
神経発生に関する方法および物質
ドーパミン作動性ニューロンの発生を促進し、ドーパミン作動性の表現型を有する神経細胞を産生させる方法。ドーパミン作動性神経細胞はパーキンソン病などの神経変性疾患に罹患している個体の治療に使用し得る。ドーパミン作動性細胞を個体の脳内に移植してもよく、かつ/またはドーパミン作動性神経の発生を個体の脳内で誘導もしくは増強させてもよい。方法は、NG4Aサブファミリーの核内受容体、たとえばNurr1を細胞内で基底レベルより高く発現させ、その細胞をWntリガンドで処理することによって増殖、自己再生、生存および/またはドーパミン作動性の誘導、分化、生存もしくはニューロンのドーパミン作動性の表現型の獲得を生じさせるあるいは増強させることを含む。細胞はアストロサイトまたはグリア細胞と共に同時培養してよく、FGF成長因子と接触させてもよい。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、神経幹細胞または神経始原細胞もしくは前駆細胞、あるいは他の幹細胞におけるニューロンとなる運命の誘導に関する。本発明は、特定のニューロン表現型の誘導および誘導の増強、詳細には中脳ドーパミン作動性ニューロン表現型の誘導および誘導の増強に関する。
【背景技術】
【0002】
パーキンソン病(PD)とは非常に多く見られる神経変性障害であり、その病因は中脳ドーパミン作動性(DA)ニューロンの選択的かつ進行性な損失によって特徴づけられる。ニューロン表現型の誘導を増強すると、パーキンソン病および他の重篤な衰弱性の神経変性障害を治療できる可能性が出てくる。
【0003】
これまでは、結果が陽性のパーキンソン病患者にヒト胎児中脳組織を移植していたが、本発明を利用する特異的細胞置換治療の開発により、このような従来手法に伴う実用的問題や倫理的問題が克服される。具体的には、本発明により移植用の細胞調製物の開発が可能となり、一方、胚組織または胚細胞を使用する必要性が低減または完全に排除される。幹細胞は、通常は廃棄される組織である臍帯から得られ得る。別の選択肢は、たとえば骨髄、血液、皮膚、眼、嗅球または嗅上皮から成人幹細胞を得ることである。
【0004】
これまでに(国際公開公報WO00/66713号およびWagner他、1999)、本発明者らの研究室において、ドーパミン作動性ニューロン表現型の誘導がNurr1を発現する細胞中で、腹側中脳の1型アストロサイト/初期グリア細胞から得られる1つまたは複数の因子の存在下で増強されることを示した。本発明は、Wnt因子がNurr1を発現する細胞のニューロン表現型の誘導を増強させることに有用であるという実験的知見に基づいている。
【0005】
詳細には、本発明者らは、DAニューロンが誕生する時点までに背側中脳よりも高いレベルでVM内において発現されている全てのWntが、神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞における増殖、自己再生、ドーパミン作動性の誘導、生存、分化および/あるいは成熟を増強させることによってドーパミン作動性ニューロンの発生を誘導させるまたは促進することに有用であることを見出した。
【0006】
本発明者らは、以下を見出した。
【0007】
Wnt-1は、ドーパミン作動性の前駆体の増殖ならびにドーパミン作動性の前駆細胞および/または幹細胞のドーパミン作動性ニューロンへの成熟を促進すること;
Wnt-7aは、ドーパミン作動性の前駆体の増殖を促進し、これらがドーパミン作動性ニューロンへと分化することを可能にすること;
Wnt-3aは、ドーパミン作動性の前駆細胞および/または幹細胞の増殖および/または自己再生を促進すること;
Wnt-2は、細胞周期離脱およびNurr1+前駆体によるドーパミン作動性ニューロン表現型の獲得を促進すること;
Wnt-5aが、神経幹細胞、前駆細胞もしくは始原細胞においてドーパミン作動性の表現型を誘導させること、およびニューロン細胞においてドーパミン作動性の誘導または分化を増強させることに最も有効であること。
【0008】
Wnt-1は、ドーパミン作動性の前駆細胞および/または幹細胞の増殖および成熟を促進することに関してWnt-3aおよびWnt-5aよりも有効である。
【0009】
特定のニューロン表現型の誘導は、遺伝的および後成的シグナルのどちらもの組込みを必要とする。発生中の中脳では、ドーパミン作動性ニューロンの誘導はオーファン核内受容体Nurr1を必要とするが(Zetterstrom他、1997; Saucedo-Cardenas他、1998; Castillo他、1998)、Nurr1の発現は神経幹細胞においてドーパミン作動性の表現型を誘導させるのに十分ではない(Wagner他、1999)。本発明者らは以前に、Nurr1と発生中の腹側中脳1型アストロサイト/初期グリア細胞由来の未知の可溶性シグナルとの組み合わせが、神経幹細胞において中脳ドーパミン作動性の表現型を誘導させるのに十分であると報告した(Wagner他、1999)。本発明者らは本明細書中で、Wnt-5aがこのようなシグナルの一部であり、またWnt-1、-2、-3a、-5aおよび-7aを含めたWntタンパク質ファミリーのメンバーが発生的に制御されており、かつ中脳ドーパミン作動性ニューロンの発生を示差的に制御することを述べる。部分的に精製したWnt-1、-2、-5a、および-7aは2つの異なる機構によりE14.5中脳DAニューロンの数を増加させたが、Wnt-3aは増加させなかった。Wnt-1および-7aは主にNurr1前駆体の増殖を増大させ、それらがドーパミン作動性ニューロンへと分化することを可能にした。Wnt-2は細胞周期離脱およびNurr1+前駆体によるドーパミン作動性ニューロン表現型の獲得を支持した。Wnt-5aは主にニューロンのDA表現型を獲得したNurr1前駆体の割合を増加させた。本発明者らの知見と一致して、Wnt-5aはNurr1を発現する中脳または皮質E13.5前駆体においてドーパミン作動性ニューロンを誘導させることに関して、中脳アストロサイト/初期グリア細胞と同様に有効であった。さらに、Frizzled8のシステインリッチドメインは、Nurr1を発現する神経前駆培養物におけるドーパミン作動性の表現型を有する細胞の増加に対する基底効果およびVM T1A-、Wnt-1またはWnt-5aに媒介される効果、ならびに神経幹細胞またはFGF-8で拡大させたNurr1+中脳ニューロスフェアに対する内因性Wntの効果を有効に遮断した。したがって、本明細書中に含めるデータは、それぞれのWntが部分的に異なる機構によってNurr1を発現する前駆細胞/幹細胞におけるDA表現型を有するニューロンの産生をそれぞれ独立して調節することの指標を提供する。
【0010】
これらの知見により、Wntリガンドが腹側中脳の神経発生中における増殖、自己再生、分化および運命決定の重要なレギュレーターとして位置づけられる。さらに、本発明者らの結果により、in vitroにおける中脳DAニューロンの大規模な産生、およびパーキンソン病などの神経変性疾患の治療における幹細胞置き換え戦略の将来の実行の道が開かれる(BjorklundおよびLindvall、2000; PriceおよびWilliams、2001; Arenas、2002; RossiおよびCattaneo、2002; Gottlieb他、2002)。
【0011】
胚性、神経性および多能性幹細胞は、ニューロン、アストロサイトおよびオリゴデンドロサイトを含めた神経細胞系列へと分化する能力を有する。さらに、幹細胞を単離し、拡大させて、脳移植用の原料物質として使用することができる(Snyder, E.Y.他、Cell 68、33-51 (1992); Rosenthal, A.、Neuron 20、169-172 (1998); Bain他、1995; Gage, F.H.他、Ann. Rev. Neurosci. 18、159-192 (1995); Okabe他、1996; Weiss, S.他、Trends Neurosci. 19、387-393 (1996); Snyder, E.Y.他、Clin. Neurosci. 3、310-316 (1996); Martinez-Serrano, A.他、Trends Neurosci. 20、530-538 (1997); McKay, R.、Science 276、66-71 (1997); Deacon他、1998; Studer, L.他、Nature Neurosci. 1、290-295 (1998); BjorklundおよびLindvall、2000; Brustle他、1999; Lee他、2000; Shuldiner他、2000および2001; Reubinoff他、2000および2001; Tropepe他、2001; Zhang他、2001; PriceおよびWilliams、2001; Arenas、2002; Bjorklund他、2002; RossiおよびCattaneo、2002; Gottlieb他、2002)。
【0012】
ほとんどの神経変性疾患はニューロン集団を冒す。さらに、損傷のほとんどが特定の神経化学的な表現型に起こる。たとえばヒトのパーキンソン病では、失われる主な細胞型は中脳ドーパミン作動性ニューロンである。神経組織の移植による特定のニューロン集団の機能置換は、神経変性疾患を治療するための魅力的な治療戦略を表している(Rosenthal, A.、Neuron 20、169-172 (1998))。別の代替方法は、幹細胞、始原細胞もしくは前駆細胞の再生を促進する、またはその発生および/もしくは動員を修復または導くために必要なシグナルを直接注入すること、あるいはこのような機能を調節する薬物を投与することであろう。
【0013】
幹細胞/始原細胞または前駆細胞は、拡大することができ、かつ特定のニューロン表現型を担うように指示できるので、移植治療用の理想的な材料である。これらの細胞では、移植用にヒト胎児組織を使用することに絡む倫理的問題および実用的問題が回避されるであろう。具体的には、移植した非自己組織の生存度は制限されており免疫系に拒絶され得る。さらに、それぞれの胎児からは少数の細胞しか提供されない。
【0014】
幹細胞または始原細胞もしくは前駆細胞において単一の特定のニューロン表現型を誘導させることは理解しにくいことが分かっている。
【発明の開示】
【0015】
本発明は、細胞中におけるドーパミン作動性ニューロン表現型の誘導を提供する。
【0016】
本発明は、ドーパミン作動性ニューロンの発生の誘導を可能とする。したがって、培養物中もしくは脳内におけるWntレベルおよび/または機能を増大させることによって、本発明は以下の誘導または促進を可能とする:ドーパミン作動性の前駆体、始原細胞もしくは幹細胞の増殖および/または自己再生;ならびに/あるいはドーパミン作動性ニューロン、前駆細胞、始原細胞または幹細胞の生存、分化および成熟の促進(ドーパミン作動性ニューロンの収量を増加させる);ならびに/あるいはin vitroまたはin vivoにおける幹細胞、始原細胞、前駆細胞もしくはニューロン細胞のニューロン性ドーパミン作動性の運命の誘導。
【0017】
本発明の任意の態様または実施形態はニューロン細胞、すなわちニューロンに適用することができ、またはそれを使用することができる。本開示における「神経細胞」はニューロン細胞であり得る。
【0018】
ドーパミン作動性ニューロンに富んだ細胞調製物を、パーキンソン病または他の障害における細胞置換治療に、ならびにドーパミン作動性ニューロンにおけるシグナル伝達現象およびin vitroにおけるドーパミン作動性ニューロンに対する薬物の効果を研究するために、たとえばハイスループットスクリーニングにおいて使用し得る。
【0019】
本発明の態様および実施形態を、添付の特許請求の範囲に示すように提供する。
【発明を実施するための最良の形態】
【0020】
一態様では、本発明は、幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞においてドーパミン作動性のニューロンとなる運命を誘導させる方法、あるいはニューロン細胞においてドーパミン作動性の誘導または分化を増強させる方法、あるいはドーパミン作動性前駆細胞もしくは始原細胞またはNurr1を発現する幹細胞を拡大させる方法であって、
細胞内でNurr1サブファミリーの核内受容体を基底レベルより高く発現させること、
および
細胞をWntリガンドで処理すること、
によってドーパミン作動性ニューロンを産生させることを含む方法を提供する。
【0021】
Nurr1サブファミリーはNR4Aサブファミリーとしても知られており、Nurr1/NR4A2、Nor1/NR4A3およびNGFI-B/NR4A1が含まれる。したがって、本発明の方法は、細胞内でたとえばNurr1/NR4A2、Nor1/NR4A3および/またはNGFI-B/NR4A1を基底レベルより高く発現させることを含み得る。好ましくは、Nurr1サブファミリーの核内受容体はNurr1である。したがって、本発明の方法は細胞内においてNurr1を基底レベルより高く発現させることを含むことが好ましい。
【0022】
本発明は、神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、ドーパミン作動性の誘導、生存、分化および/あるいは成熟を増強させることによってドーパミン作動性ニューロンの発生を誘導させるまたは促進する方法であって、
細胞内においてNurr1を基底レベルより高く発現させること、および
細胞をWntリガンドで処理すること、
によって増殖、自己再生、生存および/またはドーパミン作動性の誘導、分化、生存もしくはニューロンのドーパミン作動性の表現型の獲得を生じさせるあるいは増強させることを含む方法を提供する。
【0023】
Wntリガンドを用いた処理は、in vivo、ex vivo、または培養下であり得る。
【0024】
本発明の方法では、Wntリガンドを用いた処理とは細胞をリガンドと接触させることによりものとし得る。Wntリガンドを用いた処理とは、精製したWntリガンドおよび/もしくは組換えWntリガンドを幹細胞、始原細胞もしくは前駆細胞を含む培養物に供給すること、またはそのような細胞にin vivoで供給することによるものとし得る。Wntリガンドを用いた処理は、Wnt核酸またはタンパク質の1つもしくは複数のコピーを細胞内に導入することを含み得る。細胞を核酸で形質転換させる方法および細胞内にタンパク質を導入する方法を以下に詳述する。Wntリガンドと接触させることとは、in vivoにおいて、あるいは幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞を含む培養物中に、Wntリガンドを産生する細胞を提供することにより得る。Wntリガンドを産生する細胞は、組換え発現によってWntリガンドを産生する組換え宿主細胞であり得る。同時培養した宿主細胞をWntリガンドをコードしている核酸で形質転換させてもよく、かつ/または同時培養した細胞は導入されたWntタンパク質を含んでいてもよい。Wntタンパク質またはWntをコードしている核酸を当技術分野で利用可能な技術に従って細胞内に導入してよく、それらの例を以下に記載する。
【0025】
同時培養した細胞または宿主細胞は、別の幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞であり得る。Wntリガンドを用いた処理はまた、細胞においてWntの発現をアップレギュレーションすること、またはWntリガンドの阻害分子をダウンレギュレーションもしくは阻害することにもより得る。したがって、Wntリガンドを用いた処理は、SFRP、WIF、dkkまたはCerberusなどのWntと相互作用する分子の発現または活性を低減させることによって起こりうる(Martinez Arias他、1999; http://www.stanford.edu/〜rnusse/wntwindow.html、または任意のウェブブラウザを使用して見つけることができる)。
【0026】
Wntリガンドの供給に加えて、幹細胞または神経幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞は1型アストロサイト/グリア細胞と同時培養中であるか、あるいはin vitroまたはin vivoにおいてこのような細胞またはそれらに由来する因子と接触しているかもしれない。しかし、本発明によれば、Wntリガンドの提供は1型アストロサイトに依存しておらず、Wntリガンドを外部から供給しない神経幹細胞または神経始原細胞および1型アストロサイトを含む同時培養物は本発明の範囲に企図されておらず、またこのような同時培養を使用する方法もそうである。
【0027】
Nurr1(Law他、1992; Xing他、1997; Castillo, 1997; GenBank第S53744号、第U72345号、第U86783号)は、甲状腺ホルモン/レチノイン酸核内受容体スーパーファミリーの転写因子である。以前に国際公開公報WO00/66713号およびWagner他、1999に示されているように、神経幹細胞または神経始原細胞における基底レベルより高いNurr1の発現により、ニューロンとなる運命に向かって分化する細胞の割合が高まる。ニューロンとなる運命の誘導はin vitroまたはin vivoで実施し得る。移植前または移植後に幹細胞または神経幹細胞、始原細胞もしくは前駆細胞の分化をニューロンとなる運命に向かって誘導する能力により、成人の脳に移植した場合に移植した幹細胞がアストロサイトとなる運命に分化する傾向が改善される。Nurr1はNR4Aサブファミリーのメンバーである。Nurr1が好ましいかもしれないが、本発明の方法はNurr1に限定されず、方法は細胞内においてNR4Aサブファミリーの任意の核内受容体を基底レベルより高く発現させることを含み得る。NR4Aサブファミリーの受容体にはNurr1/NR4A2、Nor1/NR4A3およびNGFI-B/NR4A1が含まれる。したがって、本発明の方法では、NR4Aサブファミリーの核内受容体(たとえばNurr1、Nor1またはNGFI-B)は細胞内において基底レベルより高く発現され得る。たとえばNR4Aサブファミリーメンバーのアクセッション番号は以下のとおりである。
【0028】
NGF-IBタンパク質: NP775181、NP775180、NP002126
NGFI-Bヌクレオチド: NM_173158、NM_173157、NM_002135
Nor-1タンパク質: NP775292、NP775291、NP775290、NP008912、S71930、Q92570
Nor-1ヌクレオチド: NM_005413、NM_173200、NM_173199、NM_173198、NM_006981
【0029】
Wnt糖タンパク質ファミリーのメンバーは溶解性が乏しく(BradleyおよびBrown、1990および1995)、発生中の中脳で発現される(Parr他、1993)。Wntは中脳-後脳の発生(McMahonおよびBradley、1990; ThomasおよびCapecchi、1990)、神経パターン形成(KieckerおよびNiehrs、2001; Nordstrom他、2002; Houart他、2002)、前駆体の増殖(TaipaleおよびBeachy、2001; ChennおよびWalsh、2002; MegasonおよびMcMahon、2002)ならびに神経系を含めた複数の組織における運命決定(Kispert他、1998; Ross他、2000; HartmannおよびTabin、2001; Marvin他、2001; SchneiderおよびMercola、2001; TzahorおよびLassar、2001; Pandur他、2002)を調節する(Dorsky他、1998; Baker他、1999; Wilson他、2001; Garcia-Castro他、2002; Muroyama他、2002)。
【0030】
本明細書中で使用する「Wntポリペプチド」、「Wnt糖タンパク質」または「Wntリガンド」とは、細胞間の相互作用を調節する分泌タンパク質のWingless-intファミリーのメンバーをいう。Wntはショウジョウバエ(Drosophila)や線虫(Caenorhabditis elegans)からアフリカツメガエル(Xenopus)、ゼブラフィッシュおよび哺乳動物に至るまで高度に保存されている。哺乳動物で現在知られている19種のWntタンパク質は2種類の細胞表面受容体型、すなわち現在10種の受容体で形成されている7回膜貫通ドメインFrizzled受容体ファミリー、ならびに低密度リポタンパク質-受容体関連タンパク質(Low density lipoprotein-receptor related proteins、LRP)5および6、ならびにkremen1および2受容体に結合する。Wntによって伝達されるシグナルは以下の3つの既知のシグナル伝達経路によって伝達される。(1)GSK3βが阻害されるいわゆる古典的シグナル伝達経路はβ-カテニンをリン酸化せず、したがってこれは分解されず核に転位してTCFと複合体を形成し、Wnt標的遺伝子の転写を活性化させる。(2)Jnkを介する平面極性および収束-伸張経路。(3)カルシニュリンが活性化T細胞の核因子(NFAT)を脱リン酸化および活性化するイノシトール1,4,5三リン酸(IP3)/カルシウム経路(Saneyoshi他、2002)。総説には、任意の利用可能なブラウザを使用してウェブ上で見つけることができるWntホームページを参照されたい(www.stanford.edu/〜rnusse/wntwindow.html)。Wntシグナル伝達に関与している他の補助受容体には、チロシンキナーゼ受容体Ror1およびRor2(Oishi I他、2003)、触媒的に不活性な受容体チロシンキナーゼをコードしているderailed/RYK受容体ファミリー(Yoshikawa他、2003)が含まれる。
【0031】
本発明の様々な態様の一部の好ましい実施形態では、WntリガンドはWnt1リガンドである。ヒトWnt1のアミノ酸配列はGenBank参照番号Swiss proteinアクセッション番号P04628から利用可能であり、コードしている核酸は、DNAは参照番号X03072.1から、RNAはNM_005430.2から利用可能である。ヒトWnt1核酸は、以下にさらに同定した配列番号1および2を有するプライマーを用いて増幅することができる。
【0032】
本発明の様々な態様の一部の好ましい実施形態では、WntリガンドはWnt5aリガンドである。ヒトWnt5aのアミノ酸配列はGenBank参照番号Swiss proteinアクセッション番号P41221から利用可能であり、コードしている核酸は、DNAは参照番号AI634753.1、AK021503、L20861、L20861.1、U39837.1から、RNAはNM_003392から利用可能である。ヒトWnt5a核酸は、以下にさらに同定した配列番号5および6を有するプライマーを用いて増幅することができる。
【0033】
ドーパミン作動性ニューロンの産生には好ましくないが、本発明の一部の好ましい実施形態ではWnt3aリガンドを使用し得る。ドーパミン作動性ニューロンを産生させるおよび使用するための本明細書中に開示したものと類似した本発明の様々な態様および実施形態が本発明によって提供され、ここではWnt3aリガンドを使用して幹細胞/始原細胞の増殖もしくは自己再生を維持するおよび/またはそれらが他の、すなわち非ドーパミン作動性のニューロン表現型へと分化することを可能にするもしくは誘導する。本明細書中に記載した実験によって実証されるように、Wnt3aは、ドーパミン作動性ニューロンを生じさせる、Nurr-1を発現する始原体の数を減少させる。しかし、ニューロンの全体数は減少しないので他のニューロン表現型、たとえばセロトニン作動性ニューロンを含めた背側中脳表現型が生じるかもしれない。セロトニン作動性ニューロンの消失はうつ病に関連しているので、Wnt3aリガンドの使用を含む方法、および/またはWnt3aリガンド自体によって産生させたニューロンをたとえばうつ病の治療に使用し得る。
【0034】
ヒトWnt3aのアミノ酸配列はGenBank参照番号SwissProtアクセッション番号P56704から入手可能であり、コードしている核酸は、DNAは参照番号AB060284、AB060284.1、AK056278、AK056278.1から、mRNAはNM_033131から入手可能である。ヒトWnt3a核酸は、以下にさらに同定した配列番号3および4を有するプライマーを用いて増幅することができる。
【0035】
他の好ましいWntはWnt-2、Wnt-4、Wnt-7aおよびWnt-7b、特にWnt-2およびWnt-7aである。Wnt-2は、ニューロン表現型の獲得を促進するおよび/または誘導させるため、また幹細胞ならびに神経幹細胞、前駆細胞および始原細胞をDAニューロンへと分化および/または成熟させるために使用し得る。Wnt-7aは、幹細胞ならびに神経幹細胞、前駆細胞および始原細胞の増殖を促進するおよび/または誘導させることによって細胞のDAニューロンへの分化および/または成熟を促進するために使用し得る。Wnt-2およびWnt-7aのどちらかまたは両方を使用してNurr1+細胞からのTH+ニューロンの数を増加させ得る。
【0036】
Wnt-2核酸は、GenBank参照番号SwissProtアクセッション番号P09544、NCBI RefSeqタンパク質NP00382、NCBI RefSeq mRNA NM_003391およびNM_003391.1、NCBI RefSeq DNA NT_007933として寄託されており、核酸は参照番号AK056742、AK056742.1、BC029854、BC029854.1、X07876、X07876.1、AC002465、AC006326として寄託されており、以下に記載のように配列番号33および34を有するプライマーを使用して増幅することができる。Wnt-4核酸はGenBank参照番号SwissProtアクセッション番号P56705、NCBI RefSeqタンパク質NP110388、NCBI RefSeq mRNA NM_030761およびNM_030761.2、NCBI RefSeq DNA NT_004610として寄託されており、核酸は参照番号AA984007、AL031281、AY009398.1、AF009398.1、AB062766.1、BC034923.1、AF416743.1、AB061675.1、BQ891671.1、BU502468.1、BM043406.1、CB991983.1として寄託されており、以下に記載のように配列番号39および40を有するプライマーを使用して増幅することができる。Wnt-7a核酸は、GenBank参照番号SwissProtアクセッション番号000755、NCBI RefSeqタンパク質NP004616およびNP004616.2、NCBI RefSeq mRNA NM_004625およびNM_004625.2、NCBI RefSeq DNA NT_005927として寄託されており、核酸は参照番号D83175、D83175.1、BC008811、BC008811.1、U53476、U53476.1、BI823772.1、CB989433.1、BI552826.1、BI551057.1、BE740508.1として寄託されており、以下に記載のように配列番号45および46を有するプライマーを使用して増幅することができる。Wnt-7b核酸は、GenBank参照番号SwissProtアクセッション番号P56706、NCBI RefSeqタンパク質NP478679およびNP478679.1、NCBI RefSeq mRNA NM_004625およびNM_004625.2として寄託されており、核酸は参照番号AA062766、AF416743、BC034923、BM047487、BM047487.1、BU543397.1、BU541891.1、BU541105.1として寄託されており、以下に記載のように配列番号47および48を有するプライマーを使用して増幅することができる。
【0037】
野生型Wntリガンドを使用してもよく、あるいは、幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞におけるドーパミン作動性のニューロンとなる運命の増強された発生の機能が保持されている限りは、たとえば1つまたは複数のアミノ酸の付加、欠失、置換および/もしくは挿入による変異体または誘導体を使用してもよい。
【0038】
「幹細胞」とは自己再生が可能な任意の細胞型を意味し、それが胚性幹(ES)細胞である場合は個体内の全ての細胞を生み出すことができ、またはそれが多能性もしくは神経幹細胞である場合はニューロン、アストロサイトおよびオリゴデンドロサイトを含めた神経系内の全ての細胞型を生み出すことができる。幹細胞は以下のマーカーのうち1つまたは複数を発現し得る:Oct-4;Sox1-3;時期に特異的な胚性抗原(SSEA-1、-3、および-4)、ならびに腫瘍拒絶抗原TRA-1-60および-1-81(Tropepe他、2001; Xu他、2001に記載)。神経幹細胞は以下のマーカーのうち1つまたは複数を発現し得る:Nestin;p75ニューロトロフィン受容体;Notch1、SSEA-1(CapelaおよびTemple、2002)。
【0039】
「神経始原細胞」とは神経幹細胞の娘細胞または子孫細胞を意味し、幹細胞と比較してより分化した表現型および/またはより低減した分化能力を有する。前駆細胞とは、発生中にニューロンと直接的な系統関係にあってもなくても、定義された環境条件下で分化転換もしくは再分化するように、またはニューロン表現型を獲得するように誘導させられる、任意の他の細胞を意味する。好ましい実施形態では、幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞は、自発的にまたはマイトジェン(たとえばbFGF、EGFもしくは血清)を欠乏させた際にチロシンヒドロキシラーゼを発現しないあるいは有効には発現しない。
【0040】
幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞は、骨髄、皮膚、眼、鼻上皮、もしくは臍帯を含めた任意の胚性、胎児または成体組織、あるいは神経系の領域、たとえば小脳、脳室領域、脳室下領域、線条体、中脳、後脳、大脳皮質または海馬から得るか、それ由来であり得る。これらは、脊椎動物、たとえばヒトまたはウサギ、モルモット、ラット、マウスや他のげっ歯類、ネコ、イヌ、ブタ、ヒツジ、ヤギ、ウシ、ウマ、もしくは霊長類などの非ヒトであり得る哺乳動物、あるいはニワトリなどのトリから得るか、それ由来であり得る。
【0041】
本発明の好ましい実施形態では、成人の幹細胞/始原細胞/前駆細胞をin vitro、ex vivoまたはin vivoで使用する。これには、同意した成人(たとえば細胞が得られる成人)および適切な倫理委員会の承認が必要である。ヒト胚/胎児を供給源として使用する場合は、そのヒト胚は、本来ならば使用せずに破棄されるかまたは無期限に保存されるもの、特に妊娠が困難なカップルのためのIVF処理用に作製されたヒト胚である。IVFでは一般に、着床および最終的には妊娠に使用される数よりも多い数のヒト胚が作製される。このような予備の胚は通常破棄されることがある。関係する人々、特に関係する卵子ドナーおよび/または精子ドナーの適切な同意により、本来ならば破棄される胚をパーキンソン病などの重篤な神経変性障害の罹患者の利益のために倫理的に前向きな方法で使用することができる。本発明自体は、発生の任意の段階にあるヒト胚の使用に関連しない。上述のように、本発明はヒト胚に直接由来する材料を使用する必要の可能性を最小限にする一方で、ひどい病気の貴重な治療の開発を可能にする。
【0042】
一部の好ましい実施形態では、Wntリガンドと接触させ、本発明の任意の態様に従って他の方法で処理および/または使用する幹細胞または始原細胞もしくは前駆細胞は、同意した成人または適切な同意が出されている子供、たとえば障害のある患者であって、続いて本発明に従って産生させたニューロンを患者内に移植して戻すことによって処置する、かつ/あるいは内因性ドーパミン作動性ニューロンの発生もしくは機能を促進するまたは誘導させるために1つもしくは複数のWntリガンドならびに/または1つもしくは複数の1型アストロサイト/初期グリア細胞由来の因子で処置した患者から得る。
【0043】
幹細胞または始原細胞もしくは前駆細胞で誘導されるニューロンとなる運命は、未分化の表現型または原始ニューロン表現型を示し得る。これは、個体内の任意の細胞型を生み出す能力を有する全能性細胞、または複数の明確なニューロン表現型を生み出す能力を有する多能性細胞、または通常の発生中ではより限定された表現型を生み出す能力を有するがin vitroで適切な環境因子に曝した場合に他の細胞を生み出す能力を有する前駆細胞もしくは始原細胞であり得る。これは、特定のニューロンとなる運命に関連するマーカー、たとえばチロシンヒドロキシラーゼを欠いているかもしれない。
【0044】
複数の幹細胞、神経幹細胞ならびに/または始原細胞および/もしくは前駆細胞がNurr1またはNurr1サブファミリーの別の核内受容体を基底レベルより高く発現し、細胞をWntリガンドで処理する、本発明によるニューロンとなる運命を誘導させる方法では、細胞の大多数がニューロンとなる運命をとるように誘導させ得る。ニューロン細胞でドーパミン作動性の誘導または分化を増強させ得る。好ましい実施形態では、60%を超える、70%を超える、80%を超える、90%を超える幹細胞および/または始原細胞をニューロンとなる運命に誘導させ得る。
【0045】
「細胞内においてNurr1を基底レベルより高く発現させること」とは、in vivo非病的条件下において(未改変の)細胞内で発現されるレベルより高いレベルでNurr1を発現させることを意味する。同様に、細胞内においてNurr1サブファミリーの核内受容体を基底レベルより高く発現させることとは、in vivo非病的条件下において(未改変の)細胞内で発現されるレベルより高いレベルで核内受容体を発現させることを意味する。基底レベルより高い発現には、転写発現、翻訳発現、翻訳後発現、薬理学的発現、人工的アップレギュレーション発現および過剰発現が含まれる。基底レベルより高い核内受容体の発現は、本明細書中ではNurr1に関して記載する。本開示はNurr1サブファミリーの他のメンバーにも適用可能であり、このサブファミリーの他の核内受容体、たとえばNor1またはNGFI-Bと共に本発明の方法において使用し得る。したがって、Nurr1について例示しているが、本発明の方法はNurr1に限定されず任意のNurr1サブファミリーの核内受容体にまで及ぶ。
【0046】
基底レベルより高いNurr1の発現は、当業者に知られている任意の方法によって実現される。例として、基底レベルより高い発現はネイティブゲノムNurr1の調節をモジュレーションすることによって誘導し得る。これは、Nurr1のmRNAもしくはNurr1タンパク質の分解を阻害または阻止することによって、あるいはNurr1の転写および/または翻訳を増大させることによって、たとえば細胞をNurr1の転写をアップレギュレーションする線維芽細胞成長因子8(FGF8)と接触させることによって(Rosenthal, A.、(1998) Cell、93(5)、755-766)、ならびに/あるいは異種調節配列をNurr1のネイティブ調節領域内またはその隣接に導入することによって、ならびに/あるいはNurr1のネイティブ調節領域をたとえば相同組換えによってこのような異種調節配列で置き換えることによって、ならびに/あるいはNurr1の転写、翻訳もしくは機能を負に調節する、遮断するまたはダウンレギュレーションする分子、たとえばNurr2を破壊またはダウンレギュレーションすることによって(Ohkura他、(1999) Biochim Biophys Acta 14444: 69-79)行い得る。
【0047】
転写は、幹細胞、神経幹細胞、前駆細胞、始原細胞、または神経細胞に増大させたレベルの転写活性化因子を与えることによって、たとえば細胞をこのような活性化因子と接触させるか、または活性化因子をコードしている核酸を用いて細胞を形質転換させることによって、増大させ得る。あるいは、転写は、Nurr1の転写インヒビターに対するアンチセンス核酸を用いて細胞を形質転換させることによって増大させ得る。
【0048】
したがって、幹細胞、神経幹細胞、前駆細胞、始原細胞、または神経細胞においてニューロンとなる運命を誘導させるまたは誘導を増強する本発明の方法は、細胞をFGF8またはFGF20と接触させることを含み得る(Ohmachi他、2000)。
【0049】
内因性Nurr1の転写および/または翻訳を増大させることに代わってまたはそれに加えて、基底レベルより高いNurr1の発現は1つまたは複数の余分なNurr1のコピーを幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞内に導入することによって引き起こされ得る。
【0050】
したがって、さらなる態様では、本発明は、ニューロンとなる運命を誘導させるならびに/あるいは幹細胞、神経幹細胞、神経始原細胞、前駆細胞もしくは神経細胞におけるドーパミン作動性の発生の誘導を増強する、またはニューロン細胞におけるドーパミン作動性の誘導もしくは分化を増強する方法であって、細胞をWntリガンドと接触させることに加えて細胞をNurr1で形質転換させることを含む方法を提供する。
【0051】
幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞の形質転換はin vitro、in vivoまたはex vivoで実施し得る。細胞に誘導させるニューロンとなる運命は本明細書中に記載の種類のものでよく、たとえば原始ニューロン表現型を示し、特定のニューロンとなる運命に関連するマーカーが欠けていてもよい。本発明はさらに、Nurr1で形質転換させ、Wntリガンドと接触させた幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞を提供する。
【0052】
形質転換させたNurr1および/またはWntリガンドはゲノム外ベクターに含まれるか、またはゲノム内に、好ましくは安定的に組み込まれ得る。これは幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞、またはニューロン細胞においてその基底レベルより高い発現を駆動するプロモーターに機能的に連結されており、以下により詳細に記述されている。
【0053】
「機能的に連結されている」とは、同じ核酸分子の一部として結合されており、プロモーターから転写が開始されるよう適切に配置かつ配向されていることを意味する。
【0054】
細胞内に遺伝子を導入する方法は当業者に周知である。Nurr1および/またはWntリガンドがベクター上に留まるのかゲノム内に組み込まれるのかにかかわらず、幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞内にNurr1および/またはWntリガンドを導入するためにベクターを使用し得る。適切なベクターを選択するか、またはプロモーター配列、ターミネーター断片、ポリアデニル化配列、エンハンサー配列を含めた適切な調節配列を含むよう構築することができる。ベクターは必要に応じてマーカー遺伝子および他の配列を含み得る。調節配列は幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞または神経細胞内においてNurr1および/またはWntリガンドの発現を駆動し得る。たとえば、ベクターはゲノム外発現ベクターであり得、または調節配列はNurr1および/またはWntリガンドと共にゲノム内に取り込まれ得る。ベクターはプラスミドまたはウイルスベクターであり得る。
【0055】
Nurr1および/またはWntリガンドを使用者の制御下におくために、これを外因的に誘導性遺伝子プロモーターの制御下においてもよい。プロモーターにあてはめた場合の用語「誘導性」は当業者によく理解されている。要するに、誘導性プロモーターの制御下における発現が、与えた刺激に応答して「スイッチが入る」または増大される。刺激の性質はプロモーター間で異なる。一部の誘導性プロモーターは、適切な刺激が存在しない場合はわずかなもしくは検出されないレベルの発現しか引き起こさない(または全く発現させない)。刺激が存在しない場合の発現レベルがどのようであっても、任意の誘導性プロモーターによる発現は妥当な刺激の存在下で増大する。誘導性プロモーターの例はテトラサイクリンON/OFFシステムであり(Gossen他、1995)、ここでは遺伝子の発現はテトラサイクリン類似体によって調節される。
【0056】
さらなる詳細には、たとえばMolecular Cloning: a Laboratory Manual: 第3版、SambrookおよびRussell、2001、Cold Spring Harbor Laboratory Pressを参照されたい。核酸を操作するための多くの知られている技術およびプロトコル、たとえば核酸構築物の調製、突然変異誘発、シーケンシング、細胞内へのDNAの導入および遺伝子発現、ならびにタンパク質の解析がCurrent Protocols in Molecular Biology、Ausubel他編、John Wiley & Sons、1992またはそれ以降の版に詳述されている。
【0057】
抗生物質耐性または感受性遺伝子などのマーカー遺伝子を、当技術分野で周知のように目的核酸を含むクローンの同定に使用し得る。クローンはまた、結合の研究によって、たとえばサザンブロットハイブリダイゼーションによって同定またはさらに調査し得る。
【0058】
Nurr1を含むかつ/またはWntリガンドをコードしている核酸を宿主幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞のゲノムに組み込ませ得る。組込みは、形質転換させた核酸に、標準の技術に従って、ゲノムとの組換えを促進する配列を含めることによって促進され得る。組み込まれた核酸は、幹細胞、または神経幹細胞、始原細胞もしくは前駆細胞、またはニューロン細胞においてNurr1遺伝子のおよび/またはWntリガンドの発現を駆動する能力を有する調節配列を含み得る。核酸は、Nurr1および/またはWntリガンドをコードしている配列が幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞、またはニューロン細胞内におけるその発現を駆動および/または制御する能力を有する調節エレメントの制御下におかれる、ゲノム内の部位へのその組込みを指令する配列を含み得る。組み込まれた核酸は、本明細書中に記載のように、Nurr1および/またはWntリガンドを幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞、またはニューロン細胞内に形質転換させるために使用したベクター由来であり得る。
【0059】
Nurr1を含むかつ/またはWntリガンドをコードしている核酸の導入は、その核酸が直鎖状、分枝鎖状または環状であるかにかかわらず、制限なしに概して「形質転換」と称し得る。これには任意の利用可能な技術を使用し得る。適切な技術には、リン酸カルシウムトランスフェクション、DEAE-デキストラン、PEI、電気穿孔、微量注入などの機械的技法、直接的なDNAの取り込み、受容体に媒介されるDNAの移送、レトロウイルスまたは他のウイルスを用いた形質導入ならびにリポソームまたは脂質に媒介されるトランスフェクションが含まれ得る。選択した遺伝子構築物を細胞内に導入する際、当業者に周知の特定の検討事項を考慮に入れなければならない。当業者には、Nurr1および/またはWntリガンドを幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞内に導入するための具体的な形質転換方法の選択は、本発明によって重要ではなくその制限とならないことが明らかであろう。
【0060】
Nurr1および/またはWntリガンドを用いた幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞のin vivo形質転換のための適切なベクターおよび技術は当業者に周知である。適切なベクターには、アデノウイルス、アデノ関連ウイルス、パポバウイルス、ワクシニアウイルス、ヘルペスウイルス、レンチウイルスおよびレトロウイルスが含まれる。無効化したウイルスベクターは、感染性ウイルス粒子の産生に必要な遺伝子が発現されているヘルパー細胞系内で産生させ得る。適切なヘルパー細胞系は当業者に周知である。例として、Fallaux, F.J.他、(1996) Hum Gene Ther 7(2)、215-222; Willenbrink, W.他、(1994) J Virol 68(12)、8413-8417; Cosset, F.L.他、(1993) Virology 193(1)、385-395; Highkin, M.K.他、(1991) Poult Sci 70(4)、970-981; Dougherty, J.P.他、(1989) J Virol 63(7)、3209-3212; Salmons, B.他、(1989) Biochem Biophys Res Commun 159(3)、1191-1198; Sorge, J.他、(1984) Mol Cell Biol 4(9)、1730-1737; Wang, S.他、(1997) Gene Ther 4(11)、1132-1141; Moore, K.W.他、(1990) Science 248(4960)、1230-1234; Reiss, C.S.他、(1987) J Immunol 139(3)、711-714を参照されたい。ヘルパー細胞系は一般に、ウイルスゲノムをパッケージする機構によって認識される配列を欠いている。これらは、核酸を全く含まないビリオンを産生する。無傷のパッケージングシグナルと送達する遺伝子または他の配列(たとえばNurr1および/またはWnt)とを含むウイルスベクターをヘルパー細胞内にパッケージして感染性ビリオン粒子内に入れ、その後これを幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞への遺伝子の送達に使用し得る。
【0061】
