
神経変性状態を治療するためのヒト神経細胞の移植
【課題】神経変性状態を治療する方法を提供する。神経幹細胞は、ニューロン変性の部位および/または遠位に移植することができる。
【解決手段】本方法は、交換されるニューロンに対応する、特定の種類のニューロンが発生する領域から、神経幹細胞を分離することを含むことができる。本方法は、特定の種類のニューロンの増殖および/または再生に影響を及ぼす増殖因子を分泌する神経幹細胞を分離することを含むことができる。本発明では、我々は、神経回路で特定の神経伝達物質を産生する細胞を欠くことから生じるいくつかの神経変性障害を含む、そのような障害を、外因的に培養および増殖させた神経前駆体を移植することによって、治療する方法を開示し、その神経前駆体は、神経組織へ移植すると、集積することができ、神経変性に関連した症状を克服するために十分な量および十分な方法で神経伝達物質を産生することができる、ニューロンに分化する方法を開示する。
【解決手段】本方法は、交換されるニューロンに対応する、特定の種類のニューロンが発生する領域から、神経幹細胞を分離することを含むことができる。本方法は、特定の種類のニューロンの増殖および/または再生に影響を及ぼす増殖因子を分泌する神経幹細胞を分離することを含むことができる。本発明では、我々は、神経回路で特定の神経伝達物質を産生する細胞を欠くことから生じるいくつかの神経変性障害を含む、そのような障害を、外因的に培養および増殖させた神経前駆体を移植することによって、治療する方法を開示し、その神経前駆体は、神経組織へ移植すると、集積することができ、神経変性に関連した症状を克服するために十分な量および十分な方法で神経伝達物質を産生することができる、ニューロンに分化する方法を開示する。
【発明の詳細な説明】
【技術分野】
【0001】
本開示の方法は、そのような治療法にとって比類なく有益である、細胞の移植を通して疾患を治療する方法に関する。特に、本開示の方法は、神経幹細胞(NSC)を用いた神経変性状態を治療する方法を提供する。
【背景技術】
【0002】
神経変性障害は、疾患、遺伝性疾患、または外傷性もしくは虚血性の脊髄損傷もしくは脳損傷などの損傷の結果としてのニューロン変性に関わる状態によって特徴付けられる。
【0003】
四肢の骨格筋の収縮を支配する脊髄の回路には、興奮性の運動ニューロン、ならびに抑制性のGABA作動性(すなわち、GABAを産生する)介在ニューロンおよびグリシン作動性(すなわち、グリシンを産生する)介在ニューロンが関与する。運動ニューロンは、脊髄の灰白質の前角から始まる神経である。運動ニューロンの軸索は、筋線維の神経を刺激する遠心性運動線維として、脊髄の一分節から出現する。運動ニューロンによって伝導されるインパルスは、筋線維を刺激して収縮させる。γアミノ酪酸(gamma-amino butyric acid:GABA)は、哺乳動物の神経系において自然に発生される代謝物であって、電位の神経伝導を抑制または弱める神経伝達物質として作用する。GABA作動性の介在ニューロンが失われると、運動ニューロンによって引き起こされる筋収縮の抑制性調性(inhibitory tonality)の調節不全が生じる。抑制性介在ニューロンによって発揮される興奮性ニューロンに対する制御がないと、四肢の筋肉の痙攣性の抑制困難な収縮または抑制困難な硬直につながる、興奮性ニューロンの過大加熱(over-firing)が生ずる。運動ニューロンが失われると、弛緩性対麻痺が生じ、対象は、筋肉を収縮させることができず、したがって、動くことができない。
【0004】
GABA作動性の介在ニューロンが脊椎索で損傷を受ける1つの例は、胸部下行大動脈または胸腹部大動脈の一過性の遮断(cross-clamping)に関連する合併症を含む。そのような遮断は、胸部大動脈または胸腹部大動脈の動脈瘤を修復する血管手術における必要な工程である。遮断の間、脊髄の一部は血液循環を受けることができず、虚血状態となる可能性がある。虚血時間の長さによって、その後の神経変性の機能不全が、不全対麻痺、または完全に進行した痙攣性もしくは弛緩性の対麻痺として神経変性的に発現する可能性がある。
【0005】
虚血誘導性のニューロン変性につながるメカニズムは、一部しか理解されておらず、興奮性アミノ酸、プロスタグランジンおよび/または活性酸素の過剰な放出/活性が関与する可能性がある一方、一過性の虚血性障害(ischemic insult)による影響を受ける脊髄のニューロン集団は、十分明確にされている。例えば、完全に進行した痙性対麻痺の動物から採取した脊髄に関する組織病理学的分析は、小型の抑制性ニューロンの選択的な減少(selective loss)を明らかにしているが、α運動ニューロンは、あらかじめ虚血の脊髄分節内に残存する。脊髄の虚血性傷害を有するヒトの対象における同様な脊髄の神経病態について述べられている。
【0006】
対照的に、弛緩性対麻痺の動物において、汎壊死性(pan-necrotic)の神経変性による変化が、腹側の運動ニューロンだけでなく、小型の抑制性および興奮性の介在ニューロンに影響を及ぼすことが認められている。また、局所または全体的な脳虚血などで、脊髄の虚血後の神経変性の間に、局所の小膠細胞の傷害依存的な活性化、および大食細胞による浸潤などの炎症性の変化が見られる。傷害の程度によって、一般的に炎症性変化は、虚血性傷害の2日から7日後にピークに達して、その後、2週間から4週間の虚血後期間にかけて炎症性要素が次第に消失していく。
【0007】
過去20年から30年間に、動物モデルで、さまざまな物質の脊髄移植の治療可能性について評価するためにかなりの努力が払われている。したがって、細胞系または急速に分離された脊髄胎児組織が損傷部位に送達されており、さらに直接的な脊髄遺伝子治療(direct spinal gene therapy)も、機械的外傷的損傷、化学的に損傷を起こした脊髄、または進行性α運動ニューロン変性(α-motoneuronal degeneration)の遺伝子組換え動物(ALS遺伝子移植マウスまたはラット)などの、いくつかの脊髄損傷モデルにおいて、神経変性機能不全を改善するために使用されている。
【0008】
一般に、いくつかの試験では、胎児組織から生じた移植片でニューロン表現型(neuronal phenotype)の長期生存および保存が実証されているが、in vitroで増殖させた神経前駆細胞(neural precursor)から生じた移植片については実証されていない。実際に、in vitroで増殖され、機械的および化学的に損傷した脊髄へ移植した神経前駆細胞のニューロン分化および成熟は、限られたものが実証されているに過ぎない。細胞は、優先的に非ニューロン性の細胞型に分化する。この優先的な非ニューロン性分化のメカニズムは、完全には理解されていないが、以前の損傷部位での炎症誘発性サイトカイン(TNFα、TGFβなど)の局所的放出が恐らく関与すると仮定されている。
【0009】
神経変性は、特に、細胞療法についての厳しい生物学的環境を示し、確立された神経変性疾患に現れる細胞死シグナル(cell death signal)(Rothsteinら、1992年; Howlandら、2002年; Turnerら、2005年)は、移植片の生存とは同時に起こり得ないと考えられる。さらに、成人の脊髄は、再生を可能にする細胞および/またはシグナルを欠いていると見られ(Parkら、2002年)、大部分のNSC移植試験が、不良または限定的な分化を示している(Caoら、2002年; Yanら、2003年; Yanら、2004年)。
【0010】
細胞治療学(cell therapeutics)における主な問題の1つは、移植した細胞の細胞生存率が低いということである(5%未満)。現在に至るまで、移植した全ての細胞は、in vivoへの注射直後にかなりの細胞死を起こす。したがって、細胞の有効量を送達するために、最終用量を少なくとも20回注射しなければならない。その結果として、このことは、より大規模な細胞製造を必要とし、さらなる規制の障壁および経済の障壁を引き起こす。さらに、in vivoで、そのような細胞の生存率を維持することができていない。再現性のある、細胞療法の有効量の投与が実証されていないことが、政府、および米国食品医薬品局などの他の規制機関による使用の承認を妨げている。
【0011】
さらなる困難は、1つの限局部分というよりむしろ全ての神経系のような広範囲の身体、組織または器官にわたって広がる神経変性疾患および状態を治療するときに現れる。例えば、ALSでは、神経変性が、運動皮質の運動ニューロンだけでなく、脊髄全体に沿った運動ニューロンの緩やかな死(slow death)に関わる。同様に、大部分のリソソーム病で、ニューロンの破壊が脳および脊髄の大部分の部位に影響を与える。アルツハイマー病は、大部分の大脳に影響を与える。パーキンソン病およびハンチントン舞踏病などのより局所的な神経変性疾患でさえ、線条体の患部は非常に大きく、手術によって達することができる移植部位より遥かに大きい。したがって、神経変性疾患に対する細胞治療学は、より広い移植手法が必要とされることが予期される。
【0012】
したがって、神経変性状態を治療する方法の改善が必要とされる。また、一旦、移植されたならば、以前に見られた全ての制限を克服し、機能的有益性を提供する、ヒトの神経幹細胞およびヒトの神経前駆体を培養し、移植する方法の改善が必要とされる。したがって、このin vivoでの神経変性状態を治療する方法は、確固としたニューロン分化を生じさせ、さまざまな変性状態下でのニューロンの長期生存、および発生の手掛かり(developmental cue)を欠く成体組織での治療に関係したニューロン亜集団への成熟を可能にし、細胞自身の位置より広範囲の治療域を提供する。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】Rothsteinら、1992年
【非特許文献2】Howlandら、2002年
【非特許文献3】Turnerら、2005年
【非特許文献4】Parkら、2002年
【非特許文献5】Caoら、2002年
【非特許文献6】Yanら、2003年
【非特許文献7】Yanら、2004年
【発明の概要】
【発明が解決しようとする課題】
【0014】
本開示の方法は、神経変性状態を治療するための方法を含む。特に、本開示の方法は、細胞が神経変性状態を改善することができるように、in vitroで増殖させたNSC、神経前駆体または神経前駆細胞を、それを必要とする対象へ移植することを含む。一実施形態では、本開示の方法は、ドナー細胞を同定、分離、増殖および調製して、神経変性状態の治療として使用することを含む。移植されるドナー細胞は、その状態、その症状および/またはその影響の一因となる要素またはその欠如に対応するように選択することができる。
【0015】
本開示の方法の細胞は、移植直後に、ニューロンの基礎構造内に組み込まれるのに十分なニューロン量を産生して、疾患の状況または状態を改善する細胞を含む。一実施形態では、本開示の方法は、哺乳動物の中枢神経系から分離され、in vitroで増殖させた多分化能を有する神経前駆体または神経幹細胞を移植することによって、神経変性疾患または状態を治療することを含む。例えば、増殖させた神経幹細胞の移植は、痙縮、硬直、発作、麻痺またはその他の筋肉の活動亢進の症状を有するさまざまな脊髄症の形態を患う対象における歩行機能を改善するために使用することができる。
【0016】
治療法は、損傷した神経領域に、移植を通して十分な数のGABAを産生するニューロンおよび/またはグリシンを産生するニューロンに分化することができる適切な数のNSCを供給して、活動亢進の神経回路を含む欠陥のある神経回路を弱めることを含む。
【課題を解決するための手段】
【0017】
一実施形態では、本開示の方法は、運動ニューロン疾患における運動機能を回復させることを含む。運動ニューロンへ分化可能な、適切な数または治療有効量のNSCまたは神経前駆体を、変性脊髄などの神経変性の少なくとも一領域に供給し、運動機能を回復させることができる。NSCは、変性した神経筋接合部を置換することによって、その治療効果を発揮する。
【0018】
同時または代わりに、NSCは、変性組織のニューロンを保護して、より多くのニューロンをより長期間生存させる栄養分子を発現および放出することによって、その治療効果を発揮する。変性運動ニューロン疾患の対象において、NSC由来ニューロンを、腹根へ突き出させ、NSCが宿主の運動ニューロンと広範な相互関係をもつ筋肉に神経を刺激するように促進させることができる。したがって、一実施形態では、ヒトの胎児脊髄由来のNSCを、これらの細胞が宿主のニューロンとシナプス接合(synaptic contact)を形成し、運動ニューロン増殖因子を発現および放出するニューロンへ分化を起こすことができる腰髄に移植することができる。
【0019】
一実施形態では、本開示の方法は、宿主組織と同化し、1種または2種以上の増殖因子を宿主ニューロンに供給する神経幹細胞または神経前駆体を提供し、したがって、組織に存在する変性の影響からそれらを保護することを含む。本方法は、十分な数のNSCまたは神経前駆体を、少なくとも1種の増殖因子の有効量がNSCによって分泌されるように、脊髄の一領域に導入することを含む。
【0020】
一実施形態では、本開示の方法は、神経変性状態での細胞置換(cell replacement)に対する幹細胞の前臨床評価において、動物モデルを使用する方法を提供することを含む。
【0021】
一実施形態では、本開示の方法は、ニューロンへ移植するNSCの分化効率を上げることを含む。本方法は、移植後に、移植片において、20%というような、十分な数の細胞がニューロンの運命(neuronal fate)を選ぶように、未分化の状態で非常に高濃縮のNSCまたは神経前駆体を増殖することを含む。
【0022】
一実施形態では、本開示の方法は、移植するNSCまたは神経前駆体の数を増加させることなく分化細胞の数を増加させることを含む。一実施形態では、本方法は、一旦移植されると、NSCまたは神経前駆体が、in vivoで10回ほども継続して、腫瘍を生じることなく分裂するというように増殖するドナー集団を調製し、したがって、導入細胞の総数を有効に増加させることを含む。
【0023】
本開示の方法の細胞は、哺乳動物の胎児、新生児、若年者、成人および死後の組織から分離または入手することができる。本開示の方法の細胞は、中枢神経系、血液、またはニューロンに分化する他の適した幹細胞源から分離または入手することができる。また、細胞は、胚幹細胞から入手することができる。例えば、一実施形態では、本細胞は、発育する胎児の脊髄から分離された神経上皮細胞を含む。場合によっては、神経前駆細胞は、中枢神経系の特定の小領域から分離される神経前駆体である可能性がある。
【0024】
本開示の方法によれば、神経幹細胞は培養で増殖する。一実施形態では、神経前駆細胞は、培養で増殖し、分化に際してニューロンと神経膠の両方を発生することができる多分化能を有するNSCとすることができる。
【0025】
細胞は、移植時にin vitroで未分化、分化前または完全分化のいずれかとすることができる。一実施形態では、細胞は、神経系列(neural lineage)へ分化させるために導入される。本開示の方法の細胞は、炎症誘発性サイトカインおよび損傷した組織に存在するその他の環境因子の存在下における生体内で、ニューロン分化を起こすことができる。
【0026】
現在の方法を使用して、疾患、障害または状態の改善のために、本細胞を適切な部位に移植または導入することによって、神経回路を治療することができる。通常、移植は、移植した細胞の生存を補助する神経組織または非神経組織へ行われる。本開示の方法で使用されるNSC移植片は、神経変性の環境で良好に生存し、神経変性状態または疾患の発症および進行を遅らせるという形で、NSCが強力な臨床効果を発揮することができる。
【0027】
場合によっては、移植は、身体の遠位の領域へ行うことが可能で、本細胞は意図する標的へ移動することができる。したがって、本開示の方法は、ヒトNSCの部分移植(partial grafting)も含み得る。本書では、「部分移植」という用語は、神経変性の一部の領域だけ、または全体より少ない領域における増殖NSCの移植を指すことができる。例えば、脊髄の腰椎部へのヒトNSCの部分的移植。変性する運動ニューロンに対するNSCの効果の少なくとも一部は、古典的な細胞メカニズムを通して、変性する宿主の運動ニューロンへニューロトロフィンおよび栄養サイトカイン(trophic cytokine)を送達することを含む。このために、本開示の方法を使用して脊髄の腰椎部への部分移植を受けているNSCが、運動ニューロン疾患のトランスジェニック動物モデルにおいて、移植の隣接領域および移植領域から遠位の領域で生存し、広範囲なニューロン分化を起こし、運動ニューロンの生存を促進し、機能することを示している。
【0028】
したがって、本開示の方法は、痙縮、硬直または筋肉活動亢進状態を治療する方法を提供する。本方法は、少なくとも1種の神経幹細胞を哺乳動物から分離すること、in vitroで、神経幹細胞を増殖集団へ増殖させることを含む。また、本方法は、増殖集団を濃縮すること、治療有効量の増殖集団をレシピエントの脊髄の少なくとも一領域に導入することを含む。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0029】
一実施形態では、本状態は、外傷性脊椎損傷、虚血性脊椎損傷、外傷性脳損傷、脳卒中、多発性硬化症、脳性麻痺、てんかん、ハンチントン舞踏病、筋萎縮性側索硬化症、慢性虚血、遺伝性疾患またはそのいずれかの組合せに由来する。
【0030】
一実施形態では、神経幹細胞は、中枢神経系、末梢神経系、骨髄、末梢血、臍帯血および少なくとも1つの胚からなる群から選択される供給源から分離される。
【0031】
一実施形態では、哺乳動物は、発育中の哺乳動物である。
【0032】
一実施形態では、発育中の哺乳動物の在胎齢は、約6.5週から約20週までの間である。
【0033】
一実施形態では、神経幹細胞は、ヒトの胎児の脊髄から分離される。
【0034】
一実施形態では、神経幹細胞を増殖することは、血清の不存在下で、神経幹細胞を培養することを含む。
【0035】
一実施形態では、神経幹細胞を増殖することは、神経幹細胞を、少なくとも1種の増殖因子に曝露させることを含む。
【0036】
一実施形態では、増殖因子は、増殖因子はbFGF、EGF、TGF-α、aFGFおよびその組合せからなる群から選択される。
【0037】
一実施形態では、治療有効量の増殖集団は、in vivoで少なくとも1,000のGABAを産生するニューロンを発生させることができる。
【0038】
一実施形態では、治療有効量の増殖集団は、in vivoで少なくとも1,000のグリシンを産生するニューロンを発生させることができる。
【0039】
一実施形態では、増殖集団の少なくとも40%は、脊髄でニューロンを発生させることができる。
【0040】
一実施形態では、治療有効量の増殖集団を導入することは、レシピエントの脊髄の複数の領域に、治療有効量の少なくとも一部を注射することを含む。
【0041】
一実施形態では、増殖集団の少なくとも30%は、in vitroでニューロンに分化することができる。
【0042】
他の実施形態では、神経幹細胞を提供する。本神経幹細胞は、痙縮、硬直または筋肉活動亢進状態を治療することができる。神経幹細胞は、哺乳動物から分離し、in vitroで増殖集団へ増殖させる。幹細胞を含む増殖集団を濃縮して、治療有効量の増殖集団を、レシピエントの脊髄の少なくとも一領域に導入する。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0043】
本開示の方法の他の実施形態では、慢性痛を治療する方法を提供する。本方法は、少なくとも1種の神経幹細胞を哺乳動物から分離すること、in vitroで、神経幹細胞を増殖集団へ増殖させることを含む。また、本方法は、増殖集団を濃縮すること、治療有効量の増殖集団をレシピエントの脊髄の少なくとも一領域に導入することを含む。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0044】
一実施形態では、慢性疼痛は、外傷性脊椎損傷、虚血性脊椎損傷、外傷性脳損傷、脳卒中、多発性硬化症、脳性麻痺、てんかん、ハンチントン舞踏病、筋萎縮性側索硬化症、慢性虚血、遺伝性疾患またはそのいずれかの組合せに由来する。
【0045】
一実施形態では、治療有効量の増殖集団は、少なくとも1,000のGABAを産生するニューロンを発生させることができる。
【0046】
一実施形態では、治療有効量の増殖集団は、少なくとも1,000のグリシンを産生するニューロンを発生させることができる。
【0047】
一実施形態では、増殖集団の少なくとも40%は、脊髄でニューロンを発生させることができる。
【0048】
一実施形態では、治療有効量の増殖集団を導入することは、レシピエントの脊髄の複数の領域に、治療有効量の少なくとも一部を注射することを含む。
【0049】
一実施形態では、本領域は、後角を含む。
【0050】
一実施形態では、本領域は、クモ膜下腔を含む。
【0051】
さらなる実施形態では、神経幹細胞を提供する。神経幹細胞は、慢性疼痛を治療することができる。神経幹細胞は、哺乳動物から分離し、in vitroで増殖集団へ増殖させる。幹細胞を含む増殖集団を濃縮して、治療有効量の増殖集団を、レシピエントの脊髄の少なくとも一領域に導入する。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0052】
本開示の方法の他の実施形態では、運動ニューロンの変性を治療する方法を提供する。本方法は、少なくとも1種の神経幹細胞を哺乳動物から分離すること、in vitroで、神経幹細胞を増殖集団へ増殖させることを含む。また、本方法は、増殖集団を濃縮すること、治療有効量の増殖集団をレシピエントの脊髄の少なくとも一領域に導入することを含む。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0053】
一実施形態では、運動ニューロンの変性は、外傷性脊椎損傷、虚血性脊椎損傷、外傷性脳損傷、脳卒中、多発性硬化症、脳性麻痺、てんかん、ハンチントン舞踏病、筋萎縮性側索硬化症、慢性虚血、遺伝性疾患またはそのいずれかの組合せに由来する。
【0054】
一実施形態では、本方法は、少なくとも1種のニューロンのサブタイプが豊富な領域から神経幹細胞を分離することを含み、この場合、ニューロンのサブタイプは、運動障害を改善することにおいて有効な増殖因子を産生する。
【0055】
一実施形態では、増殖集団は、治療有効量の少なくとも1種の増殖因子を分泌するために十分なニューロンに分化することができる神経幹細胞量を含む。
【0056】
一実施形態では、本方法は、運動ニューロンの豊富な領域から神経幹細胞を分離することを含む。
【0057】
さらなる実施形態では、脊髄空洞症を治療することができる神経幹細胞を提供する。神経幹細胞は、哺乳動物から分離し、in vitroで増殖集団へ増殖させる。幹細胞を含む増殖集団を濃縮して、治療有効量の増殖集団を、レシピエントの脊髄の少なくとも一領域に導入する。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0058】
本開示の方法の他の実施形態では、脊髄空洞症を治療する方法を提供する。本方法は、少なくとも1種の神経幹細胞を哺乳動物から分離すること、in vitroで、神経幹細胞を増殖集団へ増殖させることを含む。また、本方法は、増殖集団を濃縮し、治療有効量の増殖集団をレシピエントの脊髄の空洞に導入することを含む。増殖集団の少なくとも20%は、レシピエントの脊髄の空洞でニューロンを発生させることができる。
【0059】
一実施形態では、脊髄空洞症は、外傷性脊椎損傷、虚血性脊椎損傷、外傷性脳損傷、脳卒中、多発性硬化症、脳性麻痺、てんかん、ハンチントン舞踏病、筋萎縮性側索硬化症、慢性虚血、遺伝性疾患またはそのいずれかの組合せに由来する。
【0060】
一実施形態では、本方法は、少なくとも1種のニューロンのサブタイプが豊富な領域から神経幹細胞を分離することを含み、この場合、ニューロンのサブタイプは、脊髄空洞症を改善することにおいて有効な増殖因子を産生する。
【0061】
一実施形態では、本方法は、運動ニューロンの豊富な領域から神経幹細胞を分離することを含む。
【0062】
一実施形態では、増殖集団は、治療有効量の少なくとも1種の増殖因子を分泌するために十分なニューロンに分化することができる神経幹細胞量を含む。
【0063】
一実施形態では、治療有効量の増殖集団は、少なくとも1,000のニューロンを発生させることができる。
【0064】
一実施形態では、増殖集団の少なくとも100,000の神経幹細胞をレシピエントの脊髄の空洞に導入する。
【0065】
なおさらなる実施形態では、脊髄空洞症を治療することができる神経幹細胞を提供する。神経幹細胞は、哺乳動物から分離し、in vitroで増殖集団へ増殖させる。幹細胞を含む増殖集団を濃縮して、治療有効量の増殖集団を、レシピエントの脊髄の空洞に導入する。増殖集団の少なくとも20%は、レシピエントの脊髄の空洞でニューロンを発生させることができる。
【0066】
本開示の方法のさらなる実施形態では、in vitroにおいて少なくとも1種の神経幹細胞を神経幹細胞の増殖集団に増殖させる方法を提供する。各神経幹細胞の増殖は、分化することなしに、細胞の倍増が30回を上回る。本方法は、中枢神経系組織から神経幹細胞を分離して、少なくとも1種の細胞外タンパク質(extracellular protein)を培養容器に供給することを含む。細胞外タンパク質は、少なくとも約10μg/mlのポリ-D-リシンおよび約1mg/mlのフィブロネクチンを含む。また、本方法は、血清の不存在下において、分離した神経幹細胞を培養容器で培養し、少なくとも1種の増殖因子を培養容器に加えることを含む。増殖因子は、増殖因子はbFGF、EGF、TGF-α、aFGFおよびその組合せからなる群から選択される。本方法は、集密に達する前に、さらに培養細胞を継代することを含む。
【0067】
一実施形態では、増殖された神経幹細胞は、ニューロンに分化することができる。
【0068】
一実施形態では、神経幹細胞を増殖することは、可溶性因子としてフィブロネクチンを培地に加えることを含む。
【0069】
一実施形態では、細胞を分離すること、および細胞を継代することは、酵素による分離を含む。
【0070】
一実施形態では、酵素による分離は、トリプシンで細胞を処置することを含む。
【0071】
一実施形態では、神経変性状態を治療するために、治療有効量の増殖集団をレシピエントの神経系の少なくとも一領域に導入する。
【0072】
したがって、既存の薬理学的方法に対する本開示の方法の利点は、栄養分子を分泌させるために、移植したNSCの能力を増強させ、最適の生物学的利用度の条件下で、変性している運動ニューロンに送達することができる、方法を提供することである。
【0073】
本発明のさらにもう1つの利点は、ヒトの胎児脊髄由来のNSCを培養および増殖して、腰髄へのNSCの良好な移植を促進する方法を提供することである。
【0074】
本開示の方法のさらなる利点は、NSC集団のより高率のニューロン分化を達成する方法を提供することを含む。
【0075】
本開示の方法のもう1つの利点は、NSCの部分移植により臨床効果を得ることを含む。
【0076】
本開示の方法の付加的な特徴および効果は、以下の本発明の詳細な説明および図において記載され、これらによって明らかとなろう。
【図面の簡単な説明】
【0077】
【図1】ヒト脊髄幹細胞の増殖を示す図である。ヒト脊髄前駆体(NSCとしても知られている)系を、7〜8週齢の死後胎児脊髄組織から単離し、約130日の正味培養期間、連続継代した。各継代の採収時に回収した細胞数を、平板培養時の最初の細胞数で割って、細胞数の増加倍数を得た。各継代時の増加倍数を掛け合わせることによって累積増加倍数(左Y軸)を得た。細胞数の倍増を各培養期間(X軸)で割って、各継代時における細胞の倍加時間(右Y軸)を算出した。この処理を3回繰り返した(連続増殖1、2、および3)。
【図2】増殖させたヒト脊髄幹細胞の形態を示す図である。(A)固定、非染色、増殖培養物、20×対物レンズの位相差図、(B)抗ネスチン抗体染色。
【図3】増殖させたヒト脊髄幹細胞から得られた分化培養物の特徴を示す図である。15〜16継代の増殖細胞を培養物中でおよそ14日間分化させ、固定し、種々のニューロン特異的抗体によって染色した。(A)TauおよびMAP2; (B)タイプ3ベータチューブリン; (C)GABA; (D)アセチルコリントランスフェラーゼ。
【図4】ヒト中脳幹細胞の増殖を示す図である。ヒト中脳前駆体(NSCとしても知られている)系を、7〜8週齢の死後胎児中脳組織から単離し、約170日の正味培養期間、連続継代した。各継代の採収時に回収した細胞数を、平板培養時の最初の細胞数で割って、細胞数の増加倍数を得た。各継代時の増加倍数を掛け合わせることによって累積増加倍数(Y軸)を得た。
