
神経系、神経変性および気分障害(disorders)の治療のための、アファニゾメノン・フロスアクアエ(AphanizomenonFlosAquae)、調製物、その抽出物およびその精製成分
本発明は、神経系、神経変性および気分病態、または疾患の予防または治療に有用な、微細藻類アファニゾメノン・フロスアクアエ(Aphanizomenon Flos Aquae Aquae Ralfs ex Born.& Flah.Var.flos aquae)(クラマス湖産AFA)の抽出物およびその精製成分を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、微細藻類アファニゾメノン・フロスアクアエ(Aphanizomenon Flos Aquae Aquae Ralfs ex Born.& Flah.Var.flos aquae)(クラマス湖産AFA)に関する。より正確には、本発明は、神経系、神経変性および気分病態(conditions)または疾患(diseases)の予防または治療に有用なクラマス湖産AFAの抽出物およびその精製成分を提供する。
【背景技術】
【0002】
発明の背景
フェニルエチルアミン(PEA)は、黒質線条体系のドーパミン作動性ニューロンにおいてフェニルアラニンの脱炭酸化により合成される内因性アミンであり、脳におけるカテコールアミン神経伝達の神経調節物質として作用しうる(1)。PEAの最も重要な作用は、カテコールアミン神経伝達を促進することである。PEAは、ドーパミンと同様アセチルコリンの放出を刺激することが知られる(2)。さらに、PEAは、ノルエピネフリン神経伝達(NE)(6)、およびセロトニン神経伝達さえも増大させる。
【0003】
近年、PEAは、その特異的なニューロン受容体とともに、自律的神経伝達物質としてもまた働きうること、また、必要な場合は、神経伝達物質を抑制することもまた可能な、真の神経調節物質として作用することが示されている(8)。
【0004】
注意および記憶の刺激;顕著な抗うつ活性を伴う気分の高揚;情動行動および性行動を含む、共感ひいては社会性の促進;空腹の抑制;薬物(substance)乱用および薬剤(drug)依存の必要性の低減といった、一連の非常に多くの効果が、このことに由来する。
【0005】
PEAと感情的な気分とのつながりは、血漿または尿中のそれ自体またはその代謝物であるPAA(フェニル酢酸)により測定される、PEAレベルの著しい低下が、うつ状態の対象において見出された、研究により確認されている(9)。
【0006】
パーキンソン病患者は、血漿中で直接測定されたPEAレベルが著しく低いことがわかっている(12)。これらの患者における、神経伝達、特にドーパミン作動性、の漸減は、黒質のドーパミン作動性ニューロンの進行性変性と関連する。
【0007】
パーキンソン病患者において、PEAレベルのこのような減少は、MAO−Bレベルの並行する上昇を伴い、このため、パーキンソン病において用いられる薬剤は、セレギリンなどのMAO−B阻害物質である(14)。さらに、ひとたび摂取されたPEAは、血液脳関門を容易に通過することができ、低用量時でさえも黒質線条体組織からのドーパミン放出を刺激することができる。これは重要な著しい特徴である。なぜなら、現在用いられる薬剤である、セレギリンは、MAO−Bおよびドーパミンの再取り込みを阻害する一方で、黒質線条体組織からのその放出に対しては作用せず、したがって、さらなるドーパミン産生の一助とはならず、まさにドーパミンの産生自体が大いに脅かされる、パーキンソン病などの病変において深刻な限界があるからである。
【0008】
アルツハイマー病は、ドーパミンの産生および再取り込み機構の変性、ならびに、時間経過とともにドーパミン作動性ニューロン数を減少させる、線条体領域のニューロンの進行性破壊、および結果として起こる、ドーパミン伝達の進行性破壊を伴う(15)。
【0009】
ADHD(注意欠陥過活動性障害)が、神経変性病変であるという事実に関する明確なデータは存在しないが、神経破壊が、小児および成人の双方において、ADHDの主要原因であることを、いくつかの研究が示そうとしてきた(19)。
【0010】
最も重要なことは、ADHDおよび学習障害に罹患した小児では、PEAレベルが顕著に低く(21)、したがって、注意(ドーパミン)および鎮静(セロトニン)の神経調節(neuromodulation)において減少する、証拠があることである。ADHDの選択薬が、PEAの、ひいては、ADHDの病因に直接関与する2つの神経伝達物質である、ドーパミンおよびノルエピネフリンの、より高い産生を刺激することによっても作用する、PEAの合成誘導体、メチルフェニデート(22)であるのはこのためである。
【0011】
アンフェタミンの使用が空腹および、結果として、体重を制御することはよく知られている。この領域におけるその使用は、その耐性をも踏まえると、時間経過とともに潜在的に極めて重度となる傾向のある、その副作用により、常に問題となってきた。このことは、空腹および体重の制御に現在用いられる主な薬剤が、ベンラファキシンおよびブプロプリオンなどの、アンフェタミン様ドーパミン作動性抗うつ剤である事実によって確認される。これらの分子は、すべてのアンフェタミンと同様、PEAの合成誘導体である。後者(ブプロプリオン)は、MAO−B酵素によるその分解が阻害される限りにおいて、強力な食欲抑制剤として作用する。
【0012】
モノアミンオキシダーゼ(MAO)AおよびBは、CNSおよび末梢組織における神経作用性アミンおよび血管作用性アミンの分解を触媒する。特に、MAO−Bは、ドーパミン作動性伝達に対するその直接および間接の関与性を踏まえると、うつ病および気分障害、パーキンソン病およびアルツハイマー病など、ドーパミンが重要な神経障害に関与する。このため、MAO−B阻害物質が、こうした神経障害の治療において用いられるのである(26)。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】1. Zhou G. et al., Platelet monoamine oxidase B and plasma β-phenylethylamine in Parkinson's disease, in J Neurol Neurosurg Psychiatry, 2001; 70:229-231, 229.
【非特許文献2】2. Ispida K. et al., β-phenylethylamine stimulates striatal acetylcoline release through activation of the AMPA glutamatergic pathway, in Biol Pharm Bull 2005 Sep.; 28(9):1626-9.
【非特許文献3】6. Paterson I.A. et al., 2-phenylethylamine: a modulator of catecholamine transmission in the mammalian central nervous system?, in Journal of Neurochemistry (1990), 55:1827-1837.
【非特許文献4】8. Mauro Federici et al., Trace Amines Depress Gabab Response In Dopaminergic Neurons By Inhibiting Girk Channels, in Molecular Pharmacology Fast Forward. Published on January 11, 2005 as doi:10.1124/mol.104.007427.
【非特許文献5】9. Gusovsky F. et al., A high pressare liquid chromatography method for plasma phenylacetic acid, a putative marker for depressive disorders, in Anal Biochem, 1985 Feb. 15; 145(1):101-5.
【非特許文献6】12. Miura Y., Plasma beta-phenylethylamine in Parkinson's disease, in Kurume Med J 2000;47(4):267-72.
【非特許文献7】14. Ebadi M. et al., Neuroprotective actions of selegiline, in J Neurosci Res 2002 Feb 1;67(3):285-289.
【非特許文献8】15. Kemppainen N. et al., Different pattern of reduction of striatal dopamine reuptake sites in Alzheimer's disease and ageing, in J Neural Transm 2001;108(7):827-36.
【非特許文献9】19. Swanson J. et al., Cognitive neuroscience of attention deficit hyperactivity disorder and hyperkinetic disorder, in Curr Opin Neurobiol. 1998 Apr;8(2):263-71.
【非特許文献10】21. Kusaga A., Decreased beta-phenylethylamine in urine of children with attention deficit hyperactivity disorder and autistic disorder, in No To Hattatsu 2002 May; 34(3):243-8
【非特許文献11】21. Matsuishi T, Yamashita Y., Neurochemical and neurotransmitter studies in patients with learning disabilities, in No To Hattatsu 1999 May;31(3):245-8.
【非特許文献12】22. Kusaga A. et al., Increased urine phenylethylamine after methylphenidate treatment in children with ADHD, in Ann Neurol 2002 Sep;52(3):372-4.
【非特許文献13】26. Yamada M. et al., Clinical Pharmacology of MAO Inhibitors: Safety and Future, in Neurotoxicology 2004; 25:215-21
【非特許文献14】26. Youdim M., et al., Therapeutic Applications of Selective and Non-Selective Inhibitors of Monoamine Oxidase A and B that do not Cause Significant Tyramine Potentiation, in Neurotoxicology 2004;25:243-50.
【発明の概要】
【0014】
発明の説明
本発明は、微細藻類アファニゾメノン・フロスアクアエ(Aphanizomenon Flos Aquae Aquae Ralfs ex Born.& Flah.Var.flos aquae)(クラマス湖産AFA)における、アルツハイマー病およびパーキンソン病、多発性硬化症、注意欠陥過活動性障害(ADHD)、自閉症、うつ病、記憶欠損および気分障害などの神経変性疾患を含む、各種の神経系の疾患、病態、機能不全または障害に対して、単独または組み合わせて有益な効果を及ぼす、物質の、同定に基づく。特に、クラマス湖産AFA微細藻類は、ドーパミン作動性およびノルアドレナリン作動性活性を特徴とする神経調節物質である、フェニルエチルアミン以外に、酵素モノアミンオキシダーゼB(MAO−B)の極めて効果的な阻害物質であることがまったく驚くべきことに示された特異的分子、すなわち:a)特異的なAFAフィトクロム;b)C−フィコシアニン(C−PC)およびフィコエリスロシアニン(PEC、その発色団であるフィコビオロビリンまたはPVBを含む)により形成されるフィコビリソームを含む、AFAフィコビリタンパク質複合体(complex)(「AFAフィコシアニン」);c)マイコスポリン様アミノ酸またはMAA、を含むことがわかっている。その藻類中に含まれるPEAが、MAO−B阻害物質により保護されない限り、MAO−B酵素による取り込みと同時に迅速に破壊されるため、この知見は極めて重要である。
【0015】
MAO−Bの選択的阻害物質として作用する同じ分子が、強力な神経保護物質としての役割をも果たし、したがって、神経学的な健康を促進するその抽出物の能力を顕著に高める。
【0016】
したがって、本発明は、クラマス湖産AFA調製物、特に、こうした活性成分に富む抽出物、あるいは、a)AFAフィトクロム;b)AFAもしくは他の任意の微細藻類中に存在する、C−フィコシアニン/フィコエリスロシアニン複合体;c)AFA中に存在するかもしくは他の任意の藻類供給源に由来する、マイコスポリン様アミノ酸ポルフィラおよびシノリン;またはd)その混合物から選択された単離および精製成分を、それを必要とする対象に投与することにより、上述の神経系の疾患、病態、機能不全または障害を予防し、制御し、または治療する方法を提供する。
【0017】
本発明に従うクラマス湖産AFA抽出物は、以下のステップにより調製されることが好ましい:
a)採取直後のAFA藻類を冷凍し解凍するか、または、出発物質が乾燥AFA粉末である場合は、水で希釈したAFA粉末を超音波で分解して、細胞を破砕する;
b)ステップa)の生成物を遠心分離し、沈殿物(細胞壁分画の大半を保持する)から上清(細胞質(cytoplasmatic)部分の大半を保持する)を分離する;
c)水溶性成分を含む上清を回収する。
【0018】
結果として得られる生成物が、PEAのほか、AFAフィトクロム、AFAフィコシアニン、およびMAAなど他の相乗作用分子をも濃縮する抽出物(「基本抽出物」と呼ぶ)である。例えば、クラマス湖産微細藻類が2〜4mg/gの範囲の天然PEA含量を有するのに対し、基本抽出物は、この濃度を、9〜11mg/gの範囲のレベルにまで上昇させる(HPLC分析)。
【0019】
抽出物を限外濾過システム、好ましくは、30,000ドルトン(Daltons)のカットオフ分子量を有する膜に通すことによりさらに精製することができる。限外濾過残留物(retentate)(抽出物A)は、AFAフィコシアニン(分子量=121,000)およびAFAフィトクロム(分子量=480,000)をともに主要な活性成分として含む。興味深いことに、MAAが、用いられるそのカットオフサイズを大きく下回る分子量を有するにもかかわらず、その残留物は、MAAの濃度も上昇させる。
【0020】
ステップa)〜c)により得られる、すなわち、限外濾過を伴わない基本抽出物は、最適量のPEA、AFAフィトクロム、AFA−PCおよびMAAを含むので、一般に好ましい。さらに、この基本抽出物はまた、低下した比率においてではあるが、その抗酸化および抗炎症特性に寄与する、クロロフィルおよびカロテンなどの物質をも含む。
【0021】
あるいは、クラマス湖産AFAの活性成分、すなわち、複合体C−フィコシアニン/フィコエリスロシアニン(C−PC/PEC)、AFAフィトクロム、およびMAAは、さらに以下に記載の通りに、単離および精製することができ、本発明に従う方法において用いることができる。
【0022】
好ましい実施形態において、クラマス湖産AFAのC−PC/PEC複合体、AFAフィトクロムおよびマイコスポリン様アミノ酸は、それを必要とする対象に対する同時または個別投与のための併用調製物として用いられ、さらに好ましい実施形態において、こうした併用調製物は、フェニルエチルアミンを追加の活性成分として含む。マイコスポリン様アミノ酸のうち、シノリンおよびポルフィラ334が、クラマス湖産AFA微細藻類中に比較的高濃度で含まれるので、特に好ましい。
【0023】
観察されたモノアミンオキシダーゼBの阻害は、ドーパミン作動性伝達を増大させ、PEAの異化を最小化することを可能とするので、特に重要である。意義深いことに、フィトクロムおよびAFAフィコシアニンがともにMAO−Bを可逆的および混合的な方法で阻害するのに対し、MAAによるMAO−B阻害は、競合的および可逆的であり、このため、3つの分子は全て、生理学的状態および副作用の不在下で高い有効性を確保する。
【0024】
さらなる態様において、本発明は、場合によってフェニルエチルアミンを添加した、AFA藻類中に存在するもしくは他の任意の藻類供給源に由来する、C−PC/PEC複合体、または単離したC−PCおよびPECの単体成分;AFAフィトクロム;AFA中に存在するもしくは他の任意の微細藻類供給源に由来する、マイコスポリン様アミノ酸ポルフィラおよびシノリン;またはその混合物から選択されることが好ましい、クラマス湖産AFA調製物、その抽出物またはその単離成分を含む栄養補助組成物または医薬組成物を意図する。好ましい実施形態において、栄養組成物は、錠剤、カプセル、飲料の形態における栄養補助食品であり、さらに好ましい実施形態において、医薬組成物は、錠剤、カプセル、小袋、シロップ、坐薬、バイアルおよび軟膏の形態であり、上記に示した神経系または神経変性の疾患もしくは病態の予防または治療に用いられることができる。本発明に従うクラマス湖産AFA液体抽出物は、それ自体で用いられるかまたは例えば凍結乾燥、噴霧乾燥などの手法により乾燥させることができる。その単離した活性成分は、当業者に知られる手法および以下の手順を用いて調合することができる。
【0025】
その活性成分の用量は、栄養補助食品としてかまたは医薬調製物としてかの、組成物の使用目的に依存するであろう。各成分の有効量は、一般に以下の範囲に含まれるであろう。
PEA=0.1〜100mg、好ましくは、5〜30mg;
フィトクロム=0.1〜1000mg、好ましくは、0.8〜10mg;
MAA=0.1〜1000mg、好ましくは、10〜100mg;
フィコシアニン=1〜2500mg、好ましくは、50〜1000mg。
【図面の簡単な説明】
【0026】
【図1】MAA試料の波長と吸光度の関係を示す図である。
【図2】MAA標準物質とMAA抽出物のクロマトグラムを示す図である。
【図3】精製済みMAAのUVスペクトルを示す図である。
【図4】AFAの細胞溶解物の成分とシネコシスチス属PCC6803のその成分との比較を示す図である。
【図5】抽出物の分光分析を示す図である。
【図6】PCB発色団の波長を示す図である。
【図7A】基本抽出物の水溶性および脂溶性成分のMAO−B阻害活性を示す図である。
【図7B】AFA水溶性抽出物のMAO−AおよびMAO−Bの阻害活性を示す図である。
【図8】水溶性抽出物のラインウィーバー−バークプロットを示す図である。
【図9】AFA−PCのMAO−AおよびMAO−Bの阻害活性を示す図である。
【図10】PCのラインウィーバー−バークプロットを示す図である。
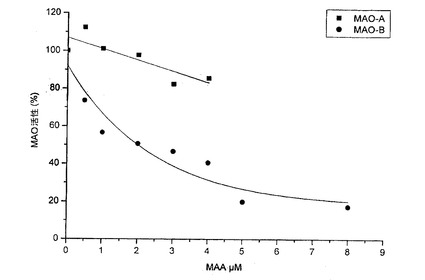
【図11】MAAによる用量依存的なMAO−B阻害を示す図である。
【図12】MAAのラインウィーバー−バークプロットを示す図である。
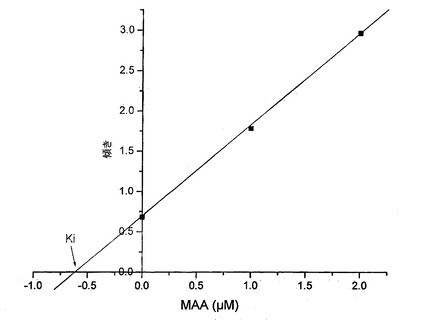
【図13】MAA濃度と傾きの関係を示す図である。
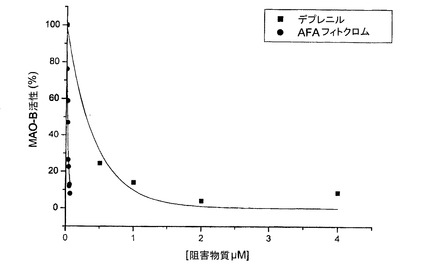
【図14】、デプレニルに関してAFAの3つの分子のMAO−B阻害活性を示す図である。
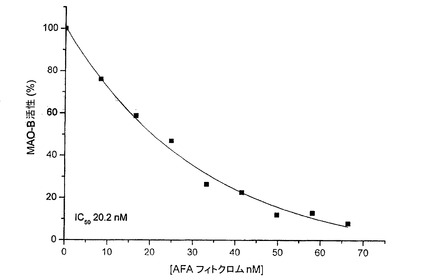
【図15】AFAフィトクロムのMAO−B活性の用量依存的な低下を示す図である。
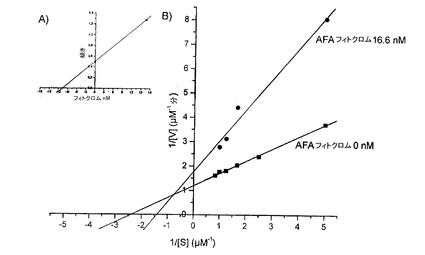
【図16】AFAフィトクロムのラインウィーバー−バークプロットを示す図である。
【図17】デプレニルおよびAFAフィトクロムのMAO−B阻害活性を示す図である。
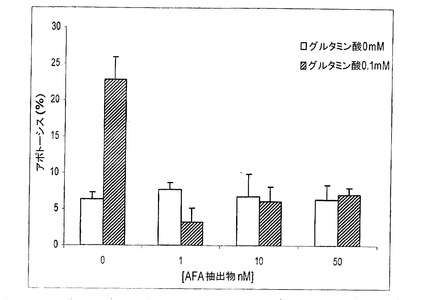
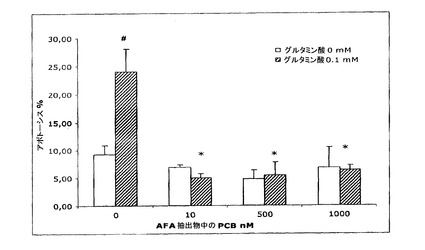
【図18】アポトーシスへのグルタミン酸とAFA抽出物の添加の影響を示す図である。
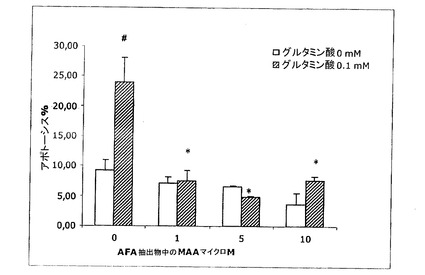
【図19】アポトーシスへのグルタミン酸とAFA抽出物のMAAの添加の影響を示す図である。
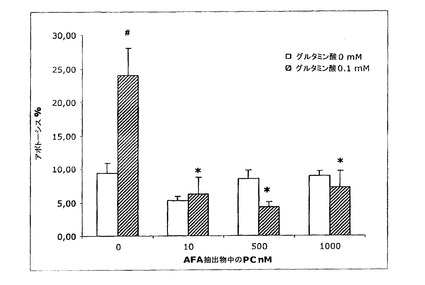
【図20】アポトーシスへのグルタミン酸とAFA抽出物のPCの添加の影響を示す図である。
