
神経化誘導に関与する新規遺伝子、およびそのタンパク質、並びにその利用方法
【課題】脊椎動物における神経誘導に関与する新規遺伝子、およびそのタンパク質を提供するとともに、これらの利用方法を提案する。
【解決手段】アフリカツメガエルのアニマル・キャップの神経誘導に関与するXMAN1遺伝子を単離し、XMAN1遺伝子およびXMAN1タンパク質の構造を決定した。このXMAN1遺伝子およびXMAN1タンパク質の機能解析を通じ、脊椎動物における神経誘導による形態形成の分子機構をより明らかにすることができる。また、XMAN1タンパク質のヒトホモログタンパク質であるヒトMAN1並びにその遺伝子も同様の利用可能性がある。さらに本発明は神経系の疾患の治療薬、治療法等への応用も可能である。
【解決手段】アフリカツメガエルのアニマル・キャップの神経誘導に関与するXMAN1遺伝子を単離し、XMAN1遺伝子およびXMAN1タンパク質の構造を決定した。このXMAN1遺伝子およびXMAN1タンパク質の機能解析を通じ、脊椎動物における神経誘導による形態形成の分子機構をより明らかにすることができる。また、XMAN1タンパク質のヒトホモログタンパク質であるヒトMAN1並びにその遺伝子も同様の利用可能性がある。さらに本発明は神経系の疾患の治療薬、治療法等への応用も可能である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、神経化誘導に関与する新規タンパク質、およびその遺伝子、並びにその利用方法に関し、より詳細には、当該タンパク質、若しくはその遺伝子を含有する神経化誘導剤、神経化誘導方法、並びに骨形成因子BMPシグナル伝達経路阻害剤に関するものである。
【背景技術】
【0002】
アフリカツメガエルの胚の胞胚期から原腸胚期にかけて動物極側に位置する、予定外胚葉組織(アニマル・キャップ)は多分化能を有しており、種々の因子に反応して外、中、内胚葉由来の様々な組織、器官に分化することが知られている。
【0003】
近年、このアニマル・キャップの性質を利用して、脊椎動物における神経誘導のメカニズムを解明するための研究が盛んに行われており、これまでの研究成果として、脊椎動物における神経誘導には、骨形成因子BMP(Bone Morphogenetic Protein)を介するシグナル伝達の経路を阻害することが重要であるとの報告も多数なされている(例えば、非特許文献1参照)。
【0004】
この骨形成因子として、ヒトでは現在までに9種類のBMP(BMP−1〜BMP−9)がクローニングされている。このBMPは骨形成ばかりでなく、受精卵から分裂増殖、細胞分化により新しい組織、器官が形成されてくる、いわゆる形態形成にも関与していることが知られている(例えば、非特許文献2参照)。
【非特許文献1】Harland, R.(2000). Neural induction, Curr. Opin. Genet. Dev. 10, pp357-62.
【非特許文献2】Hogan, BL.(1996). Bone morphogenetic proteins: multifunctional regulators of vertebrate development. Genes Dev. 10, pp1580-94.
【発明の開示】
【発明が解決しようとする課題】
【0005】
脊椎動物における神経誘導のメカニズムは、様々な因子が関与していることから非常に複雑であり、そのメカニズムは未だ十分に解明されているとはいえず、神経誘導と密接に関与していると考えられる骨形成因子BMPのシグナル伝達経路の詳細についても明らかにはなっていない。
【0006】
他方、もし脊椎動物における神経誘導に関与する未知の遺伝子や未知のタンパク質が明らかにされれば、当該遺伝子およびタンパク質の機能解析を通じ、神経誘導による形態形成の分子機構やBMPシグナル伝達経路の詳細がより一層明らかにされるといった学術的意義もさることながら、例えば神経系の異常により引き起こされる疾病の病態機序の研究や治療薬、治療法等の開発にも利用できる可能性があり、医学的領域への幅広い利用が期待できる。
【0007】
本発明は、上記の課題に鑑みなされたものであり、その目的は、脊椎動物における神経誘導に関与する新規遺伝子、およびそのタンパク質を提供するとともに、これらの利用方法を提案することにある。
【課題を解決するための手段】
【0008】
本発明者らは、鋭意検討を重ねた結果、アフリカツメガエルのアニマル・キャップを神経化させる因子をクローニングし、新規神経化誘導因子XMAN1(Xenopus MAN1)を同定した。さらに、その機能を詳細に解析することにより、この新規神経化誘導因子XMAN1が、骨形成因子BMPを介するシグナル伝達経路を阻害し神経化を誘導する効果があることを見出し、本発明を完成させるに至った。
【0009】
すなわち、本発明に係るタンパク質は、以下の(a)または(b)のタンパク質である。
(a)配列番号1に示されるアミノ酸配列からなるタンパク質。
(b)配列番号1に示されるアミノ酸配列において、1またはそれ以上のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導因子としての活性を持つタンパク質。
【0010】
上記タンパク質の具体的な一例として、アフリカツメガエル由来の神経化誘導因子XMAN1が挙げられる。具体的には、上記XMAN1タンパク質は、配列番号1に示すアミノ酸配列を有する781アミノ酸からなる、分子量約88000のタンパク質である。
【0011】
上記XMAN1タンパク質は、後述のように、骨形成因子BMPのシグナル伝達を阻害し、神経化を誘導する活性を有する。従って、XMAN1タンパク質の構造、機能を詳細に解析することにより、脊椎動物における神経化誘導のメカニズムが明らかにされるだけでなく、例えば神経系の異常により引き起こされる疾病の病態機序の研究や治療薬等にも利用できる可能性がある。
【0012】
ここで、上記「タンパク質」とは、細胞、組織などから単離精製された状態であってもよいし、タンパク質をコードする遺伝子を宿主細胞に導入して、そのタンパク質を細胞内発現させた状態であってもよい。また、本発明に係るタンパク質は、付加的なポリペプチドを含むものであってもよい。
【0013】
また、上記「1またはそれ以上のアミノ酸が置換、欠失、挿入、及び/または付加」とは、部位特異的突然変異誘発法等の公知の変異タンパク質作製法により置換、欠失、挿入、及び/または付加できる程度の数のアミノ酸が置換、欠失、挿入、及び/または付加されることを意味する。このように、上記(b)のタンパク質は、換言すれば、上記(a)のタンパク質の変異タンパク質であり、ここにいう「変異」は、主として公知の変異タンパク質作製法により人為的に導入された変異を意味するが、天然に存在する同様の変異タンパク質を単離精製したものであってもよい。また、タンパク質の精製や検出等を容易に行うために、公知のヒスチジンタグ等の付加配列を末端に含ませてもよい。
【0014】
上記のように変異を導入すれば、神経化誘導因子として高い活性を持つタンパク質や精製の容易な組替えタンパク質等を人為的に得られる可能性がある。
【0015】
また、本発明に係る遺伝子は、上記(a)または(b)のタンパク質をコードする遺伝子である。
【0016】
本発明に係る上記構成の遺伝子として、例えば、XMAN1遺伝子が挙げられる。具体的には、上記XMAN1遺伝子は、配列番号2に示す塩基配列をcDNAとして有している。
【0017】
上記XMAN1遺伝子は、後述のように、アフリカツメガエルのアニマル・キャップの多分化能を利用して、アニマル・キャップを神経化する活性(神経化誘導活性)を指標に発現クローニング法とシブ・セレクション法を用いて単離された。上記遺伝子は、アフリカツメガエルにおける神経誘導に関与する遺伝子である。従って、上記遺伝子は、XMAN1タンパク質と同様な目的に利用することができる。さらに、上記XMAN1遺伝子を用いて、他の動物から相同性を有する、より有用な遺伝子を単離することも可能である。
【0018】
なお、本発明に係る「遺伝子」には、DNAおよびRNAが含まれるものとする。また、DNAには少なくともゲノムDNA、cDNAが含まれ、RNAには、mRNAなどが含まれる。また、DNAは2本鎖のみならず、それを構成するセンス鎖、アンチセンス鎖といった1本鎖でもよい。さらに、上記「遺伝子」は、上記(a)、(b)のタンパク質をコードする配列以外に、非翻訳領域(UTR)の配列やベクター配列(発現ベクター配列を含む)などの配列を含むものであってもよい。
【0019】
また、本発明に係る神経化誘導剤は、上記(a)、(b)のタンパク質、上記(a)タンパク質(即ち、XMAN1タンパク質)のヒトホモログであるタンパク質、または、これらタンパク質(即ち、上記(a)タンパク質、上記(b)タンパク質、もしくは上記(a)タンパク質のヒトホモログ)をコードする遺伝子のいずれか一つを含有する神経化誘導剤である。
【0020】
上記(a)タンパク質のヒトホモログの具体的な一例である、ヒトMAN1タンパク質は、後述のように、上記XMAN1タンパク質と相同性が高く、神経化を誘導する活性を有していると推定され、従って、XMAN1タンパク質と同様の利用可能性があると考えられる。なお、「上記(a)タンパク質のヒトホモログ」には、ヒトMAN1タンパク質以外に、このヒトMAN1タンパク質と一部のアミノ酸配列が異なるものの、その相違部分が機能上重要な領域ではなく、実質的にヒトMAN1タンパク質と同等の機能を有するタンパク質も含まれる。
【0021】
従って、例えば、上記ヒトMAN1タンパク質をコードする遺伝子である、ヒトMAN1遺伝子(アクセッションNo.AF112299)を本発明の神経化誘導剤に用いることも可能である。
【0022】
本発明の神経化誘導剤の利用法としては、(1)脊椎動物の未分化細胞に作用させることにより、その未分化細胞を神経化することへの利用、(2)神経系の疾患を有する脊椎動物に用いることにより、その個体における神経系の疾患を治療若しくは緩和することへの利用、等が挙げられる。
【0023】
本発明に係る神経化誘導方法は、上記(a)、(b)のタンパク質、上記(a)タンパク質のヒトホモログであるタンパク質、または、これらタンパク質をコードする遺伝子のいずれか一つを用いて細胞又は組織の神経化を誘導する方法である。また、本発明に係る骨形成因子BMPシグナル伝達経路阻害剤は、上記(a)、(b)のタンパク質、上記(a)タンパク質のヒトホモログであるタンパク質、または、これらタンパク質をコードする遺伝子のいずれか一つを含有するものである。
