
真核細胞におけるエキソンスキッピングの誘導
【課題】細胞による異常型タンパク質の産生を少なくとも部分的に減少させるための方法であって、異常型タンパク質をコードするエキソンを含むmRNA前駆体を有する細胞にいわゆるエキソンスキッピングを誘導することを特徴とする方法を提供する。
【解決手段】エキソンスキッピングは、スキップされたエキソンを有さない成熟mRNAを生じ、スキップされたエキソンがアミノ酸をコードしている場合、変化した産物が得られる。エキソンスキッピングはあるエキソンのエキソン封入シグナル、例えばエキソン認識配列、を特異的に阻害することが可能な試薬を細胞に与えることによって行われる。該エキソン封入シグナルはエキソンの一部に対して相補的な核酸によって干渉することができる。医薬、例えば遺伝病の治療を目的とした医薬の調製に用いることができる上記の核酸。
【解決手段】エキソンスキッピングは、スキップされたエキソンを有さない成熟mRNAを生じ、スキップされたエキソンがアミノ酸をコードしている場合、変化した産物が得られる。エキソンスキッピングはあるエキソンのエキソン封入シグナル、例えばエキソン認識配列、を特異的に阻害することが可能な試薬を細胞に与えることによって行われる。該エキソン封入シグナルはエキソンの一部に対して相補的な核酸によって干渉することができる。医薬、例えば遺伝病の治療を目的とした医薬の調製に用いることができる上記の核酸。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アンチセンスオリゴヌクレオチドに関する。より詳細には、産生量の減少が求められる望ましくないタンパク質をコードするmRNA前駆体のエキソンの内部領域に対するアンチセンスオリゴヌクレオチドであって、14〜40個のヌクレオチドを含有し、該エキソンの有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチド、および上記アンチセンスオリゴヌクレオチドを用いたRNAスプライシングの制御方法を提供する。
【背景技術】
【0002】
急速なヒトゲノム研究の発展に伴い、専門家及び一般の人々は、病気のメカニズムの解明及び正確かつ信頼性の高い診断法に加えて、数多くの重度の遺伝病の治療法が近い将来確立されることを期待している。
【0003】
新たな見識によって投与の容易な低分子遺伝子療法が疾病(例えば代謝異常)の治療法として開発されることが期待されるが、その一方で、多くの疾病においては、上記以外の遺伝子療法、即ち、異常な遺伝子産物の修正、付加または置換、が最終的に必要となることが考えられる。
【0004】
近年、この分野における調査及び開発によって、克服しなければならないいくつかの技術的問題が浮き彫りになった。例えば、遺伝病に関わる複数の遺伝子が広範にわたること(よって治療遺伝子を投与すべき適切な反応系の選択が制限される)、治療用遺伝子の機能すべき組織への到達性(特異的なターゲッティング技術、即ち遺伝子の制限的挿入による物理的ターゲッティング、または組織特異的な親和性を有するシステムの開発による生物学的ターゲッティング、の構築が必要である)、及び投与システムの患者に対する安全性に関連する問題が挙げられる。これらの問題はある程度の相互関係があり、治療用薬剤が小さくなるほど、高効率で、ターゲッティング可能な安全性の高い投与システムの開発が容易であると一般的に結論付けることができる。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明においては、いわゆるエキソンスキッピングを細胞に誘導することによってこれらの問題に対処する。エキソンスキッピングは、スキップされたエキソンを有さない成熟mRNAを生じるので、スキップされたエキソンがアミノ酸をコードしている場合、このスキッピングは変化した産物の発現につながる。現在、エキソンスキッピングに関する技術はいわゆる「アンチセンスオリゴヌクレオチド (Anti-sense Oligonucleotide)」(AON)を使用するものであり、その研究のほとんどは、デュシェンヌ型筋ジストロフィー(Duchenne muscular dystrophy)(DMD)のモデルであるmdxマウスを利用して行われている。ジストロフィン遺伝子のエキソン23にナンセンス突然変異を有するmdxマウスは、デュシェンヌ型筋ジストロフィーの動物モデルとして利用されている。機能性ジストロフィンタンパク質の合成を排除するはずのmdx突然変異を有するにもかかわらず、稀に天然のジストロフィン陽性繊維がmdx筋組織中に観察される。このようなジストロフィン陽性繊維は、体性突然変異、または選択的スプライシングによる、天然のエキソンスキッピング機構によって発生したものと考えられる。ジストロフィンmRNA前駆体のイントロン22及び23のそれぞれの3’及び5’スプライス部位に特異的なAONはイントロン23の除去に通常関与する因子と干渉し、エキソン23もmRNAから除去することが示された(Wilton, 1999)。同様の研究において、Dunckleyら(1998)は、3’及び5’スプライス部位に特異的なAONを使ったエキソンスキッピングが思いがけない結果をもたらしうることを示した。彼らは、エキソン23のみならず、エキソン24〜29のスキッピングを観察し、その結果、エキソン22とエキソン30の結合体を有するmRNAが生成された。思いがけない22−30スプライシングバリアントの出現の根底にあるメカニズムは不明である。このようなバリアントの出現は、スプライス部位がコンセンサス配列を含んでおり、その結果、エキソンスキッピングを導くために使用したオリゴヌクレオチドによる非特異的なハイブリダイゼーションが生じたからと考えられる。オリゴヌクレオチドをスキップされるエキソン部位以外のスプライス部位にハイブリダイズさせることによって、当然のことながら、スプライシング反応の精度に容易に干渉することができる。一方、スプライシング反応が精度に欠ける理由としては、(5’及び3’スプライス部位のそれぞれのための)2個のオリゴヌクレオチドの使用が必要であるためと考えられる。1個のオリゴヌクレオチドを有するが、他のオリゴヌクレオチドを有しないmRNA前駆体は、思いがけないスプライシングバリアントとなる傾向がある。
【課題を解決するための手段】
【0006】
上記及びその他の諸問題を克服するために、本発明はRNAスプライシング反応系のmRNA前駆体を、該mRNA前駆体に含まれる少なくとも1つのエキソンが有するエキソン封入シグナル(exon inclusion signal)(EIS)を特異的に阻害することが可能な試薬と接触させ、そして該mRNA前駆体のスプライシングを行わせしめることを包含する、RNAスプライシング反応系においてmRNA前駆体のスプライシングを制御するための方法を提供する。エキソン封入シグナルに干渉することの利点は、このような因子がエキソン内に存在するということである。スキップされるエキソン内の配列に対するアンチセンスオリゴヌクレオチドを与えることによって、エキソン封入シグナルと干渉して、エキソンをスプライシング機構から効果的に遮蔽することができる。スプライシング機構がスキップすべきエキソンを認識しないことによって、最終的なmRNAからエキソンが排除される。本発明は、スプライシング機構の酵素反応(エキソンの結合)に直接干渉するものではない。このことによって本発明は、確実且つ信頼性の高い方法であると考えられる。EISはエキソンに特異的な構造であり、スプライス受容部位及びスプライス供与部位に特定の立体配置を付与するものと考えられる。このような概念においては、特定の立体配置によってスプライシング機構がエキソンを認識すると考えられている。しかしながら、本発明はこのモデルに限定されるものではない。本発明者らは、エキソンに結合可能な試薬がEISを阻害することを見出した。このような試薬は、エキソンのいかなる部位にも特異的に接触することができるにもかかわらず、EISを特異的に阻害することができる。本発明の方法で得られるmRNAそのものが有用である。例えば、mRNAに必要なエキソンが封入されるのを阻害することで、望ましくないタンパク質の産生を少なくとも部分的に減少させることができる。本発明の方法は、mRNA前駆体のスプライシングによって生じるmRNAを翻訳する工程を更に包含することが好ましい。また、mRNAが機能性タンパク質をコードしていることも好ましい。本発明の好ましい態様においては、タンパク質が2つ以上のドメインを包含し、ドメインの少なくとも1つは、mRNA前駆体に含まれるエキソンの少なくとも一部のスキッピングによって生じたmRNAにコードされている。エキソンスキッピングが有効な典型的な例としては、これに限定されるわけではないが、少なくとも2つの機能性ドメインを有する野生型構造のタンパク質であって、各ドメインは一次構造であるアミノ酸配列の個別の部分から生成されるものである。具体的には、転写因子が挙げられる。典型的な転写因子はDNA結合ドメイン及び細胞中の他のタンパク質と反応するドメインを包含する。2つのドメインの間に存在するアミノ酸の一次構造の一部をコードするエキソンのスキッピングによって、少なくとも部分的には同等の機能を発揮する、より短いタンパク質の産生につながる。従って、中間領域における不利益な突然変異(例えばフレームシフト突然変異や停止変異)はエキソンスキッピングを誘導することで少なくとも部分的に修復することが可能であり、(部分的な)機能を有する短いタンパク質を合成することができる。本発明の方法を用いると、エキソンの部分的なスキッピングを誘導することも可能である。この態様においては、試薬とエキソンの接触によって、試薬と接触したエキソンの隠蔽されたスプライス部位が活性化される。この態様は、機能性タンパク質を誘導するmRNA前駆体の操作性を拡大する。上記反応系は細胞を包含することが好ましい。細胞は試験管内(in vitro)で培養されるか、または生体内(in vivo)で増殖したものであることが好ましく、これらに限定されるわけではないが、典型的な生物の例にはヒトやマウスが包含される。
【図面の簡単な説明】
【0007】
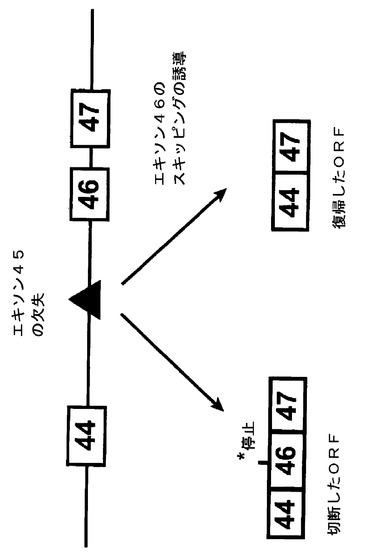
【図1】エキソン45の欠失は最も頻発するDMD突然変異の1種である。この欠失によって、エキソン44はエキソン46にスプライシングされ、翻訳読み枠は切断されてエキソン46の内部に終止コドンが発生し、その結果、ジストロフィンの欠乏が生じる。本発明者らの目的は、更なるエキソン、つまりエキソン46のスキッピングを人工的に誘導して読み枠を再構築することで、エキソン45及び46の欠失を保有する、より穏やかな疾患であるベッカー型筋ジストロフィーの患者に見られるようなわずかに短いが非常に高い機能性を有するジストロフィンタンパク質の合成を復帰させる。
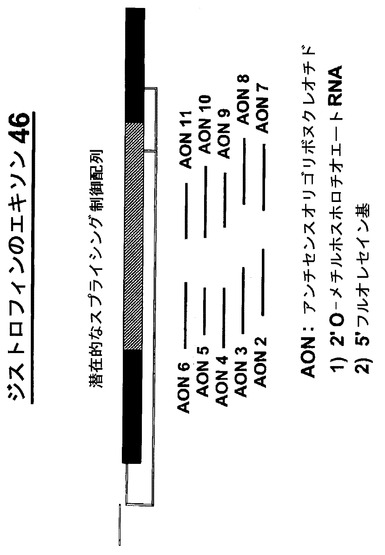
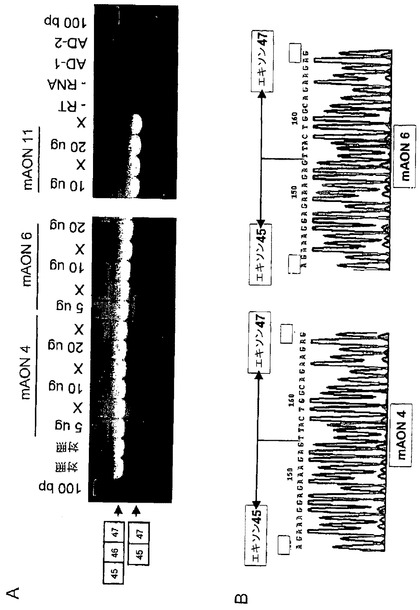
【図2】エキソン46は、プリン塩基に富んだ、mRNA前駆体のスプライシング制御を潜在的に担うと考えられる領域を有する。このマウスジストロフィンエキソン46の内部に存在するプリン塩基に富んだ領域に対して、重なり合う2’−O−メチルホスホロチオエートアンチセンスオリゴリボヌクレオチド(AON)からなるプライマー系列を設計した。これらのAONは長さと配列の両方の点で異なっている。AONの化学的修飾は、筋肉細胞内のエンドヌクレアーゼ及びRNase Hに対する抵抗性を付与する。in vitro の研究におけるトランスフェクション効率を調べるために、AONはAON陽性細胞の同定を可能にする5’フルオレセイン基を含有していた。
【図3】ターゲットとなるエキソン46のRNAに対する種々のAONの結合親和性を調べるために、ゲルシフトアッセイを行った。図にはターゲットRNAに最も高い親和性を示す5個のmAON(mAON#4,6,8,9及び11)を示した。AONがRNAに結合すると、ゲル移動度の低下した複合体が形成されるので、バンドの変化によって検出することができる。AONのターゲットに対する結合は配列特異的である。ランダムmAON、即ちエキソン46に非特異的なmAONはバンドの変化を生じなかった。
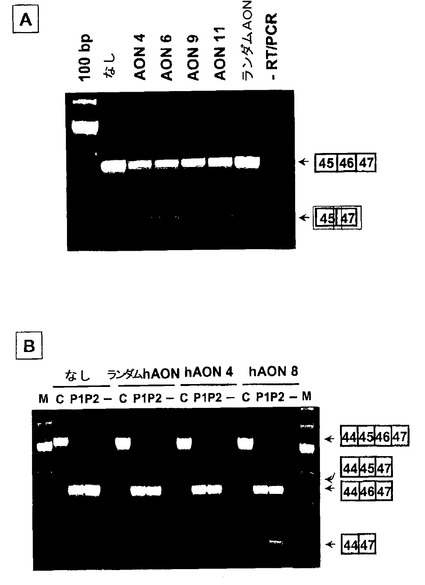
【図4】ゲルシフトアッセイで最も高い結合親和性を示したマウス特異的AON及びヒト特異的AONをそれぞれマウス培養筋管及びヒト培養筋管にトランスフェクトした。図4A:mAON#4,6,9及び11をトランスフェクトした培養マウス細胞のRT−PCR分析は、エキソン45がエキソン47に直接スプライシングされた産物に相当する大きさの部分産物を示した。ランダムAONをトランスフェクトした場合には、エキソン46のスキッピングは検出されなかった。図4B:罹患していない1人(図中の“C”)及び血縁ではない2人のDMD患者(図中の“P1”及び“P2”)のそれぞれに由来する培養ヒト筋肉細胞のRT−PCR分析は、hAON#4及びhAON#8をトランスフェクトした際に部分産物が生じることを明らかにした。対照においては、この産物はエキソン45がエキソン47にスプライシングされた産物に相当するが、一方で、患者に見られる断片の大きさは、エキソン44がエキソン47にスプライシングされた断片に相当した。トランスフェクトしなかった培養細胞及びランダムhAONでトランスフェクトした培養細胞では、エキソン46のスキッピングは検出されなかった。最も高いエキソン46スキッピング効率がhAON#8で得られた。
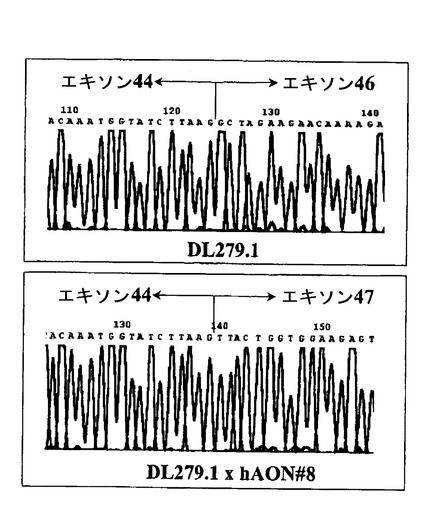
【図5】患者DL279.1(図4の“P1”に対応する)から得られたRT−PCR産物の配列データであり、このデータによって患者のエキソン45が欠失しており(上のパネル)、hAON#8のトランスフェクションによって更にエキソン46のスキッピングが起こる(下のパネル)ことを確認した。エキソン46のスキッピングは特異的であり、エキソン44はエキソン47に正確にスプライシングされ、翻訳読み枠を再構築する。
