
神経幹細胞
本発明は、出生後の被験体から神経幹細胞を取得するための組成物および方法ならびに神経学的障害を処置することにおけるその使用を提供する。一局面において、本発明は、神経幹細胞(NSC)が富化された終糸(FT)神経細胞の集団を含む組成物を提供する。別の局面において、本発明は、出生後の動物からFT−NSCを単離する方法を提供し、この方法は、該動物からFT組織を提供する工程、該FT組織を分離して、神経球を取得する工程、およびネスチン陽性NSCを回収する工程を包含する。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願
この出願は、2007年6月27日に出願された米国仮特許出願第60/937,571号(この内容は、その全体が参考として本明細書に援用される)に関する。
【0002】
発明の分野
本発明は、細胞治療の分野に関する。
【背景技術】
【0003】
神経幹細胞(NSC)および前駆体細胞は、中枢神経系(CNS)内に存在する発生的に始原の細胞であり、そこでの主要な細胞型(ニューロン、星状膠細胞および乏突起膠細胞)の全てを生じることができる。NSCは、ヒトの神経の発生および神経学的疾患の研究ならびにそのような疾患を処置するために有用である。
【0004】
NSCは、脳内の深い領域(area)(例えば、側脳室の脳室下帯および海馬の顆粒細胞層)から採取されてきた。これらの領域からNSCを採取する間、上にある脳領域を損傷する可能性は、自系NSCの採取についてのこのストラテジを危険かつ非現実的にしている。
【発明の概要】
【課題を解決するための手段】
【0005】
本発明の要旨
本発明は、容易に入手可能なNSCの集団を同定すること、ならびに既存の方法に比べて傷害の危険がはるかに少ない、それらを単離するための方法を記載することによって、これらの困難に取り組む。これらの細胞は、同種異系または自系である。異種組織の移植に起因する合併症を回避するために、NSCは、好ましくは自系である。例えば、細胞置換(cell replacement)または組織再生のための薬学的組成物は、NSCに富んだ終糸(FT)神経細胞の集団を含む。この集団は、神経球または神経球開始細胞(neurosphere initiating cell)(NS−IC)を含む。神経球は、神経幹細胞を含む細胞の凝集物または塊である。これらは、代表的に、事実上球状であり、培養において自由に浮遊する。神経球内の細胞は、培養において増殖し、一方、ニューロンおよびグリアに分化する能力を維持している。NS−ICは、長期間の神経球培養を開始することのできる細胞である。NS−ICは、ネスチン陽性であり、適切な分化条件下で、ニューロン、星状膠細胞および乏突起膠細胞に分化する能力を有する。好ましくは、この組成物は、単離されたFT細胞の集団(このうちの少なくとも10%は、NSCまたはNS−ICである)を含む。例えば、この細胞集団のうちの少なくとも30%、50%、85%、90%、99%または100%はNSCである。
【0006】
FT(必須ではない神経組織)は、被験体(例えば、神経学的傷害または他の障害に罹患しているか、これらを発症する危険を有するヒト患者)から採取される。出生後の動物からのNSCの単離は、被験体由来のFT組織を提供すること、FT組織を分離して神経球を取得すること、そしてネスチン陽性NSCを回収することによって実行される。この様式で取得された、単離されたNSC(すなわち、自系NSCの集団)を含む組成物は、この組織が取得された被験体を処置するために使用される。FT組織から取得された神経幹細胞は、FT−NSCと呼ぶ。FT−NSCの多分化能の子孫細胞を含む細胞株もまた、本発明の範囲内である。
【0007】
したがって、被験体の神経学的機能を増強または回復させる方法は、単離されたFT細胞の集団をこの被験体に投与することによって実行される。これらの細胞は、被験体に投与される(例えば、これらの細胞は、傷害もしくは損傷の部位に局所的に直接移植されるか、または罹患した部位から離れた部位に投与される)。例えば、細胞は、脳の損傷した部分に脳室内に導入されるか、または脊髄液に注入される。神経学的障害を処置する方法は、被験体からFT組織を採取する工程、FT細胞を生体外で培養して、単離されたFT−NSCの富化された集団を生成する工程、および単離されたFT−NSCの富化された集団を被験体に投与する工程を包含する。処置されるべき神経学的障害は、傷害(急性または慢性)または変性状態を含む。神経学的障害は、欠陥の起源に関係なく、脳もしくは脊髄の傷害または脳もしくは脊髄の機能の減損を含む。例えば、この被験体は、脳卒中に罹患していると診断されるか、脳卒中に罹患していると疑われる。
【0008】
上記の細胞は、疾患または傷害を通じて失われた神経組織を再構成するために使用される。神経細胞に関連する遺伝病は、遺伝子の欠損を修正するか、または疾患に対して保護するように処置する、自系もしくは同種異系の幹細胞の遺伝子改変によって処置され得る。処置されるべきCNS障害としては、神経変性疾患(例えば、アルツハイマー病、多発性硬化症(MS)、ハンティングトン病、筋萎縮性側索硬化症およびパーキンソン病)、急性脳傷害(例えば、脳卒中、頭部傷害、脳性麻痺)および他のCNS機能不全(例えば、うつ病、てんかんおよび統合失調症)が挙げられる。
【0009】
動物由来の単離されたFT−NSCを提供すること、およびFT−NSCの増殖または分化を可能にする条件下でFT−NSCを培養することによって、出生後の動物由来のFT−NSCを増殖させる方法もまた本発明の範囲内である。
【0010】
本明細書に記載の組成物は、精製または単離される。「実質的に純粋」によって、天然でそれに付随する成分から分離されている核酸、ポリペプチドまたは他の分子を意味する。代表的に、ポリペプチドは、それが、天然で関連しているタンパク質および天然に存在する有機分子を、少なくとも60重量%、70重量%、80重量%、90重量%、95重量%またはさらには99重量%含まない場合、実質的に純粋である。例えば、実質的に純粋なポリペプチドは、天然の供給源からの抽出、通常そのタンパク質を発現しない細胞における組換え核酸の発現または化学合成によって取得され得る。用語「単離された核酸」は、DNAであって、所定の核酸が由来する生物の天然に存在するゲノムにおいて、このDNAと隣接している遺伝子を含まない、DNAを意味する。したがって、用語「単離された核酸」は、クローニングされた核酸または合成核酸(RNA、RNAi、DNA)を包含する。
【0011】
有効量は、臨床上の利益を付与するのに必要とされる組成物(例えば、細胞のサンプル)の量である。この有効量は、投与の経路、被験体の年齢、体重および全身の健康に依存して変動する。薬学的組成物は、少なくとも1つの治療的または生物学的に活性な薬剤を含み、そして患者への投与に適した組成物である。このような組成物は、当該分野の周知でありかつ容認された方法によって調製される。例えば、Remington:The Science and Practice of Pharmacy,第20版(A.R.Gennaro編)Mack Publishing Co.,Easton,Pa.,2000を参照のこと。静脈内送達経路、皮下送達経路、筋肉内送達経路および腹腔内送達経路のような非経口投与は、これらの薬学的組成物を送達するために使用され得る。あるいは、化合物は、局所的に(例えば、CNS部位に直接)投与される。神経学的障害の処置のために、脳脊髄液への直接注入または脳組織への直接注射が使用される。任意の1人の患者のための投与量は、患者のサイズ、体表面積、年齢、投与される特定の核酸、性別、時間および投与の経路、全身の健康、ならびに同時に投与されている他の薬物を含む多くの要因に依存する。例えば、約5×105〜1×106個の細胞を含む、濃縮された細胞懸濁物が、部位に注射される。パーキンソン病の処置のために、これらの細胞は、脳室の壁に注射される。
【0012】
本発明の他の特徴および利点は、それらの好ましい実施形態の以下の説明および特許請求の範囲から明らかである。引用された参考文献は、参考として本明細書に援用される。
【図面の簡単な説明】
【0013】
【図1】図1Aは、腰椎および仙椎に対するFTを示す矢状の断面の図(左側のパネル)であり、胚発生の間のFTへの尾側の脊髄の脱分化を示す図(中央のパネルおよび右側のパネル)である。図1Bは、切開した17.5週のヒト胎児脊髄である。矢印は、FTを示す。スケールバー=2.5mm。図1Cは、NSCマーカーであるネスチン(赤色)およびDAPI(青色)について染色した8ヶ月齢のFTの横断面の部分からの合成画像(merged image)である。スケールバー=100μm。
【図2】図2Aは、7 DIV後の胚性ヒト(17週)FTに由来する神経球におけるネスチン発現神経幹細胞(緑色)の存在を示す、蛍光顕微鏡写真である。図2Bは、35 DIV後の出生後日数(P)5のラットの終糸に由来する神経球におけるネスチン発現神経幹細胞(赤色)の継続した存在を示す蛍光顕微鏡写真である。
【図3】図3Aは、6ヶ月齢のヒトの終糸に由来する神経球から採取された細胞を示す位相差顕微鏡写真である。図3Bは、18 DIV後に接着性培養においてβ−IIIチューブリンを発現するニューロンを示す蛍光顕微鏡写真である。細胞を、12歳のヒトの終糸に由来する神経球から採取した。図3Cは、18 DIV後の培養においてO1(緑色)を発現する乏突起膠細胞を示す蛍光顕微鏡写真である。細胞は、P6のラットの終糸から得られる神経球に由来する。細胞核を、DAPIで対比染色する(青色)。図3Dは、神経球由来の細胞の接着性培養におけるビメンチンを発現する神経前駆体細胞(neural precursor cell)(赤色)とGFAPを発現する大グリア細胞(緑色)との共在を示す蛍光顕微鏡写真である。神経球自体は、P4のラットの終糸に由来した。核を、DAPIで対比染色する(青色)。
【図4】図4Aは、GDNF、BDNFおよびCNTFとともにレチノイン酸およびソニックヘッジホッグ(sonic hedgehog)へ神経球由来細胞を曝露した後のβ−IIIチューブリン(緑色)を発現する運動ニューロンを示す蛍光顕微鏡写真である。細胞は、P7のラットの終糸から採取された神経球に由来した。図4Bは、コリンアセチルトランスフェラーゼ(赤色)の運動ニューロンの発現を示す、4(A)における細胞と同一の視野の蛍光顕微鏡写真である。図4Cは、β−IIIチューブリンを発現する運動ニューロン(赤色)を示す異なる神経球の蛍光顕微鏡写真である。図4Dは、MCN−2(運動ニューロンによって固有に発現されるマーカー)の同時発現を示す、同じ視野の細胞の蛍光顕微鏡写真である。全ての細胞は、P7のラットの終糸に由来する。これらの図は、終糸の神経球に由来する細胞の運動ニューロンへの指向性の分化を示す。
【図5】図5A〜Fは、神経前駆体細胞(NPC)、ニューロンおよび/またはグリアへ分化した神経球を示す蛍光顕微鏡写真である。a)ビメンチン(i、緑色)およびTuj−1(ii、赤色)(合成画像(iii))について染色した、単一の神経球に由来する分化した細胞。5%血清中ポリ−L−リジンおよびラミニン上で7日間平板培養した。ドナー:12歳、インビトロで18日。b)GFAP(i、赤色)およびTuj−1(ii、緑色)を発現する、単一の神経球に由来する分化した細胞。合成画像は、(iii)に示される。(a)と同一のドナーおよび条件。インビトロで17日間。c)7日間にわたって5%血清中においてFT神経球から分化したニューロンの3[H]−チミジン標識された核に対する銀粒子を示す位相差顕微鏡法(Phase microscopy)。神経球を、分化条件で培養する前に3[H]−チミジンに8時間曝露した。細胞を、Tuj−1について対比染色する(緑色)。ドナー:6ヶ月のFT、インビトロで107日間、d、e)RAおよびShh−Nで処理した後に、MNマーカーについて染色した、FT神経球から分化した細胞。d)Tuj−1(赤色)およびMNR−2(緑色)について染色した、分化した細胞。ドナー:14週の胎児、インビトロで81日間。e)Tuj−1(i、緑色)、コリンアセチルトランスフェラーゼ(ii、赤色)について染色した、分化した細胞。合成画像を、(iii)に示す。ドナー:18歳、インビトロで25日間。f)O1(緑色)について染色し、DAPIについて対比染色した、分化した細胞。ドナー:6ヶ月齢、インビトロで111日間。スケールバー:a〜f=50um。
【図6】図6Aは、FTを示すP7でのラット脊髄の尾側の面(caudal aspect)の顕微鏡写真である。矢印は、神経球を生成するために使用した構造物の部分を示す。図6Bは、(i)NSCマーカーであるネスチン(赤色)の発現;(ii)DAPI対比染色(青色);および(iii)合成画像を示す、P7のFT組織切片の蛍光顕微鏡写真である。スケールバーは、100umである。図6Cは、インビトロで10日後の、P10のラットFTに由来する神経球を示す顕微鏡写真である。スケールバーは、100umである。図6Dは、(i)ネスチン発現;(ii)DAPI対比染色、および(iii)合成画像を示す、インビトロで5日後の、P5のFTに由来する神経球の蛍光顕微鏡写真である。スケールバーは、100umである。
【図7】図7Aは、(i)NSCマーカーであるOlig2の発現(赤色);(ii)DAPI対比染色(青色);および(iii)合成画像を示す、インビトロで34日後の、P7のFTに由来する神経球の蛍光顕微鏡写真である。図7Bは、(i)ビメンチンの発現(緑色);(ii)DAPI対比染色(青色);および(iii)合成画像を示す、60 DIV後の、P7のFTに由来する神経球の蛍光顕微鏡写真である。図7Cは、(i)一部の細胞におけるSox2の発現(赤色);(ii)DAPI対比染色(青色);および(iii)合成画像を示す、30 DIV後の、P6のrFTから単離された神経球の蛍光顕微鏡写真である。図7Dは、(i)Musashi(色(COLOR))の弱い染色;(ii)DAPI対比染色(青色);および(iii)合成画像を示す、30 DIV後の、P6のFTに由来する神経球の蛍光顕微鏡写真である。図7Eは、(i)β−III−チューブリンの発現(色);(ii)GFAPの発現(COLOR);(iii)DAPI対比染色(青色);および(iv)合成画像を示す、34 DIV後の、P7のFTに由来する神経球の蛍光顕微鏡写真である。
【図8】図8Aは、(i)Pax6発現(緑色);(ii)Olig2発現(赤色);(iii)DAPI対比染色(青色);および(iv)合成画像を示す蛍光顕微鏡写真である。スケールバーは、100umである。30 DIVでのP7のラットFTに由来する細胞。データは、運動ニューロンがFT由来神経球から生成されることを示す。図8Bは、(i)Pax6発現(緑色);(ii)Olig2発現(赤色);(iii)DAPI対比染色(青色);および(iv)合成画像を示す、同様な蛍光顕微鏡写真である。スケールバーは、50umである。30 DIVでのP7のラットFTに由来する細胞。図8Cは、(i)β−IIIチューブリンの発現(緑色)(初期のニューロンのマーカー);(ii)ChAT(赤色)(コリン作用性神経伝達のマーカー);(iii)DAPI対比染色(青色);および(iv)合成画像を示す蛍光顕微鏡写真である。スケールバーは、100umである。ドナー:P6のラットFT 36 DIV。図8Dは、(i)MNR2(緑色)(運動ニューロン特異的マーカー)の発現;(ii)β−III−チューブリン(赤色)(初期のニューロンのマーカー)の発現;(iii)DAPI対比染色;および(iv)合成画像を示す蛍光顕微鏡写真である。スケールバーは、50umである。ドナー:P7のFT、30 DIV。図8Eは、(i)MRN2(緑色);(ii)GFAP(赤色)の重ならない発現;(iii)DAPI対比染色;および(iv)合成画像を示す、分離していない神経球培養物の蛍光顕微鏡写真である。これらの培養物は、ソニックヘッジホッグおよびレチノイン酸で処理されていない;正確に言えば、これらは、専ら、BDNF、CNTFおよびGDNFを含む分化培地で維持した。スケールバーは、50umである。ドナー:P7のFT、30 DIV。図8Fは、GFAP発現細胞の中の運動ニューロンの島(island)を示す、7Eと同じ条件下で培養した別の分離していない神経球の蛍光顕微鏡写真である。パネルは、(i)MRN2(緑色)の発現;(ii)GFAP(赤色)の発現;(iii)DAPI対比染色;および(iv)合成画像を示す。スケールバーは、50umである。ドナー:P7のFT、30 DIV。
【図9】図9は、レチノイン酸(RA)、ソニックヘッジホッグ(SHH)、BDNF、CNTFおよびGDNFで処理した培養物における増大した割合のMRN2発現を示す、ラットのFT NSCを使用する28個の指向性の分化実験の結果をまとめる散布図(scatter−plot)である。MNR2がまた、Shhのみで処理した神経球から分化した細胞においても発現され、一部の場合において、BDNF、CNTFおよびGDNFのみの存在下で増殖させた未処理の神経球から分化した細胞において発現されたことに留意すること。3例のうちの1例において、MNR2についての免疫染色を、いかなる特定の神経栄養因子も含まない血清で分化させた、未処理の神経球に由来する細胞のうちの40%において観察した。
【図10】図10A〜Fは、細胞型発現(cell−type expression)を特徴付けるために、細胞マーカーについて染色した神経球の蛍光顕微鏡写真である。a)ネスチン(赤色)について染色した、図1(c)と同一の8ヶ月のFTに由来する神経球、インビトロで11日間。b)ビメンチン(緑色)について染色した神経球。ドナー:6ヶ月のFT、インビトロで123日間。c)(b)と同一のドナーに由来する、2つの神経球におけるOlig−2染色(赤色)。d)10歳に由来する神経球におけるSox−2発現(赤色)。インビトロで51日間。e)(a)と同一のドナーに由来する神経球のTuj−1(i、緑色)、GFAP(ii、赤色)、合成画像(iii)。f)18歳のFT由来の神経球の位相差顕微鏡写真。インビトロで6日間。スケールバー:a〜f=100μm。画像a〜dおよびe(iii)は、DAPIで対比染色する(青色)。
【図11】図11A〜Cは、α−アマガサヘビ毒素(緑色)およびTuj−1(赤色)について染色した、得られたヒトMNおよびラット筋肉の同時培養物(co−culture)を示す神経筋接合部の蛍光顕微鏡写真である。RAおよびShh−Nで分化させた単一の神経球を、6日間、ラット筋線維と同時培養した。a)MN(赤色)およびα−アマガサヘビ毒素標識アセチルコリンレセプター(緑色)の蛍光顕微鏡写真。b)(a)に見られる神経筋接合部の共焦点顕微鏡画像。c)Imaris Surpassにおいて最大値投影(maximum intensity projection)(MIP)レンダリングに処理させた(b)の側面図。神経終末(赤色)は上にあり、標識レセプター(緑色)は下にあり、黄色は、重なった領域を示す。ドナー:6ヶ月齢のFT。スケールバー:50μm。
【図12】図12は、rFT由来神経球の不均質な分化能力を示す散布図である。この散布図は、神経球分化の14の実験における、Tuj−1(ニューロンのマーカー)およびGFAP(星状膠細胞のマーカー)の発現の変動性を示す。個々の神経球を、ポリ−L−リジンおよびラミニンでコートしたカバーガラス上にプレートし、5%血清中でそれらを培養することによって分化させた。分化した細胞を、24時間後、または7〜10日後のいずれかに免疫細胞化学によって評価した。両方の場合において、ニューロンおよび星状膠細胞の生成の変動が存在した。両方のマーカーについて二重に染色した、神経球から分化した細胞のおおよその割合は、24時間後に見出された重なりの割合と比較して、分化条件に7〜10日間の曝露した後では減少した。
【図13】図13は、MN分化をしているrFT由来神経球におけるMNR2の発現の変動を示す散布図である。この散布図は、分化した神経球におけるMNR2発現の変動性を示す。MNR2は、MN生成を同定するのに使用される複数のマーカーのうちの一つである。チャートで明らかなように、MNR2はまた、Shhのみで処理した神経球から分化した細胞においても発現され、BDNF、CNTFおよびGDNFの存在下で増殖させた、未処理の神経球に由来するものにおいても発現された。3例のうちの1例において、MNR2についての免疫染色は、いかなる特定の神経栄養因子も含まない血清中で分化させた、未処理の神経球に由来する細胞の‘40%で観察された。
【発明を実施するための形態】
【0014】
詳細な説明
本発明は、幹細胞ベースの治療において多大な進歩を表す。この分野は、治療目的のための幹細胞を獲得する、供給源および信頼のおける方法の不足に悩まされている。脳または脊髄の傷害または変性疾患に起因して失われた脳細胞(ニューロンおよびグリア)の置換において使用するための、神経幹細胞の獲得が特に困難であった。
【0015】
自系ヒト神経幹細胞のための完全に新規な供給源が同定された。FTは、脊髄の尾側末端にあり、索(cord)を尾骨に付着させる。これは、脊髄円錐から分離した、構造的に異なる解剖学的組織である。例えば、成人のヒトにおいて、この組織は、2〜3cm離れている。それは、神経系において消耗性の痕跡組織である。神経幹細胞の他の供給源とは対照的に、FTは、外科的に到達可能であり、組織は、「係留脊髄症候群」の手術の間にそれから慣例的に取得される。これは、神経幹細胞のための信頼できる供給源である。この供給源から取得された幹細胞を、組織培養中に長期の時間にわたって増殖および維持させることができ、かつニューロンおよびグリアへ分化したことが実証されている。FT NSCを、神経学的傷害または変性を罹患する患者から容易に採取し、組織培養において増殖および分化させ、続いて患者に移植して戻す。このストラテジは、非自系細胞の移植に伴う組織拒絶の問題を克服する。FTは、神経細胞置換ストラテジに適した細胞の供給源として役立つ、第一の容易に入手可能で、増殖可能な神経組織を代表する。
【0016】
FTの単離
本発明は、FT(幹細胞の存在について調査されたことのない中枢神経系の領域)からの神経幹細胞の単離を介してNSC治療の実行可能性を増大させる組成物および方法を提供する。以前の研究は、幹細胞を含むCNS内の既知の領域(これは、正常な脳機能に必須であり、外科的に到達することが困難である)からのNSCの単離および増殖に注力していた。FTは、NSCの以前に使用された供給源に対する、新規かつ魅力的な代替物である。なぜなら、FTは、出生後の哺乳動物における発達中の脊髄の、機能を有さない(non−function)レムナントを示す、組織学的に始原の構造物だからである。これらの特性は、FTを、治療上の使用のための自系NSCの魅力的な供給源にする。FTは、出生後の哺乳動物(例えば、ラットまたはヒト)、廃棄されたヒト胎児、小児から青年までのヒト(特に、係留脊髄についての外科的切除を受けた者)、および治療上の使用に神経幹細胞を必要とするあらゆる出生後の被験体から取得される。この組織を分離し、神経球を単離し、幹細胞の生存および/または増殖を促進する条件下で増殖させる。FTに由来する細胞は、発生もしくは変性状態もしくは障害、疾患、傷害もしくは外傷、感染、投薬もしくは医学手順に由来する合併症、または任意の他の自然の原因(例えば、加齢)もしくは誘発された原因(例えば、脳卒中)に起因して、神経学的機能の回復を必要とする患者に投与される。移植された細胞は、宿主の中枢または末梢の神経系に統合され、機能的な回復を促進させる。
【0017】
幹細胞
幹細胞は、大半(全てではないが)の多細胞生物に見出される細胞である。それらは、有糸細胞分裂を介して自身を更新し、そして特殊化した細胞型に分化する能力によって特徴付けられる。2つの大まかな型の哺乳動物幹細胞は、以下である:胚盤胞に見出される胚性幹細胞、および成体組織に見出される成体幹細胞。発生している胚において、幹細胞は、特殊化した胚性組織の全てに分化することができる。成体の器官において、幹細胞および前駆体細胞は、特殊化した細胞を補充する、身体のための修復システムとして働くが、血液、皮膚または腸組織のような再生性器官の正常なターンオーバーもまた維持する。
【0018】
本明細書で使用されるように、用語「幹細胞」は、自己再生し、1つより多い型の細胞に分化することのできる細胞を記載することが意図される。自己再生は、細胞分裂の多くの周期を通過するが、未分化の状態を維持する能力として本明細書で定義される。
【0019】
能力は、特殊化した細胞型に分化する能力である。最も厳密な意味において、これは、幹細胞が、全能または多能性のいずれかであること−あらゆる成熟細胞型を生じることができることを必要とするが、多分化能または単能性の前駆体細胞は、時折、幹細胞と呼ばれる。他の用語において、能力は、幹細胞の分化能力(異なる細胞型に分化する能力)を指定する。全能性幹細胞は、卵と精子細胞との融合物から生成される。受精した卵の最初の数回の分裂によって生成される細胞もまた全能である。これらの細胞は、胚および胚外の細胞型に分化することができる。多能性幹細胞(pluripotent stem cell)は、全能細胞の子孫であり、3つの胚葉のいずれかに由来する細胞に分化することができる。多分化能幹細胞(multipotent stem cell)は、密接に関連するファミリーの細胞の細胞しか生成できない(例えば、造血幹細胞は、赤血球、白血球、血小板などに分化する)。単能性細胞は、一つの細胞型しか生成できないが、非幹細胞からこれらを区別する自己再生の特性を有する(例えば、筋肉幹細胞)。
【0020】
本発明の幹細胞は、分子的および機能的な方法(molecular and functional method)によって同定される。幹細胞の実際の定義は、機能的な定義−一生にわたって組織を再生する能力である。幹細胞の特性は、単一の細胞がそれらの分化および自己再生する能力によって特徴付けられる試験管内腫瘍細胞感受性試験のような方法を使用して、インビトロで示され得る。さらに、幹細胞および幹細胞の集団は、細胞表面マーカーおよび細胞内マーカーの特有のセットに基づいて同定および単離される。
【0021】
胚性幹細胞株(ES細胞株)は、胚盤胞またはより初期の桑実胚段階の胚の内細胞塊(ICM)の胚盤葉上層組織に由来する細胞の培養物である。胚盤胞は、初期段階の胚である-ヒトにおける約4〜5日齢であり、50〜150個の細胞からなる。ES細胞は、多能性であり、発生の間に3つの一次胚葉:外胚葉、内胚葉および中胚葉の全ての派生物のもとになる。言い換えると、これらは、特定の細胞型のための十分かつ必要な刺激を与えられた場合、成体の身体の200種超の細胞型の各々に発達し得る。
【0022】
ほぼ10年間の研究の後に、胚性幹細胞を使用する認可された処置もヒト治験も存在しない。全能細胞であるES細胞は、正しい分化のために特異的なシグナルを必要とする−別の身体に直接注射された場合、ES細胞は、多くの異なる型の細胞に分化し、奇形腫(すなわち、新生物の一つの型)を生じる。移植片拒絶を回避しながら、使用可能な細胞に分化するES細胞は、胚性幹細胞の研究者達が依然として直面しているごくわずかの障害物である。多くの国が、現在、ES細胞の研究も新規なES細胞株の生産も一時停止している。
【0023】
本明細書で使用されるように、用語「成体幹細胞」とは、2つの特性:分裂し、それ自体に似た別の細胞を生成する(すなわち、自己再生)能力、およびまた分裂し、それ自体よりもより分化した細胞を生成する能力(すなわち、この細胞は、少なくとも単能性であるが、選択的に少なくとも多分化能である)を有する、発生した生物に見出される任意の細胞をいう。成体幹細胞はまた、体性幹細胞および生殖系列幹細胞としても一般的に知られている。成体幹細胞は、全ての出生後の哺乳動物において同定され得る。
【0024】
多能性の成体幹細胞は、臍帯血を含む多くの組織に見出され得る。大半の成体幹細胞は、系統が制限されており(多分化能)、一般的に、それらの組織起源によって呼ばれる(間葉幹細胞、脂肪由来幹細胞、内皮幹細胞など)。
【0025】
一局面において、自己再生を確実にするために、幹細胞は、分裂の面(plane of division)および/または細胞内要素の分裂が異なる2つの型の細胞分裂を実行する。対称的な分裂は、共に幹細胞の特性を付与された、2つの同一の娘細胞を生じる。対称的な分裂は、2つの同一の細胞を生成する細胞分裂、上皮の障壁(例えば、大脳半球の側脳室)に平行な分裂の面で生じる細胞分裂、2つの均等なサイズの細胞を生成するかまたは細胞の中心点(center point)(赤道)で生じる細胞分裂、あるいは分離に続いて同一または等価な細胞内成分/要素を有する2つの細胞を生成する細胞分裂として定義される。他方、非対称的な分裂は、1つの幹細胞および制限された自己再生能力を有する先祖細胞のみを生成する。非対称的な分裂は、先に列挙されたシナリオの全てについて非対称的である細胞分裂として定義される。娘細胞間で対称的にまたは非対称的に分裂され得る例示的な細胞内成分または要素としては、細胞内または細胞表面タンパク質;細胞骨格要素;接着結合、細胞接触または細胞接着要素;細胞内小器官;およびシグナル伝達分子が挙げられるが、これらに限定されない。前駆体は、成熟細胞に最終的に分化する前に、数回の細胞分裂を経験し得る。
【0026】
代替的な理論は、幹細胞が、それらの特定のニッチにおいて環境上のキューに起因して未分化のままであることである。幹細胞は、それらがそのニッチから離れるか、またはもはやそのようなシグナルを受け取らなくなった場合、分化する。インビトロにおいて、幹細胞のニッチを繰り返し、指向性の生体外幹細胞分化を誘導し得る。特定の局面において、幹細胞のニッチの特定の成分が過剰発現、除去、組換え、および/または合成され、所望の幹細胞応答を誘導し得る。
【0027】
神経幹細胞
本明細書で使用されるように、用語「神経幹細胞」とは、ニューロンおよびグリア細胞を生じ得る、成体神経組織に見出される幹細胞を記載することが意図される。例示的なグリア細胞としては、星状膠細胞および乏突起膠細胞が挙げられるが、これらに限定されない。ニューロン(ニューロンおよび神経細胞としても知られる)は、情報を処理し伝達する神経系内の電気的に興奮性の細胞である。ニューロンは、脊椎動物の脳、脊髄および末梢神経の中心的な成分である。十分に発達したニューロンは、永久に有糸分裂しない(amitotic)(これらは分裂しない)が;しかしながら、脳のいたるところのさらなるニューロンは、神経発生の過程を介して、脳室下帯およびやや顆粒状のゾーン(subgranular zone)で見出される神経幹細胞から生じ得る。本発明はまた、FTを含む脊髄内の神経幹細胞の供給源を提供する。
【0028】
ニューロン
ニューロンは、代表的に細胞体(soma)(すなわち、細胞体(cell body))、樹状突起(dendritic tree)および軸索から構成される。大部分の脊椎動物のニューロンは、細胞体および樹状突起に入力を受け、軸索を介して出力を伝達する。しかしながら、ニューロンのサイズ、形状および機能において、神経系および動物界を通じて大きな異質性が存在する。一部の特殊化したニューロンのサブタイプが知られており、これらとしては、かご細胞(小脳内の、拡張した、節の多い樹状突起を有するニューロン);ベッツ細胞(大型の運動ニューロン);中型の有棘ニューロン(Medium spiny neurons)(線条体における大半のニューロン);プルキンエ細胞(小脳内の巨大ニューロン、ゴルジI型多極ニューロンの1つの型);錐体細胞(三角形の細胞体を有するニューロン、ゴルジII型の1つの型);レンショー細胞(両端がα運動ニューロンに連結されたニューロン);顆粒細胞(ゴルジII型ニューロンとしての1つの型);および前角細胞(脊髄にある運動ニューロン)が挙げられるが、これらに限定されない。ニューロンのサブタイプの広範な変動に起因して、本発明の成熟ニューロンは、細胞型間の形態学的な差、分子的な差および/または機能的な差を利用する1つ以上の方法を使用して同定される。
【0029】
分化したニューロンは、他のニューロンまたは細胞に対するそれらの作用によって特徴付けられる。興奮性ニューロンは、それらの標的ニューロンを興奮させる。脳を含む中枢神経系内の興奮性ニューロンは、しばしばグルタミン酸作用性(glutamatergic)である。筋肉細胞へシナプスを形成する脊椎の運動ニューロンのような末梢神経系のニューロンは、しばしばそれらの興奮性神経伝達物質としてアセチルコリンを使用する。抑制ニューロン(inhibitory neuron)は、それらの標的ニューロンを抑制する。抑制ニューロンは、しばしば介在ニューロンである。一部の脳の構造物(新線条体、淡蒼球、小脳)の出力は、抑制性である。一次抑制性神経伝達物質は、GABAおよびグリシンである。調節性ニューロン(modulatory neuron)は、ニューロン調節(neuromodulation)と呼ばれるより複雑な効果を引き起こす。これらのニューロンは、ドーパミン、アセチルコリン、セロトニンおよびその他のような神経伝達物質を使用する。
【0030】
分化したニューロンは、それらの放電パターンによって特徴付けられる。ニューロンは、それらの電気生理学的特徴にしたがって分類される。ニューロンは、持続性または規則正しい「スパイキング(spiking)」を示す(例えば、スパイク(spike)は、活動電位の検出をいう)。一部のニューロンは、代表的に、一定で(または持続して)活性である(例えば、神経線条(neurostriatum)内の介在ニューロン)。他のニューロンは、規則正しいスパイキング(これは、少なくとも1回の刺激によって引き起こされる活動電位をいう)を示す。あるいは、またはさらに、ニューロンは、位相性の挙動またはバースト挙動を示す。バーストで発火するニューロンは、位相性と呼ばれる。一部のニューロンは、それらの迅速な発火速度に関して顕著である(例えば、一部の型の皮質の抑制介在ニューロン、淡蒼球内の細胞)。あるいは、またはさらに、一部のニューロンの活動電位は、その他のものと比べてより狭い(検出した場合、薄いスパイク(thin−spike))。例えば、前前頭皮質内の介在ニューロンは、薄いスパイクのニューロンである。
【0031】
分化したニューロンは、それらが放出する神経伝達物質によって特徴付けられる。限定しない例示的なニューロンの型は、コリン作用性ニューロン、GABA作用性ニューロン、グルタミン酸作用性ニューロン、ドーパミン作用性ニューロンおよび5−ヒドロキシトリプタミンニューロン(5−HT;セロトニン)が挙げられる。
【0032】
FT−NSCの特徴付け
さらに上記の細胞を特徴付けるために、FT−NSC株を樹立し維持した。FTに由来する神経球を、NSCマーカーであるビメンチン、CD133、Olig2およびSox2の発現について調べる。FT−NSCを、分化を促進する条件下でインビトロで培養する。分化した細胞の型を、免疫細胞化学技術および電気生理学的方法の両方を使用して同定する。ラットおよびヒトのFT−NSCを、正常なげっ歯類または脊髄外傷もしくは神経学的疾患のモデルとなるげっ歯類のいずれかのCNSに移植する。
【0033】
FT−NSCを、ラットおよびヒト(約24人のヒトドナー)から単離し、組織培養において多くの継代にわたって神経球として維持した。例えば、細胞を、15〜20回継代し、一部の細胞株を、12ヶ月超にわたり維持した。この様式で取得したFT−NSCを、免疫細胞化学によって決定されるCNSニューロンおよびグリアに分化するように誘導した。細胞は、ニューロンおよびグリアの両方(星状膠細胞と乏突起膠細胞との両方)を生じる。細胞を、誘導して分化させ、80〜100%の運動ニューロンである細胞集団を得た。
【0034】
自系幹細胞の供給源の同定
ヒトNSCの供給源を同定し、神経系の外傷および変性の症例におけるそれらの治療上の使用を研究した。自系NSCを、脊髄のFTから単離し、増殖させ、続いて、神経組織の損傷の部位に移植する。
【0035】
自系NSCの信頼できる供給源は、脊髄のFTであり、これは、脊髄の尾側末端の細長い延長部分であり、脊柱の基部にある尾骨に索を係留する(図1A)。組織学的研究は、FTが、室周囲(peri−ventricular)上衣細胞に囲まれた脊柱管(ventricular canal)ならびに種々の型のニューロンおよびグリアを封入することを示す。この局所的な環境は、NSCを生成する他のCNS領域に類似する(Alvarez−Buylla,A.およびD.A.Lim. Neuron(2004)41(5):p.683〜6;Doetsch,F. Curr Opin Genet Dev(2003)13(5):p.543〜50;Riquelme,P.A.、E.DrapeauおよびF.Doetsch.Philos Trans R Soc Lond B Biol Sci,2007)。FTは、神経系の残りと相互に連結されてなく、また身体の神経も刺激しない。本質的に、それは、神経系の消耗性のレムナントである。
【0036】
FTは、独特な発生上の由来を有する(Streeter,G.L. Am J Anat(1919)22:p.1〜12;Nievelstein,R.A.ら、Teratology(1993)48(1):p.21〜31;Kernohan,J.W. J Comp Neurol(1924)38:p.107〜125;Kunitomo,K.(1918)8:p.161〜204)。これは、胚の痕跡尾(vestigial tail)に対する神経支配(またはげっ歯類の場合、最も尾側の尾分節(tail segment)の一過的な神経支配)を発生の初期に提供する神経系のレムナントである。初期の段階において、推定上のFTは、脊髄神経節が付いた、十分に分化した脊髄である(図1B、左側)。尾が再吸収されたときに、糸の細胞は、Streeterによって「脱分化」と呼ばれる過程(図1B、右側)を経験し、上衣細胞で裏打ちされ、線維芽細胞、ニューロンおよびグリアの見かけ上ゆるく組織化された収集物で環状に囲まれた中心管を有する膠原性の構造物をもたらす。パラガングリオーマおよび始原の神経外胚葉性腫瘍は、この構造物から生じることが示され、このことは、幹細胞が存在する可能性を示唆する(Ashkenazi,E.ら、J Spinal Disord,(1998)11(6):p.540〜2;Gagliardi,F.M.ら、Childs Nerv Syst,(1993)9(1):p.3〜6;Kamalian,N.ら、J Neurol,(1987)235(1):p.56〜9;Koeller,K.K.、R.S.RosenblumおよびA.L.Morrison. Radiographics(2000)20(6):p.1721〜49)。FTは、外科的に容易に到達可能であり、脊柱にしっかりと係留され、動きに十分な自由を欠く場合、脊髄に対する緊張を軽減するために、慣例的に切開される。この状態は、「係留脊髄症候群」と呼ばれる(Bakker−Niezen,S.H.、H.A.WalderおよびJ.L.Merx.Z Kinderchir、(1984)39 Suppl 2:p.100〜3;Bode,H.ら、Klin Padiatr(1985)197(5):p.409〜14)。本発明より前に、終糸が、神経幹細胞の潜在的な供給源として採取されたことはない。
【0037】
