
相補性決定領域とは異なる分子の領域に設計された結合性を持つ合成免疫グロブリンドメイン
免疫グロブリンの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンを設計し、ある抗原のエピトープに対する前記免疫グロブリンの結合を決定するための方法(ただし、無修飾免疫グロブリンは前記エピトープには有意に結合しない)であって、
・少なくとも一つの構造ループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記構造ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、および
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ
を含む方法、ならびに修飾免疫グロブリン。
・少なくとも一つの構造ループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記構造ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、および
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ
を含む方法、ならびに修飾免疫グロブリン。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は修飾免疫グロブリンの設計および製造方法に関する。
【0002】
一般分野は、タンパク質に特異的結合性を付与することを目的とするタンパク質の設計である。より具体的に述べると、本発明に関連する改変タンパク質は、免疫グロブリン(抗体)であり、さらに具体的に述べると、免疫グロブリンの単一ドメインまたは単一ドメインの対または単一ドメインの組み合わせである。免疫グロブリンの特異的結合性は重要な特徴である。というのも、それらは他の分子(例えば抗原)との相互作用を制御し、免疫グロブリンを診断用途および治療用途に役立つものにするからである。
【背景技術】
【0003】
基本的な抗体構造を、ここではインタクトIgG1免疫グロブリンを例にとって説明する。
【0004】
二つの同一な重(H)鎖と二つの同一な軽(L)鎖とが一体となってY字状の抗体分子を形成する。重鎖はそれぞれ四つのドメインを持っている。アミノ末端の可変領域(VH)はそのYの先端部にある。これに、三つの定常ドメイン、すなわちCH1、CH2、およびカルボキシ末端のCH3(これはYの幹の根本にある)が続いている。スイッチ(switch)という短いストレッチが、重鎖可変領域と定常領域とをつなぐ。ヒンジはCH2およびCH3(Fc断片)を抗体の残りの部分(Fab断片)につなぐ。インタクト抗体分子のヒンジのタンパク質分解切断により、一つのFcと二つの同一なFab断片とを生成させることができる。軽鎖は、スイッチによって分離された二つのドメイン、すなわち可変(VL)ドメインと定常(CL)ドメインとでできている。
【0005】
ヒンジ領域にあるジスルフィド結合が、二つの重鎖をつなぐ。軽鎖は、さらなるジスルフィド結合によって、重鎖に結びつけられる。Asn結合型糖質部分は、免疫グロブリンのクラスに依存して、定常ドメイン中の異なる位置に取り付けられる。IgG1の場合は、Cys235対およびCys238対の間に形成されるヒンジ領域中の二つのジスルフィド結合が、二つの重鎖を合体させる。軽鎖は、CH1ドメイン中のCys229とCLドメイン中のCys214との間に形成される二つのさらなるジスルフィド結合によって、二つの重鎖に結びつけられる。糖質部分は各CH2のAsn306に取り付けられて、Yの幹に顕著な出っ張りを作る。
【0006】
これらの特徴は機能上重大な結果をもたらす。重鎖の可変領域(VH)と軽鎖の可変領域(VL)はどちらもYの「先端部」にあり、そこで抗原と反応するように配置される。本分子のこの先端部はアミノ酸配列のN末端が位置している側である。Yの幹は、エフェクター機能、例えば補体の活性化およびFc受容体との相互作用またはADCCおよびADCPが、効率よく媒介されるように、突き出している。そのCH2ドメインおよびCH3ドメインは、エフェクタータンパク質との相互作用が容易になるように膨らんでいる。アミノ酸配列のC末端は先端部とは反対側(これをYの「基部」と称することができる)に位置する。インタクトIgG1の構造を図1aに図解する。
【0007】
抗体にはラムダ(λ)およびカッパ(κ)と呼ばれる2タイプの軽鎖が見出される。与えられた免疫グロブリンはκ鎖を持つか、λ鎖を持つかのどちらかであり、それぞれを一つずつ持つということは決してない。λ軽鎖またはκ軽鎖を持つ抗体の間に機能的相違は見出されていない。
【0008】
主なヒト免疫グロブリンクラス単量体の構造的構成を図1bに示す。これらのクラスは、各重鎖の組成および配列が異なっている。IgMとIgEはどちらもヒンジ領域を欠くが、それぞれに余分な重鎖ドメイン(CH4)を含有している。鎖を連結するジスルフィド結合(線分)の数および位置はアイソタイプ間で異なる。また、円記号で示すN結合型糖質基の分布も異なる。
【0009】
抗体分子中の各ドメインは、二つのβシートが互いに堅く充填されて圧縮された逆平行βバレルになっている類似の構造を持つ。この保存された構造は免疫グロブリンフォールドと呼ばれる。定常ドメインの免疫グロブリンフォールドは、4ストランドシートに向かい合って充填された3ストランドシートを含有する。このフォールドは、各シートのβストランド間に生じる水素結合、内部で向かい合ったシートの残基間に生じる疎水結合、およびシート間のジスルフィド結合によって安定化される。この3ストランドシートはストランドC、FおよびGを含み、4ストランドシートはストランドA、B、E、およびDを持つ。文字A〜Gは、免疫グロブリンフォールドのアミノ酸配列に沿ったβストランドの順序を表す。
【0010】
可変ドメインのフォールドは、4ストランドおよび5ストランドの二つのシートに配置された9本のβストランドを持つ。5ストランドシートは、定常ドメインの3ストランドシートと構造的に相同であるが、余分なストランドC'およびC''を含有している。残りのストランド(A、B、C、D、E、F、G)は、定常ドメイン免疫グロブリンフォールド中の対応部分と同じトポロジーおよび類似する構造を持っている。定常ドメインと同様に、一つのジスルフィド結合が、向かい合ったシート中のストランドBとストランドFとを連結している。免疫グロブリンフォールドを、免疫グロブリンの定常ドメインおよび可変ドメインについて、図2に図解する。
【0011】
免疫グロブリン軽鎖および免疫グロブリン重鎖の可変ドメインはどちらも、三つの超可変ループ、すなわち相補性決定領域(CDR)を含有している。Vドメインの三つのCDR(CDR1、CDR2、CDR3)は、βバレルの一端に集まっている。CDRは、免疫グロブリンフォールドのβストランドB-C、C'-C''およびF-Gをつなぐループである。CDR中の残基は免疫グロブリン分子ごとに異なり、各抗体に抗原特異性を付与している。
【0012】
抗体分子の先端部にあるVLドメインおよびVHドメインは、六つのCDR(各ドメイン上に三つずつ)が協力して抗原特異的な結合のための表面(または空洞)を構築するように、密に充填される。したがって、抗体の天然抗原結合部位は、軽鎖可変ドメインのストランドB-C、C'-C''およびF-G、ならびに重鎖可変ドメインのストランドB-C、C'-C''およびF-Gをつなぐループで構成される。
【0013】
タンパク質の3D構造を設計の一助とし、そのタンパク質のコア構造を足場として使用することにより、多くのタンパク質の表面に位置するアミノ酸残基にランダム突然変異が導入されてきた。この戦略の一例は、参照により本明細書に組み入れられる以下の参考文献に、記載または概説されている:Nygren PA, Uhlen M., Curr Opin Struct Biol. (1997) 7:463-9;Binz HK, Amstutz P, Kohl A, Stumpp MT, Briand C, Forrer P, Grutter MG, Pluckthun A. Nat Biotechnol. (2004) 22:575-82;Vogt M, Skerra A. Chembiochem. (2004) 5:191-9;US 6,562,617。
【0014】
この技法の基本原理は、多くのタンパク質が、ループ、ターン、またはランダムコイルなどの構造によって相互につながれたβシートまたはαヘリックスなどの二次構造要素の特異的配置によって形成される安定なコアを持つという観察結果に基づいている。典型的には、これらのうちループ、ターンおよびランダムコイルという三つの構造要素は、タンパク質の全体構造にとって、さほど決定的ではなく、これらの構造要素中のアミノ酸残基は、多くの場合、タンパク質の全体的フォールドを破壊することなく、交換することができる。この設計原理の天然の例が、抗体のCDRである。人工的な例には、リポカリン、アンキリンおよび他のタンパク質足場が含まれる。
【0015】
ネイティブ免疫グロブリン中のCDRループでないループは、抗原結合特異性またはエピトープ結合特異性を持たないが、免疫グロブリン分子全体の正しいフォールディングおよび/またはそのエフェクターもしくは他の機能に寄与するので、本発明ではこれを構造ループと呼ぶ。
【0016】
米国特許第6,294,654号には、ヒンジ領域と可変領域の間のCH1領域にある抗体の非CDRループにペプチド抗原を組み入れることが可能な改変抗体を作製することができ、その結果生じるAbは、そのペプチド抗原がMHC IIと共にAPCの表面に提示されることによって免疫応答を生成するような形で、APCに取り込まれうることが示されている。これらの挿入ペプチドはエピトープであり、担体分子の全体的構造は重要でない。免疫グロブリンの(非CDR)ループ上にrasペプチドを置くことができること、そしてそれでもなおその免疫グロブリンは分泌されることが、実証された。免疫グロブリンが適正にフォールドされない限り免疫グロブリンが分泌されるのを防止するという、厳格な「品質管理」が細胞内には存在し、ループのアミノ酸配列を変えると、そのタンパク質は、細胞が不適正と感知してそれを分解するような構造に、フォールドされることになるかもしれない。したがって、示された例の他に、免疫グロブリンの性質を変えずに構造ループをさらに修飾することは、困難であると考えられた。
【0017】
米国特許出願公開第2004/0101905号には、標的結合部位およびFcエフェクターペプチドを含む結合分子が記載されている。Fcエフェクターペプチドは、エフェクター分子と相互作用するペプチドである。免疫グロブリン断片のCH1ドメインの非CDRループへのエフェクターペプチドの挿入が示されている。
【0018】
Fcエフェクターペプチドは、抗体の非CDRループ中に天然に存在する構造であり、それゆえに、免疫グロブリン中の異なる等価な位置に移植しても、免疫グロブリンの構造を乱さないと予想される。
【0019】
それにもかかわらず、この開示に従って非CDRループに移植されるペプチドはいずれも、選択された異なる構造環境により、不活性になる可能性が高い。
【0020】
上述したどちらの先行技術文書でも、免疫グロブリンフォールディング構造は機能および分泌にとって重要であるから、これを乱さないことは極めて大切であり、それゆえに、その構造および機能を保つべきループ中にペプチドを挿入することは困難であると、述べられている。
【0021】
米国特許出願公開第2004/0132101号および同第2005/0244403号には、抗体の構造ループにとって天然のリガンドであるエフェクターリガンドに対して、変化した結合アフィニティを持つ、突然変異型免疫グロブリンが記載されている。この文書には、免疫グロブリン分子全体にわたるさまざまな領域中の突然変異であって、抗体全体のエフェクター機能に影響を及ぼすものが、いくつか記載されている。
【0022】
従来から、既存の抗原結合部位を操作し、その結果として、新しい結合性を導入することを目的として、免疫グロブリン様足場が利用されてきたことは、他の先行技術文書によって示されている。しかし従来は、抗原結合のためにCDR領域だけが設計されてきた。言い換えると、免疫グロブリンフォールドの場合は、天然の抗原結合部位だけが、その結合アフィニティまたは結合特異性を変化させるために修飾されてきた。ファージ粒子の表面にディスプレイされるか、さまざまな原核もしくは真核発現系で可溶性に発現される、異なる形式の、そのような操作免疫グロブリン(単鎖Fv断片(scFv)またはFab断片の形で発現されることが多い)を記述している文献は、多量に存在する。この分野の主だった著者にはGreg Winter、Andreas PlueckthunおよびHennie Hoogenboomがいる。
【発明の開示】
【0023】
本発明の目的は、新しい抗原結合部位が導入された免疫グロブリン、および前記免疫グロブリンを設計し製造するための方法を提供することである。
【0024】
したがって本発明は、免疫グロブリンの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンを設計し、ある抗原のエピトープに対する前記免疫グロブリンの結合を決定するための方法(ただし、無修飾免疫グロブリンは前記エピトープには有意に結合しない)であって、
・少なくとも一つの構造ループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記構造ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、および
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ
を含む方法に関する。
【0025】
特に本発明は、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原からなる群より選択される抗原のエピトープに特異的に結合する免疫グロブリンを設計するための方法に関する。構造ループ領域の修飾により、それらのエピトープに結合するように、免疫グロブリンを設計することができる。好ましい実施形態では、免疫グロブリンが、同じ抗原のまたは異なる抗原の、互いに異なる少なくとも二つのそのようなエピトープに特異的に結合する。
【0026】
例えば本発明の方法は、少なくとも一つの第1エピトープに特異的に結合する免疫グロブリンであって、前記免疫グロブリンの少なくとも一つの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンを設計し、前記少なくとも一つのループ領域の、少なくとも一つの第2エピトープに対する特異的結合を決定する方法(ただし、エピトープは、上述した抗原の群から選択され、無修飾構造ループ領域(非CDR領域)は、前記少なくとも一つの第2エピトープには特異的に結合しない)であって、
・少なくとも一つの構造ループ領域を含む、少なくとも一つの第1エピトープに特異的に結合する免疫グロブリンをコードする核酸を用意するステップ、
・前記核酸によってコードされる前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンを前記少なくとも一つの第2エピトープと接触させるステップ、
・前記修飾免疫グロブリンが第2エピトープに特異的に結合するかどうかを決定するステップ
を含む方法に関する。
【0027】
本発明の方法は、好ましくは、少なくとも一つの構造ループ領域中の、少なくとも一つの修飾、ならびにアレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、ウイルス抗原および原虫抗原からなる群より選択される少なくとも一つの抗原に対する、前記少なくとも一つのループ領域の特異的結合を決定すること(ただし、無修飾構造ループ領域を含有する免疫グロブリンは、前記少なくとも一つの抗原には特異的に結合しない)に関する。
【0028】
本発明において、修飾の対象となる「免疫グロブリン」という用語(本明細書で使用する用語、免疫グロブリンと抗体は、相互に可換である)は、抗原、エフェクター分子/タンパク質などのエピトープに対して、単一特異性もしくは多重特異性、または多価結合性、少なくとも二つ、好ましくは少なくとも三つの特異的結合部位を示しうる。また、本発明における免疫グロブリンは、当技術分野で受け入れられている機能的断片、例えばFc、Fab、scFv、CH/CLドメインの単鎖二量体、Fv、または免疫グロブリンの他の誘導体もしくは組み合わせ、インタクト抗体の可変領域(例えばFd、Vl、Vk、Vh)および定常領域の重鎖および軽鎖のドメイン、例えばCH1、CH2、CH3、CH4、ClおよびCk、ならびに構造ループによってつながれた免疫ブロブリンドメインの二つのβストランドからなるミニドメインでもある。
【0029】
「免疫グロブリン」「修飾免疫グロブリン」または「本発明の免疫グロブリン」という用語は、免疫グロブリンの誘導体も包含すると理解される。誘導体は、本発明の1以上の免疫グロブリンの任意の組み合わせ、および/または本発明の免疫グロブリンのどのドメインまたはミニドメインが1以上の他のタンパク質(例えば他の免疫グロブリン、リガンド、足場タンパク質、酵素、毒素など)のどの位置に融合されていてもよい融合タンパク質である。本発明の免疫グロブリンの誘導体は、さまざまな化学的技法(例えば共有結合、静電相互作用、ジスルフィド結合など)で他の物質に結合することによって、取得することもできる。
【0030】
免疫グロブリンに結合される他の物質は、脂質、糖質、核酸、有機分子および無機分子またはその任意の組み合わせ(例えばPEG、プロドラッグまたは薬物)であることができる。誘導体は、同じアミノ酸配列を持つが、完全にまたは部分的に非天然アミノ酸または化学修飾アミノ酸からできている免疫グロブリンでもある。
【0031】
本発明の改変分子は、独立したタンパク質として有用であると共に、融合タンパク質または融合誘導体(最も典型的には、より大きい抗体構造の一部、または完全な抗体分子もしくはその一部、例えばFab断片、Fc断片、Fv断片などになるように融合されたもの)としても有用であるだろう。改変タンパク質を使って、単一特異性、二重特異性、三重特異性である分子、さらにはより多くの特異性を同時に保持する分子を作ることも可能だろう。そしてまた同時に、そのような分子の使用予定の要件に応じて、同時に結合する結合価を制御し、前もって選択することが可能だろう。
【0032】
本発明によれば、あらゆる種類のアレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原に対する結合領域または抗原結合部位を、与えられた抗体構造の構造ループ中に導入することができる。
【0033】
本発明において「抗原」という用語は、免疫グロブリンのCDRループ領域と相互作用することが知られている、または相互作用する能力を持つ、分子または構造を意味するものとする。先行技術の構造ループ領域は抗原とは相互作用せず、むしろ全体的構造および/またはエフェクター分子への結合に寄与する。
【0034】
本発明において「アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原」という用語は、抗体構造によって認識されうる全てのアレルゲンおよび抗原、ならびにそのような分子の断片(特に一般に「エピトープ」と呼ばれる部分構造(例えばB細胞エピトープ))を(それらが免疫に関連する限り、すなわちそれらもまた、天然抗体またはモノクローナル抗体によって認識されうる限り)包含するものとする。
【0035】
本発明において「エピトープ」という用語は、本発明の結合ドメインまたは免疫グロブリンに対する特異的結合パートナーを完全に形成するか、またはそのような特異的結合パートナーの一部であることができる分子構造を意味するものとする。
【0036】
化学的には、エピトープは、糖質、ペプチド、脂肪酸、無機物質またはその誘導体およびそれらの任意の組み合わせから構成されうる。エピトープがポリペプチドである場合、それは通常、少なくとも3個のアミノ酸、好ましくは8〜50個のアミノ酸、より好ましくは約10〜20個のアミノ酸を、そのペプチド中に含むだろう。ペプチドの長さに決定的な上限はなく、ポリペプチド配列のほぼ全長を含んでもよい。エピトープはリニアエピトープまたはコンフォメーショナルエピトープであることができる。リニアエピトープは、ポリペプチド鎖の一次配列の単一のセグメントから構成される。リニアエピトープは連続していたり、一部重複していたりすることができる。コンフォメーショナルエピトープは、ポリペプチドのフォールディングによって寄せ集められて三次元構造を形成するアミノ酸から構成され、それらのアミノ酸は、必ずしもその直線配列中で互いに隣接していない。
【0037】
具体的に述べると、エピトープは、診断に関連する分子の少なくとも一部である。すなわち、試料中のエピトープの有無は、疾患または健康状態または製造時の加工状態または環境および食品状態と、定性的または定量的に相関する。エピトープは、治療に関連する分子(すなわち、疾患の経過を変化させる、特異的結合ドメインの標的になりうる分子)の少なくとも一部であることもできる。
【0038】
好ましい「アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原」は、免疫または治療に関連することが既に証明されているか、免疫または治療に関連する能力を持つアレルゲンまたは抗原、特に、その臨床的有効性が既に試験されているものである。
【0039】
他方、本発明のもう一つの側面によれば、他の結合能力、例えば薬物などの小分子、または酵素、酵素の触媒部位、もしくは酵素基質に対する結合能力、あるいは酵素基質の遷移状態類似体に対する結合能力などを、構造ループ領域に導入することもできる。
【0040】
好ましくは、構造ループ中の新しい抗原結合部位は、無修飾免疫グロブリンにとって外来である。したがって、エフェクター分子またはFc受容体のような標的は、好ましくは、本発明の免疫グロブリンの結合分子および特異性からは除外される。
【0041】
好ましくは、構造ループ中の新しい抗原結合部位は、選択された核酸によってコードされる免疫グロブリンの置換、欠失および/または挿入によって導入される。
【0042】
本発明のもう一つの好ましい実施形態によれば、少なくとも一つのヌクレオチドの修飾が、前記核酸によってコードされる免疫グロブリンの置換、欠失および/または挿入をもたらす。
【0043】
少なくとも一つのループ領域の修飾は、1以上のアミノ酸の置換、欠失および/または挿入、好ましくは点突然変異、ループ全体の交換、より好ましくは、少なくとも2、3、4、5、6、7、8、9、10〜30個のアミノ酸の変化をもたらしうる。
【0044】
部位指定ランダム突然変異も好ましい。この方法では、ループの1以上の特定アミノ酸残基を、ランダムに生成させたインサートを使って交換するか、そのような構造ループ中に導入する。あるいは、コンビナトリアルアプローチの使用も好ましい。
【0045】
少なくとも一つのループ領域を、好ましくは、ランダム、セミランダム、または特に、部位指定ランダム突然変異誘発法によって、突然変異させるか、または修飾する。これらの方法は、本発明の免疫グロブリンの望ましい位置でアミノ酸修飾を行うために使用することができる。これらの場合、位置はランダムに選択されるか、極端に単純化した規則を使ってアミノ酸変化が加えられる。例えば全ての残基をアラニンに突然変異させることができ、これをアラニンスキャニングという。そのような方法を、より高レベルな配列多様性をスクリーニングするための選択法を使用する、より洗練された工学的アプローチと結びつけてもよい。本発明の好ましい方法は、5'-NHS-3'、5'-NNN-3'または5'-NNK-3'という配列を持つ少なくとも一つのヌクレオチド繰返し単位を含むランダムに修飾された核酸分子に関する。
【0046】
ランダムに修飾された核酸分子は、全ての既知天然アミノ酸をコードする、上述の繰返し単位を含むことができる。
【0047】
当技術分野ではよく知られているように、例えば、後述のファージディスプレイ、リボソームディスプレイ、細胞表面ディスプレイなどのディスプレイ技術をはじめとして、一定の結合特性およびアフィニティーを持つタンパク質の同定および単離に使用することができるさまざまな選択技術がある。抗体変種を生産しスクリーニングするための方法は、当技術分野では周知である。抗体の分子生物学、発現、精製およびスクリーニングの一般方法は、DuebelおよびKontermann編「Antibody Engineering」(Springer-Verlag, ハイデルベルク, 2001);ならびにHayhurstおよびGeorgiou, 2001, Curr Opin Chem Biol 5:683-689;MaynardおよびGeorgiou, 2000, Annu Rev Biomed Eng 2:339-76に記述されている。
【0048】
本発明において「構造ループ」または「非CDRループ」とは、以下のように理解すべきである。免疫グロブリンはいわゆる免疫グロブリンフォールドを持つドメインでできている。基本的には、逆平行βシートがループでつながれて、圧縮された逆平行βバレルを形成する。可変領域では、ドメインのループの一部が抗体の特異性(すなわち抗原への結合)にとって不可欠な寄与をする。これらのループはCDRループと呼ばれる。抗体ドメインの他のループは全て、むしろ、その分子の構造および/またはエフェクター機能に寄与している。これらのループを、ここでは構造ループまたは非CDRループと定義する。
【0049】
修飾免疫グロブリン(この用語には本明細書の全体にわたって常に免疫グロブリン断片が包含される)をコードする核酸分子を、宿主細胞にクローニングし、発現させ、その結合特異性についてアッセイすることができる。これらの作業は周知の手法を使って行われ、本発明に役立ちうるさまざまな方法が、「Molecular Cloning-A Laboratory Manual」第3版(Maniatis, Cold Spring Harbor Laboratory Press, ニューヨーク, 2001)および「Current Protocols in Molecular Biology」(John Wiley & Sons)に記述されている。本発明の修飾免疫グロブリンをコードする核酸は、前記免疫グロブリンを発現させるために、発現ベクターに組み入れることができる。発現ベクターは、典型的には、制御配列または調節配列、選択可能マーカー、任意の融合パートナー、および/または追加要素と作動可能に連結された(すなわち機能的な関係に置かれた)免疫グロブリンを含む。本発明の修飾免疫グロブリンは、修飾免疫グロブリンをコードする核酸を含有する核酸(好ましくは発現ベクター)で形質転換された宿主細胞を、修飾免疫グロブリンの発現を誘導しまたは引き起こすのに適当な条件下で培養することによって、生産することができる。外因性核酸分子を宿主に導入する方法は当技術分野では周知であり、使用する宿主によって異なるだろう。もちろん、修飾免疫グロブリンを発現させるための非細胞発現系または無細胞発現系も使用することができる。
【0050】
本発明の好ましい実施形態では、修飾免疫グロブリンを、発現後に精製または単離する。修飾免疫グロブリンは、当業者に知られているさまざまな方法で単離または精製することができる。標準的な精製方法には、クロマトグラフィー技法、電気泳動技法、免疫学的技法、沈降、透析、濾過、濃縮、およびクロマトフォーカシング技法などがある。多くの場合、個別の融合パートナーによって、精製を可能にすることができる。例えば、GST融合物を使用する場合にはグルタチオン樹脂を使って、Hisタグを使用する場合にはNi+2アフィニティクロマトグラフィーを使って、またflagタグを使用する場合には固定化抗flag抗体を使って、抗体を精製することができる。適切な精製技法における一般指針については、「Antibody Purification: Principles and Practice」(第3版, Scopes, Springer-Verlag, NY, 1994)を参照されたい。もちろん、本発明の修飾免疫グロブリンを、宿主の表面に、特に細菌、昆虫もしくは酵母細胞の表面に、またはファージもしくはウイルスの表面に、発現させることも可能である。
【0051】
修飾免疫グロブリンは、例えばインビトロアッセイ、インビボおよび細胞系アッセイ、ならびに選択技術を用いる方法など(ただしこれらに限定されるわけではない)、さまざまな方法を使ってスクリーニングすることができる。スクリーニング手法には自動およびハイスループットスクリーニング技術を利用することができる。スクリーニングには、融合パートナー、またはラベル、例えば酵素、免疫ラベル、同位体ラベル、もしくは小分子ラベル、例えば蛍光色素もしくは比色測定用色素または発光原分子などを利用することができる。
【0052】
好ましい実施形態では、免疫グロブリンの機能的および/または生物物理的性質を、インビトロアッセイでスクリーニングする。好ましい実施形態では、抗体を機能性(例えば反応を触媒する能力またはその標的への結合アフィニティー)について、スクリーニングする。
【0053】
アッセイには、発色ラベル、蛍光ラベル、発光ラベル、または同位体ラベルを含む(ただしこれらに限定されるわけではない)さまざまな検出方法を使用することができる。
【0054】
当技術分野では知られているように、一部のスクリーニング方法は、ライブラリーの好ましいメンバーを選択する方法である。それらの方法をここでは「選択法」と呼び、これらの方法は、本発明において、修飾免疫グロブリンのスクリーニングに役立つ。選択法を使って免疫グロブリンライブラリーをスクリーニングする場合は、好ましいライブラリーのメンバーだけが、すなわち何らかの選択基準に合致するライブラリーのメンバーだけが、増殖され、単離され、そして/または観察される。理解されるであろうが、最も適合した変種だけを観察するので、そのような方法では、ライブラリーメンバーの適合性を個別にアッセイする方法によってスクリーニングすることが可能なライブラリーよりも大きなライブラリーをスクリーニングすることができる。選択は、免疫グロブリンの表現型を、共有結合的にまたは非共有結合的に、その遺伝子型と(すなわち抗体の機能を、それをコードする核酸と)関連づける任意の方法、技法、または融合パートナーによって可能になる。例えば、選択法としてのファージディスプレイの使用は、ライブラリーメンバーを遺伝子IIIタンパク質に融合することによって可能になる。このようにして、何らかの基準(例えばその免疫グロブリンの標的に対する結合アフィニティ)を満たす修飾免疫グロブリンの選択または単離により、それをコードする核酸も選択または単離される。ひとたび単離されたら、次に、修飾免疫グロブリンをコードする遺伝子または遺伝子群を増幅することができる。この単離および増幅の工程(これをパンニングという)を繰り返して、ライブラリー中の好ましい抗体変種を濃縮させることができる。最終的には、取り付けられた核酸の核酸配列決定によって、遺伝子同定が可能になる。
【0055】
当技術分野では、本発明において免疫グロブリンライブラリーのスクリーニングに役立ちうる、さまざまな選択法が知られている。これらには、ファージディスプレイ(「Phage display of peptides and antibodies: a laboratory manual」Kayら, 1996, Academic Press, カリフォルニア州サンディエゴ, 1996;Lowmanら, 1991, Biochemistry 30:10832-10838;Smith, 1985, Science 228:1315-1317)およびその変法、例えば選択的ファージ感染(selective phage infection)(Malmborgら, 1997, J Mol Biol 273:544-551)、選択感染性ファージ(selectively infective phage)(Krebberら, 1997, J Mol Biol 268:619-630)、および遅延感染性パンニング(delayed infectivity panning)(Benharら, 2000, J Mol Biol 301:893-904)、細胞表面ディスプレイ(Witrrup, 2001, Curr Opin Biotechnol, 12:395-399)、例えば細菌(Georgiouら, 1997, Nat Biotechnol 15:29-34;Georgiouら, 1993, Trends Biotechnol 11:6-10;Leeら, 2000, Nat Biotechnol 18:645-648;Junら, 1998, Nat Biotechnol 16:576-80)、酵母(BoderおよびWittrup, 2000, Methods Enzymol 328:430-44;BoderおよびWittrup, 1997, Nat Biotechnol 15:553-557)、および哺乳動物細胞(Whitehornら, 1995, Bio/technology 13:1215-1219)でのディスプレイ、ならびにインビトロディスプレイ技術(Amstutzら, 2001, Curr Opin Biotechnol 12:400-405)、例えばポリソームディスプレイ(Mattheakisら, 1994, Proc Natl Acad Sci USA 91:9022-9026)、リボソームディスプレイ(Hanesら, 1997, Proc Natl Acad Sci USA 94:4937-4942)、mRNAディスプレイ(RobertsおよびSzostak, 1997, Proc Natl Acad Sci USA 94:12297-12302;Nemotoら, 1997, FEBS Lett 414:405-408)、およびリボソーム不活化ディスプレイ系(ribosome-inactivation display system)(Zhouら, 2002, J Am Chem Soc 124, 538-543)などがあるが、これらに限定されるわけではない。
【0056】
本発明に役立ちうる他の選択法には、ディスプレイに頼らない方法、例えばインビボ法、例えばペリプラズム発現およびサイトメトリースクリーニング(Chenら, 2001, Nat Biotechnol 19:537-542)、抗体断片再構成アッセイ(JohnssonおよびVarshavsky, 1994, Proc Natl Acad Sci USA 91:10340-10344;Pelletierら, 1998, Proc Natl Acad Sci USA 95:12141-12146)、および選択モード(Visintinら, 1999, Proc Natl Acad Sci USA 96:11723-11728)で用いられる酵母ツーハイブリッドスクリーニング(FieldsおよびSong, 1989, Nature 340:245-246)などがある(ただしこれらに限定されるわけではない)。これに代わる実施形態では、発現ベクター上の特異的配列に結合することにより、その融合パートナーおよびそれに付随するFc変種ライブラリーメンバーを、それらをコードする核酸と、共有結合的または非共有結合的に関連づける融合パートナーによって、選択が可能になる。例えば、PCT WO 00/22906;PCT WO 01/49058;PCT WO 02/04852;PCT WO 02/04853;PCT WO 02/08023;PCT WO 01/28702およびPCT WO 02/07466には、本発明に役立ちうるそのような融合パートナーおよび技法が記述されている。これに代わる実施形態として、抗体の発現が、細胞に成長上、増殖上、または生存上の利点を何かしら付与するのであれば、インビボ選択を行うことができる。
【0057】
「定方向進化」法と呼ばれる一部の選択法は、選択時に好ましい配列の交配または育種を含み、時には新しい突然変異の組み込みを伴う方法である。当業者には明らかであるだろうが、定方向進化法はライブラリー中の最も好ましい配列の同定を容易にすることができ、スクリーニングの対象となる配列の多様性を増加させることができる。当技術分野では、例えばDNAシャフリング(PCT WO 00/42561 A3;PCT WO 01/70947 A3)、エクソンシャフリング(米国特許第6,365,377号;KolkmanおよびStemmer, 2001, Nat Biotechnol 19:423-428)、ファミリーシャフリング(Crameriら, 1998, Nature 391:288-291;米国特許第6,376,246号)、RACHITT(商標)(Cocoら, 2001, Nat Biotechnol 19:354-359;PCT WO 02/06469)、STEPおよびインビトロ組換えのランダムプライミング(Zhaoら, 1998, Nat Biotechnol 16:258-261;Shaoら, 1998, Nucleic Acids Res 26:681-683)、エキソヌクレアーゼ媒介遺伝子アセンブリー(米国特許第6,352,842号;米国特許第6,361,974号)、Gene Site Saturation Mutagenesis(商標)(米国特許第6,358,709号)、Gene Reassembly(商標)(米国特許第6,358,709号)、SCRATCHY(Lutzら, 2001, Proc Natl Acad Sci USA 98:11248-11253)、DNA断片化法(Kikuchiら, Gene 236:159-167)、一本鎖DNAシャフリング(Kikuchiら, 2000, Gene 243:133-137)、およびAMEsystem(商標)定方向進化抗体工学技術(directed evolution antibody engineering technology)(Applied Molecular Evolution)(米国特許第5,824,514号;米国特許第5,817,483号;米国特許第5,814,476号;米国特許第5,763,192号;米国特許第5,723,323号)など(ただしこれらに限定されるわけではない)、本発明において抗体変種のスクリーニングに役立ちうる、さまざまな定方向進化法が知られている。
【0058】
好ましい実施形態では、1以上の細胞系アッセイまたはインビボアッセイを使って、抗体変種がスクリーニングされる。そのようなアッセイの場合、典型的には、細胞がライブラリーに属する個々の免疫グロブリンまたは免疫グロブリンのプールに曝露されるように、精製または未精製修飾免疫グロブリンが外因的に加えられる。これらのアッセイは、常にではないが典型的には、免疫グロブリンの機能、すなわち標的に結合して何らかの生化学的事象(例えばエフェクター機能、リガンド/受容体結合阻害、アポトーシスなど)を媒介するという抗体の能力に基づいている。そのようなアッセイでは、多くの場合、抗体に対する細胞の応答、例えば細胞の生残、細胞死、細胞形態の変化、または転写活性化、例えば天然遺伝子もしくはレポーター遺伝子の細胞発現などを監視することが必要になる。例えばそのようアッセイでは、ADCC、ADCP、またはCDCを誘発するという抗体変種の能力を測定することができる。一部のアッセイでは、例えば血清補体、またはエフェクター細胞、例えば末梢血単球(PBMC)、NK細胞、マクロファージなどといった、追加の細胞または成分を(すなわち標的細胞以外に)加える必要があるかもしれない。そのような追加細胞は任意の生物(好ましくはヒト、マウス、ラット、ウサギ、およびサル)に由来することができる。免疫グロブリンは、標的を発現するある種の細胞株のアポトーシスを引き起こしたり、アッセイに加えておいた免疫細胞による標的細胞への攻撃を媒介したりすることができる。細胞の死または生存能力を監視するための方法は当技術分野では知られており、色素、免疫化学試薬、細胞化学試薬、および放射性試薬の使用が含まれる。例えばカスパーゼ染色アッセイはアポトーシスの測定を可能にし、放射性基質または蛍光色素、例えばアラマーブルーなどの取り込みまたは放出は、細胞の成長または活性化の監視を可能にしうる。好ましい実施形態では、DELFIA(登録商標)EuTDA系の細胞毒性アッセイ(Perkin Elmer, マサチューセッツ州)を使用することができる。あるいは、1以上の天然細胞内成分(例えば乳酸デヒドロゲナーゼ)の放出を測定することによって、死んだ標的細胞または損傷を受けた標的細胞を監視することもできる。転写活性化も、細胞系アッセイにおいて機能をアッセイする方法として役立ちうる。この場合は、アップレギュレートされうる天然遺伝子または免疫グロブリンをアッセイすることによって、応答を監視することができる。例えば、ある種のインターロイキンの放出を測定するか、あるいはレポーターコンストラクトによって読出しを行うことができる。細胞系アッセイでは、修飾免疫グロブリンの存在に対する応答として、細胞の形態変化を測定する場合もある。そのようなアッセイのための細胞タイプは原核でも真核でもよく、当技術分野で知られるさまざまな細胞株を使用することができる。あるいは、変種をコードする核酸で形質転換またはトランスフェクトされた細胞を使って、細胞系スクリーニングを行う。すなわち細胞に抗体変種を外因的には加えない。例えばある実施形態では、細胞系スクリーニングに、細胞表面ディスプレイを利用する。細胞の表面への修飾免疫グロブリンのディスプレイを可能にする融合パートナーを使用することができる(Witrrup, 2001, Curr Opin Biotechnol, 12:395-399)。
【0059】
好ましい実施形態として、1以上の細胞系アッセイを使って、修飾免疫グロブリンの免疫原性を、実験的に決定することができる。好ましい一実施形態では、エクスビボT細胞活性化アッセイを使って、免疫原性を実験的に定量する。この方法では、一致ドナーから得た抗原提示細胞およびナイーブT細胞を、関心対象のペプチドまたは抗体全体で1回以上攻撃する。次に、いくつかの方法を使って(例えば、サイトカインの産生を監視すること、またはトリチウム化チミジンの取り込みを測定することなどによって)、T細胞活性化を検出することができる。最も好ましい実施形態では、インターフェロンγ産生をElispotアッセイを使って監視する(Schmittelら, 2000, J. Immunol. Meth., 24: 17-24)。
【0060】
本発明の修飾免疫グロブリンの生物学的性質は、細胞、組織、および生物全体を使った実験で特徴づけることができる。当技術分野では知られているように、薬物は、多くの場合、ある疾患または疾患モデルの処置に関する薬物の有効性を測定するために、または薬物の薬物動態、毒性、および他の性質を測定するために、動物(例えばマウス、ラット、ウサギ、イヌ、ネコ、ブタ、およびサルを含むが、これらに限定されるわけではない)で試験される。それらの動物は疾患モデルと呼ぶことができる。治療薬は、多くの場合、マウス(例えばヌードマウス、SCIDマウス、異種移植マウス、およびトランスジェニックマウス(ノックインおよびノックアウトを含む)を含むが、これに限定されるわけではない)で試験される。そのような実験により、治療薬として使用される抗体の潜在能力を決定するための有意義なデータを得ることができる。試験には任意の生物(好ましくは哺乳動物)を使用することができる。例えばサルは、遺伝的にヒトに類似しているので、適切な治療モデルになることができ、したがって本発明の修飾免疫グロブリンの有効性、毒性、薬物動態、または他の性質を試験するために使用することができる。最終的には、薬物として承認されるために、ヒトでの試験が要求されるので、もちろんこれらの実験も予想される。したがって本発明の修飾免疫グロブリンは、その治療有効性、毒性、免疫原性、薬物動態、および/または他の臨床的性質を決定するために、ヒトで試験することができる。
【0061】
本発明の修飾免疫グロブリンは、広範囲にわたる抗体製品に役立ちうる。ある実施形態では、本発明の抗体変種が、治療または予防に、製造または分析用途に、診断薬、工業用化合物、または研究試薬として、好ましくは治療薬として使用される。本抗体変種は、モノクローナルまたはポリクローナルである抗体組成物に役立ちうる。好ましい実施形態では、標的抗原を持つ標的細胞(例えば癌細胞)を殺すために、本発明の修飾免疫グロブリンを使用する。これに代わる実施形態では、例えばサイトカインまたはサイトカイン受容体をアンタゴナイズすることなどにより、標的抗原をブロック(block)、アンタゴナイズ(antagonize)、またはアゴナイズ(agonize)するために、本発明の修飾免疫グロブリンを使用する。もう一つの好ましい実施形態では、標的抗原をブロック、アンタゴナイズまたはアゴナイズし、標的抗原を持つ標的細胞を殺すために、本発明の修飾免疫グロブリンを使用する。もう一つの好ましい実施形態では、成長因子または成長因子受容体をブロック、アンタゴナイズまたはアゴナイズし、標的抗原を持つ標的細胞または標的抗原を必要とする標的細胞を殺すために、本発明の修飾免疫グロブリンを使用する。もう一つの好ましい実施形態では、酵素および酵素の基質をブロック、アンタゴナイズまたはアゴナイズするために、本発明の修飾免疫グロブリンを使用する。
【0062】
本発明の修飾免疫グロブリンは、さまざまな治療目的に使用することができる。好ましい実施形態では、特異的障害を処置するために、本修飾免疫グロブリンを含む抗体を患者に投与する。本発明における「患者」には、ヒトと他の動物、好ましくは哺乳動物がどちらも包含され、最も好ましくはヒトである。本明細書において「特異的障害」とは、本発明の修飾免疫グロブリンを含む医薬組成物の投与によって改善されうる障害を意味する。
【0063】
ある実施形態では、本発明の修飾免疫グロブリンが、患者に投与される唯一の治療活性剤である。あるいは、本発明の修飾免疫グロブリンは、1以上の他の治療剤(例えば細胞毒性剤、化学療法剤、サイトカイン、成長阻害剤、抗ホルモン剤、キナーゼ阻害剤、抗血管新生剤、心保護薬、または他の治療剤などであるが、これらに限定されるわけではない)と組み合わせて投与される。修飾免疫グロブリンは、1以上の他の治療レジメンと同時に投与することができる。例えば本発明の抗体変種は、化学療法、放射線療法、または化学療法と放射線療法の両方と一緒に、患者に投与することができる。一実施形態として、本発明の修飾免疫グロブリンは、本発明の抗体変種を含んでも含まなくてもよい1以上の抗体と共に投与することができる。本発明のもう一つの実施形態によれば、本発明の修飾免疫グロブリンおよび1以上の他の抗癌療法を使って、癌細胞をエクスビボで処置することができる。そのようなエクスビボ処置は、骨髄移植(特に自家骨髄移植)に役立ちうると考えられる。もちろん、本発明の抗体は、さらに他の治療技法、例えば手術などと組み合わせて使用することもできると考えられる。
【0064】
本発明の修飾免疫グロブリンとの併用投与には、さまざまな他の治療剤が役立ちうる。ある実施形態では、修飾免疫グロブリンを抗血管新生剤(血管の発達を阻止するか、血管の発達にある程度干渉する化合物)と共に投与する。抗血管新生因子は、例えば、血管新生の促進に関与する成長因子または成長因子受容体に結合する小分子またはタンパク質(例えば抗体、Fc融合物、またはサイトカイン)であることができる。本発明において好ましい抗血管新生因子は、血管内皮増殖因子(VEGF)に結合する抗体である。これに代わる実施形態では、適応免疫応答を誘導または増進する治療剤(例えばCTLA-4を標的とする抗体)と共に、修飾免疫グロブリンを投与する。これに代わる実施形態では、修飾免疫グロブリンを、チロシンキナーゼ阻害剤(チロシンキナーゼのチロシンキナーゼ活性をある程度阻害する分子)と共に投与する。これに代わる実施形態では、本発明の修飾免疫グロブリンをサイトカインと共に投与する。本明細書にいう「サイトカイン」とは、ある細胞集団が放出するタンパク質であって、別の細胞に細胞間媒介物質として作用するものの総称であり、ケモカインを包含する。
【0065】
本発明の修飾免疫グロブリンと1以上の治療活性剤とが調剤された医薬組成物が考えられる。本発明の抗体変種の製剤は、貯蔵に備えて、望ましい純度を持つ前記免疫グロブリンを、随意の医薬的に許容できる担体、賦形剤または安定剤(「Remington's Pharmaceutical Sciences」第16版, Osol, A.編, 1980)と、凍結乾燥製剤または水溶液の形で混合することにより、製造することができる。インビボ投与に使用される製剤は、好ましくは、滅菌状態にある。これは、滅菌濾過膜を通した濾過または他の方法によって、容易に達成される。本明細書に開示する修飾免疫グロブリンおよび他の治療活性剤は、免疫リポソームとして製剤化し、そして/またはマイクロカプセルに封入することもできる。
【0066】
本発明の修飾免疫グロブリンを含む医薬組成物(好ましくは滅菌水溶液の形態をとっているもの)の投与は、例えば経口投与、皮下投与、静脈内投与、鼻腔内投与、耳内投与、経皮投与、局所外用(例えばゲル剤、軟膏、ローション剤、クリーム剤など)、腹腔内投与、筋肉内投与、肺内投与(例えばAradigmから市販されているAERx(商標)吸入技術、またはInhale Therapeuticsから市販されているInhance(商標)肺送達系)、腟投与、非経口投与、直腸投与、または眼内投与など(ただしこれらに限定されるわけではない)、さまざまな方法で行うことができる。
【0067】
本明細書で使用する「特異的に結合」という用語は、不均一な分子集団における関心対象のコグネイトリガンドを決定するような結合反応を指す。したがって、指定の条件下(例えば免疫グロブリンの場合であれば免疫アッセイ条件下)で、明示された抗体は、その個別の「標的」に結合し、試料中に存在する他の分子には有意な量では結合しない。抗体のCDRに似て、修飾構造ループ領域は、抗原結合性または分子結合性のタンパク質部分であって、抗原そのものではない。
【0068】
「発現系」という用語は、所望のコード配列および制御配列を含有し、これらの配列で形質転換またはトランスフェクトされた宿主が、コードされているタンパク質を産生することができるように、それらが作動可能に連結されている核酸分子を指す。形質転換を達成するには、発現系をベクター上に含めることができるが、関連DNAを宿主染色体に組み込むこともできる。
【0069】
本発明の好ましい実施形態によれば、発現系はベクターを含む。この目的には、当技術分野で知られている任意の発現ベクターを適宜、使用することができる。
【0070】
修飾免疫グロブリンは、好ましくは、宿主中で、好ましくは細菌細胞、酵母細胞、植物細胞中で、動物細胞中で、または植物もしくは動物中で、発現される。
【0071】
例えば哺乳動物細胞(動物細胞)、植物細胞、細菌(例えば枯草菌、大腸菌)、昆虫細胞、および酵母(ピキアパストリス、サッカロミセスセレビシェ)など(ただしこれらに限定されるわけではない)、広範囲わたる種々の適当な宿主細胞を使って、修飾免疫グロブリンを発現させることができる。例えば、本発明に役立ちうる種々の細胞株は、American Type Culture Collectionから入手することができるATCC細胞株カタログに記載されている。さらにまた、本発明の免疫グロブリンを発現させるための宿主として、植物および動物も使用することができる。発現ならびにトランスフェクションベクターまたはカセットは、使用する宿主に応じて選択することができる。
【0072】
もちろん、非細胞または無細胞タンパク質発現系も、使用することができる。十分な量のタンパク質を産生するインビトロ転写/翻訳タンパク質発現プラットフォームは、細胞に基づく発現系に通例付随する面倒な上流工程および下流工程(例えば宿主細胞の形質転換、培養、または溶解)の必要性を排除する無細胞タンパク質発現の多くの利点をもたらす。
【0073】
本発明のもう一つの側面は、免疫グロブリンの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンまたはその医薬調製物を製造し、ある抗原のエピトープに対する前記免疫グロブリンの結合を決定するための方法(ただし、無修飾免疫グロブリンは前記エピトープには有意に結合しない)であって、
・少なくとも一つのループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ、および
・前記エピトープに結合する修飾免疫グロブリンを用意し、要すればそれを医薬調製物に仕上げるステップ、
を含む方法に関する。
【0074】
特に本発明は、免疫グロブリンの少なくとも一つの構造ループ領域に少なくとも一つの修飾を含む、少なくとも一つの第1分子に特異的に結合する多重特異性免疫グロブリンまたはその医薬調製物を製造し、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原からなる群より選択される少なくとも一つの第2分子に対する、前記少なくとも一つのループ領域の特異的結合を決定するための方法(ただし、無修飾構造ループ領域を含有する免疫グロブリンは、前記少なくとも一つの第2分子には特異的に結合しない)であって、
・少なくとも一つの構造ループ領域を含む、少なくとも一つの第1分子に特異的に結合する免疫グロブリンをコードする核酸を用意するステップ、
・前記核酸によってコードされる前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンを前記少なくとも一つの第2分子と接触させるステップ、
・前記修飾免疫グロブリンが第2分子に特異的に結合するかどうかを決定するステップ、および
・前記少なくとも一つの第2分子に特異的に結合する修飾免疫グロブリンを用意し、要すればそれを医薬調製物に仕上げるステップ、
を含む方法に関する。
【0075】
特異的結合対の一つのメンバー中に、一つより多い特異性を設計することが好ましい(Kuferら (2004) Trends in Biotechnology, Vol 22, p. 238-244)。
【0076】
多重特異性(例えば二特異性)モノクローナル抗体または抗体断片を作製するために、数多くの試みがなされてきた。二つの異なるポリペプチド鎖(重鎖および軽鎖)でできた二重特異性抗体を作製する際の課題の一つは、一つの細胞内で四つの異なる鎖(二つの重鎖および二つの軽鎖)を発現させる必要があり、その結果、分子のさまざまな組み合わせがいくつも生じ、それらを、その混合物中の所望の二重特異性分子から分離する必要があるということである。これらの分子の分離は、その類似性ゆえに、困難であり、多くの費用がかかる。そのような不必要な対形成の発生を最小限に抑えるために、いくつかの技法が使用されてきた(Carter (2001) Journal of Immunological Methods, vol 248, p. 7-15)。
【0077】
この課題に対する解決策の一つは、例えば互いに連結された二つのscFvのような、二つの特異性を持つ一本のポリペプチド鎖の作製、またはいわゆるダイアボディの作製である。そのような分子は、天然分子のフォールドとはかけ離れていることが示されており、その作製は困難であることがよく知られている(LeGallら (2004) Protein Engineering, Design & Selection vol 17 p.357-366)。
【0078】
現在の二重特異性抗体設計のもう一つの課題は、たとえ親抗体がそれぞれの結合パートナーに二価で結合するとしても(例えばIgG)、結果として生じる二重特異性抗体はそれぞれの結合パートナーの各々に関して一価であるという事実である。
【0079】
本発明の好ましい多重特異性分子はこれらの課題を解決する。
【0080】
二本の抗体ポリペプチド鎖の発現(Cabillyら Proc. Natl. Acad. Sci. USA 81:3273-3277 (1984))よりも達成が容易な、一本のペプチド鎖としての二重特異性分子の発現が可能である(二つの結合特異性を持つ修飾Igドメイン、実施例参照)。
【0081】
第2の特異性は分子の非可変部分に位置するという事実ゆえに、抗体様分子(すなわち2本のポリペプチド鎖でできた分子)として作製することもでき、二つの異なる重鎖または二つの異なる軽鎖は必要でない。したがって二本の鎖が間違った対を形成する可能性はない。
【0082】
本発明の抗体は、特異的結合パートナーに結合する可変領域を全体として形成する重鎖および軽鎖からなることができ、第2の特異性は、重鎖または軽鎖の構造ループのいずれかの修飾ループによって形成される。結合部位は(重鎖上、または軽鎖上、または両鎖上で)構造的に隣接しうる、1個より多い非CDRループによって形成されてもよい。
【0083】
修飾抗体または誘導体は、完全な抗体または抗体断片(例えばFab、CH1-CH2、CH2-CH3)であることができる。
【0084】
これは、結合パートナーに一価または多価で結合するか、設計によっては、異なる結合パートナーに異なる結合価で結合することさえできる。
【0085】
