
神経障害に関連したポリペプチド蓄積を阻害する方法および組成物
【課題】細胞内ポリペプチドの異常な多量体または凝集体を含む神経障害を治療する新規組成物及び方法の提供。
【解決手段】細胞内凝集体を形成するポリペプチドに結合して凝集を最小にする内部抗体、および内部抗体又はその機能的断片をコードする核酸分子を提供。細胞内ポリペプチド凝集体または複合体を形成できるポリペプチドを特異的に認識する内部抗体、またはその機能的断片を同定する方法を提供。並びに内部抗体(intrabody)を用いて神経障害に関連した望ましくない細胞内ポリペプチド複合体または凝集体の形成を阻害する方法を提供する。
【解決手段】細胞内凝集体を形成するポリペプチドに結合して凝集を最小にする内部抗体、および内部抗体又はその機能的断片をコードする核酸分子を提供。細胞内ポリペプチド凝集体または複合体を形成できるポリペプチドを特異的に認識する内部抗体、またはその機能的断片を同定する方法を提供。並びに内部抗体(intrabody)を用いて神経障害に関連した望ましくない細胞内ポリペプチド複合体または凝集体の形成を阻害する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
関連情報
本出願は、参照として本明細書に組み入れられる、1999年7月27日に提出された「神経障害に関連したポリペプチド蓄積を阻害する方法と組成物」と題する米国特許仮出願番号第60/146,047号の優先権を主張する。本明細書において引用した特許、特許出願、および参考文献は全て、その全文が参照として本明細書に組み入れられる。
【0002】
政府助成研究
本発明は、その一部を米国保健省と遺伝疾患基金の助成金(第NS38002号)の補助を受けた。
【背景技術】
【0003】
発明の背景
神経変性障害は、人類を襲う最も怖れられている疾患に属する。例えば、年齢65歳以上の10人に1人がアルツハイマー病に罹患すると推定されているが、この疾患は、記憶の喪失と、最終的に死に至る徐々に衰弱する疾患である。実際、現在のところ、北米において400万人以上の人々がこの疾患に罹患しており、現在の治療は本質的に待期的である。同様に、パーキンソン病は、北米においてほぼ100万人に罹患しており、現在、疾患の発病を遅らせる、または実質的に疾患の進行を遅らせるために利用できる治療はない。もう一つの潜伏性の神経障害はハンチントン病(HD)であり、これは現在、北米において30,000人以上が罹患している。あまり一般的ではないが、等しく衰弱するのが、新しい変異型クロイツフェルトヤコブ病のようなプリオン疾患であり、これも同様に精神の痴呆と最終的に死に至る。前頭側頭痴呆(FTD)、筋萎縮性側索硬化症(ALS)、脊髄延髄筋萎縮(SBMA、またはケネディ病)、歯状赤核淡蒼球萎縮症(DRPLA)、および脊髄小脳運動失調(例えば、SCA1〜SCA7)のような他の多くの神経変性疾患も同様に、類似の経過を示す。これらの疾患の多くの病因はなおも完全には理解されていないが、これらの疾患のほとんどが神経系の細胞において変化したポリペプチドが産生されたことが原因である可能性があるように思われる(例えば、非特許文献1、非特許文献2、非特許文献3、および非特許文献4、非特許文献5を参照のこと)。
【0004】
例えば、ハンチントン病は、主に皮質と線条体に起こる選択的な神経細胞死に関連する(非特許文献6、非特許文献7)。障害は、ハンチンチンポリペプチドにおいてポリグルタミン伸長部をコードするht遺伝子の最初のエキソンにおける伸長CAG反復配列によって引き起こされるように思われる;これは、ヒト細胞において広汎に認められる機能不明の大きい〜350 kDaのポリペプチドであるが、皮質と線条体細胞においてその最高濃度が存在する(非特許文献8)。CAG反復配列は非常に多形性であり、非罹患者の染色体では6〜39反復まで変化し、HD患者の染色体では36〜180反復まで変化する(非特許文献9、非特許文献10)。成人発症例の大多数は、40〜55単位の伸長部を有するが、伸長部が70およびそれ以上になると、疾患の若年性型を必ず引き起こす。脳において、ハンチンチンポリペプチドは、主にニューロンに認められて、主として細胞質ポリペプチドであり、その分画は、小胞および/または微小管に関連しており、このことは、これが細胞骨格接着または小胞の輸送において機能的役割を有する可能性があることを示唆している(非特許文献11、非特許文献12、非特許文献13)。ハンチンチンはまた、核にも検出され(非特許文献14、非特許文献15)、このことは、転写の調節がこのポリペプチドの可能性のある機能として除外できないことを示唆している。ハンチンチンはまた、膵臓の細胞にも認められ、この場合、ハンチンチンにおける病的に伸長した反復配列の存在は、ハンチントン病に関連した糖尿病の発症に関連している(非特許文献16、非特許文献17)。
【0005】
HDの他に、CAG/ポリグルタミン伸長部は、脊髄延髄筋萎縮症、歯状赤核淡蒼球萎縮症、ならびに1、2、3および6型脊髄小脳運動失調を含む他の少なくとも6つの遺伝性神経変性障害において認められている(非特許文献18、非特許文献19)。正常で伸長した大きさの範囲はSCA6を例外として同等であり、SCA6の場合、伸長した対立遺伝子がより小さく、変異がおそらく異なる経路によって作用する可能性がある。しかし、全ての場合において、CAG反復配列は、コード領域内に存在して、ポリグルタミン残基の鎖に翻訳される。ポリグルタミン配列を有するポリペプチドは無関係で、ほとんどの機能が不明であるが、変異は、類似のメカニズムを通じて作用する可能性がある(非特許文献20)。これらのポリペプチドは通常、広く発現されて、一般的に核および細胞質の双方に存在する。さらに、アルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆、筋萎縮性側索硬化症、脊髄延髄筋萎縮症、歯状赤核淡蒼球萎縮症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、4型脊髄小脳運動失調、5型脊髄小脳運動失調、6型脊髄小脳運動失調、および7型脊髄小脳運動失調を含む神経変性疾患はまた、神経系の細胞における変化したポリペプチドの産生にも関連している(非特許文献3)。
【0006】
したがって、神経系の細胞における変化したポリペプチドの蓄積が、細胞のサブセットの神経変性および疾患の発症の前兆となる可能性があると推測されている。
【非特許文献1】ハートレー(Hurtley, S.)、Science 282:1071(1998)
【非特許文献2】ショールソン(Shoulson, I.)、Science 282:1072〜1074(1998)
【非特許文献3】ハーディ(Hardy)ら、Science 282:1075〜1079(1998)
【非特許文献4】プライス(Price)ら、Science 282:1079〜1083(1998)
【非特許文献5】クレメント(Klement)ら、Cell 95:41〜53(1998)
【非特許文献6】ハーパー(Harper)、「ハンチントン病(Huntington's Disease)」、第22版、ソーンダース社(1991)
【非特許文献7】フォンサッテル(Vonsattel)ら、J. Neuropath. Exp. Neurol. 44:559〜577、(1985)
【非特許文献8】ハンチントン病共同研究グループ、Cell 72:971〜983(1983)
【非特許文献9】ルビンスタイン(Rubinsztein)ら、Am. J. Hum. Genet. 59:16〜22(1996)
【非特許文献10】サタシバム(Sathasivam)ら、Hum. Genet. 99:692〜695(1997)
【非特許文献11】ジフィグリア(DiFiglia)ら、Neuron 14:1075〜1081(1995)
【非特許文献12】グテクンスト(Gutekunst)ら、P.N.A.S. 92:8710〜8714(1995)
【非特許文献13】シャープ(Sharp)ら、Neuron 14:1065〜1074(1995)
【非特許文献14】フーゲベーン(Hoogeveen)ら、Hum. Mol. Genet. 2:2069〜2073(1993)
【非特許文献15】デルーイ(de Rooij)ら、Hum. Mel. Genet. 5:1093〜1098(1996)
【非特許文献16】ハールバート(Hurlbert)ら、Diabetes 48:649〜651(1999)
【非特許文献17】ファラー(Farrer, L.)、Clin. Genet. 27:62〜67(1985)
【非特許文献18】ベイテス(Bates)ら、Human Mol. Genet. 6:1633〜1637(1997)
【非特許文献19】トロッティア(Trottier)ら、Nature 378:403〜406(1995)
【非特許文献20】シソディア(Sisodia, S.)、Cell 95:1〜4(1998)
【発明の開示】
【0007】
発明の概要
細胞内ポリペプチドの蓄積を異常な多量体または凝集体として含む神経障害の衰弱する作用を治療する方法は、病態が発生する前にこれらの複合体または凝集体の形成を防止することである。
【0008】
この目的のため、これらの神経変性ポリペプチドおよびその複合体または凝集体の形成を阻害する新規組成物および方法が開発されている。
【0009】
本発明は、細胞内の病的ハンチンチンポリペプチドを内部抗体(intrabody)に接触させると、その異常な病態の特徴であるポリペプチド凝集体の形成を予防できる、という発見に基づいている。その上、そのような内部抗体は、細胞内で正常なポリペプチドを切断させるようにして凝集体の形成を阻害することができ、ターゲティングシグナルと結合すると、望ましくないポリペプチドが、例えば、ライソゾームまたはプロテアソームにおいて破壊されるように、再度ターゲティングすることができる。
【0010】
したがって、一つの局面において、本発明は、複合体を形成することができるポリペプチドをポリペプチド結合分子、例えば凝集を最小限にして、それによって細胞内凝集体の形成を減少させるようにポリペプチドに特異的に結合する内部抗体、に接触させる段階を含む、選択されたポリペプチドの細胞内凝集体の形成を阻害する方法を提供する。
【0011】
第二の局面において、本発明は、細胞内凝集体を有する危険にさらされている対象に、ポリペプチド結合分子、例えば凝集を最小限にして、それによって細胞内凝集体の形成を減少させるようにポリペプチドに特異的に結合する内部抗体、を投与することを含む、対象において選択されたポリペプチドの細胞内凝集体の形成を阻害する方法を提供する。一つの態様において、対象は神経障害のリスクを有し、好ましくはヒト対象である。もう一つの態様において、対象は、実験動物、好ましくはハンチントン病の動物モデルである。
【0012】
第三の局面において、本発明は、ポリペプチド結合分子、例えば神経障害に関連したポリペプチド凝集体または複合体を形成することができ、それによって凝集体または複合体が形成されることを阻害するポリペプチドに特異的に結合する内部抗体、を対象に投与する段階を含む、神経障害を有する、または有する可能性がある対象を治療する方法を提供する。
【0013】
第四の局面において、本発明は、細胞内ポリペプチド凝集体を形成することができるポリペプチドを提供する段階;ポリペプチドを、試験ポリペプチド結合分子、例えば、内部抗体またはその機能的断片、に接触させる段階;および試験ポリペプチド結合分子、例えば内部抗体またはその機能的断片がポリペプチドを特異的に認識する能力を決定し、それによってポリペプチド結合分子、例えば、細胞内ポリペプチド凝集体を形成することができるポリペプチドを特異的に認識する内部抗体またはその機能的断片であることが同定される段階を含む、ポリペプチド結合分子、例えば細胞内ポリペプチド凝集体または複合体を形成することができるポリペプチドを特異的に認識する内部抗体、またはその機能的断片を同定する方法を提供する。
【0014】
上記の局面の一つの態様において、本発明の内部抗体は、多価であってもよく、スペーサー領域を有してもよく、ファージディスプレイライブラリのような発現ライブラリから選択してもよく、例えば、Fv配列が含まれる。
【0015】
好ましい態様において、内部抗体は、所定のポリペプチドに対して少なくとも105 M-1、より好ましくは少なくとも106 M-1の結合親和性を有するが(解離定数、Ka)、内部抗体が多価である場合、それぞれの原子価は、ポリペプチドに対して少なくとも102 M-1、好ましくは103 M-1の結合親和性を有してもよい。一つの態様において、選択された内部抗体は、野生型ポリペプチドと比較して変異型ポリペプチドに対する親和性が高い。もう一つの態様において、選択された内部抗体は、野生型ポリペプチド、例えば野生型ポリペプチドと変異型ポリペプチドの双方に共通するポリペプチド領域に対して特異的親和性を有する。
【0016】
第五の局面において、本発明は、細胞内凝集体または複合体を形成することができるポリペプチドを提供する段階;ポリペプチドに結合する試験ポリペプチド結合分子、例えば内部抗体またはその機能的断片を提供する段階;ポリペプチドおよび内部抗体断片またはその断片を結合分子と共にインキュベートする段階;ならびに試験化合物が、ポリペプチド結合分子、例えば内部抗体またはその機能的断片の結合を変化させるか否かを決定して、内部抗体に結合するそれらの結合分子が消失すれば、それによって試験化合物が、細胞内ポリペプチド凝集体を形成することができるポリペプチドと相互作用することができると同定される段階を含む、望まない細胞内ポリペプチド凝集体または複合体を形成することができるポリペプチドを特異的に認識する化合物を同定する方法を提供する。好ましい態様において、方法は、異なる結合分子少なくとも102、103、104、105、106、107、108個および好ましくは109個またはそれ以上の多様なライブラリに適用する、および再適用することができる。
【0017】
関連する態様において、試験化合物は小さい有機分子、ペプチド、または天然物抽出物である。
【0018】
第六の局面において、本発明は、神経障害に関連した細胞内ポリペプチド凝集体を形成することができる選択されたポリペプチドに結合する、内部抗体またはその機能的断片をコードする単離核酸分子を提供する。
【0019】
一つの態様において、本発明は、それぞれ配列番号:2、配列番号:4および配列番号:6に記載したアミノ酸配列をコードする、配列番号:1または配列番号:3に記載した核酸配列、および好ましくは配列番号:5に記載した核酸配列を提供し、これらの配列は、好ましい内部抗体、またはその断片に対応する。
【0020】
もう一つの局面において、本発明の内部抗体は、配列番号:1、配列番号:3または配列番号:5の核酸分子と、ストリンジェントな条件下でハイブリダイズする核酸分子によってコードされる。好ましい態様において、核酸は哺乳類に由来する。そのような核酸はまた、ベクター、好ましくは例えばアデノウイルス、アデノ随伴ウイルス、レトロウイルス、または単純ヘルペスウイルスに由来する発現ベクターのような発現ベクター、によってコードされてもよい。その上、内部抗体またはポリペプチド内部抗体をコードするベクターは、薬学的に許容される担体を含む薬学的組成物の形であってもよい。
【0021】
さらにもう一つの態様において、本発明は、宿主細胞が好ましくは、神経起源である、または神経起源の組織の一部である、上記のベクターのいかなるものも含む宿主細胞を提供する。
【0022】
第七の局面において、本発明は、病態、例えば神経障害、に関連した細胞内ポリペプチド凝集体を形成することができる選択されたポリペプチドに結合するポリペプチド骨格のポリペプチド結合分子、好ましくは内部抗体またはその機能的断片を提供する。
【0023】
第八の局面において、本発明は、免疫が凝集体、例えば選択されたポリペプチドの細胞内凝集体の形成、を阻害するために十分な宿主抗体免疫応答を誘発する、動物、例えばヒト患者、を選択されたポリペプチドと共通のエピトープを有する免疫原によって免疫することによって、動物において、選択されたポリペプチドの細胞内凝集体の形成を阻害する方法を提供する。好ましい態様において、免疫原は、例えば、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、SCA7、および好ましくはハンチンチンのようなポリペプチドと共有するエピトープを含む、ポリペプチドをコードする発現可能な核酸ワクチン、例えばDNAワクチンである。
【0024】
関連する態様において、動物は、ハンチントン病またはハンチントン関連糖尿病(例えば、高い絶食時血糖レベルおよび/または低いインスリンレベル)を有する、または有する危険にさらされている。
【0025】
さらにもう一つの態様において、本発明は、好ましくは内部抗体が、神経障害、例えば、ハンチンチンポリペプチド、例えば、対応する野生型ハンチンチンポリペプチドと比較してさらなるグルタミン残基を有するハンチンチンポリペプチド、によって引き起こされるハンチントン病、に関連する凝集体を形成することができるポリペプチドに選択的に結合しうる、内部抗体をコードする核酸を発現するように遺伝子操作されたトランスジェニック動物を提供する。
【0026】
上記の局面のそれぞれにおいて、本発明は、好ましい態様において、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、またはSCA7、および好ましくはハンチンチン、タウ、または野生型ポリペプチドもしくは変異型ポリペプチドのいかなる断片も含むその変異型ポリペプチドのようなポリペプチドを含む細胞内凝集体の形成を阻害する方法を提供する。
【0027】
上記の局面のもう一つの態様において、ポリペプチドには、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドが含まれ、この場合、好ましくは、ポリペプチドはハンチンチンであり、より好ましくは対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有するハンチンチンポリペプチドである。
【0028】
上記の局面のなおもう一つの態様において、本発明の内部抗体は多価であってもよく、または同じもしくは異なる特異性を有する結合部位のあいだにスペーサー領域を有してもよい。
【0029】
上記の局面のさらにもう一つの態様において、本発明の内部抗体は、低分子ペプチド、ペプチド模倣抗体、抗体断片、または好ましくは内部抗体、例えばターゲティングシグナルに対応するアミノ酸配列、例えばそれによって内部抗体が標的ポリペプチドをプロテアソームに再ターゲティングさせることができるユビキチン、を含む内部抗体を含んでもよい。
