
真正細菌の蛋白質へのアルキニルアミノ酸のインビボ組込み
【課題】セレクターコドン(例えばアンバー終止コドン、4塩基以上のコドン等)に応答して成長中のポリペプチド鎖にアルキニルアミノ酸をin vivo又はin vitroで組込むための直交成分を作製する。
【解決手段】細胞で機能する第1の直交アミノアシルtRNAシンテターゼ(O−RS)を含む真正細菌細胞であって、前記O−RSがアルキニルアミノ酸である第1の非天然アミノ酸で第1の直交tRNA(O−tRNA)を優先的にアミノアシル化する。
【解決手段】細胞で機能する第1の直交アミノアシルtRNAシンテターゼ(O−RS)を含む真正細菌細胞であって、前記O−RSがアルキニルアミノ酸である第1の非天然アミノ酸で第1の直交tRNA(O−tRNA)を優先的にアミノアシル化する。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願とのクロスリファレンス)
本願は米国仮特許出願第60/612,220号(出願日2004年9月21日);米国仮特許出願第60/630,876号(出願日2004年11月24日);及び米国仮特許出願第60/634,151号(出願日2004年12月7日)の優先権と特典を主張し、各々その開示内容全体を参考資料として全目的で本明細書に組込む。
【0002】
本発明は国立衛生研究所助成番号GM62159として米国政府助成下に創出された。米国政府は本発明に所定の権利をもつことができる。
【0003】
本発明は翻訳生化学の分野に関する。本発明はアルキニルアミノ酸を蛋白質に組込む直交tRNA、直交アミノアシルtRNAシンテターゼ、及びその対を作製及び使用するための組成物及び方法に関する。本発明は前記対を使用して細胞で蛋白質を生産する方法と関連組成物にも関する。
【背景技術】
【0004】
分光プローブ、触媒助剤、又はポリマー等の非ペプチド分子で蛋白質を部位特異的に化学修飾したり、蛋白質を別の蛋白質又は他の任意部分と共有結合できるならば、蛋白質の化学的及び生物学的性質を調査及び操作するための強力な手段となる。一般的なアプローチは蛋白質の求核性表面残基(例えばリジン、ヒスチジン、又はシステインの側鎖)と外来分子の求電子基(例えばアルデヒド、α−ハロカルボキサミド、及びN−ヒドロキシスクシンイミド)のバイオコンジュゲーションを利用している(Lemineux, G. A.; Bertozzi, C. R. TIBTECH 1996, 16, 506)。
【0005】
残念ながら、修飾を標的化するために蛋白質の天然求核性ターゲットを使用するには、これらの反応の選択性が低く、蛋白質には求核性アミノ酸が複数存在するため、標識蛋白質の不均質混合物が形成されるという問題がある。更に、求核標的修飾反応には非生理的条件を要することが多いため、in vivo修飾ストラテジーを実施できないか、及び/又は蛋白質生物活性が低下する可能性がある。
【0006】
特異的標的蛋白質修飾のための新規ターゲットと新規ストラテジーを開発することが当分野で必要とされている。残念ながら、細菌からヒトに至る全公知生物は同一の20種の標準アミノ酸をコードする(稀な例外としてセレノシステイン(例えばA. Bockら, (1991), Molecular Microbiology 5:515-20参照)とピロリジン(例えばG.Srinivasanら, (2002), Science 296:1459-62参照)も挙げられる)。この特徴により、標的蛋白質修飾のための新規化学の開発に天然アミノ酸を使用することは制限される。
【0007】
この制限を解決するための1つのストラテジーは遺伝子コードを拡張し、顕著な化学的性質をもつアミノ酸を生物レパートリーに付加する方法である。このアプローチは真正細菌である大腸菌(E.coli)や他の生物のin vivo蛋白質生合成機構を使用して非天然アミノ酸を蛋白質に付加するために「直交」tRNAと対応する新規「直交」アミノアシルtRNAシンテターゼの使用により、実現可能であることが分かっている(例えばWangら, (2001), Science 292:498-500; Chinら, (2002) Journal of the American Chemical Society 124:9026-9027; Chin and Schultz, (2002), ChemBioChem 11:1135-1137; Chinら, (2002), PNAS United States of America 99:11020-11024; 及びWang and Schultz, (2002), Chem.Comm., 1-10)。国際公開WO2002/086075、発明の名称「直交tRNA−アミノアシルtRNAシンテターゼ対を作製するための方法及び組成物(METHODS AND COMPOSITIONS FOR THE PRODUCTION OF ORTHOGONAL tRNA AMINOACYL-tRNA SYNTHETASE PAIRS)」;WO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」;WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」;WO2005/019415(出願日2004年7月7日);WO2005/007870(出願日2004年7月7日);及びWO2005/007624(出願日2004年7月7日)も参照。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】国際公開WO2002/086075
【特許文献2】国際公開WO2002/085923
【特許文献3】国際公開WO2004/094593
【特許文献4】国際公開WO2005/019415
【特許文献5】国際公開WO2005/007870
【特許文献6】国際公開WO2005/007624
【非特許文献】
【0009】
【非特許文献1】Lemineux, G. A.;Bertozzi, C. R. TIBTECH 1996, 16, 506
【非特許文献2】A. Bockら, (1991), Molecular Microbiology 5:515-20
【非特許文献3】G. Srinivasanら, (2002), Science 296:1459-62
【非特許文献4】Wangら, (2001), Science 292:498-500
【非特許文献5】Chinら, (2002) Journal of the American Chemical Society 124:9026-9027
【非特許文献6】Chin and Schultz, (2002), ChemBioChem 11:1135-1137
【非特許文献7】Chinら, (2002), PNAS United States of America 99:11020-11024
【非特許文献8】Wang and Schultz,(2002), Chem. Comm., 1-10
【発明の概要】
【発明が解決しようとする課題】
【0010】
高度に特異的な標的蛋白質修飾を実施するための新規方法が当分野で必要とされている。非天然アミノ酸を大腸菌の蛋白質にin vivoで組込む直交翻訳成分の開発も当分野で必要とされており、非天然アミノ酸を規定位置に組込むことができ、非天然アミノ酸が蛋白質の他の部分と交差反応又は副反応することなしに特異的修飾のターゲットとして利用できるように顕著な化学的性質をもつことが望ましい。真正細菌蛋白質発現システムは科学研究又は治療用組換え蛋白質材料を大量に生産することができるので、当分野の上記必要は特に大腸菌で実現されることが望ましい。本発明は以下の開示から明らかなように、上記及び他の必要を満たすものである。
【課題を解決するための手段】
【0011】
本発明はセレクターコドン(例えばアンバー終止コドン、4塩基以上のコドン等)に応答して成長中のポリペプチド鎖にアルキニルアミノ酸をin vivo又はin vitroで組込むための直交成分を作製するための組成物及び方法を提供する。本発明は直交tRNA(O−tRNA)、直交アミノアシルtRNAシンテターゼ(O−RS)及びその対を提供する。これらの対は成長中のポリペプチド鎖にアルキニルアミノ酸を組込むために細胞又は無細胞系で使用することができる。アルキニルアミノ酸を含むポリペプチドはアルキン部分が[3+2]シクロ付加反応でアジド部分と容易且つ特異的に反応してトリアゾール結合を形成する結合反応で特に使用される。アルキン基はin vivo系に対して外来性であり、原則的に任意化合物にアジド基を付加できるので、本明細書に実証するようにアルキニルアミノ酸の部位特異的組込みシステムは有用な部位特異的修飾用ツールである。
【0012】
1側面において、真正細菌細胞は直交アミノアシルtRNAシンテターゼ(O−RS)を含み、前記O−RSはアルキニルアミノ酸である非天然アミノ酸で直交tRNA(O−tRNA)を優先的にアミノアシル化する。所定態様において、真正細菌細胞は大腸菌細胞である。所定側面において、O−RSはMethanococcus jannaschiiアミノアシルtRNAシンテターゼ、例えばMethanococcus jannaschiiチロシルtRNAシンテターゼから誘導される。所定態様において、O−RSを誘導するために使用されるチロシルtRNAシンテターゼは配列番号2のアミノ酸配列をもつ野生型Methanococcus jannaschiiチロシルtRNAシンテターゼである。
【0013】
所定態様において、配列番号2の野生型Methanococcus jannaschiiチロシルtRNAシンテターゼから誘導されるO−RSはコンセンサス位置の組合わせに突然変異、例えば、
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリンを含む。所定態様において、O−RSのアミノ酸配列は配列番号4、6、8、10、12、14、16、18、又はその任意保存変異体の1種を含む。O−RSを含む細胞は一般にO−RS、例えば上記任意O−RS種をコードする核酸を含む。O−RSをコードする核酸は例えば配列番号5、7、9、11、13、15、17又は19のヌクレオチド配列を含むことができる。
【0014】
所定態様において、細胞で使用されるO−tRNAはアンバーサプレッサーtRNAである。例えば、O−tRNAは配列番号1のポリヌクレオチド配列であるか又は前記配列を含む。
【0015】
所定態様において、O−RSの基質であるアルキニルアミノ酸はパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)である。細胞系は更に少なくとも1個のセレクターコドンをもつ核酸を含み、セレクターコドンはO−tRNAにより認識される。直交成分を含む細胞は更にアルキニルアミノ酸(例えばpPRO−Phe)を含むことができる。
【0016】
所定態様において、細胞は第2の直交対(即ち第2のO−tRNAと第2のO−RS)を含み、第2の対は第1の非天然アミノ酸とは異なる非天然アミノ酸に特異的であり、第2のO−tRNAは第1のO−tRNAにより認識されるセレクターコドンとは異なるセレクターコドンを認識する。
【0017】
所定側面において、直交成分を含む細胞は翻訳系を含み、前記翻訳系はO−RSとO−tRNAに加え、該当ポリペプチドをコードし、O−tRNAにより認識される少なくとも1個のセレクターコドンを含む核酸と;アルキニルアミノ酸を含むことができ、O−RSはO−tRNAにアルキニルアミノ酸を負荷することができる。
【0018】
所定側面において、本発明はポリペプチド(例えば本明細書に教示するようなO−RSポリペプチド)を提供する。これらのポリペプチドは配列番号2のMethanococcus jannaschiiチロシルアミノアシルtRNAシンテターゼから誘導することができ、以下のアミノ酸コンセンサス:
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリン
をもつことができ、前記ポリペプチドは直交tRNA(O−tRNA)をアルキニルアミノ酸で優先的にアミノアシル化することが可能なアミノアシルtRNAシンテターゼ活性をもつ。所定態様において、本発明のポリペプチドは配列番号4、6、8、10、12、14、16、18、又はその保存変異体から選択される。本発明のこのような任意O−RSポリペプチドは真正細菌細胞において直交tRNA(O−tRNA)をアルキニルアミノ酸で優先的にアミノアシル化することが可能なアミノアシルtRNAシンテターゼである。本発明は上記のような本発明の任意O−RSポリペプチドをコードするポリヌクレオチドも提供する。
【0019】
所定態様において、(本発明のO−RSをコードする)本発明のポリヌクレオチドは配列番号5、7、9、11、13、15、17及び19から選択される。本発明のO−RSをコードする本発明の任意ポリヌクレオチドはベクター(例えば発現ベクター)に組込むことができる。本発明のベクターは細胞で使用することができる。
【0020】
所定側面において、本発明は非天然アルキニルアミノ酸を含む蛋白質を真正細菌細胞で生産する方法を提供する。本方法は蛋白質の任意所望特定位置にアルキニルアミノ酸を挿入するようにプログラムすることができる。本方法は、
(a)(i)直交アミノアシルtRNAシンテターゼ(O−RS)と;
(ii)直交tRNA(O−tRNA)(O−RSはO−tRNAをアルキニルアミノ酸で優先的にアミノアシル化する)と;
(iii)蛋白質をコードする核酸(核酸はO−tRNAにより認識される少なくとも1個のセレクターコドンを含む)と;
(iv)アルキニルアミノ酸を含む真正細菌細胞を提供する段階と;
(b)細胞を増殖させる段階と;
(c)蛋白質の翻訳中に核酸によりコードされる蛋白質の特定位置(蛋白質の特定位置は核酸におけるセレクターコドンの位置に対応する)にアルキニルアミノ酸を組込むことにより、アルキニルアミノ酸を特定位置に含む蛋白質を生産する段階を含む。本方法は一般に大腸菌細胞を使用する。
【0021】
これらの方法で使用されるO−RSは一般にMethanococcus jannaschiiアミノアシルtRNAシンテターゼ、例えばMethanococcus jannaschiiチロシルtRNAシンテターゼから誘導される。所定態様において、Methanococcus jannaschiiチロシルtRNAシンテターゼは配列番号2のシンテターゼである。所定態様において、O−RSは配列番号2のMethanococcus jannaschiiチロシルtRNAシンテターゼから誘導され、O−RSは以下の突然変異:
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリンをもつアミノ酸配列をもつ。
【0022】
所定態様において、前記方法は配列番号4、6、8、10、12、14、16、18、又はその任意保存変異体から選択されるアミノ酸配列をもつO−RSを使用する。本発明の方法では、細胞はこれらのO−RSポリペプチドの任意のものをコードするポリヌクレオチドを含むことができる。例えば、配列番号5、7、9、11、13、15、17又は19のヌクレオチド配列を含むポリヌクレオチドを使用することができる。
【0023】
これらの方法の所定態様において、O−tRNAはアンバーサプレッサーtRNAであり、セレクターコドンはアンバー終止コドン(TAG)である。所定態様において、O−tRNAは配列番号1に記載のポリヌクレオチド配列を含むか又は前記配列によりコードされる。これらの方法はアルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)をもつ蛋白質を生産するために使用することができる。本発明の方法により生産された蛋白質は野生型治療用蛋白質、診断用蛋白質、産業用酵素、又はその一部のアミノ酸配列と少なくとも75%一致するアミノ酸配列を含むことができる。これらの蛋白質は場合により医薬的に許容可能なキャリヤーと結合することができる。
【0024】
所定態様において、本発明の方法により生産された蛋白質は例えば[3+2]シクロ付加反応によりトリアゾール結合を形成することによりアルキニルアミノ酸の位置を修飾することができる。
【図面の簡単な説明】
【0025】
【図1】図1Aは(1)非天然アルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(IUPAC命名法によると、2−アミノ−3−[4−(プロプ−2−イニルオキシ)フェニル]プロピオン酸とも言う)の化学構造を示す。図1Bは室温で銅の存在下でのアジドとアルキンの[3+2]シクロ付加反応による不可逆的トリアゾール形成の一般反応化学反応を示す。
【図2】野生型Methanococcus jannaschiiチロシルtRNAシンテターゼ(MjTyrRS)のヌクレオチド及びアミノ酸配列を示す。パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)tRNAシンテターゼにおける部位特異的突然変異誘発又は他の突然変異の標的アミノ酸位置(及び対応するトリプレットコドン)を四角で囲む。
【図3】野生型Methanococcus jannaschiiチロシルtRNAシンテターゼをコードするポリヌクレオチドの突然変異誘発後に同定単離した8種のパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)tRNAシンテターゼ種をまとめた表である。野生型Methanococcus jannaschiiチロシルtRNAシンテターゼとパラ−プロパルギルオキシフェニルアラニンtRNAシンテターゼ(pPRO−PheRS)において指定コドンによりコードされるアミノ酸を示す。突然変異位置のコドンも示す。突然変異体のアミノ酸位置ナンバリングは図2に示すような野生型Methanococcus jannaschiiチロシルtRNAシンテターゼのアミノ酸ナンバリングに従う。
【図4】精製Ser4→pPRO−Phe4突然変異体ミオグロビンのGelcode(登録商標)Blue(Pierce Biotechnology,Inc.)染色SDS−PAGEゲルを示す。レーン1はパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)の存在下に最小培地で培養した大腸菌で発現された蛋白質を含み;レーン2はpPRO−Pheの不在下で生成された蛋白質サンプルを示す。下部の図はミオグロビンのC末端のヘキサヒスチジンタグを検出するために抗His6抗体を使用した同一サンプル材料のウェスタンブロットを示す。
【図5】アルキニル非天然アミノ酸(Y*で示す)を含むトリプシンペブチドHGVTVLTALGY*ILKのタンデム質量スペクトルを予想フラグメントイオン質量と共に示す。矢印はペプチドに観測されたb及びyイオン系列を示す。
【図6】図6A及び6Bはアジド官能基を付けた色素の化学構造(夫々2及び3)を示す。図6Aの色素2はダンシルフルオロフォアを含み、図6Bの色素3はフルオレセインフルオロフォアを含む。
【図7】図7Aは組換えアンバーコドン(4TAG)の部位にアルキニルアミノ酸を含む突然変異体ミオグロビンと(図6A及び6Bに示すような)アジド官能基を付けた色素の[3+2]シクロ付加反応からの不可逆的トリアゾール形成の一般反応化学を示す。図7Bは分解した標識ミオグロビンの紫外線照射下の蛍光ゲルイメージングを示し、[3+2]シクロ付加反応物は色素2又は色素3と共有結合している。
【図8】図8A及び8Bアルキニル非天然アミノ酸の例の構造と名称を示す。図8Aは非天然前駆体から化学的に合成することができるアルキニル非天然アミノ酸を示す。図8Bは既存天然アミノ酸基質から潜在的に合成することができるアルキニル非天然アミノ酸を示す。
【発明を実施するための形態】
【0026】
(定義)
本発明を詳細に記載する前に、本発明は特定生物系に限定されず、当然のことながら種々のものに適用できると理解すべきである。同様に、本明細書で使用する用語は特定態様のみの記載を目的とし、限定的でないことも理解すべきである。本明細書と特許請求の範囲で使用する単数形はそうでないことが内容から明白である場合を除き、複数形も含む。従って、例えば「細胞」と言う場合には2個以上の細胞の組合せを含み、「ポリヌクレオチド」と言う場合には実際問題としてそのポリヌクレオチドの多数のコピーを含む。
【0027】
本欄及び以下に特に定義しない限り、本明細書で使用する全科学技術用語は本発明が属する分野の当業者に通常理解されている通りの意味をもつ。
【0028】
直交:本明細書で使用する「直交」なる用語は細胞又は他の翻訳系に内在する対応分子に比較して低効率で細胞の内在成分と共働するか、あるいは細胞の内在成分と共働できない分子(例えば直交tRNA(O−tRNA)及び/又は直交アミノアシルtRNAシンテターゼ(O−RS))を意味する。tRNA及びアミノアシルtRNAシンテターゼに関して直交とは、内在tRNAが内在tRNAシンテターゼと共働する能力に比較して直交tRNAが内在tRNAシンテターゼと共働できないか又は低効率(例えば20%未満、10%未満、5%未満、又は1%未満の効率)でしか共働できず、あるいは、内在tRNAシンテターゼが内在tRNAと共働する能力に比較して直交アミノアシルtRNAシンテターゼが内在tRNAと共働できないか又は低効率でしか共働できないことを意味する。直交分子は細胞内に機能的に正常な相補的内在分子をもたない。例えば、細胞中の直交tRNAがこの細胞の任意内在tRNAシンテターゼ(RS)によりアミノアシル化される効率は内在tRNAが内在RSによりアミノアシル化される効率に比較して低いか又はゼロである。別の例では、直交RSが該当細胞の任意内在tRNAをアミノアシル化する効率は内在tRNAが内在RSによりアミノアシル化される効率に比較して低いか又はゼロである。第1の直交分子と共働する第2の直交分子を細胞に導入することができる。例えば、直交tRNA/RS対は対照(例えば対応するtRNA/RS内在対、又は活性直交対(例えばチロシル直交tRNA/RS対))の効率に比較して所定の効率(例えば45%の効率、50%の効率、60%の効率、70%の効率、75%の効率、80%の効率、90%の効率、95%の効率、又は99%以上の効率)で細胞において共働する導入相補成分を含む。
【0029】
直交チロシルtRNA:本明細書で使用する直交チロシルtRNA(チロシル−O−tRNA)とは該当翻訳系に直交性のtRNAであり、tRNAは(1)天然に存在するチロシルtRNAと同一であるか又は実質的に類似しているか、(2)自然又は人工突然変異誘発により天然に存在するチロシルtRNAから誘導されるか、(3)(1)又は(2)の野生型又は突然変異体チロシルtRNA配列の配列を考慮する任意プロセスにより誘導されるか、(4)野生型又は突然変異体チロシルtRNAと相同であるか;(5)表4にチロシルtRNAシンテターゼの基質として指定する任意特定tRNAと相同であるか、あるいは(6)表4にチロシルtRNAシンテターゼの基質として指定する任意特定tRNAの保存変異体である。チロシルtRNAはアミノ酸を負荷した状態でも負荷しない状態でも存在することができる。更に当然のことながら、「チロシル−O−tRNA」は場合によりコグネイトシンテターゼによりチロシン以外のアミノ酸(例えば非天然アミノ酸パラ−プロパルギルオキシフェニルアラニン)を負荷(アミノアシル化)される。実際に、当然のことながら、本発明のチロシル−O−tRNAは翻訳中にセレクターコドンに応答して成長中のポリペプチドに天然又は人工のいずれかに拘わらずほぼ任意アミノ酸を挿入するために有利に使用される。
【0030】
直交チロシルアミノ酸シンテターゼ:本明細書で使用する直交チロシルアミノ酸シンテターゼ(チロシル−O−RS)とは該当翻訳系においてチロシル−O−tRNAをアミノ酸で優先的にアミノアシル化する酵素である。チロシル−O−RSがチロシル−O−tRNAに負荷するアミノ酸は天然又は人工のいずれかを問わずに任意アミノ酸とすることができ、本明細書では限定しない。シンテターゼは場合により天然に存在するチロシルアミノ酸シンテターゼと同一又は相同であるか、あるいは表4にO−RSとして指定するシンテターゼと同一又は相同である。例えば、O−RSは表4のチロシル−O−RSの保存変異体とすることができ、及び/又は表4のO−RSと少なくとも50%、60%、70%、80%、90%、95%、98%、99%又はそれ以上配列が一致することができる。
【0031】
コグネイト:「コグネイト」なる用語は共働する成分、例えば直交tRNAと直交アミノアシルtRNAシンテターゼを意味する。これらの成分は「相補的」であると言うこともできる。
【0032】
優先的にアミノアシル化する:本明細書で直交翻訳系に関して使用する場合に、O−RSはO−RSが発現系で任意内在tRNAに負荷するよりも効率的にO−tRNAにアミノ酸を負荷するときにコグネイトO−tRNAを「優先的にアミノアシル化する」。即ち、O−tRNAと所与の任意内在tRNAがほぼ等モル比で翻訳系に存在するとき、O−RSは内在tRNAに負荷するよりも高頻度でO−tRNAに負荷する。O−RSにより負荷されるO−tRNAとO−RSにより負荷される内在tRNAの相対比は高いことが好ましく、従って、O−tRNAと内在tRNAが等モル濃度で翻訳系に存在する場合にはO−RSはO−tRNAに排他的、又はほぼ排他的に負荷することが好ましい。O−tRNAとO−RSが等モル濃度で存在する場合にO−RSにより負荷されるO−tRNAと内在tRNAの相対比は1:1を上回り、好ましくは少なくとも約2:1、より好ましくは5:1、更に好ましくは10:1、更に好ましくは20:1、更に好ましくは50:1、更に好ましくは75:1、更に好ましくは95:1、98:1、99:1、100:1、500:1、1,000:1、5,000:1又はそれ以上である。
【0033】
(a)O−RSが内在tRNAに比較してO−tRNAを優先的にアミノアシル化するとき、及び(b)O−RSがO−tRNAを任意天然アミノ酸でアミノアシル化する場合に比較してそのアミノアシル化が非天然アミノ酸に特異的であるときにO−RSは「O−tRNAを非天然アミノ酸で優先的にアミノアシル化する」。即ち、非天然アミノ酸と天然アミノ酸がO−RSとO−tRNAを含む翻訳系に等モル量で存在するとき、O−RSは天然アミノ酸よりも高頻度で非天然アミノ酸をO−tRNAに負荷する。非天然アミノ酸を負荷されたO−tRNAと天然アミノ酸を負荷されたO−tRNAの相対比は高いことが好ましい。O−RSはO−tRNAに排他的、又はほぼ排他的に非天然アミノ酸を負荷することがより好ましい。天然アミノ酸と非天然アミノ酸の両者が等モル濃度で翻訳系に存在するとき、O−tRNAの非天然アミノ酸負荷とO−tRNAの天然アミノ酸負荷の相対比は1:1を上回り、好ましくは少なくとも約2:1、より好ましくは5:1、更に好ましくは10:1、更に好ましくは20:1、更に好ましくは50:1、更に好ましくは75:1、更に好ましくは95:1、98:1、99:1、100:1、500:1、1,000:1、5,000:1又はそれ以上である。
【0034】
セレクターコドン:「セレクターコドン」なる用語は翻訳プロセスでO−tRNAにより認識され、内在tRNAにより認識されないコドンを意味する。O−tRNAアンチコドンループはmRNA上のセレクターコドンを認識し、そのアミノ酸(例えばアルキニルアミノ酸等の非天然アミノ酸)をポリペプチドのこの部位に組込む。セレクターコドンとしては例えば終止コドン(例えばアンバー、オーカー及びオパールコドン)等のナンセンスコドン、4塩基以上のコドン、レアコドン、天然又は非天然塩基対から誘導されるコドン及び/又は同等物を挙げることができる。
【0035】
サプレッサーtRNA:サプレッサーtRNAは例えばセレクターコドンに応答してポリペプチド鎖にアミノ酸を組込むためのメカニズムを提供することにより、所与翻訳系でメッセンジャーRNA(mRNA)の読取りを変更するtRNAである。例えば、サプレッサーtRNAは例えば終止コドン(例えばアンバー、オーカー又はオパールコドン)、4塩基コドン、レアコドン等を読み飛ばすことができる。
【0036】
抑圧活性:本明細書で使用する「抑圧活性」なる用語は一般に、読み飛ばさないと翻訳終結又は誤訳(例えばフレームシフト)をもたらすコドン(例えばアンバーコドンや4塩基以上のコドンであるセレクターコドン)の翻訳読み飛ばしを行うtRNA(例えばサプレッサーtRNA)の能力を意味する。サプレッサーtRNAの抑圧活性は第2のサプレッサーtRNA又は対照系(例えばO−RSをもたない対照系)に比較して観測される翻訳読み飛ばし活性の百分率として表すことができる。
【0037】
本発明は抑圧活性を定量することが可能な種々の手段を提供する。該当セレクターコドン(例えばアンバーコドン)に対する特定O−tRNA及びO−RSの抑圧百分率とは、O−tRNA、O−RS及びセレクターコドンをもたないポジティブ対照構築物に比較して、O−RSとO−tRNAを含む該当翻訳系で発現され、そのコーディング核酸中にセレクターコドンを含む所与試験マーカー(例えばLacZ)の活性百分率を意味する。従って、例えば、セレクターコドンをもたない活性ポジティブ対照マーカー構築物が所与翻訳系において該当マーカーアッセイに関連する単位で表した観測活性Xをもつ場合には、セレクターコドンを含む試験構築物の抑圧百分率は、O−tRNAとO−RSを更に含む翻訳系で発現される以外はポジティブ対照マーカーが発現される条件とほぼ同一の環境条件下で試験マーカー構築物が示すXの百分率である。一般に、試験マーカーを発現する翻訳系は更にO−RSとO−tRNAにより認識されるアミノ酸を含む。場合により、抑圧百分率測定値は、O−tRNA、O−RS及び/又はO−tRNA及び/又はO−RSにより認識される該当アミノ酸を含まない系で試験マーカーと同一のセレクターコドンを含む「バックグラウンド」又は「ネガティブ」対照マーカー構築物に試験マーカーを比較することにより精密化することができる。このネガティブ対照は該当翻訳系におけるマーカーからのバックグラウンドシグナル効果を考慮するように抑圧百分率測定値を正規化するのに有用である。
【0038】
抑圧効率は当分野で公知の多数のアッセイの任意のものにより測定することができる。例えば、β−ガラクトシダーゼレポーターアッセイを使用することができ、例えば本発明のO−tRNAを含むプラスミドと共に修飾lacZプラスミド(構築物はlacZ核酸配列中にセレクターコドンをもつ)を適当な生物(例えば直交成分を使用することができる生物)に由来する細胞に導入する。コグネイトシンテターゼも(ポリペプチド又は発現されるとコグネイトシンテターゼをコードするポリヌクレオチドとして)導入することができる。細胞を培地で所望密度(例えばOD600=約0.5)まで増殖させ、例えばBetaFluor(登録商標)β−ガラクトシダーゼアッセイキット(Novagen)を使用してβ−ガラクトシダーゼアッセイを実施する。比較可能な対照(例えば所望位置にセレクターコドンではなく対応するセンスコドンをもつ修飾lacZ構築物から観測される値)に対するサンプルの活性百分率として抑圧百分率を計算することができる。
【0039】
翻訳系:「翻訳系」なる用語は成長中のポリペプチド鎖(蛋白質)にアミノ酸を組込む成分を意味する。翻訳系の成分としては例えばリボソーム、tRNA、シンテターゼ、mRNA等を挙げることができる。本発明のO−tRNA及び/又はO−RSは例えば非真核細胞(例えば細菌(例えば大腸菌))、又は真核細胞(例えば酵母細胞、哺乳動物細胞、植物細胞、藻類細胞、真菌細胞、昆虫細胞、及び/又は同等物)でin vitro又はin vivo翻訳系に付加するか又はその一部とすることができる。
【0040】
非天然アミノ酸:本明細書で使用する「非天然アミノ酸」なる用語は20種の標準天然アミノ酸の1種又はセレノシステインもしくはピロリジン以外の任意アミノ酸、修飾アミノ酸、及び/又はアミノ酸類似体(例えばアルキニルアミノ酸)を意味する。
【0041】
から誘導:本明細書で使用する「から誘導」なる用語は特定分子もしくは生物から単離されているか、あるいは特定分子もしくは生物を使用するか又は特定分子もしくは生物からの情報を使用して作製された成分を意味する。例えば、第2のポリペプチドから誘導されるポリペプチドは第2のポリペプチドのアミノ酸配列と同一又は実質的に同様のアミノ酸配列を含む。ポリペプチドの場合には、誘導種は例えば自然突然変異誘発、人工的特異的突然変異誘発又は人工的ランダム突然変異誘発により得ることができる。ポリペプチドを誘導するために使用される突然変異誘発は意図的に特異的でも意図的にランダムでもよい。第1のポリペプチドから誘導される別のポリペプチドを作製するためのポリペプチドの突然変異誘発は(例えばポリメラーゼの非忠実性に起因する)ランダムなイベントとすることができ、誘導されたポリペプチドの同定は偶然とすることができる。ポリペプチドの突然変異誘発は一般にポリペプチドをコードするポリヌクレオチドの操作を伴う。
【0042】
ポジティブ選択又はスクリーニングマーカー:本明細書で使用する「ポジティブ選択又はスクリーニングマーカー」なる用語は、このマーカーが存在する(例えば発現、活性化等される)と、対応する形質をもたない細胞からこの形質を含む細胞(例えばポジティブ選択マーカーをもつ細胞)を識別することができるマーカーを意味する。
【0043】
ネガティブ選択又はスクリーニングマーカー:本明細書で使用する「ネガティブ選択又はスクリーニングマーカー」なる用語は、このマーカーが存在する(例えば発現、活性化等される)と、(例えば選択性質又は形質をもつ細胞に対して)選択性質又は形質をもたない細胞を識別することができるマーカーを意味する。
【0044】
レポーター:本明細書で使用する「レポーター」なる用語は該当系のターゲット成分を同定及び/又は選択するために使用することができる成分を意味する。例えば、レポーターとしては蛋白質、例えば抗生物質耐性又は感受性を付与する酵素(例えばβ−ラクタマーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)等)、蛍光スクリーニングマーカー(例えば緑色蛍光蛋白質(例えばGFP)、YFP、EGFP、RFP等)、発光マーカー(例えばホタルルシフェラーゼ蛋白質)、アフィニティースクリーニングマーカー、又はポジティブもしくはネガティブ選択マーカー遺伝子(例えばlacZ、β−gal/lacZ(β−ガラクトシダーゼ)、ADH(アルコールデヒドロゲナーゼ)、his3、ura3、leu2、lys2等)を挙げることができる。
【0045】
真核生物:本明細書で使用する「真核生物」なる用語は真核生物界に属する生物を意味する。真核生物はその構成が一般に多細胞であり(但し、例えば酵母のように多細胞ではないものもある)、膜結合核と他の膜結合オルガネラが存在し、遺伝物質が線状であり(即ち染色体が線状)、オペロンが存在せず、イントロン、メッセージキャッピング及びポリA mRNAが存在し、更に他の生化学的特徴(例えば特徴的リボソーム構造)により一般に原核細胞から区別できる。真核生物としては例えば動物(例えば哺乳動物、昆虫、爬虫類、鳥類等)、繊毛虫、植物(例えば単子葉植物、双子葉植物、藻類等)、真菌類、酵母、鞭毛虫、微胞子虫、原生生物等が挙げられる。
【0046】
原核生物:本明細書で使用する「原核生物」なる用語はモネラ界(原核生物界とも言う)に属する生物を意味する。原核生物はその構成が単細胞であり、出芽又は分裂による無性生殖であり、膜結合核又は他の膜結合オルガネラをもたず、染色体が環状であり、オペロンが存在し、イントロン、メッセージキャッピング及びポリA mRNAが存在せず、更に他の生化学的特徴(例えば特徴的リボソーム構造)により一般に真核細胞から区別できる。原核生物は真正細菌及び古細菌亜界を含む。シアノバクテリア(藍藻類)とマイコプラズマをモネラ界の別々の分類にする場合もある。
【0047】
細菌:本明細書で使用する「細菌」及び「真正細菌」なる用語は「古細菌」から区別できる原核生物を意味する。同様に、古細菌は真核生物から区別できる原核生物を意味する。真正細菌と古細菌は多数の形態学的及び生化学的基準により区別できる。例えば、リボソームRNA配列、RNAポリメラーゼ構造、イントロンの有無、抗生物質感受性、細胞壁ペプチドグリカン及び他の細胞壁成分の有無、膜脂質構造の分岐の有無、並びにヒストン及びヒストン様蛋白質の有無を使用して生物を真正細菌又は古細菌に分類する。
【0048】
真正細菌の例としては、Escherichia coli、Thermus thermophilus及びBacillus stearothermophilusが挙げられる。古細菌の例としては、Methanococcus jannaschii(Mj)、Methanosarcina mazei(Mm)、Methanobacterium thermoautotrophicum(Mt)、Methanococcus maripaludis、Methanopyrus kandleri、Halobacterium(例えばHaloferax volcanii及びHalobacterium種NRC−1)、Archaeoglobus fulgidus(Af)、Pyrococcus furiosus(Pf)、Pyrococcus horikoshii(Ph)、Pyrobaculum aerophilum、Pyrococcus abyssi、Sulfolobus solfataricus(Ss)、Sulfolobus tokodaii、Aeuropyrum pernix(Ap)、Thermoplasma acidophilum及びThermoplasma volcaniumが挙げられる。
【0049】
保存変異体:翻訳成分に関して本明細書で使用する「保存変異体」なる用語は類似する基本成分(例えばO−tRNA又はO−RS)と同様に機能するが、参照O−tRNA又はO−RSに比較して配列に変異をもつ翻訳成分(例えば保存変異体O−tRNA又は保存変異体O−RS)を意味する。例えば、O−RSは相補的O−tRNA又は保存変異体O−tRNAを非天然アミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン等のアルキニルアミノ酸)でアミノアシル化するが、O−tRNAと保存変異体O−tRNAは同一配列をもたない。保存変異体は保存変異体が対応するO−tRNA又はO−RSに相補的である限り、例えば配列の1カ所、2カ所、3カ所、4カ所、又は5カ所以上に変異をもつことができる。
【0050】
選択又はスクリーニング物質:本明細書で使用する「選択又はスクリーニング物質」なる用語はこのような物質が存在すると、集団から所定成分を選択/スクリーニングすることができる物質を意味する。例えば、選択又はスクリーニング物質としては限定されないが、例えば栄養素、抗生物質、光波長、抗体、発現されたポリヌクレオチド等が挙げられる。選択物質は例えば濃度、強度等を変動させることができる。
【0051】
〜に応答して:本明細書で使用する「〜に応答して」なる用語は本発明のtRNAがセレクターコドンを認識し、tRNAと結合したアルキニルアミノ酸を成長中のポリペプチド鎖に組込むのを媒介するプロセスを意味する。
【0052】
コードする:本明細書で使用する「コードする」なる用語は第1の分子又は配列鎖とは異なる第2の分子又は配列鎖の生産を導くためにポリマー巨大分子又は配列鎖中の情報を使用する任意プロセスを意味する。本明細書ではこの用語を広義に使用し、種々に適用することができる。1側面では、「コードする」なる用語は新規に合成された相補的姉妹鎖をDNA依存性DNAポリメラーゼによりコードするための鋳型として2本鎖DNA分子の一方の鎖を使用する半保存的DNA複製プロセスを意味する。
【0053】
別の側面では、「コードする」なる用語は第1の分子とは異なる化学的性質をもつ第2の分子の生産を導くためにある分子中の情報を使用する任意プロセスを意味する。例えば、DNA分子は(例えばDNA依存性RNAポリメラーゼ酵素を含む転写プロセスにより)RNA分子をコードすることができる。また、RNA分子は翻訳プロセスと同様にポリペプチドをコードすることができる。翻訳プロセスについて使用する場合には、「コードする」なる用語はアミノ酸をコードするトリプレットコドンにも適用する。所定側面では、RNA分子は例えばRNA依存性DNAポリメラーゼを含む逆転写プロセスによりDNA分子をコードすることができる。別の側面では、DNA分子はポリペプチドをコードすることができ、この場合に使用する「コードする」とは当然のことながら転写プロセスと翻訳プロセスの両者を含む。
【0054】
アルキン:本明細書で使用する「アルキン」(「アセチレン」と言う場合もある)なる用語は(図1Bに示すように)2個の炭素原子間に三重結合を含み、一般構造:
【化1】

(式中、Rは任意原子又は構造である)をもつ化学構造を意味する。置換基として使用する場合には、アルキン部分を「アルキニル」基と言う。アルキニル炭素原子はsp2結合しており、2個の他の原子と結合しか形成せず、これらの結合の一方は単結合であり、第2の結合は三重結合である。例えば、アルキニルアミノ酸は2個の炭素中心間に三重結合を含むアミノ酸である。アルキニル置換基は自然界ではアミノ酸に認められないので、全アルキニルアミノ酸は非天然アミノ酸である。
【0055】
アジド:本明細書で使用する「アジド」なる用語は一般構造:
R−N=N+=N−
をもつ化学基−N3を意味する。アジド基は一般に炭素原子と結合している。
【0056】
例えば、アジド色素はアジド置換基をもつ色素分子である(例えば図6A及び6Bのアジド色素2及び3参照)。「アジド」なる用語はアジド基を含む化合物(例えば、アジ化ベンジル、アジ化ナトリウム等)を意味する。
【0057】
生理的条件下で高度に選択的に蛋白質を修飾する化学反応が大いに必要とされている(Lemineux and Bertozzi (1996) TIBTECH, 16:506)。蛋白質の選択的修飾に当分野で現在使用されている大半の反応は蛋白質アミノ酸側鎖の天然求核性残基を標的とする求核性反応パートナーと求電子性反応パートナーの間の共有結合形成、例えばα−ハロケトンとヒスチジン又はシステイン側鎖の反応を利用している。これらの場合の選択性は蛋白質の求核性残基の数とアクセシビリティにより決定される。残念ながら、天然蛋白質は反応部位の配置が不良であったり(例えば接近しにくい)、反応ターゲットを複数含む(例えばリジン、ヒスチジン及びシステイン残基)ことが多いため、修飾反応の選択性が低く、求核/求電子試薬による高度標的蛋白質修飾は困難になる。更に、修飾部位は一般にリジン、ヒスチジン及びシステインの天然求核性側鎖に限定される。他の部位の修飾は困難又は不可能になる。
【0058】
この問題の1つの解決方法は直交翻訳成分を使用して新規反応性をもつ非天然アミノ酸を蛋白質にプログラム下で部位特異的に生合成的に組込む方法である(Wang and Schultz (2002) Chem. Commun., 1:1; 及びvan Maarseveen and Back (2003) Angew. Chem., 115:6106)。本発明ではアンバーナンセンスコドンTAGに応答して細菌(例えば大腸菌)で産生された蛋白質にアルキニル含有非天然アミノ酸を遺伝的に組込む選択的蛋白質修飾のための非常に効率的な新規方法を報告する。その後、これらのアルキニルアミノ酸側鎖を特異的且つ位置選択的に修飾することができる。アルキニル基のユニークな反応化学により、蛋白質を非常に高い選択性で修飾することができる。
【0059】
細菌発現系で産生された蛋白質のユニーク部位(例えば所望部位)にアルキニル官能基を選択的に導入するために、本発明者らはアルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe;図1A参照)を遺伝的にコードする真正細菌で機能する直交RNA/アミノアシルtRNAシンテターゼ対を開発した。要約すると、本発明者らは大腸菌細胞でアンバーサプレッサーtRNAにパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)を選択的に負荷するMethanococcus janaschiiチロシルtRNAシンテターゼの新規突然変異体を同定した。これらの進化型tRNAシンテターゼ対はアルキニル基を蛋白質に部位特異的に組込むために使用することができる。
【0060】
標的蛋白質修飾
本発明ではアンバーナンセンスコドンTAGに応答して真正細菌(例えば大腸菌)で産生された蛋白質にアルキニル含有非天然アミノ酸を遺伝的に組込む選択的蛋白質修飾のための非常に効率的な方法を報告する。本明細書に記載する新規組成物と方法は直交tRNA/アミノアシルtRNAシンテターゼシステムを利用し、直交システムはMethanococcus janaschiiに由来する成分を使用し、これらの成分は該当蛋白質を産生するための真正細菌宿主系で使用される。アルキニルアミノ酸の蛋白質組込みはアルキニルアミノ酸の組込みを指示するセレクターコドンを含むように該当蛋白質をコードするポリヌクレオチドを組換えることにより任意所望位置で行われるようにプログラムすることができる。
【0061】
その後、該当蛋白質のこれらのアルキニルアミノ酸側鎖をアジド誘導体とのHuisgen[3+2]シクロ付加反応により特異的且つ位置選択的に修飾することができる(図1B参照)(Padwa,In Comprehensive Organic Synthesis; [Trost,B.M.,Ed.] Pergamon: Oxford, 1991, Vol.4, p 1069-1109; Huisgen, In 1,3-Dipolar Cycloaddition Chemistry, [Padwa,A.,Ed.] Wiley: New York, 1984; p 1-176)。この方法は求核置換ではなくシクロ付加を利用するので、蛋白質を非常に高い選択性で修飾することができる。この反応は触媒量のCu(I)塩を反応混合物に加えることにより室温で水性条件下に優れた位置選択的性(1,4>1,5)で実施できるという利点がある(Tornoeら, (2002) J. Org. Chem., 67:3057-3064; Rostovtsevら, (2002) Angew. Chem., Int. Ed., 41:2596-2599)。
【0062】
アルキニル反応性ターゲットはin vivo系に対して完全に外来性であるという利点があり、その反応化学において高度に選択性(例えばアゾ含有部分に対して高度に反応性)であり、蛋白質に関するin vitro及びin vivo両者の結合反応を可能にする比較的温和な反応条件を使用して結合することができるので、蛋白質の生物活性を維持できる。(限定的ではないが)本発明を実証するために、アルキニル部分をミオグロビンモデル蛋白質に組込んだ後に、安定なトリアゾール結合による[3+2]シクロ付加反応(図1B参照)によりアジド蛍光色素(図6A及び6B参照)とバイオコンジュゲートさせる。
【0063】
本発明はアルキニルアミノ酸とアジド部分の[3+2]シクロ付加を例証するために2種のアジド蛍光色素を使用する(実施例4参照)が、本発明はこれらの2種のアジド色素、又は任意色素もしくはラベル、又は実際に任意単一種の結合性材料の使用に限定されない。本発明のアジド含有部分はアジド誘導体であるほぼ任意分子とすることができる。このような分子としては限定されないが、色素、フルオロフォア、架橋剤、糖誘導体、ポリマー(例えばポリエチレングリコール誘導体)、光架橋剤、細胞傷害性化合物、アフィニティーラベル、ビオチン誘導体、樹脂、ビーズ、第2(又は第3以下)の蛋白質又はポリペプチド、ポリヌクレオチド(例えばDNA,RNA等)、金属キレート剤、補因子、脂肪酸、炭水化物等が挙げられる。これらのアジド分子はアルキニル基をもつ非天然アミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン)と結合することができる(図1A参照)。
【0064】
本発明は図6A及び6Bに示すアジド色素の合成について詳細に記載する。夫々実施例6及び7参照。しかし、任意該当特定蛋白質のアジド誘導体を合成することは当業者が当然に実施できる範囲内である。例えば、アジド化合物の合成方法を記載した多数の教科書やプロトコールが入手可能である。一般情報については、Patai, Saul, “The chemistry of the azido group” in The Chemistry of Functional Groups, London, New York, Interscience Publishers, 1971参照。
【0065】
別の側面では、本発明はアルキニル含有ポリペプチドとの結合反応にポリエチレングリコールの誘導体(アジド−PEG)を使用することによりペグ化ポリペプチドを作製するための組成物及び方法を提供する。アジドポリエチレングリコールの一般構造は下式:
N3−CH2−(CH2−O−CH2)n−CH2OR
により表され、式中、RはH又はCH3であり、nは例えば50〜10,000、75〜5,000、100〜2,000、100〜1,000等の整数である。本発明の各種態様において、アジドポリエチレングリコールは例えば約5,000〜約100,000Da(即ち約5kDa〜約100kDa)、約20,000〜約50,000Da、約20,000〜約10,000Da(例えば20,000Da)等の分子量をもつ。アジドポリエチレングリコールの合成技術は当業者に周知である。例えば、アジド基を含む求核性分子(例えばアジ化ナトリウム又は3−アジドプロピルアミン)と求電子基(例えば臭化物又はN−ヒドロキシスクシンイミドエステル)を含むポリエチレングリコール分子を反応させてアジドポリエチレングリコールを生成することができる。
【0066】
アジド−PEGはトリアゾール結合によりアルキニル含有蛋白質とバイオコンジュゲートした場合に本発明で利用できる。蛋白質治療薬をポリエチレングリコールで修飾(ペグ化)すると、多くの場合には蛋白質の薬物動態及び薬力学的性質を改善し、従って、効力を改善し、投薬頻度を最小限にすることができる。蛋白質治療薬のペグ化の各種利点は例えば、Deitersら,“Site-specific PEGylation of proteins containing unnatural amino acids,” Bioorganic & Medicinal Chemistry Letters 14:5743-5745 (2004)に記載及び例証されている。
【0067】
更に、エステル結合も含む非天然アルキニルアミノ酸を含むポリペプチドの作製に関連する他の利点も考えられる。例えば、アルキニルアミノ酸とエステル結合を併用することにより作製したペグ化ポリペプチドはin vivo又はin vitroエステル結合の鹸化によるポリペプチドの緩慢な遊離を可能にすることができる。また、アジド−PEG分子の代わりにポリマー支持体(アジド樹脂)を使用すると、蛋白質アフィニティー精製が可能になる。トリアゾール共有結合は非常に強力な洗浄段階を可能にし、エステルアルキニルアミノ酸の使用は塩基処理による蛋白質の遊離を可能にする。特筆すべき点として、このようなアフィニティー精製スキームは精製のために該当蛋白質に人工タグ(例えばヘキサヒスチジン)又はエピトープを付ける必要がない。使用する非天然アミノ酸に応じて、開裂段階後に原則的に野生型(天然)ポリペプチドをアフィニティー樹脂から遊離させることができる。
【0068】
エステル結合をもつ非天然アルキニルアミノ酸を合成し、蛋白質に組込むことができる(例えば、3−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノプロパン酸や4−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノブタン酸)(図8B参照)。[3+2]シクロ付加によるバイオコンジュゲーション後に、in vivo又はin vitro鹸化によりエステル結合を開裂することができ、1応用例は例えばペグ化蛋白質からのペブチド部分の緩慢な遊離である。
【0069】
所定側面において、本発明のポリペプチドはアルキニル含有ポリペプチドを含み、更にこれらのポリペプチドのコンジュゲート形を含む。例えば、所定側面において、本発明はトリアゾール結合と共有結合蛍光アジド色素を含むポリペプチドを含む(例えば図6A、6B及び7A参照)。この側面では、ポリペプチドは最初にアルキニル基を含んでおり、色素は最初にアジド基を含んでおり、[3+2]シクロ付加により両者を結合し、トリアゾール結合を形成する。別の態様では、本発明のアルキニル含有蛋白質はアジドポリエチレングリコールを含む(化学構造6参照)。
【0070】
直交tRNA/アミノアシルtRNAシンテターゼ技術
本発明の新規組成物及び方法の理解は直交tRNAと直交アミノアシルtRNAシンテターゼの対に関連する活性を理解することにより容易になる。直交tRNA及びアミノアシルtRNAシンテターゼ技術については例えば、国際公開WO2002/085923、WO2002/086075、WO204/09459、WO2005/019415、WO2005/007870及びWO2005/007624に記載されている。
【0071】
付加反応性非天然アミノ酸(例えばアルキニルアミノ酸)を遺伝コードに付加するためには、宿主翻訳機構で効率的に機能することができるが、対が翻訳系に内在性のシンテターゼ及びtRNAから独立して機能するという意味で該当翻訳系に対して「直交性」のアミノアシルtRNAシンテターゼと適切なtRNAを含む新規直交対が必要である。直交対の所望特徴としては、内在tRNAによりデコードされない特定新規コドン(例えばセレクターコドン)のみをデコード又は認識するtRNAと、そのコグネイトtRNAをただ1個の特定非天然アミノ酸で優先的にアミノアシル化(又は負荷)するアミノアシルtRNAシンテターゼが挙げられる。O−tRNAは一般に内在シンテターゼによりアミノアシル化されない。例えば大腸菌では、直交対は例えば大腸菌に存在する40種の内在tRNAのいずれとも交差反応しないアミノアシルtRNAシンテターゼと、例えば大腸菌に存在する21種の内在シンテターゼのいずれによってもアミノアシル化されない直交tRNAを含む。
【0072】
本発明は真正細菌(例えば大腸菌)の蛋白質にアルキニルアミノ酸を遺伝的にコード及び組込むための直交対を提供し、直交成分は宿主細胞の翻訳機構の内在大腸菌成分と交差反応しないが、所望非天然アミノ酸を認識し、アンバーナンセンスコドンTAGに応答して蛋白質に組込む。本発明により提供される直交成分としては、Methanococcus jannaschiiチロシルtRNAシンテターゼから誘導される直交アミノアシルtRNAシンテターゼと、突然変異体チロシルtRNACUAアンバーサプレッサーが挙げられる。この系では、突然変異体アミノアシルtRNAシンテターゼはサプレッサーtRNAをpPRO−PHEでアミノアシル化するが、20種の標準アミノ酸ではアミノアシル化しない。
【0073】
本発明はアルキニルアミノ酸を蛋白質に組込むために使用することができる付加直交tRNA−アミノアシルtRNAシンテターゼ対(例えばO−tRNA/O−RS対)の組成物と、前記対の同定及び作製方法を提供する。本発明のO−tRNAはO−tRNAにより認識されるセレクターコドンを含むポリヌクレオチドによりコードされる蛋白質へのアルキニルアミノ酸の例えばin vivoでの組込みを媒介することができる。O−tRNAのアンチコドンループはmRNA上のセレクターコドンを認識し、そのアミノ酸(例えばアルキニルアミノ酸)をポリペプチドのこの部位に組込む。本発明の直交アミノアシルtRNAシンテターゼはそのO−tRNAを特定アルキニルアミノ酸のみで優先的にアミノアシル化(又は負荷)する。
【0074】
例えば、本明細書に実証するように、修飾のために高度に選択的に標的化することができるアルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe;図1A,構造1参照)をセレクターコドン(例えばTAGコドン)に応答して真正細菌細胞(大腸菌;E.coli)の蛋白質に選択的且つ効率的に組込んだ。蛋白質に組込まれると、pPRO−Pheを細胞内で化学的に標的化することができ、例えばアジド基をもつ色素による修飾のために標的化することができる。色素分子のアジド基はアルキニルアミノ酸と反応し、色素標識のために蛋白質を高度に選択的に標的化することができる。