重鎖および軽鎖の非CDR領域中の特異的結合部位の選択および設計には、いくつかのさまざまなループを利用することができるので、上述の問題を伴わずに、2個より多い特異性を持つ抗体誘導体でさえ、設計することが可能である。
【0086】
一本のポリペプチド鎖内の特異的結合ドメインは、ペプチドリンカーを使って、またはペプチドリンカーを使わずに、つなぐことができる。
【0087】
一部の抗体クラスは、本来、多重特異性(特に二重特異性)であるとみなすことができる。すなわちそれらは、可変領域で抗原(これは通例、外来構造物であるか、癌関連構造物である)に結合し、Fc部分でFcエフェクター分子(例えばさまざまな免疫細胞上のFc受容体または補体タンパク質)に結合して、ADCC、ADCPまたはCDCなどの効果を可能にする。
【0088】
Fcエフェクター分子は、免疫グロブリン分子のFc部分(IgG1の場合、それはCH2ドメインおよびCH3ドメインからなる)によって結合される。糖鎖工学的技法により(US 6,602,684)、または直接的にFcにおけるタンパク質工学により(US 2005/0054832)、もしくは間接的にFc外での設計により(US 2005/02444403)、抗体分子のFc部分の結合を改善することによって、エフェクター機能を最適化するための方法が、いくつか記述されている。Fc受容体に対するFc領域の結合および/またはCq1などの補体タンパク質への結合が、どちらも、そのような技法によって改変されている。通常は、そのようなFcエフェクター分子に対する結合アフィニティーを改善する努力がなされる。なぜならそれはエフェクター機能の改善と相関するからである。
【0089】
本発明では、天然のFc結合領域外でFcエフェクター分子に結合する抗体を、設計することができる。「天然」Fcエフェクター分子結合に関与するループ以外の抗体ドメイン中の修飾ループを、1以上のFcエフェクター分子に結合するように、ライブラリーから選択するか、またはそのように設計することができる。そのような追加のFcエフェクター分子結合部位を持つ抗体は、一定のFcエフェクター分子またはFcエフェクター分子をディスプレイするエフェクター細胞に対して、より強いアビディティを持ち、それゆえに、糖鎖工学によって作製された抗体または他の方法で改善されたFc領域よりも、さらに強い効果を持ちうる。しかし本発明の一定の実施形態では、本発明による構造ループ中の修飾によって、修飾対象である与えられた抗体のエフェクター特徴を直接変化させず、そのまま残しておくべきである。
【0090】
抗体断片は、抗体全体と比較して一定の利点を持っている。断片は、通常は、良好な生体分布性を持ち、より容易に製造することができる。しかし抗体断片設計の大半はエフェクター機能を欠き、インビボ半減期が短い(Holliger Pら Nat Biotechnol.(2005) 23:1126-36)。
【0091】
CH1ドメインもCκまたはCλドメインもエフェクター機能を媒介せず、それが、FabがADCC、ADCPまたはCDCを示さない理由である。WO 02/44215には、抗体の抗原結合部位とFcエフェクター分子を結合するペプチドとからなる結合分子が記述されている。そのようにして、エフェクター機能を示す抗体断片を構築することができる。そのペプチドは、抗原結合も、Fcエフェクター分子に結合するというペプチドの能力も、破壊しないような位置で、その結合分子に組み入れられている。
【0092】
しかし本発明によれば、免疫グロブリンドメインの固定された足場内のランダムループ配列のライブラリーからFcエフェクター分子結合に関して選択された修飾免疫グロブリンを使って、Fcエフェクター分子への結合を達成することができる。したがって、Igドメイン足場外ではFcエフェクター分子に結合しないであろう特異的ループ配列を選択することが可能である。したがって、本発明によって得られるポリペプチドは、好ましくは、100個を越えるアミノ酸からなることができる。
【0093】
本発明のそのようなドメインの潜在的エフェクター機能を選択するために、突然変異型CH1、CκまたはCλドメインのライブラリーを、Fc受容体および/またはC1qなどの補体因子への結合に関して選択することができる。
【0094】
そのようなドメイン(例えばCH1、CH2、CH3、CH4、CκまたはCλ)からなる分子またはそのようなドメインを含有する分子のインビボ半減期を増加させるために、例えば本発明の突然変異型CH1-、CH2-、CH3-、CH4-、Cκ-またはCλ-ドメインなどのライブラリーを使って、FcRnへの結合を選択することができる。
【0095】
選択用のFcRn受容体は、それぞれの受容体を天然に発現させる細胞の表面上に用意するか、それぞれの受容体の細胞外部分の発現および精製によって用意することができる。本発明の目的には、FcRnでの第1スクリーニングによって突然変異型ドメインを選択し、それをインビトロでさらに試験し、さらには、FcRn受容体を発現させる細胞への結合により、FACS実験でさらに特徴づけることができる。例えば表面プラズモン共鳴技法などを使って、さまざまな組換えFcRn、アイソフォームおよびアロタイプへの結合のアフィニティーランキングにより、さらに特徴づけることができる。
【0096】
本発明の好ましい実施形態によれば、免疫グロブリンは、ヒトまたはネズミ由来である。
【0097】
修飾免疫グロブリンはさまざまな目的に使用することができ、特に医薬組成物では、免疫グロブリンが、好ましくはヒトまたはネズミ由来である。もちろん修飾免疫グロブリンはヒト化またはキメラ免疫グロブリンであってもよい。
【0098】
本発明のもう一つの好ましい実施形態によれば、ヒト免疫グロブリンは、IgA1、IgA2、IgD、IgE、IgG1、IgG2、IgG3、IgG4およびIgMからなる群より選択される。
【0099】
ネズミ免疫グロブリンは、好ましくは、IgA、IgD、IgE、IgG1、IgG2A、IgG2B、IgG2C、IgG3およびIgMからなる群より選択される。
【0100】
修飾免疫グロブリンは、上に挙げた免疫グロブリンクラスの一つから誘導することができる。
【0101】
免疫グロブリンは、好ましくは、免疫グロブリンの重鎖および/もしくは軽鎖またはその一部を含む。
【0102】
修飾免疫グロブリンは、重鎖および/または軽鎖、少なくとも一つの可変および/または定常ドメインを含みうる。
【0103】
本発明の免疫グロブリンは、好ましくは、免疫グロブリンの少なくとも一つの定常ドメインおよび/もしくは少なくとも一つの可変ドメインまたはその一部(ミニドメインを含む)を含む。
【0104】
定常ドメインは、定常領域のドメインとも呼ばれる免疫グロブリンの定常部分の免疫グロブリンフォールド単位である(例えばCH1、CH2、CH3、CH4、Ck、Cl)。
【0105】
可変ドメインは、可変領域のドメインとも呼ばれる免疫グロブリンの可変部分の免疫グロブリンフォールド単位である(例えばVh、Vk、Vl、Vd)。
【0106】
本発明の好ましい免疫グロブリンは、少なくとも一つのループ領域を持つ、CH1、CH2、CH3、CH4、Igk-C、Igl-C、またはその一部(ミニドメインを含む)からなる群より選択される定常ドメインからなり、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする。
【0107】
本発明のもう一つの好ましい免疫グロブリンは、少なくとも一つのループ領域を持つ、重鎖もしくは軽鎖の可変ドメイン、またはその一部(ミニドメインを含む)からなり、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする。
【0108】
好ましい実施形態によれば、定常ドメインは、CH1、CH2、CH3、CH4、Igk-C、Igl-C、またはその一部(ミニドメインを含む)の群より選択される。
【0109】
本発明の修飾免疫グロブリンは、1以上の定常領域(例えば少なくとも2個、3個、4個、5個、6個、10個のドメイン)を含みうる。修飾免疫グロブリン中に存在するドメインが1個より多い場合、これらのドメインは同じタイプであっても、異なるタイプであってもよい(例えばCH1-CH1-CH2、CH3-CH3)。もちろん、単一ドメインの順序も、どの種類であってもよい(例えばCH1-CH3-CH2、CH4-CH1-CH3-CH2)。
【0110】
免疫グロブリンのアミノ酸配列のナンバリングは全てIMGTナンバリングスキームに従う(IMGT, the international ImMunoGeneTics information system@imgt.cines.fr;http://imgt.cines.fr;Lefrancら, 1999, Nucleic Acids Res. 27: 209-212;Ruizら, 2000 Nucleic Acids Res. 28: 219-221;Lefrancら, 2001, Nucleic Acids Res. 29: 207-209;Lefrancら, 2003, Nucleic Acids Res. 31: 307-310;Lefrancら, 2005, Dev Comp Immunol 29:185-203)。
【0111】
本発明の好ましい実施形態によれば、CH1、CH2、CH3およびCH4の修飾ループ領域は、アミノ酸7〜21、アミノ酸25〜39、アミノ酸41〜81、アミノ酸83〜85、アミノ酸89〜103およびアミノ酸106〜117を含む。
【0112】
ヒト由来のIgk-CおよびIgl-Cのループ領域は、好ましくは、アミノ酸8〜18、アミノ酸27〜35、アミノ酸42〜78、アミノ酸83〜85、アミノ酸92〜100、アミノ酸108〜117およびアミノ酸123〜126を含む。
【0113】
ネズミ由来のIgk-CおよびIgl-Cのループ領域は、好ましくは、アミノ酸8〜20、アミノ酸26〜36、アミノ酸43〜79、アミノ酸83〜85、アミノ酸90〜101、アミノ酸108〜116およびアミノ酸122〜125を含む。
【0114】
ヒト由来の免疫グロブリンの可変ドメインの構造ループ領域は、好ましくは、アミノ酸8〜20、アミノ酸44〜50、アミノ酸67〜76およびアミノ酸89〜101を含む。
【0115】
本発明の好ましい実施形態によれば、ネズミ由来の免疫グロブリンの可変ドメインの構造ループ領域は、アミノ酸6〜20、アミノ酸44〜52、アミノ酸67〜76およびアミノ酸92〜101を含む。
【0116】
上に定義した各免疫グロブリンのアミノ酸領域は、修飾されるべきループ領域を含む。
【0117】
本発明の免疫グロブリンは、好ましくは、ラクダ由来である。
【0118】
ラクダ抗体は一本の重鎖だけを含み、軽鎖および重鎖からなる通常の抗体と同じ抗原アフィニティを持つ。したがってラクダ抗体は例えばヒト抗体などよりもはるかに小さく、それゆえに、密な組織に浸透して、大きなタンパク質では到達できないような抗原に到達することができる。さらにまた、ラクダの重鎖抗体は、比較的単純であり、アフィニティおよび特異性が高く、活性部位に到達しそれと相互作用する潜在能力を持つので、臨床的に有益な化合物の設計、生産および応用において、一般的な抗体よりも有利である。
【0119】
ラクダ由来の免疫グロブリンは、好ましくは、CH1、CH2およびCH3からなる群より選択される少なくとも一つの定常ドメインを含む。
【0120】
本発明の好ましい実施形態によれば、ラクダ免疫グロブリンのCH1、CH2およびCH3のループ領域は、アミノ酸8〜20、アミノ酸24〜39、アミノ酸42〜78、アミノ酸82〜85、アミノ酸91〜103およびアミノ酸108〜117を含む。
【0121】
本発明の好ましい実施形態によれば、分子に対する修飾免疫グロブリンの特異的結合は、免疫学的アッセイ、好ましくは酵素結合免疫吸着アッセイ(ELISA)、表面プラズモン共鳴アッセイ、飽和移動差核磁気共鳴分光法、転移NOE(trNOE)核磁気共鳴分光法、競合アッセイ、組織結合アッセイ、生細胞結合アッセイおよび細胞抽出物アッセイからなる群より選択される結合アッセイによって決定される。
【0122】
結合アッセイは、例えばFRET(蛍光共鳴エネルギー移動)およびBRET(生物発光共鳴エネルギー移動)に基づくアッセイ、AlphaScreen(商標)(増幅発光近接均一アッセイ(Amplified Luminescent Proximity Homogeneous Assay))、シンチレーション近接アッセイ、ELISA(酵素結合免疫吸着アッセイ)、SPR(表面プラズモン共鳴、BIACORE(登録商標)とも呼ばれている)、等温滴定熱量測定法、示差走査熱量測定、ゲル電気泳動、およびクロマトグラフィー(ゲル濾過を含む)などといった(ただしこれらに限定されるわけではない)、当技術分野で知られるさまざまな方法を使って行うことができる。これらの方法および他の方法は、何らかの融合パートナーまたはラベルを利用するものでありうる。
【0123】
修飾免疫グロブリンを、好ましくは、有機分子、酵素ラベル、放射性ラベル、着色ラベル、蛍光ラベル、発色ラベル、発光ラベル、ハプテン、ジゴキシゲニン、ビオチン、金属錯体、金属、コロイド金およびそれらの混合物からなる群より選択されるラベルにコンジュゲートする。
【0124】
修飾免疫グロブリンは、例えば結合アッセイ(例:ELISA)および結合研究などにおいて、当該コンジュゲートの簡単な検出を可能にする他の分子にコンジュゲートすることもできる。
【0125】
本発明のもう一つの側面は、少なくとも一つのループ領域を持つ、CH1、CH2、CH3、CH4、Igk-C、Igl-C、もしくはその一部(ミニドメインを含む)、またはそれらの組み合わせからなる群より選択される定常ドメインからなり、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする免疫グロブリンに関する。
【0126】
少なくとも一つの修飾抗体ドメイン(=非可変配列または構造ループによって特異的パートナーに結合するもの)を、少なくとも一つの他の結合分子(これは、抗体、抗体断片、可溶性受容体、リガンドまたはもうひとつの修飾抗体ドメインであることができる)と、分子的に組み合わせることが好ましい。
【0127】
分子は、タンパク質性分子、核酸、および糖質からなる群より選択される。
【0128】
修飾免疫グロブリンのループ領域は、どの種類の結合分子(特にタンパク質性分子、タンパク質、ペプチド、ポリペプチド、核酸、グリカン、糖質、脂質、小さい有機分子、無機分子)に特異的に結合してもよい。もちろん、修飾免疫グロブリンが少なくとも二つのループ領域を含んでいて、それらのループ領域のそれぞれが他の分子またはエピトープに特異的に結合してもよい。
【0129】
本発明の好ましい実施形態によれば、修飾構造ループ領域に結合する分子は、腫瘍関連抗原、特にEpCAM、腫瘍関連糖タンパク質72(TAG-72)、腫瘍関連抗原CA125、前立腺特異膜抗原(PSMA)、高分子量メラノーマ関連抗原(HMW-MAA)、ルイスY関連糖質を発現させる腫瘍関連抗原、癌胎児性抗原(CEA)、CEACAM5、HMFG PEM、ムチンMUC1、MUC18およびサイトケラチン腫瘍関連抗原、細菌抗原、ウイルス抗原、アレルゲン、フルオレセイン、リゾチーム、toll様受容体9、エリスロポエチン、CD2、CD3、CD3E、CD4、CD11、CD11a、CD14、CD18、CD19、CD20、CD22、CD23、CD25、CD28、CD29、CD30、CD33(p67タンパク質)、CD38、CD40、CD40L、CD52、CD54、CD56、CD80、CD147、GD3、IL-1、IL-1R、IL-2、IL-2R、IL-4、IL-5、IL-6、IL-6R、IL-8、IL-12、IL-15、IL-18、IL-23、インターフェロンα、インターフェロンβ、インターフェロンγ;TNF-α、TNFβ2、TNFα、TNFαβ、TNF-R1、TNF-RII、FasL、CD27L、CD30L、4-1BBL、TRAIL、RANKL、TWEAK、APRIL、BAFF、LIGHT、VEG1、OX40L、TRAIL受容体-1、A1アデノシン受容体、リンフォトキシンβ受容体、TACI、BAFF-R、EPO;LFA-3、ICAM-1、ICAM-3、インテグリンβ1、インテグリンβ2、インテグリンα4/β7、インテグリンα2、インテグリンα3、インテグリンα4、インテグリンα5、インテグリンα6、インテグリンαv、αVβ3インテグリン、FGFR-3、ケラチノサイト増殖因子、VLA-1、VLA-4、L-セレクチン、抗Id、E-セレクチン、HLA、HLA-DR、CTLA-4、T細胞受容体、B7-1、B7-2、VNRインテグリン、TGFβ1、TGFβ2、エオタキシン1、BLyS(Bリンパ球刺激因子)、補体C5、IgE、第VII因子、CD64、CBL、NCA90、EGFR(ErbB-1)、Her2/neu(ErbB-2)、Her3(ErbB-3)、Her4(ErbB4)、組織因子、VEGF、VEGFR、エンドセリン受容体、VLA-4、糖質、例えば血液型抗原および関連糖質、ガリリグリコシレーション(Galili Glycosylation)、ガストリン、ガストリン受容体、腫瘍関連糖質、ハプテンNP-capまたはNIP-cap、T細胞受容体α/β、E-セレクチン、ジゴキシン、胎盤アルカリホスファターゼ(PLAP)および精巣PLAP様アルカリホスファターゼ、トランスフェリン受容体、へパラナーゼI、ヒト心ミオシン、糖タンパク質IIb/IIIa(GPIIb/IIIa)、ヒトサイトメガロウイルス(HCMV)gHエンベロープ糖タンパク質、HIV gp120、HCMV、呼吸器合胞体ウイルスRSV F、RSVF Fgp、VNRインテグリン、Hep B gp120、CMV、gpIIbIIIa、HIV IIIB gp120 V3ループ、呼吸器合胞体ウイルス(RSV)Fgp、単純ヘルペスウイルス(HSV)gD糖タンパク質、HSV gB糖タンパク質、HCMV gBエンベロープ糖タンパク質、クロストリジウムパーフリンジェンス毒素およびその断片からなる群より選択される。
【0130】
本発明の修飾免疫グロブリンは、好ましくは、上に開示した分子の一つに結合しうる。これらの分子はアレルゲンも含む。
【0131】
本発明のもう一つの好ましい実施形態によれば、CH3の位置15〜17、29〜34、85.4〜85.3、92〜94、97〜98および/または108〜110のアミノ酸残基が修飾される。
【0132】
本発明の免疫グロブリンの修飾は、好ましくは、欠失、置換または挿入である。
【0133】
本発明によれば、少なくとも1個、好ましくは少なくも2、3、4、5、6、7、8、9、10および15個のアミノ酸が、削除されるか、他のアミノ酸で置換(修飾アミノ酸によるものを含む)されるか、または免疫グロブリンのループ領域中に挿入される。しかし、免疫グロブリンのループ領域中に挿入されるアミノ酸の最大数は、アミノ酸数30、好ましくは25、より好ましくは20を越えないだろう。アミノ酸の置換および挿入は、好ましくは、当技術分野で知られる方法により、本特許出願に開示するように、ランダムに行われる。
【0134】
本発明の免疫グロブリンは、ある特定の実施形態によれば、CH3領域が、EpCamが前記免疫グロブリンに結合する場合は配列番号16または配列番号18を、フルオレセインが前記免疫グロブリンに結合する場合は配列番号20を、リゾチームが前記免疫グロブリンに結合する場合は配列番号22、24、26、28、30または32を、TLR9が前記免疫グロブリンに結合する場合は配列番号34、36、38または40を、そしてリゾチームおよび/またはエリスロポエチンが前記免疫グロブリンに結合する場合は配列番号42を含むことを特徴とする。
【0135】
本発明の、ある特定の実施形態によれば、免疫グロブリンは、リゾチームおよびgp41が前記免疫グロブリンに結合する場合に、免疫グロブリンが配列番号44または配列番号46を含むことを特徴とする。
【0136】
修飾免疫グロブリンは、好ましくは、有機分子、酵素ラベル、放射性ラベル、着色ラベル、蛍光ラベル、発色ラベル、発光ラベル、ハプテン、ジゴキシゲニン、ビオチン、金属錯体、金属、コロイド金およびそれらの混合物からなる群より選択されるラベルまたはレポーター分子にコンジュゲートされる。
【0137】
上に明示したラベルにコンジュゲートされた修飾免疫グロブリンは、例えば診断方法に使用することができる。
【0138】
本発明のもう一つの側面は、能動免疫用ワクチンの製造を目的とする、本発明の免疫グロブリンまたは本発明の方法によって得ることができる免疫グロブリンの使用に関する。この場合、免疫グロブリンは抗原性原薬として、ワクチンの調剤に使用されるか、ワクチン製剤用の抗原構造を釣り上げるためまたは捕捉するために使用される。
【0139】
本発明のもう一つの側面は、免疫グロブリンのタンパク質ライブラリーの作製を目的とする、本発明の免疫グロブリンまたは本発明の方法によって得ることができる免疫グロブリンの使用に関する。
【0140】
本発明のさらにもう一つの側面は、ある分子を特異的に結合および/または検出するための方法であって、
(a)本発明の修飾免疫グロブリンまたは本発明の方法によって得ることができる修飾免疫グロブリンを、前記分子を含有すると疑われる試験試料と接触させるステップ、および
(b)特異的免疫グロブリン/分子複合体の潜在的形成を検出するステップ
を含む方法に関する。
【0141】
本発明のもう一つの側面は、ある分子を特異的に単離する方法であって、
(a)本発明の修飾免疫グロブリンまたは本発明の方法によって得ることができる修飾免疫グロブリンを、前記分子を含有する試料と接触させるステップ、
(b)形成された特異的免疫グロブリン/分子複合体を分離するステップ、および
(c)要すれば、前記複合体から分子を単離するステップ
を含む方法に関する。
【0142】
本発明の免疫グロブリンは、試料から分子を特異的に単離するために使用することができる。多重特異性免疫グロブリンを使用すれば、2以上の分子を試料から単離することができる。そのような方法で修飾免疫グロブリンを使用することはとりわけ有利である。なぜなら、単離されるべき分子に結合することができる所定の量の結合パートナー(すなわち修飾免疫グロブリン)が固定化されている均一な表面を持つマトリックスを作製することができるからである。これに対し、単一特異性結合パートナーを使用した場合には、均一なマトリックスを作製することができない。なぜなら、その単一結合パートナーはマトリックスに同じ効率では結合しないからである。
【0143】
本発明のもう一つの側面は、ある化合物を標的にターゲティングするための方法であって、
(a)前記化合物に特異的に結合する能力を持つ、本発明の修飾免疫グロブリンまたは本発明の方法によって得ることができる修飾免疫グロブリンを接触させるステップ、
(b)免疫グロブリン/化合物複合体を標的に送達するステップ、
を含む方法に関する。
【0144】
本発明の修飾免疫グロブリンは、CDRおよび/または修飾ループ領域に結合した少なくとも一つの化合物を、標的に送達するために使用することができる。そのような免疫グロブリンは、疾患の処置の過程で、治療物質を好ましい作用部位にターゲティングするために使用することができる。
【0145】
本発明のもう一つの側面は、本発明の修飾免疫グロブリンまたは本発明の方法によって得ることができる修飾免疫グロブリンを含むタンパク質ライブラリーに関する。
【0146】
前記ライブラリーを構築するための好ましい方法は、上記の説明および実施例に見出すことができる。本発明のライブラリーは、特異な分子に結合する免疫グロブリンを同定するために使用することができる。
【0147】
特に本発明は、免疫グロブリン誘導体の設計を目的とする、本発明の免疫グロブリンまたは本発明の方法によって得ることができる免疫グロブリンを含むタンパク質ライブラリーの使用に関する。少なくとも一つの修飾ループを持つ、少なくとも10、好ましくは100、より好ましくは1000、さらに好ましくは10000、さらに好ましくは100000、最も好ましくは1000000を越える変種ドメインを含む各ドメインのタンパク質ライブラリーを用いることにより、既存の免疫グロブリンを変化させて、任意のドメインまたはミニドメイン中に抗原結合部位を導入することができる。次にライブラリーを、特異的抗原への結合に関してスクリーニングする。望ましい性質に関する分子特性解析の後、選択したドメインまたはミニドメインを、遺伝子工学的技法により、それが野生型領域と置き換わるような形で、元の免疫グロブリン中にクローニングする。あるいは、ループをコードするDNAまたは突然変異させたアミノ酸をコードするDNAだけを交換して、特異的抗原に対する追加結合部位を持つ免疫グロブリンを得ることもできる。
【0148】
突然変異型抗原特異的構造ループを得るための部位の選択は、元の免疫グロブリンの構造および追加結合部位の目的に依存する。例えば元の分子が、エフェクター機能を乱さずに追加抗原結合部位を挿入される必要がある完全な免疫グロブリンである場合、修飾されるべきループは、Fcエフェクター分子に対する天然の結合パートナーであるCH2およびCH3から離れたドメインより選択されるだろう。元の免疫グロブリンがFabである場合は、軽鎖または重鎖の定常ドメインまたは各可変ドメインのループを修飾することができる。ライブラリーを作製するために、1以上のドメインの1以上の構造ループ中に突然変異を持つ突然変異型の元分子のライブラリーを調製することができる。完全な突然変異型元分子を使った選択はいくつかの利点を持ちうる。というのも、突然変異型免疫グロブリンが示すべき他の性質についても試験すれば、修飾構造ループによる抗原結合に関する選択によって、立体的に有利な修飾がもたらされるだろうからである。
【0149】
突然変異型ドメインもしくはミニドメインまたはドメインの融合分子のタンパク質ライブラリーのサイズ要件(すなわち変種タンパク質の数)は、作業課題に依存する。一般に、抗原結合部位を新規に生成させるためのライブラリーは、修飾構造ループでできた既存の改変抗原結合部位を(例えば抗原に対するアフィニティーを高めたり、抗原に対する微細特異性を変化させたりする目的で)さらに修飾するために用いられるライブラリーよりも大きい必要がある。
【0150】
本発明は、複数の免疫グロブリン(例えば定常もしくは可変ドメイン、ミニドメインおよび/またはミニドメインに含有される少なくとも一つの構造ループ領域)またはそれをコードする核酸分子を含む免疫グロブリンライブラリーまたは核酸ライブラリーにも関係する。このライブラリーは、異なる修飾を持つメンバーを含有し、その複数性(plurality)は、少なくとも一つの構造ループ領域中の修飾によって定義づけられる。核酸ライブラリーは、好ましくは、(一つのアミノ酸交換をもたらす)少なくとも10個の異なるメンバー、より好ましくは少なくとも100個、より好ましくは1000個または10000個の異なるメンバー(例えばランダム突然変異戦略またはコンビナトリアル技法によって設計されたもの)を含む。さらに多くの多様化された個別メンバーの数、例えば少なくとも1000000または少なくとも10000000なども好ましい。
【0151】
本発明のさらにもう一つの側面は、少なくとも二つの本発明ライブラリーから選択される二つの異なるドメインまたはミニドメインの組み合わせによる多重特異性免疫グロブリンの作製である。これらの選択された特異的免疫グロブリンを、ビルディングブロックのように、互いに、そして他の分子と組み合わせることにより、望ましい性質が得られるようにドメインまたはミニドメインの最適な配置を設計することができる。
【0152】
さらにまた、1以上の修飾免疫グロブリンを、あるタンパク質の考えうるさまざまな部位に、または考えうる異なる部位の全てに、そのタンパク質の構造を破壊せずに導入することもできる。そのような「ドメインシャフリング」技法により、新しいライブラリーを作り出し、それをまた望ましい性質について選択することができる。
【0153】
好ましいライブラリーは、免疫グロブリンのドメイン、ミニドメインまたはその誘導体からなる群より選択される本発明の免疫グロブリンを含有する。
【0154】
本発明の好ましい実施形態は、ある抗原に対する結合分子(抗原結合分子)であって、少なくとも一つの免疫ドメインと、その抗原に結合するように本発明に従って修飾される構造ループ領域とを含み、抗体の可変ドメインを含まない結合分子である。これは、抗体活性に使用することができる他の部分(例えば天然または修飾エフェクター領域(配列))を含んでもよいが、抗体の「天然」の結合領域は欠いている(すなわち可変ドメインを本来の位置には持っていない)。本発明のこれらの抗原結合分子は、本分子について上述した利点を持つが、抗体の特異的結合活性は持たず、その一方で、構造ループ領域中に新たに導入された特異的結合活性を持つ。
【0155】
好ましくは、本発明のこれらの抗原結合分子は、CH1、CH2、CH3、CH4、Igk-C、Igl-Cおよびそれらの組み合わせを含み、前記組み合わせは、少なくとも2個、好ましくは少なくとも4個、とりわけ少なくとも6個の定常ドメインと、本発明に従って修飾された少なくとも1個の構造ループ領域とを含む。好ましくは、これらの構造ループ領域は、本発明に従って修飾された構造ループ領域によってつながれるか、またはそのような2個の定常ドメイン間に天然に存在する構造ループ領域によってつながれる。本発明のこれらの抗原結合分子の一実施形態は、本発明に従って構造ループ中に少なくとも一つの修飾を持つ、抗体のFc領域からなる。本発明の抗原結合分子については、構造ループ中の新しい抗原結合部位がランダム突然変異技術によって(すなわちループの1以上のアミノ酸残基をランダム突然変異技法で交換することによって、またはランダムに生成させたインサートをそのような構造ループに導入することによって)導入されることも好ましい。あるいは、コンビナトリアルアプローチの使用も好ましい。
【0156】
もう一つの側面によれば、本発明は、1以上の構造ループ中に組み込まれた、無修飾免疫グロブリンにとって外来である抗原結合部位を持つ、修飾免疫グロブリンに関する。「外来」という用語は、その抗原結合部位がその免疫グロブリンのその特定領域によって天然には形成されないこと、そしてその抗原結合部位によって外来の結合パートナーは結合されるが、免疫グロブリンの天然の結合パートナーは結合されないことを意味する。これは、結合パートナー(例えばFc受容体または免疫系のエフェクター)が、無修飾免疫グロブリンにとって外来である抗原結合部位によっては、結合されないとみなされることを意味する。
【0157】
好ましくは、抗原は、病原体抗原、腫瘍関連抗原、酵素、基質、自己抗原、有機分子またはアレルゲンからなる群より選択される。より好ましい抗原は、ウイルス抗原、細菌抗原または真核生物の病原体もしくはファージに由来する抗原からなる群より選択される。好ましいウイルス抗原には、HAV-、HBV-、HCV-、HIV I-、HIV II-、パルボウイルス-、インフルエンザ-、HSV-、肝炎ウイルス、フラビウイルス、ウエストナイルウイルス、エボラウイルス、ポックスウイルス、天然痘ウイルス、麻疹ウイルス、ヘルペスウイルス、アデノウイルス、パピローマウイルス、ポリオーマウイルス、パルボウイルス、ライノウイルス、コクサッキーウイルス、ポリオウイルス、エコーウイルス、日本脳炎ウイルス、デングウイルス、ダニ媒介脳炎ウイルス、黄熱ウイルス、コロナウイルス、呼吸器合胞体ウイルス、パラインフルエンザウイルス、ラクロスウイルス、ラッサウイルス、狂犬病ウイルス、ロタウイルス抗原などがある;好ましい細菌抗原には、シュードモナス-、マイコバクテリウム-、スタフィロコッカス-、サルモネラ-、髄膜炎菌-、ボレリア-、リステリア、ナイセリア-、クロストリジウム-、エシェリキア-、レジオネラ-、バチルス-、ラクトバチルス-、ストレプトコッカス-、エンテロコッカス-、コリネバクテリウム-、ノカルジア-、ロドコッカス-、モラクセラ-、ブルセラ、カンピロバクター-、カルジオバクテリウム-、フランシセラ-、ヘリコバクター-、ヘモフィルス-、クレブシエラ-、シゲラ-、エルシニア-、ビブリオ-、クラミジア-、レプトスピラ-、リケッチア-、マイコバクテリウム-、トレポネーマ-、バルトネラ-抗原などがある。病原性真核生物の好ましい真核抗原には、ジアルジア、トキソプラズマ、シクロスポラ、クリプトスポリジウム、旋毛虫、酵母、カンジダ、アスペルギルス、クリプトコッカス、ブラストミセス、ヒストプラスマ、コクシジオイデス由来の抗原などがある。
【0158】
本発明の好ましい免疫グロブリンは少なくとも二つの抗原結合部位を含み、第1部位は第1エピトープに結合し、第2部位は第2エピトープに結合する。
【0159】
好ましい実施形態によれば、本免疫グロブリンは少なくとも二つのループ領域を含み、第1ループ領域は第1エピトープに結合し、第2ループ領域は第2エピトープに結合する。少なくとも第1ループ領域もしくは少なくとも第2ループ領域またはその両方が構造ループを含有しうる。本発明の免疫グロブリンは、本発明の必須要素(すなわち本発明に従って修飾された構造ループ領域)を含有する、機能的であることが当技術分野で知られているその断片を包含する。
【0160】
好ましくは、本発明の免疫グロブリンは、少なくとも二つの免疫グロブリンドメイン、またはその一部(ミニドメインを含む)から構成され、各ドメインは少なくとも一つの抗原結合部位を含有する。
【0161】
また、免疫グロブリンの定常領域の少なくとも一つのドメインおよび/または可変領域の少なくとも一つのドメイン、またはその一部(ミニドメインを含む)を含む、本発明の免疫グロブリンも好ましい。したがって、例えばC末端領域が修飾されている可変ドメイン、または修飾CH1領域(例えば修飾CH1ミニドメイン)に連結された可変領域は、好ましい実施形態の一つである。
【0162】
本発明の好ましい免疫グロブリンは、無修飾ドメインと少なくとも50%の相同性を持つドメインを含む。
【0163】
「相同性」という用語は、ポリペプチドがその一次、二次または三次構造中の対応する位置に同じ残基または保存された残基を持つことを示す。この用語は、相同なポリペプチドをコードする2以上のヌクレオチド配列にも及ぶ。
【0164】
「相同免疫グロブリンドメイン」とは、本明細書に開示する完全長ネイティブ配列免疫グロブリンドメイン配列または完全長免疫グロブリンドメイン配列の他の任意の断片に対して、少なくとも約50%のアミノ酸配列一致度を持つ、本発明の免疫グロブリンドメインを意味する。好ましくは、相同免疫グロブリンドメインは、ネイティブ免疫グロブリンドメイン配列、または本明細書に開示する完全長免疫グロブリンドメイン配列の他の任意の明確に定義された断片に対して、少なくとも約50%のアミノ酸配列一致度、好ましくは少なくとも約55%のアミノ酸配列一致度、より好ましくは少なくとも約60%のアミノ酸配列一致度、より好ましくは少なくとも約65%のアミノ酸配列一致度、より好ましくは少なくとも約70%のアミノ酸配列一致度、より好ましくは少なくとも約75%のアミノ酸配列一致度、より好ましくは少なくとも約80%のアミノ酸配列一致度、より好ましくは少なくとも約85%のアミノ酸配列一致度、より好ましくは少なくとも約90%のアミノ酸配列一致度、より好ましくは少なくとも約95%のアミノ酸配列一致度を持つだろう。
【0165】
本明細書で同定される免疫グロブリン配列に対する「パーセント(%)アミノ酸配列一致度」は、最大のパーセント配列一致度が得られるように配列を整列させて、必要ならばギャップを導入した後に、特定免疫グロブリンドメイン配列中のアミノ酸残基と一致する候補配列中のアミノ酸残基のパーセンテージ(保存的置換はいずれも配列一致部分とはみなさない)と定義される。パーセントアミノ酸配列一致度を決定するためのアラインメントは、当技術分野で知られるさまざまな方法により、例えばBLAST、BLAST-2、ALIGNまたはMegalign(DNASTAR)ソフトウェアなどの公に利用可能なコンピュータソフトウェアを使って達成することができる。当業者は、比較する配列の全長にわたって最大のアラインメントを得るのに必要な任意のアルゴリズムを含めて、アラインメントを判定するのに適したパラメータを決定することができる。
【0166】
%アミノ酸配列一致度の値は、後述のように、WU-BLAST-2コンピュータプログラム(Altschulら, Methods in Enzymology 266:460-480 (1996))を使って得ることができる。WU-BLAST-2検索パラメータの大半はデフォルト値に設定される。デフォルト値に設定されない値、すなわち調節可能パラメータは、以下の値に設定される:オーバーラップスパン(overlap span)=1、オーバーラップフラクション(overlap fraction)=0.125、ワード閾値(word threshold)(T)=11、およびスコア行列(scoring matrix)=BLOSUM62。WU-BLAST-2を使用する場合、%アミノ酸配列一致度値は、(a)WU-BLAST-2によって決定される、ネイティブ免疫グロブリンドメインから誘導された配列を持つ関心対象の免疫グロブリンドメインのアミノ酸配列と、関心対象の比較アミノ酸配列(すなわち、関心対象の免疫グロブリンドメインと比較される配列であり、これは、無修飾免疫グロブリンドメインであることができる)との間の、一致する同一アミノ酸残基の数を、(b)関心対象の免疫グロブリンドメインの非ランダム突然変異部分のアミノ酸残基の総数で割ることによって決定される。例えば「アミノ酸配列Bに対して少なくとも80%のアミノ酸配列一致度を持つアミノ酸配列Aを含むポリペプチド」という記述において、アミノ酸配列Aは、関心対象の比較アミノ酸配列であり、アミノ酸配列Bは、関心対象の免疫グロブリンドメインのアミノ酸配列である。
【0167】
好ましい実施形態において、本発明の免疫グロブリンは、二重特異性抗体または二重特異性単鎖抗体である。免疫グロブリンが二重特異性ドメインまたはその一部(ミニドメインを含む)を含むことは、さらに好ましい。
【0168】
本発明の免疫グロブリンは、免疫グロブリンに関して当技術分野で知られているどの目的にも使用できるが、本発明によって導入される特異性の組み合わせに依存する応用も可能にする。したがって本発明の免疫グロブリンは、好ましくは、治療および予防用途に(例えば能動免疫療法または受動免疫療法として)、製造および分析用途に、ならびに診断用途に使用される。
【0169】
本発明のもう一つの側面は、
(a)1以上の構造ループ中に組み込まれた、当該免疫グロブリンにとっては外来の抗原結合部位を持つ、修飾免疫グロブリン、および
(b)前記抗原のエピトープを含有する結合分子
を含有する、結合パートナーのキットに関する。
【0170】
本発明のこのキットのそのような結合分子は、本発明の修飾免疫グロブリンの結合特異性を同定するために使用することができる。本発明のこのキットの結合分子を使用することにより、本発明の修飾免疫グロブリンの効力を決定することができる。
【0171】
本明細書にいう効力とは、修飾分子の、その抗原に対する結合性である。結合は、品質管理目的に使用される特異性および/またはアフィニティおよび/またはアビディティに関して、定量的および/または定性的に決定することができる。
【0172】
さらにまた、本発明のキットの結合分子は、構造ループ中に異なる修飾を持つ少なくとも10、好ましくは少なくとも100、より好ましくは少なくとも1000、より好ましくは少なくとも10000、特に少なくとも100000個の免疫グロブリンからなるライブラリーより、本発明の修飾免疫グロブリンを選択するために使用することもできる。
【0173】
本発明によれば、本発明の重要な特徴の一つは、免疫グロブリンドメインの設計が、通常は抗原結合に関与しない領域で行われること、言い換えると、抗体のCDR以外の領域で行われることにある。免疫グロブリンドメインの特異的フォールドは、CDRと構造的に類似するが配列における位置が異なっている領域に、ランダム突然変異を導入することを許容することが、観察された。本発明によって同定された領域は、CDRと同様に、免疫グロブリンフォールドのβストランドをつなぐループ領域である。
【0174】
より具体的に述べると、本明細書には、ヒトIgG1 CH3ドメインのβストランドA-BおよびE-Fをつなぐループ中にランダム突然変異を導入することにより、それぞれヒトのIgG1 CH3ドメインによって通常は認識および結合されないペプチドおよびタンパク質であるToll様受容体9ペプチド(TLR-9)または鶏卵リゾチームに特異的に結合する突然変異型CH3ドメインが選択されたことを記載する。本発明者らが導入した突然変異には、野生型配列中の選択したアミノ酸残基をランダムに選択された残基で置き換える突然変異が含まれ、また、上述したループ中への余分なアミノ酸残基の挿入も含まれる。
【0175】
類推により、どの免疫グロブリンクラスの免疫グロブリンドメインでも、どの種に由来する免疫グロブリンの免疫グロブリンドメインでも、このタイプの設計を適用することができる。さらにまた、本発明において標的とする特定ループを操作することができるだけでなく、免疫グロブリンドメイン中のβストランドをつなぐ任意のループを同じように操作することができる。
【0176】
本発明では、どの生物およびどの免疫グロブリンクラスに由来する改変免疫グロブリンドメインでも、そのまま(単一ドメインとして)、またはより大きな分子の一部として、使用することができる。例えば、それらはインタクト免疫グロブリンの一部であることができる(したがって、そのインタクト免疫グロブリンは、6個のCDRによって形成されるその「通常の」抗原結合領域と、新しい改変抗原結合領域とを持つだろう)。これと同様に、多重特異性(例えば二重特異性)免疫グログロブリンを作製することができる。改変免疫グロブリンドメインは、任意の融合タンパク質の一部であることもできる。これらの改変免疫グロブリンドメインは、免疫グロブリンの使用分野全般に有用である。
【0177】
以下の免疫グロブリンのドメインは、本明細書において、免疫グロブリンドメインと理解される:
IgG、IgDおよびIgAの場合:VL、CL、VH、CH1、CH2、CH3
IgMおよびIgEの場合:VL、CL、VH、CH1、CH2、CH3、CH4。
【0178】
1.片側に、すなわちβストランドB-C、D-EもしくはF-Gをつなぐループ(多くの特許がカバーしている可変ドメインを除く「先端部」)内に、またはβストランドA-B、C-D(可変ドメインの場合はC-C'およびC''-D)もしくはE-Fをつなぐループ(「基部」)内に、ランダム突然変異が導入された単一の免疫グロブリンドメイン。単一のループまたはループの任意の組み合わせにランダム突然変異を導入することができる。残基は変化もしくは欠失させるか、または追加残基を挿入することができる。
【0179】
2.両側(先端部および基部)にランダム突然変異が導入された単一の免疫グロブリンドメイン。
【0180】
3.ランダム突然変異が導入された単一ドメインの一つを含有する任意のタンパク質、例えば
a)片側または両側にランダム突然変異が導入された「単鎖CH3」二量体(scCH3)、scCH2、scCH1/CL、
b)「基部」側に、すなわちCDRとは反対側に、ランダム突然変異が導入された単鎖Fv、
c)「基部」に、すなわちCH1およびCLドメインのC末端に、ランダム突然変異が導入されたFab断片、
d)片側または両側にランダム突然変異が導入されたFc断片(すなわちCH2-CH3からなるタンパク質)、
e)Fcの基部にランダム突然変異が導入された完全な免疫グロブリン、
f)他の適当なドメイン。
【0181】
単一ドメインの主な利点は、ラクダVH分子(「ナノ抗体(nanobody)」、www.ablynx.com参照)を奨励するために用いられる議論の全てと非常に似ている。ランダム突然変異が導入された免疫グロブリンドメインは極めて小さいタンパク質(挿入されるアミノ酸残基の数に依存して分子量約12〜15kDa)であるので、従来の抗体または抗体断片、例えばscFvおよびFabと比較して、以下の利点を持つだろう:珍しいまたは隠れたエピトープを認識すること、タンパク質標的の空洞または活性部位中に結合すること、製造が容易であること、およびその他数多くの利点。両側にランダム突然変異が導入される免疫グロブリンドメインの場合は、二価または二重特異性分子を生成させることができる。融合タンパク質の一部としての単一ドメインの主な利点は、他の任意のタンパク質上に追加の結合性を設計することができるということである。
【0182】
タンパク質の製造には任意の発現系を使用することができると考えられる。本明細書に記載する単一ドメインとの類似性を、VLがなくVHしか持たないラクダ由来の抗体に見出すことができる。これらのタンパク質では(「通常の」抗体のように6個ではなく)3個のCDRが抗原結合を担っている。
【0183】
以下の特許文献は、参照により、あたかもその全てを本明細書に記載したかのように、本明細書に組み入れられる:
US 6,294,654「Modified immunoglobulin molecule incorporating an antigen in a non-CDR loop region(非CDRループ領域に抗原を組み入れた修飾免疫グロブリン分子)」
US 5,844,094「Target binding polypeptide(標的結合性ポリペプチド)」
US 5,395,750「Methods for producing proteins which bind to predetermined antigens(所定の抗原に結合するタンパク質を製造するための方法)」
US 2004/0071690「High avidity polyvalent and polyspecific reagents(高アビディティ多価および多重特異性試薬)」
US 2004/0018508「Surrogate antibodies and methods of preparation and use thereof(代理抗体ならびにその製造および使用方法)」
US 2003/0157091「Multi-functional proteins(多機能タンパク質)」
US 2003/0148372「Method to screen phage display libraries with different ligands(異なるリガンドを使ってファージディスプレイライブラリーをスクリーニングする方法)」
US 2002/0103345「Bispecific immunologobulin-like antigen binding proteins and method of production(二重特異性免疫グロブリン様抗原結合タンパク質および製造方法)」
US 2004/0097711「Immunoglobulin superfamily proteins(免疫グロブリンスーパーファミリータンパク質)」
US 2004/0082508「Secreted proteins(分泌タンパク質)」
US 2004/0063924「Secreted proteins(分泌タンパク質)」
US 2004/0043424「Immunoglobulin superfamily proteins(免疫グロブリンスーパーファミリータンパク質)」
US 5,892,019「Production of a single-gene-encoded Immunoglobulin(単一遺伝子にコードされた免疫グロブリンの製造)」
US 5,844,094「Target binding polypeptide(標的結合ポリペプチド)」。
【0184】
以下の図面および実施例では本発明をさらに詳しく例示するが、本発明はこれらに限定されるわけではない。
図1aはインタクトIgG1の構造を表す。ドメインを矢印で示す。図1bは、主なヒト免疫グロブリンアイソタイプ単量体の構造的構成を図解している。ジスルフィド結合を線分で表し、N結合型糖質基を丸で表す。
図2は免疫グロブリンの定常ドメイン(左)および可変ドメイン(右)について、免疫グロブリンフォールドを表す。βストランドを矢印で示す。
図3は本発明の改変CH3ドメインの分子モデルを表し、ランダム突然変異を導入した部分が、溶媒接触表面によって示されている。この表面を丸で囲む。
図4は突然変異型CH3ドメインのアセンブリーに用いる断片の作製に使用したPCRを表す概略図である。PCRプライマーをそれぞれの5'−3'方向に矢印で示し、垂直線は導入した制限部位(突然変異型遺伝子のアセンブリーに使用したもの)のおよその位置を示す。PCR断片のライゲーションに備えてプライマーには以下の制限部位が含まれている:CH3LNCO:NcoI;CH3LSACおよびCH3CSAC:SacI;CH3CHINおよびCH3RHIN:HindIII;CH3RNOT:NotI。
図5は本願の免疫グロブリンドメインの使い方の例をいくつか表している。ランダム突然変異が導入された領域を星印で示す。一分子中のランダム突然変異導入領域の特異性は同一でも異なってもよい。
図6は二重特異性改変CH3ドメインの設計を表す概略図である。プライマー名を枠内に記載し、矢印はプライマーが伸長される方向を示す。斜線を引いた枠は、このコンストラクトにおいてランダム突然変異が導入された領域の相対的位置を示し、垂直線を引いた枠は、クローンC24を生成させるために導入された領域の相対的位置を示し、このクローニング手法に使用した制限部位が記載されている。
図7は二重特異性改変CH3ドメインの設計を表す概略図である。二重特異性改変CH3ドメインの基本設計のヌクレオチド配列およびその翻訳を表す。赤い配列は、二重特異性コンストラクトを生成させるためにランダム突然変異を導入した領域を示し、緑の枠はクローンC24を生成させるためにランダム突然変異を導入した領域を示す。
図8は本明細書に開示する配列の配列表である。
【実施例】
【0185】
実施例1
CH3ライブラリーの構築およびファージ表面ディスプレイ
突然変異型CH3ドメインの設計を支援するために、Brookhaven Databaseにエントリー1OQO.pdbとして公表されているIgG1 Fc断片の結晶構造を使った。
【0186】
CH3ライブラリーを構築するための基礎として使用した配列を配列番号1に示す。この配列では、第1アミノ酸がBrookhavenデータベースエントリー1oqo.pdbのA鎖のプロリン343に相当する。1oqo.pdbに含有される最後の残基は配列番号1のセリン102である。1oqo.pdbの構造の詳細な解析後に、βストランドをつなぐループを形成する残基を目視検査することにより、配列番号1の、βストランドA-Bをつなぐループの一部である残基17、18および19、ならびにβストランドE-Fをつなぐループの一部である残基71、72、73、76、および77に、ランダム突然変異を導入することにした。改変CH3ドメインの分子モデル(ランダム突然変異を導入した部分が溶媒接触表面で示されている)を図3に示す。一連のPCR反応に続いて、得られたPCR産物のライゲーションを行うことにより、改変遺伝子を作製した。ライゲーションを容易にするために、アミノ酸配列を変えずに制限部位が生じるように、配列番号1をコードするヌクレオチド配列のコドンの一部を修飾した(サイレント突然変異)。クローニングベクターpHEN1(Nucleic Acids Res. 1991 Aug 11;19(15):4133-7「Multi-subunit proteins on the surface of filamentous phage: methodologies for displaying antibody (Fab) heavy and light chains(繊維状ファージ表面上のマルチサブユニットタンパク質:抗体(Fab)重鎖および軽鎖をディスプレイするための方法論)」Hoogenboom HR, Griffiths AD, Johnson KS, Chiswell DJ, Hudson P, Winter G.)中にpelB分泌シグナルとインフレームで挿入するために、Met-Alaをコードする余分なヌクレオチド残基を配列の5'末端に取り付けて、NcoI制限部位を作った。ランダム突然変異を導入する残基については、20の天然アミノ酸の全てをコードするが、3つの停止コドンのうちの2つを回避する、コドンNNS(IUPACコード、この場合、SはCまたはGを意味する)を選択した。改変配列を配列番号2にヌクレオチド配列として記載し、配列番号3にアミノ酸配列として記載する。配列番号3中の文字Xは、ランダム突然変異が導入されたアミノ酸残基を表す。突然変異型CH3ドメインのアセンブリーに使用したPCRプライマーの配列を配列番号4〜9に示す。図4に、突然変異型遺伝子のアセンブリーのために生成させたPCR断片およびそのために使用したプライマーの概略図を示す。
【0187】
ヒトモノクローナル抗体3D6の重鎖のcDNA(Felgenhauer M, Kohl J, Rueker F.「Nucleotide sequences of the cDNAs encoding the V-regions of H- and L-chains of a human monoclonal antibody specific to HIV-1-gp41(HIV-1-gp41に特異的なヒトモノクローナル抗体のH鎖およびL鎖のV領域をコードするcDNAのヌクレオチド配列)」Nucleic Acids Res. 1990 Aug 25;18(16):4927)をPCR反応のテンプレートとして使用した。3つのPCR産物をSacIおよび/またはHindIIIでそれぞれ消化し、一つにライゲートした。そのライゲーション産物をさらにNcoIおよびNotIで消化し、前もってNcoIおよびNotIで消化しておいた表面ディスプレイファージミドベクターpHEN1中にライゲートした。選択したいくつかのクローンを制限解析およびDNA配列決定によって確認したところ、正しく挿入されたランダム突然変異配列を含むインサートを計画どおりに含有することがわかった。その後のファージ調製ステップについては、標準的なプロトコールに従った。簡単に述べると、ライゲーション混合物をエレクトロポレーションによって大腸菌TG1細胞に形質転換した。次に、ファージ粒子を大腸菌TG1細胞からヘルパーファージM13-K07を使ってレスキューした。次に、培養上清からPEG/NaClにより2段階でファージ粒子を沈殿させ、水に溶解し、パンニングによる選択に使用するか、あるいはそれらを−80℃で保存した。
実施例2
【0188】
CH3+3ライブラリーの構築
このライブラリーは、CH3ライブラリーと同じ方法で構築し、クローニングした。コンストラクトのアミノ酸配列を配列番号10に示し、対応するヌクレオチド配列を配列番号11に示す。構築に使用したプライマーは配列番号4〜7、配列番号9および配列番号12である。
実施例3
【0189】
CH3+5ライブラリーの構築
このライブラリーは、CH3ライブラリーと同じ方法で構築し、クローニングした。コンストラクトのアミノ酸配列を配列番号13に示し、対応するヌクレオチド配列を配列番号14に示す。構築に使用したプライマーは配列番号4〜7、配列番号9および配列番号15である。
実施例4
【0190】
TLR-9ペプチドによるCH3ファージライブラリーのパンニング
標準的プロトコールに従って3ラウンドのパンニングを行った。簡単に述べると、以下の方法を適用した。Maxisorp 96ウェルプレート(Nunc)をToll様受容体9(TLR-9)の配列の一部を表す合成ペプチドでコートした。ペプチドが以下の濃度で溶解している0.1M 炭酸Na緩衝液(pH9.6)200μlを各ウェルに加えた:
第1ラウンドのパンニング:1mg/ml TLR-9ペプチド;
第2ラウンドのパンニング:500μg/ml TLR-9ペプチド;
第3ラウンドのパンニング:100μg/ml TLR-9ペプチド。
【0191】
インキュベーションを37℃で1時間行った後、各ウェル200μlの2%ドライミルク(M-PBS)により、室温で1時間のブロッキングを行った。
【0192】
次に、ファージ懸濁液100μlおよび4%ドライミルク(M-PBS)100μlを加えた後、室温で、振とうしながら45分間、振とうせずに90分間インキュベートすることにより、表面ディスプレイファージライブラリーを、結合したペプチドと反応させた。
【0193】
結合していないファージ粒子を以下のように洗い流した。第1ラウンドのパンニング後:T-PBS 300μl×10、PBS 300μl×5;第2ラウンドのパンニング後:T-PBS 300μl×15、PBS 300μl×10;第3ラウンドのパンニング後:T-PBS 300μl×20、PBS 300μl×20。
【0194】
結合したファージ粒子の溶出は、各ウェル200μlの0.1Mグリシン(pH2.2)を加え、室温で振とうしながら30分間インキュベートすることによって行った。次に、2M Tris塩基60μlの添加によってファージ懸濁液を中和してから、指数増殖期の培養物10mlを、溶出させたファージ0.5mlと混合して、37℃で30分間インキュベートすることにより、大腸菌TG1細胞に感染させた。最後に、感染した細菌を、1%グルコースおよび100μg/mlアンピシリンを含むTYE培地に播種し、30℃で終夜インキュベートした。
【表1】

