
神経発生およびその調節に関連する遺伝子、方法および組成物
本開示は、神経発生、神経細胞増殖および分化を調べるための方法を提供する。具体的に、本開示は、神経発生および神経細胞増殖を調節し得る薬剤を同定する方法、神経発生および神経前駆細胞の増殖を調節する遺伝子をスクリーニングする方法、ならびに神経発生および神経増殖または分化の候補調節因子としての薬剤を同定する方法に関する。本開示はまた、神経発生を特徴づけおよび調節する薬剤を同定する方法、かかる方法により同定された薬剤、かかる薬剤で患者を治療する方法、ならびにかかる薬剤を含む組成物に関する。したがって、本方法は、健常な動物および神経系の障害において神経発生、脳の発生および機能を調節する機構の解明を可能にする。さらに、本方法は、認知障害を含む種々の神経系障害における損なわれた神経発生を予防、改善または安定化する組成物の開発を容易にする。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本願は、その全体において参照により本明細書に明示して援用される2009年9月29日に出願された米国仮特許出願第61/246,967号に対する35 U.S.C. § 119による優先権を主張する。
【0002】
配列表
本願は、ASCII形式で、EFS-Webを介して提出され、その全体において参照により本明細書に援用される配列表を含む。2010年9月28日に作成された前記ASCIIコピーは、21RE6840.txtと称し、2,011,398バイトのサイズである。
【0003】
発明の分野
本発明は、神経発生、特に中枢神経系の神経発生の活動性依存性調節に関連する遺伝子、方法および組成物に関する。より具体的には、本発明は、神経発生に関連する遺伝子の同定および操作の方法、ならびに神経発生を調節する医薬剤のスクリーニングおよび評価の方法に関する。
【背景技術】
【0004】
発明の背景
神経発生は、神経系の発生および成熟の基礎となる複雑な課程である。この過程は、細胞の増殖、生存、分化および移動の適切な時間空間的な制御に依存する。新たに生成された神経細胞は、中枢神経系の機能的細胞に分化し得、脳の神経回路に組み込まれ得る。さらに、多くの動物の脳において、新しい神経細胞は、生物の生涯を通じて継続的に生成される。例えば、神経発生は、現在、成体期を通じて、哺乳動物の脳の2つの領域:側脳室の脳室下(subventricular)帯(SVZ)および海馬の歯状回において続くことが分かっている。これらの領域では、多能性神経前駆細胞(NPC)は、分裂を続け、新しい機能的ニューロンおよびグリア細胞を生じる(Jacobs, Mol. Psychiatry 2000, 5(3): 262-9)。そのため、神経発生の調節は、CNSの領域的分化(specialization)および機能的回路を形成する特異的細胞型の確立の基礎となる。
【0005】
神経発生は、CNS生理学において重要な役割を担う。神経前駆細胞(NPC)のプールは、さらなるNPCを生じる対称分裂により増殖され得るか、または子孫の、ニューロンもしくはグリアへの最終的な分化により枯渇され得る(Butt et al., Neuron 2005, 48: 591-604;Gotz et al., Nat Rev Mol Cell Biol 2005, 6: 777-788;Huttner et al., Curr Opin Cell Biol 2005, 17: 648-657;Noctor et al., Arch Neurol 2007, 64: 639-642;Kriegstein et al., Ann Rev Neurosci 2009, 32: 149-184)。そのためNPCを活発に増殖する状態に維持することにより、前駆細胞プールを拡大することができ、マウスの大脳皮質を大きくすることができる(Chenn et al., Science 2002, 297: 365-369;Lehmann et al., Eur J Neurosci 2005, 21: 3205-3216)。逆に、増殖状態から分化状態へのNPCの成熟前の移行により、前駆細胞プールを枯渇することができ、短期間で、分化したニューロンおよびグリアの生成を増加することができるが、長期間では、前駆細胞プールの枯渇により、神経子孫のさらなる生成が制限される。NPCの生存を増加するかまたは減少することにより、皮質のサイズを変化させることもできる。(Depaepe et al., Nature 2005, 435: 1244-1250;Putz et al., Nat. Neurosci. 2005, 8: 322-331)。
【0006】
神経発生の破壊は、CNSの疾患および障害において根本的な役割を担い得る。神経細胞の数は、NPCの増殖ならびにその子孫の生存および分化により大体が決定される。これらの工程は独立して制御され得るが、神経系中で適切に機能する回路を確立するためには、これらの工程はまた、適切に調和されなければならない。実際に、ニューロンの生成およびそれらの回路への組み立てにおける誤りは、異所性、精神遅滞、自閉症スペクトラム障害、癲癇、限局性皮質形成異常を含む多くの神経学的障害を引き起こし得る。神経細胞の種類の生成における変化は、自閉症スペクトラム障害、鬱および統合失調症の基礎となり得る、脳における興奮性回路と抑制性回路との異常な割合、不均衡を引き起こし得る。
【0007】
これらの観察は、NPCの増殖を調節する機構およびNPCの子孫の運命の重要な役割を強調する。細胞増殖を制御する機構は、細胞自律的または非細胞自律的に分類され得る。細胞自律的機構に関して、細胞を「多能性(pluripotent)/多能性(multipotent)」状態へと押し進め、その状態に維持するために必要な因子の同定において、近年、大きな進歩がなされた。(Welstead et al., Curr. Opin. Genet. Dev. 2008, 18: 123-129)。
【0008】
しかしながら、インビボにおいて神経活動性に依存する非細胞自律的な機構はあまりよく分かっていない。そのため、インタクトの動物において神経発生を調節する機構を解明することは、脳の発生ならびに健常動物および神経系の障害における機能の適切な理解に重要である。このことはまた、神経系、具体的には認知障害などの神経系の障害における神経発生、具体的には損なわれた神経発生を予防、改善および/または安定化(例えば調節)するための方法および組成物の開発にも重要である。
【0009】
本発明は、活動性依存的様式における神経発生の調節に関連する標的、方法および組成物を提供することにより、当該技術分野におけるこれらのおよび他の必要性を満たす。
【発明の概要】
【0010】
発明の概要
第一の局面において、本開示は、第一の動物のインタクトの脳の領域中の神経前駆細胞と医薬剤とを接触させる工程、第一の動物および第二の対照動物を、インタクトの脳の領域において活動性を誘発し得る外的刺激に曝露する工程、ならびに第一の動物における神経前駆細胞の増殖割合および第二の動物における神経前駆細胞の増殖割合を任意の順序で測定する工程を含む方法に関し、ここで神経前駆被検細胞と神経前駆対照細胞の間の増殖の差により、神経増殖を調節し得るものとしての該医薬剤が同定される。
【0011】
特定の態様において、第一の動物および第二の動物は、両生類および哺乳類などの脊椎動物であり得る。より具体的には、第一の動物および第二の動物はアフリカツメガエルであり得、より具体的にはアフリカツメガエルのオタマジャクシであり得る。
【0012】
特定の態様において、インタクトの脳の領域は、嗅覚入力、視覚入力もしくは機械感覚入力の処理に関与し得るか、または行動出力の媒介に関与し得る。具体的な態様において、第一および第二の動物はアフリカツメガエルであり得、インタクトの脳の領域は、視蓋であり得る。インタクトの脳の領域はまた、菱脳、中脳、後脳/脊髄、網膜または鼻窩(olfactory pit)の回路を含み得る。
【0013】
特定の態様において、実験動物および対照動物中の神経前駆細胞の増殖割合を測定する工程は、第一の動物および第二の動物の視蓋中の細胞の数および型を計測することを含む。
【0014】
いくつかの態様において、神経前駆細胞と医薬剤を接触させる工程は、前記医薬剤を前記神経前駆細胞にエレクトロポレーションすることを含み得る。
【0015】
別の局面において、本開示は、神経前駆被検細胞と、前記神経前駆被検細胞中の1つ以上の遺伝子の発現を調節するのに有効な量の医薬剤とを接触させる工程、神経前駆被検細胞の増殖割合および医薬剤と接触させていない神経前駆対照細胞の増殖割合を測定する工程、ならびに神経前駆被検細胞の増殖割合および神経前駆対照細胞を任意の順序で比較する工程を含む方法に関し、ここで神経前駆被検細胞と神経前駆対照細胞の間の増殖割合の差は、神経前駆細胞の増殖の調節物質として該1つ以上の遺伝子を同定する。
【0016】
特定の態様において、神経前駆被検細胞は、第一の動物中に存在し得、神経前駆対照細胞は、第二の動物中に存在し得る。いくつかの態様において、神経前駆被検細胞および神経前駆対照細胞は、それぞれ、第一の動物および第二の動物のそれぞれの視蓋中に存在し得る。一態様において、第一の動物および第二の動物はアフリカツメガエルであり得る。
【0017】
いくつかの態様において、方法はさらに、神経前駆被検細胞および神経前駆対照細胞中に、レポーター構築物を導入する工程を含み得る。特定の態様において、レポーター構築物は、蛍光タンパク質をコードする遺伝子を含み得る。一態様において、蛍光タンパク質の発現は、空間的に、特に神経前駆細胞などの特定の細胞型に制限され得る。別の態様において、蛍光タンパク質の発現はまた、時間的に制限、例えば特定の時点後に脳の領域で生成された子孫細胞に制限され得る。他の態様において、レポーター構築物を神経前駆被検細胞に導入する工程は、細胞を、レポーター構築物をコードするプラスミドでトランスフェクトすることを含み得る。
【0018】
いくつかの態様において、神経前駆細胞の増殖割合を測定する工程は、少なくとも1つの所定時間の前および後に細胞の数および型を計測することを含み得る。
【0019】
別の態様において、方法はさらに、第一の動物および第二の動物を視覚刺激に曝露することを含み得る。
【0020】
いくつかの態様において、医薬剤は、化合物またはアンチセンスオリゴヌクレオチドを含み得る。特定の態様において、アンチセンスオリゴヌクレオチドは、siRNA、shRNAおよび/またはモルホリノを含み得る。
【0021】
本方法の別の態様において、神経前駆被検細胞中の1つ以上の遺伝子は、配列番号:1〜651、あるいはその機能的短縮体、修飾体および/または置換体から選択され得る。
【0022】
別の局面において、本開示は、神経前駆被検細胞と医薬剤を接触させる工程、神経前駆被検細胞の増殖割合および該医薬剤と接触させていない神経前駆対照細胞の増殖割合を測定する工程、神経前駆被検細胞の増殖割合と神経前駆対照細胞の増殖割合を、任意の順序で比較する工程を含む方法に関し、ここで、神経前駆被検細胞と神経前駆対照細胞の間の増殖割合の差は、増殖を調節し得るものとしての該医薬剤を同定する。
【0023】
いくつかの態様において、方法は、神経前駆被検細胞および神経前駆対照細胞にレポーター構築物を導入する工程を含み得る。特定の態様において、レポーター構築物は、蛍光タンパク質をコードする遺伝子を含み得る。いくつかの態様において、蛍光タンパク質は、神経前駆細胞中で特異的に発現し得る。
【0024】
いくつかの態様において、神経前駆細胞にレポーター構築物を導入する工程は、神経前駆細胞を、レポーター構築物をコードするプラスミドでトランスフェクトすることを含む。
【0025】
いくつかの態様において、神経前駆被検細胞と医薬剤を接触させる工程は、神経前駆被検細胞に医薬剤をエレクトロポレーションすることを含む。
【0026】
特定の態様において、方法はさらに、第一の動物および第二の動物を視覚刺激に曝露することを含み得る。
【0027】
第四の局面において、本開示は、配列番号:1〜651からなる群より選択される標的遺伝子あるいはその機能的短縮体、修飾体および/または置換体を発現する被検細胞に医薬剤を投与する工程、医薬剤を投与していない被検細胞中の標的遺伝子の発現に対して(compared with)、医薬剤を投与した被検細胞中の標的遺伝子の発現を、任意の順序で比較する工程を含む方法に関し、ここで、医薬剤を投与していない被検細胞と比較した、医薬剤を投与した被検細胞中の標的遺伝子の発現の差は、神経の増殖または分化の候補調節物質として該医薬剤を同定する。
【0028】
別の局面において、本開示は、本明細書に記載の方法により同定される医薬剤に関する。
【0029】
さらに別の局面において、本開示は、本明細書に記載される方法により同定される医薬剤を含む医薬組成物に関する。
【0030】
さらに別の局面において、本開示は、本明細書に記載される方法により同定される(identified be)化合物を投与する工程を含む、患者を治療する方法に関する。
【0031】
本発明は、神経細胞の増殖および分化の現象を調査する方法、かかる方法により同定された医薬剤、該医薬剤を含む組成物、ならびにかかる医薬剤または組成物を投与する工程を含む治療方法を含む。したがって、本開示は、神経発生の制御に関与する遺伝子を同定する方法、神経発生を特徴付けて調節するための医薬剤を同定する方法、ならびに神経系、具体的には種々の神経系の障害および/または損傷に対する調節物質および/または治療、例えば認知障害を含む神経系、具体的には神経系の障害における神経発生、具体的には損なわれた神経発生を予防、改善および/または安定化(すなわち調節)する方法および組成物を提供する。本発明はまた、本発明の方法により選択される医薬剤、およびかかる選択された医薬剤を含む医薬組成物、ならびにかかる医薬剤および組成物を患者に投与する方法であって、患者はヒト患者を含み、前記投与は患者、具体的にはヒトにおいて、神経系、具体的には認知障害を含む神経系の障害における、神経発生を調節、具体的には神経発生、具体的には損なわれた神経発生を予防、改善および/または安定化する目的のためである方法を含む。
【図面の簡単な説明】
【0032】
【図1】図1は、アフリカツメガエルオタマジャクシの透明な脳(A);脳の視蓋領域(B)ならびに視蓋葉(tectal lobe)中の細胞の増殖および分化(C)を示す。
【図2】図2は、神経前駆細胞(NPC)の種々の系統(lineage)を示す模式図である。
【図3】図3は、インタクトの脳で画像化された標識細胞の空間的および時間的解像を容易にする増殖レポーターの模式図である。
【図4】図4は、アフリカツメガエルの視蓋における24時間にわたるNPCの増殖を示す画像である。
【図5】図5は、細胞分裂ブロッカーへの曝露後の、蓋(tectal)細胞の増殖割合の減少を示すグラフである。
【図6】図6は、1日目に視覚刺激なしおよび2日目に視覚刺激ありで、細胞分裂ブロッカーの存在下または非存在下での蓋増殖割合(A);ならびに3日目に細胞分裂ブロッカーの存在下または非存在下での蓋ニューロンのパーセント(B)を示すグラフである。
【図7】図7は、対照動物と比較した、視覚刺激ありおよびなしの脱ヨード化酵素ヨードサイロニンIII型(Deiodinase iodothyronine type III)に対するモルホリノ(dio3-MO)を発現する動物における蓋増殖割合を示すグラフである。
【図8】図8は、対照動物と比較した、3日間にわたるグルタチオンS-トランスフェラーゼπ1に対するモルホリノ(GSTπ-MO)を発現する動物における蓋ニューロンおよびグリア細胞のパーセントを示すグラフである。
【図9】図9は、対照動物と比較した、3日間にわたる11の目的の遺伝子(GOI)の1つに対するモルホリノを発現する動物における、蓋増殖割合(A)ならびに蓋ニューロンおよびグリア細胞のパーセント(B)を示すグラフである。GOI:熱ショックタンパク質70(HSPA5);Ephrin受容体B-1型(EphRB1);脱ヨード化酵素ヨードサイロニンIII型(Dio3);ETSドメイン含有タンパク質Elk-4(ELK4);ウイングレスMMTV型(マウス乳癌ウイルス)組み込み部位ファミリー、メンバー7b(Wnt7b);脆弱X精神遅滞、常染色体ホモログ(FXR1);脆弱X精神遅滞タンパク質1(FMR1A);マトリックスメタロペプチダーゼ9(MMP9)、cAMP-依存型タンパク質キナーゼ触媒サブユニットα(PRKACA)、Neuritin 1-A(cpg15);グルタチオンS-トランスフェラーゼπ1(GSTπ)。
【発明を実施するための形態】
【0033】
発明の詳細な説明
そうではないと定義されない限り、本明細書に使用される全ての科学技術用語は、当業者に一般的に理解されるものと同じ意味を有する。本明細書に記載されるものと同様または同等の方法および材料を、本発明の実施または試験に使用できるが、適切な方法および材料が本明細書に記載される。本明細書で言及される全ての刊行物、特許出願、特許および他の参考文献は、その全体において参照により援用される。また、材料、方法および実施例は、例示のためだけであり、限定を意図しない。
【0034】
本発明に包含される基本的な技術を実施するための定義および方法および手段を含む、標準的な実験マニュアルが参照される。例えば、Sambrook et al., Molecular Cloning: A Laboratory Manual, 第3版、Cold Spring Harbor Laboratory Press, New York (2001);Current Protocols in Molecular Biology, Ausubel et al. (編), John Wiley & Sons, Inc., Hoboken, NJ (2010)、Current Protocols in Cell Biology, Bonifacino et al. (編), John Wiley & Sons, Inc., Hoboken, NJ (2010);Current Protocols in Immunology, Coligan et al. (編), John Wiley & Sons, Inc., Hoboken, NJ (2010);Current Protocols in Neuroscience, Gerfen et al. (編), John Wiley & Sons, Inc., Hoboken, NJ (2010);Current Protocols in Nucleic Acid Chemistry, Egli et al. (編), John Wiley and Sons, Inc., Hoboken, NJ (2010);Current Protocols in Pharmacology, Enna et al. (編), John Wiley and Sons, Inc., Hoboken, NJ (2010);およびそこに引用される種々の参考文献を参照されたい。
【0035】
定義
本開示で使用する場合、用語「約」または「およそ」は、当業者により決定される場合の特定の値について許容され得る範囲内にあることを意味し、部分的に、どのようにして値を測定または決定したか、例えば測定系または技術の限界に依存し得る。
【0036】
用語「a」、「an」および「the」は、そうではないと明確に記述されない限り、単数形および複数形の両方を意味すると理解される。したがって「a」、「an」および「the」(および適切な場合はその文法上の変形物)は、一般的に1つ以上のことをいう。
【0037】
本明細書で使用する場合、「神経発生」は、インビボ、インビトロまたはエキソビボにおける神経細胞の増殖、生存、分化および移動を含む。細胞は、中枢神経系または動物もしくはヒト内の他の場所(例えば末梢神経系)に局在し得るか、そこから得られ得るか、またはそこから由来し得る。神経発生は、正常発生中に起こるもの、および疾患、損傷または治療介入後に起こる神経再生としての神経発生を含むことが意図される。開示される発明の態様は、神経発生の非限定的な指標としての増殖または分化のいずれかの検出または測定を含む。
【0038】
「外的刺激」は、神経活動性を誘導し得る単純または複雑な細胞外刺激の任意の型を含むように広く定義される。したがって、外的刺激は、動物の視覚系への入力を含む。外的刺激はまた、嗅覚、機械感覚または視覚入力の処理および行動出力の媒介に関与するものなどの脳の他の領域への入力も含む。
【0039】
本明細書で使用する場合、用語「調節」は、遺伝子の発現、またはRNA分子もしくは同等のRNA分子、例えば非コードRNAおよび1つ以上のタンパク質もしくはタンパク質サブユニットをコードするもののレベルを変化させることを含む。「調節」はまた、調節物質の存在下で発現、レベルまたは活性が、調節物質の非存在下で観察されるものと異なるように、1つ以上の遺伝子産物(非コードRNAを含む)、タンパク質またはタンパク質サブユニットの活性を変化させることも含む。例えば、用語「調節」は、「上方調節」または「下方調節」を意味し得るが、単語「調節」の使用はこれらの定義に限定されない。調節は、発現もしくは活性の増加または減少、遺伝子産物の結合特性の変化、または生物学的活性分子の生物学的、機能的もしくは免疫学的性質の任意の他の変化であり得る。
【0040】
「神経発生の調節」または「神経発生を調節すること」は、細胞の増殖、生存、分化または移動における変化を含む。かかる変化は、インタクトの脳の領域中のものなどの細胞または細胞の集団において起こり得る。非限定的な例としては、NPC増殖に直接関連する遺伝子産物のレベルの変化などの神経発生の誘導体(または阻害剤)のレベルの増加(または減少)が挙げられる。かかる変化はまた、神経回路中の細胞分化または細胞移動の違いも含み得る。特定の態様において、神経発生を調節することは、細胞増殖および細胞運命(例えばニューロン 対 グリア)における影響のことをいう。
【0041】
用語「調節物質」、「化合物」および「医薬剤」は、本明細書において互換的に使用され得、単離された形態の薬理学的に活性な物質またはその混合物を含む。例えば、医薬剤、化合物または調節物質は、単離され構造的に画定された生成物、未知の構造の単離された生成物、いくつかの公知かつ特徴付けられた生成物の混合物、または1つ以上の生成物を含む画定されていない組成物であり得る。かかる画定されていない組成物の例としては、例えば組織試料、生物学的液体(fluid)、細胞上清、植物性調製物(vegetal preparation)等が挙げられる。医薬剤、化合物または調節物質は、ポリペプチド(またはタンパク質もしくはペプチド)、核酸、脂質、多糖、化学実体、またはそれらの混合物もしくは誘導体などの有機または無機の任意の生成物であり得る。医薬剤、化合物または調節物質は、天然または合成起源であり得、化合物(1つまたは複数)または調節物質は化合物のライブラリーを含み得る。
【0042】
「調節物質」、「化合物」または「医薬剤」は、インビボ、インビトロまたはエキソビボにおいて、該剤または試薬の非存在下の神経発生の量、程度または性質と比べて、神経性応答(neurogenic response)の量、程度または性質を増加(または減少)し得る。特定の態様において、かかる「神経性」剤による処置は、神経発生の検出または決定に使用される方法の条件下で、神経性応答の量、程度または性質を、該剤の非存在下の神経性応答の量、程度または性質と比較して、少なくとも約1%、2%、3%、4%、5%、10%、20%、40%、50%、75%、100%、200%(2倍)、300%(3倍)、400%(4倍)、500%(5倍)またはそれ以上もしくは以下だけ増加(または減少)し得る。
【0043】