【0030】
上記の局面のさらにもう一つの態様において、本発明のポリペプチド結合分子は、ターゲティングシグナルに対応し、それによって内部抗体が、特定の細胞位置に標的ポリペプチドを再ターゲティングさせることができるアミノ酸配列を含んでもよい。好ましい態様において、ターゲティングシグナルは、細胞質、核、ライソゾーム、細胞膜関連、小胞体関連、ペルオキシソーム、またはプロテオソームシグナルであり、好ましくは核またはライソゾームシグナルである。
【0031】
上記の局面のさらにもう一つの態様において、内部抗体には、配列番号:2または配列番号:4に提供されたハンチンチンに対して特異的な最小のsFv結合領域が含まれ、好ましくは、配列番号:6に提供される内部抗体α-Nt-HD-C4 sFvのアミノ酸配列を含む。もう一つの態様において、ポリペプチドをインビボまたはインビトロで内部抗体と接触させてもよい。
【0032】
したがって、本発明は、多くの長所を含むと認識される。例えば、内部抗体を、細胞内で作用して、細胞内レベルで細胞に保護を与える発現可能な遺伝子の形で投与してもよい。さらに、本発明の組成物および方法は、それらが免疫系とは独立して機能し、いかなる望ましくない免疫応答も誘発する可能性が低いように設計される。
【0033】
本発明のその他の特徴および長所は、以下の詳細な説明および請求の範囲から明らかとなるであろう。
【0034】
発明の詳細な説明
本発明の完全な範囲を明らかに理解するために、以下の定義を提供する。
【0035】
I.定義
本明細書において用いられるように、「形成の阻害」という用語には、化合物もしくは内部抗体の減少もしくは消失能、例えば、凝集体形成を阻害して、HD類似体ポリペプチドのポリペプチド切断を可能にする可溶性状態に抗体抗原複合体を維持することによって、細胞において望ましくないポリペプチドの蓄積速度を減少させる、蓄積を防止するために利用できるポリペプチドを減少させる、またはそうでなければ蓄積もしくは形成を相殺もしくは消失させる消失させることができる能力が含まれると解釈される。
【0036】
「ポリペプチド凝集体」という用語には、ポリペプチドのいかなる望ましくない、逸脱した、もしくは異常な蓄積、ポリペプチド・ポリペプチド相互作用、ポリペプチド複合体、またはポリペプチド凝集体が含まれると解釈される。この用語にはまた、特定の細胞内空間に攻撃ポリペプチドを局在させることによって、細胞に対する害を最小限にしようとして細胞が形成する可能性があるポリペプチド凝集体が含まれると解釈される。好ましい態様において、ポリペプチドは、疾患、例えば神経疾患に関連した変化したポリペプチドである。この用語には、細胞、例えば神経起源の細胞において認められる、例えば、ハンチンチン凝集体、アミロイド斑、神経原線維濃縮体、レーヴィ小体、プリオン斑、レーヴィ様小体、および望ましくないいかなる複合体または異常な蓄積または封入体も含まれると解釈される。さらになお、この用語はまた、細胞外ポリペプチド凝集体を意味してもよい。「ポリペプチド」または「複数のポリペプチド」、および「蛋白質」という用語は、明細書全体において互換的に用いられる。
【0037】
「選択されたポリペプチド」または「標的ポリペプチド」という用語は、望ましくない蓄積または凝集、例えばポリペプチド凝集体を形成するいかなるポリペプチドも含まれると解釈される。典型的に、選択または標的ポリペプチドは、対応する野生型ポリペプチドと比較して変化したポリペプチドである。
【0038】
「ポリペプチド結合分子」という用語には、例えば、望ましくない蓄積、立体構造、または凝集体を形成することができるポリペプチドに特異的に結合することができるいかなる分子も含まれる。したがって、この用語には、低分子、ペプチド、ペプチド模倣体、抗体(例えば、内因性抗体、すなわち、宿主動物の免疫系によって産生される;外来の抗体、すなわち、発現可能な核酸(複数)、外来核酸(複数)を含む細胞(複数)の形で、またはポリペプチド治療として投与される抗体、を含む)、抗体断片(例えば、Fab断片)ならびに好ましくは内部抗体、または細胞において細胞内で抗原に結合するように作用する免疫グロブリンスーパーファミリーからの類似の遺伝子操作蛋白質が含まれる。
【0039】
「内部抗体(intrabody)」という用語には、標的ポリペプチドに細胞内で特異的に結合することができるいかなる一本鎖ポリペプチド結合物質も含まれると解釈される。典型的に、内部抗体は、重鎖可変領域に結合した最小の軽鎖可変領域を含む一本鎖Fv(sFv)である。内部抗体は典型的に、ポリペプチド治療の形で投与されるが、発現可能な核酸の形、またはそのような核酸を発現する細胞の形で投与してもよい。
【0040】
「多価」という用語には、例えば、分子内において異なるsFv成分によって決定される一つまたはそれ以上の結合特異性を有する、一つ以上の結合部位を含むいかなる物質または内部抗体も含まれると解釈される。
【0041】
「スペーサー領域」という用語には、一つまたはそれ以上の機能的ドメインに、例えば一つのsFvをもう一つのsFvに、またはsFv結合部位をターゲティングシグナルに、結合させるように設計されたペプチド骨格の結合物質または内部抗体のいかなる領域も含まれると解釈される。
【0042】
「リンカー領域」という用語は、リンカーが遺伝子レベルでコードされる、VHリンカーVL、またはVLリンカーVHとして軽鎖可変領域と重鎖領域とを結合させるsFvのペプチド配列を含むと解釈される。
【0043】
「再ターゲティングする」という用語は、標的ポリペプチドに結合した場合に、結合物質または内部抗体がポリペプチドの細胞内位置を再び向け直す能力が含まれると解釈される。
【0044】
「ターゲティングシグナル」という用語は、異種ポリペプチドと結合すると、細胞内の特定の位置にポリペプチドの細胞内輸送を向けることができる、当技術分野で認識されたいかなるアミノ酸配列も含まれると解釈される。この用語には、例えば、核、細胞質、細胞膜関連、小胞体関連、ライソゾーム、ペルオキシソーム、またはプロテアソームのターゲティングシグナルが含まれると解釈される。
【0045】
「野生型ハンチンチンポリペプチド」という用語は、疾患に関連しないグルタミン残基の正常数のみを有するハンチンチンポリペプチドが含まれると解釈される。典型的に、グルタミン残基が38個未満であるいかなるハンチンチンポリペプチドも疾患に関連しておらず、これらは正常または野生型ポリペプチドであると見なしてもよい。
【0046】
「神経障害」という用語には、中枢神経系および/または末梢神経系の細胞を含むいかなる疾患または病態も含まれると解釈される。この用語には、例えば、アルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆(FTD)、筋萎縮性側索硬化症(ALS)、脊髄延髄筋萎縮症(SBMAまたはケネディ病)、歯状赤核淡蒼球萎縮症(DRPLA)、および脊髄小脳運動失調(例えば、SCA-1〜SCA-7)が含まれると解釈される。
【0047】
「発現ライブラリ」という用語には、発現可能な核酸を含むいかなるライブラリも含まれると解釈される。
【0048】
「ファージディスプレイライブラリ」という用語には、当技術分野で認識された意味が含まれると解釈され、典型的に、その発現および/または増殖がファージをパッケージングした核酸配列によって促進される、発現可能な核酸のいかなる集合体も意味する。一つの典型的な態様において、sFvは、sFv抗体結合部位(その特異的結合が表現型である)が、ファージ表面上で抗原結合に利用できるように、そしてその遺伝子型(sFvをコードするDNA)がファージ抗体遺伝子材料内に含まれるように、ファージ外膜蛋白質に融合させる。
【0049】
「Fv」という用語は、例えば、Fab、Fv、Fd、VH、またはVLのような抗体のいかなる抗原結合断片も含まれると解釈される。「sFv」という用語は一本鎖Fvであり、エピトープと相互作用することができる結合ポリペプチドを形成するために必要な最小の重鎖可変領域に結合した最小の軽鎖可変領域が含まれると解釈される。典型的に、内部抗体はFab、Fd、Fv、またはsFv領域を少なくとも一つ含む。
【0050】
「変異型ポリペプチド」という用語は、少なくとも1つのアミノ酸置換または付加、例えばグルタミン付加を有することによって、その対応する野生型ポリペプチドとは異なるいかなるポリペプチドまたはその表示が含まれると解釈される。典型的に、変異型ポリペプチドは、アミノ酸置換およびフレームシフトもしくはスプライシング部位の変化、または一つもしくはそれ以上のグルタミンの伸長によるアミノ酸鎖の変化を有する。他の場合、「変異型ポリペプチド」は、野生型分子と比較して、その結果凝集したポリペプチド種の異常な形成が起こる異常な立体構造を意味する。
【0051】
「変化に富むライブラリ」という用語は、核酸または化合物、例えば有機分子または天然物抽出物のいかなる集合体も含まれると解釈される。
【0052】
「神経起源の細胞」という用語には、中枢神経系または末梢神経系の組織に由来するいかなる細胞も含まれると解釈される。この用語には、例えばニューロン(例えば、運動神経ニューロン、知覚神経ニューロンを含む)、シュワン細胞、プルキンエ細胞、星状細胞、小膠細胞、上衣細胞、および乏突起膠細胞が含まれると解釈される。この用語にはまた、例えば、胚幹細胞または他の全能性もしくは多能性細胞もしくは組織に由来する神経細胞特徴を有するいかなる細胞も含まれると解釈される。
【0053】
「神経起源の組織試料」という用語には、中枢神経系または末梢神経系の組織に由来する多細胞試料が含まれると解釈される。この用語には、例えば、典型的に一つまたはそれ以上の異なる細胞タイプを含む全脳切片および他の組織が含まれると解釈される。
【0054】
「プロテアソーム」という用語には、その通常の技術分野で認識される意味が含まれると解釈される。典型的にプロテアソームは、細胞蛋白質を分解して、多くの蛋白質の寿命の調節に関与して、同様にペプチド源を提供することができる細胞における多数のサブユニットポリペプチド部分を意味する。
【0055】
「ユビキチン」という用語には、その通常の技術分野で認識される意味が含まれると解釈される。この用語には、異種ポリペプチド、例えばプロテアソームと融合すると、異種ポリペプチドが破壊されるようターゲティングするために適している、ヒトを含むいかなる種からのユビキチンアミノ酸配列が含まれると解釈される。
【0056】
「特異的に認識する」という用語には、分子、例えば内部抗体または結合分子の、選択されたポリペプチドに対する特異的結合能が含まれると解釈される。
【0057】
「アミロイド前駆蛋白質」、「プレセニリン1」、「プレセニリン2」「α-2マクログロブリン」、「アポリポ蛋白質」、「αシヌクレイン」、「ハンチンチン」、「プリオン蛋白質」、「タウ」、「スーパーオキシドジスムターゼ(SOD)」、「アンドロゲン受容体(AR)」、「アトロフィン1」、「アタキシン1」、「アタキシン2」、「アタキシン3」、「CACNL1A4」、および「SCA7」という用語には、その通常の技術分野で認識される意味が含まれると解釈される。
【0058】
「アルツハイマー病」、「パーキンソン病」、「ハンチントン病」、「プリオン疾患」、「前頭側頭痴呆(FTD)」、「筋萎縮性側索硬化症(ALS)」、「脊髄延髄筋萎縮症(SBMAまたはケネディ病)」、「歯状赤核淡蒼球萎縮症(DRPLA)」、「1型脊髄小脳運動失調(SCA1)」、「2型脊髄小脳運動失調(SCA)」、「3型脊髄小脳運動失調(SCA3またはマチャドジョゼフ病(MJD)」、「4型脊髄小脳運動失調(SCA4)」、「5型脊髄小脳運動失調(SCA5)」、「6型脊髄小脳運動失調(SCA6)」、および「7型脊髄小脳運動失調(SCA7)」という用語は、当技術分野で認識されるその通常の意味が含まれると解釈される。
【0059】
「選択されたポリペプチド」という用語には、細胞における望ましくない蓄積またはポリペプチド凝集体を形成するとして同定されているいかなるポリペプチドも含まれると解釈される。典型的に、ポリペプチドは疾患に関連しており、変化している、例えば、対応する野生型ポリペプチドと比較してアミノ酸置換もしくはポリグルタミン伸長を含む、または野生型ポリペプチドの対応する三次構造と比較して異常な三次構造を呈する、またはその双方である。
【0060】
「宿主細胞」という用語は、例えば、トランスフェクション、リポフェクション、または感染によって異種ポリヌクレオチド配列を組み入れることができる遺伝子操作に適した細胞が含まれると解釈される。細胞は、微生物または高等真核細胞となりうる。この用語には、最初にトランスフェクトした細胞の子孫が含まれると解釈される。好ましい態様において、細胞は神経起源の細胞である。
【0061】
「異種ポリヌクレオチドセグメント」という用語には、一つまたはそれ以上のポリペプチド、またはポリペプチドの一部もしくは断片をコードするポリヌクレオチドセグメントが含まれると解釈される。異種ポリヌクレオチドセグメントはいかなる起源に由来してもよく、例えば、真核細胞、原核細胞、ウイルス、ファージ、または合成ポリヌクレオチド断片であってもよい。
【0062】
「由来する」という用語には、記載の起源からポリヌクレオチドセグメントを(全体または一部)の単離することが含まれると解釈される。この用語には、例えば、表示のポリヌクレオチド起源に関連した配列から、または配列に基づく直接クローニング、PCR増幅、または人工的な合成が含まれると解釈される。
【0063】
「トランスジェニック動物」という用語には、動物、例えば、動物の細胞の一つまたはそれ以上、好ましくは本質的に全てがトランスジーンを含む、ヒト以外の哺乳類、例えばブタ、サル、ヤギ、または齧歯類、例えばマウスが含まれると解釈される。トランスジーンは、例えば、マイクロインジェクション、トランスフェクション、または感染、例えば組み換え型ウイルスによる感染によって、細胞の前駆体に導入することによって、細胞に直接または間接的に導入される。遺伝子操作という用語は、組み換え型DNA分子の導入に向けられる。この分子は、染色体内に組み入れてもよく、または染色体外の複製DNAであってもよい。
【0064】
「投与する」という用語は、非蛍光または経口経路、筋肉内注射、皮下/皮内注射、静脈内注射、経頬投与、経皮輸送、頭蓋内輸送、のいずれかによる輸送を含む、動物またはヒトにおける所望の位置に治療物質を輸送するために適したいかなる経路によって、動物またはヒトに治療物質を投薬、輸送、または適用することを意味すると解釈される。「投与する」という用語はさらに、治療物質がその作用を細胞において発揮できるように、治療物質を細胞に近接させることを意味すると解釈される。
【0065】
「ベクター」という用語には、それに結合しているもう一つの核酸を輸送することができる核酸分子が含まれると解釈される。一つの態様において、ベクターに結合した核酸は、ペプチド結合分子または内部抗体をコードする。
【0066】
II.ポリペプチド結合分子
一般的に、本発明は、望ましくない凝集体、立体構造、または蓄積を形成することができ、この望ましくないポリペプチド活性を減少または阻害することができるポリペプチドに特異的に結合することができる分子に関する。したがって、本発明のポリペプチド結合分子には、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体が含まれる。本発明の抗体は典型的に、ポリペプチド骨格の治療物質として投与されるが、発現可能な核酸の形で投与してもよい。または、本発明のもう一つの態様において、本発明のポリペプチド結合分子は、ポリペプチドまたはペプチド骨格抗原またはそのような抗原をコードする核酸(すなわち核酸ワクチン)のいずれかを含むワクチンの投与に反応して宿主自身によって産生された内因性の抗体である。しかし、最も好ましい態様において、本発明のポリペプチド結合分子は内部抗体である。
【0067】
内部抗体
本発明の内部抗体は、望ましくない細胞内蓄積または凝集体を形成するポリペプチドに結合することができる。一つの態様において、内部抗体は神経疾患、例えばアルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆、筋萎縮性側索硬化症、脊髄延髄筋萎縮症、歯状赤核淡蒼球萎縮症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、4型脊髄小脳運動失調、5型脊髄小脳運動失調、6型脊髄小脳運動失調、および7型脊髄小脳運動失調に関連したポリペプチドに結合する。
【0068】
本発明は、変化したポリペプチド、例えばアミロイド前駆蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、スーパーオキシドジスムターゼ(SOD)、アンドロゲン受容体(AR)、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7を含む神経疾患との関連を有するいかなるポリペプチド、野生型および/または異常な変異型にも結合することができる内部抗体のような分子を含むと解釈される。さらに、本発明はまた、伸長したポリグルタミン領域を有するいかなるポリペプチド、例えばAR、ハンチンチン、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7にも結合することができる内部抗体を含むと解釈される(例えば、ハートレー(Hurtley, S)、Science 282:1071(1998);ショールソン(Shoulson, I.)、Science 282:1072〜1074(1998);ハーディ(Hardy)ら、Science 282:1075〜1079(1998);およびプライス(Price)ら、Science 282:1079〜1083(1998)を参照のこと)。
【0069】
一つの態様において、本発明は、疾患に関連したポリペプチドを表す変化したポリペプチドを提供し、米国特許第5,132,405号;第5,091,513号;第5,084,398号;第5,525,491号;および第5,851,829号、または国際特許出願国際公開公報第99/14353号のいずれかに記載されている方法を用いること、ならびに対応する正常なポリペプチドと比較して疾患に関連する変化したポリペプチドに特異的なエピトープを認識することができる遺伝子操作された内部抗体を提供する(同様に、ザネッティ(Zanetti)ら、「抗体(The Antibodies)」、ハーウッド教育出版、4:1〜141(1997);チェン(Chen)ら、Human Gene Therapy 5:595〜601(1994);ジョーンズ(Jones)ら、Ad. Drug Delivery Reviews 31:153〜170(1998)も参照のこと)。好ましい態様において、内部抗体はハンチンチンポリペプチドに結合することができる。結合は例えば、神経細胞に蓄積して凝集してハンチントン病を引き起こす、正常および異常な全てのハンチンチンに対して特異的となりうる。
