モチーフを含まないクロマチンエレメントの共トランスフェクションと比較して、共トランスフェクトした遺伝子の強い発現を示す細胞集団の細胞数を有意に増加する。
【0113】
(実施例4)
改変されたネオマイシンホスホトランスフェラーゼをトランスフェクトされたCHO細胞を選択するために使用すると、レポーター遺伝子の発現を増強する
1つまたは2つのアミノ酸変化を有する変異ネオマイシンホスホトランスフェラーゼ遺伝子を、実施例2に記載の通り調製した。SV40初期プロモーターの調節下にある改変されたネオマイシン耐性遺伝子ならびにヒトサイトメガロウイルス(CMV)最初期プロモーターの調節下にあるレポーター遺伝子EGFPを保持するベクターをCHO細胞中にトランスフェクトした。対照として、SV40初期プロモーターの調節下にある未改変のネオマイシンホスホトランスフェラーゼ遺伝子ならびにヒトサイトメガロウイルス(CMV)最初期プロモーターの調節下にあるレポーター遺伝子EGFPを保持するベクターpEGPP-N1を、CHO細胞中にトランスフェクトした。細胞を培養し、FuGENE6でトランスフェクトし、実施例3に記載の通りG418を用いて選択した。培養後、形質転換された細胞のレポーター遺伝子発現をFACS分析により分析した。レポーター遺伝子(EGFP)発現をFACS分析により、PARTECフローサイトメーター(PartecAnalysingSystemPAS、PartecGmbH、Muenster)を用いて数値化した。蛍光イベントは、緑色蛍光タンパク質シグナルに対する530/15帯域通過フィルターを用いて得た。(計器設定:光学検出器のgain=200、増幅モード=4 decade logarithmic)。FACS分析の結果を図6のグラフに示す。グラフは細胞による蛍光発光レベル(Y軸)に対してプロットした細胞数(X軸)を示す。明確に見られるように、改変されたネオマイシンホスホトランスフェラーゼ遺伝子を選択可能なマーカーとして用いた場合に、非常に強いレポーター遺伝子活性を示す細胞の比率は有意に高い。選択可能なマーカーが未改変の耐性遺伝子である場合、レポーター遺伝子発現は低い。特に、配列番号16に示した核酸配列を有する改変されたネオマイシンホスホトランスフェラーゼを使用すると、高比率の高生産細胞を生じた。
【0114】
総括すると、改変された選択可能なマーカーの使用は、形質転換された細胞集団中の比較的強いレポーター遺伝子の発現を示す細胞の比率を増強する。
【0115】
(実施例5)
さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のATリッチ配列と改変されたネオマイシントランスフェラーゼをコードする遺伝子とのCHO細胞中への共導入は、高いレポーター遺伝子活性を示す細胞の比率を有意に増強する。
【0116】
さらなる湾曲起点モチーフを保持する、発現を増強できるクロマチンエレメント(ここでは、ヒトトポイソメラーゼIのイントロン13のATリッチ配列)(配列番号3に示した核酸配列)と、改変されたネオマイシンホスホトランスフェラーゼ(配列番号16に示した核酸配列)およびレポーター遺伝子EGPF(目的のポリヌクレオチド)を含むベクターとの共導入がレポーター遺伝子発現に与える効果を分析した。対照として、レポーター遺伝子および未改変のネオマイシンホスホトランスフェラーゼ遺伝子(配列番号17に示した核酸配列)を含むベクターをCHO細胞中にトランスフェクトするか、またはレポーター遺伝子EGFPおよび未改変のネオマイシンホスホトランスフェラーゼ遺伝子(配列番号17に示した核酸配列)と、さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のATリッチ配列をコードするDNA(配列番号3に示した核酸配列)を含むベクターとをCHO細胞中に共トランスフェクトした。これらの実験のために、細胞を培養し、FuGENE6(90ng直鎖プラスミドおよび/または300ngATリッチ配列)を用いてトランスフェクトし、実施例3に記載の通りG418培地を用いて選択した。8週間培養後、個々の細胞のレポーター遺伝子発現を、実施例3に記載の通り、FACS分析により測定した。(計器設定:光学検出器のgain=200または300、増幅モード=4decadelogarithmic)。図4(光学検出器のgain=300)および図5(光学検出器のgain=200)は発現分析の結果を示す。これらのグラフは、細胞数(X軸)および細胞による蛍光発光のレベル(Y軸)を示す。明確に見られるように、細胞が湾曲起点モチーフを含む発現を増強できるクロマチンエレメント(核酸配列配列番号3)と、改変されたネオマイシンホスホトランスフェラーゼ(配列番号13に示した核酸配列)を含むベクターとで共トランスフェクトした場合、非常に強いレポーター遺伝子活性を示す細胞の比率は、有意に高かった。従って、改変されたネオマイシンホスホトランスフェラーゼおよびクロマチンエレメントとさらなる湾曲起点モチーフとの組合わせのCHO細胞中への導入は、共導入された目的の産物(ここではレポーター遺伝子)をコードするポリヌクレオチドの発現を増強する。
【0117】
さらに、SautterおよびEnekel(突然変異体D227V、SautterおよびEnekel, Biotechnology and Bioengineering, 89, (2005) 530-538)に記載の改変されたNPT II遺伝子を含むpEGFP-N1ベクターを、さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域に対するDNA(配列番号3)とCHO細胞に共トランスフェクトした。細胞を培養し、FuGENE6でトランスフェクトし、実施例3に記載の通りG418培地を用いて選択した。0.6mg/ml G418遺伝子を補充した培地で6週間の培養後、個々の細胞の発現を、FACS分析により実施例3に記載の通り測定した(計器設定:光学検出器のgain=200、増幅モード=4decadelogarithmic)(図7)。
【0118】
前記ATリッチ領域(配列番号3)と共トランスフェクトした、記載の改変されたNPT IID227V突然変異体は、目的のポリヌクレオチドの発現を、前記ATリッチ領域(配列番号3)で共トランスフェクトした未改変のNPT遺伝子と比較して増強するが、その増加は、本発明の改変された、例えば配列番号16に示した核酸配列を有するネオマイシンホスホトランスフェラーゼを共導入した場合より低かった。
【技術分野】
【0001】
本発明は、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドに関する。さらに本発明は、前記ポリヌクレオチドを含む宿主細胞、非ヒトトランスジェニック生物、ベクターおよびキットに関する。さらに、本発明は、目的のポリヌクレオチドを発現する方法に関する。
【背景技術】
【0002】
基礎研究ならびに商業的応用の両方において、目的の産物(例えば、ポリペプチドまたは機能性RNA)をコードするポリヌクレオチドを発現するために真核細胞および原核細胞が通常用いられる。目的の産物を宿主細胞で合成するために、前記産物をコードする遺伝子(しばしばトランスジーンと呼ばれる)が宿主細胞中に、例えば、トランスフェクションまたは形質転換により導入される。次いで細胞を目的の産物を発現するのに好適な条件下で増殖する。この培養ステップは選択可能なマーカー遺伝子を使用することが多く、該遺伝子を宿主細胞中に目的の産物をコードするポリヌクレオチドと一緒に導入し、前記ポリヌクレオチドを保持する細胞を選択する。培養後、産物、例えば組換えタンパク質を適当な精製ステップにより細胞から精製することができる。
【0003】
一般に、真核細胞と原核細胞の両方を組換えタンパク質の発現に利用することができる。原核生物発現系は、大量のタンパク質を容易に生産するのに有利である。
【0004】
しかし、かかる系で生産される組換えタンパク質は、翻訳後に改変されることなく、不溶性の封入体に含まれうる。機能性になるために徹底した翻訳後改変および/またはフォールディングを経る必要があるタンパク質については、真核生物発現系を使用することが好ましい。
【0005】
目的のポリヌクレオチドによりコードされた所望の産物の合成にとって、前記ポリヌクレオチドの強くかつ信頼しうる安定な発現は、強くかつ安定な発現が費用および時間の点でより効果的であるので、非常に重要である。ポリヌクレオチドの強い発現は、前記ポリヌクレオチドの高度に転写されるクロマチン領域中への組込みおよび/または多重組込みイベントに関係することが多い。しかし、とりわけ、複雑な、いわゆる「逆方向反復(inverted repeat)」パターンと組合わせた多重組込みイベントは、トランスジーンサイレンシング、すなわち、ポリヌクレオチドの不安定なかつ低下した発現を生じがちである。
【0006】
最近の数十年間、多大な努力が、細胞系におけるトランスジーン発現の増強に向けられてきた。例えば、発現の増強は、遺伝子発現を駆動する強いプロモーターを利用することにより達成することができる。さらに、トランスジーンをある特定のクロマチンエレメントと組合わせて宿主細胞に導入すると、トランスジーンの発現を増強することができる。
【0007】
クロマチンは、真核細胞の核内部においてDNA、RNAおよびタンパク質から成る複合体である。一般に、クロマチンにはループとドメインから構成され、色々な凝縮状態で存在する。凝縮の程度が転写、複写またはDNA修復などのDNAにアクセスを必要とする過程の進行に影響を与えると思われる。一般に、比較的凝縮した構造は転写的にサイレントであると考えられ、他方、比較的脱凝縮した(オープンな)構造は効率的な遺伝子発現に欠かせないと思われる。おそらく、共トランスフェクトしたトランスジーン、すなわち目的のポリヌクレオチドの発現を増強できるクロマチンドメイン/エレメントは、クロマチン構造を脱凝縮することができる因子に対する結合部位を有する。これらのドメインの近隣の配列エレメントは、転写関係因子が容易にアクセスしうるので、より高い発現を示すと思われる。
【0008】
WO 02/24930は、哺乳動物細胞におけるトランスジーン発現を増強できる偏在性クロマチン開放性エレメント(UCOE;ubiquitous chromatin opening elements)を開示している。
【0009】
Zahn-Zabalら(J Biotechnol.、2001、87(1):29-42)は、高生産クローンの比率の増加をもたらす共トランスフェクトしたレポーター遺伝子の安定な発現を有意に増加する、ニワトリリゾチームマトリックス付着領域(MAR)を開示している。
【0010】
WO2005/040377は、タンパク質生産を増加する活性を有するDNA配列およびこれらのDNAの、真核細胞におけるタンパク質生産活性を増加するための使用を開示している。
【0011】
KalosおよびFournier(Mol Cell Biol.、1995、15(1):198-207)は、マトリックス付着領域に隣接するアポリポタンパク質B(apoB)クロマチンドメインのエレメントがトランスジーン発現に介在することを開示している。
【0012】
Klehrら(Biochemistry、1991、30(5):1264-70)は、ヒトインターフェロンβドメイン由来のヒトスカフォールド付着領域は遺伝子の発現を増強できることを示唆してる。さらに、スカフォールド付着領域(SAR)は一方向にかつ部分的に距離と無関係に、全般的なプロモーター機能を増強することを示した。
【0013】
Dangら(J Virol.、2000、74(6):2671-8)は、ヒトβインターフェロンスカフォールド付着領域(IFN-SAR)の2kb断片を挿入するとこのベクターでの遺伝子の安定な発現を引き起こすことを開示している。
【0014】
EP 1,676,916は、共トランスフェクトしたトランスジーンの発現を増強するための伸長因子1αの、5'領域の非コーディング12kb核酸配列および3'領域の4kb核酸配列を開示している。
【0015】
生産性を増加するために、増幅可能な遺伝子をトランスフェクトするベクターに対する選択可能なマーカーとして用いる遺伝子増幅細胞エンジニアリング技法を応用することができる。第1のステップで、トランスジーンと増幅可能なマーカー遺伝子を含有するベクターを宿主細胞中に導入する。選択可能でかつ増幅可能な遺伝子を増幅すると、遺伝的に連結されたトランスジーンを含有する配列は共増幅される。増幅は、選択剤の濃度を段階的に増加する、時間のかかる多段階選択プロセスにより誘導する。サブクローニングした耐性細胞を引き続いてスクリーニングすることによって、生産性の高い細胞株を同定することができる。哺乳動物細胞における遺伝子増幅のための増幅可能かつ選択可能なマーカーは、T. Omasha, Journal of Bioscience and Bioengineering (2002) 94、600-605に総括されている。
【0016】
さらに、トランスジーンの発現は、形質転換された細胞を選択するための改変された低活性の選択可能なマーカーを用いることにより増加することができる。WO/2004/050884は細胞集団中の高生産体の比率を増強する発現系を開示している。形質転換されたCHO細胞を選択するマーカーとして、改変されたネオマイシンホスホトランスフェラーゼを使用すると、高生産細胞数-対-全細胞数の比を増強した。
【先行技術文献】
【特許文献】
【0017】
【特許文献1】WO 02/24930
【特許文献2】WO2005/040377
【特許文献3】EP 1,676,916
【非特許文献】
【0018】
【非特許文献1】Zahn-Zabalら、J Biotechnol.、2001、87(1):29-42
【非特許文献2】KalosおよびFournier、Mol Cell Biol.、1995、15(1):198-207
【非特許文献3】Klehrら、Biochemistry、1991、30(5):1264-70
【非特許文献4】Dangら(J Virol.、2000、74(6):2671-8
【非特許文献5】T. Omasha, Journal of Bioscience and Bioengineering (2002) 94, 600-605
【発明の概要】
【発明が解決しようとする課題】
【0019】
しかし、宿主細胞中の目的のポリヌクレオチドの発現のレベルは、記載した技法により増加できるが、高発現はしばしば、発現のサイレンシング、すなわち、低くかつ不安定な発現をもたらす。遺伝子サイレンシング効果によって、培養初期に目的のポリヌクレオチドを強く発現する細胞クローンは、培養の過程で発現の有意な低下を示しうる。さらに、もし細胞集団における発現の不安定な細胞の比率が非常に高ければ、十分な量の所望の産物を得るために、単一の高生産クローンを、時間と費用のかかるスクリーニング手順により単離しなければならないかも知れない。
【0020】
従って、以上の欠点なしに、目的のポリヌクレオチドの発現の改善、すなわち、強くかつ安定な発現を可能にする発現系が大いに所望されている。
【0021】
従って、本発明の根底に横たわる技術的問題は、宿主細胞における遺伝子発現を改善して、高効率でかつ比較的安定な発現を達成する手段と方法を提供することにあると考えることができる。
【課題を解決するための手段】
【0022】
よって、本発明は、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドに関する。好ましいことに、かかるポリヌクレオチドは、宿主細胞中に前記ポリヌクレオチドと組合わせて導入した目的のポリヌクレオチドの発現の増強を可能にする。
【発明を実施するための形態】
【0023】
本明細書に使用した用語「ポリヌクレオチド」は核酸配列に関する。核酸配列はDNAであってもまたはRNA配列であってもよく、好ましくは核酸配列はDNA配列である。
【0024】
本発明のポリヌクレオチドは上述の核酸配列から成るかまたは上述の核酸配列を含むことが必須である。従って、これらはさらなる核酸配列も含有することができる。
【0025】
本発明のポリヌクレオチドは、好ましくは、単離された(すなわち、自然環境から単離された)ポリヌクレオチドとしてまたは遺伝的に改変された形態において提供されよう。本明細書で意味する単離されたポリヌクレオチドはまた、その自然細胞環境以外の細胞環境で存在するポリヌクレオチド、すなわち、異種ポリヌクレオチドを包含する。用語「ポリヌクレオチド」は、1本鎖ならびに2本鎖のポリヌクレオチドを包含する。さらに、グリコシル化またはメチル化ポリヌクレオチドなどの天然の改変されたポリヌクレオチドを含む化学的に改変されたポリヌクレオチド、またはビオチン化ポリヌクレオチドなどの人工の改変されたものも含まれる。
【0026】
本明細書に使用される用語「発現」は、宿主細胞内で起こる転写だけまたは転写と翻訳を意味する。用語「発現」は、好ましくは、宿主細胞中に本発明の少なくとも1つのポリペプチドと一緒に共導入された少なくとも1つの目的のポリヌクレオチドの転写または転写と翻訳に関すると解釈されるべきである。本明細書に使用される用語「共導入された」は、様々なポリヌクレオチドの宿主細胞中への、例えばトランスフェクションまたは形質転換技法による導入が、好ましくは、同じ時点で、すなわち同時に行われることを意味する。それにより、好ましくは、近傍クロマチン領域における、共導入された配列の組込みが可能になろう。