用語「阻害」、「下方調節」または「低減」は、遺伝子の発現、または1つ以上のタンパク質もしくはタンパク質サブユニットをコードするRNA分子もしくは同等のRNA分子のレベル、または1つ以上の遺伝子産物、タンパク質もしくはタンパク質サブユニットの活性を、本発明の方法において規定されるような1つ以上の調節物質(例えば、siRNA、shRNA、アンチセンスモルホリノ等)の非存在下で観察されるもの未満に減少することを含む。用語「促進」または「上方調節」は、遺伝子の発現、1つ以上のタンパク質もしくはタンパク質サブユニットをコードするRNA分子もしくは同等のRNA分子のレベル、または1つ以上の遺伝子産物、タンパク質もしくはタンパク質サブユニットの活性を、特許請求される方法において規定されるような1つ以上の調節物質の非存在下で観察されるもの未満(below)に増加することを含む。
【0044】
用語「標的遺伝子」または「目的の遺伝子(gene or interest)」は、タンパク質コード遺伝子だけではなく(not just)、非コード遺伝子を含む。かかる非コード遺伝子は、リボソームRNA(rRNA)、転移RNA(tRNA)および核内低分子RNA(snRNA)、ならびにマイクロRNA、snoRNA、siRNA、piRNAおよびncRNAをコードするものを含む。標的遺伝子はまた、複製、転写、翻訳もしくは遺伝子産物の発現に重要な他の過程を制御するポリヌクレオチド領域、または遺伝子産物をコードする領域およびそれに作動可能に連結された、発現を制御する領域の両方を含むポリヌクレオチドを含み得る。標的化遺伝子は、染色体(ゲノム)または染色体外であり得る。標的化遺伝子は細胞に対して内在するもの(endogenous)であり得るか、または外来遺伝子(トランスジーン)であり得る。外来遺伝子は、宿主ゲノムに取り込まれ得るか、またはプラスミドもしくはコスミドなどの染色体外の遺伝子構築物上に存在し得る。標的化遺伝子はまた、生物体または細胞に感染し得るウイルス、細菌、真菌または原生動物などの病原体由来であり得る。標的遺伝子は、ウイルス遺伝子およびプロウイルスの遺伝子であり得る。特定の態様において、標的遺伝子は、神経発生に重要な細胞活動性の進行に関与するかまたは関連するものである。
【0045】
用語「標的核酸」は、その発現または活性が調節される任意の核酸配列を含む。標的核酸は、DNAまたはRNAであり得る。さらに、標的遺伝子または遺伝子は、一般的に生物学的に活性である、より大きな核酸配列の断片を含み得る。
【0046】
添付の図面および実施例により図示され、それらに関連して記載される本開示のいくつかの態様、実施例をここで参照する。本明細書には特定の態様が記載されるが、記載される態様は、添付の特許請求の範囲により規定される本発明の範囲を限定することを意図しないことが理解される。反対に、本開示は、添付の特許請求の範囲に規定される発明の精神および範囲に含まれ得る変形物、改変物および均等物を包含することが意図される。さらに、本開示において、添付の特許請求の範囲に規定される発明の完全な理解を伝達するために、特定の詳細が提供される。しかしながら、これらの詳細なく、特定の態様が実施され得ることが当業者には明らかである。特定の例において、周知の方法、手順または他の具体的な詳細は、添付の特許請求の範囲に規定される発明の局面を不必要に不明瞭にすることを避けるために記載されていない。
【0047】
神経発生に関連する遺伝子の同定および特徴づけの方法
モデル系としてのアフリカツメガエル
アフリカツメガエルは、神経発生および脳の発生のインビボ研究に有利であることが明らかにされている。これらの利点にはいくつかの要因がある:
【0048】
神経発生の密な(compact)期間
カエルのオタマジャクシは、CNS発生の幼生期間の間にわたる、比較的長く利用可能な細胞増殖および分化の期間を有する。アフリカツメガエルの発生の前変態期(pre-metamorphic stage)の経過中に、細胞増殖により新しいニューロンが生成される。新たに形成されたニューロンは、その後発生中のオタマジャクシの脳の機能的な回路に組み込まれる。誕生から個々のニューロンの分化までの神経発生の流れは、アフリカツメガエルでは2〜4日の期間に捕えられ得るのに対して、哺乳動物系では1ヶ月を超える。
【0049】
この密な期間は、神経発生を含む種々の段階の詳細な調査を容易にする。この利点は、神経発生の機構が進化的に保存されているという証拠と合わせると、哺乳動物系に関連のある神経発生の基本的な機構を明らかにすることにおいてだけでなく、ヒトの神経発生的疾患を研究するための実験モデル系を提供することにおいてもツメガエル属の価値を強調する。
【0050】
利用可能な脳回路
カエルにおける神経発生および回路の形成における研究により、その後ヒトを含む哺乳動物において機能的であることが示されたいくつかの脳発生の基本的な機構が明らかにされた。CNSは、アフリカツメガエルの前後軸に沿って高度に局所化(regionalize)され、種々のCNS領域は、感覚的入力処理、情報統合、記憶、意思決定、および運動調節を含む特有の機能を有する、特定の神経回路により構成される。
【0051】
CNSの脳室層における神経前駆体の前後パターン形成は、脳機能のその後の領域分化(region specilization)の基礎となり、神経前駆体中のOtx、PaxおよびHoxなどの転写因子ファミリーの進化的に保存された発現パターン、次いで前駆体およびその神経子孫の領域的サブセットを画定する転写カスケードの活性化により確立されると考えられている(O'Leary et al., Curr. Opin. Neurobiol. 2008, 18: 90-100;Lichtneckert et al., Adv. Exp. Med. Biol. 2008, 628: 32-41)。
【0052】
実際に、Hoxファミリー転写因子は、菱脳(telecephalon)から脊髄への前後軸に沿って前駆体および細胞運命を、調節するだけでなく、ニューロン分化の際に下流の転写因子カスケードを調節することにより発生中の回路におけるニューロン間の接触を指定(specify)もする。(Dasen et al., Curr. Top. Dev. Biol. 2009, 88: 169-200;Dalla Torre di Sanguinetto et al., Curr. Opin. Neurobiol. 2008, 18: 36-43)。
【0053】
本発明の態様は、種々の脳回路および細胞型において、トランスクリプトームを分析するための方法を包含する。これらは、菱脳、中脳および後脳/脊髄、ならびに網膜および鼻窩を含む。これらの領域のそれぞれは、NPC子孫の別個の組によって特徴付けられる。例えば、菱脳は、嗅覚入力を受けて処理し、記憶および運動調節に関与する海馬および基底核に相同な領域を含む(includes regions includes regions)(Maier et al., J Chem Neuroanat, 40(1): 21-35;Brox et al., J Comp Neurol 2004, 474(4): 562-77)。中脳は、機械感覚および視覚入力を処理し(Hiramoto et al., Dev Neurobiol 2009, 69(14): 959-71;Deeg et al., J Neurophysiol 2009, 102(6): 3392-404)、菱脳および脊髄は、行動出力を媒介する(Soffe et al., J Physiol 2009, 587(Pt 20): 4829-44;Orger et al., Nat Neurosci 2008, 11(3): 327-33)。
【0054】
本発明の方法は、別個の脳領域の分析だけでなくかかる回路中の特殊化された細胞型の分析も包含する。それぞれの脳の領域における別個の回路は、領域特異的な回路に特定の性質を付与する、興奮性ニューロンおよび抑制性ニューロンと細胞特異的転写組成(composition)との特有の組合せにより構成されると考えられている。したがって、本発明の方法は、本明細書に記載された脳領域におけるGABA作動性ニューロンおよびグルタミン酸作動性ニューロンなどの同定されたニューロンのトランスクリプトームの分析を含む。
【0055】
いくつかの態様において、方法は、アフリカツメガエルの視蓋領域に関し得る。視蓋は、非哺乳類脊椎動物において主要な視覚中枢であり、中脳(または中脳(mesencephalon))の主要な構成要素を形成する対になった構造をとっており、局所的に(topographically)順序だった様式で網膜繊維からの入力を受け取る。例えば、Dingwell et al., J. Neurobiol. 2000, 44: 246-259参照。
【0056】
直接画像化
NPC増殖および分化は、前変態期(オタマジャクシ段階)のアフリカツメガエルにおいて直接的に観察され得る。図1Aは、脳を四角で示したオタマジャクシの透明な頭部を示す。図1Bは、視蓋を含む脳の領域のより詳細な図である。図1Cは、2時間および6日の時点の視蓋における神経細胞の相対的な位置(BrdUにより可視化)を示し、新たに生成された細胞は、ニューロンへと分化し、該ニューロンは脳室層から離れて移動する。(Wu et al., J. Neuroscience 1999, 19(11): 4472-4483)。
【0057】
したがって、ツメガエル属のオタマジャクシは、インビボにおける時間差画像化で分析できるので、神経前駆細胞(NPC)およびその子孫をインタクトの動物において同定および画像化し得る。これに関して、NPCは、図2に示すように、種々のモードで分裂し得る未分化の放射状グリアである。第一のモードにおいて、単一のNPCは、分裂して2つの娘NPCを形成し得る(例えば「増殖モード」)。第二のモードにおいて、単一のNPCは、分裂して1つの娘NPCおよび1つの娘ニューロンを形成し得る(例えば「混合モード」)。第3の様式において、単一のNPCは、分裂して、2つの娘ニューロンを形成し得る(「分化モード」または「最終様式」。Kriegstein et al., Ann Rev Neurosci 2009, 32: 149-184参照)。
【0058】
神経発生レポーター
かかる画像化により、形態学および発生段階に基づいた異なる細胞型の同定および分析のための方法の基礎が提供される。本願は、かかる分析を容易にするための多数の細胞レポーターの使用を包含する。例えば、かかるレポーターは、神経前駆細胞、およびGABA作動性ニューロンもしくはグルタミン酸作動性ニューロンなどの分化した細胞の異なる集団の(or)タグ化および時間差モニタリングを可能にする。
【0059】
したがって、いくつかの態様において、本方法は、分裂中のNPC細胞に特異的なレポーターを使用し得る。例えば、レポーターは、二成分(binary)Gal4-UAS(上流活性化配列)レポーター系を含み得る(Hartley et al., Proc Nat Acad Sci 2002, 99(3): 1377-1382)。図3は、2つの構成要素:Gal4ドライバー(driver)およびUAS-レポーターを含む例示的なGal4-UASレポーター系を示す。したがって、Gal4の発現を調節する配列は、UAS-レポーターの発現を指令する。
【0060】
図3に示される例において、Gal4の調節領域は、線維芽細胞増殖因子4(FGF4)遺伝子のプロモーター由来の多数のエンハンサー因子を含む。この調節領域の活性化には、増殖中のNPCにおいて発現されるが、分化した非増殖ニューロン中には有意なレベルでは存在しない内因性のsox2/oct3転写因子への結合を必要とする。そのため、この系により、増殖中のNPCにおけるUAS-レポーターの特異的な検出が可能になる。
【0061】
レポーターは、任意の目的のマーカーを含み得る。図3に示されるものなどのいくつかの態様において、レポーターは、蛍光タンパク質 (例えば、Kaede蛍光タンパク質または緑色蛍光タンパク質(GFP))、または検出もしくは可視化が可能な任意の他の適切なレポーターをコードし得る。したがって、活発に分裂する細胞は、レポーター(例えばKaedeまたはGFP)タンパク質を発現する。NPCの場合において、増殖状態にとどまっている娘細胞(例えば増殖モードまたは混合モードで分裂している(ここで1つの娘細胞はNPCである)は、例えばKaede蛍光タンパク質の発現を続ける。対照的に、最終モードのNPCの分裂は、分裂を終了し、従ってKaede蛍光タンパク質をほとんどまたは全く発現しなくなり、細胞分裂(cytokinesis)中に受け継いだ残存Kaedeタンパク質のみを含む、2つの分化したニューロンを生じる。したがって、上述のGal4-UASレポーター系は、NPCの増殖の評価基準(measure)を提供し得る。
【0062】
上述のGal4-UASレポーター系は、例示的なレポーター系である。しかしながら、任意の適切なレポーター系(例えば、一成分系または他の二成分系)を使用してもよい。他の態様において、該レポーター系は、標的化細胞における機能を調節し得る遺伝子産物を含み得る。実際に、該系は、Gal4ドライバーと協調した多数のUAS-レポーターの使用を可能にする。
【0063】
さらに、レポーターは、インビボにおける神経細胞の時間的解像を可能にするなどのさらなる性質を有し得る。1つのかかるレポーターは、図3に示される光変換可能な(photoconvertible)蛍光タンパク質Kaedeである。Kaedeタンパク質は、緑色蛍光放出スペクトルを示すが、それは、UV光、405nmレーザー、または任意の他の適切な光源のいずれかに曝露された場合光変換されて、赤色蛍光放出スペクトルを示し得る。したがって、Kaedeの光変換特性により、NPCの挙動の観察および特徴づけにおける時間的調節物質が利用可能になる。
【0064】
例えば、NPCは、Kaede蛍光エフェクター因子を有する上述のGal4-UASレポーターでトランスフェクトされ得る。レポーターを発現する増殖細胞は、緑色Kaedeタンパク質を有する娘を生じる。好ましい所定の時間(例えば、1時間、2時間、4時間、8時間、12時間、16時間、20時間、24時間、30時間、36時間、48時間、72時間、96時間、168時間、または任意の他の値もしくはその中の値の範囲)後、Kaedeタンパク質を含むNPC細胞を光源(例えば、405nmレーザー、UV光等)に曝露して、Kaedeタンパク質を緑色から赤色へと光変換し得る。新たに生じた子孫は、細胞分裂(cytokinesis)中は赤色Kaedeタンパク質を受け継ぐが、未分化なままでいるこれらのNPCは、緑色Kaedeタンパク質を生成する(Caron et al., Development 2008, 17: 107-117)。したがって、新たに生成された娘細胞は、親細胞と識別され得、緑色対赤色のKaedeタンパク質の相対的なレベルの測定により、親細胞と娘細胞の相対数を決定し得る。この測定はまた、細胞分裂(cell division)のそれぞれの型(例えば、増殖、混合、最終)の相対的な割合の評価を提供し得る。
【0065】
本方法における使用に適切なレポーター系を、当該技術分野で公知の任意の適切な技術(例えばトランスフェクション等)により、目的の細胞または組織に導入し得る。特定の態様において、レポーター系はプラスミド上に構築され得、例えばエレクトロポレーションにより細胞に導入され得る(Haas et al., Differentiation 2002, 70: 148-154)。
【0066】
マイクロアレイ分析によるディファレンシャルスクリーニング
いくつかの態様において、本方法は、神経発生、特にNPC増殖および分化に関与する遺伝子の同定を可能にする。候補遺伝子の同定は、例えば放射状グリア中および分化したニューロン中それぞれの核酸のマイクロアレイ分析により実施され得る。かかるマイクロアレイ技術は当業者に周知である。形態学に基づいて細胞を選択して、マイクロアレイ分析のための処理の前に異なる集団に分類し得る。例えば、細胞の特性に基づいて、細胞をNPCまたは分化したニューロンのいずれかに分類し得る。代替的に、細胞集団(または目的のかかる細胞集団を含む動物)を、分化を促進または抑制のいずれかする条件および/または刺激に曝露し得る。
【0067】
充分に特徴付けられたアフリカツメガエルのオタマジャクシの神経系の発生は、神経発生に関与する候補遺伝子の同定の基礎を提供した。例えば、目的の遺伝子(例えば、配列番号:1〜651)、あるいはその機能的短縮体、修飾体および/または置換体は、アフリカツメガエルにおいて、未分化NPCと比較しての分化したニューロン中で発現された遺伝子の比較マイクロアレイ分析により同定し得る(例えば以下の実施例2参照)。一態様において、アフリカツメガエルの視蓋由来の細胞を、神経発生中の異なる日、例えば1日目および5日目に回収する。1日目に回収した細胞は、高い割合の分化したニューロンを有する5日目に回収した細胞に比べて、高い割合の未分化NPCを有する。5日目に回収した細胞に比べて、1日目に回収した細胞中での発現差異(differential expression)、具体的には有意な(例えば、p<0.05、0.04、0.03、0.02、0.01以下、またはその中の任意の他の値)発現差異を示した遺伝子は、NPCの増殖および分化の制御または調節における目的の遺伝子を示す。
【0068】
別の実験プロトコルにおいて、光、嗅覚合図または機械感覚刺激などの脳の領域への入力の外的刺激を受けるかまたは受けない動物の脳の領域から細胞を回収し得る。一態様において、細胞は、暗所に維持した動物の視蓋および光に曝露した動物から回収される。典型的に、暗所および光に曝露した動物由来の細胞は、同時(例えば12時間、24時間、48時間、72時間等の後)に回収され、次いでそこに含まれる遺伝子のマイクロアレイ分析が行なわれる。本明細書で議論するように(以下の実施例1および2参照)、暗所に維持した動物から回収した細胞は、一般的に、高い割合の未分化NPCを有する(have a have)。同じ時間の間光に曝露した細胞は、一般的に、相対的に高い割合の分化したニューロンを有する。したがって、これらの2つの集団のマイクロアレイ分析により、光に曝露した動物から回収した細胞と比較して、暗所に維持した動物(例えば相対的に高い割合のNPCを有する)由来の細胞中で異なって発現される(p<0.01)遺伝子が明らかにされ得、それによりNPC増殖および分化に関与するとして、これらの遺伝子が同定される。
【0069】
例えば、本明細書に記載されるように、ツメガエル属の光への曝露は、視蓋においてNPCがニューロンに分化することを促進することが示された。したがって、光に曝露された動物は、暗所に維持された対照動物と比較して、より高い割合の分化したニューロンを示し得る。そのため、個々のツメガエル属動物を一定時間(例えば12時間、24時間、1日、2日、3日、4日、7日等)暗条件または明条件のいずれかに曝露し得、次いで、例えば視蓋由来の細胞を動物から回収してマイクロアレイ分析に供し得る。代替的に、より長い時間維持された動物は、ますます発達した視蓋を有する。したがって、24時間後に動物から回収してマイクロアレイにより分析した細胞は、例えば2日、3日、5日、7日またはそれ以上の後に動物から回収したものに比べて、より高い割合のNPCを有し得る。いずれの場合も(明対暗または初期対後期)、第一の動物集団の蓋から回収した細胞集団は、典型的により多くのNPCを有し、第二の動物由来の残りの集団は、典型的により多くの分化したニューロンを有する。したがって、比較マイクロアレイ分析により、NPCにおいて優先的に発現されるこれらの遺伝子が明らかにされ得る。かかる方法により同定される遺伝子は、本明細書に示される配列番号:1〜651を含む。これらの遺伝子は、NPCにおいて優先的に発現され得、従って神経発生(例えばNPC増殖、分化および/または生存)に関与する。
【0070】
インビボにおける候補遺伝子発現の操作
神経発生の制御において局所的環境および非細胞自律的因子の確立された役割(Peunova et al., J. Neurosci 2001, 21(22): 8809-8818;Kreigstein et al., 149-84, Cheng et al., Nat Neurosci 2009, 12(4): 399-408;Javaherian et al., Cereb Cortex 2009, 19 Supp 1: i70-77;Suh et al., Ann Rev Cell Dev Biol 2009; 25: 253-75)は、インタクトの動物において増殖活性の直接的な評価を行なって、神経発生を調節する内因性の細胞的および分子的機構が同定され得る、ツメガエル属などの実験系の重要性を強調する。
【0071】
アフリカツメガエルのCNS(例えば視蓋)中のNPC中の遺伝子発現を調節する医薬剤の使用により、目的の遺伝子がNPCの増殖に関与することが証明される。例えば、既知の遺伝子配列に基づいてモルホリノを設計して、下流に発現される(downstream expressed)目的の遺伝子の産物(例えばRNA、タンパク質)を効果的に抑制し得る。したがって、NPC増殖を制御する遺伝子の同定により、神経発生およびNPC増殖を調節し得る医薬剤のスクリーニングにおける使用のための公知の標的が提供される。
【0072】
神経発生の調節物質のスクリーニングおよび評価の方法
医薬剤
本明細書で使用する場合、医薬剤(薬物)は、薬理学的活性を有する化合物を含み、無機化合物、イオン性物質、有機化合物、有機リガンド、例えば補因子、多糖類、組み換えおよび合成ペプチド、タンパク質、ペプトイド、核酸配列、例えば遺伝子、核酸産物が挙げられる。医薬剤は、個々にスクリーニングされ得る。代替的に、神経活動性、または神経発生に関与する遺伝子の発現を調節する能力について、1つより多くの医薬剤が同時に試験され得る。医薬剤の混合物を試験する場合、記載される方法により選択された医薬剤は、(適切な場合は)別々にされ得、適切な方法(例えばクロマトグラフィー、配列決定、PCR等により同定され得る。
【0073】
コンビナトリアル化学合成または他の方法により作製された医薬剤(例えば有機化合物、組み換えもしくは合成ペプチド、ペプトイド、核酸)の大きなコンビナトリアルライブラリーが試験され得る(例えばZuckerman, R. N. et al., J. Med. Chem., 37: 2678-2685 (1994)およびそこに引用される参考文献参照;また、タグ化化合物に関連するOhlmeyer, M. H. J. et al., Proc. Nat. Acad. Sci. USA, 90:10922-10926 (1993) およびDeWitt, S. H. et al., Proc. Natl. Acad. Sci. USA, 90:6909-6913 (1993);Rutter, W. J. et al., 米国特許第5,010,175号;Huebner, V. D. et al., 米国特許第5,182,366号;ならびにGeysen, H. M., 米国特許第4,833,092号も参照)、それぞれの関連部分は、参照により本明細書に援用される。コンビナトリアルライブラリーから選択された医薬剤が特有のタグを有する場合、クロマトグラフィー法による個々の医薬剤の同定が可能である。本明細書に記載された方法に従い、神経活動性、または神経発生に関与する1つ以上の遺伝子の発現を調節し得る1つ以上の医薬剤の存在について、化学ライブラリー、微生物培養液およびファージディスプレイライブラリーも試験(スクリーニング)され得る。
【0074】
本明細書に開示される方法により、選択された特性(例えば、神経前駆細胞の増殖の調節、標的遺伝子発現の調節等)を示す化合物のスクリーニングまたは同定が可能になり得る。本明細書に開示される方法はまた、神経活性剤(neuroactive pharmaceutical agent)の構造および機能の評価または特徴づけにも使用され得る。例えば、かかる方法により、(例えば、特異性、有効性等に関しておよび/または活性を調節するために前記候補化合物の誘導体をアッセイまたはスクリーニングすることおよびかかる誘導体の活性を親非改変調節物質と比較することにより、活性の評価が可能になる。当業者に認識されるように、例えば、化学実体は、さらなる原子、官能基および/または置換基を用いた、あるいは原子もしくは基の置換を介したホモログ化(homologation)により、構造的に改変され得る。