【0070】
典型的に、本発明のそのような操作された内部抗体は、本明細書において述べたポリペプチドのいずれかにおけるエピトープに対する可変領域断片(Fv)を含む最小のsFv抗体を含む。完全な抗原認識および結合部位を含むFv、sFv、または最小の抗体断片は、堅固に非共有結合している一つの重鎖と一つの軽鎖可変ドメインとのへテロダイマーからなる、この形状において、各可変ドメインの3つの相補性決定領域は相互作用して、VH-VLダイマーの表面上の抗原結合部位を定める。集合的に、6つの相補性決定領域(CDRs)が、内部抗体に対する抗原結合特異性を付与する。
【0071】
免疫グロブリンポリペプチドの構造に関するこの知識は、望ましくない細胞内蓄積または凝集体を形成するポリペプチドと会合する結合部位(複数)を有するFab、VH、FV、Fd、または一本鎖Fv(sFv)を開発するために利用されている。これらの抗体は、細胞内で機能することを意図しており、したがって内部抗体と呼ばれる。
【0072】
選択された親和性を有する内部抗体
本発明の態様の内部抗体の結合部位は、それらが合成DNA、すなわちポリメラーゼ連鎖反応によって得られた、および/または化学合成されたオリゴヌクレオチドであってもよい複数のDNAセグメントのライゲーションから作製された、またはハイブリドーマ、成熟B細胞クローン、cDNAライブラリ、もしくは哺乳類に由来する核酸を含むファージライブラリのゲノムに由来するDNA断片のライゲーションによって作製された、組み換え型DNAである合成DNAを発現するように作製された細胞宿主において合成されるという点において生合成的である、または生合成的であってもよい。好ましい態様において、ファージライブラリは、選択された抗原に対して免疫されている(免疫ライブラリ)、または所定の抗原に対して特に免疫されていない哺乳類(無傷のライブラリ)哺乳類に由来する。一つの態様において、哺乳類は、当技術分野で既知の方法を用いてDNAワクチン接種を用いて特定の抗原に対して免疫してもよい。または、抗原特異的ファージ抗体を無傷のファージディスプレイライブラリから選択してもよい。このように、本発明の内部抗体は、予め選択した抗原物質に対する特異性と親和性を有するように特に設計される。本発明の内部抗体は、その構造が抗原認識の原因であることが知られている無傷の抗体の領域後でパターン化されているという点において抗体に似ている。
【0073】
より詳しく述べると、ポリペプチドに結合特性を付与する領域におけるこれらの内部抗体の構造は、天然の抗体のFv領域と類似である。したがって、本発明の内部抗体は、ポリペプチドの特定の計画した有用性を念頭に置いて設計してもよく、既知の親和性および特異性を有する抗体、例えば、選択されたポリペプチドに対するマウス、ラット、またはヒトモノクローナル抗体の高度可変領域のCDRs、または類似のCDRsもしくはその一部を含む。
【0074】
もう一つの態様において、本発明は、抗原プロセシングおよび他の複雑な免疫応答を必要とせずに、エクスビボで適当な内部抗体結合を選択するファージディスプレイ方法を提供する。この方法は、溶媒条件制御能と抗原提示方法を与える(例えば、溶液中で遊離で、グリセロールまたは蔗糖のような共溶媒の存在下で、磁気ビーズに結合して、またはプラスチックマイクロタイタープレートに吸着させて)。
【0075】
一つの態様において、本発明の内部抗体は、HおよびL鎖可変遺伝子が遺伝子レベルで融合されている、一本鎖Fvの形でヒト抗体の最小結合部位を利用する。そのような抗体の構築は、大きい、例えばヒトsFv結合部位の7×109個のファージライブラリから結合部位を選択することによって促進される。これらのsFv遺伝子は典型的に、M13バクテリオファージの小外膜蛋白質(遺伝子3蛋白質)に典型的にライゲーションされており、sFv遺伝子3融合体が線維状ファージの先端で示される生存ファージに再構築されている。このように、表現型(すなわち、結合部位特異性および親和性)と遺伝子型はあらゆるファージ粒子に連結している。この方法は、免疫系が存在しない場合に結合部位の遺伝子選択を行う長所を提供する(例えば、実施例4および5を参照のこと)。
【0076】
その上、数ラウンド行った後(ファージ選択、感染、および感染細胞の再増殖)、本方法は、結合試験にとって十分量の対応するsFvを産生する条件で、細菌培養において増殖させることができる選択されたファージを増幅させる長所をさらに与えた。
【0077】
より大きい調製物が望ましい場合、選択の第3ラウンド後に、ファージを遊離のsFvを産生する大腸菌株の感染によって個々のプラークから増殖させ、これを例えばいかなるエピトープまたは融合免疫タグ(例えば、mycエピトープ(すなわち9E10)またはHis6タグ)に向けられたIMACクロマトグラフィーによってインビボで精製してもよく、PBS中でスーパーロース6カラム上でのFPLCによって存在する単量体または二量体を評価して、BIAコア2000バイオセンサー装置においてストレプトアビジンコーティングマイクロチップ上で固定された選択されたポリペプチド、例えば標的ポリグルタミンバイオポリマーに対して分析してもよい。このバイオセンサー装置は、結合反応に関する結合速度定数(Ka)および解離速度定数(Kd)、ならびにKa/Kdの商から計算した結合平衡定数Kaの測定を与える。したがって、そのような物理化学研究において示され、ファージ選択において用いられる溶媒条件は、関連するポリペプチド立体構造を、例えばBIAコア測定のあいだに保存するように設計することができる。
【0078】
二機能/多機能内部抗体
さらに、本発明の内部抗体は、生物活性領域を定義するさらなるポリペプチド領域、例えばターゲティングシグナル、酵素配列、毒素、またはそこに毒素もしくは治療薬、結合ポリペプチド、酵素もしくは酵素断片、または造影剤のような、遠隔操作で検出可能な物質を結合させることができる部位をさらに含んでもよい。
【0079】
好ましい態様において、内部抗体は、酵素、例えばポリペプチドの蛋白質溶解に至りうるプロテアーゼのようなもう一つの機能的ドメインを含む。もう一つの態様において、内部抗体は、その後破壊されるようにポリペプチドをプロテアソームにターゲティングすることができるターゲティングシグナル、例えば、ユビキチンを含む。さらにもう一つの態様において、本発明の内部抗体は、再ターゲティングすることができるターゲティングシグナルを含み、本発明の内部抗体はもう一つの細胞内位置に内部抗体結合ポリペプチドを再ターゲティングすることができるターゲティングシグナルを含む。例えば、そのような場所は、例えば細胞質、核、ライソゾーム、細胞質関連、小胞体関連、ペルオキシソーム、またはプロテアソームであってもよい。さらに、本発明の内部抗体または結合分子は、異種ポリペプチドの細胞位置を変化させるために当技術分野で認識されたいかなるターゲティングシグナルを含んでもよい。
【0080】
一つの態様において、内部抗体は、特定の細胞位置にポリペプチドをターゲティングするために適した当技術分野で認識された(例えば、国際公開公報第99/14353号に記載するように、または図2に示すように)いかなるターゲティングシグナルも含むように操作してもい。
【0081】
多価内部抗体
当然のこととして、本発明の内部抗体は、一つ以上の結合部位または単一の結合部位のコピー、および他の多くの機能的領域を含んでもよい。例えば、多価内部抗体は本発明の範囲に含まれ、そのような抗体は、選択されたポリペプチド内で認められる一つまたはそれ以上のエピトープに対して親和性を有してもよい。その上、好ましい態様において、多価内部抗体は、変化したポリペプチドにおいて認められるエピトープに対する一つまたはそれ以上の親和性の他に、正常なペプチド内に認められるエピトープに対して一つまたはそれ以上の親和性を有してもよい。もう一つの態様において、内部抗体は、変化したポリペプチド、例えば疾患に関連したポリペプチドにおいて認められる一つまたはそれ以上のエピトープに対する親和性を有する。もう一つの態様において、多価内部抗体は、変化したポリペプチドに対する選択的な結合能を有し、これは正常なポリペプチドとは異なる半減期を有してもよく、異なる速度でまたは異なる細胞内空間に蓄積してもよく(または分泌される)、変化した立体構造を呈してもよく、凝集してもよく(自身で、または他のポリペプチドと共に)、他のポリペプチドとの望ましくない相互作用を形成してもよく、細胞増殖の変化または細胞死を引き起こしてもよく、および/または障害または疾患を引き起こしてもよい。
【0082】
したがって、好ましい態様において、本発明の内部抗体は、例えば、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7のようないかなる一つまたはそれ以上のポリペプチドに対する親和性を有する一つまたはそれ以上の結合部位を有する。
【0083】
もう一つの好ましい態様において、本発明の内部抗体は、例えば、アルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆(FTD)、筋萎縮性側索硬化症(ALS)、脊髄延髄筋萎縮症(SBMAまたはケネディ病)、歯状赤核淡蒼球萎縮症(DRPLA)、1型脊髄小脳運動失調(SCA1)、2型脊髄小脳運動失調(SCA2)、3型脊髄小脳運動失調(SCA3またはマチャドジョゼフ病(MJD))、4型脊髄小脳運動失調(SCA4)、5型脊髄小脳運動失調(SCA5)、6型脊髄小脳運動失調(SCA6)、および7型脊髄小脳運動失調(SCA7)のような、本質的に神経学的な障害または疾患に関係するいかなるポリペプチドにも向けられる。
【0084】
その上、本発明の内部抗体は、短いポリグルタミンに富む領域を有する正常なポリペプチドと、疾患の状態に関連する可能性がある伸長したポリグルタミンに富む領域を有するポリペプチドとを区別するように操作することができる。長さが長いか短いかが異なるに過ぎないポリグルタミンを区別するために、一つの低親和性結合部位または多数の低親和性結合部位を有する内部抗体を、変化したポリペプチドを選択的に結合させるために用いてもよい(例えば、ハストン(Huston, J.S.)ら、(1992)、Biophysical Journal 62;87〜91;ハストン(Huston, J.S.)ら、(1996)、Adv. in Prot. Chem. 49:329〜450)。このように、本発明は、最小の可能性がある種において二つまたはそれ以上の結合部位を組み入れる内部抗体のsFv領域の二価または多価型を含む(ジョージおよびハストン(George, A.J., and Huston, J.S.)、(1997)、The Antibodies 4:99〜141;ハストン(Huston, J.S.)ら、(1996)、Quarterly Journal of Nuclear Medicine 40:320〜333)。さらに、他のドメイン、例えば四量体ストレプトアビジン融合体もまた、内部抗体の、選択されたポリペプチドの特異的ターゲティング能を促進するために用いてもよい(キプリヤノフ(Kipriyanov, S.M.)ら、(1996)、Protein Engineering 9:203〜211)。
【0085】
好ましい態様において、内部抗体、例えば単価内部抗体または結合分子のKaは、105 M-1であり、より好ましくは106 M-1である。もう一つの態様において、多価内部抗体または結合分子の所定の価数のKaは、102 M-1であり、より好ましくは103 M-1である。
【0086】
上記から明らかなように、本発明は、可変領域または天然の免疫グロブリン領域の後にパターン化される結合部位(複数)を含む内部抗体の大きいファミリーを提供する。本発明の本質が、単独で、または他のポリペプチドと様々なように結合して、選択されたポリペプチド、好ましくは、例えば不適当な細胞機能を呈する変化したポリペプチドに結合することができる限り、内部抗体に結合して、本発明の態様の試薬のために用いられるいかなるポリペプチド断片の性質も本質的に無限であることは明らかであろう。
【0087】
内部抗体をコードする核酸
本発明の内部抗体はDNAレベルで設計してもよい。したがって、本明細書に記載の内部抗体をコードする核酸もまた本発明の範囲に含まれる。
【0088】
ファージディスプレイライブラリは、その発現および/または増殖がファージをパッケージングした核酸配列によって促進される、発現可能な核酸の集合体を提供する。sFvファージディスプレイライブラリにおいて、sFvは、sFv抗体結合部位(その特異的結合が表現型である)がファージ表面上での抗原結合に利用でき、その遺伝子型(sFvをコードするDNA)がファージ抗体遺伝子材料内に含まれるように、ファージ外膜蛋白質に融合される。この遺伝子型DNAは、例えば、増幅とシークエンシングを行うためと共に、sFv遺伝子を内部抗体として用いるためのその後の操作を可能にするためにSfiIおよびNotI部位での制限的切断によって容易に単離される。遺伝子は、大腸菌において発現させるためにpHEN1プラスミドにスプライシングしてもよく、ポリペプチドを本来の折り畳まれたポリペプチドとして周辺質に発現および分泌してもよい。
【0089】
本発明の内部抗体を設計できるか否かは、当該モノクローナル抗体の可変領域においてアミノ酸配列またはそれらをコードするDNAを決定することができるかに依存する。ハイブリドーマ技術は、免疫応答を産生する本質的にいかなる望ましい物質に対する抗体を産生する細胞株の産生を可能にする。次に、免疫グロブリン軽鎖および重鎖をコードするRNAをハイブリドーマの細胞質から得ることができ、mRNAの5'末端部分を用いて、その後のシークエンシングのためにcDNAを調製し、または高度可変であって隣接するフレームワーク領域(FR)のアミノ酸配列を、HおよびL鎖ならびにそのV領域断片のアミノ酸シークエンシングによって決定することができる。そのような配列分析は現在、日常的に行われている。この知識によって、抗原に結合する可能性があるFRおよびCDR配列をコードする合成遺伝子の設計が可能となる。これらの合成遺伝子は既知の技術によって、または下記に開示の技術を用いて調製し、次にこれを適した宿主に挿入して発現させ、精製する。宿主細胞に応じて、適切な立体構造を得るために再生技術を必要としてもよい。次に、様々なポリペプチドの結合能を試験し、適当な親和性を有するポリペプチドを上記のタイプの試薬に組み入れるために選択する。必要であれば、従来のカセット変異誘発または他の蛋白質操作技術を用いて、最適な結合を得るためにDNAに一点置換を行ってもよい。
【0090】
当然のこととして、当該アミノ酸配列をコードするDNAを操作、増幅、および組み換えするプロセスは一般的に当技術分野で周知であり、したがって、本明細書において詳細に記述しない。当該内部抗体領域をコードする遺伝子を同定および単離する方法は十分に理解されており、本明細書ならびに例えば米国特許第5,132,405号;第5,091,513号;第5,084,398号;第5,525,491号;第5,851,829号、および国際特許出願国際公開公報第99/14353号に記載されており、これらの参考文献は参照として本明細書に組み入れられる。
【0091】
内部抗体抗原−標的ポリペプチド
単離された標的ポリペプチド、好ましくは望ましくない細胞内蓄積または凝集を形成することが知られているポリペプチド、より好ましくは、他の場所と比較して脳または神経起源の細胞において比較的高濃度で発現されるポリペプチド、または関連する非神経障害を引き起こすポリペプチド、またはその一部もしくは断片を、ファージライブラリ選択技術の標的として、またはポリクローナル抗体およびモノクローナル抗体調製のための標準的な技術を用いて標的ポリペプチドを結合する内部抗体を産生するための免疫原として用いることができる。完全長の標的ポリペプチドを用いることができ、または本発明は、免疫原もしくはファージディスプレイ選択における標的として用いられる標的ポリペプチドの抗原性ペプチド断片、例えばハンチンチンN末端配列(例えば、残基1〜17)を提供する。好ましくは、抗原性ペプチドは、ペプチドに対する内部抗体が天然の条件で標的ポリペプチドと特異的免疫複合体を形成するように、アミノ酸残基少なくとも5個、より好ましくはアミノ酸残基少なくとも15個によって形成される標的ポリペプチドのエピトープを含む。
【0092】
または、変化したポリペプチドのいかなる数にも共通する可能性があるポリグルタミンに富む領域を示すポリペプチドを、標的抗原として(例えばハンチンチンおよびアトロフィン)用いてもよい。そのようなポリペプチドによって、ポリグルタミンに富む領域を有するいかなる数のポリペプチドも結合することができる内部抗体を作製または選択することができる。したがって、そのような内部抗体は、それぞれが最小のポリグルタミンに富む領域を形成すれば、変化したポリペプチドのいくつかの型の蓄積、凝集体形成、または細胞内位置の再ターゲティングを阻害しうるであろう。しかし、さらなる特異性が非ポリグルタミン、ポリペプチド特異的エピトープに対して付与される、独自のクラスの当該ポリペプチド、例えばハンチンチンの特異性は、好ましくは双特異的もしくは多特異的sFv抗体もしくは二抗体、またはFvの関連型を必要とする。二抗体は二価または双特異的であり、リンカーがVドメインを結合する場合に形成する一本鎖sFvの時に三価および多特異的な型は、異常なほど短く、通常10残基またはそれより少なく、好ましくは5残基またはそれより少なく、その結果可変ドメイン融合蛋白質のあいだにFv結合部位を形成する。
【0093】
選択されたポリペプチド免疫原は典型的に、ファージライブラリから内部抗体を選択するために、または適した対象(例えば、ウサギ、ヤギ、マウス、または例えばヒト患者を含む他の哺乳類)を標的ポリペプチド免疫原によって免疫することによって抗体を誘発するために用いられる。適当な免疫原調製物は、例えば組み換え発現された標的ポリペプチドまたは化学合成されたポリペプチドを含みうる。ポリペプチドはさらに、検出または精製を促進するための部分と融合してもよく、そのような部分には、免疫反応性タグ、GFP、ビオチン等が含まれる。調製物はさらに、フロイントの完全もしくは不完全アジュバントのようなアジュバント、または類似の免疫刺激物質を含みうる。
【0094】
または、免疫原は、レシピエント動物の細胞によって核酸が取り込まれて発現されるように、宿主細胞免疫応答、好ましくは液性応答を誘発するために十分量のポリペプチドが得られるように、発現可能な核酸の形で、例えばDNAワクチンとして投与してもよい。適した対象を免疫原性標的ポリペプチド調製物によって免疫すると、抗標的ポリペプチド抗体反応を誘発する。
