
真核生物細胞を改変する方法
【課題】真核生物細胞における内因性遺伝子および染色体座を改変するために、大きな相同性領域を含む標的化ベクターの使用を可能にする迅速かつ便利な方法を提供すること。
【解決手段】真核生物細胞において内因性遺伝子または染色体座を遺伝子改変する方法。この方法は、a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、真核生物において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを真核生物細胞に導入して、この細胞中の内因性遺伝子または染色体座を改変する工程;ならびにd)(c)の真核生物細胞において対立遺伝子の改変を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された真核生物細胞を同定する工程を包含する。
【解決手段】真核生物細胞において内因性遺伝子または染色体座を遺伝子改変する方法。この方法は、a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、真核生物において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを真核生物細胞に導入して、この細胞中の内因性遺伝子または染色体座を改変する工程;ならびにd)(c)の真核生物細胞において対立遺伝子の改変を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された真核生物細胞を同定する工程を包含する。
【発明の詳細な説明】
【技術分野】
【0001】
本願は、2001年2月16日に出願された米国特許実用出願番号09/784,859に対する優先権を主張する。この出願は、本明細書中に参考として援用される。本願を通じて、種々の刊行物が参照される。これらの刊行物のそれらの全体における開示は、本明細書によって本願に参考として援用される。
【0002】
(発明の分野)
本発明の分野は、真核生物細胞における内因性遺伝子および染色体座を、相同組み換えを通じて標的化し、任意の望ましい様式で改変するための大きなDNAベクターを操作および利用するための方法である。真核生物細胞に対するこれらの大きなDNA標的化ベクター(LTVECと呼ばれる)は、真核生物細胞において相同性標的化を実施することを意図する他のアプローチにより代表的に使用されるフラグメントよりも大きなクローン化されたゲノムDNAのフラグメントから獲得される。本発明の分野はさらに、LTVECが所望の内因性遺伝子または染色体座を正確に標的化および改変した真核生物細胞を検出する迅速かつ便利な方法を提供する。この分野はまた、遺伝子改変を有する生物を作製するためのこれらの細胞の使用、その生物自体、およびそれらの使用方法を包含する。
【0003】
(導入)
LTVECの使用は、現在の方法を上回る実質的な利点を提供する。例えば、これらは標的化ベクターを作製するために現在使用されているDNAフラグメントよりも大きなDNAフラグメントから獲得されるので、LTVECは、現在の技術を用いて作製される標的化ベクターよりも大きなゲノムDNAフラグメントの利用可能なライブラリー(例えば、BACおよびPACライブラリー)からより迅速かつ便利に作製され得る。さらに、より大きな改変およびより大きなゲノム領域にまたがる改変は、現在の技術を使用するよりも便利に作製され得る。さらに、本発明は、「標的化の困難な」座の標的化頻度を増大するための長い相同領域を利用し、また、これらの標的化ベクターにおいて同系DNA(存在する場合)を使用する利益を減らす。
【0004】
従って、本発明は、所定の生物の実質的に全ての内因性遺伝子および染色体座を体系的に改変するための迅速、便利、かつ効率的な方法を提供する。
【背景技術】
【0005】
(発明の背景)
相同な外因性DNAと内因性染色体配列との間の相同組み換えによる遺伝子標的化は、マウスにおいて、欠失、挿入を作製するため、変異を設計するため、遺伝子変異を修正するため、導入遺伝子を導入するため、または他の遺伝子改変を作製するために極めて価値のある方法であることが証明されている。現在の方法は、代表的に、合計10〜20kb未満の内因性DNAに対する相同領域を有する標準的な標的化ベクターを使用してマウス胚性幹(ES)細胞に所望の遺伝子改変を導入し、その後変更されたES細胞をマウス胚に注入してこれらの操作された遺伝子改変をマウス生殖系統に移す工程を包含する(非特許文献1;非特許文献2;非特許文献3;非特許文献4;非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;特許文献1(GenPharm Internationalの名の下で1998年8月4日発行))。これらの現在の方法において、標準的な標的化ベクターが所望の内因性遺伝子または染色体座を正確に標的化および改変している希なES細胞を検出することは、標的化ベクターに含まれる相同標的化配列の外側の配列情報を必要とする。首尾良い標的化のためのアッセイは、標的化ベクターの外側であり相同性アーム(定義を参照のこと)全体にまたがる配列からの標準的なサザンブロッティングまたは長いPCRを伴う(非特許文献10;非特許文献11;非特許文献12;Takara Shuzo Co.,Ltd.に対して発行された特許文献2)。従って、これらの方法を制限する大きさの考慮から、相同性アームの大きさは、合計10〜20kb未満に制限される(非特許文献13)。
【0006】
現在の方法において使用されている相同性アームよりも長い相同性アームを有する標的化ベクターを使用する能力は、極めて価値が高い。例えば、このような標的化ベクターは、大きなゲノムインサートが広範に特徴付けられ、使用前にトリミングされる必要のある現在の技術を用いて作製される標的化ベクターよりも、このような大きなゲノムインサートを含む利用可能なライブラリー(例えば、BACまたはPACライブラリー)からより迅速かつ便利に作製され得る。さらに、より大きな改変およびより大きなゲノム領域にまたがる改変は、現在の技術を使用するよりもより便利にかつより少ない工程で作製され得る。さらに、長い相同性領域の使用は、真核生物細胞における「標的化の困難な」座の標的化頻度を増大し得る。なぜならば、真核生物細胞における相同組み換えの標的化は、その標的化ベクターに含まれる総相同性に関連するようであるからである(非特許文献14)。さらに、長い相同性アームを用いて獲得された増加した標的化頻度は、これらの標的化ベクターにおいて同系DNAを使用することから得られ得る任意の潜在的な利益を減らし得る。
【0007】
非常に大きなゲノムフラグメント(例えば、BACライブラリーにおいてクローン化されたフラグメント)への正確な改変を操作する上での問題は、真核生物の内因性遺伝子または染色体座に対する大きな相同性領域を含むベクターの構築を可能にする、細菌における相同組み換えの使用を通して大部分が解決された(非特許文献15;非特許文献16;非特許文献17;非特許文献18;非特許文献19)。しかし、一旦作製されると、これらのベクターは、一般的に、相同組み換えを通じて内因性遺伝子または染色体座を改変するのに有用ではない。なぜならば相同性アームが10〜20kbよりも長い場合、希な正確な標的化事象を検出することが困難であるからである(非特許文献20)。結果として、BACゲノムフラグメントから細菌相同組み換えを使用して作製されたベクターは、標的化ベクターとして使用する前になおも広範にトリミングされなければならない(非特許文献21)。従って、真核生物細胞における内因性遺伝子および染色体座を改変するために、大きな相同性領域を含む標的化ベクターの使用を可能にする迅速かつ便利な方法論に対する必要性がなおも存在する。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第5,789,215号明細書
【特許文献2】米国特許第5,436,149号明細書
【非特許文献】
【0009】
【非特許文献1】Smithiesら、Nature、317:230−234、1985
【非特許文献2】Thomasら、Cell、51:503−512、1987
【非特許文献3】Kollerら、Proc Natl Acad Sci USA、86;8927−8931、1989
【非特許文献4】Kuhnら、Science、254:707−710、1991
【非特許文献5】Thomasら、Nature、346:847−850、1990
【非特許文献6】Schwarzbergら、Science、246:799−803、1989
【非特許文献7】Doetschmanら、Nature、330:576−578、1987
【非特許文献8】Thomsonら、Cell、5:313−321、1989
【非特許文献9】DeChiaraら、Nature、345:78−80、1990
【非特許文献10】Chengら、Nature、369:684−5、1994
【非特許文献11】FoordおよびRose、PCR Methods Appl、3:S149−61、1994
【非特許文献12】PonceおよびMicol、Nucleic Acids Res、20:623,1992
【非特許文献13】Joyner、The Practical Approach Series、293、1999
【非特許文献14】DengおよびCapecchi、Mol Cell Biol、12:3365−71、1992
【非特許文献15】Zhangら、Nat Genet、20:123−8、1998
【非特許文献16】Yangら、Nat Biotechnol、15:859−65、1997
【非特許文献17】Angrandら、Nucleic Acids Res、27:e16、1999
【非特許文献18】Muyrersら、Nucleic Acid Res、27:1555−7、1999
【非特許文献19】Narayananら、Gene Ther、6:442−7、1999
【非特許文献20】Joyner、The Practical Approach Series、293、1999
【非特許文献21】Hillら、Genomics、64:111−3、2000
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明に従って、本出願人らは、相同組み換えを通じて真核生物細胞における内因性遺伝子および染色体座を改変するために、大きな相同性領域を含む標的化ベクターの使用を可能にする新規の方法を提供する。このような方法は、上記の現在の技術の制限を克服する。さらに、当業者は、本発明の方法が、任意の真核生物(動物(例えば、マウス、ラット、他の齧歯類、またはヒト)および植物(例えば、ダイズ、トウモロコシ、およびコムギ)が挙げられるがこれらに限定されない)の任意のゲノムDNAを用いた使用に容易に適合されることを容易に認識する。
【課題を解決するための手段】
【0011】
(発明の要旨)
本発明に従って、本出願人らは、改変された内因性遺伝子または染色体座を含む真核生物細胞を作製およびスクリーニングするための新規、迅速、効率的、かつ効果的な方法を開発した。この新規の方法は、初めて、以下を組み合わせる:
1.大きなクローン化されたゲノムフラグメント中での所望の遺伝子改変を正確に操作し、それによって真核生物細胞において使用するための大きな標的化ベクター(LTVEC)を作製するための細菌相同組み換え;
2.これらの真核生物細胞における目的の内因性染色体座を改変するための、これらの細胞へのこれらのLTVECの直接的な導入;および
3.標的化配列の外側の配列情報を必要としない親対立遺伝子の対立遺伝子改変(MOA)のためのアッセイ(例えば、定量PCR)を含む、標的化された対立遺伝子が所望するように改変された希な真核生物細胞を決定するための分析。
【0012】
本発明の好ましい実施形態は、以下の工程を包含する、真核生物細胞において内因性遺伝子または染色体座を遺伝子改変する方法である:a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、真核生物において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを真核生物細胞に導入して、この細胞中の内因性遺伝子または染色体座を改変する工程;ならびにd)(c)の真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された真核生物細胞を同定する工程。
【0013】
本発明の別の実施形態は、内因性遺伝子または染色体座に対するこの遺伝子改変が、コード配列、遺伝子セグメント、または調節エレメントの欠失;コード配列、遺伝子セグメント、または調節エレメントの変更;新たなコード配列、遺伝子セグメント、または調節エレメントの挿入;条件付き対立遺伝子の作製;または1つの種由来のコード配列または遺伝子セグメントと、異なる種由来の相同またはオルソロガスな(orthologous)コード配列との置換を含む方法である。
【0014】
本発明の別の実施形態は、コード配列、遺伝子セグメント、または調節エレメントの変更が、置換、付加、または融合を含み、その融合体がエピトープタグまたは二官能性タンパク質を含む方法である。
【0015】
本発明のなお別の実施形態は、定量アッセイが、定量PCR、比較ゲノムハイブリダイゼーション、等温DNA増幅、または固定化されたプローブに対する定量ハイブリダイゼーションを含み、定量PCRが、TaqMan(登録商標)技術または分子ビーコンを用いる定量PCRを含む方法である。
【0016】
本発明の別の好ましい実施形態は、真核生物細胞が、哺乳動物胚性幹細胞であり、特に、この胚性幹細胞がマウス、ラット、または他の齧歯類の胚性幹細胞である方法である。
【0017】
本発明の別の好ましい実施形態は、内因性遺伝子または染色体座が、哺乳動物の遺伝子または染色体座、好ましくは、ヒトの遺伝子または染色体座、あるいはマウス、ラット、または他の齧歯類の遺伝子または染色体座である方法である。
【0018】
本発明のさらなる好ましい実施形態は、LTVECが20kbよりも大きなDNAフラグメント、特に100kbより大きなDNAフラグメントを収容し得る方法である。
【0019】
別の好ましい実施形態は、本発明の方法により産生される遺伝子改変された内因性遺伝子または染色体座である。
【0020】
なお別の好ましい実施形態は、本発明の方法により産生される遺伝子改変された真核生物細胞である。
【0021】
本発明の好ましい実施形態は、本発明の方法により産生される遺伝子改変された内因性遺伝子または染色体座を含む非ヒト生物である。
【0022】
本発明の方法により産生される遺伝子改変された真核生物細胞または胚性幹細胞から産生される非ヒト生物もまた好ましい。
【0023】
好ましい実施形態は、以下の工程を含む方法により産生される、遺伝子改変された内因性遺伝子または染色体座を含む、非ヒト生物である:a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、胚性幹細胞において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを胚性幹細胞に導入して、細胞中の内因性遺伝子または染色体座を改変する工程;d)(c)の胚性幹細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された胚性幹細胞を同定する工程;e)(d)の胚性幹細胞を胚盤胞に導入する工程;ならびにf)(e)の胚盤胞を妊娠のために代理母に導入する工程。
【0024】
本発明のさらなる好ましい実施形態は、以下の工程を含む方法により産生される、遺伝子改変された内因性遺伝子または染色体座を含む、非ヒト生物である:a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、真核生物において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを真核生物細胞に導入して、この細胞中の内因性遺伝子または染色体座を遺伝子改変する工程;d)(c)の真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された真核生物細胞を同定する工程;e)(d)の真核生物細胞から核を取り除く工程;f)(e)の核を卵母細胞に導入する工程;ならびにg)(f)の卵母細胞を妊娠のための代理母に導入する工程。
【0025】
なお別の好ましい実施形態は、以下の工程を含む方法により産生される、遺伝子改変された内因性遺伝子または染色体座を含む、非ヒト生物である:a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、真核生物において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを真核生物細胞に導入して、この細胞中で内因性遺伝子または染色体座を遺伝子改変する工程;d)(c)の真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された真核生物細胞を同定する工程;e)(d)の真核生物細胞と別の真核生物細胞とを融合させる工程;ならびにf)(e)の融合された真核生物細胞を妊娠のための代理母に導入する工程。
【0026】
別の好ましい実施形態において、非ヒト生物はマウス、ラット、または他の齧歯類であり;胚盤胞は、マウス、ラット、または他の齧歯類の胚盤胞であり;卵母細胞は、マウス、ラット、または他の齧歯類の卵母細胞であり;そして代理母は、マウス、ラット、または他の齧歯類である。
【0027】
別の好ましい実施形態は、胚性幹細胞が哺乳動物胚性幹細胞、好ましくは、マウス、ラット、または他の齧歯類の胚性幹細胞である方法である。
【0028】
さらなる好ましい実施形態は、非ヒト生物の産生のための、本発明の遺伝子改変された真核生物細胞の使用、特に、非ヒト生物の産生のための、本発明の遺伝子改変された胚性幹細胞の使用である。
【0029】
本発明の好ましい実施形態は、マウス胚性幹細胞において目的の内因性遺伝子または染色体座を遺伝子改変する方法である。この方法は、a)目的のDNA配列を含む20kbよりも大きなクローン化されたゲノムフラグメントを得る工程(この大きなクローン化されたDNAフラグメントは内因性遺伝子または染色体座に相同である);b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、マウス胚性幹細胞において使用するための大きな標的化ベクターを作製する工程(遺伝子改変は、コード配列、遺伝子セグメント、または調節エレメントの欠失である);c)(b)の大きな標的化ベクターをマウス胚性幹細胞に導入して、この細胞中の内因性遺伝子または染色体座を改変する工程;およびd)(c)のマウス胚性幹細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変されたマウス胚性幹細胞を同定する工程(定量アッセイは定量PCRである)を包含する。この方法により産生される遺伝子改変されたマウス胚性幹細胞;この方法により産生される遺伝子改変された内因性遺伝子または染色体座を含むマウス;および遺伝子改変されたマウス胚性幹細胞から産生されるマウスもまた好ましい。
【0030】
別の好ましい実施形態は、以下の工程を包含する方法により産生される、遺伝子改変された目的の内因性遺伝子または染色体座を含むマウスである:a)目的のDNA配列を含む20kbよりも大きなクローン化されたゲノムフラグメントを得る工程(この大きなクローン化されたDNAフラグメントは内因性遺伝子または染色体座に相同である);b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、マウス胚性幹細胞において使用するための大きな標的化ベクターを作製する工程(遺伝子改変は、コード配列、遺伝子セグメント、または調節エレメントの欠失である);c)(b)の大きな標的化ベクターをマウス胚性幹細胞に導入して、この細胞中の内因性遺伝子または染色体座を改変する工程;d)(c)のマウス胚性幹細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変されたマウス胚性幹細胞を同定する工程(定量アッセイは定量PCRである);e)(d)のマウス胚性幹細胞を胚盤胞に導入する工程;ならびにf)(e)の胚盤胞を妊娠のために代理母に導入する工程。
【0031】
マウスの産生のための、上記遺伝子改変されたマウス胚性幹細胞の使用もまた好ましい。
【0032】
本発明の1つの実施形態は、以下の工程を包含する、非ヒト真核生物細胞において、内因性免疫グロブリン可変領域遺伝子座を相同ヒト遺伝子座またはオルソログなヒト遺伝子座と、全部または一部置換する方法である:
a)ホモログヒト遺伝子座またはオルソログヒト遺伝子座を、全部または一部含む大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)のクローン化されたゲノムフラグメントを遺伝子改変し、真核生物細胞における使用のための大きな標的化ベクター(LTVEC)を作製する工程;
c)(b)のLTVECを真核生物細胞に導入して、内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
d)(c)の真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性免疫グロブリン可変領域遺伝子座が、ホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換された真核生物細胞を同定する工程。
【0033】
別の実施形態は、以下の工程をさらに包含する、非ヒト真核生物細胞において、内因性免疫グロブリン可変領域遺伝子座をホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換する方法である:
e)(a)のフラグメントと異なるホモログヒト遺伝子座またはオルソログヒト遺伝子座の一部を含む、大きなクローン化されたゲノムフラグメントを得る工程;
f)細菌相同組換えを使用して、(e)のクローン化されたゲノムフラグメントを遺伝子改変し、第2のLTVECを作製する工程;
g)(f)の第2のLTVECを工程(d)で同定された真核生物細胞に導入して、内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
h)(g)の真核生物細胞において対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、内因性免疫グロブリン可変領域遺伝子座が、ホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換された真核生物生物を同定する工程。
【0034】
上記方法の別の実施形態は、工程(e)〜(h)が、内因性免疫グロブリン可変領域遺伝子座がホモログヒト遺伝子座またはオルソログヒト遺伝子座と全部置換されるまで繰り返される方法である。
【0035】
この方法の別の実施形態は、免疫グロブリン可変遺伝子座が、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される座である方法である。
【0036】
好ましい実施形態は、定量アッセイが、定量PCR、FISH、比較ゲノムハイブリダイゼーション、等温DNA増幅、または固定されたプローブに対する定量ハイブリダイゼーションを含み、特に、定量PCRが、TaqMan(登録商標)技術または分子ビーコンを用いる定量PCRを含む方法である。
【0037】
なお別の好ましい実施形態は、以下の工程を包含する、マウス胚性幹細胞において、内因性免疫グロブリン可変領域遺伝子座をホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換する方法である:
a)ホモログヒト遺伝子座またはオルソログヒト遺伝子座を、全部または一部含む大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、胚性幹細胞における使用のための大きな標的化ベクターを作製する工程;
c)(b)の大きな標的化ベクターをマウス胚性幹細胞に導入して、細胞中の内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
d)(d)のマウス胚性幹細胞において対立遺伝子の改変(MOA)を検出するために定量PCRアッセイを使用して、内因性可変遺伝子座が、ホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換されたマウス胚性幹細胞を同定する工程。
【0038】
別の実施形態において、この方法はさらに、以下の工程を包含する:
e)(a)のフラグメントと異なるホモログヒト遺伝子座またはオルソログヒト遺伝子座の一部を含む、大きなクローン化されたゲノムフラグメントを得る工程;
f)細菌相同組換えを使用して、(e)のクローン化されたゲノムフラグメントを遺伝子改変し、胚性幹細胞における使用のための大きな標的化ベクターを作製する工程;
g)(f)の大きな標的化ベクターを工程(d)で同定されたマウス胚性幹細胞に導入して、内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
h)(g)のマウス胚性幹細胞において対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、内因性免疫グロブリン可変領域遺伝子座が、ホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換されたマウス胚性幹細胞を同定する工程。
【0039】
さらに別の好ましい実施形態は、上記工程(e)〜(h)が、内因性免疫グロブリン可変領域遺伝子座がホモログヒト遺伝子座またはオルソログヒト遺伝子座と全部置換されるまで、繰り返される、方法である。
【0040】
好ましいのはまた、上記免疫グロブリン可変遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される座を含む、方法である。
【0041】
別の好ましい実施形態は、上記の方法によって産生される遺伝子改変された免疫グロブリン可変領域遺伝子座;上記の方法によって産生される遺伝子改変された免疫グロブリン可変領域遺伝子座を含む遺伝子改変された真核生物細胞;上記の方法により遺伝子改変された免疫グロブリン可変領域遺伝子座を含む非ヒト生物;および上記の方法により遺伝子改変された免疫グロブリン可変領域遺伝子座を含むマウス胚幹細胞である。
【0042】
好ましいのはまた、マウス重鎖可変遺伝子座が、ヒト重鎖可変遺伝子座と、全部または一部置換された胚幹細胞;マウスκ軽鎖可変領域遺伝子座が、ヒトκ軽鎖可変領域遺伝子座と、全部または一部置換された胚幹細胞;マウスλ軽鎖可変領域遺伝子座が、ヒトλ軽鎖可変領域遺伝子座と、全部または一部置換された胚幹細胞;および重鎖可変領域遺伝子座および軽鎖可変領域遺伝子座が、それらのヒトホモログまたはオルソログと、全部置換された胚幹細胞である。
【0043】
別の好ましい実施形態は、上記の胚幹細胞から作製されたマウスである。
【0044】
なお別の好ましい実施形態は、上記の遺伝子改変された可変領域遺伝子座によりコードされるヒト可変領域を含む抗体;非ヒト定常領域をさらに含む抗体;およびヒト定常領域をさらに含む抗体である。
【0045】
また好ましいのは、トランスジェニックマウスであって、このマウスは、内因性マウス定常領域座を完全に作動可能に連結されたヒト重鎖可変領域座およびヒト軽鎖可変領域座を完全に含むゲノムを有し、その結果、このマウスが、抗原性刺激に応答してヒト可変領域およびマウス定常領域を含む抗体を含有する血清を産生する、トランスジェニックマウス;トランスジェニックマウスであって、このマウスは、内因性マウス定常領域座に作動可能に連結されたヒト重鎖可変領域座および/またはヒト軽鎖可変領域座を含むゲノムを有し、その結果、このマウスは、抗原性刺激に応答してヒト可変領域およびマウス定常領域を含む抗体を含有する血清を産生する、トランスジェニックマウス;トランスジェニックマウスであって、このマウスは、ホモログヒト可変座またはオルソログヒト可変座で置換された内因性免疫グロブリン可変領域座を含み、このマウスは、以下:
a)ホモログヒト可変領域座全体またはオルソログヒト可変領域座全体を含む、1つ以上の大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)のクローン化されたゲノムフラグメントを遺伝子改変し、マウス胚幹細胞における使用のための大きな標的化ベクターを作製する工程;
c)(b)の大きな標的化ベクターをマウス胚幹細胞に導入して、この細胞において内因性可変領域座全体を置換する工程;ならびに
d)(c)のマウス胚幹細胞において対立遺伝子の改変(MOA)を検出するために定量PCRアッセイを使用して、内因性可変領域座全体が、ホモログヒト可変領域座またはオルソログヒト可変領域座と置換されたマウス胚幹細胞を同定する、工程、
e)(d)のマウス胚幹細胞を胚盤胞に導入する工程;ならびに
f)(e)の胚盤胞を妊娠のために代理母に導入する工程、
を包含する、方法によって作製される、
トランスジェニックマウスである。
【0046】
別の好ましい実施形態は、上記のトランスジェニックマウスであって、免疫グロブリン可変領域遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される1つ以上の座を含む、トランスジェニックマウスである。
【0047】
また好ましいのは、マウス胚幹細胞が、上記の方法により作製されたトランスジェニックマウスに由来する、上記の方法である。
【0048】
本発明のなおさらに別の好ましい実施形態は、ヒト抗体を作製する方法であって、この方法は、以下:
a)上記のマウスを抗原性刺激に曝す工程であって、その結果、このマウスは抗原に対する抗体を産生する、工程;
b)抗体の重鎖可変領域および軽鎖可変領域をコードするDNAを単離する工程;
c)活性な抗体を発現し得る細胞中で、(b)の可変領域をコードするDNAをヒト重鎖定常領域およびヒト軽鎖定常領域をコードするDNAに作動可能に連結する工程;
d)この細胞を、ヒト抗体を発現するような条件下で増殖させる工程;ならびに
e)抗体を回収する工程、
を包含する、方法である。
【0049】
別の好ましい実施形態において、上記の細胞は、CHO細胞である。
【0050】
また好ましいのは、上記の工程(b)のDNAが、上記工程(a)中の抗原性刺激に曝されたマウスの脾臓から作製されたハイブリドーマから単離される方法である。
【0051】
また好ましいのは、DNAがPCRによって単離される、上記方法である。
【0052】
別の好ましい実施形態は、内因性免疫グロブリン可変領域遺伝子座をホモログ遺伝子座(homologous gene locus)またはオルソログ遺伝子座(orthologous gene locus)と、全部または一部置換する方法であって、この方法は、以下:
a)部位特異的組換え部位、その可変遺伝子座内の下流相同性アーム、およびその可変遺伝子座内の上流相同性アームを含む、LTVECを作製する工程であって、
ここで、この下流相同性アームは、この免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、このJセグメントを含まない、工程;
b)部位特異的組換え部位、その可変遺伝子座内の上流相同性アーム、およびその可変遺伝子座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、この上流相同性アームは、この免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECを真核生物細胞に導入する工程;
d)この可変遺伝子座中の対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、この部位特異的組換え部位がこの内因性可変領域遺伝子座に隣接する工程(c)の真核生物細胞を同定する工程;
e)このオルソログ遺伝子座またはこのホモログ遺伝子座の全部または一部に隣接する部位特異的組換え配列を含むベクターを作製する工程;ならびに
f)組換えを介して、この内因性免疫グロブリン可変領域遺伝子座がこのホモログ遺伝子座またはこのオルソログ遺伝子座と、全部または一部置換されるように、工程(d)において同定された真核生物中に(e)のベクターを導入する工程、
を包含する、方法である。
【0053】
また好ましいのは、トランスジェニックマウスであって、このマウスは、ホモログヒト免疫グロブリン可変領域座またはオルソログヒト免疫グロブリン可変領域座で置換された内因性免疫グロブリン可変領域座を含み、このマウスは、以下:
a)部位特異的組換え部位および下流相同性アームを含む、LTVECを作製する工程であって、
ここで、この下流相同性アームは、このマウス免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、このJセグメントを含まない、工程;
b)部位特異的組換え部位および上流相同性アームを含む、LTVECを作製する工程であって、
ここで、この上流相同性アームは、このマウス免疫グロブリン可変遺伝子座領域の最も遠位のマウスV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECをその真核生物細胞に導入する工程;この可変遺伝子座中の対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、この部位特異的組換え部位がこの内因性免疫グロブリン可変領域遺伝子座に隣接する工程(c)の真核生物細胞を同定する工程;
d)このオルソログ遺伝子座またはホモログ遺伝子座の全部または一部に隣接する部位特異的組換え配列を含むベクターを作製する工程;
e)組換えを介して、この内因性免疫グロブリン可変領域遺伝子座がこのホモログ遺伝子座またはオルソログ遺伝子座と、全部または一部置換されるように、工程(d)において同定されたこの真核生物細胞中に(d)のベクターを導入する工程;
f)(e)のマウス胚幹細胞を胚盤胞に導入する工程;ならびに
(f)の胚盤胞を妊娠のために代理母に導入する工程、
を包含する、方法によって作製される、トランスジェニックマウスである。
【0054】
なお別の好ましい実施形態は、真核生物細胞において、下流が部位特異的組換え部位と隣接した内因性遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、この座内の下流相同性アーム、この座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、この下流相同性アームは、この内因性遺伝子座領域の3’末端に隣接する領域を含む、工程;
b)(a)のLTVECをこの真核生物細胞内に導入する工程;ならびに
c)この内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この内因性遺伝子座の下流がこの部位特異的組換え部位と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法である。
【0055】
なお別の好ましい実施形態は、真核生物細胞において、上流が部位特異的組換え部位と隣接した内因性遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、その座内の上流相同性アーム、その座内の下流相同性アームを含むLTVECを作製する工程であって、
ここで、この上流相同性アームは、この内因性遺伝子座領域の5’末端に隣接する領域を含む、工程;
