
神経細胞の製造方法及び神経細胞分化促進剤
【課題】薬剤を用いて幹細胞を神経細胞へと迅速に分化させることを含む、安全性に優れた神経細胞を製造する方法を提供する。
【解決手段】胚様体と硫酸化阻害物質とを接触させ、該胚様体中における糖鎖の硫酸化を抑制して該胚様体の神経細胞への分化を促進することを含む、神経細胞の製造方法。
【解決手段】胚様体と硫酸化阻害物質とを接触させ、該胚様体中における糖鎖の硫酸化を抑制して該胚様体の神経細胞への分化を促進することを含む、神経細胞の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、胚様体から神経細胞を製造する方法に関する。また、本発明は前記製造方法に用いうる神経細胞分化促進剤に関する。
【背景技術】
【0002】
胚性幹細胞(ES細胞)や誘導多能性幹細胞(iPS細胞)は、生体に存在する様々な細胞に分化する能力(分化多能性)を有した状態で長期にわたり継代培養することができるという性質をもつ。したがって、細胞移植が有効な治療方法として期待される白血病、パーキンソン病等の治療にこれらの幹細胞を用いることが期待されている。
【0003】
多能性幹細胞を用いた細胞移植療法においては、多能性幹細胞を目的とする細胞又はその前駆細胞に分化誘導した上で移植する必要がある。例えば、パーキンソン病の治療にあたっては、多能性幹細胞を神経細胞又はその前駆細胞へと分化誘導し、これを患者に移植することになる。これまで多能性幹細胞の分化誘導には、主に遺伝子導入や遺伝子改変といった分子生物学的手法が報告されているが(例えば、非特許文献1)、分化誘導された細胞の安全性が懸念され、実用化には至っていない。また、上記分子生物学的手法は熟練した技術と高価な試薬類を要する一方、多能性幹細胞の分化誘導速度は比較的緩やかである。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Plos ONE December 2009 Vol.4 Issue 12 e8262
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明者らは、従来の遺伝子導入や遺伝子改変手法を用いずに、薬剤を用いて多能性幹細胞の分化誘導を行うことができれば、より簡便にしかも安価に分化誘導ができ、治療上の安全性にも優れた実用性の高い細胞を創出できると考えた。
本発明は、薬剤を用いて多能性幹細胞を神経細胞へと迅速に分化させることを含む、安全性に優れた神経細胞を製造する方法を提供することを課題とする。
また、本発明は、多能性幹細胞を安全性に優れた神経細胞へと迅速に分化誘導しうる薬剤を提供することを課題とする。
【課題を解決するための手段】
【0006】
本発明者らは上記課題に鑑み鋭意検討を行った。その結果、ES細胞やiPS細胞等の多能性幹細胞に由来する胚様体(Embryoid body:EB)と硫酸化阻害物質とを接触させると、該胚様体を神経細胞へと迅速に分化誘導できることを見い出した。本発明はこの知見に基づいてなされたものである。
【0007】
本発明の課題は下記の手段により達成された。
[1]胚様体と硫酸化阻害物質とを接触させ、該胚様体中における糖鎖の硫酸化を抑制して該胚様体の神経細胞への分化を促進することを含む、神経細胞の製造方法、
[2]硫酸化阻害物質が塩素酸塩である、[1]に記載の方法、
[3]硫酸化阻害物質を含有する培地で胚様体を培養することにより胚様体と硫酸化阻害物質とを接触させる、[1]又は[2]に記載の方法、
[4]さらに胚様体とレチノイン酸とを接触させる工程を含む、[1]〜[3]のいずれかに記載の方法、
[5]胚様体が哺乳類のES細胞又は哺乳類の人工多能性幹細胞に由来する、[1]〜[4]のいずれかに記載の方法、
[6]硫酸化阻害物質を有効成分として含有する神経細胞分化促進剤、
[7]硫酸化阻害物質が塩素酸塩である、[6]に記載の神経細胞分化促進剤、
[8]さらにレチノイン酸を含有する、[6]又は[7]に記載の神経細胞分化促進剤。
【発明の効果】
【0008】
本発明の製造方法によれば、ES細胞やiPS細胞に由来する胚様体から神経細胞を簡便に、しかも迅速に得ることができる。また、本発明の神経細胞分化促進剤は、硫酸化阻害物質を有効成分とするものであり、遺伝子のように細胞内に組み込まれて細胞分裂と共に複製される性質のものではない。したがって、細胞から神経細胞分化促進剤を容易に取り除くことができ、安全性の高い神経細胞の提供することができる。
【図面の簡単な説明】
【0009】
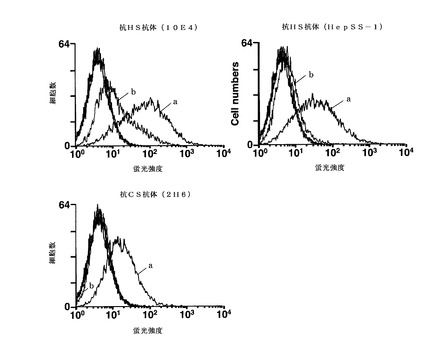
【図1】塩素酸ナトリウムとの接触から1日経過後(EB形成から3日後)のmESC由来のEBにおけるHS及びCSの硫酸化度合の変化をFACSにより分析した結果を示す図である。図1中、aは塩素酸ナトリウムと接触させていないEBのFACS分析結果を示し、bは塩素酸ナトリウムと接触させたEBのFACS分析結果を示す。a及びb以外のプロットは一次抗体としてIgMアイソタイプコントロールを用いたときの結果である。
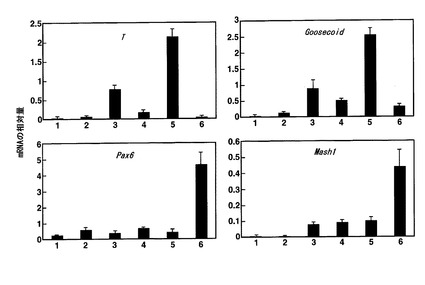
【図2】mESC由来のEBにおいて、中胚葉分化のマーカー遺伝子であるT及びGoosecoid、並びに外胚葉分化のマーカーであるPax6及びMash1のmRNA量をリアルタイムPCRで定量した結果を示すグラフである。縦軸はβアクチンのmRNAに対する相対量を示し、単位は[1000/βアクチンのmRNA量]である。横軸の1〜6の数字は、1:EB形成から0日後のEB、2:EB形成から2日後のEB、3:塩素酸ナトリウムと接触させていないEB形成から3日後のEB、4:塩素酸ナトリウムと接触させたEB形成から3日後のEB、5:塩素酸ナトリウムと接触させていないEB形成から4日後のEB、6:塩素酸ナトリウムと接触させたEB形成から4日後のEB、を示す。
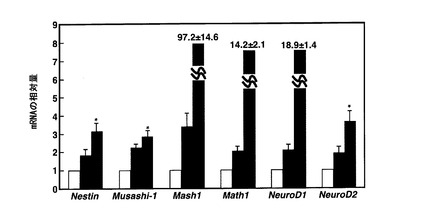
【図3】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のmESC由来のEBにおいて、神経細胞分化のマーカー遺伝子であるNestin、Musashi−1、Mash1、Math1、NeuroD1及びNeuroD2のmRNA量をリアルタイムPCRで定量した結果を示すグラフである。縦軸は塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEBにおける各遺伝子のmRNA量を1としたときの相対値を示す。グラフ中の各遺伝子のmRNA量を示す3つの棒は、左から順に、塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、レチノイン酸のみと接触させたEB、塩素酸ナトリウムとレチノイン酸の双方と接触させたEB、を示す。
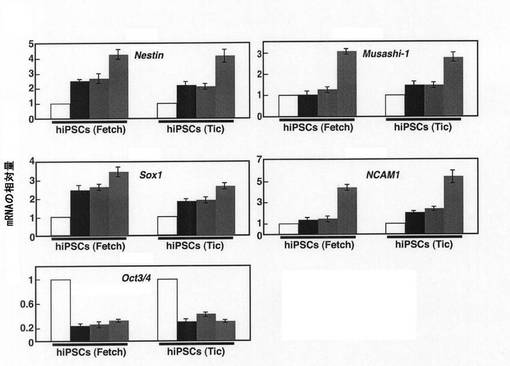
【図4】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のhiPSC由来のEBにおいて、神経細胞分化のマーカー遺伝子であるNestin、Musashi−1、Sox1及びNCAM1並びに未分化細胞のマーカーであるOct3/4のmRNA量をリアルタイムPCRで定量した結果を示すグラフである。縦軸はEB形成後0日目における各遺伝子のmRNA量を1としたときの相対値を示す。グラフ中の各遺伝子のmRNA量を示す4つの棒は、左から順に、EB形成後0日目のEB、塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB形成から5日後のEB、レチノイン酸のみと接触させたEB形成から5日後のEB、塩素酸ナトリウムとレチノイン酸の双方と接触させたEB形成から5日後のEB、を示す。
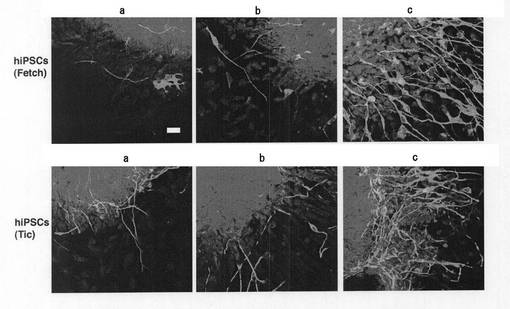
【図5】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のmESC由来のEBにおいて、抗βIII−チューブリン抗体を用いて免疫染色を行った結果を示す顕微鏡写真である。図5中、aは塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、bはレチノイン酸のみに接触させたEB、cは塩素酸ナトリウムとレチノイン酸の双方と接触させたEB、を示す。写真中の白い繊維状のものがβIII−チューブリンである。
【図6】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のhiPSC由来のEBにおいて、抗βIII−チューブリン抗体を用いて免疫染色を行った結果を示す顕微鏡写真である。図6中、aは塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、bはレチノイン酸のみに接触させたEB、cは塩素酸ナトリウムとレチノイン酸の双方と接触させたEB、を示す。写真中の白い繊維状のものがβIII−チューブリンである。
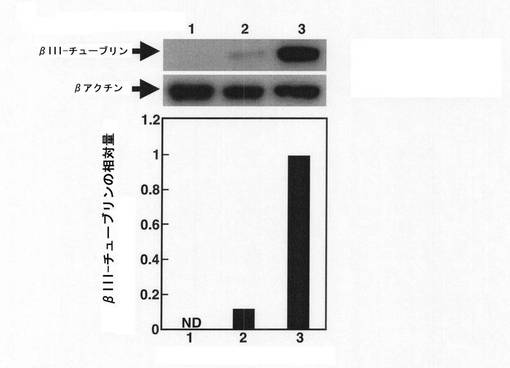
【図7】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のmESC由来のEBにおいて、抗βIII−チューブリン抗体を用いて免疫ブロットを行った結果を示す図である。縦軸は、βアクチンに対するβIII−チューブリンの相対量(βIII−チューブリンの量/βアクチンの量)を示す。横軸の1〜3の番号は、1:塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、2:レチノイン酸のみに接触させたEB、3:塩素酸ナトリウムとレチノイン酸の双方に接触させたEB、を示す。