【0095】
したがって、内部抗体は、標準的な技術を用いて得られたモノクローナル抗体に由来してもよい。より好ましくは、内部抗体は、DNAワクチン接種を用いて標的ポリペプチドに対して免疫した動物に由来する。さらにより好ましくは、内部抗体は、標的ポリペプチドまたは標的ポリペプチドをコードする発現可能なDNAによって免疫した動物に由来するファージライブラリに由来する。特定の場合において、同じ標的ポリペプチドに由来する副疾患を有する神経細胞または他の細胞によらず、標的ポリペプチドに対する抗体のインサイチュー産生によって、無傷のIgGを生成してもよく、または疾患を有する細胞に入って内部抗体として作用し、それによって標的ポリペプチドの病的な凝集に拮抗する他のクラスの抗体を生成してもよい。これらの場合、治療的抗体は、ポリペプチド免疫治療として、またはそれらが内部抗体ポリペプチドを分泌する、エクスビボで改変された細胞が患者に再投与される養子免疫細胞移入による免疫治療の形として投与してもよい(例えば、カバッツァーナカルボ(Cavazzana-Calvo, M.)ら、Science 288:669〜672)。
【0096】
さらに、内部抗体は、組み換え型組み合わせ免疫グロブリンライブラリ(例えば、内部抗体ファージディスプレイライブラリ)を標的ポリペプチドによってスクリーニングして、それによって、標的ポリペプチドに結合する免疫グロブリンライブラリメンバーを単離することによって同定および単離することができる。ファージディスプレイライブラリを作製およびスクリーニングするキットは市販されている(例えば、ファルマシア社の組み換え型ファージ抗体システム、カタログ番号27-9400-01;およびストラタジーン社のSurfZAP(商標)ファージディスプレイキット、カタログ番号240612)。さらに、特定の親和性を有する内部抗体を作製、スクリーニング、および選択するために特に用いやすい方法および試薬の例は、例えば、米国特許第5,132,405号;第5,091,513号;第5,084,398号;第5,525,491号;第5,851,829号、国際特許出願国際公開公報第99/14353号、およびシーツ(Sheets)ら、P.N.A.S. 95:6157〜6162(1998)に見ることができる。
【0097】
このように、本発明は、以下の少なくとも内部抗体の概念を(下記の実施例はさらなる証拠となるであろう)示し、それらは:1)内部抗体は遺伝子治療として輸送することができること;2)内部抗体は標的細胞に入る(免疫毒素のように)蛋白質免疫治療として輸送することができ、標的抗原との結合を通して、細胞内で機能することができること;3)内部抗体は、静脈内投与する代わりに、例えば宿主液性応答の形で免疫治療のための内部抗体蛋白質が免疫系によって作製されるように、適当な免疫によって誘発することができること;および4)異常なポリペプチド蓄積は、非常に低レベルで存在しうること、そして蛋白質・蛋白質相互作用を通じて細胞機能を有意に破壊でき、このように、内部抗体の中等度の細胞内レベルでも治療的となることである。
【0098】
III. 発現ベクターと宿主細胞
本発明のもう一つの局面は、ベクター、好ましくは内部抗体(またはその一部)をコードする核酸を含む発現ベクターに関する。一つのタイプのベクターは「プラスミド」であり、これは、その中にさらなるDNAセグメントがライゲーションされうる環状二本鎖DNAループを意味する。もう一つのタイプのベクターは、さらなるDNAセグメントをウイルスゲノムの中にライゲーションすることができるウイルスベクターである。特定のベクターは、それらが導入された宿主細胞において自律的に複製することができる(例えば、細菌の複製開始点を有する細菌ベクターおよびエピソーム哺乳類ベクター)。他のベクター(例えば、非エピソーム哺乳類ベクター)は、宿主細胞に導入されると宿主細胞のゲノムに組み入れられ、それによって宿主ゲノムと共に複製される。その上、特定のベクターは、それらが機能的に結合する遺伝子の発現を指示することができる。そのようなベクターは、本明細書において「発現ベクター」と呼ぶ。一般的に、組み換えDNA技術において用いられる発現ベクターはしばしば、プラスミドの形である。プラスミドは、最も一般的に用いられるベクターの型であるため、本明細書において、「プラスミド」および「ベクター」は互換的に用いることができる。しかし、本発明には、同等の機能を提供するウイルスベクター(例えば、複製欠損レトロウイルス、レンチウイルス、アデノウイルスおよびアデノ随伴ウイルス)のような発現ベクターのそのような他の形も含まれると解釈される。
【0099】
本発明の組み換え発現ベクターは、宿主細胞における発現に適した形で本発明の内部抗体をコードする核酸を含み、このことは、組み換え型発現ベクターが、発現される核酸配列に機能的に結合している、発現のために用いられる宿主細胞に基づいて選択される一つまたはそれ以上の調節配列を含むことを意味する。組み換え発現ベクターにおいて、「機能的に結合」とは、当該ヌクレオチド配列が調節配列(複数)に対して、ヌクレオチド配列を発現させるように結合していることを意味すると解釈される(例えば、インビトロ転写/翻訳系において、またはベクターが宿主細胞に導入される場合は宿主細胞において)。「調節配列」という用語には、プロモーター、誘導型プロモーター、エンハンサー、および他の発現制御エレメント(例えば、ポリアデニル配列)が含まれると解釈される。そのような調節配列は、例えば、ゴッデル(Goeddel)(「遺伝子発現技術(Gene Expression Technology)」Methods in Enzymology 185、アカデミック出版、サンディエゴ、カリフォルニア州(1990))に記載されている。調節配列には、多くのタイプの宿主細胞においてヌクレオチド配列の構成的発現を指示する配列、および特定の宿主細胞に限ってヌクレオチド配列の発現を指示する配列(例えば、組織特異的調節配列)が含まれる。発現ベクターの設計は、形質転換すべき宿主細胞の選択、望ましい蛋白質の発現レベル等のような要因に依存しうることは当業者によって認識されるであろう。本発明の発現ベクターは、宿主細胞に導入されて、それによって内部抗体を産生することができ、その後これをポリペプチド製剤として投与してもよい。または、内部抗体をコードする核酸を、好ましくは哺乳類、より好ましくはヒト、より好ましくは哺乳類の神経起源の細胞において発現することができるベクターに導入する。
【0100】
多くのプロモーターおよび発現ベクターが哺乳類細胞において高い発現を与えるとして当技術分野で既知である。例えば、本発明の核酸分子は哺乳類の発現ベクターを用いて発現させることができる。哺乳類発現ベクターの例には、pCDM8(シード(Seed, B.)(1987)Nature 329:840)およびpMT2PC(カウフマン(Kaufman)ら(1987)、EMBO J 6:187〜195)が含まれる。哺乳類細胞において用いる場合、発現ベクターの制御機能はしばしば、ウイルス調節エレメントによって提供される。例えば、一般的に用いられるプロモーターは、ポリオーマ、アデノウイルス2、サイトメガロウイルス、およびシミアンウイルス40に由来する。原核細胞と真核細胞の双方に適した他の発現系に関しては、サムブルック、フリッチュおよびマニアティス(Sambrook, J., Fritsh, E.F., and Maniatis, T.)、「分子のクローニング:実験マニュアル(Molecular Cloning:A Laboratry Manual)」第二版、コールドスプリングハーバー出版、コールドスプリングハーバー、(1989);アウスユベール(Ausubel)ら、(1998)、「分子生物学の現行プロトコール(Current Protocols in Molecular Biology)」、ジョンウィリー&サンズインクを参照のこと。
【0101】
もう一つの態様において、組み換え型哺乳類発現ベクターは、選択的に特定の細胞タイプにおいて核酸の発現を指示することができる(例えば、組織特異的調節エレメントを用いて核酸を発現させる)。組織特異的調節エレメントは当技術分野で既知である。適した組織特異的プロモーターの非制限的な例には、アルブミンプロモーター(肝臓特異的;ピンカート(Pinkert)ら、(1987)、Genes Dev. 1:268〜277)、リンパ様特異的プロモーター(カラムおよびイートン(Calame and Eaton)(1988)、Adv. Immunol. 43:235〜275)、特にT細胞受容体(ウィノトおよびバルチモア(Winoto and Baltimore)(1989)EMBO J. 8:729〜733)および免疫グロブリン(バネリ(Banerji)ら(1983)、Cell 33:729〜740;クイーンおよびバルチモア(Queen and Baltimore)(1983)Cell 33:741〜748)のプロモーター、ニューロン特異的プロモーター(例えば、神経細糸プロモーター;ビルネおよびラドル(Byrne and Ruddle)(1989)PNAS 86:5473〜5477)、膵臓特異的プロモーター(エドルンド(Edlund)ら、(1983)Science 230:912〜916)、および乳腺特異的プロモーター(例えば、乳清プロモーター;米国特許第4,873,316号および欧州特許第246,166号)が含まれる。発達的に調節されるプロモーター、例えばマウスホックスプロモーター(ケッセルおよびグルス(Kessel and Gruss)(1990)、Science 249:374〜379)およびαフェトプロテインプロモーター(カンペスおよびチルフマン(Campes and Tilghman)(1989)Genes Dev.3:537〜546)も同様に含まれる。
【0102】
本発明のもう一つの局面は、本発明の組み換え型発現ベクターが導入される宿主細胞に関する。「宿主細胞」および「組み換え型宿主細胞」という用語は本明細書において互換的に用いられる。そのような用語は、特定の被験細胞を意味するのみならず、そのような細胞の子孫または可能性がある子孫も意味すると理解される。変異または環境的影響によりその後の世代に特定の改変が起こる可能性があるため、そのような子孫は、実際に親細胞と同一でない可能性があるが、なおも本明細書において用いられる用語の範囲に含まれる。
【0103】
宿主細胞は、いかなる原核または真核細胞となりうる。例えば、内部抗体蛋白質は、大腸菌のような細菌細胞、昆虫細胞、酵母または哺乳類細胞(例えば、チャイニーズハムスター卵巣細胞(CHO)、哺乳類腎細胞(COS)、ハムスター腎細胞(BHK-21)、ヒト上皮細胞(293T)のような)において発現されうる。その他の適した宿主細胞は当業者に既知である。
【0104】
ベクターDNAは、従来の形質転換またはトランスフェクション技術によって原核細胞または真核細胞に導入することができる。本明細書において用いられるように、「形質転換」および「トランスフェクション」という用語は、リン酸カルシウムもしくは塩化カルシウム共沈殿、DEAEデキストラン媒介トランスフェクション、リポフェクション、または電気穿孔を含む、外来核酸(例えば、DNA)を宿主細胞に導入する当技術分野で認識された多様な技術を意味すると解釈される。好ましい態様において、内部抗体をコードする核酸は、プロタミン骨格の非ウイルスベクターを用いてトランスフェクションしてもよい(リチャードソン(Richardson, J.H.)(1995)、Trends in Biotechnology, 13:306〜310)。宿主細胞を形質転換またはトランスフェクトする適した方法は、上記のサムブルック(Sambrook)ら、ならびに例えば、本明細書において言及した他の実験マニュアルおよび参考文献に見ることができる。
【0105】
哺乳類細胞の安定なトランスフェクションに関して、用いる発現ベクターとトランスフェクション技術に応じて、外来DNAをそのゲノムに組み入れるのはごく小さい分画の細胞に過ぎない場合があることが知られている。これらの組み込み体を同定および選択するために、選択マーカー(例えば、抗生物質に対する抵抗性)をコードする遺伝子を一般的に当該遺伝子と共に宿主細胞に導入する。好ましい選択マーカーには、G418、ヒグロマイシン、およびメソトレキセートのような薬剤に対する抵抗性を付与するマーカーが含まれる。選択マーカーをコードする核酸は、内部抗体をコードするベクターと同じベクター上で宿主細胞に導入することができ、または異なるベクターに導入することができる。導入した核酸によって安定にトランスフェクトされた細胞は、薬剤選択によって同定することができる(例えば、選択マーカー遺伝子を組み入れた細胞は生存するが、他の細胞は死滅する)。
【0106】
原核または真核宿主培養細胞のような本発明の宿主細胞は、内部抗体を産生する(すなわち発現する)ために用いることができる。したがって、本発明はさらに、本発明の宿主細胞を用いて内部抗体ポリペプチドを産生する方法を提供する。一つの態様において、方法は、本発明の宿主細胞(その中に内部抗体をコードする組み換え型発現ベクターが導入される)を適した培地において、内部抗体が産生されるように培養する段階を含む。もう一つの態様において、方法は、培地または宿主細胞から内部抗体を単離することをさらに含む。
【0107】
本発明の宿主細胞はまた、ヒト以外のトランスジェニック動物を作製するために用いることができる。例えば、一つの態様において、本発明の宿主細胞は、その中に内部抗体コード配列が導入される受精卵または胚肝細胞である。次に、そのような宿主細胞を用いてその中に外来内部抗体配列がそのゲノムに導入されているヒト以外のトランスジェニック動物、または内因性Fv配列が変化している相同組み換え動物を作製することができる。そのような動物は、内部抗体の機能および/または活性を調べるために、および内部抗体/標的ポリペプチド活性の調節物質を同定および/または評価するために有用である。本明細書において用いられるように、「トランスジェニック動物」は、ヒト以外の動物、好ましくは哺乳類、より好ましくは動物の一つまたはそれ以上の細胞がトランスジーンを含む、ラットまたはマウスのような齧歯類である。トランスジェニック動物の他の例には、ヒト以外の霊長類、ヒツジ、イヌ、ウシ、ヤギ、ニワトリ、両生類等が含まれる。トランスジーンはそこからトランスジェニック動物が発生する細胞のゲノムに組み入れられ、成熟動物のゲノムに留まり、それによってトランスジェニック動物の一つまたはそれ以上の細胞タイプまたは組織においてコードされた遺伝子産物の発現を指向する外来DNAである。本明細書において用いられるように、「相同的組み換え型動物」とは、ヒト以外の動物、好ましくは哺乳類、より好ましくは内因性の遺伝子と、動物が発達する前に動物の細胞、例えば動物の胚細胞に導入された外来DNA分子とのあいだの相同的組み換えによって内因性標的ポリペプチド遺伝子が変化しているマウスである。この動物は、内部抗体を発現するようにさらに改変してもよく、または内部抗体を発現する動物と交配させることができる。より好ましくは、トランスジェニック動物は、当該ポリペプチドの一つまたはそれ以上、すなわち標的ポリペプチド、またはより好ましくは内部抗体が例えば、実施例6に記載のように条件的制御下にあるように操作される。さらに、当技術分野で認識されるいかなる技術も、本発明のトランスジェニック動物および/または相同的組み換え動物を産生するために用いてもよい。
【0108】
IV.スクリーニングアッセイ
本発明は、調節物質、すなわち標的ポリペプチド、例えばハンチンチンポリペプチドに結合する候補結合分子(例えば、ペプチド、ペプチド模倣体、低分子または他の薬剤)を同定する方法(本明細書において「スクリーニングアッセイ」と呼ぶ)を提供する。方法は、所定の疾患のモデルを表す異常を示す細胞内でインサイチューで行う。このように、標的ポリペプチドの病的な凝集をインサイチューで相殺する能力は、さらなる治療物質開発のための必要条件となる。
【0109】
特に、本発明の一つまたはそれ以上の方法は、選択されたポリペプチドの望ましくない蓄積、複合体形成、または凝集を変化させる化合物をスクリーニングするためにヒト以外の対象の細胞において内部抗体を用いることを含む。好ましい態様において、選択されるポリペプチドは、例えば、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、スーパーオキシドジスムターゼ(SOD)、アンドロゲン受容体(AR)、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、またはSCA7である。もう一つの好ましい態様において、ポリペプチドはアルツハイマー病に関連する。さらにもう一つの好ましい態様において、ポリペプチドは、ハンチンチンであり、ハンチンチンの異常型はハンチントン病に関係している。したがって、特定の疾患に関係しているポリペプチドに対して特異的な内部抗体を、細胞に基づくアッセイにおいて、インビトロで、または動物全体においてポリペプチドに接触させ、そして一つまたはそれ以上の化合物をほぼ同じ時期に投与し、内部抗体/ポリペプチド相互作用による凝集の変化をモニターする。
【0110】
インビトロにおいて、そのような変化は、結合の変化または他のいくつかの生化学パラメータの変化として測定してもよい。細胞に基づくアッセイにおいて、内部抗体/ポリペプチド相互作用の変化は、標的ポリペプチドの蓄積、複合体形成、または凝集における変化、または宿主細胞の細胞生物学における変化(例えば、細胞死レベルの変化)として測定してもよい。生物全体の意味において、内部抗体と標的ポリペプチドとのインビボ相互作用のそのような変化は、疾患の症状の変化として測定してもよい。
【0111】
一つまたは全てのアッセイを用いて、内部抗体/ポリペプチド相互作用に関して凝集の蓄積を変化させる化合物の可能性がある治療的有効性を確認してもよい。そのような化合物は、さらなる分析のために選択してもよい。理想的には、好ましい態様において、化合物は標的ポリペプチドに対する特異的結合として同定される。もう一つの好ましい態様において、化合物は正常および変異型ポリペプチドの双方を認識することができ、それらを適当なレベルで複合体を形成させることによって、正常な細胞機能と病理プロセスの排除の双方を促進することができる。なおもう一つの好ましい態様において、化合物は、野生型ポリペプチドより高い特異性を有する変異型ポリペプチドを認識することができ、またはこのように機能するように変化することができる。なおもう一つの好ましい態様において、化合物は、例えば、毒性、血液脳関門の通過能、半減期、もう一つの機能的部分との結合能、またはさらに改変を受ける能力のような特定の薬理学的特性または長所に関して選択してもよい。