【図6】hAON#8でトランスフェクトした患者DL279.1由来の培養筋肉細胞の免疫組織化学分析。タンパク質の異なる領域に対して作成した2種のジストロフィン抗体と細胞を接触させた。使用した抗体は、ターゲットであるエキソン46に近接する領域に対する抗体(ManDys-1、ex. -31-32)及びエキソン46から離れた領域に対する抗体(Dys-2、ex. -77-79)である。下のパネルは筋管中にジストロフィンタンパク質が存在しないことを示すのに対し、いずれの抗体でもジストロフィンタンパク質が検出されることから、エキソン46のhAON#8誘導性スキッピングは明らかにジストロフィンタンパク質の合成を復帰させた(上のパネル)。
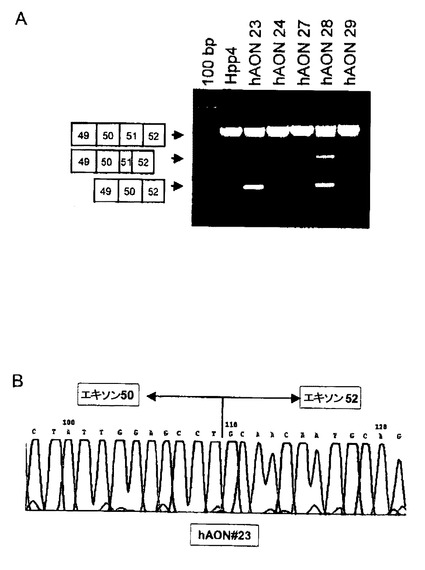
【図7】図7A:hAON#23,#24,#27,#28または#29で処理した、対照である培養ヒト筋肉細胞から単離したRNAのRT−PCR分析。エキソン50がエキソン52にスプライシングされた産物に相当する大きさの部分産物が、hAON#23及び#28で処理した細胞で検出された。これらの産物の配列を決定したところ、エキソン51が正確にスキップされていることが確認された(図7B)。さらに異常スプライシング産物をhAON#28及び#29で処理した細胞から得た。配列を決定したところ、AON処理に伴い、低い頻度で使用されるエキソン51のフレーム内の隠蔽されたスプライス部位の存在が明らかになった。AON処理によって生じた産物には復帰した読み枠からなるエキソン51の部分産物が含まれており、更なる治療価値が確認された。
【図8】図8A:エキソン29ターゲットRNAに対する種々のhAON#29の結合親和性を確認するために、ゲルシフトアッセイを行った。ハイブリダイゼーションしていないRNA(図中の「なし」)に比べると、h29AON#1,#2,#4,#6,#9,#10及び#11はゲル移動度の低い複合体を形成し、RNAに対する結合を示した。ジストロフィンエキソン19由来のランダムAONは複合体を形成しなかった。図8B:h29AON#1,#2,#4,#6,#9,#10または#11で処理した対照である培養ヒト筋肉細胞から単離したRNAのRT−PCR分析は、エキソン28がエキソン30にスプライシングされた産物に相当する大きさの部分産物の存在を明らかにした。これらの結果は、エキソン29内のERSであると考えられる領域の内側の配列に対するAON(h29AON#1,#2,#4または#6)あるいは外側の配列に対するAON(h29AON#9,#10または#11)を使ってエキソン29を特異的にスキップすることが可能であることを示す。エキソン28及び29の両方のスキッピングによって生じた更なる異常型スプライシング産物が検出された(配列データによって確認したがここには示さない)。この異常型スプライシング産物は未処理の細胞にも存在したが、これはAON処理によって促進される天然の選択的なスキッピングによるものと考えられる。ジストロフィンエキソン19から得たAON19はエキソン29のスキッピングを誘導しなかった。図8C:エキソン29の特異的スキッピングを部分配列であるRT−PCR断片の配列データによって確認した。h29AON#1で処理した細胞中のエキソン29スキッピング産物の塩基配列をここに示す。
【図9】図9A:マウス腓腹筋由来RNAのRT−PCR分析であって、RNAはmAON#4,#6または#11をそれぞれ5,10または20μg投与後2日目に単離したものである。エキソン45がエキソン47にスプライシングされた産物に相当する大きさの部分産物をすべての処理筋肉から検出した。RT−PCR反応の負の対照として“−RT”,“−RNA”,“AD−1”及び“AD−2”について分析した。図9B:mAON#4と#6(#11も実験したが、データは示さない)によって生じた部分産物の配列を決定し、エキソン46の正確なスキッピングを確認した。
【発明を実施するための形態】
【0008】
好ましい態様において、本発明は異常型タンパク質をコードするエキソンを含むmRNA前駆体を有する細胞に対して、該エキソンの少なくとも1つが有するエキソン封入シグナルを特異的に阻害することが可能な試薬を細胞に与え、そして該mRNA前駆体のスプライシングによって生じるmRNAの翻訳を行わせしめることを包含する、細胞による異常型タンパク質の産生を少なくとも部分的に減少させるための方法を提供する。
【0009】
エキソン封入シグナルを特異的に阻害することが可能ないかなる試薬も本発明に用いることができる。該試薬は、核酸またはそれと同等の機能を有する分子を含有することが好ましいが、該核酸は必ずしも一本鎖でなくてもよい。ペプチド核酸及び同程度の核酸結合特性を有する他の分子も用いることができるが、その結合特性は必ずしも結合量が同等でなくてもよい。核酸またはそれと同等の機能を有する分子は付加的な機能性を発揮するために修飾されていてもよい。例えば、2’−O−メチルオリゴリボヌクレオチドを使用することができる。このようなリボヌクレオチドは従来のオリゴヌクレオチドよりもRNase活性に対する抵抗性が高い。
【0010】
本発明の好ましい態様において、エキソン封入シグナルはエキソン認識配列(exon recognition sequence)(ERS)に特異的なアンチセンス核酸による干渉を受ける。このような配列は比較的プリン塩基に富み、スキップされるエキソンの配列情報を精査することで特定することが可能である(Tanakaら、1994, Mol. Cell Biol. 14: p. 1347-1354)。エキソン認識配列は、いわゆる弱いエキソンのmRNAへの封入を補助すると考えられる(Achselら、1996, J. Biochem. 120; p. 53-60)。このような弱いエキソンは、例えば、スプライシング機構による認識効率が低い5’及び/または3’スプライス部位を包含する。本発明においては、エキソンスキッピングはいわゆる強いエキソン、すなわち細胞のスプライシング機構によって通常効率よく認識されるエキソン、においても誘導することができることを見出した。与えられたいかなる配列からも、その配列がエキソンと推定される領域を包含するかどうかを予測し、そのエキソンが強いか弱いかを確認することが(ほとんどの場合)可能である。エキソンの強度を確認するためのいくつかの手法が存在する。有用な手法としてNetgene2スプライス部位予測サーバー(NetGene splice site prediction server)(Brunakら、1991, J. Mol. Biol. 220: p. 49-65)で検索することができる。本発明の方法によるエキソンスキッピングは(ほとんど)すべてのエキソンに誘導することが可能であり、エキソンが弱いエキソンであるか強いエキソンであるか、ならびにエキソンがERSを包含するかどうかにも依存することはない。好ましい態様において、スキッピングのターゲットとなるエキソンは強いエキソンである。他の1つの好ましい態様において、スキッピングの対象となるエキソンはERSを包含しない。
【0011】
本発明の方法は様々な用途に使用することが可能である。1つの態様において、本発明の方法は異常型タンパク質の産生を少なくとも部分的に減少させるために使用する。このようなタンパク質としては、例えば、腫瘍タンパク質またはウイルス性タンパク質が挙げられる。多くの腫瘍において、腫瘍タンパク質の存在のみならず、その相対的な発現程度が腫瘍細胞の表現型に関係している。同様に、ウイルス性タンパク質の存在のみならず、細胞中のウイルス性タンパク質の量が特定のウイルスの毒性を決定している。さらに、ウイルスの有効な増加及び蔓延のために、ウイルスの生活環における発現の時期及び細胞中の特定のウイルス性タンパク質の量的バランスによって、ウイルスが効率的にまたは非効率的に増殖するかどうかが決定される。本発明の方法を使用して細胞中の異常型タンパク質の量を減少させることで、例えば腫瘍細胞の腫瘍形成性(転移性)及び/またはウイルス感染細胞のウイルス増殖性を低下させることができる。
【0012】
好ましい態様において、本発明の方法を用いて異常型タンパク質を機能性タンパク質に変更する。1つの態様において、上記のような機能性タンパク質は、細胞に通常存在するが、処理する細胞には存在しないタンパク質の機能を発揮する。大抵の場合、機能を部分的に修復しただけでも、処理した細胞の性能が著しく向上する。向上した性能故に処理した細胞は未処理の細胞よりも選択的な優位性を示すことができ、処理の有効性を高めている。
【0013】
上述した本発明の態様は、特に欠失した遺伝子の発現を復帰させるのに適している。これはターゲットとなるエキソンの特異的なスキッピングを行い、有害な変異(典型的なものとしては停止変異またはフレームシフト点突然変異、あるいは翻訳停止をもたらす1つ以上のエキソンの欠失または挿入)を回避するか修正することによって達成される。
【0014】
遺伝子導入法に比べて、本発明の新規なスプライスシング制御型遺伝子治療(splice-modulation gene therapy)は、より小さな治療薬、これに限定されるわけではないが、典型的な例としては14〜40個のヌクレオチドからなる治療薬の投与を必要とする。好ましい態様においては、製造が容易であり、細胞による取り込みがより効果的に行われることから、14〜25個のヌクレオチドからなる分子を使用する。本発明の方法は、治療薬に付随する、効果的で安全な投与システムの設計をより柔軟に行うことを可能とする。本発明のこのような態様の重要な更なる利点は、大部分またはすべての遺伝子調節回路を未だ保持する内因性遺伝子活性の(少なくとも一部)を復帰させることで、組織特異的なアイソフォームの適切な発現レベルと合成を保障することである。
【0015】
上述の本発明の態様は、原則として、いかなる遺伝病や遺伝的疾病素質にも適用することができ、配列内部においてわずかに短いタンパク質の翻訳が完全にまたは部分的に機能している場合に限り、特定のエキソンを標的とするスキッピングによって、元々変異によって破壊された翻訳読み枠を復帰させることができる。この方法の適用に治療的価値が見出される態様としては、腫瘍抑制遺伝子の二次変異によって生じる疾病素質が挙げられ、例えば、乳ガン、結腸ガン、結節性硬化症、神経線維腫症などにおいては、活性の(部分的な)回復が二次変異による異数性の発現を排除し、腫瘍形成を防ぐ。他の好ましい態様には、遺伝子産物の欠失が直接的な疾病誘引効果を示す遺伝子の(部分的な)修復が含まれる。例えば、血友病A(血液凝固第VIII因子の欠乏によるもの)、先天的甲状腺機能不全症のいくつかの形態(チログロブリン合成欠損によるもの)、及びデュシェンヌ型筋ジストロフィー(DMD)が挙げられる。DMDは、フレームシフトによって生じるX連鎖性ジストロフィン遺伝子の欠失、重複及び停止突然変異が重度の進行性筋力低下をもたらす疾患である。通常DMDは発症すると青年期の後期または成人期の初期に死に至るが、その一方で、同じ遺伝子座にフレームシフト以外の方法で欠失または重複突然変異が生じて発症したベッカー型筋ジストロフィーは軽症であり、平均余命は35〜40年から正常人と同じである。DMDに適用する本発明の態様においては、本発明の方法によって現存する欠失領域をエキソンスキッピングによって伸張する(または現存する重複突然変異体のmRNA産物を改変する)ことができる。これは読み枠を復帰させ、配列内部においてわずかに短くなっているが機能性のタンパク質を産生するのに必要な数の近接したエキソンのスキッピングによって行う。このような欠失の誘導と同等の変異を有するBMD患者の臨床的症状が軽いことに基づき、AON療法を受けたDMD患者の症状は軽減されると考えられる。
【0016】
ジストロフィン遺伝子に生じる種々の変異が機能不全性タンパク質につながる。(総合目録としてhttp://www.dmd.nl、即ち、DMD及び関連疾患の国際的に承認されたデータベース、を参照)。機能性ジストロフィンタンパク質を産生させるために実際にスキップすべきエキソンは突然変異によって変化する。表1はスキップすることのできるエキソンの一覧であるが、本願はこれらに制限されることはない。この表においては、上述のエキソンに対して、ヒトで観察された高頻度で発生するジストロフィン遺伝子変異であって、本発明の方法で治療することができるものも示した。表に示したエキソンのスキッピングは、少なくともベッカー型筋ジスロトフィータンパク質の機能を有する変化したジストロフィンタンパク質の産生をもたらす。このように、本発明の1つの態様において、エキソン封入シグナルがヒトジストロフィン遺伝子のエキソン2,8,43,44,45,46,50,51,52または53に存在することを特徴とする方法が提供される。ある欠失/挿入変異の発生は、他の変異より高頻度で発生する。本発明の方法でエキソン46のスキッピングを誘導すると、DMDの欠失を保有する患者の約7%で治療に成功し、ジストロフィン陽性筋肉繊維がジストロフィン患者に検出されることが明らかとなった。また、本発明の方法でエキソン51のスキッピングを誘導することによって、DMDの欠失を保有する患者の約15%を治療することに成功した。この治療方法によって、該患者は少なくとも少量のジストロフィン陽性筋肉繊維を有するようになった。本発明の方法を使用したエキソン46または51のスキッピングによって、ジストロフィン遺伝子の欠失を保有する患者の約22%を治療することができる。従って、本発明の好ましい態様においては、エキソン封入シグナルはエキソン46またはエキソン51に存在する。特に好ましい態様においては、使用する試薬は以下の核酸配列を包含する:hAON#4、hAON#6、hAON#8、hAON#9、hAON#11及び/またはhAON#21〜30からなる群から選ばれる少なくとも1つの核酸配列;或いは上記hAON#の機能性領域、誘導体及び/または類似体。機能性領域、誘導体及び/または類似体は、上記hAON#と同等のエキソンスキッピング活性を有するが、本発明の方法においては、該活性量が必ずしも同等でなくてもよい。
【0017】