b)(a)のLTVECをこの真核生物に導入する工程;ならびに
c)この内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この内因性遺伝子座の上流がこの部位特異的組換え部位と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法である。
【0056】
また好ましいのは、真核生物細胞において、部位特異的組換え部位が隣接した内因性遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、この座内の下流相同性アーム、この座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、この下流相同性アームは、この内因性遺伝子座領域の3’末端に隣接する領域を含む、工程;
b)部位特異的組換え部位、その座内の上流相同性アーム、その座内の下流相同性アームを含むLTVECを作製する工程であって、
ここで、この上流相同性アームは、この内因性遺伝子座領域の5’末端に隣接する領域を含む、工程;
c)(a)および(b)のLTVECをこの真核生物細胞内に導入する工程;ならびに
d)この内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この部位特異的組換え部位がこの内因性遺伝子座と隣接する(c)の真核生物細胞を同定する工程、
を包含する、方法である。
【0057】
なお別の好ましい実施形態は、真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、その可変遺伝子座内の下流相同性アーム、その可変遺伝子座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、この下流相同性アームは、この免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、このJセグメントを含まない、工程;
b)(a)のLTVECをこの真核生物細胞内に導入する工程;ならびに
c)この可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この部位特異的組換え部位がこの内因性免疫グロブリン可変遺伝子座の下流末端と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法である。
【0058】
また好ましいのは、真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、この座内の上流相同性アーム、この座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、この上流相同性アームは、この免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
b)(a)のLTVECをこの真核生物細胞内に導入する工程;ならびに
c)この可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この部位特異的組換え部位がこの内因性免疫グロブリン可変領域遺伝子座の上流末端と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法である。
【0059】
なお別の実施形態は、真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、この座内の下流相同性アーム、この座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、この下流相同性アームは、この免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、このJセグメントを含まない、工程;
b)部位特異的組換え部位、この座内の上流相同性アーム、この座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、この上流相同性アームは、この免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECをこの真核生物細胞内に導入する工程;ならびに
d)この可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この部位特異的組換え部位がこの内因性免疫グロブリン可変領域遺伝子座と隣接する(c)の真核生物細胞を同定する工程、
を包含する、方法である。
本発明の好適な実施形態によれば、例えば以下の方法などが提供される:
(項目1)
非ヒト真核生物細胞において、内因性免疫グロブリン可変領域遺伝子座をホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換する方法であって、該方法は、以下:
a)改変改変該ホモログヒト遺伝子座または該オルソログヒト遺伝子座を全部または一部含む、大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)の該クローン化されたゲノムフラグメントを遺伝子改変し、真核生物細胞における使用のための大きな標的化ベクター(LTVEC)を作製する工程;
c)(b)の該LTVECを該真核生物細胞に導入して、該内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
d)(c)の該真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、該内因性免疫グロブリン可変領域遺伝子座が、該ホモログヒト遺伝子座または該オルソログヒト遺伝子座と、全部または一部置換された真核生物細胞を同定する、工程、
を包含する、方法。
(項目2)
項目1に記載の方法であって、該方法はさらに、以下:
e)(a)の前記フラグメントと異なるホモログヒト遺伝子座またはオルソログヒト遺伝子座の一部を含む、大きなクローン化されたゲノムフラグメントを得る工程;
f)細菌相同組換えを使用して、(e)の該クローン化されたゲノムフラグメントを遺伝子改変し、第2のLTVECを作製する工程;
g)(f)の該第2のLTVECを工程(d)で同定された前記真核生物細胞に導入して、前記内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
h)(g)の該真核生物細胞において対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該内因性免疫グロブリン可変領域遺伝子座が、該ホモログヒト遺伝子座または該オルソログヒト遺伝子座と、全部または一部置換された真核生物細胞を同定する、工程、
を包含する、方法。
(項目3)
項目2に記載の方法であって、ここで、工程(e)〜(h)は、前記内因性免疫グロブリン可変領域遺伝子座がホモログヒト遺伝子座またはオルソログヒト遺伝子座と全部置換されるまで、繰り返される、方法。
(項目4)
項目1に記載の方法であって、ここで、前記免疫グロブリン可変遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される座である、方法。
(項目5)
項目4に記載の方法であって、ここで、前記定量アッセイが、定量PCR、FISH、比較ゲノムハイブリダイゼーション、等温DNA増幅、または固定されたプローブに対する定量ハイブリダイゼーションを含む、方法。
(項目6)
前記定量PCRが、TaqMan(登録商標)技術または分子ビーコンを用いる定量PCRを含む、項目5に記載の方法。
(項目7)
マウス胚幹細胞において、内因性免疫グロブリン可変領域遺伝子座をホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換する方法であって、該方法は、以下:
a)改変該ホモログヒト遺伝子座または該オルソログヒト遺伝子座を全部または一部含む、大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)の該大きなクローン化されたゲノムフラグメントを遺伝子改変し、該胚幹細胞における使用のための大きな標的化ベクターを作製する工程;
c)(b)の該大きな標的化ベクターを胚幹細胞に導入して、該細胞中の該内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
d)(c)の該胚幹細胞において対立遺伝子の改変(MOA)を検出するために定量PCRアッセイを使用して、該内因性可変遺伝子座が、該ホモログヒト遺伝子座または該オルソログヒト遺伝子座と、全部または一部置換された胚幹細胞を同定する、工程、
を包含する、方法。
(項目8)
項目7に記載の方法であって、該方法はさらに、以下:
e)(a)の前記フラグメントと異なるホモログヒト遺伝子座またはオルソログヒト遺伝子座の一部を含む、大きなクローン化されたゲノムフラグメントを得る工程;
f)細菌相同組換えを使用して、(e)の該クローン化されたゲノムフラグメントを遺伝子改変し、前記胚幹細胞における使用のための大きな標的化ベクターを作製する工程;
g)(f)の該大きな標的化ベクターを工程(d)で同定された該胚幹細胞に導入して、前記内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
h)(g)の該マウス胚幹細胞において対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該内因性免疫グロブリン可変領域遺伝子座が、該ホモログヒト遺伝子座または該オルソログヒト遺伝子座と、全部または一部置換された胚幹細胞を同定する、工程、
を包含する、方法。
(項目9)
項目8に記載の方法であって、ここで、工程(e)〜(h)は、前記内因性免疫グロブリン可変領域遺伝子座がホモログヒト遺伝子座またはオルソログヒト遺伝子座と全部置換されるまで、繰り返される、方法。
(項目10)
項目7に記載の方法であって、ここで、前記免疫グロブリン可変遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される座を含む、方法。
(項目11)
内因性免疫グロブリン可変領域遺伝子座をホモログ遺伝子座またはオルソログ遺伝子座と、全部または一部置換する方法であって、該方法は、以下:
a)部位特異的組換え部位、該可変遺伝子座内の下流相同性アーム、該可変遺伝子座内の上流相同性アームを含む、LTVECを作製する工程であって、
ここで、該下流相同性アームは、該免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、該Jセグメントを含まない、工程;
b)部位特異的組換え部位、該可変遺伝子座内の上流相同性アーム、該可変遺伝子座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、該上流相同性アームは、該免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECを真核生物細胞に導入する工程;
d)該可変遺伝子座中の対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、該部位特異的組換え部位が該内因性可変領域遺伝子座に隣接する工程(c)の真核生物細胞を同定する工程;
e)該オルソログ遺伝子座または該ホモログ遺伝子座の全部または一部に隣接する部位特異的組換え配列を含むベクターを作製する工程;ならびに
f)組換えを介して、該内因性免疫グロブリン可変領域遺伝子座が該ホモログ遺伝子座または該オルソログ遺伝子座と、全部または一部置換されるように、工程(d)において同定された真核生物細胞中に(e)のベクターを導入する工程、
を包含する、方法。
(項目12)
前記真核生物細胞が胚幹細胞である、項目11に記載の方法。
(項目13)
項目1、4、7、8、10、11、または12に記載の方法によって産生される、遺伝子改変された免疫グロブリン可変領域遺伝子座。
(項目14)
項目1、4、7、8、11、または12に記載の方法によって産生される遺伝子改変された免疫グロブリン可変領域遺伝子座を含む、遺伝子改変された真核生物細胞。
(項目15)
項目1、4、7、8、10、または11に記載の方法によって産生される遺伝子改変された免疫グロブリン可変領域遺伝子座を含む、非ヒト生物。
(項目16)
項目7、8、10、または12に記載の方法によって作製される遺伝子改変された免疫グロブリン可変領域遺伝子座を含む、マウス胚幹細胞。
(項目17)
前記マウス重鎖可変領域座が、ヒト重鎖可変遺伝子座で全部または一部置換された、項目16に記載の胚幹細胞。
(項目18)
前記マウスκ軽鎖可変領域座が、ヒトκ軽鎖可変領域座で全部または一部置換された、項目16に記載の胚幹細胞。
(項目19)
前記マウスλ軽鎖可変領域座が、ヒトλ軽鎖可変領域座で全部または一部置換された、項目16に記載の胚幹細胞。
(項目20)
前記重鎖可変領域遺伝子座および前記軽鎖可変領域遺伝子座が、そのヒトホモログまたはヒトオルソログで全部置換された、項目16に記載の胚幹細胞。
(項目21)
項目16に記載の胚幹細胞から作製される、マウス。
(項目22)
項目17に記載の胚幹細胞から作製される、マウス。
(項目23)
項目18に記載の胚幹細胞から作製される、マウス。
(項目24)
項目19に記載の胚幹細胞から作製される、マウス。
(項目25)
項目20に記載の胚幹細胞から作製される、マウス。
(項目26)
項目11に記載の遺伝子改変された改変遺伝子座によってコードされるヒト可変領域を含む、抗体。
(項目27)
非ヒト定常領域をさらに含む、項目26に記載の抗体。
(項目28)
ヒト定常領域をさらに含む、項目26に記載の抗体。
(項目29)
トランスジェニックマウスであって、該マウスは、内因性マウス定常領域座に完全に作動可能に連結されたヒト重鎖可変領域座およびヒト軽鎖可変領域座を完全に含むゲノムを有し、その結果、該マウスは、抗原性刺激に応答してヒト可変領域およびマウス定常領域を含む抗体を含有する血清を産生する、トランスジェニックマウス。
(項目30)
トランスジェニックマウスであって、該マウスは、内因性マウス定常領域座に作動可能に連結されたヒト重鎖可変領域座および/またはヒト軽鎖可変領域座を含むゲノムを有し、その結果、該マウスは、抗原性刺激に応答してヒト可変領域およびマウス定常領域を含む抗体を含有する血清を産生する、トランスジェニックマウス。
(項目31)
トランスジェニックマウスであって、該マウスは、ホモログヒト可変領域座またはオルソログヒト可変領域座で置換された内因性免疫グロブリン可変領域座を含み、該マウスは、以下:
a)結合された場合に該ホモログヒト可変領域座または該オルソログヒト可変領域座に広がる、1つ以上の大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)の該クローン化されたゲノムフラグメントを遺伝子改変し、マウス胚幹細胞における使用のための大きな標的化ベクターを作製する工程;
c)(b)の該大きな標的化ベクターをマウス胚幹細胞に導入して、該細胞において該内因性可変領域座を置換する工程;
d)(c)の該マウス胚幹細胞において対立遺伝子の改変(MOA)を検出するために定量PCRアッセイを使用して、該内因性可変領域座が、該ホモログヒト可変領域座または該オルソログヒト可変領域座と置換されたマウス胚幹細胞を同定する、工程、
e)(d)の該マウス胚幹細胞を胚盤胞に導入する工程;ならびに
f)(e)の該胚盤胞を妊娠のために代理母に導入する工程、
を包含する、方法
によって作製される、トランスジェニックマウス。
(項目32)
トランスジェニックマウスであって、該マウスは、ホモログヒト免疫グロブリン可変領域座またはオルソログヒト免疫グロブリン可変領域座で置換された内因性免疫グロブリン可変領域座を含み、該マウスは、以下:
a)部位特異的組換え部位および下流相同性アームを含む、LTVECを作製する工程であって、
ここで、該下流相同性アームは、該マウス免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、該Jセグメントを含まない、工程;
b)部位特異的組換え部位および上流相同性アームを含む、LTVECを作製する工程であって、
ここで、該上流相同性アームは、該マウス免疫グロブリン可変遺伝子座領域の最も遠位のマウスV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECを該真核生物細胞に導入する工程;
d)該可変遺伝子座中の対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、該部位特異的組換え部位が該内因性免疫グロブリン可変領域遺伝子座に隣接する工程(c)の真核生物細胞を同定する工程;
e)該オルソログ遺伝子座または該ホモログ遺伝子座の全部または一部に隣接する部位特異的組換え配列を含むベクターを作製する工程;
f)組換えを介して、該内因性免疫グロブリン可変領域遺伝子座が該ホモログ遺伝子座または該オルソログ遺伝子座と、全部または一部置換されるように、工程(d)において同定された該真核生物中に(e)の該ベクターを導入する工程;
g)(d)の該マウス胚幹細胞を胚盤胞に導入する工程;ならびに
h)(e)の該胚盤胞を妊娠のために代理母に導入する工程、
を包含する、方法
によって作製される、トランスジェニックマウス。
(項目33)
項目30、31、または32に記載のトランスジェニックマウスであって、ここで、前記免疫グロブリン可変領域遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される1つ以上の座を含む、トランスジェニックマウス。
(項目34)
前記マウス胚幹細胞が、項目31に記載の方法によって作製されたトランスジェニックマウスに由来する、項目7、8、9、10または12に記載の方法。
(項目35)
前記マウス胚幹細胞が、項目32に記載の方法によって作製されたトランスジェニックマウスに由来する、項目7、8、9、10または12に記載の方法。
(項目36)
ヒト抗体を作製する方法であって、該方法は、以下:
a)項目26に記載のマウスを抗原性刺激に曝す工程であって、その結果、該マウスは該抗原に対する抗体を産生する、工程;
b)該抗体の重鎖可変領域および軽鎖可変領域をコードするDNAを単離する工程;
c)活性な抗体を発現し得る細胞中で、(b)の該可変領域をコードするDNAをヒト重鎖定常領域およびヒト軽鎖定常領域をコードするDNAに作動可能に連結する工程;
d)該細胞を、該ヒト抗体を発現するような条件下で増殖させる工程;ならびに
e)抗体を回収する工程、
を包含する、方法。
(項目37)
前記細胞がCHO細胞である、項目36に記載の方法。
(項目38)
工程(b)の前記DNAが、工程(a)中の抗原性刺激に曝された前記マウスの脾臓から作製されたハイブリドーマから単離される、項目36に記載の方法。
(項目39)
前記DNAがPCRによって単離される、項目31に記載の方法。
(項目40)
真核生物細胞において、下流が部位特異的組換え部位と隣接した内因性遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の下流相同性アーム、該座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、該下流相同性アームは、該内因性遺伝子座領域の3’末端に隣接する領域を含む、工程;
b)(a)の該LTVECを該真核生物細胞に導入する工程;ならびに
c)該内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該内因性遺伝子座の下流が該部位特異的組換え部位と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法。
(項目41)
真核生物細胞において、上流が部位特異的組換え部位と隣接した内因性遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の上流相同性アーム、該座内の下流相同性アームを含むLTVECを作製する工程であって、
ここで、該上流相同性アームは、該内因性遺伝子座領域の5’末端に隣接する領域を含む、工程;
b)(a)の該LTVECを該真核生物細胞に導入する工程;ならびに
c)該内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該内因性遺伝子座の上流が該部位特異的組換え部位と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法。
(項目42)
真核生物細胞において、部位特異的組換え部位が隣接した内因性遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の下流相同性アーム、該座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、該下流相同性アームは、該内因性遺伝子座領域の3’末端に隣接する領域を含む、工程;
b)部位特異的組換え部位、該座内の上流相同性アーム、該座内の下流相同性アームを含むLTVECを作製する工程であって、
ここで、該上流相同性アームは、該内因性遺伝子座領域の5’末端に隣接する領域を含む、工程;
c)(a)および(b)の該LTVECを該真核生物細胞に導入する工程;ならびに
d)該内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該部位特異的組換え部位が該内因性遺伝子座と隣接する(c)の真核生物細胞を同定する工程、
を包含する、方法。
(項目43)
真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該可変遺伝子座内の下流相同性アーム、該可変遺伝子座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、該下流相同性アームは、該免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、該Jセグメントを含まない、工程;
b)(a)の該LTVECを該真核生物に導入する工程;ならびに
c)該可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該部位特異的組換え部位が該内因性免疫グロブリン可変遺伝子座の下流末端と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法。
(項目44)
真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の上流相同性アーム、該座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、該上流相同性アームは、該免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
b)(a)の該LTVECを該真核生物に導入する工程;ならびに
c)該可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該部位特異的組換え部位が該内因性免疫グロブリン可変領域遺伝子座の上流末端と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法。
(項目45)
真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の下流相同性アーム、該座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、該下流相同性アームは、該免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、該Jセグメントを含まない、工程;
b)部位特異的組換え部位、該座内の上流相同性アーム、該座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、該上流相同性アームは、該免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)の該LTVECを該真核生物に導入する工程;ならびに
d)該可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該部位特異的組換え部位が該内因性免疫グロブリン可変領域遺伝子座と隣接する(c)の真核生物細胞を同定する工程、
を包含する、方法。
(項目46)
1つの部位特異的組換え部位が隣接する、内因性免疫グロブリン可変遺伝子座。
(項目47)
部位特異的組換え部位が隣接する、内因性免疫グロブリン可変遺伝子座。
(項目48)
1つの部位特異的組換え部位が隣接する内因性免疫グロブリン可変遺伝子座を含む、ES細胞。
(項目49)
部位特異的組換え部位が隣接する内因性免疫グロブリン可変遺伝子座を含む、ES細胞。
(項目50)
項目48または49の細胞によって作製される、マウス。
【図面の簡単な説明】
【0060】
【図1】細菌相同組換えを使用する代表的なLTVECの生成の模式図である(hb1=相同性ボックス1;hb2=相同性ボックス2;RE=制限酵素部位)。
【図2】マウス OCR10についてのドナーフラグメントおよびLTVECの模式図である(hb1=相同性ボックス1;lacZ=β−ガラクトシダーゼORF;SV40ポリA=シミアンウイルス40由来のDNAフラグメント(ポリアデニル化部位およびシグナルを含む);PGKp=マウスホスホグリセレートキナーゼ(PGK)。プロモーター、EM7=細菌プロモーター;neo=ネオマイシンホスホトランスフェラーゼ;PGK polyA=PGK遺伝子に由来し、ポリアデニル化部位およびシグナルを含む3’非翻訳領域;hb2=相同性ボックス2)。
【図3A】図3A〜3Dは、マウスOCR10 cDNA、相同性ボックス1(hb1)、相同性ボックス2(hb2)の配列であり、TaqMan(登録商標)プローブおよびプライマーを定量PCRアッセイで使用して、mOCR10 LTVECを使用して標的化されたES細胞中の対立遺伝子の改変(MOA)を検出した。 hb1:塩基対1〜211 hb2:塩基対1586〜1801 TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン3に由来している: TaqMan(登録商標)プローブ:ヌクレオチド413〜439(上の鎖) プライマー ex3−5’:ヌクレオチド390〜410(上の鎖) プライマー ex3−3’:ヌクレオチド445〜461(下の鎖) TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン4に由来している: TaqMan(登録商標)プローブ:ヌクレオチド608〜639(上の鎖) プライマー ex4−5’:ヌクレオチド586〜605(上の鎖) プライマー ex4−3’:ヌクレオチド642〜662(下の鎖)
【図3B】図3A〜3Dは、マウスOCR10 cDNA、相同性ボックス1(hb1)、相同性ボックス2(hb2)の配列であり、TaqMan(登録商標)プローブおよびプライマーを定量PCRアッセイで使用して、mOCR10 LTVECを使用して標的化されたES細胞中の対立遺伝子の改変(MOA)を検出した。 hb1:塩基対1〜211 hb2:塩基対1586〜1801 TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン3に由来している: TaqMan(登録商標)プローブ:ヌクレオチド413〜439(上の鎖) プライマー ex3−5’:ヌクレオチド390〜410(上の鎖) プライマー ex3−3’:ヌクレオチド445〜461(下の鎖) TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン4に由来している: TaqMan(登録商標)プローブ:ヌクレオチド608〜639(上の鎖) プライマー ex4−5’:ヌクレオチド586〜605(上の鎖) プライマー ex4−3’:ヌクレオチド642〜662(下の鎖)
【図3C】図3A〜3Dは、マウスOCR10 cDNA、相同性ボックス1(hb1)、相同性ボックス2(hb2)の配列であり、TaqMan(登録商標)プローブおよびプライマーを定量PCRアッセイで使用して、mOCR10 LTVECを使用して標的化されたES細胞中の対立遺伝子の改変(MOA)を検出した。 hb1:塩基対1〜211 hb2:塩基対1586〜1801 TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン3に由来している: TaqMan(登録商標)プローブ:ヌクレオチド413〜439(上の鎖) プライマー ex3−5’:ヌクレオチド390〜410(上の鎖) プライマー ex3−3’:ヌクレオチド445〜461(下の鎖) TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン4に由来している: TaqMan(登録商標)プローブ:ヌクレオチド608〜639(上の鎖) プライマー ex4−5’:ヌクレオチド586〜605(上の鎖) プライマー ex4−3’:ヌクレオチド642〜662(下の鎖)
【図3D】図3A〜3Dは、マウスOCR10 cDNA、相同性ボックス1(hb1)、相同性ボックス2(hb2)の配列であり、TaqMan(登録商標)プローブおよびプライマーを定量PCRアッセイで使用して、mOCR10 LTVECを使用して標的化されたES細胞中の対立遺伝子の改変(MOA)を検出した。 hb1:塩基対1〜211 hb2:塩基対1586〜1801 TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン3に由来している: TaqMan(登録商標)プローブ:ヌクレオチド413〜439(上の鎖) プライマー ex3−5’:ヌクレオチド390〜410(上の鎖) プライマー ex3−3’:ヌクレオチド445〜461(下の鎖) TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン4に由来している: TaqMan(登録商標)プローブ:ヌクレオチド608〜639(上の鎖) プライマー ex4−5’:ヌクレオチド586〜605(上の鎖) プライマー ex4−3’:ヌクレオチド642〜662(下の鎖)
【図4】図4A〜4Dは、マウスVDJ領域をヒトVDJ領域で置き換えて構築した2つのLTVECの模式図である。 図4A:ヒト重鎖遺伝子座のVDJ領域全体にまたがる大きなインサート(BAC)クローンを単離した。 図4B:この例において、相同性アームの供給源として、大きなインサート(BAC)クローンをマウスVDJ領域の末端から単離する。この相同性アームを使用して、2工程プロセスにおいてヒトVDJ配列の相同組換えを介して組み込んだ。 図4C〜4D:第1の工程において、LTVEC1(図4D)を、E.coliにおける細菌相同組換えにより構築する。LTVEC1は、順番に以下を含む:マウスDJ領域から上流の領域に由来する大きなマウス相同性アーム(しかし、その絶対エンドポイントは、重要ではない);ES細胞において機能的な選択マーカーをコードするカセット(この例では、PGK−ネオマイシンR);loxP部位;いくつかのV遺伝子セグメントからDJ領域全体にまたがる大きなヒトインサート;およびマウスJセグメントにすぐ隣接するが、このJセグメントを含まない領域を含むマウス相同性アーム。第2の工程において、LTVEC2(図4C)を、E.coliにおける細菌相同組換えにより構築する。LTVEC2は、順番に以下を含む:最も遠位のマウスV遺伝子セグメントに隣接するが、いかなるマウスV遺伝子セグメントも含まない領域を含む大きなマウス相同性アーム;多数の遠位ヒトV遺伝子セグメントを含む大きなインサート;変異loxP部位(LTVEC2およびLTVEC1における野生型loxP部位とは反対の配向にてloxP511といわれる)(この部位は、野生型loxPと組換わらないが、他のloxP511とは容易に組換わる);野生型loxP部位;第2の選択マーカー(この例では、PGK−ハイグロマイシンR);およびV領域に由来するマウス相同性アーム(しかし、その絶対エンドポイントは重要ではない)。
【発明を実施するための形態】
【0061】
(定義)
「標的化ベクター」は、所望の遺伝子改変に隣接する内因性染色体核酸配列に「相同(ホモログ)(homologous)」な配列を含むDNA構築物である。隣接相同性配列(「相同性アーム」と称する)は、標的化ベクターを、相同性アームと対応する内因性配列との間に存在する相同性によってゲノム内の特定の染色体位置に方向付け、「相同組換え」と称されるプロセスにより、所望の遺伝子改変を導入する。
【0062】
「相同(ホモログ)(homologous)」とは、2以上の核酸配列が、互いにハイブリダイズし得るかまたは分子間交換を受け得るに十分、同一または類似のいずれかである、2以上の核酸配列を意味する。
【0063】
「遺伝子標的化」は、標的化ベクターを使用して、相同組換えを介した内因性配列への挿入、内因性配列の欠失または内因性配列の置換による、内因性染色体座の改変である。
【0064】
「遺伝子ノックアウト」は、染色体座においてコードされた遺伝情報の破壊から生じた遺伝子改変である。
【0065】
「遺伝子ノックイン」は、染色体座においてコードされた遺伝情報を、異なるDNA配列と置換することから生じる遺伝子改変である。
【0066】
「ノックアウト生物」は、その生物の細胞の大部分が、遺伝子ノックアウトを有する生物である。
【0067】
「ノックイン生物」は、その生物の細胞の大部分が、遺伝子ノックインを有する生物である。
【0068】
「マーカー」または「選択マーカー」は、集団中の処理細胞の大部分から、選択マーカーを発現する稀なトランスフェクト細胞を単離することを可能にする選択マーカーである。このようなマーカー遺伝子としては、ネオマイシンホスホトランスフェラーゼおよびハイグロマイシンBホスホトランスフェラーゼ、または蛍光タンパク質(例えば、GFP)が挙げられるが、これらに限定されない。
【0069】
「ES細胞」は、胚幹細胞である。この細胞は、通常、胚盤胞期の胚の内部細胞塊に由来する。
【0070】
「ES細胞クローン」は、DNAの導入、引き続く選択の後の、ES細胞集団の単一細胞由来の細胞の亜集団である。
【0071】
「隣接DNA」は、参照の特定の点と同一線上でありそして隣接する、DNAのセグメントである。
【0072】
「LTVEC」は、真核生物細胞において相同標的化を行うことを目的とされた他のアプローチによって典型的に使用されるものよりも大きな、クローニングされたゲノムDNAのフラグメント由来の、真核生物細胞についての大きな標的ベクターである。
【0073】
「非ヒト生物」は、通常ヒトであることが公的に受容されない生物である。
【0074】
「対立遺伝子の改変」(MOA)は、遺伝子、またはゲノム中の染色体遺伝子座の対立遺伝子の正確なDNA配列の改変をいう。この対立遺伝子改変(MOA)としては、以下が挙げられるがこれらに限定されない:単一ヌクレオチド程度の小さなヌクレオチドのわずかな欠失、置換または挿入、または多くのキロベースにわたる目的の遺伝子または染色体遺伝子座の欠失、ならびにこれらの2つの極値の間で可能な改変のいずれかおよび全て。
【0075】
「オルソログ(orthologous)」配列は、別の種中のその配列の機能的等価物である種由来の配列をいう。
【0076】
とりわけ示される記載または実施例は、本発明に例証されることで提供される。当業者は、これらの実施例が本発明の例示の目的のみに提供され、限定の目的には含まれないことを、理解する。
【0077】
