
神経保護医薬組成物の製造のためのプロインスリンの使用、それを含む治療組成物、およびそれらの応用
本発明は、プロインスリン、好ましくはヒトプロインスリンの活性を誘発して、プログラム細胞死、好ましくは中枢および末梢神経系の神経変性病変、更に好ましくは網膜色素変性症として知られる異形変性疾患などの神経変性状態、障害または疾患の予防および治療用の医薬品または医薬組成物を製造する化合物の使用に関する。アクチベーター化合物は、化学分子、ペプチド、タンパク質、またはヌクレオチド配列からなってもよい。
【発明の詳細な説明】
【発明の背景】
【0001】
発明の分野
本発明は、生物医学分野に含まれ、更に具体的には治療化合物の開発に含まれる。本発明は、詳細には網膜色素変性症(retinitis pigmentosa)のような網膜変性疾患並びに他の神経変性疾患の治療用の医薬品の製造のためのプロインスリン分子の特定の使用に関する。
【0002】
背景技術
神経変性疾患は、遺伝性(遺伝的)、外傷性、散発性、または老年性起源の中枢神経系の様々な進行性疾患を包含する。神経変性疾患のほとんどは、それらの起源またはそれらの進行においてニューロンおよびグリア細胞のプログラム細胞死を示す。神経組織に対する損傷の不可逆的な段階を構成する前記の死の工程は、遺伝性、外傷性、散発性、または老年性であるかに関わらず疾患の主要な原因とは独立していると思われる。
【0003】
多数の増殖因子は、ニューロンやグリア細胞など様々な種類の細胞の生と死とのバランス調節に本質的な役割を果たしている。これらには、インスリン、その前駆体であるプロインスリン並びにIGF-IおよびIGF-IIなどのインスリンファミリーのものが含まれる(Varela-Nieto, I., de la Rosa, E. J., Valenciano, A. I., and Leon, Y. (2003) Cell death in the nervous system: lessons from insulin and insulin-like growth factors.Mol Neurobiol 28: 23-50)。網膜は中枢神経系の一部を構成しており、神経系の生理学的および病理学的過程の双方の研究に詳細に確認されているモデルであり、この理由により、これは本発明において用いられる細胞モデルである。網膜において検討された最も重要な病理学的過程の一つはいわゆる網膜色素変性症であり、この病状は包括的な群の遺伝性網膜障害を含んでなり、しかも世界における失明の最大原因の一つであり、概算の発生率は4,000人に1人であるからである。120を上回る関連の遺伝子座は特定されており、原因は様々であるが、いずれの場合にもプログラム細胞死による網膜ニューロン、特に光受容体の慢性的且つ進行性の喪失があり、ヒトを失明させる。現在のところ、網膜色素変性症には治療法はなく、さしあたりrdマウス、RCSラット、並びにイヌ、ネコおよびショウジョウバエでの他のモデルのような変性を起こしている動物モデルにおいて神経保護、遺伝子療法、神経修復、および生体工学的方法が行われているだけである。rd1(桿状体変性)マウスは、細胞変性を決定する分子および細胞機構を研究するための最初のモデルの一つであり、光受容体の死のアポトーシス性が確定されている(Chang, G. Q., Hao, Y., and Wong, F. (1993) Apoptosis: final common pathway of photoreceptor death in rd, rds, and rhodopsin mutant mice.Neuron 11: 595-605)。様々なrdマウスは、遺伝性の網膜ジストロフィーの治療に対する新規治療方法を試験するための理想的モデルを提供しているが、これらのマウスを用いて分子、細胞、および遺伝子の観点から光受容体の変性過程を検討することができるからである。
【0004】
遺伝子療法を介在することにより、神経変性を引き起こす突然変異遺伝子の機能的コピーの再導入に役立つ。組換えアデノウイルスまたはアデノウイルス関連のウイルスベクターにより、伸展が図られた。具体的には、新生rdマウスの桿状体特異的cGMPホスホジエステラーゼ(βPDE)のβ-サブユニットの置換により、rd表現型の組織を少なくとも6週間救済することができた(Bennett, J., Tanabe, T., Sun, D., Zeng, Y., Kjeldbye, H., Gouras, P., and Maguire, A. M. (1996) Photoreceptor cell rescue in retinal degeneration (rd) mice by in vivo gene therapy. Nat Med 2: 649-654)。しかしながら、この療法はそれぞれの親における突然変異遺伝子を明確に同定する必要があるが、これは現在のところ症例の40%で可能なだけである(Wang, D. Y., Chan, W. M., Tam, P.O., Baum, L., Lam, D. S., Chong, K. K., Fan, B. J., and Pang, CP. (2005) Gene mutations in retinosis pigmentosa and their clinical implications. Clin Chim Acta 351: 5-16)。
【0005】
新規な光受容体を開発するための神経幹細胞または前駆体の移植片は、神経修復療法の目的である。新規な光受容体は、内網膜のニューロンと適当な結合を回復しなければならない。
【0006】
増殖因子の投与により誘発される神経保護は、神経変性過程に関連した細胞死の防止を目指す。様々な投与形態が、網膜変性に罹っている数種類の動物モデルにおいて試験されている。最初の試みは、網膜変性に罹っているラットまたはマウスでの数種類の組換えタンパク質の硝子体への注射からなっていた(Faktorovich, E. G., Steinberg, R. H., Yasumura, D., Matthes, M. T., and LaVail, M. M. (1990) Photoreceptor degeneration in inherited retinal dystrophy delayed by basic fibroblast growth factor. Nature 347: 83-86、 LaVail, M. M., Unoki, K., Yasumura, D., Matthes, M. T., Yancopoulos, G. D., and Steinberg, R. H. (1992) Multiple growth factors, cytokines, and neurotrophins rescue photoreceptors from the damaging effects of constant light. Proc Nati Acad Sci U S A 89: 11249-11253)。これらの実験は、FGF2がRCSラットにおける光受容体変性を遅らせることを明らかにした(Royal Collage of Surgeon)。FGF2、FGF1、BDNF、およびCNTFなどの数種類の生存因子は、光損傷によって誘発される光受容体の死を減少させた(LaVail, M. M., Yasumura, D., Matthes, M. T., Lau-Villacorta, C, Unoki, K., Sung, CH., and Steinberg, R. H. (1998) Protection of mouse photoreceptors by survival factors in retinal degenerations. Invest Ophthalmol Vis Sci 39: 592-602)。遺伝性網膜変性に罹っているマウスモデル(Q433ter、rd、およびnrモデル、網膜色素変性症のある種のマウスモデルを表すのに用いられる用語)にCNTF類似体を硝子体内投与することによって、幾つかの変性が明らかに改善された。CNTF類似体による神経保護効果も、常染色体優性錐体-桿状体ジストロフィーのネコでの研究においてCNTFの硝子体内投与が有効な効果を有することを実証した(Chong, N. H., Alexander, R. A., Waters, L., Barnett, K. C, Bird, A. C, and Luthert, P. J. (1999) Repeated injections of a ciliary neurotrophic factor analogue leading to long-term photoreceptor survival in hereditary retinal degeneration.Invest Ophthalmol Vis Sci 40: 1298-1305)。
【0007】
もう一つの方法は、網膜変性マウスまたはラットの網膜における生存因子を発現するための遺伝子療法ベクターの使用であった。2種類のマウスモデルでは、rdおよびPrph2Rd(ペリフェリンにおける劣性突然変異) (Travis, G. H., Brennan, M. B., Danielson, P. E., Kozak, CA., and Sutcliffe, J. G. (1989) Identification of a photoreceptor-specific mRNA encoded by the gene responsible for retinal degeneration slow (rds). Nature 338: 70-73、 Connell, G., Bascom, R., Molday, L., Reid, D., Mclnnes, R. R., and Molday, R. S. (1991) Photoreceptor peripherin is the normal product of the gene responsible for retinal degeneration in the rds mouse. Proc Nati Acad Sci USA 88: 723-726)、CNTFの分泌可能な形態をコードするアデノウイルスベクターの網膜下投与により、光受容体の死を遅らせた(Cayouette, M., and Gravel, C. (1997) Adenovirus-mediated gene transfer of ciliary neurotrophic factor can prevent photoreceptor degeneration in the retinal degeneration (rd) mouse. Hum Gene Ther 8: 423- 430、 Cayouette, M., Behn, D., Sendtner, M., Lachapelle, P., and Gravel, C. (1998) Infraocular gene transfer of ciliary neurotrophic factor prevents death and increases responsiveness of rod photoreceptors in the retinal degeneration slow mouse. J Neurosci 18: 9282-9293)。一方、硝子体内投与によってほとんど活性にならない網膜におけるトランスジェニックBDNF発現は、Q344terマウスの神経変性を遅らせる(Okoye, G., Zimmer, J., Sung, J., Gehlbach, P., Deering, T., Nambu, H., Hackett, S., Melia, M., Esumi, N., Zack, D. J., and Campochiaro, P. A. (2003) Increased expression of brain-derived neurotrophic factor preserves retinal function and slows cell death from rhodopsin mutation or oxidative damage. J Neurosci 23: 4164- 4172、 Sung, CH. , Makino, C, Baylor, D., and Nathans, J. (1994) A rhodopsin gene mutation responsible for autosomal dominant retinosis pigmentosa results in a protein that is defective in localization to the photoreceptor outer segment. J Neurosci 14: 5818-5833)。これらの結果は、技術的問題を別として、硝子体内投与だけでは神経保護効果を有するには不十分である可能性があることも示している。更に、単回投与で神経保護効果をも示すCNTFは、網膜変性に罹っているPrph2Rdマウスおよび2種類のトランスジェニックラットでCNTFの分泌可能な形態をコードするAAVベクターによってその長時間発現が誘発されるときには逆効果であることが明らかにされており(Liang, F. Q., Aleman, T. S., Dejneka, N. S., Dudus, L., Fisher, K. J., Maguire, A. M., Jacobson, S. G., and Bennett, J. (2001) Long-term protection of retinal structure but not function using RAAV. CNTF in animal models of retinosis pigmentaria. Mol Ther 4: 461-472)、これは、光受容体の機能はERGによる分析によって悪化するためである。
【0008】
上記の総ての方法を考慮して、本発明は、今日まで用いられてきたシステムと比較して実際的な解決法を提供する。ニワトリ胚でのインスリンの代謝機能とは異なるインスリンの生存機能は以前に示されており、これは、プロインスリン形態でのインスリンが膵臓外形の存在以前に発生中に発現されるからである。神経系の発生中に、プロインスリンは多数の細胞過程を制御する。これは、アンチセンスオリゴヌクレオチドを用いることによるプロインスリン遺伝子またはその受容体の発現を阻害する研究において明らかにされたように、初期胚での生存因子である(Morales, A.V., Serna, J., Alarcon, C, de la Rosa, E. J., and de Pablo, F. (1997) Role of prepancreatic (pro)insulin and the insulin receptor in prevention of embryonic apoptosis. Endocrinology 138: 3967-3975)。抗体によりそれをブロックすることによっても、ニワトリ胚網膜でのアポトーシス細胞の数が増加するが(Diaz, B., Serna, J., De Pablo, F., and de la Rosa, E. J. (2000) In vivo regulation of cell death by embryonic (pro) insulin and the insulin receptor during early retinal neurogenesis. Development 127: 1641-1649)、胚への外因不活性によりアポトーシス細胞の数が減少する(Hernandez-Sanchez, C, Mansilla, A., de la Rosa, E. J., Pollerberg, G. E., Martinez- Salas, E., and de Pablo, F. (2003) Upstream AUGs in embryonic proinsulin mRNA control its low translation level. Embo J 22: 5582-5592)。ニワトリ胚から単離することができる分子は、遺伝子翻訳の第一の産物であるプロインスリンであり、その代謝活性は小さく、インスリンの活性の約5-10%である。
【0009】
慢性神経変性疾患では、ニューロンおよび/またはグリア細胞は次第に死滅する。この疾患の症状は、通常極めて僅かの細胞が死んだときに現れる。従って、比較的正常な視覚機能を維持する上で細胞生存に有利な効果的分子を決定することが重要である。それぞれが異なる病因を有する多種類の神経変性疾患があるが、それらはいずれも最終的には罹患細胞の死をもたらす。この死はプログラムされた種類のものであり、異なる種類のアポトーシスまたは非アポトーシスによる死もある。細胞が損傷前は最初のうち、良好な状態である急性損傷とは異なり、慢性疾患においては、細胞は内因性の損傷を有しており、細胞が最早損傷を維持しきれなくなり、および/または細胞が行わなければならない機能を実行することができなくなったある時点において細胞が死亡する。
【発明の概要】
【0010】
本発明の目的は、プログラム細胞死が起こる神経変性状態、障害、または疾患、好ましくは中枢および末梢神経系の神経変性疾患、更に好ましくは網膜色素変性症として知られる遺伝性変性疾患の群の予防および治療用の医薬品または医薬組成物を製造するためのプロインスリンの活性を誘発する化合物の使用(以後、本発明の誘発化合物の使用)によって構成される。本発明の好ましい態様では、上記医薬組成物は、その全身または局所への持続投与に適する。
【0011】
本発明の特定の目的は、プロインスリンの活性を誘発する化合物であって、誘発化合物がヌクレオチド配列であるものの使用(以後、本発明のプロインスリンヌクレオチド配列の使用)であり、神経保護タンパク質またはペプチドを発現させ且つ下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一方の断片、並びに
d) a)、b)および/またはc)に属するいずれか1個の配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列によって構成されるものの使用によって構成される。
【0012】
本発明の特定の態様は、プロインスリンの活性を誘発する化合物であって、ヌクレオチド配列がヒトプロインスリンをコードする配列番号1によって構成されているものの使用によって構成される。
本発明のもう一つの特定の態様は、プロインスリンの活性を誘発する化合物であって、d)のヌクレオチド配列が発現ベクターであるものの使用(以後、本発明のプロインスリン発現ベクターの使用)であって、神経保護を誘発できるプロインスリンタンパク質をコードするヌクレオチド配列または遺伝子構築物を含んでなることによって構成される。
【0013】
本発明のもう一つの特定の態様は、プロインスリンの活性を誘発する化合物であって、誘発化合物が好ましくはヒト真核細胞であるものの使用(以後、本発明のプロインスリン細胞の使用)であって、遺伝的に修飾され、且つ本発明のプロインスリンヌクレオチド配列、構築物、または発現ベクターを含んでなり、プロインスリンタンパク質を細胞外媒質に好適に発現しまたは放出できることによって構成される。
【0014】
本発明のもう一つの特定の目的は、プロインスリンの活性を誘発する化合物であって、誘発化合物がタンパク質またはペプチドであるものの使用(以後、本発明のプロインスリンタンパク質の使用)であり、神経保護活性を有し且つ下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つを含んでなるアミノ酸配列
に属する1または数個のアミノ酸配列を含んでなる化合物の使用によって構成される。
【0015】
本発明のもう一つの特定の態様は、本発明の誘発化合物であって、誘発化合物がヒトプロインスリンタンパク質(配列番号2)であるものの使用によって構成される。
【0016】
本発明のもう一つの目的は、神経退行変性をもたらす疾患、障害、または病変の治療用の医薬組成物または医薬品(以後、本発明の医薬組成物)であって、治療上有効量の本発明のプロインスリンの活性を誘発する化合物を場合によっては1種類以上の薬学上許容可能なアジュバントおよび/または担体と共に含んでなるものによって構成される。
【0017】
本発明の特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つの配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列であるものによって構成される。
【0018】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、ヌクレオチド配列がヒトプロインスリンをコードする配列番号1からなる医薬組成物によって構成される。
【0019】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、ヌクレオチド配列がヒトプロインスリン発現ベクターである医薬組成物によって構成される。
【0020】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が本発明のプロインスリン配列、遺伝子構築物、またはベクターによってコードされるタンパク質またはペプチドである医薬組成物によって構成される。
【0021】
本発明の特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発するタンパク質またはペプチドが、下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つの配列を含んでなるアミノ酸配列
に属する医薬組成物によって構成される。
【0022】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、アミノ酸配列がヒトプロインスリン(配列番号2)によって構成される医薬組成物によって構成される。
【0023】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が好ましくはヒト細胞、更に好ましくは本発明のプロインスリン配列、遺伝子構築物、または発現ベクターによって形質転換された中枢神経系細胞である医薬組成物によって構成される。
【0024】