【0027】
本明細書に使用される用語「目的のポリヌクレオチド」は核酸配列に関する。前記核酸配列はRNAまたはDNA配列であってもよく、好ましくは、DNA配列である。本発明の方法によって、目的のポリヌクレオチドは目的の産物をコードすることができる。目的の産物は、目的のポリペプチド、例えば、タンパク質、または目的のRNA、例えば、機能性のRNA、例えば、2本鎖RNA、マイクロRNA、またはsiRNAであってもよい。機能性RNAはしばしば対応する標的遺伝子をサイレントにするために使用される。好ましくは、目的のポリヌクレオチドは、当技術分野で周知でありよく記載されていて、目的のポリヌクレオチドの転写に有効である好適な調節配列(例えば、プロモーター)と機能しうる形で連結されている。
【0028】
宿主細胞における所望の産物の発現のレベルは、細胞中に存在する対応するmRNAの量または目的のポリヌクレオチドによりコードされる所望の産物の量に基づいて決定することができる。例えば、選択した配列から転写されるmRNAは、PCRによりまたはノーザンハイブリダイゼーションにより数値化することができる(Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press (1989)を参照)。ポリペプチドは様々な方法により、例えば、ポリペプチドの生物学的活性をアッセイすることより(例えば、酵素アッセイにより)、または、かかる活性に依らないアッセイ、例えば、タンパク質を認識して結合する抗体を用いるウェスタンブロット、ELISA、またはラジオイムノアッセイを利用することにより、数値化することができる。
【0029】
本明細書で使用される用語「発現を増強できるクロマチンエレメントに対する核酸配列」は、ポリヌクレオチドの発現を増強できるクロマチンエレメントまたはドメインに対する核酸配列に関する。好ましくは、この用語は、宿主細胞中に前記クロマチンエレメントとともに共導入された目的のポリヌクレオチドの発現を増強できるクロマチンエレメントに対する核酸配列に関する。目的のポリヌクレオチドの発現を増強できるクロマチンエレメント/ドメイン(例えば、MARまたは偏在性クロマチンオープニングエレメント)は当技術分野で周知であり、例えば、EP1676916、WO2005/040377、WO0224930、Dangら、Klehrら、Kalosら、Zahn-Zabalら(全て、上記引用文中に)に記載されている。さらに、クロマチンの構造に影響を与えることにより遺伝子活性を調節できる非ランダムなゲノムの分散された遺伝エレメントが、Hodgetts in Current Opinion in Gemtetics & Development (2004) 14 680-685により記載されている。WO2005/040377は、活性を増加するタンパク質生産を有する、例えば、DNA配列、および真核細胞中のタンパク質生産活性を増加するための、これらのDNA配列の使用を記載している。好ましくは、発現を増強できるクロマチンエレメントは、クロマチン中への(すなわち、染色体DNA中への)その組込みが、より脱凝縮した(「オープンな」)クロマチン構造、好ましくは、組込み部位に隣接するクロマチン領域のより脱凝縮した構造を生じさせることができる核酸エレメントである。
【0030】
以上説明したように、好ましいクロマチンエレメントはマトリックス付着活性を有するDNAエレメントを含む。このDNAエレメントは共トランスフェクトされる目的のポリヌクレオチドの発現を増強できることは理解されよう。好ましくは、クロマチンエレメントは発現を増強できるMARを含む。マトリックス付着領域はクロマチンループのベースに位置する特定のDNA配列であって、DNAを核マトリックスの繊維に固着する。マトリックス付着領域はSAR(スカフォールド付着領域)に類似し、従って、SARまたはスカフォールド/マトリックス付着領域(S/MAR)とも呼ばれる。MARを含有する配列はプロモーター活性を抑制できることは公知である(Lauberら J Biol Chem. 1997 Sep 26;272(39):24657-65.)。また、特定のMARエレメントの存在は、トランスジーン発現の平均レベルの位置誘導性変動の低下をもたらさない(Van Leeuwen ら Plant Mol Biol. 2001 Nov;47(4):543-54)。興味深いことに、マウスパーフォリン遺伝子のMAR配列は、グローバルなサイレンシング効果を果たす(Mol Cells. 2002 Feb 28;13(1):61-8)。また、MARβ、T細胞受容体βエンハンサー(Eβ)の上流に位置するマトリックス関連領域は、Eβ介在性TCR活性をサイレント化する重大な役割を果たす(Kaul-Ghanekarら Nucleic Acids Res. 2004 Sep 15;32(16):4862-75)。特定のMAR配列は、遺伝子、例えば、共導入されたトランスジーンの発現を増強することができ、複製起点として機能できることは公知である。MAR全体について明確なコンセンサス配列を規定できないが、いくつかのDNA配列モチーフが、典型的には、MAR様複製起点、TGリッチ配列、湾曲DNAモチーフ、よじれDNAモチーフ、トポイソメラーゼ部位、ATリッチ配列に見出される。これらのモチーフは当技術分野で周知であり、例えば、Singhら(Nucleic Acids Research、1997、25(7): 1419-1425)に掲げられている。本文献は開示したポリヌクレオチド配列に関する参照により本明細書にその全てが組み入れられる。さらに、S/MAR予測プログラムはゲノム配列からMARまたはSARの予測を可能にする(例えば、 Fioriniら, Biochemistry (Moscow), 2006, 71(5): 481-488。
【0031】
本明細書で言及する、発現を増強できるクロマチンエレメントは、好ましくは、配列番号28に示された核酸配列(ヒト白血球由来のゲノムDNAから増幅された;実施例1を参照)を含む。より好ましくは、クロマチンエレメントは、ヒトトポイソメラーゼIのイントロン13のATリッチMII領域を含む配列番号27に示された核酸配列(HeLa細胞由来のゲノムDNAから増幅された)を含む(GenBank L23999.1; GI: 398042; Romigら, 1994、Eur J Biochem. 221:411-419)。
【0032】
ニワトリリゾチーム5'マトリックス付着領域のMARエレメントもトランスジーンの発現を増強できることは公知である(Girodら, Biotechnol Bioeng. 2005 Jul 5;91(1):1-11)。
【0033】
本明細書で使用する「発現を増強する」は、宿主細胞中に本発明のポリヌクレオチドとともに共導入しておいた目的の産物をコードするポリヌクレオチドの転写、または転写および翻訳を含む発現を、改善または増加することを意味する。好ましくは、目的の産物の量が増加する。この増加は、より安定な発現によるかもしくはより強い発現によるかまたはそれらの両方によるものでありうる。増強された発現は、本発明によるポリヌクレオチドと一緒に宿主細胞中に共導入しておいた目的のポリヌクレオチドの発現を、参照宿主細胞中の発現と比較することにより示すことができる。好ましくは、参照宿主細胞は、本発明によるポリヌクレオチドのさらなる導入なしに、同じ目的のポリヌクレオチドを導入することにより作製した宿主細胞である。ポリペプチドの量(例えば、ウェスタンブロットにより、Elisaによりまたは酵素アッセイにより、前掲)およびRNA分子(例えば、PCRまたはノーザンブロットにより、前掲)を決定するためのならびに前記量を比較するための手段と方法は当技術分野において周知である。好ましくは、所望の産物の量は、参照量と比較して統計的に有意に増加し、さらに好ましくは、所望の産物の量は、2倍、3倍、4倍、5倍、10倍、20倍、30倍、50倍、100倍、200倍または1000倍増加する。
【0034】
本明細書で使用する用語「湾曲起点モチーフ」は、少なくとも1つの複製起点シグナル(また「複製起点シグナル」、「起点シグナル」、または「oriシグナル」とも呼ばれる)と少なくとも1つの湾曲DNAシグナルとの組合わせと解釈することとする。好ましくは、湾曲起点モチーフは、1、2、3、4、5、7、10、15、20、21、25、30、35、40、50、55、60、70、80、90、100、200、500、1000以上の複製起点シグナルおよび1、2、3、4、5、7、10、15、20、21、25、30、35、40、50、55、60、70、80、90、100、200、500、1000以上の湾曲DNAシグナルを含む。本発明による湾曲起点モチーフは異なるoriシグナルを含んでもよい。さらに、本発明による湾曲起点モチーフは異なる湾曲DNAシグナルを含んでもよい。本明細書に参照によりその全てが組み入れられるSinghら(Nucleic Acids Research, 1997, 25(7), 1419 - 1425)は、oriシグナルならびに湾曲DNAシグナルを開示している。本明細書で意味する湾曲DNAシグナルは、これらの配列を含有するDNA分子中に湾曲(内因性の)を導入できることで特徴付けられる核酸配列である。当業者にとってDNA分子中に湾曲を導入する核酸配列エレメントは公知である。例えば、湾曲は、例えば、アデニンの短いホモポリマー列(Aトラック)(4〜6個の塩基対)が、一致してらせん型スクリューで繰り返されると起こりうる。1つのらせんスクリューはおよそ10.4bpを含む(Haranら, 1994, Sequences elements responsible for DNA curvature. J Mol Biol, 244: 135-143を参照)。DNA断片の湾曲は、例えば、DNA断片の電気泳動移動度により決定するかまたは好適なコンピューターソフトウエア(例えば、Shpigelmanら, 1993, Comput Appl Biosci.;9(4):435-40に記載)を用いることにより予測することができる。湾曲DNAは、例えば、いくつかのマトリックス付着部位またはその近傍にて同定されている。湾曲DNAはまた、組換え、複製、および転写などのDNAおよびタンパク質の相互作用に関わる核プロセスにおいて重要な役割を果たしうる。DNA配列エレメント中への湾曲は、AAAANNNNNNNAAANNNNNNNAAAA(配列番号18に示した)、またはAAAANNNNNNNAAAANNNNNNNAAAA(配列番号19に示した)、またはTTTTNNNNNNNTTTNNNNNNNTTTT(配列番号20に示した)、またはTTTTNNNNNNNTTTTNNNNNNNTTTT(配列番号21に示した)[ここでNは任意のヌクレオチド、従ってA、T、CまたはGまたは前記ヌクレオチドのいずれかの化学的に改変された誘導体を表し、好ましくはNはGまたはCを表す]の核酸配列を有する湾曲DNAシグナルを含むDNAモチーフにより導入することができる。特に、湾曲はこれらのシグナルの反復配列/多量体により導入される。
【0035】
さらに、DNA配列エレメント中への湾曲は、湾曲DNAシグナル、好ましくは、湾曲DNAシグナルの多量体/反復配列、より好ましくは、おおよそ、らせんスクリューの相にある湾曲DNAシグナルの多量体/反復配列を含むDNAモチーフにより導入することができ、ここで、湾曲DNAシグナルはAAAANNNNNN(配列番号37に示した)、またはAAAANNNNNNN(配列番号38に示した)、またはAAAAANNNNN(配列番号39に示した)、またはAAAAANNNNNN(配列番号40に示した)、AAAAAANNNN(配列番号41に示した)、またはAAAAAANNNNN(配列番号42に示した)、またはTTTTNNNNNN(配列番号43に示した)、またはTTTTNNNNNNN(配列番号44に示した)、またはTTTTTNNNNN(配列番号45に示した)、またはTTTTTNNNNNN(配列番号46に示した)、またはTTTTTTNNNN(配列番号47に示した)、またはTTTTTTNNNNN(配列番号48に示した)[ここでNは任意のヌクレオチド、従ってA、T、CまたはGまたは前記ヌクレオチドのいずれかの化学的に改変された誘導体を表し、好ましくはNはGまたはCを表す]の核酸配列を有する。さらに、DNA配列中への湾曲は、湾曲DNAシグナルTTTAAA(配列番号22に示した)、およびAAATTT(配列番号23に示した)、(例えば、Singhら、前掲)、とりわけこれらのシグナルの多量体/反復配列により導入される。
【0036】
oriシグナルはoriのエレメント、好ましくはATリッチである。当業者にとってori(複製の起点)は公知である。oriはDNA複製を開始することができるDNA配列である。oriのDNA配列は、種毎に、特に原核生物と真核生物では変化しうるが、oriモチーフは一般にいくつかの共通の特性を共有する。本明細書で使用するoriシグナルは、好ましくは、真核生物の複製起点由来の配列モチーフであるが、原核生物の複製起点由来のシグナル/モチーフであってもよい。真核生物のoriはよく記載されていて、例えば、出芽酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)については起点活性(自律複製配列またはARSエレメントと呼ばれる)を与える具体的な配列が単離されている。DNA複製が核マトリックスと関係していることは公知である。また、核マトリックス付着部位、ホメオティックなタンパク質認識および結合部位、ならびに複製起点は、次のoriシグナル:ATTA(配列番号24に示した)、ATTTA(配列番号25に示した)、およびATTTTA(配列番号26に示した)、(例えば、Singhら、上記引用文中)を共有することが実証されている。
【0037】
湾曲起点モチーフの湾曲DNAシグナルと湾曲起点モチーフのoriシグナルは離れていても、すなわち、さらなる核酸配列により分離されていてもよい。さらに、湾曲DNAシグナルとoriシグナルの核酸配列は重複していてもよい、すなわち、oriシグナルのエレメント/部分であるヌクレオチドが湾曲DNAシグナルのエレメントであってもよい。さらに、湾曲DNAシグナルが1個以上のoriシグナルを含んでもよく、その逆もありうる。湾曲DNAシグナルとoriシグナルの重複する例は、例えば、配列番号1、配列番号2、配列番号29、または配列番号31に示した湾曲起点モチーフである(下記参照)。本発明による湾曲起点モチーフが、それを含有するDNA中に湾曲を導入することができるのは理解されよう。湾曲は、上記の手段と方法により決定することができる。さらに、湾曲起点モチーフはまた、様々な湾曲起点モチーフの組合わせを含みうることも理解されよう。
【0038】
本発明のある好ましい実施形態において、湾曲起点モチーフは、配列番号1に示した核酸配列ATTTTANNNNNATTTTANNNNNATTTTA、または少なくとも2つの配列番号29に示した核酸配列ATTTTANNNNNATTTTANNNNNATTTTANNNNNの反復配列(従って、前記核酸配列の少なくとも二量体をもたらす、少なくとも2倍の配列番号29に示した核酸)、配列番号2に示した核酸配列TAAAATNNNNNTAAAATNNNNNTAAAATまたは少なくとも2つの配列番号30に示した核酸配列TAAAATNNNNNTAAAATNNNNNTAAAATNNNNNの反復配列[ここで、Nは任意のヌクレオチド、すなわち、A、T、CまたはG、好ましくはCまたはG、または前記ヌクレオチドのいずれかの化学的に改変された誘導体を表す]を含む。より好ましくは、湾曲起点モチーフは、配列番号31に示した核酸配列ATTTTAGCTCGATTTTAGCTCGATTTTA、または少なくとも2つの配列番号32に示した核酸配列ATTTTAGCTCGATTTTAGCTCGATTTTAGCTCGの反復配列、または配列番号33に示した核酸配列TAAAATCGAGCTAAAATCGAGCTAAAAT、配列番号34に示したTAAAATCGAGCTAAAATCGAGCTAAAATCGAGC、または少なくとも2つの配列番号75に示した核酸配列AAAATCGAGCTAAAATCGAGCTAAAATCGAGCTの反復配列を含む。本明細書で使用する用語「少なくとも2つの反復配列」は、好ましくは、2以上の反復配列(および従って2以上のユニットまたはモノマー)、好ましくは、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7または少なくとも10、少なくとも20、少なくとも30、少なくとも40、少なくとも50、少なくとも100、少なくとも200の反復配列/ユニット/モノマーを意味する。従って、湾曲起点モチーフは、好ましくは、2倍以上、または3倍以上などの、本明細書で先に言及したそれぞれの核酸を含み、従って、好ましくは前記核酸の二量体(2つの反復配列を含む場合)、三量体(3つの反復配列を含む場合)、オリゴマー、多量体などである。本発明のクロマチンエレメント中に含まれる湾曲起点モチーフの、特に好ましい数は7である。本発明のクロマチンエレメント中に含まれる湾曲起点モチーフの、他の特に好ましい数は22である。好ましくは、前記少なくとも2つの反復配列は、正方向の反復配列(頭部から尾部への反復配列、好ましくは、反復配列の間にさらなるヌクレオチドの無いもの)である。