mRNA前駆体に含まれる1つ以上のエキソンのエキソンスキッピングを誘導することは効果的である。例えば、変異の多様性並びにエキソンの長さや変異に隣接するアミノ酸配列といった一定の性質を考慮すると、機能を復帰させるためには1つ以上のエキソンをスキップしなければならないという状況が起こりうる。DMD欠失データベースに見られるこのような状況の好ましい例としては、エキソン46〜50の欠失が挙げられるが、これに限定されるものではない。エキソン46〜50が欠失している患者は機能性ジストロフィンタンパク質を生成しない。しかしながら、エキソン45及びエキソン51のスキッピングを誘導することによって少なくとも部分的な機能を有するジストロフィンタンパク質を生成することができる。他の好ましい例としてはエキソン2の重複を有する患者が挙げられるが、これに限定されるものではない。エキソン2のEISを阻害することが可能な試薬を与えることで、エキソン2の一方または両方を部分的にスキップし、エキソン2の部分配列からなる領域または2つのエキソン2を欠失した領域の隣から野生型タンパク質を得ることが可能である。他の好ましい例として、エキソン45〜50のスキッピングが挙げられるが、これに限定されるものではない。このスキッピングにより読み枠内でベッカー様バリアントが生じる。ベッカー様バリアントはエキソン45,46,47,48,49及び/または50あるいはそれらの組み合わせに存在するいかなる変異を治療するためにも誘導することができる。本発明のその他の態様においては、mRNA前駆体に含まれる他のエキソンの有するエキソン封入シグナルを阻害する他の試薬を細胞に与える工程を更に包含することを特徴とする方法が提供される。mRNA前駆体の2つ以上の異なる遺伝子にエキソンスキッピングを誘導するために2つ以上の試薬を使用することは、当然のことながら本発明の範囲に含まれる。
【0018】
他の1つの態様において、本発明はスプライシング制御型遺伝子治療に適した試薬を選択するための方法及び選択した試薬の特異的エキソンスキッピング試薬としての有効性を予備実験で確認するための方法を提供する。目的エキソンの一部に対して相補的な試薬が、該エキソンの有するエキソン封入シグナルを特異的に阻害することが可能であることを確認するための方法であって、目的のエキソンを含むmRNA前駆体を有する細胞に被験試薬を与え、該細胞を培養して該mRNA前駆体からmRNAを生成せしめ、そして生成した該mRNAに該エキソンが存在しないことを確認することを包含する方法が提供される。好ましい態様において、該試薬が、核酸またはそれと同等の機能を有する分子を含有し、該核酸は目的エキソンの一部に対して相補的である。特異的エキソンスキッピングを誘導することが可能な試薬を本発明の方法で同定することができる。この方法には、該試薬がエキソンを含む核酸、好ましくはRNA、に比較的高い親和性で結合するかどうかを確認するための予備スクリーニングを行うことも含まれる。この結果、試薬がエキソンの有するエキソン封入シグナルを特異的に阻害することが可能であることを確認するための方法が提供され、該方法は試験管内の条件において、該核酸またはそれと同等の機能を有する分子と該目的エキソンを包含するmRNA分子との相対的な結合親和性を測定する工程を更に包含する。
【0019】
他の態様において、本発明の方法によって得られる試薬を提供する。好ましい態様においては、該試薬は核酸またはそれと同等の機能を有する分子を包含する。該試薬は、それを用いて細胞にエキソンスキッピングを誘導した場合、細胞中の異常型タンパク質の量を少なくとも部分的に減少させることが好ましく、該エキソンスキッピングによって、該細胞で機能を発揮しうるタンパク質をコードするRNAを生じることが更に好ましい。特に好ましい態様において、mRNA前駆体はジストロフィン遺伝子から誘導されたものである。機能性タンパク質は変異ジストロフィンタンパク質または正常ジストロフィンタンパク質を包含することが好ましい。また、変異ジストロフィンタンパク質が少なくともベッカー型筋ジストロフィー患者のジストロフィンタンパク質の機能を有することが好ましい。特に好ましい態様においては、使用する試薬は以下の核酸配列を包含する:hAON#4、hAON#6、hAON#8、hAON#9、hAON#11及び/またはhAON#21〜30からなる群から選ばれる少なくとも1つの核酸配列;或いは上記hAON#の機能性領域、誘導体及び/または類似体。機能性領域、誘導体及び/または類似体は、上記hAON#と同等のエキソンスキッピング活性を有するが、本発明の方法においては、該活性量が必ずしも同等でなくてもよい。
【0020】
細胞に試薬を導入するための種々の方法が当業界では知られている。特に核酸の導入方法は広範にわたって開発されている。当業者はある導入方法が本発明を実施するのに適しているかどうかを十分確認することができる。このような方法の一例は、本発明の試薬をリポソームに封入し、該リポソームを目的mRNA前駆体を有する細胞に与えることを包含するが、本発明はこの例に限定されるものではない。リポソームは核酸を細胞に導入するための運搬体として特に適している。エキソンスキッピングを誘導することのできるアンチセンス分子は、アンチセンスRNAを生成するための転写単位を包含する核酸を導入し、細胞に生成させることが可能である。適切な転写単位としては核内低分子RNAまたはtRNA転写単位が挙げられるが、これらに限定されるものではない。従って、本発明は、エキソン封入シグナルを阻害することが可能な本発明の核酸またはそれと同等の機能を有する分子を包含する核酸運搬体を更に提供する。1つの態様において、該運搬体は本発明の核酸を発現させることが可能である。一本鎖ウイルスを運搬体として利用した場合には、当然のことながら、そのようなウイルスが本発明の試薬のアンチセンス配列だけを包含するものであっても本発明の範囲に含まれる。また、このような一本鎖ウイルスの他の1つの態様においては、本発明のAONは、運搬体としてのウイルスに封入されているウイルス核に存在する核内低分子RNAまたはtRNA転写単位によってコードされている。好ましい一本鎖ウイルスはアデノ随伴ウイルスである。
【0021】
更なる1つの態様において、本発明は医薬の調製に用いる、核酸または核酸運搬体を提供する。好ましい態様において、該医薬を遺伝病の治療に使用する。さらに好ましくは、該医薬をデュシェンヌ型筋ジストロフィーの治療に使用する。
【実施例】
【0022】
エキソン45はDMDにおいて最も高頻度で欠失が起こるエキソンなので、本発明者らははじめにエキソン46の特異的なスキッピングの誘導を試みた(図1)。この誘導によってエキソン45及び46の欠失を保有するBMD患者に見られる、短いがより機能的なジストロフィンタンパク質を産生する。マウスジストロフィン遺伝子のジストロフィンmRNA前駆体のスプライシングを制御するための実験系を初めに構成した。その後、エキソン45の欠失を保有するDMD患者由来の筋肉細胞において翻訳読み枠とジストロフィン合成を復帰させることを目的として、ヒトジストロフィン遺伝子にねらいを定めた。
【0023】
mAONs及びhAONsの設計
エキソン46の内部領域に対するマウス特異的AON系列及びヒト特異的AON系列(mAON及びhAON)を設計した(図2)。使用したエキソン46の内部領域は、プリン塩基に富んだ配列を有し、エキソン46のスプライシング制御に係る推定上の機能を有すると考えられる。ゲルシフトアッセイによるAONの初期スクリーニング(下記参照)のために未修飾のDNAオリゴヌクレオチド(ベルギー国、EuroGentec社に合成を依頼)を使用した。筋肉細胞のトランスフェクション実験を実際に行う際には、2’−O−メチルホスホロチオエートアンチセンスオリゴリボヌクレオチド(ベルギー国、EuroGentec社に合成を依頼)を使用した。このような修飾されたRNAオリゴヌクレオチドはエンドヌクレアーゼ及びRNase Hに対する抵抗性を有し、高い親和性でRNAに結合することが知られている。最終的に有効であると認められ、in vitro で筋肉細胞に与えたAONの配列を下記に示す。対応するマウス特異的AONとヒト特異的AONは高い相同性があるが完全に同一ではない。
【0024】
以下に使用したAONのデオキシ体を示すが、最終的に使用した2’−O−メチルリボヌクレオチドにおいては、TをUに置き換える。
【0025】

【0026】
【0027】
ゲルシフトアッセイ
AONの有効性は、ターゲット配列に対する結合親和性に基づいて決定した。RNAフォールディングを予測するためのコンピュータシミュレーションプログラムの近年の発展にもかかわらず、設計したAONのどれが高い親和性でターゲット配列に結合するかを推測することは難しい。従って、ゲルシフトアッセイを(Bruiceら、1997に記載のプロトコールに従って)行った。エキソン46ターゲットRNA断片を、(マウスまたはヒト筋肉mRNAから、T7プロモーター配列を有するセンスプライマ−を利用して増幅した)PCR断片から32P−CTPの存在下のin vitro T7転写法で生成した。ターゲットである転写断片に対する個々のAON(0.5pmol)の結合親和性は、37℃で30分間ハイブリダイゼーションを行い、次いでポリアクリルアミド(8%)ゲル電気泳動を行うことで測定した。これらの分析をマウス特異的AON及びヒト特異的AONをスクリーニングするために行った(図3)。少なくとも5個の異なるマウス特異的AON(mAON#4、6、8、9及び11)及びそれらに対応する4個のヒト特異的AON(hAON#4、6、8及び9)において移動度の変化が観察され、ターゲットRNAに対して結合親和性を有することを示した。
【0028】
培養筋肉細胞のトランスフェクション
AONによるin vitro の筋肉細胞におけるスキッピング誘導効率を分析するために、ゲルシフトアッセイを用いてターゲットエキソンに対して最も高い結合親和性を示したエキソン46特異的AONを選択した。すべてのトランスフェクション実験において、非特異的AONをエキソン46の特異的スキッピングの負の対照として用いた。前述したように、はじめにマウス筋肉細胞の実験系を構築した。マウス筋肉細胞系であるC2C12細胞由来の(ジストロフィン発現レベルの高い)培養増殖筋芽細胞及び培養分裂後筋管の両方を使用した。後に行ったヒト由来培養筋肉細胞の実験には、罹患していない1人の筋肉生検材料及びエキソン45の欠失を保有する、血縁ではない2人のDMD患者の筋肉生検材料のそれぞれから単離した培養一次筋肉細胞を使用した。これらの異種培養物はおよそ20〜40%の筋原細胞を含んでいた。異なるAON(濃度:1μM)に対して3当量比のカチオンポリマーPEI(MBI Fermentas社製)を用い、細胞にAONをトランスフェクトした。これらの実験でトランスフェクトしたAONは5’フルオレセイン基を有し、そのため蛍光性の核を数えることでトランスフェクション効率を測定することができる。概して、60%以上の細胞が核に特異的にAONの取り込みを示した。RT−PCR分析を促進するために、トランスフェクトの24時間後にRNAzolB(オランダ国、CamPro Scientific社製)を使ってRNAを単離した。
【0029】
RT−PCR及び配列の決定
RNAの逆転写をC. therm. ポリメラーゼ(Roche社製)及びエキソン48特異的逆転写プライマーを使って行った。ジストロフィン遺伝子のエキソン46のスキッピングを検出するために、PCRを2サイクル行ってcDNAを増幅した。PCRは、エキソン44及び47に含まれるプライマー(ヒト実験系)またはエキソン45及び47に含まれるプライマー(マウス実験系)を使用したプライマーシフト増幅(nested amplification)を包含する。培養マウス筋芽細胞及び培養マウス筋管細胞では、エキソン45がエキソン47に直接スプライシングされた産物に相当する大きさの部分産物を検出した(図4)。続いて行った配列の決定によって、マウスジストロフィン転写産物におけるエキソン46の特異的なスキッピングを確認した。エキソンスキッピングの効率は個々のAONで異なり、mAON#4及び#11が最も高い効率を示した。これらの好ましい結果に基づき、培養ヒト筋肉細胞においても、ジストロフィンのスプライシングを同様に制御することに着目した実験を行った。その結果、エキソン45がエキソン47にスプライシングされた産物に相当する部分産物を対照となる筋肉細胞で検出した。興味深いことに、患者由来筋肉細胞では、エキソン44がエキソン47にスプライシングされてなる短い断片が検出された。エキソン46の特異的なスキッピングをヒトジストロフィン転写産物の配列データによって確認した。マウスとヒトの両方のジストロフィン転写産物に見られたこのようなスプライシングの制御は、トランスフェクトされていない培養細胞や非特異的AONでトランスフェクトされた培養細胞には見られなかった。
【0030】
免疫組織化学分析
ジストロフィンタンパク質の翻訳および合成を復帰させるために、エキソン45の欠失を保有する患者由来の筋肉細胞においてエキソン46のスキッピングの誘導を試みた。hAON#8でトランスフェクトした際のジストロフィン産物を検出するために、ジストロフィンタンパク質のターゲット領域に隣接するドメインと遠位のドメインのそれぞれに対して作成した2種のジストロフィンモノクローナル抗体(Mandys-1及びDys-2)を使用して、2種の患者由来培養筋肉細胞を免疫細胞化学分析に付した。蛍光分析によって、いずれの患者由来培養細胞においてもジストロフィン合成が復帰していることが明らかになった(図5)。処理したサンプルにおいては、約80%以上の筋繊維がジストロフィンに対して陽性に染色された。
【0031】
本発明者らの実験結果は、DMD患者由来筋肉細胞の内因性DMD遺伝子によるジストロフィン合成の復帰を初めて示す。これは、ジストロフィンmRNA前駆体の目的に応じたスプライシング制御を治療目的で実施するための原理を証明する。
【0032】
エキソン51のターゲットスキッピング
ジストロフィンエキソンの同時スキッピング
エキソン51のターゲットスキッピング。本発明者らは、in vitro のマウス及びヒト筋肉細胞におけるジストロフィンのエキソン46のAONによる制御の可能性について明示した。これらの知見はAONをDMDの治療用の試薬として評価する更なる研究の根拠を示した。DMDを誘発する欠失の大部分は遺伝子内の突然変異が起こりやすい2つの部位で集中的に発生し、ある1つの特定エキソンのターゲットスキッピングによって種々の突然変異を有する一連の患者の読み枠を復帰することができる(表1を参照)。エキソン51は興味深いターゲットエキソンである。このエキソンのスキッピングは、エキソン50,エキソン45〜50,エキソン48〜50,エキソン49〜50,エキソン52,又はエキソン52〜63にわたる領域の欠失を保有する患者の治療に用いることが可能であり、このような患者の合計は本発明者らのライデンデータベース(Leiden database)に登録されている総患者数の15%に達する。