本発明のもう一つの目的は、プログラム細胞死が起こるヒトを冒す中枢または末梢神経系の神経変性疾患、障害、または病変に冒されている哺乳類、好ましくはヒトの治療または予防方法において、本発明の医薬組成物の使用(以後、本発明の医薬組成物の使用)であって、上記治療組成物を上記神経変性を減少させる適量により投与することからなる使用によって構成される。
【図面の簡単な説明】
【0025】
【図1】ヒトプロインスリンのcDNAおよびプラスミドpMLC-hInsのインサートの配置図(schematic representation)。(A) プラスミドに挿入されているDNA配列の配置図。ヒトプロインスリン遺伝子のcDNAに相当。図は、3個のエキソン(エキソン1= E1、エキソン2=E2、エキソン3=E3)によって形成される462塩基対(bp)のmRNAを示す。挿入されたDNA配列は、翻訳されているものであり、347bpのORF(オープンリーディングフレーム)である。未翻訳のフランキング部分(5'UTRおよび3'UTR)は、構築物に挿入されない。翻訳されたタンパク質は、110個のアミノ酸(aa)のプレプロインスリンである。このタンパク質は、シグナルペプチド(signal pep.)、鎖B、ペプチドC、および鎖Aからなる。シグナルペプチドは除かれ、プロインスリン分子が残る。(B) プラスミドは1.A項において記載したインサートを含み、110アミノ酸プレプロインスリン(黒色の太い部分)を横紋筋ミオシン軽鎖線維の構成的筋肉プロモーターMLC1(ミオシン軽鎖)の転写制御下でコードする。このプラスミドは、rd10についてホモ接合に到達するように交雑した2系列のトランスジェニックヒトプロインスリン産生マウスを産生するのに用いられる。
【図2】P32でのマウスの12μmのクリオスタットの眼切片(cryostat eye section)であって、様々なマーカーにより変性の進行を示す桿状体および錐体の状態を観察することができる。野生型コントロールマウス(AおよびD)、プロインスリンを産生するホモ接合rd10およびトランスジェニックマウス、Proins/rd10-/-(BおよびE)、およびコントロールホモ接合rd10マウス(CおよびF)。桿状体および錐体の核が配置されている外顆粒層(ONL)をDAPIによる核染色によって見ることができる。内顆粒層(INL)には、二極性、無軸索、水平、およびミュラー細胞の核が配置されている。蛍光色素 Alexa 488で標識した凝集素であって、錐体の外側部分(上の矢印)および錐体のシナプス基部(下の矢印)を示すその反応を用いて錐体を標識する。棒線は45μmを表す。
【図3】P32におけるマウスの12μmの眼のクリオスタット切片であって、シナプス結合状態を観察することができる。野生型コントロールマウス(A)、プロインスリンを産生するホモ接合rd10およびトランスジェニックマウスProins/rd10-/- (B)、およびコントロールホモ接合rd10マウス(C)。顆粒層ONL、INL、およびガングリオン細胞層(GCL)を示す。シナプス結合が起こる網状層であり、外網状層(OPL)、および内網状層(IPL)はそれらの間に配置される。この図は、一般にシナプス結合を標識するSV2抗体による免疫組織化学的標識を示す。棒線は0.45μmを表す。
【図4】P30で行った網膜電位記録の結果。野生型マウス(wt)、コントロールRd10-/-マウス、およびProins/Rd10-/-マウスであり、いずれも系列1(L1)および系列2(L2)由来。暗順応(夜間)で記録した桿状体に生じた網膜電位応答(上列)および混合応答(中列)の例は、それぞれの動物について示される。明所視(昼間)状態で記録した錐体に生じた網膜電位応答(下列)も示す。両系列(L1およびL2)由来のProins/rd10-/-マウスで得た応答範囲が、rd10-/-マウスの応答と比較して大きいことに留意すべきである。
【図5】ヒトプロインスリンレベルと視覚パラメーターの保持との間の相関。Proins/Rd10-/-マウスにおけるヒトプロインスリンレベルを、P30において完全な網膜電位検討を行ったマウスの大腿四頭筋でP32において測定した。プロインスリンおよび様々な視覚パラメーターのそれぞれの値は、双曲線となる。網膜電位波の振幅bmax(1.5 log cd.s.m-2に応じて記録)、OP(振動電位)、bphot(1.5 log cd.s.m-2に応じて記録)、およびフリッカー応答を示す。
【図6】様々な日に行った網膜電位記録の結果。産後発生の様々な日、すなわちP35、P45、P55の野生型マウス(wt)、コントロールRd10-/-マウス、およびProins/Rd10-/-マウス。Aは、桿状体(-2,55 log cd.s.m-2)を除く暗順応状態で記録した応答、およびそれぞれの動物種および産後発生の様々な時期における混合応答(1,48 log cd.s.m-2)の例を示す。Bは、それぞれの動物種および産後発生の様々な時期において錐体に生じた(1.48 log cd.s.m-2)明所視状態で記録した応答の例を示す。
【図7】ヒトプロインスリンを皮下注射した後の網膜における存在。抽出物調製の2時間前にまたは毎日、P11とP14の間においてプロインスリンの所定量を皮下投与したC57B1/6マウスの網膜抽出物におけるヒトプロインスリンのELISAによる定量。最後の投与も抽出物調製の2時間前。
【図8】神経変性マウスの網膜組織構造に対するヒトプロインスリンの皮下投与の効果。P6と、P14またはP18との間の12時間毎にヒトプロインスリン(BおよびD)または担体(AおよびC)を投与したP14のRd1-/-マウスの網膜切片。外顆粒層における光受容体の喪失は、総ての場合にDAPIを用いる核染色によって観察することができる。ONL,外顆粒層、 INL, 内顆粒層、 GCL, ガングリオン細胞層。棒線は25μmを表す。
【図9】神経変性マウスの網膜組織構造に対するヒトプロインスリンの皮下投与の効果。P6と、P14またはP18との間の12時間毎にヒトプロインスリン(BおよびD)または担体(AおよびC)を投与したRd1-/-マウスの網膜電位図。様々な治療を施した同腹兄弟のrd突然変異マウスの網膜電位図を示す。プロインスリンを投与しても、視覚機能喪失過程を示さなかった。
【発明の具体的説明】
【0026】
ほとんどの神経変性疾患、特に網膜色素変性症には効率的および/または効果的な治療法がないことを考慮し、本発明は代替解決法を提供する。
【0027】
本発明は、プロインスリン(通常はインスリンの前駆体型として知られているインスリンファミリーの増殖因子)が、慢性神経変性過程、特に網膜色素変性症で起こる網膜神経変性過程における更なる細胞生存因子であるという事実に基づく。
【0028】
桿状体特異的サイクリックGMPホスホジエステラーゼ酵素遺伝子に劣性ホモ接合突然変異を有し、この酵素の機能を変更させ、進行性の光受容体細胞死と続いて残りの網膜細胞の種類、主に錐体の二次的変性をもたらすrd10 (Pdebrd10/rd10)およびrd1 (Pdebrd1/rd1)マウスを、網膜変性モデルとして選択した。
【0029】
ヒトプロインスリンタンパク質と、rd10突然変異に対する劣性ホモ接合体とを発現する2系列のトランスジェニックマウスProins/rd10-/-を生成した(実施例1.1)。網膜変性に罹っているこれらのProins/rd10-/-マウスは横紋筋でヒトプロインスリンを構成的に生成し(これはインスリンには処理されない)、これは血清中に検出され、グルコースの水準により膵臓の通常の制御を受けにくい。これにより、動物に用量、投与前の産物の状態(それは内因性の産生に由来するので、商業産物のようには分解しない)、並びに投与の形態および時間に依存することなく、循環しているヒトプロインスリンの水準を持続させる。これにより、その投与方法を見出すための薬物動態検討を行わなくともよくなった。
【0030】
両系統のProins/rd10-/-マウスの筋肉および血清にヒトプロインスリンが含まれることは、ヒトプロインスリンに対するELISA検出キットを用いて立証された。更に、血糖を測定し、プロインスリンが望ましくない代謝効果を持たないことが立証された。更に、ヒトプロインスリンの皮下投与によって、前記プロインスリンが神経網膜に達することができることが立証されており(網膜抽出物でのELISAキットによるプロインスリン同定)、これはプロインスリンが血液-網膜関門を通過することができることを意味する。
【0031】
網膜神経変性におけるProins/rd10-/-マウスの高プロインスリン血症の影響を、幾つかの方法により同定した。網膜の状態を最初に組織学的に観察した(実施例1)。ヒトプロインスリンについてのトランスジェニックマウスおよびrd10についてのホモ接合体では変性は遅くなり、P32においては、網膜に、一層多数の桿状体、錐体、およびシナプス結合が保持されており、網膜の状態は一層良好であることが立証された(実施例1)。
【0032】
網膜の状態が更に長時間一層良好に維持されるという事実は好都合である。実際に損傷を受けた細胞(網膜の桿状体)での細胞死の過程を遅らせることは別にして、残りの網膜は、例えば、本質的な損傷を有しておらずむしろ二次的に変性する錐体の死を防止することによって一層良好な状態に保持されており、この疾患の幾つかの症状は改善されている。この特殊な症例の場合には、桿状体が変性されていても錐体が更に長時間保持される場合には、夜間の視覚は失われるが昼間の視覚は保持され、これは、網膜色素変性症の患者の生活質を意味している。
【0033】
次いで、視覚機能を変性過程中、5種類の標準化した網膜電位図により分析し、rd10のトランスジェニックマウスコントロールと比較し、Proins/rd10-/-マウスでは、P55では視覚機能を未だ有しており、この視覚機能はrd10マウスではP35で失われ、これによって視覚応答はProins/rd10-/-マウスにおけるよりも良好であり、且つ更に長時間延長されると結論づけれることが明らかにされた(実施例2)。更に、トランスジェニックProins/rd10-/-マウスは、P30において少なくとも明所(昼間)視力では健常マウスに匹敵するERGを有する(実施例2、図4)。
【0034】
更に、本発明では、プロインスリンの単独の皮下および硝子体内投与で観察されたのとは対照的に、長時間持続する生理学的レベルでの投与形態でプロインスリンを用いると、プロインスリンにその神経保護機能を発揮させることが観察されている(実施例1.4参照)。これに関して、プロインスリンの単独においてのの皮下投与を試験したが、網膜神経変性の改善には成功せず、これは恐らく安定で長時間にわたって持続する水準が得られなかったからであろう(実施例3および4)。
【0035】
もう一つの利点は、プロインスリンはインスリンの代謝作用を持たず、従ってグルコース代謝が変更されていない患者に投与することができる点である。インスリンはイン・ビトロ研究においては抗アポトーシス活性も有するが、その通常の代謝活性は本明細書に記載したような治療についてその使用を実現できなくする。循環血清プロインスリンがこの救済を行うという事実は、それが投与を容易にするので良好な治療法であることを示す。プロインスリンは損傷したニューロンの死を防止するので、その適用は神経変性疾患の予防的および抑制的治療法となる。
【0036】
要約すれば、遺伝子療法について認可された遅延放出担体または発現ベクター上に担持されたヒトプロインスリンの他の形態の持続投与、皮下投与によって得ることもできるミオシン軽鎖プロモーターによって制御されるトランスジェニックヒトプロインスリンの発現によって得られる慢性血清プロインスリン血症水準(1−15pM)などは、遺伝的rd10マウスモデルにおいては網膜に到達して、光受容体神経変性を減少させることができる。
【0037】
従って、本発明の目的は、プログラム細胞死が起こる神経変性状態、障害、または疾患、好ましくは神経変性疾患、好ましくは中枢および末梢神経系の神経変性疾患、更に好ましくは網膜色素変性症として知られる遺伝性変性疾患の群の予防および治療用の医薬品または医薬組成物を製造するためのプロインスリンの活性を誘発する化合物の使用(以後、本発明の誘発化合物の使用)によって構成される。本発明の好ましい態様では、医薬組成物は、この医薬組成物を持続的に全身または局所投与するのに適した担体をも含んでなる。
【0038】
本発明では、上記医薬組成物が体内に定常的に放出され、標的組織に有効用量で長時間保持されるように薬学的に担持され、圧縮され、または処方されるとき、医薬組成物はその全身または局所への持続投与に適していると理解される。本発明の一つの態様においては、医薬組成物の持続投与に適する任意の担体、例えば、ポリマーまたはパッチなどこれらに限定されないものは、本発明の保護の範囲内に包含される。
【0039】
本発明で用いられているように、「プロインスリンの活性を誘発する化合物」という用語は、ヒトプロインスリンタンパク質の神経保護活性を模倣し、強度を増加し、または持続期間を延長する分子に関する。アクチベーター化合物は、化学的分子、ペプチド、タンパク質、またはヌクレオチド配列、並びに神経保護活性を有するタンパク質をコードするヌクレオチド配列を発現させる分子から構成できる。
【0040】
本発明で用いられているように、「神経保護活性」という用語は、神経変性疾患に最初に冒されている細胞および/または二次的に神経変性に冒されている細胞のプログラム細胞死の過程の減少、および/または残りの細胞の神経機能活性の増強に関する。
【0041】
本発明で用いられているように、「神経変性疾患」という用語は、とりわけ実例として且つ本発明の範囲を限定することのない下記の群、すなわち、アルツハイマー病、パーキンソン病、多発性硬化症、網膜色素変性症、レヴィー小体認知症、筋萎縮性側索硬化症、脊髄小脳性萎縮症、前頭側頭型認知症、ピック病、血管性認知症、ハンティングトン病、バッテン病、脊髄損傷、黄斑変性、および緑内障に属する疾患、障害、または病変に関する。
【0042】
従って、本発明の特定の目的は、プロインスリンの活性を誘発する化合物であって、誘発化合物が、神経保護タンパク質またはペプチドを発現させ且つ下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一方の断片、並びに
d) a)、b)、および/またはc)に属するいずれか1個の配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列によって構成されるヌクレオチド配列であるヌクレオチド配列であるものの使用(以後、本発明のプロインスリンヌクレオチド配列の使用)によって構成される。
【0043】
本発明の説明に用いられる意味において、「同様の」という用語は、例えば、1個以上のヌクレオチドの挿入、分子の末端のいずれかにおける1個以上のヌクレオチドの付加、または任意の末端または配列内部における1個以上の分子の欠失などの保存的および非保存的ヌクレオチド置換を導入することによって本明細書に示される配列に基づいて単離または構築することができ、且つヒトプロインスリンの活性を模倣することができるペプチドまたはタンパク質をコードすることができる任意のヌクレオチド配列(配列番号2)を包含することを意味する。
【0044】
本発明および当該技術の状態において記載された情報に基づいて、当業者であれば、本発明に記載したのと同様のヌクレオチド配列を続いて用いるために単離または構築することができる。
【0045】
同様のヌクレオチド配列は、通常は上記ヌクレオチド配列と実質的に相同である。本明細書で用いられる意味では、「実質的に相同」という表現は、問題のヌクレオチド配列の同一性の程度が少なくとも30%、好ましくは少なくとも85%、更に好ましくは少なくとも95%であることを意味する。用いられるヌクレオチド配列の好ましい形態は、ヒトプロインスリンヌクレオチド配列(配列番号1)およびその誘導体である。
【0046】
本発明で用いられているように、「ヌクレオチド配列」という用語は、DNA、cDNA、またはmRNA配列に関する。
【0047】
本発明の特定の態様は、プロインスリンの活性を誘発する化合物であって、ヌクレオチド配列がヒトプロインスリンをコードする配列番号1からなるものの使用によって構成される。
【0048】
d)の項に定義されているヌクレオチド配列は、プロインスリンタンパク質を発現させる遺伝子構築物および遺伝子発現ベクターに相当する。遺伝子構築物(本発明のプロインスリン遺伝子構築物)の場合には、これは、必要ならばおよび一層良好に発現したペプチドを単離、検出、または細胞の外部に分泌させるために、上記ペプチドの単離、検出、または分泌を目的として用いることができるペプチドをコードするヌクレオチド配列をも含んでなることができる。従って、本発明のもう一つの特定の目的は、本発明のプロインスリンヌクレオチド配列の他に、発現したペプチドを単離、検出、または細胞の外側へ分泌させるペプチドまたはペプチド配列をコードする任意のもう一つのヌクレオチド配列、例えば、実例として且つ本発明の範囲を限定することのないポリヒスチジン(6xHis)配列、モノクローナル抗体(例えば、その同定のための)によって認識することができるペプチド配列、またはイムノアフィニティークロマトグラフィーから生じる融合タンパク質の精製に有用な任意の他の配列、すなわち、c-myc、HA、E-tagのようなタグペプチドを含んでなる遺伝子構築物によって構成される(「抗体の使用: 実験室マニュアル」Ed Harlow and David Lane (1999). Cold Spring Harbor Laboratory Press. New York. 章: 「タンパク質のタギング」pp. 347-377)。
【0049】
上記ヌクレオチド配列および遺伝子構築物は、当業者であれば当該技術分野において広く知られている手法を用いることによって単離して得ることができる(Sambrook et al.「分子クローニング、実験室マニュアル第2版」Cold Sping Harbor Laboratory Press,ニューヨーク, 1989 vol 1-3)。上記ヌクレオチド配列は、細胞内で適当な条件において発現を調節することができる遺伝子発現ベクターに組込むことができる。
【0050】
従って、本発明のもう一つの特定の目的は、d)のヌクレオチド配列が発現ベクターであるプロインスリンの活性を誘発する化合物の使用(以後、本発明のプロインスリン発現ベクターの使用)であって、神経保護を誘発できるプロインスリンタンパク質をコードするヌクレオチド配列または遺伝子構築物を含んでなるものによって構成される。特定の態様の一例は、本発明において調製された発現ベクターの使用であって、発現が筋肉特異的プロモーターおよび配列番号1のヌクレオチド配列によって調節されるものによって構成される(実施例1参照)。
【0051】
本発明に記載のヌクレオチド配列または遺伝子構築物の他に、発現ベクターは、通常はその転写を指示するプロモーター(例えば、pT7、plac、ptrc、ptac、pBAD、5 retなど)、好ましくは組織プロモーターであって、これに転写を指示するプロモーターが操作結合するもの、並びに上記転写および適当な場合には、目的の産物の翻訳を制御し調節する他の必要なまたは適当な配列、例えば、転写開始および終結シグナル(tlt2など)、ポリアデニル化シグナル、複製起源、リボソーム結合配列(RBS)、転写レギュレーター(エンハンサー)をコードする配列、転写サイレンサー、リプレッサーなどを含んでなる。適当な発現ベクターの例は、それぞれの特殊な症例の状態および要求に従って発現プラスミド、ウイルスベクター(DNAまたはRNA)、コスミド、人工染色体などから選択することができ、目的の遺伝子または遺伝子類によりトランスフェクションまたは形質転換した細胞を選択するのに用いることができるマーカーを更に含むこともできる。ベクターの選択は、宿主細胞および実行する使用の種類によって変化する。従って、本発明の特定の態様によれば、上記ベクターはプラスミドまたはウイルスベクターである。上記ベクターは、真核細胞および微生物の形質転換に用いることができる様々なマニュアル[Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989)「分子クローニング: 実験室マニュアル, 第2版」Cold Spring Harbor Laboratory, Cold Spring Harbor,ニューヨーク]に記載されている様々な周知の方法(化学的形質転換、エレクトロポレーション、マイクロインジェクションなど)と同じ方法で当業者に知られている通常の方法によって得ることができる。一つの方法は、他の種類の療法において既に試みられているように、標的細胞に感染させる目的でレンチウイルスを用いることである(Ralph GS, Binley K, Wong LF, Azzouz M, Mazarakis ND (2006) Gene therapy for neurodegenerative and ocular diseases using lentiviral vectors. Clin Sci (Lond) 110:37-46)。
【0052】
遺伝子発現システムは、宿主細胞のゲノムに新たな遺伝子材料を組込むことができまたはできない。次に、ヌクレオチド配列、遺伝子構築物、またはプロインスリン発現ベクターを、ニューロンおよび/またはグリア変化をもたらす疾患に冒されているヒトの遺伝子療法による予防および治療の過程において、ヒト細胞、好ましくは神経変性変化に冒されているニューロンおよび/またはグリア細胞を保護するための医薬品として用いることができる。これらの遺伝子発現システムを神経変性疾患に冒されているヒトに投与すると、それらを一般的または特異的に組織細胞に導入することができ、そこでそれらが細胞ゲノムに組込まれると、プロインスリンタンパク質を発現することができ、そのタンパク質は、細胞外媒質に分泌されると、中枢神経系に到達して神経保護作用を行うことができる(実施例参照)。
【0053】
その上、これらの遺伝子発現システムを潜在的レシピエントに関して、自己または異種の人体外のヒト細胞の形質転換に用いることもでき、これらの細胞はヒトニューロンおよび/またはグリア細胞の神経保護活性を有するプロインスリンタンパク質を発現および放出するため、神経変性疾患に罹っているヒトに投与するとプロインスリンを誘発する化合物となる。
【0054】
従って、本発明のもう一つの特定の目的は、プロインスリンの活性を誘発する化合物の使用によって構成され、誘発化合物が好ましくはヒト真核細胞(以後、本発明のプロインスリン細胞)であり、これが遺伝的に修飾されており、且つ本発明のプロインスリンヌクレオチド配列、構築物、または発現ベクターを含んでなり、細胞外媒質にプロインスリンタンパク質を適当に発現または放出できる。