【0075】
アルキニルアミノ酸を蛋白質に部位特異的に組込むことができると、蛋白質の試験を容易にすることができると共に、新規特性をもつ蛋白質の開発が可能になる。例えば、アルキニル含有蛋白質の発現は特異的標識による蛋白質の試験を容易にし、酵素の触媒機能を改変し、蛋白質を他の蛋白質、小分子及び生体分子等と架橋することができる。
直交tRNA、直交アミノアシルtRNAシンテターゼ、及びその対
【0076】
1種以上の非天然アミノ酸を含む蛋白質の作製に適した翻訳系は例えば国際公開WO2002/086075、発明の名称「直交tRNA−アミノアシルtRNAシンテターゼ対を作製するための方法及び組成物(METHODS AND COMPOSITION FOR THE PRODUCTION OF ORTHOGONAL tRNA-AMINOACYL-tRNA SYNTHETASE PAIRS)」;WO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」;WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」;WO2005/019415(出願日2004年7月7日);WO2005/007870(出願日2004年7月7日);及びWO2005/007624(出願日2004年7月7日)に記載されている。これらの各出願はその開示内容全体を参考資料として本明細書に組込む。このような翻訳系は一般に直交tRNA(O−tRNA)と、直交アミノアシルtRNAシンテターゼ(O−RS)と、非天然アミノ酸(本発明ではアルキニルアミノ酸)を含む細胞(例えば大腸菌等の非真核細胞又は酵母等の真核細胞とすることができる)を含み、O−RSはO−tRNAをアルキニルアミノ酸でアミノアシル化する。本発明の直交対はO−tRNA(例えばサプレッサーtRNA、フレームシフトtRNA等)とO−RSを含む。本発明は個々の成分も提供する。
【0077】
一般に、直交対がセレクターコドンを認識し、セレクターコドンに応答してアミノ酸を負荷するとき、直交対はセレクターコドンを「抑圧」すると言う。即ち、翻訳系の(例えば細胞の)内在機構により認識されないセレクターコドンは通常翻訳されないので、非抑圧下で核酸から翻訳されるポリペプチドの生産を阻止することができる。本発明のO−tRNAはセレクターコドンを認識し、本明細書の配列表に記載するようなポリヌクレオチド配列を含むか又は前記配列によりコードされるO−tRNAの抑圧効率に比較してコグネイトシンテターゼの存在下でセレクターコドンに応答して少なくとも例えば約45%、50%、60%、75%、80%、又は90%以上の抑圧効率を含む。O−RSはO−tRNAを該当非天然アミノ酸(例えばアルキニルアミノ酸)でアミノアシル化する。細胞は例えば該当ポリペプチドをコードするポリヌクレオチドを含む核酸を介して成長中のポリペプチド鎖に非天然アミノ酸を組込むためにO−tRNA/O−RS対を使用し、ポリヌクレオチドはO−tRNAにより認識されるセレクターコドンを含む。所定の望ましい側面では、細胞は付加O−tRNA/O−RS対を含むことができ、付加O−tRNAは付加O−RSにより別の非天然アミノ酸を負荷される。例えば、O−tRNAの一方は4塩基コドンを認識することができ、他方は終止コドンを認識することができる。あるいは、複数の異なる終止コドン又は複数の異なる4塩基コドンが異なるセレクターコドンを特異的に認識することができる。
【0078】
本発明の所定態様では、直交tRNA(O−tRNA)と、直交アミノアシルtRNAシンテターゼ(O−RS)と、アルキニルアミノ酸と、該当ポリペプチドをコードするポリヌクレオチドを含む核酸を含む細胞(例えば大腸菌細胞)が提供され、ポリヌクレオチドはO−tRNAにより認識されるセレクターコドンを含む。翻訳系は無細胞系でもよく、例えば各種市販「in vitro」転写/翻訳系の任意のものを本明細書に記載するようなO−tRNA/ORS対及び非天然アミノ酸と併用することができる。
【0079】
1態様では、O−RSとO−tRNAの併用による抑圧効率はO−RSの不在下のO−tRNAの抑圧効率の例えば約5倍、10倍、15倍、20倍、又は25倍以上である。1側面では、O−RSとO−tRNAの併用による抑圧効率は本明細書の配列表に記載するような直交シンテターゼ対の抑圧効率の少なくとも例えば約35%、40%、45%、50%、60%、75%、80%、又は90%以上である。
【0080】
上述のように、本発明は場合により細胞又は他の翻訳系に複数のO−tRNA/O−RS対を含み、2種以上の非天然アミノ酸(例えばアルキニルアミノ酸と別の非天然アミノ酸)を組込むことができる。例えば、細胞は更に別の付加O−tRNA/O−RS対と第2の非天然アミノ酸を含むことができ、この付加O−tRNAは第2のセレクターコドンを認識し、この付加O−RSはO−tRNAを第2の非天然アミノ酸で優先的にアミノアシル化する。例えば、O−tRNA/O−RS対(O−tRNAは例えばアンバーセレクターコドンを認識する)を含む細胞は更に第2の直交対(例えばロイシル、リシル、グルタミル等)を含むことができる(第2のO−tRNAは別のセレクターコドン、例えばオパールコドン、4塩基コドン等を認識する)。各直交対は異なるセレクターコドンの認識を助長できるように異なる起源に由来することが望ましい。
【0081】
O−tRNA及び/又はO−RSは天然に存在するものでもよいし、例えば種々の生物の任意のものに由来するtRNAのライブラリー及び/又はRSのライブラリーを作製するか及び/又は種々の利用可能な突然変異ストラテジーの任意のものを使用することにより、天然に存在するtRNA及び/又はRSの突然変異により誘導してもよい。例えば、直交tRNA/アミノアシルtRNAシンテターゼ対を作製する1ストラテジーは例えば宿主細胞以外の起源又は多重起源に由来する(宿主に対して)異種のtRNA/シンテターゼ対を宿主細胞に導入する方法である。異種シンテターゼ候補の特性としては、例えば宿主細胞tRNAに負荷しないことが挙げられ、異種tRNA候補の特性としては、例えば宿主細胞シンテターゼによりアミノアシル化されないことが挙げられる。更に、異種tRNAは全宿主細胞シンテターゼに対して直交性である。
【0082】
直交対を作製するための第2のストラテジーはO−tRNA又はO−RSをスクリーニング及び/又は選択するための突然変異体ライブラリーを作製する方法である。これらのストラテジーを組み合わせてもよい。
直交tRNA(O−tRNA)
【0083】
本発明の直交tRNA(O−tRNA)はO−tRNAにより認識されるセレクターコドンを含むポリヌクレオチドによりコードされる蛋白質への非天然アミノ酸(例えばアルキニルアミノ酸)の例えばin vivo又はin vitroでの組込みを媒介することが望ましい。所定態様では、本発明のO−tRNAは本明細書の配列表のO−tRNA配列に記載するようなポリヌクレオチド配列を含むか又は前記配列によりコードされるO−tRNAに比較してコグネイトシンテターゼの存在下でセレクターコドンに応答して少なくとも例えば約45%、50%、60%、75%、80%、又は90%以上の抑圧効率を含む。
【0084】
抑圧効率は当分野で公知の多数のアッセイの任意のものにより測定することができる。例えば、β−ガラクトシダーゼレポーターアッセイを使用することができ、例えば本発明のO−tRNAを含むプラスミドと共に修飾lacZプラスミド(構築物はlacZ核酸配列中にセレクターコドンをもつ)を適当な生物(例えば直交成分を使用することができる生物)に由来する細胞に導入する。コグネイトシンテターゼも(ポリペプチド又は発現されるとコグネイトシンテターゼをコードするポリヌクレオチドとして)導入することができる。細胞を培地で所望密度(例えばOD600=約0.5)まで増殖させ、例えばBetaFluor(登録商標)β−ガラクトシダーゼアッセイキット(Novagen)を使用してβ−ガラクトシダーゼアッセイを実施する。比較可能な対照(例えば所望位置にセレクターコドンではなく対応するセンスコドンをもつ修飾lacZ構築物から観測される値)に対するサンプルの活性百分率として抑圧百分率を計算することができる。
【0085】
本発明のO−tRNAの例を本明細書の配列表に記載する。代表的O−tRNA及びO−RS分子の配列については本明細書の表、実施例及び図面も参照。更に、本明細書の「核酸及びポリペプチド配列と変異体」のセクションも参照。O−RS mRNA又はO−tRNA分子等のRNA分子では、所与配列又はその相補配列に対してチミン(T)がウラシル(U)で置換されている(コーディングDNAでは逆)。塩基に付加修飾を加えてもよい。
【0086】
本発明は本明細書に記載する特定O−tRNAに対応するO−tRNAの保存変異体も含む。例えば、O−tRNAの保存変異体としては、例えば本明細書の配列表に記載するような特定O−tRNAと同様に機能し、適当な自己相補性によりtRNA L形構造を維持するが、例えば本明細書の配列表、図面又は実施例に記載する配列と同一配列をもたない分子が挙げられる(更に野生型tRNA分子以外のものであることが望ましい)。本明細書の「核酸及びポリペプチド配列と変異体」のセクションも参照。
【0087】
O−tRNAを含む組成物は更に直交アミノアシルtRNAシンテターゼ(O−RS)を含むことができ、O−RSはO−tRNAを非天然アミノ酸(例えばアルキニルアミノ酸)で優先的にアミノアシル化する。所定態様では、O−tRNAを含む組成物は更に(例えばin vitro又はin vivo)翻訳系を含むことができる。該当ポリペプチドをコードし、O−tRNAにより認識されるセレクターコドンを含むポリヌクレオチドを含む核酸、又はこれらの1種以上の組み合わせも細胞に加えることができる。本明細書の「直交アミノアシルtRNAシンテターゼ」のセクションも参照。
【0088】
直交tRNA(O−tRNA)の作製方法も本発明の特徴である。本方法により作製されたO−tRNAも本発明の特徴である。本発明の所定態様では、O−tRNAは突然変異体ライブラリーを作製することにより作製することができる。突然変異体tRNAのライブラリーは当分野で公知の種々の突然変異誘発技術を使用して作製することができる。例えば、突然変異体tRNAは部位特異的突然変異、ランダム点突然変異、相同組換え、DNAシャフリング又は他の帰納的突然変異誘発法、キメラ構築又はその任意組み合わせにより作製することができる。
【0089】
tRNAの所望ループ又は領域(例えばアンチコドンループ、受容体ステム、Dアーム又はループ、可変ループ、TPCアーム又はループ、tRNA分子の他の領域又はその組合せ)の特定位置(例えば非保存位置又は保存位置)、ランダム位置又は両者の組合せに付加突然変異を導入することができる。一般に、tRNAの突然変異としてはセレクターコドンの認識を可能にするように突然変異体tRNAライブラリーの各メンバーのアンチコドンループを突然変異させる方法が挙げられる。この方法は更にO−tRNAの末端に付加配列(CCA)を付加する段階を含むことができる。一般に、O−tRNAは所望RSに対するその親和性等を維持しながら、出発材料(例えば複数のtRNA配列)に比較して所望生物に対する直交性が改善されている。
【0090】
前記方法は場合によりtRNA及び/又はアミノアシルtRNAシンテターゼの配列の類似性(及び/又は推定相同性)を分析し、特定生物について直交性であると思われるO−tRNA、O−RS及び/又はその対の潜在候補を決定する段階を含む。分析には、当分野で公知であり、本明細書に記載するコンピュータープログラム(例えばBLAST及びpileupプログラム)を使用することができる。1例では、大腸菌で使用するための潜在的直交翻訳成分を選択するためには、真正細菌生物に密接な配列類似性を示さないシンテターゼ及び/又はtRNAを選択する。
【0091】
一般に、O−tRNAは複数の潜在的O−tRNAのメンバーを含む第1の種の細胞の集団に例えばネガティブ選択を実施することにより得られる。ネガティブ選択は細胞に内在性のアミノアシルtRNAシンテターゼ(RS)によりアミノアシル化される潜在的O−tRNAライブラリーのメンバーを含む細胞を排除する。こうして、第1の種の細胞に直交性のtRNAプールが得られる。
【0092】
所定態様において、ネガティブ選択では、ネガティブ選択マーカー(例えば抗生物質耐性を付与する酵素(例えばβ−ラクタマーゼ)、検出可能な産物を付与する酵素(例えばβ−ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT))、例えば毒性物質(例えばバルナーゼ))をコードするポリヌクレオチドの(例えば機能的バルナーゼを依然として産生する)非必須位置等にセレクターコドンを導入する。場合により選択物質(例えばアンピシリン等の抗生物質)の存在下で細胞集団を増殖させることによりスクリーニング/選択を実施する。1態様では、選択物質の濃度を変動させる。
【0093】
例えば、サプレッサーtRNAの活性を測定するためには、セレクターコドンのin vivo抑圧(例えばネガティブ選択マーカー(例えばβ−ラクタマーゼ遺伝子(bla))をコードするポリヌクレオチドに導入されたナンセンス又はフレームシフト突然変異)に基づく選択システムを使用する。例えば、所定位置(例えばA184)にセレクターコドンをもつポリヌクレオチド変異体(例えばbla変異体)を構築する。細胞(例えば細菌)をこれらのポリヌクレオチドで形質転換する。内在大腸菌シンテターゼにより効率的に負荷することができない直交tRNAの場合には、抗生物質耐性(例えばアンピシリン耐性)はプラスミドで形質転換されていない細菌と同等以下のはずである。tRNAが非直交性の場合、又はtRNAに負荷することが可能な異種シンテターゼをシステムで同時発現させる場合には、より高レベルの抗生物質(例えばアンピシリン)耐性が観測される。プラスミドで形質転換されていない細胞と同等の抗生物質濃度のLB寒天プレートで増殖することができない細胞(例えば細菌)を選択する。
【0094】
毒性物質(例えばリボヌクレアーゼ又はバルナーゼ)の場合には、複数の潜在的tRNAのメンバーが内在宿主(例えば大腸菌)シンテターゼによりアミノアシル化されるとき(即ち、宿主(例えば大腸菌)シンテターゼに非直交性であるとき)には、セレクターコドンは抑圧され、生産される毒性ポリヌクレオチド産物は細胞死を生じる。直交tRNA又は非機能的tRNAを含む細胞は生存する。
【0095】
1態様では、所望生物に直交性のtRNAプールに次にポジティブ選択を実施し、セレクターコドンを例えばβ−ラクタマーゼ遺伝子等の薬剤耐性遺伝子によりコードされるポジティブ選択マーカーに配置する。ポジティブ選択は細胞に直交性のtRNAプールのメンバーをコードするか又は前記メンバーを含むポリヌクレオチドと、ポジティブ選択マーカーをコードするポリヌクレオチドと、コグネイトRSをコードするポリヌクレオチドを含む細胞で実施される。所定態様では、第2の細胞集団はネガティブ選択により排除されなかった細胞を含む。ポリヌクレオチドを細胞で発現させ、選択物質(例えばアンピシリン)の存在下で細胞を増殖させる。次に、同時発現させたコグネイトシンテターゼによりアミノアシル化され、このセレクターコドンに応答してアミノ酸を挿入することができるtRNAを選択する。一般に、これらの細胞は非機能的tRNA、又は該当シンテターゼにより効率的に認識することができないtRNAを含む細胞に比較して高い抑圧効率を示す。非機能的tRNA又は該当シンテターゼにより効率的に認識されないtRNAを含む細胞は抗生物質に感受性である。従って、(i)内在宿主(例えば大腸菌)シンテターゼの基質ではなく;(ii)該当シンテターゼによりアミノアシル化することができ;(iii)翻訳で機能的なtRNAが両者選択後に生存している。
【0096】
従って、スクリーニングされる状況に応じて同一マーカーがポジティブマーカーにもネガティブマーカーにもなる。即ち、マーカーがポジティブスクリーニングされる場合にはポジティブマーカーであり、ネガティブスクリーニングされる場合にはネガティブマーカーである。
【0097】
上記方法における選択(例えばポジティブ選択、ネガティブ選択又はポジティブ選択とネガティブ選択の両者)のストリンジェンシーは場合により選択ストリンジェンシーの変動を含む。例えば、バルナーゼは極毒性蛋白質であるので、異なる数のセレクターコドンをバルナーゼ遺伝子に導入するか及び/又は誘導プロモーターを使用することによりネガティブ選択のストリンジェンシーを制御することができる。別の例では、選択又はスクリーニング物質の濃度(例えばアンピシリン濃度)を変動させる。本発明の1側面では、初期ラウンド中の所望活性は低いと思われるのでストリンジェンシーを変動させる。即ち、初期ラウンドでは低ストリンジェンシー選択条件を適用し、後期ラウンドの選択では高ストリンジェンシー条件を適用する。所定態様では、ネガティブ選択、ポジティブ選択又はネガティブ選択とポジティブ選択の両方を複数回反復する。複数の異なるネガティブ選択マーカー、ポジティブ選択マーカー又はネガティブ選択マーカーとポジティブ選択マーカーの両方を使用することができる。所定態様では、ポジティブ選択マーカーとネガティブ選択マーカーを同一にすることができる。
【0098】
本発明ではセレクターコドンに応答して非天然アミノ酸(例えばアルキニルアミノ酸)を負荷する直交翻訳成分(例えばO−tRNA、O−RS、及びO−tRNA/O−RS対)を作製するために他の型の選択/スクリーニングを使用することもできる。例えば、ネガティブ選択マーカー、ポジティブ選択マーカー又はポジティブ選択マーカーとネガティブ選択マーカーの両方として、適切な反応体の存在下で蛍光発光するか又は発光反応を触媒するマーカーを使用することができる。別の態様では、蛍光活性化細胞ソーティング(FACS)又は発光によりマーカーの産物を検出する。場合により、マーカーはアフィニティースクリーニングマーカーを含む。Francisco, J. A. ら,(1993) Production and fluorescence-activated cell sorting of Escherichia coli expressing a functional antibody fragment on the external surface. Proc Natl Acad Sci USA. 90:10444-8も参照。
【0099】
組換え直交tRNAの他の作製方法も例えば国際出願公開WO2002/086075、発明の名称「直交tRNA−アミノアシルtRNAシンテターゼ対を作製するための方法及び組成物(METHODS AND COMPOSITIONS FOR THE PRODUCTION OF ORTHOGONAL tRNA AMINOACYL-tRNA SYNTHETASE PAIRS)」;WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」;及びWO2005/019415(出願日2004年7月7日)に記載されている。Forsterら, (2003) Programming peptidomimetic synthetases by translating genetic codes designed de novo PNAS 100 (11):6353-6357; 及びFengら, (2003), Expanding tRNA recognition of a tRNA synthetase by a single amino acid change, PNAS 100 (10):5676-5681も参照。
【0100】
直交アミノアシルtRNAシンテターゼ(O−RS)
本発明のO−RSはO−tRNAを非天然アミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン等のアルキニルアミノ酸)で優先的にin vitro又はin vivoでアミノアシル化する。本発明のO−RSはO−RSを含むポリペプチド及び/又はO−RS又はその一部をコードするポリヌクレオチドにより翻訳系(例えば細胞)に提供することができる。例えば、O−RSの1例は本明細書の配列表と実施例に記載するようなアミノ酸配列、又はその保存変異体を含む。別の例では、O−RS、又はその一部は本明細書の配列表又は実施例に記載する配列を含むアミノ酸をコードするポリヌクレオチド配列、又はその相補的ポリヌクレオチド配列によりコードされる。例えば、代表的O−RS分子の配列については下記表及び実施例参照。本明細書の「核酸及びポリペプチド配列と変異体」のセクションも参照。
【0101】
直交アミノアシルtRNAシンテターゼ(O−RS)、例えばO−tRNAと併用するためのO−RSの同定方法も本発明の特徴である。例えば、1方法は第1の種の細胞集団について選択(例えばポジティブ選択)を実施する段階を含み、前記細胞は1)複数のアミノアシルtRNAシンテターゼ(RS)のメンバー(例えば複数のRSは突然変異体RS、第1の種以外の種に由来するRS又は突然変異体RSと第1の種以外の種に由来するRSの両方を含むことができる)と;2)(例えば1種以上の種に由来する)直交tRNA(O−tRNA)と;3)(例えばポジティブ)選択マーカーをコードし、少なくとも1個のセレクターコドンを含むポリヌクレオチドを各々含む。複数のRSのメンバーを含まないか又はその量の少ない細胞に比較して抑圧効率の高い細胞について細胞を選択又はスクリーニングする。抑圧効率は当分野で公知の技術及び本明細書に記載するように測定することができる。抑圧効率の高い細胞はO−tRNAをアミノアシル化する活性RSを含む。第1の種に由来する第1組のtRNAの活性RSによる(in vitro又はin vivo)アミノアシル化レベルを第2の種に由来する第2組のtRNAの活性RSによる(in vitro又はin vivo)アミノアシル化レベルと比較する。アミノアシル化レベルは検出可能な物質(例えば標識アミノ酸又は非天然アミノ酸、例えば標識パラ−プロパルギルオキシフェニルアラニン)により測定することができる。第1組のtRNAに比較して第2組のtRNAをより効率的にアミノアシル化する活性RSを一般に選択することにより、O−tRNAと併用するための効率的(最適化)直交アミノアシルtRNAシンテターゼが得られる。この方法により同定されたO−RSも本発明の特徴である。
【0102】
アミノアシル化を測定するためには多数のアッセイの任意のものを使用することができる。これらのアッセイはin vitro又はin vivoで実施することができる。例えば、in vitroアミノアシル化アッセイは例えばHoben and Soll(1985)Methods Enzymol.113:55-59に記載されている。アミノアシル化は直交翻訳成分と共にレポーターを使用し、蛋白質をコードする少なくとも1個のセレクターコドンを含むポリヌクレオチドを発現する細胞でレポーターを検出することにより測定することもできる。WO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」;及びWO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」も参照。
【0103】
同定されたO−RSを更に操作し、所望非天然アミノ酸(例えばアルキニルアミノ酸)のみをO−tRNAに負荷し、20種の標準アミノ酸を負荷しないようにシンテターゼの基質特異性を改変することができる。非天然アミノ酸に対して基質特異性をもつ直交アミノアシルtRNAシンテターゼの作製方法は例えばシンテターゼの活性部位、シンテターゼの編集メカニズム部位、各種シンテターゼドメインを組み合わせることによる各種部位等でシンテターゼを突然変異させる段階と、選択プロセスを適用する段階を含む。ポジティブ選択後にネガティブ選択を行う併用に基づくストラテジーを使用する。ポジティブ選択では、ポジティブマーカーの非必須位置に導入したセレクターコドンが抑圧されると、細胞はポジティブ選択圧下で生存する。従って、天然及び非天然アミノ酸両者の存在下で生存細胞は直交サプレッサーtRNAに天然又は非天然アミノ酸を負荷する活性シンテターゼをコードする。ネガティブ選択では、ネガティブマーカーの非必須位置に導入したセレクターコドンが抑圧されると、天然アミノ酸特異性をもつシンテターゼは除去される。ネガティブ選択とポジティブ選択の生存細胞は直交サプレッサーtRNAを非天然アミノ酸のみでアミノアシル化(負荷)するシンテターゼをコードする。その後、これらのシンテターゼを例えばDNAシャフリング又は他の帰納的突然変異誘発法により更に突然変異誘発することができる。
【0104】
突然変異体O−RSのライブラリーは当分野で公知の種々の突然変異誘発技術を使用して作製することができる。例えば、突然変異体RSは部位特異的突然変異、ランダム点突然変異、相同組換え、DNAシャフリング又は他の帰納的突然変異誘発法、キメラ構築又はその任意組み合わせにより作製することができる。例えば、突然変異体RSのライブラリーは2種以上の他の例えばサイズとダイバーシティーの小さい「サブライブラリー」から作製することができる。RSのキメラライブラリーも本発明に含まれる。なお、場合により各種生物(例えば真正細菌又は古細菌等の微生物)に由来するtRNAシンテターゼのライブラリー(例えば天然ダイバーシティーを含むライブラリー)(例えば米国特許第6,238,884号(Shortら);米国特許第5,756,316号(Schallenbergerら);米国特許第5,783,431号(Petersenら);米国特許第5,824,485号(Thompsonら);米国特許第5,958,672号(Shortら)参照)を構築し、直交対についてスクリーニングする。
【0105】
シンテターゼにポジティブ及びネガティブ選択/スクリーニングストラテジーを実施した後、これらのシンテターゼを更に突然変異誘発することができる。例えば、O−RSをコードする核酸を単離することができ;(例えばランダム突然変異誘発、部位特異的突然変異誘発、組換え又はその任意組み合わせにより)突然変異O−RSをコードする1組のポリヌクレオチドを核酸から作製することができ;O−tRNAを非天然アミノ酸(例えばアルキニルアミノ酸)で優先的にアミノアシル化する突然変異O−RSが得られるまでこれらの個々の段階又はこれらの段階の組み合わせを繰り返すことができる。本発明の1側面では、前記段階を複数回、例えば少なくとも2回実施する。
【0106】
O−tRNA、O−RS、又はその対を作製するための本発明の方法では、付加レベルの選択/スクリーニングストリンジェンシーを使用することもできる。選択又はスクリーニングストリンジェンシーはO−RSを作製するための方法の一方又は両方の段階で変動させることができる。これは例えば選択/スクリーニング物質の使用量等の変動とすることができる。付加ラウンドのポジティブ及び/又はネガティブ選択を実施することもできる。選択又はスクリーニングは更にアミノ酸浸透率の変化、翻訳効率の変化、翻訳忠実度の変化等の1種以上を含むこともできる。一般に、1種以上の変化は蛋白質を生産するために直交tRNA−tRNAシンテターゼ対を使用する生物における1個以上の遺伝子の突然変異に基づく。
【0107】
O−RSの作製と、シンテターゼの基質特異性の改変に関するその他の一般的な詳細についてはWO2002/086075、発明の名称「直交tRNA−アミノアシルtRNAシンテターゼ対を作製するための方法及び組成物(METHODS AND COMPOSITIONS FOR THE PRODUCTION OF ORTHOGONAL tRNA AMINOACYL-tRNA SYNTHETASE PAIRS)」;及びWO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」に記載されている。
【0108】
資源及び宿主生物
本発明の直交翻訳成分(O−tRNAとO−RS)はO−tRNA/O−RS成分と宿主系が直交に作用するという条件で他の任意種に由来する宿主翻訳系で使用するために任意生物(又は生物組み合わせ)に由来することができる。O−tRNAとO−RSは同一生物に由来する必要はない。1側面では、直交成分は真正細菌宿主系で使用するために古細菌遺伝子(即ち古細菌)に由来することができる。
【0109】
例えば、直交O−tRNAは古細菌生物、例えばMethanococcus jannaschii、Methanobacterium thermoautotrophicum、Halobacterium(例えばHaloferax volcanii及びHalobacterium種NRC−1)、Archaeoglobus fulgidus、Pyrococcus furiosus、Pyrococcus horikoshii、Aeuropyrum pernix、Methanococcus maripaludis、Methanopyrus kandleri、Methanosarcina mazei(Mm)、Pyrobaculum aerophilum、Pyrococcus abyssi、Sulfolobus solfataricus(Ss)、Sulfolobus tokodaii、Thermoplasma acidophilum、Thermoplasma volcanium等の古細菌や、Escherichia coli,Thermus thermophilus,Bacillus stearothermphilus等の真正細菌に由来することができ、直交O−RSは生物又は生物組み合わせ、例えばMethanococcus jannaschii、Methanobacterium thermoautotrophicum、Halobacterium(例えばHaloferax volcanii及びHalobacterium種NRC−1)、Archaeoglobus fulgidus、Pyrococcus furiosus、Pyrococcus horikoshii、Aeuropyrum pernix、Methanococcus maripaludis、Methanopyrus kandleri、Methanosarcina mazei、Pyrobaculum aerophilum、Pyrococcus abyssi、Sulfolobus solfataricus、Sulfolobus tokodaii、Thermoplasma acidophilum、Thermoplasma volcanium等の古細菌や、Escherichia coli,Thermus thermophilus,Bacillus stearothermphilus等の真正細菌に由来することができる。1態様では、例えば植物、藻類、原生動物、真菌類、酵母、動物(例えば哺乳動物、昆虫、節足動物等)等の真核資源もO−tRNA及びO−RS資源として使用することができる。
【0110】
O−tRNA/O−RS対の個々の成分は同一生物に由来するものでも異なる生物に由来するものでもよい。1態様では、O−tRNA/O−RS対は同一生物に由来する。あるいは、O−tRNA/O−RS対のO−tRNAとO−RSは異なる生物に由来する。
【0111】
O−tRNA、O−RS又はO−tRNA/O−RS対はin vivo又はin vitroで選択又はスクリーニングすることができ、及び/又はアルキニルアミノ酸を組込んだポリペプチドを作製するために細胞(例えば真正細菌細胞)で使用することができる。使用される真正細菌細胞は限定されないが、例えばEscherichia coli、Thermus thermophilus、Bacillus stearothermphilus等である。本発明の翻訳成分を含む真正細菌細胞の組成物も本発明の特徴である。
【0112】
別の種で使用するためにある種でO−tRNA及び/又はO−RSをスクリーニングする方法については、国際出願公開WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」(出願日2004年4月16日)も参照。
【0113】
セレクターコドン
本発明のセレクターコドンは蛋白質生合成機構の遺伝コドン枠を拡張する。例えば、セレクターコドンとしては例えばユニーク3塩基コドン、ナンセンスコドン(例えばアンバーコドン(UAG)又はオパールコドン(UGA)等の終止コドン)、非天然コドン、少なくとも4塩基のコドン、レアコドン等が挙げられる。例えば1個以上、2個以上、3個以上等の多数のセレクターコドンを所望遺伝子に導入することができる。複数の異なるセレクターコドンを使用することにより、これらの異なるセレクターコドンを使用して(例えば少なくとも1種のアルキニルアミノ酸を含む)複数の非天然アミノ酸の同時部位特異的組込みを可能にする複数の直交tRNA/シンテターゼ対を使用することができる。
【0114】
1態様では、本方法はアルキニルアミノ酸を細胞にin vivo組込むために終止コドンであるセレクターコドンを使用する。例えば、終止コドンを認識し、O−RSによりアルキニルアミノ酸でアミノアシル化されるO−tRNAを作製する。このO−tRNAは天然に存在する翻訳系のアミノアシルtRNAシンテターゼにより認識されない。慣用部位特異的突然変異誘発法を使用して該当ポリペプチドをコードするポリヌクレオチドの該当部位に終止コドンを導入することができる。例えばSayers,J.R.ら,(1988),5’,3’Exonuclease in phosphorothioate−based oligonucleotide−directed mutagenesis.Nucleic Acids Res.791−802参照。O−RS、O−tRNA及び該当ポリペプチドをコードする核酸を例えばin vivoで併用すると、終止コドンに応答してアルキニルアミノ酸が組込まれ、特定位置にアルキニル活性アミノ酸を含むポリペプチドが得られる。本発明の1態様では、セレクターコドンとして使用される終止コドンはアンバーコドンUAG及び/又はオパールコドンUGAである。1例では、セレクターコドンとしてUAGとUGAの両者を使用する遺伝子コードは存在度が最も高い終結シグナルであるオーカーナンセンスコドンUAAを保存しながら22種のアミノ酸をコードすることができる。
【0115】
アルキニル活性アミノ酸のin vivo組込みは宿主細胞をさほど撹乱せずに実施することができる。例えば、大腸菌等の非真核細胞では、UAGコドンの抑圧効率はO−tRNA(例えばアンバーサプレッサーtRNA)と(UAGコドンと結合してリボソームから成長中のペプチドの放出を開始する)放出因子1(RF1)の競合に依存するので、例えばO−tRNA(例えばサプレッサーtRNA)の発現レベルを増加するか又はRF1欠損株を使用することにより抑圧効率を調節することができる。真核細胞では、UAGコドンの抑圧効率はO−tRNA(例えばアンバーサプレッサーtRNA)と(終止コドンと結合してリボソームから成長中のペプチドの放出を開始する)真核放出因子1(例えばeRF)の競合に依存するので、例えばO−tRNA(例えばサプレッサーtRNA)の発現レベルを増加することにより抑圧効率を調節することができる。更に、付加化合物、例えばジチオスレイトール(DTT)等の還元剤も加えてもよい。
【0116】
アルキニルアミノ酸はレアコドンでコードすることもできる。例えば、in vitro蛋白質合成反応でアルギニン濃度を下げると、レアアルギニンコドンAGGはアラニンでアシル化された合成tRNAによるAlaの挿入に有効であることが分かっている。例えばMaら, Biochemistry, 32:7939(1993)参照。この場合には、合成tRNAは大腸菌に少量種として存在する天然tRNAArgと競合する。更に、生物によっては全三重項コドンを使用しないものもある。Micrococcus luteusで割り当てられないコドンAGAがin vitro転写/翻訳抽出物へのアミノ酸挿入に使用されている。例えばKowal and Oliver, Nucl. Acid. Res., 25:4685 (1997)参照。本発明の成分はこれらのレアコドンをin vivo使用するために作製することができる。
【0117】
セレクターコドンは更に拡張コドン(例えば4、5、6塩基以上のコドン等の4塩基以上のコドン)も含むことができる。4塩基コドンの例としては例えばAGGA、CUAG、UAGA、CCCU等が挙げられる。5塩基コドンの例としては例えばAGGAC、CCCCU、CCCUC、CUAGA、CUACU、UAGGC等が挙げられる。本発明の方法はフレームシフト抑圧に基づく拡張コドンの使用を含む。4塩基以上のコドンは例えば1又は複数の非天然アミノ酸(例えばアルキニルアミノ酸)を同一蛋白質に挿入することができる。他の態様では、アンチコドンループは例えば少なくとも4塩基コドン、少なくとも5塩基コドン、又は少なくとも6塩基コドン又はそれ以上をデコードすることができる。4塩基コドンは256種が考えられるので、4塩基以上のコドンを使用すると同一細胞で複数の非天然アミノ酸をコードすることができる。Andersonら, (2002) Exploring the Limits of Codon and Anticodon Size,Chemistry and Biology, 9:237-244; 及びMagliery, (2001) Expanding the Genetic Code:Selection of Efficient Suppressors of Four-base Codons and Identification of “Shifty” Four-base Codons with a Library Approach in Escherichia coli, J. Mol. Biol. 307:755-769 も参照。
【0118】
例えば、in vitro生合成法を使用して非天然アミノ酸を蛋白質に組込むために4塩基コドンが使用されている。例えばMaら, (1993) Biochemistry, 32, 7939;及びHohsakaら, (1999) J. Am. Chem. Soc., 121:34 参照。2個の化学的にアシル化されたフレームシフトサプレッサーtRNAを用いて2−ナフチルアラニンとリジンのNBD誘導体をストレプトアビジンに同時にin vitroで組込むためにCGGGとAGGUが使用されている。例えばHohsakaら, (1999) J. Am. Chem. Soc., 121:12194参照。in vivo試験では、MooreらはNCUAアンチコドンをもつtRNALeu誘導体がUAGNコドン(NはU、A、G又はCであり得る)を抑圧する能力を試験し、四重項UAGAはUCUAアンチコドンをもつtRNALeuにより13〜26%の効率でデコードすることができるが、0又は−1フレームでは殆どデコードできないことを見出した。Mooreら, (2000) J. Mol. Biol., 298:195 参照。1態様では、レアコドン又はナンセンスコドンに基づく拡張コドンを本発明で使用し、他の望ましくない部位でのミスセンス読み飛ばしとフレームシフト抑圧を減らすことができる。
【0119】
所与系では、セレクターコドンは更に天然3塩基コドンの1種を含むことができ、内在系はこの天然塩基コドンを使用しない(又は殆ど使用しない)。例えば、天然3塩基コドンを認識するtRNAをもたない系、及び/又は3塩基コドンがレアコドンである系がこれに該当する。
【0120】
セレクターコドンは場合により非天然塩基対を含む。これらの非天然塩基対は更に既存遺伝子アルファベットを拡張する。塩基対が1個増えると、トリプレットコドン数は64から125に増す。第3の塩基対の性質としては安定的且つ選択的な塩基対合、高い忠実度でポリメラーゼによるDNAへの効率的な酵素組込み、及び未完成非天然塩基対の合成後の効率的な持続的プライマー伸長が挙げられる。方法と組成物に適応可能な非天然塩基対については、例えばHiraoら, (2002) An unnatural base pair for incorporating amino acid analogues into protein, Nature Biotechnology, 20:177-182 に記載されている。Wu, Y. ら,(2002) J. Am. Chem. Soc. 124:14626-14630も参照。他の関連文献は以下に挙げる。
【0121】
in vivo使用では、非天然ヌクレオシドは膜透過性であり、リン酸化され、対応する三リン酸塩を形成する。更に、増加した遺伝情報は安定しており、細胞内酵素により破壊されない。Bennerらによる従来の報告はカノニカルワトソンクリック対とは異なる水素結合パターンを利用しており、そのうちで最も注目される例はイソC:イソG対である。例えばSwitzerら, (1989)J. Am. Chem. Soc., 111:8322;及びPiccirilliら, (1990) Nature, 343:33; Kool, (2000) Curr. Opin. Chem. Biol., 4:602参照。これらの塩基は一般に天然塩基とある程度まで誤対合し、酵素複製することができない。Koolらは水素結合を塩基間の疎水性パッキング相互作用に置き換えることにより塩基対の形成を誘導できることを立証した。Kool, (2000) Curr. Opin. Chem. Biol., 4:602;及びGuckian and Kool, (1998) Angew. Chem. Int. Ed. Engl., 36, 2825参照。上記全要件を満足する非天然塩基対を開発する目的でSchultz, Romerbergらは一連の非天然疎水性塩基を体系的に合成し、試験した。PICS:PICS自己対は天然塩基対よりも安定しており、大腸菌DNAポリメラーゼIのKlenowフラグメント(KF)によりDNAに効率的に組込むことができる。例えばMcMinnら, (1999) J. Am. Chem. Soc., 121:11586;及びOgawaら, (2000) J. Am. Chem. Soc., 122:3274参照。生体機能に十分な効率と選択性でKFにより3MN:3MN自己対を合成することができる。例えばOgawaら, (2000) J. Am. Chem. Soc., 122:8803参照。しかし、どちらの塩基も後期複製用チェーンターミネーターとして作用するものである。PICS自己対を複製するために使用できる突然変異体DNAポリメラーゼが最近開発された。更に、7AI自己対も複製することができる。例えばTaeら, (2001) J. Am. Chem. Soc., 123:7439参照。Cu(II)と結合すると安定な対を形成する新規メタロ塩基対Dipic:Pyも開発された。Meggersら, (2000) J. Am. Chem. Soc., 122:10714参照。拡張コドンと非天然コドンは天然コドンに本質的に直交性であるので、本発明の方法は天然コドンに直交性のtRNAを作製するためにこの性質を利用することができる。
【0122】
翻訳バイパス系を使用してアルキニルアミノ酸を所望ポリペプチドに組込むこともできる。1翻訳バイパス系では、大きい配列が遺伝子に挿入されるが、蛋白質に翻訳されない。この配列はリボソームに配列を飛び越させて挿入の下流の翻訳を再開するための合図として機能する構造を含む。
【0123】
非天然アミノ酸
本明細書で使用する非天然アミノ酸とはセレノシステイン及び/又はピロリジンと20種の遺伝的にコードされる以下のα−アミノ酸、即ちアラニン、アルギニン、アスパラギン、アスパラギン酸、システイン、グルタミン、グルタミン酸、グリシン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、プロリン、セリン、スレオニン、トリプトフアン、チロシン、バリン以外の任意アミノ酸、修飾アミノ酸又はアミノ酸類似体を意味する。α−アミノ酸の一般構造は式I:
【化2】
により表される。
【0124】
非天然アミノ酸は一般に式Iをもつ任意構造であり、式中、R基は20種の天然アミノ酸で使用されている以外の任意置換基である。20種の天然アミノ酸の構造については、例えばL. Stryer著Biochemistry, 第3版, 1988, Freeman and Company, New York参照。なお、本発明の非天然アミノ酸は上記20種のα−アミノ酸以外の天然化合物でもよい。
【0125】
本発明の非天然アミノ酸は一般に側鎖が天然アミノ酸と異なるので、天然蛋白質と同様に他のアミノ酸(例えば天然又は非天然アミノ酸)とアミド結合を形成する。一方、非天然アミノ酸は天然アミノ酸と異なる側鎖基をもつ。
【0126】
本発明では反応性アルキニル基を含む非天然アミノ酸(例えばアジド部分と特異的且つ位置選択的に反応するアルキン部分をもつ非天然アミノ酸)が特に重要である。例えば、アルキニルアミノ酸では、式IにおけるRは任意アルキン含有構造を含む。例えば、パラ−プロパルギルオキシフェニルアラニン(略称pPRO−Phe;図1A参照)が本発明で利用される所望非天然アルキニルアミノ酸である。本発明はpPRO−Pheと直交翻訳成分の併用に限定するものではない。例えば、他の各種アルキニルアミノ酸(図8A及び8B参照)が考えられ、限定されないが、例えば、
2−アミノ−4−ペンチン酸、
2−アミノ−3−(4−エチニルフェニル)プロパン酸、
2−アミノ−3−[4−(プロプ−2−イニル)フェニル]プロパン酸、
2−アミノ−3−(プロプ−2−イニルオキシ)プロパン酸、
2−アミノ−3−(プロプ−2−イニルチオ)プロパン酸、
3−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノプロパン酸、
4−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノブタン酸が挙げられる。
【0127】
他の非天然アミノ酸では、例えば、式IにおけるRは場合によりアルキル、アリール、アシル、ヒドラジン、シアノ、ハロ、ヒドラジド、アルケニル、エーテル、硼酸、ボロン酸、ホスホ、ホスホノ、ホスフィン、エノン、イミン、エステル、ヒドロキシルアミン、アミン基等又はその任意組合せを含む。他の該当非天然アミノ酸としては限定されないが、光架橋基をもつアミノ酸、スピン標識アミノ酸、蛍光アミノ酸、金属結合性アミノ酸、金属含有アミノ酸、放射性アミノ酸、新規官能基をもつアミノ酸、他の分子と共有又は非共有的に相互作用するアミノ酸、フォトケージド及び/又は光異性化可能なアミノ酸、ビオチン又はビオチン類似体を含有するアミノ酸、ケト含有アミノ酸、グリコシル化アミノ酸、アミノ酸側鎖と結合した糖部分、ポリエチレングリコール又はポリエーテルを含むアミノ酸、重原子置換アミノ酸、化学分解性又は光分解性アミノ酸、天然アミノ酸に比較して延長側鎖(例えばポリエーテル又は例えば約5もしくは約10炭素長を上回る長鎖炭化水素等)をもつアミノ酸、炭素結合糖含有アミノ酸、アミノチオ酸含有アミノ酸、及び1個以上の毒性部分を含むアミノ酸が挙げられる。
【0128】
別の側面では、本発明は下式IV:
【化3】
により表される一般構造をもつアルキニルアミノ酸を提供する。
【0129】
この構造をもつアルキンアミノ酸は一般にR1が20種の天然アミノ酸の1種で使用される置換基であり、R2がアルキニル置換基である任意構造である。従って、この種のアルキニルアミノ酸は天然アミノ酸誘導体とみなすことができる。
【0130】
上記のように、本発明は非天然アルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)の使用に限定するものではない。実際に、真正細菌において本発明の直交翻訳系で使用することができる任意アルキニルアミノ酸が本発明の範囲に含まれる。他の各種アルキニルアミノ酸が公知であり、例えば、図8に示すアルキニルアミノ酸が挙げられる。これらのアルキニルアミノ酸構造のいくつかはpPRO−Pheと非常によく似ているので、本明細書に記載する直交tRNA及びアミノアシルtRNAシンテターゼ成分(例えば配列番号1のO−tRNAと配列番号4、6、8、10、12、14、16、18、又はその保存変異体のO−RS)を使用してこれらのアミノ酸のいくつかを真正細菌の蛋白質に組込むことができると考えられる。従って、本発明はpPRO−Pheに加えて他のアルキニルアミノ酸の組込み方法も提供する。表4(実施例9参照)に示す直交成分がpPRO−Phe以外にアルキニルアミノ酸を組込むことができるか否かに関係なく、本明細書の開示はこれらの他のアルキニルアミノ酸を組込む直交tRNA成分を構築するために十分な教示を提供するものであり、更にこれらの直交成分も本発明の範囲に含まれる。
【0131】
アルキニル基等の新規側鎖を含む非天然アミノ酸に加え、非天然アルキニルアミノ酸は場合により例えば式II及びIII:
【化4】
の構造により表されるような修飾主鎖構造も含み、上記式中、Zは一般にOH、NH2、SH、NH−R’又はS−R’を含み、XとYは同一でも異なっていてもよく、一般にS又はOであり、RとR’は場合により同一又は異なり、一般に式Iをもつ非天然アミノ酸について上記に記載したR基と同一の基及び水素から選択される。例えば、本発明の非天然アミノ酸は場合により式II及びIIIにより表されるようにアミノ又はカルボキシル基に置換を含む。この種の非天然アミノ酸としては限定されないが、例えば20種の標準天然アミノ酸に対応する側鎖又は非天然アルキニル側鎖をもつα−ヒドロキシ酸、α−チオ酸、α−アミノチオカルボキシレートが挙げられる。更に、α炭素の置換は場合によりL、D又はα,α−ジ置換アミノ酸(例えばD−グルタミン酸、D−アラニン、D−メチル−O−チロシン、アミノ酪酸等)を含む。他の代替構造としては環状アミノ酸(例えばプロリン類似体や、3、4、6、7、8及び9員環プロリン類似体)、β及びγアミノ酸(例えば置換β−アラニン及びγ−アミノ酪酸)が挙げられる。
【0132】
例えば、(所定のアルキニルアミノ酸を含む)多くの非天然アミノ酸はチロシン、セリン、システイン、アスパラギン酸、グルタミン酸等の天然アミノ酸をベースとする。例えば、以下のアルキニルアミノ酸:
2−アミノ−3−(プロプ−2−イニルオキシ)プロパン酸;
2−アミノ−3−(プロプ−2−イニルチオ)プロパン酸;
3−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノプロパン酸;及び
4−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノブタン酸
はいずれも天然アミノ酸から誘導することができる。
【0133】
チロシン類似体としてはパラ置換チロシン、オルト置換チロシン、及びメタ置換チロシンが挙げられ、置換チロシンはアルキニル基、アセチル基、ベンゾイル基、アミノ基、ヒドラジン、ヒドロキシルアミン、チオール基、カルボキシ基、イソプロピル基、メチル基、C6−C20直鎖又は分岐鎖炭化水素、飽和又は不飽和炭化水素、O−メチル基、ポリエーテル基、ニトロ基等を含む。更に、多置換アリール環も考えられる。本発明のグルタミン類似体としては限定されないが、α−ヒドロキシ誘導体、γ置換誘導体、環状誘導体及びアミド置換グルタミン誘導体が挙げられる。フェニルアラニン類似体の例としては限定されないが、パラ置換フェニルアラニン、オルト置換フェニルアラニン、及びメタ置換フェニルアラニンが挙げられ、置換基はアルキニル基、ヒドロキシ基、メトキシ基、メチル基、アリル基、アルデヒド、ニトロ、チオール基又はケト基等を含む。非天然アミノ酸の特定例としては限定されないが、p−プロパルギルオキシフェニルアラニン、3,4−ジヒドロキシ−L−フェニルアラニン(DHP)、3,4,6−トリヒドロキシ−L−フェニルアラニン、3,4,5−トリヒドロキシ−L−フェニルアラニン、4−ニトロ−フェニルアラニン、p−アセチル−L−フェニルアラニン、O−メチル−L−チロシン、L−3−(2−ナフチル)アラニン、3−メチルフェニルアラニン、O−4−アリル−L−チロシン、4−プロピル−L−チロシン、3−ニトロ−チロシン、3−チオール−チロシン、トリ−O−アセチル−GlcNAcβ−セリン、L−Dopa、フッ素化フェニルアラニン、イソプロピル−L−フェニルアラニン、p−アジド−L−フェニルアラニン、p−アシル−L−フェニルアラニン、p−ベンゾイル−L−フェニルアラニン、L−ホスホセリン、ホスホノセリン、ホスホノチロシン、p−ヨードフェニルアラニン、p−ブロモフェニルアラニン、p−アミノ−L−フェニルアラニン、及びイソプロピル−L−フェニルアラニン等が挙げられる。各種非天然アミノ酸の構造は例えば図1A、8A及び8Bに示す。公開国際出願WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」も参照。
非天然アミノ酸の化学的合成
【0134】
上記非天然アミノ酸の多くは例えばSigma(米国)やAldrich(Milwaukee, WI,米国)から市販されている。市販されていないものは場合により各種刊行物に記載されている方法や当業者に公知の標準方法を使用して合成される。有機合成技術については例えばFessendonとFessendon著Organic Chemistry(1982, 第2版, Willard Grant Press, Boston Mass.);March著Advanced Organic Chemistry(第3版, 1985, Wiley and Sons, New York);及びCareyとSundberg著Advanced Organic Chemistry(第3版, Parts A and B, 1990, Plenum Press, New York)を参照されたい。非天然アミノ酸の合成について記載しているその他の刊行物としては例えばWO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(In vivo incorporation of unnatural amino acids)」;Matsoukasら, (1995) J. Med. Chem., 38, 4660-4669;King, F. E. & Kidd, D. A. A. (1949) A New Synthesis of Glutamine and of γ-Dipeptides of Glutamic Acid from Phthylated Intermediates. J. Chem. Soc., 3315-3319;Friedman, O. M. & Chatterrji, R. (1959) Synthesis of Derivatives of Glutamine as Model Substrates for Anti-Tumor Agents. J. Am. Chem. Soc. 81, 3750-3752; Craig, J. C. ら(1988)Absolute Configuration of the Enantiomers of 7-Chloro-4[[4-(diethylamino)-1-methylbutyl]amino]quinoline (Chloroquine). J. Org. Chem. 53, 1167-1170; Azoulay, M., Vilmont, M. & Frappier, F. (1991) Glutamine analogues as Potential Antimalarials. Eur. J. Med. Chem. 26, 201-5;Koskinen, A. M. P. & Rapoport, H. (1989) Synthesis of 4-Substituted Prolines as Conformationally Constrained Amino Acid Analogues. J. Org. Chem. 54, 1859-1866; Christie, B. D. & Rapoport, H. (1985) Synthesis of Optically Pure Pipecolates from L-Asparagine. Application to the Total Synthesis of(+)-Apovincamine through Amino Acid Decarbonylation and Iminium Ion Cyclization. J. Org. Chem. 1989:1859-1866;Bartonら, (1987)Synthesis of Novel α-Amino-Acids and Derivatives Using Radical Chemistry:Synthesis of L-and D-α-Amino-Adipic Acids, L-α-aminopimelic Acid and Appropriate Unsaturated Derivatives. Tetrahedron Lett. 43:4297-4308;及びSubasingheら, (1992)Quisqualic acid analogues:synthesis of beta-heterocyclic 2-aminopropanoic acid derivatives and their activity at a novel quisqualate-sensitized site. J. Med. Chem. 35:4602-7が挙げられる。国際公開WO2004/058946、発明の名称「蛋白質アレー(PROTEIN ARRAYS)」(出願日2003年12月22日)も参照。