(発明の詳細な説明)
本出願人らは、改変された内因性遺伝子または染色体遺伝子座を含む真核生物細胞を作製およびスクリーニングするための、新規で、迅速で、能率的で、そして効率的な方法を開発した。これらの細胞において、改変は、遺伝子ノックアウト、ノックイン、点変異または大きなゲノム挿入もしくは欠失、あるいは他の改変であり得る。非限定的な例示の目的で、これらの細胞は、変更、欠失および/または挿入された遺伝子の機能を決定する目的のためのノックアウト生物またはノックイン生物(特に、ノックアウトマウスまたはノックインマウス)に有用である胚性幹細胞であり得る。
【0078】
本明細書中に記載される新規な方法は、最初に以下を組合わせる:
1.大きなクローニングされたゲノムDNAフラグメント内に所望の遺伝子改変を正確に操作し、これにより真核生物細胞(LTVEC)における使用のための大きな標的化ベクターを作製するための細菌性相同組換え;
2.これらのLTVEC細胞において対応する目的の内因性遺伝子または染色体遺伝子座を改変するための、真核生物細胞へのこれらの細胞の直接導入;および
3.標的化対立遺伝子が所望されるように改変されている、まれな真核生物細胞を決定するための分析(親対立遺伝子の対立遺伝子改変(MOA)のための定量的アッセイを含む)。
【0079】
真核生物細胞における相同組換えを首尾よく検出するための以前の方法が、LTVEC中に存在する長い相同性アームのために、本出願人らの発明のLTVECと組合わせて利用され得ないことは、強調されるべきである。真核生物細胞中の内因性遺伝子または染色体遺伝子座を、相同組換えを介して故意に改変するためにLTVECを利用することによって、まれな真核生物細胞を決定するためのアッセイの新規適用によっておそらくなされる。ここで、標的化された対立遺伝子は所望されるように改変されており、このようなアッセイとしては、親対立遺伝子の対立遺伝子改変(MOA)のための定量的アッセイ(例えば、定量的PCRまたはMOAについての他の適切な定量的アッセイを使用することによる)が挙げられる。
【0080】
現在の方法において使用されるものより長い相同性アームを有する標的化ベクターを利用する能力は、以下の理由について極めて有益である:
1.標的化ベクターは、大きなゲノム挿入物を含む利用可能なライブラリー(例えば、BACライブラリーまたはPACライブラリー)から、以前の技術を使用して作製される標的化ベクターより迅速かつ簡便に作製される。ここで、ゲノム挿入物は、広範に特徴付けられており、使用前に「整えられる(trim)」(以下に詳細に説明される)。さらに、最小配列情報が、目的の遺伝子座について知られていることが必要である。すなわち、相同性ボックス(以下に詳細に記載される)を作成するため、およびMOAについての定量的アッセイ(以下に詳細に記載される)に使用され得るプローブを作成するために必要とされる約80〜100ヌクレオチドが知られていることのみが必要である。
2.より大きな改変、ならびにより長いゲノム領域にわたる改変が、以前の技術を使用するより簡便に、そしてよりわずかな工程において作製される。例えば、本発明の方法は、そのサイズ制限のために伝統的プラスミドベースの標的化ベクターによっては収容され得ない大きな遺伝子座の正確な改変を可能にする。これはまた、1工程における複数の点での任意の所定の遺伝子座の改変(例えば、複数のエキソン遺伝子の異なるエキソンでの特定の変異の導入)を可能にして、複数の標的化ベクターを操作する必要性、およびES細胞での相同組換えのための複数回の標的化およびスクリーニングを行う必要性を緩和する。
3.相同性の長い領域(長い相同性アーム)の使用は、真核生物細胞における相同組換えの標的化が標的化ベクター内に含まれる総相同性に関連するようであるという以前の知見と一致して、真核生物細胞における「標的化しにくい(hard to target)」遺伝子座の標的化頻度を増加させる。
4.長い相同性アームを使用して得られた増加した標的化頻度は、これらの標的化ベクターにおいて同系のDNAを使用することから、たとえあったとしても、その利点を明らかに減少させる。
5.真核生物細胞を相同組換えについてスクリーニングするための定量的MOAアッセイの適用は、標的化ベクターとしてのLTVECの使用を可能にするだけでなく(利点は上記に概説した)、正確に改変された真核生物細胞を同定するための時間を代表的な7日から数時間までに減少させる。さらに、定量的MOAの適用は、改変されている、内因性遺伝子または染色体遺伝子座の外側に位置されるプローブの使用を必要としない。よって、改変された遺伝子または遺伝子座に隣接する配列を知る必要性を除去する。これは、過去に行われたスクリーニングを、真核生物細胞における相同組換え事象についてスクリーニングするためにほとんど労力を要さず、そしてかなり高い費用効果的なアプローチとなさしめる、有意な改善である。
【0081】
(方法)
本明細書中に記載されるDNAベクターを構築するために使用される技術の多くは、当業者に周知である標準的な分子生物学的技術である(例えば、Sambrook,J.,E.F.Fritsch And T.Maniatis.Molecular Cloning:A Laboratory Manual,Second Edition,Vols 1,2,and 3,1989;Current Protocols in Molecular Biology,Eds.Ausubel et al.,Greene Publ.Assoc.,Wiley Interscience,NYを参照のこと)。全てのDNA配列決定を、ABI 373A DNA配列決定機およびTaq Dideoxy Terminator Cycle Sequencing Kit(Applied Biosystems,Inc.,Foster City,CA)を使用する標準的な技術によって行った。
【0082】
(工程1.目的の遺伝子または染色体遺伝子座を含有する大きなゲノムDNAクローンを得る)
目的の遺伝子または遺伝子座を、特定の基準(例えば、詳細な構造的データまたは機能的データ)に基づいて選択し得るか、または種々のゲノム配列決定プロジェクトの努力を介して潜在的な遺伝子フラグメントが予想されるようになるような詳細な情報の非存在下で選択し得る。重要なことは、LTVECを産生させるために、目的の遺伝子の完全な配列および遺伝子構造を知って本発明の方法に適用する必要がないことを、注意するべきである。実際に、目的のゲノムクローンを得、そしてLTVECを作製する際に使用される相同性ボックス(以下に詳細に記載される)を作製し、そして定量性MOAアッセイにおける使用のためのプローブを作製するために必要とされる唯一の情報は、約80〜100ヌクレオチドである
一旦、目的の遺伝子または遺伝子座が選択されると、この遺伝子または遺伝子座を含有する大きなゲノムクローンが得られる。このクローンを、適切なDNAライブラリー(例えば、BAC、PAC、YACまたはコスミド)を標準的なハイブリダイゼーション技術またはPCR技術、あるいは当業者が精通する任意の他の方法によるスクリーニングが挙げられるがこれに限定されないいくつかの方法のいずれか1つにおいて取得し得る。
【0083】
(工程2.改変カセットへの相同性ボックス1および2の付加ならびにLTVECの作製)
相同性ボックスは、クローニングされた大きなゲノムフラグメントからLTVECを作成するために使用される細菌性相同組換えの部位を印す(図1)。相同性ボックスは、一般には、二本鎖であり、そして少なくとも40ヌクレオチド長であるDNAの短いセグメントであり、このセグメントは、「改変されるべき領域」に隣接する、このクローニングされた大きなゲノムフラグメント内の領域に相同である。相同性ボックスは、改変されたカセットに付加され、その結果、細菌中での相同組換えに続いて、この改変カセットは、改変されるべき領域に置換する(図1)。細菌性相同組換えを使用する標的化ベクターの作製技術は、種々の系において行われ得る(Yang et al.,Nat Biotechnol,15:859−65,1997;Muyrers et al.,Nucleic Acids Res,27:1555−7,1999;Angrand et
al.,Nucleic Acids Res,27:e16,1999;Narayanan et al.,Gene Ther,6:442−7,1999;Yu,et
al.,Proc Natl Acad Sci USA,97:5978−83,2000)。現在使用に好ましい技術の1つの例は、ETクローニング(Zhang et
al.,Nat Genet,20:123−8,1998;Narayanan et al.,Gene Ther,6:442−7,1999)およびこの技術の改変(Yu,et al.,Proc Natl Acad Sci USA,97:5978−83,2000)である。ETは、相同組換え反応を実施するrecE(Hall and Kolodner,Proc Natl Acad Sci USA,91:3205−9,1994)およびrecTタンパク質(Kusano et al.,Gene,138:17−25,1994)をいう。RecEは、線状二本鎖DNA(実質的にはとりわけ記載されるドナーDNAフラグメント)の一方の鎖を5’から3’に整え、よって3’一本鎖オーバーハングを有する線状二本鎖フラグメントの後ろに残すエクソヌクレアーゼである。この一本鎖オーバーハングは、一本鎖DNA(ssDNA)結合活性を有するrecTタンパク質によってコートされる(Kovall and Matthews,Science,277:1824−7,1997)。ETクローニングは、recEおよびrecTのE.coli遺伝子産物(Hall and Kolodner,Proc Natl Acad Sci USA,91:3205−9,1994;Clark et al.,Cold Spring Harb Symp Quant Biol,49:453−62,1984;Noirot and Kolodner,J
Biol Chem,273:12274−80,1998;Thresher et
al.,J Mol Biol,254:364−71,1995;Kolodner
et al.,Mol Microbiol,11:23−30,1994;Hall
et al.,J Bacteriol,175:277−87,1993)、およびバクテリオファージ(λ)タンパク質λgam(Murphy,J Bacteriol,173:5808−21,1991; Poteete et al.,J Bacteriol,170:2012−21,1988)を一過性に発現するE.coliを使用して行われる。λgamタンパク質は、recBCエクソヌクレアーゼ系による分解からドナーDNAフラグメントを保護するために必要とされ(Myers and Stahl,Annu Rev Genet,28:49−70,1994)、そしてrecBC+宿主(例えば、しばしば使用されるE.coli株DH10b)における効率的なETクローニングのために必要とされる。
【0084】
細菌性相同組換えを使用して改変および置換されるべき領域は、0ヌクレオチド(もともとの遺伝子座に挿入を作製する)〜数十キロベース(もともとの遺伝子座の欠失および/または置換を作製する)の長さにわたり得る。改変カセットに基づくと、改変は以下を生じ得る:
(a)コード配列、遺伝子セグメント、または調節エレメントの欠失;
(b)コード配列、遺伝子セグメント、または調節エレメントの変更(置換、付加、および融合(例えば、エピトープタグまたは二官能性タンパク質(例えば、GFPとの融合)の作製を含む);
(c)新たなコード領域、遺伝子セグメント、または調節エレメントの挿入(例えば、選択マーカー遺伝子またはレポーター遺伝子についてのもの)あるいは内因性転写調節下での新たな遺伝子の配置;
(d)条件付き対立遺伝子の作製(例えば、Creリコンビナーゼによって切除されるべき領域に隣接するloxP部位(Abremski and Hoess,J Biol
Chem,259:1509−14,1984)、またはFlpリコンビナーゼによって切除されるべき領域に隣接するFRT部位(Andrews et al.,Cell,40:795−803,1985;Meyer−Leon et al.,Cold Spring Harb Symp Quant Biol,49:797−804,1984;Cox,Proc Natl Acad Sci USA,80:4223−7,1983)の導入による);あるいは
(e)異なる種由来のオルソログコード配列での、ある種からのコード配列または遺伝子セグメントの置換(例えば、マウス遺伝子座をオルソログヒト遺伝子座と置換して、ここでこれらの特定の遺伝子座が「ヒト化」される)。
【0085】
これらの改変のいずれかまたは全てが、LTVECに組み込まれ得る。特定の、非限定的な例示(ここで、内因性コード配列が完全に欠失され、そして、同時にレポーター遺伝子および選択マーカーの両方と置換されている)が、以下の実施例1に提供され、以前の技術と比較して本発明の方法の利点である。
【0086】
(工程3(任意).各LTVECが正確に操作されていることの検証)
各LTVECが正確に操作されていることの検証は、以下による:
a.ドナーフラグメントを、目的の遺伝子または染色体遺伝子座へ導入することによって作製された新規な連結を検証するための診断的PCR。よって、この得られたPCRフラグメントを配列決定し、ドナーフラグメントを、目的の遺伝子または染色体遺伝子座へ導入することによって作製された新規な連結をさらに検証し得る。
b.細菌性相同組換えプロセスの間に所望の改変のみがLTVECに導入されたことを確認するための、診断的制限酵素消化。
c.ドナーフラグメントを、目的の遺伝子または染色体遺伝子座へ導入することによって作製された新規な連結を検証するための、LTVEC(特に、改変の部位にわたる領域)の直接配列決定。
【0087】
(工程4.真核生物細胞への導入のための、LTVEC DNAの精製、調製および線状化)
(a.LTVEC DNAの調製)
選択したLTVECのminiprep DNA(Sambrook,J.,E.F.Fritsch And T.Maniatis.Molecular Cloning:A Laboratory Manual,Second Edition,Vols
1,2,and3,1989;Tillett and Neilan,Biotechniques,24:568−70,572,1998;http://www.qiagen.com/literature/handbooks/plkmini/plm_399.pdf)を調製し、そしてこのminiprep LTVEC DNAを、エレクトロポレーション(Sambrook,J.,E.F.Fritsch and T.Maniatis,Molecular Cloning:A Laboratory Manual,Second Edition,Vols 1,2,and 3,1989)を使用してE.coliに再度形質転換した。この工程は、細菌性相同組換え工程に利用される組み換えタンパク質をコードするプラスミドを除くために必要である(Zhang et al.,Nat Genet,20:123−8,1998;Narayanan et al.,Gene Ther,6:442−7,1999)。(a)なぜならこれは、コピー数の多いプラスミドであって、大規模LTVEC prepにおいて得られる収量を減少し得、(b)組換えタンパク質の発現を誘導する可能性を排除するためであり;そして(c)なぜなら、これは、LTVECの物理的マッピングをあいまいにし得るからである。LTVECを真核生物細胞に導入する前に、より多量のLTVEC DNAを、標準的な方法論(http://www.qiagen.com/literature/handbooks/plk/plklow.pdf;Sambrook,J.,E.F.Fritsch And T.Maniatis.Molecular Cloning:A Laboratory Manual,Second Edition,Vols 1,2,and 3,1989;Tillett and Neilan,Biotechniques 24:568−70,572,1998)によって調製した。しかし、この工程は、組換えプロファージを利用する細菌性相同組換え法が使用される(すなわち、組換えタンパク質をコードする遺伝子が細菌染色体中に組み込まれる)場合、回避され得る(Yu,et al.,Proc Natl Acad Sci USA,97:5978−83,2000)。
【0088】
(b.LTVEC DNAの線状化)
真核生物細胞への導入のためにLTVECを調製するために、LTVECを、好ましくは、長い相同性アームに隣接する、改変された内因性遺伝子または染色体遺伝子座DNAを残す様式で線状化する。これは、めったに消化しない適切な制限酵素のいずれかを用いて、LTVEC、好ましくは、ベクターバックボーン中のを線状化することによって達成され得る。適切な制限酵素の例としては、NotI、PacI、SfiI、SrfI、SwaI、FseIなどが挙げられる。制限酵素の選択は、実験的に(すなわち、いくつかの異なる候補レアカッター(rare cutter)を試験することによって)、またはLTVECの配列が公知である場合は、配列を分析し、そして分析に基づいて適切な制限酵素を選択することによって)決定され得る。LTVECがまれな部位(例えば、CosN部位)を含むベクターバックボーンを有する状況では、次いで、このような部位を認識する酵素(例えば、λターミナーゼ(Shizuya et al.,Proc Natl Acad Sci USA,89:8794−7,1992;Becker and Gold,Proc Natl Acad Sci USA,75:4199−203,1978;Rackwitz et al.,Gene,40:259−66,1985))で切断し得る。
【0089】
(工程5.真核生物細胞へのLTVECの導入、およびLTVECの導入が首尾よく生じた場合の細胞の選択)
LTVEC DNAを、標準的な方法論(例えば、リン酸カルシウム、脂質を介するトランスフェクション、またはエレクトロポレーション(Sambrook,J.,E.F.Fritsch And T.Maniatis.Molecular Cloning:A Laboratory Manual,Second Edition,Vols 1,2,and 3,1989))を使用して真核生物細胞に導入し得る。LTVECが首尾よく導入された細胞を、選択因子に曝露することによって、LTVEC中に操作されている選択マーカー遺伝子に依存して選択し得る。非限定的な例として、選択マーカーがネオマイシンホスホトランスフェラーゼ(neo)遺伝子(Beck,et al.,Gene,19:327−36,1982)である場合、次いで、LTVECを取り込んだ細胞を、G418含有培地中で選択し得る:LTVECを有さない細胞は死ぬが、LTVECを取り込んだ細胞は、生存する(Santerre,et al.,Gene,30:147−56,1984)。他の適切な選択マーカーとしては、真核生物細胞中で活性を有する任意の薬物(例えば、ハイグロマイシンB(Santerre,et al.,Gene,30:147−56,1984;Bernard,et al.,Exp
Cell Res,158:237−43,1985;Giordano and McAllister,Gene,88:285−8,1990)、ブラスチシジン(Blasticidin)S(Izumi,et al.,Exp Cell Res,197:229−33,1991)、および当業者の精通した他のもの)が挙げられる。
【0090】
(工程6.対立遺伝子改変(MOA)についての定量的アッセイを使用する、真核生物細胞における相同組換え事象のスクリーニング)
目的の遺伝子座にLTVECを標的化することにより首尾よく改変された真核生物細胞を、目的の遺伝子座内の対立遺伝子改変を指向し得、そして相同性アーム全体にわたるアッセイに依存しない種々のアプローチを使用して同定し得る。このようなアプローチとしては、以下が挙げられるがこれらに限定されない:
(a)TaqMan(登録商標)(Lie and Petropoulos,Curr
Opin Biotechnol,9:43−8,1998)を使用する定量的PCR;
(b)分子ビーコンを使用する定量的MOAアッセイ(Tan,et al.,Chemistry,6:1107−11,2000);
(c)インサイチュハイブリダイゼーションFISHでの蛍光(Laan,et al.,Hum Genet,96:275−80,1995)または比較ゲノムハイブリダイゼーション(CGH)(Forozan,et al.,Trends Genet,13:405−9,1997;Thompson and Gray,J Cell Biochem Suppl,139−43,1993;Houldsworth and Chaganti,Am J Pathol,145:1253−60,1994);
(d)等温DNA増幅(Lizardi,et al.,Nat Genet,19:225−32,1998;Mitra and Church,Nucleic Acids Res,27:e34,1999);および
(e)固定化プローブへの定量的ハイブリダイゼーション(Southern,J.Mol.Biol.98:503,1975;Kafatos FC;Jones CW;Efstratiadis A,Nucleic Acids Res 7(6):1541−52,1979)。
【0091】
出願人は、TaqMan(登録商標)定量的PCRを使用して、標的化真核生物細胞を首尾よくスクリーニングする例示を、本明細書中に提供する。この非限定的例示において、TaqMan(登録商標)を使用して、相同組換えを受けた真核生物を同定し、ここで、二倍体ゲノム中の2つの内因性対立遺伝子のうちの1つの部分は、別の配列に置換されている。相同性アーム全体にわたる制限フラグメントの長さにおける差異が、2つの対立遺伝子のうちの1つの改変を示す伝統的な方法とは対照的に、定量的TaqMan(登録商標)法は、改変されていない対立遺伝子のコピー数の(半分だけの)減少を測定することによって一方の対立遺伝子の改変を検出する。詳細には、プローブは、改変されていない対立遺伝子を検出し、改変されている対立遺伝子を検出しない。よって、この方法は、改変の正確な特徴に依存せず、本実施例に記載される配列置換に限定されない。TaqMan(登録商標)を使用して、ゲノムDNAサンプル中のDNAテンプレートのコピー数を、特に、参照遺伝子と比較することによって定量する(Lie and Petropoulos,Curr Opin Biotechnol,9:43−8,1998)。参照遺伝子は、標的遺伝子または遺伝子座として同一のゲノムDNAにおいて定量される。よって、2つのTaqMan(登録商標)増幅(各々それぞれのプローブを有する)が行われる。一方のTaqMan(登録商標)プローブは、参照遺伝子の「Ct」(閾値サイクル)を決定し、他方のプローブは、首尾よく標的化することによって置換されている、標的化された遺伝子または遺伝子座の領域のCtを決定する。Ctは、各TaqMan(登録商標)プローブについての開始DNAの量を反映する量である(すなわち、ほとんどの配列が、閾値サイクルに達するためにPCRのより多いサイクルを必要としない)。TaqMan(登録商標)についてのテンプレート配列のコピー数を半分に減少することによって、約1Ct単位の増加を生じる。標的化遺伝子または遺伝子座の一方が相同組換えによって置換されている細胞でのTaqMan(登録商標)反応は、標的化されていない細胞由来のDNAと比較する場合、参照遺伝子についてのCtの増加することなく標的TaqMan(登録商標)反応について1Ctの増加を生じる。このことによって、LTVECを使用して真核生物細胞に目的の遺伝子の一方の対立遺伝子の改変を容易に検出し得る。
【0092】
上記の状態において、対立遺伝子改変(MOA)スクリーニングは、相同組換えを受けた細胞を同定するための、一方の対立遺伝子の改変の検出する任意の方法の使用である。標的化対立遺伝子が互いに同一(相同)であることは必要とされず、マウスの異なる2つの系統を交配することによって生じる子孫の場合のように、実際に、多型を含み得る。さらに、MOAスクリーニングによってまたカバーされる、ある特定の状況は、遺伝子(例えば、性染色体、特にY染色体上に位置するいくつか)の標的化であり、これは、細胞中に単一のコピーとして正常に存在する。この場合において、単一の標的化対立遺伝子の改変を検出する方法(例えば、定量的PCR、サザンブロッティングなど)を使用して、標的化事象を検出し得る。対立遺伝子が多型であるかまたは標的化細胞中の単一コピー中に存在する場合でさえ、本発明の方法を使用して、改変された真核生物細胞を作製し得る。
【0093】
(工程8.遺伝子改変された真核生物細胞の使用)
(a)細胞の表現型の変化が所望される場合、工程1〜7に記載された方法によって作製された、遺伝子改変された真核生物細胞を、任意のインビトロアッセイまたはインビボアッセイにおいて使用し得る。
【0094】
(b)工程1〜7に記載された方法によって作製された、遺伝子改変された真核生物細胞をまた、遺伝子改変を保有する生物を作製するために使用し得る。遺伝子改変された生物を、以下の異なる種々の技術(しかし、これらに限定されない)を使用することによって作製し得る:
1.改変された胚性幹細胞(ES細胞)(例えば、ラットES細胞およびマウスES細胞がしばしば使用される)。ES細胞を使用して、遺伝子改変されたラットまたはマウスを、標準的な胚盤胞注入技術または凝集技術(Robertson,Practical
Approach Series,254,1987;Wood et al.,Nature,365:87−9,1993;Joyner,The Practical Approach Series,293,1999)、テトラプロイド胚盤胞注入(Wang,et al.,Mech Dev,62:137−45,1997)または核移入およびクローニング(Wakayama,et al.,Proc Natl Acad Sci USA,96:14984−9,1999)によって作製し得る。他の生物(例えば、ウサギ(Wang,et al.,Mech Dev,62:137−45,1997;Schoonjans,et al.,Mol Reprod Dev,45:439−43,1996)またはニワトリ(Pain,et al.,Development,122:2339−48,1996)あるいは他の種)由来のES細胞はまた、本発明の方法を使用する遺伝子改変に従うべきである。
【0095】
2.改変されたプロトプラストは、遺伝的に改変された植物を作製するために用いられ得る(例えば、米国特許第5,350,689号「プロトプラストまたはプロトプラスト由来細胞から再生されるトウモロコシ植物およびトランスジェニックトウモロコシ植物」、米国特許第5,508,189号「培養された孔片細胞プロトプラストからの植物の再生」、およびこれらの参考文献を参照のこと)。
【0096】
3.改変された対立遺伝子を有するクローン化生物体を作製するための、改変された真核生物細胞から卵母細胞への核移入(Wakayamaら、Proc Natl Acad Sci USA,96:14984−9,1999;Baguisiら、Nat Biotechnol,17:456−61,1999;Wilmutら、Reprod Fertil Dev,10:639−43,1998;Wilmutら、Nature,385:810−3,1997;Wakayamaら、Nat Genet,24:108−9,2000;Wakayamaら、Nature,394:369−74,1998;Rideoutら、Nat Genet,24:109−10,2000;Campbellら、Nature,380:64−6,1996)。
【0097】
4.操作された染色体の移動を含む、改変された対立遺伝子を別の細胞に移入させるための細胞融合、および改変された対立遺伝子または操作された染色体を有する生物体を産生するためのこのような細胞の使用(Kuroiwaら、Nat Biotechnol,18:1086−1090,2000)。
【0098】
5.本発明の方法はまた、用いられているか、または、まだ発見されていない、他の任意のアプローチに受け入れられる。
【0099】
本発明の方法の個々の工程を実行する際に使用される多くの技術は、当業者によく知られているが、本出願人は、本発明の方法の新規性が、真核生物細胞にLTVECを直接導入して、染色体座を改変する、以前に記載されたことのない方法と、適切に改変された真核生物細胞を同定するための定量的MOAアッセイの使用とを組み合わせた、工程および技術の独自の組み合わせにあることを主張する。この新規組み合わせは、内因性の遺伝子または染色体遺伝子座の改変を有する生物を作製するための、先行技術を上回る有意な改善を示す。
【実施例】
【0100】
(実施例1:OCR10遺伝子の欠失を有するマウスES細胞の操作)
(a.mOCR10を含む大きなゲノムDNAクローンの選択)
マウスOCR10(mOCR10)遺伝子のコード配列を含んだ大きなゲノムDNAフラグメントを保有する細菌人工染色体(BAC)クローンを、PCRを使用して、整列されたマウスゲノムDNAのBACライブラリー(Incyte Genomics)をスクリーニングすることによって獲得した。このライブラリーをスクリーニングするために使用したプライマーは、mOCR10遺伝子のcDNA配列に由来した。
【0101】
使用した2つのプライマー対は、以下であった:
(a)OCR10.RAA(5’−AGCTACCAGCTGCAGATGCGGGCAG−3’)およびOCR10.PVIrc(5’−CTCCCCAGCCTGGGTCTGAAAGATGACG−3’)、これらは、102bpのDNAを増幅する;ならびに
(b)OCR10.TDY(5’−GACCTCACTTGCTACACTGACTAC−3’)およびOCR10.QETrc(5’−ACTTGTGTAGGCTGCAGAAGGTCTCTTG−3’)、これらは、1500bpのDNAを増幅する。
【0102】
このmOCR10 BACは、完全なmOCR10コード配列を含む、約180kbのゲノムDNAを含んだ。このBACクローンを使用して、LTVECを作製し、続いて、これを使用して、開始コドンがOCR10の開始コドンで正確に置換されたレポーター遺伝子を導入し、同時にこのレポーター遺伝子の後ろに、E.coliおよび哺乳動物細胞の両方において選択するのに有用な選択マーカー遺伝子を挿入しながら、mOCR10のコード領域の一部を欠失させた(図2)。レポーター遺伝子(この非限定的な例において、LacZ、この配列は、当業者に容易に入手可能である)は、E.coliのβ−ガラクトシダーゼ酵素をコードする。LacZ(その開始コドンは、mOCR10の開始コドンと同じ位置である)の挿入位置に起因して、LacZでの類似の置換が先行技術を使用して実行された他の例において観察されたように(「Gene trap strategies in ES cells」、W WurstおよびA.Gossler、Joyner、The Practical Approach Series、293、1999を参照のこと)、lacZの発現は、mOCR10の発現を模倣するはずである。LacZ遺伝子は、その発現パターンをインサイチュで明らかにし得る、単純かつ標準的な酵素アッセイが実行されるのを可能にし、従って、置換された遺伝子または染色体遺伝子座の正常な発現パターンを反映する代理アッセイを提供する。
【0103】
(b.ドナーフラグメントの構築およびLTVECの作製)
mOCR10 LTVECの構築において使用される改変カセットは、lacZ−SV40ポリA−PGKp−EM7−neo−PGKポリAカセットであり、ここでlacZは、上記のようなマーカー遺伝子であり、SV40ポリAは、シミアンウイルス40由来のフラグメントであり(Subramanianら、Prog Nucleic Acid Res Mol Biol、19:157−64、1976;Thimmappayaら、J Biol Chem、253:1613−8、1978;Dharら、Proc Natl Acad Sci U S A、71:371−5、1974;Reddyら、Science,200:494−502、1978)、そしてポリアデニル化部位およびシグナルを含み(Subramanianら、Prog Nucleic Acid Res Mol Biol、19:157−64、1976;Thimmappayaら、J Biol Chem、253:1613−8、1978;Dharら、Proc Natl Acad Sci U S A、71:371−5、1974;Reddyら、Science,200:494−502、1978)、PGKpは、マウスホスホグリセレートキナーゼ(PGK)プロモーターであり(Adraら、Gene、60:65−74、1987)(これは、哺乳動物細胞における薬物耐性遺伝子の発現を駆動するために広く使用されてきた)、EM7は、ネオマイシンホスホトランスフェラーゼ(neo)遺伝子の発現を駆動することによって、細菌中の、完成したLTVEC構築物のポジティブ選択を可能にする利点を有する、強力な細菌プロモーターであり、neoは、原核生物細胞におけるカナマイシン耐性および真核生物細胞におけるG418耐性を付与する選択マーカーであり(Beckら、Gene、19:327−36、1982)、そしてPGKポリAは、PGK遺伝子由来の3’非翻訳領域であり、ポリアデニル化部位およびシグナルを含む(Boerら、Biochem Genet、28:299−308、1990)。
【0104】
mOCR10 LTVECを構築するために、改変カセット中のLacZ遺伝子の上流に結合したmOCR10相同性ボックス1(hb1)、および改変カセット中のneo−PGKポリA配列の下流に結合したmOCR10相同性ボックス2(hb2)からなる第一のドナーフラグメント(図2)を、標準的な組換え遺伝子操作技術を使用して作製した。相同性ボックス1(hb1)は、mOCR10のオープンリーディングフレーム(mOCR10 ORF)の開始メチオニンの直ぐ上流の、211bpの非翻訳配列からなる(図3A〜3D)。相同性ボックス2(hb2)は、終止コドンで終わる、mOCR10 ORFの最後の216bpからなる(図3A〜3D)。
【0105】
続いて、細菌相同組換えを用いて(Zhangら、Nat Genet、20:123−8、1998;Angrandら、Nucleic Acid Res、27:e16、1999;Muyrersら、Nucleic Acids Res、27:1555−7、1999;Narayananら、Gene Ther、6:442−7、1999;Yuら、Proc Natl Acads Sci U S A、97:5978−83、2000)、このドナーフラグメントを使用して、mOCR10コード領域(開始メチオニンから終止コドンまで)を挿入カセットで正確に置換し、mOCR10 LTVECの構築を生じた(図2)。従って、このmOCR10 LTVECにおいて、mOCR10コード配列を、挿入カセットによって置換して、mOCR10遺伝子座における約20kbの欠失を作製し、一方で約130kbの上流相同性(上流相同性アーム)および32kbの下流相同性(下流相同性アーム)を残した。
【0106】