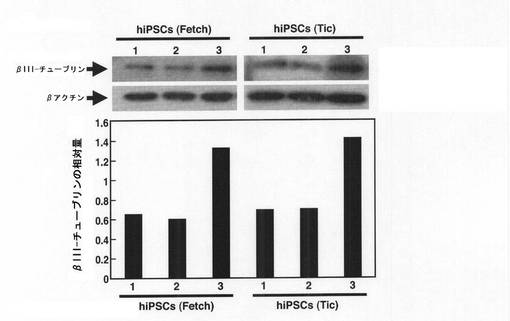
【図8】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のhiPSC由来のEBにおいて、抗βIII−チューブリン抗体を用いて免疫ブロットを行った結果を示す図である。縦軸は、βアクチンに対するβIII−チューブリンの相対量(βIII−チューブリンの量/βアクチンの量)を示す。横軸の1〜3の番号は、1:塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、2:レチノイン酸のみに接触させたEB、3:塩素酸ナトリウムとレチノイン酸の双方に接触させたEB、の結果を示す。
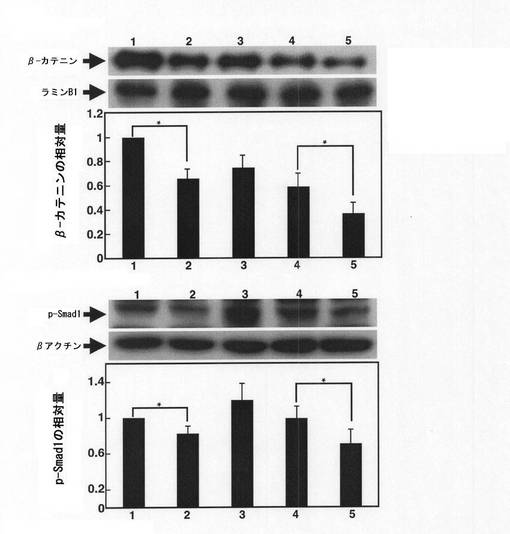
【図9】塩素酸ナトリウムとの接触から1日後(EB形成から3日後)及び塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のmESC由来のEBについて、βカテニンとp−Smad1のタンパク質量を免疫ブロットで解析した結果を示す図である。グラフの縦軸は、ラミンB1に対するβカテニンの相対量、及びβアクチンに対するp−Smad1の相対量を表わす。横軸の番号1〜5は、1:塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB形成から3日後のEB、2:塩素酸ナトリウムに接触させたEB形成から3日後のEB、3:塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB形成から5日後のEB、4:レチノイン酸のみに接触させたEB形成から5日後のEB、5:塩素酸ナトリウムとレチノイン酸の双方に接触させたEB形成から5日後のEB、の結果を示す。
【発明を実施するための形態】
【0010】
以下、本発明について、その好ましい実施態様に基づき詳細に説明する。
[本発明の製造方法]
本発明に用いる胚様体とは、多能性幹細胞を浮遊培養したときに形成されうる初期胚に似た構造をもつ球状の細胞塊である。本発明に用いる胚様体を形成する多能性幹細胞の種類に特に制限はないが、ES細胞又はiPS細胞であることが好ましい。前記多能性幹細胞は三胚葉性動物に由来する限り特に制限はない。例えば、哺乳類、鳥類等に由来する多能性幹細胞を用いることができるが、哺乳類に由来する多能性幹細胞を好適に用いることができる。前記哺乳類としては、例えば、ヒト、サル、ウシ、ヤギ、ヒツジ、ブタ、ウマ、ウサギ、ラット、マウス、モルモット等が挙げられる。
上記多能性幹細胞は、通常の方法で調製することができる。また、寄託機関等に寄託されているセルライン等の分譲により得ることもできる。
【0011】
本発明に用いる胚様体は通常の方法で調製することができる。例えば、マウスES細胞であれば、細胞吸着能の低い培養皿で、leukemia inhibitory factor(LIF)無しのESC培地等で浮遊培養することで得ることができる。また、ヒトiPS細胞であれば、細胞吸着能の低い培養皿で、bFGF無しのiPSellon培地等で浮遊培養することで得ることができる。これらの具体的な培養方法は後述の実施例で詳述する。
胚様体を形成させるための温度、CO2濃度等は通常の条件を採用することができ、例えば、37℃、5%CO2の条件下で多能性幹細胞から胚様体を形成させることができる。
本発明に用いる胚様体の調製方法に特に制限はなく、胚様体を形成しうる方法であればいずれの方法で調製したものを用いてもよい。
【0012】
本発明に用いる硫酸化阻害物質とは、細胞に取り込まれて細胞内で少なくとも糖鎖の硫酸化を抑制しうる物質である。本発明において「糖鎖の硫酸化を抑制する」とは、糖鎖の硫酸化を完全に阻害することの他、糖鎖の硫酸化を部分的に阻害することも含む概念である。「糖鎖の硫酸化を部分的に阻害する」とは、糖鎖に硫酸基が結合してはいるが、通常よりも硫酸基の数が少ない状態にすることをいう。本発明に用いる硫酸化阻害物質により、例えば、細胞膜や細胞内に存在するヘパラン硫酸(HS)やコンドロイチン硫酸(CS)といった硫酸化グリカンの硫酸基の数が減少しうる。
本発明に用いる硫酸化阻害物質は、細胞内において糖鎖の硫酸化を阻害する作用を有すれば特に制限はないが、塩素酸塩を好適に用いることができる。塩素酸塩の種類に特に制限はなく、例えば、塩素酸ナトリウム、塩素酸カリウム、塩素酸リチウム、塩素酸アンモニウム、塩素酸カルシウム等が挙げられる。本発明に用いる硫酸化阻害物質は、通常の方法で合成して得ることもできるし、市販品を用いてもよい。
【0013】
本発明において、胚様体と硫酸化阻害物質とを接触させる方法に特に制限はない。硫酸化阻害物質を含む培地で胚様体を培養することで、胚様体と硫酸化阻害物質とを容易に接触させることができる。該培地は液体培地であることが好ましい。胚様体が生存できる環境であれば、培地の種類に特に制限はなく、例えば、マウスES細胞由来の胚様体であれば上記のLIF無しのESC培地等を、ヒトiPS細胞由来の胚様体であれば上記のbFGF無しのiPSellon培地等を好適に用いることができる。
胚様体と硫酸化阻害物質との接触の開始時期は、迅速に分化誘導を促進する観点から、胚様体の形成後0〜5日後であることが好ましく、0〜4日後であることがより好ましく、0〜3日後であることがさらに好ましく、1〜2日後であることがさらに好ましく、2日後であることが特に好ましい。
胚様体と硫酸化阻害物質との接触時間に特に制限はないが、硫酸化阻害物質が細胞内に取り込まれて生理作用を発揮するために、少なくとも1日間接触させることが好ましく、1〜5日間接触させることがより好ましく、1〜4日間接触させることがさらに好ましく、1〜3日間接触させることがさらに好ましく、2〜3日間接触させることが特に好ましい。
胚様体に接触させる硫酸化阻害物質の量に特に制限はないが、硫酸化阻害物質を含む培地で胚様体を培養する場合には、培地中の硫酸化阻害物質の濃度が0.1〜500mMであることが好ましく、1〜500mMであることがより好ましく、1〜300mMであることがさらに好ましく、1〜200mMであることがさらに好ましく、10〜100mMであることが特に好ましい。
【0014】
胚様体を神経細胞へと効率的に分化誘導するために、さらに胚様体とレチノイン酸とを接触させることが好ましい。レチノイン酸は従来から多能性幹細胞の神経細胞への分化誘導に用いられている物質である。胚様体とレチノイン酸とを接触させる方法に特に制限はないが、レチノイン酸を含む培地で胚様体を培養することで、胚様体とレチノイン酸とを容易に接触させることができる。該培地は液体培地であることが好ましい。胚様体が生存できる環境であれば、培地の種類に特に制限はなく、例えば、マウスES細胞由来の胚様体であれば上記のLIF無しのESC培地等を、ヒトiPS細胞由来の胚様体であれば上記のbFGF無しのiPSellon培地等を好適に用いることができる。胚様体とレチノイン酸との接触の開始時期は、胚様体と硫酸化阻害物質との接触を開始する前であっても後であってもよく、胚様体と硫酸化阻害物質との接触の開始と同時に胚様体とレチノイン酸との接触を開始させてもよい。胚様体とレチノイン酸との接触の開始時期は、胚様体の形成後0〜5日後であることが好ましく、1〜5日後であることがより好ましく、1〜4日後であることがさらに好ましく、2〜4日後であることがさらに好ましく、3〜4日後であることが特に好ましい。
胚様体とレチノイン酸との接触時間に特に制限はないが、レチノイン酸が細胞内に取り込まれて生理作用を発揮するために、少なくとも5時間接触させることが好ましく、5〜72時間接触させることがより好ましく、12〜48時間接触させることがさらに好ましく、20〜30時間接触させることが特に好ましい。
胚様体に接触させるレチノイン酸の量に特に制限はないが、レチノイン酸を含む培地で胚様体を培養する場合には、培地中のレチノイン酸の濃度が0.1〜10μMであることが好ましく、0.5〜5μMであることがより好ましく、0.5〜2μMであることがさらに好ましい。
レチノイン酸は、生理機能を発揮させるために通常はトランス体(all−trans)が用いられる。
【0015】
胚様体は硫酸化阻害物質及び/又はレチノイン酸の存在下で神経細胞への分化誘導を継続させてもよいが、通常には、上記の好ましい接触時間硫酸化阻害物質及び/又はレチノイン酸と接触させた後、硫酸化阻害部物質及び/又はレチノイン酸との接触を終了させる。
胚様体と硫酸化阻害物質及び/又は胚様体とレチノイン酸との接触を終了するには、胚様体の培養環境中から硫酸化阻害物質やレチノイン酸を取り除けばよい。例えば、胚様体を硫酸化阻害物質やレチノイン酸を含まない培地に移して培養することで、胚様体と硫酸化阻害物質及び/又は胚様体とレチノイン酸との接触を終了させることができる。
胚様体は、硫酸化阻害物質やレチノイン酸との接触を終了させた後も、適当な培養条件で培養することで神経細胞へと分化が進む。当該培養に使用する培地に特に制限はないが、例えば、N2サプリメントを含むDMEM−F12等が挙げられる。また、培地は定期的に交換することが好ましい。
【0016】
胚様体を培養する温度、CO2濃度等は通常の条件を採用することができ、例えば、37℃、5%CO2の条件下で培養することができる。
