
神経再生ペプチド及び脳損傷治療におけるそれらの使用方法
本発明は、ニューロンの遊走、神経突起伸長、ニューロンの増殖、神経の分化及び/又はニューロンの生存を促進することができるNRP化合物又はNRPと称されるペプチドファミリーを開示し、さらに脳損傷及び神経変性疾患の治療でNRPを使用するための組成物及び方法を提供する。NRP化合物は、ニューロン及び神経芽細胞の増殖及び、急性脳損傷又は慢性神経変性疾患(例えば毒素への暴露、卒中、外傷、神経系の感染、脱髄疾患、痴呆、及び代謝異常)によって惹起される傷害領域へのニューロン及び神経芽細胞の遊走を誘導することができる。NRP化合物は、対象者又は対象細胞へ多様な手段によって直接的に(経口的、腹腔内、血管内及び患者の神経系に直接的に投与することを含む)投与することができる。NRP化合物は、治療に使用するために医薬的に許容できる投与形態に製剤化することができる。神経の再生、神経の増殖、神経の分化、神経突起伸長及び神経の生存を検出する方法を用いて、神経に対して活性を有する他の薬剤を開発することができる。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願:
本出願は、米国仮特許出願60/516,018号(2003年10月31日出願、発明の名称:神経再生ペプチド及びそれらの使用方法(”Neural Regeneration Peptides and Methods for Their Use”);発明者:Frank Sieg & Thorsten Gorba(Attorney Docket No: NRNZ 01023 US3 DBB))、米国仮特許出願60/585,041号(2004年7月2日出願、発明の名称:神経再生ペプチド:新規な種類の化学走性及びニューロン生存促進ペプチド(”Neural Regeneration Peptides: A New Class of Chemoattractive and Neuronal Survival Promoting Peptides”);発明者:Thorsten Gorba & Frank Sieg(Attorney Docket No: NRNZ 01023 US4 DBB))、及び米国仮特許出願60/616271号(2004年10月5日出願、発明の名称:神経再生ペプチド及び脳傷害治療におけるそれらの使用方法(”Neural Regeneration Peptides and Methods for Their Use in Treatment of Brain Damage”);発明者:Frank Sieg , Paul Edmond Hughes & Thorsten Gorba(Attorney Docket No: NRNZ 01023 US5 DBB))に対して優先権を主張する。上記出願の全てが参照により本明細書に含まれる。
配列リスト:
本出願は、(1)配列リストの印刷物及び(2)コンピュータ読出し可能形の配列リストを収納するディスクとして提出される配列リストを含む。本配列リストは参照により本明細書に含まれる。
【0002】
本発明は、ニューロンの遊走、増殖、生存、分化及び/又は神経突起伸長を促進するオリゴヌクレオチド及びペプチドの組成物並びにそれらの使用方法を目的とする。より具体的には、本発明は、そのようなペプチドの脳損傷及び神経変性疾患の治療における使用を目的とする。本発明はまた、神経細胞の増殖、遊走、神経突起伸長、生存及び/又は分化を検出する新規な方法を含む。
【背景技術】
【0003】
軽度から重度の外傷性脳損傷(TBI)及び限局性又は総体虚血は、障害後短時間内に重大なニューロン細胞消失及び脳機能低下を生じえる。これまでのところ、疾患によって引き起こされる頭部の損傷又は傷害の結果として脳に生じる細胞死を防止するために利用可能な治療はない。さらにまた今日まで、ニューロン機能を回復させるために利用できる治療も存在しない。慢性の神経変性疾患、例えばアルツハイマー病、パーキンソン病、ハンチントン病及び多発性硬化症に現時点で利用できる治療は対症療法のみである。疾患過程に介入するか又は細胞死を防ぐために利用可能な薬は現在のところない。
皮質-皮質下の非視床病巣は、損傷後3−7日で視床領域内のアポトーシスを生じえることはよく知られている。逆行視床変性は、視床内のアストログリア及びミクログリアの活性化を伴う(Hermann et al., 2000)。MRIのような非侵襲性技術は、非視床部の構造的病変を有するTBI患者においては視床の体積が小さいこと、及び脳室対脳比率値が増すことを明らかにした(Anderson et al., 1996)。これらの発見は、視床皮質系の興奮投射ニューロンは逆行性ニューロン細胞死に対して非常に脆弱であることを示し、したがって損傷された又は外傷を受けた視床ニューロンの救済手法の必要性を示している。
【0004】
視床内部の抑制性ニューロン回路の機能は、視床内部のニューロン活動の視床内ダウンレギュレーションにとって線条体系内部のそれと同様に必須である。ハンチントン病で生じる病変と類似する線条体病変を有する動物は、GABA-放出ポリマーマトリックスを視床に移植されたとき行動に関する所産の改善が認められることが示された(Rozas et al., 1996)。線条体内部の細胞レベルでは、カルビンジン免疫反応(“calbindin-ir”)抑制ニューロンはアクチビンAの投与によって救済できることが示された(Hughes at al., 1999)。
これまで、胎児線条体の移植を必要とする移植のみが、ハンチントン病の動物モデルにおいて運動機能の改善又は回復をもたらした(Nakao and Itakura, 2000)。ハンチントン病で傷害された視床及び線条体のGABA作動系の回復は、行動に関する所産を改善することができる(Beal et al., 1986)。
神経系発達の特徴は、前駆体細胞がそれらの正しい三次元的空間的配置へと広範囲に遊走していくことである。このような遊走は、一連の表現型分化及び未成熟ニューロンの脊椎脳への配置を促進する。およそ十兆個のニューロンを正確に結ぶために、より高次の皮質領域における層状構造の形成のような細胞性機構の構築が必要である(概説としてHatten and Heintzを参照)。
【0005】
遊走ニューロンの運動方向に対して相関関係をもつ細胞側のものは、単一遊走細胞内の一過性Ca2+変化の頻度及び大きさであろうが(Gomez and Spitzer, 1999)、ただし膜結合分子または拡散性分子によるニューロン細胞遊走の開始及び/又は実行の引き金も排除できない。
しかしながら神経突起伸長及びニューロンの遊走に必要な指示の多くは同定されている。インテグリンレセプターファミリーに属する形質膜分子は、ラミニンのような細胞外マトリックスリガンドと相互作用し、下層へのニューロン接着を開始させる(Liang and Crutcher, 1992; De Curtis and Reichardt, 1993)。インテグリン発現の制御は、遺伝子発現の調節、細胞接着、軸作成長及び細胞遊走を含む広範囲の発生プロセス及び細胞性プロセスに影響を与える。細胞遊走を促進する他のリガンドは、細胞接着分子(すなわちN-CAM;カドヘリン;TAG-1)、ラミニン様分子ネットリン-1、ニューロン-グリア接着リガンドアストロタクチン及び増殖又は神経栄養因子、例えばEGF、TGF-α、血小板活性化因子及びBDNFである(Dodd et al., 1988; Yamamoto et al., 1990; Ishii et al., 1992; Ferri and Levitt, 1995; Ganzler and Redies, 1995)。
最近、コラプシン-1(セマフォリン3A)が発見された。コラプシン-1は、一次感覚ニューロンに対する軸索誘導及び遊走パターンにおいて化学反発活性を有する(Pasterkamp et al., 2000)。対照的に、コラプシン-1は、新皮質領域の皮質先端突起を誘導するために化学誘引物質として機能する(Polleux et al., 2000)。軸索誘導に対する化学誘引作用と同様に同じような化学反発作用がスリット-1(拡散性タンパク質)によって示された(Brose et al., 2000)。
【0006】
これまでのところ、ニューロン移動の開始をもたらす連鎖事象、すなわちニューロンの付着からそれに続く遊走の開始、長距離に及ぶ遊走のプロセス(方向転換及び遊走停止シグナルを含む)は未だ説明されていない。
TGF-αが同時に投与された中脳病巣は、脳質下部帯(subventricular zone; “SVZ”)を起源とする多能性幹細胞の大規模増殖及びそれに続くこれら幹細胞の線条体への遊走をもたらす(Fallon et al., 2000)。しかしながら、SVZを起源とするニューロン前駆体の長距離遊走を防ぐために、病巣部位の極めて近傍に存在するニューロンの増殖及び遊走を活性化することが望ましい。
ニューロン遊走化学誘引物質として支質由来因子ケモカイン(SDF-1)を取り上げた報告が1件だけ存在する。胎児発現型のSDF-1は、小脳顆粒細胞を誘引して外側胚芽層から内側顆粒層へ遊走させる(Zhu et al., 2002)。それにもかかわらず、このケモカインは出生後の組織に対しては影響をもたない。脳外傷又は神経変性疾患後に成人における神経芽細胞又はニューロンの遊走行動に対して化学誘因性作用を誘導する遊走誘導因子は知られていない。
【発明の開示】
【0007】
発明の要旨
したがって、本発明の実施態様の目的は、脳損傷及び脳疾患の治療に新規なアプローチを提供することである。そのような実施態様は、神経の遊走、神経の成長、神経の増殖、神経の分化及び/又は神経の生存の1つ又は2つ以上を誘導することができるペプチドを含む。これらのペプチドは本明細書では“神経再生ペプチド”又は“NRP”と称される。他の実施態様には、脳損傷に続いて又は慢性神経変性疾患時に、1つ又は2つ以上のNRPを投与することが含まれる。“NRP”又は“NRP化合物”という用語には、NRP、NRPホモログ、PRPパラログ、NRPオルソログ及び/又はNRPアナログが含まれる。NRPは単独で、又は1つ若しくは2つ以上の他のNRPと一緒に、又は神経の成長、神経の遊走、神経の生存、神経の分化及び/又は神経の増殖を促進する他のタイプの薬剤と一緒に投与することができる。
NRP及び関連ペプチドは全般的に、所望の生物学的特性を前記分子に付与する一定のアミノ酸配列を有する。
一定のドメイン(下線を付して強調されている)を有するNRPペプチドのいくつかの実施態様は下記の表1に示されている。
【0008】
表1:神経再生ペプチド*
NRP-1: YDPEAAS-APGSGNP------CH
NRP-2KG: KDPEARR-APGSLHP------C--LAA-SCSAAG
NRP-3SF: SDSFKSQ-ARGQVPPFLGGVGCPWF
NRP-4GG: GTPGRAE-AGGQVSP------C--LAA-SCSQAYG
NRP-5RP2: RE--GRRDAPGRA--GGGG------AARSVSPSP
NRP-7SW: SEPEARR-APGRK----GGVVCASLAADW
NRP-8SG: SEVDARR-AKKSLH-------C-ILS-DTSHPRG
NRP-9SD: SEPEARRAQGGQIPSERVLSD
【0009】
いくつかの実施態様では、一般にNRPは約8から約25アミノ酸の鎖の長さを含み、約0.8から約2.7kDaの分子量を有する。さらにまた、他の実施態様では、NRPは約6.5から約10.0の等電点を有し、さらに神経の生存、神経突起伸長、ニューロンの増殖、ニューロンの分化及びニューロンの遊走から選択される成果を促進する少なくとも1つの生物学的特性を有することができる。さらにまた、NRPは、上記の表1の下線で示される1つ又は2つ以上のドメインを有することができる。いくつかの実施態様では、NRPは[A]PG[R,S]ドメインをPEドメインと一緒に有し(例えばNRP-1及びNRP-2)、また別にはPEドメインを含まない(例えばNRP-5、NRP-7)。[A]PG[R,S]ドメインの存在はNRPの生物学的特性のために望ましい。さらにまた、C-末端のGGドメインはNRPに所望の神経保護特性を付与することができる。したがって、また別の実施態様では、NRPは、[A]PG[R,S]ドメイン、[A,G]RRドメイン及びARGドメインから成る群から選択される第一のドメインを有することができ、所望の生物学的活性を有する。他の実施態様では、望ましくは、NRPは上述の第一のドメインに加えて、第一のドメインとは異なる第二のドメインを有することができる。第二のドメインはPEドメイン、[A,G]RPドメイン又はC-末端GGドメインでありえる。さらに別のある実施態様では、NRPは上述のドメインの第三のドメインを有することができる。
したがって、ある種の実施態様では、NRPは[A]PG[R,S]ドメインのみを含み、他のNRPはARGドメインのみを含み、さらに他のNRPは[G,A]RRドメインのみを含むことができる。さらにまた、他のNRPは、[A]PG[R,S]ドメイン及びPEドメインを有することができ、また他のNRPは[A]PG[R,S]ドメイン及び[G,A]ドメインを有することができる。なお別のNRPは[A]PG[R,S]ドメイン、[A,G]RRドメイン及びPEドメインを有することができる。
【0010】
NRPファミリーメンバーの遺伝子は、CAATボックス及びTATAボックスの少なくとも1つ、又はCAATボックス及びTATAボックスの両方をプロモーター領域内に一緒に含んでいる。NRPファミリーメンバー由来のオリゴヌクレオチドを用いて、NRPペプチドの発現を、そのような産生が所望される領域で高めることができる。
別の特徴では、本発明の実施態様は、頭部損傷又は慢性の神経変性疾患の結果としての脳の傷害領域のための治療方法を提供する。前記方法は、1つ又は2つ以上のNRP、NRPアナログ(構造的類似性を有するペプチドが含まれる)及び/又はNRPプロドラッグ(プロ-NRPペプチドを含む)を投与して、ニューロン又は神経芽細胞の遊走、増殖、生存及び/又は神経突起伸長を促進することによる。この治療方法は、皮質下領域の損傷と同様に新皮質の傷害を含む、軽度から重度の外傷性脳損傷(TBI)を受けた患者に特に有用であるが、ただしこれに限定されない。
ある実施態様では、NRP-2(配列番号:5)は、ゲノムクローンbA87G1(Sanger Sequencing Center)内のヒト染色体13のリバース鎖の塩基対77232−76768に位置する核酸配列によってコードされる。このペプチドはラットNRP-1のペプチドと類似の機能を有し、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走及び神経突起伸長を促進することができる。
【0011】
別の実施態様では、NRP-3(配列番号:3)は、ヒトゲノムの染色体3のリバース鎖上に、ダブルツィスト(Double Twist)アノテーションにしたがって塩基対34764−33003に位置する核酸配列によってコードされる。このNRPはまた、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
さらに別の実施態様では、NRP-4(配列番号:9)は、NCBIヒトゲノムアノテーションプロジェクトにしたがい、ヒト染色体15のフォワード鎖の塩基対21970003−21972239に位置する核酸によってコードされる。前記核酸配列から翻訳されるペプチドもヒトNRPファミリーに属する。この配列によってコードされるペプチドは、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
さらに別の実施態様では、NRP-5(配列番号:11)は、NCBIのアノテーションによって表示されているように、ヒト染色体7のリバース鎖の塩基対15047153−14824042に位置する核酸配列によってコードされる。この配列によってコードされるペプチドは、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
【0012】
NRPのまた別の実施態様は、染色体6のリバース鎖の116668725−116667697領域に位置するヒトゲノム由来のDNA配列に関するアノテーションが付されている(NCBIヒトゲノムアノテーションプロジェクトによる領域)。得られたペプチド、NRP-6(配列番号:13)は、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
NRPのさらに別の実施態様はげっ歯類で見出される。マウスのNRPは、リーディングフレーム1を用いる339の核酸から成るNCBIのアラクネ(arachne)contig_191157内に位置する核酸配列によってコードされる。オーバーラップする領域内に、フレーム3を用いるアノテーション付きNRPの29位で開始する、198の核酸の第二のORFが存在する。このORFは、切り縮められたヒトDNA修復タンパク質と高い同一性を有するタンパク質をコードする。得られたペプチド、NRP-7(配列番号:17)は、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
さらに別の実施態様はNRP-8(配列番号:20)であり、これもまた、マウスサンガー(Sanger)データベースのゲノムクローンbM344E9内のリバース鎖の核酸配列によってコードされるマウスペプチドである。前記タンパク質コード配列はアノテーションされており、塩基対5609−4052に位置する。NRP-8は、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
さらに別の実施態様はNRP-9(配列番号:28)である。前記はマウスNRP-7(配列番号:17)のラットオルソログNRPであり、ラット染色体の以下のエクソン内のリバース鎖に位置する核酸によってコードされる:7022614−7022326位に位置するエクソン1、7018685−7018651位に位置するエクソン2(NCBIデータベース)。NRP-9は、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
【0013】
また別の特徴では、本発明は、増殖誘導及び遊走誘導活性を判定するin vitroアッセイのための実施態様を含む。最近まで、長期にわたってタグのないニューロンの遊走を検出することができる、利用可能なin vitroのニューロン遊走アッセイはほとんどなかった。これらのバイオアッセイの1つは、器官型組織培養(“OTC”)として構築された嗅板周辺細胞を5日間のタイムコースでモニターする(Fueshko and Wray, 1994)。本発明のある種の実施態様では、成人の視床皮質系OTCを利用するin vitroアッセイを用いることによって、NRPを、ニューロンの遊走、ニューロンの増殖、ニューロンの分化、ニューロンの生存及び/又は神経突起伸長を誘導するそれらの能力について評価することができる。これらの実施態様は以下の理由から特に有用である:1)制御条件下では、組織培養基質上で2つの器官を互いに物理的に十分に遠く(約3から約5mm)分離させることによって両培養器官(例えば視床と皮質)の間の細胞架橋の形成を回避することができる、さらに2)出生後は、視床内のニューロン遊走は、視床の個体発生のタイムコースのために実質的に完了しているからである。これらのバイオアッセイは、したがってニューロン遊走誘導因子のスクリーニング及び同定に非常に適切であろう。
ある種の実施態様では、in vitro視床皮質系OTCアッセイは、皮質内の放射状遊走及び視床内の誘導による接線方向遊走の両方を明らかにするという利点を含む。in vitro制御条件下では、新皮質の正常な個体発生のタイムコースのために本来的な皮質内放射状遊走のみを観察することができる。
【0014】
他の実施態様では、基質に付着させた小脳の微小エクスプラントを含むin vitroバイオアッセイが提供される。これらの実施態様は、ニューロン遊走パターンに関するデータ(遊走ニューロンの数及び微小エクスプラントに対する遊走距離の定量を含む)を提供するために用いることができる。
培養からわずか2−3日後に、全体的な細胞遊走の強化と同様に小ニューロンから成る遊走鎖の発生を観察することができる。このアッセイによって得られる結果は、視床皮質系OTC内の細胞鎖誘導に類似する。
本発明の別の特徴に関する実施態様は、神経変性疾患及び脳損傷を治療又は防止するためにNRPを使用することを含む。具体的には、NRPは、損傷領域又は疾患領域近くに休止ニューロン幹細胞を欠く脳領域での使用に特に適している。
予防薬としてNRPを使用することは、折衷的外科手術、例えば冠状動脈バイパス移植法(CABG)又は脳への酸素供給が損なわれることを含む他の手術方法で有用でありえる。さらにまた、NRPは、例えば卒中、外傷又は脳若しくは脊髄への酸素付加を損なう他の損傷によって惹起される急性脳損傷の治療に有用でありえる。さらにまた、放射線療法または化学療法の前に予防的措置を実施することもできる。
【0015】
NRP化合物は、出生後の分化した神経組織内でニューロンの増殖、ニューロンの遊走、ニューロンの生存及び/又は神経突起伸長を開始させることができる。これらのペプチドは、軽度から重度の外傷性脳損傷(“TBI”)(びまん性軸索損傷、低酸素性虚血性脳障害及び頭蓋脳外傷の他の形態を含む)をもつ患者の損傷領域内のニューロン回路の改善又は修復を目的とする治療方法で利用することができる。NRP化合物は、神経系の感染(例えば一般的な細菌性髄膜炎)の治療、及び卒中(虚血性梗塞、塞栓症及び出血(例えば低血圧出血)及び他の原因によって惹起されるものを含む)の治療のために用いることができる。さらにまた、NRP化合物は、神経変性疾患(アルツハイマー病、レーヴィ小体痴呆、パーキンソン病(PD)、ハンチントン病(HD)、多発性硬化症、運動ニューロン病、筋ジストロフィー、末梢神経障害、神経系の代謝異常(糖原病を含む)、及びニューロンが傷害又は破壊される他の症状を含む)の治療に有用でありえよう。
本発明のある種の実施態様では、我々は、NRP遺伝子ファミリーのメンバーは、哺乳類の幹細胞(不死化幹細胞及び幹細胞の初代培養の両者で)で発現されることを見出した。
【0016】
他の実施態様では、我々は、NRPは、幹細胞の神経前駆細胞(神経芽細胞)への分化を促進することができることを見出した。また別の実施態様では、NRPは化学誘引物質に応答して幹細胞の遊走を刺激することができ、成熟ニューロンの形態(例えば軸索)を有する細胞への神経芽細胞の分化を促進することができ、さらに分化した神経芽細胞から神経突起(例えば軸索及び樹状突起)の成長を促進することができる。
本発明の他の特徴の実施態様は、NRPを使用して嗅覚細胞の増殖を促進することを含む。
したがって、NRPは、損傷神経細胞を修復し、神経組織の集団を再形成し、ニューロンの分化を促進し、シナプス形成(例えば神経突起伸長及び/又は神経分化)の促進に必要な過程を支援するために重要な治療的ツールとなりえる。
他の実施態様では、NRPと組み合わせた幹細胞の外科的移植を用いて、神経組織の集団を再形成することができる。幹細胞また別には神経芽細胞とNRPとを組合せることによって、神経組織の再増殖を促進することができる。そのような方法は成熟神経組織の再形成をもたらし、したがって、神経変性症状の治療に用いることができる。前記のような症状には、低酸素症/虚血、卒中、心臓バイパス移植術、アルツハイマー病、パーキンソン病、ハンチントン病、筋萎縮性側索硬化症、多発性硬化症、及び神経細胞の死又は変性を伴う他の疾患が含まれる。
本発明はその具体的な実施態様を記載することによって説明されるであろう。本発明の実施態様の他の目的、特徴及び利点は詳細な説明及び図面から明らかとなっていくであろう。
【0017】
定義
“ホモログ”という用語には、1つ又は2つ以上の遺伝子であって、前記遺伝子の配列が、進化上の関係(種間(オルソログ)又は種内(パラログ)のいずれかにおける関係)のために有意な関連を有する遺伝子が含まれる。ホモログはまた、共通の祖先のDNA配列から後世に伝えられることによって関係を有する遺伝子を含む。ホモログはまた、種の形成事象によって分離された遺伝子間の関係、又は遺伝的複製事象による遺伝子間の関係(パラログ参照)も含む。本明細書で用いられるように、“ホモログ”という用語はまた、進化上の関係により互いに関連する遺伝子産物も含む。保存的アミノ酸配列ドメインを有するNRPはホモログの例である。
“パラログ”という用語には、遺伝的重複の結果として互いに枝分かれした相同な遺伝子セットが含まれる。例えば、マウスのαグロビン及びβグロビンはパラログである。本明細書で用いられるように、“パラログ”という用語はまた、進化上の関係により互いに関連する遺伝子産物も含む。保存的アミノ酸配列ドメインを有するヒトNRPはパラログの例である。
“オルソログ”という用語には、種の形成の結果として互いに枝分かれした相同な遺伝子セットが含まれる。例えばマウス及びニワトリのαグロビン遺伝子はオルソログである。本明細書で用いられるように、“オルソログ”という用語はまた、進化の関係により互いに関連する遺伝子産物も含む。保存的アミノ酸配列ドメインを有するヒト及びマウスのNRPはホモログの例である。
“パラログペプチド”という用語には、パラログヌクレオチド配列によってコードされるペプチドが含まれる。
【0018】
“ペプチド”及び“タンパク質”という用語はアミノ酸で構成されたポリマーを含む。
“プロドラッグ”という用語には、酵素的、代謝的プロセッシング又は他のプロセッシングの結果、活性なNRP、活性なNRPアナログ又は活性なNRPパラログを生じるプロペプチドを含む分子が含まれる。
“NRP化合物”という用語には、NRP、NRPホモログ、NRPパラログ、NRPオルソログ、NRPアナログ及びNRPのプロドラッグが含まれる。
“NRP”という用語は、進化上の関係にかかわらず、神経の遊走、神経芽細胞の遊走、神経の増殖、ニューロンの分化、ニューロンの生存及び神経突起伸長の1つ又は2つ以上を含む機能を有するペプチドを含む。
アミノ酸は標準的な記号によって表される。この場合、アラニンは、“A”又は“Ala”で、アルギニンは“R”又は“Arg”で、アスパラギンは“N”又は“Asn”で、アスパラギン酸は“D”又は“Asp”で、システインは“C”又は“Cys”で、グルタミン酸は“E”又は“Glu”で、グルタミンは“Q”又は“Gln”で、グリシンは“G”又は“Gly”で、ヒスチジンは“H”又は“His”で、イソロイシンは“I”又は“Ile”で、ロイシンは“L”又は“Leu”で、リジンは“K”又は“Lys”で、メチオニンは“M”又は“Met”で、フェニルアラニンは“F”又は“Phe”で、プロリンは“P”又は“Pro”で、セリンは“S”又は“Ser”で、スレオニンは“T”又は“Thr”で、トリプトファンは“W”又は“Trp”で、チロシンは“Y”又は“Tyr”で、さらにバリンは“V”又は“Val”で表される。カルボキシ末端がアミド化されたペプチドは-NH2によって表示される。
【0019】
核酸は、アデニン(“a”で表される)、チミン(“t”で表される)、シトシン(“c”で表される)、及びグアニン(“g”で表される)を含むヌクレオチドで構成される。グアニン又はアデニンのどちらでもよいヌクレオチドは“r”で表され、チミン又はシトシンのどちらでもよいヌクレオチドは“y”で表され、さらにグアニン、アデニン、シトシン又はチミンのいずれかでありえるヌクレオチドは“n”で表される。ポリヌクレオチドはDNAでもRNAでもよく、一本鎖でも二本鎖でもよい。ポリヌクレオチドがRNAポリヌクレオチドである場合、ウラシル“u”がチミンの代用となりえる。
“疾患”は、動物のCNS又は末梢神経系の健常ではない任意の状態を含み、前記には特にパーキンソン病、レーヴィ小体痴呆、ハンチントン病、アルツハイマー病、多発性硬化症、運動ニューロン疾患、筋ジストロフィー、末梢神経障害、神経系の代謝異常(糖原病を含む)が含まれる。
“損傷”は動物の任意の急性傷害を含み、前記には特に卒中、外傷性脳損傷、低酸素症、虚血、(例えば胎盤剥離、臍帯閉塞に続く)胎児ジストレスに付随するか、又は子宮内発育遅延に付随する周産期仮死、適切な救急蘇生又は吸引の失敗に付随する周産期仮死、以下に付随する重篤なCNS障害(溺死、幼児突然死、一酸化炭素の吸入、アンモニア又は他のガス中毒、心停止、昏睡、髄膜炎、低血糖及びてんかん重積持続状態)、冠状動脈バイパス手術に伴う脳低酸素エピソード、低血圧エピソード及び高血圧危機、脳外傷及び脊椎損傷)が含まれる。
【0020】
具体的な実施態様の記載
本発明の実施態様は、神経再生ペプチド(NRP)を包含する、脳傷害の治療のための組成物及び方法を含む。NRPは、ニューロンの遊走、神経突起伸長、神経の分化、神経の生存及び/又は神経の増殖を誘導する。NRPはNRPアナログ、パラログ、オルソログ及び/又はNRPプロドラッグ、並びにヒト、マウス又は他の種の遺伝子によってコードされるペプチドでありえる。本明細書に記載されるNRPのいくつかは、本明細書に記載の遺伝子に対応する、以前に配列決定されていたオリゴヌクレオチドを基に予想されるタンパク質配列によるものである。他のペプチドは合成されたものであり、さらに少なくともいくつかはC-末端がアミド化された形態で提供される。しかしながら非アミド化型のタンパク質及びペプチドも本発明の範囲内に包含されることは理解されよう。
ラットNRP-1のヌクレオチド配列(配列番号:1)及びアミノ酸配列(配列番号:2)は以下のとおりである:
9 18 27 36
5' tat gat cca gag gcc gcc tct gcc cca gga tcg ggg
Tyr Asp Pro Glu Ala Ala Ser Ala Pro Gly Ser Gly
45
aac cct tgc cat 3' 配列番号:1
Asn Pro Cys His-NH2 配列番号:2
【0021】
しかしながら、遺伝暗号の縮退のために、多くのコドンが同じアミノ酸をコードしえる。したがって、多様な核酸配列が同じアミノ酸配列をコードすることができる。これら変型の各々は翻訳されて配列番号:2を生じることができ、したがってこれら変型のいずれも本発明の範囲内に包含される。例えば、多くの核酸配列(配列番号:1に挙げた核酸配列を含む)によってラットのNRP-1アミノ酸配列がコードされる。本発明はさらに配列番号:1のヌクレオチド配列の変種を含む。前記には前記核酸によってコードされるアミノ酸配列を保存する変種とともに、ラットのNRP-1アナログ及びNRP-1オルソログ及び/又はパラログをコードする核酸配列が含まれる。単なる例示として、DNAのための遺伝暗号にしたがって配列番号:1の変種を下記に示す(ここで各コドンは隣のコドンからスペースによって分けられ、“/”の後の核酸は“/”の前の核酸の変種を示す):
5' tat/c gat/c cca/t/c/g gag/a gcc/g/a/t gcc/g/a/t tct/a/c/g gcc/g/a/t cca/t/c/g gga/t/c/g tcg/a/t/c ggg/a/t/c aac/t cct/a/c/g tgc/t cat/c 3'
上記の配列(表示の変種を含む)は、上記の定義に従ってr、y及びnの文字を用いて表し、以下の配列を提供することができる:
5' tay gay ccn gar gcn gcn tcn gcn ccn ggn tcn ggn aay ccn tgy cay 3' 配列番号:3
【0022】
他のNRPをコードする他のヌクレオチド配列も遺伝暗号の縮退にしたがって変動しえることは理解されよう。さらにまた、DNA同様にRNAも本発明のペプチドをコードすることができ、さらに核酸がRNA核酸の場合はウラシルによってチミンを置換することができる。
ヒトの遺伝子は、鋳型としてヒトの悪液質のcDNA(米国特許5,834,192号)を用いてアノテーションが付されている。生存促進ペプチドは生存促進ペプチド(Cunningham et al., 1998)と96%を超える同一性を有し、さらにラットNRP-1は前記悪液質タンパク質と100%同一性を有し、既知の対応するcDNAとのただ1つのNRP-1ホモログである。ヒト悪液質タンパク質は染色体12の塩基対621841−625428の領域内に位置し、5つのエクソンから成る。我々は、悪液質のmRNAのスプライシング部位を、同定した染色体13上のNRPヒトパラログ(サンガー配列決定センターから入手したゲノムクローン:塩基対77232−76768)と比較し、NRP−1ヒトオルソログのコード領域をアノテーションした(このオルソログは本明細書ではNRP-2と称する)。NRP-2に関連するヌクレオチド及びアミノ酸配列は以下のとおりである:
【0023】
配列番号:4及び5
下線を付したヌクレオチド配列はシグナルペプチドを示す。
【0024】
前記タンパク質をコードするDNA配列は、鋳型として染色体12上のヒト悪液質遺伝子(米国特許5,834,192号のcDNA)のパラログ型の配列を用いるスプライシング部位分析によって予測したように4つのエクソンから成る。ゲノムクローンbA87G1の染色体地図は、正確なエクソンの位置決定のための基礎と考えられる。エクソン1はbp77232−77170の間に位置する。エクソン2はbp77088−77046の間に位置する。エクソン3はbp77036−76824の間に位置する。エクソン4は塩基対76778−76768の間に位置し、その後に翻訳終止コドンTAAが続く。翻訳されたタンパク質は110アミノ酸から成り、長さがヒトの悪液質タンパク質と同一で、ヒト悪液質タンパク質と24.5%の全体的同一性を有する。両タンパク質の細胞外局在化のためのシグナルペプチド(アミノ酸1−19)の配列比較によって、31.6%の同一性が明らかになった。重大なことには、成熟(切断)ペプチドの最初の30アミノ酸の比較によって、46.7%のアミノ酸同一性が明らかになった。さらにまた、このペプチドは、NRP-1と同様なニューロンの遊走、増殖、生存及び神経突起伸長活性を有する(図16、17及び18参照)。
NRP-1の第二のオルソログがアノテーションされている。前記は、染色体3の相補性リバース鎖の塩基対34764−33033の間(ダブルツィストヒトゲノムアノテーションプロジェクトによる領域)に位置するヒトゲノム由来のDNA配列によってコードされる。このタンパク質コード配列は、以下のように配置される5つのエクソンから成る:エクソン1、34764−34743;エクソン2、34729−34700;エクソン3、33745−33596;エクソン4、33498−33459;エクソン5、33043−33003。このヌクレオチド配列(配列番号:6)は333のヌクレオチドを有し、アミノ酸配列(配列番号:7;本明細書ではNRP-3と称される)は111のアミノ酸を有し、以下のように表示される:
【0025】
配列番号:6及び7
【0026】
これらの配列はNRPのヒト遺伝子ファミリーに属し、本明細書ではNRP-3と称される。この配列は、ヒト悪液質付随タンパク質と50%同一性及び67.2%類似性を有する。さらにまた、このヌクレオチド配列によってコードされるペプチドはNRP-1と類似の特性を有する。
第三のNRP-1オルソログがアノテーションされており、ヒト染色体15のフォワード鎖の領域21970003−21972239間に位置する、ヒトゲノム由来のDNA配列に含まれている(NCBIヒトゲノムアノテーションプロジェクトによる領域である)前記タンパク質コード配列は、以下のように配置されている6つのエクソンから成る:エクソン1、21970003−21970031;エクソン2、21970515−21970545;エクソン3、21970571−21970644;エクソン4、21970818−21970861;エクソン5、21971526−21971731;エクソン6、21972189−21972239。この遺伝子は再度配列決定され、今では下記の配列番号:48及び配列番号:49で表されている。しかしながら、遺伝子の再配列決定で、それによりコードされるNRPは変わらなかった。前記配列は、145のアミノ酸をコードする435の核酸から成る。このヌクレオチド配列(配列番号:8)及び予想されるアミノ酸配列(配列番号:9、本明細書ではNRP-4と称される)は以下のとおりである:
【0027】
配列番号:8及び9
【0028】
この配列はヒト遺伝子ファミリーNRPに属する。この配列はヒト染色体13に位置する核酸配列によってコードされるNRPと45%のアミノ酸類似性を有する。トリプレット244−246(アミノ酸82位);トリプレット391−393(アミノ酸131位)及びトリプレット421−423(アミノ酸141位)は潜在的なN-グリコシル化部位をコードする。アミノ酸118位は核酸配列内の不明確さのためにxである。ペプチド、NRP−4は、神経の増殖促進活性、神経突起伸長及びニューロンの生存促進活性を示す。オリゴヌクレオチド353−354位では、このヌクレオチドは決定されておらず、対応するアミノ酸は判明していないことに留意されたい。しかしながら、上記配列の発表の後で、正確な配列が提供され、実施例26の配列番号:48及び配列番号:49として本明細書では記載されている。配列情報におけるこの変更は、配列番号:8又は配列番号:9に由来するペプチドの神経保護作用に関連する部位に影響を与えるようには見えないことを記しておく。
ラットNRP-1のまた別のヒトオルソログ(“NRP-5”)は、リバース鎖に関するNCBIデータベースのホモ・サピエンス染色体7の作業ドラフト内に存在するDNA配列によってコードされる。このタンパク質コード配列はアノテーションを付されてあり、266のアミノ酸をコードする、全長が798の核酸を有する3つのエクソンから成る。タンパク質コードエクソンの正確な配置は以下のとおりである:エクソン1、15047153−15046815;エクソン2、14897885−14897772;エクソン3、14824386−14824042。このmRNAが発現されているというヒトEST(GenBank AW138864)での形跡がある。ヌクレオチド配列(配列番号:10)及びアミノ酸配列(配列番号:11、NRP−5)は以下のとおりである:
【0029】
配列番号:10及び11
【0030】
完全なタンパク質NRP-5は266のアミノ酸から成る。アノテーションされている翻訳されたNRPアミノ酸配列NRP-5は、染色体3に位置する分泌に関するヒトカルシウム依存アクチベータータンパク質(GenBankXP_036915)と76%の類似性を有する。さらにまた、翻訳されたヒト染色体7のNRP-5のエクソン1(339核酸)は、翻訳されたマウス5'EST(RIKENBB632392)と95.5%相同性を有する。このタンパク質は、神経突起伸長、ニューロンの生存、ニューロンの増殖及びニューロンの遊走という生物学的特性を示す、NRP-1及び他のNRPに存在するドメインを共有する。
我々は、染色体6の相補性リバース鎖の領域116668725−116667697間に位置するヒトゲノム由来のDNA配列にアノテーションを付した(NCBIヒトゲノムアノテーションプロジェクトによる領域)。このタンパク質コード配列は、以下のように配置された3つのエクソンから成る:エクソン1、116668725−116668697;エクソン2、116668333−116668305;及びエクソン3、116667872−116667697。前記配列(本明細書ではNRP-6と称される)は、78アミノ酸をコードする234の核酸から成る。この配列はNRPのヒト遺伝子ファミリーに属する。ヒトESTに対して見出された最高の相同性によって、ヒト胎盤組織から単離されたヒトcDNAクローンGenBankCS0DK001Y119と比較して核酸59−234の独自性が提示される。このクローンは3'-プライム末端から配列が決定され、924の核酸からなっている。我々のホモログは234個の核酸の後ろに終止コドンTGAを有する終端を形成するので、我々はこの遺伝子産物を対象としていない。NRPをコードするこのヌクレオチド配列(配列番号:12)及びこのペプチドのアミノ酸配列(配列番号:13、NRP-6)は以下のとおりである:
【0031】
配列番号:12及び13
【0032】
NRP-6のアミノ酸配列は、ヒト染色体13上のアノテーションを付されたNRPパラログ、NRP-2に対して14.1%の同一性及び44.9%の類似性を有する。このタンパク質は、神経突起伸長、ニューロンの生存、ニューロンの増殖及びニューロンの遊走という生物学的特性を有するNRP-1及び他のNRP(例えばNRP-2から5)に存在するドメインを共有する。
さらにまた、別のNRP-1オルソログ、マウスNRPファミリーメンバーが同定された。このマウスNRPファミリーメンバーは(ここではタンパク質2、配列番号:17と表示する;本明細書ではNRP-7と称される)は、NCBIのアラクネcontig_191157内に位置し、リーディングフレーム1を使用する339核酸から成る。オーバーラップする領域内には、フレーム3を使用する前記アノテーション付パラログの29位で開始する198核酸の第二のORFが存在する。このORFは切り縮められたヒトDNA修復タンパク質と高い同一性を有するタンパク質(ここではタンパク質1と表示する)をコードする。生物学的に活性なNRPペプチド配列:KDPEARRAPGSLHPCLAASCSAAG-NH2(配列番号:18)を使用し検索パラダイムtBLASTIを利用することによって、我々はマウスのEST RIKENデータベースでブラストヒットを得た。この5'-生成マウスESTはアクセッション番号GenBankAK012518及び以下の配列(配列番号:14)を有する:
【0033】
5'- GGCAGCCTCGAGATGGGGAAGATGGCGGCTGCTGTGGCTTCATTAGCCACGCTGGCTGCAGAGCCCAGAGAGGATGCTTTCCGGAAGCTTTTCCGCTTCTACCGGCAGAGCCGGCCGGGGACAGCGGACCTGGGAGCCGTCATCGACTTCTCAGAGGCGCACTTGGCTCGGAGCCCGAAGCCCGGCGTGCCCCAGGTAGGAAAGGAGGAGTAGTGTGTGCCAGCCTAGCGGCCGACTGGGCCACCCGAGACTGGGCCGCCTCCGGGCCGGCTTTGGAGGGAAGCCCCTGCTGGGCCTGTCCAGTGAGCTGTAATGTCGAGCGATGAGCGACCAGCTGCCTCGCTGTCCCAACGCTCTGGCCACGGCTTGTGCCTTGCCGCCATTTCCCCCAACCCACGCGGGCCACGGCTTGTGCCCTGCCGCCATTTCCCCCAACCCACGCGACCTTGCTC - 3'配列番号:14
【0034】
タンパク質1のリーディングフレーム3
オープンリーディングフレーム3(ESTの13位で開始する198個の核酸のORF)の翻訳によって、以下のタンパク質配列(配列番号:15)が明らかになった:
MGKMAAAVASLATLAAEPREDAFRKLFRFYRQSRPGTADLGAVIDFSEAHLARSPKPGVPQVGKEE 配列番号:15
この配列は、GenBankアクセッション番号Q13686のヒトアルキル化DNA修復タンパク質と82%のアミノ酸配列相同性(同一性及び化学的類似性)を有する。マウス型はC-末端が短縮されており、ヒトDNA修復タンパク質の389アミノ酸のうち66アミノ酸を有するだけである。
タンパク質2のリーディングフレーム1