【0024】
上記骨形成因子BMPは、生体内で多彩な生理作用を引き起こすことが知られているが、未だBMPのシグナル伝達経路の詳細は明らかにされていない。従って、本発明に係る阻害剤を脊椎動物の細胞に用いることにより、脊椎動物の生体内で多彩な生理作用を引き起こすBMPのシグナル伝達経路を阻害することができ、分子生物学的、細胞生物学的、発生生物学的にBMPのシグナル伝達経路の詳細を解析する上での有効な手段となる。例えば、BMPのシグナル伝達経路の研究用アッセイキットや試薬の開発を行うことができる。また、本発明に係る阻害剤は、BMPのシグナル伝達経路に関係する種々の疾患の治療薬としても利用できる可能性がある。
【発明の効果】
【0025】
以上のように、本発明のタンパク質、遺伝子、神経化誘導剤、神経化誘導方法並びにBMPシグナル伝達経路阻害剤は、脊椎動物における神経化のメカニズムの解明という基礎研究上の利用のみならず、神経系の疾患等に対する治療薬や治療法の開発等への応用が可能であるという効果を奏する。
【0026】
また、本発明は、(1)ヒト膠原病患者血清中の抗ヒトMAN1抗体の存在の有無やヒト膠原病の病態機序を調べるための診断法、診断薬の開発並びにBMPシグナル伝達経路を標的とした治療薬への利用可能性、さらには、(2)癌化を含む細胞分裂、細胞周期、遺伝子発現制御らと密接に関係している核膜の消失・生成を調べるためのアッセイ法や試薬への利用可能性を有するものである。
【発明を実施するための最良の形態】
【0027】
本発明に係るタンパク質は、脊椎動物において胚の神経化を誘導する活性を有するタンパク質である。本実施の形態では、アフリカツメガエル由来のXMAN1タンパク質を挙げて説明する。後述する実施例に示すように、このXMAN1タンパク質は、骨形成因子BMPのシグナル伝達経路を阻害し、アフリカツメガエルのアニマル・キャップの神経化を誘導する活性(神経化誘導活性)を有しているタンパク質である。上記XMAN1は、配列番号1に示すアミノ酸配列を有し、781アミノ酸からなる、分子量約88000のタンパク質である。
【0028】
上記XMAN1タンパク質は、XMAN1遺伝子にコードされている。このXMAN1遺伝子のcDNAは、配列番号2に示すように、2645塩基対(約2.65kbp)のサイズを有しており、10番目から12番目の塩基配列が開始コドン(ATG)であり、2353番目から2355番目の塩基配列が終止コドン(TGA)である。従って、上記XMAN1遺伝子は、配列番号2に示す塩基配列のうち、10番目から2352番目の塩基配列をオープンリーディングフレーム(ORF)として有しており、このORFは、2343塩基対(約2.34kbp)のサイズを有している。
【0029】
また、本発明にかかるXMAN1遺伝子としてのDNAは、動物のゲノム中に含まれる形態、すなわちイントロンなどの非コード配列を含む「ゲノム」形DNAであってもよいし、逆転写酵素やポリメラーゼを用いてmRNAを経て得られるcDNA、すなわちイントロンなどの非コード配列を含まない「転写」形DNAであってもよい。
【0030】
また、本発明にかかるXMAN1遺伝子としてのDNAには、コードされているXMAN1タンパク質の形成に悪影響を与えないか妨げない限りにおいて、他のDNA配列を含んでいてもよい。例えば、遺伝子組み換え等に用いるために、リンカーを末端に含ませることもできる。
【0031】
本発明にかかるXMAN1遺伝子のクローニング方法としては、従来公知の方法を利用することが可能であり、特に限定されるものではない。本発明においては、その目的が、従来知られていないアフリカツメガエルにおける神経誘導に関与する遺伝子をクローニングすることであったので、後述する実施例で説明するように、アフリカツメガエルのアニマル・キャップの多分化能を利用して、神経化誘導活性を指標に発現クローニング法とシブ・セレクション法とを用いて単離している。
【0032】
本発明では、アフリカツメガエル由来の上記XMAN1遺伝子を用いて、他の動物からXMAN1遺伝子と相同性を有するDNAをクローニングすることが可能である。この場合のクローニング方法としても、従来公知の方法を利用することが可能であり、特に限定されるものではない。
【0033】
具体的には、ゲノムの少なくとも一部がデータベース化されている動物の場合には、上記XMAN1遺伝子の塩基配列、若しくはアミノ酸配列に基づいて相同性のある塩基配列をデータベース中から検索すればよい。例えば、汎用されている相同性検索アルゴリズムであるBLASTによる塩基配列及びアミノ酸配列レベルの相同性検索を好適に用いることができる。
【0034】
また、ゲノムがデータベース化されていない動物の場合には、例えば、従来公知のDNAライブラリーを用いたハイブリダイゼーション法を用いることもできる。具体的には、適切なクローニング・ベクターを使用して対象となる動物からゲノムライブラリーまたはcDNAライブラリーを調製するステップと、上記XMAN1遺伝子の少なくとも一部をプローブとして用いてハイブリダイゼーションを行い、ライブラリーから上記プローブにポジティブの断片を検出するステップとを含む方法を用いることができる。このように本発明のXMAN1遺伝子のうち、少なくとも一部の領域はプローブとして有用である。プローブに用いる領域には、XMAN1遺伝子とそのヒトホモログMAN1遺伝子間で保存性の高い配列が含まれることが好ましく、例えば、PCR法にてこのような領域を増幅させてプローブとして用いることが可能である。
【0035】
本発明に係るXMAN1遺伝子の一部を改変することによって、変異型XMAN1遺伝子が得られる。この変異型XMAN1遺伝子は、動物生体内にて発現することで、野性型と比べてより優れた神経化誘導活性を有するものであってもよいし、野性型と同じ神経化誘導活性を有しているサイレント突然変異(アミノ酸配列は変わらず、塩基配列のみが変化する変異)であってもよい。
【0036】
つまり、本発明に係る変異XMAN1タンパク質は、神経化誘導因子としての活性が野生型XMAN1タンパク質と略同等であってもよいし、より優れていてもよい。
【0037】
また、変異の様式は特に限定されるものではなく、配列番号1に示されるアミノ酸配列において、1またはそれ以上のアミノ酸残基が置換、欠失、挿入及び/または付加されていればよい。
【0038】
例えば、本発明者らは、後述する実施例に示すように、種々のXMAN1欠失変異タンパク質を作製し、その機能解析を行うことにより、XMAN1タンパク質の活性発現に必要な領域として、XMAN1タンパク質のカルボキシル末端領域(C末端領域;582番目から781番目のアミノ酸残基)を同定した。このC末端領域のみで、BMPのシグナル伝達経路を阻害しアフリカツメガエルのアニマル・キャップの神経化を誘導することができる。
【0039】
つまり、本発明には、配列番号1に示すアミノ酸配列を有するXMAN1タンパク質の変異体であって、神経化誘導因子として野生型と略同等の活性を持つ変異タンパク質が含まれていてもよく、具体的には、例えば、後述する実施例に示すように、配列番号1に示されるアミノ酸配列において、アミノ末端側(N末端側)のLEMドメインを含む43個のアミノ酸が欠失された変異タンパク質ΔLEM、N末端側の449個のアミノ酸が欠失された変異タンパク質ΔTM1、N末端側の581個のアミノ酸が欠失され、C末端領域のみからなる変異タンパク質CT(582番目から781番目のアミノ酸残基)を挙げることができる。勿論、変異タンパク質はこれに限られず、神経化誘導活性の発現に重要な領域と考えられる後述のRNP1領域(701番目から708番目のアミノ酸残基)以外の領域に変異が導入され、野生型と略同等またはより優れた神経化活性誘導を有する変異タンパク質も含まれる。また、本発明には、上記変異タンパク質をコードする遺伝子も含まれる。
【0040】
このように、本発明に係るXMAN1遺伝子としては、(a)配列番号1に示されるアミノ酸配列からなるタンパク質か、(b)配列番号1に示されるアミノ酸配列において、1またはそれ以上のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導因子としての活性を有するタンパク質をコードする遺伝子であればよい。
【0041】
また、後述する実施例に示すように、相同性検索の結果、XMAN1遺伝子はヒトMAN1遺伝子と高い相同性を有している。しかしながら、本発明完成時までヒトMAN1遺伝子は機能未知であり、今回、XMAN1遺伝子は、ヒトMAN1遺伝子とは無関係にクローニングされたものである。
【0042】
さらに、後述する実施例に示すように、XMAN1タンパク質のC末端領域とヒトMAN1タンパク質のC末端領域とは、アミノ酸レベルで相同性94%を示している。従って、ヒトMAN1タンパク質もXMAN1タンパク質と同様に、神経化を誘導し、BMPのシグナル伝達経路の阻害活性を有すると推定できる。つまり、本発明は、ヒトMAN1タンパク質がBMPのシグナル伝達経路を阻害すると共に、脊椎動物において神経誘導に関与する機能を持つことを最初に明らかにしたものである。
【0043】
ヒトMAN1タンパク質は、ヒトの核内膜に局在する膜タンパク質であることが知られており(文献 Lin, F. et al. (2000).MAN1, an inner nuclear membrane protein that shares the LEM domain with lamina-associated polypeptide 2 and emerin. J. Biol. Chem. 275, pp4840-4847参照)、XMAN1タンパク質も構造上の類似性から核内膜タンパク質であると推定される。
【0044】
また、ヒト膠原病患者の血清中に、ヒトMAN1タンパク質に対する自己抗体が現れることがこれまで報告されている(文献 Paulin-Levasseur, M. et al. (1996). The MAN antigens are non-lamin constituents of the nuclear lamina in vertebrate cells. Chromosoma, 104, pp367-79参照)。ヒト膠原病は、一種の自己免疫疾患であると考えられており、その原因は未だ明らかにされておらず、治療法等も十分ではない。