【0017】
胚様体の分化には、Wnt、BMP及びFGF等を介したシグナル伝達が関与していることが知られており、Wnt及びBMPは神経細胞への分化を阻害し、中胚葉への分化を促すことも示唆されてきている(例えば、Plos ONE December 2009 Vol.4 Issue 12 e8262参照)。また、上記シグナル伝達には細胞膜に存在する硫酸化グリカンが関与しうることも報告されてきている。
本発明に用いる硫酸化阻害物質は、細胞内で糖鎖の硫酸化を阻害する作用を有する。したがって、ヘパラン硫酸やコンドロイチン硫酸等の細胞膜及び/又は細胞内に存在する硫酸化グリカンの硫酸基の数を減少させ、これによりWnt及びBMPのシグナル伝達を阻害し、胚葉体の神経細胞への分化を促進すると考えられる。
これまで、糖鎖の硫酸化に関与する特定の遺伝子の働きを抑制することで、胚様体の神経細胞への分化を誘導したことの報告はあるが(Plos ONE December 2009 Vol.4 Issue 12 e8262参照)、胚葉体と硫酸化阻害物質とを接触させるだけで、胚様体の神経細胞への分化誘導に成功した例は知られていない。
本発明により、これまでにない簡便な手法で、しかも迅速に胚様体を神経細胞へと迅速に分化させて神経細胞を得ることが可能になる。また、本発明に用いる硫酸化阻害物質は、培地から該硫酸化阻害物質を除去することで分化した神経細胞から該硫酸化阻害物質を取り除くことができる。したがって、本発明の方法に分化誘導された神経細胞は安全性に優れ、細胞移植療法等への実用化が期待される。
なお、本発明において「神経細胞」には神経細胞及び神経細胞の前駆細胞の双方が含まれる。
【0018】
[本発明の神経細胞分化促進剤]
本発明の神経細胞分化促進剤は、胚様体の神経細胞への分化を促進するための薬剤であり、有効成分として上記の硫酸化阻害物質を含有する。本発明の神経細胞分化促進剤は、上述の本発明の製造方法における硫酸化阻害物質の供給源として用いることができる。すなわち、本発明の神経細胞分化促進剤と、多能性幹細胞を浮遊培養して得られうる上記胚様体とを接触させることで、該胚様体細胞を神経細胞へと迅速に分化させることができる。
【0019】
本発明の神経細胞分化促進剤は、硫酸化阻害物質の他にレチノイン酸を含有していてもよい。レチノイン酸は、生理機能を発揮させるために通常はオールトランス体である。また、本発明の神経細胞分化促進剤はさらに栄養素、pH調整剤、防腐剤、安定剤、乳化剤等の他の成分を含有していてもよい。
【0020】
本発明の神経細胞分化促進剤の剤形に特に制限はなく、各成分が溶解又は分散した液状であっても良いし、粉末状であっても良い。また、タブレット等の錠剤の形状であってもよい。
【実施例】
【0021】
以下、実施例に基づいて本発明をさらに詳細に説明するが、本発明はその要旨を越えない限りこれらに限定されるものではない。
【0022】
[セルラインの維持]
マウスES細胞(mESC)として、R1セルライン(Proc.Natl.Acad.Sci.USA 90(1993)8424−8428に記載、自然科学研究機構 生理学研究所 分子神経生理研究部門 より供与)及びE14TG2aセルライン(Dev.Biol.121(1987)1−9に記載、東京大学大学院理学系研究科より供与)を使用した。各々の細胞を、1000U/mLの濃度でLIF(ケミコン社製)を加えたESC培地(15質量%のFBS(Hyclone社製)、1質量%のペニシリン/ストレプトマイシン(Gibco社製)、0.1mMの2−メルカプトエタノール(Gibco社製)及び0.1mMの非必須アミノ酸(Gibco社製)を添加したDMEM)中において、10μg/mLのマイトマイシン C(シグマ社製)で不活化したマウス線維芽細胞(MEFs)上で維持した。
【0023】
また、ヒトiPS細胞(hiPSC)として、MRC−hiPS_Fetch(細胞番号:NIHS0604)及びMRC−hiPS_Tic(細胞番号:JCRB1331)(いずれもExp.Cell Res.315(2009)2727−2740に記載、国立生育医療センターから入手)を使用した。各々の細胞を、10ng/mLの濃度でbFGF(和光純薬社製)を加えたiPSellon(Cell−Sight社製)中において、10μg/mLのマイトマイシン C(シグマ社製)で不活化したMEFs上で維持した。
【0024】
[胚様体の形成]
上記mESC及びhiPSCをLow Cell Binding 60mm dishes(Nunc社製)に移し、mESCについてはLIFを含まない上記のESC培地で、hiPSCについてはbFGFを含まないiPSellonでそれぞれ浮遊培養することで胚様体(EB)を形成させた。なお、hiPSCは、EB形成前にゼラチンコートされたディッシュに移してフィーダー細胞を除去した。
本発明においては、上記の浮遊培養の開始した時点をEB形成時とする。したがって、例えば、「EB形成から1日後」とは、多能性幹細胞について上記の浮遊培養を開始してから1日後を意味する。
【0025】
[胚様体の神経細胞への分化誘導]
EB形成から2日後に、培地中の最終濃度が50mMとなるように、硫酸化阻害物質としての塩素酸ナトリウム(シグマ社製)を添加した。さらにその2日後(EB形成から4日後)、培地中の最終濃度が1μMとなるようにオールトランス−レチノイン酸(以下、RAと呼ぶことがある。シグマ社製)を添加した。RAを添加してから1日経過後(EB形成から5日後)、EBをPDL/laminin−coated 60mm dishes(ベクトンディッキンソン社製)に移し、N2サプリメント(Gibco社製)を含むDMEM−F12中で培養した。この操作により培養液中から硫酸化阻害物質とRAが除かれた。
【0026】
[FACS分析]
mESCであるR1セルライン由来のEBにおいて、硫酸化阻害物質の添加によるグリカンの硫酸化度合の変化を調べるために、抗HS抗体又は抗CS抗体(いずれもマウス由来IgM)をプライマリー抗体として用いたFACS分析を行った。上記抗HS抗体としては10E4又はHepSS−1(いずれも商品名、生化学工業社製)を用い、上記抗CS抗体としては2H6(商品名、生化学工業社製)を用いた。また、上記抗体にかえてマウスIgMアイソタイプ(ケミコン社製)を用いたケースをコントロールとした。具体的な実験方法は以下のとおりである。
硫酸化阻害物質との接触から1日経過後(EB形成から3日後)の細胞塊をEDTAで処理した後、この細胞懸濁液をFACS緩衝液(0.5質量%ウシ血清アルブミン(BSA)及び0.1質量%アジ化ナトリウムを含むPBS溶液)で希釈した上記プライマリー抗体溶液中でインキュベートした。洗浄後、細胞懸濁液を上記FACS緩衝液で希釈したFITC標識した抗マウスIgM抗体(シグマ社製)溶液中でインキュベートした。洗浄後、FACSAria Cell Sorter(ベクトンディッキンソン社製)を用いてFACS分析を行った。
結果を図1に示す。
【0027】
HS及びCSは細胞の分化に重要な役割を担う硫酸化グリカンであり、mESCではEB形成後少なくとも8日目まで発現することが知られている。図1に結果から、培地に硫酸化阻害物質を加えたEB(図1中のa)では、硫酸化阻害物質を加えていないEB(図1中のb)と比較して抗HS抗体及び抗CS抗体の結合量が少なく(蛍光強度が小さく)なっており、糖鎖に結合した硫酸基の量が減少していることがわかる。図1の結果は、EBと硫酸化阻害物質の接触後わずか1日で糖鎖の硫酸化が顕著に抑制されたことを示している。
【0028】
[リアルタイムPCR分析]
mESCであるR1セルライン由来のEBにおいて、硫酸化阻害物質がEBの外胚様(神経細胞)への分化を誘導することを、特定の遺伝子のmRNAの発現レベルを指標にして調べた。具体的には、中胚葉分化のマーカーであるT及びGoosecoid、並びに外胚葉分化のマーカーであるMash1及びPax6の各遺伝子について、それらのmRNAの発現量をABI PRISM(登録商標)7700 sequence detection systemを用いたリアルタイムPCRにより定量した。J.Biol.Chem.283(2008)の第3597頁右欄第3行目〜15行目に記載の方法に基づき、下記の塩基配列のプライマーセットとプローブとを用いて定量を行った。
プライマーセット プローブ
T : 配列番号1及び配列番号2 配列番号31
Goosecoid : 配列番号3及び配列番号4 配列番号32
Mash1 : 配列番号5及び配列番号6 配列番号33
Pax6 : 配列番号7及び配列番号8 配列番号34
βアクチン :配列番号19及び配列番号20 配列番号40
なお、プローブは5’末端をレポーター色素の3FAM、3’末端をクエンチャー色素のTAMRAで標識されたものをアプライドバイオシステム社から購入した。
結果を図2に示す。
【0029】
図2の結果から、硫酸化阻害物質と接触させたEBでは、硫酸化阻害物質と接触していないEBに比べて中胚葉マーカーであるT及びGoosecoidのmRNAレベルが抑制されている一方、外胚葉マーカーであるMash1及びPax6のmRNAレベルは顕著に増加していることがわかる(図2中、横軸の番号3と4との比較、及び番号5と6との比較)。上記各遺伝子のmRNAレベルの変化は、EBと硫酸化阻害物質との接触後わずか1日で生じており(横軸の番号3と4の比較)、硫酸化阻害物質の分化誘導作用が極めて迅速であることを示している。
【0030】