【0075】
したがって、本開示は、神経活動性化合物、例えば神経細胞機能(1つまたは複数)に活性であるかまたは神経細胞機能を調節する調節物質または化合物をスクリーニング、同定、特徴づけおよび修飾するため、ならびにニューロンに活性であり得るかまたはニューロンを調節し得る化合物を同定および/または特徴づけおよび/または改善するための方法および組成物を提供する。かかる調節物質または化合物は、神経前駆細胞または神経細胞の機能および/または挙動(例えば増殖および分化)が関与し得る神経系の障害の治療に有用であり得る。したがって、特定の態様において、本方法は、神経前駆細胞(NPC)のニューロンへの分化を調節し得る化合物を同定および/または特徴づけおよび/または改善するために使用され得る。
【0076】
本明細書に記載されるような神経活性剤、化合物または調節物質はまた、細胞(具体的にはニューロンまたは神経前駆体)の1つまたは複数の機能を改変(例えば回復または修正)する能力を有する任意の化合物を含み得る。例えば、調節物質の化合物は、細胞(ニューロン)の少なくとも1つの代謝経路または生物学的もしくは機能的特性を変化させ得、ニューロンに対して活性であり、かつ具体的には神経前駆細胞(NPC)のニューロンへの分化を調節し得る化合物を同定および/または特徴づけおよび/または改善し得る。例として、本発明の生物学的に活性な化合物は、正常な表現型を、傷害されたニューロンに回復し得る、またはニューロンに対する傷害の有害な効果を少なくとも部分的に抑制し得る化合物である。特定の態様において、活性化合物は、細胞機構を抑制または活性化するその能力について、代謝経路を刺激または抑制する、生物学的特性を回復する、細胞死を防止する等のその能力について、選択され得る。
【0077】
インビボ分析に適切な医薬剤としては、例えば神経発生において遺伝子の機能の分析を可能にする、GOIのノックダウンのためのモルホリノが挙げられ得る。他の医薬剤としてはshRNA構築物が挙げられ得る。shRNA媒介ノックダウンは、MOと比較して独立したノックダウン方法を提供し、タンパク質発現の細胞型特異的操作を可能にする。ツメガエル属のニューロンにおいてshRNA-媒介ノックダウンを増強するための方法が開発され、多くのshRNA構築物の生成を能率的にするためにプラスミドカセットが利用可能である。したがって、GOIに対するshRNA構築物は、GOI発現の特異的ノックダウンについて試験され得(Chen et al., Front Neurosci 2009, 3: 63)、その後、例えばNPC増殖に対する効果について試験され得る。当然ながら、本明細書に記載される方法またはプロトコルは、細胞生存、ニューロンまたはグリアへのNPCの分化、ならびに脳領域または神経回路中の細胞の移動および集合などの神経発生の任意の他の寄与因子に対して活性な化合物をスクリーニング(または同定、特徴づけもしくは改善)するために使用され得る。
【0078】
本明細書に記載される方法およびプロトコルは、神経生存または発生に活性であり、かつNPCのニューロンへの分化を特異的に調節し得る化合物をスクリーニング(または同定、特徴づけもしくは改善)するために使用され得る。
【0079】
医薬組成物
別の局面において、本発明は、本発明の方法により記載され、同定される医薬剤、調節物質または化合物を含む医薬組成物を提供する。かかる医薬組成物は、例えば神経発生または神経細胞の機能(1つまたは複数)を調節し得るか、NPC分化および/または増殖を調節し得るか、またはニューロンに対して活性であり得るかもしくはニューロンを調節し得、かつ神経前駆細胞または神経細胞の機能および/または挙動(例えば増殖、分化等)が関与し得る神経系の障害の治療に有用であり得る、本明細書に記載される医薬剤を含み得る。
【0080】
本明細書に記載される医薬剤は、医薬組成物として調製され得、例えばヒト患者などの哺乳動物宿主に、選択された投与経路、例えば経口または非経口、静脈内、筋内、局所または皮下経路に適合された種々の形態で投与され得、かつ1つ以上の薬学的に許容され得る賦形剤を含み得る。
【0081】
細胞系アッセイ
本発明の方法はさらに、単離された細胞(例えば神経前駆細胞)または適切な培養細胞株を、1つ以上の候補化合物または調節物質と接触させる工程を含む。細胞は、医薬剤の効果、濃度、細胞集団および/または評価技術に依存して種々の時間の間接触され得る。特定の態様において、細胞は、例えば約1nM〜約1mMの範囲の候補化合物(1つまたは複数)に曝露され得る。本願から逸脱することなく、他の濃度が試験され得ることが理解されるはずである。さらに、それぞれの化合物は、いくつかの濃度で並行して試験され得る。さらに、必要な場合は、リポソーム、カチオン性脂質またはポリマー、ペネトラチン(penetratin)、Tat PDT、アデノウイルス由来のペプチド(例えばペントンまたはファイバー)もしくは他のウイルス由来のペプチド等を含む、化合物の細胞への進入を補助する種々のアジュバントおよび/またはベクターおよび/または生成物が添加され得る。接触は、生きたツメガエル属のオタマジャクシの調製物のためのインキュベーションチャンバーなどの任意の適切な支持体またはデバイス中で実施され得る。
【0082】
候補化合物のプロフィールを決定することは、いくつかの方法に従って実施され得る。特に、化合物の神経活動性を評価するために、例えば細胞数、生存、抗原の発現、特定の遺伝子の転写、および形態学的変化、例えばサイズ、神経突起成長等の種々の終点が測定され得る。
【0083】
具体的な態様において、候補化合物の神経活動性は、任意の化合物の非存在下および/または参照化合物で処理された対照神経細胞集団と比較することにより決定され得る。ニューロンの状態の決定は、種々の物理的測定値、例えば光学特性、種々の波長での蛍光、放出等を評価することによりなされ得る。ランプまたはレーザー等が備え付けられた自動化顕微鏡などの種々の装置が使用され得る。他の技術としては、冷却CCDカメラによる光検出が挙げられる。測定されたシグナルは、例えばピクセルヒストグラム、クラスター分析および形態学解析を含むソフトウェアを使用して、公知の技術に従って処理され得る。
【0084】
本発明は、また、例えば、(i)実験研究のための標的、および/または(ii)調節物質としての、および特に神経学的障害を治療するための医薬組成物の製造のような、本発明の方法によって同定され、選択され、概略(profile)され、または特徴付けられる任意の化合物(またはその誘導体)の使用に関する。
【0085】
いくつかの態様において、細胞分化モードを、細胞を調節剤と接触させることによって調節し得る。他の態様において、例えば、エレクトロポレーション、または細胞に調節剤を導入するのに適した任意の他の技術により目的の細胞に調節剤を導入することによって、細胞分化モードを調節し得る。
【0086】
特定の態様において、医薬剤(pharmaceutical agent)は、他の分裂モードに比べて、増殖モードで分裂する細胞の数または割合を増加させ得る。他の態様において、医薬剤は、増殖モードで分裂する細胞の数または割合を減少させ得る。他の態様において、医薬剤は、混合モードで分裂する細胞の数または割合を増加させ得る。あるいは、医薬剤は、混合モードで分裂する細胞の数または割合を減少させ得る。さらなる態様において、医薬剤は、分化モードで分裂する細胞の数または割合を増加させ得る。あるいは、医薬剤は、分化モードで分裂する細胞の数または割合を減少させ得る。医薬剤の組合せを、また、NPC増殖および分化に対する所望の効果を達成するために使用し得る。
【0087】
本発明の医薬剤は、NPC増殖および分化の調節に関与する公知の遺伝子標的を調節するように、選択され得る。従って、医薬剤(例えば、モルホリノ、siRNA等)は、例えば、NPC増殖および分化において調節的役割を有することが公知である標的遺伝子の翻訳の阻害または上方調節をするために構築され得るか、または選択され得る。従って、標的細胞(1つまたは複数)をかかる医薬剤と接触させること、または標的細胞(1つまたは複数)にかかる医薬剤を導入することによって、NPC挙動の調節がもたらされ得る。
【0088】
インビボアッセイ
本願で公開する方法は、単離された細胞(例えば、神経前駆細胞)または適切な培養された細胞株を、1つ以上の候補化合物または調節物質と接触させる工程をさらに含む。その効果、濃度、細胞集団、および/または評価技術に依存して、細胞を、種々の時間接触させることができる。一般的に、細胞を、1nM〜1mMの範囲の候補化合物(1つまたは複数)に曝露する。他の濃度を本願から逸脱することなく試験し得ることが理解されるはずである。さらに、各化合物を、いくつかの濃度で並行して試験し得る。さらに、必要であれば、化合物が細胞に入り込むのを補助する、リポソーム、カチオン性脂質もしくはポリマー、ペネトラチン、TatPDT、アデノウイルス由来のペプチド(ペントンまたはファイバー)または他のウイルス由来のペプチド等の種々のアジュバントおよび/またはベクターおよび/または生成物を添加できる。
【0089】
候補化合物のプロフィールを決定することは、いくつかの方法に従って実施され得る。特に、例えば、化合物の神経活性(neuro-activity)を評価するために、細胞数、生存、抗原の発現、特定の遺伝子の転写、および、例えば、サイズ、神経突起成長等の形態学的変化等の種々の終点が測定され得る。
【0090】
好ましくは、候補化合物の神経活性は、任意の化合物の非存在下の、および/または参照化合物で処置された対照神経細胞集団との比較により、測定される。ニューロンの状態の決定は、種々の物理的測定、光学的特性、種々の波長での蛍光、発光等を評価することによって行われ得る。ランプまたはレーザーを備えた自動化顕微鏡等を含む種々の装置が使用され得る。他の技術としては、冷却CCDカメラによる光検出が挙げられる。測定されたシグナルは、例えば、ピクセルヒストグラム、クラスター分析および形態学的解析を含むソフトウェアを使用する公知の技術に従って処理され得る。
【0091】
本発明は、また、(i)実験研究のための標的としての、または(ii)神経学的障害を治療するための医薬組成物の製造のための、上記に開示される方法により同定され、選択され、概略され、または特徴付けられる任意の化合物(またはその誘導体)の使用を包含する。
【0092】
いくつかの態様において、細胞分化モードを、細胞と調節剤を接触させることにより調節し得る。他の態様において、例えば、エレクトロポレーション、または細胞に調節剤を導入する任意の他の適切な技術により目的の細胞に調節剤を導入することによって、細胞分化モードを調節し得る。
【0093】
特定の態様において、医薬剤は、他の分裂モードに比べて、増殖モードで分裂する細胞の数または割合を増加し得る。他の態様において、医薬剤は、増殖モードで分裂する細胞の数または割合を減少し得る。なお他の態様において、医薬剤は、混合モードで分裂する細胞の数または割合を増加し得る。あるいは、医薬剤は、混合モードで分裂する細胞の数または割合を減少し得る。さらなる態様において、医薬剤は、分化モードで分裂する細胞の数または割合を増加し得る。あるいは、医薬剤は、分化モードで分裂する細胞の数または割合を減少し得る。医薬剤の組合せを、また、NPC増殖および分化に対する所望の効果を達成するために使用し得る。
【0094】
医薬剤は、NPC増殖および分化の調節に関与する公知の遺伝子標的を調節するように、選択され得る。従って、医薬剤(例えば、モルホリノ、siRNA等)は、NPC増殖および分化に調節的役割を有することが公知の標的遺伝子の翻訳を、例えば、阻害または上方調節するように、構築され得、または選択され得る。従って、標的細胞(1つまたは複数)とかかる医薬剤を接触させること、またはかかる医薬剤を標的細胞(1つまたは複数)に導入することは、NPC挙動の調節をもたらし得る。
【0095】
インビボアッセイ
マイクロアレイおよび/または他のゲノム分析技術により生成されたゲノムデータの解析は、オタマジャクシCNS中のNPCの増殖および分化を調節し得る遺伝子を優先させる(prioritize)ために使用され得る。従って、アンチセンスモルホリノオリゴヌクレオチド(MO)(Eisen et al., Development 2008, 135(10): 1735-43)は、目的の遺伝子(GOI)に対して生成され得、各GOIは、CNS中のBrdU取り込みを評価するための画像化を使用して、細胞増殖の効果に対するモルホリノ媒介ノックダウンによってスクリーニングされる。細胞増殖を増大または低減するGOIについて、いくつかのインビボアッセイが、視蓋における神経発生の制御におけるその機能を試験するために使用され得る(Nedivi et al, Science 1998, 281: 1863-1866; Ewald et al, J Neurosci 2008, 28(4): 850-61; Cantallopset al., Nat. Neurosci. 2000, 3: 498-503; Javaherian et al., Neuron, 2009 (in press); Van Aelst et al., Curr Opin Neurobiol 2004, 14(3): 297-304; Van Keuren-Jensen et al., Dev Neurobiol 2008, 68(11): 1315-24; Wu et al., Science 1998, 279: 222-226; Cline et al., J Physiol 2008, 586(6): 1509-17)。
【0096】
第1の例示的アッセイは、BrdU取り込みとニューロンのマーカーに対する免疫標識を組み合わせる。BrdU標識法の利点は、該方法が、非侵襲性であり、GOIノックダウンまたは過剰発現の細胞増殖に対する効果の比較的ハイスループットのスクリーニングとして使用され得ることである。BrdUとニューロン標識を組み合わせることにより、神経発生におけるGOIの定量的評価が可能になる。オタマジャクシを、BrdUを含む飼育溶液(rearing solution)に曝露し得る。この方法は、増殖細胞を効率的に標識し、BrdU曝露時間についてより良い制御を可能にする。BrdUの接近は、研究された発生期を通して変わらないこと、およびBrdUの取り込みは、DNA損傷に応答して生じないことが示された。BrdU曝露は、典型的には、全ての動物について増殖に対する潜在的概日効果を調節するために、一日の同じ時間に行われる。BrdU曝露の後、動物を、即座に固定するか、または2〜3日飼育し、その後、動物を固定し、BrdU免疫標識のために動物を処理するかのいずれかをする。
【0097】
次いで、動物を終末麻酔し(terminally anesthetize)、マイクロウェーブ固定し(Paupard et al., J. Histochem. Cytochem. 2001, 49(8): 949-956)、脳を、一次抗体を用いた抗体標識のために、全載またはビブラトーム切片(vibratome section)(Peunova et al., 8809-8818)に(in)処理される(マウス抗−BrdU、BD Bioscience;ウサギ抗NeuroD;AbcamおよびAlexafluor二次抗体(Molecular Probes))。
【0098】
第2の例示的アッセイは、Sox2.mFGF4::FP-標識NPC等の適切なレポーターまたはマーカーで標識された細胞のインビボ時間差撮影画像化法を使用する。時間差撮影インビボ画像化実験の利点は、長期的研究により、細胞増殖、細胞の運命およびこれらの事象の基礎をなす構造動力学の直接的観察が可能になることである。GOI過剰発現/ノックダウンによりインビボで神経発生がどのように変更させるかが定量され、細胞分裂速度および子孫の運命が決定され得る。
【0099】
脳室切片、全載固定脳調製物およびインビボFP標識試料中の増殖細胞を、レーザー列ならびにUVを遠赤外蛍光団に分離するためのダイクロイックミラーおよびフィルターを備える顕微鏡に搭載されたスピニングディスク共焦点(spinning disk confocal)結合物を使用して画像化し得る。シグナルは、感度の高い、高シグナル対ノイズのEMCCDカメラで捕捉される。適切な画像捕捉ソフトウェアを使用して、画像を捕捉する。二重標識実験において、ブリードスルー(bleed-through)を排除するために、画像は連続的に捕捉される。単一の蛍光団標識での調節は、ブリードスルーの非存在を確実にするためになされる。
【0100】
インビボ画像時間差撮影のために、麻酔したオタマジャクシを、特別注文の画像化チャンバに配置する。蛍光団についての適切なレーザー/フィルター設定を使用して、1μmのz幅で完全な共焦点スタック(stack)を取得する。固定された組織全載および組織切片について、視蓋(tectal lobe)の完全なzスタック(0.5μmのz間隔)を取得する。
【0101】
時間差撮影データの解析のために、放射状グリア(radial glia)とニューロンを、細胞の三次元構造に基づく形態学的基準に従って区別する。実験の時間経過の間の系統の各細胞の対称増殖性分裂;非対称神経発生分裂;最終対称神経発生分裂の数および順序、ならびに分化の段階を決定する。個々のNPCの時間差撮影画像化により、画像化実験の経過の間の、分裂細胞および分化している細胞における形態学的変化の評価を含む、細胞の運命の同定が可能になる。
【0102】
従って、特定の態様において、本発明の方法は、インビボ、より具体的には、インタクトの脳において、神経前駆細胞(NPC)のニューロンへの分化を変更する医薬剤のスクリーニング(または同定、特徴付け、もしくは改善)に有用である。本発明の方法の利点は、かかる方法が、ツメガエル属(Xenopus)オタマジャクシのインタクトの視覚系中の神経細胞集団等の、インビボの神経細胞集団を使用し得ることである。従って、インビボの神経細胞集団の使用により、化合物または調節物質の生物学的活性の予測的かつ信頼できる評価が可能になる。使用されるNPCおよびニューロンは、哺乳動物起源(例えば、齧歯類、ヒト、霊長類等)およびアフリカツメガエル(Xenopus laevis)等の両生類を含む種々の起源のものであり得る。
【0103】
医薬剤のインビボ活性を、例えば、神経発生に関連する遺伝子のスクリーニングに関して本明細書に記載されるように決定し得る。例えば、ツメガエル属オタマジャクシはインビボ時間差撮影に敏感に反応するので、神経前駆細胞およびその子孫を、インタクトの動物において画像化し得る。従って、NPC細胞をインビボで候補医薬剤と接触させた後、対照動物集団に対する試験動物集団の増殖割合を測定し得(例えば、可視化し得)、候補調節剤(modulating)の活性を、対照集団に対する、候補医薬剤で処置されたまたは候補医薬剤と接触した動物中のNPC増殖の相対速度によって決定する。従って、NPC増殖割合を変更する(例えば、増大または低下させる)医薬剤を、神経発生および神経細胞増殖の調節物質として同定する。
【0104】
神経発生の調節物質としての医薬剤のスクリーニングのためのインビトロ方法、およびインビボ遺伝子スクリーニング方法に関して本明細書に記載される条件(例えば、医薬剤の濃度、読み取り等)を、また、ここでも使用し得る。例えば、インタクト動物CNS領域(例えば、視蓋)中のNPC細胞集団を、蛍光タンパク質(FP)を発現するように、(例えば、エレクトロポレーションによって、)トランスフェクトさせ得る。インタクトの動物中のFP発現細胞を、時間t=0で画像化し得、次いで、所定の時間(例えば、24h、48h、96時間等)後に再度画像化し得、所定の時間(例えば、12時間、24時間、36時間等)の間の間隔で画像化し得る。細胞(例えば、NPCおよび分化したニューロン)の相対的型および数の変化を、所定の時間にわたって、各24時間間隔で、細胞をその形態に従って分類することによって決定し得る。形態の変化を、対照集団に対する、試験集団中のNPCおよびグリア細胞の割合として特徴付けることができる。試験動物集団中の細胞と対照動物集団中の細胞の違いは、候補剤がNPC増殖の調節物質であることを示し得る。
【0105】
特定の態様において、CNS細胞増殖に対するGOI効果のスクリーニングを、脳にGOIに対するモルホリノをエレクトロポレートしたアフリカツメガエル(X.laevis)オタマジャクシにおけるBrdU取り込みを評価するために画像化アッセイを使用して実施し得る。1〜2日後、増殖細胞を、屠殺前に2時間、BrdUに曝露することにより標識し得る。次いで、脳を、全載においてBrdUを検出するために処理し得、脳の端から端までの共焦点画像の完全なzシリーズを収集することによって画像化し得る。全載脳のかかる画像化により、細胞増殖のレベルを定量するための優れた方法が提供される。
【実施例】
【0106】
実施例
本開示は、以下の非限定的な実施例によってさらに例示される。以下の実施例は、例示のみのために理解され、添付の特許請求の範囲により規定される本発明の範囲を限定するとして解釈するべきではない。
【0107】
1.視覚刺激およびNPC増殖
概観
神経前駆細胞(NPC)の増殖および分化を、インタクトのツメガエル属オタマジャクシ中枢神経系の視覚系においてアッセイした。この実験系は、動物を視覚系刺激に曝露するか、または動物の視覚系刺激を与えない(deprive)ことによって神経活動性を操作できる。結果は、NPCの増殖割合が、12時間光/12時間暗所の条件下で飼育された動物に比べて、視覚刺激を与えられない動物において、増大することを示す。24時間視覚刺激を与えられず、その後、24時間視覚刺激を受けた動物は、視覚経験の非存在下の最初の24時間の間増殖割合が上昇し、その後、新たに生成された細胞の大部分が分化することを示した(図6)。これらのデータは、神経活動性を操作することにより、増殖割合および分化の両方が制御されることを示す:低下した神経活動性により増殖が増加し、増大した神経活動性により、前駆細胞のニューロンへの分化が誘発される。
【0108】
方法
蛍光タンパク質(FP)を発現するように、インタクトのツメガエル属オタマジャクシ中のNPCをトランスフェクトした。インタクト動物中のFP発現細胞を、共焦点顕微鏡を使用して画像化した。次いで、画像化の後、動物を、光を通さないチャンバに配置し、その結果、動物は、次の24時間にわたって視覚刺激を受けなかった。FP発現細胞を、再度画像化し、動物をチャンバに配置し、そこで、動物は、24時間視覚刺激を受けた。動物から、三度目の画像を採った。各24時間間隔の間の細胞数の変化および放射状グリア(NPC)またはニューロンとしての細胞の同一性を、それらの形態に従って決定した。24時間あたりの細胞数の変化として、およびNPCまたはニューロンの形態を有する細胞の割合として、データを表した。
【0109】
結果
視覚経験の非存在下では、細胞数は、24時間の間に、+19.9±5.8%増加した(N=12の蓋が分析された)。動物を24時間の視覚刺激に曝露した後、本発明者らは、細胞数の有意な6.3±5.4%の減少を見いだした(ウィルコクソン符号付(singed)順位検定、p=0.01)。細胞を検出するために使用された蛍光マーカーが、増殖の間のみ発現されるプロモーターにより駆動されるので、この負の割合は、細胞が増殖サイクルを離れたことを示す。視覚経験に曝露されなかった動物の別のセットは、2つの24時間の期間の間で増殖割合に有意な変化がなかったことを示した(N=7の蓋が分析された、p=0.4)。