【図21】アポトーシスへのグルタミン酸とAFA抽出物のPCBの添加の影響を示す図である。
【図22】AFAフィトクロムのバンドを示す図である。
【図23】光吸収特性を示す図である。
【図24】AFAフィトクロム量のMDA形成の用量依存的減少を示す図である。
【発明を実施するための形態】
【0027】
発明の詳細な説明
クラマス湖産藻類の典型的な固有のフィトクロムである「AFAフィトクロム」の同定
フィトクロムは、植物が光を検出するのに用い、可視スペクトルの赤色および遠赤色領域における光に対して感受性がある、光受容体、色素である。これらは、(日周期のリズム(rythms)を通して)開花の調節、発芽およびクロロフィルの合成を含む、植物における多数の異なる機能を果たす。AFAにおけるこの固有の種類のフィトクロムの存在が、光合成過程においてCフィコシアニンを補うよう他のシアノバクテリアにより通常用いられる他のフィコビリタンパク質、すなわち、アロフィコシアニン、の欠如により説明しうるので、後者(クロロフィルの合成)は、AFA藻類との関連で特に重要である。一方、クラマス湖産藻類におけるアロフィコシアニンの位置は、フィコエリスロシアニンまたはPEC(下記参照)により占められるが、特に、クラマス湖産藻類が、高い光回収能力を必要とする非熱帯環境において生育することを考えると、PECのみでは十分ではないと思われ、したがって、AFA藻類は、そのより高い必要性をフィトクロムで調和させていると思われる。
【0028】
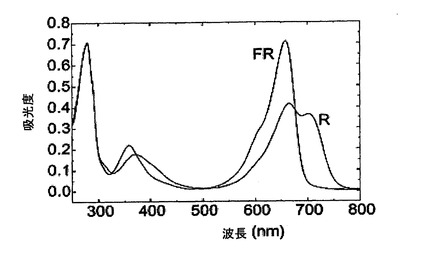
特異な構造を有するAFAフィトクロムは、本明細書において初めて記述される。長年にわたり、異なる種類のフィトクロムが植物中に見出されており、それらは、異なるフィトクロム遺伝子(例えば、コメにおいて3種類、トウモロコシにおいて6種類)を有するだけでなく、大半の場合、顕著に異なるタンパク質成分および構造を有する。これらがすべてフィトクロムであるのは、これらがすべて、フィトクロモビリンと呼ばれる、同じビリタンパク質を、光吸収発色団として、用いるからである。この発色団は、フィコシアニンの発色団であるフィコシアノビリンと類似し、4つのピロール環(テトラピロール)の開鎖からなる単一のビリン分子であることを特徴とする。より具体的に、その正常のPr状態において、このビリタンパク質は、650〜670nMの最大値で吸光するのに対し、赤色光により活性化されると、730nMの最大吸収を有するPfrに転換される。
【0029】
発見された最初のシアノバクテリアのフィトクロムは、シネコシスチス属のそれであり、植物フィトクロムとの構造的類似性がわずかであることを示した。にもかかわらず、シネコシスチス属のビリタンパク質は、赤色/遠赤色の可逆的色素タンパク質である限りにおいて、一般にフィトクロムと考えられる(48)。
【0030】
AFAフィトクロムの精製と特性解析
AFAフィトクロムは、赤色/遠赤色スペクトルにおいて吸光するその発色団として、ビリタンパク質を有する。その構造および活性を確立するため、本発明者らは、以下のプロトコールによりフィトクロムを精製した。
【0031】
・ 1gの抽出物を10mlの1Kリン酸緩衝液、pH7.0中に懸濁させる
・ その半分の体積で、1分間にわたるボルテックスを2回行う
・ 細胞を2%トリトンX100とともに35分間(35’)インキュベートする
・ 28000rpmで16〜18時間遠心分離する
・ スクロース密度段階勾配法により、上清を回収する
・ スイングアウトローターを用いて、150000gで12時間、その勾配分離物をスピンする
・ −20℃で保存する
フィトクロムが、約1Mのスクロースで可視化される、強い橙色の溶解物バンドに対応するのに対し、一方、フィコビリソームは、約0.75Mに位置する。2つのバンドのこの関係は、三量体AFA−PCの約4倍である、その藻類中に存在するフィトクロムの分子量についても、信頼できる指標を与える。後者(三量体AFA−PC)が121Kdであるので、本発明者らは、AFAフィトクロムのMWを約480Kdにおいてあらかじめ確立することができる(図22)。
【0032】
その光吸収特性について調べたところ、フィトクロムは、平衡状態を形成するPr(赤色光吸収)形態およびPfr(遠赤色光吸収)形態にそれぞれ対応する、672nMおよび694nMにおいて2つのピークを伴い吸光することを示す(図23)。
【0033】
AFA中に含まれるフィトクロムの量について、本発明者らの最初の評価は、以下の予備的な結果を与える:2mg/g(または0.2% DW)。その抽出物について、基本抽出物中では濃度が約0.5%に上昇し、抽出物B中では約1%に上昇した。これらは低濃度であるが、この分子の抗酸化/抗炎症効力は極めて強いので、ごく少量でさえも極めて重要な効果をもたらすことができる。
【0034】
抗酸化活性
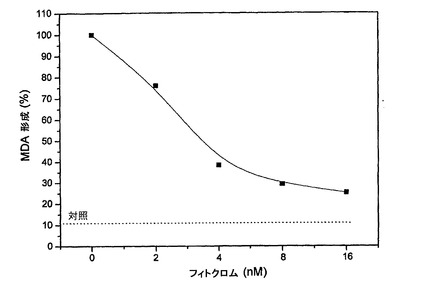
精製済みAFAフィトクロムは、極めて強力な抗酸化剤であることを示した。実際、絶対的に、今までにクラマス湖産藻類中で見出された最も強力な分子である。100μMで酸化作用物質CuCl2とともにヒト血漿試料を2時間インキュベートすると、チオバルビツール酸との反応後535nmにおける分光光度計により計測(TBA試験)される、脂質過酸化の後期副生成物(a late byproduct)である、マロンジアルデヒド(MDA)レベルの上昇が生じる。AFA藻類から抽出したAFAフィトクロム量を増加させながら(2〜16nM)100μMでCuCl2とともに37℃で2時間、血漿をインキュベートすると、MDAレベルの極めて強力な用量依存的減少が観察される(図24)。実際、わずか16nMのAFAフィトクロムにより、対照に近いMDAレベルを有する脂質過酸化のほぼ完全な阻害が得られる。意義深いことに、3.6nMのIC50が、PCBについて得られるそれ(IC50)を45倍下回る。フィトクロムが、AFA−PCよりも高い、基本抽出物の抗酸化および神経保護効果の主要原因である。
【0035】
MAAの抽出、精製および定量
本発明者らは、一般にクラマス湖産藻類として知られている、クラマス湖産の藍色植物門のアファニゾメノン・フロスアクアエにおけるMAAの存在を調べた。本発明者らの知る限り、任意のアファニゾメノン属種におけるMAAの存在についてはごく最近の報告(47)が存在するのみであり、しかしながら、こうした報告が、ポルフィラを現存のMAAとして同定するのみであるのに対し、本発明者らの研究は、ポルフィラおよびシノリン双方の2つのMAAの存在を示す。一方、藻類に関する全般的な文献に関して、今まで報告されているシアノバクテリアの大半は、その主要MAAとしてシノリンを含むのに対して、本発明者らは、シノリンに加え、アファニゾメノン・フロスアクアエ中の主要MAAとして、ポルフィラ334のまれな存在を見出した。
【0036】
【化1】
【0037】
MAAは、既に報告されているように抽出された(29)。簡潔に言うと、45℃の水浴中に2.5時間インキュベートすることにより、2mlの20%(v/v)含水メタノール(HPLCグレード)中で20mgのAFA粉末または20mgの水性抽出物を抽出した。遠心分離(5000g;米国、パロアルト、Beckman社製、GS−15R型遠心分離機)後、上清を蒸発乾燥させ、2mlの100%メタノール中に再溶解し、2〜3分間ボルテックスし、10000gで10分間遠心分離した。上清を蒸発させ、HPLCでの分析用に同じ容量の0.2%酢酸中か、または、抗酸化特性の評価用に200μlのリン酸緩衝液(PBS)中に抽出物を再溶解した。試料は、HPLC分析、または抗酸化特性試験(下記を参照のこと)にかける前に、0.2μm孔径のシリンジフィルター(イタリア、ミラノ、VWR International社製)により濾過した。
【0038】
クラマス湖産藻類のMAAは、334nmの最大吸収を有する。MAAのさらなる精製は、文献(30)に従い、Alltima C18型カラムおよびガード(イタリア、ミラノ、Alltech社製、4.6×250mm内径、5μm充填剤)を装備したHPLCシステム(日本、東京、日本分光株式会社製)を用いて行われた。検出波長は330nm、移動相は流量1.0ml分-1で0.2%酢酸であった。MAAの同定は、ドイツ、エアランゲン、フリードリヒアレクサンダー大学、Manfred Klisch博士のご厚意により提供された、ポルフィラ334、シノリンおよびパリシンを主に含む、アマノリ属種およびオバクサ属種などの標準品と吸収スペクトルおよび保持時間を比較することにより行った。試料の吸収スペクトルは、シングルビーム分光光度計(米国、パロアルト、Beckman社製、DU640型)の200〜800nmにおいて測定した。生スペクトルは、コンピュータに転送し、MAAのピーク解析のために数学的に処理された。
【0039】
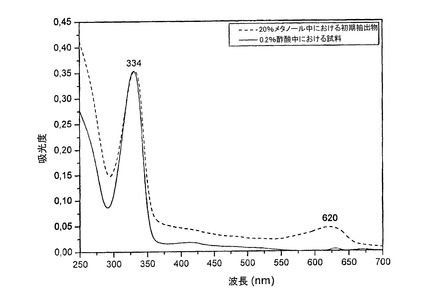
MAAは、前述のようにAFA試料およびその水性抽出物から部分的に精製された。45℃で2.5時間の20%メタノールによる試料の抽出は、この手順により、少量の光合成色素(620nmにおけるフィコシアニンなど)も抽出されたにせよ、334nmにおいて顕著なピーク(MAA)を生じた(図1の破線を参照のこと)。MAA試料は、タンパク質および塩を除去するために、100%メタノールによりさらに処理され、最終的に非極性光合成色素を除去するために、0.2%酢酸によりさらに処理された。その結果として得られる、部分的に精製されたMAAは、334nmに最大吸収を有した(図1、実線)。
【0040】
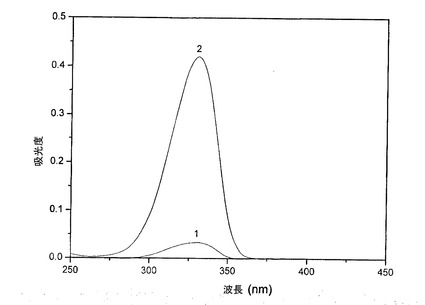
334nmにおいて吸収する化合物が、単一のMAAであるか、または1以上の混合物のMAAであるかを見出すため、HPLCによってMAAのさらなる解析および精製を行った。その試料のクロマトグラム(図2)は、シノリンおよびポルフィラ334としてそれぞれ同定された、保持時間4.2分(ピーク1)および7.6分(ピーク2)を有する2つのMAAの存在を示す。ポルフィラ334は、シノリンが少量のみで存在する(ピーク面積比1:15)ので、AFAにおける主要なMAAであると思われる。
【0041】
精製済みMAAのUVスペクトルは、334nmでその最大吸収が確認された(図3)。
【0042】
シノリンおよびポルフィラ334に対する334nmにおけるモル吸光係数が、それぞれ、44700および42300M-1cm-1であることを考慮し、本発明者らは、
a)AFA藻類について、シノリンに対して0.49mgg-1DWの濃度およびポルフィラ334に対して7.09mgg-1DWの濃度;したがって、そのMAA総含量は、0.76%藻類DWに等しい;
b)基本抽出物について、17〜21mgのMAA濃度(すなわち、1.7〜2.1% DW)
を計算した。
【0043】
全AFAが、UV曝露下で見出される最大濃度、すなわち、0.84%に近い、高構成レベルのMAA(0.76% DW)を含む(31)ので、これらは重要なデータである。また、本発明者らは、その抽出物が、全藻類よりもさらに高い濃度を有し、最大可能濃度よりもはるかに高いレベルに達していることをも見出した。
【0044】
MAA(シノリンおよびポルフィラ334)は、分子量300を有する、構造的に単純な分子である。このことは、それが最も必要とされる領域である、脳において、そのMAO−B阻害可能性を発現する能力を確かにし、これらの水溶性分子が血液脳関門を容易に通過することを可能にする。
【0045】
フィコシアニン
フィコシアニンは、8〜10%の濃度(定量については、下記を参照のこと)でその抽出物中に存在する。フィコシアニンは、特定の微細藻類ごとに特異的な特性を有するが、すべてのシアノバクテリアまたは藍藻類の、典型的な青色色素である(32)。フィコシアニンの機能的および治療的特性について、現在まで、研究は、主に、微細藻類スピルリナの特性に焦点を合わせている。スピルリナから精製したフィコシアニンは、肝臓(37)、呼吸器系(38)、および脳(39、40)などの異なる生理系での抗酸化(33)および抗炎症(34、35、36)特性を有することを示している。スピルリナから精製したPCのこうした特性は、その実質的な類似性を踏まえると、一般に、他の藻類フィコシアニンにも帰属させることができる。にもかかわらず、上述の機能的および治療的特性の説明における異なる効力が生じうる、異なる微細藻類に由来する異なるフィコシアニンには種特異的な差異が存在しうる。
【0046】
クラマス湖産藻類のフィコビリソームの構造決定および特異的特性
一般的に言うと、無傷のシアノバクテリア細胞において、フィコシアニン(PC)は、機能的形態(αβ)6でフィコビリソームの内部に存在する(41)。その細胞の破砕後、タンパク質は、分析される生体に応じて、異なる凝集状態(単量体、二量体、三量体、六量体)で見出すことができる。クラマス湖産藻類の場合、抽出物中に含まれるおよび抽出物自体から精製される両方の、PCの電気泳動解析は、タンパク質が、大半の場合、総分子量121000を有する、その三量体形態(αβ)3で見出されることを示している。単量体αβは、約40000(18500のサブユニットα+21900のサブユニットβ)の分子量である。これとは別に、スピルリナから精製したFCに関する研究の大半が、タンパク質は、スピルリナにおいて約37500の分子量を有する単量体形態αβで見出され、したがって、AFAから精製したPCと比較して異なる凝集状態を示すことを示唆する。
【0047】
AFAのフィコビリソームのクロマトグラフィー解析は、他のシアノバクテリア種において見られるように、PCのαサブユニットが1つの補欠分子族に結合するのに対し、βサブユニットは2つの補欠分子族に結合することをも示している。その補欠分子族または発色団は、フィコシアノビリン(PCB)と呼ばれ、タンパク質の青色およびその抗酸化力の両者の原因である(42)。
【0048】
AFAとスピルリナとの根本的な差異は、フィコビリソームの異なる構造に基づく。スピルリナに対して、クラマス湖産AFAのフィコビリソームは、色素アロフィコシアニンを含まず、スピルリナには欠如する構造的要素、すなわち、フィコエリスロシアニン(PEC)、に結合する色素cフィコシアニンのみを含む。FECは、今日現在、限られた数のシアノバクテリア種中のみにおいて同定されている、光合成色素である(43)。PECは、FCの化学的構造と極めて類似する化学的構造を有し、単量体および三量体を形成するために会合する2つのサブユニットαおよびβにより構成される。にもかかわらず、PCの各単量体がPCBの3分子に結合するのに対し、PECはPCBの2分子をサブユニットβに、また、フィコビオロビリン(PVB)の1分子をαサブユニットに結合させる固有の特性を有し、これが、その色素の紫色の原因となっている。
【0049】
クラマス湖産藻類のフィコビリソームが、cフィコシアニンおよびフィコエリスロシアニンの結合体により特異的に構成されると定義されるのはまったく初めてのことであり、クラマス湖産AFA藻類フィコビリソームの、この異なる性質的構造が、スピルリナからAFAを区別するさらに決定的な因子を追加する。
【0050】
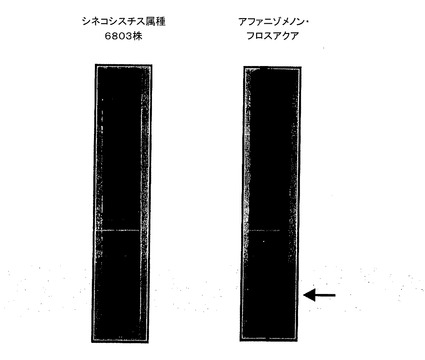
図4は、AFAの細胞溶解物の成分を、別のよく知られたシアノバクテリアである、シネコシスチス属PCC6803の、成分と比較して、言われていることを確認する。両シアノバクテリアにおいても、フィコビリソームを表す青色バンドを見ることができるが、AFA藻類におけるフィコビリソームは、より低い分子量を示し、スピルリナなど一般的な微細藻類とは異なり、AFAフィコビリソームにおいては、アロフィコシアニンではなく、フィコシアニンのみが存在することを確認する。さらに、図は、AFA中にフィコエリスロシアニンに典型的な明るい紫色のバンド(矢印により示す)もまた存在することを示し、これにより、クラマス湖産藻類フィコビリソーム中におけるその存在を提供する。
【0051】
定義を深めるため、各青色バンドは、質量分析計に接続したHPLC(RP−HPLC−ESI−MS)によりさらに解析した。保持時間の差により、フィコビリソームのタンパク質を分離し、その分子量に基づきこれを同定した。得られた結果を、以下の表に示す。第1に、本発明者らは、シネコシスチス属(表1)においては、フィコシアニン(28.2分におけるcpcAおよび28.9分におけるcpcB)およびアロフィコシアニン(30.7分におけるapcAおよび31.2分におけるapcB)がともに存在するのに対し、AFA(表2)においては、フィコシアニン(28.8分におけるcpcAおよび30.0分におけるcpcB)のみが存在することがわかる。第2に、AFAにおいては、シネコシスチス属中には存在せず、2つのビリンが付着したフィコエリスロシアニンのベータサブユニットに対応する、分子量19469のタンパク質(25分におけるpecB)が同定されている。
【0052】
【表1】
【0053】
【表2】
【0054】
この固有の構造が、全AFA−PCの、そのPCBと比べてより強力な抗酸化および抗炎症作用を説明するのに重要な要素である。抗酸化および抗炎症特性は、これらが強力な神経保護をもたらす限りにおいて、この文脈で重要となり、全PCは、神経保護の観点でもそのPCBよりも強力であり、これは、フィコビリソーム中のPCB以外の他の活性成分、すなわち、その特異的なPVB発色団を有するPECが、AFA−PCにおける最も有効な健康増進主要成分である可能性が極めて高いことを明確に示唆する。精製済みAFA−PCが、そのPCB発色団を有するC−PCだけでなく、PECおよびそのPVB発色団をも含むことは、精製の結果として得られる抽出物の分光分析を参照することにより明らかである(図5)。実際、C−PCの最大吸収は、図5の分光分析においてピークの頂点を表す、620nmである。しかし、PECの最大吸収は、αサブユニット(フィコビオロビリンまたはPVB)について566nm、およびβサブユニットの2つのPCBについてそれぞれ593nmおよび639nmであることも知られている。3つの値すべては、実際、精製済みPCの分光分析プロファイルを構成する釣り鐘型ピークに含まれている。AFA藻類におけるC−PCとPECとの間の、極めて断ち切りがたい、強い連結を考えると、これは、C−PC以外にPECも、必然的に、精製済みPC抽出物の一部であるということを裏付ける。これは、同様に、AFAに由来するPCが、大半の研究がなされてきた、スピルリナ由来のそれ(PC)を含む、他のシアノバクテリアのPCとは、構造および機能の両面で顕著に異なること、および、この差異が、共通の一部分のみ、すなわちC−Pを有し、その他は有しないことにあり、C−PCの特性がAFA−PCのC−PC成分にも帰属させることができるのに対し、AFAに由来する全PCの、C−PC/PEC複合体(その発色団であるPCBおよびPVBを含む)であることにおける、特性は、(他の任意の微細藻類中に存在する任意のC−PC/PEC複合体と同様に)もっぱらこの複合体のみに帰属するという帰結を伴うことを意味する。
【0055】
精製手法(図5)
PCは、以下の通りに、乾燥AFA抽出物から精製した。
・ 500mgの抽出物を、50mlの100mMリン酸Na緩衝液pH7.4中に懸濁させる;
・ 4℃、2500rpmで10分間(10’)遠心分離する;
・ 上清を回収し、固体の硫酸アンモニウムを50%飽和まで添加する;
・ 試料を攪拌状態に保ちつつ、4℃で60分間タンパク質を沈殿させる;
・ 4℃、10,000rpmで30分間遠心分離する;
・ 無色透明の上清を廃棄し、少量の5mMリン酸Na緩衝液pH7.4中に青色の沈殿物を再懸濁させる;
・ 同じ緩衝液に対して、4℃で一晩透析する;
・ 5mMのリン酸Na緩衝液pH7.4で平衡化したヒドロキシアパタイト製カラム内に透析済みPCを入れる;
・ イオン強度を上昇(5〜150mM)させているリン酸Na緩衝液pH7.0で試料を溶出する;
・ その分画を回収し、分光光度計により620nmおよび280nmにおける吸光度を読み取る;
・ Abs620/Abs280>4(純PCの指標)である分画をプールする;
・ 50%飽和の硫酸アンモニウムにより、4℃で1時間PCを沈殿させる;
・ 4℃、10,000rpmで30分間(30’)遠心分離する;
・ 上清を廃棄し、150mMのリン酸Na緩衝液pH7.4中にPCを再懸濁する;
・ 同じ緩衝液に対して、4℃で透析する;
・ 精製済みPCをフラスコに移し、暗所中+4℃または−20℃で保存する。
【0056】
フィコシアニンの定量
純PCのモル濃度を測定するため、本発明者らは、三量体形態(αβ)3では770000M-1cm-1に等しい、620nmにおけるモル吸光係数εを用いた。これは、1MのPC溶液が、620nmにおいて吸光値770000を有することを意味する。
【0057】
その抽出物中のPC濃度を測定するため、本発明者らは、620nmにおける比吸光度(the coefficient of specific extinction)E1%として70lg-1cm-1を用いた。これは、1%のPC(すなわち、1g/100ml)を含む溶液が、620nmにおいて70を吸収することを意味する。これらの計算によれば、その抽出物中の平均PC含量は、80〜100mg/g DW(8〜10% DW)に等しい。