【図5】増殖させたヒト中脳幹細胞のドーパミン取り込み活性を示す図である。生細胞におけるドーパミントランスポーター活性(DAT)を、ヒト中脳幹細胞系およびアッセイ時に22日または44日間分化させたそのクローンの亜系の1つから判定した。該細胞を、DAT阻害剤ノミフェンシン(10μM)の存在下(+)または不在下(-)、放射標識したドーパミンと共にインキュベートした。シンチレーションカクテル中、取り込まれなかったドーパミンを除去するために、細胞を洗浄し、すすいだ。次いで、シンチレーションカウンターを用いて、総細胞放射性(dpm)を測定した。
【図6A】ヒト中脳幹細胞系のニューロン分化およびドーパミン作動性分化の誘導に及ぼす外来因子の影響を示す図である。2つのヒト中脳幹細胞系(527RMBおよび796RMB)からの冷凍保存した神経幹細胞を解凍し、bFGFの存在下、4ウェルチャンバースライド内で、1ウェル当たり40,000細胞の密度で塗布し、6日間増殖させた。引き続き、bFGFを除去し、細胞をさらに8日間分化させた。セルトリ細胞馴化培地(SCCM、N2中、1:1に希釈)に曝露するタイミングと時間に基づき、細胞を4群に分けた。1つの群は、増殖中および分化中、SCCMに曝露し(条件1);第2群は増殖中にのみ曝露し(条件2);第3群は分化中にのみ曝露し(条件3);第4群はSCCMに曝露しなかった(対照、Cont.)。培地は1日おきに交換し、増殖期の間、マイトジェンを毎日添加した。1つの条件当たり、4つのウェルを維持し、複数のマーカーに関して染色した。分化の際、4%パラホルムアルデヒドを用いて細胞を固定し、MAP2abに対する抗体(図6A)およびチロシンヒドロキシラーゼに対する抗体(図6B)、ならびにGFAPおよびGalCを用いて免疫染色した。免疫染色された細胞を、40×対物レンズを用いてカウントすると、各ウェルで少なくとも3つのフィールドがカウントされた。いずれの条件下で維持された細胞を分析した際にも、GFAP+細胞またはGalC+細胞はほとんど検出されないか、または全く検出されず、したがって、これらの抗原は分析から除外した。
【図6B】図6Aの説明参照。
【図7】ヒト脊髄幹細胞移植によるラットの痙攣/硬直および運動欠損の減少を示す図である。腰脊髄の虚血損傷化によって痙攣ラットを作製した。1つの群(黒丸)では、培養物中で増殖させた(継代16)ヒト脊髄幹細胞を、ラット(n=9)に移植し、他方、対照の群(白丸、N=7)には、細胞のない培地のみを与えた。この試験の間(8週間)、両群に免疫抑制剤、FK506を、1日1mg/kgで投与した。個々の動物の運動協調性を1週間に1回、BBBスコアリングによって評価した。
【図8】ヒト脊髄幹細胞移植によるラットの痙攣/硬直および運動欠損の減少を示す図である。腰脊髄の虚血損傷化によって痙攣ラットを作製した。1つの群(黒丸および黒四角)では、培養物中で増殖させた(継代16)ヒト脊髄幹細胞を、ラット(n=13)に移植し、他方、対照の群(黒三角、n=6)には、細胞のない培地のみを与えた。この試験の間(12週間)、両群に免疫抑制剤、FK506を、1日3mg/kgで投与した。個々の動物の運動協調性を1週間に1回、BBBスコアリングによって評価した。
【図9A】G93A SOD1 ラットの運動ニューロン疾患重症度に及ぼすヒトNSC処置の効果を、生細胞(L、赤)移植片および死細胞(対照、C)移植片(青)の場合における進行(A〜B)ならびに臨床的および病理学的尺度のエンドポイント(C〜E)分析によって示す図である。 図9AおよびB。図9Aは、観察経過にわたる実験動物と対照動物との間の有意な区別を示すKaplan-Meierプロット (P=0.0003)であり、図9Bは、2つの群間の筋肉衰弱の2つの主要な尺度(BBBおよび傾斜平面スコア)における区別を示している(それぞれ、P=0.00168および0.00125)。
【図9B】図9Aの説明参照。
【図9C】図9C〜E。実験ラットおよび対照ラットにおける生存(C)、疾患発症までの時間(D)および運動ニューロン数(E)のエンドポイント分析。図9Cは、両群間の寿命における有意な11日の違いを示している(P=0.0005)。図9Dは、両群間の疾患発症までの時間における有意な7日の違いを示している(P=0.0001)。図9Eは、生きているNSC群と死んでいるNSC群の腰プロチュバランスにおける3,212個の細胞の違いを示している(P=0.01)。E下部の差込図は、128日齢における代表的な実験ラット(上部)と対照ラット(下部)間の運動ニューロン生存度における違いを示しており;矢印は、側方運動ニューロン群を示す。サイズバー:150μm。
【図9D】図9Cの説明参照。
【図9E】図9Cの説明参照。
【発明を実施するための形態】
【0078】
本開示の方法は、神経変性状態を治療することに関する。特に、本開示の方法は、それを必要とする対象に移植するための神経幹細胞を調製する方法を含む。移植のために細胞を調製することは、神経変性状態の治療用として商業的利用に十分な水準まで、in vitroで、特定の細胞集団を増殖することを含むことができる。一実施形態では、変性または損傷した神経領域の治療法は、神経変性状態を改善するのに十分な神経幹細胞の有効数を本領域に供給することを含む。
【0079】
本書では、神経変性状態は、ニューロンの損傷または変性が関わる、何らかの疾患または障害または症状または原因またはその影響を含む可能性がある。神経変性状態は、アレキサンダー病、アルパーズ病、アルツハイマー病、筋萎縮性側索硬化症、血管拡張性失調症、カナヴァン病、コケーン症候群、大脳皮質基底核変性症、クロイツフェルトヤコブ病、ハンチントン舞踏病、ケネディ病、クラッベ病、レヴィー小体痴呆、マシャドジョセフ病、多発性硬化症、パーキンソン病、ペリツェウスメルツバッハー病、ニーマンピック病、原発性側索硬化症、レフサム病、サンドホフ病、シルダー病、スティールリチャードソンオルゼウスキー病、脊髄癆または損傷したニューロンと関連するその他の状態を含む可能性があるが、これらに制限されない。他の神経変性状態は、外傷性脊椎損傷、虚血性脊椎損傷、脳卒中、外傷性脳損傷および遺伝性疾患を含むか、これらによって引き起こされることがある。
【0080】
本開示の方法は、神経変性状態を改善するためのNSCの使用を含む。本書では、「NSC」という用語は、神経もしくはニューロンの前駆体または神経上皮前駆体を指すこともある。NSCは、ニューロン、星状膠細胞および希突起膠細胞というCNSのいずれか3つの主な細胞の種類へ分化する能力によって機能的に定義することができる。
【0081】
一実施形態では、NSCは、各細胞がニューロン、星状膠細胞または希突起膠細胞に分化する能力を有するというように、多分化能を有する。一実施形態では、NSCは、各細胞が3種のCNSの細胞型中2種に分化する能力を有するというように、二分化能を有する。一実施形態では、NSCは、in vitroでニューロンおよび星状膠細胞の両方を発生する、少なくとも二分化能を有する細胞を含み、in vitroでニューロンを発生する、少なくとも単分化能を有する細胞を含む。
【0082】
増殖状態は、1種の細胞またはもう1種の細胞への細胞の分化の方向に影響を及ぼすことがあり、このことは細胞が1つの系譜に関係付けられていないことを示している。神経分化にとって有利に働く培養条件で、細胞の特にヒトCNS由来の細胞は、主にニューロンおよび星状膠細胞への二分化能を有し、希突起膠細胞への分化は稀である。このように、本開示の方法の分化細胞の培養は、ニューロンおよび星状膠細胞を生じさせることが可能である。一実施形態では、星状膠細胞に対するニューロンの比率は、50:50の比率に近づく可能性がある。
【0083】
本開示の方法は、感覚上皮などの哺乳動物のCNS部位に存在するNSCを得ることを含む。NSCを分離することができる他のCNS部位は、CNSの脳室帯(ventricular zone)および脳室下帯(subventricular zone)、ならびに有糸分裂後ニューロンだけでなく、有糸分裂前駆体(mitotic precursor)を含む他のCNS部位を含む。一実施形態では、本開示の方法は、哺乳動物のCNSが形成される部位に存在するNSCを使用することができる。
【0084】
一実施形態では、NSCは、所望のニューロン集団のために自然の神経原性(neurogenic)を有する領域から得る。所望の細胞集団は、神経学的状態で失われた、または不活性のような表現型と置換またはそのような表現型を補うことができる特定のニューロン表現型の細胞を含むことが可能である。
【0085】
特定の神経変性疾患または状態の治療のために有用なニューロンのサブタイプを含む、さまざまな異なるニューロンのサブタイプは、CNSの異なる部位または領域から、および胎児の発育中の異なる在胎齢にわたって、NSCを分離することによって得ることができる。CNSの異なる部位または領域、および胎児の発育中の異なる在胎齢にわたって分離されたNSCを、最適な増殖およびニューロン分化能のために使用する。哺乳動物のCNSの顕著な特徴の1つは、ニューロンのサブタイプの多様性である。例えば、NSCの単一集団は、培養で、ほんの少数の明瞭なニューロンのサブタイプを自然に発生することが可能である。さらに、胎児の特定の在胎齢から得た細胞は、培養細胞の生理的関連性(physiological relevance)を確定させる可能性がある。
【0086】
本開示の方法の実施例では、対象に移植される細胞は、損傷した神経領域のヒトの胎児の対応物に由来する。一実施形態では、NSCを、約6.5週から約20週の間の在胎齢におけるヒトの胎児のCNS領域から分離する。一実施形態では、胎児の脊髄由来の細胞を、約7週から約9週の在胎齢で分離する。分離可能な神経幹細胞集団の比率は、ドナーの年齢によって異なる可能性があることを認識すべきである。また、細胞集団の増殖能力も、ドナーの年齢によって変化する。そのようなNSCの局所的および時間的特異性は、NSCが運命制限的な前駆体として機能し、ブランク細胞(blank cell)または単一の細胞集団として機能しないことを示している。
【0087】
in vitroでのGABAを産生するニューロンを含む集団の比率は、一般に、約5%から10%で一定である。
【0088】
例えば、中脳腹側部のNSCは、同じ妊娠期間の脊髄から得られるNSCとは異なっている。特に、中脳腹側部由来のNSCは、独占的にチロシン水酸化酵素を発現するドーパミン作動性ニューロンを生じるが、一方、脊髄由来のNSCは、独占的にアセチルコリンを産生するコリン作動性ニューロンを発生する。しかし、両方の細胞型とも、同時に、より偏在性のグルタミン酸およびGABAを産生するニューロンを発生する。したがって、一実施形態では、本開示の方法は、少なくとも部分的に、チロシン水酸化酵素を発現するドーパミン作動性ニューロンの移植によって、状態を治療して改善または軽減するために、中脳腹側部からNSCを得ることを含む。本開示の方法は、少なくとも部分的に、アセチルコリンを産生するコリン作動性ニューロンの移植によって、神経変性状態を治療して改善または軽減するために、脊髄からNSCを得ることを含む。
【0089】
したがって、ドーパミン作動性ニューロンの消失によって特徴付けられるパーキンソン病などの運動障害の治療のために、本開示の方法の実施形態は、ドーパミン作動性ニューロンの神経形成がかなり高率である中脳腹側部などの領域由来のNSCの使用を含む。さらに、NSCは、ドーパミン作動性ニューロンの神経発生がかなり高率である期間のヒトの胎児発育の在胎齢で得ることができる。したがって、一実施形態では、本開示の方法は、運動障害を治療するために、約7週から約9週の在胎齢の中脳腹側部からNSCを得ることを含む。
【0090】
腹側の運動ニューロンの消失から生じる筋萎縮性側索硬化症または弛緩性対麻痺などの運動ニューロン疾患を治療するために、本開示の方法の実施形態は、腹側の運動ニューロンの神経発生がかなり高率である脊髄などの領域に由来するNSCの使用、および腹側の運動ニューロンの神経発生がかなり高率である期間のヒトの胎児発育の在胎齢に得られたNSCの使用を含む。したがって、一実施形態では、NSCは、運動ニューロン疾患を治療するために、約7週から約9週の在胎齢の脊髄から分離する。
【0091】
しかし、場合によっては、そのような局所の特異性に関する制限は、実際的な目的には、かなり幅が広いということを認識すべきである。したがって、頸椎、胸椎、腰椎および仙骨などの脊髄のさまざまな領域から得たNSCを、区別なく、移植し、対応するNSCの起源以外の部位を治療するために使用することができる。例えば、頸髄由来のNSCは、患者の腰椎部に細胞を移植することによって痙縮および/または硬直を治療するために使用することができる。
【0092】
また、NSCは、生後組織および成体組織から分離することができる。生後組織および成体組織由来のNSCは、それらの増殖および分化の特徴だけでなく、ニューロンおよび神経膠に分化するそれらの能力に関して量的に同等である。しかし、in vitroでのさまざまな生後CNSおよび成体CNSからのNSC分離の効率は、より多量のNSC集団を保有する胎児組織由来のNSCより著しく低い可能性がある。にもかかわらず、胎児由来のNSCと同様に、本開示の方法は、新生児および成人の供給源由来のNSCの少なくとも30%が、in vitroでニューロンに分化することができる。したがって、前述のように胎児由来のNSCの場合、生後組織および成体組織を使用することができるものの、胎児組織の使用が好ましい。
【0093】
さまざまなニューロンのサブタイプを、培養で増殖した胚幹細胞の操作から得ることができる。したがって、本開示の方法に基づく特定のニューロンのサブタイプは、必要に応じて、結果を改善するために他の関連のない、または所望でない細胞から分離および精製することができ、同じ神経変性状態の治療のために使用することができる。
【0094】
本開示の方法におけるNSCは、1つの部位から得て、自家移植片として、同じ対象の別の部位へ移植することができる。さらに、本開示の方法におけるNSCは、遺伝子的に同一のドナーから得て、同系移植片として移植することができる。またさらに、本開示の方法におけるNSCは、遺伝子的に同一でない同一種のメンバーから得て、同種移植片として移植することができる。代わりに、NSCは、ヒト以外の種から得て、異種移植片として移植することができる。強力な免疫抑制薬の開発に伴い、同種移植片、およびブタ起源の神経前駆細胞などの、ヒト以外の神経前駆細胞の異種移植片をヒトの対象に移植することができる。
【0095】
試料の組織は、いずれかの標準的方法によって分離することができる。一実施形態では、組織は、ピペットおよび二価陽イオンを含まない生理食塩水を使用した穏やかな機械的粉砕(trituration)によって分離し、分離細胞の懸濁液を形成する。主として、単細胞を得るための十分な分離が、局所的な過度の細胞密度を避けるために望ましい。
【0096】
NSCの成功した商用的利用のために、多くの連続的継代を通して、安定した増殖能および分化能を有する強固で、ばらつきのない培養を維持することが望ましい。前述のように、本培養法は、CNS発育の異なる領域および年齢由来のNSCにおいて、これらの異なる前駆体特性を維持するのと同時に、個々の細胞系の長期の安定した増殖を達成するために最適化させることができる。
【0097】
このために、驚くべきことに、NSC(NSC)の基質への接着を促進することが、NSCまたは前駆細胞の有糸分裂率を加速する一因となり、したがって、NSCまたは前駆細胞のより強固な培養を著しく向上させることが認められている。特に、局所的な過度の細胞密度を避けること、およびマイトジェン濃度を維持することに加えて、細胞外基質タンパク質の濃度が、NSCの長期の有糸分裂能および分化能に影響を及ぼすことが認められている。細胞外基質タンパク質は、ポリ-D-リシン、ポリ-L-リシン、ポリ-D-オルニチン、ポリ-L-オルニチン、フィブロネクチンおよびその組合せを含み得る。他の細胞外基質タンパク質は、さまざまなアイソタイプ、断片、組換え型、またはフィブロネクチン、ラミン、コラーゲンおよびそれらを組み合わせた合成模倣剤(synthetic mimetics)を含み得る。代わりに、または加えて、本開示の方法は、各個別の細胞が、細胞毒性を生じるか、細胞分化を遅らせることなく、培養の全期間において培養基質に細胞が接着するように、有効な細胞接着(cell adhesion)を促進することができる、その他の全ての適切な物質を含むことができることを認識すべきである。
【0098】
細胞外基質タンパク質は、細胞接着を促進することにおいて有効である可能性があるが、ポリ-L/D-オルニチンまたはポリ-L/D-リシンなどの異なるアミノ酸ポリマーが、各個別の細胞系に対して特定の濃度で、細胞に対して有毒である可能性がある。また、インキュベーションの継続時間は、培養皿の表面に沈着した本ポリマーの最終量に影響を及ぼし、細胞の生存度に影響する可能性がある。本開示の方法で使用されるNSCにおいて、ポリマーの濃度は、約0.1μg/mlから約1mg/mlの間の範囲内とすることができる。一実施形態では、100μg/mlのポリ-D-リシンを、中性pHの0.01M HEPES緩衝液または水で溶解し、培養容器に加える。培養容器を室温で1時間インキュベートする。次に、培養容器を水で完全にすすぎ、使用前に乾燥させる。
【0099】
また、本開示の方法は、細胞外基質タンパク質で培養容器を二重塗布することを含むことができる。一実施形態では、前述のポリ-L/D-オルニチンまたはポリ-L/D-リシンの処理後、培養容器をフィブロネクチンまたはフィブロネクチン誘導体によって処理する。一実施形態では、ヒトの血漿から調製されたフィブロネクチンタンパク質を使用する。しかし、ブタまたはウシのフィブロネクチン、組換えフィブロネクチン、フィブロネクチンタンパク質の断片、合成ペプチドおよびフィブロネクチンの他の化学的模倣剤などの、他のいかなるフィブロネクチンタンパク質の適した形態または供給源も使用することができることを認識すべきである。一実施形態では、約0.1μg/mlから約1mg/mlのフィブロネクチンを加えることができる。
【0100】
ヒトの脊髄由来のNSCの増殖を含む一実施形態では、細胞外タンパク質が培養容器に十分に結合およびコーティング可能となる期間、ポリ-D-リシン100μg/mlで培養容器を処理する。そのような時間は、約5分から約3時間までの間とすることができる。培養容器は、その後水で洗浄することができる。培養容器を空気乾燥した後に、容器を室温で約5分から数時間、約25mg/mlのフィブロネクチンで処理、または37℃で約1時間から数日間、約1mg/mlのフィブロネクチンで処理することができる。その後、フィブロネクチンを取り除き、培養容器は、少なくとも1回洗浄するか、使用するまでPBS中に保存してもよい。
【0101】
あるいは、フィブロネクチンを細胞に直接供給する可溶性因子として増殖培地に加えることができる。本実施形態では、NSCは、フィブロネクチンによる培養容器の処理に加えて、またはこの処理の代わりに、増殖培地にフィブロネクチン1μg/mlを加えることによって増殖させることができる。フィブロネクチンをコーティングした容器の貯蔵時間が比較的短いため、細胞の平面培養時に可溶性因子として付着タンパク質(attachment protein)を増殖培地に供給することは、特にNSCの大規模な商業規模の培養にとって有益である。また、この方法は、例えば、cGMPプロトコルの下で必要とされるかなり正確な状態および再現性が必要とされる神経幹細胞系を製造するため、および治療上の使用のための神経幹細胞系を製造するために有用である。
【0102】
一実施形態では、分離されたNSCを約1,000細胞/cm2から約20,000細胞/cm2の密度で、培養容器に加える。そのような密度は、培養容器における個々の細胞の平均な分散および接着をもたらし、局所の細胞の高濃度化を防止し、NSCの培養物を富化する。
【0103】
一実施形態では、NSCを血清の不存在下で増殖させる。一実施形態では、NSCの有糸分裂および分化能を不安定にさせるのに十分な血清濃度にNSCが曝露するのを避けるために、NSCを規定された無血清培地で培養する。さらに、白血病抑制因子(LIF)または毛様体神経栄養因子(ciliary neurotrophic factor:CNTF)などの特定の増殖因子に対するNSCの曝露もNSCを不安定にさせるので、避けるべきである。
【0104】
NSCの増殖を高めるために、いずれの培養過程の段階でもマイトジェンを培養に加えることができる。マイトジェンは、塩基性線維芽細胞増殖因子(bFGF)、酸性線維芽細胞増殖因子(aFGF)、上皮増殖因子(EGF)、トランスフォーミング増殖因子-α(TGFa)およびその組合せを含み得る。
【0105】
本開示の方法のNSCは、少なくとも2種の異なる培養形態で成長および増殖させることができる。培養の一形態は、一般に、クラスターと呼ばれる凝集した形態、懸濁培養と呼ばれる凝集した形態を含む。培養のもう1つの形態は、懸垂培養(adhesion culture)と呼ばれる分散した、凝集していない形態を含む。
【0106】
本開示の方法のNSCを分散する付着培養(adherent culture)において、本細胞は、最初に個々の細胞が直接基質に接触する単層を形成する。最終的に、インキュベーションの期間後、細胞は、散発的にクラスターを形成することがあり、この場合、底の層の細胞が個々に基質に付着しているのと同時に、少なくとも1つのさらなる細胞の層が底の層上に形成される。そのようなクラスター形成は、培養物が高い細胞密度で播種させたか、高い細胞密度に達することが可能なときに特に発生し、一実施形態では、NSCもしくは前駆細胞の最適な増殖、またはNSCの多分化能の最適な維持のために最小化される。本開示の方法の実施形態における分散型の付着培養では、ヒトNSCを、細胞分裂当たり約4日未満で分裂させることができる。
【0107】
分散型の付着培養のもう1つの特有の特徴は、本開示の方法のNSCがそれぞれの多分化能を保持する娘細胞を発生するように分裂することである。一実施形態では、本開示の方法におけるNSCの分散型の付着培養は、実質的な分化がない状態で、少なくとも20回の細胞倍増を行う増殖能力を含む。大部分のNSCは、それらの神経原性の潜在性を失う前に、少なくとも50回の細胞倍増を超えて増殖できる。一実施形態では、本開示の方法の分散型の付着培養で増殖されるNSCは、ニューロン分化が高められることが実証され、一実施形態では、少なくとも約30%のニューロン分化が起こる。多くの場合、NSCの少なくとも50%は、ニューロンに分化する。分散付着の培養形態が、より好ましい培養形態であるが、異なる培養法は、in vitroまたはin vivoのいずれかでの異なる分化能によって、生来異なる細胞集団の分離を可能にすることができる。
【0108】
また、本方法は、遺伝子組換えまたは支持細胞(feeder cell)の包含なしに、さまざまな供給源からのNSCのクローン分離(clonal isolation)を可能にする。したがって、非常に少数の、好ましくは1000細胞/cm2未満の細胞を、前述のように調製された細胞培養皿に播種することができる。
【0109】
NSCの播種から2、3日後、細胞は、良好に分離したコロニーを形成することができる。本コロニーは、例えば、少なくとも約250細胞から約2000細胞の所望の大きさに増殖することができる。一実施形態では、少なくとも1つの細胞のコロニーは、手で採取し、マルチウェルプレートなどの新しい細胞培養皿に個々に接種する。
【0110】
分離したクローン集団は、連続的な継代によって増殖させ、複数の神経幹細胞系を得るために使用することができる。多くのそのようなクローン細胞系が、脊髄、中脳および後脳などのヒトのCNSのさまざまな領域から分離されている。クローン細胞系は、より高率なニューロンのサブタイプなど、特定の細胞の表現型を富化するために有用である。例えば、チロシンヒドロキシラーゼを発現するドーパミン作動性ニューロン、GABA作動性ニューロン、コリン作動性ニューロンおよび他の特定の表現型のニューロンが濃縮したクローン細胞系を本開示の方法によって分離することができる。
【0111】
一実施形態では、ポリクローナルまたはモノクローナルのいずれかの神経幹細胞系を、さらに特定のサブタイプのニューロンを濃縮にするために導入することができる。中脳または脊髄のNSCからチロシンヒドロキシラーゼを発現するドーパミン作動性ニューロンおよびアセチルコリンを産生するコリン作動性ニューロンなど、特定のニューロンの有効な誘導物質を同定するために、多くの増殖因子、化学物質および天然物質(natural substance)がスクリーニングされている。因子もしくは化学物質または、その組合せを、NSCの分裂期および/または分化期に導入することができる。一実施形態では、ドーパミン作動性の表現型の神経幹細胞系を、パーキンソン病を治療するためのドナー集団としてさらに濃縮させる。
【0112】
さまざまなニューロンのサブタイプを、in vitroで所望の分化パターンを有する幹細胞の分離から得ることができる。in vitroでの結果は、実質的にin vivoで再現可能である。このことは、in vivoでの幹細胞の潜在的な有効性が、in vitroでの幹細胞の分化パターンによって予測できることを意味している。生後の生きた対象への注射後、未分化または分化前のいずれかの状態のNSCは、大体において、in vitroで観察されたin vivoの分化パターンを生じる。したがって、in vitroでチロシンヒドロキシラーゼを産生するニューロンを生じるNSCも、in vivoでチロシンヒドロキシラーゼを産生するニューロンを発生する。逆に言えば、in vitroでチロシンヒドロキシラーゼを産生するニューロンを生じないNSCは、構成的に、in vivoでチロシンヒドロキシラーゼを産生するニューロンを発生しない。
【0113】
しかし、in vitroに存在する分化の合図は、in vivoでの合図と比べて限られる。したがって、分化したニューロンのかなりの画分は、主要な神経伝達物質の表現型を発現しないことがある。求心性または遠心性ニューロンからのシグナルなどの付加的な合図、またはそのような自然のシグナルを模倣する薬剤を、NSCの有糸分裂期またはNSCの分化のいずれかの間に、分化した表現型を再構成するために使用することができる。NSCは、in vitroだけでなくin vivoに存在する合図に反応する能力を有する。したがって、一旦、虚血により損傷した脊髄に移植したならば、脊髄のNSCは、in vitroよりかなり高率のGABAを産生するニューロンを発生する。したがって、NSCは可塑性である。NSCのそのような可塑的な性質は、NSCの多分化能の性質によるものであり、それ自体として、この可塑性を、その特性の方向を変更させるために、NSC集団とさらに結合することができる、表現型を誘導する作用物質および状態を同定するために使用することができる。
【0114】
一実施形態では、そのような再プログラミング(re-programming)は、運動ニューロンの表現型の発現増強をもたらすために、脊髄由来のNSCを処理することを含む。処理の状態は、さまざまな筋細胞または神経冠細胞もしくは神経節ニューロン(ganglionic neuron)などの末梢神経系由来細胞とNSCまたはその分化細胞を共培養することを含む。また、NSCを、運動ニューロンまたは脊髄で発現または産生することが知られる分子のカクテル(cocktail)で処理して、運動ニューロン表現型のNSCの発現を高めることができる。
【0115】
ヒトの中脳におけるドーパミン作動性の表現型を増大させたNSCを誘導するために、リチウム、GDNF、BDNF、プレイオトロフィン、エリスロポエチンなどの分子、セルトリ細胞(sertoli cell)などの細胞由来の条件培地(conditioned media)、またはスクリーニングによって得た他の全ての適した化学物質もしくは細胞、ならびにその組合せによって、NSCを処理する。そのような誘導により、in vivoにおいて、移植したNSCにドーパミン作動性の表現型を発現および維持させることができる。
【0116】
一実施形態では、本開示の方法のNSCは、移植のために分化前の細胞を含むことができる。細胞の最大収量および手法の単純化のために、主に未分化細胞の集団を含む集密培養物(confluent culture)を移植のために収集する。しかし、細胞密度が増加することによって、自発的に分化をまさに始めようとする少数の細胞集団も、存在する可能性があるということを認識すべきである。
【0117】
一実施形態では、NSCの継代は、基質から細胞を収集または分離することを含む。一実施形態では、本開示の方法は、少なくとも1種の酵素を使用して基質から細胞を収集または分離することを含む。酵素処理は、NSCの細胞周期が細胞表面のマイトジェン受容体を不活性化するのに十分なほど短いとき、避けることができる。しかし、ヒトのNSCの細胞周期は、ヒトのNSCが酵素処理に対して感受性がないというほど、げっ歯類のNSCよりもかなり長い。したがって、本開示の方法では、ヒト由来のNSCを収集するために、酵素処理を使用する。ヒトのNSCは、酵素処理の存在下で、一時的にマイトジェンに不応性になることがあるが、マイトジェン受容体の不活性化を繰り返すことで、NSC比率を減少させることができる。