【0039】
さらに、湾曲起点モチーフは、核酸配列TAAAATNNNN(配列番号49に示した)、またはTAAAATNNNNN(配列番号50に示した)、またはTAAAAATNNN(配列番号51に示した)、またはTAAAAATNNNN(配列番号52に示した)、TAAAAAATNN(配列番号53に示した)、またはTAAAAAATNNN(配列番号54に示した)、またはATTTTANNNN(配列番号55に示した)、またはATTTTANNNNN(配列番号56に示した)、またはATTTTTANNN(配列番号57に示した)、またはATTTTTANNNN(配列番号58に示した)、またはATTTTTTANN(配列番号59に示した)、またはATTTTTTANNN(配列番号60に示した)の少なくとも1つの反復配列(従って、少なくとも1回の核酸配列)を含むことができる[ここでここでNは任意のヌクレオチド、従ってA、T、CまたはGまたは前記ヌクレオチドのいずれかの化学的に改変された誘導体を表し、好ましくはNはGまたはCを表す]。本明細書で使用する用語「少なくとも1つの反復配列」は、1以上の反復配列/ユニット/モノマー、好ましくは、少なくとも2、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7または少なくとも10、少なくとも20、少なくとも22、少なくとも30、少なくとも40、少なくとも50、少なくとも75、少なくとも100、少なくとも200の反復配列/ユニット/モノマーを意味する。先に記載の通り、本発明のクロマチンエレメント中に含まれるべき湾曲起点モチーフの、特に好ましい数は22である。好ましくは、前記反復配列は正方向の反復配列である(前記参照)。
【0040】
他のさらに好ましい本発明の実施形態において、湾曲起点モチーフは配列番号7または配列番号8、または配列番号9、または配列番号10に示した核酸配列を含む。
【0041】
本発明によれば、湾曲起点モチーフ用の核酸配列はクロマチンエレメントに対する核酸配列と共有結合により連結されている。当業者にとって、共有結合によって2以上の核酸配列を連結するのに使用できる技法、例えば、ライゲーションまたはクローニングなどの組換えDNA技法として知られる技法は公知である。湾曲起点モチーフが、発現を増強できるクロマチンエレメント中にさらに組み込まれることは理解されよう。それにより、「合成」クロマチンエレメントが作製される。湾曲起点モチーフ用核酸配列はクロマチンエレメント用核酸配列のいずれの位置にあってもよいことは理解されよう。好ましくは、湾曲起点モチーフは、配列番号27のヌクレオチドの1900と2200の間、より好ましくは、2000と2100の間、そして最も好ましくは、位置2031の位置に導入される。
【0042】
さらに、共トランスフェクトしたポリヌクレオチドの発現を増強できるクロマチンエレメントは、湾曲起点モチーフ用の核酸配列を、1つ以上の、例えば、2、3、4、5、10、20、30、40、50の位置にまたは100の位置に含んでもよく、好ましくは、前記ポリヌクレオチドは前記核酸配列を1つの位置に含むことが理解されよう。
【0043】
他の最も好ましい本発明の実施形態において、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドは、配列番号3にまたは配列番号4にまたは配列番号5にまたは配列番号6にまたは前記配列番号のいずれか1つの変異体に示した核酸配列を有する。かかる変異体は、好ましくは、配列番号3、4、5または6のいずれか1つと少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%または少なくとも99%同一である核酸配列を有する。2以上の核酸配列の間の同一性の程度(パーセント、%)は、好ましくは、NeedlemanおよびWunschまたはSmithおよびWatermanのアルゴリズムにより決定される。配列アラインメントを行うために、プログラムPileUp(J. Mol. Evolution., 25, 351-360, 1987, Higgins 1989, CABIOS, 5: 151-153)またはプログラムGapおよびBestFit(Needleman 1970, J. Mol. Biol. 48; 443-453 および Smith 1981, Adv. Appl. Math. 2; 482-489)が利用され、これらはGCGソフトウエアパケット(Genetics Computer Group, 575 Science Drive, Madison, Wisconsin, USA 53711, vers. 1991)の一部分である。パーセント(%)で以上に記載される配列同一性値は、好ましくは、プログラムGapを用いて、全配列領域にわたり、次の設定値、ギャップ重み:50、長さ重み:3、平均マッチ:10.000および平均ミスマッチ:0.000を用いて決定し、これは、特に断らない限り、常に配列アラインメント用の標準設定として用いることとする。さらに、前記変異体核酸配列は、好ましくは、先に規定した少なくとも1つの湾曲起点モチーフを含む。好ましくは、かかる変異体核酸配列を含む、ポリヌクレオチドは共トランスフェクトしたポリヌクレオチドの発現を増強することができる。この増強は、添付した実施例に記載のアッセイにより試験することができる。
【0044】
本発明はまた、先に規定した湾曲起点モチーフにも関する。好ましくは、前記モチーフは、共トランスフェクトしたポリヌクレオチドの発現を増強するために使われる。
【0045】
有利なことに、本発明によって、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドと目的のポリヌクレオチドとの宿主細胞中への共導入は、目的のポリヌクレオチドの発現を増強することが見出されている。例えば、ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域を含むポリヌクレオチド(Romig, 1994 Eur J Biochem. 221:411-419)と湾曲起点モチーフを、選択可能なマーカーとしてのレポーター遺伝子EGFP(Enhanced Green Fluorescent Protein;緑色蛍光が増強されたタンパク質)および未改変ネオマイシンホスホトランスフェラーゼ遺伝子を保持するClontechから入手したpEGFP-N1ベクターと一緒に、CHO細胞(Chinese Hamster Ovary;チャイニーズハムスター卵巣)中に同時トランスフェクトした。対照としてpEGFP-N1ベクターを、単独で、またはヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域に対するDNAと湾曲起点モチーフなしで組合わせて、CHO細胞中に導入した。トランスフェクション後、G418を含有する培地(0.6mg/mlG418)で細胞を培養して選択した。10日後、単一のG418耐性クローンがコロニーを形成した。全体の耐性細胞集団を用いて、FACS分析(Fluorescent Activated Cell Sorting;蛍光活性化細胞ソーティング)によりレポーター遺伝子発現の数値化を行った。対照と比較して、高発現細胞の比率、従って発現が、レポーター遺伝子と湾曲起点モチーフを含む前記ATリッチ領域を共導入することにより作製したCHO細胞において、有意に増加することを見出した。従って、共トランスフェクトした目的のポリペプチドの発現をそれ自体増強できるクロマチンエレメント中への、湾曲起点モチーフの挿入は、前記クロマチンエレメントの発現増強効果をさらに増強する。湾曲モチーフを単独で(湾曲起点モチーフの代わりに)前記クロマチンエレメントに組み込んだ場合、前記クロマチンエレメント単独(湾曲モチーフを挿入しないで)と比較すると、発現の増強が驚くほど検出されなかった。この驚く
べき発現増強効果は、例えば、配列番号3、配列番号4および配列番号5に示した核酸配列を有するポリヌクレオチドについて示された。従って、本発明による上述のポリヌクレオチドの使用は、高生産クローンを単離するための時間と費用のかかるスクリーニングステップを軽減するであろう。
【0046】
さらに、本発明は、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするか、または配列番号14、配列番号15および配列番号16のいずれか1つに示した核酸配列を含む、ポリヌクレオチドに関する。
【0047】
前記ポリヌクレオチドは、配列番号35に示したアミノ酸配列を有する未改変の(野生型)ネオマイシンホスホトランスフェラーゼと比較して、アミノ酸配列に1または2個のアミノ酸変化を有する改変されたネオマイシンホスホトランスフェラーゼII(NPT II)をコードする。具体的には、本発明の改変されたNPT IIは、それぞれRのAへの、およびRのVへのアミノ酸置換を、それぞれ配列番号11および配列番号12に記載のNPT IIの226位に対応する位置に有するか、またはそれぞれAのGへの、およびTのSへのアミノ酸置換を、それぞれ配列番号13に記載のNPT IIの224位および225位に対応する位置に有することを想定している。ネオマイシン耐性遺伝子とも呼ばれるNPT II遺伝子は、最初に大腸菌株K12のトランスポゾンTn5から単離された。NPT IIタンパク質はアミノグリコシド-アミノシクリトール抗生物質を位置特異的リン酸化により不活性化する酵素のクラスに属する。これはカナマイシンおよびG418などの様々なアミノグリコシド抗生物質に対する耐性を与える。この遺伝子は、一般に選択可能なマーカーとして、細菌、植物、動物および酵母のような様々な種類の生物を形質転換するために利用される。当技術分野では、NPT IIアミノ酸配列の改変、すなわち、アミノ酸変化がNPT IIの酵素活性を低下させうることは公知である。さらに、酵素活性の低下した突然変異NPT II変異体の形質転換への使用は、未改変のNPT II遺伝子を保持する生物よりもネオマイシンに対する耐性が低いトランスジェニック生物をもたらすことは公知である。さらに、これらの変異体を高生産細胞の選択に利用することができる(例えば、WO2004/050884を参照)。
【0048】
改変されたネオマイシンホスホトランスフェラーゼをコードする本発明のポリヌクレオチドは、融合タンパク質の1つのパートナーが先に挙げた核酸配列によりコードされているポリペプチドである融合タンパク質をコードしうることは理解されよう。かかる融合タンパク質は、さらなる部分として脂肪酸または脂質生合成経路の他の酵素、発現をモニターするためのポリペプチド(例えば、緑色、黄色、青色または赤色蛍光タンパク質、アルカリホスファターゼなど)または検出可能なマーカーとしてまたは精製のための補助的基準として役割を果たしうるいわゆる「タグ」を含んでもよい。色々な目的のためのタグが当技術分野では周知であり、それには、FLAG-タグ、6-ヒスチジン-タグ、MYC-タグなどが含まれる。
【0049】
さらに本発明は、配列番号11、配列番号12および配列番号13のいずれか1つに示されるアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16のいずれか1つに示される核酸配列を含むものである、ポリヌクレオチドによりコードされるポリペプチドに関する。
【0050】
有利なことに、本発明によれば、配列番号14、配列番号15および配列番号16のいずれか1つに示した核酸配列をCHO細胞のトランスフェクションに対する選択可能なマーカー遺伝子として含むポリヌクレオチドの使用は、特に、共導入した目的のポリヌクレオチドの発現を増強することが示されている。例えば、改変されたNPT II遺伝子およびレポーター遺伝子EGPFを含む様々なベクターをCHO細胞中に導入した。トランスフェクション後、前記細胞をG418含有培地(0.6mg/ml)で培養することにより選択した。選択可能なマーカーとして、改変されたNPT IIの代わりに未改変のネオマイシンホスホトランスフェラーゼをコードするポリヌクレオチドを用いる比較実験を実施した。高発現細胞、従ってレポーター遺伝子発現の比率は、改変されたネオマイシンホスホトランスフェラーゼを選択可能なマーカーとして用いることにより作製したトランスジェニック宿主細胞において有意に増強されることを見出した。従って、記載した本発明によるポリヌクレオチドの使用は、高生産クローンを単離するための時間と費用のかかるスクリーニングステップを軽減するであろう。
【0051】
本発明はまた、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列、または、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするポリヌクレオチドまたは配列番号14、配列番号15および配列番号16に示した核酸配列のいずれか1つに示した核酸配列、またはその組合わせを含むものである、以上説明したポリヌクレオチドを含む宿主細胞に関する。
【0052】
本明細書で使用する「宿主細胞」には、異種DNAの導入により改変することができる任意の培養可能な細胞が含まれる。好ましくは、宿主細胞は、その細胞において目的のポリヌクレオチドを安定して発現することができ、前記ポリペプチドがコードするポリペプチドを翻訳後に改変して適当なサブ細胞コンパートメントに局在化できる細胞である。適当な宿主細胞の選択はまた、検出シグナルの選択によっても影響を受けうる。本発明の宿主細胞には原核細胞および真核細胞が含まれる。原核生物にはグラム陰性またはグラム陽性生物、例えば、大腸菌または桿菌が含まれる。好ましくは、本発明によるポリヌクレオチド、特に本発明のポリヌクレオチドを含むベクター(下記参照)の増殖には原核細胞が使われることは理解されよう。形質転換のための好適な原核生物の宿主細胞には、例えば、大腸菌、枯草菌、ネズミチフス菌、ならびにシュードモナス菌、ストレプトマイセス菌、およびブドウ球菌属内の様々な他の種が含まれる。真核細胞には、限定されるものでないが、動物細胞、脊椎動物細胞、酵母細胞、植物細胞、真菌細胞、昆虫細胞(例えば、バキュロウイルスにより感染させることができるSF9細胞)、哺乳動物細胞、爬虫類動物細胞、藻類細胞および寄生生物、例えば、トリパノソーマ類の細胞が含まれる。本明細書で使用する、酵母には、厳密に分類学的な意味の酵母、すなわち、単細胞生物だけでなく、糸状真菌の酵母様多細胞真菌が含まれる。種の例には、クライベラ・ラクチス(Kluyvera lactis)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)、およびウスチラゴ・マイディス(Ustilago maydis)が含まれ、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)が好ましい。本発明の実施に使用できる他の酵母は、アカパンカビ(Neurospora crassa)、クロコウジカビ(Aspergillus niger)、アスペルギルス・ニデュランス(Aspergillus nidulans)、ピチア・パストリス(Pichia pastoris)、カンジダ・トロピカリス(Candida tropicalis)、および ハンセヌラ・ポリモルファ(Hansenula polymorpha)である。哺乳動物の宿主細胞培養系には、
COS細胞、L細胞、3T3細胞、HeLa細胞、ベビーハムスター腎(BHK)細胞、チャイニーズハムスター卵巣(CHO)細胞、胚幹細胞などの確立された細胞が含まれる。
【0053】
本発明のある好ましい実施形態において、宿主細胞は哺乳動物細胞である。さらに好ましい実施形態において、宿主細胞はCHO細胞である。CHO細胞培養系には、血清を含有するおよび/または血清を含有しない培地で培養できる全てのCHO細胞、例えば、CHO-K1、CHO-S、CHO DG44、CHO DXB11、CHO(タンパク質を含有しない)ECACC No. 00102307、CHO(dhfr-ACを含有しない)ECACC No. 05011002 細胞が含まれる。
【0054】
さらに、本発明は、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列、または、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするポリヌクレオチドまたは配列番号14、配列番号15および配列番号16に示した核酸配列のいずれか1つに示した核酸配列、またはその組合わせを含む、以上説明したポリヌクレオチドを含む、非ヒトトランスジェニック生物に関する。
【0055】