同様に、mESCであるR1セルライン由来のEBにおいて、硫酸化阻害物質がEBの神経細胞への分化を誘導していることを、硫酸化阻害物質との接触から3日後(EB形成から5日後)の細胞を用いて調べた。具体的には、神経細胞分化のマーカーであるNestin、Musashi、Mash1、Math1、NeuroD1及びNeuroD2の各遺伝子について、それらのmRNAの発現量を、下記の塩基配列のプライマーセットとプローブとを用いて上記と同様にリアルタイムPCRにより調べた。
プライマーセット プローブ
Nestin : 配列番号9及び配列番号10 配列番号35
Musashi−1:配列番号11及び配列番号12 配列番号36
Mash1 : 配列番号5及び配列番号6 配列番号33
Math1 :配列番号13及び配列番号14 配列番号37
NeuroD1 :配列番号15及び配列番号16 配列番号38
NeuroD2 :配列番号17及び配列番号18 配列番号39
なお、プローブは5’末端をレポーター色素の3FAM、3’末端をクエンチャー色素のTAMRAで標識されたものをアプライドバイオシステム社から購入した。
結果を図3に示す。
【0031】
図3の結果から、硫酸化阻害物質と接触させたEBでは、硫酸化阻害物質と接触していないEBに比べて、硫酸化阻害物質を添加して3日後(EB形成から5日後)のEBにおいて、神経細胞分化のマーカー遺伝子のmRNAレベルが顕著に増加していることがわかった。
【0032】
続いて、hiPSCであるFetch及びTic由来のEBについても上記と同様に、硫酸化阻害物質との接触から3日後(EB形成から5日後)の細胞について、神経細胞分化のマーカーであるNestin、Musashi−1、NCAM1及びSox1、並びに非分化細胞のマーカーであるOct3/4の各遺伝子のmRNAレベルをリアルタイムPCRにより調べた。なお、hiPSC由来EBのリアルタイムPCR分析には、プローブを用いずに、FastStart Universal SYBR GreenMaster(ロシュ社製)を用いた定量分析を行った。用いたプライマーセットの塩基配列は下記のとおりである。
プライマーセット
Nestin : 配列番号21及び配列番号22
Musashi−1 : 配列番号23及び配列番号24
Sox1 : 配列番号25及び配列番号26
NCAM1 : 配列番号27及び配列番号28
Oct3/4 : 配列番号29及び配列番号30
結果を図4に示す。
【0033】
図4の結果から、硫酸化阻害物質と接触させたEBでは、硫酸化阻害物質と接触していないEBに比べて、神経細胞分化のマーカー遺伝子の発現量が顕著に増加していることがわかった。すなわち、硫酸化阻害物質は、hiPSC由来のEBに対しても優れた神経細胞分化誘導能を示すことがわかる。
【0034】
[免疫染色分析]
上述のように硫酸化阻害物質と接触させたEBについて、硫酸化阻害物質との接触から3日後(EB形成から5日後)に、PLL/ラミニンコートしたガラスチャンバースライド(Iwaki社製)上に移して培養し、その2日後の細胞について神経細胞への分化を免疫染色により分析した。具体的には、上記ガラスチャンバースライド上の細胞を4%パラホルムアルデヒドで固定化し、0.1%サポニンで透過性とした。これを洗浄後ブロッキング処理を行い、抗βIII−チューブリン抗体(マウスIgG、ケミコン社製)を反応させた。さらにこれを洗浄後、FITC標識抗マウスIgG抗体(ケミコン社製)を反応させ、続いてプロピジウムヨウ化物(PI)で対比染色を行った。LSM5Pascal confocal laser scanning microscope(Carl Zeiss社製)を用いて免疫蛍光画像を得た。mESC細胞(R1セルライン)由来のEBから分化した細胞の結果を図5に、hiPSC(Fetch及びTic)から分化した細胞の結果を図6に示す。
【0035】
図5及び図6の結果から、硫酸化阻害物質と接触させたEBから分化した細胞において、神経細胞分化の指標となるβIII−チューブリンの量がタンパク質レベルで著しく増加していることがわかる(すなわち、EBが神経細胞へと構造的に大きく変化していることがわかる。)。この構造的な変化は、EBをPLL/ラミニンコートしたガラスチャンバースライド(Iwaki社製)上に移してわずか2日後(EB形成から7日後)で観察されることから、硫酸化阻害物質とEBとを接触させることによるEBの神経細胞への構造的変化は極めて迅速であるといえる。
なお、Plos ONE December 2009 Vol.4 Issue 12 e8262には、硫酸化に関与する遺伝子をノックアウトして胚様体を神経細胞へと分化させたことが記載されている。それによれば、EB形成から14日後の細胞でも、上記図5及び図6に示される量と同程度かそれ以下の量のβIII−チューブリンの発現が確認できるに過ぎない。
【0036】
[免疫ブロット分析]
上記免疫染色分析で確認したβIII−チューブリンの発現量の増加をより定量的に測定するために、免疫ブロットを実施した。上記のようにPLL/ラミニンコートしたガラスチャンバースライド(Iwaki社製)上に移して2日経過した細胞を溶解緩衝液(150mM NaCl、1%TritonX−100、1mMN3VO4、10mM NaF、プロテアーゼインヒビター(シグマ社製)を含む50mM Tris−HCl pH7.4)で溶解した。この細胞溶解液を10%SDS−PAGEにかけた後、PVDFメンブレン(ミリポア社製)にトランスファーした。このメンブレンをブロッキング後、プライマリー抗体として上記の抗βIII−チューブリン抗体(マウスIgG、ケミコン社製)を、2次抗体としてペルオキシダーゼ標識抗マウスIgG抗体(Cell signaling社製)をそれぞれ反応させた。洗浄後、メンブレン上のペルオキシダーゼをECL Plus試薬(GEヘルスケア社製)を用いて発色させた。なお、抗βアクチン抗体(シグマ社製)をβIII−チューブリンの発現量の変化を分析するためのコントロール抗体として用いた。結果を図7及び図8に示す。
【0037】
図7及び図8の結果から、硫酸化阻害物質と接触したEBでは、mESC由来のものでβIII−チューブリン量が約10倍にも上昇しており、hiPSC由来のものでもβIII−チューブリン量が2倍以上に上昇していることがわかった。なお、図7及び8のグラフは、免疫ブロットのバンド強度をNIHimageで測定した結果である。
【0038】
続いて、細胞の中胚葉分化への関与が報告されているWntシグナル及びBMPシグナルに係わるタンパク質の発現量についても免疫ブロットにより分析した。
硫酸化阻害物質との接触から1日経過後(EB形成から3日後)及び3日経過後(EB形成から5日後)のR1由来細胞を上記溶解緩衝液で溶解して細胞溶解液を調製した。また、J.Biol.Chem.283(2008)の第3596頁右欄第3行目〜18行目に記載の方法に沿って核抽出液を調製した。上記細胞溶解液はBMPシグナル伝達経路の下流で働くリン酸化Smad1(p−Smad1)の発現量の分析に用い、上記核抽出液はWintシグナル伝達経路において、核内で標的遺伝子の発現を促進する役割を果たすβ−カテニンの発現量の分析に用いた。
細胞溶解液又は核抽出液を10%SDS−PAGE後にPVDFメンブレン(ミリポア社製)にトランスファーし、このメンブレンをブロッキング処理後、プライマリー抗体である抗p−Smad1抗体(Ser463/465、ウサギIgG、セルシグナリングテクノロジー社製)又は抗βカテニン抗体(ウサギIgG、セルシグナリングテクノロジー社製)を反応させ、続いて2次抗体であるペルオキシダーゼ標識抗ウサギIgG抗体(Cell signaling社製)と反応させた。なお、細胞溶解液については抗βアクチン抗体(マウスIgG、シグマ社製)を、核抽出液については抗ラミンB1抗体(マウスIgG、Zymed社製)を、それぞれp−Smad1及びβカテニンの発現量の変化を分析するためのコントロール抗体として用いた(この場合、2次抗体としてペルオキシダーゼ標識抗マウスIgG抗体を用いた)。洗浄後、メンブレン上のペルオキシダーゼをECL Plus試薬(GEヘルスケア社製)を用いて発色させた。結果を図9に示す。
【0039】
図9の結果から、EBと硫酸化阻害物質とを接触させることで、βカテニン及びp−Smad1のいずれの発現量も減少していることがわかる。この結果は、硫酸化阻害物質により、EBを中胚葉分化へと導くWintシグナル伝達やBMPシグナル伝達が抑制され、その結果、EBの神経細胞への分化が促進されていることを示唆するものである。
【0040】
なお、上記の実験結果のうち、mESCのR1セルラインを用いた実験結果は、別のmESCのセルラインであるE14TG2aでも再現することを確認した。
【技術分野】
【0001】
本発明は、胚様体から神経細胞を製造する方法に関する。また、本発明は前記製造方法に用いうる神経細胞分化促進剤に関する。
【背景技術】
【0002】
胚性幹細胞(ES細胞)や誘導多能性幹細胞(iPS細胞)は、生体に存在する様々な細胞に分化する能力(分化多能性)を有した状態で長期にわたり継代培養することができるという性質をもつ。したがって、細胞移植が有効な治療方法として期待される白血病、パーキンソン病等の治療にこれらの幹細胞を用いることが期待されている。
【0003】