【0058】
PCB発色団の精製(図6)
・ 500mgの抽出物を、50mlの蒸留済みH2O中に懸濁させる。
・ 4℃、2500rpmで10分間(10’)遠心分離する。
・ 藍色の上清を静かに移し、1%のトリクロロ酢酸によりPCを沈殿させる。
・ 攪拌しつつ、暗所中4℃で1時間インキュベートする。
・ 4℃、10000rpmで30分間(30’)遠心分離する。
・ PCを含むペレットを回収し、メタノールで3回洗浄する。
・ 1mg/mlのHgCl2を含む10mlのメタノール中にそのペレットを再懸濁させる。
・ 暗所中42℃で20時間インキュベートし、PCからPCBを放出させる。
・ 2500rpmで10分間(10’)遠心分離し、タンパク質を除去する。
・ PCBを含む上清にβメルカプトエタノール(1μl/ml)を添加し、HgCl2を沈殿させる。
・ −20℃で24時間インキュベートする。
・ 4℃、10000rpmで30分間(30’)遠心分離し、白色の沈殿物を除去する。
・ 上清に10mlの塩化メチレン/ブタノール(2:1、v/v)を添加する。
・ 20mlの蒸留済みH2Oで洗浄し、3000rpmで10分間(10’)遠心分離する。
・ 上層を除去し、PCBを含む下層部分を回収する。
・ 15ml H2O中で、PCBを3回洗浄する。
・ 窒素下で乾燥させ、−20℃で保存する。
【0059】
クラマス湖産AFA抽出物およびそれを構成する主要な活性のある構成要素であるフィトクロム、フィコシアニンおよびMAAによるMAO−B阻害の評価
本発明者らは、特異的基質であるベンジルアミン(1mM)を用いて、基本抽出物のMAO−B阻害活性を試験した。その試験は、上記のステップa)〜c)によって取り出した(初期濃度10mg/ml)、基本抽出物の水溶性および脂溶性成分の異なる濃度とともに、MAO−B(2μg/ml)をプレインキュベートすることにより、波長250nm、30℃で分光光度計により実施した。水溶性成分に富む抽出物は、水中にその水溶性抽出物を再懸濁させ、遠心分離後に上清を回収することにより調製した。親油性成分に富む可溶性抽出物は、アセトン中にその抽出物を再懸濁させ、その後、上清を乾燥させ、ペレットを、MAO−B投与と相溶性のある溶媒である、DMSO中に再懸濁させることによって得られた。
【0060】
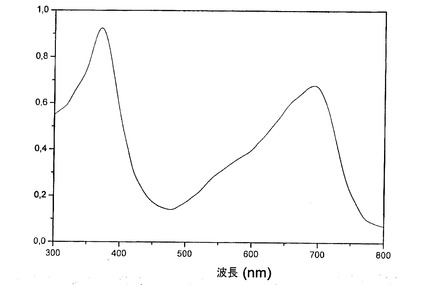
図7Aに示す通り、その水溶性分画が、MAO−Bを用量依存的な方法で阻害するのに対して、その親油性分画は、酵素を阻害しない。AFA基本抽出物の水溶性分画は、6.9μLのIC50を有する、強力な選択的MAO−B阻害物質である。そのMAO−B選択性は、4である(IC50 MAO−B/IC50 MAO−A>4.05)(図7B)。
【0061】
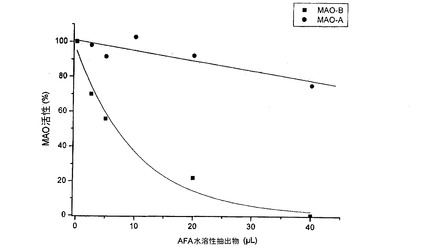
図8のラインウィーバー−バークプロットは、このような阻害が、可逆的で、競合に関して混合型であり、Vmaxの低下およびミカエリス−メンテンによるKm定数の上昇を伴うことを示す。AFA抽出物の水溶性分画の濃度に対して傾きをプロットすると、1μLの阻害定数Kiが得られる。基本抽出物の水溶性分画と比べると、この低いKi値は、MAO−B酵素に対する高い親和性を示す。
【0062】
その抽出物の阻害が可逆的であるという事実は、この阻害が、もっともらしく副作用なしに生理活性を及ぼすことを意味する。混合型競合については、競合的でもあれば非競合的でもある、異なる機能分子を含む、抽出物の複合的な性質に起因する可能性が極めて高い。その抽出物の主要活性成分は、AFAフィトクロム(0.5% DW)、フィコシアニン(8〜10% DW)、およびMAAまたはマイコスポリン様アミノ酸(1.7〜2.1% DW)であり、本発明者らは、これらを個別に、MAO−B阻害物質として試験した。
【0063】
フィコシアニンによるMAO−B阻害
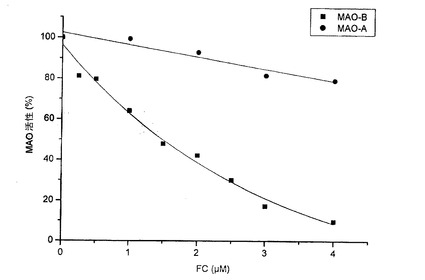
その試験は、基質としてベンジルアミンを用い、MAO−Bを、AFAから精製した各濃度のPC(0.5〜4μM)とともにプレインキュベートすることにより、波長250nm、30℃で分光光度計により実施した。図9に示す通り、AFA−PCは、1.44μMのIC50により、MAO−B活性の用量依存的な低下をもたらした。AFA−PCのMAO−B選択性は、3.5より高い(IC50MAO−B/IC50MAO−A>3.5)。
【0064】
図10におけるラインウィーバー−バークプロットは、その抽出物の場合と同様に、阻害が可逆的で、VmaxおよびKm双方の変更を伴う混合型(競合的および非競合的)であることを示す。
【0065】
PC濃度に対して傾きをプロットすることにより、本発明者らは、この場合1.06μMである、阻害定数Kiの値を得る。阻害定数は、酵素に対する阻害物質の親和性を評価する。高いKiが酵素に対する低親和性を示し、逆もまた同様である。この場合、低いKi値は、MAO−Bに対するAFA−PCの高い親和性を示す。
【0066】
MAAによるMAO−B阻害
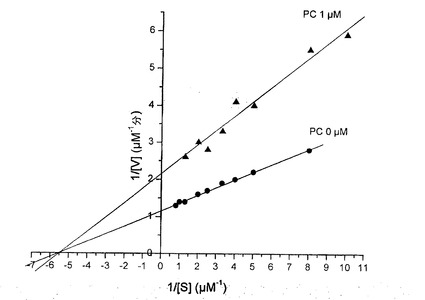
ベンジルアミン基質上におけるMAO−B活性を、20%メタノールにより基本抽出物からあらかじめ精製した、MAA濃度の上昇(0.5〜8μM)との関連で評価した。図11は、1.98μMのIC50を有する、MAAによる用量依存的なMAO−B阻害を示す。MAAのMAO−B選択性は、2より高い(IC50MAO−B/IC50MAO−A>2.02)。
【0067】
ラインウィーバー−バークプロット(図12)は、阻害が、Kmの上昇を伴うがVmaxの変化は伴わず、可逆的かつ競合的であることを示す。これは、MAAが、化学構造により、酵素の活性部位に対する連結について基質と競合することを意味する。MAA濃度に対して傾きをプロットすることにより(図13)、本発明者らは、0.585μMであり、酵素に対するとても高度の親和性を示す、阻害定数Kiの値を得る。
【0068】
AFAフィトクロムによるMAO−B阻害
その試験は、基質としてベンジルアミンを用い、MAO−Bを、各濃度の精製済みAFAフィトクロム(8.3〜66.4nM)とともにプレインキュベートすることにより、波長250nm、30℃で分光光度計により実施した。図15に示す通り、AFAフィトクロムは、20.2nMという低いIC50により、MAO−B活性の用量依存的な低下をもたらす。
【0069】
図16におけるラインウィーバー−バークプロットは、その抽出物の場合と同様に、阻害が可逆的で、VmaxおよびKm双方の変更(modification)を伴う混合型(競合的および非競合的)であることを示す。
【0070】
AFAフィトクロム濃度に対して傾きをプロットすることにより、本発明者らは、この場合10.48nMである、阻害定数Kiの値を得る。阻害定数は、酵素に対する阻害物質の親和性を評価する。高いKiが酵素に対する低親和性を示し、逆もまた同様である。この場合、極めて低いKi値は、MAO−Bに対するAFAフィトクロムの極めて高い親和性を示す。
【0071】
MAAの競合的で可逆的な作用は、これらの分子をMAO−B阻害において極めて強力にする。実際、MAO−B阻害の競合的で可逆的な特徴は、高度の有効性と生理的で副作用のない活性とを同時に確保する。この意味で、その抽出物中に含まれるMAAは、その分子量および結果としての血液脳関門を容易に通過する能力にも起因して、in vivoにおいても、MAO−B阻害に由来する治療的効果を生じるために、決定的な成分を構成する。
【0072】
MAAもさることながら、フィトクロムは、今まで知られるすべての物質で最も強力なMAO−B阻害物質であることがわかっている。MAO−B酵素に対するその極めて高い親和性、および数ナノモルの用量におけるその有効な阻害は、この分子を、それ自体において完全な治療用作用物質とするだけでなく、1つまたは複数のAFA抽出物の高い神経学的な有効性への最も重要な寄与を提供すると思われる因子にさせる。
【0073】
MAAおよびフィトクロムに関するいくつかの考察は、フィコシアニンのin vivoにおける挙動にも適用することができることを付け加えるべきであろう。本発明者らは、PCが、in vivoにおける脳に対する神経保護効果をもたらし、そのために、PCが血液脳関門を通過しうることを知っている(44)。これは、PCが、in vivoにおいて脳におけるそのMAO−B阻害活性を実現しうることを意味する。発色団の分子量は、実際、700に過ぎず、MAAの分子量とあまりかわらない。フィコシアノビリンと構造的に類似する、フィトクロムの発色団である、フィトクロモビリンについても同じことがあてはまる。
【0074】
結論として、その抽出物およびその活性成分である、AFAフィトクロム、AFA−PCおよびMAA側のMAO−Bの阻害活性は、以下の表に示す通り、分子および抽出物がともに、薬理学的物質以上の、最高レベルの活性に位置し、いずれの試験天然分子よりもはるかに優れているため、極めて重要である(45)。
【0075】
【表3】
【0076】
表が示す通り、フィコシアニンおよびMAAのみが、1μMよりもわずかに高いIC50を有し、このため、デプレニル(0.31μM)のIC50に極めて近く、考察した他の分子のIC50よりも10倍低い。一方、AFAフィトクロムは、デプレニルのIC50よりも15倍低いIC50を有する。酵素に対する阻害物質の親和性を評価する阻害定数Kiについても同じことがあてはまる。AFAフィコシアニンは、コーヒーおよびタバコの非ハーマンアルカロイドと同様に(しかし、もちろん、これら2つの物質に関連する問題なしに)約1μMのKiを有する。一方、MAAおよびAFAフィトクロムのみが、デプレニルと共に、1μMより低いKiを有し、したがって、MAO−Bに対して極めて高い親和性を有する分子である。実際、AFAフィトクロムは、セレギリン/デプレニル以外で、そのKiが数ナノモルのオーダーにある唯一の天然分子である。しかし、セレギリン/デプレニルとAFA抽出物の分子との間には、本質的な差異がある。前者が非可逆的な阻害物質であり、このため、潜在的な副作用を特徴とするのに対して、クラマス湖産AFAのMAO−B阻害分子は、すべてが可逆的であり、合成分子に関連する問題のない生理活性を特徴とする。
【0077】
図14は、デプレニルに関してAFAの3つの分子のMAO−B阻害活性を図示する。基本抽出物(および他のAFA抽出物)中における3つの分子すべての相乗作用を考えると、基本抽出物の全体のMAO−B阻害活性は極めて高い結果となる。その中に存在するPEAの高い分量をも考えると、特に重要となる事柄である。もし本発明者らが基本抽出物を、そのPC含量に基づきデプレニルと比較する場合、本発明者らは、基本抽出物が、0.05μMという低いPC用量でIC50に達し、これが、デプレニルよりも7.5倍高い(および天然物質よりも数十倍高い)効力を示すということを得る。これは、基本抽出物中に含まれるフィトクロムの効力に照らして意味を持つ。実際、7.5倍とは、PCとMAAの阻害効力の間の平均であり、それは、デプレニルよりやや低く、そして、15倍高い、フィトクロムのそれ(阻害効力)である。(図17)。これは、精製済みAFA−PCと比べて、その抽出物のより高い効力が、大部分、フィトクロムに起因することをも示す。
【0078】
さらに、その抽出物は、そのMAO−B阻害が可逆的で主に競合的であり、このため、デプレニルおよび他の合成物質などの不可逆的分子と潜在的に関連する副作用がない、生理的に作用する天然物質であることの利点を依然として維持する(46)。
【0079】
その抽出物のさらなる利点は、他の分子と完全に相乗的に作用する、強力なドーパミン作動性神経調節物質である、フェニルエチルアミンの高含量であり、この相乗活性を、本発明者らは以下のようにまとめることができる。
【0080】
・ フェニルエチルアミンまたはPEAは、黒質線条体組織からのドーパミンの放出を刺激すること、およびドーパミン自体のシナプス後の再取り込みを阻害することの両方において、二重のドーパミン作動性活性を有する;
・ フィトクロム、MAAおよびフィコシアニンは、減少したMAO−B活性が、ドーパミンを含む、神経アミンのより長い寿命を示唆する限りにおいて、強力なMAO−B阻害物質として、またドーパミン作動性伝達をも増大させる;
・ フィトクロム、MAAおよびフィコシアニンは、MAO−B阻害物質として、また、それ自体がMAO−B酵素の脱アミノ化活性の対象である、フェニルエチルアミンの寿命および活性をも延長させ、結果として、PEAによって引き起こされる、より全般的な神経調節ならびにドーパミン作動性伝達および活性に対するさらなる支援の好循環を作り出す。
・ 最後に、それ自身またはその発色団の血液脳関門を越える能力と一緒に、フィコシアニンの強力な抗酸化および抗炎症活性のほかに、フィトクロムの極めて高度な抗酸化活性、およびそれほど強力ではないが顕著なMAAの抗酸化活性が、いかなる酸化的損傷および炎症による損傷から、異なる活性分子およびより一般的にそれらが作り出す神経的好循環を保護する、神経保護もまた、もたらす。
【0081】
神経保護
本発明者らは、グルタミン酸の神経毒性効果に対する、AFA抽出物、特異的なAFA−PC、およびその発色団であるPCBのほか、MAAの神経保護特性を調べた。
【0082】
グルタミン酸は、哺乳動物の中枢神経系における主要な興奮性神経伝達物質であるが、ニューロンにおけるそのNMDAサブタイプ受容体の過剰刺激はCa2+の大量の細胞内蓄積を引き起こし、細胞死をもたらす。ミトコンドリア内のCa2+蓄積に加えて、NMDA受容体刺激後、細胞質における遊離Ca2+の一過性の増加は、一酸化窒素(NO・)または、主に初代(primary)ニューロンにおいて、そのスーパーオキシド(O2・-)反応生成物、ペルオキシ亜硝酸(ONOO-)を形成する酵素、一酸化窒素シンターゼ(NOS)のニューロンのアイソフォーム(49)を活性化する。
【0083】
ニューロンのグルタミン酸への曝露は、若干変更した方法(50)に従い実施した。
培地を除去し、ニューロンをあらかじめ37℃に温めて緩衝処理したハンクス液(5.26mM KCl、0.43mM KH2H2PO4、132.4mM NaCl、4.09mM NaHCO3、0.33mM Na2HPO4、20mMグルコース、2mM CaCl2、および20mM HEPES、pH7.4)で1回洗浄し、あらかじめ37℃に温めて緩衝処理したハンクス液中で、複数の濃度のAFA抽出物(1〜50nM)、PC(10〜1000nM)、PCB(10〜1000nM)およびMAA(1〜10μM)の不在または存在下においてプレインキュベートした。30分間のプレインキュベーション後に、Lグルタミン酸を濃縮液から100μMさらに10μMのグリシン、を示す最終濃度まで添加された。37℃で15分間ニューロンをインキュベートし、その緩衝液を吸引し、DMEMで置換し、細胞をエフェクター不在下においてさらに24時間37℃でインキュベートした。
【0084】
DNAに結合し、アポトーシスニューロン、すなわち、細胞核の断片化または凝縮化を示すニューロンの定量を可能とする、膜透過性蛍光染料である、DAPI(50)で細胞核を染色することにより、アポトーシスを評価した。簡潔に言うと、グルタミン酸曝露後の24時間、ニューロン培養物を温熱PBS(37℃)で洗浄し、室温で30分間、PBS中に4%(wt/vol)のパラホルムアルデヒドで固定した。PBSによる洗浄後、暗所内室温で10分間、細胞を3μM DAPIに曝露し、次いで、PBSで2回洗浄した。フルオレセインフィルター(330〜380の励起、30Xの倍率)を用いて、蛍光顕微鏡によりクロマチン凝縮について、細胞を記録した。総細胞核およびアポトーシス細胞核をカウントした。いずれの場合も、プロトコールデザインについて明るくない実施者により、ウェル当たり(per well)約600〜1,000個の細胞がカウントされた。個々の培養物に基づく測定は重複して(duplicate)実施し、結果は、表示された培養調製物数の平均(mean)SEM値として表す。その結果の統計学的解析は、クルスカル−ワリス検定の後、最小有意差多重範囲検定により判定した。いずれの場合も、p−0.05を有意と考えた。
【0085】
このグルタミン酸損傷試験により、本発明者らは、AFA基本抽出物、AFA−PC、そのPCB、およびMAAの神経保護能を初めて示した。図18が示す通り、培養済みニューロン細胞へのグルタミン酸の添加は、アポトーシスレベルを22.9%±3n=4(p<0.05)の比率まで上昇させたのに対し、
そのAFA基本抽出物の同時の添加は、グルタミン酸毒性に対する極めて高い保護をもたらし、既に1nMのような低量の抽出物によって、アポトーシスレベルを対照レベル(6.3±1 p>0.05)未満に低下させた(結果は、3〜8種類の異なる細胞培養物の平均±SEM。#対照群と比べたとき、有意に異なる(p<0.05);*グルタミン酸対照と比べたとき、有意に異なる(p<0.05))。MAAが与える保護については、それらはまた、より高用量の1μMで、アポトーシスレベルを対照レベル未満に低下させる(図19)結果は、3〜8種類の異なる細胞培養物の平均±SEM。#対照群と比べたとき、有意に異なる(p<0.05);*グルタミン酸対照と比べたとき、有意に異なる(p<0.05))。AFA−PCおよびPCBについて、本発明者らは、それらのアポトーシス阻害が極めて類似し、細胞培養物へのそれらの添加が、約10nMの用量で、アポトーシスの程度を対照未満に低下させるということがわかる(図20および21:結果は、3〜8種類の異なる細胞培養物の平均±SEM。#対照群と比べたとき、有意に異なる(p<0.05);*グルタミン酸対照と比べたとき、有意に異なる(p<0.05))。
【0086】
AFA−PCによる阻害の程度は、PCBの場合とほぼ等しい。おそらくその最も活性な成分と思われるPCBが、一度精製されひいてはより濃縮されると、その活性成分である全分子よりも顕著に強力であるべきことを踏まえると、これは、いささか驚くべきことである。それが実質的に同じ効力を有する事実は、全PCにおいて、PCB自体よりも実際にはさらに強力でありうる他の因子が存在することを意味する。本発明者らは、全PCが、C−PCおよびそのPCB発色団のほかに、PCBおよびPVB(フィコビオロビリン)をともにその発色団として含む、PECからもなることを知る。したがって、本発明者らは、本明細書において、精製済みPCBと全PCとの間の効力での顕著な差異を創出する因子は、正確にPEC成分、特に、極めて強い抗酸化物質であると推定される、そのPVB発色団であるということを推定することができる。
【0087】
神経保護の観点では、MAAは役割を果たすと思われるが、PCおよびPCBよりも顕著に低度である。しかし、最も強力な神経保護物質は、明らかに、全AFA抽出物であり、これは、わずか1nM(ナノモル)で細胞アポトーシスを完全に阻害することができる。これは、PCおよびPCBの効力の10倍である。これは、全AFA抽出物中に存在する多数の異なる抗酸化因子の相乗作用により確かに説明しうるが、本発明者らは、上述で、AFAフィトクロムが、今までおそらく最も強力な抗酸化物質であり、わずか16ナノモルでMDA(脂質過酸化の後期副生成物)の形成をほぼ完全に阻害しうる、ということを見たので、AFAフィトクロムが、基本抽出物のより高い効力を説明する中で重要な因子である可能性が極めて高い。こうして、本発明者らは、AFAフィトクロムが、任意のそしてすべてのフィトクロムと同様に、重要な神経保護作用物質であると結論付けることができる。
【0088】
参考文献
1. Zhou G. et al., Platelet monoamine oxidase B and plasma β-phenylethylamine in Parkinson's disease, in J Neurol Neurosurg Psychiatry, 2001; 70:229-231, 229.
2. Ispida K. et al., β-phenylethylamine stimulates striatal acetylcoline release through activation of the AMPA glutamatergic pathway, in Biol Pharm Bull 2005 Sep.; 28(9):1626-9.
3. Barroso N., Rodriguez M., Action of β-phenylethylamine and related amines on nigrostriatal dopamine neurotransmission, in European Journal of Pharmacology, 297 (1996), 195-203, 200.