【0045】
本発明を用いることにより、ヒトMAN1タンパク質の機能(BMPシグナル伝達経路阻害活性、神経化誘導活性)とヒト膠原病の病態との関係を調べるための診断薬や診断方法等を開発することが可能である。さらに、ヒト膠原病の病態機序を解明するための有効な手段となり、BMPシグナル伝達経路を標的とした治療薬を開発することも可能である。即ち、ヒト膠原病の病態機序の解明という基礎研究的な利用だけでなく、治療薬や治療法等の医学的な利用も可能である。
【0046】
また、核膜の消失・生成は、細胞分裂(癌化を含む)、細胞周期、遺伝子の発現制御と密接に関係している。この核膜の消失・生成と核内膜タンパク質であるMAN1、XMAN1のBMPシグナル伝達経路阻害活性との関連性を調べるためのアッセイ法の確立、試薬の開発等も可能である。
【0047】
以上、アフリカツメガエルの神経化誘導因子XMAN1やヒトMAN1タンパク質について、それぞれ利用可能性を述べてきたが、これら2つのタンパク質は高い相同性を有していることから、同様な機能があると推定される。従って、いずれのタンパク質、若しくはこれらのタンパク質をコードするいずれの遺伝子を用いても、本発明は、脊椎動物における神経化のメカニズム、骨形成因子BMPのシグナル伝達経路、ヒト膠原病の病態機序等の解明に役立つと考えられる。
【0048】
以下添付した図面に沿って実施例を示し、本発明の実施の形態についてさらに詳しく説明する。もちろん、本発明は以下の実施例に限定されるものではなく、細部については様々な態様が可能であることはいうまでもない。
【実施例】
【0049】
以下の実施例では、アフリカツメガエルの新規神経化誘導因子XMAN1のクローニングとその機能解析を行った結果を概略説明する。
【0050】
〔実施例1〕 新規神経化誘導因子のクローニング
アフリカツメガエルの胚の胞胚期から原腸胚期にかけての予定外胚葉組織(アニマル・キャップ)の多分化能を利用して、将来脳組織となる前方神経外胚葉から作製したcDNAライブラリーからアニマル・キャップを神経化する活性(神経化誘導活性)を有する因子を、発現クローニング法とシブ・セレクション法を用いて単離した。具体的には、アフリカツメガエルの体外受精と胚の培養は、Kay and Pengの方法(Xenopus Laevis: practical uses in cell and molecular biology, Methods in Cell Biology, Volume 36, Academic Press,1991)に従った。胚のステージは、Nieuwkoop and Faber(Normal table of Xenopus Laevis, Garland Publishing,1994)に基づいて決定した。前方神経外胚葉特異的cDNAライブラリーは、Shingaらの報告(Mech. Of Dev., Vol. 109, pp225-239, 2001)に基づいて、SuperScript Plasmid System for cDNA Synthesis and Plasmid Cloning(GibcoBRL)を用いて作製し、pCS105ベクター(カリフォルニア大学 Richard Harland博士からの供与)に導入した。シブ・セレクション法は、Lustigらの報告(Methods in Enzymology, Vol. 283, pp83-99, 1997)を参考にした。
【0051】
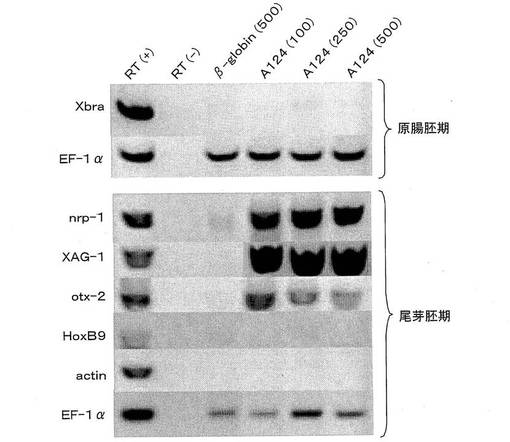
即ち、cDNAライブラリーを200クローンからなるプールに分割し、プールごとにcDNAから合成したRNAをアニマル・キャップに微量注入した。その後、アニマル・キャップにおける汎神経マーカーnrp-1の誘導の有無を、RT−PCR法により調べた。RT−PCR法は、2細胞期にmRNAを胚の動物極に微量注入し、胞胚期にアニマル・キャップを切り出した後、原腸胚期、尾芽胚期のそれぞれにおいてアニマル・キャップを回収した。RNAの抽出、cDNAの合成、PCR反応は、Wilson and Melton(Curr. Biol., Vol.4, pp676-686, 1994)に基づいて、非ラジオアイソトープ存在下で行った。以下、RT−PCR法はこの方法を用いた。なお、RT−PCR法の替わりに、in situハイブリダイゼーション法を用いることも可能である。次いで、nrp-1の発現を誘導する活性をもつプールに関して、プールサイズを順次小さくしていき、最終的に神経化誘導活性を有する単一のクローンA124を得た。
【0052】
次いで、図1(a)に示すように、2細胞期の胚の動物極にクローンA124のmRNAを微量注入した後、図1(b)に示すように、胞胚期にアニマル・キャップを切り出した。その後、図2に示すように、原腸胚期、尾芽胚期のアニマル・キャップそれぞれにおいて、種々のマーカーの発現誘導をRT−PCR法により調べた。なお、注入したクローンA124のmRNA量は、1胚あたり100p(ピコ)g、250pg、500pgである。また、β-globin(500)は、ネガティブコントロールとしてβ-globinのmRNAを500pg注入したことを示す。また、RT(+)、RT(−)は、それぞれポジティブコントロール、ネガティブコントロールを示す。EF−1αは反応に用いたcDNA量のコントロールである。
【0053】
RT−PCRの結果、中胚葉マーカーとして知られるXbra、actinの発現誘導なしに、汎神経マーカーであるnrp-1、前方神経マーカーであるXAG-1、otx-2の発現が誘導された。しかし、後方神経マーカーであるHoxB-9は発現誘導されなかった。この結果により、クローンA124にコードされている因子は、前方神経の誘導に関わる新規因子であることが示唆された。また、その神経誘導化活性は、中胚葉の誘導を介した2次的なものではないことを意味している。
【0054】
〔実施例2〕 クローンA124の構造解析
次に、クローンA124の全長を含むcDNAを単離した。具体的には、クローンA124をプローブとしてアフリカツメガエルのステージ30期の頭部cDNAライブラリーを用いてスクリーニングし、得られたポジティブクローンのうち、インサートの最も長いクローンPC11の塩基配列を決定した。しかし、このクローンPC11も開始コドンを欠いていたので、5’-Full RACE Core Set(TAKARA)を用いて、開始コドンを含む上流領域を増幅し、この遺伝子の全長を決定した。その全構造を決定したところ、図3に示すように、781アミノ酸からなる分子量約88000のタンパク質をコードしていることがわかった。
【0055】
この遺伝子の相同性検索を行ったところ、ヒトMAN1遺伝子と高い相同性を示すことがわかった。ヒトMAN1遺伝子は、910アミノ酸からなり、核内膜に存在する機能未知のタンパク質をコードする遺伝子である。
【0056】
従って、ヒトMAN1遺伝子と相同性が高かったことから、この遺伝子をXMAN1(Xenopus MAN1)と命名した。
【0057】
このXMAN1タンパク質とヒトMAN1タンパク質とは、構造上の特徴も非常に類似していることが明らかとなった。XMAN1タンパク質は、ヒトMAN1タンパク質と同様に、N末端側にクロマチンと結合すると考えられるLEMドメイン(5番目から43番目のアミノ酸残基)を有している。なお、これら2つのタンパク質間における、このLEMドメインの相同性は74%であった。
【0058】
また、XMAN1タンパク質は、疎水性アミノ酸に富む2つの膜貫通領域TM1、TM2(TM1;349番目から372番目のアミノ酸残基、TM2;497番目から519番目のアミノ酸残基)を有しており、この点でもヒトMAN1タンパク質と類似している。ヒトMAN1タンパク質はこの2つの膜貫通領域を介して核内膜に埋め込まれており、N末端とC末端は共に核の内側に存在する膜タンパク質である事が知られている。従って、XMAN1タンパク質も生体内でヒトMAN1タンパク質と同様の局在性を示すと考えられる。
【0059】
さらに、XMAN1タンパク質はC末端領域(582番目から781番目のアミノ酸残基)に、RNA結合タンパク質に多く見られるRNP2(661番目から666番目のアミノ酸残基)、RNP1(701番目から708番目のアミノ酸残基)と呼ばれる保存配列を含むRNA認識モチーフ(RRM;656番目から733番目のアミノ酸残基)を有することがわかった。また、ヒトMAN1タンパク質も非常に類似しているC末端領域を有していることがわかった。即ち、これら2つのタンパク質間における、このC末端領域の相同性は、アミノ酸レベルで94%であった。これまで、ヒトMAN1タンパク質にRRMが存在することは文献的にも指摘されておらず、今回初めてその存在を明らかにした。
【0060】
〔実施例3〕 新規神経化因子XMAN1の活性発現に必須な領域の特定
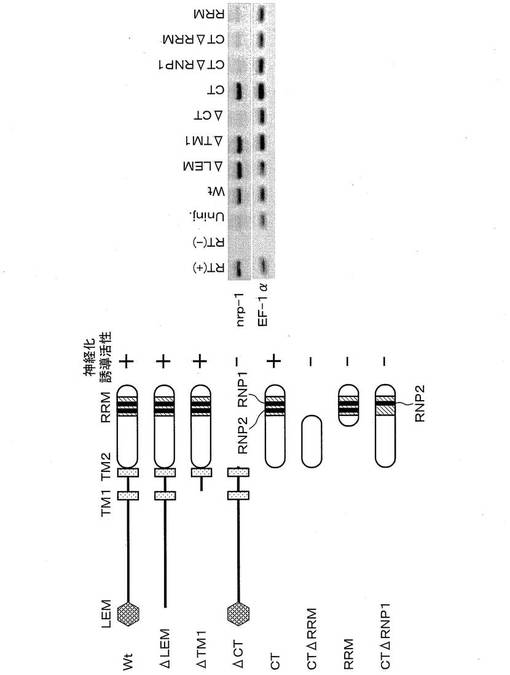
XMAN1タンパク質のどの領域に神経化誘導活性があるのかを調べるために、図4に示すように、XMAN1タンパク質の欠失変異タンパク質ΔLEM、ΔTM1、ΔCT、CTをそれぞれ作製した。欠失変異タンパク質ΔLEM、ΔTM1、ΔCTとは、野生型XMAN1タンパク質WtからそれぞれLEMドメイン、TM1領域、C末端領域を欠失させた変異タンパク質であり、欠失変異タンパク質CTとは、C末端領域のみ残存させ、この領域以外を欠失させた変異タンパク質である。なお、Δ(デルタ)とは、欠失変異体を意味する記号である。
【0061】
具体的には、XMAN1タンパク質の種々の変異タンパク質は、PCR法を用いて作製し、pCS+MTベクターに導入した。微量注入に用いたmRNAは、MEGAscript IN Vitro Transcription Kits(Ambion)を用いて合成した。
【0062】
それぞれの欠失変異タンパク質のmRNAを2細胞期の胚の動物極に微量注入し、胞胚期にアニマル・キャップを切り出した後、尾芽胚期におけるアニマル・キャップでの汎神経マーカーnrp-1の発現誘導をRT−PCR法により調べた。
【0063】