本発明によれば、非ヒトトランスジェニック生物は非ヒト動物または植物であってもよい。本明細書で意味するトランスジェニック非ヒト動物は、出生前に、例えば、胚段階に非ヒト動物または前記非ヒト動物の祖先中に導入されたトランスジーンを含有する細胞を有するものとする。トランスジーンは、細胞のゲノム中に組み込まれているDNAであり、これからトランスジェニック非ヒト動物が発生し、これが成熟非ヒト動物のゲノム中に残り、それによりトランスジェニック非ヒト動物の1以上の細胞型または組織中のコードされた遺伝子産物の発現を駆動する。好ましい非ヒトトランスジェニック動物は、マウス、ヤギ、ヒツジ、ブタ、ウシ、ウマ、魚類、および非ヒト霊長類からなる群より選択される。非ヒト農場動物は、タンパク質の大規模生産のために有用である(いわゆる「遺伝子農場(gene farm)」)。トランスジェニック非ヒト動物は、例えば、トランスジーンを受精卵母細胞の雄前核中に、例えば、マイクロインジェクションによって導入し、次いで卵母細胞を擬妊娠雌里親動物中で発生させることにより作製することができる。
【0056】
トランスジェニック動物、特にマウスなどの動物を作製する方法は当技術分野で一般的になっていて、例えば、米国特許第4,736,866号および第4,870,009号およびHogan 1986, A Laboratory Manual, Cold Spring Harbor, New York, Cold Spring Harbor Laboratoryに記載されている。トランスジェニック里親動物を用いてトランスジーンを保持するさらなる動物を育種することができる。本発明によるトランスジーンを保持するトランスジーン動物を、他のトランスジーンを保持する他の動物、例えば、テトラサイクリン依存性転写レギュレーターをコードするポリヌクレオチドを発現するトランスジェニックを育種することができる(以下にさらに詳細に考察される)。本発明はまた、本発明のベクターまたは発現制御配列を、好ましくは、先に言及したトランスジーンの形態で含む相同的組換え非ヒト動物も提供する。かかる非ヒト相同的組換え動物においては、前記核酸はゲノムの特定部位中に導入されている、すなわち、核酸分子はゲノムの内因性遺伝子または他の部分と相同的に組換えられている。前記事例において、動物は好ましくはマウスである。かかる相同的組換え動物を作製するために、好ましくは、相同的組換えが起こるゲノムの遺伝子または部分のさらなる核酸にその5'および3'末端で隣接した、導入すべき核酸をコードするDNAを含有するベクターを調製する。融合タンパク質をコードする隣接したさらなる核酸は、真核生物の遺伝子との相同的組換えが成功するのに十分な長さである。典型的には、数キロ塩基の(5'および3'末端の両方で)隣接するDNAがベクター中に含まれる(例えば、相同的組換えベクターの説明については、Thomas, 1987, Cell 51:503を参照)。ベクターを胚幹細胞株中に導入し(例えば、エレクトロポレーションにより)、導入したDNAが内因性DNAと相同的に組換えられた細胞を選択する(例えば、Li 1992, Cell 69:915を参照)。選択した細胞を次いで動物(例えば、マウス)の胚盤胞中に注射して凝縮キメラを形成する(例えば、Bradley, A., Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E.J. Robertson, 編 (IRL, Oxford, 1987) pp.113-152を参照)。キメラ胚を次いで好適な偽妊娠雌里親動物中に移植し、胚を出産させる。相同的に組換えたDNAをその生殖細胞中に保持する子孫を用いて、その全細胞が相同的に組換えられたDNAを含有する動物を育種する。これらの「生殖系列導入」動物をさらにテトラサイクリン依存性転写レギュレーターをコードする遺伝子を保持する動物と交配することができる。以上記載した相同的組換え手法だけでなく、酵素の助けをかりる部位特的組込み系が当技術分野では公知であり、本発明の調節システムの成分に適用して第2の標的DNA分子中の予め定めた位置にDNAを組み込むことができる。かかる酵素の助けをかりる組込み系の例としては、Creリコンビナーゼ-lox標的系(例えば、Baubonis 1993, Nucl. Acids Res. 21:2025-2029;およびFukushige 1992, Proc. Natl. Acad. Sci. USA 89:7905-7909に記載)およびFLPリコンビナーゼ-FRT標的系(例えば、Dang 1992, Dev. Genet. 13:367-375;およびFiering 1993、Proc. Natl. Acad. Sci. USA 90:8469-8473に記載)が挙げられる。
【0057】
本明細書で使用する用語「植物」は植物と藻類を包含する。好ましくは、この用語は多細胞陸生植物に関する。より好ましくは、多細胞陸生植物は作物植物、などの高等植物であり、トウモロコシ、キャノーラ、ダイズ、イネ、マリーゴールド、アブラナ、コムギ、または グリシン(glycine)が含まれる。原理的に、トランスジェニック植物は、Becker 1992, Plant Mol. Biol. 20:1195-1197、Bevan 1984, Nucleic Acids Res. 12:8711-8721、および“Vectors for Gene Transfer in Higher Plants” in: Transgenic Plants, Vol. 1, Engineering and Utilization, 編: Kung、およびR. Wu, Academic Press, 1993, p. 15-38に記載のように得ることができる。好ましくは、植物細胞の形質転換、従って、トランスジェニック植物の作製は、アグロバクテリウム介在性形質転換によりまたは物理的な力(例えば、「遺伝子銃」)により達成しうる。
【0058】
本発明はまた、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列、または、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16に示した核酸配列のいずれか1つに示した核酸配列、またはその組合わせを含むポリヌクレオチドを含むものである、以上に規定したポリヌクレオチドを含むベクターに関する。
【0059】
用語「ベクター」は、好ましくは、ファージ、プラスミド、ウイルスまたはレトロウイルスベクター、ならびに細菌または酵母人工染色体などの人工染色体を包含する。さらに、本用語はまた、標的化構築物のゲノムDNA中への無作為または部位指定組込みを可能にする標的化構築物に関する。かかる標的構築物は、好ましくは、以下に詳細に記載する相同的なまたは異種組換えに十分な長さのDNAを含む。本発明のポリヌクレオチドを包含するベクターは、好ましくは、さらに宿主における繁殖および/または選択のための選択可能なマーカーを含む。ベクターは、宿主細胞中に当技術分野で周知の様々な技法により組み込むことができる。例えば、プラスミドベクターはリン酸カルシウム沈降物または塩化ルビジウム沈降物などの沈降物で、または荷電脂質との複合体で、またはフラーレンなどの炭素に基づくクラスターで導入することができる。あるいは、プラスミドベクターは熱ショックまたはエレクトロポレーション技法により導入することができる。ベクターがウイルスであれば、適当なパッケージング細胞株を用いてin vitroでパックした後、宿主細胞に適用してもよい。レトロウイルスベクターは複製コンピテントまたは複製欠陥性であってもよい。後者の場合、ウイルス繁殖は一般に補完性宿主細胞/細胞でのみ起こりうる。
【0060】
より好ましくは、本発明のベクターにおいて、本発明によるポリヌクレオチドは、原核生物または真核細胞、特に真核細胞またはそれらの単離された画分における発現または繁殖を可能にする発現制御配列と機能しうる形で連結されている。前記ポリヌクレオチドの発現は、ポリヌクレオチドの、好ましくは、翻訳可能なmRNA中への転写を含む。真核細胞、好ましくは哺乳動物の細胞における発現を保証する調節エレメントは当技術分野で周知である。これらは、好ましくは、転写の開始を保証する調節配列および、任意に、転写の終結および転写物の安定化を保証するポリ-Aシグナルを含む。さらなる調節エレメントには、転写ならびに翻訳エンハンサーが含まれてもよい。原核生物宿主細胞における発現を可能にすると思われる調節エレメントとしては、例えば、大腸菌のlac、trpまたはtacプロモーターが含まれ、真核生物宿主細胞における発現を可能にする調節エレメントの例は、酵母のAOX1またはGAL1プロモーターまたは哺乳動物および他の動物細胞のCMV-、SV40-、RSV-プロモーター(ラウス肉腫ウイルス)、CMV-エンハンサー、SV40-エンハンサーまたはグロビン-イントロンである。好適な発現制御配列は当技術分野で周知である。転写の開始に関わるエレメントに加えて、かかる調節エレメントはポリヌクレオチドの下流にSV40-ポリA部位またはtk-ポリA部位などの転写終結シグナルを含んでもよい。この関係において、好適な発現ベクターとしては、Okayama-Berg cDNA発現ベクターpcDV1(Pharmacia)、pBluescript(Stratagene)、pCDM8、pRc/CMV、pcDNA1、pcDNA3(Invitrogene)またはpSPORT1(GIBCO BRL)などが当技術分野において公知である。好ましくは、前記ベクターは発現ベクターおよび遺伝子導入または標的化ベクターである。レトロウイルス、ワクシニアウイルス、アデノ随伴ウイルス、ヘルペスウイルス、またはウシパピローマウイルスなどのウイルスから誘導される発現ベクターを、本発明のポリヌクレオチドまたはベクターを標的化細胞集団中への送達するのに利用することができる当業者に周知の方法を、構築物組換えウイルスベクターを構築するために利用することができる;例えば、Sambrook, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (1989) N.Y. および Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience、N.Y. (1994)に記載の技法を参照されたい。
【0061】
本発明はまた、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列、または、配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするポリヌクレオチドまたは配列番号14、配列番号15および配列番号16に示した核酸配列のいずれか1つに示した核酸配列、またはその組合わせを含むものである、以上説明したポリヌクレオチドを含む、キットに関する。
【0062】
本発明のポリヌクレオチドは、本発明の方法を行うのに適応したキットとして、宿主細胞における宿主細胞の核酸の発現に必要な他の成分と一緒に提供することができる。キットの他の成分は、好ましくは、発現のために利用する宿主細胞および/またはポリヌクレオチドの細胞中への導入のために用いる物質である。
【0063】
本明細書で使用する用語「キット」は、上述した本発明の化合物、手段または試薬のコレクションを意味する。キットの成分は、別のバイアルに含まれていてもよいし(すなわち、別の部品のキットとして)または単一バイアルで提供されてもよい。さらに、本発明のキットは本明細書で以上言及した方法を実施するために使われることは理解されよう。好ましくは、以上言及した方法を実施するために直ぐ使用できる方式で提供されると想定している。さらに、本キットは好ましくは、前記方法を実行するための取扱説明書を含む。取扱説明書は、紙または電子形式のユーザーマニュアルで提供することができる。例えば、マニュアルは、本発明のキットを用いて上述の方法を行ったときに得られる結果の解釈に対する指示書を含むものである。
【0064】
以上記載した用語の定義と説明は、以下の実施形態に対して必要な変更を加えて適用される。
【0065】
本発明はまた、少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)宿主細胞中に
(i)発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチド、および
(ii)少なくとも1つの目的のポリヌクレオチド
を導入するステップ、ならびに
(b)前記宿主細胞を培養するステップ
を含んでなる前記方法に関する。
【0066】
本発明による方法においては、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドを用いて少なくとも1つの目的のポリヌクレオチドを発現する。好ましくは、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドを用いて少なくとも1つの目的のポリヌクレオチドの発現を増強する。本明細書で使用する用語「少なくとも1つの目的のポリヌクレオチド」は、好ましくは、1以上の、例えば、少なくとも2、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7または少なくとも10、または少なくとも20の目的のポリヌクレオチドを意味し、好ましくは用語「少なくとも1つの目的のポリヌクレオチド」は1つの目的のポリヌクレオチドを意味する。好ましくは、本発明の方法のポリヌクレオチドおよび目的のポリヌクレオチドは宿主細胞中に共導入(同時に導入)される。本発明のポリヌクレオチドは、宿主細胞中にポリヌクレオチドを導入するのに好適ないずれの技法により宿主細胞中に導入してもよく、かかる技法は当技術分野で周知でありかつよく記載されている。例えば、ポリヌクレオチドはエレクトロポレーションにより、または、マイクロインジェクションにより、リン酸カルシウム沈降物または塩化ルビジウム沈降物などの沈降物で、または脂質に基づくトランスフェクションにより、または荷電脂質との複合体で、またはフラーレンなどの炭素に基づくクラスターで導入することができる。さらに、ポリペプチドは、レトロウイルスなどのウイルスを用いることにより組み込むことができる。本発明のポリヌクレオチドは。いずれの宿主細胞中に導入してもよく、好ましくは、本出願の文脈で記述したいずれの宿主細胞中に導入してもよい。より好ましくは、ポリヌクレオチドはCHO細胞中に脂質に基づくトランスフェクションにより導入される。宿主細胞中に導入されるべき目的のポリヌクレオチドおよび本発明の方法によるポリヌクレオチドは同じ核酸分子、例えば、DNAまたはRNAベクター上に、または異なる核酸分子上に位置してもよい。好ましくは、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドと目的のポリヌクレオチドは異なる核酸分子上に存在する。さらに、宿主細胞中に導入すべきポリヌクレオチドは、直鎖または環状型で存在してもよい。核酸分子、特に、DNA分子に関する用語「直鎖」または用語「環状」は当業者により理解されている。好ましくは、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドは、直鎖または環状型で存在し、目的のポリヌクレオチドまたはいずれかの他のポリヌクレオチド(例えば、マーカーをコードするポリヌクレオチド)は環状型、例えば、プラスミドで存在する。より好ましくは、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドは直鎖型で存在し、目的のポリヌクレオチドまたはいずれかの他のポリヌクレオチド(例えば、マーカーをコードするポリヌクレオチド)は直鎖型、例えば、直鎖プラスミドで存在する。
【0067】
本発明の方法によると、好ましくは、参照宿主細胞における目的のポリヌクレオチドの発現と比較して、目的のポリヌクレオチドの発現が増強されるであろうことは理解されよう。かかる参照細胞は、本発明のポリヌクレオチドを導入せずに目的のポリヌクレオチドを導入すること(例えば、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドなしに、目的のヌクレオチドをトランスフェクトすること)により作製することができる。発現の増強は本明細書に先に記載した手段と方法により示すことができる。
【0068】