323もの核酸をもつ長いORFがEST配列のフレーム1内に見出すことができる。続いて、我々は、マウスゲノム内の前記323核酸のORFの5'末端をアノテーションし、NCBIデータベースのマウスアラクネcontig_191157配列の23970と24374との間に位置する新規な遺伝子を発見した。このタンパク質コード配列は、113アミノ酸をコードする全長が339核酸を有する2つのエクソンから成る。エクソン1の位置は23970−23990であり、エクソン2の位置は24057−24374である。ラットNRP-1のこのマウスNRPオルソログのヌクレオチド配列(配列番号:16)及びアミノ酸配列(配列番号:17、NRP-7)は以下のとおりである:
【0035】
配列番号:16及び17
【0036】
発現された完全なNRP-7のアミノ酸配列は113のアミノ酸を含む(配列番号:17)。NRP-7のまた別のバージョンはまた別のスプライス型であり、配列番号:16の21位の後にさらに別の66ヌクレオチドを含み(配列番号:35)、前記は135アミノ酸を有する長型のNRP-7(“NRP-7ロング”)(配列番号:36)を生じる。
【0037】
【0038】
タンパク質機能プログラムツールSMARTは28アミノ酸から成る単一ペプチド配列を予想する。このタンパク質は、ヒト染色体13のNRPオルソログに対して13.6%同一性及び23.6%類似性を有し、さらにNRP-1に類似するニューロン生存、遊走、増殖及び成長活性を有する。
第二のマウスNRPファミリーメンバーは、マウスサンガーデータベースのゲノムクローンbM344E9内の相補性リバース鎖に位置する。生物学的に活性なNRPペプチド配列:KDPEARRAPGSLHPCLAASCSAAG-NH2(配列番号:18)を使用し検索プログラムtBLASTNを利用することによって、我々は、マウスゲノムサンガーデーターベースでゲノムクローンbM344E9内に類似性をもつ領域を得た。このタンパク質コード配列はアノテーションを付されてあり、5つのエクソンから成っている。前記は、141アミノ酸をコードする全長が423個の核酸である。コードエクソンの配置は以下のとおりである:エクソン1、5609−5596;エクソン2、5502−5489;エクソン3、5398−5283;エクソン4、5243−5229;及びエクソン5、5215−4952。ラットNRP-1のマウスオルソログ(本明細書ではNRP-8と称される)のコードヌクレオチド配列(配列番号:19)及びアミノ酸配列(配列番号:20)は以下のとおりである:
【0039】
配列番号:19及び20
【0040】
NRP-8の発現されたアミノ酸配列は141のアミノ酸残基を含む。112−114位のアスパラギン残基は、N-グリコシル化コンセンサス配列が存在することによりN-グリコシル化されると推定される。この新規なマウスNRP-1オルソログ、NRP-10は、染色体13に位置するヒトNRPオルソログ(NRP-2)に対して35.5%相同性を、さらにNCBIのアラクネcontigに位置するマウスNRP-1オルソログに対して28.9%の相同性を有する。さらにまた、このペプチドは、NRP-1又は他のNRPペプチドに存在するドメインに類似するアミノ酸配列ドメインを含み、さらにこのペプチドは、ニューロンの遊走、増殖、生存及び/又は神経突起伸長の促進を含む生物学的特性を有する。
NRP-9(マウスNRP-7のラットオルソログ)のヌクレオチド配列(配列番号:27)及びアミノ酸配列(配列番号:28)は以下のとおりである:
【0041】
配列番号:27及び28
【0042】
上述のNRP化合物の他に、我々は、NRPの発現にまた有用でありえる、NRP様ペプチドドメインを有する他の遺伝子を同定した。これらには、マイコバクテリウム及び腫瘍細胞に由来する遺伝子が含まれる。最近刊行された論文では、コンセンサス配列(PE_PGRS)(これは我々が提唱する配列(PGR/S)と類似している)を含むヒト型結核菌のPEマルチ遺伝子ファミリーが開示された。彼らはまた、これらタンパク質は宿主においてで前記細菌によって放出され、細菌の生存を促進するとも述べている。彼らが前記論文で提供した、PE_PGRSコンセンサス配列が見出される例が存在する。ヒト型結核菌のRv1818c遺伝子産物のアミノ酸配列(配列番号:21)は以下に示される:
【0043】
msfvvtipea laavatdlag igstigtana aaavptttvl aaaadevsaa maalfsghaq ayqalsaqaa lfheqfvral tagagsyaaa eaasaapleg vldvinapal allgrplign gangapgtga nggdggilig nggaggsgaa gmpggnggaa glfgnggagg aggnvasgta gfggaggagg llygaggagg aggragggvg giggaggagg nggllfgagg aggvgglaad agdggaggdg glffgvggag gaggtgtnvt ggaggaggng gllfgaggvg gvggdgvafl
gtapggpgga ggagglfgvg gaggaggigl vgnggaggsg gsallwgdgg aggaggvgst tggaggaggn agllvgagga ggagalggga tgvggaggng gtagllfgag gaggfgfgga ggagglggka gligdggdgg aggngtgakg gdggagggai lvgnggnggn agsgtpngsa gtggaggllg kngmnglp 配列番号:21
【0044】
エプスタイン-バーウイルス核抗原1のアミノ酸配列(配列番号:22)は以下に示される:
【0045】
msdegpgtgp gnglgekgdt sgpegsggsg pqrrggdnhg rgrgrgrgrg ggrpgapggs
gsgprhrdgv rrpqkrpsci gckgthggtg agagaggaga ggagagggag agggaggagg
aggagaggga gagggaggag gagagggaga gggaggagag ggaggaggag agggagaggg
aggagaggga ggaggagagg gagaggagga ggagaggaga gggaggagga gaggagagga
gaggagagga ggagaggagg agaggaggag agggaggaga gggaggagag gaggagagga
ggagaggagg agagggagag gagaggggrg rggsggrgrg gsggrgrggs ggrrgrgrer
arggsrerar grgrgrgekr prspssqsss sgspprrppp grrpffhpvg eadyfeyhqe
ggpdgepdvp pgaieqgpad dpgegpstgp rgqgdggrrk kggwfgkhrg qggsnpkfen
iaeglralla rshverttde gtwvagvfvy ggsktslynl rrgtalaipq crltplsrlp
fgmapgpgpq pgplresivc yfmvflqthi faevlkdaik dlvmtkpapt cnirvtvcsf
ddgvdlppwf ppmvegaaae gddgddgdeg gdgdegeegq e 配列番号:22
(M.J. Brennan and G. Delogu, The PE multigene family: a “molecular mantra” for mycobacteria. Trends in Microbiology 5: 246-249 (2002)より)
【0046】
NRP-1からNRP-9の完全な配列を用いる必要がないことは理解されよう。それよりはむしろ、本発明の実施態様にしたがって約8つのアミノ酸のペプチドフラグメントを用いることができる。本明細書で同定したコンセンサス配列ドメインが与えられるならば、合成ペプチドを作り上げるか、又は天然に存在するNRPを切り縮めて、生物学的に活性なペプチド部分を得ることができる。発現ペプチドを短縮する方法(例えば合成DNAを用いる)又は酵素的に改変する方法は当業界では公知である。
本発明のある実施態様は、配列KDPEARRAPGSLHPCLAASCSAAG-NH2(NRP-2セグメントKG;配列番号:18)を含むNRP-2(配列番号:5)の24merフラグメントである。
本発明のまた別の実施態様は、配列KDPEARRAPGSLHPCLAAS-NH2(NRP-2セグメントKS;配列番号:23)を含むNRP-2(配列番号:5)の19merフラグメントである。
本発明のさらに別の実施態様は、配列SEPEARRAPGRKGGVVCASLAADW-NH2(NRP-7セグメントSW;配列番号:24)を含むNRP-7(配列番号:17又は配列番号:36)の24mer型である。
本発明のさらに別の実施態様は、配列SDSFKSQARGO-NH2(NRP-3セグメントSQ;配列番号:25)を含む(NRP-3(配列番号:7)のアミノ酸11−23の間に位置する)11merペプチドである。
【0047】
本発明のまた別の実施態様は、配列GTPGRAEAGGQ-NH2(NRP-4セグメントGQ;配列番号:26)を含む(アノテーションを付されたNRP-4(配列番号:9)のアミノ酸22−32の間に位置する)11merペプチドである。
本発明のまた別の実施態様は、配列GTPGRAEAGGQVSPCLAASCSQAYG-NH2(NRP-4セグメントGQ;配列番号:29)を含むNRP-4(配列番号:9)の25merフラグメントである。
本発明のさらに別の実施態様は、配列REGRRDAPGRAGG-NH2(NRP-5セグメントRG;配列番号:30)を含むNRP-5(配列番号:11)の13merフラグメントである。
本発明のさらに別の実施態様は、配列SEVDARRAKKSLHCILSDTSHPRG-NH2(NRP-8セグメントSG;配列番号:31)を含むNRP-8(配列番号:20)の24merフラグメントである。
本発明のまた別の実施態様は、配列SDSFKSQARGQVPPFLGGVGCPWF-NH2(NRP-3セグメントSF;配列番号:32)を含むNRP-3(配列番号:7)の24merフラグメントである。
本発明のまた別の実施態様は、配列REGRRDAP-NH2(NRP-5 RP;配列番号:33)を含むNRP-5(配列番号:11)の8merフラグメントである。
本発明のさらに別の実施態様は、配列SEPEARRAQGGQIPSERVLSD-NH2(NRP-9セグメントSD;配列番号:34)を含む21merフラグメントで、前記配列はNRP-9(配列番号:28)のアミノ酸残基88-108の間に位置している。
本発明のまた別の実施態様は、配列PGRAEAGGQ-NH2(NRP-4セグメントPQ;配列番号:43)を含む9merペプチドで、前記配列はアノテーションを付されたNRP-4(配列番号:9)のアミノ酸24-32の間に位置している。さらに別の実施態様が本明細書の他の箇所に記載されている。
【0048】
NRP化合物の使用
したがって、本発明は、NRP、NRPによってコードされるペプチド、NRPのホモログ、オルソログ又はパラログ、NRPのアナログ及びNRPのプロドラッグに関する実施態様を含み、ここでNRPのプロドラッグは、酵素的に、代謝的に又は他の態様で改変されてNRP、NRPホモログ、NRPパラログ、NRPオルソログ又はNRPアナログとなることができる分子である。そのような分子は包括的に“NRP化合物”又は“NRP”と称される。NRP化合物はヌクレオチド配列によってコードされてもよい。前記ヌクレオチド配列はDNAでもRNAでもよく、さらに一本鎖でも二本鎖でもよい。本発明には本出願に開示された配列そのものと同様に、前記配列に対して相補的な配列も含まれることは理解されよう。さらにまた、NRPのまた別のスプライス形が存在しえることは理解されよう。そのような事例では、NRP RNAのまた別のスプライス形、並びにそれらがコードすることができるタンパク質及びペプチドもまた本発明の部分であると考えられる。
上記で示したように、本発明の実施態様は、NRPはニューロン及び神経芽細胞の増殖、遊走、分化、神経突起の生成を誘導することができ、さらにニューロンを神経損傷によってもたらされる障害から保護することができるという、本発明者らの驚くべき発見を基にしている。神経細胞の増殖及び急性脳損傷または慢性神経変性疾患によって生じた傷害領域への遊走は、神経機能の改善をもたらすことができる。さらにまた、NRP化合物を用いて、脳組織が変性するか、変性又は死のリスクを有するか、又は既に死んでいる、多様な異常及び症状を治療することができる。
細胞はまた、トランスフェクションの後、NRPオリゴヌクレオチドを使用してNRPの産生を刺激することができる。いくつかの事例では、トランスフェクションは複製可能なビヒクルとして行うことができ、さらに他の事例では、NRPオリゴヌクレオチドは裸のDNAとして導入することもできる。
【0049】
NRPで治療できる異常及び症状
NRP化合物が有用でありえる異常及び症状には以下が含まれる:
中枢神経系の感染(細菌、菌類、スピロヘータ、寄生虫感染を含む)及び類肉腫(化膿性感染、急性細菌性髄膜炎、軟膜炎を含む);
脳血管性疾患(卒中、虚血性発作、アテローム性硬化症血栓、ラクーン(lacune)、塞栓症、高血圧性出血、動脈瘤破裂、血管奇形、一過性虚血発作、頭蓋内出血、特発性くも膜下出血、高血圧性脳障害、脳動脈の炎症性疾患、例えば心不全(おそらく冠状動脈バイパス手術から生じる)によって惹起される灌流の低下、及び脳血管障害の他の形態を含む);
脳頭蓋外傷(頭蓋骨基底部骨折及び頭蓋内神経損傷を含む)、頚動脈洞フィステル、気脳症、気瘤及び鼻漏、脳挫傷、外傷性脳内出血、小児の急性脳腫脹;
脱髄性疾患(視神経脊髄炎、急性播種性脳脊髄炎、急性及び亜急性壊死性出血性脳炎、シルダーびまん性脳硬化症、及び末梢神経障害に併発する多発性硬化症を含む);
神経系の変性疾患(1つ又は2つ以上の進行性痴呆の症状、びまん性脳萎縮、非アルツハイマー型のびまん性皮質萎縮、レーヴィ小体痴呆、ピック病、前側頭性痴呆、視床変性、非ハンチントン型舞踏病及び痴呆、皮質脊髄変性(ヤコブ病)、痴呆-パーキンソン病-筋萎縮性側索硬化症合併症(Guamaninaら)を含む);
神経系の後天的代謝異常(錯乱、昏迷又は昏睡-虚血-低酸素症、低血糖症、高血糖症、高炭酸ガス症、肝不全及びライ症候群の1つ又は2つ以上を含む症状として提示される代謝性疾患、進行性錐体外路症候群として提示される代謝性疾患、小脳性運動失調、高体温、セリアック-スプルー病として提示される代謝性疾患、精神病又は痴呆を引き起こす代謝性疾患(クッシング病及びステロイド脳障害を含む)、甲状腺精神病及び甲状腺機能低下、膵臓性脳障害を含む);
栄養欠乏による神経系の疾患;
アルコール及びアルコール中毒;
薬物及び他の化学物質(アヘン誘導体及び合成鎮痛剤、沈静睡眠薬、興奮薬、向精神薬、細菌毒素、植物毒素、蛇咬傷及び刺創毒、重金属、工業毒物、抗腫瘍剤及び免疫抑制剤、サリドマイド、アミノグリコシド系抗生物質(聴器毒)及びペニシリン誘導体(痙攣)、心保護剤(β-ブロッカー、ジギタリス誘導体及びアミオダロンを含む)による神経系の異常。
【0050】
上記のリストによって示したように、本発明の組成物及び方法は、ヒトの神経の損傷及び疾患の治療に有用でありえる。さらに一般的には、本発明の組成物及び方法は、急性脳損傷(びまん性軸索損傷、周産期低酸素-虚血性損傷、外傷性脳損傷、卒中、虚血性梗塞、塞栓症及び高血圧性出血;CNS毒物への暴露、脳神経系の感染(例えば細菌性髄膜炎);代謝性疾患(例えば低酸素-虚血性脳障害、末梢神経障害及び糖原病を含むようなもの)を含むが、ただしこれらに限定されない)の結果として神経の傷害をもつか、又は慢性神経損傷若しくは神経変性疾患(多発性硬化症、レーヴィ小体痴呆、アルツハイマー病、パーキンソン病及びハンチントン病を含むが、ただしこれらに限定されない)に罹患している人間の患者の治療で有用である。そのような疾患又は損傷をもつ患者は、神経突起伸長と同様にニューロンの増殖及び遊走を開始させることができる治療プロトコルによって大きな利益を得ることができる。
さらに一般的には、本発明は、外傷、毒物への暴露、仮死又は低酸素-虚血による損傷後の傷害領域へのニューロン及び神経芽細胞の遊走の誘導に適用される。
NRP化合物(NRP-1、そのオルソログ、アナログ、パラログ及び同定されたNRPペプチドドメインを含むプロドラッグを含む)を用いて、ニューロン及び神経芽細胞の遊走を促進することができる。
【0051】
しかしながら、NRPを有利に用いることができるが、他の形態のNRP化合物の投与を排除しようとするものではない。例えば、ヒトパラログ型又はNRPのペプチドフラグメントもNRPの代わりに投与することができる。例示すれば、NRP及び担体を含むNRPのプロドラッグ型(NRP及び前記担体は患者の体内での切断又は消化に対し感受性を有する結合によって結合されてある)を投与することによって、NRPのCNS内の有効量を増加させることができる。投与後に切断又は消化されてNRPを放出する、任意の適切な結合を利用することができる。
また別の適切な治療方法は、中枢神経系の内部でNRP又はNRPのアナログ、パラログ若しくはプロペプチドを活性な形で発現させることができる細胞株である移植物、又は前記細胞株を含む移植物を通して、患者のNRPレベルを増加させる。
NRPは、医薬又は医薬調製物の部分として投与することができる。前記は、医薬的に適切な任意の担体、アジュバント又は賦形剤とNRP化合物を一緒にすることを必要とする。さらにまた、NRP化合物は他の非NRP性神経保護薬剤、増殖薬剤又は他の薬剤とともに用いてもよい。もちろんのこと、担体、アジュバント又は賦形剤の選択は用いられる投与ルートに一般的に左右される。
投与ルートは広く変動しえる。NRPは種々の方法で、すなわち腹腔内、静脈内又は脳室内に投与することができる。末梢適用も、中枢神経系との直接的干渉がないので選択方法でありえる。
【0052】
当業界で公知のいずれの末梢投与ルートも用いることができる。これらには、非経口ルート、例えば末梢循環への注射、皮下、眼窩内、眼内、くも膜下槽内輸液(徐放性装置又はミニポンプ、例えば浸透圧ポンプ又は皮膚パッチを用いる)、移植物、エーロゾル、吸入、乱切法、腹腔内、嚢内、筋肉内、鼻内、経口、頬内、肺、直腸又は膣ルートが含まれる。組成物は、人間又は他の哺乳動物へ、上述の神経学的疾患に対する治療を提供するために治療的に有効な量(例えば患者の病変を排除又は減少させる量)で非経口的に投与するために製剤化することができる。
ある投与ルートは、皮下注射(例えば0.9%塩化ナトリウムに溶解)及び経口投与(例えばカプセルで)を含む。
場合によってNRP化合物を直接患者のCNSに投与することが所望されることもまた理解されよう。前記は、適切な任意の直接投与ルートによって実施することができる。具体例には、側方脳室注射又は外科的に挿入した患者の脳の側方脳室へのシャントによる投与が含まれる。
【0053】
NRP投与量の決定
投与されるべきNRPの有効量の決定は当業者の技術範囲内であり、当業者には日常的な作業であろう。ある種の実施態様では、使用されるNRPの量は、本明細書に記載のアッセイ系を用いてin vitro試験で概算することができる。投与される最終量は、投与ルート、使用されるNRP、及び治療されるべき神経学的疾患または症状の性質に左右されるであろう。適切な用量範囲は、例えば体重100g当たり約0.01mgから約1mg、また別には体重100g当たりNRPの約0.06μgから約0.6mgで、この場合前記用量は中枢に投与される。
医薬に内包するためには、NRPは、通常の方法、例えばMerrtfieldら(J. Am. Chem. Soc. 15:2149-2154 (1963))の段階的固相合成法によって直接合成することができる。そのようなペプチド合成方法は当業界では公知であり、例えば以下の文献に記載されている:Fields and Colowick, 1997, Solid Phase Peptide Synthesis (Methods in Enzymology, vol.289), Academic Press, San Diego, CA。また別には、合成は市販のペプチド合成装置、例えばアプライドバイオシステムモデル(Applied Biosystems model)430Aを含むことができる。
一般的な提案として、非経口的に投与される1回分のNRP-1(配列番号:2)の医薬的に有効な総量は、用量応答曲線によって測定可能な範囲内にある。ある範囲は体重100g当たり約0.06mgから約0.6mgである。例えば、血中NRP-1(配列番号:2)は、治療される哺乳動物の体液中で測定され、用量を決定することができる。また別には、NRP-1(配列番号:2)化合物の量を増加さえながら患者に投与し、前記患者のNRP-1(配列番号:2)の血清レベルをチェックすることができる。使用されるべきNRP-1(配列番号:2)の量は、これらのNRP-1(配列番号:2)の血清レベルを基準にしてモル基準で算出することができる。
【0054】
特に、本化合物の適切な投与量を決定するある方法は、生物学的流体(例えば体液又は血液)中でのNRPレベルを測定することを必要とする。そのようなレベルの測定は任意の方法(RIA及びELISAを含む)実施することができる。NRPレベルを測定した後、単回投与又は複数回投与を用いて前記流体を化合物と接触させる。この接触工程の後で、前記流体中のNRPレベルを再度測定する。流体のNRPレベルが、前記分子が投与される目的について所望の有効性を生じるさせるに十分な量低下した場合、前記分子の投与量を調整して最大の効能を生じさせることができる。この方法はin vitroでもin vivoでも実施することができる。この方法はin vitroで、例えば哺乳動物から前記流体を抜き取り、NRPレベルを測定し、本明細書の化合物が単回又は複数回投与で哺乳動物に投与し(すなわち、接触工程は哺乳動物への投与によって達成される)、NRPレベルを前記哺乳動物から抜き取った流体で再度測定した後で実施することができる。
NRP化合物は、徐放系によって適切に投与することができる。徐放性組成物の適切な例には、形状物品(例えばフィルム又はマイクロカプセル)としての形状をもつ半透性ポリマーマトリックスが含まれる。徐放性マトリックスにはポリペプチドラクチド(米国特許3,773,919号、EO58,481号)、ポリ(2-ヒドロキシエチルメタクリレート)(Langer et al., 1981)、エチレンビニルアセテート(Langer et al., 上掲書)、又はポリ-D-(-)-3-ヒドロキシ酪酸(EP133,988号)が含まれる。徐放性組成物にはまたリポソーム結合化合物が含まれる。本化合物を含有するリポソームは当業者に公知の方法によって製造される。前記が以下によって実施される:DE3,218,121号;Hwang et al., 1980;EP52,322号;EP36,676号;EP88,046号;EP143,949号;EP142,641号;日本国特許出願83-118008、米国特許2,485,045号及び4,544,545号、並びにEP102,324号。いくつかの実施態様では、リポソームは小さな(約200オングストローム、或いは約200オングストロームから800オングストローム)単一薄層型で、その場合、脂質含有量は約30molパーセントコレステロールを超え、この選択した割合はもっとも有効な治療のために調整される。本明細書(上記及び下記)に引用される米国特許は全て参照により本明細書に含まれる。
【0055】
例えばWO95/32003(1995年11月30日公開)に記載された複合技術による、非PEG化ペプチドより寿命が長いPEG化ペプチドもまた用いることができる。
非経口的投与のためには、投与用量は約0.01から約1mg/100g体重、また別には体重100g当たり約0.06μgから約0.6mgのNRP化合物である。いくつかの実施態様では、本化合物は、一般的には所望の純度の各化合物を混合し、 医薬的に又は非経口的に許容可能な担体(すなわち用いられる投与量及び濃度で受容者にとって無毒なもの)であって、製剤の他の成分と適合するものとともに注射可能なユニット投薬形で製剤化することができる。例えば、製剤は好ましくは、酸化剤及びポリペプチドにとって有害であることが知られている他の化合物を含まない。上記の投与用量は限定的なものではないことは理解されよう。上記範囲を外れる他の用量も当業者は決定することができる。
いくつかの実施態様では、化合物は液体担体、又は微細に分割した固体担体、又はその両者と均一に及び密接に接触させることによって製造することができる。続いて、所望する場合は、前記生成物を所望の製剤に成型することができる。いくつかの実施態様では、前記担体は非経口的担体であり、また別には受容者の血液と等張な溶液である。そのような担体としての媒体の例は、水、食塩水、リンゲル溶液、緩衝溶液、及びデキストロース溶液が含まれる。非水性の媒体(例えば固定油およびオレイン酸エチル)も本明細書では有用である。
【0056】
担体は適切には微量の添加物、例えば等張性及び化学的安定性を強化する物質を含む。そのような物質は、望ましくは、用いられる投与量及び濃度で受容者に無毒であり、例示すれば以下を含む:緩衝液(例えばリン酸、クエン酸、コハク酸、酢酸及び他の有機酸又はそれらの塩);抗酸化剤(例えばアスコルビン酸);低分子量(10残基未満)ポリペプチド(例えばポリアルギニン)又はトリペプチド;タンパク質(例えば血清アルブミン、ゼラチン又は免疫グロブリン);親水性ポリマー(例えばポリビニルピロリドン);グリシン;アミノ酸(例えばグルタミン酸、アスパラギン酸、ヒスチジン、又はアルギニン);単糖類、二糖類、及び他の炭水化物(セルロース若しくはその誘導体、グルコース、マンノース、トレハロース、又はデキストロリン);キレート剤(例えばEDTA);糖アルコール(例えばマンニトール又はソルビトール);対イオン(例えばナトリウム);非イオン性界面活性剤(例えばポリソルベート、ポロキサマー又はポリエチレングリコール(PEG));及び/又は中性塩(例えばNaCl、KCl、MgCl2、CaCl2など)。
NRP化合物は、望ましくは上記のような媒体中で約4.5から約8のpHで製剤化することができる。前述のビヒクル、担体又は安定化剤のある種のものの使用は化合物の塩の形成をもたらすことは理解されよう。最終的な調製物は安定な液体又は凍結乾燥固体でもよい。
【0057】
他の実施態様ではアジュバントを用いることができる。錠剤、カプセルなどに取り込むことができる典型的なアジュバントは、結合剤(例えばアラビアゴム、ともろこしデンプン又はゼラチン);ビヒクル(例えば微晶質セルロース、);崩壊剤(例えばトウモロコシデンプン又はアルギン酸);滑沢剤(例えばステアリン酸マグネシウム);甘味剤(例えばシュクロース又はラクトース);香料(例えばペパーミント、ヒメコウジ、又はチェリー)である。投薬形態がカプセルであるとき、上記の材料の他に、液状担体(例えば脂肪油)もまた含むことができる。種々のタイプの他の材料をコーティングとして、又は投薬ユニットの物理的形態の改変物質として用いることができる。シロップ又はエリキシルは、活性化合物、甘味料(例えばシュクロース)、保存料(例えばプロピルパラベン)、着色剤及び香料(例えばチェリー)を含むことができる。注射用滅菌組成物は、通常の製薬慣行にしたがって製剤化することができる。例えば、活性化合物の媒体(例えば水又は天然に存在する植物油(例えばごま油、ピーナツ油又は綿実油)又は合成脂肪性媒体(例えばオレイン酸エチル)など)中の溶解液又は懸濁液を所望することができる。緩衝剤、保存料、抗酸化剤などを容認されている製薬慣行にしたがって取り入れることができる。
望ましくは、治療的投与に用いられるNRP化合物は滅菌することができる。滅菌は、滅菌ろ過膜(例えば約0.2ミクロンのポアサイズを有する膜)でろ過することによって容易に実施することができる。治療用組成物は一般的には、滅菌アクセスポートを有する容器(例えば静脈内溶液バッグ又は皮下注射針によって刺し通すことができる栓を有するバイアル)に入れることができる。
他の実施態様では、NRP化合物は、単位用量又は多用量容器(例えば封入アンプル又はバイアル)に、水溶液又は再構成用の凍結乾燥製剤として保存することができる。凍結乾燥製剤の例として、10mLのバイアルに化合物の5mL滅菌ろ過水溶液(0.01%w/v)を充填し、得られた混合物を凍結乾燥する。輸液溶液は、制菌水又は他の適切な溶媒を用いて凍結乾燥化合物を再構成することによって調製することができる。
【0058】
遺伝子治療
本発明の他の実施態様では、治療方法は、器官治療のための遺伝子治療を含み、前記はNRP化合物をコードする核酸を用いる。一般的には、遺伝子治療は器官のNRPレベルを増加させる(又は過剰発現させる)ために用いることができる。ヌクレオチド配列の例には、配列番号:1、3、4、6、8、10、12、16、19、27若しくは35、又はNRPのコンセンサスドメイン及び生物学的特性を有するペプチドをコードするその部分が含まれる。プロNRPをコードする他の配列も用いることができることは理解されよう(プロNRPは切断に際して生物学的に活性なNRPを生じることができる)。
生物にNRPをコードする配列をトランスフェクトするために任意の適切な方法を用いることができる。例えば、in vivo及びex vivoでの方法を用いることができる。in vivoデリバリーのためには、核酸(単独又はベクター、リポソーム、沈殿物と結合されている)を生物(例えば人間の患者)に直接、いくつかの実施態様では、NRP化合物の発現が所望される部位に注射することができる。ex vivo治療のためには、生物の細胞を取り出し、これらの細胞に核酸を導入し、この改変した細胞を前記生物に直接投与するか、又は例えば多孔性膜に被包化してこれを前記患者に移植する(例えば米国特許4,892,538号及び5,283,187号を参照されたい)。
我々は、本明細書で培養細胞がNRPを発現することができること、及びこれらNRP-発現細胞を有害な傷害に感受性を有するニューロンとインキュベートするとき、NRPが発現され、培養液中に分泌され、ニューロンを有害な傷害から保護することができることを示した。この驚くべき発見は、遺伝子移入及び細胞移植によって神経変性を処置する治療方法を支持する。
核酸を生細胞に移入するために利用可能な多様な技術が存在する。これらの技術は、核酸が培養細胞にin vitroで移入されるか、又は目的の宿主の細胞にin vivoで移入されるかにしたがって変動しえる。核酸の哺乳細胞へのin vitro移入に適した技術には、リポソームの使用、エレクトロポレーション、マイクロインジェクション、細胞融合、DEAE-デキストラン、リン酸カルシウム沈殿方法などが含まれる。遺伝子のex vivoデリバリーに一般的に用いられるベクターはレトロウイルスである。
【0059】
ある種の実施態様では、in vivo核酸移入技術には、例えばウイルスベクター(例えばアデノウイルス、単純ヘルペスI型ウイルス、又はアデノ付随ウイルス)によるトランスフェクション及び脂質利用系が含まれる。脂質仲介遺伝子移入に有用な資質は、N-[1-(2,3-ジオレイルオキシ)プロピル]-N,N,N-トリメチルアンモニウム(DOTMA)、ジオレオイルファチジルエタノールアミン(DOPE)及び3-β[N-(N',N'-ジメチルアミノエタン)カルボモイル]コレステロール(DC-Chol)である。いくつかの状況では、核酸含有ベクターを標的細胞に誘導する薬剤とともに核酸源を提供することが所望されえる。そのような“標的誘導”分子には、細胞表面膜タンパク質又は標的細胞に特異的な抗体、標的細胞上のレセプターに対するリガンドなどが含まれる。リポソームが用いられる場合は、エンドサイトーシス関連細胞表面膜タンパク質と結合するタンパク質を、標的誘導及び/又は取り込み促進に用いることができる。そのようなタンパク質の例には、特別な細胞タイプに親和性を有するカプシドタンパク質及びそのフラグメント、サイクリングにおいてインターナリゼーションを受けるタンパク質に対する抗体、及び細胞内局在化を標的とし細胞内半減期を強化するタンパク質が含まれる。他の実施態様では、レセプター仲介エンドサイトーシスを用いることができる。そのような方法は、例えば以下に記載されている:Wu et al., 1987;及びWanger et al., 1990。これまでに知られている遺伝子マーキング及び遺伝子治療プロトコルの総論のためには文献(Anderson, 1992)を参照されたい。さらにWO93/2563号及び前記に引用されている参考文献もまた参照されたい。
キットもまた本発明の範囲内に意図される。典型的なキットは、NRP製剤のための容器(いくつかの実施態様ではバイアル)を含み、前記容器は、医薬的に許容できる緩衝液中の1つ又は2つ以上のNRP化合物及び、使用者に前記医薬製剤の使用を指示する使用説明(例えば製品挿入物又はラベル)を含む。
【0060】
実施例
以下の実施例は、本発明のある種の実施態様を具体的に説明するために提供される。他の実施態様も考案することが可能で、前記もなお本発明の範囲内であることは容易に理解されよう。これら他の実施態様のいずれも本発明の範囲内であると考えられる。
【0061】
実施例1:NRP遺伝子の同定
ニューロンの生存、増殖及び分化に対して有効性を有する哺乳動物の遊走誘導因子を同定するために、我々は、ヒト悪液質関連タンパク質のcDNA及びコードされたその16mer悪液質フラグメントを鋳型として用いラット及びマウスのゲノムをスクリーニングし、新規なNRPホモログを同定した。
NRP遺伝子の同定は種々の細胞源(in vivo組織、神経幹細胞培養)の全RNAを入手することを含む。RNAはロシュ全RNA単離キット(Roche Total RNA Isolation Kit)を用いて抽出した。相補性DNA(cDNA)は、スーパースクリプトRT IIを用いて合成し、続いて以下のプライマーを用いマウスNRP遺伝子及びベータアクチンのマルチプレックスPCR増幅を実施した:mouFS NRPフォワードプライマー:5' AACGGAATGAATCGAAACCC 3'(配列番号:37);mouFS Revプライマー:5' CGCTCGACATTACAGCTCA 3'(配列番号:38);マウスβ-アクチンFwd:5' GAAAGGGTGTAAAACGCAGC 3'(配列番号:39);マウスβ-アクチンRev:5' GGTACCACCATGTACCCAGG 3'(配列番号:40)。正しいサイズを有するフラグメントをゲル精製し、直接pGEMベクターにクローニングし、大腸菌(E. coli, DH5α)のコンピテント株を形質転換した。形質転換細胞コロニーをNRP遺伝子フラグメントの存在についてスクリーニングし、陽性コロニー由来のプラスミドの配列を決定した。NRP遺伝子をコードする遺伝子の構造は図1Aに示されている。
【0062】
NRPタンパク質ドメインの性状決定:NRPは、セクレトームP(SecretomeP)サーバー(Technical University of Denmark)を用いて前記タンパク質配列を解析すると極めて有意なスコアを有するので、おそらくFGF-1、FGF-2のように非古典的経路で分泌されるのであろう。陽電荷アミノ酸残基の数及びN-末端NRP内の全体的な原子の数が多いことがこの予想につながった(Bendsten et al., 2004)。
単一のトレフォイルドメイン配列モチーフがマウスNRP配列内に予想され、このドメインはヒトpS2タンパク質(Jakowlew et al., 1984)と52.5%の相同性を有する(図1D)。推定的NRPトレフォイルドメインは、前記のトレフォイルドメイン(データベースSMART)のコンセンサス配列内の15の保存アミノ酸のうちの9個が類似又は同一である。この60%コンセンサス値はまたトレフォイル因子ファミリーのメンバーとしてNRPを受け入れるための閾値でもある。トレフォイルドメインは、タンパク質-タンパク質相互作用に関与し、シクロオキシゲナーゼ2(COX-2)及びトロンボキサンA2レセプター(TXA2-R)活性化経路を介して作用すると示唆されてきた(Rodrigues et al., 2003)。ヒトPs2タンパク質はトレフォイル因子ファミリー1(TFF-1)に属し、REK1/2経路でのシグナリング作用によって乳がん細胞の化学誘引(Prest et al., 2002)における関係が考えられている(Graness et al., 2002)。
ニューロンの生存、増殖、遊走及び分化を促進するマウスNRP(NRP-7セグメントSW)ドメインはトレフォイルドメインからC-末端側に位置し、さらにラットNRP-9についてはC-末端に位置する(図1Bのアラインメントされた21mer及び24mer配列を参照されたい)。
【0063】
NRP、悪液質関連タンパク質及びSDF-1間の相同性:NRPとSDF-1αとの間の驚くほど類似する生物学的活性にもかかわらず、両ペプチドは、32.6%相同性という中程度の類似性を示しただけである(図1E)。
成熟SDF-1の最初の9アミノ酸は残存化学誘引活性を示し、これはCDF-1αよりほぼ100倍低い(Loetscher et al., 1998)。マウス24merペプチド(NRP-7セグメントSW;配列番号:24)のN-末端11アミノ酸は、成熟SDF-1の開始部と50%を超える相同性でアラインメントされる。この関連性を示すタンパク質ドメインによって、悪液質タンパク質、NRP及びSDF-1αは、ニューロンの生存、増殖、遊走及び分化にとって重要な生物学的に活性なドメインを共有することが示される。前記生物学的に活性な領域は、悪液質関連タンパク質及びSDF-1については成熟タンパク質のN-末端に配置されるが、NRPについてはこの領域はトレフォイルドメインからC-末端側に位置する。
【0064】
実施例2:小脳微小エクスプラント:神経保護及びニューロン増殖方法
薄層状の小脳皮質をP3/4、P7/8ラットの子供から摘出し、125μmのポアサイズを有する金属紗を通してすり潰して均一サイズの微小エクスプラントを得た。61gで3分間遠心後、ペレットをスタートV(StartV)培養液(Biochrom)に懸濁し、前記懸濁液を6ウェルプレートのポリ-D-リジン被覆カバースリップに播種し、3時間インキュベートして接着させ、その後ウェル当たり1mLのスタートVを添加した。以前に記載したように、グルタメート/3-NP及びNRPも添加した。前記エクスプラントを5%CO2及び100%湿度下で34℃で48−72時間培養した。増殖速度測定のために、BrdUを培養開始時に加え、顕微鏡視野当たりの細胞を48−72時間後に数えた。
【0065】
結果:
(a)神経保護:ナノモル濃度のNRP-4セグメントGQ(配列番号:26)は重篤な損傷後50%の生存率をもたらした(図2)。
(b)ニューロン増殖:NRP-7セグメントSW(配列番号:24)は、これらの培養で増殖速度を200%以上に高めた(図3)。小脳の微小エクスプラントの増殖細胞は、ビヒクル単独の作用と比較して、興奮毒性(excitotoxic)及び酸化性ストレスに対して耐性を示した。
【0066】
実施例3:接触走性遊走アッセイ
マウス神経幹細胞、EGF依存性不死化マウス神経幹細胞MEB5及び野生型PC-12細胞に対する遊走誘導/化学誘引活性について、NRPを下記のように接触走性遊走アッセイでテストした。
方法:
最初のNRPコーティング:12μmのポアサイズを有するトランスウェルプレート(Corning)のコントロールウェルをBSA/PBS媒体の1.5mLで被覆した。残りのプレートを1−100ng/mLの範囲の種々の濃度のNRP(10μg/mLのBSAを含むPBSで調製)を用いて被覆した。続いて、このプレートを37℃で1時間インキュベートして被覆した。続いてウェルを1mLの滅菌PBSで2回洗い流した。
細胞外マトリックスコーティング:MEB-5細胞にはラミニン(10μg/mL)、マウス初代幹細胞にはPDL(50μg/mL)+マトリゲル、及びPC-12細胞にはフィブロネクチン(25μg/mL)+マトリゲルを細胞のための細胞外マトリックス(ECM)として用いた。全てのECM化合物はPBSで希釈した。ウェル当たり1.5mLのECMを室温で2時間インキュベートした。続いてウェルを1mLの血清非含有培養液(例えばNB/B27)で1回洗浄し、続いて1mLのPBSで洗浄した。
インサートのコーティング:5μg/mLのPDL/PLL混合物(PBS中)を用いてインサートを被覆した。続いて前記インサートをMiliQ水で洗浄した。
培養液への移動及び細胞の播種:適切な培養液を12ウェルプレートに移した(MBE-5細胞:DMEM高グルコース+N2(Invitrogenの増殖培養液補充物)+10ng/ビオチン+2mMのL-グルタミン;初代幹細胞:EuroclonのNSA培養液;及びPC-12細胞:NB/B27倍溶液)。続いてプレートを37℃、5%CO2でインキュベートし、1−2x105細胞を播種した。プレートを1−2日in vitroで固定した(DIV)。
固定:インサートを廃棄し、ウェルをPFAの連続希釈(0.4、1.2、3及び4%)で各希釈について3−5分固定した。ウェルを洗浄し、PFAの連続希釈(0.4、1.2、3及び4%)中で各希釈について3−5分保存した。ウェルを計測までPBS中で洗浄した。神経突起伸長を示し、底部の小室へ移動した全ての細胞を遊走細胞として数えた。
【0067】
結果:
10ng/mLのNRP-4GG(配列番号:29)は、BSA-ビヒクル単独と比較して195%を超えるMEB細胞を培養皿の底へ遊走させた(図4A)。NRP-4GG(配列番号:29)ペプチドの存在下では、BSA-ビヒクル単独と比較して93.7%を超えるE14細胞が遊走した(図4B)。NRP-2KS(配列番号:23)の存在下では、BSA-ビヒクル単独と比較して109%を超えるMEB-5細胞が底部へ遊走した(図4B)。NRP-7SW(配列番号:24)処理ウェルでは、BSA-ビヒクル単独と比較して35%を超えるE14細胞密度が得られた(図4D)。NRP-4GG(配列番号:29)ペプチドの存在下では、BSA-ビヒクル単独と比較して80.8%を超えるPC-12細胞が底部へ遊走した(図4E)。NRP-7SW(配列番号:24)は、BSA-ビヒクル単独と比較して333%を超えるPC-12細胞を遊走させた(図4F)。
【0068】
実施例4:神経幹細胞培養及び軸索伸長についての分化アッセイ
NSA幹細胞培養液はEuroclone(Italy)から購入した。Neurobasal、DMEM/F12培養液、N2及びB27補充物は全てライフテクノロジー(Life Technology)から得た。抗βIII-チュブリン抗体はシグマ(Sigma)から購入した。Cy3複合ヤギ抗マウス抗体はアマーシャム(Amersham)から購入し、Syto21はモレキュラープローブ(Molecular Probes)から購入した。NRP-2KS(配列番号:23)及びNRP-7SW(配列番号:24)を用いた。ヒト組換えエリスロポエチン(EPO)はR&D システムズから購入した。
神経幹細胞培養:
神経幹細胞はE15 C3Hマウスの前脳に由来し、文献(Gritti et al., 2001)に記載されているように、20ng/mLのEGF及び10ng/mLのbFGFの存在下でニューロスフェアとして培養した。簡単に記せば、時期を定めた妊娠マウスを犠牲にし、胎児を取り出した。無菌条件下で、脳を取り出し前脳を切り出した。すり潰して組織を分離し、75gで10分遠心して沈殿させ、非被覆組織培養フラスコで増殖因子を含むNSA培養液中に細胞を播種した。継代のためにニューロスフェアをすり潰し、単一細胞として播種した。