具体的には、尾芽胚期のアニマル・キャップからRNAを単離し、公知の手法に従いcDNAを合成して、PCR法による増幅を行った。なお、Uninj,はRNAを注入していないアニマル・キャップを示す。そのRT−PCRの結果、欠失変異タンパク質ΔLEM、ΔTM1、CTは野生型タンパク質Wtと略同等のnrp-1の発現を誘導した。しかし、欠失変異タンパク質ΔCTは、ほとんどnrp-1の発現を誘導しなかった。従って、XMAN1タンパク質のC末端領域が神経化誘導活性に必須であることがわかった。このC末端領域は、ヒトMAN1タンパク質のC末端領域と高い相同性を示しており、進化上もこの領域が重要な機能を有していることが示唆された。従って、このXMAN1タンパク質のC末端領域を神経化誘導活性ドメインCTと呼ぶこととした。
【0064】
さらに、この神経化誘導活性ドメインCTについて、欠失変異タンパク質CTΔRRM、RRM、CTΔRNP1を作製して詳細に検討した。欠失変異タンパク質CTΔRRM、CTΔRNP1は、神経化誘導活性ドメインCTからそれぞれRRM、RNP1を欠失させた変異タンパク質である。欠失変異タンパク質RRMは、神経化誘導活性ドメインCTのRRMのみ残存させて、N末端からRRMまでの領域を欠失させた変異タンパク質である。
【0065】
これらの欠失変異体のmRNAを用いて、上記と同様の方法でnrp-1の誘導を指標に、神経化誘導活性ドメインの機能に重要な部位を調べた結果、図4に示すように、欠失変異タンパク質CTΔRRM、RRM、CTΔRNP1は、いずれもnrp-1の発現を誘導しなかった。この結果から、RRM、特にその中のRNP1配列が神経化誘導活性に重要な役割を果たしていることがわかった。これまで、ヒトMAN1タンパク質にRRMの存在のみならず、その機能についても全くわかっていなかったが、今回初めてその存在意義を示すことができた。
【0066】
〔実施例4〕 神経活性化ドメインによるBMPシグナル伝達経路の阻害
さらに、XMAN1タンパク質は骨形成因子BMPのシグナル伝達経路を阻害することにより、アニマル・キャップを神経化するのか否か検討した。BMPのシグナルは細胞表面のBMPI型受容体、BMPII型受容体の2種類の受容体と細胞内因子Smad(特にSmad1,4,5,8)とを介して核に伝えられた後、そこで標的因子の発現誘導、活性化を通して多様な生理作用を引き起こすと考えられている。これら、BMPシグナル伝達経路の直接的な標的因子として、アフリカツメガエルではXmsx-1、Xhox-3、Xvent-2等が知られている。
【0067】
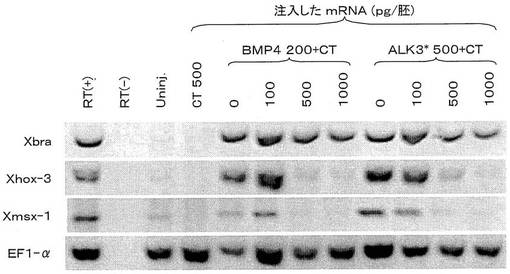
そこで、2細胞期の胚の動物極にBMP−4、若しくは活性化型BMPI型受容体(ALK3*)のmRNAを単独で微量注入し、胞胚期にアニマル・キャップを切り出した後、原腸胚期に種々のマーカーの発現誘導をRT−PCR法により調べた。具体的には、原腸胚期のアニマル・キャップからRNAを単離し、公知の手法に従いcDNAを合成して、PCR法による増幅を行った。なお、図5に示すように、注入したBMP−4のmRNA量は、胚1個あたり、200pg、ALK3*は500pgであり、CTは注入していない。コントロールとして、CTのmRNAのみを500pg注入したものを示した。そのRT−PCRの結果は図5に示すように、BMP−4またはALK3*のmRNAのみを注入した場合、標的因子Xmsx-1、Xhox-3の発現は誘導された。また、CTのmRNAを単独で注入した場合、標的因子Xmsx-1、Xhox-3の発現は誘導されなかった。なお、活性化型BMPI型受容体(ALK3*)とは、BMPを受容しなくても活性型として機能し、恒常的にシグナルを細胞内に伝達する受容体である。
【0068】
しかし、同様にBMP−4のmRNAと、若しくはALK3*のmRNAと神経化誘導活性ドメインCTのmRNAとを一緒に微量注入した場合(注入したBMP−4、ALK3*のmRNA量はそれぞれ、胚1個あたり、200pg、500pg、またCTのmRNA量はそれぞれ、100,500,1000pgである。)、標的因子Xmsx-1、Xhox-3の発現誘導は、注入したCTのmRNA量に依存して著しく阻害された。なお、中胚葉マーカーXbraの発現誘導はCTのmRNAの注入量によって、ほとんど変化しなかった。これらの結果から、神経化誘導活性ドメインCTがBMPのシグナル伝達経路を受容体より下流で阻害することが示された。
【0069】
〔実施例5〕 新規神経化因子XMAN1によるBMP標的因子の転写抑制
さらに、XMAN1がBMPの標的因子の発現誘導を転写レベルで抑制しているか否かをルシフェラーゼ・アッセイ法を用いて調べた。
【0070】
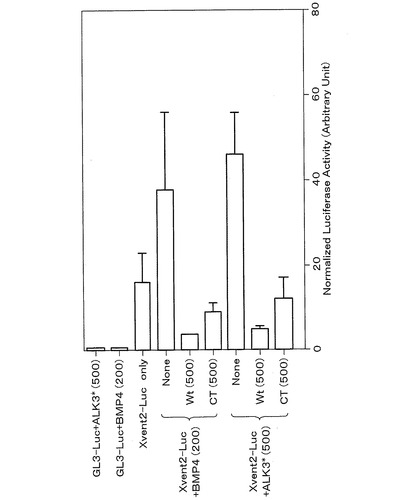
具体的には、ルシフェラーゼ活性は、Dual Luciferase Reporter Assay System(Promega)を用いて測定した。まず、BMPの標的因子の一つ、Xvent-2のプロモーターの下流にリポーター遺伝子としてfireflyルシフェラーゼ遺伝子を組み込んだプラスミドDNA(Xvent2-Luc、カリフォルニア大学 Ken Cho博士からの供与)を用いた。その後、このプラスミドDNA(Xvent2-Luc)を100pg、Renillaルシフェラーゼ遺伝子を持つコントロールプラスミドDNA(pRL-TK)を2pg、BMP−4のmRNA並びにWt、CTのいずれか1つのmRNAを混合して、2細胞期の胚の動物極に微量注入した。胞胚期にアニマル・キャップを切り出した後、原腸胚期に5個ずつ、3本のチューブに集め、各チューブに50μlのルシフェラーゼ反応液を加えて、fireflyルシフェラーゼ活性を測定した。その後、Gro & Stop溶液を加えてRenillaルシフェラーゼ活性を測定した。測定には、ルミノメーター(Berthold)を用いた。Fireflyルシフェラーゼ活性値をRenillaルシフェラーゼ活性値で割った値を補正値とし、3本のチューブの平均値を算出した。また、BMP−4の替わりにALK3*を用いて、同様にルシフェラーゼ活性を測定した。なお、注入したBMP−4のmRNA量は、1胚あたり200pg、ALK3*は500pg、XMAN1のmRNAは野生型Wt、欠失変異体CTともに500pgである。また、コントロールとして、BMP−4のmRNAと、若しくはALK3*のmRNAと、ルシフェラーゼ遺伝子をXvent-2のプロモーターと異なるプロモーター(GL-3)の下流に組み込んだプラスミドDNA(GL3-Luc)とを注入した場合、Xvent2-Lucのみを注入した場合を示す。
【0071】
これらのルシフェラーゼ・アッセイの結果、図6に示すように、BMP−4若しくはALK3*のmRNAとXvent2-Lucとを注入した場合、ルシフェラーゼ活性は上昇した。しかし、この活性上昇は、野生型XMAN1タンパク質Wt、神経化誘導活性ドメインCTにより著しく抑制された。従って、XMAN1タンパク質、特にその神経化誘導活性ドメインCTは、BMPの標的因子の転写活性化を抑制することがわかった。
【0072】
なお、BMP−4若しくはALK3*非存在下でも、ある程度のルシフェラーゼ活性が見られるのは、アニマル・キャップ内に存在する内在性のBMPの働きによるものと考えられる。
【産業上の利用可能性】
【0073】
本発明は、(1)ヒト膠原病患者血清中の抗ヒトMAN1抗体の存在の有無やヒト膠原病の病態機序を調べるための診断法、診断薬の開発並びにBMPシグナル伝達経路を標的とした治療薬への利用可能性、さらには、(2)癌化を含む細胞分裂、細胞周期、遺伝子発現制御らと密接に関係している核膜の消失・生成を調べるためのアッセイ法や試薬への利用可能性を有するものである。
【図面の簡単な説明】
【0074】
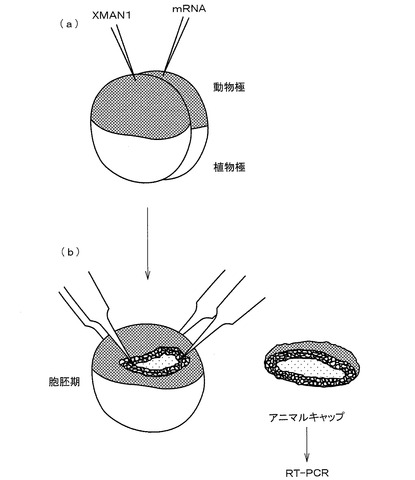
【図1】(a)は、本発明の実施例におけるアフリカツメガエルの2細胞期の胚へのmRNAの注入操作を示した概略説明図であり、(b)は、胞胚期におけるアニマル・キャップの切り出し操作を示した概略説明図である。
【図2】図1で切り出したアニマル・キャップの原腸胚期、尾芽胚期における各種マーカーの発現誘導をRT−PCR法にて調べた結果を示す電気泳動図である。
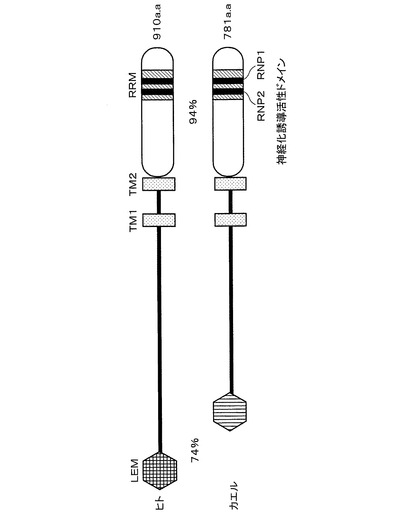
【図3】本発明の実施例におけるヒトMAN1並びにXMAN1タンパク質の各構造を概略的に示す模式図である。
【図4】XMAN1の各種欠失変異タンパク質を示す概略模式図、並びに各種変異mRNAを注入したアニマル・キャップの尾芽胚期における各種マーカーの発現誘導をRT−PCR法にて調べた結果を示す電気泳動図である。
【図5】各種mRNAを注入したアニマル・キャップの原腸胚期における各種マーカーの発現誘導をRT−PCR法にて調べた結果を示す電気泳動図である。
【図6】XMAN1の各種変異タンパク質によるBMPの標的因子の転写活性の抑制を、ルシフェラーゼ・アッセイ法により調べた結果を示すグラフである。
【技術分野】
【0001】
本発明は、神経化誘導に関与する新規タンパク質、およびその遺伝子、並びにその利用方法に関し、より詳細には、当該タンパク質、若しくはその遺伝子を含有する神経化誘導剤、神経化誘導方法、並びに骨形成因子BMPシグナル伝達経路阻害剤に関するものである。
【背景技術】
【0002】
アフリカツメガエルの胚の胞胚期から原腸胚期にかけて動物極側に位置する、予定外胚葉組織(アニマル・キャップ)は多分化能を有しており、種々の因子に反応して外、中、内胚葉由来の様々な組織、器官に分化することが知られている。
【0003】
近年、このアニマル・キャップの性質を利用して、脊椎動物における神経誘導のメカニズムを解明するための研究が盛んに行われており、これまでの研究成果として、脊椎動物における神経誘導には、骨形成因子BMP(Bone Morphogenetic Protein)を介するシグナル伝達の経路を阻害することが重要であるとの報告も多数なされている(例えば、非特許文献1参照)。
【0004】
この骨形成因子として、ヒトでは現在までに9種類のBMP(BMP−1〜BMP−9)がクローニングされている。このBMPは骨形成ばかりでなく、受精卵から分裂増殖、細胞分化により新しい組織、器官が形成されてくる、いわゆる形態形成にも関与していることが知られている(例えば、非特許文献2参照)。
【非特許文献1】Harland, R.(2000). Neural induction, Curr. Opin. Genet. Dev. 10, pp357-62.