【0055】
これらの細胞は、当業者に知られている遺伝子工学技術によって上記ヌクレオチド配列により形質転換、感染、またはトランスフェクションすることができる[Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989).「分子クローニング:実験室マニュアル, 第2版」Cold Spring Harbor Laboratory]。生物薬剤学的手段および遺伝子療法の方法は当業者に十分知られており、過度の努力することなく本発明に記載の常法を用いて開発することができる。
【0056】
もう一つの特定の態様は、様々な細胞株、好ましくは中枢神経系、更に好ましくはヒト組織を再生する細胞として用いることができるニューロンからヒトプロインスリンヌクレオチド配列(配列番号1)によって形質転換したヒト細胞の使用である。
【0057】
更に、本発明のもう一つの特定の目的は、プロインスリンの活性を誘発する化合物であって、誘発化合物がタンパク質またはペプチドであるものの使用(以後、本発明のプロインスリンタンパク質の使用)であり、神経保護活性を有し且つ下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つを含んでなるアミノ酸配列
に属する1または数個のアミノ酸配列を含んでなる化合物の使用によって構成される。
【0058】
本明細書で用いられる意味において、「同様の」という用語は、例えば、1個以上のアミノ酸の挿入、分子の末端のいずれかにおける1個以上のアミノ酸の付加、または任意の末端もしくは配列内部における1個以上のアミノ酸の欠失などの保存的および非保存的アミノ酸置換を導入し、且つヒトプロインスリンの活性を模倣することによって本明細書に示される配列に基づいて単離または構築することができる任意のアミノ酸配列を包含することを意味する。
【0059】
本発明に記載の情報に基づいて、当業者であれば、本発明に記載したのと同様のアミノ酸配列を単離または構築することができる。
【0060】
同様のアミノ酸配列は、通常は上記アミノ酸配列に実質的に相同である。本明細書で用いられる意味では、「実質的に相同」という表現は、問題のアミノ酸配列の同一性の程度が少なくとも30%、好ましくは少なくとも85%、更に好ましくは少なくとも95%であることを意味する。
【0061】
本発明のもう一つの特定の態様は、本発明の誘発化合物の使用であって、誘発化合物がヒトプロインスリンタンパク質(配列番号2)であるものによって構成される。
【0062】
本発明のもう一つの目的は、神経退行変性をもたらす疾患、障害、または病変の治療用の医薬組成物または医薬品(以後、本発明の医薬組成物)であって、治療上有効量の本発明のプロインスリンの活性を誘発する化合物を場合によってはプロインスリンの活性を持続的に誘発する化合物の全身または局所投与に適当な1種類以上の薬学上許容可能なアジュバントおよび/または担体と共に含んでなる医薬組成物または医薬品によって構成される。
【0063】
上記組成物で用いることができる薬学上許容可能なアジュバントおよび担体は、当業者に知られており、且つ治療組成物の製造に一般に用いられるアジュバントおよび担体である。
【0064】
本明細書で用いられる意味において、「治療上有効量」という表現は、所望な効果を生じるように計算された神経保護を発揮することができ、通常は、特に患者の年齢、患者の容体、変化または障害の重篤度、並びに投与の経路および頻度などの化合物の特徴によって決定される薬剤または化合物の量に関する。
【0065】
もう一つの特定の態様においては、上記治療組成物は、薬学上許容可能な希釈剤中で固形形態または水性懸濁液の形態に調製される。本発明によって提供される治療組成物は任意の適当な投与方法によって投与することができ、そのために上記組成物は選択された投与方法に適する投薬形態に処方される。特定の態様においては、本発明によって提供される治療組成物は、非経口、経口、鼻内吸入、腹腔内、皮下などに投与される。医薬品の投与のための様々な投薬形態およびそれらを得るのに必要な賦形剤の総説は、例えば、"Tratado de Farmacia Galenica", C. Fauli i Trillo, 1993, Luzan 5, S. A. Ediciones, Madridに見出すことができる。
【0066】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、神経保護薬剤または化合物が下記の群、すなわちプロインスリン活性を有するタンパク質またはペプチドを発現させるプロインスリン配列、遺伝子構築物、または発現ベクターに属する医薬組成物によって構成される。
【0067】
本発明の特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が、下記の群
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列の任意の一つの断片、並びに
d) a)、b)、および/またはc)に属する任意の一つの配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列である医薬組成物によって構成される。
【0068】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、ヌクレオチド配列がヒトプロインスリンをコードする配列番号1を含んでなる医薬組成物によって構成される。
【0069】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、ヌクレオチド配列がヒトプロインスリン発現ベクターであることを特徴とする医薬組成物によって構成される。
【0070】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が本発明のプロインスリン配列、遺伝子構築物、またはベクターによってコードされるタンパク質またはペプチドであることを特徴とする医薬組成物によって構成される。
【0071】
本発明の特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発するタンパク質またはペプチドが下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属する任意の一つの配列を含んでなるアミノ酸配列
に属することを特徴とする医薬組成物によって構成される。
【0072】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、アミノ酸配列がヒトプロインスリン(配列番号2)によって構成されることを特徴とする医薬組成物によって構成される。
【0073】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が、好ましくはヒト細胞、好ましくは本発明のプロインスリン配列、構築物、または発現ベクターによって形質転換された中枢神経系細胞であることを特徴とする医薬組成物によって構成される。
【0074】
本発明のもう一つの目的は、プログラム細胞死が起こるヒトを冒す中枢または末梢神経系の神経変性疾患、障害、または病変に冒されている哺乳類、好ましくはヒトの治療または予防方法において、本発明の医薬組成物の使用(以後、本発明の医薬組成物の使用)であって、上記治療組成物を、上記神経変性を減少させる適量で投与することからなる使用によって構成される。
【0075】
本発明の医薬組成物は、単独でまたは他の医薬化合物と共に治療方法に用いることができる。
【0076】
本発明のもう一つの特定の態様は、下記の群、すなわち、アルツハイマー病、パーキンソン病、多発性硬化症、レヴィー小体認知症、筋萎縮性側索硬化症、脊髄小脳性萎縮症、前頭側頭型認知症、ピック病、血管性認知症、ハンティングトン病、バッテン病、および脊髄損傷に属する神経変性疾患の治療方法における本発明の医薬組成物の使用によって構成される。
【0077】
本発明のもう一つの特定の態様は、下記の群、すなわち、網膜色素変性症、黄斑変性、および緑内障に属する神経変性疾患の治療方法における本発明の医薬組成物の使用によって構成される。
【実施例】
【0078】
本明細書において提供される下記の具体例は、本発明の性質を説明するためのに役立つ。これらの実施例は説明目的のためにのみ包含されており、本明細書に記載の本発明が限定されると解釈すべきではない。
【0079】
実施例1. ヒトプロインスリンは、トランスジェニックProins/rd10-/-マウスの網膜桿状体の死を防止し、シナプス結合を保持することができる。
1.1. rd10突然変異(Proins/rd10-/-)についてヒトプロインスリンおよびホモ接合体を産生する2種類のトランスジェニック系列の産生
横紋筋ミオシン軽鎖(MLC1)マウスの構成的プロモーターの制御下において、遺伝的背景が50% C57B1/6および50% SJLの数系列のトランスジェニックヒトプロインスリン産生マウスを得た後、それらをホモ接合rd10マウス(100% C57B1/6)と連続的に交雑させた。従って、遺伝的背景は95% C57B1/6を上回る比率に到達するまで均質化され、Proins/rd10-/-マウスを得た。これは6回目の戻し交雑によって達成され、二つの主系列L1およびL2を得た(L2動物は下記の実施例で用い、一方、図4のP30におけるERGに記載の結果もL1動物を用いて行い、プロインスリン血症および血糖分析も両方で行った)。
【0080】
本発明では、野生型マウスは、コントロールとして用いられるマウスと理解されている。それは変化しない。それは市販のものであり、その名称はJackson laboratories製のC57B1/6Jである。
【0081】
rd1マウスは、桿状体特異的サイクリックGMPホスホジエステラーゼ遺伝子に突然変異を有する市販マウスと理解されており、その商品名はJackson laboratories製のPdebrd1/rd1である。
【0082】
rd10マウスは、桿状体特異的サイクリックGMPホスホジエステラーゼ遺伝子に突然変異を有する市販マウスと理解されており、その商品名はJackson laboratories製のPdebrd10/rd10である。
【0083】
Proins/rd10-/-マウスは、rd10マウスと、トランスジェニックヒトプロインスリン産生マウスとを横紋筋プロモーターMLC1下において交雑させることによって生成するマウスであると理解されている。これらのマウスに導入された構築物は、軽鎖ミオシン筋肉プロモーターによって制御されているヒトプロインスリンタンパク質のcDNAを有し、その発現は構成的である(図1B, 配列番号1)。発現の変動性が観察され、これはトランスジェニックマウスの異なる浸透度による可能性がある。これは、血清に見出されるヒトプロインスリンの産生と相関している。
【0084】
トランスジェニックマウスの産生に用いた構築物は、サイズが6.4kbのプラスミド(pMLC-hIns)からなっていた。発現は、ミオシン軽鎖構成的筋肉プロモーター(MLC)によって伝達される。ヒトプロインスリン遺伝子のcDNAを、二つのEcoRI部位間でクローニングする(図1B)。
【0085】
ジェノタイピング ゲノムDNAは、Miller et al., 1988において記載された手法に従って組織から得た(Miller, S. A., Dykes, D. D. and Polesky, H. F. (1988) A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res, 16, 1215.)。離乳マウスの尾を、プロテイナーゼK(Roche Diagnostics, マンハイム, 独国)0.3mgを有するリーシス緩衝液(Tris- HCl 40 mM, pH 8.0, EDTA 20 mM, SDS 0.5%、およびNaCl 200 mM)0.5ml中で消化した。DNAを沈澱させたならば、これをHindIII酵素(Roche)で開裂させた。ゲノムDNA 10μgを用いて、これを最終容積50μlのHindIII酵素により37℃において一晩消化した。それらを、長さ12cmの1%アガロースゲルのそれぞれの個々のウェルに装填し、良好なサイズ分離を行った。導入前に、下記の溶液、すなわち、最初は脱プリン溶液 (0.5M HCl)で15分間、次いで変性溶液(0.5 N NaOHおよび1.5 M NaCl)で30分間、最後に中和溶液(0.5 M Tris-HCl, pH 8)で30分間で、ゲルを調製した。DNA断片化はこの処理によって行われ、それによって導入が促進される。ナイロン膜(Schleicher & Schuell BioSciencie, 米国)を用いて、DNAをStratalinkerオーブン(Stratagene, La Jolla, カリフォルニア, 米国)中でUV線によって膜に固定した。湿式導入は、室温において一晩行った。
【0086】
ジェノタイピングには、構築物に挿入されたヒトインスリンcDNA配列に対してdCTPベース中の32P-放射能標識プローブを用いた。プローブを作製するための鋳型は、プラスミド自身からEcoRI (Roche)で消化して得た。
【0087】
構築物をEcoRIで消化した後に放出されたインサートを、DNA抽出キット(Millipore)によって精製した。プローブの標識は、[32P]dCTPの存在下においてRandom Primer Kit (Stratagene)を用いて行った。次いで、プローブをMicrospin G25カラム(Amersham Pharmacia Biotech)で精製した。
【0088】
膜を、50%ホルムアミド、1 x Denhardt (0.02% Ficol, 0.02 % ポリビニルピロリドン、および0.02% BSA)、1% SDS、5 x SSC (0.15 M NaClおよび15 mMクエン酸ナトリウム, pH 7.2)、および0.1 mg/mlサケ精子DNAを含む溶液で65℃で少なくとも2時間予備ハイブリダイゼーションした後、予備ハイブリダイゼーション溶液で65℃において一晩ハイブリダイゼーションし、これにプローブ1.5 x 106 cpm/mlを加えた。2x SSC で室温にて15分間の2回の洗浄、2 x SSCおよび1% SDSで30分間の更に1回の洗浄、並びに2 x SSCおよび0.1% SDSで62.5℃での最終洗浄を行った。
【0089】
フィルターを洗浄したならば、乾燥させることなくGLAD型プラスチックに包み、2個の増幅スクリーン(Genescreen plus, DuPont)と18x24 cmのKodak Biomax MS型写真フィルム(Eastman Kodak Company, ロチェスター, ニューヨーク, 米国)との間に露光した。シャープなシグナルを得るには、3-4日の露出時間が必要であった。ヒトプロインスリンcDNAに対して生成したプローブは、ゲノムDNAゲルに1.5 Kbのバンドを検出した。
【0090】
rd10マウスは、桿状体特異的サイクリックGMPホスホジエステラーゼ6酵素遺伝のエキソン13に位置する点突然変異を有する。野生型マウスでは、その位置にCfoI酵素による開裂部位がある。突然変異体の場合には、酵素開裂部位は消失している。これにより、内因性ジェノタイピング技術により制御することができる。
【0091】
ゲノムPCRは、次のプライマー3'-CTTTCTATTCTCTGTCAGCAAAGC-5' (オリゴA, 配列番号3)および3'-CATGAGTAGGGTAAACATGGTCTG-5' (オリゴB, 配列番号4)を用いて行い、97 bpの断片を増幅した。次に、PCR生成物を、CfoI酵素(Roche)により37℃において2時間消化した。これを、次に3% Metaphorアガロースゲル中で分画した。野生型マウスは、消化後に二つの54および43bpのバンドを示した。ホモ接合rd10マウスは1本の97bpのバンドを生じたが、これは、酵素がアンプリコンを開裂しなかったからであり、またヘテロ接合rd10/+マウスは3本のバンド(対立遺伝子の一方は開裂され、他方は開裂しない)を示した。この遺伝子では、個体間の変動性の種類は見出されず、これは、変性が標準的パターンに従い、通常はこれを有する総ての個体について同様に実現されることを意味する。
【0092】
1.2. トランスジェニックマウスの血糖およびプロインスリンレベルの測定
総てのマウスの血糖を、12時間の絶食後に試験ストリップ(Accu-Chek, Roche)で測定した。マウスの正常レベルは、通常は100-200 mg/dlである。Proins/rd10-/-二重突然変異体では、80-150 mg/dlの変動が見られ、この変動はプロインスリン血症の水準とは直接相関しなかった。
【0093】
市販のELISAアッセイ(Lineo Research, ミズーリー, 米国)を用いて、トランスジェニックマウスによるヒトプロインスリン産生レベルを検出し、これは特異的にヒトプロインスリンを検出する。
【0094】
トランスジェニックおよびコントロールマウスの筋肉および血清を分析した。血清の場合には、総て製造業者の推奨に従ったが、筋肉の場合には、事前にリーシス緩衝液(50 mM Tris-HCl pH 7.0, 100 mM NaClおよび0.1% Triton)によるタンパク質抽出およびBCAキット(Pierce, Rockford, イリノイ, 米国)によるその定量が必要であった。
【0095】
筋肉抽出物で検出されたヒトプロインスリンは常に極めて高く、タンパク質の量によって補正しなければならなかった。血清で検出されたヒトプロインスリンは、1−15pMであった。この濃度は、製造業者の指示に従って20μlの容積で測定した。測定は、P30で組織的に行った。これは、トランスジェニックProins/rd10-/-マウスが筋肉でヒトプロインスリンを産生し、これが血液循環に流れ込み、網膜神経に到達することができることを示唆していた。
【0096】
1.3. トランスジェニックマウスの網膜切片のクリオスタットでの免疫染色
網膜組織構造をP32マウスで分析し(図2)、変性に罹っていない野生型マウス(AおよびD)と、変性に罹っているProins/rd10-/-マウス(BおよびE)およびrd10-/-マウス(CおよびF)とを比較した。変性がrd10-/-マウスでは極めて進行しているP32では、カラム当たりの光受容体細胞の数、錐体の状態およびアバンダンス、並びにシナプスの状態を分析した(図3)。このようにして、総体的な網膜の状態および変性の進行を、様々なマーカーで明らかにしようとした。
【0097】
総ての組織はTissue-tec (Sakura Finetek Europe B.U. オランダ国)に埋設し、ドライアイスで冷凍して、処理まで-80℃で保管した。12μmの厚さのクリオスタット切片を作製した。これらをポリ-L-リシンをコーティングしたスライド(Fisher Biotech, ピッツバーグ, 米国)に集め、使用するときまで-80℃に保管した。
【0098】
染色を行う総ての場合に、スライドをフリーザーから取り出した後、室温において30分間放置し、4%パラホルムアルデヒド(PFA)で20分間固定した後、PBSで洗浄した。
【0099】
残りの錐体を標識することを決定したが、これらの錐体が突然変異を全く行わずに二次的に変性するので、網膜の状態およびその保持のアイディアを得ることができるからである。次に、変性の進行を知るには、外顆粒層(ONL)の厚みに残っている細胞の列の数を数えることで十分であった。蛍光色素Alexa 488で標識した凝集素(Molecular Probes, Eugene, オレゴン, 米国)を、0.1% BSAを含むPBS溶液中で2時間用いて、錐体を標識した。 1 mM MgCl2および1 mM CaCl2を含む溶液で洗浄した。錐体の外側部分およびその軸索末端は、このようにして検出した。切片をDAPIを含む固定媒質で固定し、核を対比染色した。
【0100】
変性のない野生型コントロールマウスは、ONLに8-12列の核を有する(図2D)。OPLに外側部分およびそのシナプスボタンを支持している錐体の数は、OPLで観察される凝集素染色による極めて典型的なものである(図2A)。コントロールrd10-/-マウスでは、ONLにおける桿状体の列の数は極めて小さく1-2列であり、この点における変性が極めて高いからである(図2F)。極めて驚くべきことは、錐体が初めは突然変異によって影響を受けていないにも拘わらず、この時点で既に変性し始めていることである(図2C)。錐体の外側部分はほとんど現れず、そのシナプスボタンの数はかなり減少していることが観察された。Proins/rd10-/-マウスにおいては、5-6列の桿状体核がこの症例で観察され、これは最大のプロインスリン血症が検出されたマウスの1匹であった(図2E)。錐体の状態は極めて良好であり、実際に、それは野生型マウスに類似していた。良好な錐体が保存されていることを示唆しているOPLにおけるその外側部分およびおよびシナプスボタンが見える(図2B)。血中プロインスリンレベルと相関するONL保存における変化が観察された。
【0101】
更に、ニューロンのシナプス結合が起こる網状層(図3)の状態を分析したが、プロインスリンを用いるこの治療が変性死を防止または遅延させるだけでなく、細胞が内因性損傷を有しまたは二次的に変化しているか否か、またはそれらが機能的であるか否かに拘わらず細胞を生きたままにしておこうとするものであるので、分析した。SV2 (シナプス小胞2)抗体による染色のため、クリオスタット切片を4% PFAで固定した後、0.1% Triton x-100で浸透させ10% NGS(正常なヤギ血清)で1時間ブロックした。1:50の希釈倍率のSV2抗体をシナプス小胞のタンパク質に結合し、従って網膜の網状層(外網状層、OPLおよび内側網状層、IPL)を標識する。これを、ブロック溶液中で4℃において一晩インキュベーションした。Alexa 488に接合した二次抗体(1/200)を用いるインキュベーションは、室温において1時間行った。対応するPBS洗浄の後、切片をDAPIを含む固定媒質で固定し、核を対比染色した。
【0102】