内因性Nurr1および/またはWntの転写および/または翻訳を増大させることに代わってまたはそれに加えて、基底レベルより高いNurr1および/またはWntの発現は1つまたは複数の余分なNurr1および/またはWntタンパク質のコピーを、微量注入または細胞透過ペプチド、すなわちTAT、トランスポータン、アンテナペディアペネトラチン(penetratin)ペプチド(Lindsay、2002)を含めた他の担体系もしくはタンパク質送達系によって幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞内に導入することによって引き起こされ得る。
【0062】
さらなる態様では、本発明は、幹細胞、神経幹細胞または始原細胞もしくは前駆細胞、またはニューロン細胞において特定のニューロンとなる運命を誘導させる方法であって、幹細胞または始原細胞またはニューロン細胞がNurr1を基底レベルより高く発現し、細胞をWntリガンドおよび任意選択で1型アストロサイト/グリア細胞によって供給されるまたはそれ由来である1つまたは複数の因子と接触させることを含む方法を提供する。この因子または複数の因子は、幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞を1型アストロサイト/グリア細胞と同時培養するあるいは接触させることによって提供し得る。方法はin vitroまたはin vivoで起こり得る。基底レベルより高くNurr1および/またはWntを発現する幹細胞または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞は、Nurr1および/またはWntを用いた細胞の形質転換によって産生させ得る。
【0063】
因子または複数の因子は不死化したアストロサイト/グリア細胞によって供給されるまたはそれ由来であり得る。因子または複数の因子はグリア細胞系、たとえばアストロサイトもしくは放射状グリアもしくは未熟なグリア中脳細胞系によって供給されるまたはそれ由来であり得る。細胞系は同種細胞集団を提供する。
【0064】
本発明の重要な態様は、ドーパミン作動性ニューロンは、幹細胞または始原細胞もしくは前駆細胞からin vitroで、細胞において基底レベルより高くNurr1を発現させること、および細胞を腹側中脳の1型アストロサイト/初期グリア細胞によって供給されるまたはそれ由来である1つまたは複数の因子と接触させることを含む方法によって産生させることができるが、ドーパミン作動性の運命の誘導はWntリガンドと接触させることによって増強または促進されるという知見に基づいている。
【0065】
本発明は多数のドーパミン作動性ニューロンの産生を可能にする。これらのドーパミン作動性ニューロンは、パーキンソン病において変性するまたは損傷もしくは損失される細胞と置き換わる原料物質として使用し得る。
【0066】
好ましくは、Nurr1を基底レベルより高く発現する細胞は、Wntリガンドと接触させた場合に有糸分裂する。
【0067】
本発明の方法では、細胞を以下から選択された1つまたは複数の物質とさらに接触させ得る:塩基性線維芽細胞成長因子(bFGF);上皮成長因子(EGF);およびレチノイドX受容体(RXR)の活性化因子、たとえば合成レチノイド類似体SR11237(Gendimenico, G.J.他、(1994) J Invest Dermatol 102(5)、676-80)、9-シスレチノールもしくはドコサヘキサン酸(DHA)またはLG849(Mata de Urquiza他、2000)。実験によって以下に実証するように、これらの物質のうち1つまたは複数を用いた本発明による細胞の処理を使用して、ドーパミン作動性の運命をとる幹細胞、始原細胞もしくは前駆細胞の割合を増加させるか、またはニューロン細胞においてドーパミン作動性の誘導もしくは分化を増強させ得る。本発明によるニューロン細胞においてドーパミン作動性の運命を誘導させることまたはドーパミン作動性の誘導もしくは分化を増強させる方法は、細胞をFGF成長因子ファミリーのメンバー、たとえばFGF4、FGF8またはFGF20と接触させることを含み得る。
【0068】
有利には、細胞を上記物質のうち2つ以上と接触させ得る。本発明者らは、bFGFまたはEGFおよびSR11237の有益な効果が飽和用量で付加的であることを思いがけず見出した。この知見は、これらの物質が異なる機構によって作用するかもしれないことを示唆している。
【0069】
ドーパミン作動性の表現型を誘導させる方法は、Wntリガンドおよび任意選択で腹側中脳の1型アストロサイト/グリア細胞によって供給されるまたはそれ由来である1つまたは複数のさらなる因子と接触させる前に、たとえば腹側中脳の1型アストロサイト/グリア細胞もしくはそれらに由来する因子と接触させるまたは同時培養する前に、幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞またはニューロン細胞をbFGFおよび/またはEGFで前処理することを含み得る。
【0070】
任意選択の前処理ステップは、既に国際公開公報WO00/66713号およびWagner他、(1999)に報告されている本発明者らによる2つの思いがけない知見から生じる。(i)Nurr1を基底レベルより高く発現しており、高い増殖を示す神経幹細胞系では、腹側中脳の1型アストロサイト/グリア細胞と同時培養した際にドーパミン作動性の運命の誘導の増強が実証されたこと;および(ii)血清を含まない培地(SFM)中でbFGFまたはEGFを処理した後、Nurr1を基底レベルより高く発現するほとんどの幹細胞系の基底増殖がSFMのみに継代した後に上昇し続けたこと。
【0071】
ドーパミン作動性の表現型を誘導させる方法は、幹細胞または神経幹細胞、始原細胞もしくは前駆細胞をFGF成長因子ファミリーのメンバー、たとえばFGF2、FGF4、FGF8またはFGF20で前処理することを含み得る。
【0072】
前処理の代わりにまたは前処理と共に、Wnt処理と同時に追加の因子を用いて細胞を処理し得る。
【0073】
幹細胞、神経幹細胞または始原細胞もしくは前駆細胞においてニューロンとなる運命が誘導される、あるいはニューロン細胞においてドーパミン作動性の誘導または分化が増強されている本発明による方法は、ニューロンとなる運命のマーカーを検出することを含み得る。b-チューブリンIII(TuJ1)はニューロンとなる運命のマーカーの1つである(Menezes, J.R.他、(1994) J Neurosci 14(9)、5399-5416)。他のニューロンマーカーには神経フィラメントおよびMAP2が含まれる。特定のニューロン表現型を誘導させる場合は、マーカーはその表現型に特異的であるべきである。ドーパミン作動性の運命には、チロシンヒドロキシラーゼ(TH)、ドーパミントランスポーター(DAT)およびドーパミン受容体の発現を、たとえば免疫反応性またはin situハイブリダイゼーションによって検出し得る。チロシンヒドロキシラーゼはDA細胞の主要なマーカーである。ドーパミンおよび代謝物の含有量および/または放出は、たとえば高圧液体クロマトグラフィー(HPLC)によって検出し得る(Cooper, J.R.他、The Biochemical Basis of Neuropharmacology、第7版、(1996) Oxford University Press)。ドーパミンβヒドロキシラーゼおよびGABAまたはGADが存在しないこと(TH/ドーパミン/DATの存在下)もドーパミン作動性の運命の指標である。さらなるマーカーには、2型アルデヒドデヒドロゲナーゼ(ADH-2)、GIRK2、Lmx1bおよびPtx3が含まれる。
【0074】
マーカーの検出は当業者に知られている任意の方法に従って実施し得る。検出方法には、マーカーをコードしている核酸配列に結合する能力を有する特異的な結合メンバーであって、その配列とハイブリダイズ可能な核酸プローブ、または核酸配列もしくはそれにコードされているポリペプチドに対する特異性を有する免疫グロブリン/抗体ドメインを含み、特異的な結合メンバーの配列もしくはポリペプチドへの結合が検出可能であるように標識されている特異的な結合メンバーを使用し得る。「特異的な結合メンバー」はあるマーカーに対して特有の特異性を有し、通常の条件下では他の種類に優先してそのマーカーに結合する。あるいは、マーカーが特定のmRNAである場合は、特異的なオリゴヌクレオチドプライマーへの結合およびたとえばポリメラーゼ連鎖反応における増幅によって検出し得る。
【0075】
核酸プローブおよびプライマーはストリンジェントな条件下でマーカーとハイブリダイズし得る。たとえば約80〜90%同一であるマーカー配列を検出するための適切な条件には、42℃、0.25MのNa2HPO4、pH7.2、6.5%のSDS、10%の硫酸デキストラン中で終夜のハイブリダイゼーション、および55℃、0.1×SSC、0.1%のSDS中での最終洗浄が含まれる。約90%より高い同一性のマーカー配列の検出では、適切な条件には、65℃、0.25MのNa2HPO4、pH7.2、6.5%のSDS、10%の硫酸デキストラン中で終夜のハイブリダイゼーション、および60℃、0.1×SSC、0.1%のSDS中での最終洗浄が含まれる。
【0076】
さらなる態様では、本発明は、本明細書中に開示した方法のうち任意の1つに従って産生させたニューロンを提供する。このニューロンは原始ニューロン表現型を有し得る。これは、複数の明確なニューロン表現型を生み出す能力を有し得る。このニューロンは特定のニューロン表現型を有し得、この表現型はNurr1を基底レベルより高く発現する幹細胞、神経幹細胞、始原細胞、前駆細胞もしくは神経細胞と接触させた因子もしくは複数の因子が供給されたまたはそれが由来するWntリガンドおよび/またはアストロサイト/グリア細胞の種類、ならびに/あるいは幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞を同時培養したまたは接触させたアストロサイト/グリア細胞の種類によって影響を受ける。好ましい実施形態では、ニューロンはドーパミン作動性の表現型を有する。
【0077】
ニューロンは、ニューロンにおいて分子の発現を駆動する能力を有するプロモーターに機能的に連結されている、神経保護または神経再生特性を有する分子をコードしている核酸を含み得る。プロモーターは、損傷を与える全ての過剰発現を阻止し得るように誘導性プロモーター、たとえばTetONキメラプロモーターであり得る。プロモーターは特定のニューロン表現型、たとえばTHプロモーターまたはNurr1プロモーターと関連し得る。
【0078】
コードされた分子は、その発現がその環境に依存せずにニューロンを与えるように、すなわちその生存が1つもしくは複数の因子の存在、またはたとえばそれを移植する神経環境における条件に依存しないようであり得る。例として、ニューロンは、ニューロンにおいて分子の発現を駆動する能力を有するプロモーターに機能的に連結されている、以下に記載の神経保護または神経再生分子のうち1つまたは複数をコードしている核酸を含み得る。
【0079】
それに追加してまたはその代わりに、コードされた分子の発現はそのニューロンを取り囲む細胞環境の神経保護または神経再生において機能し得る。このようにして、ニューロンは神経保護および神経再生特性を有する分子を送達するために組み合わせた細胞および遺伝子治療手法において使用し得る。
【0080】
神経保護および神経再生特性を有する分子の例には以下のものが含まれる。
【0081】
(i)神経変性を補償および阻止することができる神経栄養因子。一例はグリア由来の神経栄養成長因子(GDNF)であり、これは強力な神経生存因子であり、ドーパミン作動性ニューロンからの発芽を促進し、チロシンヒドロキシラーゼの発現を増大させる(Tomac他、(1995) Nature、373、335-339; Arenas他、(1995) Neuron、15、1465-1473)。軸索伸長GDNFを増強することにより、GDNFはニューロンがその局所的環境を神経支配する能力を増大させ得る。GDNFファミリーの他の神経栄養分子には、ニューチュリン(Neurturin)、ペルセフィン(Persephin)およびアルテミン(Artemin)が含まれる。ノイロトロピンファミリーの神経栄養分子には、神経成長因子(NGF)、脳由来神経栄養因子(BDNF)、ならびにノイロトロピン-3、-4/5および-6が含まれる。神経栄養活性を有する他の因子には、FGFファミリーのメンバー、たとえばFGF2、4、8および20;Wnt-1、-2、-5a、-3aおよび7aを含めたWntファミリーのメンバー;BMP2、4、5および7、nodal、アクチビンおよびGDFを含めたBMPファミリーのメンバー;ならびにTGFα/βファミリーのメンバーが含まれる。
【0082】
(ii)抗アポトーシス分子。細胞死において中心的な役割を果たすBcl2。Bcl2の過剰発現はニューロンを自然に発生する細胞死および虚血から保護する(Martinou他、(1994) Neuron、1017-1030)。ニューロンに特異的な別の抗アポトーシス分子はBclX-Lである。
【0083】
(iii)神経支配およびその環境と連結を形成することにおいてニューロンを補助する軸索再生および/または伸長および/または誘導分子、たとえばエフリン。エフリンは、チロシンキナーゼ受容体を活性化する能力を有する膜結合リガンドのクラスを定義する。エフリンは神経発生に関係づけられている(Irving他、(1996) Dev. Biol.、173、26-38; Krull他、(1997) Curr. Biol. 7、571-580; Frisen他、(1998) Neuron、20、235-243; Gao他、(1996) PNAS、93、11161-11166; Torres他、(1998) Neuron、21、1453-1463; Winslow他、(1995) Neuron、14、973-981; Yue他、(1999) J Neurosci 19(6)、2090-2101)。
【0084】
(iv)転写因子、たとえばホメオボックスドメインタンパク質Ptx3(Smidt, M.P.他、(1997) Proc Natl Acad Sci USA、94(24)、13305-13310)、Lmx1b、Pax2、Pax5、Pax8、またはengrailed 1もしくは2(WurstおよびBally-Cuif、2001; RhinnおよびBrand、2001)、あるいは塩基性ヘリックス-ループ-ヘリックスファミリーの神経性遺伝子。
【0085】
本発明によるニューロンまたは本発明で使用するためのニューロンは、1つまたは複数の他の細胞型、たとえば幹細胞、神経幹細胞、前駆細胞もしくは始原細胞を実質的に含まなくてもよい。ニューロンは、抗体および磁気ビーズによる細胞外エピトープの認識に基づいた方法または蛍光活性化細胞分取(FACS)を含めた、当業者に知られている任意の技術を用いて神経幹細胞または始原細胞から分離し得る。例として、幹細胞、神経幹細胞、前駆細胞もしくは始原細胞上に見つかるがニューロン上には見つからない分子の細胞外領域に対する抗体を使用し得る。このような分子には、Notch1、CD133、SSEA1、プロミニン(prominin)1/2、RPTPβ/ホスホキャン(phosphocan)、TIS21およびグリア細胞系由来の神経栄養因子受容体GFRαまたはNCAMが含まれる。抗体に結合した幹細胞は補体に曝すことによって溶解するか、またはたとえば磁気選別によって分離し得る(Johansson他、(1999) Cell、96、25-34)。ニューロンの対象レシピエントに対して異種である抗体を使用する場合は、このような細胞選別手順を回避する全てのたとえば幹細胞、神経幹細胞または始原細胞もしくは前駆細胞を異種抗体で標識し、これらはレシピエントの免疫系の第一の標的となる。あるいは、所望の表現型を獲得する細胞は、細胞外エピトープに対する抗体、または細胞型に特異的なプロモーターの制御下における、蛍光タンパク質を含めた導入遺伝子の発現によっても分離することができる。例として、ドーパミン作動性ニューロンはTH、DAT、Ptx3またはドーパミン作動性ニューロンに特異的に使用される他のプロモーターの制御下で発現される蛍光タンパク質を用いて単離することができる。
【0086】
本発明の方法は、神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において所望の表現型を高めるために追加の陰性または陽性選択方法を含み得る。
【0087】
DAニューロンを富化するために陰性選択を使用し得る。非DAニューロン用の選択的神経毒素、たとえば5-7-ジヒドロキシトリプタミン(セロトニン作動性ニューロンを排除するため)、またはサポニンもしくは毒素に結合した抗体、または補体の追加後に、たとえばGABAトランスポーターに対する抗体(GABA作動性ニューロンを排除するため)を使用し得る。本発明の方法は、たとえば細胞を含むin vitro培養物に陰性選択剤を加えることによって、または陰性選択剤の存在下で細胞を培養することによって、神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を陰性選択剤で、好ましくはin vitroで処理するあるいはそれと接触させることをさらに含み得る。陰性選択剤は所望の細胞型(または複数の細胞型)以外の細胞型に対する選択を行う。たとえば、本発明がドーパミン作動性ニューロン表現型を促進する、増強させるまたは誘導させることに関係する場合、陰性選択剤はDAニューロンならびに幹細胞および神経幹細胞、前駆細胞および始原細胞などのDAニューロンへ発生する細胞以外の細胞に対する選択を行い得る。したがって、陰性選択剤は非DAニューロンなどの非DA表現型を有する分化細胞に対する選択を行い得る。陰性選択剤は、所望の細胞型(または複数の細胞型)以外の細胞の増殖を低減もしくは阻止するかつ/または死滅させ得る。陰性選択剤は、DAニューロン以外のニューロン集団を減少させる選択的神経毒素であり得る。たとえば、陰性選択剤は5-7-ジヒドロキシトリプタミンであり得る(セロトニン作動性ニューロンを減少させるため)。陰性選択剤は非DAニューロンに特異的な抗体または抗体フラグメントであってよく、この抗体または抗体フラグメント(たとえばscFvまたはFab)はサポニンまたは毒素に結合している。たとえば、抗体はGABAトランスポーターに特異的であり得る(GABA作動性ニューロンを減少させるため)。
【0088】
本発明の方法では、神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞を抗酸化剤(たとえばアスコルビン酸)、低酸素圧および/または低酸素誘導因子(たとえばHIFまたはエリスロポイエチン)の存在下で増殖させ得る。
【0089】
本発明はさらに、様々な態様および実施形態中で、Wntリガンド、またはWntリガンドをコードしている核酸、または可溶性frizzed関連タンパク質もしくはdikkopfaもしくはWIFのWnt阻害活性を阻害もしくは遮断する合成Wntリガンド類似体、またはタンパク質、核酸もしくは合成拮抗剤、またはWntの下流にある1つもしくは複数のシグナル伝達構成要素を阻害、遮断、増強、切り替えもしくはモジュレーションするように作用するタンパク質、核酸もしくは合成薬物から選択された物質の使用であって、たとえばパーキンソン症候群またはパーキンソン病に罹患している個体の治療において、内因的または外因的に供給されたもののどちらかの幹細胞、始原細胞もしくは前駆細胞、またはニューロン細胞に作用することによって脳内でドーパミン作動性ニューロンの発生を誘導させる、促進するもしくは増強させるため、ならびに/あるいはドーパミン作動性ニューロンの消失を阻害もしくは阻止する、または生存、表現型の分化もしくは成熟、神経突起生成もしくはシナプス形成、機能的出力を促進するための、Wntリガンドもしくはそれをコードしている核酸または他の前記の物質を個体に投与することを含む治療方法における使用を提供する。Wntリガンドもしくはそれをコードしている核酸または他の前記の物質は、たとえば製薬上許容される賦形剤または担体を含む任意の適切な組成物中で投与し得、神経変性障害、パーキンソン症候群またはパーキンソン病を治療するための医薬の製造に使用し得る。Wntリガンドもしくはそれをコードしている核酸は、中枢神経系および/もしくは脳に投与するか、またはそれを標的とし得る。
【0090】
本発明は様々な態様で、本明細書中で開示した方法のうち任意の1つに従って産生させたニューロンだけでなく、このようなニューロン、幹細胞、始原細胞もしくは前駆細胞および/またはWntリガンドを含む医薬組成物、医薬、薬物または他の組成物、医療の方法におけるこのようなニューロン、幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞および/あるいはWntリガンドあるいは組成物の使用、たとえばパーキンソン病または他の(たとえば神経変性性)疾患を治療する(予防的処置を含み得る)ためのこのようなニューロン、幹細胞、始原細胞、前駆細胞またはニューロン細胞および/あるいはWntリガンドあるいは組成物を患者に投与することを含む方法、たとえばパーキンソン病または他の(たとえば神経変性疾患)を治療するための投与用組成物の製造におけるこのようなニューロンまたは細胞および/またはWntリガンドの使用、ならびにこのようなニューロンまたは細胞および/またはWntリガンドを製薬上許容される賦形剤、ビヒクルまたは担体、および任意選択で1つまたは複数の他の成分、たとえば神経保護分子、神経再生分子、レチノイド、成長因子、アストロサイト/グリア細胞、抗アポトーシス因子、あるいは幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞または宿主脳内において遺伝子の発現を調節する因子を混合することを含む医薬組成物を作製する方法にまで及ぶ。このような任意選択の成分はその環境に依存せずに、すなわちその生存がその環境における1つもしくは複数の因子または条件の存在に依存しないように、ニューロンを与え得る。例として、医薬組成物を作製する方法はニューロンを発生中の腹側中脳で見つかる1つまたは複数の因子と混合することを含み得る。ニューロンをGDNFおよび/またはニューチュリン(NTN)と混合し得る。
【0091】
本発明は、本発明に従って産生させたニューロン、幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞および/あるいはWntリガンド、ならびに1つまたは複数の追加の成分を含む組成物を提供する。本発明による医薬組成物および本発明に従って使用する医薬組成物は、ニューロンまたは細胞に加えて、製薬上許容される賦形剤、担体、緩衝剤、保存剤、安定化剤、抗酸化剤または当業者に周知の他の材料を含み得る。このような材料は無毒性であるべきであり、ニューロンの活性を妨害するべきでない。担体または他の材料の正確な性質は投与経路に依存するだろう。組成物は、1つまたは複数の神経保護分子、神経再生分子、レチノイド、成長因子、アストロサイト/グリア細胞、あるいは幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞において遺伝子の発現を調節する因子を含み得る。このような物質は、上述のように、その環境に依存せずにニューロンを与え得る。
【0092】
液状医薬組成物は一般に、水、石油、動物性もしくは植物性油、鉱油または合成油などの液状担体を含む。生理食塩水溶液、組織もしくは細胞培養培地、デキストロースまたは他の糖溶液あるいはエチレングリコール、プロピレングリコールもしくはポリエチレングリコールなどのグリコールを含め得る。
【0093】
組成物は、発熱物質を含まず適切なpH、等張性および安定性を有する、非経口的に許容される水溶液の形態であり得る。当業者は、たとえば塩化ナトリウム、リンゲル液、または乳酸化リンゲル液などの等張ビヒクルを用いて適切な溶液を十分に調製することができる。人工脳脊髄液を用いて組成物を調製し得る。
【0094】
本発明は、医療方法、特にニューロン細胞の変性、損傷、損失、または障害に関連する病状の治療における本発明に従って産生させたニューロンおよび/またはWntリガンドの使用にまで及ぶ。さらに、本発明は、特定の表現型のニューロンの産生、損傷、または損失に関連する状態、疾患または障害の治療における、その表現型のニューロンおよび/またはWntリガンドの使用を提供し得る。より詳細には、本発明はヒトパーキンソン病の治療におけるドーパミン作動性ニューロンおよび/またはWntリガンドの使用を提供する。本発明は神経変性疾患(たとえばパーキンソン病)を治療するための材料および方法に特に関するが、それに限定はされない。例として、本発明は、脊髄および/もしくは大脳皮質、またはNurr1+細胞を含む神経系の他の領域の変性または損傷の治療にまで及ぶ。
【0095】
投与した細胞が2つ以上の明確なニューロン表現型を生み出す能力を有する幹細胞、始原細胞または前駆細胞である治療方法では、ニューロン、細胞および/もしくはWntリガンドまたは組成物を、細胞を所望する特定のニューロンとなる運命へと分化するよう指示するアストロサイト/グリア細胞を含む領域内に導入し得る。細胞および/もしくはWntリガンドまたは組成物をたとえば腹側中脳に注射し、それらはここで1型アストロサイト/グリア細胞と相互作用し得、ドーパミン作動性の表現型をとるように誘導させ得る。あるいはまたはそれに加えて、移植した組成物は、ニューロンまたは細胞を上述のように特定のニューロンとなる運命に向かうその発生を指示する1つまたは複数の因子、たとえば1型アストロサイト/グリア細胞と組み合わせて含み得る。
【0096】
細胞は、当技術分野で知られている任意の技法によって患者内に移植し得る(たとえばLindvall, O.、(1998) Mov. Disord. 13、Suppl. 1:83-7; Freed, C.R.他、(1997) Cell Transplant、6、201-202; Kordower他、(1995) New England Journal of Medicine、332、1118-1124; Freed, C.R.、(1992) New England Journal of Medicine、327、1549-1555)。
【0097】
本発明による組成物の投与量は、好ましくは「予防上有効量」または「治療上有効量」(状況次第とはいえ、予防は治療と見なされ得る)であり、これは、個体において利点を示すのに十分である。投与する実際の量、ならびに投与速度および間隔は、治療するものの性質および重篤度に依存するだろう。治療の処方、たとえば用量の決定などは、一般医および他の医師の責任範囲内にある。
【0098】
組成物は単独でまたは他の治療と組み合わせて、治療する状況に応じて同時にまたは連続的に投与し得る。
【0099】
本明細書中に提供する方法は、初代細胞を用いてin vivoもしくはin vitroで、または細胞系を原料物質として用いて実施し得る。in vitroで拡大させた細胞の利点は、産生させ得るニューロンの数に実質上制限がないことである。
【0100】
移植した細胞の免疫学的拒絶に関連する可能性のある不利点を寛解させるために、幹細胞または始原細胞もしくは前駆細胞を患者から単離して所望する表現型に誘導させ得る。その後、細胞を患者に移植し得る。有利なことに、多数の免疫適合性のあるニューロン細胞を産生させ得るように、単離した幹細胞または始原細胞もしくは前駆細胞を用いて細胞系を確立させ得る。さらなる選択肢は、広範囲の免疫学的適合性を網羅する細胞バンクを確立することであり、そこから個体患者に適切な選択をすることができる。一個体由来の幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞を、それらまたはそれらの子孫を第2の個体へと導入した場合に拒絶が寛解されるように改変し得る。例として、ドナー細胞における1つまたは複数のMHC対立遺伝子をレシピエントのそれと、たとえば相同組換えによって置き換え得る。
【0101】
不死化癌遺伝子を有する細胞系由来の細胞を使用して患者に移植する場合は、細胞を患者に移植する前にCRE-locP系を用いて癌遺伝子を除去し得る(Westerman, K.A.他、Proc. Natl. Acad Sci. USA 93、8971 (1996))。患者の体温では不活性である不死化癌遺伝子を使用し得る。
【0102】
さらなる態様では、本発明は、神経変性疾患の治療に使用する物質についてスクリーニングする方法における、本発明に従って産生させた細胞またはニューロンの使用にまで及ぶ。ニューロンはドーパミン作動性ニューロンであり得る。神経変性疾患はパーキンソン症候群またはパーキンソン病であり得る。物質は神経保護分子および/もしくは神経再生分子ならびに/または発生的可溶性シグナルならびに/または腹側中脳の1型アストロサイトもしくはグリア細胞由来の因子もしくは複数の因子であり得る。方法は、in vitroまたはin vivoで実施し得る。
【0103】
この方法は、以下を含み得る。
【0104】
(i)本発明のニューロンを前記ニューロンに対する毒素で処理すること;
(ii)ニューロンを毒素から分離すること;
(iii)処理したニューロンを1つまたは複数の試験物質と接触させること;
(iv)ニューロンが毒素から回復する能力を決定すること;
(v)ニューロンが毒素から回復する前記能力を、試験物質(または複数の試験物質)と接触させない場合におけるそのニューロンまたは同一のニューロンが毒素から回復する能力と比較すること。
【0105】
この方法は、以下を含み得る。
【0106】
(i)1つまたは複数の試験物質存在下で本発明のニューロンをそのニューロンに対する毒素で処理すること;
(ii)ニューロンが毒素に耐性を示す能力を決定すること;
(iii)ニューロンが毒素に耐性を示す前記能力を、試験物質(または複数の試験物質)と接触させない場合におけるそのニューロンまたは同一のニューロンが毒素に耐性を示す能力と比較すること。
【0107】
毒素は6-ヒドロキシドーパミン、5,7-ジヒドロキシトリプタミンまたは1-メチル-4-フェニル-1,2,3,6-テトラヒドロピリジン(MPTP)、ラクタシスチンを含めたプロテアソームインヒビター、またはロテノンを含めた殺虫剤であり得、これらは全てカテコールアミン作動性ニューロンの死をもたらし、パーキンソン病の特徴が実験的に再現される。ニューロンが毒素から回復するまたは毒素に耐性を示す能力は、当業者に知られている任意の方法、たとえば細胞生存度をモニターすること(たとえば、たとえばTUNEL技法による細胞計数によって)、形態をモニターすること(たとえば発芽、軸索伸長および/または枝分かれ)、および/または生化学をモニターすること(たとえばTH活性、たとえば神経伝達物質の取り込み/放出/含有量)によって決定し得る。
【0108】
本発明において使用し得る幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞にはC17.2(Snyder, E.Y.他、Cell 68、33-51 (1992))およびH6ヒト細胞系(Flax他、Nature Biotech 16 (1998))が含まれる。さらなる例がGage他、1995、およびGotlieb、2002)に記載されている。
【0109】
本議論は神経幹細胞または神経始原細胞もしくは前駆細胞に関して行ったが、本明細書中で提供する方法は他の幹細胞、始原細胞または前駆細胞におけるニューロンとなる運命の誘導に応用し得る。このような細胞の例には、非神経系に関連する幹細胞が含まれる。これらの方法は間質もしくは造血幹細胞および/または表皮由来の増殖性細胞に応用し得る。造血細胞は血液または骨髄生検から採取し得る。間質細胞は骨髄生検から採取し得る。上皮細胞は皮膚生検、またはたとえば口腔粘膜を掻取ることによって採取し得る。ニューロン表現型はこれら幹細胞、始原細胞または前駆細胞の生理的なin vivoでの運命ではないので、誘導過程は分化転換、または脱分化および神経再分化と呼ぶことができる。このような細胞をニューロンとなる運命へと誘導させる方法は、非ニューロン表現型に関連する遺伝子に対するアンチセンスレギュレーターの使用、すなわちこのような細胞がニューロンでない運命に向かって分化することを抑制かつ/または逆転させることを含み得る。
【0110】
本発明の方法は神経運命に傾倒していない幹細胞に適用し得る。これは、異なる発生系に関連する2つ以上の娘幹細胞を生み出す能力を有する幹細胞に適用し得る。このような幹細胞の例は胚性幹細胞、造血幹細胞、表皮由来の増殖性細胞、および神経幹細胞である。
【0111】
上述のように、本開示は、基底レベルより高いNurr1の発現と組み合わせてWntリガンドおよび/または腹側中脳の1型アストロサイトもしくはグリア細胞由来の1つもしくは複数の因子を必要とする方法によって、幹細胞または始原細胞もしくは前駆細胞からドーパミン作動性ニューロンを作製することができることを実証している。
【0112】
様々なさらなる態様において、本発明は、神経幹細胞または始原細胞もしくは前駆細胞においてドーパミン作動性の運命の誘導を増強させる因子または複数の因子、あるいはNurr1を基底レベルより高く発現し、Wntリガンドおよびこれによって同定する因子または複数の因子で処理したニューロン細胞においてドーパミン作動性の誘導または分化を増強させる因子または複数の因子の、アッセイならびにスクリーニング方法の供給に関する。
【0113】
本発明は、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞において、単独でまたは組み合わせて、ドーパミン作動性の運命の誘導を増強、増大または強化する能力を有する因子もしくは複数の因子のスクリーニング方法を提供する。本発明のさらなる態様は、このような幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞においてドーパミン作動性の運命の誘導を増強させる因子もしくは複数の因子のスクリーニングまたは探索および/もしくは獲得/同定における、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞の使用を提供する。
【0114】
スクリーニング方法は以下を含み得る。
【0115】
(a)試験物質を、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞と接触させることであり、この接触により試験物質と細胞との間の相互作用が生じ得;および
(b)試験物質と細胞との間の相互作用を決定すること。
【0116】
スクリーニング方法は、試験物質を、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞由来の膜画分、可溶性画分または核画分と接触させ、試験物質と画分との間の相互作用を決定することを含み得る。このような画分の調製は当業者の能力範囲内に十分ある。
【0117】
結合または相互作用は任意数の当技術分野で知られている定性的または定量的な技法によって決定し得る。試験物質と幹細胞または始原細胞またはニューロン細胞との間の相互作用は、どちらか一方を検出可能な標識で標識し、これを、固体支持体上に、たとえば固体支持体に結合した抗体を使用して、またはBiacoreシステムを含めたそれ自体で知られている他の技術によって固定していてもよいもう一方と接触させることによって調査し得る。
【0118】
スクリーニング方法は、1つまたは複数の試験物質の存在下で幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞を培養すること、およびたとえば本明細書中に記載のようにドーパミン作動性の表現型のマーカーを検出することによってドーパミン作動性の表現型への分化について細胞を解析することを含み得る。チロシンヒドロキシラーゼ(TH)はドーパミン作動性の表現型のマーカーの1つである。
【0119】
本発明に従ってスクリーニングを行った全ての物質は天然または合成化合物であり得る。
【0120】
スクリーニング方法は、腹側中脳の1型アストロサイトまたは初期グリア細胞を、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞においてドーパミン作動性の運命を誘導させる能力のない神経細胞(たとえばアストロサイト)と比較することを含み得る。比較は、たとえば腹側中脳の発生中の1型アストロサイトまたは初期グリア細胞と他の神経位置由来の1型アストロサイトまたは初期グリアとの間であり得る。
【0121】
アストロサイトまたは初期グリア細胞を含むスクリーニング方法では、不死化アストロサイトまたは不死化グリア細胞を使用し得る。これは、アストロサイト細胞系またはグリア細胞系、たとえばアストロサイトまたはグリア中脳細胞系を含み得る。このような細胞系は同種細胞集団を提供する。
【0122】
スクリーニング方法では、細胞間の表現型の差を解析するための任意の知られている方法を使用し得、これはDNA、mRNA、cDNAまたはポリペプチドのレベルであり得る。示差的スクリーニングおよび遺伝子スクリーニングがこのような技法のうちの2つである。本明細書中に記載した任意のスクリーニング方法で同定した物質を、本明細書中に記載した任意の他のスクリーニング方法の試験物質として使用し得る。
【0123】
スクリーニング方法では、核発現アレイ、たとえばマウスcDNA発現アレイを使用し得る。この手法では、様々な核酸分子のアレイをフィルター、石英または別の表面上に、たとえば核酸をフィルターに架橋結合させるよって配列させる。試験溶液または抽出物を得て、その中にある核酸をたとえば蛍光によって標識する。その後、溶液または抽出物をフィルターまたは遺伝子チップに施用する。試験核酸とフィルターまたは遺伝子チップ上の核酸とのハイブリダイゼーションを決定し、対照溶液で得られたハイブリダイゼーションと比較する。試験試料と対照試料とで得られたハイブリダイゼーションの差は、異なる核酸含有の指標である。核酸アレイについてのさらなる情報には、任意の利用可能なウェブブラウザを使用して検索可能なClontechウェブサイト(たとえばwww.clontech.com)またはAffymetrixウェブサイト(たとえばwww.affymetrix.com)を参照されたい。
【0124】
スクリーニング方法は、本明細書中では基底レベルより高く発現されたNurr1に関して記載しているが、本開示はNurr1サブファミリーの全ての核内受容体、たとえばNor-1およびNGFI-Bにも及ぶ。したがって、Nurr1は例として記載し、限定するものではない。本発明の任意の方法において、Nurr1または任意の他の受容体、たとえばNor-1やNGFI-Bを含めたNurr1サブファミリーの核内受容体は、細胞において基底レベルより高く発現され得る。
【0125】
スクリーニング方法では、幹細胞または始原細胞もしくは前駆細胞または神経細胞をWntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞または始原細胞もしくは前駆細胞または神経細胞と比較することを含み得る。これは、たとえばNurr1、ならびに/あるいは、幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞において増殖および/もしくは自己再生および/もしくは分化および/もしくは生存を増強させるかつ/あるいはドーパミン作動性の運命の獲得や誘導を促進するおよび/もしくはドーパミン作動性ニューロンの発生を誘導させる、ならびに/またはWntリガンドの存在下でNurr1を基底レベルより高く発現するニューロン細胞においてドーパミン作動性の誘導もしくは分化を増強させる、因子もしくは複数の因子の標的遺伝子を同定するためである。標的遺伝子(もしくは複数の遺伝子)および/または因子(もしくは複数の因子)が同定された後は、これらを単離および/または精製および/またはクローニングし、さらなる方法で使用し得る。
【0126】
スクリーニング方法は、物質もしくは複数の物質を混合物から精製および/または単離することを含み得る。この方法は、混合物の1つまたは複数の画分が、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞または神経細胞と相互作用する能力、たとえばこのような幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞に結合するならびに/または増殖および/もしくは自己再生を促進するならびに/またはそれにおいてドーパミン作動性の表現型もしくは運命の誘導、獲得、分化または発生を増強させる能力を決定することを含み得る。精製および/または単離には、当業者に知られている任意の方法を使用し得る。
【0127】
スクリーニング方法では、試験物質をコードしている核酸に機能的に連結されている誘導性プロモーターを使用し得る。このような構築物を宿主細胞内に取り込ませ、許容条件および非許容条件下におけるその細胞の1つまたは複数の特性を決定して比較する。決定された特性は、宿主細胞が、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞においてドーパミン作動性の表現型を誘導させる能力であるかもしれない。許容条件と非許容条件とにおける宿主細胞の能力の差は、試験物質が、単独でまたは組み合わせて、幹細胞、神経幹細胞または始原細胞もしくは前駆細胞において、増殖および/または自己再生および/またはドーパミン作動性の運命の誘導および/またはドーパミン作動性の分化、生存もしくは発生を増強させる、あるいはWntリガンドの存在下でNurr1を基底レベルより高く発現するニューロン細胞においてドーパミン作動性の誘導または分化を増強させる能力を有するかもしれないことを示している。
【0128】
本発明の任意のスクリーニング方法の詳細な様式は、ルーチンな技術および知識を用いて、当業者によって変形させ得る。
【0129】
本発明によって提供するいずれか1つの方法によって同定した因子もしくは複数の因子を単離および/または精製し、かつ/またはさらに調査し得る。これを製造することもできる。
【0130】
様々なさらなる態様では、本発明は、本明細書中で開示したいずれか1つの方法によって同定した因子;このような因子を含む医薬組成物、医薬、薬物または他の組成物(この組成物は、Nurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロンおよびWntリガンドを含み得る);Wntリガンドの存在下で基底レベルより高くNurr1を発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞由来のドーパミン作動性ニューロンの誘導および/または表現型の分化もしくは成熟および/または生存および/または神経突起生成および/またはシナプス形成および/または機能的出力を増強させるためのこのような因子の使用;医療方法であって、たとえばドーパミン作動性ニューロンの変性、損傷、損失、もしくは障害またはそれに影響を与えることに関連する症状を治療する(予防的処置を含み得る)、たとえばパーキンソン病または別の神経変性疾患を治療するためにこのような因子または組成物を患者に投与することを含む方法における、このような因子または組成物の使用;たとえばパーキンソン病など(たとえば神経変性疾患)を治療するために投与する組成物、医薬または薬物の製造におけるこのような因子の使用;ならびに、このような因子を製薬上許容される賦形剤、ビヒクルまたは担体、および任意選択で他の成分と混合することを含む、医薬組成物を作製する方法をさらに提供する。
【0131】
関連の態様では、本発明は、幹細胞、神経幹細胞、前駆細胞もしくは始原細胞においてドーパミン作動性の運命を誘導させる、またはNurr1を基底レベルより高く発現するニューロン細胞においてドーパミン作動性の誘導もしくは分化を増強させる、Wntリガンドの能力をモジュレーションする物質のスクリーニング方法を提供する。
【0132】
したがって、この方法では、Nurr1を基底レベルより高く発現する幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞において増殖、自己再生、ドーパミン作動性の発生、分化、成熟および/またはドーパミン作動性の運命の獲得を誘導させる、Wntリガンドの能力をモジュレーションする物質をスクリーニングし得る。
【0133】
このような方法は以下のうちの1つまたは複数を含み得る。
【0134】
(i)Wntリガンドおよび1つまたは複数の試験物質の存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞を提供すること;
(ii)ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答するこのような細胞の割合を解析すること;
(iii) ドーパミン作動性の運命をとるこのような細胞の割合を、1つまたは複数の試験物質を存在させない同等の反応培地および条件における、ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答するこのような細胞の数と、比較すること。処理した細胞と未処理の細胞との間のドーパミン作動性ニューロンの割合の差は、関連する1つまたは複数の試験物質のモジュレーション効果の指標である。
【0135】
このようなスクリーニング方法は以下を含み得る。
【0136】
(i)Nurr1を基底レベルより高く発現する幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞を、1つまたは複数の試験物質の存在下でWntリガンドと接触させること;
(ii)ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答する幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞の割合を分析すること;
(iii)ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答する幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞の割合を、1つまたは複数の試験物質を存在させない同等の反応条件における、ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答する幹細胞、前駆細胞、始原細胞または神経細胞の数と比較すること。