LTVECが、先行技術を使用して作製された標的化ベクターよりも、入手可能なBACライブラリーからより迅速かつ簡便に作製され得る(なぜなら、単一の細菌相同組換え工程のみが必要であり、必要な配列情報は、相同性ボックスを作製するために必要な配列情報のみであるからである)ことに注意することが重要である。対照的に、細菌相同組換えを使用して標的化ベクターを作製するための先行技術のアプローチは、ES細胞への導入の前に、大きな標的化ベクターが「トリミング」される必要がある(Hillら、Genomics、64:111−3、2000)。先行技術のアプローチによって利用されるスクリーニング方法に対応するために、十分短い相同性アームを作製することが必要なので、このトリミングは必要とされる。Hillらの方法の1つの主な欠点は、2つのさらなる相同組換え工程が、単にトリミングのために必要とされることである(1つの工程は、改変された遺伝子座の上流領域をトリミングし、1つの工程は、改変された遺伝子座の下流領域をトリミングする)。これを行うために、実質的により多くの配列情報(トリミング部位にわたる配列情報を含む)が必要とされる。
【0107】
さらに、上記の実施例によって例示される別の明らかな利点は、mOCR10遺伝子にわたる非常に大きな欠失(約20kb)が、単一工程において容易に作製され得ることである。対照的に、先行技術を使用して同じ作業を達成することは、いくつかの工程を必要とし得、そしてCreリコンビナーゼを使用して、真核生物細胞において改変された遺伝子座の導入後にloxP部位が隣接する配列を除去するために、loxP部位を有するコード配列の上流および下流の領域を作製する工程を含み得る。このことは、1工程では達成不能であり得、従って、異なる選択マーカーを使用する2つの標的化ベクターの構築、およびES細胞における2つの連続する標的化事象(1つの事象は、コード配列の上流領域にloxP部位を導入すること、そして他方の事象は、コード配列の下流領域にloxP部位を導入すること)を必要とし得る。相同組換えの達成頻度は、比較的短い相同性アームが隣接する大きな欠失を含む標的化ベクターを使用する場合、低い可能性があるので、大きな欠失の作製は、しばしば、真核生物細胞における先行の標的化技術を使用すると、低い効率で生じることもさらに注意されるべきである。本発明の方法(以下を参照のこと)を使用して得られる高い効率は、真核生物細胞における相同組換えの割合を増大させる、LTVEC中に存在する非常に長い相同性アームに起因する。
【0108】
(c.mOCR10 LTVEC DNAの確認、調製およびES細胞への導入)
挿入カセットおよび相同性配列の連結部の周辺の配列を、DNA配列決定によって確認した。mOCR10 LTVECのサイズを、制限分析、その後のパルスフィールドゲル電気泳動(PFGE)によって確認した(Cantorら、Annu Rev Biophys Chem、17:287−304、1988;SchwartzおよびCantor、Cell、37:67−75、1984)。mOCR10 LTVECの標準的なラージスケールのプラスミド調製を行って、プラスミドDNAを制限酵素NotI(これは、mOCR10 LTVECのベクター骨格を切断する)で消化して、直鎖状DNAを作製した。続いて、線状化されたDNAを、エレクトロポレーションによって、マウスES細胞に導入した(Robertson、Practical Approach Series、254、1987;Joyner、The Practical Approach Series、293、1999;Sambrookら、Sambrook,J.、E.F.FritschおよびT.Maniatis、Molecular Cloning:A Laboratory Manual、第二版、1巻、2巻および3巻、1989)。mOCR10 LTVECで首尾よくトランスフェクトされたES細胞を、標準的な選択方法を使用して、G418含有培地中で選択した(Robertson、Practical Approach Series、254、1987;Joyner、The Practical Approach Series、293、1999)。
【0109】
(d.対立遺伝子の定量的改変(MOA)アッセイを使用する、標的化ES細胞クローンの同定)
2つの内因性mOCR10遺伝子のうち1つが改変カセット配列によって置換されたES細胞を同定するため、個々のES細胞クローン由来のDNAを、標準的なTaqMan(登録商標)方法論を、記載されたように使用する定量的PCRによって分析した(Applied Biosystems、TaqMan(登録商標)Universal PCR Master Mix、カタログ番号P/N4304437;http://www.pebiodocs.com/pebiodocs/04304449.pdfもまた参照のこと)。使用したプライマーおよびTaqMan(登録商標)プローブは、図3A〜3Dに記載の通りである。合計69個の個々のES細胞クローンをスクリーニングし、そして3個のクローンが陽性として、すなわち、内因性mOCR10コード配列の1つが、上記の改変カセットによって置換されたクローンとして、同定された。
【0110】
MOAアプローチのいくつかの利点が明らかである:
(i)改変されている遺伝子座の外側のプローブの使用を必要としない。従って、改変された遺伝子座に隣接する配列を知る必要を排除する。
【0111】
(ii)以前の選り抜きの方法であった従来のサザンブロット方法論(Robertson、Practical Approach Series、254、1987;Joyner、The Practical Approach Series、293、1999)と比較して、実行するために必要な時間が非常に短い。従って、正確に改変された細胞を同定するための時間を、代表的には数日間からほんの数時間に減少させる。
【0112】
これは、過去にスクリーニングが実行された方法における有意な改善であり、この方法を、真核生物細胞における相同組換え事象についてスクリーニングするための、労力集約がかなり少なく、かつコスト効率のよいアプローチにしている。
【0113】
本発明の方法のなお別の利点は、難しい遺伝子座を標的化する能力に起因してもまた、先行技術より優れていることである。先行技術を使用すると、特定の遺伝子座についての首尾よい標的化頻度は、2000の組み込み事象のうち1、おそらくそれより低い頻度でさえあり得ることが示された。本発明の方法を使用して、本出願人は、このような難しい遺伝子座が、長い相同性アーム(すなわち、先行技術によって可能な長さよりも長い)を含むLTVECを使用して、より効率的に標的化され得ることを実証した。上記の非限定的な実施例が実証するように、本出願人は、OCR10遺伝子座(この遺伝子座は、従来技術を使用する標的化が困難であることが以前に証明された)を標的化した。本発明の方法を使用して、本出願人は、mOCR10 LTVEC(160kbより長い相同性アームを含み、20kbの欠失を導入する)が組み込まれた69個のES細胞クローンのうち3個において、首尾よい標的化を得たが、一方、ES細胞標的化の先行技術(Joyner、The Practical Approach Series、293、1999)を使用すると、10〜20kbより短い相同性アームを有し、一方でまた15kb未満の欠失を導入する、プラスミドベースのベクターを使用して、ベクターの600を超える構成要素のうち、標的化事象は同定されなかったことを示した。これらのデータは明らかに、本発明の方法が、先行技術よりも優れていることを実証する。
【0114】
(実施例2:LTVECが標的化ベクターとして使用される場合の、増大した標的化頻度および同質遺伝子DNAを使用する必要性の撤廃)
上記のように、長い相同性アームを使用して得られた増大した標的化頻度は、存在する場合、LTVECを構築する際の、標的化される真核生物細胞のDNAと同質遺伝子の(すなわち、このDNAの配列と同一の)ゲノムDNAを使用することに由来する利点を低減するはずである。この仮説を試験するため、本出願人は、標的化されるべき真核生物細胞と同じマウス副系統に由来するゲノムDNA(おそらく同質遺伝子)を使用して、多数のLTVECを構築し、そして標的化されるべき真核生物細胞と異なるマウス副系統に由来するゲノムDNA(おそらく非同質遺伝子)を使用して、多数の他のLTVECを構築した。LTVECの2つの組は、1〜13%の範囲の類似の標的化頻度を示した(表1)。このことは、LTVECを使用した首尾よい標的化の割合が、遺伝子同質性に依存しないことを示す。
【0115】
(表1)
(BACクローンベクターを用いる遺伝子標的化の概要)
【0116】
【表1】

要約すると、ES細胞における相同組換え事象についてのMOAスクリーニングと組合わせた、LTVECを作製し、これらを標的ベクターとして直接使用するアプローチは、遺伝子改変した遺伝子座を操作するための新規の方法を作成する。この方法は、迅速で、安価であり、そして使用の際に、以前の冗長な、時間のかかる方法よりも有意な改善を示す。従って、以前の方法論により必要とされる時間および費用のほんの少しで、生物体のゲノム中の本質的に任意のおよび全ての遺伝子の、迅速な大スケールのインビボでの機能的ゲノム分析の可能性を開く。
【0117】
(実施例3:キメラ抗体およびヒト抗体を産生するためのLTVECの使用)
(a.導入)
抗体は、2つの鎖(軽鎖および重鎖)から構成され、これらの鎖の各々は、2つのドメイン(可変ドメインおよび定常ドメイン)から構成される。抗体タンパク質の可変領域は、抗体のN末端部分であり、この部分は抗原に結合する。重鎖可変ドメインは、重鎖可変遺伝子の遺伝子座のDNAによりコードされ、この遺伝子座は、可変(V)遺伝子セグメント、多様性(D)遺伝子セグメント、および連結(J)遺伝子セグメントから構成される。軽鎖可変ドメインは、軽鎖可変遺伝子の遺伝子座(κおよびλ)のDNAによりコードされ、この遺伝子座は、可変(V)遺伝子セグメントおよび連結(J)遺伝子セグメントから構成される。
【0118】
B細胞の初期の発達の間の可変領域(VDJ/VJ)遺伝子の再構成は、最初の機構であり、これにより、免疫系は、遭遇し得る莫大な数の抗原を認識し得る抗体を産生する。本質的に、B細胞発達の間のDNA再構成を通じて、莫大なレパートリーの可変(VDJ/VJ)領域の配列をアセンブルし、次いで、これを、定常(C)領域に連結して、完全な重鎖および軽鎖を作製し、この重鎖および軽鎖をアセンブルして抗体を形成する。機能的抗体をアセンブルした後、第2のリンパ器官を生じる体細胞過剰変異は、生物体が抗体のアフィニティーを選択および最適化し得るさらなる多様性を導入する。
【0119】
非ヒト種における種々の抗原に対する抗体の産生は、ヒト治療剤として用いられ得る抗体の大規模産生についての大きな見込みを最初に提供した。しかし、種の違いは、外来の抗体を不活化し、そしてアレルギー反応を引き起こすヒトによる抗体の産生を導く。抗体を「ヒト化」し、それにより、ヒトにおいて外来物として認識される可能性が低い抗体を作製する試みが、引き続いてなされた。初めに、この手順は、マウス由来の抗体の抗原結合部位を、ヒト抗体の定常領域と組合わせ、それにより、ヒトにおいて免疫原性が低い組換え抗体を作製することに関した。開発された第2のアプローチは、ファージディスプレーであり、これにより、ヒトV領域は、ファージディスプレーライブラリーへとクローン化され、そして適切な結合特性を有する領域が、ヒト定常領域に連結され、ヒト抗体を作製する。しかし、この技術は、B細胞において天然で生じる、抗体発達およびアフィニティーの成熟の不足により、制限される。
【0120】
より最近では、内因性遺伝子をマウスでノックアウトし、そして遺伝子を、ヒト対応物と交換して、完全にヒト抗体を産生している。残念なことに、これらの構築物の使用は、B細胞における抗体の発達および最適化における、内因性定常領域の重要性が強調されている。完全なヒト抗体を産生するマウスは、免疫応答が低下されている。このことは、マウスの対応物と比べてアフィニティーが低下されている、完全なヒト構築物を有するトランスジェニックマウスにより産生されるヒト抗体に起因し得る。低下したアフィニティーは、B細胞成熟および生存をもたらし得る。従って、マウスまたは他の生物体においてヒト化抗体を産生するより称賛される方法(この方法において、マウスの内因性の可変領域および定常領域をノックアウトし、そしてヒト対応物と交換する)は、最適な抗体を生じ得ない。
【0121】
キメラ抗体(ヒト可変領域(VDJ/VJ)とマウス定常領域とをB細胞成熟の間に利用し、その後、この抗体は、そのマウス定常領域をそのヒト対応物で置換されるよう操作される)の使用が、示唆されている(1998年6月23日発行の米国特許第5,770,429号)。しかし、このようなキメラを作製するために現在まで存在している唯一の方法論は、トランススイッチであり、この方法論において、そのキメラの形成は、重鎖においてのみ生じるほんの稀な事象である。従来、トランスジェニック動物において、セグメント群をコードする可変遺伝子全体をヒト遺伝子で大規模に置換し、それにより重鎖および軽鎖の両方でキメラを作製する機構は、存在しない。本明細書中に開示される本出願人の方法論を使用すると、キメラ抗体が作製され、その後、このキメラ抗体は、標準的方法論を介して、高親和性ヒト抗体を作製するように変化され得る。
【0122】
(b.簡単な説明)
ヒト可変領域(VDJ/VJ)およびマウス定常領域を含むハイブリッド抗体を産生する、トランスジェニックマウスが、作製される。これは、そのマウス可変領域(VDJ/VJ)遺伝子群をそれらのヒト対応物で直接インサイチュにて置換することによって、達成される。生じるハイブリッド免疫グロブリン座は、そのハイブリッド抗体を産生するように、B細胞発生の間に天然の再配列プロセスを受ける。
【0123】
その後、完全にヒト化した抗体が、そのマウス定常領域を所望のヒト対応物で置換することによって、作製される。このアプローチは、以前の方法(例えば、マウスモノクローナル抗体の「ヒト化」またはHuMabマウスにおける完全にヒト化した抗体の産生)よりもかなり効率良く、治療抗体を生じる。さらに、この方法は、以前の方法が失敗してきた多くの抗原に対する治療抗体の産生に成功する。このマウスは、ヒト(VDJ/VJ)−マウス定常領域である抗体を産生し、このことは、完全にヒト化した抗体を産生する以前に利用可能であったHuMAbマウスを越える、以下の利点を有する。この新規なマウスにより産生される抗体は、そのマウスのB細胞レセプター複合体の他の成分(適切なB細胞分化に必要なシグナル伝達成分(例えば、IgaおよびIgb)を含む)と、より効率良く相互作用するマウスFc領域を保持する。さらに、このマウスFc領域は、マウス細胞、補体分子などの上のFcレセプターとそのマウスFc領域との相互作用において、ヒトFc領域よりも特異的である。これらの相互作用は、強力かつ特異的な免疫応答のため、B細胞の増殖および成熟のため、ならびに抗体の親和性成熟のために、重要である。
【0124】
ヒトV−D−J/V−J領域によるそのマウス座の等価な領域の直接置換が存在するので、適切な転写、組換え、および/またはクラススイッチに必要な配列のすべてが、インタクトなままである。例えば、そのマウス免疫グロブリン重鎖イントロンエンハンサー(Em)は、V−D−J組換えおよびB細胞発生の初期段階の間の重鎖遺伝子発現に重要であることが示されているが[Ronia,D.,Berru,M.およびShulman,M.,J.Mol.Cell Biol 19:7031〜7040(1999)]、その免疫グロブリン重鎖3’エンハンサー領域は、クラススイッチ[Pan,Q.,Petit−Frere,C.,Stavnezer,J.およびHammarstrom,L.,Eur J Immunol 30:1019−1029(2000)]ならびにB細胞分化の後期段階での重鎖遺伝子発現のために重要であるようである[Ong,J.,Stevens,S.Roeder,R.G.,およびEckhardt,L.A.,J Immunol 160:4896−4903(1998)]。その転写制御エレメントのこれらの様々であるが重要な機能を考慮すると、これらの配列をインタクトなまま維持することが、望ましい。
【0125】
正常なB細胞分化過程の間に免疫グロブリン座で生じる必要な組換え事象は、これらの座が不適切な染色体に挿入されるかまたは現在利用可能なマウスにおけるように多コピーで挿入される場合に、異常で非生産的な免疫グロブリン再配列の頻度を増加し得る。生産的免疫グロブリン再配列の減少により、従って、特定のB細胞発生段階での適切なシグナル伝達により、異常な細胞が排除される。初期発生段階でのB細胞数の減少は、全体的最終B細胞集団を減少させ、そのマウスの免疫応答を非常に制限する。(現在利用可能なマウスにおける、変異免疫グロブリン座と対照的に、ヒトトランスジェニック座が、重鎖および軽鎖について異なる染色体位置に組み込まれている)唯一のキメラの重鎖座または軽鎖座のみが存在するので、非生産的再配列または治療上不適切なキメラ抗体を生じ得る座のトランススプライシングもトランス再配列も存在しない(Willers,J.,Kolb,C.およびWeiler,E.Immunobiology 200:150−164(2000);Fujieda,S.,Lin,Y.Q.,Saxon,A.およびZhang,K.,J Immunol 157:3450−3459(1996))。
【0126】
真のマウス染色体免疫グロブリン座中へのヒトV−D−J領域またはV−J領域の置換は、実質的により安定であり、現在利用可能なマウスと比較して、子孫への伝達率が増加し、そしてB細胞遺伝子型のモザイク現象が減少する(Tomizuka,K.,Shinohara,T.,Yoshida,H.,Uejima,H.,Ohguma,A.,Tanaka,S.,Sato,K.,Oshimura,M.およびIshida,I.,Proc Natl Acad Sci(USA)97:722−727(2000))。さらに、インビボでの真のマウス座へのヒト可変領域(VDJ/VJ)の導入は、適切な時期の組換え事象に重要であることが以前に示されている、染色体接近性の適切な全体調節を維持する(Haines,B.B.およびBrodeur,P.H.,Eur J Immunol 28:4228−4235(1998))。
【0127】
ヒト抗体のうちの約1/3は、λ軽鎖を含み、対して、マウス抗体のうちの1/20のみがλ軽鎖を含む。従って、マウスλ軽鎖V−J配列を、ヒト座由来のλ軽鎖V−J配列で置換することは、抗体のレパートリーを増加し、そして真のヒト免疫応答とより密接に一致するために役立ち、従って、治療上有用な抗体を含む可能性を増加させる。
【0128】
ヒト配列を真マウス免疫グロブリン座中に組み込むさらなる利点は、その挿入部位に変異原性破壊を生じ得、そして生存するホモ接合体マウスの単離を阻止し得る、新規な組み込み部位に導入されないことである。これは、繁殖中のマウス群の作製および維持を非常に簡単にする。
【0129】
以下は、上記の利点のすべてを備える抗体を作製するための新規な方法を提供する。当業者は、本明細書中に記載される一般的方法が、等価な結果を生じるように改変され得ることを、認識する。
【0130】
(C.材料および方法)
マウス重鎖可変領域(VDJ)をヒト対応物で正確に置換することは、以下の例における相同組換えと部位特異的組換えの組み合わせを使用して例証され、この例は、2段階プロセスを使用する。当業者は、マウス座を、ヒトホモログ座またはオルソログヒト座で置換することが、1つ以上の工程で達成され得ることを、認識する。従って、本発明は、マウス座の全体または一部を、相同組換えを介して、各組み込みにより置換することを企図する。
【0131】
ヒト重鎖座のVDJ領域全体に広がる大インサート(BAC)クローンを、単離する(図4A)。この領域全体の配列は、以下のGenBankファイル(AB019437、AB019438、AB019439、AB019440、AB019441、X97051、およびX54713)にて入手可能である。この例において、大インサート(BAC)クローンは、相同性アームの供給源として、マウスVDJ領域の末端から単離し(図4B)、この末端を、2段階プロセスにおけるヒトVDJ配列の相同組換えを介して直接組み込みに使用する。
【0132】
第1段階において、LTVEC1(図4D)を、E.coliにおける細菌相同組換えにより構築する。LTVEC1は、順番に、マウスDJ領域から上流にある領域に由来する大マウス相同性アーム(しかし、その絶対的端部は重要ではない);ES細胞において機能する選択マーカー(この例においてはPGK−ネオマイシンR)をコードするカセット;loxP部位;いくつかのV遺伝子セグメントからDJ領域全体にまで広がる、大ヒトインサート;およびマウスJセグメントとすぐ近接するがこのセグメントは含まない領域を含む、マウス相同性アーム;を含む。この下流アームの5’末端およびloxP部位の配置は、置換されるべき領域の3’末端を、この座中に規定する。マウスES細胞を、標準的技術(例えば、エレクトロポレーション)によって、線状化LTVEC1で形質転換する。LTVEC1の直接導入は、内因性可変遺伝子座の改変を生じるので、ネオマイシン耐性コロニーを、MOAアッセイを使用して正確な標的化についてスクリーニングし得る。これらの標的化ES細胞は、ハイブリッド重鎖を含む抗体を産生するマウスを生じ得る。しかし、このマウス可変セグメントの残りを排除するその後の段階を続行することが、好ましい。
【0133】
第2段階において、LTVEC2(図4C)を、E.coli中にて、細菌相同組換えによって構築する。LTVEC2は、順番に、最も遠位のマウスV遺伝子セグメントと近接するがいかなるマウスV遺伝子セグメントも含まない、領域を含む大マウス相同性アーム;多数の遠位ヒトV遺伝子セグメントを含む大インサート;(LTVEC2およびLTVEC1中の野生型loxP部位と反対の方向にある)lox511と呼ばれる変異体loxP部位[Hoess,R.H.,Wierzbicki,A.およびAbremski,K.,Nucleic Acids Res.14:2287−2300(1986)](この部位は、野生型loxP部位と組換わらないが、他のlox511部位と容易に組み換わる);野生型loxP部位;第2選択マーカー(この例においてはPGK−ピューロマイシンR)およびV領域に由来する、マウス相同性アーム(しかし、その絶対的端部は重要ではない);を含む。この上流相同性アームの3’末端およびloxP部位の配置は、置換されるべき領域の5’末端を、この座中に規定する。その後、LTVEC1で正確に標的化されたマウスES細胞を、標準的技術によって、線状化LTVEC2で形質転換し、そしてハイグロマイシン耐性コロニーを、内因性可変遺伝子座における改変について、MOAアッセイを使用して正確な標的化についてスクリーニングする。この形質転換から生じる正確に標的化されたES細胞を、本明細書中で、以後、「二重標的化ES細胞」と呼ぶ。
【0134】
その後の、この二重標的化ES細胞におけるCREリコンビナーゼの一過性発現は、マウスV領域の残りの欠如を生じる。あるいは、この二重標的化ES細胞を、キメラマウスの作製のために、宿主胚盤胞中に注入し得る。生じたキメラマウスを、発生初期にCREリコンビナーゼを発現するマウスと交配させると、子孫F1において、マウスV領域の残りの欠如を生じる。この後者の選択肢は、ハイブリッド重鎖座が、生殖細胞系を通過する可能性を増加させる。なぜなら、これは、より少ない世代の間ES細胞を培養する工程を包含するからである。
【0135】
LTVEC2中にlox511を含めると、ハイブリッド座中へのさらなるヒトV遺伝子セグメントの挿入が可能である。1つのアプローチは、多くのさらなるヒトV遺伝子セグメントを含む大ゲノムDNAクローンをlox511部位およびloxP部位と隣接するように、細菌相同組換えを使用することである。このような改変大ゲノムDNAクローンを、CREリコンビナーゼを一過性発現するプラスミドとともに、二重標的化ES細胞中に同時形質転換すると、カセット交換によって、このさらなるV遺伝子セグメントの導入を生じる(Bethke,B.およびSauer,B.,Nucleic Acids
Res.25:2828−2834(1997))。
【0136】
さらなるV遺伝子セグメントを組み込むための第2のアプローチは、多くのさらなるヒトV遺伝子セグメントを含む大ゲノムDNAクローンを、例えば、LTVEC2中に含まれる同じマウス相同性アームを使用して、マウス座中に独立して標的化することである。この場合、このさらなるヒトV遺伝子セグメントは、lox511部位およびloxP部位と隣接しており、その標的化ES細胞を、マウスを作製するために使用する。二重標的化ES細胞由来のマウスおよびこのさらなるV遺伝子セグメントを含むES細胞由来のマウスを、減数分裂の間にCREリコンビナーゼの発現を指向する第3のマウスと交配する。減数分裂対合の間のその2つの組換え座の近接性によって、他の系(Herault,Y.,Rassoulzadegan,M.,Cuzin.F.およびDuboule,D.,Nature Genetics 20:381−384(1998))において観察されたような、高頻度のCRE誘導性染色体内組換えが生じる。
【0137】
別のアプローチは、上記に概略したものと類似するが、ヒトLTVEC1およびLTVEC2とともにloxP部位およびlox511部位を導入するのではなく、loxP部位およびlox511部位を、マウスLTVEC上に導入し、その後、CREを使用して、隣接するloxP部位およびlox511部位を介するカセット交換によって、そのヒトx座中に特異的に標的化する。下記に概略する方法論は、このLTVEC技術をどのように使用して、任意の非ヒト動物において目的の任意の内因性遺伝子の末端に、隣接部位特異的組換え部位を配置し得るかを、示す。
【0138】
マウスLTVEC1は、J領域の下流かつそのJ領域に近接して、細菌組換えにより挿入されたカセットを含む。このカセットは、loxP部位および細菌選択マーカー/哺乳動物選択マーカー(例えば、ハイグロマイシン耐性)を含む。LTVEC1は、順番に、マウスDJ領域から上流にある(が、この可変遺伝子座内にある)領域由来の大相同性アーム(しかし、その絶対的末端は、重要ではない);ES細胞において機能する選択マーカー(この例においてはPGK−ハイグロマイシンR)をコードするカセット;loxP部位;およびマウスJセグメントとすぐ近接するがこのJセグメントを含まない、相同性アーム;を含む。この下流相同性アームの5’末端およびloxP部位の配置は、置換されるべき領域の3’末端を、この座中に規定する。カセット挿入部位にある内因性可変遺伝子の3’末端の改変は、MOAアッセイにより、ES細胞中での正確に挿入されたLTVEC1の欠失を可能にする。薬物耐性マーカーに、FRT部位が隣接する。FRT部位の導入は、ES細胞中においてFLPによるか、または生殖細胞系能力を有する細胞においてFLPを発現するマウスと生じるマウスとの交配のいずれかによる、残りすべての薬物耐性マーカーの除去を可能にする。
【0139】
LTVEC2を、その座の最も遠位のV領域の上流にカセットを挿入するような細菌組換えによって、構築する。このカセットは、lox511部位および細菌選択マーカー/哺乳動物選択マーカー(例えば、ネオマイシン耐性)を含む。LTVEC2は、順番に、最も遠位にあるマウスV遺伝子セグメントと近接する領域を含むがいかなるマウスV遺伝子セグメントも含まない、大相同性アーム;LTVEC2およびLTVEC1中の野生型loxP部位とは反対の方向にあるlox511部位;野生型loxP部位;第2選択マーカー(この例においてはPGK−ネオマイシンR);およびV領域に由来する(従って、その可変遺伝子座内にある)マウス相同性アーム(しかし、その絶対的端部は重要ではない);を含む。この上流相同性アームの3’末端およびloxP部位の配置は、置換されるべき領域の5’末端を、この座中に規定する。カセット挿入部位にある内因性可変遺伝子の5’末端の改変は、MOAアッセイにより、ES細胞中での正確に挿入されたLTVEC2の欠失を可能にする。これらのLTVECを、一緒にか連続して、標準的技術を使用してES細胞中に導入し、MOAアッセイを使用して、正確な標的化についてスクリーニングする。
【0140】
VDJ/VJ領域の一部または全体を含むヒトBACを、細菌組換えにより、そのヒト配列とlox511部位およびloxP部位が隣接するカセット群を挿入するように改変する。その上流カセットを、マウス改変領域を置換する領域のすぐ上流に挿入する。この上流カセットは、順番に、lox511部位、その後、細菌選択マーカー/哺乳動物選択マーカー(例えば、ピューロマイシン耐性)を含む。その下流カセットを、J領域の下流に近接して挿入する。この下流カセットは、順番に、loxP部位、その後、細菌選択マーカー(例えば、スペクチノマイシン耐性)を含む。
【0141】
ライブラリーから単離される単一BAC上に存在するよりも大きなヒト可変領域を挿入するために、いくつかの方法を使用し得る。これらのうちのいくつかを、以下に記載する。
【0142】
loxP部位およびlox511部位を、細菌組換えによって、重複するBAC上に別個に挿入し得、これらのBACは、ES細胞中に形質転換した場合、互いに組み換わる。この場合、上流BACは、1つのカセットを有し、このカセットは、マウス可変領域を置換する領域のすぐ上流で組み換わり、lox511部位、その後ろに細菌選択マーカー/哺乳動物選択マーカー(例えば、ネオマイシン耐性)を有する。下流BACは、1つのカセットを有し、このカセットは、J領域のすぐ下流でこのJ領域と近接して組み換わり、細菌選択マーカー/哺乳動物選択マーカー(例えば、ピューロマイシン耐性)の後ろに、loxP部位を含む。これらの2つのBACが重複していない場合、重複するホモロジーによりこの上流BACおよび下流BACと関連するさらなるBACを、このスキーム中に組み込む。これらを、細菌組換えによって、細菌選択マーカー/哺乳動物選択マーカー(例えば、ピューロマイシン耐性)を含むように改変する。これらの上流BACおよび下流BACを、ネオマイシン耐性マーカーおよびハイグロマイシン耐性マーカーを保有する、loxPカセットおよびlox511カセットを含むように改変する。
【0143】
ヒトBACを、可変領域と隣接するlox511組換え部位および可変領域と隣接するloxP組換え部位を含むES細胞株中に、CREリコンビナーゼとともに同時形質転換する。重複するBACを使用する場合、それらのBACの間で相同組換えが生じて、より大きなDNAフラグメントが生じ、そして隣接するloxP部位およびlox511部位が、この大フラグメントを、マウス座中に標的化する。細胞を、ピューロマイシン耐性について選択し、そしてそのマウス可変領域の置換についてスクリーニングする。あるいは、このマウス配列を、まず、その2つのloxP部位を介して欠失させ得、その後、そのヒト配列を、残りのlox511部位およびloxP部位を介して導入し得る。
【0144】
LTVEC1が、第3の部位特異的組換え部位(例えば、lox2272(Anal Biochem 2001 Mar15;290(2):260−71))を、細菌耐性遺伝子/哺乳動物耐性遺伝子(例えば、ピューロマイシン耐性)のすぐ下流に含む場合、第4のBACを挿入し得、LTVECを作製し得る。このLTVECは、順番に、ピューロマイシン耐性遺伝子、loxP遺伝子、およびlox2272遺伝子、その後ろに、ヒト配列、を含む。このBACを、マウス免疫グロブリン座中に組み込んだ後、lox511部位/lox2272部位は、第2回目のカセット交換におけるレシピンエトとして役立ち得、この第2回目のカセット交換において、ピューロマイシン耐性遺伝子が、ヒト免疫グロブリン座可変領域のさらなる上流部分、およびlox511部位およびlox2272部位が隣接する異なる細菌耐性遺伝子/哺乳動物耐性遺伝子によって、置換される。
【0145】
このヒト可変領域のより大きなストレッチを導入するための別の方法は、稀な制限エンドヌクレアーゼ切断部位を使用して、複数のBACに由来する配列をインビトロで合わせることである。これは、細菌相同組換えを使用して、最も下流のBACの最後のJのすぐ下流にloxPおよびスペクチノマイシン耐性遺伝子を挿入すること、そして下流のBACのヒト配列の上流に、第2の細菌選択マーカーおよび稀なI−Ceu1部位を挿入することによって、達成する。lox511部位および細菌選択マーカー/哺乳動物選択マーカー(例えば、ピューロマイシン耐性)を、第1のBAC中の配列から上流の、ヒト可変領域の領域を含む第2のBACの上流末端に、挿入する。I−Ceu1部位を、第2のBACの下流末端に挿入する。両方のBACをI−Ceu1およびNot1(これは、両方の改変BACのベクター部分中で独特である)で消化した後、この2つのBACを連結し、そして組換え体を、ピューロマイシン耐性およびスペクチノマイシン耐性について細菌中で選択する。生じるより大きなBACは、順番に、lox511部位、上流ヒト配列、I−Ceu1部位、下流ヒト配列、loxP部位、およびスペクチノマイシン耐性遺伝子を含む。lox511部位とloxP部位との間の領域を、上記のようなカセット交換およびピューロマイシン選択によって、マウス免疫グロブリン座中に挿入する。
【0146】
ヒト可変領域のより大きなストレッチを挿入するための第3の方法は、上記のように複数のBACに由来する配列を合わせることであるが、制限消化/連結の代わりに、細菌相同組換えを使用する。組換え体BACについての同じ選択を、細菌において適用するが、但し、2つのBACのうちの1つを消化し、消化後にその末端を、他の「レシピンエト」BACと相同であるように設計し、そしてそのレシピエントBACを、細菌株中で、細菌相同組換えを許容するように改変する。
【0147】
ヒト可変/マウス定常モノクローナル抗体産生マウスを作製する際の最後の段階は、軽鎖λ座および軽鎖κ座上での等価な可変領域置換を実施し、そして同じマウスにおいてともにホモ接合性であるように3つすべてのハイブリッド座を交配することである。生じるトランスジェニックマウスは、完全に内因性マウス定常領域に作動可能に連結した完全にヒトの重鎖および軽鎖の可変遺伝子座を含むゲノムを有し、そのマウスは、抗原性刺激に応答して、ヒト可変領域およびマウス定常領域を含む抗体を含む血清を産生するようになっている。その後、そのようなマウスを、ヒト抗体の可変領域をコードするDNAの供給源として使用し得る。標準的組換え技術を使用して、この抗体の重鎖および軽鎖の可変領域をコードするDNAを、ヒト重鎖定常領域およびヒト軽鎖定常領域をコードするDNAと、細胞(例えば、CHO細胞)中で作動可能に連結し、この細胞は、活性な抗体を発現可能である。この細胞を、完全なヒト抗体を発現するに適切な条件下で増殖させ、その後、この完全なヒト抗体を回収する。可変領域をコードする配列を、例えば、PCR増幅またはcDNAクローニングによって単離し得る。好ましい実施形態において、ヒト可変領域免疫グロブリン座のいくつかまたはすべてを含むトランスジェニックマウスから作製したハイブリドーマ(KohlerおよびMilstein,Eur.J.Immunol.,6:511−519(1976))を、ヒト可変領域をコードするDNAの供給源として使用する。
【0148】
要約すると、LTVECを作製し、そのLTVECを、ES細胞における相同組換え事象についてMOAスクリーニングと組み合わせて、標的化ベクターとして直接使用するアプローチは、遺伝子改変された座を操作するための新規な方法を生じる。この方法は、迅速かつ安価である。そしてこの方法は、以前に使用された冗漫で時間がかかる方法を超える有意な改善を示す。従って、この方法は、以前の方法論により必要とされた時間および価格の何分の1かで、生物のゲノム中の本質的に任意かつすべての遺伝子の迅速な、大規模の、インビボでの機能的なゲノム分析の可能性を開く。
【0149】
上記の発明は、説明および例としていくらか詳細に記載されているが、添付の特許請求の範囲の趣旨からも範囲からも逸脱することなく、本発明の教示に対して特定の変化および改変がなされ得ることが、当業者にとって容易に明らかである。
【技術分野】
【0001】