【0112】
当該結合分子はまた、組織切片培養、例えば動物、例えば、神経疾患(例えば、AD、HD、SCA疾患等)のトランスジェニック動物モデルからの臓器型切片培養を用いてアッセイしてもよい。好ましい態様において、HDまたはSCA1を有するマウスを用いる。さらに、前述のいかなる実験動物も、興奮毒のような化合物または酸化的障害を引き起こすことが知られている化合物に対する感受性に関して、野生型対照と平行して調べることができる。神経変性、例えば、線条体のHD様変性、または他の神経毒性作用を誘導するために適している当技術分野で記載の多くの化合物を、インビボで、またはそのような動物に由来する臓器型切片培養を用いて試験することができる(例えば表1、および例えば、ボウリング(Bowling)ら、Life Sci. 56:1151〜1171(1995);ジャンセル(Jansel)ら、Brain Res. 532:351〜354(1990);ロススタイン(Rothstein)ら、P.N.A.S. 90:6591〜6595(1993);およびシュワルツ(Schwarcz)ら、Life Sci. 35:19〜32(1963)を参照のこと)。短期間の暴露の後に野生型細胞または組織に対して明白な毒性を示さない用量を用いることによって、通常誘発される初期の臨床下病態を中和することによる疾患を有する細胞の異なる脆弱性を確認することができる。
【0113】
好ましい態様において、表1に記載した化合物を用いて、変異型ニューロン、野生型ニューロン、または内部抗体もしくは結合分子を有するニューロンのいずれかにおいて異なる作用を誘発してもよい。
【0114】
【表1】

【0115】
一つの態様において、培養は、本明細書に記述した方法を用いて調製することができ、2週間成熟させた。例えばHDまたはSCA1トランスジーンのいずれかの存在による認識可能な病態は、この時点で最小であるはずであるが、切片におけるニューロンは、なおもリスクが増加している可能性がある。次に、上記に示した化合物を加えて、結果を形態学的にモニターすることができる。
【0116】
したがって、このアッセイ系は、低分子治療物質をスクリーニングするために理想的である。一つの態様において、特異的結合に対する構造的基礎を発見するために、ポリグルタミン領域の伸長した反復配列に特異的なsFv結合部位(例えば、相補性決定領域、CDRs)におけるアミノ酸配列の差の比較を行う。特に、ハンチンチンポリペプチドに対して特異的なsFvのH3 CDRループは、内部抗体/ポリペプチド結合相互作用のほとんどの原因であり、したがってH3ペプチド模倣体をこの系を用いてスクリーニングして、無傷の内部抗体の代わりに用いてもよい。次に、この情報を用いて、立体構造エピトープのような異常なポリグルタミン領域と相互作用する他の結合分子を同定することができ、これらを例えばペプチドファージディスプレイライブラリ(ニューイングランドバイオラブス社)を用いて適当な類似体を発見するためにスクリーニングしてもよい。
【0117】
好ましい態様において、上記のアッセイを用いてポリペプチドの蓄積または凝集に対して阻害作用を有する候補結合分子を同定する。もう一つの態様において、候補化合物は、内部抗体の生物活性を模倣する、例えば、凝集を防止してポリペプチドを再ターゲティングしてもよい。
【0118】
本発明の試験化合物は、以下を含む当技術分野で既知の組み合わせライブラリ法を含む様々なアプローチのいずれかを用いて得ることができる:生物ライブラリ;空間的に位置特定可能な平行な固相または液相ライブラリ;脱重畳積分を必要とする合成ライブラリ法;「1ビーズ1化合物」ライブラリ法;およびアフィニティクロマトグラフィー選択を用いる合成ライブラリ法。生物ライブラリアプローチはペプチドライブラリに限定されるが、他の4つのアプローチはペプチド、非ペプチドオリゴマーまたは低分子化合物ライブラリに適用できる(ラム(Lam, K.S.)(1997)Anticancer Drug Des. 12:145)。
【0119】
分子ライブラリを合成するための方法の例は、例えば、デウィット(DeWitt)ら(1993)Proc. Natl. Acad. Sci. USA 90:6909;エルブ(Erb)ら(1994、Proc. Natl. Acad. Sci. USA91:11422);ズッカーマン(Zuckermann)ら(1994、J. Med. Chem. 37:2678);チョー(Cho)ら(1993)Science 261:1303;カレル(Carell)ら(1994)Angew. Chem. Int. Ed. Engl. 33:2059;カレル(Carell)ら(1994)Angrew. Chem. Int. Ed. Engl. 33:2061;およびギャロップ(Gallop)ら(1994)J. Med. Chem. 37:1233に見ることができる。
【0120】
化合物のライブラリは、溶液(例えば、ホーテン(Houghten)、(1992)、Biotechniques 13:412〜421)、またはビーズ(ラム(Lam)、(1991)、Nature 354:82〜84)、チップ(フォドー(Fodor)(1993)、Nature 364:555〜556)、細菌(ラドナー(Ladner)、米国特許第5,223,409号)、胞子(ラドナー(Ladner)、米国特許第5,223,409号)、プラスミド(カル(Cull)ら、(1992)、Proc. Natl. Acad. Sci. USA 89:1865〜1869)、またはファージ(スコットおよびスミス(Scott and Smith)(1990)Science 249:386〜390;デブリン(Devlin)、(1990)、Science 249:404〜406;チャーラ(Cwirla)ら(1990)Proc. Natl. Acad. Sci. USA 87:6378〜6382;フェリチ(Felici)(1991)、J. Mol. Biol. 222:301〜310;およびラドナー(Ladner)、上記)に存在してもよい。
【0121】
一つの態様において、アッセイは、その中で標的ポリペプチド、例えばハンチンチンと内部抗体とを発現する細胞を試験化合物に接触させて、試験化合物がポリペプチド/内部抗体相互作用を変化させるか否かを決定する、細胞に基づくアッセイである。例えば、細胞は、哺乳類起源または酵母細胞となりうる。試験化合物がポリペプチドに結合するか否かを決定することは、例えば試験化合物とポリペプチドとの結合が複合体における標識化合物の検出によって決定されうるように、放射性同位元素または酵素標識に化合物をカップリングさせることによって行うことができる。例えば、試験化合物は、125I、35S、14C、3H、または32Pによって直接または間接的に標識することができ、放射性同位元素は、放射線放出の直接計数またはシンチレーション計数によって検出することができる。または、試験化合物は、例えば、ホースラディッシュペルオキシダーゼ、アルカリホスファターゼ、またはルシフェラーゼによって酵素標識することができ、酵素標識は、適当な基質の産物への変換を測定することによって検出される。
【0122】
いかなる相互作用体も標識せずに、試験化合物が標的ポリペプチドと相互作用できるか否かを決定することも本発明の範囲内である。例えば、マイクロフィジオメーターを用いて、試験化合物または標的ポリペプチドのいずれも標識せずに、標的ポリペプチドと試験化合物との相互作用を検出することができる(マッコネル(McConnell, H.M.)ら、(1992)、Science 257:1906〜1912)。さらにもう一つの態様において、本発明のアッセイは、標的ポリペプチドと内部抗体を試験化合物に接触させ、試験化合物がポリペプチド/内部抗体相互作用を変化させるか否かを決定する細胞不含アッセイである。試験化合物と標的ポリペプチドとの結合は、直接または間接的に決定することができる。候補化合物が標的ポリペプチドに結合するか否かを決定することは、リアルタイム生体分子相互作用分析(BIA)のような技術を用いて行うことができる(シェランダーおよびウルバニクスキー(Sjolander, S. and Urbaniczky, C.)、(1991)、Anal. Chem. 63:2338〜2345およびスザボ(Szabo)ら(1995)、Curr. Opin. Struct. Biol. 5:699〜705)。本明細書において用いられるように、「BIA」は、いかなる相互作用も標識せずにリアルタイムで生体特異的相互作用を調べる技術である(例えば、BIAcoa(商標))。光学現象表面プラズモン共鳴(SPR)の変化は、生体分子間のリアルタイム反応の指標として用いることができる。
【0123】
化合物および天然抽出物の試験ライブラリの多くの薬剤スクリーニングプログラムにおいて、所定の期間において調べる化合物数を最大限にするために高処理量アッセイが望ましい。精製、または半精製蛋白質を用いて行ってもよいアッセイのような細胞不含系において行われるアッセイは、しばしばその中で試験化合物によって媒介される分子標的の変化を迅速に開始させて比較的容易に検出するために作製することができる「一次」スクリーニングと呼ばれる。その上、試験化合物の細胞毒性および/または生物学的利用性の影響は、一般的にインビトロ系では無視することができ、アッセイはその代わりに上流または下流のエレメントによる結合親和性の変化として現れる可能性がある分子標的上の薬剤の影響に主に重点が置かれる。したがって、本発明の例としてのスクリーニングアッセイにおいて、当該化合物を標的ポリペプチドに接触させ、標的ポリペプチドの蓄積または凝集の定量は望ましくない複合体または凝集体の形成を阻害する化合物の有効性を評価することによって決定される。化合物の有効性は、試験化合物の様々な濃度を用いて得られるデータから用量反応曲線を作製することによって評価することができる。その上、対照アッセイも同様に、比較の基準値を提供するために行うことができる。
【0124】
本発明の上記のアッセイ方法の一つ以上の態様において、複合体型と非複合体との分離を促進するため、またはアッセイの自動化に適合させるために標的ポリペプチドを固定することが望ましいかもしれない。試験化合物の標的ポリペプチドへの結合は、反応物質を含むために適したいかなる容器においても行うことができる。そのような容器の例には、マイクロタイタープレート、試験管、および微量遠心管が含まれる。一つの態様において、一つまたはそれ以上の蛋白質をマトリクスに結合させるドメインを付加する融合蛋白質を提供することができる。例えば、グルタチオンSトランスフェラーゼ/標的ポリペプチド融合蛋白質をグルタチオンセファロースビーズ(シグマケミカル社、セントルイス、ミズーリ州)またはグルタチオン誘導体化マイクロタイタープレートに吸着させることができ、次にこれを試験化合物または試験化合物および内部抗体と組み合わせて、複合体形成を行うための条件(例えば、イオン強度およびpHの生理的条件)でインキュベートする。インキュベーション後、ビーズまたはマイクロタイタープレートウェルを洗浄して、いかなる未結合成分も除去して、ビーズの場合にはマトリクスを固定して、複合体を例えば上記のように直接または間接的に測定する。または、複合体は、マトリクスから解離させることができ、標的ポリペプチド結合または活性のレベルを標準的な技術を用いて決定することができる。
【0125】
そのようなスクリーニング方法によって選択された化合物の試験は、モデルの階層性を用いて、トランスフェクト細胞モデルから開始して、適当な正常または変異体マウスからの脳切片について行い、次にウイルスを形質導入したマウスまたは疾患のトランスジェニックマウスモデルにおいて試験を行って、ハンチントンのような所定の神経疾患に対する関連を調べてもよい。これらの平均によって、試験化合物を臨床試験で利益が得られる可能性がある少数の化合物まで狭めてもよい。
【0126】
マトリクス上に蛋白質を固定する他の技術はまた、本発明のスクリーニングアッセイにおいて用いることができる。例えば、いずれかの標的ポリペプチドをビオチンおよびストレプトアビジンの結合を利用して固定することができる。ビオチン結合標的ポリペプチドまたは内部抗体は、当技術分野で既知の技術を用いて(例えば、ビオチン結合キット、ピアスケミカル社、ロックフォード、イリノイ州)ビオチンNHS(Nヒドロキシスクシニミド)から調製して、ストレプトアビジンをコーティングした96ウェルプレート(ピアスケミカル社)のウェルに固定することができる。
【0127】
本発明のさらにもう一つの局面において、標的ポリペプチドまたは内部抗体結合領域は、二ハイブリッドアッセイまたは三ハイブリッドアッセイにおいて、標的ポリペプチドおよび/または内部抗体に結合または相互作用する他の蛋白質また化合物を同定するための「おとり蛋白質」として用いることができる(例えば、米国特許第5,283,317号;ゼルボス(Zervos)ら(1993)、Cell 72:223〜232;マデュラ(Madura)ら、(1993)、J. Biol. Chem. 268:12046〜12054;バーテル(Bartel)ら(1993)、Biotechniques 14:920〜924;イワブチ(Iwabuchi)ら、(1993)、Oncogene 8:1693〜1696;およびブレント(Brent)、国際公開公報第94/10300号を参照のこと)。
【0128】
本発明はさらに、上記のスクリーニングアッセイによって同定される新規物質およびこれらのアッセイを用いることによってそのような物質を産生するためのプロセスに関する。したがって、一つの態様において、本発明は、上記のスクリーニングアッセイ(例えば、細胞に基づくアッセイまたは細胞不含アッセイ)のいずれか一つの段階を含む方法によって得ることができる化合物または物質を含む。例えば、一つの態様において、本発明には、本明細書に記載の方法のいずれかによって得ることができる化合物または物質が含まれる。
【0129】
したがって、物質、例えば適当な動物モデルにおいて本明細書に記載のように同定された化合物または内部抗体をさらに用いることも本発明の範囲内である。例えば、本明細書において記述されたように同定された物質を動物モデルにおいて用いて、そのような物質による治療の有効性、毒性、または副作用を決定することができる。または、本明細書において同定された物質を動物モデルにおいて用いて、そのような物質の作用機序を決定することができる。さらに、適当であると思われる場合には、そのような物質をヒト対象、好ましくは神経障害の危険にさらされている対象に投与してもよい。
【0130】
本発明はまた、本明細書に記載したいかなる神経障害も診断、予後を決定、および治療するために、上記のスクリーニングアッセイによって同定された新規物質を用いることに関する。したがって、本明細書に記載のいかなる神経障害の診断、予後、または治療において用いられる薬剤または薬学的組成物の設計、処方、合成、製造、および/または産生においてそのような物質を用いることは本発明の範囲内である。
【0131】
V.使用方法
予防的方法
一つの局面において、本発明は、ポリペプチドの蓄積またはそのような複合体もしくは凝集体の形成を阻害する物質を対象に投与することによって、対象において、神経ポリペプチドの異常または望ましくない細胞内蓄積または凝集に関連した疾患または病態を予防する方法を提供する。好ましい態様において、疾患または病態は神経疾患(例えば、ハーディ(Hardy)ら、Science 282:1075〜1079(1998)から抜粋した表2を参照のこと)である。そのような異常または望ましくないポリペプチド活性によって引き起こされる、またはそれらに関係する神経疾患の危険にさらされている対象は、例えば、当技術分野で認識された診断または予後的技法によって同定することができる。したがって、陽性診断が示されると、本明細書に記載した適当な化合物または内部抗体のいかなる投与も、神経疾患または障害が予防される、その重症度が減少する、またはその進行もしくは発症が遅れるように、異常に特徴的な症状が発現する前に予防的物質として投与してもよい。
【0132】
常染色体優性原発性神経変性疾患
【表2】
Ch、染色体;PrP、プリオン蛋白質;T、タングル;LB、レーヴィ小体;+、存在する;AD、アルツハイマー病;PD、パーキンソン病;HD、ハンチントン病;SOD;スーパーオキシドジスムターゼ。
* SBMAは技術的には常染色体優性ではないが、その細胞での作用機序からみておそらく優性である。
【0133】
治療的方法
本発明のもう一つの局面は、標的ポリペプチド、好ましくは神経細胞において発現されたポリペプチドの異常な発現または活性を特徴とする疾患または障害に罹患した人を治療する方法に関する。
【0134】
したがって、本発明は、中枢または末梢神経系の細胞に関連する疾患または病態を治療する方法を提供する。例えば、本発明は、以下の細胞のいずれかの異常な細胞増殖、分化、または細胞死に至る可能性がある異常なポリペプチド蓄積または凝集に関連した神経系の病変を治療する方法を提供する:ニューロン(例えば、運動神経ニューロン、知覚神経ニューロン)、シュワン細胞、プルキンエ細胞、星状細胞、小膠細胞、上衣細胞、乏突起膠細胞または他のいかなるタイプの神経細胞。神経系の障害には、アルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆(FTD)、筋萎縮性側索硬化症(ALS)、脊髄延髄筋萎縮症(SBMAまたはケネディ病)、歯状赤核淡蒼球萎縮症(DRPLA)、1型脊髄小脳運動失調(SCA1)、2型脊髄小脳運動失調(SCA2)、3型脊髄小脳運動失調(SCA3またはマチャドジョゼフ病(MJD))、4型脊髄小脳運動失調(SCA4)、5型脊髄小脳運動失調(SCA5)、6型脊髄小脳運動失調(SCA6)、および7型脊髄小脳運動失調(SCA7)が含まれるが、これらに限定しない。
【0135】
望ましくない細胞内ポリペプチド複合体/蓄積の形成の阻害
一つの態様において、本発明は、望ましくない細胞内ポリペプチド凝集または蓄積の形成を阻害する方法を提供する。特に、本発明の内部抗体は、ポリペプチドに特異的に結合することによって望ましくないポリペプチドが複合体または凝集体を蓄積または形成することを阻害するように設計する。いくつかの状況において、ポリペプチドに対して内部抗体が単に特異的結合しただけで、ポリペプチドを含む複合体または凝集体の形成を阻害するために十分であるかもしれない。そのような内部抗体は、標的ポリペプチドに対して親和性を有する一つまたはそれ以上の原子価、すなわち結合部位を有し、例えば、立体的な妨害によって、または蛋白質をより不安定にする変化した立体構造を誘導することによって、および/またはポリペプチド複合体がポリペプチドの蓄積または凝集に至る相互作用を形成できないようにする、例えばその溶解性を増加させる、または凝集体を形成する臨界濃度を増加させることによって、ポリペプチドの蓄積または凝集を阻害してもよい。