【0118】
一実施形態では、細胞の収集に際し、短時間の遠心分離によって細胞を濃縮する。細胞を、生理食塩水、緩衝生理食塩水などの最終的、臨床的に使用可能な溶液でさらに洗浄および再懸濁するか、代わりに、保存溶液またはハイバーネーション「冬眠」溶液(hibernation solution)で再懸濁することができる。代わりに、細胞を、培地+ジメチルスルホキシド、または他の全ての適切な凍結保護物質などの寒剤で再懸濁し、保存のために凍結させることができる。
【0119】
ハイバーネーション溶液は、長期間の間、生きた細胞の生存度を維持するために配合する。一実施形態では、保存溶液は、すぐに使用できる剤形で生きた細胞を移植手術現場へ送り、すぐに使用するために適応される可能性がある。また、遠く離れた場所に、生きた細胞を送るのに適した状態は、約0℃から約20℃の範囲で少なくとも24時間、安定した温度を維持することができる断熱装置を含む。約0℃から約8℃の間で、約24時間から約48時間保存した生きた細胞は、疾患または状態の治療のために移植可能である。
【0120】
一実施形態では、細胞は、前述の臨床的に使用可能なハイバーネーションまたは凍結液などの溶液中で濃縮させることができる。一実施形態では、細胞を、細胞を投与するための細胞密度と同一または異なってもよい適切な細胞密度に濃縮する。一実施形態では、投与のための細胞密度を、注射部位、注射部位の神経変性状態、有益な効果のための必要最小量、および毒性副作用に対する考慮などの要因によって、約1,000/μlから約1,000,000/μlまで変えることができる。一実施形態では、本開示の方法は、約5,000/μlから約50,000/μlまでの細胞密度で細胞を注射することを含む。
【0121】
増殖した細胞を治療部位に送るために懸濁させた媒質の量を、本書では注射量(injection volume)と呼ぶことがある。注射量は、注射部位および組織の変質状況に依存する。より具体的には、注射量の下限は、細胞の密集する傾向だけでなく、細胞密度の高い粘性懸濁液に対する実際的な液体の処理方法によって決定することができる。注射量の上限は、実際的な手術時間だけでなく、宿主組織を損傷させるのを避けるために必要な、注射量によって発揮される圧縮力の限界によって決定することができる。
【0122】
公知の方法を使用したドナー細胞の細胞生存が低いことから、有効な治療をするために、比較的小さな領域に多量の細胞を送達することが必要とされてきた。しかし、注射量は、宿主組織に対して発揮される静水圧であって、多量の注射量に関連して注射時間が長引くと、手術のリスクが増大する。さらに、ドナー細胞の過剰注射は、宿主の実質組織の圧迫およびその後の損傷につながる。量の制約を補う試みにおいて、公知の方法は、注射のために、細胞密度の高い懸濁液の調製を必要としている。しかし、高い細胞密度は、移植した細胞の密集化を促進し、細胞の移動または拡散を抑制し、制限領域を越えた有効な治療を妨げ、宿主組織へ途切れのない統合(seamless integration)を損なう。
【0123】
対照的に、本開示の方法によって調製された細胞のin vivoでの生存が改善される結果として、注射ごとに必要となる細胞数がより少なくなる。実際、注射時から6カ月後に、最高で3から4倍の注射細胞数が生存していることが明らかにされており、本開示の方法を使用した著明な定量的生存(quantitative survival)が実証されている。また、定量的な生存により、所望の細胞量の再現性のある投与が実現できる。したがって、一実施形態では、細胞を約1,000から約200,000細胞/μlの密度に濃縮する。一実施形態では、約5,000から約50,000細胞/μlを有効な移植のために使用している。他の実施形態では、約10,000から約30,000細胞/μlを使用する。一実施形態では、注射部位当たり約100/μl未満の注射量で懸濁された細胞を治療領域へ送達することができる。例えば、脊髄路に沿って両側的に複数の注射が行われる可能性ある、ヒトの神経変性状態の対象に対する治療では、0.1の注射量および注射部位当たり約100/μlを使用することができる。
【0124】
所望の領域に細胞を注射するのに適したいかなる器具も、本開示の方法で使用することができる。一実施形態では、ほぼ一定の流速で、ある時間にわたって、マイクロリッター以下の量を送ることができる注射器を使用する。針もしくは柔軟な管、または他のあらゆる適した転送器具を通して、細胞をこの器具に充填することができる。
【0125】
一実施形態では、神経変性状態の治療における所望の注射部位は、脊髄の少なくとも一領域を含む。一実施形態では、細胞を、脊髄の頸椎、胸椎または腰椎部などの、脊髄の少なくとも1カ所の特定の分節または部位に移植する。例えば、腰椎部では、わずか5対の神経根が椎骨の脊椎管と交差し、各対の神経根は、各腰椎の高さで脊椎から出て、広い領域に分布している。脊髄の腰椎部における神経根の密度が低いことにより、腰椎部は、特に細胞の安全な注射部位としてよく適している。一実施形態では、細胞を脊髄の軟部組織(parenchyma)の中間帯に移植する。
【0126】
一実施形態では、細胞を約5から約50の部位に注射する。一実施形態では、細胞を脊髄の各側の約10から約30の部位に注射する。少なくとも2カ所の部位は、約100μから約5000μの間隔を離すことができる。一実施形態では、注射部位間の間隔は、約400μから約600μである。注射部位の間隔は、脊髄分節の全体を通して発生する、実質的に中断されておらず、連続したドナー細胞の存在に基づいて、さらにラットまたはブタなどの動物モデルで、約2から3カ月の生存を達成することを示した平均注射量に基づいて決定することができる。一実施形態では、痙縮/硬直などの症状を治療するために、または運動ニューロンの生存に有効な、少なくともいくつかの腰椎部の長さにわたり、脊髄正中の両側に沿って細胞を注射する。ヒトにおける注射の実数値は、動物モデルの結果より推定することができる。
【0127】
一実施形態では、標的の注射部位は、脊髄の灰白質である。灰白質内で、ラミナ(lamina)の特定の高さにNSCを注入するように注射針の先端を配置することができる。例えば、痙縮/硬直を治療するためにGABA/グリシンを産生するニューロンを送達することは、ラミナV〜VIIを取り囲む領域にNSCを送達することである。あるいは、神経障害性疼痛または慢性疼痛を治療するために、頸椎から腰椎までのさまざまな脊髄分節の灰白質後角またはその近接にNSCを送達することができる。あるいは、ALSなどの運動ニューロン疾患を治療するために、頸椎から腰椎までのさまざまな脊髄分節の灰白質前角またはその近接にNSCを送達することができる。
【0128】
本開示の方法の細胞は、in vivoで多数のニューロンを発生させることができる。NSCが移植前に明白に分化前ではないとき、NSCを分化する前に、in vivoで最高で2つから4つの細胞分裂まで増殖させ、それによって、さらに有効なドナー細胞数を増加させることができる。分化すると、ニューロンは特定の神経伝達物質を分泌する。さらに、このニューロンは、in vivoで移植片を取り囲む環境(milieu)に対して、異なる状態に有益な増殖因子、酵素、およびその他のタンパク質または物質を分泌する。したがって、さまざまな状態が本開示の方法によって治療できるのは、移植した細胞がin vivoで多数のニューロンを発生する能力によるためと、神経変性状態が、ニューロン由来の要素などの要素が失われていることが原因であるか、これらを結果的に失ったことによるためである。したがって、増殖因子、酵素およびその他のタンパク質などのようなニューロン由来の要素を欠いていることによりCNS組織の変性を患う対象を、本開示の方法によって効果的に治療することができる。
【0129】
移植したニューロンによって分泌される増殖因子、酵素、およびその他のタンパク質または物質に反応する状態は、テイサックス病、ニーマンピック病、バッテン病、クラッベ病、運動失調、およびその他の状態などの、遺伝性リソソーム病を含む。
【0130】
さらに、治療に関する本開示の方法は、in vitroで増殖して、損傷もしくは変性したニューロンを交換し、他のニューロンに対して抑制効果もしくは刺激効果をもたらし、および/またはニューロンの再生に寄与する栄養因子を放出することができる、細胞を移植することを含む。
【0131】
実施形態は、さらなる運動ニューロンを、損傷または変性したニューロンの代替として供給することを含む。例えば、本開示の方法は、空洞化を埋めるために、脊髄の空洞内に十分な神経の基礎構造を供給することを含む。神経の基礎構造は、外傷性脊髄損傷、遺伝性疾患または他のあらゆる原因から生じる脊髄空洞症に関連する空洞の拡大を減速させることができるならば十分である。また、十分な神経基礎構造を提供することは、脊髄が変性することにより生じるさらなる合併症の軽減に役立つことを認識すべきである。
【0132】
全てのNSCが既知の疾患に対して治療的に作用するわけではない。異なる疾患で影響を受けるニューロン集団の種類は、異なる可能性がある。したがって、治療的に有効なNSCのドナー集団は、失われた神経の要素を交換することに寄与する。例えば、痙縮、発作、運動障害およびその他の筋肉活動亢進障害の治療は、GABAまたはグリシンを産生する抑制性ニューロンに分化することができる治療効果のある細胞量を供給することを含み得る。異なるNSC集団は、分化したニューロン表現型を調べることによってin vitroで評価することができる。それから、in vitroでの分化パターンを、in vivoでの適切な表現型を産生する細胞の有効性を予測するために用いる。これは、適切な神経伝達物質の表現型に関してだけでなく、ニューロンの適切な形態、遊走およびその他の表現型の特性に関しても予測される。
【0133】
一実施形態では、症状の病因と関連して損傷および破壊したニューロンのサブタイプに対応するニューロンのサブタイプを産生することができるNSCを移植する。例えば、対象の興奮性回路(excitatory circuit)の活動亢進は、脊髄の外傷、胸部/胸腹部の大動脈手術、脳卒中、てんかん、脳外傷、ハンチントン舞踏病、膀胱性尿失禁、便通の亢進、および損傷または遺伝的症状から生じるその他の制御不能の筋肉収縮による、遺伝的症状またはニューロンの損傷によって引き起こされる可能性がある。痙縮、発作またはその他の活動亢進は、多くの異なる病因の起源による脊髄とは対照的に脳で生じる。例えば、焦点てんかんは、GABAの発揮する回路に対する調性制御(tonal control)が失われることによる、非制御の活動亢進から発症すると考えられる。このため、本開示の方法は、in vitroで増殖されたNSCの移植によって、影響を受けた領域にGABAまたはグリシンなどの抑制性神経伝達物質(inhibitory neurotransmitter)を供給することを含む。例えば、痙縮、発作およびその他の活動亢進の場合、痙縮、発作およびその他のニューロンの活動亢進に関連する少なくとも1つの活動亢進の神経回路を弱めるために、GABAまたはグリシンを産生するニューロンなどの抑制性ニューロンに分化することができる多くのNSCを、移植されるin vivoで発生させる。したがって、本開示の方法は、てんかんおよび同様の発作の状態を治療するために適用することができる。
【0134】
また、本開示の方法は、不全麻痺、完全麻痺、痙縮、硬直またはその他全ての脳虚血によって生じる運動、言語もしくは認識症状を治療するために適用することができる。脳虚血は、脳における脳卒中発作、または脳への血液循環がかなりの時間、中断される心臓発作により生じる可能性がある。したがって、前述の脊髄虚血と類似している。一部の脳卒中の対象は、記憶喪失、完全麻痺または不全麻痺などの他の障害に加えて、中枢の起源の発作を発症する。また、これらの脳虚血による障害は、海馬および/または他の脳領域における抑制性介在ニューロンの選択的な減少による可能性がある。したがって、本開示の方法は、不全麻痺、完全麻痺、痙縮、またはその他の運動、言語もしくは認識症状を患う脳卒中の対象を治療するために適用することができる。
【0135】
不全麻痺、弛緩性対麻痺、およびALS、外傷性脊椎損傷、虚血性傷害または遺伝性疾患に起因する筋収縮の制御不能などの、筋収縮の制御不能に関連するその他の状態の場合、本開示の方法は、運動ニューロンの消失を遅らせるために、十分な栄養的影響を発揮するニューロンの移植を提供することを含む。特に、本開示の方法は、最適の生物学的利用度の条件下で、変性する運動ニューロンを送達することができる栄養分子を分泌する移植NSCの能力を促進する。そのような栄養分子は、スーパーオキシドジスムターゼ(SOD1)などのエキソサイトーシスにより分泌されたスーパーオキシドジスムターゼ、リソソーム酵素、および細胞産生の抗酸化物質などの非タンパク質性分子を含む。移植した細胞によって分泌されるその他の栄養因子は、グリア細胞株由来神経栄養因子(GDNF)、脳由来神経栄養因子(BDNF)、血管上皮増殖因子(VEGF)、プレイオトロフィン、血管内皮増殖因子(VEGF)、エリスロポエチン、ミッドカイン、インスリン、インスリン様増殖因子1(IGF-1)およびインスリン様増殖因子2(IGF-2)、または他の全ての有益な栄養要素を含むことができる。
【0136】
広範囲にわたる神経変性状態を治療するために、本開示の方法の能力に寄与するもう1つの要因は、存在するニューロン線維に沿って広範囲に移動するNSC分化細胞の能力を含む。移植した細胞の移動は、ドナーニューロンおよび/または神経膠の全体的な分布および統合、ならびにそのような細胞によって分泌される治療要素の全体的または分散した供給に結び付く。
【0137】
細胞の広範な移動は、それを必要とする対象の神経系および身体の全体を通して、主要な治療用のタンパク質および物質の全体的および安定した送達を可能にする。したがって、本開示の方法の細胞は、治療用のタンパク質および物質のための有効な送達媒体である。そのような送達の目的のために、本開示の方法は、腸、筋肉、血管内系および皮下部位を含む神経系以外の領域に加えて、CNS実質、脳室、硬膜下腔、クモ膜下腔、硬膜外腔、末梢神経系部位を含む神経系内のさまざまな部位へ細胞を移植することを含む。
【実施例】
【0138】
(実施例1:ヒト脊髄神経幹細胞/前駆細胞の増殖)
在胎齢およそ7〜8.5週の少なくとも1つのドナーからの脊髄を得る。機械的粉砕を用いて、Ca++およびMg++を含まないリン酸緩衝生理食塩水中、該脊髄の単一の近接組織を解離させる。次いで、得られた細胞懸濁液を、ポリ-L-オルニチンまたはポリ-D-リシンおよびヒトフィブロネクチンまたは他の細胞外基質タンパク質によって予備コーティングした組織培養プレートに播種する。組織培養処理したプレートまたはフラスコを、100μg/mlのポリ-D-リシンと共に、室温で1時間インキュベートした。次いでそれらを水で3回洗浄し、乾燥した。次にそれらを25mg/mlと共に、室温で5分間インキュベートした。時には、10mg/mlのフィブロネクチンを、室温で1時間用いた。時には、1mg/mlのフィブロネクチンを、37℃で18時間用いた。N2(DMEM/F12 + インスリン、トランスフェリン、セレニウム、プトレシン、およびプロゲステロン)からなる培養培地に、1種のヒト組換え塩基性線維芽細胞成長因子(bFGF)を添加した。一実施形態において、0.1ng/ml〜100ng/mlの範囲を使用することができる。一実施形態において、最適には、10ng/mlのbFGFが使用される。
【0139】
得られた最初の培養物は、単層中、有糸分裂後ニューロンおよび増殖性NSCからなる。引き続き、およそ5日からおよそ20日培養後、分裂性のネスチン陽性NSCは、非分裂性ニューロンまたは緩徐分裂性神経膠より優勢となる。これらの培養条件下、NSCを増殖のために選択的に促進させる。増殖NSC集団をトリプシン使用などのマイルドな酵素処置によって継代接種する。該細胞を、無血清または実質的に無血清の培地中で培養する。低濃度の血清は該細胞により忍容され得るが、血清はNSCのグリア分化を促進するLIFおよびCNTFなどの多くのサイトカイン類を含有しているため、該細胞を血清に曝露することは避けることが最良である。したがって、継代接種の間、血清ではなく、トリプシン阻害剤などの特定の酵素阻害剤を添加することによって、使用酵素を抑制する。各継代接種時に、採収した細胞数をカウントし、さらに増殖させるために、フラクションを再播種する。図1に示されるように、本発明の方法を用いて、ヒトNSCを、それらの増殖性および分化性を維持しつつ、個体数が1018倍超増加する増殖をさせることができる。該細胞は再現可能に増殖できる。図1に示されるように、該細胞の連続的継代を3回繰り返したところ、再現性のある細胞増殖曲線および細胞倍増時間が得られた。増殖の間、ほとんど全ての細胞が有糸分裂神経上皮細胞のin vivoマーカーであるネスチンを発現し、タイプ3ベータチューブリンおよびGFAPなどの分化ニューロンおよび神経膠の抗原を有さない。また、細胞は、committed neuronal progenitorの可能性があるPSA-NCAMに関する免疫染色、希突起膠細胞のマーカーであるO4およびGalC、ならびに放射神経膠のマーカーであるRC2でも陰性である。したがって、免疫染色によって判定したところ、NSCは長期増殖期間にわたって、それらの抗原プロフィルの発現を安定に保持する。形態およびネスチン発現の例はそれぞれ図2Aおよび2Bに示されている。
【0140】
(実施例2:ヒト脊髄神経幹細胞/前駆細胞の分化)
NSCの増殖中、任意の時点で、培養物中のbFGFなどのマイトジェンを取り出すことによって、培養物を分化させることができる。マイトジェン除去後、約1〜3日以内に、引き続きNSCの分化が起こり、別な異種細胞形態が明らかである。分化のおよそ4〜7日までに、MAP2c、タウ、およびタイプIIIベータチューブリンなどのニューロン特異的抗原を免疫染色によって可視化することができる。およそ12〜14日までに、細胞内タンパク質輸送の明らかな局在化と共に、培養物全体にわたる、長形、索状の軸索性過程が明らかになる。およそ28日までに、シナプシンおよびシナプトフィジンなどのシナプスタンパク質は軸索終末内へ局在化し、斑点状の染色として現れる。該ニューロンの長期成熟をさらに促進するために、星状膠細胞のフィーダー層をさらに供給することができる。図3に示されるように、ヒト脊髄NSCの分化によって、ニューロンと神経膠との混合培養物が生成され、混合物中、ニューロンはタウ、MAP2ab(A)およびタイプ3ベータチューブリン(B)などのニューロン特異的抗原を強く発現し、該培養物のおよそ50%を含む。図3Bに示されるように、該培養物は、数センチメートル伸長する長く束ねられた軸索ケーブルを自然に生成させる。図3Cに示されるように、ニューロンのかなりの部分がGABA作動性である。コリン作動性運動ニューロンもまた該培養物中に存在する(図3D)。培養物中かなりのGABAニューロンが存在することにより、一定の回路におけるGABA産生の減少に起因する種々の神経学的状態の治療に対するヒト脊髄NSCの有用性が予想される。同様に、コリン作動性ニューロンの存在により、ヒト脊髄NSCが運動ニューロン分化の能力を有することが実証され、運動ニューロンの漸進的変性に起因する種々の運動ニューロン疾患の治療に対する有用性が予想される。治療のためには、NSCをさらなる表現型増強条件の有無で増殖させ、採収し、欠損神経領域に注射する。
【0141】
(実施例3:ヒト中脳神経幹細胞/前駆細胞の増殖)
在胎齢7〜8.5週胎児の1つの中脳組織を得る。実施例1に記載したとおり、中脳組織からNSCを得る。該細胞を、160日の正味培養期間にわたって連続的に継代し、得られた増殖を図4に示す。増殖期間を通して、NSCはそれらの多分化能および神経形成能、ならびに、ドーパミン作動性ニューロンをもたらす分化能を安定して保持する。ドーパミン作動性ニューロンは、チロシンヒドロキシラーゼ(TH)およびドーパミントランスポーター(DAT)のニューロン発現によって評価される。
【0142】
DAT発現はドーパミン産生ニューロンのマーカーである。ニューロン中のDAT発現は、培養物中の分化ニューロンのシナプス膜を越えて放射標識ドーパミンを輸送するその機能を測定することによって評価できる。分化したヒト中脳NSCおよび単クローン誘導したヒト中脳NSCからの培養物中のDAT機能は、放射標識ドーパミン取り込みアッセイによって評価される(図5)。該アッセイの結果により、ヒト中脳NSCの強い機能的ドーパミン作動活性が示される。それはさらに、NSCの単離モノクローナル集団によって、ドーパミン作動性表現型を増加することができ、これが特に、分化の際にドーパミン作動性ニューロンを高い割合へと増加させる傾向があることを示している(図5)。
【0143】
増加したドーパミン作動性ニューロンを生成するNSCは、パーキンソン病の治療に特に有用である。同様に、特定の表現型に関する分化を増強させるために、組織からの単離時点で予めプログラミングされたNSCを用いて、アルツハイマー病の治療に有用な前脳コリン作動性ニューロン、ALSなどの運動ニューロン疾患の治療に有用な脊髄コリン作動性ニューロン、うつ病の治療に有用なセロトニン作動性ニューロン、癲癇およびハンチントン舞踏病の治療に有用なGABA作動性ニューロンなどの他の特定の所望ニューロンを単離することができる。
【0144】
(実施例4:ヒト中脳神経幹細胞/前駆細胞の分化)
ヒト中脳NSC/前駆体は、実施例2に記載されるとおり分化させることができる。NSCの有糸分裂期の間、またはそれらの分化の間、種々の外来因子によって該培養物を処置することによって、所望の表現型の割合を高くすることができる。ヒト中脳NSCからドーパミン作動性表現型を増加させることができるこのような因子の例は、図6Aおよび6Bに示されるとおり、セルトリ細胞からの馴化培地によって実証される。
【0145】
図6Aおよび6Bに示される試験において、凍結保存された神経幹細胞を解凍し、マイトジェンの存在下、4ウェルチャンバースライド中、1ウェル当たり40,000細胞の密度で塗布し、6日間増殖させ、マイトジェンを除去して細胞を8日間分化させた。セルトリ細胞馴化培地(SCCM、N2a中1:1に希釈)への曝露のタイミング及び時間に基づいて、細胞を4つの群に分けた。1つの群は、増殖時および分化時、SCCMに曝露し(条件1);第2の群は、増殖時のみ曝露し(条件2);第3の群は、分化時のみ曝露し(条件3);第4の群は、SCCMに曝露しなかった(対照またはcont.)。培地は1日おきに交換し、マイトジェンは増殖期の間、毎日添加した。1条件当たり、4つのウェルを維持して複数マーカーに関して染色させ、3つの細胞系:796MB、527MB、および566SCを試験した。脊髄由来の566SC細胞は測定可能なTH陽性ニューロンを有さず、図は省略した。
【0146】
分化の際、4%パラホルムアルデヒドを用いて細胞を固定し、MAP2[MAP2abサブタイプを認識するAP20クローン(Sigma)]、ニューロン特異的βチューブリン[TuJ1(Covance)]、およびチロシンヒドロキシラーゼ(Pel-Freez)に対する抗体、ならびにGFAP(Dako)およびGalC(Chemicon)を用いて免疫染色した。免疫染色した細胞を、40×対物レンズを用いてカウントし、各ウェルに関して少なくとも3つの視野をカウントした。いずれの条件下で維持された細胞の分析の際にも、GFAP+またはGalC+細胞は、ほとんど、または全く検出されず、したがって、これらの抗原は分析から除外した。また、566SC細胞は、記載された条件下で維持した後、高濃度過ぎて定量化できなかったので、最終分析に含めなかった。
【0147】
この分析によって、外来因子による処置によって、ヒト中脳NSCが影響されることが示され、ドーパミン作動性ニューロンをさらに濃縮するのに有用なタンパク質因子または化学物質を探索することができることが示される。したがって、パーキンソン病などの特定の適応症の治療を目的として特に有用な集団を得るために、ヒトNSCを使用することにより、新規の合成/天然化学物質およびタンパク質因子を効果的にスクリーンすることができる。
【0148】
(実施例5:ヒト脊髄神経幹細胞/前駆細胞の移植によるラットにおける痙攣および硬直の処置)
一過性脊髄虚血を誘導するために、先にTaira(1996)に記載された方法を用いる。Sprague Dawley(SD)ラットをハロタン(1.5%)で麻酔する。2 Fr Fogarty(登録商標)カテーテルを左大腿動脈を通して、左鎖骨下動脈のレベルまで下行胸椎大動脈に通す。大動脈閉塞のレベル下の遠位動脈圧(DBP)を測定するために、ポリエチレン(PE-50)カテーテルによって、尾動脈にカニューレ挿入する。
【0149】
0.05mlの生理食塩水を用いて、動脈内バルーンカテーテルの膨張時に脊髄虚血を誘導する。動脈閉塞時の全身低血圧は、PE-50カテーテルによりカニューレ挿入した頚動脈からの動脈血(10.5〜11cc)の一部抜き取りにより再現する。およそ40mmHgの全身低血圧をこの方法によって誘導することができる。閉塞の有効性は、尾動脈で測定されるDBPの即時的および持続的低下によって証明される。脊髄虚血誘導のおよそ10分後、バルーンを収縮させ、頚動脈から抜き取った血液を再注入する。動脈血圧が安定化したら(再流動後約20〜30分以内)、動脈ラインを除去し、傷を閉じる。
【0150】
脊髄虚血の誘導後、運動機能の回復を、改変21ポイントオープンフィールドロコモータースケール(BBBスケール)を用い、およそ2日間隔で評価する。0〜4のBBBスコアを有する動物のみを、移植試験のための実験群用に選択する。
【0151】
虚血損傷の約7〜21日後、0〜4のBBBスコアを有する痙攣ラットを大気中、1.5〜2%ハロタンで麻酔し、脊髄ユニット装置内に入れる。次いで、Thl1〜L2脊椎の部分的椎弓切除を実施する。80〜100μmの先端直径を有するガラスキャピラリーを圧力制御微量注射器に接続する。1回の注射当たり5,000、10,000、15,000または20,000のヒト神経幹細胞/前駆細胞を含有する0.5μlの細胞懸濁液をラットに注射する。脊髄の各々の側(左および右)に、L2〜L6曝露セグメント間に均一に分布した、合計6〜8回の注射を、各ラットに投与する。注射の中心を、中央灰白質内(V層〜VII層)に標的化する(L3レベルにおける脊髄の背側表面からの距離:1mm)。移植後、切開部を3%H2O2およびペニシリン/ストレプトマイシン混合物で洗浄し、2層で閉じる。次いでラットを回復させる。
【0152】
脊髄移植の3日前に、全動物にFK-506 (Prograf;フジサワ;1mg/kg;i.p.)による免疫抑制処置を開始する。移植後、全動物は全生存期間の間、免疫抑制処置を毎日受ける。これら移植片の免疫拒絶は、FK-506によって効果的に防ぐことができる。該ラットはおよそ2または7週間生存する(各時点でn=5)。
【0153】
生存期間の最後に、ラットをペントバルビタール(40mg/kg;i.p.)によって麻酔し、ヘパリン化生理食塩水で1〜2分間、引き続き0.1Mのリン酸緩衝液(PB)中4%パラホルムアルデヒドによって経噴門潅流する。脊髄を切開し、同一固定液中、4℃で一晩後固定する。後固定後、脊髄組織を段階的蔗糖液(10、20および30%)中で合計3日間凍結保護する。次いで、凍結した冠状、側矢状または水平状の脊髄切片(10〜30μm)を切断する。免疫組織化学では、遊離浮遊切片(30μm)を、5%正常ヤギ血清(NGS)、0.2%トリトンX100(TX)を含有するPBS、0.1M(pH=7.4)中に、室温で2時間入れ、非特異的タンパク質活性を阻止する。この後、種々のヒト一次特異的抗体と共に、4℃で一晩インキュベーションする。
【0154】
一次抗体と共にインキュベーションした後、切片をPBS中で3回洗浄し、蛍光マーカーが結合したヤギ二次抗ウサギ抗体またはマウス抗体(Alexa 488または594;4μl/ml;Molecular Probes)と共にインキュベートする。ブロッキング調製物および抗体調製物は全て、0.1MのPBS/0.2%TX/5%NGS中で作製する。二重標識実験では、種々の種からの一次抗体を同時に適用し、引き続いて、種々の蛍光マーカーに結合させた二次抗体を適用する。対照実験では一次抗体を省略する。一般的な核染色では、最終二次抗体溶液にDAPI(3μl/ml)を添加する。染色後、切片を室温で乾燥し、Prolong 抗退色キット(Molecular Probes)で覆う。
【0155】
Leica 蛍光顕微鏡を用いてスライドを分析する。Olympusデジタルカメラで画像(512×512ピクセル)をキャプチャーし、Adobe Photoshop 5.5(Adobe Systems、マウンテンビュー、カリフォルニア州)によって処理する。二重染色切片における種々の抗体の同時局在化を確認するために、Nikon顕微鏡に据えつけた測光CCDを含むDelta Vision 逆応答顕微鏡システム(Applied Precision社)によって画像をキャプチャーする。一般に、0.1μmまたは0.2μm離れた60個の光学切片を取る。使用されるレンズは20×、40×および60×(NA 1.3)である。データセットを逆応答させ、Silicon Graphics Octane ワークステーション上、SoftWorxソフトウェア(Applied Precision社)を用いて分析する。