4. Dyck L.E., Release of monoamines from striatal slices by phenelzine and β-phenylethylamine, in Prog Neuropsychopharmacol Biol Psychiatry, 1983, 7:797-800; Philips S.R., Robson A.M., In vivo release of endogenous dopamine from rat caudate nucleus by phenylethylamine, in Neuropharmacology 1983, 22:1297-1301; Raitieri m., et al., Effect of sympathomimetic amines on the synaptosomal transport of noradrenaline, dopamine and 5-hydroxytryptamine, in Eur J Pharmacol 1977, 41:133-143.
5. Janssen P.A.J, et al., Does phenylethylamine act as an endogenous amphetamine in some patients?, in International Journal of Neuropsychopharmacology 1999, 2: 229-240, 232.
6. Paterson I.A. et al., 2-phenylethylamine: a modulator of catecholamine transmission in the mammalian central nervous system?, in Journal of Neurochemistry (1990), 55:1827-1837.
7. Sabelli H.C., Javaid I.J., Phenylethylamine Modulation of Affect: Therapeutic and Diagnostic Implications, in Journal of Neuropsychiatry (1995), 7(1):6-14, 7.
8. Mauro Federici et al., Trace Amines Depress Gabab Response In Dopaminergic Neurons By Inhibiting Girk Channels, in Molecular Pharmacology Fast Forward. Published on January 11, 2005 as doi:10.1124/mol.104.007427.
9. Gusovsky F. et al., A high pressare liquid chromatography method for plasma phenylacetic acid, a putative marker for depressive disorders, in Anal Biochem, 1985 Feb. 15; 145(1):101-5. この研究では、うつ状態の患者は、対照群の536.18+/−54.99ng/mlに対して、327.64+/−45.44ng/mlの血漿中PAAレベルを有する。別の研究では、うつ状態の患者の尿において、うつではない患者の104+/−23mg/dieに対して、66+/−23mg/dieの平均PAAであることがわかった。参照:Sabelli HC. et al., Urinary phenylacetic acid in panic disorder with and without depression, in Acta Psychiatr Scand 1990 Jul;82(1):14-6.
10. Szabo A. et al., Phenylethylamine, a possible link to the antidepressant effects of exercise?, in Br J Sports Med 2001 Oct; 35(5):3423.
11. Sabelli H et al., Sustained antidepressant effect of PEA replacement, in J Neuropsychiatry Clin Neurosci, 8(2): 168-71.
12. Miura Y., Plasma beta-phenylethylamine in Parkinson's disease, in Kurume Med J 2000;47(4):267-72.
13. Ibid.,
14. Ebadi M. et al., Neuroprotective actions of selegiline, in J Neurosci Res 2002 Feb 1;67(3):285-289.
15. Kemppainen N. et al., Different pattern of reduction of striatal dopamine reuptake sites in Alzheimer's disease and ageing, in J Neural Transm 2001;108(7):827-36.
16. Knoll J., (-)Deprenyl (Selegiline): past, present and future, in Neurobiology (Bp) 2000;8(2):179-99.
17. Knoll J., The pharmacological basis of the beneficial effects of (-)deprenyl (selegiline) in Parkinson's and Alzheimer's diseases, in J Neural Transm Suppl 1993;40:69-91.
18. Rimbau V., et al., Protective effects of C-phycocyanin against kainic acid-induced neuronal damage in rat hippocampus, in Neurosci Lett 1999 Dec 3;276(2):75-8. この研究では、フィコシアニンは微細藻類スピルリナから用いられている。クラマス湖産藻類からのフィコシアニンは、異なっており、高い抗酸化活性を持っている。参照:Benedetti S., Scoglio S., Canestrari F., et al., Antioxydant properties of a novel phycocyanin extract from the blue-green alga Aphanizomenon Flos Aquae, in Life Sciences, 75 (2004): 2353-2362.
19. Swanson J. et al., Cognitive neuroscience of attention deficit hyperactivity disorder and hyperkinetic disorder, in Curr Opin Neurobiol. 1998 Apr;8(2):263-71.
20. Citazione solo di Benedetti et al. LifeScience; o menzione del parallelo brevetto? Attendere l'anno provisional in attesa di effettuare studi sulla neuroprotezione?
21. Kusaga A., Decreased beta-phenylethylamine in urine of children with attention deficit hyperactivity disorder and autistic disorder, in No To Hattatsu 2002 May; 34(3):243-8; Matsuishi T, Yamashita Y., Neurochemical and neurotransmitter studies in patients with learning disabilities, in No To Hattatsu 1999 May;31(3):245-8.
22. Kusaga A. et al., Increased urine phenylethylamine after methylphenidate treatment in children with ADHD, in Ann Neurol 2002 Sep;52(3):372-4.
23. Jain AK,. Et al., Bupropion SR vs. placebo for weight loss in obese patients with depressive symptoms, in Obes Res. 2002 Oct;10(10):1049-56.
24. Rudolph et al., A randomized, placebo-controlled, dose-response trial of venlafaxine hydrochloride in the treatment of major depression, in J Clin Psychiatry (1998); 59(3):116-22.
25. PEAは、熱によって非常に損傷を受けやすい脂溶性分子である。これは、凍結乾燥などの、高い温度を用いた乾燥方法が、通常PEAの低濃度を有することを意味する。PEAの最も高い含有量は、Refractance Window(登録商標) methodで乾燥された藻類に見られる。この基本抽出物が実現されるのは、この種類の藻類由来であるからである。
26. Yamada M. et al., Clinical Pharmacology of MAO Inhibitors: Safety and Future, in Neurotoxicology 2004; 25:215-21; Youdim M., et al., Therapeutic Applications of Selective and Non-Selective Inhibitors of Monoamine Oxidase A and B that do not Cause Significant Tyramine Potentiation, in Neurotoxicology 2004;25:243-50.
27. Groniger A et al., Photoprotective compounds in cyanobacteria, phytoplankton and macroalgae-a database, in J Photochem Photobiol B. 2000 Nov;58(2-3):115-22.
28. Suh HJ et al., Mycosporine glycine protects biological systems against photodynamic damage by quenching singlet oxygen with a high efficiency, in Photochem Photobiol. 2003 Aug;78(2):109-13.
29. Groniger A et al., Photoprotective compounds in cyanobacteria, phytoplankton and macroalgae-a database, in J Photochem Photobiol B. 2000 Nov;58(2-3):115-22.
30. Sinha RP et al., Induction of mycosporine-like amino acids (MAAs) in cyanobacteria by solar ultraviolet-B radiation, in J Photochem Photobiol B. 2001 Jul; 60(2-3):129-35.
31. Garcia-Pichel F et al., Occurrence of UV-Absorbing, Mycosporine-Like Compounds among Cyanobacterial Isolates and an Estimate of Their Screening Capacity, in Appl Environ Microbiol. 1993 Jan;59(1):163-169.
32. Glazer A.N., Phycobiliproteins, in Methods Enzymol, 1988, 167: 291303.
33. Bhat V.B., et al., C-phycocyanin: a potent peroxyl radical scavenger in vivo and in vitro, in Biochem Biophys Res Commun., 2000; 275(1):20-25; Romay, C. et al., Antioxidant and antinflammatory properties of Cphycocyanin from blue-green algae, in Inflamm Res, 1998, Jan.; 47(1): 3641.
34. Reddy C.M., et al., Selective Inhibition of cyclooxygenase-2 by Cphycocyanin, in Biochem Biophys Res Commun. 2000; 277(3): 599-603.
35. Gonzales R., et al., Anti-infl ammatory activity of phycocyanin extract in acetic acid induced colitis in rats, in Pharmacol Res, 1999; 39(1): 55-9.
36. Gonzales R., et al., Anti-infl ammatory activity of phycocyanin extract in acetic acid induced colitis in rats, in Pharmacol Res, 1999; 39(1): 55-9.
37. Vadiraja BB. et al., Hepatoprotective effect of Cphycocyanin:protection for carbon tetrachloride and R-(+)-pulegone-mediated hepatotoxicty in rats, in Biochem Biophys Res Commun, 1998; 249(2):428-31.
38. Romay C., et al., Phycocyanin extract reduces leukotriene B4 levels in arachidonic induced mouse-ear infl ammation test, in J Pharm Pharmacol. 1999,51(5):641-42. Come e noto, il leucotriene B4 e uno dei fattori principalmente responsabili di patologie respiratorie quali asma e allergie.
39. Rimbau V., et al., Protective effects of C-phycocyanin against kainic acid-induced neuronal damage in rat hippocampus, in Neurosci Lett 1999, 276(2):75-8.
40. Rimbau V. et al., C-phycocyanin protects cerebellar granule cells from low potassium/serum deprivation-induced apoptosis, in Naunyn Schmiedebergs Arch Pharmacol 2001; 364(2): 96-104.
41. Glazer A.N., Phycobilisomes, in Methods Enzymol 1988, 167;304-312.
42. Hirata T., et al., Antioxidant avtivities of phycocyanobilin prepared from Spirulina platensis, in J Appl Phycol 2000, 12:435-439.
43. Fuglistaller P., et al., Isolation and characterization of phycoerythrocyanin and chromatic adptation of the thermophilic cyanobacterium Mastigocladus laminosus, in Arch Microbiol 1981, 129:268274.
44. Rimbau V., et al., Protective effects of C-phycocyanin against kainic acid-induced neuronal damage in rat hippocampus, in Neurosci Lett 1999, 276(2):75-8.
45. この表に記載されているデータは、以下の研究を引用したものである: Magyar K. et al., Pharmacological aspects of (-)-deprenyl, in Curr Med Chem, 2004 Aug, 11(15):2017-31; Hou et al., Monoamine oxidase B (MAO-B) inhibition by active principles from Uncaria rhyncophylla, in Journal of Ethnopharmacology 100 (2005) 216-220; Herraiz T, Chaparro C., Human monoamine oxidase is inhibited by tobacco smoke: β-carboline alkaloids act as potent and revesible inhibitors, in Biochemical and Biophysical Research Communications 326 (2005) 378-386; Kong LD et al., Inihibition MAO-A and B by some plantderived alkaloids, phenols and anthraquinones, in Journal of Ethnopharmacology 91 (2004) 351-355.
46. Yoshida S. et al., Fluorinated phenylcyclopropylamines. Part 3: Inhibition of monoamine oxidase A and B, in Bioorganic & Medicinal Chemistry 12 (2004) 2645-2652.
47. Torres A. et al., Porphyra-334, a potential natural source for UVA protective sunscreens, in Photochem. Photobiol. Sci. 5 (2006) 432-435.
48. Hughes J, Lamparter T., Prokaryotes and Phytochrome. The Connection to Chromophores and Signaling, in Plant Physiology, December 1999, Vol. 121, pp. 1059-1068.
49. Garthwaite et al., Endothelium-derived relaxing factor release on activation of NMDA receptors suggests role as intercellular messenger in the brain, in Nature. 1988 Nov 24;336(6197):385-8.
50. Delgado-Esteban M. et al., D-Glucose prevents glutathione oxidation and mitochondrial damage after glutamate receptor stimulation in rat cortical primary neurone, in J Neurochem. 2000 Oct;75(4):1618-24.