【0033】
ジストロフィンのエキソン51内に存在するプリン塩基に富んだ種々の領域に対するヒト特異的AON10種(hAON#21〜30、下記参照)を設計した。このようなプリン塩基に富んだ領域は、エキソンのスプライシング反応の制御を担うと推定される因子、すなわちスプライシング反応の際にエキソンの除去を誘導するために遮蔽を試みた領域の存在を示唆した。すべての実験をエキソン46のスキッピングに用いたプロトコール(上記参照)に従って行った。ターゲットRNAに高い結合親和性を有するhAONを確認するために、ゲルシフトアッセイを行った。最も高い親和性を示す5種のhAONを選択した。エキソン51のスキッピングの可能性をin vitro で分析するために、これらのhAONを対照となるヒト筋肉細胞にトランスフェクトした。トランスフェクションの24時間後にRNAを単離し、エキソン53または65に特異的な逆転写用プライマーを使ってcDNAを作製した。ターゲット領域のPCRによる増幅は、エキソン51に隣接する種々のプライマーを組み合わせて行った。RT−PCR及び配列の決定によって、エキソン51の特異的なスキッピングが誘導されていたことがヒトジストロフィン転写産物から明らかとなった。続いて、エキソンのスキッピングを誘導することが示されている2種のhAON(#23及び#29)を、上記の突然変異の1つを有するDMD患者由来の6種の培養筋肉細胞にトランスフェクトした。これらの培養細胞におけるエキソン51のスキッピングをRT−PCR及び配列の決定によって確認した(図7)。更に重要なことに、ジストロフィンタンパク質の異なる部分に対して作成した複数の抗体を用いた免疫組織化学分析によると、すべての分析結果がエキソン51のスキッピングによってジストロフィンタンパク質の合成が復帰したことを示していた。
【0034】
【0035】
複数のジストロフィンエキソンの同時スキッピング。
欠失変異に加えて1つのエキソン、例えばエキソン46またはエキソン51、のスキッピングを行うことで、多種にわたるDMD突然変異の読み枠を復帰させることができる。この方法が適用可能な突然変異の範囲は、1つ以上のエキソンを同時にスキッピングすることによって拡大することができる。例えば、エキソン46からエキソン50の欠失を有するDMD患者において、欠失領域に隣接したエキソン45及び51の両方のスキッピングを行うだけで翻訳読み枠の再構成を可能にした。
【0036】
ERS非依存性のエキソンスキッピング。
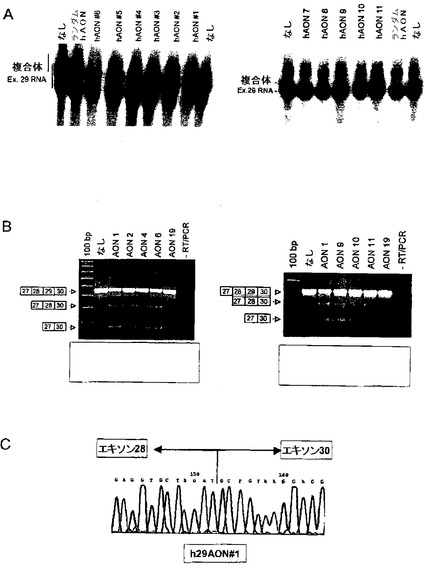
2人のベッカー型筋ジストロフィー患者に見られるエキソン29の突然変異はエキソン29のスキッピングを生じた(Ginjaarら、2000, EJHG, vol. 8: p. 793-796)。AONを用いた突然変異部位ターゲッティングによってエキソン29のスキッピングを誘導する可能性について検討した。突然変異はERS活性と関連しうるプリン塩基に富んだ領域に位置している。ERSであると考えられる領域の内側(h29AON#1〜h29AON#6)及び外側(h29AON#7〜h29AON#11)に対するAON系列を設計した(下記参照)。ターゲットRNAに対して高い親和性を示すAONを同定するために、ゲルシフトアッセイを(前述のように)行った(図8)。続いて、h29AON#1,#2,#4,#6,#9,#10及び#11を対照である培養ヒト筋管にPEIトランスフェクション試薬を用いてトランスフェクトした。トランスフェクションの24時間後にRNAを単離し、エキソン31に特異的な逆転写プライマーを使ってcDNAの逆転写を行った。ターゲット領域のPCRによる増幅はエキソン29に隣接する種々のプライマーを組み合わせて行った。このRT−PCR及びそれに続く配列の決定(図8BとC)によって、ヒトジストロフィン転写産物においてエキソン29のスキッピングが誘導されたことが明らかになった。しかしながら、エキソン29のスキッピングを促進するこれらのAONは、AONが結合すると考えられるERSの内側と外側の両方の配列に基づいて設計されたものだった(h29AON#1,#2,#4,#6,#9及び#11)。これらの結果から、エキソン29のスキッピングはエキソン29がERSを含むか含まないかには依存せず、従ってAONのエキソン29に対する結合はERSよりもエキソン封入シグナルを不活性化すると考えられる。ここに証明したERS非依存性エキソンスキッピングは、本発明の治療方法の適応範囲全体をERSを含まないエキソンにまで広げる可能性がある。
【0037】
【0038】
【0039】
マウス筋肉組織におけるin vivo のAON誘導性エキソン46スキッピング。
培養筋肉細胞を用いた実験で見込みのある結果が得られたので、次にin vivo で種々のマウスジストロフィンエキソン46に特異的なAONを試験した。試験はポリエチレンイミン(PEI)に結合させたマウスジストロフィンエキソン46に特異的AONを対照となるマウスの腓腹筋に筋肉注射することで行った。in vitro のマウス筋肉細胞における有効性が既に明らかとなっているmAON#4,#6及び#11は、RT−PCR及び配列の決定によってin vivo の筋肉組織でエキソン46のスキッピングを誘導することが判明した(図9)。in vivo のエキソン46のスキッピングは投与量に依存しており、20μg/筋肉/日の2日間にわたる注射で最も高い効率(最大10%)を示した。
【先行技術文献】
【非特許文献】
【0040】
【非特許文献1】Achsel et al., 1996, J. Biochem. 120: p. 53-60.
【非特許文献2】Bruice T.W. and Lima, W.F. (1997) Biochemistry 36 (16): p. 5004-5019.
【非特許文献3】Brunak et al., 1991, J. Mol. Biol. 220: p. 49-65.
【非特許文献4】Dunckley, MG et al, (1998) Human molecular genetics 7: p. 1083-1090.
【非特許文献5】Ginjaar et al., 2000, EJHG, vol. 8, p. 793-796.
【非特許文献6】Mann et al., 2001, PNAS vol. 98, p. 42-47.
【非特許文献7】Tanaka et al., 1994, Mol. Cell. Biol. 14: p. 1347-1354.
【非特許文献8】Wilton SD et al., (1999) Neuromuscular disorders 9: p. 330-338.
【0041】
デュシェンヌ型筋ジストロフィー及び関連疾患の詳細及び背景はウェブサイトhttp://www.dmd.nlで検索することができる。
【0042】
【表1】
【配列表フリーテキスト】
【0043】
配列番号1: マウス特異的AONであるmAON#2
配列番号2: マウス特異的AONであるmAON#3
配列番号3: マウス特異的AONであるmAON#4
配列番号4: マウス特異的AONであるmAON#5
配列番号5: マウス特異的AONであるmAON#6
配列番号6: マウス特異的AONであるmAON#7
配列番号7: マウス特異的AONであるmAON#8
配列番号8: マウス特異的AONであるmAON#9
配列番号9: マウス特異的AONであるmAON#10
配列番号10: マウス特異的AONであるmAON#11
配列番号11: ヒト特異的AONであるhAON#4
配列番号12: ヒト特異的AONであるhAON#6
配列番号13: ヒト特異的AONであるhAON#8
配列番号14: ヒト特異的AONであるhAON#9
配列番号15: ヒト特異的AONであるhAON#11
配列番号16: エキソン51特異的なhAON#21
配列番号17: エキソン51特異的なhAON#22
配列番号18: エキソン51特異的なhAON#23
配列番号19: エキソン51特異的なhAON#24
配列番号20: エキソン51特異的なhAON#25
配列番号21: エキソン51特異的なhAON#26
配列番号22: エキソン51特異的なhAON#27
配列番号23: エキソン51特異的なhAON#28
配列番号24: エキソン51特異的なhAON#29
配列番号25: エキソン51特異的なhAON#30
配列番号26: ヒト特異的AONであるh29AON#1
配列番号27: ヒト特異的AONであるh29AON#2
配列番号28: ヒト特異的AONであるh29AON#3
配列番号29: ヒト特異的AONであるh29AON#4
配列番号30: ヒト特異的AONであるh29AON#5
配列番号31: ヒト特異的AONであるh29AON#6
配列番号32: ヒト特異的AONであるh29AON#7
配列番号33: ヒト特異的AONであるh29AON#8
配列番号34: ヒト特異的AONであるh29AON#9
配列番号35: ヒト特異的AONであるh29AON#10
配列番号36: ヒト特異的AONであるh29AON#11
配列番号38: hAON#8で処理した細胞に見られる、エクソン44がエクソン47にスプライシングしたものに相当するサイズの短縮産物
配列番号39: hAON#23で処理した細胞に見られる、エクソン50がエクソン52にスプライシングしたものに相当するサイズの短縮産物
配列番号40: h29AON#1で処理した細胞に見られる、エクソン28がエクソン30にスプライシングしたものに相当するサイズの短縮産物
配列番号41: mAON#4で処理したマウスに見られる、エクソン45がエクソン47にスプライシングしたものに相当するサイズの短縮産物
配列番号42: mAON#6で処理したマウスに見られる、エクソン45がエクソン47にスプライシングしたものに相当するサイズの短縮産物
【技術分野】
【0001】
本発明は、アンチセンスオリゴヌクレオチドに関する。より詳細には、産生量の減少が求められる望ましくないタンパク質をコードするmRNA前駆体のエキソンの内部領域に対するアンチセンスオリゴヌクレオチドであって、14〜40個のヌクレオチドを含有し、該エキソンの有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチド、および上記アンチセンスオリゴヌクレオチドを用いたRNAスプライシングの制御方法を提供する。
【背景技術】
【0002】
急速なヒトゲノム研究の発展に伴い、専門家及び一般の人々は、病気のメカニズムの解明及び正確かつ信頼性の高い診断法に加えて、数多くの重度の遺伝病の治療法が近い将来確立されることを期待している。
【0003】
新たな見識によって投与の容易な低分子遺伝子療法が疾病(例えば代謝異常)の治療法として開発されることが期待されるが、その一方で、多くの疾病においては、上記以外の遺伝子療法、即ち、異常な遺伝子産物の修正、付加または置換、が最終的に必要となることが考えられる。
【0004】
近年、この分野における調査及び開発によって、克服しなければならないいくつかの技術的問題が浮き彫りになった。例えば、遺伝病に関わる複数の遺伝子が広範にわたること(よって治療遺伝子を投与すべき適切な反応系の選択が制限される)、治療用遺伝子の機能すべき組織への到達性(特異的なターゲッティング技術、即ち遺伝子の制限的挿入による物理的ターゲッティング、または組織特異的な親和性を有するシステムの開発による生物学的ターゲッティング、の構築が必要である)、及び投与システムの患者に対する安全性に関連する問題が挙げられる。これらの問題はある程度の相互関係があり、治療用薬剤が小さくなるほど、高効率で、ターゲッティング可能な安全性の高い投与システムの開発が容易であると一般的に結論付けることができる。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明においては、いわゆるエキソンスキッピングを細胞に誘導することによってこれらの問題に対処する。エキソンスキッピングは、スキップされたエキソンを有さない成熟mRNAを生じるので、スキップされたエキソンがアミノ酸をコードしている場合、このスキッピングは変化した産物の発現につながる。現在、エキソンスキッピングに関する技術はいわゆる「アンチセンスオリゴヌクレオチド (Anti-sense Oligonucleotide)」(AON)を使用するものであり、その研究のほとんどは、デュシェンヌ型筋ジストロフィー(Duchenne muscular dystrophy)(DMD)のモデルであるmdxマウスを利用して行われている。ジストロフィン遺伝子のエキソン23にナンセンス突然変異を有するmdxマウスは、デュシェンヌ型筋ジストロフィーの動物モデルとして利用されている。機能性ジストロフィンタンパク質の合成を排除するはずのmdx突然変異を有するにもかかわらず、稀に天然のジストロフィン陽性繊維がmdx筋組織中に観察される。このようなジストロフィン陽性繊維は、体性突然変異、または選択的スプライシングによる、天然のエキソンスキッピング機構によって発生したものと考えられる。ジストロフィンmRNA前駆体のイントロン22及び23のそれぞれの3’及び5’スプライス部位に特異的なAONはイントロン23の除去に通常関与する因子と干渉し、エキソン23もmRNAから除去することが示された(Wilton, 1999)。同様の研究において、Dunckleyら(1998)は、3’及び5’スプライス部位に特異的なAONを使ったエキソンスキッピングが思いがけない結果をもたらしうることを示した。彼らは、エキソン23のみならず、エキソン24〜29のスキッピングを観察し、その結果、エキソン22とエキソン30の結合体を有するmRNAが生成された。思いがけない22−30スプライシングバリアントの出現の根底にあるメカニズムは不明である。このようなバリアントの出現は、スプライス部位がコンセンサス配列を含んでおり、その結果、エキソンスキッピングを導くために使用したオリゴヌクレオチドによる非特異的なハイブリダイゼーションが生じたからと考えられる。オリゴヌクレオチドをスキップされるエキソン部位以外のスプライス部位にハイブリダイズさせることによって、当然のことながら、スプライシング反応の精度に容易に干渉することができる。一方、スプライシング反応が精度に欠ける理由としては、(5’及び3’スプライス部位のそれぞれのための)2個のオリゴヌクレオチドの使用が必要であるためと考えられる。1個のオリゴヌクレオチドを有するが、他のオリゴヌクレオチドを有しないmRNA前駆体は、思いがけないスプライシングバリアントとなる傾向がある。