【0110】
視覚刺激に曝露されなかった動物の脳において、定常速度(steady rate)の神経発生があった。視覚刺激の非存在下では、新しいニューロンの割合が、第1の24時間の間隔の間に32.9±11.3%増え、これは、第2の24時間間隔の間の30.1±9.4%の増加からは有意ではなかった(N=7蓋、ウィルコクソン符号付順位検定p=.73)。対照的に、最後の24時間の間隔の間に視覚刺激に曝露された動物の神経発生割合は、第1の24時間の間隔中の79.4±22.9%から、第2の24時間の間隔の間の18.0±9.4%まで有意に減少した(N=12蓋、p=.05)。
【0111】
24時間の視覚経験の非存在下では、グリアの外観を喪失する細胞の割合は、第1の24時間の間に17.1±9.7%の低下、および最後の24時間間隔の間に44.9±10.6%の減少を示す(N=7蓋、p=0.18)。24時間視覚を与えず、その後24時間視覚刺激に曝露された動物由来の細胞がグリアの外観を喪失する割合は、視覚刺激に曝露した後、有意に増大する(前では17.1±9.7%喪失、および後では56.0±10.0%喪失、p=.04)。
【0112】
これらの結果を合わせると、視覚経験が細胞増殖を中断させること、および視覚経験がグリア前駆体がニューロンに分化する割合が上昇することと一致する。
【0113】
2.視覚経験ありなしの調節剤およびNPC増殖
背景および方法
NPC検出
これらの実験は、分裂細胞におけるタンパク質発現を駆動する、細胞型特異的蛍光レポーター系を利用した(図3参照)。レポーターは、線維芽細胞増殖因子4(FGF4)の上流調節因子(regulatory element)の6つのリピートからなる。生体内の(endogenous)Sox2/oct3転写因子は、FGF4調節因子に結合し、Gal4の発現を駆動(drive)し、該Gal4は、次いで、UAS蛍光タンパク質を駆動する。FGF4、Sox2およびoct3は、各々、増殖細胞中で発現し、生体内のSox2/oct3転写因子に依存して、増殖細胞内でレポーター発現のGal4促進性特異性を駆動する。UAS蛍光タンパク質を別の構築物として発現させ;このレポーターは、このレポーター系に調節性および特異性を付加した。
【0114】
上記の蛍光レポーターを発現する蓋細胞を、種々の視覚経験を有するオタマジャクシから、および種々の時間の間、該構築物を発現したオタマジャクシから、採取した。次いで、これらの細胞由来のRNAを処理し、マイクロアレイ比較を行った。さらなる分析のために選択された、同定された目的の遺伝子を、以下の表1にまとめる:
【表1】
表1−マイクロアレイ分析を介して同定された目的の遺伝子
【0115】
種々の時期(age)のオタマジャクシの間(より多くの集団の増殖細胞 対 より多くの集団の最終的に分化した細胞)、および、増殖 対 最終分化の相対レベルにも偏りを加え得る、種々の視覚経験を有するオタマジャクシの間で、比較を行った。複数のマイクロアレイ比較において、その発現に有意な変化を示した遺伝子を優先させた。
【0116】
上記のレポーターを使用して、オタマジャクシの視蓋の増殖細胞において蛍光タンパク質Kaedeを駆動(drive)した。Kaedeの蛍光放出スペクトルは、実験に一時的な制御を加えたUV光または405nmのレーザー源への曝露の後、緑から赤に光転換(photoconvert)され得る。オタマジャクシをレポーターでトランスフェクトした24〜36時間後、全てのKaede発現細胞は、該タンパク質の赤形態に光転換された。類似の方法を使用した公開されたデータ(Caron et al., Development 2008, 135:3259-3269)に基づくと、新たに生じた子孫は、細胞質分裂の間に赤Kaedeを受け継ぐが、新しい緑Kaedeタンパク質を合成し続ける。例えば、Caronら(同書)は、緑 対 赤の相対レベルによって新たに生成された細胞を同定した。
【0117】
組織中に複数で標識された細胞を確実に生じる十分に確立された方法である、エレクトロポレーションにより、視蓋の細胞を増殖レポーターでトランスフェクトした(Haas et al., Differentiation 2002, 70: 148-154)。Gal4駆動体(driver)プラスミド(0.5μg/μl)およびUAS−kaedeプラスミド(0.5μg/μl)を、視蓋の脳室(ventricle)に注射し、次いで、電圧パルスを組織に印加し、該プラスミドを蓋の細胞に駆動した。これらのプラスミドの構築物を、候補遺伝子の翻訳を阻害するように設計されたアンチセンスモルホリノオリゴヌクレオチドと共に共エレクトロポレートした(Eisen et al., Development 2008, 135: 1735-1743)。モルホリノを、0.1mMでエレクトロポレートし、リサミン蛍光タグにより可視化した。周囲光からのUV光によるKaedeの転換を防止するために、動物を暗所に維持した。24〜36時間後、オタマジャクシを麻酔し、各動物の視蓋の端から端までの完全なz−スタックを、3日の連続する日について、各日に収集した。蓋の第1の画像化の後、全ての動物を、第2の画像化まで24時間、暗所に戻した。そのときに、0.2Hzで1秒間光を放つLED光(567nmを放出する。これは、Kaedeタンパク質を転換する波長を超えている)のアレイを有するチャンバーからなる増強された視覚環境に、数匹の動物を曝露した。この視覚刺激は、蓋のニューロンにおけるシナプス駆動を増強することが示されており、シナプス可塑性に有意な変化を生じる(Sin et al., Nature 2002, 419: 475-480)。
【0118】
データ取得および解析
第1日に使用した取得設定を、ピクセル値が第3日までに飽和に達することから保護するために使用した。選択した設定を、実験の間ずっと使用した。取得したz−スタックからの3D情報を使用するVolocityソフトウェア(Improvision, Perkin Elmer)を使用し、対象物の強度およびサイズの標準偏差に基づいて、標識された細胞の細胞体を同定し、かつ選択した。次いで、同定した対象物は、実験者が検証し、細胞形態に基づいて細胞型(グリア、ニューロン、または不確定)に帰属させた。各視蓋から、典型的に、15〜45細胞がトランスフェクトされた。24時間あたりの細胞数の変化として、パーセント増殖を計算した。これらの測定を、平均±s.e.m.として報告する。
【0119】
対照動物、細胞増殖ブロッカーを有する動物、および24時間の増強された視覚刺激を受けた動物における増殖の分析を報告する。マイクロアレイから同定された2つの候補(Dio3およびGstπ)をアンチセンスモルホリノでノックダウンした実験の結果もまた報告する。さらに、視覚経験がDio3モルホリノノックダウンの存在下での増殖に影響することを測定するための実験からの結果を報告する。使用した実験条件を、以下の表2にまとめる:
【表2】
表2−モルホリノノックダウン実験条件
【0120】
結果および考察
Sox2レポーター構築物の発現は、放射状グリアが、ツメガエル属視蓋中の主要な神経前駆細胞であることを示した(図4)。分化した細胞は、もはや、Sox2/oct3を産生し、gal4−UAS−Kaede構築物を駆動しないので、ニューロンは、分化後、緑Kaedeを発現し続けなかった。NPC分裂の大部分は、最後の分裂であることが見出された。
【0121】
細胞分裂ブロッカー
アフィジコリン(150μM)およびヒドロキシ尿素(2%DMSO中20mM)を、細胞に加え、細胞分裂をブロックし、これらの細胞の増殖割合を、DMSOのみを受けた対照動物と比較した。この処理は、ツメガエル属の増殖を中断させる(Harris et al., Neuron 1991, 6: 499-515)。DMSOのみに曝露された対照動物において、24時間で16.4±4.2%の細胞数(N=8視蓋)の平均増加が観察された。アフィジコリンおよびヒドロキシ尿素は、細胞増殖をブロックした(細胞数の増加:0.41±6.28%(N=10、p=0.1、マン−ホイットニー;図5参照)。
【0122】
NPC運命の視覚経験依存性調節
視覚経験の非存在下では、細胞数は、24時間の間に19.9±5.8%増加した(N=12視蓋)。24時間の視覚刺激に動物を曝露した後、6.3±5.4%の細胞数の有意な減少が見出された(p=0.01)。この負の割合は、細胞が増殖サイクルを離れ、徐々に成長している樹状突起分枝(dendritic arbor)中での赤Kaedeレポーターの希釈により、もはや検出可能ではないことを示す。Kaedeレポーターは、増殖の間のみ発現され、従って、最終的に分化した細胞は、増殖モードの間にレポーターを産生したことのみを示す。24時間の視覚刺激を受けなかった対照動物は、2つの24時間の期間の間で増殖割合に有意な差を示さなかった(10.3±7.2%および−0.6±3.5%;N=12分析された視蓋、p=0.23;図6A)。
【0123】
視覚刺激への曝露が蓋中の細胞の運命に影響するかどうかを試験するために、24時間の視覚刺激有り無しの後の標識された集団中のニューロンの割合を比較した。ニューロンの形態を有する細胞の割合は、対照動物に比べると視覚刺激に曝露された動物において、より大きく(75.7±4.1% 対 60.1±4.8%、p=0.02、マン−ホイットニー;図6B)、このことは、視覚刺激が、NPC子孫の分化を推進することを示した。
【0124】
Dio3−モルホリノ発現および神経発生
脱ヨード酵素ヨードサイロニンIII型は、甲状腺ホルモンの活性形態(T3)からヨウ素を除去しそれを効率的に不活性化する、甲状腺ホルモン経路中の酵素である。マイクロアレイ分析は、Dio3発現が活動中の前駆細胞中で増大したことを示唆した。アフリカツメガエル中のT3レベルは、変態期前では低いが、T3受容体の存在により、NPC中で検出され、T3レベルの相対的変化が増殖に影響し得ることを示唆した。増大した増殖は、変態期のアフリカツメガエルオタマジャクシ中の甲状腺ホルモンの上昇および受容体活性化と相関する(Denver et al., Dev Biol 2009, 326: 155-168)。従って、モルホリノ発現を用いたDio3のノックダウンにより、T3レベルが上昇し、従って、増殖が増大するはずである。
【0125】
Dio3に対するモルホリノ(Dio3−MO)でトランスフェクトした動物は、第1の24時間の期間において11.4±17.3%、および第2の24時間の期間の間に33.9±22.2%の細胞数の増加を示した(N=4、図7)。対照的に、対照動物は、第1の24時間の期間に最大の増殖の増大を示し、次いで、続く24時間の期間の間にほとんど増殖を示さない。クローンのようである細胞の異常なクラスターが、Dio3−MOでトランスフェクトした動物の蓋内に見出され、これは、移動のための正常な時間を有さない増殖の異常なパターンと一致した。
【0126】
Dio3−MOを発現する数匹の動物をまた、第2の24時間の期間の間に、視覚刺激に曝露した(図7)。視覚経験により、Dio3−MOでトランスフェクトした動物中(N=5)の増殖の割合が減少することが見出されたが、この相違は、増殖割合の大きな変動のために有意とはみなされなかった。
【0127】
GSTπのノックダウンにより神経分化が促進される
グルタチオンS−トランスフェラーゼπ1(GSTP1)は、多くの疎水性化合物および親電子性化合物と還元グルタチオンとのコンジュゲーションを触媒することにより解毒において重要な役割を果たす、タンパク質のグルタチオンS−トランスフェラーゼファミリーのメンバーである。GST−π1は、癌に対する感受性において役割を果たすと考えられている。GST−π1は、神経前駆細胞中で上方調節されていることが観察され、従って、GST−πのノックダウンは、増殖割合を低減すると予測される。
【0128】
GSTπに対するモルホリノを発現するオタマジャクシの蓋において、顕著な結果を観察した。第1日までのニューロンの数でさえ、対照動物で観察されたニューロンの数より有意に多かった(図8)。試験動物中のGSTπ−MOを発現する蓋細胞の、対照動物(42.0±4.7%)に比べて有意により大きな集団(7.27±4.7%、N=7視蓋)が、第1日までにニューロンに分化した(P=0.002、マン−ホイットニー)。第2日までに、対照集団内の49.2±5.7%に比べて、試験動物の細胞の86.4±3.7%がニューロンに分化した(p=0.001)。このニューロンの割合は、ニューロンの数が第3の期間の間に試験動物において増加することが観察されなかったので、最大を示し得(86.8±1.0%)、対照動物内のニューロンの割合より有意に大きい(60.1±16.6%;p=0.004)。
【0129】
ニューロンの全割合の増大により、神経前駆体の集団が、GSTπ−MOの発現と共に減少することが示される。放射状グリアの割合は、GSTπ−MOを発現する動物および対照動物において3日の期間にわたり減少する(図8)。GSTπ−MOを発現する脳由来の細胞の割合を対照動物と比較することにより、第1日までに有意に少ないグリアが存在し(13.3±4.6%および41.5±4.8%;p=0.005)、これらの相違は、次の2日の間続く(8.4±2.0%および17.4±3.5%、p=0.02;4.8±1.6%および13.3±4.6%、p=0.02)ことが示された。小さな割合の細胞は、放射状グリアまたはニューロンに分類することはできなかった。
【0130】
脆弱X関連遺伝子
Fmr1A。脆弱X精神遅滞タンパク質1は、核から細胞質へのmRNAトラフィッキング(trafficking)およびニューロン内の局所タンパク質翻訳を制御すると考えられているmRNA結合タンパク質である。マイクロアレイデータにより、分化したニューロンに比べると、NPC中のFMR1および82kD FMRP相互作用タンパク質、増殖誘導遺伝子1(AKA核脆弱X精神遅滞タンパク質相互作用タンパク質)の発現は低いことが示唆された。ニューロンの増殖におけるFMRPおよび関連するタンパク質の潜在的な役割は、完全には明らかではない。1つの研究により、FMR1が、NPC増殖を増大させ、分化を変更することが示されたが(Castren et al., Proc Natl Acad Sci USA 2005, 102: 17834-17839)、別の研究(another)により、FMR1はNPCの分化のみを変更することが示された(Bhattacharyya et al., Stem Cells Dev 2008 17: 107-117)。これらの研究の間の考え得る不一致は、これらの研究の各々が、異なる供給源のインビトロ細胞を使用しているという事実のためであり得る。インビトロ研究は、NPCの増殖におけるFMR1および関連する遺伝子の役割を明確にし得る。
【0131】
FXR1.脆弱X精神遅滞、常染色体ホモログ遺伝子は、機能的に類似するタンパク質FMR1およびFXR2と相互作用する。マイクロアレイデータに基づくと、FXR1をノックダウンすることは、分化を阻害することにより増殖を増大し得る。
【0132】
FMR1AおよびFXR1に対するモルホリノは、蓋におけるNPC増殖を減少させるようである。定性的観察があった。
【技術分野】
【0001】
関連出願
本願は、その全体において参照により本明細書に明示して援用される2009年9月29日に出願された米国仮特許出願第61/246,967号に対する35 U.S.C. § 119による優先権を主張する。
【0002】
配列表
本願は、ASCII形式で、EFS-Webを介して提出され、その全体において参照により本明細書に援用される配列表を含む。2010年9月28日に作成された前記ASCIIコピーは、21RE6840.txtと称し、2,011,398バイトのサイズである。
【0003】
発明の分野
本発明は、神経発生、特に中枢神経系の神経発生の活動性依存性調節に関連する遺伝子、方法および組成物に関する。より具体的には、本発明は、神経発生に関連する遺伝子の同定および操作の方法、ならびに神経発生を調節する医薬剤のスクリーニングおよび評価の方法に関する。
【背景技術】
【0004】
発明の背景
神経発生は、神経系の発生および成熟の基礎となる複雑な課程である。この過程は、細胞の増殖、生存、分化および移動の適切な時間空間的な制御に依存する。新たに生成された神経細胞は、中枢神経系の機能的細胞に分化し得、脳の神経回路に組み込まれ得る。さらに、多くの動物の脳において、新しい神経細胞は、生物の生涯を通じて継続的に生成される。例えば、神経発生は、現在、成体期を通じて、哺乳動物の脳の2つの領域:側脳室の脳室下(subventricular)帯(SVZ)および海馬の歯状回において続くことが分かっている。これらの領域では、多能性神経前駆細胞(NPC)は、分裂を続け、新しい機能的ニューロンおよびグリア細胞を生じる(Jacobs, Mol. Psychiatry 2000, 5(3): 262-9)。そのため、神経発生の調節は、CNSの領域的分化(specialization)および機能的回路を形成する特異的細胞型の確立の基礎となる。
【0005】
神経発生は、CNS生理学において重要な役割を担う。神経前駆細胞(NPC)のプールは、さらなるNPCを生じる対称分裂により増殖され得るか、または子孫の、ニューロンもしくはグリアへの最終的な分化により枯渇され得る(Butt et al., Neuron 2005, 48: 591-604;Gotz et al., Nat Rev Mol Cell Biol 2005, 6: 777-788;Huttner et al., Curr Opin Cell Biol 2005, 17: 648-657;Noctor et al., Arch Neurol 2007, 64: 639-642;Kriegstein et al., Ann Rev Neurosci 2009, 32: 149-184)。そのためNPCを活発に増殖する状態に維持することにより、前駆細胞プールを拡大することができ、マウスの大脳皮質を大きくすることができる(Chenn et al., Science 2002, 297: 365-369;Lehmann et al., Eur J Neurosci 2005, 21: 3205-3216)。逆に、増殖状態から分化状態へのNPCの成熟前の移行により、前駆細胞プールを枯渇することができ、短期間で、分化したニューロンおよびグリアの生成を増加することができるが、長期間では、前駆細胞プールの枯渇により、神経子孫のさらなる生成が制限される。NPCの生存を増加するかまたは減少することにより、皮質のサイズを変化させることもできる。(Depaepe et al., Nature 2005, 435: 1244-1250;Putz et al., Nat. Neurosci. 2005, 8: 322-331)。
【0006】
神経発生の破壊は、CNSの疾患および障害において根本的な役割を担い得る。神経細胞の数は、NPCの増殖ならびにその子孫の生存および分化により大体が決定される。これらの工程は独立して制御され得るが、神経系中で適切に機能する回路を確立するためには、これらの工程はまた、適切に調和されなければならない。実際に、ニューロンの生成およびそれらの回路への組み立てにおける誤りは、異所性、精神遅滞、自閉症スペクトラム障害、癲癇、限局性皮質形成異常を含む多くの神経学的障害を引き起こし得る。神経細胞の種類の生成における変化は、自閉症スペクトラム障害、鬱および統合失調症の基礎となり得る、脳における興奮性回路と抑制性回路との異常な割合、不均衡を引き起こし得る。
【0007】
これらの観察は、NPCの増殖を調節する機構およびNPCの子孫の運命の重要な役割を強調する。細胞増殖を制御する機構は、細胞自律的または非細胞自律的に分類され得る。細胞自律的機構に関して、細胞を「多能性(pluripotent)/多能性(multipotent)」状態へと押し進め、その状態に維持するために必要な因子の同定において、近年、大きな進歩がなされた。(Welstead et al., Curr. Opin. Genet. Dev. 2008, 18: 123-129)。
【0008】
しかしながら、インビボにおいて神経活動性に依存する非細胞自律的な機構はあまりよく分かっていない。そのため、インタクトの動物において神経発生を調節する機構を解明することは、脳の発生ならびに健常動物および神経系の障害における機能の適切な理解に重要である。このことはまた、神経系、具体的には認知障害などの神経系の障害における神経発生、具体的には損なわれた神経発生を予防、改善および/または安定化(例えば調節)するための方法および組成物の開発にも重要である。
【0009】
本発明は、活動性依存的様式における神経発生の調節に関連する標的、方法および組成物を提供することにより、当該技術分野におけるこれらのおよび他の必要性を満たす。
【発明の概要】
【0010】
発明の概要
第一の局面において、本開示は、第一の動物のインタクトの脳の領域中の神経前駆細胞と医薬剤とを接触させる工程、第一の動物および第二の対照動物を、インタクトの脳の領域において活動性を誘発し得る外的刺激に曝露する工程、ならびに第一の動物における神経前駆細胞の増殖割合および第二の動物における神経前駆細胞の増殖割合を任意の順序で測定する工程を含む方法に関し、ここで神経前駆被検細胞と神経前駆対照細胞の間の増殖の差により、神経増殖を調節し得るものとしての該医薬剤が同定される。
【0011】
特定の態様において、第一の動物および第二の動物は、両生類および哺乳類などの脊椎動物であり得る。より具体的には、第一の動物および第二の動物はアフリカツメガエルであり得、より具体的にはアフリカツメガエルのオタマジャクシであり得る。
【0012】
特定の態様において、インタクトの脳の領域は、嗅覚入力、視覚入力もしくは機械感覚入力の処理に関与し得るか、または行動出力の媒介に関与し得る。具体的な態様において、第一および第二の動物はアフリカツメガエルであり得、インタクトの脳の領域は、視蓋であり得る。インタクトの脳の領域はまた、菱脳、中脳、後脳/脊髄、網膜または鼻窩(olfactory pit)の回路を含み得る。
【0013】
特定の態様において、実験動物および対照動物中の神経前駆細胞の増殖割合を測定する工程は、第一の動物および第二の動物の視蓋中の細胞の数および型を計測することを含む。
【0014】
いくつかの態様において、神経前駆細胞と医薬剤を接触させる工程は、前記医薬剤を前記神経前駆細胞にエレクトロポレーションすることを含み得る。
【0015】
別の局面において、本開示は、神経前駆被検細胞と、前記神経前駆被検細胞中の1つ以上の遺伝子の発現を調節するのに有効な量の医薬剤とを接触させる工程、神経前駆被検細胞の増殖割合および医薬剤と接触させていない神経前駆対照細胞の増殖割合を測定する工程、ならびに神経前駆被検細胞の増殖割合および神経前駆対照細胞を任意の順序で比較する工程を含む方法に関し、ここで神経前駆被検細胞と神経前駆対照細胞の間の増殖割合の差は、神経前駆細胞の増殖の調節物質として該1つ以上の遺伝子を同定する。
【0016】
特定の態様において、神経前駆被検細胞は、第一の動物中に存在し得、神経前駆対照細胞は、第二の動物中に存在し得る。いくつかの態様において、神経前駆被検細胞および神経前駆対照細胞は、それぞれ、第一の動物および第二の動物のそれぞれの視蓋中に存在し得る。一態様において、第一の動物および第二の動物はアフリカツメガエルであり得る。
【0017】
いくつかの態様において、方法はさらに、神経前駆被検細胞および神経前駆対照細胞中に、レポーター構築物を導入する工程を含み得る。特定の態様において、レポーター構築物は、蛍光タンパク質をコードする遺伝子を含み得る。一態様において、蛍光タンパク質の発現は、空間的に、特に神経前駆細胞などの特定の細胞型に制限され得る。別の態様において、蛍光タンパク質の発現はまた、時間的に制限、例えば特定の時点後に脳の領域で生成された子孫細胞に制限され得る。他の態様において、レポーター構築物を神経前駆被検細胞に導入する工程は、細胞を、レポーター構築物をコードするプラスミドでトランスフェクトすることを含み得る。