多能性幹細胞を用いた細胞移植療法においては、多能性幹細胞を目的とする細胞又はその前駆細胞に分化誘導した上で移植する必要がある。例えば、パーキンソン病の治療にあたっては、多能性幹細胞を神経細胞又はその前駆細胞へと分化誘導し、これを患者に移植することになる。これまで多能性幹細胞の分化誘導には、主に遺伝子導入や遺伝子改変といった分子生物学的手法が報告されているが(例えば、非特許文献1)、分化誘導された細胞の安全性が懸念され、実用化には至っていない。また、上記分子生物学的手法は熟練した技術と高価な試薬類を要する一方、多能性幹細胞の分化誘導速度は比較的緩やかである。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Plos ONE December 2009 Vol.4 Issue 12 e8262
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明者らは、従来の遺伝子導入や遺伝子改変手法を用いずに、薬剤を用いて多能性幹細胞の分化誘導を行うことができれば、より簡便にしかも安価に分化誘導ができ、治療上の安全性にも優れた実用性の高い細胞を創出できると考えた。
本発明は、薬剤を用いて多能性幹細胞を神経細胞へと迅速に分化させることを含む、安全性に優れた神経細胞を製造する方法を提供することを課題とする。
また、本発明は、多能性幹細胞を安全性に優れた神経細胞へと迅速に分化誘導しうる薬剤を提供することを課題とする。
【課題を解決するための手段】
【0006】
本発明者らは上記課題に鑑み鋭意検討を行った。その結果、ES細胞やiPS細胞等の多能性幹細胞に由来する胚様体(Embryoid body:EB)と硫酸化阻害物質とを接触させると、該胚様体を神経細胞へと迅速に分化誘導できることを見い出した。本発明はこの知見に基づいてなされたものである。
【0007】
本発明の課題は下記の手段により達成された。
[1]胚様体と硫酸化阻害物質とを接触させ、該胚様体中における糖鎖の硫酸化を抑制して該胚様体の神経細胞への分化を促進することを含む、神経細胞の製造方法、
[2]硫酸化阻害物質が塩素酸塩である、[1]に記載の方法、
[3]硫酸化阻害物質を含有する培地で胚様体を培養することにより胚様体と硫酸化阻害物質とを接触させる、[1]又は[2]に記載の方法、
[4]さらに胚様体とレチノイン酸とを接触させる工程を含む、[1]〜[3]のいずれかに記載の方法、
[5]胚様体が哺乳類のES細胞又は哺乳類の人工多能性幹細胞に由来する、[1]〜[4]のいずれかに記載の方法、
[6]硫酸化阻害物質を有効成分として含有する神経細胞分化促進剤、
[7]硫酸化阻害物質が塩素酸塩である、[6]に記載の神経細胞分化促進剤、
[8]さらにレチノイン酸を含有する、[6]又は[7]に記載の神経細胞分化促進剤。
【発明の効果】
【0008】
本発明の製造方法によれば、ES細胞やiPS細胞に由来する胚様体から神経細胞を簡便に、しかも迅速に得ることができる。また、本発明の神経細胞分化促進剤は、硫酸化阻害物質を有効成分とするものであり、遺伝子のように細胞内に組み込まれて細胞分裂と共に複製される性質のものではない。したがって、細胞から神経細胞分化促進剤を容易に取り除くことができ、安全性の高い神経細胞の提供することができる。
【図面の簡単な説明】
【0009】
【図1】塩素酸ナトリウムとの接触から1日経過後(EB形成から3日後)のmESC由来のEBにおけるHS及びCSの硫酸化度合の変化をFACSにより分析した結果を示す図である。図1中、aは塩素酸ナトリウムと接触させていないEBのFACS分析結果を示し、bは塩素酸ナトリウムと接触させたEBのFACS分析結果を示す。a及びb以外のプロットは一次抗体としてIgMアイソタイプコントロールを用いたときの結果である。
【図2】mESC由来のEBにおいて、中胚葉分化のマーカー遺伝子であるT及びGoosecoid、並びに外胚葉分化のマーカーであるPax6及びMash1のmRNA量をリアルタイムPCRで定量した結果を示すグラフである。縦軸はβアクチンのmRNAに対する相対量を示し、単位は[1000/βアクチンのmRNA量]である。横軸の1〜6の数字は、1:EB形成から0日後のEB、2:EB形成から2日後のEB、3:塩素酸ナトリウムと接触させていないEB形成から3日後のEB、4:塩素酸ナトリウムと接触させたEB形成から3日後のEB、5:塩素酸ナトリウムと接触させていないEB形成から4日後のEB、6:塩素酸ナトリウムと接触させたEB形成から4日後のEB、を示す。
【図3】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のmESC由来のEBにおいて、神経細胞分化のマーカー遺伝子であるNestin、Musashi−1、Mash1、Math1、NeuroD1及びNeuroD2のmRNA量をリアルタイムPCRで定量した結果を示すグラフである。縦軸は塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEBにおける各遺伝子のmRNA量を1としたときの相対値を示す。グラフ中の各遺伝子のmRNA量を示す3つの棒は、左から順に、塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、レチノイン酸のみと接触させたEB、塩素酸ナトリウムとレチノイン酸の双方と接触させたEB、を示す。
【図4】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のhiPSC由来のEBにおいて、神経細胞分化のマーカー遺伝子であるNestin、Musashi−1、Sox1及びNCAM1並びに未分化細胞のマーカーであるOct3/4のmRNA量をリアルタイムPCRで定量した結果を示すグラフである。縦軸はEB形成後0日目における各遺伝子のmRNA量を1としたときの相対値を示す。グラフ中の各遺伝子のmRNA量を示す4つの棒は、左から順に、EB形成後0日目のEB、塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB形成から5日後のEB、レチノイン酸のみと接触させたEB形成から5日後のEB、塩素酸ナトリウムとレチノイン酸の双方と接触させたEB形成から5日後のEB、を示す。
【図5】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のmESC由来のEBにおいて、抗βIII−チューブリン抗体を用いて免疫染色を行った結果を示す顕微鏡写真である。図5中、aは塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、bはレチノイン酸のみに接触させたEB、cは塩素酸ナトリウムとレチノイン酸の双方と接触させたEB、を示す。写真中の白い繊維状のものがβIII−チューブリンである。
【図6】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のhiPSC由来のEBにおいて、抗βIII−チューブリン抗体を用いて免疫染色を行った結果を示す顕微鏡写真である。図6中、aは塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、bはレチノイン酸のみに接触させたEB、cは塩素酸ナトリウムとレチノイン酸の双方と接触させたEB、を示す。写真中の白い繊維状のものがβIII−チューブリンである。
【図7】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のmESC由来のEBにおいて、抗βIII−チューブリン抗体を用いて免疫ブロットを行った結果を示す図である。縦軸は、βアクチンに対するβIII−チューブリンの相対量(βIII−チューブリンの量/βアクチンの量)を示す。横軸の1〜3の番号は、1:塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、2:レチノイン酸のみに接触させたEB、3:塩素酸ナトリウムとレチノイン酸の双方に接触させたEB、を示す。
【図8】塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のhiPSC由来のEBにおいて、抗βIII−チューブリン抗体を用いて免疫ブロットを行った結果を示す図である。縦軸は、βアクチンに対するβIII−チューブリンの相対量(βIII−チューブリンの量/βアクチンの量)を示す。横軸の1〜3の番号は、1:塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB、2:レチノイン酸のみに接触させたEB、3:塩素酸ナトリウムとレチノイン酸の双方に接触させたEB、の結果を示す。
【図9】塩素酸ナトリウムとの接触から1日後(EB形成から3日後)及び塩素酸ナトリウムとの接触から3日後(EB形成から5日後)のmESC由来のEBについて、βカテニンとp−Smad1のタンパク質量を免疫ブロットで解析した結果を示す図である。グラフの縦軸は、ラミンB1に対するβカテニンの相対量、及びβアクチンに対するp−Smad1の相対量を表わす。横軸の番号1〜5は、1:塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB形成から3日後のEB、2:塩素酸ナトリウムに接触させたEB形成から3日後のEB、3:塩素酸ナトリウムとレチノイン酸のいずれにも接触させていないEB形成から5日後のEB、4:レチノイン酸のみに接触させたEB形成から5日後のEB、5:塩素酸ナトリウムとレチノイン酸の双方に接触させたEB形成から5日後のEB、の結果を示す。
【発明を実施するための形態】
【0010】
以下、本発明について、その好ましい実施態様に基づき詳細に説明する。
[本発明の製造方法]