非天然アミノ酸の細胞取込み
【0135】
細胞による非天然アミノ酸取り込みは例えば蛋白質に組込むように非天然アミノ酸を設計及び選択する場合に一般に考慮される問題の1つである。例えば、α−アミノ酸は電荷密度が高いため、これらの化合物は細胞に浸透しにくいと思われる。天然アミノ酸は異なる程度のアミノ酸特異性を示すことが多い一連の蛋白質輸送システムにより細胞に取り込まれる。非天然アミノ酸が細胞に取り込まれる場合にはどの非天然アミノ酸が取り込まれるかを判断する迅速なスクリーニングを実施することができる。例えば国際公開WO2004/058946、発明の名称「蛋白質アレー(PROTEIN ARRAYS)」(出願日2003年12月22日);及びLiu and Schultz(1999)Progress toward the evolution of an organism with an expanded genetic code. PNAS 96:4780-4785における毒性アッセイ参照。取込みは種々のアッセイで容易に分析されるが、細胞取込み経路に利用可能な非天然アミノ酸を設計する代替方法は、アミノ酸をin vivo生産する生合成経路を提供する方法である。
【0136】
非天然アミノ酸の生合成
細胞にはアミノ酸と他の化合物を生産するために多数の生合成経路が元々存在している。特定非天然アミノ酸の生合成法は自然界(例えば細胞中)には存在しないと思われるが、本発明はこのような方法を提供する。例えば、非天然アミノ酸の生合成経路は場合により新規酵素を付加するか又は既存宿主細胞経路を改変することにより宿主細胞で作製される。付加新規酵素は場合により天然酵素又は人工的に進化させた酵素である。例えば、(WO2002/085923、前出の実施例に記載されているような)p−アミノフェニルアラニンの生合成は他の生物に由来する公知酵素の組合せの付加に依存している。これらの酵素の遺伝子はこれらの遺伝子を含むプラスミドで細胞を形質転換することにより細胞に導入することができる。これらの遺伝子は細胞で発現されると、所望化合物を合成するための酵素経路を提供する。場合により付加される酵素種の例は下記実施例に記載する。その他の酵素配列は例えばGenbankに登録されている。人工的に進化させた酵素も場合により同様に細胞に付加する。このように、非天然アミノ酸を生産するように細胞機構と細胞資源を操作する。
【0137】
実際に、生合成経路で使用する新規酵素を生産するため又は非天然アミノを生産するように既存経路をin vitroもしくはin vivoで進化させるためには種々の方法の任意のものを使用することができる。非天然アミノ酸を生産するため(又は、実際に新規基質特異性もしくは他の該当活性をもつようにシンテターゼを進化させるため)には、酵素又は他の生合成経路成分を進化させる方法として入手可能な多数の方法を本発明に適用することができる。例えば、非天然アミノ酸を生産(又は新規シンテターゼを生産)するように新規酵素及び/又は前記酵素の経路をin vitro又はin vivoで開発するためには、場合によりDNAシャフリングを使用する。例えばStemmer(1994)Rapid evolution of a protein in vitro by DNA shuffling, Nature 370(4): 389-391;及びStemmer, (1994)DNA shuffling by random fragmentation and reassembly:In vitro recombination for molecular evolution, Proc. Natl. Acad. Sci. USA, 91:10747-10751参照。関連アプローチは所望特性をもつ酵素を迅速に進化させるために関連(例えば相同)遺伝子のファミリーをシャフリングする。このような「ファミリー遺伝子シャフリング」法の1例はCrameriら(1998)“DNA shuffling of a family of genes from diverse species accelerates directed evolution, ”Nature, 391 (6664) :288-291に記載されている。(生合成経路成分であるか又はシンテターゼであるかを問わずに)新規酵素は例えばOstermeierら(1999)“A combinatorial approach to hybrid enzymes independent of DNA homology”, Nature Biotech 17:1205に記載されているような「ハイブリッド酵素作製用インクリメンタルトランケーション(incremental truncation for the creation of hybrid enzymes:ITCHY)として知られるDNA組換え法を使用して作製することもできる。このアプローチは1種以上のin vitro又はin vivo組換え法の基質として使用することができる酵素又は他の経路変異体のライブラリーを作製するために使用することもできる。Ostermeierら(1999)“Combinatorial Protein Engineering by Incremental Truncation,” Proc. Natl. Acad. Sci. USA, 96:3562-67, 及び Ostermeierら(1999)“Incremental Truncation as a Strategy in the Engineering of Novel Biocatalysts, ” Biological and Medicinal Chemistry, 7:2139-44も参照。別のアプローチは例えば非天然アミノ酸(又は新規シンテターゼ)の生産に関連する生合成反応を触媒する能力について選択する酵素又は他の経路変異体のライブラリーを作製するために指数的集合突然変異誘発を使用する。このアプローチでは、該当配列中の小群の残基を並行してランダム化し、機能的蛋白質をもたらすアミノ酸を各変異位置で同定する。非天然アミノ酸(又は新規シンテターゼ)の生産用新規酵素を作製するように本発明に応用することができるこのような方法の例はDelegrave & Youvan(1993)Biotechnology Research 11:1548-1552に記載されている。更に別のアプローチでは、例えばArkin and Youvan(1992)“Optimizing nucleotide mixtures to encode specific subsets of amino acids for semi-random mutagenesis”Biotechnology 10:297-300;又はReidhaar-Olsonら(1991)“Random mutagenesis of protein sequences using oligonucleotide cassettes”Methods Enzymol. 208:564-86の一般突然変異誘発法を使用することにより、酵素及び/又は経路成分操作にドープ又は縮重オリゴヌクレオチドを使用するランダム又は半ランダム突然変異誘発を使用することができる。ポリヌクレオチドリアセンブリと部位飽和突然変異誘発を使用する更に別のアプローチ(「非確率論的」突然変異誘発と呼ぶことが多い)を使用すると、酵素及び/又は経路成分を作製した後に、(例えば非天然アミノ酸のin vivo生産のための)1種以上のシンテターゼ又は生合成経路機能を実施する能力についてスクリーニングすることができる。例えばShort「遺伝子ワクチン及び酵素の非確率論的作製(NON-STOCHASTIC GENERATION OF GENETIC VACCINES AND ENZYMES)」WO00/46344参照。
【0138】
このような突然変異法の代替方法は生物の全ゲノムを組換え、得られた子孫を特定経路機能について選択する方法である(「全ゲノムシャフリング」と呼ぶことが多い)。このアプローチは、例えばゲノム組換えと非天然アミノ酸(又はその中間体)の生産能に関する生物(例えば大腸菌又は他の細胞)の選択により本発明に適用することができる。例えば、非天然アミノ酸をin vivo生産するように細胞で既存及び/又は新規経路を進化させるための経路設計には、以下の刊行物に教示されている方法を適用することができる:Patnaikら(2002)“Genome shuffling of lactobacillus for improved acid tolerance”Nature Biotechnology, 20(7):707-712;及びZhangら(2002)“Genome shuffling leads to rapid phenotypic improvement in bacteria”Nature, February 7, 415(6872):644-646。
【0139】
例えば所望化合物を生産するための他の生物及び代謝経路組換え技術も利用可能であり、同様に非天然アミノ酸の生産に適用することができる。有用な経路組換えアプローチを教示している刊行物の例としては、Nakamura and White (2003) “Metabolic engineering for the microbial production of 1, 3 propanediol”Curr. Opin. Biotechnol. 14(5):454-9; Berryら(2002)“Application of Metabolic Engineering to improve both the production and use of Biotech Indigo”J. Industrial Microbiology and Biotechnology 28:127-133; Bantaら(2002)“Optimizing an artificial metabolic pathway:Engineering the cofactor specificity of Corynebacterium 2, 5-diketo-D-gluconic acid reductase for use in vitamin C biosynthesis” Biochemistry, 41(20), 6226-36; Selivonovaら(2001)“Rapid Evolution of Novel Traits in Microorganisms”Applied and Environmental Microbiology, 67:3645、及び他の多数の文献が挙げられる。
【0140】
使用する方法に関係なく、一般に本発明の組換え生合成経路を使用して生産される非天然アミノ酸は効率的な蛋白質生合成に十分な濃度(例えば天然細胞量)で生産されるが、他の細胞内アミノ酸濃度を有意に変化させたり細胞資源を枯渇させる程ではない。このようにin vivo生産される典型的な濃度は約10mM〜約0.05mMである。特定経路に所望される酵素を生産するように細胞を組換え、非天然アミノ酸を作製したら、場合によりin vivo選択を使用してリボソーム蛋白質合成と細胞増殖の両方に適合するように非天然アミノ酸の生産を更に最適化させる。
【0141】
パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)を組込むための直交成分
本発明はセレクターコドン(例えばアンバー終止コドン、ナンセンスコドン、4塩基以上のコドン等)に応答して成長中のポリペプチド鎖にアルキニルアミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe))を例えばin vivoで組込むための直交成分を作製する組成物及び方法を提供する。例えば、本発明は直交tRNA(O−tRNA)、直交アミノアシルtRNAシンテターゼ(O−RS)及びその対を提供する。これらの対は成長中のポリペプチド鎖にpPRO−Pheを組込むために使用することができる。
【0142】
本発明の組成物は直交アミノアシルtRNAシンテターゼ(O−RS)を含み、O−RSはO−tRNAをpPRO−Pheで優先的にアミノアシル化する。所定態様では、O−RSは配列番号4、6、8、10、12、14、16もしくは18を含むアミノ酸配列又はその保存変異体を含む。本発明の所定態様では、O−RSは内在tRNAよりもO−tRNAを優先的にpPRO−Phe等のアルキニルアミノ酸でアミノアシル化し、O−RSはO−tRNAを優先し、pPRO−Pheを負荷されるO−tRNAとpPRO−Pheを負荷される内在tRNAの比は1:1を上回り、O−RSはO−tRNAに排他的又はほぼ排他的に負荷することがより好ましい。
【0143】
O−RSを含む組成物は場合により更に直交tRNA(O−tRNA)を含むことができ、O−tRNAはセレクターコドンを認識する。一般に、本発明のO−tRNAは下記配列表(例えば配列番号1)及び実施例に記載するようなポリヌクレオチド配列を含むか又は前記配列によりコードされるO−tRNAの抑圧効率に比較してコグネイトシンテターゼの存在下でセレクターコドンに応答して少なくとも例えば約45%、50%、60%、75%、80%、又は90%以上の抑圧効率を含む。1態様では、O−RSとO−tRNAの併用による抑圧効率はO−RSの不在下のO−tRNAの抑圧効率の例えば5倍、10倍、15倍、20倍、25倍又はそれ以上である。1側面では、O−RSとO−tRNAの併用による抑圧効率はMethanococcus jannaschiiに由来する直交チロシルtRNAシンテターゼ対の抑圧効率の少なくとも45%である。
【0144】
O−tRNAを含む組成物は場合により細胞(例えば大腸菌細胞等の真正細菌細胞)、及び/又は翻訳系を含むことができる。
【0145】
本発明は翻訳系を含む細胞(例えば真正細菌細胞)も提供し、前記翻訳系は直交tRNA(O−tRNA)と;直交アミノアシルtRNAシンテターゼ(O−RS)と;アルキニルアミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe))を含む。一般に、O−RSは内在tRNAよりもO−tRNAを優先的にpPRO−Phe等のアルキニルアミノ酸でアミノアシル化し、O−RSはO−tRNAを優先し、pPRO−Pheを負荷されるO−tRNAとpPRO−Pheを負荷される内在tRNAの比は1:1を上回り、O−RSはO−tRNAに排他的又はほぼ排他的に負荷することがより好ましい。O−tRNAは第1のセレクターコドンを認識し、O−RSはO−tRNAをpPRO−Pheで優先的にアミノアシル化する。1態様では、O−tRNAは配列番号1に記載のポリヌクレオチド配列又はその相補的ポリヌクレオチド配列を含むか又は前記配列によりコードされる。1態様では、O−RSは配列番号4、6、8、10、12、14、16もしくは18の任意1種に記載のアミノ酸配列又はその保存変異体を含む。
【0146】
本発明の細胞は場合により異なる付加O−tRNA/O−RS対と第2の非天然アミノ酸を更に含むことができ、例えばこのO−tRNAは第2のセレクターコドンを認識し、このO−RSは対応するO−tRNAを第2の非天然アミノ酸で優先的にアミノアシル化し、第2のアミノ酸はpPRO−Pheと異なる。場合により、本発明の細胞は該当ポリペプチドをコードするポリヌクレオチドを含む核酸を含み、前記ポリヌクレオチドはO−tRNAにより認識されるセレクターコドンを含む。
【0147】
所定態様では、本発明の細胞は直交−tRNA(O−tRNA)と、直交アミノアシルtRNAシンテターゼ(O−RS)と、アルキニルアミノ酸(例えばpPRO−Phe)と、該当ポリペプチドをコードするポリヌクレオチドを含む核酸を含む真正細菌細胞(例えば大腸菌細胞)であり、前記ポリヌクレオチドはO−tRNAにより認識されるセレクターコドンを含む。本発明の所定態様では、O−RSはO−RSが任意内在tRNAをアミノアシル化する効率よりも高い効率でO−tRNAを優先的にアミノアシル化する。
【0148】
本発明の所定態様では、本発明のO−tRNAは本明細書の配列表(例えば配列番号1)及び実施例に記載するようなポリヌクレオチド配列、又はその相補的ポリヌクレオチド配列を含むか又は前記配列によりコードされる。本発明の所定態様では、O−RSは本明細書の配列表に記載するようなアミノ酸配列、又はその保存変異体を含む。1態様では、O−RS又はその一部は本明細書の配列又は実施例に記載するようなアミノ酸をコードするポリヌクレオチド配列、又はその相補的ポリヌクレオチド配列によりコードされる。
【0149】
本発明のO−tRNA及び/又はO−RSは種々の生物(例えば真核及び/又は非真核生物)の任意のものに由来することができる。
【0150】
ポリヌクレオチドも本発明の特徴である。本発明のポリヌクレオチドは本明細書の配列表に記載するようなポリペプチドをコードするヌクレオチド配列を含む人工(例えば人為的に作製され、天然には存在しない)ポリヌクレオチドを含むか、及び/又は前記ポリヌクレオチド配列に相補的であるか又は前記配列をコードする。本発明のポリヌクレオチドは更に核酸の実質的に全長にわたって高ストリンジェント条件下で上記ポリヌクレオチドとハイブリダイズする核酸も含むことができる。本発明のポリヌクレオチドは更に天然に存在するtRNA又は対応するコーディング核酸のポリヌクレオチドと例えば少なくとも75%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%又はそれ以上一致するポリヌクレオチドを含み(但し、本発明のポリヌクレオチドは天然に存在するtRNA又は対応するコーディング核酸以外のものである)、前記tRNAはセレクターコドン(例えば4塩基コドン)を認識する。上記任意ポリヌクレオチドと例えば少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%又はそれ以上一致する人工ポリヌクレオチド及び/又は上記任意ポリヌクレオチドの保存変異体を含むポリヌクレオチドも本発明のポリヌクレオチドに含まれる。
【0151】
本発明のポリヌクレオチドを含むベクターも本発明の特徴である。例えば、本発明のベクターはプラスミド、コスミド、ファージ、ウイルス、発現ベクター及び/又は同等物を含むことができる。本発明のベクターを含む細胞も本発明の特徴である。
【0152】
O−tRNA/O−RS対の成分の作製方法も本発明の特徴である。これらの方法により作製された成分も本発明の特徴である。例えば、細胞に対して直交性である少なくとも1種のtRNA(O−tRNA)の作製方法は突然変異体tRNAのライブラリーを作製する段階と;セレクターコドンを認識できるように突然変異体tRNAのライブラリーの各メンバーのアンチコドンループを突然変異させることにより、潜在的O−tRNAのライブラリーを提供する段階と、潜在的O−tRNAのライブラリーのメンバーを含む第1の種の第1の細胞集団についてネガティブ選択を実施する段階を含む。ネガティブ選択は細胞に内在性のアミノアシルtRNAシンテターゼ(RS)によりアミノアシル化される潜在的O−tRNAライブラリーのメンバーを含む細胞を排除する。こうして、第1の種の細胞に対して直交性のtRNAプールが得られ、それにより少なくとも1種のO−tRNAが得られる。本発明の方法により作製されたO−tRNAも提供する。
【0153】
所定態様では、前記方法は更に第1の種の第2の細胞集団についてポジティブ選択を実施する段階を含み、前記細胞は第1の種の細胞に対して直交性のtRNAプールのメンバーと、コグネイトアミノアシルtRNAシンテターゼと、ポジティブ選択マーカーを含む。ポジティブ選択を使用し、コグネイトアミノアシルtRNAシンテターゼによりアミノアシル化され、ポジティブ選択マーカーの存在下で所望応答を示すtRNAプールのメンバーを含む細胞について細胞を選択又はスクリーニングすることにより、O−tRNAを得る。所定態様では、第2の細胞集団はネガティブ選択により排除されなかった細胞を含む。
【0154】
アルキニルアミノ酸をO−tRNAに負荷する直交アミノアシルtRNAシンテターゼの同定方法も提供する。例えば、方法は第1の種の細胞集団に選択を実施する段階を含み、前記細胞は、1)複数のアミノアシルtRNAシンテターゼ(RS)(例えば複数のRSは突然変異体RS、第1の種以外の種に由来するRS、又は突然変異体RSと第1の種以外の種に由来するRSの両方を含むことができる)のメンバーと;2)(例えば1種以上の種に由来する)直交−tRNA(O−tRNA)と;3)ポジティブ選択マーカーをコードし、少なくとも1個のセレクターコドンを含むポリヌクレオチドを各々含む。
【0155】
複数のRSのメンバーを含まないか又はその量の少ない細胞に比較して抑圧効率の高い細胞について細胞(例えば宿主細胞)を選択又はスクリーニングする。これらの選択/スクリーニングした細胞はO−tRNAをアミノアシル化する活性RSを含む。この方法により同定された直交アミノアシルtRNAシンテターゼも本発明の特徴である。
【0156】
パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)を特定位置にもつ蛋白質を細胞(例えば大腸菌細胞等の真正細菌細胞)で生産する方法も本発明の特徴である。例えば、1方法は、少なくとも1個のセレクターコドンを含み、蛋白質をコードする核酸を含む細胞を適当な培地で増殖させる段階と、pPRを提供する段階と、少なくとも1個のセレクターコドンによる核酸の翻訳中に蛋白質の特定位置にpPRを組込むことにより、蛋白質を生産する段階を含む。細胞は更に、細胞で機能し、セレクターコドンを認識する直交tRNAと;O−tRNAをpPRO−Pheで優先的にアミノアシル化する直交アミノアシルtRNAシンテターゼ(O−RS)を含む。この方法により生産された蛋白質も本発明の特徴である。
【0157】
本発明は例えばpPRO−Pheを組込んだ蛋白質を含有する組成物も提供する。所定態様では、蛋白質は公知蛋白質(例えば治療用蛋白質、診断用蛋白質、産業用酵素、又はその部分)のアミノ酸配列と少なくとも75%一致するアミノ酸配列を含む。場合により、組成物は医薬的に許容可能なキャリヤーを含有する。
【0158】
核酸及びポリペプチド配列と変異体
上記及び下記に記載するように、本発明は例えばO−tRNA及びO−RSをコードするポリヌクレオチド配列と、ポリペプチドアミノ酸配列(例えばO−RS)と、前記配列を含む例えば組成物、系及び方法を提供する。前記配列(例えばO−tRNA及びO−RSアミノ酸及びヌクレオチド配列)の例を本明細書に開示する(表4、例えば配列番号5、7、9、11、13、15、17及び19参照)。しかし、当業者に自明の通り、本発明は本明細書、例えば実施例や配列表に開示する配列に限定されない。当業者に自明の通り、本発明は例えば本明細書に記載する機能をもつ(例えばO−tRNA又はO−RSをコードする)多数の関連配列も提供する。
【0159】
O−tRNAをpPRO−Pheでアミノアシル化することが可能なO−RS種の構築と分析を実施例1に記載する。同実施例は単離された8種のO−RS種について記載する(図3及び実施例9参照)。これらのアミノ酸配列から明らかなように、8種の突然変異体O−RSクローンのアミノ酸置換には部分的コンセンサス傾向が認められる。2個以上のクローンの結合ポケットに以下のアミノ酸の少なくとも2種が認められた:Ala32,Pro107/Gln107,Ala158,Ile159,及びAla162/Pro162(配列番号21参照)。突然変異Tyr32→Ala32及びAsp158→Ala158の結果、Tyr32、Asp158及び天然基質チロシン間の水素結合が失われ、その結合が弱まると思われる。小さい主に疎水性の側鎖が存在するならば、pPRO−Pheの結合を助長すると予想される。これらのコンセンサス傾向により、pPRO−Pheを組込むために真正細菌宿主細胞系で配列番号1のO−tRNAと直交系で共働すると予想される付加O−RS種の設計が可能になる。これらのコンセンサス傾向は以下のように予測することができる。
【表1】
【0160】
従って、これらのコンセンサス傾向に基づき、実験により同定された8種のpPRO−PheRS種(即ちpPRO−PheRS−1〜pPRO−PheRS−8)で表されない少なくとも4種の付加直交pPRO−Pheシンテターゼ(pPRO−PheRS−con1〜pPRO−PheRS−con4)を合理的に設計することができる。これらは以下の通りである。
【表2】
【0161】
本発明はポリペプチド(O−RS)とポリヌクレオチド(例えばO−tRNA、O−RS又はその部分をコードするポリヌクレオチド、アミノアシルtRNAシンテターゼクローンを単離するために使用されるオリゴヌクレオチド等)を提供する。本発明のポリヌクレオチドとしては、1個以上のセレクターコドンをもつ本発明の該当蛋白質又はポリペプチドをコードするものが挙げられる。更に、本発明のポリヌクレオチドとしては、例えば配列番号5、7、9、11、13、15、17又は19に記載のヌクレオチド配列を含むポリヌクレオチド;そのポリヌクレオチド配列に相補的であるか又は前記配列をコードするポリヌクレオチドが挙げられる。本発明のポリヌクレオチドは更に配列番号4、6、8、10、12、14、16又は18を含むアミノ酸配列をコードする任意ポリヌクレオチドを含む。本発明のポリヌクレオチドは更に本発明のポリペプチドをコードするポリヌクレオチドを含む。同様に、核酸の実質的に全長にわたって高ストリンジェント条件下で上記ポリヌクレオチドとハイブリダイズする(天然ポリヌクレオチド以外の)人工核酸も本発明のポリヌクレオチドである。1態様では、組成物は本発明のポリペプチドと賦形剤(例えば緩衝液、水、医薬的に許容可能な賦形剤等)を含有する。本発明は本発明のポリペプチドに対して特異的に免疫反応性の抗体又は抗血清も提供する。人工ポリヌクレオチドとは人為的に作製され、天然には存在しないポリヌクレオチドである。
【0162】
本発明のポリヌクレオチドは天然に存在するtRNAのポリヌクレオチドと例えば少なくとも75%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%又はそれ以上一致する人工ポリヌクレオチド(但し、天然に存在するtRNA以外のもの)も含む。ポリヌクレオチドは天然に存在するtRNAのポリヌクレオチドと例えば少なくとも75%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%又はそれ以上一致する(但し100%は一致しない)人工ポリヌクレオチドも含む。
【0163】
所定態様では、ベクター(例えばプラスミド、コスミド、ファージ、ウイルス等)に本発明のポリヌクレオチドを組込む。1態様では、ベクターは発現ベクターである。別の態様では、発現ベクターは本発明のポリヌクレオチドの1種以上と機能的に連結されたプロモーターを含む。別の態様では、本発明のポリヌクレオチドを組込んだベクターを細胞に導入する。
【0164】
同様に当業者に自明の通り、開示配列の多数の変異体も本発明に含まれる。例えば、機能的に同一配列となる開示配列の保存変異体も本発明に含まれる。少なくとも1種の開示配列とハイブリダイスする核酸ポリヌクレオチド配列の変異体も本発明に含むものとする。例えば標準配列比較法により本明細書に開示する配列のユニークサブ配列であると判断される配列も本発明に含まれる。
【0165】
保存変異
遺伝コードの縮重により、「サイレント置換」(即ちコードされるポリペプチドに変化を生じない核酸配列の置換)はアミノ酸をコードする全核酸配列の暗黙の特徴である。同様に、「保存アミノ酸置換」はアミノ酸配列中の1又は数個のアミノ酸を高度に類似する特性をもつ別のアミノ酸で置換するものであり、このような置換も開示構築物と高度に類似することが容易に認められる。各開示配列のこのような保存変異は本発明の特徴である。
【0166】
特定核酸配列の「保存変異」とは同一又は本質的に同一のアミノ酸配列をコードする核酸を意味し、核酸がアミノ酸配列をコードしない場合には本質的に同一の配列を意味する。当業者に自明の通り、コードされる配列中の単一アミノ酸又は低百分率(一般に5%未満、より一般には4%、2%又は1%未満)のアミノ酸を置換、付加又は欠失させる個々の置換、欠失又は付加の結果としてアミノ酸を欠失するか、アミノ酸が付加されるか、又はアミノ酸が化学的に類似するアミノ酸で置換される場合には、これらの変異は「保存修飾変異」である。従って、本発明のポリペプチド配列の「保存変異」としては、ポリペプチド配列のアミノ酸の低百分率、一般に5%未満、より一般には2%又は1%未満が同一保存置換基のアミノ酸で置換される場合が挙げられる。最後に、非機能的配列の付加のように核酸分子のコードされる活性を変えない配列の付加も基本核酸の保存変異である。
【0167】
機能的に類似するアミノ酸を示す保存置換表は当分野で周知であり、このようなアミノ酸では、あるアミノ酸残基が同様の化学的性質(例えば芳香族側鎖又は正荷電側鎖)をもつ別のアミノ酸残基に置換しているため、ポリペプチド分子の機能的性質は実質的に変化しない。化学的性質が類似する天然アミノ酸を含む代表的グループを以下に示すが、これらのグループ内の置換は「保存置換」である。
【表3】
【0168】
核酸ハイブリダイゼーション
本発明の核酸の保存変異体を含めて本発明の核酸を同定するためには比較ハイブリダイゼーションを使用することができ、この比較ハイブリダイゼーション法は本発明の核酸を識別する1好適方法である。更に、高、超高及び超々高ストリンジェンシー条件下で配列番号5、7、9、11、13、15、17及び19により表される核酸とハイブリダイズするターゲット核酸も本発明の特徴である。このような核酸の例としては所与核酸配列と比較して1又は数個のサイレント又は保存核酸置換をもつものが挙げられる。
【0169】
試験核酸が完全にマッチする相補的ターゲットに比較して少なくとも50%の割合でプローブとハイブリダイズする場合、即ちマッチしないターゲット核酸の任意のものとのハイブリダイゼーションに観測されるシグナル対ノイズ比の少なくとも約5倍〜10倍で完全にマッチするプローブが完全にマッチする相補的ターゲットと結合する条件下におけるプローブとターゲットのハイブリダイゼーションに比較してシグナル対ノイズ比が少なくとも1/2である場合に試験核酸はプローブ核酸と特異的にハイブリダイズすると言う。
【0170】
核酸は一般に溶液中で会合するときに「ハイブリダイズ」する。核酸は水素結合、溶媒排除、塩基スタッキング等の種々の十分に特性決定された物理化学的力によりハイブリダイズする。核酸ハイブリダイゼーションの詳しい手引きはTijssen(1993)Laboratory Techniques in Biochemistry and Molecular Biology--Hybridization with Nucleic Acid Probes part I chapter 2, “Overview of principles of hybridization and the strategy of nucleic acid probe assays,”(Elsevier, New York) 及び Current Protocols in Molecular Biology, Ausubelら編, Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (2004年補遺)(「Ausubel」)に記載されている。Hames and Higgins(1995)Gene Probes 1 IRL Press at Oxford University Press, Oxford, England, (Hames and Higgins 1)及びHames and Higgins (1995) Gene Probes 2 IRL Press at Oxford University Press, Oxford, England (Hames and Higgins 2) はオリゴヌクレオチドを含むDNAとRNAの合成、標識、検出及び定量について詳細に記載している。
【0171】
サザン又はノーザンブロットで100個を上回る相補的残基をもつ相補的核酸のハイブリダイゼーションをフィルター上で行うためのストリンジェントハイブリダイゼーション条件の1例は、50%ホルマリンにヘパリン1mgを加え、42℃で一晩ハイブリダイゼーションを実施する。ストリンジェント洗浄条件の1例は65℃、0.2×SSCで15分間洗浄する(SSC緩衝液の説明についてはSambrook,前出参照)。多くの場合には高ストリンジェンシー洗浄の前に低ストリンジェンシー洗浄を実施してバックグラウンドプローブシグナルを除去する。低ストリンジェンシー洗浄の1例は40℃、2×SSCで15分間である。一般に、シグナル対ノイズ比が特定ハイブリダイゼーションアッセイで無関係プローブに観測される比の5倍(以上)である場合に特異的ハイブリダイゼーションが検出されたとみなす。
【0172】
サザン及びノーザンハイブリダイゼーション等の核酸ハイブリダイゼーション実験において「ストリンジェントハイブリダイゼーション洗浄条件」は配列依存性であり、各種環境パラメーターにより異なる。核酸ハイブリダイゼーションの詳しい手引きはTijssen (1993), 前出やHames and Higgins 1及び2に記載されている。ストリンジェントハイブリダイゼーション及び洗浄条件は任意試験核酸について経験により容易に決定することができる。例えば、高ストリンジェントハイブリダイゼーション及び洗浄条件を決定するには、一連の選択基準に合致するまで(例えばハイブリダイゼーション又は洗浄における温度上昇、塩濃度低下、界面活性剤濃度増加及び/又はホルマリン等の有機溶媒濃度増加により)ハイブリダイゼーション及び洗浄条件を徐々に増加する。例えば、高ストリンジェントハイブリダイゼーション及び洗浄条件では、マッチしないターゲットとプローブのハイブリダイゼーションに観測されるシグナル対ノイズ比の少なくとも約5倍でプローブが完全にマッチする相補的ターゲットと結合するまでハイブリダイゼーション及び洗浄条件を徐々に増加する。
【0173】
「超ストリンジェント」条件は特定プローブの熱融点(Tm)に等しくなるように選択される。Tmは(規定イオン強度及びpH下で)試験配列の50%が完全にマッチするプローブとハイブリダイズする温度である。本発明の目的には、一般に規定イオン強度及びpHで特定配列のTmよりも約5℃低くなるように「高ストリンジェント」ハイブリダイゼーション及び洗浄条件を選択する。
【0174】
「超高ストリンジェンシー」ハイブリダイゼーション及び洗浄条件は完全にマッチする相補的ターゲット核酸とプローブの結合に観測されるシグナル対ノイズ比がマッチしないターゲット核酸の任意のものとのハイブリダイゼーションに観測されるシグナル対ノイズ比の少なくとも10倍になるまでハイブリダイゼーション及び洗浄条件のストリンジェンシーを増加する条件である。完全にマッチする相補的ターゲット核酸のシグナル対ノイズ比の少なくとも1/2で前記条件下にプローブとハイブリダイズする場合にターゲット核酸は超高ストリンジェンシー条件下でプローブと結合すると言う。
【0175】
同様に、該当ハイブリダイゼーションアッセイのハイブリダイゼーション及び/又は洗浄条件を徐々に増加することにより更に高レベルのストリンジェンシーを決定することもできる。例えば、完全にマッチする相補的ターゲット核酸とプローブの結合に観測されるシグナル対ノイズ比がマッチしないターゲット核酸の任意のものとのハイブリダイゼーションに観測されるシグナル対ノイズ比の少なくとも10倍、20倍、50倍、100倍又は500倍以上になるまでハイブリダイゼーション及び洗浄条件のストリンジェンシーを増加する条件である。完全にマッチする相補的ターゲット核酸のシグナル対ノイズ比の少なくとも1/2で前記条件下にプローブとハイブリダイズする場合にターゲット核酸は超々高ストリンジェンシー条件下でプローブと結合すると言う。
【0176】
ストリンジェント条件下で相互にハイブリダイズしない核酸でも、これらの核酸によりコードされるポリペプチドが実質的に同一である場合には実質的に同一である。これは、例えば遺伝コードに許容される最大コドン縮重を使用して核酸のコピーを作製する場合に該当する。
【0177】
ユニークサブ配列
1側面では、本発明は本明細書に開示するO−tRNA及びO−RSの配列から選択される核酸中にユニークサブ配列を含む核酸を提供する。ユニークサブ配列は任意公知O−tRNA及びO−RS核酸配列に対応する核酸に比較してユニークである。例えばデフォルトパラメーターに設定したBLASTを使用してアラインメントを実施することができる。任意ユニークサブ配列は例えば本発明の核酸を同定するためのプローブとして有用である。
【0178】
同様に、本発明は本明細書に開示するO−RSの配列から選択されるポリペプチド中にユニークサブ配列を含むポリペプチドを含む。この場合には、ユニークサブ配列は任意公知ポリペプチド配列に対応するポリペプチドに比較してユニークである。
【0179】
本発明はO−RSの配列から選択されるポリペプチド中のユニークサブ配列をコードするユニークコーディングオリゴヌクレオチドとストリンジェント条件下でハイブリダイズするターゲット核酸も提供し、この場合には、ユニークサブ配列は対照ポリペプチド(例えば本発明のシンテターゼを例えば突然変異により誘導した元の親配列)の任意のものに対応するポリペプチドに比較してユニークである。ユニーク配列は上記のように決定する。
【0180】
配列比較、一致度及び相同度
2種以上の核酸又はポリペプチド配列に関して「一致」又は「一致度」百分率なる用語は2種以上の配列又はサブ配列を最大限に対応するように対比及び整列させ、以下に記載する配列比較アルゴリズム(又は当業者に入手可能な他のアルゴリズム)の1種を使用するか又は目視により測定した場合に相互に同一であるか又は同一のアミノ酸残基もしくはヌクレオチドの百分率が特定値であることを意味する。
【0181】
2種以上の核酸又はポリペプチド(例えばO−tRNAもしくはO−RSをコードするDNA又はO−RSのアミノ酸配列)に関して「実質的に一致」なる用語は2種以上の配列又はサブ配列を最大限に対応するように対比及び整列させ、配列比較アルゴリズムを使用するか又は目視により測定した場合にヌクレオチド又はアミノ酸残基一致度が少なくとも約60%、約80%、約90〜95%、約98%、約99%又はそれ以上であることを意味する。このような「実質的に一致」する配列は一般に実際の起源が記載されていなくても「相同」であるとみなす。少なくとも約50残基長の配列の領域、より好ましくは少なくとも約100残基長の領域にわたって「実質的一致」が存在していることが好ましく、少なくとも約150残基又は比較する2配列の全長にわたって配列が実質的に一致していることが最も好ましい。
【0182】
蛋白質及び/又は蛋白質配列は共通の祖先蛋白質又は蛋白質配列から天然又は人工的に誘導される場合に「相同」である。同様に、核酸及び/又は核酸配列は共通の祖先核酸又は核酸配列から天然又は人工的に誘導される場合に相同である。例えば、1個以上のセレクターコドンを含むように任意天然核酸を利用可能な任意突然変異誘発法により改変することができる。この突然変異誘発した核酸は発現されると、1種以上の非天然アミノ酸(例えばアルキニルアミノ酸)を含むポリペプチドをコードすることができる。突然変異法は当然のことながら更に1個以上の標準コドンを変異させ、得られる突然変異体蛋白質の1個以上の標準アミノ酸も変異させることができる。相同性は一般に2種以上の核酸又は蛋白質(又はその配列)間の配列類似度から推定される。相同性の判定に有用な配列間類似度の厳密な百分率は該当核酸及び蛋白質により異なるが、通常は25%程度の低い配列類似度を使用して相同性を判定する。例えば30%、40%、50%、60%、70%、80%、90%、95%又は99%以上のより高レベルの配列類似度を使用して相同性を判定することもできる。配列類似度百分率の決定方法(例えばデフォルトパラメーターを使用するBLASTP及びBLASTN)は本明細書に記載し、一般に入手可能である。
【0183】
配列比較及び相同性判定には、一般にある配列を参照配列としてこれに試験配列を比較する。配列比較アルゴリズムを使用する場合には、試験配列と参照配列をコンピューターに入力し、必要に応じてサブ配列座標を指定し、配列アルゴリズムプログラムパラメーターを指定する。こうすると、配列比較アルゴリズムは指定プログラムパラメーターに基づいて参照配列に対して試験配列の配列一致度百分率を計算する。
【0184】
比較のための最適な配列アラインメントは例えばSmith & Waterman, Adv. Appl. Math. 2:482(1981)の局所相同性アルゴリズム、Needleman & Wunsch, J. Mol. Biol. 48:443(1970)の相同性アラインメントアルゴリズム、Pearson & Lipman, Proc. Natl. Acad. Sci. USA 85:2444(1988)の類似性探索法、これらのアルゴリズムのコンピューターソフトウェア(Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WIのGAP、BESTFIT、FASTA及びTFASTA)、又は目視(一般にCurrent Protocols in Molecular Biology, Ausubelら編, Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (2004年補遺)参照)により実施することができる。
【0185】
配列一致度及び配列類似度百分率を決定するのに適したアルゴリズムの1例はAltschulら, J. Mol. Biol. 215:403-410(1990) に記載されているBLASTアルゴリズムである。BLAST分析を実施するためのソフトウェアはNational Center for Biotechnology Information(www. ncbi. nlm. nih. gov/)から公共入手可能である。このアルゴリズムはデータベース配列中の同一長さの単語と整列した場合に所定の正の閾値スコアTと一致するか又はこれを満足するクエリー配列中の長さWの短い単語を識別することによりまず高スコア配列対(HSP)を識別する。Tを隣接単語スコア閾値と言う(Altschulら,前出)。これらの初期隣接単語ヒットをシードとして検索を開始し、これらの単語を含むもっと長いHSPを探索する。次に、累積アラインメントスコアを増加できる限り、単語ヒットを各配列に沿って両方向に延長する。ヌクレオチド配列の場合にはパラメーターM(1対のマッチ残基のリウォードスコア、常に>0)及びN(ミスマッチ残基のペナルティースコア、常に<0)を使用して累積スコアを計算する。アミノ酸配列の場合には、スコアリングマトリクスを使用して累積スコアを計算する。累積アラインメントスコアがその最大到達値から量Xだけ低下するか、累積スコアが1カ所以上の負スコア残基アラインメントの累積によりゼロ以下になるか、又はどちらかの配列の末端に達したら各方向の単語ヒットの延長を停止する。BLASTアルゴリズムパラメーターW、T及びXはアラインメントの感度と速度を決定する。BLASTNプログラム(ヌクレオチド配列用)は語長(W)11、期待値(E)10、カットオフ100、M=5、N=4、及び両鎖の比較をデフォルトとして使用する。アミノ酸配列用として、BLASTPプログラムは語長(W)3、期待値(E)10、及びBLOSUM62スコアリングマトリクスをデフォルトとして使用する(Henikoff & Henikoff (1989) Proc. Natl. Acad. Sci. USA 89:10915参照)。
【0186】
配列一致度百分率の計算に加え、BLASTアルゴリズムは2配列間の類似性の統計分析も実施する(例えばKarlin & Altschul, Proc. Natl. Acad. Sci. USA 90:5873-5787 (1993)参照)。BLASTアルゴリズムにより提供される類似性の1尺度は2種のヌクレオチド又はアミノ酸配列間に偶然にマッチが起こる確率を示す最小合計確率(P(N))である。例えば、試験核酸を参照核酸に比較した場合の最小合計確率が約0.1未満、より好ましくは約0.01未満、最も好ましくは約0.001未満である場合に核酸は参照核酸に類似しているとみなす。
【0187】
突然変異誘発及び他の分子生物学技術
本発明のポリヌクレオチドとポリペプチド及び本発明で使用されるポリヌクレオチドとポリペプチドは分子生物学技術を使用して操作することができる。分子生物学技術について記載している一般教科書としてはBerger and Kimmel, Guide to Molecular Cloning Techniques, Methods in Enzymology volume 152 Academic Press, Inc., San Diego, CA (Berger);Sambrookら, Molecular Cloning-A Laboratory Manual (第3版), Vol. 1-3, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 2001 (「Sambrook」)及びCurrent Protocols in Molecular Biology, F. M. Ausubelら編, Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (2004年補遺) (「Ausubel」))が挙げられる。これらの教科書は突然変異誘発法、ベクターの使用、プロモーター及び他の多くの関連事項について記載しており、アルキニルアミノ酸(例えばpPRO−Phe)、直交tRNA、直交シンテターゼ及びその対を含む蛋白質を生産するためのセレクターコドンを含む遺伝子の作製についても記載している。
【0188】
例えばtRNA分子を突然変異させるため、tRNAのライブラリーを作製するため、シンテターゼのライブラリーを作製するため、アルキニルアミノ酸をコードするセレクターコドンを該当蛋白質又はポリペプチドに挿入するために、本発明では各種突然変異誘発法を使用する。これらの方法としては限定されないが、部位特異的、ランダム点突然変異誘発、相同組換え、DNAシャフリング又は他の帰納的突然変異誘発法、キメラ構築、ウラシル含有鋳型を使用する突然変異誘発、オリゴヌクレオチド特異的突然変異誘発、ホスホロチオエート修飾DNA突然変異誘発、ギャップデュプレクスDNAを使用する突然変異誘発等、又はその任意組み合わせが挙げられる。他の適切な方法としては点ミスマッチ修復、修復欠損宿主株を使用する突然変異誘発、制限−選択及び制限−精製、欠失突然変異誘発、完全遺伝子合成による突然変異誘発、2本鎖切断修復等が挙げられる。例えばキメラ構築物を使用する突然変異誘発も本発明に含まれる。1態様では、天然分子又は改変もしくは突然変異させた天然分子の既知情報(例えば配列、配列比較、物性、結晶構造等)に基づいて突然変異誘発を行うことができる。
【0189】
本発明のポリヌクレオチド又は本発明のポリヌクレオチドを組込んだ構築物(例えばクローニングベクター又は発現ベクター等の本発明のベクター)で宿主細胞を遺伝子組換え(例えば形質転換、形質導入又はトランスフェクション)する。例えば、直交tRNA、直交tRNAシンテターゼ、及び修飾すべき蛋白質のコーディング領域を所望宿主細胞で機能的な遺伝子発現制御エレメントに機能的に連結する。典型的なベクターは転写及び翻訳ターミネーターと、転写及び翻訳開始配列と、特定ターゲット核酸の発現の調節に有用なプロモーターを含む。ベクターは場合により少なくとも1個の独立ターミネーター配列と、真核生物又は原核生物又は両者(例えばシャトルベクター)でカセットの複製を可能にする配列と、原核系と真核系の両者の選択マーカーを含む包括的発現カセットを含む。ベクターは原核生物、真核生物、又は好ましくは両者での複製及び/又は組込みに適している。Giliman & Smith, Gene 8:81 (1979); Robertsら, Nature, 328:731 (1987); Schneider, B. ら, Protein Expr. Purif. 6435:10(1995); Ausubel, Sambrook, Berger(いずれも前出)参照。ベクターは例えばプラスミド、細菌、ウイルス、裸のポリヌクレオチド又はポリヌクレオチドコンジュゲートの形態とすることができる。ベクターはエレクトロポレーション(Fromら, Proc. Natl. Acad. Sci. USA 82, 5824 (1985))、ウイルスベクターによる感染、核酸を小ビーズもしくは粒子のマトリックスに埋込むか又は表面に付着させて小粒子形態で高速射入する方法(Kleinら, Nature 327, 70-73 (1987))、及び/又は同等方法等の標準方法により細胞及び/又は微生物に導入する。
【0190】
クローニングに有用な細菌とバクテリオファージのカタログは例えばATCCから入手でき、例えばATCCから刊行されたThe ATCC Catalogue of Bacteria and Bacteriophage (1996) Ghernaら(編)が挙げられる。その他のシーケンシング、クローニング及び分子生物学の他の側面の基本手順と基礎理論事項もSambrook(前出)、Ausubel(前出)、及びWatsonら(1992) Recombinant DNA Second Edition, Scientific American Books (New York)に記載されている。更に、Midland Certified Reagent Company (Midland, TX;mcrc. com)、The Great American Gene Company(Ramona, CA, www.genco.comで販売)、ExpressGen Inc.(Chicago, IL, www.expressgen.comで販売)、Operon Technologies Inc. (Alameda, CA)及び他の多数の企業等の各種販売会社からほぼ任意核酸(及び標準又は標準外を問わずほぼ任意標識核酸)をオーダーメード又は標準注文することができる。
【0191】
遺伝子組換えした宿主細胞は例えばスクリーニング段階、プロモーター活性化又は形質転換細胞選択等の操作に合うように適宜改変した慣用栄養培地で培養することができる。これらの細胞は場合によりトランスジェニック生物で培養することができる。(例えば後期核酸単離のための)例えば細胞単離及び培養に関する他の有用な文献としてはFreshney (1994) Culture of Animal Cells, a Manual of Basic Technique, 第3版, Wiley-Liss, New Yorkとその引用文献; Payneら(1992) Plant Cell and Tissue Culture in Liquid Systems John Wiley & Sons, Inc. New York, NY; Gamborg and Phillips (eds) (1995) Plant Cell, Tissue and Organ Culture; Fundamental Methods Springer Lab Manual, Springer-Verlag (Berlin Heidelberg New York) 及び Atlas and Parks (eds) The Handbook of Microbiological Media (1993) CRC Press, Boca Raton, FLが挙げられる。
【0192】
該当蛋白質及びポリペプチド
アルキニルアミノ酸の1つの大きな利点は(限定されないが)各種小分子、生体分子又は他の蛋白質等の任意のものと蛋白質を架橋又は結合するためにアルキニルアミノ酸を含む蛋白質を使用できる点である。少なくとも1個のアルキニルアミノ酸を含む該当蛋白質又はポリペプチドは本発明の特徴である。本発明は本発明の組成物と方法を使用して生産された少なくとも1個のアルキニルアミノ酸を含むポリペプチド又は蛋白質も含む。賦形剤(例えば医薬的に許容可能な賦形剤)も蛋白質に添加することができる。場合により、本発明の蛋白質は単一アミノ酸位置又は複数の位置に(アルキニルアミノ酸残基のその後の可能な修飾に加えて)翻訳後修飾を含むことができるか、あるいは蛋白質は複数の異なる型の修飾をもつことができる。
【0193】
アルキニルアミノ酸を特定位置にもつ蛋白質を細胞で生産する方法も本発明の特徴である。例えば、1方法は、少なくとも1個のセレクターコドンを含み且つ蛋白質をコードする核酸を含む細胞を適当な培地で増殖させる段階と、アルキニルアミノ酸を提供する段階を含み;細胞は更に、細胞で機能し、セレクターコドンを認識する直交tRNA(O−tRNA)と;O−tRNAをアルキニルアミノ酸で優先的にアミノアシル化する直交アミノアシルtRNAシンテターゼ(O−RS)を含む。この方法により生産された蛋白質も本発明の特徴である。
【0194】
所定態様では、O−RSは発現系において内在tRNAよりもコグネイトO−tRNAのアミノアシル化を優先する。O−tRNAとO−RSが等モル濃度で存在する場合にO−RSにより負荷されるO−tRNAと内在tRNAの相対比は1:1を上回り、好ましくは少なくとも約2:1、より好ましくは5:1、更に好ましくは10:1、更に好ましくは20:1、更に好ましくは50:1、更に好ましくは75:1、更に好ましくは95:1、98:1、99:1、100:1、500:1、1,000:1、5,000:1又はそれ以上である。
【0195】
本発明はアルキニルアミノ酸を組込んだ蛋白質を含有する組成物も提供する。所定態様では、蛋白質は治療用蛋白質、診断用蛋白質、産業用酵素、又はその部分のアミノ酸配列と少なくとも75%一致するアミノ酸配列を含む。
【0196】
本発明の組成物と本発明の方法により作製された組成物は場合により細胞に導入する。その後、本発明のO−tRNA/O−RS対又は個々の成分を宿主系の翻訳機構で使用することができ、その結果として、アルキニルアミノ酸は蛋白質に組込まれる。国際公開WO2004/094593、出願日2004年4月16日、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」、及びWO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」はこのプロセスを記載しており、参考資料として本明細書に組込む。例えば、O−tRNA/O−RS対を宿主(例えば大腸菌細胞)に導入すると、前記対はセレクターコドンに応答してパラ−プロパルギルオキシフェニルアラニン等のアルキニルアミノ酸のin vivo蛋白質組込みを誘導する。系に付加されるパラ−プロパルギルオキシフェニルアラニンは外部から増殖培地に添加することができる合成アミノ酸(例えばフェニルアラニンやチロシンの誘導体)である。場合により、本発明の組成物はin vitro翻訳系に導入してもよいし、in vivo系に導入してもよい。