【0156】
立体解析学的で偏りのない系統的なサンプリングを用いて、ヒト核NUMA抗体に対して免疫反応性の移植ニューロンの総数を予測する。L2〜L6脊髄セグメントから採取した、先に染色した各々10番目の切片を、fractionator sampling schemeを適用後、立体解析学的定量化のために使用する。アパーチャー数値1.3を有する100×油浸漬対物鏡を用い、Leica DMLB顕微鏡によって、視像(1μmの厚さ)を得る。デジタルカメラ(Olympus)、および、StagePro制御電動式Zステージ(Media Cybernetics)に付属のImageProソフトウェア(Media Cybernetics)を用いて、視像をキャプチャーする。移植細胞の総数は、fractionator formula:N=Q×1/hsf×1/asf×1/ssf(式中、Nは陽性核の総数、Qはカウントした細胞の合計、hsfは高さのサンプリング分数、asfは面積のサンプリング分数、およびssfはスライスサンプリング分数である)を適用して算出する。
【0157】
虚血脊髄における移植したヒトNSCの三次元再構成視像を提供するために、連続的に切断した脊髄から採取した、先に保存した画像を使用する。三次元再構成のために、平均して約60〜100の連続切片を用いる。第1の工程で、Ellipseソフトウェアを用いて、連続画像のスタックを開き、特注開発されたAllignモジュール(VidiTo、SK)を用いてアラインさせる。アラインメントの方法は、全ての連続脊髄画像における2つの形態学的参照点を規定すること(第1の点:中央カナルの中心;第2の点:背角の内側境界)、および引き続いて、全画像のコンピュータ処理アラインメントからなる。次いで背角と腹角の境界を確認するために、Laminar Mapsモジュール(Ellipse)を用いて先にアラインした画像のスタック上にラインを引く。最後に、3-Dコンストラクター(Media Cybernetics)を用いた三次元再構成のために、先にアラインし、層状マップ標識した画像のスタックを用いる。
【0158】
ラット星状膠細胞上ヒト脊髄NSCの2週間から3週間の培養により、培養物の大多数において、ニューロン表現型の時間依存性の成熟および発達が示された。これは、NSEまたはMOCに対するヒト特異的抗体による染色によって確認される。十分に発達した軸索樹状突起間樹を有する多数のニューロンもまた確認される。NSE陽性ニューロンの大多数(85〜90%)はGABA陽性である。軸索および樹状突起におけるシナプトフィジンの発現が、いくつかのMOC陽性ニューロンにおいて見られる。
【0159】
虚血損傷の21日後に移植した場合、細胞の強健な生存が見られる。これは、全てがヒト特異的な抗体のNUMA、MOC、またはNSEに対して免疫反応性である、明白に確認される両側性の移植片として発現する。移植された脊髄セグメントから採取した水平切片の分析により、個々の注射部位間にNUMA陽性細胞の明らかな移動が示される。NUMA陽性細胞の大多数は、MOC免疫反応性との同時局在化を示す。
【0160】
共焦点顕微鏡によって分析されたNUMAおよびGABA抗体による脊髄切片の二重標識化により、平均25〜35%のGABA陽性細胞が明らかになる。7週間生存時の全ての移植動物に、GABA作動性表現型の一貫した発現が見られる。
【0161】
同時点(すなわち、移植後7週目)でのシナプトフィジンおよびNUMA抗体による二重染色により、移植片内に高密度のシナプトフィジン陽性網状構造が示される。
【0162】
NUMA陽性細胞はGFAP抗体による同時局在化を時々示すのみとなる。これらの細胞は移植片周囲に典型的に局在化する。
【0163】
NUMA陽性細胞の立体解析学的予測により、個々の移植片内に平均75,460±5,697の残留移植細胞が示される。これは、もともと注射されたものよりも平均3〜3.6倍多い細胞である。細胞周期抗原のKi67は活性有糸分裂細胞のマーカーである。移植後2週間目または7週間目における脊髄切片の染色により、移植後わずか2週間目で、hKi67免疫反応性が示される。生存7週間目では、Ki67陽性細胞は時折存在するのみである(1〜2細胞/10切片)。これらの結果により、移植したヒト脊髄NSCおよびそれらの後代は、最初の2週間は平均約3回の細胞倍増に相当する数で増殖し、次いで分裂終了となり、安定して組み込まれることが示される。
【0164】
MOC抗体およびDAPIにより染色した40μmの厚さの連続脊髄切片(総数150〜200切片)から取った画像を用いることによって、移植したL3〜L5セグメントのボリューム再構成を実施する。三次元移植片再構成により、灰白質の領域内に分布した、体軸方向に方向付けられたMOC陽性移植片が十分に認識されることが示される。図7および図8に示されるように、ヒト脊髄NSCの機能的効果は、該細胞を移植し、BBBスコアリングによって、運動利益の回復を測定することによって評価される。
【0165】
本試験において見られた挙動回復の程度に関して、移植後、3つの主要な群が見られた。第1のものは、もっとも強健な回復および歩行能力を示した動物(BBB<16)、第2のものは、下肢の3つの関節全ての活動的な運動性において改善を示したが、立つことはできなかった動物(BBBは8付近)、そして、第3の群は、何も回復を示さなかった動物(すなわち、非レスポンダー)。移植片に対する応答性の違いの理由は明らかではないが、阻害困難な主要求心性神経および/またはα運動ニューロンに関して、移植片位置のわずかな違いがある役割を演じていると我々は推測している。さらに、本試験では、動物がわずか3カ月しか生存しなかったことに注意すべきである。移植後の長期的生存および身体的回復の継続は、高程度の機能的回復に関連している可能性が高いと我々は推測している。それにもかかわらず、処置群とは対照的に、培地のみを注射された動物ではいずれも有意な回復は見られなかった。
【0166】
(実施例6:ヒト脊髄NSCの移植による運動ニューロン疾患の処置)
開示された方法のNSCは、強力で顕著な臨床的利益と生物学的利益の双方を与える。そのため、開示された方法により、ALSなどの中枢神経系全体に散在した状態、ならびに上記の脊髄虚血などの特定の領域に局在化した状態の双方の処置が可能になる。ALSにおいて、腰髄における移植では、セグメントの運動器官の他の重要な部分、すなわち、呼吸運動を担っている頚髄運動ニューロンカラムが除外され得るが、脊髄にNSCを移植する開示された方法は、髄全体にわたって宿主の運動ニューロンに広範囲の作用が生じ得るCSF内への、移植した細胞からのBDNFおよびGDNFならびに他の因子の放出を促進する。
【0167】
驚くべきことに、神経変性脊髄環境の腰髄セグメント内へのヒトNSCの部分的移植片は、生存し、大量に神経分化し、運動ニューロンの生存を促進して移植部位と他の場所の双方で機能することが判明した。NSCは、ヒトALS(筋萎縮性側索硬化症)のモデルであるSOD1 G93Aラットの症状の発症を有意に遅らせ、それらの寿命を延ばす。
【0168】
SOD1 G93Aラットは、神経病理学およびALSの特に攻撃的な形態の臨床的症状の包括的モデルである[(Nagaiら、2001年;Howlandら、2002年)]。ニューロンの広範な分化が生じ、引き続き、分化したニューロンが宿主ニューロンとシナプス接触を形成し、GDNFおよびBDNFを発現し放出するSOD1 G93Aラットおよびマウスの腰髄内に、ヒト胎児脊髄のNSCを移植することができる。例えば、劇症性運動ニューロン疾患を特徴とするラットSOD1 G93Aモデルを、該疾患におけるNSCの有益な作用を試験し実証するために用いることができる。このため、開示された方法で使用されたNSC移植片は、神経変性環境において十分に生存し、強力な臨床的効果を発揮する。これらの効果の少なくとも一部は、運動ニューロン成長因子を発現し放出するこれらの移植片の能力に関連している。したがって、腰突起に限定された制限された移植スケジュールにも関わらず、開示された方法の移植NSCは、劇症性運動ニューロン疾患の発症および進行を遅らせ、これらの動物の寿命を10日超延ばす。
【0169】
移植前に、線維芽細胞成長因子(FGF-2)を含有する無血清培地において、8週在胎齢の死後ヒト胎児の脊髄組織からのヒトNSC(NSI-566RSC)を約10〜12継代増殖させる(Joheら、1996年)。これらの細胞の結果をヒト核抗原(HNu)に対する抗体によって確実に追跡する(Yanら、2003年)。これらの細胞を用いる全ての外科的操作は、本明細書に参照として組み込まれている、Johns Hopkins Medical Institutionsの動物の管理および利用委員会によって承認されたプロトコルに従って、ガス麻酔(エンフルラン:酸素:亜酸化窒素=1:33:66)および無菌的方法を用いて実施される。
【0170】
顕微鏡先導下、Kopf脊髄定位ユニットに乗せた雌雄混合の9週齢(220〜300g)SOD1 G93Aラットの腰プロチュバランス(L4 & L5)内に、生きているNSCまたは死んでいるNSCを移植する。死細胞は、移植前に、凍結と解凍の反復により調製する。細胞懸濁液は、サイラスティックチューブを介して、10μlのHamilton微量注射器に連結したプルド面取りガラスマイクロピペットを用い、腹側角の両側の腹側角を目標に(注射部位1つ当たり5×104 NSC、片側に4つの注射部位)、およそ8回の注射によって、無菌条件下、送達される。非処置動物またはシクロスポリンを受けている動物で移植片生存が1カ月を超えないことを示しているパイロットデータに基づき、免疫拒絶を防ぐために、全ラットにFK-506(1mg/kg i.p.)を与えた。
【0171】
ラットを、運動の強さに関して試験し、週に2回体重を量る。運動の強さ試験は、Basso、BeattieおよびBresnahan(BBB)運動等級評価(Bassoら、1995年)、および傾斜平面評価(RivlinおよびTator、1977年)を含んだ。BBBスコア試験では、動物をオープンスペースで、約4分間または5分間試験する。全ての運動機能を記録し、スケールに従って評価する。傾斜平面試験では、ラットを傾斜平面マットの上に置き、ラットの位置が約5秒間安定化できる最大の箇所まで角度を調整する。次いでこの角度を、その動物の傾斜平面スコアとして記録する。BBBおよび傾斜平面スコアを、hoc試験後、MANOVA、引き続き、Fisher LSDによって解析する。疾患の発症は、体重が急激に減少し始める点として定める。移植タイプ(生細胞移植片または死細胞移植片)の効果としての疾患経過は、(スチューデントのt検定によって)2つの群間の発症時の年齢および死亡時の年齢を比較することにより、ならびにKaplan-Meier生存解析、引き続いて長期ランク試験により解析する。
【0172】
ラットのBBBスコア(下記参照)が3未満で、1つの関節のみが動くか、または全く動きがなく、その動物が瀕死状態と考えられる段階で、潅流-固定により安楽死させる。
【0173】
4%の中性緩衝パラホルムアルデヒドによって潅流させた動物からの組織を調製する。付着根を有する胸-腰脊髄セグメントおよび腰神経を、同一の固定剤中、さらに4時間浸漬することによってさらに固定する。移植領域全体およびその上下の縁1mmを含むブロックを冷凍保護し、さらなる処置のために冷凍する。L3-S1根を、全体を着けた調製物として別々に処理するか、またはガラスピペットの熱凝固端を用いて細根を分離した後に処理する。ブロックを横断面または正中面(35μm)で切片化する。NSC生存および分化を、多くの場合、ヒト特異的マーカー、HNuと他の細胞マーカーとを組み合わせた二重標識免疫蛍光によって試験し、Yanら、2004年に記載されているとおりに実施する。
【0174】
NSC分化を試験するために、HNu(+)細胞の総数をカウントする非立体解析学的方法、ならびに我々の免疫蛍光調製物からランダムに選択した高強力(100×)フィールド上のHNuおよび表現型マーカーによって二重標識した細胞を用いる。移植領域にわたっておよそ1mm離れている6つの切片の各々における1つのフィールドを、各動物から用いる。各場合から、カウントした6つのフィールド全てからのHNu(+)の数および二重標識のプロフィルをプールし、1つの実験プロトコルごとに群分けする。各処置群(一群当たりn=6)に関して、単一および二重標識細胞の平均数を出す。
【0175】
生細胞または死細胞を移植したラット(各n=4)における運動ニューロンの生存を評価するために、128日齢で殺処置したラットの組織を評価する。立体解析学的要件(Yanら、2004年)に関して各ラットからのL3-S1部位における6つの切片おきにサンプリングし、明確な核および神経細胞体の直径が>35μmを有する多極細胞として同定されたα運動ニューロンを、Yanら、2004年に記載されたとおり光学精留塔によってカウントする。生細胞を移植した動物対死細胞を移植した動物間の違いを、スチューデントのt検定によって解析する。
【0176】
運動神経栄養因子のELISA判定のため、ガス麻酔下、動物の第4脳室から25G注射器によってCSFをサンプリングする。移植部位およびそれらに隣接する領域を含有する組織サンプルを、1mmの厚さの新鮮脊髄スライスから横断切開する。CSFまたは組織サンプルを処理し、総タンパク質を、Shengら、2003年に記載されたとおりに先ず測定する。E-Max免疫アッセイシステム(Promega、マジソン、ウィスコンシン州)を用いて、CSFおよび脊髄サンプル中のGDNFおよびBDNFの濃度を測定する。TMB-クロモゲンの吸光度を450nmで読み取る。生移植片、移植片隣接領域および死細胞移植片からのサンプル間における濃度の分散を、一元配置ANOVA、引き続き、チューキーの多比較hoc後検定によって解析する。生細胞移植動物対死細胞移植動物間のCFS濃度の差を、スチューデントのt検定によって解析する。
【0177】
運動神経栄養因子をウェスタンブロッティングするために、ELISAに関して調製したCSFまたは脊髄からのタンパク質サンプルを、分子量マーカーと共に電気泳動させ、ニトロセルロース膜に移す。ブロットを、5%のロバ血清を含有するTBS、pH7.4中でブロックし、次いで、先ずGDNF抗体およびBDNF抗体(1:500;一晩、4℃)中、次いでHRP結合ロバ抗ヤギIgG(GDNFに関して)および抗ウサギIgG(BDNFに関して)(1:2000;Jackson ImmunoResearch)(1hr、RT)中でインキュベートする。抗体は全て、5%のロバ血清を含有するTBS中で希釈する。ブロットをSuperSignal Chemiluminescent Substrate(Pierce)によって展開し、Kodak-XARフィルム(Eastman Kodak、ロチェスター、ニューヨーク)に曝露する。次いで、ブロットをストリップし、βアクチン抗体(1:500、Sigma)およびHRP結合ロバ抗マウスIgG(1:10000、Jackson ImmunoResearch)によって再ブロットする。免疫反応性のバンドをBio-Rad Quantity Oneソフトウェア(Bio-Rad Laboratories、ヘルキュールス、カリフォルニア州)によって解析する。バンド密度比(GDNFまたはBDNF:βアクチン)を動物ごとに算出し、群の平均値を、ELISA実験の場合のように、統計解析に入れる。
【0178】
移植後22週目のSOD1 G93Aラットの脊髄におけるヒトNSCを、ヒト特異的HNu抗体を用いる免疫染色によって同定する。HNu(+)細胞は、腹角(A)内で生存し、それらの大多数が、微小管関連エピトープであるTUJ-1などのニューロン系列マーカーによって染色されることが示される。ヒトNSCは、それらのヒト核タンパク質(HNu)識別特性によって同定され、それらの表現型の結果は、HNuに関する二重免疫細胞化学、ならびに神経前駆細胞、ニューロン細胞および膠細胞に対して特異的なエピトープによって追跡される。SOD1 G93Aラットの実験の最後に、ヒトNSCは、強健な移植および優れた長期生存を示す。HNu(+)細胞の大多数(70.4±6.4%)は、TUJ-1のそれらの同時局在化に基づいたニューロン系列へと分化する。HNu(+)細胞のおよそ5分の1(19.2±5.6%)がネスチンと同時局在化し、ごくわずかな(1.3±0.9%)HNu(+)細胞がGFAPに関して陽性であった。
【0179】
宿主の循環内に組み込まれるヒトNSCの能力を、移植片/宿主細胞に関する神経細胞体マーカーおよび宿主または移植片終末に対して選択的なマーカーによって試験する。移植片の出所を確定するためにHNuに関して、ニューロン分化を確定するためにTUJ-1に関し、ラットおよびマウスのエピトープを認識するが、ヒトのエピトープは認識しない前シナプスタンパク質Bassoon(BSN)に関してはモノクローカル(monoclocal)抗体に関して、切片を染色する。柔組織の場における多数のHNu(+)細胞、TUJ-1(+)細胞は、ラット出所であるシナプスボタンに接触することが分かる。
【0180】
共焦点顕微鏡において、HNu(+)核およびTUJ-1 (+)細胞質を有するNSC由来ニューロン細胞は、ラット終末に接触する。反対に、TUJ-1およびヒト特異的シナプトフィジンによって染色された調製物は、宿主のニューロン、特に大型および小型の運動ニューロンに対する小型ボタンの高密度の終末フィールドを現す。宿主の運動ニューロンは、多数の移植片由来のボタンに接触する。HNuおよびヒトNF-70に関して染色された横断切片は、移植片を左方に残し、前索の灰白質に沿って優先的に進む移植片由来の多数の軸索を示す。ChAT免疫反応性を有する細胞/方法を用いて、腹角における白質から灰質を描写する。神経フィラメントエピトープNF70に対するヒト特異的抗体によって標識化された多数の軸索が移植部位に関して見られ、多くのヒトNSCがプロジェクションニューロンへと分化することを実証し;これらの軸索は腹角の白質を優先させることを示している。
【0181】
SOD1 G93Aラットの腰髄内へのNSC移植片は、寿命を延ばし、運動ニューロンの死滅ならびに疾患の発症および進行を遅らせる。生細胞(L)移植片および死細胞(対照、C)移植片を用いた場合における臨床的および病理学的尺度の進行分析は図9に示されている。生きているNSCを移植した動物は、Kaplan-Meier分析とエンドポイント分析の双方で有意な生存の増加を示した。Kaplan-Meierプロット(図9A)は、観察の過程を通して、実験動物と対照動物との間の有意な区別を示す(P=0.0003)。BBBオープンフィールドおよび傾斜平面試験スコア(図9B)の時間プロットは、死んでいるNSCを受けた動物に比較して、生きているNSCを移植した動物では、筋肉衰弱における進行が有意により遅いことを示している。
【0182】
Tgラットの腰プロチュバランス(L3〜S1)における運動ニューロン生存に及ぼすNSCの効果を、生きているNSCまたは死んでいるNSCを与え、128日齢で安楽死させた小群の動物で調べる。死んでいるNSCを移植した動物では、平均寿命が138日であり、一方、生きているNSCを移植したラットは149日間生きる。このように、寿命における有意な11日の違いが、実験ラットと対照ラットとの間に生じる(P=0.0005)。死細胞を与えた動物では、疾患発症までの時間の平均は115日であり、生きているNSCを移植した動物では、122日である。これら両群間には、疾患発症までの時間において、有意な7日の違いが見られる(P=0.0001)。
【0183】
立体解析学的に見積もったα運動ニューロンの数は、生いているNSCを与えた動物では6,418、死んでいるNSCを移植したラットでは3,206であり、すなわち、同年齢の対照動物に比較して、実験動物の腰プロチュバランスには2倍多いニューロンが存在する。生きているNSC群と死んでいるNSC群との間の腰プロチュバランスにおける3,212個の細胞の違いが、128日齢における代表的な実験ラットと対照ラットに見られる(P=0.01)。
【0184】
ヒトNSCによってもたらされる変性運動ニューロンに対する神経保護の可能性のある機構としては、哺乳動物の運動ニューロンに対する古典的な栄養効果を有する2つのペプチドの発現および放出が挙げられる[BDNFおよびGDNF](Hendersonら、1994年;Koliatsosら、1993年)。移植を行ったSOD1 G93Aラットの脊髄におけるGDNF およびBDNFの発現および放出を判定する。ウェスタンブロッティングおよびELISAによって、BDNFおよびGDNFに関して、脊髄調製物およびCSFサンプルを評価する。生細胞を移植したラット(L1およびL2)および死細胞を移植した動物(C)の柔組織およびCSFにおけるGDNF濃度をELISAによって判定する。L1は、移植部位全体の濃度を表し、一方、L2は1セグメント上または下の組織における濃度を示す。群間の差異は有意であり、これはL1またはL2とC群間の大きな違いによって生じる。
【0185】
実験(生細胞、L)群と対照(死細胞、C)群との間のCSF濃度の違いは、t検定によっても有意である。ELISAは、生きているNSC移植動物の脊髄における移植部位では0.912±0.050pg/μgの濃度、1セグメント離れた部位では0.819±0.115pg/μgを示した。死んでいるNSCを移植した動物では、これらの濃度は、移植片を含有する脊髄セグメントにおいて、0.368±0.026 pg/μgであった。CSFでは、GDNF濃度は、実験動物で0.027±0.012 pg/μlであり、0.006±0.002 pg/μlであった。これらのデータは、生きているNSCを用いた動物のCSFにおいて、脊髄におけるGDNFの発現および放出が3倍増加し、GDNFの分泌が5倍増加することを示す。
【0186】
また、ウェスタンブロッティングは、生きているNSCを移植した動物におけるより高いノーマライズされたGDNF濃度を示す。GDNFウェスタンブロッティングによって、16kDaタンパク質の検出による増加のELISAパターンが確認される。また、ウェスタンブロッティングは、生細胞移植片では0.860±0.007、死細胞移植片では0.708±0.052のノーマライズされたGDNF濃度を示す。
【0187】
実験ラットおよび対照の実質組織およびCSFにおけるBDNFのELISA染色を判定する。ELISA 分析により、実験動物において、移植部位(L1)では0.086±0.014 pg/μg、移植片から1セグメント離れた部位(L2)では0.054±0.009 pg/μgの濃度が示される。対照ラットでは、移植含有セグメントにおけるBDNF濃度は、0.010±0.003 pg/μgであった。実験と対照のCSF濃度間の違いは有意である。CSFでは、BDNF濃度が、実験動物では0.041±0.013 pg/μl、対照では0.010±0.008 pg/μlである。これらの所見は、実験動物において、脊髄のBDNF濃度は8倍増加し、CSFのBDNF濃度は4倍増加することを示している。まとめると、ELISAデータは、生きているNSCを用いて移植した動物では、特にCSFでは、BDNFに比較してGDNFのより広範な分泌を示唆している。
【0188】
免疫化学もまた、移植したHNu(+)細胞の大多数がGDNFを発現していることを明らかにしている。生移植片を有する動物におけるGDNF源は移植した細胞自体である。HNu(赤)およびGDNF(緑)に関する二重染色調製物は、移植NSCの細胞質内GDNF免疫反応性の存在度を示す。生移植片を受ける動物では、宿主運動ニューロン内の分泌小胞に類似した丸い細胞質構造内に強力なGDNF免疫反応性が存在する。
【0189】
GDNFおよびヒトシナプトフィジン(後者は移植片由来の終末を標識するため)によって染色した宿主運動ニューロン全体の共焦点顕微鏡により、小胞構造内にはGDNFの局在化が示されたが、例示の宿主運動ニューロンの表面上に位置する移植片由来の終末内には示されない。ヒトシナプトフィジン抗体(宿主の運動ニューロンを神経刺激している全ての移植片終末を標識するため)およびGDNFによって染色した切片は、宿主の運動ニューロンに接触しているボタン内の2つのタンパク質の同時局在化の欠如を示す。
【0190】
生移植片を用いたラットは、移植片に源を発し、中央カナルの内および周囲の構造を神経刺激している経路の確立を示す。宿主運動ニューロンに接触している終末におけるGDNFタンパク質の欠如とは対照的に、中央カナルを神経刺激しているNSC由来軸索終末の大多数は、GDNF免疫反応性と同時局在化する。ヒトシナプトフィジンおよびGDNFの広範な同時局在化が中央カナルにおける上衣細胞を神経刺激している移植片由来終末内に見られる。これらの解剖学的パターンは、GDNFが、移植片を逆行性輸送によって神経刺激し、経シナプス輸送によってこれらのニューロンに送達されない宿主運動ニューロン終末によって恐らく取り込まれることを示している(Rindら、2005年)。
【0191】
SOD1 G93Aラットの腹角における進行性変性過程に対する移植NSCの明らかな抵抗性は、特に有望である。本明細書に報告されたNSCの生存性で広範囲の分化により、SOD1 G93Aに存在する運動ニューロンに関与する炎症性/興奮毒性のシグナル伝達(Rothsteinら、1992年;Howlandら、2002年;Turnerら、2005年)が細胞に明らかな毒性を有さないことの強い指標である。この因子単独で、運動ニューロン変性疾患における運動機能を回復させるために移植片を用いる将来の細胞戦略に対する楽観主義を高める。
【0192】
(実施例7:ヒト脊髄神経幹細胞/前駆細胞の移植による外傷性脊髄損傷の処置、脊髄空洞症の処置)
増殖させたヒト脊髄幹細胞を、免疫抑制した成体メスSprague-Dawleyまたは免疫不全胸腺欠損ヌードマウスのいずれかに注射する。移植の1カ月前に、両群にC4-5挫傷損傷を作製する。移植片レシピエント(n=24)は、移植後60〜150日生存した。ヒト由来のNSCは、ニューロン、星状膠細胞および希突起膠細胞からなる大型の細胞凝集体を形成した。これらの移植片は、各損傷をいつも完全に満たした。未熟な外観のNeuN+/ヒト核+ニューロンは、50%のドナー細胞集団を含むことが多かった。これらのニューロンは、移植部位から少なくとも2cm距離で、灰質と白質の双方を介してヒト神経フィラメント+処理を送達した。宿主ニューロンと移植片ニューロンの双方の近位に、強いヒト特異的シナプトフィジン免疫反応性が認められ、非反応性と思われるGFAP+細胞はドナーニューロンと並列した。さらに、これらの移植片は、宿主源から生じると思われるTH+および5HT+線維の増殖を支持した。したがって、このNSC系は、脊髄内灰白質の修復に有利である主要な胎児CNS様性質を有していると思われる。
【0193】
当然のことながら、本明細書に記載された好ましい本実施形態に対する種々の変更および改変が当業者には明らかであろう。このような変更および改変は、本開示の方法の精神および範囲を逸脱することなく、また、その意図された利益を減ずることなく行うことができる。したがって、このような変更および改変は、添付の請求項に含まれることが意図されている。
【技術分野】
【0001】
本開示の方法は、そのような治療法にとって比類なく有益である、細胞の移植を通して疾患を治療する方法に関する。特に、本開示の方法は、神経幹細胞(NSC)を用いた神経変性状態を治療する方法を提供する。
【背景技術】
【0002】
神経変性障害は、疾患、遺伝性疾患、または外傷性もしくは虚血性の脊髄損傷もしくは脳損傷などの損傷の結果としてのニューロン変性に関わる状態によって特徴付けられる。
【0003】
四肢の骨格筋の収縮を支配する脊髄の回路には、興奮性の運動ニューロン、ならびに抑制性のGABA作動性(すなわち、GABAを産生する)介在ニューロンおよびグリシン作動性(すなわち、グリシンを産生する)介在ニューロンが関与する。運動ニューロンは、脊髄の灰白質の前角から始まる神経である。運動ニューロンの軸索は、筋線維の神経を刺激する遠心性運動線維として、脊髄の一分節から出現する。運動ニューロンによって伝導されるインパルスは、筋線維を刺激して収縮させる。γアミノ酪酸(gamma-amino butyric acid:GABA)は、哺乳動物の神経系において自然に発生される代謝物であって、電位の神経伝導を抑制または弱める神経伝達物質として作用する。GABA作動性の介在ニューロンが失われると、運動ニューロンによって引き起こされる筋収縮の抑制性調性(inhibitory tonality)の調節不全が生じる。抑制性介在ニューロンによって発揮される興奮性ニューロンに対する制御がないと、四肢の筋肉の痙攣性の抑制困難な収縮または抑制困難な硬直につながる、興奮性ニューロンの過大加熱(over-firing)が生ずる。運動ニューロンが失われると、弛緩性対麻痺が生じ、対象は、筋肉を収縮させることができず、したがって、動くことができない。
【0004】