【0137】
このようなスクリーニング方法は、同等のもしくは同一の非ヒト動物においてin vivoで、またはin vitroもしくは培養中で、細胞上で実施し得る。
【0138】
Wntまたは誘導活性をモジュレーションする物質を同定した後、物質をさらに調査し得る。これは、医薬、医薬組成物または薬物などの組成物の調製、すなわち製造または配合において製造および/または使用し得る。そのモジュレーション活性について試験した全ての物質は天然または合成化合物であり得る。
【0139】
以降、本発明の態様および実施形態を、例として、添付の図面を参照しながら例示する。さらなる態様および実施形態は当業者には明らかであろう。本明細書中で言及した全ての文献は、本明細書中に参考として組み込まれる。
【0140】
実験
本明細書中に参照により特に組み込まれるのは、1型アストロサイトまたはグリア細胞の存在下、ドーパミン作動性前駆体の増殖および/または自己再生、および、Nurr1を発現している幹細胞、神経幹細胞、前駆細胞または始原細胞におけるドーパミン作動性ニューロンの誘導を示し、そしてかかる細胞を、FGF(たとえばFGF8)またはレチノイドなどの追加の因子と接触させたときに得られた追加の結果を示しているWO OO/66713に記載された実験結果である。
【0141】
Wnt、Nurr1およびチロシンヒドロキシラーゼ(TH)の発現を発生中の腹側中脳で検討し、E10.5〜P1の背側中脳(DM)および大脳皮質と比較した。リアルタイムRT-PCRおよびISHは、THのメッセージがマウスにおいてE11.5ではっきり検出され、ラット腹側中脳においてE11.5で最初に検出されたことを示したが(図1aおよび図1b)、これは、Nurr1の発現の一日後に起こり(図1c)、ラットVMにおいてE16まで続くドーパミン作動性ニューロンの誕生(Fosterら、1988)と一致するものである。Wnt-1およびWnt-5aは、マウス腹側中脳において最高の発現レベルを有するWntであり(図1dおよび図1e)、ラットにおいてE11.5でピークとなった(図1a)。同様のピークは、背側中脳においてE11.5に、Wnt-1およびWnt-3aについて検出されたが、Wnt-5aについては検出されず、これは、腹側部の発生におけるWnt-5aの支配的な役割(Saneyoshiら、2002)と整合するものである。さらに、精製した1型アストロサイトにおけるWntの発現のリアルタイムRT-PCR分析は、Wnt-5aが腹側中脳のアストロサイトに発現しており、発現レベルは、腹側中脳での方が、背側中脳または大脳皮質のアストロサイトにおけるより有意に高かったことを示した(図1f)。
【0142】
発生中の中脳において高レベルで発現されていたWnt-1、3aおよび5aのmRNA発現に加え、リアルタイムRT-PCRによりなされたより詳細な分析は、Wnt-2および-7aのmRNAが中脳において中レベルで発現し、Wnt-7bおよびWnt-16が中脳において低レベルで発現されていたことを明らかにした。これらのリガンドはいずれも、腹側中脳での方が、背側中脳よりも高いレベルで発現しており、これは、Wnt-2、-7a、-7b、および-16の腹側中脳の発生における可能な役割を示唆するものである。本発明者らはまた、Wnt-4、-6、-1Ob、および-11が中脳において低レベルで発現していたことを見出した。Wnt-3、-13および-2bは、中脳において極めて低レベルで発現しており、Wnt-5b、-8a/d、-1Oa、および-15は、中脳においてバックグラウンドレベルで検出された。
【0143】
VMにおけるDAニューロンの誕生は、ラットにおいて胚11.5日(E11.5)に起こることが知られている。本発明者らは、Wntタンパク質の全ファミリーの発現パターンを発生中のラット脳において検討し、そして、本発明者らの研究を、DAニューロン誕生時までにVMにおいて有意なレベルで発現しているWntに広げた。分析したWntの中で、Wnt-2、-4、-7a、-7bは、E11.5において、VMでの方がDMよりも高いレベルの発現を示し、および/またはE11.5のDAニューロン誕生時にVMにおいてピークとなった(図8A、8B、8Cおよび8D)。これらのうち、Wnt-2およびWnt-7aは、VMにおいて、より高いレベルで発現していた(それぞれ図8Aおよび8B)。これに対し、Wnt-5b、-8a/8d、-10a、および-15の発現は、分析した脳領域のいずれにおいても観察されなかった。Wnt-2b/13、Wnt-3、Wnt-6、およびWnt-1Ob、およびWnt-11の転写産物は、主として背側中脳領域に限定されていたが、低レベルであった(図9A、9B、9Cおよび9D)。低レベルではあったが、Wnt-16の転写産物は、胚発生の間中両方の領域で検出された(図9E)。これらのデータは、緊密に関連した細胞外シグナル伝達分子のファミリーの間での、動的な発現のパターンを明らかにするものである。さらに、これらの遺伝子の空間的および時間的発現は、Wntファミリーのメンバーが、胚発生の間中、全く別様に機能することを示唆するものである。したがって、ここで提示された発現プロファイルは、Wnt-2、Wnt-4、Wnt-7a、およびWnt-7bの、DAニューロンの発生における可能な役割を示したものである。
【0144】
本発明者らは、Wntの標準的な経路の中心的なシグナル伝達構成要素であるβ-カテニンの発現を、E10.5のマウスにおけるオーファン核内受容体Nurr1の発現により特徴付けられるDA前駆細胞において検討した。腹側中脳のドーパミン作動性前駆体が高レベルのβ-カテニンを発現していることが見出された。二重免疫組織化学は、β-カテニンがE10.5のマウスにおいて、Nurr1と同じドメインに発現していたことを示したが、これはWntのシグナル伝達とβ-カテニンの安定化とが、in vivoの正常な発生の最中に、VM DA前駆細胞で生じることを示すものである。
【0145】
Wntリガンドを分泌するように操作された安定した線維芽細胞の細胞系(Shimizuら、1997)からの部分的に精製された馴化培地は、サイズ排除に基づくろ過により作製した。連続2回の精製の後、当初のCMに比べて約250〜1500倍の濃度の増大が得られた。部分的に精製されたWntリガンドの個々のロットは、互いに、合わせたウェスタンブロット産物のバンドの密度に基づいて正規化し、任意単位として表現され、ここで、1単位は、正規化された部分的に精製された産物の1μlに等しい。E14.5のラットから得られた腹側中脳の培養物は、その後、増大する濃度の部分的に精製されたWnt-1、-3a、-5a、または対照(CP)培地で3日間処置した(図2A)。Wnt-1または5aによる処置は、TH+細胞数の強い用量反応性の増加をもたらしたものの、Wnt-3aまたはCPによる処置は、100単位までの用量で効果をもたらさず、対照培地は、50単位(μl)までわずかな効果をもたらしたか、または効果をもたらさなかったが、このことは、Wnt-1または-5aによる処置で観察されたTH+細胞数の増加が、これらのリガンドに特異的であることを示唆するものである。興味深いことに、10単位のWnt-1または-5aの効果はin vitroで早くも1日後には検出され、3日目のCPおよびWnt-3aより高く、そして7日後に減少し始めた(図2B)。本発明者らの結果は、部分的に精製されたWntが少なくとも3日間は完全に安定しており、そしてその効果が1週間にわたって維持されたことを示したものである。さらに、TH+細胞数のこれらの増加は、これらが、GDNF、FGF-2またはFGF-8を含む既知のドーパミン栄養分子(dopaminotrophic molecule)により3日後にもたらされる増加を超えており、そしてNurr1発現神経幹細胞のための誘導シグナルを含むVM T1A馴化培地の効果におよそ等しいことから、生物学的に意味があるものと考えられた(図2C)。
【0146】
Wnt-2およびWnt-7aを過剰発現している線維芽細胞細胞系からの馴化培地は、Wnt-1およびWnt-5aの部分的に精製された培地と同様の結果を与え、Wnt-2およびWnt-7aの両方ともE14.5の腹側中脳前駆体培養物におけるTH+ニューロンの数を、対照線維芽細胞馴化培地に比べて増加させた。部分的に精製されたWntによる処置により、TH免疫反応性の増加がWnt-2およびWnt-7aの両方について観察された(図10A)。Wnt-7aによる処置は、対照および他のWntに比べて小幅なTH+の増加をもたらした。並行して行った実験において、Wnt-2による処置は、TH+ニューロンの約2倍の増加をもたらした。これらのデータは、Wnt-2およびWnt-7aによる処置が、VM前駆体培養物においてTH+ニューロンの数を増加させることを示している。Wnt-2およびWnt-7aの同様の効果が、THおよびNurr1マーカーによる二重免疫組織化学を用いた場合に観察されたが(図10B)、これは、これらのWntにより発揮される効果がNurr1発現前駆体に対するものであることを示すものである。
【0147】
したがって、本発明者らの結果は、糖タンパク質のWntファミリーのメンバーが、始原細胞から発生するドーパミン作動性ニューロンの数を極めて有効に調節していることを示している。さらに、本発明者らのデータは、Wntリガンドが、TH細胞数の調節について重複する効果を有していることを示しており、これは、Wntのいくつかの機能がある程度冗長であり得ることを示唆するものである。
【0148】
Wntリガンドによる処置はまた、大きな球形の増殖性クラスターの出現を誘導した。これらのクラスターは最初は小さく、培養期間中大きさを増した。5細胞径より大きく、1個またはそれ以上のTH+ニューロンを含むTH+ニューロンのクラスターを、in vitroで様々な時間に計数した。注目すべきは、10単位のWnt-1および-5aによる処置はin vitroで7日後に観察されたこれらのクラスターの数の3〜8倍の増加をもたらしたが、Wnt-3aはこれをもたらさなかったことである(図2Eおよび図2F)。さらに、Wntリガンドに反応して出現したクラスターは、より大きく、大部分がほとんど全てTH+ニューロンからなっていた。一方、対照およびWnt-3aにより処置された培養物は、一般的により小さなスフェアを含んでいた。興味深いことに、実質的に全てのクラスターと、培養物中の顕著な数の単離細胞とが、固定の前のBrdUの急激なパルスの後にBrdU陽性であった(図2f)。したがって、有糸分裂能のあるスフェアは、前駆体(および/またはコミットメントは少ないが、依然増殖性の)細胞のTH陽性細胞への誘導または分化からの、ドーパミン作動性前駆体の細胞周期への動員に由来する可能性がある。
【0149】
Wnt-1の欠失(McMahonおよびBradley、1990; ThomasおよびCapecchi、1990)またはWnt受容体LRP6の欠失(Pinsonら、2000)が中脳-後脳接合部全体の欠損をもたらし、そしてWnt-1ヌル変異体がドーパミン作動性ニューロンを欠損している(DanielianおよびMcMahon、1996)ことから、本発明者らはWntリガンドがVM前駆体培養物においてTH+ニューロンの数を増加させる可能性を調べた。Wntは、神経系における細胞運命決定の制御(Dorskyら、1998; Bakerら、1999; Wilsonら、2001; Garcia-Castroら、2002; Muroyamaら、2002)、細胞増殖(ChennおよびWalsh、2002; MegasonおよびMcMahon、2002)および分化(Hallら、2000; PatapoutianおよびReichardt、2000; Krylovaら、2002)に必須の役割を担っていることが知られており、これは、複数のメカニズムがVM前駆細胞におけるドーパミン作動性ニューロンの発生の調節に関与し得ることを示唆するものである。
【0150】
本発明者らは、リアルタイムRT-PCRにより、Wnt-5aではなく(Kioussiら、2002)、Wnt-1による細胞周期の進行を媒介する(MegasonおよびMcMahon、2002)β-カテニン(TetsuおよびMcCormick、1999; Shtutmanら、1999)の標的であるサイクリンD1、またはその他のサイクリンの発現が、Wnt-1、-5aおよびVM-T1Aによって調節されているかどうか検討した(図3Aおよび図7A)。サイクリンD2のmRNAは、これらの処置のいずれにも影響されなかったが、サイクリンD1のmRNAは、Wnt-5aまたはVM T1AによってではなくWnt-1によりアップレギュレーションされ、そしてサイクリンD3は、Wnt-5aによってではなくWnt-1によりアップレギュレーションされた。本発明者らの結果はWnt-1が、細胞周期の進行を転写レベルで調節していることを示したものである。同様に、細胞周期インヒビターの分析は、Wnt-5aではなくWnt-1がp27およびp57のmRNAをダウンレギュレーションしたが(図7Bおよび7C)、いずれもp21のmRNAの発現に影響しなかったことを明らかにした。したがって、本発明者らの結果は、Wnt-1が、G1-Sの進行を転写レベルで調節することにより、Nurr1が誘導する細胞周期の停止に拮抗するよう作用し得ることを示唆するものである(Castroら、2001)。
【0151】
Wntの標的遺伝子は種々の生物系で同定されており、多くの細胞周期レギュレーターを包含する。Wnt-2およびWnt-7aによる処置は、重要な細胞周期レギュレーターの発現における明確な差異をもたらした(図11)。驚くべきことに、Wnt-2による処置によって、前駆体培養物は、サイクリンD1のmRNAレベルにおける若干のアップレギュレーションを示した(図11A)。予想通り、Wnt-7aはサイクリンD1の転写レベルを増加させたが(図11A)、これは、本明細書に記載されたBrdUの取り込みの増大したレベルの知見と整合するものである。サイクリンD2はWnt-7aにより調節されなかったが、Wnt-2による処置はサイクリンD2のmRNAレベルの増加を導いた(図11B)。サイクリンD3の調節は、Wnt-7aまたはWnt-2aによる処置のいずれにおいても観察されなかった(図11C)。興味深いことに、Wnt-7aによる処置は、細胞周期インヒビターp27およびp57の発現を減少させた(それぞれ、図11Dおよび11E)。これらのデータは、Wnt-7aがそのドーパミン作動性前駆体に対する機能を、本明細書に記載されたWnt-1のものと同様のメカニズムにより発揮することを示すものである。これに対して、p27およびp57のmRNAレベルにおける増加は、Wnt-2による処置の後に観察された(それぞれ、図11Dおよび11E)。これらのデータは、Wnt-2は前駆体培養物におけるTH+ニューロンの数を増加させるが、Wnt-5aのものと別異のメカニズムによってこれを行ない、そして、これはG1-Sの進行の転写レベルでの負の調節を伴うことを示唆するものである。
【0152】
本発明者らは、Wntが、VM培養物におけるDA前駆細胞の増殖を調節するかどうか検討した。興味深いことに、本発明者らは、新生DAニューロン(Nurr1+/TH+細胞)においてBrdUの取り込みを観察できなかったが、これは、増殖している細胞は、実験の最後の6時間の標識期間中TH+細胞に分化しないことを示すものである。本発明者らは次に、WntがVM前駆体培養物において増殖を調節するかどうか、および、中脳におけるNurr1発現ニューロン前駆体(Nurr1+/BrdU+細胞)がWntリガンドの潜在的な標的であるかどうかを検討した。VM前駆体培養物におけるBrdUの取り込みは、Wnt-5aによる処置によってではなく、Wnt-1および-3aによる処置によって増加したが(図7D)、これは、Wnt-1および-3aがマイトジェンとして作用し得、培養物における全体的な前駆体の増殖を促進しうることを示すものである。Wnt-1および-3aの効果は、Nurr1およびBrdU二重陽性前駆細胞をin vitroで1日後に検討した場合にさらに大きかった(3倍増)。部分的に精製されたWnt-5aも、Nurr1+集団の有糸分裂を増加させたが、程度はそれ程大きくなかった(図3B)。in vitroの1日目におけるNurr1およびBrdUに対する二重免疫組織化学は、Wnt-1および-3aは、有糸分裂を3倍増加させたが、Wnt-5a、およびWnt-5aを発現し、Nurr1発現前駆体にドーパミン作動性表現型を誘導するVM1型アストロサイトからの馴化培地は、Nurr1+集団において有糸分裂を2倍増加させたことを示した(図3B)。この知見は、Wnt-1によるサイクリンD1発現の増強と合わせて、Wnt-1が、有糸分裂/増殖を調節することによりTH+細胞数を主として増強させることを示したものである。驚くべきことに、Wnt-3aは、Nurr1陽性細胞の有糸分裂を増強することについてWnt-1と同程度有効であったものの、TH陽性細胞数をわずかに増加させるか、または増加させなかったが、このことは、増加した有糸分裂は、腹側中脳におけるWntの活性のほんの一要素にすぎないことを示唆するものである。Wnt-1およびWnt-3aによって誘導される増殖の明確な増加は、β-カテニンを安定化するこれらの能力(Shimizuら、1997)、幹細胞および神経前駆体において細胞周期リエントリー(reentry)を促進するβ-カテニンの役割(TaipaleおよびBeachy、2001; ChennおよびWalsh、2002)、および造血幹細胞の自己再生におけるWnt-3aの役割(Reyaら、2003)とよく相関する。したがって、Wnt-1の、前駆体の増殖およびG1-Sの進行を制御する遺伝子の調節に対する効果は、Wnt-1が、前駆細胞における有糸分裂/増殖/自己再生を調節することにより、主としてTH+細胞数を増強させることを示すものである。しかしながら、Wnt-3aは、Nurr1+細胞における有糸分裂を増強することについてWnt-1と同程度に有効であったものの、Wnt-1と異なり、DAニューロンの数に影響を与えないか、または数を減少させたが、このことは、自己再生および/または前駆体集団の維持におけるWnt-3aの特異的な役割を示唆するものである。
【0153】
BrdU(有糸分裂のS期にある細胞のマーカー)の取り込みを評価する実験において、BrdU+細胞数の増加が、Wnt-7aによる処置の後でも観察された(図10C)。実際、Wnt-7aによる処置は、上記のWnt-1と同様のBrdU+の増加をもたらした。これに対し、BrdUの取り込みの若干の減少が、Wnt-2による処置の後で認められた。これらのデータは、Wnt-7aが前駆体集団を拡大することによりTH+細胞数を増加させる一方、Wnt-2の効果は、別個のメカニズムを介して起こり、そして前駆体の増殖の減少を伴うことを示すものである。
【0154】
本発明者らはまた、Wnt-1の増殖効果が特異的であり、そしてWntインヒビターであるFrizzled-8のシステインリッチドメイン(Fz8-CRD)(35、36)によりブロックされ得るかどうかを検討した。Fz8-CRDを過剰発現している線維芽細胞からの部分的に精製されたCMを対照およびWnt-1で処置された培養物に加え、そしてTH+ニューロンの数を検討した(図7Dおよび7E)。Fz8-CRDは、両方の条件においてTH+ニューロンの数を減少させたが、これは、内因性のWntが培養物中のDAニューロンの発生に必要であり、そして、WntのTH+細胞への効果が特異的であることを示すものである。
【0155】
したがって、まとめると、全ての結果は、Wntの効果が特異的であり、そしてVMにおけるTH+かつ増殖性の細胞の数がWntによって独立して調節されていることを示していた。さらに、これらの結果は、増大した有糸分裂/増殖/自己再生が、腹側中脳におけるWntの活性のほんの一要素にすぎないことを示唆したものである。
【0156】
本発明者らはまた、Wntリガンドの効果がより多くのニューロンの数をもたらすかどうか、およびこれらの効果が増殖性クラスター内部または外部のドーパミン作動性ニューロンに特異的かどうかを問うた。本発明者らは、Wnt-1または-3aが、TuJ1+細胞を含む増殖性クラスターの数を、対照、Wnt-5aまたはVM T1Aに比べて10〜12倍増加させたことを見出した(図3d)。Wnt-1および-3aの両方の、ニューロンを発生させることができる増殖性クラスターの数を増加させる能力にもかかわらず、Wnt-1のみが培養物中のTuJ1+ニューロンの総数を増加させた(図3f)。さらに、Wnt-1は、TH+ニューロン(図3g)、またはドーパミン作動性ニューロンを含むVMクラスターの比率を増加させず(クラスターの約30%)、そしてWnt-3aは、ドーパミン作動性ニューロン(図3g)およびクラスター(図3e)の比率を10%未満に減少させさえした。本発明者らの知見は、Wnt-1およびWnt-3aが、全てのVMニューロンの数を、増殖および増殖性クラスターの数を増加させることにより増加させることを示したものである。Wnt-1はVMニューロンの総数を、増殖的なメカニズムにより非選択的に増加させる一方、Wnt-3aは、クラスターとして増殖しているニューロン前駆細胞の増殖を増加させ、これらのTH+細胞への分化を抑制したが、これは、これらが、ドーパミン作動性前駆体の自己再生において役割を果たし得ることを示唆するものである。Wnt-1および-3aに対して、Wnt-5aおよびVM T1Aは、TuJ1+ニューロン(図3f)またはニューロンを含む増殖性クラスターの数を増加させなかったものの(図3d)、代わりにドーパミン作動性クラスターおよびドーパミン作動性ニューロンについて、2倍を超えて選択的に富化したが(それぞれ図3eおよび3g)、これは、Wnt-5aおよびVM T1Aが、VM培養物におけるドーパミン作動性表現型の獲得を指示または促進することに関与し得ることを示すものである。Wntの、クラスター外のTuJ1+ニューロンに対する効果は、Wnt-3aを除きクラスター内のニューロンに対するものと極めて類似しており、Wnt-3aはTuJ1+ニューロンの数を増加させなかったが(図3f)、これは、Wnt-3aがクラスター内の前駆体の増殖および/または自己再生を調節し、これを行うことによりドーパミン作動性表現型を獲得するニューロンの比率を減少させていることを示すものである(図3g)。
【0157】
したがって、本発明者らの結果は、Wntが、VMにおける相異なる機能:前駆体の増殖および/または自己再生、ニューロンの数、および/またはドーパミン作動性表現型を獲得するニューロンの比率、を独立かつ示差的に調節していることを示唆したものである。さらに、Wnt-1、-5aおよびVM T1Aの、ドーパミン作動性ニューロンおよび細胞クラスターの数に対する同様の効果にもかかわらず、本発明者らの結果は、Wnt-1およびWnt-5aが、ドーパミン作動性細胞系統に、2つの部分的に相異なるメカニズムを介して作用することを示すものである。Wnt-1はドーパミン作動性ニューロンの比率ではなく、細胞の増殖を増大させる一方、Wnt-5aおよびVM T1Aは、増殖にはあまり作用しないが、その代わり、ドーパミン作動性ニューロンの比率を増殖性クラスターの内部および外部で、追加のメカニズムにより増大させる。
【0158】
VM T1Aがドーパミン作動性誘導シグナルの供給源であるという前提のもと(Wagnerら、1999)、本発明者らは、WntまたはVM T1Aによる処置が、腹側中脳のNurr1+ニューロン前駆体(Nurr1+/TH-細胞)のドーパミン作動性ニューロン(Nurr1+/TH+細胞)への変換に影響を与えるかどうか検討した。対照条件においては、全Nurr1陽性細胞の約50%がTHを発現したが、Wnt-3aによる処置はこの集団を30%に減少させる。これに対し、Wnt-1はNurr1+/TH+の比率を70%まで増加させ、そしてWnt-5aまたはVM T1Aはこの比率を90%まで増加させた(図4A)。これらの結果は、ADH-2/TH二重免疫染色により得られたものと極めて類似していたが、これは、ADH2もドーパミン作動性前駆体およびニューロンを標識したからである(Wagnerら、1999)。したがって、本発明者らのデータは、Wnt-5aが、Wnt-1とは異なり、主にNurr1発現前駆体にドーパミン作動性表現型を誘導することにより、ドーパミン作動性ニューロンの数を増加させるモデルを示唆したものである。このモデルを試験するために、本発明者らは、VM以外の脳構造に由来するNurr1陽性/TH陰性細胞も、WntまたはVM T1Aのいずれかによりドーパミン作動性表現型を獲得するよう誘導されうるかどうかを検討した。Nurr1+前駆細胞に富む、E13.5の大脳皮質からの初代培養物を、種々のWntリガンドに暴露するか、またはP1のVMもしくは(対照としての)皮質のアストロサイトと共に共培養した。Wnt-3aによる処置または皮質のT1Aとの共培養のもとでは、THを共発現したNurr1陽性の皮質細胞は実質的に存在しなかった。しかしながら、Wnt-1、Wnt-5a、またはVM T1Aによる処置は、THを発現したNurr1陽性細胞の比率の有意な増加を誘導した(図4B)。最も劇的な効果は、Wnt-5aによって誘導され、これは、わずか3日間で、二重Nurr1/TH+細胞の数を、対照に比べて>40倍、そしてWnt-1に比べて>4倍増加させたが、この効果は増殖のみでは説明できないものである。さらに、Wnt-5aが腹側部のドーパミン作動性表現型の獲得を促進するという知見は、先に報告された、Wnt-5aの、非古典的な(non-canonical)Wntシグナル伝達を介した腹側部の運命を促進する役割(Saneyoshiら、2002)とよく相関する。
【0159】
本発明者らはまた、Wnt-5aによる処置が、中脳ドーパミン作動性ニューロンに特徴的な遺伝子の発現を調節することにより、前駆体培養物においてより分化した中脳DA表現型の獲得を促進するかどうか検討した。本発明者らはまず、ビコイド(bicoid)関連ホメオドメイン遺伝子Ptx3(Smidtら、1997)の発現を、リアルタイムRT-PCRにより検討し、そしてWnt-1ではなく、Wnt-5aによる処置がPtx3のmRNAを増大させたことを見出した(図6A)。この知見は、Ptx3がDAニューロンの発生に必要であり(Van Den Munckhof Pら、2003; Nunes Iら、2003)、そして同じファミリーの別のホメオドメイン遺伝子であるPtx2が、Wntシグナル伝達により直接調節されていることから(Kioussi Cら、2002; Baek SHら、2003)、極めて興味深いものである。本発明者らは次に、癌原遺伝子c-ret(Trupp Mら、1996)ならびに2種類の他のGDNF補助受容体、GFRα1(AiraksinenおよびSaarma、2002)およびNCAM(Paratchaら、2003)の発現も、Wntにより調節されているかどうか検討した。興味深いことに、c-retおよびNCAMともWntシグナル伝達により調節され(Zheng Sら、1996; Conacci-Sorrellら、2002)、そしてこれらのリガンドであるGDNFは、DAニューロンの出生後の発生に必要である(Granholmら、2000)。本発明者らは、分析した全てのGDNF受容体が、Wnt-1およびWnt-5aにより示差的に調節されていることを見出した。c-retの発現は、Wnt-1ではなWnt-5aによってアップレギュレーションされる一方(図6B)、GFRα1およびNCAMの発現は、Wnt-5aにより維持され、そしてWnt-1により抑制された(図6Cおよび6D)。したがって、本発明者らの結果は、Wnt-5aが、中脳DAニューロンの2つの弁別的な特徴であるPtx3およびGDNF受容体の発現を増大させることにより、前駆細胞におけるDA表現型の獲得を促進すること、ならびにこれらの分化および生存を増強することに用いられ得ることを示すものである。したがって、Wnt-5aは、DAニューロンの数を増加させることができる。
【0160】
Wnt-5aの生物学的活性が、測定したパラメーターの全てにおいてVM T1Aとほぼ同一であったという知見(図1、図2、図3および図4)、およびVM T1AがWnt-5aを発現するという知見(図1F)は、VM T1Aの効果が、一部アストロサイトによるWntの分泌によりうることを示したものであり、本発明者らにWntの神経幹細胞培養物における機能を検討することを促した。本発明者らは、したがって、Wntの効果をブロックするFrizzled-8のシステインリッチドメイン(Fz8-CRD)(Hsiehら、1999)を過剰発現する線維芽細胞からの馴化培地が、VM T1AまたはWnt-5aの、ドーパミン作動性ニューロンの誘導に対する活性をブロックすることができるかどうか検討した。本発明者らはFz8-CRDの、VM T1Aと共培養しているNurr1発現神経幹細胞(Nurr1-c17.2-c42、Wagnerら、1999)への添加が、ドーパミン作動性ニューロンの誘導を部分的にブロックしたことを見出した。さらに、Fz8-CRDは、発生中のマウス脳(Yeら、1998)およびNurr1発現マウス胚性幹細胞に由来するニューロスフェア(Kimら、2002)を包含する、アストロサイトを含有する調製物において、ドーパミン作動性ニューロンをShhとの組み合わせで誘導することが知られている因子であるFGF8により拡大させたニューロスフェアとして増殖させた、VM神経幹細胞におけるドーパミン作動性ニューロンの数を減少させた(図5a)。同様に、E14.5のVM前駆体のFz8-CRDによる共処置は、未処置(図5b)およびVM T1A(図5c)またはWnt-5aにより処置した培養物(図5d)におけるNurr1+前駆体からのドーパミン作動性ニューロンの誘導をブロックした。したがって本発明者らの結果は、Wntが発生に関する他のシグナルと協力して作用し、そして部分的に前駆細胞/神経幹細胞培養物におけるニューロンドーパミン作動性表現型の獲得に必要であることを示すものである。したがって、本発明者らの研究は、Wntが腹側中脳における神経発生の重大かつ順次的な2つの側面である、前駆体の増殖およびドーパミン作動性表現型の獲得の、本質的なレギュレーターであるモデルを示唆するものである(図5F)。このように、Wntは、VM前駆体の運命決定を調節するものであると思われる。Wnt-1は、前駆体の増殖の増大により神経発生を促進し、かつDAおよび非DA VMニューロンの両方に影響を与えた。Wnt-3aは、Nurr1+前駆体の増殖または維持および/または自己再生を促進し、かつDAニューロンの数を減少させた。これに対し、Wnt-5aは弱いマイトジェンであり、Ptx3およびGDNF受容体の発現を増大させ、かつNurr1発現前駆体におけるDA表現型の獲得を効果的に促進した。
【0161】
これらの結果は、ドーパミン作動性前駆体が、Wntに極めて特異的な様式で応答することを明確に示している。Wnt-1、-3a、および-5aは、中脳DAニューロンの発生を、一部重複するメカニズムにより示差的に調節しており、これは、DA前駆体の増殖を促進すること(Wnt-1≧Wnt-3a>Wnt-5a)、これらの分化を抑制すること(Wnt-3a)、神経発生を拡大すること(Wnt-1)、および中脳ドーパミン作動性表現型の獲得を促進すること(DA前駆体のDAニューロンへの分化を促進すること)(Wnt-5a>Wnt-1)を包含する。Wnt-1と同様に、Wnt-7aは前駆体集団を拡大し、そして細胞周期をG1-S移行において促進することによりTH+細胞の数を増加させた。Wnt-2は、前駆体の低減した増殖および細胞周期のG1での停止を伴う、異なるメカニズムにより、DAニューロンの数を増加させた。
【0162】
神経幹細胞における中脳ドーパミン作動性表現型の獲得を調節することに関するWntの役割について、本発明者らは、アストロサイト由来のシグナルが、中脳ドーパミン作動性ニューロンのものを包含する、Nurr1発現前駆体における組織特異的なニューロン表現型を誘導することにより、神経発生において決定的な役割を果たすことを先に示した(Wagnerら、1999、WO00/66713)。興味深いことに、海馬のアストロサイトの、成人における神経発生への同様の役割が最近報告された(Songら、2002)。本明細書で、本発明者らは、アストロサイト由来のシグナルが、Nurr1発現前駆体に対して部分的に重複する相異なる機能を発揮するWntリガンドのファミリーのメンバーを含むことを示した。Wnt-1は、ニューロン前駆体の増殖を増大させることにより神経発生を促進し、かつ全てのVMニューロンに影響を与えた。Wnt-3aは、Nurr1+前駆体の増殖および/または自己再生を促進し、かつドーパミン作動性ニューロンの数を減少させた。これに対し、Wnt-5aはマイトジェンとしてはWnt-1および-3aより有効ではなかったが、Nurr1発現前駆体においてドーパミン作動性表現型を誘導することにおいて最も有効であった。さらに、Fz8 CRDが、Nurr1発現前駆体ばかりでなくFGF8により拡大したニューロスフェアにおいてもドーパミン作動性ニューロンの誘導をブロックしたという知見は、Wntがドーパミン作動性ニューロンの誘導に必要であることを示しており、FGF8の誘導効果がWntによって媒介されている可能性があることを示すものである。この可能性は、FGF8の、発生中の中脳-後脳においてWntの発現およびオーガナイザー活性の両方を誘導する既知の能力と合致するものである(総説として、WurstおよびBally-Cuif、2001;およびRhinnおよびBrand、2001を参照)。
【0163】
Wntを、ドーパミン作動性前駆体/幹細胞の拡大および中脳ドーパミン作動性ニューロンの誘導における重要な細胞外分子プレーヤーとして同定する本発明者らの結果は、パーキンソン病における細胞置換のための、幹細胞に由来するドーパミン作動性ニューロンの有効かつ大規模な産生への扉を開くものである(BjorklundおよびLindvall、2000; PriceおよびWilliams、2001; Arenas、2002; RossiおよびCattaneo、2002; Gottliebら、2002)。したがって、Wntリガンドおよび腹側中脳アストロサイトまたは初期グリア細胞に由来する追加のシグナルは、腹側中脳のドーパミン作動性ニューロンを誘導するのに用いることができる。本発明は、さらに、有効な幹細胞治療の将来的な改善に必要な追加の成分の同定を提供する。
【0164】
神経変性疾患の治療
本発明のニューロンの、神経変性疾患を治療する能力の確認は、パーキンソン病のin vivoモデルを用いて得る。ドーパミン神経毒素である、6-ヒドロキシドーパミン(6-OHDA)またはMPTPは、ニューロンにより特異的に取り込まれ、そして酸化ストレスならびにドーパミン作動性およびノルアドレナリン作動性ニューロンの消失を引き起こす。また、ラクタシスチンを包含するプロテアソームインヒビター、またはロテノンを包含する殺虫剤の注入が、中脳ドーパミン作動性ニューロンの死を引き起こし、そして実験的にパーキンソン病の特徴を再現することも知られている。
【0165】
Nurr1サブファミリーの核内受容体、たとえばNurr1を発現する細胞であって、in vitroでドーパミン作動性ニューロンに分化したものを、6-OHDAまたはMPTPまたはラクタシスチンまたはロテノンで処置したマウスもしくはその他の非ヒト動物の、黒質および/または線条体に外科的に移植する。これらの細胞の、統合および完全に分化する能力は、LacZおよびEGFPなどのレポーター遺伝子を発現する細胞において、電気生理学的および/または形態学的技法により評価する。神経化学的技法は、カテコールアミンの含量および放出の測定を含む。形態学的分析は、チロシンヒドロキシラーゼ、ドーパミントランスポーターおよびドーパミン受容体、Ptx3、Lmx1bおよびADH-2を包含する、ドーパミン作動性ニューロンに特徴的なマーカー遺伝子の発現、およびシナプス接合の形成を研究することを含み得る。
【0166】
未分化のNurr1+細胞がin vivoでドーパミン作動性の表現型に向けて自発的に分化する能力は、かかる細胞を、軸索を切断した、または6-OHDAもしくはMPTPPもしくはラクタシスチンもしくはロテノンにより処置した動物に、線条体内または黒質内移植することにより評価した。ドーパミン作動性表現型、分化および統合は、上記のとおりに検出した。
【0167】
線条体および/または黒質内に移植されたNurr1+細胞に由来するドーパミン作動性ニューロンの、片側性の毒素(たとえば6-OHDA)による処置によって誘導された運動非対称性、またはMPPの全身投与によって誘導された運動異常のいずれかを救済する能力は、アポモルフィンおよびアンフェタミン試験における旋回行動の評価(Schwarting、R. K.ら、(1996)Progress in Neurobiology、50(2-3)、275〜331)、および/または、階段試験、足踏み試験もしくはシリンダー試験(cylinder test)における熟練した足使いの能力(performance in skilled paw usage)によって確認する。
【0168】
in vivoで基底レベルを超えるNurr1を発現する、宿主由来の内因性の幹細胞、神経幹細胞、始原細胞もしくは前駆細胞または神経細胞を、Wntリガンドのin vivoでの投与後における、ドーパミン作動性ニューロンへ分化するこれらの能力について検討することができる。分析は、上記のとおり、形態学的、生化学的および行動学的レベルでの評価を含んでもよい。腹側中脳のアストロサイト/グリア細胞またはこれらに由来する因子の能力は、上記のとおり、Wntリガンド投与と共に分析してもよい。
【0169】
概括
Wntは、細胞増殖、自己再生、運命決定および分化を調節する糖タンパク質のファミリーである。本発明者らの結果は、β-カテニンがNurr1+DA前駆細胞で発現していること、ならびにWnt-1、-3aおよび-5aがVMにおいて高レベルで存在しており、そして発生の最中に示差的に調節されていることを示すものである。部分的に精製されたWntは、VMの発生を弁別的に調節した:Wnt-3aはNurr1+前駆細胞の増殖および/または自己再生を促進したが、TH+ニューロンの数を増加させなかった。代わりに、Wnt-1および-5aは前駆体培養物におけるラット中脳のDAニューロンの数を、2つの相異なるメカニズムによって増加させた。Wnt-1は、主にNurr1+前駆体の増殖を増大させ、サイクリンD1およびD3をアップレギュレーションし、かつp27およびp57のmRNAをダウンレギュレーションした。これに対し、Wnt-5aは、主としてニューロンDA表現型を獲得するNurr1+前駆体の比率を増加させ、これは、Ptx3およびc-retのmRNAのアップレギュレーションを含んだ。さらに、Frizzled8(Wntインヒビター)の可溶性システインリッチドメインは、Wnt-1およびWnt-5aの、増殖および前駆体培養物におけるDA表現型の獲得に対する効果をブロックし、そしてまた、内因性Wntの、Nurr1発現神経幹細胞およびFGF8拡大VMニューロスフェア培養物におけるドーパミン作動性表現型の獲得に対する効果をもブロックした。
【0170】
本明細書に記載された結果は、発生中のラット中脳における、Wntタンパク質の全ファミリーの複雑な空間的および時間的発現パターンを明らかにするものである。Wnt-1、Wnt-3aおよびWnt-5aに加えて、この広範な発現分析は、Wnt-2、Wnt-4、Wnt-7aおよびWnt-7bを、ドーパミン作動性ニューロンの生成における役割を有する候補として同定した。この研究に記載されている知見は、Wnt-7aが、ドーパミン作動性前駆体の増殖を促進し、そしてこれらのドーパミン作動性ニューロンへの分化を可能にすることにより、Wnt-1と同様の様式で作用することを示すものである。Wnt-7aによる処置での、増大したBrdUの取り込み、増大したサイクリンD1の発現、および低減したcdkインヒビターの発現という本発明者らの観察は、これを強く支持するものである。また、この調査は、Wnt-2による処置の結果が、低減したレベルのBrdUの取り込み、サイクリンD2の発現の増大、および細胞周期インヒビターp27およびp57の発現の大幅な増大であることを明らかにした。これらのデータは、Wnt-2が、研究した他のWntに比べ、細胞周期離脱およびNurr1+前駆体によるドーパミン作動性ニューロン表現型の獲得を有利にする、新規なメカニズムを介して作用することを示すものである。これらの知見は、Wntによる類似しているが、相異なる作用メカニズムを浮き彫りにし、そしてWntが、増殖、自己再生および幹細胞/前駆細胞のDAニューロンへの分化の重要なレギュレーターであることを示すものである。さらに本発明者らは、分析した全てのWntが、前駆細胞/幹細胞の増殖の維持およびニューロンドーパミン作動性表現型の獲得に関してユニークな活性プロファイルを示すことを見出したが、これは、Wntファミリーの全てのメンバーがニューロン変性の処置のための魅力ある標的となり得ることを示唆するものである。
【0171】
したがって、幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞の増殖および分化ポテンシャル、Nurr1などの選択遺伝子、未成熟グリア細胞またはアストロサイト、およびWntを利用する本明細書に記載した方法は、神経変性疾患の治療において、所望の神経化学的表現型のニューロンの作製を提供する(たとえばニューロン移植の供給原料として)。中脳ドーパミン作動性ニューロンの誘導は、パーキンソン病を治療する細胞置換戦略に用いることができる。
【0172】
方法
in situハイブリダイゼーション(ISH)
雄および雌の野生型CD-1マウス(25〜35g、Charles River、Uppsala)は、欧州共同体(86/609/EEC)、神経科学学会(1985年1月)のガイドラインに従って飼育、給餌および処置し、そして全ての実験は、地域の倫理委員会により承認を受けた。胚10.5および11.5日からのマウスを取り出し、O.C.T.中に-70℃で迅速に凍結した。胚全体にわたる連続矢状切片(14μm厚)を、ガラス製顕微鏡スライド(StarFrost、KnittelGlaser)上に回収した。ISHは、新鮮な凍結組織に対し、35S標識リボプローブを用いて先に記載されたとおりに行った(Truppら、1997)。mRNAレベルを保全するため、切片を15分間、氷冷4%PFAにて固定し、そしてPBSで2回すすいだ。組織を、0.2MのHCl中で12分間除タンパクし、0.1Mのトリエタノールアミン中の0.25%無水酢酸で20分間アセチル化し、そして増加する濃度のエタノール中で脱水した。スライドを、加湿チャンバー内で16時間、54℃で200μlのハイブリダイゼーションカクテル中の106cpmのプローブと共にインキュベートした。全ての洗浄は、62℃で行なった。1×SSC中の15分間の最初の2回の洗浄、50%ホルムアミド/0.5×SSC中の30分間および1×SSC中の15分間であった。引き続き、37℃での30分間のRNase処置(40μg/ml)および1×SSC中の15分間の2回の洗浄がなされ、その後に、エタノール中での脱水および風乾を行なった。クロロホルムは省略し、脱水時間は30秒〜1分に短縮した。その後、スライドを水で1:1に希釈したNTB-2感光乳剤(Eastman Kodak、Rochester、NY)中に浸し、4℃で6〜8週間感光させ、D19(Eastman Kodak)で現像し、AL-4(Agfa Gevaert、Kista、Sweden)で固定し、そしてチオニンで対比染色した。
【0173】
免疫組織化学
免疫組織化学は、4%パラホルムアルデヒド(PFA)を最後に添加した(postfixed)スライド上で行なった。インキュベーションは、4℃で一晩、希釈バッファー(1%のウシ血清アルブミン、BSA、および0.3%のTriton-X100を含有するリン酸緩衝生理食塩水、PBS)中のマウス抗β-カテニン、1:250(BD Transduction Lab.)およびウサギ抗Nurr1、1:200(Santa Cruz Biotech.)と共に行なった。0.2%Tween-20/PBSでの洗浄に引き続き、切片を希釈バッファー中30分間ブロッキングし、これに2次抗体(全てJacksonからの、Cy2ウマ抗マウスIgGもしくはウマ抗ウサギIgG、またはローダミンウマ抗マウスIgG)、1:200による2時間インキュベーションを続けた。
【0174】
リアルタイムRT-PCRおよび遺伝子発現の定量
マウスおよびヒトWnt1、Wnt3a、Wnt5a、Frizzled8、Ptx3、サイクリンD1およびサイクリンD2およびチロシンヒドロキシラーゼについてのものを含むGenbankのcDNA配列を、プライマーの設計のためにPrimer Express 1.0(PE Applied Biosystems、Foster City、CA、USA)およびPrimer 3(http://www-genome.wi.mit.edu/cgi-bin/primer/primer3_www.cgi)に用いた。以下のオリゴヌクレオチドを用いた:
QuantumRNA古典18S内部標準(米国オースチン、Ambion);
mWnt1フォワード - 5'-CTTCGGCAAGATCGTCAACC-3'(配列番号1);
mWnt1リバース - 5'-GCGAAGATGAACGCTGTTTCT-3'(配列番号2);
mWnt3aフォワード - 5'-GAACGCGACCTGGTCTACTACG-3'(配列番号3);
mWnt3aリバース - 5'-GTTAGGTTCGCAGAAGTTGGGT-3'(配列番号4);
mWnt5aフォワード - 5'-AATAACCCTGTTCAGATGTCA-3'(配列番号5);
mWnt5aリバース - 5'-TACTGCATGTGGTCCTGATA-3'(配列番号6);
チロシンヒドロキシラーゼフォワード - 5'-AGTACTTTGTGCGCTTCGAGGTG-3'(配列番号7);
チロシンヒドロキシラーゼリバース - 5'-CTTGGGAACCAGGGAACCTTG-3'(配列番号8);
Fz8フォワード - 5'-TTGGAAGTGACCTCGCTCCTAG-3'(配列番号9);
Fz8リバース - 5'-GGTTGGGCATGTAAGTGTAGTTGT-3'(配列番号10);
Ptx3フォワード - 5'-AGGGTGGACTCCTACAGATTGG-3'(配列番号11);
Ptx3リバース - 5'-CCGATCCCAGATATTGAAGCC-3'(配列番号12);