二つの網膜網状層、外網状層(OPL)、および内網状層(IPL)において、発現が観察された(図3)。脳に突出しているニューロン、ガングリオン細胞を有する二極性介在ニューロン結合が起こる内網状層は、3例においてかなりの染色と良好な状態を示した。差異は、光受容体細胞と二極性介在ニューロンとの結合が主として位置している外網状層にあった。OPLは、野生型コントロールマウス(図3A)およびトランスジェニックProins/rd10-/-マウス(図3B)においてはかなり画定して、濃かった。コントロールrd10マウスはIPLでは染色を維持したが、OPLは全く明確に保存されず、ほとんど染色されなかった(図3C)。このコントロールrd10-/-マウスにおいては、組織化の欠如によりおよび残りのニューロンが光受容体を有するシナプスを喪失しているという事実により、核層間にシナプス投影の試みさえ見られた。これは、Proins/rd10-/-マウスの網膜の機能状態が、治療なしのrd10-/-マウスより良好であることを示唆している。
【0103】
これは、トランスジェニック Proins/rd10-/-マウスでは、ヒトプロインスリンが治療なしの変性状態よりシナプス結合を長く保持することができることを示していた。これは、変性による細胞死を遅らせることの他に、それらを良好に保ち且つそれらの生物学的機能を行うことができることを含んでいる。
【0104】
rd10-/-マウスと比較して、Proins/rd10-/-マウスにおいて組織学的に観察された改善点は、次の網膜電位記録で一層明らかである(図4、5、および6)。
【0105】
更に、神経保護効果でのプロインスリンレベルの値を分析するため、網膜および他の神経系の部分の神経変性疾患の遺伝子モデル並びに老衰関連のもので見られる慢性および進行性損傷以外のモデルにより分析を行った。従って、予備検討を行い、網膜ガングリオン細胞死を急性損傷(視神経の切断)によって成熟ラットに引き起こし、ヒトプロインスリンの皮下投与を行った。この治療は、この典型的な急性損傷で必然的瞬間までガングリオン細胞死を僅かに遅延させた(データーは示さず)。ヒトプロインスリンは、これらのrdマウスの治療として皮下および硝子体内注射によっても投与したが(図8および9)、皮下投与したプロインスリンは網膜神経に到達することができたにも拘わらず、変性は停止しなかった(図7)。これらの結果は、血清プロインスリンは、持続および伸張レベルが達成されるときにのみ、中枢神経系の損傷部分に到達して神経保護作用を発揮することができることを示している。
【0106】
実施例2. ヒトプロインスリンはコントロールおよび疾病マウスと比較してトランスジェニック Proins/rd10-/-マウスでERGによって評価した視覚機能を向上させる。
マウスは、網膜電位の検討のため12時間毎の明-暗サイクルを常に保持したが、マウスを一晩中暗闇に順化させた。情報目的には、錐体は明所(昼間)視覚に関与しており、桿状体は暗所(夜間)視覚に関与する。マウスを弱い赤色光下において、ケタミン(95mg/kg)およびキシラジン(5mg/kg)を含む溶液の腹腔内投与により麻酔した。瞳孔を、1%トロピカミド(Colircusi Tropicamida, Alcon Cusi, SA, E1 Masnou, バルセロナ, スペイン国)を含む溶液1滴で散大した。記録電極は、マウスの眼に置いたレンズである。参照電極は口に付け、接地電極は尾に付けた。麻酔した動物をファラデーケージに入れ、総ての暗所実験は完全な暗闇で行った。このようにして、ガンツフェルトスティミュレーターによって生じた低強度光のフラッシュによって誘発された網膜電位応答を記録して、桿状体のみで生じた応答(桿状体応答)または暗闇への順化での錐体と桿状体とで生じた応答(混合応答)記録することができた。用いた光刺激の強度は、-4 - 1.52 log cd.s.m-2の値に設定した。光強度は、眼のレベルの光度計(Mavo Monitor USB)で測定した。64回の応答の最大値を、それぞれの刺激強度について平均した。光刺激の間隔は強度に応じて変化し、従って低強度刺激(-4 log cd.s.m-2)については、刺激間時間を10秒間に設定し、高強度刺激(1.52 log cd.s.m-2)については、60秒間に設定した。動物を、錐体の単独応答を記録するために明所条件に順化させた。これらの条件下では、閃光の間隔は1秒間に設定した。
【0107】
網膜からの電気シグナルをグラス増幅器(CP511 AC増幅器, Grass Instruments, クインシー, マサチューセッツ)で0.3-1000Hzの間で増幅し、濾過した。シグナルを数値化した(PC-card ADI instruments, カリフォルニア)。記録をコンピューターに保管して、後で解析した。
【0108】
桿状体性応答を暗所順化状態で記録した後、強度が-4 - -1.52 log cd.s.m-2の閃光を当てた。錐体および桿状体によって生じた混合応答を記録した後、-1.52 - 0.48 log cd.s.m-2の強度の閃光を当てた。振動電位も、100 - 1000Hzの電気フィルターを適用することによって単離した。錐体性応答は、(30cd.m-2のバックグラウンド光を記録して)光順化状態で記録した後、強度が-0.52 - 2 log cd.s.m-2の閃光を当てた。フリッカー応答(30 Hz)を光順化状態で記録した後、1.48 log cd.s.m-2を加えた。
【0109】
図4は、野生型(WT)、コントロールrd10-/-および系列1および系列2由来のP30で得たProins/rd10-/-マウスの、暗所に順化(暗順応状態)および光に順化(明所視状態)した状態で記録した。
【0110】
図5は、より大きな群またはマウスでのインスリン間網膜電位応答とプロインスリンレベルとの相関を解析した結果を示す。この相関は、用量-応答の関係を示唆しており、ヒトプロインスリンを用いる薬理学的方法が実行可能な有効性を有することを支持している。
【0111】
更に、図6は、産後発生の様々な時期(P35、P45、およびP55)に得られた系列2由来の野生型(wt)、コントロールrd10、およびProins/rd10-/-マウスに対応する網膜電位記録を示す。暗順応(暗所への順応)および明所視(光への順応)状態で得た記録を、別々に示す。野生型マウスがどのようにP35で広範囲の暗順応(桿状体)および明所視(錐体)網膜電位応答を生じるかが観察された。応答は、桿状体および錐体で生じた応答のいずれでも、コントロールrd10-/-マウスではP35でゼロであった。Proins/rd10-/-マウスは、P35で極めて有意な明所視および暗順応網膜電位応答を保持し、P55まである程度の応答の保持を達成した。このように、視覚応答はトランスジェニックProins/rd10-/-マウスで一層良好であり、更に長時間伸張することが観察された。
【0112】
実施例3: プロインスリンのrd1マウスの網膜の硝子体内投与による効果
下記の実施例3および4を実施するため、rd10マウスと同じC57BL/6遺伝的背景並びに同じ桿状体特異的サイクリックGMPホスホジエステラーゼ遺伝子を除く様々な突然変異を共有するrd1型マウスを用いた。
【0113】
プロインスリンのイン・ビボでのrd1マウス網膜の硝子体内投与の効果、具体的には、P13におけるrd1マウスのヒトプロインスリンを1μg/μlの濃度で1μgの硝子体内投与に関して。右目にプロインスリンを投与し、左目に担体を投与した。投与は単回投与であり、効果を24または48時間後にTUNELによって平面的におよび切片で固定した網膜で分析した。後者では、外顆粒層に残っている光受容体の数も評価した。TUNEL(TdTによるdUTPニックエンド標識)法を用いて、細胞死を検出した。末端トランスフェラーゼ酵素(TdT)は、フルオレセイン標識したヌクレオチド(dUTP)をDNAの遊離の3'末端に付加し、これによりプログラム細胞死中に起こるDNAの断片化を検出することができる。Promega製TUNELキットを用いた。手法は、細胞、断片、または組織で行った。組織の性質による透過性化(permeabilization)工程後、組織をPBSで洗浄し、キット溶液を用いて室温において30分間予備インキュベーションした。反応混合物を製造者取扱説明書に従い調製し、その反応を37℃において1時間行った。反応を2 x SSC溶液で室温にて15分間停止した。これをPBSにより洗浄し、ベクタシールドで固定した。平面的に固定された網膜での細胞死を検出するため、眼の残りの要素から切り離した全網膜神経を、取扱いの際にコントラストが良好になるように黒色ニトロセルロース膜(Sartorius, ゲッティンゲン, 独国)上で顕微鏡に固定した。これは、通常は光受容体層を上にして置き、ニトロセルロースに精巧な切開用ピンセットの助けを借りて付着させた。平面的に取り付けた網膜を、24穴プレートに0.1 M リン酸緩衝液, pH 7.1中で4%(w/v)PFAで4℃において一晩固定した。翌日、それらをPBSおよびBSA(30 mg/ml PBS)で洗浄した。浸透化はPBS中1% (w/v) Triton X-100を用いて(4回、それぞれ30分間)、および予めPBSで十分に洗浄することによってTriton X-100残留物を除去してコラゲナーゼおよびプロテイナーゼKで行った。コラゲナーゼ(20 U/ml, Sigma)を37℃において1時間作用させた後、プロテイナーゼK(20μg/ml, Promega)を37℃において15分間作用させた。次に、網膜を少なくとも2時間再固定する必要があった。これをPBSおよびBSA (30 mg/ml PBS)で十分に洗浄し、TUNEL反応を上記の方法で行った。平面的に固定した網膜を共焦点顕微鏡法(Leica TCS-SP2-A0BS)によって分析した。
【0114】
網膜層中の細胞死の位置を確認するため、上記のような網膜切片についてTUNELを行い、常に0.1 M リン酸緩衝液(pH 7.1)中4%(w/v)PFAに固定し、30%(w/v)スクロース/10 mMリン酸緩衝液(pH 7.1)で凍結防止した。切片のTUNEL標識には、平面的に固定した網膜より浸透を低くする必要がある。従って、BGT [100 mMグリシン、3 mg/ml BSA、0.25%(w/v)Triton X-100、PBS中]を用いて2シリーズの浸透をそれぞれ15分間行った。それらをPBSにより洗浄し、TUNEL反応を上記のように行った。スライドの総ての切片をカバーするため、相当する試薬50μlを加え、それらをパラフィルムでカバーした。
【0115】
いずれの定量(切片のTUNEL、平面的に固定した網膜のTUNEL、切片の光受容体)でも、プロインスリンと担体による治療の間には有意差は見られなかった。
【0116】
実施例4. プロインスリンのrd1マウスでの皮下注射による効果
更に長時間の治療を行うために、新たな投与経路である皮下注射を試験した。糖尿病患者にインスリン投与の目的で一般に用いられるこの経路は、硝子体内注射より多少外傷が少ない。更に、この方法でのヒトプロインスリン投与は、成熟ラットにおいて視神経を切断した後の網膜ガングリオン細胞の変性に予め正の遅延を生じた。多数のプロトコルを行い、ヒトプロインスリンの薬理学的供給を最適にした(表)。
【0117】
【表1】

【0118】
プロインスリンが皮下注射によって網膜に到達するか否かを、最初に測定した。その目的のため、プロインスリンを皮下投与したマウスでELISAによって検討を行い、ヒトプロインスリンを検出した(図7)(Lineo Research, ミズーリー, 米国)。
【0119】
この分析法は血清プロインスリンを検出するように設計されているので、筋肉および網膜抽出物用に改良しなければならなかった。トランスジェニックマウスによるヒトプロインスリン産生レベルを検出するため、筋肉および網膜、並びに血清抽出物を分析した。血清の場合には、マウスの眼の涙嚢部分から血液をパスツールピペットで採取した。これをエッペンドルフ試験管に移し、室温において2時間血液凝固させ、1,300 gで15分間遠心分離して血清を集め、これを使用まで凍結した。組織の場合には、タンパク質抽出をリーシス緩衝液[50 mM Tris-HCl, pH 7、100 mM NaClおよび0.1%(w/v) Triton X-100]で行い、BCAキット(Pierce)による抽出タンパク質の定量を行った。網膜神経は、それぞれ60μlの容積で抽出した。後脚の筋肉を、得られた筋肉片に従って200-300μlの容積で抽出した。
【0120】
血清の場合には、製造業者の推奨に従い、常に20μlで二つの試料を用いた。組織の場合には、抽出物20μlを二つ作製し、場合によっては精度を上げるため連続希釈を用いた。網膜および筋肉抽出物データは、タンパク質の量によって補正した。
【0121】
P14の野生型マウスの網膜抽出物におけるヒトプロインスリンは、用量依存的な皮下投与の2時間後にのみ同定することができた。1.6μgの用量では、0.72pMの網膜濃縮を生じたが、5μgの用量では3.62pMとなった。P11とP14の間に1.6μgを毎日投与することによって、更に高い、恐らくは蓄積された、レベルの3.56pMを生じることができた。従って、この方法によって、末梢の外来プロインスリンが網膜神経に到達する能力および少なくとも網膜における一層高いプロインスリンレベルを達成する反復治療の有効性が確かめられた。
【0122】
神経変性における光受容体および網膜電位図(表、図8、および9)によって測定された視覚機能に対する様々な間隔での反復治療の効果を実証した。P8およびP15の間の1.6μgの1日用量を最初に投与した。この場合には、組織学的レベルにおいては保護効果はなかった。プロインスリンを変性過程が開始する前に投与すれば効果は一層明白になり得るという信念により救済を一層速く開始するため、治療をP6およびP14の間に試験したが、光受容体の保持はこの時点においては得られなかった。組織学的回復を試み且つ網膜電位図による視覚機能を評価するため、P6およびP14の間にヒトプロインスリンを1.6μgの1日投与を2回行った。幾匹かの動物はP18まで保持されたが、プロインスリン用量はこの第二の期間では2.5μgに増加した。この治療は、組織学的レベル(図8)または機能レベル(図9)で何ら保護効果を持たなかった。
【0123】
投与されたプロインスリンは網膜に到達することができ、それに対する応答を可能にする総ての要素が明らかに配置されているという事実にも拘わらず、最もありふれた結果は、組織学的回復の欠如およびERGによる視覚機能の欠如(図8および図9)であった。
【発明の背景】
【0001】
発明の分野
本発明は、生物医学分野に含まれ、更に具体的には治療化合物の開発に含まれる。本発明は、詳細には網膜色素変性症(retinitis pigmentosa)のような網膜変性疾患並びに他の神経変性疾患の治療用の医薬品の製造のためのプロインスリン分子の特定の使用に関する。
【0002】
背景技術
神経変性疾患は、遺伝性(遺伝的)、外傷性、散発性、または老年性起源の中枢神経系の様々な進行性疾患を包含する。神経変性疾患のほとんどは、それらの起源またはそれらの進行においてニューロンおよびグリア細胞のプログラム細胞死を示す。神経組織に対する損傷の不可逆的な段階を構成する前記の死の工程は、遺伝性、外傷性、散発性、または老年性であるかに関わらず疾患の主要な原因とは独立していると思われる。
【0003】
多数の増殖因子は、ニューロンやグリア細胞など様々な種類の細胞の生と死とのバランス調節に本質的な役割を果たしている。これらには、インスリン、その前駆体であるプロインスリン並びにIGF-IおよびIGF-IIなどのインスリンファミリーのものが含まれる(Varela-Nieto, I., de la Rosa, E. J., Valenciano, A. I., and Leon, Y. (2003) Cell death in the nervous system: lessons from insulin and insulin-like growth factors.Mol Neurobiol 28: 23-50)。網膜は中枢神経系の一部を構成しており、神経系の生理学的および病理学的過程の双方の研究に詳細に確認されているモデルであり、この理由により、これは本発明において用いられる細胞モデルである。網膜において検討された最も重要な病理学的過程の一つはいわゆる網膜色素変性症であり、この病状は包括的な群の遺伝性網膜障害を含んでなり、しかも世界における失明の最大原因の一つであり、概算の発生率は4,000人に1人であるからである。120を上回る関連の遺伝子座は特定されており、原因は様々であるが、いずれの場合にもプログラム細胞死による網膜ニューロン、特に光受容体の慢性的且つ進行性の喪失があり、ヒトを失明させる。現在のところ、網膜色素変性症には治療法はなく、さしあたりrdマウス、RCSラット、並びにイヌ、ネコおよびショウジョウバエでの他のモデルのような変性を起こしている動物モデルにおいて神経保護、遺伝子療法、神経修復、および生体工学的方法が行われているだけである。rd1(桿状体変性)マウスは、細胞変性を決定する分子および細胞機構を研究するための最初のモデルの一つであり、光受容体の死のアポトーシス性が確定されている(Chang, G. Q., Hao, Y., and Wong, F. (1993) Apoptosis: final common pathway of photoreceptor death in rd, rds, and rhodopsin mutant mice.Neuron 11: 595-605)。様々なrdマウスは、遺伝性の網膜ジストロフィーの治療に対する新規治療方法を試験するための理想的モデルを提供しているが、これらのマウスを用いて分子、細胞、および遺伝子の観点から光受容体の変性過程を検討することができるからである。
【0004】
遺伝子療法を介在することにより、神経変性を引き起こす突然変異遺伝子の機能的コピーの再導入に役立つ。組換えアデノウイルスまたはアデノウイルス関連のウイルスベクターにより、伸展が図られた。具体的には、新生rdマウスの桿状体特異的cGMPホスホジエステラーゼ(βPDE)のβ-サブユニットの置換により、rd表現型の組織を少なくとも6週間救済することができた(Bennett, J., Tanabe, T., Sun, D., Zeng, Y., Kjeldbye, H., Gouras, P., and Maguire, A. M. (1996) Photoreceptor cell rescue in retinal degeneration (rd) mice by in vivo gene therapy. Nat Med 2: 649-654)。しかしながら、この療法はそれぞれの親における突然変異遺伝子を明確に同定する必要があるが、これは現在のところ症例の40%で可能なだけである(Wang, D. Y., Chan, W. M., Tam, P.O., Baum, L., Lam, D. S., Chong, K. K., Fan, B. J., and Pang, CP. (2005) Gene mutations in retinosis pigmentosa and their clinical implications. Clin Chim Acta 351: 5-16)。
【0005】
新規な光受容体を開発するための神経幹細胞または前駆体の移植片は、神経修復療法の目的である。新規な光受容体は、内網膜のニューロンと適当な結合を回復しなければならない。
【0006】
増殖因子の投与により誘発される神経保護は、神経変性過程に関連した細胞死の防止を目指す。様々な投与形態が、網膜変性に罹っている数種類の動物モデルにおいて試験されている。最初の試みは、網膜変性に罹っているラットまたはマウスでの数種類の組換えタンパク質の硝子体への注射からなっていた(Faktorovich, E. G., Steinberg, R. H., Yasumura, D., Matthes, M. T., and LaVail, M. M. (1990) Photoreceptor degeneration in inherited retinal dystrophy delayed by basic fibroblast growth factor. Nature 347: 83-86、 LaVail, M. M., Unoki, K., Yasumura, D., Matthes, M. T., Yancopoulos, G. D., and Steinberg, R. H. (1992) Multiple growth factors, cytokines, and neurotrophins rescue photoreceptors from the damaging effects of constant light. Proc Nati Acad Sci U S A 89: 11249-11253)。これらの実験は、FGF2がRCSラットにおける光受容体変性を遅らせることを明らかにした(Royal Collage of Surgeon)。FGF2、FGF1、BDNF、およびCNTFなどの数種類の生存因子は、光損傷によって誘発される光受容体の死を減少させた(LaVail, M. M., Yasumura, D., Matthes, M. T., Lau-Villacorta, C, Unoki, K., Sung, CH., and Steinberg, R. H. (1998) Protection of mouse photoreceptors by survival factors in retinal degenerations. Invest Ophthalmol Vis Sci 39: 592-602)。遺伝性網膜変性に罹っているマウスモデル(Q433ter、rd、およびnrモデル、網膜色素変性症のある種のマウスモデルを表すのに用いられる用語)にCNTF類似体を硝子体内投与することによって、幾つかの変性が明らかに改善された。CNTF類似体による神経保護効果も、常染色体優性錐体-桿状体ジストロフィーのネコでの研究においてCNTFの硝子体内投与が有効な効果を有することを実証した(Chong, N. H., Alexander, R. A., Waters, L., Barnett, K. C, Bird, A. C, and Luthert, P. J. (1999) Repeated injections of a ciliary neurotrophic factor analogue leading to long-term photoreceptor survival in hereditary retinal degeneration.Invest Ophthalmol Vis Sci 40: 1298-1305)。