FTは、以下の理由に関してNSCの安全かつ信頼できる供給源である。これは、外科的に到達可能であり、消耗性の神経組織である。ヒト組織は、胎児組織から、そして脊髄の外科的な解放(untethering)に続いてFTの生検を行う小児神経外科センターから容易に入手可能である。NSCの自家移植は、神経系への異種移植に伴う重大な問題であることが実証されている、免疫学的拒絶を回避する明確な利点を有する(Barker,R.A.およびH.Widner. NeuroRx,(2004)1(4):p.472〜81;Linazasoro,G. Neurologia(2003)18(2):p.74〜100)。NSCは、哺乳動物のCNSで同定されているが、現行の供給源は、外科的に到達するのが困難であり、これは、代表的に、正常な機能に重要な領域(例えば、脊髄および前脳の側脳室)に由来する(Alvarez−Buylla,A.、D.G.HerreraおよびH.Wichterle. Prog Brain Res,(2000)127:p.1〜11;Alvarez−Buylla,A.、B.SeriおよびF.Doetsch. Brain Res Bull,(2002)57(6):p.751〜8)。これらの領域の外科的な破壊は、顕著な(profound)神経学的な欠損をもたらす。FT由来のNSCは、一生にわたって採取され得、自系NSCのための供給源として役立ち得、それによって、免疫学的拒絶の問題を回避する。単離、培養、増殖、分化に続いて、自系FT−NCSは、神経学的な障害または傷害の処置のために、被験体に移植される。
【0038】
主要な組織は、動物から取得される(例えば、げっ歯類(ラット、マウス)、またはヒト(例えば、胎児および出生後のヒト)FT)。例えば、胎児ヒト組織は、廃棄された胎児から取得される。若年のヒトFTは、係留された索の切除のための手術の間に取得される。加えて、FTは、出生後のラットから取得される。出生後ラットFTは、ヒトFTと同じ組織学的特徴を有し、このクラスのNSCのさらなる特徴付けのための、容易に入手可能で、大規模の組織の供給であるという利点を有する。
【0039】
胎児から成人までの種々な年齢の、死後のヒト材料由来の組織切片におけるFT−NSC局在は、種々の段階において存在するNSCの数に関する情報を提供し、より正確な切開をもたらす。FTを、切開し、固定し、低温槽上で切片にする(section)。NSCを、ネスチンおよびSox2のようなNSCマーカーに対して指向される抗体を用いて組織切片を染色することによって同定する(Bazan,E.ら、Histol Histopathol,(2004)19(4):p.1261〜75)。
【0040】
NSCの培養
本発明のNSCを、被験体への移植に先立って、維持および増殖のために培養する。
【0041】
FTを、当該分野で公知の培地および増殖因子を含む種々の条件(例えば、N2、EGFおよびbFGF+ヘパリンを含むDMEM/F12)を使用して、組織培養に置く(Rajan,P.およびE.Snyder. Methods Enzymol,(2006)419:p.23〜52;Vescovi,A.L.およびE.Y.Snyder. Brain Pathol,(1999)9(3):p.569〜98)。神経球を単離し、継代する。これらは、非限定的な、先に検討した免疫学的なマーカーを使用して同定する。
【0042】
分離のための手順は、抗体でコートされた磁気ビーズを使用する磁気分離、固体マトリクス(例えば、プレート)に付着させた抗体を使用するアフィニティクロマトグラフィおよびパニング、または他の便利な技術を含む。他の分離技術は、多色チャネル、小角および鈍角光散乱検出チャネル、インピーダンスチャネルなどのような種々の程度の洗練を有し得る蛍光励起セルソーターを含む。死細胞は、例えば、死細胞と結合する色素(ヨウ化プロピジウム[PI]、LDS)を使用する選択による標準的な方法を使用して排除される。選択された細胞の生存度に過度に有害ではない、あらゆる技術が使用され得る。
【0043】
NSC株を樹立し、移植/回復研究のために増殖され、ヒトNSCの場合、治療用の細胞株の供給源として発達させる。ヒトNSCのための培養条件は、増殖速度および細胞の収量を最大化するために最適化される。培地、増殖因子、酵素などの操作による。細胞株の増殖速度は、生成された神経球の数を計数すること、および生存細胞の数を計数することによって測定される(Cardozo,D.L. Neuroscience(1993)56(2):p.409〜21)。
【0044】
FT−NSCは、ニューロンおよびグリアに分化する
個々の神経球を単離し、ニューロンおよびグリアへの分化を促進する種々の条件下でインビトロで平板培養する(Reynolds,B.A.、W.TetzlaffおよびS.Weiss. J Neurosci,(1992)12(11):p.4565〜74;Reynolds,B.A.およびS.Weiss. Science,(1992)255(5052):p.1707〜10)。分化をさらに特徴付けるために、神経球を他の細胞と同時培養し、分化条件への曝露の前に、脂肪親和性膜染料(lipophilic membrane stain)(例えば、DiI)、カルボキシフルオレセインまたはBrdUのような試薬で予め標識する。この追跡ストラテジは、生成されるニューロンおよびグリアの供給源として神経球を確実に樹立する。3つの基本的なアプローチが、これらの細胞を特徴付けるために使用される:a.血清の存在下で接着基質上で平板培養すること;b.特定の表現型を促進することが知られている規定された試薬(例えば、運動ニューロンを生成するために、レチノイン酸+ソニックヘッジホッグタンパク質を使用すること)での前処理に続いて、接着基質上で平板培養すること;ならびにc.標識された神経球と筋肉のような標的組織に由来する培養細胞またはCNSの異なる領域に由来する細胞との同時培養。同時培養データは、FT−NSCの運命に対する標的組織の影響を確立し、FT神経球が、筋肉細胞およびニューロンとのシナプスの相互作用を形成することを確認する。
【0045】
神経球由来の細胞型は、ニューロンおよびグリアのサブタイプに特異的な抗体ならびに未熟なニューロンのサブタイプと成熟ニューロンのサブタイプとの間を区別する抗体を含む、ニューロンおよびグリアに対して指向される抗体を使用して同定される。これらとしては、以下が挙げられる:神経−特異的エノラーゼおよび神経−チューブリンIII(ニューロン);ビメンチン(神経前駆体);GFAP(星状膠細胞);O1(乏突起膠細胞);コリンアセチルトランスフェラーゼおよびMNR2(運動ニューロン)(Gage,F.H.、J.RayおよびLJ.Fisher. Annu Rev Neurosci,(1995)18:p.159〜92;Schwartz,P.H.ら、J Neurosci Res,(2003)74(6):p.838〜51;Schwartz,P.H.ら、Stem Cells,(2005)23(9):p.1286〜94;Wichterle,H.ら、Cell,(2002)110(3):p.385〜97)。
【0046】
運動ニューロンの表現型を促進する試薬に曝露した神経球を、筋肉細胞(筋管)と同時培養する。得られたニューロンを、神経筋接合部に結合する標識されたアマガサヘビ毒素で培養物を染色することによって、筋肉細胞とのシナプスを確立するそれらの能力について試験する。機能的な運動ニューロンを、電子顕微鏡法を使用して、さらに評価し、機能しているシナプスのシナプス前の要素およびシナプス後の要素(例えば、シナプス小胞、シナプス後密度)の存在を同定する。
【0047】
神経球に由来する形態学的に同定可能なニューロンを、細胞が、ニューロンに典型的な生理的特徴を有するか否かを決定する、標準的な電気生理学的技術(静止電位、活動電位を生成する能力、および添加された神経伝達物質に対する応答を測定すること)を使用して特徴付ける。筋肉の同時培養の場合に、分化したニューロンを、視覚的方法および電気生理学的方法の両方で筋肉の単収縮応答について評価する(Cardozo,D.L.およびB.P.Bean. J Neurophysiol,(1995)74(3):p.1137〜48;Elkes,D.A.ら、Neuron,(1997)19(1): p.165〜74)。
【0048】
インビボ移植、統合(integration)および機能的な回復実験
FT−NSC株を、げっ歯類宿主組織へ再統合するそれらの能力ならびにヒト被験体および脊髄外傷と神経変性とについてのげっ歯類モデルにおける機能的な回復を生成するそれらの能力について評価する。当該分野で認識される腰部脊髄外傷についてのげっ歯類モデルを、FT−NSCの評価について使用する。これらのモデルは、欠損の程度および機能的な回復の程度に特異的な測定基準を含む(Karimi−Abdolrezaee,S.ら、J Neurosci,(2006)26(13):p.3377〜89;Kimura,H.ら、Neurol Res,(2005)27(8):p.812〜9;Nakamura,M.ら、J Neurosci Res,(2005)81(4):p.457〜68;Vroemen,M.ら、Eur J Neurosci,(2003)18(4):p.743〜51)。GFP標識されたFT−NSCを、脊髄外傷を経験したラットの傷害部位に移植する。動物を機能的な回復について試験し、その後屠殺し、死後の脊髄を、移植した細胞の統合について組織学的に調べる。
【0049】
FT−NSC細胞の再統合を、以下のように評価する。FT−NSC株を、全てのCNS細胞が緑色蛍光タンパク質(GFP)で標識されているトランスジェニックラットから樹立する(Dombrowski,M.A.ら、Brain Res,(2006)1125(1):p.1〜8)。これらのGFP標識されたFT−NSCを濃縮し、正常なラットの脊髄に移植する。移植された細胞が生存し、宿主組織に統合する能力を、標準的な組織学的方法を使用して死後に調べる。
【0050】
ヒト由来FT−NSC株を、ヒト被験体および免疫不全マウス系統(NOD−SCID)への移植によって評価する。移植した細胞を、SC101およびSC121のようなヒト特異的抗原についての染色によって同定する(Anderson,AJ.、B.J.CummingsおよびCW.Cotman. Exp Neurol,(1994)125(2):p.286〜95;Cummings,B.J.ら、Proc Natl Acad Sci U S A,(2005)102(39):p.14069〜74)。
【0051】
細胞をまた、神経変性疾患についてのげっ歯類モデルにおいても評価する(Kitamura,Y.ら、Jpn J Pharmacol,(2000)84(3):p.237〜43;Orth,M.およびS.J.Tabrizi.(2003)18(7):p.729〜37;Springer,W.およびP.J.Kahle. Curr Neurol Neurosci Rep,(2006)6(5):p.432〜6)。動物を、機能的な回復について試験し、そして死後に、宿主組織への移植した細胞の統合について試験する。
【0052】
神経変性疾患および神経外傷を処置するためのFT−NSCの使用
自系FT NSCは、パーキンソン病および他の神経変性疾患ならびに外傷性脳傷害の処置に有用である。研究は、ドーパミン産生細胞が、マウスパーキンソン病(Parkinson’s)モデルおよびヒトパーキンソン病患者の両方の線条に統合し得、この疾患の症状を緩和し得ることを示唆する。本発明以前には、NSCの豊富かつ容易に入手可能な供給源の欠如は、個々の移植片において比較的少数の細胞が使用されていたことを意味する。さらに、以前の移植片は、中枢神経系の「免疫学的に特権のある(immunoprivileged)」性質にも拘らず、拒絶をもたらす免疫反応を引き起こす、異種細胞を使用した。本発明は、これらの問題の両方を解決する:全ての患者はFTを有し、したがって、全ての患者は、増殖可能であり、かつ適切な条件下で、自家移植のための特定の細胞型(ドーパミンニューロンおよびコリン作用性運動ニューロンが挙げられる)を効率よく生成する、自系NSCの準備のできた供給源を有する。FT−NSCは、自系細胞の利用可能性に対する根本的な拘束を排除するので、それらの治療上の使用に対する唯一の制限は、生じる細胞型の多様性である。例えば、FT由来乏突起膠細胞を使用して、多発性硬化症のような脱髄疾患を処置する一方で、FT由来運動ニューロンは、筋萎縮性側索硬化症のような運動ニューロンの喪失の疾患または脊髄傷害に適用可能である。
【0053】
ヒト患者からの細胞の採取
FT細胞を、公知の方法を使用して患者から採取する。例えば、手術を使用して、「係留脊髄症候群」の症例においてなされるように、FTを切断および切開する。第二に、少量の組織を、針穿刺吸引(needle aspiration)によって採取する。これらの細胞に関してたくさんの数の継代が可能であること、そして単回の注射に必要とされるのが比較的少数(500,000〜1,000,000)であることを考慮して、単一のサンプルは、複数回の注射に十分な細胞をもたらし得る。蛍光検査(fluoroscopic)または他の誘導を使用して、複数のサンプルを、単一の手順の間に採取することができる。
【0054】
出生後組織をTCSの外科的な標本から取得したが、同じ実験の全てをまた、胎児由来HuFT NSCおよび出生後のラット終糸を使用して実行した。この結果は、全ての年齢において、終糸内にNSCが存在することを一貫して確認した。実際に、正常な78歳のHuFTの免疫染色は、ネスチン陽性細胞の証拠を示し、この主張をさらに支持した。18歳までのHuFT由来のNSCの単離および分化は、これらの幹細胞が、成人期に残存する可能性を示唆する。
【0055】
FTは、バイオセイフティー、組織適合性およびその除去に続いて何の欠損も存在しないという利点を付与する(profer)、自系、消耗性かつ到達可能なNSCのための未開発の資源である。この組織供給源由来のNSCは、神経系の外傷および変性の処置に有用である。
【0056】
以下の試薬および方法を使用して、以下の実施例に記載されたデータを生成した。
【0057】
インビトロ分化
非特異的分化のために、単一の神経球を、可視化のための解剖顕微鏡の補助を使用して単離し、1% N2、1% ペニシリン−ストレプトマイシン、および5〜10% ウシ胎児血清(Gibco)を含むDMEM/F12培地中で96穴の培養ディッシュ(Corning)の個々のウェルにおいて、ポリ−L−リジン(0.01%、Sigma)およびラミニン(20mg/ml、Sigma)でコートしたカバーガラス上で平板培養した。培地を、実験の残りの間、変更しなかった。免疫細胞化学のために、カバーガラスを2〜10日後に処理した。分化した細胞が増殖性細胞に由来したことを確認するために、神経球を、トリチウム化チミジン(Cepko labから贈呈、培地1mlあたり5mL)で8時間インキュベートした。続いて、神経球を、視覚的に単離し、3回、幹細胞培地で洗浄し、次に、上記のように分化させた。運動ニューロンへの指向性の分化は、まず、4〜5日間のレチノイン酸(RA、2mM、Sigma)およびソニックヘッジホッグ(Shh−N 500〜100nM、R&D systems製、またはHh−Ag1.3、Curis)での神経球の処理を含んだ。この処理を、以前に記載した幹細胞培地中で行った。その後、個々の神経球を単離し、1% N2、1% ペニシリン−ストレプトマイシン、5% ウマ血清(Gibco)、CNTF(25ng/ml、Sigma)、GDNF(25ng/ml、Sigma)およびBDNF(50ng/ml)を含むDMEM/F12培地中で7〜10日間、96穴の培養ディッシュ(Corning)の個々のウェルにおいて、ポリ−L−オルニチン(0.01%、Sigma)、I型コラーゲン(0.01%、Sigma)およびラミニン(20mg/ml、Sigma)でコートされたカバーガラス上で平板培養した。3つの型の対照実験を実行した。神経球を、RAを含まずShh−Nのみで処理した。神経球を、RAおよびShh−Nで処理しないが、BDNF、CNTFおよびGDNFの存在下で増殖させた。最後に、一部の神経球は、Shh−NでもRAでも処理せず、それらを、BDNFもCNTFもGDNFも存在しない中で培養したが、上記のように、5〜10%血清中で非特異的分化を受けた。また、先に記載したように、対照も、コートしたカバーガラスを含む96穴の培養ディッシュの個々のウェル中で7〜10日間培養した。その後、全てのカバーガラスを免疫細胞化学のために処理した。
【0058】
神経筋接合部の形成の存在を証明するために、個々の神経球を、RA(2mM)およびShh−N(1000nM)で4〜6日間処理し、続いて、先に記載した、CNTF、BDNFおよびGDNFを含むMNの増殖および生存のための分化培地中で、筋肉培養物上で平板培養した。対照培養物は、筋肉培養物上で平板培養した未処理の神経球を有したか、または全く神経球を有さなかった。21日後、培養物を、蛍光αアマガサヘビ毒素(2.5mg/ml、alexa fluor 488で標識)で2.5時間インキュベートした。その後、これらを洗浄し、固定し、免疫細胞化学(ニューロンのマーカーBTIII)のために処理した。
【0059】
抗体
ネスチンに対するウサギポリクローナル抗血清(1:400)、ChATに対するヤギポリクローナル抗体(1:100)およびニューロン特異的エノラーゼに対するマウスモノクローナル抗体(1:1000)を、Chemiconから取得した。ウサギポリクローナルSox2(1:1000)は、Sigma Abcamに由来する。ビメンチンに対するマウスモノクローナル抗体は、Cepko laboratoryから贈呈された。マウスモノクローナルCD133(1:1000)は、Miltenyi Biotecから購入した。GFAPに対するウサギポリクローナル抗体(1:1000)は、Dakoに由来し、GFAPに対するマウスモノクローナル(1:1000)は、Sigmaに由来した。βチューブリンIIIに対するウサギポリクローナルは、Covanceに由来した。Tuj 1(1:1000)およびNeu−N(1:1000)に対するマウスモノクローナル抗体も、Olig−2に対する抗体と同様に使用した。NICHDの賛助の下に開発され、The University of Iowa,Department of Biological Sciences,Iowa City,IA 52242によって維持された、神経フィラメント、MNR2、Lim3およびIs1−1、ならびにPax6に対するモノクローナル抗体を、Developmental Studies Hybridoma Bankから購入した。AF 488結合体化ロバ抗ウサギIgG、AF 488結合体化ロバ抗マウスIgG、AF 568結合体化ロバ抗ヤギIgG、AF 488結合体化ヤギ抗マウスIgG、およびAF 568結合体化ヤギ抗ウサギIgGは、InvitrogenのAlexa Fluor productsから購入した二次抗体であった。全て1:1000で使用した。
【0060】
免疫細胞化学
免疫細胞化学を、カバーガラスに付着した全神経球または分化した神経球を使用して実行した。カバーガラスを、4% ホルムアルデヒド(PBS中、pH7.2)で20〜30分間、固定し、次に、PBSでそれぞれ10分間、3回洗浄した。抗体希釈物を、ブロッキング溶液(PBS、0.2% ウシ血清アルブミン中、10% 正常ヤギ血清、10% アイシングラス、0.3% Triton X)で調製し、一次抗体を、カバーガラスを用いて、それら各々の希釈物で一晩(8時間)インキュベートした。このことの後、PBSで3回洗浄し、その後、適切な二次抗体で4時間インキュベートした。PBSでの3回のさらなる洗浄の後、カバーガラスをDapi(0.03mg/ml)で30分間インキュベートした。その後、カバーガラスを、もう一度3回(それぞれ10分間)洗浄し、それから封入剤としてVectashieldを使用してガラススライド上に載せた。このスライドを、Zeiss顕微鏡写真機(photomicroscope)を使用して免疫蛍光検査のために視覚化した。特定のマーカーについての細胞染色のおおよその割合を、4〜5 20×視野の平均カウントによって決定した。Dapiを、細胞の総数を計数するためのマーカーとして使用した。細胞の数が極めて多い(>500)場合、またはそれらが、一部の視野で共に塊になるが、他の視野では存在しない場合、Dapiに対するマーカー陽性細胞のパーセンテージを見積もった。
【0061】
免疫細胞化学を使用して、インビトロでのHuFT由来の未分化神経球における種々のNSC、神経前駆体細胞(NPC)、ニューロンおよびグリアのマーカーの存在を証明した。全ての神経球(n=13)は、NSCマーカーであるネスチンについて陽性に染色した。より小さな神経球(<100ミクロン)において、100%の細胞がネスチンを発現した。しかしながら、より大きな神経球において、それらのコアは、ネスチン陰性のようであった。このコアは、おそらく細胞の壊死性の塊である。神経球(n=33)はまた、神経前駆体マーカーであるビメンチン、CD133(n=18)、Olig2(n=17)およびSox2(n=17)について陽性である細胞を含んだ。NPCマーカーの発現は、神経球間で変動した。試験した全ての神経球(n=30)はまた、ニューロンのマーカーであるBTIIIおよびグリアのマーカーであるGFAPを示差的に発現し、おそらく分化に際して観察された変動したパターンを先触れした。
【実施例】
【0062】
実施例1:FT組織からの神経球の生成
29の実験において、FTを、P4〜P19齢のラットから切開し、培養した。神経球は、29個の培養物のうち、26個で生成された(89.5%)。個々の培養物を、最大18回まで継代し、3つの細胞株を樹立し、凍結させた。ヒトFT組織を、4つの胎児および6ヶ月齢〜18歳の12の出生後手術から単離した。神経球は、16個の培養物のうち、14個で生成され(87.5%)、これらの培養物を、最大6回継代した。神経球をもたらした最も年長の組織ドナーは、18歳である。図2A〜Dは、FTおよび尾方の脊髄ならびにこれらから得られた神経球を示す。20個を超えるヒトまたはラット神経球を、ネスチン免疫反応性について染色した(図2、CおよびD)。試験した全ての神経球はネスチン陽性であった。図3A〜Dは、FT神経球からニューロンおよびグリアへの細胞の分化を示す。ラットおよびヒト由来の個々の神経球を、血清存在下でポリ−l−リジン、ラミニンおよびコラーゲンを含む種々の接着基質上で平板培養した。全ての場合において、FT−NSCは、免疫細胞化学的判断基準によって決定されるように、ニューロンおよびグリア(星状膠細胞および乏突起膠細胞を含む)に分化した。
【0063】
単一の神経球を、レチノイン酸およびソニックヘッジホッグタンパク質とともにインキュベートし、血清および神経栄養因子の存在下で接着基質上で平板培養した。FT−NSCは、形態学的に同定可能なニューロンに分化し、MNR2、LIM−3、ISL−Iおよびコリンアセチルトランスフェラーゼを含む運動ニューロンマーカーに対して染色した(図4A〜D)。DiIまたはカルボキシフルオレセインで予め標識した神経球は、初代ラット筋肉細胞と同時培養した場合にニューロンに分化した(運動ニューロンまたはそれらが神経筋接合部を形成したか否かの証明に関して)。
【0064】
実施例2:ヒトFTの培養ならびに得られたNSCの増殖および継代
14〜21週齢のヒト胎児組織を、妊娠の人工的な終結の後に取得した。脊髄を迅速に切開し、氷冷ハンクス溶液中に置いた。その後、顕微鏡での視覚化の下で、ヒトFTを同定し、切開した。ヒトFTの周囲の脊髄神経根を、時折切開し、陰性対照として別個に培養した。6ヶ月齢〜18歳のヒト出生後組織を、係留された索の解放(TCSのための慣例的な神経外科手順)を受けた小児から取得した。これらの場合において、ヒトFTを、顕微鏡の補助を使用して神経外科医が視覚的に同定し、取り出しに先立って、その正体を、電気生理学的試験を用いて確認した。この組織を、氷冷ハンクス溶液中で、手術室から研究室に移した。
【0065】
一度、胎児または出生後のFT組織を取得したら、それを、標準的な培地(例えば、DMEM/F12(1:1,Gibco)、1% N2処方物(Gibco)、1% ペニシリン−ストレプトマイシン溶液(Gibco)、EGF(20ng/ml、Gibco)、bGFG(20ng/ml、Gibco)、LIF(10ng/ml)および3mMカルシウムをともなうII型コラゲナーゼ 100U/ml(Gibco))を含む培養ディッシュ(Corning)に移し、ピンセットおよび外科用メスを用いて掻き裂いた。FGFは、安定性のために、8mg/mlのヘパリン(Sigma)を含む溶液中で調製した。培養物は、加湿したインキュベーター中に、37℃、5% CO2で維持した。24時間後、その組織を、コラゲナーゼで部分的に消化し、さらなる分離のために先端熱加工したピペットで機械的に粉砕した。初代幹細胞の増殖を、3〜5日後にインビトロで検出し、そして未分化細胞の球体の形成により特徴付けた。
【0066】
組織を、4つの胚供給源および17の出生後供給源から取得した。インビトロでの3〜4日間の後、神経球は、胚培養物のうちの100%および出生後培養物のうちの82%で観察された。神経球は、球状、自由に浮遊する、NSCの異種起源の凝集物であり、これは、種々のニューロンおよびグリアに分化する能力を維持しながら、培養において増殖する。観察された神経球の数は、初代培養物あたり1個から50個を超える神経球の範囲で変動し、ドナーの年齢と相関しなかった。増殖および自己再生についてのそれらの能力を示すために、それらを、最大10回まで継代し、最大6ヶ月までそれらをインビトロで維持した。8つの株を凍結し、神経球の首尾良い回復について試験した。
【0067】
継代頻度は、初代培養物の間で変動した。一部の培養物は迅速に増殖し、10〜14日ごとに継代することを必要とし、他のものは、3〜4週間ごとに継代することしか必要としなかった。神経球を、5〜7分間、1×AccumaxTM(Innovative Cell Technologies)を用いて分離し、その後、機械的に粉砕して、神経球の部分的な分離を達成した。遠心分離(10分間、1000rpm)後、細胞を、新鮮培地と馴化培地の1:1の組み合わせ中に再懸濁した。これらの継代の間に、神経球を単一の細胞に分離した場合に、死滅率は高く、時折100%であったことに注意すべきである。
【0068】
実施例3:ニューロンおよびグリアへのヒトFT−NSCの自発的な分化
一部の神経球は、培養容器に接着し、そして培地からのいずれの因子の添加も除去もなく自発的に分化するようだった。種々のドナーに由来する単一の神経球は、LIF、bFGFおよびEGFの除去の後に血清の存在下で神経前駆体細胞(NPC)、ニューロンおよびグリアに成功裏に分化した。個々の神経球を、ポリリジンおよびラミニンでコートしたカバーガラスに2〜10日間、これらの分化条件で平板培養した。全ての場合(n=50の実験)において、神経球は、免疫細胞化学で同定されるNPC、ニューロンおよび/またはグリアの変動した組み合わせ(assortment)を生成した(図5 AおよびB)。これらとしては、ニューロン特異的エノラーゼ、神経フィラメント、neu−nおよびβチューブリンIII(ニューロンのマーカー)、GFAP(星状膠細胞のマーカー)、O1(成熟乏突起膠細胞のマーカー)、mushashi、ビメンチンおよびsox−2(NPCマーカー)が挙げられる(図5 AおよびB)。
【0069】
これらの分化実験は、インビトロでの細胞の組成および分化能力における神経球の異質性の観察と一致して、使用したドナー供給源(6ヶ月齢および12歳)内およびそれらの間の両方で不均質な神経球の能力を明らかにした。観察された染色パターンもまた、細胞塊が特定のマーカーを発現すること、またはより均一に分散した細胞が種々のマーカーを発現することのいずれかをともなって、変動した。48時間を超えて分化させた場合、高い割合の細胞(約79%、n=8、SEM 0.08)が、ニューロンのマーカーおよびグリアのマーカーの両方について二重染色された。しかしながら、分化条件で7〜10日後(n=7)、神経球あたりのニューロンおよびグリアのマーカーについて二重染色した細胞の平均の割合は、約23%(SEM 0.09)に低下した。さらに、7日後に、分化した細胞を、ニューロンのマーカーおよびグリアのマーカーの両方について染色した、これらの実験において、神経球の能力のおおよそ半分が、ニューロン優位であるかまたはグリア優位であるかのいずれかのようであり、二重染色は最小であるかまたは存在しなかった。他の場合において、ニューロンおよびグリアの両方の変動する割合を生成し、部分的に二重染色された。この変動性の能力は、供給源にかかわらず神経球で一貫し、ドナーの年齢には関係しないようだった。NPCマーカーの染色の一貫性は、7〜10日後でさえも分化した細胞の約98%で観察された(n=21、SEM 0.01)。これらの細胞は、頻繁にニューロンのマーカーまたはグリアのマーカーを同時発現した。このことは、ニューロンおよびグリアのマーカーの二重染色(分化時間の増大とともに低下する)と組み合わせて、生成された細胞が未熟なニューロンおよびグリアを表すことを示す。
【0070】
ヒトFT神経球は、インビトロで増殖し、これを継代した。これらの増殖性神経球は、NPC、ニューロンおよびグリアの収集物に分化した。分化したニューロンおよびグリアが増殖性細胞に由来することを確認するために、単一の神経球を、トリチウム化チミジンで8時間処理した(n=4の実験)。その後、これらの神経球を、トリチウム化チミジンを含む環境から取り出し、先に記載したように7日間分化させた。4つの場合全てにおいて、生じたニューロンおよびグリアのうち33〜63%が、それらの核にトリチウム化チミジンの証拠を有し、8時間の曝露ウインドウ(exposure window)が、細胞周期のS期にある増殖性の神経球細胞を捕捉したことを示した。これらの細胞は、S期にある間に放射性ヌクレオチド標識を取り込み、続いてニューロンおよびグリアに分化した(図5C)。
【0071】
同様に、ラットFT神経球は増殖し、これをインビトロで継代することができ、これらの増殖性の神経球は、NPC、ニューロンおよびグリアの収集物に分化する。これらの分化した細胞が増殖性細胞に由来することを証明するために、トリチウム化チミジンを使用して細胞を標識した。5つの実験において、神経球をトリチウム化チミジンで8時間処理した。その後、これらを、トリチウム化チミジンを含む環境から取り出し、洗浄し、上記の標準的な条件において7日間にわたって分化させ、BTIIIおよびGFAPについて染色した。5つの場合全てにおいて、生じたニューロンおよびグリアのうちの27〜90%が、それらの核にトリチウム化チミジンの証拠を有した。これらのデータは、8時間の曝露ウインドウが、細胞周期の「S」期にあるNSCの一部のパーセンテージを捕捉し、これらの増殖性細胞がこの時間の間に放射性ヌクレオチド標識を取り込んだことを示す。これらの細胞は、続いてニューロンおよびグリアに分化し、それらの核内で明白なトリチウム化チミジンによって同定された。
【0072】
実施例4:ヒトFT−NSCの運動ニューロンへの指向性の分化R
本発明以前には、運動ニューロンを生成する出生後の神経球についての先行する報告はなかった。以前に記載されたWichterleの、胚性幹細胞の運動ニューロン(MN)への指向性の分化の方法の変形を用いることで、単離され本明細書中に記載されたようなHuFT由来神経球を、MNに分化するように一貫して誘導した。最初に、単一の神経球をレチノイン酸およびソニックヘッジホッグ(Shh−Nタンパク質またはHh−Ag1.3として知られるShhシグナル伝達の特異的小分子アゴニスト)で、運動ニューロン前駆体(MNP)の誘導のために処理した。次に、これらの個々の神経球を、血清ならびにMNの成長および生存を支持することが公知の三つの神経栄養因子の存在下で、接着基質上で培養した。その三つの神経栄養因子は、毛様体由来神経栄養因子(CNTF)、脳由来神経栄養因子(BDNF)、および神経膠由来神経栄養因子(GDNF)である。これらの分化条件での7〜10日の後、MN特異的マーカーの存在について免疫細胞化学により細胞を分析した。これらは、motor neuron restricted−2(MNR2)、Islet 1(Isl1)、Lim3およびコリンアセチルトランスフェラーゼ(ChAT)を含んだ。MNPの最終分割の間に最初に発現されるMNR2は、MNの同一性にかかわる決定因子である。Isl1およびLim3は、MNの発達に関連するホメオボックス転写因子である。Isl1は、全てのクラスのMNにより発現される。分化した神経球は、MNPにおいて発現されることが公知の二種類の前駆体マーカー(転写因子のOlig−2およびPax6)について試験した。全ての実験において(n=16)、ニューロンの異なる部分が、上記(図5Dおよび5E)のMNまたはMNPマーカーを発現した。この変動性は、単一のマーカー(例えば、MNR2)の使用においてさえ持続する。体細胞性の運動ニューロンにより選択的かつ一貫して発現されるホメオボックスドメインタンパク質であるホメオボックス9(HB9)、およびPax6の存在は、RT−PCRにより確認した。
【0073】
過去の研究において、胚性幹細胞(ESC)は、Hh−Ag1.3およびRAに曝露した場合、機能性の運動ニューロンに容易に分化する。アゴニストは、実際のペプチドよりも、より効果があることが公知であり、そしてMNの生成のためのペプチドに優先して用いられた。本発明者らのデータは、この観察と矛盾がなかった。RAおよびHh−Ag1.3への初期の曝露後HuFT神経球が分化した両方の実験において、100%のニューロンがMNR2を発現した。RAおよびShh−Nでの処理後生成した5〜40%のMNR2陽性細胞と比較して、この観察はHh−Ag1.3が、HuFT神経球のMN分化のための、より有力な薬剤であることを示した。Shh−N濃度を増加させることは、結果を変化させないようだった。Shh−N単独ではまた、Shh−NおよびRAで処理したものと同様の範囲でMNR2陽性細胞を生成した。GDNF、CDNFおよびBDNFの存在下で培養された未処理神経球はまた、変動する割合のMNR2陽性細胞を一貫して生成した(図5C)。多数の例において、MNR2について免疫染色された細胞は、一緒にかたまりになる傾向があった。指向性のMN分化のためのRAおよびShh−Nの使用は、CDNF、BDNFおよびGDNFの存在下で神経球を単に分化させることよりも顕著には優れていないようだった(図5D)。さらに3つの実験の1つにおいて、血清のみにおいて増殖した未処理神経球由来の細胞の約40%が、MNR2について陽性に染色された。神経球の公知の異質性、および発生的に意図される本来のHuFTの機能と組み合わせて、この観察は、RAの腹方化作用および誘導的なShh−Nシグナル伝達を必要とすることなく、MNに分化するいくらかのHuFT NSCの本質的な能力を示した。Hh−Ag1.3の有効な作用は、全てのHuFT神経球由来細胞をMNに指向させるようである。RAおよびShhで処理した、ならびに/またはCNTF、GDNFおよびBDNFを含む培地中で培養されたHuFT由来NSCは、インビトロで筋繊維と神経筋シナプスを形成することができた(n=16)。
【0074】
実施例5:EGF+FGF+LIFに応答して自己再生するラット−FT由来神経球の単離と性格づけ
ラットFT(rFT)において可能性のあるNSCに、いずれかの特定のニッチまたは特異的な場所があるか否かを決定するため、組織を切開し、そしてホルムアルデヒドで固定した組織の複数の切片をNSCマーカーであるネスチンについて染色した(図6Aおよび6B)。これらの試料の免疫組織化学が、明白な分布パターンをともなわない散在する個別的なネスチン陽性細胞を示した。
【0075】
インビトロでのNSCの単離は、コラゲナーゼで分離した一次組織を、bFGF(20ng/ml)、EGF(20ng/ml)およびヒトLIF(10ng/ml)を含む標準的な幹細胞培地(DMEM、F12、N2補充)中で培養することを含む。先行する研究は、これらの分裂促進因子を、おそらく付加的な効果をともなう、NSC増殖に対する好結果の刺激剤として同定した。インビトロで3〜4日後、神経球が、34の初代培養のうち31で観察された。これらの神経球は、初めは自由に浮遊しており、そしてその球状構造、位相での明るい外観、一定の細胞膜、および回折リングにより同定された(図6C)。初代培養当たりの神経球の数は、10から40より多くまで変化した。この数は、ドナーの年齢と相関していないようだった。それらの増殖および自己再生の能力を示すため、神経球を分離し、そして二次球体を生成するよう19回まで継代し、そしてインビトロで7ヶ月まで維持した。
【0076】
初期の考察とは反対に、神経球は、NSCの同種集団ではないが、おそらく異なる能力を持つ、異なるNSCおよび神経前駆体細胞(NPC)の不均質な収集物であることが示された。この不均質な性質を考慮して、rFT由来神経球は、種々のNSC、NPC、神経およびグリアのマーカーの発現を決定する免疫細胞化学を用いて特徴付けられた(表1)。特に、神経球は、NSCマーカーであるネスチン(n=9)、NPCマーカーであるSox2(n=8)、ビメンチン(n=6)、Olig−2(n=3)、およびMusashi(n=4)、ニューロン特異的マーカーであるβ−チューブリンIII(BT III、n=12)、ならびに星状膠細胞マーカーであるグリア繊維酸性タンパク(GFAP、n=12)について染色した。
【0077】
9例全てにおいて、変化する割合での細胞が、ネスチンについて陽性であった。9例のうち4例で、神経球中の100%の細胞が、このNSCマーカーを発現し、これはNSの年齢またはサイズに相関しないようであった(図2D)。神経前駆体マーカーについての染色は変化した。3つの実験全てにおいて、より強い染色を示すいくらかの領域をともなって、神経球細胞の100%が、Olig−2について陽性に染まった(図7A)。Sox2およびビメンチンに関して、全ての神経球が、これらのマーカーについて陽性に染まるいくらかの割合の細胞を有したが(図7Bおよび7C)、このパーセンテージは、Sox2で40から100%、ビメンチンで33から100%で変化した。Musashiの染色は弱く、時折のホットスポットをともなった(図7D)。全ての神経球がまた、BT IIIおよびGFAPを発現し、この発現は、いくらかの神経球では均一であったが、他では違った(図7E)。示差的発現の後者の例では、BT IIIまたはGFAPのどちらかが周縁部で発現し、他のマーカーは、逆に神経球の中心部で発現した。
【0078】
実施例6:運動ニューロン(MN)を生成するRFT由来神経球の指向性の分化
rFT由来神経球からのMNの生成を一貫して誘導するために、単一の神経球をレチノイン酸(RA)およびソニックヘッジホッグ(Shh−Nタンパク質、またはHh−Ag1.3と呼ばれるShhシグナル伝達の特異的小分子アゴニスト)で、MN前駆体を誘導するために4〜5日間処理した。これらの処理された個々の神経球は、その後、血清ならびに、MNの成長および生存を支持することが公知の三つの神経栄養因子の存在下で、接着基質上で平板培養した。用いた神経栄養因子は、毛様体由来神経栄養因子(CNTF)、脳由来神経栄養因子(BDNF)、および神経膠由来神経栄養因子(GDNF)である。これらの条件において処理した神経球を7〜10日間分化させた後、細胞をMN特異的マーカーの存在について免疫細胞化学により分析した(図8、図9)。用いたマーカーは、motor neuron restricted−2(MNR2)、Islet 1(Isl1)、Lim3およびコリンアセチルトランスフェラーゼ(ChAT)であった。MNR2、Isl1およびLim3は、Wichterleらにより、MNSへの胚様体の規定された分化を記載する彼らのもともとの論文において用いられた。Pax6およびOlig2は、MN前駆体細胞を同定するためのマーカーとして用いた。