【課題を解決するための手段】
【0006】
上記及びその他の諸問題を克服するために、本発明はRNAスプライシング反応系のmRNA前駆体を、該mRNA前駆体に含まれる少なくとも1つのエキソンが有するエキソン封入シグナル(exon inclusion signal)(EIS)を特異的に阻害することが可能な試薬と接触させ、そして該mRNA前駆体のスプライシングを行わせしめることを包含する、RNAスプライシング反応系においてmRNA前駆体のスプライシングを制御するための方法を提供する。エキソン封入シグナルに干渉することの利点は、このような因子がエキソン内に存在するということである。スキップされるエキソン内の配列に対するアンチセンスオリゴヌクレオチドを与えることによって、エキソン封入シグナルと干渉して、エキソンをスプライシング機構から効果的に遮蔽することができる。スプライシング機構がスキップすべきエキソンを認識しないことによって、最終的なmRNAからエキソンが排除される。本発明は、スプライシング機構の酵素反応(エキソンの結合)に直接干渉するものではない。このことによって本発明は、確実且つ信頼性の高い方法であると考えられる。EISはエキソンに特異的な構造であり、スプライス受容部位及びスプライス供与部位に特定の立体配置を付与するものと考えられる。このような概念においては、特定の立体配置によってスプライシング機構がエキソンを認識すると考えられている。しかしながら、本発明はこのモデルに限定されるものではない。本発明者らは、エキソンに結合可能な試薬がEISを阻害することを見出した。このような試薬は、エキソンのいかなる部位にも特異的に接触することができるにもかかわらず、EISを特異的に阻害することができる。本発明の方法で得られるmRNAそのものが有用である。例えば、mRNAに必要なエキソンが封入されるのを阻害することで、望ましくないタンパク質の産生を少なくとも部分的に減少させることができる。本発明の方法は、mRNA前駆体のスプライシングによって生じるmRNAを翻訳する工程を更に包含することが好ましい。また、mRNAが機能性タンパク質をコードしていることも好ましい。本発明の好ましい態様においては、タンパク質が2つ以上のドメインを包含し、ドメインの少なくとも1つは、mRNA前駆体に含まれるエキソンの少なくとも一部のスキッピングによって生じたmRNAにコードされている。エキソンスキッピングが有効な典型的な例としては、これに限定されるわけではないが、少なくとも2つの機能性ドメインを有する野生型構造のタンパク質であって、各ドメインは一次構造であるアミノ酸配列の個別の部分から生成されるものである。具体的には、転写因子が挙げられる。典型的な転写因子はDNA結合ドメイン及び細胞中の他のタンパク質と反応するドメインを包含する。2つのドメインの間に存在するアミノ酸の一次構造の一部をコードするエキソンのスキッピングによって、少なくとも部分的には同等の機能を発揮する、より短いタンパク質の産生につながる。従って、中間領域における不利益な突然変異(例えばフレームシフト突然変異や停止変異)はエキソンスキッピングを誘導することで少なくとも部分的に修復することが可能であり、(部分的な)機能を有する短いタンパク質を合成することができる。本発明の方法を用いると、エキソンの部分的なスキッピングを誘導することも可能である。この態様においては、試薬とエキソンの接触によって、試薬と接触したエキソンの隠蔽されたスプライス部位が活性化される。この態様は、機能性タンパク質を誘導するmRNA前駆体の操作性を拡大する。上記反応系は細胞を包含することが好ましい。細胞は試験管内(in vitro)で培養されるか、または生体内(in vivo)で増殖したものであることが好ましく、これらに限定されるわけではないが、典型的な生物の例にはヒトやマウスが包含される。
【図面の簡単な説明】
【0007】
【図1】エキソン45の欠失は最も頻発するDMD突然変異の1種である。この欠失によって、エキソン44はエキソン46にスプライシングされ、翻訳読み枠は切断されてエキソン46の内部に終止コドンが発生し、その結果、ジストロフィンの欠乏が生じる。本発明者らの目的は、更なるエキソン、つまりエキソン46のスキッピングを人工的に誘導して読み枠を再構築することで、エキソン45及び46の欠失を保有する、より穏やかな疾患であるベッカー型筋ジストロフィーの患者に見られるようなわずかに短いが非常に高い機能性を有するジストロフィンタンパク質の合成を復帰させる。
【図2】エキソン46は、プリン塩基に富んだ、mRNA前駆体のスプライシング制御を潜在的に担うと考えられる領域を有する。このマウスジストロフィンエキソン46の内部に存在するプリン塩基に富んだ領域に対して、重なり合う2’−O−メチルホスホロチオエートアンチセンスオリゴリボヌクレオチド(AON)からなるプライマー系列を設計した。これらのAONは長さと配列の両方の点で異なっている。AONの化学的修飾は、筋肉細胞内のエンドヌクレアーゼ及びRNase Hに対する抵抗性を付与する。in vitro の研究におけるトランスフェクション効率を調べるために、AONはAON陽性細胞の同定を可能にする5’フルオレセイン基を含有していた。
【図3】ターゲットとなるエキソン46のRNAに対する種々のAONの結合親和性を調べるために、ゲルシフトアッセイを行った。図にはターゲットRNAに最も高い親和性を示す5個のmAON(mAON#4,6,8,9及び11)を示した。AONがRNAに結合すると、ゲル移動度の低下した複合体が形成されるので、バンドの変化によって検出することができる。AONのターゲットに対する結合は配列特異的である。ランダムmAON、即ちエキソン46に非特異的なmAONはバンドの変化を生じなかった。
【図4】ゲルシフトアッセイで最も高い結合親和性を示したマウス特異的AON及びヒト特異的AONをそれぞれマウス培養筋管及びヒト培養筋管にトランスフェクトした。図4A:mAON#4,6,9及び11をトランスフェクトした培養マウス細胞のRT−PCR分析は、エキソン45がエキソン47に直接スプライシングされた産物に相当する大きさの部分産物を示した。ランダムAONをトランスフェクトした場合には、エキソン46のスキッピングは検出されなかった。図4B:罹患していない1人(図中の“C”)及び血縁ではない2人のDMD患者(図中の“P1”及び“P2”)のそれぞれに由来する培養ヒト筋肉細胞のRT−PCR分析は、hAON#4及びhAON#8をトランスフェクトした際に部分産物が生じることを明らかにした。対照においては、この産物はエキソン45がエキソン47にスプライシングされた産物に相当するが、一方で、患者に見られる断片の大きさは、エキソン44がエキソン47にスプライシングされた断片に相当した。トランスフェクトしなかった培養細胞及びランダムhAONでトランスフェクトした培養細胞では、エキソン46のスキッピングは検出されなかった。最も高いエキソン46スキッピング効率がhAON#8で得られた。
【図5】患者DL279.1(図4の“P1”に対応する)から得られたRT−PCR産物の配列データであり、このデータによって患者のエキソン45が欠失しており(上のパネル)、hAON#8のトランスフェクションによって更にエキソン46のスキッピングが起こる(下のパネル)ことを確認した。エキソン46のスキッピングは特異的であり、エキソン44はエキソン47に正確にスプライシングされ、翻訳読み枠を再構築する。
【図6】hAON#8でトランスフェクトした患者DL279.1由来の培養筋肉細胞の免疫組織化学分析。タンパク質の異なる領域に対して作成した2種のジストロフィン抗体と細胞を接触させた。使用した抗体は、ターゲットであるエキソン46に近接する領域に対する抗体(ManDys-1、ex. -31-32)及びエキソン46から離れた領域に対する抗体(Dys-2、ex. -77-79)である。下のパネルは筋管中にジストロフィンタンパク質が存在しないことを示すのに対し、いずれの抗体でもジストロフィンタンパク質が検出されることから、エキソン46のhAON#8誘導性スキッピングは明らかにジストロフィンタンパク質の合成を復帰させた(上のパネル)。
【図7】図7A:hAON#23,#24,#27,#28または#29で処理した、対照である培養ヒト筋肉細胞から単離したRNAのRT−PCR分析。エキソン50がエキソン52にスプライシングされた産物に相当する大きさの部分産物が、hAON#23及び#28で処理した細胞で検出された。これらの産物の配列を決定したところ、エキソン51が正確にスキップされていることが確認された(図7B)。さらに異常スプライシング産物をhAON#28及び#29で処理した細胞から得た。配列を決定したところ、AON処理に伴い、低い頻度で使用されるエキソン51のフレーム内の隠蔽されたスプライス部位の存在が明らかになった。AON処理によって生じた産物には復帰した読み枠からなるエキソン51の部分産物が含まれており、更なる治療価値が確認された。
【図8】図8A:エキソン29ターゲットRNAに対する種々のhAON#29の結合親和性を確認するために、ゲルシフトアッセイを行った。ハイブリダイゼーションしていないRNA(図中の「なし」)に比べると、h29AON#1,#2,#4,#6,#9,#10及び#11はゲル移動度の低い複合体を形成し、RNAに対する結合を示した。ジストロフィンエキソン19由来のランダムAONは複合体を形成しなかった。図8B:h29AON#1,#2,#4,#6,#9,#10または#11で処理した対照である培養ヒト筋肉細胞から単離したRNAのRT−PCR分析は、エキソン28がエキソン30にスプライシングされた産物に相当する大きさの部分産物の存在を明らかにした。これらの結果は、エキソン29内のERSであると考えられる領域の内側の配列に対するAON(h29AON#1,#2,#4または#6)あるいは外側の配列に対するAON(h29AON#9,#10または#11)を使ってエキソン29を特異的にスキップすることが可能であることを示す。エキソン28及び29の両方のスキッピングによって生じた更なる異常型スプライシング産物が検出された(配列データによって確認したがここには示さない)。この異常型スプライシング産物は未処理の細胞にも存在したが、これはAON処理によって促進される天然の選択的なスキッピングによるものと考えられる。ジストロフィンエキソン19から得たAON19はエキソン29のスキッピングを誘導しなかった。図8C:エキソン29の特異的スキッピングを部分配列であるRT−PCR断片の配列データによって確認した。h29AON#1で処理した細胞中のエキソン29スキッピング産物の塩基配列をここに示す。
【図9】図9A:マウス腓腹筋由来RNAのRT−PCR分析であって、RNAはmAON#4,#6または#11をそれぞれ5,10または20μg投与後2日目に単離したものである。エキソン45がエキソン47にスプライシングされた産物に相当する大きさの部分産物をすべての処理筋肉から検出した。RT−PCR反応の負の対照として“−RT”,“−RNA”,“AD−1”及び“AD−2”について分析した。図9B:mAON#4と#6(#11も実験したが、データは示さない)によって生じた部分産物の配列を決定し、エキソン46の正確なスキッピングを確認した。
【発明を実施するための形態】
【0008】
好ましい態様において、本発明は異常型タンパク質をコードするエキソンを含むmRNA前駆体を有する細胞に対して、該エキソンの少なくとも1つが有するエキソン封入シグナルを特異的に阻害することが可能な試薬を細胞に与え、そして該mRNA前駆体のスプライシングによって生じるmRNAの翻訳を行わせしめることを包含する、細胞による異常型タンパク質の産生を少なくとも部分的に減少させるための方法を提供する。
【0009】
エキソン封入シグナルを特異的に阻害することが可能ないかなる試薬も本発明に用いることができる。該試薬は、核酸またはそれと同等の機能を有する分子を含有することが好ましいが、該核酸は必ずしも一本鎖でなくてもよい。ペプチド核酸及び同程度の核酸結合特性を有する他の分子も用いることができるが、その結合特性は必ずしも結合量が同等でなくてもよい。核酸またはそれと同等の機能を有する分子は付加的な機能性を発揮するために修飾されていてもよい。例えば、2’−O−メチルオリゴリボヌクレオチドを使用することができる。このようなリボヌクレオチドは従来のオリゴヌクレオチドよりもRNase活性に対する抵抗性が高い。
【0010】
本発明の好ましい態様において、エキソン封入シグナルはエキソン認識配列(exon recognition sequence)(ERS)に特異的なアンチセンス核酸による干渉を受ける。このような配列は比較的プリン塩基に富み、スキップされるエキソンの配列情報を精査することで特定することが可能である(Tanakaら、1994, Mol. Cell Biol. 14: p. 1347-1354)。エキソン認識配列は、いわゆる弱いエキソンのmRNAへの封入を補助すると考えられる(Achselら、1996, J. Biochem. 120; p. 53-60)。このような弱いエキソンは、例えば、スプライシング機構による認識効率が低い5’及び/または3’スプライス部位を包含する。本発明においては、エキソンスキッピングはいわゆる強いエキソン、すなわち細胞のスプライシング機構によって通常効率よく認識されるエキソン、においても誘導することができることを見出した。与えられたいかなる配列からも、その配列がエキソンと推定される領域を包含するかどうかを予測し、そのエキソンが強いか弱いかを確認することが(ほとんどの場合)可能である。エキソンの強度を確認するためのいくつかの手法が存在する。有用な手法としてNetgene2スプライス部位予測サーバー(NetGene splice site prediction server)(Brunakら、1991, J. Mol. Biol. 220: p. 49-65)で検索することができる。本発明の方法によるエキソンスキッピングは(ほとんど)すべてのエキソンに誘導することが可能であり、エキソンが弱いエキソンであるか強いエキソンであるか、ならびにエキソンがERSを包含するかどうかにも依存することはない。好ましい態様において、スキッピングのターゲットとなるエキソンは強いエキソンである。他の1つの好ましい態様において、スキッピングの対象となるエキソンはERSを包含しない。