【0136】
したがって、本発明は、本明細書に記載の内部抗体のいずれかを、単独、またはもう一つの内部抗体、または望ましくない細胞内ポリペプチド蓄積もしくは凝集を形成することが知られているポリペプチドをターゲティングするために適した他の治療物質と組み合わせて、対象に投与する方法を提供する。一つの態様において、対象は実験動物であってもよい。好ましい態様において、本発明の方法は、ヒト患者を治療するために適している。もう一つの好ましい態様において、内部抗体または結合分子の投与は、アルツハイマー病の治療または治癒のためである。なおもう一つの好ましい態様において、内部抗体または結合分子の投与は、ハンチントン病の治療のためである。本発明の一つの態様において、内部抗体を投与する方法は、ポリペプチドとしてである。本発明の好ましい態様において、内部抗体は、下記の章に記載するように、遺伝子治療の形で発現可能な核酸として投与される。
【0137】
遺伝子治療
本発明の内部抗体またはペプチド骨格の結合分子は、遺伝子治療として投与した場合に、本明細書に記載のいかなる神経疾患の治療においても特に有用である。一般的アプローチは、導入された遺伝子材料によってコードされる一つまたはそれ以上の遺伝子産物が、望ましくないポリペプチド蓄積または凝集を阻害するために細胞において産生されるように、内部抗体またはペプチド骨格の結合分子をコードする核酸を細胞に導入することを含む。
【0138】
内部抗体またはペプチド骨格の結合分子をコードする本発明の核酸分子は、核酸によってコードされ、ベクターに挿入され、遺伝子治療ベクターとして用いることができる。遺伝子治療ベクターは、例えば静脈内注射、局所投与(例えば、米国特許第5,328,470号を参照のこと)または定位注入(例えば、チェン(Chen)ら、(1994)、PNAS 91:3054〜3057)によって対象に輸送することができる。遺伝子治療ベクターの薬学的調製物は、許容される希釈剤において遺伝子治療ベクターを含みうる、またはその中で遺伝子輸送媒体が埋め込まれている徐放性マトリクスを含みうる。または、組み換え型細胞から完全な遺伝子輸送ベクター、例えばレトロウイルスベクターを無傷で産生することができる場合、薬学的調製物は、遺伝子輸送系を産生する一つまたはそれ以上の細胞を含みうる。薬学的組成物は容器に含めることができる。遺伝子治療ベクターは、典型的に、内因性の転写因子に反応する構成的な調節エレメントを利用する。好ましい態様において、内部抗体またはペプチド結合分子をコードする遺伝子治療ベクターは、アデノウイルス、アデノ随伴ウイルス、レトロウイルス、または単純ヘルペスウイルスであるウイルスに由来する発現ベクターである(例えば、デュリング(During)ら、Mol. Med. Today 4:485〜93(1998);デュリング(During)ら、Nat. Med. 4:1131〜5(1998);デュリング(During)ら、Gene Ther. 5:820〜7(1998);カプリット(Kaplitt)ら、Pediatr. Neurosurg. 28:3〜14(1998);フリース(Freese)ら、Epilepsia 38:759〜66(1997);オコナー(O'Connor)ら、Exp. Neurol. 148:167〜78(1997);デュリング(During)ら、Exp. Neurol. 144:74〜81(1997);フリース(Freese)ら、Mov. Disord. 11:469〜88(1996);デュリング(During)、Lancet 348:618(1996);フリース(Freese)ら、J. Clin. Endocrinol. Metab. 81:2404〜4(1996);デュリング(During)ら、Clin. Neurosci. 3:292〜300(1995〜6);デュリング(During)ら、Science 266:1399〜403(1994);およびカプリット(Kaplitt)ら、Nat. Genet. 8:148〜54(1994)を参照のこと)。
【0139】
非ウイルス遺伝子輸送媒体はまた、本発明の治療的プラスミドの細胞特異的輸送を行うための手段である。これらは、DNA複合体形成剤とカップリング、または融合した従来の抗体または一本鎖Fv抗体である(ウエレク(Uherek)ら、J. Biol. Chem. 273:8835〜8841(1998);フォスター(Foster)ら、Human Gene Therapy 8:719〜727(1997);チェン(Chen)ら、Gene Therapy 2:116〜123(1995)を参照のこと)。このクラスの遺伝子輸送媒体はまた、リポソームにカップリングした抗体またはその断片が含まれる(ヒュアン(Huang)ら、米国特許第4,925,661号/第4,957,735号/第6,008,202号)。
【0140】
遺伝子治療のアプローチに関する検討に関しては、アンダーソン(Anderson, W.F.)(1992)Science 256:808〜813;ミラー(Miller, A.D.)(1992)Nature 357:455〜460;フリードマン(Friedmann, T.)(1989)Science 244:1275〜1281;およびクーノヤー(Cournoyer, D.)ら(1990)Curr. Opin. Biotech. 1:196〜208を参照のこと。
【0141】
細胞タイプ、遺伝子、および遺伝子治療の方法に関する詳しい記述は、例えば、ウィルソン(Wilson, J.M.)ら(1988)Proc. Natl. Acad. Sci. USA 85:3014〜3018;アーメンターノ(Armentano, D.)ら(1990)Proc. Natl. Acad. Sci. USA 87:6141〜6145;ウルフ(Wolff, J.A.)ら(1990)Science 247:1465〜1468;チョウデュリー(Chowdhury, J.R.)ら、(1991)Science 254:1802〜1805;フェリー(Ferry, N.)ら(1991)Proc. Natl. Acad. Sci. USA 88:8377〜8381;ウィルソン(Wilson, J.M.)ら(1992)J. Biol. Chem. 267:963〜967;クアンチン(Quantin, B.)ら(1992)Proc. Natl. Acad. Sci. USA 89:2581〜2584;ダイ(Dai, Y.)ら(1992)Proc. Natl. Acad. Sci. USA 89:10892〜10895;バン・ビューセケム(van Beusechem, V.W.)ら(1992)Proc. Natl. Acad. Sci. USA 89:7640〜7644;ローゼンフェルド(Rosenfeld, M.A.)ら(1992)Cell 68:143〜155;ケイ(Kay, M.A.)ら、(1992)Human Gene Therapy 3:641〜647;クリスチャーノ(Christiano, R.J.)ら(1993)Proc. Natl. Acad. Sci. USA 90:2122〜2126;ヒュ(Hwu, P.)ら(1993)J. Immunol. 150:4104〜4115;およびヘルツおよびジェラルド(Herz, J. and Gerard, R.D.)(1993)Proc. Natl. Acad. Sci. USA 90:2812〜2816;バコード・レーヴィ(Bachoud-Levi)ら(1998)Progress in Brain Research, 117:511〜524;ローウェンスタイン(Lowenstein)ら(1998)Progress in Brain Research 117:485〜501;ウェイヤーブロック(Weyerbrock)ら(1999)Current Opinion in Oncology 11:168〜173;カルパチ(Karpati)ら(1996)Trends Neurosci. 19:49〜54;スケイパー(Skaper)ら(1998)Mol. and Cell Neurosci. 12:179〜193;スール(Suhr)ら(1999)Arch. Neurol. 56:287〜292;ラビノヴィッツ(Rabinowitz)ら(1998)Current Opinion in Biotechnology 9:470〜475;タイラー(Tyler)ら(1999)P.N.A.S. 96:7053〜7058を参照のこと。
【0142】
VI.薬学的組成物
本発明の内部抗体は、投与に適した薬学的組成物に組み入れることができる。そのような組成物は典型的に、内部抗体と薬学的に許容される担体とを含む。本明細書において用いられるように、「薬学的に許容される担体」という用語には、例えばポリペプチド、核酸、ペプチド骨格の結合分子、または低分子の形での内部抗体または結合分子の薬学的な投与に適合する、いかなるおよび全ての溶媒、分散媒体、コーティング、抗菌および抗真菌剤、等張および吸収遅延剤等も含まれると解釈される。そのような媒体および物質を薬学的に活性な物質に用いることは、当技術分野で周知である。従来の培地または物質が内部抗体と不適合性である場合を除き、組成物におけるその使用が意図される。補助活性化合物もまた、組成物に組み入れることができる。
【0143】
本発明の薬学的組成物は、その意図する投与経路と適合するように製剤化される。投与経路の例には、非経口、例えば静脈内、皮内、くも膜下、皮下、経口(例えば吸入)、経皮(局所)、経粘膜、および直腸投与が含まれる。非経口、皮内、または皮下投与に用いられる溶液または懸濁液は、以下の成分を含みうる:注射用水、生理食塩液、固定油、ポリエチレングリコール、グリセリン、プロピレングリコール、または他の合成溶媒のような滅菌希釈剤;ベンジルアルコールまたはメチルパラベンのような抗菌剤;アスコルビン酸または重炭酸ナトリウムのような抗酸化剤;エチレンジアミン四酢酸のようなキレート化剤;酢酸塩、クエン酸塩またはリン酸塩のような緩衝剤および塩化ナトリウムまたはデキストロースのような等張性調節剤。pHは、塩酸または水酸化ナトリウムのような酸または塩基によって調節することができる。非経口調製物は、アンプル、使い捨てシリンジ、またはガラスもしくはプラスチック製の多用量バイアルに封入することができる。
【0144】
注射可能な用途に適した薬学的組成物には、滅菌水溶液(水溶性の場合)または分散剤および滅菌注射溶液または分散液の即時調製用の滅菌粉末が含まれる。静脈内投与の場合、適した担体には、生理食塩液、静菌水、CremophorEL(商標)(BASF、パルシッパニー、ニュージャージー州)またはリン酸緩衝生理食塩液(PBS)が含まれる。全ての場合において、組成物は滅菌でなければならず、容易なシリンジ操作性が存在する程度に流動性でなければならない。これは製造および保存条件で安定でなければならず、細菌および真菌のような微生物の混入作用に対して保護されなければならない。担体は、例えば、水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコール、液体ポリエチレングリコール等)、およびその適した混合物を含む溶媒または分散媒体となりうる。適切な流動性は、例えば、レシチンのようなコーティングを用いることによって、分散剤の場合には必要な粒子径を維持することによって、そして界面活性剤を利用することによって維持することができる。微生物の作用の予防は、様々な抗菌剤および抗真菌剤、例えばパラベン、クロロブタノール、フェノール、アスコルビン酸、チメロサル等によって行うことができる。多くの場合において、等張剤、例えば糖、マンニトール、ソルビトールのような多価アルコール、塩化ナトリウムを組成物に含めることが好ましいであろう。注射可能組成物の持続的な吸収は、吸収を遅らせる物質、例えばモノステアリン酸アルミニウムおよびゼラチンを組成物に含めることによって得ることができる。
【0145】
滅菌注射溶液は、先に列挙した成分の一つまたは組み合わせと共に適当な溶媒に内部抗体の必要量を組み入れて、必要に応じてその後濾過滅菌することによって調製することができる。一般的に、分散剤は、基礎となる分散媒体と先に列挙した必要な他の成分とを含む滅菌媒体に活性化合物を組み入れることによって調製される。滅菌注射溶液を調製するための滅菌粉末の場合、調製の好ましい方法は、先に濾過滅菌したその溶液から活性成分プラスさらに望ましい成分の粉末を生じる真空乾燥および凍結乾燥である。
【0146】
経口組成物は一般的に、不活性希釈剤または食用担体を含む。それらは、ゼラチンカプセルに封入することができ、または錠剤に圧縮することができる。経口治療的投与の目的の場合、活性化合物を賦形剤と共に組み入れて、錠剤、トローチ剤、またはカプセル剤の形で用いることができる。経口組成物はまた、液体担体中の化合物が経口適用され、ヒューッと音を立てて喀出されるか、または飲み込まれる、口腔洗浄剤として用いるための液体担体を用いて調製することができる。薬学的に適合性の結合物質、および/またはアジュバント材料は組成物の一部として含めることができる。錠剤、丸剤、カプセル剤、トローチ剤等は、以下の成分のいかなるものも、または類似の性質の以下の化合物を含みうる:微結晶セルロース、トラガカントガム、またはゼラチンのような結合剤;デンプンまたは乳糖のような賦形剤;アルギン酸、プリモゲル、またはコーンスターチのような崩壊剤;ステアリン酸マグネシウムまたはステローツのような潤滑剤;コロイド二酸化珪素のような滑り剤;蔗糖またはサッカリンのような甘味料;またはペパーミント、サリチル酸メチル、またはオレンジ香料のような着香料。
【0147】
吸入によって投与するために、内部抗体は、適した噴射剤、例えば二酸化炭素のような気体を含む圧縮容器もしくはディスペンサー、またはネブライザーからエアロゾルスプレーの形で輸送することができる。
【0148】
全身投与はまた、経粘膜または経皮手段によって行うことができる。経粘膜または経皮投与の場合、浸透すべき障壁に対して適当な浸透剤を製剤に用いる。そのような浸透剤は、一般的に当技術分野で既知であり、例えば、経粘膜投与の場合、洗浄剤、胆汁酸塩、およびフシジン酸誘導体が含まれる。経粘膜投与は、点鼻スプレーまたは坐剤を利用することによって行うことができる。経皮投与の場合、活性化合物を、当技術分野で一般的に既知の軟膏、軟膏、ゲル、またはクリームに製剤化する。
【0149】
内部抗体はまた、坐剤(例えば、カカオバターおよび他のグリセリドのような従来の坐剤基剤と共に)の形で、または直腸輸送のための固定浣腸の形で調製することができる。
【0150】
一つの態様において、内部抗体は、インプラントおよび微量封入輸送系を含む徐放製剤のような体内からの急速な排泄に対して内部抗体を保護する担体と共に調製される。エチレン酢酸ビニル、多価無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、およびポリ乳酸のような生体分解性の生体適合性のポリマーを用いることができる。そのような製剤を調製する方法は、当業者に明らかであろう。材料はまた、アルザコーポレーションおよびノバファーマシューティカルズインクから購入することができる。リポソーム懸濁液(ウイルス抗原に対するモノクローナル抗体によって感染細胞にターゲティングされるリポソームを含む)もまた、薬学的に許容される担体として用いることができる。これらは、例えば、米国特許第4,522,811号に記載されるように、当業者に既知の方法に従って調製することができる。
【0151】
投与を容易にするため、そして用量を均一にするために、経口または非経口組成物を単位投与剤形に処方することは特に都合がよい。本明細書において用いられる単位投与剤形は、治療すべき対象の単位用量として適している物理的に個別の単位を意味する;各単位は、必要な薬学的担体に関連して望ましい治療効果を生じるように計算された内部抗体の既定量を含む。本発明の単位投与剤形の仕様は、内部抗体の独自の特徴、得られる特定の治療効果、および対象を治療するためにそのような内部抗体を合成する技術分野に固有の制限によって影響を受け、直接左右される。
【0152】
そのような内部抗体の毒性および治療効果は、例えば、LD50(集団の50%に対して致死的な用量)およびED50(集団の50%において治療的に有効な用量)を決定するために、細胞培養または実験動物において標準的な薬学的技法によって決定することができる。毒性と治療効果の用量比が治療指数であり、これはLD50/ED50の比として表すことができる。大きい治療指数を示す内部抗体が好ましい。毒性副作用を示す内部抗体を用いてもよいが、非感染細胞に対する起こりうる障害を最小限にして、それによって副作用を減少させるために、罹患組織部位にそのような内部抗体をターゲティングする輸送システムを設計する場合には注意しなければならない。
【0153】
細胞培養アッセイおよび動物実験から得られたデータは、ヒトにおいて用いられる用量範囲を決定するために用いることができる。そのような化合物の用量は、ほとんど、または全く毒性を示さないED50を含む循環中の濃度範囲内に存在する。用量は、用いる投与剤形および利用する投与経路に応じてこの用量範囲内で変化してもよい。本発明の方法において用いられる内部抗体に関して、治療的有効量は、細胞培養アッセイからまず推定することができる。用量は、細胞培養において決定したIC50(すなわち、症状の半最大阻害が得られる試験内部抗体の濃度)を含む循環中の血漿濃度範囲が得られるように動物モデルにおいて処方してもよい。そのような情報を用いて、ヒトにおいて有用な用量をより正確に決定することができる。血漿レベルは、例えば、高速液体クロマトグラフィーによって測定してもよい。
【0154】
本発明の内部抗体をコードする遺伝子をベクターに挿入して、遺伝子治療ベクターとして用いることができる。遺伝子治療ベクターは、上記の章において記載したように、対象に輸送することができる。
【0155】
薬学的組成物は、投与説明書と共に、容器、パック、またはディスペンサーに含めることができる。
【0156】
本発明は、制限的に解釈してはならない以下の実施例によってさらに説明する。
【0157】
実施例
実施例全体を通じて、特に明記していない限り、以下の材料と方法を用いた。
【0158】
材料と方法