【0079】
MNR2は、運動ニューロン前駆体の最終分裂の間に最初に発現され、そしてMNの同一性にかかわる決定因子である。Isl1およびLim3は、MNの発達に関連する二つのホメオボックス転写因子である。Isl1は、全てのクラスのMNにより発現される。全ての実験において(n=25)、種々の割合の分化したニューロンが、上に記載したMNまたはMNPマーカーを発現した(図8)。この変動性は、MNR2のような単一のマーカーの使用においても持続する(図9)。Hh−Ag1.3を使用した9つの実験において、分化したニューロンの95〜100%が、MNマーカー(例えば、MNR2、Isl1、Lim3およびChAT)を発現した。このアゴニストは、実際のペプチドよりも、より有効であり、そしてMNの生成について、Shh−Nを優先して用いられてきた。そのデータは、Shh−Nで処理した神経球は、そのうちほんの20〜40%がMNマーカーを発現する分化したニューロンを生じるという観察と一致する。Shh−N濃度の増加は、結果を変化させないようであった。
【0080】
三つの形態の対照実験を実施した。第一番目は、神経球をRAなしで、Shh−Nで処理し(n=8)、そしてその後それらを、血清ならびにBDNF、CNTFおよびGDNFを含む培地中で分化させることを含んだ。第二番目は、未処理の神経球を、血清および3つのノイロトロピンを含む培地中で培養することを含んだ(n=8)。そして第三番目は、未処理の神経球を、特定の神経栄養のサポートはない、血清を含む培地中で分化させることを含む(n=3)。初めの二つの条件は、変動する割合のMNR2陽性細胞を一貫して生成した(図8Eおよび8F)。複数の例において、MNR2について免疫染色した細胞は、島において互いに塊になる傾向があり、そしてグリアにより取り巻かれることが、時折言及された。指向性および一貫したMNの生成のためのRAおよびShh−Nの使用は、BDNF、CNTFおよびGDNFの存在下において神経球を単に分化させることよりも優れてはいないことが明らかとなった。しかしながら、神経球を、神経栄養のサポートなしで、血清中で培養した場合、MNの生成は一貫性がなく、3つの実験の1つのみで、約40%の細胞がMNR2を発現した。神経球の公知の異質性、および発生的に意図される本来のFTの機能と組み合わせて、これらのデータは、RAの尾方化作用または外因性の腹方化Shh−Nシグナル伝達を必要とすることなく、MNに分化するいくらかのrFT NSCの本質的な能力を示す。しかしながらHh−Ag1.3の使用は、大半だが全てではない神経球由来細胞がMNを生成するように指向した点において、MNの収量を増加させるために有益であることが見出された。
【0081】
実施例7:FT由来神経球は、多分化能性であり、ニューロンおよびグリアに分化する
いくらかの神経球は、培養容器に接着し、そして培地からのいずれの因子の添加も除去もなく自発的に分化する。rFT由来神経球が、ニューロンおよびグリアに分化するために必要とされる条件を決定するために研究を実行した。bFGF、EGFおよびLIFの除去後、34の実験において単一の神経球を、ポリ−L−リジンおよび/もしくはラミニンコートしたカバーガラスならびに/または5〜10%の血清への曝露の種々の組み合わせ上で平板培養した。その神経球をこれらの分化条件に7日間供した。接着基質単独または血清単独のどちらかの使用が、形態学的分化を開始するために十分であったが、血清の添加は、より迅速な分化を結果として生じた。全ての例において、分化した神経球は、BTIII、神経フィラメント、O1、およびGFAPを含むニューロンのマーカーおよびグリアのマーカーを発現した(表1および2)。
【0082】
一度これらの条件が確立されると、次の65の実験が、rFT神経球の分化の能力をさらに特徴付けた。これらの実験において、その分化条件は、3つの増殖因子全ての除去、5〜10%の血清による培地の補充、ならびにポリ−L−リジンおよびラミニンでコートしたカバーガラス上での単一神経球の平板培養を含んだ。これらの実験のうちの30において、神経球は、24時間を越えて分化し、そして35の実験において、神経球は7〜10日間を越えて分化した。哺乳動物CNSの他の領域から培養された神経球の報告された不均一さと一致して、rFT由来神経球は、さまざまな分化能力を有した。
【0083】
その変動性にもかかわらず、全ての実験において、各神経球由来のいくらかの割合のNPC、ニューロンおよび/またはグリアが、免疫細胞化学マーカーを用いて同定された。これらは、ニューロン特異的エノラーゼ(NSE)、NeuN、BTIII、GFAP、O1、Musashi、ビメンチンおよびSox2を含んだ(表1)。24時間を越えて分化した神経球(n=9)は、ニューロンのマーカーおよびグリアのマーカーの両方について二重染色された総細胞の高い割合を有した(約70%、表3)。この割合は、神経球が7〜10日間を越えて分化した後、平均約14.5%まで減少した(n=13、表3)。時折、7日間の分化の後、神経球由来の細胞は、ニューロンのマーカーまたはグリアのマーカーのどちらかを主に発現した。しかしながら、ほとんどの例において、明白な優勢は観察されなかった。14の実験における同じ二つのマーカーの使用にもかかわらず、分化した神経球は、BTIIIおよびGFAP発現のさまざまなアレイを示した。この変化は、同一の供給源由来、および異なるrFT供給源の間由来の両方の神経球において持続していた。
【0084】
分化した神経球の間の染色パターンもまた変化する。しばしば、神経球由来の細胞の塊は、一つの特定のマーカーについて全て染色陽性であり、そして他の領域における細胞は異なるマーカーを発現する。しかしながら、より頻繁に、異なるマーカーについて染まる細胞が、一緒に点在する。
【0085】
【表1−1】
【0086】
【表1−2】
【0087】
【表2】
【0088】
【表3】
実施例8:FT−NSCはインビトロで運動ニューロンを生成する
神経幹細胞(NSC)は、中枢神経系(CNS)における未分化細胞であって自己再生が可能であり、そしてニューロンおよびグリアに分化するように誘導され得る。哺乳動物のNSCの現用の供給源は、正常な機能にとって大切であり、外科的に到達することが困難であるCNSの領域に制限されている。これは、ヒト疾患におけるそれらの治療上の能力を制限する。以前に調べられておらず、犠牲にしてもよく、そして容易に到達することができる、脊髄の尾方末端の組織である終糸(FT)が、哺乳動物における多分化能の神経球の供給源であることが、予期せず発見された。本研究では、単離し、そしてこれらの細胞の能力を特徴付けるためにラットモデルを用いた。ラットFT(rFT)由来の神経球は、上皮細胞成長因子(EGF)、塩基性線維芽細胞成長因子(bFGF)、および白血病抑制因子(LIF)の組み合わせによるインビトロでの増殖に耐えられる。増殖している細胞は、血清への曝露により神経前駆体細胞、ニューロン、星状膠細胞およびグリアへと分化するように誘導された神経球を形成した。種々の神経栄養因子と組み合わせてソニックヘッジホッグ(Shh)およびレチノイン酸(RA)を用いる指向性の分化をとおして、rFT由来神経球は、運動ニューロン(MN)をインビトロで生じた。
【0089】
多分化能NSCの存在が、ラットからヒトまでの範囲の種において成体の哺乳動物CNSの複数の領域において示された。これらの領域は、嗅球、脳室の上衣下裏打ち、海馬、小脳、脊髄および網膜を含む。哺乳動物NSCの現用の供給源は、それらが、正常な機能にとって重要であり、そして到達することが困難な領域から取得されることから、ヒト疾患における移植治療のためには理想的ではない。これらの領域の外科的破壊は、自系NSCを採取するためにそれらを用いることを非現実的にする、甚大な神経学的な欠損を導いた。
【0090】
FTは、自系多分化細胞の供給源として優れた候補である。それは、容易に到達することができ、そして犠牲にしてもよい、成体において持続する組織である点において、現在利用可能な供給源を凌駕する明確な利点を提供する。本発明の方法は、FTを自家置換療法のために用いることにより、免疫学的な問題を回避する。
【0091】
発生の初期に、FTは、胚の予定運命の尾への神経支配(あるいはげっ歯類において、最も尾側の尾分節の一時的な神経支配)を与える。成体において、それは痕跡のレムナントである。いくらかのヒトは、動かせる尾をもって生まれるが、しかしながら、尾の異常な残存は、おそらく発生過程の欠陥を示す。ヒトの尾の胚発生は、最初に3.5〜5mmの段階(約4週)で検出される。11〜15mmの段階(約7週)では、尾骨領域はより進んだ発達を示し、脊髄が上衣領域、マント領域および境界領域に分化し、そしてそれぞれの脊髄神経節からそれに入るよく発達した脊髄根を有す。この時点では、この領域が脊髄のより頭方の部分のような成体の状態へと完全に分化し始めないという、組織学的証拠はない。
【0092】
発生が続くと、脊髄の尾骨/尾の部分は再吸収され、そして細胞は、Streeterにより「脱分化」と名付けられた(Streeter,G.L.1919.Am J Anat 22:1−12)過程をうける。30mmの段階(約9.9週)までに、脊髄の尾骨領域は、顕著に変化した。尾骨の脊髄組織は初期の胚の型に戻り、線維芽細胞、ニューロンおよびグリアのゆるく組織化された収集物によって囲まれた上衣細胞により裏打ちされた狭い中心管をもつ膠原性の構造を、結果として生じる(Streeter,G.L.1919.Am J Anat 22:1−12)。最後の三つの尾骨神経節が消失するように、境界領域およびマント領域は、完全に消失する。再吸収は、尾から吻への方向で生じ、そして結果として生じる構造が、FT(脊髄を尾骨に固定する脊髄の尾方末端の細長い延長部分)として成体中に残る。
【0093】
通常、脱分化した出生後のFTは、残りの神経系と相互連結せず、身体の神経を刺激しない。それは偽りなく痕跡のレムナントである。ヒトにおいて、脊椎に固くつながれ、動作の十分な自由を欠く(係留脊髄症候群として公知の状態である)例において、脊髄に対する緊張を軽減するために、それは慣例的に外科的に切開される。FT細胞は、初期の胚細胞型に似ており、脊髄の残部に存在する複数の細胞型に再分化する能力を維持する。
【0094】
ヒトおよびラットの両方において、FTは、脊柱管を囲む膠原性の構造である。室周囲の上衣細胞ならびにゆるく組織化された線維芽細胞、ニューロンおよびグリアの収集物が、その管を囲む。ラットにおいてFTニューロンは、通常よりも、より小さいものとして記載され、そして拘束および分化の初期段階においてニューロンを示す。パラガングリオーマおよび他の初期の神経外胚葉性腫瘍は、成体FTから生じ、NSCが存在することを再び示唆する。FTは、多分化細胞の供給源である。ラットモデルの使用は、制御された環境中での、これらの細胞の全身性の制限のない研究を可能にする。げっ歯類では、FTは、最も尾側の尾分節の一時的な神経支配を与える。
【0095】
細胞培養
rFTの培養および得られた神経球
初代培養:全ての手順は、滅菌条件下で実施した。P2〜P11齢の出生後のラット(雄および雌、SD系(Sprague Dawley)、チャールズ・リバー(Charles River))を、イソフルラン(Abbott)で麻酔し、そして頚椎脱臼により殺した。脊柱を、氷冷したHanks溶液中で迅速に切開した。顕微鏡での可視化のもと、rFTを同定し切開した。各切開は、細胞死を最小にするため、5分未満で実施した。rFTの周囲の脊髄神経根を時折切開し、陰性対照として別個に培養した。rFT(通常、培養ディッシュ当たり、3つの同胞rFT)をプールし、そして幹細胞培地(SCM)(Weissら、1996.J Neurosci 16:7599−7609;Carpenterら、1999.Exp Neurol 158:265−278;Liら、2005a.Biochem Biophys Res Commun 326:425−434;Kimら、2006.Exp Neurol 199:222−235)を含む培養ディッシュ(Corning)中に移した。この培地は、DMEM/F12(1:1、Gibco)、1% N2処方物(Gibco)、1% ペニシリン−ストレプトマイシン溶液(Gibco)、EGF(20ng/ml、Gibco)、bFGF(20ng/ml、Gibco)、およびLIF(10ng/ml)から作製した。FGFは、安定性のために、8mg/mlのヘパリン(Sigma)を含む溶液中で調製した。
【0096】
組織を分離するため、3mMのカルシウム(Gibco)をともなうII型コラゲナーゼ 100U/ml(Gibco)をSCMに加えた。切開した組織をその後、このコラゲナーゼを含む培地に移し、そしてピンセットおよび外科用メスを用いて掻き裂いた。培養物は、加湿したインキュベーター中に、37℃、5% CO2で維持した。24時間後、その組織を、さらなる分離のために先端熱加工したピペットで機械的に粉砕し、そしてコラゲナーゼ含有SCM中に残した。初代幹細胞の増殖を、3〜5日後にインビトロで検出し、そして未分化細胞の球体の形成(Reynolds,B.A. and Weiss,S.1992.Science 255:1707−1710)により特徴付けた。
【0097】
培養の継代:培養物は、2〜3週毎に継代した。神経球を、5〜8分間、1×AccumaxTM(Innovative Cell Technologies)を用いて分離し、そしてその後、神経球の部分的分離を達成するために機械的に粉砕した。初期の実験において、神経球を完全に分離した場合、細胞が生存しても少数だった。酵素的分離後、細胞を遠心分離し(1000rpmで10分間)、そして新鮮培地と馴化培地の1:1の組み合わせ中に再懸濁した。
【0098】
ラット筋肉の培養
P0〜P7のラットを殺し、そして近位の肢の筋肉を、氷冷したHBSS中で迅速に切開した。その組織を、穏やかに別々に掻き裂き、そしてその後、DMEM/F12(1:1)、1% N2補充物および1% ペニシリン−ストレプトマイシンからなる培地を含む培養ディッシュに移した。3mMのカルシウムをともなうII型コラゲナーゼ(100U/ml)を、筋肉繊維を単一の細胞に分散させるために、この培地に加えた。ディッシュは、37℃、5% CO2でインキュベーター中に24時間置いた。24時間後、組織を完全に分離するために、先端熱加工したパスツールピペットで培養物を粉砕した。培養物をその後、1000rpmで5分間遠心分離した。そのペレットを2回洗浄し、その後、DMEM/F12 1:1、1% N2補充物、1% ペニシリン−ストレプトマイシン、10% ウシ胎児血清を含む培地中に再懸濁した。シス−ヒドロキシプロリン(100μg/ml)を、線維芽細胞の増殖を抑制するために平板培養培地に加えた。細胞を、ポリ−L−リジン(0.01%)およびラミニン(20μg/ml)でコートしたカバーガラス上で、約106細胞/mlの密度で平板培養した。
【0099】
細胞分化
インビトロ分化
血清をもちいての非特異的分化:単一の神経球を、可視化のための解剖顕微鏡を用いて単離し、そして96穴の培養ディッシュ(Corning)の個々のウエル中で、1% N2、1% ペニシリン−ストレプトマイシン、および5〜10% ウシ胎児血清(Gibco)を含むDMEM/F12培地において、ポリ−L−リジン(0.01%)およびラミニン(20μg/ml)でコートしたカバーガラス上で平板培養した。培地は、実験の残りの間、変更しなかった。カバーガラスを、24時間後、または7〜10日後に免疫細胞化学のために処理した。
【0100】
トリチウム化したチミジンとのインキュベーション:チミジン標識実験を、Cepko laboratory(Dyer,M.A. and Cepko,C.L.2000.Nat Neurosci 3:873−880)のプロトコールを用いて実施した。神経球を、SCM中で8時間、3Hチミジン(NEN、5μCi/ml;89Ci/mmol)とインキュベートした。個々の神経球を単離し、SCM中で3回洗浄し、そして上述のように血清中で分化させた。分化後、カバーガラスを免疫細胞化学のために処理した。カバーガラスをスライド上に載せる前に、乳化剤のオイルをカバーガラスに加え、そして暗室に2日間置いた。乳化剤のオイルを除去し、そしてカバーガラスを水で洗浄した。現像液を4分間加えた。続いて、その現像液を吸引し、そしてそのカバーガラスを4% パラホルムアルデヒドで20分間固定した。その後、カバーガラスを水で洗浄し、Vectashieldを用いてスライド上に載せ、そして蛍光顕微鏡使用法下で(免疫細胞化学について)および明視野顕微鏡使用法下で(トリチウム化チミジンの取り込みについて)可視化した。
【0101】
指向性の分化:神経球を、レチノイン酸(RA、2mM、Sigma)およびソニックヘッジホッグ(Shh)タンパク質(Shh−N 400〜1000nM、R&D systemsより入手)、またはソニックヘッジホッグシグナル伝達の小分子アゴニスト(Hh−Ag1.3、Curis)で、当該分野で認識される方法(Wichterleら、2002.Cell 110:385−397;Soundararajanら、2006.J Neurosci 26:3256−3268)の変形を用いて4〜5日間、処理した。この処理は、SCM中で実施した。個々の神経球をその後単離し、そして96穴の培養ディッシュ(Corning)の個々のウエル中で、1% N2、1% ペニシリン−ストレプトマイシン、5% ウマ血清(Gibco)、CNTF(25ng/ml、Sigma)、GDNF(25ng/ml、Sigma)、およびBDNF(50ng/ml)を含むDMEM/F12培地において、ポリ−L−オルニチン(0.01%、Sigma)、I型コラーゲン(0.01%)およびラミニン(20mg/ml)でコートしたカバーガラス上で7〜10日間、平板培養した。四つの条件を用いた:(1)神経球を上記のように処理した;(2)RAなしで上記のように処理した;(3)ShhもRAもなしで上記のように処理した;(4)ShhもRAも三つのノイロトロピンもなしで、血清単独を用いて。カバーガラスをその後、免疫細胞化学のために処理した。
【0102】
神経筋接合部の形成:個々の神経球を、RA(2mM)およびShh−N(600〜1000nM)で、4〜6日間処理し、そして続いて上記の、MN成長および生存のための分化培地中の筋培養物上で平板培養した。二つの型の対照培養物を用いた:1)筋細胞単独および2)未処理の神経球をその上で平板培養した筋細胞。6〜21日後、培養物を蛍光α−アマガサヘビ毒素(2μg/ml、Molecular Probes alexa fluor 488)と2.5時間インキュベートした。それらをその後、洗浄し、固定し、そして免疫細胞化学(ニューロンマーカーTUJ−1)のために処理した。単一の神経球を、同時培養に先立って、1時間、1μMのジ−I(Di−I)またはジ−D(Di−D)で標識した。
【0103】
細胞マーカー
抗体:ネスチンに対するヤギポリクローナル抗血清(1:50)(R&D systemsより入手)。ChATに対するヤギポリクローナル抗体(1:100)およびニューロン特異的エノラーゼに対するマウスモノクローナル抗体(1:1000)はChemiconから入手した。Sox2に対するウサギポリクローナル抗体(1:1000)は、SigmaおよびAbcamから入手した。ビメンチンに対するマウスモノクロ−ナル抗体はZymedから入手した。GFAPに対するウサギポリクローナル抗体(1:1000)はDakoから入手し、GFAPに対するマウスモノクローナル抗体(1:1000)はSigmaから入手した。β−チューブリンIII(Tuj−1)に対するウサギポリクローナル抗体はCovanceから入手した。Tuj−1に対するマウスモノクローナル抗体(1:1000)およびNeu−Nに対するマウスモノクロ−ナル抗体(1:1000)はCovanceから入手した。Olig−2に対するマウスモノクローナル抗体は、使用に先立って前希釈した。NICHDの援助のもとに開発され、University of Iowa,Department of Biological Sciences,Iowa City,IA 52242によって維持された、神経フィラメント;MNR2、Lim3およびIsl−1;ならびにPax6に対するモノクローナル抗体は、Developmental Studies Hybridoma Bankから入手した。AF488結合体化ロバ抗ウサギIgG、AF488結合体化ロバ抗マウスIgG、AF568結合体化ロバ抗ヤギIgG、AF488結合体化ヤギ抗マウスIgG、およびAF568結合体化ヤギ抗ウサギIgGは、InvitrogenからのAlexa Fluor productsより入手し、全て1:1000で使用した。
【0104】
免疫細胞化学:はカバーガラスに付着した全神経球または分化した神経球で実施した。カバーガラスを、(PBS、pH7.4中の)4%ホルムアルデヒド中で20〜30分間固定し、その後PBS中で各10分で3回洗浄した。抗体希釈液をブロッキング溶液(PBS中0.2% ウシ血清アルブミンにおいて、10% 正常ヤギ血清、10% アイシングラス、0.3% Triton X)中で調製し、そして一次抗体をそのカバーガラスと一晩(8時間)インキュベートした。その後、適切な二次抗体との4時間のインキュベーションの前に、PBS中で3回洗浄した。PBS中での追加の3回の洗浄後、カバーガラスをDapi(0.03mg/ml)中で30分間インキュベートした。カバーガラスを、その後、3回洗浄し(各10分間)、そしてVectashield中にスライドガラス上に載せた。そのスライドは、Zeiss写真用顕微鏡または共焦点顕微鏡を用いて、免疫蛍光検査のために可視化した。特定のマーカーについての細胞染色のおおよその割合を、4〜5の20×視野の平均カウントにより決定した。細胞カウントは、Dapiを用いる核染色に基づく。
【0105】
rFT由来神経球の単離および特徴付け
単離:rFTから単離した細胞を、コラゲナーゼで分離し、そして、bFGF(20ng/ml)、EGF(20ng/ml)およびヒトLIF(10ng/ml)を含む標準的な幹細胞培地(DMEM、F12、N2補充物)中で培養した。インビトロで3〜4日後(DIV)、34の初代培養物中31において、神経球が観察された。これらの神経球は、最初は自由に浮遊し、そしてそれらの球状構造、位相での明るい外観、および一定の細胞膜により同定した。神経球は、やがてより大きな神経球へと成長する、3〜4の丸い細胞のより小さな塊として、初めは出現する。これらの、より大きな神経球のサイズは、サイズにおいて約50um未満〜1mm超までの広い範囲であった。30umより小さい細胞塊は、神経球としてカウントしなかった。初代培養物当たりの神経球の数は、約30から、50を超えるまで変化した。この数はドナーのラットの年齢とは相関しないようだった。増殖および自己再生の能力を示すため、神経球を分離し、そして19回まで継代した。これらの培養物は、インビトロで7ヶ月まで維持した。12の培養物を凍結し、そして二つを生存能力について試験し、成功裡に回復した。
【0106】
特徴付け:神経球は、NSCの均質な集団ではなく、むしろ異なる分化能力をもつ異なるNSCおよび神経前駆体細胞(NPC)の不均質な収集物である。rFT由来神経球を、種々の神経幹細胞(NSC)マーカー、神経前駆体細胞(NPC)マーカー、ニューロンのマーカーおよびグリアのマーカー(表1)の発現を決定するために、免疫細胞化学を用いて特徴付けた。特に、神経球を、NSCマーカーのネスチン(n=9);NPCマーカーのSox2(n=8)、ビメンチン(n=6)、Olig−2(n=3)、およびMusashi(n=4);ニューロン特異的マーカーのβ−チューブリンIII(Tuj−1、n=12);ならびに星状膠細胞のマーカーのグリア線維酸性タンパク(GFAP、n=12)について染色した。
【0107】
全ての例において、変動する割合の細胞が、ネスチンについて陽性であった。9例中4例において、神経球中の100%の細胞が、ネスチン+であった。このネスチン染色の存在は、神経球の培養における時間とは相関しないようであった。加えて、固定された全載(whole mount)(n=1)および組織切片(n=2)をネスチンについて染色した。免疫組織化学が、ネスチン+細胞を示した。神経前駆体のマーカーの染色は変化した。三つの実験のうちの三つにおいて、神経球内の100%の細胞が、Olig−2について陽性に染色され、いくらかの領域は、より強い染色を示した。全ての神経球が、Sox−2およびビメンチンについて陽性に染まる、ある割合の細胞を有したけれども、このパーセンテージは、Sox−2については40から100%まで変化し、そしてビメンチンについては33から100%まで変化した。Musashiの染色は弱く、時折の高強度の染色の塊をともなった。
【0108】
Tuj−1+およびGFAP+細胞は、全ての神経球中に存在した(n=12)。どの神経球も、両方のマーカーについて陽性のいくらかの細胞を含み、そしてこの比は神経球の間で大きく変化した。異なるマーカーを発現する細胞の空間的な塊形成がある。この塊形成は、ほとんどの神経球において明瞭であるが、そのパターンは変化した。
【0109】
ニューロンおよびグリアへの分化
いくらかの神経球は、培養容器に接着し、そして培地へのいずれの因子の付加も除去もなしに、ニューロンおよびグリアの形態学的特徴を有する細胞へ自発的に分化する。rFT由来神経球をニューロンおよびグリアに分化させるために必要とされる条件を決定した。bFGF、EGFおよびLIFの除去後、単一の神経球を、表2に示した接着基質±5〜10%のウシ胎児血清への曝露の7種類の異なる組み合わせで処理したカバーガラス上に平板培養した。各条件につき5回の実験を実施した。7日後、培養物をTuj−1、神経フィラメント、O1、GFAPおよびネスチンについて染色した。接着基質単独または血清単独のどちらかの使用が、形態学的な分化を開始するために十分であったが、血清の付加が、より迅速な分化を結果として生じた。全ての例において、Tuj−1、神経フィラメント、O1、およびGFAPを含むニューロンのマーカーまたはグリアのマーカーのどちらかを発現する神経球に由来する細胞を検出した。
【0110】
全ての続いて起こる分化の実験(n=65)は、3つの増殖因子全ての除去、5〜10%ウシ胎児血清の培地への補充、およびポリ−L−リジンおよびラミニンの両方でコートしたカバーガラス上への単一神経球の平板培養により、実施した。細胞を、これらの条件で24時間(n=30)または7〜10日間(n=35)培養し、そして培養物を免疫細胞化学のためにその後固定した。これらの実験において用いられた神経球のサイズの広い分布を考慮すると、得られた分化した細胞の数は、神経球当たり約50細胞未満から、5000細胞超の範囲であり、最初に平板培養した神経球のサイズと相関する。より大きい神経球(通常100umより大きい)は、5000より多い分化した細胞を生成することができる。
【0111】
神経球が、NPC、ニューロン、星状膠細胞および乏突起膠細胞を産生する能力があるか否かを決定した。これらの細胞型を同定するために用いた免疫細胞化学マーカーは、ニューロン特異的エノラーゼ(NSE)、NeuN、Tuj−1、GFAP、O1、Musashi、ビメンチンおよびSox2を含んだ(表1および表3)。各例において、単一の神経球から得られた細胞を、これらのマーカーの二つについて二重染色した。これらの実験由来のデータは、rFT由来神経球が変動した分化能力を有することを示した。24時間を超えて分化した神経球(n=9)は、ニューロンのマーカーおよびグリアのマーカーの両方について二重に染まった細胞の高い割合を有した(表3)。Tuj−1およびGFAP染色の例において、69±14%(n=5)の細胞がその二つのマーカーについて二重に染まった。7〜10日後、ニューロンのマーカーおよびグリアのマーカーの両方について二重に染まった細胞の割合は、顕著に減少した(表3)。Tuj−1およびGFAP染色の例において、わずかに13±10%(n=9)の細胞が、その二つのマーカーについて二重に染まった。Tuj−1およびGFAPの変動する発現が、24時間後の分化を7〜10日後の分化と比べた14の実験において観察された。これらのrFT由来細胞集団中に存在するTuj−1+およびGFAP+の変動する割合は、各神経球の不均質な分化能力を反映する。この変動は、ラットの年齢とは拘わりなく、両方を同一の供給源から得た神経球の間でなされた比較において、異なるrFT供給源に由来する場合と同様に持続した。
【0112】
まれな場合において、7日間の分化の後、単一の神経球由来の85%を超える細胞が、ニューロンのマーカーまたはグリアのマーカーのどちらかを発現した(n=2)。しかしながら、ほとんどの例において、明白な優勢は観察されず、そしてニューロンの細胞およびグリア細胞の両方の変動する割合は、単一の神経球の分化から見出された。NPCマーカーの染色は、分化条件に7〜10日間曝露した後でさえ持続した(表3)。実際、その染色は、使用した全てのNPCマーカーについての24時間での染色と比較して、7〜10日後にわずかに増加するようである。
【0113】
分化した細胞が、増殖性細胞に由来することを立証するため、本発明者らは、活発に分裂している細胞をトリチウム化したチミジン(3H)で標識した。神経球を、3Hで8時間処理した(n=5)。その神経球を、その後洗浄し、上述の標準的な条件において、7日間を超えて分化させた。得られた細胞を、Tuj−1およびGFAPについて染色した。5例全てにおいて、ニューロンおよびグリアとして免疫学的に同定された細胞の27〜90%が、それらの核中に取り込まれたトリチウム化したチミジンを有した(図12)。この結果は、得られた細胞が、活発に分裂している細胞の子孫であったことを示す。
【0114】
RFT由来神経球は運動ニューロン(MN)を生成する
全ての実験において(n=25)、種々の割合の分化した細胞が、上述のMNマーカーまたは運動ニューロン前駆体マーカーを発現した。図12は、MNR−2、Tuj−1およびGFAPについての染色に基づいて、運動ニューロン、ニューロンおよびグリアの割合を示す。
【0115】
Shh−Nで処理した神経球は、分化したニューロンを生じ、その20〜40%のみがMNマーカーであるMNR−2、Isl1、Lim3およびChATを発現した(n=14)。Shh−N濃度を400nMから1000nMまで増加させることは、その結果を変化させないようだった。Hh−Ag1.3が用いられた場合(n=9、1.5μM)、分化したニューロンの95〜100%がMNマーカーを発現した。この結果は、これらの濃度において、Hh−Ag1.3アゴニストが、実際のShh−Nペプチドよりも、FT由来神経球からのMNの生成のために、より有効であり得ることを示唆した。
【0116】
FTからMNを生成するために、どの因子が本質的であるかを決定するため、分化条件を変化させた:(1)神経球を、RAなしで、Shh−Nで処理し(n=8)、そしてそれらを血清ならびにBDNF、CNTFおよびGDNFを含む培地中で分化させた;(2)未処理の神経球を、血清および3つのノイロトロピンを含む培地中で培養した(n=8);(3)未処理の神経球を、神経栄養因子の付加なしで、血清を含む培地中で分化させた(n=3)。図13に示したように、(1)および(2)の条件において、神経球は、変動する割合のMNR2+細胞(5〜67%)を一貫して生成した。
【0117】
条件(3)において、MNの生成は一貫性がなかった。3例中1例において、神経球から得られた細胞の40%はMNR2を発現し、そして3例中2例において、細胞はMNR2+ではなかった。MNの指向されかつ一貫した生成のための、RAおよびShh−Nの使用は、BDNF、CNTFおよびGDNFの存在下において神経球を単に分化させることよりも優れているとは立証しなかった。しかしながら、上述し、そして図13に示したように、Hh−Ag1.3は、MNの収量を増大させるうえで有益である。FTが脊髄の痕跡のレムナントであることを考慮して、これらの結果は、RAの尾方化作用も外因性の腹方化Shh−Nシグナル伝達もなしに、いくらかのrFT NSCをMNに分化させる能力を示す。
【0118】
この研究の結果は、多分化能幹細胞が出生後のrFT中に存在することを示す。これらの細胞は、NSCの二つの基本的な特性を示す:それらは、自己再生/増殖でき、ならびにニューロン、星状膠細胞および乏突起膠細胞を含む複数の細胞型へ分化できる。MNを生成するFTの能力は、ALSのような神経変性疾患に対して、特に治療上重要であり得る。
【0119】
NSCのニッチとしてのFT
本発明の方法を、先に記載された、例えば側脳室の脳室下帯(SVZ)のようなNSCのためのCNSのニッチの多くの特性を、FTの組織学的環境が有することを決定するために用いた。SVZにおける細胞構成は、A型(神経芽細胞)、B型(ゆっくりと増殖するGFAP+神経形成星状膠細胞)、C型(中間前駆体細胞)およびE型(上衣)細胞からなる。この系において、E型細胞は脳室を裏打ちし、そして、脳室に接触するためにE細胞の間を縫うように続くB細胞によって、時折置き換えられる。B型細胞は、E細胞の脳室下側に向かって位置し、吻側移動経路(Rostral Migratory Stream)として公知の経路に沿って嗅球へ移動するA細胞をさや状に覆う。C型細胞は、A細胞の鎖に沿って分散する。
【0120】
これらの研究は、FTにおける細胞型が、上衣細胞、神経芽細胞、星状膠細胞様細胞、小膠細胞、乏突起膠細胞、ニューロン、線維芽細胞、脂肪細胞および神経節細胞を含むことを示した。FT細胞は、脊柱管を裏打ちする上衣細胞の周りにゆるく組織化される。FTの上衣細胞は、広範囲なモザイクまたはサイズの変化する環を形成する脳室の裏打ち上のロゼットとして、しばしば伸長する。理論によって拘束されることを望まないが、本発明は、FTの上衣細胞は、SVZのE細胞に類似するという驚くべき発見に基づく。さらに、SVZのB細胞と同様に、FT中のいくらかのGFAP+星状膠細胞様細胞は、上衣細胞の間に互いに入り込む突起を有する。さらに、これらの突起の構造は、SVZ中でA細胞をさや状に覆うB細胞に類似する。本発明の方法を用いて実施した研究からのデータは、FTの頭方の部分に見出されるニューロンが、ラットおよびヒトの両方における形態において神経芽細胞に類似することを示す。それらは、しばしば神経線維の管に沿って存在し、そして時折、FTの外側縁に伸長する。本発明の特定の実施形態において、これらの細胞は、SVZのA細胞に相似性であると考慮される。
【0121】
腫瘍形成
CNSの腫瘍は、神経組織のニッチ近くにしばしば見出される。脊髄およびFT内の腫瘍は、全てのCNSの腫瘍の4〜10%を構成する。パラガングリオーマおよび原始神経外胚葉性腫瘍(例えば、脳室上衣腫)は、終糸に対する親和性を有する。パラガングリオーマは、神経内分泌腫瘍であり、そして脳室上衣腫は、中心管の上衣細胞から生じる腫瘍である。それらは、全てのグリアの脊髄腫瘍の60%の原因であり、そして成体における最も一般的な髄内脊髄性新生物である;粘液乳頭型脳室上衣腫(FTにおける上衣のグリアの腫瘍)は、脳室上衣腫の13%を構成し、そしてFTに対する明瞭な偏好を有する(Koeller,K.K.et al.2000.Radiographics 20:1721−1749)。
【0122】
細胞の同一性は分化プロセスの前に決定される
24時間と7〜10日間で分化した単一の神経球におけるニューロンのマーカーおよびグリアのマーカーの発現を対比した。24時間の分化プロセス後、ほとんどの細胞が、ニューロンのマーカーおよびグリアのマーカーの両方を発現した。一つの局面において、この結果は、分化プロセスの初期における未解明の細胞運命を反映する。7〜10日後、単一の神経球に由来するほとんどの細胞は、ニューロンのマーカーまたはグリアのマーカーのどちらかを発現し、非常に少数の細胞が両方について二重に染色される。この結果は、ドナーの年齢とともには変化しない。24時間後の相対的に不明瞭な細胞同一性と比べて、細胞は時間とともに、より拘束された細胞運命を発現する。
【0123】
細胞同一性は、分化プロセスの前に決定される。分化前に染色した神経球は、同一の処理にもかかわらずTuj−1+およびGFAP+細胞の異なるパターンを示した。神経球内のいくらかの細胞が、両方のマーカーについて二重に染まったが、ほとんどの細胞は、Tuj−1またはGFAPのどちらかの一つのマーカーのみを発現した。同一のマーカーについて陽性の細胞は、空間的に一緒に塊を形成する傾向がある。
【0124】
神経球分化の前に取得されたニューロンの特徴またはグリアの特徴は、各神経球の分化能力を予測する。分化の初期段階の間の一過的な二重染色は、細胞運命が未決定であるよりもむしろあいまいである分化経路に沿った点を示す。分化前の神経球細胞の初期の染色パターン(GFAP+またはTuj−1+)は、よりニューロンまたはグリアの運命へと神経球を操るためには、最初とは異なる条件でその神経球を培養することを必要とすることを示す。
【0125】
神経前駆体細胞(NPC)マーカーは、7〜10日で持続する
NPCマーカーの発現は、分化の24時間後に高く、そして7〜10日後でさらに高かった。ほとんどの細胞は、二重に染まらなくなり、そしてより成熟した表現型を示すマーカー(すなわち、ニューロンまたはグリア細胞マーカー)を発現することを考慮すると、これは驚きであった。
【0126】
その持続、およびNPC染色におけるわずかな増加は、二つの機構に帰する。一つの説明は、時間にそった細胞の代謝活性における変化を含む。一局面において、分化プロセスの初期に、いくらかの細胞は、迅速なタンパク質ターンオーバーをともなって高度に代謝的に活性であり、NPCマーカーの検出を妨げる。数日の分化の後、ターンオーバー速度が低下することから、そのタンパク質レベルが増加し、それゆえ免疫細胞化学を介してのタンパク質検出を可能とする。代替的な説明は、7〜10日の分化後、細胞は系統特異的なNPCであり、そしてそれゆえ、ニューロンまたはグリア特異的マーカーに加えてNPCマーカーを発現するというものである。
【0127】
RFT神経球はMNを生成する本質的な能力を有する
RFT由来神経球は、インビトロで胚性幹細胞をMNに分化させるために用いられたRAおよびShhへの暴露の有りおよび無しでMNを生成した。胎仔身体における吻側神経前駆体は、RA(尾方化シグナル)に応答して脊髄の位置の同一性を獲得し、そしてその後、Shhの腹方化シグナルに応答して運動ニューロン前駆体の同一性を獲得する。
【0128】
BDNF、CNTFおよびGDNFは、MNの成長および生存を支持することが公知のノイロトロピンである。血清、BDNF、CNTF、およびGDNFの存在下で分化させる前に、RAおよびShh−Nで処理したRFT神経球は、20〜40%のMNを生成した。RA処理もShh−N処理もなく、BDNF、CNTF、およびGDNFを有する血清中で平板培養した神経球は、5〜67%のMNを生成し、rFT神経球をShh−NおよびRAで処理することは、MNを生成するためにBDNF、CNTF、およびGDNFの存在下で、それらを平板培養するよりも、より効果的ではないことを示した。RAおよびShhは、胚性幹細胞の分化をMNへ指向させるために重要であるが、胚の発生の間にすでに、ある程度まで尾方化され腹方化されていてもよい出生後rFTから得られたNSCとは、これらのシグナルは関連するものでなくともよい。