実施例5
【0195】
TLR-9に対して選択されたCH3突然変異体の選ばれたクローンの可溶性発現のためのクローニング
3ラウンドのパンニングによって選択されたファージから、ミディプレップによってファージミドDNAを単離した。突然変異型CH3領域をコードするDNAを、PCRによってバッチ増幅し、ベクターpNOTBAD/Myc-His(これは、大腸菌発現ベクターpBAD/Myc-His(Invitrogen)に、クローニングを容易にするためのNotI制限部位が挿入されているものである)に、NcoI-NotIクローニングした。ライゲートしたコンストラクトをエレクトロポレーションによって大腸菌LMG194細胞(Invitrogen)に形質転換し、1%グルコースおよびアンピシリンを含むTYE培地にて、30℃で終夜成長させた。選択したクローンを、アンピシリンを含む2×YT培地200μlに接種し、30℃で終夜成長させ、L-アラビノースを最終濃度が0.1%になるように加えることによって誘導した。16℃で終夜発現させた後、細胞を遠心分離によって収集し、ペリプラズム抽出物を調製するために、100μlのホウ酸Na緩衝液(pH8.0)により、4℃で終夜処理した。50μlのペリプラズム抽出物をELISA(下記参照)に使用した。
実施例6
【0196】
TLR-9に対して選択されたCH3突然変異体のELISA
選択したクローンをTLR-9ペプチドへの特異的結合についてELISAでアッセイした。
コーティング:マイクロタイタープレート(NUNC, Maxisorp)、各ウェル100μl、20μg TLR-9ペプチド/ml-0.1M炭酸Na緩衝液(pH9.6)、37℃で1時間;
洗浄:PBS 200μl×3;
ブロッキング:1%BSA-PBS、室温で1時間;
洗浄:PBS 200μl×3
ペリプラズム抽出物結合:ペリプラズム抽出物50μl、2%BSA-PBS 50μl、室温で終夜;
洗浄:PBS 200μl×3;
一次抗体:抗His4(Qiagen)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
二次抗体:ヤギ抗マウス*HRP(SIGMA)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
検出:クエン酸Na/リン酸緩衝液(pH4.5)中の3mg/ml OPD、30%H2O2 0.4μl;
停止:3M H2SO4 100ml;
吸光度の読取り:492/620nm。
【0197】
この最初の予備ELISAで高いシグナルを与えたクローンを、上記と同じ条件下に、20mlの体積で培養した。そのペリプラズム抽出物を、上述のように、培養体積の1/20の体積に単離し、確認のためにELISA(上述したもの)で試験した。
【表2】
実施例7
【0198】
鶏卵リゾチームによるCH3およびCH3+5ファージライブラリーのパンニング
3ラウンドのパンニングを行った。鶏卵リゾチームが以下の濃度で溶解しているPBS 200μlを各ウェルに加えることにより、Maxisorp 96ウェルプレート(Nunc)を鶏卵リゾチームでコートした:
第1ラウンドのパンニング:2mg/ml HEL;
第2ラウンドのパンニング:1mg/ml HEL;
第3ラウンドのパンニング:1mg/ml HEL。
【0199】
インキュベーションを37℃で1時間行った後、各ウェル200μlの2%ドライミルク(M-PBS)により、室温で1時間のブロッキングを行った。
【0200】
次に、ファージ懸濁液100μlおよび4%ドライミルク(M-PBS)100μlを加えた後、室温で、振とうしながら45分間、振とうせずに90分間インキュベートすることにより、表面ディスプレイファージライブラリーを、結合した鶏卵リゾチームと反応させた。
【0201】
結合していないファージ粒子を以下のように洗い流した。
第1ラウンドのパンニング後:T-PBS 300μl×10、PBS 300μl×5;
第2ラウンドのパンニング後:T-PBS 300μl×15、PBS 300μl×10;
第3ラウンドのパンニング後:T-PBS 300μl×20、PBS 300μl×20。
【0202】
結合したファージ粒子の溶出は、各ウェル200μlの0.1Mグリシン(pH2.2)を加え、室温で振とうしながら30分間インキュベートすることによって行った。次に、2M Tris塩基60μlの添加によってファージ懸濁液を中和してから、指数増殖期の培養物10mlを、溶出させたファージ0.5mlと混合して、37℃で30分間インキュベートすることにより、大腸菌TG1細胞に感染させた。最後に、感染した細菌を、1%グルコースおよび100μg/mlアンピシリンを含むTYE培地に播種し、30℃で終夜インキュベートした。
【表3】
【表4】
実施例8
【0203】
実施例7の選ばれたクローンの可溶性発現のためのクローニング
可溶性発現のための、選ばれたクローンのクローニングは、TLR-9に対して選択されたCH3突然変異体について上述したとおりに行った。
実施例9
【0204】
実施例7の選ばれたクローンの可溶性発現
選ばれたクローンの可溶性発現は、TLR-9に対して選択されたCH3突然変異体について上述したとおりに行った。ペリプラズム抽出物を予備ELISAで試験した(プロトコールは実施例10参照)。
【0205】
この最初の予備ELISAで高いシグナルを与えたクローンを、上記と同じ条件下に、20mlの体積で培養した。そのペリプラズム抽出物を、上述のように、培養体積の1/20の体積に単離し、確認のためにELISA(実施例10に記載)で試験した。
実施例10
【0206】
鶏卵リゾチームに対して選択されたCH3突然変異体のELISA
コーティング:マイクロタイタープレート(NUNC, Maxisorp)、各ウェル100μl、PBS中の100μg鶏卵リゾチーム/ml、37℃で1時間;
洗浄:PBS 200μl×3;
ブロッキング:1%BSA-PBS、室温で1時間;
洗浄:PBS 200μl×3;
ペリプラズム抽出物結合:ペリプラズム抽出物50μl、2%BSA-PBS 50μl、室温で終夜;
洗浄:PBS 200μl×3;
一次抗体:抗His4(Qiagen)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
二次抗体:ヤギ抗マウス*HRP(SIGMA)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
検出:クエン酸Na/リン酸緩衝液(pH4.5)中の3mg/ml OPD、30%H2O2 0.4μl;
停止:3M H2SO4 100ml;
吸光度の読取り:492/620nm。
【表5】
【表6】
【0207】
鶏卵リゾチームは抗his4抗体と反応するので、比較的高いバックグラウンドが観察された。
【表7】
実施例11
【0208】
CLライブラリー
Fab断片の結晶構造の目視検査(ヒトモノクローナル抗体3D6のFabの構造を使用する:RSCB Protein Data Bank(http://www.rcsb.org/pdb/)エントリー1DFB.PDB(He XMら Proc Natl Acad Sci U S A. 1992 Aug 1;89(15):7154-8))ならびにこのタンパク質の二次構造および三次構造のコンピュータ支援解析(例えばこの目的にはProtein Explorerを使用する(http://molvis.sdsc.edu/protexpl/frntdoor.htm))により、CLドメイン足場のβストランドをつなぐループ領域中に位置する残基を同定することができる。これらの残基は、アミノ酸8〜18、アミノ酸27〜35、アミノ酸42〜78、アミノ酸83〜85、アミノ酸92〜100、アミノ酸108〜117およびアミノ酸123〜126を含む(IMGTナンバリングシステム(Lefranc MPら Nucleic Acids Res. 2005 Jan 1;33 (Database issue):D593-7;Lefranc MPら Dev Comp Immunol. 2005;29(3):185-203)によるナンバリング)。
【0209】
より具体的に述べると、ヒトCLドメイン(配列番号48)内で、残基11、12、14〜18、および92〜95にランダム突然変異を導入する。ランダム突然変異の導入は、関連コドンの位置がヌクレオチド配列5'-NNS-3'(これは、潜在的に20の天然アミノ酸の全てをコードすると共に、3つの停止コドンのうちの2つを回避する配列である)でコードされるPCRプライマーを使って、コード配列をPCR増幅することにより、達成される。ライブラリーインサートは、2回の別個のPCR反応によって増幅され、その二つのPCR断片が、HpyCH4IV制限部位(PCRプライマーによりサイレント突然変異として導入されるもの)により、一つにライゲートされる。さらに、ファージディスプレイベクターpHEN(Hoogenboom HRら Nucleic Acids Res. 1991 Aug 11;19(15):4133-7)へのクローニングのために、制限エンドヌクレアーゼ部位NcoIおよびNotIも、プライマーによってそれぞれ与えられる。ファージディスプレイの場合、CLドメインのC末端システインは含まれないが、修飾CLクローンを例えばFab断片の構築などに使用する時には、後から付加することができる。
【0210】
PCR増幅のテンプレートとしては、ヒトモノクローナル抗体の軽鎖全体をインサートとして含有するpRcCMV-3D6LC(Rueker Fら Ann N Y Acad Sci. 1991 Dec 27;646:212-9)などのプラスミドを使用する。
【0211】
CLドメインの位置92と位置95の間に挿入された追加残基を含有するCL+3(配列番号50、51)およびCL+5(配列番号52、53)ライブラリーの場合は、プライマーCLRHPYの代わりにプライマーCLRHPY3およびCLRHPY5をそれぞれ使用する。
【0212】
pHEN1のNcoI部位にクローニングされる(これにより、コンストラクトのN末端にpelBリーダー配列が取り付けられることになる)、PCRおよびライゲーションの最終産物のヌクレオチド配列およびアミノ酸配列を、以下に示す(配列番号48、49):
【化1】
【0213】
CLライブラリー用のプライマーリスト:
cllnco:5'-cttaccatgg ccgtggctgc accatctgtc ttcatcttcc cgccatctnn snnscagnns nnsnnsnnsn nsgcctctgt tgtgtgc-3'(配列番号56)
cllhpy:5'-tgacaacgtc agggtgctgc tgaggc-3'(配列番号57)
clrhpy:5'-tcagaacgtt gnnsnnsnns nnstacgaga aacacaaagt c-3'(配列番号58)
clrhpy3:5'-tcagaacgtt gnnsnnsnns nnsnnsnnsn nstacgagaa acacaaagtc-3'(配列番号59)
clrhpy5:5'-tcagaacgtt gnnsnnsnns nnsnnsnnsn nsnnsnnsta cgagaaacac aaagtc-3'(配列番号60)
clrnot:5'-catcgcggcc gcctctcccc tgttgaagct c-3'(配列番号61)
【0214】
選ばれたいくつかのライブラリークローン(ファージミドベクターpHEN1中にクローニングされた突然変異型CLドメイン)を制限解析およびDNA配列決定に付して、それらが正しく挿入されたランダム突然変異配列を含むインサートを計画どおりに含有することを確認する。その後のファージ調製ステップについては、標準的なプロトコールに従う。簡単に述べると、ライゲーション混合物をエレクトロポレーションによって大腸菌TG1細胞に形質転換する。次に、ファージ粒子を大腸菌TG1細胞からヘルパーファージM13-K07を使ってレスキューする。次に、培養上清からPEG/NaClにより2段階でファージ粒子を沈殿させ、水に溶解し、パンニングによる選択に使用するか、あるいはそれらを−80℃で保存することができる。
実施例12
【0215】
CH1ライブラリー
Fab断片の結晶構造(ヒトモノクローナル抗体3D6のFabの構造を使用する:RSCB Protein Data Bankエントリー1DFB.PDB)の目視検査、ならびにこのタンパク質の二次構造および三次構造のコンピュータ支援解析(この目的にはProtein Explorerを使用する)により、CH1ドメイン足場のβストランドをつなぐループ領域中に位置する残基を同定することができる。これらの残基は、アミノ酸7〜21、アミノ酸25〜39、アミノ酸41〜81、アミノ酸83〜85、アミノ酸89〜103およびアミノ酸106〜117を含む(IMGTナンバリングシステムによるナンバリング)。
【0216】
より具体的に述べると、ヒトCH1ドメイン内で、残基12〜19および93〜100にランダム突然変異を導入する(配列番号54、55)。ランダム突然変異の導入は、関連コドンの位置がヌクレオチド配列5'-NNS-3'(これは、潜在的に20の天然アミノ酸の全てをコードすると共に、3つの停止コドンのうちの2つを回避する配列である)でコードされるPCRプライマーを使って、コード配列をPCR増幅することにより、達成される。ライブラリーインサートは、2回の別個のPCR反応によって増幅され、その二つのPCR断片が、BstEII制限部位(CH1ドメイン中に天然に存在するもの)により、一つにライゲートされる。さらに、ファージディスプレイベクターpHENへのクローニングのために、制限エンドヌクレアーゼ部位NcoIおよびNotIも、プライマーによってそれぞれ与えられる。ファージディスプレイの場合、CH1ドメインのC末端システインは含まれないが、修飾CH1クローンを例えばFab断片の構築などに使用する時には、後から付加することができる。
【0217】
PCR増幅のテンプレートとしては、ヒトモノクローナル抗体の重鎖全体をインサートとして含有するpRcCMV-3D6HCなどのプラスミドを使用する。
【0218】
pHEN1のNcoI部位にクローニングされる(これにより、コンストラクトのN末端にpelBリーダー配列が取り付けられることになる)、PCRおよびライゲーションの最終産物のヌクレオチド配列およびアミノ酸配列を、以下に示す(配列番号54、55):
【化2】
【0219】
CH1ライブラリー用のプライマーリスト:
CH1LNCO:5'-acgtccatgg ccgcctccac caagggccca tcggtcttcc ccctggcacc ctcctccnns nnsnnsnnsn nsnnsnnsnn sgccctgggc tgcctggtc-3'(配列番号62)
CH1LBST:5'-ggcacggtca ccacgctgct gag-3'(配列番号63)
CH1RBST:5'-agcgtggtga ccgtgcccnn snnsnnsnns nnsnnsnnsa cctacatctg caacgtgaat c-3'(配列番号64)
CH1RNOT:5'-catagcggcc gcagatttgg gctcaacttt cttgtc-3'(配列番号65)
【0220】
選ばれたいくつかのライブラリークローン(ファージミドベクターpHEN1中にクローニングされた突然変異型CH1ドメイン)を制限解析およびDNA配列決定に付して、それらが正しく挿入されたランダム突然変異配列を含むインサートを計画どおりに含有することを確認する。その後のファージ調製ステップについては、標準的なプロトコールに従う。簡単に述べると、ライゲーション混合物をエレクトロポレーションによって大腸菌TG1細胞に形質転換する。次に、ファージ粒子を大腸菌TG1細胞からヘルパーファージM13-K07を使ってレスキューする。次に、培養上清からPEG/NaClにより2段階でファージ粒子を沈殿させ、水に溶解し、パンニングによる選択に使用するか、あるいはそれらを−80℃で保存することができる。
実施例13
【0221】
鶏卵リゾチーム(HEL)によるCH1ファージライブラリーのパンニング
CH1ファージライブラリー(実施例12参照)を使って3ラウンドのパンニングを行う。鶏卵リゾチームが以下の濃度で溶解しているPBS 200μlを各ウェルに加えることにより、Maxisorp 96ウェルプレート(Nunc)を鶏卵リゾチームでコートする:
第1ラウンドのパンニング:2mg/ml HEL;
第2ラウンドのパンニング:1mg/ml HEL;
第3ラウンドのパンニング:1mg/ml HEL。
【0222】
インキュベーションを37℃で1時間行った後、各ウェル200μlの2%ドライミルク(M-PBS)により、室温で1時間のブロッキングを行う。
【0223】
次に、ファージ懸濁液100μlおよび4%ドライミルク(M-PBS)100μlを加えた後、室温で、振とうしながら45分間、振とうせずに90分間インキュベートすることにより、表面ディスプレイファージライブラリーを、結合した鶏卵リゾチームと反応させる。
【0224】
結合していないファージ粒子を以下のように洗い流す:
第1ラウンドのパンニング:T-PBS 300μl×10、PBS 300μl×5
第2ラウンドのパンニング:T-PBS 300μl×15、PBS 300μl×10
第3ラウンドのパンニング:T-PBS 300μl×20、PBS 300μl×20。
【0225】
結合したファージ粒子の溶出は、各ウェル200μlの0.1Mグリシン(pH2.2)を加え、室温で振とうしながら30分間インキュベートすることによって行う。次に、2M Tris塩基60μlの添加によってファージ懸濁液を中和してから、指数増殖期の培養物10mlを、溶出させたファージ0.5mlと混合して、37℃で30分間インキュベートすることにより、大腸菌TG1細胞に感染させる。最後に、感染した細菌を、1%グルコースおよび100μg/mlアンピシリンを含むTYE培地に播種し、30℃で終夜インキュベートする。
【0226】
リゾチームに対して選択されたCH1突然変異体の選ばれたクローンの可溶性発現のためのクローニング
3ラウンドのパンニングによって選択されるファージから、ミディプレップによってファージミドDNAを単離する。突然変異型CH1ドメインをコードするDNAを、PCRによってバッチ増幅し、ベクターpNOTBAD/Myc-His(これは、大腸菌発現ベクターpBAD/Myc-His(Invitrogen)に、クローニングを容易にするためのNotI制限部位が挿入されているものである)に、NcoI-NotIクローニングする。ライゲートしたコンストラクトをエレクトロポレーションによって大腸菌LMG194細胞(Invitrogen)に形質転換し、1%グルコースおよびアンピシリンを含むTYE培地にて、30℃で終夜成長させる。選択したクローンを、アンピシリンを含む2×YT培地200μlに接種し、30℃で終夜成長させ、L-アラビノースを最終濃度が0.1%になるように加えることによって誘導する。16℃で終夜発現させた後、細胞を遠心分離によって収集し、ペリプラズム抽出物を調製するために、100μlのホウ酸Na緩衝液(pH8.0)により、4℃で終夜処理する。50μlのペリプラズム抽出物をELISAに使用する。
【0227】
この最初の予備ELISAで高いシグナルを与えるクローンを、上記と同じ条件下に、20mlの体積で培養する。そのペリプラズム抽出物を、上述のように、培養体積の1/20の体積に単離し、確認のためにELISA(後述)で試験する。
【0228】
鶏卵リゾチームに対して選択されたCH1突然変異体のELISA
コーティング:マイクロタイタープレート(NUNC, Maxisorp)、各ウェル100μl、PBS中の100μg鶏卵リゾチーム/ml、37℃で1時間;
洗浄:PBS 200μl×3;
ブロッキング:1%BSA-PBS、室温で1時間;
洗浄:PBS 200μl×3;
ペリプラズム抽出物結合:ペリプラズム抽出物50μl、2%BSA-PBS 50μl、室温で終夜;
洗浄:PBS 200μl×3;
一次抗体:抗His4(Qiagen)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
二次抗体:ヤギ抗マウス*HRP(SIGMA)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
検出:クエン酸Na/リン酸緩衝液(pH4.5)中の3mg/ml OPD、30%H2O2 0.4μl;
停止:3M H2SO4 100ml;
吸光度の読取り:492/620nm。
【0229】
ELISAシグナルがバックグラウンドシグナルの少なくとも3倍である場合に、そのクローンを陽性であると解釈する。
実施例14
【0230】
鶏卵リゾチーム(HEL)によるCLファージライブラリーのパンニング
CLファージライブラリー(実施例11参照)を使って3ラウンドのパンニングを行う。鶏卵リゾチームが以下の濃度で溶解しているPBS 200μlを各ウェルに加えることにより、Maxisorp 96ウェルプレート(Nunc)を鶏卵リゾチームでコートする:
第1ラウンドのパンニング:2mg/ml HEL;
第2ラウンドのパンニング:1mg/ml HEL;
第3ラウンドのパンニング:1mg/ml HEL。
【0231】
インキュベーションを37℃で1時間行った後、各ウェル200μlの2%ドライミルク(M-PBS)により、室温で1時間のブロッキングを行う。
【0232】
次に、ファージ懸濁液100μlおよび4%ドライミルク(M-PBS)100μlを加えた後、室温で、振とうしながら45分間、振とうせずに90分間インキュベートすることにより、表面ディスプレイファージライブラリーを、結合した鶏卵リゾチームと反応させる。
【0233】
結合していないファージ粒子を以下のように洗い流す:
第1ラウンドのパンニング:T-PBS 300μl×10、PBS 300μl×5;
第2ラウンドのパンニング:T-PBS 300μl×15、PBS 300μl×10;
第3ラウンドのパンニング:T-PBS 300μl×20、PBS 300μl×20。
【0234】
結合したファージ粒子の溶出は、各ウェル200μlの0.1Mグリシン(pH2.2)を加え、室温で振とうしながら30分間インキュベートすることによって行う。次に、2M Tris塩基60μlの添加によってファージ懸濁液を中和してから、指数増殖期の培養物10mlを、溶出させたファージ0.5mlと混合して、37℃で30分間インキュベートすることにより、大腸菌TG1細胞に感染させる。最後に、感染した細菌を、1%グルコースおよび100μg/mlアンピシリンを含むTYE培地に播種し、30℃で終夜インキュベートする。
【0235】
リゾチームに対して選択されたCL突然変異体の選ばれたクローンの可溶性発現のためのクローニング
3ラウンドのパンニングによって選択されるファージから、ミディプレップによってファージミドDNAを単離する。突然変異型CLドメインをコードするDNAを、PCRによってバッチ増幅し、ベクターpNOTBAD/Myc-His(これは、大腸菌発現ベクターpBAD/Myc-His(Invitrogen)に、クローニングを容易にするためのNotI制限部位が挿入されているものである)に、NcoI-NotIクローニングする。ライゲートしたコンストラクトをエレクトロポレーションによって大腸菌LMG194細胞(Invitrogen)に形質転換し、1%グルコースおよびアンピシリンを含むTYE培地にて、30℃で終夜成長させる。選択したクローンを、アンピシリンを含む2×YT培地200μlに接種し、30℃で終夜成長させ、L-アラビノースを最終濃度が0.1%になるように加えることによって誘導する。16℃で終夜発現させた後、細胞を遠心分離によって収集し、ペリプラズム抽出物を調製するために、100μlのホウ酸Na緩衝液(pH8.0)により、4℃で終夜処理する。50μlのペリプラズム抽出物をELISAに使用する。
【0236】
この最初の予備ELISAで高いシグナルを与えるクローンを、上記と同じ条件下に、20mlの体積で培養する。そのペリプラズム抽出物を、上述のように、培養体積の1/20の体積に単離し、確認のためにELISA(後述)で試験する。
【0237】
鶏卵リゾチームに対して選択されたCL突然変異体のELISA
コーティング:マイクロタイタープレート(NUNC, Maxisorp)、各ウェル100μl、PBS中の100μg鶏卵リゾチーム/ml、37℃で1時間;
洗浄:PBS 200μl×3;
ブロッキング:1%BSA-PBS、室温で1時間;
洗浄:PBS 200μl×3;
ペリプラズム抽出物結合:ペリプラズム抽出物50μl、2%BSA-PBS 50μl、室温で終夜;
洗浄:PBS 200μl×3;
一次抗体:抗His4(Qiagen)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
二次抗体:ヤギ抗マウス*HRP(SIGMA)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
検出:クエン酸Na/リン酸緩衝液(pH4.5)中の3mg/ml OPD、30%H2O2 0.4μl;
停止:3M H2SO4 100ml;
吸光度の読取り:492/620nm。
【0238】
ELISAシグナルがバックグラウンドシグナルの少なくとも3倍である場合に、そのクローンを陽性であると解釈する。
実施例15
【0239】
両側にランダム突然変異が導入された免疫グロブリンドメインの構築(二重特異性改変CH3ドメイン)
この実施例では二つの結合特異性を持つ改変免疫グロブリンドメインを記述する。
【0240】
この改変免疫グロブリンドメインの設計には以下の戦略を含めた:
・リゾチームに特異的に結合するCH3+5ライブラリー由来の改変CH3ドメイン・クローンC24(実施例10参照)を、出発点として使用した。
・免疫グロブリンフォールドのβストランドをつないでいて、クローンC24の生成時に突然変異させた残基に対してドメインの反対側に存在する残基を、ランダム突然変異を導入する対象として、この修飾CH3ドメイン中に同定した。
・CH3、CH3+3およびCH3+5ライブラリーについて上述したものと同様の手法で、これらの残基のランダム突然変異導入と、この改変免疫グロブリンドメインの合成とを可能にするPCRプライマーを設計した。
【0241】
ランダム突然変異が導入された位置を含有する四つのPCR産物をライゲートし、完全長インサートをPCRによって増幅した。次に、それらをpHEN-1中にNcoI-NotI部位を利用してクローニングし、大腸菌TG-1細胞に形質転換することにより、約108コロニーのライブラリーを構築した。ランダムに選んだ20個のコロニーを配列決定したところ、ランダム突然変異を導入した位置には、独立して突然変異が導入されていることがわかった。また、「野生型」(C24)配列は観察されなかった。標準的なプロトコールに従ってファージライブラリーを作製し、6.32×1010TU/mlというファージ力価を得た。
【0242】
二重特異性を試験するために、組換えヒトエリスロポエチン(rhEPO)を第2抗体として選択し、一方、最初に設計した鶏卵リゾチームに対する特異性は、そのコンストラクトが保っているものと予想した。rhEPO反応性ファージを4ラウンドのパンニングによって選択した。突然変異導入後も鶏卵リゾチームを結合するC24クローンの集団が保たれるように、rhEPOによる第1ラウンドの選択後に、鶏卵リゾチーム(PBS中、1mg/ml)によるファージ集団のパンニングを1ラウンド行った。その後のパンニングでは、0.1M炭酸Na緩衝液(pH9.6)中の濃度を減らしながら、200μlのrhEPOを、マイクロタイタープレート(Maxisorp, Nunc)の5ウェルにコートした(下記の表参照)。2%M-PBSでブロッキングを行った後、ブロッキング剤中のファージを、室温で2時間結合させた。T-PBSで20回、PBSで20回洗浄した後、0.1Mグリシン(pH2.2)で溶出させ、2M Trisで中和した。溶出させたファージを直ちに使用して、指数増殖期のTG-1を感染させた。感染した細胞をアンピシリン含有培地で選択した。ヘルパーファージM13-K07の重複感染によってファージ粒子を培養上清からレスキューし、PEGで濃縮し、もう1ラウンドのパンニングに使用した。各ラウンドのパンニング後に、インプットファージ数およびアウトプットファージ数を、大腸菌の形質転換単位として決定した(表8)。
【表8】
【0243】
結果として生じたコロニーをプレートから掻き落とし、アンピシリンを含む2×YTで培養し、そのプラスミドDNAをミディプレップで単離した。インサートをPCRで増幅した後、ベクターpNOTBAD中にサブクローニングし、大腸菌E104株に形質転換した。4×72コロニーを、アンピシリンを含む2×YT培地 200μlで培養し、翌日、0.1%L-アラビノースで誘導した。16℃で24時間発現させた後、それらを、200μlのホウ酸Na緩衝液(pH8.0)により、4℃で6時間溶解し、ペリプラズム抽出物をELISAに使用した。
【0244】
ELISAのために、Maxisorpプレートを、PBS中の鶏卵リゾチーム(20μg/ml)または0.1M炭酸Na緩衝液(pH9.6)中のrhEPOで、それぞれ37℃で1時間コートした。1%BSA-PBSによるブロッキングの後、同じブロッキング剤中のペリプラズム抽出物を終夜結合させた。抗His-(4)抗体およびHRP(鶏卵リゾチーム検出用)またはAP(rhEPO検出用)とコンジュゲートしたヤギ抗マウスIgG抗体を使って結合を明らかにした。OPD変換の呈色反応(HRP)は1.25M H2SO4で停止させた後に492/620nmで読取り、pNPP変換(AP)は405/620nmで読み取った。有望な吸光度値を持つ14個のクローンを、20ml規模での発現用に選択した。16℃で24時間のアラビノース誘導後に、細胞を収集し、1mlのホウ酸Na緩衝液中、4℃で終夜溶解し、その溶解物をELISAに使用した。ELISAは4重に並行して上述のように行い、ペリプラズム抽出物を含まないウェルおよび抗原を含まないウェルを陰性対照として使用した。配列番号42、43のクローンを使って結果(表9)を得た。
【表9】
実施例16
【0245】
改変CH3ドメインはFab様形式の二重特異性をもたらす
この実施例で使用するコンストラクトでは、抗体のVL鎖およびVH鎖がどちらも改変CH3ドメインに融合される。
【0246】
HIV-1のgp41上のエピトープを認識するヒトモノクローナル抗体3D6(He XMら Proc Natl Acad Sci U S A. 1992 89:7154-8;Kohl Jら Ann N Y Acad Sci. 1991 646:106-14;Felgenhauer Mら Nucleic Acids Res. 1990 18:4927)のVLおよびVH領域を、鶏卵リゾチームに特異的に結合する改変CH3ドメイン・クローン24の融合パートナーとして使用した。
【0247】
ジスルフィド結合によるVL-CH3/VH-CH3二量体の形成を促進するために、C24配列のC末端に残基Ser-Cysを付加した。
【0248】
二つの鎖3D6VL-C24および3D6VH-C24のそれぞれヌクレオチド配列およびアミノ酸配列を、それぞれ配列番号47、46および配列番号45、44に示す。
【0249】
コード領域の増幅を可能にすると同時に、コード領域を一つにライゲートする際に使用した制限部位(サイレント突然変異)を導入するプライマーを、設計した。遺伝子を発現させるためにピキアパストリス発現系を選択した。コンストラクトを適切なピキアパストリス発現ベクターにクローニングした。すなわち、3D6VL-C24をpPIC9K(最終名:pPIC9K3LC)にクローニングし、3D6VH-C24(最終名:pPICZ3HC)をpPICZalphaAにクローニングした。コンストラクトpPICZ3HCをBglIIで線状化し、ピキアパストリスGS115に形質転換し、形質転換体をゼオシン含有固形培地上で選択した。次に、形質転換体の一つを、SalI線状化コンストラクトpPIC9K3LCの宿主細胞として使用した。次に、二重形質転換体をRDB培地上で選択した。
【0250】
クローンをYPG 30mlに接種し、OD600=10になるまで成長させた後、BMMY培地中の1%メタノールを加えることによって誘導した。この誘導を16℃で36時間続けた。上清を遠心分離によって取り出し、約10倍濃縮した。組換えタンパク質の存在を、抗His(4)抗体を用いるウェスタンブロットによって確認し、約50〜100μg/l-初期培養物の濃度であると見積った。
【0251】
最初の機能試験は、10倍濃縮上清で行った。まず、Maxisorpプレートのウェルを、PBS中20μg/mlの鶏卵リゾチームまたは0.1M炭酸Na緩衝液(pH9.6)中20μg/mlの抗体3D6のエピトープで、それぞれ37℃で1時間コートした。3D6エピトープは組換え生産されたGST融合タンパク質の形で使用した。1%BSA-PBSによるブロッキング後に、濃縮された上清を同じブロッキング剤中で終夜結合させた。抗His(4)抗体およびHRPにコンジュゲートしたヤギ抗マウス抗体を使って、結合を明らかにし、OPD変換がもたらす呈色反応として492/620nmで可視化した(表10)。
【表10】
【図面の簡単な説明】
【0252】
【図1】図1aはインタクトIgG1の構造を表す。ドメインを矢印で示す。図1bは、主なヒト免疫グロブリンアイソタイプ単量体の構造的構成を図解している。ジスルフィド結合を線分で表し、N結合型糖質基を丸で表す。
【図2】免疫グロブリンの定常ドメイン(左)および可変ドメイン(右)について、免疫グロブリンフォールドを表す。βストランドを矢印で示す。
【図3】本発明の改変CH3ドメインの分子モデルを表し、ランダム突然変異を導入した部分が、溶媒接触表面によって示されている。この表面を丸で囲む。
【図4】突然変異型CH3ドメインのアセンブリーに用いる断片の作製に使用したPCRを表す概略図である。PCRプライマーをそれぞれの5'−3'方向に矢印で示し、垂直線は導入した制限部位(突然変異型遺伝子のアセンブリーに使用したもの)のおよその位置を示す。PCR断片のライゲーションに備えてプライマーには以下の制限部位が含まれている:CH3LNCO:NcoI;CH3LSACおよびCH3CSAC:SacI;CH3CHINおよびCH3RHIN:HindIII;CH3RNOT:NotI。
【図5】本願の免疫グロブリンドメインの使い方の例をいくつか表している。ランダム突然変異が導入された領域を星印で示す。一分子中のランダム突然変異導入領域の特異性は同一でも異なってもよい。
【図6】二重特異性改変CH3ドメインの設計を表す概略図である。プライマー名を枠内に記載し、矢印はプライマーが伸長される方向を示す。斜線を引いた枠は、このコンストラクトにおいてランダム突然変異が導入された領域の相対的位置を示し、垂直線を引いた枠は、クローンC24を生成させるために導入された領域の相対的位置を示し、このクローニング手法に使用した制限部位が記載されている。
【図7】二重特異性改変CH3ドメインの設計を表す概略図である。二重特異性改変CH3ドメインの基本設計のヌクレオチド配列およびその翻訳を表す。赤い配列は、二重特異性コンストラクトを生成させるためにランダム突然変異を導入した領域を示し、緑の枠はクローンC24を生成させるためにランダム突然変異を導入した領域を示す。
【図8】本明細書に開示する配列の配列表である。
【図1a】
【図1b】
【技術分野】
【0001】
本発明は修飾免疫グロブリンの設計および製造方法に関する。
【0002】
一般分野は、タンパク質に特異的結合性を付与することを目的とするタンパク質の設計である。より具体的に述べると、本発明に関連する改変タンパク質は、免疫グロブリン(抗体)であり、さらに具体的に述べると、免疫グロブリンの単一ドメインまたは単一ドメインの対または単一ドメインの組み合わせである。免疫グロブリンの特異的結合性は重要な特徴である。というのも、それらは他の分子(例えば抗原)との相互作用を制御し、免疫グロブリンを診断用途および治療用途に役立つものにするからである。
【背景技術】
【0003】
基本的な抗体構造を、ここではインタクトIgG1免疫グロブリンを例にとって説明する。
【0004】
二つの同一な重(H)鎖と二つの同一な軽(L)鎖とが一体となってY字状の抗体分子を形成する。重鎖はそれぞれ四つのドメインを持っている。アミノ末端の可変領域(VH)はそのYの先端部にある。これに、三つの定常ドメイン、すなわちCH1、CH2、およびカルボキシ末端のCH3(これはYの幹の根本にある)が続いている。スイッチ(switch)という短いストレッチが、重鎖可変領域と定常領域とをつなぐ。ヒンジはCH2およびCH3(Fc断片)を抗体の残りの部分(Fab断片)につなぐ。インタクト抗体分子のヒンジのタンパク質分解切断により、一つのFcと二つの同一なFab断片とを生成させることができる。軽鎖は、スイッチによって分離された二つのドメイン、すなわち可変(VL)ドメインと定常(CL)ドメインとでできている。
【0005】
ヒンジ領域にあるジスルフィド結合が、二つの重鎖をつなぐ。軽鎖は、さらなるジスルフィド結合によって、重鎖に結びつけられる。Asn結合型糖質部分は、免疫グロブリンのクラスに依存して、定常ドメイン中の異なる位置に取り付けられる。IgG1の場合は、Cys235対およびCys238対の間に形成されるヒンジ領域中の二つのジスルフィド結合が、二つの重鎖を合体させる。軽鎖は、CH1ドメイン中のCys229とCLドメイン中のCys214との間に形成される二つのさらなるジスルフィド結合によって、二つの重鎖に結びつけられる。糖質部分は各CH2のAsn306に取り付けられて、Yの幹に顕著な出っ張りを作る。
【0006】
これらの特徴は機能上重大な結果をもたらす。重鎖の可変領域(VH)と軽鎖の可変領域(VL)はどちらもYの「先端部」にあり、そこで抗原と反応するように配置される。本分子のこの先端部はアミノ酸配列のN末端が位置している側である。Yの幹は、エフェクター機能、例えば補体の活性化およびFc受容体との相互作用またはADCCおよびADCPが、効率よく媒介されるように、突き出している。そのCH2ドメインおよびCH3ドメインは、エフェクタータンパク質との相互作用が容易になるように膨らんでいる。アミノ酸配列のC末端は先端部とは反対側(これをYの「基部」と称することができる)に位置する。インタクトIgG1の構造を図1aに図解する。
【0007】
抗体にはラムダ(λ)およびカッパ(κ)と呼ばれる2タイプの軽鎖が見出される。与えられた免疫グロブリンはκ鎖を持つか、λ鎖を持つかのどちらかであり、それぞれを一つずつ持つということは決してない。λ軽鎖またはκ軽鎖を持つ抗体の間に機能的相違は見出されていない。
【0008】
主なヒト免疫グロブリンクラス単量体の構造的構成を図1bに示す。これらのクラスは、各重鎖の組成および配列が異なっている。IgMとIgEはどちらもヒンジ領域を欠くが、それぞれに余分な重鎖ドメイン(CH4)を含有している。鎖を連結するジスルフィド結合(線分)の数および位置はアイソタイプ間で異なる。また、円記号で示すN結合型糖質基の分布も異なる。
【0009】
抗体分子中の各ドメインは、二つのβシートが互いに堅く充填されて圧縮された逆平行βバレルになっている類似の構造を持つ。この保存された構造は免疫グロブリンフォールドと呼ばれる。定常ドメインの免疫グロブリンフォールドは、4ストランドシートに向かい合って充填された3ストランドシートを含有する。このフォールドは、各シートのβストランド間に生じる水素結合、内部で向かい合ったシートの残基間に生じる疎水結合、およびシート間のジスルフィド結合によって安定化される。この3ストランドシートはストランドC、FおよびGを含み、4ストランドシートはストランドA、B、E、およびDを持つ。文字A〜Gは、免疫グロブリンフォールドのアミノ酸配列に沿ったβストランドの順序を表す。
【0010】
可変ドメインのフォールドは、4ストランドおよび5ストランドの二つのシートに配置された9本のβストランドを持つ。5ストランドシートは、定常ドメインの3ストランドシートと構造的に相同であるが、余分なストランドC'およびC''を含有している。残りのストランド(A、B、C、D、E、F、G)は、定常ドメイン免疫グロブリンフォールド中の対応部分と同じトポロジーおよび類似する構造を持っている。定常ドメインと同様に、一つのジスルフィド結合が、向かい合ったシート中のストランドBとストランドFとを連結している。免疫グロブリンフォールドを、免疫グロブリンの定常ドメインおよび可変ドメインについて、図2に図解する。
【0011】
免疫グロブリン軽鎖および免疫グロブリン重鎖の可変ドメインはどちらも、三つの超可変ループ、すなわち相補性決定領域(CDR)を含有している。Vドメインの三つのCDR(CDR1、CDR2、CDR3)は、βバレルの一端に集まっている。CDRは、免疫グロブリンフォールドのβストランドB-C、C'-C''およびF-Gをつなぐループである。CDR中の残基は免疫グロブリン分子ごとに異なり、各抗体に抗原特異性を付与している。
【0012】
抗体分子の先端部にあるVLドメインおよびVHドメインは、六つのCDR(各ドメイン上に三つずつ)が協力して抗原特異的な結合のための表面(または空洞)を構築するように、密に充填される。したがって、抗体の天然抗原結合部位は、軽鎖可変ドメインのストランドB-C、C'-C''およびF-G、ならびに重鎖可変ドメインのストランドB-C、C'-C''およびF-Gをつなぐループで構成される。
【0013】
タンパク質の3D構造を設計の一助とし、そのタンパク質のコア構造を足場として使用することにより、多くのタンパク質の表面に位置するアミノ酸残基にランダム突然変異が導入されてきた。この戦略の一例は、参照により本明細書に組み入れられる以下の参考文献に、記載または概説されている:Nygren PA, Uhlen M., Curr Opin Struct Biol. (1997) 7:463-9;Binz HK, Amstutz P, Kohl A, Stumpp MT, Briand C, Forrer P, Grutter MG, Pluckthun A. Nat Biotechnol. (2004) 22:575-82;Vogt M, Skerra A. Chembiochem. (2004) 5:191-9;US 6,562,617。
【0014】
この技法の基本原理は、多くのタンパク質が、ループ、ターン、またはランダムコイルなどの構造によって相互につながれたβシートまたはαヘリックスなどの二次構造要素の特異的配置によって形成される安定なコアを持つという観察結果に基づいている。典型的には、これらのうちループ、ターンおよびランダムコイルという三つの構造要素は、タンパク質の全体構造にとって、さほど決定的ではなく、これらの構造要素中のアミノ酸残基は、多くの場合、タンパク質の全体的フォールドを破壊することなく、交換することができる。この設計原理の天然の例が、抗体のCDRである。人工的な例には、リポカリン、アンキリンおよび他のタンパク質足場が含まれる。
【0015】
ネイティブ免疫グロブリン中のCDRループでないループは、抗原結合特異性またはエピトープ結合特異性を持たないが、免疫グロブリン分子全体の正しいフォールディングおよび/またはそのエフェクターもしくは他の機能に寄与するので、本発明ではこれを構造ループと呼ぶ。
【0016】
米国特許第6,294,654号には、ヒンジ領域と可変領域の間のCH1領域にある抗体の非CDRループにペプチド抗原を組み入れることが可能な改変抗体を作製することができ、その結果生じるAbは、そのペプチド抗原がMHC IIと共にAPCの表面に提示されることによって免疫応答を生成するような形で、APCに取り込まれうることが示されている。これらの挿入ペプチドはエピトープであり、担体分子の全体的構造は重要でない。免疫グロブリンの(非CDR)ループ上にrasペプチドを置くことができること、そしてそれでもなおその免疫グロブリンは分泌されることが、実証された。免疫グロブリンが適正にフォールドされない限り免疫グロブリンが分泌されるのを防止するという、厳格な「品質管理」が細胞内には存在し、ループのアミノ酸配列を変えると、そのタンパク質は、細胞が不適正と感知してそれを分解するような構造に、フォールドされることになるかもしれない。したがって、示された例の他に、免疫グロブリンの性質を変えずに構造ループをさらに修飾することは、困難であると考えられた。
【0017】
米国特許出願公開第2004/0101905号には、標的結合部位およびFcエフェクターペプチドを含む結合分子が記載されている。Fcエフェクターペプチドは、エフェクター分子と相互作用するペプチドである。免疫グロブリン断片のCH1ドメインの非CDRループへのエフェクターペプチドの挿入が示されている。
【0018】
Fcエフェクターペプチドは、抗体の非CDRループ中に天然に存在する構造であり、それゆえに、免疫グロブリン中の異なる等価な位置に移植しても、免疫グロブリンの構造を乱さないと予想される。
【0019】
それにもかかわらず、この開示に従って非CDRループに移植されるペプチドはいずれも、選択された異なる構造環境により、不活性になる可能性が高い。
【0020】
上述したどちらの先行技術文書でも、免疫グロブリンフォールディング構造は機能および分泌にとって重要であるから、これを乱さないことは極めて大切であり、それゆえに、その構造および機能を保つべきループ中にペプチドを挿入することは困難であると、述べられている。
【0021】
米国特許出願公開第2004/0132101号および同第2005/0244403号には、抗体の構造ループにとって天然のリガンドであるエフェクターリガンドに対して、変化した結合アフィニティを持つ、突然変異型免疫グロブリンが記載されている。この文書には、免疫グロブリン分子全体にわたるさまざまな領域中の突然変異であって、抗体全体のエフェクター機能に影響を及ぼすものが、いくつか記載されている。
【0022】
従来から、既存の抗原結合部位を操作し、その結果として、新しい結合性を導入することを目的として、免疫グロブリン様足場が利用されてきたことは、他の先行技術文書によって示されている。しかし従来は、抗原結合のためにCDR領域だけが設計されてきた。言い換えると、免疫グロブリンフォールドの場合は、天然の抗原結合部位だけが、その結合アフィニティまたは結合特異性を変化させるために修飾されてきた。ファージ粒子の表面にディスプレイされるか、さまざまな原核もしくは真核発現系で可溶性に発現される、異なる形式の、そのような操作免疫グロブリン(単鎖Fv断片(scFv)またはFab断片の形で発現されることが多い)を記述している文献は、多量に存在する。この分野の主だった著者にはGreg Winter、Andreas PlueckthunおよびHennie Hoogenboomがいる。
【発明の開示】
【0023】
本発明の目的は、新しい抗原結合部位が導入された免疫グロブリン、および前記免疫グロブリンを設計し製造するための方法を提供することである。
【0024】
したがって本発明は、免疫グロブリンの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンを設計し、ある抗原のエピトープに対する前記免疫グロブリンの結合を決定するための方法(ただし、無修飾免疫グロブリンは前記エピトープには有意に結合しない)であって、
・少なくとも一つの構造ループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記構造ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、および
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ
を含む方法に関する。
【0025】
特に本発明は、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原からなる群より選択される抗原のエピトープに特異的に結合する免疫グロブリンを設計するための方法に関する。構造ループ領域の修飾により、それらのエピトープに結合するように、免疫グロブリンを設計することができる。好ましい実施形態では、免疫グロブリンが、同じ抗原のまたは異なる抗原の、互いに異なる少なくとも二つのそのようなエピトープに特異的に結合する。
【0026】
例えば本発明の方法は、少なくとも一つの第1エピトープに特異的に結合する免疫グロブリンであって、前記免疫グロブリンの少なくとも一つの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンを設計し、前記少なくとも一つのループ領域の、少なくとも一つの第2エピトープに対する特異的結合を決定する方法(ただし、エピトープは、上述した抗原の群から選択され、無修飾構造ループ領域(非CDR領域)は、前記少なくとも一つの第2エピトープには特異的に結合しない)であって、
・少なくとも一つの構造ループ領域を含む、少なくとも一つの第1エピトープに特異的に結合する免疫グロブリンをコードする核酸を用意するステップ、
・前記核酸によってコードされる前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンを前記少なくとも一つの第2エピトープと接触させるステップ、
・前記修飾免疫グロブリンが第2エピトープに特異的に結合するかどうかを決定するステップ
を含む方法に関する。
【0027】
本発明の方法は、好ましくは、少なくとも一つの構造ループ領域中の、少なくとも一つの修飾、ならびにアレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、ウイルス抗原および原虫抗原からなる群より選択される少なくとも一つの抗原に対する、前記少なくとも一つのループ領域の特異的結合を決定すること(ただし、無修飾構造ループ領域を含有する免疫グロブリンは、前記少なくとも一つの抗原には特異的に結合しない)に関する。
【0028】
本発明において、修飾の対象となる「免疫グロブリン」という用語(本明細書で使用する用語、免疫グロブリンと抗体は、相互に可換である)は、抗原、エフェクター分子/タンパク質などのエピトープに対して、単一特異性もしくは多重特異性、または多価結合性、少なくとも二つ、好ましくは少なくとも三つの特異的結合部位を示しうる。また、本発明における免疫グロブリンは、当技術分野で受け入れられている機能的断片、例えばFc、Fab、scFv、CH/CLドメインの単鎖二量体、Fv、または免疫グロブリンの他の誘導体もしくは組み合わせ、インタクト抗体の可変領域(例えばFd、Vl、Vk、Vh)および定常領域の重鎖および軽鎖のドメイン、例えばCH1、CH2、CH3、CH4、ClおよびCk、ならびに構造ループによってつながれた免疫ブロブリンドメインの二つのβストランドからなるミニドメインでもある。
【0029】
「免疫グロブリン」「修飾免疫グロブリン」または「本発明の免疫グロブリン」という用語は、免疫グロブリンの誘導体も包含すると理解される。誘導体は、本発明の1以上の免疫グロブリンの任意の組み合わせ、および/または本発明の免疫グロブリンのどのドメインまたはミニドメインが1以上の他のタンパク質(例えば他の免疫グロブリン、リガンド、足場タンパク質、酵素、毒素など)のどの位置に融合されていてもよい融合タンパク質である。本発明の免疫グロブリンの誘導体は、さまざまな化学的技法(例えば共有結合、静電相互作用、ジスルフィド結合など)で他の物質に結合することによって、取得することもできる。
【0030】
免疫グロブリンに結合される他の物質は、脂質、糖質、核酸、有機分子および無機分子またはその任意の組み合わせ(例えばPEG、プロドラッグまたは薬物)であることができる。誘導体は、同じアミノ酸配列を持つが、完全にまたは部分的に非天然アミノ酸または化学修飾アミノ酸からできている免疫グロブリンでもある。
【0031】
本発明の改変分子は、独立したタンパク質として有用であると共に、融合タンパク質または融合誘導体(最も典型的には、より大きい抗体構造の一部、または完全な抗体分子もしくはその一部、例えばFab断片、Fc断片、Fv断片などになるように融合されたもの)としても有用であるだろう。改変タンパク質を使って、単一特異性、二重特異性、三重特異性である分子、さらにはより多くの特異性を同時に保持する分子を作ることも可能だろう。そしてまた同時に、そのような分子の使用予定の要件に応じて、同時に結合する結合価を制御し、前もって選択することが可能だろう。
【0032】
本発明によれば、あらゆる種類のアレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原に対する結合領域または抗原結合部位を、与えられた抗体構造の構造ループ中に導入することができる。
【0033】
本発明において「抗原」という用語は、免疫グロブリンのCDRループ領域と相互作用することが知られている、または相互作用する能力を持つ、分子または構造を意味するものとする。先行技術の構造ループ領域は抗原とは相互作用せず、むしろ全体的構造および/またはエフェクター分子への結合に寄与する。
【0034】
本発明において「アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原」という用語は、抗体構造によって認識されうる全てのアレルゲンおよび抗原、ならびにそのような分子の断片(特に一般に「エピトープ」と呼ばれる部分構造(例えばB細胞エピトープ))を(それらが免疫に関連する限り、すなわちそれらもまた、天然抗体またはモノクローナル抗体によって認識されうる限り)包含するものとする。
【0035】
本発明において「エピトープ」という用語は、本発明の結合ドメインまたは免疫グロブリンに対する特異的結合パートナーを完全に形成するか、またはそのような特異的結合パートナーの一部であることができる分子構造を意味するものとする。
【0036】
化学的には、エピトープは、糖質、ペプチド、脂肪酸、無機物質またはその誘導体およびそれらの任意の組み合わせから構成されうる。エピトープがポリペプチドである場合、それは通常、少なくとも3個のアミノ酸、好ましくは8〜50個のアミノ酸、より好ましくは約10〜20個のアミノ酸を、そのペプチド中に含むだろう。ペプチドの長さに決定的な上限はなく、ポリペプチド配列のほぼ全長を含んでもよい。エピトープはリニアエピトープまたはコンフォメーショナルエピトープであることができる。リニアエピトープは、ポリペプチド鎖の一次配列の単一のセグメントから構成される。リニアエピトープは連続していたり、一部重複していたりすることができる。コンフォメーショナルエピトープは、ポリペプチドのフォールディングによって寄せ集められて三次元構造を形成するアミノ酸から構成され、それらのアミノ酸は、必ずしもその直線配列中で互いに隣接していない。
【0037】
具体的に述べると、エピトープは、診断に関連する分子の少なくとも一部である。すなわち、試料中のエピトープの有無は、疾患または健康状態または製造時の加工状態または環境および食品状態と、定性的または定量的に相関する。エピトープは、治療に関連する分子(すなわち、疾患の経過を変化させる、特異的結合ドメインの標的になりうる分子)の少なくとも一部であることもできる。
【0038】
好ましい「アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原」は、免疫または治療に関連することが既に証明されているか、免疫または治療に関連する能力を持つアレルゲンまたは抗原、特に、その臨床的有効性が既に試験されているものである。
【0039】
他方、本発明のもう一つの側面によれば、他の結合能力、例えば薬物などの小分子、または酵素、酵素の触媒部位、もしくは酵素基質に対する結合能力、あるいは酵素基質の遷移状態類似体に対する結合能力などを、構造ループ領域に導入することもできる。