【非特許文献2】Hogan, BL.(1996). Bone morphogenetic proteins: multifunctional regulators of vertebrate development. Genes Dev. 10, pp1580-94.
【発明の開示】
【発明が解決しようとする課題】
【0005】
脊椎動物における神経誘導のメカニズムは、様々な因子が関与していることから非常に複雑であり、そのメカニズムは未だ十分に解明されているとはいえず、神経誘導と密接に関与していると考えられる骨形成因子BMPのシグナル伝達経路の詳細についても明らかにはなっていない。
【0006】
他方、もし脊椎動物における神経誘導に関与する未知の遺伝子や未知のタンパク質が明らかにされれば、当該遺伝子およびタンパク質の機能解析を通じ、神経誘導による形態形成の分子機構やBMPシグナル伝達経路の詳細がより一層明らかにされるといった学術的意義もさることながら、例えば神経系の異常により引き起こされる疾病の病態機序の研究や治療薬、治療法等の開発にも利用できる可能性があり、医学的領域への幅広い利用が期待できる。
【0007】
本発明は、上記の課題に鑑みなされたものであり、その目的は、脊椎動物における神経誘導に関与する新規遺伝子、およびそのタンパク質を提供するとともに、これらの利用方法を提案することにある。
【課題を解決するための手段】
【0008】
本発明者らは、鋭意検討を重ねた結果、アフリカツメガエルのアニマル・キャップを神経化させる因子をクローニングし、新規神経化誘導因子XMAN1(Xenopus MAN1)を同定した。さらに、その機能を詳細に解析することにより、この新規神経化誘導因子XMAN1が、骨形成因子BMPを介するシグナル伝達経路を阻害し神経化を誘導する効果があることを見出し、本発明を完成させるに至った。
【0009】
すなわち、本発明に係るタンパク質は、以下の(a)または(b)のタンパク質である。
(a)配列番号1に示されるアミノ酸配列からなるタンパク質。
(b)配列番号1に示されるアミノ酸配列において、1またはそれ以上のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導因子としての活性を持つタンパク質。
【0010】
上記タンパク質の具体的な一例として、アフリカツメガエル由来の神経化誘導因子XMAN1が挙げられる。具体的には、上記XMAN1タンパク質は、配列番号1に示すアミノ酸配列を有する781アミノ酸からなる、分子量約88000のタンパク質である。
【0011】
上記XMAN1タンパク質は、後述のように、骨形成因子BMPのシグナル伝達を阻害し、神経化を誘導する活性を有する。従って、XMAN1タンパク質の構造、機能を詳細に解析することにより、脊椎動物における神経化誘導のメカニズムが明らかにされるだけでなく、例えば神経系の異常により引き起こされる疾病の病態機序の研究や治療薬等にも利用できる可能性がある。
【0012】
ここで、上記「タンパク質」とは、細胞、組織などから単離精製された状態であってもよいし、タンパク質をコードする遺伝子を宿主細胞に導入して、そのタンパク質を細胞内発現させた状態であってもよい。また、本発明に係るタンパク質は、付加的なポリペプチドを含むものであってもよい。
【0013】
また、上記「1またはそれ以上のアミノ酸が置換、欠失、挿入、及び/または付加」とは、部位特異的突然変異誘発法等の公知の変異タンパク質作製法により置換、欠失、挿入、及び/または付加できる程度の数のアミノ酸が置換、欠失、挿入、及び/または付加されることを意味する。このように、上記(b)のタンパク質は、換言すれば、上記(a)のタンパク質の変異タンパク質であり、ここにいう「変異」は、主として公知の変異タンパク質作製法により人為的に導入された変異を意味するが、天然に存在する同様の変異タンパク質を単離精製したものであってもよい。また、タンパク質の精製や検出等を容易に行うために、公知のヒスチジンタグ等の付加配列を末端に含ませてもよい。
【0014】
上記のように変異を導入すれば、神経化誘導因子として高い活性を持つタンパク質や精製の容易な組替えタンパク質等を人為的に得られる可能性がある。
【0015】
また、本発明に係る遺伝子は、上記(a)または(b)のタンパク質をコードする遺伝子である。
【0016】
本発明に係る上記構成の遺伝子として、例えば、XMAN1遺伝子が挙げられる。具体的には、上記XMAN1遺伝子は、配列番号2に示す塩基配列をcDNAとして有している。
【0017】
上記XMAN1遺伝子は、後述のように、アフリカツメガエルのアニマル・キャップの多分化能を利用して、アニマル・キャップを神経化する活性(神経化誘導活性)を指標に発現クローニング法とシブ・セレクション法を用いて単離された。上記遺伝子は、アフリカツメガエルにおける神経誘導に関与する遺伝子である。従って、上記遺伝子は、XMAN1タンパク質と同様な目的に利用することができる。さらに、上記XMAN1遺伝子を用いて、他の動物から相同性を有する、より有用な遺伝子を単離することも可能である。
【0018】
なお、本発明に係る「遺伝子」には、DNAおよびRNAが含まれるものとする。また、DNAには少なくともゲノムDNA、cDNAが含まれ、RNAには、mRNAなどが含まれる。また、DNAは2本鎖のみならず、それを構成するセンス鎖、アンチセンス鎖といった1本鎖でもよい。さらに、上記「遺伝子」は、上記(a)、(b)のタンパク質をコードする配列以外に、非翻訳領域(UTR)の配列やベクター配列(発現ベクター配列を含む)などの配列を含むものであってもよい。
【0019】
また、本発明に係る神経化誘導剤は、上記(a)、(b)のタンパク質、上記(a)タンパク質(即ち、XMAN1タンパク質)のヒトホモログであるタンパク質、または、これらタンパク質(即ち、上記(a)タンパク質、上記(b)タンパク質、もしくは上記(a)タンパク質のヒトホモログ)をコードする遺伝子のいずれか一つを含有する神経化誘導剤である。
【0020】
上記(a)タンパク質のヒトホモログの具体的な一例である、ヒトMAN1タンパク質は、後述のように、上記XMAN1タンパク質と相同性が高く、神経化を誘導する活性を有していると推定され、従って、XMAN1タンパク質と同様の利用可能性があると考えられる。なお、「上記(a)タンパク質のヒトホモログ」には、ヒトMAN1タンパク質以外に、このヒトMAN1タンパク質と一部のアミノ酸配列が異なるものの、その相違部分が機能上重要な領域ではなく、実質的にヒトMAN1タンパク質と同等の機能を有するタンパク質も含まれる。
【0021】
従って、例えば、上記ヒトMAN1タンパク質をコードする遺伝子である、ヒトMAN1遺伝子(アクセッションNo.AF112299)を本発明の神経化誘導剤に用いることも可能である。
【0022】
本発明の神経化誘導剤の利用法としては、(1)脊椎動物の未分化細胞に作用させることにより、その未分化細胞を神経化することへの利用、(2)神経系の疾患を有する脊椎動物に用いることにより、その個体における神経系の疾患を治療若しくは緩和することへの利用、等が挙げられる。
【0023】
本発明に係る神経化誘導方法は、上記(a)、(b)のタンパク質、上記(a)タンパク質のヒトホモログであるタンパク質、または、これらタンパク質をコードする遺伝子のいずれか一つを用いて細胞又は組織の神経化を誘導する方法である。また、本発明に係る骨形成因子BMPシグナル伝達経路阻害剤は、上記(a)、(b)のタンパク質、上記(a)タンパク質のヒトホモログであるタンパク質、または、これらタンパク質をコードする遺伝子のいずれか一つを含有するものである。
【0024】
上記骨形成因子BMPは、生体内で多彩な生理作用を引き起こすことが知られているが、未だBMPのシグナル伝達経路の詳細は明らかにされていない。従って、本発明に係る阻害剤を脊椎動物の細胞に用いることにより、脊椎動物の生体内で多彩な生理作用を引き起こすBMPのシグナル伝達経路を阻害することができ、分子生物学的、細胞生物学的、発生生物学的にBMPのシグナル伝達経路の詳細を解析する上での有効な手段となる。例えば、BMPのシグナル伝達経路の研究用アッセイキットや試薬の開発を行うことができる。また、本発明に係る阻害剤は、BMPのシグナル伝達経路に関係する種々の疾患の治療薬としても利用できる可能性がある。
【発明の効果】
【0025】
以上のように、本発明のタンパク質、遺伝子、神経化誘導剤、神経化誘導方法並びにBMPシグナル伝達経路阻害剤は、脊椎動物における神経化のメカニズムの解明という基礎研究上の利用のみならず、神経系の疾患等に対する治療薬や治療法の開発等への応用が可能であるという効果を奏する。
【0026】
また、本発明は、(1)ヒト膠原病患者血清中の抗ヒトMAN1抗体の存在の有無やヒト膠原病の病態機序を調べるための診断法、診断薬の開発並びにBMPシグナル伝達経路を標的とした治療薬への利用可能性、さらには、(2)癌化を含む細胞分裂、細胞周期、遺伝子発現制御らと密接に関係している核膜の消失・生成を調べるためのアッセイ法や試薬への利用可能性を有するものである。
【発明を実施するための最良の形態】
【0027】
本発明に係るタンパク質は、脊椎動物において胚の神経化を誘導する活性を有するタンパク質である。本実施の形態では、アフリカツメガエル由来のXMAN1タンパク質を挙げて説明する。後述する実施例に示すように、このXMAN1タンパク質は、骨形成因子BMPのシグナル伝達経路を阻害し、アフリカツメガエルのアニマル・キャップの神経化を誘導する活性(神経化誘導活性)を有しているタンパク質である。上記XMAN1は、配列番号1に示すアミノ酸配列を有し、781アミノ酸からなる、分子量約88000のタンパク質である。
【0028】
上記XMAN1タンパク質は、XMAN1遺伝子にコードされている。このXMAN1遺伝子のcDNAは、配列番号2に示すように、2645塩基対(約2.65kbp)のサイズを有しており、10番目から12番目の塩基配列が開始コドン(ATG)であり、2353番目から2355番目の塩基配列が終止コドン(TGA)である。従って、上記XMAN1遺伝子は、配列番号2に示す塩基配列のうち、10番目から2352番目の塩基配列をオープンリーディングフレーム(ORF)として有しており、このORFは、2343塩基対(約2.34kbp)のサイズを有している。
【0029】
また、本発明にかかるXMAN1遺伝子としてのDNAは、動物のゲノム中に含まれる形態、すなわちイントロンなどの非コード配列を含む「ゲノム」形DNAであってもよいし、逆転写酵素やポリメラーゼを用いてmRNAを経て得られるcDNA、すなわちイントロンなどの非コード配列を含まない「転写」形DNAであってもよい。
【0030】