【0018】
いくつかの態様において、神経前駆細胞の増殖割合を測定する工程は、少なくとも1つの所定時間の前および後に細胞の数および型を計測することを含み得る。
【0019】
別の態様において、方法はさらに、第一の動物および第二の動物を視覚刺激に曝露することを含み得る。
【0020】
いくつかの態様において、医薬剤は、化合物またはアンチセンスオリゴヌクレオチドを含み得る。特定の態様において、アンチセンスオリゴヌクレオチドは、siRNA、shRNAおよび/またはモルホリノを含み得る。
【0021】
本方法の別の態様において、神経前駆被検細胞中の1つ以上の遺伝子は、配列番号:1〜651、あるいはその機能的短縮体、修飾体および/または置換体から選択され得る。
【0022】
別の局面において、本開示は、神経前駆被検細胞と医薬剤を接触させる工程、神経前駆被検細胞の増殖割合および該医薬剤と接触させていない神経前駆対照細胞の増殖割合を測定する工程、神経前駆被検細胞の増殖割合と神経前駆対照細胞の増殖割合を、任意の順序で比較する工程を含む方法に関し、ここで、神経前駆被検細胞と神経前駆対照細胞の間の増殖割合の差は、増殖を調節し得るものとしての該医薬剤を同定する。
【0023】
いくつかの態様において、方法は、神経前駆被検細胞および神経前駆対照細胞にレポーター構築物を導入する工程を含み得る。特定の態様において、レポーター構築物は、蛍光タンパク質をコードする遺伝子を含み得る。いくつかの態様において、蛍光タンパク質は、神経前駆細胞中で特異的に発現し得る。
【0024】
いくつかの態様において、神経前駆細胞にレポーター構築物を導入する工程は、神経前駆細胞を、レポーター構築物をコードするプラスミドでトランスフェクトすることを含む。
【0025】
いくつかの態様において、神経前駆被検細胞と医薬剤を接触させる工程は、神経前駆被検細胞に医薬剤をエレクトロポレーションすることを含む。
【0026】
特定の態様において、方法はさらに、第一の動物および第二の動物を視覚刺激に曝露することを含み得る。
【0027】
第四の局面において、本開示は、配列番号:1〜651からなる群より選択される標的遺伝子あるいはその機能的短縮体、修飾体および/または置換体を発現する被検細胞に医薬剤を投与する工程、医薬剤を投与していない被検細胞中の標的遺伝子の発現に対して(compared with)、医薬剤を投与した被検細胞中の標的遺伝子の発現を、任意の順序で比較する工程を含む方法に関し、ここで、医薬剤を投与していない被検細胞と比較した、医薬剤を投与した被検細胞中の標的遺伝子の発現の差は、神経の増殖または分化の候補調節物質として該医薬剤を同定する。
【0028】
別の局面において、本開示は、本明細書に記載の方法により同定される医薬剤に関する。
【0029】
さらに別の局面において、本開示は、本明細書に記載される方法により同定される医薬剤を含む医薬組成物に関する。
【0030】
さらに別の局面において、本開示は、本明細書に記載される方法により同定される(identified be)化合物を投与する工程を含む、患者を治療する方法に関する。
【0031】
本発明は、神経細胞の増殖および分化の現象を調査する方法、かかる方法により同定された医薬剤、該医薬剤を含む組成物、ならびにかかる医薬剤または組成物を投与する工程を含む治療方法を含む。したがって、本開示は、神経発生の制御に関与する遺伝子を同定する方法、神経発生を特徴付けて調節するための医薬剤を同定する方法、ならびに神経系、具体的には種々の神経系の障害および/または損傷に対する調節物質および/または治療、例えば認知障害を含む神経系、具体的には神経系の障害における神経発生、具体的には損なわれた神経発生を予防、改善および/または安定化(すなわち調節)する方法および組成物を提供する。本発明はまた、本発明の方法により選択される医薬剤、およびかかる選択された医薬剤を含む医薬組成物、ならびにかかる医薬剤および組成物を患者に投与する方法であって、患者はヒト患者を含み、前記投与は患者、具体的にはヒトにおいて、神経系、具体的には認知障害を含む神経系の障害における、神経発生を調節、具体的には神経発生、具体的には損なわれた神経発生を予防、改善および/または安定化する目的のためである方法を含む。
【図面の簡単な説明】
【0032】
【図1】図1は、アフリカツメガエルオタマジャクシの透明な脳(A);脳の視蓋領域(B)ならびに視蓋葉(tectal lobe)中の細胞の増殖および分化(C)を示す。
【図2】図2は、神経前駆細胞(NPC)の種々の系統(lineage)を示す模式図である。
【図3】図3は、インタクトの脳で画像化された標識細胞の空間的および時間的解像を容易にする増殖レポーターの模式図である。
【図4】図4は、アフリカツメガエルの視蓋における24時間にわたるNPCの増殖を示す画像である。
【図5】図5は、細胞分裂ブロッカーへの曝露後の、蓋(tectal)細胞の増殖割合の減少を示すグラフである。
【図6】図6は、1日目に視覚刺激なしおよび2日目に視覚刺激ありで、細胞分裂ブロッカーの存在下または非存在下での蓋増殖割合(A);ならびに3日目に細胞分裂ブロッカーの存在下または非存在下での蓋ニューロンのパーセント(B)を示すグラフである。
【図7】図7は、対照動物と比較した、視覚刺激ありおよびなしの脱ヨード化酵素ヨードサイロニンIII型(Deiodinase iodothyronine type III)に対するモルホリノ(dio3-MO)を発現する動物における蓋増殖割合を示すグラフである。
【図8】図8は、対照動物と比較した、3日間にわたるグルタチオンS-トランスフェラーゼπ1に対するモルホリノ(GSTπ-MO)を発現する動物における蓋ニューロンおよびグリア細胞のパーセントを示すグラフである。
【図9】図9は、対照動物と比較した、3日間にわたる11の目的の遺伝子(GOI)の1つに対するモルホリノを発現する動物における、蓋増殖割合(A)ならびに蓋ニューロンおよびグリア細胞のパーセント(B)を示すグラフである。GOI:熱ショックタンパク質70(HSPA5);Ephrin受容体B-1型(EphRB1);脱ヨード化酵素ヨードサイロニンIII型(Dio3);ETSドメイン含有タンパク質Elk-4(ELK4);ウイングレスMMTV型(マウス乳癌ウイルス)組み込み部位ファミリー、メンバー7b(Wnt7b);脆弱X精神遅滞、常染色体ホモログ(FXR1);脆弱X精神遅滞タンパク質1(FMR1A);マトリックスメタロペプチダーゼ9(MMP9)、cAMP-依存型タンパク質キナーゼ触媒サブユニットα(PRKACA)、Neuritin 1-A(cpg15);グルタチオンS-トランスフェラーゼπ1(GSTπ)。
【発明を実施するための形態】
【0033】
発明の詳細な説明
そうではないと定義されない限り、本明細書に使用される全ての科学技術用語は、当業者に一般的に理解されるものと同じ意味を有する。本明細書に記載されるものと同様または同等の方法および材料を、本発明の実施または試験に使用できるが、適切な方法および材料が本明細書に記載される。本明細書で言及される全ての刊行物、特許出願、特許および他の参考文献は、その全体において参照により援用される。また、材料、方法および実施例は、例示のためだけであり、限定を意図しない。
【0034】
本発明に包含される基本的な技術を実施するための定義および方法および手段を含む、標準的な実験マニュアルが参照される。例えば、Sambrook et al., Molecular Cloning: A Laboratory Manual, 第3版、Cold Spring Harbor Laboratory Press, New York (2001);Current Protocols in Molecular Biology, Ausubel et al. (編), John Wiley & Sons, Inc., Hoboken, NJ (2010)、Current Protocols in Cell Biology, Bonifacino et al. (編), John Wiley & Sons, Inc., Hoboken, NJ (2010);Current Protocols in Immunology, Coligan et al. (編), John Wiley & Sons, Inc., Hoboken, NJ (2010);Current Protocols in Neuroscience, Gerfen et al. (編), John Wiley & Sons, Inc., Hoboken, NJ (2010);Current Protocols in Nucleic Acid Chemistry, Egli et al. (編), John Wiley and Sons, Inc., Hoboken, NJ (2010);Current Protocols in Pharmacology, Enna et al. (編), John Wiley and Sons, Inc., Hoboken, NJ (2010);およびそこに引用される種々の参考文献を参照されたい。
【0035】
定義
本開示で使用する場合、用語「約」または「およそ」は、当業者により決定される場合の特定の値について許容され得る範囲内にあることを意味し、部分的に、どのようにして値を測定または決定したか、例えば測定系または技術の限界に依存し得る。
【0036】
用語「a」、「an」および「the」は、そうではないと明確に記述されない限り、単数形および複数形の両方を意味すると理解される。したがって「a」、「an」および「the」(および適切な場合はその文法上の変形物)は、一般的に1つ以上のことをいう。
【0037】
本明細書で使用する場合、「神経発生」は、インビボ、インビトロまたはエキソビボにおける神経細胞の増殖、生存、分化および移動を含む。細胞は、中枢神経系または動物もしくはヒト内の他の場所(例えば末梢神経系)に局在し得るか、そこから得られ得るか、またはそこから由来し得る。神経発生は、正常発生中に起こるもの、および疾患、損傷または治療介入後に起こる神経再生としての神経発生を含むことが意図される。開示される発明の態様は、神経発生の非限定的な指標としての増殖または分化のいずれかの検出または測定を含む。
【0038】
「外的刺激」は、神経活動性を誘導し得る単純または複雑な細胞外刺激の任意の型を含むように広く定義される。したがって、外的刺激は、動物の視覚系への入力を含む。外的刺激はまた、嗅覚、機械感覚または視覚入力の処理および行動出力の媒介に関与するものなどの脳の他の領域への入力も含む。
【0039】
本明細書で使用する場合、用語「調節」は、遺伝子の発現、またはRNA分子もしくは同等のRNA分子、例えば非コードRNAおよび1つ以上のタンパク質もしくはタンパク質サブユニットをコードするもののレベルを変化させることを含む。「調節」はまた、調節物質の存在下で発現、レベルまたは活性が、調節物質の非存在下で観察されるものと異なるように、1つ以上の遺伝子産物(非コードRNAを含む)、タンパク質またはタンパク質サブユニットの活性を変化させることも含む。例えば、用語「調節」は、「上方調節」または「下方調節」を意味し得るが、単語「調節」の使用はこれらの定義に限定されない。調節は、発現もしくは活性の増加または減少、遺伝子産物の結合特性の変化、または生物学的活性分子の生物学的、機能的もしくは免疫学的性質の任意の他の変化であり得る。
【0040】
「神経発生の調節」または「神経発生を調節すること」は、細胞の増殖、生存、分化または移動における変化を含む。かかる変化は、インタクトの脳の領域中のものなどの細胞または細胞の集団において起こり得る。非限定的な例としては、NPC増殖に直接関連する遺伝子産物のレベルの変化などの神経発生の誘導体(または阻害剤)のレベルの増加(または減少)が挙げられる。かかる変化はまた、神経回路中の細胞分化または細胞移動の違いも含み得る。特定の態様において、神経発生を調節することは、細胞増殖および細胞運命(例えばニューロン 対 グリア)における影響のことをいう。
【0041】
用語「調節物質」、「化合物」および「医薬剤」は、本明細書において互換的に使用され得、単離された形態の薬理学的に活性な物質またはその混合物を含む。例えば、医薬剤、化合物または調節物質は、単離され構造的に画定された生成物、未知の構造の単離された生成物、いくつかの公知かつ特徴付けられた生成物の混合物、または1つ以上の生成物を含む画定されていない組成物であり得る。かかる画定されていない組成物の例としては、例えば組織試料、生物学的液体(fluid)、細胞上清、植物性調製物(vegetal preparation)等が挙げられる。医薬剤、化合物または調節物質は、ポリペプチド(またはタンパク質もしくはペプチド)、核酸、脂質、多糖、化学実体、またはそれらの混合物もしくは誘導体などの有機または無機の任意の生成物であり得る。医薬剤、化合物または調節物質は、天然または合成起源であり得、化合物(1つまたは複数)または調節物質は化合物のライブラリーを含み得る。
【0042】
「調節物質」、「化合物」または「医薬剤」は、インビボ、インビトロまたはエキソビボにおいて、該剤または試薬の非存在下の神経発生の量、程度または性質と比べて、神経性応答(neurogenic response)の量、程度または性質を増加(または減少)し得る。特定の態様において、かかる「神経性」剤による処置は、神経発生の検出または決定に使用される方法の条件下で、神経性応答の量、程度または性質を、該剤の非存在下の神経性応答の量、程度または性質と比較して、少なくとも約1%、2%、3%、4%、5%、10%、20%、40%、50%、75%、100%、200%(2倍)、300%(3倍)、400%(4倍)、500%(5倍)またはそれ以上もしくは以下だけ増加(または減少)し得る。
【0043】
用語「阻害」、「下方調節」または「低減」は、遺伝子の発現、または1つ以上のタンパク質もしくはタンパク質サブユニットをコードするRNA分子もしくは同等のRNA分子のレベル、または1つ以上の遺伝子産物、タンパク質もしくはタンパク質サブユニットの活性を、本発明の方法において規定されるような1つ以上の調節物質(例えば、siRNA、shRNA、アンチセンスモルホリノ等)の非存在下で観察されるもの未満に減少することを含む。用語「促進」または「上方調節」は、遺伝子の発現、1つ以上のタンパク質もしくはタンパク質サブユニットをコードするRNA分子もしくは同等のRNA分子のレベル、または1つ以上の遺伝子産物、タンパク質もしくはタンパク質サブユニットの活性を、特許請求される方法において規定されるような1つ以上の調節物質の非存在下で観察されるもの未満(below)に増加することを含む。
【0044】
用語「標的遺伝子」または「目的の遺伝子(gene or interest)」は、タンパク質コード遺伝子だけではなく(not just)、非コード遺伝子を含む。かかる非コード遺伝子は、リボソームRNA(rRNA)、転移RNA(tRNA)および核内低分子RNA(snRNA)、ならびにマイクロRNA、snoRNA、siRNA、piRNAおよびncRNAをコードするものを含む。標的遺伝子はまた、複製、転写、翻訳もしくは遺伝子産物の発現に重要な他の過程を制御するポリヌクレオチド領域、または遺伝子産物をコードする領域およびそれに作動可能に連結された、発現を制御する領域の両方を含むポリヌクレオチドを含み得る。標的化遺伝子は、染色体(ゲノム)または染色体外であり得る。標的化遺伝子は細胞に対して内在するもの(endogenous)であり得るか、または外来遺伝子(トランスジーン)であり得る。外来遺伝子は、宿主ゲノムに取り込まれ得るか、またはプラスミドもしくはコスミドなどの染色体外の遺伝子構築物上に存在し得る。標的化遺伝子はまた、生物体または細胞に感染し得るウイルス、細菌、真菌または原生動物などの病原体由来であり得る。標的遺伝子は、ウイルス遺伝子およびプロウイルスの遺伝子であり得る。特定の態様において、標的遺伝子は、神経発生に重要な細胞活動性の進行に関与するかまたは関連するものである。
【0045】
用語「標的核酸」は、その発現または活性が調節される任意の核酸配列を含む。標的核酸は、DNAまたはRNAであり得る。さらに、標的遺伝子または遺伝子は、一般的に生物学的に活性である、より大きな核酸配列の断片を含み得る。
【0046】
添付の図面および実施例により図示され、それらに関連して記載される本開示のいくつかの態様、実施例をここで参照する。本明細書には特定の態様が記載されるが、記載される態様は、添付の特許請求の範囲により規定される本発明の範囲を限定することを意図しないことが理解される。反対に、本開示は、添付の特許請求の範囲に規定される発明の精神および範囲に含まれ得る変形物、改変物および均等物を包含することが意図される。さらに、本開示において、添付の特許請求の範囲に規定される発明の完全な理解を伝達するために、特定の詳細が提供される。しかしながら、これらの詳細なく、特定の態様が実施され得ることが当業者には明らかである。特定の例において、周知の方法、手順または他の具体的な詳細は、添付の特許請求の範囲に規定される発明の局面を不必要に不明瞭にすることを避けるために記載されていない。
【0047】
神経発生に関連する遺伝子の同定および特徴づけの方法
モデル系としてのアフリカツメガエル
アフリカツメガエルは、神経発生および脳の発生のインビボ研究に有利であることが明らかにされている。これらの利点にはいくつかの要因がある:
【0048】
神経発生の密な(compact)期間
カエルのオタマジャクシは、CNS発生の幼生期間の間にわたる、比較的長く利用可能な細胞増殖および分化の期間を有する。アフリカツメガエルの発生の前変態期(pre-metamorphic stage)の経過中に、細胞増殖により新しいニューロンが生成される。新たに形成されたニューロンは、その後発生中のオタマジャクシの脳の機能的な回路に組み込まれる。誕生から個々のニューロンの分化までの神経発生の流れは、アフリカツメガエルでは2〜4日の期間に捕えられ得るのに対して、哺乳動物系では1ヶ月を超える。
【0049】
この密な期間は、神経発生を含む種々の段階の詳細な調査を容易にする。この利点は、神経発生の機構が進化的に保存されているという証拠と合わせると、哺乳動物系に関連のある神経発生の基本的な機構を明らかにすることにおいてだけでなく、ヒトの神経発生的疾患を研究するための実験モデル系を提供することにおいてもツメガエル属の価値を強調する。
【0050】
利用可能な脳回路
カエルにおける神経発生および回路の形成における研究により、その後ヒトを含む哺乳動物において機能的であることが示されたいくつかの脳発生の基本的な機構が明らかにされた。CNSは、アフリカツメガエルの前後軸に沿って高度に局所化(regionalize)され、種々のCNS領域は、感覚的入力処理、情報統合、記憶、意思決定、および運動調節を含む特有の機能を有する、特定の神経回路により構成される。
【0051】
CNSの脳室層における神経前駆体の前後パターン形成は、脳機能のその後の領域分化(region specilization)の基礎となり、神経前駆体中のOtx、PaxおよびHoxなどの転写因子ファミリーの進化的に保存された発現パターン、次いで前駆体およびその神経子孫の領域的サブセットを画定する転写カスケードの活性化により確立されると考えられている(O'Leary et al., Curr. Opin. Neurobiol. 2008, 18: 90-100;Lichtneckert et al., Adv. Exp. Med. Biol. 2008, 628: 32-41)。
【0052】
実際に、Hoxファミリー転写因子は、菱脳(telecephalon)から脊髄への前後軸に沿って前駆体および細胞運命を、調節するだけでなく、ニューロン分化の際に下流の転写因子カスケードを調節することにより発生中の回路におけるニューロン間の接触を指定(specify)もする。(Dasen et al., Curr. Top. Dev. Biol. 2009, 88: 169-200;Dalla Torre di Sanguinetto et al., Curr. Opin. Neurobiol. 2008, 18: 36-43)。
【0053】
本発明の態様は、種々の脳回路および細胞型において、トランスクリプトームを分析するための方法を包含する。これらは、菱脳、中脳および後脳/脊髄、ならびに網膜および鼻窩を含む。これらの領域のそれぞれは、NPC子孫の別個の組によって特徴付けられる。例えば、菱脳は、嗅覚入力を受けて処理し、記憶および運動調節に関与する海馬および基底核に相同な領域を含む(includes regions includes regions)(Maier et al., J Chem Neuroanat, 40(1): 21-35;Brox et al., J Comp Neurol 2004, 474(4): 562-77)。中脳は、機械感覚および視覚入力を処理し(Hiramoto et al., Dev Neurobiol 2009, 69(14): 959-71;Deeg et al., J Neurophysiol 2009, 102(6): 3392-404)、菱脳および脊髄は、行動出力を媒介する(Soffe et al., J Physiol 2009, 587(Pt 20): 4829-44;Orger et al., Nat Neurosci 2008, 11(3): 327-33)。
【0054】
本発明の方法は、別個の脳領域の分析だけでなくかかる回路中の特殊化された細胞型の分析も包含する。それぞれの脳の領域における別個の回路は、領域特異的な回路に特定の性質を付与する、興奮性ニューロンおよび抑制性ニューロンと細胞特異的転写組成(composition)との特有の組合せにより構成されると考えられている。したがって、本発明の方法は、本明細書に記載された脳領域におけるGABA作動性ニューロンおよびグルタミン酸作動性ニューロンなどの同定されたニューロンのトランスクリプトームの分析を含む。
【0055】
いくつかの態様において、方法は、アフリカツメガエルの視蓋領域に関し得る。視蓋は、非哺乳類脊椎動物において主要な視覚中枢であり、中脳(または中脳(mesencephalon))の主要な構成要素を形成する対になった構造をとっており、局所的に(topographically)順序だった様式で網膜繊維からの入力を受け取る。例えば、Dingwell et al., J. Neurobiol. 2000, 44: 246-259参照。
【0056】
直接画像化
NPC増殖および分化は、前変態期(オタマジャクシ段階)のアフリカツメガエルにおいて直接的に観察され得る。図1Aは、脳を四角で示したオタマジャクシの透明な頭部を示す。図1Bは、視蓋を含む脳の領域のより詳細な図である。図1Cは、2時間および6日の時点の視蓋における神経細胞の相対的な位置(BrdUにより可視化)を示し、新たに生成された細胞は、ニューロンへと分化し、該ニューロンは脳室層から離れて移動する。(Wu et al., J. Neuroscience 1999, 19(11): 4472-4483)。
【0057】
したがって、ツメガエル属のオタマジャクシは、インビボにおける時間差画像化で分析できるので、神経前駆細胞(NPC)およびその子孫をインタクトの動物において同定および画像化し得る。これに関して、NPCは、図2に示すように、種々のモードで分裂し得る未分化の放射状グリアである。第一のモードにおいて、単一のNPCは、分裂して2つの娘NPCを形成し得る(例えば「増殖モード」)。第二のモードにおいて、単一のNPCは、分裂して1つの娘NPCおよび1つの娘ニューロンを形成し得る(例えば「混合モード」)。第3の様式において、単一のNPCは、分裂して、2つの娘ニューロンを形成し得る(「分化モード」または「最終様式」。Kriegstein et al., Ann Rev Neurosci 2009, 32: 149-184参照)。