Shingoら(2001)がエリスロポエチン(EPO)について記載したように、神経幹細胞の対称的分裂(2つの幹細胞、両者とも幹細胞へ)から非対称的分裂(1つの幹細胞、1つのニューロン前駆細胞又は神経芽細胞へ)へのニューロスフェア内での細胞のシフトをNRPが誘導することができるか否かを決定するために、ニューロスフェアをすり潰して分離させ、以下を補充したNSA培養液中で6ウェルプレートに200,000細胞/ウェルの密度で拡張(expansion)させるために播種した:EGFのみ;EGF+種々の濃度のNRP;又はEGF+10U/mLのEPO。1日おきに半分の培養液を新しいNRP化合物を含む培養液で交換した。7日後にニューロスフェアの細胞をプレート培養した。EGFの存在下では、細胞は対称的分裂を保持する。EGFの除去とともに、細胞は分化を開始する(非対称的細胞分裂)。続いて、下記に記載するように細胞を分化アッセイに付したが、ただしbFGFは培養液に最初の24時間は添加せず、NRP又は他の化合物を含まないコントロール培養液で7日間分化させた。NRPに全く暴露されなかった細胞では、神経芽細胞の数は、NRP又はEPOに暴露された細胞の場合よりも少なかった。したがって、NRPは未分化幹細胞の神経芽細胞への分化を増進することができる。
分化アッセイ:
十分なサイズに達したとき、ニューロスフェアをすり潰して分離し、Nuncの24ウェルプレートに入れた、ラミニンを被覆した直径13mmのカバースリップ上に、ウェル当たり200,000細胞の密度でプレート培養した。プレート培養の培養液は、N2を補充したDMEM/F12及びB27と2mMグルタミンを補充したNeurobasalの1:1混合物であった。プレート培養後の生存を強化するために培養液には2ng/mLのbFGFが含まれていた。24時間後に、培養液をニューロン分化促進培養液(N2を補充したDMEM/F12及びB27と2mMグルタミンを補充したNeurobasalの1:3混合物)と交換し、さらにコントロールを除いてテスト化合物又はIGF-1を同時に加えた。新しい化合物を含む培養液を1日おきに交換した。プレート培養の7日後に、分化細胞を100ng/mLのSyto21を含む分化培養液で20分インキュベートし、生細胞の核を標識した。続いてそれらを4%のパラホルムアミドで固定し、マウス抗βIIIチュブリン抗体およびヤギ抗マウスCy3-複合二次抗体で免疫染色した。
ニューロン分化及び軸索伸長の定量:
染色後、カバースリップを組織培養プレートから取り出し、イムノフルア(Immunofluore)蛍光マウント媒体を用いてカバースリップ上にマウントした。全細胞に対するニューロンの百分率を解析するために、アクソイビジョン(Axoivision)ソフトを搭載したツァイスのアクシオフォト顕微鏡を用い、20xの倍率でウェル当たり任意の2視野を、赤色(チュブリン染色ニューロン)及び緑色(Syto21染色核)蛍光チャネルで調べた。視野当たりのニューロン及び核を数え、全細胞数に対するニューロンの百分率を決定した。結果は図5A−5Hに示されている。
【0069】
結果:
広い濃度範囲にわたって、全生細胞に対してニューロン細胞数を分化アッセイ内で正規化したとき、NRP-7セグメントSWは、分化培養液中のラミニン上でプレート培養したNSCから生じるニューロンの子孫細胞の百分率を増加させた。10pMの濃度で、βIII-チュブリン陽性ニューロンでは最大2倍の増加が観察され(図5E)、一方、軸索全長の成長における最大の増加活性はより高いナノモル濃度範囲にあった(図5A)。IGF-1は軸索伸長促進の陽性コントロールとして用いられ(Ishii et al., 1993)、NRPと同様に有効あったが、より低い濃度で用いたときは能力は劣っていた(図5A)。ラットに21merのNRP-9をNSCの分化中にBrdUとともに24時間投与したとき、増殖速度の増加を観察することができ(図5H)、このことは、アッセイでのニューロンの百分率の増加は、少なくとも部分的にはニューロン前駆細胞の増殖に起因する可能性を高める。
NRP-7SW及びNRP-2KSは、IGF-1と同様な有効性で分化神経前駆細胞の軸索伸長促進活性を有していたが、IGF-1よりもはるかに高い能力を有していた(図5A及び5B)。
NRP-2KSについてのこれらのデータの定量は図5C(ビヒクルによる処理)及び図5D(NRP-2KS処理)で例として示されている。
広い濃度範囲にわたって、全生細胞に対してニューロン細胞数を分化アッセイ内で正規化したとき、NRP-2KSは、分化培養液中のラミニン上でプレート培養したNSCから生じるニューロンの子孫細胞の百分率を増加させた。10nMの濃度で、βIII-チュブリン陽性ニューロンでは最大2倍の増加が観察された(図5F)。NSCの分化中にBrdUとともに24時間NRP-2KSを投与したとき、増殖速度の増加を観察することができ(図5G)、このことは、アッセイでのニューロンの百分率の増加は、少なくとも部分的にはニューロン前駆細胞の増殖に起因する可能性を高める。
【0070】
実施例5:in vivoで神経集団の再形成を促進するためのNRPの使用
本明細書に開示した発見を考慮して、我々は、NRPが神経組織の集団再形成を動物で促進することができるか否かを決定するためのin vivo実験を実施する。OEG細胞を損傷(脊髄損傷のような損傷)後の神経組織の集団再形成のために細胞供給源として評価した。NRPをOEG細胞培養に添加するか、及び/又は脊髄の移植操作においてOEG細胞とともに同時投与する。脊髄損傷をもつ患者の損傷部位での外科手術のために準備を整え、脊椎管には当業界で公知の方法を用いてアクセスし、損傷領域を特定する。OEG細胞及びNRPを損傷部位に移植し、場合によって更にNRP供給源を局所的に提供する。
【0071】
実施例6:NRPの発現
方法:
NRP遺伝子発現の分析:アノテーションを付されたマウスのNRP遺伝子の発現はRT-PCR、ノザンブロット及びin situハイブリダイゼーションによって確認した。
in situハイブリダイゼーション及びノザンブロット:マウスNRP(NRP-7(配列番号:35))の固有のコード領域(alkBホモログ遺伝子転写開始部位の上流)を包含する88merのオリゴヌクレオチドをpGEM7Zf(-)(Promega, Madison, WI, USA)のBamHIとEcoRI部位の間にクローン化した。適切な5'オーバーハング(下線部)も与えられた相補的な合成オリゴヌクレオチド;mfsNRP.S88、センス鎖(Invitrogen)(下記に示される:5' AATTCGGAATGAATCGAAACCCTGGAGTCGTGACCCCGGAAGAACCTGCCAGAGCCGGAATTTCGAGTTCTGCTTCCGGGCCAAACTG(配列番号:41))及び、mfsNRP.AS88、アンチセンス鎖(Invitrogen)(下記に示される:5' GATCCAGTTTGGCCCGGAAGCAGAACTCGAAATTCCGGCTCGGCAGGTTCTTCCGGGGTCACGACTCCAGGGTTTCGATTCATTCCG(配列番号:42))を変性させ、さらに、100mMのNaCl、0.1mMのEDTA(Sigma)、20mMのHEPES(pH7.9)中で各々48μMでオリゴヌクレオチドを95℃2分加熱してアニールさせ、1℃/分で60℃に冷却し、60℃でさらに1.5時間維持した。1μLのアニールさせた88mer及び300ngのQiaexを抽出し(Qiagen)、ゲル精製し、BamHI(Roche Diagnostics Ltd., Auckland, New Zealand)EcoRI(New England Biolabs Inc.)二重消化したpGEM7Zf(-)を4℃で19時間0.2ユニットのT4リガーゼ(Epicentre, Madison, WI, USA)で連結した。コンピテントDH10B(Invitrogen)で調製したクローンを、ClaI(Roche)耐性プラスミドの存在及びEcoRI/BamHI二重消化時の88merの切り出しについてスクリーニングした。センス及びアンチセンスRNAプローブの合成のための鋳型は、それぞれ20UのBamHI又はEcoRIを用い、10μgのDNA(JetStar Maxi Kit(Genomed)を用いて調製)を完全に消化することによって調製した。コンサート・ラピッド・ゲル抽出系(Concert Rapid Gel Extraction System)(Invvitrogen)を用いて鋳型をゲル精製した。
リボプローブを転写し、DIGノザンスターターキット(Roche)を用いてノザンブロットを実施した。周産期の星状細胞又はpUSE-myc-mNRPをトランスフェクトしたHEK細胞の全RNAの10μgを1.2%のホルムアルデヒドRNAゲルで0.16−1.77kbのRNAラダーとともに分離し、SSC中で陽性に荷電したナイロン膜(Roche)に移した。マーカーレーンを切り出しメチレンブルーで染色した。DIG標識NRPプローブを52℃で一晩ハイブリダイズさせた。ストリンジェントな洗浄及びDIG抗体とのインキュベーションの後、バイオイメージングシステム(UVP)を用いてCDP*ルミネッセンスによってシグナルを検出した。全脳をE15マウスから摘出し、4%PFAで3時間固定し、20%シュクロースで一晩凍結に対して保護し、ティシュテク(Tissuetek)OCT媒体に包埋し、-80℃で保存した。前記凍結保護した脳をPLL被覆スライド上で14μmの切片に切断し、8μg/malのプロテアーゼKで8分処理し、4%PFAで5分間後染色し、さらに45℃でDIG標識NRPプローブ(88merプローブ配列)(1:100希釈)で、センスコントロールとともに一晩ハイブリダイズした。抗DIG抗体で標識し、NBT/BCIPでシグナルを発色させた後、切片をネスチン/GFAP/b-チュブリンで二重標識し、蛍光二次抗体で可視化した。
【0072】
結果:
PCRを用いて、NRPメッセージがE15マウス胎児の脳の種々の領域で検出されたが、興味深いことには発現レベルは培養されているマウスの神経幹細胞ではるかに高かった(図6A)。神経幹細胞特異的発現は、マウスNRP mRNAに対するin situハイブリダイゼーション及び幹細胞/前駆細胞特異的中間体フィラメントネスチンによる二重染色によってさらに立証された(Lendahl et al., 1990)(図6E−6H)。NRPメッセージ(mRNA)がNSCに由来するニューロン又はグリア子孫細胞で維持されているか否かを判定するために、未分化NSCにおける発現レベルをCNTFにより星状細胞に分化させたNSC及びBDNFによってニューロンへ分化させたNSCと比較した。NRP発現レベルはCNTF処理細胞でのみ顕著に増加し、星状細胞系列は分泌タンパク質の主要な供給であることを示した(図6B)。E19マウスのいくつかの組織における発現レベルを比較し、肺を除いて非神経組織での発現は低く、存在しないこともあることが明らかになった(図6C)。星状細胞での比較的高い発現レベルを基にして、周産期星状細胞培養由来のRNAでのノザンブロットを試み、それによってほぼ0.8及び1.2kbのサイズの2つの転写物が明らかになった。転写は翻訳開始点の上流-114及び-509から始まるので、これは予想される完全長の転写物に一致する(図6D)。E15マウス胎児の前脳の切片のin situハイブリダイゼーションによって、辺縁ゾーン及びサブプレートにおける強いシグナルが明らかになり、皮質プレートの弱い標識によって補完されていた(図6I)。マウスNRP mRNAは、ネスチン免疫反応性と同時局在を示した。この段階で、GFAP又はMAP2の同時標識は観察されなかった(データは示されていない)。この染色パターンは、放射状グリア様細胞における発現を示し、前記細胞は未成熟な星状細胞への形質転換を経ている(Steindler & Laywell, 2003)。
【0073】
実施例7:小脳微小エクスプラントII−NRP-2セグメントSW(配列番号:24及びSDF-1)の神経保護の比較
方法:
小脳微小エクスプラントを、実施例9で述べたようにグルタメート/3-NP及びNRPの他にSDF-1も加えて調製した。
結果:
非保護小脳微小エクスプラントの0.5mMの3-NP/グルタメートによる48時間の処理で生じた損傷は75−92%の範囲の細胞死であった。図7AはNRP-7SWは極めて顕著な神経保護を100fMから100nMの濃度範囲にわたって付与し、100pMでほぼ50%の損傷からの回復を示した。比較すれば、10nMのヒトSDF-1は30%未満の神経保護を付与し、狭い用量範囲の有効性を示した(図7B)。
16merの悪液質フラグメントはより低い有効性を示し、ニューロン保護及びニューロンの増殖誘導については10倍低い潜在能力を有していた(データは示されていない)。
【0074】
実施例8:ニューロン遊走の誘導:NRP-9セグメントSD(配列番号:34)、NRP-2セグメントSW(配列番号:24)及びSDF-1の比較
接触遊走アッセイのための方法:
接触遊走アッセイは、ラットNRP-9SD(配列番号:34)(4nM)及びマウスNRP-7SW(配列番号:24)(0.4nM)をボイデン(Boyden)チャンバー中の誘引物質として用い、実施例10の記載にしたがって実施した。
視床皮質系-皮質培養の方法
ロングエヴァンス(Long Evans)ラットの新生児(P0)由来の後頭皮質及び視床背面を、Paxinos37のP0アトラスにしたがって切り出した。後頭皮質は冠状に、視床背面は前方から組織ショッパー(McIlwain)で350μmの厚さの切片にし、これらを直ちに0.65%のD-グルコース(Merck)を補充したゲイのバランス塩類溶液(Gey's Balanced Salt Solution(GBSS))に移し、7℃で1時間回復させた。垂直の向きを有する視床切片を立体顕微鏡下で選別し、皮質組織と少なくとも3mm離して一緒にカバースリップ上に配置した。この事例では、視床は手綱核が皮質層VIに向き合う方向に置かれている。これらの切片は、10μLのトロンビン(25U/mL、ICN)で凝固させたニワトリ血漿(Cocalico)の10μLを用いて血漿凝塊中でカバースリップに接着させた。カバースリップをローラーチューブ(Nunc)内に置き、0.75半人工培養液(2/4基準培養液イーグル、1/4ハンクスバランス塩類溶液、1/4不活化ウマ血清、2mMのL-グルタミン及び0.65%D-グルコース)を供給した。培養物をローラーチューブのインキュベーターで36℃で20日までin vitroで維持し(DIV)、ラットの悪液質関連NRPを含む培養液は3日毎に交換した。
ラット胎児の前脳のOTCは記載したように調製した。簡単に記せば、妊娠E17期の妊娠ラットを犠牲にし、迅速に胎児を取り出した。前脳の冠状切片を400μmの厚さに切り、3cmのペトリ皿(1mLのDMEM/F12、6.5mg/mLのグルコース、0.1mMグルタミン及び10%FCSを含む)中のミリセルPICM ORG50メンブレン(Millipore)上に静置した。その後、培養液を1mLのNeurobasal/B27、6.5mg/mLグルコース、0.1mMグルタミンに交換した。前記には10μLのPBS、NRP又はSDF-1が表示の濃度で添加され、さらに均一なサイズの小さなDiI(1,1'-ジオクトデシシル-3,3,3',3-テトラメチルインドカルボシアニン)結晶を、立体顕微鏡下で内側神経節隆起に静置した(n=10−15)。24時間培養した後、総数又は神経節隆起及び皮質内に分散したニューロンを倒立顕微鏡下で数えた。
【0075】
結果:
NRP-9SD(配列番号:34)(4nM)及びマウスNRP-7SW(配列番号:24)(0.4nM)は、マウス神経幹細胞に対してNRPの実質的な化学誘引活性を示した。コントロール条件の細胞数のほぼ4倍の細胞がNRP-9SDによって誘引されたが、10分の1未満の量のNRP-7SWによってなお2倍を超えるNSCが培養皿の底に誘引された。比較すれば、100nMのSDF-1被覆は、0.4nMのマウスNRP-7SWの場合と同様な数の細胞を誘引した(図36A)。それらの代表的な形態的表現型が図8Bに示されている。
神経節隆起及び皮質原基を含む矢状方向の胎児脳切片を用いるOTCアッセイでは、OTCに投与されたNRP-7SWは、遊走するニューロン前駆体細胞の数を5倍増強させた(図8C)。ヒトSDF-1は同様な数の細胞を誘導して内側神経節隆起から皮質原基へ遊走させるが、接触走性遊走アッセイの場合のように、63アミノ酸の長さを有する成熟ヒトSDF-1はNRPよりはるかに能力は低い。
【0076】
実施例9:組換えNRPの発現及び活性のアッセイ
HEK293-発現mfsNRPの調製:
HEK293細胞にN-末端及びC-末端にタグを付したmNRP融合タンパク質発現構築物(それぞれpUSE-Flag-mNRP2及びpUSE-myc-mNRP11)を25kDaポリエチレニミンを用いてトランスフェクトした(Boussif et al., 1995)。使用したNRP配列は配列番号:35であった。pEGFP-N1(Clontech)との並行トランスフェクションもまた実施した。簡単に記せば、各構築物をPEIと6、9及び12対1PEIの比で、5%グルコース中に10ng/μLの DNAでDNAホスフェートと一緒にした。36mmウェルで2mLの増殖培養液(10%ウシ胎児血清補充DMEM)中で70%コンフルエントのHEK239細胞に、ウェル当たり200μLのトランスフェクション混合物とともに4.5時間インキュベートすることによってトランスフェクトした。この後、2xの抗生物質/抗マイコプラズマミックスを含む2mLの増殖培養液を各ウェルに添加した。次の日、1mL/ウェルのトリプシン-EDTAを用いて培養を増殖培養液(800μg/mLのジェネチシンを含む)に1:15に分割した。選別は、培養液を週に2回交換することによって17日間維持した。コンフルエンスに達したときに、1:10に分割した細胞の部分で選別を継続し、残りは10%DMSOを含む増殖培養液中で凍結した。さらにまた、800μg/mLのジェネチシン選別から1:2分割を13日目に調製した。安定なトランスフェクタントを発現されたタグ付タンパク質の検出のために採集した。
HEK293細胞でのMyc-NRPの発現はウェスタンブロットで確認された:
ウェスタンブロットを実施して、HEK293細胞でのMyc-NRPの発現を確認した。細胞溶解物を変性条件下で可溶化し、SDSポリアクリルアミド電気泳動(PAGE)を用いてタンパク質を分離した。発現された組換えマウスMyc-NRPは抗Mycモノクローナル抗体によって検出された。
【0077】
結果:
HEK細胞にトランスフェクトされた、c-mycプロモーターの制御下での完全長組み換えマウスNRPの生成により、アンチセンス88bpのNRP cRNAプローブを用いたとき、予想された0.8kbサイズの転写物が明らかになった(図9A)。Myc-NRP-HEK細胞によって発現された16.5kDaの組換えタンパク質生成物は、レムリSDSゲルで20kDaのマーカーサイズへ泳動した(図9B)。
NRP遺伝子によってコードされた、完全なNRP遺伝子組換え発現産物の測定された生物学的活性は、テストしたNRP合成ペプチドのそれと同じであった。小脳微小エクスプラントとの同時培養アッセイでは、Myc-NRP-HEK細胞は、種々の細胞濃度で播種したとき実質的な神経保護作用を付与し、Myc-NRP-HEK細胞数5000で、回復値はMAP-2陽性ニューロンの51%であった(図9C)。空ベクターのみを含むコントロールMyc-HEKは、酸化性/興奮毒性ストレスの後MAP-2陽性細胞の回復を全く示さなかった。添加したHEK細胞は、神経保護について釣鐘型の用量応答曲線を示した。
Myc-NRP-HEK細胞は、マウスのNSCを用いて接触走性遊走アッセイでテストしたとき、化学誘引ニューロン遊走誘導活性を示した。マウスNRPを発現しているMyc-NRP-HEK細胞をボイデンチャンバーの底に播種し、マウスNCSをインサートに播種したとき、24時間後に検出された神経突起伸長を示すニューロン細胞集団はコントロールMyc-HEK細胞を用いた同じアッセイと比較して2倍増加した(図9D)。
実施例10:NRPの作用の可能なメカニズム:
最近のデータは、SDF-1(CXCL12)及びそのレセプターCXCR4は、免疫系及び神経系で同様な作用を有する可能性を示唆している。すなわち、それらは細胞の移動、ニューロン及びリンパ球の増殖、可塑性(神経突起伸長及び分化)並びに生存を調節することができる(Vlahakis et al., 2002 and Lazarini et al., 2003)。CXCL12に対する同様な依存は協調的なホメオスタシスによる相互作用を支持することができるかもしれないが、CXCR4レセプター結合の結果として、HIV感染及びそれに続く神経障害罹患のように炎症性プロセスに対する固有の脆弱性を構成するかもしれない(Keswani et al., 2003)。HIV及び関連ウイルスは、リンパ球レセプターCD-4の他にコレセプターを標的細胞への感染に必要とする。HIVに使用されるコレセプターの1つは、G-タンパク質結合ケモカインレセプターCXCR4である。SDF-1とCXCR4との相互作用はCD4-リンパ球へのHIVの侵入を阻止することができる。さらにまた、CXCR4-レセプターと拮抗することによって癌細胞の移動/侵襲を阻害することにより、動物モデルで癌の転移を阻止することができることが知られている(Rubin et al., 2003;Liang et al., 2004)。
これまでのところ、SDF-1は、CXCR4レセプターと結合する唯一の公知リガンドである。我々は、ここで、NRPはCXCR4レセプターに対する新規なクラスのリガンドでありえること、さらに生物学的活性(例えば化学誘引及びニューロン生存)はCXCR4レセプターの活性化によって発揮できることを示すいくつかの証拠を提供する。したがって、CXCR4レセプターに対するNRPの拮抗作用が存在しえる可能性も排除しないが、NRPはCXCR4レセプターのためのアゴニストであることができる。同様な観察がSDF-1についても得られた。すなわち、単一アミノ酸置換がCXCR4に対するSDF-1作用に拮抗することができる(Tudan et al., WO0185196)。しかしながら、これはNRPの作用のただ1つの可能なメカニズムではなく、他のメカニズムもここに記載したこの観察を説明しえることは理解されよう。
【0078】
方法:
小脳微小エクスプラントを、グルタミン/3-NP及びNRPの他に、SDF-1、ワートマンニン及びPD98059(両者ともCalbiochem)を添加しながら実施例9で述べたように調製した。
NRPの生存促進及び遊走誘導活性に対するCXCR4レセプター中和抗体の作用
CXCR4レセプター(フュージンレセプター)に対して作製した中和抗体1μg/mLを72時間適用することによって、NRPで処理した小脳微小エクスプラント内のニューロンの生存及びニューロンの遊走パターンは顕著に低下した。1μg/mLの抗体濃度で、NRP-9 SD(配列番号:34)の神経保護及び/又は遊走促進作用は失われた(図10A)。同様な阻害は、ヒトNRP-2KS(配列番号:23)(図10B)又はマウスNRP-7SW(配列番号:24)(データは示されていない)についても観察された。
NRPの化学遊走作用に対するCXCR4レセプター中和抗体の作用
培養皿に被覆された10ng/mLのNRPの化学誘引作用は、ニューロン幹細胞株MEB-5をCXCR4に対する中和抗体で1.5時間プレ-インキュベートすることによって阻止された。NRP-9SD(配列番号:34)ペプチド単独で観察された遊走と比較して、極めてわずかな細胞が遊走した(図10C)。
NRPの作用はERK1/2及びAktリン酸化によって媒介され得る
神経保護及び/又は遊走作用を発揮するためにNRPによって利用されるシグナリング経路を解明するために、我々は、MEK阻害剤PD98509及びホスファチジルイノシトール3-キナーゼ(PI-3K)阻害剤ワートマンニンを用いてMAPK又はAktリン酸化を阻害した。
NRP媒介神経保護及び化学誘引の作用メカニズムにおける、MAPK及びakt活性の上流の調節因子PI-3K(Aktのリン酸化)の役割を知るために、我々は、小脳微小エクスプラント及び接触走性遊走アッセイにおける有効性についてMAPKキナーゼ(MEK)阻害剤PD98509をテストした。ビヒクル及びPD98509による又はPD98509なしの損傷治療によって前記阻害剤による軽度の毒性のみが示され、一方、阻害剤とNRP-9SD(配列番号:34)による同時処理によってNRPの神経保護活性は失われた(図11A)。Aktリン酸化のワートマンニン(100nM)による阻害は、ビヒクル及びNRP無しの損傷治療後には有意な影響を示さなかったが、ERK1/2のリン酸化の阻害とは同様であり、NRPによって生じる神経保護は失われた(図11B)。接触走性遊走アッセイでは、遊走NSCでほぼ3倍の増加がPD98509(0.1pMから100pM)でほぼ完全に阻止され、BSA被覆ウェルでの基礎遊走に対しては有意な影響はなかった(図11C)。神経保護とは対照的に、NRPの化学誘引遊走活性はワートマンニンによるPI-3Kの阻害によっては抑制されなかった(データは示されていない)。底部の区画へ遊走した細胞数の低下がニューロン生存の障害によるものであったという可能性を排除するために、インサートをSyto2で生細胞について染色し、上部表面を分析した。生存細胞における顕著な相違は見出されなかった。
【0079】
実施例11:in vivoでのNRP有効性
材料と方法:
in vivoでのNRPの有効性をテストするために、低酸素虚血損傷(HI)に暴露したラットで実験を行った。成獣ラット(50日齢、ウィスターラット、250−300g、雄)を用いた。改変したレヴィン(Levine)のモデル調製及び実験方法を用いた(Rice et al.,1981, Ann. Neurol. 9:131-141;Guan et al., 1993, Cereb. Blood Flow Metab. 13(4):609-16)。簡単に記せば、これらの方法は、側脳室にカニューレを埋め込んだ動物での一側性頚動脈結紮により誘導したHI損傷及びそれに続く吸入性仮死から成る。ハロタン麻酔下で定位脳手術により、誘導カニューレを硬膜の最頂部上の正中線より右へ1.5mm及び耳間ゼロ平面に対して前方7.5mmの位置に設置した。カニューレ設置2日後に右頚動脈を二重結紮した。麻酔から1時間の回復時間後に、各ラットを、湿度(90±5%)及び温度(31℃±0.5℃)が制御されているインキュベーターにさらに1時間置き、続いて各ラットを低酸素(6%酸素)に10分間暴露した。処置前に動物をインキュベーターでさらに2時間維持した。
低酸素虚血損傷の後、ラットを、生理食塩水に溶解したNRP-4セグメントPQ(PGRAEAGGQ;配列番号:43)の5nM(n=12)、50nM(n=12)若しくは500nM(n=12)又はビヒクル(n=10)(通常の生理食塩水)の脳室内(icv)投与により処置した。
組織学的試験を低酸素虚血後5日のラットで実施した。ラットを過剰のペントバルビタールで殺し、通常の生理食塩水、その後10%のホルマリンを心臓経由で灌流させた。脳は処理前最低2日間同じ固定液中に維持し、標準的なパラフィン包埋術を用いた。
厚さ8μmの冠状切片を線条体、大脳皮質及び海馬から切り出し、チオニン及び酸性フクシンで染色した。組織損傷の重篤度は、線条体、皮質並びに海馬のCA1-2、CA3、CA4及び歯状回でスコアを決定した。組織傷害はニューロンの消失(好酸性(赤色)細胞質及び凝縮核)、広範囲壊死及び細胞性反応として特定した。組織傷害は以下のスコアシステムを用いてスコアを決定した:0:組織が組織傷害を示さなかった;1:<5%の組織が傷害された;2:<50%の組織が傷害された;3:>50%の組織が傷害された;及び4:>95%の組織が傷害された。
結果と結論:
この実験の結果は図12に示されている。ヒト染色体15の9merフラグメント、NRP-4セグメントPQ(PGRAEAGGQ-NH2;配列番号:43;低酸素症後2時間で9ng/icv)によって、損傷後5日の全ての解析した脳内で100%の神経保護が付与された。
【0080】
実施例12:嗅球鞘性グリア(“OEG”)細胞の成長促進
NRPが神経組織の再ミエリン化に有用でありえるか否かを決定するために、我々は、ヒト嗅球鞘性(ensheating)グリア(OEG)細胞を用いて実験を行った。OEG細胞は、傷害組織(例えば脊椎の組織)の集団再形成のための細胞の供給集団として評価されている。OEG細胞は、当業界で公知の方法を用いてヒトの鼻粘膜の掻爬検体から得た。細胞は制御条件下及びNRP-7SW(配列番号:24)の存在下で培養液で増殖させた。
制御条件下では、OEG細胞の正常な増殖速度は72時間内で約25%である。我々は、NRP-7SW(配列番号:24)の存在下では、OEG細胞は72時間にわたって約50%の速度で増殖することを見いだした。これらの結果は、NRPはOEG細胞の増殖を強化することができ、それによって神経細胞移植方法において有用で細胞増殖を促進し、傷害組織に神経機能を取り戻すことができることを示している(図13)。
【0081】
実施例13:NRP-5セグメントRGペプチドアナログの神経保護活性
NRP-5セグメントRG関連ペプチドのアミノ酸配列における変更が、NRP-5セグメントRG(配列番号:30)の活性と異なる活性を有するペプチドを生成するか否かを決定するために、我々は種々のアミノ酸置換を有する合成ペプチドを作製した。我々は、本明細書に開示した小脳微小エクスプラントアッセイで置換NRP-5RGペプチドをテストした。
アミノ酸配列REGRRDAPGRAGG-NH2(配列番号:30)を有するアミド化ペプチドのN-末端の最初の5つのアミノ酸内でのアミノ酸置換によって、配列REAAADAPGRAGG-NH2(配列番号:44)及びAAARRDAPGRAGG-NH2(配列番号:45)を有するペプチドを作製した。
PGRドメインにおけるアミノ酸置換はアミノ酸配列REGRRDAAAAAGG-NH2(配列番号:47)を有するペプチドを生じ、C-末端GGドメインの置換はアミノ酸配列REGRRDAPGRAAA-NH2(配列番号:46)を有するペプチドを生じた。上記の置換ペプチドの各々をテストし、その結果をNRP-5セグメントRG(配列番号:30;“NRP-5RG”)を用いて得られた結果と比較した。
図14Aは、配列番号:44は、最大効果が約1pMの濃度で観察される神経保護活性を有していた。右側の棒線(“標準13mer”と表示されている)はNRP-5RG(配列番号:30)を示す。NRP-5RGのアミノ酸3−5の置換はペプチドの活性プロフィルを有意には変化させなかった(図42A)。
図14Bは、配列番号:45が神経保護活性を有し、最大効果は約1pMで観察されることを示している。右側の棒線(「原型配列」と表示されている)はNRP-5RG(配列番号:30)を示す。AAARRDAPGRAGG-NH2(配列番号:45)を生じる1−3位の置換は、原型配列NRP-5RGよりもいっそう高い神経保護活性をもたらした。この結果は、NRP-5RGよりも高いこのアナログの安定性プロフィルによるものであろう。ただし他の仮説もこの観察の説明になりえる。
図42Cは、配列番号:46がNRP-5RP(配列番号:30;右の棒線;“標準13mer”)よりも、又は配列番号:44又は配列番号:45(それぞれ図14A及び14B)よりも低い効果を有することを示している。実際、統計的に有意な効果は濃度100nMで観察されたに過ぎない。原型配列(配列番号:30)は20%高い神経保護を生じ、配列番号:46よりも106から107倍高い潜在能力を有する。
図14Dは、配列番号:47はある種の濃度(例えば0.1pM、10pM及び100nM)ではある程度の神経保護作用を有するが、他の濃度ではNRP-5RG(配列番号:30;「標準」と標識された右の棒線)について観察されたものと比較して活性はなかったことを示す。
我々はこれらの実験から、PGR及びC-末端GGドメインは、NRP-5RG(配列番号:30)の活性の維持のために有用であると結論した。我々はまた、NRPのアミド化C-末端は神経保護作用を生じるために十分ではないと結論した。なぜならば、C-末端アミド化NRP(配列番号:46)は、何らかの神経保護作用があるとしても、大半の濃度でその作用はほとんどなかったからである。さらにまた、NRPの内部における変更は、たとえ内部のアミノ酸のいずれもC-末端アミド基を持っていないとしても、活性に影響を与えた。
【0082】
実施例14:NRP-5RGの増殖誘導及び化学誘引活性
また別の実験セットで、我々は、本明細書に記載した胎児の小脳微小エクスプラントにおける神経細胞増殖の誘導に対するNRP-5RGの作用を決定した。図15は、NRP-5RGが増殖誘導活性を有し、最大活性は100pMの濃度で観察されることを示している。ある程度の作用は1mPで、さらに0.1pMの濃度でさえ観察されるが、これらの作用は統計的には有意ではなかった。
さらに別の実験で、我々は、本明細書に記載した接触走性遊走アッセイでNRP-5RGの化学誘引作用を調べた。図16は、NRP-5RGが化学誘引性であり、コントロール(BSA含有)培養液に暴露された細胞と比較して42.1%強い作用を有することを示している。
【0083】
実施例15:NRPは酸化性ストレスに応答して神経細胞を保護した
この実験シリーズでは、我々は、NRP-4セグメントPQ(“NRP-4PQ”)は酸化性損傷から小脳微小エクスプラントを保護することができるか否かを調べた。我々は、ビヒクル、0.1mM過酸化水素、又はビヒクル中の種々の濃度のNRP-4PQに細胞を暴露した。図17は、0.1mMの過酸化水素に対する酸化性ストレスの48時間後のNRP-4PQの神経保護作用を示す。NRP-4PQ処理に付随する神経保護は、過酸化物を投与されなかったコントロールのエクスプラントの神経保護すら超えていた。神経突起成長のこの有効性強化のメカニズムは明確ではないが、前記は、微小エクスプラント自体を作製又は維持することに付随するストレスを軽減する可能性がある。
これらの発見は酸化的ストレスが役割を演じる多くの条件と関係がある。例えば、酸化的ストレスは慢性のCNS損傷及び疾患の全てに付随する。NRP作用による酸化的ストレス仲介神経毒性の抑制は、多くのCNS損傷又は疾患に対して非常に有益でありえる。
【0084】
実施例16:リン酸化NRP-7SWによって仲介される神経保護
我々は、一連の実験を実施して、N-末端アミノ酸セリンのリン酸化がNRP-7セグメントSW(「NRP-7SW」)の神経保護作用を変化させるか否かを決定した。NRP-7SWのリン酸化によってNRP-7PSWが生成される。セリンはin vivo条件下ではリン酸化される(NetPhos 2.0 Server−Technical University of Denmark)という高い確率予想(0.9)のために、我々はN-末端セリンの役割の仮説をたてた。小脳微小エクスプラントを0.5mMの3-NP/グルタメートで損傷させ、NRP-7PSWで処理した。
図18は、NRP-7PSWは神経保護作用を持つことを示す実験結果を示している。0.1pMという低い濃度で顕著な作用が観察され、前記濃度は、テストした他の濃度よりも高い神経保護活性(44.2%の神経保護)を示した。使用した最低の濃度(0.1pM)がかなりの神経保護作用を示したので、より低濃度でも神経保護作用が示されるであろう。
【0085】
実施例17:NRP-9SDによる分化の誘導後のNSCの増殖強化
E15マウスのNSC P10細胞をラミニン上に播種し、NRP-9SD(配列番号:34)の存在下で3日間培養し、培養期間の最後の48時間にはBrdUの存在下で培養した。各条件について、2つの別個のウェルで4視野を数え、増殖細胞数を決定した。
結果:
NRP-9SDは、分化中のマウスNSC前駆体細胞の増殖速度を高めた(図19)。この生物学的活性は100fMから1nMの広い用量範囲で観察された。
【0086】
実施例18:NRP IIの発現
in situハイブリダイゼーション:
全脳をE15及びE17マウスから摘出し、4%パラホルムアルデヒド(PFA)で3時間固定し、20%シュクロースで一晩凍結保護し、ティシュテック(Tissuetek)OCT(Sakura finetek)で包埋し-80℃で保存した。この凍結保護脳を14μmの厚さの切片に切り出し、PLL被覆スライドに静置し、8μg/malのプロテイナーゼKで8分処理し、4%PFAで5分後固定し、さらにDIG-標識NRPセンス及びアンチセンスプローブで45℃で一晩ハイブリダイズさせた(実施例13に記載の88merプローブ配列;センス鎖は配列番号:41、アンチセンス鎖は配列番号:42;1:100希釈)。抗DIG抗体での標識及びNBT/BCIPによるシグナルの発色後、切片をネスチン/GFAP/βIII-チュブリン及びヴィメンチンの1:100希釈(abcam, マウスモノクローナル[RV202], ab8978-1)で二重標識し、二次蛍光抗体を用いて可視化した。センスコントロールは陰性のままであった。
結果:
図20A及び20Bは上述の動物の脳の蛍光顕微鏡写真を示す。マウスのフレームシフトNRPメッセージと放射状グリアマーカー、ヴィメンチンとの同時局在は、NRP発現は神経上皮幹細胞-放射状グリア-星状細胞系列で維持されていることを示した(図20A及び20B)。このことは神経幹細胞の重要な機能を指摘している。なぜならば、放射状グリアは発生中に多くの脳領域でニューロンを生じることが示され(TE Anthony, C. Klein, G. Fishell, N. Heinz, 2004 ”Radial glial cells serve as neuronal progenitors in all regions of the central nervous system.” Neuron, 41:881-890)、成獣の脳室下ゾーンの星状細胞は神経幹細胞であることが示された(F. Doetsch, I. Caile, DA Lim, JM Garcia-Verdugo, A. Alvarez-Buylla, 1999 ”Subventricular zone astrocytes are neural stem cells in adult mammalian brain”, Cell 97:703-706”)からである。
これらの結果から我々は、NRP mRNAはマウスの脳で生成されると結論した。
【0087】
実施例19:配列番号:8及び配列番号:9の再配列決定
2004年8月20日に、配列番号:8及び配列番号:9に対応する更改配列の報告が刊行された(NBCI, NT_026446)。新しく同定された配列を基に、我々は、ヌクレオチドNRP配列は75アミノ酸を有するペプチドをコードする225ヌクレオチドから成るとアノテーションを付した。NRP(配列番号:8)をコードする染色体15上の以前にアノテーションを付された配列にしたがって、前記配列内で以下を変更した:168位はGからCに変更し、それ以外エクソン1−4は(NRPの生物学的に活性な配列を含めて)変更なし。179位から先へ向かって新規にアノテーションを付されたエクソン5が始まる。222位から先へ向かってエクソン6が始まる。
【0088】
全アミノ酸配列を一文字コードで記す:
>humchrom15NRPエクソン1 2 3 4 5 6
MAVVLLAPFGDISQEITKVGTGTPGRAEAGGQVSPCLAASCSQAYGAILAHCNLCLPGSSDLPASASQSARLQVD
配列番号:49
【0089】
実施例20:卒中の治療
患者は卒中の徴候を示している。卒中の診断が為され、続いて医師は、NRP化合物を患者に静脈内に、或いは脳室内に直接又は患者の脳の罹患部分に直接投与する。NRP化合物は本明細書に開示したようにペプチド又はタンパク質であり、医薬的に許容できる形態(所望する場合にはビヒクル、緩衝剤及び安定剤を含む)で投与される。NRPによる処置は、卒中に付随する神経変性を減少させ、さらに予想される症状の悪化を少なくとも部分的に遅らせるか、停止させるか、又は少なくとも部分的に回復させる。
【0090】
実施例21:NRPの予防的使用
患者は心脈管機能不全と診断され、冠状動脈バイパス(CABG)手術が指示される。CABG手術は脳灌流の低下を伴い、脳灌流の低下は低酸素性又は虚血性脳損傷をもたらしえる。そのような低酸素症及び虚血の悪影響を減少させるために、患者はNRP化合物で前処置される。NRP化合物は患者に医薬的に許容できる形態(所望する場合にはビヒクル及び/又は安定剤を含む)で投与される。投与ルートには静脈内、脳内、または脳室経由投与が含まれる。所望の場合は、マルチルート投与を用いることができる。CABG手術を受ける患者の前処置は、CABG手術に伴う神経変性を減少させ、患者は、NRP前処置を施されずにCABG手術を受けた患者と比較して、術後の神経学的損失が減少する。
本発明をその具体的な実施態様に関して記載した。当業者は、本出願の開示及び教示を取り入れた他の実施態様を面倒な実験なしに開発することができるであろう。それらの実施態様はいずれも本発明の部分と考えられる。本明細書に引用した全ての参考文献は引用により本明細書に含まれるものとする。
【0091】
参考文献
Akerblom, IE, and Murry, LE (1996). Human cachexia associated protein. US patent 5,834,192.
Anderson, CV, Wood, DM, Bigler, ED, and Blatter, DD (1996). Lesion volume, injury severity, and thalamic integrity following head injury. J. Neurotrauma 13: 35-40.
Anderson, WF (1992) Human gene therapy. Science 256: 808-813.
Bach et al., (1995) Insulin like growth factor binding proteins. Diabetes Reviews 3: 38-61.
Baldwin, ME, Roufail, S, Halford, MM, Alitalo, K, Stacker, SA and Achen, MG (2001). Multiple forms of mouse vascular endothelial growth factor-D are generated by RNA splicing and proteolysis. J Biol Chem 276: 44307-44314.
Beal, MF, Kowall, NW, Ellison, DW, Mazurek, MF, Swartz, KJ, and Martin, JB (1986). Replication of the neurochemical characteristics of Huntington's disease by quinolinic acid. Nature 321: 168-171.
Bolz, J, Novak, N, and Staiger, V (1992). Formation of specific afferent connections on organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12:3054-3070.
Brose, K, and Tessier-Lavigne, M (2000). Slit proteins: key regulators of axon guidance, axonal branching, and cell migration. Curr. Opin. Neurobiol. 10: 95-102.