また、本発明にかかるXMAN1遺伝子としてのDNAには、コードされているXMAN1タンパク質の形成に悪影響を与えないか妨げない限りにおいて、他のDNA配列を含んでいてもよい。例えば、遺伝子組み換え等に用いるために、リンカーを末端に含ませることもできる。
【0031】
本発明にかかるXMAN1遺伝子のクローニング方法としては、従来公知の方法を利用することが可能であり、特に限定されるものではない。本発明においては、その目的が、従来知られていないアフリカツメガエルにおける神経誘導に関与する遺伝子をクローニングすることであったので、後述する実施例で説明するように、アフリカツメガエルのアニマル・キャップの多分化能を利用して、神経化誘導活性を指標に発現クローニング法とシブ・セレクション法とを用いて単離している。
【0032】
本発明では、アフリカツメガエル由来の上記XMAN1遺伝子を用いて、他の動物からXMAN1遺伝子と相同性を有するDNAをクローニングすることが可能である。この場合のクローニング方法としても、従来公知の方法を利用することが可能であり、特に限定されるものではない。
【0033】
具体的には、ゲノムの少なくとも一部がデータベース化されている動物の場合には、上記XMAN1遺伝子の塩基配列、若しくはアミノ酸配列に基づいて相同性のある塩基配列をデータベース中から検索すればよい。例えば、汎用されている相同性検索アルゴリズムであるBLASTによる塩基配列及びアミノ酸配列レベルの相同性検索を好適に用いることができる。
【0034】
また、ゲノムがデータベース化されていない動物の場合には、例えば、従来公知のDNAライブラリーを用いたハイブリダイゼーション法を用いることもできる。具体的には、適切なクローニング・ベクターを使用して対象となる動物からゲノムライブラリーまたはcDNAライブラリーを調製するステップと、上記XMAN1遺伝子の少なくとも一部をプローブとして用いてハイブリダイゼーションを行い、ライブラリーから上記プローブにポジティブの断片を検出するステップとを含む方法を用いることができる。このように本発明のXMAN1遺伝子のうち、少なくとも一部の領域はプローブとして有用である。プローブに用いる領域には、XMAN1遺伝子とそのヒトホモログMAN1遺伝子間で保存性の高い配列が含まれることが好ましく、例えば、PCR法にてこのような領域を増幅させてプローブとして用いることが可能である。
【0035】
本発明に係るXMAN1遺伝子の一部を改変することによって、変異型XMAN1遺伝子が得られる。この変異型XMAN1遺伝子は、動物生体内にて発現することで、野性型と比べてより優れた神経化誘導活性を有するものであってもよいし、野性型と同じ神経化誘導活性を有しているサイレント突然変異(アミノ酸配列は変わらず、塩基配列のみが変化する変異)であってもよい。
【0036】
つまり、本発明に係る変異XMAN1タンパク質は、神経化誘導因子としての活性が野生型XMAN1タンパク質と略同等であってもよいし、より優れていてもよい。
【0037】
また、変異の様式は特に限定されるものではなく、配列番号1に示されるアミノ酸配列において、1またはそれ以上のアミノ酸残基が置換、欠失、挿入及び/または付加されていればよい。
【0038】
例えば、本発明者らは、後述する実施例に示すように、種々のXMAN1欠失変異タンパク質を作製し、その機能解析を行うことにより、XMAN1タンパク質の活性発現に必要な領域として、XMAN1タンパク質のカルボキシル末端領域(C末端領域;582番目から781番目のアミノ酸残基)を同定した。このC末端領域のみで、BMPのシグナル伝達経路を阻害しアフリカツメガエルのアニマル・キャップの神経化を誘導することができる。
【0039】
つまり、本発明には、配列番号1に示すアミノ酸配列を有するXMAN1タンパク質の変異体であって、神経化誘導因子として野生型と略同等の活性を持つ変異タンパク質が含まれていてもよく、具体的には、例えば、後述する実施例に示すように、配列番号1に示されるアミノ酸配列において、アミノ末端側(N末端側)のLEMドメインを含む43個のアミノ酸が欠失された変異タンパク質ΔLEM、N末端側の449個のアミノ酸が欠失された変異タンパク質ΔTM1、N末端側の581個のアミノ酸が欠失され、C末端領域のみからなる変異タンパク質CT(582番目から781番目のアミノ酸残基)を挙げることができる。勿論、変異タンパク質はこれに限られず、神経化誘導活性の発現に重要な領域と考えられる後述のRNP1領域(701番目から708番目のアミノ酸残基)以外の領域に変異が導入され、野生型と略同等またはより優れた神経化活性誘導を有する変異タンパク質も含まれる。また、本発明には、上記変異タンパク質をコードする遺伝子も含まれる。
【0040】
このように、本発明に係るXMAN1遺伝子としては、(a)配列番号1に示されるアミノ酸配列からなるタンパク質か、(b)配列番号1に示されるアミノ酸配列において、1またはそれ以上のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導因子としての活性を有するタンパク質をコードする遺伝子であればよい。
【0041】
また、後述する実施例に示すように、相同性検索の結果、XMAN1遺伝子はヒトMAN1遺伝子と高い相同性を有している。しかしながら、本発明完成時までヒトMAN1遺伝子は機能未知であり、今回、XMAN1遺伝子は、ヒトMAN1遺伝子とは無関係にクローニングされたものである。
【0042】
さらに、後述する実施例に示すように、XMAN1タンパク質のC末端領域とヒトMAN1タンパク質のC末端領域とは、アミノ酸レベルで相同性94%を示している。従って、ヒトMAN1タンパク質もXMAN1タンパク質と同様に、神経化を誘導し、BMPのシグナル伝達経路の阻害活性を有すると推定できる。つまり、本発明は、ヒトMAN1タンパク質がBMPのシグナル伝達経路を阻害すると共に、脊椎動物において神経誘導に関与する機能を持つことを最初に明らかにしたものである。
【0043】
ヒトMAN1タンパク質は、ヒトの核内膜に局在する膜タンパク質であることが知られており(文献 Lin, F. et al. (2000).MAN1, an inner nuclear membrane protein that shares the LEM domain with lamina-associated polypeptide 2 and emerin. J. Biol. Chem. 275, pp4840-4847参照)、XMAN1タンパク質も構造上の類似性から核内膜タンパク質であると推定される。
【0044】
また、ヒト膠原病患者の血清中に、ヒトMAN1タンパク質に対する自己抗体が現れることがこれまで報告されている(文献 Paulin-Levasseur, M. et al. (1996). The MAN antigens are non-lamin constituents of the nuclear lamina in vertebrate cells. Chromosoma, 104, pp367-79参照)。ヒト膠原病は、一種の自己免疫疾患であると考えられており、その原因は未だ明らかにされておらず、治療法等も十分ではない。
【0045】
本発明を用いることにより、ヒトMAN1タンパク質の機能(BMPシグナル伝達経路阻害活性、神経化誘導活性)とヒト膠原病の病態との関係を調べるための診断薬や診断方法等を開発することが可能である。さらに、ヒト膠原病の病態機序を解明するための有効な手段となり、BMPシグナル伝達経路を標的とした治療薬を開発することも可能である。即ち、ヒト膠原病の病態機序の解明という基礎研究的な利用だけでなく、治療薬や治療法等の医学的な利用も可能である。
【0046】
また、核膜の消失・生成は、細胞分裂(癌化を含む)、細胞周期、遺伝子の発現制御と密接に関係している。この核膜の消失・生成と核内膜タンパク質であるMAN1、XMAN1のBMPシグナル伝達経路阻害活性との関連性を調べるためのアッセイ法の確立、試薬の開発等も可能である。
【0047】
以上、アフリカツメガエルの神経化誘導因子XMAN1やヒトMAN1タンパク質について、それぞれ利用可能性を述べてきたが、これら2つのタンパク質は高い相同性を有していることから、同様な機能があると推定される。従って、いずれのタンパク質、若しくはこれらのタンパク質をコードするいずれの遺伝子を用いても、本発明は、脊椎動物における神経化のメカニズム、骨形成因子BMPのシグナル伝達経路、ヒト膠原病の病態機序等の解明に役立つと考えられる。
【0048】
以下添付した図面に沿って実施例を示し、本発明の実施の形態についてさらに詳しく説明する。もちろん、本発明は以下の実施例に限定されるものではなく、細部については様々な態様が可能であることはいうまでもない。
【実施例】
【0049】
以下の実施例では、アフリカツメガエルの新規神経化誘導因子XMAN1のクローニングとその機能解析を行った結果を概略説明する。
【0050】
〔実施例1〕 新規神経化誘導因子のクローニング
アフリカツメガエルの胚の胞胚期から原腸胚期にかけての予定外胚葉組織(アニマル・キャップ)の多分化能を利用して、将来脳組織となる前方神経外胚葉から作製したcDNAライブラリーからアニマル・キャップを神経化する活性(神経化誘導活性)を有する因子を、発現クローニング法とシブ・セレクション法を用いて単離した。具体的には、アフリカツメガエルの体外受精と胚の培養は、Kay and Pengの方法(Xenopus Laevis: practical uses in cell and molecular biology, Methods in Cell Biology, Volume 36, Academic Press,1991)に従った。胚のステージは、Nieuwkoop and Faber(Normal table of Xenopus Laevis, Garland Publishing,1994)に基づいて決定した。前方神経外胚葉特異的cDNAライブラリーは、Shingaらの報告(Mech. Of Dev., Vol. 109, pp225-239, 2001)に基づいて、SuperScript Plasmid System for cDNA Synthesis and Plasmid Cloning(GibcoBRL)を用いて作製し、pCS105ベクター(カリフォルニア大学 Richard Harland博士からの供与)に導入した。シブ・セレクション法は、Lustigらの報告(Methods in Enzymology, Vol. 283, pp83-99, 1997)を参考にした。
【0051】