【技術分野】
【0001】
本発明は、微細藻類アファニゾメノン・フロスアクアエ(Aphanizomenon Flos Aquae Aquae Ralfs ex Born.& Flah.Var.flos aquae)(クラマス湖産AFA)に関する。より正確には、本発明は、神経系、神経変性および気分病態(conditions)または疾患(diseases)の予防または治療に有用なクラマス湖産AFAの抽出物およびその精製成分を提供する。
【背景技術】
【0002】
発明の背景
フェニルエチルアミン(PEA)は、黒質線条体系のドーパミン作動性ニューロンにおいてフェニルアラニンの脱炭酸化により合成される内因性アミンであり、脳におけるカテコールアミン神経伝達の神経調節物質として作用しうる(1)。PEAの最も重要な作用は、カテコールアミン神経伝達を促進することである。PEAは、ドーパミンと同様アセチルコリンの放出を刺激することが知られる(2)。さらに、PEAは、ノルエピネフリン神経伝達(NE)(6)、およびセロトニン神経伝達さえも増大させる。
【0003】
近年、PEAは、その特異的なニューロン受容体とともに、自律的神経伝達物質としてもまた働きうること、また、必要な場合は、神経伝達物質を抑制することもまた可能な、真の神経調節物質として作用することが示されている(8)。
【0004】
注意および記憶の刺激;顕著な抗うつ活性を伴う気分の高揚;情動行動および性行動を含む、共感ひいては社会性の促進;空腹の抑制;薬物(substance)乱用および薬剤(drug)依存の必要性の低減といった、一連の非常に多くの効果が、このことに由来する。
【0005】
PEAと感情的な気分とのつながりは、血漿または尿中のそれ自体またはその代謝物であるPAA(フェニル酢酸)により測定される、PEAレベルの著しい低下が、うつ状態の対象において見出された、研究により確認されている(9)。
【0006】
パーキンソン病患者は、血漿中で直接測定されたPEAレベルが著しく低いことがわかっている(12)。これらの患者における、神経伝達、特にドーパミン作動性、の漸減は、黒質のドーパミン作動性ニューロンの進行性変性と関連する。
【0007】
パーキンソン病患者において、PEAレベルのこのような減少は、MAO−Bレベルの並行する上昇を伴い、このため、パーキンソン病において用いられる薬剤は、セレギリンなどのMAO−B阻害物質である(14)。さらに、ひとたび摂取されたPEAは、血液脳関門を容易に通過することができ、低用量時でさえも黒質線条体組織からのドーパミン放出を刺激することができる。これは重要な著しい特徴である。なぜなら、現在用いられる薬剤である、セレギリンは、MAO−Bおよびドーパミンの再取り込みを阻害する一方で、黒質線条体組織からのその放出に対しては作用せず、したがって、さらなるドーパミン産生の一助とはならず、まさにドーパミンの産生自体が大いに脅かされる、パーキンソン病などの病変において深刻な限界があるからである。
【0008】
アルツハイマー病は、ドーパミンの産生および再取り込み機構の変性、ならびに、時間経過とともにドーパミン作動性ニューロン数を減少させる、線条体領域のニューロンの進行性破壊、および結果として起こる、ドーパミン伝達の進行性破壊を伴う(15)。
【0009】
ADHD(注意欠陥過活動性障害)が、神経変性病変であるという事実に関する明確なデータは存在しないが、神経破壊が、小児および成人の双方において、ADHDの主要原因であることを、いくつかの研究が示そうとしてきた(19)。
【0010】
最も重要なことは、ADHDおよび学習障害に罹患した小児では、PEAレベルが顕著に低く(21)、したがって、注意(ドーパミン)および鎮静(セロトニン)の神経調節(neuromodulation)において減少する、証拠があることである。ADHDの選択薬が、PEAの、ひいては、ADHDの病因に直接関与する2つの神経伝達物質である、ドーパミンおよびノルエピネフリンの、より高い産生を刺激することによっても作用する、PEAの合成誘導体、メチルフェニデート(22)であるのはこのためである。
【0011】
アンフェタミンの使用が空腹および、結果として、体重を制御することはよく知られている。この領域におけるその使用は、その耐性をも踏まえると、時間経過とともに潜在的に極めて重度となる傾向のある、その副作用により、常に問題となってきた。このことは、空腹および体重の制御に現在用いられる主な薬剤が、ベンラファキシンおよびブプロプリオンなどの、アンフェタミン様ドーパミン作動性抗うつ剤である事実によって確認される。これらの分子は、すべてのアンフェタミンと同様、PEAの合成誘導体である。後者(ブプロプリオン)は、MAO−B酵素によるその分解が阻害される限りにおいて、強力な食欲抑制剤として作用する。
【0012】
モノアミンオキシダーゼ(MAO)AおよびBは、CNSおよび末梢組織における神経作用性アミンおよび血管作用性アミンの分解を触媒する。特に、MAO−Bは、ドーパミン作動性伝達に対するその直接および間接の関与性を踏まえると、うつ病および気分障害、パーキンソン病およびアルツハイマー病など、ドーパミンが重要な神経障害に関与する。このため、MAO−B阻害物質が、こうした神経障害の治療において用いられるのである(26)。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】1. Zhou G. et al., Platelet monoamine oxidase B and plasma β-phenylethylamine in Parkinson's disease, in J Neurol Neurosurg Psychiatry, 2001; 70:229-231, 229.
【非特許文献2】2. Ispida K. et al., β-phenylethylamine stimulates striatal acetylcoline release through activation of the AMPA glutamatergic pathway, in Biol Pharm Bull 2005 Sep.; 28(9):1626-9.
【非特許文献3】6. Paterson I.A. et al., 2-phenylethylamine: a modulator of catecholamine transmission in the mammalian central nervous system?, in Journal of Neurochemistry (1990), 55:1827-1837.
【非特許文献4】8. Mauro Federici et al., Trace Amines Depress Gabab Response In Dopaminergic Neurons By Inhibiting Girk Channels, in Molecular Pharmacology Fast Forward. Published on January 11, 2005 as doi:10.1124/mol.104.007427.
【非特許文献5】9. Gusovsky F. et al., A high pressare liquid chromatography method for plasma phenylacetic acid, a putative marker for depressive disorders, in Anal Biochem, 1985 Feb. 15; 145(1):101-5.
【非特許文献6】12. Miura Y., Plasma beta-phenylethylamine in Parkinson's disease, in Kurume Med J 2000;47(4):267-72.
【非特許文献7】14. Ebadi M. et al., Neuroprotective actions of selegiline, in J Neurosci Res 2002 Feb 1;67(3):285-289.
【非特許文献8】15. Kemppainen N. et al., Different pattern of reduction of striatal dopamine reuptake sites in Alzheimer's disease and ageing, in J Neural Transm 2001;108(7):827-36.
【非特許文献9】19. Swanson J. et al., Cognitive neuroscience of attention deficit hyperactivity disorder and hyperkinetic disorder, in Curr Opin Neurobiol. 1998 Apr;8(2):263-71.
【非特許文献10】21. Kusaga A., Decreased beta-phenylethylamine in urine of children with attention deficit hyperactivity disorder and autistic disorder, in No To Hattatsu 2002 May; 34(3):243-8
【非特許文献11】21. Matsuishi T, Yamashita Y., Neurochemical and neurotransmitter studies in patients with learning disabilities, in No To Hattatsu 1999 May;31(3):245-8.
【非特許文献12】22. Kusaga A. et al., Increased urine phenylethylamine after methylphenidate treatment in children with ADHD, in Ann Neurol 2002 Sep;52(3):372-4.
【非特許文献13】26. Yamada M. et al., Clinical Pharmacology of MAO Inhibitors: Safety and Future, in Neurotoxicology 2004; 25:215-21
【非特許文献14】26. Youdim M., et al., Therapeutic Applications of Selective and Non-Selective Inhibitors of Monoamine Oxidase A and B that do not Cause Significant Tyramine Potentiation, in Neurotoxicology 2004;25:243-50.
【発明の概要】
【0014】
発明の説明
本発明は、微細藻類アファニゾメノン・フロスアクアエ(Aphanizomenon Flos Aquae Aquae Ralfs ex Born.& Flah.Var.flos aquae)(クラマス湖産AFA)における、アルツハイマー病およびパーキンソン病、多発性硬化症、注意欠陥過活動性障害(ADHD)、自閉症、うつ病、記憶欠損および気分障害などの神経変性疾患を含む、各種の神経系の疾患、病態、機能不全または障害に対して、単独または組み合わせて有益な効果を及ぼす、物質の、同定に基づく。特に、クラマス湖産AFA微細藻類は、ドーパミン作動性およびノルアドレナリン作動性活性を特徴とする神経調節物質である、フェニルエチルアミン以外に、酵素モノアミンオキシダーゼB(MAO−B)の極めて効果的な阻害物質であることがまったく驚くべきことに示された特異的分子、すなわち:a)特異的なAFAフィトクロム;b)C−フィコシアニン(C−PC)およびフィコエリスロシアニン(PEC、その発色団であるフィコビオロビリンまたはPVBを含む)により形成されるフィコビリソームを含む、AFAフィコビリタンパク質複合体(complex)(「AFAフィコシアニン」);c)マイコスポリン様アミノ酸またはMAA、を含むことがわかっている。その藻類中に含まれるPEAが、MAO−B阻害物質により保護されない限り、MAO−B酵素による取り込みと同時に迅速に破壊されるため、この知見は極めて重要である。
【0015】
MAO−Bの選択的阻害物質として作用する同じ分子が、強力な神経保護物質としての役割をも果たし、したがって、神経学的な健康を促進するその抽出物の能力を顕著に高める。
【0016】
したがって、本発明は、クラマス湖産AFA調製物、特に、こうした活性成分に富む抽出物、あるいは、a)AFAフィトクロム;b)AFAもしくは他の任意の微細藻類中に存在する、C−フィコシアニン/フィコエリスロシアニン複合体;c)AFA中に存在するかもしくは他の任意の藻類供給源に由来する、マイコスポリン様アミノ酸ポルフィラおよびシノリン;またはd)その混合物から選択された単離および精製成分を、それを必要とする対象に投与することにより、上述の神経系の疾患、病態、機能不全または障害を予防し、制御し、または治療する方法を提供する。
【0017】
本発明に従うクラマス湖産AFA抽出物は、以下のステップにより調製されることが好ましい:
a)採取直後のAFA藻類を冷凍し解凍するか、または、出発物質が乾燥AFA粉末である場合は、水で希釈したAFA粉末を超音波で分解して、細胞を破砕する;
b)ステップa)の生成物を遠心分離し、沈殿物(細胞壁分画の大半を保持する)から上清(細胞質(cytoplasmatic)部分の大半を保持する)を分離する;
c)水溶性成分を含む上清を回収する。
【0018】
結果として得られる生成物が、PEAのほか、AFAフィトクロム、AFAフィコシアニン、およびMAAなど他の相乗作用分子をも濃縮する抽出物(「基本抽出物」と呼ぶ)である。例えば、クラマス湖産微細藻類が2〜4mg/gの範囲の天然PEA含量を有するのに対し、基本抽出物は、この濃度を、9〜11mg/gの範囲のレベルにまで上昇させる(HPLC分析)。
【0019】
抽出物を限外濾過システム、好ましくは、30,000ドルトン(Daltons)のカットオフ分子量を有する膜に通すことによりさらに精製することができる。限外濾過残留物(retentate)(抽出物A)は、AFAフィコシアニン(分子量=121,000)およびAFAフィトクロム(分子量=480,000)をともに主要な活性成分として含む。興味深いことに、MAAが、用いられるそのカットオフサイズを大きく下回る分子量を有するにもかかわらず、その残留物は、MAAの濃度も上昇させる。
【0020】
ステップa)〜c)により得られる、すなわち、限外濾過を伴わない基本抽出物は、最適量のPEA、AFAフィトクロム、AFA−PCおよびMAAを含むので、一般に好ましい。さらに、この基本抽出物はまた、低下した比率においてではあるが、その抗酸化および抗炎症特性に寄与する、クロロフィルおよびカロテンなどの物質をも含む。
【0021】
あるいは、クラマス湖産AFAの活性成分、すなわち、複合体C−フィコシアニン/フィコエリスロシアニン(C−PC/PEC)、AFAフィトクロム、およびMAAは、さらに以下に記載の通りに、単離および精製することができ、本発明に従う方法において用いることができる。
【0022】
好ましい実施形態において、クラマス湖産AFAのC−PC/PEC複合体、AFAフィトクロムおよびマイコスポリン様アミノ酸は、それを必要とする対象に対する同時または個別投与のための併用調製物として用いられ、さらに好ましい実施形態において、こうした併用調製物は、フェニルエチルアミンを追加の活性成分として含む。マイコスポリン様アミノ酸のうち、シノリンおよびポルフィラ334が、クラマス湖産AFA微細藻類中に比較的高濃度で含まれるので、特に好ましい。
【0023】
観察されたモノアミンオキシダーゼBの阻害は、ドーパミン作動性伝達を増大させ、PEAの異化を最小化することを可能とするので、特に重要である。意義深いことに、フィトクロムおよびAFAフィコシアニンがともにMAO−Bを可逆的および混合的な方法で阻害するのに対し、MAAによるMAO−B阻害は、競合的および可逆的であり、このため、3つの分子は全て、生理学的状態および副作用の不在下で高い有効性を確保する。
【0024】
さらなる態様において、本発明は、場合によってフェニルエチルアミンを添加した、AFA藻類中に存在するもしくは他の任意の藻類供給源に由来する、C−PC/PEC複合体、または単離したC−PCおよびPECの単体成分;AFAフィトクロム;AFA中に存在するもしくは他の任意の微細藻類供給源に由来する、マイコスポリン様アミノ酸ポルフィラおよびシノリン;またはその混合物から選択されることが好ましい、クラマス湖産AFA調製物、その抽出物またはその単離成分を含む栄養補助組成物または医薬組成物を意図する。好ましい実施形態において、栄養組成物は、錠剤、カプセル、飲料の形態における栄養補助食品であり、さらに好ましい実施形態において、医薬組成物は、錠剤、カプセル、小袋、シロップ、坐薬、バイアルおよび軟膏の形態であり、上記に示した神経系または神経変性の疾患もしくは病態の予防または治療に用いられることができる。本発明に従うクラマス湖産AFA液体抽出物は、それ自体で用いられるかまたは例えば凍結乾燥、噴霧乾燥などの手法により乾燥させることができる。その単離した活性成分は、当業者に知られる手法および以下の手順を用いて調合することができる。
【0025】
その活性成分の用量は、栄養補助食品としてかまたは医薬調製物としてかの、組成物の使用目的に依存するであろう。各成分の有効量は、一般に以下の範囲に含まれるであろう。
PEA=0.1〜100mg、好ましくは、5〜30mg;
フィトクロム=0.1〜1000mg、好ましくは、0.8〜10mg;
MAA=0.1〜1000mg、好ましくは、10〜100mg;
フィコシアニン=1〜2500mg、好ましくは、50〜1000mg。
【図面の簡単な説明】
【0026】
【図1】MAA試料の波長と吸光度の関係を示す図である。
【図2】MAA標準物質とMAA抽出物のクロマトグラムを示す図である。
【図3】精製済みMAAのUVスペクトルを示す図である。
【図4】AFAの細胞溶解物の成分とシネコシスチス属PCC6803のその成分との比較を示す図である。
【図5】抽出物の分光分析を示す図である。
【図6】PCB発色団の波長を示す図である。
【図7A】基本抽出物の水溶性および脂溶性成分のMAO−B阻害活性を示す図である。
【図7B】AFA水溶性抽出物のMAO−AおよびMAO−Bの阻害活性を示す図である。
【図8】水溶性抽出物のラインウィーバー−バークプロットを示す図である。
【図9】AFA−PCのMAO−AおよびMAO−Bの阻害活性を示す図である。
【図10】PCのラインウィーバー−バークプロットを示す図である。
【図11】MAAによる用量依存的なMAO−B阻害を示す図である。
【図12】MAAのラインウィーバー−バークプロットを示す図である。
【図13】MAA濃度と傾きの関係を示す図である。
【図14】、デプレニルに関してAFAの3つの分子のMAO−B阻害活性を示す図である。
【図15】AFAフィトクロムのMAO−B活性の用量依存的な低下を示す図である。
【図16】AFAフィトクロムのラインウィーバー−バークプロットを示す図である。
【図17】デプレニルおよびAFAフィトクロムのMAO−B阻害活性を示す図である。
【図18】アポトーシスへのグルタミン酸とAFA抽出物の添加の影響を示す図である。
【図19】アポトーシスへのグルタミン酸とAFA抽出物のMAAの添加の影響を示す図である。
【図20】アポトーシスへのグルタミン酸とAFA抽出物のPCの添加の影響を示す図である。
【図21】アポトーシスへのグルタミン酸とAFA抽出物のPCBの添加の影響を示す図である。
【図22】AFAフィトクロムのバンドを示す図である。
【図23】光吸収特性を示す図である。
【図24】AFAフィトクロム量のMDA形成の用量依存的減少を示す図である。
【発明を実施するための形態】
【0027】
発明の詳細な説明
クラマス湖産藻類の典型的な固有のフィトクロムである「AFAフィトクロム」の同定