GABA作動性の介在ニューロンが脊椎索で損傷を受ける1つの例は、胸部下行大動脈または胸腹部大動脈の一過性の遮断(cross-clamping)に関連する合併症を含む。そのような遮断は、胸部大動脈または胸腹部大動脈の動脈瘤を修復する血管手術における必要な工程である。遮断の間、脊髄の一部は血液循環を受けることができず、虚血状態となる可能性がある。虚血時間の長さによって、その後の神経変性の機能不全が、不全対麻痺、または完全に進行した痙攣性もしくは弛緩性の対麻痺として神経変性的に発現する可能性がある。
【0005】
虚血誘導性のニューロン変性につながるメカニズムは、一部しか理解されておらず、興奮性アミノ酸、プロスタグランジンおよび/または活性酸素の過剰な放出/活性が関与する可能性がある一方、一過性の虚血性障害(ischemic insult)による影響を受ける脊髄のニューロン集団は、十分明確にされている。例えば、完全に進行した痙性対麻痺の動物から採取した脊髄に関する組織病理学的分析は、小型の抑制性ニューロンの選択的な減少(selective loss)を明らかにしているが、α運動ニューロンは、あらかじめ虚血の脊髄分節内に残存する。脊髄の虚血性傷害を有するヒトの対象における同様な脊髄の神経病態について述べられている。
【0006】
対照的に、弛緩性対麻痺の動物において、汎壊死性(pan-necrotic)の神経変性による変化が、腹側の運動ニューロンだけでなく、小型の抑制性および興奮性の介在ニューロンに影響を及ぼすことが認められている。また、局所または全体的な脳虚血などで、脊髄の虚血後の神経変性の間に、局所の小膠細胞の傷害依存的な活性化、および大食細胞による浸潤などの炎症性の変化が見られる。傷害の程度によって、一般的に炎症性変化は、虚血性傷害の2日から7日後にピークに達して、その後、2週間から4週間の虚血後期間にかけて炎症性要素が次第に消失していく。
【0007】
過去20年から30年間に、動物モデルで、さまざまな物質の脊髄移植の治療可能性について評価するためにかなりの努力が払われている。したがって、細胞系または急速に分離された脊髄胎児組織が損傷部位に送達されており、さらに直接的な脊髄遺伝子治療(direct spinal gene therapy)も、機械的外傷的損傷、化学的に損傷を起こした脊髄、または進行性α運動ニューロン変性(α-motoneuronal degeneration)の遺伝子組換え動物(ALS遺伝子移植マウスまたはラット)などの、いくつかの脊髄損傷モデルにおいて、神経変性機能不全を改善するために使用されている。
【0008】
一般に、いくつかの試験では、胎児組織から生じた移植片でニューロン表現型(neuronal phenotype)の長期生存および保存が実証されているが、in vitroで増殖させた神経前駆細胞(neural precursor)から生じた移植片については実証されていない。実際に、in vitroで増殖され、機械的および化学的に損傷した脊髄へ移植した神経前駆細胞のニューロン分化および成熟は、限られたものが実証されているに過ぎない。細胞は、優先的に非ニューロン性の細胞型に分化する。この優先的な非ニューロン性分化のメカニズムは、完全には理解されていないが、以前の損傷部位での炎症誘発性サイトカイン(TNFα、TGFβなど)の局所的放出が恐らく関与すると仮定されている。
【0009】
神経変性は、特に、細胞療法についての厳しい生物学的環境を示し、確立された神経変性疾患に現れる細胞死シグナル(cell death signal)(Rothsteinら、1992年; Howlandら、2002年; Turnerら、2005年)は、移植片の生存とは同時に起こり得ないと考えられる。さらに、成人の脊髄は、再生を可能にする細胞および/またはシグナルを欠いていると見られ(Parkら、2002年)、大部分のNSC移植試験が、不良または限定的な分化を示している(Caoら、2002年; Yanら、2003年; Yanら、2004年)。
【0010】
細胞治療学(cell therapeutics)における主な問題の1つは、移植した細胞の細胞生存率が低いということである(5%未満)。現在に至るまで、移植した全ての細胞は、in vivoへの注射直後にかなりの細胞死を起こす。したがって、細胞の有効量を送達するために、最終用量を少なくとも20回注射しなければならない。その結果として、このことは、より大規模な細胞製造を必要とし、さらなる規制の障壁および経済の障壁を引き起こす。さらに、in vivoで、そのような細胞の生存率を維持することができていない。再現性のある、細胞療法の有効量の投与が実証されていないことが、政府、および米国食品医薬品局などの他の規制機関による使用の承認を妨げている。
【0011】
さらなる困難は、1つの限局部分というよりむしろ全ての神経系のような広範囲の身体、組織または器官にわたって広がる神経変性疾患および状態を治療するときに現れる。例えば、ALSでは、神経変性が、運動皮質の運動ニューロンだけでなく、脊髄全体に沿った運動ニューロンの緩やかな死(slow death)に関わる。同様に、大部分のリソソーム病で、ニューロンの破壊が脳および脊髄の大部分の部位に影響を与える。アルツハイマー病は、大部分の大脳に影響を与える。パーキンソン病およびハンチントン舞踏病などのより局所的な神経変性疾患でさえ、線条体の患部は非常に大きく、手術によって達することができる移植部位より遥かに大きい。したがって、神経変性疾患に対する細胞治療学は、より広い移植手法が必要とされることが予期される。
【0012】
したがって、神経変性状態を治療する方法の改善が必要とされる。また、一旦、移植されたならば、以前に見られた全ての制限を克服し、機能的有益性を提供する、ヒトの神経幹細胞およびヒトの神経前駆体を培養し、移植する方法の改善が必要とされる。したがって、このin vivoでの神経変性状態を治療する方法は、確固としたニューロン分化を生じさせ、さまざまな変性状態下でのニューロンの長期生存、および発生の手掛かり(developmental cue)を欠く成体組織での治療に関係したニューロン亜集団への成熟を可能にし、細胞自身の位置より広範囲の治療域を提供する。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】Rothsteinら、1992年
【非特許文献2】Howlandら、2002年
【非特許文献3】Turnerら、2005年
【非特許文献4】Parkら、2002年
【非特許文献5】Caoら、2002年
【非特許文献6】Yanら、2003年
【非特許文献7】Yanら、2004年
【発明の概要】
【発明が解決しようとする課題】
【0014】
本開示の方法は、神経変性状態を治療するための方法を含む。特に、本開示の方法は、細胞が神経変性状態を改善することができるように、in vitroで増殖させたNSC、神経前駆体または神経前駆細胞を、それを必要とする対象へ移植することを含む。一実施形態では、本開示の方法は、ドナー細胞を同定、分離、増殖および調製して、神経変性状態の治療として使用することを含む。移植されるドナー細胞は、その状態、その症状および/またはその影響の一因となる要素またはその欠如に対応するように選択することができる。
【0015】
本開示の方法の細胞は、移植直後に、ニューロンの基礎構造内に組み込まれるのに十分なニューロン量を産生して、疾患の状況または状態を改善する細胞を含む。一実施形態では、本開示の方法は、哺乳動物の中枢神経系から分離され、in vitroで増殖させた多分化能を有する神経前駆体または神経幹細胞を移植することによって、神経変性疾患または状態を治療することを含む。例えば、増殖させた神経幹細胞の移植は、痙縮、硬直、発作、麻痺またはその他の筋肉の活動亢進の症状を有するさまざまな脊髄症の形態を患う対象における歩行機能を改善するために使用することができる。
【0016】
治療法は、損傷した神経領域に、移植を通して十分な数のGABAを産生するニューロンおよび/またはグリシンを産生するニューロンに分化することができる適切な数のNSCを供給して、活動亢進の神経回路を含む欠陥のある神経回路を弱めることを含む。
【課題を解決するための手段】
【0017】
一実施形態では、本開示の方法は、運動ニューロン疾患における運動機能を回復させることを含む。運動ニューロンへ分化可能な、適切な数または治療有効量のNSCまたは神経前駆体を、変性脊髄などの神経変性の少なくとも一領域に供給し、運動機能を回復させることができる。NSCは、変性した神経筋接合部を置換することによって、その治療効果を発揮する。
【0018】
同時または代わりに、NSCは、変性組織のニューロンを保護して、より多くのニューロンをより長期間生存させる栄養分子を発現および放出することによって、その治療効果を発揮する。変性運動ニューロン疾患の対象において、NSC由来ニューロンを、腹根へ突き出させ、NSCが宿主の運動ニューロンと広範な相互関係をもつ筋肉に神経を刺激するように促進させることができる。したがって、一実施形態では、ヒトの胎児脊髄由来のNSCを、これらの細胞が宿主のニューロンとシナプス接合(synaptic contact)を形成し、運動ニューロン増殖因子を発現および放出するニューロンへ分化を起こすことができる腰髄に移植することができる。
【0019】
一実施形態では、本開示の方法は、宿主組織と同化し、1種または2種以上の増殖因子を宿主ニューロンに供給する神経幹細胞または神経前駆体を提供し、したがって、組織に存在する変性の影響からそれらを保護することを含む。本方法は、十分な数のNSCまたは神経前駆体を、少なくとも1種の増殖因子の有効量がNSCによって分泌されるように、脊髄の一領域に導入することを含む。
【0020】
一実施形態では、本開示の方法は、神経変性状態での細胞置換(cell replacement)に対する幹細胞の前臨床評価において、動物モデルを使用する方法を提供することを含む。
【0021】
一実施形態では、本開示の方法は、ニューロンへ移植するNSCの分化効率を上げることを含む。本方法は、移植後に、移植片において、20%というような、十分な数の細胞がニューロンの運命(neuronal fate)を選ぶように、未分化の状態で非常に高濃縮のNSCまたは神経前駆体を増殖することを含む。
【0022】
一実施形態では、本開示の方法は、移植するNSCまたは神経前駆体の数を増加させることなく分化細胞の数を増加させることを含む。一実施形態では、本方法は、一旦移植されると、NSCまたは神経前駆体が、in vivoで10回ほども継続して、腫瘍を生じることなく分裂するというように増殖するドナー集団を調製し、したがって、導入細胞の総数を有効に増加させることを含む。
【0023】
本開示の方法の細胞は、哺乳動物の胎児、新生児、若年者、成人および死後の組織から分離または入手することができる。本開示の方法の細胞は、中枢神経系、血液、またはニューロンに分化する他の適した幹細胞源から分離または入手することができる。また、細胞は、胚幹細胞から入手することができる。例えば、一実施形態では、本細胞は、発育する胎児の脊髄から分離された神経上皮細胞を含む。場合によっては、神経前駆細胞は、中枢神経系の特定の小領域から分離される神経前駆体である可能性がある。
【0024】
本開示の方法によれば、神経幹細胞は培養で増殖する。一実施形態では、神経前駆細胞は、培養で増殖し、分化に際してニューロンと神経膠の両方を発生することができる多分化能を有するNSCとすることができる。
【0025】
細胞は、移植時にin vitroで未分化、分化前または完全分化のいずれかとすることができる。一実施形態では、細胞は、神経系列(neural lineage)へ分化させるために導入される。本開示の方法の細胞は、炎症誘発性サイトカインおよび損傷した組織に存在するその他の環境因子の存在下における生体内で、ニューロン分化を起こすことができる。
【0026】
現在の方法を使用して、疾患、障害または状態の改善のために、本細胞を適切な部位に移植または導入することによって、神経回路を治療することができる。通常、移植は、移植した細胞の生存を補助する神経組織または非神経組織へ行われる。本開示の方法で使用されるNSC移植片は、神経変性の環境で良好に生存し、神経変性状態または疾患の発症および進行を遅らせるという形で、NSCが強力な臨床効果を発揮することができる。
【0027】
場合によっては、移植は、身体の遠位の領域へ行うことが可能で、本細胞は意図する標的へ移動することができる。したがって、本開示の方法は、ヒトNSCの部分移植(partial grafting)も含み得る。本書では、「部分移植」という用語は、神経変性の一部の領域だけ、または全体より少ない領域における増殖NSCの移植を指すことができる。例えば、脊髄の腰椎部へのヒトNSCの部分的移植。変性する運動ニューロンに対するNSCの効果の少なくとも一部は、古典的な細胞メカニズムを通して、変性する宿主の運動ニューロンへニューロトロフィンおよび栄養サイトカイン(trophic cytokine)を送達することを含む。このために、本開示の方法を使用して脊髄の腰椎部への部分移植を受けているNSCが、運動ニューロン疾患のトランスジェニック動物モデルにおいて、移植の隣接領域および移植領域から遠位の領域で生存し、広範囲なニューロン分化を起こし、運動ニューロンの生存を促進し、機能することを示している。
【0028】
したがって、本開示の方法は、痙縮、硬直または筋肉活動亢進状態を治療する方法を提供する。本方法は、少なくとも1種の神経幹細胞を哺乳動物から分離すること、in vitroで、神経幹細胞を増殖集団へ増殖させることを含む。また、本方法は、増殖集団を濃縮すること、治療有効量の増殖集団をレシピエントの脊髄の少なくとも一領域に導入することを含む。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0029】
一実施形態では、本状態は、外傷性脊椎損傷、虚血性脊椎損傷、外傷性脳損傷、脳卒中、多発性硬化症、脳性麻痺、てんかん、ハンチントン舞踏病、筋萎縮性側索硬化症、慢性虚血、遺伝性疾患またはそのいずれかの組合せに由来する。
【0030】
一実施形態では、神経幹細胞は、中枢神経系、末梢神経系、骨髄、末梢血、臍帯血および少なくとも1つの胚からなる群から選択される供給源から分離される。
【0031】
一実施形態では、哺乳動物は、発育中の哺乳動物である。
【0032】
一実施形態では、発育中の哺乳動物の在胎齢は、約6.5週から約20週までの間である。
【0033】
一実施形態では、神経幹細胞は、ヒトの胎児の脊髄から分離される。
【0034】
一実施形態では、神経幹細胞を増殖することは、血清の不存在下で、神経幹細胞を培養することを含む。
【0035】
一実施形態では、神経幹細胞を増殖することは、神経幹細胞を、少なくとも1種の増殖因子に曝露させることを含む。
【0036】
一実施形態では、増殖因子は、増殖因子はbFGF、EGF、TGF-α、aFGFおよびその組合せからなる群から選択される。
【0037】
一実施形態では、治療有効量の増殖集団は、in vivoで少なくとも1,000のGABAを産生するニューロンを発生させることができる。
【0038】
一実施形態では、治療有効量の増殖集団は、in vivoで少なくとも1,000のグリシンを産生するニューロンを発生させることができる。
【0039】
一実施形態では、増殖集団の少なくとも40%は、脊髄でニューロンを発生させることができる。
【0040】
一実施形態では、治療有効量の増殖集団を導入することは、レシピエントの脊髄の複数の領域に、治療有効量の少なくとも一部を注射することを含む。
【0041】
一実施形態では、増殖集団の少なくとも30%は、in vitroでニューロンに分化することができる。
【0042】
他の実施形態では、神経幹細胞を提供する。本神経幹細胞は、痙縮、硬直または筋肉活動亢進状態を治療することができる。神経幹細胞は、哺乳動物から分離し、in vitroで増殖集団へ増殖させる。幹細胞を含む増殖集団を濃縮して、治療有効量の増殖集団を、レシピエントの脊髄の少なくとも一領域に導入する。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0043】
本開示の方法の他の実施形態では、慢性痛を治療する方法を提供する。本方法は、少なくとも1種の神経幹細胞を哺乳動物から分離すること、in vitroで、神経幹細胞を増殖集団へ増殖させることを含む。また、本方法は、増殖集団を濃縮すること、治療有効量の増殖集団をレシピエントの脊髄の少なくとも一領域に導入することを含む。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0044】
一実施形態では、慢性疼痛は、外傷性脊椎損傷、虚血性脊椎損傷、外傷性脳損傷、脳卒中、多発性硬化症、脳性麻痺、てんかん、ハンチントン舞踏病、筋萎縮性側索硬化症、慢性虚血、遺伝性疾患またはそのいずれかの組合せに由来する。
【0045】
一実施形態では、治療有効量の増殖集団は、少なくとも1,000のGABAを産生するニューロンを発生させることができる。
【0046】
一実施形態では、治療有効量の増殖集団は、少なくとも1,000のグリシンを産生するニューロンを発生させることができる。
【0047】
一実施形態では、増殖集団の少なくとも40%は、脊髄でニューロンを発生させることができる。
【0048】
一実施形態では、治療有効量の増殖集団を導入することは、レシピエントの脊髄の複数の領域に、治療有効量の少なくとも一部を注射することを含む。
【0049】
一実施形態では、本領域は、後角を含む。
【0050】
一実施形態では、本領域は、クモ膜下腔を含む。
【0051】
さらなる実施形態では、神経幹細胞を提供する。神経幹細胞は、慢性疼痛を治療することができる。神経幹細胞は、哺乳動物から分離し、in vitroで増殖集団へ増殖させる。幹細胞を含む増殖集団を濃縮して、治療有効量の増殖集団を、レシピエントの脊髄の少なくとも一領域に導入する。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0052】
本開示の方法の他の実施形態では、運動ニューロンの変性を治療する方法を提供する。本方法は、少なくとも1種の神経幹細胞を哺乳動物から分離すること、in vitroで、神経幹細胞を増殖集団へ増殖させることを含む。また、本方法は、増殖集団を濃縮すること、治療有効量の増殖集団をレシピエントの脊髄の少なくとも一領域に導入することを含む。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0053】
一実施形態では、運動ニューロンの変性は、外傷性脊椎損傷、虚血性脊椎損傷、外傷性脳損傷、脳卒中、多発性硬化症、脳性麻痺、てんかん、ハンチントン舞踏病、筋萎縮性側索硬化症、慢性虚血、遺伝性疾患またはそのいずれかの組合せに由来する。
【0054】
一実施形態では、本方法は、少なくとも1種のニューロンのサブタイプが豊富な領域から神経幹細胞を分離することを含み、この場合、ニューロンのサブタイプは、運動障害を改善することにおいて有効な増殖因子を産生する。
【0055】
一実施形態では、増殖集団は、治療有効量の少なくとも1種の増殖因子を分泌するために十分なニューロンに分化することができる神経幹細胞量を含む。
【0056】
一実施形態では、本方法は、運動ニューロンの豊富な領域から神経幹細胞を分離することを含む。
【0057】
さらなる実施形態では、脊髄空洞症を治療することができる神経幹細胞を提供する。神経幹細胞は、哺乳動物から分離し、in vitroで増殖集団へ増殖させる。幹細胞を含む増殖集団を濃縮して、治療有効量の増殖集団を、レシピエントの脊髄の少なくとも一領域に導入する。増殖集団の少なくとも20%は、レシピエントの脊髄でニューロンを発生させることができる。
【0058】
本開示の方法の他の実施形態では、脊髄空洞症を治療する方法を提供する。本方法は、少なくとも1種の神経幹細胞を哺乳動物から分離すること、in vitroで、神経幹細胞を増殖集団へ増殖させることを含む。また、本方法は、増殖集団を濃縮し、治療有効量の増殖集団をレシピエントの脊髄の空洞に導入することを含む。増殖集団の少なくとも20%は、レシピエントの脊髄の空洞でニューロンを発生させることができる。
【0059】
一実施形態では、脊髄空洞症は、外傷性脊椎損傷、虚血性脊椎損傷、外傷性脳損傷、脳卒中、多発性硬化症、脳性麻痺、てんかん、ハンチントン舞踏病、筋萎縮性側索硬化症、慢性虚血、遺伝性疾患またはそのいずれかの組合せに由来する。
【0060】
一実施形態では、本方法は、少なくとも1種のニューロンのサブタイプが豊富な領域から神経幹細胞を分離することを含み、この場合、ニューロンのサブタイプは、脊髄空洞症を改善することにおいて有効な増殖因子を産生する。
【0061】
一実施形態では、本方法は、運動ニューロンの豊富な領域から神経幹細胞を分離することを含む。
【0062】
一実施形態では、増殖集団は、治療有効量の少なくとも1種の増殖因子を分泌するために十分なニューロンに分化することができる神経幹細胞量を含む。
【0063】
一実施形態では、治療有効量の増殖集団は、少なくとも1,000のニューロンを発生させることができる。
【0064】
一実施形態では、増殖集団の少なくとも100,000の神経幹細胞をレシピエントの脊髄の空洞に導入する。
【0065】
なおさらなる実施形態では、脊髄空洞症を治療することができる神経幹細胞を提供する。神経幹細胞は、哺乳動物から分離し、in vitroで増殖集団へ増殖させる。幹細胞を含む増殖集団を濃縮して、治療有効量の増殖集団を、レシピエントの脊髄の空洞に導入する。増殖集団の少なくとも20%は、レシピエントの脊髄の空洞でニューロンを発生させることができる。
【0066】
本開示の方法のさらなる実施形態では、in vitroにおいて少なくとも1種の神経幹細胞を神経幹細胞の増殖集団に増殖させる方法を提供する。各神経幹細胞の増殖は、分化することなしに、細胞の倍増が30回を上回る。本方法は、中枢神経系組織から神経幹細胞を分離して、少なくとも1種の細胞外タンパク質(extracellular protein)を培養容器に供給することを含む。細胞外タンパク質は、少なくとも約10μg/mlのポリ-D-リシンおよび約1mg/mlのフィブロネクチンを含む。また、本方法は、血清の不存在下において、分離した神経幹細胞を培養容器で培養し、少なくとも1種の増殖因子を培養容器に加えることを含む。増殖因子は、増殖因子はbFGF、EGF、TGF-α、aFGFおよびその組合せからなる群から選択される。本方法は、集密に達する前に、さらに培養細胞を継代することを含む。
【0067】
一実施形態では、増殖された神経幹細胞は、ニューロンに分化することができる。
【0068】
一実施形態では、神経幹細胞を増殖することは、可溶性因子としてフィブロネクチンを培地に加えることを含む。
【0069】
一実施形態では、細胞を分離すること、および細胞を継代することは、酵素による分離を含む。
【0070】
一実施形態では、酵素による分離は、トリプシンで細胞を処置することを含む。
【0071】
一実施形態では、神経変性状態を治療するために、治療有効量の増殖集団をレシピエントの神経系の少なくとも一領域に導入する。
【0072】
したがって、既存の薬理学的方法に対する本開示の方法の利点は、栄養分子を分泌させるために、移植したNSCの能力を増強させ、最適の生物学的利用度の条件下で、変性している運動ニューロンに送達することができる、方法を提供することである。
【0073】
本発明のさらにもう1つの利点は、ヒトの胎児脊髄由来のNSCを培養および増殖して、腰髄へのNSCの良好な移植を促進する方法を提供することである。
【0074】
本開示の方法のさらなる利点は、NSC集団のより高率のニューロン分化を達成する方法を提供することを含む。
【0075】
本開示の方法のもう1つの利点は、NSCの部分移植により臨床効果を得ることを含む。
【0076】
本開示の方法の付加的な特徴および効果は、以下の本発明の詳細な説明および図において記載され、これらによって明らかとなろう。
【図面の簡単な説明】
【0077】
【図1】ヒト脊髄幹細胞の増殖を示す図である。ヒト脊髄前駆体(NSCとしても知られている)系を、7〜8週齢の死後胎児脊髄組織から単離し、約130日の正味培養期間、連続継代した。各継代の採収時に回収した細胞数を、平板培養時の最初の細胞数で割って、細胞数の増加倍数を得た。各継代時の増加倍数を掛け合わせることによって累積増加倍数(左Y軸)を得た。細胞数の倍増を各培養期間(X軸)で割って、各継代時における細胞の倍加時間(右Y軸)を算出した。この処理を3回繰り返した(連続増殖1、2、および3)。
【図2】増殖させたヒト脊髄幹細胞の形態を示す図である。(A)固定、非染色、増殖培養物、20×対物レンズの位相差図、(B)抗ネスチン抗体染色。
【図3】増殖させたヒト脊髄幹細胞から得られた分化培養物の特徴を示す図である。15〜16継代の増殖細胞を培養物中でおよそ14日間分化させ、固定し、種々のニューロン特異的抗体によって染色した。(A)TauおよびMAP2; (B)タイプ3ベータチューブリン; (C)GABA; (D)アセチルコリントランスフェラーゼ。
【図4】ヒト中脳幹細胞の増殖を示す図である。ヒト中脳前駆体(NSCとしても知られている)系を、7〜8週齢の死後胎児中脳組織から単離し、約170日の正味培養期間、連続継代した。各継代の採収時に回収した細胞数を、平板培養時の最初の細胞数で割って、細胞数の増加倍数を得た。各継代時の増加倍数を掛け合わせることによって累積増加倍数(Y軸)を得た。
【図5】増殖させたヒト中脳幹細胞のドーパミン取り込み活性を示す図である。生細胞におけるドーパミントランスポーター活性(DAT)を、ヒト中脳幹細胞系およびアッセイ時に22日または44日間分化させたそのクローンの亜系の1つから判定した。該細胞を、DAT阻害剤ノミフェンシン(10μM)の存在下(+)または不在下(-)、放射標識したドーパミンと共にインキュベートした。シンチレーションカクテル中、取り込まれなかったドーパミンを除去するために、細胞を洗浄し、すすいだ。次いで、シンチレーションカウンターを用いて、総細胞放射性(dpm)を測定した。
【図6A】ヒト中脳幹細胞系のニューロン分化およびドーパミン作動性分化の誘導に及ぼす外来因子の影響を示す図である。2つのヒト中脳幹細胞系(527RMBおよび796RMB)からの冷凍保存した神経幹細胞を解凍し、bFGFの存在下、4ウェルチャンバースライド内で、1ウェル当たり40,000細胞の密度で塗布し、6日間増殖させた。引き続き、bFGFを除去し、細胞をさらに8日間分化させた。セルトリ細胞馴化培地(SCCM、N2中、1:1に希釈)に曝露するタイミングと時間に基づき、細胞を4群に分けた。1つの群は、増殖中および分化中、SCCMに曝露し(条件1);第2群は増殖中にのみ曝露し(条件2);第3群は分化中にのみ曝露し(条件3);第4群はSCCMに曝露しなかった(対照、Cont.)。培地は1日おきに交換し、増殖期の間、マイトジェンを毎日添加した。1つの条件当たり、4つのウェルを維持し、複数のマーカーに関して染色した。分化の際、4%パラホルムアルデヒドを用いて細胞を固定し、MAP2abに対する抗体(図6A)およびチロシンヒドロキシラーゼに対する抗体(図6B)、ならびにGFAPおよびGalCを用いて免疫染色した。免疫染色された細胞を、40×対物レンズを用いてカウントすると、各ウェルで少なくとも3つのフィールドがカウントされた。いずれの条件下で維持された細胞を分析した際にも、GFAP+細胞またはGalC+細胞はほとんど検出されないか、または全く検出されず、したがって、これらの抗原は分析から除外した。
【図6B】図6Aの説明参照。
【図7】ヒト脊髄幹細胞移植によるラットの痙攣/硬直および運動欠損の減少を示す図である。腰脊髄の虚血損傷化によって痙攣ラットを作製した。1つの群(黒丸)では、培養物中で増殖させた(継代16)ヒト脊髄幹細胞を、ラット(n=9)に移植し、他方、対照の群(白丸、N=7)には、細胞のない培地のみを与えた。この試験の間(8週間)、両群に免疫抑制剤、FK506を、1日1mg/kgで投与した。個々の動物の運動協調性を1週間に1回、BBBスコアリングによって評価した。
【図8】ヒト脊髄幹細胞移植によるラットの痙攣/硬直および運動欠損の減少を示す図である。腰脊髄の虚血損傷化によって痙攣ラットを作製した。1つの群(黒丸および黒四角)では、培養物中で増殖させた(継代16)ヒト脊髄幹細胞を、ラット(n=13)に移植し、他方、対照の群(黒三角、n=6)には、細胞のない培地のみを与えた。この試験の間(12週間)、両群に免疫抑制剤、FK506を、1日3mg/kgで投与した。個々の動物の運動協調性を1週間に1回、BBBスコアリングによって評価した。
【図9A】G93A SOD1 ラットの運動ニューロン疾患重症度に及ぼすヒトNSC処置の効果を、生細胞(L、赤)移植片および死細胞(対照、C)移植片(青)の場合における進行(A〜B)ならびに臨床的および病理学的尺度のエンドポイント(C〜E)分析によって示す図である。 図9AおよびB。図9Aは、観察経過にわたる実験動物と対照動物との間の有意な区別を示すKaplan-Meierプロット (P=0.0003)であり、図9Bは、2つの群間の筋肉衰弱の2つの主要な尺度(BBBおよび傾斜平面スコア)における区別を示している(それぞれ、P=0.00168および0.00125)。