【0197】
本発明の細胞は非天然アミノ酸を組込んだ蛋白質を大量の有用な量で合成することができる。1側面では、組成物は場合により、アルキニルアミノ酸を組込んだ蛋白質を例えば少なくとも10μg、少なくとも50μg、少なくとも75μg、少なくとも100μg、少なくとも200μg、少なくとも250μg、少なくとも500μg、少なくとも1mg、少なくとも10mg以上、又はそれ以上、あるいはin vivo蛋白質生産方法で達成可能な量で含有する(組換え蛋白質生産及び精製に関する詳細は本明細書に記載する)。別の側面では、蛋白質は場合により例えば細胞溶解液、緩衝液、医薬緩衝液、又は他の懸濁液中(例えば約1nl〜約100Lの任意の容量中)に例えば少なくとも10μg蛋白質/l、少なくとも50μg蛋白質/l、少なくとも75μg蛋白質/l、少なくとも100μg蛋白質/l、少なくとも200μg蛋白質/l、少なくとも250μg蛋白質/l、少なくとも500μg蛋白質/l、少なくとも1mg蛋白質/l、又は少なくとも10mg蛋白質/l以上の濃度で組成物中に存在する。少なくとも1種のアルキニルアミノ酸を組込んだ蛋白質を細胞で大量(例えば他の方法、例えばin vitro翻訳で一般に可能な量よりも多量)に生産することも本発明の特徴である。
【0198】
アルキニルアミノ酸の組込みは例えば寸法、酸性度、求核性、水素結合、疎水性、プロテアーゼ標的部位接近性、(例えば蛋白質アレーのための)部分へのターゲット等を変化させるように例えば蛋白質構造及び/又は機能の変化を調整するために実施することができる。アルキニルアミノ酸を組込んだ蛋白質は触媒性又は物性を強化するか又は全く新規にすることができる。例えば、アルキニルアミノ酸を蛋白質に組込むことにより、場合により毒性、生体分布、構造的性質、分光学的性質、化学及び/又は光化学的性質、触媒能、半減期(例えば血清半減期)、他の分子との(例えば共有又は非共有)反応性等の性質を改変する。少なくとも1種のアルキニルアミノ酸を組込んだ蛋白質を含有する組成物は例えば新規治療薬、診断薬、触媒酵素、産業用酵素、結合蛋白質(例えば抗体)、及び例えば蛋白質構造と機能の研究に有用である。例えばDougherty, (2000) Unnatural Amino Acids as Probes of Protein Structure and Function, Current Opinion in Chemical Biology, 4:645-652参照。
【0199】
本発明の1側面では、組成物は少なくとも1個、例えば少なくとも2個、少なくとも3個、少なくとも4個、少なくとも5個、少なくとも6個、少なくとも7個、少なくとも8個、少なくとも9個、又は少なくとも10個以上の非天然アミノ酸(例えばアルキニルアミノ酸及び/又は他の非天然アミノ酸)を組込んだ少なくとも1種の蛋白質を含有する。非天然アミノ酸は同一でも異なっていてもよく、例えば1、2、3、4、5、6、7、8、9、又は10種以上の異なる非天然アミノ酸を含む1、2、3、4、5、6、7、8、9、又は10個以上の異なる部位が蛋白質に存在することができる。別の側面では、組成物は蛋白質に存在する特定アミノ酸の全部よりは少ないが少なくとも1個がアルキニルアミノ酸で置換された蛋白質を含有する。2個以上の非天然アミノ酸を組込んだ所与蛋白質では、非天然アミノ酸は同一でも異なっていてもよい(例えば蛋白質は2個以上の異なる型の非天然アミノ酸を組込んでもよいし、2個の同一非天然アミノ酸を組込んでもよい)。3個以上の非天然アミノ酸を組込んだ所与蛋白質では、非天然アミノ酸は同一でも異なっていてもよいし、同一種の複数の非天然アミノ酸と少なくとも1個の別の非天然アミノ酸の組み合わせでもよい。
【0200】
本明細書に記載する組成物と方法を使用してアルキニルアミノ酸を含むほぼ任意蛋白質(又はその部分)(及び例えば1個以上のセレクターコドンを含む対応する任意コーディング核酸)を生産することができる。数十万種の公知蛋白質を列挙するまでもなく、例えば該当翻訳系に1個以上の適当なセレクターコドンを含むように入手可能な任意突然変異法を調整することにより、1種以上の非天然アミノ酸を組込むように公知蛋白質の任意のものを改変することができる。公知蛋白質の一般的な配列寄託機関としてはGenBank、EMBL、DDBJ及びNCBIが挙げられる。他の寄託機関もインターネットを検索することにより容易に確認できる。
【0201】
一般に、蛋白質は入手可能な任意蛋白質(例えば治療用蛋白質、診断用蛋白質、産業用酵素、又はその部分等)と例えば少なくとも60%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも90%、少なくとも95%、又は少なくとも99%以上一致しており、1種以上の非天然アミノ酸を含む。1種以上のアルキニルアミノ酸を組込むように改変することができる治療用、診断用、及び他の蛋白質の例としては限定されないが、国際公開WO2004/094593、出願日2004年4月16日、発明の名称「真核遺伝コードの拡張(Expanding the Eukaryotic Genetic Code)」;及びWO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」に記載されているものが挙げられる。1種以上のアルキニルアミノ酸を組込むように改変することができる治療用、診断用、及び他の蛋白質の例としては限定されないが、例えばα1アンチトリプシン、アンジオスタチン、抗血友病因子、抗体(抗体の詳細については後述する)、アポリポ蛋白質、アポ蛋白質、心房性ナトリウム利尿因子、心房性ナトリウム利尿ポリペプチド、心房性ペプチド、C−X−Cケモカイン(例えばT39765、NAP−2、ENA−78、Gro−a、Gro−b、Gro−c、IP−10、GCP−2、NAP−4、SDF−1、PF−4、MIG)、カルシトニン、CCケモカイン(例えば単球化学誘引蛋白質−1、単球化学誘引蛋白質−2、単球化学誘引蛋白質−3、単球炎症性蛋白質−1α、単球炎症性蛋白質−1β、RANTES、I309、R83915、R91733、HCC1、T58847、D31065、T64262)、CD40リガンド、Cキットリガンド、コラーゲン、コロニー刺激因子(CSF)、補体因子5a、補体阻害剤、補体受容体1、サイトカイン(例えば上皮好中球活性化ペプチド−78、GROα/MGSA、GROβ、GROγ、MIP−1α、MIP−1δ、MCP−1)、表皮増殖因子(EGF)、エリスロポエチン(「EPO」)、表皮剥離毒素A及びB、IX因子、VII因子、VIII因子、X因子、繊維芽細胞増殖因子(FGF)、フィブリノーゲン、フィブロネクチン、G−CSF、GM−CSF、グルコセレブロシダーゼ、ゴナドトロピン、増殖因子、ヘッジホッグ蛋白質(例えばソニック、インディアン、デザート)、ヘモグロビン、肝細胞増殖因子(HGF)、ヒルジン、ヒト血清アルブミン、インスリン、インスリン様増殖因子(IGF)、インターフェロン(例えばIFN−α、IFN−β、IFN−γ)、インターロイキン(例えばIL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12等)、ケラチノサイト増殖因子(KGF)、ラクトフェリン、白血病阻害因子、ルシフェラーゼ、ニュールチュリン、好中球阻害因子(NIF)、オンコスタチンM、骨形成蛋白質、副甲状腺ホルモン、PD−ECSF、PDGF、ペプチドホルモン(例えばヒト成長ホルモン)、プレイオトロピン、プロテインA、プロテインG、発熱外毒素A、B及びC、リラキシン、レニン、SCF、可溶性補体受容体I、可溶性I−CAM1、可溶性インターロイキン受容体(IL−1、2、3、4、5、6、7、9、10、11、12、13、14、15)、可溶性TNF受容体、ソマトメジン、ソマトスタチン、ソマトトロピン、ストレプトキナーゼ、スーパー抗原即ちブドウ球菌エンテロトキシン(SEA、SEB、SEC1、SEC2、SEC3、SED、SEE)、スーパーオキシドジスムターゼ(SOD)、毒素性ショック症候群毒素(TSST−1)、チモシンα1、組織プラスミノーゲンアクチベーター、腫瘍壊死因子β(TNFβ)、腫瘍壊死因子受容体(TNFR)、腫瘍壊死因子α(TNFα)、血管内皮増殖因子(VEGEF)、ウロキナーゼ及び他の多数の蛋白質が挙げられる。
【0202】
本明細書に記載するアルキニルアミノ酸のin vivo組込み用組成物及び方法を使用して生産することができる蛋白質の1類としては転写モジュレーター又はその部分が挙げられる。転写モジュレーターの例としては細胞増殖、分化、制御等を調節する遺伝子及び転写モジュレーター蛋白質が挙げられる。転写モジュレーターは原核生物、ウイルス及び真核生物(例えば真菌、植物、酵母、昆虫、及び哺乳動物を含む動物)に存在し、広範な治療ターゲットを提供する。当然のことながら、発現及び転写アクチベーターは例えば受容体との結合、シグナル伝達カスケードの刺激、転写因子の発現調節、プロモーターやエンハンサーとの結合、プロモーターやエンハンサーと結合する蛋白質との結合、DNA巻き戻し、プレmRNAスプライシング、RNAポリアデニル化及びRNA分解等の多数のメカニズムにより転写を調節する。
【0203】
本発明の蛋白質(例えば1種以上のアルキニルアミノ酸を組込んだ蛋白質)の1類としてはサイトカイン、炎症性分子、増殖因子、その受容体、及び腫瘍遺伝子産物等の生体活性蛋白質が挙げられ、例えばインターロイキン(例えばIL−1、IL−2、IL−8等)、インターフェロン、FGF、IGF−I、IGF−II、FGF、PDGF、TNF、TGF−α、TGF−β、EGF、KGF、SCF/c−キット、CD40L/CD40、VLA−4/VCAM−1、ICAM−1/LFA−1及びヒアルリン/CD44;シグナル伝達分子及び対応する腫瘍遺伝子産物(例えばMos、Ras、Raf及びMet);並びに転写アクチベーター及びサプレッサー(例えばp53、Tat、Fos、Myc、Jun、Myb、Rel、及びステロイドホルモン受容体(例えばエストロゲン、プロゲステロン、テストステロン、アルドステロン、LDL受容体リガンド及びコルチコステロン))が挙げられる。
【0204】
少なくとも1種のアルキニルアミノ酸を組込んだ酵素(例えば産業用酵素)又はその部分も本発明により提供される。酵素の例としては限定されないが、例えばアミダーゼ、アミノ酸ラセマーゼ、アシラーゼ、デハロゲナーゼ、ジオキシゲナーゼ、ジアリールプロパンペルオキシダーゼ、エピメラーゼ、エポキシドヒドロラーゼ、エステラーゼ、イソメラーゼ、キナーゼ、グルコースイソメラーゼ、グリコシダーゼ、グリコシルトランスフェラーゼ、ハロペルオキシダーゼ、モノオキシゲナーゼ(例えばp450類)、リパーゼ、リグニンペルオキシダーゼ、ニトリルヒドラターゼ、ニトリラーゼ、プロテアーゼ、ホスファターゼ、スブチリシン、トランスアミナーゼ及びヌクレアーゼが挙げられる。
【0205】
これらの蛋白質の多くは市販されており(例えばSigma BioSciences 2003カタログ及び価格表参照)、対応する蛋白質配列と遺伝子及び、一般に多くのその変異体も周知である(例えばGenbank参照)。例えば1種以上の該当治療、診断又は酵素特性に関して蛋白質を改変するように本発明に従って1種以上のアルキニルアミノ酸を挿入することにより前記蛋白質の任意のものを改変することができる。治療関連特性の例としては血清半減期、貯蔵半減期、安定性、免疫原性、治療活性、(例えば非天然アミノ酸(例えばアルキニルアミノ酸)へのレポーター基(例えばラベル又はラベル結合部位)の付加による)検出性、LD50又は他の副作用の低減、胃管を経由して体内に導入できること(例えば経口利用性)等が挙げられる。診断特性の例としては貯蔵半減期、安定性、診断活性、検出性等が挙げられる。該当酵素特性の例としては貯蔵半減期、安定性、酵素活性、産生能等が挙げられる。
【0206】
他の各種蛋白質も本発明の組成物と方法を使用して1種以上のアルキニルアミノ酸を含むように改変することができる。例えば、本発明は例えば感染性真菌(例えばAspergillus、Candida種);細菌、特に病原細菌モデルとして利用できる大腸菌や医学的に重要な細菌(例えばStaphylococci(例えばaureus)又はStreptococci(例えばpneumoniae));原生動物(例えば胞子虫類(例えばPlasmodia)、根足虫類(例えばEntamoeba)及び鞭毛虫類(Trypanosoma、Leishmania、Trichomonas、Giardia等));ウイルス(例えば(+)RNAウイルス(例えばポックスウイルス(例えばワクシニア)、ピコルナウイルス(例えばポリオ)、トガウイルス(例えば風疹)、フラビウイルス(例えばHCV)、及びコロナウイルス)、(−)RNAウイルス(例えばラブドウイルス(例えばVSV)、パラミクソウイルス(例えばRSV)、オルトミクソウイルス(例えばインフルエンザ)、ブンヤウイルス及びアレナウイルス)、dsDNAウイルス(例えばレオウイルス)、RNA→DNAウイルス(即ちレトロウイルス、例えばHIV及びHTLV)、及び所定のDNA→RNAウイルス(例えばB型肝炎))に由来する蛋白質において、1種以上のワクチン蛋白質中の1種以上の天然アミノ酸をアルキニルアミノ酸で置換することができる。
【0207】
昆虫耐性蛋白質(例えばCry蛋白質)、澱粉及び脂質産生酵素、植物及び昆虫毒素、毒素耐性蛋白質、マイコトキシン解毒蛋白質、植物成長酵素(例えばリブロース1,5−ビスリン酸カルボキシラーゼ/オキシゲナーゼ、「RUBISCO」)、リポキシゲナーゼ(LOX)及びホスホエノールピルビン酸(PEP)カルボキシラーゼ等の農業関連蛋白質もアルキニルアミノ酸修飾の適切なターゲットである。
【0208】
所定態様では、本発明の方法及び/又は組成物における該当蛋白質又はポリペプチド(又はその部分)は核酸によりコードされる。一般に、核酸は少なくとも1個のセレクターコドン、少なくとも2個のセレクターコドン、少なくとも3個のセレクターコドン、少なくとも4個のセレクターコドン、少なくとも5個のセレクターコドン、少なくとも6個のセレクターコドン、少なくとも7個のセレクターコドン、少なくとも8個のセレクターコドン、少なくとも9個のセレクターコドン、10個以上のセレクターコドンを含む。
【0209】
該当蛋白質又はポリペプチドをコードする遺伝子はアルキニルアミノ酸を組込むための1個以上のセレクターコドンを付加するように本明細書の「突然変異誘発及び他の分子生物学技術」のセクションに記載する当業者に周知の方法を使用して突然変異誘発することができる。例えば、1個以上のセレクターコドンを付加するように該当蛋白質の核酸を突然変異誘発し、1種以上のアルキニルアミノ酸を挿入できるようにする。本発明は例えば少なくとも1種のアルキニルアミノ酸を組込んだ任意蛋白質のこのような任意変異体(例えば突然変異体、変形)を含む。同様に、本発明は対応する核酸、即ち1種以上のアルキニルアミノ酸をコードする1個以上のセレクターコドンを含む任意核酸も含む。
【0210】
アルキニルアミノ酸を組込んだ蛋白質を生産するためには、直交tRNA/RS対によるアルキニルアミノ酸のin vivo組込みに適した宿主細胞及び生物を使用することができる。直交tRNA、直交tRNAシンテターゼ、及び修飾すべき蛋白質をコードするベクターを発現する1種以上のベクターで宿主細胞を遺伝子組換え(例えば形質転換、形質導入又はトランスフェクション)する。これらの成分の各々を同一ベクターに配置してもよいし、各々別々のベクターに配置してもよいし、2成分を第1のベクターに配置し、第3の成分を第2のベクターに配置してもよい。ベクターは例えばプラスミド、細菌、ウイルス、裸のポリヌクレオチド又はポリヌクレオチドコンジュゲートの形態とすることができる。
【0211】
免疫反応性によるポリペプチドの定義
本発明のポリペプチドは種々の新規ポリペプチド配列(例えば本発明の翻訳系で合成される蛋白質の場合にはアルキニルアミノ酸を含み、例えば新規シンテターゼの場合には標準アミノ酸の新規配列を含むポリペプチド)を提供するので、ポリペプチドは例えばイムノアッセイで認識することができる新規構造特徴も提供する。本発明のポリペプチドに特異的に結合する抗血清の作製及び前記抗血清と結合するポリペプチドも本発明の特徴である。本明細書で使用する「抗体」なる用語は限定されないが、検体(抗原)と特異的に結合してこれを認識する1又は複数の免疫グロブリン遺伝子又はそのフラグメントにより実質的にコードされるポリペプチドを意味する。例としてはポリクローナル、モノクローナル、キメラ、及び1本鎖抗体等が挙げられる。Fabフラグメントやファージディスプレイを含む発現ライブラリーにより生産されるフラグメント等の免疫グロブリンフラグメントも本明細書で使用する「抗体」なる用語に含まれる。抗体構造及び用語については、例えばPaul, Fundamental Immunology, 4th Ed., 1999, Raven Press, New York参照。
【0212】
イムノアッセイ用抗血清を作製するためには、免疫原性ポリペプチドの1種以上を本明細書に記載するように作製し、精製する。例えば、組換え蛋白質を組換え細胞で生産することができる。(マウスの仮想遺伝子一致により結果の再現性が高いのでこのアッセイで使用する)近交系マウスに標準マウス免疫プロトコールに従って標準アジュバント(例えばフロイントアジュバント)と共に免疫原性蛋白質を免疫する(抗体作製、イムノアッセイフォーマット及び特異的免疫反応性を測定するために使用可能な条件の標準解説については例えばHarlow and Lane (1988) Antibodies. A Laboratory Manual, Cold Spring Harbor Publications, New York 参照)。蛋白質、抗体、抗血清等に関するその他の詳細は国際公開WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」;WO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」;WO2004/035605、発明の名称「糖蛋白質合成(GLYCOPROTEIN SYNTHESIS)」;及びWO2004/058946、発明の名称「蛋白質アレー(PROTEIN ARRAYS)」に記載されている。
【0213】
O−tRNA及びO−RS及びO−tRNA/O−RS対の使用
本発明の組成物と本発明の方法により作製された組成物は場合により細胞に導入する。本発明のO−tRNA/O−RS対又は個々の成分をその後、宿主系の翻訳機構で使用することができ、その結果として、アルキニルアミノ酸は蛋白質に組込まれる。国際公開WO2002/085923、発明者Schultzら、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」はこのプロセスを記載しており、参考資料として本明細書に組込む。例えば、O−tRNA/O−RS対を宿主(例えば大腸菌細胞)に導入すると、前記対はセレクターコドン(例えばアンバーナンセンスコドン)に応答して外部から増殖培地に添加することができるアルキニルアミノ酸のin vivo蛋白質(例えばミオグロビン試験蛋白質又は治療用蛋白質)組込みを誘導する。場合により、本発明の組成物はin vitro翻訳系に導入してもよいし、細胞内in vivo系に導入してもよい。アルキニルアミノ酸を組込んだ蛋白質は多様な用途の任意のものに使用することができる。最も特筆すべき点として、蛋白質に組込まれたアルキニル部分は例えば、他の蛋白質、小分子(例えばラベルや色素)及び/又は生体分子との架橋をはじめとする多様な修飾の任意のもののターゲットとして利用することができる。これらの修飾により、アルキニルアミノ酸の組込みの結果、治療用蛋白質を改善することができ、更に酵素の触媒機能を改変又は改善するために使用することもできる。所定側面では、アルキニルアミノ酸の蛋白質組込みとその後の修飾は蛋白質構造、他の蛋白質との相互作用等の研究を助長することができる。
【0214】
キット
キットも本発明の特徴である。例えば、少なくとも1種のアルキニルアミノ酸を組込んだ蛋白質を細胞で生産するためのキットが提供され、前記キットはO−tRNAをコードするポリヌクレオチド配列、及び/又はO−tRNA、及び/又はO−RSをコードするポリヌクレオチド配列、及び/又はO−RSを収容する容器を含む。1態様では、キットは更にアルキニルアミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン)を含む。別の態様では、キットは更に蛋白質を生産するための説明書を含む。
【0215】
以下、実施例により本発明を例証するが、これらの実施例により発明を限定するものではない。特許請求の範囲に記載する発明の範囲から逸脱せずに変更可能な種々の非必須パラメーターが当業者に自明である。
【実施例1】
【0216】
アルキニルアミノ酸を大腸菌の蛋白質に組込むための直交tRNA/シンテターゼ対の進化
直交tRNA−シンテターゼ対の特異性を進化させることにより、本発明者らは原核生物と真核生物の両者でナンセンス及びフレームシフトコドンに応答して多数の非天然アミノ酸を蛋白質に選択的且つ効率的に組込んでいる(Andersonら(2004) Proc. Natl. Acad. Sci. U.S.A., 101:1566; Alfontaら(2003) J. Am. Chem. Soc., 125:14662; Wangら(2003) Proc. Natl. Acad. Sci. U.S.A., 100:56; Chinら(2003) Science 301:964; Chinら(2002) Proc. Natl. Acad. Sci., 99:11020; 及び Wang ら(2001) Science 292:498)。本発明は大腸菌翻訳機構を使用して反応性アルキニル部分を含むアミノ酸を蛋白質に生合成的に組込むための組成物と方法を提供する。大腸菌翻訳成分を使用する生合成はin vivo(例えば大腸菌細胞中)で実施することもできるし、粗細胞抽出液又は精製翻訳成分を使用してin vitroで実施することもできる。蛋白質に組込まれるアルキニル基はアジド含有部分と容易且つ特異的に結合するので、蛋白質修飾/操作の有用なターゲットとなる。
【0217】
アルキニル基とアジド基の化学反応(図1Bに示す)は蛋白質に存在する全内在官能基の化学反応に対して完全に直交性である。それらのユニークな反応性の1例は[3+2]シクロ付加による不可逆的トリアゾール形成である(図1B;及びPadwa, In Comprehensive Organic Synthesis; [Trost, B. M., Ed.] Pergamon: Oxford, 1991, Vol. 4, p 1069; Huisgen, In 1,3-Dipolar Cycloaddition Chemistry, [Padwa, A., Ed.] Wiley:New York, 1984;p1参照)。この反応は室温にて水性媒体中(生物サンプルを修飾するために十分温和な条件)で銅(I)の存在下に実施する場合には、完全に位置選択的に進行し(Rostovtsevら(2002)Angew. Chem. Int. Ed., 41:2596)、アルキニル官能基とアジド官能基が導入された蛋白質を選択的に修飾するために使用することができる(Deitersら(2003) J. Am. Chem. Soc., 125:11182; Wangら(2003) J. Am. Chem. Soc., 125:3192; Link and Tirrell (2003) J. Am. Chem. Soc., 125:11164)。
【0218】
本明細書に記載する発明はアルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(略称pPRO−Phe;IUPAC命名法によると、2−アミノ−3−[4−(プロプ−2−イニルオキシ)プロピオン酸とも言う;構造は図1Aに示し、化学構造1と称する)を大腸菌宿主系に選択的に組込むMethanococcus jannaschii成分から誘導される直交tRNA/tRNAシンテターゼ対を提供する。本実施例はpPRO−Pheが本明細書に記載する新規直交tRNA及びtRNAシンテターゼ試薬を使用して大腸菌で発現される蛋白質に選択的に組込まれることを実証する。
【0219】
本実施例ではM. jannaschiiチロシルtRNA/tRNAシンテターゼ対(MjTyrRS/tRNACUA)から誘導される直交tRNA/シンテターゼ対の進化について報告し、直交対は標準(即ち天然)アミノ酸に対して親和性がないか又は非常に低い。誘導される直交tRNAシンテターゼはアンバーサプレッサーtRNACUAにpPRO−Pheを選択的に負荷し、更に、アミノアシル化されたtRNA(即ち「負荷されたtRNA」)は転写産物中で遭遇したTAGアンバー終止コドン(セレクターコドン)に応答してpPRO−Pheを組込むために内在大腸菌翻訳系により基質として使用される。このtRNA/シンテターゼ対の直交性(Steer and Schimmel (1999) Biol. Chem., 274:35601)により、tRNAもシンテターゼも内在大腸菌tRNA又はシンテターゼと交差反応せず、非天然アミノ酸はアンバーナンセンスコドンTAGのみに応答して遊離する。
【0220】
野生型M. jannaschiiチロシルtRNAシンテターゼの突然変異誘発により〜107個の異なるM. jannaschiiチロシルtRNAシンテターゼから成るライブラリーを作製した。MjTyrRSライブラリーを作製するために、5個の突然変異標的位置をアラニンコドンに変換した。pBR322から誘導したカナマイシン耐性をもつプラスミドpBK−JYRSで大腸菌GlnRSプロモーター及びターミネーターの制御下にMjTyrRS遺伝子を発現させた。残基Tyr32、Glu107、Asp158、Ile159、及びLeu162を部位特異的突然変異誘発によりAlaで置換し、プラスミドpBK−JYA5を得た。突然変異部位にNNK(N=A+T+G+C及びK=G+T)をもつ8個のオリゴヌクレオチドを使用してAla5MjTyrRS突然変異体(pBK−JYA5)をPCR増幅し、NdeI−PstIで消化したpBK−JYA5にライゲーションし、MjTyrRSライブラリーを作製した。ライゲーションしたベクターを大腸菌DH10Bコンピテント細胞に形質転換し、1.6×109コロニー形成単位のライブラリーを得た。
【0221】
野生型M. jannaschiiチロシルtRNAシンテターゼ分子のポリヌクレオチド及びアミノ酸配列を図2に示し、更に夫々配列番号3及び2にも示す。突然変異誘発はBacillus stearothermophilusに由来する相同チロシルtRNAシンテターゼの結晶構造に基づいて5個の活性部位残基(Tyr32,Glu107,Asp158,Ile159,及びLeu162)をランダム化することから構成した。
【0222】
突然変異誘発後、シンテターゼプールを次にポジティブ及びネガティブ選択ラウンドに付した。ポジティブ選択はクロラムフェニコールアセチルトランスフェラーゼ(CAT)遺伝子の許容部位(Asp112)におけるアンバー終止コドンの抑圧に基づく。MjTyrRS突然変異体ライブラリーと、突然変異CAT遺伝子と、同時発現させたMjアンバーサプレッサーtRNACUAを含む大腸菌細胞を最小培地でpPRO−Phe(1mM)とクロラムフェニコール(80μg/mL)の存在下に増殖させると、tRNACUAを内在アミノ酸又はpPRO−Pheでアミノアシル化する突然変異体シンテターゼを含む細胞のみが生存する。次に、許容部位に3個のアンバー突然変異(Gln2,Asp44,Gly65)をもつ毒性蛋白質バルナーゼをコードする突然変異遺伝子を含む細胞にシンテターゼライブラリー遺伝子を形質転換した。バルナーゼレポーターをもつベクターにはサプレッサーtRNAも加えた。これらの細胞をpPRO−Pheの不在下で増殖させ、内在アミノ酸を基質として受容することが可能なシンテターゼを逆選択した。3ラウンドの選択後、pPRO−Pheの有無に対する増殖率依存性について96個のクローンをスクリーニングし、8個の候補クローンを同定し、配列決定した。これらの単離クローンに認められたアミノ酸置換を図3に示す。8個のクローンのポリヌクレオチド配列とアミノ酸配列も配列番号4〜19に示す。
【0223】
8個の突然変異体O−RSクローンのアミノ酸置換にはコンセンサス傾向が認められる。クローンの殆どの結合ポケットには以下のアミノ酸:Ala32,Pro107/Gln107,Ala158,Ile159,及びAla162/Pro162が高頻度で認められた。突然変異Tyr32→Ala32及びAsp158→Ala158の結果、Tyr32、Asp158及び天然基質チロシン間の水素結合が失われ、その結合が弱まると思われる。小さい主に疎水性の側鎖が存在するならば、pPRO−Pheの結合を助長すると予想される。クローンの1個(pPRO−PheRS−1)には更にLeu110→Phe110突然変異も認められた。
【0224】
シンテターゼpPRO−PheRS−1を更に特性決定するために選択した。このシンテターゼはクロラムフェニコール耐性を大腸菌に付与し、pPRO−Pheの存在下と不在下のIC50は夫々110及び5μg/mLであった。pPRO−Pheの有無によるクロラムフェニコール耐性の大きな差異は非天然アミノ酸pRPO−Pheに対するpPRO−PheRS−1の実質的なin vivo特異性を示唆している。
【実施例2】
【0225】
大腸菌の蛋白質へのアルキニルアミノ酸の部位特異的組込み
突然変異体アンバーサプレッサーtRNACUAとpPRO−PheRS−1の直交対を大腸菌で使用し、多数の構造、力学、及び蛋白質フォールディング研究の対象となっているモノマー153残基ヘム含有蛋白質であるマッコウクジラミオグロビンにpPRO−Pheを選択的に組込んだ(Reedy and Gibney (2004) Chem. Rev., 104:617とその引用文献; Uzawaら(2004) Proc. Natl. Acad. Sci. U.S.A., 101:1171とその引用文献;Wright,and Baldwin in Frontiers in Molecular Biology: Mechanisms of Protein Folding, [Pain, R., ed.] Oxford University Press, London, 2000, pp.309)。
【0226】
アルキニル修飾ミオグロビンを作製するために、ミオグロビンオープンリーディングフレームの4番目のコドン(Ser4)をTAG(アンバー終止)に突然変異させ、C末端6×His(ヘキサヒスチジン)タグをオープンリーディングフレームに付加した。突然変異体蛋白質を発現させるために、(アラビノースプロモーターとrrnBターミネーターの制御下の突然変異体マッコウクジラミオグロビン遺伝子と;lppプロモーターとrrnCターミネーターの制御下のチロシルtRNACUAと;テトラサイクリン耐性遺伝子をコードする)プラスミドpBAD/JYAMB−4TAGを(突然変異体シンテターゼとカナマイシン耐性遺伝子をコードする)pBKベクターと共にDH10B大腸菌に同時形質転換した。テトラサイクリン(25mg/L)とカナマイシン(30mg/L)を補充したLuria−Bertani培地(5mL)で細胞を増幅させ、リン酸緩衝液で洗浄し、適当な抗生物質、pPRO−Phe(1mM)、及びアラビノース(0.002%)を加えた液体グリセロール最小培地(0.3mMロイシンを補充)500mLを接種するために使用した。細胞を飽和まで増殖させた後、遠心により回収した。Ni−アフィニティークロマトグラフィーを使用して蛋白質を精製した処、Ni−アフィニティークロマトグラフィーによる精製後に収量2mg/Lが得られ、SDS−PAGE/Gelcode(登録商標)Blue染色(Pierce Biotechnology,Inc.)により90%均質であると推定された。総収量〜1mgの突然変異体ミオグロビンが得られた。
【0227】
こうして作製した蛋白質をGelcode(登録商標)Blue染色と抗His6抗体を使用するウェスタンブロッティングの両者で図4,レーン1に可視化する。pPRO−Pheの不在下では、染色又は(抗His6抗体を使用する)ウェスタンブロッティング後にミオグロビンは認められず、進化型シンテターゼの高い選択性が示唆された(図4,レーン2参照)。
【実施例3】
【0228】
大腸菌の蛋白質へのアルキニルアミノ酸組込みの質量分析による確認
突然変異体ミオグロビンのアンバー終止コドンの部位に組込まれたアミノ酸の実体を更に確認するために、ミオグロビンのトリプシン消化物を液体クロマトグラフィー/タンデム質量分析に付した。この実験で使用した突然変異体ミオグロビンは74位に組換えアンバー終止コドンを含むものを使用した。この位置のpPRO−Pheの組込み(pPRO−Phe74)を試験し、74位突然変異を調べた。ミオグロビン−74TAG突然変異体はLC MS/MS分析の特性が改善されているので、先に記載したSer4→TAG(アンバーコドン)の代わりに使用した。
【0229】
真正細菌発現後に、ニッケルアフィニティーカラムを使用してミオグロビン74TAGを精製した。SDS−PAGEゲルのGelcode(登録商標)Blue染色により蛋白質バンドを可視化した。突然変異体ミオグロビンに対応するゲルバンドをポリアクリルアミドゲルから切り出し、1.5mm立方体にスライスし、ほぼ従来の記載通りにトリプシン加水分解した(Shevchenkoら(1996) Anal. Chem., 68:850-858)。
【0230】
非天然アミノ酸を含むトリプシンペプチドをLCQイオントラップ質量分析計でナノフロー逆相HPLC/μESI/MSにより分析した。Nanospray HPLC(Agilent 1100シリーズ)を取付たFinnigan LCQ Decaイオントラップ質量分析計(Thermo Finnigan)で液体クロマトグラフィータンデム質量分析(LC−MS/MS)を実施した。
【0231】
非天然アミノ酸(Y*で示す)を含むペプチドHGVTVLTALGY*ILK(配列番号X)の単一及び二重荷電イオンに対応する前駆体イオンをイオントラップ質量分析計で分離分画した。この分析の結果を図5に示す。フラグメントイオン質量は明確に帰属させることができ、pPRO−Pheの部位特異的組込みが確認された。LC MS/MS試験によると、この位置に天然アミノ酸の組込みは示唆されず、進化型シンテターゼの高い選択性が確認された。
【実施例4】
【0232】
[3+2]シクロ付加によるアルキニルアミノ酸を含む蛋白質の修飾
アルキニル官能基を含む蛋白質は[3+2]シクロ付加反応の使用による修飾の有効なターゲットとすることができる。本実施例は2個の異なるアジド含有色素分子によるアルキニルミオグロビンの修飾について記載する。本実施例で使用した突然変異体ミオグロビンは実施例1に記載したように4番目のコドンにpPRO−Pheを組込んだ(Ser4→pPRO−Phe4)。
【0233】
実施例2に記載したようにSer4→pPRQ−Phe4ミオグロビンを大腸菌で作製した後、夫々ダンシル及びフルオレセインフルオロフォアを含むアジド官能基を付けた色素2又は3で修飾した(図6A及び6Bに示す;Deitersら(2003) J. Am. Chem. Soc., 725:11782; Wangら(2003) J. Am. Chem. Soc., 125:3192; Link and Tirrell (2003) J. Am. Chem. Soc., 125:11164も参照)。[3+2]シクロ付加修飾反応を図7Aに示す。
【0234】
シクロ付加反応のために、CuSO4(H2O中50mMストック溶液;最終反応容量中1mM)1μL、色素2又は3(EtOH中50mM)2μL、トリス(1−ベンジル−1H−[1,2,3]トリアゾール−4−イルメチル)アミン(DMSO中50mM)2μL、及び(還元剤として)銅線1mg又はトリス(カルボキシエチル)ホスフィン(H2O中100mM)1μLを0.1Mリン酸緩衝液(pH=8)中精製突然変異体ミオグロビン(〜0.5mg/mL)45μLに加えた。室温で8時間又は4℃で一晩後にH2O 450μLを加え、透析膜(10kDaカットオフ)を通して混合物を遠心した。上清を2×500μLリン酸緩衝液で遠心洗浄後、溶液を容量50μLにした。
【0235】
還元剤として銅線又はトリス(カルボキシエチル)ホスフィン(2mM)を使用すると、一般に同様の標識効率が得られた。従来の知見(Wangら(2003) J. Am. Chem. Soc., 125:3192)とは対照的に、リガンドトリス(1−ベンジル−1H−[1,2,3]トリアゾール−4−イルメチル)アミンの有無はこれらの反応の結果に実質的な影響がなかった。次に蛍光標識蛋白質(Blake (2001) Curr. Opin. Pharmacol., 1:533; Woutersら(2001) Trends in Cell Biology 11:203; Zachariasら(2000) Curr. Opin. Neurobiol., 20:416)20μLのサンプルをSDS−PAGEにより分析し、ゲル内イメージングした。ダンシル色素2(λex=337nm,λem=506nm)で修飾した突然変異体ミオグロビンは360±30nmでEagle Eyeデンシトメーター(Stratagene)によりゲル内イメージングした。フルオレセイン色素3(λex=495nm,λem=516nm)の結合は450±30nmでStorm Phosphorimager(Molecular Dynamics)により可視化した。この蛍光イメージングの結果を図7Bに示す。突然変異体ミオグロビンは色素2及び3のいずれでも有効に標識される。3で標識したミオグロビンのA280/A495値の比較によると、標識効率は〜75%であった(Wangら(2003)J.Am.Chem.Soc.,125:3192参照)。このバイオコンジュゲーションの選択性は野生型ミオグロビンと2又は3の反応が認められなかったという事実により確認された(結果は示さず)。
【0236】
本実施例の記載はアルキニルアミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン)を生物(例えば大腸菌)の蛋白質に有効かつ選択的に組込むことができることを実証するものである。これらのアミノ酸はその後、蛋白質内で例えばアジド部分を使用する[3+2]シクロ付加による結合の化学的ターゲットとすることができ、更に、この標的修飾は高度に特異的且つ位置選択的である。アルキニルアミノ酸を蛋白質に部位特異的に組込むことができると、蛋白質結合又は修飾が所望される任意蛋白質の試験で有用なツールとなる。
【実施例5】
【0237】
非天然アルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニンの合成
非天然アルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(略称pPRO−Phe;図1A参照,化合物1)を市販N−Boc−チロシンから3段階で合成し(Deitersら(2003) J. Am. Chem. Soc., 125:11782; Wangら(2003) J. Am. Chem. Soc., 125:3192; Link and Tirrell (2003) J. Am. Chem. Soc., 125:11164参照)、総収率81%であった。
【0238】
ステップ1
N−tert−ブトキシカルボニル−チロシン(2g,7mmol,1当量)とK2CO3(3g,21mmol,3当量)を無水DMF(15mL)に懸濁した。臭化プロパルギル(2.1mL,21mmol,3当量,80%トルエン溶液)をゆっくりと加え、反応混合物を18時間室温で撹拌した。水(75mL)とEt2O(50mL)を加え、層分離し、水相をEt2O(2×50mL)で抽出した。有機層を合わせて乾燥し(MgSO4)、溶媒を減圧除去した。下記構造及び名称の生成物(4)が黄色油状物(2.3g,91%)として得られ、それ以上精製せずに次段階で使用した。
【化5】
【0239】
ステップ2
塩化アセチル(7mL)を0℃のメタノール(60mL)に注意深く加え、無水HClのMeOH中5M溶液を得た。前段階の生成物(化合物4;2g,5.6mmol)を加え、周囲温度まで昇温させながら反応混合物を4時間撹拌した。揮発分を減圧除去後に黄色がかった固体(下記構造及び名称の化合物5;1.6g,98%)が得られ、次段階で直接使用した。
【化6】
【0240】
ステップ3
前段階(5)からのプロパルギルエステル(1.6g,5.5mmol)を2N NaOH水溶液(14mL)とMeOH(10mL)の混合物に溶かした。1.5時間室温で撹拌後、濃HClを加えることによりpHを7に調整した。水(20mL)を加え、混合物を4℃に一晩維持した。沈殿を濾過し、氷冷H2Oで洗浄し、減圧乾燥すると、pPRO−Phe(1)1.23g(90%)が白色固体として得られた。1H NMR(400MHz,D2O;D2O中カリウム塩として)δ7.20(d,7=8.8Hz,2H),6.99(d,J=8.8Hz,2H),4.75(s,2H),3.50(dd,J=5.6,7.2Hz,1H),2.95(dd,J=5.6,13.6Hz,1H),2.82(dd,J=7.2,13.6Hz,1H);13C NMR(100MHz,D2O)δ181.3,164.9,155.6,131.4,130.7,115.3,57.3,56.1,39.3;HRMS(CI)m/z 220.0969[C12H13NO3(M+1)必要値220.0968]。
【化7】
【実施例6】
【0241】
アジド色素2の合成
アジド色素2(図6A参照;化合物2)を以下のプロトコールに従って合成した。3−アジドプロピルアミン(371mg,3.71mmol,3当量)(Carboniら(1993)Org.Chem.,58:3736−3741に従って合成)を塩化ダンシル(500mg,1.85mmol,1当量)とトリエチルアミン(258μL,1.85mmol,1当量)の0℃のCH2Cl2(10mL)溶液に加えた。1時間撹拌後、反応混合物を室温まで昇温させ、更に1時間撹拌した。揮発分を減圧除去し、粗生成物をシリカゲルクロマトグラフィー(Et2O/ヘキサン=1:1)により精製すると、2(548mg,89%)が黄色油状物として得られた。1H NMR(400MHz,CDCl3)δ8.55(d,J=8.4Hz,1H),8.29(d,J=8.8Hz,1H),8.23(dd,J=1.2,7.2Hz,1H),7.56−7.49(comp,2H),7.18(d,J=7.6Hz,1H),5.24(br s,1H),3.21(t,J=6.4Hz,2H),2.95(dt,J=6.4Hz,2H),2.89(s,6H),1.62(quin,J=6.4Hz,2H);13C NMR(100MHz,CDCl3)δ134.3,130.4,129.7,129.4,128.4,123.3,118.8,115.3,48.6,45.4,40.6,28.7(13C NMRスペクトルに4級炭素原子の全シグナルは認められない);HRMS(CI)m/z 334.1336[C15H20N5O2S(M+1)必要値334.1332]。
【化8】
【実施例7】
【0242】
アジド色素3の合成
アジド色素3(図6B参照;化合物3)を以下のプロトコールに従って合成した。EDCI(83mg,0.43mmol,1当量)をフルオレセインアミン(150mg,0.43mmol,1当量)と4−(3−アジドプロピルカルバモイル)酪酸(92mg,0.43,1当量)の室温のピリジン(2mL)溶液に加えた。(4−(3−アジドプロピルカルバモイル)酪酸は3−アジドプロピルアミンをグルタル酸無水物と反応させることにより合成した。)懸濁液を一晩撹拌し、反応混合物をH2O(15mL)に注入した。濃HClを加えることにより溶液を酸性化した(pH<2)。1時間撹拌後、沈殿を濾別し、1N HCl(3×3mL)で洗浄し、少量のEtOAcに溶かした。ヘキサンを加えると、3がオレンジ色結晶として沈殿したので、採取し、減圧乾燥した(200mg,86%)。1H NMR(400MHz,CD3OD)δ8.65(s,1H),8.15(d,J=8.4Hz,1H),7.61−7.51(comp,2H),7.40(d,J=8.4Hz,1H),7.35(br s,2H),7.22−7.14(comp,2H),6.85−6.56(comp,3H),3.40−3.24(comp,4H),2.54(t,J=7.2Hz,2H),2.39−2.30(comp,2H),2.10−1.99(comp,2H),1.82−1.72(comp,2H);13C NMR(100MHz,CD3OD)δ175.7,174.4,172.4,167.9,160.8,143.0,134.3,132.9,131.8,129.6,124.4,123.3,121.1,118.5 103.5,50.2,38.0,37.2,36.2,29.8,22.9;4 HRMS(CI)m/z 544.1835[C28H25N5O7(M+1)必要値544.1827]。
【化9】
【実施例8】
【0243】
アルキニルアミノ酸の大腸菌組込み用の代表的O−RS及びO−tRNA
代表的O−tRNAとしては配列番号1が挙げられる(実施例9,表4参照)。代表的O−RSとしては配列番号4、6、8、10、12、14、16及び18に記載のアミノ酸配列が挙げられる(図3及び実施例9,表4参照)。
【0244】
O−RS又はその部分をコードするポリヌクレオチドの例としては配列番号4、6、8、10、12、14、16及び18を含む任意アミノ酸配列をコードする任意ポリヌクレオチドが挙げられる。例えば、配列番号5、7、9、11、13、15、17及び19に記載のポリヌクレオチドが代表的O−RSをコードする。
【0245】
当然のことながら、本明細書に記載する実施例及び態様は例証の目的に過ぎず、これらの記載に鑑みて種々の変形又は変更が当業者に示唆され、このような変形又は変更も本願の精神及び範囲と特許請求の範囲に含むものとする。
【0246】
以上、明確に理解できるように本発明を多少詳細に記載したが、本発明の真の範囲を逸脱することなく形態や細部に種々の変更が可能であることは以上の開示から当業者に自明である。例えば、上記全技術及び装置は種々に組合せて使用することができる。本明細書に引用した全刊行物、特許、特許出願、及び/又は他の文献はその開示内容全体を全目的で参考資料として組込み、各刊行物、特許、特許出願、及び/又は他の文献を全目的で参考資料として組込むと個々に記載しているものとして扱う。
【実施例9】
【0247】
ヌクレオチド及びアミノ酸配列
本実施例は夫々各種ポリヌクレオチド及びポリペプチドのヌクレオチド及びアミノ酸配列を示す。下表4に記載する配列は例示に過ぎず、表4に記載する配列に本発明を限定するものではない。
【表4−1】
【表4−2】
【表4−3】
【表4−4】
【表4−5】
【表4−6】
【表4−7】
【表4−8】
【表4−9】
【表4−10】
【技術分野】
【0001】
(関連出願とのクロスリファレンス)
本願は米国仮特許出願第60/612,220号(出願日2004年9月21日);米国仮特許出願第60/630,876号(出願日2004年11月24日);及び米国仮特許出願第60/634,151号(出願日2004年12月7日)の優先権と特典を主張し、各々その開示内容全体を参考資料として全目的で本明細書に組込む。
【0002】
本発明は国立衛生研究所助成番号GM62159として米国政府助成下に創出された。米国政府は本発明に所定の権利をもつことができる。
【0003】
本発明は翻訳生化学の分野に関する。本発明はアルキニルアミノ酸を蛋白質に組込む直交tRNA、直交アミノアシルtRNAシンテターゼ、及びその対を作製及び使用するための組成物及び方法に関する。本発明は前記対を使用して細胞で蛋白質を生産する方法と関連組成物にも関する。
【背景技術】
【0004】
分光プローブ、触媒助剤、又はポリマー等の非ペプチド分子で蛋白質を部位特異的に化学修飾したり、蛋白質を別の蛋白質又は他の任意部分と共有結合できるならば、蛋白質の化学的及び生物学的性質を調査及び操作するための強力な手段となる。一般的なアプローチは蛋白質の求核性表面残基(例えばリジン、ヒスチジン、又はシステインの側鎖)と外来分子の求電子基(例えばアルデヒド、α−ハロカルボキサミド、及びN−ヒドロキシスクシンイミド)のバイオコンジュゲーションを利用している(Lemineux, G. A.; Bertozzi, C. R. TIBTECH 1996, 16, 506)。
【0005】
残念ながら、修飾を標的化するために蛋白質の天然求核性ターゲットを使用するには、これらの反応の選択性が低く、蛋白質には求核性アミノ酸が複数存在するため、標識蛋白質の不均質混合物が形成されるという問題がある。更に、求核標的修飾反応には非生理的条件を要することが多いため、in vivo修飾ストラテジーを実施できないか、及び/又は蛋白質生物活性が低下する可能性がある。
【0006】
特異的標的蛋白質修飾のための新規ターゲットと新規ストラテジーを開発することが当分野で必要とされている。残念ながら、細菌からヒトに至る全公知生物は同一の20種の標準アミノ酸をコードする(稀な例外としてセレノシステイン(例えばA. Bockら, (1991), Molecular Microbiology 5:515-20参照)とピロリジン(例えばG.Srinivasanら, (2002), Science 296:1459-62参照)も挙げられる)。この特徴により、標的蛋白質修飾のための新規化学の開発に天然アミノ酸を使用することは制限される。
【0007】
この制限を解決するための1つのストラテジーは遺伝子コードを拡張し、顕著な化学的性質をもつアミノ酸を生物レパートリーに付加する方法である。このアプローチは真正細菌である大腸菌(E.coli)や他の生物のin vivo蛋白質生合成機構を使用して非天然アミノ酸を蛋白質に付加するために「直交」tRNAと対応する新規「直交」アミノアシルtRNAシンテターゼの使用により、実現可能であることが分かっている(例えばWangら, (2001), Science 292:498-500; Chinら, (2002) Journal of the American Chemical Society 124:9026-9027; Chin and Schultz, (2002), ChemBioChem 11:1135-1137; Chinら, (2002), PNAS United States of America 99:11020-11024; 及びWang and Schultz, (2002), Chem.Comm., 1-10)。国際公開WO2002/086075、発明の名称「直交tRNA−アミノアシルtRNAシンテターゼ対を作製するための方法及び組成物(METHODS AND COMPOSITIONS FOR THE PRODUCTION OF ORTHOGONAL tRNA AMINOACYL-tRNA SYNTHETASE PAIRS)」;WO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」;WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」;WO2005/019415(出願日2004年7月7日);WO2005/007870(出願日2004年7月7日);及びWO2005/007624(出願日2004年7月7日)も参照。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】国際公開WO2002/086075
【特許文献2】国際公開WO2002/085923
【特許文献3】国際公開WO2004/094593
【特許文献4】国際公開WO2005/019415
【特許文献5】国際公開WO2005/007870
【特許文献6】国際公開WO2005/007624
【非特許文献】
【0009】
【非特許文献1】Lemineux, G. A.;Bertozzi, C. R. TIBTECH 1996, 16, 506
【非特許文献2】A. Bockら, (1991), Molecular Microbiology 5:515-20
【非特許文献3】G. Srinivasanら, (2002), Science 296:1459-62
【非特許文献4】Wangら, (2001), Science 292:498-500
【非特許文献5】Chinら, (2002) Journal of the American Chemical Society 124:9026-9027
【非特許文献6】Chin and Schultz, (2002), ChemBioChem 11:1135-1137
【非特許文献7】Chinら, (2002), PNAS United States of America 99:11020-11024
【非特許文献8】Wang and Schultz,(2002), Chem. Comm., 1-10
【発明の概要】
【発明が解決しようとする課題】
【0010】
高度に特異的な標的蛋白質修飾を実施するための新規方法が当分野で必要とされている。非天然アミノ酸を大腸菌の蛋白質にin vivoで組込む直交翻訳成分の開発も当分野で必要とされており、非天然アミノ酸を規定位置に組込むことができ、非天然アミノ酸が蛋白質の他の部分と交差反応又は副反応することなしに特異的修飾のターゲットとして利用できるように顕著な化学的性質をもつことが望ましい。真正細菌蛋白質発現システムは科学研究又は治療用組換え蛋白質材料を大量に生産することができるので、当分野の上記必要は特に大腸菌で実現されることが望ましい。本発明は以下の開示から明らかなように、上記及び他の必要を満たすものである。
【課題を解決するための手段】
【0011】
本発明はセレクターコドン(例えばアンバー終止コドン、4塩基以上のコドン等)に応答して成長中のポリペプチド鎖にアルキニルアミノ酸をin vivo又はin vitroで組込むための直交成分を作製するための組成物及び方法を提供する。本発明は直交tRNA(O−tRNA)、直交アミノアシルtRNAシンテターゼ(O−RS)及びその対を提供する。これらの対は成長中のポリペプチド鎖にアルキニルアミノ酸を組込むために細胞又は無細胞系で使用することができる。アルキニルアミノ酸を含むポリペプチドはアルキン部分が[3+2]シクロ付加反応でアジド部分と容易且つ特異的に反応してトリアゾール結合を形成する結合反応で特に使用される。アルキン基はin vivo系に対して外来性であり、原則的に任意化合物にアジド基を付加できるので、本明細書に実証するようにアルキニルアミノ酸の部位特異的組込みシステムは有用な部位特異的修飾用ツールである。
【0012】
1側面において、真正細菌細胞は直交アミノアシルtRNAシンテターゼ(O−RS)を含み、前記O−RSはアルキニルアミノ酸である非天然アミノ酸で直交tRNA(O−tRNA)を優先的にアミノアシル化する。所定態様において、真正細菌細胞は大腸菌細胞である。所定側面において、O−RSはMethanococcus jannaschiiアミノアシルtRNAシンテターゼ、例えばMethanococcus jannaschiiチロシルtRNAシンテターゼから誘導される。所定態様において、O−RSを誘導するために使用されるチロシルtRNAシンテターゼは配列番号2のアミノ酸配列をもつ野生型Methanococcus jannaschiiチロシルtRNAシンテターゼである。
【0013】
所定態様において、配列番号2の野生型Methanococcus jannaschiiチロシルtRNAシンテターゼから誘導されるO−RSはコンセンサス位置の組合わせに突然変異、例えば、
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリンを含む。所定態様において、O−RSのアミノ酸配列は配列番号4、6、8、10、12、14、16、18、又はその任意保存変異体の1種を含む。O−RSを含む細胞は一般にO−RS、例えば上記任意O−RS種をコードする核酸を含む。O−RSをコードする核酸は例えば配列番号5、7、9、11、13、15、17又は19のヌクレオチド配列を含むことができる。