【0007】
もう一つの方法は、網膜変性マウスまたはラットの網膜における生存因子を発現するための遺伝子療法ベクターの使用であった。2種類のマウスモデルでは、rdおよびPrph2Rd(ペリフェリンにおける劣性突然変異) (Travis, G. H., Brennan, M. B., Danielson, P. E., Kozak, CA., and Sutcliffe, J. G. (1989) Identification of a photoreceptor-specific mRNA encoded by the gene responsible for retinal degeneration slow (rds). Nature 338: 70-73、 Connell, G., Bascom, R., Molday, L., Reid, D., Mclnnes, R. R., and Molday, R. S. (1991) Photoreceptor peripherin is the normal product of the gene responsible for retinal degeneration in the rds mouse. Proc Nati Acad Sci USA 88: 723-726)、CNTFの分泌可能な形態をコードするアデノウイルスベクターの網膜下投与により、光受容体の死を遅らせた(Cayouette, M., and Gravel, C. (1997) Adenovirus-mediated gene transfer of ciliary neurotrophic factor can prevent photoreceptor degeneration in the retinal degeneration (rd) mouse. Hum Gene Ther 8: 423- 430、 Cayouette, M., Behn, D., Sendtner, M., Lachapelle, P., and Gravel, C. (1998) Infraocular gene transfer of ciliary neurotrophic factor prevents death and increases responsiveness of rod photoreceptors in the retinal degeneration slow mouse. J Neurosci 18: 9282-9293)。一方、硝子体内投与によってほとんど活性にならない網膜におけるトランスジェニックBDNF発現は、Q344terマウスの神経変性を遅らせる(Okoye, G., Zimmer, J., Sung, J., Gehlbach, P., Deering, T., Nambu, H., Hackett, S., Melia, M., Esumi, N., Zack, D. J., and Campochiaro, P. A. (2003) Increased expression of brain-derived neurotrophic factor preserves retinal function and slows cell death from rhodopsin mutation or oxidative damage. J Neurosci 23: 4164- 4172、 Sung, CH. , Makino, C, Baylor, D., and Nathans, J. (1994) A rhodopsin gene mutation responsible for autosomal dominant retinosis pigmentosa results in a protein that is defective in localization to the photoreceptor outer segment. J Neurosci 14: 5818-5833)。これらの結果は、技術的問題を別として、硝子体内投与だけでは神経保護効果を有するには不十分である可能性があることも示している。更に、単回投与で神経保護効果をも示すCNTFは、網膜変性に罹っているPrph2Rdマウスおよび2種類のトランスジェニックラットでCNTFの分泌可能な形態をコードするAAVベクターによってその長時間発現が誘発されるときには逆効果であることが明らかにされており(Liang, F. Q., Aleman, T. S., Dejneka, N. S., Dudus, L., Fisher, K. J., Maguire, A. M., Jacobson, S. G., and Bennett, J. (2001) Long-term protection of retinal structure but not function using RAAV. CNTF in animal models of retinosis pigmentaria. Mol Ther 4: 461-472)、これは、光受容体の機能はERGによる分析によって悪化するためである。
【0008】
上記の総ての方法を考慮して、本発明は、今日まで用いられてきたシステムと比較して実際的な解決法を提供する。ニワトリ胚でのインスリンの代謝機能とは異なるインスリンの生存機能は以前に示されており、これは、プロインスリン形態でのインスリンが膵臓外形の存在以前に発生中に発現されるからである。神経系の発生中に、プロインスリンは多数の細胞過程を制御する。これは、アンチセンスオリゴヌクレオチドを用いることによるプロインスリン遺伝子またはその受容体の発現を阻害する研究において明らかにされたように、初期胚での生存因子である(Morales, A.V., Serna, J., Alarcon, C, de la Rosa, E. J., and de Pablo, F. (1997) Role of prepancreatic (pro)insulin and the insulin receptor in prevention of embryonic apoptosis. Endocrinology 138: 3967-3975)。抗体によりそれをブロックすることによっても、ニワトリ胚網膜でのアポトーシス細胞の数が増加するが(Diaz, B., Serna, J., De Pablo, F., and de la Rosa, E. J. (2000) In vivo regulation of cell death by embryonic (pro) insulin and the insulin receptor during early retinal neurogenesis. Development 127: 1641-1649)、胚への外因不活性によりアポトーシス細胞の数が減少する(Hernandez-Sanchez, C, Mansilla, A., de la Rosa, E. J., Pollerberg, G. E., Martinez- Salas, E., and de Pablo, F. (2003) Upstream AUGs in embryonic proinsulin mRNA control its low translation level. Embo J 22: 5582-5592)。ニワトリ胚から単離することができる分子は、遺伝子翻訳の第一の産物であるプロインスリンであり、その代謝活性は小さく、インスリンの活性の約5-10%である。
【0009】
慢性神経変性疾患では、ニューロンおよび/またはグリア細胞は次第に死滅する。この疾患の症状は、通常極めて僅かの細胞が死んだときに現れる。従って、比較的正常な視覚機能を維持する上で細胞生存に有利な効果的分子を決定することが重要である。それぞれが異なる病因を有する多種類の神経変性疾患があるが、それらはいずれも最終的には罹患細胞の死をもたらす。この死はプログラムされた種類のものであり、異なる種類のアポトーシスまたは非アポトーシスによる死もある。細胞が損傷前は最初のうち、良好な状態である急性損傷とは異なり、慢性疾患においては、細胞は内因性の損傷を有しており、細胞が最早損傷を維持しきれなくなり、および/または細胞が行わなければならない機能を実行することができなくなったある時点において細胞が死亡する。
【発明の概要】
【0010】
本発明の目的は、プログラム細胞死が起こる神経変性状態、障害、または疾患、好ましくは中枢および末梢神経系の神経変性疾患、更に好ましくは網膜色素変性症として知られる遺伝性変性疾患の群の予防および治療用の医薬品または医薬組成物を製造するためのプロインスリンの活性を誘発する化合物の使用(以後、本発明の誘発化合物の使用)によって構成される。本発明の好ましい態様では、上記医薬組成物は、その全身または局所への持続投与に適する。
【0011】
本発明の特定の目的は、プロインスリンの活性を誘発する化合物であって、誘発化合物がヌクレオチド配列であるものの使用(以後、本発明のプロインスリンヌクレオチド配列の使用)であり、神経保護タンパク質またはペプチドを発現させ且つ下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一方の断片、並びに
d) a)、b)および/またはc)に属するいずれか1個の配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列によって構成されるものの使用によって構成される。
【0012】
本発明の特定の態様は、プロインスリンの活性を誘発する化合物であって、ヌクレオチド配列がヒトプロインスリンをコードする配列番号1によって構成されているものの使用によって構成される。
本発明のもう一つの特定の態様は、プロインスリンの活性を誘発する化合物であって、d)のヌクレオチド配列が発現ベクターであるものの使用(以後、本発明のプロインスリン発現ベクターの使用)であって、神経保護を誘発できるプロインスリンタンパク質をコードするヌクレオチド配列または遺伝子構築物を含んでなることによって構成される。
【0013】
本発明のもう一つの特定の態様は、プロインスリンの活性を誘発する化合物であって、誘発化合物が好ましくはヒト真核細胞であるものの使用(以後、本発明のプロインスリン細胞の使用)であって、遺伝的に修飾され、且つ本発明のプロインスリンヌクレオチド配列、構築物、または発現ベクターを含んでなり、プロインスリンタンパク質を細胞外媒質に好適に発現しまたは放出できることによって構成される。
【0014】
本発明のもう一つの特定の目的は、プロインスリンの活性を誘発する化合物であって、誘発化合物がタンパク質またはペプチドであるものの使用(以後、本発明のプロインスリンタンパク質の使用)であり、神経保護活性を有し且つ下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つを含んでなるアミノ酸配列
に属する1または数個のアミノ酸配列を含んでなる化合物の使用によって構成される。
【0015】
本発明のもう一つの特定の態様は、本発明の誘発化合物であって、誘発化合物がヒトプロインスリンタンパク質(配列番号2)であるものの使用によって構成される。
【0016】
本発明のもう一つの目的は、神経退行変性をもたらす疾患、障害、または病変の治療用の医薬組成物または医薬品(以後、本発明の医薬組成物)であって、治療上有効量の本発明のプロインスリンの活性を誘発する化合物を場合によっては1種類以上の薬学上許容可能なアジュバントおよび/または担体と共に含んでなるものによって構成される。
【0017】
本発明の特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つの配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列であるものによって構成される。
【0018】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、ヌクレオチド配列がヒトプロインスリンをコードする配列番号1からなる医薬組成物によって構成される。
【0019】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、ヌクレオチド配列がヒトプロインスリン発現ベクターである医薬組成物によって構成される。
【0020】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が本発明のプロインスリン配列、遺伝子構築物、またはベクターによってコードされるタンパク質またはペプチドである医薬組成物によって構成される。
【0021】
本発明の特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発するタンパク質またはペプチドが、下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つの配列を含んでなるアミノ酸配列
に属する医薬組成物によって構成される。
【0022】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、アミノ酸配列がヒトプロインスリン(配列番号2)によって構成される医薬組成物によって構成される。
【0023】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が好ましくはヒト細胞、更に好ましくは本発明のプロインスリン配列、遺伝子構築物、または発現ベクターによって形質転換された中枢神経系細胞である医薬組成物によって構成される。
【0024】
本発明のもう一つの目的は、プログラム細胞死が起こるヒトを冒す中枢または末梢神経系の神経変性疾患、障害、または病変に冒されている哺乳類、好ましくはヒトの治療または予防方法において、本発明の医薬組成物の使用(以後、本発明の医薬組成物の使用)であって、上記治療組成物を上記神経変性を減少させる適量により投与することからなる使用によって構成される。
【図面の簡単な説明】
【0025】
【図1】ヒトプロインスリンのcDNAおよびプラスミドpMLC-hInsのインサートの配置図(schematic representation)。(A) プラスミドに挿入されているDNA配列の配置図。ヒトプロインスリン遺伝子のcDNAに相当。図は、3個のエキソン(エキソン1= E1、エキソン2=E2、エキソン3=E3)によって形成される462塩基対(bp)のmRNAを示す。挿入されたDNA配列は、翻訳されているものであり、347bpのORF(オープンリーディングフレーム)である。未翻訳のフランキング部分(5'UTRおよび3'UTR)は、構築物に挿入されない。翻訳されたタンパク質は、110個のアミノ酸(aa)のプレプロインスリンである。このタンパク質は、シグナルペプチド(signal pep.)、鎖B、ペプチドC、および鎖Aからなる。シグナルペプチドは除かれ、プロインスリン分子が残る。(B) プラスミドは1.A項において記載したインサートを含み、110アミノ酸プレプロインスリン(黒色の太い部分)を横紋筋ミオシン軽鎖線維の構成的筋肉プロモーターMLC1(ミオシン軽鎖)の転写制御下でコードする。このプラスミドは、rd10についてホモ接合に到達するように交雑した2系列のトランスジェニックヒトプロインスリン産生マウスを産生するのに用いられる。
【図2】P32でのマウスの12μmのクリオスタットの眼切片(cryostat eye section)であって、様々なマーカーにより変性の進行を示す桿状体および錐体の状態を観察することができる。野生型コントロールマウス(AおよびD)、プロインスリンを産生するホモ接合rd10およびトランスジェニックマウス、Proins/rd10-/-(BおよびE)、およびコントロールホモ接合rd10マウス(CおよびF)。桿状体および錐体の核が配置されている外顆粒層(ONL)をDAPIによる核染色によって見ることができる。内顆粒層(INL)には、二極性、無軸索、水平、およびミュラー細胞の核が配置されている。蛍光色素 Alexa 488で標識した凝集素であって、錐体の外側部分(上の矢印)および錐体のシナプス基部(下の矢印)を示すその反応を用いて錐体を標識する。棒線は45μmを表す。
【図3】P32におけるマウスの12μmの眼のクリオスタット切片であって、シナプス結合状態を観察することができる。野生型コントロールマウス(A)、プロインスリンを産生するホモ接合rd10およびトランスジェニックマウスProins/rd10-/- (B)、およびコントロールホモ接合rd10マウス(C)。顆粒層ONL、INL、およびガングリオン細胞層(GCL)を示す。シナプス結合が起こる網状層であり、外網状層(OPL)、および内網状層(IPL)はそれらの間に配置される。この図は、一般にシナプス結合を標識するSV2抗体による免疫組織化学的標識を示す。棒線は0.45μmを表す。
【図4】P30で行った網膜電位記録の結果。野生型マウス(wt)、コントロールRd10-/-マウス、およびProins/Rd10-/-マウスであり、いずれも系列1(L1)および系列2(L2)由来。暗順応(夜間)で記録した桿状体に生じた網膜電位応答(上列)および混合応答(中列)の例は、それぞれの動物について示される。明所視(昼間)状態で記録した錐体に生じた網膜電位応答(下列)も示す。両系列(L1およびL2)由来のProins/rd10-/-マウスで得た応答範囲が、rd10-/-マウスの応答と比較して大きいことに留意すべきである。
【図5】ヒトプロインスリンレベルと視覚パラメーターの保持との間の相関。Proins/Rd10-/-マウスにおけるヒトプロインスリンレベルを、P30において完全な網膜電位検討を行ったマウスの大腿四頭筋でP32において測定した。プロインスリンおよび様々な視覚パラメーターのそれぞれの値は、双曲線となる。網膜電位波の振幅bmax(1.5 log cd.s.m-2に応じて記録)、OP(振動電位)、bphot(1.5 log cd.s.m-2に応じて記録)、およびフリッカー応答を示す。
【図6】様々な日に行った網膜電位記録の結果。産後発生の様々な日、すなわちP35、P45、P55の野生型マウス(wt)、コントロールRd10-/-マウス、およびProins/Rd10-/-マウス。Aは、桿状体(-2,55 log cd.s.m-2)を除く暗順応状態で記録した応答、およびそれぞれの動物種および産後発生の様々な時期における混合応答(1,48 log cd.s.m-2)の例を示す。Bは、それぞれの動物種および産後発生の様々な時期において錐体に生じた(1.48 log cd.s.m-2)明所視状態で記録した応答の例を示す。
【図7】ヒトプロインスリンを皮下注射した後の網膜における存在。抽出物調製の2時間前にまたは毎日、P11とP14の間においてプロインスリンの所定量を皮下投与したC57B1/6マウスの網膜抽出物におけるヒトプロインスリンのELISAによる定量。最後の投与も抽出物調製の2時間前。
【図8】神経変性マウスの網膜組織構造に対するヒトプロインスリンの皮下投与の効果。P6と、P14またはP18との間の12時間毎にヒトプロインスリン(BおよびD)または担体(AおよびC)を投与したP14のRd1-/-マウスの網膜切片。外顆粒層における光受容体の喪失は、総ての場合にDAPIを用いる核染色によって観察することができる。ONL,外顆粒層、 INL, 内顆粒層、 GCL, ガングリオン細胞層。棒線は25μmを表す。
【図9】神経変性マウスの網膜組織構造に対するヒトプロインスリンの皮下投与の効果。P6と、P14またはP18との間の12時間毎にヒトプロインスリン(BおよびD)または担体(AおよびC)を投与したRd1-/-マウスの網膜電位図。様々な治療を施した同腹兄弟のrd突然変異マウスの網膜電位図を示す。プロインスリンを投与しても、視覚機能喪失過程を示さなかった。
【発明の具体的説明】
【0026】
ほとんどの神経変性疾患、特に網膜色素変性症には効率的および/または効果的な治療法がないことを考慮し、本発明は代替解決法を提供する。
【0027】
本発明は、プロインスリン(通常はインスリンの前駆体型として知られているインスリンファミリーの増殖因子)が、慢性神経変性過程、特に網膜色素変性症で起こる網膜神経変性過程における更なる細胞生存因子であるという事実に基づく。
【0028】
桿状体特異的サイクリックGMPホスホジエステラーゼ酵素遺伝子に劣性ホモ接合突然変異を有し、この酵素の機能を変更させ、進行性の光受容体細胞死と続いて残りの網膜細胞の種類、主に錐体の二次的変性をもたらすrd10 (Pdebrd10/rd10)およびrd1 (Pdebrd1/rd1)マウスを、網膜変性モデルとして選択した。
【0029】
ヒトプロインスリンタンパク質と、rd10突然変異に対する劣性ホモ接合体とを発現する2系列のトランスジェニックマウスProins/rd10-/-を生成した(実施例1.1)。網膜変性に罹っているこれらのProins/rd10-/-マウスは横紋筋でヒトプロインスリンを構成的に生成し(これはインスリンには処理されない)、これは血清中に検出され、グルコースの水準により膵臓の通常の制御を受けにくい。これにより、動物に用量、投与前の産物の状態(それは内因性の産生に由来するので、商業産物のようには分解しない)、並びに投与の形態および時間に依存することなく、循環しているヒトプロインスリンの水準を持続させる。これにより、その投与方法を見出すための薬物動態検討を行わなくともよくなった。
【0030】
両系統のProins/rd10-/-マウスの筋肉および血清にヒトプロインスリンが含まれることは、ヒトプロインスリンに対するELISA検出キットを用いて立証された。更に、血糖を測定し、プロインスリンが望ましくない代謝効果を持たないことが立証された。更に、ヒトプロインスリンの皮下投与によって、前記プロインスリンが神経網膜に達することができることが立証されており(網膜抽出物でのELISAキットによるプロインスリン同定)、これはプロインスリンが血液-網膜関門を通過することができることを意味する。
【0031】
網膜神経変性におけるProins/rd10-/-マウスの高プロインスリン血症の影響を、幾つかの方法により同定した。網膜の状態を最初に組織学的に観察した(実施例1)。ヒトプロインスリンについてのトランスジェニックマウスおよびrd10についてのホモ接合体では変性は遅くなり、P32においては、網膜に、一層多数の桿状体、錐体、およびシナプス結合が保持されており、網膜の状態は一層良好であることが立証された(実施例1)。