即ち、cDNAライブラリーを200クローンからなるプールに分割し、プールごとにcDNAから合成したRNAをアニマル・キャップに微量注入した。その後、アニマル・キャップにおける汎神経マーカーnrp-1の誘導の有無を、RT−PCR法により調べた。RT−PCR法は、2細胞期にmRNAを胚の動物極に微量注入し、胞胚期にアニマル・キャップを切り出した後、原腸胚期、尾芽胚期のそれぞれにおいてアニマル・キャップを回収した。RNAの抽出、cDNAの合成、PCR反応は、Wilson and Melton(Curr. Biol., Vol.4, pp676-686, 1994)に基づいて、非ラジオアイソトープ存在下で行った。以下、RT−PCR法はこの方法を用いた。なお、RT−PCR法の替わりに、in situハイブリダイゼーション法を用いることも可能である。次いで、nrp-1の発現を誘導する活性をもつプールに関して、プールサイズを順次小さくしていき、最終的に神経化誘導活性を有する単一のクローンA124を得た。
【0052】
次いで、図1(a)に示すように、2細胞期の胚の動物極にクローンA124のmRNAを微量注入した後、図1(b)に示すように、胞胚期にアニマル・キャップを切り出した。その後、図2に示すように、原腸胚期、尾芽胚期のアニマル・キャップそれぞれにおいて、種々のマーカーの発現誘導をRT−PCR法により調べた。なお、注入したクローンA124のmRNA量は、1胚あたり100p(ピコ)g、250pg、500pgである。また、β-globin(500)は、ネガティブコントロールとしてβ-globinのmRNAを500pg注入したことを示す。また、RT(+)、RT(−)は、それぞれポジティブコントロール、ネガティブコントロールを示す。EF−1αは反応に用いたcDNA量のコントロールである。
【0053】
RT−PCRの結果、中胚葉マーカーとして知られるXbra、actinの発現誘導なしに、汎神経マーカーであるnrp-1、前方神経マーカーであるXAG-1、otx-2の発現が誘導された。しかし、後方神経マーカーであるHoxB-9は発現誘導されなかった。この結果により、クローンA124にコードされている因子は、前方神経の誘導に関わる新規因子であることが示唆された。また、その神経誘導化活性は、中胚葉の誘導を介した2次的なものではないことを意味している。
【0054】
〔実施例2〕 クローンA124の構造解析
次に、クローンA124の全長を含むcDNAを単離した。具体的には、クローンA124をプローブとしてアフリカツメガエルのステージ30期の頭部cDNAライブラリーを用いてスクリーニングし、得られたポジティブクローンのうち、インサートの最も長いクローンPC11の塩基配列を決定した。しかし、このクローンPC11も開始コドンを欠いていたので、5’-Full RACE Core Set(TAKARA)を用いて、開始コドンを含む上流領域を増幅し、この遺伝子の全長を決定した。その全構造を決定したところ、図3に示すように、781アミノ酸からなる分子量約88000のタンパク質をコードしていることがわかった。
【0055】
この遺伝子の相同性検索を行ったところ、ヒトMAN1遺伝子と高い相同性を示すことがわかった。ヒトMAN1遺伝子は、910アミノ酸からなり、核内膜に存在する機能未知のタンパク質をコードする遺伝子である。
【0056】
従って、ヒトMAN1遺伝子と相同性が高かったことから、この遺伝子をXMAN1(Xenopus MAN1)と命名した。
【0057】
このXMAN1タンパク質とヒトMAN1タンパク質とは、構造上の特徴も非常に類似していることが明らかとなった。XMAN1タンパク質は、ヒトMAN1タンパク質と同様に、N末端側にクロマチンと結合すると考えられるLEMドメイン(5番目から43番目のアミノ酸残基)を有している。なお、これら2つのタンパク質間における、このLEMドメインの相同性は74%であった。
【0058】
また、XMAN1タンパク質は、疎水性アミノ酸に富む2つの膜貫通領域TM1、TM2(TM1;349番目から372番目のアミノ酸残基、TM2;497番目から519番目のアミノ酸残基)を有しており、この点でもヒトMAN1タンパク質と類似している。ヒトMAN1タンパク質はこの2つの膜貫通領域を介して核内膜に埋め込まれており、N末端とC末端は共に核の内側に存在する膜タンパク質である事が知られている。従って、XMAN1タンパク質も生体内でヒトMAN1タンパク質と同様の局在性を示すと考えられる。
【0059】
さらに、XMAN1タンパク質はC末端領域(582番目から781番目のアミノ酸残基)に、RNA結合タンパク質に多く見られるRNP2(661番目から666番目のアミノ酸残基)、RNP1(701番目から708番目のアミノ酸残基)と呼ばれる保存配列を含むRNA認識モチーフ(RRM;656番目から733番目のアミノ酸残基)を有することがわかった。また、ヒトMAN1タンパク質も非常に類似しているC末端領域を有していることがわかった。即ち、これら2つのタンパク質間における、このC末端領域の相同性は、アミノ酸レベルで94%であった。これまで、ヒトMAN1タンパク質にRRMが存在することは文献的にも指摘されておらず、今回初めてその存在を明らかにした。
【0060】
〔実施例3〕 新規神経化因子XMAN1の活性発現に必須な領域の特定
XMAN1タンパク質のどの領域に神経化誘導活性があるのかを調べるために、図4に示すように、XMAN1タンパク質の欠失変異タンパク質ΔLEM、ΔTM1、ΔCT、CTをそれぞれ作製した。欠失変異タンパク質ΔLEM、ΔTM1、ΔCTとは、野生型XMAN1タンパク質WtからそれぞれLEMドメイン、TM1領域、C末端領域を欠失させた変異タンパク質であり、欠失変異タンパク質CTとは、C末端領域のみ残存させ、この領域以外を欠失させた変異タンパク質である。なお、Δ(デルタ)とは、欠失変異体を意味する記号である。
【0061】
具体的には、XMAN1タンパク質の種々の変異タンパク質は、PCR法を用いて作製し、pCS+MTベクターに導入した。微量注入に用いたmRNAは、MEGAscript IN Vitro Transcription Kits(Ambion)を用いて合成した。
【0062】
それぞれの欠失変異タンパク質のmRNAを2細胞期の胚の動物極に微量注入し、胞胚期にアニマル・キャップを切り出した後、尾芽胚期におけるアニマル・キャップでの汎神経マーカーnrp-1の発現誘導をRT−PCR法により調べた。
【0063】
具体的には、尾芽胚期のアニマル・キャップからRNAを単離し、公知の手法に従いcDNAを合成して、PCR法による増幅を行った。なお、Uninj,はRNAを注入していないアニマル・キャップを示す。そのRT−PCRの結果、欠失変異タンパク質ΔLEM、ΔTM1、CTは野生型タンパク質Wtと略同等のnrp-1の発現を誘導した。しかし、欠失変異タンパク質ΔCTは、ほとんどnrp-1の発現を誘導しなかった。従って、XMAN1タンパク質のC末端領域が神経化誘導活性に必須であることがわかった。このC末端領域は、ヒトMAN1タンパク質のC末端領域と高い相同性を示しており、進化上もこの領域が重要な機能を有していることが示唆された。従って、このXMAN1タンパク質のC末端領域を神経化誘導活性ドメインCTと呼ぶこととした。
【0064】
さらに、この神経化誘導活性ドメインCTについて、欠失変異タンパク質CTΔRRM、RRM、CTΔRNP1を作製して詳細に検討した。欠失変異タンパク質CTΔRRM、CTΔRNP1は、神経化誘導活性ドメインCTからそれぞれRRM、RNP1を欠失させた変異タンパク質である。欠失変異タンパク質RRMは、神経化誘導活性ドメインCTのRRMのみ残存させて、N末端からRRMまでの領域を欠失させた変異タンパク質である。
【0065】
これらの欠失変異体のmRNAを用いて、上記と同様の方法でnrp-1の誘導を指標に、神経化誘導活性ドメインの機能に重要な部位を調べた結果、図4に示すように、欠失変異タンパク質CTΔRRM、RRM、CTΔRNP1は、いずれもnrp-1の発現を誘導しなかった。この結果から、RRM、特にその中のRNP1配列が神経化誘導活性に重要な役割を果たしていることがわかった。これまで、ヒトMAN1タンパク質にRRMの存在のみならず、その機能についても全くわかっていなかったが、今回初めてその存在意義を示すことができた。
【0066】
〔実施例4〕 神経活性化ドメインによるBMPシグナル伝達経路の阻害
さらに、XMAN1タンパク質は骨形成因子BMPのシグナル伝達経路を阻害することにより、アニマル・キャップを神経化するのか否か検討した。BMPのシグナルは細胞表面のBMPI型受容体、BMPII型受容体の2種類の受容体と細胞内因子Smad(特にSmad1,4,5,8)とを介して核に伝えられた後、そこで標的因子の発現誘導、活性化を通して多様な生理作用を引き起こすと考えられている。これら、BMPシグナル伝達経路の直接的な標的因子として、アフリカツメガエルではXmsx-1、Xhox-3、Xvent-2等が知られている。
【0067】
そこで、2細胞期の胚の動物極にBMP−4、若しくは活性化型BMPI型受容体(ALK3*)のmRNAを単独で微量注入し、胞胚期にアニマル・キャップを切り出した後、原腸胚期に種々のマーカーの発現誘導をRT−PCR法により調べた。具体的には、原腸胚期のアニマル・キャップからRNAを単離し、公知の手法に従いcDNAを合成して、PCR法による増幅を行った。なお、図5に示すように、注入したBMP−4のmRNA量は、胚1個あたり、200pg、ALK3*は500pgであり、CTは注入していない。コントロールとして、CTのmRNAのみを500pg注入したものを示した。そのRT−PCRの結果は図5に示すように、BMP−4またはALK3*のmRNAのみを注入した場合、標的因子Xmsx-1、Xhox-3の発現は誘導された。また、CTのmRNAを単独で注入した場合、標的因子Xmsx-1、Xhox-3の発現は誘導されなかった。なお、活性化型BMPI型受容体(ALK3*)とは、BMPを受容しなくても活性型として機能し、恒常的にシグナルを細胞内に伝達する受容体である。
【0068】
しかし、同様にBMP−4のmRNAと、若しくはALK3*のmRNAと神経化誘導活性ドメインCTのmRNAとを一緒に微量注入した場合(注入したBMP−4、ALK3*のmRNA量はそれぞれ、胚1個あたり、200pg、500pg、またCTのmRNA量はそれぞれ、100,500,1000pgである。)、標的因子Xmsx-1、Xhox-3の発現誘導は、注入したCTのmRNA量に依存して著しく阻害された。なお、中胚葉マーカーXbraの発現誘導はCTのmRNAの注入量によって、ほとんど変化しなかった。これらの結果から、神経化誘導活性ドメインCTがBMPのシグナル伝達経路を受容体より下流で阻害することが示された。
【0069】
〔実施例5〕 新規神経化因子XMAN1によるBMP標的因子の転写抑制
さらに、XMAN1がBMPの標的因子の発現誘導を転写レベルで抑制しているか否かをルシフェラーゼ・アッセイ法を用いて調べた。
【0070】