本明細書で使用する用語「前記宿主細胞を培養する」は、好適な条件下で細胞を培養する方法および手段に関する。細胞を培養するための好適な条件は当技術分野で周知であり、よく記載されている。好ましくは、培養は、細胞が増殖しまたは成長し、目的のポリヌクレオチドを発現することを可能にする。例えば、培養に用いる培地は最小限の成長速度を可能にする培地(最小培地)であってもまたはさらに高い成長速度を可能にする培地であってもよい。さらに、培地の組成は生物/宿主 細胞の亜集団を選択することを可能にしてもよい。好ましくは、選択によって、導入された選択可能なマーカー遺伝子を保持しかつ十分な量の選択可能なマーカー遺伝子を発現する宿主細胞だけが、増殖することができる。選択は、選択剤、例えば、ある特定の抗生物質を、培養に用いる培地に加えることにより、または物質、例えば、ある特定の栄養分を培地に加えないことにより達成することができる。本発明の方法によって用いることができる選択剤は、酵素ネオマイシンホスホトランスフェラーゼにより不活性化されるアミノグリコシド抗生物質、好ましくはジェネティシン(G418)である。従って、上述の方法の関係で使用することができる選択可能なマーカー遺伝子は、ネオマイシンホスホトランスフェラーゼ、ハイグロマイシンホスホトランスフェラーゼ、ピューロマイシン-N-アセチル-トランスフェラーゼ、ジヒドロ葉酸レダクターゼ、グルタミンシンテターゼ、アスパラギン酸トランスカルバミラーゼ、メタロチオネイン、アデノシンデアミナーゼ、アデニル酸デアミナーゼ、UMPシンテターゼ、P-糖タンパク質(p-gp、MDR)、アスパラギンシンテターゼまたはオルニチンデカルボキシラーゼである。本明細書で以下に記載するように、マーカー遺伝子は、選択能力の低い選択可能なマーカータンパク質をコードする遺伝子であってもよい。先に記載の通り、宿主はCHO細胞であることが好ましい。好ましくは、CHO細胞を、L-グルタミン(例えば、PAA Laboratories GmbHより供給される)と10%ウシ胎児血清(FBS)を加えたHamのF-12培地において37℃にて、5%CO2雰囲気中で増殖する。サブコンフルエントな培養物(70〜80%)を1:4〜1:10に分割して、すなわち、0.25%トリプシン/EDTA(PAA Laboratories GmbH)を用いて1〜2x10,000細胞/cm2にて接種した。細胞の培養には、単離するステップおよび引き続いての個々の細胞、特に高発現する細胞を繁殖するステップなどのさらなるステップが含まれることは理解されよう。
【0069】
有利なことに、本発明の上述の方法によって、発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチドと目的のポリペプチドとの宿主細胞中への共導入は、目的のポリヌクレオチドの発現を増強することが見出されている。例えば、ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域(Romig,1994, Eur J Biochem. 221:411-419)およびさらなる湾曲起点モチーフを含むポリヌクレオチドを、レポーター遺伝子EGFP(Enhanced Green Fluorescent Protein;増強緑色蛍光タンパク質)およびNPT耐性遺伝子を含むpEGFP-N1ベクター(Clontech)と一緒に、CHO細胞(Chinese Hamster Ovary)中に共トランスフェクト(すなわち共導入)した。対照として、(a)ベクターpEGFP-N1単独、または(b)ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域のDNAと組合わせた、さらなる湾曲起点モチーフを伴わないベクターをCHO細胞中に導入した。G418(0.6mg/ml)による選択を含む培養ステップの後に、個々の細胞のレポーター遺伝子発現をFACS分析(Fluorescent Activated Cell Sorting)により数値化した。対照と比較して、高発現細胞の比率、従ってその発現は、レポーター遺伝子およびさらなる湾曲起点モチーフを含む前記ATリッチ領域を共導入することにより作製したCHO細胞において、有意に増加することが見出された。従って、目的のポリヌクレオチド(ここではレポーター遺伝子)および湾曲起点モチーフを含む、発現を増強できる前記ATリッチ領域をコードするDNAの共導入は、目的のポリヌクレオチドの高くかつ安定な発現を可能にする。従って、共トランスフェクトした目的のポリペプチドの発現をそれ自体で増強できるクロマチンエレメント中への、湾曲起点モチーフの挿入は、さらに前記クロマチンエレメントの発現増強効果を増強する。
【0070】
有意に増強しかつ比較的安定な発現の驚くべき効果は、例えば、配列番号3および配列番号4に示した核酸配列を有するポリヌクレオチドについて、レポーター遺伝子による共トランスフェクションと耐性細胞の選択の後に示された。従って、本発明の方法は単一の高生産クローンに対する時間と費用のかかるスクリーニングステップを軽減しうる。
【0071】
本発明はまた、少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)宿主細胞中に
(i)配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16のいずれか1つに示した核酸配列を含む、少なくとも1つのポリヌクレオチド、および
(ii)少なくとも1つの目的のポリヌクレオチド
を導入するステップ、ならびに
(b)前記宿主細胞を培養するステップ
を含んでなる前記方法に関する。
【0072】
上述の方法は、少なくとも1つの目的のポリヌクレオチドおよび少なくとも1つの改変されたネオマイシンホスホトランスフェラーゼ遺伝子をコードする本発明によるポリヌクレオチドの宿主細胞中への共導入を含んでなる。用語「宿主細胞中に導入する」、「宿主細胞」、「少なくとも1つの目的のポリヌクレオチド」は既に本明細書において先に記載されている。上述の方法の文脈において前記宿主細胞を培養するとは、宿主細胞を好適な培養条件下でインキュベートすることを意味する(前記参照)。上述の方法の場合、選択可能なマーカー遺伝子はネオマイシンホスホトランスフェラーゼであり、選択剤は、酵素ネオマイシンホスホトランスフェラーゼにより不活性化されるアミノグリコシド抗生物質であってもよい。好ましくは、選択剤はG418である。
【0073】
上述の方法によれば、宿主細胞における目的のポリヌクレオチドの発現は参照宿主細胞と比較して増強されるであろうことは理解されよう。好ましくは、上述の方法の文脈において、参照宿主細胞は、未改変のNPT II遺伝子と共導入された対応する目的のポリヌクレオチドを発現する宿主細胞である。
【0074】
さらに、上述の方法の文脈において、NPT IIをコードするポリヌクレオチドおよび目的のポリヌクレオチドは、宿主細胞中に導入される前に、同じ核酸分子上にまたは異なる核酸分子上に位置してもよいことは理解されよう。好ましくは、選択可能なマーカーをコードするポリヌクレオチドと目的のポリヌクレオチドは1分子上に存在する。
【0075】
有利なことに、本発明によれば、本明細書の随所に記載されるNPT II突然変異体ポリペプチドをコードするポリヌクレオチド(すなわち、配列番号11、配列番号12および配列番号13のいずれかの1つに示したアミノ酸配列を含むポリペプチドをコードするか、または配列番号14、配列番号15および配列番号16のいずれかの1つに示した核酸配列を含むものであるポリヌクレオチド)の、CHO細胞を形質転換するための選択可能なマーカー遺伝子としての使用は、共導入したレポーター遺伝子の発現を増強することが見出されている。上述の核酸配列を含むDNA配列をEGPFレポーター遺伝子と共にCHO細胞中に共導入した。トランスフェクション後、細胞をG418を含有する培地で増殖した。比較実験を行い、ここでは改変されたネオマイシンホスホトランスフェラーゼをコードする本発明によるポリヌクレオチドの代わりに、未改変のNPT II遺伝子(配列番号17に示した核酸配列)を選択可能なマーカー遺伝子として用いた。未改変のマーカーでトランスフェクトした宿主細胞の大部分と比較して、改変されたNPT IIでトランスフェクトした宿主細胞の大部分においてレポーター遺伝子発現が有意に増強されることが見出された。従って、本発明による上述の方法は、単一の高生産クローンに対する時間と費用のかかるスクリーニングステップを軽減しうる。
【0076】
本発明はまた、少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)宿主細胞中に
(i)少なくとも1つの、低下した選択能力を有する選択可能なマーカーをコードするポリヌクレオチド、および
(ii)少なくとも1つの、遺伝子発現を増強することができる核酸配列を含む、ポリヌクレオチド、および
(iii)少なくとも1つの目的のポリヌクレオチドを導入するステップ
を導入するステップ、ならびに
(b)前記宿主細胞を培養するステップ
を含んでなる前記方法に関する。
【0077】
用語「遺伝子発現を増強できる核酸配列」は、共トランスフェクトした目的のポリヌクレオチドの発現を増強できるエンハンサー、MAR、SARまたはUCOE配列などの共トランスフェクトした目的のポリヌクレオチドの発現を増強できるいずれかの核酸配列を含み、好ましくは、遺伝子発現を増強できる核酸配列を含むポリヌクレオチドは発現を増強できるクロマチンエレメントに対する核酸配列を含むポリヌクレオチドであり、より好ましくは、遺伝子発現を増強できる核酸配列を含むポリヌクレオチドは本明細書に先に記載した発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものである先に説明したポリヌクレオチドであり、最も好ましくは、遺伝子発現を増強できる核酸配列を含むポリヌクレオチドは配列番号3に、配列番号4に、配列番号5に、または配列番号6に示した核酸配列を含むものであるポリヌクレオチドである。上述の方法の文脈において、遺伝子発現を増強できる核酸配列を含む、ポリヌクレオチドはさらなる湾曲起点モチーフを含んでもまたは含まなくてもよいことは理解されよう。
【0078】
本明細書で使用する用語「低下した選択能力を有する選択可能なマーカー」は、その選択能力がそれぞれの未改変のマーカー(例えば、野生型)と比較して低下した、遺伝的に改変された選択可能なマーカーに関する。WO 2004/050884は、共トランスフェクトした目的のポリヌクレオチドの発現を増強する、低下した選択能力を有するマーカーを開示している。低下した選択能力のもとでは、マーカータンパク質の酵素活性が低下していることは理解されよう。酵素活性は、マーカー遺伝子タンパク質のアミノ酸配列を改変することにより、例えば、前記マーカータンパク質をコードするポリヌクレオチド配列を変異させることにより低下させることができる。酵素活性は、当技術分野で周知でありかつよく記載された酵素アッセイにより決定することができる。全ての酵素アッセイは、好適な基質の消費または産物の生産を経時的に測定する。基質および産物の濃度を測定する多数の色々な方法が存在し、多くの酵素をいくつかの異なる方法でアッセイすることができる。本明細書で意味する酵素アッセイは、次の手法により行うことができる:初期速度実験、進行曲線実験、過渡的動力学実験、緩和実験。基質の消費または産物の生産は、分光学的手段、蛍光測定的手段、熱量測定的手段または化学発光的手段、放射線測定的手段およびクロマトグラフィ的手段により決定することができる。アッセイは連続的に(生産と消費を連続的に測定する)または非連続的に(サンプルを間隔をおいて採取する)行うことができる。酵素活性を比較するために、第1の酵素(例えば、改変されたマーカータンパク質)の活性を第2の酵素(例えば、未改変のマーカータンパク質)の酵素活性と、同じ条件下で比較する。酵素は精製されていてもまたは細胞抽出物の成分であってもよい。好ましくは、上述の方法の文脈において低下した選択能力を有する選択可能なマーカーの酵素活性は、未改変の選択可能なマーカーの酵素活性と比較すると、1〜80%低下、より好ましくは1〜50%低下、最も好ましくは1〜20%低下している。選択能力の低下は、野生型マーカー遺伝子の核酸配列を改変して、対応するアミノ酸配列がそれぞれの野生型マーカーのアミノ酸配列と比較して少なくとも1つのアミノ酸変化があるようにすることにより達成することができる。本明細書で使用する用語「少なくとも1つのアミノ酸変化」は、好ましくは、1以上、例えば、少なくとも2、少なくとも3、少なくとも4、少なくとも5、少なくとも6、少なくとも7または少なくとも10、少なくとも20、少なくとも30、少なくとも40、少なくとも50、または少なくとも100のアミノ酸変化を意味する。本発明の文脈において、選択能力の低下はまた、他の発現制御配列と比較して選択可能なマーカー遺伝子の発現を低下させる発現制御配列、例えば、プロモーターを用いることにより引き起こしてもよい。本発明の上述の方法によると、選択可能なマーカー遺伝子は、ネオマイシンホスホトランスフェラーゼ、ハイグロマイシンホスホトランスフェラーゼ、ピューロマイシン-N-アセチル-トランスフェラーゼ、ジヒドロ葉酸レダクターゼ、グルタミンシンテターゼ、アスパラギン酸トランスカルバミラーゼ、メタロチオネイン、アデノシンデアミナーゼ、アデニル酸デアミナーゼ、UMPシンテターゼ、P-糖タンパク質(p-gp、MDR)、アスパラギンシンテターゼまたはオルニチンデカルボキシラーゼであってもよい。
【0079】
上述の方法のある好ましい実施形態において、選択可能なマーカーをコードするポリヌクレオチドは、改変されたネオマイシンホスホトランスフェラーゼをコードするポリヌクレオチドである。上述の方法で用いることができる改変されたネオマイシンホスホトランスフェラーゼ遺伝子は、例えば、WO 2004/050884に記載されていて、これは本明細書に参照によりその全てが組み入れられる。
【0080】
上述の方法の他のさらに好ましい実施形態において、低下した選択能力を有する選択可能なマーカーをコードするポリヌクレオチドは、配列番号11、配列番号12および配列番号13のいずれかの1つに示したアミノ酸配列を含むポリペプチドをコードするか、または配列番号14、配列番号15および配列番号16のいずれかの1つに示した核酸配列を含むものであるポリヌクレオチドである。最も好ましくは、前記ポリヌクレオチドは配列番号13に示したアミノ酸配列を含むポリペプチドをコードするポリヌクレオチド、または配列番号16に示した核酸配列を含むものであるポリヌクレオチドである。また、本発明の方法のための低下した選択能力を有する選択可能なマーカーをコードするポリヌクレオチドと考えられるのは、野生型NPT II(配列番号35を参照)と比較して、野生型NPT IIポリペプチドのアミノ酸182位にてEがDへ、190位にてDがGへ、208位にてDがGへ、227位にてDがGへ、182位にてEがGへ、91位にてWがAへ、198位にてVがGへ、227位にてDがAへ、227位にてDがVへ、261位にてDがGへ、261位にてDがNへ、または240位にてFがIへのアミノ酸置換を有する、改変されたNPT IIポリヌクレオチドをコードするポリヌクレオチドである(これらは全て、Sauter, BIOTECHNOLOGY AND BIOENGINEERING, VOL. 89, NO. 5, 2005により開示されている)。さらに好ましいのは、野生型NPT II(配列番号35を参照)と比較して、位置にてVがMへ、182位にてEがDへ、205位にてGがEへ、210位にてGがAへ、211位においてRがPへ、211位においてRがKへ、211位においてRがHへのアミノ酸置換を有する、改変されたNPT IIポリペプチドをコードするポリヌクレオチドである。
【0081】
有利なことに、本発明によって、改変されたNPT II選択可能なマーカー遺伝子をコードするポリヌクレオチド、および遺伝子発現を増強できる核酸配列を含むポリヌクレオチド、および目的のポリヌクレオチドの宿主細胞中への共導入は、目的のポリヌクレオチドの発現を有意に増強することが見出されている。例えば、配列番号13に示したアミノ酸配列を含む改変されたNPT IIマーカーをコードするポリヌクレオチド、ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域(Romig、1994、Eur J Biochem. 221:411-419)およびさらなる湾曲起点モチーフを含むポリヌクレオチド(例えば、配列番号3に示した核酸配列)およびレポーター遺伝子EGFPをCHO(チャイニーズハムスター卵巣)細胞中に共導入した。対照として、a)EGFP遺伝子および未改変のNPT II遺伝子を保持するベクターpEGFP-N1、またはb)湾曲起点モチーフを含むヒトトポイソメラーゼIのイントロン13のMII領域の対応するATリッチ領域に対するDNA(配列番号3)と組合わせたpEGFP-N1ベクターを、CHO細胞中に導入した。さらに、SautterおよびEnekel(変異体D227V、Sautter および Enekel, Biotechnology and Bioengineering, 89, (2005) 530-538)により記載されている改変されたNPT II遺伝子を、さらなる湾曲起点モチーフを伴うヒトトポイソメラーゼIのイントロン13のMII領域の対応するATリッチ領域に対するDNA(配列番号3)と組合わせて、レポーター遺伝子とCHO細胞に共導入した。トランスフェクション後、耐性細胞を、G418を含有する培地における培養中に選択した。個々のG418耐性宿主細胞のレポーター遺伝子発現をFACS分析(Fluorescent Activated Cell Sorting)により数値化した。対照と比較して、高い発現細胞の比率、従って発現が、EGFP遺伝子、前記ATリッチ領域および湾曲起点モチーフを含むポリヌクレオチド、ならびに改変されたNPT II選択可能なマーカー遺伝子をコードするポリヌクレオチドを共導入することにより作製したCHO細胞において、有意に増加したことを見出した。さらに、改変されたNPT II D227V突然変異体の使用は、目的のポリヌクレオチドの発現を未改変のNPT遺伝子の使用と比較して増強する、しかしながら、改変されたNPT II D227V遺伝子の使用は、例えば配列番号13に示したアミノ酸配列を含む改変されたNPT IIマーカーより有意に低い遺伝子発現レベルをもたらした。従って、改変されたNPT IIをコードするポリヌクレオチドのような低下した選択能力を有する選択可能なマーカーをコードする少なくとも1つのポリヌクレオチド、発現を増強できるクロマチンエレメントに対する第1の核酸配列と少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含む先に規定したポリヌクレオチドのような遺伝子発現を増強できる核酸配列を含む少なくとも1つのポリヌクレオチド、および目的のポリヌクレオチドを宿主細胞中へ共導入すること、ならびに、前記細胞を好適な選択剤を用いて培養することは、目的のポリヌクレオチドの発現を有意に増強する。さらに、この効果はCHO細胞に対してだけでなく、爬虫類細胞(毒ヘビ細胞株VH2)に対しても示された。本発明のポリヌクレオチドの宿主細胞中への組合わせた導入の発現に与える増強効果は、個々の成分単独での効果、または個々の効果の合計よりもより大きく、従って、前記組合わせた導入は予想外の相乗効果を示すものである。例えば、配列番号13に示したアミノ酸配列を有する改変されたNPT IIマーカーを用いると、G418での選択後に、大部分が高生産クローンである耐性クローンをもたらすが、低生産クローンの大部分は選択過程で失われた。