【0032】
網膜の状態が更に長時間一層良好に維持されるという事実は好都合である。実際に損傷を受けた細胞(網膜の桿状体)での細胞死の過程を遅らせることは別にして、残りの網膜は、例えば、本質的な損傷を有しておらずむしろ二次的に変性する錐体の死を防止することによって一層良好な状態に保持されており、この疾患の幾つかの症状は改善されている。この特殊な症例の場合には、桿状体が変性されていても錐体が更に長時間保持される場合には、夜間の視覚は失われるが昼間の視覚は保持され、これは、網膜色素変性症の患者の生活質を意味している。
【0033】
次いで、視覚機能を変性過程中、5種類の標準化した網膜電位図により分析し、rd10のトランスジェニックマウスコントロールと比較し、Proins/rd10-/-マウスでは、P55では視覚機能を未だ有しており、この視覚機能はrd10マウスではP35で失われ、これによって視覚応答はProins/rd10-/-マウスにおけるよりも良好であり、且つ更に長時間延長されると結論づけれることが明らかにされた(実施例2)。更に、トランスジェニックProins/rd10-/-マウスは、P30において少なくとも明所(昼間)視力では健常マウスに匹敵するERGを有する(実施例2、図4)。
【0034】
更に、本発明では、プロインスリンの単独の皮下および硝子体内投与で観察されたのとは対照的に、長時間持続する生理学的レベルでの投与形態でプロインスリンを用いると、プロインスリンにその神経保護機能を発揮させることが観察されている(実施例1.4参照)。これに関して、プロインスリンの単独においてのの皮下投与を試験したが、網膜神経変性の改善には成功せず、これは恐らく安定で長時間にわたって持続する水準が得られなかったからであろう(実施例3および4)。
【0035】
もう一つの利点は、プロインスリンはインスリンの代謝作用を持たず、従ってグルコース代謝が変更されていない患者に投与することができる点である。インスリンはイン・ビトロ研究においては抗アポトーシス活性も有するが、その通常の代謝活性は本明細書に記載したような治療についてその使用を実現できなくする。循環血清プロインスリンがこの救済を行うという事実は、それが投与を容易にするので良好な治療法であることを示す。プロインスリンは損傷したニューロンの死を防止するので、その適用は神経変性疾患の予防的および抑制的治療法となる。
【0036】
要約すれば、遺伝子療法について認可された遅延放出担体または発現ベクター上に担持されたヒトプロインスリンの他の形態の持続投与、皮下投与によって得ることもできるミオシン軽鎖プロモーターによって制御されるトランスジェニックヒトプロインスリンの発現によって得られる慢性血清プロインスリン血症水準(1−15pM)などは、遺伝的rd10マウスモデルにおいては網膜に到達して、光受容体神経変性を減少させることができる。
【0037】
従って、本発明の目的は、プログラム細胞死が起こる神経変性状態、障害、または疾患、好ましくは神経変性疾患、好ましくは中枢および末梢神経系の神経変性疾患、更に好ましくは網膜色素変性症として知られる遺伝性変性疾患の群の予防および治療用の医薬品または医薬組成物を製造するためのプロインスリンの活性を誘発する化合物の使用(以後、本発明の誘発化合物の使用)によって構成される。本発明の好ましい態様では、医薬組成物は、この医薬組成物を持続的に全身または局所投与するのに適した担体をも含んでなる。
【0038】
本発明では、上記医薬組成物が体内に定常的に放出され、標的組織に有効用量で長時間保持されるように薬学的に担持され、圧縮され、または処方されるとき、医薬組成物はその全身または局所への持続投与に適していると理解される。本発明の一つの態様においては、医薬組成物の持続投与に適する任意の担体、例えば、ポリマーまたはパッチなどこれらに限定されないものは、本発明の保護の範囲内に包含される。
【0039】
本発明で用いられているように、「プロインスリンの活性を誘発する化合物」という用語は、ヒトプロインスリンタンパク質の神経保護活性を模倣し、強度を増加し、または持続期間を延長する分子に関する。アクチベーター化合物は、化学的分子、ペプチド、タンパク質、またはヌクレオチド配列、並びに神経保護活性を有するタンパク質をコードするヌクレオチド配列を発現させる分子から構成できる。
【0040】
本発明で用いられているように、「神経保護活性」という用語は、神経変性疾患に最初に冒されている細胞および/または二次的に神経変性に冒されている細胞のプログラム細胞死の過程の減少、および/または残りの細胞の神経機能活性の増強に関する。
【0041】
本発明で用いられているように、「神経変性疾患」という用語は、とりわけ実例として且つ本発明の範囲を限定することのない下記の群、すなわち、アルツハイマー病、パーキンソン病、多発性硬化症、網膜色素変性症、レヴィー小体認知症、筋萎縮性側索硬化症、脊髄小脳性萎縮症、前頭側頭型認知症、ピック病、血管性認知症、ハンティングトン病、バッテン病、脊髄損傷、黄斑変性、および緑内障に属する疾患、障害、または病変に関する。
【0042】
従って、本発明の特定の目的は、プロインスリンの活性を誘発する化合物であって、誘発化合物が、神経保護タンパク質またはペプチドを発現させ且つ下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一方の断片、並びに
d) a)、b)、および/またはc)に属するいずれか1個の配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列によって構成されるヌクレオチド配列であるヌクレオチド配列であるものの使用(以後、本発明のプロインスリンヌクレオチド配列の使用)によって構成される。
【0043】
本発明の説明に用いられる意味において、「同様の」という用語は、例えば、1個以上のヌクレオチドの挿入、分子の末端のいずれかにおける1個以上のヌクレオチドの付加、または任意の末端または配列内部における1個以上の分子の欠失などの保存的および非保存的ヌクレオチド置換を導入することによって本明細書に示される配列に基づいて単離または構築することができ、且つヒトプロインスリンの活性を模倣することができるペプチドまたはタンパク質をコードすることができる任意のヌクレオチド配列(配列番号2)を包含することを意味する。
【0044】
本発明および当該技術の状態において記載された情報に基づいて、当業者であれば、本発明に記載したのと同様のヌクレオチド配列を続いて用いるために単離または構築することができる。
【0045】
同様のヌクレオチド配列は、通常は上記ヌクレオチド配列と実質的に相同である。本明細書で用いられる意味では、「実質的に相同」という表現は、問題のヌクレオチド配列の同一性の程度が少なくとも30%、好ましくは少なくとも85%、更に好ましくは少なくとも95%であることを意味する。用いられるヌクレオチド配列の好ましい形態は、ヒトプロインスリンヌクレオチド配列(配列番号1)およびその誘導体である。
【0046】
本発明で用いられているように、「ヌクレオチド配列」という用語は、DNA、cDNA、またはmRNA配列に関する。
【0047】
本発明の特定の態様は、プロインスリンの活性を誘発する化合物であって、ヌクレオチド配列がヒトプロインスリンをコードする配列番号1からなるものの使用によって構成される。
【0048】
d)の項に定義されているヌクレオチド配列は、プロインスリンタンパク質を発現させる遺伝子構築物および遺伝子発現ベクターに相当する。遺伝子構築物(本発明のプロインスリン遺伝子構築物)の場合には、これは、必要ならばおよび一層良好に発現したペプチドを単離、検出、または細胞の外部に分泌させるために、上記ペプチドの単離、検出、または分泌を目的として用いることができるペプチドをコードするヌクレオチド配列をも含んでなることができる。従って、本発明のもう一つの特定の目的は、本発明のプロインスリンヌクレオチド配列の他に、発現したペプチドを単離、検出、または細胞の外側へ分泌させるペプチドまたはペプチド配列をコードする任意のもう一つのヌクレオチド配列、例えば、実例として且つ本発明の範囲を限定することのないポリヒスチジン(6xHis)配列、モノクローナル抗体(例えば、その同定のための)によって認識することができるペプチド配列、またはイムノアフィニティークロマトグラフィーから生じる融合タンパク質の精製に有用な任意の他の配列、すなわち、c-myc、HA、E-tagのようなタグペプチドを含んでなる遺伝子構築物によって構成される(「抗体の使用: 実験室マニュアル」Ed Harlow and David Lane (1999). Cold Spring Harbor Laboratory Press. New York. 章: 「タンパク質のタギング」pp. 347-377)。
【0049】
上記ヌクレオチド配列および遺伝子構築物は、当業者であれば当該技術分野において広く知られている手法を用いることによって単離して得ることができる(Sambrook et al.「分子クローニング、実験室マニュアル第2版」Cold Sping Harbor Laboratory Press,ニューヨーク, 1989 vol 1-3)。上記ヌクレオチド配列は、細胞内で適当な条件において発現を調節することができる遺伝子発現ベクターに組込むことができる。
【0050】
従って、本発明のもう一つの特定の目的は、d)のヌクレオチド配列が発現ベクターであるプロインスリンの活性を誘発する化合物の使用(以後、本発明のプロインスリン発現ベクターの使用)であって、神経保護を誘発できるプロインスリンタンパク質をコードするヌクレオチド配列または遺伝子構築物を含んでなるものによって構成される。特定の態様の一例は、本発明において調製された発現ベクターの使用であって、発現が筋肉特異的プロモーターおよび配列番号1のヌクレオチド配列によって調節されるものによって構成される(実施例1参照)。
【0051】
本発明に記載のヌクレオチド配列または遺伝子構築物の他に、発現ベクターは、通常はその転写を指示するプロモーター(例えば、pT7、plac、ptrc、ptac、pBAD、5 retなど)、好ましくは組織プロモーターであって、これに転写を指示するプロモーターが操作結合するもの、並びに上記転写および適当な場合には、目的の産物の翻訳を制御し調節する他の必要なまたは適当な配列、例えば、転写開始および終結シグナル(tlt2など)、ポリアデニル化シグナル、複製起源、リボソーム結合配列(RBS)、転写レギュレーター(エンハンサー)をコードする配列、転写サイレンサー、リプレッサーなどを含んでなる。適当な発現ベクターの例は、それぞれの特殊な症例の状態および要求に従って発現プラスミド、ウイルスベクター(DNAまたはRNA)、コスミド、人工染色体などから選択することができ、目的の遺伝子または遺伝子類によりトランスフェクションまたは形質転換した細胞を選択するのに用いることができるマーカーを更に含むこともできる。ベクターの選択は、宿主細胞および実行する使用の種類によって変化する。従って、本発明の特定の態様によれば、上記ベクターはプラスミドまたはウイルスベクターである。上記ベクターは、真核細胞および微生物の形質転換に用いることができる様々なマニュアル[Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989)「分子クローニング: 実験室マニュアル, 第2版」Cold Spring Harbor Laboratory, Cold Spring Harbor,ニューヨーク]に記載されている様々な周知の方法(化学的形質転換、エレクトロポレーション、マイクロインジェクションなど)と同じ方法で当業者に知られている通常の方法によって得ることができる。一つの方法は、他の種類の療法において既に試みられているように、標的細胞に感染させる目的でレンチウイルスを用いることである(Ralph GS, Binley K, Wong LF, Azzouz M, Mazarakis ND (2006) Gene therapy for neurodegenerative and ocular diseases using lentiviral vectors. Clin Sci (Lond) 110:37-46)。
【0052】
遺伝子発現システムは、宿主細胞のゲノムに新たな遺伝子材料を組込むことができまたはできない。次に、ヌクレオチド配列、遺伝子構築物、またはプロインスリン発現ベクターを、ニューロンおよび/またはグリア変化をもたらす疾患に冒されているヒトの遺伝子療法による予防および治療の過程において、ヒト細胞、好ましくは神経変性変化に冒されているニューロンおよび/またはグリア細胞を保護するための医薬品として用いることができる。これらの遺伝子発現システムを神経変性疾患に冒されているヒトに投与すると、それらを一般的または特異的に組織細胞に導入することができ、そこでそれらが細胞ゲノムに組込まれると、プロインスリンタンパク質を発現することができ、そのタンパク質は、細胞外媒質に分泌されると、中枢神経系に到達して神経保護作用を行うことができる(実施例参照)。
【0053】
その上、これらの遺伝子発現システムを潜在的レシピエントに関して、自己または異種の人体外のヒト細胞の形質転換に用いることもでき、これらの細胞はヒトニューロンおよび/またはグリア細胞の神経保護活性を有するプロインスリンタンパク質を発現および放出するため、神経変性疾患に罹っているヒトに投与するとプロインスリンを誘発する化合物となる。
【0054】
従って、本発明のもう一つの特定の目的は、プロインスリンの活性を誘発する化合物の使用によって構成され、誘発化合物が好ましくはヒト真核細胞(以後、本発明のプロインスリン細胞)であり、これが遺伝的に修飾されており、且つ本発明のプロインスリンヌクレオチド配列、構築物、または発現ベクターを含んでなり、細胞外媒質にプロインスリンタンパク質を適当に発現または放出できる。
【0055】
これらの細胞は、当業者に知られている遺伝子工学技術によって上記ヌクレオチド配列により形質転換、感染、またはトランスフェクションすることができる[Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989).「分子クローニング:実験室マニュアル, 第2版」Cold Spring Harbor Laboratory]。生物薬剤学的手段および遺伝子療法の方法は当業者に十分知られており、過度の努力することなく本発明に記載の常法を用いて開発することができる。
【0056】
もう一つの特定の態様は、様々な細胞株、好ましくは中枢神経系、更に好ましくはヒト組織を再生する細胞として用いることができるニューロンからヒトプロインスリンヌクレオチド配列(配列番号1)によって形質転換したヒト細胞の使用である。
【0057】
更に、本発明のもう一つの特定の目的は、プロインスリンの活性を誘発する化合物であって、誘発化合物がタンパク質またはペプチドであるものの使用(以後、本発明のプロインスリンタンパク質の使用)であり、神経保護活性を有し且つ下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つを含んでなるアミノ酸配列
に属する1または数個のアミノ酸配列を含んでなる化合物の使用によって構成される。
【0058】
本明細書で用いられる意味において、「同様の」という用語は、例えば、1個以上のアミノ酸の挿入、分子の末端のいずれかにおける1個以上のアミノ酸の付加、または任意の末端もしくは配列内部における1個以上のアミノ酸の欠失などの保存的および非保存的アミノ酸置換を導入し、且つヒトプロインスリンの活性を模倣することによって本明細書に示される配列に基づいて単離または構築することができる任意のアミノ酸配列を包含することを意味する。
【0059】
本発明に記載の情報に基づいて、当業者であれば、本発明に記載したのと同様のアミノ酸配列を単離または構築することができる。
【0060】
同様のアミノ酸配列は、通常は上記アミノ酸配列に実質的に相同である。本明細書で用いられる意味では、「実質的に相同」という表現は、問題のアミノ酸配列の同一性の程度が少なくとも30%、好ましくは少なくとも85%、更に好ましくは少なくとも95%であることを意味する。
【0061】
本発明のもう一つの特定の態様は、本発明の誘発化合物の使用であって、誘発化合物がヒトプロインスリンタンパク質(配列番号2)であるものによって構成される。
【0062】
本発明のもう一つの目的は、神経退行変性をもたらす疾患、障害、または病変の治療用の医薬組成物または医薬品(以後、本発明の医薬組成物)であって、治療上有効量の本発明のプロインスリンの活性を誘発する化合物を場合によってはプロインスリンの活性を持続的に誘発する化合物の全身または局所投与に適当な1種類以上の薬学上許容可能なアジュバントおよび/または担体と共に含んでなる医薬組成物または医薬品によって構成される。
【0063】
上記組成物で用いることができる薬学上許容可能なアジュバントおよび担体は、当業者に知られており、且つ治療組成物の製造に一般に用いられるアジュバントおよび担体である。
【0064】
本明細書で用いられる意味において、「治療上有効量」という表現は、所望な効果を生じるように計算された神経保護を発揮することができ、通常は、特に患者の年齢、患者の容体、変化または障害の重篤度、並びに投与の経路および頻度などの化合物の特徴によって決定される薬剤または化合物の量に関する。
【0065】
もう一つの特定の態様においては、上記治療組成物は、薬学上許容可能な希釈剤中で固形形態または水性懸濁液の形態に調製される。本発明によって提供される治療組成物は任意の適当な投与方法によって投与することができ、そのために上記組成物は選択された投与方法に適する投薬形態に処方される。特定の態様においては、本発明によって提供される治療組成物は、非経口、経口、鼻内吸入、腹腔内、皮下などに投与される。医薬品の投与のための様々な投薬形態およびそれらを得るのに必要な賦形剤の総説は、例えば、"Tratado de Farmacia Galenica", C. Fauli i Trillo, 1993, Luzan 5, S. A. Ediciones, Madridに見出すことができる。
【0066】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、神経保護薬剤または化合物が下記の群、すなわちプロインスリン活性を有するタンパク質またはペプチドを発現させるプロインスリン配列、遺伝子構築物、または発現ベクターに属する医薬組成物によって構成される。
【0067】
本発明の特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が、下記の群
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列の任意の一つの断片、並びに
d) a)、b)、および/またはc)に属する任意の一つの配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列である医薬組成物によって構成される。
【0068】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、ヌクレオチド配列がヒトプロインスリンをコードする配列番号1を含んでなる医薬組成物によって構成される。
【0069】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、ヌクレオチド配列がヒトプロインスリン発現ベクターであることを特徴とする医薬組成物によって構成される。
【0070】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が本発明のプロインスリン配列、遺伝子構築物、またはベクターによってコードされるタンパク質またはペプチドであることを特徴とする医薬組成物によって構成される。
【0071】
本発明の特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発するタンパク質またはペプチドが下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属する任意の一つの配列を含んでなるアミノ酸配列
に属することを特徴とする医薬組成物によって構成される。
【0072】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、アミノ酸配列がヒトプロインスリン(配列番号2)によって構成されることを特徴とする医薬組成物によって構成される。
【0073】
本発明のもう一つの特定の態様は、本発明の医薬組成物であって、プロインスリンの活性を誘発する化合物が、好ましくはヒト細胞、好ましくは本発明のプロインスリン配列、構築物、または発現ベクターによって形質転換された中枢神経系細胞であることを特徴とする医薬組成物によって構成される。
【0074】
本発明のもう一つの目的は、プログラム細胞死が起こるヒトを冒す中枢または末梢神経系の神経変性疾患、障害、または病変に冒されている哺乳類、好ましくはヒトの治療または予防方法において、本発明の医薬組成物の使用(以後、本発明の医薬組成物の使用)であって、上記治療組成物を、上記神経変性を減少させる適量で投与することからなる使用によって構成される。
【0075】
本発明の医薬組成物は、単独でまたは他の医薬化合物と共に治療方法に用いることができる。
【0076】
本発明のもう一つの特定の態様は、下記の群、すなわち、アルツハイマー病、パーキンソン病、多発性硬化症、レヴィー小体認知症、筋萎縮性側索硬化症、脊髄小脳性萎縮症、前頭側頭型認知症、ピック病、血管性認知症、ハンティングトン病、バッテン病、および脊髄損傷に属する神経変性疾患の治療方法における本発明の医薬組成物の使用によって構成される。
【0077】
本発明のもう一つの特定の態様は、下記の群、すなわち、網膜色素変性症、黄斑変性、および緑内障に属する神経変性疾患の治療方法における本発明の医薬組成物の使用によって構成される。
【実施例】
【0078】
本明細書において提供される下記の具体例は、本発明の性質を説明するためのに役立つ。これらの実施例は説明目的のためにのみ包含されており、本明細書に記載の本発明が限定されると解釈すべきではない。
【0079】
実施例1. ヒトプロインスリンは、トランスジェニックProins/rd10-/-マウスの網膜桿状体の死を防止し、シナプス結合を保持することができる。