【0040】
好ましくは、構造ループ中の新しい抗原結合部位は、無修飾免疫グロブリンにとって外来である。したがって、エフェクター分子またはFc受容体のような標的は、好ましくは、本発明の免疫グロブリンの結合分子および特異性からは除外される。
【0041】
好ましくは、構造ループ中の新しい抗原結合部位は、選択された核酸によってコードされる免疫グロブリンの置換、欠失および/または挿入によって導入される。
【0042】
本発明のもう一つの好ましい実施形態によれば、少なくとも一つのヌクレオチドの修飾が、前記核酸によってコードされる免疫グロブリンの置換、欠失および/または挿入をもたらす。
【0043】
少なくとも一つのループ領域の修飾は、1以上のアミノ酸の置換、欠失および/または挿入、好ましくは点突然変異、ループ全体の交換、より好ましくは、少なくとも2、3、4、5、6、7、8、9、10〜30個のアミノ酸の変化をもたらしうる。
【0044】
部位指定ランダム突然変異も好ましい。この方法では、ループの1以上の特定アミノ酸残基を、ランダムに生成させたインサートを使って交換するか、そのような構造ループ中に導入する。あるいは、コンビナトリアルアプローチの使用も好ましい。
【0045】
少なくとも一つのループ領域を、好ましくは、ランダム、セミランダム、または特に、部位指定ランダム突然変異誘発法によって、突然変異させるか、または修飾する。これらの方法は、本発明の免疫グロブリンの望ましい位置でアミノ酸修飾を行うために使用することができる。これらの場合、位置はランダムに選択されるか、極端に単純化した規則を使ってアミノ酸変化が加えられる。例えば全ての残基をアラニンに突然変異させることができ、これをアラニンスキャニングという。そのような方法を、より高レベルな配列多様性をスクリーニングするための選択法を使用する、より洗練された工学的アプローチと結びつけてもよい。本発明の好ましい方法は、5'-NHS-3'、5'-NNN-3'または5'-NNK-3'という配列を持つ少なくとも一つのヌクレオチド繰返し単位を含むランダムに修飾された核酸分子に関する。
【0046】
ランダムに修飾された核酸分子は、全ての既知天然アミノ酸をコードする、上述の繰返し単位を含むことができる。
【0047】
当技術分野ではよく知られているように、例えば、後述のファージディスプレイ、リボソームディスプレイ、細胞表面ディスプレイなどのディスプレイ技術をはじめとして、一定の結合特性およびアフィニティーを持つタンパク質の同定および単離に使用することができるさまざまな選択技術がある。抗体変種を生産しスクリーニングするための方法は、当技術分野では周知である。抗体の分子生物学、発現、精製およびスクリーニングの一般方法は、DuebelおよびKontermann編「Antibody Engineering」(Springer-Verlag, ハイデルベルク, 2001);ならびにHayhurstおよびGeorgiou, 2001, Curr Opin Chem Biol 5:683-689;MaynardおよびGeorgiou, 2000, Annu Rev Biomed Eng 2:339-76に記述されている。
【0048】
本発明において「構造ループ」または「非CDRループ」とは、以下のように理解すべきである。免疫グロブリンはいわゆる免疫グロブリンフォールドを持つドメインでできている。基本的には、逆平行βシートがループでつながれて、圧縮された逆平行βバレルを形成する。可変領域では、ドメインのループの一部が抗体の特異性(すなわち抗原への結合)にとって不可欠な寄与をする。これらのループはCDRループと呼ばれる。抗体ドメインの他のループは全て、むしろ、その分子の構造および/またはエフェクター機能に寄与している。これらのループを、ここでは構造ループまたは非CDRループと定義する。
【0049】
修飾免疫グロブリン(この用語には本明細書の全体にわたって常に免疫グロブリン断片が包含される)をコードする核酸分子を、宿主細胞にクローニングし、発現させ、その結合特異性についてアッセイすることができる。これらの作業は周知の手法を使って行われ、本発明に役立ちうるさまざまな方法が、「Molecular Cloning-A Laboratory Manual」第3版(Maniatis, Cold Spring Harbor Laboratory Press, ニューヨーク, 2001)および「Current Protocols in Molecular Biology」(John Wiley & Sons)に記述されている。本発明の修飾免疫グロブリンをコードする核酸は、前記免疫グロブリンを発現させるために、発現ベクターに組み入れることができる。発現ベクターは、典型的には、制御配列または調節配列、選択可能マーカー、任意の融合パートナー、および/または追加要素と作動可能に連結された(すなわち機能的な関係に置かれた)免疫グロブリンを含む。本発明の修飾免疫グロブリンは、修飾免疫グロブリンをコードする核酸を含有する核酸(好ましくは発現ベクター)で形質転換された宿主細胞を、修飾免疫グロブリンの発現を誘導しまたは引き起こすのに適当な条件下で培養することによって、生産することができる。外因性核酸分子を宿主に導入する方法は当技術分野では周知であり、使用する宿主によって異なるだろう。もちろん、修飾免疫グロブリンを発現させるための非細胞発現系または無細胞発現系も使用することができる。
【0050】
本発明の好ましい実施形態では、修飾免疫グロブリンを、発現後に精製または単離する。修飾免疫グロブリンは、当業者に知られているさまざまな方法で単離または精製することができる。標準的な精製方法には、クロマトグラフィー技法、電気泳動技法、免疫学的技法、沈降、透析、濾過、濃縮、およびクロマトフォーカシング技法などがある。多くの場合、個別の融合パートナーによって、精製を可能にすることができる。例えば、GST融合物を使用する場合にはグルタチオン樹脂を使って、Hisタグを使用する場合にはNi+2アフィニティクロマトグラフィーを使って、またflagタグを使用する場合には固定化抗flag抗体を使って、抗体を精製することができる。適切な精製技法における一般指針については、「Antibody Purification: Principles and Practice」(第3版, Scopes, Springer-Verlag, NY, 1994)を参照されたい。もちろん、本発明の修飾免疫グロブリンを、宿主の表面に、特に細菌、昆虫もしくは酵母細胞の表面に、またはファージもしくはウイルスの表面に、発現させることも可能である。
【0051】
修飾免疫グロブリンは、例えばインビトロアッセイ、インビボおよび細胞系アッセイ、ならびに選択技術を用いる方法など(ただしこれらに限定されるわけではない)、さまざまな方法を使ってスクリーニングすることができる。スクリーニング手法には自動およびハイスループットスクリーニング技術を利用することができる。スクリーニングには、融合パートナー、またはラベル、例えば酵素、免疫ラベル、同位体ラベル、もしくは小分子ラベル、例えば蛍光色素もしくは比色測定用色素または発光原分子などを利用することができる。
【0052】
好ましい実施形態では、免疫グロブリンの機能的および/または生物物理的性質を、インビトロアッセイでスクリーニングする。好ましい実施形態では、抗体を機能性(例えば反応を触媒する能力またはその標的への結合アフィニティー)について、スクリーニングする。
【0053】
アッセイには、発色ラベル、蛍光ラベル、発光ラベル、または同位体ラベルを含む(ただしこれらに限定されるわけではない)さまざまな検出方法を使用することができる。
【0054】
当技術分野では知られているように、一部のスクリーニング方法は、ライブラリーの好ましいメンバーを選択する方法である。それらの方法をここでは「選択法」と呼び、これらの方法は、本発明において、修飾免疫グロブリンのスクリーニングに役立つ。選択法を使って免疫グロブリンライブラリーをスクリーニングする場合は、好ましいライブラリーのメンバーだけが、すなわち何らかの選択基準に合致するライブラリーのメンバーだけが、増殖され、単離され、そして/または観察される。理解されるであろうが、最も適合した変種だけを観察するので、そのような方法では、ライブラリーメンバーの適合性を個別にアッセイする方法によってスクリーニングすることが可能なライブラリーよりも大きなライブラリーをスクリーニングすることができる。選択は、免疫グロブリンの表現型を、共有結合的にまたは非共有結合的に、その遺伝子型と(すなわち抗体の機能を、それをコードする核酸と)関連づける任意の方法、技法、または融合パートナーによって可能になる。例えば、選択法としてのファージディスプレイの使用は、ライブラリーメンバーを遺伝子IIIタンパク質に融合することによって可能になる。このようにして、何らかの基準(例えばその免疫グロブリンの標的に対する結合アフィニティ)を満たす修飾免疫グロブリンの選択または単離により、それをコードする核酸も選択または単離される。ひとたび単離されたら、次に、修飾免疫グロブリンをコードする遺伝子または遺伝子群を増幅することができる。この単離および増幅の工程(これをパンニングという)を繰り返して、ライブラリー中の好ましい抗体変種を濃縮させることができる。最終的には、取り付けられた核酸の核酸配列決定によって、遺伝子同定が可能になる。
【0055】
当技術分野では、本発明において免疫グロブリンライブラリーのスクリーニングに役立ちうる、さまざまな選択法が知られている。これらには、ファージディスプレイ(「Phage display of peptides and antibodies: a laboratory manual」Kayら, 1996, Academic Press, カリフォルニア州サンディエゴ, 1996;Lowmanら, 1991, Biochemistry 30:10832-10838;Smith, 1985, Science 228:1315-1317)およびその変法、例えば選択的ファージ感染(selective phage infection)(Malmborgら, 1997, J Mol Biol 273:544-551)、選択感染性ファージ(selectively infective phage)(Krebberら, 1997, J Mol Biol 268:619-630)、および遅延感染性パンニング(delayed infectivity panning)(Benharら, 2000, J Mol Biol 301:893-904)、細胞表面ディスプレイ(Witrrup, 2001, Curr Opin Biotechnol, 12:395-399)、例えば細菌(Georgiouら, 1997, Nat Biotechnol 15:29-34;Georgiouら, 1993, Trends Biotechnol 11:6-10;Leeら, 2000, Nat Biotechnol 18:645-648;Junら, 1998, Nat Biotechnol 16:576-80)、酵母(BoderおよびWittrup, 2000, Methods Enzymol 328:430-44;BoderおよびWittrup, 1997, Nat Biotechnol 15:553-557)、および哺乳動物細胞(Whitehornら, 1995, Bio/technology 13:1215-1219)でのディスプレイ、ならびにインビトロディスプレイ技術(Amstutzら, 2001, Curr Opin Biotechnol 12:400-405)、例えばポリソームディスプレイ(Mattheakisら, 1994, Proc Natl Acad Sci USA 91:9022-9026)、リボソームディスプレイ(Hanesら, 1997, Proc Natl Acad Sci USA 94:4937-4942)、mRNAディスプレイ(RobertsおよびSzostak, 1997, Proc Natl Acad Sci USA 94:12297-12302;Nemotoら, 1997, FEBS Lett 414:405-408)、およびリボソーム不活化ディスプレイ系(ribosome-inactivation display system)(Zhouら, 2002, J Am Chem Soc 124, 538-543)などがあるが、これらに限定されるわけではない。
【0056】
本発明に役立ちうる他の選択法には、ディスプレイに頼らない方法、例えばインビボ法、例えばペリプラズム発現およびサイトメトリースクリーニング(Chenら, 2001, Nat Biotechnol 19:537-542)、抗体断片再構成アッセイ(JohnssonおよびVarshavsky, 1994, Proc Natl Acad Sci USA 91:10340-10344;Pelletierら, 1998, Proc Natl Acad Sci USA 95:12141-12146)、および選択モード(Visintinら, 1999, Proc Natl Acad Sci USA 96:11723-11728)で用いられる酵母ツーハイブリッドスクリーニング(FieldsおよびSong, 1989, Nature 340:245-246)などがある(ただしこれらに限定されるわけではない)。これに代わる実施形態では、発現ベクター上の特異的配列に結合することにより、その融合パートナーおよびそれに付随するFc変種ライブラリーメンバーを、それらをコードする核酸と、共有結合的または非共有結合的に関連づける融合パートナーによって、選択が可能になる。例えば、PCT WO 00/22906;PCT WO 01/49058;PCT WO 02/04852;PCT WO 02/04853;PCT WO 02/08023;PCT WO 01/28702およびPCT WO 02/07466には、本発明に役立ちうるそのような融合パートナーおよび技法が記述されている。これに代わる実施形態として、抗体の発現が、細胞に成長上、増殖上、または生存上の利点を何かしら付与するのであれば、インビボ選択を行うことができる。
【0057】
「定方向進化」法と呼ばれる一部の選択法は、選択時に好ましい配列の交配または育種を含み、時には新しい突然変異の組み込みを伴う方法である。当業者には明らかであるだろうが、定方向進化法はライブラリー中の最も好ましい配列の同定を容易にすることができ、スクリーニングの対象となる配列の多様性を増加させることができる。当技術分野では、例えばDNAシャフリング(PCT WO 00/42561 A3;PCT WO 01/70947 A3)、エクソンシャフリング(米国特許第6,365,377号;KolkmanおよびStemmer, 2001, Nat Biotechnol 19:423-428)、ファミリーシャフリング(Crameriら, 1998, Nature 391:288-291;米国特許第6,376,246号)、RACHITT(商標)(Cocoら, 2001, Nat Biotechnol 19:354-359;PCT WO 02/06469)、STEPおよびインビトロ組換えのランダムプライミング(Zhaoら, 1998, Nat Biotechnol 16:258-261;Shaoら, 1998, Nucleic Acids Res 26:681-683)、エキソヌクレアーゼ媒介遺伝子アセンブリー(米国特許第6,352,842号;米国特許第6,361,974号)、Gene Site Saturation Mutagenesis(商標)(米国特許第6,358,709号)、Gene Reassembly(商標)(米国特許第6,358,709号)、SCRATCHY(Lutzら, 2001, Proc Natl Acad Sci USA 98:11248-11253)、DNA断片化法(Kikuchiら, Gene 236:159-167)、一本鎖DNAシャフリング(Kikuchiら, 2000, Gene 243:133-137)、およびAMEsystem(商標)定方向進化抗体工学技術(directed evolution antibody engineering technology)(Applied Molecular Evolution)(米国特許第5,824,514号;米国特許第5,817,483号;米国特許第5,814,476号;米国特許第5,763,192号;米国特許第5,723,323号)など(ただしこれらに限定されるわけではない)、本発明において抗体変種のスクリーニングに役立ちうる、さまざまな定方向進化法が知られている。
【0058】
好ましい実施形態では、1以上の細胞系アッセイまたはインビボアッセイを使って、抗体変種がスクリーニングされる。そのようなアッセイの場合、典型的には、細胞がライブラリーに属する個々の免疫グロブリンまたは免疫グロブリンのプールに曝露されるように、精製または未精製修飾免疫グロブリンが外因的に加えられる。これらのアッセイは、常にではないが典型的には、免疫グロブリンの機能、すなわち標的に結合して何らかの生化学的事象(例えばエフェクター機能、リガンド/受容体結合阻害、アポトーシスなど)を媒介するという抗体の能力に基づいている。そのようなアッセイでは、多くの場合、抗体に対する細胞の応答、例えば細胞の生残、細胞死、細胞形態の変化、または転写活性化、例えば天然遺伝子もしくはレポーター遺伝子の細胞発現などを監視することが必要になる。例えばそのようアッセイでは、ADCC、ADCP、またはCDCを誘発するという抗体変種の能力を測定することができる。一部のアッセイでは、例えば血清補体、またはエフェクター細胞、例えば末梢血単球(PBMC)、NK細胞、マクロファージなどといった、追加の細胞または成分を(すなわち標的細胞以外に)加える必要があるかもしれない。そのような追加細胞は任意の生物(好ましくはヒト、マウス、ラット、ウサギ、およびサル)に由来することができる。免疫グロブリンは、標的を発現するある種の細胞株のアポトーシスを引き起こしたり、アッセイに加えておいた免疫細胞による標的細胞への攻撃を媒介したりすることができる。細胞の死または生存能力を監視するための方法は当技術分野では知られており、色素、免疫化学試薬、細胞化学試薬、および放射性試薬の使用が含まれる。例えばカスパーゼ染色アッセイはアポトーシスの測定を可能にし、放射性基質または蛍光色素、例えばアラマーブルーなどの取り込みまたは放出は、細胞の成長または活性化の監視を可能にしうる。好ましい実施形態では、DELFIA(登録商標)EuTDA系の細胞毒性アッセイ(Perkin Elmer, マサチューセッツ州)を使用することができる。あるいは、1以上の天然細胞内成分(例えば乳酸デヒドロゲナーゼ)の放出を測定することによって、死んだ標的細胞または損傷を受けた標的細胞を監視することもできる。転写活性化も、細胞系アッセイにおいて機能をアッセイする方法として役立ちうる。この場合は、アップレギュレートされうる天然遺伝子または免疫グロブリンをアッセイすることによって、応答を監視することができる。例えば、ある種のインターロイキンの放出を測定するか、あるいはレポーターコンストラクトによって読出しを行うことができる。細胞系アッセイでは、修飾免疫グロブリンの存在に対する応答として、細胞の形態変化を測定する場合もある。そのようなアッセイのための細胞タイプは原核でも真核でもよく、当技術分野で知られるさまざまな細胞株を使用することができる。あるいは、変種をコードする核酸で形質転換またはトランスフェクトされた細胞を使って、細胞系スクリーニングを行う。すなわち細胞に抗体変種を外因的には加えない。例えばある実施形態では、細胞系スクリーニングに、細胞表面ディスプレイを利用する。細胞の表面への修飾免疫グロブリンのディスプレイを可能にする融合パートナーを使用することができる(Witrrup, 2001, Curr Opin Biotechnol, 12:395-399)。
【0059】
好ましい実施形態として、1以上の細胞系アッセイを使って、修飾免疫グロブリンの免疫原性を、実験的に決定することができる。好ましい一実施形態では、エクスビボT細胞活性化アッセイを使って、免疫原性を実験的に定量する。この方法では、一致ドナーから得た抗原提示細胞およびナイーブT細胞を、関心対象のペプチドまたは抗体全体で1回以上攻撃する。次に、いくつかの方法を使って(例えば、サイトカインの産生を監視すること、またはトリチウム化チミジンの取り込みを測定することなどによって)、T細胞活性化を検出することができる。最も好ましい実施形態では、インターフェロンγ産生をElispotアッセイを使って監視する(Schmittelら, 2000, J. Immunol. Meth., 24: 17-24)。
【0060】
本発明の修飾免疫グロブリンの生物学的性質は、細胞、組織、および生物全体を使った実験で特徴づけることができる。当技術分野では知られているように、薬物は、多くの場合、ある疾患または疾患モデルの処置に関する薬物の有効性を測定するために、または薬物の薬物動態、毒性、および他の性質を測定するために、動物(例えばマウス、ラット、ウサギ、イヌ、ネコ、ブタ、およびサルを含むが、これらに限定されるわけではない)で試験される。それらの動物は疾患モデルと呼ぶことができる。治療薬は、多くの場合、マウス(例えばヌードマウス、SCIDマウス、異種移植マウス、およびトランスジェニックマウス(ノックインおよびノックアウトを含む)を含むが、これに限定されるわけではない)で試験される。そのような実験により、治療薬として使用される抗体の潜在能力を決定するための有意義なデータを得ることができる。試験には任意の生物(好ましくは哺乳動物)を使用することができる。例えばサルは、遺伝的にヒトに類似しているので、適切な治療モデルになることができ、したがって本発明の修飾免疫グロブリンの有効性、毒性、薬物動態、または他の性質を試験するために使用することができる。最終的には、薬物として承認されるために、ヒトでの試験が要求されるので、もちろんこれらの実験も予想される。したがって本発明の修飾免疫グロブリンは、その治療有効性、毒性、免疫原性、薬物動態、および/または他の臨床的性質を決定するために、ヒトで試験することができる。
【0061】
本発明の修飾免疫グロブリンは、広範囲にわたる抗体製品に役立ちうる。ある実施形態では、本発明の抗体変種が、治療または予防に、製造または分析用途に、診断薬、工業用化合物、または研究試薬として、好ましくは治療薬として使用される。本抗体変種は、モノクローナルまたはポリクローナルである抗体組成物に役立ちうる。好ましい実施形態では、標的抗原を持つ標的細胞(例えば癌細胞)を殺すために、本発明の修飾免疫グロブリンを使用する。これに代わる実施形態では、例えばサイトカインまたはサイトカイン受容体をアンタゴナイズすることなどにより、標的抗原をブロック(block)、アンタゴナイズ(antagonize)、またはアゴナイズ(agonize)するために、本発明の修飾免疫グロブリンを使用する。もう一つの好ましい実施形態では、標的抗原をブロック、アンタゴナイズまたはアゴナイズし、標的抗原を持つ標的細胞を殺すために、本発明の修飾免疫グロブリンを使用する。もう一つの好ましい実施形態では、成長因子または成長因子受容体をブロック、アンタゴナイズまたはアゴナイズし、標的抗原を持つ標的細胞または標的抗原を必要とする標的細胞を殺すために、本発明の修飾免疫グロブリンを使用する。もう一つの好ましい実施形態では、酵素および酵素の基質をブロック、アンタゴナイズまたはアゴナイズするために、本発明の修飾免疫グロブリンを使用する。
【0062】
本発明の修飾免疫グロブリンは、さまざまな治療目的に使用することができる。好ましい実施形態では、特異的障害を処置するために、本修飾免疫グロブリンを含む抗体を患者に投与する。本発明における「患者」には、ヒトと他の動物、好ましくは哺乳動物がどちらも包含され、最も好ましくはヒトである。本明細書において「特異的障害」とは、本発明の修飾免疫グロブリンを含む医薬組成物の投与によって改善されうる障害を意味する。
【0063】
ある実施形態では、本発明の修飾免疫グロブリンが、患者に投与される唯一の治療活性剤である。あるいは、本発明の修飾免疫グロブリンは、1以上の他の治療剤(例えば細胞毒性剤、化学療法剤、サイトカイン、成長阻害剤、抗ホルモン剤、キナーゼ阻害剤、抗血管新生剤、心保護薬、または他の治療剤などであるが、これらに限定されるわけではない)と組み合わせて投与される。修飾免疫グロブリンは、1以上の他の治療レジメンと同時に投与することができる。例えば本発明の抗体変種は、化学療法、放射線療法、または化学療法と放射線療法の両方と一緒に、患者に投与することができる。一実施形態として、本発明の修飾免疫グロブリンは、本発明の抗体変種を含んでも含まなくてもよい1以上の抗体と共に投与することができる。本発明のもう一つの実施形態によれば、本発明の修飾免疫グロブリンおよび1以上の他の抗癌療法を使って、癌細胞をエクスビボで処置することができる。そのようなエクスビボ処置は、骨髄移植(特に自家骨髄移植)に役立ちうると考えられる。もちろん、本発明の抗体は、さらに他の治療技法、例えば手術などと組み合わせて使用することもできると考えられる。
【0064】
本発明の修飾免疫グロブリンとの併用投与には、さまざまな他の治療剤が役立ちうる。ある実施形態では、修飾免疫グロブリンを抗血管新生剤(血管の発達を阻止するか、血管の発達にある程度干渉する化合物)と共に投与する。抗血管新生因子は、例えば、血管新生の促進に関与する成長因子または成長因子受容体に結合する小分子またはタンパク質(例えば抗体、Fc融合物、またはサイトカイン)であることができる。本発明において好ましい抗血管新生因子は、血管内皮増殖因子(VEGF)に結合する抗体である。これに代わる実施形態では、適応免疫応答を誘導または増進する治療剤(例えばCTLA-4を標的とする抗体)と共に、修飾免疫グロブリンを投与する。これに代わる実施形態では、修飾免疫グロブリンを、チロシンキナーゼ阻害剤(チロシンキナーゼのチロシンキナーゼ活性をある程度阻害する分子)と共に投与する。これに代わる実施形態では、本発明の修飾免疫グロブリンをサイトカインと共に投与する。本明細書にいう「サイトカイン」とは、ある細胞集団が放出するタンパク質であって、別の細胞に細胞間媒介物質として作用するものの総称であり、ケモカインを包含する。
【0065】
本発明の修飾免疫グロブリンと1以上の治療活性剤とが調剤された医薬組成物が考えられる。本発明の抗体変種の製剤は、貯蔵に備えて、望ましい純度を持つ前記免疫グロブリンを、随意の医薬的に許容できる担体、賦形剤または安定剤(「Remington's Pharmaceutical Sciences」第16版, Osol, A.編, 1980)と、凍結乾燥製剤または水溶液の形で混合することにより、製造することができる。インビボ投与に使用される製剤は、好ましくは、滅菌状態にある。これは、滅菌濾過膜を通した濾過または他の方法によって、容易に達成される。本明細書に開示する修飾免疫グロブリンおよび他の治療活性剤は、免疫リポソームとして製剤化し、そして/またはマイクロカプセルに封入することもできる。
【0066】
本発明の修飾免疫グロブリンを含む医薬組成物(好ましくは滅菌水溶液の形態をとっているもの)の投与は、例えば経口投与、皮下投与、静脈内投与、鼻腔内投与、耳内投与、経皮投与、局所外用(例えばゲル剤、軟膏、ローション剤、クリーム剤など)、腹腔内投与、筋肉内投与、肺内投与(例えばAradigmから市販されているAERx(商標)吸入技術、またはInhale Therapeuticsから市販されているInhance(商標)肺送達系)、腟投与、非経口投与、直腸投与、または眼内投与など(ただしこれらに限定されるわけではない)、さまざまな方法で行うことができる。
【0067】
本明細書で使用する「特異的に結合」という用語は、不均一な分子集団における関心対象のコグネイトリガンドを決定するような結合反応を指す。したがって、指定の条件下(例えば免疫グロブリンの場合であれば免疫アッセイ条件下)で、明示された抗体は、その個別の「標的」に結合し、試料中に存在する他の分子には有意な量では結合しない。抗体のCDRに似て、修飾構造ループ領域は、抗原結合性または分子結合性のタンパク質部分であって、抗原そのものではない。
【0068】
「発現系」という用語は、所望のコード配列および制御配列を含有し、これらの配列で形質転換またはトランスフェクトされた宿主が、コードされているタンパク質を産生することができるように、それらが作動可能に連結されている核酸分子を指す。形質転換を達成するには、発現系をベクター上に含めることができるが、関連DNAを宿主染色体に組み込むこともできる。
【0069】
本発明の好ましい実施形態によれば、発現系はベクターを含む。この目的には、当技術分野で知られている任意の発現ベクターを適宜、使用することができる。
【0070】
修飾免疫グロブリンは、好ましくは、宿主中で、好ましくは細菌細胞、酵母細胞、植物細胞中で、動物細胞中で、または植物もしくは動物中で、発現される。
【0071】
例えば哺乳動物細胞(動物細胞)、植物細胞、細菌(例えば枯草菌、大腸菌)、昆虫細胞、および酵母(ピキアパストリス、サッカロミセスセレビシェ)など(ただしこれらに限定されるわけではない)、広範囲わたる種々の適当な宿主細胞を使って、修飾免疫グロブリンを発現させることができる。例えば、本発明に役立ちうる種々の細胞株は、American Type Culture Collectionから入手することができるATCC細胞株カタログに記載されている。さらにまた、本発明の免疫グロブリンを発現させるための宿主として、植物および動物も使用することができる。発現ならびにトランスフェクションベクターまたはカセットは、使用する宿主に応じて選択することができる。
【0072】
もちろん、非細胞または無細胞タンパク質発現系も、使用することができる。十分な量のタンパク質を産生するインビトロ転写/翻訳タンパク質発現プラットフォームは、細胞に基づく発現系に通例付随する面倒な上流工程および下流工程(例えば宿主細胞の形質転換、培養、または溶解)の必要性を排除する無細胞タンパク質発現の多くの利点をもたらす。
【0073】
本発明のもう一つの側面は、免疫グロブリンの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンまたはその医薬調製物を製造し、ある抗原のエピトープに対する前記免疫グロブリンの結合を決定するための方法(ただし、無修飾免疫グロブリンは前記エピトープには有意に結合しない)であって、
・少なくとも一つのループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ、および
・前記エピトープに結合する修飾免疫グロブリンを用意し、要すればそれを医薬調製物に仕上げるステップ、
を含む方法に関する。
【0074】
特に本発明は、免疫グロブリンの少なくとも一つの構造ループ領域に少なくとも一つの修飾を含む、少なくとも一つの第1分子に特異的に結合する多重特異性免疫グロブリンまたはその医薬調製物を製造し、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、原虫抗原およびウイルス抗原からなる群より選択される少なくとも一つの第2分子に対する、前記少なくとも一つのループ領域の特異的結合を決定するための方法(ただし、無修飾構造ループ領域を含有する免疫グロブリンは、前記少なくとも一つの第2分子には特異的に結合しない)であって、
・少なくとも一つの構造ループ領域を含む、少なくとも一つの第1分子に特異的に結合する免疫グロブリンをコードする核酸を用意するステップ、
・前記核酸によってコードされる前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンを前記少なくとも一つの第2分子と接触させるステップ、
・前記修飾免疫グロブリンが第2分子に特異的に結合するかどうかを決定するステップ、および
・前記少なくとも一つの第2分子に特異的に結合する修飾免疫グロブリンを用意し、要すればそれを医薬調製物に仕上げるステップ、
を含む方法に関する。
【0075】
特異的結合対の一つのメンバー中に、一つより多い特異性を設計することが好ましい(Kuferら (2004) Trends in Biotechnology, Vol 22, p. 238-244)。
【0076】
多重特異性(例えば二特異性)モノクローナル抗体または抗体断片を作製するために、数多くの試みがなされてきた。二つの異なるポリペプチド鎖(重鎖および軽鎖)でできた二重特異性抗体を作製する際の課題の一つは、一つの細胞内で四つの異なる鎖(二つの重鎖および二つの軽鎖)を発現させる必要があり、その結果、分子のさまざまな組み合わせがいくつも生じ、それらを、その混合物中の所望の二重特異性分子から分離する必要があるということである。これらの分子の分離は、その類似性ゆえに、困難であり、多くの費用がかかる。そのような不必要な対形成の発生を最小限に抑えるために、いくつかの技法が使用されてきた(Carter (2001) Journal of Immunological Methods, vol 248, p. 7-15)。
【0077】
この課題に対する解決策の一つは、例えば互いに連結された二つのscFvのような、二つの特異性を持つ一本のポリペプチド鎖の作製、またはいわゆるダイアボディの作製である。そのような分子は、天然分子のフォールドとはかけ離れていることが示されており、その作製は困難であることがよく知られている(LeGallら (2004) Protein Engineering, Design & Selection vol 17 p.357-366)。
【0078】
現在の二重特異性抗体設計のもう一つの課題は、たとえ親抗体がそれぞれの結合パートナーに二価で結合するとしても(例えばIgG)、結果として生じる二重特異性抗体はそれぞれの結合パートナーの各々に関して一価であるという事実である。
【0079】
本発明の好ましい多重特異性分子はこれらの課題を解決する。
【0080】
二本の抗体ポリペプチド鎖の発現(Cabillyら Proc. Natl. Acad. Sci. USA 81:3273-3277 (1984))よりも達成が容易な、一本のペプチド鎖としての二重特異性分子の発現が可能である(二つの結合特異性を持つ修飾Igドメイン、実施例参照)。
【0081】
第2の特異性は分子の非可変部分に位置するという事実ゆえに、抗体様分子(すなわち2本のポリペプチド鎖でできた分子)として作製することもでき、二つの異なる重鎖または二つの異なる軽鎖は必要でない。したがって二本の鎖が間違った対を形成する可能性はない。
【0082】
本発明の抗体は、特異的結合パートナーに結合する可変領域を全体として形成する重鎖および軽鎖からなることができ、第2の特異性は、重鎖または軽鎖の構造ループのいずれかの修飾ループによって形成される。結合部位は(重鎖上、または軽鎖上、または両鎖上で)構造的に隣接しうる、1個より多い非CDRループによって形成されてもよい。
【0083】
修飾抗体または誘導体は、完全な抗体または抗体断片(例えばFab、CH1-CH2、CH2-CH3)であることができる。
【0084】
これは、結合パートナーに一価または多価で結合するか、設計によっては、異なる結合パートナーに異なる結合価で結合することさえできる。
【0085】
重鎖および軽鎖の非CDR領域中の特異的結合部位の選択および設計には、いくつかのさまざまなループを利用することができるので、上述の問題を伴わずに、2個より多い特異性を持つ抗体誘導体でさえ、設計することが可能である。
【0086】
一本のポリペプチド鎖内の特異的結合ドメインは、ペプチドリンカーを使って、またはペプチドリンカーを使わずに、つなぐことができる。
【0087】
一部の抗体クラスは、本来、多重特異性(特に二重特異性)であるとみなすことができる。すなわちそれらは、可変領域で抗原(これは通例、外来構造物であるか、癌関連構造物である)に結合し、Fc部分でFcエフェクター分子(例えばさまざまな免疫細胞上のFc受容体または補体タンパク質)に結合して、ADCC、ADCPまたはCDCなどの効果を可能にする。
【0088】
Fcエフェクター分子は、免疫グロブリン分子のFc部分(IgG1の場合、それはCH2ドメインおよびCH3ドメインからなる)によって結合される。糖鎖工学的技法により(US 6,602,684)、または直接的にFcにおけるタンパク質工学により(US 2005/0054832)、もしくは間接的にFc外での設計により(US 2005/02444403)、抗体分子のFc部分の結合を改善することによって、エフェクター機能を最適化するための方法が、いくつか記述されている。Fc受容体に対するFc領域の結合および/またはCq1などの補体タンパク質への結合が、どちらも、そのような技法によって改変されている。通常は、そのようなFcエフェクター分子に対する結合アフィニティーを改善する努力がなされる。なぜならそれはエフェクター機能の改善と相関するからである。
【0089】
本発明では、天然のFc結合領域外でFcエフェクター分子に結合する抗体を、設計することができる。「天然」Fcエフェクター分子結合に関与するループ以外の抗体ドメイン中の修飾ループを、1以上のFcエフェクター分子に結合するように、ライブラリーから選択するか、またはそのように設計することができる。そのような追加のFcエフェクター分子結合部位を持つ抗体は、一定のFcエフェクター分子またはFcエフェクター分子をディスプレイするエフェクター細胞に対して、より強いアビディティを持ち、それゆえに、糖鎖工学によって作製された抗体または他の方法で改善されたFc領域よりも、さらに強い効果を持ちうる。しかし本発明の一定の実施形態では、本発明による構造ループ中の修飾によって、修飾対象である与えられた抗体のエフェクター特徴を直接変化させず、そのまま残しておくべきである。
【0090】
抗体断片は、抗体全体と比較して一定の利点を持っている。断片は、通常は、良好な生体分布性を持ち、より容易に製造することができる。しかし抗体断片設計の大半はエフェクター機能を欠き、インビボ半減期が短い(Holliger Pら Nat Biotechnol.(2005) 23:1126-36)。
【0091】
CH1ドメインもCκまたはCλドメインもエフェクター機能を媒介せず、それが、FabがADCC、ADCPまたはCDCを示さない理由である。WO 02/44215には、抗体の抗原結合部位とFcエフェクター分子を結合するペプチドとからなる結合分子が記述されている。そのようにして、エフェクター機能を示す抗体断片を構築することができる。そのペプチドは、抗原結合も、Fcエフェクター分子に結合するというペプチドの能力も、破壊しないような位置で、その結合分子に組み入れられている。
【0092】
しかし本発明によれば、免疫グロブリンドメインの固定された足場内のランダムループ配列のライブラリーからFcエフェクター分子結合に関して選択された修飾免疫グロブリンを使って、Fcエフェクター分子への結合を達成することができる。したがって、Igドメイン足場外ではFcエフェクター分子に結合しないであろう特異的ループ配列を選択することが可能である。したがって、本発明によって得られるポリペプチドは、好ましくは、100個を越えるアミノ酸からなることができる。
【0093】
本発明のそのようなドメインの潜在的エフェクター機能を選択するために、突然変異型CH1、CκまたはCλドメインのライブラリーを、Fc受容体および/またはC1qなどの補体因子への結合に関して選択することができる。
【0094】
そのようなドメイン(例えばCH1、CH2、CH3、CH4、CκまたはCλ)からなる分子またはそのようなドメインを含有する分子のインビボ半減期を増加させるために、例えば本発明の突然変異型CH1-、CH2-、CH3-、CH4-、Cκ-またはCλ-ドメインなどのライブラリーを使って、FcRnへの結合を選択することができる。
【0095】
選択用のFcRn受容体は、それぞれの受容体を天然に発現させる細胞の表面上に用意するか、それぞれの受容体の細胞外部分の発現および精製によって用意することができる。本発明の目的には、FcRnでの第1スクリーニングによって突然変異型ドメインを選択し、それをインビトロでさらに試験し、さらには、FcRn受容体を発現させる細胞への結合により、FACS実験でさらに特徴づけることができる。例えば表面プラズモン共鳴技法などを使って、さまざまな組換えFcRn、アイソフォームおよびアロタイプへの結合のアフィニティーランキングにより、さらに特徴づけることができる。
【0096】
本発明の好ましい実施形態によれば、免疫グロブリンは、ヒトまたはネズミ由来である。
【0097】
修飾免疫グロブリンはさまざまな目的に使用することができ、特に医薬組成物では、免疫グロブリンが、好ましくはヒトまたはネズミ由来である。もちろん修飾免疫グロブリンはヒト化またはキメラ免疫グロブリンであってもよい。
【0098】
本発明のもう一つの好ましい実施形態によれば、ヒト免疫グロブリンは、IgA1、IgA2、IgD、IgE、IgG1、IgG2、IgG3、IgG4およびIgMからなる群より選択される。
【0099】
ネズミ免疫グロブリンは、好ましくは、IgA、IgD、IgE、IgG1、IgG2A、IgG2B、IgG2C、IgG3およびIgMからなる群より選択される。
【0100】
修飾免疫グロブリンは、上に挙げた免疫グロブリンクラスの一つから誘導することができる。
【0101】
免疫グロブリンは、好ましくは、免疫グロブリンの重鎖および/もしくは軽鎖またはその一部を含む。
【0102】
修飾免疫グロブリンは、重鎖および/または軽鎖、少なくとも一つの可変および/または定常ドメインを含みうる。
【0103】
本発明の免疫グロブリンは、好ましくは、免疫グロブリンの少なくとも一つの定常ドメインおよび/もしくは少なくとも一つの可変ドメインまたはその一部(ミニドメインを含む)を含む。
【0104】
定常ドメインは、定常領域のドメインとも呼ばれる免疫グロブリンの定常部分の免疫グロブリンフォールド単位である(例えばCH1、CH2、CH3、CH4、Ck、Cl)。
【0105】
可変ドメインは、可変領域のドメインとも呼ばれる免疫グロブリンの可変部分の免疫グロブリンフォールド単位である(例えばVh、Vk、Vl、Vd)。
【0106】
本発明の好ましい免疫グロブリンは、少なくとも一つのループ領域を持つ、CH1、CH2、CH3、CH4、Igk-C、Igl-C、またはその一部(ミニドメインを含む)からなる群より選択される定常ドメインからなり、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする。
【0107】
本発明のもう一つの好ましい免疫グロブリンは、少なくとも一つのループ領域を持つ、重鎖もしくは軽鎖の可変ドメイン、またはその一部(ミニドメインを含む)からなり、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする。
【0108】
好ましい実施形態によれば、定常ドメインは、CH1、CH2、CH3、CH4、Igk-C、Igl-C、またはその一部(ミニドメインを含む)の群より選択される。
【0109】
本発明の修飾免疫グロブリンは、1以上の定常領域(例えば少なくとも2個、3個、4個、5個、6個、10個のドメイン)を含みうる。修飾免疫グロブリン中に存在するドメインが1個より多い場合、これらのドメインは同じタイプであっても、異なるタイプであってもよい(例えばCH1-CH1-CH2、CH3-CH3)。もちろん、単一ドメインの順序も、どの種類であってもよい(例えばCH1-CH3-CH2、CH4-CH1-CH3-CH2)。
【0110】
免疫グロブリンのアミノ酸配列のナンバリングは全てIMGTナンバリングスキームに従う(IMGT, the international ImMunoGeneTics information system@imgt.cines.fr;http://imgt.cines.fr;Lefrancら, 1999, Nucleic Acids Res. 27: 209-212;Ruizら, 2000 Nucleic Acids Res. 28: 219-221;Lefrancら, 2001, Nucleic Acids Res. 29: 207-209;Lefrancら, 2003, Nucleic Acids Res. 31: 307-310;Lefrancら, 2005, Dev Comp Immunol 29:185-203)。
【0111】
本発明の好ましい実施形態によれば、CH1、CH2、CH3およびCH4の修飾ループ領域は、アミノ酸7〜21、アミノ酸25〜39、アミノ酸41〜81、アミノ酸83〜85、アミノ酸89〜103およびアミノ酸106〜117を含む。
【0112】
ヒト由来のIgk-CおよびIgl-Cのループ領域は、好ましくは、アミノ酸8〜18、アミノ酸27〜35、アミノ酸42〜78、アミノ酸83〜85、アミノ酸92〜100、アミノ酸108〜117およびアミノ酸123〜126を含む。
【0113】
ネズミ由来のIgk-CおよびIgl-Cのループ領域は、好ましくは、アミノ酸8〜20、アミノ酸26〜36、アミノ酸43〜79、アミノ酸83〜85、アミノ酸90〜101、アミノ酸108〜116およびアミノ酸122〜125を含む。
【0114】
ヒト由来の免疫グロブリンの可変ドメインの構造ループ領域は、好ましくは、アミノ酸8〜20、アミノ酸44〜50、アミノ酸67〜76およびアミノ酸89〜101を含む。
【0115】
本発明の好ましい実施形態によれば、ネズミ由来の免疫グロブリンの可変ドメインの構造ループ領域は、アミノ酸6〜20、アミノ酸44〜52、アミノ酸67〜76およびアミノ酸92〜101を含む。
【0116】
上に定義した各免疫グロブリンのアミノ酸領域は、修飾されるべきループ領域を含む。
【0117】
本発明の免疫グロブリンは、好ましくは、ラクダ由来である。
【0118】
ラクダ抗体は一本の重鎖だけを含み、軽鎖および重鎖からなる通常の抗体と同じ抗原アフィニティを持つ。したがってラクダ抗体は例えばヒト抗体などよりもはるかに小さく、それゆえに、密な組織に浸透して、大きなタンパク質では到達できないような抗原に到達することができる。さらにまた、ラクダの重鎖抗体は、比較的単純であり、アフィニティおよび特異性が高く、活性部位に到達しそれと相互作用する潜在能力を持つので、臨床的に有益な化合物の設計、生産および応用において、一般的な抗体よりも有利である。
【0119】
ラクダ由来の免疫グロブリンは、好ましくは、CH1、CH2およびCH3からなる群より選択される少なくとも一つの定常ドメインを含む。
【0120】
本発明の好ましい実施形態によれば、ラクダ免疫グロブリンのCH1、CH2およびCH3のループ領域は、アミノ酸8〜20、アミノ酸24〜39、アミノ酸42〜78、アミノ酸82〜85、アミノ酸91〜103およびアミノ酸108〜117を含む。
【0121】
本発明の好ましい実施形態によれば、分子に対する修飾免疫グロブリンの特異的結合は、免疫学的アッセイ、好ましくは酵素結合免疫吸着アッセイ(ELISA)、表面プラズモン共鳴アッセイ、飽和移動差核磁気共鳴分光法、転移NOE(trNOE)核磁気共鳴分光法、競合アッセイ、組織結合アッセイ、生細胞結合アッセイおよび細胞抽出物アッセイからなる群より選択される結合アッセイによって決定される。
【0122】
結合アッセイは、例えばFRET(蛍光共鳴エネルギー移動)およびBRET(生物発光共鳴エネルギー移動)に基づくアッセイ、AlphaScreen(商標)(増幅発光近接均一アッセイ(Amplified Luminescent Proximity Homogeneous Assay))、シンチレーション近接アッセイ、ELISA(酵素結合免疫吸着アッセイ)、SPR(表面プラズモン共鳴、BIACORE(登録商標)とも呼ばれている)、等温滴定熱量測定法、示差走査熱量測定、ゲル電気泳動、およびクロマトグラフィー(ゲル濾過を含む)などといった(ただしこれらに限定されるわけではない)、当技術分野で知られるさまざまな方法を使って行うことができる。これらの方法および他の方法は、何らかの融合パートナーまたはラベルを利用するものでありうる。
【0123】
修飾免疫グロブリンを、好ましくは、有機分子、酵素ラベル、放射性ラベル、着色ラベル、蛍光ラベル、発色ラベル、発光ラベル、ハプテン、ジゴキシゲニン、ビオチン、金属錯体、金属、コロイド金およびそれらの混合物からなる群より選択されるラベルにコンジュゲートする。
【0124】
修飾免疫グロブリンは、例えば結合アッセイ(例:ELISA)および結合研究などにおいて、当該コンジュゲートの簡単な検出を可能にする他の分子にコンジュゲートすることもできる。
【0125】
本発明のもう一つの側面は、少なくとも一つのループ領域を持つ、CH1、CH2、CH3、CH4、Igk-C、Igl-C、もしくはその一部(ミニドメインを含む)、またはそれらの組み合わせからなる群より選択される定常ドメインからなり、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする免疫グロブリンに関する。
【0126】
少なくとも一つの修飾抗体ドメイン(=非可変配列または構造ループによって特異的パートナーに結合するもの)を、少なくとも一つの他の結合分子(これは、抗体、抗体断片、可溶性受容体、リガンドまたはもうひとつの修飾抗体ドメインであることができる)と、分子的に組み合わせることが好ましい。
【0127】
分子は、タンパク質性分子、核酸、および糖質からなる群より選択される。
【0128】
修飾免疫グロブリンのループ領域は、どの種類の結合分子(特にタンパク質性分子、タンパク質、ペプチド、ポリペプチド、核酸、グリカン、糖質、脂質、小さい有機分子、無機分子)に特異的に結合してもよい。もちろん、修飾免疫グロブリンが少なくとも二つのループ領域を含んでいて、それらのループ領域のそれぞれが他の分子またはエピトープに特異的に結合してもよい。
【0129】
本発明の好ましい実施形態によれば、修飾構造ループ領域に結合する分子は、腫瘍関連抗原、特にEpCAM、腫瘍関連糖タンパク質72(TAG-72)、腫瘍関連抗原CA125、前立腺特異膜抗原(PSMA)、高分子量メラノーマ関連抗原(HMW-MAA)、ルイスY関連糖質を発現させる腫瘍関連抗原、癌胎児性抗原(CEA)、CEACAM5、HMFG PEM、ムチンMUC1、MUC18およびサイトケラチン腫瘍関連抗原、細菌抗原、ウイルス抗原、アレルゲン、フルオレセイン、リゾチーム、toll様受容体9、エリスロポエチン、CD2、CD3、CD3E、CD4、CD11、CD11a、CD14、CD18、CD19、CD20、CD22、CD23、CD25、CD28、CD29、CD30、CD33(p67タンパク質)、CD38、CD40、CD40L、CD52、CD54、CD56、CD80、CD147、GD3、IL-1、IL-1R、IL-2、IL-2R、IL-4、IL-5、IL-6、IL-6R、IL-8、IL-12、IL-15、IL-18、IL-23、インターフェロンα、インターフェロンβ、インターフェロンγ;TNF-α、TNFβ2、TNFα、TNFαβ、TNF-R1、TNF-RII、FasL、CD27L、CD30L、4-1BBL、TRAIL、RANKL、TWEAK、APRIL、BAFF、LIGHT、VEG1、OX40L、TRAIL受容体-1、A1アデノシン受容体、リンフォトキシンβ受容体、TACI、BAFF-R、EPO;LFA-3、ICAM-1、ICAM-3、インテグリンβ1、インテグリンβ2、インテグリンα4/β7、インテグリンα2、インテグリンα3、インテグリンα4、インテグリンα5、インテグリンα6、インテグリンαv、αVβ3インテグリン、FGFR-3、ケラチノサイト増殖因子、VLA-1、VLA-4、L-セレクチン、抗Id、E-セレクチン、HLA、HLA-DR、CTLA-4、T細胞受容体、B7-1、B7-2、VNRインテグリン、TGFβ1、TGFβ2、エオタキシン1、BLyS(Bリンパ球刺激因子)、補体C5、IgE、第VII因子、CD64、CBL、NCA90、EGFR(ErbB-1)、Her2/neu(ErbB-2)、Her3(ErbB-3)、Her4(ErbB4)、組織因子、VEGF、VEGFR、エンドセリン受容体、VLA-4、糖質、例えば血液型抗原および関連糖質、ガリリグリコシレーション(Galili Glycosylation)、ガストリン、ガストリン受容体、腫瘍関連糖質、ハプテンNP-capまたはNIP-cap、T細胞受容体α/β、E-セレクチン、ジゴキシン、胎盤アルカリホスファターゼ(PLAP)および精巣PLAP様アルカリホスファターゼ、トランスフェリン受容体、へパラナーゼI、ヒト心ミオシン、糖タンパク質IIb/IIIa(GPIIb/IIIa)、ヒトサイトメガロウイルス(HCMV)gHエンベロープ糖タンパク質、HIV gp120、HCMV、呼吸器合胞体ウイルスRSV F、RSVF Fgp、VNRインテグリン、Hep B gp120、CMV、gpIIbIIIa、HIV IIIB gp120 V3ループ、呼吸器合胞体ウイルス(RSV)Fgp、単純ヘルペスウイルス(HSV)gD糖タンパク質、HSV gB糖タンパク質、HCMV gBエンベロープ糖タンパク質、クロストリジウムパーフリンジェンス毒素およびその断片からなる群より選択される。
【0130】
本発明の修飾免疫グロブリンは、好ましくは、上に開示した分子の一つに結合しうる。これらの分子はアレルゲンも含む。
【0131】
本発明のもう一つの好ましい実施形態によれば、CH3の位置15〜17、29〜34、85.4〜85.3、92〜94、97〜98および/または108〜110のアミノ酸残基が修飾される。
【0132】
本発明の免疫グロブリンの修飾は、好ましくは、欠失、置換または挿入である。
【0133】
本発明によれば、少なくとも1個、好ましくは少なくも2、3、4、5、6、7、8、9、10および15個のアミノ酸が、削除されるか、他のアミノ酸で置換(修飾アミノ酸によるものを含む)されるか、または免疫グロブリンのループ領域中に挿入される。しかし、免疫グロブリンのループ領域中に挿入されるアミノ酸の最大数は、アミノ酸数30、好ましくは25、より好ましくは20を越えないだろう。アミノ酸の置換および挿入は、好ましくは、当技術分野で知られる方法により、本特許出願に開示するように、ランダムに行われる。
【0134】
本発明の免疫グロブリンは、ある特定の実施形態によれば、CH3領域が、EpCamが前記免疫グロブリンに結合する場合は配列番号16または配列番号18を、フルオレセインが前記免疫グロブリンに結合する場合は配列番号20を、リゾチームが前記免疫グロブリンに結合する場合は配列番号22、24、26、28、30または32を、TLR9が前記免疫グロブリンに結合する場合は配列番号34、36、38または40を、そしてリゾチームおよび/またはエリスロポエチンが前記免疫グロブリンに結合する場合は配列番号42を含むことを特徴とする。
【0135】
本発明の、ある特定の実施形態によれば、免疫グロブリンは、リゾチームおよびgp41が前記免疫グロブリンに結合する場合に、免疫グロブリンが配列番号44または配列番号46を含むことを特徴とする。
【0136】
修飾免疫グロブリンは、好ましくは、有機分子、酵素ラベル、放射性ラベル、着色ラベル、蛍光ラベル、発色ラベル、発光ラベル、ハプテン、ジゴキシゲニン、ビオチン、金属錯体、金属、コロイド金およびそれらの混合物からなる群より選択されるラベルまたはレポーター分子にコンジュゲートされる。
【0137】
上に明示したラベルにコンジュゲートされた修飾免疫グロブリンは、例えば診断方法に使用することができる。
【0138】
本発明のもう一つの側面は、能動免疫用ワクチンの製造を目的とする、本発明の免疫グロブリンまたは本発明の方法によって得ることができる免疫グロブリンの使用に関する。この場合、免疫グロブリンは抗原性原薬として、ワクチンの調剤に使用されるか、ワクチン製剤用の抗原構造を釣り上げるためまたは捕捉するために使用される。