フィトクロムは、植物が光を検出するのに用い、可視スペクトルの赤色および遠赤色領域における光に対して感受性がある、光受容体、色素である。これらは、(日周期のリズム(rythms)を通して)開花の調節、発芽およびクロロフィルの合成を含む、植物における多数の異なる機能を果たす。AFAにおけるこの固有の種類のフィトクロムの存在が、光合成過程においてCフィコシアニンを補うよう他のシアノバクテリアにより通常用いられる他のフィコビリタンパク質、すなわち、アロフィコシアニン、の欠如により説明しうるので、後者(クロロフィルの合成)は、AFA藻類との関連で特に重要である。一方、クラマス湖産藻類におけるアロフィコシアニンの位置は、フィコエリスロシアニンまたはPEC(下記参照)により占められるが、特に、クラマス湖産藻類が、高い光回収能力を必要とする非熱帯環境において生育することを考えると、PECのみでは十分ではないと思われ、したがって、AFA藻類は、そのより高い必要性をフィトクロムで調和させていると思われる。
【0028】
特異な構造を有するAFAフィトクロムは、本明細書において初めて記述される。長年にわたり、異なる種類のフィトクロムが植物中に見出されており、それらは、異なるフィトクロム遺伝子(例えば、コメにおいて3種類、トウモロコシにおいて6種類)を有するだけでなく、大半の場合、顕著に異なるタンパク質成分および構造を有する。これらがすべてフィトクロムであるのは、これらがすべて、フィトクロモビリンと呼ばれる、同じビリタンパク質を、光吸収発色団として、用いるからである。この発色団は、フィコシアニンの発色団であるフィコシアノビリンと類似し、4つのピロール環(テトラピロール)の開鎖からなる単一のビリン分子であることを特徴とする。より具体的に、その正常のPr状態において、このビリタンパク質は、650〜670nMの最大値で吸光するのに対し、赤色光により活性化されると、730nMの最大吸収を有するPfrに転換される。
【0029】
発見された最初のシアノバクテリアのフィトクロムは、シネコシスチス属のそれであり、植物フィトクロムとの構造的類似性がわずかであることを示した。にもかかわらず、シネコシスチス属のビリタンパク質は、赤色/遠赤色の可逆的色素タンパク質である限りにおいて、一般にフィトクロムと考えられる(48)。
【0030】
AFAフィトクロムの精製と特性解析
AFAフィトクロムは、赤色/遠赤色スペクトルにおいて吸光するその発色団として、ビリタンパク質を有する。その構造および活性を確立するため、本発明者らは、以下のプロトコールによりフィトクロムを精製した。
【0031】
・ 1gの抽出物を10mlの1Kリン酸緩衝液、pH7.0中に懸濁させる
・ その半分の体積で、1分間にわたるボルテックスを2回行う
・ 細胞を2%トリトンX100とともに35分間(35’)インキュベートする
・ 28000rpmで16〜18時間遠心分離する
・ スクロース密度段階勾配法により、上清を回収する
・ スイングアウトローターを用いて、150000gで12時間、その勾配分離物をスピンする
・ −20℃で保存する
フィトクロムが、約1Mのスクロースで可視化される、強い橙色の溶解物バンドに対応するのに対し、一方、フィコビリソームは、約0.75Mに位置する。2つのバンドのこの関係は、三量体AFA−PCの約4倍である、その藻類中に存在するフィトクロムの分子量についても、信頼できる指標を与える。後者(三量体AFA−PC)が121Kdであるので、本発明者らは、AFAフィトクロムのMWを約480Kdにおいてあらかじめ確立することができる(図22)。
【0032】
その光吸収特性について調べたところ、フィトクロムは、平衡状態を形成するPr(赤色光吸収)形態およびPfr(遠赤色光吸収)形態にそれぞれ対応する、672nMおよび694nMにおいて2つのピークを伴い吸光することを示す(図23)。
【0033】
AFA中に含まれるフィトクロムの量について、本発明者らの最初の評価は、以下の予備的な結果を与える:2mg/g(または0.2% DW)。その抽出物について、基本抽出物中では濃度が約0.5%に上昇し、抽出物B中では約1%に上昇した。これらは低濃度であるが、この分子の抗酸化/抗炎症効力は極めて強いので、ごく少量でさえも極めて重要な効果をもたらすことができる。
【0034】
抗酸化活性
精製済みAFAフィトクロムは、極めて強力な抗酸化剤であることを示した。実際、絶対的に、今までにクラマス湖産藻類中で見出された最も強力な分子である。100μMで酸化作用物質CuCl2とともにヒト血漿試料を2時間インキュベートすると、チオバルビツール酸との反応後535nmにおける分光光度計により計測(TBA試験)される、脂質過酸化の後期副生成物(a late byproduct)である、マロンジアルデヒド(MDA)レベルの上昇が生じる。AFA藻類から抽出したAFAフィトクロム量を増加させながら(2〜16nM)100μMでCuCl2とともに37℃で2時間、血漿をインキュベートすると、MDAレベルの極めて強力な用量依存的減少が観察される(図24)。実際、わずか16nMのAFAフィトクロムにより、対照に近いMDAレベルを有する脂質過酸化のほぼ完全な阻害が得られる。意義深いことに、3.6nMのIC50が、PCBについて得られるそれ(IC50)を45倍下回る。フィトクロムが、AFA−PCよりも高い、基本抽出物の抗酸化および神経保護効果の主要原因である。
【0035】
MAAの抽出、精製および定量
本発明者らは、一般にクラマス湖産藻類として知られている、クラマス湖産の藍色植物門のアファニゾメノン・フロスアクアエにおけるMAAの存在を調べた。本発明者らの知る限り、任意のアファニゾメノン属種におけるMAAの存在についてはごく最近の報告(47)が存在するのみであり、しかしながら、こうした報告が、ポルフィラを現存のMAAとして同定するのみであるのに対し、本発明者らの研究は、ポルフィラおよびシノリン双方の2つのMAAの存在を示す。一方、藻類に関する全般的な文献に関して、今まで報告されているシアノバクテリアの大半は、その主要MAAとしてシノリンを含むのに対して、本発明者らは、シノリンに加え、アファニゾメノン・フロスアクアエ中の主要MAAとして、ポルフィラ334のまれな存在を見出した。
【0036】
【化1】
【0037】
MAAは、既に報告されているように抽出された(29)。簡潔に言うと、45℃の水浴中に2.5時間インキュベートすることにより、2mlの20%(v/v)含水メタノール(HPLCグレード)中で20mgのAFA粉末または20mgの水性抽出物を抽出した。遠心分離(5000g;米国、パロアルト、Beckman社製、GS−15R型遠心分離機)後、上清を蒸発乾燥させ、2mlの100%メタノール中に再溶解し、2〜3分間ボルテックスし、10000gで10分間遠心分離した。上清を蒸発させ、HPLCでの分析用に同じ容量の0.2%酢酸中か、または、抗酸化特性の評価用に200μlのリン酸緩衝液(PBS)中に抽出物を再溶解した。試料は、HPLC分析、または抗酸化特性試験(下記を参照のこと)にかける前に、0.2μm孔径のシリンジフィルター(イタリア、ミラノ、VWR International社製)により濾過した。
【0038】
クラマス湖産藻類のMAAは、334nmの最大吸収を有する。MAAのさらなる精製は、文献(30)に従い、Alltima C18型カラムおよびガード(イタリア、ミラノ、Alltech社製、4.6×250mm内径、5μm充填剤)を装備したHPLCシステム(日本、東京、日本分光株式会社製)を用いて行われた。検出波長は330nm、移動相は流量1.0ml分-1で0.2%酢酸であった。MAAの同定は、ドイツ、エアランゲン、フリードリヒアレクサンダー大学、Manfred Klisch博士のご厚意により提供された、ポルフィラ334、シノリンおよびパリシンを主に含む、アマノリ属種およびオバクサ属種などの標準品と吸収スペクトルおよび保持時間を比較することにより行った。試料の吸収スペクトルは、シングルビーム分光光度計(米国、パロアルト、Beckman社製、DU640型)の200〜800nmにおいて測定した。生スペクトルは、コンピュータに転送し、MAAのピーク解析のために数学的に処理された。
【0039】
MAAは、前述のようにAFA試料およびその水性抽出物から部分的に精製された。45℃で2.5時間の20%メタノールによる試料の抽出は、この手順により、少量の光合成色素(620nmにおけるフィコシアニンなど)も抽出されたにせよ、334nmにおいて顕著なピーク(MAA)を生じた(図1の破線を参照のこと)。MAA試料は、タンパク質および塩を除去するために、100%メタノールによりさらに処理され、最終的に非極性光合成色素を除去するために、0.2%酢酸によりさらに処理された。その結果として得られる、部分的に精製されたMAAは、334nmに最大吸収を有した(図1、実線)。
【0040】
334nmにおいて吸収する化合物が、単一のMAAであるか、または1以上の混合物のMAAであるかを見出すため、HPLCによってMAAのさらなる解析および精製を行った。その試料のクロマトグラム(図2)は、シノリンおよびポルフィラ334としてそれぞれ同定された、保持時間4.2分(ピーク1)および7.6分(ピーク2)を有する2つのMAAの存在を示す。ポルフィラ334は、シノリンが少量のみで存在する(ピーク面積比1:15)ので、AFAにおける主要なMAAであると思われる。
【0041】
精製済みMAAのUVスペクトルは、334nmでその最大吸収が確認された(図3)。
【0042】
シノリンおよびポルフィラ334に対する334nmにおけるモル吸光係数が、それぞれ、44700および42300M-1cm-1であることを考慮し、本発明者らは、
a)AFA藻類について、シノリンに対して0.49mgg-1DWの濃度およびポルフィラ334に対して7.09mgg-1DWの濃度;したがって、そのMAA総含量は、0.76%藻類DWに等しい;
b)基本抽出物について、17〜21mgのMAA濃度(すなわち、1.7〜2.1% DW)
を計算した。
【0043】
全AFAが、UV曝露下で見出される最大濃度、すなわち、0.84%に近い、高構成レベルのMAA(0.76% DW)を含む(31)ので、これらは重要なデータである。また、本発明者らは、その抽出物が、全藻類よりもさらに高い濃度を有し、最大可能濃度よりもはるかに高いレベルに達していることをも見出した。
【0044】
MAA(シノリンおよびポルフィラ334)は、分子量300を有する、構造的に単純な分子である。このことは、それが最も必要とされる領域である、脳において、そのMAO−B阻害可能性を発現する能力を確かにし、これらの水溶性分子が血液脳関門を容易に通過することを可能にする。
【0045】
フィコシアニン
フィコシアニンは、8〜10%の濃度(定量については、下記を参照のこと)でその抽出物中に存在する。フィコシアニンは、特定の微細藻類ごとに特異的な特性を有するが、すべてのシアノバクテリアまたは藍藻類の、典型的な青色色素である(32)。フィコシアニンの機能的および治療的特性について、現在まで、研究は、主に、微細藻類スピルリナの特性に焦点を合わせている。スピルリナから精製したフィコシアニンは、肝臓(37)、呼吸器系(38)、および脳(39、40)などの異なる生理系での抗酸化(33)および抗炎症(34、35、36)特性を有することを示している。スピルリナから精製したPCのこうした特性は、その実質的な類似性を踏まえると、一般に、他の藻類フィコシアニンにも帰属させることができる。にもかかわらず、上述の機能的および治療的特性の説明における異なる効力が生じうる、異なる微細藻類に由来する異なるフィコシアニンには種特異的な差異が存在しうる。
【0046】
クラマス湖産藻類のフィコビリソームの構造決定および特異的特性
一般的に言うと、無傷のシアノバクテリア細胞において、フィコシアニン(PC)は、機能的形態(αβ)6でフィコビリソームの内部に存在する(41)。その細胞の破砕後、タンパク質は、分析される生体に応じて、異なる凝集状態(単量体、二量体、三量体、六量体)で見出すことができる。クラマス湖産藻類の場合、抽出物中に含まれるおよび抽出物自体から精製される両方の、PCの電気泳動解析は、タンパク質が、大半の場合、総分子量121000を有する、その三量体形態(αβ)3で見出されることを示している。単量体αβは、約40000(18500のサブユニットα+21900のサブユニットβ)の分子量である。これとは別に、スピルリナから精製したFCに関する研究の大半が、タンパク質は、スピルリナにおいて約37500の分子量を有する単量体形態αβで見出され、したがって、AFAから精製したPCと比較して異なる凝集状態を示すことを示唆する。
【0047】
AFAのフィコビリソームのクロマトグラフィー解析は、他のシアノバクテリア種において見られるように、PCのαサブユニットが1つの補欠分子族に結合するのに対し、βサブユニットは2つの補欠分子族に結合することをも示している。その補欠分子族または発色団は、フィコシアノビリン(PCB)と呼ばれ、タンパク質の青色およびその抗酸化力の両者の原因である(42)。
【0048】
AFAとスピルリナとの根本的な差異は、フィコビリソームの異なる構造に基づく。スピルリナに対して、クラマス湖産AFAのフィコビリソームは、色素アロフィコシアニンを含まず、スピルリナには欠如する構造的要素、すなわち、フィコエリスロシアニン(PEC)、に結合する色素cフィコシアニンのみを含む。FECは、今日現在、限られた数のシアノバクテリア種中のみにおいて同定されている、光合成色素である(43)。PECは、FCの化学的構造と極めて類似する化学的構造を有し、単量体および三量体を形成するために会合する2つのサブユニットαおよびβにより構成される。にもかかわらず、PCの各単量体がPCBの3分子に結合するのに対し、PECはPCBの2分子をサブユニットβに、また、フィコビオロビリン(PVB)の1分子をαサブユニットに結合させる固有の特性を有し、これが、その色素の紫色の原因となっている。
【0049】
クラマス湖産藻類のフィコビリソームが、cフィコシアニンおよびフィコエリスロシアニンの結合体により特異的に構成されると定義されるのはまったく初めてのことであり、クラマス湖産AFA藻類フィコビリソームの、この異なる性質的構造が、スピルリナからAFAを区別するさらに決定的な因子を追加する。
【0050】
図4は、AFAの細胞溶解物の成分を、別のよく知られたシアノバクテリアである、シネコシスチス属PCC6803の、成分と比較して、言われていることを確認する。両シアノバクテリアにおいても、フィコビリソームを表す青色バンドを見ることができるが、AFA藻類におけるフィコビリソームは、より低い分子量を示し、スピルリナなど一般的な微細藻類とは異なり、AFAフィコビリソームにおいては、アロフィコシアニンではなく、フィコシアニンのみが存在することを確認する。さらに、図は、AFA中にフィコエリスロシアニンに典型的な明るい紫色のバンド(矢印により示す)もまた存在することを示し、これにより、クラマス湖産藻類フィコビリソーム中におけるその存在を提供する。
【0051】
定義を深めるため、各青色バンドは、質量分析計に接続したHPLC(RP−HPLC−ESI−MS)によりさらに解析した。保持時間の差により、フィコビリソームのタンパク質を分離し、その分子量に基づきこれを同定した。得られた結果を、以下の表に示す。第1に、本発明者らは、シネコシスチス属(表1)においては、フィコシアニン(28.2分におけるcpcAおよび28.9分におけるcpcB)およびアロフィコシアニン(30.7分におけるapcAおよび31.2分におけるapcB)がともに存在するのに対し、AFA(表2)においては、フィコシアニン(28.8分におけるcpcAおよび30.0分におけるcpcB)のみが存在することがわかる。第2に、AFAにおいては、シネコシスチス属中には存在せず、2つのビリンが付着したフィコエリスロシアニンのベータサブユニットに対応する、分子量19469のタンパク質(25分におけるpecB)が同定されている。
【0052】
【表1】
【0053】
【表2】
【0054】
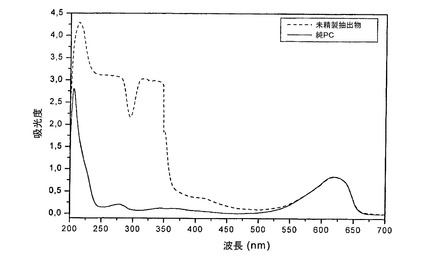
この固有の構造が、全AFA−PCの、そのPCBと比べてより強力な抗酸化および抗炎症作用を説明するのに重要な要素である。抗酸化および抗炎症特性は、これらが強力な神経保護をもたらす限りにおいて、この文脈で重要となり、全PCは、神経保護の観点でもそのPCBよりも強力であり、これは、フィコビリソーム中のPCB以外の他の活性成分、すなわち、その特異的なPVB発色団を有するPECが、AFA−PCにおける最も有効な健康増進主要成分である可能性が極めて高いことを明確に示唆する。精製済みAFA−PCが、そのPCB発色団を有するC−PCだけでなく、PECおよびそのPVB発色団をも含むことは、精製の結果として得られる抽出物の分光分析を参照することにより明らかである(図5)。実際、C−PCの最大吸収は、図5の分光分析においてピークの頂点を表す、620nmである。しかし、PECの最大吸収は、αサブユニット(フィコビオロビリンまたはPVB)について566nm、およびβサブユニットの2つのPCBについてそれぞれ593nmおよび639nmであることも知られている。3つの値すべては、実際、精製済みPCの分光分析プロファイルを構成する釣り鐘型ピークに含まれている。AFA藻類におけるC−PCとPECとの間の、極めて断ち切りがたい、強い連結を考えると、これは、C−PC以外にPECも、必然的に、精製済みPC抽出物の一部であるということを裏付ける。これは、同様に、AFAに由来するPCが、大半の研究がなされてきた、スピルリナ由来のそれ(PC)を含む、他のシアノバクテリアのPCとは、構造および機能の両面で顕著に異なること、および、この差異が、共通の一部分のみ、すなわちC−Pを有し、その他は有しないことにあり、C−PCの特性がAFA−PCのC−PC成分にも帰属させることができるのに対し、AFAに由来する全PCの、C−PC/PEC複合体(その発色団であるPCBおよびPVBを含む)であることにおける、特性は、(他の任意の微細藻類中に存在する任意のC−PC/PEC複合体と同様に)もっぱらこの複合体のみに帰属するという帰結を伴うことを意味する。
【0055】
精製手法(図5)
PCは、以下の通りに、乾燥AFA抽出物から精製した。
・ 500mgの抽出物を、50mlの100mMリン酸Na緩衝液pH7.4中に懸濁させる;
・ 4℃、2500rpmで10分間(10’)遠心分離する;
・ 上清を回収し、固体の硫酸アンモニウムを50%飽和まで添加する;
・ 試料を攪拌状態に保ちつつ、4℃で60分間タンパク質を沈殿させる;
・ 4℃、10,000rpmで30分間遠心分離する;
・ 無色透明の上清を廃棄し、少量の5mMリン酸Na緩衝液pH7.4中に青色の沈殿物を再懸濁させる;
・ 同じ緩衝液に対して、4℃で一晩透析する;
・ 5mMのリン酸Na緩衝液pH7.4で平衡化したヒドロキシアパタイト製カラム内に透析済みPCを入れる;
・ イオン強度を上昇(5〜150mM)させているリン酸Na緩衝液pH7.0で試料を溶出する;
・ その分画を回収し、分光光度計により620nmおよび280nmにおける吸光度を読み取る;
・ Abs620/Abs280>4(純PCの指標)である分画をプールする;
・ 50%飽和の硫酸アンモニウムにより、4℃で1時間PCを沈殿させる;
・ 4℃、10,000rpmで30分間(30’)遠心分離する;
・ 上清を廃棄し、150mMのリン酸Na緩衝液pH7.4中にPCを再懸濁する;
・ 同じ緩衝液に対して、4℃で透析する;
・ 精製済みPCをフラスコに移し、暗所中+4℃または−20℃で保存する。
【0056】
フィコシアニンの定量
純PCのモル濃度を測定するため、本発明者らは、三量体形態(αβ)3では770000M-1cm-1に等しい、620nmにおけるモル吸光係数εを用いた。これは、1MのPC溶液が、620nmにおいて吸光値770000を有することを意味する。
【0057】