【図9B】図9Aの説明参照。
【図9C】図9C〜E。実験ラットおよび対照ラットにおける生存(C)、疾患発症までの時間(D)および運動ニューロン数(E)のエンドポイント分析。図9Cは、両群間の寿命における有意な11日の違いを示している(P=0.0005)。図9Dは、両群間の疾患発症までの時間における有意な7日の違いを示している(P=0.0001)。図9Eは、生きているNSC群と死んでいるNSC群の腰プロチュバランスにおける3,212個の細胞の違いを示している(P=0.01)。E下部の差込図は、128日齢における代表的な実験ラット(上部)と対照ラット(下部)間の運動ニューロン生存度における違いを示しており;矢印は、側方運動ニューロン群を示す。サイズバー:150μm。
【図9D】図9Cの説明参照。
【図9E】図9Cの説明参照。
【発明を実施するための形態】
【0078】
本開示の方法は、神経変性状態を治療することに関する。特に、本開示の方法は、それを必要とする対象に移植するための神経幹細胞を調製する方法を含む。移植のために細胞を調製することは、神経変性状態の治療用として商業的利用に十分な水準まで、in vitroで、特定の細胞集団を増殖することを含むことができる。一実施形態では、変性または損傷した神経領域の治療法は、神経変性状態を改善するのに十分な神経幹細胞の有効数を本領域に供給することを含む。
【0079】
本書では、神経変性状態は、ニューロンの損傷または変性が関わる、何らかの疾患または障害または症状または原因またはその影響を含む可能性がある。神経変性状態は、アレキサンダー病、アルパーズ病、アルツハイマー病、筋萎縮性側索硬化症、血管拡張性失調症、カナヴァン病、コケーン症候群、大脳皮質基底核変性症、クロイツフェルトヤコブ病、ハンチントン舞踏病、ケネディ病、クラッベ病、レヴィー小体痴呆、マシャドジョセフ病、多発性硬化症、パーキンソン病、ペリツェウスメルツバッハー病、ニーマンピック病、原発性側索硬化症、レフサム病、サンドホフ病、シルダー病、スティールリチャードソンオルゼウスキー病、脊髄癆または損傷したニューロンと関連するその他の状態を含む可能性があるが、これらに制限されない。他の神経変性状態は、外傷性脊椎損傷、虚血性脊椎損傷、脳卒中、外傷性脳損傷および遺伝性疾患を含むか、これらによって引き起こされることがある。
【0080】
本開示の方法は、神経変性状態を改善するためのNSCの使用を含む。本書では、「NSC」という用語は、神経もしくはニューロンの前駆体または神経上皮前駆体を指すこともある。NSCは、ニューロン、星状膠細胞および希突起膠細胞というCNSのいずれか3つの主な細胞の種類へ分化する能力によって機能的に定義することができる。
【0081】
一実施形態では、NSCは、各細胞がニューロン、星状膠細胞または希突起膠細胞に分化する能力を有するというように、多分化能を有する。一実施形態では、NSCは、各細胞が3種のCNSの細胞型中2種に分化する能力を有するというように、二分化能を有する。一実施形態では、NSCは、in vitroでニューロンおよび星状膠細胞の両方を発生する、少なくとも二分化能を有する細胞を含み、in vitroでニューロンを発生する、少なくとも単分化能を有する細胞を含む。
【0082】
増殖状態は、1種の細胞またはもう1種の細胞への細胞の分化の方向に影響を及ぼすことがあり、このことは細胞が1つの系譜に関係付けられていないことを示している。神経分化にとって有利に働く培養条件で、細胞の特にヒトCNS由来の細胞は、主にニューロンおよび星状膠細胞への二分化能を有し、希突起膠細胞への分化は稀である。このように、本開示の方法の分化細胞の培養は、ニューロンおよび星状膠細胞を生じさせることが可能である。一実施形態では、星状膠細胞に対するニューロンの比率は、50:50の比率に近づく可能性がある。
【0083】
本開示の方法は、感覚上皮などの哺乳動物のCNS部位に存在するNSCを得ることを含む。NSCを分離することができる他のCNS部位は、CNSの脳室帯(ventricular zone)および脳室下帯(subventricular zone)、ならびに有糸分裂後ニューロンだけでなく、有糸分裂前駆体(mitotic precursor)を含む他のCNS部位を含む。一実施形態では、本開示の方法は、哺乳動物のCNSが形成される部位に存在するNSCを使用することができる。
【0084】
一実施形態では、NSCは、所望のニューロン集団のために自然の神経原性(neurogenic)を有する領域から得る。所望の細胞集団は、神経学的状態で失われた、または不活性のような表現型と置換またはそのような表現型を補うことができる特定のニューロン表現型の細胞を含むことが可能である。
【0085】
特定の神経変性疾患または状態の治療のために有用なニューロンのサブタイプを含む、さまざまな異なるニューロンのサブタイプは、CNSの異なる部位または領域から、および胎児の発育中の異なる在胎齢にわたって、NSCを分離することによって得ることができる。CNSの異なる部位または領域、および胎児の発育中の異なる在胎齢にわたって分離されたNSCを、最適な増殖およびニューロン分化能のために使用する。哺乳動物のCNSの顕著な特徴の1つは、ニューロンのサブタイプの多様性である。例えば、NSCの単一集団は、培養で、ほんの少数の明瞭なニューロンのサブタイプを自然に発生することが可能である。さらに、胎児の特定の在胎齢から得た細胞は、培養細胞の生理的関連性(physiological relevance)を確定させる可能性がある。
【0086】
本開示の方法の実施例では、対象に移植される細胞は、損傷した神経領域のヒトの胎児の対応物に由来する。一実施形態では、NSCを、約6.5週から約20週の間の在胎齢におけるヒトの胎児のCNS領域から分離する。一実施形態では、胎児の脊髄由来の細胞を、約7週から約9週の在胎齢で分離する。分離可能な神経幹細胞集団の比率は、ドナーの年齢によって異なる可能性があることを認識すべきである。また、細胞集団の増殖能力も、ドナーの年齢によって変化する。そのようなNSCの局所的および時間的特異性は、NSCが運命制限的な前駆体として機能し、ブランク細胞(blank cell)または単一の細胞集団として機能しないことを示している。
【0087】
in vitroでのGABAを産生するニューロンを含む集団の比率は、一般に、約5%から10%で一定である。
【0088】
例えば、中脳腹側部のNSCは、同じ妊娠期間の脊髄から得られるNSCとは異なっている。特に、中脳腹側部由来のNSCは、独占的にチロシン水酸化酵素を発現するドーパミン作動性ニューロンを生じるが、一方、脊髄由来のNSCは、独占的にアセチルコリンを産生するコリン作動性ニューロンを発生する。しかし、両方の細胞型とも、同時に、より偏在性のグルタミン酸およびGABAを産生するニューロンを発生する。したがって、一実施形態では、本開示の方法は、少なくとも部分的に、チロシン水酸化酵素を発現するドーパミン作動性ニューロンの移植によって、状態を治療して改善または軽減するために、中脳腹側部からNSCを得ることを含む。本開示の方法は、少なくとも部分的に、アセチルコリンを産生するコリン作動性ニューロンの移植によって、神経変性状態を治療して改善または軽減するために、脊髄からNSCを得ることを含む。
【0089】
したがって、ドーパミン作動性ニューロンの消失によって特徴付けられるパーキンソン病などの運動障害の治療のために、本開示の方法の実施形態は、ドーパミン作動性ニューロンの神経形成がかなり高率である中脳腹側部などの領域由来のNSCの使用を含む。さらに、NSCは、ドーパミン作動性ニューロンの神経発生がかなり高率である期間のヒトの胎児発育の在胎齢で得ることができる。したがって、一実施形態では、本開示の方法は、運動障害を治療するために、約7週から約9週の在胎齢の中脳腹側部からNSCを得ることを含む。
【0090】
腹側の運動ニューロンの消失から生じる筋萎縮性側索硬化症または弛緩性対麻痺などの運動ニューロン疾患を治療するために、本開示の方法の実施形態は、腹側の運動ニューロンの神経発生がかなり高率である脊髄などの領域に由来するNSCの使用、および腹側の運動ニューロンの神経発生がかなり高率である期間のヒトの胎児発育の在胎齢に得られたNSCの使用を含む。したがって、一実施形態では、NSCは、運動ニューロン疾患を治療するために、約7週から約9週の在胎齢の脊髄から分離する。
【0091】
しかし、場合によっては、そのような局所の特異性に関する制限は、実際的な目的には、かなり幅が広いということを認識すべきである。したがって、頸椎、胸椎、腰椎および仙骨などの脊髄のさまざまな領域から得たNSCを、区別なく、移植し、対応するNSCの起源以外の部位を治療するために使用することができる。例えば、頸髄由来のNSCは、患者の腰椎部に細胞を移植することによって痙縮および/または硬直を治療するために使用することができる。
【0092】
また、NSCは、生後組織および成体組織から分離することができる。生後組織および成体組織由来のNSCは、それらの増殖および分化の特徴だけでなく、ニューロンおよび神経膠に分化するそれらの能力に関して量的に同等である。しかし、in vitroでのさまざまな生後CNSおよび成体CNSからのNSC分離の効率は、より多量のNSC集団を保有する胎児組織由来のNSCより著しく低い可能性がある。にもかかわらず、胎児由来のNSCと同様に、本開示の方法は、新生児および成人の供給源由来のNSCの少なくとも30%が、in vitroでニューロンに分化することができる。したがって、前述のように胎児由来のNSCの場合、生後組織および成体組織を使用することができるものの、胎児組織の使用が好ましい。
【0093】
さまざまなニューロンのサブタイプを、培養で増殖した胚幹細胞の操作から得ることができる。したがって、本開示の方法に基づく特定のニューロンのサブタイプは、必要に応じて、結果を改善するために他の関連のない、または所望でない細胞から分離および精製することができ、同じ神経変性状態の治療のために使用することができる。
【0094】
本開示の方法におけるNSCは、1つの部位から得て、自家移植片として、同じ対象の別の部位へ移植することができる。さらに、本開示の方法におけるNSCは、遺伝子的に同一のドナーから得て、同系移植片として移植することができる。またさらに、本開示の方法におけるNSCは、遺伝子的に同一でない同一種のメンバーから得て、同種移植片として移植することができる。代わりに、NSCは、ヒト以外の種から得て、異種移植片として移植することができる。強力な免疫抑制薬の開発に伴い、同種移植片、およびブタ起源の神経前駆細胞などの、ヒト以外の神経前駆細胞の異種移植片をヒトの対象に移植することができる。
【0095】
試料の組織は、いずれかの標準的方法によって分離することができる。一実施形態では、組織は、ピペットおよび二価陽イオンを含まない生理食塩水を使用した穏やかな機械的粉砕(trituration)によって分離し、分離細胞の懸濁液を形成する。主として、単細胞を得るための十分な分離が、局所的な過度の細胞密度を避けるために望ましい。
【0096】
NSCの成功した商用的利用のために、多くの連続的継代を通して、安定した増殖能および分化能を有する強固で、ばらつきのない培養を維持することが望ましい。前述のように、本培養法は、CNS発育の異なる領域および年齢由来のNSCにおいて、これらの異なる前駆体特性を維持するのと同時に、個々の細胞系の長期の安定した増殖を達成するために最適化させることができる。
【0097】
このために、驚くべきことに、NSC(NSC)の基質への接着を促進することが、NSCまたは前駆細胞の有糸分裂率を加速する一因となり、したがって、NSCまたは前駆細胞のより強固な培養を著しく向上させることが認められている。特に、局所的な過度の細胞密度を避けること、およびマイトジェン濃度を維持することに加えて、細胞外基質タンパク質の濃度が、NSCの長期の有糸分裂能および分化能に影響を及ぼすことが認められている。細胞外基質タンパク質は、ポリ-D-リシン、ポリ-L-リシン、ポリ-D-オルニチン、ポリ-L-オルニチン、フィブロネクチンおよびその組合せを含み得る。他の細胞外基質タンパク質は、さまざまなアイソタイプ、断片、組換え型、またはフィブロネクチン、ラミン、コラーゲンおよびそれらを組み合わせた合成模倣剤(synthetic mimetics)を含み得る。代わりに、または加えて、本開示の方法は、各個別の細胞が、細胞毒性を生じるか、細胞分化を遅らせることなく、培養の全期間において培養基質に細胞が接着するように、有効な細胞接着(cell adhesion)を促進することができる、その他の全ての適切な物質を含むことができることを認識すべきである。
【0098】
細胞外基質タンパク質は、細胞接着を促進することにおいて有効である可能性があるが、ポリ-L/D-オルニチンまたはポリ-L/D-リシンなどの異なるアミノ酸ポリマーが、各個別の細胞系に対して特定の濃度で、細胞に対して有毒である可能性がある。また、インキュベーションの継続時間は、培養皿の表面に沈着した本ポリマーの最終量に影響を及ぼし、細胞の生存度に影響する可能性がある。本開示の方法で使用されるNSCにおいて、ポリマーの濃度は、約0.1μg/mlから約1mg/mlの間の範囲内とすることができる。一実施形態では、100μg/mlのポリ-D-リシンを、中性pHの0.01M HEPES緩衝液または水で溶解し、培養容器に加える。培養容器を室温で1時間インキュベートする。次に、培養容器を水で完全にすすぎ、使用前に乾燥させる。
【0099】
また、本開示の方法は、細胞外基質タンパク質で培養容器を二重塗布することを含むことができる。一実施形態では、前述のポリ-L/D-オルニチンまたはポリ-L/D-リシンの処理後、培養容器をフィブロネクチンまたはフィブロネクチン誘導体によって処理する。一実施形態では、ヒトの血漿から調製されたフィブロネクチンタンパク質を使用する。しかし、ブタまたはウシのフィブロネクチン、組換えフィブロネクチン、フィブロネクチンタンパク質の断片、合成ペプチドおよびフィブロネクチンの他の化学的模倣剤などの、他のいかなるフィブロネクチンタンパク質の適した形態または供給源も使用することができることを認識すべきである。一実施形態では、約0.1μg/mlから約1mg/mlのフィブロネクチンを加えることができる。
【0100】
ヒトの脊髄由来のNSCの増殖を含む一実施形態では、細胞外タンパク質が培養容器に十分に結合およびコーティング可能となる期間、ポリ-D-リシン100μg/mlで培養容器を処理する。そのような時間は、約5分から約3時間までの間とすることができる。培養容器は、その後水で洗浄することができる。培養容器を空気乾燥した後に、容器を室温で約5分から数時間、約25mg/mlのフィブロネクチンで処理、または37℃で約1時間から数日間、約1mg/mlのフィブロネクチンで処理することができる。その後、フィブロネクチンを取り除き、培養容器は、少なくとも1回洗浄するか、使用するまでPBS中に保存してもよい。
【0101】
あるいは、フィブロネクチンを細胞に直接供給する可溶性因子として増殖培地に加えることができる。本実施形態では、NSCは、フィブロネクチンによる培養容器の処理に加えて、またはこの処理の代わりに、増殖培地にフィブロネクチン1μg/mlを加えることによって増殖させることができる。フィブロネクチンをコーティングした容器の貯蔵時間が比較的短いため、細胞の平面培養時に可溶性因子として付着タンパク質(attachment protein)を増殖培地に供給することは、特にNSCの大規模な商業規模の培養にとって有益である。また、この方法は、例えば、cGMPプロトコルの下で必要とされるかなり正確な状態および再現性が必要とされる神経幹細胞系を製造するため、および治療上の使用のための神経幹細胞系を製造するために有用である。
【0102】
一実施形態では、分離されたNSCを約1,000細胞/cm2から約20,000細胞/cm2の密度で、培養容器に加える。そのような密度は、培養容器における個々の細胞の平均な分散および接着をもたらし、局所の細胞の高濃度化を防止し、NSCの培養物を富化する。
【0103】
一実施形態では、NSCを血清の不存在下で増殖させる。一実施形態では、NSCの有糸分裂および分化能を不安定にさせるのに十分な血清濃度にNSCが曝露するのを避けるために、NSCを規定された無血清培地で培養する。さらに、白血病抑制因子(LIF)または毛様体神経栄養因子(ciliary neurotrophic factor:CNTF)などの特定の増殖因子に対するNSCの曝露もNSCを不安定にさせるので、避けるべきである。
【0104】
NSCの増殖を高めるために、いずれの培養過程の段階でもマイトジェンを培養に加えることができる。マイトジェンは、塩基性線維芽細胞増殖因子(bFGF)、酸性線維芽細胞増殖因子(aFGF)、上皮増殖因子(EGF)、トランスフォーミング増殖因子-α(TGFa)およびその組合せを含み得る。
【0105】
本開示の方法のNSCは、少なくとも2種の異なる培養形態で成長および増殖させることができる。培養の一形態は、一般に、クラスターと呼ばれる凝集した形態、懸濁培養と呼ばれる凝集した形態を含む。培養のもう1つの形態は、懸垂培養(adhesion culture)と呼ばれる分散した、凝集していない形態を含む。
【0106】
本開示の方法のNSCを分散する付着培養(adherent culture)において、本細胞は、最初に個々の細胞が直接基質に接触する単層を形成する。最終的に、インキュベーションの期間後、細胞は、散発的にクラスターを形成することがあり、この場合、底の層の細胞が個々に基質に付着しているのと同時に、少なくとも1つのさらなる細胞の層が底の層上に形成される。そのようなクラスター形成は、培養物が高い細胞密度で播種させたか、高い細胞密度に達することが可能なときに特に発生し、一実施形態では、NSCもしくは前駆細胞の最適な増殖、またはNSCの多分化能の最適な維持のために最小化される。本開示の方法の実施形態における分散型の付着培養では、ヒトNSCを、細胞分裂当たり約4日未満で分裂させることができる。
【0107】
分散型の付着培養のもう1つの特有の特徴は、本開示の方法のNSCがそれぞれの多分化能を保持する娘細胞を発生するように分裂することである。一実施形態では、本開示の方法におけるNSCの分散型の付着培養は、実質的な分化がない状態で、少なくとも20回の細胞倍増を行う増殖能力を含む。大部分のNSCは、それらの神経原性の潜在性を失う前に、少なくとも50回の細胞倍増を超えて増殖できる。一実施形態では、本開示の方法の分散型の付着培養で増殖されるNSCは、ニューロン分化が高められることが実証され、一実施形態では、少なくとも約30%のニューロン分化が起こる。多くの場合、NSCの少なくとも50%は、ニューロンに分化する。分散付着の培養形態が、より好ましい培養形態であるが、異なる培養法は、in vitroまたはin vivoのいずれかでの異なる分化能によって、生来異なる細胞集団の分離を可能にすることができる。
【0108】
また、本方法は、遺伝子組換えまたは支持細胞(feeder cell)の包含なしに、さまざまな供給源からのNSCのクローン分離(clonal isolation)を可能にする。したがって、非常に少数の、好ましくは1000細胞/cm2未満の細胞を、前述のように調製された細胞培養皿に播種することができる。
【0109】
NSCの播種から2、3日後、細胞は、良好に分離したコロニーを形成することができる。本コロニーは、例えば、少なくとも約250細胞から約2000細胞の所望の大きさに増殖することができる。一実施形態では、少なくとも1つの細胞のコロニーは、手で採取し、マルチウェルプレートなどの新しい細胞培養皿に個々に接種する。
【0110】
分離したクローン集団は、連続的な継代によって増殖させ、複数の神経幹細胞系を得るために使用することができる。多くのそのようなクローン細胞系が、脊髄、中脳および後脳などのヒトのCNSのさまざまな領域から分離されている。クローン細胞系は、より高率なニューロンのサブタイプなど、特定の細胞の表現型を富化するために有用である。例えば、チロシンヒドロキシラーゼを発現するドーパミン作動性ニューロン、GABA作動性ニューロン、コリン作動性ニューロンおよび他の特定の表現型のニューロンが濃縮したクローン細胞系を本開示の方法によって分離することができる。
【0111】
一実施形態では、ポリクローナルまたはモノクローナルのいずれかの神経幹細胞系を、さらに特定のサブタイプのニューロンを濃縮にするために導入することができる。中脳または脊髄のNSCからチロシンヒドロキシラーゼを発現するドーパミン作動性ニューロンおよびアセチルコリンを産生するコリン作動性ニューロンなど、特定のニューロンの有効な誘導物質を同定するために、多くの増殖因子、化学物質および天然物質(natural substance)がスクリーニングされている。因子もしくは化学物質または、その組合せを、NSCの分裂期および/または分化期に導入することができる。一実施形態では、ドーパミン作動性の表現型の神経幹細胞系を、パーキンソン病を治療するためのドナー集団としてさらに濃縮させる。
【0112】
さまざまなニューロンのサブタイプを、in vitroで所望の分化パターンを有する幹細胞の分離から得ることができる。in vitroでの結果は、実質的にin vivoで再現可能である。このことは、in vivoでの幹細胞の潜在的な有効性が、in vitroでの幹細胞の分化パターンによって予測できることを意味している。生後の生きた対象への注射後、未分化または分化前のいずれかの状態のNSCは、大体において、in vitroで観察されたin vivoの分化パターンを生じる。したがって、in vitroでチロシンヒドロキシラーゼを産生するニューロンを生じるNSCも、in vivoでチロシンヒドロキシラーゼを産生するニューロンを発生する。逆に言えば、in vitroでチロシンヒドロキシラーゼを産生するニューロンを生じないNSCは、構成的に、in vivoでチロシンヒドロキシラーゼを産生するニューロンを発生しない。
【0113】
しかし、in vitroに存在する分化の合図は、in vivoでの合図と比べて限られる。したがって、分化したニューロンのかなりの画分は、主要な神経伝達物質の表現型を発現しないことがある。求心性または遠心性ニューロンからのシグナルなどの付加的な合図、またはそのような自然のシグナルを模倣する薬剤を、NSCの有糸分裂期またはNSCの分化のいずれかの間に、分化した表現型を再構成するために使用することができる。NSCは、in vitroだけでなくin vivoに存在する合図に反応する能力を有する。したがって、一旦、虚血により損傷した脊髄に移植したならば、脊髄のNSCは、in vitroよりかなり高率のGABAを産生するニューロンを発生する。したがって、NSCは可塑性である。NSCのそのような可塑的な性質は、NSCの多分化能の性質によるものであり、それ自体として、この可塑性を、その特性の方向を変更させるために、NSC集団とさらに結合することができる、表現型を誘導する作用物質および状態を同定するために使用することができる。
【0114】
一実施形態では、そのような再プログラミング(re-programming)は、運動ニューロンの表現型の発現増強をもたらすために、脊髄由来のNSCを処理することを含む。処理の状態は、さまざまな筋細胞または神経冠細胞もしくは神経節ニューロン(ganglionic neuron)などの末梢神経系由来細胞とNSCまたはその分化細胞を共培養することを含む。また、NSCを、運動ニューロンまたは脊髄で発現または産生することが知られる分子のカクテル(cocktail)で処理して、運動ニューロン表現型のNSCの発現を高めることができる。
【0115】
ヒトの中脳におけるドーパミン作動性の表現型を増大させたNSCを誘導するために、リチウム、GDNF、BDNF、プレイオトロフィン、エリスロポエチンなどの分子、セルトリ細胞(sertoli cell)などの細胞由来の条件培地(conditioned media)、またはスクリーニングによって得た他の全ての適した化学物質もしくは細胞、ならびにその組合せによって、NSCを処理する。そのような誘導により、in vivoにおいて、移植したNSCにドーパミン作動性の表現型を発現および維持させることができる。
【0116】
一実施形態では、本開示の方法のNSCは、移植のために分化前の細胞を含むことができる。細胞の最大収量および手法の単純化のために、主に未分化細胞の集団を含む集密培養物(confluent culture)を移植のために収集する。しかし、細胞密度が増加することによって、自発的に分化をまさに始めようとする少数の細胞集団も、存在する可能性があるということを認識すべきである。
【0117】
一実施形態では、NSCの継代は、基質から細胞を収集または分離することを含む。一実施形態では、本開示の方法は、少なくとも1種の酵素を使用して基質から細胞を収集または分離することを含む。酵素処理は、NSCの細胞周期が細胞表面のマイトジェン受容体を不活性化するのに十分なほど短いとき、避けることができる。しかし、ヒトのNSCの細胞周期は、ヒトのNSCが酵素処理に対して感受性がないというほど、げっ歯類のNSCよりもかなり長い。したがって、本開示の方法では、ヒト由来のNSCを収集するために、酵素処理を使用する。ヒトのNSCは、酵素処理の存在下で、一時的にマイトジェンに不応性になることがあるが、マイトジェン受容体の不活性化を繰り返すことで、NSC比率を減少させることができる。
【0118】
一実施形態では、細胞の収集に際し、短時間の遠心分離によって細胞を濃縮する。細胞を、生理食塩水、緩衝生理食塩水などの最終的、臨床的に使用可能な溶液でさらに洗浄および再懸濁するか、代わりに、保存溶液またはハイバーネーション「冬眠」溶液(hibernation solution)で再懸濁することができる。代わりに、細胞を、培地+ジメチルスルホキシド、または他の全ての適切な凍結保護物質などの寒剤で再懸濁し、保存のために凍結させることができる。
【0119】
ハイバーネーション溶液は、長期間の間、生きた細胞の生存度を維持するために配合する。一実施形態では、保存溶液は、すぐに使用できる剤形で生きた細胞を移植手術現場へ送り、すぐに使用するために適応される可能性がある。また、遠く離れた場所に、生きた細胞を送るのに適した状態は、約0℃から約20℃の範囲で少なくとも24時間、安定した温度を維持することができる断熱装置を含む。約0℃から約8℃の間で、約24時間から約48時間保存した生きた細胞は、疾患または状態の治療のために移植可能である。
【0120】
一実施形態では、細胞は、前述の臨床的に使用可能なハイバーネーションまたは凍結液などの溶液中で濃縮させることができる。一実施形態では、細胞を、細胞を投与するための細胞密度と同一または異なってもよい適切な細胞密度に濃縮する。一実施形態では、投与のための細胞密度を、注射部位、注射部位の神経変性状態、有益な効果のための必要最小量、および毒性副作用に対する考慮などの要因によって、約1,000/μlから約1,000,000/μlまで変えることができる。一実施形態では、本開示の方法は、約5,000/μlから約50,000/μlまでの細胞密度で細胞を注射することを含む。
【0121】
増殖した細胞を治療部位に送るために懸濁させた媒質の量を、本書では注射量(injection volume)と呼ぶことがある。注射量は、注射部位および組織の変質状況に依存する。より具体的には、注射量の下限は、細胞の密集する傾向だけでなく、細胞密度の高い粘性懸濁液に対する実際的な液体の処理方法によって決定することができる。注射量の上限は、実際的な手術時間だけでなく、宿主組織を損傷させるのを避けるために必要な、注射量によって発揮される圧縮力の限界によって決定することができる。
【0122】
公知の方法を使用したドナー細胞の細胞生存が低いことから、有効な治療をするために、比較的小さな領域に多量の細胞を送達することが必要とされてきた。しかし、注射量は、宿主組織に対して発揮される静水圧であって、多量の注射量に関連して注射時間が長引くと、手術のリスクが増大する。さらに、ドナー細胞の過剰注射は、宿主の実質組織の圧迫およびその後の損傷につながる。量の制約を補う試みにおいて、公知の方法は、注射のために、細胞密度の高い懸濁液の調製を必要としている。しかし、高い細胞密度は、移植した細胞の密集化を促進し、細胞の移動または拡散を抑制し、制限領域を越えた有効な治療を妨げ、宿主組織へ途切れのない統合(seamless integration)を損なう。
【0123】
対照的に、本開示の方法によって調製された細胞のin vivoでの生存が改善される結果として、注射ごとに必要となる細胞数がより少なくなる。実際、注射時から6カ月後に、最高で3から4倍の注射細胞数が生存していることが明らかにされており、本開示の方法を使用した著明な定量的生存(quantitative survival)が実証されている。また、定量的な生存により、所望の細胞量の再現性のある投与が実現できる。したがって、一実施形態では、細胞を約1,000から約200,000細胞/μlの密度に濃縮する。一実施形態では、約5,000から約50,000細胞/μlを有効な移植のために使用している。他の実施形態では、約10,000から約30,000細胞/μlを使用する。一実施形態では、注射部位当たり約100/μl未満の注射量で懸濁された細胞を治療領域へ送達することができる。例えば、脊髄路に沿って両側的に複数の注射が行われる可能性ある、ヒトの神経変性状態の対象に対する治療では、0.1の注射量および注射部位当たり約100/μlを使用することができる。