本願は、2001年2月16日に出願された米国特許実用出願番号09/784,859に対する優先権を主張する。この出願は、本明細書中に参考として援用される。本願を通じて、種々の刊行物が参照される。これらの刊行物のそれらの全体における開示は、本明細書によって本願に参考として援用される。
【0002】
(発明の分野)
本発明の分野は、真核生物細胞における内因性遺伝子および染色体座を、相同組み換えを通じて標的化し、任意の望ましい様式で改変するための大きなDNAベクターを操作および利用するための方法である。真核生物細胞に対するこれらの大きなDNA標的化ベクター(LTVECと呼ばれる)は、真核生物細胞において相同性標的化を実施することを意図する他のアプローチにより代表的に使用されるフラグメントよりも大きなクローン化されたゲノムDNAのフラグメントから獲得される。本発明の分野はさらに、LTVECが所望の内因性遺伝子または染色体座を正確に標的化および改変した真核生物細胞を検出する迅速かつ便利な方法を提供する。この分野はまた、遺伝子改変を有する生物を作製するためのこれらの細胞の使用、その生物自体、およびそれらの使用方法を包含する。
【0003】
(導入)
LTVECの使用は、現在の方法を上回る実質的な利点を提供する。例えば、これらは標的化ベクターを作製するために現在使用されているDNAフラグメントよりも大きなDNAフラグメントから獲得されるので、LTVECは、現在の技術を用いて作製される標的化ベクターよりも大きなゲノムDNAフラグメントの利用可能なライブラリー(例えば、BACおよびPACライブラリー)からより迅速かつ便利に作製され得る。さらに、より大きな改変およびより大きなゲノム領域にまたがる改変は、現在の技術を使用するよりも便利に作製され得る。さらに、本発明は、「標的化の困難な」座の標的化頻度を増大するための長い相同領域を利用し、また、これらの標的化ベクターにおいて同系DNA(存在する場合)を使用する利益を減らす。
【0004】
従って、本発明は、所定の生物の実質的に全ての内因性遺伝子および染色体座を体系的に改変するための迅速、便利、かつ効率的な方法を提供する。
【背景技術】
【0005】
(発明の背景)
相同な外因性DNAと内因性染色体配列との間の相同組み換えによる遺伝子標的化は、マウスにおいて、欠失、挿入を作製するため、変異を設計するため、遺伝子変異を修正するため、導入遺伝子を導入するため、または他の遺伝子改変を作製するために極めて価値のある方法であることが証明されている。現在の方法は、代表的に、合計10〜20kb未満の内因性DNAに対する相同領域を有する標準的な標的化ベクターを使用してマウス胚性幹(ES)細胞に所望の遺伝子改変を導入し、その後変更されたES細胞をマウス胚に注入してこれらの操作された遺伝子改変をマウス生殖系統に移す工程を包含する(非特許文献1;非特許文献2;非特許文献3;非特許文献4;非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;特許文献1(GenPharm Internationalの名の下で1998年8月4日発行))。これらの現在の方法において、標準的な標的化ベクターが所望の内因性遺伝子または染色体座を正確に標的化および改変している希なES細胞を検出することは、標的化ベクターに含まれる相同標的化配列の外側の配列情報を必要とする。首尾良い標的化のためのアッセイは、標的化ベクターの外側であり相同性アーム(定義を参照のこと)全体にまたがる配列からの標準的なサザンブロッティングまたは長いPCRを伴う(非特許文献10;非特許文献11;非特許文献12;Takara Shuzo Co.,Ltd.に対して発行された特許文献2)。従って、これらの方法を制限する大きさの考慮から、相同性アームの大きさは、合計10〜20kb未満に制限される(非特許文献13)。
【0006】
現在の方法において使用されている相同性アームよりも長い相同性アームを有する標的化ベクターを使用する能力は、極めて価値が高い。例えば、このような標的化ベクターは、大きなゲノムインサートが広範に特徴付けられ、使用前にトリミングされる必要のある現在の技術を用いて作製される標的化ベクターよりも、このような大きなゲノムインサートを含む利用可能なライブラリー(例えば、BACまたはPACライブラリー)からより迅速かつ便利に作製され得る。さらに、より大きな改変およびより大きなゲノム領域にまたがる改変は、現在の技術を使用するよりもより便利にかつより少ない工程で作製され得る。さらに、長い相同性領域の使用は、真核生物細胞における「標的化の困難な」座の標的化頻度を増大し得る。なぜならば、真核生物細胞における相同組み換えの標的化は、その標的化ベクターに含まれる総相同性に関連するようであるからである(非特許文献14)。さらに、長い相同性アームを用いて獲得された増加した標的化頻度は、これらの標的化ベクターにおいて同系DNAを使用することから得られ得る任意の潜在的な利益を減らし得る。
【0007】
非常に大きなゲノムフラグメント(例えば、BACライブラリーにおいてクローン化されたフラグメント)への正確な改変を操作する上での問題は、真核生物の内因性遺伝子または染色体座に対する大きな相同性領域を含むベクターの構築を可能にする、細菌における相同組み換えの使用を通して大部分が解決された(非特許文献15;非特許文献16;非特許文献17;非特許文献18;非特許文献19)。しかし、一旦作製されると、これらのベクターは、一般的に、相同組み換えを通じて内因性遺伝子または染色体座を改変するのに有用ではない。なぜならば相同性アームが10〜20kbよりも長い場合、希な正確な標的化事象を検出することが困難であるからである(非特許文献20)。結果として、BACゲノムフラグメントから細菌相同組み換えを使用して作製されたベクターは、標的化ベクターとして使用する前になおも広範にトリミングされなければならない(非特許文献21)。従って、真核生物細胞における内因性遺伝子および染色体座を改変するために、大きな相同性領域を含む標的化ベクターの使用を可能にする迅速かつ便利な方法論に対する必要性がなおも存在する。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第5,789,215号明細書
【特許文献2】米国特許第5,436,149号明細書
【非特許文献】
【0009】
【非特許文献1】Smithiesら、Nature、317:230−234、1985
【非特許文献2】Thomasら、Cell、51:503−512、1987
【非特許文献3】Kollerら、Proc Natl Acad Sci USA、86;8927−8931、1989
【非特許文献4】Kuhnら、Science、254:707−710、1991
【非特許文献5】Thomasら、Nature、346:847−850、1990
【非特許文献6】Schwarzbergら、Science、246:799−803、1989
【非特許文献7】Doetschmanら、Nature、330:576−578、1987
【非特許文献8】Thomsonら、Cell、5:313−321、1989
【非特許文献9】DeChiaraら、Nature、345:78−80、1990
【非特許文献10】Chengら、Nature、369:684−5、1994
【非特許文献11】FoordおよびRose、PCR Methods Appl、3:S149−61、1994
【非特許文献12】PonceおよびMicol、Nucleic Acids Res、20:623,1992
【非特許文献13】Joyner、The Practical Approach Series、293、1999
【非特許文献14】DengおよびCapecchi、Mol Cell Biol、12:3365−71、1992
【非特許文献15】Zhangら、Nat Genet、20:123−8、1998
【非特許文献16】Yangら、Nat Biotechnol、15:859−65、1997
【非特許文献17】Angrandら、Nucleic Acids Res、27:e16、1999
【非特許文献18】Muyrersら、Nucleic Acid Res、27:1555−7、1999
【非特許文献19】Narayananら、Gene Ther、6:442−7、1999
【非特許文献20】Joyner、The Practical Approach Series、293、1999
【非特許文献21】Hillら、Genomics、64:111−3、2000
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明に従って、本出願人らは、相同組み換えを通じて真核生物細胞における内因性遺伝子および染色体座を改変するために、大きな相同性領域を含む標的化ベクターの使用を可能にする新規の方法を提供する。このような方法は、上記の現在の技術の制限を克服する。さらに、当業者は、本発明の方法が、任意の真核生物(動物(例えば、マウス、ラット、他の齧歯類、またはヒト)および植物(例えば、ダイズ、トウモロコシ、およびコムギ)が挙げられるがこれらに限定されない)の任意のゲノムDNAを用いた使用に容易に適合されることを容易に認識する。
【課題を解決するための手段】
【0011】
(発明の要旨)
本発明に従って、本出願人らは、改変された内因性遺伝子または染色体座を含む真核生物細胞を作製およびスクリーニングするための新規、迅速、効率的、かつ効果的な方法を開発した。この新規の方法は、初めて、以下を組み合わせる:
1.大きなクローン化されたゲノムフラグメント中での所望の遺伝子改変を正確に操作し、それによって真核生物細胞において使用するための大きな標的化ベクター(LTVEC)を作製するための細菌相同組み換え;
2.これらの真核生物細胞における目的の内因性染色体座を改変するための、これらの細胞へのこれらのLTVECの直接的な導入;および
3.標的化配列の外側の配列情報を必要としない親対立遺伝子の対立遺伝子改変(MOA)のためのアッセイ(例えば、定量PCR)を含む、標的化された対立遺伝子が所望するように改変された希な真核生物細胞を決定するための分析。
【0012】
本発明の好ましい実施形態は、以下の工程を包含する、真核生物細胞において内因性遺伝子または染色体座を遺伝子改変する方法である:a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、真核生物において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを真核生物細胞に導入して、この細胞中の内因性遺伝子または染色体座を改変する工程;ならびにd)(c)の真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された真核生物細胞を同定する工程。
【0013】
本発明の別の実施形態は、内因性遺伝子または染色体座に対するこの遺伝子改変が、コード配列、遺伝子セグメント、または調節エレメントの欠失;コード配列、遺伝子セグメント、または調節エレメントの変更;新たなコード配列、遺伝子セグメント、または調節エレメントの挿入;条件付き対立遺伝子の作製;または1つの種由来のコード配列または遺伝子セグメントと、異なる種由来の相同またはオルソロガスな(orthologous)コード配列との置換を含む方法である。
【0014】
本発明の別の実施形態は、コード配列、遺伝子セグメント、または調節エレメントの変更が、置換、付加、または融合を含み、その融合体がエピトープタグまたは二官能性タンパク質を含む方法である。
【0015】
本発明のなお別の実施形態は、定量アッセイが、定量PCR、比較ゲノムハイブリダイゼーション、等温DNA増幅、または固定化されたプローブに対する定量ハイブリダイゼーションを含み、定量PCRが、TaqMan(登録商標)技術または分子ビーコンを用いる定量PCRを含む方法である。
【0016】
本発明の別の好ましい実施形態は、真核生物細胞が、哺乳動物胚性幹細胞であり、特に、この胚性幹細胞がマウス、ラット、または他の齧歯類の胚性幹細胞である方法である。
【0017】
本発明の別の好ましい実施形態は、内因性遺伝子または染色体座が、哺乳動物の遺伝子または染色体座、好ましくは、ヒトの遺伝子または染色体座、あるいはマウス、ラット、または他の齧歯類の遺伝子または染色体座である方法である。
【0018】
本発明のさらなる好ましい実施形態は、LTVECが20kbよりも大きなDNAフラグメント、特に100kbより大きなDNAフラグメントを収容し得る方法である。
【0019】
別の好ましい実施形態は、本発明の方法により産生される遺伝子改変された内因性遺伝子または染色体座である。
【0020】
なお別の好ましい実施形態は、本発明の方法により産生される遺伝子改変された真核生物細胞である。
【0021】
本発明の好ましい実施形態は、本発明の方法により産生される遺伝子改変された内因性遺伝子または染色体座を含む非ヒト生物である。
【0022】
本発明の方法により産生される遺伝子改変された真核生物細胞または胚性幹細胞から産生される非ヒト生物もまた好ましい。
【0023】
好ましい実施形態は、以下の工程を含む方法により産生される、遺伝子改変された内因性遺伝子または染色体座を含む、非ヒト生物である:a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、胚性幹細胞において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを胚性幹細胞に導入して、細胞中の内因性遺伝子または染色体座を改変する工程;d)(c)の胚性幹細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された胚性幹細胞を同定する工程;e)(d)の胚性幹細胞を胚盤胞に導入する工程;ならびにf)(e)の胚盤胞を妊娠のために代理母に導入する工程。
【0024】
本発明のさらなる好ましい実施形態は、以下の工程を含む方法により産生される、遺伝子改変された内因性遺伝子または染色体座を含む、非ヒト生物である:a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、真核生物において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを真核生物細胞に導入して、この細胞中の内因性遺伝子または染色体座を遺伝子改変する工程;d)(c)の真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された真核生物細胞を同定する工程;e)(d)の真核生物細胞から核を取り除く工程;f)(e)の核を卵母細胞に導入する工程;ならびにg)(f)の卵母細胞を妊娠のための代理母に導入する工程。
【0025】
なお別の好ましい実施形態は、以下の工程を含む方法により産生される、遺伝子改変された内因性遺伝子または染色体座を含む、非ヒト生物である:a)目的のDNA配列を含む大きなクローン化されたゲノムフラグメントを得る工程;b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、真核生物において使用するための大きな標的化ベクター(LTVEC)を作製する工程;c)(b)のLTVECを真核生物細胞に導入して、この細胞中で内因性遺伝子または染色体座を遺伝子改変する工程;d)(c)の真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変された真核生物細胞を同定する工程;e)(d)の真核生物細胞と別の真核生物細胞とを融合させる工程;ならびにf)(e)の融合された真核生物細胞を妊娠のための代理母に導入する工程。
【0026】
別の好ましい実施形態において、非ヒト生物はマウス、ラット、または他の齧歯類であり;胚盤胞は、マウス、ラット、または他の齧歯類の胚盤胞であり;卵母細胞は、マウス、ラット、または他の齧歯類の卵母細胞であり;そして代理母は、マウス、ラット、または他の齧歯類である。
【0027】
別の好ましい実施形態は、胚性幹細胞が哺乳動物胚性幹細胞、好ましくは、マウス、ラット、または他の齧歯類の胚性幹細胞である方法である。
【0028】
さらなる好ましい実施形態は、非ヒト生物の産生のための、本発明の遺伝子改変された真核生物細胞の使用、特に、非ヒト生物の産生のための、本発明の遺伝子改変された胚性幹細胞の使用である。
【0029】
本発明の好ましい実施形態は、マウス胚性幹細胞において目的の内因性遺伝子または染色体座を遺伝子改変する方法である。この方法は、a)目的のDNA配列を含む20kbよりも大きなクローン化されたゲノムフラグメントを得る工程(この大きなクローン化されたDNAフラグメントは内因性遺伝子または染色体座に相同である);b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、マウス胚性幹細胞において使用するための大きな標的化ベクターを作製する工程(遺伝子改変は、コード配列、遺伝子セグメント、または調節エレメントの欠失である);c)(b)の大きな標的化ベクターをマウス胚性幹細胞に導入して、この細胞中の内因性遺伝子または染色体座を改変する工程;およびd)(c)のマウス胚性幹細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変されたマウス胚性幹細胞を同定する工程(定量アッセイは定量PCRである)を包含する。この方法により産生される遺伝子改変されたマウス胚性幹細胞;この方法により産生される遺伝子改変された内因性遺伝子または染色体座を含むマウス;および遺伝子改変されたマウス胚性幹細胞から産生されるマウスもまた好ましい。
【0030】
別の好ましい実施形態は、以下の工程を包含する方法により産生される、遺伝子改変された目的の内因性遺伝子または染色体座を含むマウスである:a)目的のDNA配列を含む20kbよりも大きなクローン化されたゲノムフラグメントを得る工程(この大きなクローン化されたDNAフラグメントは内因性遺伝子または染色体座に相同である);b)細菌相同組み換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、マウス胚性幹細胞において使用するための大きな標的化ベクターを作製する工程(遺伝子改変は、コード配列、遺伝子セグメント、または調節エレメントの欠失である);c)(b)の大きな標的化ベクターをマウス胚性幹細胞に導入して、この細胞中の内因性遺伝子または染色体座を改変する工程;d)(c)のマウス胚性幹細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性遺伝子または染色体座が遺伝子改変されたマウス胚性幹細胞を同定する工程(定量アッセイは定量PCRである);e)(d)のマウス胚性幹細胞を胚盤胞に導入する工程;ならびにf)(e)の胚盤胞を妊娠のために代理母に導入する工程。
【0031】
マウスの産生のための、上記遺伝子改変されたマウス胚性幹細胞の使用もまた好ましい。
【0032】
本発明の1つの実施形態は、以下の工程を包含する、非ヒト真核生物細胞において、内因性免疫グロブリン可変領域遺伝子座を相同ヒト遺伝子座またはオルソログなヒト遺伝子座と、全部または一部置換する方法である:
a)ホモログヒト遺伝子座またはオルソログヒト遺伝子座を、全部または一部含む大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)のクローン化されたゲノムフラグメントを遺伝子改変し、真核生物細胞における使用のための大きな標的化ベクター(LTVEC)を作製する工程;
c)(b)のLTVECを真核生物細胞に導入して、内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
d)(c)の真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、内因性免疫グロブリン可変領域遺伝子座が、ホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換された真核生物細胞を同定する工程。
【0033】
別の実施形態は、以下の工程をさらに包含する、非ヒト真核生物細胞において、内因性免疫グロブリン可変領域遺伝子座をホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換する方法である:
e)(a)のフラグメントと異なるホモログヒト遺伝子座またはオルソログヒト遺伝子座の一部を含む、大きなクローン化されたゲノムフラグメントを得る工程;
f)細菌相同組換えを使用して、(e)のクローン化されたゲノムフラグメントを遺伝子改変し、第2のLTVECを作製する工程;
g)(f)の第2のLTVECを工程(d)で同定された真核生物細胞に導入して、内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
h)(g)の真核生物細胞において対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、内因性免疫グロブリン可変領域遺伝子座が、ホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換された真核生物生物を同定する工程。
【0034】
上記方法の別の実施形態は、工程(e)〜(h)が、内因性免疫グロブリン可変領域遺伝子座がホモログヒト遺伝子座またはオルソログヒト遺伝子座と全部置換されるまで繰り返される方法である。
【0035】
この方法の別の実施形態は、免疫グロブリン可変遺伝子座が、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される座である方法である。
【0036】
好ましい実施形態は、定量アッセイが、定量PCR、FISH、比較ゲノムハイブリダイゼーション、等温DNA増幅、または固定されたプローブに対する定量ハイブリダイゼーションを含み、特に、定量PCRが、TaqMan(登録商標)技術または分子ビーコンを用いる定量PCRを含む方法である。
【0037】
なお別の好ましい実施形態は、以下の工程を包含する、マウス胚性幹細胞において、内因性免疫グロブリン可変領域遺伝子座をホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換する方法である:
a)ホモログヒト遺伝子座またはオルソログヒト遺伝子座を、全部または一部含む大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)の大きなクローン化されたゲノムフラグメントを遺伝子改変し、胚性幹細胞における使用のための大きな標的化ベクターを作製する工程;
c)(b)の大きな標的化ベクターをマウス胚性幹細胞に導入して、細胞中の内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
d)(d)のマウス胚性幹細胞において対立遺伝子の改変(MOA)を検出するために定量PCRアッセイを使用して、内因性可変遺伝子座が、ホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換されたマウス胚性幹細胞を同定する工程。
【0038】
別の実施形態において、この方法はさらに、以下の工程を包含する:
e)(a)のフラグメントと異なるホモログヒト遺伝子座またはオルソログヒト遺伝子座の一部を含む、大きなクローン化されたゲノムフラグメントを得る工程;
f)細菌相同組換えを使用して、(e)のクローン化されたゲノムフラグメントを遺伝子改変し、胚性幹細胞における使用のための大きな標的化ベクターを作製する工程;
g)(f)の大きな標的化ベクターを工程(d)で同定されたマウス胚性幹細胞に導入して、内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
h)(g)のマウス胚性幹細胞において対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、内因性免疫グロブリン可変領域遺伝子座が、ホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換されたマウス胚性幹細胞を同定する工程。
【0039】
さらに別の好ましい実施形態は、上記工程(e)〜(h)が、内因性免疫グロブリン可変領域遺伝子座がホモログヒト遺伝子座またはオルソログヒト遺伝子座と全部置換されるまで、繰り返される、方法である。
【0040】
好ましいのはまた、上記免疫グロブリン可変遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される座を含む、方法である。
【0041】
別の好ましい実施形態は、上記の方法によって産生される遺伝子改変された免疫グロブリン可変領域遺伝子座;上記の方法によって産生される遺伝子改変された免疫グロブリン可変領域遺伝子座を含む遺伝子改変された真核生物細胞;上記の方法により遺伝子改変された免疫グロブリン可変領域遺伝子座を含む非ヒト生物;および上記の方法により遺伝子改変された免疫グロブリン可変領域遺伝子座を含むマウス胚幹細胞である。
【0042】
好ましいのはまた、マウス重鎖可変遺伝子座が、ヒト重鎖可変遺伝子座と、全部または一部置換された胚幹細胞;マウスκ軽鎖可変領域遺伝子座が、ヒトκ軽鎖可変領域遺伝子座と、全部または一部置換された胚幹細胞;マウスλ軽鎖可変領域遺伝子座が、ヒトλ軽鎖可変領域遺伝子座と、全部または一部置換された胚幹細胞;および重鎖可変領域遺伝子座および軽鎖可変領域遺伝子座が、それらのヒトホモログまたはオルソログと、全部置換された胚幹細胞である。
【0043】
別の好ましい実施形態は、上記の胚幹細胞から作製されたマウスである。
【0044】
なお別の好ましい実施形態は、上記の遺伝子改変された可変領域遺伝子座によりコードされるヒト可変領域を含む抗体;非ヒト定常領域をさらに含む抗体;およびヒト定常領域をさらに含む抗体である。
【0045】
また好ましいのは、トランスジェニックマウスであって、このマウスは、内因性マウス定常領域座を完全に作動可能に連結されたヒト重鎖可変領域座およびヒト軽鎖可変領域座を完全に含むゲノムを有し、その結果、このマウスが、抗原性刺激に応答してヒト可変領域およびマウス定常領域を含む抗体を含有する血清を産生する、トランスジェニックマウス;トランスジェニックマウスであって、このマウスは、内因性マウス定常領域座に作動可能に連結されたヒト重鎖可変領域座および/またはヒト軽鎖可変領域座を含むゲノムを有し、その結果、このマウスは、抗原性刺激に応答してヒト可変領域およびマウス定常領域を含む抗体を含有する血清を産生する、トランスジェニックマウス;トランスジェニックマウスであって、このマウスは、ホモログヒト可変座またはオルソログヒト可変座で置換された内因性免疫グロブリン可変領域座を含み、このマウスは、以下:
a)ホモログヒト可変領域座全体またはオルソログヒト可変領域座全体を含む、1つ以上の大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)のクローン化されたゲノムフラグメントを遺伝子改変し、マウス胚幹細胞における使用のための大きな標的化ベクターを作製する工程;
c)(b)の大きな標的化ベクターをマウス胚幹細胞に導入して、この細胞において内因性可変領域座全体を置換する工程;ならびに
d)(c)のマウス胚幹細胞において対立遺伝子の改変(MOA)を検出するために定量PCRアッセイを使用して、内因性可変領域座全体が、ホモログヒト可変領域座またはオルソログヒト可変領域座と置換されたマウス胚幹細胞を同定する、工程、
e)(d)のマウス胚幹細胞を胚盤胞に導入する工程;ならびに
f)(e)の胚盤胞を妊娠のために代理母に導入する工程、
を包含する、方法によって作製される、
トランスジェニックマウスである。
【0046】
別の好ましい実施形態は、上記のトランスジェニックマウスであって、免疫グロブリン可変領域遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される1つ以上の座を含む、トランスジェニックマウスである。
【0047】
また好ましいのは、マウス胚幹細胞が、上記の方法により作製されたトランスジェニックマウスに由来する、上記の方法である。
【0048】
本発明のなおさらに別の好ましい実施形態は、ヒト抗体を作製する方法であって、この方法は、以下:
a)上記のマウスを抗原性刺激に曝す工程であって、その結果、このマウスは抗原に対する抗体を産生する、工程;
b)抗体の重鎖可変領域および軽鎖可変領域をコードするDNAを単離する工程;
c)活性な抗体を発現し得る細胞中で、(b)の可変領域をコードするDNAをヒト重鎖定常領域およびヒト軽鎖定常領域をコードするDNAに作動可能に連結する工程;
d)この細胞を、ヒト抗体を発現するような条件下で増殖させる工程;ならびに
e)抗体を回収する工程、
を包含する、方法である。
【0049】
別の好ましい実施形態において、上記の細胞は、CHO細胞である。
【0050】
また好ましいのは、上記の工程(b)のDNAが、上記工程(a)中の抗原性刺激に曝されたマウスの脾臓から作製されたハイブリドーマから単離される方法である。
【0051】
また好ましいのは、DNAがPCRによって単離される、上記方法である。
【0052】
別の好ましい実施形態は、内因性免疫グロブリン可変領域遺伝子座をホモログ遺伝子座(homologous gene locus)またはオルソログ遺伝子座(orthologous gene locus)と、全部または一部置換する方法であって、この方法は、以下:
a)部位特異的組換え部位、その可変遺伝子座内の下流相同性アーム、およびその可変遺伝子座内の上流相同性アームを含む、LTVECを作製する工程であって、
ここで、この下流相同性アームは、この免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、このJセグメントを含まない、工程;
b)部位特異的組換え部位、その可変遺伝子座内の上流相同性アーム、およびその可変遺伝子座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、この上流相同性アームは、この免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECを真核生物細胞に導入する工程;
d)この可変遺伝子座中の対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、この部位特異的組換え部位がこの内因性可変領域遺伝子座に隣接する工程(c)の真核生物細胞を同定する工程;
e)このオルソログ遺伝子座またはこのホモログ遺伝子座の全部または一部に隣接する部位特異的組換え配列を含むベクターを作製する工程;ならびに
f)組換えを介して、この内因性免疫グロブリン可変領域遺伝子座がこのホモログ遺伝子座またはこのオルソログ遺伝子座と、全部または一部置換されるように、工程(d)において同定された真核生物中に(e)のベクターを導入する工程、
を包含する、方法である。
【0053】
また好ましいのは、トランスジェニックマウスであって、このマウスは、ホモログヒト免疫グロブリン可変領域座またはオルソログヒト免疫グロブリン可変領域座で置換された内因性免疫グロブリン可変領域座を含み、このマウスは、以下:
a)部位特異的組換え部位および下流相同性アームを含む、LTVECを作製する工程であって、
ここで、この下流相同性アームは、このマウス免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、このJセグメントを含まない、工程;
b)部位特異的組換え部位および上流相同性アームを含む、LTVECを作製する工程であって、
ここで、この上流相同性アームは、このマウス免疫グロブリン可変遺伝子座領域の最も遠位のマウスV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECをその真核生物細胞に導入する工程;この可変遺伝子座中の対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、この部位特異的組換え部位がこの内因性免疫グロブリン可変領域遺伝子座に隣接する工程(c)の真核生物細胞を同定する工程;
d)このオルソログ遺伝子座またはホモログ遺伝子座の全部または一部に隣接する部位特異的組換え配列を含むベクターを作製する工程;
e)組換えを介して、この内因性免疫グロブリン可変領域遺伝子座がこのホモログ遺伝子座またはオルソログ遺伝子座と、全部または一部置換されるように、工程(d)において同定されたこの真核生物細胞中に(d)のベクターを導入する工程;
f)(e)のマウス胚幹細胞を胚盤胞に導入する工程;ならびに
(f)の胚盤胞を妊娠のために代理母に導入する工程、
を包含する、方法によって作製される、トランスジェニックマウスである。
【0054】
なお別の好ましい実施形態は、真核生物細胞において、下流が部位特異的組換え部位と隣接した内因性遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、この座内の下流相同性アーム、この座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、この下流相同性アームは、この内因性遺伝子座領域の3’末端に隣接する領域を含む、工程;
b)(a)のLTVECをこの真核生物細胞内に導入する工程;ならびに
c)この内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この内因性遺伝子座の下流がこの部位特異的組換え部位と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法である。
【0055】
なお別の好ましい実施形態は、真核生物細胞において、上流が部位特異的組換え部位と隣接した内因性遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、その座内の上流相同性アーム、その座内の下流相同性アームを含むLTVECを作製する工程であって、
ここで、この上流相同性アームは、この内因性遺伝子座領域の5’末端に隣接する領域を含む、工程;
b)(a)のLTVECをこの真核生物に導入する工程;ならびに
c)この内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この内因性遺伝子座の上流がこの部位特異的組換え部位と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法である。
【0056】
また好ましいのは、真核生物細胞において、部位特異的組換え部位が隣接した内因性遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、この座内の下流相同性アーム、この座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、この下流相同性アームは、この内因性遺伝子座領域の3’末端に隣接する領域を含む、工程;
b)部位特異的組換え部位、その座内の上流相同性アーム、その座内の下流相同性アームを含むLTVECを作製する工程であって、
ここで、この上流相同性アームは、この内因性遺伝子座領域の5’末端に隣接する領域を含む、工程;
c)(a)および(b)のLTVECをこの真核生物細胞内に導入する工程;ならびに
d)この内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この部位特異的組換え部位がこの内因性遺伝子座と隣接する(c)の真核生物細胞を同定する工程、
を包含する、方法である。
【0057】