【0082】
従って、上述の方法は、高生産細胞を単離するための効果的かつ迅速な手順を可能にし、費用と時間のかかる遺伝子増幅およびスクリーニングステップの必要性を軽減する。
【0083】
さらに、前記のように作製した細胞は、目的のポリヌクレオチドの驚くべき安定な発現を示した。単一クローンを、前記の通り[改変されたNPT IIマーカー(例えば、配列番号13に示したアミノ酸配列を有する)をコードするポリヌクレオチド、ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域および湾曲起点モチーフを含む、例えば、配列番号3に示した核酸配列を有するポリヌクレオチドを共トランスフェクトし、選択圧力下で培養することによって]作製した細胞から成る耐性コロニーから単離し、そして8週間選択圧力なしに連続培養した。驚くべきことに、選択圧力なしの8週間培養後のFACS分析は、試験したクローンの50%が変わらないレポーター遺伝子発現を示す一方、クローンの他の50%は、8週間培養の開始時のレポーター発現と比較してわずかに低下したレポーター遺伝子発現を示した(開始時の発現の50%〜90%)。従って、上述の方法は、有利なことに、選択可能なマーカー剤の不在のもとですら強くかつ安定な発現をするクローンの時間効率的な生産を可能にする。
【0084】
本発明の方法のある好ましい実施形態においては、宿主細胞を無血清培地または血清含有培地にて培養することができる。より好ましくは、宿主細胞を無血清培地で培養する。宿主細胞を無血清培地で培養することは、血清含有培地での培養から生じる感染のリスクを軽減できるので有利である。さらに、無血清培地の使用は、特に大規模な手法にとって、さらに費用効果が高い。用語「血清」の意味は当業者によく理解されている。本明細書で使用する用語「無血清培地」は、いずれの型の血清も、例えば、ウシ胎児血清または血清から分離されるタンパク質などのいずれの血清成分も含まない、任意の宿主細胞を培養するための増殖培地を意味する。対照的に、本明細書で意味する血清含有培地は、血清または血清から分離されるタンパク質などの血清成分を含有する培地に関する。
【0085】
本発明の方法の他の実施形態において、目的のポリヌクレオチドは、ヒト化抗体などの免疫グロブリン、治療タンパク質、膜タンパク質および酵素、ホルモン、イオンチャネルタンパク質およびプロテアーゼからなる群より選択されるポリペプチドをコードするものである。
【0086】
さらに、本発明は、目的のポリペプチドを製造する方法であって、上述の方法のいずれかのステップ、および目的のポリヌクレオチドがコードするポリペプチドを得るさらなるステップを含んでなる前記方法に関する。
【0087】
本発明の方法の他のさらに好ましい実施形態において、宿主細胞は動物細胞、植物細胞、酵母細胞、真菌細胞および藻類細胞である。より好ましくは、前記細胞は哺乳動物細胞または脊椎動物細胞である。最も好ましくは、先に記載した通り、宿主細胞はCHO細胞である。本発明で想定されるのはまた、爬虫類細胞、好ましくは毒ヘビ細胞、好ましくは細胞株VH2(Viper Russells心臓、ECACC No. 90102539)である。
【0088】
さらに、本発明は、発現を増強できるクロマチンエレメントに対する第1の核酸配列と少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列とを含むポリヌクレオチドおよび/または配列番号11、配列番号12および配列番号13のいずれかの1つに示したアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16のいずれかの1つに示した核酸配列を含む、ポリヌクレオチドを含むものである医薬組成物に関する。さらに、医薬組成物は治療効果を有する産物をコードする目的のポリヌクレオチドを含んでもよい。
【0089】
本明細書で使用する用語「医薬組成物」は、本発明の化合物および任意に1種以上の製薬上許容される担体を含む。本発明の化合物は製薬上許容される塩として製剤することができる。許容される塩には、酢酸塩、塩酸塩、硫酸塩、塩化物などが含まれる。医薬組成物は、好ましくは、局所にまたは全身に投与される。薬物投与用に好都合な、好適な投与の経路は、経口、静脈内または非経口投与、ならびに吸入である。しかし、化合物の性質と作用様式に応じて、医薬組成物を他の経路により投与することもできる。例えば、ポリヌクレオチド化合物は、遺伝子治療手法で、ウイルスベクターまたはウイルスまたはリポソームを用いることにより投与することができる。
【0090】
化合物は、好ましくは、薬物を標準医薬品担体と組合わせることにより調製した慣用の投与剤形で、慣用の手順によって投与される。これらの手順は、所望の調製に適当なように、成分を混合、造粒および圧縮または溶解することに関わりうる。製薬上許容される担体または希釈剤の形態と特性は、組み合わせるべき活性成分の量、投与の経路および他の周知の変数により規定されることは理解されるであろう。
【0091】
担体は、製剤の他成分と共存しうるおよびそのレシピエントに有害でないという意味で、許容されなければならない。使用される医薬品担体は、例えば、固体、ゲルまたは液体でありうる。固体担体の例は、ラクトース、白土、スクロース、タルク、ゼラチン、寒天、ペクチン、アカシア、ステアリン酸マグネシウム、ステアリン酸などである。液体担体の例は、リン酸緩衝生理食塩水溶液、シロップ、ピーナッツ油およびオリーブ油などの油、水、乳濁液、様々なタイプの湿潤剤、無菌溶液などである。同様に、担体または希釈剤は、モノステアリン酸グリセリルまたはジステアリン酸グリセリル単独でまたはワックス併用でなどの当技術分野で周知の時間遅延材料を含んでもよい。前記好適な担体には、前記および他の当技術分野で周知の担体が含まれ、例えば、Remington's Pharmaceutical Sciences, Mack Publishing Company, Easton, Pennsylvaniaを参照されたい。
【0092】
希釈剤は、組合わせの生物学的物性に影響を与えないように選択される。かかる希釈剤の例は、蒸留水、生理食塩水、リンゲル液、ブドウ糖液、およびハンク(Hank)液である。さらに、医薬組成物または製剤はまた、他の担体、アジュバント、または無毒性、無治療性、無免疫原性の安定剤などを含んでもよい。
【0093】
治療上有効な用量は、本明細書で言及した疾患に随伴する症候群または症状を予防、改善または治療する本発明の医薬組成物で使われる化合物の量を意味する。かかる化合物の治療効力および毒性は、細胞培養または実験動物における標準医薬品手順、例えば、ED50(集団の50%にとって治療上有効な用量)およびLD50(集団の50%が死に至る用量)により決定することができる。治療と毒性効果の間の用量比が治療指数であり、比LD50/ED50で表される。
【0094】
投薬レジメンは担当医によりおよび他の臨床的因子;好ましくは以上記載した方法のいずれか1つによって決定されるであろう。医療業界では周知のように、いずれかの一患者に対する投薬は、患者サイズ、身体表面積、年齢、投与する特定の化合物、性別、時間および投与の経路、一般的な健康状態、および同時に投与される他の薬物を含む多くの因子に依存する。進行状態は定期的な評価を行うことによりモニターすることができる。
【0095】
本明細書で意味する医薬組成物および製剤は少なくとも1回、本明細書に挙げた疾患または症状を治療または改善または予防するために投与される。しかし、前記医薬組成物を1回以上、例えば毎日1〜4回、日数に限定されることなく、投与することができる。
【0096】
さらに、本発明は、遺伝子治療に用いる医薬組成物を製造するためのポリヌクレオチドの使用であって、該ポリヌクレオチドが、発現を増強できるクロマチンエレメントに対する第1の核酸配列と少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列とを含むポリヌクレオチドならびに/または配列番号11、配列番号12および配列番号13のいずれかの1つに示したアミノ酸配列を含むポリペプチドをコードするかもしくは配列番号14、配列番号15および配列番号16のいずれかの1つに示した核酸配列を含む、ポリヌクレオチドを含むものである前記使用に関する。
【0097】
本明細書で使用する用語「遺伝子治療」は、少なくとも1つの本発明のポリヌクレオチドをin vivoで、または宿主細胞中へのin vitro導入と次いで被験者(例えば、ヒトまたは非ヒト動物)中への再移植により被験者の宿主細胞中に導入する治療の方法として理解されよう。好ましくは、本発明のポリヌクレオチドを遺伝子治療に用いる宿主細胞中に、好ましくは治療効果を有する目的の産物をコードする目的のポリヌクレオチドと一緒に共導入する。本明細書で使用する用語「遺伝子治療」は、宿主細胞遺伝子の機能の欠如を補償するために目的のポリヌクレオチドを細胞中に導入することを意味してもよい。さらに、異常な遺伝子を、相同的組換えを介して機能性遺伝子により置換えることができる。さらに、調節、すなわち、特定の遺伝子の発現を、例えば、調節RNAにより、例えばRNAi手法で改変することができる。好ましくは、導入は、当技術分野で周知の技法により行われる。より好ましくは、導入は、ウイルスベクターにより行われる。この目的のためには、ウイルスベクターを、ウイルス疾患を生じる遺伝子を取り除いて上述した本発明のポリヌクレオチドを挿入することにより、遺伝的に改変することができる。ウイルスベクターには、限定されるものでないが、レトロウイルス、アデノウイルス、アデノ随伴ウイルスが含まれる。上述したポリヌクレオチドは、裸のDNAによるかまたはリポソームの使用によるトランスフェクションなどの非ウイルス的な方法により送達することもできる。遺伝子療法により治療される疾患は、限定されるものでないが、癌、神経疾患、代謝性疾患、免疫不全疾患でありうる。非ウイルス遺伝子療法に対する出現中のベクターおよび標的化方法は、Lavigne および Gorecki(Expert Opinion of Emerging Drugs (2006) 11、541-557)に記載されている。
【0098】
本明細書に引用される全ての参照は、それらの全開示内容および本明細書に具体的に記載した開示内容に関する参照により、本明細書に組み入れられる。
【図面の簡単な説明】
【0099】
【図1】配列番号3に示した、挿入された湾曲起点モチーフを含むヒトトポイソメラーゼのイントロン13のATリッチ領域の核酸配列である。湾曲起点モチーフに下線が引かれている。
【図2】配列番号4に示した、挿入された湾曲起点モチーフを含むヒトトポイソメラーゼのイントロン13のATリッチ領域の核酸配列である。湾曲起点モチーフに下線が引かれている。
【図3】次のトランスフェクション後のCHO細胞におけるレポーター遺伝子EGFPの発現である:環状pEGFP-N1 100ng単独(「o」)によるトランスフェクション、または環状pEGFP-N1ベクター(Clontech)100ngと配列番号27に示した核酸配列を含むヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域を含むものである直鎖DNA 100ngとの共トランスフェクション(「X」)、または環状pEGFP-N1ベクター(Clontech)100ngと配列番号28に示した核酸配列を含むヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域をコードする直鎖DNA 100ngとの共トランスフェクション(黒丸)、または環状pEGFP-N1ベクター(Clontech)100 ngと配列番号3に示した湾曲起点モチーフを含むヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域を含む直鎖DNA 100ngとの共トランスフェクション(黒四角)。ベクターpEGPF-N1は、EGPFレポーター遺伝子および野生型NPT II遺伝子を含む。およそ8週間の選択圧力(0.6 mg/ml G418)下の培養後、個々の細胞のレポーター遺伝子発現をFACS分析(蛍光検出の感度:4 decades、gain 300)により数値化した。レポーター遺伝子発現が比較的強い細胞の比率は、細胞を湾曲起点モチーフを含む発現を増強できるクロマチンエレメントを用いて共トランスフェクトした場合に、pEGPF-N1単独を用いてまたはpEGPF-N1と湾曲起点モチーフなしの発現を増強できるクロマチンエレメントを用いてトランスフェクトした細胞と比較して、有意に増強する。
【図4】次のトランスフェクション後のCHO細胞におけるEGFPの発現である:直鎖pEGFP-N1単独のDNA 90ngを用いたトランスフェクション(「o」)、または直鎖pEGFP-N1 90ngとヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域およびさらなる湾曲起点モチーフ(配列番号3に示した核酸配列)DNA 300ngとの共トランスフェクション(黒四角)、またはEGFPレポーター遺伝子および配列番号16に示した核酸配列を含む改変されたネオマイシンホスホトランスフェラーゼを保持するベクター(pAG)90ngとヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域およびさらなる配列番号3に示した核酸配列をもつ湾曲起点モチーフをコードするDNA 300ngとの共トランスフェクション(黒丸)。およそ8週間の選択圧力(0.6 mg/ml G418)下の培養後、個々の細胞のレポーター遺伝子発現をFACS分析(計器設定:光学検出器のgain=300、増幅モード=4 decade logarithmic)により数値化した。強いレポーター遺伝子発現を示す細胞の比率は、レポーター遺伝子をCHO細胞中に、さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のATリッチ領域をコードするDNAと組合わせて導入した場合、pEGFP-N1を単独でトランスフェクトした場合より有意に高かった。さらに、強いレポーター遺伝子発現をもつ細胞の比率は、湾曲起点モチーフを含むヒトトポイソメラーゼIのイントロン13のATリッチ領域と、改変されたネオマイシンホスホトランスフェラーゼ遺伝子(未改変のネオマイシンホスホトランスフェラーゼの代わりに、pEGFP-N1プラスミド上に局在する)をコードするDNAとの両方をCHO細胞中に導入した場合に、最高であった。配列番号16に示した核酸配列を有する改変されたネオマイシンホスホトランスフェラーゼと湾曲起点モチーフを含むATリッチ領域(配列番号3に示した核酸配列)とを用いてトランスフェクトした細胞における安定な発現は予想外に高く、蛍光検出の範囲は、低発現細胞と高発現細胞を同時に検出するには不十分であった。低発現細胞と高発現細胞の間の蛍光強度の差は104のファクターより大きいので、細胞を低感度で再測定した(図5:計器設定:光学検出器のgain=200、増幅モード=4 decade logarithmic)。