【0124】
所望の領域に細胞を注射するのに適したいかなる器具も、本開示の方法で使用することができる。一実施形態では、ほぼ一定の流速で、ある時間にわたって、マイクロリッター以下の量を送ることができる注射器を使用する。針もしくは柔軟な管、または他のあらゆる適した転送器具を通して、細胞をこの器具に充填することができる。
【0125】
一実施形態では、神経変性状態の治療における所望の注射部位は、脊髄の少なくとも一領域を含む。一実施形態では、細胞を、脊髄の頸椎、胸椎または腰椎部などの、脊髄の少なくとも1カ所の特定の分節または部位に移植する。例えば、腰椎部では、わずか5対の神経根が椎骨の脊椎管と交差し、各対の神経根は、各腰椎の高さで脊椎から出て、広い領域に分布している。脊髄の腰椎部における神経根の密度が低いことにより、腰椎部は、特に細胞の安全な注射部位としてよく適している。一実施形態では、細胞を脊髄の軟部組織(parenchyma)の中間帯に移植する。
【0126】
一実施形態では、細胞を約5から約50の部位に注射する。一実施形態では、細胞を脊髄の各側の約10から約30の部位に注射する。少なくとも2カ所の部位は、約100μから約5000μの間隔を離すことができる。一実施形態では、注射部位間の間隔は、約400μから約600μである。注射部位の間隔は、脊髄分節の全体を通して発生する、実質的に中断されておらず、連続したドナー細胞の存在に基づいて、さらにラットまたはブタなどの動物モデルで、約2から3カ月の生存を達成することを示した平均注射量に基づいて決定することができる。一実施形態では、痙縮/硬直などの症状を治療するために、または運動ニューロンの生存に有効な、少なくともいくつかの腰椎部の長さにわたり、脊髄正中の両側に沿って細胞を注射する。ヒトにおける注射の実数値は、動物モデルの結果より推定することができる。
【0127】
一実施形態では、標的の注射部位は、脊髄の灰白質である。灰白質内で、ラミナ(lamina)の特定の高さにNSCを注入するように注射針の先端を配置することができる。例えば、痙縮/硬直を治療するためにGABA/グリシンを産生するニューロンを送達することは、ラミナV〜VIIを取り囲む領域にNSCを送達することである。あるいは、神経障害性疼痛または慢性疼痛を治療するために、頸椎から腰椎までのさまざまな脊髄分節の灰白質後角またはその近接にNSCを送達することができる。あるいは、ALSなどの運動ニューロン疾患を治療するために、頸椎から腰椎までのさまざまな脊髄分節の灰白質前角またはその近接にNSCを送達することができる。
【0128】
本開示の方法の細胞は、in vivoで多数のニューロンを発生させることができる。NSCが移植前に明白に分化前ではないとき、NSCを分化する前に、in vivoで最高で2つから4つの細胞分裂まで増殖させ、それによって、さらに有効なドナー細胞数を増加させることができる。分化すると、ニューロンは特定の神経伝達物質を分泌する。さらに、このニューロンは、in vivoで移植片を取り囲む環境(milieu)に対して、異なる状態に有益な増殖因子、酵素、およびその他のタンパク質または物質を分泌する。したがって、さまざまな状態が本開示の方法によって治療できるのは、移植した細胞がin vivoで多数のニューロンを発生する能力によるためと、神経変性状態が、ニューロン由来の要素などの要素が失われていることが原因であるか、これらを結果的に失ったことによるためである。したがって、増殖因子、酵素およびその他のタンパク質などのようなニューロン由来の要素を欠いていることによりCNS組織の変性を患う対象を、本開示の方法によって効果的に治療することができる。
【0129】
移植したニューロンによって分泌される増殖因子、酵素、およびその他のタンパク質または物質に反応する状態は、テイサックス病、ニーマンピック病、バッテン病、クラッベ病、運動失調、およびその他の状態などの、遺伝性リソソーム病を含む。
【0130】
さらに、治療に関する本開示の方法は、in vitroで増殖して、損傷もしくは変性したニューロンを交換し、他のニューロンに対して抑制効果もしくは刺激効果をもたらし、および/またはニューロンの再生に寄与する栄養因子を放出することができる、細胞を移植することを含む。
【0131】
実施形態は、さらなる運動ニューロンを、損傷または変性したニューロンの代替として供給することを含む。例えば、本開示の方法は、空洞化を埋めるために、脊髄の空洞内に十分な神経の基礎構造を供給することを含む。神経の基礎構造は、外傷性脊髄損傷、遺伝性疾患または他のあらゆる原因から生じる脊髄空洞症に関連する空洞の拡大を減速させることができるならば十分である。また、十分な神経基礎構造を提供することは、脊髄が変性することにより生じるさらなる合併症の軽減に役立つことを認識すべきである。
【0132】
全てのNSCが既知の疾患に対して治療的に作用するわけではない。異なる疾患で影響を受けるニューロン集団の種類は、異なる可能性がある。したがって、治療的に有効なNSCのドナー集団は、失われた神経の要素を交換することに寄与する。例えば、痙縮、発作、運動障害およびその他の筋肉活動亢進障害の治療は、GABAまたはグリシンを産生する抑制性ニューロンに分化することができる治療効果のある細胞量を供給することを含み得る。異なるNSC集団は、分化したニューロン表現型を調べることによってin vitroで評価することができる。それから、in vitroでの分化パターンを、in vivoでの適切な表現型を産生する細胞の有効性を予測するために用いる。これは、適切な神経伝達物質の表現型に関してだけでなく、ニューロンの適切な形態、遊走およびその他の表現型の特性に関しても予測される。
【0133】
一実施形態では、症状の病因と関連して損傷および破壊したニューロンのサブタイプに対応するニューロンのサブタイプを産生することができるNSCを移植する。例えば、対象の興奮性回路(excitatory circuit)の活動亢進は、脊髄の外傷、胸部/胸腹部の大動脈手術、脳卒中、てんかん、脳外傷、ハンチントン舞踏病、膀胱性尿失禁、便通の亢進、および損傷または遺伝的症状から生じるその他の制御不能の筋肉収縮による、遺伝的症状またはニューロンの損傷によって引き起こされる可能性がある。痙縮、発作またはその他の活動亢進は、多くの異なる病因の起源による脊髄とは対照的に脳で生じる。例えば、焦点てんかんは、GABAの発揮する回路に対する調性制御(tonal control)が失われることによる、非制御の活動亢進から発症すると考えられる。このため、本開示の方法は、in vitroで増殖されたNSCの移植によって、影響を受けた領域にGABAまたはグリシンなどの抑制性神経伝達物質(inhibitory neurotransmitter)を供給することを含む。例えば、痙縮、発作およびその他の活動亢進の場合、痙縮、発作およびその他のニューロンの活動亢進に関連する少なくとも1つの活動亢進の神経回路を弱めるために、GABAまたはグリシンを産生するニューロンなどの抑制性ニューロンに分化することができる多くのNSCを、移植されるin vivoで発生させる。したがって、本開示の方法は、てんかんおよび同様の発作の状態を治療するために適用することができる。
【0134】
また、本開示の方法は、不全麻痺、完全麻痺、痙縮、硬直またはその他全ての脳虚血によって生じる運動、言語もしくは認識症状を治療するために適用することができる。脳虚血は、脳における脳卒中発作、または脳への血液循環がかなりの時間、中断される心臓発作により生じる可能性がある。したがって、前述の脊髄虚血と類似している。一部の脳卒中の対象は、記憶喪失、完全麻痺または不全麻痺などの他の障害に加えて、中枢の起源の発作を発症する。また、これらの脳虚血による障害は、海馬および/または他の脳領域における抑制性介在ニューロンの選択的な減少による可能性がある。したがって、本開示の方法は、不全麻痺、完全麻痺、痙縮、またはその他の運動、言語もしくは認識症状を患う脳卒中の対象を治療するために適用することができる。
【0135】
不全麻痺、弛緩性対麻痺、およびALS、外傷性脊椎損傷、虚血性傷害または遺伝性疾患に起因する筋収縮の制御不能などの、筋収縮の制御不能に関連するその他の状態の場合、本開示の方法は、運動ニューロンの消失を遅らせるために、十分な栄養的影響を発揮するニューロンの移植を提供することを含む。特に、本開示の方法は、最適の生物学的利用度の条件下で、変性する運動ニューロンを送達することができる栄養分子を分泌する移植NSCの能力を促進する。そのような栄養分子は、スーパーオキシドジスムターゼ(SOD1)などのエキソサイトーシスにより分泌されたスーパーオキシドジスムターゼ、リソソーム酵素、および細胞産生の抗酸化物質などの非タンパク質性分子を含む。移植した細胞によって分泌されるその他の栄養因子は、グリア細胞株由来神経栄養因子(GDNF)、脳由来神経栄養因子(BDNF)、血管上皮増殖因子(VEGF)、プレイオトロフィン、血管内皮増殖因子(VEGF)、エリスロポエチン、ミッドカイン、インスリン、インスリン様増殖因子1(IGF-1)およびインスリン様増殖因子2(IGF-2)、または他の全ての有益な栄養要素を含むことができる。
【0136】
広範囲にわたる神経変性状態を治療するために、本開示の方法の能力に寄与するもう1つの要因は、存在するニューロン線維に沿って広範囲に移動するNSC分化細胞の能力を含む。移植した細胞の移動は、ドナーニューロンおよび/または神経膠の全体的な分布および統合、ならびにそのような細胞によって分泌される治療要素の全体的または分散した供給に結び付く。
【0137】
細胞の広範な移動は、それを必要とする対象の神経系および身体の全体を通して、主要な治療用のタンパク質および物質の全体的および安定した送達を可能にする。したがって、本開示の方法の細胞は、治療用のタンパク質および物質のための有効な送達媒体である。そのような送達の目的のために、本開示の方法は、腸、筋肉、血管内系および皮下部位を含む神経系以外の領域に加えて、CNS実質、脳室、硬膜下腔、クモ膜下腔、硬膜外腔、末梢神経系部位を含む神経系内のさまざまな部位へ細胞を移植することを含む。
【実施例】
【0138】
(実施例1:ヒト脊髄神経幹細胞/前駆細胞の増殖)
在胎齢およそ7〜8.5週の少なくとも1つのドナーからの脊髄を得る。機械的粉砕を用いて、Ca++およびMg++を含まないリン酸緩衝生理食塩水中、該脊髄の単一の近接組織を解離させる。次いで、得られた細胞懸濁液を、ポリ-L-オルニチンまたはポリ-D-リシンおよびヒトフィブロネクチンまたは他の細胞外基質タンパク質によって予備コーティングした組織培養プレートに播種する。組織培養処理したプレートまたはフラスコを、100μg/mlのポリ-D-リシンと共に、室温で1時間インキュベートした。次いでそれらを水で3回洗浄し、乾燥した。次にそれらを25mg/mlと共に、室温で5分間インキュベートした。時には、10mg/mlのフィブロネクチンを、室温で1時間用いた。時には、1mg/mlのフィブロネクチンを、37℃で18時間用いた。N2(DMEM/F12 + インスリン、トランスフェリン、セレニウム、プトレシン、およびプロゲステロン)からなる培養培地に、1種のヒト組換え塩基性線維芽細胞成長因子(bFGF)を添加した。一実施形態において、0.1ng/ml〜100ng/mlの範囲を使用することができる。一実施形態において、最適には、10ng/mlのbFGFが使用される。
【0139】
得られた最初の培養物は、単層中、有糸分裂後ニューロンおよび増殖性NSCからなる。引き続き、およそ5日からおよそ20日培養後、分裂性のネスチン陽性NSCは、非分裂性ニューロンまたは緩徐分裂性神経膠より優勢となる。これらの培養条件下、NSCを増殖のために選択的に促進させる。増殖NSC集団をトリプシン使用などのマイルドな酵素処置によって継代接種する。該細胞を、無血清または実質的に無血清の培地中で培養する。低濃度の血清は該細胞により忍容され得るが、血清はNSCのグリア分化を促進するLIFおよびCNTFなどの多くのサイトカイン類を含有しているため、該細胞を血清に曝露することは避けることが最良である。したがって、継代接種の間、血清ではなく、トリプシン阻害剤などの特定の酵素阻害剤を添加することによって、使用酵素を抑制する。各継代接種時に、採収した細胞数をカウントし、さらに増殖させるために、フラクションを再播種する。図1に示されるように、本発明の方法を用いて、ヒトNSCを、それらの増殖性および分化性を維持しつつ、個体数が1018倍超増加する増殖をさせることができる。該細胞は再現可能に増殖できる。図1に示されるように、該細胞の連続的継代を3回繰り返したところ、再現性のある細胞増殖曲線および細胞倍増時間が得られた。増殖の間、ほとんど全ての細胞が有糸分裂神経上皮細胞のin vivoマーカーであるネスチンを発現し、タイプ3ベータチューブリンおよびGFAPなどの分化ニューロンおよび神経膠の抗原を有さない。また、細胞は、committed neuronal progenitorの可能性があるPSA-NCAMに関する免疫染色、希突起膠細胞のマーカーであるO4およびGalC、ならびに放射神経膠のマーカーであるRC2でも陰性である。したがって、免疫染色によって判定したところ、NSCは長期増殖期間にわたって、それらの抗原プロフィルの発現を安定に保持する。形態およびネスチン発現の例はそれぞれ図2Aおよび2Bに示されている。
【0140】
(実施例2:ヒト脊髄神経幹細胞/前駆細胞の分化)
NSCの増殖中、任意の時点で、培養物中のbFGFなどのマイトジェンを取り出すことによって、培養物を分化させることができる。マイトジェン除去後、約1〜3日以内に、引き続きNSCの分化が起こり、別な異種細胞形態が明らかである。分化のおよそ4〜7日までに、MAP2c、タウ、およびタイプIIIベータチューブリンなどのニューロン特異的抗原を免疫染色によって可視化することができる。およそ12〜14日までに、細胞内タンパク質輸送の明らかな局在化と共に、培養物全体にわたる、長形、索状の軸索性過程が明らかになる。およそ28日までに、シナプシンおよびシナプトフィジンなどのシナプスタンパク質は軸索終末内へ局在化し、斑点状の染色として現れる。該ニューロンの長期成熟をさらに促進するために、星状膠細胞のフィーダー層をさらに供給することができる。図3に示されるように、ヒト脊髄NSCの分化によって、ニューロンと神経膠との混合培養物が生成され、混合物中、ニューロンはタウ、MAP2ab(A)およびタイプ3ベータチューブリン(B)などのニューロン特異的抗原を強く発現し、該培養物のおよそ50%を含む。図3Bに示されるように、該培養物は、数センチメートル伸長する長く束ねられた軸索ケーブルを自然に生成させる。図3Cに示されるように、ニューロンのかなりの部分がGABA作動性である。コリン作動性運動ニューロンもまた該培養物中に存在する(図3D)。培養物中かなりのGABAニューロンが存在することにより、一定の回路におけるGABA産生の減少に起因する種々の神経学的状態の治療に対するヒト脊髄NSCの有用性が予想される。同様に、コリン作動性ニューロンの存在により、ヒト脊髄NSCが運動ニューロン分化の能力を有することが実証され、運動ニューロンの漸進的変性に起因する種々の運動ニューロン疾患の治療に対する有用性が予想される。治療のためには、NSCをさらなる表現型増強条件の有無で増殖させ、採収し、欠損神経領域に注射する。
【0141】
(実施例3:ヒト中脳神経幹細胞/前駆細胞の増殖)
在胎齢7〜8.5週胎児の1つの中脳組織を得る。実施例1に記載したとおり、中脳組織からNSCを得る。該細胞を、160日の正味培養期間にわたって連続的に継代し、得られた増殖を図4に示す。増殖期間を通して、NSCはそれらの多分化能および神経形成能、ならびに、ドーパミン作動性ニューロンをもたらす分化能を安定して保持する。ドーパミン作動性ニューロンは、チロシンヒドロキシラーゼ(TH)およびドーパミントランスポーター(DAT)のニューロン発現によって評価される。
【0142】
DAT発現はドーパミン産生ニューロンのマーカーである。ニューロン中のDAT発現は、培養物中の分化ニューロンのシナプス膜を越えて放射標識ドーパミンを輸送するその機能を測定することによって評価できる。分化したヒト中脳NSCおよび単クローン誘導したヒト中脳NSCからの培養物中のDAT機能は、放射標識ドーパミン取り込みアッセイによって評価される(図5)。該アッセイの結果により、ヒト中脳NSCの強い機能的ドーパミン作動活性が示される。それはさらに、NSCの単離モノクローナル集団によって、ドーパミン作動性表現型を増加することができ、これが特に、分化の際にドーパミン作動性ニューロンを高い割合へと増加させる傾向があることを示している(図5)。
【0143】
増加したドーパミン作動性ニューロンを生成するNSCは、パーキンソン病の治療に特に有用である。同様に、特定の表現型に関する分化を増強させるために、組織からの単離時点で予めプログラミングされたNSCを用いて、アルツハイマー病の治療に有用な前脳コリン作動性ニューロン、ALSなどの運動ニューロン疾患の治療に有用な脊髄コリン作動性ニューロン、うつ病の治療に有用なセロトニン作動性ニューロン、癲癇およびハンチントン舞踏病の治療に有用なGABA作動性ニューロンなどの他の特定の所望ニューロンを単離することができる。
【0144】
(実施例4:ヒト中脳神経幹細胞/前駆細胞の分化)
ヒト中脳NSC/前駆体は、実施例2に記載されるとおり分化させることができる。NSCの有糸分裂期の間、またはそれらの分化の間、種々の外来因子によって該培養物を処置することによって、所望の表現型の割合を高くすることができる。ヒト中脳NSCからドーパミン作動性表現型を増加させることができるこのような因子の例は、図6Aおよび6Bに示されるとおり、セルトリ細胞からの馴化培地によって実証される。
【0145】
図6Aおよび6Bに示される試験において、凍結保存された神経幹細胞を解凍し、マイトジェンの存在下、4ウェルチャンバースライド中、1ウェル当たり40,000細胞の密度で塗布し、6日間増殖させ、マイトジェンを除去して細胞を8日間分化させた。セルトリ細胞馴化培地(SCCM、N2a中1:1に希釈)への曝露のタイミング及び時間に基づいて、細胞を4つの群に分けた。1つの群は、増殖時および分化時、SCCMに曝露し(条件1);第2の群は、増殖時のみ曝露し(条件2);第3の群は、分化時のみ曝露し(条件3);第4の群は、SCCMに曝露しなかった(対照またはcont.)。培地は1日おきに交換し、マイトジェンは増殖期の間、毎日添加した。1条件当たり、4つのウェルを維持して複数マーカーに関して染色させ、3つの細胞系:796MB、527MB、および566SCを試験した。脊髄由来の566SC細胞は測定可能なTH陽性ニューロンを有さず、図は省略した。
【0146】
分化の際、4%パラホルムアルデヒドを用いて細胞を固定し、MAP2[MAP2abサブタイプを認識するAP20クローン(Sigma)]、ニューロン特異的βチューブリン[TuJ1(Covance)]、およびチロシンヒドロキシラーゼ(Pel-Freez)に対する抗体、ならびにGFAP(Dako)およびGalC(Chemicon)を用いて免疫染色した。免疫染色した細胞を、40×対物レンズを用いてカウントし、各ウェルに関して少なくとも3つの視野をカウントした。いずれの条件下で維持された細胞の分析の際にも、GFAP+またはGalC+細胞は、ほとんど、または全く検出されず、したがって、これらの抗原は分析から除外した。また、566SC細胞は、記載された条件下で維持した後、高濃度過ぎて定量化できなかったので、最終分析に含めなかった。
【0147】
この分析によって、外来因子による処置によって、ヒト中脳NSCが影響されることが示され、ドーパミン作動性ニューロンをさらに濃縮するのに有用なタンパク質因子または化学物質を探索することができることが示される。したがって、パーキンソン病などの特定の適応症の治療を目的として特に有用な集団を得るために、ヒトNSCを使用することにより、新規の合成/天然化学物質およびタンパク質因子を効果的にスクリーンすることができる。
【0148】
(実施例5:ヒト脊髄神経幹細胞/前駆細胞の移植によるラットにおける痙攣および硬直の処置)
一過性脊髄虚血を誘導するために、先にTaira(1996)に記載された方法を用いる。Sprague Dawley(SD)ラットをハロタン(1.5%)で麻酔する。2 Fr Fogarty(登録商標)カテーテルを左大腿動脈を通して、左鎖骨下動脈のレベルまで下行胸椎大動脈に通す。大動脈閉塞のレベル下の遠位動脈圧(DBP)を測定するために、ポリエチレン(PE-50)カテーテルによって、尾動脈にカニューレ挿入する。
【0149】
0.05mlの生理食塩水を用いて、動脈内バルーンカテーテルの膨張時に脊髄虚血を誘導する。動脈閉塞時の全身低血圧は、PE-50カテーテルによりカニューレ挿入した頚動脈からの動脈血(10.5〜11cc)の一部抜き取りにより再現する。およそ40mmHgの全身低血圧をこの方法によって誘導することができる。閉塞の有効性は、尾動脈で測定されるDBPの即時的および持続的低下によって証明される。脊髄虚血誘導のおよそ10分後、バルーンを収縮させ、頚動脈から抜き取った血液を再注入する。動脈血圧が安定化したら(再流動後約20〜30分以内)、動脈ラインを除去し、傷を閉じる。
【0150】
脊髄虚血の誘導後、運動機能の回復を、改変21ポイントオープンフィールドロコモータースケール(BBBスケール)を用い、およそ2日間隔で評価する。0〜4のBBBスコアを有する動物のみを、移植試験のための実験群用に選択する。
【0151】
虚血損傷の約7〜21日後、0〜4のBBBスコアを有する痙攣ラットを大気中、1.5〜2%ハロタンで麻酔し、脊髄ユニット装置内に入れる。次いで、Thl1〜L2脊椎の部分的椎弓切除を実施する。80〜100μmの先端直径を有するガラスキャピラリーを圧力制御微量注射器に接続する。1回の注射当たり5,000、10,000、15,000または20,000のヒト神経幹細胞/前駆細胞を含有する0.5μlの細胞懸濁液をラットに注射する。脊髄の各々の側(左および右)に、L2〜L6曝露セグメント間に均一に分布した、合計6〜8回の注射を、各ラットに投与する。注射の中心を、中央灰白質内(V層〜VII層)に標的化する(L3レベルにおける脊髄の背側表面からの距離:1mm)。移植後、切開部を3%H2O2およびペニシリン/ストレプトマイシン混合物で洗浄し、2層で閉じる。次いでラットを回復させる。
【0152】
脊髄移植の3日前に、全動物にFK-506 (Prograf;フジサワ;1mg/kg;i.p.)による免疫抑制処置を開始する。移植後、全動物は全生存期間の間、免疫抑制処置を毎日受ける。これら移植片の免疫拒絶は、FK-506によって効果的に防ぐことができる。該ラットはおよそ2または7週間生存する(各時点でn=5)。
【0153】
生存期間の最後に、ラットをペントバルビタール(40mg/kg;i.p.)によって麻酔し、ヘパリン化生理食塩水で1〜2分間、引き続き0.1Mのリン酸緩衝液(PB)中4%パラホルムアルデヒドによって経噴門潅流する。脊髄を切開し、同一固定液中、4℃で一晩後固定する。後固定後、脊髄組織を段階的蔗糖液(10、20および30%)中で合計3日間凍結保護する。次いで、凍結した冠状、側矢状または水平状の脊髄切片(10〜30μm)を切断する。免疫組織化学では、遊離浮遊切片(30μm)を、5%正常ヤギ血清(NGS)、0.2%トリトンX100(TX)を含有するPBS、0.1M(pH=7.4)中に、室温で2時間入れ、非特異的タンパク質活性を阻止する。この後、種々のヒト一次特異的抗体と共に、4℃で一晩インキュベーションする。
【0154】
一次抗体と共にインキュベーションした後、切片をPBS中で3回洗浄し、蛍光マーカーが結合したヤギ二次抗ウサギ抗体またはマウス抗体(Alexa 488または594;4μl/ml;Molecular Probes)と共にインキュベートする。ブロッキング調製物および抗体調製物は全て、0.1MのPBS/0.2%TX/5%NGS中で作製する。二重標識実験では、種々の種からの一次抗体を同時に適用し、引き続いて、種々の蛍光マーカーに結合させた二次抗体を適用する。対照実験では一次抗体を省略する。一般的な核染色では、最終二次抗体溶液にDAPI(3μl/ml)を添加する。染色後、切片を室温で乾燥し、Prolong 抗退色キット(Molecular Probes)で覆う。
【0155】
Leica 蛍光顕微鏡を用いてスライドを分析する。Olympusデジタルカメラで画像(512×512ピクセル)をキャプチャーし、Adobe Photoshop 5.5(Adobe Systems、マウンテンビュー、カリフォルニア州)によって処理する。二重染色切片における種々の抗体の同時局在化を確認するために、Nikon顕微鏡に据えつけた測光CCDを含むDelta Vision 逆応答顕微鏡システム(Applied Precision社)によって画像をキャプチャーする。一般に、0.1μmまたは0.2μm離れた60個の光学切片を取る。使用されるレンズは20×、40×および60×(NA 1.3)である。データセットを逆応答させ、Silicon Graphics Octane ワークステーション上、SoftWorxソフトウェア(Applied Precision社)を用いて分析する。
【0156】
立体解析学的で偏りのない系統的なサンプリングを用いて、ヒト核NUMA抗体に対して免疫反応性の移植ニューロンの総数を予測する。L2〜L6脊髄セグメントから採取した、先に染色した各々10番目の切片を、fractionator sampling schemeを適用後、立体解析学的定量化のために使用する。アパーチャー数値1.3を有する100×油浸漬対物鏡を用い、Leica DMLB顕微鏡によって、視像(1μmの厚さ)を得る。デジタルカメラ(Olympus)、および、StagePro制御電動式Zステージ(Media Cybernetics)に付属のImageProソフトウェア(Media Cybernetics)を用いて、視像をキャプチャーする。移植細胞の総数は、fractionator formula:N=Q×1/hsf×1/asf×1/ssf(式中、Nは陽性核の総数、Qはカウントした細胞の合計、hsfは高さのサンプリング分数、asfは面積のサンプリング分数、およびssfはスライスサンプリング分数である)を適用して算出する。
【0157】
虚血脊髄における移植したヒトNSCの三次元再構成視像を提供するために、連続的に切断した脊髄から採取した、先に保存した画像を使用する。三次元再構成のために、平均して約60〜100の連続切片を用いる。第1の工程で、Ellipseソフトウェアを用いて、連続画像のスタックを開き、特注開発されたAllignモジュール(VidiTo、SK)を用いてアラインさせる。アラインメントの方法は、全ての連続脊髄画像における2つの形態学的参照点を規定すること(第1の点:中央カナルの中心;第2の点:背角の内側境界)、および引き続いて、全画像のコンピュータ処理アラインメントからなる。次いで背角と腹角の境界を確認するために、Laminar Mapsモジュール(Ellipse)を用いて先にアラインした画像のスタック上にラインを引く。最後に、3-Dコンストラクター(Media Cybernetics)を用いた三次元再構成のために、先にアラインし、層状マップ標識した画像のスタックを用いる。
【0158】
ラット星状膠細胞上ヒト脊髄NSCの2週間から3週間の培養により、培養物の大多数において、ニューロン表現型の時間依存性の成熟および発達が示された。これは、NSEまたはMOCに対するヒト特異的抗体による染色によって確認される。十分に発達した軸索樹状突起間樹を有する多数のニューロンもまた確認される。NSE陽性ニューロンの大多数(85〜90%)はGABA陽性である。軸索および樹状突起におけるシナプトフィジンの発現が、いくつかのMOC陽性ニューロンにおいて見られる。
【0159】
虚血損傷の21日後に移植した場合、細胞の強健な生存が見られる。これは、全てがヒト特異的な抗体のNUMA、MOC、またはNSEに対して免疫反応性である、明白に確認される両側性の移植片として発現する。移植された脊髄セグメントから採取した水平切片の分析により、個々の注射部位間にNUMA陽性細胞の明らかな移動が示される。NUMA陽性細胞の大多数は、MOC免疫反応性との同時局在化を示す。
【0160】
共焦点顕微鏡によって分析されたNUMAおよびGABA抗体による脊髄切片の二重標識化により、平均25〜35%のGABA陽性細胞が明らかになる。7週間生存時の全ての移植動物に、GABA作動性表現型の一貫した発現が見られる。
【0161】
同時点(すなわち、移植後7週目)でのシナプトフィジンおよびNUMA抗体による二重染色により、移植片内に高密度のシナプトフィジン陽性網状構造が示される。