【0129】
FTにおけるいくらかのNSCは、血清以外の因子を必要とすることなくMNへと分化する能力を持ち得る。FT由来神経球を血清単独で平板培養した場合、1/3は、40%のMNを生成し、時折のMN発現は、特定の介入なしに起こり得ることを示した。一局面において、FTは、胎仔の尾への神経支配を与える(または、げっ歯類の例においては、最も尾側の尾分節の一過的な神経支配を与える)、脊髄の部分の痕跡のレムナントであることから、このことが起こる。FT「脱分化」の発生上のプロセスがヒトで欠けている場合、新生児は、動かせる尾をもって生まれ得、持続するMN神経支配を示唆する。この状況において、FTから単離されたNSCは、MNのような脊髄に固有の細胞型を生成する能力を有し得る。
【0130】
rFT由来神経球からのMN収量の増加
Shh−Nに加えて、Shhシグナル伝達はまた、小分子アゴニストのHh−Ag1.3により活性化される。胚性幹細胞(ESC)からのMNの生成のために、研究は300〜500nMのShh−Nまたは1〜2μMのHh−Ag1.3を使用した(Wichterle,H.ら、 2002. Cell 110:385−397;Harper,J.M.ら、 2004. Proc Natl Acad Sci U S A 101: 7123−7128;Miles,G.B.ら、 2004. J Neurosci 24:7848−7858;Li,X.J.ら、 2005. Nat Biotechnol 23: 215−221 ;Soundararajan,P.ら、 2006. J Neurosci 26:3256−3268). Wichterleらは、これらの濃度においてShh−NおよびHh−Ag1.3で同一の結果を報告したが、ほとんどの研究は、ESCからMNを生成するために1μMのHh−Ag1.3を使用した。rFT神経球において、Hh−Ag1.3は、Shh−Nと比較した場合、生成されたMNの収量の増加において特に有効であった。Hh−Ag1.3を用いた場合、100%近くのMNが生成され、一方、Shh−Nが加えられた場合、わずか20〜40%のMNが生成した。Hh−Ag1.3は、ESC神経球に比較してrFT神経球におけるMN収量の増加のために選択的に有効のようである。これらの結果は、rFT神経球の一つの予期せぬ、そして優れた特性を強調する。
【0131】
FT神経球由来のGFAP+細胞
GFAPを星状膠細胞のマーカーとして用いたが、しかしながらGFAPは、星状膠細胞様成体幹細胞についてのマーカーでもある。成体哺乳動物において、神経形成星状膠細胞は、側脳室のSVZ、および海馬中の歯状回のやや顆粒状のゾーンにおいてインビボで同定された。神経形成星状膠細胞を、非神経形成星状膠細胞の非常に大きい集団から区別する特徴およびマーカーは、未解明である。rFTから分化したGFAP+細胞は、神経形成および/または非神経形成星状膠細胞である。分化実験において、細胞は、時折GFAPおよびニューロンのマーカーについて二重染色される。指向性のMN分化をうけた神経球由来の細胞は、時折MNR2およびGFAPの両方を発現する。GFAPが、単に非神経形成星状膠細胞マーカーである場合、GFAPと運動ニューロンマーカーの同時発現は驚きである。なぜならば、GFAPはまた、星状膠細胞様成体NSCについてのマーカーであり、二重染色された細胞は、MNの細胞運命に拘束された神経形成星状膠細胞を示し得る。
【0132】
RFT神経球は増殖し、インビトロで継代され得、そしてNPC、ニューロンおよびグリアの収集物に分化する。哺乳動物CNS内の多分化能細胞の発見は、外傷、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症(ALS)、および多発性硬化症を含む多くの現在治療不能のCNS疾患の治療の可能性のためにすばらしい意味を有した。多分化能細胞の供給源としてのFTの発見は、これらの神経学的疾患の自家移植療法の分野における新しい可能性を開く。
【0133】
実施例9:出生後のヒト終糸は、運動ニューロンを生成することが可能な自系多分化能神経球の供給源である
本発明の方法を、年齢18歳までのドナーからヒトNSCを単離するために用いた。これらの細胞は、培養の時間の延長した期間にわたって増殖した神経球を生じた。その神経球を、ニューロンおよびグリアに分化するように誘導した。さらに、それらを、インビトロで横紋筋を刺激することが可能な運動ニューロンを形成するように誘導した。これは、到達可能および消耗可能の両方である多分化能CNS細胞の最初のヒトの供給源であり、そして出生後の組織から得られたヒト神経球由来の運動ニューロンの最初の報告である。本発明は、免疫学的拒絶を回避する自系細胞に基づく移植療法を提供する。
【0134】
ヒトにおいて消耗可能な自系NSCの供給源を探した。その独特の発生歴ならびにパラガングリオーマおよび神経外胚葉性腫瘍を生成するその性質のために、FTを焦点として選択した。FTは、脊柱の基部にある尾骨に脊髄を固定する脊髄の尾方末端の細長い延長(成体で約15cm)である(図1A)。それは、発生初期に胎仔の痕跡尾への神経支配を与えるか、または尾のある脊椎動物において最も尾側の尾分節の一過的な神経支配を与える神経系のレムナントである。初期の段階において、予定運命のFTは、三つの追加の脊髄神経節(C3〜C5)をもつ分化した脊髄である(図1A)。尾が再吸収される時、FTの細胞は、Streeterにより「脱分化」と名付けられたプロセスにより初期の胚段階への逆戻りをうける(図1A、右図)。 その結果は、消失する尾側の点へと狭まり、上衣細胞により裏打ちされ、そして見たところゆるく組織化された線維芽細胞、脂肪細胞、繊毛のない上衣細胞のロゼット、神経芽細胞、ニューロンおよびグリアの収集物で環状に囲まれた中心管を有する、細長い構造物である。この局所的環境は、NSCを生成する他のCNS領域の特性の多くを有する。FTは、外科的に接近可能であり、そして脊髄が動作の十分な自由を欠く「係留脊髄症候群」(TCS)の例において脊髄上で牽引力を軽減するために、慣例的に外科的に切断される。
【0135】
FT組織は、ヒト胎児および出生後の手術より得た。FTは、選択的に人工中絶された14週〜21週齢の胎児から切開した(図1B)。年齢が6ヶ月から18歳の出生後組織を、TCSの神経外科例から得た。組織供給源に関する不明瞭さはなく、全ての外科的FT試料を硬膜鞘内から取得し、そしてFTの同一性を、切除前に臨床的電気生理学を用いて確認した。切片は、3つの出生後のFTから作製し、そしてNSCのマーカーのネスチンの存在について試験した。年齢が8ヶ月〜5歳の3つのFT試料の免疫組織化学が、ネスチン+上衣細胞ならびに分散した神経前駆体細胞の存在を示した。(図1C)。
【0136】
増殖している細胞の浮遊凝集物である神経球を、FTから単離した。組織は、4つの胎児および17つの出生後ドナーから得た。初代組織は、酵素的に分離し、そして神経球の成長を促進する標準的な条件を用いて培養した。インビトロで3〜4日後(DIV)、本発明者らは、胎児の100%および出生後培養の82%に神経球を観察した(図10F)。その神経球は、細胞の小さい塊としてスタートし、そして直径で約25μmから500μm超の球体に成長した。単離した神経球の数は、初代培養物当たり約1から50を超える神経球で変化し、そしてこの存在量は、ドナーの年齢に依存しなかった。それらの増殖および自己再生の能力を示すため、神経球を成功裏に分離し、そして10回まで継代し、そして試みた最長の期間である6ヶ月間インビトロで維持した。8つの培養物を凍結し、そして一つを生存能力について試験し、そして成功裏に回復した。
【0137】
神経球、ならびにニューロンおよびグリアを生成するそれらの能力を特徴付けるため、神経球を種々の免疫細胞学的マーカーの発現について試験した。免疫細胞化学を単一の神経球で実施し、そして各アッセイについて、神経球は、1以上のドナーに由来した。神経球を、種々の神経の幹細胞(NSC)、神経前駆体細胞(NPC)、ニューロンのマーカーおよびグリアのマーカーについて試験した。全ての神経球はネスチン+(n=13)(図10A)であった。より小さい神経球(100μm未満)においては、100%の細胞がネスチン+であったが、一方、より大きな神経球においては、中心部はネスチン−のようであった。試験した全ての神経球はまた、NPCマーカーのビメンチン(n=33)、CD133(n=18)、Olig−2(n=17)およびSox−2(n=17)(図10B〜D)に対して陽性の細胞を含んだ。NPC+細胞の発現パターンおよび割合は、神経球の間で変化した。本発明者らは、神経のタンパク質であるβ−チューブリンIIIを認識するTuj−1、および星状膠細胞マーカーのGFAPについて42の神経球を染色した(図10E)。Tuj−1+およびGFAP+細胞は、全ての神経球中に存在した。加えて、各神経球は、両方のマーカーについて陽性のいくらかの細胞を含み、そしてこの二重に陽性の割合は著しく変化した。図10Eで示したように、異なるマーカーを発現する細胞の空間的な塊形成があった。ほとんどの神経球について、塊形成は明白であったが、一方でそのパターンは変化した。
【0138】
分化した細胞型を生成する神経球の能力を試験するため、単一の神経球を、5%ウシ胎児血清で増殖因子を置換した上記の培地を用いて、ポリ−L−リジンおよびラミニンでコートしたカバーガラス上に平板培養した。その神経球は、年齢が6ヶ月および12歳の2人のドナーから得た。個々の神経球から得た培養物を、平板培養2〜10日後に免疫細胞化学を用いて試験した。分化した細胞が、増殖している細胞に由来したことを確認するために、単一の神経球を、トリチウム化したチミジンと8時間インキュベーションした(n=4)。その神経球をその後、分化させ、そして7日後に試験した。全ての例において、結果として生じるニューロンおよびグリアの顕著な割合(33〜63%)が、それらの核に取り込まれた放射性のヌクレオチドを有した(図5C)。
【0139】
培養物を、ニューロンのマーカーであるニューロン特異的エノラーゼ(n=4)、およびTuj−1(n=19);星状膠細胞マーカーであるGFAP(n=11);乏突起膠細胞マーカーであるO1(n=5);ならびにNPCマーカーであるビメンチン(n=9)、CD133(n=4)、Olig−2(n=3)およびSox−2(n=8)に対する抗体で染色した。各培養物から得た分化した細胞の数は、約50細胞より少ないから5000細胞超までの範囲であって、それは平板培養した神経球のサイズと相関した(n=50)。各神経球により生成された細胞型の割合には大きな変動性があり、それは神経球のサイズまたはドナーの年齢とは相関しなかった。平板培養の2日後、神経球から得た細胞の79+/−8%(n=8)が、ニューロンのマーカーおよびGFAPまたはO1のどちらかについて二重に染まった(図5B)。7〜10日後、二重に染まったものは、23+/−9%(n=7)に減少し、そして培養物の約半分において、ニューロンまたは星状膠細胞のどちらかが優位を占め、神経球の変動する能力を示唆した。分化後10日でネスチン+細胞はなかったが(n=3)、しかしながら細胞の98%が、ニューロンのマーカーまたはグリアのマーカーに加えてNPCマーカーについて陽性で残り(n=21)(図5A)、その細胞が、まだ十分に分化していないことを示した。
【0140】
FTは、脊髄の痕跡の部分を示すことから、本発明者らは、FTの神経球が、脊髄の外傷またはMNの変成の例における細胞置換ストラテジにおいて用いられ得る、脊髄運動ニューロン(MN)を生成することが可能かどうかを決定した。MN前駆体を生成するために、単一の神経球を、2μMのレチノイン酸および0.4〜1μMのソニックヘッジホッグタンパク質(Shh−N)で4〜6日処理した(Wichterle,H.ら、 2002. Cell 110:385−97)。神経球をその後、5%のウマ血清ならびにMNの成長および生存を促進することが公知の3つの神経栄養因子:毛様体由来神経栄養因子、脳由来神経栄養因子、および神経膠由来神経栄養因子(Zurn,A.D.ら、 1996. J Neurosci Res 44:133−41)の存在下で接着基質上で平板培養した。7〜10日後、各神経球により生成されたMNの割合を、MNマーカーのMotor Neuron Restricted−2(MNR−2)(Jessel,T.M. Nat Rev Genet 1:20−9)についての免疫細胞化学を用いて決定した。Shh−Nで処理した神経球は20+/−12%のMNR−2+細胞(n=6)を生成し、そしてShh−N濃度を増加させることは、MNの割合に影響しないようだった(図5D)。興味深いことに、小分子Shh−NアゴニストのHh−Ag1.3(1.5μM)(Frank− Kamenetsky,Mら、 2002. J Biol 1:10;Harper,J.M.ら、 2004. PNAS USA 101:7123−8)を処理に含めた場合、100%の細胞がMNR−2を発現し(n=2)、Hh−Ag1.3が、MN分化の誘導において、より有効であり得ることを示唆した。未処理の神経球を3つの神経栄養因子と平板培養した場合(n=4)、本発明者らは、5〜50%のMNR2+細胞、および血清単独において平板培養した2/3の未処理神経球が陽性細胞を生じたことを検出し(1%および40%)、いくらかの神経球は、加えられた因子の非存在下でMNを生成し得ることを示唆した。MNの同一性を確認するため、本発明者らは追加のMNマーカーについて試験した:Lim−3(n=3)、Islet−1(n=1)、およびコリンアセチルトランスフェラーゼ(n=2)(図5E)(Oda,Y. and Nakanishi,I. 2000. Histol Histopathol 15,825−34;Arber,S.ら、1999. Neuron 23,659−74;Pfaff,S. L.ら、1996. Cell 84,309−20;Tsuchida,T.ら、 1994. Cell 79,957−70)。全ての例において、その結果は、染まった細胞の割合に関して、MNR−2についての結果と類似した。さらに4/4の神経球は、RT/PCR(Pfaff,S.L.ら、 1996. Cell 84:309−20)により決定されたように体細胞の運動ニューロンによって選択的に発現されるホメオボックスタンパク質のHomeobox−9を発現した。
【0141】
免疫細胞化学の基準によりMNとして特徴付けられた細胞が、筋肉を刺激することが可能であるか決定するため、神経球を出生後ラット由来の横紋筋培養物に加えた。神経球を、上述のようにレチノイン酸およびShh−Nで処理し、そしてその後、単一の神経球を各筋肉培養物に加えた。同時培養中のニューロンが、平板培養した神経球に由来したか決定するため、4つの神経球を、平板培養の前に脂肪親和性カルボシアニン色素のDiDで2.5時間プレインキュベーションした。全ての例において、本発明者らは、ニューロンの形態学的な特徴を有するDiD+細胞を検出した。位相差顕微鏡法により、または神経球の付加なしに筋肉培養物におけるTuj−1染色により検出できるニューロンはなかった(n=12)。神経球と6〜21日の同時培養後、神経筋接合部でのニコチン酸アセチルコリンレセプターの塊形成を検出するために、培養物を、蛍光α−アマガサヘビ毒素とインキュベートした。全ての同時培養物は、この基準による神経筋接合部の証拠を示した(n=18)。図11Bは、神経筋接合部および刺激を与えるニューロンを示すために、α−アマガサヘビ毒素およびTuj−1の両方について染色した培養物を示す。筋繊維のみを含む対照培養物は、神経筋接合部を含まなかった(n=12)。
【0142】
ほとんどのFTは、TCSの外科的試料から得たが、実質的に区別できない結果を、中絶した胎児由来のFTをもちいて、および出生後のラット由来のFTでの広範囲な実験から得た。これらの結果は、FT中の多分化能細胞の存在が、正常の条件を反映することを示す。FTは、神経系の外傷または変性の例における使用のための、自系の、消耗可能な、到達可能な多分化能細胞の供給源である。年齢が18歳までのドナーからの、これらの細胞の単離および分化は、それらが成人期まで持続することを示す。
【0143】
HuFTからの神経球の単離および分化
ヒト胎児組織(14〜21週齢)を、ヒト小児組織でと同じように、氷冷したHanks緩衝液中で切開した(図1b)。その組織をDMEM/F12(1:1、Gibco)中で、II型コラゲナーゼ 100U/ml(Gibco)で分散し、そして標準的な幹細胞培地のDMEM/F12、1% N2処方物(Gibco)、1%ペニシリン−ストレプトマイシン溶液(Gibco)、EGF(20ng/ml、Gibco) bGFG(20ng/ml、Gibco)、LIF(10ng/ml)(Weiss,S.ら、 1996. J Neurosci 16,7599−609)中で維持した。2〜4週毎に、神経球を、Accumax(Innovative Cell Technologies)での分離後継代した。神経球の分化を、増殖因子の除去、培地への5〜10%血清の付加およびポリリジンおよび/またはラミニンでコートしたカバーガラスでの平板培養により誘導した。MNへの分化のために、神経球をRA(2uM、Sigma)および0.4〜1μMのShh−N(R&D systems)または1.5μMのHh−Ag1.3(Curis)で4〜6日で処理し、その後、ポリ−L−オルニチン、ラミニンおよびコラーゲンでコートしたカバーガラス上で、BDNF、 CNTFおよびGDNF(Sigma)を含む神経基本培地(Wichterle,H.ら、 2002. Cell 110,385−97)中で7〜10日間、平板培養した。
【0144】
他の実施形態
本発明をその詳細な明細とともに記載したが、先述の明細は説明を意図するものであって、付録の特許請求の範囲により規定される本発明の範囲を制限するものではない。多の局面、利点、および改変は、以下の特許請求の範囲内にある。
【技術分野】
【0001】
関連出願
この出願は、2007年6月27日に出願された米国仮特許出願第60/937,571号(この内容は、その全体が参考として本明細書に援用される)に関する。
【0002】
発明の分野
本発明は、細胞治療の分野に関する。
【背景技術】
【0003】
神経幹細胞(NSC)および前駆体細胞は、中枢神経系(CNS)内に存在する発生的に始原の細胞であり、そこでの主要な細胞型(ニューロン、星状膠細胞および乏突起膠細胞)の全てを生じることができる。NSCは、ヒトの神経の発生および神経学的疾患の研究ならびにそのような疾患を処置するために有用である。
【0004】
NSCは、脳内の深い領域(area)(例えば、側脳室の脳室下帯および海馬の顆粒細胞層)から採取されてきた。これらの領域からNSCを採取する間、上にある脳領域を損傷する可能性は、自系NSCの採取についてのこのストラテジを危険かつ非現実的にしている。
【発明の概要】
【課題を解決するための手段】
【0005】
本発明の要旨
本発明は、容易に入手可能なNSCの集団を同定すること、ならびに既存の方法に比べて傷害の危険がはるかに少ない、それらを単離するための方法を記載することによって、これらの困難に取り組む。これらの細胞は、同種異系または自系である。異種組織の移植に起因する合併症を回避するために、NSCは、好ましくは自系である。例えば、細胞置換(cell replacement)または組織再生のための薬学的組成物は、NSCに富んだ終糸(FT)神経細胞の集団を含む。この集団は、神経球または神経球開始細胞(neurosphere initiating cell)(NS−IC)を含む。神経球は、神経幹細胞を含む細胞の凝集物または塊である。これらは、代表的に、事実上球状であり、培養において自由に浮遊する。神経球内の細胞は、培養において増殖し、一方、ニューロンおよびグリアに分化する能力を維持している。NS−ICは、長期間の神経球培養を開始することのできる細胞である。NS−ICは、ネスチン陽性であり、適切な分化条件下で、ニューロン、星状膠細胞および乏突起膠細胞に分化する能力を有する。好ましくは、この組成物は、単離されたFT細胞の集団(このうちの少なくとも10%は、NSCまたはNS−ICである)を含む。例えば、この細胞集団のうちの少なくとも30%、50%、85%、90%、99%または100%はNSCである。
【0006】
FT(必須ではない神経組織)は、被験体(例えば、神経学的傷害または他の障害に罹患しているか、これらを発症する危険を有するヒト患者)から採取される。出生後の動物からのNSCの単離は、被験体由来のFT組織を提供すること、FT組織を分離して神経球を取得すること、そしてネスチン陽性NSCを回収することによって実行される。この様式で取得された、単離されたNSC(すなわち、自系NSCの集団)を含む組成物は、この組織が取得された被験体を処置するために使用される。FT組織から取得された神経幹細胞は、FT−NSCと呼ぶ。FT−NSCの多分化能の子孫細胞を含む細胞株もまた、本発明の範囲内である。
【0007】
したがって、被験体の神経学的機能を増強または回復させる方法は、単離されたFT細胞の集団をこの被験体に投与することによって実行される。これらの細胞は、被験体に投与される(例えば、これらの細胞は、傷害もしくは損傷の部位に局所的に直接移植されるか、または罹患した部位から離れた部位に投与される)。例えば、細胞は、脳の損傷した部分に脳室内に導入されるか、または脊髄液に注入される。神経学的障害を処置する方法は、被験体からFT組織を採取する工程、FT細胞を生体外で培養して、単離されたFT−NSCの富化された集団を生成する工程、および単離されたFT−NSCの富化された集団を被験体に投与する工程を包含する。処置されるべき神経学的障害は、傷害(急性または慢性)または変性状態を含む。神経学的障害は、欠陥の起源に関係なく、脳もしくは脊髄の傷害または脳もしくは脊髄の機能の減損を含む。例えば、この被験体は、脳卒中に罹患していると診断されるか、脳卒中に罹患していると疑われる。
【0008】
上記の細胞は、疾患または傷害を通じて失われた神経組織を再構成するために使用される。神経細胞に関連する遺伝病は、遺伝子の欠損を修正するか、または疾患に対して保護するように処置する、自系もしくは同種異系の幹細胞の遺伝子改変によって処置され得る。処置されるべきCNS障害としては、神経変性疾患(例えば、アルツハイマー病、多発性硬化症(MS)、ハンティングトン病、筋萎縮性側索硬化症およびパーキンソン病)、急性脳傷害(例えば、脳卒中、頭部傷害、脳性麻痺)および他のCNS機能不全(例えば、うつ病、てんかんおよび統合失調症)が挙げられる。
【0009】
動物由来の単離されたFT−NSCを提供すること、およびFT−NSCの増殖または分化を可能にする条件下でFT−NSCを培養することによって、出生後の動物由来のFT−NSCを増殖させる方法もまた本発明の範囲内である。
【0010】
本明細書に記載の組成物は、精製または単離される。「実質的に純粋」によって、天然でそれに付随する成分から分離されている核酸、ポリペプチドまたは他の分子を意味する。代表的に、ポリペプチドは、それが、天然で関連しているタンパク質および天然に存在する有機分子を、少なくとも60重量%、70重量%、80重量%、90重量%、95重量%またはさらには99重量%含まない場合、実質的に純粋である。例えば、実質的に純粋なポリペプチドは、天然の供給源からの抽出、通常そのタンパク質を発現しない細胞における組換え核酸の発現または化学合成によって取得され得る。用語「単離された核酸」は、DNAであって、所定の核酸が由来する生物の天然に存在するゲノムにおいて、このDNAと隣接している遺伝子を含まない、DNAを意味する。したがって、用語「単離された核酸」は、クローニングされた核酸または合成核酸(RNA、RNAi、DNA)を包含する。
【0011】
有効量は、臨床上の利益を付与するのに必要とされる組成物(例えば、細胞のサンプル)の量である。この有効量は、投与の経路、被験体の年齢、体重および全身の健康に依存して変動する。薬学的組成物は、少なくとも1つの治療的または生物学的に活性な薬剤を含み、そして患者への投与に適した組成物である。このような組成物は、当該分野の周知でありかつ容認された方法によって調製される。例えば、Remington:The Science and Practice of Pharmacy,第20版(A.R.Gennaro編)Mack Publishing Co.,Easton,Pa.,2000を参照のこと。静脈内送達経路、皮下送達経路、筋肉内送達経路および腹腔内送達経路のような非経口投与は、これらの薬学的組成物を送達するために使用され得る。あるいは、化合物は、局所的に(例えば、CNS部位に直接)投与される。神経学的障害の処置のために、脳脊髄液への直接注入または脳組織への直接注射が使用される。任意の1人の患者のための投与量は、患者のサイズ、体表面積、年齢、投与される特定の核酸、性別、時間および投与の経路、全身の健康、ならびに同時に投与されている他の薬物を含む多くの要因に依存する。例えば、約5×105〜1×106個の細胞を含む、濃縮された細胞懸濁物が、部位に注射される。パーキンソン病の処置のために、これらの細胞は、脳室の壁に注射される。
【0012】
本発明の他の特徴および利点は、それらの好ましい実施形態の以下の説明および特許請求の範囲から明らかである。引用された参考文献は、参考として本明細書に援用される。
【図面の簡単な説明】
【0013】
【図1】図1Aは、腰椎および仙椎に対するFTを示す矢状の断面の図(左側のパネル)であり、胚発生の間のFTへの尾側の脊髄の脱分化を示す図(中央のパネルおよび右側のパネル)である。図1Bは、切開した17.5週のヒト胎児脊髄である。矢印は、FTを示す。スケールバー=2.5mm。図1Cは、NSCマーカーであるネスチン(赤色)およびDAPI(青色)について染色した8ヶ月齢のFTの横断面の部分からの合成画像(merged image)である。スケールバー=100μm。
【図2】図2Aは、7 DIV後の胚性ヒト(17週)FTに由来する神経球におけるネスチン発現神経幹細胞(緑色)の存在を示す、蛍光顕微鏡写真である。図2Bは、35 DIV後の出生後日数(P)5のラットの終糸に由来する神経球におけるネスチン発現神経幹細胞(赤色)の継続した存在を示す蛍光顕微鏡写真である。
【図3】図3Aは、6ヶ月齢のヒトの終糸に由来する神経球から採取された細胞を示す位相差顕微鏡写真である。図3Bは、18 DIV後に接着性培養においてβ−IIIチューブリンを発現するニューロンを示す蛍光顕微鏡写真である。細胞を、12歳のヒトの終糸に由来する神経球から採取した。図3Cは、18 DIV後の培養においてO1(緑色)を発現する乏突起膠細胞を示す蛍光顕微鏡写真である。細胞は、P6のラットの終糸から得られる神経球に由来する。細胞核を、DAPIで対比染色する(青色)。図3Dは、神経球由来の細胞の接着性培養におけるビメンチンを発現する神経前駆体細胞(neural precursor cell)(赤色)とGFAPを発現する大グリア細胞(緑色)との共在を示す蛍光顕微鏡写真である。神経球自体は、P4のラットの終糸に由来した。核を、DAPIで対比染色する(青色)。
【図4】図4Aは、GDNF、BDNFおよびCNTFとともにレチノイン酸およびソニックヘッジホッグ(sonic hedgehog)へ神経球由来細胞を曝露した後のβ−IIIチューブリン(緑色)を発現する運動ニューロンを示す蛍光顕微鏡写真である。細胞は、P7のラットの終糸から採取された神経球に由来した。図4Bは、コリンアセチルトランスフェラーゼ(赤色)の運動ニューロンの発現を示す、4(A)における細胞と同一の視野の蛍光顕微鏡写真である。図4Cは、β−IIIチューブリンを発現する運動ニューロン(赤色)を示す異なる神経球の蛍光顕微鏡写真である。図4Dは、MCN−2(運動ニューロンによって固有に発現されるマーカー)の同時発現を示す、同じ視野の細胞の蛍光顕微鏡写真である。全ての細胞は、P7のラットの終糸に由来する。これらの図は、終糸の神経球に由来する細胞の運動ニューロンへの指向性の分化を示す。
【図5】図5A〜Fは、神経前駆体細胞(NPC)、ニューロンおよび/またはグリアへ分化した神経球を示す蛍光顕微鏡写真である。a)ビメンチン(i、緑色)およびTuj−1(ii、赤色)(合成画像(iii))について染色した、単一の神経球に由来する分化した細胞。5%血清中ポリ−L−リジンおよびラミニン上で7日間平板培養した。ドナー:12歳、インビトロで18日。b)GFAP(i、赤色)およびTuj−1(ii、緑色)を発現する、単一の神経球に由来する分化した細胞。合成画像は、(iii)に示される。(a)と同一のドナーおよび条件。インビトロで17日間。c)7日間にわたって5%血清中においてFT神経球から分化したニューロンの3[H]−チミジン標識された核に対する銀粒子を示す位相差顕微鏡法(Phase microscopy)。神経球を、分化条件で培養する前に3[H]−チミジンに8時間曝露した。細胞を、Tuj−1について対比染色する(緑色)。ドナー:6ヶ月のFT、インビトロで107日間、d、e)RAおよびShh−Nで処理した後に、MNマーカーについて染色した、FT神経球から分化した細胞。d)Tuj−1(赤色)およびMNR−2(緑色)について染色した、分化した細胞。ドナー:14週の胎児、インビトロで81日間。e)Tuj−1(i、緑色)、コリンアセチルトランスフェラーゼ(ii、赤色)について染色した、分化した細胞。合成画像を、(iii)に示す。ドナー:18歳、インビトロで25日間。f)O1(緑色)について染色し、DAPIについて対比染色した、分化した細胞。ドナー:6ヶ月齢、インビトロで111日間。スケールバー:a〜f=50um。
【図6】図6Aは、FTを示すP7でのラット脊髄の尾側の面(caudal aspect)の顕微鏡写真である。矢印は、神経球を生成するために使用した構造物の部分を示す。図6Bは、(i)NSCマーカーであるネスチン(赤色)の発現;(ii)DAPI対比染色(青色);および(iii)合成画像を示す、P7のFT組織切片の蛍光顕微鏡写真である。スケールバーは、100umである。図6Cは、インビトロで10日後の、P10のラットFTに由来する神経球を示す顕微鏡写真である。スケールバーは、100umである。図6Dは、(i)ネスチン発現;(ii)DAPI対比染色、および(iii)合成画像を示す、インビトロで5日後の、P5のFTに由来する神経球の蛍光顕微鏡写真である。スケールバーは、100umである。
【図7】図7Aは、(i)NSCマーカーであるOlig2の発現(赤色);(ii)DAPI対比染色(青色);および(iii)合成画像を示す、インビトロで34日後の、P7のFTに由来する神経球の蛍光顕微鏡写真である。図7Bは、(i)ビメンチンの発現(緑色);(ii)DAPI対比染色(青色);および(iii)合成画像を示す、60 DIV後の、P7のFTに由来する神経球の蛍光顕微鏡写真である。図7Cは、(i)一部の細胞におけるSox2の発現(赤色);(ii)DAPI対比染色(青色);および(iii)合成画像を示す、30 DIV後の、P6のrFTから単離された神経球の蛍光顕微鏡写真である。図7Dは、(i)Musashi(色(COLOR))の弱い染色;(ii)DAPI対比染色(青色);および(iii)合成画像を示す、30 DIV後の、P6のFTに由来する神経球の蛍光顕微鏡写真である。図7Eは、(i)β−III−チューブリンの発現(色);(ii)GFAPの発現(COLOR);(iii)DAPI対比染色(青色);および(iv)合成画像を示す、34 DIV後の、P7のFTに由来する神経球の蛍光顕微鏡写真である。
【図8】図8Aは、(i)Pax6発現(緑色);(ii)Olig2発現(赤色);(iii)DAPI対比染色(青色);および(iv)合成画像を示す蛍光顕微鏡写真である。スケールバーは、100umである。30 DIVでのP7のラットFTに由来する細胞。データは、運動ニューロンがFT由来神経球から生成されることを示す。図8Bは、(i)Pax6発現(緑色);(ii)Olig2発現(赤色);(iii)DAPI対比染色(青色);および(iv)合成画像を示す、同様な蛍光顕微鏡写真である。スケールバーは、50umである。30 DIVでのP7のラットFTに由来する細胞。図8Cは、(i)β−IIIチューブリンの発現(緑色)(初期のニューロンのマーカー);(ii)ChAT(赤色)(コリン作用性神経伝達のマーカー);(iii)DAPI対比染色(青色);および(iv)合成画像を示す蛍光顕微鏡写真である。スケールバーは、100umである。ドナー:P6のラットFT 36 DIV。図8Dは、(i)MNR2(緑色)(運動ニューロン特異的マーカー)の発現;(ii)β−III−チューブリン(赤色)(初期のニューロンのマーカー)の発現;(iii)DAPI対比染色;および(iv)合成画像を示す蛍光顕微鏡写真である。スケールバーは、50umである。ドナー:P7のFT、30 DIV。図8Eは、(i)MRN2(緑色);(ii)GFAP(赤色)の重ならない発現;(iii)DAPI対比染色;および(iv)合成画像を示す、分離していない神経球培養物の蛍光顕微鏡写真である。これらの培養物は、ソニックヘッジホッグおよびレチノイン酸で処理されていない;正確に言えば、これらは、専ら、BDNF、CNTFおよびGDNFを含む分化培地で維持した。スケールバーは、50umである。ドナー:P7のFT、30 DIV。図8Fは、GFAP発現細胞の中の運動ニューロンの島(island)を示す、7Eと同じ条件下で培養した別の分離していない神経球の蛍光顕微鏡写真である。パネルは、(i)MRN2(緑色)の発現;(ii)GFAP(赤色)の発現;(iii)DAPI対比染色;および(iv)合成画像を示す。スケールバーは、50umである。ドナー:P7のFT、30 DIV。
【図9】図9は、レチノイン酸(RA)、ソニックヘッジホッグ(SHH)、BDNF、CNTFおよびGDNFで処理した培養物における増大した割合のMRN2発現を示す、ラットのFT NSCを使用する28個の指向性の分化実験の結果をまとめる散布図(scatter−plot)である。MNR2がまた、Shhのみで処理した神経球から分化した細胞においても発現され、一部の場合において、BDNF、CNTFおよびGDNFのみの存在下で増殖させた未処理の神経球から分化した細胞において発現されたことに留意すること。3例のうちの1例において、MNR2についての免疫染色を、いかなる特定の神経栄養因子も含まない血清で分化させた、未処理の神経球に由来する細胞のうちの40%において観察した。
【図10】図10A〜Fは、細胞型発現(cell−type expression)を特徴付けるために、細胞マーカーについて染色した神経球の蛍光顕微鏡写真である。a)ネスチン(赤色)について染色した、図1(c)と同一の8ヶ月のFTに由来する神経球、インビトロで11日間。b)ビメンチン(緑色)について染色した神経球。ドナー:6ヶ月のFT、インビトロで123日間。c)(b)と同一のドナーに由来する、2つの神経球におけるOlig−2染色(赤色)。d)10歳に由来する神経球におけるSox−2発現(赤色)。インビトロで51日間。e)(a)と同一のドナーに由来する神経球のTuj−1(i、緑色)、GFAP(ii、赤色)、合成画像(iii)。f)18歳のFT由来の神経球の位相差顕微鏡写真。インビトロで6日間。スケールバー:a〜f=100μm。画像a〜dおよびe(iii)は、DAPIで対比染色する(青色)。
【図11】図11A〜Cは、α−アマガサヘビ毒素(緑色)およびTuj−1(赤色)について染色した、得られたヒトMNおよびラット筋肉の同時培養物(co−culture)を示す神経筋接合部の蛍光顕微鏡写真である。RAおよびShh−Nで分化させた単一の神経球を、6日間、ラット筋線維と同時培養した。a)MN(赤色)およびα−アマガサヘビ毒素標識アセチルコリンレセプター(緑色)の蛍光顕微鏡写真。b)(a)に見られる神経筋接合部の共焦点顕微鏡画像。c)Imaris Surpassにおいて最大値投影(maximum intensity projection)(MIP)レンダリングに処理させた(b)の側面図。神経終末(赤色)は上にあり、標識レセプター(緑色)は下にあり、黄色は、重なった領域を示す。ドナー:6ヶ月齢のFT。スケールバー:50μm。
【図12】図12は、rFT由来神経球の不均質な分化能力を示す散布図である。この散布図は、神経球分化の14の実験における、Tuj−1(ニューロンのマーカー)およびGFAP(星状膠細胞のマーカー)の発現の変動性を示す。個々の神経球を、ポリ−L−リジンおよびラミニンでコートしたカバーガラス上にプレートし、5%血清中でそれらを培養することによって分化させた。分化した細胞を、24時間後、または7〜10日後のいずれかに免疫細胞化学によって評価した。両方の場合において、ニューロンおよび星状膠細胞の生成の変動が存在した。両方のマーカーについて二重に染色した、神経球から分化した細胞のおおよその割合は、24時間後に見出された重なりの割合と比較して、分化条件に7〜10日間の曝露した後では減少した。
【図13】図13は、MN分化をしているrFT由来神経球におけるMNR2の発現の変動を示す散布図である。この散布図は、分化した神経球におけるMNR2発現の変動性を示す。MNR2は、MN生成を同定するのに使用される複数のマーカーのうちの一つである。チャートで明らかなように、MNR2はまた、Shhのみで処理した神経球から分化した細胞においても発現され、BDNF、CNTFおよびGDNFの存在下で増殖させた、未処理の神経球に由来するものにおいても発現された。3例のうちの1例において、MNR2についての免疫染色は、いかなる特定の神経栄養因子も含まない血清中で分化させた、未処理の神経球に由来する細胞の‘40%で観察された。
【発明を実施するための形態】
【0014】
詳細な説明
本発明は、幹細胞ベースの治療において多大な進歩を表す。この分野は、治療目的のための幹細胞を獲得する、供給源および信頼のおける方法の不足に悩まされている。脳または脊髄の傷害または変性疾患に起因して失われた脳細胞(ニューロンおよびグリア)の置換において使用するための、神経幹細胞の獲得が特に困難であった。
【0015】
自系ヒト神経幹細胞のための完全に新規な供給源が同定された。FTは、脊髄の尾側末端にあり、索(cord)を尾骨に付着させる。これは、脊髄円錐から分離した、構造的に異なる解剖学的組織である。例えば、成人のヒトにおいて、この組織は、2〜3cm離れている。それは、神経系において消耗性の痕跡組織である。神経幹細胞の他の供給源とは対照的に、FTは、外科的に到達可能であり、組織は、「係留脊髄症候群」の手術の間にそれから慣例的に取得される。これは、神経幹細胞のための信頼できる供給源である。この供給源から取得された幹細胞を、組織培養中に長期の時間にわたって増殖および維持させることができ、かつニューロンおよびグリアへ分化したことが実証されている。FT NSCを、神経学的傷害または変性を罹患する患者から容易に採取し、組織培養において増殖および分化させ、続いて患者に移植して戻す。このストラテジは、非自系細胞の移植に伴う組織拒絶の問題を克服する。FTは、神経細胞置換ストラテジに適した細胞の供給源として役立つ、第一の容易に入手可能で、増殖可能な神経組織を代表する。
【0016】
FTの単離