その抽出物中のPC濃度を測定するため、本発明者らは、620nmにおける比吸光度(the coefficient of specific extinction)E1%として70lg-1cm-1を用いた。これは、1%のPC(すなわち、1g/100ml)を含む溶液が、620nmにおいて70を吸収することを意味する。これらの計算によれば、その抽出物中の平均PC含量は、80〜100mg/g DW(8〜10% DW)に等しい。
【0058】
PCB発色団の精製(図6)
・ 500mgの抽出物を、50mlの蒸留済みH2O中に懸濁させる。
・ 4℃、2500rpmで10分間(10’)遠心分離する。
・ 藍色の上清を静かに移し、1%のトリクロロ酢酸によりPCを沈殿させる。
・ 攪拌しつつ、暗所中4℃で1時間インキュベートする。
・ 4℃、10000rpmで30分間(30’)遠心分離する。
・ PCを含むペレットを回収し、メタノールで3回洗浄する。
・ 1mg/mlのHgCl2を含む10mlのメタノール中にそのペレットを再懸濁させる。
・ 暗所中42℃で20時間インキュベートし、PCからPCBを放出させる。
・ 2500rpmで10分間(10’)遠心分離し、タンパク質を除去する。
・ PCBを含む上清にβメルカプトエタノール(1μl/ml)を添加し、HgCl2を沈殿させる。
・ −20℃で24時間インキュベートする。
・ 4℃、10000rpmで30分間(30’)遠心分離し、白色の沈殿物を除去する。
・ 上清に10mlの塩化メチレン/ブタノール(2:1、v/v)を添加する。
・ 20mlの蒸留済みH2Oで洗浄し、3000rpmで10分間(10’)遠心分離する。
・ 上層を除去し、PCBを含む下層部分を回収する。
・ 15ml H2O中で、PCBを3回洗浄する。
・ 窒素下で乾燥させ、−20℃で保存する。
【0059】
クラマス湖産AFA抽出物およびそれを構成する主要な活性のある構成要素であるフィトクロム、フィコシアニンおよびMAAによるMAO−B阻害の評価
本発明者らは、特異的基質であるベンジルアミン(1mM)を用いて、基本抽出物のMAO−B阻害活性を試験した。その試験は、上記のステップa)〜c)によって取り出した(初期濃度10mg/ml)、基本抽出物の水溶性および脂溶性成分の異なる濃度とともに、MAO−B(2μg/ml)をプレインキュベートすることにより、波長250nm、30℃で分光光度計により実施した。水溶性成分に富む抽出物は、水中にその水溶性抽出物を再懸濁させ、遠心分離後に上清を回収することにより調製した。親油性成分に富む可溶性抽出物は、アセトン中にその抽出物を再懸濁させ、その後、上清を乾燥させ、ペレットを、MAO−B投与と相溶性のある溶媒である、DMSO中に再懸濁させることによって得られた。
【0060】
図7Aに示す通り、その水溶性分画が、MAO−Bを用量依存的な方法で阻害するのに対して、その親油性分画は、酵素を阻害しない。AFA基本抽出物の水溶性分画は、6.9μLのIC50を有する、強力な選択的MAO−B阻害物質である。そのMAO−B選択性は、4である(IC50 MAO−B/IC50 MAO−A>4.05)(図7B)。
【0061】
図8のラインウィーバー−バークプロットは、このような阻害が、可逆的で、競合に関して混合型であり、Vmaxの低下およびミカエリス−メンテンによるKm定数の上昇を伴うことを示す。AFA抽出物の水溶性分画の濃度に対して傾きをプロットすると、1μLの阻害定数Kiが得られる。基本抽出物の水溶性分画と比べると、この低いKi値は、MAO−B酵素に対する高い親和性を示す。
【0062】
その抽出物の阻害が可逆的であるという事実は、この阻害が、もっともらしく副作用なしに生理活性を及ぼすことを意味する。混合型競合については、競合的でもあれば非競合的でもある、異なる機能分子を含む、抽出物の複合的な性質に起因する可能性が極めて高い。その抽出物の主要活性成分は、AFAフィトクロム(0.5% DW)、フィコシアニン(8〜10% DW)、およびMAAまたはマイコスポリン様アミノ酸(1.7〜2.1% DW)であり、本発明者らは、これらを個別に、MAO−B阻害物質として試験した。
【0063】
フィコシアニンによるMAO−B阻害
その試験は、基質としてベンジルアミンを用い、MAO−Bを、AFAから精製した各濃度のPC(0.5〜4μM)とともにプレインキュベートすることにより、波長250nm、30℃で分光光度計により実施した。図9に示す通り、AFA−PCは、1.44μMのIC50により、MAO−B活性の用量依存的な低下をもたらした。AFA−PCのMAO−B選択性は、3.5より高い(IC50MAO−B/IC50MAO−A>3.5)。
【0064】
図10におけるラインウィーバー−バークプロットは、その抽出物の場合と同様に、阻害が可逆的で、VmaxおよびKm双方の変更を伴う混合型(競合的および非競合的)であることを示す。
【0065】
PC濃度に対して傾きをプロットすることにより、本発明者らは、この場合1.06μMである、阻害定数Kiの値を得る。阻害定数は、酵素に対する阻害物質の親和性を評価する。高いKiが酵素に対する低親和性を示し、逆もまた同様である。この場合、低いKi値は、MAO−Bに対するAFA−PCの高い親和性を示す。
【0066】
MAAによるMAO−B阻害
ベンジルアミン基質上におけるMAO−B活性を、20%メタノールにより基本抽出物からあらかじめ精製した、MAA濃度の上昇(0.5〜8μM)との関連で評価した。図11は、1.98μMのIC50を有する、MAAによる用量依存的なMAO−B阻害を示す。MAAのMAO−B選択性は、2より高い(IC50MAO−B/IC50MAO−A>2.02)。
【0067】
ラインウィーバー−バークプロット(図12)は、阻害が、Kmの上昇を伴うがVmaxの変化は伴わず、可逆的かつ競合的であることを示す。これは、MAAが、化学構造により、酵素の活性部位に対する連結について基質と競合することを意味する。MAA濃度に対して傾きをプロットすることにより(図13)、本発明者らは、0.585μMであり、酵素に対するとても高度の親和性を示す、阻害定数Kiの値を得る。
【0068】
AFAフィトクロムによるMAO−B阻害
その試験は、基質としてベンジルアミンを用い、MAO−Bを、各濃度の精製済みAFAフィトクロム(8.3〜66.4nM)とともにプレインキュベートすることにより、波長250nm、30℃で分光光度計により実施した。図15に示す通り、AFAフィトクロムは、20.2nMという低いIC50により、MAO−B活性の用量依存的な低下をもたらす。
【0069】
図16におけるラインウィーバー−バークプロットは、その抽出物の場合と同様に、阻害が可逆的で、VmaxおよびKm双方の変更(modification)を伴う混合型(競合的および非競合的)であることを示す。
【0070】
AFAフィトクロム濃度に対して傾きをプロットすることにより、本発明者らは、この場合10.48nMである、阻害定数Kiの値を得る。阻害定数は、酵素に対する阻害物質の親和性を評価する。高いKiが酵素に対する低親和性を示し、逆もまた同様である。この場合、極めて低いKi値は、MAO−Bに対するAFAフィトクロムの極めて高い親和性を示す。
【0071】
MAAの競合的で可逆的な作用は、これらの分子をMAO−B阻害において極めて強力にする。実際、MAO−B阻害の競合的で可逆的な特徴は、高度の有効性と生理的で副作用のない活性とを同時に確保する。この意味で、その抽出物中に含まれるMAAは、その分子量および結果としての血液脳関門を容易に通過する能力にも起因して、in vivoにおいても、MAO−B阻害に由来する治療的効果を生じるために、決定的な成分を構成する。
【0072】
MAAもさることながら、フィトクロムは、今まで知られるすべての物質で最も強力なMAO−B阻害物質であることがわかっている。MAO−B酵素に対するその極めて高い親和性、および数ナノモルの用量におけるその有効な阻害は、この分子を、それ自体において完全な治療用作用物質とするだけでなく、1つまたは複数のAFA抽出物の高い神経学的な有効性への最も重要な寄与を提供すると思われる因子にさせる。
【0073】
MAAおよびフィトクロムに関するいくつかの考察は、フィコシアニンのin vivoにおける挙動にも適用することができることを付け加えるべきであろう。本発明者らは、PCが、in vivoにおける脳に対する神経保護効果をもたらし、そのために、PCが血液脳関門を通過しうることを知っている(44)。これは、PCが、in vivoにおいて脳におけるそのMAO−B阻害活性を実現しうることを意味する。発色団の分子量は、実際、700に過ぎず、MAAの分子量とあまりかわらない。フィコシアノビリンと構造的に類似する、フィトクロムの発色団である、フィトクロモビリンについても同じことがあてはまる。
【0074】
結論として、その抽出物およびその活性成分である、AFAフィトクロム、AFA−PCおよびMAA側のMAO−Bの阻害活性は、以下の表に示す通り、分子および抽出物がともに、薬理学的物質以上の、最高レベルの活性に位置し、いずれの試験天然分子よりもはるかに優れているため、極めて重要である(45)。
【0075】
【表3】
【0076】
表が示す通り、フィコシアニンおよびMAAのみが、1μMよりもわずかに高いIC50を有し、このため、デプレニル(0.31μM)のIC50に極めて近く、考察した他の分子のIC50よりも10倍低い。一方、AFAフィトクロムは、デプレニルのIC50よりも15倍低いIC50を有する。酵素に対する阻害物質の親和性を評価する阻害定数Kiについても同じことがあてはまる。AFAフィコシアニンは、コーヒーおよびタバコの非ハーマンアルカロイドと同様に(しかし、もちろん、これら2つの物質に関連する問題なしに)約1μMのKiを有する。一方、MAAおよびAFAフィトクロムのみが、デプレニルと共に、1μMより低いKiを有し、したがって、MAO−Bに対して極めて高い親和性を有する分子である。実際、AFAフィトクロムは、セレギリン/デプレニル以外で、そのKiが数ナノモルのオーダーにある唯一の天然分子である。しかし、セレギリン/デプレニルとAFA抽出物の分子との間には、本質的な差異がある。前者が非可逆的な阻害物質であり、このため、潜在的な副作用を特徴とするのに対して、クラマス湖産AFAのMAO−B阻害分子は、すべてが可逆的であり、合成分子に関連する問題のない生理活性を特徴とする。
【0077】
図14は、デプレニルに関してAFAの3つの分子のMAO−B阻害活性を図示する。基本抽出物(および他のAFA抽出物)中における3つの分子すべての相乗作用を考えると、基本抽出物の全体のMAO−B阻害活性は極めて高い結果となる。その中に存在するPEAの高い分量をも考えると、特に重要となる事柄である。もし本発明者らが基本抽出物を、そのPC含量に基づきデプレニルと比較する場合、本発明者らは、基本抽出物が、0.05μMという低いPC用量でIC50に達し、これが、デプレニルよりも7.5倍高い(および天然物質よりも数十倍高い)効力を示すということを得る。これは、基本抽出物中に含まれるフィトクロムの効力に照らして意味を持つ。実際、7.5倍とは、PCとMAAの阻害効力の間の平均であり、それは、デプレニルよりやや低く、そして、15倍高い、フィトクロムのそれ(阻害効力)である。(図17)。これは、精製済みAFA−PCと比べて、その抽出物のより高い効力が、大部分、フィトクロムに起因することをも示す。
【0078】
さらに、その抽出物は、そのMAO−B阻害が可逆的で主に競合的であり、このため、デプレニルおよび他の合成物質などの不可逆的分子と潜在的に関連する副作用がない、生理的に作用する天然物質であることの利点を依然として維持する(46)。
【0079】
その抽出物のさらなる利点は、他の分子と完全に相乗的に作用する、強力なドーパミン作動性神経調節物質である、フェニルエチルアミンの高含量であり、この相乗活性を、本発明者らは以下のようにまとめることができる。
【0080】
・ フェニルエチルアミンまたはPEAは、黒質線条体組織からのドーパミンの放出を刺激すること、およびドーパミン自体のシナプス後の再取り込みを阻害することの両方において、二重のドーパミン作動性活性を有する;
・ フィトクロム、MAAおよびフィコシアニンは、減少したMAO−B活性が、ドーパミンを含む、神経アミンのより長い寿命を示唆する限りにおいて、強力なMAO−B阻害物質として、またドーパミン作動性伝達をも増大させる;
・ フィトクロム、MAAおよびフィコシアニンは、MAO−B阻害物質として、また、それ自体がMAO−B酵素の脱アミノ化活性の対象である、フェニルエチルアミンの寿命および活性をも延長させ、結果として、PEAによって引き起こされる、より全般的な神経調節ならびにドーパミン作動性伝達および活性に対するさらなる支援の好循環を作り出す。
・ 最後に、それ自身またはその発色団の血液脳関門を越える能力と一緒に、フィコシアニンの強力な抗酸化および抗炎症活性のほかに、フィトクロムの極めて高度な抗酸化活性、およびそれほど強力ではないが顕著なMAAの抗酸化活性が、いかなる酸化的損傷および炎症による損傷から、異なる活性分子およびより一般的にそれらが作り出す神経的好循環を保護する、神経保護もまた、もたらす。
【0081】
神経保護
本発明者らは、グルタミン酸の神経毒性効果に対する、AFA抽出物、特異的なAFA−PC、およびその発色団であるPCBのほか、MAAの神経保護特性を調べた。
【0082】
グルタミン酸は、哺乳動物の中枢神経系における主要な興奮性神経伝達物質であるが、ニューロンにおけるそのNMDAサブタイプ受容体の過剰刺激はCa2+の大量の細胞内蓄積を引き起こし、細胞死をもたらす。ミトコンドリア内のCa2+蓄積に加えて、NMDA受容体刺激後、細胞質における遊離Ca2+の一過性の増加は、一酸化窒素(NO・)または、主に初代(primary)ニューロンにおいて、そのスーパーオキシド(O2・-)反応生成物、ペルオキシ亜硝酸(ONOO-)を形成する酵素、一酸化窒素シンターゼ(NOS)のニューロンのアイソフォーム(49)を活性化する。
【0083】
ニューロンのグルタミン酸への曝露は、若干変更した方法(50)に従い実施した。
培地を除去し、ニューロンをあらかじめ37℃に温めて緩衝処理したハンクス液(5.26mM KCl、0.43mM KH2H2PO4、132.4mM NaCl、4.09mM NaHCO3、0.33mM Na2HPO4、20mMグルコース、2mM CaCl2、および20mM HEPES、pH7.4)で1回洗浄し、あらかじめ37℃に温めて緩衝処理したハンクス液中で、複数の濃度のAFA抽出物(1〜50nM)、PC(10〜1000nM)、PCB(10〜1000nM)およびMAA(1〜10μM)の不在または存在下においてプレインキュベートした。30分間のプレインキュベーション後に、Lグルタミン酸を濃縮液から100μMさらに10μMのグリシン、を示す最終濃度まで添加された。37℃で15分間ニューロンをインキュベートし、その緩衝液を吸引し、DMEMで置換し、細胞をエフェクター不在下においてさらに24時間37℃でインキュベートした。
【0084】
DNAに結合し、アポトーシスニューロン、すなわち、細胞核の断片化または凝縮化を示すニューロンの定量を可能とする、膜透過性蛍光染料である、DAPI(50)で細胞核を染色することにより、アポトーシスを評価した。簡潔に言うと、グルタミン酸曝露後の24時間、ニューロン培養物を温熱PBS(37℃)で洗浄し、室温で30分間、PBS中に4%(wt/vol)のパラホルムアルデヒドで固定した。PBSによる洗浄後、暗所内室温で10分間、細胞を3μM DAPIに曝露し、次いで、PBSで2回洗浄した。フルオレセインフィルター(330〜380の励起、30Xの倍率)を用いて、蛍光顕微鏡によりクロマチン凝縮について、細胞を記録した。総細胞核およびアポトーシス細胞核をカウントした。いずれの場合も、プロトコールデザインについて明るくない実施者により、ウェル当たり(per well)約600〜1,000個の細胞がカウントされた。個々の培養物に基づく測定は重複して(duplicate)実施し、結果は、表示された培養調製物数の平均(mean)SEM値として表す。その結果の統計学的解析は、クルスカル−ワリス検定の後、最小有意差多重範囲検定により判定した。いずれの場合も、p−0.05を有意と考えた。
【0085】
このグルタミン酸損傷試験により、本発明者らは、AFA基本抽出物、AFA−PC、そのPCB、およびMAAの神経保護能を初めて示した。図18が示す通り、培養済みニューロン細胞へのグルタミン酸の添加は、アポトーシスレベルを22.9%±3n=4(p<0.05)の比率まで上昇させたのに対し、
そのAFA基本抽出物の同時の添加は、グルタミン酸毒性に対する極めて高い保護をもたらし、既に1nMのような低量の抽出物によって、アポトーシスレベルを対照レベル(6.3±1 p>0.05)未満に低下させた(結果は、3〜8種類の異なる細胞培養物の平均±SEM。#対照群と比べたとき、有意に異なる(p<0.05);*グルタミン酸対照と比べたとき、有意に異なる(p<0.05))。MAAが与える保護については、それらはまた、より高用量の1μMで、アポトーシスレベルを対照レベル未満に低下させる(図19)結果は、3〜8種類の異なる細胞培養物の平均±SEM。#対照群と比べたとき、有意に異なる(p<0.05);*グルタミン酸対照と比べたとき、有意に異なる(p<0.05))。AFA−PCおよびPCBについて、本発明者らは、それらのアポトーシス阻害が極めて類似し、細胞培養物へのそれらの添加が、約10nMの用量で、アポトーシスの程度を対照未満に低下させるということがわかる(図20および21:結果は、3〜8種類の異なる細胞培養物の平均±SEM。#対照群と比べたとき、有意に異なる(p<0.05);*グルタミン酸対照と比べたとき、有意に異なる(p<0.05))。
【0086】
AFA−PCによる阻害の程度は、PCBの場合とほぼ等しい。おそらくその最も活性な成分と思われるPCBが、一度精製されひいてはより濃縮されると、その活性成分である全分子よりも顕著に強力であるべきことを踏まえると、これは、いささか驚くべきことである。それが実質的に同じ効力を有する事実は、全PCにおいて、PCB自体よりも実際にはさらに強力でありうる他の因子が存在することを意味する。本発明者らは、全PCが、C−PCおよびそのPCB発色団のほかに、PCBおよびPVB(フィコビオロビリン)をともにその発色団として含む、PECからもなることを知る。したがって、本発明者らは、本明細書において、精製済みPCBと全PCとの間の効力での顕著な差異を創出する因子は、正確にPEC成分、特に、極めて強い抗酸化物質であると推定される、そのPVB発色団であるということを推定することができる。
【0087】
神経保護の観点では、MAAは役割を果たすと思われるが、PCおよびPCBよりも顕著に低度である。しかし、最も強力な神経保護物質は、明らかに、全AFA抽出物であり、これは、わずか1nM(ナノモル)で細胞アポトーシスを完全に阻害することができる。これは、PCおよびPCBの効力の10倍である。これは、全AFA抽出物中に存在する多数の異なる抗酸化因子の相乗作用により確かに説明しうるが、本発明者らは、上述で、AFAフィトクロムが、今までおそらく最も強力な抗酸化物質であり、わずか16ナノモルでMDA(脂質過酸化の後期副生成物)の形成をほぼ完全に阻害しうる、ということを見たので、AFAフィトクロムが、基本抽出物のより高い効力を説明する中で重要な因子である可能性が極めて高い。こうして、本発明者らは、AFAフィトクロムが、任意のそしてすべてのフィトクロムと同様に、重要な神経保護作用物質であると結論付けることができる。
【0088】
参考文献
1. Zhou G. et al., Platelet monoamine oxidase B and plasma β-phenylethylamine in Parkinson's disease, in J Neurol Neurosurg Psychiatry, 2001; 70:229-231, 229.