Cote, F, Do, TH, Laflamme, L, Gallo, J-M, and Gallo-Payet, N (1999). Activation of the AT2 receptor of angiotensin II induces neurite outgrowth and cell migration in microexplant cultures of the cerebellum. J. Biol. Chem. 274: 31686-31692.
Cunningham, TJ, Hodge, L, Speicher, D, Reim, D, Tyler-Polsz, C, Levitt, P, Eagleson, K, Kennedy, S, and Wang, Y (1998). Identification of a survival promoting peptide in medium conditioned byoxidatively stressed cell lined of nervous system origin. J. Neurosci. 18: 7047-7060.
De Curtis, I, and Reichardt, LF (1993). Function and spatial distribution in developing chick retina of the laminin receptor a6b1 and its isoforms. Development 118: 377-388.
Dodd, J, Morton, SB, Karagogeos, D, Yamamoto, M, and Jessell, TM (1988). Spatial regulation of axonal glycoprotein expression on subsets of embryonic spinal neurons. Neuron 1: 105-116.
Dyke, MW, Bianchi-Scarra, G, and Musso, M (2001). Characterization of a triplex DNA-binding protein encoded by an alternative reading frame of loricrin. Eur. J. Biochem 268: 225-234.
Fallon, J, Reid, S, Kinyamu, R, Opole, I, Opole, R, Baratta, J, Korc, M, Endo, TL, Duong, A, Nguyen, G, Karkehabadhi, M, Twardzik, D, and Loughlin, S (2000). In vivo induction of massive proliferation, directed migration, and differentiation of neural cells in the adult mammalian brain. PNAS 97: 14686-1491.
Ferri, RT, and Levitt, P (1995). Regulation of regional differences in the differentiation of cerebral cortical neurons by EGF family-matrix interactions. Development 121: 1151-1160.
Fueshko, S, and Wray S (1994). LHRH cells migrate on peripherin fibers in embryonic olfactory explant cultures: an in vitro model for neurophilic neuronal migration. Dev. Biol. 166: 331-348.
Gahwiler, BH (1981). Organotypic monolayer cultures of nervous tissue. J. Neurosci. Methods 4: 329-342.
Ganzler, SI, and Redies, C (1995). R-cadherin expression during nucleus formation in chicken forebrain. J. Neurosci. 15: 57-72.
Gomez, TM, and Spitzer, NC (1999). In vivo regulation of axon extension and pathfinding by growth-cone calcium transients. Nature 397: 350-355.
Gulyas, AI, Hajos, N, and Freund, TF (1996). Interneurons containing calretinin are specialized to control other neurons in the rat hippocampus. J. Neurosci. 16: 3397-3411.
Guth, S, Tange, TO, Kellenberger, E, Valcarcel, J (2001). Dual function for U2AF35 in AG-dependent pre-mRNA splicing. Mol Cell Biol 21: 7673-7681.
Hatten, ME, and Heintz, N (1999). Neurogenesis and migration In Fundamental Neuroscience, (R: Zigmond, ed.), pp 451-479, Academic Press, San Diego.
Hermann, DM, Mies, G, Hata, R, and Hossmann, KA (2000). Microglial and astrocytic reactions prior to onset of thalamic cell death after traumatic lesion of the rat sensorimotor cortex. Acta Neuropathol (Berl) 99: 147-153.
Hughes, PE, Alexi, T, Williams, CE, Clark, RG, and Gluckman, PD (1999). Administration of recombinant human Activin-A has powerful neurotrophic effects on select striatal phenotypes in the quinolinic acid lesion model of Huntington's disease. Neuroscience 92: 197-209.
Hwang et al., (1980). Hepatic Uptake and degredation of unilamellar sphingomyelin/cholesterol liposomes: a kinetic study. Proc. of the Natl. Acad. Of Sciences USA 77: 4030-4034.
Ishii, N, Wadsworth, WG, Stern, BD, Culotti, JG, and Hedgecock, EM (1992). UNC-6, a laminin-related protein, guides cell and pioneer axon migrations in C. elegans. Neuron 9: 873-881.
Langer et al., (1981) Biocompatibility of polymeric delivery systems for macromolecules. J. Biomed. Mater. Res. 15: 27-277.
Liang, S, and Crutcher, KA (1992). Neuronal migration in vitro. Dev. Brain Res. 66: 127-132.
Lu, Q., Sun, E., Klein, R. S. and Flanagan I. G.(2001). Ephrin-b reverse signalling is mediated by a novel PDZ-RGS protein and selectively inhibits G-protein coupled in chemoattraction. Cell 105: 69-79.
Nakao, N, and Itakura, T (2000). Fetal tissue transplants in animal models of Huntington's disease: the effects on damaged neuronal circuitry and behavioural deficits. Prog. Neurobiol. 61: 313-338.
Obst, K, and Wahle, P (1995). Areal differences of NPY mRNA expressing neurons are established in the late postnatal rat visual cortex in vivo, but not in organotypic cultures. Eur. J. Neurosci. 7: 2139-2158.
Pasterkamp, RJ, Giger, RJ, Baker, RE, Hermens, WT, and Verhaagen J (2000). Ectopic adenoviral vector-directed expression of Sema3A in organotypic spinal cord explants inhibits growth of primary sensory afferents. Dev. Biol. 220: 129-141.
Paxinos, G, Toerk, I, Tecott, LH, and Valentino, KL (1991). Atlas of the Developing Brain. Academic Press: San Diego.
Polleux, F, Morrow, T, and Ghosh, A (2000). Semaphorin3A is a chemoattractant for cortical dendrites. Nature 404: 567-573.
Rozas, G, Liste, I, Lopez-Martin, E, Guerra, Mj, Kokaia, M, and Labandeira-Garcia, JL (1996). Intrathalamic implants of GABA-releasing polymer matrices reduce motor impairments in rats with excitotoxically lesioned striata. Exp. Neurol. 142: 323-330.
Sieg, F, Obst, K, Gorba, T, Riederer, B. Pape, H-C, and Wahle, P (1998). Postnatal expression pattern of calcium-binding proteins in organotypic thalamic cultures and in the dorsal thalamus in vivo. Dev. Brain Res. 110: 83-95.
Stoppini, L, Buchs, P-A, and Muller, D (1991). A simple method for organotypic cultures of nervous tissue. J. Neurosci. Methods 37: 173-182.
Van der Flier, A, Kuikman, I, Kramer, D, Geerts, D, Kreft, M, Takafuta, T, Shapiro, SS and Sonnenberg, A (2002). Different splice variants of filamin-B affect myogenesis, subcellular distribution, and determine binding to integrin β subunits. J Cell Biol 156: 361-376.
Wagener, R, Kobbe, B, Aszodi, A, Aeschlimann, D, and Paulsson M (2001). Characterization of the mouse matrilin-4 gene: A 5' antiparallel overlap with the gene encoding the transcription factor RBP-L. Genomics 76: 89-98.
Wagner et al., (1990) Transferrin-polycation conjugates as carriers for DNA uptake into cells. Proc. Natl. Acad. Sci. USA. 87: 3410-3414.
Wu et al., (1987) Receptor-mediated in vitro gene transformation by a soluble DNA carrier system. J. Biol. Chem. 262: 4429-4432.
Yamamoto M, Hassinger, L, and Crandall, JE (1990). Ultrastructural localization of stage-specific neurite-associated proteins in the developing rat cerebral and cerebellar cortices. J. Neurocytol. 19: 619-627.
Zhu, Y, Yu, T, Zhang, X-C, Nagasawa, T, Wu, JY, Rao, Y (2002). Role of the chemokine SDF-1 as the meningeal attractant for embryonic cerebellar neurons. Nature Neurosci. 5: 719-720.
【0092】
工業的応用性
本発明の実施態様は、神経変性又は神経細胞死を含む症状の治療に使用される組成物の製造に有用な神経再生ペプチド(NRP)のための遺伝子及びペプチドを含む。NRPによる治療は、ニューロンの生存、ニューロンの遊走、ニューロンの分化、神経突起伸長及び/又はニューロンの増殖を増加させる。アルツハイマー病、パーキンソン病、低酸素症、卒中及び冠状動脈バイパス手術のような症状は、本発明の組成物及び方法によって有効に治療されえる。
【図面の簡単な説明】
【0093】
【図1A】図1Aは、イントロンを有するNRP-7(配列番号:35)をコードする遺伝子の構造を、DNA修復タンパク質をコードする既知遺伝子の2bpフレームシフトとしてのNRP遺伝子の存在とともに示している。2つのCpGアイランドを含むプロモーター領域が強調されている(一方のCpGアイランドはNRPについて予想されたものであり、下流のもう一方はDNA修復タンパク質について予想されたものである)。NRP-7(配列番号:35)遺伝子産物を得るためのプライマーの位置が表示されている。
【図1B】図1Bは、マウスNRP(NRP-7ロング)とラットNRP(NRP-9)オルソログとの間の72.2%相同性を示している。赤い数字は、生物学的に活性なNRPドメイン内の対応する相同性を示し、他方、青い線は推定的なN-グリコシル化部位を示す。
【図1C】図1Cは、ヒト悪液質関連タンパク質とマウスNRP(NRP-7ロング)とのアラインメントを示す。両配列の全体にわたるロイシン、グリシン及びプロリンアミノ酸残基の保存に注目されたい。全体的な相同性は34.4%である。悪液質関連タンパク質の生物学的に活性なドメインは20位のシグナルペプチドの直後に位置し、一方マウスNRP(NRP-7ロング)の活性ドメインはアミノ酸83位で開始する。
【図1D】図1Dは、マウスNRPフラグメントとヒトトレホイルタンパク質Ps2との間の全体的な相同性は52.5%であることを示す。トレホイル因子ファミリー(TFF)のコンセンサス配列を規定する15アミノ酸残基のうち9アミノ酸がNRPに存在し、それによってトレホイル因子ドメインがNRPに存在することが確認される。
【図1E】図1Eは、マウスSDF-1αとマウスNRP(NRP-7ロング)タンパク質配列とのアラインメントを示す。両ニューロン化学誘引分子間には32.6%の中等度の相同性が存在する。
【図2】3-NP/グルタメートを用いた興奮毒性(excitotoxic)/酸化性損傷後のNRP-4セグメントGQ(配列番号:26)による生存誘導を示す。
【図3】3-NP/グルタメートを用いて損傷されたニューロンにおけるNRP-7セグメントSW(配列番号:24)による増殖誘導を示す。
【図4A】コーティングに10ng/mLのNRP-4GG(配列番号:29)ペプチドを用いたマウスNEB-5細胞による接触走性(haptotactic)遊走アッセイを示す。200,000個のMEB-5細胞を用いた。
【図4B】コーティングに10ng/mLのNRP-4 GG(配列番号:29)ペプチドを用いた初代マウス幹細胞による遊走アッセイの結果を示す。200,000細胞を播種した。
【図4C】1ng/mLのNRP-2KS(配列番号:23)ペプチドコーティングを用いたマウスNEB-5細胞による遊走アッセイの結果を示す。200,000細胞を播種した。
【図4D】10ng/mLのNRP-7SW(配列番号:24)コーティングを用いた初代マウス幹細胞による遊走アッセイを示す。400,000細胞を播種した。
【図4E】50ng/mLのNRP-4GG(配列番号:29)コーティングを用いた野生型PC-12細胞(200,000細胞/ウェル)による遊走アッセイを示す。
【図4F】野生型PC-12細胞及び1ng/mLのNRP-7SW(配列番号:24)コーティングとそれに続くマトリゲル/PDLコーティングを用いた遊走アッセイを示す。100,000細胞を播種した。
【図5A】図5A及び5Bは、NRPの存在下で7日間分化させた神経幹細胞の各培養ウェルでもっとも長い5本の軸索の中間の長さを示すグラフである。図5AはNRP-7SW(配列番号:24)及びIGF-1の影響を示す。
【図5B】図5A及び5Bは、NRPの存在下で7日間分化させた神経幹細胞の各培養ウェルでもっとも長い5本の軸索の中間の長さを示すグラフである。図5BはNRP-2KS(配列番号:23)、IGF-1又は分化培養液単独の影響を示す。コントロール培養液との有意差が、2-テール(2-tailed)スチューデントのt-検定によって決定されたとおり観察された。**p<0.01;***p<0.001
【図5C】図5C及び5Dは分化培養液単独の場合の最も長い軸索伸長の代表例を示す。
【図5D】図5C及び5Dは分化培養液単独の場合の最も長い軸索伸長の代表例を示す。図5Dは10nMのNRP-7 SW(配列番号:24)の存在下における最長の軸索の成長を示し、図33Dはコントロール条件を示す。
【図5E】図5E及び5Fは、ニューロンの分化を経たNSC細胞におけるNRP-7SW(配列番号:24)及びNRP-2KS(配列番号:23)の増加に対する影響を示すグラフである。
【図5F】図5E及び5Fは、ニューロンの分化を経たNSC細胞におけるNRP-7SW(配列番号:24)及びNRP-2KS(配列番号:23)の増加に対する影響を示すグラフである。
【図5G】ナノモル濃度のNRP-2KS(配列番号:23)が、マウス前脳の神経幹細胞由来の多能性幹細胞を犠牲にしてニューロン前駆体の生成を促進することを示している。
【図5H】分化実験スキームの開始前のNRP-9セグメントSD(配列番号:34)の未分化NSCへの投与を示す。NRP処理下ではBrdU陽性細胞のアップレギュレーションが生じるが、全体的な細胞数には明白な変化はない。
【図6A】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。 図6Aは、マウスNRPの発現は、E15マウスの胎児の脳組織の皮質(Ctx)、線条体(Str)及び嗅球(OB)で検出されたことを示す。しかしながら、そのレベルは、急性的に単離して不死化したマウス神経幹細胞と比較してはるかに低かった。
【図6B】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Bは、CNTFによる神経幹細胞の星状細胞への分化は、未分化幹細胞又はBDNFによるニューロンの分化と比較してNRP mRNA発現を顕著に増加させることを示す。
【図6C】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Cは、種々のE19マウス胎児組織の分析を示すゲルである。前記は、周産期前脳に由来する胎児の皮質、星状細胞系分化NSC及び星状細胞培養と比較して、肺を除きマウスのNRPの発現は非神経組織では低いか又は存在しないことを示している。
【図6D】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Dは、DNA修復タンパク質配列とオーバーラップしない88bpプローブによるノザンブロットハイブリダイゼーションを示す。前記によって、周産期星状細胞培養に由来するRNAで、サイズがほぼ0.8及び1.2kbの2つの別個のmRNAが検出される。
【図6E】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Eは、NSCを標識する、マウスNRPアンチセンスプローブによるin situハイブリダイゼーションを示す。
【図6F】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Fは、センスコントロールにおける特異的なシグナルの欠如を示す。
【図6G】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。 図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Gは、NSCにおけるNRP mRNA発現を示す。
【図6H】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Hは、図6Gで示されたNRP mRNA発現と同じ部位にネスチンが分布することを示す。
【図6I】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Iは、E15マウス前脳切片、脳室帯、皮質原基(特にサブプレート及び辺縁帯)の冠状のNRP mRNAのin situハイブリダイゼーションを示し、皮質プレートではハイブリダイゼーションの強度は低下する。
【図6J】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Jは、サブプレートから辺縁帯までの全長に広がるネスチン陽性細胞を示す。
【図6K】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Kは、図6Jのネスチン発現と同じ部位でのマウスNRPメッセージの同時発現を示す。
【図6L】図6Lは、損傷の非存在下及びミトコンドリア毒素3-ニトロプロピオン酸により惹起された損傷の6時間後におけるNT-2細胞のNRP-2(配列番号:4)遺伝子産物の発現を示す。遺伝子発現は、細胞が1時間以上3-NPで処理されたとき実質的に減少した。
【図7A】図7A及び7Bは、NRP-7セグメントSWによるニューロンの生存及び増殖誘導を示す。図7Aは、0.1pMから100nMの広い用量範囲にわたるNRP-7SW(配列番号:24)の神経保護活性に対する影響を示す。
【図7B】図7A及び7Bは、NRP-7セグメントSWによるニューロンの生存及び増殖誘導を示す。図7Bは、SDF-1はNRPと比較してほんの限定的な神経保護作用を有したことを示す。
【図8A】図8A及び図8Bは、NRPによるニューロン遊走誘導実験の結果を示す。図8Aは、ラット及びマウスの合成ペプチド由来NRP(NRP-9SD及びNRP-7SW)は、接触走性遊走アッセイで示されるように、ニューロン幹細胞(“NSC”)の誘引において化学誘引特性を表すことを示すグラフである。NRP-7SWの有効性はSDF-1のそれと類似したが、NRPはより高い潜在能力を示した。SDF-1とNRP-7SWの作用の規模は類似しているように見えるが、SDF-1の量(100nM)はNRP-7SWの量(0.2nM)よりも多い。したがってNRP-7SWはSDF-1よりも約500倍強力である。
【図8B】図8A及び図8Bは、NRPによるニューロン遊走誘導実験の結果を示す。図8Bは、皮質原基に向かって遊走する、中央神経節隆起由来の神経前駆体細胞の移動を示す脳OTCの顕微鏡写真である。
【図8C】1nMの濃度でNRP-7SWの最大効果を示すOTCアッセイの定量のグラフを示す。一方、SDF-1の能力は前記よりも低かった。スチューデントのt-検定を統計分析に用いた(*p<0.05、**p<0.01、***p<0.001;N=6)。
【図9A】図9A−9Dは、完全長のマウス組換えNRP-7ロング(配列番号:35)の発現実験及び機能実験の結果を示す。図9Aは、サイトメガロウイルス(“CMV”)プロモーターの制御下では、マウスNRP-7ロング(配列番号:35)遺伝子の0.8kbシグナルは、NRP遺伝子特異的88bpのcRNAプローブによって検出したとき、HEK細胞で極めて過剰発現されることを示すノザンブロットを示す。
【図9B】図9A−9Dは、完全長のマウス組換えNRP-7ロング(配列番号:35)の発現実験及び機能実験の結果を示す。図9Bは、NRP-7ロング(配列番号:36)がHEK細胞によって発現され、20kDaの分子量で移動することを示すウェスタンブロットである。
【図9C】図9A−9Dは、完全長のマウス組換えNRP-7ロング(配列番号:35)の発現実験及び機能実験の結果を示す。図9Cは、組換えMyc-NRP-7ロング(配列番号:35)−HEK細胞は、酸化性/興奮毒性損傷を受けた小脳微小エクスプラントに対し51%の神経保護を提供することを示すグラフである。
【図9D】図9A−9Dは、完全長のマウス組換えNRP-7ロング(配列番号:35)の発現実験及び機能実験の結果を示す。図9Dは、Myc-NRP-7ロング(配列番号:36)−HEK細胞は、培養皿の底に播種したとき、接触走性遊走アッセイでNSCを誘引するための化学誘引活性をもつことを示すグラフである。Myc−HEKコントロール実験と比較して、2倍以上の数の細胞が、インサートから培養皿の底へ誘引された。スチューデントt-検定を統計分析に適用した(微小エクスプラントについては***p<0.001;N=4、さらに接触走性遊走アッセイについては***p<0.001;N=6)。
【図10A】図10A−10Cは、CSCR4及びNRPに対する抗体を用いた、P4-小脳エクスプラントに関する実験の結果を示す。図10Aは、神経突起伸長に関するNRP-9SD(配列番号:34)の阻害を示すグラフである。
【図10B】図10A−10Cは、CSCR4及びNRPに対する抗体を用いた、P4-小脳エクスプラントに関する実験の結果を示す。図10Bは、神経突起伸長に関するNRP-2KS(配列番号:23)の阻害を示す。培養皿に被覆した10ng/mLのNRPの化学誘引作用は、ニューロン幹細胞株MEB-5をCXCR4に対する中和抗体と予備インキュベートすることによって完全に阻害された。
【図10C】図10A−10Cは、CSCR4及びNRPに対する抗体を用いた、P4-小脳エクスプラントに関する実験の結果を示す。図10Cは、CXCR4に対する抗体は、NRP-9SD(配列番号:34)だけで処理した細胞と比較してNRP-9SDの化学誘引作用を低下させたことを示す。
【図11A】図11A−11Cは、NRPの神経保護誘導及び遊走誘導活性に中心的に関与する可能なシグナリング連鎖事象を提案する、MAPK阻害物質を用いたNRPの作用に対する実験の結果を示す。図11Aは、MAPK(MEK)-阻害物質のPD98509は、種々のNRP濃度範囲にわたってラットのNRP-9SD(配列番号:34)の神経保護活性を完全に阻害することを示すグラフである。
【図11B】図11A−11Cは、NRPの神経保護誘導及び遊走誘導活性に中心的に関与する可能なシグナリング連鎖事象を提案する、MAPK阻害物質を用いたNRPの作用に対する実験の結果を示す。図11Bは、PD98509のように、PIK-3阻害物質のワートマンニンは、NRP-9SD(配列番号:34)の神経保護活性を阻害することを示すグラフである。
【図11C】図11A−11Cは、NRPの神経保護誘導及び遊走誘導活性に中心的に関与する可能なシグナリング連鎖事象を提案する、MAPK阻害物質を用いたNRPの作用に対する実験の結果を示す。図11Cは、NRP-9SD(4nM)によって誘導される遊走NSC数は15μMのPD98509によって阻害され、一方、BSA被覆ウェルにおける基礎遊走レベルは、MEK阻害物質によって顕著には変化しないことを示すグラフである(*p<0.05、**p<0.01、***p<0.001;N=8)。
【図12A】低酸素症の2時間後にICV投与したとき、NRP-4セグメントPQ(配列番号:43)は、損傷の5日後に分析した脳領域の全てにおいて実質的な神経保護を提供した実験の結果を示す。
【図12B】低酸素症の2時間後にICV投与したNRP-5セグメントRG(配列番号:30)は、低酸素障害の5日後に分析した脳領域の全てにおいて実質的な神経保護を提供した実験の結果を示す。
【図13】NRP-7セグメントSW(配列番号:24)は、ヒト成人嗅球鞘性グリア(ensheating glia)の初代培養の増殖を強化することを示す実験の結果である。
【図14A】図14A−14Dは、4つのNRP-5 RGアナログの神経保護活性の原型NRP-5RG配列に対する実験結果を示す。
【図14B】図14A−14Dは、4つのNRP-5 RGアナログの神経保護活性の原型NRP-5RG配列に対する実験結果を示す。
【図14C】図14A−14Dは、4つのNRP-5 RGアナログの神経保護活性の原型NRP-5RG配列に対する実験結果を示す。
【図14D】図14A−14Dは、4つのNRP-5 RGアナログの神経保護活性の原型NRP-5RG配列に対する実験結果を示す。
【図15】胎児小脳細胞におけるNRP-5 RGの増殖誘導作用のグラフを示す。
【図16】マウスNSC及び化学誘引物質としてNRP-5 RGを用いた接触走性遊走アッセイの結果を示すグラフである。
【図17】酸化ストレス(0.1mMの過酸化水素)の48時間後のNRP-4 PQの神経保護作用の結果を示すグラフである。
【図18】リン酸化NRP-7 SW(NRP-7 PSW)の神経保護作用を示すグラフである。
【図19】BrDU染色核に対するNRP-9SDの作用を示す実験のグラフである。
【図20A】図20Aおよび20Bは、神経上皮幹細胞−放射状グリア−星状細胞系列内におけるNRP-7(配列番号:35)の遺伝子発現の顕微鏡写真を示す。 図20Aは、E17マウス脳の大脳皮質におけるマウスフレームシフトNRP転写物の発現を示す。88merの特異的プローブを用い、凍結切片でのin situハイブリダイゼーションによって検出された。
【図20B】図20Aおよび20Bは、神経上皮幹細胞−放射状グリア−星状細胞系列内におけるNRP-7(配列番号:35)の遺伝子発現の顕微鏡写真を示す。 図20Bは、同じ切片で抗ビメンチン抗体を用いた光学顕微鏡法を示す。マウスフレームシフトメッセージとビメンチン中間体のフィラメントタンパク質の高度な同時局在が示されている。
【技術分野】
【0001】
関連出願:
本出願は、米国仮特許出願60/516,018号(2003年10月31日出願、発明の名称:神経再生ペプチド及びそれらの使用方法(”Neural Regeneration Peptides and Methods for Their Use”);発明者:Frank Sieg & Thorsten Gorba(Attorney Docket No: NRNZ 01023 US3 DBB))、米国仮特許出願60/585,041号(2004年7月2日出願、発明の名称:神経再生ペプチド:新規な種類の化学走性及びニューロン生存促進ペプチド(”Neural Regeneration Peptides: A New Class of Chemoattractive and Neuronal Survival Promoting Peptides”);発明者:Thorsten Gorba & Frank Sieg(Attorney Docket No: NRNZ 01023 US4 DBB))、及び米国仮特許出願60/616271号(2004年10月5日出願、発明の名称:神経再生ペプチド及び脳傷害治療におけるそれらの使用方法(”Neural Regeneration Peptides and Methods for Their Use in Treatment of Brain Damage”);発明者:Frank Sieg , Paul Edmond Hughes & Thorsten Gorba(Attorney Docket No: NRNZ 01023 US5 DBB))に対して優先権を主張する。上記出願の全てが参照により本明細書に含まれる。
配列リスト:
本出願は、(1)配列リストの印刷物及び(2)コンピュータ読出し可能形の配列リストを収納するディスクとして提出される配列リストを含む。本配列リストは参照により本明細書に含まれる。
【0002】
本発明は、ニューロンの遊走、増殖、生存、分化及び/又は神経突起伸長を促進するオリゴヌクレオチド及びペプチドの組成物並びにそれらの使用方法を目的とする。より具体的には、本発明は、そのようなペプチドの脳損傷及び神経変性疾患の治療における使用を目的とする。本発明はまた、神経細胞の増殖、遊走、神経突起伸長、生存及び/又は分化を検出する新規な方法を含む。
【背景技術】
【0003】
軽度から重度の外傷性脳損傷(TBI)及び限局性又は総体虚血は、障害後短時間内に重大なニューロン細胞消失及び脳機能低下を生じえる。これまでのところ、疾患によって引き起こされる頭部の損傷又は傷害の結果として脳に生じる細胞死を防止するために利用可能な治療はない。さらにまた今日まで、ニューロン機能を回復させるために利用できる治療も存在しない。慢性の神経変性疾患、例えばアルツハイマー病、パーキンソン病、ハンチントン病及び多発性硬化症に現時点で利用できる治療は対症療法のみである。疾患過程に介入するか又は細胞死を防ぐために利用可能な薬は現在のところない。
皮質-皮質下の非視床病巣は、損傷後3−7日で視床領域内のアポトーシスを生じえることはよく知られている。逆行視床変性は、視床内のアストログリア及びミクログリアの活性化を伴う(Hermann et al., 2000)。MRIのような非侵襲性技術は、非視床部の構造的病変を有するTBI患者においては視床の体積が小さいこと、及び脳室対脳比率値が増すことを明らかにした(Anderson et al., 1996)。これらの発見は、視床皮質系の興奮投射ニューロンは逆行性ニューロン細胞死に対して非常に脆弱であることを示し、したがって損傷された又は外傷を受けた視床ニューロンの救済手法の必要性を示している。
【0004】
視床内部の抑制性ニューロン回路の機能は、視床内部のニューロン活動の視床内ダウンレギュレーションにとって線条体系内部のそれと同様に必須である。ハンチントン病で生じる病変と類似する線条体病変を有する動物は、GABA-放出ポリマーマトリックスを視床に移植されたとき行動に関する所産の改善が認められることが示された(Rozas et al., 1996)。線条体内部の細胞レベルでは、カルビンジン免疫反応(“calbindin-ir”)抑制ニューロンはアクチビンAの投与によって救済できることが示された(Hughes at al., 1999)。
これまで、胎児線条体の移植を必要とする移植のみが、ハンチントン病の動物モデルにおいて運動機能の改善又は回復をもたらした(Nakao and Itakura, 2000)。ハンチントン病で傷害された視床及び線条体のGABA作動系の回復は、行動に関する所産を改善することができる(Beal et al., 1986)。
神経系発達の特徴は、前駆体細胞がそれらの正しい三次元的空間的配置へと広範囲に遊走していくことである。このような遊走は、一連の表現型分化及び未成熟ニューロンの脊椎脳への配置を促進する。およそ十兆個のニューロンを正確に結ぶために、より高次の皮質領域における層状構造の形成のような細胞性機構の構築が必要である(概説としてHatten and Heintzを参照)。
【0005】
遊走ニューロンの運動方向に対して相関関係をもつ細胞側のものは、単一遊走細胞内の一過性Ca2+変化の頻度及び大きさであろうが(Gomez and Spitzer, 1999)、ただし膜結合分子または拡散性分子によるニューロン細胞遊走の開始及び/又は実行の引き金も排除できない。
しかしながら神経突起伸長及びニューロンの遊走に必要な指示の多くは同定されている。インテグリンレセプターファミリーに属する形質膜分子は、ラミニンのような細胞外マトリックスリガンドと相互作用し、下層へのニューロン接着を開始させる(Liang and Crutcher, 1992; De Curtis and Reichardt, 1993)。インテグリン発現の制御は、遺伝子発現の調節、細胞接着、軸作成長及び細胞遊走を含む広範囲の発生プロセス及び細胞性プロセスに影響を与える。細胞遊走を促進する他のリガンドは、細胞接着分子(すなわちN-CAM;カドヘリン;TAG-1)、ラミニン様分子ネットリン-1、ニューロン-グリア接着リガンドアストロタクチン及び増殖又は神経栄養因子、例えばEGF、TGF-α、血小板活性化因子及びBDNFである(Dodd et al., 1988; Yamamoto et al., 1990; Ishii et al., 1992; Ferri and Levitt, 1995; Ganzler and Redies, 1995)。
最近、コラプシン-1(セマフォリン3A)が発見された。コラプシン-1は、一次感覚ニューロンに対する軸索誘導及び遊走パターンにおいて化学反発活性を有する(Pasterkamp et al., 2000)。対照的に、コラプシン-1は、新皮質領域の皮質先端突起を誘導するために化学誘引物質として機能する(Polleux et al., 2000)。軸索誘導に対する化学誘引作用と同様に同じような化学反発作用がスリット-1(拡散性タンパク質)によって示された(Brose et al., 2000)。
【0006】
これまでのところ、ニューロン移動の開始をもたらす連鎖事象、すなわちニューロンの付着からそれに続く遊走の開始、長距離に及ぶ遊走のプロセス(方向転換及び遊走停止シグナルを含む)は未だ説明されていない。
TGF-αが同時に投与された中脳病巣は、脳質下部帯(subventricular zone; “SVZ”)を起源とする多能性幹細胞の大規模増殖及びそれに続くこれら幹細胞の線条体への遊走をもたらす(Fallon et al., 2000)。しかしながら、SVZを起源とするニューロン前駆体の長距離遊走を防ぐために、病巣部位の極めて近傍に存在するニューロンの増殖及び遊走を活性化することが望ましい。
ニューロン遊走化学誘引物質として支質由来因子ケモカイン(SDF-1)を取り上げた報告が1件だけ存在する。胎児発現型のSDF-1は、小脳顆粒細胞を誘引して外側胚芽層から内側顆粒層へ遊走させる(Zhu et al., 2002)。それにもかかわらず、このケモカインは出生後の組織に対しては影響をもたない。脳外傷又は神経変性疾患後に成人における神経芽細胞又はニューロンの遊走行動に対して化学誘因性作用を誘導する遊走誘導因子は知られていない。
【発明の開示】
【0007】
発明の要旨
したがって、本発明の実施態様の目的は、脳損傷及び脳疾患の治療に新規なアプローチを提供することである。そのような実施態様は、神経の遊走、神経の成長、神経の増殖、神経の分化及び/又は神経の生存の1つ又は2つ以上を誘導することができるペプチドを含む。これらのペプチドは本明細書では“神経再生ペプチド”又は“NRP”と称される。他の実施態様には、脳損傷に続いて又は慢性神経変性疾患時に、1つ又は2つ以上のNRPを投与することが含まれる。“NRP”又は“NRP化合物”という用語には、NRP、NRPホモログ、PRPパラログ、NRPオルソログ及び/又はNRPアナログが含まれる。NRPは単独で、又は1つ若しくは2つ以上の他のNRPと一緒に、又は神経の成長、神経の遊走、神経の生存、神経の分化及び/又は神経の増殖を促進する他のタイプの薬剤と一緒に投与することができる。
NRP及び関連ペプチドは全般的に、所望の生物学的特性を前記分子に付与する一定のアミノ酸配列を有する。
一定のドメイン(下線を付して強調されている)を有するNRPペプチドのいくつかの実施態様は下記の表1に示されている。
【0008】
表1:神経再生ペプチド*
NRP-1: YDPEAAS-APGSGNP------CH
NRP-2KG: KDPEARR-APGSLHP------C--LAA-SCSAAG
NRP-3SF: SDSFKSQ-ARGQVPPFLGGVGCPWF
NRP-4GG: GTPGRAE-AGGQVSP------C--LAA-SCSQAYG
NRP-5RP2: RE--GRRDAPGRA--GGGG------AARSVSPSP
NRP-7SW: SEPEARR-APGRK----GGVVCASLAADW
NRP-8SG: SEVDARR-AKKSLH-------C-ILS-DTSHPRG
NRP-9SD: SEPEARRAQGGQIPSERVLSD
【0009】
いくつかの実施態様では、一般にNRPは約8から約25アミノ酸の鎖の長さを含み、約0.8から約2.7kDaの分子量を有する。さらにまた、他の実施態様では、NRPは約6.5から約10.0の等電点を有し、さらに神経の生存、神経突起伸長、ニューロンの増殖、ニューロンの分化及びニューロンの遊走から選択される成果を促進する少なくとも1つの生物学的特性を有することができる。さらにまた、NRPは、上記の表1の下線で示される1つ又は2つ以上のドメインを有することができる。いくつかの実施態様では、NRPは[A]PG[R,S]ドメインをPEドメインと一緒に有し(例えばNRP-1及びNRP-2)、また別にはPEドメインを含まない(例えばNRP-5、NRP-7)。[A]PG[R,S]ドメインの存在はNRPの生物学的特性のために望ましい。さらにまた、C-末端のGGドメインはNRPに所望の神経保護特性を付与することができる。したがって、また別の実施態様では、NRPは、[A]PG[R,S]ドメイン、[A,G]RRドメイン及びARGドメインから成る群から選択される第一のドメインを有することができ、所望の生物学的活性を有する。他の実施態様では、望ましくは、NRPは上述の第一のドメインに加えて、第一のドメインとは異なる第二のドメインを有することができる。第二のドメインはPEドメイン、[A,G]RPドメイン又はC-末端GGドメインでありえる。さらに別のある実施態様では、NRPは上述のドメインの第三のドメインを有することができる。
したがって、ある種の実施態様では、NRPは[A]PG[R,S]ドメインのみを含み、他のNRPはARGドメインのみを含み、さらに他のNRPは[G,A]RRドメインのみを含むことができる。さらにまた、他のNRPは、[A]PG[R,S]ドメイン及びPEドメインを有することができ、また他のNRPは[A]PG[R,S]ドメイン及び[G,A]ドメインを有することができる。なお別のNRPは[A]PG[R,S]ドメイン、[A,G]RRドメイン及びPEドメインを有することができる。