【0058】
神経発生レポーター
かかる画像化により、形態学および発生段階に基づいた異なる細胞型の同定および分析のための方法の基礎が提供される。本願は、かかる分析を容易にするための多数の細胞レポーターの使用を包含する。例えば、かかるレポーターは、神経前駆細胞、およびGABA作動性ニューロンもしくはグルタミン酸作動性ニューロンなどの分化した細胞の異なる集団の(or)タグ化および時間差モニタリングを可能にする。
【0059】
したがって、いくつかの態様において、本方法は、分裂中のNPC細胞に特異的なレポーターを使用し得る。例えば、レポーターは、二成分(binary)Gal4-UAS(上流活性化配列)レポーター系を含み得る(Hartley et al., Proc Nat Acad Sci 2002, 99(3): 1377-1382)。図3は、2つの構成要素:Gal4ドライバー(driver)およびUAS-レポーターを含む例示的なGal4-UASレポーター系を示す。したがって、Gal4の発現を調節する配列は、UAS-レポーターの発現を指令する。
【0060】
図3に示される例において、Gal4の調節領域は、線維芽細胞増殖因子4(FGF4)遺伝子のプロモーター由来の多数のエンハンサー因子を含む。この調節領域の活性化には、増殖中のNPCにおいて発現されるが、分化した非増殖ニューロン中には有意なレベルでは存在しない内因性のsox2/oct3転写因子への結合を必要とする。そのため、この系により、増殖中のNPCにおけるUAS-レポーターの特異的な検出が可能になる。
【0061】
レポーターは、任意の目的のマーカーを含み得る。図3に示されるものなどのいくつかの態様において、レポーターは、蛍光タンパク質 (例えば、Kaede蛍光タンパク質または緑色蛍光タンパク質(GFP))、または検出もしくは可視化が可能な任意の他の適切なレポーターをコードし得る。したがって、活発に分裂する細胞は、レポーター(例えばKaedeまたはGFP)タンパク質を発現する。NPCの場合において、増殖状態にとどまっている娘細胞(例えば増殖モードまたは混合モードで分裂している(ここで1つの娘細胞はNPCである)は、例えばKaede蛍光タンパク質の発現を続ける。対照的に、最終モードのNPCの分裂は、分裂を終了し、従ってKaede蛍光タンパク質をほとんどまたは全く発現しなくなり、細胞分裂(cytokinesis)中に受け継いだ残存Kaedeタンパク質のみを含む、2つの分化したニューロンを生じる。したがって、上述のGal4-UASレポーター系は、NPCの増殖の評価基準(measure)を提供し得る。
【0062】
上述のGal4-UASレポーター系は、例示的なレポーター系である。しかしながら、任意の適切なレポーター系(例えば、一成分系または他の二成分系)を使用してもよい。他の態様において、該レポーター系は、標的化細胞における機能を調節し得る遺伝子産物を含み得る。実際に、該系は、Gal4ドライバーと協調した多数のUAS-レポーターの使用を可能にする。
【0063】
さらに、レポーターは、インビボにおける神経細胞の時間的解像を可能にするなどのさらなる性質を有し得る。1つのかかるレポーターは、図3に示される光変換可能な(photoconvertible)蛍光タンパク質Kaedeである。Kaedeタンパク質は、緑色蛍光放出スペクトルを示すが、それは、UV光、405nmレーザー、または任意の他の適切な光源のいずれかに曝露された場合光変換されて、赤色蛍光放出スペクトルを示し得る。したがって、Kaedeの光変換特性により、NPCの挙動の観察および特徴づけにおける時間的調節物質が利用可能になる。
【0064】
例えば、NPCは、Kaede蛍光エフェクター因子を有する上述のGal4-UASレポーターでトランスフェクトされ得る。レポーターを発現する増殖細胞は、緑色Kaedeタンパク質を有する娘を生じる。好ましい所定の時間(例えば、1時間、2時間、4時間、8時間、12時間、16時間、20時間、24時間、30時間、36時間、48時間、72時間、96時間、168時間、または任意の他の値もしくはその中の値の範囲)後、Kaedeタンパク質を含むNPC細胞を光源(例えば、405nmレーザー、UV光等)に曝露して、Kaedeタンパク質を緑色から赤色へと光変換し得る。新たに生じた子孫は、細胞分裂(cytokinesis)中は赤色Kaedeタンパク質を受け継ぐが、未分化なままでいるこれらのNPCは、緑色Kaedeタンパク質を生成する(Caron et al., Development 2008, 17: 107-117)。したがって、新たに生成された娘細胞は、親細胞と識別され得、緑色対赤色のKaedeタンパク質の相対的なレベルの測定により、親細胞と娘細胞の相対数を決定し得る。この測定はまた、細胞分裂(cell division)のそれぞれの型(例えば、増殖、混合、最終)の相対的な割合の評価を提供し得る。
【0065】
本方法における使用に適切なレポーター系を、当該技術分野で公知の任意の適切な技術(例えばトランスフェクション等)により、目的の細胞または組織に導入し得る。特定の態様において、レポーター系はプラスミド上に構築され得、例えばエレクトロポレーションにより細胞に導入され得る(Haas et al., Differentiation 2002, 70: 148-154)。
【0066】
マイクロアレイ分析によるディファレンシャルスクリーニング
いくつかの態様において、本方法は、神経発生、特にNPC増殖および分化に関与する遺伝子の同定を可能にする。候補遺伝子の同定は、例えば放射状グリア中および分化したニューロン中それぞれの核酸のマイクロアレイ分析により実施され得る。かかるマイクロアレイ技術は当業者に周知である。形態学に基づいて細胞を選択して、マイクロアレイ分析のための処理の前に異なる集団に分類し得る。例えば、細胞の特性に基づいて、細胞をNPCまたは分化したニューロンのいずれかに分類し得る。代替的に、細胞集団(または目的のかかる細胞集団を含む動物)を、分化を促進または抑制のいずれかする条件および/または刺激に曝露し得る。
【0067】
充分に特徴付けられたアフリカツメガエルのオタマジャクシの神経系の発生は、神経発生に関与する候補遺伝子の同定の基礎を提供した。例えば、目的の遺伝子(例えば、配列番号:1〜651)、あるいはその機能的短縮体、修飾体および/または置換体は、アフリカツメガエルにおいて、未分化NPCと比較しての分化したニューロン中で発現された遺伝子の比較マイクロアレイ分析により同定し得る(例えば以下の実施例2参照)。一態様において、アフリカツメガエルの視蓋由来の細胞を、神経発生中の異なる日、例えば1日目および5日目に回収する。1日目に回収した細胞は、高い割合の分化したニューロンを有する5日目に回収した細胞に比べて、高い割合の未分化NPCを有する。5日目に回収した細胞に比べて、1日目に回収した細胞中での発現差異(differential expression)、具体的には有意な(例えば、p<0.05、0.04、0.03、0.02、0.01以下、またはその中の任意の他の値)発現差異を示した遺伝子は、NPCの増殖および分化の制御または調節における目的の遺伝子を示す。
【0068】
別の実験プロトコルにおいて、光、嗅覚合図または機械感覚刺激などの脳の領域への入力の外的刺激を受けるかまたは受けない動物の脳の領域から細胞を回収し得る。一態様において、細胞は、暗所に維持した動物の視蓋および光に曝露した動物から回収される。典型的に、暗所および光に曝露した動物由来の細胞は、同時(例えば12時間、24時間、48時間、72時間等の後)に回収され、次いでそこに含まれる遺伝子のマイクロアレイ分析が行なわれる。本明細書で議論するように(以下の実施例1および2参照)、暗所に維持した動物から回収した細胞は、一般的に、高い割合の未分化NPCを有する(have a have)。同じ時間の間光に曝露した細胞は、一般的に、相対的に高い割合の分化したニューロンを有する。したがって、これらの2つの集団のマイクロアレイ分析により、光に曝露した動物から回収した細胞と比較して、暗所に維持した動物(例えば相対的に高い割合のNPCを有する)由来の細胞中で異なって発現される(p<0.01)遺伝子が明らかにされ得、それによりNPC増殖および分化に関与するとして、これらの遺伝子が同定される。
【0069】
例えば、本明細書に記載されるように、ツメガエル属の光への曝露は、視蓋においてNPCがニューロンに分化することを促進することが示された。したがって、光に曝露された動物は、暗所に維持された対照動物と比較して、より高い割合の分化したニューロンを示し得る。そのため、個々のツメガエル属動物を一定時間(例えば12時間、24時間、1日、2日、3日、4日、7日等)暗条件または明条件のいずれかに曝露し得、次いで、例えば視蓋由来の細胞を動物から回収してマイクロアレイ分析に供し得る。代替的に、より長い時間維持された動物は、ますます発達した視蓋を有する。したがって、24時間後に動物から回収してマイクロアレイにより分析した細胞は、例えば2日、3日、5日、7日またはそれ以上の後に動物から回収したものに比べて、より高い割合のNPCを有し得る。いずれの場合も(明対暗または初期対後期)、第一の動物集団の蓋から回収した細胞集団は、典型的により多くのNPCを有し、第二の動物由来の残りの集団は、典型的により多くの分化したニューロンを有する。したがって、比較マイクロアレイ分析により、NPCにおいて優先的に発現されるこれらの遺伝子が明らかにされ得る。かかる方法により同定される遺伝子は、本明細書に示される配列番号:1〜651を含む。これらの遺伝子は、NPCにおいて優先的に発現され得、従って神経発生(例えばNPC増殖、分化および/または生存)に関与する。
【0070】
インビボにおける候補遺伝子発現の操作
神経発生の制御において局所的環境および非細胞自律的因子の確立された役割(Peunova et al., J. Neurosci 2001, 21(22): 8809-8818;Kreigstein et al., 149-84, Cheng et al., Nat Neurosci 2009, 12(4): 399-408;Javaherian et al., Cereb Cortex 2009, 19 Supp 1: i70-77;Suh et al., Ann Rev Cell Dev Biol 2009; 25: 253-75)は、インタクトの動物において増殖活性の直接的な評価を行なって、神経発生を調節する内因性の細胞的および分子的機構が同定され得る、ツメガエル属などの実験系の重要性を強調する。
【0071】
アフリカツメガエルのCNS(例えば視蓋)中のNPC中の遺伝子発現を調節する医薬剤の使用により、目的の遺伝子がNPCの増殖に関与することが証明される。例えば、既知の遺伝子配列に基づいてモルホリノを設計して、下流に発現される(downstream expressed)目的の遺伝子の産物(例えばRNA、タンパク質)を効果的に抑制し得る。したがって、NPC増殖を制御する遺伝子の同定により、神経発生およびNPC増殖を調節し得る医薬剤のスクリーニングにおける使用のための公知の標的が提供される。
【0072】
神経発生の調節物質のスクリーニングおよび評価の方法
医薬剤
本明細書で使用する場合、医薬剤(薬物)は、薬理学的活性を有する化合物を含み、無機化合物、イオン性物質、有機化合物、有機リガンド、例えば補因子、多糖類、組み換えおよび合成ペプチド、タンパク質、ペプトイド、核酸配列、例えば遺伝子、核酸産物が挙げられる。医薬剤は、個々にスクリーニングされ得る。代替的に、神経活動性、または神経発生に関与する遺伝子の発現を調節する能力について、1つより多くの医薬剤が同時に試験され得る。医薬剤の混合物を試験する場合、記載される方法により選択された医薬剤は、(適切な場合は)別々にされ得、適切な方法(例えばクロマトグラフィー、配列決定、PCR等により同定され得る。
【0073】
コンビナトリアル化学合成または他の方法により作製された医薬剤(例えば有機化合物、組み換えもしくは合成ペプチド、ペプトイド、核酸)の大きなコンビナトリアルライブラリーが試験され得る(例えばZuckerman, R. N. et al., J. Med. Chem., 37: 2678-2685 (1994)およびそこに引用される参考文献参照;また、タグ化化合物に関連するOhlmeyer, M. H. J. et al., Proc. Nat. Acad. Sci. USA, 90:10922-10926 (1993) およびDeWitt, S. H. et al., Proc. Natl. Acad. Sci. USA, 90:6909-6913 (1993);Rutter, W. J. et al., 米国特許第5,010,175号;Huebner, V. D. et al., 米国特許第5,182,366号;ならびにGeysen, H. M., 米国特許第4,833,092号も参照)、それぞれの関連部分は、参照により本明細書に援用される。コンビナトリアルライブラリーから選択された医薬剤が特有のタグを有する場合、クロマトグラフィー法による個々の医薬剤の同定が可能である。本明細書に記載された方法に従い、神経活動性、または神経発生に関与する1つ以上の遺伝子の発現を調節し得る1つ以上の医薬剤の存在について、化学ライブラリー、微生物培養液およびファージディスプレイライブラリーも試験(スクリーニング)され得る。
【0074】
本明細書に開示される方法により、選択された特性(例えば、神経前駆細胞の増殖の調節、標的遺伝子発現の調節等)を示す化合物のスクリーニングまたは同定が可能になり得る。本明細書に開示される方法はまた、神経活性剤(neuroactive pharmaceutical agent)の構造および機能の評価または特徴づけにも使用され得る。例えば、かかる方法により、(例えば、特異性、有効性等に関しておよび/または活性を調節するために前記候補化合物の誘導体をアッセイまたはスクリーニングすることおよびかかる誘導体の活性を親非改変調節物質と比較することにより、活性の評価が可能になる。当業者に認識されるように、例えば、化学実体は、さらなる原子、官能基および/または置換基を用いた、あるいは原子もしくは基の置換を介したホモログ化(homologation)により、構造的に改変され得る。
【0075】
したがって、本開示は、神経活動性化合物、例えば神経細胞機能(1つまたは複数)に活性であるかまたは神経細胞機能を調節する調節物質または化合物をスクリーニング、同定、特徴づけおよび修飾するため、ならびにニューロンに活性であり得るかまたはニューロンを調節し得る化合物を同定および/または特徴づけおよび/または改善するための方法および組成物を提供する。かかる調節物質または化合物は、神経前駆細胞または神経細胞の機能および/または挙動(例えば増殖および分化)が関与し得る神経系の障害の治療に有用であり得る。したがって、特定の態様において、本方法は、神経前駆細胞(NPC)のニューロンへの分化を調節し得る化合物を同定および/または特徴づけおよび/または改善するために使用され得る。
【0076】
本明細書に記載されるような神経活性剤、化合物または調節物質はまた、細胞(具体的にはニューロンまたは神経前駆体)の1つまたは複数の機能を改変(例えば回復または修正)する能力を有する任意の化合物を含み得る。例えば、調節物質の化合物は、細胞(ニューロン)の少なくとも1つの代謝経路または生物学的もしくは機能的特性を変化させ得、ニューロンに対して活性であり、かつ具体的には神経前駆細胞(NPC)のニューロンへの分化を調節し得る化合物を同定および/または特徴づけおよび/または改善し得る。例として、本発明の生物学的に活性な化合物は、正常な表現型を、傷害されたニューロンに回復し得る、またはニューロンに対する傷害の有害な効果を少なくとも部分的に抑制し得る化合物である。特定の態様において、活性化合物は、細胞機構を抑制または活性化するその能力について、代謝経路を刺激または抑制する、生物学的特性を回復する、細胞死を防止する等のその能力について、選択され得る。
【0077】
インビボ分析に適切な医薬剤としては、例えば神経発生において遺伝子の機能の分析を可能にする、GOIのノックダウンのためのモルホリノが挙げられ得る。他の医薬剤としてはshRNA構築物が挙げられ得る。shRNA媒介ノックダウンは、MOと比較して独立したノックダウン方法を提供し、タンパク質発現の細胞型特異的操作を可能にする。ツメガエル属のニューロンにおいてshRNA-媒介ノックダウンを増強するための方法が開発され、多くのshRNA構築物の生成を能率的にするためにプラスミドカセットが利用可能である。したがって、GOIに対するshRNA構築物は、GOI発現の特異的ノックダウンについて試験され得(Chen et al., Front Neurosci 2009, 3: 63)、その後、例えばNPC増殖に対する効果について試験され得る。当然ながら、本明細書に記載される方法またはプロトコルは、細胞生存、ニューロンまたはグリアへのNPCの分化、ならびに脳領域または神経回路中の細胞の移動および集合などの神経発生の任意の他の寄与因子に対して活性な化合物をスクリーニング(または同定、特徴づけもしくは改善)するために使用され得る。
【0078】
本明細書に記載される方法およびプロトコルは、神経生存または発生に活性であり、かつNPCのニューロンへの分化を特異的に調節し得る化合物をスクリーニング(または同定、特徴づけもしくは改善)するために使用され得る。
【0079】
医薬組成物
別の局面において、本発明は、本発明の方法により記載され、同定される医薬剤、調節物質または化合物を含む医薬組成物を提供する。かかる医薬組成物は、例えば神経発生または神経細胞の機能(1つまたは複数)を調節し得るか、NPC分化および/または増殖を調節し得るか、またはニューロンに対して活性であり得るかもしくはニューロンを調節し得、かつ神経前駆細胞または神経細胞の機能および/または挙動(例えば増殖、分化等)が関与し得る神経系の障害の治療に有用であり得る、本明細書に記載される医薬剤を含み得る。
【0080】
本明細書に記載される医薬剤は、医薬組成物として調製され得、例えばヒト患者などの哺乳動物宿主に、選択された投与経路、例えば経口または非経口、静脈内、筋内、局所または皮下経路に適合された種々の形態で投与され得、かつ1つ以上の薬学的に許容され得る賦形剤を含み得る。
【0081】
細胞系アッセイ
本発明の方法はさらに、単離された細胞(例えば神経前駆細胞)または適切な培養細胞株を、1つ以上の候補化合物または調節物質と接触させる工程を含む。細胞は、医薬剤の効果、濃度、細胞集団および/または評価技術に依存して種々の時間の間接触され得る。特定の態様において、細胞は、例えば約1nM〜約1mMの範囲の候補化合物(1つまたは複数)に曝露され得る。本願から逸脱することなく、他の濃度が試験され得ることが理解されるはずである。さらに、それぞれの化合物は、いくつかの濃度で並行して試験され得る。さらに、必要な場合は、リポソーム、カチオン性脂質またはポリマー、ペネトラチン(penetratin)、Tat PDT、アデノウイルス由来のペプチド(例えばペントンまたはファイバー)もしくは他のウイルス由来のペプチド等を含む、化合物の細胞への進入を補助する種々のアジュバントおよび/またはベクターおよび/または生成物が添加され得る。接触は、生きたツメガエル属のオタマジャクシの調製物のためのインキュベーションチャンバーなどの任意の適切な支持体またはデバイス中で実施され得る。
【0082】
候補化合物のプロフィールを決定することは、いくつかの方法に従って実施され得る。特に、化合物の神経活動性を評価するために、例えば細胞数、生存、抗原の発現、特定の遺伝子の転写、および形態学的変化、例えばサイズ、神経突起成長等の種々の終点が測定され得る。
【0083】
具体的な態様において、候補化合物の神経活動性は、任意の化合物の非存在下および/または参照化合物で処理された対照神経細胞集団と比較することにより決定され得る。ニューロンの状態の決定は、種々の物理的測定値、例えば光学特性、種々の波長での蛍光、放出等を評価することによりなされ得る。ランプまたはレーザー等が備え付けられた自動化顕微鏡などの種々の装置が使用され得る。他の技術としては、冷却CCDカメラによる光検出が挙げられる。測定されたシグナルは、例えばピクセルヒストグラム、クラスター分析および形態学解析を含むソフトウェアを使用して、公知の技術に従って処理され得る。
【0084】
本発明は、また、例えば、(i)実験研究のための標的、および/または(ii)調節物質としての、および特に神経学的障害を治療するための医薬組成物の製造のような、本発明の方法によって同定され、選択され、概略(profile)され、または特徴付けられる任意の化合物(またはその誘導体)の使用に関する。
【0085】
いくつかの態様において、細胞分化モードを、細胞を調節剤と接触させることによって調節し得る。他の態様において、例えば、エレクトロポレーション、または細胞に調節剤を導入するのに適した任意の他の技術により目的の細胞に調節剤を導入することによって、細胞分化モードを調節し得る。
【0086】
特定の態様において、医薬剤(pharmaceutical agent)は、他の分裂モードに比べて、増殖モードで分裂する細胞の数または割合を増加させ得る。他の態様において、医薬剤は、増殖モードで分裂する細胞の数または割合を減少させ得る。他の態様において、医薬剤は、混合モードで分裂する細胞の数または割合を増加させ得る。あるいは、医薬剤は、混合モードで分裂する細胞の数または割合を減少させ得る。さらなる態様において、医薬剤は、分化モードで分裂する細胞の数または割合を増加させ得る。あるいは、医薬剤は、分化モードで分裂する細胞の数または割合を減少させ得る。医薬剤の組合せを、また、NPC増殖および分化に対する所望の効果を達成するために使用し得る。
【0087】
本発明の医薬剤は、NPC増殖および分化の調節に関与する公知の遺伝子標的を調節するように、選択され得る。従って、医薬剤(例えば、モルホリノ、siRNA等)は、例えば、NPC増殖および分化において調節的役割を有することが公知である標的遺伝子の翻訳の阻害または上方調節をするために構築され得るか、または選択され得る。従って、標的細胞(1つまたは複数)をかかる医薬剤と接触させること、または標的細胞(1つまたは複数)にかかる医薬剤を導入することによって、NPC挙動の調節がもたらされ得る。
【0088】
インビボアッセイ
本願で公開する方法は、単離された細胞(例えば、神経前駆細胞)または適切な培養された細胞株を、1つ以上の候補化合物または調節物質と接触させる工程をさらに含む。その効果、濃度、細胞集団、および/または評価技術に依存して、細胞を、種々の時間接触させることができる。一般的に、細胞を、1nM〜1mMの範囲の候補化合物(1つまたは複数)に曝露する。他の濃度を本願から逸脱することなく試験し得ることが理解されるはずである。さらに、各化合物を、いくつかの濃度で並行して試験し得る。さらに、必要であれば、化合物が細胞に入り込むのを補助する、リポソーム、カチオン性脂質もしくはポリマー、ペネトラチン、TatPDT、アデノウイルス由来のペプチド(ペントンまたはファイバー)または他のウイルス由来のペプチド等の種々のアジュバントおよび/またはベクターおよび/または生成物を添加できる。
【0089】
候補化合物のプロフィールを決定することは、いくつかの方法に従って実施され得る。特に、例えば、化合物の神経活性(neuro-activity)を評価するために、細胞数、生存、抗原の発現、特定の遺伝子の転写、および、例えば、サイズ、神経突起成長等の形態学的変化等の種々の終点が測定され得る。