一般的に、本発明の実践は、特に明記していない限り、化学、分子生物学、組み換え型DNA技術、PCR技術、免疫学(特に、例えば抗体技術)および必要な細胞培養または動物飼育技術に関する従来の技術を用い、これらは当業者の範囲内であり、文献に説明されている。例えばサムブルック、フリッチュおよびマニアティス(Sambrook, J., Fritsh, E.F., and Maniatis, T.)、「分子のクローニング(Molecular Cloning)」、コールドスプリングハーバー研究所出版(1989);「DNAクローニング(DNA Cloning)」、第1巻および2巻(グロバー(D.N. Glover)編、1985);「PCRハンドブック:核酸化学の現行プロトコール(PCR Handbook Current Protocols in Mucleic Acid Chemistry)」、ビューケージ(Beaucage)編、ジョンウィリー&サンズ(1999);「抗体操作プロトコール(分子生物学の方法)(Antibody Engineering Protocols)(Methods in Molecular Biology)」、510、ポール(Paul, S.)、ヒュマナ出版(1996);「抗体の操作:実践アプローチ(Antibody Engineering:A Practical Approach(実践アプローチシリーズ、169)」、マッカファーティ(McCafferty)編、アール出版(1996);「抗体:実験マニュアル(Antibodies:A Laboratory Manual)」、ハーロウ(Harlow)ら、C.S.H.J.出版、(1999);「分子生物学の現行プロトコール(Current Protocols in Molecular Biology)」、アウスユベール(Ausubel)ら、編、ジョンウィリー&サンズ(1992);「哺乳類細胞大量培養技術(Large-Scale Mammalian Cell Culture Technology)」、ルビニエッキ(Lubiniecki, A.)編、マーセルデッカー出版(1990);ならびに「マウス胚の操作(Manipulating the Mouse Embryo)」、ホーガン(Hogan)ら、C.S.H.L.出版、(1994)を参照のこと。
【0159】
GST融合蛋白質およびGFP融合蛋白質の構築
全てのプラスミド構築に関して標準的な技術を用いた(例えば、アウスユベール(Ausubel)ら、およびサムブルック(Sambrook)ら、上記を参照のこと)。
【0160】
DRPLA構築物は、オノデラ(Onodera)らから得るか、またはオノデラ(Onodera)ら(FEBS letters(1996)399:135〜139)が記述したように誘導する。
【0161】
HD構築物、例えばHD-Q25およびHD-Q104 cDNAは、pcDNA-HD17およびpcDNA-HD14(カザンツェフ(Kazantsev)らからの寄贈、Proc. Natl. Acad. Sci. USA 96:11404〜11409(1999))から、BamHI部位を含むHDセンスプライマー
およびc-mycアンチセンスプライマー
を用いるPCR(95℃で1分間、50℃で1分間、および72℃で1分間を25サイクル)によって増幅した。PCR産物は、プロテナーゼK(最終濃度1mg/ml)によって37℃で30分間処理してから、QIAEX IIキット(キアゲン社)を用いてゲル精製を行った。次に、精製したHD-Q25およびQ104 PCR産物をXmaIおよびBamHI制限酵素によって消化してpGEXベクターに個別にライゲーションした。HD-Q25およびQ104 PCR産物はまた、BamHIおよびSmaI制限酵素によって消化して、BamHIおよびNruIによって消化したpQBi25c3ベクター(QUANTUMバイオテクノロジー社、モントリオール、ケベック州、カナダ)に個別にライゲーションした。
【0162】
プラスミドpQBi-25c3-HD-Q(-)は、pQBi-25c3-HD-Q104をEcoRIおよびHindIIIによって消化することによって調製した。酵素を75℃で10分不活化した後、消化した産物をdNTPの存在下でクレノウポリメラーゼによって30℃で30分処理して、平滑末端DNAを生じた。次に、消化したベクターをゲル精製して、自己ライゲーションした。
【0163】
プラスミドpGEX-HD-42QおよびHD-65Qは以下のように調製した:42Qおよび65QをコードするcDNAセグメントを、pBSK-42QおよびpBSK-65QからHindIIIおよびXmaIによる消化によって単離した。ゲル精製を行った後、cDNAセグメントを、同じ制限酵素によって消化してHD-25Q cDNAコード配列を置換したpGEX-HD 25Qにクローニングした。本発明の発現された実験ポリペプチドの関連するアミノ酸配列を下記の表3に要約する。
【0164】
実験ポリペプチドのアミノ酸配列
【表3】
略語:DRPLA、歯状赤核淡蒼球萎縮症(アトロフィン1);HD、ハンチントン病(ハンチンチン);GST、グルタチオントランスフェラーゼ;GFP、緑色蛍光蛋白質。アトロフィン1およびハンチンチン蛋白質配列には下線を示す。隣接する配列は発現ベクターの一部である。GSTとGFPのポリペプチド配列は当技術分野において認識されており、例えばGFP:プラスミドpQB125-fC3(カタログ番号AFP2133)、カンタムバイオテクノロジーズ社(1801 de Maissonneuve Bvld. West Montreal、Quebec, h3H 159、カナダ);GST:プラスミドpGEX-4T-3(カタログ番号27-4583-01)、ファルマシア社(ゲンバンクアクセッション番号U13855号);およびオノデラ(Onodera)ら(1996)「伸長したポリグルタミンドメイン蛋白質の大腸菌における毒性(Toxicity of expanded polyglutamine-domain proteins in Escherichia coli)」)に記載のGST-DRPLA構築物を参照のこと。
【0165】
蛋白質発現と精製
N-HD-C4 sFvからのVHリンカーVL cDNAコード配列は、NcoIおよびNotI制限酵素による消化によってpHEN-1ベクターから切除した。ゲル精製を行った後、cDNAを、同じ制限酵素によって先に消化されたpSYN1ベクターにライゲーションした。次に、ライゲーションしたプラスミドを電気穿孔によってDH5αコンピテント細菌に導入して、増幅されたプラスミドDNAをQIAプレップスピンミニプレップキット(キアゲンインク、バレンシア、カリフォルニア州)によって精製した。N-HD-C4 sFvは、以下のように大腸菌株TG1において発現させた:形質転換体を100 μg/mlアンピシリンと1%グルコースを含む2×TY培地10 mlに接種した。37℃で一晩インキュベートした後、培養を100 μg/mlアンピシリンと0.1%グルコースを含む2XTY培地1Lに接種して、Abs600 nm=1が得られるまで37℃で強く攪拌した。次に、IPTGを加えて最終濃度0.1 nMとして、培養をRTで200 rpmで一晩インキュベートした。遠心によって細胞を回収して、pH 8.0のPBS 30 mlに浮遊させた。ライソザイムを最終濃度1mg/mlとなるように加えて、混合物をRTで5分インキュベートした。次に、細菌溶解物を氷中で超音波処理して、染色体DNAを切断して、25,000×gで4℃で30分間遠心して細菌の破片および他の不溶性材料を除去した。上清を、予め洗浄したNi-NTA樹脂(キアゲン社)の20 mMイミダゾールを含むPBS溶液1mlと共に4℃で2時間インキュベートした。次に、樹脂をPBS 30 mlによって洗浄した後、35 mMイミダゾールを含むPBS 10 mlおよび40 mMイミダゾールを含むPBS 10 mlによって洗浄した。次に、sFv分子を、250 mMイミダゾールを含むPBSによって1ml分画に溶出した。当該分画をプールしてPBS中で透析した。
【0166】
GST融合蛋白質は、以下のように大腸菌株BL21において発現させた:新鮮な形質転換体を、100 μg/mlアンピシリンと0.1%グルコースを含むLB培地1Lに接種して、Abs600 nm=1が得られるまで37℃で強く攪拌した。次に、IPTGを最終濃度0.1 nMとなるように加えて、培養を200 rpmで3時間インキュベートした。遠心によって細胞を回収して、pH 8.0のPBS 30 mlに浮遊させた。ライソザイムを最終濃度1mg/mlとなるように加えて、混合物をRTで5分インキュベートした。次に、細菌溶解物を氷中で超音波処理して、25,000×gで4℃で30分間遠心した。上清を、予め洗浄したグルタチオンセファロース4Bビーズ(ファルマシア社)のPBS溶液1mlと共に4℃で2時間インキュベートした。次に、樹脂をPBS 50 mlによって洗浄した後、蛋白質を10 mM還元グルタチオンを含む50 mMトリス、pH 8.0によって1ml分画に溶出した。当該分画をプールしてPBS中で透析した。
【0167】
免疫精製実験
N-HD-C4 sFv約3μgをGST-DRPLA-Q35、GST-HD-Q25、またはGST-HD-Q42のPBS溶液のいずれか4μg(最終容量200 μl)と共に混合して、RTで1時間揺り動かした。グルタチオンセファロース4Bビーズ(50%スラリー、PBSで洗浄済み)20 μlを加えて、RTでさらに30分インキュベートして、sFv/GST融合蛋白質複合体を捕獲した。ビーズを12,000×gで10分間遠心することによって回収して、PBS1mlで2回洗浄した。複合体を、10 mM還元グルタチオンを含むpH 8.0の50 mMトリス10 μlと共に室温で10分間溶出した。次に、試験管を12,000×gで1分間遠心して、可溶性複合体を含む上清を新しい試験管に移し、2×SDS試料緩衝液の等量と混合して、95℃に加熱してからSDS-PAGEにローディングした。
【0168】
COS7細胞の同時トランスフェクションと免疫蛍光
COS7細胞を6ウェルプレートのカバーガラス(細胞105個/ウェル)上で増殖させて、スーパーフェクトトランスフェクション試薬(キアゲン社)5μlを用いて、N-HD-C4 sFvまたは陰性対照sFv(HAタグおよび核配置配列(NLS)を含む)のいずれかを含むプラスミドと、GFP標的化融合蛋白質を含むプラスミドを3:1の比率(標的に対するsFv)で同時トランスフェクトした。トランスフェクションの24時間後、細胞を以下のように免疫蛍光のために調製した:細胞を4%パラホルムアルデヒドによって固定して、0.2%トライトンX100によって透過させた。二次抗体の非特異的結合を防止するために、細胞を10%正常ヤギ血清の3%BSA溶液によってブロッキングした。次に、予め吸収させたポリクローナルウサギ抗HA IgGを加えてsFv抗体を検出し、結合したIgGをローダミン結合ヤギ抗ウサギIgG抗体(ピアス社)によって顕色させた。
【0169】
内部抗体
再ターゲティングする内部抗体は、以下のように構築した。簡単に説明すると、プラスミドpcDNA3.1(+)(インビトロゲン社、カールスバッド、カリフォルニア州)を、HA免疫タグを表すインフルエンザ血液凝集素エピトープ(YPYDVPDYA(配列番号:43))と、SV40核ターゲティング配列(TPPLLLRLV(配列番号:44))、ライソゾームターゲティングシグナル、またはターゲティングシグナルなし(ターゲティングシグナルが存在しなければsFvは細胞質に残される)のいずれか、およびハンチンチンに特異的なC4 sFv(抗HD-C4)、または無関係なsFv対照をコードするように改変した。sFv内部抗体構築物の発現は、サイトメガロウイルス(CMV)プロモーターの制御下であった。
【0170】
速度論的結合分析
精製scFvクローンC4のKdは、BIAコア2000(ビアコアAG社、アップサラ、スウェーデン)において表面プラズモン共鳴を用いて決定した。BIAコアフローセルにおいて、ビオチン結合HDペプチド(1M NaClおよび50 mM NaOH中で250 nM)の約50共鳴単位(RU)を、1M NaClの50 mM NaOH溶液を連続的に1分間3回注入して予め条件を設定したストレプトアビジンセンサーチップにカップリングさせた。カップリングしたペプチドのこの量によって、結合した内部抗体の最大量40〜80 RUが得られた。内部抗体が結合した後の表面を再生するために、1M NaClを含む50 mM NaOH5μlを注入して、基準値に回復させた。濃度範囲60〜100 nMで5μl/分の連続的な流れの下で結合を測定した。konはln(dR/dt)/t対濃度のプロットから決定し、式中Rは反応であり、tは時間である(カールッソン(Karlsson)ら、J. Immunol. Methods 145:229〜240(1991))。koffは、流速20 μl/分を用いることによって分析した内部抗体の最高濃度でのセンサーグラムの解離部分から決定した。Kdはkoff/konとして計算した。
【0171】
脳組織培養
臓器型切片培養は、公表された方法に軽微な改変を加えて(1)(14)、P12〜P14小脳、大脳皮質、または線条体から作製した。脳をマウスから摘出して、高マグネシウム(1.8 mM CaCl2、および2.4 mM MgCl2)HEPES緩衝ハンクス生理食塩液(HBHS)の4℃のビーカーに入れた。次に脳をブロックにして、プレキシグラス(厚さ0.3 cm)の1cm2片上で2%アガロースブロックの隣にSuperGlue(登録商標)を用いて載せる。歯科用ワックスを切片チャンバー組織ホルダー上のプラスチックに載せる。載せた後、組織をビブラトームの切片チャンバーに置く。ビブラトームの水槽は、冷却した4℃の高マグネシウムHBHSを含む。温度は切片チャンバーに接続したペルティエ熱電流冷蔵システム(FHC、ブランズウィック、メイン州)によって維持する。厚さ300、または400 μmの切片を当該組織から切断する。新しい各切片を作製すると、それを切片の回収が終了するまで4℃の増殖培地に入れた。一つの調製物からの切片を封入したフィルター単位に入れて、高マグネシウム人工脳脊髄液(12 mM NaCl、0.33 mM KCl、0.12 mM NaH2PO4、2.5 mM NaHCO3、1mMデキストロース、2.4 mM MgCl2、1.8 mM CaCl2;全ての物質をSigma(登録商標)、セントルイス、ミズーリ州から得た)中で37℃で1時間インキュベートした。溶液には95%O2と5%CO2を絶えず通気した。空気培地界面で増殖した切片を0.4 μmのTranswell(登録商標)フィルター(Corning Costar(登録商標)、ケンブリッジ、マサチューセッツ州)で維持した。増殖培地は、25%熱不活化ウシ血清、25%HBHS、重炭酸塩を含まない50%MEM、4mM Lグルタミン、30 mM Dグルコース、50 mM重炭酸ナトリウム、および12.5 mM HEPES(全ての物質をシグマ社、セントルイス、ミズーリ州から得た)を含む。温度は33℃で5%CO2で維持した。培地は、週に3回交換した。
【0172】
脳組織培養のトランスフェクション
プラスミドを、バイオラド社からのバイオリスティックPDS-1000/He粒子輸送システム、「遺伝子銃」を用いて神経組織に導入した。プラスミドは、金またはタングステンからなるミクロ担体上にコーティングした。次に、ミクロ担体をマクロ担体にローディングした。マクロ担体は、高圧ヘリウムと、停止スクリーン方向の部分的真空とによって加速された。停止スクリーンは、マクロ担体を停止させるが、微小担体を標的細胞に向けさせ、浸透させる。臓器型培養を衝突のために1%寒天を含むペトリ皿に入れ、その後培養を増殖チャンバーに戻した。典型的に、厚さ300〜400 μmの切片を用いた。
【0173】
顕微鏡
標準的な倒立および直立蛍光顕微鏡を用いた。しかしさらに、これまでに記載されているようにレーザー共焦点顕微鏡を適用してもよい(例えば、ベッカー(Becker)ら、IEEE Transactions on Biomed. Eng. 45:105〜118(1998);ターナー(Turner)ら、Neuron 4:833〜845(1990);およびターナー(Turner)ら、International Rev. Exp. Path. 36:53〜72(1996))を参照のこと。
【0174】
一本鎖Fvファージライブラリ構築およびアミノ末端HDペプチドに対して特異的なヒトsFv抗体の選択
グルタミン残基の数が変化しており、GFPと融合したN末端ハンチンチンペプチドに対して免疫したマウスの脾臓からの一本鎖Fvライブラリ構築は、以下のように実施してもよい。簡単に説明するとVHおよびVL遺伝子に対応するcDNAは、スーパースクリプトII RNアーゼH逆転写酵素(ギブコBRL社)とプライマーを用いて、ガンマ重鎖とカッパ軽鎖のそれぞれ、可変および定常領域のあいだの接合部に対して免疫したマウスの総脾臓RNAから作製することができる。VHおよびVL遺伝子は、例えば、PCR 25サイクル(94℃を1分間、55℃を1分間、72℃を2分間)によって、cDNAからPfuポリメラーゼ(ストラタジーン社)およびVHプライマー[VH1 BACK(34)およびVH1 FOR2(49)、およびVLプライマーVK2BACKおよびVK4FOR(10)]によってそれぞれ増幅する。リンカーDNAは、反応をふさぐためにクレノウポリメラーゼおよびデオキシヌクレオチド三リン酸(dNTPs)を用いて、以下のプライマーの等モル比を混合することによって調製してもよい。
【0175】
次に、VHおよびVL PCR産物と二本鎖リンカーDNAをアガロースゲル上で精製し、VHおよびVL産物各1μgとDNAリンカー300 ngとをプライマーを加えずにPCR反応混合物50 μl中で混合した。7サイクル(94℃で2分、72℃で4分)後、VH1 BACK-SfiI(SfiI制限部位を有するVH1 BACK)およびVK4FOR-NotI(NotI制限部位を有するVK4FOR)プライマー各25 pmolを反応混合物に加えて、20サイクル(94℃で1.5分、および72℃で2.5分)増幅する。ゲル精製後、集合したVHリンカーVL PCR産物をSfiIおよびNotIによって十分に消化して、pHEN1ベクターにクローニングする。ライゲーション混合物の少量をTG1コンピテント細胞に電気穿孔する。ライブラリの多様性はVH1 BACKとVH4FORプライマーによる組み換え型コロニーのPCRスクリーニングの後、BstNI制限酵素による消化によって分析してもよい(グッソウ(Gussow)ら、Nucleic Acid Res. 17:4000(1989))。次に、ファージライブラリを、100 mg/mlアンピシリンおよび1%グルコースを含む2×TY培地50 mlにコロニーを採取して、37℃で45分振とうさせ、ヘルパーファージを加えて、ファージを救出することによって増幅させることができる。
【0176】