本発明に用いる胚様体とは、多能性幹細胞を浮遊培養したときに形成されうる初期胚に似た構造をもつ球状の細胞塊である。本発明に用いる胚様体を形成する多能性幹細胞の種類に特に制限はないが、ES細胞又はiPS細胞であることが好ましい。前記多能性幹細胞は三胚葉性動物に由来する限り特に制限はない。例えば、哺乳類、鳥類等に由来する多能性幹細胞を用いることができるが、哺乳類に由来する多能性幹細胞を好適に用いることができる。前記哺乳類としては、例えば、ヒト、サル、ウシ、ヤギ、ヒツジ、ブタ、ウマ、ウサギ、ラット、マウス、モルモット等が挙げられる。
上記多能性幹細胞は、通常の方法で調製することができる。また、寄託機関等に寄託されているセルライン等の分譲により得ることもできる。
【0011】
本発明に用いる胚様体は通常の方法で調製することができる。例えば、マウスES細胞であれば、細胞吸着能の低い培養皿で、leukemia inhibitory factor(LIF)無しのESC培地等で浮遊培養することで得ることができる。また、ヒトiPS細胞であれば、細胞吸着能の低い培養皿で、bFGF無しのiPSellon培地等で浮遊培養することで得ることができる。これらの具体的な培養方法は後述の実施例で詳述する。
胚様体を形成させるための温度、CO2濃度等は通常の条件を採用することができ、例えば、37℃、5%CO2の条件下で多能性幹細胞から胚様体を形成させることができる。
本発明に用いる胚様体の調製方法に特に制限はなく、胚様体を形成しうる方法であればいずれの方法で調製したものを用いてもよい。
【0012】
本発明に用いる硫酸化阻害物質とは、細胞に取り込まれて細胞内で少なくとも糖鎖の硫酸化を抑制しうる物質である。本発明において「糖鎖の硫酸化を抑制する」とは、糖鎖の硫酸化を完全に阻害することの他、糖鎖の硫酸化を部分的に阻害することも含む概念である。「糖鎖の硫酸化を部分的に阻害する」とは、糖鎖に硫酸基が結合してはいるが、通常よりも硫酸基の数が少ない状態にすることをいう。本発明に用いる硫酸化阻害物質により、例えば、細胞膜や細胞内に存在するヘパラン硫酸(HS)やコンドロイチン硫酸(CS)といった硫酸化グリカンの硫酸基の数が減少しうる。
本発明に用いる硫酸化阻害物質は、細胞内において糖鎖の硫酸化を阻害する作用を有すれば特に制限はないが、塩素酸塩を好適に用いることができる。塩素酸塩の種類に特に制限はなく、例えば、塩素酸ナトリウム、塩素酸カリウム、塩素酸リチウム、塩素酸アンモニウム、塩素酸カルシウム等が挙げられる。本発明に用いる硫酸化阻害物質は、通常の方法で合成して得ることもできるし、市販品を用いてもよい。
【0013】
本発明において、胚様体と硫酸化阻害物質とを接触させる方法に特に制限はない。硫酸化阻害物質を含む培地で胚様体を培養することで、胚様体と硫酸化阻害物質とを容易に接触させることができる。該培地は液体培地であることが好ましい。胚様体が生存できる環境であれば、培地の種類に特に制限はなく、例えば、マウスES細胞由来の胚様体であれば上記のLIF無しのESC培地等を、ヒトiPS細胞由来の胚様体であれば上記のbFGF無しのiPSellon培地等を好適に用いることができる。
胚様体と硫酸化阻害物質との接触の開始時期は、迅速に分化誘導を促進する観点から、胚様体の形成後0〜5日後であることが好ましく、0〜4日後であることがより好ましく、0〜3日後であることがさらに好ましく、1〜2日後であることがさらに好ましく、2日後であることが特に好ましい。
胚様体と硫酸化阻害物質との接触時間に特に制限はないが、硫酸化阻害物質が細胞内に取り込まれて生理作用を発揮するために、少なくとも1日間接触させることが好ましく、1〜5日間接触させることがより好ましく、1〜4日間接触させることがさらに好ましく、1〜3日間接触させることがさらに好ましく、2〜3日間接触させることが特に好ましい。
胚様体に接触させる硫酸化阻害物質の量に特に制限はないが、硫酸化阻害物質を含む培地で胚様体を培養する場合には、培地中の硫酸化阻害物質の濃度が0.1〜500mMであることが好ましく、1〜500mMであることがより好ましく、1〜300mMであることがさらに好ましく、1〜200mMであることがさらに好ましく、10〜100mMであることが特に好ましい。
【0014】
胚様体を神経細胞へと効率的に分化誘導するために、さらに胚様体とレチノイン酸とを接触させることが好ましい。レチノイン酸は従来から多能性幹細胞の神経細胞への分化誘導に用いられている物質である。胚様体とレチノイン酸とを接触させる方法に特に制限はないが、レチノイン酸を含む培地で胚様体を培養することで、胚様体とレチノイン酸とを容易に接触させることができる。該培地は液体培地であることが好ましい。胚様体が生存できる環境であれば、培地の種類に特に制限はなく、例えば、マウスES細胞由来の胚様体であれば上記のLIF無しのESC培地等を、ヒトiPS細胞由来の胚様体であれば上記のbFGF無しのiPSellon培地等を好適に用いることができる。胚様体とレチノイン酸との接触の開始時期は、胚様体と硫酸化阻害物質との接触を開始する前であっても後であってもよく、胚様体と硫酸化阻害物質との接触の開始と同時に胚様体とレチノイン酸との接触を開始させてもよい。胚様体とレチノイン酸との接触の開始時期は、胚様体の形成後0〜5日後であることが好ましく、1〜5日後であることがより好ましく、1〜4日後であることがさらに好ましく、2〜4日後であることがさらに好ましく、3〜4日後であることが特に好ましい。
胚様体とレチノイン酸との接触時間に特に制限はないが、レチノイン酸が細胞内に取り込まれて生理作用を発揮するために、少なくとも5時間接触させることが好ましく、5〜72時間接触させることがより好ましく、12〜48時間接触させることがさらに好ましく、20〜30時間接触させることが特に好ましい。
胚様体に接触させるレチノイン酸の量に特に制限はないが、レチノイン酸を含む培地で胚様体を培養する場合には、培地中のレチノイン酸の濃度が0.1〜10μMであることが好ましく、0.5〜5μMであることがより好ましく、0.5〜2μMであることがさらに好ましい。
レチノイン酸は、生理機能を発揮させるために通常はトランス体(all−trans)が用いられる。
【0015】
胚様体は硫酸化阻害物質及び/又はレチノイン酸の存在下で神経細胞への分化誘導を継続させてもよいが、通常には、上記の好ましい接触時間硫酸化阻害物質及び/又はレチノイン酸と接触させた後、硫酸化阻害部物質及び/又はレチノイン酸との接触を終了させる。
胚様体と硫酸化阻害物質及び/又は胚様体とレチノイン酸との接触を終了するには、胚様体の培養環境中から硫酸化阻害物質やレチノイン酸を取り除けばよい。例えば、胚様体を硫酸化阻害物質やレチノイン酸を含まない培地に移して培養することで、胚様体と硫酸化阻害物質及び/又は胚様体とレチノイン酸との接触を終了させることができる。
胚様体は、硫酸化阻害物質やレチノイン酸との接触を終了させた後も、適当な培養条件で培養することで神経細胞へと分化が進む。当該培養に使用する培地に特に制限はないが、例えば、N2サプリメントを含むDMEM−F12等が挙げられる。また、培地は定期的に交換することが好ましい。
【0016】
胚様体を培養する温度、CO2濃度等は通常の条件を採用することができ、例えば、37℃、5%CO2の条件下で培養することができる。
【0017】
胚様体の分化には、Wnt、BMP及びFGF等を介したシグナル伝達が関与していることが知られており、Wnt及びBMPは神経細胞への分化を阻害し、中胚葉への分化を促すことも示唆されてきている(例えば、Plos ONE December 2009 Vol.4 Issue 12 e8262参照)。また、上記シグナル伝達には細胞膜に存在する硫酸化グリカンが関与しうることも報告されてきている。
本発明に用いる硫酸化阻害物質は、細胞内で糖鎖の硫酸化を阻害する作用を有する。したがって、ヘパラン硫酸やコンドロイチン硫酸等の細胞膜及び/又は細胞内に存在する硫酸化グリカンの硫酸基の数を減少させ、これによりWnt及びBMPのシグナル伝達を阻害し、胚葉体の神経細胞への分化を促進すると考えられる。
これまで、糖鎖の硫酸化に関与する特定の遺伝子の働きを抑制することで、胚様体の神経細胞への分化を誘導したことの報告はあるが(Plos ONE December 2009 Vol.4 Issue 12 e8262参照)、胚葉体と硫酸化阻害物質とを接触させるだけで、胚様体の神経細胞への分化誘導に成功した例は知られていない。
本発明により、これまでにない簡便な手法で、しかも迅速に胚様体を神経細胞へと迅速に分化させて神経細胞を得ることが可能になる。また、本発明に用いる硫酸化阻害物質は、培地から該硫酸化阻害物質を除去することで分化した神経細胞から該硫酸化阻害物質を取り除くことができる。したがって、本発明の方法に分化誘導された神経細胞は安全性に優れ、細胞移植療法等への実用化が期待される。
なお、本発明において「神経細胞」には神経細胞及び神経細胞の前駆細胞の双方が含まれる。
【0018】
[本発明の神経細胞分化促進剤]
本発明の神経細胞分化促進剤は、胚様体の神経細胞への分化を促進するための薬剤であり、有効成分として上記の硫酸化阻害物質を含有する。本発明の神経細胞分化促進剤は、上述の本発明の製造方法における硫酸化阻害物質の供給源として用いることができる。すなわち、本発明の神経細胞分化促進剤と、多能性幹細胞を浮遊培養して得られうる上記胚様体とを接触させることで、該胚様体細胞を神経細胞へと迅速に分化させることができる。
【0019】
本発明の神経細胞分化促進剤は、硫酸化阻害物質の他にレチノイン酸を含有していてもよい。レチノイン酸は、生理機能を発揮させるために通常はオールトランス体である。また、本発明の神経細胞分化促進剤はさらに栄養素、pH調整剤、防腐剤、安定剤、乳化剤等の他の成分を含有していてもよい。