【0139】
本発明のもう一つの側面は、免疫グロブリンのタンパク質ライブラリーの作製を目的とする、本発明の免疫グロブリンまたは本発明の方法によって得ることができる免疫グロブリンの使用に関する。
【0140】
本発明のさらにもう一つの側面は、ある分子を特異的に結合および/または検出するための方法であって、
(a)本発明の修飾免疫グロブリンまたは本発明の方法によって得ることができる修飾免疫グロブリンを、前記分子を含有すると疑われる試験試料と接触させるステップ、および
(b)特異的免疫グロブリン/分子複合体の潜在的形成を検出するステップ
を含む方法に関する。
【0141】
本発明のもう一つの側面は、ある分子を特異的に単離する方法であって、
(a)本発明の修飾免疫グロブリンまたは本発明の方法によって得ることができる修飾免疫グロブリンを、前記分子を含有する試料と接触させるステップ、
(b)形成された特異的免疫グロブリン/分子複合体を分離するステップ、および
(c)要すれば、前記複合体から分子を単離するステップ
を含む方法に関する。
【0142】
本発明の免疫グロブリンは、試料から分子を特異的に単離するために使用することができる。多重特異性免疫グロブリンを使用すれば、2以上の分子を試料から単離することができる。そのような方法で修飾免疫グロブリンを使用することはとりわけ有利である。なぜなら、単離されるべき分子に結合することができる所定の量の結合パートナー(すなわち修飾免疫グロブリン)が固定化されている均一な表面を持つマトリックスを作製することができるからである。これに対し、単一特異性結合パートナーを使用した場合には、均一なマトリックスを作製することができない。なぜなら、その単一結合パートナーはマトリックスに同じ効率では結合しないからである。
【0143】
本発明のもう一つの側面は、ある化合物を標的にターゲティングするための方法であって、
(a)前記化合物に特異的に結合する能力を持つ、本発明の修飾免疫グロブリンまたは本発明の方法によって得ることができる修飾免疫グロブリンを接触させるステップ、
(b)免疫グロブリン/化合物複合体を標的に送達するステップ、
を含む方法に関する。
【0144】
本発明の修飾免疫グロブリンは、CDRおよび/または修飾ループ領域に結合した少なくとも一つの化合物を、標的に送達するために使用することができる。そのような免疫グロブリンは、疾患の処置の過程で、治療物質を好ましい作用部位にターゲティングするために使用することができる。
【0145】
本発明のもう一つの側面は、本発明の修飾免疫グロブリンまたは本発明の方法によって得ることができる修飾免疫グロブリンを含むタンパク質ライブラリーに関する。
【0146】
前記ライブラリーを構築するための好ましい方法は、上記の説明および実施例に見出すことができる。本発明のライブラリーは、特異な分子に結合する免疫グロブリンを同定するために使用することができる。
【0147】
特に本発明は、免疫グロブリン誘導体の設計を目的とする、本発明の免疫グロブリンまたは本発明の方法によって得ることができる免疫グロブリンを含むタンパク質ライブラリーの使用に関する。少なくとも一つの修飾ループを持つ、少なくとも10、好ましくは100、より好ましくは1000、さらに好ましくは10000、さらに好ましくは100000、最も好ましくは1000000を越える変種ドメインを含む各ドメインのタンパク質ライブラリーを用いることにより、既存の免疫グロブリンを変化させて、任意のドメインまたはミニドメイン中に抗原結合部位を導入することができる。次にライブラリーを、特異的抗原への結合に関してスクリーニングする。望ましい性質に関する分子特性解析の後、選択したドメインまたはミニドメインを、遺伝子工学的技法により、それが野生型領域と置き換わるような形で、元の免疫グロブリン中にクローニングする。あるいは、ループをコードするDNAまたは突然変異させたアミノ酸をコードするDNAだけを交換して、特異的抗原に対する追加結合部位を持つ免疫グロブリンを得ることもできる。
【0148】
突然変異型抗原特異的構造ループを得るための部位の選択は、元の免疫グロブリンの構造および追加結合部位の目的に依存する。例えば元の分子が、エフェクター機能を乱さずに追加抗原結合部位を挿入される必要がある完全な免疫グロブリンである場合、修飾されるべきループは、Fcエフェクター分子に対する天然の結合パートナーであるCH2およびCH3から離れたドメインより選択されるだろう。元の免疫グロブリンがFabである場合は、軽鎖または重鎖の定常ドメインまたは各可変ドメインのループを修飾することができる。ライブラリーを作製するために、1以上のドメインの1以上の構造ループ中に突然変異を持つ突然変異型の元分子のライブラリーを調製することができる。完全な突然変異型元分子を使った選択はいくつかの利点を持ちうる。というのも、突然変異型免疫グロブリンが示すべき他の性質についても試験すれば、修飾構造ループによる抗原結合に関する選択によって、立体的に有利な修飾がもたらされるだろうからである。
【0149】
突然変異型ドメインもしくはミニドメインまたはドメインの融合分子のタンパク質ライブラリーのサイズ要件(すなわち変種タンパク質の数)は、作業課題に依存する。一般に、抗原結合部位を新規に生成させるためのライブラリーは、修飾構造ループでできた既存の改変抗原結合部位を(例えば抗原に対するアフィニティーを高めたり、抗原に対する微細特異性を変化させたりする目的で)さらに修飾するために用いられるライブラリーよりも大きい必要がある。
【0150】
本発明は、複数の免疫グロブリン(例えば定常もしくは可変ドメイン、ミニドメインおよび/またはミニドメインに含有される少なくとも一つの構造ループ領域)またはそれをコードする核酸分子を含む免疫グロブリンライブラリーまたは核酸ライブラリーにも関係する。このライブラリーは、異なる修飾を持つメンバーを含有し、その複数性(plurality)は、少なくとも一つの構造ループ領域中の修飾によって定義づけられる。核酸ライブラリーは、好ましくは、(一つのアミノ酸交換をもたらす)少なくとも10個の異なるメンバー、より好ましくは少なくとも100個、より好ましくは1000個または10000個の異なるメンバー(例えばランダム突然変異戦略またはコンビナトリアル技法によって設計されたもの)を含む。さらに多くの多様化された個別メンバーの数、例えば少なくとも1000000または少なくとも10000000なども好ましい。
【0151】
本発明のさらにもう一つの側面は、少なくとも二つの本発明ライブラリーから選択される二つの異なるドメインまたはミニドメインの組み合わせによる多重特異性免疫グロブリンの作製である。これらの選択された特異的免疫グロブリンを、ビルディングブロックのように、互いに、そして他の分子と組み合わせることにより、望ましい性質が得られるようにドメインまたはミニドメインの最適な配置を設計することができる。
【0152】
さらにまた、1以上の修飾免疫グロブリンを、あるタンパク質の考えうるさまざまな部位に、または考えうる異なる部位の全てに、そのタンパク質の構造を破壊せずに導入することもできる。そのような「ドメインシャフリング」技法により、新しいライブラリーを作り出し、それをまた望ましい性質について選択することができる。
【0153】
好ましいライブラリーは、免疫グロブリンのドメイン、ミニドメインまたはその誘導体からなる群より選択される本発明の免疫グロブリンを含有する。
【0154】
本発明の好ましい実施形態は、ある抗原に対する結合分子(抗原結合分子)であって、少なくとも一つの免疫ドメインと、その抗原に結合するように本発明に従って修飾される構造ループ領域とを含み、抗体の可変ドメインを含まない結合分子である。これは、抗体活性に使用することができる他の部分(例えば天然または修飾エフェクター領域(配列))を含んでもよいが、抗体の「天然」の結合領域は欠いている(すなわち可変ドメインを本来の位置には持っていない)。本発明のこれらの抗原結合分子は、本分子について上述した利点を持つが、抗体の特異的結合活性は持たず、その一方で、構造ループ領域中に新たに導入された特異的結合活性を持つ。
【0155】
好ましくは、本発明のこれらの抗原結合分子は、CH1、CH2、CH3、CH4、Igk-C、Igl-Cおよびそれらの組み合わせを含み、前記組み合わせは、少なくとも2個、好ましくは少なくとも4個、とりわけ少なくとも6個の定常ドメインと、本発明に従って修飾された少なくとも1個の構造ループ領域とを含む。好ましくは、これらの構造ループ領域は、本発明に従って修飾された構造ループ領域によってつながれるか、またはそのような2個の定常ドメイン間に天然に存在する構造ループ領域によってつながれる。本発明のこれらの抗原結合分子の一実施形態は、本発明に従って構造ループ中に少なくとも一つの修飾を持つ、抗体のFc領域からなる。本発明の抗原結合分子については、構造ループ中の新しい抗原結合部位がランダム突然変異技術によって(すなわちループの1以上のアミノ酸残基をランダム突然変異技法で交換することによって、またはランダムに生成させたインサートをそのような構造ループに導入することによって)導入されることも好ましい。あるいは、コンビナトリアルアプローチの使用も好ましい。
【0156】
もう一つの側面によれば、本発明は、1以上の構造ループ中に組み込まれた、無修飾免疫グロブリンにとって外来である抗原結合部位を持つ、修飾免疫グロブリンに関する。「外来」という用語は、その抗原結合部位がその免疫グロブリンのその特定領域によって天然には形成されないこと、そしてその抗原結合部位によって外来の結合パートナーは結合されるが、免疫グロブリンの天然の結合パートナーは結合されないことを意味する。これは、結合パートナー(例えばFc受容体または免疫系のエフェクター)が、無修飾免疫グロブリンにとって外来である抗原結合部位によっては、結合されないとみなされることを意味する。
【0157】
好ましくは、抗原は、病原体抗原、腫瘍関連抗原、酵素、基質、自己抗原、有機分子またはアレルゲンからなる群より選択される。より好ましい抗原は、ウイルス抗原、細菌抗原または真核生物の病原体もしくはファージに由来する抗原からなる群より選択される。好ましいウイルス抗原には、HAV-、HBV-、HCV-、HIV I-、HIV II-、パルボウイルス-、インフルエンザ-、HSV-、肝炎ウイルス、フラビウイルス、ウエストナイルウイルス、エボラウイルス、ポックスウイルス、天然痘ウイルス、麻疹ウイルス、ヘルペスウイルス、アデノウイルス、パピローマウイルス、ポリオーマウイルス、パルボウイルス、ライノウイルス、コクサッキーウイルス、ポリオウイルス、エコーウイルス、日本脳炎ウイルス、デングウイルス、ダニ媒介脳炎ウイルス、黄熱ウイルス、コロナウイルス、呼吸器合胞体ウイルス、パラインフルエンザウイルス、ラクロスウイルス、ラッサウイルス、狂犬病ウイルス、ロタウイルス抗原などがある;好ましい細菌抗原には、シュードモナス-、マイコバクテリウム-、スタフィロコッカス-、サルモネラ-、髄膜炎菌-、ボレリア-、リステリア、ナイセリア-、クロストリジウム-、エシェリキア-、レジオネラ-、バチルス-、ラクトバチルス-、ストレプトコッカス-、エンテロコッカス-、コリネバクテリウム-、ノカルジア-、ロドコッカス-、モラクセラ-、ブルセラ、カンピロバクター-、カルジオバクテリウム-、フランシセラ-、ヘリコバクター-、ヘモフィルス-、クレブシエラ-、シゲラ-、エルシニア-、ビブリオ-、クラミジア-、レプトスピラ-、リケッチア-、マイコバクテリウム-、トレポネーマ-、バルトネラ-抗原などがある。病原性真核生物の好ましい真核抗原には、ジアルジア、トキソプラズマ、シクロスポラ、クリプトスポリジウム、旋毛虫、酵母、カンジダ、アスペルギルス、クリプトコッカス、ブラストミセス、ヒストプラスマ、コクシジオイデス由来の抗原などがある。
【0158】
本発明の好ましい免疫グロブリンは少なくとも二つの抗原結合部位を含み、第1部位は第1エピトープに結合し、第2部位は第2エピトープに結合する。
【0159】
好ましい実施形態によれば、本免疫グロブリンは少なくとも二つのループ領域を含み、第1ループ領域は第1エピトープに結合し、第2ループ領域は第2エピトープに結合する。少なくとも第1ループ領域もしくは少なくとも第2ループ領域またはその両方が構造ループを含有しうる。本発明の免疫グロブリンは、本発明の必須要素(すなわち本発明に従って修飾された構造ループ領域)を含有する、機能的であることが当技術分野で知られているその断片を包含する。
【0160】
好ましくは、本発明の免疫グロブリンは、少なくとも二つの免疫グロブリンドメイン、またはその一部(ミニドメインを含む)から構成され、各ドメインは少なくとも一つの抗原結合部位を含有する。
【0161】
また、免疫グロブリンの定常領域の少なくとも一つのドメインおよび/または可変領域の少なくとも一つのドメイン、またはその一部(ミニドメインを含む)を含む、本発明の免疫グロブリンも好ましい。したがって、例えばC末端領域が修飾されている可変ドメイン、または修飾CH1領域(例えば修飾CH1ミニドメイン)に連結された可変領域は、好ましい実施形態の一つである。
【0162】
本発明の好ましい免疫グロブリンは、無修飾ドメインと少なくとも50%の相同性を持つドメインを含む。
【0163】
「相同性」という用語は、ポリペプチドがその一次、二次または三次構造中の対応する位置に同じ残基または保存された残基を持つことを示す。この用語は、相同なポリペプチドをコードする2以上のヌクレオチド配列にも及ぶ。
【0164】
「相同免疫グロブリンドメイン」とは、本明細書に開示する完全長ネイティブ配列免疫グロブリンドメイン配列または完全長免疫グロブリンドメイン配列の他の任意の断片に対して、少なくとも約50%のアミノ酸配列一致度を持つ、本発明の免疫グロブリンドメインを意味する。好ましくは、相同免疫グロブリンドメインは、ネイティブ免疫グロブリンドメイン配列、または本明細書に開示する完全長免疫グロブリンドメイン配列の他の任意の明確に定義された断片に対して、少なくとも約50%のアミノ酸配列一致度、好ましくは少なくとも約55%のアミノ酸配列一致度、より好ましくは少なくとも約60%のアミノ酸配列一致度、より好ましくは少なくとも約65%のアミノ酸配列一致度、より好ましくは少なくとも約70%のアミノ酸配列一致度、より好ましくは少なくとも約75%のアミノ酸配列一致度、より好ましくは少なくとも約80%のアミノ酸配列一致度、より好ましくは少なくとも約85%のアミノ酸配列一致度、より好ましくは少なくとも約90%のアミノ酸配列一致度、より好ましくは少なくとも約95%のアミノ酸配列一致度を持つだろう。
【0165】
本明細書で同定される免疫グロブリン配列に対する「パーセント(%)アミノ酸配列一致度」は、最大のパーセント配列一致度が得られるように配列を整列させて、必要ならばギャップを導入した後に、特定免疫グロブリンドメイン配列中のアミノ酸残基と一致する候補配列中のアミノ酸残基のパーセンテージ(保存的置換はいずれも配列一致部分とはみなさない)と定義される。パーセントアミノ酸配列一致度を決定するためのアラインメントは、当技術分野で知られるさまざまな方法により、例えばBLAST、BLAST-2、ALIGNまたはMegalign(DNASTAR)ソフトウェアなどの公に利用可能なコンピュータソフトウェアを使って達成することができる。当業者は、比較する配列の全長にわたって最大のアラインメントを得るのに必要な任意のアルゴリズムを含めて、アラインメントを判定するのに適したパラメータを決定することができる。
【0166】
%アミノ酸配列一致度の値は、後述のように、WU-BLAST-2コンピュータプログラム(Altschulら, Methods in Enzymology 266:460-480 (1996))を使って得ることができる。WU-BLAST-2検索パラメータの大半はデフォルト値に設定される。デフォルト値に設定されない値、すなわち調節可能パラメータは、以下の値に設定される:オーバーラップスパン(overlap span)=1、オーバーラップフラクション(overlap fraction)=0.125、ワード閾値(word threshold)(T)=11、およびスコア行列(scoring matrix)=BLOSUM62。WU-BLAST-2を使用する場合、%アミノ酸配列一致度値は、(a)WU-BLAST-2によって決定される、ネイティブ免疫グロブリンドメインから誘導された配列を持つ関心対象の免疫グロブリンドメインのアミノ酸配列と、関心対象の比較アミノ酸配列(すなわち、関心対象の免疫グロブリンドメインと比較される配列であり、これは、無修飾免疫グロブリンドメインであることができる)との間の、一致する同一アミノ酸残基の数を、(b)関心対象の免疫グロブリンドメインの非ランダム突然変異部分のアミノ酸残基の総数で割ることによって決定される。例えば「アミノ酸配列Bに対して少なくとも80%のアミノ酸配列一致度を持つアミノ酸配列Aを含むポリペプチド」という記述において、アミノ酸配列Aは、関心対象の比較アミノ酸配列であり、アミノ酸配列Bは、関心対象の免疫グロブリンドメインのアミノ酸配列である。
【0167】
好ましい実施形態において、本発明の免疫グロブリンは、二重特異性抗体または二重特異性単鎖抗体である。免疫グロブリンが二重特異性ドメインまたはその一部(ミニドメインを含む)を含むことは、さらに好ましい。
【0168】
本発明の免疫グロブリンは、免疫グロブリンに関して当技術分野で知られているどの目的にも使用できるが、本発明によって導入される特異性の組み合わせに依存する応用も可能にする。したがって本発明の免疫グロブリンは、好ましくは、治療および予防用途に(例えば能動免疫療法または受動免疫療法として)、製造および分析用途に、ならびに診断用途に使用される。
【0169】
本発明のもう一つの側面は、
(a)1以上の構造ループ中に組み込まれた、当該免疫グロブリンにとっては外来の抗原結合部位を持つ、修飾免疫グロブリン、および
(b)前記抗原のエピトープを含有する結合分子
を含有する、結合パートナーのキットに関する。
【0170】
本発明のこのキットのそのような結合分子は、本発明の修飾免疫グロブリンの結合特異性を同定するために使用することができる。本発明のこのキットの結合分子を使用することにより、本発明の修飾免疫グロブリンの効力を決定することができる。
【0171】
本明細書にいう効力とは、修飾分子の、その抗原に対する結合性である。結合は、品質管理目的に使用される特異性および/またはアフィニティおよび/またはアビディティに関して、定量的および/または定性的に決定することができる。
【0172】
さらにまた、本発明のキットの結合分子は、構造ループ中に異なる修飾を持つ少なくとも10、好ましくは少なくとも100、より好ましくは少なくとも1000、より好ましくは少なくとも10000、特に少なくとも100000個の免疫グロブリンからなるライブラリーより、本発明の修飾免疫グロブリンを選択するために使用することもできる。
【0173】
本発明によれば、本発明の重要な特徴の一つは、免疫グロブリンドメインの設計が、通常は抗原結合に関与しない領域で行われること、言い換えると、抗体のCDR以外の領域で行われることにある。免疫グロブリンドメインの特異的フォールドは、CDRと構造的に類似するが配列における位置が異なっている領域に、ランダム突然変異を導入することを許容することが、観察された。本発明によって同定された領域は、CDRと同様に、免疫グロブリンフォールドのβストランドをつなぐループ領域である。
【0174】
より具体的に述べると、本明細書には、ヒトIgG1 CH3ドメインのβストランドA-BおよびE-Fをつなぐループ中にランダム突然変異を導入することにより、それぞれヒトのIgG1 CH3ドメインによって通常は認識および結合されないペプチドおよびタンパク質であるToll様受容体9ペプチド(TLR-9)または鶏卵リゾチームに特異的に結合する突然変異型CH3ドメインが選択されたことを記載する。本発明者らが導入した突然変異には、野生型配列中の選択したアミノ酸残基をランダムに選択された残基で置き換える突然変異が含まれ、また、上述したループ中への余分なアミノ酸残基の挿入も含まれる。
【0175】
類推により、どの免疫グロブリンクラスの免疫グロブリンドメインでも、どの種に由来する免疫グロブリンの免疫グロブリンドメインでも、このタイプの設計を適用することができる。さらにまた、本発明において標的とする特定ループを操作することができるだけでなく、免疫グロブリンドメイン中のβストランドをつなぐ任意のループを同じように操作することができる。
【0176】
本発明では、どの生物およびどの免疫グロブリンクラスに由来する改変免疫グロブリンドメインでも、そのまま(単一ドメインとして)、またはより大きな分子の一部として、使用することができる。例えば、それらはインタクト免疫グロブリンの一部であることができる(したがって、そのインタクト免疫グロブリンは、6個のCDRによって形成されるその「通常の」抗原結合領域と、新しい改変抗原結合領域とを持つだろう)。これと同様に、多重特異性(例えば二重特異性)免疫グログロブリンを作製することができる。改変免疫グロブリンドメインは、任意の融合タンパク質の一部であることもできる。これらの改変免疫グロブリンドメインは、免疫グロブリンの使用分野全般に有用である。
【0177】
以下の免疫グロブリンのドメインは、本明細書において、免疫グロブリンドメインと理解される:
IgG、IgDおよびIgAの場合:VL、CL、VH、CH1、CH2、CH3
IgMおよびIgEの場合:VL、CL、VH、CH1、CH2、CH3、CH4。
【0178】
1.片側に、すなわちβストランドB-C、D-EもしくはF-Gをつなぐループ(多くの特許がカバーしている可変ドメインを除く「先端部」)内に、またはβストランドA-B、C-D(可変ドメインの場合はC-C'およびC''-D)もしくはE-Fをつなぐループ(「基部」)内に、ランダム突然変異が導入された単一の免疫グロブリンドメイン。単一のループまたはループの任意の組み合わせにランダム突然変異を導入することができる。残基は変化もしくは欠失させるか、または追加残基を挿入することができる。
【0179】
2.両側(先端部および基部)にランダム突然変異が導入された単一の免疫グロブリンドメイン。
【0180】
3.ランダム突然変異が導入された単一ドメインの一つを含有する任意のタンパク質、例えば
a)片側または両側にランダム突然変異が導入された「単鎖CH3」二量体(scCH3)、scCH2、scCH1/CL、
b)「基部」側に、すなわちCDRとは反対側に、ランダム突然変異が導入された単鎖Fv、
c)「基部」に、すなわちCH1およびCLドメインのC末端に、ランダム突然変異が導入されたFab断片、
d)片側または両側にランダム突然変異が導入されたFc断片(すなわちCH2-CH3からなるタンパク質)、
e)Fcの基部にランダム突然変異が導入された完全な免疫グロブリン、
f)他の適当なドメイン。
【0181】
単一ドメインの主な利点は、ラクダVH分子(「ナノ抗体(nanobody)」、www.ablynx.com参照)を奨励するために用いられる議論の全てと非常に似ている。ランダム突然変異が導入された免疫グロブリンドメインは極めて小さいタンパク質(挿入されるアミノ酸残基の数に依存して分子量約12〜15kDa)であるので、従来の抗体または抗体断片、例えばscFvおよびFabと比較して、以下の利点を持つだろう:珍しいまたは隠れたエピトープを認識すること、タンパク質標的の空洞または活性部位中に結合すること、製造が容易であること、およびその他数多くの利点。両側にランダム突然変異が導入される免疫グロブリンドメインの場合は、二価または二重特異性分子を生成させることができる。融合タンパク質の一部としての単一ドメインの主な利点は、他の任意のタンパク質上に追加の結合性を設計することができるということである。
【0182】
タンパク質の製造には任意の発現系を使用することができると考えられる。本明細書に記載する単一ドメインとの類似性を、VLがなくVHしか持たないラクダ由来の抗体に見出すことができる。これらのタンパク質では(「通常の」抗体のように6個ではなく)3個のCDRが抗原結合を担っている。
【0183】
以下の特許文献は、参照により、あたかもその全てを本明細書に記載したかのように、本明細書に組み入れられる:
US 6,294,654「Modified immunoglobulin molecule incorporating an antigen in a non-CDR loop region(非CDRループ領域に抗原を組み入れた修飾免疫グロブリン分子)」
US 5,844,094「Target binding polypeptide(標的結合性ポリペプチド)」
US 5,395,750「Methods for producing proteins which bind to predetermined antigens(所定の抗原に結合するタンパク質を製造するための方法)」
US 2004/0071690「High avidity polyvalent and polyspecific reagents(高アビディティ多価および多重特異性試薬)」
US 2004/0018508「Surrogate antibodies and methods of preparation and use thereof(代理抗体ならびにその製造および使用方法)」
US 2003/0157091「Multi-functional proteins(多機能タンパク質)」
US 2003/0148372「Method to screen phage display libraries with different ligands(異なるリガンドを使ってファージディスプレイライブラリーをスクリーニングする方法)」
US 2002/0103345「Bispecific immunologobulin-like antigen binding proteins and method of production(二重特異性免疫グロブリン様抗原結合タンパク質および製造方法)」
US 2004/0097711「Immunoglobulin superfamily proteins(免疫グロブリンスーパーファミリータンパク質)」
US 2004/0082508「Secreted proteins(分泌タンパク質)」
US 2004/0063924「Secreted proteins(分泌タンパク質)」
US 2004/0043424「Immunoglobulin superfamily proteins(免疫グロブリンスーパーファミリータンパク質)」
US 5,892,019「Production of a single-gene-encoded Immunoglobulin(単一遺伝子にコードされた免疫グロブリンの製造)」
US 5,844,094「Target binding polypeptide(標的結合ポリペプチド)」。
【0184】
以下の図面および実施例では本発明をさらに詳しく例示するが、本発明はこれらに限定されるわけではない。
図1aはインタクトIgG1の構造を表す。ドメインを矢印で示す。図1bは、主なヒト免疫グロブリンアイソタイプ単量体の構造的構成を図解している。ジスルフィド結合を線分で表し、N結合型糖質基を丸で表す。
図2は免疫グロブリンの定常ドメイン(左)および可変ドメイン(右)について、免疫グロブリンフォールドを表す。βストランドを矢印で示す。
図3は本発明の改変CH3ドメインの分子モデルを表し、ランダム突然変異を導入した部分が、溶媒接触表面によって示されている。この表面を丸で囲む。
図4は突然変異型CH3ドメインのアセンブリーに用いる断片の作製に使用したPCRを表す概略図である。PCRプライマーをそれぞれの5'−3'方向に矢印で示し、垂直線は導入した制限部位(突然変異型遺伝子のアセンブリーに使用したもの)のおよその位置を示す。PCR断片のライゲーションに備えてプライマーには以下の制限部位が含まれている:CH3LNCO:NcoI;CH3LSACおよびCH3CSAC:SacI;CH3CHINおよびCH3RHIN:HindIII;CH3RNOT:NotI。
図5は本願の免疫グロブリンドメインの使い方の例をいくつか表している。ランダム突然変異が導入された領域を星印で示す。一分子中のランダム突然変異導入領域の特異性は同一でも異なってもよい。
図6は二重特異性改変CH3ドメインの設計を表す概略図である。プライマー名を枠内に記載し、矢印はプライマーが伸長される方向を示す。斜線を引いた枠は、このコンストラクトにおいてランダム突然変異が導入された領域の相対的位置を示し、垂直線を引いた枠は、クローンC24を生成させるために導入された領域の相対的位置を示し、このクローニング手法に使用した制限部位が記載されている。
図7は二重特異性改変CH3ドメインの設計を表す概略図である。二重特異性改変CH3ドメインの基本設計のヌクレオチド配列およびその翻訳を表す。赤い配列は、二重特異性コンストラクトを生成させるためにランダム突然変異を導入した領域を示し、緑の枠はクローンC24を生成させるためにランダム突然変異を導入した領域を示す。
図8は本明細書に開示する配列の配列表である。
【実施例】
【0185】
実施例1
CH3ライブラリーの構築およびファージ表面ディスプレイ
突然変異型CH3ドメインの設計を支援するために、Brookhaven Databaseにエントリー1OQO.pdbとして公表されているIgG1 Fc断片の結晶構造を使った。
【0186】
CH3ライブラリーを構築するための基礎として使用した配列を配列番号1に示す。この配列では、第1アミノ酸がBrookhavenデータベースエントリー1oqo.pdbのA鎖のプロリン343に相当する。1oqo.pdbに含有される最後の残基は配列番号1のセリン102である。1oqo.pdbの構造の詳細な解析後に、βストランドをつなぐループを形成する残基を目視検査することにより、配列番号1の、βストランドA-Bをつなぐループの一部である残基17、18および19、ならびにβストランドE-Fをつなぐループの一部である残基71、72、73、76、および77に、ランダム突然変異を導入することにした。改変CH3ドメインの分子モデル(ランダム突然変異を導入した部分が溶媒接触表面で示されている)を図3に示す。一連のPCR反応に続いて、得られたPCR産物のライゲーションを行うことにより、改変遺伝子を作製した。ライゲーションを容易にするために、アミノ酸配列を変えずに制限部位が生じるように、配列番号1をコードするヌクレオチド配列のコドンの一部を修飾した(サイレント突然変異)。クローニングベクターpHEN1(Nucleic Acids Res. 1991 Aug 11;19(15):4133-7「Multi-subunit proteins on the surface of filamentous phage: methodologies for displaying antibody (Fab) heavy and light chains(繊維状ファージ表面上のマルチサブユニットタンパク質:抗体(Fab)重鎖および軽鎖をディスプレイするための方法論)」Hoogenboom HR, Griffiths AD, Johnson KS, Chiswell DJ, Hudson P, Winter G.)中にpelB分泌シグナルとインフレームで挿入するために、Met-Alaをコードする余分なヌクレオチド残基を配列の5'末端に取り付けて、NcoI制限部位を作った。ランダム突然変異を導入する残基については、20の天然アミノ酸の全てをコードするが、3つの停止コドンのうちの2つを回避する、コドンNNS(IUPACコード、この場合、SはCまたはGを意味する)を選択した。改変配列を配列番号2にヌクレオチド配列として記載し、配列番号3にアミノ酸配列として記載する。配列番号3中の文字Xは、ランダム突然変異が導入されたアミノ酸残基を表す。突然変異型CH3ドメインのアセンブリーに使用したPCRプライマーの配列を配列番号4〜9に示す。図4に、突然変異型遺伝子のアセンブリーのために生成させたPCR断片およびそのために使用したプライマーの概略図を示す。
【0187】
ヒトモノクローナル抗体3D6の重鎖のcDNA(Felgenhauer M, Kohl J, Rueker F.「Nucleotide sequences of the cDNAs encoding the V-regions of H- and L-chains of a human monoclonal antibody specific to HIV-1-gp41(HIV-1-gp41に特異的なヒトモノクローナル抗体のH鎖およびL鎖のV領域をコードするcDNAのヌクレオチド配列)」Nucleic Acids Res. 1990 Aug 25;18(16):4927)をPCR反応のテンプレートとして使用した。3つのPCR産物をSacIおよび/またはHindIIIでそれぞれ消化し、一つにライゲートした。そのライゲーション産物をさらにNcoIおよびNotIで消化し、前もってNcoIおよびNotIで消化しておいた表面ディスプレイファージミドベクターpHEN1中にライゲートした。選択したいくつかのクローンを制限解析およびDNA配列決定によって確認したところ、正しく挿入されたランダム突然変異配列を含むインサートを計画どおりに含有することがわかった。その後のファージ調製ステップについては、標準的なプロトコールに従った。簡単に述べると、ライゲーション混合物をエレクトロポレーションによって大腸菌TG1細胞に形質転換した。次に、ファージ粒子を大腸菌TG1細胞からヘルパーファージM13-K07を使ってレスキューした。次に、培養上清からPEG/NaClにより2段階でファージ粒子を沈殿させ、水に溶解し、パンニングによる選択に使用するか、あるいはそれらを−80℃で保存した。
実施例2
【0188】
CH3+3ライブラリーの構築
このライブラリーは、CH3ライブラリーと同じ方法で構築し、クローニングした。コンストラクトのアミノ酸配列を配列番号10に示し、対応するヌクレオチド配列を配列番号11に示す。構築に使用したプライマーは配列番号4〜7、配列番号9および配列番号12である。
実施例3
【0189】
CH3+5ライブラリーの構築
このライブラリーは、CH3ライブラリーと同じ方法で構築し、クローニングした。コンストラクトのアミノ酸配列を配列番号13に示し、対応するヌクレオチド配列を配列番号14に示す。構築に使用したプライマーは配列番号4〜7、配列番号9および配列番号15である。
実施例4
【0190】
TLR-9ペプチドによるCH3ファージライブラリーのパンニング
標準的プロトコールに従って3ラウンドのパンニングを行った。簡単に述べると、以下の方法を適用した。Maxisorp 96ウェルプレート(Nunc)をToll様受容体9(TLR-9)の配列の一部を表す合成ペプチドでコートした。ペプチドが以下の濃度で溶解している0.1M 炭酸Na緩衝液(pH9.6)200μlを各ウェルに加えた:
第1ラウンドのパンニング:1mg/ml TLR-9ペプチド;
第2ラウンドのパンニング:500μg/ml TLR-9ペプチド;
第3ラウンドのパンニング:100μg/ml TLR-9ペプチド。
【0191】
インキュベーションを37℃で1時間行った後、各ウェル200μlの2%ドライミルク(M-PBS)により、室温で1時間のブロッキングを行った。
【0192】
次に、ファージ懸濁液100μlおよび4%ドライミルク(M-PBS)100μlを加えた後、室温で、振とうしながら45分間、振とうせずに90分間インキュベートすることにより、表面ディスプレイファージライブラリーを、結合したペプチドと反応させた。
【0193】
結合していないファージ粒子を以下のように洗い流した。第1ラウンドのパンニング後:T-PBS 300μl×10、PBS 300μl×5;第2ラウンドのパンニング後:T-PBS 300μl×15、PBS 300μl×10;第3ラウンドのパンニング後:T-PBS 300μl×20、PBS 300μl×20。
【0194】
結合したファージ粒子の溶出は、各ウェル200μlの0.1Mグリシン(pH2.2)を加え、室温で振とうしながら30分間インキュベートすることによって行った。次に、2M Tris塩基60μlの添加によってファージ懸濁液を中和してから、指数増殖期の培養物10mlを、溶出させたファージ0.5mlと混合して、37℃で30分間インキュベートすることにより、大腸菌TG1細胞に感染させた。最後に、感染した細菌を、1%グルコースおよび100μg/mlアンピシリンを含むTYE培地に播種し、30℃で終夜インキュベートした。
【表1】
実施例5
【0195】
TLR-9に対して選択されたCH3突然変異体の選ばれたクローンの可溶性発現のためのクローニング
3ラウンドのパンニングによって選択されたファージから、ミディプレップによってファージミドDNAを単離した。突然変異型CH3領域をコードするDNAを、PCRによってバッチ増幅し、ベクターpNOTBAD/Myc-His(これは、大腸菌発現ベクターpBAD/Myc-His(Invitrogen)に、クローニングを容易にするためのNotI制限部位が挿入されているものである)に、NcoI-NotIクローニングした。ライゲートしたコンストラクトをエレクトロポレーションによって大腸菌LMG194細胞(Invitrogen)に形質転換し、1%グルコースおよびアンピシリンを含むTYE培地にて、30℃で終夜成長させた。選択したクローンを、アンピシリンを含む2×YT培地200μlに接種し、30℃で終夜成長させ、L-アラビノースを最終濃度が0.1%になるように加えることによって誘導した。16℃で終夜発現させた後、細胞を遠心分離によって収集し、ペリプラズム抽出物を調製するために、100μlのホウ酸Na緩衝液(pH8.0)により、4℃で終夜処理した。50μlのペリプラズム抽出物をELISA(下記参照)に使用した。
実施例6
【0196】
TLR-9に対して選択されたCH3突然変異体のELISA
選択したクローンをTLR-9ペプチドへの特異的結合についてELISAでアッセイした。
コーティング:マイクロタイタープレート(NUNC, Maxisorp)、各ウェル100μl、20μg TLR-9ペプチド/ml-0.1M炭酸Na緩衝液(pH9.6)、37℃で1時間;
洗浄:PBS 200μl×3;
ブロッキング:1%BSA-PBS、室温で1時間;
洗浄:PBS 200μl×3
ペリプラズム抽出物結合:ペリプラズム抽出物50μl、2%BSA-PBS 50μl、室温で終夜;
洗浄:PBS 200μl×3;
一次抗体:抗His4(Qiagen)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
二次抗体:ヤギ抗マウス*HRP(SIGMA)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
検出:クエン酸Na/リン酸緩衝液(pH4.5)中の3mg/ml OPD、30%H2O2 0.4μl;
停止:3M H2SO4 100ml;
吸光度の読取り:492/620nm。
【0197】
この最初の予備ELISAで高いシグナルを与えたクローンを、上記と同じ条件下に、20mlの体積で培養した。そのペリプラズム抽出物を、上述のように、培養体積の1/20の体積に単離し、確認のためにELISA(上述したもの)で試験した。
【表2】
実施例7
【0198】
鶏卵リゾチームによるCH3およびCH3+5ファージライブラリーのパンニング
3ラウンドのパンニングを行った。鶏卵リゾチームが以下の濃度で溶解しているPBS 200μlを各ウェルに加えることにより、Maxisorp 96ウェルプレート(Nunc)を鶏卵リゾチームでコートした:
第1ラウンドのパンニング:2mg/ml HEL;
第2ラウンドのパンニング:1mg/ml HEL;
第3ラウンドのパンニング:1mg/ml HEL。
【0199】
インキュベーションを37℃で1時間行った後、各ウェル200μlの2%ドライミルク(M-PBS)により、室温で1時間のブロッキングを行った。
【0200】
次に、ファージ懸濁液100μlおよび4%ドライミルク(M-PBS)100μlを加えた後、室温で、振とうしながら45分間、振とうせずに90分間インキュベートすることにより、表面ディスプレイファージライブラリーを、結合した鶏卵リゾチームと反応させた。
【0201】
結合していないファージ粒子を以下のように洗い流した。
第1ラウンドのパンニング後:T-PBS 300μl×10、PBS 300μl×5;
第2ラウンドのパンニング後:T-PBS 300μl×15、PBS 300μl×10;
第3ラウンドのパンニング後:T-PBS 300μl×20、PBS 300μl×20。
【0202】
結合したファージ粒子の溶出は、各ウェル200μlの0.1Mグリシン(pH2.2)を加え、室温で振とうしながら30分間インキュベートすることによって行った。次に、2M Tris塩基60μlの添加によってファージ懸濁液を中和してから、指数増殖期の培養物10mlを、溶出させたファージ0.5mlと混合して、37℃で30分間インキュベートすることにより、大腸菌TG1細胞に感染させた。最後に、感染した細菌を、1%グルコースおよび100μg/mlアンピシリンを含むTYE培地に播種し、30℃で終夜インキュベートした。
【表3】
【表4】
実施例8
【0203】
実施例7の選ばれたクローンの可溶性発現のためのクローニング
可溶性発現のための、選ばれたクローンのクローニングは、TLR-9に対して選択されたCH3突然変異体について上述したとおりに行った。
実施例9
【0204】
実施例7の選ばれたクローンの可溶性発現
選ばれたクローンの可溶性発現は、TLR-9に対して選択されたCH3突然変異体について上述したとおりに行った。ペリプラズム抽出物を予備ELISAで試験した(プロトコールは実施例10参照)。
【0205】
この最初の予備ELISAで高いシグナルを与えたクローンを、上記と同じ条件下に、20mlの体積で培養した。そのペリプラズム抽出物を、上述のように、培養体積の1/20の体積に単離し、確認のためにELISA(実施例10に記載)で試験した。
実施例10
【0206】
鶏卵リゾチームに対して選択されたCH3突然変異体のELISA
コーティング:マイクロタイタープレート(NUNC, Maxisorp)、各ウェル100μl、PBS中の100μg鶏卵リゾチーム/ml、37℃で1時間;
洗浄:PBS 200μl×3;
ブロッキング:1%BSA-PBS、室温で1時間;
洗浄:PBS 200μl×3;
ペリプラズム抽出物結合:ペリプラズム抽出物50μl、2%BSA-PBS 50μl、室温で終夜;
洗浄:PBS 200μl×3;
一次抗体:抗His4(Qiagen)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
二次抗体:ヤギ抗マウス*HRP(SIGMA)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
検出:クエン酸Na/リン酸緩衝液(pH4.5)中の3mg/ml OPD、30%H2O2 0.4μl;
停止:3M H2SO4 100ml;
吸光度の読取り:492/620nm。
【表5】
【表6】
【0207】
鶏卵リゾチームは抗his4抗体と反応するので、比較的高いバックグラウンドが観察された。
【表7】
実施例11
【0208】
CLライブラリー
Fab断片の結晶構造の目視検査(ヒトモノクローナル抗体3D6のFabの構造を使用する:RSCB Protein Data Bank(http://www.rcsb.org/pdb/)エントリー1DFB.PDB(He XMら Proc Natl Acad Sci U S A. 1992 Aug 1;89(15):7154-8))ならびにこのタンパク質の二次構造および三次構造のコンピュータ支援解析(例えばこの目的にはProtein Explorerを使用する(http://molvis.sdsc.edu/protexpl/frntdoor.htm))により、CLドメイン足場のβストランドをつなぐループ領域中に位置する残基を同定することができる。これらの残基は、アミノ酸8〜18、アミノ酸27〜35、アミノ酸42〜78、アミノ酸83〜85、アミノ酸92〜100、アミノ酸108〜117およびアミノ酸123〜126を含む(IMGTナンバリングシステム(Lefranc MPら Nucleic Acids Res. 2005 Jan 1;33 (Database issue):D593-7;Lefranc MPら Dev Comp Immunol. 2005;29(3):185-203)によるナンバリング)。
【0209】
より具体的に述べると、ヒトCLドメイン(配列番号48)内で、残基11、12、14〜18、および92〜95にランダム突然変異を導入する。ランダム突然変異の導入は、関連コドンの位置がヌクレオチド配列5'-NNS-3'(これは、潜在的に20の天然アミノ酸の全てをコードすると共に、3つの停止コドンのうちの2つを回避する配列である)でコードされるPCRプライマーを使って、コード配列をPCR増幅することにより、達成される。ライブラリーインサートは、2回の別個のPCR反応によって増幅され、その二つのPCR断片が、HpyCH4IV制限部位(PCRプライマーによりサイレント突然変異として導入されるもの)により、一つにライゲートされる。さらに、ファージディスプレイベクターpHEN(Hoogenboom HRら Nucleic Acids Res. 1991 Aug 11;19(15):4133-7)へのクローニングのために、制限エンドヌクレアーゼ部位NcoIおよびNotIも、プライマーによってそれぞれ与えられる。ファージディスプレイの場合、CLドメインのC末端システインは含まれないが、修飾CLクローンを例えばFab断片の構築などに使用する時には、後から付加することができる。
【0210】
PCR増幅のテンプレートとしては、ヒトモノクローナル抗体の軽鎖全体をインサートとして含有するpRcCMV-3D6LC(Rueker Fら Ann N Y Acad Sci. 1991 Dec 27;646:212-9)などのプラスミドを使用する。
【0211】
CLドメインの位置92と位置95の間に挿入された追加残基を含有するCL+3(配列番号50、51)およびCL+5(配列番号52、53)ライブラリーの場合は、プライマーCLRHPYの代わりにプライマーCLRHPY3およびCLRHPY5をそれぞれ使用する。
【0212】
pHEN1のNcoI部位にクローニングされる(これにより、コンストラクトのN末端にpelBリーダー配列が取り付けられることになる)、PCRおよびライゲーションの最終産物のヌクレオチド配列およびアミノ酸配列を、以下に示す(配列番号48、49):
【化1】
【0213】
CLライブラリー用のプライマーリスト:
cllnco:5'-cttaccatgg ccgtggctgc accatctgtc ttcatcttcc cgccatctnn snnscagnns nnsnnsnnsn nsgcctctgt tgtgtgc-3'(配列番号56)
cllhpy:5'-tgacaacgtc agggtgctgc tgaggc-3'(配列番号57)
clrhpy:5'-tcagaacgtt gnnsnnsnns nnstacgaga aacacaaagt c-3'(配列番号58)
clrhpy3:5'-tcagaacgtt gnnsnnsnns nnsnnsnnsn nstacgagaa acacaaagtc-3'(配列番号59)
clrhpy5:5'-tcagaacgtt gnnsnnsnns nnsnnsnnsn nsnnsnnsta cgagaaacac aaagtc-3'(配列番号60)
clrnot:5'-catcgcggcc gcctctcccc tgttgaagct c-3'(配列番号61)
【0214】
選ばれたいくつかのライブラリークローン(ファージミドベクターpHEN1中にクローニングされた突然変異型CLドメイン)を制限解析およびDNA配列決定に付して、それらが正しく挿入されたランダム突然変異配列を含むインサートを計画どおりに含有することを確認する。その後のファージ調製ステップについては、標準的なプロトコールに従う。簡単に述べると、ライゲーション混合物をエレクトロポレーションによって大腸菌TG1細胞に形質転換する。次に、ファージ粒子を大腸菌TG1細胞からヘルパーファージM13-K07を使ってレスキューする。次に、培養上清からPEG/NaClにより2段階でファージ粒子を沈殿させ、水に溶解し、パンニングによる選択に使用するか、あるいはそれらを−80℃で保存することができる。
実施例12
【0215】
CH1ライブラリー
Fab断片の結晶構造(ヒトモノクローナル抗体3D6のFabの構造を使用する:RSCB Protein Data Bankエントリー1DFB.PDB)の目視検査、ならびにこのタンパク質の二次構造および三次構造のコンピュータ支援解析(この目的にはProtein Explorerを使用する)により、CH1ドメイン足場のβストランドをつなぐループ領域中に位置する残基を同定することができる。これらの残基は、アミノ酸7〜21、アミノ酸25〜39、アミノ酸41〜81、アミノ酸83〜85、アミノ酸89〜103およびアミノ酸106〜117を含む(IMGTナンバリングシステムによるナンバリング)。
【0216】
より具体的に述べると、ヒトCH1ドメイン内で、残基12〜19および93〜100にランダム突然変異を導入する(配列番号54、55)。ランダム突然変異の導入は、関連コドンの位置がヌクレオチド配列5'-NNS-3'(これは、潜在的に20の天然アミノ酸の全てをコードすると共に、3つの停止コドンのうちの2つを回避する配列である)でコードされるPCRプライマーを使って、コード配列をPCR増幅することにより、達成される。ライブラリーインサートは、2回の別個のPCR反応によって増幅され、その二つのPCR断片が、BstEII制限部位(CH1ドメイン中に天然に存在するもの)により、一つにライゲートされる。さらに、ファージディスプレイベクターpHENへのクローニングのために、制限エンドヌクレアーゼ部位NcoIおよびNotIも、プライマーによってそれぞれ与えられる。ファージディスプレイの場合、CH1ドメインのC末端システインは含まれないが、修飾CH1クローンを例えばFab断片の構築などに使用する時には、後から付加することができる。
【0217】
PCR増幅のテンプレートとしては、ヒトモノクローナル抗体の重鎖全体をインサートとして含有するpRcCMV-3D6HCなどのプラスミドを使用する。
【0218】
pHEN1のNcoI部位にクローニングされる(これにより、コンストラクトのN末端にpelBリーダー配列が取り付けられることになる)、PCRおよびライゲーションの最終産物のヌクレオチド配列およびアミノ酸配列を、以下に示す(配列番号54、55):
【化2】
【0219】
CH1ライブラリー用のプライマーリスト:
CH1LNCO:5'-acgtccatgg ccgcctccac caagggccca tcggtcttcc ccctggcacc ctcctccnns nnsnnsnnsn nsnnsnnsnn sgccctgggc tgcctggtc-3'(配列番号62)
CH1LBST:5'-ggcacggtca ccacgctgct gag-3'(配列番号63)
CH1RBST:5'-agcgtggtga ccgtgcccnn snnsnnsnns nnsnnsnnsa cctacatctg caacgtgaat c-3'(配列番号64)
CH1RNOT:5'-catagcggcc gcagatttgg gctcaacttt cttgtc-3'(配列番号65)
【0220】
選ばれたいくつかのライブラリークローン(ファージミドベクターpHEN1中にクローニングされた突然変異型CH1ドメイン)を制限解析およびDNA配列決定に付して、それらが正しく挿入されたランダム突然変異配列を含むインサートを計画どおりに含有することを確認する。その後のファージ調製ステップについては、標準的なプロトコールに従う。簡単に述べると、ライゲーション混合物をエレクトロポレーションによって大腸菌TG1細胞に形質転換する。次に、ファージ粒子を大腸菌TG1細胞からヘルパーファージM13-K07を使ってレスキューする。次に、培養上清からPEG/NaClにより2段階でファージ粒子を沈殿させ、水に溶解し、パンニングによる選択に使用するか、あるいはそれらを−80℃で保存することができる。
実施例13
【0221】
鶏卵リゾチーム(HEL)によるCH1ファージライブラリーのパンニング
CH1ファージライブラリー(実施例12参照)を使って3ラウンドのパンニングを行う。鶏卵リゾチームが以下の濃度で溶解しているPBS 200μlを各ウェルに加えることにより、Maxisorp 96ウェルプレート(Nunc)を鶏卵リゾチームでコートする:
第1ラウンドのパンニング:2mg/ml HEL;
第2ラウンドのパンニング:1mg/ml HEL;
第3ラウンドのパンニング:1mg/ml HEL。
【0222】
インキュベーションを37℃で1時間行った後、各ウェル200μlの2%ドライミルク(M-PBS)により、室温で1時間のブロッキングを行う。
【0223】
次に、ファージ懸濁液100μlおよび4%ドライミルク(M-PBS)100μlを加えた後、室温で、振とうしながら45分間、振とうせずに90分間インキュベートすることにより、表面ディスプレイファージライブラリーを、結合した鶏卵リゾチームと反応させる。
【0224】
結合していないファージ粒子を以下のように洗い流す:
第1ラウンドのパンニング:T-PBS 300μl×10、PBS 300μl×5
第2ラウンドのパンニング:T-PBS 300μl×15、PBS 300μl×10
第3ラウンドのパンニング:T-PBS 300μl×20、PBS 300μl×20。
【0225】
結合したファージ粒子の溶出は、各ウェル200μlの0.1Mグリシン(pH2.2)を加え、室温で振とうしながら30分間インキュベートすることによって行う。次に、2M Tris塩基60μlの添加によってファージ懸濁液を中和してから、指数増殖期の培養物10mlを、溶出させたファージ0.5mlと混合して、37℃で30分間インキュベートすることにより、大腸菌TG1細胞に感染させる。最後に、感染した細菌を、1%グルコースおよび100μg/mlアンピシリンを含むTYE培地に播種し、30℃で終夜インキュベートする。
【0226】
リゾチームに対して選択されたCH1突然変異体の選ばれたクローンの可溶性発現のためのクローニング
3ラウンドのパンニングによって選択されるファージから、ミディプレップによってファージミドDNAを単離する。突然変異型CH1ドメインをコードするDNAを、PCRによってバッチ増幅し、ベクターpNOTBAD/Myc-His(これは、大腸菌発現ベクターpBAD/Myc-His(Invitrogen)に、クローニングを容易にするためのNotI制限部位が挿入されているものである)に、NcoI-NotIクローニングする。ライゲートしたコンストラクトをエレクトロポレーションによって大腸菌LMG194細胞(Invitrogen)に形質転換し、1%グルコースおよびアンピシリンを含むTYE培地にて、30℃で終夜成長させる。選択したクローンを、アンピシリンを含む2×YT培地200μlに接種し、30℃で終夜成長させ、L-アラビノースを最終濃度が0.1%になるように加えることによって誘導する。16℃で終夜発現させた後、細胞を遠心分離によって収集し、ペリプラズム抽出物を調製するために、100μlのホウ酸Na緩衝液(pH8.0)により、4℃で終夜処理する。50μlのペリプラズム抽出物をELISAに使用する。