【0010】
NRPファミリーメンバーの遺伝子は、CAATボックス及びTATAボックスの少なくとも1つ、又はCAATボックス及びTATAボックスの両方をプロモーター領域内に一緒に含んでいる。NRPファミリーメンバー由来のオリゴヌクレオチドを用いて、NRPペプチドの発現を、そのような産生が所望される領域で高めることができる。
別の特徴では、本発明の実施態様は、頭部損傷又は慢性の神経変性疾患の結果としての脳の傷害領域のための治療方法を提供する。前記方法は、1つ又は2つ以上のNRP、NRPアナログ(構造的類似性を有するペプチドが含まれる)及び/又はNRPプロドラッグ(プロ-NRPペプチドを含む)を投与して、ニューロン又は神経芽細胞の遊走、増殖、生存及び/又は神経突起伸長を促進することによる。この治療方法は、皮質下領域の損傷と同様に新皮質の傷害を含む、軽度から重度の外傷性脳損傷(TBI)を受けた患者に特に有用であるが、ただしこれに限定されない。
ある実施態様では、NRP-2(配列番号:5)は、ゲノムクローンbA87G1(Sanger Sequencing Center)内のヒト染色体13のリバース鎖の塩基対77232−76768に位置する核酸配列によってコードされる。このペプチドはラットNRP-1のペプチドと類似の機能を有し、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走及び神経突起伸長を促進することができる。
【0011】
別の実施態様では、NRP-3(配列番号:3)は、ヒトゲノムの染色体3のリバース鎖上に、ダブルツィスト(Double Twist)アノテーションにしたがって塩基対34764−33003に位置する核酸配列によってコードされる。このNRPはまた、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
さらに別の実施態様では、NRP-4(配列番号:9)は、NCBIヒトゲノムアノテーションプロジェクトにしたがい、ヒト染色体15のフォワード鎖の塩基対21970003−21972239に位置する核酸によってコードされる。前記核酸配列から翻訳されるペプチドもヒトNRPファミリーに属する。この配列によってコードされるペプチドは、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
さらに別の実施態様では、NRP-5(配列番号:11)は、NCBIのアノテーションによって表示されているように、ヒト染色体7のリバース鎖の塩基対15047153−14824042に位置する核酸配列によってコードされる。この配列によってコードされるペプチドは、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
【0012】
NRPのまた別の実施態様は、染色体6のリバース鎖の116668725−116667697領域に位置するヒトゲノム由来のDNA配列に関するアノテーションが付されている(NCBIヒトゲノムアノテーションプロジェクトによる領域)。得られたペプチド、NRP-6(配列番号:13)は、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
NRPのさらに別の実施態様はげっ歯類で見出される。マウスのNRPは、リーディングフレーム1を用いる339の核酸から成るNCBIのアラクネ(arachne)contig_191157内に位置する核酸配列によってコードされる。オーバーラップする領域内に、フレーム3を用いるアノテーション付きNRPの29位で開始する、198の核酸の第二のORFが存在する。このORFは、切り縮められたヒトDNA修復タンパク質と高い同一性を有するタンパク質をコードする。得られたペプチド、NRP-7(配列番号:17)は、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
さらに別の実施態様はNRP-8(配列番号:20)であり、これもまた、マウスサンガー(Sanger)データベースのゲノムクローンbM344E9内のリバース鎖の核酸配列によってコードされるマウスペプチドである。前記タンパク質コード配列はアノテーションされており、塩基対5609−4052に位置する。NRP-8は、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
さらに別の実施態様はNRP-9(配列番号:28)である。前記はマウスNRP-7(配列番号:17)のラットオルソログNRPであり、ラット染色体の以下のエクソン内のリバース鎖に位置する核酸によってコードされる:7022614−7022326位に位置するエクソン1、7018685−7018651位に位置するエクソン2(NCBIデータベース)。NRP-9は、ニューロンの生存、ニューロンの分化、ニューロンの増殖、ニューロンの遊走又は神経突起伸長を促進することができる。
【0013】
また別の特徴では、本発明は、増殖誘導及び遊走誘導活性を判定するin vitroアッセイのための実施態様を含む。最近まで、長期にわたってタグのないニューロンの遊走を検出することができる、利用可能なin vitroのニューロン遊走アッセイはほとんどなかった。これらのバイオアッセイの1つは、器官型組織培養(“OTC”)として構築された嗅板周辺細胞を5日間のタイムコースでモニターする(Fueshko and Wray, 1994)。本発明のある種の実施態様では、成人の視床皮質系OTCを利用するin vitroアッセイを用いることによって、NRPを、ニューロンの遊走、ニューロンの増殖、ニューロンの分化、ニューロンの生存及び/又は神経突起伸長を誘導するそれらの能力について評価することができる。これらの実施態様は以下の理由から特に有用である:1)制御条件下では、組織培養基質上で2つの器官を互いに物理的に十分に遠く(約3から約5mm)分離させることによって両培養器官(例えば視床と皮質)の間の細胞架橋の形成を回避することができる、さらに2)出生後は、視床内のニューロン遊走は、視床の個体発生のタイムコースのために実質的に完了しているからである。これらのバイオアッセイは、したがってニューロン遊走誘導因子のスクリーニング及び同定に非常に適切であろう。
ある種の実施態様では、in vitro視床皮質系OTCアッセイは、皮質内の放射状遊走及び視床内の誘導による接線方向遊走の両方を明らかにするという利点を含む。in vitro制御条件下では、新皮質の正常な個体発生のタイムコースのために本来的な皮質内放射状遊走のみを観察することができる。
【0014】
他の実施態様では、基質に付着させた小脳の微小エクスプラントを含むin vitroバイオアッセイが提供される。これらの実施態様は、ニューロン遊走パターンに関するデータ(遊走ニューロンの数及び微小エクスプラントに対する遊走距離の定量を含む)を提供するために用いることができる。
培養からわずか2−3日後に、全体的な細胞遊走の強化と同様に小ニューロンから成る遊走鎖の発生を観察することができる。このアッセイによって得られる結果は、視床皮質系OTC内の細胞鎖誘導に類似する。
本発明の別の特徴に関する実施態様は、神経変性疾患及び脳損傷を治療又は防止するためにNRPを使用することを含む。具体的には、NRPは、損傷領域又は疾患領域近くに休止ニューロン幹細胞を欠く脳領域での使用に特に適している。
予防薬としてNRPを使用することは、折衷的外科手術、例えば冠状動脈バイパス移植法(CABG)又は脳への酸素供給が損なわれることを含む他の手術方法で有用でありえる。さらにまた、NRPは、例えば卒中、外傷又は脳若しくは脊髄への酸素付加を損なう他の損傷によって惹起される急性脳損傷の治療に有用でありえる。さらにまた、放射線療法または化学療法の前に予防的措置を実施することもできる。
【0015】
NRP化合物は、出生後の分化した神経組織内でニューロンの増殖、ニューロンの遊走、ニューロンの生存及び/又は神経突起伸長を開始させることができる。これらのペプチドは、軽度から重度の外傷性脳損傷(“TBI”)(びまん性軸索損傷、低酸素性虚血性脳障害及び頭蓋脳外傷の他の形態を含む)をもつ患者の損傷領域内のニューロン回路の改善又は修復を目的とする治療方法で利用することができる。NRP化合物は、神経系の感染(例えば一般的な細菌性髄膜炎)の治療、及び卒中(虚血性梗塞、塞栓症及び出血(例えば低血圧出血)及び他の原因によって惹起されるものを含む)の治療のために用いることができる。さらにまた、NRP化合物は、神経変性疾患(アルツハイマー病、レーヴィ小体痴呆、パーキンソン病(PD)、ハンチントン病(HD)、多発性硬化症、運動ニューロン病、筋ジストロフィー、末梢神経障害、神経系の代謝異常(糖原病を含む)、及びニューロンが傷害又は破壊される他の症状を含む)の治療に有用でありえよう。
本発明のある種の実施態様では、我々は、NRP遺伝子ファミリーのメンバーは、哺乳類の幹細胞(不死化幹細胞及び幹細胞の初代培養の両者で)で発現されることを見出した。
【0016】
他の実施態様では、我々は、NRPは、幹細胞の神経前駆細胞(神経芽細胞)への分化を促進することができることを見出した。また別の実施態様では、NRPは化学誘引物質に応答して幹細胞の遊走を刺激することができ、成熟ニューロンの形態(例えば軸索)を有する細胞への神経芽細胞の分化を促進することができ、さらに分化した神経芽細胞から神経突起(例えば軸索及び樹状突起)の成長を促進することができる。
本発明の他の特徴の実施態様は、NRPを使用して嗅覚細胞の増殖を促進することを含む。
したがって、NRPは、損傷神経細胞を修復し、神経組織の集団を再形成し、ニューロンの分化を促進し、シナプス形成(例えば神経突起伸長及び/又は神経分化)の促進に必要な過程を支援するために重要な治療的ツールとなりえる。
他の実施態様では、NRPと組み合わせた幹細胞の外科的移植を用いて、神経組織の集団を再形成することができる。幹細胞また別には神経芽細胞とNRPとを組合せることによって、神経組織の再増殖を促進することができる。そのような方法は成熟神経組織の再形成をもたらし、したがって、神経変性症状の治療に用いることができる。前記のような症状には、低酸素症/虚血、卒中、心臓バイパス移植術、アルツハイマー病、パーキンソン病、ハンチントン病、筋萎縮性側索硬化症、多発性硬化症、及び神経細胞の死又は変性を伴う他の疾患が含まれる。
本発明はその具体的な実施態様を記載することによって説明されるであろう。本発明の実施態様の他の目的、特徴及び利点は詳細な説明及び図面から明らかとなっていくであろう。
【0017】
定義
“ホモログ”という用語には、1つ又は2つ以上の遺伝子であって、前記遺伝子の配列が、進化上の関係(種間(オルソログ)又は種内(パラログ)のいずれかにおける関係)のために有意な関連を有する遺伝子が含まれる。ホモログはまた、共通の祖先のDNA配列から後世に伝えられることによって関係を有する遺伝子を含む。ホモログはまた、種の形成事象によって分離された遺伝子間の関係、又は遺伝的複製事象による遺伝子間の関係(パラログ参照)も含む。本明細書で用いられるように、“ホモログ”という用語はまた、進化上の関係により互いに関連する遺伝子産物も含む。保存的アミノ酸配列ドメインを有するNRPはホモログの例である。
“パラログ”という用語には、遺伝的重複の結果として互いに枝分かれした相同な遺伝子セットが含まれる。例えば、マウスのαグロビン及びβグロビンはパラログである。本明細書で用いられるように、“パラログ”という用語はまた、進化上の関係により互いに関連する遺伝子産物も含む。保存的アミノ酸配列ドメインを有するヒトNRPはパラログの例である。
“オルソログ”という用語には、種の形成の結果として互いに枝分かれした相同な遺伝子セットが含まれる。例えばマウス及びニワトリのαグロビン遺伝子はオルソログである。本明細書で用いられるように、“オルソログ”という用語はまた、進化の関係により互いに関連する遺伝子産物も含む。保存的アミノ酸配列ドメインを有するヒト及びマウスのNRPはホモログの例である。
“パラログペプチド”という用語には、パラログヌクレオチド配列によってコードされるペプチドが含まれる。
【0018】
“ペプチド”及び“タンパク質”という用語はアミノ酸で構成されたポリマーを含む。
“プロドラッグ”という用語には、酵素的、代謝的プロセッシング又は他のプロセッシングの結果、活性なNRP、活性なNRPアナログ又は活性なNRPパラログを生じるプロペプチドを含む分子が含まれる。
“NRP化合物”という用語には、NRP、NRPホモログ、NRPパラログ、NRPオルソログ、NRPアナログ及びNRPのプロドラッグが含まれる。
“NRP”という用語は、進化上の関係にかかわらず、神経の遊走、神経芽細胞の遊走、神経の増殖、ニューロンの分化、ニューロンの生存及び神経突起伸長の1つ又は2つ以上を含む機能を有するペプチドを含む。
アミノ酸は標準的な記号によって表される。この場合、アラニンは、“A”又は“Ala”で、アルギニンは“R”又は“Arg”で、アスパラギンは“N”又は“Asn”で、アスパラギン酸は“D”又は“Asp”で、システインは“C”又は“Cys”で、グルタミン酸は“E”又は“Glu”で、グルタミンは“Q”又は“Gln”で、グリシンは“G”又は“Gly”で、ヒスチジンは“H”又は“His”で、イソロイシンは“I”又は“Ile”で、ロイシンは“L”又は“Leu”で、リジンは“K”又は“Lys”で、メチオニンは“M”又は“Met”で、フェニルアラニンは“F”又は“Phe”で、プロリンは“P”又は“Pro”で、セリンは“S”又は“Ser”で、スレオニンは“T”又は“Thr”で、トリプトファンは“W”又は“Trp”で、チロシンは“Y”又は“Tyr”で、さらにバリンは“V”又は“Val”で表される。カルボキシ末端がアミド化されたペプチドは-NH2によって表示される。
【0019】
核酸は、アデニン(“a”で表される)、チミン(“t”で表される)、シトシン(“c”で表される)、及びグアニン(“g”で表される)を含むヌクレオチドで構成される。グアニン又はアデニンのどちらでもよいヌクレオチドは“r”で表され、チミン又はシトシンのどちらでもよいヌクレオチドは“y”で表され、さらにグアニン、アデニン、シトシン又はチミンのいずれかでありえるヌクレオチドは“n”で表される。ポリヌクレオチドはDNAでもRNAでもよく、一本鎖でも二本鎖でもよい。ポリヌクレオチドがRNAポリヌクレオチドである場合、ウラシル“u”がチミンの代用となりえる。
“疾患”は、動物のCNS又は末梢神経系の健常ではない任意の状態を含み、前記には特にパーキンソン病、レーヴィ小体痴呆、ハンチントン病、アルツハイマー病、多発性硬化症、運動ニューロン疾患、筋ジストロフィー、末梢神経障害、神経系の代謝異常(糖原病を含む)が含まれる。
“損傷”は動物の任意の急性傷害を含み、前記には特に卒中、外傷性脳損傷、低酸素症、虚血、(例えば胎盤剥離、臍帯閉塞に続く)胎児ジストレスに付随するか、又は子宮内発育遅延に付随する周産期仮死、適切な救急蘇生又は吸引の失敗に付随する周産期仮死、以下に付随する重篤なCNS障害(溺死、幼児突然死、一酸化炭素の吸入、アンモニア又は他のガス中毒、心停止、昏睡、髄膜炎、低血糖及びてんかん重積持続状態)、冠状動脈バイパス手術に伴う脳低酸素エピソード、低血圧エピソード及び高血圧危機、脳外傷及び脊椎損傷)が含まれる。
【0020】
具体的な実施態様の記載
本発明の実施態様は、神経再生ペプチド(NRP)を包含する、脳傷害の治療のための組成物及び方法を含む。NRPは、ニューロンの遊走、神経突起伸長、神経の分化、神経の生存及び/又は神経の増殖を誘導する。NRPはNRPアナログ、パラログ、オルソログ及び/又はNRPプロドラッグ、並びにヒト、マウス又は他の種の遺伝子によってコードされるペプチドでありえる。本明細書に記載されるNRPのいくつかは、本明細書に記載の遺伝子に対応する、以前に配列決定されていたオリゴヌクレオチドを基に予想されるタンパク質配列によるものである。他のペプチドは合成されたものであり、さらに少なくともいくつかはC-末端がアミド化された形態で提供される。しかしながら非アミド化型のタンパク質及びペプチドも本発明の範囲内に包含されることは理解されよう。
ラットNRP-1のヌクレオチド配列(配列番号:1)及びアミノ酸配列(配列番号:2)は以下のとおりである:
9 18 27 36
5' tat gat cca gag gcc gcc tct gcc cca gga tcg ggg
Tyr Asp Pro Glu Ala Ala Ser Ala Pro Gly Ser Gly
45
aac cct tgc cat 3' 配列番号:1
Asn Pro Cys His-NH2 配列番号:2
【0021】
しかしながら、遺伝暗号の縮退のために、多くのコドンが同じアミノ酸をコードしえる。したがって、多様な核酸配列が同じアミノ酸配列をコードすることができる。これら変型の各々は翻訳されて配列番号:2を生じることができ、したがってこれら変型のいずれも本発明の範囲内に包含される。例えば、多くの核酸配列(配列番号:1に挙げた核酸配列を含む)によってラットのNRP-1アミノ酸配列がコードされる。本発明はさらに配列番号:1のヌクレオチド配列の変種を含む。前記には前記核酸によってコードされるアミノ酸配列を保存する変種とともに、ラットのNRP-1アナログ及びNRP-1オルソログ及び/又はパラログをコードする核酸配列が含まれる。単なる例示として、DNAのための遺伝暗号にしたがって配列番号:1の変種を下記に示す(ここで各コドンは隣のコドンからスペースによって分けられ、“/”の後の核酸は“/”の前の核酸の変種を示す):
5' tat/c gat/c cca/t/c/g gag/a gcc/g/a/t gcc/g/a/t tct/a/c/g gcc/g/a/t cca/t/c/g gga/t/c/g tcg/a/t/c ggg/a/t/c aac/t cct/a/c/g tgc/t cat/c 3'
上記の配列(表示の変種を含む)は、上記の定義に従ってr、y及びnの文字を用いて表し、以下の配列を提供することができる:
5' tay gay ccn gar gcn gcn tcn gcn ccn ggn tcn ggn aay ccn tgy cay 3' 配列番号:3
【0022】
他のNRPをコードする他のヌクレオチド配列も遺伝暗号の縮退にしたがって変動しえることは理解されよう。さらにまた、DNA同様にRNAも本発明のペプチドをコードすることができ、さらに核酸がRNA核酸の場合はウラシルによってチミンを置換することができる。
ヒトの遺伝子は、鋳型としてヒトの悪液質のcDNA(米国特許5,834,192号)を用いてアノテーションが付されている。生存促進ペプチドは生存促進ペプチド(Cunningham et al., 1998)と96%を超える同一性を有し、さらにラットNRP-1は前記悪液質タンパク質と100%同一性を有し、既知の対応するcDNAとのただ1つのNRP-1ホモログである。ヒト悪液質タンパク質は染色体12の塩基対621841−625428の領域内に位置し、5つのエクソンから成る。我々は、悪液質のmRNAのスプライシング部位を、同定した染色体13上のNRPヒトパラログ(サンガー配列決定センターから入手したゲノムクローン:塩基対77232−76768)と比較し、NRP−1ヒトオルソログのコード領域をアノテーションした(このオルソログは本明細書ではNRP-2と称する)。NRP-2に関連するヌクレオチド及びアミノ酸配列は以下のとおりである:
【0023】
配列番号:4及び5
下線を付したヌクレオチド配列はシグナルペプチドを示す。
【0024】
前記タンパク質をコードするDNA配列は、鋳型として染色体12上のヒト悪液質遺伝子(米国特許5,834,192号のcDNA)のパラログ型の配列を用いるスプライシング部位分析によって予測したように4つのエクソンから成る。ゲノムクローンbA87G1の染色体地図は、正確なエクソンの位置決定のための基礎と考えられる。エクソン1はbp77232−77170の間に位置する。エクソン2はbp77088−77046の間に位置する。エクソン3はbp77036−76824の間に位置する。エクソン4は塩基対76778−76768の間に位置し、その後に翻訳終止コドンTAAが続く。翻訳されたタンパク質は110アミノ酸から成り、長さがヒトの悪液質タンパク質と同一で、ヒト悪液質タンパク質と24.5%の全体的同一性を有する。両タンパク質の細胞外局在化のためのシグナルペプチド(アミノ酸1−19)の配列比較によって、31.6%の同一性が明らかになった。重大なことには、成熟(切断)ペプチドの最初の30アミノ酸の比較によって、46.7%のアミノ酸同一性が明らかになった。さらにまた、このペプチドは、NRP-1と同様なニューロンの遊走、増殖、生存及び神経突起伸長活性を有する(図16、17及び18参照)。
NRP-1の第二のオルソログがアノテーションされている。前記は、染色体3の相補性リバース鎖の塩基対34764−33033の間(ダブルツィストヒトゲノムアノテーションプロジェクトによる領域)に位置するヒトゲノム由来のDNA配列によってコードされる。このタンパク質コード配列は、以下のように配置される5つのエクソンから成る:エクソン1、34764−34743;エクソン2、34729−34700;エクソン3、33745−33596;エクソン4、33498−33459;エクソン5、33043−33003。このヌクレオチド配列(配列番号:6)は333のヌクレオチドを有し、アミノ酸配列(配列番号:7;本明細書ではNRP-3と称される)は111のアミノ酸を有し、以下のように表示される:
【0025】
配列番号:6及び7
【0026】
これらの配列はNRPのヒト遺伝子ファミリーに属し、本明細書ではNRP-3と称される。この配列は、ヒト悪液質付随タンパク質と50%同一性及び67.2%類似性を有する。さらにまた、このヌクレオチド配列によってコードされるペプチドはNRP-1と類似の特性を有する。
第三のNRP-1オルソログがアノテーションされており、ヒト染色体15のフォワード鎖の領域21970003−21972239間に位置する、ヒトゲノム由来のDNA配列に含まれている(NCBIヒトゲノムアノテーションプロジェクトによる領域である)前記タンパク質コード配列は、以下のように配置されている6つのエクソンから成る:エクソン1、21970003−21970031;エクソン2、21970515−21970545;エクソン3、21970571−21970644;エクソン4、21970818−21970861;エクソン5、21971526−21971731;エクソン6、21972189−21972239。この遺伝子は再度配列決定され、今では下記の配列番号:48及び配列番号:49で表されている。しかしながら、遺伝子の再配列決定で、それによりコードされるNRPは変わらなかった。前記配列は、145のアミノ酸をコードする435の核酸から成る。このヌクレオチド配列(配列番号:8)及び予想されるアミノ酸配列(配列番号:9、本明細書ではNRP-4と称される)は以下のとおりである:
【0027】
配列番号:8及び9
【0028】
この配列はヒト遺伝子ファミリーNRPに属する。この配列はヒト染色体13に位置する核酸配列によってコードされるNRPと45%のアミノ酸類似性を有する。トリプレット244−246(アミノ酸82位);トリプレット391−393(アミノ酸131位)及びトリプレット421−423(アミノ酸141位)は潜在的なN-グリコシル化部位をコードする。アミノ酸118位は核酸配列内の不明確さのためにxである。ペプチド、NRP−4は、神経の増殖促進活性、神経突起伸長及びニューロンの生存促進活性を示す。オリゴヌクレオチド353−354位では、このヌクレオチドは決定されておらず、対応するアミノ酸は判明していないことに留意されたい。しかしながら、上記配列の発表の後で、正確な配列が提供され、実施例26の配列番号:48及び配列番号:49として本明細書では記載されている。配列情報におけるこの変更は、配列番号:8又は配列番号:9に由来するペプチドの神経保護作用に関連する部位に影響を与えるようには見えないことを記しておく。
ラットNRP-1のまた別のヒトオルソログ(“NRP-5”)は、リバース鎖に関するNCBIデータベースのホモ・サピエンス染色体7の作業ドラフト内に存在するDNA配列によってコードされる。このタンパク質コード配列はアノテーションを付されてあり、266のアミノ酸をコードする、全長が798の核酸を有する3つのエクソンから成る。タンパク質コードエクソンの正確な配置は以下のとおりである:エクソン1、15047153−15046815;エクソン2、14897885−14897772;エクソン3、14824386−14824042。このmRNAが発現されているというヒトEST(GenBank AW138864)での形跡がある。ヌクレオチド配列(配列番号:10)及びアミノ酸配列(配列番号:11、NRP−5)は以下のとおりである:
【0029】
配列番号:10及び11
【0030】
完全なタンパク質NRP-5は266のアミノ酸から成る。アノテーションされている翻訳されたNRPアミノ酸配列NRP-5は、染色体3に位置する分泌に関するヒトカルシウム依存アクチベータータンパク質(GenBankXP_036915)と76%の類似性を有する。さらにまた、翻訳されたヒト染色体7のNRP-5のエクソン1(339核酸)は、翻訳されたマウス5'EST(RIKENBB632392)と95.5%相同性を有する。このタンパク質は、神経突起伸長、ニューロンの生存、ニューロンの増殖及びニューロンの遊走という生物学的特性を示す、NRP-1及び他のNRPに存在するドメインを共有する。
我々は、染色体6の相補性リバース鎖の領域116668725−116667697間に位置するヒトゲノム由来のDNA配列にアノテーションを付した(NCBIヒトゲノムアノテーションプロジェクトによる領域)。このタンパク質コード配列は、以下のように配置された3つのエクソンから成る:エクソン1、116668725−116668697;エクソン2、116668333−116668305;及びエクソン3、116667872−116667697。前記配列(本明細書ではNRP-6と称される)は、78アミノ酸をコードする234の核酸から成る。この配列はNRPのヒト遺伝子ファミリーに属する。ヒトESTに対して見出された最高の相同性によって、ヒト胎盤組織から単離されたヒトcDNAクローンGenBankCS0DK001Y119と比較して核酸59−234の独自性が提示される。このクローンは3'-プライム末端から配列が決定され、924の核酸からなっている。我々のホモログは234個の核酸の後ろに終止コドンTGAを有する終端を形成するので、我々はこの遺伝子産物を対象としていない。NRPをコードするこのヌクレオチド配列(配列番号:12)及びこのペプチドのアミノ酸配列(配列番号:13、NRP-6)は以下のとおりである:
【0031】
配列番号:12及び13
【0032】
NRP-6のアミノ酸配列は、ヒト染色体13上のアノテーションを付されたNRPパラログ、NRP-2に対して14.1%の同一性及び44.9%の類似性を有する。このタンパク質は、神経突起伸長、ニューロンの生存、ニューロンの増殖及びニューロンの遊走という生物学的特性を有するNRP-1及び他のNRP(例えばNRP-2から5)に存在するドメインを共有する。
さらにまた、別のNRP-1オルソログ、マウスNRPファミリーメンバーが同定された。このマウスNRPファミリーメンバーは(ここではタンパク質2、配列番号:17と表示する;本明細書ではNRP-7と称される)は、NCBIのアラクネcontig_191157内に位置し、リーディングフレーム1を使用する339核酸から成る。オーバーラップする領域内には、フレーム3を使用する前記アノテーション付パラログの29位で開始する198核酸の第二のORFが存在する。このORFは切り縮められたヒトDNA修復タンパク質と高い同一性を有するタンパク質(ここではタンパク質1と表示する)をコードする。生物学的に活性なNRPペプチド配列:KDPEARRAPGSLHPCLAASCSAAG-NH2(配列番号:18)を使用し検索パラダイムtBLASTIを利用することによって、我々はマウスのEST RIKENデータベースでブラストヒットを得た。この5'-生成マウスESTはアクセッション番号GenBankAK012518及び以下の配列(配列番号:14)を有する:
【0033】
5'- GGCAGCCTCGAGATGGGGAAGATGGCGGCTGCTGTGGCTTCATTAGCCACGCTGGCTGCAGAGCCCAGAGAGGATGCTTTCCGGAAGCTTTTCCGCTTCTACCGGCAGAGCCGGCCGGGGACAGCGGACCTGGGAGCCGTCATCGACTTCTCAGAGGCGCACTTGGCTCGGAGCCCGAAGCCCGGCGTGCCCCAGGTAGGAAAGGAGGAGTAGTGTGTGCCAGCCTAGCGGCCGACTGGGCCACCCGAGACTGGGCCGCCTCCGGGCCGGCTTTGGAGGGAAGCCCCTGCTGGGCCTGTCCAGTGAGCTGTAATGTCGAGCGATGAGCGACCAGCTGCCTCGCTGTCCCAACGCTCTGGCCACGGCTTGTGCCTTGCCGCCATTTCCCCCAACCCACGCGGGCCACGGCTTGTGCCCTGCCGCCATTTCCCCCAACCCACGCGACCTTGCTC - 3'配列番号:14
【0034】
タンパク質1のリーディングフレーム3
オープンリーディングフレーム3(ESTの13位で開始する198個の核酸のORF)の翻訳によって、以下のタンパク質配列(配列番号:15)が明らかになった:
MGKMAAAVASLATLAAEPREDAFRKLFRFYRQSRPGTADLGAVIDFSEAHLARSPKPGVPQVGKEE 配列番号:15
この配列は、GenBankアクセッション番号Q13686のヒトアルキル化DNA修復タンパク質と82%のアミノ酸配列相同性(同一性及び化学的類似性)を有する。マウス型はC-末端が短縮されており、ヒトDNA修復タンパク質の389アミノ酸のうち66アミノ酸を有するだけである。
タンパク質2のリーディングフレーム1
323もの核酸をもつ長いORFがEST配列のフレーム1内に見出すことができる。続いて、我々は、マウスゲノム内の前記323核酸のORFの5'末端をアノテーションし、NCBIデータベースのマウスアラクネcontig_191157配列の23970と24374との間に位置する新規な遺伝子を発見した。このタンパク質コード配列は、113アミノ酸をコードする全長が339核酸を有する2つのエクソンから成る。エクソン1の位置は23970−23990であり、エクソン2の位置は24057−24374である。ラットNRP-1のこのマウスNRPオルソログのヌクレオチド配列(配列番号:16)及びアミノ酸配列(配列番号:17、NRP-7)は以下のとおりである:
【0035】
配列番号:16及び17
【0036】
発現された完全なNRP-7のアミノ酸配列は113のアミノ酸を含む(配列番号:17)。NRP-7のまた別のバージョンはまた別のスプライス型であり、配列番号:16の21位の後にさらに別の66ヌクレオチドを含み(配列番号:35)、前記は135アミノ酸を有する長型のNRP-7(“NRP-7ロング”)(配列番号:36)を生じる。
【0037】
【0038】
タンパク質機能プログラムツールSMARTは28アミノ酸から成る単一ペプチド配列を予想する。このタンパク質は、ヒト染色体13のNRPオルソログに対して13.6%同一性及び23.6%類似性を有し、さらにNRP-1に類似するニューロン生存、遊走、増殖及び成長活性を有する。
第二のマウスNRPファミリーメンバーは、マウスサンガーデータベースのゲノムクローンbM344E9内の相補性リバース鎖に位置する。生物学的に活性なNRPペプチド配列:KDPEARRAPGSLHPCLAASCSAAG-NH2(配列番号:18)を使用し検索プログラムtBLASTNを利用することによって、我々は、マウスゲノムサンガーデーターベースでゲノムクローンbM344E9内に類似性をもつ領域を得た。このタンパク質コード配列はアノテーションを付されてあり、5つのエクソンから成っている。前記は、141アミノ酸をコードする全長が423個の核酸である。コードエクソンの配置は以下のとおりである:エクソン1、5609−5596;エクソン2、5502−5489;エクソン3、5398−5283;エクソン4、5243−5229;及びエクソン5、5215−4952。ラットNRP-1のマウスオルソログ(本明細書ではNRP-8と称される)のコードヌクレオチド配列(配列番号:19)及びアミノ酸配列(配列番号:20)は以下のとおりである:
【0039】
配列番号:19及び20
【0040】
NRP-8の発現されたアミノ酸配列は141のアミノ酸残基を含む。112−114位のアスパラギン残基は、N-グリコシル化コンセンサス配列が存在することによりN-グリコシル化されると推定される。この新規なマウスNRP-1オルソログ、NRP-10は、染色体13に位置するヒトNRPオルソログ(NRP-2)に対して35.5%相同性を、さらにNCBIのアラクネcontigに位置するマウスNRP-1オルソログに対して28.9%の相同性を有する。さらにまた、このペプチドは、NRP-1又は他のNRPペプチドに存在するドメインに類似するアミノ酸配列ドメインを含み、さらにこのペプチドは、ニューロンの遊走、増殖、生存及び/又は神経突起伸長の促進を含む生物学的特性を有する。
NRP-9(マウスNRP-7のラットオルソログ)のヌクレオチド配列(配列番号:27)及びアミノ酸配列(配列番号:28)は以下のとおりである:
【0041】
配列番号:27及び28
【0042】
上述のNRP化合物の他に、我々は、NRPの発現にまた有用でありえる、NRP様ペプチドドメインを有する他の遺伝子を同定した。これらには、マイコバクテリウム及び腫瘍細胞に由来する遺伝子が含まれる。最近刊行された論文では、コンセンサス配列(PE_PGRS)(これは我々が提唱する配列(PGR/S)と類似している)を含むヒト型結核菌のPEマルチ遺伝子ファミリーが開示された。彼らはまた、これらタンパク質は宿主においてで前記細菌によって放出され、細菌の生存を促進するとも述べている。彼らが前記論文で提供した、PE_PGRSコンセンサス配列が見出される例が存在する。ヒト型結核菌のRv1818c遺伝子産物のアミノ酸配列(配列番号:21)は以下に示される:
【0043】
msfvvtipea laavatdlag igstigtana aaavptttvl aaaadevsaa maalfsghaq ayqalsaqaa lfheqfvral tagagsyaaa eaasaapleg vldvinapal allgrplign gangapgtga nggdggilig nggaggsgaa gmpggnggaa glfgnggagg aggnvasgta gfggaggagg llygaggagg aggragggvg giggaggagg nggllfgagg aggvgglaad agdggaggdg glffgvggag gaggtgtnvt ggaggaggng gllfgaggvg gvggdgvafl
gtapggpgga ggagglfgvg gaggaggigl vgnggaggsg gsallwgdgg aggaggvgst tggaggaggn agllvgagga ggagalggga tgvggaggng gtagllfgag gaggfgfgga ggagglggka gligdggdgg aggngtgakg gdggagggai lvgnggnggn agsgtpngsa gtggaggllg kngmnglp 配列番号:21
【0044】
エプスタイン-バーウイルス核抗原1のアミノ酸配列(配列番号:22)は以下に示される:
【0045】
msdegpgtgp gnglgekgdt sgpegsggsg pqrrggdnhg rgrgrgrgrg ggrpgapggs
gsgprhrdgv rrpqkrpsci gckgthggtg agagaggaga ggagagggag agggaggagg
aggagaggga gagggaggag gagagggaga gggaggagag ggaggaggag agggagaggg
aggagaggga ggaggagagg gagaggagga ggagaggaga gggaggagga gaggagagga
gaggagagga ggagaggagg agaggaggag agggaggaga gggaggagag gaggagagga
ggagaggagg agagggagag gagaggggrg rggsggrgrg gsggrgrggs ggrrgrgrer
arggsrerar grgrgrgekr prspssqsss sgspprrppp grrpffhpvg eadyfeyhqe
ggpdgepdvp pgaieqgpad dpgegpstgp rgqgdggrrk kggwfgkhrg qggsnpkfen
iaeglralla rshverttde gtwvagvfvy ggsktslynl rrgtalaipq crltplsrlp
fgmapgpgpq pgplresivc yfmvflqthi faevlkdaik dlvmtkpapt cnirvtvcsf
ddgvdlppwf ppmvegaaae gddgddgdeg gdgdegeegq e 配列番号:22
(M.J. Brennan and G. Delogu, The PE multigene family: a “molecular mantra” for mycobacteria. Trends in Microbiology 5: 246-249 (2002)より)
【0046】
NRP-1からNRP-9の完全な配列を用いる必要がないことは理解されよう。それよりはむしろ、本発明の実施態様にしたがって約8つのアミノ酸のペプチドフラグメントを用いることができる。本明細書で同定したコンセンサス配列ドメインが与えられるならば、合成ペプチドを作り上げるか、又は天然に存在するNRPを切り縮めて、生物学的に活性なペプチド部分を得ることができる。発現ペプチドを短縮する方法(例えば合成DNAを用いる)又は酵素的に改変する方法は当業界では公知である。
本発明のある実施態様は、配列KDPEARRAPGSLHPCLAASCSAAG-NH2(NRP-2セグメントKG;配列番号:18)を含むNRP-2(配列番号:5)の24merフラグメントである。
本発明のまた別の実施態様は、配列KDPEARRAPGSLHPCLAAS-NH2(NRP-2セグメントKS;配列番号:23)を含むNRP-2(配列番号:5)の19merフラグメントである。
本発明のさらに別の実施態様は、配列SEPEARRAPGRKGGVVCASLAADW-NH2(NRP-7セグメントSW;配列番号:24)を含むNRP-7(配列番号:17又は配列番号:36)の24mer型である。
本発明のさらに別の実施態様は、配列SDSFKSQARGO-NH2(NRP-3セグメントSQ;配列番号:25)を含む(NRP-3(配列番号:7)のアミノ酸11−23の間に位置する)11merペプチドである。
【0047】
本発明のまた別の実施態様は、配列GTPGRAEAGGQ-NH2(NRP-4セグメントGQ;配列番号:26)を含む(アノテーションを付されたNRP-4(配列番号:9)のアミノ酸22−32の間に位置する)11merペプチドである。
本発明のまた別の実施態様は、配列GTPGRAEAGGQVSPCLAASCSQAYG-NH2(NRP-4セグメントGQ;配列番号:29)を含むNRP-4(配列番号:9)の25merフラグメントである。
本発明のさらに別の実施態様は、配列REGRRDAPGRAGG-NH2(NRP-5セグメントRG;配列番号:30)を含むNRP-5(配列番号:11)の13merフラグメントである。
本発明のさらに別の実施態様は、配列SEVDARRAKKSLHCILSDTSHPRG-NH2(NRP-8セグメントSG;配列番号:31)を含むNRP-8(配列番号:20)の24merフラグメントである。
本発明のまた別の実施態様は、配列SDSFKSQARGQVPPFLGGVGCPWF-NH2(NRP-3セグメントSF;配列番号:32)を含むNRP-3(配列番号:7)の24merフラグメントである。
本発明のまた別の実施態様は、配列REGRRDAP-NH2(NRP-5 RP;配列番号:33)を含むNRP-5(配列番号:11)の8merフラグメントである。
本発明のさらに別の実施態様は、配列SEPEARRAQGGQIPSERVLSD-NH2(NRP-9セグメントSD;配列番号:34)を含む21merフラグメントで、前記配列はNRP-9(配列番号:28)のアミノ酸残基88-108の間に位置している。
本発明のまた別の実施態様は、配列PGRAEAGGQ-NH2(NRP-4セグメントPQ;配列番号:43)を含む9merペプチドで、前記配列はアノテーションを付されたNRP-4(配列番号:9)のアミノ酸24-32の間に位置している。さらに別の実施態様が本明細書の他の箇所に記載されている。
【0048】
NRP化合物の使用
したがって、本発明は、NRP、NRPによってコードされるペプチド、NRPのホモログ、オルソログ又はパラログ、NRPのアナログ及びNRPのプロドラッグに関する実施態様を含み、ここでNRPのプロドラッグは、酵素的に、代謝的に又は他の態様で改変されてNRP、NRPホモログ、NRPパラログ、NRPオルソログ又はNRPアナログとなることができる分子である。そのような分子は包括的に“NRP化合物”又は“NRP”と称される。NRP化合物はヌクレオチド配列によってコードされてもよい。前記ヌクレオチド配列はDNAでもRNAでもよく、さらに一本鎖でも二本鎖でもよい。本発明には本出願に開示された配列そのものと同様に、前記配列に対して相補的な配列も含まれることは理解されよう。さらにまた、NRPのまた別のスプライス形が存在しえることは理解されよう。そのような事例では、NRP RNAのまた別のスプライス形、並びにそれらがコードすることができるタンパク質及びペプチドもまた本発明の部分であると考えられる。
上記で示したように、本発明の実施態様は、NRPはニューロン及び神経芽細胞の増殖、遊走、分化、神経突起の生成を誘導することができ、さらにニューロンを神経損傷によってもたらされる障害から保護することができるという、本発明者らの驚くべき発見を基にしている。神経細胞の増殖及び急性脳損傷または慢性神経変性疾患によって生じた傷害領域への遊走は、神経機能の改善をもたらすことができる。さらにまた、NRP化合物を用いて、脳組織が変性するか、変性又は死のリスクを有するか、又は既に死んでいる、多様な異常及び症状を治療することができる。
細胞はまた、トランスフェクションの後、NRPオリゴヌクレオチドを使用してNRPの産生を刺激することができる。いくつかの事例では、トランスフェクションは複製可能なビヒクルとして行うことができ、さらに他の事例では、NRPオリゴヌクレオチドは裸のDNAとして導入することもできる。
【0049】
NRPで治療できる異常及び症状
NRP化合物が有用でありえる異常及び症状には以下が含まれる:
中枢神経系の感染(細菌、菌類、スピロヘータ、寄生虫感染を含む)及び類肉腫(化膿性感染、急性細菌性髄膜炎、軟膜炎を含む);
脳血管性疾患(卒中、虚血性発作、アテローム性硬化症血栓、ラクーン(lacune)、塞栓症、高血圧性出血、動脈瘤破裂、血管奇形、一過性虚血発作、頭蓋内出血、特発性くも膜下出血、高血圧性脳障害、脳動脈の炎症性疾患、例えば心不全(おそらく冠状動脈バイパス手術から生じる)によって惹起される灌流の低下、及び脳血管障害の他の形態を含む);
脳頭蓋外傷(頭蓋骨基底部骨折及び頭蓋内神経損傷を含む)、頚動脈洞フィステル、気脳症、気瘤及び鼻漏、脳挫傷、外傷性脳内出血、小児の急性脳腫脹;
脱髄性疾患(視神経脊髄炎、急性播種性脳脊髄炎、急性及び亜急性壊死性出血性脳炎、シルダーびまん性脳硬化症、及び末梢神経障害に併発する多発性硬化症を含む);