【0011】
本発明の方法は様々な用途に使用することが可能である。1つの態様において、本発明の方法は異常型タンパク質の産生を少なくとも部分的に減少させるために使用する。このようなタンパク質としては、例えば、腫瘍タンパク質またはウイルス性タンパク質が挙げられる。多くの腫瘍において、腫瘍タンパク質の存在のみならず、その相対的な発現程度が腫瘍細胞の表現型に関係している。同様に、ウイルス性タンパク質の存在のみならず、細胞中のウイルス性タンパク質の量が特定のウイルスの毒性を決定している。さらに、ウイルスの有効な増加及び蔓延のために、ウイルスの生活環における発現の時期及び細胞中の特定のウイルス性タンパク質の量的バランスによって、ウイルスが効率的にまたは非効率的に増殖するかどうかが決定される。本発明の方法を使用して細胞中の異常型タンパク質の量を減少させることで、例えば腫瘍細胞の腫瘍形成性(転移性)及び/またはウイルス感染細胞のウイルス増殖性を低下させることができる。
【0012】
好ましい態様において、本発明の方法を用いて異常型タンパク質を機能性タンパク質に変更する。1つの態様において、上記のような機能性タンパク質は、細胞に通常存在するが、処理する細胞には存在しないタンパク質の機能を発揮する。大抵の場合、機能を部分的に修復しただけでも、処理した細胞の性能が著しく向上する。向上した性能故に処理した細胞は未処理の細胞よりも選択的な優位性を示すことができ、処理の有効性を高めている。
【0013】
上述した本発明の態様は、特に欠失した遺伝子の発現を復帰させるのに適している。これはターゲットとなるエキソンの特異的なスキッピングを行い、有害な変異(典型的なものとしては停止変異またはフレームシフト点突然変異、あるいは翻訳停止をもたらす1つ以上のエキソンの欠失または挿入)を回避するか修正することによって達成される。
【0014】
遺伝子導入法に比べて、本発明の新規なスプライスシング制御型遺伝子治療(splice-modulation gene therapy)は、より小さな治療薬、これに限定されるわけではないが、典型的な例としては14〜40個のヌクレオチドからなる治療薬の投与を必要とする。好ましい態様においては、製造が容易であり、細胞による取り込みがより効果的に行われることから、14〜25個のヌクレオチドからなる分子を使用する。本発明の方法は、治療薬に付随する、効果的で安全な投与システムの設計をより柔軟に行うことを可能とする。本発明のこのような態様の重要な更なる利点は、大部分またはすべての遺伝子調節回路を未だ保持する内因性遺伝子活性の(少なくとも一部)を復帰させることで、組織特異的なアイソフォームの適切な発現レベルと合成を保障することである。
【0015】
上述の本発明の態様は、原則として、いかなる遺伝病や遺伝的疾病素質にも適用することができ、配列内部においてわずかに短いタンパク質の翻訳が完全にまたは部分的に機能している場合に限り、特定のエキソンを標的とするスキッピングによって、元々変異によって破壊された翻訳読み枠を復帰させることができる。この方法の適用に治療的価値が見出される態様としては、腫瘍抑制遺伝子の二次変異によって生じる疾病素質が挙げられ、例えば、乳ガン、結腸ガン、結節性硬化症、神経線維腫症などにおいては、活性の(部分的な)回復が二次変異による異数性の発現を排除し、腫瘍形成を防ぐ。他の好ましい態様には、遺伝子産物の欠失が直接的な疾病誘引効果を示す遺伝子の(部分的な)修復が含まれる。例えば、血友病A(血液凝固第VIII因子の欠乏によるもの)、先天的甲状腺機能不全症のいくつかの形態(チログロブリン合成欠損によるもの)、及びデュシェンヌ型筋ジストロフィー(DMD)が挙げられる。DMDは、フレームシフトによって生じるX連鎖性ジストロフィン遺伝子の欠失、重複及び停止突然変異が重度の進行性筋力低下をもたらす疾患である。通常DMDは発症すると青年期の後期または成人期の初期に死に至るが、その一方で、同じ遺伝子座にフレームシフト以外の方法で欠失または重複突然変異が生じて発症したベッカー型筋ジストロフィーは軽症であり、平均余命は35〜40年から正常人と同じである。DMDに適用する本発明の態様においては、本発明の方法によって現存する欠失領域をエキソンスキッピングによって伸張する(または現存する重複突然変異体のmRNA産物を改変する)ことができる。これは読み枠を復帰させ、配列内部においてわずかに短くなっているが機能性のタンパク質を産生するのに必要な数の近接したエキソンのスキッピングによって行う。このような欠失の誘導と同等の変異を有するBMD患者の臨床的症状が軽いことに基づき、AON療法を受けたDMD患者の症状は軽減されると考えられる。
【0016】
ジストロフィン遺伝子に生じる種々の変異が機能不全性タンパク質につながる。(総合目録としてhttp://www.dmd.nl、即ち、DMD及び関連疾患の国際的に承認されたデータベース、を参照)。機能性ジストロフィンタンパク質を産生させるために実際にスキップすべきエキソンは突然変異によって変化する。表1はスキップすることのできるエキソンの一覧であるが、本願はこれらに制限されることはない。この表においては、上述のエキソンに対して、ヒトで観察された高頻度で発生するジストロフィン遺伝子変異であって、本発明の方法で治療することができるものも示した。表に示したエキソンのスキッピングは、少なくともベッカー型筋ジスロトフィータンパク質の機能を有する変化したジストロフィンタンパク質の産生をもたらす。このように、本発明の1つの態様において、エキソン封入シグナルがヒトジストロフィン遺伝子のエキソン2,8,43,44,45,46,50,51,52または53に存在することを特徴とする方法が提供される。ある欠失/挿入変異の発生は、他の変異より高頻度で発生する。本発明の方法でエキソン46のスキッピングを誘導すると、DMDの欠失を保有する患者の約7%で治療に成功し、ジストロフィン陽性筋肉繊維がジストロフィン患者に検出されることが明らかとなった。また、本発明の方法でエキソン51のスキッピングを誘導することによって、DMDの欠失を保有する患者の約15%を治療することに成功した。この治療方法によって、該患者は少なくとも少量のジストロフィン陽性筋肉繊維を有するようになった。本発明の方法を使用したエキソン46または51のスキッピングによって、ジストロフィン遺伝子の欠失を保有する患者の約22%を治療することができる。従って、本発明の好ましい態様においては、エキソン封入シグナルはエキソン46またはエキソン51に存在する。特に好ましい態様においては、使用する試薬は以下の核酸配列を包含する:hAON#4、hAON#6、hAON#8、hAON#9、hAON#11及び/またはhAON#21〜30からなる群から選ばれる少なくとも1つの核酸配列;或いは上記hAON#の機能性領域、誘導体及び/または類似体。機能性領域、誘導体及び/または類似体は、上記hAON#と同等のエキソンスキッピング活性を有するが、本発明の方法においては、該活性量が必ずしも同等でなくてもよい。
【0017】
mRNA前駆体に含まれる1つ以上のエキソンのエキソンスキッピングを誘導することは効果的である。例えば、変異の多様性並びにエキソンの長さや変異に隣接するアミノ酸配列といった一定の性質を考慮すると、機能を復帰させるためには1つ以上のエキソンをスキップしなければならないという状況が起こりうる。DMD欠失データベースに見られるこのような状況の好ましい例としては、エキソン46〜50の欠失が挙げられるが、これに限定されるものではない。エキソン46〜50が欠失している患者は機能性ジストロフィンタンパク質を生成しない。しかしながら、エキソン45及びエキソン51のスキッピングを誘導することによって少なくとも部分的な機能を有するジストロフィンタンパク質を生成することができる。他の好ましい例としてはエキソン2の重複を有する患者が挙げられるが、これに限定されるものではない。エキソン2のEISを阻害することが可能な試薬を与えることで、エキソン2の一方または両方を部分的にスキップし、エキソン2の部分配列からなる領域または2つのエキソン2を欠失した領域の隣から野生型タンパク質を得ることが可能である。他の好ましい例として、エキソン45〜50のスキッピングが挙げられるが、これに限定されるものではない。このスキッピングにより読み枠内でベッカー様バリアントが生じる。ベッカー様バリアントはエキソン45,46,47,48,49及び/または50あるいはそれらの組み合わせに存在するいかなる変異を治療するためにも誘導することができる。本発明のその他の態様においては、mRNA前駆体に含まれる他のエキソンの有するエキソン封入シグナルを阻害する他の試薬を細胞に与える工程を更に包含することを特徴とする方法が提供される。mRNA前駆体の2つ以上の異なる遺伝子にエキソンスキッピングを誘導するために2つ以上の試薬を使用することは、当然のことながら本発明の範囲に含まれる。
【0018】
他の1つの態様において、本発明はスプライシング制御型遺伝子治療に適した試薬を選択するための方法及び選択した試薬の特異的エキソンスキッピング試薬としての有効性を予備実験で確認するための方法を提供する。目的エキソンの一部に対して相補的な試薬が、該エキソンの有するエキソン封入シグナルを特異的に阻害することが可能であることを確認するための方法であって、目的のエキソンを含むmRNA前駆体を有する細胞に被験試薬を与え、該細胞を培養して該mRNA前駆体からmRNAを生成せしめ、そして生成した該mRNAに該エキソンが存在しないことを確認することを包含する方法が提供される。好ましい態様において、該試薬が、核酸またはそれと同等の機能を有する分子を含有し、該核酸は目的エキソンの一部に対して相補的である。特異的エキソンスキッピングを誘導することが可能な試薬を本発明の方法で同定することができる。この方法には、該試薬がエキソンを含む核酸、好ましくはRNA、に比較的高い親和性で結合するかどうかを確認するための予備スクリーニングを行うことも含まれる。この結果、試薬がエキソンの有するエキソン封入シグナルを特異的に阻害することが可能であることを確認するための方法が提供され、該方法は試験管内の条件において、該核酸またはそれと同等の機能を有する分子と該目的エキソンを包含するmRNA分子との相対的な結合親和性を測定する工程を更に包含する。
【0019】
他の態様において、本発明の方法によって得られる試薬を提供する。好ましい態様においては、該試薬は核酸またはそれと同等の機能を有する分子を包含する。該試薬は、それを用いて細胞にエキソンスキッピングを誘導した場合、細胞中の異常型タンパク質の量を少なくとも部分的に減少させることが好ましく、該エキソンスキッピングによって、該細胞で機能を発揮しうるタンパク質をコードするRNAを生じることが更に好ましい。特に好ましい態様において、mRNA前駆体はジストロフィン遺伝子から誘導されたものである。機能性タンパク質は変異ジストロフィンタンパク質または正常ジストロフィンタンパク質を包含することが好ましい。また、変異ジストロフィンタンパク質が少なくともベッカー型筋ジストロフィー患者のジストロフィンタンパク質の機能を有することが好ましい。特に好ましい態様においては、使用する試薬は以下の核酸配列を包含する:hAON#4、hAON#6、hAON#8、hAON#9、hAON#11及び/またはhAON#21〜30からなる群から選ばれる少なくとも1つの核酸配列;或いは上記hAON#の機能性領域、誘導体及び/または類似体。機能性領域、誘導体及び/または類似体は、上記hAON#と同等のエキソンスキッピング活性を有するが、本発明の方法においては、該活性量が必ずしも同等でなくてもよい。
【0020】
細胞に試薬を導入するための種々の方法が当業界では知られている。特に核酸の導入方法は広範にわたって開発されている。当業者はある導入方法が本発明を実施するのに適しているかどうかを十分確認することができる。このような方法の一例は、本発明の試薬をリポソームに封入し、該リポソームを目的mRNA前駆体を有する細胞に与えることを包含するが、本発明はこの例に限定されるものではない。リポソームは核酸を細胞に導入するための運搬体として特に適している。エキソンスキッピングを誘導することのできるアンチセンス分子は、アンチセンスRNAを生成するための転写単位を包含する核酸を導入し、細胞に生成させることが可能である。適切な転写単位としては核内低分子RNAまたはtRNA転写単位が挙げられるが、これらに限定されるものではない。従って、本発明は、エキソン封入シグナルを阻害することが可能な本発明の核酸またはそれと同等の機能を有する分子を包含する核酸運搬体を更に提供する。1つの態様において、該運搬体は本発明の核酸を発現させることが可能である。一本鎖ウイルスを運搬体として利用した場合には、当然のことながら、そのようなウイルスが本発明の試薬のアンチセンス配列だけを包含するものであっても本発明の範囲に含まれる。また、このような一本鎖ウイルスの他の1つの態様においては、本発明のAONは、運搬体としてのウイルスに封入されているウイルス核に存在する核内低分子RNAまたはtRNA転写単位によってコードされている。好ましい一本鎖ウイルスはアデノ随伴ウイルスである。
【0021】
更なる1つの態様において、本発明は医薬の調製に用いる、核酸または核酸運搬体を提供する。好ましい態様において、該医薬を遺伝病の治療に使用する。さらに好ましくは、該医薬をデュシェンヌ型筋ジストロフィーの治療に使用する。
【実施例】
【0022】
エキソン45はDMDにおいて最も高頻度で欠失が起こるエキソンなので、本発明者らははじめにエキソン46の特異的なスキッピングの誘導を試みた(図1)。この誘導によってエキソン45及び46の欠失を保有するBMD患者に見られる、短いがより機能的なジストロフィンタンパク質を産生する。マウスジストロフィン遺伝子のジストロフィンmRNA前駆体のスプライシングを制御するための実験系を初めに構成した。その後、エキソン45の欠失を保有するDMD患者由来の筋肉細胞において翻訳読み枠とジストロフィン合成を復帰させることを目的として、ヒトジストロフィン遺伝子にねらいを定めた。
【0023】
mAONs及びhAONsの設計