サイクリンD1フォワード - 5'-ACCCTGACACCAATCTCCTCAAC-3'(配列番号13);
サイクリンD1リバース - 5'-GTAAGATACGGAGGGCGCACAG-3'(配列番号14);
サイクリンD2フォワード - 5'-ACTGATGTGGATTGTCTCAAAGCCT-3'(配列番号15);
サイクリンD2リバース - 5'-CGTTATGCTGCTCTTGACGGAA-3'(配列番号16);
C-retフォワード - 5'-ATGCACAATTACAGGCTGGTTCT-3'(配列番号17)
C-retリバース: 5'-GTCATTGACCAGGACTACTAGCTGC-3'(配列番号18)
NCAMフォワード 5'-CACTTCGTGTTCAGGACTTCAGC-3'(配列番号19)
NCAMリバース 5'-GGACGAAAATGACAATGAGGATG-3'(配列番号20)
GFRa1フォワード: 5'-GTCTGAGAATGAGATCCCCACAC-3'(配列番号21)
GFRa1リバース: 5'-ACACATTGGATTTCAGCTTCTGAG-3'(配列番号22)
サイクリンD3フォワード: 5'-GGCTATGAACTACCTGGATCGCTA-3'(配列番号23)
サイクリンD3リバース: 5'-ACGGTACCTAGAAGCTGCAATTG-3'(配列番号24)
p21フォワード - 5'-AGCAAAGTGTGCCGTTGTCTCT-3'(配列番号25)
p21リバース - 5'-TCTCCGTGACGAAGTCAAAGTTC-3'(配列番号26)
p27フォワード - 5'-TTAATTGGGTCTCAGGCAAACTCT-3'(配列番号27)
p27リバース - 5'-CTAACCCAGCCTGATTGTCTGAC-3'(配列番号28)
p57フォワード - 5'-GAGGACCAGAACCGCTGGGACTT-3'(配列番号29)
p57リバース - 5'-ACTCGCTGTCCACCTCCATCCA-3'(配列番号30)
GDNFフォワード: 5'-TTTCGATATTGTAGCGGTTCCTGT-3'(配列番号31)
GDNFリバース: 5'-GCCTACCTTGTCACTTGTTAGCCT-3'(配列番号32)
Wnt2フォワード: 5'-AACGTCCCTCTCGGTGGAATC-3'(配列番号33)
Wnt2リバース: 5'-TGTACCACCATGAAGAGCTGACC-3'(配列番号34)
Wnt2b/13フォワード: 5'-CCACCCGGACTGATCTTGTCTACT-3'(配列番号35)
Wnt2b/13リバース: 5'-GGAACCTGAAGCCTTGTCCAA-3'(配列番号36)
Wnt3フォワード: 5'-CAGCGTAGCAGAAGGTGTGAAG-3'(配列番号37)
Wnt3リバース: 5'-ATGGCCAGGCTGTCATCTATG-3'(配列番号38)
Wnt4フォワード: 5'-GCTGTACCTGGCCAAGCTGTC-3'(配列番号39)
Wnt4リバース: 5'-TGGATCAGGCCTTTGAGTTTCTC-3'(配列番号40)
Wnt5bフォワード: 5'-GCCGAGCTCTCATGAACCTACAG-3'(配列番号41)
Wnt5bリバース: 5'-GGCGACATCAGCCATCTTATACAC-3'(配列番号42)
Wnt6フォワード: 5'-GCGGAGACGATGTGGACTTC-3'(配列番号43)
Wnt6リバース: 5'-ATGCACGGATATCTCCACGG-3'(配列番号44)
Wnt7aフォワード: 5'-TGCGTGCCAGTCGAAACAAG-3'(配列番号45)
Wnt7aリバース: 5'-GATATACACCAGGTCAGTGTCCATGG-3'(配列番号46)
Wnt7bフォワード:5'-GCCAACATCATCTGCAACAAGA-3'(配列番号47)
Wnt7bリバース: 5'-CCGATCACAATGATGGCATC-3'(配列番号48)
Wnt8a/8dフォワード: 5'-CAGCGACAACGTGGAGTTCG-3'(配列番号49)
Wnt8a/8dリバース: 5'-CATCCTTCCCTTTCTCCAAACTG-3'(配列番号50)
Wnt1Oaフォワード: 5'-CCACTCCGACCTGGTCTACTTTG-3'(配列番号51)
Wnt1Oaリバース: 5'-TGCTGCTCTTATTGCACAGGC-3'(配列番号52)
Wnt1Obフォワード: 5'-ACGACATGGACTTCGGAGAGAAGT-3'(配列番号53)
Wnt1Obリバース: 5'-CATTCTCGCCTGGATGTCCC-3'(配列番号54)
Wnt11フォワード: 5'-CAAGTTTTCCGATGCTCCTATGAA-3'(配列番号55)
Wnt11リバース: 5'-TTGTGTAGACGCATCAGTTTATTGG-3'(配列番号56)
Wnt15フォワード: 5'-CTGTTCGTACCTGTTGGAAGCA-3'(配列番号57)
Wnt15リバース: 5'-CAGCCGTGTCATAGCGTAGCT-3'(配列番号58)
Wnt16フォワード: 5'-ACCCCCATTCTCAAGGATGACTT-3'(配列番号59)
Wnt16リバース: 5'-CAGTTTCTTGTTCTCCACGCAGTA-3'(配列番号60)
【0175】
18Sを除き、残りの全てのプライマーはEurogentec、Seraing、BelgiumおよびDNA Technology A/S、Aarhus、Denmarkから購入した。
【0176】
全RNAを、P1のラットの腹側中脳、背側中脳および皮質に由来するコンフルエントなT1Aアストロサイト培養物、ならびにE10.5、E11.5、E13.5、E15.5およびP1の腹側中脳および背側中脳、およびE14、E16、E18およびP1の皮質から切除した組織から、RNeasy抽出キット(Qiagen、Hilden、Germany)を用いて単離した。全RNAはまた、Wntで3日間in vitroにて処置したE14.5のVM前駆体培養物(100mmディッシュに細胞750万個)からも抽出した(3重測定)。逆転写のために、1μgの全RNAを最初に1単位のRQ1 RnaseフリーDNAse(Promega、Madison、USA)で40分間処置した。DNAseは、1μlのEDTA 0.02Mの添加と、65℃での10分間のインキュベーションとにより不活化した。0.5μgのランダムプライマー(Life Technologies、Grand Island、NY、USA)を次に添加し、そして混合物を70℃で10分間インキュベートした。次に、各試料を2本のチューブ、cDNA反応チューブと陰性対照チューブとに均等に分割した。次に、1×First-Strand Buffer(Life Technologies、Grand Island、NY、USA)、0.01MのDTT(Life Technologies、Grand Island、NY、USA)および0.5mMのdNTPS(Promega、Madison、USA)を含有するマスターミックスを、cDNAおよびRT-チューブの両方に添加し、そして25℃で10分間インキュベートし、その後、42℃での2分間のインキュベーションを続けた。200単位のSuperscript II逆転写酵素(Life Technologies、Grand Island、NY、USA)を、次に、cDNAチューブのみに加え、全ての試料を42℃で50分間インキュベートした。Superscript IIは、70℃での10分間のインキュベーションにより不活化した。次に、cDNAおよびRT-の両方を、さらなる分析のために10倍に希釈した。リアルタイムPCRは1μlの1:10に希釈したcDNAおよびRT-と共に、総容量25μlで、三重で行なった。各PCR反応は、1×PCRバッファー(Life Technologies、Grand Island、NY、USA)、3mMのMgCl2(Life Technologies、Grand Island、NY、USA)、0.2mMのdNTPs(Promega、Madison、USA)、0.3μMの各フォワードおよびリバースプライマー、1単位のPlatinum Taq DNAポリメラーゼ(Life Technologies、Grand Island、NY、USA)および1×SYBR Green(Molecular Probes、Leiden、The Netherlands)で構成された。PCRは、94℃で2分間、その後94℃で30秒間、60℃で30〜45秒間、72℃で45〜60秒間を35〜40サイクルおよび80℃で15秒間(SYBR Green検出について)、ABI PRISM 5700検出システム(PE Applied Biosystems、Foster City、CA、USA)で行なった。他のアニーリング温度は、Wnt5aについての54℃、p27についての57℃、サイクリンD1についての62℃、サイクリンD2についての61℃およびp57についての65℃を含めた。SYBR Greenシグナルが、ユニークかつ特異的なアンプリコンに一致することを確認するために、各ランの後、各PCR産物について融解曲線を得た。PCR産物の特異性は、シーケンシングによって検証した。
【0177】
標準曲線は、96ウェルプレートのリアルタイムPCRランごとに、逆転写されたRNAまたは各プローブの対象となる配列を含むプラスミドの3倍連続希釈を用いて作成した。得られた標準曲線のプロットを、次に、Cts(所定のテンプレートを設定した蛍光閾値まで増幅するのに必要なPCRサイクル数)を、所定の試料の最初のテンプレートの任意量に変換するのに用いた。
【0178】
発現レベルは、各試料のRT-値を対応するRT+値から減じ、そしてこの数値を、各試料について並行するアッセイで得たハウスキーピング遺伝子18Sの値で除すことにより得た。
【0179】
結果の統計学的分析は、一元配置ANOVAで行なった。種々の発生段階が、最も早い発生段階と異なった特定の時点を、有意な相互作用が生じた場合にのみ同定するために、フィッシャーの制約付最小有意差(protected least significant difference)を事後的に用いた。全ての試験について、有意性はp<0.05のレベルと仮定した(*p<0.05; **p<0.001; ***p<0.0001)。
【0180】
特定の遺伝子についての各アッセイは、3重で、2回または3回反復した。18Sのアッセイは、各試料について、アッセイの開始時、およびアッセイの途中に1回または2回、試料の完全性を検証するために実行した。PCRプライマーの特異性は、プライマー配列のBLASTラン(BLAST run)により決定した。全てのPCR産物は、アンプリコンのサイズを検証するためにゲルに流した。PCR産物の特異性は、無作為試料中のアンプリコンのシーケンシングによって決定した。
【0181】
前駆体の培養および処置
適時に交配させた(timed-mated)Sprague-Dawleyラットから得たE13.5〜14.5の胚からの腹側中脳および大脳皮質を切除し、機械的に解離し、ポリ-D-リジンでコートされた12または24ウェルプレートに、規定無血清培地(defined, serum-free medium)(インスリン(10ng/ml)、トランスフェリン(100μg/ml)、プトレシン(100μM)、プロゲステロン(20nM)、セレン(30nM)、グルコース(6mg/ml)、およびウシ血清アルブミン(1mg/ml)を含有するF12とDMEMとの1:1混合物からなるN2)中、最終密度1×105細胞/cm2でプレーティングした。精製した1型アストロサイトは、P1のラットのVMまたはCTXに由来する混合グリア培養物から、標準的なプロトコル(Wagnerら、1999)にしたがって得た。振盪し、12ウェルプレートに再プレーティングした後、アストロサイトを15%ウシ胎児血清含有培地中でコンフルエントまで増殖させ、N2培地に変え、そのときに新鮮に切除したVMまたはCTX細胞を、アストロサイトの上に1×105細胞/cm2の密度でプレーティングした。固定の4〜6時間前に添加した5-ブロモデオキシウリジン(BrdU)を除き、全ての因子は、培養の開始時に一度に加えた。培養物は、加湿された、C02 5%、空気95%のインキュベータ内で37℃にて維持し、そして所定時間後、4%パラホルムアルデヒドで45分間、免疫細胞化学分析の前に固定した。
【0182】
ニューロスフェアの培養
E13.5のラット胚からの腹側中脳を切除し、そして一緒にプールし、N2無血清培地に再懸濁し、機械的に解離し、そして、事前にポリ-D-リジン(Sigma、Stockholm、Sweden)でコートした24ウェルプレート(BD Bioscience、Erembodegem、Belgium)に、最終密度100〜125×103細胞/cm2でプレーティングした。細胞は、20ng/mlのFGF8b(R&D Systems、Minneapolis、USA)および8μg/mlのヘパリン(Sigma、Stockholm、Sweden)の存在下で増殖させ、そして7〜10日後に、高いスフェア密度で、部分的に精製された馴化培地、およびタンパク質125〜250μg/mlのFz8CRDが添加されたN2に再プレーティングした。ニューロスフェアは、その後5〜7日間分化させた。固定および免疫組織化学分析は、上述のとおりに行なった。
【0183】
Wnt馴化培地の調製、特性決定および精製
ヘマグルチニンタグ付Wnt-1a、3a、または5aを安定的に過剰発現するB1A線維芽細胞系(Shimizu H.、1997)を、1OOμg/mlのG-418が添加された標準的な完全培地(DMEM+10%FBS)中で増殖させた。馴化培地の回収のために、細胞を完全倍地中に低密度で再プレーティングし、そして50〜70%コンフルエントに到達させ、その時点で細胞を洗浄し、培地を無血清N2(1OμMの酪酸ナトリウムを含む)と24時間置換した。次に、シスター(sister)フラスコからの馴化培地を採取し、ロットとしてプールし、-80℃で2カ月までの間保存した。この回収および保存のルーチンを用いた場合、新鮮なCMと比べ活性の喪失は観察されなかった。T1A CMは、培地をin vitroで3日後に回収することを除き、同様の手順で採取した。濃縮のため、CMの個々のロットを室温で解凍し、80mlのアリコートに分割し、Centricon-Plus 80カラム(Millipore)にローディングし、そして製造者の指示に従い、遠心分離により濃縮した。濃縮に続き、アリコートを再プールし、そして、タンパク質含量の決定およびウェスタンブロット分析のための試料を取ってから、-80℃で凍結させた。簡単に述べると、20μgのタンパク質を10%ポリアクリルアミドミニゲルにローディングし、変性条件下150Vで約30分間流した。PVDF膜(Hybond P、Amersham Pharmacia Biotech、Uppsala、Sweden)へのドライエレクトロブロットトランスファー、およびトリス緩衝生理食塩水中の3%ウシ血清アルブミンとX%のTriton-X100とからなるブロッキングバッファー(TBST)中での30分間のプレインキュベーションの後、ブロットをTBST中1:1000に希釈したマウス抗ヘマグルチニン(Babco)と共に一晩4℃でインキュベートした。洗浄後、ブロットを、TBST中1:1000に希釈したアルカリホスファターゼ結合ヤギ抗マウスIgG(Santa Cruz)と共に、室温で1時間インキュベートした。ブロットをAmersham ECF試薬で視覚化し、そして青色蛍光をMolecular Devices Storm 840 phosphoimagerで定量した。
【0184】
Fz8CRD馴化培地の調製、特性決定および精製
B1A線維芽細胞系を、mFz8CRD-IgG/pRK5およびpIRES puro 2(Clontech、BD Bioscience、Erembodegem、Belgium)プラスミドで、Lipofectamine Plus試薬(Invitrogen、Lidingo、Sweden)を用い、製造者の指示に従って共トランスフェクトした。ピューロマイシン耐性(1μg/ml Sigma、Stockholm、Sweden)について選択し、クローンを単離し、そして増殖させた。Fz8CRD 700xを過剰発現するクローン(リアルタイムRT-PCR分析)を同定した。Fz8CRD馴化培地の採取および部分精製を上述のとおりに行なった。行なった全てのブロッキング実験において、Fz8CRDおよびB1Aの対照部分精製馴化培地を、125〜250μg/mlの最終濃度で用いた。
【0185】
免疫細胞化学分析
固定した培養物を、1%ウシ血清アルブミンおよび0.3%Triton-X100を含有するリン酸緩衝生理食塩水(PBS)で適切に希釈した以下の抗体の1つと共にインキュベートした:マウス抗BrdU、1:50(DAKO、Denmark)、マウス抗βチューブリン、III型(TuJ1)、1:250(Sigma)、マウス抗TH、1:1000(Incstar、USA)、ウサギ抗TH、1:250(PelFreeze)、ウサギ抗Nurr1、1:2000(Dr. T. Perlmann、Swedenからの寄贈)、ウサギ抗Nurr1、1:1000(Santa Cruz、USA)またはマウス抗Nurr1、1:250(BD Transduction Laboratories、USA)、ウサギ抗Adh2、1:4000(Dr. R. Lindahl、South Carolinaからの寄贈)。インキュベーションは、4℃で一晩か、または室温で1時間行なった。両方の方法とも同様の結果をもたらした。洗浄後、培養物を1〜3時間、同じ希釈バッファー中の適切な2次抗体(ビオチン化1:500; CY2-、FITC-、またはローダミン結合ウマ抗マウスIgG 1:100;またはヤギ抗ウサギIgG 1:100;全てVector、USAから)と共にインキュベートした。明視野免疫染色は、Vector Laboratory ABC免疫ペルオキシダーゼキットによりNovaRed(赤)、またはAEC(赤)、SGもしくは3-3'ジアミノベンジジン四塩酸塩(DAB 0.5mg/ml)/塩化ニッケル(1.6mg/ml)(灰/黒)、またはVIP(紫)基質のいずれかを用いて視覚化した。二重染色は、記載のとおりに順次的な単一染色により行なった。染色の順序は、BrdU二重標識についてのみ重要であり、この場合、BrdU手技は常に2番目に行なった。用いた1次または2次抗体のいずれかを欠いた対照実験は、用いた抗体ペアの間に交差反応性がわずかにあるか、またはないことを示した。写真はZeiss Axioplan 100M顕微鏡で得、そして浜松ホトニクス社製カメラC4742-95(QED画像化ソフト付)に集めた。
【0186】
データ分析および統計
明瞭に染色された細胞のみを陽性細胞として計数した。スフェアは、これらが1個またはそれ以上の陽性細胞を含む場合に陽性とみなした。個々の細胞についての定量的免疫細胞化学のデータは、他に別記のない限り、盲検化された観察者により、3〜4回の独立した実験からの、条件あたりそれぞれ3〜4ウェルにおける10〜20の重複しない視野(細胞)からの、またはウェル全体における(クラスター)得られた計測数の平均値および標準誤差を表す。各変数について、初期の統計学的比較は、用量、時点、処置および/または領域の、複数の要因によるグローバルANOVA(global ANOVA)により行い、処置と他の任意の変数との間に有意な相互作用が存在する場合は、データを、別個の時間、用量または領域にさらに分割した。フィッシャーの制約付最小有意差は、処置が対照から(または相互に)異なった特定のポイントを、要因である処置と他の変数との間に有意な相互作用が生じたときのみ同定するために、事後的に用いた。全ての試験についての有意性は、p<0.05のレベルを前提とした。結果部分における表示を簡略化するために、個々の統計分析の結果、ならびに行なった分析の種類は、個々の図の凡例中に表示する。
【0187】
参考文献
【図面の簡単な説明】
【0188】
【図1】図1は、Wntリガンドが発生中の中脳で示差的に発現していることを示す。図1aは、Wnt-1が腹側中脳および背側中脳で高レベルに発現されているが、Wnt-3aの発現が背側中脳で、およびWnt-5aが腹側中脳で優勢であることを明らかにしたリアルタイムPCR分析の結果を示す。図1b、図1c、図1dおよび図1eは、腹側中脳内において、Wnt-1(図1d)およびWnt-5a発現(図1e)のドメインが、ドーパミン作動性前駆体およびドーパミン作動性ニューロンを標識するNurr1(図1c)およびチロシンヒドロキシラーゼ(TH)(図1b)のものと合致することを示したin situハイブリダイゼーションの結果を示す。図1fは、出生後1日の腹側中脳(VM)から単離した1型アストロサイトが、背側中脳(DM)または大脳皮質(CC)よりも有意に高いレベルのWnt-5a mRNAを発現することを実証するin situハイブリダイゼーションの結果を示す。各脳領域について、フィッシャーの事後検定を用いた一元配置ANOVAによりE10.5に対して比較した場合、*p<0.05; **p<0.001; ***p<0.0001である。
【図2】図2は、ラットのE14.5の腹側中脳(VM)培養物において、Wnt-1およびWnt-5aは、ドーパミン作動性ニューロンの数を増加させ(図2a、図2bおよび図2c)、ドーパミン作動性ニューロン(図2eおよび図2f)を含むクラスターを増殖させたが、Wnt-3aはこれらをしなかったことを示す。図2aおよび図2eは用量依存性を示し、図2bおよび図2fは経時変化分析を示し、そして図2cはWntリガンドの効果の、対照(N2)、グリア細胞系由来神経栄養因子(GDNF)、線維芽細胞成長因子2および8(FGF-2および-8)、VM1型アストロサイト(T1A)および対照精製培地(control purified media)(CP)との比較を示す。結果は、部分的に精製されたWntは、活性があり、安定であり、および培養中のドーパミン作動性ニューロンの数の増加に関して、VM T1Aと同程度に有効であり得ることを示している。図2bおよびc:フィッシャーの事後検定を用いた一元配置ANOVAによりN2およびCPと比較した場合、それぞれ*、**p<0.01である。図2e:フィッシャーの事後検定を用いた一元配置ANOVAによりCPと比較した場合、*p<0.01である。チロシンヒドロキシラーゼ(TH)の免疫染色培養物は、Wnt-1およびWnt-5aが、ドーパミン作動性クラスターの外部または内部でのドーパミン作動性ニューロンの数の極めて劇的な増加を誘導することを示した。視野は3.14mm<SUP>2であり、ウェルは4cm2である。
【図3】図3は、Wnt-1が前駆体の増殖およびニューロンの総数を増大させたが、Wnt-5aとは異なり、ドーパミン作動性表現型を獲得したドーパミン作動性前駆体の比率は増大させなかったことを示す。図3aは、Wnt-1は、サイクリンD1のmRNAの発現を増大させたが、Wnt-5aはそれをしなかったことを示す。図3bは、Wnt-1およびWnt-3aが、BrdUを取り込んだNurr1免疫染色細胞の比率の3倍の増加を誘導したが、Wnt-5aおよび腹側中脳の1型アストロサイト(VM T1A)からの馴化培地は、Nurr1+細胞におけるBrdUの取り込みをより少ない程度で増加させたことを示す。図3dは、Wnt-1およびWnt-3aが、Tuj1陽性ニューロンを含む増殖性クラスターの数を増大させたことを示す。図3eは、ニューロンを含む増殖性クラスターにおけるドーパミン作動性ニューロンの比率は、Wnt-1処置の後では変化せず、Wnt-3aにより減少し、Wnt-5aおよびVM T1Aでの処置により増大したことを示す。図3fは、Wnt-1のみが、増殖性クラスター外の個々のTuJ1陽性ニューロンの数を増加させたことを示す。図3gは、クラスター外のニューロン数の増加にもかかわらず、Wnt-1は、ドーパミン作動性ニューロンの比率を増大させなかったことを示す。一方、Tuj1陽性ニューロンの数を変化させなかった処置は、ドーパミン作動性ニューロンの比率を減少(Wnt-3a)、または増大させた(Wnt-5aまたはVM T1A)。フィッシャーの事後検定を用いた一元配置ANOVAにより対照精製培地(CP)と比較した場合、*p<0.05; **p<0.001; ***p<0.0001である。
【図4】図4は、Wnt-5aが、Nurr1+細胞においてドーパミン作動性表現型を誘導するのに最も有効な因子であることを示す。図4aは、Wnt-5aまたはVM T1A処置が、ドーパミン作動性表現型を獲得したVMにおけるNurr1発現細胞の比率を、対照条件における50%から90%に増大させたことを明らかにした、チロシンヒドロキシラーゼ(TH)とNurr1との二重免疫組織化学の結果を示す。一方、増殖を誘導するのに最も効果的な2因子、Wnt-1およびWnt-3aは、Wnt-5aより有効ではないか(Wnt-1)、またはドーパミン作動性細胞の比率を50%から30%に減少させさえした(Wnt-3a)。図4bは、同様に、Wnt-5aおよびVM T1Aが、Nurr1を発現したE13.5の皮質前駆体培養物におけるドーパミン作動性表現型の獲得を促進するのに最も効果的な処置であったことを示す。Wnt-1はWnt-5aよりはるかに低い効果を有し、そしてWnt-3aまたは皮質1型アストロサイト(CTX T1A)は、THを発現するNurr1+細胞の比率を変化させなかった。フィッシャーの事後検定を用いた一元配置ANOVAにより対照精製培地(CP)と比較した場合、*p<0.05; **p<0.001; ***p<0.0001である。
【図5】図5は、Wntシグナル伝達がドーパミン作動性ニューロンの発生に必要であることを説明する。図5aは、FGF8で増殖させたE13.5のVMニューロスフェアが、5〜7日でグリアおよびニューロン系統に分化し、スフェアの12%にドーパミン作動性ニューロンを生じさせたことを示す。これに対し、培養培地へのFz8 CRDの添加は、ドーパミン作動性ニューロンを含むニューロスフェアの数を、対照(CM=CP)に比べ減少させた。同様に、E14.5のVM前駆体培養物のFz8 CRD過剰発現線維芽細胞からの馴化培地による処置(図5b、図5cおよび図5d)は、チロシンヒドロキシラーゼの発現を獲得したNurr1免疫反応性細胞の比率を、対照条件にて(CM、図5b)、または腹側中脳の1型アストロサイトからの馴化培地(VM T1A、図5c)もしくはWnt-5a(図5d)による処置の後に減少させた。フィッシャーの事後検定を用いた一元配置ANOVAにより対照(N2)または馴化培地(CM)と比較した場合、*p<0.05; ***p<0.0001であり、VM T1AまたはWnt-5aと比較した場合、#p<0.05; ###p<0.0001である。図5fは、Wnt-1、-3aおよび-5aが、VMドーパミン作動性ニューロンの発生を調節するメカニズムのモデルである。Wnt1は、恐らく中脳-終脳のオーガナイザーに由来し、Nurr1発現前駆体の増殖を制御し、そして、VMニューロンの数を増加させる。Wnt-5aは、VMアストロサイトによって発現され、Nurr1発現前駆体におけるドーパミン作動性表現型の誘導を調節することによりVMドーパミン作動性ニューロンの数を特異的に増加させる。最後に、Wnt-3aは、主に背側中脳で発現され、Nurr1発現前駆体の増殖および/または自己再生を増進させ、そしてドーパミン作動性表現型を獲得するニューロンの比率を減少させた。疑問符は、Wnt-1およびWnt-3aの正確な細胞供給源が不明であることを示す。矢印の大きさは効果の強度に相関することに留意されたい。
【図6】図6は、Wntがドーパミン作動性ニューロンの発生を、前駆体の増殖およびDA表現型の獲得を調節することにより、示差的に制御することを示す。リアルタイムRT-PCRで評価したところ、Wnt-5aは、in vitroで3日目に、Ptx3のmRNA(A)およびc-ret(B)の発現をアップレギュレーションし、GFRα1(C)およびNCAM(D)の発現を維持したが、Wnt-1はこれらをしなかった。
【図7】図7は、Wnt-1がサイクリンD1および細胞周期インヒビターp27およびp57の発現を調節し、VM前駆体の増殖を増大させ、そしてTHニューロンの数を特異的に増加させることを示す。リアルタイムRT-PCRは、in vitroで3日目に、Wnt-1は、サイクリンD3のmRNAの発現を増加させ(A)および細胞周期インヒビターp27(B)およびp57(C)の発現を減少させたが、Wnt-5aがこれらをしなかったことを示した。Dに示すように、Wnt-1およびWnt-3aは、in vitroで3日目に、VM前駆体の増殖を増大させたが、Wnt-5aはこれをしなかった。Eに示すように、Wnt遮断試薬である部分的に精製されたFz8-CRDの増大する単位は、in vitroで3日後、VM培養物におけるTH+ニューロンの数を用量依存的に減少させたが、これは内因性WntがDAの発生に必要であることを示している。Fに示すように、Wnt-1によるTH+ニューロンの数の増加は、Fz8-CRDにより部分的にブロックされたが、これは、Wntの効果が特異的であることを示唆している。統計学:フィッシャーの事後検定を用いた一元配置ANOVAにより対照と比較した場合、・p<0.05;*p<0.01;**p<0.001;***p<0.0001であり、そしてWnt処置のみと比較した場合、###p<O.0001である(n=3〜5、Fz8-CRDについては例外的に、n=2〜3)。濃度:10単位/μlのWntまたはCP、および5単位/μlのFz8-CRD。視野は3.14mm<SUP>2である。
【図8】図8は、発生中の中脳におけるWntの発現を示す。リアルタイムRT-PCR分析は、Wnt-2(8a)、Wnt-4(8b)、Wnt-7a(8c)、およびWnt-7b(8d)の転写産物が、ドーパミン作動性ニューロン誕生時の腹側中脳領域で優勢であることを明らかにした。
【図9】図9は、発生中のCNSにおけるWntの発現を示す。リアルタイムRT-PCR分析は、Wnt-3(9A)、Wnt-6(9B)、Wnt-1Ob(9C)、Wnt-11(9D)、およびWnt-16(9E)の発現が、背側中脳領域およびCNSのその他の領域に、より特異的であることを示した。
【図10】図10は、Wnt-2およびWnt-7aが、ラットのE14.5の腹側部前駆体培養物におけるドーパミン作動性ニューロンの数を増加させたことを示す。Wnt-2およびWnt-7aによる処置は、培養物(10A)およびNurr1発現前駆体(10B)におけるチロシンヒドロキシラーゼ陽性ニューロンの総数を両方とも増加させた。Wnt-7aは、腹側部前駆体の増殖を増大させたが(10C)、Wnt-2による処置は、初代培養物におけるBrdUの取り込み量の減少をもたらした(10C)。
【図11】図11は、細胞周期レギュレーターへのWntの種々の効果を示す。Wnt-7aは、サイクリンD1のmRNAを増加させたが、cdkインヒビターp27およびp57をダウンレギュレーションした(11A、11Dおよび11E)。Wnt-2による処置では、サイクリンD2の発現の増加と共に、p27およびp57のmRNAの増加が観察された(11B、11Dおよび11E)。
【技術分野】
【0001】
本発明は、神経幹細胞または神経始原細胞もしくは前駆細胞、あるいは他の幹細胞におけるニューロンとなる運命の誘導に関する。本発明は、特定のニューロン表現型の誘導および誘導の増強、詳細には中脳ドーパミン作動性ニューロン表現型の誘導および誘導の増強に関する。
【背景技術】
【0002】
パーキンソン病(PD)とは非常に多く見られる神経変性障害であり、その病因は中脳ドーパミン作動性(DA)ニューロンの選択的かつ進行性な損失によって特徴づけられる。ニューロン表現型の誘導を増強すると、パーキンソン病および他の重篤な衰弱性の神経変性障害を治療できる可能性が出てくる。
【0003】
これまでは、結果が陽性のパーキンソン病患者にヒト胎児中脳組織を移植していたが、本発明を利用する特異的細胞置換治療の開発により、このような従来手法に伴う実用的問題や倫理的問題が克服される。具体的には、本発明により移植用の細胞調製物の開発が可能となり、一方、胚組織または胚細胞を使用する必要性が低減または完全に排除される。幹細胞は、通常は廃棄される組織である臍帯から得られ得る。別の選択肢は、たとえば骨髄、血液、皮膚、眼、嗅球または嗅上皮から成人幹細胞を得ることである。
【0004】
これまでに(国際公開公報WO00/66713号およびWagner他、1999)、本発明者らの研究室において、ドーパミン作動性ニューロン表現型の誘導がNurr1を発現する細胞中で、腹側中脳の1型アストロサイト/初期グリア細胞から得られる1つまたは複数の因子の存在下で増強されることを示した。本発明は、Wnt因子がNurr1を発現する細胞のニューロン表現型の誘導を増強させることに有用であるという実験的知見に基づいている。
【0005】
詳細には、本発明者らは、DAニューロンが誕生する時点までに背側中脳よりも高いレベルでVM内において発現されている全てのWntが、神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞における増殖、自己再生、ドーパミン作動性の誘導、生存、分化および/あるいは成熟を増強させることによってドーパミン作動性ニューロンの発生を誘導させるまたは促進することに有用であることを見出した。
【0006】
本発明者らは、以下を見出した。
【0007】
Wnt-1は、ドーパミン作動性の前駆体の増殖ならびにドーパミン作動性の前駆細胞および/または幹細胞のドーパミン作動性ニューロンへの成熟を促進すること;
Wnt-7aは、ドーパミン作動性の前駆体の増殖を促進し、これらがドーパミン作動性ニューロンへと分化することを可能にすること;
Wnt-3aは、ドーパミン作動性の前駆細胞および/または幹細胞の増殖および/または自己再生を促進すること;
Wnt-2は、細胞周期離脱およびNurr1+前駆体によるドーパミン作動性ニューロン表現型の獲得を促進すること;
Wnt-5aが、神経幹細胞、前駆細胞もしくは始原細胞においてドーパミン作動性の表現型を誘導させること、およびニューロン細胞においてドーパミン作動性の誘導または分化を増強させることに最も有効であること。
【0008】
Wnt-1は、ドーパミン作動性の前駆細胞および/または幹細胞の増殖および成熟を促進することに関してWnt-3aおよびWnt-5aよりも有効である。
【0009】
特定のニューロン表現型の誘導は、遺伝的および後成的シグナルのどちらもの組込みを必要とする。発生中の中脳では、ドーパミン作動性ニューロンの誘導はオーファン核内受容体Nurr1を必要とするが(Zetterstrom他、1997; Saucedo-Cardenas他、1998; Castillo他、1998)、Nurr1の発現は神経幹細胞においてドーパミン作動性の表現型を誘導させるのに十分ではない(Wagner他、1999)。本発明者らは以前に、Nurr1と発生中の腹側中脳1型アストロサイト/初期グリア細胞由来の未知の可溶性シグナルとの組み合わせが、神経幹細胞において中脳ドーパミン作動性の表現型を誘導させるのに十分であると報告した(Wagner他、1999)。本発明者らは本明細書中で、Wnt-5aがこのようなシグナルの一部であり、またWnt-1、-2、-3a、-5aおよび-7aを含めたWntタンパク質ファミリーのメンバーが発生的に制御されており、かつ中脳ドーパミン作動性ニューロンの発生を示差的に制御することを述べる。部分的に精製したWnt-1、-2、-5a、および-7aは2つの異なる機構によりE14.5中脳DAニューロンの数を増加させたが、Wnt-3aは増加させなかった。Wnt-1および-7aは主にNurr1前駆体の増殖を増大させ、それらがドーパミン作動性ニューロンへと分化することを可能にした。Wnt-2は細胞周期離脱およびNurr1+前駆体によるドーパミン作動性ニューロン表現型の獲得を支持した。Wnt-5aは主にニューロンのDA表現型を獲得したNurr1前駆体の割合を増加させた。本発明者らの知見と一致して、Wnt-5aはNurr1を発現する中脳または皮質E13.5前駆体においてドーパミン作動性ニューロンを誘導させることに関して、中脳アストロサイト/初期グリア細胞と同様に有効であった。さらに、Frizzled8のシステインリッチドメインは、Nurr1を発現する神経前駆培養物におけるドーパミン作動性の表現型を有する細胞の増加に対する基底効果およびVM T1A-、Wnt-1またはWnt-5aに媒介される効果、ならびに神経幹細胞またはFGF-8で拡大させたNurr1+中脳ニューロスフェアに対する内因性Wntの効果を有効に遮断した。したがって、本明細書中に含めるデータは、それぞれのWntが部分的に異なる機構によってNurr1を発現する前駆細胞/幹細胞におけるDA表現型を有するニューロンの産生をそれぞれ独立して調節することの指標を提供する。
【0010】
これらの知見により、Wntリガンドが腹側中脳の神経発生中における増殖、自己再生、分化および運命決定の重要なレギュレーターとして位置づけられる。さらに、本発明者らの結果により、in vitroにおける中脳DAニューロンの大規模な産生、およびパーキンソン病などの神経変性疾患の治療における幹細胞置き換え戦略の将来の実行の道が開かれる(BjorklundおよびLindvall、2000; PriceおよびWilliams、2001; Arenas、2002; RossiおよびCattaneo、2002; Gottlieb他、2002)。
【0011】
胚性、神経性および多能性幹細胞は、ニューロン、アストロサイトおよびオリゴデンドロサイトを含めた神経細胞系列へと分化する能力を有する。さらに、幹細胞を単離し、拡大させて、脳移植用の原料物質として使用することができる(Snyder, E.Y.他、Cell 68、33-51 (1992); Rosenthal, A.、Neuron 20、169-172 (1998); Bain他、1995; Gage, F.H.他、Ann. Rev. Neurosci. 18、159-192 (1995); Okabe他、1996; Weiss, S.他、Trends Neurosci. 19、387-393 (1996); Snyder, E.Y.他、Clin. Neurosci. 3、310-316 (1996); Martinez-Serrano, A.他、Trends Neurosci. 20、530-538 (1997); McKay, R.、Science 276、66-71 (1997); Deacon他、1998; Studer, L.他、Nature Neurosci. 1、290-295 (1998); BjorklundおよびLindvall、2000; Brustle他、1999; Lee他、2000; Shuldiner他、2000および2001; Reubinoff他、2000および2001; Tropepe他、2001; Zhang他、2001; PriceおよびWilliams、2001; Arenas、2002; Bjorklund他、2002; RossiおよびCattaneo、2002; Gottlieb他、2002)。
【0012】
ほとんどの神経変性疾患はニューロン集団を冒す。さらに、損傷のほとんどが特定の神経化学的な表現型に起こる。たとえばヒトのパーキンソン病では、失われる主な細胞型は中脳ドーパミン作動性ニューロンである。神経組織の移植による特定のニューロン集団の機能置換は、神経変性疾患を治療するための魅力的な治療戦略を表している(Rosenthal, A.、Neuron 20、169-172 (1998))。別の代替方法は、幹細胞、始原細胞もしくは前駆細胞の再生を促進する、またはその発生および/もしくは動員を修復または導くために必要なシグナルを直接注入すること、あるいはこのような機能を調節する薬物を投与することであろう。
【0013】
幹細胞/始原細胞または前駆細胞は、拡大することができ、かつ特定のニューロン表現型を担うように指示できるので、移植治療用の理想的な材料である。これらの細胞では、移植用にヒト胎児組織を使用することに絡む倫理的問題および実用的問題が回避されるであろう。具体的には、移植した非自己組織の生存度は制限されており免疫系に拒絶され得る。さらに、それぞれの胎児からは少数の細胞しか提供されない。
【0014】
幹細胞または始原細胞もしくは前駆細胞において単一の特定のニューロン表現型を誘導させることは理解しにくいことが分かっている。
【発明の開示】
【0015】
本発明は、細胞中におけるドーパミン作動性ニューロン表現型の誘導を提供する。
【0016】
本発明は、ドーパミン作動性ニューロンの発生の誘導を可能とする。したがって、培養物中もしくは脳内におけるWntレベルおよび/または機能を増大させることによって、本発明は以下の誘導または促進を可能とする:ドーパミン作動性の前駆体、始原細胞もしくは幹細胞の増殖および/または自己再生;ならびに/あるいはドーパミン作動性ニューロン、前駆細胞、始原細胞または幹細胞の生存、分化および成熟の促進(ドーパミン作動性ニューロンの収量を増加させる);ならびに/あるいはin vitroまたはin vivoにおける幹細胞、始原細胞、前駆細胞もしくはニューロン細胞のニューロン性ドーパミン作動性の運命の誘導。
【0017】
本発明の任意の態様または実施形態はニューロン細胞、すなわちニューロンに適用することができ、またはそれを使用することができる。本開示における「神経細胞」はニューロン細胞であり得る。
【0018】
ドーパミン作動性ニューロンに富んだ細胞調製物を、パーキンソン病または他の障害における細胞置換治療に、ならびにドーパミン作動性ニューロンにおけるシグナル伝達現象およびin vitroにおけるドーパミン作動性ニューロンに対する薬物の効果を研究するために、たとえばハイスループットスクリーニングにおいて使用し得る。
【0019】
本発明の態様および実施形態を、添付の特許請求の範囲に示すように提供する。
【発明を実施するための最良の形態】
【0020】
一態様では、本発明は、幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞においてドーパミン作動性のニューロンとなる運命を誘導させる方法、あるいはニューロン細胞においてドーパミン作動性の誘導または分化を増強させる方法、あるいはドーパミン作動性前駆細胞もしくは始原細胞またはNurr1を発現する幹細胞を拡大させる方法であって、
細胞内でNurr1サブファミリーの核内受容体を基底レベルより高く発現させること、
および
細胞をWntリガンドで処理すること、
によってドーパミン作動性ニューロンを産生させることを含む方法を提供する。
【0021】
Nurr1サブファミリーはNR4Aサブファミリーとしても知られており、Nurr1/NR4A2、Nor1/NR4A3およびNGFI-B/NR4A1が含まれる。したがって、本発明の方法は、細胞内でたとえばNurr1/NR4A2、Nor1/NR4A3および/またはNGFI-B/NR4A1を基底レベルより高く発現させることを含み得る。好ましくは、Nurr1サブファミリーの核内受容体はNurr1である。したがって、本発明の方法は細胞内においてNurr1を基底レベルより高く発現させることを含むことが好ましい。
【0022】
本発明は、神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、ドーパミン作動性の誘導、生存、分化および/あるいは成熟を増強させることによってドーパミン作動性ニューロンの発生を誘導させるまたは促進する方法であって、
細胞内においてNurr1を基底レベルより高く発現させること、および
細胞をWntリガンドで処理すること、
によって増殖、自己再生、生存および/またはドーパミン作動性の誘導、分化、生存もしくはニューロンのドーパミン作動性の表現型の獲得を生じさせるあるいは増強させることを含む方法を提供する。
【0023】
Wntリガンドを用いた処理は、in vivo、ex vivo、または培養下であり得る。
【0024】
本発明の方法では、Wntリガンドを用いた処理とは細胞をリガンドと接触させることによりものとし得る。Wntリガンドを用いた処理とは、精製したWntリガンドおよび/もしくは組換えWntリガンドを幹細胞、始原細胞もしくは前駆細胞を含む培養物に供給すること、またはそのような細胞にin vivoで供給することによるものとし得る。Wntリガンドを用いた処理は、Wnt核酸またはタンパク質の1つもしくは複数のコピーを細胞内に導入することを含み得る。細胞を核酸で形質転換させる方法および細胞内にタンパク質を導入する方法を以下に詳述する。Wntリガンドと接触させることとは、in vivoにおいて、あるいは幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞を含む培養物中に、Wntリガンドを産生する細胞を提供することにより得る。Wntリガンドを産生する細胞は、組換え発現によってWntリガンドを産生する組換え宿主細胞であり得る。同時培養した宿主細胞をWntリガンドをコードしている核酸で形質転換させてもよく、かつ/または同時培養した細胞は導入されたWntタンパク質を含んでいてもよい。Wntタンパク質またはWntをコードしている核酸を当技術分野で利用可能な技術に従って細胞内に導入してよく、それらの例を以下に記載する。
【0025】
同時培養した細胞または宿主細胞は、別の幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞であり得る。Wntリガンドを用いた処理はまた、細胞においてWntの発現をアップレギュレーションすること、またはWntリガンドの阻害分子をダウンレギュレーションもしくは阻害することにもより得る。したがって、Wntリガンドを用いた処理は、SFRP、WIF、dkkまたはCerberusなどのWntと相互作用する分子の発現または活性を低減させることによって起こりうる(Martinez Arias他、1999; http://www.stanford.edu/〜rnusse/wntwindow.html、または任意のウェブブラウザを使用して見つけることができる)。
【0026】