【0014】
所定態様において、細胞で使用されるO−tRNAはアンバーサプレッサーtRNAである。例えば、O−tRNAは配列番号1のポリヌクレオチド配列であるか又は前記配列を含む。
【0015】
所定態様において、O−RSの基質であるアルキニルアミノ酸はパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)である。細胞系は更に少なくとも1個のセレクターコドンをもつ核酸を含み、セレクターコドンはO−tRNAにより認識される。直交成分を含む細胞は更にアルキニルアミノ酸(例えばpPRO−Phe)を含むことができる。
【0016】
所定態様において、細胞は第2の直交対(即ち第2のO−tRNAと第2のO−RS)を含み、第2の対は第1の非天然アミノ酸とは異なる非天然アミノ酸に特異的であり、第2のO−tRNAは第1のO−tRNAにより認識されるセレクターコドンとは異なるセレクターコドンを認識する。
【0017】
所定側面において、直交成分を含む細胞は翻訳系を含み、前記翻訳系はO−RSとO−tRNAに加え、該当ポリペプチドをコードし、O−tRNAにより認識される少なくとも1個のセレクターコドンを含む核酸と;アルキニルアミノ酸を含むことができ、O−RSはO−tRNAにアルキニルアミノ酸を負荷することができる。
【0018】
所定側面において、本発明はポリペプチド(例えば本明細書に教示するようなO−RSポリペプチド)を提供する。これらのポリペプチドは配列番号2のMethanococcus jannaschiiチロシルアミノアシルtRNAシンテターゼから誘導することができ、以下のアミノ酸コンセンサス:
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリン
をもつことができ、前記ポリペプチドは直交tRNA(O−tRNA)をアルキニルアミノ酸で優先的にアミノアシル化することが可能なアミノアシルtRNAシンテターゼ活性をもつ。所定態様において、本発明のポリペプチドは配列番号4、6、8、10、12、14、16、18、又はその保存変異体から選択される。本発明のこのような任意O−RSポリペプチドは真正細菌細胞において直交tRNA(O−tRNA)をアルキニルアミノ酸で優先的にアミノアシル化することが可能なアミノアシルtRNAシンテターゼである。本発明は上記のような本発明の任意O−RSポリペプチドをコードするポリヌクレオチドも提供する。
【0019】
所定態様において、(本発明のO−RSをコードする)本発明のポリヌクレオチドは配列番号5、7、9、11、13、15、17及び19から選択される。本発明のO−RSをコードする本発明の任意ポリヌクレオチドはベクター(例えば発現ベクター)に組込むことができる。本発明のベクターは細胞で使用することができる。
【0020】
所定側面において、本発明は非天然アルキニルアミノ酸を含む蛋白質を真正細菌細胞で生産する方法を提供する。本方法は蛋白質の任意所望特定位置にアルキニルアミノ酸を挿入するようにプログラムすることができる。本方法は、
(a)(i)直交アミノアシルtRNAシンテターゼ(O−RS)と;
(ii)直交tRNA(O−tRNA)(O−RSはO−tRNAをアルキニルアミノ酸で優先的にアミノアシル化する)と;
(iii)蛋白質をコードする核酸(核酸はO−tRNAにより認識される少なくとも1個のセレクターコドンを含む)と;
(iv)アルキニルアミノ酸を含む真正細菌細胞を提供する段階と;
(b)細胞を増殖させる段階と;
(c)蛋白質の翻訳中に核酸によりコードされる蛋白質の特定位置(蛋白質の特定位置は核酸におけるセレクターコドンの位置に対応する)にアルキニルアミノ酸を組込むことにより、アルキニルアミノ酸を特定位置に含む蛋白質を生産する段階を含む。本方法は一般に大腸菌細胞を使用する。
【0021】
これらの方法で使用されるO−RSは一般にMethanococcus jannaschiiアミノアシルtRNAシンテターゼ、例えばMethanococcus jannaschiiチロシルtRNAシンテターゼから誘導される。所定態様において、Methanococcus jannaschiiチロシルtRNAシンテターゼは配列番号2のシンテターゼである。所定態様において、O−RSは配列番号2のMethanococcus jannaschiiチロシルtRNAシンテターゼから誘導され、O−RSは以下の突然変異:
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリンをもつアミノ酸配列をもつ。
【0022】
所定態様において、前記方法は配列番号4、6、8、10、12、14、16、18、又はその任意保存変異体から選択されるアミノ酸配列をもつO−RSを使用する。本発明の方法では、細胞はこれらのO−RSポリペプチドの任意のものをコードするポリヌクレオチドを含むことができる。例えば、配列番号5、7、9、11、13、15、17又は19のヌクレオチド配列を含むポリヌクレオチドを使用することができる。
【0023】
これらの方法の所定態様において、O−tRNAはアンバーサプレッサーtRNAであり、セレクターコドンはアンバー終止コドン(TAG)である。所定態様において、O−tRNAは配列番号1に記載のポリヌクレオチド配列を含むか又は前記配列によりコードされる。これらの方法はアルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)をもつ蛋白質を生産するために使用することができる。本発明の方法により生産された蛋白質は野生型治療用蛋白質、診断用蛋白質、産業用酵素、又はその一部のアミノ酸配列と少なくとも75%一致するアミノ酸配列を含むことができる。これらの蛋白質は場合により医薬的に許容可能なキャリヤーと結合することができる。
【0024】
所定態様において、本発明の方法により生産された蛋白質は例えば[3+2]シクロ付加反応によりトリアゾール結合を形成することによりアルキニルアミノ酸の位置を修飾することができる。
【図面の簡単な説明】
【0025】
【図1】図1Aは(1)非天然アルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(IUPAC命名法によると、2−アミノ−3−[4−(プロプ−2−イニルオキシ)フェニル]プロピオン酸とも言う)の化学構造を示す。図1Bは室温で銅の存在下でのアジドとアルキンの[3+2]シクロ付加反応による不可逆的トリアゾール形成の一般反応化学反応を示す。
【図2】野生型Methanococcus jannaschiiチロシルtRNAシンテターゼ(MjTyrRS)のヌクレオチド及びアミノ酸配列を示す。パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)tRNAシンテターゼにおける部位特異的突然変異誘発又は他の突然変異の標的アミノ酸位置(及び対応するトリプレットコドン)を四角で囲む。
【図3】野生型Methanococcus jannaschiiチロシルtRNAシンテターゼをコードするポリヌクレオチドの突然変異誘発後に同定単離した8種のパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)tRNAシンテターゼ種をまとめた表である。野生型Methanococcus jannaschiiチロシルtRNAシンテターゼとパラ−プロパルギルオキシフェニルアラニンtRNAシンテターゼ(pPRO−PheRS)において指定コドンによりコードされるアミノ酸を示す。突然変異位置のコドンも示す。突然変異体のアミノ酸位置ナンバリングは図2に示すような野生型Methanococcus jannaschiiチロシルtRNAシンテターゼのアミノ酸ナンバリングに従う。
【図4】精製Ser4→pPRO−Phe4突然変異体ミオグロビンのGelcode(登録商標)Blue(Pierce Biotechnology,Inc.)染色SDS−PAGEゲルを示す。レーン1はパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)の存在下に最小培地で培養した大腸菌で発現された蛋白質を含み;レーン2はpPRO−Pheの不在下で生成された蛋白質サンプルを示す。下部の図はミオグロビンのC末端のヘキサヒスチジンタグを検出するために抗His6抗体を使用した同一サンプル材料のウェスタンブロットを示す。
【図5】アルキニル非天然アミノ酸(Y*で示す)を含むトリプシンペブチドHGVTVLTALGY*ILKのタンデム質量スペクトルを予想フラグメントイオン質量と共に示す。矢印はペプチドに観測されたb及びyイオン系列を示す。
【図6】図6A及び6Bはアジド官能基を付けた色素の化学構造(夫々2及び3)を示す。図6Aの色素2はダンシルフルオロフォアを含み、図6Bの色素3はフルオレセインフルオロフォアを含む。
【図7】図7Aは組換えアンバーコドン(4TAG)の部位にアルキニルアミノ酸を含む突然変異体ミオグロビンと(図6A及び6Bに示すような)アジド官能基を付けた色素の[3+2]シクロ付加反応からの不可逆的トリアゾール形成の一般反応化学を示す。図7Bは分解した標識ミオグロビンの紫外線照射下の蛍光ゲルイメージングを示し、[3+2]シクロ付加反応物は色素2又は色素3と共有結合している。
【図8】図8A及び8Bアルキニル非天然アミノ酸の例の構造と名称を示す。図8Aは非天然前駆体から化学的に合成することができるアルキニル非天然アミノ酸を示す。図8Bは既存天然アミノ酸基質から潜在的に合成することができるアルキニル非天然アミノ酸を示す。
【発明を実施するための形態】
【0026】
(定義)
本発明を詳細に記載する前に、本発明は特定生物系に限定されず、当然のことながら種々のものに適用できると理解すべきである。同様に、本明細書で使用する用語は特定態様のみの記載を目的とし、限定的でないことも理解すべきである。本明細書と特許請求の範囲で使用する単数形はそうでないことが内容から明白である場合を除き、複数形も含む。従って、例えば「細胞」と言う場合には2個以上の細胞の組合せを含み、「ポリヌクレオチド」と言う場合には実際問題としてそのポリヌクレオチドの多数のコピーを含む。
【0027】
本欄及び以下に特に定義しない限り、本明細書で使用する全科学技術用語は本発明が属する分野の当業者に通常理解されている通りの意味をもつ。
【0028】
直交:本明細書で使用する「直交」なる用語は細胞又は他の翻訳系に内在する対応分子に比較して低効率で細胞の内在成分と共働するか、あるいは細胞の内在成分と共働できない分子(例えば直交tRNA(O−tRNA)及び/又は直交アミノアシルtRNAシンテターゼ(O−RS))を意味する。tRNA及びアミノアシルtRNAシンテターゼに関して直交とは、内在tRNAが内在tRNAシンテターゼと共働する能力に比較して直交tRNAが内在tRNAシンテターゼと共働できないか又は低効率(例えば20%未満、10%未満、5%未満、又は1%未満の効率)でしか共働できず、あるいは、内在tRNAシンテターゼが内在tRNAと共働する能力に比較して直交アミノアシルtRNAシンテターゼが内在tRNAと共働できないか又は低効率でしか共働できないことを意味する。直交分子は細胞内に機能的に正常な相補的内在分子をもたない。例えば、細胞中の直交tRNAがこの細胞の任意内在tRNAシンテターゼ(RS)によりアミノアシル化される効率は内在tRNAが内在RSによりアミノアシル化される効率に比較して低いか又はゼロである。別の例では、直交RSが該当細胞の任意内在tRNAをアミノアシル化する効率は内在tRNAが内在RSによりアミノアシル化される効率に比較して低いか又はゼロである。第1の直交分子と共働する第2の直交分子を細胞に導入することができる。例えば、直交tRNA/RS対は対照(例えば対応するtRNA/RS内在対、又は活性直交対(例えばチロシル直交tRNA/RS対))の効率に比較して所定の効率(例えば45%の効率、50%の効率、60%の効率、70%の効率、75%の効率、80%の効率、90%の効率、95%の効率、又は99%以上の効率)で細胞において共働する導入相補成分を含む。
【0029】
直交チロシルtRNA:本明細書で使用する直交チロシルtRNA(チロシル−O−tRNA)とは該当翻訳系に直交性のtRNAであり、tRNAは(1)天然に存在するチロシルtRNAと同一であるか又は実質的に類似しているか、(2)自然又は人工突然変異誘発により天然に存在するチロシルtRNAから誘導されるか、(3)(1)又は(2)の野生型又は突然変異体チロシルtRNA配列の配列を考慮する任意プロセスにより誘導されるか、(4)野生型又は突然変異体チロシルtRNAと相同であるか;(5)表4にチロシルtRNAシンテターゼの基質として指定する任意特定tRNAと相同であるか、あるいは(6)表4にチロシルtRNAシンテターゼの基質として指定する任意特定tRNAの保存変異体である。チロシルtRNAはアミノ酸を負荷した状態でも負荷しない状態でも存在することができる。更に当然のことながら、「チロシル−O−tRNA」は場合によりコグネイトシンテターゼによりチロシン以外のアミノ酸(例えば非天然アミノ酸パラ−プロパルギルオキシフェニルアラニン)を負荷(アミノアシル化)される。実際に、当然のことながら、本発明のチロシル−O−tRNAは翻訳中にセレクターコドンに応答して成長中のポリペプチドに天然又は人工のいずれかに拘わらずほぼ任意アミノ酸を挿入するために有利に使用される。
【0030】
直交チロシルアミノ酸シンテターゼ:本明細書で使用する直交チロシルアミノ酸シンテターゼ(チロシル−O−RS)とは該当翻訳系においてチロシル−O−tRNAをアミノ酸で優先的にアミノアシル化する酵素である。チロシル−O−RSがチロシル−O−tRNAに負荷するアミノ酸は天然又は人工のいずれかを問わずに任意アミノ酸とすることができ、本明細書では限定しない。シンテターゼは場合により天然に存在するチロシルアミノ酸シンテターゼと同一又は相同であるか、あるいは表4にO−RSとして指定するシンテターゼと同一又は相同である。例えば、O−RSは表4のチロシル−O−RSの保存変異体とすることができ、及び/又は表4のO−RSと少なくとも50%、60%、70%、80%、90%、95%、98%、99%又はそれ以上配列が一致することができる。
【0031】
コグネイト:「コグネイト」なる用語は共働する成分、例えば直交tRNAと直交アミノアシルtRNAシンテターゼを意味する。これらの成分は「相補的」であると言うこともできる。
【0032】
優先的にアミノアシル化する:本明細書で直交翻訳系に関して使用する場合に、O−RSはO−RSが発現系で任意内在tRNAに負荷するよりも効率的にO−tRNAにアミノ酸を負荷するときにコグネイトO−tRNAを「優先的にアミノアシル化する」。即ち、O−tRNAと所与の任意内在tRNAがほぼ等モル比で翻訳系に存在するとき、O−RSは内在tRNAに負荷するよりも高頻度でO−tRNAに負荷する。O−RSにより負荷されるO−tRNAとO−RSにより負荷される内在tRNAの相対比は高いことが好ましく、従って、O−tRNAと内在tRNAが等モル濃度で翻訳系に存在する場合にはO−RSはO−tRNAに排他的、又はほぼ排他的に負荷することが好ましい。O−tRNAとO−RSが等モル濃度で存在する場合にO−RSにより負荷されるO−tRNAと内在tRNAの相対比は1:1を上回り、好ましくは少なくとも約2:1、より好ましくは5:1、更に好ましくは10:1、更に好ましくは20:1、更に好ましくは50:1、更に好ましくは75:1、更に好ましくは95:1、98:1、99:1、100:1、500:1、1,000:1、5,000:1又はそれ以上である。
【0033】
(a)O−RSが内在tRNAに比較してO−tRNAを優先的にアミノアシル化するとき、及び(b)O−RSがO−tRNAを任意天然アミノ酸でアミノアシル化する場合に比較してそのアミノアシル化が非天然アミノ酸に特異的であるときにO−RSは「O−tRNAを非天然アミノ酸で優先的にアミノアシル化する」。即ち、非天然アミノ酸と天然アミノ酸がO−RSとO−tRNAを含む翻訳系に等モル量で存在するとき、O−RSは天然アミノ酸よりも高頻度で非天然アミノ酸をO−tRNAに負荷する。非天然アミノ酸を負荷されたO−tRNAと天然アミノ酸を負荷されたO−tRNAの相対比は高いことが好ましい。O−RSはO−tRNAに排他的、又はほぼ排他的に非天然アミノ酸を負荷することがより好ましい。天然アミノ酸と非天然アミノ酸の両者が等モル濃度で翻訳系に存在するとき、O−tRNAの非天然アミノ酸負荷とO−tRNAの天然アミノ酸負荷の相対比は1:1を上回り、好ましくは少なくとも約2:1、より好ましくは5:1、更に好ましくは10:1、更に好ましくは20:1、更に好ましくは50:1、更に好ましくは75:1、更に好ましくは95:1、98:1、99:1、100:1、500:1、1,000:1、5,000:1又はそれ以上である。
【0034】
セレクターコドン:「セレクターコドン」なる用語は翻訳プロセスでO−tRNAにより認識され、内在tRNAにより認識されないコドンを意味する。O−tRNAアンチコドンループはmRNA上のセレクターコドンを認識し、そのアミノ酸(例えばアルキニルアミノ酸等の非天然アミノ酸)をポリペプチドのこの部位に組込む。セレクターコドンとしては例えば終止コドン(例えばアンバー、オーカー及びオパールコドン)等のナンセンスコドン、4塩基以上のコドン、レアコドン、天然又は非天然塩基対から誘導されるコドン及び/又は同等物を挙げることができる。
【0035】
サプレッサーtRNA:サプレッサーtRNAは例えばセレクターコドンに応答してポリペプチド鎖にアミノ酸を組込むためのメカニズムを提供することにより、所与翻訳系でメッセンジャーRNA(mRNA)の読取りを変更するtRNAである。例えば、サプレッサーtRNAは例えば終止コドン(例えばアンバー、オーカー又はオパールコドン)、4塩基コドン、レアコドン等を読み飛ばすことができる。
【0036】
抑圧活性:本明細書で使用する「抑圧活性」なる用語は一般に、読み飛ばさないと翻訳終結又は誤訳(例えばフレームシフト)をもたらすコドン(例えばアンバーコドンや4塩基以上のコドンであるセレクターコドン)の翻訳読み飛ばしを行うtRNA(例えばサプレッサーtRNA)の能力を意味する。サプレッサーtRNAの抑圧活性は第2のサプレッサーtRNA又は対照系(例えばO−RSをもたない対照系)に比較して観測される翻訳読み飛ばし活性の百分率として表すことができる。
【0037】
本発明は抑圧活性を定量することが可能な種々の手段を提供する。該当セレクターコドン(例えばアンバーコドン)に対する特定O−tRNA及びO−RSの抑圧百分率とは、O−tRNA、O−RS及びセレクターコドンをもたないポジティブ対照構築物に比較して、O−RSとO−tRNAを含む該当翻訳系で発現され、そのコーディング核酸中にセレクターコドンを含む所与試験マーカー(例えばLacZ)の活性百分率を意味する。従って、例えば、セレクターコドンをもたない活性ポジティブ対照マーカー構築物が所与翻訳系において該当マーカーアッセイに関連する単位で表した観測活性Xをもつ場合には、セレクターコドンを含む試験構築物の抑圧百分率は、O−tRNAとO−RSを更に含む翻訳系で発現される以外はポジティブ対照マーカーが発現される条件とほぼ同一の環境条件下で試験マーカー構築物が示すXの百分率である。一般に、試験マーカーを発現する翻訳系は更にO−RSとO−tRNAにより認識されるアミノ酸を含む。場合により、抑圧百分率測定値は、O−tRNA、O−RS及び/又はO−tRNA及び/又はO−RSにより認識される該当アミノ酸を含まない系で試験マーカーと同一のセレクターコドンを含む「バックグラウンド」又は「ネガティブ」対照マーカー構築物に試験マーカーを比較することにより精密化することができる。このネガティブ対照は該当翻訳系におけるマーカーからのバックグラウンドシグナル効果を考慮するように抑圧百分率測定値を正規化するのに有用である。
【0038】
抑圧効率は当分野で公知の多数のアッセイの任意のものにより測定することができる。例えば、β−ガラクトシダーゼレポーターアッセイを使用することができ、例えば本発明のO−tRNAを含むプラスミドと共に修飾lacZプラスミド(構築物はlacZ核酸配列中にセレクターコドンをもつ)を適当な生物(例えば直交成分を使用することができる生物)に由来する細胞に導入する。コグネイトシンテターゼも(ポリペプチド又は発現されるとコグネイトシンテターゼをコードするポリヌクレオチドとして)導入することができる。細胞を培地で所望密度(例えばOD600=約0.5)まで増殖させ、例えばBetaFluor(登録商標)β−ガラクトシダーゼアッセイキット(Novagen)を使用してβ−ガラクトシダーゼアッセイを実施する。比較可能な対照(例えば所望位置にセレクターコドンではなく対応するセンスコドンをもつ修飾lacZ構築物から観測される値)に対するサンプルの活性百分率として抑圧百分率を計算することができる。
【0039】
翻訳系:「翻訳系」なる用語は成長中のポリペプチド鎖(蛋白質)にアミノ酸を組込む成分を意味する。翻訳系の成分としては例えばリボソーム、tRNA、シンテターゼ、mRNA等を挙げることができる。本発明のO−tRNA及び/又はO−RSは例えば非真核細胞(例えば細菌(例えば大腸菌))、又は真核細胞(例えば酵母細胞、哺乳動物細胞、植物細胞、藻類細胞、真菌細胞、昆虫細胞、及び/又は同等物)でin vitro又はin vivo翻訳系に付加するか又はその一部とすることができる。
【0040】
非天然アミノ酸:本明細書で使用する「非天然アミノ酸」なる用語は20種の標準天然アミノ酸の1種又はセレノシステインもしくはピロリジン以外の任意アミノ酸、修飾アミノ酸、及び/又はアミノ酸類似体(例えばアルキニルアミノ酸)を意味する。
【0041】
から誘導:本明細書で使用する「から誘導」なる用語は特定分子もしくは生物から単離されているか、あるいは特定分子もしくは生物を使用するか又は特定分子もしくは生物からの情報を使用して作製された成分を意味する。例えば、第2のポリペプチドから誘導されるポリペプチドは第2のポリペプチドのアミノ酸配列と同一又は実質的に同様のアミノ酸配列を含む。ポリペプチドの場合には、誘導種は例えば自然突然変異誘発、人工的特異的突然変異誘発又は人工的ランダム突然変異誘発により得ることができる。ポリペプチドを誘導するために使用される突然変異誘発は意図的に特異的でも意図的にランダムでもよい。第1のポリペプチドから誘導される別のポリペプチドを作製するためのポリペプチドの突然変異誘発は(例えばポリメラーゼの非忠実性に起因する)ランダムなイベントとすることができ、誘導されたポリペプチドの同定は偶然とすることができる。ポリペプチドの突然変異誘発は一般にポリペプチドをコードするポリヌクレオチドの操作を伴う。
【0042】
ポジティブ選択又はスクリーニングマーカー:本明細書で使用する「ポジティブ選択又はスクリーニングマーカー」なる用語は、このマーカーが存在する(例えば発現、活性化等される)と、対応する形質をもたない細胞からこの形質を含む細胞(例えばポジティブ選択マーカーをもつ細胞)を識別することができるマーカーを意味する。
【0043】
ネガティブ選択又はスクリーニングマーカー:本明細書で使用する「ネガティブ選択又はスクリーニングマーカー」なる用語は、このマーカーが存在する(例えば発現、活性化等される)と、(例えば選択性質又は形質をもつ細胞に対して)選択性質又は形質をもたない細胞を識別することができるマーカーを意味する。
【0044】
レポーター:本明細書で使用する「レポーター」なる用語は該当系のターゲット成分を同定及び/又は選択するために使用することができる成分を意味する。例えば、レポーターとしては蛋白質、例えば抗生物質耐性又は感受性を付与する酵素(例えばβ−ラクタマーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)等)、蛍光スクリーニングマーカー(例えば緑色蛍光蛋白質(例えばGFP)、YFP、EGFP、RFP等)、発光マーカー(例えばホタルルシフェラーゼ蛋白質)、アフィニティースクリーニングマーカー、又はポジティブもしくはネガティブ選択マーカー遺伝子(例えばlacZ、β−gal/lacZ(β−ガラクトシダーゼ)、ADH(アルコールデヒドロゲナーゼ)、his3、ura3、leu2、lys2等)を挙げることができる。
【0045】
真核生物:本明細書で使用する「真核生物」なる用語は真核生物界に属する生物を意味する。真核生物はその構成が一般に多細胞であり(但し、例えば酵母のように多細胞ではないものもある)、膜結合核と他の膜結合オルガネラが存在し、遺伝物質が線状であり(即ち染色体が線状)、オペロンが存在せず、イントロン、メッセージキャッピング及びポリA mRNAが存在し、更に他の生化学的特徴(例えば特徴的リボソーム構造)により一般に原核細胞から区別できる。真核生物としては例えば動物(例えば哺乳動物、昆虫、爬虫類、鳥類等)、繊毛虫、植物(例えば単子葉植物、双子葉植物、藻類等)、真菌類、酵母、鞭毛虫、微胞子虫、原生生物等が挙げられる。
【0046】
原核生物:本明細書で使用する「原核生物」なる用語はモネラ界(原核生物界とも言う)に属する生物を意味する。原核生物はその構成が単細胞であり、出芽又は分裂による無性生殖であり、膜結合核又は他の膜結合オルガネラをもたず、染色体が環状であり、オペロンが存在し、イントロン、メッセージキャッピング及びポリA mRNAが存在せず、更に他の生化学的特徴(例えば特徴的リボソーム構造)により一般に真核細胞から区別できる。原核生物は真正細菌及び古細菌亜界を含む。シアノバクテリア(藍藻類)とマイコプラズマをモネラ界の別々の分類にする場合もある。
【0047】
細菌:本明細書で使用する「細菌」及び「真正細菌」なる用語は「古細菌」から区別できる原核生物を意味する。同様に、古細菌は真核生物から区別できる原核生物を意味する。真正細菌と古細菌は多数の形態学的及び生化学的基準により区別できる。例えば、リボソームRNA配列、RNAポリメラーゼ構造、イントロンの有無、抗生物質感受性、細胞壁ペプチドグリカン及び他の細胞壁成分の有無、膜脂質構造の分岐の有無、並びにヒストン及びヒストン様蛋白質の有無を使用して生物を真正細菌又は古細菌に分類する。
【0048】
真正細菌の例としては、Escherichia coli、Thermus thermophilus及びBacillus stearothermophilusが挙げられる。古細菌の例としては、Methanococcus jannaschii(Mj)、Methanosarcina mazei(Mm)、Methanobacterium thermoautotrophicum(Mt)、Methanococcus maripaludis、Methanopyrus kandleri、Halobacterium(例えばHaloferax volcanii及びHalobacterium種NRC−1)、Archaeoglobus fulgidus(Af)、Pyrococcus furiosus(Pf)、Pyrococcus horikoshii(Ph)、Pyrobaculum aerophilum、Pyrococcus abyssi、Sulfolobus solfataricus(Ss)、Sulfolobus tokodaii、Aeuropyrum pernix(Ap)、Thermoplasma acidophilum及びThermoplasma volcaniumが挙げられる。
【0049】
保存変異体:翻訳成分に関して本明細書で使用する「保存変異体」なる用語は類似する基本成分(例えばO−tRNA又はO−RS)と同様に機能するが、参照O−tRNA又はO−RSに比較して配列に変異をもつ翻訳成分(例えば保存変異体O−tRNA又は保存変異体O−RS)を意味する。例えば、O−RSは相補的O−tRNA又は保存変異体O−tRNAを非天然アミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン等のアルキニルアミノ酸)でアミノアシル化するが、O−tRNAと保存変異体O−tRNAは同一配列をもたない。保存変異体は保存変異体が対応するO−tRNA又はO−RSに相補的である限り、例えば配列の1カ所、2カ所、3カ所、4カ所、又は5カ所以上に変異をもつことができる。
【0050】
選択又はスクリーニング物質:本明細書で使用する「選択又はスクリーニング物質」なる用語はこのような物質が存在すると、集団から所定成分を選択/スクリーニングすることができる物質を意味する。例えば、選択又はスクリーニング物質としては限定されないが、例えば栄養素、抗生物質、光波長、抗体、発現されたポリヌクレオチド等が挙げられる。選択物質は例えば濃度、強度等を変動させることができる。
【0051】
〜に応答して:本明細書で使用する「〜に応答して」なる用語は本発明のtRNAがセレクターコドンを認識し、tRNAと結合したアルキニルアミノ酸を成長中のポリペプチド鎖に組込むのを媒介するプロセスを意味する。
【0052】
コードする:本明細書で使用する「コードする」なる用語は第1の分子又は配列鎖とは異なる第2の分子又は配列鎖の生産を導くためにポリマー巨大分子又は配列鎖中の情報を使用する任意プロセスを意味する。本明細書ではこの用語を広義に使用し、種々に適用することができる。1側面では、「コードする」なる用語は新規に合成された相補的姉妹鎖をDNA依存性DNAポリメラーゼによりコードするための鋳型として2本鎖DNA分子の一方の鎖を使用する半保存的DNA複製プロセスを意味する。
【0053】
別の側面では、「コードする」なる用語は第1の分子とは異なる化学的性質をもつ第2の分子の生産を導くためにある分子中の情報を使用する任意プロセスを意味する。例えば、DNA分子は(例えばDNA依存性RNAポリメラーゼ酵素を含む転写プロセスにより)RNA分子をコードすることができる。また、RNA分子は翻訳プロセスと同様にポリペプチドをコードすることができる。翻訳プロセスについて使用する場合には、「コードする」なる用語はアミノ酸をコードするトリプレットコドンにも適用する。所定側面では、RNA分子は例えばRNA依存性DNAポリメラーゼを含む逆転写プロセスによりDNA分子をコードすることができる。別の側面では、DNA分子はポリペプチドをコードすることができ、この場合に使用する「コードする」とは当然のことながら転写プロセスと翻訳プロセスの両者を含む。
【0054】
アルキン:本明細書で使用する「アルキン」(「アセチレン」と言う場合もある)なる用語は(図1Bに示すように)2個の炭素原子間に三重結合を含み、一般構造:
【化1】
(式中、Rは任意原子又は構造である)をもつ化学構造を意味する。置換基として使用する場合には、アルキン部分を「アルキニル」基と言う。アルキニル炭素原子はsp2結合しており、2個の他の原子と結合しか形成せず、これらの結合の一方は単結合であり、第2の結合は三重結合である。例えば、アルキニルアミノ酸は2個の炭素中心間に三重結合を含むアミノ酸である。アルキニル置換基は自然界ではアミノ酸に認められないので、全アルキニルアミノ酸は非天然アミノ酸である。
【0055】
アジド:本明細書で使用する「アジド」なる用語は一般構造:
R−N=N+=N−
をもつ化学基−N3を意味する。アジド基は一般に炭素原子と結合している。
【0056】
例えば、アジド色素はアジド置換基をもつ色素分子である(例えば図6A及び6Bのアジド色素2及び3参照)。「アジド」なる用語はアジド基を含む化合物(例えば、アジ化ベンジル、アジ化ナトリウム等)を意味する。
【0057】
生理的条件下で高度に選択的に蛋白質を修飾する化学反応が大いに必要とされている(Lemineux and Bertozzi (1996) TIBTECH, 16:506)。蛋白質の選択的修飾に当分野で現在使用されている大半の反応は蛋白質アミノ酸側鎖の天然求核性残基を標的とする求核性反応パートナーと求電子性反応パートナーの間の共有結合形成、例えばα−ハロケトンとヒスチジン又はシステイン側鎖の反応を利用している。これらの場合の選択性は蛋白質の求核性残基の数とアクセシビリティにより決定される。残念ながら、天然蛋白質は反応部位の配置が不良であったり(例えば接近しにくい)、反応ターゲットを複数含む(例えばリジン、ヒスチジン及びシステイン残基)ことが多いため、修飾反応の選択性が低く、求核/求電子試薬による高度標的蛋白質修飾は困難になる。更に、修飾部位は一般にリジン、ヒスチジン及びシステインの天然求核性側鎖に限定される。他の部位の修飾は困難又は不可能になる。
【0058】
この問題の1つの解決方法は直交翻訳成分を使用して新規反応性をもつ非天然アミノ酸を蛋白質にプログラム下で部位特異的に生合成的に組込む方法である(Wang and Schultz (2002) Chem. Commun., 1:1; 及びvan Maarseveen and Back (2003) Angew. Chem., 115:6106)。本発明ではアンバーナンセンスコドンTAGに応答して細菌(例えば大腸菌)で産生された蛋白質にアルキニル含有非天然アミノ酸を遺伝的に組込む選択的蛋白質修飾のための非常に効率的な新規方法を報告する。その後、これらのアルキニルアミノ酸側鎖を特異的且つ位置選択的に修飾することができる。アルキニル基のユニークな反応化学により、蛋白質を非常に高い選択性で修飾することができる。
【0059】
細菌発現系で産生された蛋白質のユニーク部位(例えば所望部位)にアルキニル官能基を選択的に導入するために、本発明者らはアルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe;図1A参照)を遺伝的にコードする真正細菌で機能する直交RNA/アミノアシルtRNAシンテターゼ対を開発した。要約すると、本発明者らは大腸菌細胞でアンバーサプレッサーtRNAにパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)を選択的に負荷するMethanococcus janaschiiチロシルtRNAシンテターゼの新規突然変異体を同定した。これらの進化型tRNAシンテターゼ対はアルキニル基を蛋白質に部位特異的に組込むために使用することができる。
【0060】
標的蛋白質修飾
本発明ではアンバーナンセンスコドンTAGに応答して真正細菌(例えば大腸菌)で産生された蛋白質にアルキニル含有非天然アミノ酸を遺伝的に組込む選択的蛋白質修飾のための非常に効率的な方法を報告する。本明細書に記載する新規組成物と方法は直交tRNA/アミノアシルtRNAシンテターゼシステムを利用し、直交システムはMethanococcus janaschiiに由来する成分を使用し、これらの成分は該当蛋白質を産生するための真正細菌宿主系で使用される。アルキニルアミノ酸の蛋白質組込みはアルキニルアミノ酸の組込みを指示するセレクターコドンを含むように該当蛋白質をコードするポリヌクレオチドを組換えることにより任意所望位置で行われるようにプログラムすることができる。
【0061】
その後、該当蛋白質のこれらのアルキニルアミノ酸側鎖をアジド誘導体とのHuisgen[3+2]シクロ付加反応により特異的且つ位置選択的に修飾することができる(図1B参照)(Padwa,In Comprehensive Organic Synthesis; [Trost,B.M.,Ed.] Pergamon: Oxford, 1991, Vol.4, p 1069-1109; Huisgen, In 1,3-Dipolar Cycloaddition Chemistry, [Padwa,A.,Ed.] Wiley: New York, 1984; p 1-176)。この方法は求核置換ではなくシクロ付加を利用するので、蛋白質を非常に高い選択性で修飾することができる。この反応は触媒量のCu(I)塩を反応混合物に加えることにより室温で水性条件下に優れた位置選択的性(1,4>1,5)で実施できるという利点がある(Tornoeら, (2002) J. Org. Chem., 67:3057-3064; Rostovtsevら, (2002) Angew. Chem., Int. Ed., 41:2596-2599)。
【0062】
アルキニル反応性ターゲットはin vivo系に対して完全に外来性であるという利点があり、その反応化学において高度に選択性(例えばアゾ含有部分に対して高度に反応性)であり、蛋白質に関するin vitro及びin vivo両者の結合反応を可能にする比較的温和な反応条件を使用して結合することができるので、蛋白質の生物活性を維持できる。(限定的ではないが)本発明を実証するために、アルキニル部分をミオグロビンモデル蛋白質に組込んだ後に、安定なトリアゾール結合による[3+2]シクロ付加反応(図1B参照)によりアジド蛍光色素(図6A及び6B参照)とバイオコンジュゲートさせる。
【0063】
本発明はアルキニルアミノ酸とアジド部分の[3+2]シクロ付加を例証するために2種のアジド蛍光色素を使用する(実施例4参照)が、本発明はこれらの2種のアジド色素、又は任意色素もしくはラベル、又は実際に任意単一種の結合性材料の使用に限定されない。本発明のアジド含有部分はアジド誘導体であるほぼ任意分子とすることができる。このような分子としては限定されないが、色素、フルオロフォア、架橋剤、糖誘導体、ポリマー(例えばポリエチレングリコール誘導体)、光架橋剤、細胞傷害性化合物、アフィニティーラベル、ビオチン誘導体、樹脂、ビーズ、第2(又は第3以下)の蛋白質又はポリペプチド、ポリヌクレオチド(例えばDNA,RNA等)、金属キレート剤、補因子、脂肪酸、炭水化物等が挙げられる。これらのアジド分子はアルキニル基をもつ非天然アミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン)と結合することができる(図1A参照)。
【0064】
本発明は図6A及び6Bに示すアジド色素の合成について詳細に記載する。夫々実施例6及び7参照。しかし、任意該当特定蛋白質のアジド誘導体を合成することは当業者が当然に実施できる範囲内である。例えば、アジド化合物の合成方法を記載した多数の教科書やプロトコールが入手可能である。一般情報については、Patai, Saul, “The chemistry of the azido group” in The Chemistry of Functional Groups, London, New York, Interscience Publishers, 1971参照。
【0065】
別の側面では、本発明はアルキニル含有ポリペプチドとの結合反応にポリエチレングリコールの誘導体(アジド−PEG)を使用することによりペグ化ポリペプチドを作製するための組成物及び方法を提供する。アジドポリエチレングリコールの一般構造は下式:
N3−CH2−(CH2−O−CH2)n−CH2OR
により表され、式中、RはH又はCH3であり、nは例えば50〜10,000、75〜5,000、100〜2,000、100〜1,000等の整数である。本発明の各種態様において、アジドポリエチレングリコールは例えば約5,000〜約100,000Da(即ち約5kDa〜約100kDa)、約20,000〜約50,000Da、約20,000〜約10,000Da(例えば20,000Da)等の分子量をもつ。アジドポリエチレングリコールの合成技術は当業者に周知である。例えば、アジド基を含む求核性分子(例えばアジ化ナトリウム又は3−アジドプロピルアミン)と求電子基(例えば臭化物又はN−ヒドロキシスクシンイミドエステル)を含むポリエチレングリコール分子を反応させてアジドポリエチレングリコールを生成することができる。
【0066】
アジド−PEGはトリアゾール結合によりアルキニル含有蛋白質とバイオコンジュゲートした場合に本発明で利用できる。蛋白質治療薬をポリエチレングリコールで修飾(ペグ化)すると、多くの場合には蛋白質の薬物動態及び薬力学的性質を改善し、従って、効力を改善し、投薬頻度を最小限にすることができる。蛋白質治療薬のペグ化の各種利点は例えば、Deitersら,“Site-specific PEGylation of proteins containing unnatural amino acids,” Bioorganic & Medicinal Chemistry Letters 14:5743-5745 (2004)に記載及び例証されている。
【0067】
更に、エステル結合も含む非天然アルキニルアミノ酸を含むポリペプチドの作製に関連する他の利点も考えられる。例えば、アルキニルアミノ酸とエステル結合を併用することにより作製したペグ化ポリペプチドはin vivo又はin vitroエステル結合の鹸化によるポリペプチドの緩慢な遊離を可能にすることができる。また、アジド−PEG分子の代わりにポリマー支持体(アジド樹脂)を使用すると、蛋白質アフィニティー精製が可能になる。トリアゾール共有結合は非常に強力な洗浄段階を可能にし、エステルアルキニルアミノ酸の使用は塩基処理による蛋白質の遊離を可能にする。特筆すべき点として、このようなアフィニティー精製スキームは精製のために該当蛋白質に人工タグ(例えばヘキサヒスチジン)又はエピトープを付ける必要がない。使用する非天然アミノ酸に応じて、開裂段階後に原則的に野生型(天然)ポリペプチドをアフィニティー樹脂から遊離させることができる。
【0068】
エステル結合をもつ非天然アルキニルアミノ酸を合成し、蛋白質に組込むことができる(例えば、3−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノプロパン酸や4−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノブタン酸)(図8B参照)。[3+2]シクロ付加によるバイオコンジュゲーション後に、in vivo又はin vitro鹸化によりエステル結合を開裂することができ、1応用例は例えばペグ化蛋白質からのペブチド部分の緩慢な遊離である。
【0069】
所定側面において、本発明のポリペプチドはアルキニル含有ポリペプチドを含み、更にこれらのポリペプチドのコンジュゲート形を含む。例えば、所定側面において、本発明はトリアゾール結合と共有結合蛍光アジド色素を含むポリペプチドを含む(例えば図6A、6B及び7A参照)。この側面では、ポリペプチドは最初にアルキニル基を含んでおり、色素は最初にアジド基を含んでおり、[3+2]シクロ付加により両者を結合し、トリアゾール結合を形成する。別の態様では、本発明のアルキニル含有蛋白質はアジドポリエチレングリコールを含む(化学構造6参照)。
【0070】
直交tRNA/アミノアシルtRNAシンテターゼ技術
本発明の新規組成物及び方法の理解は直交tRNAと直交アミノアシルtRNAシンテターゼの対に関連する活性を理解することにより容易になる。直交tRNA及びアミノアシルtRNAシンテターゼ技術については例えば、国際公開WO2002/085923、WO2002/086075、WO204/09459、WO2005/019415、WO2005/007870及びWO2005/007624に記載されている。
【0071】
付加反応性非天然アミノ酸(例えばアルキニルアミノ酸)を遺伝コードに付加するためには、宿主翻訳機構で効率的に機能することができるが、対が翻訳系に内在性のシンテターゼ及びtRNAから独立して機能するという意味で該当翻訳系に対して「直交性」のアミノアシルtRNAシンテターゼと適切なtRNAを含む新規直交対が必要である。直交対の所望特徴としては、内在tRNAによりデコードされない特定新規コドン(例えばセレクターコドン)のみをデコード又は認識するtRNAと、そのコグネイトtRNAをただ1個の特定非天然アミノ酸で優先的にアミノアシル化(又は負荷)するアミノアシルtRNAシンテターゼが挙げられる。O−tRNAは一般に内在シンテターゼによりアミノアシル化されない。例えば大腸菌では、直交対は例えば大腸菌に存在する40種の内在tRNAのいずれとも交差反応しないアミノアシルtRNAシンテターゼと、例えば大腸菌に存在する21種の内在シンテターゼのいずれによってもアミノアシル化されない直交tRNAを含む。
【0072】
本発明は真正細菌(例えば大腸菌)の蛋白質にアルキニルアミノ酸を遺伝的にコード及び組込むための直交対を提供し、直交成分は宿主細胞の翻訳機構の内在大腸菌成分と交差反応しないが、所望非天然アミノ酸を認識し、アンバーナンセンスコドンTAGに応答して蛋白質に組込む。本発明により提供される直交成分としては、Methanococcus jannaschiiチロシルtRNAシンテターゼから誘導される直交アミノアシルtRNAシンテターゼと、突然変異体チロシルtRNACUAアンバーサプレッサーが挙げられる。この系では、突然変異体アミノアシルtRNAシンテターゼはサプレッサーtRNAをpPRO−PHEでアミノアシル化するが、20種の標準アミノ酸ではアミノアシル化しない。
【0073】
本発明はアルキニルアミノ酸を蛋白質に組込むために使用することができる付加直交tRNA−アミノアシルtRNAシンテターゼ対(例えばO−tRNA/O−RS対)の組成物と、前記対の同定及び作製方法を提供する。本発明のO−tRNAはO−tRNAにより認識されるセレクターコドンを含むポリヌクレオチドによりコードされる蛋白質へのアルキニルアミノ酸の例えばin vivoでの組込みを媒介することができる。O−tRNAのアンチコドンループはmRNA上のセレクターコドンを認識し、そのアミノ酸(例えばアルキニルアミノ酸)をポリペプチドのこの部位に組込む。本発明の直交アミノアシルtRNAシンテターゼはそのO−tRNAを特定アルキニルアミノ酸のみで優先的にアミノアシル化(又は負荷)する。
【0074】
例えば、本明細書に実証するように、修飾のために高度に選択的に標的化することができるアルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe;図1A,構造1参照)をセレクターコドン(例えばTAGコドン)に応答して真正細菌細胞(大腸菌;E.coli)の蛋白質に選択的且つ効率的に組込んだ。蛋白質に組込まれると、pPRO−Pheを細胞内で化学的に標的化することができ、例えばアジド基をもつ色素による修飾のために標的化することができる。色素分子のアジド基はアルキニルアミノ酸と反応し、色素標識のために蛋白質を高度に選択的に標的化することができる。
【0075】
アルキニルアミノ酸を蛋白質に部位特異的に組込むことができると、蛋白質の試験を容易にすることができると共に、新規特性をもつ蛋白質の開発が可能になる。例えば、アルキニル含有蛋白質の発現は特異的標識による蛋白質の試験を容易にし、酵素の触媒機能を改変し、蛋白質を他の蛋白質、小分子及び生体分子等と架橋することができる。
直交tRNA、直交アミノアシルtRNAシンテターゼ、及びその対
【0076】
1種以上の非天然アミノ酸を含む蛋白質の作製に適した翻訳系は例えば国際公開WO2002/086075、発明の名称「直交tRNA−アミノアシルtRNAシンテターゼ対を作製するための方法及び組成物(METHODS AND COMPOSITION FOR THE PRODUCTION OF ORTHOGONAL tRNA-AMINOACYL-tRNA SYNTHETASE PAIRS)」;WO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」;WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」;WO2005/019415(出願日2004年7月7日);WO2005/007870(出願日2004年7月7日);及びWO2005/007624(出願日2004年7月7日)に記載されている。これらの各出願はその開示内容全体を参考資料として本明細書に組込む。このような翻訳系は一般に直交tRNA(O−tRNA)と、直交アミノアシルtRNAシンテターゼ(O−RS)と、非天然アミノ酸(本発明ではアルキニルアミノ酸)を含む細胞(例えば大腸菌等の非真核細胞又は酵母等の真核細胞とすることができる)を含み、O−RSはO−tRNAをアルキニルアミノ酸でアミノアシル化する。本発明の直交対はO−tRNA(例えばサプレッサーtRNA、フレームシフトtRNA等)とO−RSを含む。本発明は個々の成分も提供する。
【0077】
一般に、直交対がセレクターコドンを認識し、セレクターコドンに応答してアミノ酸を負荷するとき、直交対はセレクターコドンを「抑圧」すると言う。即ち、翻訳系の(例えば細胞の)内在機構により認識されないセレクターコドンは通常翻訳されないので、非抑圧下で核酸から翻訳されるポリペプチドの生産を阻止することができる。本発明のO−tRNAはセレクターコドンを認識し、本明細書の配列表に記載するようなポリヌクレオチド配列を含むか又は前記配列によりコードされるO−tRNAの抑圧効率に比較してコグネイトシンテターゼの存在下でセレクターコドンに応答して少なくとも例えば約45%、50%、60%、75%、80%、又は90%以上の抑圧効率を含む。O−RSはO−tRNAを該当非天然アミノ酸(例えばアルキニルアミノ酸)でアミノアシル化する。細胞は例えば該当ポリペプチドをコードするポリヌクレオチドを含む核酸を介して成長中のポリペプチド鎖に非天然アミノ酸を組込むためにO−tRNA/O−RS対を使用し、ポリヌクレオチドはO−tRNAにより認識されるセレクターコドンを含む。所定の望ましい側面では、細胞は付加O−tRNA/O−RS対を含むことができ、付加O−tRNAは付加O−RSにより別の非天然アミノ酸を負荷される。例えば、O−tRNAの一方は4塩基コドンを認識することができ、他方は終止コドンを認識することができる。あるいは、複数の異なる終止コドン又は複数の異なる4塩基コドンが異なるセレクターコドンを特異的に認識することができる。
【0078】
本発明の所定態様では、直交tRNA(O−tRNA)と、直交アミノアシルtRNAシンテターゼ(O−RS)と、アルキニルアミノ酸と、該当ポリペプチドをコードするポリヌクレオチドを含む核酸を含む細胞(例えば大腸菌細胞)が提供され、ポリヌクレオチドはO−tRNAにより認識されるセレクターコドンを含む。翻訳系は無細胞系でもよく、例えば各種市販「in vitro」転写/翻訳系の任意のものを本明細書に記載するようなO−tRNA/ORS対及び非天然アミノ酸と併用することができる。
【0079】
1態様では、O−RSとO−tRNAの併用による抑圧効率はO−RSの不在下のO−tRNAの抑圧効率の例えば約5倍、10倍、15倍、20倍、又は25倍以上である。1側面では、O−RSとO−tRNAの併用による抑圧効率は本明細書の配列表に記載するような直交シンテターゼ対の抑圧効率の少なくとも例えば約35%、40%、45%、50%、60%、75%、80%、又は90%以上である。
【0080】
上述のように、本発明は場合により細胞又は他の翻訳系に複数のO−tRNA/O−RS対を含み、2種以上の非天然アミノ酸(例えばアルキニルアミノ酸と別の非天然アミノ酸)を組込むことができる。例えば、細胞は更に別の付加O−tRNA/O−RS対と第2の非天然アミノ酸を含むことができ、この付加O−tRNAは第2のセレクターコドンを認識し、この付加O−RSはO−tRNAを第2の非天然アミノ酸で優先的にアミノアシル化する。例えば、O−tRNA/O−RS対(O−tRNAは例えばアンバーセレクターコドンを認識する)を含む細胞は更に第2の直交対(例えばロイシル、リシル、グルタミル等)を含むことができる(第2のO−tRNAは別のセレクターコドン、例えばオパールコドン、4塩基コドン等を認識する)。各直交対は異なるセレクターコドンの認識を助長できるように異なる起源に由来することが望ましい。
【0081】
O−tRNA及び/又はO−RSは天然に存在するものでもよいし、例えば種々の生物の任意のものに由来するtRNAのライブラリー及び/又はRSのライブラリーを作製するか及び/又は種々の利用可能な突然変異ストラテジーの任意のものを使用することにより、天然に存在するtRNA及び/又はRSの突然変異により誘導してもよい。例えば、直交tRNA/アミノアシルtRNAシンテターゼ対を作製する1ストラテジーは例えば宿主細胞以外の起源又は多重起源に由来する(宿主に対して)異種のtRNA/シンテターゼ対を宿主細胞に導入する方法である。異種シンテターゼ候補の特性としては、例えば宿主細胞tRNAに負荷しないことが挙げられ、異種tRNA候補の特性としては、例えば宿主細胞シンテターゼによりアミノアシル化されないことが挙げられる。更に、異種tRNAは全宿主細胞シンテターゼに対して直交性である。
【0082】
直交対を作製するための第2のストラテジーはO−tRNA又はO−RSをスクリーニング及び/又は選択するための突然変異体ライブラリーを作製する方法である。これらのストラテジーを組み合わせてもよい。
直交tRNA(O−tRNA)
【0083】
本発明の直交tRNA(O−tRNA)はO−tRNAにより認識されるセレクターコドンを含むポリヌクレオチドによりコードされる蛋白質への非天然アミノ酸(例えばアルキニルアミノ酸)の例えばin vivo又はin vitroでの組込みを媒介することが望ましい。所定態様では、本発明のO−tRNAは本明細書の配列表のO−tRNA配列に記載するようなポリヌクレオチド配列を含むか又は前記配列によりコードされるO−tRNAに比較してコグネイトシンテターゼの存在下でセレクターコドンに応答して少なくとも例えば約45%、50%、60%、75%、80%、又は90%以上の抑圧効率を含む。
【0084】
抑圧効率は当分野で公知の多数のアッセイの任意のものにより測定することができる。例えば、β−ガラクトシダーゼレポーターアッセイを使用することができ、例えば本発明のO−tRNAを含むプラスミドと共に修飾lacZプラスミド(構築物はlacZ核酸配列中にセレクターコドンをもつ)を適当な生物(例えば直交成分を使用することができる生物)に由来する細胞に導入する。コグネイトシンテターゼも(ポリペプチド又は発現されるとコグネイトシンテターゼをコードするポリヌクレオチドとして)導入することができる。細胞を培地で所望密度(例えばOD600=約0.5)まで増殖させ、例えばBetaFluor(登録商標)β−ガラクトシダーゼアッセイキット(Novagen)を使用してβ−ガラクトシダーゼアッセイを実施する。比較可能な対照(例えば所望位置にセレクターコドンではなく対応するセンスコドンをもつ修飾lacZ構築物から観測される値)に対するサンプルの活性百分率として抑圧百分率を計算することができる。
【0085】
本発明のO−tRNAの例を本明細書の配列表に記載する。代表的O−tRNA及びO−RS分子の配列については本明細書の表、実施例及び図面も参照。更に、本明細書の「核酸及びポリペプチド配列と変異体」のセクションも参照。O−RS mRNA又はO−tRNA分子等のRNA分子では、所与配列又はその相補配列に対してチミン(T)がウラシル(U)で置換されている(コーディングDNAでは逆)。塩基に付加修飾を加えてもよい。
【0086】
本発明は本明細書に記載する特定O−tRNAに対応するO−tRNAの保存変異体も含む。例えば、O−tRNAの保存変異体としては、例えば本明細書の配列表に記載するような特定O−tRNAと同様に機能し、適当な自己相補性によりtRNA L形構造を維持するが、例えば本明細書の配列表、図面又は実施例に記載する配列と同一配列をもたない分子が挙げられる(更に野生型tRNA分子以外のものであることが望ましい)。本明細書の「核酸及びポリペプチド配列と変異体」のセクションも参照。
【0087】
O−tRNAを含む組成物は更に直交アミノアシルtRNAシンテターゼ(O−RS)を含むことができ、O−RSはO−tRNAを非天然アミノ酸(例えばアルキニルアミノ酸)で優先的にアミノアシル化する。所定態様では、O−tRNAを含む組成物は更に(例えばin vitro又はin vivo)翻訳系を含むことができる。該当ポリペプチドをコードし、O−tRNAにより認識されるセレクターコドンを含むポリヌクレオチドを含む核酸、又はこれらの1種以上の組み合わせも細胞に加えることができる。本明細書の「直交アミノアシルtRNAシンテターゼ」のセクションも参照。
【0088】