2. Ispida K. et al., β-phenylethylamine stimulates striatal acetylcoline release through activation of the AMPA glutamatergic pathway, in Biol Pharm Bull 2005 Sep.; 28(9):1626-9.
3. Barroso N., Rodriguez M., Action of β-phenylethylamine and related amines on nigrostriatal dopamine neurotransmission, in European Journal of Pharmacology, 297 (1996), 195-203, 200.
4. Dyck L.E., Release of monoamines from striatal slices by phenelzine and β-phenylethylamine, in Prog Neuropsychopharmacol Biol Psychiatry, 1983, 7:797-800; Philips S.R., Robson A.M., In vivo release of endogenous dopamine from rat caudate nucleus by phenylethylamine, in Neuropharmacology 1983, 22:1297-1301; Raitieri m., et al., Effect of sympathomimetic amines on the synaptosomal transport of noradrenaline, dopamine and 5-hydroxytryptamine, in Eur J Pharmacol 1977, 41:133-143.
5. Janssen P.A.J, et al., Does phenylethylamine act as an endogenous amphetamine in some patients?, in International Journal of Neuropsychopharmacology 1999, 2: 229-240, 232.
6. Paterson I.A. et al., 2-phenylethylamine: a modulator of catecholamine transmission in the mammalian central nervous system?, in Journal of Neurochemistry (1990), 55:1827-1837.
7. Sabelli H.C., Javaid I.J., Phenylethylamine Modulation of Affect: Therapeutic and Diagnostic Implications, in Journal of Neuropsychiatry (1995), 7(1):6-14, 7.
8. Mauro Federici et al., Trace Amines Depress Gabab Response In Dopaminergic Neurons By Inhibiting Girk Channels, in Molecular Pharmacology Fast Forward. Published on January 11, 2005 as doi:10.1124/mol.104.007427.
9. Gusovsky F. et al., A high pressare liquid chromatography method for plasma phenylacetic acid, a putative marker for depressive disorders, in Anal Biochem, 1985 Feb. 15; 145(1):101-5. この研究では、うつ状態の患者は、対照群の536.18+/−54.99ng/mlに対して、327.64+/−45.44ng/mlの血漿中PAAレベルを有する。別の研究では、うつ状態の患者の尿において、うつではない患者の104+/−23mg/dieに対して、66+/−23mg/dieの平均PAAであることがわかった。参照:Sabelli HC. et al., Urinary phenylacetic acid in panic disorder with and without depression, in Acta Psychiatr Scand 1990 Jul;82(1):14-6.
10. Szabo A. et al., Phenylethylamine, a possible link to the antidepressant effects of exercise?, in Br J Sports Med 2001 Oct; 35(5):3423.
11. Sabelli H et al., Sustained antidepressant effect of PEA replacement, in J Neuropsychiatry Clin Neurosci, 8(2): 168-71.
12. Miura Y., Plasma beta-phenylethylamine in Parkinson's disease, in Kurume Med J 2000;47(4):267-72.
13. Ibid.,
14. Ebadi M. et al., Neuroprotective actions of selegiline, in J Neurosci Res 2002 Feb 1;67(3):285-289.
15. Kemppainen N. et al., Different pattern of reduction of striatal dopamine reuptake sites in Alzheimer's disease and ageing, in J Neural Transm 2001;108(7):827-36.
16. Knoll J., (-)Deprenyl (Selegiline): past, present and future, in Neurobiology (Bp) 2000;8(2):179-99.
17. Knoll J., The pharmacological basis of the beneficial effects of (-)deprenyl (selegiline) in Parkinson's and Alzheimer's diseases, in J Neural Transm Suppl 1993;40:69-91.
18. Rimbau V., et al., Protective effects of C-phycocyanin against kainic acid-induced neuronal damage in rat hippocampus, in Neurosci Lett 1999 Dec 3;276(2):75-8. この研究では、フィコシアニンは微細藻類スピルリナから用いられている。クラマス湖産藻類からのフィコシアニンは、異なっており、高い抗酸化活性を持っている。参照:Benedetti S., Scoglio S., Canestrari F., et al., Antioxydant properties of a novel phycocyanin extract from the blue-green alga Aphanizomenon Flos Aquae, in Life Sciences, 75 (2004): 2353-2362.
19. Swanson J. et al., Cognitive neuroscience of attention deficit hyperactivity disorder and hyperkinetic disorder, in Curr Opin Neurobiol. 1998 Apr;8(2):263-71.
20. Citazione solo di Benedetti et al. LifeScience; o menzione del parallelo brevetto? Attendere l'anno provisional in attesa di effettuare studi sulla neuroprotezione?
21. Kusaga A., Decreased beta-phenylethylamine in urine of children with attention deficit hyperactivity disorder and autistic disorder, in No To Hattatsu 2002 May; 34(3):243-8; Matsuishi T, Yamashita Y., Neurochemical and neurotransmitter studies in patients with learning disabilities, in No To Hattatsu 1999 May;31(3):245-8.
22. Kusaga A. et al., Increased urine phenylethylamine after methylphenidate treatment in children with ADHD, in Ann Neurol 2002 Sep;52(3):372-4.
23. Jain AK,. Et al., Bupropion SR vs. placebo for weight loss in obese patients with depressive symptoms, in Obes Res. 2002 Oct;10(10):1049-56.
24. Rudolph et al., A randomized, placebo-controlled, dose-response trial of venlafaxine hydrochloride in the treatment of major depression, in J Clin Psychiatry (1998); 59(3):116-22.
25. PEAは、熱によって非常に損傷を受けやすい脂溶性分子である。これは、凍結乾燥などの、高い温度を用いた乾燥方法が、通常PEAの低濃度を有することを意味する。PEAの最も高い含有量は、Refractance Window(登録商標) methodで乾燥された藻類に見られる。この基本抽出物が実現されるのは、この種類の藻類由来であるからである。
26. Yamada M. et al., Clinical Pharmacology of MAO Inhibitors: Safety and Future, in Neurotoxicology 2004; 25:215-21; Youdim M., et al., Therapeutic Applications of Selective and Non-Selective Inhibitors of Monoamine Oxidase A and B that do not Cause Significant Tyramine Potentiation, in Neurotoxicology 2004;25:243-50.
27. Groniger A et al., Photoprotective compounds in cyanobacteria, phytoplankton and macroalgae-a database, in J Photochem Photobiol B. 2000 Nov;58(2-3):115-22.
28. Suh HJ et al., Mycosporine glycine protects biological systems against photodynamic damage by quenching singlet oxygen with a high efficiency, in Photochem Photobiol. 2003 Aug;78(2):109-13.
29. Groniger A et al., Photoprotective compounds in cyanobacteria, phytoplankton and macroalgae-a database, in J Photochem Photobiol B. 2000 Nov;58(2-3):115-22.
30. Sinha RP et al., Induction of mycosporine-like amino acids (MAAs) in cyanobacteria by solar ultraviolet-B radiation, in J Photochem Photobiol B. 2001 Jul; 60(2-3):129-35.
31. Garcia-Pichel F et al., Occurrence of UV-Absorbing, Mycosporine-Like Compounds among Cyanobacterial Isolates and an Estimate of Their Screening Capacity, in Appl Environ Microbiol. 1993 Jan;59(1):163-169.
32. Glazer A.N., Phycobiliproteins, in Methods Enzymol, 1988, 167: 291303.
33. Bhat V.B., et al., C-phycocyanin: a potent peroxyl radical scavenger in vivo and in vitro, in Biochem Biophys Res Commun., 2000; 275(1):20-25; Romay, C. et al., Antioxidant and antinflammatory properties of Cphycocyanin from blue-green algae, in Inflamm Res, 1998, Jan.; 47(1): 3641.
34. Reddy C.M., et al., Selective Inhibition of cyclooxygenase-2 by Cphycocyanin, in Biochem Biophys Res Commun. 2000; 277(3): 599-603.
35. Gonzales R., et al., Anti-infl ammatory activity of phycocyanin extract in acetic acid induced colitis in rats, in Pharmacol Res, 1999; 39(1): 55-9.
36. Gonzales R., et al., Anti-infl ammatory activity of phycocyanin extract in acetic acid induced colitis in rats, in Pharmacol Res, 1999; 39(1): 55-9.
37. Vadiraja BB. et al., Hepatoprotective effect of Cphycocyanin:protection for carbon tetrachloride and R-(+)-pulegone-mediated hepatotoxicty in rats, in Biochem Biophys Res Commun, 1998; 249(2):428-31.
38. Romay C., et al., Phycocyanin extract reduces leukotriene B4 levels in arachidonic induced mouse-ear infl ammation test, in J Pharm Pharmacol. 1999,51(5):641-42. Come e noto, il leucotriene B4 e uno dei fattori principalmente responsabili di patologie respiratorie quali asma e allergie.
39. Rimbau V., et al., Protective effects of C-phycocyanin against kainic acid-induced neuronal damage in rat hippocampus, in Neurosci Lett 1999, 276(2):75-8.
40. Rimbau V. et al., C-phycocyanin protects cerebellar granule cells from low potassium/serum deprivation-induced apoptosis, in Naunyn Schmiedebergs Arch Pharmacol 2001; 364(2): 96-104.
41. Glazer A.N., Phycobilisomes, in Methods Enzymol 1988, 167;304-312.
42. Hirata T., et al., Antioxidant avtivities of phycocyanobilin prepared from Spirulina platensis, in J Appl Phycol 2000, 12:435-439.
43. Fuglistaller P., et al., Isolation and characterization of phycoerythrocyanin and chromatic adptation of the thermophilic cyanobacterium Mastigocladus laminosus, in Arch Microbiol 1981, 129:268274.
44. Rimbau V., et al., Protective effects of C-phycocyanin against kainic acid-induced neuronal damage in rat hippocampus, in Neurosci Lett 1999, 276(2):75-8.
45. この表に記載されているデータは、以下の研究を引用したものである: Magyar K. et al., Pharmacological aspects of (-)-deprenyl, in Curr Med Chem, 2004 Aug, 11(15):2017-31; Hou et al., Monoamine oxidase B (MAO-B) inhibition by active principles from Uncaria rhyncophylla, in Journal of Ethnopharmacology 100 (2005) 216-220; Herraiz T, Chaparro C., Human monoamine oxidase is inhibited by tobacco smoke: β-carboline alkaloids act as potent and revesible inhibitors, in Biochemical and Biophysical Research Communications 326 (2005) 378-386; Kong LD et al., Inihibition MAO-A and B by some plantderived alkaloids, phenols and anthraquinones, in Journal of Ethnopharmacology 91 (2004) 351-355.
46. Yoshida S. et al., Fluorinated phenylcyclopropylamines. Part 3: Inhibition of monoamine oxidase A and B, in Bioorganic & Medicinal Chemistry 12 (2004) 2645-2652.
47. Torres A. et al., Porphyra-334, a potential natural source for UVA protective sunscreens, in Photochem. Photobiol. Sci. 5 (2006) 432-435.
48. Hughes J, Lamparter T., Prokaryotes and Phytochrome. The Connection to Chromophores and Signaling, in Plant Physiology, December 1999, Vol. 121, pp. 1059-1068.
49. Garthwaite et al., Endothelium-derived relaxing factor release on activation of NMDA receptors suggests role as intercellular messenger in the brain, in Nature. 1988 Nov 24;336(6197):385-8.
50. Delgado-Esteban M. et al., D-Glucose prevents glutathione oxidation and mitochondrial damage after glutamate receptor stimulation in rat cortical primary neurone, in J Neurochem. 2000 Oct;75(4):1618-24.
【特許請求の範囲】
【請求項1】
神経系の疾患、病態、機能不全または障害の予防、制御または治療用の組成物の製造のための、微細藻類アファニゾメノン・フロスアクアエ(Aphanizomenon Flos Aquae Aquae Ralfs ex Born.& Flah.Var.flos aquae)(クラマス湖産AFA)の調製物、その抽出物、または、C−フィコシアニン/フィコエリスロシアニン複合体(C−PC/PEC)、その個別の発色団であるフィコシアノビリン(PCB)およびフィコビオロビリン(PVB)、AFAフィトクロムおよびマイコスポリン様アミノ酸(MAA)、もしくはその混合物から選択されるその単離成分の使用。
【請求項2】
前記神経系の疾患、病態、機能不全または障害が、以下のうちの1つである:アルツハイマー病、パーキンソン病、多発性硬化症、注意欠陥過活動性障害(ADHD)、自閉症、うつ病、記憶欠損および気分障害、請求項1に記載の使用。
【請求項3】
以下のステップ:
a)採取直後のAFA藻類を冷凍し解凍するか、または、出発物質が乾燥AFA粉末である場合は、水で希釈したAFA粉末を超音波で分解して、細胞を破砕する;
b)ステップa)の生成物を遠心分離し、沈殿物から上清を分離する;
c)水溶性成分を含む上清を回収する
により調製される、AFA抽出物の、請求項1から2に記載の使用。
【請求項4】
前記上清を、30,000ドルトンのカットオフ分子量を有する限外濾過膜を用いる限外濾過にかけることにより、クラマス湖産AFA抽出物がさらに精製される、請求項3に記載の使用。
【請求項5】
前記単離成分が、シノリンおよびポルフィラ334から選択されるマイコスポリン様アミノ酸である、請求項1に記載の使用。
【化1】
【請求項6】
前記単離成分が、フィコシアニン/フィコエリスロシアニン複合体(C−PC/PEC)またはその単独の部分成分(subcomponent)、すなわち、C−PCまたはPECである、請求項1に記載の使用。
【請求項7】
前記単離成分が、フィトクロムである、請求項1に記載の使用。
【請求項8】
前記単離成分の混合物が、マイコスポリン様アミノ酸、フィコシアニン、フィコエリスロシアニン、およびフィトクロムからなる、請求項1に記載の使用。
【請求項9】
前記混合物が、フェニルエチルアミン(PEA)をさらに含む、請求項8に記載の使用。
【請求項10】
前記組成物がヒトへの投与に適する、請求項1に記載の使用。
【請求項11】
クラマス湖産AFA調製物、その抽出物またはその単離成分が、薬剤的に許容できる媒体(vehicles)または賦形剤(excipients)と共に調合される、請求項1に記載の使用。
【請求項12】
フィトクロム、フィコシアニン、フィコエリスロシアニンおよびマイコスポリン様アミノ酸またはその混合物から選択されるクラマス湖産AFAの単離成分を対象に投与することを含む、それを必要とする対象における酵素モノアミンオキシダーゼBの阻害方法。
【請求項13】
前記対象が、カテコールアミン関連の神経系または神経変性疾患に罹患したヒト対象である、請求項12に記載の方法。
【請求項1】
神経系の疾患、病態、機能不全または障害の予防、制御または治療用の組成物の製造のための、微細藻類アファニゾメノン・フロスアクアエ(Aphanizomenon Flos Aquae Aquae Ralfs ex Born.& Flah.Var.flos aquae)(クラマス湖産AFA)の調製物、その抽出物、または、C−フィコシアニン/フィコエリスロシアニン複合体(C−PC/PEC)、その個別の発色団であるフィコシアノビリン(PCB)およびフィコビオロビリン(PVB)、AFAフィトクロムおよびマイコスポリン様アミノ酸(MAA)、もしくはその混合物から選択されるその単離成分の使用。
【請求項2】
前記神経系の疾患、病態、機能不全または障害が、以下のうちの1つである:アルツハイマー病、パーキンソン病、多発性硬化症、注意欠陥過活動性障害(ADHD)、自閉症、うつ病、記憶欠損および気分障害、請求項1に記載の使用。
【請求項3】
以下のステップ:
a)採取直後のAFA藻類を冷凍し解凍するか、または、出発物質が乾燥AFA粉末である場合は、水で希釈したAFA粉末を超音波で分解して、細胞を破砕する;
b)ステップa)の生成物を遠心分離し、沈殿物から上清を分離する;
c)水溶性成分を含む上清を回収する
により調製される、AFA抽出物の、請求項1から2に記載の使用。
【請求項4】
前記上清を、30,000ドルトンのカットオフ分子量を有する限外濾過膜を用いる限外濾過にかけることにより、クラマス湖産AFA抽出物がさらに精製される、請求項3に記載の使用。
【請求項5】
前記単離成分が、シノリンおよびポルフィラ334から選択されるマイコスポリン様アミノ酸である、請求項1に記載の使用。
【化1】
【請求項6】
前記単離成分が、フィコシアニン/フィコエリスロシアニン複合体(C−PC/PEC)またはその単独の部分成分(subcomponent)、すなわち、C−PCまたはPECである、請求項1に記載の使用。
【請求項7】
前記単離成分が、フィトクロムである、請求項1に記載の使用。
【請求項8】
前記単離成分の混合物が、マイコスポリン様アミノ酸、フィコシアニン、フィコエリスロシアニン、およびフィトクロムからなる、請求項1に記載の使用。
【請求項9】
前記混合物が、フェニルエチルアミン(PEA)をさらに含む、請求項8に記載の使用。
【請求項10】
前記組成物がヒトへの投与に適する、請求項1に記載の使用。
【請求項11】
クラマス湖産AFA調製物、その抽出物またはその単離成分が、薬剤的に許容できる媒体(vehicles)または賦形剤(excipients)と共に調合される、請求項1に記載の使用。
【請求項12】
フィトクロム、フィコシアニン、フィコエリスロシアニンおよびマイコスポリン様アミノ酸またはその混合物から選択されるクラマス湖産AFAの単離成分を対象に投与することを含む、それを必要とする対象における酵素モノアミンオキシダーゼBの阻害方法。
【請求項13】
前記対象が、カテコールアミン関連の神経系または神経変性疾患に罹患したヒト対象である、請求項12に記載の方法。
【図1】

【図2】

【図3】
【図4】
【図5】
【図6】
【図7A】

【図7B】
【図8】

【図9】
【図10】
【図11】
【図12】

【図13】
【図14】

【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】

【図23】
【図24】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【公表番号】特表2009−541388(P2009−541388A)
【公表日】平成21年11月26日(2009.11.26)
【国際特許分類】
【出願番号】特願2009−516969(P2009−516969)
【出願日】平成19年6月26日(2007.6.26)
【国際出願番号】PCT/EP2007/005622
【国際公開番号】WO2008/000430
【国際公開日】平成20年1月3日(2008.1.3)
【出願人】(508375343)ヌトラテック ソシエタ ア レスポンサビリタ リミタータ (2)
【氏名又は名称原語表記】NUTRATEC S.R.L.
【Fターム(参考)】
【公表日】平成21年11月26日(2009.11.26)
【国際特許分類】
【出願日】平成19年6月26日(2007.6.26)
【国際出願番号】PCT/EP2007/005622
【国際公開番号】WO2008/000430
【国際公開日】平成20年1月3日(2008.1.3)
【出願人】(508375343)ヌトラテック ソシエタ ア レスポンサビリタ リミタータ (2)
【氏名又は名称原語表記】NUTRATEC S.R.L.
【Fターム(参考)】
[ Back to top ]