なお別の好ましい実施形態は、真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、その可変遺伝子座内の下流相同性アーム、その可変遺伝子座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、この下流相同性アームは、この免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、このJセグメントを含まない、工程;
b)(a)のLTVECをこの真核生物細胞内に導入する工程;ならびに
c)この可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この部位特異的組換え部位がこの内因性免疫グロブリン可変遺伝子座の下流末端と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法である。
【0058】
また好ましいのは、真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、この座内の上流相同性アーム、この座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、この上流相同性アームは、この免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
b)(a)のLTVECをこの真核生物細胞内に導入する工程;ならびに
c)この可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この部位特異的組換え部位がこの内因性免疫グロブリン可変領域遺伝子座の上流末端と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法である。
【0059】
なお別の実施形態は、真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、この方法は、以下:
a)部位特異的組換え部位、この座内の下流相同性アーム、この座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、この下流相同性アームは、この免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、このJセグメントを含まない、工程;
b)部位特異的組換え部位、この座内の上流相同性アーム、この座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、この上流相同性アームは、この免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECをこの真核生物細胞内に導入する工程;ならびに
d)この可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、この部位特異的組換え部位がこの内因性免疫グロブリン可変領域遺伝子座と隣接する(c)の真核生物細胞を同定する工程、
を包含する、方法である。
本発明の好適な実施形態によれば、例えば以下の方法などが提供される:
(項目1)
非ヒト真核生物細胞において、内因性免疫グロブリン可変領域遺伝子座をホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換する方法であって、該方法は、以下:
a)改変改変該ホモログヒト遺伝子座または該オルソログヒト遺伝子座を全部または一部含む、大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)の該クローン化されたゲノムフラグメントを遺伝子改変し、真核生物細胞における使用のための大きな標的化ベクター(LTVEC)を作製する工程;
c)(b)の該LTVECを該真核生物細胞に導入して、該内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
d)(c)の該真核生物細胞において対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、該内因性免疫グロブリン可変領域遺伝子座が、該ホモログヒト遺伝子座または該オルソログヒト遺伝子座と、全部または一部置換された真核生物細胞を同定する、工程、
を包含する、方法。
(項目2)
項目1に記載の方法であって、該方法はさらに、以下:
e)(a)の前記フラグメントと異なるホモログヒト遺伝子座またはオルソログヒト遺伝子座の一部を含む、大きなクローン化されたゲノムフラグメントを得る工程;
f)細菌相同組換えを使用して、(e)の該クローン化されたゲノムフラグメントを遺伝子改変し、第2のLTVECを作製する工程;
g)(f)の該第2のLTVECを工程(d)で同定された前記真核生物細胞に導入して、前記内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
h)(g)の該真核生物細胞において対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該内因性免疫グロブリン可変領域遺伝子座が、該ホモログヒト遺伝子座または該オルソログヒト遺伝子座と、全部または一部置換された真核生物細胞を同定する、工程、
を包含する、方法。
(項目3)
項目2に記載の方法であって、ここで、工程(e)〜(h)は、前記内因性免疫グロブリン可変領域遺伝子座がホモログヒト遺伝子座またはオルソログヒト遺伝子座と全部置換されるまで、繰り返される、方法。
(項目4)
項目1に記載の方法であって、ここで、前記免疫グロブリン可変遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される座である、方法。
(項目5)
項目4に記載の方法であって、ここで、前記定量アッセイが、定量PCR、FISH、比較ゲノムハイブリダイゼーション、等温DNA増幅、または固定されたプローブに対する定量ハイブリダイゼーションを含む、方法。
(項目6)
前記定量PCRが、TaqMan(登録商標)技術または分子ビーコンを用いる定量PCRを含む、項目5に記載の方法。
(項目7)
マウス胚幹細胞において、内因性免疫グロブリン可変領域遺伝子座をホモログヒト遺伝子座またはオルソログヒト遺伝子座と、全部または一部置換する方法であって、該方法は、以下:
a)改変該ホモログヒト遺伝子座または該オルソログヒト遺伝子座を全部または一部含む、大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)の該大きなクローン化されたゲノムフラグメントを遺伝子改変し、該胚幹細胞における使用のための大きな標的化ベクターを作製する工程;
c)(b)の該大きな標的化ベクターを胚幹細胞に導入して、該細胞中の該内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
d)(c)の該胚幹細胞において対立遺伝子の改変(MOA)を検出するために定量PCRアッセイを使用して、該内因性可変遺伝子座が、該ホモログヒト遺伝子座または該オルソログヒト遺伝子座と、全部または一部置換された胚幹細胞を同定する、工程、
を包含する、方法。
(項目8)
項目7に記載の方法であって、該方法はさらに、以下:
e)(a)の前記フラグメントと異なるホモログヒト遺伝子座またはオルソログヒト遺伝子座の一部を含む、大きなクローン化されたゲノムフラグメントを得る工程;
f)細菌相同組換えを使用して、(e)の該クローン化されたゲノムフラグメントを遺伝子改変し、前記胚幹細胞における使用のための大きな標的化ベクターを作製する工程;
g)(f)の該大きな標的化ベクターを工程(d)で同定された該胚幹細胞に導入して、前記内因性免疫グロブリン可変遺伝子座を、全部または一部置換する工程;ならびに
h)(g)の該マウス胚幹細胞において対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該内因性免疫グロブリン可変領域遺伝子座が、該ホモログヒト遺伝子座または該オルソログヒト遺伝子座と、全部または一部置換された胚幹細胞を同定する、工程、
を包含する、方法。
(項目9)
項目8に記載の方法であって、ここで、工程(e)〜(h)は、前記内因性免疫グロブリン可変領域遺伝子座がホモログヒト遺伝子座またはオルソログヒト遺伝子座と全部置換されるまで、繰り返される、方法。
(項目10)
項目7に記載の方法であって、ここで、前記免疫グロブリン可変遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される座を含む、方法。
(項目11)
内因性免疫グロブリン可変領域遺伝子座をホモログ遺伝子座またはオルソログ遺伝子座と、全部または一部置換する方法であって、該方法は、以下:
a)部位特異的組換え部位、該可変遺伝子座内の下流相同性アーム、該可変遺伝子座内の上流相同性アームを含む、LTVECを作製する工程であって、
ここで、該下流相同性アームは、該免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、該Jセグメントを含まない、工程;
b)部位特異的組換え部位、該可変遺伝子座内の上流相同性アーム、該可変遺伝子座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、該上流相同性アームは、該免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECを真核生物細胞に導入する工程;
d)該可変遺伝子座中の対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、該部位特異的組換え部位が該内因性可変領域遺伝子座に隣接する工程(c)の真核生物細胞を同定する工程;
e)該オルソログ遺伝子座または該ホモログ遺伝子座の全部または一部に隣接する部位特異的組換え配列を含むベクターを作製する工程;ならびに
f)組換えを介して、該内因性免疫グロブリン可変領域遺伝子座が該ホモログ遺伝子座または該オルソログ遺伝子座と、全部または一部置換されるように、工程(d)において同定された真核生物細胞中に(e)のベクターを導入する工程、
を包含する、方法。
(項目12)
前記真核生物細胞が胚幹細胞である、項目11に記載の方法。
(項目13)
項目1、4、7、8、10、11、または12に記載の方法によって産生される、遺伝子改変された免疫グロブリン可変領域遺伝子座。
(項目14)
項目1、4、7、8、11、または12に記載の方法によって産生される遺伝子改変された免疫グロブリン可変領域遺伝子座を含む、遺伝子改変された真核生物細胞。
(項目15)
項目1、4、7、8、10、または11に記載の方法によって産生される遺伝子改変された免疫グロブリン可変領域遺伝子座を含む、非ヒト生物。
(項目16)
項目7、8、10、または12に記載の方法によって作製される遺伝子改変された免疫グロブリン可変領域遺伝子座を含む、マウス胚幹細胞。
(項目17)
前記マウス重鎖可変領域座が、ヒト重鎖可変遺伝子座で全部または一部置換された、項目16に記載の胚幹細胞。
(項目18)
前記マウスκ軽鎖可変領域座が、ヒトκ軽鎖可変領域座で全部または一部置換された、項目16に記載の胚幹細胞。
(項目19)
前記マウスλ軽鎖可変領域座が、ヒトλ軽鎖可変領域座で全部または一部置換された、項目16に記載の胚幹細胞。
(項目20)
前記重鎖可変領域遺伝子座および前記軽鎖可変領域遺伝子座が、そのヒトホモログまたはヒトオルソログで全部置換された、項目16に記載の胚幹細胞。
(項目21)
項目16に記載の胚幹細胞から作製される、マウス。
(項目22)
項目17に記載の胚幹細胞から作製される、マウス。
(項目23)
項目18に記載の胚幹細胞から作製される、マウス。
(項目24)
項目19に記載の胚幹細胞から作製される、マウス。
(項目25)
項目20に記載の胚幹細胞から作製される、マウス。
(項目26)
項目11に記載の遺伝子改変された改変遺伝子座によってコードされるヒト可変領域を含む、抗体。
(項目27)
非ヒト定常領域をさらに含む、項目26に記載の抗体。
(項目28)
ヒト定常領域をさらに含む、項目26に記載の抗体。
(項目29)
トランスジェニックマウスであって、該マウスは、内因性マウス定常領域座に完全に作動可能に連結されたヒト重鎖可変領域座およびヒト軽鎖可変領域座を完全に含むゲノムを有し、その結果、該マウスは、抗原性刺激に応答してヒト可変領域およびマウス定常領域を含む抗体を含有する血清を産生する、トランスジェニックマウス。
(項目30)
トランスジェニックマウスであって、該マウスは、内因性マウス定常領域座に作動可能に連結されたヒト重鎖可変領域座および/またはヒト軽鎖可変領域座を含むゲノムを有し、その結果、該マウスは、抗原性刺激に応答してヒト可変領域およびマウス定常領域を含む抗体を含有する血清を産生する、トランスジェニックマウス。
(項目31)
トランスジェニックマウスであって、該マウスは、ホモログヒト可変領域座またはオルソログヒト可変領域座で置換された内因性免疫グロブリン可変領域座を含み、該マウスは、以下:
a)結合された場合に該ホモログヒト可変領域座または該オルソログヒト可変領域座に広がる、1つ以上の大きなクローン化されたゲノムフラグメントを得る工程;
b)細菌相同組換えを使用して、(a)の該クローン化されたゲノムフラグメントを遺伝子改変し、マウス胚幹細胞における使用のための大きな標的化ベクターを作製する工程;
c)(b)の該大きな標的化ベクターをマウス胚幹細胞に導入して、該細胞において該内因性可変領域座を置換する工程;
d)(c)の該マウス胚幹細胞において対立遺伝子の改変(MOA)を検出するために定量PCRアッセイを使用して、該内因性可変領域座が、該ホモログヒト可変領域座または該オルソログヒト可変領域座と置換されたマウス胚幹細胞を同定する、工程、
e)(d)の該マウス胚幹細胞を胚盤胞に導入する工程;ならびに
f)(e)の該胚盤胞を妊娠のために代理母に導入する工程、
を包含する、方法
によって作製される、トランスジェニックマウス。
(項目32)
トランスジェニックマウスであって、該マウスは、ホモログヒト免疫グロブリン可変領域座またはオルソログヒト免疫グロブリン可変領域座で置換された内因性免疫グロブリン可変領域座を含み、該マウスは、以下:
a)部位特異的組換え部位および下流相同性アームを含む、LTVECを作製する工程であって、
ここで、該下流相同性アームは、該マウス免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、該Jセグメントを含まない、工程;
b)部位特異的組換え部位および上流相同性アームを含む、LTVECを作製する工程であって、
ここで、該上流相同性アームは、該マウス免疫グロブリン可変遺伝子座領域の最も遠位のマウスV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)のLTVECを該真核生物細胞に導入する工程;
d)該可変遺伝子座中の対立遺伝子の改変(MOA)を検出するために定量アッセイを使用して、該部位特異的組換え部位が該内因性免疫グロブリン可変領域遺伝子座に隣接する工程(c)の真核生物細胞を同定する工程;
e)該オルソログ遺伝子座または該ホモログ遺伝子座の全部または一部に隣接する部位特異的組換え配列を含むベクターを作製する工程;
f)組換えを介して、該内因性免疫グロブリン可変領域遺伝子座が該ホモログ遺伝子座または該オルソログ遺伝子座と、全部または一部置換されるように、工程(d)において同定された該真核生物中に(e)の該ベクターを導入する工程;
g)(d)の該マウス胚幹細胞を胚盤胞に導入する工程;ならびに
h)(e)の該胚盤胞を妊娠のために代理母に導入する工程、
を包含する、方法
によって作製される、トランスジェニックマウス。
(項目33)
項目30、31、または32に記載のトランスジェニックマウスであって、ここで、前記免疫グロブリン可変領域遺伝子座は、以下:
a)κ軽鎖の可変遺伝子座;
b)λ軽鎖の可変遺伝子座;および
c)重鎖の可変遺伝子座、
からなる群より選択される1つ以上の座を含む、トランスジェニックマウス。
(項目34)
前記マウス胚幹細胞が、項目31に記載の方法によって作製されたトランスジェニックマウスに由来する、項目7、8、9、10または12に記載の方法。
(項目35)
前記マウス胚幹細胞が、項目32に記載の方法によって作製されたトランスジェニックマウスに由来する、項目7、8、9、10または12に記載の方法。
(項目36)
ヒト抗体を作製する方法であって、該方法は、以下:
a)項目26に記載のマウスを抗原性刺激に曝す工程であって、その結果、該マウスは該抗原に対する抗体を産生する、工程;
b)該抗体の重鎖可変領域および軽鎖可変領域をコードするDNAを単離する工程;
c)活性な抗体を発現し得る細胞中で、(b)の該可変領域をコードするDNAをヒト重鎖定常領域およびヒト軽鎖定常領域をコードするDNAに作動可能に連結する工程;
d)該細胞を、該ヒト抗体を発現するような条件下で増殖させる工程;ならびに
e)抗体を回収する工程、
を包含する、方法。
(項目37)
前記細胞がCHO細胞である、項目36に記載の方法。
(項目38)
工程(b)の前記DNAが、工程(a)中の抗原性刺激に曝された前記マウスの脾臓から作製されたハイブリドーマから単離される、項目36に記載の方法。
(項目39)
前記DNAがPCRによって単離される、項目31に記載の方法。
(項目40)
真核生物細胞において、下流が部位特異的組換え部位と隣接した内因性遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の下流相同性アーム、該座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、該下流相同性アームは、該内因性遺伝子座領域の3’末端に隣接する領域を含む、工程;
b)(a)の該LTVECを該真核生物細胞に導入する工程;ならびに
c)該内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該内因性遺伝子座の下流が該部位特異的組換え部位と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法。
(項目41)
真核生物細胞において、上流が部位特異的組換え部位と隣接した内因性遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の上流相同性アーム、該座内の下流相同性アームを含むLTVECを作製する工程であって、
ここで、該上流相同性アームは、該内因性遺伝子座領域の5’末端に隣接する領域を含む、工程;
b)(a)の該LTVECを該真核生物細胞に導入する工程;ならびに
c)該内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該内因性遺伝子座の上流が該部位特異的組換え部位と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法。
(項目42)
真核生物細胞において、部位特異的組換え部位が隣接した内因性遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の下流相同性アーム、該座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、該下流相同性アームは、該内因性遺伝子座領域の3’末端に隣接する領域を含む、工程;
b)部位特異的組換え部位、該座内の上流相同性アーム、該座内の下流相同性アームを含むLTVECを作製する工程であって、
ここで、該上流相同性アームは、該内因性遺伝子座領域の5’末端に隣接する領域を含む、工程;
c)(a)および(b)の該LTVECを該真核生物細胞に導入する工程;ならびに
d)該内因性遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該部位特異的組換え部位が該内因性遺伝子座と隣接する(c)の真核生物細胞を同定する工程、
を包含する、方法。
(項目43)
真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該可変遺伝子座内の下流相同性アーム、該可変遺伝子座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、該下流相同性アームは、該免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、該Jセグメントを含まない、工程;
b)(a)の該LTVECを該真核生物に導入する工程;ならびに
c)該可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該部位特異的組換え部位が該内因性免疫グロブリン可変遺伝子座の下流末端と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法。
(項目44)
真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の上流相同性アーム、該座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、該上流相同性アームは、該免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
b)(a)の該LTVECを該真核生物に導入する工程;ならびに
c)該可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該部位特異的組換え部位が該内因性免疫グロブリン可変領域遺伝子座の上流末端と隣接する(b)の真核生物細胞を同定する工程、
を包含する、方法。
(項目45)
真核生物細胞において、部位特異的組換え部位が隣接した内因性免疫グロブリン可変遺伝子座を作製する方法であって、該方法は、以下:
a)部位特異的組換え部位、該座内の下流相同性アーム、該座内の上流相同性アームを含むLTVECを作製する工程であって、
ここで、該下流相同性アームは、該免疫グロブリン可変遺伝子座領域のJセグメントにすぐ隣接する領域を含むが、該Jセグメントを含まない、工程;
b)部位特異的組換え部位、該座内の上流相同性アーム、該座内の下流相同性アームを含む、LTVECを作製する工程であって、
ここで、該上流相同性アームは、該免疫グロブリン可変遺伝子座領域の最も遠位のV遺伝子セグメントに隣接する領域を含むが、いかなるV遺伝子セグメントも含まない、工程;
c)(a)および(b)の該LTVECを該真核生物に導入する工程;ならびに
d)該可変遺伝子座における対立遺伝子の改変(MOA)を検出するための定量アッセイを使用して、該部位特異的組換え部位が該内因性免疫グロブリン可変領域遺伝子座と隣接する(c)の真核生物細胞を同定する工程、
を包含する、方法。
(項目46)
1つの部位特異的組換え部位が隣接する、内因性免疫グロブリン可変遺伝子座。
(項目47)
部位特異的組換え部位が隣接する、内因性免疫グロブリン可変遺伝子座。
(項目48)
1つの部位特異的組換え部位が隣接する内因性免疫グロブリン可変遺伝子座を含む、ES細胞。
(項目49)
部位特異的組換え部位が隣接する内因性免疫グロブリン可変遺伝子座を含む、ES細胞。
(項目50)
項目48または49の細胞によって作製される、マウス。
【図面の簡単な説明】
【0060】
【図1】細菌相同組換えを使用する代表的なLTVECの生成の模式図である(hb1=相同性ボックス1;hb2=相同性ボックス2;RE=制限酵素部位)。
【図2】マウス OCR10についてのドナーフラグメントおよびLTVECの模式図である(hb1=相同性ボックス1;lacZ=β−ガラクトシダーゼORF;SV40ポリA=シミアンウイルス40由来のDNAフラグメント(ポリアデニル化部位およびシグナルを含む);PGKp=マウスホスホグリセレートキナーゼ(PGK)。プロモーター、EM7=細菌プロモーター;neo=ネオマイシンホスホトランスフェラーゼ;PGK polyA=PGK遺伝子に由来し、ポリアデニル化部位およびシグナルを含む3’非翻訳領域;hb2=相同性ボックス2)。
【図3A】図3A〜3Dは、マウスOCR10 cDNA、相同性ボックス1(hb1)、相同性ボックス2(hb2)の配列であり、TaqMan(登録商標)プローブおよびプライマーを定量PCRアッセイで使用して、mOCR10 LTVECを使用して標的化されたES細胞中の対立遺伝子の改変(MOA)を検出した。 hb1:塩基対1〜211 hb2:塩基対1586〜1801 TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン3に由来している: TaqMan(登録商標)プローブ:ヌクレオチド413〜439(上の鎖) プライマー ex3−5’:ヌクレオチド390〜410(上の鎖) プライマー ex3−3’:ヌクレオチド445〜461(下の鎖) TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン4に由来している: TaqMan(登録商標)プローブ:ヌクレオチド608〜639(上の鎖) プライマー ex4−5’:ヌクレオチド586〜605(上の鎖) プライマー ex4−3’:ヌクレオチド642〜662(下の鎖)
【図3B】図3A〜3Dは、マウスOCR10 cDNA、相同性ボックス1(hb1)、相同性ボックス2(hb2)の配列であり、TaqMan(登録商標)プローブおよびプライマーを定量PCRアッセイで使用して、mOCR10 LTVECを使用して標的化されたES細胞中の対立遺伝子の改変(MOA)を検出した。 hb1:塩基対1〜211 hb2:塩基対1586〜1801 TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン3に由来している: TaqMan(登録商標)プローブ:ヌクレオチド413〜439(上の鎖) プライマー ex3−5’:ヌクレオチド390〜410(上の鎖) プライマー ex3−3’:ヌクレオチド445〜461(下の鎖) TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン4に由来している: TaqMan(登録商標)プローブ:ヌクレオチド608〜639(上の鎖) プライマー ex4−5’:ヌクレオチド586〜605(上の鎖) プライマー ex4−3’:ヌクレオチド642〜662(下の鎖)
【図3C】図3A〜3Dは、マウスOCR10 cDNA、相同性ボックス1(hb1)、相同性ボックス2(hb2)の配列であり、TaqMan(登録商標)プローブおよびプライマーを定量PCRアッセイで使用して、mOCR10 LTVECを使用して標的化されたES細胞中の対立遺伝子の改変(MOA)を検出した。 hb1:塩基対1〜211 hb2:塩基対1586〜1801 TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン3に由来している: TaqMan(登録商標)プローブ:ヌクレオチド413〜439(上の鎖) プライマー ex3−5’:ヌクレオチド390〜410(上の鎖) プライマー ex3−3’:ヌクレオチド445〜461(下の鎖) TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン4に由来している: TaqMan(登録商標)プローブ:ヌクレオチド608〜639(上の鎖) プライマー ex4−5’:ヌクレオチド586〜605(上の鎖) プライマー ex4−3’:ヌクレオチド642〜662(下の鎖)
【図3D】図3A〜3Dは、マウスOCR10 cDNA、相同性ボックス1(hb1)、相同性ボックス2(hb2)の配列であり、TaqMan(登録商標)プローブおよびプライマーを定量PCRアッセイで使用して、mOCR10 LTVECを使用して標的化されたES細胞中の対立遺伝子の改変(MOA)を検出した。 hb1:塩基対1〜211 hb2:塩基対1586〜1801 TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン3に由来している: TaqMan(登録商標)プローブ:ヌクレオチド413〜439(上の鎖) プライマー ex3−5’:ヌクレオチド390〜410(上の鎖) プライマー ex3−3’:ヌクレオチド445〜461(下の鎖) TaqMan(登録商標)プローブおよび対応するPCRプライマーセットは、mOCR10エキソン4に由来している: TaqMan(登録商標)プローブ:ヌクレオチド608〜639(上の鎖) プライマー ex4−5’:ヌクレオチド586〜605(上の鎖) プライマー ex4−3’:ヌクレオチド642〜662(下の鎖)
【図4】図4A〜4Dは、マウスVDJ領域をヒトVDJ領域で置き換えて構築した2つのLTVECの模式図である。 図4A:ヒト重鎖遺伝子座のVDJ領域全体にまたがる大きなインサート(BAC)クローンを単離した。 図4B:この例において、相同性アームの供給源として、大きなインサート(BAC)クローンをマウスVDJ領域の末端から単離する。この相同性アームを使用して、2工程プロセスにおいてヒトVDJ配列の相同組換えを介して組み込んだ。 図4C〜4D:第1の工程において、LTVEC1(図4D)を、E.coliにおける細菌相同組換えにより構築する。LTVEC1は、順番に以下を含む:マウスDJ領域から上流の領域に由来する大きなマウス相同性アーム(しかし、その絶対エンドポイントは、重要ではない);ES細胞において機能的な選択マーカーをコードするカセット(この例では、PGK−ネオマイシンR);loxP部位;いくつかのV遺伝子セグメントからDJ領域全体にまたがる大きなヒトインサート;およびマウスJセグメントにすぐ隣接するが、このJセグメントを含まない領域を含むマウス相同性アーム。第2の工程において、LTVEC2(図4C)を、E.coliにおける細菌相同組換えにより構築する。LTVEC2は、順番に以下を含む:最も遠位のマウスV遺伝子セグメントに隣接するが、いかなるマウスV遺伝子セグメントも含まない領域を含む大きなマウス相同性アーム;多数の遠位ヒトV遺伝子セグメントを含む大きなインサート;変異loxP部位(LTVEC2およびLTVEC1における野生型loxP部位とは反対の配向にてloxP511といわれる)(この部位は、野生型loxPと組換わらないが、他のloxP511とは容易に組換わる);野生型loxP部位;第2の選択マーカー(この例では、PGK−ハイグロマイシンR);およびV領域に由来するマウス相同性アーム(しかし、その絶対エンドポイントは重要ではない)。
【発明を実施するための形態】
【0061】
(定義)
「標的化ベクター」は、所望の遺伝子改変に隣接する内因性染色体核酸配列に「相同(ホモログ)(homologous)」な配列を含むDNA構築物である。隣接相同性配列(「相同性アーム」と称する)は、標的化ベクターを、相同性アームと対応する内因性配列との間に存在する相同性によってゲノム内の特定の染色体位置に方向付け、「相同組換え」と称されるプロセスにより、所望の遺伝子改変を導入する。
【0062】
「相同(ホモログ)(homologous)」とは、2以上の核酸配列が、互いにハイブリダイズし得るかまたは分子間交換を受け得るに十分、同一または類似のいずれかである、2以上の核酸配列を意味する。
【0063】
「遺伝子標的化」は、標的化ベクターを使用して、相同組換えを介した内因性配列への挿入、内因性配列の欠失または内因性配列の置換による、内因性染色体座の改変である。
【0064】
「遺伝子ノックアウト」は、染色体座においてコードされた遺伝情報の破壊から生じた遺伝子改変である。
【0065】
「遺伝子ノックイン」は、染色体座においてコードされた遺伝情報を、異なるDNA配列と置換することから生じる遺伝子改変である。
【0066】
「ノックアウト生物」は、その生物の細胞の大部分が、遺伝子ノックアウトを有する生物である。
【0067】
「ノックイン生物」は、その生物の細胞の大部分が、遺伝子ノックインを有する生物である。
【0068】
「マーカー」または「選択マーカー」は、集団中の処理細胞の大部分から、選択マーカーを発現する稀なトランスフェクト細胞を単離することを可能にする選択マーカーである。このようなマーカー遺伝子としては、ネオマイシンホスホトランスフェラーゼおよびハイグロマイシンBホスホトランスフェラーゼ、または蛍光タンパク質(例えば、GFP)が挙げられるが、これらに限定されない。
【0069】
「ES細胞」は、胚幹細胞である。この細胞は、通常、胚盤胞期の胚の内部細胞塊に由来する。
【0070】
「ES細胞クローン」は、DNAの導入、引き続く選択の後の、ES細胞集団の単一細胞由来の細胞の亜集団である。
【0071】
「隣接DNA」は、参照の特定の点と同一線上でありそして隣接する、DNAのセグメントである。
【0072】
「LTVEC」は、真核生物細胞において相同標的化を行うことを目的とされた他のアプローチによって典型的に使用されるものよりも大きな、クローニングされたゲノムDNAのフラグメント由来の、真核生物細胞についての大きな標的ベクターである。
【0073】
「非ヒト生物」は、通常ヒトであることが公的に受容されない生物である。
【0074】
「対立遺伝子の改変」(MOA)は、遺伝子、またはゲノム中の染色体遺伝子座の対立遺伝子の正確なDNA配列の改変をいう。この対立遺伝子改変(MOA)としては、以下が挙げられるがこれらに限定されない:単一ヌクレオチド程度の小さなヌクレオチドのわずかな欠失、置換または挿入、または多くのキロベースにわたる目的の遺伝子または染色体遺伝子座の欠失、ならびにこれらの2つの極値の間で可能な改変のいずれかおよび全て。
【0075】
「オルソログ(orthologous)」配列は、別の種中のその配列の機能的等価物である種由来の配列をいう。
【0076】
とりわけ示される記載または実施例は、本発明に例証されることで提供される。当業者は、これらの実施例が本発明の例示の目的のみに提供され、限定の目的には含まれないことを、理解する。
【0077】
(発明の詳細な説明)
本出願人らは、改変された内因性遺伝子または染色体遺伝子座を含む真核生物細胞を作製およびスクリーニングするための、新規で、迅速で、能率的で、そして効率的な方法を開発した。これらの細胞において、改変は、遺伝子ノックアウト、ノックイン、点変異または大きなゲノム挿入もしくは欠失、あるいは他の改変であり得る。非限定的な例示の目的で、これらの細胞は、変更、欠失および/または挿入された遺伝子の機能を決定する目的のためのノックアウト生物またはノックイン生物(特に、ノックアウトマウスまたはノックインマウス)に有用である胚性幹細胞であり得る。
【0078】
本明細書中に記載される新規な方法は、最初に以下を組合わせる:
1.大きなクローニングされたゲノムDNAフラグメント内に所望の遺伝子改変を正確に操作し、これにより真核生物細胞(LTVEC)における使用のための大きな標的化ベクターを作製するための細菌性相同組換え;
2.これらのLTVEC細胞において対応する目的の内因性遺伝子または染色体遺伝子座を改変するための、真核生物細胞へのこれらの細胞の直接導入;および
3.標的化対立遺伝子が所望されるように改変されている、まれな真核生物細胞を決定するための分析(親対立遺伝子の対立遺伝子改変(MOA)のための定量的アッセイを含む)。
【0079】
真核生物細胞における相同組換えを首尾よく検出するための以前の方法が、LTVEC中に存在する長い相同性アームのために、本出願人らの発明のLTVECと組合わせて利用され得ないことは、強調されるべきである。真核生物細胞中の内因性遺伝子または染色体遺伝子座を、相同組換えを介して故意に改変するためにLTVECを利用することによって、まれな真核生物細胞を決定するためのアッセイの新規適用によっておそらくなされる。ここで、標的化された対立遺伝子は所望されるように改変されており、このようなアッセイとしては、親対立遺伝子の対立遺伝子改変(MOA)のための定量的アッセイ(例えば、定量的PCRまたはMOAについての他の適切な定量的アッセイを使用することによる)が挙げられる。