Wntリガンドの供給に加えて、幹細胞または神経幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞は1型アストロサイト/グリア細胞と同時培養中であるか、あるいはin vitroまたはin vivoにおいてこのような細胞またはそれらに由来する因子と接触しているかもしれない。しかし、本発明によれば、Wntリガンドの提供は1型アストロサイトに依存しておらず、Wntリガンドを外部から供給しない神経幹細胞または神経始原細胞および1型アストロサイトを含む同時培養物は本発明の範囲に企図されておらず、またこのような同時培養を使用する方法もそうである。
【0027】
Nurr1(Law他、1992; Xing他、1997; Castillo, 1997; GenBank第S53744号、第U72345号、第U86783号)は、甲状腺ホルモン/レチノイン酸核内受容体スーパーファミリーの転写因子である。以前に国際公開公報WO00/66713号およびWagner他、1999に示されているように、神経幹細胞または神経始原細胞における基底レベルより高いNurr1の発現により、ニューロンとなる運命に向かって分化する細胞の割合が高まる。ニューロンとなる運命の誘導はin vitroまたはin vivoで実施し得る。移植前または移植後に幹細胞または神経幹細胞、始原細胞もしくは前駆細胞の分化をニューロンとなる運命に向かって誘導する能力により、成人の脳に移植した場合に移植した幹細胞がアストロサイトとなる運命に分化する傾向が改善される。Nurr1はNR4Aサブファミリーのメンバーである。Nurr1が好ましいかもしれないが、本発明の方法はNurr1に限定されず、方法は細胞内においてNR4Aサブファミリーの任意の核内受容体を基底レベルより高く発現させることを含み得る。NR4Aサブファミリーの受容体にはNurr1/NR4A2、Nor1/NR4A3およびNGFI-B/NR4A1が含まれる。したがって、本発明の方法では、NR4Aサブファミリーの核内受容体(たとえばNurr1、Nor1またはNGFI-B)は細胞内において基底レベルより高く発現され得る。たとえばNR4Aサブファミリーメンバーのアクセッション番号は以下のとおりである。
【0028】
NGF-IBタンパク質: NP775181、NP775180、NP002126
NGFI-Bヌクレオチド: NM_173158、NM_173157、NM_002135
Nor-1タンパク質: NP775292、NP775291、NP775290、NP008912、S71930、Q92570
Nor-1ヌクレオチド: NM_005413、NM_173200、NM_173199、NM_173198、NM_006981
【0029】
Wnt糖タンパク質ファミリーのメンバーは溶解性が乏しく(BradleyおよびBrown、1990および1995)、発生中の中脳で発現される(Parr他、1993)。Wntは中脳-後脳の発生(McMahonおよびBradley、1990; ThomasおよびCapecchi、1990)、神経パターン形成(KieckerおよびNiehrs、2001; Nordstrom他、2002; Houart他、2002)、前駆体の増殖(TaipaleおよびBeachy、2001; ChennおよびWalsh、2002; MegasonおよびMcMahon、2002)ならびに神経系を含めた複数の組織における運命決定(Kispert他、1998; Ross他、2000; HartmannおよびTabin、2001; Marvin他、2001; SchneiderおよびMercola、2001; TzahorおよびLassar、2001; Pandur他、2002)を調節する(Dorsky他、1998; Baker他、1999; Wilson他、2001; Garcia-Castro他、2002; Muroyama他、2002)。
【0030】
本明細書中で使用する「Wntポリペプチド」、「Wnt糖タンパク質」または「Wntリガンド」とは、細胞間の相互作用を調節する分泌タンパク質のWingless-intファミリーのメンバーをいう。Wntはショウジョウバエ(Drosophila)や線虫(Caenorhabditis elegans)からアフリカツメガエル(Xenopus)、ゼブラフィッシュおよび哺乳動物に至るまで高度に保存されている。哺乳動物で現在知られている19種のWntタンパク質は2種類の細胞表面受容体型、すなわち現在10種の受容体で形成されている7回膜貫通ドメインFrizzled受容体ファミリー、ならびに低密度リポタンパク質-受容体関連タンパク質(Low density lipoprotein-receptor related proteins、LRP)5および6、ならびにkremen1および2受容体に結合する。Wntによって伝達されるシグナルは以下の3つの既知のシグナル伝達経路によって伝達される。(1)GSK3βが阻害されるいわゆる古典的シグナル伝達経路はβ-カテニンをリン酸化せず、したがってこれは分解されず核に転位してTCFと複合体を形成し、Wnt標的遺伝子の転写を活性化させる。(2)Jnkを介する平面極性および収束-伸張経路。(3)カルシニュリンが活性化T細胞の核因子(NFAT)を脱リン酸化および活性化するイノシトール1,4,5三リン酸(IP3)/カルシウム経路(Saneyoshi他、2002)。総説には、任意の利用可能なブラウザを使用してウェブ上で見つけることができるWntホームページを参照されたい(www.stanford.edu/〜rnusse/wntwindow.html)。Wntシグナル伝達に関与している他の補助受容体には、チロシンキナーゼ受容体Ror1およびRor2(Oishi I他、2003)、触媒的に不活性な受容体チロシンキナーゼをコードしているderailed/RYK受容体ファミリー(Yoshikawa他、2003)が含まれる。
【0031】
本発明の様々な態様の一部の好ましい実施形態では、WntリガンドはWnt1リガンドである。ヒトWnt1のアミノ酸配列はGenBank参照番号Swiss proteinアクセッション番号P04628から利用可能であり、コードしている核酸は、DNAは参照番号X03072.1から、RNAはNM_005430.2から利用可能である。ヒトWnt1核酸は、以下にさらに同定した配列番号1および2を有するプライマーを用いて増幅することができる。
【0032】
本発明の様々な態様の一部の好ましい実施形態では、WntリガンドはWnt5aリガンドである。ヒトWnt5aのアミノ酸配列はGenBank参照番号Swiss proteinアクセッション番号P41221から利用可能であり、コードしている核酸は、DNAは参照番号AI634753.1、AK021503、L20861、L20861.1、U39837.1から、RNAはNM_003392から利用可能である。ヒトWnt5a核酸は、以下にさらに同定した配列番号5および6を有するプライマーを用いて増幅することができる。
【0033】
ドーパミン作動性ニューロンの産生には好ましくないが、本発明の一部の好ましい実施形態ではWnt3aリガンドを使用し得る。ドーパミン作動性ニューロンを産生させるおよび使用するための本明細書中に開示したものと類似した本発明の様々な態様および実施形態が本発明によって提供され、ここではWnt3aリガンドを使用して幹細胞/始原細胞の増殖もしくは自己再生を維持するおよび/またはそれらが他の、すなわち非ドーパミン作動性のニューロン表現型へと分化することを可能にするもしくは誘導する。本明細書中に記載した実験によって実証されるように、Wnt3aは、ドーパミン作動性ニューロンを生じさせる、Nurr-1を発現する始原体の数を減少させる。しかし、ニューロンの全体数は減少しないので他のニューロン表現型、たとえばセロトニン作動性ニューロンを含めた背側中脳表現型が生じるかもしれない。セロトニン作動性ニューロンの消失はうつ病に関連しているので、Wnt3aリガンドの使用を含む方法、および/またはWnt3aリガンド自体によって産生させたニューロンをたとえばうつ病の治療に使用し得る。
【0034】
ヒトWnt3aのアミノ酸配列はGenBank参照番号SwissProtアクセッション番号P56704から入手可能であり、コードしている核酸は、DNAは参照番号AB060284、AB060284.1、AK056278、AK056278.1から、mRNAはNM_033131から入手可能である。ヒトWnt3a核酸は、以下にさらに同定した配列番号3および4を有するプライマーを用いて増幅することができる。
【0035】
他の好ましいWntはWnt-2、Wnt-4、Wnt-7aおよびWnt-7b、特にWnt-2およびWnt-7aである。Wnt-2は、ニューロン表現型の獲得を促進するおよび/または誘導させるため、また幹細胞ならびに神経幹細胞、前駆細胞および始原細胞をDAニューロンへと分化および/または成熟させるために使用し得る。Wnt-7aは、幹細胞ならびに神経幹細胞、前駆細胞および始原細胞の増殖を促進するおよび/または誘導させることによって細胞のDAニューロンへの分化および/または成熟を促進するために使用し得る。Wnt-2およびWnt-7aのどちらかまたは両方を使用してNurr1+細胞からのTH+ニューロンの数を増加させ得る。
【0036】
Wnt-2核酸は、GenBank参照番号SwissProtアクセッション番号P09544、NCBI RefSeqタンパク質NP00382、NCBI RefSeq mRNA NM_003391およびNM_003391.1、NCBI RefSeq DNA NT_007933として寄託されており、核酸は参照番号AK056742、AK056742.1、BC029854、BC029854.1、X07876、X07876.1、AC002465、AC006326として寄託されており、以下に記載のように配列番号33および34を有するプライマーを使用して増幅することができる。Wnt-4核酸はGenBank参照番号SwissProtアクセッション番号P56705、NCBI RefSeqタンパク質NP110388、NCBI RefSeq mRNA NM_030761およびNM_030761.2、NCBI RefSeq DNA NT_004610として寄託されており、核酸は参照番号AA984007、AL031281、AY009398.1、AF009398.1、AB062766.1、BC034923.1、AF416743.1、AB061675.1、BQ891671.1、BU502468.1、BM043406.1、CB991983.1として寄託されており、以下に記載のように配列番号39および40を有するプライマーを使用して増幅することができる。Wnt-7a核酸は、GenBank参照番号SwissProtアクセッション番号000755、NCBI RefSeqタンパク質NP004616およびNP004616.2、NCBI RefSeq mRNA NM_004625およびNM_004625.2、NCBI RefSeq DNA NT_005927として寄託されており、核酸は参照番号D83175、D83175.1、BC008811、BC008811.1、U53476、U53476.1、BI823772.1、CB989433.1、BI552826.1、BI551057.1、BE740508.1として寄託されており、以下に記載のように配列番号45および46を有するプライマーを使用して増幅することができる。Wnt-7b核酸は、GenBank参照番号SwissProtアクセッション番号P56706、NCBI RefSeqタンパク質NP478679およびNP478679.1、NCBI RefSeq mRNA NM_004625およびNM_004625.2として寄託されており、核酸は参照番号AA062766、AF416743、BC034923、BM047487、BM047487.1、BU543397.1、BU541891.1、BU541105.1として寄託されており、以下に記載のように配列番号47および48を有するプライマーを使用して増幅することができる。
【0037】
野生型Wntリガンドを使用してもよく、あるいは、幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞におけるドーパミン作動性のニューロンとなる運命の増強された発生の機能が保持されている限りは、たとえば1つまたは複数のアミノ酸の付加、欠失、置換および/もしくは挿入による変異体または誘導体を使用してもよい。
【0038】
「幹細胞」とは自己再生が可能な任意の細胞型を意味し、それが胚性幹(ES)細胞である場合は個体内の全ての細胞を生み出すことができ、またはそれが多能性もしくは神経幹細胞である場合はニューロン、アストロサイトおよびオリゴデンドロサイトを含めた神経系内の全ての細胞型を生み出すことができる。幹細胞は以下のマーカーのうち1つまたは複数を発現し得る:Oct-4;Sox1-3;時期に特異的な胚性抗原(SSEA-1、-3、および-4)、ならびに腫瘍拒絶抗原TRA-1-60および-1-81(Tropepe他、2001; Xu他、2001に記載)。神経幹細胞は以下のマーカーのうち1つまたは複数を発現し得る:Nestin;p75ニューロトロフィン受容体;Notch1、SSEA-1(CapelaおよびTemple、2002)。
【0039】
「神経始原細胞」とは神経幹細胞の娘細胞または子孫細胞を意味し、幹細胞と比較してより分化した表現型および/またはより低減した分化能力を有する。前駆細胞とは、発生中にニューロンと直接的な系統関係にあってもなくても、定義された環境条件下で分化転換もしくは再分化するように、またはニューロン表現型を獲得するように誘導させられる、任意の他の細胞を意味する。好ましい実施形態では、幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞は、自発的にまたはマイトジェン(たとえばbFGF、EGFもしくは血清)を欠乏させた際にチロシンヒドロキシラーゼを発現しないあるいは有効には発現しない。
【0040】
幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞は、骨髄、皮膚、眼、鼻上皮、もしくは臍帯を含めた任意の胚性、胎児または成体組織、あるいは神経系の領域、たとえば小脳、脳室領域、脳室下領域、線条体、中脳、後脳、大脳皮質または海馬から得るか、それ由来であり得る。これらは、脊椎動物、たとえばヒトまたはウサギ、モルモット、ラット、マウスや他のげっ歯類、ネコ、イヌ、ブタ、ヒツジ、ヤギ、ウシ、ウマ、もしくは霊長類などの非ヒトであり得る哺乳動物、あるいはニワトリなどのトリから得るか、それ由来であり得る。
【0041】
本発明の好ましい実施形態では、成人の幹細胞/始原細胞/前駆細胞をin vitro、ex vivoまたはin vivoで使用する。これには、同意した成人(たとえば細胞が得られる成人)および適切な倫理委員会の承認が必要である。ヒト胚/胎児を供給源として使用する場合は、そのヒト胚は、本来ならば使用せずに破棄されるかまたは無期限に保存されるもの、特に妊娠が困難なカップルのためのIVF処理用に作製されたヒト胚である。IVFでは一般に、着床および最終的には妊娠に使用される数よりも多い数のヒト胚が作製される。このような予備の胚は通常破棄されることがある。関係する人々、特に関係する卵子ドナーおよび/または精子ドナーの適切な同意により、本来ならば破棄される胚をパーキンソン病などの重篤な神経変性障害の罹患者の利益のために倫理的に前向きな方法で使用することができる。本発明自体は、発生の任意の段階にあるヒト胚の使用に関連しない。上述のように、本発明はヒト胚に直接由来する材料を使用する必要の可能性を最小限にする一方で、ひどい病気の貴重な治療の開発を可能にする。
【0042】
一部の好ましい実施形態では、Wntリガンドと接触させ、本発明の任意の態様に従って他の方法で処理および/または使用する幹細胞または始原細胞もしくは前駆細胞は、同意した成人または適切な同意が出されている子供、たとえば障害のある患者であって、続いて本発明に従って産生させたニューロンを患者内に移植して戻すことによって処置する、かつ/あるいは内因性ドーパミン作動性ニューロンの発生もしくは機能を促進するまたは誘導させるために1つもしくは複数のWntリガンドならびに/または1つもしくは複数の1型アストロサイト/初期グリア細胞由来の因子で処置した患者から得る。
【0043】
幹細胞または始原細胞もしくは前駆細胞で誘導されるニューロンとなる運命は、未分化の表現型または原始ニューロン表現型を示し得る。これは、個体内の任意の細胞型を生み出す能力を有する全能性細胞、または複数の明確なニューロン表現型を生み出す能力を有する多能性細胞、または通常の発生中ではより限定された表現型を生み出す能力を有するがin vitroで適切な環境因子に曝した場合に他の細胞を生み出す能力を有する前駆細胞もしくは始原細胞であり得る。これは、特定のニューロンとなる運命に関連するマーカー、たとえばチロシンヒドロキシラーゼを欠いているかもしれない。
【0044】
複数の幹細胞、神経幹細胞ならびに/または始原細胞および/もしくは前駆細胞がNurr1またはNurr1サブファミリーの別の核内受容体を基底レベルより高く発現し、細胞をWntリガンドで処理する、本発明によるニューロンとなる運命を誘導させる方法では、細胞の大多数がニューロンとなる運命をとるように誘導させ得る。ニューロン細胞でドーパミン作動性の誘導または分化を増強させ得る。好ましい実施形態では、60%を超える、70%を超える、80%を超える、90%を超える幹細胞および/または始原細胞をニューロンとなる運命に誘導させ得る。
【0045】
「細胞内においてNurr1を基底レベルより高く発現させること」とは、in vivo非病的条件下において(未改変の)細胞内で発現されるレベルより高いレベルでNurr1を発現させることを意味する。同様に、細胞内においてNurr1サブファミリーの核内受容体を基底レベルより高く発現させることとは、in vivo非病的条件下において(未改変の)細胞内で発現されるレベルより高いレベルで核内受容体を発現させることを意味する。基底レベルより高い発現には、転写発現、翻訳発現、翻訳後発現、薬理学的発現、人工的アップレギュレーション発現および過剰発現が含まれる。基底レベルより高い核内受容体の発現は、本明細書中ではNurr1に関して記載する。本開示はNurr1サブファミリーの他のメンバーにも適用可能であり、このサブファミリーの他の核内受容体、たとえばNor1またはNGFI-Bと共に本発明の方法において使用し得る。したがって、Nurr1について例示しているが、本発明の方法はNurr1に限定されず任意のNurr1サブファミリーの核内受容体にまで及ぶ。
【0046】
基底レベルより高いNurr1の発現は、当業者に知られている任意の方法によって実現される。例として、基底レベルより高い発現はネイティブゲノムNurr1の調節をモジュレーションすることによって誘導し得る。これは、Nurr1のmRNAもしくはNurr1タンパク質の分解を阻害または阻止することによって、あるいはNurr1の転写および/または翻訳を増大させることによって、たとえば細胞をNurr1の転写をアップレギュレーションする線維芽細胞成長因子8(FGF8)と接触させることによって(Rosenthal, A.、(1998) Cell、93(5)、755-766)、ならびに/あるいは異種調節配列をNurr1のネイティブ調節領域内またはその隣接に導入することによって、ならびに/あるいはNurr1のネイティブ調節領域をたとえば相同組換えによってこのような異種調節配列で置き換えることによって、ならびに/あるいはNurr1の転写、翻訳もしくは機能を負に調節する、遮断するまたはダウンレギュレーションする分子、たとえばNurr2を破壊またはダウンレギュレーションすることによって(Ohkura他、(1999) Biochim Biophys Acta 14444: 69-79)行い得る。
【0047】
転写は、幹細胞、神経幹細胞、前駆細胞、始原細胞、または神経細胞に増大させたレベルの転写活性化因子を与えることによって、たとえば細胞をこのような活性化因子と接触させるか、または活性化因子をコードしている核酸を用いて細胞を形質転換させることによって、増大させ得る。あるいは、転写は、Nurr1の転写インヒビターに対するアンチセンス核酸を用いて細胞を形質転換させることによって増大させ得る。
【0048】
したがって、幹細胞、神経幹細胞、前駆細胞、始原細胞、または神経細胞においてニューロンとなる運命を誘導させるまたは誘導を増強する本発明の方法は、細胞をFGF8またはFGF20と接触させることを含み得る(Ohmachi他、2000)。
【0049】
内因性Nurr1の転写および/または翻訳を増大させることに代わってまたはそれに加えて、基底レベルより高いNurr1の発現は1つまたは複数の余分なNurr1のコピーを幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞内に導入することによって引き起こされ得る。
【0050】
したがって、さらなる態様では、本発明は、ニューロンとなる運命を誘導させるならびに/あるいは幹細胞、神経幹細胞、神経始原細胞、前駆細胞もしくは神経細胞におけるドーパミン作動性の発生の誘導を増強する、またはニューロン細胞におけるドーパミン作動性の誘導もしくは分化を増強する方法であって、細胞をWntリガンドと接触させることに加えて細胞をNurr1で形質転換させることを含む方法を提供する。
【0051】
幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞の形質転換はin vitro、in vivoまたはex vivoで実施し得る。細胞に誘導させるニューロンとなる運命は本明細書中に記載の種類のものでよく、たとえば原始ニューロン表現型を示し、特定のニューロンとなる運命に関連するマーカーが欠けていてもよい。本発明はさらに、Nurr1で形質転換させ、Wntリガンドと接触させた幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞を提供する。
【0052】
形質転換させたNurr1および/またはWntリガンドはゲノム外ベクターに含まれるか、またはゲノム内に、好ましくは安定的に組み込まれ得る。これは幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞、またはニューロン細胞においてその基底レベルより高い発現を駆動するプロモーターに機能的に連結されており、以下により詳細に記述されている。
【0053】
「機能的に連結されている」とは、同じ核酸分子の一部として結合されており、プロモーターから転写が開始されるよう適切に配置かつ配向されていることを意味する。
【0054】
細胞内に遺伝子を導入する方法は当業者に周知である。Nurr1および/またはWntリガンドがベクター上に留まるのかゲノム内に組み込まれるのかにかかわらず、幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞内にNurr1および/またはWntリガンドを導入するためにベクターを使用し得る。適切なベクターを選択するか、またはプロモーター配列、ターミネーター断片、ポリアデニル化配列、エンハンサー配列を含めた適切な調節配列を含むよう構築することができる。ベクターは必要に応じてマーカー遺伝子および他の配列を含み得る。調節配列は幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞または神経細胞内においてNurr1および/またはWntリガンドの発現を駆動し得る。たとえば、ベクターはゲノム外発現ベクターであり得、または調節配列はNurr1および/またはWntリガンドと共にゲノム内に取り込まれ得る。ベクターはプラスミドまたはウイルスベクターであり得る。
【0055】
Nurr1および/またはWntリガンドを使用者の制御下におくために、これを外因的に誘導性遺伝子プロモーターの制御下においてもよい。プロモーターにあてはめた場合の用語「誘導性」は当業者によく理解されている。要するに、誘導性プロモーターの制御下における発現が、与えた刺激に応答して「スイッチが入る」または増大される。刺激の性質はプロモーター間で異なる。一部の誘導性プロモーターは、適切な刺激が存在しない場合はわずかなもしくは検出されないレベルの発現しか引き起こさない(または全く発現させない)。刺激が存在しない場合の発現レベルがどのようであっても、任意の誘導性プロモーターによる発現は妥当な刺激の存在下で増大する。誘導性プロモーターの例はテトラサイクリンON/OFFシステムであり(Gossen他、1995)、ここでは遺伝子の発現はテトラサイクリン類似体によって調節される。
【0056】
さらなる詳細には、たとえばMolecular Cloning: a Laboratory Manual: 第3版、SambrookおよびRussell、2001、Cold Spring Harbor Laboratory Pressを参照されたい。核酸を操作するための多くの知られている技術およびプロトコル、たとえば核酸構築物の調製、突然変異誘発、シーケンシング、細胞内へのDNAの導入および遺伝子発現、ならびにタンパク質の解析がCurrent Protocols in Molecular Biology、Ausubel他編、John Wiley & Sons、1992またはそれ以降の版に詳述されている。
【0057】
抗生物質耐性または感受性遺伝子などのマーカー遺伝子を、当技術分野で周知のように目的核酸を含むクローンの同定に使用し得る。クローンはまた、結合の研究によって、たとえばサザンブロットハイブリダイゼーションによって同定またはさらに調査し得る。
【0058】
Nurr1を含むかつ/またはWntリガンドをコードしている核酸を宿主幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞のゲノムに組み込ませ得る。組込みは、形質転換させた核酸に、標準の技術に従って、ゲノムとの組換えを促進する配列を含めることによって促進され得る。組み込まれた核酸は、幹細胞、または神経幹細胞、始原細胞もしくは前駆細胞、またはニューロン細胞においてNurr1遺伝子のおよび/またはWntリガンドの発現を駆動する能力を有する調節配列を含み得る。核酸は、Nurr1および/またはWntリガンドをコードしている配列が幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞、またはニューロン細胞内におけるその発現を駆動および/または制御する能力を有する調節エレメントの制御下におかれる、ゲノム内の部位へのその組込みを指令する配列を含み得る。組み込まれた核酸は、本明細書中に記載のように、Nurr1および/またはWntリガンドを幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞、またはニューロン細胞内に形質転換させるために使用したベクター由来であり得る。
【0059】
Nurr1を含むかつ/またはWntリガンドをコードしている核酸の導入は、その核酸が直鎖状、分枝鎖状または環状であるかにかかわらず、制限なしに概して「形質転換」と称し得る。これには任意の利用可能な技術を使用し得る。適切な技術には、リン酸カルシウムトランスフェクション、DEAE-デキストラン、PEI、電気穿孔、微量注入などの機械的技法、直接的なDNAの取り込み、受容体に媒介されるDNAの移送、レトロウイルスまたは他のウイルスを用いた形質導入ならびにリポソームまたは脂質に媒介されるトランスフェクションが含まれ得る。選択した遺伝子構築物を細胞内に導入する際、当業者に周知の特定の検討事項を考慮に入れなければならない。当業者には、Nurr1および/またはWntリガンドを幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞内に導入するための具体的な形質転換方法の選択は、本発明によって重要ではなくその制限とならないことが明らかであろう。
【0060】
Nurr1および/またはWntリガンドを用いた幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞のin vivo形質転換のための適切なベクターおよび技術は当業者に周知である。適切なベクターには、アデノウイルス、アデノ関連ウイルス、パポバウイルス、ワクシニアウイルス、ヘルペスウイルス、レンチウイルスおよびレトロウイルスが含まれる。無効化したウイルスベクターは、感染性ウイルス粒子の産生に必要な遺伝子が発現されているヘルパー細胞系内で産生させ得る。適切なヘルパー細胞系は当業者に周知である。例として、Fallaux, F.J.他、(1996) Hum Gene Ther 7(2)、215-222; Willenbrink, W.他、(1994) J Virol 68(12)、8413-8417; Cosset, F.L.他、(1993) Virology 193(1)、385-395; Highkin, M.K.他、(1991) Poult Sci 70(4)、970-981; Dougherty, J.P.他、(1989) J Virol 63(7)、3209-3212; Salmons, B.他、(1989) Biochem Biophys Res Commun 159(3)、1191-1198; Sorge, J.他、(1984) Mol Cell Biol 4(9)、1730-1737; Wang, S.他、(1997) Gene Ther 4(11)、1132-1141; Moore, K.W.他、(1990) Science 248(4960)、1230-1234; Reiss, C.S.他、(1987) J Immunol 139(3)、711-714を参照されたい。ヘルパー細胞系は一般に、ウイルスゲノムをパッケージする機構によって認識される配列を欠いている。これらは、核酸を全く含まないビリオンを産生する。無傷のパッケージングシグナルと送達する遺伝子または他の配列(たとえばNurr1および/またはWnt)とを含むウイルスベクターをヘルパー細胞内にパッケージして感染性ビリオン粒子内に入れ、その後これを幹細胞、または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞への遺伝子の送達に使用し得る。
【0061】
内因性Nurr1および/またはWntの転写および/または翻訳を増大させることに代わってまたはそれに加えて、基底レベルより高いNurr1および/またはWntの発現は1つまたは複数の余分なNurr1および/またはWntタンパク質のコピーを、微量注入または細胞透過ペプチド、すなわちTAT、トランスポータン、アンテナペディアペネトラチン(penetratin)ペプチド(Lindsay、2002)を含めた他の担体系もしくはタンパク質送達系によって幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞内に導入することによって引き起こされ得る。
【0062】
さらなる態様では、本発明は、幹細胞、神経幹細胞または始原細胞もしくは前駆細胞、またはニューロン細胞において特定のニューロンとなる運命を誘導させる方法であって、幹細胞または始原細胞またはニューロン細胞がNurr1を基底レベルより高く発現し、細胞をWntリガンドおよび任意選択で1型アストロサイト/グリア細胞によって供給されるまたはそれ由来である1つまたは複数の因子と接触させることを含む方法を提供する。この因子または複数の因子は、幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞を1型アストロサイト/グリア細胞と同時培養するあるいは接触させることによって提供し得る。方法はin vitroまたはin vivoで起こり得る。基底レベルより高くNurr1および/またはWntを発現する幹細胞または神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞は、Nurr1および/またはWntを用いた細胞の形質転換によって産生させ得る。
【0063】
因子または複数の因子は不死化したアストロサイト/グリア細胞によって供給されるまたはそれ由来であり得る。因子または複数の因子はグリア細胞系、たとえばアストロサイトもしくは放射状グリアもしくは未熟なグリア中脳細胞系によって供給されるまたはそれ由来であり得る。細胞系は同種細胞集団を提供する。
【0064】
本発明の重要な態様は、ドーパミン作動性ニューロンは、幹細胞または始原細胞もしくは前駆細胞からin vitroで、細胞において基底レベルより高くNurr1を発現させること、および細胞を腹側中脳の1型アストロサイト/初期グリア細胞によって供給されるまたはそれ由来である1つまたは複数の因子と接触させることを含む方法によって産生させることができるが、ドーパミン作動性の運命の誘導はWntリガンドと接触させることによって増強または促進されるという知見に基づいている。
【0065】
本発明は多数のドーパミン作動性ニューロンの産生を可能にする。これらのドーパミン作動性ニューロンは、パーキンソン病において変性するまたは損傷もしくは損失される細胞と置き換わる原料物質として使用し得る。
【0066】
好ましくは、Nurr1を基底レベルより高く発現する細胞は、Wntリガンドと接触させた場合に有糸分裂する。
【0067】
本発明の方法では、細胞を以下から選択された1つまたは複数の物質とさらに接触させ得る:塩基性線維芽細胞成長因子(bFGF);上皮成長因子(EGF);およびレチノイドX受容体(RXR)の活性化因子、たとえば合成レチノイド類似体SR11237(Gendimenico, G.J.他、(1994) J Invest Dermatol 102(5)、676-80)、9-シスレチノールもしくはドコサヘキサン酸(DHA)またはLG849(Mata de Urquiza他、2000)。実験によって以下に実証するように、これらの物質のうち1つまたは複数を用いた本発明による細胞の処理を使用して、ドーパミン作動性の運命をとる幹細胞、始原細胞もしくは前駆細胞の割合を増加させるか、またはニューロン細胞においてドーパミン作動性の誘導もしくは分化を増強させ得る。本発明によるニューロン細胞においてドーパミン作動性の運命を誘導させることまたはドーパミン作動性の誘導もしくは分化を増強させる方法は、細胞をFGF成長因子ファミリーのメンバー、たとえばFGF4、FGF8またはFGF20と接触させることを含み得る。
【0068】
有利には、細胞を上記物質のうち2つ以上と接触させ得る。本発明者らは、bFGFまたはEGFおよびSR11237の有益な効果が飽和用量で付加的であることを思いがけず見出した。この知見は、これらの物質が異なる機構によって作用するかもしれないことを示唆している。
【0069】
ドーパミン作動性の表現型を誘導させる方法は、Wntリガンドおよび任意選択で腹側中脳の1型アストロサイト/グリア細胞によって供給されるまたはそれ由来である1つまたは複数のさらなる因子と接触させる前に、たとえば腹側中脳の1型アストロサイト/グリア細胞もしくはそれらに由来する因子と接触させるまたは同時培養する前に、幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞またはニューロン細胞をbFGFおよび/またはEGFで前処理することを含み得る。
【0070】
任意選択の前処理ステップは、既に国際公開公報WO00/66713号およびWagner他、(1999)に報告されている本発明者らによる2つの思いがけない知見から生じる。(i)Nurr1を基底レベルより高く発現しており、高い増殖を示す神経幹細胞系では、腹側中脳の1型アストロサイト/グリア細胞と同時培養した際にドーパミン作動性の運命の誘導の増強が実証されたこと;および(ii)血清を含まない培地(SFM)中でbFGFまたはEGFを処理した後、Nurr1を基底レベルより高く発現するほとんどの幹細胞系の基底増殖がSFMのみに継代した後に上昇し続けたこと。
【0071】
ドーパミン作動性の表現型を誘導させる方法は、幹細胞または神経幹細胞、始原細胞もしくは前駆細胞をFGF成長因子ファミリーのメンバー、たとえばFGF2、FGF4、FGF8またはFGF20で前処理することを含み得る。
【0072】
前処理の代わりにまたは前処理と共に、Wnt処理と同時に追加の因子を用いて細胞を処理し得る。
【0073】
幹細胞、神経幹細胞または始原細胞もしくは前駆細胞においてニューロンとなる運命が誘導される、あるいはニューロン細胞においてドーパミン作動性の誘導または分化が増強されている本発明による方法は、ニューロンとなる運命のマーカーを検出することを含み得る。b-チューブリンIII(TuJ1)はニューロンとなる運命のマーカーの1つである(Menezes, J.R.他、(1994) J Neurosci 14(9)、5399-5416)。他のニューロンマーカーには神経フィラメントおよびMAP2が含まれる。特定のニューロン表現型を誘導させる場合は、マーカーはその表現型に特異的であるべきである。ドーパミン作動性の運命には、チロシンヒドロキシラーゼ(TH)、ドーパミントランスポーター(DAT)およびドーパミン受容体の発現を、たとえば免疫反応性またはin situハイブリダイゼーションによって検出し得る。チロシンヒドロキシラーゼはDA細胞の主要なマーカーである。ドーパミンおよび代謝物の含有量および/または放出は、たとえば高圧液体クロマトグラフィー(HPLC)によって検出し得る(Cooper, J.R.他、The Biochemical Basis of Neuropharmacology、第7版、(1996) Oxford University Press)。ドーパミンβヒドロキシラーゼおよびGABAまたはGADが存在しないこと(TH/ドーパミン/DATの存在下)もドーパミン作動性の運命の指標である。さらなるマーカーには、2型アルデヒドデヒドロゲナーゼ(ADH-2)、GIRK2、Lmx1bおよびPtx3が含まれる。
【0074】
マーカーの検出は当業者に知られている任意の方法に従って実施し得る。検出方法には、マーカーをコードしている核酸配列に結合する能力を有する特異的な結合メンバーであって、その配列とハイブリダイズ可能な核酸プローブ、または核酸配列もしくはそれにコードされているポリペプチドに対する特異性を有する免疫グロブリン/抗体ドメインを含み、特異的な結合メンバーの配列もしくはポリペプチドへの結合が検出可能であるように標識されている特異的な結合メンバーを使用し得る。「特異的な結合メンバー」はあるマーカーに対して特有の特異性を有し、通常の条件下では他の種類に優先してそのマーカーに結合する。あるいは、マーカーが特定のmRNAである場合は、特異的なオリゴヌクレオチドプライマーへの結合およびたとえばポリメラーゼ連鎖反応における増幅によって検出し得る。
【0075】
核酸プローブおよびプライマーはストリンジェントな条件下でマーカーとハイブリダイズし得る。たとえば約80〜90%同一であるマーカー配列を検出するための適切な条件には、42℃、0.25MのNa2HPO4、pH7.2、6.5%のSDS、10%の硫酸デキストラン中で終夜のハイブリダイゼーション、および55℃、0.1×SSC、0.1%のSDS中での最終洗浄が含まれる。約90%より高い同一性のマーカー配列の検出では、適切な条件には、65℃、0.25MのNa2HPO4、pH7.2、6.5%のSDS、10%の硫酸デキストラン中で終夜のハイブリダイゼーション、および60℃、0.1×SSC、0.1%のSDS中での最終洗浄が含まれる。
【0076】
さらなる態様では、本発明は、本明細書中に開示した方法のうち任意の1つに従って産生させたニューロンを提供する。このニューロンは原始ニューロン表現型を有し得る。これは、複数の明確なニューロン表現型を生み出す能力を有し得る。このニューロンは特定のニューロン表現型を有し得、この表現型はNurr1を基底レベルより高く発現する幹細胞、神経幹細胞、始原細胞、前駆細胞もしくは神経細胞と接触させた因子もしくは複数の因子が供給されたまたはそれが由来するWntリガンドおよび/またはアストロサイト/グリア細胞の種類、ならびに/あるいは幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞を同時培養したまたは接触させたアストロサイト/グリア細胞の種類によって影響を受ける。好ましい実施形態では、ニューロンはドーパミン作動性の表現型を有する。
【0077】
ニューロンは、ニューロンにおいて分子の発現を駆動する能力を有するプロモーターに機能的に連結されている、神経保護または神経再生特性を有する分子をコードしている核酸を含み得る。プロモーターは、損傷を与える全ての過剰発現を阻止し得るように誘導性プロモーター、たとえばTetONキメラプロモーターであり得る。プロモーターは特定のニューロン表現型、たとえばTHプロモーターまたはNurr1プロモーターと関連し得る。
【0078】
コードされた分子は、その発現がその環境に依存せずにニューロンを与えるように、すなわちその生存が1つもしくは複数の因子の存在、またはたとえばそれを移植する神経環境における条件に依存しないようであり得る。例として、ニューロンは、ニューロンにおいて分子の発現を駆動する能力を有するプロモーターに機能的に連結されている、以下に記載の神経保護または神経再生分子のうち1つまたは複数をコードしている核酸を含み得る。
【0079】
それに追加してまたはその代わりに、コードされた分子の発現はそのニューロンを取り囲む細胞環境の神経保護または神経再生において機能し得る。このようにして、ニューロンは神経保護および神経再生特性を有する分子を送達するために組み合わせた細胞および遺伝子治療手法において使用し得る。
【0080】
神経保護および神経再生特性を有する分子の例には以下のものが含まれる。
【0081】
(i)神経変性を補償および阻止することができる神経栄養因子。一例はグリア由来の神経栄養成長因子(GDNF)であり、これは強力な神経生存因子であり、ドーパミン作動性ニューロンからの発芽を促進し、チロシンヒドロキシラーゼの発現を増大させる(Tomac他、(1995) Nature、373、335-339; Arenas他、(1995) Neuron、15、1465-1473)。軸索伸長GDNFを増強することにより、GDNFはニューロンがその局所的環境を神経支配する能力を増大させ得る。GDNFファミリーの他の神経栄養分子には、ニューチュリン(Neurturin)、ペルセフィン(Persephin)およびアルテミン(Artemin)が含まれる。ノイロトロピンファミリーの神経栄養分子には、神経成長因子(NGF)、脳由来神経栄養因子(BDNF)、ならびにノイロトロピン-3、-4/5および-6が含まれる。神経栄養活性を有する他の因子には、FGFファミリーのメンバー、たとえばFGF2、4、8および20;Wnt-1、-2、-5a、-3aおよび7aを含めたWntファミリーのメンバー;BMP2、4、5および7、nodal、アクチビンおよびGDFを含めたBMPファミリーのメンバー;ならびにTGFα/βファミリーのメンバーが含まれる。
【0082】
(ii)抗アポトーシス分子。細胞死において中心的な役割を果たすBcl2。Bcl2の過剰発現はニューロンを自然に発生する細胞死および虚血から保護する(Martinou他、(1994) Neuron、1017-1030)。ニューロンに特異的な別の抗アポトーシス分子はBclX-Lである。
【0083】
(iii)神経支配およびその環境と連結を形成することにおいてニューロンを補助する軸索再生および/または伸長および/または誘導分子、たとえばエフリン。エフリンは、チロシンキナーゼ受容体を活性化する能力を有する膜結合リガンドのクラスを定義する。エフリンは神経発生に関係づけられている(Irving他、(1996) Dev. Biol.、173、26-38; Krull他、(1997) Curr. Biol. 7、571-580; Frisen他、(1998) Neuron、20、235-243; Gao他、(1996) PNAS、93、11161-11166; Torres他、(1998) Neuron、21、1453-1463; Winslow他、(1995) Neuron、14、973-981; Yue他、(1999) J Neurosci 19(6)、2090-2101)。
【0084】
(iv)転写因子、たとえばホメオボックスドメインタンパク質Ptx3(Smidt, M.P.他、(1997) Proc Natl Acad Sci USA、94(24)、13305-13310)、Lmx1b、Pax2、Pax5、Pax8、またはengrailed 1もしくは2(WurstおよびBally-Cuif、2001; RhinnおよびBrand、2001)、あるいは塩基性ヘリックス-ループ-ヘリックスファミリーの神経性遺伝子。
【0085】
本発明によるニューロンまたは本発明で使用するためのニューロンは、1つまたは複数の他の細胞型、たとえば幹細胞、神経幹細胞、前駆細胞もしくは始原細胞を実質的に含まなくてもよい。ニューロンは、抗体および磁気ビーズによる細胞外エピトープの認識に基づいた方法または蛍光活性化細胞分取(FACS)を含めた、当業者に知られている任意の技術を用いて神経幹細胞または始原細胞から分離し得る。例として、幹細胞、神経幹細胞、前駆細胞もしくは始原細胞上に見つかるがニューロン上には見つからない分子の細胞外領域に対する抗体を使用し得る。このような分子には、Notch1、CD133、SSEA1、プロミニン(prominin)1/2、RPTPβ/ホスホキャン(phosphocan)、TIS21およびグリア細胞系由来の神経栄養因子受容体GFRαまたはNCAMが含まれる。抗体に結合した幹細胞は補体に曝すことによって溶解するか、またはたとえば磁気選別によって分離し得る(Johansson他、(1999) Cell、96、25-34)。ニューロンの対象レシピエントに対して異種である抗体を使用する場合は、このような細胞選別手順を回避する全てのたとえば幹細胞、神経幹細胞または始原細胞もしくは前駆細胞を異種抗体で標識し、これらはレシピエントの免疫系の第一の標的となる。あるいは、所望の表現型を獲得する細胞は、細胞外エピトープに対する抗体、または細胞型に特異的なプロモーターの制御下における、蛍光タンパク質を含めた導入遺伝子の発現によっても分離することができる。例として、ドーパミン作動性ニューロンはTH、DAT、Ptx3またはドーパミン作動性ニューロンに特異的に使用される他のプロモーターの制御下で発現される蛍光タンパク質を用いて単離することができる。
【0086】