1.1. rd10突然変異(Proins/rd10-/-)についてヒトプロインスリンおよびホモ接合体を産生する2種類のトランスジェニック系列の産生
横紋筋ミオシン軽鎖(MLC1)マウスの構成的プロモーターの制御下において、遺伝的背景が50% C57B1/6および50% SJLの数系列のトランスジェニックヒトプロインスリン産生マウスを得た後、それらをホモ接合rd10マウス(100% C57B1/6)と連続的に交雑させた。従って、遺伝的背景は95% C57B1/6を上回る比率に到達するまで均質化され、Proins/rd10-/-マウスを得た。これは6回目の戻し交雑によって達成され、二つの主系列L1およびL2を得た(L2動物は下記の実施例で用い、一方、図4のP30におけるERGに記載の結果もL1動物を用いて行い、プロインスリン血症および血糖分析も両方で行った)。
【0080】
本発明では、野生型マウスは、コントロールとして用いられるマウスと理解されている。それは変化しない。それは市販のものであり、その名称はJackson laboratories製のC57B1/6Jである。
【0081】
rd1マウスは、桿状体特異的サイクリックGMPホスホジエステラーゼ遺伝子に突然変異を有する市販マウスと理解されており、その商品名はJackson laboratories製のPdebrd1/rd1である。
【0082】
rd10マウスは、桿状体特異的サイクリックGMPホスホジエステラーゼ遺伝子に突然変異を有する市販マウスと理解されており、その商品名はJackson laboratories製のPdebrd10/rd10である。
【0083】
Proins/rd10-/-マウスは、rd10マウスと、トランスジェニックヒトプロインスリン産生マウスとを横紋筋プロモーターMLC1下において交雑させることによって生成するマウスであると理解されている。これらのマウスに導入された構築物は、軽鎖ミオシン筋肉プロモーターによって制御されているヒトプロインスリンタンパク質のcDNAを有し、その発現は構成的である(図1B, 配列番号1)。発現の変動性が観察され、これはトランスジェニックマウスの異なる浸透度による可能性がある。これは、血清に見出されるヒトプロインスリンの産生と相関している。
【0084】
トランスジェニックマウスの産生に用いた構築物は、サイズが6.4kbのプラスミド(pMLC-hIns)からなっていた。発現は、ミオシン軽鎖構成的筋肉プロモーター(MLC)によって伝達される。ヒトプロインスリン遺伝子のcDNAを、二つのEcoRI部位間でクローニングする(図1B)。
【0085】
ジェノタイピング ゲノムDNAは、Miller et al., 1988において記載された手法に従って組織から得た(Miller, S. A., Dykes, D. D. and Polesky, H. F. (1988) A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res, 16, 1215.)。離乳マウスの尾を、プロテイナーゼK(Roche Diagnostics, マンハイム, 独国)0.3mgを有するリーシス緩衝液(Tris- HCl 40 mM, pH 8.0, EDTA 20 mM, SDS 0.5%、およびNaCl 200 mM)0.5ml中で消化した。DNAを沈澱させたならば、これをHindIII酵素(Roche)で開裂させた。ゲノムDNA 10μgを用いて、これを最終容積50μlのHindIII酵素により37℃において一晩消化した。それらを、長さ12cmの1%アガロースゲルのそれぞれの個々のウェルに装填し、良好なサイズ分離を行った。導入前に、下記の溶液、すなわち、最初は脱プリン溶液 (0.5M HCl)で15分間、次いで変性溶液(0.5 N NaOHおよび1.5 M NaCl)で30分間、最後に中和溶液(0.5 M Tris-HCl, pH 8)で30分間で、ゲルを調製した。DNA断片化はこの処理によって行われ、それによって導入が促進される。ナイロン膜(Schleicher & Schuell BioSciencie, 米国)を用いて、DNAをStratalinkerオーブン(Stratagene, La Jolla, カリフォルニア, 米国)中でUV線によって膜に固定した。湿式導入は、室温において一晩行った。
【0086】
ジェノタイピングには、構築物に挿入されたヒトインスリンcDNA配列に対してdCTPベース中の32P-放射能標識プローブを用いた。プローブを作製するための鋳型は、プラスミド自身からEcoRI (Roche)で消化して得た。
【0087】
構築物をEcoRIで消化した後に放出されたインサートを、DNA抽出キット(Millipore)によって精製した。プローブの標識は、[32P]dCTPの存在下においてRandom Primer Kit (Stratagene)を用いて行った。次いで、プローブをMicrospin G25カラム(Amersham Pharmacia Biotech)で精製した。
【0088】
膜を、50%ホルムアミド、1 x Denhardt (0.02% Ficol, 0.02 % ポリビニルピロリドン、および0.02% BSA)、1% SDS、5 x SSC (0.15 M NaClおよび15 mMクエン酸ナトリウム, pH 7.2)、および0.1 mg/mlサケ精子DNAを含む溶液で65℃で少なくとも2時間予備ハイブリダイゼーションした後、予備ハイブリダイゼーション溶液で65℃において一晩ハイブリダイゼーションし、これにプローブ1.5 x 106 cpm/mlを加えた。2x SSC で室温にて15分間の2回の洗浄、2 x SSCおよび1% SDSで30分間の更に1回の洗浄、並びに2 x SSCおよび0.1% SDSで62.5℃での最終洗浄を行った。
【0089】
フィルターを洗浄したならば、乾燥させることなくGLAD型プラスチックに包み、2個の増幅スクリーン(Genescreen plus, DuPont)と18x24 cmのKodak Biomax MS型写真フィルム(Eastman Kodak Company, ロチェスター, ニューヨーク, 米国)との間に露光した。シャープなシグナルを得るには、3-4日の露出時間が必要であった。ヒトプロインスリンcDNAに対して生成したプローブは、ゲノムDNAゲルに1.5 Kbのバンドを検出した。
【0090】
rd10マウスは、桿状体特異的サイクリックGMPホスホジエステラーゼ6酵素遺伝のエキソン13に位置する点突然変異を有する。野生型マウスでは、その位置にCfoI酵素による開裂部位がある。突然変異体の場合には、酵素開裂部位は消失している。これにより、内因性ジェノタイピング技術により制御することができる。
【0091】
ゲノムPCRは、次のプライマー3'-CTTTCTATTCTCTGTCAGCAAAGC-5' (オリゴA, 配列番号3)および3'-CATGAGTAGGGTAAACATGGTCTG-5' (オリゴB, 配列番号4)を用いて行い、97 bpの断片を増幅した。次に、PCR生成物を、CfoI酵素(Roche)により37℃において2時間消化した。これを、次に3% Metaphorアガロースゲル中で分画した。野生型マウスは、消化後に二つの54および43bpのバンドを示した。ホモ接合rd10マウスは1本の97bpのバンドを生じたが、これは、酵素がアンプリコンを開裂しなかったからであり、またヘテロ接合rd10/+マウスは3本のバンド(対立遺伝子の一方は開裂され、他方は開裂しない)を示した。この遺伝子では、個体間の変動性の種類は見出されず、これは、変性が標準的パターンに従い、通常はこれを有する総ての個体について同様に実現されることを意味する。
【0092】
1.2. トランスジェニックマウスの血糖およびプロインスリンレベルの測定
総てのマウスの血糖を、12時間の絶食後に試験ストリップ(Accu-Chek, Roche)で測定した。マウスの正常レベルは、通常は100-200 mg/dlである。Proins/rd10-/-二重突然変異体では、80-150 mg/dlの変動が見られ、この変動はプロインスリン血症の水準とは直接相関しなかった。
【0093】
市販のELISAアッセイ(Lineo Research, ミズーリー, 米国)を用いて、トランスジェニックマウスによるヒトプロインスリン産生レベルを検出し、これは特異的にヒトプロインスリンを検出する。
【0094】
トランスジェニックおよびコントロールマウスの筋肉および血清を分析した。血清の場合には、総て製造業者の推奨に従ったが、筋肉の場合には、事前にリーシス緩衝液(50 mM Tris-HCl pH 7.0, 100 mM NaClおよび0.1% Triton)によるタンパク質抽出およびBCAキット(Pierce, Rockford, イリノイ, 米国)によるその定量が必要であった。
【0095】
筋肉抽出物で検出されたヒトプロインスリンは常に極めて高く、タンパク質の量によって補正しなければならなかった。血清で検出されたヒトプロインスリンは、1−15pMであった。この濃度は、製造業者の指示に従って20μlの容積で測定した。測定は、P30で組織的に行った。これは、トランスジェニックProins/rd10-/-マウスが筋肉でヒトプロインスリンを産生し、これが血液循環に流れ込み、網膜神経に到達することができることを示唆していた。
【0096】
1.3. トランスジェニックマウスの網膜切片のクリオスタットでの免疫染色
網膜組織構造をP32マウスで分析し(図2)、変性に罹っていない野生型マウス(AおよびD)と、変性に罹っているProins/rd10-/-マウス(BおよびE)およびrd10-/-マウス(CおよびF)とを比較した。変性がrd10-/-マウスでは極めて進行しているP32では、カラム当たりの光受容体細胞の数、錐体の状態およびアバンダンス、並びにシナプスの状態を分析した(図3)。このようにして、総体的な網膜の状態および変性の進行を、様々なマーカーで明らかにしようとした。
【0097】
総ての組織はTissue-tec (Sakura Finetek Europe B.U. オランダ国)に埋設し、ドライアイスで冷凍して、処理まで-80℃で保管した。12μmの厚さのクリオスタット切片を作製した。これらをポリ-L-リシンをコーティングしたスライド(Fisher Biotech, ピッツバーグ, 米国)に集め、使用するときまで-80℃に保管した。
【0098】
染色を行う総ての場合に、スライドをフリーザーから取り出した後、室温において30分間放置し、4%パラホルムアルデヒド(PFA)で20分間固定した後、PBSで洗浄した。
【0099】
残りの錐体を標識することを決定したが、これらの錐体が突然変異を全く行わずに二次的に変性するので、網膜の状態およびその保持のアイディアを得ることができるからである。次に、変性の進行を知るには、外顆粒層(ONL)の厚みに残っている細胞の列の数を数えることで十分であった。蛍光色素Alexa 488で標識した凝集素(Molecular Probes, Eugene, オレゴン, 米国)を、0.1% BSAを含むPBS溶液中で2時間用いて、錐体を標識した。 1 mM MgCl2および1 mM CaCl2を含む溶液で洗浄した。錐体の外側部分およびその軸索末端は、このようにして検出した。切片をDAPIを含む固定媒質で固定し、核を対比染色した。
【0100】
変性のない野生型コントロールマウスは、ONLに8-12列の核を有する(図2D)。OPLに外側部分およびそのシナプスボタンを支持している錐体の数は、OPLで観察される凝集素染色による極めて典型的なものである(図2A)。コントロールrd10-/-マウスでは、ONLにおける桿状体の列の数は極めて小さく1-2列であり、この点における変性が極めて高いからである(図2F)。極めて驚くべきことは、錐体が初めは突然変異によって影響を受けていないにも拘わらず、この時点で既に変性し始めていることである(図2C)。錐体の外側部分はほとんど現れず、そのシナプスボタンの数はかなり減少していることが観察された。Proins/rd10-/-マウスにおいては、5-6列の桿状体核がこの症例で観察され、これは最大のプロインスリン血症が検出されたマウスの1匹であった(図2E)。錐体の状態は極めて良好であり、実際に、それは野生型マウスに類似していた。良好な錐体が保存されていることを示唆しているOPLにおけるその外側部分およびおよびシナプスボタンが見える(図2B)。血中プロインスリンレベルと相関するONL保存における変化が観察された。
【0101】
更に、ニューロンのシナプス結合が起こる網状層(図3)の状態を分析したが、プロインスリンを用いるこの治療が変性死を防止または遅延させるだけでなく、細胞が内因性損傷を有しまたは二次的に変化しているか否か、またはそれらが機能的であるか否かに拘わらず細胞を生きたままにしておこうとするものであるので、分析した。SV2 (シナプス小胞2)抗体による染色のため、クリオスタット切片を4% PFAで固定した後、0.1% Triton x-100で浸透させ10% NGS(正常なヤギ血清)で1時間ブロックした。1:50の希釈倍率のSV2抗体をシナプス小胞のタンパク質に結合し、従って網膜の網状層(外網状層、OPLおよび内側網状層、IPL)を標識する。これを、ブロック溶液中で4℃において一晩インキュベーションした。Alexa 488に接合した二次抗体(1/200)を用いるインキュベーションは、室温において1時間行った。対応するPBS洗浄の後、切片をDAPIを含む固定媒質で固定し、核を対比染色した。
【0102】
二つの網膜網状層、外網状層(OPL)、および内網状層(IPL)において、発現が観察された(図3)。脳に突出しているニューロン、ガングリオン細胞を有する二極性介在ニューロン結合が起こる内網状層は、3例においてかなりの染色と良好な状態を示した。差異は、光受容体細胞と二極性介在ニューロンとの結合が主として位置している外網状層にあった。OPLは、野生型コントロールマウス(図3A)およびトランスジェニックProins/rd10-/-マウス(図3B)においてはかなり画定して、濃かった。コントロールrd10マウスはIPLでは染色を維持したが、OPLは全く明確に保存されず、ほとんど染色されなかった(図3C)。このコントロールrd10-/-マウスにおいては、組織化の欠如によりおよび残りのニューロンが光受容体を有するシナプスを喪失しているという事実により、核層間にシナプス投影の試みさえ見られた。これは、Proins/rd10-/-マウスの網膜の機能状態が、治療なしのrd10-/-マウスより良好であることを示唆している。
【0103】
これは、トランスジェニック Proins/rd10-/-マウスでは、ヒトプロインスリンが治療なしの変性状態よりシナプス結合を長く保持することができることを示していた。これは、変性による細胞死を遅らせることの他に、それらを良好に保ち且つそれらの生物学的機能を行うことができることを含んでいる。
【0104】
rd10-/-マウスと比較して、Proins/rd10-/-マウスにおいて組織学的に観察された改善点は、次の網膜電位記録で一層明らかである(図4、5、および6)。
【0105】
更に、神経保護効果でのプロインスリンレベルの値を分析するため、網膜および他の神経系の部分の神経変性疾患の遺伝子モデル並びに老衰関連のもので見られる慢性および進行性損傷以外のモデルにより分析を行った。従って、予備検討を行い、網膜ガングリオン細胞死を急性損傷(視神経の切断)によって成熟ラットに引き起こし、ヒトプロインスリンの皮下投与を行った。この治療は、この典型的な急性損傷で必然的瞬間までガングリオン細胞死を僅かに遅延させた(データーは示さず)。ヒトプロインスリンは、これらのrdマウスの治療として皮下および硝子体内注射によっても投与したが(図8および9)、皮下投与したプロインスリンは網膜神経に到達することができたにも拘わらず、変性は停止しなかった(図7)。これらの結果は、血清プロインスリンは、持続および伸張レベルが達成されるときにのみ、中枢神経系の損傷部分に到達して神経保護作用を発揮することができることを示している。
【0106】
実施例2. ヒトプロインスリンはコントロールおよび疾病マウスと比較してトランスジェニック Proins/rd10-/-マウスでERGによって評価した視覚機能を向上させる。
マウスは、網膜電位の検討のため12時間毎の明-暗サイクルを常に保持したが、マウスを一晩中暗闇に順化させた。情報目的には、錐体は明所(昼間)視覚に関与しており、桿状体は暗所(夜間)視覚に関与する。マウスを弱い赤色光下において、ケタミン(95mg/kg)およびキシラジン(5mg/kg)を含む溶液の腹腔内投与により麻酔した。瞳孔を、1%トロピカミド(Colircusi Tropicamida, Alcon Cusi, SA, E1 Masnou, バルセロナ, スペイン国)を含む溶液1滴で散大した。記録電極は、マウスの眼に置いたレンズである。参照電極は口に付け、接地電極は尾に付けた。麻酔した動物をファラデーケージに入れ、総ての暗所実験は完全な暗闇で行った。このようにして、ガンツフェルトスティミュレーターによって生じた低強度光のフラッシュによって誘発された網膜電位応答を記録して、桿状体のみで生じた応答(桿状体応答)または暗闇への順化での錐体と桿状体とで生じた応答(混合応答)記録することができた。用いた光刺激の強度は、-4 - 1.52 log cd.s.m-2の値に設定した。光強度は、眼のレベルの光度計(Mavo Monitor USB)で測定した。64回の応答の最大値を、それぞれの刺激強度について平均した。光刺激の間隔は強度に応じて変化し、従って低強度刺激(-4 log cd.s.m-2)については、刺激間時間を10秒間に設定し、高強度刺激(1.52 log cd.s.m-2)については、60秒間に設定した。動物を、錐体の単独応答を記録するために明所条件に順化させた。これらの条件下では、閃光の間隔は1秒間に設定した。
【0107】
網膜からの電気シグナルをグラス増幅器(CP511 AC増幅器, Grass Instruments, クインシー, マサチューセッツ)で0.3-1000Hzの間で増幅し、濾過した。シグナルを数値化した(PC-card ADI instruments, カリフォルニア)。記録をコンピューターに保管して、後で解析した。
【0108】
桿状体性応答を暗所順化状態で記録した後、強度が-4 - -1.52 log cd.s.m-2の閃光を当てた。錐体および桿状体によって生じた混合応答を記録した後、-1.52 - 0.48 log cd.s.m-2の強度の閃光を当てた。振動電位も、100 - 1000Hzの電気フィルターを適用することによって単離した。錐体性応答は、(30cd.m-2のバックグラウンド光を記録して)光順化状態で記録した後、強度が-0.52 - 2 log cd.s.m-2の閃光を当てた。フリッカー応答(30 Hz)を光順化状態で記録した後、1.48 log cd.s.m-2を加えた。
【0109】
図4は、野生型(WT)、コントロールrd10-/-および系列1および系列2由来のP30で得たProins/rd10-/-マウスの、暗所に順化(暗順応状態)および光に順化(明所視状態)した状態で記録した。
【0110】
図5は、より大きな群またはマウスでのインスリン間網膜電位応答とプロインスリンレベルとの相関を解析した結果を示す。この相関は、用量-応答の関係を示唆しており、ヒトプロインスリンを用いる薬理学的方法が実行可能な有効性を有することを支持している。
【0111】
更に、図6は、産後発生の様々な時期(P35、P45、およびP55)に得られた系列2由来の野生型(wt)、コントロールrd10、およびProins/rd10-/-マウスに対応する網膜電位記録を示す。暗順応(暗所への順応)および明所視(光への順応)状態で得た記録を、別々に示す。野生型マウスがどのようにP35で広範囲の暗順応(桿状体)および明所視(錐体)網膜電位応答を生じるかが観察された。応答は、桿状体および錐体で生じた応答のいずれでも、コントロールrd10-/-マウスではP35でゼロであった。Proins/rd10-/-マウスは、P35で極めて有意な明所視および暗順応網膜電位応答を保持し、P55まである程度の応答の保持を達成した。このように、視覚応答はトランスジェニックProins/rd10-/-マウスで一層良好であり、更に長時間伸張することが観察された。
【0112】
実施例3: プロインスリンのrd1マウスの網膜の硝子体内投与による効果
下記の実施例3および4を実施するため、rd10マウスと同じC57BL/6遺伝的背景並びに同じ桿状体特異的サイクリックGMPホスホジエステラーゼ遺伝子を除く様々な突然変異を共有するrd1型マウスを用いた。
【0113】
プロインスリンのイン・ビボでのrd1マウス網膜の硝子体内投与の効果、具体的には、P13におけるrd1マウスのヒトプロインスリンを1μg/μlの濃度で1μgの硝子体内投与に関して。右目にプロインスリンを投与し、左目に担体を投与した。投与は単回投与であり、効果を24または48時間後にTUNELによって平面的におよび切片で固定した網膜で分析した。後者では、外顆粒層に残っている光受容体の数も評価した。TUNEL(TdTによるdUTPニックエンド標識)法を用いて、細胞死を検出した。末端トランスフェラーゼ酵素(TdT)は、フルオレセイン標識したヌクレオチド(dUTP)をDNAの遊離の3'末端に付加し、これによりプログラム細胞死中に起こるDNAの断片化を検出することができる。Promega製TUNELキットを用いた。手法は、細胞、断片、または組織で行った。組織の性質による透過性化(permeabilization)工程後、組織をPBSで洗浄し、キット溶液を用いて室温において30分間予備インキュベーションした。反応混合物を製造者取扱説明書に従い調製し、その反応を37℃において1時間行った。反応を2 x SSC溶液で室温にて15分間停止した。これをPBSにより洗浄し、ベクタシールドで固定した。平面的に固定された網膜での細胞死を検出するため、眼の残りの要素から切り離した全網膜神経を、取扱いの際にコントラストが良好になるように黒色ニトロセルロース膜(Sartorius, ゲッティンゲン, 独国)上で顕微鏡に固定した。これは、通常は光受容体層を上にして置き、ニトロセルロースに精巧な切開用ピンセットの助けを借りて付着させた。平面的に取り付けた網膜を、24穴プレートに0.1 M リン酸緩衝液, pH 7.1中で4%(w/v)PFAで4℃において一晩固定した。翌日、それらをPBSおよびBSA(30 mg/ml PBS)で洗浄した。浸透化はPBS中1% (w/v) Triton X-100を用いて(4回、それぞれ30分間)、および予めPBSで十分に洗浄することによってTriton X-100残留物を除去してコラゲナーゼおよびプロテイナーゼKで行った。コラゲナーゼ(20 U/ml, Sigma)を37℃において1時間作用させた後、プロテイナーゼK(20μg/ml, Promega)を37℃において15分間作用させた。次に、網膜を少なくとも2時間再固定する必要があった。これをPBSおよびBSA (30 mg/ml PBS)で十分に洗浄し、TUNEL反応を上記の方法で行った。平面的に固定した網膜を共焦点顕微鏡法(Leica TCS-SP2-A0BS)によって分析した。