【0080】
現在の方法において使用されるものより長い相同性アームを有する標的化ベクターを利用する能力は、以下の理由について極めて有益である:
1.標的化ベクターは、大きなゲノム挿入物を含む利用可能なライブラリー(例えば、BACライブラリーまたはPACライブラリー)から、以前の技術を使用して作製される標的化ベクターより迅速かつ簡便に作製される。ここで、ゲノム挿入物は、広範に特徴付けられており、使用前に「整えられる(trim)」(以下に詳細に説明される)。さらに、最小配列情報が、目的の遺伝子座について知られていることが必要である。すなわち、相同性ボックス(以下に詳細に記載される)を作成するため、およびMOAについての定量的アッセイ(以下に詳細に記載される)に使用され得るプローブを作成するために必要とされる約80〜100ヌクレオチドが知られていることのみが必要である。
2.より大きな改変、ならびにより長いゲノム領域にわたる改変が、以前の技術を使用するより簡便に、そしてよりわずかな工程において作製される。例えば、本発明の方法は、そのサイズ制限のために伝統的プラスミドベースの標的化ベクターによっては収容され得ない大きな遺伝子座の正確な改変を可能にする。これはまた、1工程における複数の点での任意の所定の遺伝子座の改変(例えば、複数のエキソン遺伝子の異なるエキソンでの特定の変異の導入)を可能にして、複数の標的化ベクターを操作する必要性、およびES細胞での相同組換えのための複数回の標的化およびスクリーニングを行う必要性を緩和する。
3.相同性の長い領域(長い相同性アーム)の使用は、真核生物細胞における相同組換えの標的化が標的化ベクター内に含まれる総相同性に関連するようであるという以前の知見と一致して、真核生物細胞における「標的化しにくい(hard to target)」遺伝子座の標的化頻度を増加させる。
4.長い相同性アームを使用して得られた増加した標的化頻度は、これらの標的化ベクターにおいて同系のDNAを使用することから、たとえあったとしても、その利点を明らかに減少させる。
5.真核生物細胞を相同組換えについてスクリーニングするための定量的MOAアッセイの適用は、標的化ベクターとしてのLTVECの使用を可能にするだけでなく(利点は上記に概説した)、正確に改変された真核生物細胞を同定するための時間を代表的な7日から数時間までに減少させる。さらに、定量的MOAの適用は、改変されている、内因性遺伝子または染色体遺伝子座の外側に位置されるプローブの使用を必要としない。よって、改変された遺伝子または遺伝子座に隣接する配列を知る必要性を除去する。これは、過去に行われたスクリーニングを、真核生物細胞における相同組換え事象についてスクリーニングするためにほとんど労力を要さず、そしてかなり高い費用効果的なアプローチとなさしめる、有意な改善である。
【0081】
(方法)
本明細書中に記載されるDNAベクターを構築するために使用される技術の多くは、当業者に周知である標準的な分子生物学的技術である(例えば、Sambrook,J.,E.F.Fritsch And T.Maniatis.Molecular Cloning:A Laboratory Manual,Second Edition,Vols 1,2,and 3,1989;Current Protocols in Molecular Biology,Eds.Ausubel et al.,Greene Publ.Assoc.,Wiley Interscience,NYを参照のこと)。全てのDNA配列決定を、ABI 373A DNA配列決定機およびTaq Dideoxy Terminator Cycle Sequencing Kit(Applied Biosystems,Inc.,Foster City,CA)を使用する標準的な技術によって行った。
【0082】
(工程1.目的の遺伝子または染色体遺伝子座を含有する大きなゲノムDNAクローンを得る)
目的の遺伝子または遺伝子座を、特定の基準(例えば、詳細な構造的データまたは機能的データ)に基づいて選択し得るか、または種々のゲノム配列決定プロジェクトの努力を介して潜在的な遺伝子フラグメントが予想されるようになるような詳細な情報の非存在下で選択し得る。重要なことは、LTVECを産生させるために、目的の遺伝子の完全な配列および遺伝子構造を知って本発明の方法に適用する必要がないことを、注意するべきである。実際に、目的のゲノムクローンを得、そしてLTVECを作製する際に使用される相同性ボックス(以下に詳細に記載される)を作製し、そして定量性MOAアッセイにおける使用のためのプローブを作製するために必要とされる唯一の情報は、約80〜100ヌクレオチドである
一旦、目的の遺伝子または遺伝子座が選択されると、この遺伝子または遺伝子座を含有する大きなゲノムクローンが得られる。このクローンを、適切なDNAライブラリー(例えば、BAC、PAC、YACまたはコスミド)を標準的なハイブリダイゼーション技術またはPCR技術、あるいは当業者が精通する任意の他の方法によるスクリーニングが挙げられるがこれに限定されないいくつかの方法のいずれか1つにおいて取得し得る。
【0083】
(工程2.改変カセットへの相同性ボックス1および2の付加ならびにLTVECの作製)
相同性ボックスは、クローニングされた大きなゲノムフラグメントからLTVECを作成するために使用される細菌性相同組換えの部位を印す(図1)。相同性ボックスは、一般には、二本鎖であり、そして少なくとも40ヌクレオチド長であるDNAの短いセグメントであり、このセグメントは、「改変されるべき領域」に隣接する、このクローニングされた大きなゲノムフラグメント内の領域に相同である。相同性ボックスは、改変されたカセットに付加され、その結果、細菌中での相同組換えに続いて、この改変カセットは、改変されるべき領域に置換する(図1)。細菌性相同組換えを使用する標的化ベクターの作製技術は、種々の系において行われ得る(Yang et al.,Nat Biotechnol,15:859−65,1997;Muyrers et al.,Nucleic Acids Res,27:1555−7,1999;Angrand et
al.,Nucleic Acids Res,27:e16,1999;Narayanan et al.,Gene Ther,6:442−7,1999;Yu,et
al.,Proc Natl Acad Sci USA,97:5978−83,2000)。現在使用に好ましい技術の1つの例は、ETクローニング(Zhang et
al.,Nat Genet,20:123−8,1998;Narayanan et al.,Gene Ther,6:442−7,1999)およびこの技術の改変(Yu,et al.,Proc Natl Acad Sci USA,97:5978−83,2000)である。ETは、相同組換え反応を実施するrecE(Hall and Kolodner,Proc Natl Acad Sci USA,91:3205−9,1994)およびrecTタンパク質(Kusano et al.,Gene,138:17−25,1994)をいう。RecEは、線状二本鎖DNA(実質的にはとりわけ記載されるドナーDNAフラグメント)の一方の鎖を5’から3’に整え、よって3’一本鎖オーバーハングを有する線状二本鎖フラグメントの後ろに残すエクソヌクレアーゼである。この一本鎖オーバーハングは、一本鎖DNA(ssDNA)結合活性を有するrecTタンパク質によってコートされる(Kovall and Matthews,Science,277:1824−7,1997)。ETクローニングは、recEおよびrecTのE.coli遺伝子産物(Hall and Kolodner,Proc Natl Acad Sci USA,91:3205−9,1994;Clark et al.,Cold Spring Harb Symp Quant Biol,49:453−62,1984;Noirot and Kolodner,J
Biol Chem,273:12274−80,1998;Thresher et
al.,J Mol Biol,254:364−71,1995;Kolodner
et al.,Mol Microbiol,11:23−30,1994;Hall
et al.,J Bacteriol,175:277−87,1993)、およびバクテリオファージ(λ)タンパク質λgam(Murphy,J Bacteriol,173:5808−21,1991; Poteete et al.,J Bacteriol,170:2012−21,1988)を一過性に発現するE.coliを使用して行われる。λgamタンパク質は、recBCエクソヌクレアーゼ系による分解からドナーDNAフラグメントを保護するために必要とされ(Myers and Stahl,Annu Rev Genet,28:49−70,1994)、そしてrecBC+宿主(例えば、しばしば使用されるE.coli株DH10b)における効率的なETクローニングのために必要とされる。
【0084】
細菌性相同組換えを使用して改変および置換されるべき領域は、0ヌクレオチド(もともとの遺伝子座に挿入を作製する)〜数十キロベース(もともとの遺伝子座の欠失および/または置換を作製する)の長さにわたり得る。改変カセットに基づくと、改変は以下を生じ得る:
(a)コード配列、遺伝子セグメント、または調節エレメントの欠失;
(b)コード配列、遺伝子セグメント、または調節エレメントの変更(置換、付加、および融合(例えば、エピトープタグまたは二官能性タンパク質(例えば、GFPとの融合)の作製を含む);
(c)新たなコード領域、遺伝子セグメント、または調節エレメントの挿入(例えば、選択マーカー遺伝子またはレポーター遺伝子についてのもの)あるいは内因性転写調節下での新たな遺伝子の配置;
(d)条件付き対立遺伝子の作製(例えば、Creリコンビナーゼによって切除されるべき領域に隣接するloxP部位(Abremski and Hoess,J Biol
Chem,259:1509−14,1984)、またはFlpリコンビナーゼによって切除されるべき領域に隣接するFRT部位(Andrews et al.,Cell,40:795−803,1985;Meyer−Leon et al.,Cold Spring Harb Symp Quant Biol,49:797−804,1984;Cox,Proc Natl Acad Sci USA,80:4223−7,1983)の導入による);あるいは
(e)異なる種由来のオルソログコード配列での、ある種からのコード配列または遺伝子セグメントの置換(例えば、マウス遺伝子座をオルソログヒト遺伝子座と置換して、ここでこれらの特定の遺伝子座が「ヒト化」される)。
【0085】
これらの改変のいずれかまたは全てが、LTVECに組み込まれ得る。特定の、非限定的な例示(ここで、内因性コード配列が完全に欠失され、そして、同時にレポーター遺伝子および選択マーカーの両方と置換されている)が、以下の実施例1に提供され、以前の技術と比較して本発明の方法の利点である。
【0086】
(工程3(任意).各LTVECが正確に操作されていることの検証)
各LTVECが正確に操作されていることの検証は、以下による:
a.ドナーフラグメントを、目的の遺伝子または染色体遺伝子座へ導入することによって作製された新規な連結を検証するための診断的PCR。よって、この得られたPCRフラグメントを配列決定し、ドナーフラグメントを、目的の遺伝子または染色体遺伝子座へ導入することによって作製された新規な連結をさらに検証し得る。
b.細菌性相同組換えプロセスの間に所望の改変のみがLTVECに導入されたことを確認するための、診断的制限酵素消化。
c.ドナーフラグメントを、目的の遺伝子または染色体遺伝子座へ導入することによって作製された新規な連結を検証するための、LTVEC(特に、改変の部位にわたる領域)の直接配列決定。
【0087】
(工程4.真核生物細胞への導入のための、LTVEC DNAの精製、調製および線状化)
(a.LTVEC DNAの調製)
選択したLTVECのminiprep DNA(Sambrook,J.,E.F.Fritsch And T.Maniatis.Molecular Cloning:A Laboratory Manual,Second Edition,Vols
1,2,and3,1989;Tillett and Neilan,Biotechniques,24:568−70,572,1998;http://www.qiagen.com/literature/handbooks/plkmini/plm_399.pdf)を調製し、そしてこのminiprep LTVEC DNAを、エレクトロポレーション(Sambrook,J.,E.F.Fritsch and T.Maniatis,Molecular Cloning:A Laboratory Manual,Second Edition,Vols 1,2,and 3,1989)を使用してE.coliに再度形質転換した。この工程は、細菌性相同組換え工程に利用される組み換えタンパク質をコードするプラスミドを除くために必要である(Zhang et al.,Nat Genet,20:123−8,1998;Narayanan et al.,Gene Ther,6:442−7,1999)。(a)なぜならこれは、コピー数の多いプラスミドであって、大規模LTVEC prepにおいて得られる収量を減少し得、(b)組換えタンパク質の発現を誘導する可能性を排除するためであり;そして(c)なぜなら、これは、LTVECの物理的マッピングをあいまいにし得るからである。LTVECを真核生物細胞に導入する前に、より多量のLTVEC DNAを、標準的な方法論(http://www.qiagen.com/literature/handbooks/plk/plklow.pdf;Sambrook,J.,E.F.Fritsch And T.Maniatis.Molecular Cloning:A Laboratory Manual,Second Edition,Vols 1,2,and 3,1989;Tillett and Neilan,Biotechniques 24:568−70,572,1998)によって調製した。しかし、この工程は、組換えプロファージを利用する細菌性相同組換え法が使用される(すなわち、組換えタンパク質をコードする遺伝子が細菌染色体中に組み込まれる)場合、回避され得る(Yu,et al.,Proc Natl Acad Sci USA,97:5978−83,2000)。
【0088】
(b.LTVEC DNAの線状化)
真核生物細胞への導入のためにLTVECを調製するために、LTVECを、好ましくは、長い相同性アームに隣接する、改変された内因性遺伝子または染色体遺伝子座DNAを残す様式で線状化する。これは、めったに消化しない適切な制限酵素のいずれかを用いて、LTVEC、好ましくは、ベクターバックボーン中のを線状化することによって達成され得る。適切な制限酵素の例としては、NotI、PacI、SfiI、SrfI、SwaI、FseIなどが挙げられる。制限酵素の選択は、実験的に(すなわち、いくつかの異なる候補レアカッター(rare cutter)を試験することによって)、またはLTVECの配列が公知である場合は、配列を分析し、そして分析に基づいて適切な制限酵素を選択することによって)決定され得る。LTVECがまれな部位(例えば、CosN部位)を含むベクターバックボーンを有する状況では、次いで、このような部位を認識する酵素(例えば、λターミナーゼ(Shizuya et al.,Proc Natl Acad Sci USA,89:8794−7,1992;Becker and Gold,Proc Natl Acad Sci USA,75:4199−203,1978;Rackwitz et al.,Gene,40:259−66,1985))で切断し得る。
【0089】
(工程5.真核生物細胞へのLTVECの導入、およびLTVECの導入が首尾よく生じた場合の細胞の選択)
LTVEC DNAを、標準的な方法論(例えば、リン酸カルシウム、脂質を介するトランスフェクション、またはエレクトロポレーション(Sambrook,J.,E.F.Fritsch And T.Maniatis.Molecular Cloning:A Laboratory Manual,Second Edition,Vols 1,2,and 3,1989))を使用して真核生物細胞に導入し得る。LTVECが首尾よく導入された細胞を、選択因子に曝露することによって、LTVEC中に操作されている選択マーカー遺伝子に依存して選択し得る。非限定的な例として、選択マーカーがネオマイシンホスホトランスフェラーゼ(neo)遺伝子(Beck,et al.,Gene,19:327−36,1982)である場合、次いで、LTVECを取り込んだ細胞を、G418含有培地中で選択し得る:LTVECを有さない細胞は死ぬが、LTVECを取り込んだ細胞は、生存する(Santerre,et al.,Gene,30:147−56,1984)。他の適切な選択マーカーとしては、真核生物細胞中で活性を有する任意の薬物(例えば、ハイグロマイシンB(Santerre,et al.,Gene,30:147−56,1984;Bernard,et al.,Exp
Cell Res,158:237−43,1985;Giordano and McAllister,Gene,88:285−8,1990)、ブラスチシジン(Blasticidin)S(Izumi,et al.,Exp Cell Res,197:229−33,1991)、および当業者の精通した他のもの)が挙げられる。
【0090】
(工程6.対立遺伝子改変(MOA)についての定量的アッセイを使用する、真核生物細胞における相同組換え事象のスクリーニング)
目的の遺伝子座にLTVECを標的化することにより首尾よく改変された真核生物細胞を、目的の遺伝子座内の対立遺伝子改変を指向し得、そして相同性アーム全体にわたるアッセイに依存しない種々のアプローチを使用して同定し得る。このようなアプローチとしては、以下が挙げられるがこれらに限定されない:
(a)TaqMan(登録商標)(Lie and Petropoulos,Curr
Opin Biotechnol,9:43−8,1998)を使用する定量的PCR;
(b)分子ビーコンを使用する定量的MOAアッセイ(Tan,et al.,Chemistry,6:1107−11,2000);
(c)インサイチュハイブリダイゼーションFISHでの蛍光(Laan,et al.,Hum Genet,96:275−80,1995)または比較ゲノムハイブリダイゼーション(CGH)(Forozan,et al.,Trends Genet,13:405−9,1997;Thompson and Gray,J Cell Biochem Suppl,139−43,1993;Houldsworth and Chaganti,Am J Pathol,145:1253−60,1994);
(d)等温DNA増幅(Lizardi,et al.,Nat Genet,19:225−32,1998;Mitra and Church,Nucleic Acids Res,27:e34,1999);および
(e)固定化プローブへの定量的ハイブリダイゼーション(Southern,J.Mol.Biol.98:503,1975;Kafatos FC;Jones CW;Efstratiadis A,Nucleic Acids Res 7(6):1541−52,1979)。
【0091】
出願人は、TaqMan(登録商標)定量的PCRを使用して、標的化真核生物細胞を首尾よくスクリーニングする例示を、本明細書中に提供する。この非限定的例示において、TaqMan(登録商標)を使用して、相同組換えを受けた真核生物を同定し、ここで、二倍体ゲノム中の2つの内因性対立遺伝子のうちの1つの部分は、別の配列に置換されている。相同性アーム全体にわたる制限フラグメントの長さにおける差異が、2つの対立遺伝子のうちの1つの改変を示す伝統的な方法とは対照的に、定量的TaqMan(登録商標)法は、改変されていない対立遺伝子のコピー数の(半分だけの)減少を測定することによって一方の対立遺伝子の改変を検出する。詳細には、プローブは、改変されていない対立遺伝子を検出し、改変されている対立遺伝子を検出しない。よって、この方法は、改変の正確な特徴に依存せず、本実施例に記載される配列置換に限定されない。TaqMan(登録商標)を使用して、ゲノムDNAサンプル中のDNAテンプレートのコピー数を、特に、参照遺伝子と比較することによって定量する(Lie and Petropoulos,Curr Opin Biotechnol,9:43−8,1998)。参照遺伝子は、標的遺伝子または遺伝子座として同一のゲノムDNAにおいて定量される。よって、2つのTaqMan(登録商標)増幅(各々それぞれのプローブを有する)が行われる。一方のTaqMan(登録商標)プローブは、参照遺伝子の「Ct」(閾値サイクル)を決定し、他方のプローブは、首尾よく標的化することによって置換されている、標的化された遺伝子または遺伝子座の領域のCtを決定する。Ctは、各TaqMan(登録商標)プローブについての開始DNAの量を反映する量である(すなわち、ほとんどの配列が、閾値サイクルに達するためにPCRのより多いサイクルを必要としない)。TaqMan(登録商標)についてのテンプレート配列のコピー数を半分に減少することによって、約1Ct単位の増加を生じる。標的化遺伝子または遺伝子座の一方が相同組換えによって置換されている細胞でのTaqMan(登録商標)反応は、標的化されていない細胞由来のDNAと比較する場合、参照遺伝子についてのCtの増加することなく標的TaqMan(登録商標)反応について1Ctの増加を生じる。このことによって、LTVECを使用して真核生物細胞に目的の遺伝子の一方の対立遺伝子の改変を容易に検出し得る。
【0092】
上記の状態において、対立遺伝子改変(MOA)スクリーニングは、相同組換えを受けた細胞を同定するための、一方の対立遺伝子の改変の検出する任意の方法の使用である。標的化対立遺伝子が互いに同一(相同)であることは必要とされず、マウスの異なる2つの系統を交配することによって生じる子孫の場合のように、実際に、多型を含み得る。さらに、MOAスクリーニングによってまたカバーされる、ある特定の状況は、遺伝子(例えば、性染色体、特にY染色体上に位置するいくつか)の標的化であり、これは、細胞中に単一のコピーとして正常に存在する。この場合において、単一の標的化対立遺伝子の改変を検出する方法(例えば、定量的PCR、サザンブロッティングなど)を使用して、標的化事象を検出し得る。対立遺伝子が多型であるかまたは標的化細胞中の単一コピー中に存在する場合でさえ、本発明の方法を使用して、改変された真核生物細胞を作製し得る。
【0093】
(工程8.遺伝子改変された真核生物細胞の使用)
(a)細胞の表現型の変化が所望される場合、工程1〜7に記載された方法によって作製された、遺伝子改変された真核生物細胞を、任意のインビトロアッセイまたはインビボアッセイにおいて使用し得る。
【0094】
(b)工程1〜7に記載された方法によって作製された、遺伝子改変された真核生物細胞をまた、遺伝子改変を保有する生物を作製するために使用し得る。遺伝子改変された生物を、以下の異なる種々の技術(しかし、これらに限定されない)を使用することによって作製し得る:
1.改変された胚性幹細胞(ES細胞)(例えば、ラットES細胞およびマウスES細胞がしばしば使用される)。ES細胞を使用して、遺伝子改変されたラットまたはマウスを、標準的な胚盤胞注入技術または凝集技術(Robertson,Practical
Approach Series,254,1987;Wood et al.,Nature,365:87−9,1993;Joyner,The Practical Approach Series,293,1999)、テトラプロイド胚盤胞注入(Wang,et al.,Mech Dev,62:137−45,1997)または核移入およびクローニング(Wakayama,et al.,Proc Natl Acad Sci USA,96:14984−9,1999)によって作製し得る。他の生物(例えば、ウサギ(Wang,et al.,Mech Dev,62:137−45,1997;Schoonjans,et al.,Mol Reprod Dev,45:439−43,1996)またはニワトリ(Pain,et al.,Development,122:2339−48,1996)あるいは他の種)由来のES細胞はまた、本発明の方法を使用する遺伝子改変に従うべきである。
【0095】
2.改変されたプロトプラストは、遺伝的に改変された植物を作製するために用いられ得る(例えば、米国特許第5,350,689号「プロトプラストまたはプロトプラスト由来細胞から再生されるトウモロコシ植物およびトランスジェニックトウモロコシ植物」、米国特許第5,508,189号「培養された孔片細胞プロトプラストからの植物の再生」、およびこれらの参考文献を参照のこと)。
【0096】
3.改変された対立遺伝子を有するクローン化生物体を作製するための、改変された真核生物細胞から卵母細胞への核移入(Wakayamaら、Proc Natl Acad Sci USA,96:14984−9,1999;Baguisiら、Nat Biotechnol,17:456−61,1999;Wilmutら、Reprod Fertil Dev,10:639−43,1998;Wilmutら、Nature,385:810−3,1997;Wakayamaら、Nat Genet,24:108−9,2000;Wakayamaら、Nature,394:369−74,1998;Rideoutら、Nat Genet,24:109−10,2000;Campbellら、Nature,380:64−6,1996)。
【0097】
4.操作された染色体の移動を含む、改変された対立遺伝子を別の細胞に移入させるための細胞融合、および改変された対立遺伝子または操作された染色体を有する生物体を産生するためのこのような細胞の使用(Kuroiwaら、Nat Biotechnol,18:1086−1090,2000)。
【0098】
5.本発明の方法はまた、用いられているか、または、まだ発見されていない、他の任意のアプローチに受け入れられる。
【0099】
本発明の方法の個々の工程を実行する際に使用される多くの技術は、当業者によく知られているが、本出願人は、本発明の方法の新規性が、真核生物細胞にLTVECを直接導入して、染色体座を改変する、以前に記載されたことのない方法と、適切に改変された真核生物細胞を同定するための定量的MOAアッセイの使用とを組み合わせた、工程および技術の独自の組み合わせにあることを主張する。この新規組み合わせは、内因性の遺伝子または染色体遺伝子座の改変を有する生物を作製するための、先行技術を上回る有意な改善を示す。
【実施例】
【0100】
(実施例1:OCR10遺伝子の欠失を有するマウスES細胞の操作)
(a.mOCR10を含む大きなゲノムDNAクローンの選択)
マウスOCR10(mOCR10)遺伝子のコード配列を含んだ大きなゲノムDNAフラグメントを保有する細菌人工染色体(BAC)クローンを、PCRを使用して、整列されたマウスゲノムDNAのBACライブラリー(Incyte Genomics)をスクリーニングすることによって獲得した。このライブラリーをスクリーニングするために使用したプライマーは、mOCR10遺伝子のcDNA配列に由来した。
【0101】
使用した2つのプライマー対は、以下であった:
(a)OCR10.RAA(5’−AGCTACCAGCTGCAGATGCGGGCAG−3’)およびOCR10.PVIrc(5’−CTCCCCAGCCTGGGTCTGAAAGATGACG−3’)、これらは、102bpのDNAを増幅する;ならびに
(b)OCR10.TDY(5’−GACCTCACTTGCTACACTGACTAC−3’)およびOCR10.QETrc(5’−ACTTGTGTAGGCTGCAGAAGGTCTCTTG−3’)、これらは、1500bpのDNAを増幅する。
【0102】
このmOCR10 BACは、完全なmOCR10コード配列を含む、約180kbのゲノムDNAを含んだ。このBACクローンを使用して、LTVECを作製し、続いて、これを使用して、開始コドンがOCR10の開始コドンで正確に置換されたレポーター遺伝子を導入し、同時にこのレポーター遺伝子の後ろに、E.coliおよび哺乳動物細胞の両方において選択するのに有用な選択マーカー遺伝子を挿入しながら、mOCR10のコード領域の一部を欠失させた(図2)。レポーター遺伝子(この非限定的な例において、LacZ、この配列は、当業者に容易に入手可能である)は、E.coliのβ−ガラクトシダーゼ酵素をコードする。LacZ(その開始コドンは、mOCR10の開始コドンと同じ位置である)の挿入位置に起因して、LacZでの類似の置換が先行技術を使用して実行された他の例において観察されたように(「Gene trap strategies in ES cells」、W WurstおよびA.Gossler、Joyner、The Practical Approach Series、293、1999を参照のこと)、lacZの発現は、mOCR10の発現を模倣するはずである。LacZ遺伝子は、その発現パターンをインサイチュで明らかにし得る、単純かつ標準的な酵素アッセイが実行されるのを可能にし、従って、置換された遺伝子または染色体遺伝子座の正常な発現パターンを反映する代理アッセイを提供する。
【0103】
(b.ドナーフラグメントの構築およびLTVECの作製)
mOCR10 LTVECの構築において使用される改変カセットは、lacZ−SV40ポリA−PGKp−EM7−neo−PGKポリAカセットであり、ここでlacZは、上記のようなマーカー遺伝子であり、SV40ポリAは、シミアンウイルス40由来のフラグメントであり(Subramanianら、Prog Nucleic Acid Res Mol Biol、19:157−64、1976;Thimmappayaら、J Biol Chem、253:1613−8、1978;Dharら、Proc Natl Acad Sci U S A、71:371−5、1974;Reddyら、Science,200:494−502、1978)、そしてポリアデニル化部位およびシグナルを含み(Subramanianら、Prog Nucleic Acid Res Mol Biol、19:157−64、1976;Thimmappayaら、J Biol Chem、253:1613−8、1978;Dharら、Proc Natl Acad Sci U S A、71:371−5、1974;Reddyら、Science,200:494−502、1978)、PGKpは、マウスホスホグリセレートキナーゼ(PGK)プロモーターであり(Adraら、Gene、60:65−74、1987)(これは、哺乳動物細胞における薬物耐性遺伝子の発現を駆動するために広く使用されてきた)、EM7は、ネオマイシンホスホトランスフェラーゼ(neo)遺伝子の発現を駆動することによって、細菌中の、完成したLTVEC構築物のポジティブ選択を可能にする利点を有する、強力な細菌プロモーターであり、neoは、原核生物細胞におけるカナマイシン耐性および真核生物細胞におけるG418耐性を付与する選択マーカーであり(Beckら、Gene、19:327−36、1982)、そしてPGKポリAは、PGK遺伝子由来の3’非翻訳領域であり、ポリアデニル化部位およびシグナルを含む(Boerら、Biochem Genet、28:299−308、1990)。
【0104】
mOCR10 LTVECを構築するために、改変カセット中のLacZ遺伝子の上流に結合したmOCR10相同性ボックス1(hb1)、および改変カセット中のneo−PGKポリA配列の下流に結合したmOCR10相同性ボックス2(hb2)からなる第一のドナーフラグメント(図2)を、標準的な組換え遺伝子操作技術を使用して作製した。相同性ボックス1(hb1)は、mOCR10のオープンリーディングフレーム(mOCR10 ORF)の開始メチオニンの直ぐ上流の、211bpの非翻訳配列からなる(図3A〜3D)。相同性ボックス2(hb2)は、終止コドンで終わる、mOCR10 ORFの最後の216bpからなる(図3A〜3D)。
【0105】
続いて、細菌相同組換えを用いて(Zhangら、Nat Genet、20:123−8、1998;Angrandら、Nucleic Acid Res、27:e16、1999;Muyrersら、Nucleic Acids Res、27:1555−7、1999;Narayananら、Gene Ther、6:442−7、1999;Yuら、Proc Natl Acads Sci U S A、97:5978−83、2000)、このドナーフラグメントを使用して、mOCR10コード領域(開始メチオニンから終止コドンまで)を挿入カセットで正確に置換し、mOCR10 LTVECの構築を生じた(図2)。従って、このmOCR10 LTVECにおいて、mOCR10コード配列を、挿入カセットによって置換して、mOCR10遺伝子座における約20kbの欠失を作製し、一方で約130kbの上流相同性(上流相同性アーム)および32kbの下流相同性(下流相同性アーム)を残した。
【0106】
LTVECが、先行技術を使用して作製された標的化ベクターよりも、入手可能なBACライブラリーからより迅速かつ簡便に作製され得る(なぜなら、単一の細菌相同組換え工程のみが必要であり、必要な配列情報は、相同性ボックスを作製するために必要な配列情報のみであるからである)ことに注意することが重要である。対照的に、細菌相同組換えを使用して標的化ベクターを作製するための先行技術のアプローチは、ES細胞への導入の前に、大きな標的化ベクターが「トリミング」される必要がある(Hillら、Genomics、64:111−3、2000)。先行技術のアプローチによって利用されるスクリーニング方法に対応するために、十分短い相同性アームを作製することが必要なので、このトリミングは必要とされる。Hillらの方法の1つの主な欠点は、2つのさらなる相同組換え工程が、単にトリミングのために必要とされることである(1つの工程は、改変された遺伝子座の上流領域をトリミングし、1つの工程は、改変された遺伝子座の下流領域をトリミングする)。これを行うために、実質的により多くの配列情報(トリミング部位にわたる配列情報を含む)が必要とされる。
【0107】
さらに、上記の実施例によって例示される別の明らかな利点は、mOCR10遺伝子にわたる非常に大きな欠失(約20kb)が、単一工程において容易に作製され得ることである。対照的に、先行技術を使用して同じ作業を達成することは、いくつかの工程を必要とし得、そしてCreリコンビナーゼを使用して、真核生物細胞において改変された遺伝子座の導入後にloxP部位が隣接する配列を除去するために、loxP部位を有するコード配列の上流および下流の領域を作製する工程を含み得る。このことは、1工程では達成不能であり得、従って、異なる選択マーカーを使用する2つの標的化ベクターの構築、およびES細胞における2つの連続する標的化事象(1つの事象は、コード配列の上流領域にloxP部位を導入すること、そして他方の事象は、コード配列の下流領域にloxP部位を導入すること)を必要とし得る。相同組換えの達成頻度は、比較的短い相同性アームが隣接する大きな欠失を含む標的化ベクターを使用する場合、低い可能性があるので、大きな欠失の作製は、しばしば、真核生物細胞における先行の標的化技術を使用すると、低い効率で生じることもさらに注意されるべきである。本発明の方法(以下を参照のこと)を使用して得られる高い効率は、真核生物細胞における相同組換えの割合を増大させる、LTVEC中に存在する非常に長い相同性アームに起因する。