【0090】
好ましくは、候補化合物の神経活性は、任意の化合物の非存在下の、および/または参照化合物で処置された対照神経細胞集団との比較により、測定される。ニューロンの状態の決定は、種々の物理的測定、光学的特性、種々の波長での蛍光、発光等を評価することによって行われ得る。ランプまたはレーザーを備えた自動化顕微鏡等を含む種々の装置が使用され得る。他の技術としては、冷却CCDカメラによる光検出が挙げられる。測定されたシグナルは、例えば、ピクセルヒストグラム、クラスター分析および形態学的解析を含むソフトウェアを使用する公知の技術に従って処理され得る。
【0091】
本発明は、また、(i)実験研究のための標的としての、または(ii)神経学的障害を治療するための医薬組成物の製造のための、上記に開示される方法により同定され、選択され、概略され、または特徴付けられる任意の化合物(またはその誘導体)の使用を包含する。
【0092】
いくつかの態様において、細胞分化モードを、細胞と調節剤を接触させることにより調節し得る。他の態様において、例えば、エレクトロポレーション、または細胞に調節剤を導入する任意の他の適切な技術により目的の細胞に調節剤を導入することによって、細胞分化モードを調節し得る。
【0093】
特定の態様において、医薬剤は、他の分裂モードに比べて、増殖モードで分裂する細胞の数または割合を増加し得る。他の態様において、医薬剤は、増殖モードで分裂する細胞の数または割合を減少し得る。なお他の態様において、医薬剤は、混合モードで分裂する細胞の数または割合を増加し得る。あるいは、医薬剤は、混合モードで分裂する細胞の数または割合を減少し得る。さらなる態様において、医薬剤は、分化モードで分裂する細胞の数または割合を増加し得る。あるいは、医薬剤は、分化モードで分裂する細胞の数または割合を減少し得る。医薬剤の組合せを、また、NPC増殖および分化に対する所望の効果を達成するために使用し得る。
【0094】
医薬剤は、NPC増殖および分化の調節に関与する公知の遺伝子標的を調節するように、選択され得る。従って、医薬剤(例えば、モルホリノ、siRNA等)は、NPC増殖および分化に調節的役割を有することが公知の標的遺伝子の翻訳を、例えば、阻害または上方調節するように、構築され得、または選択され得る。従って、標的細胞(1つまたは複数)とかかる医薬剤を接触させること、またはかかる医薬剤を標的細胞(1つまたは複数)に導入することは、NPC挙動の調節をもたらし得る。
【0095】
インビボアッセイ
マイクロアレイおよび/または他のゲノム分析技術により生成されたゲノムデータの解析は、オタマジャクシCNS中のNPCの増殖および分化を調節し得る遺伝子を優先させる(prioritize)ために使用され得る。従って、アンチセンスモルホリノオリゴヌクレオチド(MO)(Eisen et al., Development 2008, 135(10): 1735-43)は、目的の遺伝子(GOI)に対して生成され得、各GOIは、CNS中のBrdU取り込みを評価するための画像化を使用して、細胞増殖の効果に対するモルホリノ媒介ノックダウンによってスクリーニングされる。細胞増殖を増大または低減するGOIについて、いくつかのインビボアッセイが、視蓋における神経発生の制御におけるその機能を試験するために使用され得る(Nedivi et al, Science 1998, 281: 1863-1866; Ewald et al, J Neurosci 2008, 28(4): 850-61; Cantallopset al., Nat. Neurosci. 2000, 3: 498-503; Javaherian et al., Neuron, 2009 (in press); Van Aelst et al., Curr Opin Neurobiol 2004, 14(3): 297-304; Van Keuren-Jensen et al., Dev Neurobiol 2008, 68(11): 1315-24; Wu et al., Science 1998, 279: 222-226; Cline et al., J Physiol 2008, 586(6): 1509-17)。
【0096】
第1の例示的アッセイは、BrdU取り込みとニューロンのマーカーに対する免疫標識を組み合わせる。BrdU標識法の利点は、該方法が、非侵襲性であり、GOIノックダウンまたは過剰発現の細胞増殖に対する効果の比較的ハイスループットのスクリーニングとして使用され得ることである。BrdUとニューロン標識を組み合わせることにより、神経発生におけるGOIの定量的評価が可能になる。オタマジャクシを、BrdUを含む飼育溶液(rearing solution)に曝露し得る。この方法は、増殖細胞を効率的に標識し、BrdU曝露時間についてより良い制御を可能にする。BrdUの接近は、研究された発生期を通して変わらないこと、およびBrdUの取り込みは、DNA損傷に応答して生じないことが示された。BrdU曝露は、典型的には、全ての動物について増殖に対する潜在的概日効果を調節するために、一日の同じ時間に行われる。BrdU曝露の後、動物を、即座に固定するか、または2〜3日飼育し、その後、動物を固定し、BrdU免疫標識のために動物を処理するかのいずれかをする。
【0097】
次いで、動物を終末麻酔し(terminally anesthetize)、マイクロウェーブ固定し(Paupard et al., J. Histochem. Cytochem. 2001, 49(8): 949-956)、脳を、一次抗体を用いた抗体標識のために、全載またはビブラトーム切片(vibratome section)(Peunova et al., 8809-8818)に(in)処理される(マウス抗−BrdU、BD Bioscience;ウサギ抗NeuroD;AbcamおよびAlexafluor二次抗体(Molecular Probes))。
【0098】
第2の例示的アッセイは、Sox2.mFGF4::FP-標識NPC等の適切なレポーターまたはマーカーで標識された細胞のインビボ時間差撮影画像化法を使用する。時間差撮影インビボ画像化実験の利点は、長期的研究により、細胞増殖、細胞の運命およびこれらの事象の基礎をなす構造動力学の直接的観察が可能になることである。GOI過剰発現/ノックダウンによりインビボで神経発生がどのように変更させるかが定量され、細胞分裂速度および子孫の運命が決定され得る。
【0099】
脳室切片、全載固定脳調製物およびインビボFP標識試料中の増殖細胞を、レーザー列ならびにUVを遠赤外蛍光団に分離するためのダイクロイックミラーおよびフィルターを備える顕微鏡に搭載されたスピニングディスク共焦点(spinning disk confocal)結合物を使用して画像化し得る。シグナルは、感度の高い、高シグナル対ノイズのEMCCDカメラで捕捉される。適切な画像捕捉ソフトウェアを使用して、画像を捕捉する。二重標識実験において、ブリードスルー(bleed-through)を排除するために、画像は連続的に捕捉される。単一の蛍光団標識での調節は、ブリードスルーの非存在を確実にするためになされる。
【0100】
インビボ画像時間差撮影のために、麻酔したオタマジャクシを、特別注文の画像化チャンバに配置する。蛍光団についての適切なレーザー/フィルター設定を使用して、1μmのz幅で完全な共焦点スタック(stack)を取得する。固定された組織全載および組織切片について、視蓋(tectal lobe)の完全なzスタック(0.5μmのz間隔)を取得する。
【0101】
時間差撮影データの解析のために、放射状グリア(radial glia)とニューロンを、細胞の三次元構造に基づく形態学的基準に従って区別する。実験の時間経過の間の系統の各細胞の対称増殖性分裂;非対称神経発生分裂;最終対称神経発生分裂の数および順序、ならびに分化の段階を決定する。個々のNPCの時間差撮影画像化により、画像化実験の経過の間の、分裂細胞および分化している細胞における形態学的変化の評価を含む、細胞の運命の同定が可能になる。
【0102】
従って、特定の態様において、本発明の方法は、インビボ、より具体的には、インタクトの脳において、神経前駆細胞(NPC)のニューロンへの分化を変更する医薬剤のスクリーニング(または同定、特徴付け、もしくは改善)に有用である。本発明の方法の利点は、かかる方法が、ツメガエル属(Xenopus)オタマジャクシのインタクトの視覚系中の神経細胞集団等の、インビボの神経細胞集団を使用し得ることである。従って、インビボの神経細胞集団の使用により、化合物または調節物質の生物学的活性の予測的かつ信頼できる評価が可能になる。使用されるNPCおよびニューロンは、哺乳動物起源(例えば、齧歯類、ヒト、霊長類等)およびアフリカツメガエル(Xenopus laevis)等の両生類を含む種々の起源のものであり得る。
【0103】
医薬剤のインビボ活性を、例えば、神経発生に関連する遺伝子のスクリーニングに関して本明細書に記載されるように決定し得る。例えば、ツメガエル属オタマジャクシはインビボ時間差撮影に敏感に反応するので、神経前駆細胞およびその子孫を、インタクトの動物において画像化し得る。従って、NPC細胞をインビボで候補医薬剤と接触させた後、対照動物集団に対する試験動物集団の増殖割合を測定し得(例えば、可視化し得)、候補調節剤(modulating)の活性を、対照集団に対する、候補医薬剤で処置されたまたは候補医薬剤と接触した動物中のNPC増殖の相対速度によって決定する。従って、NPC増殖割合を変更する(例えば、増大または低下させる)医薬剤を、神経発生および神経細胞増殖の調節物質として同定する。
【0104】
神経発生の調節物質としての医薬剤のスクリーニングのためのインビトロ方法、およびインビボ遺伝子スクリーニング方法に関して本明細書に記載される条件(例えば、医薬剤の濃度、読み取り等)を、また、ここでも使用し得る。例えば、インタクト動物CNS領域(例えば、視蓋)中のNPC細胞集団を、蛍光タンパク質(FP)を発現するように、(例えば、エレクトロポレーションによって、)トランスフェクトさせ得る。インタクトの動物中のFP発現細胞を、時間t=0で画像化し得、次いで、所定の時間(例えば、24h、48h、96時間等)後に再度画像化し得、所定の時間(例えば、12時間、24時間、36時間等)の間の間隔で画像化し得る。細胞(例えば、NPCおよび分化したニューロン)の相対的型および数の変化を、所定の時間にわたって、各24時間間隔で、細胞をその形態に従って分類することによって決定し得る。形態の変化を、対照集団に対する、試験集団中のNPCおよびグリア細胞の割合として特徴付けることができる。試験動物集団中の細胞と対照動物集団中の細胞の違いは、候補剤がNPC増殖の調節物質であることを示し得る。
【0105】
特定の態様において、CNS細胞増殖に対するGOI効果のスクリーニングを、脳にGOIに対するモルホリノをエレクトロポレートしたアフリカツメガエル(X.laevis)オタマジャクシにおけるBrdU取り込みを評価するために画像化アッセイを使用して実施し得る。1〜2日後、増殖細胞を、屠殺前に2時間、BrdUに曝露することにより標識し得る。次いで、脳を、全載においてBrdUを検出するために処理し得、脳の端から端までの共焦点画像の完全なzシリーズを収集することによって画像化し得る。全載脳のかかる画像化により、細胞増殖のレベルを定量するための優れた方法が提供される。
【実施例】
【0106】
実施例
本開示は、以下の非限定的な実施例によってさらに例示される。以下の実施例は、例示のみのために理解され、添付の特許請求の範囲により規定される本発明の範囲を限定するとして解釈するべきではない。
【0107】
1.視覚刺激およびNPC増殖
概観
神経前駆細胞(NPC)の増殖および分化を、インタクトのツメガエル属オタマジャクシ中枢神経系の視覚系においてアッセイした。この実験系は、動物を視覚系刺激に曝露するか、または動物の視覚系刺激を与えない(deprive)ことによって神経活動性を操作できる。結果は、NPCの増殖割合が、12時間光/12時間暗所の条件下で飼育された動物に比べて、視覚刺激を与えられない動物において、増大することを示す。24時間視覚刺激を与えられず、その後、24時間視覚刺激を受けた動物は、視覚経験の非存在下の最初の24時間の間増殖割合が上昇し、その後、新たに生成された細胞の大部分が分化することを示した(図6)。これらのデータは、神経活動性を操作することにより、増殖割合および分化の両方が制御されることを示す:低下した神経活動性により増殖が増加し、増大した神経活動性により、前駆細胞のニューロンへの分化が誘発される。
【0108】
方法
蛍光タンパク質(FP)を発現するように、インタクトのツメガエル属オタマジャクシ中のNPCをトランスフェクトした。インタクト動物中のFP発現細胞を、共焦点顕微鏡を使用して画像化した。次いで、画像化の後、動物を、光を通さないチャンバに配置し、その結果、動物は、次の24時間にわたって視覚刺激を受けなかった。FP発現細胞を、再度画像化し、動物をチャンバに配置し、そこで、動物は、24時間視覚刺激を受けた。動物から、三度目の画像を採った。各24時間間隔の間の細胞数の変化および放射状グリア(NPC)またはニューロンとしての細胞の同一性を、それらの形態に従って決定した。24時間あたりの細胞数の変化として、およびNPCまたはニューロンの形態を有する細胞の割合として、データを表した。
【0109】
結果
視覚経験の非存在下では、細胞数は、24時間の間に、+19.9±5.8%増加した(N=12の蓋が分析された)。動物を24時間の視覚刺激に曝露した後、本発明者らは、細胞数の有意な6.3±5.4%の減少を見いだした(ウィルコクソン符号付(singed)順位検定、p=0.01)。細胞を検出するために使用された蛍光マーカーが、増殖の間のみ発現されるプロモーターにより駆動されるので、この負の割合は、細胞が増殖サイクルを離れたことを示す。視覚経験に曝露されなかった動物の別のセットは、2つの24時間の期間の間で増殖割合に有意な変化がなかったことを示した(N=7の蓋が分析された、p=0.4)。
【0110】
視覚刺激に曝露されなかった動物の脳において、定常速度(steady rate)の神経発生があった。視覚刺激の非存在下では、新しいニューロンの割合が、第1の24時間の間隔の間に32.9±11.3%増え、これは、第2の24時間間隔の間の30.1±9.4%の増加からは有意ではなかった(N=7蓋、ウィルコクソン符号付順位検定p=.73)。対照的に、最後の24時間の間隔の間に視覚刺激に曝露された動物の神経発生割合は、第1の24時間の間隔中の79.4±22.9%から、第2の24時間の間隔の間の18.0±9.4%まで有意に減少した(N=12蓋、p=.05)。
【0111】
24時間の視覚経験の非存在下では、グリアの外観を喪失する細胞の割合は、第1の24時間の間に17.1±9.7%の低下、および最後の24時間間隔の間に44.9±10.6%の減少を示す(N=7蓋、p=0.18)。24時間視覚を与えず、その後24時間視覚刺激に曝露された動物由来の細胞がグリアの外観を喪失する割合は、視覚刺激に曝露した後、有意に増大する(前では17.1±9.7%喪失、および後では56.0±10.0%喪失、p=.04)。
【0112】
これらの結果を合わせると、視覚経験が細胞増殖を中断させること、および視覚経験がグリア前駆体がニューロンに分化する割合が上昇することと一致する。
【0113】
2.視覚経験ありなしの調節剤およびNPC増殖
背景および方法
NPC検出
これらの実験は、分裂細胞におけるタンパク質発現を駆動する、細胞型特異的蛍光レポーター系を利用した(図3参照)。レポーターは、線維芽細胞増殖因子4(FGF4)の上流調節因子(regulatory element)の6つのリピートからなる。生体内の(endogenous)Sox2/oct3転写因子は、FGF4調節因子に結合し、Gal4の発現を駆動(drive)し、該Gal4は、次いで、UAS蛍光タンパク質を駆動する。FGF4、Sox2およびoct3は、各々、増殖細胞中で発現し、生体内のSox2/oct3転写因子に依存して、増殖細胞内でレポーター発現のGal4促進性特異性を駆動する。UAS蛍光タンパク質を別の構築物として発現させ;このレポーターは、このレポーター系に調節性および特異性を付加した。
【0114】
上記の蛍光レポーターを発現する蓋細胞を、種々の視覚経験を有するオタマジャクシから、および種々の時間の間、該構築物を発現したオタマジャクシから、採取した。次いで、これらの細胞由来のRNAを処理し、マイクロアレイ比較を行った。さらなる分析のために選択された、同定された目的の遺伝子を、以下の表1にまとめる:
【表1】
表1−マイクロアレイ分析を介して同定された目的の遺伝子
【0115】
種々の時期(age)のオタマジャクシの間(より多くの集団の増殖細胞 対 より多くの集団の最終的に分化した細胞)、および、増殖 対 最終分化の相対レベルにも偏りを加え得る、種々の視覚経験を有するオタマジャクシの間で、比較を行った。複数のマイクロアレイ比較において、その発現に有意な変化を示した遺伝子を優先させた。
【0116】
上記のレポーターを使用して、オタマジャクシの視蓋の増殖細胞において蛍光タンパク質Kaedeを駆動(drive)した。Kaedeの蛍光放出スペクトルは、実験に一時的な制御を加えたUV光または405nmのレーザー源への曝露の後、緑から赤に光転換(photoconvert)され得る。オタマジャクシをレポーターでトランスフェクトした24〜36時間後、全てのKaede発現細胞は、該タンパク質の赤形態に光転換された。類似の方法を使用した公開されたデータ(Caron et al., Development 2008, 135:3259-3269)に基づくと、新たに生じた子孫は、細胞質分裂の間に赤Kaedeを受け継ぐが、新しい緑Kaedeタンパク質を合成し続ける。例えば、Caronら(同書)は、緑 対 赤の相対レベルによって新たに生成された細胞を同定した。
【0117】
組織中に複数で標識された細胞を確実に生じる十分に確立された方法である、エレクトロポレーションにより、視蓋の細胞を増殖レポーターでトランスフェクトした(Haas et al., Differentiation 2002, 70: 148-154)。Gal4駆動体(driver)プラスミド(0.5μg/μl)およびUAS−kaedeプラスミド(0.5μg/μl)を、視蓋の脳室(ventricle)に注射し、次いで、電圧パルスを組織に印加し、該プラスミドを蓋の細胞に駆動した。これらのプラスミドの構築物を、候補遺伝子の翻訳を阻害するように設計されたアンチセンスモルホリノオリゴヌクレオチドと共に共エレクトロポレートした(Eisen et al., Development 2008, 135: 1735-1743)。モルホリノを、0.1mMでエレクトロポレートし、リサミン蛍光タグにより可視化した。周囲光からのUV光によるKaedeの転換を防止するために、動物を暗所に維持した。24〜36時間後、オタマジャクシを麻酔し、各動物の視蓋の端から端までの完全なz−スタックを、3日の連続する日について、各日に収集した。蓋の第1の画像化の後、全ての動物を、第2の画像化まで24時間、暗所に戻した。そのときに、0.2Hzで1秒間光を放つLED光(567nmを放出する。これは、Kaedeタンパク質を転換する波長を超えている)のアレイを有するチャンバーからなる増強された視覚環境に、数匹の動物を曝露した。この視覚刺激は、蓋のニューロンにおけるシナプス駆動を増強することが示されており、シナプス可塑性に有意な変化を生じる(Sin et al., Nature 2002, 419: 475-480)。
【0118】
データ取得および解析
第1日に使用した取得設定を、ピクセル値が第3日までに飽和に達することから保護するために使用した。選択した設定を、実験の間ずっと使用した。取得したz−スタックからの3D情報を使用するVolocityソフトウェア(Improvision, Perkin Elmer)を使用し、対象物の強度およびサイズの標準偏差に基づいて、標識された細胞の細胞体を同定し、かつ選択した。次いで、同定した対象物は、実験者が検証し、細胞形態に基づいて細胞型(グリア、ニューロン、または不確定)に帰属させた。各視蓋から、典型的に、15〜45細胞がトランスフェクトされた。24時間あたりの細胞数の変化として、パーセント増殖を計算した。これらの測定を、平均±s.e.m.として報告する。
【0119】
対照動物、細胞増殖ブロッカーを有する動物、および24時間の増強された視覚刺激を受けた動物における増殖の分析を報告する。マイクロアレイから同定された2つの候補(Dio3およびGstπ)をアンチセンスモルホリノでノックダウンした実験の結果もまた報告する。さらに、視覚経験がDio3モルホリノノックダウンの存在下での増殖に影響することを測定するための実験からの結果を報告する。使用した実験条件を、以下の表2にまとめる:
【表2】
表2−モルホリノノックダウン実験条件
【0120】
結果および考察
Sox2レポーター構築物の発現は、放射状グリアが、ツメガエル属視蓋中の主要な神経前駆細胞であることを示した(図4)。分化した細胞は、もはや、Sox2/oct3を産生し、gal4−UAS−Kaede構築物を駆動しないので、ニューロンは、分化後、緑Kaedeを発現し続けなかった。NPC分裂の大部分は、最後の分裂であることが見出された。
【0121】
細胞分裂ブロッカー
アフィジコリン(150μM)およびヒドロキシ尿素(2%DMSO中20mM)を、細胞に加え、細胞分裂をブロックし、これらの細胞の増殖割合を、DMSOのみを受けた対照動物と比較した。この処理は、ツメガエル属の増殖を中断させる(Harris et al., Neuron 1991, 6: 499-515)。DMSOのみに曝露された対照動物において、24時間で16.4±4.2%の細胞数(N=8視蓋)の平均増加が観察された。アフィジコリンおよびヒドロキシ尿素は、細胞増殖をブロックした(細胞数の増加:0.41±6.28%(N=10、p=0.1、マン−ホイットニー;図5参照)。
【0122】
NPC運命の視覚経験依存性調節
視覚経験の非存在下では、細胞数は、24時間の間に19.9±5.8%増加した(N=12視蓋)。24時間の視覚刺激に動物を曝露した後、6.3±5.4%の細胞数の有意な減少が見出された(p=0.01)。この負の割合は、細胞が増殖サイクルを離れ、徐々に成長している樹状突起分枝(dendritic arbor)中での赤Kaedeレポーターの希釈により、もはや検出可能ではないことを示す。Kaedeレポーターは、増殖の間のみ発現され、従って、最終的に分化した細胞は、増殖モードの間にレポーターを産生したことのみを示す。24時間の視覚刺激を受けなかった対照動物は、2つの24時間の期間の間で増殖割合に有意な差を示さなかった(10.3±7.2%および−0.6±3.5%;N=12分析された視蓋、p=0.23;図6A)。
【0123】
視覚刺激への曝露が蓋中の細胞の運命に影響するかどうかを試験するために、24時間の視覚刺激有り無しの後の標識された集団中のニューロンの割合を比較した。ニューロンの形態を有する細胞の割合は、対照動物に比べると視覚刺激に曝露された動物において、より大きく(75.7±4.1% 対 60.1±4.8%、p=0.02、マン−ホイットニー;図6B)、このことは、視覚刺激が、NPC子孫の分化を推進することを示した。
【0124】
Dio3−モルホリノ発現および神経発生