本発明の方法は、神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において所望の表現型を高めるために追加の陰性または陽性選択方法を含み得る。
【0087】
DAニューロンを富化するために陰性選択を使用し得る。非DAニューロン用の選択的神経毒素、たとえば5-7-ジヒドロキシトリプタミン(セロトニン作動性ニューロンを排除するため)、またはサポニンもしくは毒素に結合した抗体、または補体の追加後に、たとえばGABAトランスポーターに対する抗体(GABA作動性ニューロンを排除するため)を使用し得る。本発明の方法は、たとえば細胞を含むin vitro培養物に陰性選択剤を加えることによって、または陰性選択剤の存在下で細胞を培養することによって、神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を陰性選択剤で、好ましくはin vitroで処理するあるいはそれと接触させることをさらに含み得る。陰性選択剤は所望の細胞型(または複数の細胞型)以外の細胞型に対する選択を行う。たとえば、本発明がドーパミン作動性ニューロン表現型を促進する、増強させるまたは誘導させることに関係する場合、陰性選択剤はDAニューロンならびに幹細胞および神経幹細胞、前駆細胞および始原細胞などのDAニューロンへ発生する細胞以外の細胞に対する選択を行い得る。したがって、陰性選択剤は非DAニューロンなどの非DA表現型を有する分化細胞に対する選択を行い得る。陰性選択剤は、所望の細胞型(または複数の細胞型)以外の細胞の増殖を低減もしくは阻止するかつ/または死滅させ得る。陰性選択剤は、DAニューロン以外のニューロン集団を減少させる選択的神経毒素であり得る。たとえば、陰性選択剤は5-7-ジヒドロキシトリプタミンであり得る(セロトニン作動性ニューロンを減少させるため)。陰性選択剤は非DAニューロンに特異的な抗体または抗体フラグメントであってよく、この抗体または抗体フラグメント(たとえばscFvまたはFab)はサポニンまたは毒素に結合している。たとえば、抗体はGABAトランスポーターに特異的であり得る(GABA作動性ニューロンを減少させるため)。
【0088】
本発明の方法では、神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞を抗酸化剤(たとえばアスコルビン酸)、低酸素圧および/または低酸素誘導因子(たとえばHIFまたはエリスロポイエチン)の存在下で増殖させ得る。
【0089】
本発明はさらに、様々な態様および実施形態中で、Wntリガンド、またはWntリガンドをコードしている核酸、または可溶性frizzed関連タンパク質もしくはdikkopfaもしくはWIFのWnt阻害活性を阻害もしくは遮断する合成Wntリガンド類似体、またはタンパク質、核酸もしくは合成拮抗剤、またはWntの下流にある1つもしくは複数のシグナル伝達構成要素を阻害、遮断、増強、切り替えもしくはモジュレーションするように作用するタンパク質、核酸もしくは合成薬物から選択された物質の使用であって、たとえばパーキンソン症候群またはパーキンソン病に罹患している個体の治療において、内因的または外因的に供給されたもののどちらかの幹細胞、始原細胞もしくは前駆細胞、またはニューロン細胞に作用することによって脳内でドーパミン作動性ニューロンの発生を誘導させる、促進するもしくは増強させるため、ならびに/あるいはドーパミン作動性ニューロンの消失を阻害もしくは阻止する、または生存、表現型の分化もしくは成熟、神経突起生成もしくはシナプス形成、機能的出力を促進するための、Wntリガンドもしくはそれをコードしている核酸または他の前記の物質を個体に投与することを含む治療方法における使用を提供する。Wntリガンドもしくはそれをコードしている核酸または他の前記の物質は、たとえば製薬上許容される賦形剤または担体を含む任意の適切な組成物中で投与し得、神経変性障害、パーキンソン症候群またはパーキンソン病を治療するための医薬の製造に使用し得る。Wntリガンドもしくはそれをコードしている核酸は、中枢神経系および/もしくは脳に投与するか、またはそれを標的とし得る。
【0090】
本発明は様々な態様で、本明細書中で開示した方法のうち任意の1つに従って産生させたニューロンだけでなく、このようなニューロン、幹細胞、始原細胞もしくは前駆細胞および/またはWntリガンドを含む医薬組成物、医薬、薬物または他の組成物、医療の方法におけるこのようなニューロン、幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞および/あるいはWntリガンドあるいは組成物の使用、たとえばパーキンソン病または他の(たとえば神経変性性)疾患を治療する(予防的処置を含み得る)ためのこのようなニューロン、幹細胞、始原細胞、前駆細胞またはニューロン細胞および/あるいはWntリガンドあるいは組成物を患者に投与することを含む方法、たとえばパーキンソン病または他の(たとえば神経変性疾患)を治療するための投与用組成物の製造におけるこのようなニューロンまたは細胞および/またはWntリガンドの使用、ならびにこのようなニューロンまたは細胞および/またはWntリガンドを製薬上許容される賦形剤、ビヒクルまたは担体、および任意選択で1つまたは複数の他の成分、たとえば神経保護分子、神経再生分子、レチノイド、成長因子、アストロサイト/グリア細胞、抗アポトーシス因子、あるいは幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞または宿主脳内において遺伝子の発現を調節する因子を混合することを含む医薬組成物を作製する方法にまで及ぶ。このような任意選択の成分はその環境に依存せずに、すなわちその生存がその環境における1つもしくは複数の因子または条件の存在に依存しないように、ニューロンを与え得る。例として、医薬組成物を作製する方法はニューロンを発生中の腹側中脳で見つかる1つまたは複数の因子と混合することを含み得る。ニューロンをGDNFおよび/またはニューチュリン(NTN)と混合し得る。
【0091】
本発明は、本発明に従って産生させたニューロン、幹細胞、始原細胞もしくは前駆細胞またはニューロン細胞および/あるいはWntリガンド、ならびに1つまたは複数の追加の成分を含む組成物を提供する。本発明による医薬組成物および本発明に従って使用する医薬組成物は、ニューロンまたは細胞に加えて、製薬上許容される賦形剤、担体、緩衝剤、保存剤、安定化剤、抗酸化剤または当業者に周知の他の材料を含み得る。このような材料は無毒性であるべきであり、ニューロンの活性を妨害するべきでない。担体または他の材料の正確な性質は投与経路に依存するだろう。組成物は、1つまたは複数の神経保護分子、神経再生分子、レチノイド、成長因子、アストロサイト/グリア細胞、あるいは幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞において遺伝子の発現を調節する因子を含み得る。このような物質は、上述のように、その環境に依存せずにニューロンを与え得る。
【0092】
液状医薬組成物は一般に、水、石油、動物性もしくは植物性油、鉱油または合成油などの液状担体を含む。生理食塩水溶液、組織もしくは細胞培養培地、デキストロースまたは他の糖溶液あるいはエチレングリコール、プロピレングリコールもしくはポリエチレングリコールなどのグリコールを含め得る。
【0093】
組成物は、発熱物質を含まず適切なpH、等張性および安定性を有する、非経口的に許容される水溶液の形態であり得る。当業者は、たとえば塩化ナトリウム、リンゲル液、または乳酸化リンゲル液などの等張ビヒクルを用いて適切な溶液を十分に調製することができる。人工脳脊髄液を用いて組成物を調製し得る。
【0094】
本発明は、医療方法、特にニューロン細胞の変性、損傷、損失、または障害に関連する病状の治療における本発明に従って産生させたニューロンおよび/またはWntリガンドの使用にまで及ぶ。さらに、本発明は、特定の表現型のニューロンの産生、損傷、または損失に関連する状態、疾患または障害の治療における、その表現型のニューロンおよび/またはWntリガンドの使用を提供し得る。より詳細には、本発明はヒトパーキンソン病の治療におけるドーパミン作動性ニューロンおよび/またはWntリガンドの使用を提供する。本発明は神経変性疾患(たとえばパーキンソン病)を治療するための材料および方法に特に関するが、それに限定はされない。例として、本発明は、脊髄および/もしくは大脳皮質、またはNurr1+細胞を含む神経系の他の領域の変性または損傷の治療にまで及ぶ。
【0095】
投与した細胞が2つ以上の明確なニューロン表現型を生み出す能力を有する幹細胞、始原細胞または前駆細胞である治療方法では、ニューロン、細胞および/もしくはWntリガンドまたは組成物を、細胞を所望する特定のニューロンとなる運命へと分化するよう指示するアストロサイト/グリア細胞を含む領域内に導入し得る。細胞および/もしくはWntリガンドまたは組成物をたとえば腹側中脳に注射し、それらはここで1型アストロサイト/グリア細胞と相互作用し得、ドーパミン作動性の表現型をとるように誘導させ得る。あるいはまたはそれに加えて、移植した組成物は、ニューロンまたは細胞を上述のように特定のニューロンとなる運命に向かうその発生を指示する1つまたは複数の因子、たとえば1型アストロサイト/グリア細胞と組み合わせて含み得る。
【0096】
細胞は、当技術分野で知られている任意の技法によって患者内に移植し得る(たとえばLindvall, O.、(1998) Mov. Disord. 13、Suppl. 1:83-7; Freed, C.R.他、(1997) Cell Transplant、6、201-202; Kordower他、(1995) New England Journal of Medicine、332、1118-1124; Freed, C.R.、(1992) New England Journal of Medicine、327、1549-1555)。
【0097】
本発明による組成物の投与量は、好ましくは「予防上有効量」または「治療上有効量」(状況次第とはいえ、予防は治療と見なされ得る)であり、これは、個体において利点を示すのに十分である。投与する実際の量、ならびに投与速度および間隔は、治療するものの性質および重篤度に依存するだろう。治療の処方、たとえば用量の決定などは、一般医および他の医師の責任範囲内にある。
【0098】
組成物は単独でまたは他の治療と組み合わせて、治療する状況に応じて同時にまたは連続的に投与し得る。
【0099】
本明細書中に提供する方法は、初代細胞を用いてin vivoもしくはin vitroで、または細胞系を原料物質として用いて実施し得る。in vitroで拡大させた細胞の利点は、産生させ得るニューロンの数に実質上制限がないことである。
【0100】
移植した細胞の免疫学的拒絶に関連する可能性のある不利点を寛解させるために、幹細胞または始原細胞もしくは前駆細胞を患者から単離して所望する表現型に誘導させ得る。その後、細胞を患者に移植し得る。有利なことに、多数の免疫適合性のあるニューロン細胞を産生させ得るように、単離した幹細胞または始原細胞もしくは前駆細胞を用いて細胞系を確立させ得る。さらなる選択肢は、広範囲の免疫学的適合性を網羅する細胞バンクを確立することであり、そこから個体患者に適切な選択をすることができる。一個体由来の幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞を、それらまたはそれらの子孫を第2の個体へと導入した場合に拒絶が寛解されるように改変し得る。例として、ドナー細胞における1つまたは複数のMHC対立遺伝子をレシピエントのそれと、たとえば相同組換えによって置き換え得る。
【0101】
不死化癌遺伝子を有する細胞系由来の細胞を使用して患者に移植する場合は、細胞を患者に移植する前にCRE-locP系を用いて癌遺伝子を除去し得る(Westerman, K.A.他、Proc. Natl. Acad Sci. USA 93、8971 (1996))。患者の体温では不活性である不死化癌遺伝子を使用し得る。
【0102】
さらなる態様では、本発明は、神経変性疾患の治療に使用する物質についてスクリーニングする方法における、本発明に従って産生させた細胞またはニューロンの使用にまで及ぶ。ニューロンはドーパミン作動性ニューロンであり得る。神経変性疾患はパーキンソン症候群またはパーキンソン病であり得る。物質は神経保護分子および/もしくは神経再生分子ならびに/または発生的可溶性シグナルならびに/または腹側中脳の1型アストロサイトもしくはグリア細胞由来の因子もしくは複数の因子であり得る。方法は、in vitroまたはin vivoで実施し得る。
【0103】
この方法は、以下を含み得る。
【0104】
(i)本発明のニューロンを前記ニューロンに対する毒素で処理すること;
(ii)ニューロンを毒素から分離すること;
(iii)処理したニューロンを1つまたは複数の試験物質と接触させること;
(iv)ニューロンが毒素から回復する能力を決定すること;
(v)ニューロンが毒素から回復する前記能力を、試験物質(または複数の試験物質)と接触させない場合におけるそのニューロンまたは同一のニューロンが毒素から回復する能力と比較すること。
【0105】
この方法は、以下を含み得る。
【0106】
(i)1つまたは複数の試験物質存在下で本発明のニューロンをそのニューロンに対する毒素で処理すること;
(ii)ニューロンが毒素に耐性を示す能力を決定すること;
(iii)ニューロンが毒素に耐性を示す前記能力を、試験物質(または複数の試験物質)と接触させない場合におけるそのニューロンまたは同一のニューロンが毒素に耐性を示す能力と比較すること。
【0107】
毒素は6-ヒドロキシドーパミン、5,7-ジヒドロキシトリプタミンまたは1-メチル-4-フェニル-1,2,3,6-テトラヒドロピリジン(MPTP)、ラクタシスチンを含めたプロテアソームインヒビター、またはロテノンを含めた殺虫剤であり得、これらは全てカテコールアミン作動性ニューロンの死をもたらし、パーキンソン病の特徴が実験的に再現される。ニューロンが毒素から回復するまたは毒素に耐性を示す能力は、当業者に知られている任意の方法、たとえば細胞生存度をモニターすること(たとえば、たとえばTUNEL技法による細胞計数によって)、形態をモニターすること(たとえば発芽、軸索伸長および/または枝分かれ)、および/または生化学をモニターすること(たとえばTH活性、たとえば神経伝達物質の取り込み/放出/含有量)によって決定し得る。
【0108】
本発明において使用し得る幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞にはC17.2(Snyder, E.Y.他、Cell 68、33-51 (1992))およびH6ヒト細胞系(Flax他、Nature Biotech 16 (1998))が含まれる。さらなる例がGage他、1995、およびGotlieb、2002)に記載されている。
【0109】
本議論は神経幹細胞または神経始原細胞もしくは前駆細胞に関して行ったが、本明細書中で提供する方法は他の幹細胞、始原細胞または前駆細胞におけるニューロンとなる運命の誘導に応用し得る。このような細胞の例には、非神経系に関連する幹細胞が含まれる。これらの方法は間質もしくは造血幹細胞および/または表皮由来の増殖性細胞に応用し得る。造血細胞は血液または骨髄生検から採取し得る。間質細胞は骨髄生検から採取し得る。上皮細胞は皮膚生検、またはたとえば口腔粘膜を掻取ることによって採取し得る。ニューロン表現型はこれら幹細胞、始原細胞または前駆細胞の生理的なin vivoでの運命ではないので、誘導過程は分化転換、または脱分化および神経再分化と呼ぶことができる。このような細胞をニューロンとなる運命へと誘導させる方法は、非ニューロン表現型に関連する遺伝子に対するアンチセンスレギュレーターの使用、すなわちこのような細胞がニューロンでない運命に向かって分化することを抑制かつ/または逆転させることを含み得る。
【0110】
本発明の方法は神経運命に傾倒していない幹細胞に適用し得る。これは、異なる発生系に関連する2つ以上の娘幹細胞を生み出す能力を有する幹細胞に適用し得る。このような幹細胞の例は胚性幹細胞、造血幹細胞、表皮由来の増殖性細胞、および神経幹細胞である。
【0111】
上述のように、本開示は、基底レベルより高いNurr1の発現と組み合わせてWntリガンドおよび/または腹側中脳の1型アストロサイトもしくはグリア細胞由来の1つもしくは複数の因子を必要とする方法によって、幹細胞または始原細胞もしくは前駆細胞からドーパミン作動性ニューロンを作製することができることを実証している。
【0112】
様々なさらなる態様において、本発明は、神経幹細胞または始原細胞もしくは前駆細胞においてドーパミン作動性の運命の誘導を増強させる因子または複数の因子、あるいはNurr1を基底レベルより高く発現し、Wntリガンドおよびこれによって同定する因子または複数の因子で処理したニューロン細胞においてドーパミン作動性の誘導または分化を増強させる因子または複数の因子の、アッセイならびにスクリーニング方法の供給に関する。
【0113】
本発明は、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞において、単独でまたは組み合わせて、ドーパミン作動性の運命の誘導を増強、増大または強化する能力を有する因子もしくは複数の因子のスクリーニング方法を提供する。本発明のさらなる態様は、このような幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞においてドーパミン作動性の運命の誘導を増強させる因子もしくは複数の因子のスクリーニングまたは探索および/もしくは獲得/同定における、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞の使用を提供する。
【0114】
スクリーニング方法は以下を含み得る。
【0115】
(a)試験物質を、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞と接触させることであり、この接触により試験物質と細胞との間の相互作用が生じ得;および
(b)試験物質と細胞との間の相互作用を決定すること。
【0116】
スクリーニング方法は、試験物質を、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞由来の膜画分、可溶性画分または核画分と接触させ、試験物質と画分との間の相互作用を決定することを含み得る。このような画分の調製は当業者の能力範囲内に十分ある。
【0117】
結合または相互作用は任意数の当技術分野で知られている定性的または定量的な技法によって決定し得る。試験物質と幹細胞または始原細胞またはニューロン細胞との間の相互作用は、どちらか一方を検出可能な標識で標識し、これを、固体支持体上に、たとえば固体支持体に結合した抗体を使用して、またはBiacoreシステムを含めたそれ自体で知られている他の技術によって固定していてもよいもう一方と接触させることによって調査し得る。
【0118】
スクリーニング方法は、1つまたは複数の試験物質の存在下で幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロン細胞を培養すること、およびたとえば本明細書中に記載のようにドーパミン作動性の表現型のマーカーを検出することによってドーパミン作動性の表現型への分化について細胞を解析することを含み得る。チロシンヒドロキシラーゼ(TH)はドーパミン作動性の表現型のマーカーの1つである。
【0119】
本発明に従ってスクリーニングを行った全ての物質は天然または合成化合物であり得る。
【0120】
スクリーニング方法は、腹側中脳の1型アストロサイトまたは初期グリア細胞を、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞においてドーパミン作動性の運命を誘導させる能力のない神経細胞(たとえばアストロサイト)と比較することを含み得る。比較は、たとえば腹側中脳の発生中の1型アストロサイトまたは初期グリア細胞と他の神経位置由来の1型アストロサイトまたは初期グリアとの間であり得る。
【0121】
アストロサイトまたは初期グリア細胞を含むスクリーニング方法では、不死化アストロサイトまたは不死化グリア細胞を使用し得る。これは、アストロサイト細胞系またはグリア細胞系、たとえばアストロサイトまたはグリア中脳細胞系を含み得る。このような細胞系は同種細胞集団を提供する。
【0122】
スクリーニング方法では、細胞間の表現型の差を解析するための任意の知られている方法を使用し得、これはDNA、mRNA、cDNAまたはポリペプチドのレベルであり得る。示差的スクリーニングおよび遺伝子スクリーニングがこのような技法のうちの2つである。本明細書中に記載した任意のスクリーニング方法で同定した物質を、本明細書中に記載した任意の他のスクリーニング方法の試験物質として使用し得る。
【0123】
スクリーニング方法では、核発現アレイ、たとえばマウスcDNA発現アレイを使用し得る。この手法では、様々な核酸分子のアレイをフィルター、石英または別の表面上に、たとえば核酸をフィルターに架橋結合させるよって配列させる。試験溶液または抽出物を得て、その中にある核酸をたとえば蛍光によって標識する。その後、溶液または抽出物をフィルターまたは遺伝子チップに施用する。試験核酸とフィルターまたは遺伝子チップ上の核酸とのハイブリダイゼーションを決定し、対照溶液で得られたハイブリダイゼーションと比較する。試験試料と対照試料とで得られたハイブリダイゼーションの差は、異なる核酸含有の指標である。核酸アレイについてのさらなる情報には、任意の利用可能なウェブブラウザを使用して検索可能なClontechウェブサイト(たとえばwww.clontech.com)またはAffymetrixウェブサイト(たとえばwww.affymetrix.com)を参照されたい。
【0124】
スクリーニング方法は、本明細書中では基底レベルより高く発現されたNurr1に関して記載しているが、本開示はNurr1サブファミリーの全ての核内受容体、たとえばNor-1およびNGFI-Bにも及ぶ。したがって、Nurr1は例として記載し、限定するものではない。本発明の任意の方法において、Nurr1または任意の他の受容体、たとえばNor-1やNGFI-Bを含めたNurr1サブファミリーの核内受容体は、細胞において基底レベルより高く発現され得る。
【0125】
スクリーニング方法では、幹細胞または始原細胞もしくは前駆細胞または神経細胞をWntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞または始原細胞もしくは前駆細胞または神経細胞と比較することを含み得る。これは、たとえばNurr1、ならびに/あるいは、幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞において増殖および/もしくは自己再生および/もしくは分化および/もしくは生存を増強させるかつ/あるいはドーパミン作動性の運命の獲得や誘導を促進するおよび/もしくはドーパミン作動性ニューロンの発生を誘導させる、ならびに/またはWntリガンドの存在下でNurr1を基底レベルより高く発現するニューロン細胞においてドーパミン作動性の誘導もしくは分化を増強させる、因子もしくは複数の因子の標的遺伝子を同定するためである。標的遺伝子(もしくは複数の遺伝子)および/または因子(もしくは複数の因子)が同定された後は、これらを単離および/または精製および/またはクローニングし、さらなる方法で使用し得る。
【0126】
スクリーニング方法は、物質もしくは複数の物質を混合物から精製および/または単離することを含み得る。この方法は、混合物の1つまたは複数の画分が、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞または神経始原細胞もしくは前駆細胞または神経細胞と相互作用する能力、たとえばこのような幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞に結合するならびに/または増殖および/もしくは自己再生を促進するならびに/またはそれにおいてドーパミン作動性の表現型もしくは運命の誘導、獲得、分化または発生を増強させる能力を決定することを含み得る。精製および/または単離には、当業者に知られている任意の方法を使用し得る。
【0127】
スクリーニング方法では、試験物質をコードしている核酸に機能的に連結されている誘導性プロモーターを使用し得る。このような構築物を宿主細胞内に取り込ませ、許容条件および非許容条件下におけるその細胞の1つまたは複数の特性を決定して比較する。決定された特性は、宿主細胞が、Wntリガンドの存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞においてドーパミン作動性の表現型を誘導させる能力であるかもしれない。許容条件と非許容条件とにおける宿主細胞の能力の差は、試験物質が、単独でまたは組み合わせて、幹細胞、神経幹細胞または始原細胞もしくは前駆細胞において、増殖および/または自己再生および/またはドーパミン作動性の運命の誘導および/またはドーパミン作動性の分化、生存もしくは発生を増強させる、あるいはWntリガンドの存在下でNurr1を基底レベルより高く発現するニューロン細胞においてドーパミン作動性の誘導または分化を増強させる能力を有するかもしれないことを示している。
【0128】
本発明の任意のスクリーニング方法の詳細な様式は、ルーチンな技術および知識を用いて、当業者によって変形させ得る。
【0129】
本発明によって提供するいずれか1つの方法によって同定した因子もしくは複数の因子を単離および/または精製し、かつ/またはさらに調査し得る。これを製造することもできる。
【0130】
様々なさらなる態様では、本発明は、本明細書中で開示したいずれか1つの方法によって同定した因子;このような因子を含む医薬組成物、医薬、薬物または他の組成物(この組成物は、Nurr1を基底レベルより高く発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞またはニューロンおよびWntリガンドを含み得る);Wntリガンドの存在下で基底レベルより高くNurr1を発現する幹細胞、神経幹細胞または始原細胞もしくは前駆細胞由来のドーパミン作動性ニューロンの誘導および/または表現型の分化もしくは成熟および/または生存および/または神経突起生成および/またはシナプス形成および/または機能的出力を増強させるためのこのような因子の使用;医療方法であって、たとえばドーパミン作動性ニューロンの変性、損傷、損失、もしくは障害またはそれに影響を与えることに関連する症状を治療する(予防的処置を含み得る)、たとえばパーキンソン病または別の神経変性疾患を治療するためにこのような因子または組成物を患者に投与することを含む方法における、このような因子または組成物の使用;たとえばパーキンソン病など(たとえば神経変性疾患)を治療するために投与する組成物、医薬または薬物の製造におけるこのような因子の使用;ならびに、このような因子を製薬上許容される賦形剤、ビヒクルまたは担体、および任意選択で他の成分と混合することを含む、医薬組成物を作製する方法をさらに提供する。
【0131】
関連の態様では、本発明は、幹細胞、神経幹細胞、前駆細胞もしくは始原細胞においてドーパミン作動性の運命を誘導させる、またはNurr1を基底レベルより高く発現するニューロン細胞においてドーパミン作動性の誘導もしくは分化を増強させる、Wntリガンドの能力をモジュレーションする物質のスクリーニング方法を提供する。
【0132】
したがって、この方法では、Nurr1を基底レベルより高く発現する幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞において増殖、自己再生、ドーパミン作動性の発生、分化、成熟および/またはドーパミン作動性の運命の獲得を誘導させる、Wntリガンドの能力をモジュレーションする物質をスクリーニングし得る。
【0133】
このような方法は以下のうちの1つまたは複数を含み得る。
【0134】
(i)Wntリガンドおよび1つまたは複数の試験物質の存在下でNurr1を基底レベルより高く発現する幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞を提供すること;
(ii)ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答するこのような細胞の割合を解析すること;
(iii) ドーパミン作動性の運命をとるこのような細胞の割合を、1つまたは複数の試験物質を存在させない同等の反応培地および条件における、ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答するこのような細胞の数と、比較すること。処理した細胞と未処理の細胞との間のドーパミン作動性ニューロンの割合の差は、関連する1つまたは複数の試験物質のモジュレーション効果の指標である。
【0135】
このようなスクリーニング方法は以下を含み得る。
【0136】
(i)Nurr1を基底レベルより高く発現する幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞を、1つまたは複数の試験物質の存在下でWntリガンドと接触させること;
(ii)ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答する幹細胞、神経幹細胞、前駆細胞もしくは始原細胞またはニューロン細胞の割合を分析すること;
(iii)ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答する幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞の割合を、1つまたは複数の試験物質を存在させない同等の反応条件における、ドーパミン作動性の運命もしくは表現型をとる、かつ/またはWntに応答する幹細胞、前駆細胞、始原細胞または神経細胞の数と比較すること。
【0137】
このようなスクリーニング方法は、同等のもしくは同一の非ヒト動物においてin vivoで、またはin vitroもしくは培養中で、細胞上で実施し得る。
【0138】
Wntまたは誘導活性をモジュレーションする物質を同定した後、物質をさらに調査し得る。これは、医薬、医薬組成物または薬物などの組成物の調製、すなわち製造または配合において製造および/または使用し得る。そのモジュレーション活性について試験した全ての物質は天然または合成化合物であり得る。
【0139】
以降、本発明の態様および実施形態を、例として、添付の図面を参照しながら例示する。さらなる態様および実施形態は当業者には明らかであろう。本明細書中で言及した全ての文献は、本明細書中に参考として組み込まれる。
【0140】
実験
本明細書中に参照により特に組み込まれるのは、1型アストロサイトまたはグリア細胞の存在下、ドーパミン作動性前駆体の増殖および/または自己再生、および、Nurr1を発現している幹細胞、神経幹細胞、前駆細胞または始原細胞におけるドーパミン作動性ニューロンの誘導を示し、そしてかかる細胞を、FGF(たとえばFGF8)またはレチノイドなどの追加の因子と接触させたときに得られた追加の結果を示しているWO OO/66713に記載された実験結果である。
【0141】
Wnt、Nurr1およびチロシンヒドロキシラーゼ(TH)の発現を発生中の腹側中脳で検討し、E10.5〜P1の背側中脳(DM)および大脳皮質と比較した。リアルタイムRT-PCRおよびISHは、THのメッセージがマウスにおいてE11.5ではっきり検出され、ラット腹側中脳においてE11.5で最初に検出されたことを示したが(図1aおよび図1b)、これは、Nurr1の発現の一日後に起こり(図1c)、ラットVMにおいてE16まで続くドーパミン作動性ニューロンの誕生(Fosterら、1988)と一致するものである。Wnt-1およびWnt-5aは、マウス腹側中脳において最高の発現レベルを有するWntであり(図1dおよび図1e)、ラットにおいてE11.5でピークとなった(図1a)。同様のピークは、背側中脳においてE11.5に、Wnt-1およびWnt-3aについて検出されたが、Wnt-5aについては検出されず、これは、腹側部の発生におけるWnt-5aの支配的な役割(Saneyoshiら、2002)と整合するものである。さらに、精製した1型アストロサイトにおけるWntの発現のリアルタイムRT-PCR分析は、Wnt-5aが腹側中脳のアストロサイトに発現しており、発現レベルは、腹側中脳での方が、背側中脳または大脳皮質のアストロサイトにおけるより有意に高かったことを示した(図1f)。
【0142】
発生中の中脳において高レベルで発現されていたWnt-1、3aおよび5aのmRNA発現に加え、リアルタイムRT-PCRによりなされたより詳細な分析は、Wnt-2および-7aのmRNAが中脳において中レベルで発現し、Wnt-7bおよびWnt-16が中脳において低レベルで発現されていたことを明らかにした。これらのリガンドはいずれも、腹側中脳での方が、背側中脳よりも高いレベルで発現しており、これは、Wnt-2、-7a、-7b、および-16の腹側中脳の発生における可能な役割を示唆するものである。本発明者らはまた、Wnt-4、-6、-1Ob、および-11が中脳において低レベルで発現していたことを見出した。Wnt-3、-13および-2bは、中脳において極めて低レベルで発現しており、Wnt-5b、-8a/d、-1Oa、および-15は、中脳においてバックグラウンドレベルで検出された。
【0143】
VMにおけるDAニューロンの誕生は、ラットにおいて胚11.5日(E11.5)に起こることが知られている。本発明者らは、Wntタンパク質の全ファミリーの発現パターンを発生中のラット脳において検討し、そして、本発明者らの研究を、DAニューロン誕生時までにVMにおいて有意なレベルで発現しているWntに広げた。分析したWntの中で、Wnt-2、-4、-7a、-7bは、E11.5において、VMでの方がDMよりも高いレベルの発現を示し、および/またはE11.5のDAニューロン誕生時にVMにおいてピークとなった(図8A、8B、8Cおよび8D)。これらのうち、Wnt-2およびWnt-7aは、VMにおいて、より高いレベルで発現していた(それぞれ図8Aおよび8B)。これに対し、Wnt-5b、-8a/8d、-10a、および-15の発現は、分析した脳領域のいずれにおいても観察されなかった。Wnt-2b/13、Wnt-3、Wnt-6、およびWnt-1Ob、およびWnt-11の転写産物は、主として背側中脳領域に限定されていたが、低レベルであった(図9A、9B、9Cおよび9D)。低レベルではあったが、Wnt-16の転写産物は、胚発生の間中両方の領域で検出された(図9E)。これらのデータは、緊密に関連した細胞外シグナル伝達分子のファミリーの間での、動的な発現のパターンを明らかにするものである。さらに、これらの遺伝子の空間的および時間的発現は、Wntファミリーのメンバーが、胚発生の間中、全く別様に機能することを示唆するものである。したがって、ここで提示された発現プロファイルは、Wnt-2、Wnt-4、Wnt-7a、およびWnt-7bの、DAニューロンの発生における可能な役割を示したものである。
【0144】
本発明者らは、Wntの標準的な経路の中心的なシグナル伝達構成要素であるβ-カテニンの発現を、E10.5のマウスにおけるオーファン核内受容体Nurr1の発現により特徴付けられるDA前駆細胞において検討した。腹側中脳のドーパミン作動性前駆体が高レベルのβ-カテニンを発現していることが見出された。二重免疫組織化学は、β-カテニンがE10.5のマウスにおいて、Nurr1と同じドメインに発現していたことを示したが、これはWntのシグナル伝達とβ-カテニンの安定化とが、in vivoの正常な発生の最中に、VM DA前駆細胞で生じることを示すものである。
【0145】
Wntリガンドを分泌するように操作された安定した線維芽細胞の細胞系(Shimizuら、1997)からの部分的に精製された馴化培地は、サイズ排除に基づくろ過により作製した。連続2回の精製の後、当初のCMに比べて約250〜1500倍の濃度の増大が得られた。部分的に精製されたWntリガンドの個々のロットは、互いに、合わせたウェスタンブロット産物のバンドの密度に基づいて正規化し、任意単位として表現され、ここで、1単位は、正規化された部分的に精製された産物の1μlに等しい。E14.5のラットから得られた腹側中脳の培養物は、その後、増大する濃度の部分的に精製されたWnt-1、-3a、-5a、または対照(CP)培地で3日間処置した(図2A)。Wnt-1または5aによる処置は、TH+細胞数の強い用量反応性の増加をもたらしたものの、Wnt-3aまたはCPによる処置は、100単位までの用量で効果をもたらさず、対照培地は、50単位(μl)までわずかな効果をもたらしたか、または効果をもたらさなかったが、このことは、Wnt-1または-5aによる処置で観察されたTH+細胞数の増加が、これらのリガンドに特異的であることを示唆するものである。興味深いことに、10単位のWnt-1または-5aの効果はin vitroで早くも1日後には検出され、3日目のCPおよびWnt-3aより高く、そして7日後に減少し始めた(図2B)。本発明者らの結果は、部分的に精製されたWntが少なくとも3日間は完全に安定しており、そしてその効果が1週間にわたって維持されたことを示したものである。さらに、TH+細胞数のこれらの増加は、これらが、GDNF、FGF-2またはFGF-8を含む既知のドーパミン栄養分子(dopaminotrophic molecule)により3日後にもたらされる増加を超えており、そしてNurr1発現神経幹細胞のための誘導シグナルを含むVM T1A馴化培地の効果におよそ等しいことから、生物学的に意味があるものと考えられた(図2C)。
【0146】
Wnt-2およびWnt-7aを過剰発現している線維芽細胞細胞系からの馴化培地は、Wnt-1およびWnt-5aの部分的に精製された培地と同様の結果を与え、Wnt-2およびWnt-7aの両方ともE14.5の腹側中脳前駆体培養物におけるTH+ニューロンの数を、対照線維芽細胞馴化培地に比べて増加させた。部分的に精製されたWntによる処置により、TH免疫反応性の増加がWnt-2およびWnt-7aの両方について観察された(図10A)。Wnt-7aによる処置は、対照および他のWntに比べて小幅なTH+の増加をもたらした。並行して行った実験において、Wnt-2による処置は、TH+ニューロンの約2倍の増加をもたらした。これらのデータは、Wnt-2およびWnt-7aによる処置が、VM前駆体培養物においてTH+ニューロンの数を増加させることを示している。Wnt-2およびWnt-7aの同様の効果が、THおよびNurr1マーカーによる二重免疫組織化学を用いた場合に観察されたが(図10B)、これは、これらのWntにより発揮される効果がNurr1発現前駆体に対するものであることを示すものである。
【0147】
したがって、本発明者らの結果は、糖タンパク質のWntファミリーのメンバーが、始原細胞から発生するドーパミン作動性ニューロンの数を極めて有効に調節していることを示している。さらに、本発明者らのデータは、Wntリガンドが、TH細胞数の調節について重複する効果を有していることを示しており、これは、Wntのいくつかの機能がある程度冗長であり得ることを示唆するものである。
【0148】
Wntリガンドによる処置はまた、大きな球形の増殖性クラスターの出現を誘導した。これらのクラスターは最初は小さく、培養期間中大きさを増した。5細胞径より大きく、1個またはそれ以上のTH+ニューロンを含むTH+ニューロンのクラスターを、in vitroで様々な時間に計数した。注目すべきは、10単位のWnt-1および-5aによる処置はin vitroで7日後に観察されたこれらのクラスターの数の3〜8倍の増加をもたらしたが、Wnt-3aはこれをもたらさなかったことである(図2Eおよび図2F)。さらに、Wntリガンドに反応して出現したクラスターは、より大きく、大部分がほとんど全てTH+ニューロンからなっていた。一方、対照およびWnt-3aにより処置された培養物は、一般的により小さなスフェアを含んでいた。興味深いことに、実質的に全てのクラスターと、培養物中の顕著な数の単離細胞とが、固定の前のBrdUの急激なパルスの後にBrdU陽性であった(図2f)。したがって、有糸分裂能のあるスフェアは、前駆体(および/またはコミットメントは少ないが、依然増殖性の)細胞のTH陽性細胞への誘導または分化からの、ドーパミン作動性前駆体の細胞周期への動員に由来する可能性がある。
【0149】
Wnt-1の欠失(McMahonおよびBradley、1990; ThomasおよびCapecchi、1990)またはWnt受容体LRP6の欠失(Pinsonら、2000)が中脳-後脳接合部全体の欠損をもたらし、そしてWnt-1ヌル変異体がドーパミン作動性ニューロンを欠損している(DanielianおよびMcMahon、1996)ことから、本発明者らはWntリガンドがVM前駆体培養物においてTH+ニューロンの数を増加させる可能性を調べた。Wntは、神経系における細胞運命決定の制御(Dorskyら、1998; Bakerら、1999; Wilsonら、2001; Garcia-Castroら、2002; Muroyamaら、2002)、細胞増殖(ChennおよびWalsh、2002; MegasonおよびMcMahon、2002)および分化(Hallら、2000; PatapoutianおよびReichardt、2000; Krylovaら、2002)に必須の役割を担っていることが知られており、これは、複数のメカニズムがVM前駆細胞におけるドーパミン作動性ニューロンの発生の調節に関与し得ることを示唆するものである。
【0150】
本発明者らは、リアルタイムRT-PCRにより、Wnt-5aではなく(Kioussiら、2002)、Wnt-1による細胞周期の進行を媒介する(MegasonおよびMcMahon、2002)β-カテニン(TetsuおよびMcCormick、1999; Shtutmanら、1999)の標的であるサイクリンD1、またはその他のサイクリンの発現が、Wnt-1、-5aおよびVM-T1Aによって調節されているかどうか検討した(図3Aおよび図7A)。サイクリンD2のmRNAは、これらの処置のいずれにも影響されなかったが、サイクリンD1のmRNAは、Wnt-5aまたはVM T1AによってではなくWnt-1によりアップレギュレーションされ、そしてサイクリンD3は、Wnt-5aによってではなくWnt-1によりアップレギュレーションされた。本発明者らの結果はWnt-1が、細胞周期の進行を転写レベルで調節していることを示したものである。同様に、細胞周期インヒビターの分析は、Wnt-5aではなくWnt-1がp27およびp57のmRNAをダウンレギュレーションしたが(図7Bおよび7C)、いずれもp21のmRNAの発現に影響しなかったことを明らかにした。したがって、本発明者らの結果は、Wnt-1が、G1-Sの進行を転写レベルで調節することにより、Nurr1が誘導する細胞周期の停止に拮抗するよう作用し得ることを示唆するものである(Castroら、2001)。
【0151】
Wntの標的遺伝子は種々の生物系で同定されており、多くの細胞周期レギュレーターを包含する。Wnt-2およびWnt-7aによる処置は、重要な細胞周期レギュレーターの発現における明確な差異をもたらした(図11)。驚くべきことに、Wnt-2による処置によって、前駆体培養物は、サイクリンD1のmRNAレベルにおける若干のアップレギュレーションを示した(図11A)。予想通り、Wnt-7aはサイクリンD1の転写レベルを増加させたが(図11A)、これは、本明細書に記載されたBrdUの取り込みの増大したレベルの知見と整合するものである。サイクリンD2はWnt-7aにより調節されなかったが、Wnt-2による処置はサイクリンD2のmRNAレベルの増加を導いた(図11B)。サイクリンD3の調節は、Wnt-7aまたはWnt-2aによる処置のいずれにおいても観察されなかった(図11C)。興味深いことに、Wnt-7aによる処置は、細胞周期インヒビターp27およびp57の発現を減少させた(それぞれ、図11Dおよび11E)。これらのデータは、Wnt-7aがそのドーパミン作動性前駆体に対する機能を、本明細書に記載されたWnt-1のものと同様のメカニズムにより発揮することを示すものである。これに対して、p27およびp57のmRNAレベルにおける増加は、Wnt-2による処置の後に観察された(それぞれ、図11Dおよび11E)。これらのデータは、Wnt-2は前駆体培養物におけるTH+ニューロンの数を増加させるが、Wnt-5aのものと別異のメカニズムによってこれを行ない、そして、これはG1-Sの進行の転写レベルでの負の調節を伴うことを示唆するものである。
【0152】