【0020】
本発明の神経細胞分化促進剤の剤形に特に制限はなく、各成分が溶解又は分散した液状であっても良いし、粉末状であっても良い。また、タブレット等の錠剤の形状であってもよい。
【実施例】
【0021】
以下、実施例に基づいて本発明をさらに詳細に説明するが、本発明はその要旨を越えない限りこれらに限定されるものではない。
【0022】
[セルラインの維持]
マウスES細胞(mESC)として、R1セルライン(Proc.Natl.Acad.Sci.USA 90(1993)8424−8428に記載、自然科学研究機構 生理学研究所 分子神経生理研究部門 より供与)及びE14TG2aセルライン(Dev.Biol.121(1987)1−9に記載、東京大学大学院理学系研究科より供与)を使用した。各々の細胞を、1000U/mLの濃度でLIF(ケミコン社製)を加えたESC培地(15質量%のFBS(Hyclone社製)、1質量%のペニシリン/ストレプトマイシン(Gibco社製)、0.1mMの2−メルカプトエタノール(Gibco社製)及び0.1mMの非必須アミノ酸(Gibco社製)を添加したDMEM)中において、10μg/mLのマイトマイシン C(シグマ社製)で不活化したマウス線維芽細胞(MEFs)上で維持した。
【0023】
また、ヒトiPS細胞(hiPSC)として、MRC−hiPS_Fetch(細胞番号:NIHS0604)及びMRC−hiPS_Tic(細胞番号:JCRB1331)(いずれもExp.Cell Res.315(2009)2727−2740に記載、国立生育医療センターから入手)を使用した。各々の細胞を、10ng/mLの濃度でbFGF(和光純薬社製)を加えたiPSellon(Cell−Sight社製)中において、10μg/mLのマイトマイシン C(シグマ社製)で不活化したMEFs上で維持した。
【0024】
[胚様体の形成]
上記mESC及びhiPSCをLow Cell Binding 60mm dishes(Nunc社製)に移し、mESCについてはLIFを含まない上記のESC培地で、hiPSCについてはbFGFを含まないiPSellonでそれぞれ浮遊培養することで胚様体(EB)を形成させた。なお、hiPSCは、EB形成前にゼラチンコートされたディッシュに移してフィーダー細胞を除去した。
本発明においては、上記の浮遊培養の開始した時点をEB形成時とする。したがって、例えば、「EB形成から1日後」とは、多能性幹細胞について上記の浮遊培養を開始してから1日後を意味する。
【0025】
[胚様体の神経細胞への分化誘導]
EB形成から2日後に、培地中の最終濃度が50mMとなるように、硫酸化阻害物質としての塩素酸ナトリウム(シグマ社製)を添加した。さらにその2日後(EB形成から4日後)、培地中の最終濃度が1μMとなるようにオールトランス−レチノイン酸(以下、RAと呼ぶことがある。シグマ社製)を添加した。RAを添加してから1日経過後(EB形成から5日後)、EBをPDL/laminin−coated 60mm dishes(ベクトンディッキンソン社製)に移し、N2サプリメント(Gibco社製)を含むDMEM−F12中で培養した。この操作により培養液中から硫酸化阻害物質とRAが除かれた。
【0026】
[FACS分析]
mESCであるR1セルライン由来のEBにおいて、硫酸化阻害物質の添加によるグリカンの硫酸化度合の変化を調べるために、抗HS抗体又は抗CS抗体(いずれもマウス由来IgM)をプライマリー抗体として用いたFACS分析を行った。上記抗HS抗体としては10E4又はHepSS−1(いずれも商品名、生化学工業社製)を用い、上記抗CS抗体としては2H6(商品名、生化学工業社製)を用いた。また、上記抗体にかえてマウスIgMアイソタイプ(ケミコン社製)を用いたケースをコントロールとした。具体的な実験方法は以下のとおりである。
硫酸化阻害物質との接触から1日経過後(EB形成から3日後)の細胞塊をEDTAで処理した後、この細胞懸濁液をFACS緩衝液(0.5質量%ウシ血清アルブミン(BSA)及び0.1質量%アジ化ナトリウムを含むPBS溶液)で希釈した上記プライマリー抗体溶液中でインキュベートした。洗浄後、細胞懸濁液を上記FACS緩衝液で希釈したFITC標識した抗マウスIgM抗体(シグマ社製)溶液中でインキュベートした。洗浄後、FACSAria Cell Sorter(ベクトンディッキンソン社製)を用いてFACS分析を行った。
結果を図1に示す。
【0027】
HS及びCSは細胞の分化に重要な役割を担う硫酸化グリカンであり、mESCではEB形成後少なくとも8日目まで発現することが知られている。図1に結果から、培地に硫酸化阻害物質を加えたEB(図1中のa)では、硫酸化阻害物質を加えていないEB(図1中のb)と比較して抗HS抗体及び抗CS抗体の結合量が少なく(蛍光強度が小さく)なっており、糖鎖に結合した硫酸基の量が減少していることがわかる。図1の結果は、EBと硫酸化阻害物質の接触後わずか1日で糖鎖の硫酸化が顕著に抑制されたことを示している。
【0028】
[リアルタイムPCR分析]
mESCであるR1セルライン由来のEBにおいて、硫酸化阻害物質がEBの外胚様(神経細胞)への分化を誘導することを、特定の遺伝子のmRNAの発現レベルを指標にして調べた。具体的には、中胚葉分化のマーカーであるT及びGoosecoid、並びに外胚葉分化のマーカーであるMash1及びPax6の各遺伝子について、それらのmRNAの発現量をABI PRISM(登録商標)7700 sequence detection systemを用いたリアルタイムPCRにより定量した。J.Biol.Chem.283(2008)の第3597頁右欄第3行目〜15行目に記載の方法に基づき、下記の塩基配列のプライマーセットとプローブとを用いて定量を行った。
プライマーセット プローブ
T : 配列番号1及び配列番号2 配列番号31
Goosecoid : 配列番号3及び配列番号4 配列番号32
Mash1 : 配列番号5及び配列番号6 配列番号33
Pax6 : 配列番号7及び配列番号8 配列番号34
βアクチン :配列番号19及び配列番号20 配列番号40
なお、プローブは5’末端をレポーター色素の3FAM、3’末端をクエンチャー色素のTAMRAで標識されたものをアプライドバイオシステム社から購入した。
結果を図2に示す。
【0029】
図2の結果から、硫酸化阻害物質と接触させたEBでは、硫酸化阻害物質と接触していないEBに比べて中胚葉マーカーであるT及びGoosecoidのmRNAレベルが抑制されている一方、外胚葉マーカーであるMash1及びPax6のmRNAレベルは顕著に増加していることがわかる(図2中、横軸の番号3と4との比較、及び番号5と6との比較)。上記各遺伝子のmRNAレベルの変化は、EBと硫酸化阻害物質との接触後わずか1日で生じており(横軸の番号3と4の比較)、硫酸化阻害物質の分化誘導作用が極めて迅速であることを示している。
【0030】
同様に、mESCであるR1セルライン由来のEBにおいて、硫酸化阻害物質がEBの神経細胞への分化を誘導していることを、硫酸化阻害物質との接触から3日後(EB形成から5日後)の細胞を用いて調べた。具体的には、神経細胞分化のマーカーであるNestin、Musashi、Mash1、Math1、NeuroD1及びNeuroD2の各遺伝子について、それらのmRNAの発現量を、下記の塩基配列のプライマーセットとプローブとを用いて上記と同様にリアルタイムPCRにより調べた。
プライマーセット プローブ
Nestin : 配列番号9及び配列番号10 配列番号35
Musashi−1:配列番号11及び配列番号12 配列番号36
Mash1 : 配列番号5及び配列番号6 配列番号33
Math1 :配列番号13及び配列番号14 配列番号37
NeuroD1 :配列番号15及び配列番号16 配列番号38
NeuroD2 :配列番号17及び配列番号18 配列番号39
なお、プローブは5’末端をレポーター色素の3FAM、3’末端をクエンチャー色素のTAMRAで標識されたものをアプライドバイオシステム社から購入した。
結果を図3に示す。
【0031】
図3の結果から、硫酸化阻害物質と接触させたEBでは、硫酸化阻害物質と接触していないEBに比べて、硫酸化阻害物質を添加して3日後(EB形成から5日後)のEBにおいて、神経細胞分化のマーカー遺伝子のmRNAレベルが顕著に増加していることがわかった。
【0032】
続いて、hiPSCであるFetch及びTic由来のEBについても上記と同様に、硫酸化阻害物質との接触から3日後(EB形成から5日後)の細胞について、神経細胞分化のマーカーであるNestin、Musashi−1、NCAM1及びSox1、並びに非分化細胞のマーカーであるOct3/4の各遺伝子のmRNAレベルをリアルタイムPCRにより調べた。なお、hiPSC由来EBのリアルタイムPCR分析には、プローブを用いずに、FastStart Universal SYBR GreenMaster(ロシュ社製)を用いた定量分析を行った。用いたプライマーセットの塩基配列は下記のとおりである。
プライマーセット
Nestin : 配列番号21及び配列番号22
Musashi−1 : 配列番号23及び配列番号24
Sox1 : 配列番号25及び配列番号26
NCAM1 : 配列番号27及び配列番号28
Oct3/4 : 配列番号29及び配列番号30
結果を図4に示す。
【0033】