脱ヨード酵素ヨードサイロニンIII型は、甲状腺ホルモンの活性形態(T3)からヨウ素を除去しそれを効率的に不活性化する、甲状腺ホルモン経路中の酵素である。マイクロアレイ分析は、Dio3発現が活動中の前駆細胞中で増大したことを示唆した。アフリカツメガエル中のT3レベルは、変態期前では低いが、T3受容体の存在により、NPC中で検出され、T3レベルの相対的変化が増殖に影響し得ることを示唆した。増大した増殖は、変態期のアフリカツメガエルオタマジャクシ中の甲状腺ホルモンの上昇および受容体活性化と相関する(Denver et al., Dev Biol 2009, 326: 155-168)。従って、モルホリノ発現を用いたDio3のノックダウンにより、T3レベルが上昇し、従って、増殖が増大するはずである。
【0125】
Dio3に対するモルホリノ(Dio3−MO)でトランスフェクトした動物は、第1の24時間の期間において11.4±17.3%、および第2の24時間の期間の間に33.9±22.2%の細胞数の増加を示した(N=4、図7)。対照的に、対照動物は、第1の24時間の期間に最大の増殖の増大を示し、次いで、続く24時間の期間の間にほとんど増殖を示さない。クローンのようである細胞の異常なクラスターが、Dio3−MOでトランスフェクトした動物の蓋内に見出され、これは、移動のための正常な時間を有さない増殖の異常なパターンと一致した。
【0126】
Dio3−MOを発現する数匹の動物をまた、第2の24時間の期間の間に、視覚刺激に曝露した(図7)。視覚経験により、Dio3−MOでトランスフェクトした動物中(N=5)の増殖の割合が減少することが見出されたが、この相違は、増殖割合の大きな変動のために有意とはみなされなかった。
【0127】
GSTπのノックダウンにより神経分化が促進される
グルタチオンS−トランスフェラーゼπ1(GSTP1)は、多くの疎水性化合物および親電子性化合物と還元グルタチオンとのコンジュゲーションを触媒することにより解毒において重要な役割を果たす、タンパク質のグルタチオンS−トランスフェラーゼファミリーのメンバーである。GST−π1は、癌に対する感受性において役割を果たすと考えられている。GST−π1は、神経前駆細胞中で上方調節されていることが観察され、従って、GST−πのノックダウンは、増殖割合を低減すると予測される。
【0128】
GSTπに対するモルホリノを発現するオタマジャクシの蓋において、顕著な結果を観察した。第1日までのニューロンの数でさえ、対照動物で観察されたニューロンの数より有意に多かった(図8)。試験動物中のGSTπ−MOを発現する蓋細胞の、対照動物(42.0±4.7%)に比べて有意により大きな集団(7.27±4.7%、N=7視蓋)が、第1日までにニューロンに分化した(P=0.002、マン−ホイットニー)。第2日までに、対照集団内の49.2±5.7%に比べて、試験動物の細胞の86.4±3.7%がニューロンに分化した(p=0.001)。このニューロンの割合は、ニューロンの数が第3の期間の間に試験動物において増加することが観察されなかったので、最大を示し得(86.8±1.0%)、対照動物内のニューロンの割合より有意に大きい(60.1±16.6%;p=0.004)。
【0129】
ニューロンの全割合の増大により、神経前駆体の集団が、GSTπ−MOの発現と共に減少することが示される。放射状グリアの割合は、GSTπ−MOを発現する動物および対照動物において3日の期間にわたり減少する(図8)。GSTπ−MOを発現する脳由来の細胞の割合を対照動物と比較することにより、第1日までに有意に少ないグリアが存在し(13.3±4.6%および41.5±4.8%;p=0.005)、これらの相違は、次の2日の間続く(8.4±2.0%および17.4±3.5%、p=0.02;4.8±1.6%および13.3±4.6%、p=0.02)ことが示された。小さな割合の細胞は、放射状グリアまたはニューロンに分類することはできなかった。
【0130】
脆弱X関連遺伝子
Fmr1A。脆弱X精神遅滞タンパク質1は、核から細胞質へのmRNAトラフィッキング(trafficking)およびニューロン内の局所タンパク質翻訳を制御すると考えられているmRNA結合タンパク質である。マイクロアレイデータにより、分化したニューロンに比べると、NPC中のFMR1および82kD FMRP相互作用タンパク質、増殖誘導遺伝子1(AKA核脆弱X精神遅滞タンパク質相互作用タンパク質)の発現は低いことが示唆された。ニューロンの増殖におけるFMRPおよび関連するタンパク質の潜在的な役割は、完全には明らかではない。1つの研究により、FMR1が、NPC増殖を増大させ、分化を変更することが示されたが(Castren et al., Proc Natl Acad Sci USA 2005, 102: 17834-17839)、別の研究(another)により、FMR1はNPCの分化のみを変更することが示された(Bhattacharyya et al., Stem Cells Dev 2008 17: 107-117)。これらの研究の間の考え得る不一致は、これらの研究の各々が、異なる供給源のインビトロ細胞を使用しているという事実のためであり得る。インビトロ研究は、NPCの増殖におけるFMR1および関連する遺伝子の役割を明確にし得る。
【0131】
FXR1.脆弱X精神遅滞、常染色体ホモログ遺伝子は、機能的に類似するタンパク質FMR1およびFXR2と相互作用する。マイクロアレイデータに基づくと、FXR1をノックダウンすることは、分化を阻害することにより増殖を増大し得る。
【0132】
FMR1AおよびFXR1に対するモルホリノは、蓋におけるNPC増殖を減少させるようである。定性的観察があった。
【特許請求の範囲】
【請求項1】
a)第1の動物のインタクトの脳領域中の神経前駆細胞を医薬剤と接触させる工程;
b)該第1の動物および第2の対照動物を視覚刺激に曝露する工程;ならびに
c)該第1の動物における該神経前駆細胞の増殖割合および該第2の動物における神経前駆細胞の増殖割合を測定する工程
を含む方法であって、該神経前駆被検細胞と該神経前駆対照細胞の間の増殖割合の相違により、神経増殖を調節し得る医薬剤として該医薬剤が同定される、方法。
【請求項2】
該インタクトの脳領域が視蓋を含む、請求項1記載の方法。
【請求項3】
該インタクトの脳領域が、嗅覚的入力、視覚的入力、および機械的感覚的(mechnosensory)入力から選択される群の1つ以上の処理に関与しているか、または行動的出力の媒介に関与している、請求項1記載の方法。
【請求項4】
該インタクトの脳領域が、終脳、中脳、菱脳/脊髄、網膜、または鼻窩の回路を含む、請求項1記載の方法。
【請求項5】
該第1の動物および該第2の動物が、アフリカツメガエルである、請求項1記載の方法。
【請求項6】
測定する工程が、該第1の動物および該第2の動物の視蓋中の細胞の数および型を数えることを含む、請求項1記載の方法。
【請求項7】
該神経前駆細胞を医薬剤と接触させる工程が、該医薬剤を該神経前駆細胞にエレクトロポレートすることを含む、請求項1記載の方法。
【請求項8】
a)神経前駆被検細胞を、該神経前駆被検細胞中での1つ以上の遺伝子の発現を調節するのに有効な量の医薬剤と接触させる工程;
b)該神経前駆被検細胞の増殖割合、および該医薬剤と接触されなかった神経前駆対照細胞の増殖割合を測定する工程;ならびに
c)該神経前駆被検細胞の増殖割合と該神経前駆対照細胞の増殖割合を比較する工程
を含む方法であって、該神経前駆被検細胞と該神経前駆対照細胞の間の増殖割合の相違により、該1つ以上の遺伝子が神経前駆細胞の増殖の調節物質として同定される、方法。
【請求項9】
該神経前駆被検細胞が第1の動物中にあり、該神経前駆対照細胞が第2の動物中にある、請求項8記載の方法。
【請求項10】
該神経前駆被検細胞および該神経前駆対照細胞が、それぞれ、該第1の動物および該第2の動物の各々のインタクトの脳領域中にある、請求項9記載の方法。
【請求項11】
該インタクトの脳領域が、嗅覚的入力、視覚的入力、もしくは機械的感覚的入力の処理に関与しているか、または行動的出力の媒介に関与している、請求項10記載の方法。
【請求項12】
該インタクトの脳領域が、終脳、中脳、菱脳/脊髄、網膜、または鼻窩の回路を含む、請求項10記載の方法。
【請求項13】
該第1の動物および該第2の動物が、アフリカツメガエルである、請求項10記載の方法。
【請求項14】
レポーター構築物を、該神経前駆被検細胞および該神経前駆対照細胞に導入する工程をさらに含む、請求項8記載の方法。
【請求項15】
該レポーター構築物が、蛍光タンパク質をコードする遺伝子を含む、請求項14記載の方法。
【請求項16】
蛍光タンパク質が、神経前駆細胞中で特異的に発現する、請求項15記載の方法。
【請求項17】
導入する工程が、該レポーター構築物をコードするプラスミドでトランスフェクトすることを含む、請求項14記載の方法。
【請求項18】
測定する工程が、少なくとも1つの所定の期間の前および後に、細胞の数および型を数えることを含む、請求項8記載の方法。
【請求項19】
該神経前駆被検細胞を医薬剤と接触させる工程が、該医薬剤を該神経前駆被検細胞にエレクトロポレートすることを含む、請求項8記載の方法。
【請求項20】
該第1の動物および該第2の動物を視覚刺激に曝露する工程をさらに含む、請求項9記載の方法。
【請求項21】
該医薬剤が、化合物またはアンチセンスオリゴヌクレオチドを含む、請求項8記載の方法。
【請求項22】
該アンチセンスオリゴヌクレオチドが、SiRNAおよびshRNAおよび/またはモルホリノを含む、請求項21記載の方法。
【請求項23】
該1つ以上の遺伝子が、配列番号:1〜651、あるいはその機能的短縮体(functional truncation)、改変体および/または置換体から選択される、請求項8記載の方法。
【請求項24】
a)神経前駆被検細胞を医薬剤と接触させる工程;
b)該神経前駆被検細胞の増殖割合、および該医薬剤と接触されなかった神経前駆対照細胞の増殖割合を測定する工程;ならびに
c)該神経前駆被検細胞の増殖割合および該神経前駆対照細胞の増殖割合を比較する工程
を含む方法であって、該神経前駆被検細胞と該神経前駆対照細胞の間の増殖割合の相違により、増殖を調節し得る医薬剤として該医薬剤が同定される、方法。
【請求項25】
該神経前駆被検細胞が第1の動物中にあり、該神経前駆対照細胞が第2の動物中にある、請求項24記載の方法。
【請求項26】
該神経前駆被検細胞および該神経前駆対照細胞が、該第1の動物および該第2の動物の各々のインタクトの脳領域中にある、請求項25記載の方法。
【請求項27】
該インタクトの脳領域が、嗅覚的入力、視覚的入力、もしくは機械的感覚的入力の処理に関与しているか、または行動的出力の媒介に関与している、請求項26記載の方法。
【請求項28】
該インタクトの脳領域が、終脳、中脳、菱脳/脊髄、網膜、または鼻窩の回路を含む、請求項26記載の方法。
【請求項29】
該第1の動物および該第2の動物が、アフリカツメガエルである、請求項25記載の方法。
【請求項30】
レポーター構築物を、該神経前駆被検細胞および該神経前駆対照細胞に導入する工程をさらに含む、請求項24記載の方法。
【請求項31】
該レポーター構築物が、蛍光タンパク質をコードする遺伝子を含む、請求項30記載の方法。
【請求項32】
蛍光タンパク質が、神経前駆細胞中で特異的に発現する、請求項31記載の方法。
【請求項33】
導入する工程が、該レポーター構築物をコードするプラスミドでトランスフェクトすることを含む、請求項30記載の方法。
【請求項34】
測定する工程が、少なくとも1つの所定の期間の前および後に、細胞の数および型を数えることを含む、請求項24記載の方法。
【請求項35】
該神経前駆被検細胞を医薬剤と接触させる工程が、該医薬剤を該神経前駆被検細胞にエレクトロポレートすることを含む、請求項24記載の方法。
【請求項36】
該第1の動物および該第2の動物を視覚刺激に曝露する工程をさらに含む、請求項24記載の方法。
【請求項37】
a)医薬剤を、配列番号:1〜651からなる群より選択される標的遺伝子あるいはその機能的短縮体、改変体および/または置換体を発現する被検細胞に投与する工程、ならびに
b)該医薬剤を投与された被検細胞における標的遺伝子の発現を、該医薬剤と接触させていない被検細胞における標的遺伝子の発現と(compared with)比較する工程
を含む方法であって、該医薬剤を投与されていない被検細胞と比較しての、該医薬剤を投与された被検細胞中の標的遺伝子の発現における相違により、神経増殖または神経分化の候補調節物質として該医薬剤が同定される、方法。
【請求項38】
該被検細胞および対照細胞が、神経前駆細胞である、請求項37記載の方法。
【請求項39】
インタクトの脳領域において神経発生の該候補調節物質を評価する工程をさらに含む、請求項37記載の方法。
【請求項40】
該インタクトの脳領域が、アフリカツメガエルの視蓋である、請求項39記載の方法。
【請求項41】
請求項1記載の方法によって同定された医薬剤。
【請求項42】
請求項8記載の方法によって同定された医薬剤。
【請求項43】
請求項24記載の方法によって同定された医薬剤。
【請求項44】
請求項37記載の方法によって同定された医薬剤。
【請求項45】
請求項38記載の医薬剤を患者に投与する工程を含む方法。
【請求項46】
請求項39記載の医薬剤を患者に投与する工程を含む方法。
【請求項47】
請求項40記載の医薬剤を患者に投与する工程を含む方法。
【請求項48】
請求項41記載の医薬剤を患者に投与する工程を含む方法。
【請求項49】
請求項38記載の医薬剤を含む医薬組成物。
【請求項50】
請求項39記載の医薬剤を含む医薬組成物。
【請求項51】
請求項40記載の医薬剤を含む医薬組成物。
【請求項52】
請求項41記載の医薬剤を含む医薬組成物。
【請求項1】
a)第1の動物のインタクトの脳領域中の神経前駆細胞を医薬剤と接触させる工程;
b)該第1の動物および第2の対照動物を視覚刺激に曝露する工程;ならびに
c)該第1の動物における該神経前駆細胞の増殖割合および該第2の動物における神経前駆細胞の増殖割合を測定する工程
を含む方法であって、該神経前駆被検細胞と該神経前駆対照細胞の間の増殖割合の相違により、神経増殖を調節し得る医薬剤として該医薬剤が同定される、方法。
【請求項2】
該インタクトの脳領域が視蓋を含む、請求項1記載の方法。
【請求項3】
該インタクトの脳領域が、嗅覚的入力、視覚的入力、および機械的感覚的(mechnosensory)入力から選択される群の1つ以上の処理に関与しているか、または行動的出力の媒介に関与している、請求項1記載の方法。
【請求項4】
該インタクトの脳領域が、終脳、中脳、菱脳/脊髄、網膜、または鼻窩の回路を含む、請求項1記載の方法。
【請求項5】
該第1の動物および該第2の動物が、アフリカツメガエルである、請求項1記載の方法。
【請求項6】
測定する工程が、該第1の動物および該第2の動物の視蓋中の細胞の数および型を数えることを含む、請求項1記載の方法。
【請求項7】
該神経前駆細胞を医薬剤と接触させる工程が、該医薬剤を該神経前駆細胞にエレクトロポレートすることを含む、請求項1記載の方法。
【請求項8】
a)神経前駆被検細胞を、該神経前駆被検細胞中での1つ以上の遺伝子の発現を調節するのに有効な量の医薬剤と接触させる工程;
b)該神経前駆被検細胞の増殖割合、および該医薬剤と接触されなかった神経前駆対照細胞の増殖割合を測定する工程;ならびに
c)該神経前駆被検細胞の増殖割合と該神経前駆対照細胞の増殖割合を比較する工程
を含む方法であって、該神経前駆被検細胞と該神経前駆対照細胞の間の増殖割合の相違により、該1つ以上の遺伝子が神経前駆細胞の増殖の調節物質として同定される、方法。
【請求項9】
該神経前駆被検細胞が第1の動物中にあり、該神経前駆対照細胞が第2の動物中にある、請求項8記載の方法。
【請求項10】
該神経前駆被検細胞および該神経前駆対照細胞が、それぞれ、該第1の動物および該第2の動物の各々のインタクトの脳領域中にある、請求項9記載の方法。
【請求項11】
該インタクトの脳領域が、嗅覚的入力、視覚的入力、もしくは機械的感覚的入力の処理に関与しているか、または行動的出力の媒介に関与している、請求項10記載の方法。
【請求項12】
該インタクトの脳領域が、終脳、中脳、菱脳/脊髄、網膜、または鼻窩の回路を含む、請求項10記載の方法。
【請求項13】
該第1の動物および該第2の動物が、アフリカツメガエルである、請求項10記載の方法。
【請求項14】
レポーター構築物を、該神経前駆被検細胞および該神経前駆対照細胞に導入する工程をさらに含む、請求項8記載の方法。
【請求項15】
該レポーター構築物が、蛍光タンパク質をコードする遺伝子を含む、請求項14記載の方法。
【請求項16】
蛍光タンパク質が、神経前駆細胞中で特異的に発現する、請求項15記載の方法。
【請求項17】
導入する工程が、該レポーター構築物をコードするプラスミドでトランスフェクトすることを含む、請求項14記載の方法。
【請求項18】
測定する工程が、少なくとも1つの所定の期間の前および後に、細胞の数および型を数えることを含む、請求項8記載の方法。
【請求項19】
該神経前駆被検細胞を医薬剤と接触させる工程が、該医薬剤を該神経前駆被検細胞にエレクトロポレートすることを含む、請求項8記載の方法。
【請求項20】
該第1の動物および該第2の動物を視覚刺激に曝露する工程をさらに含む、請求項9記載の方法。
【請求項21】
該医薬剤が、化合物またはアンチセンスオリゴヌクレオチドを含む、請求項8記載の方法。
【請求項22】
該アンチセンスオリゴヌクレオチドが、SiRNAおよびshRNAおよび/またはモルホリノを含む、請求項21記載の方法。
【請求項23】
該1つ以上の遺伝子が、配列番号:1〜651、あるいはその機能的短縮体(functional truncation)、改変体および/または置換体から選択される、請求項8記載の方法。
【請求項24】
a)神経前駆被検細胞を医薬剤と接触させる工程;
b)該神経前駆被検細胞の増殖割合、および該医薬剤と接触されなかった神経前駆対照細胞の増殖割合を測定する工程;ならびに
c)該神経前駆被検細胞の増殖割合および該神経前駆対照細胞の増殖割合を比較する工程
を含む方法であって、該神経前駆被検細胞と該神経前駆対照細胞の間の増殖割合の相違により、増殖を調節し得る医薬剤として該医薬剤が同定される、方法。
【請求項25】
該神経前駆被検細胞が第1の動物中にあり、該神経前駆対照細胞が第2の動物中にある、請求項24記載の方法。
【請求項26】
該神経前駆被検細胞および該神経前駆対照細胞が、該第1の動物および該第2の動物の各々のインタクトの脳領域中にある、請求項25記載の方法。
【請求項27】
該インタクトの脳領域が、嗅覚的入力、視覚的入力、もしくは機械的感覚的入力の処理に関与しているか、または行動的出力の媒介に関与している、請求項26記載の方法。
【請求項28】
該インタクトの脳領域が、終脳、中脳、菱脳/脊髄、網膜、または鼻窩の回路を含む、請求項26記載の方法。
【請求項29】
該第1の動物および該第2の動物が、アフリカツメガエルである、請求項25記載の方法。
【請求項30】
レポーター構築物を、該神経前駆被検細胞および該神経前駆対照細胞に導入する工程をさらに含む、請求項24記載の方法。
【請求項31】
該レポーター構築物が、蛍光タンパク質をコードする遺伝子を含む、請求項30記載の方法。
【請求項32】
蛍光タンパク質が、神経前駆細胞中で特異的に発現する、請求項31記載の方法。
【請求項33】
導入する工程が、該レポーター構築物をコードするプラスミドでトランスフェクトすることを含む、請求項30記載の方法。
【請求項34】
測定する工程が、少なくとも1つの所定の期間の前および後に、細胞の数および型を数えることを含む、請求項24記載の方法。
【請求項35】
該神経前駆被検細胞を医薬剤と接触させる工程が、該医薬剤を該神経前駆被検細胞にエレクトロポレートすることを含む、請求項24記載の方法。
【請求項36】
該第1の動物および該第2の動物を視覚刺激に曝露する工程をさらに含む、請求項24記載の方法。
【請求項37】
a)医薬剤を、配列番号:1〜651からなる群より選択される標的遺伝子あるいはその機能的短縮体、改変体および/または置換体を発現する被検細胞に投与する工程、ならびに
b)該医薬剤を投与された被検細胞における標的遺伝子の発現を、該医薬剤と接触させていない被検細胞における標的遺伝子の発現と(compared with)比較する工程
を含む方法であって、該医薬剤を投与されていない被検細胞と比較しての、該医薬剤を投与された被検細胞中の標的遺伝子の発現における相違により、神経増殖または神経分化の候補調節物質として該医薬剤が同定される、方法。
【請求項38】
該被検細胞および対照細胞が、神経前駆細胞である、請求項37記載の方法。
【請求項39】
インタクトの脳領域において神経発生の該候補調節物質を評価する工程をさらに含む、請求項37記載の方法。
【請求項40】
該インタクトの脳領域が、アフリカツメガエルの視蓋である、請求項39記載の方法。
【請求項41】
請求項1記載の方法によって同定された医薬剤。
【請求項42】
請求項8記載の方法によって同定された医薬剤。
【請求項43】
請求項24記載の方法によって同定された医薬剤。
【請求項44】
請求項37記載の方法によって同定された医薬剤。
【請求項45】
請求項38記載の医薬剤を患者に投与する工程を含む方法。
【請求項46】
請求項39記載の医薬剤を患者に投与する工程を含む方法。
【請求項47】
請求項40記載の医薬剤を患者に投与する工程を含む方法。
【請求項48】
請求項41記載の医薬剤を患者に投与する工程を含む方法。
【請求項49】
請求項38記載の医薬剤を含む医薬組成物。
【請求項50】
請求項39記載の医薬剤を含む医薬組成物。
【請求項51】
請求項40記載の医薬剤を含む医薬組成物。
【請求項52】
請求項41記載の医薬剤を含む医薬組成物。
【図1】

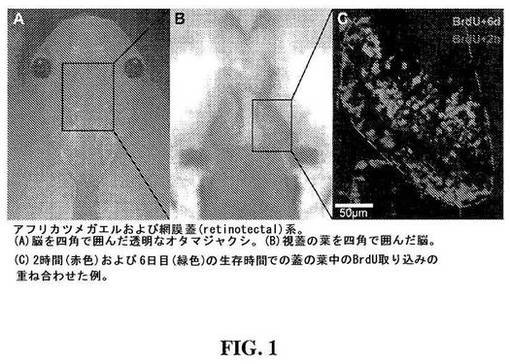
【図2】
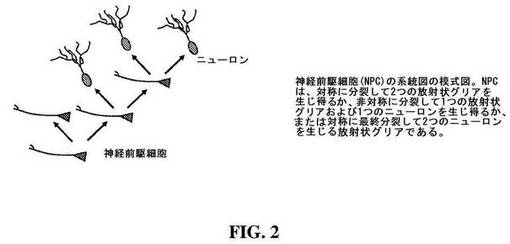
【図3】
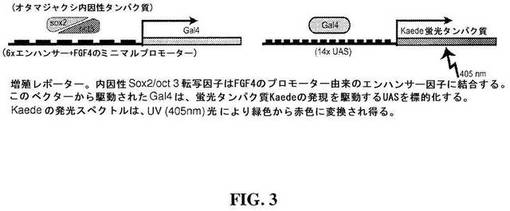
【図4】
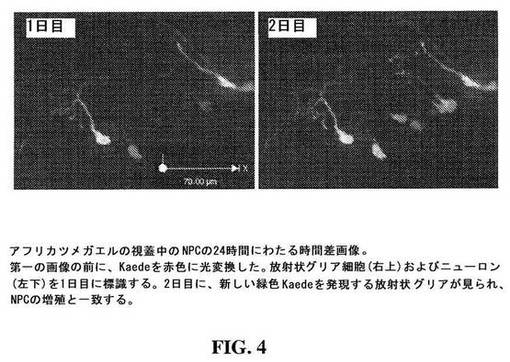
【図5】
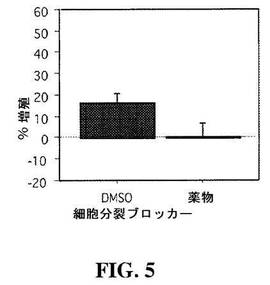
【図6】
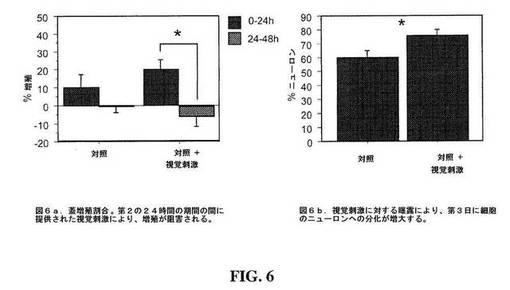
【図7】
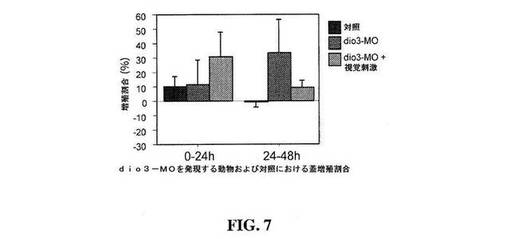
【図8】
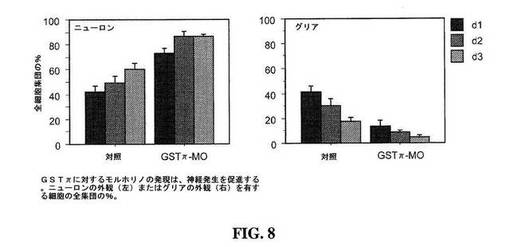
【図9】
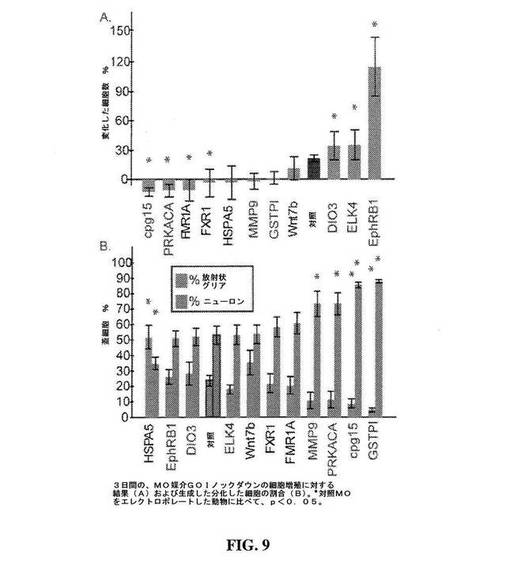
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2013−505738(P2013−505738A)
【公表日】平成25年2月21日(2013.2.21)
【国際特許分類】
【出願番号】特願2012−532268(P2012−532268)
【出願日】平成22年9月29日(2010.9.29)
【国際出願番号】PCT/US2010/050695
【国際公開番号】WO2011/041393
【国際公開日】平成23年4月7日(2011.4.7)
【出願人】(512081063)ダート ニューロサイエンス (ケイマン) エルティーディー (1)
【Fターム(参考)】
【公表日】平成25年2月21日(2013.2.21)
【国際特許分類】
【出願日】平成22年9月29日(2010.9.29)
【国際出願番号】PCT/US2010/050695
【国際公開番号】WO2011/041393
【国際公開日】平成23年4月7日(2011.4.7)
【出願人】(512081063)ダート ニューロサイエンス (ケイマン) エルティーディー (1)
【Fターム(参考)】
[ Back to top ]