【図5】図4の凡例に示したようにトランスフェクトしかつ選択したCHO細胞の集団である。蛍光検出の感度(4 decades)は200のgainで測定した。pAGおよびATリッチ領域(配列番号3に示した核酸配列)を用いて共トランスフェクトしたCHO細胞(「O」)の大部分は、pEGFP-N1およびATリッチ領域(配列番号3に示した核酸配列)を用いて共トランスフェクトしたCHO細胞(「X」)よりおよそ10倍高い発現を示した。従って、さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のATリッチ領域に対するDNAと、改変されたネオマイシンホスホトランスフェラーゼ遺伝子をコードするDNAとの両方をCHO細胞に導入した場合に、強いレポーター遺伝子発現をもつ細胞の比率が有意に増強された。
【図6】図3の凡例に示したようにレポーター遺伝子EGFPを様々なネオマイシンホスホトランスフェラーゼ耐性遺伝子(NPT II)と組合わせてトランスフェクトし、選択した後のCHO細胞におけるEGFPの発現である。次のネオマイシンホスホトランスフェラーゼ耐性遺伝子の共トランスフェクトしたレポーター遺伝子の発現に与える効果を分析した:それぞれ、配列番号17(X)に示した核酸配列を含む未改変のNPT II遺伝子、ならびに配列番号14(O)、配列番号15(黒丸)、配列番号16(黒四角)に示した核酸配列を含む改変されたNPT II遺伝子に対する結果である。個々の細胞のレポーター遺伝子発現は、十分な量のG418を含有する選択培地で8週間培養後にFACS分析により測定した。レポーター遺伝子の比較的強い発現を示す細胞の比率は、改変されたネオマイシンホスホトランスフェラーゼでトランスフェクトした細胞については、野生型ネオマイシンホスホトランスフェラーゼでトランスフェクトした細胞と比較して有意に高かった。レポーター遺伝子の比較的強い発現を示す細胞の比率は、配列番号16に示した核酸配列を有する改変されたネオマイシンホスホトランスフェラーゼでトランスフェクトした細胞が最高であった。
【図7】レポーター遺伝子EGFPを、改変されたネオマイシンホスホトランスフェラーゼ耐性遺伝子D227V(配列番号36)およびさらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域に対するDNA(配列番号3)と組合わせて共トランスフェクトした後のCHO細胞におけるEGFPの発現である(O)。対照として、未改変のネオマイシンホスホトランスフェラーゼ(pEGFP-N1)を、湾曲起点モチーフを伴うヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域に対するDNA(配列番号3)を用いて共トランスフェクトした(X)。実施例3に記載の通り、細胞を培養し、FuGENE6でトランスフェクトし、G418培地で選択した。0.6mg/ml G418を補充した培地における6週間の培養後、個々の細胞の遺伝子発現を、実施例3に記載の通りFACS分析により測定した。(計器設定:光学検出器のgain=200、増幅モード=4decadelogarithmic)(図7)。ATリッチ領域(配列番号3)と共トランスフェクトした記載の改変されたNPT IID227V突然変異体は、ATリッチ領域(配列番号3)と共トランスフェクトした未改変のNPT遺伝子と比較して、目的のポリヌクレオチドの発現を増強するが、その増加は、例えば、配列番号16に示した核酸配列を有する本発明の改変されたネオマイシンホスホトランスフェラーゼの共導入に対してより低かった。
【0100】
以下の実施例は単に本発明を説明するものである。これらは、どういう場合でも、本発明の範囲を限定すると解釈してはならない。
【実施例】
【0101】
(実施例1)
湾曲起点モチーフをもつMAR配列の作製
ヒト白血球由来およびHeLa細胞由来のゲノムDNAを、「血液および細胞培養DNAキット」(QIAGEN GmbH, Hilden, Germany)を用いて、製造業者取扱説明書に従い精製した。ヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域(Romig, Eur J Biochem. 221:411-419)を、2回の逐次PCR反応(ネストPCR)によって増幅し、ここで、テンプレートとしてHeLa細胞からまたはヒト血液から抽出したゲノムDNAのおよそ200ngを、ポリメラーゼとしてPfuポリメラーゼを使用し、そして次のプライマーを用いた:
プライマー1(フォワード):5'-CCAGCTAACTGCTATACACAATAGACATTTGTGC(配列番号61)
プライマー2(リバース):5'-CTGCCAACAGTAATTAGCACATGAATTCACTTCAGGTAAC(配列番号62)
第2のPCRについては、
プライマー3(フォワード):5'-GTGCCTGACTTGAACTGCAATGGG(配列番号63)
プライマー4(リバース):5'-CAGTAATTAGCACATGAATTCAC(配列番号64)。
【0102】
下線部は、後のクローニングステップで使用するEcoRI制限部位の配列である。
【0103】
第1のPCR反応は次の通り実施した:30サイクルの94℃における2分間変性、50℃における35秒間のアニーリングおよび72℃における2分間の伸長、その後に72℃における5分間の最後の伸長。
【0104】
第2のPCR反応は第1のPCRの一部分をテンプレートとして用いて、次の通り実施した:30サイクルの94℃における1分間変性、50℃における35秒間のアニーリングおよび72℃における2分間の伸長、その後に72℃における5分間の最後の伸長。
【0105】
以下のステップで、PCR産物をベクターpPCR-Script-Amp-SK(+)(Stratagene、LaJolla、USA)のSrfI部位(SrfIは平滑末端を作製する)中に、PCR-Script(登録商標)Ampクローニングキットを用いることにより製造業者の取扱説明書に従ってライゲートした。ライゲーションの後、ライゲーション産物を、コンピテント大腸菌細胞(XL-GoldKan、PCR-Script(登録商標)Ampクローニングキット)中に、製造業者取扱説明書に従って形質転換した。形質転換の後、細胞を、首尾よく形質転換した細胞を選択できるように、およそ20時間、37℃にて、100μg/mlアンピシリンを補充したLuria-ブロス-培地においてインキュベートした。次いで、得られるPCR断片を保持するベクターの2.9kb BamH1/EcoR1 血液ATリッチ核酸断片および2.9kb BamH1/EcoR1 HeLa ATリッチ核酸断片を、ベクターpUC18のBamH1/EcoR1制限部位中にクローニングした。クローニングしたPCR産物を、「血液ATリッチ核酸」(起源として、血液から抽出したゲノムDNAから増幅した;配列番号28に示した核酸配列)および「HeLaATリッチ核酸」(起源として、HeLa細胞から抽出したゲノムDNAから増幅した;配列番号27に示した核酸配列)と名付けた。両方のATリッチ配列は、高い配列同一性を示すがいくつかの位置で配列差を有する(613位、1079〜1082位、1128位、1160-1163位、1176位、1188位、1197位、1205位、1227位、1236位、1333位、1905位、1919位、1931位、1950位、1959位、1975位、2191位、2234位、2495位、2507位、2634位、2652位、2733位、2762位および2768位)。部位指定変異誘発によって、制限エンドヌクレアーゼAvrIIに対する制限部位を、HeLaおよび血液ATリッチ核酸配列の両方の中に、QuickChange(登録商標)部位特異的変異誘発キット(Stratagene)を用いて製造業者の取扱説明書に従い導入した。部位指定変異誘発に用いたプライマーの核酸配列は次の通りである:
プライマーavr_sense ggtcttgtatctgcctaggagatacaagaggtgctc(配列番号65)
プライマーavr_asense gagcacctcttgtatctcctaggcagatacaagacc(配列番号66)
湾曲起点モチーフを、GeneartAG(Regensburg)により、Nalezkovaaら(2005)Protein Expression and Purification 39、296-306に記載の方法を用いて合成した。
【0106】
湾曲起点モチーフを含むAvrII断片を、両方向に、新しく作製したHeLaおよび血液AT-リッチ配列のAvrII制限部位中にクローニングした。4種の得られる構築物の核酸配列を配列番号3、配列番号4、配列番号5および配列番号6に示す。
【0107】
(実施例2)
改変されたネオマイシンホスホトランスフェラーゼII(NPT II)をコードするポリヌクレオチドの作製
1つまたは2つのアミノ酸変化を有する改変されたNPT IIタンパク質をコードするNPT IIポリヌクレオチドを、部位特異的変異誘発により、未改変のNPT II(野生型)遺伝子をテンプレートとして保持するベクターpEGFP-N1を用いて作製した。次のプライマー対を部位特異的変異誘発に用いて、QuickChange(登録商標)部位特異的変異誘発キット(Stratagene)を製造業者の取扱説明書に従って使用した。
【0108】
NPT II R226A突然変異体(配列番号14に示した核酸配列を得る)について
R226A_sense:catagcgttggctaccgctgatatcgctgaagagcttggc(配列番号67)
R226A_asense:gccaagctcttcagcgatatcagcggtagccaacgctatg(配列番号68)
R226V突然変異体(配列番号15に示した核酸配列を得る)について
R226V_sense:catagcgttggctaccgttgatatcgctgaagagcttggc(配列番号69)
R226V_asense:gccaagctcttcagcgatatcaacggtagccaacgctatg(配列番号70)
NPT II A224G;T225S二重突然変異体(配列番号16に示した核酸配列を得る)について
A224G_sense:ctatcaggacatagcgttgggatcccgtgatattgctgaagagc(配列番号71)
A224G_asense:gctcttcagcaatatcacgggatcccaacgctatgtcctgatag(配列番号72)
FortheNPT IID227V突然変異体(配列番号36に示した核酸配列を得る)
D227V_sense:ggacatagcgttggctacacgcgttattgctgaagagcttgg(配列番号73)
D227V_asense:ccaagctcttcagcaataacgcgtgtagccaacgctatgtcc(配列番号74)
得られるベクターはEGFP遺伝子と改変されたネオマイシンホスホトランスフェラーゼII遺伝子の両方を含む。
【0109】
(実施例3)
さらなる湾曲起点モチーフを伴うヒトトポイソメラーゼIのイントロン13のATリッチ領域とレポーター遺伝子とのCHO細胞中への共導入は、高いレポーター遺伝子活性を示す細胞の比率を有意に増強する
ヒトトポイソメラーゼIのイントロン13のATリッチ領域と湾曲起点モチーフを含むポリヌクレオチドの、共トランスフェクトしたレポーター遺伝子の発現に与える効果を分析した。この理由で、レポーター遺伝子5EGFPおよび未改変ネオマイシンホスホトランスフェラーゼ遺伝子を保持するpEGFP-N1ベクター100ngをCHO細胞中に次のMAR構築物(各100ng)と一緒にトランスフェクトした。
【0110】
配列番号28に示したATリッチ核酸(血液ATリッチ配列)
配列番号27に示したATリッチ核酸(HeLaATリッチ配列)
配列番号3に示したATリッチ核酸(挿入された湾曲起点モチーフを伴うATリッチ配列)
対照として、pEGFP-N1ベクターを単独でトランスフェクトした。
【0111】
CHO-K1細胞株を、10%FBS(PAALaboratoriesGmbH)を補充したL-グルタミン(PAALaboratoriesGmbH)とHamF-12培地で培養した。EGFPを発現する安定なCHO細胞のプールを、FuGENE6(Roche)でのトランスフェクションにより、製造業者取扱説明書に従いかつ1:4のDNA:FuGENE比(μgDNA:μlFuGENE6)で得た。細胞を24ウエルプレートで100〜200000細胞/ウエルにて接種し、一晩付着させた。100ngのpEGFP-N1を100ngの直鎖ATリッチ配列と共トランスフェクトした。直鎖ATリッチ配列(配列番号3、配列番号27、配列番号28)は制限酵素消化およびゲル溶出によるゲル電気泳動後に単離した。48時間後、培地を0.6mg/ml G418(G418硫酸塩、Calbiochem)を補充した培地と変更した。引き続いて、培地を0.6mg/ml G418(G418硫酸塩、Calbiochem)を補充した培地と変更し、クローンのプールを8週間の選択後に試験した。レポーター遺伝子(EGFP)発現、すなわち、個々の細胞の蛍光を、FACS分析によりPARTECフローサイトメーター(PartecAnalysingSystemPAS、PartecGmbH、Muenster)を用いて数値化した。蛍光イベントは、緑色蛍光タンパク質シグナルに対する530/15帯域通過フィルターを用いて得た。(計器設定:光学検出器のgain=300、増幅モード=4decadelogarithmic)。
【0112】
レポーター遺伝子シグナルの強度は発現を示し、すなわち、強い蛍光シグナルは高いレポーター遺伝子発現を示す。特定の蛍光を有する個々の細胞の数(イベントとも呼ぶ)を蛍光のレベルに対してプロットした。様々なトランスフェクション実験の結果を図3に示す。GPF遺伝子を保持するベクターをCHO細胞中に単独でトランスフェクトした場合、大多数の細胞は比較的低いレポーター遺伝子発現を示した。そのベクターに加えてトポイソメラーゼ遺伝子のATリッチ領域(配列番号28に示した「血液ATリッチ」構築物、または配列番号27に示した「HeLaATリッチ」構築物)をコードするDNAを細胞中にトランスフェクトした場合、比較的高いレポーター遺伝子発現を示す細胞の比率はベクター単独のトランスフェクションと比較して有意に増強された。予想外なこととして、ATリッチ配列を共トランスフェクトしたほとんど全ての細胞がEGFP発現を示した。おそらく、遺伝子サイレンシングを示したのは非常に少量の細胞だけであった。対照的に、EGFP-N1ベクター単独でトランスフェクトしたG418耐性細胞プールのほぼ全ての細胞がEGFP発現を示さなかった。従って、遺伝子サイレンシングを示さなかったのは、EGFP-N1ベクター単独でトランスフェクトした細胞のうちの非常に少量だけであり、この細胞集団はFACS分析で101〜102の蛍光強度で検出された。pEGFP-N1ベクターに加えて、さらなる核酸配列番号3に示した湾曲起点モチーフをもつヒトトポイソメラーゼのATリッチ領域をCHO細胞中にトランスフェクトした場合、レポーター遺伝子発現は最高であった。発現は、配列番号28で共トランスフェクトした細胞と比較して、およそ10〜20のファクターで増強された(共トランスフェクトした配列番号3)。対応する結果はまた、湾曲起点が反対方向にあるATリッチ構築物についても観察された(配列番号4に示した核酸配列)。従って、クロマチンエレメント(単独で発現を増強できる)とさらなる湾曲起点モチーフとのトランスフェクションは、レポーター遺伝子およびさらなる湾曲起点モチーフを含まないクロマチンエレメントの共トランスフェクションと比較して、共トランスフェクトした遺伝子の強い発現を示す細胞集団の細胞数を有意に増加する。
【0113】
(実施例4)
改変されたネオマイシンホスホトランスフェラーゼをトランスフェクトされたCHO細胞を選択するために使用すると、レポーター遺伝子の発現を増強する
1つまたは2つのアミノ酸変化を有する変異ネオマイシンホスホトランスフェラーゼ遺伝子を、実施例2に記載の通り調製した。SV40初期プロモーターの調節下にある改変されたネオマイシン耐性遺伝子ならびにヒトサイトメガロウイルス(CMV)最初期プロモーターの調節下にあるレポーター遺伝子EGFPを保持するベクターをCHO細胞中にトランスフェクトした。対照として、SV40初期プロモーターの調節下にある未改変のネオマイシンホスホトランスフェラーゼ遺伝子ならびにヒトサイトメガロウイルス(CMV)最初期プロモーターの調節下にあるレポーター遺伝子EGFPを保持するベクターpEGPP-N1を、CHO細胞中にトランスフェクトした。細胞を培養し、FuGENE6でトランスフェクトし、実施例3に記載の通りG418を用いて選択した。培養後、形質転換された細胞のレポーター遺伝子発現をFACS分析により分析した。レポーター遺伝子(EGFP)発現をFACS分析により、PARTECフローサイトメーター(PartecAnalysingSystemPAS、PartecGmbH、Muenster)を用いて数値化した。蛍光イベントは、緑色蛍光タンパク質シグナルに対する530/15帯域通過フィルターを用いて得た。(計器設定:光学検出器のgain=200、増幅モード=4 decade logarithmic)。FACS分析の結果を図6のグラフに示す。グラフは細胞による蛍光発光レベル(Y軸)に対してプロットした細胞数(X軸)を示す。明確に見られるように、改変されたネオマイシンホスホトランスフェラーゼ遺伝子を選択可能なマーカーとして用いた場合に、非常に強いレポーター遺伝子活性を示す細胞の比率は有意に高い。選択可能なマーカーが未改変の耐性遺伝子である場合、レポーター遺伝子発現は低い。特に、配列番号16に示した核酸配列を有する改変されたネオマイシンホスホトランスフェラーゼを使用すると、高比率の高生産細胞を生じた。