具体的には、ルシフェラーゼ活性は、Dual Luciferase Reporter Assay System(Promega)を用いて測定した。まず、BMPの標的因子の一つ、Xvent-2のプロモーターの下流にリポーター遺伝子としてfireflyルシフェラーゼ遺伝子を組み込んだプラスミドDNA(Xvent2-Luc、カリフォルニア大学 Ken Cho博士からの供与)を用いた。その後、このプラスミドDNA(Xvent2-Luc)を100pg、Renillaルシフェラーゼ遺伝子を持つコントロールプラスミドDNA(pRL-TK)を2pg、BMP−4のmRNA並びにWt、CTのいずれか1つのmRNAを混合して、2細胞期の胚の動物極に微量注入した。胞胚期にアニマル・キャップを切り出した後、原腸胚期に5個ずつ、3本のチューブに集め、各チューブに50μlのルシフェラーゼ反応液を加えて、fireflyルシフェラーゼ活性を測定した。その後、Gro & Stop溶液を加えてRenillaルシフェラーゼ活性を測定した。測定には、ルミノメーター(Berthold)を用いた。Fireflyルシフェラーゼ活性値をRenillaルシフェラーゼ活性値で割った値を補正値とし、3本のチューブの平均値を算出した。また、BMP−4の替わりにALK3*を用いて、同様にルシフェラーゼ活性を測定した。なお、注入したBMP−4のmRNA量は、1胚あたり200pg、ALK3*は500pg、XMAN1のmRNAは野生型Wt、欠失変異体CTともに500pgである。また、コントロールとして、BMP−4のmRNAと、若しくはALK3*のmRNAと、ルシフェラーゼ遺伝子をXvent-2のプロモーターと異なるプロモーター(GL-3)の下流に組み込んだプラスミドDNA(GL3-Luc)とを注入した場合、Xvent2-Lucのみを注入した場合を示す。
【0071】
これらのルシフェラーゼ・アッセイの結果、図6に示すように、BMP−4若しくはALK3*のmRNAとXvent2-Lucとを注入した場合、ルシフェラーゼ活性は上昇した。しかし、この活性上昇は、野生型XMAN1タンパク質Wt、神経化誘導活性ドメインCTにより著しく抑制された。従って、XMAN1タンパク質、特にその神経化誘導活性ドメインCTは、BMPの標的因子の転写活性化を抑制することがわかった。
【0072】
なお、BMP−4若しくはALK3*非存在下でも、ある程度のルシフェラーゼ活性が見られるのは、アニマル・キャップ内に存在する内在性のBMPの働きによるものと考えられる。
【産業上の利用可能性】
【0073】
本発明は、(1)ヒト膠原病患者血清中の抗ヒトMAN1抗体の存在の有無やヒト膠原病の病態機序を調べるための診断法、診断薬の開発並びにBMPシグナル伝達経路を標的とした治療薬への利用可能性、さらには、(2)癌化を含む細胞分裂、細胞周期、遺伝子発現制御らと密接に関係している核膜の消失・生成を調べるためのアッセイ法や試薬への利用可能性を有するものである。
【図面の簡単な説明】
【0074】
【図1】(a)は、本発明の実施例におけるアフリカツメガエルの2細胞期の胚へのmRNAの注入操作を示した概略説明図であり、(b)は、胞胚期におけるアニマル・キャップの切り出し操作を示した概略説明図である。
【図2】図1で切り出したアニマル・キャップの原腸胚期、尾芽胚期における各種マーカーの発現誘導をRT−PCR法にて調べた結果を示す電気泳動図である。
【図3】本発明の実施例におけるヒトMAN1並びにXMAN1タンパク質の各構造を概略的に示す模式図である。
【図4】XMAN1の各種欠失変異タンパク質を示す概略模式図、並びに各種変異mRNAを注入したアニマル・キャップの尾芽胚期における各種マーカーの発現誘導をRT−PCR法にて調べた結果を示す電気泳動図である。
【図5】各種mRNAを注入したアニマル・キャップの原腸胚期における各種マーカーの発現誘導をRT−PCR法にて調べた結果を示す電気泳動図である。
【図6】XMAN1の各種変異タンパク質によるBMPの標的因子の転写活性の抑制を、ルシフェラーゼ・アッセイ法により調べた結果を示すグラフである。
【特許請求の範囲】
【請求項1】
以下の(1)〜(2)の何れか1つのタンパク質。
(1)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列からなるタンパク質、ならびに、
(2)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質。
【請求項2】
請求項1に記載のタンパク質をコードする遺伝子。
【請求項3】
配列番号4に示される塩基配列の2140番目から2742番目の塩基配列からなる遺伝子。
【請求項4】
以下の(1)〜(4)に記載のタンパク質、および該タンパク質をコードする遺伝子からなる群より選択される、少なくとも一つの物質を含有する神経化誘導剤。
(1)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列からなるタンパク質、
(2)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質、
(3)配列番号3に示されるアミノ酸配列からなるタンパク質、ならびに、
(4)配列番号3に示されるアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質。
【請求項5】
以下の(1)〜(4)に記載のタンパク質、および該タンパク質をコードする遺伝子からなる群より選択される少なくとも一つの物質を用いて細胞又は組織の神経化を誘導する方法。
(1)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列からなるタンパク質、
(2)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質、
(3)配列番号3に示されるアミノ酸配列からなるタンパク質、ならびに、
(4)配列番号3に示されるアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質。
【請求項6】
以下の(1)〜(4)に記載のタンパク質、および該タンパク質をコードする遺伝子からなる群より選択される少なくとも一つの物質を含有する骨形成因子BMPシグナル伝達経路阻害剤。
(1)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列からなるタンパク質、
(2)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質、
(3)配列番号3に示されるアミノ酸配列からなるタンパク質、ならびに、
(4)配列番号3に示されるアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質。
【請求項1】
以下の(1)〜(2)の何れか1つのタンパク質。
(1)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列からなるタンパク質、ならびに、
(2)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質。
【請求項2】
請求項1に記載のタンパク質をコードする遺伝子。
【請求項3】
配列番号4に示される塩基配列の2140番目から2742番目の塩基配列からなる遺伝子。
【請求項4】
以下の(1)〜(4)に記載のタンパク質、および該タンパク質をコードする遺伝子からなる群より選択される、少なくとも一つの物質を含有する神経化誘導剤。
(1)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列からなるタンパク質、
(2)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質、
(3)配列番号3に示されるアミノ酸配列からなるタンパク質、ならびに、
(4)配列番号3に示されるアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質。
【請求項5】
以下の(1)〜(4)に記載のタンパク質、および該タンパク質をコードする遺伝子からなる群より選択される少なくとも一つの物質を用いて細胞又は組織の神経化を誘導する方法。
(1)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列からなるタンパク質、
(2)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質、
(3)配列番号3に示されるアミノ酸配列からなるタンパク質、ならびに、
(4)配列番号3に示されるアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質。
【請求項6】
以下の(1)〜(4)に記載のタンパク質、および該タンパク質をコードする遺伝子からなる群より選択される少なくとも一つの物質を含有する骨形成因子BMPシグナル伝達経路阻害剤。
(1)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列からなるタンパク質、
(2)配列番号3に示されるアミノ酸配列の712番目から911番目のアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質、
(3)配列番号3に示されるアミノ酸配列からなるタンパク質、ならびに、
(4)配列番号3に示されるアミノ酸配列において、1または数個のアミノ酸が置換、欠失、挿入、及び/または付加されたアミノ酸配列からなり、かつ神経化誘導活性を持つタンパク質。
【図1】


【図3】
【図6】
【図2】
【図4】
【図5】
【図3】
【図6】
【図2】
【図4】
【図5】
【公開番号】特開2009−34101(P2009−34101A)
【公開日】平成21年2月19日(2009.2.19)
【国際特許分類】
【出願番号】特願2008−176903(P2008−176903)
【出願日】平成20年7月7日(2008.7.7)
【分割の表示】特願2002−116378(P2002−116378)の分割
【原出願日】平成14年4月18日(2002.4.18)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
【公開日】平成21年2月19日(2009.2.19)
【国際特許分類】
【出願日】平成20年7月7日(2008.7.7)
【分割の表示】特願2002−116378(P2002−116378)の分割
【原出願日】平成14年4月18日(2002.4.18)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
[ Back to top ]