【0114】
網膜層中の細胞死の位置を確認するため、上記のような網膜切片についてTUNELを行い、常に0.1 M リン酸緩衝液(pH 7.1)中4%(w/v)PFAに固定し、30%(w/v)スクロース/10 mMリン酸緩衝液(pH 7.1)で凍結防止した。切片のTUNEL標識には、平面的に固定した網膜より浸透を低くする必要がある。従って、BGT [100 mMグリシン、3 mg/ml BSA、0.25%(w/v)Triton X-100、PBS中]を用いて2シリーズの浸透をそれぞれ15分間行った。それらをPBSにより洗浄し、TUNEL反応を上記のように行った。スライドの総ての切片をカバーするため、相当する試薬50μlを加え、それらをパラフィルムでカバーした。
【0115】
いずれの定量(切片のTUNEL、平面的に固定した網膜のTUNEL、切片の光受容体)でも、プロインスリンと担体による治療の間には有意差は見られなかった。
【0116】
実施例4. プロインスリンのrd1マウスでの皮下注射による効果
更に長時間の治療を行うために、新たな投与経路である皮下注射を試験した。糖尿病患者にインスリン投与の目的で一般に用いられるこの経路は、硝子体内注射より多少外傷が少ない。更に、この方法でのヒトプロインスリン投与は、成熟ラットにおいて視神経を切断した後の網膜ガングリオン細胞の変性に予め正の遅延を生じた。多数のプロトコルを行い、ヒトプロインスリンの薬理学的供給を最適にした(表)。
【0117】
【表1】
【0118】
プロインスリンが皮下注射によって網膜に到達するか否かを、最初に測定した。その目的のため、プロインスリンを皮下投与したマウスでELISAによって検討を行い、ヒトプロインスリンを検出した(図7)(Lineo Research, ミズーリー, 米国)。
【0119】
この分析法は血清プロインスリンを検出するように設計されているので、筋肉および網膜抽出物用に改良しなければならなかった。トランスジェニックマウスによるヒトプロインスリン産生レベルを検出するため、筋肉および網膜、並びに血清抽出物を分析した。血清の場合には、マウスの眼の涙嚢部分から血液をパスツールピペットで採取した。これをエッペンドルフ試験管に移し、室温において2時間血液凝固させ、1,300 gで15分間遠心分離して血清を集め、これを使用まで凍結した。組織の場合には、タンパク質抽出をリーシス緩衝液[50 mM Tris-HCl, pH 7、100 mM NaClおよび0.1%(w/v) Triton X-100]で行い、BCAキット(Pierce)による抽出タンパク質の定量を行った。網膜神経は、それぞれ60μlの容積で抽出した。後脚の筋肉を、得られた筋肉片に従って200-300μlの容積で抽出した。
【0120】
血清の場合には、製造業者の推奨に従い、常に20μlで二つの試料を用いた。組織の場合には、抽出物20μlを二つ作製し、場合によっては精度を上げるため連続希釈を用いた。網膜および筋肉抽出物データは、タンパク質の量によって補正した。
【0121】
P14の野生型マウスの網膜抽出物におけるヒトプロインスリンは、用量依存的な皮下投与の2時間後にのみ同定することができた。1.6μgの用量では、0.72pMの網膜濃縮を生じたが、5μgの用量では3.62pMとなった。P11とP14の間に1.6μgを毎日投与することによって、更に高い、恐らくは蓄積された、レベルの3.56pMを生じることができた。従って、この方法によって、末梢の外来プロインスリンが網膜神経に到達する能力および少なくとも網膜における一層高いプロインスリンレベルを達成する反復治療の有効性が確かめられた。
【0122】
神経変性における光受容体および網膜電位図(表、図8、および9)によって測定された視覚機能に対する様々な間隔での反復治療の効果を実証した。P8およびP15の間の1.6μgの1日用量を最初に投与した。この場合には、組織学的レベルにおいては保護効果はなかった。プロインスリンを変性過程が開始する前に投与すれば効果は一層明白になり得るという信念により救済を一層速く開始するため、治療をP6およびP14の間に試験したが、光受容体の保持はこの時点においては得られなかった。組織学的回復を試み且つ網膜電位図による視覚機能を評価するため、P6およびP14の間にヒトプロインスリンを1.6μgの1日投与を2回行った。幾匹かの動物はP18まで保持されたが、プロインスリン用量はこの第二の期間では2.5μgに増加した。この治療は、組織学的レベル(図8)または機能レベル(図9)で何ら保護効果を持たなかった。
【0123】
投与されたプロインスリンは網膜に到達することができ、それに対する応答を可能にする総ての要素が明らかに配置されているという事実にも拘わらず、最もありふれた結果は、組織学的回復の欠如およびERGによる視覚機能の欠如(図8および図9)であった。
【特許請求の範囲】
【請求項1】
プログラム細胞死が起こる神経変性状態、障害、もしくは疾患の予防もしくは治療用の医薬品または医薬組成物を製造するための、プロインスリンの活性を誘発する化合物の使用。
【請求項2】
前記神経変性疾患が網膜色素変性症であることを特徴とする、請求項1に記載の誘発化合物の使用。
【請求項3】
前記医薬組成物がその持続的な全身または局所投与に適するものであることを特徴とする、請求項1または2に記載の化合物の使用。
【請求項4】
前記誘発化合物が、神経保護タンパク質またはペプチドを発現させ、且つ下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一方の断片、並びに
d) a)、b)、および/またはc)に属するいずれか1個の配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列によって構成されるヌクレオチド配列であることを特徴とする、請求項1−3のいずれか一項に記載の誘発化合物の使用。
【請求項5】
前記ヌクレオチド配列がヒトプロインスリンをコードする配列番号1によって構成されることを特徴とする、請求項4に記載のプロインスリンの活性を誘発化合物の使用。
【請求項6】
前記d)のヌクレオチド配列がプロインスリンヌクレオチド配列(配列番号1)を含んでなる遺伝子構築物によって構成されることを特徴とする、請求項4に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項7】
前記d)のヌクレオチド配列が、神経保護を誘発できるプロインスリンタンパク質をコードするヌクレオチド配列または遺伝子構築物を含んでなる発現ベクターによって構成されることを特徴とする、請求項4に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項8】
前記発現ベクターが配列番号1のヌクレオチド配列と、組織特異的、好ましくは筋肉特異的プロモーターとを含むことを特徴とする、請求項7に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項9】
前記誘発化合物が、遺伝的に修飾され、且つプロインスリンヌクレオチド配列、構築物、または発現ベクターを含んでなり、プロインスリンタンパク質を細胞外媒質に好適に発現しまたは放出できるヒト真核細胞であることを特徴とする、請求項1−3のいずれか一項に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項10】
前記真核細胞がヒトプロインスリンヌクレオチド配列(配列番号1)によって形質転換したヒト細胞であることを特徴とする、請求項9に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項11】
前記誘発化合物が神経保護活性を有し、下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つを含んでなるアミノ酸配列
に属する1または数個のアミノ酸配列を含んでなるタンパク質またはペプチドであることを特徴とする、請求項1−3のいずれか一項に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項12】
前記誘発化合物がヒトプロインスリンタンパク質(配列番号2)であることを特徴とする、請求項11に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項13】
治療上有効量のプロインスリンの活性を誘発する化合物を、前記誘発する化合物の全身または局所持続への投与の保持に適当な1種類以上の薬学上許容可能なアジュバントおよび/または担体と共に含んでなることを特徴とする、神経退行変性をもたらす疾患、障害、もしくは病変の治療用医薬組成物または医薬品。
【請求項14】
前記プロインスリンの活性を誘発する化合物が、下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つの配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列であることを特徴とする、請求項13に記載の医薬組成物。
【請求項15】
前記ヌクレオチド配列がヒトプロインスリンをコードする配列番号1によって構成されることを特徴とする、請求項14に記載の医薬組成物。
【請求項16】
前記d)のヌクレオチド配列がプロインスリンヌクレオチド配列(配列番号1)を含んでなる遺伝子構築物によって構成されることを特徴とする、請求項14に記載の医薬組成物。
【請求項17】
前記d)のヌクレオチド配列が、神経保護を誘発できるプロインスリンタンパク質をコードするヌクレオチド配列または遺伝子構築物を含んでなる発現ベクターであることを特徴とする、請求項14に記載の医薬組成物。
【請求項18】
前記発現ベクターが、配列番号1のヌクレオチド配列と、組織特異的、好ましくは筋肉特異的プロモーターとを含むことを特徴とする、請求項17に記載の医薬組成物。
【請求項19】
前記プロインスリンの活性を誘発する化合物が、下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つの配列を含んでなるアミノ酸配列
に属するタンパク質またはペプチドであることを特徴とする、請求項13に記載の医薬組成物。
【請求項20】
前記アミノ酸配列がヒトプロインスリンによって構成されるもの(配列番号2)であることを特徴とする、請求項19に記載の医薬組成物。
【請求項21】
前記プロインスリンの活性を誘発する化合物がヒト細胞、好ましくはプロインスリン配列、構築物、または発現ベクターによって形質転換された中枢神経系細胞であることを特徴とする、請求項13に記載の医薬組成物。
【請求項22】
前記真核細胞がヒトプロインスリンヌクレオチド配列(配列番号1)によって形質転換されたヒト細胞であることを特徴とする、請求項21に記載の医薬組成物。
【請求項23】
プログラム細胞死が起こる神経変性疾患、障害、または病変に罹っているヒトの、前記治療組成物を前記神経変性を減少させる適当用量で投与することからなる治療または予防方法における、請求項13−22のいずれか一項に記載の医薬組成物の使用。
【請求項24】
前記神経変性疾患が下記の群、すなわちアルツハイマー病、パーキンソン病、多発性硬化症、レヴィー小体認知症、筋萎縮性側索硬化症、脊髄小脳性萎縮症、前頭側頭型認知症、ピック病、血管性認知症、ハンティングトン病、バッテン病、および脊髄損傷に属することを特徴とする、請求項23に記載の医薬組成物の使用。
【請求項25】
神経変性疾患が下記の群、すなわち網膜色素変性症、黄斑変性、および緑内障に属することを特徴とする、請求項23に記載の医薬組成物の使用。
【請求項1】
プログラム細胞死が起こる神経変性状態、障害、もしくは疾患の予防もしくは治療用の医薬品または医薬組成物を製造するための、プロインスリンの活性を誘発する化合物の使用。
【請求項2】
前記神経変性疾患が網膜色素変性症であることを特徴とする、請求項1に記載の誘発化合物の使用。
【請求項3】
前記医薬組成物がその持続的な全身または局所投与に適するものであることを特徴とする、請求項1または2に記載の化合物の使用。
【請求項4】
前記誘発化合物が、神経保護タンパク質またはペプチドを発現させ、且つ下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一方の断片、並びに
d) a)、b)、および/またはc)に属するいずれか1個の配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列によって構成されるヌクレオチド配列であることを特徴とする、請求項1−3のいずれか一項に記載の誘発化合物の使用。
【請求項5】
前記ヌクレオチド配列がヒトプロインスリンをコードする配列番号1によって構成されることを特徴とする、請求項4に記載のプロインスリンの活性を誘発化合物の使用。
【請求項6】
前記d)のヌクレオチド配列がプロインスリンヌクレオチド配列(配列番号1)を含んでなる遺伝子構築物によって構成されることを特徴とする、請求項4に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項7】
前記d)のヌクレオチド配列が、神経保護を誘発できるプロインスリンタンパク質をコードするヌクレオチド配列または遺伝子構築物を含んでなる発現ベクターによって構成されることを特徴とする、請求項4に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項8】
前記発現ベクターが配列番号1のヌクレオチド配列と、組織特異的、好ましくは筋肉特異的プロモーターとを含むことを特徴とする、請求項7に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項9】
前記誘発化合物が、遺伝的に修飾され、且つプロインスリンヌクレオチド配列、構築物、または発現ベクターを含んでなり、プロインスリンタンパク質を細胞外媒質に好適に発現しまたは放出できるヒト真核細胞であることを特徴とする、請求項1−3のいずれか一項に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項10】
前記真核細胞がヒトプロインスリンヌクレオチド配列(配列番号1)によって形質転換したヒト細胞であることを特徴とする、請求項9に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項11】
前記誘発化合物が神経保護活性を有し、下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つを含んでなるアミノ酸配列
に属する1または数個のアミノ酸配列を含んでなるタンパク質またはペプチドであることを特徴とする、請求項1−3のいずれか一項に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項12】
前記誘発化合物がヒトプロインスリンタンパク質(配列番号2)であることを特徴とする、請求項11に記載のプロインスリンの活性を誘発する化合物の使用。
【請求項13】
治療上有効量のプロインスリンの活性を誘発する化合物を、前記誘発する化合物の全身または局所持続への投与の保持に適当な1種類以上の薬学上許容可能なアジュバントおよび/または担体と共に含んでなることを特徴とする、神経退行変性をもたらす疾患、障害、もしくは病変の治療用医薬組成物または医薬品。
【請求項14】
前記プロインスリンの活性を誘発する化合物が、下記の群:
a) ヒトプロインスリンをコードするヌクレオチド配列(配列番号1)によって構成されるヌクレオチド配列、
b) a)の配列と同様のヌクレオチド配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つの配列を含んでなるヌクレオチド配列
に属する1または数個のヌクレオチド配列であることを特徴とする、請求項13に記載の医薬組成物。
【請求項15】
前記ヌクレオチド配列がヒトプロインスリンをコードする配列番号1によって構成されることを特徴とする、請求項14に記載の医薬組成物。
【請求項16】
前記d)のヌクレオチド配列がプロインスリンヌクレオチド配列(配列番号1)を含んでなる遺伝子構築物によって構成されることを特徴とする、請求項14に記載の医薬組成物。
【請求項17】
前記d)のヌクレオチド配列が、神経保護を誘発できるプロインスリンタンパク質をコードするヌクレオチド配列または遺伝子構築物を含んでなる発現ベクターであることを特徴とする、請求項14に記載の医薬組成物。
【請求項18】
前記発現ベクターが、配列番号1のヌクレオチド配列と、組織特異的、好ましくは筋肉特異的プロモーターとを含むことを特徴とする、請求項17に記載の医薬組成物。
【請求項19】
前記プロインスリンの活性を誘発する化合物が、下記の群:
a) ヒトプロインスリンアミノ酸配列(配列番号2)によって構成されるアミノ酸配列、
b) a)の配列と同様のアミノ酸配列、
c) a)およびb)の配列のいずれか一つの断片、並びに
d) a)、b)、および/またはc)に属するいずれか一つの配列を含んでなるアミノ酸配列
に属するタンパク質またはペプチドであることを特徴とする、請求項13に記載の医薬組成物。
【請求項20】
前記アミノ酸配列がヒトプロインスリンによって構成されるもの(配列番号2)であることを特徴とする、請求項19に記載の医薬組成物。
【請求項21】
前記プロインスリンの活性を誘発する化合物がヒト細胞、好ましくはプロインスリン配列、構築物、または発現ベクターによって形質転換された中枢神経系細胞であることを特徴とする、請求項13に記載の医薬組成物。
【請求項22】
前記真核細胞がヒトプロインスリンヌクレオチド配列(配列番号1)によって形質転換されたヒト細胞であることを特徴とする、請求項21に記載の医薬組成物。
【請求項23】
プログラム細胞死が起こる神経変性疾患、障害、または病変に罹っているヒトの、前記治療組成物を前記神経変性を減少させる適当用量で投与することからなる治療または予防方法における、請求項13−22のいずれか一項に記載の医薬組成物の使用。
【請求項24】
前記神経変性疾患が下記の群、すなわちアルツハイマー病、パーキンソン病、多発性硬化症、レヴィー小体認知症、筋萎縮性側索硬化症、脊髄小脳性萎縮症、前頭側頭型認知症、ピック病、血管性認知症、ハンティングトン病、バッテン病、および脊髄損傷に属することを特徴とする、請求項23に記載の医薬組成物の使用。
【請求項25】
神経変性疾患が下記の群、すなわち網膜色素変性症、黄斑変性、および緑内障に属することを特徴とする、請求項23に記載の医薬組成物の使用。
【図1】

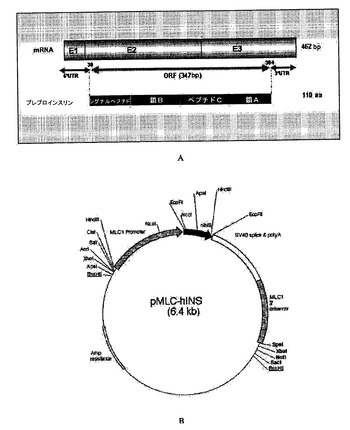
【図2】
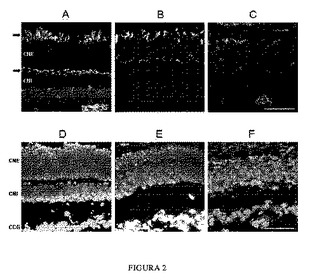
【図3】
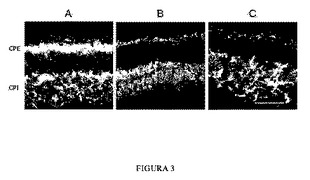
【図4】
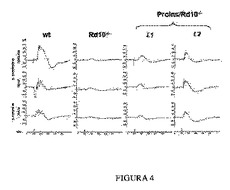
【図5】
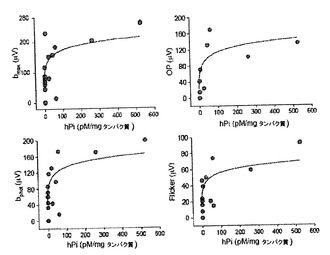
【図6】
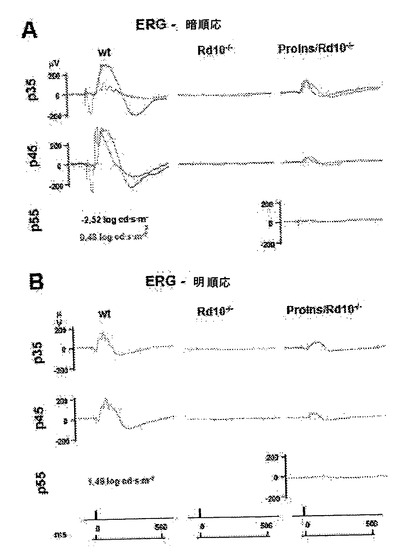
【図7】
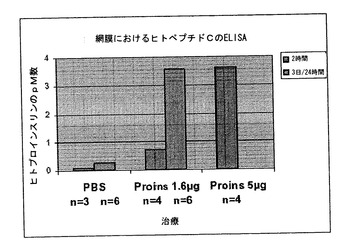
【図8】
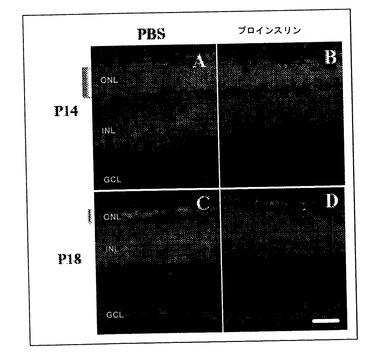
【図9】
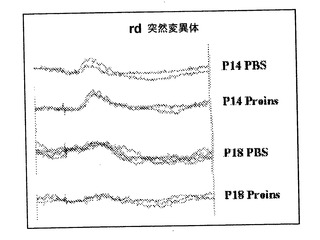
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2009−537612(P2009−537612A)
【公表日】平成21年10月29日(2009.10.29)
【国際特許分類】
【出願番号】特願2009−511534(P2009−511534)
【出願日】平成19年5月21日(2007.5.21)
【国際出願番号】PCT/ES2007/070097
【国際公開番号】WO2007/135220
【国際公開日】平成19年11月29日(2007.11.29)
【出願人】(593005895)コンセホ・スペリオール・デ・インベスティガシオネス・シエンティフィカス (67)
【氏名又は名称原語表記】CONSEJO SUPERIOR DE INVESTIGACIONES CIENTIFICAS
【出願人】(508348473)ウニベルシダッド、デ、アルカラ、デ、エナレス (1)
【氏名又は名称原語表記】UNIVERSIDAD DE ALCALA DE HENARES
【出願人】(508348484)ウニベルシダッド、アウトノマ、デ、バルセロナ (1)
【氏名又は名称原語表記】UNIVERSIDAD AUTONOMA DE BARCELONA
【Fターム(参考)】
【公表日】平成21年10月29日(2009.10.29)
【国際特許分類】
【出願日】平成19年5月21日(2007.5.21)
【国際出願番号】PCT/ES2007/070097
【国際公開番号】WO2007/135220
【国際公開日】平成19年11月29日(2007.11.29)
【出願人】(593005895)コンセホ・スペリオール・デ・インベスティガシオネス・シエンティフィカス (67)
【氏名又は名称原語表記】CONSEJO SUPERIOR DE INVESTIGACIONES CIENTIFICAS
【出願人】(508348473)ウニベルシダッド、デ、アルカラ、デ、エナレス (1)
【氏名又は名称原語表記】UNIVERSIDAD DE ALCALA DE HENARES
【出願人】(508348484)ウニベルシダッド、アウトノマ、デ、バルセロナ (1)
【氏名又は名称原語表記】UNIVERSIDAD AUTONOMA DE BARCELONA
【Fターム(参考)】
[ Back to top ]