【0114】
総括すると、改変された選択可能なマーカーの使用は、形質転換された細胞集団中の比較的強いレポーター遺伝子の発現を示す細胞の比率を増強する。
【0115】
(実施例5)
さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のATリッチ配列と改変されたネオマイシントランスフェラーゼをコードする遺伝子とのCHO細胞中への共導入は、高いレポーター遺伝子活性を示す細胞の比率を有意に増強する。
【0116】
さらなる湾曲起点モチーフを保持する、発現を増強できるクロマチンエレメント(ここでは、ヒトトポイソメラーゼIのイントロン13のATリッチ配列)(配列番号3に示した核酸配列)と、改変されたネオマイシンホスホトランスフェラーゼ(配列番号16に示した核酸配列)およびレポーター遺伝子EGPF(目的のポリヌクレオチド)を含むベクターとの共導入がレポーター遺伝子発現に与える効果を分析した。対照として、レポーター遺伝子および未改変のネオマイシンホスホトランスフェラーゼ遺伝子(配列番号17に示した核酸配列)を含むベクターをCHO細胞中にトランスフェクトするか、またはレポーター遺伝子EGFPおよび未改変のネオマイシンホスホトランスフェラーゼ遺伝子(配列番号17に示した核酸配列)と、さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のATリッチ配列をコードするDNA(配列番号3に示した核酸配列)を含むベクターとをCHO細胞中に共トランスフェクトした。これらの実験のために、細胞を培養し、FuGENE6(90ng直鎖プラスミドおよび/または300ngATリッチ配列)を用いてトランスフェクトし、実施例3に記載の通りG418培地を用いて選択した。8週間培養後、個々の細胞のレポーター遺伝子発現を、実施例3に記載の通り、FACS分析により測定した。(計器設定:光学検出器のgain=200または300、増幅モード=4decadelogarithmic)。図4(光学検出器のgain=300)および図5(光学検出器のgain=200)は発現分析の結果を示す。これらのグラフは、細胞数(X軸)および細胞による蛍光発光のレベル(Y軸)を示す。明確に見られるように、細胞が湾曲起点モチーフを含む発現を増強できるクロマチンエレメント(核酸配列配列番号3)と、改変されたネオマイシンホスホトランスフェラーゼ(配列番号13に示した核酸配列)を含むベクターとで共トランスフェクトした場合、非常に強いレポーター遺伝子活性を示す細胞の比率は、有意に高かった。従って、改変されたネオマイシンホスホトランスフェラーゼおよびクロマチンエレメントとさらなる湾曲起点モチーフとの組合わせのCHO細胞中への導入は、共導入された目的の産物(ここではレポーター遺伝子)をコードするポリヌクレオチドの発現を増強する。
【0117】
さらに、SautterおよびEnekel(突然変異体D227V、SautterおよびEnekel, Biotechnology and Bioengineering, 89, (2005) 530-538)に記載の改変されたNPT II遺伝子を含むpEGFP-N1ベクターを、さらなる湾曲起点モチーフをもつヒトトポイソメラーゼIのイントロン13のMII領域のATリッチ領域に対するDNA(配列番号3)とCHO細胞に共トランスフェクトした。細胞を培養し、FuGENE6でトランスフェクトし、実施例3に記載の通りG418培地を用いて選択した。0.6mg/ml G418遺伝子を補充した培地で6週間の培養後、個々の細胞の発現を、FACS分析により実施例3に記載の通り測定した(計器設定:光学検出器のgain=200、増幅モード=4decadelogarithmic)(図7)。
【0118】
前記ATリッチ領域(配列番号3)と共トランスフェクトした、記載の改変されたNPT IID227V突然変異体は、目的のポリヌクレオチドの発現を、前記ATリッチ領域(配列番号3)で共トランスフェクトした未改変のNPT遺伝子と比較して増強するが、その増加は、本発明の改変された、例えば配列番号16に示した核酸配列を有するネオマイシンホスホトランスフェラーゼを共導入した場合より低かった。
【特許請求の範囲】
【請求項1】
発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチド。
【請求項2】
クロマチンエレメントがMARエレメントである、請求項1に記載のポリヌクレオチド。
【請求項3】
湾曲起点モチーフが配列番号1、2、31もしくは33、または配列番号29の核酸配列の少なくとも2つの反復配列、または配列番号30の核酸配列の少なくとも2つの反復配列、または配列番号32の核酸配列の少なくとも2つの反復配列または配列番号34の核酸配列の少なくとも2つの反復配列に示される核酸を含む、請求項1に記載のポリヌクレオチド。
【請求項4】
湾曲起点モチーフが配列番号7、または配列番号8、または配列番号9、または配列番号10の配列を含む、請求項1に記載のポリヌクレオチド。
【請求項5】
ポリヌクレオチドが配列番号3、または配列番号4、または配列番号5、または配列番号6、または配列番号3、4、5または6のうちのいずれか1つと少なくとも70%同一である変異体の配列を含みかつ少なくとも1つの湾曲起点モチーフを含むものである、請求項1に記載のポリヌクレオチド。
【請求項6】
配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16のいずれか1つに示した核酸配列を含むものであるポリヌクレオチド。
【請求項7】
請求項6に記載のポリヌクレオチドがコードするポリペプチド。
【請求項8】
請求項1〜6のいずれか1項に記載のポリヌクレオチドまたはその組合わせを含む宿主細胞。
【請求項9】
請求項1〜6のいずれか1項に記載のポリヌクレオチドまたはその組合わせを含むベクター。
【請求項10】
請求項1〜6のいずれか1項に記載のポリヌクレオチドまたはその組合わせを含む非ヒトトランスジェニック生物。
【請求項11】
請求項1〜6のいずれか1項に記載のポリヌクレオチドまたはその組合わせを含むキット。
【請求項12】
少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)動物宿主細胞中に、
(i)請求項1〜5に記載のいずれか1つのポリヌクレオチドまたはその組合わせ、および
(ii)目的のポリペプチドをコードする少なくとも1つのポリヌクレオチド
を導入するステップ、ならびに
(b)前記細胞を培養するステップ
を含んでなる前記方法。
【請求項13】
少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)動物宿主細胞中に、
(i)請求項6に記載のポリヌクレオチド、および
(ii)目的のポリペプチドをコードする少なくとも1つのポリヌクレオチド
を導入するステップ、ならびに
(b)前記細胞を培養するステップ
を含んでなる前記方法。
【請求項14】
少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)動物宿主細胞中に、
(i)低下した選択能力を有する選択可能なマーカー遺伝子をコードするポリヌクレオチド、および
(ii)遺伝子発現を増強できる核酸配列を含むポリヌクレオチド、および
(iii)目的のポリペプチドをコードする少なくとも1つのポリヌクレオチド
を導入するステップ、ならびに
(b)前記細胞を培養するステップ
を含んでなる前記方法。
【請求項15】
低下した選択能力を有する選択可能なマーカー遺伝子をコードするポリヌクレオチドが改変されたネオマイシンホスホトランスフェラーゼをコードするポリヌクレオチドである、請求項14に記載の方法。
【請求項16】
改変されたネオマイシンホスホトランスフェラーゼをコードするポリヌクレオチドが請求項6に記載のポリヌクレオチドである、請求項14〜15のいずれか1項に記載の方法。
【請求項17】
遺伝子発現を増強できる核酸配列を含むポリヌクレオチドが請求項1〜5のいずれか1項に記載のポリヌクレオチドである、請求項14〜16のいずれか1項に記載の方法。
【請求項18】
培養を好適な条件下で行いかつ宿主細胞を選択剤を用いて選択することができる、請求項12〜17のいずれか1項に記載の方法。
【請求項19】
宿主細胞を無血清または血清含有培地で培養する、請求項12〜18のいずれか1項に記載の方法。
【請求項20】
ポリペプチドが免疫グロブリン、治療タンパク質、膜タンパク質および酵素からなる群より選択される、請求項12〜19のいずれか1項に記載の方法。
【請求項21】
目的のポリペプチドを製造する方法であって、請求項12〜20のいずれか1項に記載のステップおよびポリヌクレオチドがコードする目的のポリペプチドを得るステップを含んでなる前記方法。
【請求項22】
前記宿主細胞が爬虫類細胞、好ましくは毒ヘビ細胞、または哺乳動物細胞、好ましくはCHO細胞である、請求項8に記載の宿主細胞または請求項12〜21のいずれか1項に記載の方法。
【請求項23】
請求項1〜5のいずれか1項に記載のポリヌクレオチドおよび/または請求項6に記載のポリヌクレオチドを含む医薬組成物。
【請求項24】
遺伝子治療に用いる医薬組成物を製造するための、請求項1〜5のいずれか1項に記載のポリヌクレオチドおよび/または請求項6に記載のポリヌクレオチドの使用。
【請求項25】
配列番号7、または配列番号8、または配列番号9、または配列番号10の配列を含む湾曲起点モチーフ。
【請求項1】
発現を増強できるクロマチンエレメントに対する第1の核酸配列および少なくとも1つの、湾曲起点モチーフを含む第2の核酸配列を含むものであるポリヌクレオチド。
【請求項2】
クロマチンエレメントがMARエレメントである、請求項1に記載のポリヌクレオチド。
【請求項3】
湾曲起点モチーフが配列番号1、2、31もしくは33、または配列番号29の核酸配列の少なくとも2つの反復配列、または配列番号30の核酸配列の少なくとも2つの反復配列、または配列番号32の核酸配列の少なくとも2つの反復配列または配列番号34の核酸配列の少なくとも2つの反復配列に示される核酸を含む、請求項1に記載のポリヌクレオチド。
【請求項4】
湾曲起点モチーフが配列番号7、または配列番号8、または配列番号9、または配列番号10の配列を含む、請求項1に記載のポリヌクレオチド。
【請求項5】
ポリヌクレオチドが配列番号3、または配列番号4、または配列番号5、または配列番号6、または配列番号3、4、5または6のうちのいずれか1つと少なくとも70%同一である変異体の配列を含みかつ少なくとも1つの湾曲起点モチーフを含むものである、請求項1に記載のポリヌクレオチド。
【請求項6】
配列番号11、配列番号12および配列番号13のいずれか1つに示したアミノ酸配列を含むポリペプチドをコードするかまたは配列番号14、配列番号15および配列番号16のいずれか1つに示した核酸配列を含むものであるポリヌクレオチド。
【請求項7】
請求項6に記載のポリヌクレオチドがコードするポリペプチド。
【請求項8】
請求項1〜6のいずれか1項に記載のポリヌクレオチドまたはその組合わせを含む宿主細胞。
【請求項9】
請求項1〜6のいずれか1項に記載のポリヌクレオチドまたはその組合わせを含むベクター。
【請求項10】
請求項1〜6のいずれか1項に記載のポリヌクレオチドまたはその組合わせを含む非ヒトトランスジェニック生物。
【請求項11】
請求項1〜6のいずれか1項に記載のポリヌクレオチドまたはその組合わせを含むキット。
【請求項12】
少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)動物宿主細胞中に、
(i)請求項1〜5に記載のいずれか1つのポリヌクレオチドまたはその組合わせ、および
(ii)目的のポリペプチドをコードする少なくとも1つのポリヌクレオチド
を導入するステップ、ならびに
(b)前記細胞を培養するステップ
を含んでなる前記方法。
【請求項13】
少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)動物宿主細胞中に、
(i)請求項6に記載のポリヌクレオチド、および
(ii)目的のポリペプチドをコードする少なくとも1つのポリヌクレオチド
を導入するステップ、ならびに
(b)前記細胞を培養するステップ
を含んでなる前記方法。
【請求項14】
少なくとも1つの目的のポリヌクレオチドを発現する方法であって、
(a)動物宿主細胞中に、
(i)低下した選択能力を有する選択可能なマーカー遺伝子をコードするポリヌクレオチド、および
(ii)遺伝子発現を増強できる核酸配列を含むポリヌクレオチド、および
(iii)目的のポリペプチドをコードする少なくとも1つのポリヌクレオチド
を導入するステップ、ならびに
(b)前記細胞を培養するステップ
を含んでなる前記方法。
【請求項15】
低下した選択能力を有する選択可能なマーカー遺伝子をコードするポリヌクレオチドが改変されたネオマイシンホスホトランスフェラーゼをコードするポリヌクレオチドである、請求項14に記載の方法。
【請求項16】
改変されたネオマイシンホスホトランスフェラーゼをコードするポリヌクレオチドが請求項6に記載のポリヌクレオチドである、請求項14〜15のいずれか1項に記載の方法。
【請求項17】
遺伝子発現を増強できる核酸配列を含むポリヌクレオチドが請求項1〜5のいずれか1項に記載のポリヌクレオチドである、請求項14〜16のいずれか1項に記載の方法。
【請求項18】
培養を好適な条件下で行いかつ宿主細胞を選択剤を用いて選択することができる、請求項12〜17のいずれか1項に記載の方法。
【請求項19】
宿主細胞を無血清または血清含有培地で培養する、請求項12〜18のいずれか1項に記載の方法。
【請求項20】
ポリペプチドが免疫グロブリン、治療タンパク質、膜タンパク質および酵素からなる群より選択される、請求項12〜19のいずれか1項に記載の方法。
【請求項21】
目的のポリペプチドを製造する方法であって、請求項12〜20のいずれか1項に記載のステップおよびポリヌクレオチドがコードする目的のポリペプチドを得るステップを含んでなる前記方法。
【請求項22】
前記宿主細胞が爬虫類細胞、好ましくは毒ヘビ細胞、または哺乳動物細胞、好ましくはCHO細胞である、請求項8に記載の宿主細胞または請求項12〜21のいずれか1項に記載の方法。
【請求項23】
請求項1〜5のいずれか1項に記載のポリヌクレオチドおよび/または請求項6に記載のポリヌクレオチドを含む医薬組成物。
【請求項24】
遺伝子治療に用いる医薬組成物を製造するための、請求項1〜5のいずれか1項に記載のポリヌクレオチドおよび/または請求項6に記載のポリヌクレオチドの使用。
【請求項25】
配列番号7、または配列番号8、または配列番号9、または配列番号10の配列を含む湾曲起点モチーフ。
【図1】



【図2】

【図3】
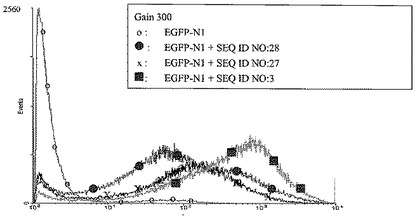
【図4】
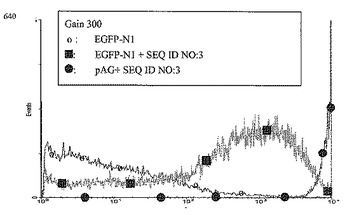
【図5】
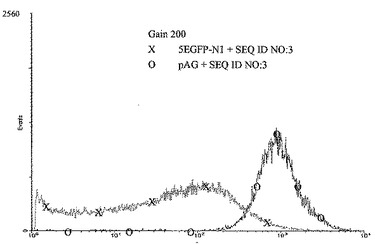
【図6】
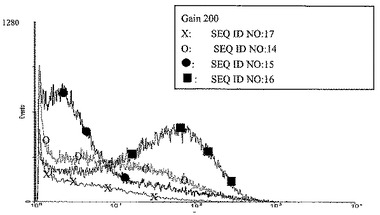
【図7】
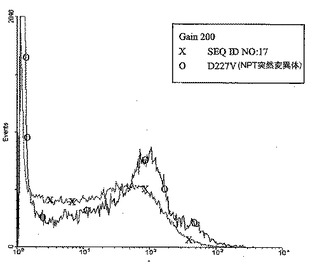
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2010−522549(P2010−522549A)
【公表日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願番号】特願2010−500290(P2010−500290)
【出願日】平成20年3月28日(2008.3.28)
【国際出願番号】PCT/EP2008/053716
【国際公開番号】WO2008/116931
【国際公開日】平成20年10月2日(2008.10.2)
【出願人】(509267270)
【出願人】(509267281)
【Fターム(参考)】
【公表日】平成22年7月8日(2010.7.8)
【国際特許分類】
【出願日】平成20年3月28日(2008.3.28)
【国際出願番号】PCT/EP2008/053716
【国際公開番号】WO2008/116931
【国際公開日】平成20年10月2日(2008.10.2)
【出願人】(509267270)
【出願人】(509267281)
【Fターム(参考)】
[ Back to top ]