神経系の変性疾患(1つ又は2つ以上の進行性痴呆の症状、びまん性脳萎縮、非アルツハイマー型のびまん性皮質萎縮、レーヴィ小体痴呆、ピック病、前側頭性痴呆、視床変性、非ハンチントン型舞踏病及び痴呆、皮質脊髄変性(ヤコブ病)、痴呆-パーキンソン病-筋萎縮性側索硬化症合併症(Guamaninaら)を含む);
神経系の後天的代謝異常(錯乱、昏迷又は昏睡-虚血-低酸素症、低血糖症、高血糖症、高炭酸ガス症、肝不全及びライ症候群の1つ又は2つ以上を含む症状として提示される代謝性疾患、進行性錐体外路症候群として提示される代謝性疾患、小脳性運動失調、高体温、セリアック-スプルー病として提示される代謝性疾患、精神病又は痴呆を引き起こす代謝性疾患(クッシング病及びステロイド脳障害を含む)、甲状腺精神病及び甲状腺機能低下、膵臓性脳障害を含む);
栄養欠乏による神経系の疾患;
アルコール及びアルコール中毒;
薬物及び他の化学物質(アヘン誘導体及び合成鎮痛剤、沈静睡眠薬、興奮薬、向精神薬、細菌毒素、植物毒素、蛇咬傷及び刺創毒、重金属、工業毒物、抗腫瘍剤及び免疫抑制剤、サリドマイド、アミノグリコシド系抗生物質(聴器毒)及びペニシリン誘導体(痙攣)、心保護剤(β-ブロッカー、ジギタリス誘導体及びアミオダロンを含む)による神経系の異常。
【0050】
上記のリストによって示したように、本発明の組成物及び方法は、ヒトの神経の損傷及び疾患の治療に有用でありえる。さらに一般的には、本発明の組成物及び方法は、急性脳損傷(びまん性軸索損傷、周産期低酸素-虚血性損傷、外傷性脳損傷、卒中、虚血性梗塞、塞栓症及び高血圧性出血;CNS毒物への暴露、脳神経系の感染(例えば細菌性髄膜炎);代謝性疾患(例えば低酸素-虚血性脳障害、末梢神経障害及び糖原病を含むようなもの)を含むが、ただしこれらに限定されない)の結果として神経の傷害をもつか、又は慢性神経損傷若しくは神経変性疾患(多発性硬化症、レーヴィ小体痴呆、アルツハイマー病、パーキンソン病及びハンチントン病を含むが、ただしこれらに限定されない)に罹患している人間の患者の治療で有用である。そのような疾患又は損傷をもつ患者は、神経突起伸長と同様にニューロンの増殖及び遊走を開始させることができる治療プロトコルによって大きな利益を得ることができる。
さらに一般的には、本発明は、外傷、毒物への暴露、仮死又は低酸素-虚血による損傷後の傷害領域へのニューロン及び神経芽細胞の遊走の誘導に適用される。
NRP化合物(NRP-1、そのオルソログ、アナログ、パラログ及び同定されたNRPペプチドドメインを含むプロドラッグを含む)を用いて、ニューロン及び神経芽細胞の遊走を促進することができる。
【0051】
しかしながら、NRPを有利に用いることができるが、他の形態のNRP化合物の投与を排除しようとするものではない。例えば、ヒトパラログ型又はNRPのペプチドフラグメントもNRPの代わりに投与することができる。例示すれば、NRP及び担体を含むNRPのプロドラッグ型(NRP及び前記担体は患者の体内での切断又は消化に対し感受性を有する結合によって結合されてある)を投与することによって、NRPのCNS内の有効量を増加させることができる。投与後に切断又は消化されてNRPを放出する、任意の適切な結合を利用することができる。
また別の適切な治療方法は、中枢神経系の内部でNRP又はNRPのアナログ、パラログ若しくはプロペプチドを活性な形で発現させることができる細胞株である移植物、又は前記細胞株を含む移植物を通して、患者のNRPレベルを増加させる。
NRPは、医薬又は医薬調製物の部分として投与することができる。前記は、医薬的に適切な任意の担体、アジュバント又は賦形剤とNRP化合物を一緒にすることを必要とする。さらにまた、NRP化合物は他の非NRP性神経保護薬剤、増殖薬剤又は他の薬剤とともに用いてもよい。もちろんのこと、担体、アジュバント又は賦形剤の選択は用いられる投与ルートに一般的に左右される。
投与ルートは広く変動しえる。NRPは種々の方法で、すなわち腹腔内、静脈内又は脳室内に投与することができる。末梢適用も、中枢神経系との直接的干渉がないので選択方法でありえる。
【0052】
当業界で公知のいずれの末梢投与ルートも用いることができる。これらには、非経口ルート、例えば末梢循環への注射、皮下、眼窩内、眼内、くも膜下槽内輸液(徐放性装置又はミニポンプ、例えば浸透圧ポンプ又は皮膚パッチを用いる)、移植物、エーロゾル、吸入、乱切法、腹腔内、嚢内、筋肉内、鼻内、経口、頬内、肺、直腸又は膣ルートが含まれる。組成物は、人間又は他の哺乳動物へ、上述の神経学的疾患に対する治療を提供するために治療的に有効な量(例えば患者の病変を排除又は減少させる量)で非経口的に投与するために製剤化することができる。
ある投与ルートは、皮下注射(例えば0.9%塩化ナトリウムに溶解)及び経口投与(例えばカプセルで)を含む。
場合によってNRP化合物を直接患者のCNSに投与することが所望されることもまた理解されよう。前記は、適切な任意の直接投与ルートによって実施することができる。具体例には、側方脳室注射又は外科的に挿入した患者の脳の側方脳室へのシャントによる投与が含まれる。
【0053】
NRP投与量の決定
投与されるべきNRPの有効量の決定は当業者の技術範囲内であり、当業者には日常的な作業であろう。ある種の実施態様では、使用されるNRPの量は、本明細書に記載のアッセイ系を用いてin vitro試験で概算することができる。投与される最終量は、投与ルート、使用されるNRP、及び治療されるべき神経学的疾患または症状の性質に左右されるであろう。適切な用量範囲は、例えば体重100g当たり約0.01mgから約1mg、また別には体重100g当たりNRPの約0.06μgから約0.6mgで、この場合前記用量は中枢に投与される。
医薬に内包するためには、NRPは、通常の方法、例えばMerrtfieldら(J. Am. Chem. Soc. 15:2149-2154 (1963))の段階的固相合成法によって直接合成することができる。そのようなペプチド合成方法は当業界では公知であり、例えば以下の文献に記載されている:Fields and Colowick, 1997, Solid Phase Peptide Synthesis (Methods in Enzymology, vol.289), Academic Press, San Diego, CA。また別には、合成は市販のペプチド合成装置、例えばアプライドバイオシステムモデル(Applied Biosystems model)430Aを含むことができる。
一般的な提案として、非経口的に投与される1回分のNRP-1(配列番号:2)の医薬的に有効な総量は、用量応答曲線によって測定可能な範囲内にある。ある範囲は体重100g当たり約0.06mgから約0.6mgである。例えば、血中NRP-1(配列番号:2)は、治療される哺乳動物の体液中で測定され、用量を決定することができる。また別には、NRP-1(配列番号:2)化合物の量を増加さえながら患者に投与し、前記患者のNRP-1(配列番号:2)の血清レベルをチェックすることができる。使用されるべきNRP-1(配列番号:2)の量は、これらのNRP-1(配列番号:2)の血清レベルを基準にしてモル基準で算出することができる。
【0054】
特に、本化合物の適切な投与量を決定するある方法は、生物学的流体(例えば体液又は血液)中でのNRPレベルを測定することを必要とする。そのようなレベルの測定は任意の方法(RIA及びELISAを含む)実施することができる。NRPレベルを測定した後、単回投与又は複数回投与を用いて前記流体を化合物と接触させる。この接触工程の後で、前記流体中のNRPレベルを再度測定する。流体のNRPレベルが、前記分子が投与される目的について所望の有効性を生じるさせるに十分な量低下した場合、前記分子の投与量を調整して最大の効能を生じさせることができる。この方法はin vitroでもin vivoでも実施することができる。この方法はin vitroで、例えば哺乳動物から前記流体を抜き取り、NRPレベルを測定し、本明細書の化合物が単回又は複数回投与で哺乳動物に投与し(すなわち、接触工程は哺乳動物への投与によって達成される)、NRPレベルを前記哺乳動物から抜き取った流体で再度測定した後で実施することができる。
NRP化合物は、徐放系によって適切に投与することができる。徐放性組成物の適切な例には、形状物品(例えばフィルム又はマイクロカプセル)としての形状をもつ半透性ポリマーマトリックスが含まれる。徐放性マトリックスにはポリペプチドラクチド(米国特許3,773,919号、EO58,481号)、ポリ(2-ヒドロキシエチルメタクリレート)(Langer et al., 1981)、エチレンビニルアセテート(Langer et al., 上掲書)、又はポリ-D-(-)-3-ヒドロキシ酪酸(EP133,988号)が含まれる。徐放性組成物にはまたリポソーム結合化合物が含まれる。本化合物を含有するリポソームは当業者に公知の方法によって製造される。前記が以下によって実施される:DE3,218,121号;Hwang et al., 1980;EP52,322号;EP36,676号;EP88,046号;EP143,949号;EP142,641号;日本国特許出願83-118008、米国特許2,485,045号及び4,544,545号、並びにEP102,324号。いくつかの実施態様では、リポソームは小さな(約200オングストローム、或いは約200オングストロームから800オングストローム)単一薄層型で、その場合、脂質含有量は約30molパーセントコレステロールを超え、この選択した割合はもっとも有効な治療のために調整される。本明細書(上記及び下記)に引用される米国特許は全て参照により本明細書に含まれる。
【0055】
例えばWO95/32003(1995年11月30日公開)に記載された複合技術による、非PEG化ペプチドより寿命が長いPEG化ペプチドもまた用いることができる。
非経口的投与のためには、投与用量は約0.01から約1mg/100g体重、また別には体重100g当たり約0.06μgから約0.6mgのNRP化合物である。いくつかの実施態様では、本化合物は、一般的には所望の純度の各化合物を混合し、 医薬的に又は非経口的に許容可能な担体(すなわち用いられる投与量及び濃度で受容者にとって無毒なもの)であって、製剤の他の成分と適合するものとともに注射可能なユニット投薬形で製剤化することができる。例えば、製剤は好ましくは、酸化剤及びポリペプチドにとって有害であることが知られている他の化合物を含まない。上記の投与用量は限定的なものではないことは理解されよう。上記範囲を外れる他の用量も当業者は決定することができる。
いくつかの実施態様では、化合物は液体担体、又は微細に分割した固体担体、又はその両者と均一に及び密接に接触させることによって製造することができる。続いて、所望する場合は、前記生成物を所望の製剤に成型することができる。いくつかの実施態様では、前記担体は非経口的担体であり、また別には受容者の血液と等張な溶液である。そのような担体としての媒体の例は、水、食塩水、リンゲル溶液、緩衝溶液、及びデキストロース溶液が含まれる。非水性の媒体(例えば固定油およびオレイン酸エチル)も本明細書では有用である。
【0056】
担体は適切には微量の添加物、例えば等張性及び化学的安定性を強化する物質を含む。そのような物質は、望ましくは、用いられる投与量及び濃度で受容者に無毒であり、例示すれば以下を含む:緩衝液(例えばリン酸、クエン酸、コハク酸、酢酸及び他の有機酸又はそれらの塩);抗酸化剤(例えばアスコルビン酸);低分子量(10残基未満)ポリペプチド(例えばポリアルギニン)又はトリペプチド;タンパク質(例えば血清アルブミン、ゼラチン又は免疫グロブリン);親水性ポリマー(例えばポリビニルピロリドン);グリシン;アミノ酸(例えばグルタミン酸、アスパラギン酸、ヒスチジン、又はアルギニン);単糖類、二糖類、及び他の炭水化物(セルロース若しくはその誘導体、グルコース、マンノース、トレハロース、又はデキストロリン);キレート剤(例えばEDTA);糖アルコール(例えばマンニトール又はソルビトール);対イオン(例えばナトリウム);非イオン性界面活性剤(例えばポリソルベート、ポロキサマー又はポリエチレングリコール(PEG));及び/又は中性塩(例えばNaCl、KCl、MgCl2、CaCl2など)。
NRP化合物は、望ましくは上記のような媒体中で約4.5から約8のpHで製剤化することができる。前述のビヒクル、担体又は安定化剤のある種のものの使用は化合物の塩の形成をもたらすことは理解されよう。最終的な調製物は安定な液体又は凍結乾燥固体でもよい。
【0057】
他の実施態様ではアジュバントを用いることができる。錠剤、カプセルなどに取り込むことができる典型的なアジュバントは、結合剤(例えばアラビアゴム、ともろこしデンプン又はゼラチン);ビヒクル(例えば微晶質セルロース、);崩壊剤(例えばトウモロコシデンプン又はアルギン酸);滑沢剤(例えばステアリン酸マグネシウム);甘味剤(例えばシュクロース又はラクトース);香料(例えばペパーミント、ヒメコウジ、又はチェリー)である。投薬形態がカプセルであるとき、上記の材料の他に、液状担体(例えば脂肪油)もまた含むことができる。種々のタイプの他の材料をコーティングとして、又は投薬ユニットの物理的形態の改変物質として用いることができる。シロップ又はエリキシルは、活性化合物、甘味料(例えばシュクロース)、保存料(例えばプロピルパラベン)、着色剤及び香料(例えばチェリー)を含むことができる。注射用滅菌組成物は、通常の製薬慣行にしたがって製剤化することができる。例えば、活性化合物の媒体(例えば水又は天然に存在する植物油(例えばごま油、ピーナツ油又は綿実油)又は合成脂肪性媒体(例えばオレイン酸エチル)など)中の溶解液又は懸濁液を所望することができる。緩衝剤、保存料、抗酸化剤などを容認されている製薬慣行にしたがって取り入れることができる。
望ましくは、治療的投与に用いられるNRP化合物は滅菌することができる。滅菌は、滅菌ろ過膜(例えば約0.2ミクロンのポアサイズを有する膜)でろ過することによって容易に実施することができる。治療用組成物は一般的には、滅菌アクセスポートを有する容器(例えば静脈内溶液バッグ又は皮下注射針によって刺し通すことができる栓を有するバイアル)に入れることができる。
他の実施態様では、NRP化合物は、単位用量又は多用量容器(例えば封入アンプル又はバイアル)に、水溶液又は再構成用の凍結乾燥製剤として保存することができる。凍結乾燥製剤の例として、10mLのバイアルに化合物の5mL滅菌ろ過水溶液(0.01%w/v)を充填し、得られた混合物を凍結乾燥する。輸液溶液は、制菌水又は他の適切な溶媒を用いて凍結乾燥化合物を再構成することによって調製することができる。
【0058】
遺伝子治療
本発明の他の実施態様では、治療方法は、器官治療のための遺伝子治療を含み、前記はNRP化合物をコードする核酸を用いる。一般的には、遺伝子治療は器官のNRPレベルを増加させる(又は過剰発現させる)ために用いることができる。ヌクレオチド配列の例には、配列番号:1、3、4、6、8、10、12、16、19、27若しくは35、又はNRPのコンセンサスドメイン及び生物学的特性を有するペプチドをコードするその部分が含まれる。プロNRPをコードする他の配列も用いることができることは理解されよう(プロNRPは切断に際して生物学的に活性なNRPを生じることができる)。
生物にNRPをコードする配列をトランスフェクトするために任意の適切な方法を用いることができる。例えば、in vivo及びex vivoでの方法を用いることができる。in vivoデリバリーのためには、核酸(単独又はベクター、リポソーム、沈殿物と結合されている)を生物(例えば人間の患者)に直接、いくつかの実施態様では、NRP化合物の発現が所望される部位に注射することができる。ex vivo治療のためには、生物の細胞を取り出し、これらの細胞に核酸を導入し、この改変した細胞を前記生物に直接投与するか、又は例えば多孔性膜に被包化してこれを前記患者に移植する(例えば米国特許4,892,538号及び5,283,187号を参照されたい)。
我々は、本明細書で培養細胞がNRPを発現することができること、及びこれらNRP-発現細胞を有害な傷害に感受性を有するニューロンとインキュベートするとき、NRPが発現され、培養液中に分泌され、ニューロンを有害な傷害から保護することができることを示した。この驚くべき発見は、遺伝子移入及び細胞移植によって神経変性を処置する治療方法を支持する。
核酸を生細胞に移入するために利用可能な多様な技術が存在する。これらの技術は、核酸が培養細胞にin vitroで移入されるか、又は目的の宿主の細胞にin vivoで移入されるかにしたがって変動しえる。核酸の哺乳細胞へのin vitro移入に適した技術には、リポソームの使用、エレクトロポレーション、マイクロインジェクション、細胞融合、DEAE-デキストラン、リン酸カルシウム沈殿方法などが含まれる。遺伝子のex vivoデリバリーに一般的に用いられるベクターはレトロウイルスである。
【0059】
ある種の実施態様では、in vivo核酸移入技術には、例えばウイルスベクター(例えばアデノウイルス、単純ヘルペスI型ウイルス、又はアデノ付随ウイルス)によるトランスフェクション及び脂質利用系が含まれる。脂質仲介遺伝子移入に有用な資質は、N-[1-(2,3-ジオレイルオキシ)プロピル]-N,N,N-トリメチルアンモニウム(DOTMA)、ジオレオイルファチジルエタノールアミン(DOPE)及び3-β[N-(N',N'-ジメチルアミノエタン)カルボモイル]コレステロール(DC-Chol)である。いくつかの状況では、核酸含有ベクターを標的細胞に誘導する薬剤とともに核酸源を提供することが所望されえる。そのような“標的誘導”分子には、細胞表面膜タンパク質又は標的細胞に特異的な抗体、標的細胞上のレセプターに対するリガンドなどが含まれる。リポソームが用いられる場合は、エンドサイトーシス関連細胞表面膜タンパク質と結合するタンパク質を、標的誘導及び/又は取り込み促進に用いることができる。そのようなタンパク質の例には、特別な細胞タイプに親和性を有するカプシドタンパク質及びそのフラグメント、サイクリングにおいてインターナリゼーションを受けるタンパク質に対する抗体、及び細胞内局在化を標的とし細胞内半減期を強化するタンパク質が含まれる。他の実施態様では、レセプター仲介エンドサイトーシスを用いることができる。そのような方法は、例えば以下に記載されている:Wu et al., 1987;及びWanger et al., 1990。これまでに知られている遺伝子マーキング及び遺伝子治療プロトコルの総論のためには文献(Anderson, 1992)を参照されたい。さらにWO93/2563号及び前記に引用されている参考文献もまた参照されたい。
キットもまた本発明の範囲内に意図される。典型的なキットは、NRP製剤のための容器(いくつかの実施態様ではバイアル)を含み、前記容器は、医薬的に許容できる緩衝液中の1つ又は2つ以上のNRP化合物及び、使用者に前記医薬製剤の使用を指示する使用説明(例えば製品挿入物又はラベル)を含む。
【0060】
実施例
以下の実施例は、本発明のある種の実施態様を具体的に説明するために提供される。他の実施態様も考案することが可能で、前記もなお本発明の範囲内であることは容易に理解されよう。これら他の実施態様のいずれも本発明の範囲内であると考えられる。
【0061】
実施例1:NRP遺伝子の同定
ニューロンの生存、増殖及び分化に対して有効性を有する哺乳動物の遊走誘導因子を同定するために、我々は、ヒト悪液質関連タンパク質のcDNA及びコードされたその16mer悪液質フラグメントを鋳型として用いラット及びマウスのゲノムをスクリーニングし、新規なNRPホモログを同定した。
NRP遺伝子の同定は種々の細胞源(in vivo組織、神経幹細胞培養)の全RNAを入手することを含む。RNAはロシュ全RNA単離キット(Roche Total RNA Isolation Kit)を用いて抽出した。相補性DNA(cDNA)は、スーパースクリプトRT IIを用いて合成し、続いて以下のプライマーを用いマウスNRP遺伝子及びベータアクチンのマルチプレックスPCR増幅を実施した:mouFS NRPフォワードプライマー:5' AACGGAATGAATCGAAACCC 3'(配列番号:37);mouFS Revプライマー:5' CGCTCGACATTACAGCTCA 3'(配列番号:38);マウスβ-アクチンFwd:5' GAAAGGGTGTAAAACGCAGC 3'(配列番号:39);マウスβ-アクチンRev:5' GGTACCACCATGTACCCAGG 3'(配列番号:40)。正しいサイズを有するフラグメントをゲル精製し、直接pGEMベクターにクローニングし、大腸菌(E. coli, DH5α)のコンピテント株を形質転換した。形質転換細胞コロニーをNRP遺伝子フラグメントの存在についてスクリーニングし、陽性コロニー由来のプラスミドの配列を決定した。NRP遺伝子をコードする遺伝子の構造は図1Aに示されている。
【0062】
NRPタンパク質ドメインの性状決定:NRPは、セクレトームP(SecretomeP)サーバー(Technical University of Denmark)を用いて前記タンパク質配列を解析すると極めて有意なスコアを有するので、おそらくFGF-1、FGF-2のように非古典的経路で分泌されるのであろう。陽電荷アミノ酸残基の数及びN-末端NRP内の全体的な原子の数が多いことがこの予想につながった(Bendsten et al., 2004)。
単一のトレフォイルドメイン配列モチーフがマウスNRP配列内に予想され、このドメインはヒトpS2タンパク質(Jakowlew et al., 1984)と52.5%の相同性を有する(図1D)。推定的NRPトレフォイルドメインは、前記のトレフォイルドメイン(データベースSMART)のコンセンサス配列内の15の保存アミノ酸のうちの9個が類似又は同一である。この60%コンセンサス値はまたトレフォイル因子ファミリーのメンバーとしてNRPを受け入れるための閾値でもある。トレフォイルドメインは、タンパク質-タンパク質相互作用に関与し、シクロオキシゲナーゼ2(COX-2)及びトロンボキサンA2レセプター(TXA2-R)活性化経路を介して作用すると示唆されてきた(Rodrigues et al., 2003)。ヒトPs2タンパク質はトレフォイル因子ファミリー1(TFF-1)に属し、REK1/2経路でのシグナリング作用によって乳がん細胞の化学誘引(Prest et al., 2002)における関係が考えられている(Graness et al., 2002)。
ニューロンの生存、増殖、遊走及び分化を促進するマウスNRP(NRP-7セグメントSW)ドメインはトレフォイルドメインからC-末端側に位置し、さらにラットNRP-9についてはC-末端に位置する(図1Bのアラインメントされた21mer及び24mer配列を参照されたい)。
【0063】
NRP、悪液質関連タンパク質及びSDF-1間の相同性:NRPとSDF-1αとの間の驚くほど類似する生物学的活性にもかかわらず、両ペプチドは、32.6%相同性という中程度の類似性を示しただけである(図1E)。
成熟SDF-1の最初の9アミノ酸は残存化学誘引活性を示し、これはCDF-1αよりほぼ100倍低い(Loetscher et al., 1998)。マウス24merペプチド(NRP-7セグメントSW;配列番号:24)のN-末端11アミノ酸は、成熟SDF-1の開始部と50%を超える相同性でアラインメントされる。この関連性を示すタンパク質ドメインによって、悪液質タンパク質、NRP及びSDF-1αは、ニューロンの生存、増殖、遊走及び分化にとって重要な生物学的に活性なドメインを共有することが示される。前記生物学的に活性な領域は、悪液質関連タンパク質及びSDF-1については成熟タンパク質のN-末端に配置されるが、NRPについてはこの領域はトレフォイルドメインからC-末端側に位置する。
【0064】
実施例2:小脳微小エクスプラント:神経保護及びニューロン増殖方法
薄層状の小脳皮質をP3/4、P7/8ラットの子供から摘出し、125μmのポアサイズを有する金属紗を通してすり潰して均一サイズの微小エクスプラントを得た。61gで3分間遠心後、ペレットをスタートV(StartV)培養液(Biochrom)に懸濁し、前記懸濁液を6ウェルプレートのポリ-D-リジン被覆カバースリップに播種し、3時間インキュベートして接着させ、その後ウェル当たり1mLのスタートVを添加した。以前に記載したように、グルタメート/3-NP及びNRPも添加した。前記エクスプラントを5%CO2及び100%湿度下で34℃で48−72時間培養した。増殖速度測定のために、BrdUを培養開始時に加え、顕微鏡視野当たりの細胞を48−72時間後に数えた。
【0065】
結果:
(a)神経保護:ナノモル濃度のNRP-4セグメントGQ(配列番号:26)は重篤な損傷後50%の生存率をもたらした(図2)。
(b)ニューロン増殖:NRP-7セグメントSW(配列番号:24)は、これらの培養で増殖速度を200%以上に高めた(図3)。小脳の微小エクスプラントの増殖細胞は、ビヒクル単独の作用と比較して、興奮毒性(excitotoxic)及び酸化性ストレスに対して耐性を示した。
【0066】
実施例3:接触走性遊走アッセイ
マウス神経幹細胞、EGF依存性不死化マウス神経幹細胞MEB5及び野生型PC-12細胞に対する遊走誘導/化学誘引活性について、NRPを下記のように接触走性遊走アッセイでテストした。
方法:
最初のNRPコーティング:12μmのポアサイズを有するトランスウェルプレート(Corning)のコントロールウェルをBSA/PBS媒体の1.5mLで被覆した。残りのプレートを1−100ng/mLの範囲の種々の濃度のNRP(10μg/mLのBSAを含むPBSで調製)を用いて被覆した。続いて、このプレートを37℃で1時間インキュベートして被覆した。続いてウェルを1mLの滅菌PBSで2回洗い流した。
細胞外マトリックスコーティング:MEB-5細胞にはラミニン(10μg/mL)、マウス初代幹細胞にはPDL(50μg/mL)+マトリゲル、及びPC-12細胞にはフィブロネクチン(25μg/mL)+マトリゲルを細胞のための細胞外マトリックス(ECM)として用いた。全てのECM化合物はPBSで希釈した。ウェル当たり1.5mLのECMを室温で2時間インキュベートした。続いてウェルを1mLの血清非含有培養液(例えばNB/B27)で1回洗浄し、続いて1mLのPBSで洗浄した。
インサートのコーティング:5μg/mLのPDL/PLL混合物(PBS中)を用いてインサートを被覆した。続いて前記インサートをMiliQ水で洗浄した。
培養液への移動及び細胞の播種:適切な培養液を12ウェルプレートに移した(MBE-5細胞:DMEM高グルコース+N2(Invitrogenの増殖培養液補充物)+10ng/ビオチン+2mMのL-グルタミン;初代幹細胞:EuroclonのNSA培養液;及びPC-12細胞:NB/B27倍溶液)。続いてプレートを37℃、5%CO2でインキュベートし、1−2x105細胞を播種した。プレートを1−2日in vitroで固定した(DIV)。
固定:インサートを廃棄し、ウェルをPFAの連続希釈(0.4、1.2、3及び4%)で各希釈について3−5分固定した。ウェルを洗浄し、PFAの連続希釈(0.4、1.2、3及び4%)中で各希釈について3−5分保存した。ウェルを計測までPBS中で洗浄した。神経突起伸長を示し、底部の小室へ移動した全ての細胞を遊走細胞として数えた。
【0067】
結果:
10ng/mLのNRP-4GG(配列番号:29)は、BSA-ビヒクル単独と比較して195%を超えるMEB細胞を培養皿の底へ遊走させた(図4A)。NRP-4GG(配列番号:29)ペプチドの存在下では、BSA-ビヒクル単独と比較して93.7%を超えるE14細胞が遊走した(図4B)。NRP-2KS(配列番号:23)の存在下では、BSA-ビヒクル単独と比較して109%を超えるMEB-5細胞が底部へ遊走した(図4B)。NRP-7SW(配列番号:24)処理ウェルでは、BSA-ビヒクル単独と比較して35%を超えるE14細胞密度が得られた(図4D)。NRP-4GG(配列番号:29)ペプチドの存在下では、BSA-ビヒクル単独と比較して80.8%を超えるPC-12細胞が底部へ遊走した(図4E)。NRP-7SW(配列番号:24)は、BSA-ビヒクル単独と比較して333%を超えるPC-12細胞を遊走させた(図4F)。
【0068】
実施例4:神経幹細胞培養及び軸索伸長についての分化アッセイ
NSA幹細胞培養液はEuroclone(Italy)から購入した。Neurobasal、DMEM/F12培養液、N2及びB27補充物は全てライフテクノロジー(Life Technology)から得た。抗βIII-チュブリン抗体はシグマ(Sigma)から購入した。Cy3複合ヤギ抗マウス抗体はアマーシャム(Amersham)から購入し、Syto21はモレキュラープローブ(Molecular Probes)から購入した。NRP-2KS(配列番号:23)及びNRP-7SW(配列番号:24)を用いた。ヒト組換えエリスロポエチン(EPO)はR&D システムズから購入した。
神経幹細胞培養:
神経幹細胞はE15 C3Hマウスの前脳に由来し、文献(Gritti et al., 2001)に記載されているように、20ng/mLのEGF及び10ng/mLのbFGFの存在下でニューロスフェアとして培養した。簡単に記せば、時期を定めた妊娠マウスを犠牲にし、胎児を取り出した。無菌条件下で、脳を取り出し前脳を切り出した。すり潰して組織を分離し、75gで10分遠心して沈殿させ、非被覆組織培養フラスコで増殖因子を含むNSA培養液中に細胞を播種した。継代のためにニューロスフェアをすり潰し、単一細胞として播種した。
Shingoら(2001)がエリスロポエチン(EPO)について記載したように、神経幹細胞の対称的分裂(2つの幹細胞、両者とも幹細胞へ)から非対称的分裂(1つの幹細胞、1つのニューロン前駆細胞又は神経芽細胞へ)へのニューロスフェア内での細胞のシフトをNRPが誘導することができるか否かを決定するために、ニューロスフェアをすり潰して分離させ、以下を補充したNSA培養液中で6ウェルプレートに200,000細胞/ウェルの密度で拡張(expansion)させるために播種した:EGFのみ;EGF+種々の濃度のNRP;又はEGF+10U/mLのEPO。1日おきに半分の培養液を新しいNRP化合物を含む培養液で交換した。7日後にニューロスフェアの細胞をプレート培養した。EGFの存在下では、細胞は対称的分裂を保持する。EGFの除去とともに、細胞は分化を開始する(非対称的細胞分裂)。続いて、下記に記載するように細胞を分化アッセイに付したが、ただしbFGFは培養液に最初の24時間は添加せず、NRP又は他の化合物を含まないコントロール培養液で7日間分化させた。NRPに全く暴露されなかった細胞では、神経芽細胞の数は、NRP又はEPOに暴露された細胞の場合よりも少なかった。したがって、NRPは未分化幹細胞の神経芽細胞への分化を増進することができる。
分化アッセイ:
十分なサイズに達したとき、ニューロスフェアをすり潰して分離し、Nuncの24ウェルプレートに入れた、ラミニンを被覆した直径13mmのカバースリップ上に、ウェル当たり200,000細胞の密度でプレート培養した。プレート培養の培養液は、N2を補充したDMEM/F12及びB27と2mMグルタミンを補充したNeurobasalの1:1混合物であった。プレート培養後の生存を強化するために培養液には2ng/mLのbFGFが含まれていた。24時間後に、培養液をニューロン分化促進培養液(N2を補充したDMEM/F12及びB27と2mMグルタミンを補充したNeurobasalの1:3混合物)と交換し、さらにコントロールを除いてテスト化合物又はIGF-1を同時に加えた。新しい化合物を含む培養液を1日おきに交換した。プレート培養の7日後に、分化細胞を100ng/mLのSyto21を含む分化培養液で20分インキュベートし、生細胞の核を標識した。続いてそれらを4%のパラホルムアミドで固定し、マウス抗βIIIチュブリン抗体およびヤギ抗マウスCy3-複合二次抗体で免疫染色した。
ニューロン分化及び軸索伸長の定量:
染色後、カバースリップを組織培養プレートから取り出し、イムノフルア(Immunofluore)蛍光マウント媒体を用いてカバースリップ上にマウントした。全細胞に対するニューロンの百分率を解析するために、アクソイビジョン(Axoivision)ソフトを搭載したツァイスのアクシオフォト顕微鏡を用い、20xの倍率でウェル当たり任意の2視野を、赤色(チュブリン染色ニューロン)及び緑色(Syto21染色核)蛍光チャネルで調べた。視野当たりのニューロン及び核を数え、全細胞数に対するニューロンの百分率を決定した。結果は図5A−5Hに示されている。
【0069】
結果:
広い濃度範囲にわたって、全生細胞に対してニューロン細胞数を分化アッセイ内で正規化したとき、NRP-7セグメントSWは、分化培養液中のラミニン上でプレート培養したNSCから生じるニューロンの子孫細胞の百分率を増加させた。10pMの濃度で、βIII-チュブリン陽性ニューロンでは最大2倍の増加が観察され(図5E)、一方、軸索全長の成長における最大の増加活性はより高いナノモル濃度範囲にあった(図5A)。IGF-1は軸索伸長促進の陽性コントロールとして用いられ(Ishii et al., 1993)、NRPと同様に有効あったが、より低い濃度で用いたときは能力は劣っていた(図5A)。ラットに21merのNRP-9をNSCの分化中にBrdUとともに24時間投与したとき、増殖速度の増加を観察することができ(図5H)、このことは、アッセイでのニューロンの百分率の増加は、少なくとも部分的にはニューロン前駆細胞の増殖に起因する可能性を高める。
NRP-7SW及びNRP-2KSは、IGF-1と同様な有効性で分化神経前駆細胞の軸索伸長促進活性を有していたが、IGF-1よりもはるかに高い能力を有していた(図5A及び5B)。
NRP-2KSについてのこれらのデータの定量は図5C(ビヒクルによる処理)及び図5D(NRP-2KS処理)で例として示されている。
広い濃度範囲にわたって、全生細胞に対してニューロン細胞数を分化アッセイ内で正規化したとき、NRP-2KSは、分化培養液中のラミニン上でプレート培養したNSCから生じるニューロンの子孫細胞の百分率を増加させた。10nMの濃度で、βIII-チュブリン陽性ニューロンでは最大2倍の増加が観察された(図5F)。NSCの分化中にBrdUとともに24時間NRP-2KSを投与したとき、増殖速度の増加を観察することができ(図5G)、このことは、アッセイでのニューロンの百分率の増加は、少なくとも部分的にはニューロン前駆細胞の増殖に起因する可能性を高める。
【0070】
実施例5:in vivoで神経集団の再形成を促進するためのNRPの使用
本明細書に開示した発見を考慮して、我々は、NRPが神経組織の集団再形成を動物で促進することができるか否かを決定するためのin vivo実験を実施する。OEG細胞を損傷(脊髄損傷のような損傷)後の神経組織の集団再形成のために細胞供給源として評価した。NRPをOEG細胞培養に添加するか、及び/又は脊髄の移植操作においてOEG細胞とともに同時投与する。脊髄損傷をもつ患者の損傷部位での外科手術のために準備を整え、脊椎管には当業界で公知の方法を用いてアクセスし、損傷領域を特定する。OEG細胞及びNRPを損傷部位に移植し、場合によって更にNRP供給源を局所的に提供する。
【0071】
実施例6:NRPの発現
方法:
NRP遺伝子発現の分析:アノテーションを付されたマウスのNRP遺伝子の発現はRT-PCR、ノザンブロット及びin situハイブリダイゼーションによって確認した。
in situハイブリダイゼーション及びノザンブロット:マウスNRP(NRP-7(配列番号:35))の固有のコード領域(alkBホモログ遺伝子転写開始部位の上流)を包含する88merのオリゴヌクレオチドをpGEM7Zf(-)(Promega, Madison, WI, USA)のBamHIとEcoRI部位の間にクローン化した。適切な5'オーバーハング(下線部)も与えられた相補的な合成オリゴヌクレオチド;mfsNRP.S88、センス鎖(Invitrogen)(下記に示される:5' AATTCGGAATGAATCGAAACCCTGGAGTCGTGACCCCGGAAGAACCTGCCAGAGCCGGAATTTCGAGTTCTGCTTCCGGGCCAAACTG(配列番号:41))及び、mfsNRP.AS88、アンチセンス鎖(Invitrogen)(下記に示される:5' GATCCAGTTTGGCCCGGAAGCAGAACTCGAAATTCCGGCTCGGCAGGTTCTTCCGGGGTCACGACTCCAGGGTTTCGATTCATTCCG(配列番号:42))を変性させ、さらに、100mMのNaCl、0.1mMのEDTA(Sigma)、20mMのHEPES(pH7.9)中で各々48μMでオリゴヌクレオチドを95℃2分加熱してアニールさせ、1℃/分で60℃に冷却し、60℃でさらに1.5時間維持した。1μLのアニールさせた88mer及び300ngのQiaexを抽出し(Qiagen)、ゲル精製し、BamHI(Roche Diagnostics Ltd., Auckland, New Zealand)EcoRI(New England Biolabs Inc.)二重消化したpGEM7Zf(-)を4℃で19時間0.2ユニットのT4リガーゼ(Epicentre, Madison, WI, USA)で連結した。コンピテントDH10B(Invitrogen)で調製したクローンを、ClaI(Roche)耐性プラスミドの存在及びEcoRI/BamHI二重消化時の88merの切り出しについてスクリーニングした。センス及びアンチセンスRNAプローブの合成のための鋳型は、それぞれ20UのBamHI又はEcoRIを用い、10μgのDNA(JetStar Maxi Kit(Genomed)を用いて調製)を完全に消化することによって調製した。コンサート・ラピッド・ゲル抽出系(Concert Rapid Gel Extraction System)(Invvitrogen)を用いて鋳型をゲル精製した。
リボプローブを転写し、DIGノザンスターターキット(Roche)を用いてノザンブロットを実施した。周産期の星状細胞又はpUSE-myc-mNRPをトランスフェクトしたHEK細胞の全RNAの10μgを1.2%のホルムアルデヒドRNAゲルで0.16−1.77kbのRNAラダーとともに分離し、SSC中で陽性に荷電したナイロン膜(Roche)に移した。マーカーレーンを切り出しメチレンブルーで染色した。DIG標識NRPプローブを52℃で一晩ハイブリダイズさせた。ストリンジェントな洗浄及びDIG抗体とのインキュベーションの後、バイオイメージングシステム(UVP)を用いてCDP*ルミネッセンスによってシグナルを検出した。全脳をE15マウスから摘出し、4%PFAで3時間固定し、20%シュクロースで一晩凍結に対して保護し、ティシュテク(Tissuetek)OCT媒体に包埋し、-80℃で保存した。前記凍結保護した脳をPLL被覆スライド上で14μmの切片に切断し、8μg/malのプロテアーゼKで8分処理し、4%PFAで5分間後染色し、さらに45℃でDIG標識NRPプローブ(88merプローブ配列)(1:100希釈)で、センスコントロールとともに一晩ハイブリダイズした。抗DIG抗体で標識し、NBT/BCIPでシグナルを発色させた後、切片をネスチン/GFAP/b-チュブリンで二重標識し、蛍光二次抗体で可視化した。
【0072】
結果:
PCRを用いて、NRPメッセージがE15マウス胎児の脳の種々の領域で検出されたが、興味深いことには発現レベルは培養されているマウスの神経幹細胞ではるかに高かった(図6A)。神経幹細胞特異的発現は、マウスNRP mRNAに対するin situハイブリダイゼーション及び幹細胞/前駆細胞特異的中間体フィラメントネスチンによる二重染色によってさらに立証された(Lendahl et al., 1990)(図6E−6H)。NRPメッセージ(mRNA)がNSCに由来するニューロン又はグリア子孫細胞で維持されているか否かを判定するために、未分化NSCにおける発現レベルをCNTFにより星状細胞に分化させたNSC及びBDNFによってニューロンへ分化させたNSCと比較した。NRP発現レベルはCNTF処理細胞でのみ顕著に増加し、星状細胞系列は分泌タンパク質の主要な供給であることを示した(図6B)。E19マウスのいくつかの組織における発現レベルを比較し、肺を除いて非神経組織での発現は低く、存在しないこともあることが明らかになった(図6C)。星状細胞での比較的高い発現レベルを基にして、周産期星状細胞培養由来のRNAでのノザンブロットを試み、それによってほぼ0.8及び1.2kbのサイズの2つの転写物が明らかになった。転写は翻訳開始点の上流-114及び-509から始まるので、これは予想される完全長の転写物に一致する(図6D)。E15マウス胎児の前脳の切片のin situハイブリダイゼーションによって、辺縁ゾーン及びサブプレートにおける強いシグナルが明らかになり、皮質プレートの弱い標識によって補完されていた(図6I)。マウスNRP mRNAは、ネスチン免疫反応性と同時局在を示した。この段階で、GFAP又はMAP2の同時標識は観察されなかった(データは示されていない)。この染色パターンは、放射状グリア様細胞における発現を示し、前記細胞は未成熟な星状細胞への形質転換を経ている(Steindler & Laywell, 2003)。
【0073】
実施例7:小脳微小エクスプラントII−NRP-2セグメントSW(配列番号:24及びSDF-1)の神経保護の比較
方法:
小脳微小エクスプラントを、実施例9で述べたようにグルタメート/3-NP及びNRPの他にSDF-1も加えて調製した。
結果:
非保護小脳微小エクスプラントの0.5mMの3-NP/グルタメートによる48時間の処理で生じた損傷は75−92%の範囲の細胞死であった。図7AはNRP-7SWは極めて顕著な神経保護を100fMから100nMの濃度範囲にわたって付与し、100pMでほぼ50%の損傷からの回復を示した。比較すれば、10nMのヒトSDF-1は30%未満の神経保護を付与し、狭い用量範囲の有効性を示した(図7B)。
16merの悪液質フラグメントはより低い有効性を示し、ニューロン保護及びニューロンの増殖誘導については10倍低い潜在能力を有していた(データは示されていない)。
【0074】
実施例8:ニューロン遊走の誘導:NRP-9セグメントSD(配列番号:34)、NRP-2セグメントSW(配列番号:24)及びSDF-1の比較
接触遊走アッセイのための方法:
接触遊走アッセイは、ラットNRP-9SD(配列番号:34)(4nM)及びマウスNRP-7SW(配列番号:24)(0.4nM)をボイデン(Boyden)チャンバー中の誘引物質として用い、実施例10の記載にしたがって実施した。
視床皮質系-皮質培養の方法
ロングエヴァンス(Long Evans)ラットの新生児(P0)由来の後頭皮質及び視床背面を、Paxinos37のP0アトラスにしたがって切り出した。後頭皮質は冠状に、視床背面は前方から組織ショッパー(McIlwain)で350μmの厚さの切片にし、これらを直ちに0.65%のD-グルコース(Merck)を補充したゲイのバランス塩類溶液(Gey's Balanced Salt Solution(GBSS))に移し、7℃で1時間回復させた。垂直の向きを有する視床切片を立体顕微鏡下で選別し、皮質組織と少なくとも3mm離して一緒にカバースリップ上に配置した。この事例では、視床は手綱核が皮質層VIに向き合う方向に置かれている。これらの切片は、10μLのトロンビン(25U/mL、ICN)で凝固させたニワトリ血漿(Cocalico)の10μLを用いて血漿凝塊中でカバースリップに接着させた。カバースリップをローラーチューブ(Nunc)内に置き、0.75半人工培養液(2/4基準培養液イーグル、1/4ハンクスバランス塩類溶液、1/4不活化ウマ血清、2mMのL-グルタミン及び0.65%D-グルコース)を供給した。培養物をローラーチューブのインキュベーターで36℃で20日までin vitroで維持し(DIV)、ラットの悪液質関連NRPを含む培養液は3日毎に交換した。