【0108】
(c.mOCR10 LTVEC DNAの確認、調製およびES細胞への導入)
挿入カセットおよび相同性配列の連結部の周辺の配列を、DNA配列決定によって確認した。mOCR10 LTVECのサイズを、制限分析、その後のパルスフィールドゲル電気泳動(PFGE)によって確認した(Cantorら、Annu Rev Biophys Chem、17:287−304、1988;SchwartzおよびCantor、Cell、37:67−75、1984)。mOCR10 LTVECの標準的なラージスケールのプラスミド調製を行って、プラスミドDNAを制限酵素NotI(これは、mOCR10 LTVECのベクター骨格を切断する)で消化して、直鎖状DNAを作製した。続いて、線状化されたDNAを、エレクトロポレーションによって、マウスES細胞に導入した(Robertson、Practical Approach Series、254、1987;Joyner、The Practical Approach Series、293、1999;Sambrookら、Sambrook,J.、E.F.FritschおよびT.Maniatis、Molecular Cloning:A Laboratory Manual、第二版、1巻、2巻および3巻、1989)。mOCR10 LTVECで首尾よくトランスフェクトされたES細胞を、標準的な選択方法を使用して、G418含有培地中で選択した(Robertson、Practical Approach Series、254、1987;Joyner、The Practical Approach Series、293、1999)。
【0109】
(d.対立遺伝子の定量的改変(MOA)アッセイを使用する、標的化ES細胞クローンの同定)
2つの内因性mOCR10遺伝子のうち1つが改変カセット配列によって置換されたES細胞を同定するため、個々のES細胞クローン由来のDNAを、標準的なTaqMan(登録商標)方法論を、記載されたように使用する定量的PCRによって分析した(Applied Biosystems、TaqMan(登録商標)Universal PCR Master Mix、カタログ番号P/N4304437;http://www.pebiodocs.com/pebiodocs/04304449.pdfもまた参照のこと)。使用したプライマーおよびTaqMan(登録商標)プローブは、図3A〜3Dに記載の通りである。合計69個の個々のES細胞クローンをスクリーニングし、そして3個のクローンが陽性として、すなわち、内因性mOCR10コード配列の1つが、上記の改変カセットによって置換されたクローンとして、同定された。
【0110】
MOAアプローチのいくつかの利点が明らかである:
(i)改変されている遺伝子座の外側のプローブの使用を必要としない。従って、改変された遺伝子座に隣接する配列を知る必要を排除する。
【0111】
(ii)以前の選り抜きの方法であった従来のサザンブロット方法論(Robertson、Practical Approach Series、254、1987;Joyner、The Practical Approach Series、293、1999)と比較して、実行するために必要な時間が非常に短い。従って、正確に改変された細胞を同定するための時間を、代表的には数日間からほんの数時間に減少させる。
【0112】
これは、過去にスクリーニングが実行された方法における有意な改善であり、この方法を、真核生物細胞における相同組換え事象についてスクリーニングするための、労力集約がかなり少なく、かつコスト効率のよいアプローチにしている。
【0113】
本発明の方法のなお別の利点は、難しい遺伝子座を標的化する能力に起因してもまた、先行技術より優れていることである。先行技術を使用すると、特定の遺伝子座についての首尾よい標的化頻度は、2000の組み込み事象のうち1、おそらくそれより低い頻度でさえあり得ることが示された。本発明の方法を使用して、本出願人は、このような難しい遺伝子座が、長い相同性アーム(すなわち、先行技術によって可能な長さよりも長い)を含むLTVECを使用して、より効率的に標的化され得ることを実証した。上記の非限定的な実施例が実証するように、本出願人は、OCR10遺伝子座(この遺伝子座は、従来技術を使用する標的化が困難であることが以前に証明された)を標的化した。本発明の方法を使用して、本出願人は、mOCR10 LTVEC(160kbより長い相同性アームを含み、20kbの欠失を導入する)が組み込まれた69個のES細胞クローンのうち3個において、首尾よい標的化を得たが、一方、ES細胞標的化の先行技術(Joyner、The Practical Approach Series、293、1999)を使用すると、10〜20kbより短い相同性アームを有し、一方でまた15kb未満の欠失を導入する、プラスミドベースのベクターを使用して、ベクターの600を超える構成要素のうち、標的化事象は同定されなかったことを示した。これらのデータは明らかに、本発明の方法が、先行技術よりも優れていることを実証する。
【0114】
(実施例2:LTVECが標的化ベクターとして使用される場合の、増大した標的化頻度および同質遺伝子DNAを使用する必要性の撤廃)
上記のように、長い相同性アームを使用して得られた増大した標的化頻度は、存在する場合、LTVECを構築する際の、標的化される真核生物細胞のDNAと同質遺伝子の(すなわち、このDNAの配列と同一の)ゲノムDNAを使用することに由来する利点を低減するはずである。この仮説を試験するため、本出願人は、標的化されるべき真核生物細胞と同じマウス副系統に由来するゲノムDNA(おそらく同質遺伝子)を使用して、多数のLTVECを構築し、そして標的化されるべき真核生物細胞と異なるマウス副系統に由来するゲノムDNA(おそらく非同質遺伝子)を使用して、多数の他のLTVECを構築した。LTVECの2つの組は、1〜13%の範囲の類似の標的化頻度を示した(表1)。このことは、LTVECを使用した首尾よい標的化の割合が、遺伝子同質性に依存しないことを示す。
【0115】
(表1)
(BACクローンベクターを用いる遺伝子標的化の概要)
【0116】
【表1】
要約すると、ES細胞における相同組換え事象についてのMOAスクリーニングと組合わせた、LTVECを作製し、これらを標的ベクターとして直接使用するアプローチは、遺伝子改変した遺伝子座を操作するための新規の方法を作成する。この方法は、迅速で、安価であり、そして使用の際に、以前の冗長な、時間のかかる方法よりも有意な改善を示す。従って、以前の方法論により必要とされる時間および費用のほんの少しで、生物体のゲノム中の本質的に任意のおよび全ての遺伝子の、迅速な大スケールのインビボでの機能的ゲノム分析の可能性を開く。
【0117】
(実施例3:キメラ抗体およびヒト抗体を産生するためのLTVECの使用)
(a.導入)
抗体は、2つの鎖(軽鎖および重鎖)から構成され、これらの鎖の各々は、2つのドメイン(可変ドメインおよび定常ドメイン)から構成される。抗体タンパク質の可変領域は、抗体のN末端部分であり、この部分は抗原に結合する。重鎖可変ドメインは、重鎖可変遺伝子の遺伝子座のDNAによりコードされ、この遺伝子座は、可変(V)遺伝子セグメント、多様性(D)遺伝子セグメント、および連結(J)遺伝子セグメントから構成される。軽鎖可変ドメインは、軽鎖可変遺伝子の遺伝子座(κおよびλ)のDNAによりコードされ、この遺伝子座は、可変(V)遺伝子セグメントおよび連結(J)遺伝子セグメントから構成される。
【0118】
B細胞の初期の発達の間の可変領域(VDJ/VJ)遺伝子の再構成は、最初の機構であり、これにより、免疫系は、遭遇し得る莫大な数の抗原を認識し得る抗体を産生する。本質的に、B細胞発達の間のDNA再構成を通じて、莫大なレパートリーの可変(VDJ/VJ)領域の配列をアセンブルし、次いで、これを、定常(C)領域に連結して、完全な重鎖および軽鎖を作製し、この重鎖および軽鎖をアセンブルして抗体を形成する。機能的抗体をアセンブルした後、第2のリンパ器官を生じる体細胞過剰変異は、生物体が抗体のアフィニティーを選択および最適化し得るさらなる多様性を導入する。
【0119】
非ヒト種における種々の抗原に対する抗体の産生は、ヒト治療剤として用いられ得る抗体の大規模産生についての大きな見込みを最初に提供した。しかし、種の違いは、外来の抗体を不活化し、そしてアレルギー反応を引き起こすヒトによる抗体の産生を導く。抗体を「ヒト化」し、それにより、ヒトにおいて外来物として認識される可能性が低い抗体を作製する試みが、引き続いてなされた。初めに、この手順は、マウス由来の抗体の抗原結合部位を、ヒト抗体の定常領域と組合わせ、それにより、ヒトにおいて免疫原性が低い組換え抗体を作製することに関した。開発された第2のアプローチは、ファージディスプレーであり、これにより、ヒトV領域は、ファージディスプレーライブラリーへとクローン化され、そして適切な結合特性を有する領域が、ヒト定常領域に連結され、ヒト抗体を作製する。しかし、この技術は、B細胞において天然で生じる、抗体発達およびアフィニティーの成熟の不足により、制限される。
【0120】
より最近では、内因性遺伝子をマウスでノックアウトし、そして遺伝子を、ヒト対応物と交換して、完全にヒト抗体を産生している。残念なことに、これらの構築物の使用は、B細胞における抗体の発達および最適化における、内因性定常領域の重要性が強調されている。完全なヒト抗体を産生するマウスは、免疫応答が低下されている。このことは、マウスの対応物と比べてアフィニティーが低下されている、完全なヒト構築物を有するトランスジェニックマウスにより産生されるヒト抗体に起因し得る。低下したアフィニティーは、B細胞成熟および生存をもたらし得る。従って、マウスまたは他の生物体においてヒト化抗体を産生するより称賛される方法(この方法において、マウスの内因性の可変領域および定常領域をノックアウトし、そしてヒト対応物と交換する)は、最適な抗体を生じ得ない。
【0121】
キメラ抗体(ヒト可変領域(VDJ/VJ)とマウス定常領域とをB細胞成熟の間に利用し、その後、この抗体は、そのマウス定常領域をそのヒト対応物で置換されるよう操作される)の使用が、示唆されている(1998年6月23日発行の米国特許第5,770,429号)。しかし、このようなキメラを作製するために現在まで存在している唯一の方法論は、トランススイッチであり、この方法論において、そのキメラの形成は、重鎖においてのみ生じるほんの稀な事象である。従来、トランスジェニック動物において、セグメント群をコードする可変遺伝子全体をヒト遺伝子で大規模に置換し、それにより重鎖および軽鎖の両方でキメラを作製する機構は、存在しない。本明細書中に開示される本出願人の方法論を使用すると、キメラ抗体が作製され、その後、このキメラ抗体は、標準的方法論を介して、高親和性ヒト抗体を作製するように変化され得る。
【0122】
(b.簡単な説明)
ヒト可変領域(VDJ/VJ)およびマウス定常領域を含むハイブリッド抗体を産生する、トランスジェニックマウスが、作製される。これは、そのマウス可変領域(VDJ/VJ)遺伝子群をそれらのヒト対応物で直接インサイチュにて置換することによって、達成される。生じるハイブリッド免疫グロブリン座は、そのハイブリッド抗体を産生するように、B細胞発生の間に天然の再配列プロセスを受ける。
【0123】
その後、完全にヒト化した抗体が、そのマウス定常領域を所望のヒト対応物で置換することによって、作製される。このアプローチは、以前の方法(例えば、マウスモノクローナル抗体の「ヒト化」またはHuMabマウスにおける完全にヒト化した抗体の産生)よりもかなり効率良く、治療抗体を生じる。さらに、この方法は、以前の方法が失敗してきた多くの抗原に対する治療抗体の産生に成功する。このマウスは、ヒト(VDJ/VJ)−マウス定常領域である抗体を産生し、このことは、完全にヒト化した抗体を産生する以前に利用可能であったHuMAbマウスを越える、以下の利点を有する。この新規なマウスにより産生される抗体は、そのマウスのB細胞レセプター複合体の他の成分(適切なB細胞分化に必要なシグナル伝達成分(例えば、IgaおよびIgb)を含む)と、より効率良く相互作用するマウスFc領域を保持する。さらに、このマウスFc領域は、マウス細胞、補体分子などの上のFcレセプターとそのマウスFc領域との相互作用において、ヒトFc領域よりも特異的である。これらの相互作用は、強力かつ特異的な免疫応答のため、B細胞の増殖および成熟のため、ならびに抗体の親和性成熟のために、重要である。
【0124】
ヒトV−D−J/V−J領域によるそのマウス座の等価な領域の直接置換が存在するので、適切な転写、組換え、および/またはクラススイッチに必要な配列のすべてが、インタクトなままである。例えば、そのマウス免疫グロブリン重鎖イントロンエンハンサー(Em)は、V−D−J組換えおよびB細胞発生の初期段階の間の重鎖遺伝子発現に重要であることが示されているが[Ronia,D.,Berru,M.およびShulman,M.,J.Mol.Cell Biol 19:7031〜7040(1999)]、その免疫グロブリン重鎖3’エンハンサー領域は、クラススイッチ[Pan,Q.,Petit−Frere,C.,Stavnezer,J.およびHammarstrom,L.,Eur J Immunol 30:1019−1029(2000)]ならびにB細胞分化の後期段階での重鎖遺伝子発現のために重要であるようである[Ong,J.,Stevens,S.Roeder,R.G.,およびEckhardt,L.A.,J Immunol 160:4896−4903(1998)]。その転写制御エレメントのこれらの様々であるが重要な機能を考慮すると、これらの配列をインタクトなまま維持することが、望ましい。
【0125】
正常なB細胞分化過程の間に免疫グロブリン座で生じる必要な組換え事象は、これらの座が不適切な染色体に挿入されるかまたは現在利用可能なマウスにおけるように多コピーで挿入される場合に、異常で非生産的な免疫グロブリン再配列の頻度を増加し得る。生産的免疫グロブリン再配列の減少により、従って、特定のB細胞発生段階での適切なシグナル伝達により、異常な細胞が排除される。初期発生段階でのB細胞数の減少は、全体的最終B細胞集団を減少させ、そのマウスの免疫応答を非常に制限する。(現在利用可能なマウスにおける、変異免疫グロブリン座と対照的に、ヒトトランスジェニック座が、重鎖および軽鎖について異なる染色体位置に組み込まれている)唯一のキメラの重鎖座または軽鎖座のみが存在するので、非生産的再配列または治療上不適切なキメラ抗体を生じ得る座のトランススプライシングもトランス再配列も存在しない(Willers,J.,Kolb,C.およびWeiler,E.Immunobiology 200:150−164(2000);Fujieda,S.,Lin,Y.Q.,Saxon,A.およびZhang,K.,J Immunol 157:3450−3459(1996))。
【0126】
真のマウス染色体免疫グロブリン座中へのヒトV−D−J領域またはV−J領域の置換は、実質的により安定であり、現在利用可能なマウスと比較して、子孫への伝達率が増加し、そしてB細胞遺伝子型のモザイク現象が減少する(Tomizuka,K.,Shinohara,T.,Yoshida,H.,Uejima,H.,Ohguma,A.,Tanaka,S.,Sato,K.,Oshimura,M.およびIshida,I.,Proc Natl Acad Sci(USA)97:722−727(2000))。さらに、インビボでの真のマウス座へのヒト可変領域(VDJ/VJ)の導入は、適切な時期の組換え事象に重要であることが以前に示されている、染色体接近性の適切な全体調節を維持する(Haines,B.B.およびBrodeur,P.H.,Eur J Immunol 28:4228−4235(1998))。
【0127】
ヒト抗体のうちの約1/3は、λ軽鎖を含み、対して、マウス抗体のうちの1/20のみがλ軽鎖を含む。従って、マウスλ軽鎖V−J配列を、ヒト座由来のλ軽鎖V−J配列で置換することは、抗体のレパートリーを増加し、そして真のヒト免疫応答とより密接に一致するために役立ち、従って、治療上有用な抗体を含む可能性を増加させる。
【0128】
ヒト配列を真マウス免疫グロブリン座中に組み込むさらなる利点は、その挿入部位に変異原性破壊を生じ得、そして生存するホモ接合体マウスの単離を阻止し得る、新規な組み込み部位に導入されないことである。これは、繁殖中のマウス群の作製および維持を非常に簡単にする。
【0129】
以下は、上記の利点のすべてを備える抗体を作製するための新規な方法を提供する。当業者は、本明細書中に記載される一般的方法が、等価な結果を生じるように改変され得ることを、認識する。
【0130】
(C.材料および方法)
マウス重鎖可変領域(VDJ)をヒト対応物で正確に置換することは、以下の例における相同組換えと部位特異的組換えの組み合わせを使用して例証され、この例は、2段階プロセスを使用する。当業者は、マウス座を、ヒトホモログ座またはオルソログヒト座で置換することが、1つ以上の工程で達成され得ることを、認識する。従って、本発明は、マウス座の全体または一部を、相同組換えを介して、各組み込みにより置換することを企図する。
【0131】
ヒト重鎖座のVDJ領域全体に広がる大インサート(BAC)クローンを、単離する(図4A)。この領域全体の配列は、以下のGenBankファイル(AB019437、AB019438、AB019439、AB019440、AB019441、X97051、およびX54713)にて入手可能である。この例において、大インサート(BAC)クローンは、相同性アームの供給源として、マウスVDJ領域の末端から単離し(図4B)、この末端を、2段階プロセスにおけるヒトVDJ配列の相同組換えを介して直接組み込みに使用する。
【0132】
第1段階において、LTVEC1(図4D)を、E.coliにおける細菌相同組換えにより構築する。LTVEC1は、順番に、マウスDJ領域から上流にある領域に由来する大マウス相同性アーム(しかし、その絶対的端部は重要ではない);ES細胞において機能する選択マーカー(この例においてはPGK−ネオマイシンR)をコードするカセット;loxP部位;いくつかのV遺伝子セグメントからDJ領域全体にまで広がる、大ヒトインサート;およびマウスJセグメントとすぐ近接するがこのセグメントは含まない領域を含む、マウス相同性アーム;を含む。この下流アームの5’末端およびloxP部位の配置は、置換されるべき領域の3’末端を、この座中に規定する。マウスES細胞を、標準的技術(例えば、エレクトロポレーション)によって、線状化LTVEC1で形質転換する。LTVEC1の直接導入は、内因性可変遺伝子座の改変を生じるので、ネオマイシン耐性コロニーを、MOAアッセイを使用して正確な標的化についてスクリーニングし得る。これらの標的化ES細胞は、ハイブリッド重鎖を含む抗体を産生するマウスを生じ得る。しかし、このマウス可変セグメントの残りを排除するその後の段階を続行することが、好ましい。
【0133】
第2段階において、LTVEC2(図4C)を、E.coli中にて、細菌相同組換えによって構築する。LTVEC2は、順番に、最も遠位のマウスV遺伝子セグメントと近接するがいかなるマウスV遺伝子セグメントも含まない、領域を含む大マウス相同性アーム;多数の遠位ヒトV遺伝子セグメントを含む大インサート;(LTVEC2およびLTVEC1中の野生型loxP部位と反対の方向にある)lox511と呼ばれる変異体loxP部位[Hoess,R.H.,Wierzbicki,A.およびAbremski,K.,Nucleic Acids Res.14:2287−2300(1986)](この部位は、野生型loxP部位と組換わらないが、他のlox511部位と容易に組み換わる);野生型loxP部位;第2選択マーカー(この例においてはPGK−ピューロマイシンR)およびV領域に由来する、マウス相同性アーム(しかし、その絶対的端部は重要ではない);を含む。この上流相同性アームの3’末端およびloxP部位の配置は、置換されるべき領域の5’末端を、この座中に規定する。その後、LTVEC1で正確に標的化されたマウスES細胞を、標準的技術によって、線状化LTVEC2で形質転換し、そしてハイグロマイシン耐性コロニーを、内因性可変遺伝子座における改変について、MOAアッセイを使用して正確な標的化についてスクリーニングする。この形質転換から生じる正確に標的化されたES細胞を、本明細書中で、以後、「二重標的化ES細胞」と呼ぶ。
【0134】
その後の、この二重標的化ES細胞におけるCREリコンビナーゼの一過性発現は、マウスV領域の残りの欠如を生じる。あるいは、この二重標的化ES細胞を、キメラマウスの作製のために、宿主胚盤胞中に注入し得る。生じたキメラマウスを、発生初期にCREリコンビナーゼを発現するマウスと交配させると、子孫F1において、マウスV領域の残りの欠如を生じる。この後者の選択肢は、ハイブリッド重鎖座が、生殖細胞系を通過する可能性を増加させる。なぜなら、これは、より少ない世代の間ES細胞を培養する工程を包含するからである。
【0135】
LTVEC2中にlox511を含めると、ハイブリッド座中へのさらなるヒトV遺伝子セグメントの挿入が可能である。1つのアプローチは、多くのさらなるヒトV遺伝子セグメントを含む大ゲノムDNAクローンをlox511部位およびloxP部位と隣接するように、細菌相同組換えを使用することである。このような改変大ゲノムDNAクローンを、CREリコンビナーゼを一過性発現するプラスミドとともに、二重標的化ES細胞中に同時形質転換すると、カセット交換によって、このさらなるV遺伝子セグメントの導入を生じる(Bethke,B.およびSauer,B.,Nucleic Acids
Res.25:2828−2834(1997))。
【0136】
さらなるV遺伝子セグメントを組み込むための第2のアプローチは、多くのさらなるヒトV遺伝子セグメントを含む大ゲノムDNAクローンを、例えば、LTVEC2中に含まれる同じマウス相同性アームを使用して、マウス座中に独立して標的化することである。この場合、このさらなるヒトV遺伝子セグメントは、lox511部位およびloxP部位と隣接しており、その標的化ES細胞を、マウスを作製するために使用する。二重標的化ES細胞由来のマウスおよびこのさらなるV遺伝子セグメントを含むES細胞由来のマウスを、減数分裂の間にCREリコンビナーゼの発現を指向する第3のマウスと交配する。減数分裂対合の間のその2つの組換え座の近接性によって、他の系(Herault,Y.,Rassoulzadegan,M.,Cuzin.F.およびDuboule,D.,Nature Genetics 20:381−384(1998))において観察されたような、高頻度のCRE誘導性染色体内組換えが生じる。
【0137】
別のアプローチは、上記に概略したものと類似するが、ヒトLTVEC1およびLTVEC2とともにloxP部位およびlox511部位を導入するのではなく、loxP部位およびlox511部位を、マウスLTVEC上に導入し、その後、CREを使用して、隣接するloxP部位およびlox511部位を介するカセット交換によって、そのヒトx座中に特異的に標的化する。下記に概略する方法論は、このLTVEC技術をどのように使用して、任意の非ヒト動物において目的の任意の内因性遺伝子の末端に、隣接部位特異的組換え部位を配置し得るかを、示す。
【0138】
マウスLTVEC1は、J領域の下流かつそのJ領域に近接して、細菌組換えにより挿入されたカセットを含む。このカセットは、loxP部位および細菌選択マーカー/哺乳動物選択マーカー(例えば、ハイグロマイシン耐性)を含む。LTVEC1は、順番に、マウスDJ領域から上流にある(が、この可変遺伝子座内にある)領域由来の大相同性アーム(しかし、その絶対的末端は、重要ではない);ES細胞において機能する選択マーカー(この例においてはPGK−ハイグロマイシンR)をコードするカセット;loxP部位;およびマウスJセグメントとすぐ近接するがこのJセグメントを含まない、相同性アーム;を含む。この下流相同性アームの5’末端およびloxP部位の配置は、置換されるべき領域の3’末端を、この座中に規定する。カセット挿入部位にある内因性可変遺伝子の3’末端の改変は、MOAアッセイにより、ES細胞中での正確に挿入されたLTVEC1の欠失を可能にする。薬物耐性マーカーに、FRT部位が隣接する。FRT部位の導入は、ES細胞中においてFLPによるか、または生殖細胞系能力を有する細胞においてFLPを発現するマウスと生じるマウスとの交配のいずれかによる、残りすべての薬物耐性マーカーの除去を可能にする。
【0139】
LTVEC2を、その座の最も遠位のV領域の上流にカセットを挿入するような細菌組換えによって、構築する。このカセットは、lox511部位および細菌選択マーカー/哺乳動物選択マーカー(例えば、ネオマイシン耐性)を含む。LTVEC2は、順番に、最も遠位にあるマウスV遺伝子セグメントと近接する領域を含むがいかなるマウスV遺伝子セグメントも含まない、大相同性アーム;LTVEC2およびLTVEC1中の野生型loxP部位とは反対の方向にあるlox511部位;野生型loxP部位;第2選択マーカー(この例においてはPGK−ネオマイシンR);およびV領域に由来する(従って、その可変遺伝子座内にある)マウス相同性アーム(しかし、その絶対的端部は重要ではない);を含む。この上流相同性アームの3’末端およびloxP部位の配置は、置換されるべき領域の5’末端を、この座中に規定する。カセット挿入部位にある内因性可変遺伝子の5’末端の改変は、MOAアッセイにより、ES細胞中での正確に挿入されたLTVEC2の欠失を可能にする。これらのLTVECを、一緒にか連続して、標準的技術を使用してES細胞中に導入し、MOAアッセイを使用して、正確な標的化についてスクリーニングする。
【0140】
VDJ/VJ領域の一部または全体を含むヒトBACを、細菌組換えにより、そのヒト配列とlox511部位およびloxP部位が隣接するカセット群を挿入するように改変する。その上流カセットを、マウス改変領域を置換する領域のすぐ上流に挿入する。この上流カセットは、順番に、lox511部位、その後、細菌選択マーカー/哺乳動物選択マーカー(例えば、ピューロマイシン耐性)を含む。その下流カセットを、J領域の下流に近接して挿入する。この下流カセットは、順番に、loxP部位、その後、細菌選択マーカー(例えば、スペクチノマイシン耐性)を含む。
【0141】
ライブラリーから単離される単一BAC上に存在するよりも大きなヒト可変領域を挿入するために、いくつかの方法を使用し得る。これらのうちのいくつかを、以下に記載する。
【0142】
loxP部位およびlox511部位を、細菌組換えによって、重複するBAC上に別個に挿入し得、これらのBACは、ES細胞中に形質転換した場合、互いに組み換わる。この場合、上流BACは、1つのカセットを有し、このカセットは、マウス可変領域を置換する領域のすぐ上流で組み換わり、lox511部位、その後ろに細菌選択マーカー/哺乳動物選択マーカー(例えば、ネオマイシン耐性)を有する。下流BACは、1つのカセットを有し、このカセットは、J領域のすぐ下流でこのJ領域と近接して組み換わり、細菌選択マーカー/哺乳動物選択マーカー(例えば、ピューロマイシン耐性)の後ろに、loxP部位を含む。これらの2つのBACが重複していない場合、重複するホモロジーによりこの上流BACおよび下流BACと関連するさらなるBACを、このスキーム中に組み込む。これらを、細菌組換えによって、細菌選択マーカー/哺乳動物選択マーカー(例えば、ピューロマイシン耐性)を含むように改変する。これらの上流BACおよび下流BACを、ネオマイシン耐性マーカーおよびハイグロマイシン耐性マーカーを保有する、loxPカセットおよびlox511カセットを含むように改変する。
【0143】
ヒトBACを、可変領域と隣接するlox511組換え部位および可変領域と隣接するloxP組換え部位を含むES細胞株中に、CREリコンビナーゼとともに同時形質転換する。重複するBACを使用する場合、それらのBACの間で相同組換えが生じて、より大きなDNAフラグメントが生じ、そして隣接するloxP部位およびlox511部位が、この大フラグメントを、マウス座中に標的化する。細胞を、ピューロマイシン耐性について選択し、そしてそのマウス可変領域の置換についてスクリーニングする。あるいは、このマウス配列を、まず、その2つのloxP部位を介して欠失させ得、その後、そのヒト配列を、残りのlox511部位およびloxP部位を介して導入し得る。
【0144】
LTVEC1が、第3の部位特異的組換え部位(例えば、lox2272(Anal Biochem 2001 Mar15;290(2):260−71))を、細菌耐性遺伝子/哺乳動物耐性遺伝子(例えば、ピューロマイシン耐性)のすぐ下流に含む場合、第4のBACを挿入し得、LTVECを作製し得る。このLTVECは、順番に、ピューロマイシン耐性遺伝子、loxP遺伝子、およびlox2272遺伝子、その後ろに、ヒト配列、を含む。このBACを、マウス免疫グロブリン座中に組み込んだ後、lox511部位/lox2272部位は、第2回目のカセット交換におけるレシピンエトとして役立ち得、この第2回目のカセット交換において、ピューロマイシン耐性遺伝子が、ヒト免疫グロブリン座可変領域のさらなる上流部分、およびlox511部位およびlox2272部位が隣接する異なる細菌耐性遺伝子/哺乳動物耐性遺伝子によって、置換される。
【0145】
このヒト可変領域のより大きなストレッチを導入するための別の方法は、稀な制限エンドヌクレアーゼ切断部位を使用して、複数のBACに由来する配列をインビトロで合わせることである。これは、細菌相同組換えを使用して、最も下流のBACの最後のJのすぐ下流にloxPおよびスペクチノマイシン耐性遺伝子を挿入すること、そして下流のBACのヒト配列の上流に、第2の細菌選択マーカーおよび稀なI−Ceu1部位を挿入することによって、達成する。lox511部位および細菌選択マーカー/哺乳動物選択マーカー(例えば、ピューロマイシン耐性)を、第1のBAC中の配列から上流の、ヒト可変領域の領域を含む第2のBACの上流末端に、挿入する。I−Ceu1部位を、第2のBACの下流末端に挿入する。両方のBACをI−Ceu1およびNot1(これは、両方の改変BACのベクター部分中で独特である)で消化した後、この2つのBACを連結し、そして組換え体を、ピューロマイシン耐性およびスペクチノマイシン耐性について細菌中で選択する。生じるより大きなBACは、順番に、lox511部位、上流ヒト配列、I−Ceu1部位、下流ヒト配列、loxP部位、およびスペクチノマイシン耐性遺伝子を含む。lox511部位とloxP部位との間の領域を、上記のようなカセット交換およびピューロマイシン選択によって、マウス免疫グロブリン座中に挿入する。
【0146】
ヒト可変領域のより大きなストレッチを挿入するための第3の方法は、上記のように複数のBACに由来する配列を合わせることであるが、制限消化/連結の代わりに、細菌相同組換えを使用する。組換え体BACについての同じ選択を、細菌において適用するが、但し、2つのBACのうちの1つを消化し、消化後にその末端を、他の「レシピンエト」BACと相同であるように設計し、そしてそのレシピエントBACを、細菌株中で、細菌相同組換えを許容するように改変する。
【0147】
ヒト可変/マウス定常モノクローナル抗体産生マウスを作製する際の最後の段階は、軽鎖λ座および軽鎖κ座上での等価な可変領域置換を実施し、そして同じマウスにおいてともにホモ接合性であるように3つすべてのハイブリッド座を交配することである。生じるトランスジェニックマウスは、完全に内因性マウス定常領域に作動可能に連結した完全にヒトの重鎖および軽鎖の可変遺伝子座を含むゲノムを有し、そのマウスは、抗原性刺激に応答して、ヒト可変領域およびマウス定常領域を含む抗体を含む血清を産生するようになっている。その後、そのようなマウスを、ヒト抗体の可変領域をコードするDNAの供給源として使用し得る。標準的組換え技術を使用して、この抗体の重鎖および軽鎖の可変領域をコードするDNAを、ヒト重鎖定常領域およびヒト軽鎖定常領域をコードするDNAと、細胞(例えば、CHO細胞)中で作動可能に連結し、この細胞は、活性な抗体を発現可能である。この細胞を、完全なヒト抗体を発現するに適切な条件下で増殖させ、その後、この完全なヒト抗体を回収する。可変領域をコードする配列を、例えば、PCR増幅またはcDNAクローニングによって単離し得る。好ましい実施形態において、ヒト可変領域免疫グロブリン座のいくつかまたはすべてを含むトランスジェニックマウスから作製したハイブリドーマ(KohlerおよびMilstein,Eur.J.Immunol.,6:511−519(1976))を、ヒト可変領域をコードするDNAの供給源として使用する。
【0148】
要約すると、LTVECを作製し、そのLTVECを、ES細胞における相同組換え事象についてMOAスクリーニングと組み合わせて、標的化ベクターとして直接使用するアプローチは、遺伝子改変された座を操作するための新規な方法を生じる。この方法は、迅速かつ安価である。そしてこの方法は、以前に使用された冗漫で時間がかかる方法を超える有意な改善を示す。従って、この方法は、以前の方法論により必要とされた時間および価格の何分の1かで、生物のゲノム中の本質的に任意かつすべての遺伝子の迅速な、大規模の、インビボでの機能的なゲノム分析の可能性を開く。
【0149】
上記の発明は、説明および例としていくらか詳細に記載されているが、添付の特許請求の範囲の趣旨からも範囲からも逸脱することなく、本発明の教示に対して特定の変化および改変がなされ得ることが、当業者にとって容易に明らかである。
【特許請求の範囲】
【請求項1】
本明細書中に記載の発明。
【請求項1】
本明細書中に記載の発明。
【図1】

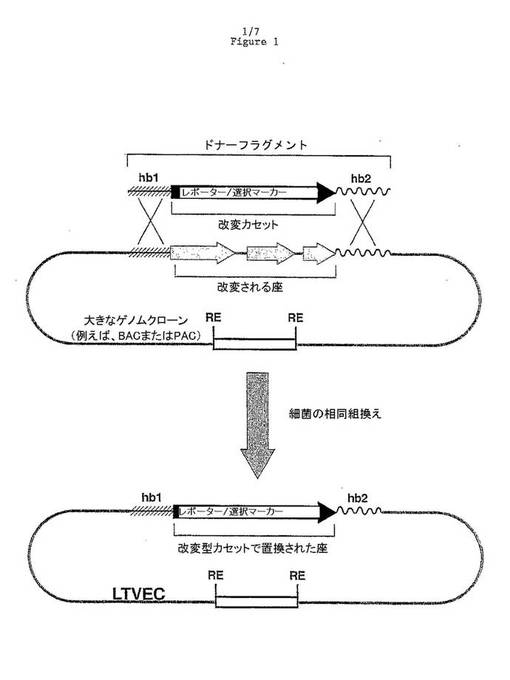
【図2】
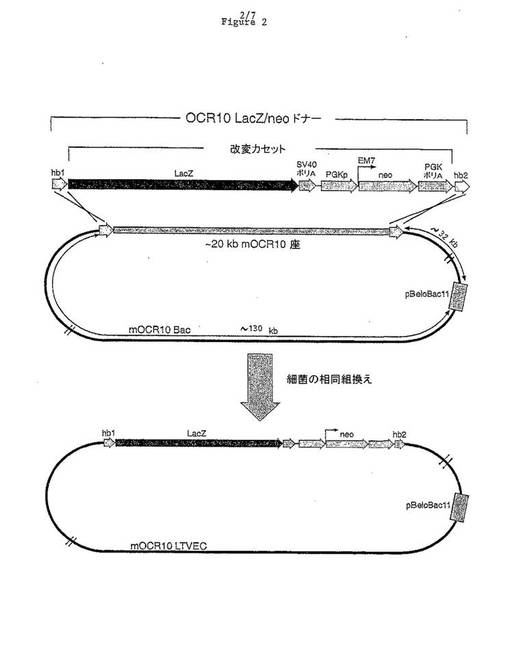
【図3A】

【図3B】

【図3C】

【図3D】

【図4】
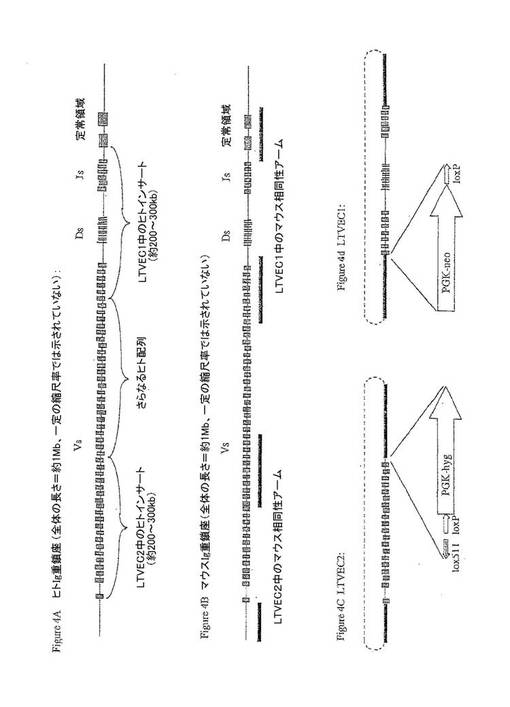
【図2】
【図3A】
【図3B】
【図3C】
【図3D】
【図4】
【公開番号】特開2012−95670(P2012−95670A)
【公開日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願番号】特願2012−32592(P2012−32592)
【出願日】平成24年2月17日(2012.2.17)
【分割の表示】特願2009−177054(P2009−177054)の分割
【原出願日】平成14年2月15日(2002.2.15)
【出願人】(597160510)リジェネロン・ファーマシューティカルズ・インコーポレイテッド (50)
【氏名又は名称原語表記】REGENERON PHARMACEUTICALS, INC.
【Fターム(参考)】
【公開日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願日】平成24年2月17日(2012.2.17)
【分割の表示】特願2009−177054(P2009−177054)の分割
【原出願日】平成14年2月15日(2002.2.15)
【出願人】(597160510)リジェネロン・ファーマシューティカルズ・インコーポレイテッド (50)
【氏名又は名称原語表記】REGENERON PHARMACEUTICALS, INC.
【Fターム(参考)】
[ Back to top ]