本発明は、FT(幹細胞の存在について調査されたことのない中枢神経系の領域)からの神経幹細胞の単離を介してNSC治療の実行可能性を増大させる組成物および方法を提供する。以前の研究は、幹細胞を含むCNS内の既知の領域(これは、正常な脳機能に必須であり、外科的に到達することが困難である)からのNSCの単離および増殖に注力していた。FTは、NSCの以前に使用された供給源に対する、新規かつ魅力的な代替物である。なぜなら、FTは、出生後の哺乳動物における発達中の脊髄の、機能を有さない(non−function)レムナントを示す、組織学的に始原の構造物だからである。これらの特性は、FTを、治療上の使用のための自系NSCの魅力的な供給源にする。FTは、出生後の哺乳動物(例えば、ラットまたはヒト)、廃棄されたヒト胎児、小児から青年までのヒト(特に、係留脊髄についての外科的切除を受けた者)、および治療上の使用に神経幹細胞を必要とするあらゆる出生後の被験体から取得される。この組織を分離し、神経球を単離し、幹細胞の生存および/または増殖を促進する条件下で増殖させる。FTに由来する細胞は、発生もしくは変性状態もしくは障害、疾患、傷害もしくは外傷、感染、投薬もしくは医学手順に由来する合併症、または任意の他の自然の原因(例えば、加齢)もしくは誘発された原因(例えば、脳卒中)に起因して、神経学的機能の回復を必要とする患者に投与される。移植された細胞は、宿主の中枢または末梢の神経系に統合され、機能的な回復を促進させる。
【0017】
幹細胞
幹細胞は、大半(全てではないが)の多細胞生物に見出される細胞である。それらは、有糸細胞分裂を介して自身を更新し、そして特殊化した細胞型に分化する能力によって特徴付けられる。2つの大まかな型の哺乳動物幹細胞は、以下である:胚盤胞に見出される胚性幹細胞、および成体組織に見出される成体幹細胞。発生している胚において、幹細胞は、特殊化した胚性組織の全てに分化することができる。成体の器官において、幹細胞および前駆体細胞は、特殊化した細胞を補充する、身体のための修復システムとして働くが、血液、皮膚または腸組織のような再生性器官の正常なターンオーバーもまた維持する。
【0018】
本明細書で使用されるように、用語「幹細胞」は、自己再生し、1つより多い型の細胞に分化することのできる細胞を記載することが意図される。自己再生は、細胞分裂の多くの周期を通過するが、未分化の状態を維持する能力として本明細書で定義される。
【0019】
能力は、特殊化した細胞型に分化する能力である。最も厳密な意味において、これは、幹細胞が、全能または多能性のいずれかであること−あらゆる成熟細胞型を生じることができることを必要とするが、多分化能または単能性の前駆体細胞は、時折、幹細胞と呼ばれる。他の用語において、能力は、幹細胞の分化能力(異なる細胞型に分化する能力)を指定する。全能性幹細胞は、卵と精子細胞との融合物から生成される。受精した卵の最初の数回の分裂によって生成される細胞もまた全能である。これらの細胞は、胚および胚外の細胞型に分化することができる。多能性幹細胞(pluripotent stem cell)は、全能細胞の子孫であり、3つの胚葉のいずれかに由来する細胞に分化することができる。多分化能幹細胞(multipotent stem cell)は、密接に関連するファミリーの細胞の細胞しか生成できない(例えば、造血幹細胞は、赤血球、白血球、血小板などに分化する)。単能性細胞は、一つの細胞型しか生成できないが、非幹細胞からこれらを区別する自己再生の特性を有する(例えば、筋肉幹細胞)。
【0020】
本発明の幹細胞は、分子的および機能的な方法(molecular and functional method)によって同定される。幹細胞の実際の定義は、機能的な定義−一生にわたって組織を再生する能力である。幹細胞の特性は、単一の細胞がそれらの分化および自己再生する能力によって特徴付けられる試験管内腫瘍細胞感受性試験のような方法を使用して、インビトロで示され得る。さらに、幹細胞および幹細胞の集団は、細胞表面マーカーおよび細胞内マーカーの特有のセットに基づいて同定および単離される。
【0021】
胚性幹細胞株(ES細胞株)は、胚盤胞またはより初期の桑実胚段階の胚の内細胞塊(ICM)の胚盤葉上層組織に由来する細胞の培養物である。胚盤胞は、初期段階の胚である-ヒトにおける約4〜5日齢であり、50〜150個の細胞からなる。ES細胞は、多能性であり、発生の間に3つの一次胚葉:外胚葉、内胚葉および中胚葉の全ての派生物のもとになる。言い換えると、これらは、特定の細胞型のための十分かつ必要な刺激を与えられた場合、成体の身体の200種超の細胞型の各々に発達し得る。
【0022】
ほぼ10年間の研究の後に、胚性幹細胞を使用する認可された処置もヒト治験も存在しない。全能細胞であるES細胞は、正しい分化のために特異的なシグナルを必要とする−別の身体に直接注射された場合、ES細胞は、多くの異なる型の細胞に分化し、奇形腫(すなわち、新生物の一つの型)を生じる。移植片拒絶を回避しながら、使用可能な細胞に分化するES細胞は、胚性幹細胞の研究者達が依然として直面しているごくわずかの障害物である。多くの国が、現在、ES細胞の研究も新規なES細胞株の生産も一時停止している。
【0023】
本明細書で使用されるように、用語「成体幹細胞」とは、2つの特性:分裂し、それ自体に似た別の細胞を生成する(すなわち、自己再生)能力、およびまた分裂し、それ自体よりもより分化した細胞を生成する能力(すなわち、この細胞は、少なくとも単能性であるが、選択的に少なくとも多分化能である)を有する、発生した生物に見出される任意の細胞をいう。成体幹細胞はまた、体性幹細胞および生殖系列幹細胞としても一般的に知られている。成体幹細胞は、全ての出生後の哺乳動物において同定され得る。
【0024】
多能性の成体幹細胞は、臍帯血を含む多くの組織に見出され得る。大半の成体幹細胞は、系統が制限されており(多分化能)、一般的に、それらの組織起源によって呼ばれる(間葉幹細胞、脂肪由来幹細胞、内皮幹細胞など)。
【0025】
一局面において、自己再生を確実にするために、幹細胞は、分裂の面(plane of division)および/または細胞内要素の分裂が異なる2つの型の細胞分裂を実行する。対称的な分裂は、共に幹細胞の特性を付与された、2つの同一の娘細胞を生じる。対称的な分裂は、2つの同一の細胞を生成する細胞分裂、上皮の障壁(例えば、大脳半球の側脳室)に平行な分裂の面で生じる細胞分裂、2つの均等なサイズの細胞を生成するかまたは細胞の中心点(center point)(赤道)で生じる細胞分裂、あるいは分離に続いて同一または等価な細胞内成分/要素を有する2つの細胞を生成する細胞分裂として定義される。他方、非対称的な分裂は、1つの幹細胞および制限された自己再生能力を有する先祖細胞のみを生成する。非対称的な分裂は、先に列挙されたシナリオの全てについて非対称的である細胞分裂として定義される。娘細胞間で対称的にまたは非対称的に分裂され得る例示的な細胞内成分または要素としては、細胞内または細胞表面タンパク質;細胞骨格要素;接着結合、細胞接触または細胞接着要素;細胞内小器官;およびシグナル伝達分子が挙げられるが、これらに限定されない。前駆体は、成熟細胞に最終的に分化する前に、数回の細胞分裂を経験し得る。
【0026】
代替的な理論は、幹細胞が、それらの特定のニッチにおいて環境上のキューに起因して未分化のままであることである。幹細胞は、それらがそのニッチから離れるか、またはもはやそのようなシグナルを受け取らなくなった場合、分化する。インビトロにおいて、幹細胞のニッチを繰り返し、指向性の生体外幹細胞分化を誘導し得る。特定の局面において、幹細胞のニッチの特定の成分が過剰発現、除去、組換え、および/または合成され、所望の幹細胞応答を誘導し得る。
【0027】
神経幹細胞
本明細書で使用されるように、用語「神経幹細胞」とは、ニューロンおよびグリア細胞を生じ得る、成体神経組織に見出される幹細胞を記載することが意図される。例示的なグリア細胞としては、星状膠細胞および乏突起膠細胞が挙げられるが、これらに限定されない。ニューロン(ニューロンおよび神経細胞としても知られる)は、情報を処理し伝達する神経系内の電気的に興奮性の細胞である。ニューロンは、脊椎動物の脳、脊髄および末梢神経の中心的な成分である。十分に発達したニューロンは、永久に有糸分裂しない(amitotic)(これらは分裂しない)が;しかしながら、脳のいたるところのさらなるニューロンは、神経発生の過程を介して、脳室下帯およびやや顆粒状のゾーン(subgranular zone)で見出される神経幹細胞から生じ得る。本発明はまた、FTを含む脊髄内の神経幹細胞の供給源を提供する。
【0028】
ニューロン
ニューロンは、代表的に細胞体(soma)(すなわち、細胞体(cell body))、樹状突起(dendritic tree)および軸索から構成される。大部分の脊椎動物のニューロンは、細胞体および樹状突起に入力を受け、軸索を介して出力を伝達する。しかしながら、ニューロンのサイズ、形状および機能において、神経系および動物界を通じて大きな異質性が存在する。一部の特殊化したニューロンのサブタイプが知られており、これらとしては、かご細胞(小脳内の、拡張した、節の多い樹状突起を有するニューロン);ベッツ細胞(大型の運動ニューロン);中型の有棘ニューロン(Medium spiny neurons)(線条体における大半のニューロン);プルキンエ細胞(小脳内の巨大ニューロン、ゴルジI型多極ニューロンの1つの型);錐体細胞(三角形の細胞体を有するニューロン、ゴルジII型の1つの型);レンショー細胞(両端がα運動ニューロンに連結されたニューロン);顆粒細胞(ゴルジII型ニューロンとしての1つの型);および前角細胞(脊髄にある運動ニューロン)が挙げられるが、これらに限定されない。ニューロンのサブタイプの広範な変動に起因して、本発明の成熟ニューロンは、細胞型間の形態学的な差、分子的な差および/または機能的な差を利用する1つ以上の方法を使用して同定される。
【0029】
分化したニューロンは、他のニューロンまたは細胞に対するそれらの作用によって特徴付けられる。興奮性ニューロンは、それらの標的ニューロンを興奮させる。脳を含む中枢神経系内の興奮性ニューロンは、しばしばグルタミン酸作用性(glutamatergic)である。筋肉細胞へシナプスを形成する脊椎の運動ニューロンのような末梢神経系のニューロンは、しばしばそれらの興奮性神経伝達物質としてアセチルコリンを使用する。抑制ニューロン(inhibitory neuron)は、それらの標的ニューロンを抑制する。抑制ニューロンは、しばしば介在ニューロンである。一部の脳の構造物(新線条体、淡蒼球、小脳)の出力は、抑制性である。一次抑制性神経伝達物質は、GABAおよびグリシンである。調節性ニューロン(modulatory neuron)は、ニューロン調節(neuromodulation)と呼ばれるより複雑な効果を引き起こす。これらのニューロンは、ドーパミン、アセチルコリン、セロトニンおよびその他のような神経伝達物質を使用する。
【0030】
分化したニューロンは、それらの放電パターンによって特徴付けられる。ニューロンは、それらの電気生理学的特徴にしたがって分類される。ニューロンは、持続性または規則正しい「スパイキング(spiking)」を示す(例えば、スパイク(spike)は、活動電位の検出をいう)。一部のニューロンは、代表的に、一定で(または持続して)活性である(例えば、神経線条(neurostriatum)内の介在ニューロン)。他のニューロンは、規則正しいスパイキング(これは、少なくとも1回の刺激によって引き起こされる活動電位をいう)を示す。あるいは、またはさらに、ニューロンは、位相性の挙動またはバースト挙動を示す。バーストで発火するニューロンは、位相性と呼ばれる。一部のニューロンは、それらの迅速な発火速度に関して顕著である(例えば、一部の型の皮質の抑制介在ニューロン、淡蒼球内の細胞)。あるいは、またはさらに、一部のニューロンの活動電位は、その他のものと比べてより狭い(検出した場合、薄いスパイク(thin−spike))。例えば、前前頭皮質内の介在ニューロンは、薄いスパイクのニューロンである。
【0031】
分化したニューロンは、それらが放出する神経伝達物質によって特徴付けられる。限定しない例示的なニューロンの型は、コリン作用性ニューロン、GABA作用性ニューロン、グルタミン酸作用性ニューロン、ドーパミン作用性ニューロンおよび5−ヒドロキシトリプタミンニューロン(5−HT;セロトニン)が挙げられる。
【0032】
FT−NSCの特徴付け
さらに上記の細胞を特徴付けるために、FT−NSC株を樹立し維持した。FTに由来する神経球を、NSCマーカーであるビメンチン、CD133、Olig2およびSox2の発現について調べる。FT−NSCを、分化を促進する条件下でインビトロで培養する。分化した細胞の型を、免疫細胞化学技術および電気生理学的方法の両方を使用して同定する。ラットおよびヒトのFT−NSCを、正常なげっ歯類または脊髄外傷もしくは神経学的疾患のモデルとなるげっ歯類のいずれかのCNSに移植する。
【0033】
FT−NSCを、ラットおよびヒト(約24人のヒトドナー)から単離し、組織培養において多くの継代にわたって神経球として維持した。例えば、細胞を、15〜20回継代し、一部の細胞株を、12ヶ月超にわたり維持した。この様式で取得したFT−NSCを、免疫細胞化学によって決定されるCNSニューロンおよびグリアに分化するように誘導した。細胞は、ニューロンおよびグリアの両方(星状膠細胞と乏突起膠細胞との両方)を生じる。細胞を、誘導して分化させ、80〜100%の運動ニューロンである細胞集団を得た。
【0034】
自系幹細胞の供給源の同定
ヒトNSCの供給源を同定し、神経系の外傷および変性の症例におけるそれらの治療上の使用を研究した。自系NSCを、脊髄のFTから単離し、増殖させ、続いて、神経組織の損傷の部位に移植する。
【0035】
自系NSCの信頼できる供給源は、脊髄のFTであり、これは、脊髄の尾側末端の細長い延長部分であり、脊柱の基部にある尾骨に索を係留する(図1A)。組織学的研究は、FTが、室周囲(peri−ventricular)上衣細胞に囲まれた脊柱管(ventricular canal)ならびに種々の型のニューロンおよびグリアを封入することを示す。この局所的な環境は、NSCを生成する他のCNS領域に類似する(Alvarez−Buylla,A.およびD.A.Lim. Neuron(2004)41(5):p.683〜6;Doetsch,F. Curr Opin Genet Dev(2003)13(5):p.543〜50;Riquelme,P.A.、E.DrapeauおよびF.Doetsch.Philos Trans R Soc Lond B Biol Sci,2007)。FTは、神経系の残りと相互に連結されてなく、また身体の神経も刺激しない。本質的に、それは、神経系の消耗性のレムナントである。
【0036】
FTは、独特な発生上の由来を有する(Streeter,G.L. Am J Anat(1919)22:p.1〜12;Nievelstein,R.A.ら、Teratology(1993)48(1):p.21〜31;Kernohan,J.W. J Comp Neurol(1924)38:p.107〜125;Kunitomo,K.(1918)8:p.161〜204)。これは、胚の痕跡尾(vestigial tail)に対する神経支配(またはげっ歯類の場合、最も尾側の尾分節(tail segment)の一過的な神経支配)を発生の初期に提供する神経系のレムナントである。初期の段階において、推定上のFTは、脊髄神経節が付いた、十分に分化した脊髄である(図1B、左側)。尾が再吸収されたときに、糸の細胞は、Streeterによって「脱分化」と呼ばれる過程(図1B、右側)を経験し、上衣細胞で裏打ちされ、線維芽細胞、ニューロンおよびグリアの見かけ上ゆるく組織化された収集物で環状に囲まれた中心管を有する膠原性の構造物をもたらす。パラガングリオーマおよび始原の神経外胚葉性腫瘍は、この構造物から生じることが示され、このことは、幹細胞が存在する可能性を示唆する(Ashkenazi,E.ら、J Spinal Disord,(1998)11(6):p.540〜2;Gagliardi,F.M.ら、Childs Nerv Syst,(1993)9(1):p.3〜6;Kamalian,N.ら、J Neurol,(1987)235(1):p.56〜9;Koeller,K.K.、R.S.RosenblumおよびA.L.Morrison. Radiographics(2000)20(6):p.1721〜49)。FTは、外科的に容易に到達可能であり、脊柱にしっかりと係留され、動きに十分な自由を欠く場合、脊髄に対する緊張を軽減するために、慣例的に切開される。この状態は、「係留脊髄症候群」と呼ばれる(Bakker−Niezen,S.H.、H.A.WalderおよびJ.L.Merx.Z Kinderchir、(1984)39 Suppl 2:p.100〜3;Bode,H.ら、Klin Padiatr(1985)197(5):p.409〜14)。本発明より前に、終糸が、神経幹細胞の潜在的な供給源として採取されたことはない。
【0037】
FTは、以下の理由に関してNSCの安全かつ信頼できる供給源である。これは、外科的に到達可能であり、消耗性の神経組織である。ヒト組織は、胎児組織から、そして脊髄の外科的な解放(untethering)に続いてFTの生検を行う小児神経外科センターから容易に入手可能である。NSCの自家移植は、神経系への異種移植に伴う重大な問題であることが実証されている、免疫学的拒絶を回避する明確な利点を有する(Barker,R.A.およびH.Widner. NeuroRx,(2004)1(4):p.472〜81;Linazasoro,G. Neurologia(2003)18(2):p.74〜100)。NSCは、哺乳動物のCNSで同定されているが、現行の供給源は、外科的に到達するのが困難であり、これは、代表的に、正常な機能に重要な領域(例えば、脊髄および前脳の側脳室)に由来する(Alvarez−Buylla,A.、D.G.HerreraおよびH.Wichterle. Prog Brain Res,(2000)127:p.1〜11;Alvarez−Buylla,A.、B.SeriおよびF.Doetsch. Brain Res Bull,(2002)57(6):p.751〜8)。これらの領域の外科的な破壊は、顕著な(profound)神経学的な欠損をもたらす。FT由来のNSCは、一生にわたって採取され得、自系NSCのための供給源として役立ち得、それによって、免疫学的拒絶の問題を回避する。単離、培養、増殖、分化に続いて、自系FT−NCSは、神経学的な障害または傷害の処置のために、被験体に移植される。
【0038】
主要な組織は、動物から取得される(例えば、げっ歯類(ラット、マウス)、またはヒト(例えば、胎児および出生後のヒト)FT)。例えば、胎児ヒト組織は、廃棄された胎児から取得される。若年のヒトFTは、係留された索の切除のための手術の間に取得される。加えて、FTは、出生後のラットから取得される。出生後ラットFTは、ヒトFTと同じ組織学的特徴を有し、このクラスのNSCのさらなる特徴付けのための、容易に入手可能で、大規模の組織の供給であるという利点を有する。
【0039】
胎児から成人までの種々な年齢の、死後のヒト材料由来の組織切片におけるFT−NSC局在は、種々の段階において存在するNSCの数に関する情報を提供し、より正確な切開をもたらす。FTを、切開し、固定し、低温槽上で切片にする(section)。NSCを、ネスチンおよびSox2のようなNSCマーカーに対して指向される抗体を用いて組織切片を染色することによって同定する(Bazan,E.ら、Histol Histopathol,(2004)19(4):p.1261〜75)。
【0040】
NSCの培養
本発明のNSCを、被験体への移植に先立って、維持および増殖のために培養する。
【0041】
FTを、当該分野で公知の培地および増殖因子を含む種々の条件(例えば、N2、EGFおよびbFGF+ヘパリンを含むDMEM/F12)を使用して、組織培養に置く(Rajan,P.およびE.Snyder. Methods Enzymol,(2006)419:p.23〜52;Vescovi,A.L.およびE.Y.Snyder. Brain Pathol,(1999)9(3):p.569〜98)。神経球を単離し、継代する。これらは、非限定的な、先に検討した免疫学的なマーカーを使用して同定する。
【0042】
分離のための手順は、抗体でコートされた磁気ビーズを使用する磁気分離、固体マトリクス(例えば、プレート)に付着させた抗体を使用するアフィニティクロマトグラフィおよびパニング、または他の便利な技術を含む。他の分離技術は、多色チャネル、小角および鈍角光散乱検出チャネル、インピーダンスチャネルなどのような種々の程度の洗練を有し得る蛍光励起セルソーターを含む。死細胞は、例えば、死細胞と結合する色素(ヨウ化プロピジウム[PI]、LDS)を使用する選択による標準的な方法を使用して排除される。選択された細胞の生存度に過度に有害ではない、あらゆる技術が使用され得る。
【0043】
NSC株を樹立し、移植/回復研究のために増殖され、ヒトNSCの場合、治療用の細胞株の供給源として発達させる。ヒトNSCのための培養条件は、増殖速度および細胞の収量を最大化するために最適化される。培地、増殖因子、酵素などの操作による。細胞株の増殖速度は、生成された神経球の数を計数すること、および生存細胞の数を計数することによって測定される(Cardozo,D.L. Neuroscience(1993)56(2):p.409〜21)。
【0044】
FT−NSCは、ニューロンおよびグリアに分化する
個々の神経球を単離し、ニューロンおよびグリアへの分化を促進する種々の条件下でインビトロで平板培養する(Reynolds,B.A.、W.TetzlaffおよびS.Weiss. J Neurosci,(1992)12(11):p.4565〜74;Reynolds,B.A.およびS.Weiss. Science,(1992)255(5052):p.1707〜10)。分化をさらに特徴付けるために、神経球を他の細胞と同時培養し、分化条件への曝露の前に、脂肪親和性膜染料(lipophilic membrane stain)(例えば、DiI)、カルボキシフルオレセインまたはBrdUのような試薬で予め標識する。この追跡ストラテジは、生成されるニューロンおよびグリアの供給源として神経球を確実に樹立する。3つの基本的なアプローチが、これらの細胞を特徴付けるために使用される:a.血清の存在下で接着基質上で平板培養すること;b.特定の表現型を促進することが知られている規定された試薬(例えば、運動ニューロンを生成するために、レチノイン酸+ソニックヘッジホッグタンパク質を使用すること)での前処理に続いて、接着基質上で平板培養すること;ならびにc.標識された神経球と筋肉のような標的組織に由来する培養細胞またはCNSの異なる領域に由来する細胞との同時培養。同時培養データは、FT−NSCの運命に対する標的組織の影響を確立し、FT神経球が、筋肉細胞およびニューロンとのシナプスの相互作用を形成することを確認する。
【0045】
神経球由来の細胞型は、ニューロンおよびグリアのサブタイプに特異的な抗体ならびに未熟なニューロンのサブタイプと成熟ニューロンのサブタイプとの間を区別する抗体を含む、ニューロンおよびグリアに対して指向される抗体を使用して同定される。これらとしては、以下が挙げられる:神経−特異的エノラーゼおよび神経−チューブリンIII(ニューロン);ビメンチン(神経前駆体);GFAP(星状膠細胞);O1(乏突起膠細胞);コリンアセチルトランスフェラーゼおよびMNR2(運動ニューロン)(Gage,F.H.、J.RayおよびLJ.Fisher. Annu Rev Neurosci,(1995)18:p.159〜92;Schwartz,P.H.ら、J Neurosci Res,(2003)74(6):p.838〜51;Schwartz,P.H.ら、Stem Cells,(2005)23(9):p.1286〜94;Wichterle,H.ら、Cell,(2002)110(3):p.385〜97)。
【0046】
運動ニューロンの表現型を促進する試薬に曝露した神経球を、筋肉細胞(筋管)と同時培養する。得られたニューロンを、神経筋接合部に結合する標識されたアマガサヘビ毒素で培養物を染色することによって、筋肉細胞とのシナプスを確立するそれらの能力について試験する。機能的な運動ニューロンを、電子顕微鏡法を使用して、さらに評価し、機能しているシナプスのシナプス前の要素およびシナプス後の要素(例えば、シナプス小胞、シナプス後密度)の存在を同定する。
【0047】
神経球に由来する形態学的に同定可能なニューロンを、細胞が、ニューロンに典型的な生理的特徴を有するか否かを決定する、標準的な電気生理学的技術(静止電位、活動電位を生成する能力、および添加された神経伝達物質に対する応答を測定すること)を使用して特徴付ける。筋肉の同時培養の場合に、分化したニューロンを、視覚的方法および電気生理学的方法の両方で筋肉の単収縮応答について評価する(Cardozo,D.L.およびB.P.Bean. J Neurophysiol,(1995)74(3):p.1137〜48;Elkes,D.A.ら、Neuron,(1997)19(1): p.165〜74)。
【0048】
インビボ移植、統合(integration)および機能的な回復実験
FT−NSC株を、げっ歯類宿主組織へ再統合するそれらの能力ならびにヒト被験体および脊髄外傷と神経変性とについてのげっ歯類モデルにおける機能的な回復を生成するそれらの能力について評価する。当該分野で認識される腰部脊髄外傷についてのげっ歯類モデルを、FT−NSCの評価について使用する。これらのモデルは、欠損の程度および機能的な回復の程度に特異的な測定基準を含む(Karimi−Abdolrezaee,S.ら、J Neurosci,(2006)26(13):p.3377〜89;Kimura,H.ら、Neurol Res,(2005)27(8):p.812〜9;Nakamura,M.ら、J Neurosci Res,(2005)81(4):p.457〜68;Vroemen,M.ら、Eur J Neurosci,(2003)18(4):p.743〜51)。GFP標識されたFT−NSCを、脊髄外傷を経験したラットの傷害部位に移植する。動物を機能的な回復について試験し、その後屠殺し、死後の脊髄を、移植した細胞の統合について組織学的に調べる。
【0049】
FT−NSC細胞の再統合を、以下のように評価する。FT−NSC株を、全てのCNS細胞が緑色蛍光タンパク質(GFP)で標識されているトランスジェニックラットから樹立する(Dombrowski,M.A.ら、Brain Res,(2006)1125(1):p.1〜8)。これらのGFP標識されたFT−NSCを濃縮し、正常なラットの脊髄に移植する。移植された細胞が生存し、宿主組織に統合する能力を、標準的な組織学的方法を使用して死後に調べる。
【0050】
ヒト由来FT−NSC株を、ヒト被験体および免疫不全マウス系統(NOD−SCID)への移植によって評価する。移植した細胞を、SC101およびSC121のようなヒト特異的抗原についての染色によって同定する(Anderson,AJ.、B.J.CummingsおよびCW.Cotman. Exp Neurol,(1994)125(2):p.286〜95;Cummings,B.J.ら、Proc Natl Acad Sci U S A,(2005)102(39):p.14069〜74)。
【0051】
細胞をまた、神経変性疾患についてのげっ歯類モデルにおいても評価する(Kitamura,Y.ら、Jpn J Pharmacol,(2000)84(3):p.237〜43;Orth,M.およびS.J.Tabrizi.(2003)18(7):p.729〜37;Springer,W.およびP.J.Kahle. Curr Neurol Neurosci Rep,(2006)6(5):p.432〜6)。動物を、機能的な回復について試験し、そして死後に、宿主組織への移植した細胞の統合について試験する。
【0052】
神経変性疾患および神経外傷を処置するためのFT−NSCの使用
自系FT NSCは、パーキンソン病および他の神経変性疾患ならびに外傷性脳傷害の処置に有用である。研究は、ドーパミン産生細胞が、マウスパーキンソン病(Parkinson’s)モデルおよびヒトパーキンソン病患者の両方の線条に統合し得、この疾患の症状を緩和し得ることを示唆する。本発明以前には、NSCの豊富かつ容易に入手可能な供給源の欠如は、個々の移植片において比較的少数の細胞が使用されていたことを意味する。さらに、以前の移植片は、中枢神経系の「免疫学的に特権のある(immunoprivileged)」性質にも拘らず、拒絶をもたらす免疫反応を引き起こす、異種細胞を使用した。本発明は、これらの問題の両方を解決する:全ての患者はFTを有し、したがって、全ての患者は、増殖可能であり、かつ適切な条件下で、自家移植のための特定の細胞型(ドーパミンニューロンおよびコリン作用性運動ニューロンが挙げられる)を効率よく生成する、自系NSCの準備のできた供給源を有する。FT−NSCは、自系細胞の利用可能性に対する根本的な拘束を排除するので、それらの治療上の使用に対する唯一の制限は、生じる細胞型の多様性である。例えば、FT由来乏突起膠細胞を使用して、多発性硬化症のような脱髄疾患を処置する一方で、FT由来運動ニューロンは、筋萎縮性側索硬化症のような運動ニューロンの喪失の疾患または脊髄傷害に適用可能である。
【0053】
ヒト患者からの細胞の採取
FT細胞を、公知の方法を使用して患者から採取する。例えば、手術を使用して、「係留脊髄症候群」の症例においてなされるように、FTを切断および切開する。第二に、少量の組織を、針穿刺吸引(needle aspiration)によって採取する。これらの細胞に関してたくさんの数の継代が可能であること、そして単回の注射に必要とされるのが比較的少数(500,000〜1,000,000)であることを考慮して、単一のサンプルは、複数回の注射に十分な細胞をもたらし得る。蛍光検査(fluoroscopic)または他の誘導を使用して、複数のサンプルを、単一の手順の間に採取することができる。
【0054】
出生後組織をTCSの外科的な標本から取得したが、同じ実験の全てをまた、胎児由来HuFT NSCおよび出生後のラット終糸を使用して実行した。この結果は、全ての年齢において、終糸内にNSCが存在することを一貫して確認した。実際に、正常な78歳のHuFTの免疫染色は、ネスチン陽性細胞の証拠を示し、この主張をさらに支持した。18歳までのHuFT由来のNSCの単離および分化は、これらの幹細胞が、成人期に残存する可能性を示唆する。
【0055】
FTは、バイオセイフティー、組織適合性およびその除去に続いて何の欠損も存在しないという利点を付与する(profer)、自系、消耗性かつ到達可能なNSCのための未開発の資源である。この組織供給源由来のNSCは、神経系の外傷および変性の処置に有用である。
【0056】
以下の試薬および方法を使用して、以下の実施例に記載されたデータを生成した。
【0057】
インビトロ分化
非特異的分化のために、単一の神経球を、可視化のための解剖顕微鏡の補助を使用して単離し、1% N2、1% ペニシリン−ストレプトマイシン、および5〜10% ウシ胎児血清(Gibco)を含むDMEM/F12培地中で96穴の培養ディッシュ(Corning)の個々のウェルにおいて、ポリ−L−リジン(0.01%、Sigma)およびラミニン(20mg/ml、Sigma)でコートしたカバーガラス上で平板培養した。培地を、実験の残りの間、変更しなかった。免疫細胞化学のために、カバーガラスを2〜10日後に処理した。分化した細胞が増殖性細胞に由来したことを確認するために、神経球を、トリチウム化チミジン(Cepko labから贈呈、培地1mlあたり5mL)で8時間インキュベートした。続いて、神経球を、視覚的に単離し、3回、幹細胞培地で洗浄し、次に、上記のように分化させた。運動ニューロンへの指向性の分化は、まず、4〜5日間のレチノイン酸(RA、2mM、Sigma)およびソニックヘッジホッグ(Shh−N 500〜100nM、R&D systems製、またはHh−Ag1.3、Curis)での神経球の処理を含んだ。この処理を、以前に記載した幹細胞培地中で行った。その後、個々の神経球を単離し、1% N2、1% ペニシリン−ストレプトマイシン、5% ウマ血清(Gibco)、CNTF(25ng/ml、Sigma)、GDNF(25ng/ml、Sigma)およびBDNF(50ng/ml)を含むDMEM/F12培地中で7〜10日間、96穴の培養ディッシュ(Corning)の個々のウェルにおいて、ポリ−L−オルニチン(0.01%、Sigma)、I型コラーゲン(0.01%、Sigma)およびラミニン(20mg/ml、Sigma)でコートされたカバーガラス上で平板培養した。3つの型の対照実験を実行した。神経球を、RAを含まずShh−Nのみで処理した。神経球を、RAおよびShh−Nで処理しないが、BDNF、CNTFおよびGDNFの存在下で増殖させた。最後に、一部の神経球は、Shh−NでもRAでも処理せず、それらを、BDNFもCNTFもGDNFも存在しない中で培養したが、上記のように、5〜10%血清中で非特異的分化を受けた。また、先に記載したように、対照も、コートしたカバーガラスを含む96穴の培養ディッシュの個々のウェル中で7〜10日間培養した。その後、全てのカバーガラスを免疫細胞化学のために処理した。
【0058】
神経筋接合部の形成の存在を証明するために、個々の神経球を、RA(2mM)およびShh−N(1000nM)で4〜6日間処理し、続いて、先に記載した、CNTF、BDNFおよびGDNFを含むMNの増殖および生存のための分化培地中で、筋肉培養物上で平板培養した。対照培養物は、筋肉培養物上で平板培養した未処理の神経球を有したか、または全く神経球を有さなかった。21日後、培養物を、蛍光αアマガサヘビ毒素(2.5mg/ml、alexa fluor 488で標識)で2.5時間インキュベートした。その後、これらを洗浄し、固定し、免疫細胞化学(ニューロンのマーカーBTIII)のために処理した。
【0059】
抗体
ネスチンに対するウサギポリクローナル抗血清(1:400)、ChATに対するヤギポリクローナル抗体(1:100)およびニューロン特異的エノラーゼに対するマウスモノクローナル抗体(1:1000)を、Chemiconから取得した。ウサギポリクローナルSox2(1:1000)は、Sigma Abcamに由来する。ビメンチンに対するマウスモノクローナル抗体は、Cepko laboratoryから贈呈された。マウスモノクローナルCD133(1:1000)は、Miltenyi Biotecから購入した。GFAPに対するウサギポリクローナル抗体(1:1000)は、Dakoに由来し、GFAPに対するマウスモノクローナル(1:1000)は、Sigmaに由来した。βチューブリンIIIに対するウサギポリクローナルは、Covanceに由来した。Tuj 1(1:1000)およびNeu−N(1:1000)に対するマウスモノクローナル抗体も、Olig−2に対する抗体と同様に使用した。NICHDの賛助の下に開発され、The University of Iowa,Department of Biological Sciences,Iowa City,IA 52242によって維持された、神経フィラメント、MNR2、Lim3およびIs1−1、ならびにPax6に対するモノクローナル抗体を、Developmental Studies Hybridoma Bankから購入した。AF 488結合体化ロバ抗ウサギIgG、AF 488結合体化ロバ抗マウスIgG、AF 568結合体化ロバ抗ヤギIgG、AF 488結合体化ヤギ抗マウスIgG、およびAF 568結合体化ヤギ抗ウサギIgGは、InvitrogenのAlexa Fluor productsから購入した二次抗体であった。全て1:1000で使用した。
【0060】
免疫細胞化学
免疫細胞化学を、カバーガラスに付着した全神経球または分化した神経球を使用して実行した。カバーガラスを、4% ホルムアルデヒド(PBS中、pH7.2)で20〜30分間、固定し、次に、PBSでそれぞれ10分間、3回洗浄した。抗体希釈物を、ブロッキング溶液(PBS、0.2% ウシ血清アルブミン中、10% 正常ヤギ血清、10% アイシングラス、0.3% Triton X)で調製し、一次抗体を、カバーガラスを用いて、それら各々の希釈物で一晩(8時間)インキュベートした。このことの後、PBSで3回洗浄し、その後、適切な二次抗体で4時間インキュベートした。PBSでの3回のさらなる洗浄の後、カバーガラスをDapi(0.03mg/ml)で30分間インキュベートした。その後、カバーガラスを、もう一度3回(それぞれ10分間)洗浄し、それから封入剤としてVectashieldを使用してガラススライド上に載せた。このスライドを、Zeiss顕微鏡写真機(photomicroscope)を使用して免疫蛍光検査のために視覚化した。特定のマーカーについての細胞染色のおおよその割合を、4〜5 20×視野の平均カウントによって決定した。Dapiを、細胞の総数を計数するためのマーカーとして使用した。細胞の数が極めて多い(>500)場合、またはそれらが、一部の視野で共に塊になるが、他の視野では存在しない場合、Dapiに対するマーカー陽性細胞のパーセンテージを見積もった。
【0061】
免疫細胞化学を使用して、インビトロでのHuFT由来の未分化神経球における種々のNSC、神経前駆体細胞(NPC)、ニューロンおよびグリアのマーカーの存在を証明した。全ての神経球(n=13)は、NSCマーカーであるネスチンについて陽性に染色した。より小さな神経球(<100ミクロン)において、100%の細胞がネスチンを発現した。しかしながら、より大きな神経球において、それらのコアは、ネスチン陰性のようであった。このコアは、おそらく細胞の壊死性の塊である。神経球(n=33)はまた、神経前駆体マーカーであるビメンチン、CD133(n=18)、Olig2(n=17)およびSox2(n=17)について陽性である細胞を含んだ。NPCマーカーの発現は、神経球間で変動した。試験した全ての神経球(n=30)はまた、ニューロンのマーカーであるBTIIIおよびグリアのマーカーであるGFAPを示差的に発現し、おそらく分化に際して観察された変動したパターンを先触れした。
【実施例】
【0062】
実施例1:FT組織からの神経球の生成
29の実験において、FTを、P4〜P19齢のラットから切開し、培養した。神経球は、29個の培養物のうち、26個で生成された(89.5%)。個々の培養物を、最大18回まで継代し、3つの細胞株を樹立し、凍結させた。ヒトFT組織を、4つの胎児および6ヶ月齢〜18歳の12の出生後手術から単離した。神経球は、16個の培養物のうち、14個で生成され(87.5%)、これらの培養物を、最大6回継代した。神経球をもたらした最も年長の組織ドナーは、18歳である。図2A〜Dは、FTおよび尾方の脊髄ならびにこれらから得られた神経球を示す。20個を超えるヒトまたはラット神経球を、ネスチン免疫反応性について染色した(図2、CおよびD)。試験した全ての神経球はネスチン陽性であった。図3A〜Dは、FT神経球からニューロンおよびグリアへの細胞の分化を示す。ラットおよびヒト由来の個々の神経球を、血清存在下でポリ−l−リジン、ラミニンおよびコラーゲンを含む種々の接着基質上で平板培養した。全ての場合において、FT−NSCは、免疫細胞化学的判断基準によって決定されるように、ニューロンおよびグリア(星状膠細胞および乏突起膠細胞を含む)に分化した。