図4の結果から、硫酸化阻害物質と接触させたEBでは、硫酸化阻害物質と接触していないEBに比べて、神経細胞分化のマーカー遺伝子の発現量が顕著に増加していることがわかった。すなわち、硫酸化阻害物質は、hiPSC由来のEBに対しても優れた神経細胞分化誘導能を示すことがわかる。
【0034】
[免疫染色分析]
上述のように硫酸化阻害物質と接触させたEBについて、硫酸化阻害物質との接触から3日後(EB形成から5日後)に、PLL/ラミニンコートしたガラスチャンバースライド(Iwaki社製)上に移して培養し、その2日後の細胞について神経細胞への分化を免疫染色により分析した。具体的には、上記ガラスチャンバースライド上の細胞を4%パラホルムアルデヒドで固定化し、0.1%サポニンで透過性とした。これを洗浄後ブロッキング処理を行い、抗βIII−チューブリン抗体(マウスIgG、ケミコン社製)を反応させた。さらにこれを洗浄後、FITC標識抗マウスIgG抗体(ケミコン社製)を反応させ、続いてプロピジウムヨウ化物(PI)で対比染色を行った。LSM5Pascal confocal laser scanning microscope(Carl Zeiss社製)を用いて免疫蛍光画像を得た。mESC細胞(R1セルライン)由来のEBから分化した細胞の結果を図5に、hiPSC(Fetch及びTic)から分化した細胞の結果を図6に示す。
【0035】
図5及び図6の結果から、硫酸化阻害物質と接触させたEBから分化した細胞において、神経細胞分化の指標となるβIII−チューブリンの量がタンパク質レベルで著しく増加していることがわかる(すなわち、EBが神経細胞へと構造的に大きく変化していることがわかる。)。この構造的な変化は、EBをPLL/ラミニンコートしたガラスチャンバースライド(Iwaki社製)上に移してわずか2日後(EB形成から7日後)で観察されることから、硫酸化阻害物質とEBとを接触させることによるEBの神経細胞への構造的変化は極めて迅速であるといえる。
なお、Plos ONE December 2009 Vol.4 Issue 12 e8262には、硫酸化に関与する遺伝子をノックアウトして胚様体を神経細胞へと分化させたことが記載されている。それによれば、EB形成から14日後の細胞でも、上記図5及び図6に示される量と同程度かそれ以下の量のβIII−チューブリンの発現が確認できるに過ぎない。
【0036】
[免疫ブロット分析]
上記免疫染色分析で確認したβIII−チューブリンの発現量の増加をより定量的に測定するために、免疫ブロットを実施した。上記のようにPLL/ラミニンコートしたガラスチャンバースライド(Iwaki社製)上に移して2日経過した細胞を溶解緩衝液(150mM NaCl、1%TritonX−100、1mMN3VO4、10mM NaF、プロテアーゼインヒビター(シグマ社製)を含む50mM Tris−HCl pH7.4)で溶解した。この細胞溶解液を10%SDS−PAGEにかけた後、PVDFメンブレン(ミリポア社製)にトランスファーした。このメンブレンをブロッキング後、プライマリー抗体として上記の抗βIII−チューブリン抗体(マウスIgG、ケミコン社製)を、2次抗体としてペルオキシダーゼ標識抗マウスIgG抗体(Cell signaling社製)をそれぞれ反応させた。洗浄後、メンブレン上のペルオキシダーゼをECL Plus試薬(GEヘルスケア社製)を用いて発色させた。なお、抗βアクチン抗体(シグマ社製)をβIII−チューブリンの発現量の変化を分析するためのコントロール抗体として用いた。結果を図7及び図8に示す。
【0037】
図7及び図8の結果から、硫酸化阻害物質と接触したEBでは、mESC由来のものでβIII−チューブリン量が約10倍にも上昇しており、hiPSC由来のものでもβIII−チューブリン量が2倍以上に上昇していることがわかった。なお、図7及び8のグラフは、免疫ブロットのバンド強度をNIHimageで測定した結果である。
【0038】
続いて、細胞の中胚葉分化への関与が報告されているWntシグナル及びBMPシグナルに係わるタンパク質の発現量についても免疫ブロットにより分析した。
硫酸化阻害物質との接触から1日経過後(EB形成から3日後)及び3日経過後(EB形成から5日後)のR1由来細胞を上記溶解緩衝液で溶解して細胞溶解液を調製した。また、J.Biol.Chem.283(2008)の第3596頁右欄第3行目〜18行目に記載の方法に沿って核抽出液を調製した。上記細胞溶解液はBMPシグナル伝達経路の下流で働くリン酸化Smad1(p−Smad1)の発現量の分析に用い、上記核抽出液はWintシグナル伝達経路において、核内で標的遺伝子の発現を促進する役割を果たすβ−カテニンの発現量の分析に用いた。
細胞溶解液又は核抽出液を10%SDS−PAGE後にPVDFメンブレン(ミリポア社製)にトランスファーし、このメンブレンをブロッキング処理後、プライマリー抗体である抗p−Smad1抗体(Ser463/465、ウサギIgG、セルシグナリングテクノロジー社製)又は抗βカテニン抗体(ウサギIgG、セルシグナリングテクノロジー社製)を反応させ、続いて2次抗体であるペルオキシダーゼ標識抗ウサギIgG抗体(Cell signaling社製)と反応させた。なお、細胞溶解液については抗βアクチン抗体(マウスIgG、シグマ社製)を、核抽出液については抗ラミンB1抗体(マウスIgG、Zymed社製)を、それぞれp−Smad1及びβカテニンの発現量の変化を分析するためのコントロール抗体として用いた(この場合、2次抗体としてペルオキシダーゼ標識抗マウスIgG抗体を用いた)。洗浄後、メンブレン上のペルオキシダーゼをECL Plus試薬(GEヘルスケア社製)を用いて発色させた。結果を図9に示す。
【0039】
図9の結果から、EBと硫酸化阻害物質とを接触させることで、βカテニン及びp−Smad1のいずれの発現量も減少していることがわかる。この結果は、硫酸化阻害物質により、EBを中胚葉分化へと導くWintシグナル伝達やBMPシグナル伝達が抑制され、その結果、EBの神経細胞への分化が促進されていることを示唆するものである。
【0040】
なお、上記の実験結果のうち、mESCのR1セルラインを用いた実験結果は、別のmESCのセルラインであるE14TG2aでも再現することを確認した。
【特許請求の範囲】
【請求項1】
胚様体と硫酸化阻害物質とを接触させ、該胚様体中における糖鎖の硫酸化を抑制して該胚様体の神経細胞への分化を促進することを含む、神経細胞の製造方法。
【請求項2】
硫酸化阻害物質が塩素酸塩である、請求項1に記載の製造方法。
【請求項3】
硫酸化阻害物質を含有する培地で胚様体を培養することにより胚様体と硫酸化阻害物質とを接触させる、請求項1又は2に記載の製造方法。
【請求項4】
さらに胚様体とレチノイン酸とを接触させる工程を含む、請求項1〜3のいずれか1項に記載の製造方法。
【請求項5】
胚様体が哺乳類のES細胞又は哺乳類の人工多能性幹細胞に由来する、請求項1〜4のいずれか1項に記載の製造方法。
【請求項6】
硫酸化阻害物質を有効成分として含有する神経細胞分化促進剤。
【請求項7】
硫酸化阻害物質が塩素酸塩である、請求項6に記載の神経細胞分化促進剤。
【請求項8】
さらにレチノイン酸を含有する、請求項6又は7に記載の神経細胞分化促進剤。
【請求項1】
胚様体と硫酸化阻害物質とを接触させ、該胚様体中における糖鎖の硫酸化を抑制して該胚様体の神経細胞への分化を促進することを含む、神経細胞の製造方法。
【請求項2】
硫酸化阻害物質が塩素酸塩である、請求項1に記載の製造方法。
【請求項3】
硫酸化阻害物質を含有する培地で胚様体を培養することにより胚様体と硫酸化阻害物質とを接触させる、請求項1又は2に記載の製造方法。
【請求項4】
さらに胚様体とレチノイン酸とを接触させる工程を含む、請求項1〜3のいずれか1項に記載の製造方法。
【請求項5】
胚様体が哺乳類のES細胞又は哺乳類の人工多能性幹細胞に由来する、請求項1〜4のいずれか1項に記載の製造方法。
【請求項6】
硫酸化阻害物質を有効成分として含有する神経細胞分化促進剤。
【請求項7】
硫酸化阻害物質が塩素酸塩である、請求項6に記載の神経細胞分化促進剤。
【請求項8】
さらにレチノイン酸を含有する、請求項6又は7に記載の神経細胞分化促進剤。
【図1】


【図2】
【図3】
【図4】
【図5】

【図6】
【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2012−75380(P2012−75380A)
【公開日】平成24年4月19日(2012.4.19)
【国際特許分類】
【出願番号】特願2010−223472(P2010−223472)
【出願日】平成22年10月1日(2010.10.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 掲載年月日:平成22年9月25日 掲載アドレス:http://www.sciencedirect.com/science/journal/0006291X
【出願人】(598123138)学校法人 創価大学 (49)
【Fターム(参考)】
【公開日】平成24年4月19日(2012.4.19)
【国際特許分類】
【出願日】平成22年10月1日(2010.10.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 掲載年月日:平成22年9月25日 掲載アドレス:http://www.sciencedirect.com/science/journal/0006291X
【出願人】(598123138)学校法人 創価大学 (49)
【Fターム(参考)】
[ Back to top ]