ラット胎児の前脳のOTCは記載したように調製した。簡単に記せば、妊娠E17期の妊娠ラットを犠牲にし、迅速に胎児を取り出した。前脳の冠状切片を400μmの厚さに切り、3cmのペトリ皿(1mLのDMEM/F12、6.5mg/mLのグルコース、0.1mMグルタミン及び10%FCSを含む)中のミリセルPICM ORG50メンブレン(Millipore)上に静置した。その後、培養液を1mLのNeurobasal/B27、6.5mg/mLグルコース、0.1mMグルタミンに交換した。前記には10μLのPBS、NRP又はSDF-1が表示の濃度で添加され、さらに均一なサイズの小さなDiI(1,1'-ジオクトデシシル-3,3,3',3-テトラメチルインドカルボシアニン)結晶を、立体顕微鏡下で内側神経節隆起に静置した(n=10−15)。24時間培養した後、総数又は神経節隆起及び皮質内に分散したニューロンを倒立顕微鏡下で数えた。
【0075】
結果:
NRP-9SD(配列番号:34)(4nM)及びマウスNRP-7SW(配列番号:24)(0.4nM)は、マウス神経幹細胞に対してNRPの実質的な化学誘引活性を示した。コントロール条件の細胞数のほぼ4倍の細胞がNRP-9SDによって誘引されたが、10分の1未満の量のNRP-7SWによってなお2倍を超えるNSCが培養皿の底に誘引された。比較すれば、100nMのSDF-1被覆は、0.4nMのマウスNRP-7SWの場合と同様な数の細胞を誘引した(図36A)。それらの代表的な形態的表現型が図8Bに示されている。
神経節隆起及び皮質原基を含む矢状方向の胎児脳切片を用いるOTCアッセイでは、OTCに投与されたNRP-7SWは、遊走するニューロン前駆体細胞の数を5倍増強させた(図8C)。ヒトSDF-1は同様な数の細胞を誘導して内側神経節隆起から皮質原基へ遊走させるが、接触走性遊走アッセイの場合のように、63アミノ酸の長さを有する成熟ヒトSDF-1はNRPよりはるかに能力は低い。
【0076】
実施例9:組換えNRPの発現及び活性のアッセイ
HEK293-発現mfsNRPの調製:
HEK293細胞にN-末端及びC-末端にタグを付したmNRP融合タンパク質発現構築物(それぞれpUSE-Flag-mNRP2及びpUSE-myc-mNRP11)を25kDaポリエチレニミンを用いてトランスフェクトした(Boussif et al., 1995)。使用したNRP配列は配列番号:35であった。pEGFP-N1(Clontech)との並行トランスフェクションもまた実施した。簡単に記せば、各構築物をPEIと6、9及び12対1PEIの比で、5%グルコース中に10ng/μLの DNAでDNAホスフェートと一緒にした。36mmウェルで2mLの増殖培養液(10%ウシ胎児血清補充DMEM)中で70%コンフルエントのHEK239細胞に、ウェル当たり200μLのトランスフェクション混合物とともに4.5時間インキュベートすることによってトランスフェクトした。この後、2xの抗生物質/抗マイコプラズマミックスを含む2mLの増殖培養液を各ウェルに添加した。次の日、1mL/ウェルのトリプシン-EDTAを用いて培養を増殖培養液(800μg/mLのジェネチシンを含む)に1:15に分割した。選別は、培養液を週に2回交換することによって17日間維持した。コンフルエンスに達したときに、1:10に分割した細胞の部分で選別を継続し、残りは10%DMSOを含む増殖培養液中で凍結した。さらにまた、800μg/mLのジェネチシン選別から1:2分割を13日目に調製した。安定なトランスフェクタントを発現されたタグ付タンパク質の検出のために採集した。
HEK293細胞でのMyc-NRPの発現はウェスタンブロットで確認された:
ウェスタンブロットを実施して、HEK293細胞でのMyc-NRPの発現を確認した。細胞溶解物を変性条件下で可溶化し、SDSポリアクリルアミド電気泳動(PAGE)を用いてタンパク質を分離した。発現された組換えマウスMyc-NRPは抗Mycモノクローナル抗体によって検出された。
【0077】
結果:
HEK細胞にトランスフェクトされた、c-mycプロモーターの制御下での完全長組み換えマウスNRPの生成により、アンチセンス88bpのNRP cRNAプローブを用いたとき、予想された0.8kbサイズの転写物が明らかになった(図9A)。Myc-NRP-HEK細胞によって発現された16.5kDaの組換えタンパク質生成物は、レムリSDSゲルで20kDaのマーカーサイズへ泳動した(図9B)。
NRP遺伝子によってコードされた、完全なNRP遺伝子組換え発現産物の測定された生物学的活性は、テストしたNRP合成ペプチドのそれと同じであった。小脳微小エクスプラントとの同時培養アッセイでは、Myc-NRP-HEK細胞は、種々の細胞濃度で播種したとき実質的な神経保護作用を付与し、Myc-NRP-HEK細胞数5000で、回復値はMAP-2陽性ニューロンの51%であった(図9C)。空ベクターのみを含むコントロールMyc-HEKは、酸化性/興奮毒性ストレスの後MAP-2陽性細胞の回復を全く示さなかった。添加したHEK細胞は、神経保護について釣鐘型の用量応答曲線を示した。
Myc-NRP-HEK細胞は、マウスのNSCを用いて接触走性遊走アッセイでテストしたとき、化学誘引ニューロン遊走誘導活性を示した。マウスNRPを発現しているMyc-NRP-HEK細胞をボイデンチャンバーの底に播種し、マウスNCSをインサートに播種したとき、24時間後に検出された神経突起伸長を示すニューロン細胞集団はコントロールMyc-HEK細胞を用いた同じアッセイと比較して2倍増加した(図9D)。
実施例10:NRPの作用の可能なメカニズム:
最近のデータは、SDF-1(CXCL12)及びそのレセプターCXCR4は、免疫系及び神経系で同様な作用を有する可能性を示唆している。すなわち、それらは細胞の移動、ニューロン及びリンパ球の増殖、可塑性(神経突起伸長及び分化)並びに生存を調節することができる(Vlahakis et al., 2002 and Lazarini et al., 2003)。CXCL12に対する同様な依存は協調的なホメオスタシスによる相互作用を支持することができるかもしれないが、CXCR4レセプター結合の結果として、HIV感染及びそれに続く神経障害罹患のように炎症性プロセスに対する固有の脆弱性を構成するかもしれない(Keswani et al., 2003)。HIV及び関連ウイルスは、リンパ球レセプターCD-4の他にコレセプターを標的細胞への感染に必要とする。HIVに使用されるコレセプターの1つは、G-タンパク質結合ケモカインレセプターCXCR4である。SDF-1とCXCR4との相互作用はCD4-リンパ球へのHIVの侵入を阻止することができる。さらにまた、CXCR4-レセプターと拮抗することによって癌細胞の移動/侵襲を阻害することにより、動物モデルで癌の転移を阻止することができることが知られている(Rubin et al., 2003;Liang et al., 2004)。
これまでのところ、SDF-1は、CXCR4レセプターと結合する唯一の公知リガンドである。我々は、ここで、NRPはCXCR4レセプターに対する新規なクラスのリガンドでありえること、さらに生物学的活性(例えば化学誘引及びニューロン生存)はCXCR4レセプターの活性化によって発揮できることを示すいくつかの証拠を提供する。したがって、CXCR4レセプターに対するNRPの拮抗作用が存在しえる可能性も排除しないが、NRPはCXCR4レセプターのためのアゴニストであることができる。同様な観察がSDF-1についても得られた。すなわち、単一アミノ酸置換がCXCR4に対するSDF-1作用に拮抗することができる(Tudan et al., WO0185196)。しかしながら、これはNRPの作用のただ1つの可能なメカニズムではなく、他のメカニズムもここに記載したこの観察を説明しえることは理解されよう。
【0078】
方法:
小脳微小エクスプラントを、グルタミン/3-NP及びNRPの他に、SDF-1、ワートマンニン及びPD98059(両者ともCalbiochem)を添加しながら実施例9で述べたように調製した。
NRPの生存促進及び遊走誘導活性に対するCXCR4レセプター中和抗体の作用
CXCR4レセプター(フュージンレセプター)に対して作製した中和抗体1μg/mLを72時間適用することによって、NRPで処理した小脳微小エクスプラント内のニューロンの生存及びニューロンの遊走パターンは顕著に低下した。1μg/mLの抗体濃度で、NRP-9 SD(配列番号:34)の神経保護及び/又は遊走促進作用は失われた(図10A)。同様な阻害は、ヒトNRP-2KS(配列番号:23)(図10B)又はマウスNRP-7SW(配列番号:24)(データは示されていない)についても観察された。
NRPの化学遊走作用に対するCXCR4レセプター中和抗体の作用
培養皿に被覆された10ng/mLのNRPの化学誘引作用は、ニューロン幹細胞株MEB-5をCXCR4に対する中和抗体で1.5時間プレ-インキュベートすることによって阻止された。NRP-9SD(配列番号:34)ペプチド単独で観察された遊走と比較して、極めてわずかな細胞が遊走した(図10C)。
NRPの作用はERK1/2及びAktリン酸化によって媒介され得る
神経保護及び/又は遊走作用を発揮するためにNRPによって利用されるシグナリング経路を解明するために、我々は、MEK阻害剤PD98509及びホスファチジルイノシトール3-キナーゼ(PI-3K)阻害剤ワートマンニンを用いてMAPK又はAktリン酸化を阻害した。
NRP媒介神経保護及び化学誘引の作用メカニズムにおける、MAPK及びakt活性の上流の調節因子PI-3K(Aktのリン酸化)の役割を知るために、我々は、小脳微小エクスプラント及び接触走性遊走アッセイにおける有効性についてMAPKキナーゼ(MEK)阻害剤PD98509をテストした。ビヒクル及びPD98509による又はPD98509なしの損傷治療によって前記阻害剤による軽度の毒性のみが示され、一方、阻害剤とNRP-9SD(配列番号:34)による同時処理によってNRPの神経保護活性は失われた(図11A)。Aktリン酸化のワートマンニン(100nM)による阻害は、ビヒクル及びNRP無しの損傷治療後には有意な影響を示さなかったが、ERK1/2のリン酸化の阻害とは同様であり、NRPによって生じる神経保護は失われた(図11B)。接触走性遊走アッセイでは、遊走NSCでほぼ3倍の増加がPD98509(0.1pMから100pM)でほぼ完全に阻止され、BSA被覆ウェルでの基礎遊走に対しては有意な影響はなかった(図11C)。神経保護とは対照的に、NRPの化学誘引遊走活性はワートマンニンによるPI-3Kの阻害によっては抑制されなかった(データは示されていない)。底部の区画へ遊走した細胞数の低下がニューロン生存の障害によるものであったという可能性を排除するために、インサートをSyto2で生細胞について染色し、上部表面を分析した。生存細胞における顕著な相違は見出されなかった。
【0079】
実施例11:in vivoでのNRP有効性
材料と方法:
in vivoでのNRPの有効性をテストするために、低酸素虚血損傷(HI)に暴露したラットで実験を行った。成獣ラット(50日齢、ウィスターラット、250−300g、雄)を用いた。改変したレヴィン(Levine)のモデル調製及び実験方法を用いた(Rice et al.,1981, Ann. Neurol. 9:131-141;Guan et al., 1993, Cereb. Blood Flow Metab. 13(4):609-16)。簡単に記せば、これらの方法は、側脳室にカニューレを埋め込んだ動物での一側性頚動脈結紮により誘導したHI損傷及びそれに続く吸入性仮死から成る。ハロタン麻酔下で定位脳手術により、誘導カニューレを硬膜の最頂部上の正中線より右へ1.5mm及び耳間ゼロ平面に対して前方7.5mmの位置に設置した。カニューレ設置2日後に右頚動脈を二重結紮した。麻酔から1時間の回復時間後に、各ラットを、湿度(90±5%)及び温度(31℃±0.5℃)が制御されているインキュベーターにさらに1時間置き、続いて各ラットを低酸素(6%酸素)に10分間暴露した。処置前に動物をインキュベーターでさらに2時間維持した。
低酸素虚血損傷の後、ラットを、生理食塩水に溶解したNRP-4セグメントPQ(PGRAEAGGQ;配列番号:43)の5nM(n=12)、50nM(n=12)若しくは500nM(n=12)又はビヒクル(n=10)(通常の生理食塩水)の脳室内(icv)投与により処置した。
組織学的試験を低酸素虚血後5日のラットで実施した。ラットを過剰のペントバルビタールで殺し、通常の生理食塩水、その後10%のホルマリンを心臓経由で灌流させた。脳は処理前最低2日間同じ固定液中に維持し、標準的なパラフィン包埋術を用いた。
厚さ8μmの冠状切片を線条体、大脳皮質及び海馬から切り出し、チオニン及び酸性フクシンで染色した。組織損傷の重篤度は、線条体、皮質並びに海馬のCA1-2、CA3、CA4及び歯状回でスコアを決定した。組織傷害はニューロンの消失(好酸性(赤色)細胞質及び凝縮核)、広範囲壊死及び細胞性反応として特定した。組織傷害は以下のスコアシステムを用いてスコアを決定した:0:組織が組織傷害を示さなかった;1:<5%の組織が傷害された;2:<50%の組織が傷害された;3:>50%の組織が傷害された;及び4:>95%の組織が傷害された。
結果と結論:
この実験の結果は図12に示されている。ヒト染色体15の9merフラグメント、NRP-4セグメントPQ(PGRAEAGGQ-NH2;配列番号:43;低酸素症後2時間で9ng/icv)によって、損傷後5日の全ての解析した脳内で100%の神経保護が付与された。
【0080】
実施例12:嗅球鞘性グリア(“OEG”)細胞の成長促進
NRPが神経組織の再ミエリン化に有用でありえるか否かを決定するために、我々は、ヒト嗅球鞘性(ensheating)グリア(OEG)細胞を用いて実験を行った。OEG細胞は、傷害組織(例えば脊椎の組織)の集団再形成のための細胞の供給集団として評価されている。OEG細胞は、当業界で公知の方法を用いてヒトの鼻粘膜の掻爬検体から得た。細胞は制御条件下及びNRP-7SW(配列番号:24)の存在下で培養液で増殖させた。
制御条件下では、OEG細胞の正常な増殖速度は72時間内で約25%である。我々は、NRP-7SW(配列番号:24)の存在下では、OEG細胞は72時間にわたって約50%の速度で増殖することを見いだした。これらの結果は、NRPはOEG細胞の増殖を強化することができ、それによって神経細胞移植方法において有用で細胞増殖を促進し、傷害組織に神経機能を取り戻すことができることを示している(図13)。
【0081】
実施例13:NRP-5セグメントRGペプチドアナログの神経保護活性
NRP-5セグメントRG関連ペプチドのアミノ酸配列における変更が、NRP-5セグメントRG(配列番号:30)の活性と異なる活性を有するペプチドを生成するか否かを決定するために、我々は種々のアミノ酸置換を有する合成ペプチドを作製した。我々は、本明細書に開示した小脳微小エクスプラントアッセイで置換NRP-5RGペプチドをテストした。
アミノ酸配列REGRRDAPGRAGG-NH2(配列番号:30)を有するアミド化ペプチドのN-末端の最初の5つのアミノ酸内でのアミノ酸置換によって、配列REAAADAPGRAGG-NH2(配列番号:44)及びAAARRDAPGRAGG-NH2(配列番号:45)を有するペプチドを作製した。
PGRドメインにおけるアミノ酸置換はアミノ酸配列REGRRDAAAAAGG-NH2(配列番号:47)を有するペプチドを生じ、C-末端GGドメインの置換はアミノ酸配列REGRRDAPGRAAA-NH2(配列番号:46)を有するペプチドを生じた。上記の置換ペプチドの各々をテストし、その結果をNRP-5セグメントRG(配列番号:30;“NRP-5RG”)を用いて得られた結果と比較した。
図14Aは、配列番号:44は、最大効果が約1pMの濃度で観察される神経保護活性を有していた。右側の棒線(“標準13mer”と表示されている)はNRP-5RG(配列番号:30)を示す。NRP-5RGのアミノ酸3−5の置換はペプチドの活性プロフィルを有意には変化させなかった(図42A)。
図14Bは、配列番号:45が神経保護活性を有し、最大効果は約1pMで観察されることを示している。右側の棒線(「原型配列」と表示されている)はNRP-5RG(配列番号:30)を示す。AAARRDAPGRAGG-NH2(配列番号:45)を生じる1−3位の置換は、原型配列NRP-5RGよりもいっそう高い神経保護活性をもたらした。この結果は、NRP-5RGよりも高いこのアナログの安定性プロフィルによるものであろう。ただし他の仮説もこの観察の説明になりえる。
図42Cは、配列番号:46がNRP-5RP(配列番号:30;右の棒線;“標準13mer”)よりも、又は配列番号:44又は配列番号:45(それぞれ図14A及び14B)よりも低い効果を有することを示している。実際、統計的に有意な効果は濃度100nMで観察されたに過ぎない。原型配列(配列番号:30)は20%高い神経保護を生じ、配列番号:46よりも106から107倍高い潜在能力を有する。
図14Dは、配列番号:47はある種の濃度(例えば0.1pM、10pM及び100nM)ではある程度の神経保護作用を有するが、他の濃度ではNRP-5RG(配列番号:30;「標準」と標識された右の棒線)について観察されたものと比較して活性はなかったことを示す。
我々はこれらの実験から、PGR及びC-末端GGドメインは、NRP-5RG(配列番号:30)の活性の維持のために有用であると結論した。我々はまた、NRPのアミド化C-末端は神経保護作用を生じるために十分ではないと結論した。なぜならば、C-末端アミド化NRP(配列番号:46)は、何らかの神経保護作用があるとしても、大半の濃度でその作用はほとんどなかったからである。さらにまた、NRPの内部における変更は、たとえ内部のアミノ酸のいずれもC-末端アミド基を持っていないとしても、活性に影響を与えた。
【0082】
実施例14:NRP-5RGの増殖誘導及び化学誘引活性
また別の実験セットで、我々は、本明細書に記載した胎児の小脳微小エクスプラントにおける神経細胞増殖の誘導に対するNRP-5RGの作用を決定した。図15は、NRP-5RGが増殖誘導活性を有し、最大活性は100pMの濃度で観察されることを示している。ある程度の作用は1mPで、さらに0.1pMの濃度でさえ観察されるが、これらの作用は統計的には有意ではなかった。
さらに別の実験で、我々は、本明細書に記載した接触走性遊走アッセイでNRP-5RGの化学誘引作用を調べた。図16は、NRP-5RGが化学誘引性であり、コントロール(BSA含有)培養液に暴露された細胞と比較して42.1%強い作用を有することを示している。
【0083】
実施例15:NRPは酸化性ストレスに応答して神経細胞を保護した
この実験シリーズでは、我々は、NRP-4セグメントPQ(“NRP-4PQ”)は酸化性損傷から小脳微小エクスプラントを保護することができるか否かを調べた。我々は、ビヒクル、0.1mM過酸化水素、又はビヒクル中の種々の濃度のNRP-4PQに細胞を暴露した。図17は、0.1mMの過酸化水素に対する酸化性ストレスの48時間後のNRP-4PQの神経保護作用を示す。NRP-4PQ処理に付随する神経保護は、過酸化物を投与されなかったコントロールのエクスプラントの神経保護すら超えていた。神経突起成長のこの有効性強化のメカニズムは明確ではないが、前記は、微小エクスプラント自体を作製又は維持することに付随するストレスを軽減する可能性がある。
これらの発見は酸化的ストレスが役割を演じる多くの条件と関係がある。例えば、酸化的ストレスは慢性のCNS損傷及び疾患の全てに付随する。NRP作用による酸化的ストレス仲介神経毒性の抑制は、多くのCNS損傷又は疾患に対して非常に有益でありえる。
【0084】
実施例16:リン酸化NRP-7SWによって仲介される神経保護
我々は、一連の実験を実施して、N-末端アミノ酸セリンのリン酸化がNRP-7セグメントSW(「NRP-7SW」)の神経保護作用を変化させるか否かを決定した。NRP-7SWのリン酸化によってNRP-7PSWが生成される。セリンはin vivo条件下ではリン酸化される(NetPhos 2.0 Server−Technical University of Denmark)という高い確率予想(0.9)のために、我々はN-末端セリンの役割の仮説をたてた。小脳微小エクスプラントを0.5mMの3-NP/グルタメートで損傷させ、NRP-7PSWで処理した。
図18は、NRP-7PSWは神経保護作用を持つことを示す実験結果を示している。0.1pMという低い濃度で顕著な作用が観察され、前記濃度は、テストした他の濃度よりも高い神経保護活性(44.2%の神経保護)を示した。使用した最低の濃度(0.1pM)がかなりの神経保護作用を示したので、より低濃度でも神経保護作用が示されるであろう。
【0085】
実施例17:NRP-9SDによる分化の誘導後のNSCの増殖強化
E15マウスのNSC P10細胞をラミニン上に播種し、NRP-9SD(配列番号:34)の存在下で3日間培養し、培養期間の最後の48時間にはBrdUの存在下で培養した。各条件について、2つの別個のウェルで4視野を数え、増殖細胞数を決定した。
結果:
NRP-9SDは、分化中のマウスNSC前駆体細胞の増殖速度を高めた(図19)。この生物学的活性は100fMから1nMの広い用量範囲で観察された。
【0086】
実施例18:NRP IIの発現
in situハイブリダイゼーション:
全脳をE15及びE17マウスから摘出し、4%パラホルムアルデヒド(PFA)で3時間固定し、20%シュクロースで一晩凍結保護し、ティシュテック(Tissuetek)OCT(Sakura finetek)で包埋し-80℃で保存した。この凍結保護脳を14μmの厚さの切片に切り出し、PLL被覆スライドに静置し、8μg/malのプロテイナーゼKで8分処理し、4%PFAで5分後固定し、さらにDIG-標識NRPセンス及びアンチセンスプローブで45℃で一晩ハイブリダイズさせた(実施例13に記載の88merプローブ配列;センス鎖は配列番号:41、アンチセンス鎖は配列番号:42;1:100希釈)。抗DIG抗体での標識及びNBT/BCIPによるシグナルの発色後、切片をネスチン/GFAP/βIII-チュブリン及びヴィメンチンの1:100希釈(abcam, マウスモノクローナル[RV202], ab8978-1)で二重標識し、二次蛍光抗体を用いて可視化した。センスコントロールは陰性のままであった。
結果:
図20A及び20Bは上述の動物の脳の蛍光顕微鏡写真を示す。マウスのフレームシフトNRPメッセージと放射状グリアマーカー、ヴィメンチンとの同時局在は、NRP発現は神経上皮幹細胞-放射状グリア-星状細胞系列で維持されていることを示した(図20A及び20B)。このことは神経幹細胞の重要な機能を指摘している。なぜならば、放射状グリアは発生中に多くの脳領域でニューロンを生じることが示され(TE Anthony, C. Klein, G. Fishell, N. Heinz, 2004 ”Radial glial cells serve as neuronal progenitors in all regions of the central nervous system.” Neuron, 41:881-890)、成獣の脳室下ゾーンの星状細胞は神経幹細胞であることが示された(F. Doetsch, I. Caile, DA Lim, JM Garcia-Verdugo, A. Alvarez-Buylla, 1999 ”Subventricular zone astrocytes are neural stem cells in adult mammalian brain”, Cell 97:703-706”)からである。
これらの結果から我々は、NRP mRNAはマウスの脳で生成されると結論した。
【0087】
実施例19:配列番号:8及び配列番号:9の再配列決定
2004年8月20日に、配列番号:8及び配列番号:9に対応する更改配列の報告が刊行された(NBCI, NT_026446)。新しく同定された配列を基に、我々は、ヌクレオチドNRP配列は75アミノ酸を有するペプチドをコードする225ヌクレオチドから成るとアノテーションを付した。NRP(配列番号:8)をコードする染色体15上の以前にアノテーションを付された配列にしたがって、前記配列内で以下を変更した:168位はGからCに変更し、それ以外エクソン1−4は(NRPの生物学的に活性な配列を含めて)変更なし。179位から先へ向かって新規にアノテーションを付されたエクソン5が始まる。222位から先へ向かってエクソン6が始まる。
【0088】
全アミノ酸配列を一文字コードで記す:
>humchrom15NRPエクソン1 2 3 4 5 6
MAVVLLAPFGDISQEITKVGTGTPGRAEAGGQVSPCLAASCSQAYGAILAHCNLCLPGSSDLPASASQSARLQVD
配列番号:49
【0089】
実施例20:卒中の治療
患者は卒中の徴候を示している。卒中の診断が為され、続いて医師は、NRP化合物を患者に静脈内に、或いは脳室内に直接又は患者の脳の罹患部分に直接投与する。NRP化合物は本明細書に開示したようにペプチド又はタンパク質であり、医薬的に許容できる形態(所望する場合にはビヒクル、緩衝剤及び安定剤を含む)で投与される。NRPによる処置は、卒中に付随する神経変性を減少させ、さらに予想される症状の悪化を少なくとも部分的に遅らせるか、停止させるか、又は少なくとも部分的に回復させる。
【0090】
実施例21:NRPの予防的使用
患者は心脈管機能不全と診断され、冠状動脈バイパス(CABG)手術が指示される。CABG手術は脳灌流の低下を伴い、脳灌流の低下は低酸素性又は虚血性脳損傷をもたらしえる。そのような低酸素症及び虚血の悪影響を減少させるために、患者はNRP化合物で前処置される。NRP化合物は患者に医薬的に許容できる形態(所望する場合にはビヒクル及び/又は安定剤を含む)で投与される。投与ルートには静脈内、脳内、または脳室経由投与が含まれる。所望の場合は、マルチルート投与を用いることができる。CABG手術を受ける患者の前処置は、CABG手術に伴う神経変性を減少させ、患者は、NRP前処置を施されずにCABG手術を受けた患者と比較して、術後の神経学的損失が減少する。
本発明をその具体的な実施態様に関して記載した。当業者は、本出願の開示及び教示を取り入れた他の実施態様を面倒な実験なしに開発することができるであろう。それらの実施態様はいずれも本発明の部分と考えられる。本明細書に引用した全ての参考文献は引用により本明細書に含まれるものとする。
【0091】
参考文献
Akerblom, IE, and Murry, LE (1996). Human cachexia associated protein. US patent 5,834,192.
Anderson, CV, Wood, DM, Bigler, ED, and Blatter, DD (1996). Lesion volume, injury severity, and thalamic integrity following head injury. J. Neurotrauma 13: 35-40.
Anderson, WF (1992) Human gene therapy. Science 256: 808-813.
Bach et al., (1995) Insulin like growth factor binding proteins. Diabetes Reviews 3: 38-61.
Baldwin, ME, Roufail, S, Halford, MM, Alitalo, K, Stacker, SA and Achen, MG (2001). Multiple forms of mouse vascular endothelial growth factor-D are generated by RNA splicing and proteolysis. J Biol Chem 276: 44307-44314.
Beal, MF, Kowall, NW, Ellison, DW, Mazurek, MF, Swartz, KJ, and Martin, JB (1986). Replication of the neurochemical characteristics of Huntington's disease by quinolinic acid. Nature 321: 168-171.
Bolz, J, Novak, N, and Staiger, V (1992). Formation of specific afferent connections on organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12:3054-3070.
Brose, K, and Tessier-Lavigne, M (2000). Slit proteins: key regulators of axon guidance, axonal branching, and cell migration. Curr. Opin. Neurobiol. 10: 95-102.
Cote, F, Do, TH, Laflamme, L, Gallo, J-M, and Gallo-Payet, N (1999). Activation of the AT2 receptor of angiotensin II induces neurite outgrowth and cell migration in microexplant cultures of the cerebellum. J. Biol. Chem. 274: 31686-31692.
Cunningham, TJ, Hodge, L, Speicher, D, Reim, D, Tyler-Polsz, C, Levitt, P, Eagleson, K, Kennedy, S, and Wang, Y (1998). Identification of a survival promoting peptide in medium conditioned byoxidatively stressed cell lined of nervous system origin. J. Neurosci. 18: 7047-7060.
De Curtis, I, and Reichardt, LF (1993). Function and spatial distribution in developing chick retina of the laminin receptor a6b1 and its isoforms. Development 118: 377-388.
Dodd, J, Morton, SB, Karagogeos, D, Yamamoto, M, and Jessell, TM (1988). Spatial regulation of axonal glycoprotein expression on subsets of embryonic spinal neurons. Neuron 1: 105-116.
Dyke, MW, Bianchi-Scarra, G, and Musso, M (2001). Characterization of a triplex DNA-binding protein encoded by an alternative reading frame of loricrin. Eur. J. Biochem 268: 225-234.
Fallon, J, Reid, S, Kinyamu, R, Opole, I, Opole, R, Baratta, J, Korc, M, Endo, TL, Duong, A, Nguyen, G, Karkehabadhi, M, Twardzik, D, and Loughlin, S (2000). In vivo induction of massive proliferation, directed migration, and differentiation of neural cells in the adult mammalian brain. PNAS 97: 14686-1491.
Ferri, RT, and Levitt, P (1995). Regulation of regional differences in the differentiation of cerebral cortical neurons by EGF family-matrix interactions. Development 121: 1151-1160.
Fueshko, S, and Wray S (1994). LHRH cells migrate on peripherin fibers in embryonic olfactory explant cultures: an in vitro model for neurophilic neuronal migration. Dev. Biol. 166: 331-348.
Gahwiler, BH (1981). Organotypic monolayer cultures of nervous tissue. J. Neurosci. Methods 4: 329-342.
Ganzler, SI, and Redies, C (1995). R-cadherin expression during nucleus formation in chicken forebrain. J. Neurosci. 15: 57-72.
Gomez, TM, and Spitzer, NC (1999). In vivo regulation of axon extension and pathfinding by growth-cone calcium transients. Nature 397: 350-355.
Gulyas, AI, Hajos, N, and Freund, TF (1996). Interneurons containing calretinin are specialized to control other neurons in the rat hippocampus. J. Neurosci. 16: 3397-3411.
Guth, S, Tange, TO, Kellenberger, E, Valcarcel, J (2001). Dual function for U2AF35 in AG-dependent pre-mRNA splicing. Mol Cell Biol 21: 7673-7681.
Hatten, ME, and Heintz, N (1999). Neurogenesis and migration In Fundamental Neuroscience, (R: Zigmond, ed.), pp 451-479, Academic Press, San Diego.
Hermann, DM, Mies, G, Hata, R, and Hossmann, KA (2000). Microglial and astrocytic reactions prior to onset of thalamic cell death after traumatic lesion of the rat sensorimotor cortex. Acta Neuropathol (Berl) 99: 147-153.
Hughes, PE, Alexi, T, Williams, CE, Clark, RG, and Gluckman, PD (1999). Administration of recombinant human Activin-A has powerful neurotrophic effects on select striatal phenotypes in the quinolinic acid lesion model of Huntington's disease. Neuroscience 92: 197-209.
Hwang et al., (1980). Hepatic Uptake and degredation of unilamellar sphingomyelin/cholesterol liposomes: a kinetic study. Proc. of the Natl. Acad. Of Sciences USA 77: 4030-4034.
Ishii, N, Wadsworth, WG, Stern, BD, Culotti, JG, and Hedgecock, EM (1992). UNC-6, a laminin-related protein, guides cell and pioneer axon migrations in C. elegans. Neuron 9: 873-881.
Langer et al., (1981) Biocompatibility of polymeric delivery systems for macromolecules. J. Biomed. Mater. Res. 15: 27-277.
Liang, S, and Crutcher, KA (1992). Neuronal migration in vitro. Dev. Brain Res. 66: 127-132.
Lu, Q., Sun, E., Klein, R. S. and Flanagan I. G.(2001). Ephrin-b reverse signalling is mediated by a novel PDZ-RGS protein and selectively inhibits G-protein coupled in chemoattraction. Cell 105: 69-79.
Nakao, N, and Itakura, T (2000). Fetal tissue transplants in animal models of Huntington's disease: the effects on damaged neuronal circuitry and behavioural deficits. Prog. Neurobiol. 61: 313-338.
Obst, K, and Wahle, P (1995). Areal differences of NPY mRNA expressing neurons are established in the late postnatal rat visual cortex in vivo, but not in organotypic cultures. Eur. J. Neurosci. 7: 2139-2158.
Pasterkamp, RJ, Giger, RJ, Baker, RE, Hermens, WT, and Verhaagen J (2000). Ectopic adenoviral vector-directed expression of Sema3A in organotypic spinal cord explants inhibits growth of primary sensory afferents. Dev. Biol. 220: 129-141.
Paxinos, G, Toerk, I, Tecott, LH, and Valentino, KL (1991). Atlas of the Developing Brain. Academic Press: San Diego.
Polleux, F, Morrow, T, and Ghosh, A (2000). Semaphorin3A is a chemoattractant for cortical dendrites. Nature 404: 567-573.
Rozas, G, Liste, I, Lopez-Martin, E, Guerra, Mj, Kokaia, M, and Labandeira-Garcia, JL (1996). Intrathalamic implants of GABA-releasing polymer matrices reduce motor impairments in rats with excitotoxically lesioned striata. Exp. Neurol. 142: 323-330.
Sieg, F, Obst, K, Gorba, T, Riederer, B. Pape, H-C, and Wahle, P (1998). Postnatal expression pattern of calcium-binding proteins in organotypic thalamic cultures and in the dorsal thalamus in vivo. Dev. Brain Res. 110: 83-95.
Stoppini, L, Buchs, P-A, and Muller, D (1991). A simple method for organotypic cultures of nervous tissue. J. Neurosci. Methods 37: 173-182.
Van der Flier, A, Kuikman, I, Kramer, D, Geerts, D, Kreft, M, Takafuta, T, Shapiro, SS and Sonnenberg, A (2002). Different splice variants of filamin-B affect myogenesis, subcellular distribution, and determine binding to integrin β subunits. J Cell Biol 156: 361-376.
Wagener, R, Kobbe, B, Aszodi, A, Aeschlimann, D, and Paulsson M (2001). Characterization of the mouse matrilin-4 gene: A 5' antiparallel overlap with the gene encoding the transcription factor RBP-L. Genomics 76: 89-98.
Wagner et al., (1990) Transferrin-polycation conjugates as carriers for DNA uptake into cells. Proc. Natl. Acad. Sci. USA. 87: 3410-3414.
Wu et al., (1987) Receptor-mediated in vitro gene transformation by a soluble DNA carrier system. J. Biol. Chem. 262: 4429-4432.
Yamamoto M, Hassinger, L, and Crandall, JE (1990). Ultrastructural localization of stage-specific neurite-associated proteins in the developing rat cerebral and cerebellar cortices. J. Neurocytol. 19: 619-627.
Zhu, Y, Yu, T, Zhang, X-C, Nagasawa, T, Wu, JY, Rao, Y (2002). Role of the chemokine SDF-1 as the meningeal attractant for embryonic cerebellar neurons. Nature Neurosci. 5: 719-720.