直交tRNA(O−tRNA)の作製方法も本発明の特徴である。本方法により作製されたO−tRNAも本発明の特徴である。本発明の所定態様では、O−tRNAは突然変異体ライブラリーを作製することにより作製することができる。突然変異体tRNAのライブラリーは当分野で公知の種々の突然変異誘発技術を使用して作製することができる。例えば、突然変異体tRNAは部位特異的突然変異、ランダム点突然変異、相同組換え、DNAシャフリング又は他の帰納的突然変異誘発法、キメラ構築又はその任意組み合わせにより作製することができる。
【0089】
tRNAの所望ループ又は領域(例えばアンチコドンループ、受容体ステム、Dアーム又はループ、可変ループ、TPCアーム又はループ、tRNA分子の他の領域又はその組合せ)の特定位置(例えば非保存位置又は保存位置)、ランダム位置又は両者の組合せに付加突然変異を導入することができる。一般に、tRNAの突然変異としてはセレクターコドンの認識を可能にするように突然変異体tRNAライブラリーの各メンバーのアンチコドンループを突然変異させる方法が挙げられる。この方法は更にO−tRNAの末端に付加配列(CCA)を付加する段階を含むことができる。一般に、O−tRNAは所望RSに対するその親和性等を維持しながら、出発材料(例えば複数のtRNA配列)に比較して所望生物に対する直交性が改善されている。
【0090】
前記方法は場合によりtRNA及び/又はアミノアシルtRNAシンテターゼの配列の類似性(及び/又は推定相同性)を分析し、特定生物について直交性であると思われるO−tRNA、O−RS及び/又はその対の潜在候補を決定する段階を含む。分析には、当分野で公知であり、本明細書に記載するコンピュータープログラム(例えばBLAST及びpileupプログラム)を使用することができる。1例では、大腸菌で使用するための潜在的直交翻訳成分を選択するためには、真正細菌生物に密接な配列類似性を示さないシンテターゼ及び/又はtRNAを選択する。
【0091】
一般に、O−tRNAは複数の潜在的O−tRNAのメンバーを含む第1の種の細胞の集団に例えばネガティブ選択を実施することにより得られる。ネガティブ選択は細胞に内在性のアミノアシルtRNAシンテターゼ(RS)によりアミノアシル化される潜在的O−tRNAライブラリーのメンバーを含む細胞を排除する。こうして、第1の種の細胞に直交性のtRNAプールが得られる。
【0092】
所定態様において、ネガティブ選択では、ネガティブ選択マーカー(例えば抗生物質耐性を付与する酵素(例えばβ−ラクタマーゼ)、検出可能な産物を付与する酵素(例えばβ−ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT))、例えば毒性物質(例えばバルナーゼ))をコードするポリヌクレオチドの(例えば機能的バルナーゼを依然として産生する)非必須位置等にセレクターコドンを導入する。場合により選択物質(例えばアンピシリン等の抗生物質)の存在下で細胞集団を増殖させることによりスクリーニング/選択を実施する。1態様では、選択物質の濃度を変動させる。
【0093】
例えば、サプレッサーtRNAの活性を測定するためには、セレクターコドンのin vivo抑圧(例えばネガティブ選択マーカー(例えばβ−ラクタマーゼ遺伝子(bla))をコードするポリヌクレオチドに導入されたナンセンス又はフレームシフト突然変異)に基づく選択システムを使用する。例えば、所定位置(例えばA184)にセレクターコドンをもつポリヌクレオチド変異体(例えばbla変異体)を構築する。細胞(例えば細菌)をこれらのポリヌクレオチドで形質転換する。内在大腸菌シンテターゼにより効率的に負荷することができない直交tRNAの場合には、抗生物質耐性(例えばアンピシリン耐性)はプラスミドで形質転換されていない細菌と同等以下のはずである。tRNAが非直交性の場合、又はtRNAに負荷することが可能な異種シンテターゼをシステムで同時発現させる場合には、より高レベルの抗生物質(例えばアンピシリン)耐性が観測される。プラスミドで形質転換されていない細胞と同等の抗生物質濃度のLB寒天プレートで増殖することができない細胞(例えば細菌)を選択する。
【0094】
毒性物質(例えばリボヌクレアーゼ又はバルナーゼ)の場合には、複数の潜在的tRNAのメンバーが内在宿主(例えば大腸菌)シンテターゼによりアミノアシル化されるとき(即ち、宿主(例えば大腸菌)シンテターゼに非直交性であるとき)には、セレクターコドンは抑圧され、生産される毒性ポリヌクレオチド産物は細胞死を生じる。直交tRNA又は非機能的tRNAを含む細胞は生存する。
【0095】
1態様では、所望生物に直交性のtRNAプールに次にポジティブ選択を実施し、セレクターコドンを例えばβ−ラクタマーゼ遺伝子等の薬剤耐性遺伝子によりコードされるポジティブ選択マーカーに配置する。ポジティブ選択は細胞に直交性のtRNAプールのメンバーをコードするか又は前記メンバーを含むポリヌクレオチドと、ポジティブ選択マーカーをコードするポリヌクレオチドと、コグネイトRSをコードするポリヌクレオチドを含む細胞で実施される。所定態様では、第2の細胞集団はネガティブ選択により排除されなかった細胞を含む。ポリヌクレオチドを細胞で発現させ、選択物質(例えばアンピシリン)の存在下で細胞を増殖させる。次に、同時発現させたコグネイトシンテターゼによりアミノアシル化され、このセレクターコドンに応答してアミノ酸を挿入することができるtRNAを選択する。一般に、これらの細胞は非機能的tRNA、又は該当シンテターゼにより効率的に認識することができないtRNAを含む細胞に比較して高い抑圧効率を示す。非機能的tRNA又は該当シンテターゼにより効率的に認識されないtRNAを含む細胞は抗生物質に感受性である。従って、(i)内在宿主(例えば大腸菌)シンテターゼの基質ではなく;(ii)該当シンテターゼによりアミノアシル化することができ;(iii)翻訳で機能的なtRNAが両者選択後に生存している。
【0096】
従って、スクリーニングされる状況に応じて同一マーカーがポジティブマーカーにもネガティブマーカーにもなる。即ち、マーカーがポジティブスクリーニングされる場合にはポジティブマーカーであり、ネガティブスクリーニングされる場合にはネガティブマーカーである。
【0097】
上記方法における選択(例えばポジティブ選択、ネガティブ選択又はポジティブ選択とネガティブ選択の両者)のストリンジェンシーは場合により選択ストリンジェンシーの変動を含む。例えば、バルナーゼは極毒性蛋白質であるので、異なる数のセレクターコドンをバルナーゼ遺伝子に導入するか及び/又は誘導プロモーターを使用することによりネガティブ選択のストリンジェンシーを制御することができる。別の例では、選択又はスクリーニング物質の濃度(例えばアンピシリン濃度)を変動させる。本発明の1側面では、初期ラウンド中の所望活性は低いと思われるのでストリンジェンシーを変動させる。即ち、初期ラウンドでは低ストリンジェンシー選択条件を適用し、後期ラウンドの選択では高ストリンジェンシー条件を適用する。所定態様では、ネガティブ選択、ポジティブ選択又はネガティブ選択とポジティブ選択の両方を複数回反復する。複数の異なるネガティブ選択マーカー、ポジティブ選択マーカー又はネガティブ選択マーカーとポジティブ選択マーカーの両方を使用することができる。所定態様では、ポジティブ選択マーカーとネガティブ選択マーカーを同一にすることができる。
【0098】
本発明ではセレクターコドンに応答して非天然アミノ酸(例えばアルキニルアミノ酸)を負荷する直交翻訳成分(例えばO−tRNA、O−RS、及びO−tRNA/O−RS対)を作製するために他の型の選択/スクリーニングを使用することもできる。例えば、ネガティブ選択マーカー、ポジティブ選択マーカー又はポジティブ選択マーカーとネガティブ選択マーカーの両方として、適切な反応体の存在下で蛍光発光するか又は発光反応を触媒するマーカーを使用することができる。別の態様では、蛍光活性化細胞ソーティング(FACS)又は発光によりマーカーの産物を検出する。場合により、マーカーはアフィニティースクリーニングマーカーを含む。Francisco, J. A. ら,(1993) Production and fluorescence-activated cell sorting of Escherichia coli expressing a functional antibody fragment on the external surface. Proc Natl Acad Sci USA. 90:10444-8も参照。
【0099】
組換え直交tRNAの他の作製方法も例えば国際出願公開WO2002/086075、発明の名称「直交tRNA−アミノアシルtRNAシンテターゼ対を作製するための方法及び組成物(METHODS AND COMPOSITIONS FOR THE PRODUCTION OF ORTHOGONAL tRNA AMINOACYL-tRNA SYNTHETASE PAIRS)」;WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」;及びWO2005/019415(出願日2004年7月7日)に記載されている。Forsterら, (2003) Programming peptidomimetic synthetases by translating genetic codes designed de novo PNAS 100 (11):6353-6357; 及びFengら, (2003), Expanding tRNA recognition of a tRNA synthetase by a single amino acid change, PNAS 100 (10):5676-5681も参照。
【0100】
直交アミノアシルtRNAシンテターゼ(O−RS)
本発明のO−RSはO−tRNAを非天然アミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン等のアルキニルアミノ酸)で優先的にin vitro又はin vivoでアミノアシル化する。本発明のO−RSはO−RSを含むポリペプチド及び/又はO−RS又はその一部をコードするポリヌクレオチドにより翻訳系(例えば細胞)に提供することができる。例えば、O−RSの1例は本明細書の配列表と実施例に記載するようなアミノ酸配列、又はその保存変異体を含む。別の例では、O−RS、又はその一部は本明細書の配列表又は実施例に記載する配列を含むアミノ酸をコードするポリヌクレオチド配列、又はその相補的ポリヌクレオチド配列によりコードされる。例えば、代表的O−RS分子の配列については下記表及び実施例参照。本明細書の「核酸及びポリペプチド配列と変異体」のセクションも参照。
【0101】
直交アミノアシルtRNAシンテターゼ(O−RS)、例えばO−tRNAと併用するためのO−RSの同定方法も本発明の特徴である。例えば、1方法は第1の種の細胞集団について選択(例えばポジティブ選択)を実施する段階を含み、前記細胞は1)複数のアミノアシルtRNAシンテターゼ(RS)のメンバー(例えば複数のRSは突然変異体RS、第1の種以外の種に由来するRS又は突然変異体RSと第1の種以外の種に由来するRSの両方を含むことができる)と;2)(例えば1種以上の種に由来する)直交tRNA(O−tRNA)と;3)(例えばポジティブ)選択マーカーをコードし、少なくとも1個のセレクターコドンを含むポリヌクレオチドを各々含む。複数のRSのメンバーを含まないか又はその量の少ない細胞に比較して抑圧効率の高い細胞について細胞を選択又はスクリーニングする。抑圧効率は当分野で公知の技術及び本明細書に記載するように測定することができる。抑圧効率の高い細胞はO−tRNAをアミノアシル化する活性RSを含む。第1の種に由来する第1組のtRNAの活性RSによる(in vitro又はin vivo)アミノアシル化レベルを第2の種に由来する第2組のtRNAの活性RSによる(in vitro又はin vivo)アミノアシル化レベルと比較する。アミノアシル化レベルは検出可能な物質(例えば標識アミノ酸又は非天然アミノ酸、例えば標識パラ−プロパルギルオキシフェニルアラニン)により測定することができる。第1組のtRNAに比較して第2組のtRNAをより効率的にアミノアシル化する活性RSを一般に選択することにより、O−tRNAと併用するための効率的(最適化)直交アミノアシルtRNAシンテターゼが得られる。この方法により同定されたO−RSも本発明の特徴である。
【0102】
アミノアシル化を測定するためには多数のアッセイの任意のものを使用することができる。これらのアッセイはin vitro又はin vivoで実施することができる。例えば、in vitroアミノアシル化アッセイは例えばHoben and Soll(1985)Methods Enzymol.113:55-59に記載されている。アミノアシル化は直交翻訳成分と共にレポーターを使用し、蛋白質をコードする少なくとも1個のセレクターコドンを含むポリヌクレオチドを発現する細胞でレポーターを検出することにより測定することもできる。WO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」;及びWO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」も参照。
【0103】
同定されたO−RSを更に操作し、所望非天然アミノ酸(例えばアルキニルアミノ酸)のみをO−tRNAに負荷し、20種の標準アミノ酸を負荷しないようにシンテターゼの基質特異性を改変することができる。非天然アミノ酸に対して基質特異性をもつ直交アミノアシルtRNAシンテターゼの作製方法は例えばシンテターゼの活性部位、シンテターゼの編集メカニズム部位、各種シンテターゼドメインを組み合わせることによる各種部位等でシンテターゼを突然変異させる段階と、選択プロセスを適用する段階を含む。ポジティブ選択後にネガティブ選択を行う併用に基づくストラテジーを使用する。ポジティブ選択では、ポジティブマーカーの非必須位置に導入したセレクターコドンが抑圧されると、細胞はポジティブ選択圧下で生存する。従って、天然及び非天然アミノ酸両者の存在下で生存細胞は直交サプレッサーtRNAに天然又は非天然アミノ酸を負荷する活性シンテターゼをコードする。ネガティブ選択では、ネガティブマーカーの非必須位置に導入したセレクターコドンが抑圧されると、天然アミノ酸特異性をもつシンテターゼは除去される。ネガティブ選択とポジティブ選択の生存細胞は直交サプレッサーtRNAを非天然アミノ酸のみでアミノアシル化(負荷)するシンテターゼをコードする。その後、これらのシンテターゼを例えばDNAシャフリング又は他の帰納的突然変異誘発法により更に突然変異誘発することができる。
【0104】
突然変異体O−RSのライブラリーは当分野で公知の種々の突然変異誘発技術を使用して作製することができる。例えば、突然変異体RSは部位特異的突然変異、ランダム点突然変異、相同組換え、DNAシャフリング又は他の帰納的突然変異誘発法、キメラ構築又はその任意組み合わせにより作製することができる。例えば、突然変異体RSのライブラリーは2種以上の他の例えばサイズとダイバーシティーの小さい「サブライブラリー」から作製することができる。RSのキメラライブラリーも本発明に含まれる。なお、場合により各種生物(例えば真正細菌又は古細菌等の微生物)に由来するtRNAシンテターゼのライブラリー(例えば天然ダイバーシティーを含むライブラリー)(例えば米国特許第6,238,884号(Shortら);米国特許第5,756,316号(Schallenbergerら);米国特許第5,783,431号(Petersenら);米国特許第5,824,485号(Thompsonら);米国特許第5,958,672号(Shortら)参照)を構築し、直交対についてスクリーニングする。
【0105】
シンテターゼにポジティブ及びネガティブ選択/スクリーニングストラテジーを実施した後、これらのシンテターゼを更に突然変異誘発することができる。例えば、O−RSをコードする核酸を単離することができ;(例えばランダム突然変異誘発、部位特異的突然変異誘発、組換え又はその任意組み合わせにより)突然変異O−RSをコードする1組のポリヌクレオチドを核酸から作製することができ;O−tRNAを非天然アミノ酸(例えばアルキニルアミノ酸)で優先的にアミノアシル化する突然変異O−RSが得られるまでこれらの個々の段階又はこれらの段階の組み合わせを繰り返すことができる。本発明の1側面では、前記段階を複数回、例えば少なくとも2回実施する。
【0106】
O−tRNA、O−RS、又はその対を作製するための本発明の方法では、付加レベルの選択/スクリーニングストリンジェンシーを使用することもできる。選択又はスクリーニングストリンジェンシーはO−RSを作製するための方法の一方又は両方の段階で変動させることができる。これは例えば選択/スクリーニング物質の使用量等の変動とすることができる。付加ラウンドのポジティブ及び/又はネガティブ選択を実施することもできる。選択又はスクリーニングは更にアミノ酸浸透率の変化、翻訳効率の変化、翻訳忠実度の変化等の1種以上を含むこともできる。一般に、1種以上の変化は蛋白質を生産するために直交tRNA−tRNAシンテターゼ対を使用する生物における1個以上の遺伝子の突然変異に基づく。
【0107】
O−RSの作製と、シンテターゼの基質特異性の改変に関するその他の一般的な詳細についてはWO2002/086075、発明の名称「直交tRNA−アミノアシルtRNAシンテターゼ対を作製するための方法及び組成物(METHODS AND COMPOSITIONS FOR THE PRODUCTION OF ORTHOGONAL tRNA AMINOACYL-tRNA SYNTHETASE PAIRS)」;及びWO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」に記載されている。
【0108】
資源及び宿主生物
本発明の直交翻訳成分(O−tRNAとO−RS)はO−tRNA/O−RS成分と宿主系が直交に作用するという条件で他の任意種に由来する宿主翻訳系で使用するために任意生物(又は生物組み合わせ)に由来することができる。O−tRNAとO−RSは同一生物に由来する必要はない。1側面では、直交成分は真正細菌宿主系で使用するために古細菌遺伝子(即ち古細菌)に由来することができる。
【0109】
例えば、直交O−tRNAは古細菌生物、例えばMethanococcus jannaschii、Methanobacterium thermoautotrophicum、Halobacterium(例えばHaloferax volcanii及びHalobacterium種NRC−1)、Archaeoglobus fulgidus、Pyrococcus furiosus、Pyrococcus horikoshii、Aeuropyrum pernix、Methanococcus maripaludis、Methanopyrus kandleri、Methanosarcina mazei(Mm)、Pyrobaculum aerophilum、Pyrococcus abyssi、Sulfolobus solfataricus(Ss)、Sulfolobus tokodaii、Thermoplasma acidophilum、Thermoplasma volcanium等の古細菌や、Escherichia coli,Thermus thermophilus,Bacillus stearothermphilus等の真正細菌に由来することができ、直交O−RSは生物又は生物組み合わせ、例えばMethanococcus jannaschii、Methanobacterium thermoautotrophicum、Halobacterium(例えばHaloferax volcanii及びHalobacterium種NRC−1)、Archaeoglobus fulgidus、Pyrococcus furiosus、Pyrococcus horikoshii、Aeuropyrum pernix、Methanococcus maripaludis、Methanopyrus kandleri、Methanosarcina mazei、Pyrobaculum aerophilum、Pyrococcus abyssi、Sulfolobus solfataricus、Sulfolobus tokodaii、Thermoplasma acidophilum、Thermoplasma volcanium等の古細菌や、Escherichia coli,Thermus thermophilus,Bacillus stearothermphilus等の真正細菌に由来することができる。1態様では、例えば植物、藻類、原生動物、真菌類、酵母、動物(例えば哺乳動物、昆虫、節足動物等)等の真核資源もO−tRNA及びO−RS資源として使用することができる。
【0110】
O−tRNA/O−RS対の個々の成分は同一生物に由来するものでも異なる生物に由来するものでもよい。1態様では、O−tRNA/O−RS対は同一生物に由来する。あるいは、O−tRNA/O−RS対のO−tRNAとO−RSは異なる生物に由来する。
【0111】
O−tRNA、O−RS又はO−tRNA/O−RS対はin vivo又はin vitroで選択又はスクリーニングすることができ、及び/又はアルキニルアミノ酸を組込んだポリペプチドを作製するために細胞(例えば真正細菌細胞)で使用することができる。使用される真正細菌細胞は限定されないが、例えばEscherichia coli、Thermus thermophilus、Bacillus stearothermphilus等である。本発明の翻訳成分を含む真正細菌細胞の組成物も本発明の特徴である。
【0112】
別の種で使用するためにある種でO−tRNA及び/又はO−RSをスクリーニングする方法については、国際出願公開WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」(出願日2004年4月16日)も参照。
【0113】
セレクターコドン
本発明のセレクターコドンは蛋白質生合成機構の遺伝コドン枠を拡張する。例えば、セレクターコドンとしては例えばユニーク3塩基コドン、ナンセンスコドン(例えばアンバーコドン(UAG)又はオパールコドン(UGA)等の終止コドン)、非天然コドン、少なくとも4塩基のコドン、レアコドン等が挙げられる。例えば1個以上、2個以上、3個以上等の多数のセレクターコドンを所望遺伝子に導入することができる。複数の異なるセレクターコドンを使用することにより、これらの異なるセレクターコドンを使用して(例えば少なくとも1種のアルキニルアミノ酸を含む)複数の非天然アミノ酸の同時部位特異的組込みを可能にする複数の直交tRNA/シンテターゼ対を使用することができる。
【0114】
1態様では、本方法はアルキニルアミノ酸を細胞にin vivo組込むために終止コドンであるセレクターコドンを使用する。例えば、終止コドンを認識し、O−RSによりアルキニルアミノ酸でアミノアシル化されるO−tRNAを作製する。このO−tRNAは天然に存在する翻訳系のアミノアシルtRNAシンテターゼにより認識されない。慣用部位特異的突然変異誘発法を使用して該当ポリペプチドをコードするポリヌクレオチドの該当部位に終止コドンを導入することができる。例えばSayers,J.R.ら,(1988),5’,3’Exonuclease in phosphorothioate−based oligonucleotide−directed mutagenesis.Nucleic Acids Res.791−802参照。O−RS、O−tRNA及び該当ポリペプチドをコードする核酸を例えばin vivoで併用すると、終止コドンに応答してアルキニルアミノ酸が組込まれ、特定位置にアルキニル活性アミノ酸を含むポリペプチドが得られる。本発明の1態様では、セレクターコドンとして使用される終止コドンはアンバーコドンUAG及び/又はオパールコドンUGAである。1例では、セレクターコドンとしてUAGとUGAの両者を使用する遺伝子コードは存在度が最も高い終結シグナルであるオーカーナンセンスコドンUAAを保存しながら22種のアミノ酸をコードすることができる。
【0115】
アルキニル活性アミノ酸のin vivo組込みは宿主細胞をさほど撹乱せずに実施することができる。例えば、大腸菌等の非真核細胞では、UAGコドンの抑圧効率はO−tRNA(例えばアンバーサプレッサーtRNA)と(UAGコドンと結合してリボソームから成長中のペプチドの放出を開始する)放出因子1(RF1)の競合に依存するので、例えばO−tRNA(例えばサプレッサーtRNA)の発現レベルを増加するか又はRF1欠損株を使用することにより抑圧効率を調節することができる。真核細胞では、UAGコドンの抑圧効率はO−tRNA(例えばアンバーサプレッサーtRNA)と(終止コドンと結合してリボソームから成長中のペプチドの放出を開始する)真核放出因子1(例えばeRF)の競合に依存するので、例えばO−tRNA(例えばサプレッサーtRNA)の発現レベルを増加することにより抑圧効率を調節することができる。更に、付加化合物、例えばジチオスレイトール(DTT)等の還元剤も加えてもよい。
【0116】
アルキニルアミノ酸はレアコドンでコードすることもできる。例えば、in vitro蛋白質合成反応でアルギニン濃度を下げると、レアアルギニンコドンAGGはアラニンでアシル化された合成tRNAによるAlaの挿入に有効であることが分かっている。例えばMaら, Biochemistry, 32:7939(1993)参照。この場合には、合成tRNAは大腸菌に少量種として存在する天然tRNAArgと競合する。更に、生物によっては全三重項コドンを使用しないものもある。Micrococcus luteusで割り当てられないコドンAGAがin vitro転写/翻訳抽出物へのアミノ酸挿入に使用されている。例えばKowal and Oliver, Nucl. Acid. Res., 25:4685 (1997)参照。本発明の成分はこれらのレアコドンをin vivo使用するために作製することができる。
【0117】
セレクターコドンは更に拡張コドン(例えば4、5、6塩基以上のコドン等の4塩基以上のコドン)も含むことができる。4塩基コドンの例としては例えばAGGA、CUAG、UAGA、CCCU等が挙げられる。5塩基コドンの例としては例えばAGGAC、CCCCU、CCCUC、CUAGA、CUACU、UAGGC等が挙げられる。本発明の方法はフレームシフト抑圧に基づく拡張コドンの使用を含む。4塩基以上のコドンは例えば1又は複数の非天然アミノ酸(例えばアルキニルアミノ酸)を同一蛋白質に挿入することができる。他の態様では、アンチコドンループは例えば少なくとも4塩基コドン、少なくとも5塩基コドン、又は少なくとも6塩基コドン又はそれ以上をデコードすることができる。4塩基コドンは256種が考えられるので、4塩基以上のコドンを使用すると同一細胞で複数の非天然アミノ酸をコードすることができる。Andersonら, (2002) Exploring the Limits of Codon and Anticodon Size,Chemistry and Biology, 9:237-244; 及びMagliery, (2001) Expanding the Genetic Code:Selection of Efficient Suppressors of Four-base Codons and Identification of “Shifty” Four-base Codons with a Library Approach in Escherichia coli, J. Mol. Biol. 307:755-769 も参照。
【0118】
例えば、in vitro生合成法を使用して非天然アミノ酸を蛋白質に組込むために4塩基コドンが使用されている。例えばMaら, (1993) Biochemistry, 32, 7939;及びHohsakaら, (1999) J. Am. Chem. Soc., 121:34 参照。2個の化学的にアシル化されたフレームシフトサプレッサーtRNAを用いて2−ナフチルアラニンとリジンのNBD誘導体をストレプトアビジンに同時にin vitroで組込むためにCGGGとAGGUが使用されている。例えばHohsakaら, (1999) J. Am. Chem. Soc., 121:12194参照。in vivo試験では、MooreらはNCUAアンチコドンをもつtRNALeu誘導体がUAGNコドン(NはU、A、G又はCであり得る)を抑圧する能力を試験し、四重項UAGAはUCUAアンチコドンをもつtRNALeuにより13〜26%の効率でデコードすることができるが、0又は−1フレームでは殆どデコードできないことを見出した。Mooreら, (2000) J. Mol. Biol., 298:195 参照。1態様では、レアコドン又はナンセンスコドンに基づく拡張コドンを本発明で使用し、他の望ましくない部位でのミスセンス読み飛ばしとフレームシフト抑圧を減らすことができる。
【0119】
所与系では、セレクターコドンは更に天然3塩基コドンの1種を含むことができ、内在系はこの天然塩基コドンを使用しない(又は殆ど使用しない)。例えば、天然3塩基コドンを認識するtRNAをもたない系、及び/又は3塩基コドンがレアコドンである系がこれに該当する。
【0120】
セレクターコドンは場合により非天然塩基対を含む。これらの非天然塩基対は更に既存遺伝子アルファベットを拡張する。塩基対が1個増えると、トリプレットコドン数は64から125に増す。第3の塩基対の性質としては安定的且つ選択的な塩基対合、高い忠実度でポリメラーゼによるDNAへの効率的な酵素組込み、及び未完成非天然塩基対の合成後の効率的な持続的プライマー伸長が挙げられる。方法と組成物に適応可能な非天然塩基対については、例えばHiraoら, (2002) An unnatural base pair for incorporating amino acid analogues into protein, Nature Biotechnology, 20:177-182 に記載されている。Wu, Y. ら,(2002) J. Am. Chem. Soc. 124:14626-14630も参照。他の関連文献は以下に挙げる。
【0121】
in vivo使用では、非天然ヌクレオシドは膜透過性であり、リン酸化され、対応する三リン酸塩を形成する。更に、増加した遺伝情報は安定しており、細胞内酵素により破壊されない。Bennerらによる従来の報告はカノニカルワトソンクリック対とは異なる水素結合パターンを利用しており、そのうちで最も注目される例はイソC:イソG対である。例えばSwitzerら, (1989)J. Am. Chem. Soc., 111:8322;及びPiccirilliら, (1990) Nature, 343:33; Kool, (2000) Curr. Opin. Chem. Biol., 4:602参照。これらの塩基は一般に天然塩基とある程度まで誤対合し、酵素複製することができない。Koolらは水素結合を塩基間の疎水性パッキング相互作用に置き換えることにより塩基対の形成を誘導できることを立証した。Kool, (2000) Curr. Opin. Chem. Biol., 4:602;及びGuckian and Kool, (1998) Angew. Chem. Int. Ed. Engl., 36, 2825参照。上記全要件を満足する非天然塩基対を開発する目的でSchultz, Romerbergらは一連の非天然疎水性塩基を体系的に合成し、試験した。PICS:PICS自己対は天然塩基対よりも安定しており、大腸菌DNAポリメラーゼIのKlenowフラグメント(KF)によりDNAに効率的に組込むことができる。例えばMcMinnら, (1999) J. Am. Chem. Soc., 121:11586;及びOgawaら, (2000) J. Am. Chem. Soc., 122:3274参照。生体機能に十分な効率と選択性でKFにより3MN:3MN自己対を合成することができる。例えばOgawaら, (2000) J. Am. Chem. Soc., 122:8803参照。しかし、どちらの塩基も後期複製用チェーンターミネーターとして作用するものである。PICS自己対を複製するために使用できる突然変異体DNAポリメラーゼが最近開発された。更に、7AI自己対も複製することができる。例えばTaeら, (2001) J. Am. Chem. Soc., 123:7439参照。Cu(II)と結合すると安定な対を形成する新規メタロ塩基対Dipic:Pyも開発された。Meggersら, (2000) J. Am. Chem. Soc., 122:10714参照。拡張コドンと非天然コドンは天然コドンに本質的に直交性であるので、本発明の方法は天然コドンに直交性のtRNAを作製するためにこの性質を利用することができる。
【0122】
翻訳バイパス系を使用してアルキニルアミノ酸を所望ポリペプチドに組込むこともできる。1翻訳バイパス系では、大きい配列が遺伝子に挿入されるが、蛋白質に翻訳されない。この配列はリボソームに配列を飛び越させて挿入の下流の翻訳を再開するための合図として機能する構造を含む。
【0123】
非天然アミノ酸
本明細書で使用する非天然アミノ酸とはセレノシステイン及び/又はピロリジンと20種の遺伝的にコードされる以下のα−アミノ酸、即ちアラニン、アルギニン、アスパラギン、アスパラギン酸、システイン、グルタミン、グルタミン酸、グリシン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、プロリン、セリン、スレオニン、トリプトフアン、チロシン、バリン以外の任意アミノ酸、修飾アミノ酸又はアミノ酸類似体を意味する。α−アミノ酸の一般構造は式I:
【化2】
により表される。
【0124】
非天然アミノ酸は一般に式Iをもつ任意構造であり、式中、R基は20種の天然アミノ酸で使用されている以外の任意置換基である。20種の天然アミノ酸の構造については、例えばL. Stryer著Biochemistry, 第3版, 1988, Freeman and Company, New York参照。なお、本発明の非天然アミノ酸は上記20種のα−アミノ酸以外の天然化合物でもよい。
【0125】
本発明の非天然アミノ酸は一般に側鎖が天然アミノ酸と異なるので、天然蛋白質と同様に他のアミノ酸(例えば天然又は非天然アミノ酸)とアミド結合を形成する。一方、非天然アミノ酸は天然アミノ酸と異なる側鎖基をもつ。
【0126】
本発明では反応性アルキニル基を含む非天然アミノ酸(例えばアジド部分と特異的且つ位置選択的に反応するアルキン部分をもつ非天然アミノ酸)が特に重要である。例えば、アルキニルアミノ酸では、式IにおけるRは任意アルキン含有構造を含む。例えば、パラ−プロパルギルオキシフェニルアラニン(略称pPRO−Phe;図1A参照)が本発明で利用される所望非天然アルキニルアミノ酸である。本発明はpPRO−Pheと直交翻訳成分の併用に限定するものではない。例えば、他の各種アルキニルアミノ酸(図8A及び8B参照)が考えられ、限定されないが、例えば、
2−アミノ−4−ペンチン酸、
2−アミノ−3−(4−エチニルフェニル)プロパン酸、
2−アミノ−3−[4−(プロプ−2−イニル)フェニル]プロパン酸、
2−アミノ−3−(プロプ−2−イニルオキシ)プロパン酸、
2−アミノ−3−(プロプ−2−イニルチオ)プロパン酸、
3−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノプロパン酸、
4−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノブタン酸が挙げられる。
【0127】
他の非天然アミノ酸では、例えば、式IにおけるRは場合によりアルキル、アリール、アシル、ヒドラジン、シアノ、ハロ、ヒドラジド、アルケニル、エーテル、硼酸、ボロン酸、ホスホ、ホスホノ、ホスフィン、エノン、イミン、エステル、ヒドロキシルアミン、アミン基等又はその任意組合せを含む。他の該当非天然アミノ酸としては限定されないが、光架橋基をもつアミノ酸、スピン標識アミノ酸、蛍光アミノ酸、金属結合性アミノ酸、金属含有アミノ酸、放射性アミノ酸、新規官能基をもつアミノ酸、他の分子と共有又は非共有的に相互作用するアミノ酸、フォトケージド及び/又は光異性化可能なアミノ酸、ビオチン又はビオチン類似体を含有するアミノ酸、ケト含有アミノ酸、グリコシル化アミノ酸、アミノ酸側鎖と結合した糖部分、ポリエチレングリコール又はポリエーテルを含むアミノ酸、重原子置換アミノ酸、化学分解性又は光分解性アミノ酸、天然アミノ酸に比較して延長側鎖(例えばポリエーテル又は例えば約5もしくは約10炭素長を上回る長鎖炭化水素等)をもつアミノ酸、炭素結合糖含有アミノ酸、アミノチオ酸含有アミノ酸、及び1個以上の毒性部分を含むアミノ酸が挙げられる。
【0128】
別の側面では、本発明は下式IV:
【化3】
により表される一般構造をもつアルキニルアミノ酸を提供する。
【0129】
この構造をもつアルキンアミノ酸は一般にR1が20種の天然アミノ酸の1種で使用される置換基であり、R2がアルキニル置換基である任意構造である。従って、この種のアルキニルアミノ酸は天然アミノ酸誘導体とみなすことができる。
【0130】
上記のように、本発明は非天然アルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)の使用に限定するものではない。実際に、真正細菌において本発明の直交翻訳系で使用することができる任意アルキニルアミノ酸が本発明の範囲に含まれる。他の各種アルキニルアミノ酸が公知であり、例えば、図8に示すアルキニルアミノ酸が挙げられる。これらのアルキニルアミノ酸構造のいくつかはpPRO−Pheと非常によく似ているので、本明細書に記載する直交tRNA及びアミノアシルtRNAシンテターゼ成分(例えば配列番号1のO−tRNAと配列番号4、6、8、10、12、14、16、18、又はその保存変異体のO−RS)を使用してこれらのアミノ酸のいくつかを真正細菌の蛋白質に組込むことができると考えられる。従って、本発明はpPRO−Pheに加えて他のアルキニルアミノ酸の組込み方法も提供する。表4(実施例9参照)に示す直交成分がpPRO−Phe以外にアルキニルアミノ酸を組込むことができるか否かに関係なく、本明細書の開示はこれらの他のアルキニルアミノ酸を組込む直交tRNA成分を構築するために十分な教示を提供するものであり、更にこれらの直交成分も本発明の範囲に含まれる。
【0131】
アルキニル基等の新規側鎖を含む非天然アミノ酸に加え、非天然アルキニルアミノ酸は場合により例えば式II及びIII:
【化4】
の構造により表されるような修飾主鎖構造も含み、上記式中、Zは一般にOH、NH2、SH、NH−R’又はS−R’を含み、XとYは同一でも異なっていてもよく、一般にS又はOであり、RとR’は場合により同一又は異なり、一般に式Iをもつ非天然アミノ酸について上記に記載したR基と同一の基及び水素から選択される。例えば、本発明の非天然アミノ酸は場合により式II及びIIIにより表されるようにアミノ又はカルボキシル基に置換を含む。この種の非天然アミノ酸としては限定されないが、例えば20種の標準天然アミノ酸に対応する側鎖又は非天然アルキニル側鎖をもつα−ヒドロキシ酸、α−チオ酸、α−アミノチオカルボキシレートが挙げられる。更に、α炭素の置換は場合によりL、D又はα,α−ジ置換アミノ酸(例えばD−グルタミン酸、D−アラニン、D−メチル−O−チロシン、アミノ酪酸等)を含む。他の代替構造としては環状アミノ酸(例えばプロリン類似体や、3、4、6、7、8及び9員環プロリン類似体)、β及びγアミノ酸(例えば置換β−アラニン及びγ−アミノ酪酸)が挙げられる。
【0132】
例えば、(所定のアルキニルアミノ酸を含む)多くの非天然アミノ酸はチロシン、セリン、システイン、アスパラギン酸、グルタミン酸等の天然アミノ酸をベースとする。例えば、以下のアルキニルアミノ酸:
2−アミノ−3−(プロプ−2−イニルオキシ)プロパン酸;
2−アミノ−3−(プロプ−2−イニルチオ)プロパン酸;
3−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノプロパン酸;及び
4−[(プロプ−2−イニルオキシ)カルボニル]−2−アミノブタン酸
はいずれも天然アミノ酸から誘導することができる。
【0133】
チロシン類似体としてはパラ置換チロシン、オルト置換チロシン、及びメタ置換チロシンが挙げられ、置換チロシンはアルキニル基、アセチル基、ベンゾイル基、アミノ基、ヒドラジン、ヒドロキシルアミン、チオール基、カルボキシ基、イソプロピル基、メチル基、C6−C20直鎖又は分岐鎖炭化水素、飽和又は不飽和炭化水素、O−メチル基、ポリエーテル基、ニトロ基等を含む。更に、多置換アリール環も考えられる。本発明のグルタミン類似体としては限定されないが、α−ヒドロキシ誘導体、γ置換誘導体、環状誘導体及びアミド置換グルタミン誘導体が挙げられる。フェニルアラニン類似体の例としては限定されないが、パラ置換フェニルアラニン、オルト置換フェニルアラニン、及びメタ置換フェニルアラニンが挙げられ、置換基はアルキニル基、ヒドロキシ基、メトキシ基、メチル基、アリル基、アルデヒド、ニトロ、チオール基又はケト基等を含む。非天然アミノ酸の特定例としては限定されないが、p−プロパルギルオキシフェニルアラニン、3,4−ジヒドロキシ−L−フェニルアラニン(DHP)、3,4,6−トリヒドロキシ−L−フェニルアラニン、3,4,5−トリヒドロキシ−L−フェニルアラニン、4−ニトロ−フェニルアラニン、p−アセチル−L−フェニルアラニン、O−メチル−L−チロシン、L−3−(2−ナフチル)アラニン、3−メチルフェニルアラニン、O−4−アリル−L−チロシン、4−プロピル−L−チロシン、3−ニトロ−チロシン、3−チオール−チロシン、トリ−O−アセチル−GlcNAcβ−セリン、L−Dopa、フッ素化フェニルアラニン、イソプロピル−L−フェニルアラニン、p−アジド−L−フェニルアラニン、p−アシル−L−フェニルアラニン、p−ベンゾイル−L−フェニルアラニン、L−ホスホセリン、ホスホノセリン、ホスホノチロシン、p−ヨードフェニルアラニン、p−ブロモフェニルアラニン、p−アミノ−L−フェニルアラニン、及びイソプロピル−L−フェニルアラニン等が挙げられる。各種非天然アミノ酸の構造は例えば図1A、8A及び8Bに示す。公開国際出願WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」も参照。
非天然アミノ酸の化学的合成
【0134】
上記非天然アミノ酸の多くは例えばSigma(米国)やAldrich(Milwaukee, WI,米国)から市販されている。市販されていないものは場合により各種刊行物に記載されている方法や当業者に公知の標準方法を使用して合成される。有機合成技術については例えばFessendonとFessendon著Organic Chemistry(1982, 第2版, Willard Grant Press, Boston Mass.);March著Advanced Organic Chemistry(第3版, 1985, Wiley and Sons, New York);及びCareyとSundberg著Advanced Organic Chemistry(第3版, Parts A and B, 1990, Plenum Press, New York)を参照されたい。非天然アミノ酸の合成について記載しているその他の刊行物としては例えばWO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(In vivo incorporation of unnatural amino acids)」;Matsoukasら, (1995) J. Med. Chem., 38, 4660-4669;King, F. E. & Kidd, D. A. A. (1949) A New Synthesis of Glutamine and of γ-Dipeptides of Glutamic Acid from Phthylated Intermediates. J. Chem. Soc., 3315-3319;Friedman, O. M. & Chatterrji, R. (1959) Synthesis of Derivatives of Glutamine as Model Substrates for Anti-Tumor Agents. J. Am. Chem. Soc. 81, 3750-3752; Craig, J. C. ら(1988)Absolute Configuration of the Enantiomers of 7-Chloro-4[[4-(diethylamino)-1-methylbutyl]amino]quinoline (Chloroquine). J. Org. Chem. 53, 1167-1170; Azoulay, M., Vilmont, M. & Frappier, F. (1991) Glutamine analogues as Potential Antimalarials. Eur. J. Med. Chem. 26, 201-5;Koskinen, A. M. P. & Rapoport, H. (1989) Synthesis of 4-Substituted Prolines as Conformationally Constrained Amino Acid Analogues. J. Org. Chem. 54, 1859-1866; Christie, B. D. & Rapoport, H. (1985) Synthesis of Optically Pure Pipecolates from L-Asparagine. Application to the Total Synthesis of(+)-Apovincamine through Amino Acid Decarbonylation and Iminium Ion Cyclization. J. Org. Chem. 1989:1859-1866;Bartonら, (1987)Synthesis of Novel α-Amino-Acids and Derivatives Using Radical Chemistry:Synthesis of L-and D-α-Amino-Adipic Acids, L-α-aminopimelic Acid and Appropriate Unsaturated Derivatives. Tetrahedron Lett. 43:4297-4308;及びSubasingheら, (1992)Quisqualic acid analogues:synthesis of beta-heterocyclic 2-aminopropanoic acid derivatives and their activity at a novel quisqualate-sensitized site. J. Med. Chem. 35:4602-7が挙げられる。国際公開WO2004/058946、発明の名称「蛋白質アレー(PROTEIN ARRAYS)」(出願日2003年12月22日)も参照。
非天然アミノ酸の細胞取込み
【0135】
細胞による非天然アミノ酸取り込みは例えば蛋白質に組込むように非天然アミノ酸を設計及び選択する場合に一般に考慮される問題の1つである。例えば、α−アミノ酸は電荷密度が高いため、これらの化合物は細胞に浸透しにくいと思われる。天然アミノ酸は異なる程度のアミノ酸特異性を示すことが多い一連の蛋白質輸送システムにより細胞に取り込まれる。非天然アミノ酸が細胞に取り込まれる場合にはどの非天然アミノ酸が取り込まれるかを判断する迅速なスクリーニングを実施することができる。例えば国際公開WO2004/058946、発明の名称「蛋白質アレー(PROTEIN ARRAYS)」(出願日2003年12月22日);及びLiu and Schultz(1999)Progress toward the evolution of an organism with an expanded genetic code. PNAS 96:4780-4785における毒性アッセイ参照。取込みは種々のアッセイで容易に分析されるが、細胞取込み経路に利用可能な非天然アミノ酸を設計する代替方法は、アミノ酸をin vivo生産する生合成経路を提供する方法である。
【0136】
非天然アミノ酸の生合成
細胞にはアミノ酸と他の化合物を生産するために多数の生合成経路が元々存在している。特定非天然アミノ酸の生合成法は自然界(例えば細胞中)には存在しないと思われるが、本発明はこのような方法を提供する。例えば、非天然アミノ酸の生合成経路は場合により新規酵素を付加するか又は既存宿主細胞経路を改変することにより宿主細胞で作製される。付加新規酵素は場合により天然酵素又は人工的に進化させた酵素である。例えば、(WO2002/085923、前出の実施例に記載されているような)p−アミノフェニルアラニンの生合成は他の生物に由来する公知酵素の組合せの付加に依存している。これらの酵素の遺伝子はこれらの遺伝子を含むプラスミドで細胞を形質転換することにより細胞に導入することができる。これらの遺伝子は細胞で発現されると、所望化合物を合成するための酵素経路を提供する。場合により付加される酵素種の例は下記実施例に記載する。その他の酵素配列は例えばGenbankに登録されている。人工的に進化させた酵素も場合により同様に細胞に付加する。このように、非天然アミノ酸を生産するように細胞機構と細胞資源を操作する。
【0137】
実際に、生合成経路で使用する新規酵素を生産するため又は非天然アミノを生産するように既存経路をin vitroもしくはin vivoで進化させるためには種々の方法の任意のものを使用することができる。非天然アミノ酸を生産するため(又は、実際に新規基質特異性もしくは他の該当活性をもつようにシンテターゼを進化させるため)には、酵素又は他の生合成経路成分を進化させる方法として入手可能な多数の方法を本発明に適用することができる。例えば、非天然アミノ酸を生産(又は新規シンテターゼを生産)するように新規酵素及び/又は前記酵素の経路をin vitro又はin vivoで開発するためには、場合によりDNAシャフリングを使用する。例えばStemmer(1994)Rapid evolution of a protein in vitro by DNA shuffling, Nature 370(4): 389-391;及びStemmer, (1994)DNA shuffling by random fragmentation and reassembly:In vitro recombination for molecular evolution, Proc. Natl. Acad. Sci. USA, 91:10747-10751参照。関連アプローチは所望特性をもつ酵素を迅速に進化させるために関連(例えば相同)遺伝子のファミリーをシャフリングする。このような「ファミリー遺伝子シャフリング」法の1例はCrameriら(1998)“DNA shuffling of a family of genes from diverse species accelerates directed evolution, ”Nature, 391 (6664) :288-291に記載されている。(生合成経路成分であるか又はシンテターゼであるかを問わずに)新規酵素は例えばOstermeierら(1999)“A combinatorial approach to hybrid enzymes independent of DNA homology”, Nature Biotech 17:1205に記載されているような「ハイブリッド酵素作製用インクリメンタルトランケーション(incremental truncation for the creation of hybrid enzymes:ITCHY)として知られるDNA組換え法を使用して作製することもできる。このアプローチは1種以上のin vitro又はin vivo組換え法の基質として使用することができる酵素又は他の経路変異体のライブラリーを作製するために使用することもできる。Ostermeierら(1999)“Combinatorial Protein Engineering by Incremental Truncation,” Proc. Natl. Acad. Sci. USA, 96:3562-67, 及び Ostermeierら(1999)“Incremental Truncation as a Strategy in the Engineering of Novel Biocatalysts, ” Biological and Medicinal Chemistry, 7:2139-44も参照。別のアプローチは例えば非天然アミノ酸(又は新規シンテターゼ)の生産に関連する生合成反応を触媒する能力について選択する酵素又は他の経路変異体のライブラリーを作製するために指数的集合突然変異誘発を使用する。このアプローチでは、該当配列中の小群の残基を並行してランダム化し、機能的蛋白質をもたらすアミノ酸を各変異位置で同定する。非天然アミノ酸(又は新規シンテターゼ)の生産用新規酵素を作製するように本発明に応用することができるこのような方法の例はDelegrave & Youvan(1993)Biotechnology Research 11:1548-1552に記載されている。更に別のアプローチでは、例えばArkin and Youvan(1992)“Optimizing nucleotide mixtures to encode specific subsets of amino acids for semi-random mutagenesis”Biotechnology 10:297-300;又はReidhaar-Olsonら(1991)“Random mutagenesis of protein sequences using oligonucleotide cassettes”Methods Enzymol. 208:564-86の一般突然変異誘発法を使用することにより、酵素及び/又は経路成分操作にドープ又は縮重オリゴヌクレオチドを使用するランダム又は半ランダム突然変異誘発を使用することができる。ポリヌクレオチドリアセンブリと部位飽和突然変異誘発を使用する更に別のアプローチ(「非確率論的」突然変異誘発と呼ぶことが多い)を使用すると、酵素及び/又は経路成分を作製した後に、(例えば非天然アミノ酸のin vivo生産のための)1種以上のシンテターゼ又は生合成経路機能を実施する能力についてスクリーニングすることができる。例えばShort「遺伝子ワクチン及び酵素の非確率論的作製(NON-STOCHASTIC GENERATION OF GENETIC VACCINES AND ENZYMES)」WO00/46344参照。
【0138】
このような突然変異法の代替方法は生物の全ゲノムを組換え、得られた子孫を特定経路機能について選択する方法である(「全ゲノムシャフリング」と呼ぶことが多い)。このアプローチは、例えばゲノム組換えと非天然アミノ酸(又はその中間体)の生産能に関する生物(例えば大腸菌又は他の細胞)の選択により本発明に適用することができる。例えば、非天然アミノ酸をin vivo生産するように細胞で既存及び/又は新規経路を進化させるための経路設計には、以下の刊行物に教示されている方法を適用することができる:Patnaikら(2002)“Genome shuffling of lactobacillus for improved acid tolerance”Nature Biotechnology, 20(7):707-712;及びZhangら(2002)“Genome shuffling leads to rapid phenotypic improvement in bacteria”Nature, February 7, 415(6872):644-646。