【0227】
この最初の予備ELISAで高いシグナルを与えるクローンを、上記と同じ条件下に、20mlの体積で培養する。そのペリプラズム抽出物を、上述のように、培養体積の1/20の体積に単離し、確認のためにELISA(後述)で試験する。
【0228】
鶏卵リゾチームに対して選択されたCH1突然変異体のELISA
コーティング:マイクロタイタープレート(NUNC, Maxisorp)、各ウェル100μl、PBS中の100μg鶏卵リゾチーム/ml、37℃で1時間;
洗浄:PBS 200μl×3;
ブロッキング:1%BSA-PBS、室温で1時間;
洗浄:PBS 200μl×3;
ペリプラズム抽出物結合:ペリプラズム抽出物50μl、2%BSA-PBS 50μl、室温で終夜;
洗浄:PBS 200μl×3;
一次抗体:抗His4(Qiagen)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
二次抗体:ヤギ抗マウス*HRP(SIGMA)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
検出:クエン酸Na/リン酸緩衝液(pH4.5)中の3mg/ml OPD、30%H2O2 0.4μl;
停止:3M H2SO4 100ml;
吸光度の読取り:492/620nm。
【0229】
ELISAシグナルがバックグラウンドシグナルの少なくとも3倍である場合に、そのクローンを陽性であると解釈する。
実施例14
【0230】
鶏卵リゾチーム(HEL)によるCLファージライブラリーのパンニング
CLファージライブラリー(実施例11参照)を使って3ラウンドのパンニングを行う。鶏卵リゾチームが以下の濃度で溶解しているPBS 200μlを各ウェルに加えることにより、Maxisorp 96ウェルプレート(Nunc)を鶏卵リゾチームでコートする:
第1ラウンドのパンニング:2mg/ml HEL;
第2ラウンドのパンニング:1mg/ml HEL;
第3ラウンドのパンニング:1mg/ml HEL。
【0231】
インキュベーションを37℃で1時間行った後、各ウェル200μlの2%ドライミルク(M-PBS)により、室温で1時間のブロッキングを行う。
【0232】
次に、ファージ懸濁液100μlおよび4%ドライミルク(M-PBS)100μlを加えた後、室温で、振とうしながら45分間、振とうせずに90分間インキュベートすることにより、表面ディスプレイファージライブラリーを、結合した鶏卵リゾチームと反応させる。
【0233】
結合していないファージ粒子を以下のように洗い流す:
第1ラウンドのパンニング:T-PBS 300μl×10、PBS 300μl×5;
第2ラウンドのパンニング:T-PBS 300μl×15、PBS 300μl×10;
第3ラウンドのパンニング:T-PBS 300μl×20、PBS 300μl×20。
【0234】
結合したファージ粒子の溶出は、各ウェル200μlの0.1Mグリシン(pH2.2)を加え、室温で振とうしながら30分間インキュベートすることによって行う。次に、2M Tris塩基60μlの添加によってファージ懸濁液を中和してから、指数増殖期の培養物10mlを、溶出させたファージ0.5mlと混合して、37℃で30分間インキュベートすることにより、大腸菌TG1細胞に感染させる。最後に、感染した細菌を、1%グルコースおよび100μg/mlアンピシリンを含むTYE培地に播種し、30℃で終夜インキュベートする。
【0235】
リゾチームに対して選択されたCL突然変異体の選ばれたクローンの可溶性発現のためのクローニング
3ラウンドのパンニングによって選択されるファージから、ミディプレップによってファージミドDNAを単離する。突然変異型CLドメインをコードするDNAを、PCRによってバッチ増幅し、ベクターpNOTBAD/Myc-His(これは、大腸菌発現ベクターpBAD/Myc-His(Invitrogen)に、クローニングを容易にするためのNotI制限部位が挿入されているものである)に、NcoI-NotIクローニングする。ライゲートしたコンストラクトをエレクトロポレーションによって大腸菌LMG194細胞(Invitrogen)に形質転換し、1%グルコースおよびアンピシリンを含むTYE培地にて、30℃で終夜成長させる。選択したクローンを、アンピシリンを含む2×YT培地200μlに接種し、30℃で終夜成長させ、L-アラビノースを最終濃度が0.1%になるように加えることによって誘導する。16℃で終夜発現させた後、細胞を遠心分離によって収集し、ペリプラズム抽出物を調製するために、100μlのホウ酸Na緩衝液(pH8.0)により、4℃で終夜処理する。50μlのペリプラズム抽出物をELISAに使用する。
【0236】
この最初の予備ELISAで高いシグナルを与えるクローンを、上記と同じ条件下に、20mlの体積で培養する。そのペリプラズム抽出物を、上述のように、培養体積の1/20の体積に単離し、確認のためにELISA(後述)で試験する。
【0237】
鶏卵リゾチームに対して選択されたCL突然変異体のELISA
コーティング:マイクロタイタープレート(NUNC, Maxisorp)、各ウェル100μl、PBS中の100μg鶏卵リゾチーム/ml、37℃で1時間;
洗浄:PBS 200μl×3;
ブロッキング:1%BSA-PBS、室温で1時間;
洗浄:PBS 200μl×3;
ペリプラズム抽出物結合:ペリプラズム抽出物50μl、2%BSA-PBS 50μl、室温で終夜;
洗浄:PBS 200μl×3;
一次抗体:抗His4(Qiagen)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
二次抗体:ヤギ抗マウス*HRP(SIGMA)、1%BSA-PBSに1:1000、室温で90分間、各ウェル100μl;
洗浄:PBS 200μl×3;
検出:クエン酸Na/リン酸緩衝液(pH4.5)中の3mg/ml OPD、30%H2O2 0.4μl;
停止:3M H2SO4 100ml;
吸光度の読取り:492/620nm。
【0238】
ELISAシグナルがバックグラウンドシグナルの少なくとも3倍である場合に、そのクローンを陽性であると解釈する。
実施例15
【0239】
両側にランダム突然変異が導入された免疫グロブリンドメインの構築(二重特異性改変CH3ドメイン)
この実施例では二つの結合特異性を持つ改変免疫グロブリンドメインを記述する。
【0240】
この改変免疫グロブリンドメインの設計には以下の戦略を含めた:
・リゾチームに特異的に結合するCH3+5ライブラリー由来の改変CH3ドメイン・クローンC24(実施例10参照)を、出発点として使用した。
・免疫グロブリンフォールドのβストランドをつないでいて、クローンC24の生成時に突然変異させた残基に対してドメインの反対側に存在する残基を、ランダム突然変異を導入する対象として、この修飾CH3ドメイン中に同定した。
・CH3、CH3+3およびCH3+5ライブラリーについて上述したものと同様の手法で、これらの残基のランダム突然変異導入と、この改変免疫グロブリンドメインの合成とを可能にするPCRプライマーを設計した。
【0241】
ランダム突然変異が導入された位置を含有する四つのPCR産物をライゲートし、完全長インサートをPCRによって増幅した。次に、それらをpHEN-1中にNcoI-NotI部位を利用してクローニングし、大腸菌TG-1細胞に形質転換することにより、約108コロニーのライブラリーを構築した。ランダムに選んだ20個のコロニーを配列決定したところ、ランダム突然変異を導入した位置には、独立して突然変異が導入されていることがわかった。また、「野生型」(C24)配列は観察されなかった。標準的なプロトコールに従ってファージライブラリーを作製し、6.32×1010TU/mlというファージ力価を得た。
【0242】
二重特異性を試験するために、組換えヒトエリスロポエチン(rhEPO)を第2抗体として選択し、一方、最初に設計した鶏卵リゾチームに対する特異性は、そのコンストラクトが保っているものと予想した。rhEPO反応性ファージを4ラウンドのパンニングによって選択した。突然変異導入後も鶏卵リゾチームを結合するC24クローンの集団が保たれるように、rhEPOによる第1ラウンドの選択後に、鶏卵リゾチーム(PBS中、1mg/ml)によるファージ集団のパンニングを1ラウンド行った。その後のパンニングでは、0.1M炭酸Na緩衝液(pH9.6)中の濃度を減らしながら、200μlのrhEPOを、マイクロタイタープレート(Maxisorp, Nunc)の5ウェルにコートした(下記の表参照)。2%M-PBSでブロッキングを行った後、ブロッキング剤中のファージを、室温で2時間結合させた。T-PBSで20回、PBSで20回洗浄した後、0.1Mグリシン(pH2.2)で溶出させ、2M Trisで中和した。溶出させたファージを直ちに使用して、指数増殖期のTG-1を感染させた。感染した細胞をアンピシリン含有培地で選択した。ヘルパーファージM13-K07の重複感染によってファージ粒子を培養上清からレスキューし、PEGで濃縮し、もう1ラウンドのパンニングに使用した。各ラウンドのパンニング後に、インプットファージ数およびアウトプットファージ数を、大腸菌の形質転換単位として決定した(表8)。
【表8】
【0243】
結果として生じたコロニーをプレートから掻き落とし、アンピシリンを含む2×YTで培養し、そのプラスミドDNAをミディプレップで単離した。インサートをPCRで増幅した後、ベクターpNOTBAD中にサブクローニングし、大腸菌E104株に形質転換した。4×72コロニーを、アンピシリンを含む2×YT培地 200μlで培養し、翌日、0.1%L-アラビノースで誘導した。16℃で24時間発現させた後、それらを、200μlのホウ酸Na緩衝液(pH8.0)により、4℃で6時間溶解し、ペリプラズム抽出物をELISAに使用した。
【0244】
ELISAのために、Maxisorpプレートを、PBS中の鶏卵リゾチーム(20μg/ml)または0.1M炭酸Na緩衝液(pH9.6)中のrhEPOで、それぞれ37℃で1時間コートした。1%BSA-PBSによるブロッキングの後、同じブロッキング剤中のペリプラズム抽出物を終夜結合させた。抗His-(4)抗体およびHRP(鶏卵リゾチーム検出用)またはAP(rhEPO検出用)とコンジュゲートしたヤギ抗マウスIgG抗体を使って結合を明らかにした。OPD変換の呈色反応(HRP)は1.25M H2SO4で停止させた後に492/620nmで読取り、pNPP変換(AP)は405/620nmで読み取った。有望な吸光度値を持つ14個のクローンを、20ml規模での発現用に選択した。16℃で24時間のアラビノース誘導後に、細胞を収集し、1mlのホウ酸Na緩衝液中、4℃で終夜溶解し、その溶解物をELISAに使用した。ELISAは4重に並行して上述のように行い、ペリプラズム抽出物を含まないウェルおよび抗原を含まないウェルを陰性対照として使用した。配列番号42、43のクローンを使って結果(表9)を得た。
【表9】
実施例16
【0245】
改変CH3ドメインはFab様形式の二重特異性をもたらす
この実施例で使用するコンストラクトでは、抗体のVL鎖およびVH鎖がどちらも改変CH3ドメインに融合される。
【0246】
HIV-1のgp41上のエピトープを認識するヒトモノクローナル抗体3D6(He XMら Proc Natl Acad Sci U S A. 1992 89:7154-8;Kohl Jら Ann N Y Acad Sci. 1991 646:106-14;Felgenhauer Mら Nucleic Acids Res. 1990 18:4927)のVLおよびVH領域を、鶏卵リゾチームに特異的に結合する改変CH3ドメイン・クローン24の融合パートナーとして使用した。
【0247】
ジスルフィド結合によるVL-CH3/VH-CH3二量体の形成を促進するために、C24配列のC末端に残基Ser-Cysを付加した。
【0248】
二つの鎖3D6VL-C24および3D6VH-C24のそれぞれヌクレオチド配列およびアミノ酸配列を、それぞれ配列番号47、46および配列番号45、44に示す。
【0249】
コード領域の増幅を可能にすると同時に、コード領域を一つにライゲートする際に使用した制限部位(サイレント突然変異)を導入するプライマーを、設計した。遺伝子を発現させるためにピキアパストリス発現系を選択した。コンストラクトを適切なピキアパストリス発現ベクターにクローニングした。すなわち、3D6VL-C24をpPIC9K(最終名:pPIC9K3LC)にクローニングし、3D6VH-C24(最終名:pPICZ3HC)をpPICZalphaAにクローニングした。コンストラクトpPICZ3HCをBglIIで線状化し、ピキアパストリスGS115に形質転換し、形質転換体をゼオシン含有固形培地上で選択した。次に、形質転換体の一つを、SalI線状化コンストラクトpPIC9K3LCの宿主細胞として使用した。次に、二重形質転換体をRDB培地上で選択した。
【0250】
クローンをYPG 30mlに接種し、OD600=10になるまで成長させた後、BMMY培地中の1%メタノールを加えることによって誘導した。この誘導を16℃で36時間続けた。上清を遠心分離によって取り出し、約10倍濃縮した。組換えタンパク質の存在を、抗His(4)抗体を用いるウェスタンブロットによって確認し、約50〜100μg/l-初期培養物の濃度であると見積った。
【0251】
最初の機能試験は、10倍濃縮上清で行った。まず、Maxisorpプレートのウェルを、PBS中20μg/mlの鶏卵リゾチームまたは0.1M炭酸Na緩衝液(pH9.6)中20μg/mlの抗体3D6のエピトープで、それぞれ37℃で1時間コートした。3D6エピトープは組換え生産されたGST融合タンパク質の形で使用した。1%BSA-PBSによるブロッキング後に、濃縮された上清を同じブロッキング剤中で終夜結合させた。抗His(4)抗体およびHRPにコンジュゲートしたヤギ抗マウス抗体を使って、結合を明らかにし、OPD変換がもたらす呈色反応として492/620nmで可視化した(表10)。
【表10】
【図面の簡単な説明】
【0252】
【図1】図1aはインタクトIgG1の構造を表す。ドメインを矢印で示す。図1bは、主なヒト免疫グロブリンアイソタイプ単量体の構造的構成を図解している。ジスルフィド結合を線分で表し、N結合型糖質基を丸で表す。
【図2】免疫グロブリンの定常ドメイン(左)および可変ドメイン(右)について、免疫グロブリンフォールドを表す。βストランドを矢印で示す。
【図3】本発明の改変CH3ドメインの分子モデルを表し、ランダム突然変異を導入した部分が、溶媒接触表面によって示されている。この表面を丸で囲む。
【図4】突然変異型CH3ドメインのアセンブリーに用いる断片の作製に使用したPCRを表す概略図である。PCRプライマーをそれぞれの5'−3'方向に矢印で示し、垂直線は導入した制限部位(突然変異型遺伝子のアセンブリーに使用したもの)のおよその位置を示す。PCR断片のライゲーションに備えてプライマーには以下の制限部位が含まれている:CH3LNCO:NcoI;CH3LSACおよびCH3CSAC:SacI;CH3CHINおよびCH3RHIN:HindIII;CH3RNOT:NotI。
【図5】本願の免疫グロブリンドメインの使い方の例をいくつか表している。ランダム突然変異が導入された領域を星印で示す。一分子中のランダム突然変異導入領域の特異性は同一でも異なってもよい。
【図6】二重特異性改変CH3ドメインの設計を表す概略図である。プライマー名を枠内に記載し、矢印はプライマーが伸長される方向を示す。斜線を引いた枠は、このコンストラクトにおいてランダム突然変異が導入された領域の相対的位置を示し、垂直線を引いた枠は、クローンC24を生成させるために導入された領域の相対的位置を示し、このクローニング手法に使用した制限部位が記載されている。
【図7】二重特異性改変CH3ドメインの設計を表す概略図である。二重特異性改変CH3ドメインの基本設計のヌクレオチド配列およびその翻訳を表す。赤い配列は、二重特異性コンストラクトを生成させるためにランダム突然変異を導入した領域を示し、緑の枠はクローンC24を生成させるためにランダム突然変異を導入した領域を示す。
【図8】本明細書に開示する配列の配列表である。
【図1a】
【図1b】
【特許請求の範囲】
【請求項1】
免疫グロブリンの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンを設計し、ある抗原のエピトープに対する前記免疫グロブリンの結合を決定するための方法(ただし、無修飾免疫グロブリンは前記エピトープには有意に結合しない)であって、
・少なくとも一つの構造ループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記構造ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、および
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ
を含む方法。
【請求項2】
前記免疫グロブリンが少なくとも二つの異なるエピトープに特異的に結合する、請求項1に記載の方法。
【請求項3】
前記免疫グロブリンの少なくとも一つの構造ループ領域に少なくとも一つの修飾を含み、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、ウイルス抗原および原虫抗原からなる群より選択される少なくとも一つの分子に対する前記少なくとも一つのループ領域の特異的結合を決定する(ただし、無修飾構造ループ領域を含有する免疫グロブリンは、前記少なくとも一つの分子に特異的には結合しない)、請求項1または請求項2のいずれか一項に記載の方法。
【請求項4】
免疫グロブリンの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンまたはその医薬調製物を製造し、ある抗原のエピトープに対する前記免疫グロブリンの結合を決定するための方法(ただし、無修飾免疫グロブリンは前記エピトープには有意に結合しない)であって、
・少なくとも一つのループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ、および
・前記エピトープに結合する修飾免疫グロブリンを用意し、要すればそれを医薬調製物に仕上げるステップ、
を含む方法。
【請求項5】
免疫グロブリンの少なくとも一つの構造ループ領域に少なくとも一つの修飾を含む、少なくとも一つの第1分子に特異的に結合する多重特異性免疫グロブリン、またはその医薬調製物を製造し、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、ウイルス抗原および原虫抗原からなる群より選択される少なくとも一つの第2分子に対する、前記少なくとも一つのループ領域の特異的結合を決定するための方法(ただし、無修飾構造ループ領域を含有する免疫グロブリンは、前記少なくとも一つの第2分子には特異的に結合しない)であって、
・少なくとも一つの構造ループ領域を含む、少なくとも一つの第1分子に特異的に結合する免疫グロブリンをコードする核酸を用意するステップ、
・前記核酸によってコードされる前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンを前記少なくとも一つの第2分子と接触させるステップ、
・前記修飾免疫グロブリンが第2分子に特異的に結合するかどうかを決定するステップ、および
・前記少なくとも一つの第2分子に特異的に結合する修飾免疫グロブリンを用意し、要すればそれを医薬調製物に仕上げるステップ、
を含む方法。
【請求項6】
免疫グロブリンがヒトまたはネズミ由来であることを特徴とする、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
ヒト免疫グロブリンがIgA1、IgA2、IgD、IgE、IgG1、IgG2、IgG3、IgG4およびIgMからなる群より選択されることを特徴とする、請求項6に記載の方法。
【請求項8】
ネズミ免疫グロブリンがIgA、IgD、IgE、IgG1、IgG2A、IgG2B、IgG2C、IgG3およびIgMからなる群より選択されることを特徴とする、請求項6に記載の方法。
【請求項9】
免疫グロブリンが免疫グロブリンの重鎖および/もしくは軽鎖またはその一部を含むことを特徴とする、請求項7または8に記載の方法。
【請求項10】
免疫グロブリンが、免疫グロブリンの少なくとも一つの定常ドメインおよび/もしくは少なくとも一つの可変ドメイン、またはその一部(ミニドメインを含む)を含むことを特徴とする、請求項7〜9のいずれか一項に記載の方法。
【請求項11】
定常ドメインがCH1、CH2、CH3、CH4、Igk-C、Igl-Cおよびそれらの組み合わせの群より選択されることを特徴とする、請求項10に記載の方法。
【請求項12】
CH1、CH2、CH3およびCH4の修飾ループ領域が、アミノ酸7〜21、アミノ酸25〜39、アミノ酸41〜81、アミノ酸83〜85、アミノ酸89〜103およびアミノ酸106〜117を含むことを特徴とする、請求項11に記載の方法。.
【請求項13】
ヒト由来のIgk-CおよびIgl-Cのループ領域が、アミノ酸8〜18、アミノ酸27〜35、アミノ酸42〜78、アミノ酸83〜85、アミノ酸92〜100、アミノ酸108〜117およびアミノ酸123〜126を含むことを特徴とする、請求項11に記載の方法。
【請求項14】
ネズミ由来のIgk-CおよびIgl-Cのループ領域が、アミノ酸8〜20、アミノ酸26〜36、アミノ酸43〜79、アミノ酸83〜85、アミノ酸90〜101、アミノ酸108〜116およびアミノ酸122〜125を含むことを特徴とする、請求項11に記載の方法。
【請求項15】
ヒト由来の免疫グロブリンの可変ドメインの構造ループ領域が、アミノ酸8〜20、アミノ酸44〜50、アミノ酸67〜76およびアミノ酸89〜101を含むことを特徴とする、請求項10に記載の方法。
【請求項16】
ネズミ由来の免疫グロブリンの可変ドメインの構造ループ領域が、アミノ酸6〜20、アミノ酸44〜52、アミノ酸67〜76およびアミノ酸92〜101を含むことを特徴とする、請求項10に記載の方法。
【請求項17】
免疫グロブリンがラクダ由来であることを特徴とする、請求項1〜5のいずれか一項に記載の方法。
【請求項18】
免疫グロブリンがCH1、CH2およびCH3からなる群より選択される少なくとも一つの定常ドメインを含むことを特徴とする、請求項17に記載の方法。
【請求項19】
CH1、CH2およびCH3のループ領域が、アミノ酸8〜20、アミノ酸24〜39、アミノ酸42〜78、アミノ酸82〜85、アミノ酸91〜103およびアミノ酸108〜117を含むことを特徴とする、請求項18に記載の方法。.
【請求項20】
少なくとも一つのヌクレオチドの修飾が、前記核酸によってコードされる免疫グロブリンの置換、欠失および/または挿入をもたらす、請求項1〜19のいずれか一項に記載の方法。
【請求項21】
少なくとも一つのループ領域のアミノ酸が部位指定ランダム突然変異によって修飾されることを特徴とする、請求項1〜20のいずれか一項に記載の方法。
【請求項22】
ランダムに修飾された核酸分子が、5'-NNS-3'、5'-NNN-3'または5'-NNK-3'という配列を持つ少なくとも一つのヌクレオチド繰返し単位を含むことを特徴とする、請求項21に記載の方法。
【請求項23】
発現系がベクターを含むことを特徴とする、請求項1〜22のいずれか一項に記載の方法。
【請求項24】
修飾免疫グロブリンが、宿主中で、好ましくは細菌、酵母、植物細胞中で、動物細胞中で、または植物もしくは動物中で発現されることを特徴とする、請求項1〜23のいずれか一項に記載の方法。
【請求項25】
分子に対する修飾免疫グロブリンの特異的結合が、免疫学的アッセイ、好ましくは酵素結合免疫吸着アッセイ(ELISA)、表面プラズモン共鳴アッセイ、飽和移動差核磁気共鳴分光法、転移NOE(trNOE)核磁気共鳴分光法、競合アッセイ、組織結合アッセイ、生細胞結合アッセイおよび細胞抽出物アッセイからなる群より選択される結合アッセイによって決定されることを特徴とする、請求項1〜24のいずれか一項に記載の方法。
【請求項26】
修飾免疫グロブリンが、有機分子、酵素ラベル、放射性ラベル、着色ラベル、蛍光ラベル、発色ラベル、発光ラベル、ハプテン、ジゴキシゲニン、ビオチン、金属錯体、金属、コロイド金およびそれらの混合物からなる群より選択されるラベルにコンジュゲートされることを特徴とする、請求項1〜25のいずれか一項に記載の方法。
【請求項27】
少なくとも一つのループ領域を持つ、CH1、CH2、CH3、CH4、Igk-C、Igl-C、またはその一部(ミニドメインを含む)からなる群より選択される定常ドメインからなる免疫グロブリンであって、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする免疫グロブリン。
【請求項28】
少なくとも一つのループ領域を持つ、重鎖もしくは軽鎖の可変ドメインまたはその一部(ミニドメインを含む)からなる免疫グロブリンであって、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする免疫グロブリン。
【請求項29】
抗原がタンパク質性分子、核酸、および糖質からなる群より選択されることを特徴とする、請求項27または28に記載の免疫グロブリン。
【請求項30】
抗原が、腫瘍関連抗原、特にEpCAM、腫瘍関連糖タンパク質72(TAG-72)、腫瘍関連抗原CA125、前立腺特異膜抗原(PSMA)、高分子量メラノーマ関連抗原(HMW-MAA)、ルイスY関連糖質を発現させる腫瘍関連抗原、癌胎児性抗原(CEA)、CEACAM5、HMFG PEM、ムチンMUC1、MUC18およびサイトケラチン腫瘍関連抗原、細菌抗原、ウイルス抗原、アレルゲン、フルオレセイン、リゾチーム、toll様受容体9、エリスロポエチン、CD2、CD3、CD3E、CD4、CD11、CD11a、CD14、CD18、CD19、CD20、CD22、CD23、CD25、CD28、CD29、CD30、CD33(p67タンパク質)、CD38、CD40、CD40L、CD52、CD54、CD56、CD80、CD147、GD3、IL-1、IL-1R、IL-2、IL-2R、IL-4、IL-5、IL-6、IL-6R、IL-8、IL-12、IL-15、IL-18、IL-23、インターフェロンα、インターフェロンβ、インターフェロンγ;TNF-α、TNFβ2、TNFα、TNFαβ、TNF-R1、TNF-RII、FasL、CD27L、CD30L、4-1BBL、TRAIL、RANKL、TWEAK、APRIL、BAFF、LIGHT、VEG1、OX40L、TRAIL受容体-1、A1アデノシン受容体、リンフォトキシンβ受容体、TACI、BAFF-R、EPO;LFA-3、ICAM-1、ICAM-3、インテグリンβ1、インテグリンβ2、インテグリンα4/β7、インテグリンα2、インテグリンα3、インテグリンα4、インテグリンα5、インテグリンα6、インテグリンαV、αVβ3インテグリン、FGFR-3、ケラチノサイト増殖因子、VLA-1、VLA-4、L-セレクチン、抗Id、E-セレクチン、HLA、HLA-DR、CTLA-4、T細胞受容体、B7-1、B7-2、VNRインテグリン、TGFβ1、TGFβ2、エオタキシン1、BLyS(Bリンパ球刺激因子)、補体C5、IgE、第VII因子、CD64、CBL、NCA90、EGFR(ErbB-1)、Her2/neu(ErbB-2)、Her3(ErbB-3)、Her4(ErbB4)、組織因子、VEGF、VEGFR、エンドセリン受容体、VLA-4、糖質、例えば血液型抗原および関連糖質、ガリリグリコシレーション(Galili Glycosylation)、ガストリン、ガストリン受容体、腫瘍関連糖質、ハプテンNP-capまたはNIP-cap、T細胞受容体α/β、E-セレクチン、ジゴキシン、胎盤アルカリホスファターゼ(PLAP)および精巣PLAP様アルカリホスファターゼ、トランスフェリン受容体、へパラナーゼI、ヒト心ミオシン、糖タンパク質IIb/IIIa(GPIIb/IIIa)、ヒトサイトメガロウイルス(HCMV)gHエンベロープ糖タンパク質、HIV gp120、HCMV、呼吸器合胞体ウイルスRSV F、RSVF Fgp、VNRインテグリン、Hep B gp120、CMV、gpIIbIIIa、HIV IIIB gp120 V3ループ、呼吸器合胞体ウイルス(RSV)Fgp、単純ヘルペスウイルス(HSV)gD糖タンパク質、HSV gB糖タンパク質、HCMV gBエンベロープ糖タンパク質、クロストリジウムパーフリンジェンス毒素およびその断片からなる群より選択されることを特徴とする、請求項27〜29のいずれか一項に記載の免疫グロブリン。
【請求項31】
CH3の位置15〜17、29〜34、85.4〜85.3、92〜94、97〜98および/または108〜110のアミノ酸残基が修飾されることを特徴とする、請求項27〜30のいずれか一項に記載の免疫グロブリン。
【請求項32】
修飾が欠失、置換、挿入またはそれらの組み合わせであることを特徴とする、請求項27〜31のいずれか一項に記載の免疫グロブリン。
【請求項33】
CH3領域が、EpCamが前記免疫グロブリンに結合する場合は配列番号16または配列番号18を、フルオレセインが前記免疫グロブリンに結合する場合は配列番号20を、リゾチームが前記免疫グロブリンに結合する場合は配列番号22、24、26、28、30または32を、TLR9が前記免疫グロブリンに結合する場合は配列番号34、36、38または40を、そしてリゾチームおよび/またはエリスロポエチンが前記免疫グロブリンに結合する場合は配列番号42を含むことを特徴とする、請求項27〜32のいずれか一項に記載の免疫グロブリン。
【請求項34】
リゾチームおよびgp41が前記免疫グロブリンに結合する場合に配列番号44または配列番号46を含むことを特徴とする、請求項27〜33のいずれか一項に記載の免疫グロブリン。
【請求項35】
修飾免疫グロブリンが、有機分子、酵素ラベル、放射性ラベル、着色ラベル、蛍光ラベル、発色ラベル、発光ラベル、ハプテン、ジゴキシゲニン、ビオチン、金属錯体、金属、コロイド金およびそれらの混合物からなる群より選択されるラベルまたはレポーター分子にコンジュゲートされることを特徴とする、請求項27〜34のいずれか一項に記載の免疫グロブリン。
【請求項36】
能動免疫用ワクチンの製造を目的とする、請求項27〜35のいずれか一項に記載の免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる免疫グロブリンの使用。
【請求項37】
免疫グロブリンのタンパク質ライブラリーの作製を目的とする、請求項27〜35のいずれか一項に記載の免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる免疫グロブリンの使用。
【請求項38】
請求項27〜35のいずれか一項に記載の方法によって得ることができる免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる免疫グロブリンを含むタンパク質ライブラリー。
【請求項39】
ある分子を特異的に結合および/または検出するための方法であって、
(a)請求項27〜35のいずれか一項に記載の修飾免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる修飾免疫グロブリンを、前記分子を含有すると疑われる試験試料と接触させるステップ、および
(b)特異的免疫グロブリン/分子複合体の潜在的形成を検出するステップ
を含む方法。
【請求項40】
ある分子を特異的に単離する方法であって、
(a)請求項27〜35のいずれか一項に記載の修飾免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる修飾免疫グロブリンを、前記分子を含有する試料と接触させるステップ、
(b)形成された特異的免疫グロブリン/分子複合体を分離するステップ、および
(c)要すれば、前記複合体から分子を単離するステップ
を含む方法。
【請求項41】
ある化合物を標的にターゲティングするための方法であって、
(a)前記化合物に特異的に結合する、請求項27〜35のいずれか一項に記載の修飾免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる修飾免疫グロブリンを、接触させるステップ、
(b)特異的免疫グロブリン/化合物複合体を標的に送達するステップ、
を含む方法。
【請求項42】
1以上の構造ループ中に組み込まれた、無修飾免疫グロブリンにとって外来である抗原結合部位を持つ、修飾免疫グロブリン。
【請求項43】
前記抗原が、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、ウイルス抗原および原虫抗原からなる群より選択される、請求項42に記載の免疫グロブリン。
【請求項44】
少なくとも二つの抗原結合部位を含み、第1部位は第1エピトープに結合し、第2部位は第2エピトープに結合する、請求項42または43に記載の免疫グロブリン。
【請求項45】
少なくとも二つのループ領域を含み、第1ループ領域は第1エピトープに結合し、第2ループ領域は第2エピトープに結合する、請求項42〜44のいずれか一項に記載の免疫グロブリン。
【請求項46】
前記ループ領域が構造ループを含有する、請求項45に記載の免疫グロブリン。
【請求項47】
少なくとも二つの免疫グロブリン、またはその一部(ミニドメインを含む)から構成され、各ドメインが少なくとも一つの抗原結合部位を含有する、請求項42〜46のいずれか一項に記載の免疫グロブリン。
【請求項48】
免疫グロブリンの定常領域の少なくとも一つのドメインおよび/もしくは可変領域の少なくとも一つのドメイン、またはその一部(ミニドメインを含む)を含む、請求項42〜47のいずれか一項に記載の免疫グロブリン。
【請求項49】
無修飾ドメインと少なくとも50%の相同性を持つドメインを含む、請求項42〜48のいずれか一項に記載の免疫グロブリン。
【請求項50】
二重特異性抗体または二重特異性単鎖抗体である、請求項42〜49のいずれか一項に記載の免疫グロブリン。
【請求項51】
前記免疫グロブリンが二重特異性ドメインまたはその一部(ミニドメインを含む)を含む、請求項42〜50のいずれか一項に記載の免疫グロブリン。
【請求項52】
治療および予防用である、請求項42〜51のいずれか一項に記載の免疫グロブリン。
【請求項53】
製造および分析用である、請求項42〜51のいずれか一項に記載の免疫グロブリン。
【請求項54】
診断用である、請求項42〜51のいずれか一項に記載の免疫グロブリン。
【請求項55】
構造ループ中に異なる修飾を持つ、少なくとも100個の、請求項27〜35および42〜54のいずれか一項に記載の免疫グロブリンからなるライブラリー。
【請求項56】
前記免疫グロブリンが、免疫グロブリンのドメイン、ミニドメインまたはその誘導体からなる群より選択される免疫グロブリンである、請求項55に記載のライブラリー。
【請求項57】
(a)1以上の構造ループ中に組み込まれた、当該免疫グロブリンにとって外来の抗原結合部位を持つ、修飾免疫グロブリン、および
(b)前記抗原のエピトープを含有する結合分子
を含有する、結合パートナーのキット。
【請求項58】
前記修飾免疫グロブリンの結合特異性を同定するための、請求項57に記載のキットの結合分子の使用。
【請求項59】
前記修飾免疫グロブリンの効力を決定するための、請求項57に記載のキットの結合分子の使用。
【請求項60】
構造ループ中に異なる修飾を持つ少なくとも100個の免疫グロブリンからなるライブラリーから前記修飾免疫グロブリンを選択するための、請求項57に記載のキットの結合分子の使用。
【請求項1】
免疫グロブリンの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンを設計し、ある抗原のエピトープに対する前記免疫グロブリンの結合を決定するための方法(ただし、無修飾免疫グロブリンは前記エピトープには有意に結合しない)であって、
・少なくとも一つの構造ループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記構造ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、および
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ
を含む方法。
【請求項2】
前記免疫グロブリンが少なくとも二つの異なるエピトープに特異的に結合する、請求項1に記載の方法。
【請求項3】
前記免疫グロブリンの少なくとも一つの構造ループ領域に少なくとも一つの修飾を含み、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、ウイルス抗原および原虫抗原からなる群より選択される少なくとも一つの分子に対する前記少なくとも一つのループ領域の特異的結合を決定する(ただし、無修飾構造ループ領域を含有する免疫グロブリンは、前記少なくとも一つの分子に特異的には結合しない)、請求項1または請求項2のいずれか一項に記載の方法。
【請求項4】
免疫グロブリンの構造ループ領域に少なくとも一つの修飾を含む免疫グロブリンまたはその医薬調製物を製造し、ある抗原のエピトープに対する前記免疫グロブリンの結合を決定するための方法(ただし、無修飾免疫グロブリンは前記エピトープには有意に結合しない)であって、
・少なくとも一つのループ領域を含む免疫グロブリンをコードする核酸を用意するステップ、
・前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンをエピトープと接触させるステップ、
・前記修飾免疫グロブリンが前記エピトープに結合するかどうかを決定するステップ、および
・前記エピトープに結合する修飾免疫グロブリンを用意し、要すればそれを医薬調製物に仕上げるステップ、
を含む方法。
【請求項5】
免疫グロブリンの少なくとも一つの構造ループ領域に少なくとも一つの修飾を含む、少なくとも一つの第1分子に特異的に結合する多重特異性免疫グロブリン、またはその医薬調製物を製造し、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、ウイルス抗原および原虫抗原からなる群より選択される少なくとも一つの第2分子に対する、前記少なくとも一つのループ領域の特異的結合を決定するための方法(ただし、無修飾構造ループ領域を含有する免疫グロブリンは、前記少なくとも一つの第2分子には特異的に結合しない)であって、
・少なくとも一つの構造ループ領域を含む、少なくとも一つの第1分子に特異的に結合する免疫グロブリンをコードする核酸を用意するステップ、
・前記核酸によってコードされる前記ループ領域の少なくとも一つの、少なくとも一つのヌクレオチド残基を修飾するステップ、
・前記修飾核酸を発現系に移すステップ、
・前記修飾免疫グロブリンを発現させるステップ、
・発現させた修飾免疫グロブリンを前記少なくとも一つの第2分子と接触させるステップ、
・前記修飾免疫グロブリンが第2分子に特異的に結合するかどうかを決定するステップ、および
・前記少なくとも一つの第2分子に特異的に結合する修飾免疫グロブリンを用意し、要すればそれを医薬調製物に仕上げるステップ、
を含む方法。
【請求項6】
免疫グロブリンがヒトまたはネズミ由来であることを特徴とする、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
ヒト免疫グロブリンがIgA1、IgA2、IgD、IgE、IgG1、IgG2、IgG3、IgG4およびIgMからなる群より選択されることを特徴とする、請求項6に記載の方法。
【請求項8】
ネズミ免疫グロブリンがIgA、IgD、IgE、IgG1、IgG2A、IgG2B、IgG2C、IgG3およびIgMからなる群より選択されることを特徴とする、請求項6に記載の方法。
【請求項9】
免疫グロブリンが免疫グロブリンの重鎖および/もしくは軽鎖またはその一部を含むことを特徴とする、請求項7または8に記載の方法。
【請求項10】
免疫グロブリンが、免疫グロブリンの少なくとも一つの定常ドメインおよび/もしくは少なくとも一つの可変ドメイン、またはその一部(ミニドメインを含む)を含むことを特徴とする、請求項7〜9のいずれか一項に記載の方法。
【請求項11】
定常ドメインがCH1、CH2、CH3、CH4、Igk-C、Igl-Cおよびそれらの組み合わせの群より選択されることを特徴とする、請求項10に記載の方法。
【請求項12】
CH1、CH2、CH3およびCH4の修飾ループ領域が、アミノ酸7〜21、アミノ酸25〜39、アミノ酸41〜81、アミノ酸83〜85、アミノ酸89〜103およびアミノ酸106〜117を含むことを特徴とする、請求項11に記載の方法。.
【請求項13】
ヒト由来のIgk-CおよびIgl-Cのループ領域が、アミノ酸8〜18、アミノ酸27〜35、アミノ酸42〜78、アミノ酸83〜85、アミノ酸92〜100、アミノ酸108〜117およびアミノ酸123〜126を含むことを特徴とする、請求項11に記載の方法。
【請求項14】
ネズミ由来のIgk-CおよびIgl-Cのループ領域が、アミノ酸8〜20、アミノ酸26〜36、アミノ酸43〜79、アミノ酸83〜85、アミノ酸90〜101、アミノ酸108〜116およびアミノ酸122〜125を含むことを特徴とする、請求項11に記載の方法。
【請求項15】
ヒト由来の免疫グロブリンの可変ドメインの構造ループ領域が、アミノ酸8〜20、アミノ酸44〜50、アミノ酸67〜76およびアミノ酸89〜101を含むことを特徴とする、請求項10に記載の方法。
【請求項16】
ネズミ由来の免疫グロブリンの可変ドメインの構造ループ領域が、アミノ酸6〜20、アミノ酸44〜52、アミノ酸67〜76およびアミノ酸92〜101を含むことを特徴とする、請求項10に記載の方法。
【請求項17】
免疫グロブリンがラクダ由来であることを特徴とする、請求項1〜5のいずれか一項に記載の方法。
【請求項18】
免疫グロブリンがCH1、CH2およびCH3からなる群より選択される少なくとも一つの定常ドメインを含むことを特徴とする、請求項17に記載の方法。
【請求項19】
CH1、CH2およびCH3のループ領域が、アミノ酸8〜20、アミノ酸24〜39、アミノ酸42〜78、アミノ酸82〜85、アミノ酸91〜103およびアミノ酸108〜117を含むことを特徴とする、請求項18に記載の方法。.
【請求項20】
少なくとも一つのヌクレオチドの修飾が、前記核酸によってコードされる免疫グロブリンの置換、欠失および/または挿入をもたらす、請求項1〜19のいずれか一項に記載の方法。
【請求項21】
少なくとも一つのループ領域のアミノ酸が部位指定ランダム突然変異によって修飾されることを特徴とする、請求項1〜20のいずれか一項に記載の方法。
【請求項22】
ランダムに修飾された核酸分子が、5'-NNS-3'、5'-NNN-3'または5'-NNK-3'という配列を持つ少なくとも一つのヌクレオチド繰返し単位を含むことを特徴とする、請求項21に記載の方法。
【請求項23】
発現系がベクターを含むことを特徴とする、請求項1〜22のいずれか一項に記載の方法。
【請求項24】
修飾免疫グロブリンが、宿主中で、好ましくは細菌、酵母、植物細胞中で、動物細胞中で、または植物もしくは動物中で発現されることを特徴とする、請求項1〜23のいずれか一項に記載の方法。
【請求項25】
分子に対する修飾免疫グロブリンの特異的結合が、免疫学的アッセイ、好ましくは酵素結合免疫吸着アッセイ(ELISA)、表面プラズモン共鳴アッセイ、飽和移動差核磁気共鳴分光法、転移NOE(trNOE)核磁気共鳴分光法、競合アッセイ、組織結合アッセイ、生細胞結合アッセイおよび細胞抽出物アッセイからなる群より選択される結合アッセイによって決定されることを特徴とする、請求項1〜24のいずれか一項に記載の方法。
【請求項26】
修飾免疫グロブリンが、有機分子、酵素ラベル、放射性ラベル、着色ラベル、蛍光ラベル、発色ラベル、発光ラベル、ハプテン、ジゴキシゲニン、ビオチン、金属錯体、金属、コロイド金およびそれらの混合物からなる群より選択されるラベルにコンジュゲートされることを特徴とする、請求項1〜25のいずれか一項に記載の方法。
【請求項27】
少なくとも一つのループ領域を持つ、CH1、CH2、CH3、CH4、Igk-C、Igl-C、またはその一部(ミニドメインを含む)からなる群より選択される定常ドメインからなる免疫グロブリンであって、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする免疫グロブリン。
【請求項28】
少なくとも一つのループ領域を持つ、重鎖もしくは軽鎖の可変ドメインまたはその一部(ミニドメインを含む)からなる免疫グロブリンであって、前記少なくとも一つのループ領域が、少なくとも一つの修飾ループ領域を形成する少なくとも一つのアミノ酸修飾を含み、前記少なくとも一つの修飾ループ領域が、ある抗原の少なくとも一つのエピトープに特異的に結合することを特徴とする免疫グロブリン。
【請求項29】
抗原がタンパク質性分子、核酸、および糖質からなる群より選択されることを特徴とする、請求項27または28に記載の免疫グロブリン。
【請求項30】
抗原が、腫瘍関連抗原、特にEpCAM、腫瘍関連糖タンパク質72(TAG-72)、腫瘍関連抗原CA125、前立腺特異膜抗原(PSMA)、高分子量メラノーマ関連抗原(HMW-MAA)、ルイスY関連糖質を発現させる腫瘍関連抗原、癌胎児性抗原(CEA)、CEACAM5、HMFG PEM、ムチンMUC1、MUC18およびサイトケラチン腫瘍関連抗原、細菌抗原、ウイルス抗原、アレルゲン、フルオレセイン、リゾチーム、toll様受容体9、エリスロポエチン、CD2、CD3、CD3E、CD4、CD11、CD11a、CD14、CD18、CD19、CD20、CD22、CD23、CD25、CD28、CD29、CD30、CD33(p67タンパク質)、CD38、CD40、CD40L、CD52、CD54、CD56、CD80、CD147、GD3、IL-1、IL-1R、IL-2、IL-2R、IL-4、IL-5、IL-6、IL-6R、IL-8、IL-12、IL-15、IL-18、IL-23、インターフェロンα、インターフェロンβ、インターフェロンγ;TNF-α、TNFβ2、TNFα、TNFαβ、TNF-R1、TNF-RII、FasL、CD27L、CD30L、4-1BBL、TRAIL、RANKL、TWEAK、APRIL、BAFF、LIGHT、VEG1、OX40L、TRAIL受容体-1、A1アデノシン受容体、リンフォトキシンβ受容体、TACI、BAFF-R、EPO;LFA-3、ICAM-1、ICAM-3、インテグリンβ1、インテグリンβ2、インテグリンα4/β7、インテグリンα2、インテグリンα3、インテグリンα4、インテグリンα5、インテグリンα6、インテグリンαV、αVβ3インテグリン、FGFR-3、ケラチノサイト増殖因子、VLA-1、VLA-4、L-セレクチン、抗Id、E-セレクチン、HLA、HLA-DR、CTLA-4、T細胞受容体、B7-1、B7-2、VNRインテグリン、TGFβ1、TGFβ2、エオタキシン1、BLyS(Bリンパ球刺激因子)、補体C5、IgE、第VII因子、CD64、CBL、NCA90、EGFR(ErbB-1)、Her2/neu(ErbB-2)、Her3(ErbB-3)、Her4(ErbB4)、組織因子、VEGF、VEGFR、エンドセリン受容体、VLA-4、糖質、例えば血液型抗原および関連糖質、ガリリグリコシレーション(Galili Glycosylation)、ガストリン、ガストリン受容体、腫瘍関連糖質、ハプテンNP-capまたはNIP-cap、T細胞受容体α/β、E-セレクチン、ジゴキシン、胎盤アルカリホスファターゼ(PLAP)および精巣PLAP様アルカリホスファターゼ、トランスフェリン受容体、へパラナーゼI、ヒト心ミオシン、糖タンパク質IIb/IIIa(GPIIb/IIIa)、ヒトサイトメガロウイルス(HCMV)gHエンベロープ糖タンパク質、HIV gp120、HCMV、呼吸器合胞体ウイルスRSV F、RSVF Fgp、VNRインテグリン、Hep B gp120、CMV、gpIIbIIIa、HIV IIIB gp120 V3ループ、呼吸器合胞体ウイルス(RSV)Fgp、単純ヘルペスウイルス(HSV)gD糖タンパク質、HSV gB糖タンパク質、HCMV gBエンベロープ糖タンパク質、クロストリジウムパーフリンジェンス毒素およびその断片からなる群より選択されることを特徴とする、請求項27〜29のいずれか一項に記載の免疫グロブリン。
【請求項31】
CH3の位置15〜17、29〜34、85.4〜85.3、92〜94、97〜98および/または108〜110のアミノ酸残基が修飾されることを特徴とする、請求項27〜30のいずれか一項に記載の免疫グロブリン。
【請求項32】
修飾が欠失、置換、挿入またはそれらの組み合わせであることを特徴とする、請求項27〜31のいずれか一項に記載の免疫グロブリン。
【請求項33】
CH3領域が、EpCamが前記免疫グロブリンに結合する場合は配列番号16または配列番号18を、フルオレセインが前記免疫グロブリンに結合する場合は配列番号20を、リゾチームが前記免疫グロブリンに結合する場合は配列番号22、24、26、28、30または32を、TLR9が前記免疫グロブリンに結合する場合は配列番号34、36、38または40を、そしてリゾチームおよび/またはエリスロポエチンが前記免疫グロブリンに結合する場合は配列番号42を含むことを特徴とする、請求項27〜32のいずれか一項に記載の免疫グロブリン。
【請求項34】
リゾチームおよびgp41が前記免疫グロブリンに結合する場合に配列番号44または配列番号46を含むことを特徴とする、請求項27〜33のいずれか一項に記載の免疫グロブリン。
【請求項35】
修飾免疫グロブリンが、有機分子、酵素ラベル、放射性ラベル、着色ラベル、蛍光ラベル、発色ラベル、発光ラベル、ハプテン、ジゴキシゲニン、ビオチン、金属錯体、金属、コロイド金およびそれらの混合物からなる群より選択されるラベルまたはレポーター分子にコンジュゲートされることを特徴とする、請求項27〜34のいずれか一項に記載の免疫グロブリン。
【請求項36】
能動免疫用ワクチンの製造を目的とする、請求項27〜35のいずれか一項に記載の免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる免疫グロブリンの使用。
【請求項37】
免疫グロブリンのタンパク質ライブラリーの作製を目的とする、請求項27〜35のいずれか一項に記載の免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる免疫グロブリンの使用。
【請求項38】
請求項27〜35のいずれか一項に記載の方法によって得ることができる免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる免疫グロブリンを含むタンパク質ライブラリー。
【請求項39】
ある分子を特異的に結合および/または検出するための方法であって、
(a)請求項27〜35のいずれか一項に記載の修飾免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる修飾免疫グロブリンを、前記分子を含有すると疑われる試験試料と接触させるステップ、および
(b)特異的免疫グロブリン/分子複合体の潜在的形成を検出するステップ
を含む方法。
【請求項40】
ある分子を特異的に単離する方法であって、
(a)請求項27〜35のいずれか一項に記載の修飾免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる修飾免疫グロブリンを、前記分子を含有する試料と接触させるステップ、
(b)形成された特異的免疫グロブリン/分子複合体を分離するステップ、および
(c)要すれば、前記複合体から分子を単離するステップ
を含む方法。
【請求項41】
ある化合物を標的にターゲティングするための方法であって、
(a)前記化合物に特異的に結合する、請求項27〜35のいずれか一項に記載の修飾免疫グロブリンまたは請求項1〜26のいずれか一項に記載の方法によって得ることができる修飾免疫グロブリンを、接触させるステップ、
(b)特異的免疫グロブリン/化合物複合体を標的に送達するステップ、
を含む方法。
【請求項42】
1以上の構造ループ中に組み込まれた、無修飾免疫グロブリンにとって外来である抗原結合部位を持つ、修飾免疫グロブリン。
【請求項43】
前記抗原が、アレルゲン、腫瘍関連抗原、自己抗原、酵素、細菌抗原、真菌抗原、ウイルス抗原および原虫抗原からなる群より選択される、請求項42に記載の免疫グロブリン。
【請求項44】
少なくとも二つの抗原結合部位を含み、第1部位は第1エピトープに結合し、第2部位は第2エピトープに結合する、請求項42または43に記載の免疫グロブリン。
【請求項45】
少なくとも二つのループ領域を含み、第1ループ領域は第1エピトープに結合し、第2ループ領域は第2エピトープに結合する、請求項42〜44のいずれか一項に記載の免疫グロブリン。
【請求項46】
前記ループ領域が構造ループを含有する、請求項45に記載の免疫グロブリン。
【請求項47】
少なくとも二つの免疫グロブリン、またはその一部(ミニドメインを含む)から構成され、各ドメインが少なくとも一つの抗原結合部位を含有する、請求項42〜46のいずれか一項に記載の免疫グロブリン。
【請求項48】
免疫グロブリンの定常領域の少なくとも一つのドメインおよび/もしくは可変領域の少なくとも一つのドメイン、またはその一部(ミニドメインを含む)を含む、請求項42〜47のいずれか一項に記載の免疫グロブリン。
【請求項49】
無修飾ドメインと少なくとも50%の相同性を持つドメインを含む、請求項42〜48のいずれか一項に記載の免疫グロブリン。
【請求項50】
二重特異性抗体または二重特異性単鎖抗体である、請求項42〜49のいずれか一項に記載の免疫グロブリン。
【請求項51】
前記免疫グロブリンが二重特異性ドメインまたはその一部(ミニドメインを含む)を含む、請求項42〜50のいずれか一項に記載の免疫グロブリン。
【請求項52】
治療および予防用である、請求項42〜51のいずれか一項に記載の免疫グロブリン。
【請求項53】
製造および分析用である、請求項42〜51のいずれか一項に記載の免疫グロブリン。
【請求項54】
診断用である、請求項42〜51のいずれか一項に記載の免疫グロブリン。
【請求項55】
構造ループ中に異なる修飾を持つ、少なくとも100個の、請求項27〜35および42〜54のいずれか一項に記載の免疫グロブリンからなるライブラリー。
【請求項56】
前記免疫グロブリンが、免疫グロブリンのドメイン、ミニドメインまたはその誘導体からなる群より選択される免疫グロブリンである、請求項55に記載のライブラリー。
【請求項57】
(a)1以上の構造ループ中に組み込まれた、当該免疫グロブリンにとって外来の抗原結合部位を持つ、修飾免疫グロブリン、および
(b)前記抗原のエピトープを含有する結合分子
を含有する、結合パートナーのキット。
【請求項58】
前記修飾免疫グロブリンの結合特異性を同定するための、請求項57に記載のキットの結合分子の使用。
【請求項59】
前記修飾免疫グロブリンの効力を決定するための、請求項57に記載のキットの結合分子の使用。
【請求項60】
構造ループ中に異なる修飾を持つ少なくとも100個の免疫グロブリンからなるライブラリーから前記修飾免疫グロブリンを選択するための、請求項57に記載のキットの結合分子の使用。
【図2】