【0092】
工業的応用性
本発明の実施態様は、神経変性又は神経細胞死を含む症状の治療に使用される組成物の製造に有用な神経再生ペプチド(NRP)のための遺伝子及びペプチドを含む。NRPによる治療は、ニューロンの生存、ニューロンの遊走、ニューロンの分化、神経突起伸長及び/又はニューロンの増殖を増加させる。アルツハイマー病、パーキンソン病、低酸素症、卒中及び冠状動脈バイパス手術のような症状は、本発明の組成物及び方法によって有効に治療されえる。
【図面の簡単な説明】
【0093】
【図1A】図1Aは、イントロンを有するNRP-7(配列番号:35)をコードする遺伝子の構造を、DNA修復タンパク質をコードする既知遺伝子の2bpフレームシフトとしてのNRP遺伝子の存在とともに示している。2つのCpGアイランドを含むプロモーター領域が強調されている(一方のCpGアイランドはNRPについて予想されたものであり、下流のもう一方はDNA修復タンパク質について予想されたものである)。NRP-7(配列番号:35)遺伝子産物を得るためのプライマーの位置が表示されている。
【図1B】図1Bは、マウスNRP(NRP-7ロング)とラットNRP(NRP-9)オルソログとの間の72.2%相同性を示している。赤い数字は、生物学的に活性なNRPドメイン内の対応する相同性を示し、他方、青い線は推定的なN-グリコシル化部位を示す。
【図1C】図1Cは、ヒト悪液質関連タンパク質とマウスNRP(NRP-7ロング)とのアラインメントを示す。両配列の全体にわたるロイシン、グリシン及びプロリンアミノ酸残基の保存に注目されたい。全体的な相同性は34.4%である。悪液質関連タンパク質の生物学的に活性なドメインは20位のシグナルペプチドの直後に位置し、一方マウスNRP(NRP-7ロング)の活性ドメインはアミノ酸83位で開始する。
【図1D】図1Dは、マウスNRPフラグメントとヒトトレホイルタンパク質Ps2との間の全体的な相同性は52.5%であることを示す。トレホイル因子ファミリー(TFF)のコンセンサス配列を規定する15アミノ酸残基のうち9アミノ酸がNRPに存在し、それによってトレホイル因子ドメインがNRPに存在することが確認される。
【図1E】図1Eは、マウスSDF-1αとマウスNRP(NRP-7ロング)タンパク質配列とのアラインメントを示す。両ニューロン化学誘引分子間には32.6%の中等度の相同性が存在する。
【図2】3-NP/グルタメートを用いた興奮毒性(excitotoxic)/酸化性損傷後のNRP-4セグメントGQ(配列番号:26)による生存誘導を示す。
【図3】3-NP/グルタメートを用いて損傷されたニューロンにおけるNRP-7セグメントSW(配列番号:24)による増殖誘導を示す。
【図4A】コーティングに10ng/mLのNRP-4GG(配列番号:29)ペプチドを用いたマウスNEB-5細胞による接触走性(haptotactic)遊走アッセイを示す。200,000個のMEB-5細胞を用いた。
【図4B】コーティングに10ng/mLのNRP-4 GG(配列番号:29)ペプチドを用いた初代マウス幹細胞による遊走アッセイの結果を示す。200,000細胞を播種した。
【図4C】1ng/mLのNRP-2KS(配列番号:23)ペプチドコーティングを用いたマウスNEB-5細胞による遊走アッセイの結果を示す。200,000細胞を播種した。
【図4D】10ng/mLのNRP-7SW(配列番号:24)コーティングを用いた初代マウス幹細胞による遊走アッセイを示す。400,000細胞を播種した。
【図4E】50ng/mLのNRP-4GG(配列番号:29)コーティングを用いた野生型PC-12細胞(200,000細胞/ウェル)による遊走アッセイを示す。
【図4F】野生型PC-12細胞及び1ng/mLのNRP-7SW(配列番号:24)コーティングとそれに続くマトリゲル/PDLコーティングを用いた遊走アッセイを示す。100,000細胞を播種した。
【図5A】図5A及び5Bは、NRPの存在下で7日間分化させた神経幹細胞の各培養ウェルでもっとも長い5本の軸索の中間の長さを示すグラフである。図5AはNRP-7SW(配列番号:24)及びIGF-1の影響を示す。
【図5B】図5A及び5Bは、NRPの存在下で7日間分化させた神経幹細胞の各培養ウェルでもっとも長い5本の軸索の中間の長さを示すグラフである。図5BはNRP-2KS(配列番号:23)、IGF-1又は分化培養液単独の影響を示す。コントロール培養液との有意差が、2-テール(2-tailed)スチューデントのt-検定によって決定されたとおり観察された。**p<0.01;***p<0.001
【図5C】図5C及び5Dは分化培養液単独の場合の最も長い軸索伸長の代表例を示す。
【図5D】図5C及び5Dは分化培養液単独の場合の最も長い軸索伸長の代表例を示す。図5Dは10nMのNRP-7 SW(配列番号:24)の存在下における最長の軸索の成長を示し、図33Dはコントロール条件を示す。
【図5E】図5E及び5Fは、ニューロンの分化を経たNSC細胞におけるNRP-7SW(配列番号:24)及びNRP-2KS(配列番号:23)の増加に対する影響を示すグラフである。
【図5F】図5E及び5Fは、ニューロンの分化を経たNSC細胞におけるNRP-7SW(配列番号:24)及びNRP-2KS(配列番号:23)の増加に対する影響を示すグラフである。
【図5G】ナノモル濃度のNRP-2KS(配列番号:23)が、マウス前脳の神経幹細胞由来の多能性幹細胞を犠牲にしてニューロン前駆体の生成を促進することを示している。
【図5H】分化実験スキームの開始前のNRP-9セグメントSD(配列番号:34)の未分化NSCへの投与を示す。NRP処理下ではBrdU陽性細胞のアップレギュレーションが生じるが、全体的な細胞数には明白な変化はない。
【図6A】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。 図6Aは、マウスNRPの発現は、E15マウスの胎児の脳組織の皮質(Ctx)、線条体(Str)及び嗅球(OB)で検出されたことを示す。しかしながら、そのレベルは、急性的に単離して不死化したマウス神経幹細胞と比較してはるかに低かった。
【図6B】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Bは、CNTFによる神経幹細胞の星状細胞への分化は、未分化幹細胞又はBDNFによるニューロンの分化と比較してNRP mRNA発現を顕著に増加させることを示す。
【図6C】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Cは、種々のE19マウス胎児組織の分析を示すゲルである。前記は、周産期前脳に由来する胎児の皮質、星状細胞系分化NSC及び星状細胞培養と比較して、肺を除きマウスのNRPの発現は非神経組織では低いか又は存在しないことを示している。
【図6D】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Dは、DNA修復タンパク質配列とオーバーラップしない88bpプローブによるノザンブロットハイブリダイゼーションを示す。前記によって、周産期星状細胞培養に由来するRNAで、サイズがほぼ0.8及び1.2kbの2つの別個のmRNAが検出される。
【図6E】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Eは、NSCを標識する、マウスNRPアンチセンスプローブによるin situハイブリダイゼーションを示す。
【図6F】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Fは、センスコントロールにおける特異的なシグナルの欠如を示す。
【図6G】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。 図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Gは、NSCにおけるNRP mRNA発現を示す。
【図6H】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Hは、図6Gで示されたNRP mRNA発現と同じ部位にネスチンが分布することを示す。
【図6I】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Iは、E15マウス前脳切片、脳室帯、皮質原基(特にサブプレート及び辺縁帯)の冠状のNRP mRNAのin situハイブリダイゼーションを示し、皮質プレートではハイブリダイゼーションの強度は低下する。
【図6J】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Jは、サブプレートから辺縁帯までの全長に広がるネスチン陽性細胞を示す。
【図6K】図6A−6KはNSC内及びマウス胎児組織でのNRPの遺伝子発現を示す。図6A−6Cは、マルチプレックスPCRでは、マウスNRP(上部バンド、412bp)の発現レベルはβ-アクチン発現(下部バンド、260bp)と比較して準定量的であることを示すゲルの写真である。図6Kは、図6Jのネスチン発現と同じ部位でのマウスNRPメッセージの同時発現を示す。
【図6L】図6Lは、損傷の非存在下及びミトコンドリア毒素3-ニトロプロピオン酸により惹起された損傷の6時間後におけるNT-2細胞のNRP-2(配列番号:4)遺伝子産物の発現を示す。遺伝子発現は、細胞が1時間以上3-NPで処理されたとき実質的に減少した。
【図7A】図7A及び7Bは、NRP-7セグメントSWによるニューロンの生存及び増殖誘導を示す。図7Aは、0.1pMから100nMの広い用量範囲にわたるNRP-7SW(配列番号:24)の神経保護活性に対する影響を示す。
【図7B】図7A及び7Bは、NRP-7セグメントSWによるニューロンの生存及び増殖誘導を示す。図7Bは、SDF-1はNRPと比較してほんの限定的な神経保護作用を有したことを示す。
【図8A】図8A及び図8Bは、NRPによるニューロン遊走誘導実験の結果を示す。図8Aは、ラット及びマウスの合成ペプチド由来NRP(NRP-9SD及びNRP-7SW)は、接触走性遊走アッセイで示されるように、ニューロン幹細胞(“NSC”)の誘引において化学誘引特性を表すことを示すグラフである。NRP-7SWの有効性はSDF-1のそれと類似したが、NRPはより高い潜在能力を示した。SDF-1とNRP-7SWの作用の規模は類似しているように見えるが、SDF-1の量(100nM)はNRP-7SWの量(0.2nM)よりも多い。したがってNRP-7SWはSDF-1よりも約500倍強力である。
【図8B】図8A及び図8Bは、NRPによるニューロン遊走誘導実験の結果を示す。図8Bは、皮質原基に向かって遊走する、中央神経節隆起由来の神経前駆体細胞の移動を示す脳OTCの顕微鏡写真である。
【図8C】1nMの濃度でNRP-7SWの最大効果を示すOTCアッセイの定量のグラフを示す。一方、SDF-1の能力は前記よりも低かった。スチューデントのt-検定を統計分析に用いた(*p<0.05、**p<0.01、***p<0.001;N=6)。
【図9A】図9A−9Dは、完全長のマウス組換えNRP-7ロング(配列番号:35)の発現実験及び機能実験の結果を示す。図9Aは、サイトメガロウイルス(“CMV”)プロモーターの制御下では、マウスNRP-7ロング(配列番号:35)遺伝子の0.8kbシグナルは、NRP遺伝子特異的88bpのcRNAプローブによって検出したとき、HEK細胞で極めて過剰発現されることを示すノザンブロットを示す。
【図9B】図9A−9Dは、完全長のマウス組換えNRP-7ロング(配列番号:35)の発現実験及び機能実験の結果を示す。図9Bは、NRP-7ロング(配列番号:36)がHEK細胞によって発現され、20kDaの分子量で移動することを示すウェスタンブロットである。
【図9C】図9A−9Dは、完全長のマウス組換えNRP-7ロング(配列番号:35)の発現実験及び機能実験の結果を示す。図9Cは、組換えMyc-NRP-7ロング(配列番号:35)−HEK細胞は、酸化性/興奮毒性損傷を受けた小脳微小エクスプラントに対し51%の神経保護を提供することを示すグラフである。
【図9D】図9A−9Dは、完全長のマウス組換えNRP-7ロング(配列番号:35)の発現実験及び機能実験の結果を示す。図9Dは、Myc-NRP-7ロング(配列番号:36)−HEK細胞は、培養皿の底に播種したとき、接触走性遊走アッセイでNSCを誘引するための化学誘引活性をもつことを示すグラフである。Myc−HEKコントロール実験と比較して、2倍以上の数の細胞が、インサートから培養皿の底へ誘引された。スチューデントt-検定を統計分析に適用した(微小エクスプラントについては***p<0.001;N=4、さらに接触走性遊走アッセイについては***p<0.001;N=6)。
【図10A】図10A−10Cは、CSCR4及びNRPに対する抗体を用いた、P4-小脳エクスプラントに関する実験の結果を示す。図10Aは、神経突起伸長に関するNRP-9SD(配列番号:34)の阻害を示すグラフである。
【図10B】図10A−10Cは、CSCR4及びNRPに対する抗体を用いた、P4-小脳エクスプラントに関する実験の結果を示す。図10Bは、神経突起伸長に関するNRP-2KS(配列番号:23)の阻害を示す。培養皿に被覆した10ng/mLのNRPの化学誘引作用は、ニューロン幹細胞株MEB-5をCXCR4に対する中和抗体と予備インキュベートすることによって完全に阻害された。
【図10C】図10A−10Cは、CSCR4及びNRPに対する抗体を用いた、P4-小脳エクスプラントに関する実験の結果を示す。図10Cは、CXCR4に対する抗体は、NRP-9SD(配列番号:34)だけで処理した細胞と比較してNRP-9SDの化学誘引作用を低下させたことを示す。
【図11A】図11A−11Cは、NRPの神経保護誘導及び遊走誘導活性に中心的に関与する可能なシグナリング連鎖事象を提案する、MAPK阻害物質を用いたNRPの作用に対する実験の結果を示す。図11Aは、MAPK(MEK)-阻害物質のPD98509は、種々のNRP濃度範囲にわたってラットのNRP-9SD(配列番号:34)の神経保護活性を完全に阻害することを示すグラフである。
【図11B】図11A−11Cは、NRPの神経保護誘導及び遊走誘導活性に中心的に関与する可能なシグナリング連鎖事象を提案する、MAPK阻害物質を用いたNRPの作用に対する実験の結果を示す。図11Bは、PD98509のように、PIK-3阻害物質のワートマンニンは、NRP-9SD(配列番号:34)の神経保護活性を阻害することを示すグラフである。
【図11C】図11A−11Cは、NRPの神経保護誘導及び遊走誘導活性に中心的に関与する可能なシグナリング連鎖事象を提案する、MAPK阻害物質を用いたNRPの作用に対する実験の結果を示す。図11Cは、NRP-9SD(4nM)によって誘導される遊走NSC数は15μMのPD98509によって阻害され、一方、BSA被覆ウェルにおける基礎遊走レベルは、MEK阻害物質によって顕著には変化しないことを示すグラフである(*p<0.05、**p<0.01、***p<0.001;N=8)。
【図12A】低酸素症の2時間後にICV投与したとき、NRP-4セグメントPQ(配列番号:43)は、損傷の5日後に分析した脳領域の全てにおいて実質的な神経保護を提供した実験の結果を示す。
【図12B】低酸素症の2時間後にICV投与したNRP-5セグメントRG(配列番号:30)は、低酸素障害の5日後に分析した脳領域の全てにおいて実質的な神経保護を提供した実験の結果を示す。
【図13】NRP-7セグメントSW(配列番号:24)は、ヒト成人嗅球鞘性グリア(ensheating glia)の初代培養の増殖を強化することを示す実験の結果である。
【図14A】図14A−14Dは、4つのNRP-5 RGアナログの神経保護活性の原型NRP-5RG配列に対する実験結果を示す。
【図14B】図14A−14Dは、4つのNRP-5 RGアナログの神経保護活性の原型NRP-5RG配列に対する実験結果を示す。
【図14C】図14A−14Dは、4つのNRP-5 RGアナログの神経保護活性の原型NRP-5RG配列に対する実験結果を示す。
【図14D】図14A−14Dは、4つのNRP-5 RGアナログの神経保護活性の原型NRP-5RG配列に対する実験結果を示す。
【図15】胎児小脳細胞におけるNRP-5 RGの増殖誘導作用のグラフを示す。
【図16】マウスNSC及び化学誘引物質としてNRP-5 RGを用いた接触走性遊走アッセイの結果を示すグラフである。
【図17】酸化ストレス(0.1mMの過酸化水素)の48時間後のNRP-4 PQの神経保護作用の結果を示すグラフである。
【図18】リン酸化NRP-7 SW(NRP-7 PSW)の神経保護作用を示すグラフである。
【図19】BrDU染色核に対するNRP-9SDの作用を示す実験のグラフである。
【図20A】図20Aおよび20Bは、神経上皮幹細胞−放射状グリア−星状細胞系列内におけるNRP-7(配列番号:35)の遺伝子発現の顕微鏡写真を示す。 図20Aは、E17マウス脳の大脳皮質におけるマウスフレームシフトNRP転写物の発現を示す。88merの特異的プローブを用い、凍結切片でのin situハイブリダイゼーションによって検出された。
【図20B】図20Aおよび20Bは、神経上皮幹細胞−放射状グリア−星状細胞系列内におけるNRP-7(配列番号:35)の遺伝子発現の顕微鏡写真を示す。 図20Bは、同じ切片で抗ビメンチン抗体を用いた光学顕微鏡法を示す。マウスフレームシフトメッセージとビメンチン中間体のフィラメントタンパク質の高度な同時局在が示されている。
【特許請求の範囲】
【請求項1】
配列番号:29、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:36、配列番号:44及び配列番号:45から成る群から選択されるアミノ酸配列を有する神経再生ペプチド(NRP)。
【請求項2】
配列番号:27、配列番号:35及び配列番号:48から成る群から選択される、NRPをコードするオリゴヌクレオチド配列。
【請求項3】
オリゴヌクレオチド配列が遺伝暗号の重複性のためによってのみ請求項2のオリゴヌクレオチド配列と異なる、NRPをコードするオリゴヌクレオチド。
【請求項4】
神経組織に請求項1記載の少なくとも1つのNRPの医薬的に有効な量をデリバーする工程を含む、神経の再生を促進する方法。
【請求項5】
神経の再生が、神経の生存、神経の増殖、神経の遊走、神経突起伸長及び神経の分化から成る群から選択される、請求項4記載の方法。
【請求項6】
NRPが静脈内にデリバーされる、請求項4記載の方法。
【請求項7】
NRPが請求項2のオリゴヌクレオチドによってコードされる、請求項4記載の方法。
【請求項8】
NRPが、請求項2のオリゴヌクレオチド又は遺伝暗号の重複性のためによってのみ請求項2のオリゴヌクレオチドの配列と異なる配列を有するオリゴヌクレオチドによってコードされる、請求項4記載の方法。
【請求項9】
NRPをコードするオリゴヌクレオチドであって、前記オリゴヌクレオチドがその配列全体にわたって請求項2記載のオリゴヌクレオチド配列と少なくとも85%の配列同一性を有する、前記NRPをコードするオリゴヌクレオチド。
【請求項10】
NRPを発現することができる、請求項2記載のオリゴヌクレオチド配列を含む発現ベクター。
【請求項11】
プラスミドである、請求項10記載の発現ベクター。
【請求項12】
ウイルスベクターである、請求項10記載の発現ベクター。
【請求項13】
請求項10記載の発現ベクターを含む宿主細胞。
【請求項14】
細胞が原核細胞である、請求項13記載の宿主細胞。
【請求項15】
細胞が真核細胞である、請求項13記載の宿主細胞。
【請求項16】
“t”が“u”により置換されている、請求項2記載のオリゴヌクレオチド配列。
【請求項17】
配列番号:27、配列番号:35及び配列番号:48から成る群から選択されるオリゴヌクレオチドと相補的なオリゴヌクレオチド。
【請求項18】
少なくとも15塩基対の長さであり、さらにNRPをコードするゲノムDNAとストリンジェントな条件下でハイブリダイズする、請求項17記載のオリゴヌクレオチド。
【請求項19】
配列番号:28、配列番号:29、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:36、配列番号:43、配列番号:44、配列番号:45、配列番号:46、配列番号:47及び配列番号:49から成る群から選択されるアミノ酸配列と配列全体にわたって少なくとも80%のアミノ酸配列同一性を有するアミノ酸配列を含むポリペプチド。
【請求項20】
神経組織が動物の脳幹神経節に存在する、請求項4記載の方法。
【請求項21】
神経組織が、パーキンソン病、アルツハイマー病、低酸素症、虚血、卒中、冠状動脈バイパス手術、外傷又はハンチントン病により変性のリスクを有する、請求項20記載の方法。
【請求項22】
神経変性を特徴とする神経学的症状を生物で治療する方法であって、請求項1のNRPの薬理学的に有効な量を前記生物の罹患神経に投与する工程を含む、前記神経学的症状の治療方法。
【請求項23】
神経学的症状が、パーキンソン病、アルツハイマー病、低酸素症、虚血、卒中、冠状動脈バイパス手術、外傷及びハンチントン病から成る群から選択される、請求項22記載の方法。
【請求項24】
請求項1記載のNRPの有効量をニューロンに投与することを含む、ニューロン増殖及びニューロン遊走の少なくとも1つをニューロンにおいて誘導する方法。
【請求項25】
請求項1記載のNRPの有効量をニューロンに投与することを含む、ニューロンにおいて神経突起伸長を促進する方法。
【請求項26】
請求項1記載のNRPの有効量をニューロンに投与することを含む、ニューロンの生存を促進する方法。
【請求項27】
ニューロンが生きている動物の内部に存在する、請求項24〜26のいずれか1項記載の方法。
【請求項28】
ニューロンが動物の体内に存在しない、請求項24〜26のいずれか1項に記載の方法。
【請求項29】
ニューロンが動物の中枢神経系内に存在する、請求項24〜26のいずれか1項に記載の方法。
【請求項30】
脊椎損傷を有する患者に請求項1記載のNRPの有効量を投与することを含む、脊椎損傷を有する患者の治療方法。
【請求項31】
アミノ酸配列が、少なくとも1つの保存的アミノ酸置換のために、配列番号:28、配列番号:29、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:36、配列番号:43、配列番号:44、配列番号:45、配列番号:46、配列番号:47又は配列番号:49のアミノ酸配列と異なる、請求項1記載のNRP。
【請求項32】
ニューロンの障害を持つ対象者に神経再生ペプチド(NRP)の医薬的に有効な量を投与することを含む、幹細胞のニューロン表現型への分化を促進する方法。
【請求項33】
NRPが、配列番号:2、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:17、配列番号:18、配列番号:20、配列番号:21、配列番号:22、配列番号:23、配列番号:24、配列番号:25、配列番号:26、配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:46、配列番号:47及び配列番号:49から成る群から選択される、請求項32記載の方法。
【請求項34】
ニューロンの障害をもつ対象者に神経再生ペプチド(NRP)の医薬的に有効な量を投与することを含む、幹細胞で軸索の成長を促進する方法。
【請求項35】
NRPが、配列番号:2、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:17、配列番号:18、配列番号:20、配列番号:21、配列番号:22、配列番号:23、配列番号:24、配列番号:25、配列番号:26、配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44及び配列番号:45から成る群から選択される、請求項34記載の方法。
【請求項36】
神経増殖ペプチド(NRP)の医薬的に有効な量を投与することを含む、神経芽細胞の増殖を促進する方法。
【請求項37】
NRPが、配列番号:2、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:17、配列番号:18、配列番号:20、配列番号:21、配列番号:22、配列番号:23、配列番号:24、配列番号:25、配列番号:26、配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される、請求項36記載の方法。
【請求項38】
幹細胞に神経再生ペプチド(NRP)の医薬的に有効な量を投与することを含む、幹細胞の遊走を誘導する方法。
【請求項39】
NRPが、配列番号:2、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:17、配列番号:18、配列番号:20、配列番号:21、配列番号:22、配列番号:23、配列番号:24、配列番号:25、配列番号:26、配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される、請求項38記載の方法。
【請求項40】
配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される神経再生ペプチドの医薬的に有効な量を対象者に投与することを含む、治療の必要がある患者で神経変性症状を治療する方法。
【請求項41】
神経芽細胞集団を医薬的に有効な量のNRPと組み合わせて移植し、前記神経芽細胞が増殖し、分化し及び/又は神経突起を形成する工程を含む、哺乳動物の神経系の一部分の集団を再成する方法。
【請求項42】
医療処置に付随する神経変性を減少させる方法であって、前記方法が、前記医療処置の前に対象者に医薬的に有効な量の神経再生ペプチド(NRP)を投与することを含む、前記神経変性を減少させる方法。
【請求項43】
医療処置が、外科手術、化学療法及び放射線療法からなる群から選択される、請求項42記載の方法。
【請求項44】
医療処置が冠状動脈バイパス手術である、請求項42記載の方法。
【請求項45】
症状が、感染、脳血管障害、頭蓋脳外傷、脱髄疾患、痴呆、代謝異常、栄養失調、アルコール中毒、末梢神経障害、薬剤誘導神経傷害及び化学物質誘導神経傷害に付随する症状から成る群から選択される、請求項40記載の方法。
【請求項46】
配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される神経再生ペプチドの医薬的に有効な量を、低酸素症又は虚血の患者に投与することを含む、低酸素症又は神経の虚血に付随する神経変性を治療する方法。
【請求項47】
低酸素症又は虚血が心不全又は卒中に付随する、請求項46記載の方法。
【請求項48】
低酸素症又は虚血が外傷性脳損傷に付随する、請求項46記載の方法。
【請求項49】
配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される少なくとも1つの神経再生ペプチド、及び医薬的に許容できる媒体を含む医薬組成物。
【請求項50】
神経変性が細菌、菌類又はウイルス感染に付随する、請求項46記載の方法。
【請求項51】
本明細書に実質的に記載された神経再生ペプチド。
【請求項52】
本明細書に実質的に記載された神経再生ペプチドを使用する方法。
【請求項1】
配列番号:29、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:36、配列番号:44及び配列番号:45から成る群から選択されるアミノ酸配列を有する神経再生ペプチド(NRP)。
【請求項2】
配列番号:27、配列番号:35及び配列番号:48から成る群から選択される、NRPをコードするオリゴヌクレオチド配列。
【請求項3】
オリゴヌクレオチド配列が遺伝暗号の重複性のためによってのみ請求項2のオリゴヌクレオチド配列と異なる、NRPをコードするオリゴヌクレオチド。
【請求項4】
神経組織に請求項1記載の少なくとも1つのNRPの医薬的に有効な量をデリバーする工程を含む、神経の再生を促進する方法。
【請求項5】
神経の再生が、神経の生存、神経の増殖、神経の遊走、神経突起伸長及び神経の分化から成る群から選択される、請求項4記載の方法。
【請求項6】
NRPが静脈内にデリバーされる、請求項4記載の方法。
【請求項7】
NRPが請求項2のオリゴヌクレオチドによってコードされる、請求項4記載の方法。
【請求項8】
NRPが、請求項2のオリゴヌクレオチド又は遺伝暗号の重複性のためによってのみ請求項2のオリゴヌクレオチドの配列と異なる配列を有するオリゴヌクレオチドによってコードされる、請求項4記載の方法。
【請求項9】
NRPをコードするオリゴヌクレオチドであって、前記オリゴヌクレオチドがその配列全体にわたって請求項2記載のオリゴヌクレオチド配列と少なくとも85%の配列同一性を有する、前記NRPをコードするオリゴヌクレオチド。
【請求項10】
NRPを発現することができる、請求項2記載のオリゴヌクレオチド配列を含む発現ベクター。
【請求項11】
プラスミドである、請求項10記載の発現ベクター。
【請求項12】
ウイルスベクターである、請求項10記載の発現ベクター。
【請求項13】
請求項10記載の発現ベクターを含む宿主細胞。
【請求項14】
細胞が原核細胞である、請求項13記載の宿主細胞。
【請求項15】
細胞が真核細胞である、請求項13記載の宿主細胞。
【請求項16】
“t”が“u”により置換されている、請求項2記載のオリゴヌクレオチド配列。
【請求項17】
配列番号:27、配列番号:35及び配列番号:48から成る群から選択されるオリゴヌクレオチドと相補的なオリゴヌクレオチド。
【請求項18】
少なくとも15塩基対の長さであり、さらにNRPをコードするゲノムDNAとストリンジェントな条件下でハイブリダイズする、請求項17記載のオリゴヌクレオチド。
【請求項19】
配列番号:28、配列番号:29、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:36、配列番号:43、配列番号:44、配列番号:45、配列番号:46、配列番号:47及び配列番号:49から成る群から選択されるアミノ酸配列と配列全体にわたって少なくとも80%のアミノ酸配列同一性を有するアミノ酸配列を含むポリペプチド。
【請求項20】
神経組織が動物の脳幹神経節に存在する、請求項4記載の方法。
【請求項21】
神経組織が、パーキンソン病、アルツハイマー病、低酸素症、虚血、卒中、冠状動脈バイパス手術、外傷又はハンチントン病により変性のリスクを有する、請求項20記載の方法。
【請求項22】
神経変性を特徴とする神経学的症状を生物で治療する方法であって、請求項1のNRPの薬理学的に有効な量を前記生物の罹患神経に投与する工程を含む、前記神経学的症状の治療方法。
【請求項23】
神経学的症状が、パーキンソン病、アルツハイマー病、低酸素症、虚血、卒中、冠状動脈バイパス手術、外傷及びハンチントン病から成る群から選択される、請求項22記載の方法。
【請求項24】
請求項1記載のNRPの有効量をニューロンに投与することを含む、ニューロン増殖及びニューロン遊走の少なくとも1つをニューロンにおいて誘導する方法。
【請求項25】
請求項1記載のNRPの有効量をニューロンに投与することを含む、ニューロンにおいて神経突起伸長を促進する方法。
【請求項26】
請求項1記載のNRPの有効量をニューロンに投与することを含む、ニューロンの生存を促進する方法。
【請求項27】
ニューロンが生きている動物の内部に存在する、請求項24〜26のいずれか1項記載の方法。
【請求項28】
ニューロンが動物の体内に存在しない、請求項24〜26のいずれか1項に記載の方法。
【請求項29】
ニューロンが動物の中枢神経系内に存在する、請求項24〜26のいずれか1項に記載の方法。
【請求項30】
脊椎損傷を有する患者に請求項1記載のNRPの有効量を投与することを含む、脊椎損傷を有する患者の治療方法。
【請求項31】
アミノ酸配列が、少なくとも1つの保存的アミノ酸置換のために、配列番号:28、配列番号:29、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:36、配列番号:43、配列番号:44、配列番号:45、配列番号:46、配列番号:47又は配列番号:49のアミノ酸配列と異なる、請求項1記載のNRP。
【請求項32】
ニューロンの障害を持つ対象者に神経再生ペプチド(NRP)の医薬的に有効な量を投与することを含む、幹細胞のニューロン表現型への分化を促進する方法。
【請求項33】
NRPが、配列番号:2、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:17、配列番号:18、配列番号:20、配列番号:21、配列番号:22、配列番号:23、配列番号:24、配列番号:25、配列番号:26、配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:46、配列番号:47及び配列番号:49から成る群から選択される、請求項32記載の方法。
【請求項34】
ニューロンの障害をもつ対象者に神経再生ペプチド(NRP)の医薬的に有効な量を投与することを含む、幹細胞で軸索の成長を促進する方法。
【請求項35】
NRPが、配列番号:2、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:17、配列番号:18、配列番号:20、配列番号:21、配列番号:22、配列番号:23、配列番号:24、配列番号:25、配列番号:26、配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44及び配列番号:45から成る群から選択される、請求項34記載の方法。
【請求項36】
神経増殖ペプチド(NRP)の医薬的に有効な量を投与することを含む、神経芽細胞の増殖を促進する方法。
【請求項37】
NRPが、配列番号:2、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:17、配列番号:18、配列番号:20、配列番号:21、配列番号:22、配列番号:23、配列番号:24、配列番号:25、配列番号:26、配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される、請求項36記載の方法。
【請求項38】
幹細胞に神経再生ペプチド(NRP)の医薬的に有効な量を投与することを含む、幹細胞の遊走を誘導する方法。
【請求項39】
NRPが、配列番号:2、配列番号:5、配列番号:7、配列番号:9、配列番号:11、配列番号:13、配列番号:17、配列番号:18、配列番号:20、配列番号:21、配列番号:22、配列番号:23、配列番号:24、配列番号:25、配列番号:26、配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される、請求項38記載の方法。
【請求項40】
配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される神経再生ペプチドの医薬的に有効な量を対象者に投与することを含む、治療の必要がある患者で神経変性症状を治療する方法。
【請求項41】
神経芽細胞集団を医薬的に有効な量のNRPと組み合わせて移植し、前記神経芽細胞が増殖し、分化し及び/又は神経突起を形成する工程を含む、哺乳動物の神経系の一部分の集団を再成する方法。
【請求項42】
医療処置に付随する神経変性を減少させる方法であって、前記方法が、前記医療処置の前に対象者に医薬的に有効な量の神経再生ペプチド(NRP)を投与することを含む、前記神経変性を減少させる方法。
【請求項43】
医療処置が、外科手術、化学療法及び放射線療法からなる群から選択される、請求項42記載の方法。
【請求項44】
医療処置が冠状動脈バイパス手術である、請求項42記載の方法。
【請求項45】
症状が、感染、脳血管障害、頭蓋脳外傷、脱髄疾患、痴呆、代謝異常、栄養失調、アルコール中毒、末梢神経障害、薬剤誘導神経傷害及び化学物質誘導神経傷害に付随する症状から成る群から選択される、請求項40記載の方法。
【請求項46】
配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される神経再生ペプチドの医薬的に有効な量を、低酸素症又は虚血の患者に投与することを含む、低酸素症又は神経の虚血に付随する神経変性を治療する方法。
【請求項47】
低酸素症又は虚血が心不全又は卒中に付随する、請求項46記載の方法。
【請求項48】
低酸素症又は虚血が外傷性脳損傷に付随する、請求項46記載の方法。
【請求項49】
配列番号:28、配列番号:20、配列番号:30、配列番号:31、配列番号:32、配列番号:33、配列番号:34、配列番号:35、配列番号:43、配列番号:44、配列番号:45、配列番号:45、配列番号:46及び配列番号:49から成る群から選択される少なくとも1つの神経再生ペプチド、及び医薬的に許容できる媒体を含む医薬組成物。
【請求項50】
神経変性が細菌、菌類又はウイルス感染に付随する、請求項46記載の方法。
【請求項51】
本明細書に実質的に記載された神経再生ペプチド。
【請求項52】
本明細書に実質的に記載された神経再生ペプチドを使用する方法。
【図1A】

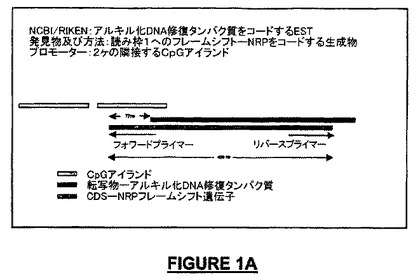
【図1B】

【図1C】

【図1D】

【図1E】

【図2】
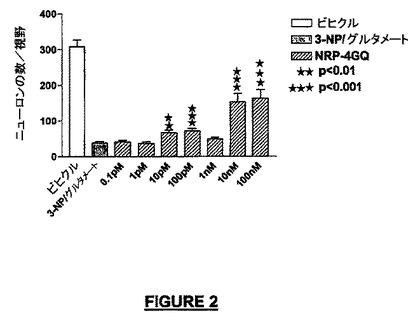
【図3】
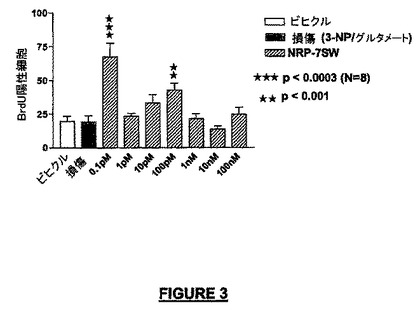
【図4A】
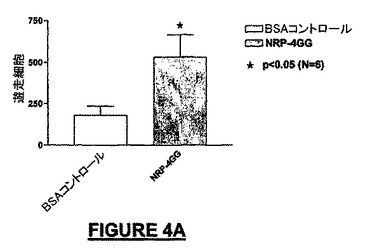
【図4B】
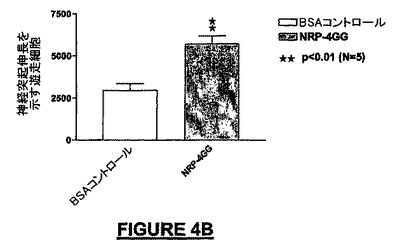
【図4C】
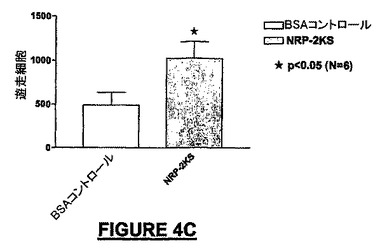
【図4D】
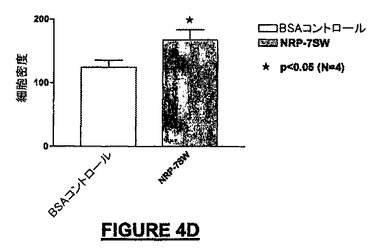
【図4E】
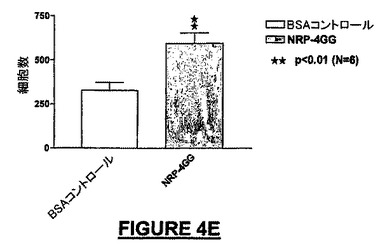
【図4F】
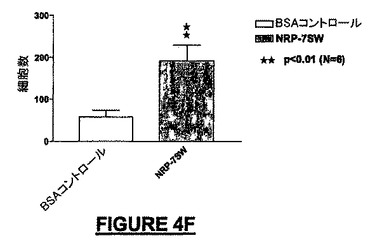
【図5A】
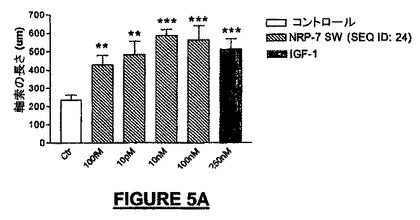
【図5B】
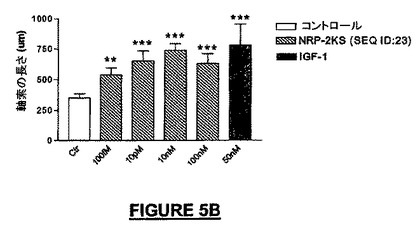
【図5C】

【図5D】

【図5E】
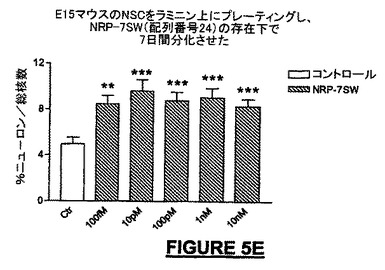
【図5F】
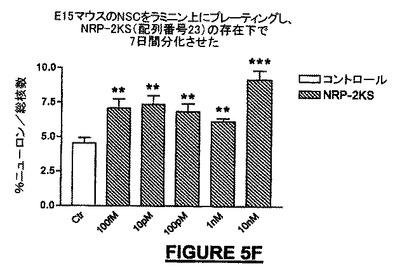
【図5G】
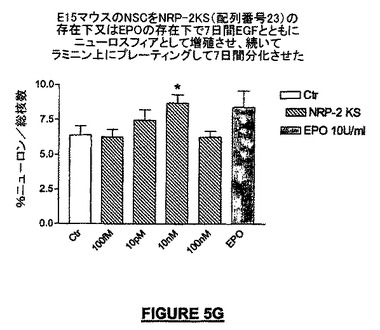
【図5H】
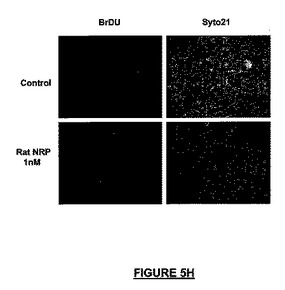
【図6A】
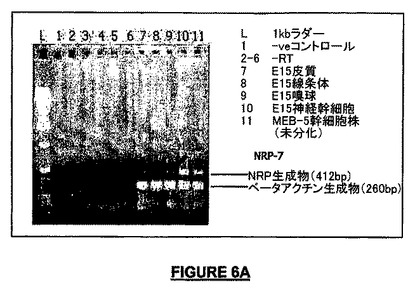
【図6B】
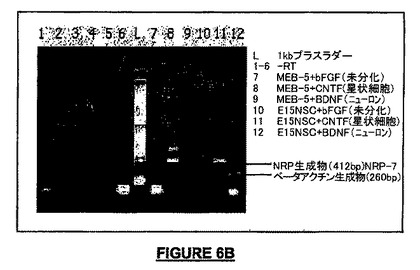
【図6C】

【図6D】
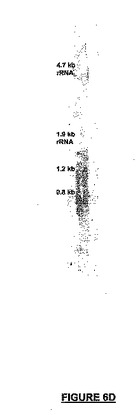
【図6E−K】
【図6L】
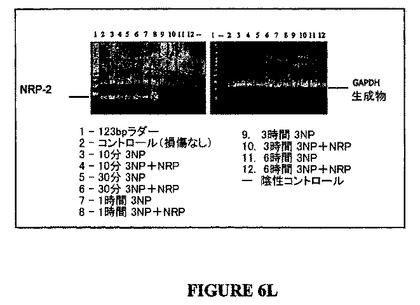
【図7A】
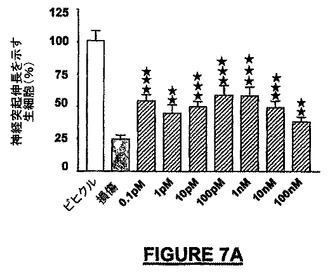
【図7B】
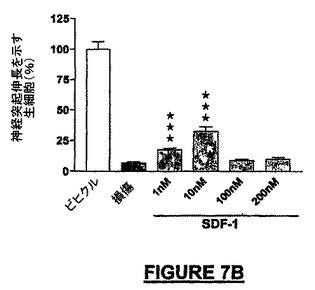
【図8A】
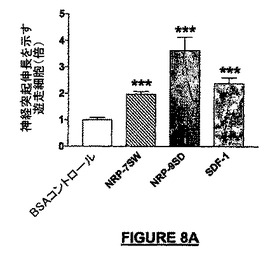
【図8B】

【図8C】
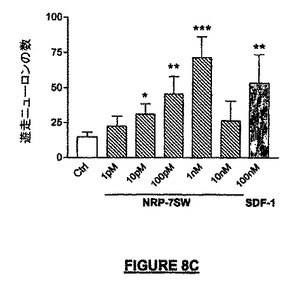
【図9A】

【図9B】
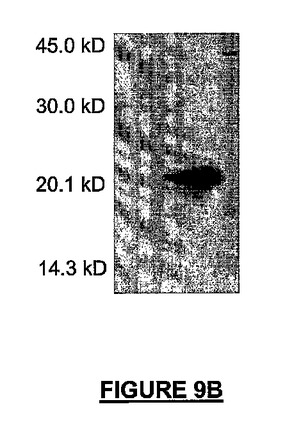
【図9C】
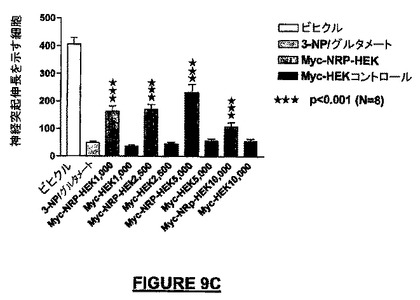
【図9D】
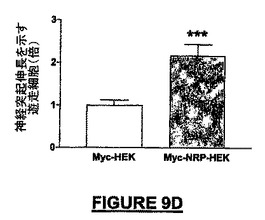
【図10A】
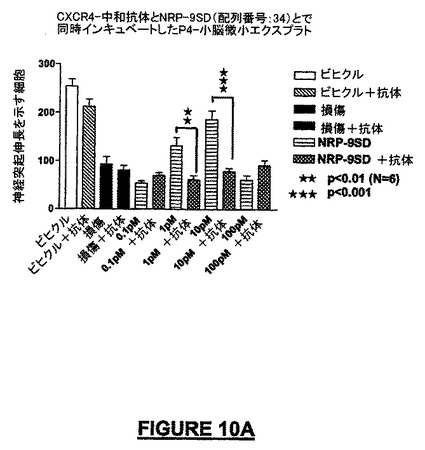
【図10B】
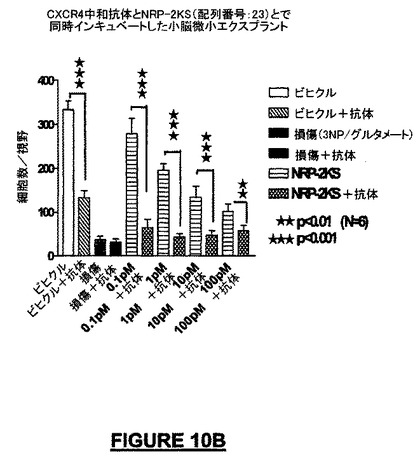
【図10C】
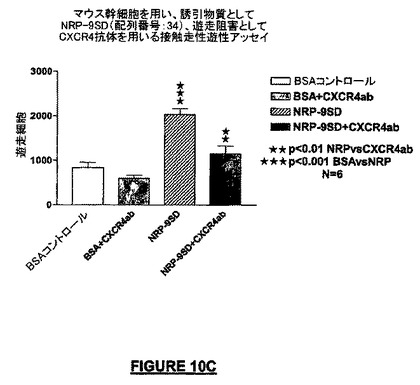
【図11A】
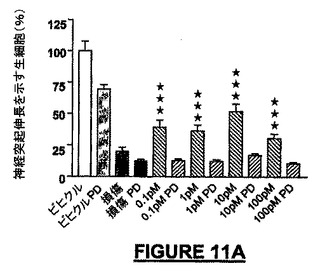
【図11B】
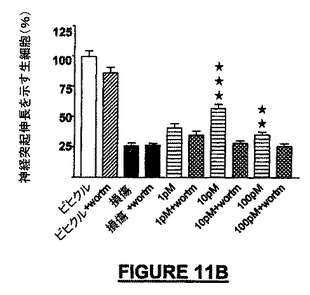
【図11C】
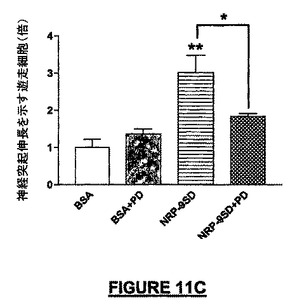
【図12A】
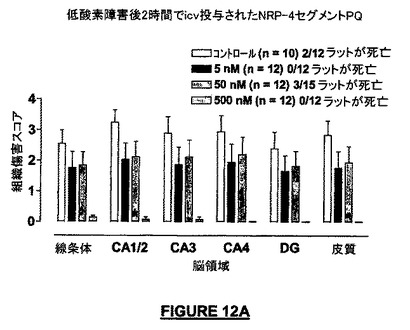
【図12B】
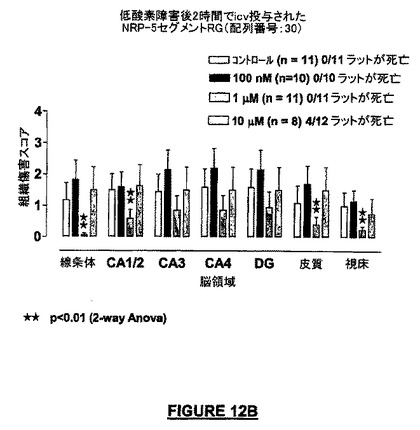
【図13】
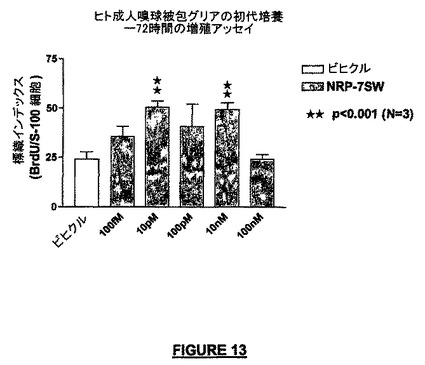
【図14A】
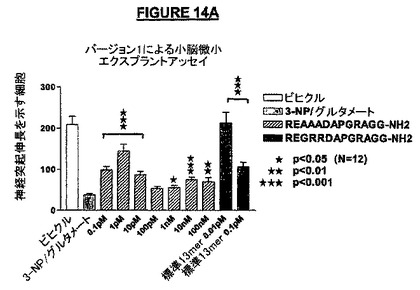
【図14B】
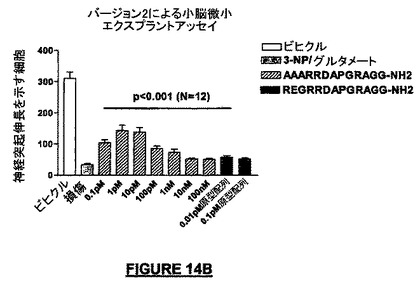
【図14C】
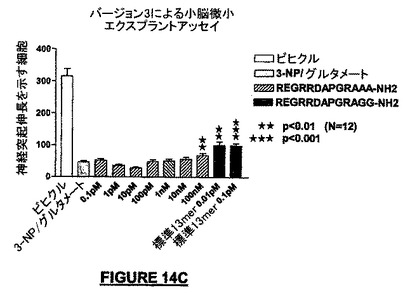
【図14D】
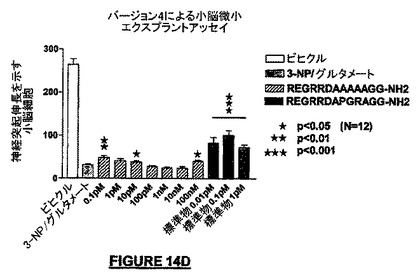
【図15】
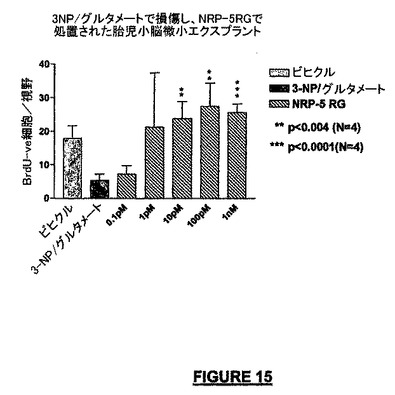
【図16】
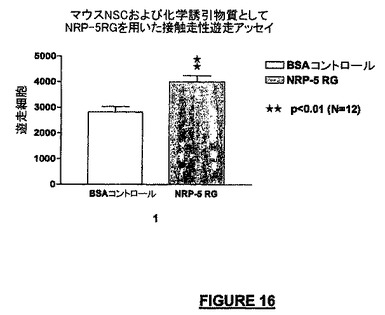
【図17】
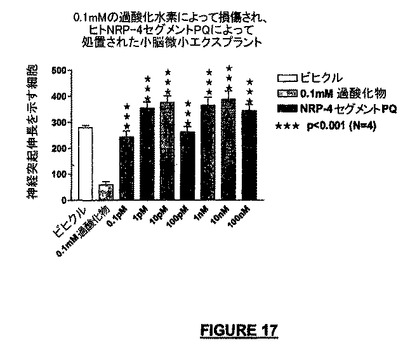
【図18】
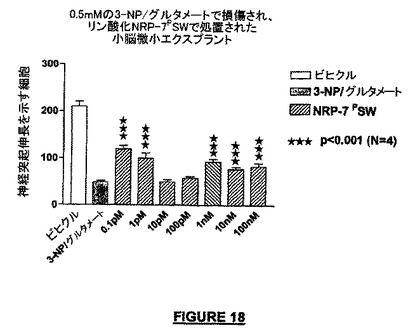
【図19】
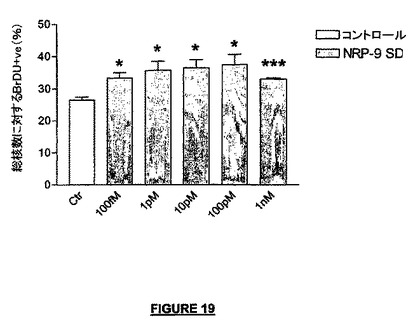
【図20A】

【図20B】

【図1B】
【図1C】
【図1D】
【図1E】
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図5H】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E−K】
【図6L】
【図7A】
【図7B】
【図8A】
【図8B】
【図8C】
【図9A】
【図9B】
【図9C】
【図9D】
【図10A】
【図10B】
【図10C】
【図11A】
【図11B】
【図11C】
【図12A】
【図12B】
【図13】
【図14A】
【図14B】
【図14C】
【図14D】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20A】
【図20B】
【公表番号】特表2007−511210(P2007−511210A)
【公表日】平成19年5月10日(2007.5.10)
【国際特許分類】
【出願番号】特願2006−538375(P2006−538375)
【出願日】平成16年11月1日(2004.11.1)
【国際出願番号】PCT/US2004/036203
【国際公開番号】WO2005/042561
【国際公開日】平成17年5月12日(2005.5.12)
【出願人】(505353744)ニューレン ファーマシューティカルズ リミテッド (9)
【Fターム(参考)】
【公表日】平成19年5月10日(2007.5.10)
【国際特許分類】
【出願日】平成16年11月1日(2004.11.1)
【国際出願番号】PCT/US2004/036203
【国際公開番号】WO2005/042561
【国際公開日】平成17年5月12日(2005.5.12)
【出願人】(505353744)ニューレン ファーマシューティカルズ リミテッド (9)
【Fターム(参考)】
[ Back to top ]