【0063】
単一の神経球を、レチノイン酸およびソニックヘッジホッグタンパク質とともにインキュベートし、血清および神経栄養因子の存在下で接着基質上で平板培養した。FT−NSCは、形態学的に同定可能なニューロンに分化し、MNR2、LIM−3、ISL−Iおよびコリンアセチルトランスフェラーゼを含む運動ニューロンマーカーに対して染色した(図4A〜D)。DiIまたはカルボキシフルオレセインで予め標識した神経球は、初代ラット筋肉細胞と同時培養した場合にニューロンに分化した(運動ニューロンまたはそれらが神経筋接合部を形成したか否かの証明に関して)。
【0064】
実施例2:ヒトFTの培養ならびに得られたNSCの増殖および継代
14〜21週齢のヒト胎児組織を、妊娠の人工的な終結の後に取得した。脊髄を迅速に切開し、氷冷ハンクス溶液中に置いた。その後、顕微鏡での視覚化の下で、ヒトFTを同定し、切開した。ヒトFTの周囲の脊髄神経根を、時折切開し、陰性対照として別個に培養した。6ヶ月齢〜18歳のヒト出生後組織を、係留された索の解放(TCSのための慣例的な神経外科手順)を受けた小児から取得した。これらの場合において、ヒトFTを、顕微鏡の補助を使用して神経外科医が視覚的に同定し、取り出しに先立って、その正体を、電気生理学的試験を用いて確認した。この組織を、氷冷ハンクス溶液中で、手術室から研究室に移した。
【0065】
一度、胎児または出生後のFT組織を取得したら、それを、標準的な培地(例えば、DMEM/F12(1:1,Gibco)、1% N2処方物(Gibco)、1% ペニシリン−ストレプトマイシン溶液(Gibco)、EGF(20ng/ml、Gibco)、bGFG(20ng/ml、Gibco)、LIF(10ng/ml)および3mMカルシウムをともなうII型コラゲナーゼ 100U/ml(Gibco))を含む培養ディッシュ(Corning)に移し、ピンセットおよび外科用メスを用いて掻き裂いた。FGFは、安定性のために、8mg/mlのヘパリン(Sigma)を含む溶液中で調製した。培養物は、加湿したインキュベーター中に、37℃、5% CO2で維持した。24時間後、その組織を、コラゲナーゼで部分的に消化し、さらなる分離のために先端熱加工したピペットで機械的に粉砕した。初代幹細胞の増殖を、3〜5日後にインビトロで検出し、そして未分化細胞の球体の形成により特徴付けた。
【0066】
組織を、4つの胚供給源および17の出生後供給源から取得した。インビトロでの3〜4日間の後、神経球は、胚培養物のうちの100%および出生後培養物のうちの82%で観察された。神経球は、球状、自由に浮遊する、NSCの異種起源の凝集物であり、これは、種々のニューロンおよびグリアに分化する能力を維持しながら、培養において増殖する。観察された神経球の数は、初代培養物あたり1個から50個を超える神経球の範囲で変動し、ドナーの年齢と相関しなかった。増殖および自己再生についてのそれらの能力を示すために、それらを、最大10回まで継代し、最大6ヶ月までそれらをインビトロで維持した。8つの株を凍結し、神経球の首尾良い回復について試験した。
【0067】
継代頻度は、初代培養物の間で変動した。一部の培養物は迅速に増殖し、10〜14日ごとに継代することを必要とし、他のものは、3〜4週間ごとに継代することしか必要としなかった。神経球を、5〜7分間、1×AccumaxTM(Innovative Cell Technologies)を用いて分離し、その後、機械的に粉砕して、神経球の部分的な分離を達成した。遠心分離(10分間、1000rpm)後、細胞を、新鮮培地と馴化培地の1:1の組み合わせ中に再懸濁した。これらの継代の間に、神経球を単一の細胞に分離した場合に、死滅率は高く、時折100%であったことに注意すべきである。
【0068】
実施例3:ニューロンおよびグリアへのヒトFT−NSCの自発的な分化
一部の神経球は、培養容器に接着し、そして培地からのいずれの因子の添加も除去もなく自発的に分化するようだった。種々のドナーに由来する単一の神経球は、LIF、bFGFおよびEGFの除去の後に血清の存在下で神経前駆体細胞(NPC)、ニューロンおよびグリアに成功裏に分化した。個々の神経球を、ポリリジンおよびラミニンでコートしたカバーガラスに2〜10日間、これらの分化条件で平板培養した。全ての場合(n=50の実験)において、神経球は、免疫細胞化学で同定されるNPC、ニューロンおよび/またはグリアの変動した組み合わせ(assortment)を生成した(図5 AおよびB)。これらとしては、ニューロン特異的エノラーゼ、神経フィラメント、neu−nおよびβチューブリンIII(ニューロンのマーカー)、GFAP(星状膠細胞のマーカー)、O1(成熟乏突起膠細胞のマーカー)、mushashi、ビメンチンおよびsox−2(NPCマーカー)が挙げられる(図5 AおよびB)。
【0069】
これらの分化実験は、インビトロでの細胞の組成および分化能力における神経球の異質性の観察と一致して、使用したドナー供給源(6ヶ月齢および12歳)内およびそれらの間の両方で不均質な神経球の能力を明らかにした。観察された染色パターンもまた、細胞塊が特定のマーカーを発現すること、またはより均一に分散した細胞が種々のマーカーを発現することのいずれかをともなって、変動した。48時間を超えて分化させた場合、高い割合の細胞(約79%、n=8、SEM 0.08)が、ニューロンのマーカーおよびグリアのマーカーの両方について二重染色された。しかしながら、分化条件で7〜10日後(n=7)、神経球あたりのニューロンおよびグリアのマーカーについて二重染色した細胞の平均の割合は、約23%(SEM 0.09)に低下した。さらに、7日後に、分化した細胞を、ニューロンのマーカーおよびグリアのマーカーの両方について染色した、これらの実験において、神経球の能力のおおよそ半分が、ニューロン優位であるかまたはグリア優位であるかのいずれかのようであり、二重染色は最小であるかまたは存在しなかった。他の場合において、ニューロンおよびグリアの両方の変動する割合を生成し、部分的に二重染色された。この変動性の能力は、供給源にかかわらず神経球で一貫し、ドナーの年齢には関係しないようだった。NPCマーカーの染色の一貫性は、7〜10日後でさえも分化した細胞の約98%で観察された(n=21、SEM 0.01)。これらの細胞は、頻繁にニューロンのマーカーまたはグリアのマーカーを同時発現した。このことは、ニューロンおよびグリアのマーカーの二重染色(分化時間の増大とともに低下する)と組み合わせて、生成された細胞が未熟なニューロンおよびグリアを表すことを示す。
【0070】
ヒトFT神経球は、インビトロで増殖し、これを継代した。これらの増殖性神経球は、NPC、ニューロンおよびグリアの収集物に分化した。分化したニューロンおよびグリアが増殖性細胞に由来することを確認するために、単一の神経球を、トリチウム化チミジンで8時間処理した(n=4の実験)。その後、これらの神経球を、トリチウム化チミジンを含む環境から取り出し、先に記載したように7日間分化させた。4つの場合全てにおいて、生じたニューロンおよびグリアのうち33〜63%が、それらの核にトリチウム化チミジンの証拠を有し、8時間の曝露ウインドウ(exposure window)が、細胞周期のS期にある増殖性の神経球細胞を捕捉したことを示した。これらの細胞は、S期にある間に放射性ヌクレオチド標識を取り込み、続いてニューロンおよびグリアに分化した(図5C)。
【0071】
同様に、ラットFT神経球は増殖し、これをインビトロで継代することができ、これらの増殖性の神経球は、NPC、ニューロンおよびグリアの収集物に分化する。これらの分化した細胞が増殖性細胞に由来することを証明するために、トリチウム化チミジンを使用して細胞を標識した。5つの実験において、神経球をトリチウム化チミジンで8時間処理した。その後、これらを、トリチウム化チミジンを含む環境から取り出し、洗浄し、上記の標準的な条件において7日間にわたって分化させ、BTIIIおよびGFAPについて染色した。5つの場合全てにおいて、生じたニューロンおよびグリアのうちの27〜90%が、それらの核にトリチウム化チミジンの証拠を有した。これらのデータは、8時間の曝露ウインドウが、細胞周期の「S」期にあるNSCの一部のパーセンテージを捕捉し、これらの増殖性細胞がこの時間の間に放射性ヌクレオチド標識を取り込んだことを示す。これらの細胞は、続いてニューロンおよびグリアに分化し、それらの核内で明白なトリチウム化チミジンによって同定された。
【0072】
実施例4:ヒトFT−NSCの運動ニューロンへの指向性の分化R
本発明以前には、運動ニューロンを生成する出生後の神経球についての先行する報告はなかった。以前に記載されたWichterleの、胚性幹細胞の運動ニューロン(MN)への指向性の分化の方法の変形を用いることで、単離され本明細書中に記載されたようなHuFT由来神経球を、MNに分化するように一貫して誘導した。最初に、単一の神経球をレチノイン酸およびソニックヘッジホッグ(Shh−Nタンパク質またはHh−Ag1.3として知られるShhシグナル伝達の特異的小分子アゴニスト)で、運動ニューロン前駆体(MNP)の誘導のために処理した。次に、これらの個々の神経球を、血清ならびにMNの成長および生存を支持することが公知の三つの神経栄養因子の存在下で、接着基質上で培養した。その三つの神経栄養因子は、毛様体由来神経栄養因子(CNTF)、脳由来神経栄養因子(BDNF)、および神経膠由来神経栄養因子(GDNF)である。これらの分化条件での7〜10日の後、MN特異的マーカーの存在について免疫細胞化学により細胞を分析した。これらは、motor neuron restricted−2(MNR2)、Islet 1(Isl1)、Lim3およびコリンアセチルトランスフェラーゼ(ChAT)を含んだ。MNPの最終分割の間に最初に発現されるMNR2は、MNの同一性にかかわる決定因子である。Isl1およびLim3は、MNの発達に関連するホメオボックス転写因子である。Isl1は、全てのクラスのMNにより発現される。分化した神経球は、MNPにおいて発現されることが公知の二種類の前駆体マーカー(転写因子のOlig−2およびPax6)について試験した。全ての実験において(n=16)、ニューロンの異なる部分が、上記(図5Dおよび5E)のMNまたはMNPマーカーを発現した。この変動性は、単一のマーカー(例えば、MNR2)の使用においてさえ持続する。体細胞性の運動ニューロンにより選択的かつ一貫して発現されるホメオボックスドメインタンパク質であるホメオボックス9(HB9)、およびPax6の存在は、RT−PCRにより確認した。
【0073】
過去の研究において、胚性幹細胞(ESC)は、Hh−Ag1.3およびRAに曝露した場合、機能性の運動ニューロンに容易に分化する。アゴニストは、実際のペプチドよりも、より効果があることが公知であり、そしてMNの生成のためのペプチドに優先して用いられた。本発明者らのデータは、この観察と矛盾がなかった。RAおよびHh−Ag1.3への初期の曝露後HuFT神経球が分化した両方の実験において、100%のニューロンがMNR2を発現した。RAおよびShh−Nでの処理後生成した5〜40%のMNR2陽性細胞と比較して、この観察はHh−Ag1.3が、HuFT神経球のMN分化のための、より有力な薬剤であることを示した。Shh−N濃度を増加させることは、結果を変化させないようだった。Shh−N単独ではまた、Shh−NおよびRAで処理したものと同様の範囲でMNR2陽性細胞を生成した。GDNF、CDNFおよびBDNFの存在下で培養された未処理神経球はまた、変動する割合のMNR2陽性細胞を一貫して生成した(図5C)。多数の例において、MNR2について免疫染色された細胞は、一緒にかたまりになる傾向があった。指向性のMN分化のためのRAおよびShh−Nの使用は、CDNF、BDNFおよびGDNFの存在下で神経球を単に分化させることよりも顕著には優れていないようだった(図5D)。さらに3つの実験の1つにおいて、血清のみにおいて増殖した未処理神経球由来の細胞の約40%が、MNR2について陽性に染色された。神経球の公知の異質性、および発生的に意図される本来のHuFTの機能と組み合わせて、この観察は、RAの腹方化作用および誘導的なShh−Nシグナル伝達を必要とすることなく、MNに分化するいくらかのHuFT NSCの本質的な能力を示した。Hh−Ag1.3の有効な作用は、全てのHuFT神経球由来細胞をMNに指向させるようである。RAおよびShhで処理した、ならびに/またはCNTF、GDNFおよびBDNFを含む培地中で培養されたHuFT由来NSCは、インビトロで筋繊維と神経筋シナプスを形成することができた(n=16)。
【0074】
実施例5:EGF+FGF+LIFに応答して自己再生するラット−FT由来神経球の単離と性格づけ
ラットFT(rFT)において可能性のあるNSCに、いずれかの特定のニッチまたは特異的な場所があるか否かを決定するため、組織を切開し、そしてホルムアルデヒドで固定した組織の複数の切片をNSCマーカーであるネスチンについて染色した(図6Aおよび6B)。これらの試料の免疫組織化学が、明白な分布パターンをともなわない散在する個別的なネスチン陽性細胞を示した。
【0075】
インビトロでのNSCの単離は、コラゲナーゼで分離した一次組織を、bFGF(20ng/ml)、EGF(20ng/ml)およびヒトLIF(10ng/ml)を含む標準的な幹細胞培地(DMEM、F12、N2補充)中で培養することを含む。先行する研究は、これらの分裂促進因子を、おそらく付加的な効果をともなう、NSC増殖に対する好結果の刺激剤として同定した。インビトロで3〜4日後、神経球が、34の初代培養のうち31で観察された。これらの神経球は、初めは自由に浮遊しており、そしてその球状構造、位相での明るい外観、一定の細胞膜、および回折リングにより同定された(図6C)。初代培養当たりの神経球の数は、10から40より多くまで変化した。この数は、ドナーの年齢と相関していないようだった。それらの増殖および自己再生の能力を示すため、神経球を分離し、そして二次球体を生成するよう19回まで継代し、そしてインビトロで7ヶ月まで維持した。
【0076】
初期の考察とは反対に、神経球は、NSCの同種集団ではないが、おそらく異なる能力を持つ、異なるNSCおよび神経前駆体細胞(NPC)の不均質な収集物であることが示された。この不均質な性質を考慮して、rFT由来神経球は、種々のNSC、NPC、神経およびグリアのマーカーの発現を決定する免疫細胞化学を用いて特徴付けられた(表1)。特に、神経球は、NSCマーカーであるネスチン(n=9)、NPCマーカーであるSox2(n=8)、ビメンチン(n=6)、Olig−2(n=3)、およびMusashi(n=4)、ニューロン特異的マーカーであるβ−チューブリンIII(BT III、n=12)、ならびに星状膠細胞マーカーであるグリア繊維酸性タンパク(GFAP、n=12)について染色した。
【0077】
9例全てにおいて、変化する割合での細胞が、ネスチンについて陽性であった。9例のうち4例で、神経球中の100%の細胞が、このNSCマーカーを発現し、これはNSの年齢またはサイズに相関しないようであった(図2D)。神経前駆体マーカーについての染色は変化した。3つの実験全てにおいて、より強い染色を示すいくらかの領域をともなって、神経球細胞の100%が、Olig−2について陽性に染まった(図7A)。Sox2およびビメンチンに関して、全ての神経球が、これらのマーカーについて陽性に染まるいくらかの割合の細胞を有したが(図7Bおよび7C)、このパーセンテージは、Sox2で40から100%、ビメンチンで33から100%で変化した。Musashiの染色は弱く、時折のホットスポットをともなった(図7D)。全ての神経球がまた、BT IIIおよびGFAPを発現し、この発現は、いくらかの神経球では均一であったが、他では違った(図7E)。示差的発現の後者の例では、BT IIIまたはGFAPのどちらかが周縁部で発現し、他のマーカーは、逆に神経球の中心部で発現した。
【0078】
実施例6:運動ニューロン(MN)を生成するRFT由来神経球の指向性の分化
rFT由来神経球からのMNの生成を一貫して誘導するために、単一の神経球をレチノイン酸(RA)およびソニックヘッジホッグ(Shh−Nタンパク質、またはHh−Ag1.3と呼ばれるShhシグナル伝達の特異的小分子アゴニスト)で、MN前駆体を誘導するために4〜5日間処理した。これらの処理された個々の神経球は、その後、血清ならびに、MNの成長および生存を支持することが公知の三つの神経栄養因子の存在下で、接着基質上で平板培養した。用いた神経栄養因子は、毛様体由来神経栄養因子(CNTF)、脳由来神経栄養因子(BDNF)、および神経膠由来神経栄養因子(GDNF)である。これらの条件において処理した神経球を7〜10日間分化させた後、細胞をMN特異的マーカーの存在について免疫細胞化学により分析した(図8、図9)。用いたマーカーは、motor neuron restricted−2(MNR2)、Islet 1(Isl1)、Lim3およびコリンアセチルトランスフェラーゼ(ChAT)であった。MNR2、Isl1およびLim3は、Wichterleらにより、MNSへの胚様体の規定された分化を記載する彼らのもともとの論文において用いられた。Pax6およびOlig2は、MN前駆体細胞を同定するためのマーカーとして用いた。
【0079】
MNR2は、運動ニューロン前駆体の最終分裂の間に最初に発現され、そしてMNの同一性にかかわる決定因子である。Isl1およびLim3は、MNの発達に関連する二つのホメオボックス転写因子である。Isl1は、全てのクラスのMNにより発現される。全ての実験において(n=25)、種々の割合の分化したニューロンが、上に記載したMNまたはMNPマーカーを発現した(図8)。この変動性は、MNR2のような単一のマーカーの使用においても持続する(図9)。Hh−Ag1.3を使用した9つの実験において、分化したニューロンの95〜100%が、MNマーカー(例えば、MNR2、Isl1、Lim3およびChAT)を発現した。このアゴニストは、実際のペプチドよりも、より有効であり、そしてMNの生成について、Shh−Nを優先して用いられてきた。そのデータは、Shh−Nで処理した神経球は、そのうちほんの20〜40%がMNマーカーを発現する分化したニューロンを生じるという観察と一致する。Shh−N濃度の増加は、結果を変化させないようであった。
【0080】
三つの形態の対照実験を実施した。第一番目は、神経球をRAなしで、Shh−Nで処理し(n=8)、そしてその後それらを、血清ならびにBDNF、CNTFおよびGDNFを含む培地中で分化させることを含んだ。第二番目は、未処理の神経球を、血清および3つのノイロトロピンを含む培地中で培養することを含んだ(n=8)。そして第三番目は、未処理の神経球を、特定の神経栄養のサポートはない、血清を含む培地中で分化させることを含む(n=3)。初めの二つの条件は、変動する割合のMNR2陽性細胞を一貫して生成した(図8Eおよび8F)。複数の例において、MNR2について免疫染色した細胞は、島において互いに塊になる傾向があり、そしてグリアにより取り巻かれることが、時折言及された。指向性および一貫したMNの生成のためのRAおよびShh−Nの使用は、BDNF、CNTFおよびGDNFの存在下において神経球を単に分化させることよりも優れてはいないことが明らかとなった。しかしながら、神経球を、神経栄養のサポートなしで、血清中で培養した場合、MNの生成は一貫性がなく、3つの実験の1つのみで、約40%の細胞がMNR2を発現した。神経球の公知の異質性、および発生的に意図される本来のFTの機能と組み合わせて、これらのデータは、RAの尾方化作用または外因性の腹方化Shh−Nシグナル伝達を必要とすることなく、MNに分化するいくらかのrFT NSCの本質的な能力を示す。しかしながらHh−Ag1.3の使用は、大半だが全てではない神経球由来細胞がMNを生成するように指向した点において、MNの収量を増加させるために有益であることが見出された。
【0081】
実施例7:FT由来神経球は、多分化能性であり、ニューロンおよびグリアに分化する
いくらかの神経球は、培養容器に接着し、そして培地からのいずれの因子の添加も除去もなく自発的に分化する。rFT由来神経球が、ニューロンおよびグリアに分化するために必要とされる条件を決定するために研究を実行した。bFGF、EGFおよびLIFの除去後、34の実験において単一の神経球を、ポリ−L−リジンおよび/もしくはラミニンコートしたカバーガラスならびに/または5〜10%の血清への曝露の種々の組み合わせ上で平板培養した。その神経球をこれらの分化条件に7日間供した。接着基質単独または血清単独のどちらかの使用が、形態学的分化を開始するために十分であったが、血清の添加は、より迅速な分化を結果として生じた。全ての例において、分化した神経球は、BTIII、神経フィラメント、O1、およびGFAPを含むニューロンのマーカーおよびグリアのマーカーを発現した(表1および2)。
【0082】
一度これらの条件が確立されると、次の65の実験が、rFT神経球の分化の能力をさらに特徴付けた。これらの実験において、その分化条件は、3つの増殖因子全ての除去、5〜10%の血清による培地の補充、ならびにポリ−L−リジンおよびラミニンでコートしたカバーガラス上での単一神経球の平板培養を含んだ。これらの実験のうちの30において、神経球は、24時間を越えて分化し、そして35の実験において、神経球は7〜10日間を越えて分化した。哺乳動物CNSの他の領域から培養された神経球の報告された不均一さと一致して、rFT由来神経球は、さまざまな分化能力を有した。
【0083】
その変動性にもかかわらず、全ての実験において、各神経球由来のいくらかの割合のNPC、ニューロンおよび/またはグリアが、免疫細胞化学マーカーを用いて同定された。これらは、ニューロン特異的エノラーゼ(NSE)、NeuN、BTIII、GFAP、O1、Musashi、ビメンチンおよびSox2を含んだ(表1)。24時間を越えて分化した神経球(n=9)は、ニューロンのマーカーおよびグリアのマーカーの両方について二重染色された総細胞の高い割合を有した(約70%、表3)。この割合は、神経球が7〜10日間を越えて分化した後、平均約14.5%まで減少した(n=13、表3)。時折、7日間の分化の後、神経球由来の細胞は、ニューロンのマーカーまたはグリアのマーカーのどちらかを主に発現した。しかしながら、ほとんどの例において、明白な優勢は観察されなかった。14の実験における同じ二つのマーカーの使用にもかかわらず、分化した神経球は、BTIIIおよびGFAP発現のさまざまなアレイを示した。この変化は、同一の供給源由来、および異なるrFT供給源の間由来の両方の神経球において持続していた。
【0084】
分化した神経球の間の染色パターンもまた変化する。しばしば、神経球由来の細胞の塊は、一つの特定のマーカーについて全て染色陽性であり、そして他の領域における細胞は異なるマーカーを発現する。しかしながら、より頻繁に、異なるマーカーについて染まる細胞が、一緒に点在する。
【0085】
【表1−1】
【0086】
【表1−2】
【0087】
【表2】
【0088】
【表3】
実施例8:FT−NSCはインビトロで運動ニューロンを生成する
神経幹細胞(NSC)は、中枢神経系(CNS)における未分化細胞であって自己再生が可能であり、そしてニューロンおよびグリアに分化するように誘導され得る。哺乳動物のNSCの現用の供給源は、正常な機能にとって大切であり、外科的に到達することが困難であるCNSの領域に制限されている。これは、ヒト疾患におけるそれらの治療上の能力を制限する。以前に調べられておらず、犠牲にしてもよく、そして容易に到達することができる、脊髄の尾方末端の組織である終糸(FT)が、哺乳動物における多分化能の神経球の供給源であることが、予期せず発見された。本研究では、単離し、そしてこれらの細胞の能力を特徴付けるためにラットモデルを用いた。ラットFT(rFT)由来の神経球は、上皮細胞成長因子(EGF)、塩基性線維芽細胞成長因子(bFGF)、および白血病抑制因子(LIF)の組み合わせによるインビトロでの増殖に耐えられる。増殖している細胞は、血清への曝露により神経前駆体細胞、ニューロン、星状膠細胞およびグリアへと分化するように誘導された神経球を形成した。種々の神経栄養因子と組み合わせてソニックヘッジホッグ(Shh)およびレチノイン酸(RA)を用いる指向性の分化をとおして、rFT由来神経球は、運動ニューロン(MN)をインビトロで生じた。
【0089】
多分化能NSCの存在が、ラットからヒトまでの範囲の種において成体の哺乳動物CNSの複数の領域において示された。これらの領域は、嗅球、脳室の上衣下裏打ち、海馬、小脳、脊髄および網膜を含む。哺乳動物NSCの現用の供給源は、それらが、正常な機能にとって重要であり、そして到達することが困難な領域から取得されることから、ヒト疾患における移植治療のためには理想的ではない。これらの領域の外科的破壊は、自系NSCを採取するためにそれらを用いることを非現実的にする、甚大な神経学的な欠損を導いた。
【0090】
FTは、自系多分化細胞の供給源として優れた候補である。それは、容易に到達することができ、そして犠牲にしてもよい、成体において持続する組織である点において、現在利用可能な供給源を凌駕する明確な利点を提供する。本発明の方法は、FTを自家置換療法のために用いることにより、免疫学的な問題を回避する。
【0091】
発生の初期に、FTは、胚の予定運命の尾への神経支配(あるいはげっ歯類において、最も尾側の尾分節の一時的な神経支配)を与える。成体において、それは痕跡のレムナントである。いくらかのヒトは、動かせる尾をもって生まれるが、しかしながら、尾の異常な残存は、おそらく発生過程の欠陥を示す。ヒトの尾の胚発生は、最初に3.5〜5mmの段階(約4週)で検出される。11〜15mmの段階(約7週)では、尾骨領域はより進んだ発達を示し、脊髄が上衣領域、マント領域および境界領域に分化し、そしてそれぞれの脊髄神経節からそれに入るよく発達した脊髄根を有す。この時点では、この領域が脊髄のより頭方の部分のような成体の状態へと完全に分化し始めないという、組織学的証拠はない。
【0092】
発生が続くと、脊髄の尾骨/尾の部分は再吸収され、そして細胞は、Streeterにより「脱分化」と名付けられた(Streeter,G.L.1919.Am J Anat 22:1−12)過程をうける。30mmの段階(約9.9週)までに、脊髄の尾骨領域は、顕著に変化した。尾骨の脊髄組織は初期の胚の型に戻り、線維芽細胞、ニューロンおよびグリアのゆるく組織化された収集物によって囲まれた上衣細胞により裏打ちされた狭い中心管をもつ膠原性の構造を、結果として生じる(Streeter,G.L.1919.Am J Anat 22:1−12)。最後の三つの尾骨神経節が消失するように、境界領域およびマント領域は、完全に消失する。再吸収は、尾から吻への方向で生じ、そして結果として生じる構造が、FT(脊髄を尾骨に固定する脊髄の尾方末端の細長い延長部分)として成体中に残る。
【0093】
通常、脱分化した出生後のFTは、残りの神経系と相互連結せず、身体の神経を刺激しない。それは偽りなく痕跡のレムナントである。ヒトにおいて、脊椎に固くつながれ、動作の十分な自由を欠く(係留脊髄症候群として公知の状態である)例において、脊髄に対する緊張を軽減するために、それは慣例的に外科的に切開される。FT細胞は、初期の胚細胞型に似ており、脊髄の残部に存在する複数の細胞型に再分化する能力を維持する。
【0094】
ヒトおよびラットの両方において、FTは、脊柱管を囲む膠原性の構造である。室周囲の上衣細胞ならびにゆるく組織化された線維芽細胞、ニューロンおよびグリアの収集物が、その管を囲む。ラットにおいてFTニューロンは、通常よりも、より小さいものとして記載され、そして拘束および分化の初期段階においてニューロンを示す。パラガングリオーマおよび他の初期の神経外胚葉性腫瘍は、成体FTから生じ、NSCが存在することを再び示唆する。FTは、多分化細胞の供給源である。ラットモデルの使用は、制御された環境中での、これらの細胞の全身性の制限のない研究を可能にする。げっ歯類では、FTは、最も尾側の尾分節の一時的な神経支配を与える。
【0095】
細胞培養
rFTの培養および得られた神経球
初代培養:全ての手順は、滅菌条件下で実施した。P2〜P11齢の出生後のラット(雄および雌、SD系(Sprague Dawley)、チャールズ・リバー(Charles River))を、イソフルラン(Abbott)で麻酔し、そして頚椎脱臼により殺した。脊柱を、氷冷したHanks溶液中で迅速に切開した。顕微鏡での可視化のもと、rFTを同定し切開した。各切開は、細胞死を最小にするため、5分未満で実施した。rFTの周囲の脊髄神経根を時折切開し、陰性対照として別個に培養した。rFT(通常、培養ディッシュ当たり、3つの同胞rFT)をプールし、そして幹細胞培地(SCM)(Weissら、1996.J Neurosci 16:7599−7609;Carpenterら、1999.Exp Neurol 158:265−278;Liら、2005a.Biochem Biophys Res Commun 326:425−434;Kimら、2006.Exp Neurol 199:222−235)を含む培養ディッシュ(Corning)中に移した。この培地は、DMEM/F12(1:1、Gibco)、1% N2処方物(Gibco)、1% ペニシリン−ストレプトマイシン溶液(Gibco)、EGF(20ng/ml、Gibco)、bFGF(20ng/ml、Gibco)、およびLIF(10ng/ml)から作製した。FGFは、安定性のために、8mg/mlのヘパリン(Sigma)を含む溶液中で調製した。
【0096】
組織を分離するため、3mMのカルシウム(Gibco)をともなうII型コラゲナーゼ 100U/ml(Gibco)をSCMに加えた。切開した組織をその後、このコラゲナーゼを含む培地に移し、そしてピンセットおよび外科用メスを用いて掻き裂いた。培養物は、加湿したインキュベーター中に、37℃、5% CO2で維持した。24時間後、その組織を、さらなる分離のために先端熱加工したピペットで機械的に粉砕し、そしてコラゲナーゼ含有SCM中に残した。初代幹細胞の増殖を、3〜5日後にインビトロで検出し、そして未分化細胞の球体の形成(Reynolds,B.A. and Weiss,S.1992.Science 255:1707−1710)により特徴付けた。
【0097】
培養の継代:培養物は、2〜3週毎に継代した。神経球を、5〜8分間、1×AccumaxTM(Innovative Cell Technologies)を用いて分離し、そしてその後、神経球の部分的分離を達成するために機械的に粉砕した。初期の実験において、神経球を完全に分離した場合、細胞が生存しても少数だった。酵素的分離後、細胞を遠心分離し(1000rpmで10分間)、そして新鮮培地と馴化培地の1:1の組み合わせ中に再懸濁した。
【0098】
ラット筋肉の培養
P0〜P7のラットを殺し、そして近位の肢の筋肉を、氷冷したHBSS中で迅速に切開した。その組織を、穏やかに別々に掻き裂き、そしてその後、DMEM/F12(1:1)、1% N2補充物および1% ペニシリン−ストレプトマイシンからなる培地を含む培養ディッシュに移した。3mMのカルシウムをともなうII型コラゲナーゼ(100U/ml)を、筋肉繊維を単一の細胞に分散させるために、この培地に加えた。ディッシュは、37℃、5% CO2でインキュベーター中に24時間置いた。24時間後、組織を完全に分離するために、先端熱加工したパスツールピペットで培養物を粉砕した。培養物をその後、1000rpmで5分間遠心分離した。そのペレットを2回洗浄し、その後、DMEM/F12 1:1、1% N2補充物、1% ペニシリン−ストレプトマイシン、10% ウシ胎児血清を含む培地中に再懸濁した。シス−ヒドロキシプロリン(100μg/ml)を、線維芽細胞の増殖を抑制するために平板培養培地に加えた。細胞を、ポリ−L−リジン(0.01%)およびラミニン(20μg/ml)でコートしたカバーガラス上で、約106細胞/mlの密度で平板培養した。
【0099】
細胞分化
インビトロ分化
血清をもちいての非特異的分化:単一の神経球を、可視化のための解剖顕微鏡を用いて単離し、そして96穴の培養ディッシュ(Corning)の個々のウエル中で、1% N2、1% ペニシリン−ストレプトマイシン、および5〜10% ウシ胎児血清(Gibco)を含むDMEM/F12培地において、ポリ−L−リジン(0.01%)およびラミニン(20μg/ml)でコートしたカバーガラス上で平板培養した。培地は、実験の残りの間、変更しなかった。カバーガラスを、24時間後、または7〜10日後に免疫細胞化学のために処理した。
【0100】
トリチウム化したチミジンとのインキュベーション:チミジン標識実験を、Cepko laboratory(Dyer,M.A. and Cepko,C.L.2000.Nat Neurosci 3:873−880)のプロトコールを用いて実施した。神経球を、SCM中で8時間、3Hチミジン(NEN、5μCi/ml;89Ci/mmol)とインキュベートした。個々の神経球を単離し、SCM中で3回洗浄し、そして上述のように血清中で分化させた。分化後、カバーガラスを免疫細胞化学のために処理した。カバーガラスをスライド上に載せる前に、乳化剤のオイルをカバーガラスに加え、そして暗室に2日間置いた。乳化剤のオイルを除去し、そしてカバーガラスを水で洗浄した。現像液を4分間加えた。続いて、その現像液を吸引し、そしてそのカバーガラスを4% パラホルムアルデヒドで20分間固定した。その後、カバーガラスを水で洗浄し、Vectashieldを用いてスライド上に載せ、そして蛍光顕微鏡使用法下で(免疫細胞化学について)および明視野顕微鏡使用法下で(トリチウム化チミジンの取り込みについて)可視化した。
【0101】
指向性の分化:神経球を、レチノイン酸(RA、2mM、Sigma)およびソニックヘッジホッグ(Shh)タンパク質(Shh−N 400〜1000nM、R&D systemsより入手)、またはソニックヘッジホッグシグナル伝達の小分子アゴニスト(Hh−Ag1.3、Curis)で、当該分野で認識される方法(Wichterleら、2002.Cell 110:385−397;Soundararajanら、2006.J Neurosci 26:3256−3268)の変形を用いて4〜5日間、処理した。この処理は、SCM中で実施した。個々の神経球をその後単離し、そして96穴の培養ディッシュ(Corning)の個々のウエル中で、1% N2、1% ペニシリン−ストレプトマイシン、5% ウマ血清(Gibco)、CNTF(25ng/ml、Sigma)、GDNF(25ng/ml、Sigma)、およびBDNF(50ng/ml)を含むDMEM/F12培地において、ポリ−L−オルニチン(0.01%、Sigma)、I型コラーゲン(0.01%)およびラミニン(20mg/ml)でコートしたカバーガラス上で7〜10日間、平板培養した。四つの条件を用いた:(1)神経球を上記のように処理した;(2)RAなしで上記のように処理した;(3)ShhもRAもなしで上記のように処理した;(4)ShhもRAも三つのノイロトロピンもなしで、血清単独を用いて。カバーガラスをその後、免疫細胞化学のために処理した。
【0102】
神経筋接合部の形成:個々の神経球を、RA(2mM)およびShh−N(600〜1000nM)で、4〜6日間処理し、そして続いて上記の、MN成長および生存のための分化培地中の筋培養物上で平板培養した。二つの型の対照培養物を用いた:1)筋細胞単独および2)未処理の神経球をその上で平板培養した筋細胞。6〜21日後、培養物を蛍光α−アマガサヘビ毒素(2μg/ml、Molecular Probes alexa fluor 488)と2.5時間インキュベートした。それらをその後、洗浄し、固定し、そして免疫細胞化学(ニューロンマーカーTUJ−1)のために処理した。単一の神経球を、同時培養に先立って、1時間、1μMのジ−I(Di−I)またはジ−D(Di−D)で標識した。
【0103】
細胞マーカー
抗体:ネスチンに対するヤギポリクローナル抗血清(1:50)(R&D systemsより入手)。ChATに対するヤギポリクローナル抗体(1:100)およびニューロン特異的エノラーゼに対するマウスモノクローナル抗体(1:1000)はChemiconから入手した。Sox2に対するウサギポリクローナル抗体(1:1000)は、SigmaおよびAbcamから入手した。ビメンチンに対するマウスモノクロ−ナル抗体はZymedから入手した。GFAPに対するウサギポリクローナル抗体(1:1000)はDakoから入手し、GFAPに対するマウスモノクローナル抗体(1:1000)はSigmaから入手した。β−チューブリンIII(Tuj−1)に対するウサギポリクローナル抗体はCovanceから入手した。Tuj−1に対するマウスモノクローナル抗体(1:1000)およびNeu−Nに対するマウスモノクロ−ナル抗体(1:1000)はCovanceから入手した。Olig−2に対するマウスモノクローナル抗体は、使用に先立って前希釈した。NICHDの援助のもとに開発され、University of Iowa,Department of Biological Sciences,Iowa City,IA 52242によって維持された、神経フィラメント;MNR2、Lim3およびIsl−1;ならびにPax6に対するモノクローナル抗体は、Developmental Studies Hybridoma Bankから入手した。AF488結合体化ロバ抗ウサギIgG、AF488結合体化ロバ抗マウスIgG、AF568結合体化ロバ抗ヤギIgG、AF488結合体化ヤギ抗マウスIgG、およびAF568結合体化ヤギ抗ウサギIgGは、InvitrogenからのAlexa Fluor productsより入手し、全て1:1000で使用した。
【0104】
免疫細胞化学:はカバーガラスに付着した全神経球または分化した神経球で実施した。カバーガラスを、(PBS、pH7.4中の)4%ホルムアルデヒド中で20〜30分間固定し、その後PBS中で各10分で3回洗浄した。抗体希釈液をブロッキング溶液(PBS中0.2% ウシ血清アルブミンにおいて、10% 正常ヤギ血清、10% アイシングラス、0.3% Triton X)中で調製し、そして一次抗体をそのカバーガラスと一晩(8時間)インキュベートした。その後、適切な二次抗体との4時間のインキュベーションの前に、PBS中で3回洗浄した。PBS中での追加の3回の洗浄後、カバーガラスをDapi(0.03mg/ml)中で30分間インキュベートした。カバーガラスを、その後、3回洗浄し(各10分間)、そしてVectashield中にスライドガラス上に載せた。そのスライドは、Zeiss写真用顕微鏡または共焦点顕微鏡を用いて、免疫蛍光検査のために可視化した。特定のマーカーについての細胞染色のおおよその割合を、4〜5の20×視野の平均カウントにより決定した。細胞カウントは、Dapiを用いる核染色に基づく。
【0105】
rFT由来神経球の単離および特徴付け