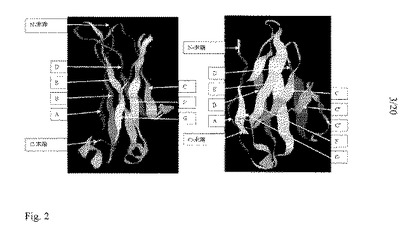
【図3】
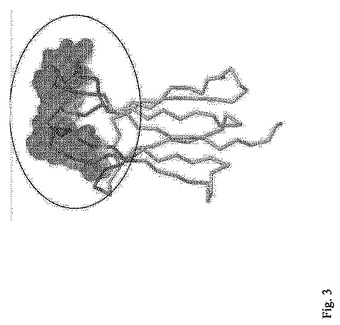
【図4】
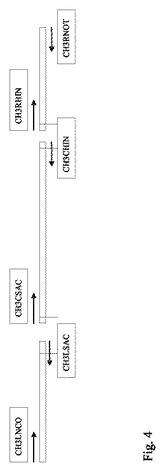
【図5】
【図6】
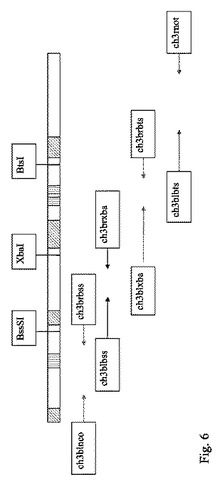
【図7】