エキソン46の内部領域に対するマウス特異的AON系列及びヒト特異的AON系列(mAON及びhAON)を設計した(図2)。使用したエキソン46の内部領域は、プリン塩基に富んだ配列を有し、エキソン46のスプライシング制御に係る推定上の機能を有すると考えられる。ゲルシフトアッセイによるAONの初期スクリーニング(下記参照)のために未修飾のDNAオリゴヌクレオチド(ベルギー国、EuroGentec社に合成を依頼)を使用した。筋肉細胞のトランスフェクション実験を実際に行う際には、2’−O−メチルホスホロチオエートアンチセンスオリゴリボヌクレオチド(ベルギー国、EuroGentec社に合成を依頼)を使用した。このような修飾されたRNAオリゴヌクレオチドはエンドヌクレアーゼ及びRNase Hに対する抵抗性を有し、高い親和性でRNAに結合することが知られている。最終的に有効であると認められ、in vitro で筋肉細胞に与えたAONの配列を下記に示す。対応するマウス特異的AONとヒト特異的AONは高い相同性があるが完全に同一ではない。
【0024】
以下に使用したAONのデオキシ体を示すが、最終的に使用した2’−O−メチルリボヌクレオチドにおいては、TをUに置き換える。
【0025】
【0026】
【0027】
ゲルシフトアッセイ
AONの有効性は、ターゲット配列に対する結合親和性に基づいて決定した。RNAフォールディングを予測するためのコンピュータシミュレーションプログラムの近年の発展にもかかわらず、設計したAONのどれが高い親和性でターゲット配列に結合するかを推測することは難しい。従って、ゲルシフトアッセイを(Bruiceら、1997に記載のプロトコールに従って)行った。エキソン46ターゲットRNA断片を、(マウスまたはヒト筋肉mRNAから、T7プロモーター配列を有するセンスプライマ−を利用して増幅した)PCR断片から32P−CTPの存在下のin vitro T7転写法で生成した。ターゲットである転写断片に対する個々のAON(0.5pmol)の結合親和性は、37℃で30分間ハイブリダイゼーションを行い、次いでポリアクリルアミド(8%)ゲル電気泳動を行うことで測定した。これらの分析をマウス特異的AON及びヒト特異的AONをスクリーニングするために行った(図3)。少なくとも5個の異なるマウス特異的AON(mAON#4、6、8、9及び11)及びそれらに対応する4個のヒト特異的AON(hAON#4、6、8及び9)において移動度の変化が観察され、ターゲットRNAに対して結合親和性を有することを示した。
【0028】
培養筋肉細胞のトランスフェクション
AONによるin vitro の筋肉細胞におけるスキッピング誘導効率を分析するために、ゲルシフトアッセイを用いてターゲットエキソンに対して最も高い結合親和性を示したエキソン46特異的AONを選択した。すべてのトランスフェクション実験において、非特異的AONをエキソン46の特異的スキッピングの負の対照として用いた。前述したように、はじめにマウス筋肉細胞の実験系を構築した。マウス筋肉細胞系であるC2C12細胞由来の(ジストロフィン発現レベルの高い)培養増殖筋芽細胞及び培養分裂後筋管の両方を使用した。後に行ったヒト由来培養筋肉細胞の実験には、罹患していない1人の筋肉生検材料及びエキソン45の欠失を保有する、血縁ではない2人のDMD患者の筋肉生検材料のそれぞれから単離した培養一次筋肉細胞を使用した。これらの異種培養物はおよそ20〜40%の筋原細胞を含んでいた。異なるAON(濃度:1μM)に対して3当量比のカチオンポリマーPEI(MBI Fermentas社製)を用い、細胞にAONをトランスフェクトした。これらの実験でトランスフェクトしたAONは5’フルオレセイン基を有し、そのため蛍光性の核を数えることでトランスフェクション効率を測定することができる。概して、60%以上の細胞が核に特異的にAONの取り込みを示した。RT−PCR分析を促進するために、トランスフェクトの24時間後にRNAzolB(オランダ国、CamPro Scientific社製)を使ってRNAを単離した。
【0029】
RT−PCR及び配列の決定
RNAの逆転写をC. therm. ポリメラーゼ(Roche社製)及びエキソン48特異的逆転写プライマーを使って行った。ジストロフィン遺伝子のエキソン46のスキッピングを検出するために、PCRを2サイクル行ってcDNAを増幅した。PCRは、エキソン44及び47に含まれるプライマー(ヒト実験系)またはエキソン45及び47に含まれるプライマー(マウス実験系)を使用したプライマーシフト増幅(nested amplification)を包含する。培養マウス筋芽細胞及び培養マウス筋管細胞では、エキソン45がエキソン47に直接スプライシングされた産物に相当する大きさの部分産物を検出した(図4)。続いて行った配列の決定によって、マウスジストロフィン転写産物におけるエキソン46の特異的なスキッピングを確認した。エキソンスキッピングの効率は個々のAONで異なり、mAON#4及び#11が最も高い効率を示した。これらの好ましい結果に基づき、培養ヒト筋肉細胞においても、ジストロフィンのスプライシングを同様に制御することに着目した実験を行った。その結果、エキソン45がエキソン47にスプライシングされた産物に相当する部分産物を対照となる筋肉細胞で検出した。興味深いことに、患者由来筋肉細胞では、エキソン44がエキソン47にスプライシングされてなる短い断片が検出された。エキソン46の特異的なスキッピングをヒトジストロフィン転写産物の配列データによって確認した。マウスとヒトの両方のジストロフィン転写産物に見られたこのようなスプライシングの制御は、トランスフェクトされていない培養細胞や非特異的AONでトランスフェクトされた培養細胞には見られなかった。
【0030】
免疫組織化学分析
ジストロフィンタンパク質の翻訳および合成を復帰させるために、エキソン45の欠失を保有する患者由来の筋肉細胞においてエキソン46のスキッピングの誘導を試みた。hAON#8でトランスフェクトした際のジストロフィン産物を検出するために、ジストロフィンタンパク質のターゲット領域に隣接するドメインと遠位のドメインのそれぞれに対して作成した2種のジストロフィンモノクローナル抗体(Mandys-1及びDys-2)を使用して、2種の患者由来培養筋肉細胞を免疫細胞化学分析に付した。蛍光分析によって、いずれの患者由来培養細胞においてもジストロフィン合成が復帰していることが明らかになった(図5)。処理したサンプルにおいては、約80%以上の筋繊維がジストロフィンに対して陽性に染色された。
【0031】
本発明者らの実験結果は、DMD患者由来筋肉細胞の内因性DMD遺伝子によるジストロフィン合成の復帰を初めて示す。これは、ジストロフィンmRNA前駆体の目的に応じたスプライシング制御を治療目的で実施するための原理を証明する。
【0032】
エキソン51のターゲットスキッピング
ジストロフィンエキソンの同時スキッピング
エキソン51のターゲットスキッピング。本発明者らは、in vitro のマウス及びヒト筋肉細胞におけるジストロフィンのエキソン46のAONによる制御の可能性について明示した。これらの知見はAONをDMDの治療用の試薬として評価する更なる研究の根拠を示した。DMDを誘発する欠失の大部分は遺伝子内の突然変異が起こりやすい2つの部位で集中的に発生し、ある1つの特定エキソンのターゲットスキッピングによって種々の突然変異を有する一連の患者の読み枠を復帰することができる(表1を参照)。エキソン51は興味深いターゲットエキソンである。このエキソンのスキッピングは、エキソン50,エキソン45〜50,エキソン48〜50,エキソン49〜50,エキソン52,又はエキソン52〜63にわたる領域の欠失を保有する患者の治療に用いることが可能であり、このような患者の合計は本発明者らのライデンデータベース(Leiden database)に登録されている総患者数の15%に達する。
【0033】
ジストロフィンのエキソン51内に存在するプリン塩基に富んだ種々の領域に対するヒト特異的AON10種(hAON#21〜30、下記参照)を設計した。このようなプリン塩基に富んだ領域は、エキソンのスプライシング反応の制御を担うと推定される因子、すなわちスプライシング反応の際にエキソンの除去を誘導するために遮蔽を試みた領域の存在を示唆した。すべての実験をエキソン46のスキッピングに用いたプロトコール(上記参照)に従って行った。ターゲットRNAに高い結合親和性を有するhAONを確認するために、ゲルシフトアッセイを行った。最も高い親和性を示す5種のhAONを選択した。エキソン51のスキッピングの可能性をin vitro で分析するために、これらのhAONを対照となるヒト筋肉細胞にトランスフェクトした。トランスフェクションの24時間後にRNAを単離し、エキソン53または65に特異的な逆転写用プライマーを使ってcDNAを作製した。ターゲット領域のPCRによる増幅は、エキソン51に隣接する種々のプライマーを組み合わせて行った。RT−PCR及び配列の決定によって、エキソン51の特異的なスキッピングが誘導されていたことがヒトジストロフィン転写産物から明らかとなった。続いて、エキソンのスキッピングを誘導することが示されている2種のhAON(#23及び#29)を、上記の突然変異の1つを有するDMD患者由来の6種の培養筋肉細胞にトランスフェクトした。これらの培養細胞におけるエキソン51のスキッピングをRT−PCR及び配列の決定によって確認した(図7)。更に重要なことに、ジストロフィンタンパク質の異なる部分に対して作成した複数の抗体を用いた免疫組織化学分析によると、すべての分析結果がエキソン51のスキッピングによってジストロフィンタンパク質の合成が復帰したことを示していた。
【0034】
【0035】
複数のジストロフィンエキソンの同時スキッピング。
欠失変異に加えて1つのエキソン、例えばエキソン46またはエキソン51、のスキッピングを行うことで、多種にわたるDMD突然変異の読み枠を復帰させることができる。この方法が適用可能な突然変異の範囲は、1つ以上のエキソンを同時にスキッピングすることによって拡大することができる。例えば、エキソン46からエキソン50の欠失を有するDMD患者において、欠失領域に隣接したエキソン45及び51の両方のスキッピングを行うだけで翻訳読み枠の再構成を可能にした。
【0036】
ERS非依存性のエキソンスキッピング。
2人のベッカー型筋ジストロフィー患者に見られるエキソン29の突然変異はエキソン29のスキッピングを生じた(Ginjaarら、2000, EJHG, vol. 8: p. 793-796)。AONを用いた突然変異部位ターゲッティングによってエキソン29のスキッピングを誘導する可能性について検討した。突然変異はERS活性と関連しうるプリン塩基に富んだ領域に位置している。ERSであると考えられる領域の内側(h29AON#1〜h29AON#6)及び外側(h29AON#7〜h29AON#11)に対するAON系列を設計した(下記参照)。ターゲットRNAに対して高い親和性を示すAONを同定するために、ゲルシフトアッセイを(前述のように)行った(図8)。続いて、h29AON#1,#2,#4,#6,#9,#10及び#11を対照である培養ヒト筋管にPEIトランスフェクション試薬を用いてトランスフェクトした。トランスフェクションの24時間後にRNAを単離し、エキソン31に特異的な逆転写プライマーを使ってcDNAの逆転写を行った。ターゲット領域のPCRによる増幅はエキソン29に隣接する種々のプライマーを組み合わせて行った。このRT−PCR及びそれに続く配列の決定(図8BとC)によって、ヒトジストロフィン転写産物においてエキソン29のスキッピングが誘導されたことが明らかになった。しかしながら、エキソン29のスキッピングを促進するこれらのAONは、AONが結合すると考えられるERSの内側と外側の両方の配列に基づいて設計されたものだった(h29AON#1,#2,#4,#6,#9及び#11)。これらの結果から、エキソン29のスキッピングはエキソン29がERSを含むか含まないかには依存せず、従ってAONのエキソン29に対する結合はERSよりもエキソン封入シグナルを不活性化すると考えられる。ここに証明したERS非依存性エキソンスキッピングは、本発明の治療方法の適応範囲全体をERSを含まないエキソンにまで広げる可能性がある。
【0037】
【0038】
【0039】
マウス筋肉組織におけるin vivo のAON誘導性エキソン46スキッピング。
培養筋肉細胞を用いた実験で見込みのある結果が得られたので、次にin vivo で種々のマウスジストロフィンエキソン46に特異的なAONを試験した。試験はポリエチレンイミン(PEI)に結合させたマウスジストロフィンエキソン46に特異的AONを対照となるマウスの腓腹筋に筋肉注射することで行った。in vitro のマウス筋肉細胞における有効性が既に明らかとなっているmAON#4,#6及び#11は、RT−PCR及び配列の決定によってin vivo の筋肉組織でエキソン46のスキッピングを誘導することが判明した(図9)。in vivo のエキソン46のスキッピングは投与量に依存しており、20μg/筋肉/日の2日間にわたる注射で最も高い効率(最大10%)を示した。
【先行技術文献】
【非特許文献】
【0040】
【非特許文献1】Achsel et al., 1996, J. Biochem. 120: p. 53-60.
【非特許文献2】Bruice T.W. and Lima, W.F. (1997) Biochemistry 36 (16): p. 5004-5019.
【非特許文献3】Brunak et al., 1991, J. Mol. Biol. 220: p. 49-65.
【非特許文献4】Dunckley, MG et al, (1998) Human molecular genetics 7: p. 1083-1090.
【非特許文献5】Ginjaar et al., 2000, EJHG, vol. 8, p. 793-796.
【非特許文献6】Mann et al., 2001, PNAS vol. 98, p. 42-47.
【非特許文献7】Tanaka et al., 1994, Mol. Cell. Biol. 14: p. 1347-1354.
【非特許文献8】Wilton SD et al., (1999) Neuromuscular disorders 9: p. 330-338.