本発明者らは、Wntが、VM培養物におけるDA前駆細胞の増殖を調節するかどうか検討した。興味深いことに、本発明者らは、新生DAニューロン(Nurr1+/TH+細胞)においてBrdUの取り込みを観察できなかったが、これは、増殖している細胞は、実験の最後の6時間の標識期間中TH+細胞に分化しないことを示すものである。本発明者らは次に、WntがVM前駆体培養物において増殖を調節するかどうか、および、中脳におけるNurr1発現ニューロン前駆体(Nurr1+/BrdU+細胞)がWntリガンドの潜在的な標的であるかどうかを検討した。VM前駆体培養物におけるBrdUの取り込みは、Wnt-5aによる処置によってではなく、Wnt-1および-3aによる処置によって増加したが(図7D)、これは、Wnt-1および-3aがマイトジェンとして作用し得、培養物における全体的な前駆体の増殖を促進しうることを示すものである。Wnt-1および-3aの効果は、Nurr1およびBrdU二重陽性前駆細胞をin vitroで1日後に検討した場合にさらに大きかった(3倍増)。部分的に精製されたWnt-5aも、Nurr1+集団の有糸分裂を増加させたが、程度はそれ程大きくなかった(図3B)。in vitroの1日目におけるNurr1およびBrdUに対する二重免疫組織化学は、Wnt-1および-3aは、有糸分裂を3倍増加させたが、Wnt-5a、およびWnt-5aを発現し、Nurr1発現前駆体にドーパミン作動性表現型を誘導するVM1型アストロサイトからの馴化培地は、Nurr1+集団において有糸分裂を2倍増加させたことを示した(図3B)。この知見は、Wnt-1によるサイクリンD1発現の増強と合わせて、Wnt-1が、有糸分裂/増殖を調節することによりTH+細胞数を主として増強させることを示したものである。驚くべきことに、Wnt-3aは、Nurr1陽性細胞の有糸分裂を増強することについてWnt-1と同程度有効であったものの、TH陽性細胞数をわずかに増加させるか、または増加させなかったが、このことは、増加した有糸分裂は、腹側中脳におけるWntの活性のほんの一要素にすぎないことを示唆するものである。Wnt-1およびWnt-3aによって誘導される増殖の明確な増加は、β-カテニンを安定化するこれらの能力(Shimizuら、1997)、幹細胞および神経前駆体において細胞周期リエントリー(reentry)を促進するβ-カテニンの役割(TaipaleおよびBeachy、2001; ChennおよびWalsh、2002)、および造血幹細胞の自己再生におけるWnt-3aの役割(Reyaら、2003)とよく相関する。したがって、Wnt-1の、前駆体の増殖およびG1-Sの進行を制御する遺伝子の調節に対する効果は、Wnt-1が、前駆細胞における有糸分裂/増殖/自己再生を調節することにより、主としてTH+細胞数を増強させることを示すものである。しかしながら、Wnt-3aは、Nurr1+細胞における有糸分裂を増強することについてWnt-1と同程度に有効であったものの、Wnt-1と異なり、DAニューロンの数に影響を与えないか、または数を減少させたが、このことは、自己再生および/または前駆体集団の維持におけるWnt-3aの特異的な役割を示唆するものである。
【0153】
BrdU(有糸分裂のS期にある細胞のマーカー)の取り込みを評価する実験において、BrdU+細胞数の増加が、Wnt-7aによる処置の後でも観察された(図10C)。実際、Wnt-7aによる処置は、上記のWnt-1と同様のBrdU+の増加をもたらした。これに対し、BrdUの取り込みの若干の減少が、Wnt-2による処置の後で認められた。これらのデータは、Wnt-7aが前駆体集団を拡大することによりTH+細胞数を増加させる一方、Wnt-2の効果は、別個のメカニズムを介して起こり、そして前駆体の増殖の減少を伴うことを示すものである。
【0154】
本発明者らはまた、Wnt-1の増殖効果が特異的であり、そしてWntインヒビターであるFrizzled-8のシステインリッチドメイン(Fz8-CRD)(35、36)によりブロックされ得るかどうかを検討した。Fz8-CRDを過剰発現している線維芽細胞からの部分的に精製されたCMを対照およびWnt-1で処置された培養物に加え、そしてTH+ニューロンの数を検討した(図7Dおよび7E)。Fz8-CRDは、両方の条件においてTH+ニューロンの数を減少させたが、これは、内因性のWntが培養物中のDAニューロンの発生に必要であり、そして、WntのTH+細胞への効果が特異的であることを示すものである。
【0155】
したがって、まとめると、全ての結果は、Wntの効果が特異的であり、そしてVMにおけるTH+かつ増殖性の細胞の数がWntによって独立して調節されていることを示していた。さらに、これらの結果は、増大した有糸分裂/増殖/自己再生が、腹側中脳におけるWntの活性のほんの一要素にすぎないことを示唆したものである。
【0156】
本発明者らはまた、Wntリガンドの効果がより多くのニューロンの数をもたらすかどうか、およびこれらの効果が増殖性クラスター内部または外部のドーパミン作動性ニューロンに特異的かどうかを問うた。本発明者らは、Wnt-1または-3aが、TuJ1+細胞を含む増殖性クラスターの数を、対照、Wnt-5aまたはVM T1Aに比べて10〜12倍増加させたことを見出した(図3d)。Wnt-1および-3aの両方の、ニューロンを発生させることができる増殖性クラスターの数を増加させる能力にもかかわらず、Wnt-1のみが培養物中のTuJ1+ニューロンの総数を増加させた(図3f)。さらに、Wnt-1は、TH+ニューロン(図3g)、またはドーパミン作動性ニューロンを含むVMクラスターの比率を増加させず(クラスターの約30%)、そしてWnt-3aは、ドーパミン作動性ニューロン(図3g)およびクラスター(図3e)の比率を10%未満に減少させさえした。本発明者らの知見は、Wnt-1およびWnt-3aが、全てのVMニューロンの数を、増殖および増殖性クラスターの数を増加させることにより増加させることを示したものである。Wnt-1はVMニューロンの総数を、増殖的なメカニズムにより非選択的に増加させる一方、Wnt-3aは、クラスターとして増殖しているニューロン前駆細胞の増殖を増加させ、これらのTH+細胞への分化を抑制したが、これは、これらが、ドーパミン作動性前駆体の自己再生において役割を果たし得ることを示唆するものである。Wnt-1および-3aに対して、Wnt-5aおよびVM T1Aは、TuJ1+ニューロン(図3f)またはニューロンを含む増殖性クラスターの数を増加させなかったものの(図3d)、代わりにドーパミン作動性クラスターおよびドーパミン作動性ニューロンについて、2倍を超えて選択的に富化したが(それぞれ図3eおよび3g)、これは、Wnt-5aおよびVM T1Aが、VM培養物におけるドーパミン作動性表現型の獲得を指示または促進することに関与し得ることを示すものである。Wntの、クラスター外のTuJ1+ニューロンに対する効果は、Wnt-3aを除きクラスター内のニューロンに対するものと極めて類似しており、Wnt-3aはTuJ1+ニューロンの数を増加させなかったが(図3f)、これは、Wnt-3aがクラスター内の前駆体の増殖および/または自己再生を調節し、これを行うことによりドーパミン作動性表現型を獲得するニューロンの比率を減少させていることを示すものである(図3g)。
【0157】
したがって、本発明者らの結果は、Wntが、VMにおける相異なる機能:前駆体の増殖および/または自己再生、ニューロンの数、および/またはドーパミン作動性表現型を獲得するニューロンの比率、を独立かつ示差的に調節していることを示唆したものである。さらに、Wnt-1、-5aおよびVM T1Aの、ドーパミン作動性ニューロンおよび細胞クラスターの数に対する同様の効果にもかかわらず、本発明者らの結果は、Wnt-1およびWnt-5aが、ドーパミン作動性細胞系統に、2つの部分的に相異なるメカニズムを介して作用することを示すものである。Wnt-1はドーパミン作動性ニューロンの比率ではなく、細胞の増殖を増大させる一方、Wnt-5aおよびVM T1Aは、増殖にはあまり作用しないが、その代わり、ドーパミン作動性ニューロンの比率を増殖性クラスターの内部および外部で、追加のメカニズムにより増大させる。
【0158】
VM T1Aがドーパミン作動性誘導シグナルの供給源であるという前提のもと(Wagnerら、1999)、本発明者らは、WntまたはVM T1Aによる処置が、腹側中脳のNurr1+ニューロン前駆体(Nurr1+/TH-細胞)のドーパミン作動性ニューロン(Nurr1+/TH+細胞)への変換に影響を与えるかどうか検討した。対照条件においては、全Nurr1陽性細胞の約50%がTHを発現したが、Wnt-3aによる処置はこの集団を30%に減少させる。これに対し、Wnt-1はNurr1+/TH+の比率を70%まで増加させ、そしてWnt-5aまたはVM T1Aはこの比率を90%まで増加させた(図4A)。これらの結果は、ADH-2/TH二重免疫染色により得られたものと極めて類似していたが、これは、ADH2もドーパミン作動性前駆体およびニューロンを標識したからである(Wagnerら、1999)。したがって、本発明者らのデータは、Wnt-5aが、Wnt-1とは異なり、主にNurr1発現前駆体にドーパミン作動性表現型を誘導することにより、ドーパミン作動性ニューロンの数を増加させるモデルを示唆したものである。このモデルを試験するために、本発明者らは、VM以外の脳構造に由来するNurr1陽性/TH陰性細胞も、WntまたはVM T1Aのいずれかによりドーパミン作動性表現型を獲得するよう誘導されうるかどうかを検討した。Nurr1+前駆細胞に富む、E13.5の大脳皮質からの初代培養物を、種々のWntリガンドに暴露するか、またはP1のVMもしくは(対照としての)皮質のアストロサイトと共に共培養した。Wnt-3aによる処置または皮質のT1Aとの共培養のもとでは、THを共発現したNurr1陽性の皮質細胞は実質的に存在しなかった。しかしながら、Wnt-1、Wnt-5a、またはVM T1Aによる処置は、THを発現したNurr1陽性細胞の比率の有意な増加を誘導した(図4B)。最も劇的な効果は、Wnt-5aによって誘導され、これは、わずか3日間で、二重Nurr1/TH+細胞の数を、対照に比べて>40倍、そしてWnt-1に比べて>4倍増加させたが、この効果は増殖のみでは説明できないものである。さらに、Wnt-5aが腹側部のドーパミン作動性表現型の獲得を促進するという知見は、先に報告された、Wnt-5aの、非古典的な(non-canonical)Wntシグナル伝達を介した腹側部の運命を促進する役割(Saneyoshiら、2002)とよく相関する。
【0159】
本発明者らはまた、Wnt-5aによる処置が、中脳ドーパミン作動性ニューロンに特徴的な遺伝子の発現を調節することにより、前駆体培養物においてより分化した中脳DA表現型の獲得を促進するかどうか検討した。本発明者らはまず、ビコイド(bicoid)関連ホメオドメイン遺伝子Ptx3(Smidtら、1997)の発現を、リアルタイムRT-PCRにより検討し、そしてWnt-1ではなく、Wnt-5aによる処置がPtx3のmRNAを増大させたことを見出した(図6A)。この知見は、Ptx3がDAニューロンの発生に必要であり(Van Den Munckhof Pら、2003; Nunes Iら、2003)、そして同じファミリーの別のホメオドメイン遺伝子であるPtx2が、Wntシグナル伝達により直接調節されていることから(Kioussi Cら、2002; Baek SHら、2003)、極めて興味深いものである。本発明者らは次に、癌原遺伝子c-ret(Trupp Mら、1996)ならびに2種類の他のGDNF補助受容体、GFRα1(AiraksinenおよびSaarma、2002)およびNCAM(Paratchaら、2003)の発現も、Wntにより調節されているかどうか検討した。興味深いことに、c-retおよびNCAMともWntシグナル伝達により調節され(Zheng Sら、1996; Conacci-Sorrellら、2002)、そしてこれらのリガンドであるGDNFは、DAニューロンの出生後の発生に必要である(Granholmら、2000)。本発明者らは、分析した全てのGDNF受容体が、Wnt-1およびWnt-5aにより示差的に調節されていることを見出した。c-retの発現は、Wnt-1ではなWnt-5aによってアップレギュレーションされる一方(図6B)、GFRα1およびNCAMの発現は、Wnt-5aにより維持され、そしてWnt-1により抑制された(図6Cおよび6D)。したがって、本発明者らの結果は、Wnt-5aが、中脳DAニューロンの2つの弁別的な特徴であるPtx3およびGDNF受容体の発現を増大させることにより、前駆細胞におけるDA表現型の獲得を促進すること、ならびにこれらの分化および生存を増強することに用いられ得ることを示すものである。したがって、Wnt-5aは、DAニューロンの数を増加させることができる。
【0160】
Wnt-5aの生物学的活性が、測定したパラメーターの全てにおいてVM T1Aとほぼ同一であったという知見(図1、図2、図3および図4)、およびVM T1AがWnt-5aを発現するという知見(図1F)は、VM T1Aの効果が、一部アストロサイトによるWntの分泌によりうることを示したものであり、本発明者らにWntの神経幹細胞培養物における機能を検討することを促した。本発明者らは、したがって、Wntの効果をブロックするFrizzled-8のシステインリッチドメイン(Fz8-CRD)(Hsiehら、1999)を過剰発現する線維芽細胞からの馴化培地が、VM T1AまたはWnt-5aの、ドーパミン作動性ニューロンの誘導に対する活性をブロックすることができるかどうか検討した。本発明者らはFz8-CRDの、VM T1Aと共培養しているNurr1発現神経幹細胞(Nurr1-c17.2-c42、Wagnerら、1999)への添加が、ドーパミン作動性ニューロンの誘導を部分的にブロックしたことを見出した。さらに、Fz8-CRDは、発生中のマウス脳(Yeら、1998)およびNurr1発現マウス胚性幹細胞に由来するニューロスフェア(Kimら、2002)を包含する、アストロサイトを含有する調製物において、ドーパミン作動性ニューロンをShhとの組み合わせで誘導することが知られている因子であるFGF8により拡大させたニューロスフェアとして増殖させた、VM神経幹細胞におけるドーパミン作動性ニューロンの数を減少させた(図5a)。同様に、E14.5のVM前駆体のFz8-CRDによる共処置は、未処置(図5b)およびVM T1A(図5c)またはWnt-5aにより処置した培養物(図5d)におけるNurr1+前駆体からのドーパミン作動性ニューロンの誘導をブロックした。したがって本発明者らの結果は、Wntが発生に関する他のシグナルと協力して作用し、そして部分的に前駆細胞/神経幹細胞培養物におけるニューロンドーパミン作動性表現型の獲得に必要であることを示すものである。したがって、本発明者らの研究は、Wntが腹側中脳における神経発生の重大かつ順次的な2つの側面である、前駆体の増殖およびドーパミン作動性表現型の獲得の、本質的なレギュレーターであるモデルを示唆するものである(図5F)。このように、Wntは、VM前駆体の運命決定を調節するものであると思われる。Wnt-1は、前駆体の増殖の増大により神経発生を促進し、かつDAおよび非DA VMニューロンの両方に影響を与えた。Wnt-3aは、Nurr1+前駆体の増殖または維持および/または自己再生を促進し、かつDAニューロンの数を減少させた。これに対し、Wnt-5aは弱いマイトジェンであり、Ptx3およびGDNF受容体の発現を増大させ、かつNurr1発現前駆体におけるDA表現型の獲得を効果的に促進した。
【0161】
これらの結果は、ドーパミン作動性前駆体が、Wntに極めて特異的な様式で応答することを明確に示している。Wnt-1、-3a、および-5aは、中脳DAニューロンの発生を、一部重複するメカニズムにより示差的に調節しており、これは、DA前駆体の増殖を促進すること(Wnt-1≧Wnt-3a>Wnt-5a)、これらの分化を抑制すること(Wnt-3a)、神経発生を拡大すること(Wnt-1)、および中脳ドーパミン作動性表現型の獲得を促進すること(DA前駆体のDAニューロンへの分化を促進すること)(Wnt-5a>Wnt-1)を包含する。Wnt-1と同様に、Wnt-7aは前駆体集団を拡大し、そして細胞周期をG1-S移行において促進することによりTH+細胞の数を増加させた。Wnt-2は、前駆体の低減した増殖および細胞周期のG1での停止を伴う、異なるメカニズムにより、DAニューロンの数を増加させた。
【0162】
神経幹細胞における中脳ドーパミン作動性表現型の獲得を調節することに関するWntの役割について、本発明者らは、アストロサイト由来のシグナルが、中脳ドーパミン作動性ニューロンのものを包含する、Nurr1発現前駆体における組織特異的なニューロン表現型を誘導することにより、神経発生において決定的な役割を果たすことを先に示した(Wagnerら、1999、WO00/66713)。興味深いことに、海馬のアストロサイトの、成人における神経発生への同様の役割が最近報告された(Songら、2002)。本明細書で、本発明者らは、アストロサイト由来のシグナルが、Nurr1発現前駆体に対して部分的に重複する相異なる機能を発揮するWntリガンドのファミリーのメンバーを含むことを示した。Wnt-1は、ニューロン前駆体の増殖を増大させることにより神経発生を促進し、かつ全てのVMニューロンに影響を与えた。Wnt-3aは、Nurr1+前駆体の増殖および/または自己再生を促進し、かつドーパミン作動性ニューロンの数を減少させた。これに対し、Wnt-5aはマイトジェンとしてはWnt-1および-3aより有効ではなかったが、Nurr1発現前駆体においてドーパミン作動性表現型を誘導することにおいて最も有効であった。さらに、Fz8 CRDが、Nurr1発現前駆体ばかりでなくFGF8により拡大したニューロスフェアにおいてもドーパミン作動性ニューロンの誘導をブロックしたという知見は、Wntがドーパミン作動性ニューロンの誘導に必要であることを示しており、FGF8の誘導効果がWntによって媒介されている可能性があることを示すものである。この可能性は、FGF8の、発生中の中脳-後脳においてWntの発現およびオーガナイザー活性の両方を誘導する既知の能力と合致するものである(総説として、WurstおよびBally-Cuif、2001;およびRhinnおよびBrand、2001を参照)。
【0163】
Wntを、ドーパミン作動性前駆体/幹細胞の拡大および中脳ドーパミン作動性ニューロンの誘導における重要な細胞外分子プレーヤーとして同定する本発明者らの結果は、パーキンソン病における細胞置換のための、幹細胞に由来するドーパミン作動性ニューロンの有効かつ大規模な産生への扉を開くものである(BjorklundおよびLindvall、2000; PriceおよびWilliams、2001; Arenas、2002; RossiおよびCattaneo、2002; Gottliebら、2002)。したがって、Wntリガンドおよび腹側中脳アストロサイトまたは初期グリア細胞に由来する追加のシグナルは、腹側中脳のドーパミン作動性ニューロンを誘導するのに用いることができる。本発明は、さらに、有効な幹細胞治療の将来的な改善に必要な追加の成分の同定を提供する。
【0164】
神経変性疾患の治療
本発明のニューロンの、神経変性疾患を治療する能力の確認は、パーキンソン病のin vivoモデルを用いて得る。ドーパミン神経毒素である、6-ヒドロキシドーパミン(6-OHDA)またはMPTPは、ニューロンにより特異的に取り込まれ、そして酸化ストレスならびにドーパミン作動性およびノルアドレナリン作動性ニューロンの消失を引き起こす。また、ラクタシスチンを包含するプロテアソームインヒビター、またはロテノンを包含する殺虫剤の注入が、中脳ドーパミン作動性ニューロンの死を引き起こし、そして実験的にパーキンソン病の特徴を再現することも知られている。
【0165】
Nurr1サブファミリーの核内受容体、たとえばNurr1を発現する細胞であって、in vitroでドーパミン作動性ニューロンに分化したものを、6-OHDAまたはMPTPまたはラクタシスチンまたはロテノンで処置したマウスもしくはその他の非ヒト動物の、黒質および/または線条体に外科的に移植する。これらの細胞の、統合および完全に分化する能力は、LacZおよびEGFPなどのレポーター遺伝子を発現する細胞において、電気生理学的および/または形態学的技法により評価する。神経化学的技法は、カテコールアミンの含量および放出の測定を含む。形態学的分析は、チロシンヒドロキシラーゼ、ドーパミントランスポーターおよびドーパミン受容体、Ptx3、Lmx1bおよびADH-2を包含する、ドーパミン作動性ニューロンに特徴的なマーカー遺伝子の発現、およびシナプス接合の形成を研究することを含み得る。
【0166】
未分化のNurr1+細胞がin vivoでドーパミン作動性の表現型に向けて自発的に分化する能力は、かかる細胞を、軸索を切断した、または6-OHDAもしくはMPTPPもしくはラクタシスチンもしくはロテノンにより処置した動物に、線条体内または黒質内移植することにより評価した。ドーパミン作動性表現型、分化および統合は、上記のとおりに検出した。
【0167】
線条体および/または黒質内に移植されたNurr1+細胞に由来するドーパミン作動性ニューロンの、片側性の毒素(たとえば6-OHDA)による処置によって誘導された運動非対称性、またはMPPの全身投与によって誘導された運動異常のいずれかを救済する能力は、アポモルフィンおよびアンフェタミン試験における旋回行動の評価(Schwarting、R. K.ら、(1996)Progress in Neurobiology、50(2-3)、275〜331)、および/または、階段試験、足踏み試験もしくはシリンダー試験(cylinder test)における熟練した足使いの能力(performance in skilled paw usage)によって確認する。
【0168】
in vivoで基底レベルを超えるNurr1を発現する、宿主由来の内因性の幹細胞、神経幹細胞、始原細胞もしくは前駆細胞または神経細胞を、Wntリガンドのin vivoでの投与後における、ドーパミン作動性ニューロンへ分化するこれらの能力について検討することができる。分析は、上記のとおり、形態学的、生化学的および行動学的レベルでの評価を含んでもよい。腹側中脳のアストロサイト/グリア細胞またはこれらに由来する因子の能力は、上記のとおり、Wntリガンド投与と共に分析してもよい。
【0169】
概括
Wntは、細胞増殖、自己再生、運命決定および分化を調節する糖タンパク質のファミリーである。本発明者らの結果は、β-カテニンがNurr1+DA前駆細胞で発現していること、ならびにWnt-1、-3aおよび-5aがVMにおいて高レベルで存在しており、そして発生の最中に示差的に調節されていることを示すものである。部分的に精製されたWntは、VMの発生を弁別的に調節した:Wnt-3aはNurr1+前駆細胞の増殖および/または自己再生を促進したが、TH+ニューロンの数を増加させなかった。代わりに、Wnt-1および-5aは前駆体培養物におけるラット中脳のDAニューロンの数を、2つの相異なるメカニズムによって増加させた。Wnt-1は、主にNurr1+前駆体の増殖を増大させ、サイクリンD1およびD3をアップレギュレーションし、かつp27およびp57のmRNAをダウンレギュレーションした。これに対し、Wnt-5aは、主としてニューロンDA表現型を獲得するNurr1+前駆体の比率を増加させ、これは、Ptx3およびc-retのmRNAのアップレギュレーションを含んだ。さらに、Frizzled8(Wntインヒビター)の可溶性システインリッチドメインは、Wnt-1およびWnt-5aの、増殖および前駆体培養物におけるDA表現型の獲得に対する効果をブロックし、そしてまた、内因性Wntの、Nurr1発現神経幹細胞およびFGF8拡大VMニューロスフェア培養物におけるドーパミン作動性表現型の獲得に対する効果をもブロックした。
【0170】
本明細書に記載された結果は、発生中のラット中脳における、Wntタンパク質の全ファミリーの複雑な空間的および時間的発現パターンを明らかにするものである。Wnt-1、Wnt-3aおよびWnt-5aに加えて、この広範な発現分析は、Wnt-2、Wnt-4、Wnt-7aおよびWnt-7bを、ドーパミン作動性ニューロンの生成における役割を有する候補として同定した。この研究に記載されている知見は、Wnt-7aが、ドーパミン作動性前駆体の増殖を促進し、そしてこれらのドーパミン作動性ニューロンへの分化を可能にすることにより、Wnt-1と同様の様式で作用することを示すものである。Wnt-7aによる処置での、増大したBrdUの取り込み、増大したサイクリンD1の発現、および低減したcdkインヒビターの発現という本発明者らの観察は、これを強く支持するものである。また、この調査は、Wnt-2による処置の結果が、低減したレベルのBrdUの取り込み、サイクリンD2の発現の増大、および細胞周期インヒビターp27およびp57の発現の大幅な増大であることを明らかにした。これらのデータは、Wnt-2が、研究した他のWntに比べ、細胞周期離脱およびNurr1+前駆体によるドーパミン作動性ニューロン表現型の獲得を有利にする、新規なメカニズムを介して作用することを示すものである。これらの知見は、Wntによる類似しているが、相異なる作用メカニズムを浮き彫りにし、そしてWntが、増殖、自己再生および幹細胞/前駆細胞のDAニューロンへの分化の重要なレギュレーターであることを示すものである。さらに本発明者らは、分析した全てのWntが、前駆細胞/幹細胞の増殖の維持およびニューロンドーパミン作動性表現型の獲得に関してユニークな活性プロファイルを示すことを見出したが、これは、Wntファミリーの全てのメンバーがニューロン変性の処置のための魅力ある標的となり得ることを示唆するものである。
【0171】
したがって、幹細胞、神経幹細胞、前駆細胞、始原細胞または神経細胞の増殖および分化ポテンシャル、Nurr1などの選択遺伝子、未成熟グリア細胞またはアストロサイト、およびWntを利用する本明細書に記載した方法は、神経変性疾患の治療において、所望の神経化学的表現型のニューロンの作製を提供する(たとえばニューロン移植の供給原料として)。中脳ドーパミン作動性ニューロンの誘導は、パーキンソン病を治療する細胞置換戦略に用いることができる。
【0172】
方法
in situハイブリダイゼーション(ISH)
雄および雌の野生型CD-1マウス(25〜35g、Charles River、Uppsala)は、欧州共同体(86/609/EEC)、神経科学学会(1985年1月)のガイドラインに従って飼育、給餌および処置し、そして全ての実験は、地域の倫理委員会により承認を受けた。胚10.5および11.5日からのマウスを取り出し、O.C.T.中に-70℃で迅速に凍結した。胚全体にわたる連続矢状切片(14μm厚)を、ガラス製顕微鏡スライド(StarFrost、KnittelGlaser)上に回収した。ISHは、新鮮な凍結組織に対し、35S標識リボプローブを用いて先に記載されたとおりに行った(Truppら、1997)。mRNAレベルを保全するため、切片を15分間、氷冷4%PFAにて固定し、そしてPBSで2回すすいだ。組織を、0.2MのHCl中で12分間除タンパクし、0.1Mのトリエタノールアミン中の0.25%無水酢酸で20分間アセチル化し、そして増加する濃度のエタノール中で脱水した。スライドを、加湿チャンバー内で16時間、54℃で200μlのハイブリダイゼーションカクテル中の106cpmのプローブと共にインキュベートした。全ての洗浄は、62℃で行なった。1×SSC中の15分間の最初の2回の洗浄、50%ホルムアミド/0.5×SSC中の30分間および1×SSC中の15分間であった。引き続き、37℃での30分間のRNase処置(40μg/ml)および1×SSC中の15分間の2回の洗浄がなされ、その後に、エタノール中での脱水および風乾を行なった。クロロホルムは省略し、脱水時間は30秒〜1分に短縮した。その後、スライドを水で1:1に希釈したNTB-2感光乳剤(Eastman Kodak、Rochester、NY)中に浸し、4℃で6〜8週間感光させ、D19(Eastman Kodak)で現像し、AL-4(Agfa Gevaert、Kista、Sweden)で固定し、そしてチオニンで対比染色した。
【0173】
免疫組織化学
免疫組織化学は、4%パラホルムアルデヒド(PFA)を最後に添加した(postfixed)スライド上で行なった。インキュベーションは、4℃で一晩、希釈バッファー(1%のウシ血清アルブミン、BSA、および0.3%のTriton-X100を含有するリン酸緩衝生理食塩水、PBS)中のマウス抗β-カテニン、1:250(BD Transduction Lab.)およびウサギ抗Nurr1、1:200(Santa Cruz Biotech.)と共に行なった。0.2%Tween-20/PBSでの洗浄に引き続き、切片を希釈バッファー中30分間ブロッキングし、これに2次抗体(全てJacksonからの、Cy2ウマ抗マウスIgGもしくはウマ抗ウサギIgG、またはローダミンウマ抗マウスIgG)、1:200による2時間インキュベーションを続けた。
【0174】
リアルタイムRT-PCRおよび遺伝子発現の定量
マウスおよびヒトWnt1、Wnt3a、Wnt5a、Frizzled8、Ptx3、サイクリンD1およびサイクリンD2およびチロシンヒドロキシラーゼについてのものを含むGenbankのcDNA配列を、プライマーの設計のためにPrimer Express 1.0(PE Applied Biosystems、Foster City、CA、USA)およびPrimer 3(http://www-genome.wi.mit.edu/cgi-bin/primer/primer3_www.cgi)に用いた。以下のオリゴヌクレオチドを用いた:
QuantumRNA古典18S内部標準(米国オースチン、Ambion);
mWnt1フォワード - 5'-CTTCGGCAAGATCGTCAACC-3'(配列番号1);
mWnt1リバース - 5'-GCGAAGATGAACGCTGTTTCT-3'(配列番号2);
mWnt3aフォワード - 5'-GAACGCGACCTGGTCTACTACG-3'(配列番号3);
mWnt3aリバース - 5'-GTTAGGTTCGCAGAAGTTGGGT-3'(配列番号4);
mWnt5aフォワード - 5'-AATAACCCTGTTCAGATGTCA-3'(配列番号5);
mWnt5aリバース - 5'-TACTGCATGTGGTCCTGATA-3'(配列番号6);
チロシンヒドロキシラーゼフォワード - 5'-AGTACTTTGTGCGCTTCGAGGTG-3'(配列番号7);
チロシンヒドロキシラーゼリバース - 5'-CTTGGGAACCAGGGAACCTTG-3'(配列番号8);
Fz8フォワード - 5'-TTGGAAGTGACCTCGCTCCTAG-3'(配列番号9);
Fz8リバース - 5'-GGTTGGGCATGTAAGTGTAGTTGT-3'(配列番号10);
Ptx3フォワード - 5'-AGGGTGGACTCCTACAGATTGG-3'(配列番号11);
Ptx3リバース - 5'-CCGATCCCAGATATTGAAGCC-3'(配列番号12);
サイクリンD1フォワード - 5'-ACCCTGACACCAATCTCCTCAAC-3'(配列番号13);
サイクリンD1リバース - 5'-GTAAGATACGGAGGGCGCACAG-3'(配列番号14);
サイクリンD2フォワード - 5'-ACTGATGTGGATTGTCTCAAAGCCT-3'(配列番号15);
サイクリンD2リバース - 5'-CGTTATGCTGCTCTTGACGGAA-3'(配列番号16);
C-retフォワード - 5'-ATGCACAATTACAGGCTGGTTCT-3'(配列番号17)
C-retリバース: 5'-GTCATTGACCAGGACTACTAGCTGC-3'(配列番号18)
NCAMフォワード 5'-CACTTCGTGTTCAGGACTTCAGC-3'(配列番号19)
NCAMリバース 5'-GGACGAAAATGACAATGAGGATG-3'(配列番号20)
GFRa1フォワード: 5'-GTCTGAGAATGAGATCCCCACAC-3'(配列番号21)
GFRa1リバース: 5'-ACACATTGGATTTCAGCTTCTGAG-3'(配列番号22)
サイクリンD3フォワード: 5'-GGCTATGAACTACCTGGATCGCTA-3'(配列番号23)
サイクリンD3リバース: 5'-ACGGTACCTAGAAGCTGCAATTG-3'(配列番号24)
p21フォワード - 5'-AGCAAAGTGTGCCGTTGTCTCT-3'(配列番号25)
p21リバース - 5'-TCTCCGTGACGAAGTCAAAGTTC-3'(配列番号26)
p27フォワード - 5'-TTAATTGGGTCTCAGGCAAACTCT-3'(配列番号27)
p27リバース - 5'-CTAACCCAGCCTGATTGTCTGAC-3'(配列番号28)
p57フォワード - 5'-GAGGACCAGAACCGCTGGGACTT-3'(配列番号29)
p57リバース - 5'-ACTCGCTGTCCACCTCCATCCA-3'(配列番号30)
GDNFフォワード: 5'-TTTCGATATTGTAGCGGTTCCTGT-3'(配列番号31)
GDNFリバース: 5'-GCCTACCTTGTCACTTGTTAGCCT-3'(配列番号32)
Wnt2フォワード: 5'-AACGTCCCTCTCGGTGGAATC-3'(配列番号33)
Wnt2リバース: 5'-TGTACCACCATGAAGAGCTGACC-3'(配列番号34)
Wnt2b/13フォワード: 5'-CCACCCGGACTGATCTTGTCTACT-3'(配列番号35)
Wnt2b/13リバース: 5'-GGAACCTGAAGCCTTGTCCAA-3'(配列番号36)
Wnt3フォワード: 5'-CAGCGTAGCAGAAGGTGTGAAG-3'(配列番号37)
Wnt3リバース: 5'-ATGGCCAGGCTGTCATCTATG-3'(配列番号38)
Wnt4フォワード: 5'-GCTGTACCTGGCCAAGCTGTC-3'(配列番号39)
Wnt4リバース: 5'-TGGATCAGGCCTTTGAGTTTCTC-3'(配列番号40)
Wnt5bフォワード: 5'-GCCGAGCTCTCATGAACCTACAG-3'(配列番号41)
Wnt5bリバース: 5'-GGCGACATCAGCCATCTTATACAC-3'(配列番号42)
Wnt6フォワード: 5'-GCGGAGACGATGTGGACTTC-3'(配列番号43)
Wnt6リバース: 5'-ATGCACGGATATCTCCACGG-3'(配列番号44)
Wnt7aフォワード: 5'-TGCGTGCCAGTCGAAACAAG-3'(配列番号45)
Wnt7aリバース: 5'-GATATACACCAGGTCAGTGTCCATGG-3'(配列番号46)
Wnt7bフォワード:5'-GCCAACATCATCTGCAACAAGA-3'(配列番号47)
Wnt7bリバース: 5'-CCGATCACAATGATGGCATC-3'(配列番号48)
Wnt8a/8dフォワード: 5'-CAGCGACAACGTGGAGTTCG-3'(配列番号49)
Wnt8a/8dリバース: 5'-CATCCTTCCCTTTCTCCAAACTG-3'(配列番号50)
Wnt1Oaフォワード: 5'-CCACTCCGACCTGGTCTACTTTG-3'(配列番号51)
Wnt1Oaリバース: 5'-TGCTGCTCTTATTGCACAGGC-3'(配列番号52)
Wnt1Obフォワード: 5'-ACGACATGGACTTCGGAGAGAAGT-3'(配列番号53)
Wnt1Obリバース: 5'-CATTCTCGCCTGGATGTCCC-3'(配列番号54)
Wnt11フォワード: 5'-CAAGTTTTCCGATGCTCCTATGAA-3'(配列番号55)
Wnt11リバース: 5'-TTGTGTAGACGCATCAGTTTATTGG-3'(配列番号56)
Wnt15フォワード: 5'-CTGTTCGTACCTGTTGGAAGCA-3'(配列番号57)
Wnt15リバース: 5'-CAGCCGTGTCATAGCGTAGCT-3'(配列番号58)
Wnt16フォワード: 5'-ACCCCCATTCTCAAGGATGACTT-3'(配列番号59)
Wnt16リバース: 5'-CAGTTTCTTGTTCTCCACGCAGTA-3'(配列番号60)
【0175】
18Sを除き、残りの全てのプライマーはEurogentec、Seraing、BelgiumおよびDNA Technology A/S、Aarhus、Denmarkから購入した。
【0176】
全RNAを、P1のラットの腹側中脳、背側中脳および皮質に由来するコンフルエントなT1Aアストロサイト培養物、ならびにE10.5、E11.5、E13.5、E15.5およびP1の腹側中脳および背側中脳、およびE14、E16、E18およびP1の皮質から切除した組織から、RNeasy抽出キット(Qiagen、Hilden、Germany)を用いて単離した。全RNAはまた、Wntで3日間in vitroにて処置したE14.5のVM前駆体培養物(100mmディッシュに細胞750万個)からも抽出した(3重測定)。逆転写のために、1μgの全RNAを最初に1単位のRQ1 RnaseフリーDNAse(Promega、Madison、USA)で40分間処置した。DNAseは、1μlのEDTA 0.02Mの添加と、65℃での10分間のインキュベーションとにより不活化した。0.5μgのランダムプライマー(Life Technologies、Grand Island、NY、USA)を次に添加し、そして混合物を70℃で10分間インキュベートした。次に、各試料を2本のチューブ、cDNA反応チューブと陰性対照チューブとに均等に分割した。次に、1×First-Strand Buffer(Life Technologies、Grand Island、NY、USA)、0.01MのDTT(Life Technologies、Grand Island、NY、USA)および0.5mMのdNTPS(Promega、Madison、USA)を含有するマスターミックスを、cDNAおよびRT-チューブの両方に添加し、そして25℃で10分間インキュベートし、その後、42℃での2分間のインキュベーションを続けた。200単位のSuperscript II逆転写酵素(Life Technologies、Grand Island、NY、USA)を、次に、cDNAチューブのみに加え、全ての試料を42℃で50分間インキュベートした。Superscript IIは、70℃での10分間のインキュベーションにより不活化した。次に、cDNAおよびRT-の両方を、さらなる分析のために10倍に希釈した。リアルタイムPCRは1μlの1:10に希釈したcDNAおよびRT-と共に、総容量25μlで、三重で行なった。各PCR反応は、1×PCRバッファー(Life Technologies、Grand Island、NY、USA)、3mMのMgCl2(Life Technologies、Grand Island、NY、USA)、0.2mMのdNTPs(Promega、Madison、USA)、0.3μMの各フォワードおよびリバースプライマー、1単位のPlatinum Taq DNAポリメラーゼ(Life Technologies、Grand Island、NY、USA)および1×SYBR Green(Molecular Probes、Leiden、The Netherlands)で構成された。PCRは、94℃で2分間、その後94℃で30秒間、60℃で30〜45秒間、72℃で45〜60秒間を35〜40サイクルおよび80℃で15秒間(SYBR Green検出について)、ABI PRISM 5700検出システム(PE Applied Biosystems、Foster City、CA、USA)で行なった。他のアニーリング温度は、Wnt5aについての54℃、p27についての57℃、サイクリンD1についての62℃、サイクリンD2についての61℃およびp57についての65℃を含めた。SYBR Greenシグナルが、ユニークかつ特異的なアンプリコンに一致することを確認するために、各ランの後、各PCR産物について融解曲線を得た。PCR産物の特異性は、シーケンシングによって検証した。
【0177】
標準曲線は、96ウェルプレートのリアルタイムPCRランごとに、逆転写されたRNAまたは各プローブの対象となる配列を含むプラスミドの3倍連続希釈を用いて作成した。得られた標準曲線のプロットを、次に、Cts(所定のテンプレートを設定した蛍光閾値まで増幅するのに必要なPCRサイクル数)を、所定の試料の最初のテンプレートの任意量に変換するのに用いた。
【0178】
発現レベルは、各試料のRT-値を対応するRT+値から減じ、そしてこの数値を、各試料について並行するアッセイで得たハウスキーピング遺伝子18Sの値で除すことにより得た。
【0179】
結果の統計学的分析は、一元配置ANOVAで行なった。種々の発生段階が、最も早い発生段階と異なった特定の時点を、有意な相互作用が生じた場合にのみ同定するために、フィッシャーの制約付最小有意差(protected least significant difference)を事後的に用いた。全ての試験について、有意性はp<0.05のレベルと仮定した(*p<0.05; **p<0.001; ***p<0.0001)。
【0180】
特定の遺伝子についての各アッセイは、3重で、2回または3回反復した。18Sのアッセイは、各試料について、アッセイの開始時、およびアッセイの途中に1回または2回、試料の完全性を検証するために実行した。PCRプライマーの特異性は、プライマー配列のBLASTラン(BLAST run)により決定した。全てのPCR産物は、アンプリコンのサイズを検証するためにゲルに流した。PCR産物の特異性は、無作為試料中のアンプリコンのシーケンシングによって決定した。
【0181】
前駆体の培養および処置
適時に交配させた(timed-mated)Sprague-Dawleyラットから得たE13.5〜14.5の胚からの腹側中脳および大脳皮質を切除し、機械的に解離し、ポリ-D-リジンでコートされた12または24ウェルプレートに、規定無血清培地(defined, serum-free medium)(インスリン(10ng/ml)、トランスフェリン(100μg/ml)、プトレシン(100μM)、プロゲステロン(20nM)、セレン(30nM)、グルコース(6mg/ml)、およびウシ血清アルブミン(1mg/ml)を含有するF12とDMEMとの1:1混合物からなるN2)中、最終密度1×105細胞/cm2でプレーティングした。精製した1型アストロサイトは、P1のラットのVMまたはCTXに由来する混合グリア培養物から、標準的なプロトコル(Wagnerら、1999)にしたがって得た。振盪し、12ウェルプレートに再プレーティングした後、アストロサイトを15%ウシ胎児血清含有培地中でコンフルエントまで増殖させ、N2培地に変え、そのときに新鮮に切除したVMまたはCTX細胞を、アストロサイトの上に1×105細胞/cm2の密度でプレーティングした。固定の4〜6時間前に添加した5-ブロモデオキシウリジン(BrdU)を除き、全ての因子は、培養の開始時に一度に加えた。培養物は、加湿された、C02 5%、空気95%のインキュベータ内で37℃にて維持し、そして所定時間後、4%パラホルムアルデヒドで45分間、免疫細胞化学分析の前に固定した。
【0182】
ニューロスフェアの培養
E13.5のラット胚からの腹側中脳を切除し、そして一緒にプールし、N2無血清培地に再懸濁し、機械的に解離し、そして、事前にポリ-D-リジン(Sigma、Stockholm、Sweden)でコートした24ウェルプレート(BD Bioscience、Erembodegem、Belgium)に、最終密度100〜125×103細胞/cm2でプレーティングした。細胞は、20ng/mlのFGF8b(R&D Systems、Minneapolis、USA)および8μg/mlのヘパリン(Sigma、Stockholm、Sweden)の存在下で増殖させ、そして7〜10日後に、高いスフェア密度で、部分的に精製された馴化培地、およびタンパク質125〜250μg/mlのFz8CRDが添加されたN2に再プレーティングした。ニューロスフェアは、その後5〜7日間分化させた。固定および免疫組織化学分析は、上述のとおりに行なった。
【0183】
Wnt馴化培地の調製、特性決定および精製
ヘマグルチニンタグ付Wnt-1a、3a、または5aを安定的に過剰発現するB1A線維芽細胞系(Shimizu H.、1997)を、1OOμg/mlのG-418が添加された標準的な完全培地(DMEM+10%FBS)中で増殖させた。馴化培地の回収のために、細胞を完全倍地中に低密度で再プレーティングし、そして50〜70%コンフルエントに到達させ、その時点で細胞を洗浄し、培地を無血清N2(1OμMの酪酸ナトリウムを含む)と24時間置換した。次に、シスター(sister)フラスコからの馴化培地を採取し、ロットとしてプールし、-80℃で2カ月までの間保存した。この回収および保存のルーチンを用いた場合、新鮮なCMと比べ活性の喪失は観察されなかった。T1A CMは、培地をin vitroで3日後に回収することを除き、同様の手順で採取した。濃縮のため、CMの個々のロットを室温で解凍し、80mlのアリコートに分割し、Centricon-Plus 80カラム(Millipore)にローディングし、そして製造者の指示に従い、遠心分離により濃縮した。濃縮に続き、アリコートを再プールし、そして、タンパク質含量の決定およびウェスタンブロット分析のための試料を取ってから、-80℃で凍結させた。簡単に述べると、20μgのタンパク質を10%ポリアクリルアミドミニゲルにローディングし、変性条件下150Vで約30分間流した。PVDF膜(Hybond P、Amersham Pharmacia Biotech、Uppsala、Sweden)へのドライエレクトロブロットトランスファー、およびトリス緩衝生理食塩水中の3%ウシ血清アルブミンとX%のTriton-X100とからなるブロッキングバッファー(TBST)中での30分間のプレインキュベーションの後、ブロットをTBST中1:1000に希釈したマウス抗ヘマグルチニン(Babco)と共に一晩4℃でインキュベートした。洗浄後、ブロットを、TBST中1:1000に希釈したアルカリホスファターゼ結合ヤギ抗マウスIgG(Santa Cruz)と共に、室温で1時間インキュベートした。ブロットをAmersham ECF試薬で視覚化し、そして青色蛍光をMolecular Devices Storm 840 phosphoimagerで定量した。
【0184】
Fz8CRD馴化培地の調製、特性決定および精製
B1A線維芽細胞系を、mFz8CRD-IgG/pRK5およびpIRES puro 2(Clontech、BD Bioscience、Erembodegem、Belgium)プラスミドで、Lipofectamine Plus試薬(Invitrogen、Lidingo、Sweden)を用い、製造者の指示に従って共トランスフェクトした。ピューロマイシン耐性(1μg/ml Sigma、Stockholm、Sweden)について選択し、クローンを単離し、そして増殖させた。Fz8CRD 700xを過剰発現するクローン(リアルタイムRT-PCR分析)を同定した。Fz8CRD馴化培地の採取および部分精製を上述のとおりに行なった。行なった全てのブロッキング実験において、Fz8CRDおよびB1Aの対照部分精製馴化培地を、125〜250μg/mlの最終濃度で用いた。