単離:rFTから単離した細胞を、コラゲナーゼで分離し、そして、bFGF(20ng/ml)、EGF(20ng/ml)およびヒトLIF(10ng/ml)を含む標準的な幹細胞培地(DMEM、F12、N2補充物)中で培養した。インビトロで3〜4日後(DIV)、34の初代培養物中31において、神経球が観察された。これらの神経球は、最初は自由に浮遊し、そしてそれらの球状構造、位相での明るい外観、および一定の細胞膜により同定した。神経球は、やがてより大きな神経球へと成長する、3〜4の丸い細胞のより小さな塊として、初めは出現する。これらの、より大きな神経球のサイズは、サイズにおいて約50um未満〜1mm超までの広い範囲であった。30umより小さい細胞塊は、神経球としてカウントしなかった。初代培養物当たりの神経球の数は、約30から、50を超えるまで変化した。この数はドナーのラットの年齢とは相関しないようだった。増殖および自己再生の能力を示すため、神経球を分離し、そして19回まで継代した。これらの培養物は、インビトロで7ヶ月まで維持した。12の培養物を凍結し、そして二つを生存能力について試験し、成功裡に回復した。
【0106】
特徴付け:神経球は、NSCの均質な集団ではなく、むしろ異なる分化能力をもつ異なるNSCおよび神経前駆体細胞(NPC)の不均質な収集物である。rFT由来神経球を、種々の神経幹細胞(NSC)マーカー、神経前駆体細胞(NPC)マーカー、ニューロンのマーカーおよびグリアのマーカー(表1)の発現を決定するために、免疫細胞化学を用いて特徴付けた。特に、神経球を、NSCマーカーのネスチン(n=9);NPCマーカーのSox2(n=8)、ビメンチン(n=6)、Olig−2(n=3)、およびMusashi(n=4);ニューロン特異的マーカーのβ−チューブリンIII(Tuj−1、n=12);ならびに星状膠細胞のマーカーのグリア線維酸性タンパク(GFAP、n=12)について染色した。
【0107】
全ての例において、変動する割合の細胞が、ネスチンについて陽性であった。9例中4例において、神経球中の100%の細胞が、ネスチン+であった。このネスチン染色の存在は、神経球の培養における時間とは相関しないようであった。加えて、固定された全載(whole mount)(n=1)および組織切片(n=2)をネスチンについて染色した。免疫組織化学が、ネスチン+細胞を示した。神経前駆体のマーカーの染色は変化した。三つの実験のうちの三つにおいて、神経球内の100%の細胞が、Olig−2について陽性に染色され、いくらかの領域は、より強い染色を示した。全ての神経球が、Sox−2およびビメンチンについて陽性に染まる、ある割合の細胞を有したけれども、このパーセンテージは、Sox−2については40から100%まで変化し、そしてビメンチンについては33から100%まで変化した。Musashiの染色は弱く、時折の高強度の染色の塊をともなった。
【0108】
Tuj−1+およびGFAP+細胞は、全ての神経球中に存在した(n=12)。どの神経球も、両方のマーカーについて陽性のいくらかの細胞を含み、そしてこの比は神経球の間で大きく変化した。異なるマーカーを発現する細胞の空間的な塊形成がある。この塊形成は、ほとんどの神経球において明瞭であるが、そのパターンは変化した。
【0109】
ニューロンおよびグリアへの分化
いくらかの神経球は、培養容器に接着し、そして培地へのいずれの因子の付加も除去もなしに、ニューロンおよびグリアの形態学的特徴を有する細胞へ自発的に分化する。rFT由来神経球をニューロンおよびグリアに分化させるために必要とされる条件を決定した。bFGF、EGFおよびLIFの除去後、単一の神経球を、表2に示した接着基質±5〜10%のウシ胎児血清への曝露の7種類の異なる組み合わせで処理したカバーガラス上に平板培養した。各条件につき5回の実験を実施した。7日後、培養物をTuj−1、神経フィラメント、O1、GFAPおよびネスチンについて染色した。接着基質単独または血清単独のどちらかの使用が、形態学的な分化を開始するために十分であったが、血清の付加が、より迅速な分化を結果として生じた。全ての例において、Tuj−1、神経フィラメント、O1、およびGFAPを含むニューロンのマーカーまたはグリアのマーカーのどちらかを発現する神経球に由来する細胞を検出した。
【0110】
全ての続いて起こる分化の実験(n=65)は、3つの増殖因子全ての除去、5〜10%ウシ胎児血清の培地への補充、およびポリ−L−リジンおよびラミニンの両方でコートしたカバーガラス上への単一神経球の平板培養により、実施した。細胞を、これらの条件で24時間(n=30)または7〜10日間(n=35)培養し、そして培養物を免疫細胞化学のためにその後固定した。これらの実験において用いられた神経球のサイズの広い分布を考慮すると、得られた分化した細胞の数は、神経球当たり約50細胞未満から、5000細胞超の範囲であり、最初に平板培養した神経球のサイズと相関する。より大きい神経球(通常100umより大きい)は、5000より多い分化した細胞を生成することができる。
【0111】
神経球が、NPC、ニューロン、星状膠細胞および乏突起膠細胞を産生する能力があるか否かを決定した。これらの細胞型を同定するために用いた免疫細胞化学マーカーは、ニューロン特異的エノラーゼ(NSE)、NeuN、Tuj−1、GFAP、O1、Musashi、ビメンチンおよびSox2を含んだ(表1および表3)。各例において、単一の神経球から得られた細胞を、これらのマーカーの二つについて二重染色した。これらの実験由来のデータは、rFT由来神経球が変動した分化能力を有することを示した。24時間を超えて分化した神経球(n=9)は、ニューロンのマーカーおよびグリアのマーカーの両方について二重に染まった細胞の高い割合を有した(表3)。Tuj−1およびGFAP染色の例において、69±14%(n=5)の細胞がその二つのマーカーについて二重に染まった。7〜10日後、ニューロンのマーカーおよびグリアのマーカーの両方について二重に染まった細胞の割合は、顕著に減少した(表3)。Tuj−1およびGFAP染色の例において、わずかに13±10%(n=9)の細胞が、その二つのマーカーについて二重に染まった。Tuj−1およびGFAPの変動する発現が、24時間後の分化を7〜10日後の分化と比べた14の実験において観察された。これらのrFT由来細胞集団中に存在するTuj−1+およびGFAP+の変動する割合は、各神経球の不均質な分化能力を反映する。この変動は、ラットの年齢とは拘わりなく、両方を同一の供給源から得た神経球の間でなされた比較において、異なるrFT供給源に由来する場合と同様に持続した。
【0112】
まれな場合において、7日間の分化の後、単一の神経球由来の85%を超える細胞が、ニューロンのマーカーまたはグリアのマーカーのどちらかを発現した(n=2)。しかしながら、ほとんどの例において、明白な優勢は観察されず、そしてニューロンの細胞およびグリア細胞の両方の変動する割合は、単一の神経球の分化から見出された。NPCマーカーの染色は、分化条件に7〜10日間曝露した後でさえ持続した(表3)。実際、その染色は、使用した全てのNPCマーカーについての24時間での染色と比較して、7〜10日後にわずかに増加するようである。
【0113】
分化した細胞が、増殖性細胞に由来することを立証するため、本発明者らは、活発に分裂している細胞をトリチウム化したチミジン(3H)で標識した。神経球を、3Hで8時間処理した(n=5)。その神経球を、その後洗浄し、上述の標準的な条件において、7日間を超えて分化させた。得られた細胞を、Tuj−1およびGFAPについて染色した。5例全てにおいて、ニューロンおよびグリアとして免疫学的に同定された細胞の27〜90%が、それらの核中に取り込まれたトリチウム化したチミジンを有した(図12)。この結果は、得られた細胞が、活発に分裂している細胞の子孫であったことを示す。
【0114】
RFT由来神経球は運動ニューロン(MN)を生成する
全ての実験において(n=25)、種々の割合の分化した細胞が、上述のMNマーカーまたは運動ニューロン前駆体マーカーを発現した。図12は、MNR−2、Tuj−1およびGFAPについての染色に基づいて、運動ニューロン、ニューロンおよびグリアの割合を示す。
【0115】
Shh−Nで処理した神経球は、分化したニューロンを生じ、その20〜40%のみがMNマーカーであるMNR−2、Isl1、Lim3およびChATを発現した(n=14)。Shh−N濃度を400nMから1000nMまで増加させることは、その結果を変化させないようだった。Hh−Ag1.3が用いられた場合(n=9、1.5μM)、分化したニューロンの95〜100%がMNマーカーを発現した。この結果は、これらの濃度において、Hh−Ag1.3アゴニストが、実際のShh−Nペプチドよりも、FT由来神経球からのMNの生成のために、より有効であり得ることを示唆した。
【0116】
FTからMNを生成するために、どの因子が本質的であるかを決定するため、分化条件を変化させた:(1)神経球を、RAなしで、Shh−Nで処理し(n=8)、そしてそれらを血清ならびにBDNF、CNTFおよびGDNFを含む培地中で分化させた;(2)未処理の神経球を、血清および3つのノイロトロピンを含む培地中で培養した(n=8);(3)未処理の神経球を、神経栄養因子の付加なしで、血清を含む培地中で分化させた(n=3)。図13に示したように、(1)および(2)の条件において、神経球は、変動する割合のMNR2+細胞(5〜67%)を一貫して生成した。
【0117】
条件(3)において、MNの生成は一貫性がなかった。3例中1例において、神経球から得られた細胞の40%はMNR2を発現し、そして3例中2例において、細胞はMNR2+ではなかった。MNの指向されかつ一貫した生成のための、RAおよびShh−Nの使用は、BDNF、CNTFおよびGDNFの存在下において神経球を単に分化させることよりも優れているとは立証しなかった。しかしながら、上述し、そして図13に示したように、Hh−Ag1.3は、MNの収量を増大させるうえで有益である。FTが脊髄の痕跡のレムナントであることを考慮して、これらの結果は、RAの尾方化作用も外因性の腹方化Shh−Nシグナル伝達もなしに、いくらかのrFT NSCをMNに分化させる能力を示す。
【0118】
この研究の結果は、多分化能幹細胞が出生後のrFT中に存在することを示す。これらの細胞は、NSCの二つの基本的な特性を示す:それらは、自己再生/増殖でき、ならびにニューロン、星状膠細胞および乏突起膠細胞を含む複数の細胞型へ分化できる。MNを生成するFTの能力は、ALSのような神経変性疾患に対して、特に治療上重要であり得る。
【0119】
NSCのニッチとしてのFT
本発明の方法を、先に記載された、例えば側脳室の脳室下帯(SVZ)のようなNSCのためのCNSのニッチの多くの特性を、FTの組織学的環境が有することを決定するために用いた。SVZにおける細胞構成は、A型(神経芽細胞)、B型(ゆっくりと増殖するGFAP+神経形成星状膠細胞)、C型(中間前駆体細胞)およびE型(上衣)細胞からなる。この系において、E型細胞は脳室を裏打ちし、そして、脳室に接触するためにE細胞の間を縫うように続くB細胞によって、時折置き換えられる。B型細胞は、E細胞の脳室下側に向かって位置し、吻側移動経路(Rostral Migratory Stream)として公知の経路に沿って嗅球へ移動するA細胞をさや状に覆う。C型細胞は、A細胞の鎖に沿って分散する。
【0120】
これらの研究は、FTにおける細胞型が、上衣細胞、神経芽細胞、星状膠細胞様細胞、小膠細胞、乏突起膠細胞、ニューロン、線維芽細胞、脂肪細胞および神経節細胞を含むことを示した。FT細胞は、脊柱管を裏打ちする上衣細胞の周りにゆるく組織化される。FTの上衣細胞は、広範囲なモザイクまたはサイズの変化する環を形成する脳室の裏打ち上のロゼットとして、しばしば伸長する。理論によって拘束されることを望まないが、本発明は、FTの上衣細胞は、SVZのE細胞に類似するという驚くべき発見に基づく。さらに、SVZのB細胞と同様に、FT中のいくらかのGFAP+星状膠細胞様細胞は、上衣細胞の間に互いに入り込む突起を有する。さらに、これらの突起の構造は、SVZ中でA細胞をさや状に覆うB細胞に類似する。本発明の方法を用いて実施した研究からのデータは、FTの頭方の部分に見出されるニューロンが、ラットおよびヒトの両方における形態において神経芽細胞に類似することを示す。それらは、しばしば神経線維の管に沿って存在し、そして時折、FTの外側縁に伸長する。本発明の特定の実施形態において、これらの細胞は、SVZのA細胞に相似性であると考慮される。
【0121】
腫瘍形成
CNSの腫瘍は、神経組織のニッチ近くにしばしば見出される。脊髄およびFT内の腫瘍は、全てのCNSの腫瘍の4〜10%を構成する。パラガングリオーマおよび原始神経外胚葉性腫瘍(例えば、脳室上衣腫)は、終糸に対する親和性を有する。パラガングリオーマは、神経内分泌腫瘍であり、そして脳室上衣腫は、中心管の上衣細胞から生じる腫瘍である。それらは、全てのグリアの脊髄腫瘍の60%の原因であり、そして成体における最も一般的な髄内脊髄性新生物である;粘液乳頭型脳室上衣腫(FTにおける上衣のグリアの腫瘍)は、脳室上衣腫の13%を構成し、そしてFTに対する明瞭な偏好を有する(Koeller,K.K.et al.2000.Radiographics 20:1721−1749)。
【0122】
細胞の同一性は分化プロセスの前に決定される
24時間と7〜10日間で分化した単一の神経球におけるニューロンのマーカーおよびグリアのマーカーの発現を対比した。24時間の分化プロセス後、ほとんどの細胞が、ニューロンのマーカーおよびグリアのマーカーの両方を発現した。一つの局面において、この結果は、分化プロセスの初期における未解明の細胞運命を反映する。7〜10日後、単一の神経球に由来するほとんどの細胞は、ニューロンのマーカーまたはグリアのマーカーのどちらかを発現し、非常に少数の細胞が両方について二重に染色される。この結果は、ドナーの年齢とともには変化しない。24時間後の相対的に不明瞭な細胞同一性と比べて、細胞は時間とともに、より拘束された細胞運命を発現する。
【0123】
細胞同一性は、分化プロセスの前に決定される。分化前に染色した神経球は、同一の処理にもかかわらずTuj−1+およびGFAP+細胞の異なるパターンを示した。神経球内のいくらかの細胞が、両方のマーカーについて二重に染まったが、ほとんどの細胞は、Tuj−1またはGFAPのどちらかの一つのマーカーのみを発現した。同一のマーカーについて陽性の細胞は、空間的に一緒に塊を形成する傾向がある。
【0124】
神経球分化の前に取得されたニューロンの特徴またはグリアの特徴は、各神経球の分化能力を予測する。分化の初期段階の間の一過的な二重染色は、細胞運命が未決定であるよりもむしろあいまいである分化経路に沿った点を示す。分化前の神経球細胞の初期の染色パターン(GFAP+またはTuj−1+)は、よりニューロンまたはグリアの運命へと神経球を操るためには、最初とは異なる条件でその神経球を培養することを必要とすることを示す。
【0125】
神経前駆体細胞(NPC)マーカーは、7〜10日で持続する
NPCマーカーの発現は、分化の24時間後に高く、そして7〜10日後でさらに高かった。ほとんどの細胞は、二重に染まらなくなり、そしてより成熟した表現型を示すマーカー(すなわち、ニューロンまたはグリア細胞マーカー)を発現することを考慮すると、これは驚きであった。
【0126】
その持続、およびNPC染色におけるわずかな増加は、二つの機構に帰する。一つの説明は、時間にそった細胞の代謝活性における変化を含む。一局面において、分化プロセスの初期に、いくらかの細胞は、迅速なタンパク質ターンオーバーをともなって高度に代謝的に活性であり、NPCマーカーの検出を妨げる。数日の分化の後、ターンオーバー速度が低下することから、そのタンパク質レベルが増加し、それゆえ免疫細胞化学を介してのタンパク質検出を可能とする。代替的な説明は、7〜10日の分化後、細胞は系統特異的なNPCであり、そしてそれゆえ、ニューロンまたはグリア特異的マーカーに加えてNPCマーカーを発現するというものである。
【0127】
RFT神経球はMNを生成する本質的な能力を有する
RFT由来神経球は、インビトロで胚性幹細胞をMNに分化させるために用いられたRAおよびShhへの暴露の有りおよび無しでMNを生成した。胎仔身体における吻側神経前駆体は、RA(尾方化シグナル)に応答して脊髄の位置の同一性を獲得し、そしてその後、Shhの腹方化シグナルに応答して運動ニューロン前駆体の同一性を獲得する。
【0128】
BDNF、CNTFおよびGDNFは、MNの成長および生存を支持することが公知のノイロトロピンである。血清、BDNF、CNTF、およびGDNFの存在下で分化させる前に、RAおよびShh−Nで処理したRFT神経球は、20〜40%のMNを生成した。RA処理もShh−N処理もなく、BDNF、CNTF、およびGDNFを有する血清中で平板培養した神経球は、5〜67%のMNを生成し、rFT神経球をShh−NおよびRAで処理することは、MNを生成するためにBDNF、CNTF、およびGDNFの存在下で、それらを平板培養するよりも、より効果的ではないことを示した。RAおよびShhは、胚性幹細胞の分化をMNへ指向させるために重要であるが、胚の発生の間にすでに、ある程度まで尾方化され腹方化されていてもよい出生後rFTから得られたNSCとは、これらのシグナルは関連するものでなくともよい。
【0129】
FTにおけるいくらかのNSCは、血清以外の因子を必要とすることなくMNへと分化する能力を持ち得る。FT由来神経球を血清単独で平板培養した場合、1/3は、40%のMNを生成し、時折のMN発現は、特定の介入なしに起こり得ることを示した。一局面において、FTは、胎仔の尾への神経支配を与える(または、げっ歯類の例においては、最も尾側の尾分節の一過的な神経支配を与える)、脊髄の部分の痕跡のレムナントであることから、このことが起こる。FT「脱分化」の発生上のプロセスがヒトで欠けている場合、新生児は、動かせる尾をもって生まれ得、持続するMN神経支配を示唆する。この状況において、FTから単離されたNSCは、MNのような脊髄に固有の細胞型を生成する能力を有し得る。
【0130】
rFT由来神経球からのMN収量の増加
Shh−Nに加えて、Shhシグナル伝達はまた、小分子アゴニストのHh−Ag1.3により活性化される。胚性幹細胞(ESC)からのMNの生成のために、研究は300〜500nMのShh−Nまたは1〜2μMのHh−Ag1.3を使用した(Wichterle,H.ら、 2002. Cell 110:385−397;Harper,J.M.ら、 2004. Proc Natl Acad Sci U S A 101: 7123−7128;Miles,G.B.ら、 2004. J Neurosci 24:7848−7858;Li,X.J.ら、 2005. Nat Biotechnol 23: 215−221 ;Soundararajan,P.ら、 2006. J Neurosci 26:3256−3268). Wichterleらは、これらの濃度においてShh−NおよびHh−Ag1.3で同一の結果を報告したが、ほとんどの研究は、ESCからMNを生成するために1μMのHh−Ag1.3を使用した。rFT神経球において、Hh−Ag1.3は、Shh−Nと比較した場合、生成されたMNの収量の増加において特に有効であった。Hh−Ag1.3を用いた場合、100%近くのMNが生成され、一方、Shh−Nが加えられた場合、わずか20〜40%のMNが生成した。Hh−Ag1.3は、ESC神経球に比較してrFT神経球におけるMN収量の増加のために選択的に有効のようである。これらの結果は、rFT神経球の一つの予期せぬ、そして優れた特性を強調する。
【0131】
FT神経球由来のGFAP+細胞
GFAPを星状膠細胞のマーカーとして用いたが、しかしながらGFAPは、星状膠細胞様成体幹細胞についてのマーカーでもある。成体哺乳動物において、神経形成星状膠細胞は、側脳室のSVZ、および海馬中の歯状回のやや顆粒状のゾーンにおいてインビボで同定された。神経形成星状膠細胞を、非神経形成星状膠細胞の非常に大きい集団から区別する特徴およびマーカーは、未解明である。rFTから分化したGFAP+細胞は、神経形成および/または非神経形成星状膠細胞である。分化実験において、細胞は、時折GFAPおよびニューロンのマーカーについて二重染色される。指向性のMN分化をうけた神経球由来の細胞は、時折MNR2およびGFAPの両方を発現する。GFAPが、単に非神経形成星状膠細胞マーカーである場合、GFAPと運動ニューロンマーカーの同時発現は驚きである。なぜならば、GFAPはまた、星状膠細胞様成体NSCについてのマーカーであり、二重染色された細胞は、MNの細胞運命に拘束された神経形成星状膠細胞を示し得る。
【0132】
RFT神経球は増殖し、インビトロで継代され得、そしてNPC、ニューロンおよびグリアの収集物に分化する。哺乳動物CNS内の多分化能細胞の発見は、外傷、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症(ALS)、および多発性硬化症を含む多くの現在治療不能のCNS疾患の治療の可能性のためにすばらしい意味を有した。多分化能細胞の供給源としてのFTの発見は、これらの神経学的疾患の自家移植療法の分野における新しい可能性を開く。
【0133】
実施例9:出生後のヒト終糸は、運動ニューロンを生成することが可能な自系多分化能神経球の供給源である
本発明の方法を、年齢18歳までのドナーからヒトNSCを単離するために用いた。これらの細胞は、培養の時間の延長した期間にわたって増殖した神経球を生じた。その神経球を、ニューロンおよびグリアに分化するように誘導した。さらに、それらを、インビトロで横紋筋を刺激することが可能な運動ニューロンを形成するように誘導した。これは、到達可能および消耗可能の両方である多分化能CNS細胞の最初のヒトの供給源であり、そして出生後の組織から得られたヒト神経球由来の運動ニューロンの最初の報告である。本発明は、免疫学的拒絶を回避する自系細胞に基づく移植療法を提供する。
【0134】
ヒトにおいて消耗可能な自系NSCの供給源を探した。その独特の発生歴ならびにパラガングリオーマおよび神経外胚葉性腫瘍を生成するその性質のために、FTを焦点として選択した。FTは、脊柱の基部にある尾骨に脊髄を固定する脊髄の尾方末端の細長い延長(成体で約15cm)である(図1A)。それは、発生初期に胎仔の痕跡尾への神経支配を与えるか、または尾のある脊椎動物において最も尾側の尾分節の一過的な神経支配を与える神経系のレムナントである。初期の段階において、予定運命のFTは、三つの追加の脊髄神経節(C3〜C5)をもつ分化した脊髄である(図1A)。尾が再吸収される時、FTの細胞は、Streeterにより「脱分化」と名付けられたプロセスにより初期の胚段階への逆戻りをうける(図1A、右図)。 その結果は、消失する尾側の点へと狭まり、上衣細胞により裏打ちされ、そして見たところゆるく組織化された線維芽細胞、脂肪細胞、繊毛のない上衣細胞のロゼット、神経芽細胞、ニューロンおよびグリアの収集物で環状に囲まれた中心管を有する、細長い構造物である。この局所的環境は、NSCを生成する他のCNS領域の特性の多くを有する。FTは、外科的に接近可能であり、そして脊髄が動作の十分な自由を欠く「係留脊髄症候群」(TCS)の例において脊髄上で牽引力を軽減するために、慣例的に外科的に切断される。
【0135】
FT組織は、ヒト胎児および出生後の手術より得た。FTは、選択的に人工中絶された14週〜21週齢の胎児から切開した(図1B)。年齢が6ヶ月から18歳の出生後組織を、TCSの神経外科例から得た。組織供給源に関する不明瞭さはなく、全ての外科的FT試料を硬膜鞘内から取得し、そしてFTの同一性を、切除前に臨床的電気生理学を用いて確認した。切片は、3つの出生後のFTから作製し、そしてNSCのマーカーのネスチンの存在について試験した。年齢が8ヶ月〜5歳の3つのFT試料の免疫組織化学が、ネスチン+上衣細胞ならびに分散した神経前駆体細胞の存在を示した。(図1C)。
【0136】
増殖している細胞の浮遊凝集物である神経球を、FTから単離した。組織は、4つの胎児および17つの出生後ドナーから得た。初代組織は、酵素的に分離し、そして神経球の成長を促進する標準的な条件を用いて培養した。インビトロで3〜4日後(DIV)、本発明者らは、胎児の100%および出生後培養の82%に神経球を観察した(図10F)。その神経球は、細胞の小さい塊としてスタートし、そして直径で約25μmから500μm超の球体に成長した。単離した神経球の数は、初代培養物当たり約1から50を超える神経球で変化し、そしてこの存在量は、ドナーの年齢に依存しなかった。それらの増殖および自己再生の能力を示すため、神経球を成功裏に分離し、そして10回まで継代し、そして試みた最長の期間である6ヶ月間インビトロで維持した。8つの培養物を凍結し、そして一つを生存能力について試験し、そして成功裏に回復した。
【0137】
神経球、ならびにニューロンおよびグリアを生成するそれらの能力を特徴付けるため、神経球を種々の免疫細胞学的マーカーの発現について試験した。免疫細胞化学を単一の神経球で実施し、そして各アッセイについて、神経球は、1以上のドナーに由来した。神経球を、種々の神経の幹細胞(NSC)、神経前駆体細胞(NPC)、ニューロンのマーカーおよびグリアのマーカーについて試験した。全ての神経球はネスチン+(n=13)(図10A)であった。より小さい神経球(100μm未満)においては、100%の細胞がネスチン+であったが、一方、より大きな神経球においては、中心部はネスチン−のようであった。試験した全ての神経球はまた、NPCマーカーのビメンチン(n=33)、CD133(n=18)、Olig−2(n=17)およびSox−2(n=17)(図10B〜D)に対して陽性の細胞を含んだ。NPC+細胞の発現パターンおよび割合は、神経球の間で変化した。本発明者らは、神経のタンパク質であるβ−チューブリンIIIを認識するTuj−1、および星状膠細胞マーカーのGFAPについて42の神経球を染色した(図10E)。Tuj−1+およびGFAP+細胞は、全ての神経球中に存在した。加えて、各神経球は、両方のマーカーについて陽性のいくらかの細胞を含み、そしてこの二重に陽性の割合は著しく変化した。図10Eで示したように、異なるマーカーを発現する細胞の空間的な塊形成があった。ほとんどの神経球について、塊形成は明白であったが、一方でそのパターンは変化した。
【0138】
分化した細胞型を生成する神経球の能力を試験するため、単一の神経球を、5%ウシ胎児血清で増殖因子を置換した上記の培地を用いて、ポリ−L−リジンおよびラミニンでコートしたカバーガラス上に平板培養した。その神経球は、年齢が6ヶ月および12歳の2人のドナーから得た。個々の神経球から得た培養物を、平板培養2〜10日後に免疫細胞化学を用いて試験した。分化した細胞が、増殖している細胞に由来したことを確認するために、単一の神経球を、トリチウム化したチミジンと8時間インキュベーションした(n=4)。その神経球をその後、分化させ、そして7日後に試験した。全ての例において、結果として生じるニューロンおよびグリアの顕著な割合(33〜63%)が、それらの核に取り込まれた放射性のヌクレオチドを有した(図5C)。
【0139】
培養物を、ニューロンのマーカーであるニューロン特異的エノラーゼ(n=4)、およびTuj−1(n=19);星状膠細胞マーカーであるGFAP(n=11);乏突起膠細胞マーカーであるO1(n=5);ならびにNPCマーカーであるビメンチン(n=9)、CD133(n=4)、Olig−2(n=3)およびSox−2(n=8)に対する抗体で染色した。各培養物から得た分化した細胞の数は、約50細胞より少ないから5000細胞超までの範囲であって、それは平板培養した神経球のサイズと相関した(n=50)。各神経球により生成された細胞型の割合には大きな変動性があり、それは神経球のサイズまたはドナーの年齢とは相関しなかった。平板培養の2日後、神経球から得た細胞の79+/−8%(n=8)が、ニューロンのマーカーおよびGFAPまたはO1のどちらかについて二重に染まった(図5B)。7〜10日後、二重に染まったものは、23+/−9%(n=7)に減少し、そして培養物の約半分において、ニューロンまたは星状膠細胞のどちらかが優位を占め、神経球の変動する能力を示唆した。分化後10日でネスチン+細胞はなかったが(n=3)、しかしながら細胞の98%が、ニューロンのマーカーまたはグリアのマーカーに加えてNPCマーカーについて陽性で残り(n=21)(図5A)、その細胞が、まだ十分に分化していないことを示した。
【0140】
FTは、脊髄の痕跡の部分を示すことから、本発明者らは、FTの神経球が、脊髄の外傷またはMNの変成の例における細胞置換ストラテジにおいて用いられ得る、脊髄運動ニューロン(MN)を生成することが可能かどうかを決定した。MN前駆体を生成するために、単一の神経球を、2μMのレチノイン酸および0.4〜1μMのソニックヘッジホッグタンパク質(Shh−N)で4〜6日処理した(Wichterle,H.ら、 2002. Cell 110:385−97)。神経球をその後、5%のウマ血清ならびにMNの成長および生存を促進することが公知の3つの神経栄養因子:毛様体由来神経栄養因子、脳由来神経栄養因子、および神経膠由来神経栄養因子(Zurn,A.D.ら、 1996. J Neurosci Res 44:133−41)の存在下で接着基質上で平板培養した。7〜10日後、各神経球により生成されたMNの割合を、MNマーカーのMotor Neuron Restricted−2(MNR−2)(Jessel,T.M. Nat Rev Genet 1:20−9)についての免疫細胞化学を用いて決定した。Shh−Nで処理した神経球は20+/−12%のMNR−2+細胞(n=6)を生成し、そしてShh−N濃度を増加させることは、MNの割合に影響しないようだった(図5D)。興味深いことに、小分子Shh−NアゴニストのHh−Ag1.3(1.5μM)(Frank− Kamenetsky,Mら、 2002. J Biol 1:10;Harper,J.M.ら、 2004. PNAS USA 101:7123−8)を処理に含めた場合、100%の細胞がMNR−2を発現し(n=2)、Hh−Ag1.3が、MN分化の誘導において、より有効であり得ることを示唆した。未処理の神経球を3つの神経栄養因子と平板培養した場合(n=4)、本発明者らは、5〜50%のMNR2+細胞、および血清単独において平板培養した2/3の未処理神経球が陽性細胞を生じたことを検出し(1%および40%)、いくらかの神経球は、加えられた因子の非存在下でMNを生成し得ることを示唆した。MNの同一性を確認するため、本発明者らは追加のMNマーカーについて試験した:Lim−3(n=3)、Islet−1(n=1)、およびコリンアセチルトランスフェラーゼ(n=2)(図5E)(Oda,Y. and Nakanishi,I. 2000. Histol Histopathol 15,825−34;Arber,S.ら、1999. Neuron 23,659−74;Pfaff,S. L.ら、1996. Cell 84,309−20;Tsuchida,T.ら、 1994. Cell 79,957−70)。全ての例において、その結果は、染まった細胞の割合に関して、MNR−2についての結果と類似した。さらに4/4の神経球は、RT/PCR(Pfaff,S.L.ら、 1996. Cell 84:309−20)により決定されたように体細胞の運動ニューロンによって選択的に発現されるホメオボックスタンパク質のHomeobox−9を発現した。
【0141】
免疫細胞化学の基準によりMNとして特徴付けられた細胞が、筋肉を刺激することが可能であるか決定するため、神経球を出生後ラット由来の横紋筋培養物に加えた。神経球を、上述のようにレチノイン酸およびShh−Nで処理し、そしてその後、単一の神経球を各筋肉培養物に加えた。同時培養中のニューロンが、平板培養した神経球に由来したか決定するため、4つの神経球を、平板培養の前に脂肪親和性カルボシアニン色素のDiDで2.5時間プレインキュベーションした。全ての例において、本発明者らは、ニューロンの形態学的な特徴を有するDiD+細胞を検出した。位相差顕微鏡法により、または神経球の付加なしに筋肉培養物におけるTuj−1染色により検出できるニューロンはなかった(n=12)。神経球と6〜21日の同時培養後、神経筋接合部でのニコチン酸アセチルコリンレセプターの塊形成を検出するために、培養物を、蛍光α−アマガサヘビ毒素とインキュベートした。全ての同時培養物は、この基準による神経筋接合部の証拠を示した(n=18)。図11Bは、神経筋接合部および刺激を与えるニューロンを示すために、α−アマガサヘビ毒素およびTuj−1の両方について染色した培養物を示す。筋繊維のみを含む対照培養物は、神経筋接合部を含まなかった(n=12)。
【0142】
ほとんどのFTは、TCSの外科的試料から得たが、実質的に区別できない結果を、中絶した胎児由来のFTをもちいて、および出生後のラット由来のFTでの広範囲な実験から得た。これらの結果は、FT中の多分化能細胞の存在が、正常の条件を反映することを示す。FTは、神経系の外傷または変性の例における使用のための、自系の、消耗可能な、到達可能な多分化能細胞の供給源である。年齢が18歳までのドナーからの、これらの細胞の単離および分化は、それらが成人期まで持続することを示す。
【0143】
HuFTからの神経球の単離および分化
ヒト胎児組織(14〜21週齢)を、ヒト小児組織でと同じように、氷冷したHanks緩衝液中で切開した(図1b)。その組織をDMEM/F12(1:1、Gibco)中で、II型コラゲナーゼ 100U/ml(Gibco)で分散し、そして標準的な幹細胞培地のDMEM/F12、1% N2処方物(Gibco)、1%ペニシリン−ストレプトマイシン溶液(Gibco)、EGF(20ng/ml、Gibco) bGFG(20ng/ml、Gibco)、LIF(10ng/ml)(Weiss,S.ら、 1996. J Neurosci 16,7599−609)中で維持した。2〜4週毎に、神経球を、Accumax(Innovative Cell Technologies)での分離後継代した。神経球の分化を、増殖因子の除去、培地への5〜10%血清の付加およびポリリジンおよび/またはラミニンでコートしたカバーガラスでの平板培養により誘導した。MNへの分化のために、神経球をRA(2uM、Sigma)および0.4〜1μMのShh−N(R&D systems)または1.5μMのHh−Ag1.3(Curis)で4〜6日で処理し、その後、ポリ−L−オルニチン、ラミニンおよびコラーゲンでコートしたカバーガラス上で、BDNF、 CNTFおよびGDNF(Sigma)を含む神経基本培地(Wichterle,H.ら、 2002. Cell 110,385−97)中で7〜10日間、平板培養した。
【0144】
他の実施形態
本発明をその詳細な明細とともに記載したが、先述の明細は説明を意図するものであって、付録の特許請求の範囲により規定される本発明の範囲を制限するものではない。多の局面、利点、および改変は、以下の特許請求の範囲内にある。
【特許請求の範囲】
【請求項1】
神経幹細胞(NSC)が富化された終糸(FT)神経細胞の集団を含む組成物。
【請求項2】
前記集団が、神経球または神経球開始幹細胞を含む、請求項1に記載の組成物。
【請求項3】
少なくとも10%の神経幹細胞を含む、単離されたFT細胞の集団を含む組成物。
【請求項4】
少なくとも30%の神経幹細胞を含む、単離されたFT細胞の集団を含む組成物。
【請求項5】
少なくとも90%の神経幹細胞を含む、単離されたFT細胞の集団を含む組成物。
【請求項6】
出生後の動物からFT−NSCを単離する方法であって、該方法は、該動物からFT組織を提供する工程、該FT組織を分離して、神経球を取得する工程、およびネスチン陽性NSCを回収する工程を包含する、方法。
【請求項7】
請求項6に記載の方法によって取得された、単離されたNSCを含む組成物。
【請求項8】
被験体の神経学的機能を増強または回復させる方法であって、該方法は、単離されたFT細胞の集団を該被験体に投与する工程を包含する、方法。
【請求項9】
神経学的障害を処置する方法であって、該方法は、被験体からFT組織を採取する工程、FT細胞を生体外で培養して、単離されたFT−NSCの富化された集団を生成する工程、および単離されたFT−NSCの該富化された集団を該被験体に投与する工程を包含する、方法。
【請求項10】
前記神経学的障害が、傷害または変性状態を含む、請求項9に記載の方法。
【請求項11】
前記神経学的障害が、脳もしくは脊髄の傷害または脳もしくは脊髄の機能の減損を含む、請求項9に記載の方法。
【請求項12】
前記被験体が、脳卒中に罹患していると診断されるか、または脳卒中に罹患していると疑われる、請求項9に記載の方法。
【請求項13】
FT−NSC由来の多分化能の子孫細胞を含む細胞株。
【請求項14】
出生後の動物由来のFT−NSCを増殖させる方法であって、該方法は、該動物由来の単離されたFT−NSCを提供する工程、および該FT−NSCの増殖または分化を可能にする条件下で、該FT−NSCを培養する工程を包含する、方法。
【請求項15】
前記FT組織が、針穿刺吸引によって取得される、請求項6に記載の方法。
【請求項1】
神経幹細胞(NSC)が富化された終糸(FT)神経細胞の集団を含む組成物。
【請求項2】
前記集団が、神経球または神経球開始幹細胞を含む、請求項1に記載の組成物。
【請求項3】
少なくとも10%の神経幹細胞を含む、単離されたFT細胞の集団を含む組成物。
【請求項4】
少なくとも30%の神経幹細胞を含む、単離されたFT細胞の集団を含む組成物。
【請求項5】
少なくとも90%の神経幹細胞を含む、単離されたFT細胞の集団を含む組成物。
【請求項6】
出生後の動物からFT−NSCを単離する方法であって、該方法は、該動物からFT組織を提供する工程、該FT組織を分離して、神経球を取得する工程、およびネスチン陽性NSCを回収する工程を包含する、方法。
【請求項7】
請求項6に記載の方法によって取得された、単離されたNSCを含む組成物。
【請求項8】
被験体の神経学的機能を増強または回復させる方法であって、該方法は、単離されたFT細胞の集団を該被験体に投与する工程を包含する、方法。
【請求項9】
神経学的障害を処置する方法であって、該方法は、被験体からFT組織を採取する工程、FT細胞を生体外で培養して、単離されたFT−NSCの富化された集団を生成する工程、および単離されたFT−NSCの該富化された集団を該被験体に投与する工程を包含する、方法。
【請求項10】
前記神経学的障害が、傷害または変性状態を含む、請求項9に記載の方法。
【請求項11】
前記神経学的障害が、脳もしくは脊髄の傷害または脳もしくは脊髄の機能の減損を含む、請求項9に記載の方法。
【請求項12】
前記被験体が、脳卒中に罹患していると診断されるか、または脳卒中に罹患していると疑われる、請求項9に記載の方法。
【請求項13】
FT−NSC由来の多分化能の子孫細胞を含む細胞株。
【請求項14】
出生後の動物由来のFT−NSCを増殖させる方法であって、該方法は、該動物由来の単離されたFT−NSCを提供する工程、および該FT−NSCの増殖または分化を可能にする条件下で、該FT−NSCを培養する工程を包含する、方法。
【請求項15】
前記FT組織が、針穿刺吸引によって取得される、請求項6に記載の方法。
【図1】