【0162】
NUMA陽性細胞はGFAP抗体による同時局在化を時々示すのみとなる。これらの細胞は移植片周囲に典型的に局在化する。
【0163】
NUMA陽性細胞の立体解析学的予測により、個々の移植片内に平均75,460±5,697の残留移植細胞が示される。これは、もともと注射されたものよりも平均3〜3.6倍多い細胞である。細胞周期抗原のKi67は活性有糸分裂細胞のマーカーである。移植後2週間目または7週間目における脊髄切片の染色により、移植後わずか2週間目で、hKi67免疫反応性が示される。生存7週間目では、Ki67陽性細胞は時折存在するのみである(1〜2細胞/10切片)。これらの結果により、移植したヒト脊髄NSCおよびそれらの後代は、最初の2週間は平均約3回の細胞倍増に相当する数で増殖し、次いで分裂終了となり、安定して組み込まれることが示される。
【0164】
MOC抗体およびDAPIにより染色した40μmの厚さの連続脊髄切片(総数150〜200切片)から取った画像を用いることによって、移植したL3〜L5セグメントのボリューム再構成を実施する。三次元移植片再構成により、灰白質の領域内に分布した、体軸方向に方向付けられたMOC陽性移植片が十分に認識されることが示される。図7および図8に示されるように、ヒト脊髄NSCの機能的効果は、該細胞を移植し、BBBスコアリングによって、運動利益の回復を測定することによって評価される。
【0165】
本試験において見られた挙動回復の程度に関して、移植後、3つの主要な群が見られた。第1のものは、もっとも強健な回復および歩行能力を示した動物(BBB<16)、第2のものは、下肢の3つの関節全ての活動的な運動性において改善を示したが、立つことはできなかった動物(BBBは8付近)、そして、第3の群は、何も回復を示さなかった動物(すなわち、非レスポンダー)。移植片に対する応答性の違いの理由は明らかではないが、阻害困難な主要求心性神経および/またはα運動ニューロンに関して、移植片位置のわずかな違いがある役割を演じていると我々は推測している。さらに、本試験では、動物がわずか3カ月しか生存しなかったことに注意すべきである。移植後の長期的生存および身体的回復の継続は、高程度の機能的回復に関連している可能性が高いと我々は推測している。それにもかかわらず、処置群とは対照的に、培地のみを注射された動物ではいずれも有意な回復は見られなかった。
【0166】
(実施例6:ヒト脊髄NSCの移植による運動ニューロン疾患の処置)
開示された方法のNSCは、強力で顕著な臨床的利益と生物学的利益の双方を与える。そのため、開示された方法により、ALSなどの中枢神経系全体に散在した状態、ならびに上記の脊髄虚血などの特定の領域に局在化した状態の双方の処置が可能になる。ALSにおいて、腰髄における移植では、セグメントの運動器官の他の重要な部分、すなわち、呼吸運動を担っている頚髄運動ニューロンカラムが除外され得るが、脊髄にNSCを移植する開示された方法は、髄全体にわたって宿主の運動ニューロンに広範囲の作用が生じ得るCSF内への、移植した細胞からのBDNFおよびGDNFならびに他の因子の放出を促進する。
【0167】
驚くべきことに、神経変性脊髄環境の腰髄セグメント内へのヒトNSCの部分的移植片は、生存し、大量に神経分化し、運動ニューロンの生存を促進して移植部位と他の場所の双方で機能することが判明した。NSCは、ヒトALS(筋萎縮性側索硬化症)のモデルであるSOD1 G93Aラットの症状の発症を有意に遅らせ、それらの寿命を延ばす。
【0168】
SOD1 G93Aラットは、神経病理学およびALSの特に攻撃的な形態の臨床的症状の包括的モデルである[(Nagaiら、2001年;Howlandら、2002年)]。ニューロンの広範な分化が生じ、引き続き、分化したニューロンが宿主ニューロンとシナプス接触を形成し、GDNFおよびBDNFを発現し放出するSOD1 G93Aラットおよびマウスの腰髄内に、ヒト胎児脊髄のNSCを移植することができる。例えば、劇症性運動ニューロン疾患を特徴とするラットSOD1 G93Aモデルを、該疾患におけるNSCの有益な作用を試験し実証するために用いることができる。このため、開示された方法で使用されたNSC移植片は、神経変性環境において十分に生存し、強力な臨床的効果を発揮する。これらの効果の少なくとも一部は、運動ニューロン成長因子を発現し放出するこれらの移植片の能力に関連している。したがって、腰突起に限定された制限された移植スケジュールにも関わらず、開示された方法の移植NSCは、劇症性運動ニューロン疾患の発症および進行を遅らせ、これらの動物の寿命を10日超延ばす。
【0169】
移植前に、線維芽細胞成長因子(FGF-2)を含有する無血清培地において、8週在胎齢の死後ヒト胎児の脊髄組織からのヒトNSC(NSI-566RSC)を約10〜12継代増殖させる(Joheら、1996年)。これらの細胞の結果をヒト核抗原(HNu)に対する抗体によって確実に追跡する(Yanら、2003年)。これらの細胞を用いる全ての外科的操作は、本明細書に参照として組み込まれている、Johns Hopkins Medical Institutionsの動物の管理および利用委員会によって承認されたプロトコルに従って、ガス麻酔(エンフルラン:酸素:亜酸化窒素=1:33:66)および無菌的方法を用いて実施される。
【0170】
顕微鏡先導下、Kopf脊髄定位ユニットに乗せた雌雄混合の9週齢(220〜300g)SOD1 G93Aラットの腰プロチュバランス(L4 & L5)内に、生きているNSCまたは死んでいるNSCを移植する。死細胞は、移植前に、凍結と解凍の反復により調製する。細胞懸濁液は、サイラスティックチューブを介して、10μlのHamilton微量注射器に連結したプルド面取りガラスマイクロピペットを用い、腹側角の両側の腹側角を目標に(注射部位1つ当たり5×104 NSC、片側に4つの注射部位)、およそ8回の注射によって、無菌条件下、送達される。非処置動物またはシクロスポリンを受けている動物で移植片生存が1カ月を超えないことを示しているパイロットデータに基づき、免疫拒絶を防ぐために、全ラットにFK-506(1mg/kg i.p.)を与えた。
【0171】
ラットを、運動の強さに関して試験し、週に2回体重を量る。運動の強さ試験は、Basso、BeattieおよびBresnahan(BBB)運動等級評価(Bassoら、1995年)、および傾斜平面評価(RivlinおよびTator、1977年)を含んだ。BBBスコア試験では、動物をオープンスペースで、約4分間または5分間試験する。全ての運動機能を記録し、スケールに従って評価する。傾斜平面試験では、ラットを傾斜平面マットの上に置き、ラットの位置が約5秒間安定化できる最大の箇所まで角度を調整する。次いでこの角度を、その動物の傾斜平面スコアとして記録する。BBBおよび傾斜平面スコアを、hoc試験後、MANOVA、引き続き、Fisher LSDによって解析する。疾患の発症は、体重が急激に減少し始める点として定める。移植タイプ(生細胞移植片または死細胞移植片)の効果としての疾患経過は、(スチューデントのt検定によって)2つの群間の発症時の年齢および死亡時の年齢を比較することにより、ならびにKaplan-Meier生存解析、引き続いて長期ランク試験により解析する。
【0172】
ラットのBBBスコア(下記参照)が3未満で、1つの関節のみが動くか、または全く動きがなく、その動物が瀕死状態と考えられる段階で、潅流-固定により安楽死させる。
【0173】
4%の中性緩衝パラホルムアルデヒドによって潅流させた動物からの組織を調製する。付着根を有する胸-腰脊髄セグメントおよび腰神経を、同一の固定剤中、さらに4時間浸漬することによってさらに固定する。移植領域全体およびその上下の縁1mmを含むブロックを冷凍保護し、さらなる処置のために冷凍する。L3-S1根を、全体を着けた調製物として別々に処理するか、またはガラスピペットの熱凝固端を用いて細根を分離した後に処理する。ブロックを横断面または正中面(35μm)で切片化する。NSC生存および分化を、多くの場合、ヒト特異的マーカー、HNuと他の細胞マーカーとを組み合わせた二重標識免疫蛍光によって試験し、Yanら、2004年に記載されているとおりに実施する。
【0174】
NSC分化を試験するために、HNu(+)細胞の総数をカウントする非立体解析学的方法、ならびに我々の免疫蛍光調製物からランダムに選択した高強力(100×)フィールド上のHNuおよび表現型マーカーによって二重標識した細胞を用いる。移植領域にわたっておよそ1mm離れている6つの切片の各々における1つのフィールドを、各動物から用いる。各場合から、カウントした6つのフィールド全てからのHNu(+)の数および二重標識のプロフィルをプールし、1つの実験プロトコルごとに群分けする。各処置群(一群当たりn=6)に関して、単一および二重標識細胞の平均数を出す。
【0175】
生細胞または死細胞を移植したラット(各n=4)における運動ニューロンの生存を評価するために、128日齢で殺処置したラットの組織を評価する。立体解析学的要件(Yanら、2004年)に関して各ラットからのL3-S1部位における6つの切片おきにサンプリングし、明確な核および神経細胞体の直径が>35μmを有する多極細胞として同定されたα運動ニューロンを、Yanら、2004年に記載されたとおり光学精留塔によってカウントする。生細胞を移植した動物対死細胞を移植した動物間の違いを、スチューデントのt検定によって解析する。
【0176】
運動神経栄養因子のELISA判定のため、ガス麻酔下、動物の第4脳室から25G注射器によってCSFをサンプリングする。移植部位およびそれらに隣接する領域を含有する組織サンプルを、1mmの厚さの新鮮脊髄スライスから横断切開する。CSFまたは組織サンプルを処理し、総タンパク質を、Shengら、2003年に記載されたとおりに先ず測定する。E-Max免疫アッセイシステム(Promega、マジソン、ウィスコンシン州)を用いて、CSFおよび脊髄サンプル中のGDNFおよびBDNFの濃度を測定する。TMB-クロモゲンの吸光度を450nmで読み取る。生移植片、移植片隣接領域および死細胞移植片からのサンプル間における濃度の分散を、一元配置ANOVA、引き続き、チューキーの多比較hoc後検定によって解析する。生細胞移植動物対死細胞移植動物間のCFS濃度の差を、スチューデントのt検定によって解析する。
【0177】
運動神経栄養因子をウェスタンブロッティングするために、ELISAに関して調製したCSFまたは脊髄からのタンパク質サンプルを、分子量マーカーと共に電気泳動させ、ニトロセルロース膜に移す。ブロットを、5%のロバ血清を含有するTBS、pH7.4中でブロックし、次いで、先ずGDNF抗体およびBDNF抗体(1:500;一晩、4℃)中、次いでHRP結合ロバ抗ヤギIgG(GDNFに関して)および抗ウサギIgG(BDNFに関して)(1:2000;Jackson ImmunoResearch)(1hr、RT)中でインキュベートする。抗体は全て、5%のロバ血清を含有するTBS中で希釈する。ブロットをSuperSignal Chemiluminescent Substrate(Pierce)によって展開し、Kodak-XARフィルム(Eastman Kodak、ロチェスター、ニューヨーク)に曝露する。次いで、ブロットをストリップし、βアクチン抗体(1:500、Sigma)およびHRP結合ロバ抗マウスIgG(1:10000、Jackson ImmunoResearch)によって再ブロットする。免疫反応性のバンドをBio-Rad Quantity Oneソフトウェア(Bio-Rad Laboratories、ヘルキュールス、カリフォルニア州)によって解析する。バンド密度比(GDNFまたはBDNF:βアクチン)を動物ごとに算出し、群の平均値を、ELISA実験の場合のように、統計解析に入れる。
【0178】
移植後22週目のSOD1 G93Aラットの脊髄におけるヒトNSCを、ヒト特異的HNu抗体を用いる免疫染色によって同定する。HNu(+)細胞は、腹角(A)内で生存し、それらの大多数が、微小管関連エピトープであるTUJ-1などのニューロン系列マーカーによって染色されることが示される。ヒトNSCは、それらのヒト核タンパク質(HNu)識別特性によって同定され、それらの表現型の結果は、HNuに関する二重免疫細胞化学、ならびに神経前駆細胞、ニューロン細胞および膠細胞に対して特異的なエピトープによって追跡される。SOD1 G93Aラットの実験の最後に、ヒトNSCは、強健な移植および優れた長期生存を示す。HNu(+)細胞の大多数(70.4±6.4%)は、TUJ-1のそれらの同時局在化に基づいたニューロン系列へと分化する。HNu(+)細胞のおよそ5分の1(19.2±5.6%)がネスチンと同時局在化し、ごくわずかな(1.3±0.9%)HNu(+)細胞がGFAPに関して陽性であった。
【0179】
宿主の循環内に組み込まれるヒトNSCの能力を、移植片/宿主細胞に関する神経細胞体マーカーおよび宿主または移植片終末に対して選択的なマーカーによって試験する。移植片の出所を確定するためにHNuに関して、ニューロン分化を確定するためにTUJ-1に関し、ラットおよびマウスのエピトープを認識するが、ヒトのエピトープは認識しない前シナプスタンパク質Bassoon(BSN)に関してはモノクローカル(monoclocal)抗体に関して、切片を染色する。柔組織の場における多数のHNu(+)細胞、TUJ-1(+)細胞は、ラット出所であるシナプスボタンに接触することが分かる。
【0180】
共焦点顕微鏡において、HNu(+)核およびTUJ-1 (+)細胞質を有するNSC由来ニューロン細胞は、ラット終末に接触する。反対に、TUJ-1およびヒト特異的シナプトフィジンによって染色された調製物は、宿主のニューロン、特に大型および小型の運動ニューロンに対する小型ボタンの高密度の終末フィールドを現す。宿主の運動ニューロンは、多数の移植片由来のボタンに接触する。HNuおよびヒトNF-70に関して染色された横断切片は、移植片を左方に残し、前索の灰白質に沿って優先的に進む移植片由来の多数の軸索を示す。ChAT免疫反応性を有する細胞/方法を用いて、腹角における白質から灰質を描写する。神経フィラメントエピトープNF70に対するヒト特異的抗体によって標識化された多数の軸索が移植部位に関して見られ、多くのヒトNSCがプロジェクションニューロンへと分化することを実証し;これらの軸索は腹角の白質を優先させることを示している。
【0181】
SOD1 G93Aラットの腰髄内へのNSC移植片は、寿命を延ばし、運動ニューロンの死滅ならびに疾患の発症および進行を遅らせる。生細胞(L)移植片および死細胞(対照、C)移植片を用いた場合における臨床的および病理学的尺度の進行分析は図9に示されている。生きているNSCを移植した動物は、Kaplan-Meier分析とエンドポイント分析の双方で有意な生存の増加を示した。Kaplan-Meierプロット(図9A)は、観察の過程を通して、実験動物と対照動物との間の有意な区別を示す(P=0.0003)。BBBオープンフィールドおよび傾斜平面試験スコア(図9B)の時間プロットは、死んでいるNSCを受けた動物に比較して、生きているNSCを移植した動物では、筋肉衰弱における進行が有意により遅いことを示している。
【0182】
Tgラットの腰プロチュバランス(L3〜S1)における運動ニューロン生存に及ぼすNSCの効果を、生きているNSCまたは死んでいるNSCを与え、128日齢で安楽死させた小群の動物で調べる。死んでいるNSCを移植した動物では、平均寿命が138日であり、一方、生きているNSCを移植したラットは149日間生きる。このように、寿命における有意な11日の違いが、実験ラットと対照ラットとの間に生じる(P=0.0005)。死細胞を与えた動物では、疾患発症までの時間の平均は115日であり、生きているNSCを移植した動物では、122日である。これら両群間には、疾患発症までの時間において、有意な7日の違いが見られる(P=0.0001)。
【0183】
立体解析学的に見積もったα運動ニューロンの数は、生いているNSCを与えた動物では6,418、死んでいるNSCを移植したラットでは3,206であり、すなわち、同年齢の対照動物に比較して、実験動物の腰プロチュバランスには2倍多いニューロンが存在する。生きているNSC群と死んでいるNSC群との間の腰プロチュバランスにおける3,212個の細胞の違いが、128日齢における代表的な実験ラットと対照ラットに見られる(P=0.01)。
【0184】
ヒトNSCによってもたらされる変性運動ニューロンに対する神経保護の可能性のある機構としては、哺乳動物の運動ニューロンに対する古典的な栄養効果を有する2つのペプチドの発現および放出が挙げられる[BDNFおよびGDNF](Hendersonら、1994年;Koliatsosら、1993年)。移植を行ったSOD1 G93Aラットの脊髄におけるGDNF およびBDNFの発現および放出を判定する。ウェスタンブロッティングおよびELISAによって、BDNFおよびGDNFに関して、脊髄調製物およびCSFサンプルを評価する。生細胞を移植したラット(L1およびL2)および死細胞を移植した動物(C)の柔組織およびCSFにおけるGDNF濃度をELISAによって判定する。L1は、移植部位全体の濃度を表し、一方、L2は1セグメント上または下の組織における濃度を示す。群間の差異は有意であり、これはL1またはL2とC群間の大きな違いによって生じる。
【0185】
実験(生細胞、L)群と対照(死細胞、C)群との間のCSF濃度の違いは、t検定によっても有意である。ELISAは、生きているNSC移植動物の脊髄における移植部位では0.912±0.050pg/μgの濃度、1セグメント離れた部位では0.819±0.115pg/μgを示した。死んでいるNSCを移植した動物では、これらの濃度は、移植片を含有する脊髄セグメントにおいて、0.368±0.026 pg/μgであった。CSFでは、GDNF濃度は、実験動物で0.027±0.012 pg/μlであり、0.006±0.002 pg/μlであった。これらのデータは、生きているNSCを用いた動物のCSFにおいて、脊髄におけるGDNFの発現および放出が3倍増加し、GDNFの分泌が5倍増加することを示す。
【0186】
また、ウェスタンブロッティングは、生きているNSCを移植した動物におけるより高いノーマライズされたGDNF濃度を示す。GDNFウェスタンブロッティングによって、16kDaタンパク質の検出による増加のELISAパターンが確認される。また、ウェスタンブロッティングは、生細胞移植片では0.860±0.007、死細胞移植片では0.708±0.052のノーマライズされたGDNF濃度を示す。
【0187】
実験ラットおよび対照の実質組織およびCSFにおけるBDNFのELISA染色を判定する。ELISA 分析により、実験動物において、移植部位(L1)では0.086±0.014 pg/μg、移植片から1セグメント離れた部位(L2)では0.054±0.009 pg/μgの濃度が示される。対照ラットでは、移植含有セグメントにおけるBDNF濃度は、0.010±0.003 pg/μgであった。実験と対照のCSF濃度間の違いは有意である。CSFでは、BDNF濃度が、実験動物では0.041±0.013 pg/μl、対照では0.010±0.008 pg/μlである。これらの所見は、実験動物において、脊髄のBDNF濃度は8倍増加し、CSFのBDNF濃度は4倍増加することを示している。まとめると、ELISAデータは、生きているNSCを用いて移植した動物では、特にCSFでは、BDNFに比較してGDNFのより広範な分泌を示唆している。
【0188】
免疫化学もまた、移植したHNu(+)細胞の大多数がGDNFを発現していることを明らかにしている。生移植片を有する動物におけるGDNF源は移植した細胞自体である。HNu(赤)およびGDNF(緑)に関する二重染色調製物は、移植NSCの細胞質内GDNF免疫反応性の存在度を示す。生移植片を受ける動物では、宿主運動ニューロン内の分泌小胞に類似した丸い細胞質構造内に強力なGDNF免疫反応性が存在する。
【0189】
GDNFおよびヒトシナプトフィジン(後者は移植片由来の終末を標識するため)によって染色した宿主運動ニューロン全体の共焦点顕微鏡により、小胞構造内にはGDNFの局在化が示されたが、例示の宿主運動ニューロンの表面上に位置する移植片由来の終末内には示されない。ヒトシナプトフィジン抗体(宿主の運動ニューロンを神経刺激している全ての移植片終末を標識するため)およびGDNFによって染色した切片は、宿主の運動ニューロンに接触しているボタン内の2つのタンパク質の同時局在化の欠如を示す。
【0190】
生移植片を用いたラットは、移植片に源を発し、中央カナルの内および周囲の構造を神経刺激している経路の確立を示す。宿主運動ニューロンに接触している終末におけるGDNFタンパク質の欠如とは対照的に、中央カナルを神経刺激しているNSC由来軸索終末の大多数は、GDNF免疫反応性と同時局在化する。ヒトシナプトフィジンおよびGDNFの広範な同時局在化が中央カナルにおける上衣細胞を神経刺激している移植片由来終末内に見られる。これらの解剖学的パターンは、GDNFが、移植片を逆行性輸送によって神経刺激し、経シナプス輸送によってこれらのニューロンに送達されない宿主運動ニューロン終末によって恐らく取り込まれることを示している(Rindら、2005年)。
【0191】
SOD1 G93Aラットの腹角における進行性変性過程に対する移植NSCの明らかな抵抗性は、特に有望である。本明細書に報告されたNSCの生存性で広範囲の分化により、SOD1 G93Aに存在する運動ニューロンに関与する炎症性/興奮毒性のシグナル伝達(Rothsteinら、1992年;Howlandら、2002年;Turnerら、2005年)が細胞に明らかな毒性を有さないことの強い指標である。この因子単独で、運動ニューロン変性疾患における運動機能を回復させるために移植片を用いる将来の細胞戦略に対する楽観主義を高める。
【0192】
(実施例7:ヒト脊髄神経幹細胞/前駆細胞の移植による外傷性脊髄損傷の処置、脊髄空洞症の処置)
増殖させたヒト脊髄幹細胞を、免疫抑制した成体メスSprague-Dawleyまたは免疫不全胸腺欠損ヌードマウスのいずれかに注射する。移植の1カ月前に、両群にC4-5挫傷損傷を作製する。移植片レシピエント(n=24)は、移植後60〜150日生存した。ヒト由来のNSCは、ニューロン、星状膠細胞および希突起膠細胞からなる大型の細胞凝集体を形成した。これらの移植片は、各損傷をいつも完全に満たした。未熟な外観のNeuN+/ヒト核+ニューロンは、50%のドナー細胞集団を含むことが多かった。これらのニューロンは、移植部位から少なくとも2cm距離で、灰質と白質の双方を介してヒト神経フィラメント+処理を送達した。宿主ニューロンと移植片ニューロンの双方の近位に、強いヒト特異的シナプトフィジン免疫反応性が認められ、非反応性と思われるGFAP+細胞はドナーニューロンと並列した。さらに、これらの移植片は、宿主源から生じると思われるTH+および5HT+線維の増殖を支持した。したがって、このNSC系は、脊髄内灰白質の修復に有利である主要な胎児CNS様性質を有していると思われる。
【0193】
当然のことながら、本明細書に記載された好ましい本実施形態に対する種々の変更および改変が当業者には明らかであろう。このような変更および改変は、本開示の方法の精神および範囲を逸脱することなく、また、その意図された利益を減ずることなく行うことができる。したがって、このような変更および改変は、添付の請求項に含まれることが意図されている。
【特許請求の範囲】
【請求項1】
ヒト神経幹細胞を提供する工程;
5分から3時間の間、0.1 μg/mLから1 mg/mLのポリ-D-リシン溶液とともに培養容器をインキュベートする工程;
培養容器を洗浄し、乾燥して、予備コーティングした培養容器を形成する工程;
血清の不存在下で、前記予備コーティングした培養容器中に神経幹細胞を播種する工程;
予備コーティングした培養容器にフィブロネクチン溶液および少なくとも一つのマイトジェンを添加し、血清の不存在下で神経幹細胞を培養する工程;
少なくとも一つのマイトジェンとともに神経幹細胞をさらに培養する工程;ならびに
集密になる前に、培養した神経幹細胞を継代接種する工程
を含む、ヒト神経幹細胞を培養する方法。
【請求項2】
前記の増殖神経幹細胞が、ニューロンに分化することができる、請求項1に記載の方法。
【請求項3】
前記神経幹細胞を増殖することが、可溶性因子としてフィブロネクチンを培養培地に加えることを含む、請求項1に記載の方法。
【請求項4】
前記細胞を分離すること、および前記細胞を継代することが、酵素による分離を含む、請求項1に記載の方法。
【請求項5】
前記酵素による分離が、トリプシンで前記細胞を処置することを含む、請求項4に記載の方法。
【請求項6】
神経変性状態を治療するために、治療有効量の前記増殖集団をレシピエント神経系の少なくとも一領域に導入する、請求項1に記載の方法。
【請求項1】
ヒト神経幹細胞を提供する工程;
5分から3時間の間、0.1 μg/mLから1 mg/mLのポリ-D-リシン溶液とともに培養容器をインキュベートする工程;
培養容器を洗浄し、乾燥して、予備コーティングした培養容器を形成する工程;
血清の不存在下で、前記予備コーティングした培養容器中に神経幹細胞を播種する工程;
予備コーティングした培養容器にフィブロネクチン溶液および少なくとも一つのマイトジェンを添加し、血清の不存在下で神経幹細胞を培養する工程;
少なくとも一つのマイトジェンとともに神経幹細胞をさらに培養する工程;ならびに
集密になる前に、培養した神経幹細胞を継代接種する工程
を含む、ヒト神経幹細胞を培養する方法。
【請求項2】
前記の増殖神経幹細胞が、ニューロンに分化することができる、請求項1に記載の方法。
【請求項3】
前記神経幹細胞を増殖することが、可溶性因子としてフィブロネクチンを培養培地に加えることを含む、請求項1に記載の方法。
【請求項4】
前記細胞を分離すること、および前記細胞を継代することが、酵素による分離を含む、請求項1に記載の方法。
【請求項5】
前記酵素による分離が、トリプシンで前記細胞を処置することを含む、請求項4に記載の方法。
【請求項6】
神経変性状態を治療するために、治療有効量の前記増殖集団をレシピエント神経系の少なくとも一領域に導入する、請求項1に記載の方法。
【図1】


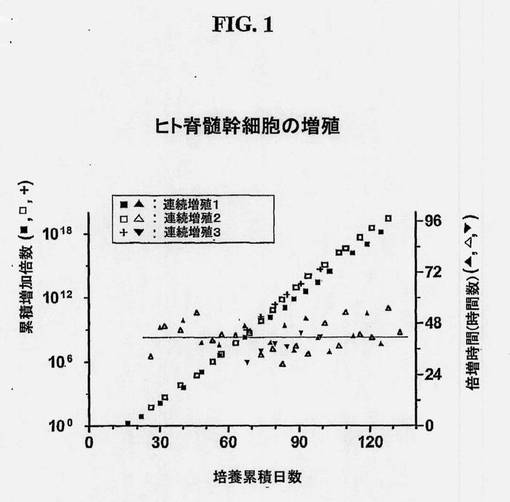
【図2】
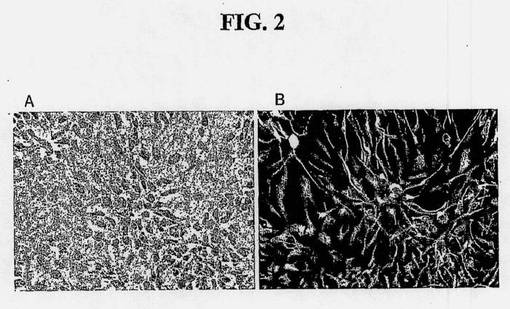
【図3】
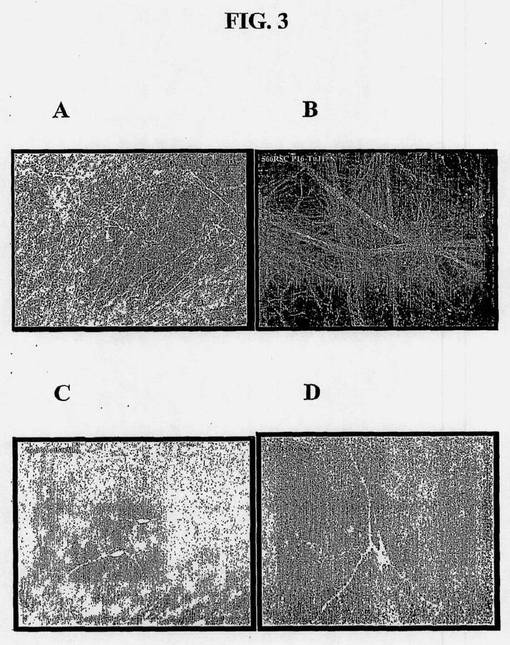
【図4】
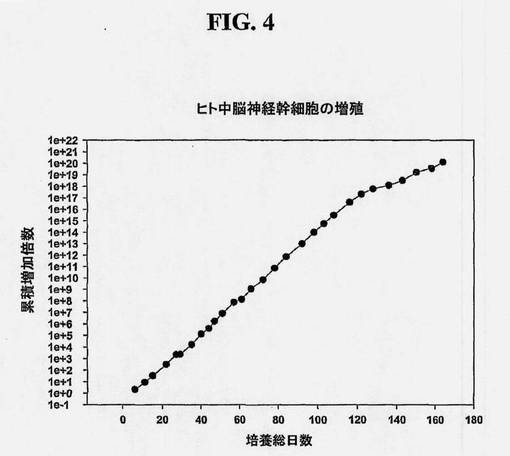
【図5】
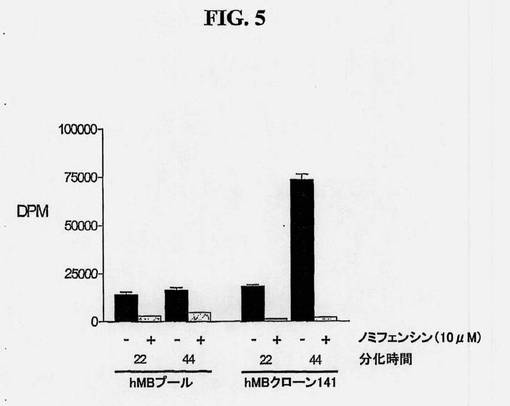
【図6A】
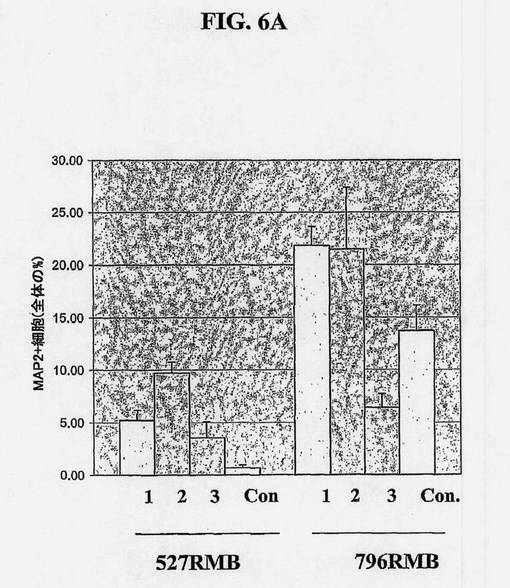
【図6B】
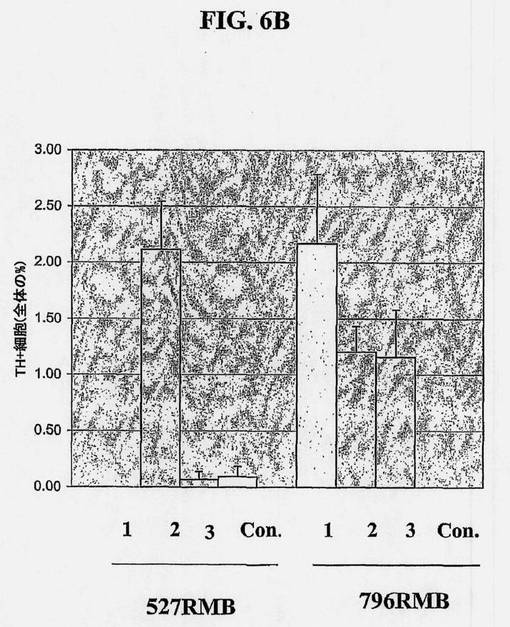
【図7】
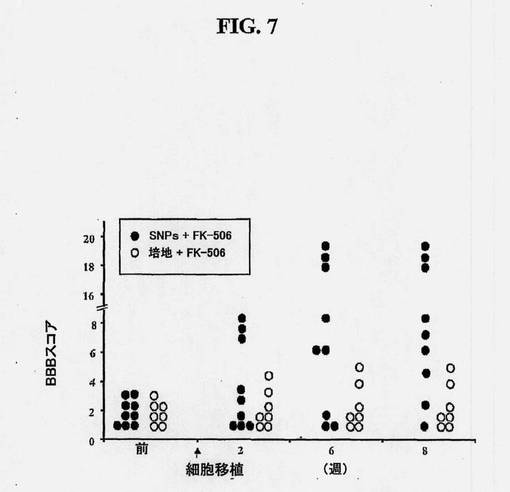
【図8】
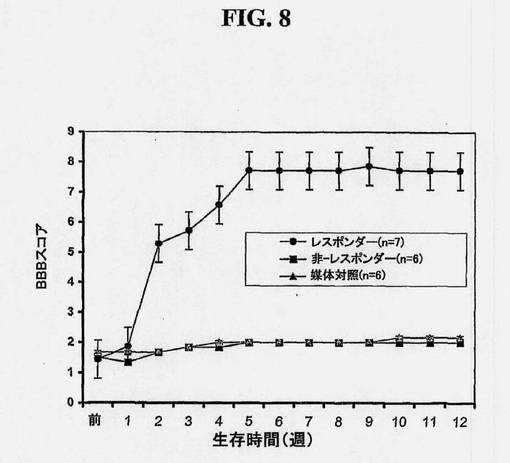
【図9A】
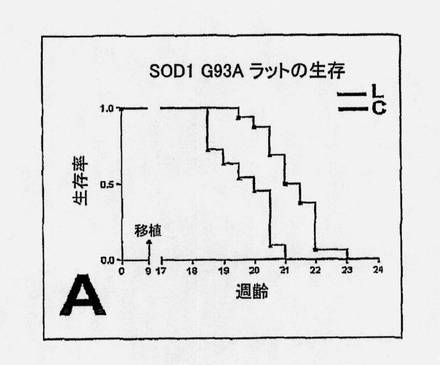
【図9B】
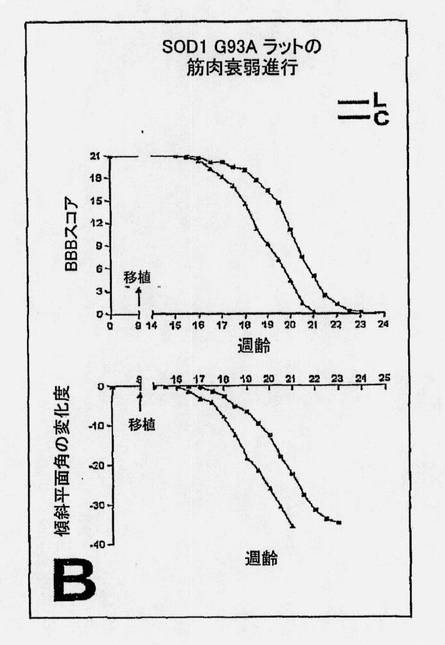
【図9C】
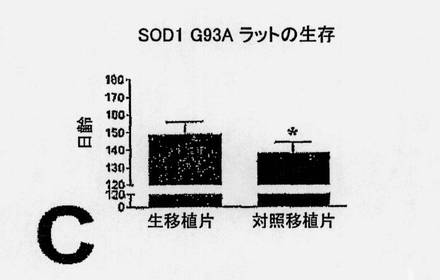
【図9D】
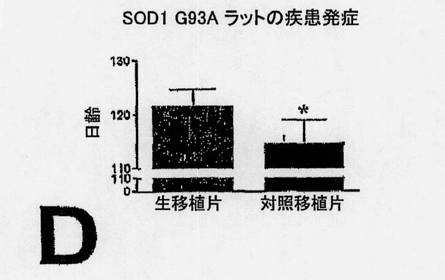
【図9E】
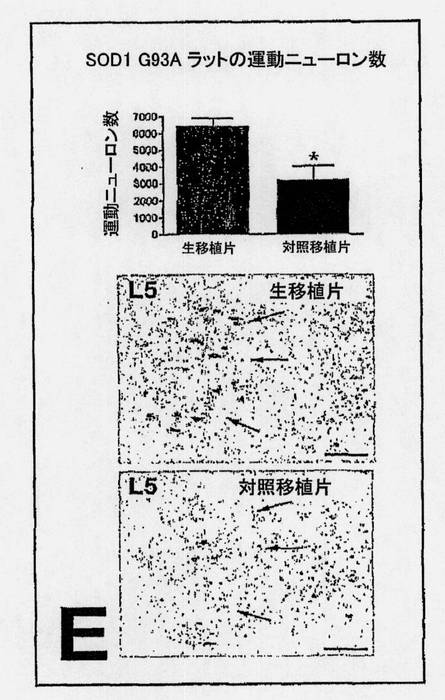
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図7】
【図8】
【図9A】
【図9B】
【図9C】
【図9D】
【図9E】
【公開番号】特開2012−95667(P2012−95667A)
【公開日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願番号】特願2012−20641(P2012−20641)
【出願日】平成24年2月2日(2012.2.2)
【分割の表示】特願2007−543219(P2007−543219)の分割
【原出願日】平成17年11月17日(2005.11.17)
【出願人】(507161466)ニューラルステム・インコーポレーテッド (2)
【Fターム(参考)】
【公開日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願日】平成24年2月2日(2012.2.2)
【分割の表示】特願2007−543219(P2007−543219)の分割
【原出願日】平成17年11月17日(2005.11.17)
【出願人】(507161466)ニューラルステム・インコーポレーテッド (2)
【Fターム(参考)】
[ Back to top ]