【図8−1】

【図8−2】

【図8−3】

【図8−4】

【図8−5】

【図8−6】

【図8−7】

【図8−8】

【図8−9】

【図8−10】

【図8−11】

【図8−12】

【図3】
【図4】
【図5】
【図6】
【図7】
【図8−1】
【図8−2】
【図8−3】
【図8−4】
【図8−5】
【図8−6】
【図8−7】
【図8−8】
【図8−9】
【図8−10】
【図8−11】
【図8−12】
【公表番号】特表2008−526809(P2008−526809A)
【公表日】平成20年7月24日(2008.7.24)
【国際特許分類】
【出願番号】特願2007−549891(P2007−549891)
【出願日】平成18年1月5日(2006.1.5)
【国際出願番号】PCT/EP2006/050059
【国際公開番号】WO2006/072620
【国際公開日】平成18年7月13日(2006.7.13)
【出願人】(507227832)エフ−シュタール・ビオテヒノロギシェ・フォルシュングス−ウント・エントヴィックルングスゲゼルシャフト・ミット・ベシュレンクテル・ハフツング (4)
【氏名又は名称原語表記】F−STAR BIOTECHNOLOGISCHE FORSCHUNGS− UND ENTWICKLUNGSGES.M.B.H.
【Fターム(参考)】
【公表日】平成20年7月24日(2008.7.24)
【国際特許分類】
【出願日】平成18年1月5日(2006.1.5)
【国際出願番号】PCT/EP2006/050059
【国際公開番号】WO2006/072620
【国際公開日】平成18年7月13日(2006.7.13)
【出願人】(507227832)エフ−シュタール・ビオテヒノロギシェ・フォルシュングス−ウント・エントヴィックルングスゲゼルシャフト・ミット・ベシュレンクテル・ハフツング (4)
【氏名又は名称原語表記】F−STAR BIOTECHNOLOGISCHE FORSCHUNGS− UND ENTWICKLUNGSGES.M.B.H.
【Fターム(参考)】
[ Back to top ]