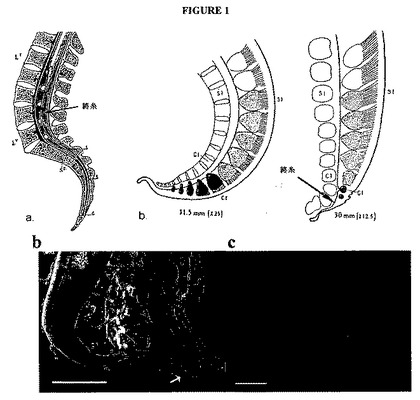
【図2A】

【図2B】

【図3A】

【図3B】

【図3C】

【図3D】

【図4A】
【図4B】

【図4C】

【図4D】

【図5】
【図6】
【図7a】

【図7b】

【図7c】

【図7d】

【図7e】

【図8a】

【図8b】

【図8c】

【図8d】

【図8e】

【図8f】

【図9】
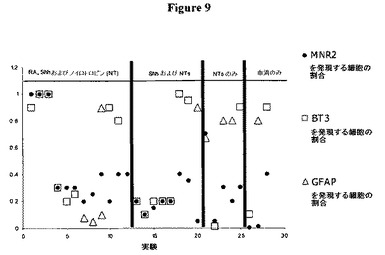
【図10】
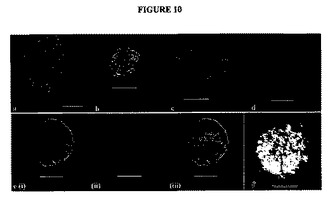
【図11】
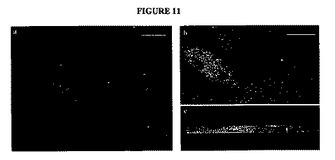
【図12】
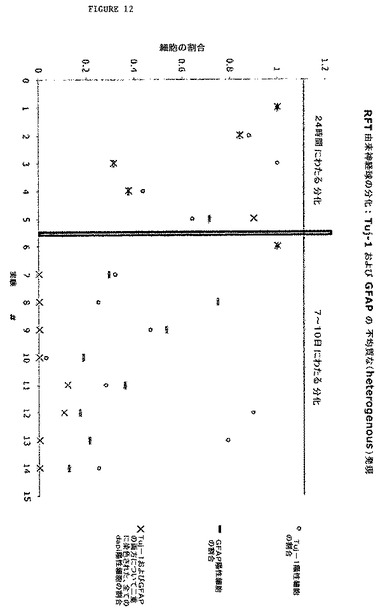
【図13】
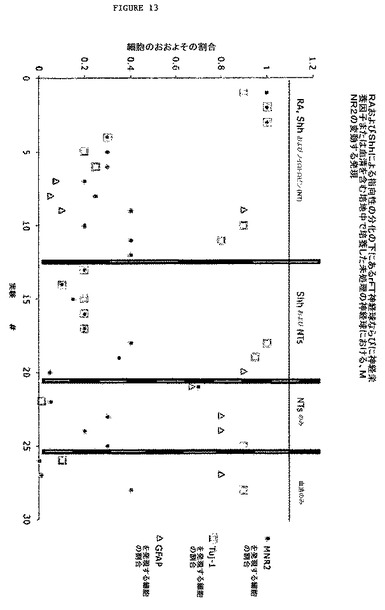
【図2A】
【図2B】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
【図4B】
【図4C】
【図4D】
【図5】
【図6】
【図7a】
【図7b】
【図7c】
【図7d】
【図7e】
【図8a】
【図8b】
【図8c】
【図8d】
【図8e】
【図8f】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2010−531663(P2010−531663A)
【公表日】平成22年9月30日(2010.9.30)
【国際特許分類】
【出願番号】特願2010−514835(P2010−514835)
【出願日】平成20年6月27日(2008.6.27)
【国際出願番号】PCT/US2008/008069
【国際公開番号】WO2009/002559
【国際公開日】平成20年12月31日(2008.12.31)
【出願人】(502072134)プレジデント アンド フェロウズ オブ ハーバード カレッジ (92)
【氏名又は名称原語表記】President and Fellows of Harvard College
【Fターム(参考)】
【公表日】平成22年9月30日(2010.9.30)
【国際特許分類】
【出願日】平成20年6月27日(2008.6.27)
【国際出願番号】PCT/US2008/008069
【国際公開番号】WO2009/002559
【国際公開日】平成20年12月31日(2008.12.31)
【出願人】(502072134)プレジデント アンド フェロウズ オブ ハーバード カレッジ (92)
【氏名又は名称原語表記】President and Fellows of Harvard College
【Fターム(参考)】
[ Back to top ]