【0041】
デュシェンヌ型筋ジストロフィー及び関連疾患の詳細及び背景はウェブサイトhttp://www.dmd.nlで検索することができる。
【0042】
【表1】
【配列表フリーテキスト】
【0043】
配列番号1: マウス特異的AONであるmAON#2
配列番号2: マウス特異的AONであるmAON#3
配列番号3: マウス特異的AONであるmAON#4
配列番号4: マウス特異的AONであるmAON#5
配列番号5: マウス特異的AONであるmAON#6
配列番号6: マウス特異的AONであるmAON#7
配列番号7: マウス特異的AONであるmAON#8
配列番号8: マウス特異的AONであるmAON#9
配列番号9: マウス特異的AONであるmAON#10
配列番号10: マウス特異的AONであるmAON#11
配列番号11: ヒト特異的AONであるhAON#4
配列番号12: ヒト特異的AONであるhAON#6
配列番号13: ヒト特異的AONであるhAON#8
配列番号14: ヒト特異的AONであるhAON#9
配列番号15: ヒト特異的AONであるhAON#11
配列番号16: エキソン51特異的なhAON#21
配列番号17: エキソン51特異的なhAON#22
配列番号18: エキソン51特異的なhAON#23
配列番号19: エキソン51特異的なhAON#24
配列番号20: エキソン51特異的なhAON#25
配列番号21: エキソン51特異的なhAON#26
配列番号22: エキソン51特異的なhAON#27
配列番号23: エキソン51特異的なhAON#28
配列番号24: エキソン51特異的なhAON#29
配列番号25: エキソン51特異的なhAON#30
配列番号26: ヒト特異的AONであるh29AON#1
配列番号27: ヒト特異的AONであるh29AON#2
配列番号28: ヒト特異的AONであるh29AON#3
配列番号29: ヒト特異的AONであるh29AON#4
配列番号30: ヒト特異的AONであるh29AON#5
配列番号31: ヒト特異的AONであるh29AON#6
配列番号32: ヒト特異的AONであるh29AON#7
配列番号33: ヒト特異的AONであるh29AON#8
配列番号34: ヒト特異的AONであるh29AON#9
配列番号35: ヒト特異的AONであるh29AON#10
配列番号36: ヒト特異的AONであるh29AON#11
配列番号38: hAON#8で処理した細胞に見られる、エクソン44がエクソン47にスプライシングしたものに相当するサイズの短縮産物
配列番号39: hAON#23で処理した細胞に見られる、エクソン50がエクソン52にスプライシングしたものに相当するサイズの短縮産物
配列番号40: h29AON#1で処理した細胞に見られる、エクソン28がエクソン30にスプライシングしたものに相当するサイズの短縮産物
配列番号41: mAON#4で処理したマウスに見られる、エクソン45がエクソン47にスプライシングしたものに相当するサイズの短縮産物
配列番号42: mAON#6で処理したマウスに見られる、エクソン45がエクソン47にスプライシングしたものに相当するサイズの短縮産物
【特許請求の範囲】
【請求項1】
産生量の減少が求められる望ましくないタンパク質をコードするmRNA前駆体のエキソンの内部領域に対するアンチセンスオリゴヌクレオチドであって、14〜40個のヌクレオチドを含有し、該エキソンの有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチド。
【請求項2】
該望ましくないタンパク質の産生を減少させるための医薬の製造用である、請求項1に記載のアンチセンスオリゴヌクレオチド。
【請求項3】
請求項1または2に記載のアンチセンスオリゴヌクレオチドまたはその相補鎖を包含する核酸運搬体。
【請求項4】
アンチセンスオリゴヌクレオチドを放出することが可能な、請求項3に記載の核酸運搬体。
【請求項5】
RNAスプライシング反応を実施可能な細胞による望ましくないタンパク質の細胞内産生の減少を目的として、該細胞内におけるmRNA前駆体のスプライシングを制御するためのin vitroの方法であって、該細胞内のmRNA前駆体を、該mRNA前駆体に含まれる少なくとも1つのエキソンの有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチドと接触させることを包含し、該アンチセンスオリゴヌクレオチドは、14〜40個のヌクレオチドを含有し、該少なくとも1つのエキソンの内部領域に対するものであることを特徴とする方法。
【請求項6】
該mRNAが機能性タンパク質をコードしていることを特徴とする、請求項5に記載の方法。
【請求項7】
該タンパク質が2つ以上のドメインを包含し、該ドメインの少なくとも1つは、該mRNA前駆体に含まれるエキソンの少なくとも一部のスキッピングによって生じたmRNAにコードされていることを特徴とする、請求項5または6に記載の方法。
【請求項8】
該接触によって、アンチセンスオリゴヌクレオチドと接触したエキソンの隠蔽されたスプライス部位が活性化されることを特徴とする、請求項5〜7のいずれかに記載の方法。
【請求項9】
該エキソン封入シグナルがエキソン認識配列を包含することを特徴とする、請求項5〜8のいずれかに記載の方法。
【請求項10】
該エキソン封入シグナルが強いスプライス供与部位/受容部位対を有するエキソンに存在することを特徴とする、請求項5〜9のいずれかに記載の方法。
【請求項11】
該アンチセンスオリゴヌクレオチドが、15〜25個のヌクレオチドを含有することを特徴とする、請求項5〜10のいずれかに記載の方法。
【請求項12】
該mRNA前駆体に含まれる他のエキソンの有するエキソン封入シグナルを阻害しうる別のアンチセンスオリゴヌクレオチドを、該細胞内の該mRNA前駆体と接触させる工程を更に包含することを特徴とする、請求項5〜11のいずれかに記載の方法。
【請求項13】
異常型タンパク質をコードするエキソンを含むmRNA前駆体を有する細胞による異常型タンパク質の産生を少なくとも部分的に減少させるためのin vitroの方法であって、
該エキソンの少なくとも1つが有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチドであって、該少なくとも1つのエキソンの内部領域に対するものであり、14〜40個のヌクレオチドを含有するアンチセンスオリゴヌクレオチドを該細胞に与え、そして
該mRNA前駆体のスプライシングによって生じるmRNAの翻訳を行わせしめる
ことを包含する方法。
【請求項14】
該エキソン封入シグナルがエキソン認識配列を包含することを特徴とする、請求項13に記載の方法。
【請求項15】
該エキソン封入シグナルが強いスプライス供与部位/受容部位対を有するエキソンに存在することを特徴とする、請求項13または14に記載の方法。
【請求項16】
該アンチセンスオリゴヌクレオチドが、15〜25個のヌクレオチドを含有することを特徴とする、請求項13〜15のいずれかに記載の方法。
【請求項17】
該mRNA前駆体に含まれる他のエキソンの有するエキソン封入シグナルを阻害しうる別のアンチセンスオリゴヌクレオチドを該細胞に与える工程を更に包含することを特徴とする、請求項13〜16のいずれかに記載の方法。
【請求項18】
目的エキソンの一部に対して相補的な核酸が、該エキソンの有するエキソン封入シグナルを特異的に阻害することが可能であることを確認するためのin vitroの方法であって、
目的のエキソンを含むmRNA前駆体を有する細胞に被験核酸を与え、
該細胞を培養して該mRNA前駆体からmRNAを生成せしめ、そして
生成した該mRNAに該エキソンが存在しないことを確認する
ことを包含する方法。
【請求項19】
該目的エキソンを包含するRNA分子と該核酸との相対的な結合親和性をin vitroで測定する工程を更に包含する、請求項18に記載の方法。
【請求項20】
請求項1または2のアンチセンスオリゴヌクレオチドを導入した、ヒト以外の動物。
【請求項21】
ヒトタンパク質をコードする核酸を更に包含する、請求項20に記載のヒト以外の動物。
【請求項22】
該ヒト以外の動物は、自らが有する該ヒトタンパク質の相同体をコードする遺伝子にサイレント突然変異が導入されていることを特徴とする、請求項21に記載のヒト以外の動物。
【請求項23】
mRNA前駆体に含まれる少なくとも1つのエキソンの有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチドを用いた、細胞内の望ましくないタンパク質の産生を減少させるための医薬の製造方法であって、該アンチセンスオリゴヌクレオチドが、該少なくとも1つのエキソンの内部領域に対するものであり、14〜40個のヌクレオチドを含有することを特徴とする方法。
【請求項24】
該望ましくないタンパク質が、野生型構造の時に少なくとも2つの機能性ドメインを有し、各ドメインは一次構造であるアミノ酸配列の個別の部分から生成されたものであることを特徴とする、請求項23に記載の方法。
【請求項25】
該望ましくないタンパク質が、異常型タンパク質であることを特徴とする、請求項23または24に記載の方法。
【請求項26】
該異常型タンパク質が、腫瘍タンパク質またはウイルス性タンパク質であることを特徴とする、請求項25に記載の方法。
【請求項27】
該医薬が、遺伝病の治療用または遺伝的疾病素質の改善用の医薬であることを特徴とする、請求項23〜26のいずれかに記載の方法。
【請求項28】
該医薬が、腫瘍およびウイルス感染からなる群より選ばれる少なくとも1種の治療を目的とした医薬であることを特徴とする、請求項23〜27のいずれかに記載の方法。
【請求項29】
該医薬が、乳ガン、結腸ガン、結節性硬化症、神経線維腫症、血友病Aおよび先天的甲状腺機能不全症からなる群より選ばれる少なくとも1種の治療を目的とした医薬であることを特徴とする、請求項27または28のいずれかに記載の方法。
【請求項1】
産生量の減少が求められる望ましくないタンパク質をコードするmRNA前駆体のエキソンの内部領域に対するアンチセンスオリゴヌクレオチドであって、14〜40個のヌクレオチドを含有し、該エキソンの有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチド。
【請求項2】
該望ましくないタンパク質の産生を減少させるための医薬の製造用である、請求項1に記載のアンチセンスオリゴヌクレオチド。
【請求項3】
請求項1または2に記載のアンチセンスオリゴヌクレオチドまたはその相補鎖を包含する核酸運搬体。
【請求項4】
アンチセンスオリゴヌクレオチドを放出することが可能な、請求項3に記載の核酸運搬体。
【請求項5】
RNAスプライシング反応を実施可能な細胞による望ましくないタンパク質の細胞内産生の減少を目的として、該細胞内におけるmRNA前駆体のスプライシングを制御するためのin vitroの方法であって、該細胞内のmRNA前駆体を、該mRNA前駆体に含まれる少なくとも1つのエキソンの有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチドと接触させることを包含し、該アンチセンスオリゴヌクレオチドは、14〜40個のヌクレオチドを含有し、該少なくとも1つのエキソンの内部領域に対するものであることを特徴とする方法。
【請求項6】
該mRNAが機能性タンパク質をコードしていることを特徴とする、請求項5に記載の方法。
【請求項7】
該タンパク質が2つ以上のドメインを包含し、該ドメインの少なくとも1つは、該mRNA前駆体に含まれるエキソンの少なくとも一部のスキッピングによって生じたmRNAにコードされていることを特徴とする、請求項5または6に記載の方法。
【請求項8】
該接触によって、アンチセンスオリゴヌクレオチドと接触したエキソンの隠蔽されたスプライス部位が活性化されることを特徴とする、請求項5〜7のいずれかに記載の方法。
【請求項9】
該エキソン封入シグナルがエキソン認識配列を包含することを特徴とする、請求項5〜8のいずれかに記載の方法。
【請求項10】
該エキソン封入シグナルが強いスプライス供与部位/受容部位対を有するエキソンに存在することを特徴とする、請求項5〜9のいずれかに記載の方法。
【請求項11】
該アンチセンスオリゴヌクレオチドが、15〜25個のヌクレオチドを含有することを特徴とする、請求項5〜10のいずれかに記載の方法。
【請求項12】
該mRNA前駆体に含まれる他のエキソンの有するエキソン封入シグナルを阻害しうる別のアンチセンスオリゴヌクレオチドを、該細胞内の該mRNA前駆体と接触させる工程を更に包含することを特徴とする、請求項5〜11のいずれかに記載の方法。
【請求項13】
異常型タンパク質をコードするエキソンを含むmRNA前駆体を有する細胞による異常型タンパク質の産生を少なくとも部分的に減少させるためのin vitroの方法であって、
該エキソンの少なくとも1つが有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチドであって、該少なくとも1つのエキソンの内部領域に対するものであり、14〜40個のヌクレオチドを含有するアンチセンスオリゴヌクレオチドを該細胞に与え、そして
該mRNA前駆体のスプライシングによって生じるmRNAの翻訳を行わせしめる
ことを包含する方法。
【請求項14】
該エキソン封入シグナルがエキソン認識配列を包含することを特徴とする、請求項13に記載の方法。
【請求項15】
該エキソン封入シグナルが強いスプライス供与部位/受容部位対を有するエキソンに存在することを特徴とする、請求項13または14に記載の方法。
【請求項16】
該アンチセンスオリゴヌクレオチドが、15〜25個のヌクレオチドを含有することを特徴とする、請求項13〜15のいずれかに記載の方法。
【請求項17】
該mRNA前駆体に含まれる他のエキソンの有するエキソン封入シグナルを阻害しうる別のアンチセンスオリゴヌクレオチドを該細胞に与える工程を更に包含することを特徴とする、請求項13〜16のいずれかに記載の方法。
【請求項18】
目的エキソンの一部に対して相補的な核酸が、該エキソンの有するエキソン封入シグナルを特異的に阻害することが可能であることを確認するためのin vitroの方法であって、
目的のエキソンを含むmRNA前駆体を有する細胞に被験核酸を与え、
該細胞を培養して該mRNA前駆体からmRNAを生成せしめ、そして
生成した該mRNAに該エキソンが存在しないことを確認する
ことを包含する方法。
【請求項19】
該目的エキソンを包含するRNA分子と該核酸との相対的な結合親和性をin vitroで測定する工程を更に包含する、請求項18に記載の方法。
【請求項20】
請求項1または2のアンチセンスオリゴヌクレオチドを導入した、ヒト以外の動物。
【請求項21】
ヒトタンパク質をコードする核酸を更に包含する、請求項20に記載のヒト以外の動物。
【請求項22】
該ヒト以外の動物は、自らが有する該ヒトタンパク質の相同体をコードする遺伝子にサイレント突然変異が導入されていることを特徴とする、請求項21に記載のヒト以外の動物。
【請求項23】
mRNA前駆体に含まれる少なくとも1つのエキソンの有するエキソン封入シグナルを特異的に阻害することが可能なアンチセンスオリゴヌクレオチドを用いた、細胞内の望ましくないタンパク質の産生を減少させるための医薬の製造方法であって、該アンチセンスオリゴヌクレオチドが、該少なくとも1つのエキソンの内部領域に対するものであり、14〜40個のヌクレオチドを含有することを特徴とする方法。
【請求項24】
該望ましくないタンパク質が、野生型構造の時に少なくとも2つの機能性ドメインを有し、各ドメインは一次構造であるアミノ酸配列の個別の部分から生成されたものであることを特徴とする、請求項23に記載の方法。
【請求項25】
該望ましくないタンパク質が、異常型タンパク質であることを特徴とする、請求項23または24に記載の方法。
【請求項26】
該異常型タンパク質が、腫瘍タンパク質またはウイルス性タンパク質であることを特徴とする、請求項25に記載の方法。
【請求項27】
該医薬が、遺伝病の治療用または遺伝的疾病素質の改善用の医薬であることを特徴とする、請求項23〜26のいずれかに記載の方法。
【請求項28】
該医薬が、腫瘍およびウイルス感染からなる群より選ばれる少なくとも1種の治療を目的とした医薬であることを特徴とする、請求項23〜27のいずれかに記載の方法。
【請求項29】
該医薬が、乳ガン、結腸ガン、結節性硬化症、神経線維腫症、血友病Aおよび先天的甲状腺機能不全症からなる群より選ばれる少なくとも1種の治療を目的とした医薬であることを特徴とする、請求項27または28のいずれかに記載の方法。
【図1】

【図2】
【図3】

【図4】
【図5】
【図6】

【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2011−200235(P2011−200235A)
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願番号】特願2011−98952(P2011−98952)
【出願日】平成23年4月27日(2011.4.27)
【分割の表示】特願2002−529499(P2002−529499)の分割
【原出願日】平成13年9月21日(2001.9.21)
【出願人】(503107439)
【Fターム(参考)】
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願日】平成23年4月27日(2011.4.27)
【分割の表示】特願2002−529499(P2002−529499)の分割
【原出願日】平成13年9月21日(2001.9.21)
【出願人】(503107439)
【Fターム(参考)】
[ Back to top ]