【0185】
免疫細胞化学分析
固定した培養物を、1%ウシ血清アルブミンおよび0.3%Triton-X100を含有するリン酸緩衝生理食塩水(PBS)で適切に希釈した以下の抗体の1つと共にインキュベートした:マウス抗BrdU、1:50(DAKO、Denmark)、マウス抗βチューブリン、III型(TuJ1)、1:250(Sigma)、マウス抗TH、1:1000(Incstar、USA)、ウサギ抗TH、1:250(PelFreeze)、ウサギ抗Nurr1、1:2000(Dr. T. Perlmann、Swedenからの寄贈)、ウサギ抗Nurr1、1:1000(Santa Cruz、USA)またはマウス抗Nurr1、1:250(BD Transduction Laboratories、USA)、ウサギ抗Adh2、1:4000(Dr. R. Lindahl、South Carolinaからの寄贈)。インキュベーションは、4℃で一晩か、または室温で1時間行なった。両方の方法とも同様の結果をもたらした。洗浄後、培養物を1〜3時間、同じ希釈バッファー中の適切な2次抗体(ビオチン化1:500; CY2-、FITC-、またはローダミン結合ウマ抗マウスIgG 1:100;またはヤギ抗ウサギIgG 1:100;全てVector、USAから)と共にインキュベートした。明視野免疫染色は、Vector Laboratory ABC免疫ペルオキシダーゼキットによりNovaRed(赤)、またはAEC(赤)、SGもしくは3-3'ジアミノベンジジン四塩酸塩(DAB 0.5mg/ml)/塩化ニッケル(1.6mg/ml)(灰/黒)、またはVIP(紫)基質のいずれかを用いて視覚化した。二重染色は、記載のとおりに順次的な単一染色により行なった。染色の順序は、BrdU二重標識についてのみ重要であり、この場合、BrdU手技は常に2番目に行なった。用いた1次または2次抗体のいずれかを欠いた対照実験は、用いた抗体ペアの間に交差反応性がわずかにあるか、またはないことを示した。写真はZeiss Axioplan 100M顕微鏡で得、そして浜松ホトニクス社製カメラC4742-95(QED画像化ソフト付)に集めた。
【0186】
データ分析および統計
明瞭に染色された細胞のみを陽性細胞として計数した。スフェアは、これらが1個またはそれ以上の陽性細胞を含む場合に陽性とみなした。個々の細胞についての定量的免疫細胞化学のデータは、他に別記のない限り、盲検化された観察者により、3〜4回の独立した実験からの、条件あたりそれぞれ3〜4ウェルにおける10〜20の重複しない視野(細胞)からの、またはウェル全体における(クラスター)得られた計測数の平均値および標準誤差を表す。各変数について、初期の統計学的比較は、用量、時点、処置および/または領域の、複数の要因によるグローバルANOVA(global ANOVA)により行い、処置と他の任意の変数との間に有意な相互作用が存在する場合は、データを、別個の時間、用量または領域にさらに分割した。フィッシャーの制約付最小有意差は、処置が対照から(または相互に)異なった特定のポイントを、要因である処置と他の変数との間に有意な相互作用が生じたときのみ同定するために、事後的に用いた。全ての試験についての有意性は、p<0.05のレベルを前提とした。結果部分における表示を簡略化するために、個々の統計分析の結果、ならびに行なった分析の種類は、個々の図の凡例中に表示する。
【0187】
参考文献
【図面の簡単な説明】
【0188】
【図1】図1は、Wntリガンドが発生中の中脳で示差的に発現していることを示す。図1aは、Wnt-1が腹側中脳および背側中脳で高レベルに発現されているが、Wnt-3aの発現が背側中脳で、およびWnt-5aが腹側中脳で優勢であることを明らかにしたリアルタイムPCR分析の結果を示す。図1b、図1c、図1dおよび図1eは、腹側中脳内において、Wnt-1(図1d)およびWnt-5a発現(図1e)のドメインが、ドーパミン作動性前駆体およびドーパミン作動性ニューロンを標識するNurr1(図1c)およびチロシンヒドロキシラーゼ(TH)(図1b)のものと合致することを示したin situハイブリダイゼーションの結果を示す。図1fは、出生後1日の腹側中脳(VM)から単離した1型アストロサイトが、背側中脳(DM)または大脳皮質(CC)よりも有意に高いレベルのWnt-5a mRNAを発現することを実証するin situハイブリダイゼーションの結果を示す。各脳領域について、フィッシャーの事後検定を用いた一元配置ANOVAによりE10.5に対して比較した場合、*p<0.05; **p<0.001; ***p<0.0001である。
【図2】図2は、ラットのE14.5の腹側中脳(VM)培養物において、Wnt-1およびWnt-5aは、ドーパミン作動性ニューロンの数を増加させ(図2a、図2bおよび図2c)、ドーパミン作動性ニューロン(図2eおよび図2f)を含むクラスターを増殖させたが、Wnt-3aはこれらをしなかったことを示す。図2aおよび図2eは用量依存性を示し、図2bおよび図2fは経時変化分析を示し、そして図2cはWntリガンドの効果の、対照(N2)、グリア細胞系由来神経栄養因子(GDNF)、線維芽細胞成長因子2および8(FGF-2および-8)、VM1型アストロサイト(T1A)および対照精製培地(control purified media)(CP)との比較を示す。結果は、部分的に精製されたWntは、活性があり、安定であり、および培養中のドーパミン作動性ニューロンの数の増加に関して、VM T1Aと同程度に有効であり得ることを示している。図2bおよびc:フィッシャーの事後検定を用いた一元配置ANOVAによりN2およびCPと比較した場合、それぞれ*、**p<0.01である。図2e:フィッシャーの事後検定を用いた一元配置ANOVAによりCPと比較した場合、*p<0.01である。チロシンヒドロキシラーゼ(TH)の免疫染色培養物は、Wnt-1およびWnt-5aが、ドーパミン作動性クラスターの外部または内部でのドーパミン作動性ニューロンの数の極めて劇的な増加を誘導することを示した。視野は3.14mm<SUP>2であり、ウェルは4cm2である。
【図3】図3は、Wnt-1が前駆体の増殖およびニューロンの総数を増大させたが、Wnt-5aとは異なり、ドーパミン作動性表現型を獲得したドーパミン作動性前駆体の比率は増大させなかったことを示す。図3aは、Wnt-1は、サイクリンD1のmRNAの発現を増大させたが、Wnt-5aはそれをしなかったことを示す。図3bは、Wnt-1およびWnt-3aが、BrdUを取り込んだNurr1免疫染色細胞の比率の3倍の増加を誘導したが、Wnt-5aおよび腹側中脳の1型アストロサイト(VM T1A)からの馴化培地は、Nurr1+細胞におけるBrdUの取り込みをより少ない程度で増加させたことを示す。図3dは、Wnt-1およびWnt-3aが、Tuj1陽性ニューロンを含む増殖性クラスターの数を増大させたことを示す。図3eは、ニューロンを含む増殖性クラスターにおけるドーパミン作動性ニューロンの比率は、Wnt-1処置の後では変化せず、Wnt-3aにより減少し、Wnt-5aおよびVM T1Aでの処置により増大したことを示す。図3fは、Wnt-1のみが、増殖性クラスター外の個々のTuJ1陽性ニューロンの数を増加させたことを示す。図3gは、クラスター外のニューロン数の増加にもかかわらず、Wnt-1は、ドーパミン作動性ニューロンの比率を増大させなかったことを示す。一方、Tuj1陽性ニューロンの数を変化させなかった処置は、ドーパミン作動性ニューロンの比率を減少(Wnt-3a)、または増大させた(Wnt-5aまたはVM T1A)。フィッシャーの事後検定を用いた一元配置ANOVAにより対照精製培地(CP)と比較した場合、*p<0.05; **p<0.001; ***p<0.0001である。
【図4】図4は、Wnt-5aが、Nurr1+細胞においてドーパミン作動性表現型を誘導するのに最も有効な因子であることを示す。図4aは、Wnt-5aまたはVM T1A処置が、ドーパミン作動性表現型を獲得したVMにおけるNurr1発現細胞の比率を、対照条件における50%から90%に増大させたことを明らかにした、チロシンヒドロキシラーゼ(TH)とNurr1との二重免疫組織化学の結果を示す。一方、増殖を誘導するのに最も効果的な2因子、Wnt-1およびWnt-3aは、Wnt-5aより有効ではないか(Wnt-1)、またはドーパミン作動性細胞の比率を50%から30%に減少させさえした(Wnt-3a)。図4bは、同様に、Wnt-5aおよびVM T1Aが、Nurr1を発現したE13.5の皮質前駆体培養物におけるドーパミン作動性表現型の獲得を促進するのに最も効果的な処置であったことを示す。Wnt-1はWnt-5aよりはるかに低い効果を有し、そしてWnt-3aまたは皮質1型アストロサイト(CTX T1A)は、THを発現するNurr1+細胞の比率を変化させなかった。フィッシャーの事後検定を用いた一元配置ANOVAにより対照精製培地(CP)と比較した場合、*p<0.05; **p<0.001; ***p<0.0001である。
【図5】図5は、Wntシグナル伝達がドーパミン作動性ニューロンの発生に必要であることを説明する。図5aは、FGF8で増殖させたE13.5のVMニューロスフェアが、5〜7日でグリアおよびニューロン系統に分化し、スフェアの12%にドーパミン作動性ニューロンを生じさせたことを示す。これに対し、培養培地へのFz8 CRDの添加は、ドーパミン作動性ニューロンを含むニューロスフェアの数を、対照(CM=CP)に比べ減少させた。同様に、E14.5のVM前駆体培養物のFz8 CRD過剰発現線維芽細胞からの馴化培地による処置(図5b、図5cおよび図5d)は、チロシンヒドロキシラーゼの発現を獲得したNurr1免疫反応性細胞の比率を、対照条件にて(CM、図5b)、または腹側中脳の1型アストロサイトからの馴化培地(VM T1A、図5c)もしくはWnt-5a(図5d)による処置の後に減少させた。フィッシャーの事後検定を用いた一元配置ANOVAにより対照(N2)または馴化培地(CM)と比較した場合、*p<0.05; ***p<0.0001であり、VM T1AまたはWnt-5aと比較した場合、#p<0.05; ###p<0.0001である。図5fは、Wnt-1、-3aおよび-5aが、VMドーパミン作動性ニューロンの発生を調節するメカニズムのモデルである。Wnt1は、恐らく中脳-終脳のオーガナイザーに由来し、Nurr1発現前駆体の増殖を制御し、そして、VMニューロンの数を増加させる。Wnt-5aは、VMアストロサイトによって発現され、Nurr1発現前駆体におけるドーパミン作動性表現型の誘導を調節することによりVMドーパミン作動性ニューロンの数を特異的に増加させる。最後に、Wnt-3aは、主に背側中脳で発現され、Nurr1発現前駆体の増殖および/または自己再生を増進させ、そしてドーパミン作動性表現型を獲得するニューロンの比率を減少させた。疑問符は、Wnt-1およびWnt-3aの正確な細胞供給源が不明であることを示す。矢印の大きさは効果の強度に相関することに留意されたい。
【図6】図6は、Wntがドーパミン作動性ニューロンの発生を、前駆体の増殖およびDA表現型の獲得を調節することにより、示差的に制御することを示す。リアルタイムRT-PCRで評価したところ、Wnt-5aは、in vitroで3日目に、Ptx3のmRNA(A)およびc-ret(B)の発現をアップレギュレーションし、GFRα1(C)およびNCAM(D)の発現を維持したが、Wnt-1はこれらをしなかった。
【図7】図7は、Wnt-1がサイクリンD1および細胞周期インヒビターp27およびp57の発現を調節し、VM前駆体の増殖を増大させ、そしてTHニューロンの数を特異的に増加させることを示す。リアルタイムRT-PCRは、in vitroで3日目に、Wnt-1は、サイクリンD3のmRNAの発現を増加させ(A)および細胞周期インヒビターp27(B)およびp57(C)の発現を減少させたが、Wnt-5aがこれらをしなかったことを示した。Dに示すように、Wnt-1およびWnt-3aは、in vitroで3日目に、VM前駆体の増殖を増大させたが、Wnt-5aはこれをしなかった。Eに示すように、Wnt遮断試薬である部分的に精製されたFz8-CRDの増大する単位は、in vitroで3日後、VM培養物におけるTH+ニューロンの数を用量依存的に減少させたが、これは内因性WntがDAの発生に必要であることを示している。Fに示すように、Wnt-1によるTH+ニューロンの数の増加は、Fz8-CRDにより部分的にブロックされたが、これは、Wntの効果が特異的であることを示唆している。統計学:フィッシャーの事後検定を用いた一元配置ANOVAにより対照と比較した場合、・p<0.05;*p<0.01;**p<0.001;***p<0.0001であり、そしてWnt処置のみと比較した場合、###p<O.0001である(n=3〜5、Fz8-CRDについては例外的に、n=2〜3)。濃度:10単位/μlのWntまたはCP、および5単位/μlのFz8-CRD。視野は3.14mm<SUP>2である。
【図8】図8は、発生中の中脳におけるWntの発現を示す。リアルタイムRT-PCR分析は、Wnt-2(8a)、Wnt-4(8b)、Wnt-7a(8c)、およびWnt-7b(8d)の転写産物が、ドーパミン作動性ニューロン誕生時の腹側中脳領域で優勢であることを明らかにした。
【図9】図9は、発生中のCNSにおけるWntの発現を示す。リアルタイムRT-PCR分析は、Wnt-3(9A)、Wnt-6(9B)、Wnt-1Ob(9C)、Wnt-11(9D)、およびWnt-16(9E)の発現が、背側中脳領域およびCNSのその他の領域に、より特異的であることを示した。
【図10】図10は、Wnt-2およびWnt-7aが、ラットのE14.5の腹側部前駆体培養物におけるドーパミン作動性ニューロンの数を増加させたことを示す。Wnt-2およびWnt-7aによる処置は、培養物(10A)およびNurr1発現前駆体(10B)におけるチロシンヒドロキシラーゼ陽性ニューロンの総数を両方とも増加させた。Wnt-7aは、腹側部前駆体の増殖を増大させたが(10C)、Wnt-2による処置は、初代培養物におけるBrdUの取り込み量の減少をもたらした(10C)。
【図11】図11は、細胞周期レギュレーターへのWntの種々の効果を示す。Wnt-7aは、サイクリンD1のmRNAを増加させたが、cdkインヒビターp27およびp57をダウンレギュレーションした(11A、11Dおよび11E)。Wnt-2による処置では、サイクリンD2の発現の増加と共に、p27およびp57のmRNAの増加が観察された(11B、11Dおよび11E)。
【特許請求の範囲】
【請求項1】
神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、ドーパミン作動性の誘導、生存、分化および/もしくは成熟を増強させることによってドーパミン作動性ニューロンの発生を誘導させるまたは促進する方法であって、
前記細胞内でNurr1サブファミリーの核内受容体を基底レベルより高く発現させ、
前記細胞をWntリガンドで処理することによって、
増殖、自己再生、生存および/もしくはドーパミン作動性の誘導、分化、生存またはニューロンのドーパミン作動性の表現型の獲得を起こさせるかあるいは増強させることを含む方法。
【請求項2】
前記核内受容体がNurr1である、請求項1に記載の方法。
【請求項3】
前記核内受容体がNor1またはNGFI-Bである、請求項1に記載の方法。
【請求項4】
細胞をNurr1のDNAで形質転換させることまたは前記細胞内にNurr1のRNAを導入することによってNurr1を基底レベルより高く発現させることを含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
前記細胞内にNurr1タンパク質を導入することによってNurr1を基底レベルより高く発現させることを含む、請求項1〜3のいずれか1項に記載の方法。
【請求項6】
前記細胞内にNurr1タンパク質を保存することによってNurr1を基底レベルより高く発現させることを含む、請求項1〜3のいずれか1項に記載の方法。
【請求項7】
前記WntリガンドがWnt1リガンドである、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
前記WntリガンドがWnt5aリガンドである、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
前記WntリガンドがWnt3aである、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
前記WntリガンドがWnt-2である、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記WntリガンドがWnt-4である、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記WntリガンドがWnt-7aである、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
前記WntリガンドがWnt-7bである、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくはニューロン細胞をWnt-1またはWnt-5a以外のWntリガンドまたは追加のWntリガンドで処理する、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞が、前記Wntリガンドで処理した場合に、有糸分裂性であり、かつ/または自己再生する能力を有する、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞をさらにFGF成長因子ファミリーのメンバーと接触させる、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞をレチノイドもしくはレチノイド誘導体、レチノイドX受容体(RXR)の活性化因子、レチノイド酸受容体(RAR)のリプレッサー、9-シスレチナール、DHA、SR11237、またはLG849と接触させる、請求項1〜16のいずれか1項に記載の方法。
【請求項18】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞をWntリガンドで処理する前にあるいはそれと同時に、前記細胞をbFGFおよび/またはEGFおよび/またはFGF-8および/またはLIFおよび/またはShhで処理する、請求項1〜17のいずれか1項に記載の方法。
【請求項19】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞を抗酸化剤、アスコルビン酸、低酸素圧または低酸素誘導因子の存在下で増殖させる、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞が腹側中脳のアストロサイトまたは初期グリア細胞の存在下で増殖および/または分化する、請求項1〜19のいずれか1項に記載の方法。
【請求項21】
前記Wntリガンドを前記細胞を含むin vitro培養物に加える、請求項1〜20のいずれか1項に記載の方法。
【請求項22】
Wntリガンドが前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞と同時培養した細胞からの発現によって産生され、その同時培養した細胞は1型アストロサイトもしくは初期グリア細胞以外の細胞であるか、または前記Wntリガンドをコードしている核酸で形質転換させた宿主細胞もしくは導入したWntタンパク質を含む細胞である、請求項21に記載の方法。
【請求項23】
前記同時培養した、1型アストロサイトもしくは初期グリア細胞以外の細胞または宿主細胞が別の幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞である、請求項22に記載の方法。
【請求項24】
前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞が、前記Wntリガンドをそれをコードしている核酸から発現するように遺伝子操作されている、請求項21に記載の方法。
【請求項25】
Wntリガンドタンパク質を前記細胞内に導入する、請求項21に記載の方法。
【請求項26】
前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を、初期グリア細胞、または場合によっては腹側中脳由来の1型アストロサイトとさらに同時培養することを含む、請求項1〜25のいずれか1項に記載の方法。
【請求項27】
前記1型アストロサイトが不死化されているか、または前記腹側中脳以外の領域のアストロサイト細胞系由来である、請求項26に記載の方法。
【請求項28】
前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を非ドーパミン作動性ニューロンに対して選択する陰性選択剤とさらに接触させることを含む、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
ニューロンを1つまたは複数の追加の成分を含む組成物に配合することをさらに含む、請求項1〜28のいずれか1項に記載の方法。
【請求項30】
前記組成物が製薬上許容される賦形剤を含む、請求項29に記載の方法。
【請求項31】
前記組成物を個体に投与することをさらに含む、請求項30に記載の方法。
【請求項32】
前記ニューロンを個体の脳内に移植する、請求項31に記載の方法。
【請求項33】
Nurr1の発現を増大させるために前記細胞を個体内でin situで処理する、請求項1〜20のいずれか1項に記載の方法。
【請求項34】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞を個体内でin situでWntリガンドを用いて処理する、請求項1〜20のいずれか1項に記載の方法。
【請求項35】
前記Wntリガンドをコードしている核酸を前記細胞内に導入する、請求項34に記載の方法。
【請求項36】
Wntリガンドタンパク質を前記細胞内に導入する、請求項34に記載の方法。
【請求項37】
Nurr1の発現を基底レベルより高いレベルまで上昇させるために前記細胞を個体内でin situで処理する、請求項34に記載の方法。
【請求項38】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞が前記個体にとって内因性である、請求項33〜37のいずれか1項に記載の方法。
【請求項39】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞が、前記個体への移植によって外因的に供給される、請求項33〜37のいずれか1項に記載の方法。
【請求項40】
前記個体がパーキンソン病、パーキンソン症候群、ニューロンの消失または神経変性疾患に罹患している、請求項31〜39のいずれか1項に記載の方法。
【請求項41】
個体を治療するための医薬の製造において前記方法に従って産生させたニューロンの使用をさらに含む、請求項1〜30のいずれか1項に記載の方法。
【請求項42】
前記医薬が個体の脳内へ移植するためのものである、請求項41に記載の方法。
【請求項43】
前記個体がパーキンソン病、パーキンソン症候群、ニューロンの消失または神経変性疾患に罹患している、請求項42に記載の方法。
【請求項44】
請求項1〜28のいずれか1項の記載に従って産生させたドーパミン作動性ニューロン。
【請求項45】
神経変性疾患の治療に使用するための物質のスクリーニング方法における、請求項44に記載のドーパミン作動性ニューロンの使用。
【請求項46】
(i)ドーパミン作動性ニューロンを前記ドーパミン作動性ニューロンの毒素で処理すること;
(ii)前記ドーパミン作動性ニューロンを前記毒素から分離すること;
(iii)前記処理したドーパミン作動性ニューロンを1つまたは複数の試験物質と接触させること;
(iv)前記ドーパミン作動性ニューロンが前記毒素から回復する能力を決定すること;
(v)前記ドーパミン作動性ニューロンが前記毒素から回復する前記能力を、前記1つまたは複数の試験物質と接触させない場合にドーパミン作動性ニューロンが前記毒素から回復する能力と比較すること、
をさらに含む、請求項1〜28のいずれか1項に記載の方法。
【請求項47】
(i)1つまたは複数の試験物質の存在下でドーパミン作動性ニューロンを前記ドーパミン作動性ニューロンの毒素で処理すること;
(ii)前記ドーパミン作動性ニューロンが前記毒素に耐性を示す能力を決定すること;
(iii)前記ドーパミン作動性ニューロンが前記毒素に耐性を示す前記能力を、前記1つまたは複数の試験物質と接触させない場合にドーパミン作動性ニューロンが前記毒素に耐性を示す能力と比較すること、
をさらに含む、請求項1〜28のいずれか1項に記載の方法。
【請求項48】
ドーパミン作動性ニューロンが前記毒素から回復するまたはそれに耐性を示す能力を向上させる物質を、1つまたは複数の追加の成分を含む組成物に配合することをさらに含む、請求項46または請求項47に記載の方法。
【請求項49】
前記組成物が製薬上許容される賦形剤を含む、請求項48に記載の方法。
【請求項50】
前記組成物を個体に投与することをさらに含む、請求項49に記載の方法。
【請求項51】
前記個体がパーキンソン病、パーキンソン症候群、ニューロンの消失または神経変性疾患に罹患している、請求項50に記載の方法。
【請求項52】
単独でまたは組み合わせて、Nurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化、もしくは成熟を増強させる因子もしくは複数の因子を得る方法であって、
(a)1つまたは複数の試験物質を存在させてまたは存在させずに、Nurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞をWntリガンドで処理すること;ならびに
(b)前記細胞の増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化、もしくは成熟を決定し、前記1つまたは複数の試験物質を存在させた場合または存在させない場合の前記増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化もしくは成熟の程度を比較することによって、前記因子または複数の因子を得ること、
を含む方法。
【請求項53】
前記Wntリガンドを前記細胞を含むin vitro培養物に加えることによって前記細胞を前記Wntリガンドで処理する、請求項52に記載の方法。
【請求項54】
前記Wntリガンドをコードしている核酸を前記細胞内に導入することによって前記細胞を前記Wntリガンドで処理する、請求項52に記載の方法。
【請求項55】
Wntリガンドタンパク質を前記細胞内に導入することによって前記細胞を前記Wntリガンドで処理する、請求項52に記載の方法。
【請求項56】
1型アストロサイトもしくは初期グリア細胞以外の細胞、または前記Wntリガンドをコードしている核酸で形質転換させた宿主細胞もしくは導入したWntタンパク質を含む細胞と同時培養することによって前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を前記Wntリガンドで処理する、請求項53に記載の方法。
【請求項57】
前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を初期グリア細胞または場合によっては腹側中脳由来の1型アストロサイトと同時培養することをさらに含む、請求項52〜56のいずれか1項に記載の方法。
【請求項58】
Nurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化もしくは成熟を増強させる能力を有する因子もしくは複数の因子が、単離かつ/または精製された形態で提供される、請求項52〜57のいずれか1項に記載の方法。
【請求項59】
Nurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化もしくは成熟を増強させる能力を有する因子もしくは複数の因子を、1つまたは複数の追加の成分を含む組成物に配合する、請求項56〜58のいずれか1項に記載の方法。
【請求項60】
前記組成物がNurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を含む、請求項59に記載の方法。
【請求項61】
前記組成物がWntリガンドを含む、請求項60に記載の方法。
【請求項62】
前記WntリガンドがWnt1リガンドである、請求項61に記載の方法。
【請求項63】
前記WntリガンドがWnt5aリガンドである、請求項61に記載の方法。
【請求項64】
前記組成物が製薬上許容される賦形剤を含む、請求項59〜63のいずれか1項に記載の方法。
【請求項65】
前記組成物を個体に投与することをさらに含む、請求項64に記載の方法。
【請求項66】
前記組成物を前記個体の脳内に移植する、請求項65に記載の方法。
【請求項67】
前記個体がパーキンソン病、パーキンソン症候群、ニューロンの消失または神経変性疾患に罹患している、請求項66に記載の方法。
【請求項1】
神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、ドーパミン作動性の誘導、生存、分化および/もしくは成熟を増強させることによってドーパミン作動性ニューロンの発生を誘導させるまたは促進する方法であって、
前記細胞内でNurr1サブファミリーの核内受容体を基底レベルより高く発現させ、
前記細胞をWntリガンドで処理することによって、
増殖、自己再生、生存および/もしくはドーパミン作動性の誘導、分化、生存またはニューロンのドーパミン作動性の表現型の獲得を起こさせるかあるいは増強させることを含む方法。
【請求項2】
前記核内受容体がNurr1である、請求項1に記載の方法。
【請求項3】
前記核内受容体がNor1またはNGFI-Bである、請求項1に記載の方法。
【請求項4】
細胞をNurr1のDNAで形質転換させることまたは前記細胞内にNurr1のRNAを導入することによってNurr1を基底レベルより高く発現させることを含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
前記細胞内にNurr1タンパク質を導入することによってNurr1を基底レベルより高く発現させることを含む、請求項1〜3のいずれか1項に記載の方法。
【請求項6】
前記細胞内にNurr1タンパク質を保存することによってNurr1を基底レベルより高く発現させることを含む、請求項1〜3のいずれか1項に記載の方法。
【請求項7】
前記WntリガンドがWnt1リガンドである、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
前記WntリガンドがWnt5aリガンドである、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
前記WntリガンドがWnt3aである、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
前記WntリガンドがWnt-2である、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
前記WntリガンドがWnt-4である、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記WntリガンドがWnt-7aである、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
前記WntリガンドがWnt-7bである、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくはニューロン細胞をWnt-1またはWnt-5a以外のWntリガンドまたは追加のWntリガンドで処理する、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞が、前記Wntリガンドで処理した場合に、有糸分裂性であり、かつ/または自己再生する能力を有する、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞をさらにFGF成長因子ファミリーのメンバーと接触させる、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞をレチノイドもしくはレチノイド誘導体、レチノイドX受容体(RXR)の活性化因子、レチノイド酸受容体(RAR)のリプレッサー、9-シスレチナール、DHA、SR11237、またはLG849と接触させる、請求項1〜16のいずれか1項に記載の方法。
【請求項18】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞をWntリガンドで処理する前にあるいはそれと同時に、前記細胞をbFGFおよび/またはEGFおよび/またはFGF-8および/またはLIFおよび/またはShhで処理する、請求項1〜17のいずれか1項に記載の方法。
【請求項19】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞を抗酸化剤、アスコルビン酸、低酸素圧または低酸素誘導因子の存在下で増殖させる、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞が腹側中脳のアストロサイトまたは初期グリア細胞の存在下で増殖および/または分化する、請求項1〜19のいずれか1項に記載の方法。
【請求項21】
前記Wntリガンドを前記細胞を含むin vitro培養物に加える、請求項1〜20のいずれか1項に記載の方法。
【請求項22】
Wntリガンドが前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞と同時培養した細胞からの発現によって産生され、その同時培養した細胞は1型アストロサイトもしくは初期グリア細胞以外の細胞であるか、または前記Wntリガンドをコードしている核酸で形質転換させた宿主細胞もしくは導入したWntタンパク質を含む細胞である、請求項21に記載の方法。
【請求項23】
前記同時培養した、1型アストロサイトもしくは初期グリア細胞以外の細胞または宿主細胞が別の幹細胞、神経幹細胞、始原細胞、前駆細胞または神経細胞である、請求項22に記載の方法。
【請求項24】
前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞が、前記Wntリガンドをそれをコードしている核酸から発現するように遺伝子操作されている、請求項21に記載の方法。
【請求項25】
Wntリガンドタンパク質を前記細胞内に導入する、請求項21に記載の方法。
【請求項26】
前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を、初期グリア細胞、または場合によっては腹側中脳由来の1型アストロサイトとさらに同時培養することを含む、請求項1〜25のいずれか1項に記載の方法。
【請求項27】
前記1型アストロサイトが不死化されているか、または前記腹側中脳以外の領域のアストロサイト細胞系由来である、請求項26に記載の方法。
【請求項28】
前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を非ドーパミン作動性ニューロンに対して選択する陰性選択剤とさらに接触させることを含む、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
ニューロンを1つまたは複数の追加の成分を含む組成物に配合することをさらに含む、請求項1〜28のいずれか1項に記載の方法。
【請求項30】
前記組成物が製薬上許容される賦形剤を含む、請求項29に記載の方法。
【請求項31】
前記組成物を個体に投与することをさらに含む、請求項30に記載の方法。
【請求項32】
前記ニューロンを個体の脳内に移植する、請求項31に記載の方法。
【請求項33】
Nurr1の発現を増大させるために前記細胞を個体内でin situで処理する、請求項1〜20のいずれか1項に記載の方法。
【請求項34】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞を個体内でin situでWntリガンドを用いて処理する、請求項1〜20のいずれか1項に記載の方法。
【請求項35】
前記Wntリガンドをコードしている核酸を前記細胞内に導入する、請求項34に記載の方法。
【請求項36】
Wntリガンドタンパク質を前記細胞内に導入する、請求項34に記載の方法。
【請求項37】
Nurr1の発現を基底レベルより高いレベルまで上昇させるために前記細胞を個体内でin situで処理する、請求項34に記載の方法。
【請求項38】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞が前記個体にとって内因性である、請求項33〜37のいずれか1項に記載の方法。
【請求項39】
前記神経幹細胞、始原細胞もしくは前駆細胞または他の幹細胞もしくは神経細胞が、前記個体への移植によって外因的に供給される、請求項33〜37のいずれか1項に記載の方法。
【請求項40】
前記個体がパーキンソン病、パーキンソン症候群、ニューロンの消失または神経変性疾患に罹患している、請求項31〜39のいずれか1項に記載の方法。
【請求項41】
個体を治療するための医薬の製造において前記方法に従って産生させたニューロンの使用をさらに含む、請求項1〜30のいずれか1項に記載の方法。
【請求項42】
前記医薬が個体の脳内へ移植するためのものである、請求項41に記載の方法。
【請求項43】
前記個体がパーキンソン病、パーキンソン症候群、ニューロンの消失または神経変性疾患に罹患している、請求項42に記載の方法。
【請求項44】
請求項1〜28のいずれか1項の記載に従って産生させたドーパミン作動性ニューロン。
【請求項45】
神経変性疾患の治療に使用するための物質のスクリーニング方法における、請求項44に記載のドーパミン作動性ニューロンの使用。
【請求項46】
(i)ドーパミン作動性ニューロンを前記ドーパミン作動性ニューロンの毒素で処理すること;
(ii)前記ドーパミン作動性ニューロンを前記毒素から分離すること;
(iii)前記処理したドーパミン作動性ニューロンを1つまたは複数の試験物質と接触させること;
(iv)前記ドーパミン作動性ニューロンが前記毒素から回復する能力を決定すること;
(v)前記ドーパミン作動性ニューロンが前記毒素から回復する前記能力を、前記1つまたは複数の試験物質と接触させない場合にドーパミン作動性ニューロンが前記毒素から回復する能力と比較すること、
をさらに含む、請求項1〜28のいずれか1項に記載の方法。
【請求項47】
(i)1つまたは複数の試験物質の存在下でドーパミン作動性ニューロンを前記ドーパミン作動性ニューロンの毒素で処理すること;
(ii)前記ドーパミン作動性ニューロンが前記毒素に耐性を示す能力を決定すること;
(iii)前記ドーパミン作動性ニューロンが前記毒素に耐性を示す前記能力を、前記1つまたは複数の試験物質と接触させない場合にドーパミン作動性ニューロンが前記毒素に耐性を示す能力と比較すること、
をさらに含む、請求項1〜28のいずれか1項に記載の方法。
【請求項48】
ドーパミン作動性ニューロンが前記毒素から回復するまたはそれに耐性を示す能力を向上させる物質を、1つまたは複数の追加の成分を含む組成物に配合することをさらに含む、請求項46または請求項47に記載の方法。
【請求項49】
前記組成物が製薬上許容される賦形剤を含む、請求項48に記載の方法。
【請求項50】
前記組成物を個体に投与することをさらに含む、請求項49に記載の方法。
【請求項51】
前記個体がパーキンソン病、パーキンソン症候群、ニューロンの消失または神経変性疾患に罹患している、請求項50に記載の方法。
【請求項52】
単独でまたは組み合わせて、Nurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化、もしくは成熟を増強させる因子もしくは複数の因子を得る方法であって、
(a)1つまたは複数の試験物質を存在させてまたは存在させずに、Nurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞をWntリガンドで処理すること;ならびに
(b)前記細胞の増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化、もしくは成熟を決定し、前記1つまたは複数の試験物質を存在させた場合または存在させない場合の前記増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化もしくは成熟の程度を比較することによって、前記因子または複数の因子を得ること、
を含む方法。
【請求項53】
前記Wntリガンドを前記細胞を含むin vitro培養物に加えることによって前記細胞を前記Wntリガンドで処理する、請求項52に記載の方法。
【請求項54】
前記Wntリガンドをコードしている核酸を前記細胞内に導入することによって前記細胞を前記Wntリガンドで処理する、請求項52に記載の方法。
【請求項55】
Wntリガンドタンパク質を前記細胞内に導入することによって前記細胞を前記Wntリガンドで処理する、請求項52に記載の方法。
【請求項56】
1型アストロサイトもしくは初期グリア細胞以外の細胞、または前記Wntリガンドをコードしている核酸で形質転換させた宿主細胞もしくは導入したWntタンパク質を含む細胞と同時培養することによって前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を前記Wntリガンドで処理する、請求項53に記載の方法。
【請求項57】
前記神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を初期グリア細胞または場合によっては腹側中脳由来の1型アストロサイトと同時培養することをさらに含む、請求項52〜56のいずれか1項に記載の方法。
【請求項58】
Nurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化もしくは成熟を増強させる能力を有する因子もしくは複数の因子が、単離かつ/または精製された形態で提供される、請求項52〜57のいずれか1項に記載の方法。
【請求項59】
Nurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞において増殖、自己再生、生存および/またはドーパミン作動性の発生、誘導、分化もしくは成熟を増強させる能力を有する因子もしくは複数の因子を、1つまたは複数の追加の成分を含む組成物に配合する、請求項56〜58のいずれか1項に記載の方法。
【請求項60】
前記組成物がNurr1を基底レベルより高く発現する神経幹細胞、始原細胞もしくは前駆細胞、または他の幹細胞もしくは神経細胞を含む、請求項59に記載の方法。
【請求項61】
前記組成物がWntリガンドを含む、請求項60に記載の方法。
【請求項62】
前記WntリガンドがWnt1リガンドである、請求項61に記載の方法。
【請求項63】
前記WntリガンドがWnt5aリガンドである、請求項61に記載の方法。
【請求項64】
前記組成物が製薬上許容される賦形剤を含む、請求項59〜63のいずれか1項に記載の方法。
【請求項65】
前記組成物を個体に投与することをさらに含む、請求項64に記載の方法。
【請求項66】
前記組成物を前記個体の脳内に移植する、請求項65に記載の方法。
【請求項67】
前記個体がパーキンソン病、パーキンソン症候群、ニューロンの消失または神経変性疾患に罹患している、請求項66に記載の方法。
【図1】

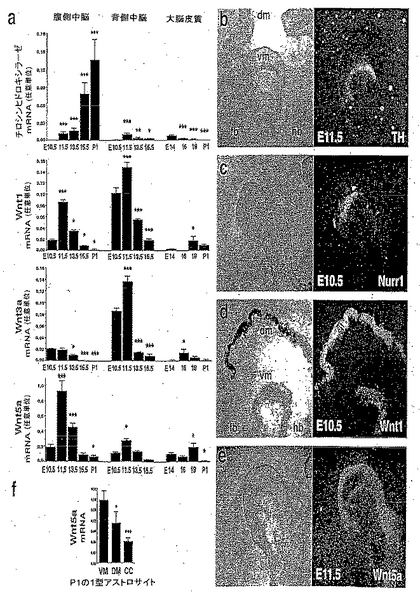
【図2】
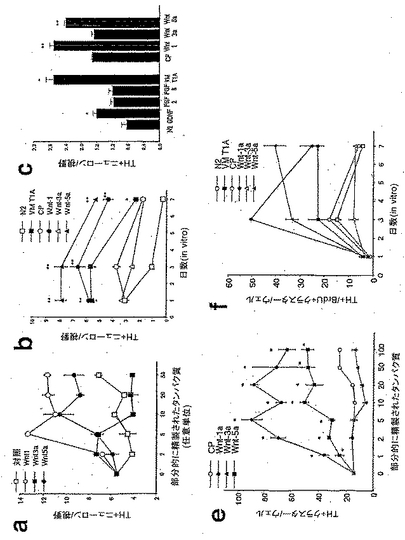
【図3】
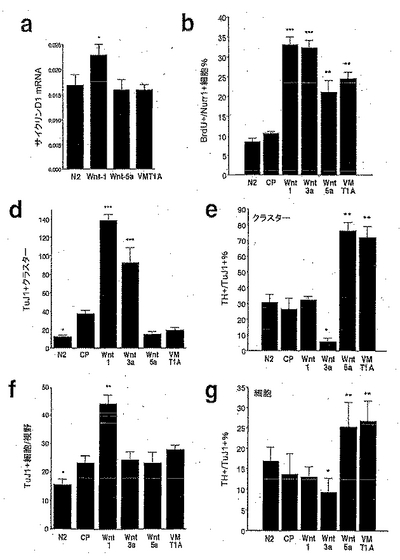
【図4】
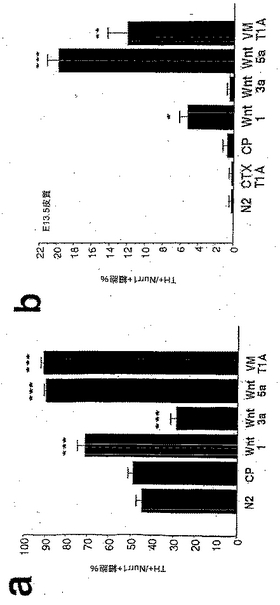
【図5】
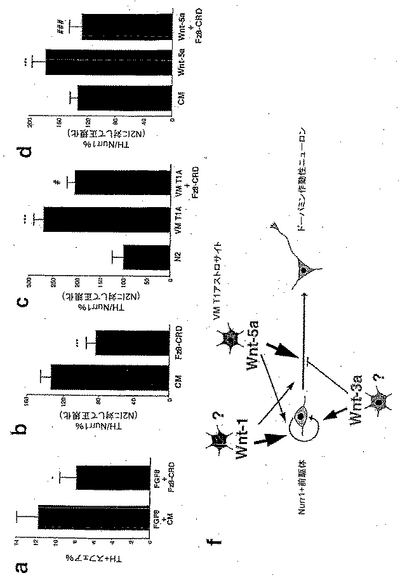
【図6】
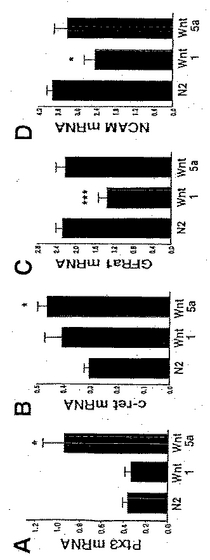
【図7】
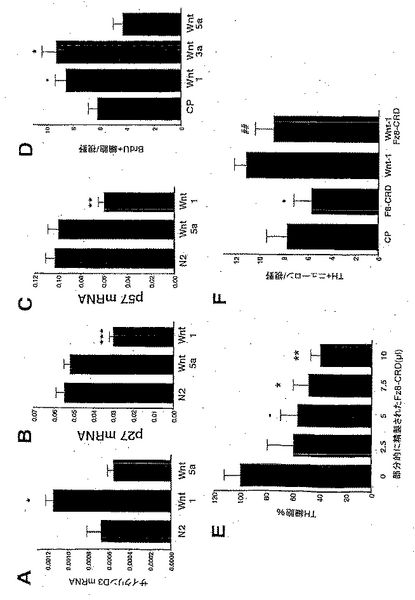
【図8】
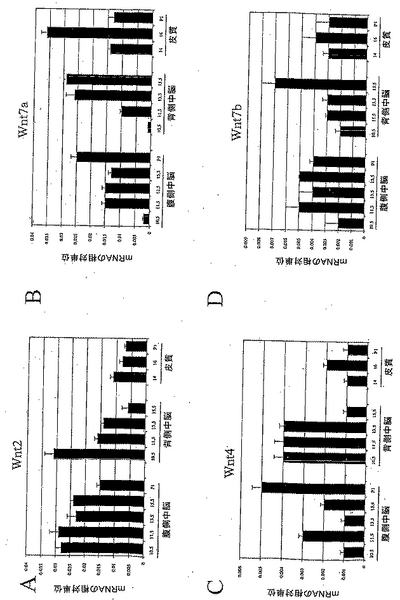
【図9】
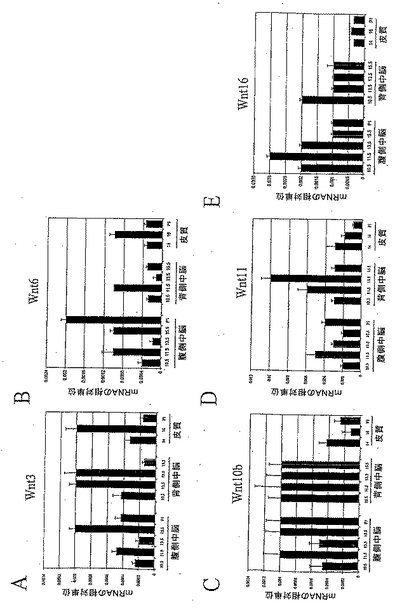
【図10】
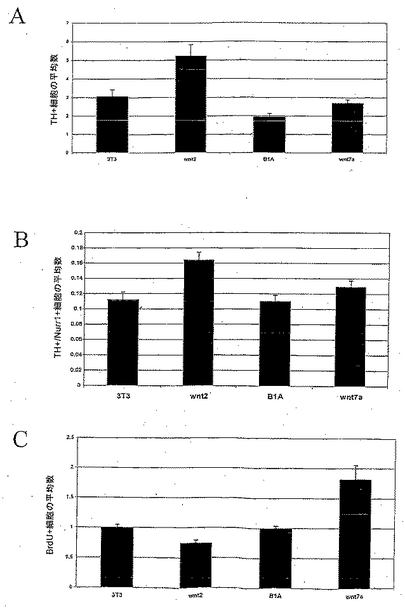
【図11】
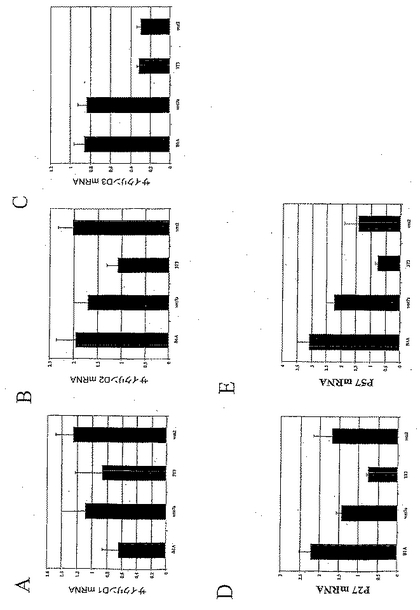
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2006−509497(P2006−509497A)
【公表日】平成18年3月23日(2006.3.23)
【国際特許分類】
【出願番号】特願2004−539370(P2004−539370)
【出願日】平成15年9月24日(2003.9.24)
【国際出願番号】PCT/IB2003/004598
【国際公開番号】WO2004/029229
【国際公開日】平成16年4月8日(2004.4.8)
【出願人】(505107077)ニューロ セラピューティクス エービー (4)
【Fターム(参考)】
【公表日】平成18年3月23日(2006.3.23)
【国際特許分類】
【出願日】平成15年9月24日(2003.9.24)
【国際出願番号】PCT/IB2003/004598
【国際公開番号】WO2004/029229
【国際公開日】平成16年4月8日(2004.4.8)
【出願人】(505107077)ニューロ セラピューティクス エービー (4)
【Fターム(参考)】
[ Back to top ]