【0139】
例えば所望化合物を生産するための他の生物及び代謝経路組換え技術も利用可能であり、同様に非天然アミノ酸の生産に適用することができる。有用な経路組換えアプローチを教示している刊行物の例としては、Nakamura and White (2003) “Metabolic engineering for the microbial production of 1, 3 propanediol”Curr. Opin. Biotechnol. 14(5):454-9; Berryら(2002)“Application of Metabolic Engineering to improve both the production and use of Biotech Indigo”J. Industrial Microbiology and Biotechnology 28:127-133; Bantaら(2002)“Optimizing an artificial metabolic pathway:Engineering the cofactor specificity of Corynebacterium 2, 5-diketo-D-gluconic acid reductase for use in vitamin C biosynthesis” Biochemistry, 41(20), 6226-36; Selivonovaら(2001)“Rapid Evolution of Novel Traits in Microorganisms”Applied and Environmental Microbiology, 67:3645、及び他の多数の文献が挙げられる。
【0140】
使用する方法に関係なく、一般に本発明の組換え生合成経路を使用して生産される非天然アミノ酸は効率的な蛋白質生合成に十分な濃度(例えば天然細胞量)で生産されるが、他の細胞内アミノ酸濃度を有意に変化させたり細胞資源を枯渇させる程ではない。このようにin vivo生産される典型的な濃度は約10mM〜約0.05mMである。特定経路に所望される酵素を生産するように細胞を組換え、非天然アミノ酸を作製したら、場合によりin vivo選択を使用してリボソーム蛋白質合成と細胞増殖の両方に適合するように非天然アミノ酸の生産を更に最適化させる。
【0141】
パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)を組込むための直交成分
本発明はセレクターコドン(例えばアンバー終止コドン、ナンセンスコドン、4塩基以上のコドン等)に応答して成長中のポリペプチド鎖にアルキニルアミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe))を例えばin vivoで組込むための直交成分を作製する組成物及び方法を提供する。例えば、本発明は直交tRNA(O−tRNA)、直交アミノアシルtRNAシンテターゼ(O−RS)及びその対を提供する。これらの対は成長中のポリペプチド鎖にpPRO−Pheを組込むために使用することができる。
【0142】
本発明の組成物は直交アミノアシルtRNAシンテターゼ(O−RS)を含み、O−RSはO−tRNAをpPRO−Pheで優先的にアミノアシル化する。所定態様では、O−RSは配列番号4、6、8、10、12、14、16もしくは18を含むアミノ酸配列又はその保存変異体を含む。本発明の所定態様では、O−RSは内在tRNAよりもO−tRNAを優先的にpPRO−Phe等のアルキニルアミノ酸でアミノアシル化し、O−RSはO−tRNAを優先し、pPRO−Pheを負荷されるO−tRNAとpPRO−Pheを負荷される内在tRNAの比は1:1を上回り、O−RSはO−tRNAに排他的又はほぼ排他的に負荷することがより好ましい。
【0143】
O−RSを含む組成物は場合により更に直交tRNA(O−tRNA)を含むことができ、O−tRNAはセレクターコドンを認識する。一般に、本発明のO−tRNAは下記配列表(例えば配列番号1)及び実施例に記載するようなポリヌクレオチド配列を含むか又は前記配列によりコードされるO−tRNAの抑圧効率に比較してコグネイトシンテターゼの存在下でセレクターコドンに応答して少なくとも例えば約45%、50%、60%、75%、80%、又は90%以上の抑圧効率を含む。1態様では、O−RSとO−tRNAの併用による抑圧効率はO−RSの不在下のO−tRNAの抑圧効率の例えば5倍、10倍、15倍、20倍、25倍又はそれ以上である。1側面では、O−RSとO−tRNAの併用による抑圧効率はMethanococcus jannaschiiに由来する直交チロシルtRNAシンテターゼ対の抑圧効率の少なくとも45%である。
【0144】
O−tRNAを含む組成物は場合により細胞(例えば大腸菌細胞等の真正細菌細胞)、及び/又は翻訳系を含むことができる。
【0145】
本発明は翻訳系を含む細胞(例えば真正細菌細胞)も提供し、前記翻訳系は直交tRNA(O−tRNA)と;直交アミノアシルtRNAシンテターゼ(O−RS)と;アルキニルアミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン(pPRO−Phe))を含む。一般に、O−RSは内在tRNAよりもO−tRNAを優先的にpPRO−Phe等のアルキニルアミノ酸でアミノアシル化し、O−RSはO−tRNAを優先し、pPRO−Pheを負荷されるO−tRNAとpPRO−Pheを負荷される内在tRNAの比は1:1を上回り、O−RSはO−tRNAに排他的又はほぼ排他的に負荷することがより好ましい。O−tRNAは第1のセレクターコドンを認識し、O−RSはO−tRNAをpPRO−Pheで優先的にアミノアシル化する。1態様では、O−tRNAは配列番号1に記載のポリヌクレオチド配列又はその相補的ポリヌクレオチド配列を含むか又は前記配列によりコードされる。1態様では、O−RSは配列番号4、6、8、10、12、14、16もしくは18の任意1種に記載のアミノ酸配列又はその保存変異体を含む。
【0146】
本発明の細胞は場合により異なる付加O−tRNA/O−RS対と第2の非天然アミノ酸を更に含むことができ、例えばこのO−tRNAは第2のセレクターコドンを認識し、このO−RSは対応するO−tRNAを第2の非天然アミノ酸で優先的にアミノアシル化し、第2のアミノ酸はpPRO−Pheと異なる。場合により、本発明の細胞は該当ポリペプチドをコードするポリヌクレオチドを含む核酸を含み、前記ポリヌクレオチドはO−tRNAにより認識されるセレクターコドンを含む。
【0147】
所定態様では、本発明の細胞は直交−tRNA(O−tRNA)と、直交アミノアシルtRNAシンテターゼ(O−RS)と、アルキニルアミノ酸(例えばpPRO−Phe)と、該当ポリペプチドをコードするポリヌクレオチドを含む核酸を含む真正細菌細胞(例えば大腸菌細胞)であり、前記ポリヌクレオチドはO−tRNAにより認識されるセレクターコドンを含む。本発明の所定態様では、O−RSはO−RSが任意内在tRNAをアミノアシル化する効率よりも高い効率でO−tRNAを優先的にアミノアシル化する。
【0148】
本発明の所定態様では、本発明のO−tRNAは本明細書の配列表(例えば配列番号1)及び実施例に記載するようなポリヌクレオチド配列、又はその相補的ポリヌクレオチド配列を含むか又は前記配列によりコードされる。本発明の所定態様では、O−RSは本明細書の配列表に記載するようなアミノ酸配列、又はその保存変異体を含む。1態様では、O−RS又はその一部は本明細書の配列又は実施例に記載するようなアミノ酸をコードするポリヌクレオチド配列、又はその相補的ポリヌクレオチド配列によりコードされる。
【0149】
本発明のO−tRNA及び/又はO−RSは種々の生物(例えば真核及び/又は非真核生物)の任意のものに由来することができる。
【0150】
ポリヌクレオチドも本発明の特徴である。本発明のポリヌクレオチドは本明細書の配列表に記載するようなポリペプチドをコードするヌクレオチド配列を含む人工(例えば人為的に作製され、天然には存在しない)ポリヌクレオチドを含むか、及び/又は前記ポリヌクレオチド配列に相補的であるか又は前記配列をコードする。本発明のポリヌクレオチドは更に核酸の実質的に全長にわたって高ストリンジェント条件下で上記ポリヌクレオチドとハイブリダイズする核酸も含むことができる。本発明のポリヌクレオチドは更に天然に存在するtRNA又は対応するコーディング核酸のポリヌクレオチドと例えば少なくとも75%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%又はそれ以上一致するポリヌクレオチドを含み(但し、本発明のポリヌクレオチドは天然に存在するtRNA又は対応するコーディング核酸以外のものである)、前記tRNAはセレクターコドン(例えば4塩基コドン)を認識する。上記任意ポリヌクレオチドと例えば少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%又はそれ以上一致する人工ポリヌクレオチド及び/又は上記任意ポリヌクレオチドの保存変異体を含むポリヌクレオチドも本発明のポリヌクレオチドに含まれる。
【0151】
本発明のポリヌクレオチドを含むベクターも本発明の特徴である。例えば、本発明のベクターはプラスミド、コスミド、ファージ、ウイルス、発現ベクター及び/又は同等物を含むことができる。本発明のベクターを含む細胞も本発明の特徴である。
【0152】
O−tRNA/O−RS対の成分の作製方法も本発明の特徴である。これらの方法により作製された成分も本発明の特徴である。例えば、細胞に対して直交性である少なくとも1種のtRNA(O−tRNA)の作製方法は突然変異体tRNAのライブラリーを作製する段階と;セレクターコドンを認識できるように突然変異体tRNAのライブラリーの各メンバーのアンチコドンループを突然変異させることにより、潜在的O−tRNAのライブラリーを提供する段階と、潜在的O−tRNAのライブラリーのメンバーを含む第1の種の第1の細胞集団についてネガティブ選択を実施する段階を含む。ネガティブ選択は細胞に内在性のアミノアシルtRNAシンテターゼ(RS)によりアミノアシル化される潜在的O−tRNAライブラリーのメンバーを含む細胞を排除する。こうして、第1の種の細胞に対して直交性のtRNAプールが得られ、それにより少なくとも1種のO−tRNAが得られる。本発明の方法により作製されたO−tRNAも提供する。
【0153】
所定態様では、前記方法は更に第1の種の第2の細胞集団についてポジティブ選択を実施する段階を含み、前記細胞は第1の種の細胞に対して直交性のtRNAプールのメンバーと、コグネイトアミノアシルtRNAシンテターゼと、ポジティブ選択マーカーを含む。ポジティブ選択を使用し、コグネイトアミノアシルtRNAシンテターゼによりアミノアシル化され、ポジティブ選択マーカーの存在下で所望応答を示すtRNAプールのメンバーを含む細胞について細胞を選択又はスクリーニングすることにより、O−tRNAを得る。所定態様では、第2の細胞集団はネガティブ選択により排除されなかった細胞を含む。
【0154】
アルキニルアミノ酸をO−tRNAに負荷する直交アミノアシルtRNAシンテターゼの同定方法も提供する。例えば、方法は第1の種の細胞集団に選択を実施する段階を含み、前記細胞は、1)複数のアミノアシルtRNAシンテターゼ(RS)(例えば複数のRSは突然変異体RS、第1の種以外の種に由来するRS、又は突然変異体RSと第1の種以外の種に由来するRSの両方を含むことができる)のメンバーと;2)(例えば1種以上の種に由来する)直交−tRNA(O−tRNA)と;3)ポジティブ選択マーカーをコードし、少なくとも1個のセレクターコドンを含むポリヌクレオチドを各々含む。
【0155】
複数のRSのメンバーを含まないか又はその量の少ない細胞に比較して抑圧効率の高い細胞について細胞(例えば宿主細胞)を選択又はスクリーニングする。これらの選択/スクリーニングした細胞はO−tRNAをアミノアシル化する活性RSを含む。この方法により同定された直交アミノアシルtRNAシンテターゼも本発明の特徴である。
【0156】
パラ−プロパルギルオキシフェニルアラニン(pPRO−Phe)を特定位置にもつ蛋白質を細胞(例えば大腸菌細胞等の真正細菌細胞)で生産する方法も本発明の特徴である。例えば、1方法は、少なくとも1個のセレクターコドンを含み、蛋白質をコードする核酸を含む細胞を適当な培地で増殖させる段階と、pPRを提供する段階と、少なくとも1個のセレクターコドンによる核酸の翻訳中に蛋白質の特定位置にpPRを組込むことにより、蛋白質を生産する段階を含む。細胞は更に、細胞で機能し、セレクターコドンを認識する直交tRNAと;O−tRNAをpPRO−Pheで優先的にアミノアシル化する直交アミノアシルtRNAシンテターゼ(O−RS)を含む。この方法により生産された蛋白質も本発明の特徴である。
【0157】
本発明は例えばpPRO−Pheを組込んだ蛋白質を含有する組成物も提供する。所定態様では、蛋白質は公知蛋白質(例えば治療用蛋白質、診断用蛋白質、産業用酵素、又はその部分)のアミノ酸配列と少なくとも75%一致するアミノ酸配列を含む。場合により、組成物は医薬的に許容可能なキャリヤーを含有する。
【0158】
核酸及びポリペプチド配列と変異体
上記及び下記に記載するように、本発明は例えばO−tRNA及びO−RSをコードするポリヌクレオチド配列と、ポリペプチドアミノ酸配列(例えばO−RS)と、前記配列を含む例えば組成物、系及び方法を提供する。前記配列(例えばO−tRNA及びO−RSアミノ酸及びヌクレオチド配列)の例を本明細書に開示する(表4、例えば配列番号5、7、9、11、13、15、17及び19参照)。しかし、当業者に自明の通り、本発明は本明細書、例えば実施例や配列表に開示する配列に限定されない。当業者に自明の通り、本発明は例えば本明細書に記載する機能をもつ(例えばO−tRNA又はO−RSをコードする)多数の関連配列も提供する。
【0159】
O−tRNAをpPRO−Pheでアミノアシル化することが可能なO−RS種の構築と分析を実施例1に記載する。同実施例は単離された8種のO−RS種について記載する(図3及び実施例9参照)。これらのアミノ酸配列から明らかなように、8種の突然変異体O−RSクローンのアミノ酸置換には部分的コンセンサス傾向が認められる。2個以上のクローンの結合ポケットに以下のアミノ酸の少なくとも2種が認められた:Ala32,Pro107/Gln107,Ala158,Ile159,及びAla162/Pro162(配列番号21参照)。突然変異Tyr32→Ala32及びAsp158→Ala158の結果、Tyr32、Asp158及び天然基質チロシン間の水素結合が失われ、その結合が弱まると思われる。小さい主に疎水性の側鎖が存在するならば、pPRO−Pheの結合を助長すると予想される。これらのコンセンサス傾向により、pPRO−Pheを組込むために真正細菌宿主細胞系で配列番号1のO−tRNAと直交系で共働すると予想される付加O−RS種の設計が可能になる。これらのコンセンサス傾向は以下のように予測することができる。
【表1】
【0160】
従って、これらのコンセンサス傾向に基づき、実験により同定された8種のpPRO−PheRS種(即ちpPRO−PheRS−1〜pPRO−PheRS−8)で表されない少なくとも4種の付加直交pPRO−Pheシンテターゼ(pPRO−PheRS−con1〜pPRO−PheRS−con4)を合理的に設計することができる。これらは以下の通りである。
【表2】
【0161】
本発明はポリペプチド(O−RS)とポリヌクレオチド(例えばO−tRNA、O−RS又はその部分をコードするポリヌクレオチド、アミノアシルtRNAシンテターゼクローンを単離するために使用されるオリゴヌクレオチド等)を提供する。本発明のポリヌクレオチドとしては、1個以上のセレクターコドンをもつ本発明の該当蛋白質又はポリペプチドをコードするものが挙げられる。更に、本発明のポリヌクレオチドとしては、例えば配列番号5、7、9、11、13、15、17又は19に記載のヌクレオチド配列を含むポリヌクレオチド;そのポリヌクレオチド配列に相補的であるか又は前記配列をコードするポリヌクレオチドが挙げられる。本発明のポリヌクレオチドは更に配列番号4、6、8、10、12、14、16又は18を含むアミノ酸配列をコードする任意ポリヌクレオチドを含む。本発明のポリヌクレオチドは更に本発明のポリペプチドをコードするポリヌクレオチドを含む。同様に、核酸の実質的に全長にわたって高ストリンジェント条件下で上記ポリヌクレオチドとハイブリダイズする(天然ポリヌクレオチド以外の)人工核酸も本発明のポリヌクレオチドである。1態様では、組成物は本発明のポリペプチドと賦形剤(例えば緩衝液、水、医薬的に許容可能な賦形剤等)を含有する。本発明は本発明のポリペプチドに対して特異的に免疫反応性の抗体又は抗血清も提供する。人工ポリヌクレオチドとは人為的に作製され、天然には存在しないポリヌクレオチドである。
【0162】
本発明のポリヌクレオチドは天然に存在するtRNAのポリヌクレオチドと例えば少なくとも75%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%又はそれ以上一致する人工ポリヌクレオチド(但し、天然に存在するtRNA以外のもの)も含む。ポリヌクレオチドは天然に存在するtRNAのポリヌクレオチドと例えば少なくとも75%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%又はそれ以上一致する(但し100%は一致しない)人工ポリヌクレオチドも含む。
【0163】
所定態様では、ベクター(例えばプラスミド、コスミド、ファージ、ウイルス等)に本発明のポリヌクレオチドを組込む。1態様では、ベクターは発現ベクターである。別の態様では、発現ベクターは本発明のポリヌクレオチドの1種以上と機能的に連結されたプロモーターを含む。別の態様では、本発明のポリヌクレオチドを組込んだベクターを細胞に導入する。
【0164】
同様に当業者に自明の通り、開示配列の多数の変異体も本発明に含まれる。例えば、機能的に同一配列となる開示配列の保存変異体も本発明に含まれる。少なくとも1種の開示配列とハイブリダイスする核酸ポリヌクレオチド配列の変異体も本発明に含むものとする。例えば標準配列比較法により本明細書に開示する配列のユニークサブ配列であると判断される配列も本発明に含まれる。
【0165】
保存変異
遺伝コードの縮重により、「サイレント置換」(即ちコードされるポリペプチドに変化を生じない核酸配列の置換)はアミノ酸をコードする全核酸配列の暗黙の特徴である。同様に、「保存アミノ酸置換」はアミノ酸配列中の1又は数個のアミノ酸を高度に類似する特性をもつ別のアミノ酸で置換するものであり、このような置換も開示構築物と高度に類似することが容易に認められる。各開示配列のこのような保存変異は本発明の特徴である。
【0166】
特定核酸配列の「保存変異」とは同一又は本質的に同一のアミノ酸配列をコードする核酸を意味し、核酸がアミノ酸配列をコードしない場合には本質的に同一の配列を意味する。当業者に自明の通り、コードされる配列中の単一アミノ酸又は低百分率(一般に5%未満、より一般には4%、2%又は1%未満)のアミノ酸を置換、付加又は欠失させる個々の置換、欠失又は付加の結果としてアミノ酸を欠失するか、アミノ酸が付加されるか、又はアミノ酸が化学的に類似するアミノ酸で置換される場合には、これらの変異は「保存修飾変異」である。従って、本発明のポリペプチド配列の「保存変異」としては、ポリペプチド配列のアミノ酸の低百分率、一般に5%未満、より一般には2%又は1%未満が同一保存置換基のアミノ酸で置換される場合が挙げられる。最後に、非機能的配列の付加のように核酸分子のコードされる活性を変えない配列の付加も基本核酸の保存変異である。
【0167】
機能的に類似するアミノ酸を示す保存置換表は当分野で周知であり、このようなアミノ酸では、あるアミノ酸残基が同様の化学的性質(例えば芳香族側鎖又は正荷電側鎖)をもつ別のアミノ酸残基に置換しているため、ポリペプチド分子の機能的性質は実質的に変化しない。化学的性質が類似する天然アミノ酸を含む代表的グループを以下に示すが、これらのグループ内の置換は「保存置換」である。
【表3】
【0168】
核酸ハイブリダイゼーション
本発明の核酸の保存変異体を含めて本発明の核酸を同定するためには比較ハイブリダイゼーションを使用することができ、この比較ハイブリダイゼーション法は本発明の核酸を識別する1好適方法である。更に、高、超高及び超々高ストリンジェンシー条件下で配列番号5、7、9、11、13、15、17及び19により表される核酸とハイブリダイズするターゲット核酸も本発明の特徴である。このような核酸の例としては所与核酸配列と比較して1又は数個のサイレント又は保存核酸置換をもつものが挙げられる。
【0169】
試験核酸が完全にマッチする相補的ターゲットに比較して少なくとも50%の割合でプローブとハイブリダイズする場合、即ちマッチしないターゲット核酸の任意のものとのハイブリダイゼーションに観測されるシグナル対ノイズ比の少なくとも約5倍〜10倍で完全にマッチするプローブが完全にマッチする相補的ターゲットと結合する条件下におけるプローブとターゲットのハイブリダイゼーションに比較してシグナル対ノイズ比が少なくとも1/2である場合に試験核酸はプローブ核酸と特異的にハイブリダイズすると言う。
【0170】
核酸は一般に溶液中で会合するときに「ハイブリダイズ」する。核酸は水素結合、溶媒排除、塩基スタッキング等の種々の十分に特性決定された物理化学的力によりハイブリダイズする。核酸ハイブリダイゼーションの詳しい手引きはTijssen(1993)Laboratory Techniques in Biochemistry and Molecular Biology--Hybridization with Nucleic Acid Probes part I chapter 2, “Overview of principles of hybridization and the strategy of nucleic acid probe assays,”(Elsevier, New York) 及び Current Protocols in Molecular Biology, Ausubelら編, Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (2004年補遺)(「Ausubel」)に記載されている。Hames and Higgins(1995)Gene Probes 1 IRL Press at Oxford University Press, Oxford, England, (Hames and Higgins 1)及びHames and Higgins (1995) Gene Probes 2 IRL Press at Oxford University Press, Oxford, England (Hames and Higgins 2) はオリゴヌクレオチドを含むDNAとRNAの合成、標識、検出及び定量について詳細に記載している。
【0171】
サザン又はノーザンブロットで100個を上回る相補的残基をもつ相補的核酸のハイブリダイゼーションをフィルター上で行うためのストリンジェントハイブリダイゼーション条件の1例は、50%ホルマリンにヘパリン1mgを加え、42℃で一晩ハイブリダイゼーションを実施する。ストリンジェント洗浄条件の1例は65℃、0.2×SSCで15分間洗浄する(SSC緩衝液の説明についてはSambrook,前出参照)。多くの場合には高ストリンジェンシー洗浄の前に低ストリンジェンシー洗浄を実施してバックグラウンドプローブシグナルを除去する。低ストリンジェンシー洗浄の1例は40℃、2×SSCで15分間である。一般に、シグナル対ノイズ比が特定ハイブリダイゼーションアッセイで無関係プローブに観測される比の5倍(以上)である場合に特異的ハイブリダイゼーションが検出されたとみなす。
【0172】
サザン及びノーザンハイブリダイゼーション等の核酸ハイブリダイゼーション実験において「ストリンジェントハイブリダイゼーション洗浄条件」は配列依存性であり、各種環境パラメーターにより異なる。核酸ハイブリダイゼーションの詳しい手引きはTijssen (1993), 前出やHames and Higgins 1及び2に記載されている。ストリンジェントハイブリダイゼーション及び洗浄条件は任意試験核酸について経験により容易に決定することができる。例えば、高ストリンジェントハイブリダイゼーション及び洗浄条件を決定するには、一連の選択基準に合致するまで(例えばハイブリダイゼーション又は洗浄における温度上昇、塩濃度低下、界面活性剤濃度増加及び/又はホルマリン等の有機溶媒濃度増加により)ハイブリダイゼーション及び洗浄条件を徐々に増加する。例えば、高ストリンジェントハイブリダイゼーション及び洗浄条件では、マッチしないターゲットとプローブのハイブリダイゼーションに観測されるシグナル対ノイズ比の少なくとも約5倍でプローブが完全にマッチする相補的ターゲットと結合するまでハイブリダイゼーション及び洗浄条件を徐々に増加する。
【0173】
「超ストリンジェント」条件は特定プローブの熱融点(Tm)に等しくなるように選択される。Tmは(規定イオン強度及びpH下で)試験配列の50%が完全にマッチするプローブとハイブリダイズする温度である。本発明の目的には、一般に規定イオン強度及びpHで特定配列のTmよりも約5℃低くなるように「高ストリンジェント」ハイブリダイゼーション及び洗浄条件を選択する。
【0174】
「超高ストリンジェンシー」ハイブリダイゼーション及び洗浄条件は完全にマッチする相補的ターゲット核酸とプローブの結合に観測されるシグナル対ノイズ比がマッチしないターゲット核酸の任意のものとのハイブリダイゼーションに観測されるシグナル対ノイズ比の少なくとも10倍になるまでハイブリダイゼーション及び洗浄条件のストリンジェンシーを増加する条件である。完全にマッチする相補的ターゲット核酸のシグナル対ノイズ比の少なくとも1/2で前記条件下にプローブとハイブリダイズする場合にターゲット核酸は超高ストリンジェンシー条件下でプローブと結合すると言う。
【0175】
同様に、該当ハイブリダイゼーションアッセイのハイブリダイゼーション及び/又は洗浄条件を徐々に増加することにより更に高レベルのストリンジェンシーを決定することもできる。例えば、完全にマッチする相補的ターゲット核酸とプローブの結合に観測されるシグナル対ノイズ比がマッチしないターゲット核酸の任意のものとのハイブリダイゼーションに観測されるシグナル対ノイズ比の少なくとも10倍、20倍、50倍、100倍又は500倍以上になるまでハイブリダイゼーション及び洗浄条件のストリンジェンシーを増加する条件である。完全にマッチする相補的ターゲット核酸のシグナル対ノイズ比の少なくとも1/2で前記条件下にプローブとハイブリダイズする場合にターゲット核酸は超々高ストリンジェンシー条件下でプローブと結合すると言う。
【0176】
ストリンジェント条件下で相互にハイブリダイズしない核酸でも、これらの核酸によりコードされるポリペプチドが実質的に同一である場合には実質的に同一である。これは、例えば遺伝コードに許容される最大コドン縮重を使用して核酸のコピーを作製する場合に該当する。
【0177】
ユニークサブ配列
1側面では、本発明は本明細書に開示するO−tRNA及びO−RSの配列から選択される核酸中にユニークサブ配列を含む核酸を提供する。ユニークサブ配列は任意公知O−tRNA及びO−RS核酸配列に対応する核酸に比較してユニークである。例えばデフォルトパラメーターに設定したBLASTを使用してアラインメントを実施することができる。任意ユニークサブ配列は例えば本発明の核酸を同定するためのプローブとして有用である。
【0178】
同様に、本発明は本明細書に開示するO−RSの配列から選択されるポリペプチド中にユニークサブ配列を含むポリペプチドを含む。この場合には、ユニークサブ配列は任意公知ポリペプチド配列に対応するポリペプチドに比較してユニークである。
【0179】
本発明はO−RSの配列から選択されるポリペプチド中のユニークサブ配列をコードするユニークコーディングオリゴヌクレオチドとストリンジェント条件下でハイブリダイズするターゲット核酸も提供し、この場合には、ユニークサブ配列は対照ポリペプチド(例えば本発明のシンテターゼを例えば突然変異により誘導した元の親配列)の任意のものに対応するポリペプチドに比較してユニークである。ユニーク配列は上記のように決定する。
【0180】
配列比較、一致度及び相同度
2種以上の核酸又はポリペプチド配列に関して「一致」又は「一致度」百分率なる用語は2種以上の配列又はサブ配列を最大限に対応するように対比及び整列させ、以下に記載する配列比較アルゴリズム(又は当業者に入手可能な他のアルゴリズム)の1種を使用するか又は目視により測定した場合に相互に同一であるか又は同一のアミノ酸残基もしくはヌクレオチドの百分率が特定値であることを意味する。
【0181】
2種以上の核酸又はポリペプチド(例えばO−tRNAもしくはO−RSをコードするDNA又はO−RSのアミノ酸配列)に関して「実質的に一致」なる用語は2種以上の配列又はサブ配列を最大限に対応するように対比及び整列させ、配列比較アルゴリズムを使用するか又は目視により測定した場合にヌクレオチド又はアミノ酸残基一致度が少なくとも約60%、約80%、約90〜95%、約98%、約99%又はそれ以上であることを意味する。このような「実質的に一致」する配列は一般に実際の起源が記載されていなくても「相同」であるとみなす。少なくとも約50残基長の配列の領域、より好ましくは少なくとも約100残基長の領域にわたって「実質的一致」が存在していることが好ましく、少なくとも約150残基又は比較する2配列の全長にわたって配列が実質的に一致していることが最も好ましい。
【0182】
蛋白質及び/又は蛋白質配列は共通の祖先蛋白質又は蛋白質配列から天然又は人工的に誘導される場合に「相同」である。同様に、核酸及び/又は核酸配列は共通の祖先核酸又は核酸配列から天然又は人工的に誘導される場合に相同である。例えば、1個以上のセレクターコドンを含むように任意天然核酸を利用可能な任意突然変異誘発法により改変することができる。この突然変異誘発した核酸は発現されると、1種以上の非天然アミノ酸(例えばアルキニルアミノ酸)を含むポリペプチドをコードすることができる。突然変異法は当然のことながら更に1個以上の標準コドンを変異させ、得られる突然変異体蛋白質の1個以上の標準アミノ酸も変異させることができる。相同性は一般に2種以上の核酸又は蛋白質(又はその配列)間の配列類似度から推定される。相同性の判定に有用な配列間類似度の厳密な百分率は該当核酸及び蛋白質により異なるが、通常は25%程度の低い配列類似度を使用して相同性を判定する。例えば30%、40%、50%、60%、70%、80%、90%、95%又は99%以上のより高レベルの配列類似度を使用して相同性を判定することもできる。配列類似度百分率の決定方法(例えばデフォルトパラメーターを使用するBLASTP及びBLASTN)は本明細書に記載し、一般に入手可能である。
【0183】
配列比較及び相同性判定には、一般にある配列を参照配列としてこれに試験配列を比較する。配列比較アルゴリズムを使用する場合には、試験配列と参照配列をコンピューターに入力し、必要に応じてサブ配列座標を指定し、配列アルゴリズムプログラムパラメーターを指定する。こうすると、配列比較アルゴリズムは指定プログラムパラメーターに基づいて参照配列に対して試験配列の配列一致度百分率を計算する。
【0184】
比較のための最適な配列アラインメントは例えばSmith & Waterman, Adv. Appl. Math. 2:482(1981)の局所相同性アルゴリズム、Needleman & Wunsch, J. Mol. Biol. 48:443(1970)の相同性アラインメントアルゴリズム、Pearson & Lipman, Proc. Natl. Acad. Sci. USA 85:2444(1988)の類似性探索法、これらのアルゴリズムのコンピューターソフトウェア(Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WIのGAP、BESTFIT、FASTA及びTFASTA)、又は目視(一般にCurrent Protocols in Molecular Biology, Ausubelら編, Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (2004年補遺)参照)により実施することができる。
【0185】
配列一致度及び配列類似度百分率を決定するのに適したアルゴリズムの1例はAltschulら, J. Mol. Biol. 215:403-410(1990) に記載されているBLASTアルゴリズムである。BLAST分析を実施するためのソフトウェアはNational Center for Biotechnology Information(www. ncbi. nlm. nih. gov/)から公共入手可能である。このアルゴリズムはデータベース配列中の同一長さの単語と整列した場合に所定の正の閾値スコアTと一致するか又はこれを満足するクエリー配列中の長さWの短い単語を識別することによりまず高スコア配列対(HSP)を識別する。Tを隣接単語スコア閾値と言う(Altschulら,前出)。これらの初期隣接単語ヒットをシードとして検索を開始し、これらの単語を含むもっと長いHSPを探索する。次に、累積アラインメントスコアを増加できる限り、単語ヒットを各配列に沿って両方向に延長する。ヌクレオチド配列の場合にはパラメーターM(1対のマッチ残基のリウォードスコア、常に>0)及びN(ミスマッチ残基のペナルティースコア、常に<0)を使用して累積スコアを計算する。アミノ酸配列の場合には、スコアリングマトリクスを使用して累積スコアを計算する。累積アラインメントスコアがその最大到達値から量Xだけ低下するか、累積スコアが1カ所以上の負スコア残基アラインメントの累積によりゼロ以下になるか、又はどちらかの配列の末端に達したら各方向の単語ヒットの延長を停止する。BLASTアルゴリズムパラメーターW、T及びXはアラインメントの感度と速度を決定する。BLASTNプログラム(ヌクレオチド配列用)は語長(W)11、期待値(E)10、カットオフ100、M=5、N=4、及び両鎖の比較をデフォルトとして使用する。アミノ酸配列用として、BLASTPプログラムは語長(W)3、期待値(E)10、及びBLOSUM62スコアリングマトリクスをデフォルトとして使用する(Henikoff & Henikoff (1989) Proc. Natl. Acad. Sci. USA 89:10915参照)。
【0186】
配列一致度百分率の計算に加え、BLASTアルゴリズムは2配列間の類似性の統計分析も実施する(例えばKarlin & Altschul, Proc. Natl. Acad. Sci. USA 90:5873-5787 (1993)参照)。BLASTアルゴリズムにより提供される類似性の1尺度は2種のヌクレオチド又はアミノ酸配列間に偶然にマッチが起こる確率を示す最小合計確率(P(N))である。例えば、試験核酸を参照核酸に比較した場合の最小合計確率が約0.1未満、より好ましくは約0.01未満、最も好ましくは約0.001未満である場合に核酸は参照核酸に類似しているとみなす。
【0187】
突然変異誘発及び他の分子生物学技術
本発明のポリヌクレオチドとポリペプチド及び本発明で使用されるポリヌクレオチドとポリペプチドは分子生物学技術を使用して操作することができる。分子生物学技術について記載している一般教科書としてはBerger and Kimmel, Guide to Molecular Cloning Techniques, Methods in Enzymology volume 152 Academic Press, Inc., San Diego, CA (Berger);Sambrookら, Molecular Cloning-A Laboratory Manual (第3版), Vol. 1-3, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 2001 (「Sambrook」)及びCurrent Protocols in Molecular Biology, F. M. Ausubelら編, Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (2004年補遺) (「Ausubel」))が挙げられる。これらの教科書は突然変異誘発法、ベクターの使用、プロモーター及び他の多くの関連事項について記載しており、アルキニルアミノ酸(例えばpPRO−Phe)、直交tRNA、直交シンテターゼ及びその対を含む蛋白質を生産するためのセレクターコドンを含む遺伝子の作製についても記載している。
【0188】
例えばtRNA分子を突然変異させるため、tRNAのライブラリーを作製するため、シンテターゼのライブラリーを作製するため、アルキニルアミノ酸をコードするセレクターコドンを該当蛋白質又はポリペプチドに挿入するために、本発明では各種突然変異誘発法を使用する。これらの方法としては限定されないが、部位特異的、ランダム点突然変異誘発、相同組換え、DNAシャフリング又は他の帰納的突然変異誘発法、キメラ構築、ウラシル含有鋳型を使用する突然変異誘発、オリゴヌクレオチド特異的突然変異誘発、ホスホロチオエート修飾DNA突然変異誘発、ギャップデュプレクスDNAを使用する突然変異誘発等、又はその任意組み合わせが挙げられる。他の適切な方法としては点ミスマッチ修復、修復欠損宿主株を使用する突然変異誘発、制限−選択及び制限−精製、欠失突然変異誘発、完全遺伝子合成による突然変異誘発、2本鎖切断修復等が挙げられる。例えばキメラ構築物を使用する突然変異誘発も本発明に含まれる。1態様では、天然分子又は改変もしくは突然変異させた天然分子の既知情報(例えば配列、配列比較、物性、結晶構造等)に基づいて突然変異誘発を行うことができる。
【0189】
本発明のポリヌクレオチド又は本発明のポリヌクレオチドを組込んだ構築物(例えばクローニングベクター又は発現ベクター等の本発明のベクター)で宿主細胞を遺伝子組換え(例えば形質転換、形質導入又はトランスフェクション)する。例えば、直交tRNA、直交tRNAシンテターゼ、及び修飾すべき蛋白質のコーディング領域を所望宿主細胞で機能的な遺伝子発現制御エレメントに機能的に連結する。典型的なベクターは転写及び翻訳ターミネーターと、転写及び翻訳開始配列と、特定ターゲット核酸の発現の調節に有用なプロモーターを含む。ベクターは場合により少なくとも1個の独立ターミネーター配列と、真核生物又は原核生物又は両者(例えばシャトルベクター)でカセットの複製を可能にする配列と、原核系と真核系の両者の選択マーカーを含む包括的発現カセットを含む。ベクターは原核生物、真核生物、又は好ましくは両者での複製及び/又は組込みに適している。Giliman & Smith, Gene 8:81 (1979); Robertsら, Nature, 328:731 (1987); Schneider, B. ら, Protein Expr. Purif. 6435:10(1995); Ausubel, Sambrook, Berger(いずれも前出)参照。ベクターは例えばプラスミド、細菌、ウイルス、裸のポリヌクレオチド又はポリヌクレオチドコンジュゲートの形態とすることができる。ベクターはエレクトロポレーション(Fromら, Proc. Natl. Acad. Sci. USA 82, 5824 (1985))、ウイルスベクターによる感染、核酸を小ビーズもしくは粒子のマトリックスに埋込むか又は表面に付着させて小粒子形態で高速射入する方法(Kleinら, Nature 327, 70-73 (1987))、及び/又は同等方法等の標準方法により細胞及び/又は微生物に導入する。
【0190】
クローニングに有用な細菌とバクテリオファージのカタログは例えばATCCから入手でき、例えばATCCから刊行されたThe ATCC Catalogue of Bacteria and Bacteriophage (1996) Ghernaら(編)が挙げられる。その他のシーケンシング、クローニング及び分子生物学の他の側面の基本手順と基礎理論事項もSambrook(前出)、Ausubel(前出)、及びWatsonら(1992) Recombinant DNA Second Edition, Scientific American Books (New York)に記載されている。更に、Midland Certified Reagent Company (Midland, TX;mcrc. com)、The Great American Gene Company(Ramona, CA, www.genco.comで販売)、ExpressGen Inc.(Chicago, IL, www.expressgen.comで販売)、Operon Technologies Inc. (Alameda, CA)及び他の多数の企業等の各種販売会社からほぼ任意核酸(及び標準又は標準外を問わずほぼ任意標識核酸)をオーダーメード又は標準注文することができる。
【0191】
遺伝子組換えした宿主細胞は例えばスクリーニング段階、プロモーター活性化又は形質転換細胞選択等の操作に合うように適宜改変した慣用栄養培地で培養することができる。これらの細胞は場合によりトランスジェニック生物で培養することができる。(例えば後期核酸単離のための)例えば細胞単離及び培養に関する他の有用な文献としてはFreshney (1994) Culture of Animal Cells, a Manual of Basic Technique, 第3版, Wiley-Liss, New Yorkとその引用文献; Payneら(1992) Plant Cell and Tissue Culture in Liquid Systems John Wiley & Sons, Inc. New York, NY; Gamborg and Phillips (eds) (1995) Plant Cell, Tissue and Organ Culture; Fundamental Methods Springer Lab Manual, Springer-Verlag (Berlin Heidelberg New York) 及び Atlas and Parks (eds) The Handbook of Microbiological Media (1993) CRC Press, Boca Raton, FLが挙げられる。
【0192】
該当蛋白質及びポリペプチド
アルキニルアミノ酸の1つの大きな利点は(限定されないが)各種小分子、生体分子又は他の蛋白質等の任意のものと蛋白質を架橋又は結合するためにアルキニルアミノ酸を含む蛋白質を使用できる点である。少なくとも1個のアルキニルアミノ酸を含む該当蛋白質又はポリペプチドは本発明の特徴である。本発明は本発明の組成物と方法を使用して生産された少なくとも1個のアルキニルアミノ酸を含むポリペプチド又は蛋白質も含む。賦形剤(例えば医薬的に許容可能な賦形剤)も蛋白質に添加することができる。場合により、本発明の蛋白質は単一アミノ酸位置又は複数の位置に(アルキニルアミノ酸残基のその後の可能な修飾に加えて)翻訳後修飾を含むことができるか、あるいは蛋白質は複数の異なる型の修飾をもつことができる。
【0193】
アルキニルアミノ酸を特定位置にもつ蛋白質を細胞で生産する方法も本発明の特徴である。例えば、1方法は、少なくとも1個のセレクターコドンを含み且つ蛋白質をコードする核酸を含む細胞を適当な培地で増殖させる段階と、アルキニルアミノ酸を提供する段階を含み;細胞は更に、細胞で機能し、セレクターコドンを認識する直交tRNA(O−tRNA)と;O−tRNAをアルキニルアミノ酸で優先的にアミノアシル化する直交アミノアシルtRNAシンテターゼ(O−RS)を含む。この方法により生産された蛋白質も本発明の特徴である。
【0194】
所定態様では、O−RSは発現系において内在tRNAよりもコグネイトO−tRNAのアミノアシル化を優先する。O−tRNAとO−RSが等モル濃度で存在する場合にO−RSにより負荷されるO−tRNAと内在tRNAの相対比は1:1を上回り、好ましくは少なくとも約2:1、より好ましくは5:1、更に好ましくは10:1、更に好ましくは20:1、更に好ましくは50:1、更に好ましくは75:1、更に好ましくは95:1、98:1、99:1、100:1、500:1、1,000:1、5,000:1又はそれ以上である。
【0195】
本発明はアルキニルアミノ酸を組込んだ蛋白質を含有する組成物も提供する。所定態様では、蛋白質は治療用蛋白質、診断用蛋白質、産業用酵素、又はその部分のアミノ酸配列と少なくとも75%一致するアミノ酸配列を含む。
【0196】
本発明の組成物と本発明の方法により作製された組成物は場合により細胞に導入する。その後、本発明のO−tRNA/O−RS対又は個々の成分を宿主系の翻訳機構で使用することができ、その結果として、アルキニルアミノ酸は蛋白質に組込まれる。国際公開WO2004/094593、出願日2004年4月16日、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」、及びWO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」はこのプロセスを記載しており、参考資料として本明細書に組込む。例えば、O−tRNA/O−RS対を宿主(例えば大腸菌細胞)に導入すると、前記対はセレクターコドンに応答してパラ−プロパルギルオキシフェニルアラニン等のアルキニルアミノ酸のin vivo蛋白質組込みを誘導する。系に付加されるパラ−プロパルギルオキシフェニルアラニンは外部から増殖培地に添加することができる合成アミノ酸(例えばフェニルアラニンやチロシンの誘導体)である。場合により、本発明の組成物はin vitro翻訳系に導入してもよいし、in vivo系に導入してもよい。
【0197】
本発明の細胞は非天然アミノ酸を組込んだ蛋白質を大量の有用な量で合成することができる。1側面では、組成物は場合により、アルキニルアミノ酸を組込んだ蛋白質を例えば少なくとも10μg、少なくとも50μg、少なくとも75μg、少なくとも100μg、少なくとも200μg、少なくとも250μg、少なくとも500μg、少なくとも1mg、少なくとも10mg以上、又はそれ以上、あるいはin vivo蛋白質生産方法で達成可能な量で含有する(組換え蛋白質生産及び精製に関する詳細は本明細書に記載する)。別の側面では、蛋白質は場合により例えば細胞溶解液、緩衝液、医薬緩衝液、又は他の懸濁液中(例えば約1nl〜約100Lの任意の容量中)に例えば少なくとも10μg蛋白質/l、少なくとも50μg蛋白質/l、少なくとも75μg蛋白質/l、少なくとも100μg蛋白質/l、少なくとも200μg蛋白質/l、少なくとも250μg蛋白質/l、少なくとも500μg蛋白質/l、少なくとも1mg蛋白質/l、又は少なくとも10mg蛋白質/l以上の濃度で組成物中に存在する。少なくとも1種のアルキニルアミノ酸を組込んだ蛋白質を細胞で大量(例えば他の方法、例えばin vitro翻訳で一般に可能な量よりも多量)に生産することも本発明の特徴である。
【0198】
アルキニルアミノ酸の組込みは例えば寸法、酸性度、求核性、水素結合、疎水性、プロテアーゼ標的部位接近性、(例えば蛋白質アレーのための)部分へのターゲット等を変化させるように例えば蛋白質構造及び/又は機能の変化を調整するために実施することができる。アルキニルアミノ酸を組込んだ蛋白質は触媒性又は物性を強化するか又は全く新規にすることができる。例えば、アルキニルアミノ酸を蛋白質に組込むことにより、場合により毒性、生体分布、構造的性質、分光学的性質、化学及び/又は光化学的性質、触媒能、半減期(例えば血清半減期)、他の分子との(例えば共有又は非共有)反応性等の性質を改変する。少なくとも1種のアルキニルアミノ酸を組込んだ蛋白質を含有する組成物は例えば新規治療薬、診断薬、触媒酵素、産業用酵素、結合蛋白質(例えば抗体)、及び例えば蛋白質構造と機能の研究に有用である。例えばDougherty, (2000) Unnatural Amino Acids as Probes of Protein Structure and Function, Current Opinion in Chemical Biology, 4:645-652参照。
【0199】
本発明の1側面では、組成物は少なくとも1個、例えば少なくとも2個、少なくとも3個、少なくとも4個、少なくとも5個、少なくとも6個、少なくとも7個、少なくとも8個、少なくとも9個、又は少なくとも10個以上の非天然アミノ酸(例えばアルキニルアミノ酸及び/又は他の非天然アミノ酸)を組込んだ少なくとも1種の蛋白質を含有する。非天然アミノ酸は同一でも異なっていてもよく、例えば1、2、3、4、5、6、7、8、9、又は10種以上の異なる非天然アミノ酸を含む1、2、3、4、5、6、7、8、9、又は10個以上の異なる部位が蛋白質に存在することができる。別の側面では、組成物は蛋白質に存在する特定アミノ酸の全部よりは少ないが少なくとも1個がアルキニルアミノ酸で置換された蛋白質を含有する。2個以上の非天然アミノ酸を組込んだ所与蛋白質では、非天然アミノ酸は同一でも異なっていてもよい(例えば蛋白質は2個以上の異なる型の非天然アミノ酸を組込んでもよいし、2個の同一非天然アミノ酸を組込んでもよい)。3個以上の非天然アミノ酸を組込んだ所与蛋白質では、非天然アミノ酸は同一でも異なっていてもよいし、同一種の複数の非天然アミノ酸と少なくとも1個の別の非天然アミノ酸の組み合わせでもよい。
【0200】
本明細書に記載する組成物と方法を使用してアルキニルアミノ酸を含むほぼ任意蛋白質(又はその部分)(及び例えば1個以上のセレクターコドンを含む対応する任意コーディング核酸)を生産することができる。数十万種の公知蛋白質を列挙するまでもなく、例えば該当翻訳系に1個以上の適当なセレクターコドンを含むように入手可能な任意突然変異法を調整することにより、1種以上の非天然アミノ酸を組込むように公知蛋白質の任意のものを改変することができる。公知蛋白質の一般的な配列寄託機関としてはGenBank、EMBL、DDBJ及びNCBIが挙げられる。他の寄託機関もインターネットを検索することにより容易に確認できる。
【0201】
一般に、蛋白質は入手可能な任意蛋白質(例えば治療用蛋白質、診断用蛋白質、産業用酵素、又はその部分等)と例えば少なくとも60%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも90%、少なくとも95%、又は少なくとも99%以上一致しており、1種以上の非天然アミノ酸を含む。1種以上のアルキニルアミノ酸を組込むように改変することができる治療用、診断用、及び他の蛋白質の例としては限定されないが、国際公開WO2004/094593、出願日2004年4月16日、発明の名称「真核遺伝コードの拡張(Expanding the Eukaryotic Genetic Code)」;及びWO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」に記載されているものが挙げられる。1種以上のアルキニルアミノ酸を組込むように改変することができる治療用、診断用、及び他の蛋白質の例としては限定されないが、例えばα1アンチトリプシン、アンジオスタチン、抗血友病因子、抗体(抗体の詳細については後述する)、アポリポ蛋白質、アポ蛋白質、心房性ナトリウム利尿因子、心房性ナトリウム利尿ポリペプチド、心房性ペプチド、C−X−Cケモカイン(例えばT39765、NAP−2、ENA−78、Gro−a、Gro−b、Gro−c、IP−10、GCP−2、NAP−4、SDF−1、PF−4、MIG)、カルシトニン、CCケモカイン(例えば単球化学誘引蛋白質−1、単球化学誘引蛋白質−2、単球化学誘引蛋白質−3、単球炎症性蛋白質−1α、単球炎症性蛋白質−1β、RANTES、I309、R83915、R91733、HCC1、T58847、D31065、T64262)、CD40リガンド、Cキットリガンド、コラーゲン、コロニー刺激因子(CSF)、補体因子5a、補体阻害剤、補体受容体1、サイトカイン(例えば上皮好中球活性化ペプチド−78、GROα/MGSA、GROβ、GROγ、MIP−1α、MIP−1δ、MCP−1)、表皮増殖因子(EGF)、エリスロポエチン(「EPO」)、表皮剥離毒素A及びB、IX因子、VII因子、VIII因子、X因子、繊維芽細胞増殖因子(FGF)、フィブリノーゲン、フィブロネクチン、G−CSF、GM−CSF、グルコセレブロシダーゼ、ゴナドトロピン、増殖因子、ヘッジホッグ蛋白質(例えばソニック、インディアン、デザート)、ヘモグロビン、肝細胞増殖因子(HGF)、ヒルジン、ヒト血清アルブミン、インスリン、インスリン様増殖因子(IGF)、インターフェロン(例えばIFN−α、IFN−β、IFN−γ)、インターロイキン(例えばIL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12等)、ケラチノサイト増殖因子(KGF)、ラクトフェリン、白血病阻害因子、ルシフェラーゼ、ニュールチュリン、好中球阻害因子(NIF)、オンコスタチンM、骨形成蛋白質、副甲状腺ホルモン、PD−ECSF、PDGF、ペプチドホルモン(例えばヒト成長ホルモン)、プレイオトロピン、プロテインA、プロテインG、発熱外毒素A、B及びC、リラキシン、レニン、SCF、可溶性補体受容体I、可溶性I−CAM1、可溶性インターロイキン受容体(IL−1、2、3、4、5、6、7、9、10、11、12、13、14、15)、可溶性TNF受容体、ソマトメジン、ソマトスタチン、ソマトトロピン、ストレプトキナーゼ、スーパー抗原即ちブドウ球菌エンテロトキシン(SEA、SEB、SEC1、SEC2、SEC3、SED、SEE)、スーパーオキシドジスムターゼ(SOD)、毒素性ショック症候群毒素(TSST−1)、チモシンα1、組織プラスミノーゲンアクチベーター、腫瘍壊死因子β(TNFβ)、腫瘍壊死因子受容体(TNFR)、腫瘍壊死因子α(TNFα)、血管内皮増殖因子(VEGEF)、ウロキナーゼ及び他の多数の蛋白質が挙げられる。
【0202】
本明細書に記載するアルキニルアミノ酸のin vivo組込み用組成物及び方法を使用して生産することができる蛋白質の1類としては転写モジュレーター又はその部分が挙げられる。転写モジュレーターの例としては細胞増殖、分化、制御等を調節する遺伝子及び転写モジュレーター蛋白質が挙げられる。転写モジュレーターは原核生物、ウイルス及び真核生物(例えば真菌、植物、酵母、昆虫、及び哺乳動物を含む動物)に存在し、広範な治療ターゲットを提供する。当然のことながら、発現及び転写アクチベーターは例えば受容体との結合、シグナル伝達カスケードの刺激、転写因子の発現調節、プロモーターやエンハンサーとの結合、プロモーターやエンハンサーと結合する蛋白質との結合、DNA巻き戻し、プレmRNAスプライシング、RNAポリアデニル化及びRNA分解等の多数のメカニズムにより転写を調節する。
【0203】
本発明の蛋白質(例えば1種以上のアルキニルアミノ酸を組込んだ蛋白質)の1類としてはサイトカイン、炎症性分子、増殖因子、その受容体、及び腫瘍遺伝子産物等の生体活性蛋白質が挙げられ、例えばインターロイキン(例えばIL−1、IL−2、IL−8等)、インターフェロン、FGF、IGF−I、IGF−II、FGF、PDGF、TNF、TGF−α、TGF−β、EGF、KGF、SCF/c−キット、CD40L/CD40、VLA−4/VCAM−1、ICAM−1/LFA−1及びヒアルリン/CD44;シグナル伝達分子及び対応する腫瘍遺伝子産物(例えばMos、Ras、Raf及びMet);並びに転写アクチベーター及びサプレッサー(例えばp53、Tat、Fos、Myc、Jun、Myb、Rel、及びステロイドホルモン受容体(例えばエストロゲン、プロゲステロン、テストステロン、アルドステロン、LDL受容体リガンド及びコルチコステロン))が挙げられる。
【0204】
少なくとも1種のアルキニルアミノ酸を組込んだ酵素(例えば産業用酵素)又はその部分も本発明により提供される。酵素の例としては限定されないが、例えばアミダーゼ、アミノ酸ラセマーゼ、アシラーゼ、デハロゲナーゼ、ジオキシゲナーゼ、ジアリールプロパンペルオキシダーゼ、エピメラーゼ、エポキシドヒドロラーゼ、エステラーゼ、イソメラーゼ、キナーゼ、グルコースイソメラーゼ、グリコシダーゼ、グリコシルトランスフェラーゼ、ハロペルオキシダーゼ、モノオキシゲナーゼ(例えばp450類)、リパーゼ、リグニンペルオキシダーゼ、ニトリルヒドラターゼ、ニトリラーゼ、プロテアーゼ、ホスファターゼ、スブチリシン、トランスアミナーゼ及びヌクレアーゼが挙げられる。
【0205】
これらの蛋白質の多くは市販されており(例えばSigma BioSciences 2003カタログ及び価格表参照)、対応する蛋白質配列と遺伝子及び、一般に多くのその変異体も周知である(例えばGenbank参照)。例えば1種以上の該当治療、診断又は酵素特性に関して蛋白質を改変するように本発明に従って1種以上のアルキニルアミノ酸を挿入することにより前記蛋白質の任意のものを改変することができる。治療関連特性の例としては血清半減期、貯蔵半減期、安定性、免疫原性、治療活性、(例えば非天然アミノ酸(例えばアルキニルアミノ酸)へのレポーター基(例えばラベル又はラベル結合部位)の付加による)検出性、LD50又は他の副作用の低減、胃管を経由して体内に導入できること(例えば経口利用性)等が挙げられる。診断特性の例としては貯蔵半減期、安定性、診断活性、検出性等が挙げられる。該当酵素特性の例としては貯蔵半減期、安定性、酵素活性、産生能等が挙げられる。
【0206】
他の各種蛋白質も本発明の組成物と方法を使用して1種以上のアルキニルアミノ酸を含むように改変することができる。例えば、本発明は例えば感染性真菌(例えばAspergillus、Candida種);細菌、特に病原細菌モデルとして利用できる大腸菌や医学的に重要な細菌(例えばStaphylococci(例えばaureus)又はStreptococci(例えばpneumoniae));原生動物(例えば胞子虫類(例えばPlasmodia)、根足虫類(例えばEntamoeba)及び鞭毛虫類(Trypanosoma、Leishmania、Trichomonas、Giardia等));ウイルス(例えば(+)RNAウイルス(例えばポックスウイルス(例えばワクシニア)、ピコルナウイルス(例えばポリオ)、トガウイルス(例えば風疹)、フラビウイルス(例えばHCV)、及びコロナウイルス)、(−)RNAウイルス(例えばラブドウイルス(例えばVSV)、パラミクソウイルス(例えばRSV)、オルトミクソウイルス(例えばインフルエンザ)、ブンヤウイルス及びアレナウイルス)、dsDNAウイルス(例えばレオウイルス)、RNA→DNAウイルス(即ちレトロウイルス、例えばHIV及びHTLV)、及び所定のDNA→RNAウイルス(例えばB型肝炎))に由来する蛋白質において、1種以上のワクチン蛋白質中の1種以上の天然アミノ酸をアルキニルアミノ酸で置換することができる。
【0207】
昆虫耐性蛋白質(例えばCry蛋白質)、澱粉及び脂質産生酵素、植物及び昆虫毒素、毒素耐性蛋白質、マイコトキシン解毒蛋白質、植物成長酵素(例えばリブロース1,5−ビスリン酸カルボキシラーゼ/オキシゲナーゼ、「RUBISCO」)、リポキシゲナーゼ(LOX)及びホスホエノールピルビン酸(PEP)カルボキシラーゼ等の農業関連蛋白質もアルキニルアミノ酸修飾の適切なターゲットである。
【0208】
所定態様では、本発明の方法及び/又は組成物における該当蛋白質又はポリペプチド(又はその部分)は核酸によりコードされる。一般に、核酸は少なくとも1個のセレクターコドン、少なくとも2個のセレクターコドン、少なくとも3個のセレクターコドン、少なくとも4個のセレクターコドン、少なくとも5個のセレクターコドン、少なくとも6個のセレクターコドン、少なくとも7個のセレクターコドン、少なくとも8個のセレクターコドン、少なくとも9個のセレクターコドン、10個以上のセレクターコドンを含む。
【0209】
該当蛋白質又はポリペプチドをコードする遺伝子はアルキニルアミノ酸を組込むための1個以上のセレクターコドンを付加するように本明細書の「突然変異誘発及び他の分子生物学技術」のセクションに記載する当業者に周知の方法を使用して突然変異誘発することができる。例えば、1個以上のセレクターコドンを付加するように該当蛋白質の核酸を突然変異誘発し、1種以上のアルキニルアミノ酸を挿入できるようにする。本発明は例えば少なくとも1種のアルキニルアミノ酸を組込んだ任意蛋白質のこのような任意変異体(例えば突然変異体、変形)を含む。同様に、本発明は対応する核酸、即ち1種以上のアルキニルアミノ酸をコードする1個以上のセレクターコドンを含む任意核酸も含む。
【0210】
アルキニルアミノ酸を組込んだ蛋白質を生産するためには、直交tRNA/RS対によるアルキニルアミノ酸のin vivo組込みに適した宿主細胞及び生物を使用することができる。直交tRNA、直交tRNAシンテターゼ、及び修飾すべき蛋白質をコードするベクターを発現する1種以上のベクターで宿主細胞を遺伝子組換え(例えば形質転換、形質導入又はトランスフェクション)する。これらの成分の各々を同一ベクターに配置してもよいし、各々別々のベクターに配置してもよいし、2成分を第1のベクターに配置し、第3の成分を第2のベクターに配置してもよい。ベクターは例えばプラスミド、細菌、ウイルス、裸のポリヌクレオチド又はポリヌクレオチドコンジュゲートの形態とすることができる。
【0211】
免疫反応性によるポリペプチドの定義
本発明のポリペプチドは種々の新規ポリペプチド配列(例えば本発明の翻訳系で合成される蛋白質の場合にはアルキニルアミノ酸を含み、例えば新規シンテターゼの場合には標準アミノ酸の新規配列を含むポリペプチド)を提供するので、ポリペプチドは例えばイムノアッセイで認識することができる新規構造特徴も提供する。本発明のポリペプチドに特異的に結合する抗血清の作製及び前記抗血清と結合するポリペプチドも本発明の特徴である。本明細書で使用する「抗体」なる用語は限定されないが、検体(抗原)と特異的に結合してこれを認識する1又は複数の免疫グロブリン遺伝子又はそのフラグメントにより実質的にコードされるポリペプチドを意味する。例としてはポリクローナル、モノクローナル、キメラ、及び1本鎖抗体等が挙げられる。Fabフラグメントやファージディスプレイを含む発現ライブラリーにより生産されるフラグメント等の免疫グロブリンフラグメントも本明細書で使用する「抗体」なる用語に含まれる。抗体構造及び用語については、例えばPaul, Fundamental Immunology, 4th Ed., 1999, Raven Press, New York参照。
【0212】
イムノアッセイ用抗血清を作製するためには、免疫原性ポリペプチドの1種以上を本明細書に記載するように作製し、精製する。例えば、組換え蛋白質を組換え細胞で生産することができる。(マウスの仮想遺伝子一致により結果の再現性が高いのでこのアッセイで使用する)近交系マウスに標準マウス免疫プロトコールに従って標準アジュバント(例えばフロイントアジュバント)と共に免疫原性蛋白質を免疫する(抗体作製、イムノアッセイフォーマット及び特異的免疫反応性を測定するために使用可能な条件の標準解説については例えばHarlow and Lane (1988) Antibodies. A Laboratory Manual, Cold Spring Harbor Publications, New York 参照)。蛋白質、抗体、抗血清等に関するその他の詳細は国際公開WO2004/094593、発明の名称「真核遺伝コードの拡張(EXPANDING THE EUKARYOTIC GENETIC CODE)」;WO2002/085923、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」;WO2004/035605、発明の名称「糖蛋白質合成(GLYCOPROTEIN SYNTHESIS)」;及びWO2004/058946、発明の名称「蛋白質アレー(PROTEIN ARRAYS)」に記載されている。
【0213】
O−tRNA及びO−RS及びO−tRNA/O−RS対の使用
本発明の組成物と本発明の方法により作製された組成物は場合により細胞に導入する。本発明のO−tRNA/O−RS対又は個々の成分をその後、宿主系の翻訳機構で使用することができ、その結果として、アルキニルアミノ酸は蛋白質に組込まれる。国際公開WO2002/085923、発明者Schultzら、発明の名称「非天然アミノ酸のインビボ組込み(IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS)」はこのプロセスを記載しており、参考資料として本明細書に組込む。例えば、O−tRNA/O−RS対を宿主(例えば大腸菌細胞)に導入すると、前記対はセレクターコドン(例えばアンバーナンセンスコドン)に応答して外部から増殖培地に添加することができるアルキニルアミノ酸のin vivo蛋白質(例えばミオグロビン試験蛋白質又は治療用蛋白質)組込みを誘導する。場合により、本発明の組成物はin vitro翻訳系に導入してもよいし、細胞内in vivo系に導入してもよい。アルキニルアミノ酸を組込んだ蛋白質は多様な用途の任意のものに使用することができる。最も特筆すべき点として、蛋白質に組込まれたアルキニル部分は例えば、他の蛋白質、小分子(例えばラベルや色素)及び/又は生体分子との架橋をはじめとする多様な修飾の任意のもののターゲットとして利用することができる。これらの修飾により、アルキニルアミノ酸の組込みの結果、治療用蛋白質を改善することができ、更に酵素の触媒機能を改変又は改善するために使用することもできる。所定側面では、アルキニルアミノ酸の蛋白質組込みとその後の修飾は蛋白質構造、他の蛋白質との相互作用等の研究を助長することができる。
【0214】
キット
キットも本発明の特徴である。例えば、少なくとも1種のアルキニルアミノ酸を組込んだ蛋白質を細胞で生産するためのキットが提供され、前記キットはO−tRNAをコードするポリヌクレオチド配列、及び/又はO−tRNA、及び/又はO−RSをコードするポリヌクレオチド配列、及び/又はO−RSを収容する容器を含む。1態様では、キットは更にアルキニルアミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン)を含む。別の態様では、キットは更に蛋白質を生産するための説明書を含む。
【0215】
以下、実施例により本発明を例証するが、これらの実施例により発明を限定するものではない。特許請求の範囲に記載する発明の範囲から逸脱せずに変更可能な種々の非必須パラメーターが当業者に自明である。
【実施例1】
【0216】
アルキニルアミノ酸を大腸菌の蛋白質に組込むための直交tRNA/シンテターゼ対の進化
直交tRNA−シンテターゼ対の特異性を進化させることにより、本発明者らは原核生物と真核生物の両者でナンセンス及びフレームシフトコドンに応答して多数の非天然アミノ酸を蛋白質に選択的且つ効率的に組込んでいる(Andersonら(2004) Proc. Natl. Acad. Sci. U.S.A., 101:1566; Alfontaら(2003) J. Am. Chem. Soc., 125:14662; Wangら(2003) Proc. Natl. Acad. Sci. U.S.A., 100:56; Chinら(2003) Science 301:964; Chinら(2002) Proc. Natl. Acad. Sci., 99:11020; 及び Wang ら(2001) Science 292:498)。本発明は大腸菌翻訳機構を使用して反応性アルキニル部分を含むアミノ酸を蛋白質に生合成的に組込むための組成物と方法を提供する。大腸菌翻訳成分を使用する生合成はin vivo(例えば大腸菌細胞中)で実施することもできるし、粗細胞抽出液又は精製翻訳成分を使用してin vitroで実施することもできる。蛋白質に組込まれるアルキニル基はアジド含有部分と容易且つ特異的に結合するので、蛋白質修飾/操作の有用なターゲットとなる。
【0217】
アルキニル基とアジド基の化学反応(図1Bに示す)は蛋白質に存在する全内在官能基の化学反応に対して完全に直交性である。それらのユニークな反応性の1例は[3+2]シクロ付加による不可逆的トリアゾール形成である(図1B;及びPadwa, In Comprehensive Organic Synthesis; [Trost, B. M., Ed.] Pergamon: Oxford, 1991, Vol. 4, p 1069; Huisgen, In 1,3-Dipolar Cycloaddition Chemistry, [Padwa, A., Ed.] Wiley:New York, 1984;p1参照)。この反応は室温にて水性媒体中(生物サンプルを修飾するために十分温和な条件)で銅(I)の存在下に実施する場合には、完全に位置選択的に進行し(Rostovtsevら(2002)Angew. Chem. Int. Ed., 41:2596)、アルキニル官能基とアジド官能基が導入された蛋白質を選択的に修飾するために使用することができる(Deitersら(2003) J. Am. Chem. Soc., 125:11182; Wangら(2003) J. Am. Chem. Soc., 125:3192; Link and Tirrell (2003) J. Am. Chem. Soc., 125:11164)。
【0218】
本明細書に記載する発明はアルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(略称pPRO−Phe;IUPAC命名法によると、2−アミノ−3−[4−(プロプ−2−イニルオキシ)プロピオン酸とも言う;構造は図1Aに示し、化学構造1と称する)を大腸菌宿主系に選択的に組込むMethanococcus jannaschii成分から誘導される直交tRNA/tRNAシンテターゼ対を提供する。本実施例はpPRO−Pheが本明細書に記載する新規直交tRNA及びtRNAシンテターゼ試薬を使用して大腸菌で発現される蛋白質に選択的に組込まれることを実証する。
【0219】
本実施例ではM. jannaschiiチロシルtRNA/tRNAシンテターゼ対(MjTyrRS/tRNACUA)から誘導される直交tRNA/シンテターゼ対の進化について報告し、直交対は標準(即ち天然)アミノ酸に対して親和性がないか又は非常に低い。誘導される直交tRNAシンテターゼはアンバーサプレッサーtRNACUAにpPRO−Pheを選択的に負荷し、更に、アミノアシル化されたtRNA(即ち「負荷されたtRNA」)は転写産物中で遭遇したTAGアンバー終止コドン(セレクターコドン)に応答してpPRO−Pheを組込むために内在大腸菌翻訳系により基質として使用される。このtRNA/シンテターゼ対の直交性(Steer and Schimmel (1999) Biol. Chem., 274:35601)により、tRNAもシンテターゼも内在大腸菌tRNA又はシンテターゼと交差反応せず、非天然アミノ酸はアンバーナンセンスコドンTAGのみに応答して遊離する。
【0220】
野生型M. jannaschiiチロシルtRNAシンテターゼの突然変異誘発により〜107個の異なるM. jannaschiiチロシルtRNAシンテターゼから成るライブラリーを作製した。MjTyrRSライブラリーを作製するために、5個の突然変異標的位置をアラニンコドンに変換した。pBR322から誘導したカナマイシン耐性をもつプラスミドpBK−JYRSで大腸菌GlnRSプロモーター及びターミネーターの制御下にMjTyrRS遺伝子を発現させた。残基Tyr32、Glu107、Asp158、Ile159、及びLeu162を部位特異的突然変異誘発によりAlaで置換し、プラスミドpBK−JYA5を得た。突然変異部位にNNK(N=A+T+G+C及びK=G+T)をもつ8個のオリゴヌクレオチドを使用してAla5MjTyrRS突然変異体(pBK−JYA5)をPCR増幅し、NdeI−PstIで消化したpBK−JYA5にライゲーションし、MjTyrRSライブラリーを作製した。ライゲーションしたベクターを大腸菌DH10Bコンピテント細胞に形質転換し、1.6×109コロニー形成単位のライブラリーを得た。
【0221】
野生型M. jannaschiiチロシルtRNAシンテターゼ分子のポリヌクレオチド及びアミノ酸配列を図2に示し、更に夫々配列番号3及び2にも示す。突然変異誘発はBacillus stearothermophilusに由来する相同チロシルtRNAシンテターゼの結晶構造に基づいて5個の活性部位残基(Tyr32,Glu107,Asp158,Ile159,及びLeu162)をランダム化することから構成した。
【0222】
突然変異誘発後、シンテターゼプールを次にポジティブ及びネガティブ選択ラウンドに付した。ポジティブ選択はクロラムフェニコールアセチルトランスフェラーゼ(CAT)遺伝子の許容部位(Asp112)におけるアンバー終止コドンの抑圧に基づく。MjTyrRS突然変異体ライブラリーと、突然変異CAT遺伝子と、同時発現させたMjアンバーサプレッサーtRNACUAを含む大腸菌細胞を最小培地でpPRO−Phe(1mM)とクロラムフェニコール(80μg/mL)の存在下に増殖させると、tRNACUAを内在アミノ酸又はpPRO−Pheでアミノアシル化する突然変異体シンテターゼを含む細胞のみが生存する。次に、許容部位に3個のアンバー突然変異(Gln2,Asp44,Gly65)をもつ毒性蛋白質バルナーゼをコードする突然変異遺伝子を含む細胞にシンテターゼライブラリー遺伝子を形質転換した。バルナーゼレポーターをもつベクターにはサプレッサーtRNAも加えた。これらの細胞をpPRO−Pheの不在下で増殖させ、内在アミノ酸を基質として受容することが可能なシンテターゼを逆選択した。3ラウンドの選択後、pPRO−Pheの有無に対する増殖率依存性について96個のクローンをスクリーニングし、8個の候補クローンを同定し、配列決定した。これらの単離クローンに認められたアミノ酸置換を図3に示す。8個のクローンのポリヌクレオチド配列とアミノ酸配列も配列番号4〜19に示す。
【0223】
8個の突然変異体O−RSクローンのアミノ酸置換にはコンセンサス傾向が認められる。クローンの殆どの結合ポケットには以下のアミノ酸:Ala32,Pro107/Gln107,Ala158,Ile159,及びAla162/Pro162が高頻度で認められた。突然変異Tyr32→Ala32及びAsp158→Ala158の結果、Tyr32、Asp158及び天然基質チロシン間の水素結合が失われ、その結合が弱まると思われる。小さい主に疎水性の側鎖が存在するならば、pPRO−Pheの結合を助長すると予想される。クローンの1個(pPRO−PheRS−1)には更にLeu110→Phe110突然変異も認められた。
【0224】
シンテターゼpPRO−PheRS−1を更に特性決定するために選択した。このシンテターゼはクロラムフェニコール耐性を大腸菌に付与し、pPRO−Pheの存在下と不在下のIC50は夫々110及び5μg/mLであった。pPRO−Pheの有無によるクロラムフェニコール耐性の大きな差異は非天然アミノ酸pRPO−Pheに対するpPRO−PheRS−1の実質的なin vivo特異性を示唆している。
【実施例2】
【0225】
大腸菌の蛋白質へのアルキニルアミノ酸の部位特異的組込み
突然変異体アンバーサプレッサーtRNACUAとpPRO−PheRS−1の直交対を大腸菌で使用し、多数の構造、力学、及び蛋白質フォールディング研究の対象となっているモノマー153残基ヘム含有蛋白質であるマッコウクジラミオグロビンにpPRO−Pheを選択的に組込んだ(Reedy and Gibney (2004) Chem. Rev., 104:617とその引用文献; Uzawaら(2004) Proc. Natl. Acad. Sci. U.S.A., 101:1171とその引用文献;Wright,and Baldwin in Frontiers in Molecular Biology: Mechanisms of Protein Folding, [Pain, R., ed.] Oxford University Press, London, 2000, pp.309)。
【0226】
アルキニル修飾ミオグロビンを作製するために、ミオグロビンオープンリーディングフレームの4番目のコドン(Ser4)をTAG(アンバー終止)に突然変異させ、C末端6×His(ヘキサヒスチジン)タグをオープンリーディングフレームに付加した。突然変異体蛋白質を発現させるために、(アラビノースプロモーターとrrnBターミネーターの制御下の突然変異体マッコウクジラミオグロビン遺伝子と;lppプロモーターとrrnCターミネーターの制御下のチロシルtRNACUAと;テトラサイクリン耐性遺伝子をコードする)プラスミドpBAD/JYAMB−4TAGを(突然変異体シンテターゼとカナマイシン耐性遺伝子をコードする)pBKベクターと共にDH10B大腸菌に同時形質転換した。テトラサイクリン(25mg/L)とカナマイシン(30mg/L)を補充したLuria−Bertani培地(5mL)で細胞を増幅させ、リン酸緩衝液で洗浄し、適当な抗生物質、pPRO−Phe(1mM)、及びアラビノース(0.002%)を加えた液体グリセロール最小培地(0.3mMロイシンを補充)500mLを接種するために使用した。細胞を飽和まで増殖させた後、遠心により回収した。Ni−アフィニティークロマトグラフィーを使用して蛋白質を精製した処、Ni−アフィニティークロマトグラフィーによる精製後に収量2mg/Lが得られ、SDS−PAGE/Gelcode(登録商標)Blue染色(Pierce Biotechnology,Inc.)により90%均質であると推定された。総収量〜1mgの突然変異体ミオグロビンが得られた。
【0227】
こうして作製した蛋白質をGelcode(登録商標)Blue染色と抗His6抗体を使用するウェスタンブロッティングの両者で図4,レーン1に可視化する。pPRO−Pheの不在下では、染色又は(抗His6抗体を使用する)ウェスタンブロッティング後にミオグロビンは認められず、進化型シンテターゼの高い選択性が示唆された(図4,レーン2参照)。
【実施例3】
【0228】
大腸菌の蛋白質へのアルキニルアミノ酸組込みの質量分析による確認
突然変異体ミオグロビンのアンバー終止コドンの部位に組込まれたアミノ酸の実体を更に確認するために、ミオグロビンのトリプシン消化物を液体クロマトグラフィー/タンデム質量分析に付した。この実験で使用した突然変異体ミオグロビンは74位に組換えアンバー終止コドンを含むものを使用した。この位置のpPRO−Pheの組込み(pPRO−Phe74)を試験し、74位突然変異を調べた。ミオグロビン−74TAG突然変異体はLC MS/MS分析の特性が改善されているので、先に記載したSer4→TAG(アンバーコドン)の代わりに使用した。
【0229】
真正細菌発現後に、ニッケルアフィニティーカラムを使用してミオグロビン74TAGを精製した。SDS−PAGEゲルのGelcode(登録商標)Blue染色により蛋白質バンドを可視化した。突然変異体ミオグロビンに対応するゲルバンドをポリアクリルアミドゲルから切り出し、1.5mm立方体にスライスし、ほぼ従来の記載通りにトリプシン加水分解した(Shevchenkoら(1996) Anal. Chem., 68:850-858)。
【0230】
非天然アミノ酸を含むトリプシンペプチドをLCQイオントラップ質量分析計でナノフロー逆相HPLC/μESI/MSにより分析した。Nanospray HPLC(Agilent 1100シリーズ)を取付たFinnigan LCQ Decaイオントラップ質量分析計(Thermo Finnigan)で液体クロマトグラフィータンデム質量分析(LC−MS/MS)を実施した。
【0231】
非天然アミノ酸(Y*で示す)を含むペプチドHGVTVLTALGY*ILK(配列番号X)の単一及び二重荷電イオンに対応する前駆体イオンをイオントラップ質量分析計で分離分画した。この分析の結果を図5に示す。フラグメントイオン質量は明確に帰属させることができ、pPRO−Pheの部位特異的組込みが確認された。LC MS/MS試験によると、この位置に天然アミノ酸の組込みは示唆されず、進化型シンテターゼの高い選択性が確認された。
【実施例4】
【0232】
[3+2]シクロ付加によるアルキニルアミノ酸を含む蛋白質の修飾
アルキニル官能基を含む蛋白質は[3+2]シクロ付加反応の使用による修飾の有効なターゲットとすることができる。本実施例は2個の異なるアジド含有色素分子によるアルキニルミオグロビンの修飾について記載する。本実施例で使用した突然変異体ミオグロビンは実施例1に記載したように4番目のコドンにpPRO−Pheを組込んだ(Ser4→pPRO−Phe4)。
【0233】
実施例2に記載したようにSer4→pPRQ−Phe4ミオグロビンを大腸菌で作製した後、夫々ダンシル及びフルオレセインフルオロフォアを含むアジド官能基を付けた色素2又は3で修飾した(図6A及び6Bに示す;Deitersら(2003) J. Am. Chem. Soc., 725:11782; Wangら(2003) J. Am. Chem. Soc., 125:3192; Link and Tirrell (2003) J. Am. Chem. Soc., 125:11164も参照)。[3+2]シクロ付加修飾反応を図7Aに示す。
【0234】
シクロ付加反応のために、CuSO4(H2O中50mMストック溶液;最終反応容量中1mM)1μL、色素2又は3(EtOH中50mM)2μL、トリス(1−ベンジル−1H−[1,2,3]トリアゾール−4−イルメチル)アミン(DMSO中50mM)2μL、及び(還元剤として)銅線1mg又はトリス(カルボキシエチル)ホスフィン(H2O中100mM)1μLを0.1Mリン酸緩衝液(pH=8)中精製突然変異体ミオグロビン(〜0.5mg/mL)45μLに加えた。室温で8時間又は4℃で一晩後にH2O 450μLを加え、透析膜(10kDaカットオフ)を通して混合物を遠心した。上清を2×500μLリン酸緩衝液で遠心洗浄後、溶液を容量50μLにした。
【0235】
還元剤として銅線又はトリス(カルボキシエチル)ホスフィン(2mM)を使用すると、一般に同様の標識効率が得られた。従来の知見(Wangら(2003) J. Am. Chem. Soc., 125:3192)とは対照的に、リガンドトリス(1−ベンジル−1H−[1,2,3]トリアゾール−4−イルメチル)アミンの有無はこれらの反応の結果に実質的な影響がなかった。次に蛍光標識蛋白質(Blake (2001) Curr. Opin. Pharmacol., 1:533; Woutersら(2001) Trends in Cell Biology 11:203; Zachariasら(2000) Curr. Opin. Neurobiol., 20:416)20μLのサンプルをSDS−PAGEにより分析し、ゲル内イメージングした。ダンシル色素2(λex=337nm,λem=506nm)で修飾した突然変異体ミオグロビンは360±30nmでEagle Eyeデンシトメーター(Stratagene)によりゲル内イメージングした。フルオレセイン色素3(λex=495nm,λem=516nm)の結合は450±30nmでStorm Phosphorimager(Molecular Dynamics)により可視化した。この蛍光イメージングの結果を図7Bに示す。突然変異体ミオグロビンは色素2及び3のいずれでも有効に標識される。3で標識したミオグロビンのA280/A495値の比較によると、標識効率は〜75%であった(Wangら(2003)J.Am.Chem.Soc.,125:3192参照)。このバイオコンジュゲーションの選択性は野生型ミオグロビンと2又は3の反応が認められなかったという事実により確認された(結果は示さず)。
【0236】
本実施例の記載はアルキニルアミノ酸(例えばパラ−プロパルギルオキシフェニルアラニン)を生物(例えば大腸菌)の蛋白質に有効かつ選択的に組込むことができることを実証するものである。これらのアミノ酸はその後、蛋白質内で例えばアジド部分を使用する[3+2]シクロ付加による結合の化学的ターゲットとすることができ、更に、この標的修飾は高度に特異的且つ位置選択的である。アルキニルアミノ酸を蛋白質に部位特異的に組込むことができると、蛋白質結合又は修飾が所望される任意蛋白質の試験で有用なツールとなる。
【実施例5】
【0237】
非天然アルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニンの合成
非天然アルキニルアミノ酸パラ−プロパルギルオキシフェニルアラニン(略称pPRO−Phe;図1A参照,化合物1)を市販N−Boc−チロシンから3段階で合成し(Deitersら(2003) J. Am. Chem. Soc., 125:11782; Wangら(2003) J. Am. Chem. Soc., 125:3192; Link and Tirrell (2003) J. Am. Chem. Soc., 125:11164参照)、総収率81%であった。
【0238】
ステップ1
N−tert−ブトキシカルボニル−チロシン(2g,7mmol,1当量)とK2CO3(3g,21mmol,3当量)を無水DMF(15mL)に懸濁した。臭化プロパルギル(2.1mL,21mmol,3当量,80%トルエン溶液)をゆっくりと加え、反応混合物を18時間室温で撹拌した。水(75mL)とEt2O(50mL)を加え、層分離し、水相をEt2O(2×50mL)で抽出した。有機層を合わせて乾燥し(MgSO4)、溶媒を減圧除去した。下記構造及び名称の生成物(4)が黄色油状物(2.3g,91%)として得られ、それ以上精製せずに次段階で使用した。
【化5】
【0239】
ステップ2
塩化アセチル(7mL)を0℃のメタノール(60mL)に注意深く加え、無水HClのMeOH中5M溶液を得た。前段階の生成物(化合物4;2g,5.6mmol)を加え、周囲温度まで昇温させながら反応混合物を4時間撹拌した。揮発分を減圧除去後に黄色がかった固体(下記構造及び名称の化合物5;1.6g,98%)が得られ、次段階で直接使用した。
【化6】
【0240】
ステップ3
前段階(5)からのプロパルギルエステル(1.6g,5.5mmol)を2N NaOH水溶液(14mL)とMeOH(10mL)の混合物に溶かした。1.5時間室温で撹拌後、濃HClを加えることによりpHを7に調整した。水(20mL)を加え、混合物を4℃に一晩維持した。沈殿を濾過し、氷冷H2Oで洗浄し、減圧乾燥すると、pPRO−Phe(1)1.23g(90%)が白色固体として得られた。1H NMR(400MHz,D2O;D2O中カリウム塩として)δ7.20(d,7=8.8Hz,2H),6.99(d,J=8.8Hz,2H),4.75(s,2H),3.50(dd,J=5.6,7.2Hz,1H),2.95(dd,J=5.6,13.6Hz,1H),2.82(dd,J=7.2,13.6Hz,1H);13C NMR(100MHz,D2O)δ181.3,164.9,155.6,131.4,130.7,115.3,57.3,56.1,39.3;HRMS(CI)m/z 220.0969[C12H13NO3(M+1)必要値220.0968]。
【化7】
【実施例6】
【0241】
アジド色素2の合成
アジド色素2(図6A参照;化合物2)を以下のプロトコールに従って合成した。3−アジドプロピルアミン(371mg,3.71mmol,3当量)(Carboniら(1993)Org.Chem.,58:3736−3741に従って合成)を塩化ダンシル(500mg,1.85mmol,1当量)とトリエチルアミン(258μL,1.85mmol,1当量)の0℃のCH2Cl2(10mL)溶液に加えた。1時間撹拌後、反応混合物を室温まで昇温させ、更に1時間撹拌した。揮発分を減圧除去し、粗生成物をシリカゲルクロマトグラフィー(Et2O/ヘキサン=1:1)により精製すると、2(548mg,89%)が黄色油状物として得られた。1H NMR(400MHz,CDCl3)δ8.55(d,J=8.4Hz,1H),8.29(d,J=8.8Hz,1H),8.23(dd,J=1.2,7.2Hz,1H),7.56−7.49(comp,2H),7.18(d,J=7.6Hz,1H),5.24(br s,1H),3.21(t,J=6.4Hz,2H),2.95(dt,J=6.4Hz,2H),2.89(s,6H),1.62(quin,J=6.4Hz,2H);13C NMR(100MHz,CDCl3)δ134.3,130.4,129.7,129.4,128.4,123.3,118.8,115.3,48.6,45.4,40.6,28.7(13C NMRスペクトルに4級炭素原子の全シグナルは認められない);HRMS(CI)m/z 334.1336[C15H20N5O2S(M+1)必要値334.1332]。
【化8】
【実施例7】
【0242】
アジド色素3の合成
アジド色素3(図6B参照;化合物3)を以下のプロトコールに従って合成した。EDCI(83mg,0.43mmol,1当量)をフルオレセインアミン(150mg,0.43mmol,1当量)と4−(3−アジドプロピルカルバモイル)酪酸(92mg,0.43,1当量)の室温のピリジン(2mL)溶液に加えた。(4−(3−アジドプロピルカルバモイル)酪酸は3−アジドプロピルアミンをグルタル酸無水物と反応させることにより合成した。)懸濁液を一晩撹拌し、反応混合物をH2O(15mL)に注入した。濃HClを加えることにより溶液を酸性化した(pH<2)。1時間撹拌後、沈殿を濾別し、1N HCl(3×3mL)で洗浄し、少量のEtOAcに溶かした。ヘキサンを加えると、3がオレンジ色結晶として沈殿したので、採取し、減圧乾燥した(200mg,86%)。1H NMR(400MHz,CD3OD)δ8.65(s,1H),8.15(d,J=8.4Hz,1H),7.61−7.51(comp,2H),7.40(d,J=8.4Hz,1H),7.35(br s,2H),7.22−7.14(comp,2H),6.85−6.56(comp,3H),3.40−3.24(comp,4H),2.54(t,J=7.2Hz,2H),2.39−2.30(comp,2H),2.10−1.99(comp,2H),1.82−1.72(comp,2H);13C NMR(100MHz,CD3OD)δ175.7,174.4,172.4,167.9,160.8,143.0,134.3,132.9,131.8,129.6,124.4,123.3,121.1,118.5 103.5,50.2,38.0,37.2,36.2,29.8,22.9;4 HRMS(CI)m/z 544.1835[C28H25N5O7(M+1)必要値544.1827]。
【化9】
【実施例8】
【0243】
アルキニルアミノ酸の大腸菌組込み用の代表的O−RS及びO−tRNA
代表的O−tRNAとしては配列番号1が挙げられる(実施例9,表4参照)。代表的O−RSとしては配列番号4、6、8、10、12、14、16及び18に記載のアミノ酸配列が挙げられる(図3及び実施例9,表4参照)。
【0244】
O−RS又はその部分をコードするポリヌクレオチドの例としては配列番号4、6、8、10、12、14、16及び18を含む任意アミノ酸配列をコードする任意ポリヌクレオチドが挙げられる。例えば、配列番号5、7、9、11、13、15、17及び19に記載のポリヌクレオチドが代表的O−RSをコードする。
【0245】
当然のことながら、本明細書に記載する実施例及び態様は例証の目的に過ぎず、これらの記載に鑑みて種々の変形又は変更が当業者に示唆され、このような変形又は変更も本願の精神及び範囲と特許請求の範囲に含むものとする。
【0246】
以上、明確に理解できるように本発明を多少詳細に記載したが、本発明の真の範囲を逸脱することなく形態や細部に種々の変更が可能であることは以上の開示から当業者に自明である。例えば、上記全技術及び装置は種々に組合せて使用することができる。本明細書に引用した全刊行物、特許、特許出願、及び/又は他の文献はその開示内容全体を全目的で参考資料として組込み、各刊行物、特許、特許出願、及び/又は他の文献を全目的で参考資料として組込むと個々に記載しているものとして扱う。
【実施例9】
【0247】
ヌクレオチド及びアミノ酸配列
本実施例は夫々各種ポリヌクレオチド及びポリペプチドのヌクレオチド及びアミノ酸配列を示す。下表4に記載する配列は例示に過ぎず、表4に記載する配列に本発明を限定するものではない。
【表4−1】
【表4−2】
【表4−3】
【表4−4】
【表4−5】
【表4−6】
【表4−7】
【表4−8】
【表4−9】
【表4−10】
【特許請求の範囲】
【請求項1】
細胞で機能する第1の直交アミノアシルtRNAシンテターゼ(O−RS)を含む真正細菌細胞であって、前記O−RSが、アルキニルアミノ酸である第1の非天然アミノ酸で第1の直交tRNA(O−tRNA)を優先的にアミノアシル化する、細胞。
【請求項2】
真正細菌細胞が大腸菌細胞である、請求項1に記載の細胞。
【請求項3】
前記O−RSが、メタノコッカス・ヤナシイ(メタノコッカス・ヤナシイ(Methanococcus jannaschii))アミノアシルtRNAシンテターゼから誘導される、請求項1に記載の細胞。
【請求項4】
前記O−RSが、メタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導される、請求項1に記載の細胞。
【請求項5】
前記O−RSが、配列番号2のアミノ酸配列を有する野生型メタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導される、請求項1に記載の細胞。
【請求項6】
前記O−RSが、配列番号2のアミノ酸配列を有する野生型メタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導されると共に、
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリン
を含むアミノ酸配列を有する、請求項1に記載の細胞。
【請求項7】
前記O−RSが、配列番号4、6、8、10、12、14、16、18、及びその保存変異体から選択されるアミノ酸配列を含む、請求項1に記載の細胞。
【請求項8】
前記細胞が、前記O−RSをコードするポリヌクレオチドを含み、
前記O−RSが、配列番号4、6、8、10、12、14、16、18、及びその保存変異体から選択されるアミノ酸配列を含む、請求項1に記載の細胞。
【請求項9】
前記ポリヌクレオチドが、配列番号5、7、9、11、13、15、17又は19のヌクレオチド配列から選択される、請求項8に記載の細胞。
【請求項10】
前記O−tRNAが、アンバーサプレッサーtRNAである、請求項1に記載の細胞。
【請求項11】
前記O−tRNAが、配列番号1に記載のポリヌクレオチド配列を含むか、又は前記配列によりコードされる、請求項1に記載の細胞。
【請求項12】
前記アルキニルアミノ酸が、パラ−プロパルギルオキシフェニルアラニンである、請求項1に記載の細胞。
【請求項13】
少なくとも1個のセレクターコドンを含む核酸を含み、前記セレクターコドンが前記第1のO−tRNAにより認識される、請求項1に記載の細胞。
【請求項14】
第2のO−RS及び第2のO−tRNAを含み、
前記第2のO−RSが、前記第1の非天然アミノ酸とは異なる第2の非天然アミノ酸で、前記第2のO−tRNAを優先的にアミノアシル化し、
前記第2のO−tRNAが、前記第1のO−tRNAにより認識されるセレクターコドンとは異なるセレクターコドンを認識する、請求項13に記載の細胞。
【請求項15】
前記アルキニルアミノ酸を含む、請求項1に記載の細胞。
【請求項16】
前記アルキニルアミノ酸が、パラ−プロパルギルオキシフェニルアラニンである、請求項15に記載の細胞。
【請求項17】
翻訳系を含む、請求項1に記載の細胞。
【請求項18】
前記翻訳系が、
(a)前記O−RSと;
(b)前記O−tRNAと;
(c)目的のポリペプチドをコードし、前記O−tRNAにより認識される少なくとも1個のセレクターコドンを含む、核酸と;
(d)アルキニルアミノ酸とを含み、
前記O−RSが、前記O−tRNAに前記アルキニルアミノ酸を負荷し得る、請求項17に記載の細胞。
【請求項19】
配列番号2のメタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルアミノアシルtRNAシンテターゼから誘導されるポリペプチドであって、
前記ポリペプチドが、
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリン
を含むアミノ酸配列を有し、
前記ポリペプチドが、直交tRNA(O−tRNA)をアルキニルアミノ酸で優先的にアミノアシル化し得るアミノアシルtRNAシンテターゼである、ポリペプチド。
【請求項20】
配列番号4、6、8、10、12、14、16、18に記載のアミノ酸配列又はその保存変異体を含むポリペプチド。
【請求項21】
前記ポリペプチドが、真正細菌細胞において直交tRNA(O−tRNA)をアルキニルアミノ酸で優先的にアミノアシル化し得るアミノアシルtRNAシンテターゼである、請求項20に記載のポリペプチド。
【請求項22】
請求項19、20又は21に記載のポリペプチドをコードするポリヌクレオチド。
【請求項23】
前記ポリヌクレオチドが、配列番号5、7、9、11、13、15、17及び19から選択される、請求項22に記載のポリヌクレオチド。
【請求項24】
請求項22に記載のポリヌクレオチドを含むベクター。
【請求項25】
ベクターが発現ベクターである、請求項24に記載のベクター。
【請求項26】
請求項24に記載のベクターを含む細胞。
【請求項27】
特定位置に非天然アルキニルアミノ酸を含む蛋白質を真正細菌細胞で生産する方法であって、前記方法が、
(a)真正細菌細胞を提供し、前記真正細菌細胞は、
(i)直交アミノアシルtRNAシンテターゼ(O−RS)と、
(ii)直交tRNA(O−tRNA)であって、前記O−RSにより前記アルキニルアミノ酸で優先的にアミノアシル化される、O−tRNAと、
(iii)前記蛋白質をコードする核酸であって、前記O−tRNAにより認識される少なくとも1個のセレクターコドンを含む核酸と、
(iv)アルキニルアミノ酸とを含み、
(b)前記細胞を増殖させ、
(c)前記核酸によりコードされる蛋白質の翻訳中に、前記核酸における前記セレクターコドンの位置に対応する特定位置に前記アルキニルアミノ酸を組込むことにより、前記アルキニルアミノ酸を前記特定位置に含む蛋白質を生産することを含む、方法。
【請求項28】
前記真正細菌細胞が大腸菌細胞である、請求項27に記載の方法。
【請求項29】
前記O−RSが、メタノコッカス・ヤナシイ(Methanococcus jannaschii)アミノアシルtRNAシンテターゼから誘導される、請求項27に記載の方法。
【請求項30】
前記O−RSが、メタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導される、請求項27に記載の方法。
【請求項31】
前記O−RSが、配列番号2のメタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導される、請求項27に記載の方法。
【請求項32】
前記O−RSが、配列番号2のメタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導され、前記O−RSが、
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリン
を含むアミノ酸配列を有する、請求項27に記載の方法。
【請求項33】
前記O−RSが、配列番号4、6、8、10、12、14、16、18、及びその保存変異体から選択されるアミノ酸配列を含む、請求項27に記載の方法。
【請求項34】
前記細胞が、配列番号4、6、8、10、12、14、16、18、及びその保存変異体から選択されるアミノ酸配列を含む前記O−RSをコードするポリヌクレオチドを含む、請求項27に記載の方法。
【請求項35】
前記ポリヌクレオチドが、配列番号5、7、9、11、13、15、17又は19のヌクレオチド配列から選択される、請求項34に記載の方法。
【請求項36】
前記O−tRNAがアンバーサプレッサーtRNAであり、前記セレクターコドンがアンバー終止コドン(TAG)である、請求項27に記載の方法。
【請求項37】
前記O−tRNAが、配列番号1に記載のポリヌクレオチド配列を含むか、又は前記配列によりコードされる、請求項27に記載の方法。
【請求項38】
前記アルキニルアミノ酸が、パラ−プロパルギルオキシフェニルアラニンである、請求項27に記載の方法。
【請求項39】
前記蛋白質が、野生型治療用蛋白質、診断用蛋白質、産業用酵素、又はその一部のアミノ酸配列と少なくとも75%の同一性を有するアミノ酸配列を含む、請求項27に記載の方法。
【請求項40】
前記蛋白質が、医薬的に許容可能なキャリヤーと結合している、請求項39に記載の方法。
【請求項41】
前記蛋白質の前記特定位置を修飾する、請求項27に記載の方法。
【請求項42】
前記蛋白質が、前記特定位置にトリアゾール結合を含む請求項41に記載の方法。
【請求項1】
細胞で機能する第1の直交アミノアシルtRNAシンテターゼ(O−RS)を含む真正細菌細胞であって、前記O−RSが、アルキニルアミノ酸である第1の非天然アミノ酸で第1の直交tRNA(O−tRNA)を優先的にアミノアシル化する、細胞。
【請求項2】
真正細菌細胞が大腸菌細胞である、請求項1に記載の細胞。
【請求項3】
前記O−RSが、メタノコッカス・ヤナシイ(メタノコッカス・ヤナシイ(Methanococcus jannaschii))アミノアシルtRNAシンテターゼから誘導される、請求項1に記載の細胞。
【請求項4】
前記O−RSが、メタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導される、請求項1に記載の細胞。
【請求項5】
前記O−RSが、配列番号2のアミノ酸配列を有する野生型メタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導される、請求項1に記載の細胞。
【請求項6】
前記O−RSが、配列番号2のアミノ酸配列を有する野生型メタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導されると共に、
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリン
を含むアミノ酸配列を有する、請求項1に記載の細胞。
【請求項7】
前記O−RSが、配列番号4、6、8、10、12、14、16、18、及びその保存変異体から選択されるアミノ酸配列を含む、請求項1に記載の細胞。
【請求項8】
前記細胞が、前記O−RSをコードするポリヌクレオチドを含み、
前記O−RSが、配列番号4、6、8、10、12、14、16、18、及びその保存変異体から選択されるアミノ酸配列を含む、請求項1に記載の細胞。
【請求項9】
前記ポリヌクレオチドが、配列番号5、7、9、11、13、15、17又は19のヌクレオチド配列から選択される、請求項8に記載の細胞。
【請求項10】
前記O−tRNAが、アンバーサプレッサーtRNAである、請求項1に記載の細胞。
【請求項11】
前記O−tRNAが、配列番号1に記載のポリヌクレオチド配列を含むか、又は前記配列によりコードされる、請求項1に記載の細胞。
【請求項12】
前記アルキニルアミノ酸が、パラ−プロパルギルオキシフェニルアラニンである、請求項1に記載の細胞。
【請求項13】
少なくとも1個のセレクターコドンを含む核酸を含み、前記セレクターコドンが前記第1のO−tRNAにより認識される、請求項1に記載の細胞。
【請求項14】
第2のO−RS及び第2のO−tRNAを含み、
前記第2のO−RSが、前記第1の非天然アミノ酸とは異なる第2の非天然アミノ酸で、前記第2のO−tRNAを優先的にアミノアシル化し、
前記第2のO−tRNAが、前記第1のO−tRNAにより認識されるセレクターコドンとは異なるセレクターコドンを認識する、請求項13に記載の細胞。
【請求項15】
前記アルキニルアミノ酸を含む、請求項1に記載の細胞。
【請求項16】
前記アルキニルアミノ酸が、パラ−プロパルギルオキシフェニルアラニンである、請求項15に記載の細胞。
【請求項17】
翻訳系を含む、請求項1に記載の細胞。
【請求項18】
前記翻訳系が、
(a)前記O−RSと;
(b)前記O−tRNAと;
(c)目的のポリペプチドをコードし、前記O−tRNAにより認識される少なくとも1個のセレクターコドンを含む、核酸と;
(d)アルキニルアミノ酸とを含み、
前記O−RSが、前記O−tRNAに前記アルキニルアミノ酸を負荷し得る、請求項17に記載の細胞。
【請求項19】
配列番号2のメタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルアミノアシルtRNAシンテターゼから誘導されるポリペプチドであって、
前記ポリペプチドが、
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリン
を含むアミノ酸配列を有し、
前記ポリペプチドが、直交tRNA(O−tRNA)をアルキニルアミノ酸で優先的にアミノアシル化し得るアミノアシルtRNAシンテターゼである、ポリペプチド。
【請求項20】
配列番号4、6、8、10、12、14、16、18に記載のアミノ酸配列又はその保存変異体を含むポリペプチド。
【請求項21】
前記ポリペプチドが、真正細菌細胞において直交tRNA(O−tRNA)をアルキニルアミノ酸で優先的にアミノアシル化し得るアミノアシルtRNAシンテターゼである、請求項20に記載のポリペプチド。
【請求項22】
請求項19、20又は21に記載のポリペプチドをコードするポリヌクレオチド。
【請求項23】
前記ポリヌクレオチドが、配列番号5、7、9、11、13、15、17及び19から選択される、請求項22に記載のポリヌクレオチド。
【請求項24】
請求項22に記載のポリヌクレオチドを含むベクター。
【請求項25】
ベクターが発現ベクターである、請求項24に記載のベクター。
【請求項26】
請求項24に記載のベクターを含む細胞。
【請求項27】
特定位置に非天然アルキニルアミノ酸を含む蛋白質を真正細菌細胞で生産する方法であって、前記方法が、
(a)真正細菌細胞を提供し、前記真正細菌細胞は、
(i)直交アミノアシルtRNAシンテターゼ(O−RS)と、
(ii)直交tRNA(O−tRNA)であって、前記O−RSにより前記アルキニルアミノ酸で優先的にアミノアシル化される、O−tRNAと、
(iii)前記蛋白質をコードする核酸であって、前記O−tRNAにより認識される少なくとも1個のセレクターコドンを含む核酸と、
(iv)アルキニルアミノ酸とを含み、
(b)前記細胞を増殖させ、
(c)前記核酸によりコードされる蛋白質の翻訳中に、前記核酸における前記セレクターコドンの位置に対応する特定位置に前記アルキニルアミノ酸を組込むことにより、前記アルキニルアミノ酸を前記特定位置に含む蛋白質を生産することを含む、方法。
【請求項28】
前記真正細菌細胞が大腸菌細胞である、請求項27に記載の方法。
【請求項29】
前記O−RSが、メタノコッカス・ヤナシイ(Methanococcus jannaschii)アミノアシルtRNAシンテターゼから誘導される、請求項27に記載の方法。
【請求項30】
前記O−RSが、メタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導される、請求項27に記載の方法。
【請求項31】
前記O−RSが、配列番号2のメタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導される、請求項27に記載の方法。
【請求項32】
前記O−RSが、配列番号2のメタノコッカス・ヤナシイ(Methanococcus jannaschii)チロシルtRNAシンテターゼから誘導され、前記O−RSが、
(a)アミノ酸32位にアラニン;
(b)アミノ酸107位にプロリン又はグルタミン;
(c)アミノ酸158位にアラニン;及び
(d)アミノ酸162位にアラニン又はプロリン
を含むアミノ酸配列を有する、請求項27に記載の方法。
【請求項33】
前記O−RSが、配列番号4、6、8、10、12、14、16、18、及びその保存変異体から選択されるアミノ酸配列を含む、請求項27に記載の方法。
【請求項34】
前記細胞が、配列番号4、6、8、10、12、14、16、18、及びその保存変異体から選択されるアミノ酸配列を含む前記O−RSをコードするポリヌクレオチドを含む、請求項27に記載の方法。
【請求項35】
前記ポリヌクレオチドが、配列番号5、7、9、11、13、15、17又は19のヌクレオチド配列から選択される、請求項34に記載の方法。
【請求項36】
前記O−tRNAがアンバーサプレッサーtRNAであり、前記セレクターコドンがアンバー終止コドン(TAG)である、請求項27に記載の方法。
【請求項37】
前記O−tRNAが、配列番号1に記載のポリヌクレオチド配列を含むか、又は前記配列によりコードされる、請求項27に記載の方法。
【請求項38】
前記アルキニルアミノ酸が、パラ−プロパルギルオキシフェニルアラニンである、請求項27に記載の方法。
【請求項39】
前記蛋白質が、野生型治療用蛋白質、診断用蛋白質、産業用酵素、又はその一部のアミノ酸配列と少なくとも75%の同一性を有するアミノ酸配列を含む、請求項27に記載の方法。
【請求項40】
前記蛋白質が、医薬的に許容可能なキャリヤーと結合している、請求項39に記載の方法。
【請求項41】
前記蛋白質の前記特定位置を修飾する、請求項27に記載の方法。
【請求項42】
前記蛋白質が、前記特定位置にトリアゾール結合を含む請求項41に記載の方法。
【図1】

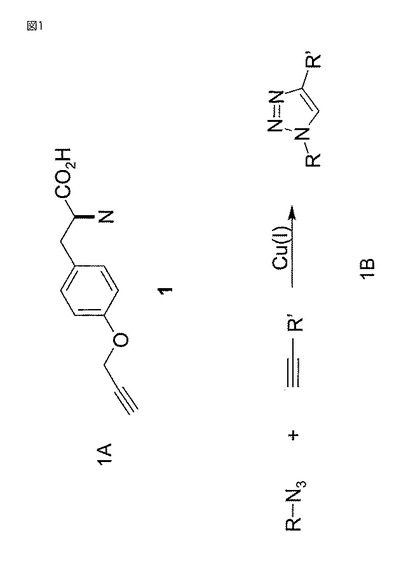
【図2】

【図3】
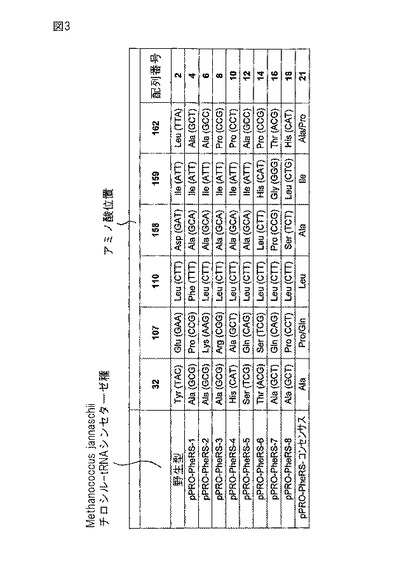
【図5】
【図6】
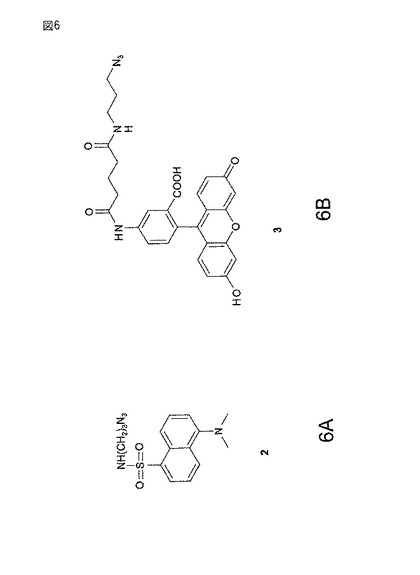
【図8】
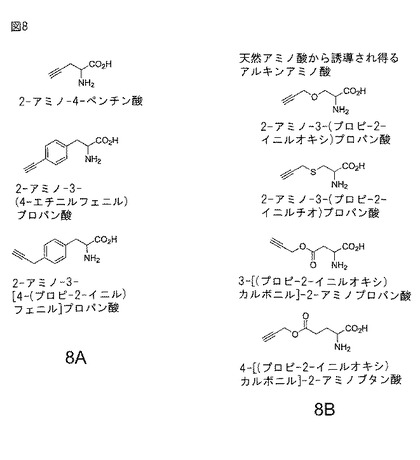
【図4】
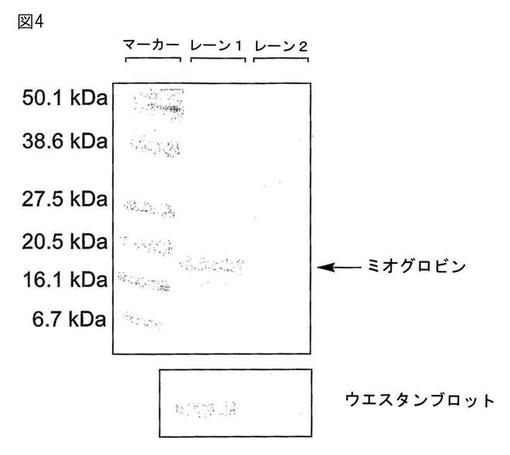
【図7】
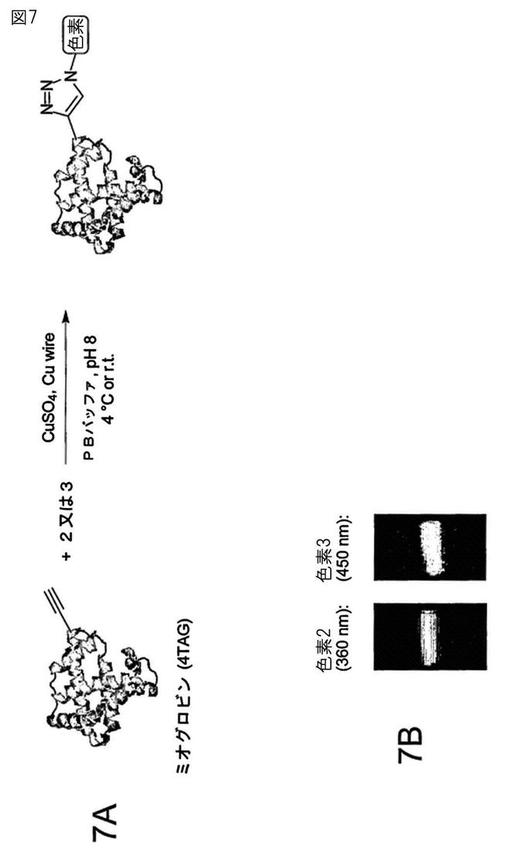
【図2】
【図3】
【図5】
【図6】
【図8】
【図4】
【図7】
【公開番号】特開2013−9682(P2013−9682A)
【公開日】平成25年1月17日(2013.1.17)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−205802(P2012−205802)
【出願日】平成24年9月19日(2012.9.19)
【分割の表示】特願2007−533599(P2007−533599)の分割
【原出願日】平成17年9月20日(2005.9.20)
【出願人】(399038620)ザ スクリプス リサーチ インスティチュート (51)
【Fターム(参考)】
【公開日】平成25年1月17日(2013.1.17)
【国際特許分類】
【出願番号】特願2012−205802(P2012−205802)
【出願日】平成24年9月19日(2012.9.19)
【分割の表示】特願2007−533599(P2007−533599)の分割
【原出願日】平成17年9月20日(2005.9.20)
【出願人】(399038620)ザ スクリプス リサーチ インスティチュート (51)
【Fターム(参考)】
[ Back to top ]