ビオチン標識抗原は、例えばeアミノ基またはリジン残基と非特異的に反応するNHSビオチン(ピアスケミカル社)を用いて調製することができる。所定のライブラリに関して、少量をその対応する抗原で平衡にしてもよい。次に、ストレプトアビジンコーティング磁気ビーズを加えて、ストレプトアビジンを抗原ファージ抗体複合体に結合させ、これをライブラリ溶液からのビーズの磁石による分離によって単離する;次にビーズを洗浄する。結合したsFvファージをpH 2の酸性溶液によってビーズから溶出し、直ちに中和して、感染細菌によって救出する。救出されたファージは、増幅して別のラウンドの選択のために用いることができる。
【0177】
選択は、例えばCOS-7細胞に発現された融合蛋白質を用いて、HDに特異的な隣接エピトープとポリグルタミン反復配列の長さとの組み合わせを有する当初の構築物から行ってもよい。後者は、異種ポリグルタミン蛋白質(例えば、DRPLAを用いて)に対する二回目の選択によって同定することができ、前者はグルタミン(Gln)の25コピー以上を含まない蛋白質のイソ型との交叉反応性によって同定することができる。
【0178】
実施例2に記載するように、選択の4ラウンドは、各ラウンドで抗原濃度を減少させることによって(100 nM、50 nM、10 nM、1nM)実施した。第3ラウンドの後、細菌クローンをスクリーニングして、N末端HD配列に対して特異的なsFv配列を分泌するクローンを単離する。スクリーニングはELISA法によって行った。分泌されたsFvを含む細菌の上清を、陰性対照として2%ウシ血清アルブミン(BSA)またはビオチン結合抗原を含む2%BSA(0.1 μg/ウェル、最終濃度)のいずれかと共に、予めブロッキングしたマイクロタイタープレートにおいて4℃で一晩インキュベートした。次に、混合物をストレプトアビジンコーティング(0.1μg/ウェル、最終濃度)したマイクロタイタープレートに移した。一晩インキュベートした後、プレートを洗浄して、マウス抗myc IgG(9E10、0.1 μg/ウェル最終濃度)を加えた。結合したIgGを、ヤギ抗マウスIgGに結合したアルカリホスファターゼによって検出した。
【0179】
さらなる特徴付けのために、可溶性sFv蛋白質を精製する必要があった。したがって、選択されたクローンのVHリンカーVLをコードするcDNAをpHENからpSAYN-1ベクターに移した。pSYN-1ベクターは、6×His尾部を有する蛋白質を発現させ、これによってNi-NTAカラムでの迅速な精製が容易となる。これらのsFv分子の結合特異性は、以下のようなアフィニティ精製によって調べることができる。
【0180】
各クローン約3μgを、GST-DRPLA-Q35、GST-HD-Q25、またはGST-HD-Q42のいずれか4μgと共に室温で1時間溶液中でインキュベートした。次に、グルタチオンセファロース4B(ファルマシア社)の底面容積10 μlを各試験管に加えて、GST融合蛋白質/sFv複合体を沈殿させた。30分インキュベートした後、試験管を遠心して、沈殿物をPBS1mlによって3回洗浄した。10 mM還元グルタチオンを沈殿物に加えることによって、複合体をグルタチオンセファロースビーズから溶出した。
【0181】
遠心後、溶出した複合体を含む上清を新しい試験管に移して、等量の2×SDSローディング緩衝液と共に混合した。試料を沸騰させて、蛋白質を12%SDS-PAGE上で分離した。蛋白質のバンドは、クーマシーブルー染色によって可視化して、および/またはsFv分子をイムノブロット法によって検出するためにニトロセルロースメンブレンに転写した。
【0182】
実施例1
細胞内ポリペプチド凝集の効果をアッセイする方法
本実施例において、細胞内ポリペプチド凝集をアッセイする方法および細胞内ポリペプチド凝集が細胞死に至ることを証明する方法を提供する。
【0183】
細胞内ポリペプチド凝集の影響を調べるために、ハンチントン病患者において認められるポリペプチドを表すハンチンチンポリペプチドを発現する細胞と比較して、正常なハンチンチンポリペプチドを表すポリペプチドを発現する細胞を調べた。霊長類細胞(COS-7)に正常なポリペプチドまたはグルタミン残基47、72、もしくは104個を有する疾患に関連した変化したハンチンチンポリペプチドを表すモデルハンチンチンGFP融合ポリペプチドをコードするプラスミド構築物をトランスフェクトさせた。24時間のトランスフェクション後、試験蛋白質のそれぞれに融合させたGFPタグによって示されるように、大量の細胞がハンチンチンポリペプチドを発現する。観察は3回の時点で記録した(表4を参照のこと)。
【0184】
【表4】
【0185】
48時間の時点での例を図3に示す。蛍光顕微鏡を用いて、グルタミン残基104個を有するハンチンチン-GFPポリペプチドを発現する細胞(図3D)は、大きい輝く凝集体を有することが認められた。最も多くの細胞死は、これらの細胞に認められた。グルタミン残基がより少ない(72Q)モデルハンチンチンポリペプチドを発現する細胞では、極めて明るくより多様な凝集体を認め、細胞死は多少少なかった。グルタミン残基47個を有するモデルハンチンチンポリペプチドを発現する細胞では、蛍光強度は多少散在していた。正常範囲のグルタミン残基(25)を有するモデルハンチンチンポリペプチドを発現する細胞は、ポリペプチド凝集の証拠を認めず散在した蛍光を示した。
【0186】
このように、この実験は、変化したハンチンチンポリペプチド、細胞内凝集および細胞死が一致することを証明する。したがって、このアッセイ系は、ハンチントン病の病態の範囲をインビトロで再現し、細胞内ポリペプチド凝集およびポリペプチド凝集体媒介細胞死を変化させる治療をスクリーニングするために用いることができる(実施例2を参照のこと)。
【0187】
実施例2
神経ポリペプチドに対して結合特異性を有する内部抗体を操作および選択する方法
本実施例において、選択したポリペプチドに対して親和性を有する内部抗体を同定および選択する方法を示す。
【0188】
ハンチンチンポリペプチドを含む細胞内ポリペプチド凝集体の形成を阻害することができる内部抗体を作製するために、ハンチンチンのN末端アミノ鎖残基17個(Nt-HD)に対応するビオチン結合ペプチドを、この配列に対して特異的なsFv分子を示すファージを捕獲するための抗原として作製した。簡単に説明すると、異なるクローン109個を含むヒトsFvファージライブラリを、タフツ大学(ボストン、マサチューセッツ州)の蛋白質中央施設で合成されたペプチドを用いてビオチン結合Nt-HDと共にインキュベートした。次に、ストレプトアビジンコーティング磁気ビーズを加えて、関連するsFvファージ抗体と複合体を形成したストレプトアビジンを単離して洗浄した。結合したsFvファージを酸によって溶出して、中和し、感染細菌によって救出した。次に、救出したファージを増幅して、もう一つのラウンドの選択に用いた。
【0189】
選択の4ラウンドは、各ラウンドで抗原の濃度を減少させて(すなわち、100 nM、50 nM、10 nM、1nM)行った。第3ラウンドの後、細菌クローンをスクリーニングして、Nt-HD配列に対して特異的なsFv分子を分泌するクローンを単離した。スクリーニングはELISA法を用いて実施した。クローン90個をスクリーニングしたところ、クローン20個がビオチン結合Nt-HDペプチドに対して高い結合(陰性対照の結合より少なくとも10倍)を示した。sFv種の結合特異性を調べるために、ELISA法においてOD 0.2を示したクローン8個を、大規模sFv発現のためにクローン20個から選択した。sFv分子を含む細菌上清はストレプトアビジン、GST-HD-42Q、GST-HD-65Q、およびGST-DRPLA-35Q(陰性対照として、オノデラ(Onodera)ら(33))をコーティングしたマイクロタイタープレート上でELISA法によってアッセイした。陽性対照に関しては、細菌上清をビオチン結合N末端HDペプチドと共に溶液中でインキュベートしてから、ストレプトアビジンをコーティングしたマイクロタイタープレートに移した。結合したsFvを、α-9E10 IgGの後、アルカリホスファターゼ結合ヤギ抗マウスIgGによって検出した。この実験から、Nt-HD-隣接ペプチド配列を含む抗原と選択的に反応するクローン1個(α-Nt-HD-C4 sFvと命名する)を同定した。
【0190】
さらに特徴を調べるために、可溶性sFv蛋白質を精製する必要があった。したがって、選択されたクローンのVHリンカーVLをコードするcDNAをpHEN-1ベクターからpSYN-1ベクターに移入した。pSYN-1ベクターは、蛋白質の発現を可能にして、ヘキサHis尾部に融合したsFvの周辺質への分泌を可能にし、これによって迅速な精製が促進される。したがって、α-Nt-HD-C4 sFvの2つのサブクローンを細菌において発現させ、得られたポリペプチドをNi-NTAカラムにおいて精製した(Qiagen(登録商標))。
【0191】
選択したsFvクローンの結合特異性は、以下のように、完全な範囲の抗原に対するアフィニティ結合によって調べた:(a)図9および10の実験結果に対応する試料数を用いて、α-Nt-HD-C4 sFvクローン1(レーン2〜4)、またはα-Nt-HD-C4 sFvクローン2(レーン5〜7)約3μgを以下の抗原4μgと共に混合して、溶液中で室温で1時間インキュベートした:(レーン2および5)GST-DRPLA-35Q(レーン3および6)GST-HD-25Q、(レーン4および7)GST-HD-42Q;(b)グルタチオンセファロース4B(ファルマシア社)の底面容量10 μlを各試験管に加えて全てのGST含有蛋白質複合体を結合させた;ビーズを10 mMグルタチオン溶液によって洗浄し、これによってGST抗原またはsFv抗原複合体を溶出して、これらの溶出物をSDS-PAGE法によって分析した。蛋白質のバンドはクーマシーブルー染色(図9)によって可視化し、同様にイムノブロット法によってsFv分子を検出するためにニトロセルロースメンブレンに転写した(図10)。いずれのサブクローンもHD隣接ペプチドのみに結合する所望の特異性を示した。
【0192】
例えば、図9は、Nt-HD配列を含む試料に限って(すなわちGST-HD-25QおよびGST-HD-42Q)、α-Nt-HD-C4 sFvに対応するバンドが他の蛋白質とほぼ同じ強度で存在することを示している。イムノブロット分析はまた、双方の抗C4 sFvクローンがN末端HD配列を含む試料中に存在するが、GST-DRPLA-Q35を含む試料では検出されないことを確認した(図10)。
【0193】
本発明の内部抗体の正確な結合特性の特徴を調べるために、固定した抗原に結合するsFvのさらに定性的および定量的試験をインビトロで実施した。C4 sFv抗原結合特性はELISAアッセイによってモニターし、同様にC4 sFvがN末端HD配列に特異的に結合することを確認した(図11A)。定量的結合実験に関して、大量輸送の人為的結果が無視できる条件で速度論の測定を行うために、C末端ビオチンを有するHDペプチド(1〜17位)を非常に低濃度でストレプトアビジンコーティングBIAコアセンサーチップ上にローディングした。結合と解離の速度論は、BIAコア2000においてC4 sFv抗体について決定した(図11B)。Kdは、単純な生体分子相互作用に関して1:1ラングミュアモデルを用いて測定したKaおよびKd速度定数に基づいて7.9 nMであると計算された(BIA評価3.0ソフトウェア、バイアコアインターナショナルAB、アップサラ、スウェーデン)(図11B、右上の挿入図)。対応する結合親和性(Ka)1.3×108 M-1は、この大きいファージディスプレイライブラリから厳密な抗原に対して選択された抗体に典型的で、これは特に短いペプチド配列に対してかなり高い親和性sFvであることを示している(シーツ(Sheets)ら、Proc. Natl. Acad. Sci. USA 95:6157〜6162)。これらの結果は、HD(1〜17位)ペプチドが、見かけの結合定数を有意に減少させる広範囲の非構造類似体よりむしろ、C4 sFv複合部位によって認識される比較的堅固なエピトープを形成する可能性があることを示唆している。
【0194】
これらの結果は、ハンチンチンのN末端配列のような特定のエピトープに対して特異的であるα-Nt-HD-C4 sFv内部抗体分子を同定および選択できることを示している。したがって、選択されたポリペプチドポリペプチドに対して独自の特異的N末端またはC末端エピトープを検出するいかなる内部抗体を選択してもよい。その上、上記の方法を用いて、ポリグルタミンに富むドメインに対して特異的な内部抗体を作製することができる。そのような内部抗体は、ポリグルタミンに富む領域を有するいかなる当該ポリペプチド、例えば、ハンチンチン、アタキシン、アトロフィン等にも結合しうるであろう。最後に、これらの方法は、選択されたポリペプチドに対して二重または多重の特異性を有する多価内部抗体の操作を提供する。多価内部抗体は、例えば標的ポリペプチドの標的ポリグルタミンに富む領域と共に、ポリペプチドの他のN末端および/またはC末端隣接エピトープに結合することができるであろう。
【0195】
実施例3
病的ハンチンチンポリペプチドを発現する哺乳類細胞におけるポリペプチド凝集を阻害する方法
本実施例において、哺乳類細胞において凝集体の形成を阻害して、細胞内ポリペプチドを再ターゲティングする方法を示す。
【0196】
内部抗体が細胞内ポリペプチドに特異的に結合して、ポリペプチド凝集を阻害できることを証明するために、哺乳類細胞(COS-7)に、GFPに融合したモデルハンチンチンポリペプチドをコードする第一のプラスミドと、モデルハンチンチンポリペプチドに特異的に結合する内部抗体をコードする第二のプラスミドとを同時トランスフェクトさせた。アッセイは、双方の標的抗原、すなわちモデルハンチンチンポリペプチドと内部抗体を可視化できるように設計する。特に、ハンチンチンポリペプチドは、ハンチンチンGFP融合ポリペプチドのGFPドメインが放出する蛍光を検出することによって可視化され、調べる内部抗体は、内部抗体に特異的に結合することができるローダミン結合抗体を用いて可視化される。
【0197】
したがって、図5A〜Hに示す実験結果は、蛍光顕微鏡を用いて、内部抗体が細胞においてハンチンチンポリペプチドに特異的に結合してこれを再ターゲティングできることを示している。図5Aおよび5Bにおいて、細胞の分布をまず、位相差光学顕微鏡を用いて示す。図5C、5E、および5Gにおいて、モデルハンチンチンGFP融合ポリペプチド(HD-25Q-GFP)(図5E)と、核ターゲティングシグナルをさらに含むモデルハンチンチンポリペプチドに特異的に結合する内部抗体(α-Nt-HD-C4 sFv-NLS)とを同時発現する細胞は、核におけるローダミンおよびGFP染色の集合によって示されるように、ハンチンチンポリペプチドを核に分布するよう再ターゲティングできることが認められた(図5G)。対照的に、無関係な蛋白質に結合する内部抗体を発現する細胞は、ハンチンチンポリペプチドの分布を再ターゲティングすることができない(図5D、5F、およびFH)。
【0198】
これらの実験を拡大して、上記のように試験した内部抗体が同様に、ハンチントン病に関連するポリグルタミン反復配列を有する変化したハンチンチンポリペプチドに対しても同じ有効性を有すること、そして細胞内凝集体を形成する傾向が最も高いことを示した。このように、哺乳類細胞に、ポリグルタミン104個を有するモデル変化ハンチンチンポリペプチド(pHD-104Q-GFP)をコードするプラスミドと、ハンチンチン特異的内部抗体(α-Nt-HD-C4 sFv)または無関係なポリペプチドに対して結合特異性を有する内部抗体のいずれかをコードする第二のプラスミドとをトランスフェクトさせた。図7A〜Hは、蛍光顕微鏡を用いて、ハンチンチン特異的内部抗体のみがモデル変化ハンチンチンポリペプチド(すなわち、グルタミン反復配列104個を有する)の細胞分布を核に特異的に再ターゲティングできるが(図7C、7E、および7G)、無関係な内部抗体は作用を示さないことを示している(図7D、7F、および7H)。
【0199】
ハンチンチン特異的内部抗体(α-Nt-HD-C4 sFv)の特異性をさらに証明するために、哺乳類細胞に、ポリグルタミンに富み、凝集して神経疾患(DRPLA)を引き起こすことができるもう一つの標的ポリペプチドを同時トランスフェクトさせた。モデルDRPLA標的ポリペプチドをGFPドメインと融合させて、トランスフェクトした細胞における標的ポリペプチドの発現および細胞内位置を追跡した。さらに、一つはハンチンチンに特異的であり、もう一つは無関係なポリペプチドに特異的な内部抗体である異なる2つの内部抗体を、個々にDRPLA標的ポリペプチドと共に同時トランスフェクトした。グルタミンに富むDRPLAポリペプチド(GFP-DRPLA-35Q)(図6C〜G)と、ハンチンチンポリペプチド(α-Nt-HD-C4 sFv;図6D、6F)またはもう一つの無関係なポリペプチド(陰性対照sFv;図6C、6E、および6G)のいずれかに対する内部抗体とを同時発現する細胞(COS-7)は、DRPLAポリペプチドの細胞分布に変化を示さない。
【0200】
さらに、ハンチンチン特異的内部抗体が細胞内ポリペプチド凝集体の形成を防止できるのみならず、異なる細胞の位置に望ましくないポリペプチドを再ターゲティングしうることを確立する実験を実施した。哺乳類細胞に、ポリグルタミン含有量が最も高く(グルタミン残基72または104個)、凝集して疾患を誘発する傾向が大きい、変化したハンチンチンポリペプチドをコードするプラスミドを、ターゲティングシグナルなし、核ターゲティングシグナル、またはライソゾームターゲティングシグナルのいずれかを有するハンチンチン特異的内部抗体の一つと共にトランスフェクトした。
【0201】
蛍光顕微鏡を用いて調べた3つの内部抗体のそれぞれは、対照(図8A、8E)と比較して細胞内ポリペプチド凝集体の形成を阻害することが示された(8B〜8D、8F〜7H)。これは、ポリペプチド凝集体によって放出された明るいGFPシグナルの減少によって証明される。さらに、同様に、核ターゲティングシグナル(図8C、8Gを参照)またはライソゾームターゲティングシグナル(図8D、8G)のいずれかを有する内部抗体と接触させると、モデルハンチンチンポリペプチドの再ターゲティングが得られることも証明された。これは、核ターゲティングシグナルを有する内部抗体を発現するそれらの細胞の核にGFPシグナルが蓄積することによって示された(図8C、8G)。ライソゾームターゲティングシグナルを有する内部抗体をトランスフェクトした細胞では、細胞質全体により散在するシグナルを認めた(図8D、8H)。
【0202】
さらになお、本発明の内部抗体がHDの細胞内凝集の形成を阻害できるか否かを調べるさらにもう一つの試験において、抗HD C4 sFv内部抗体およびHD-ポリQ-GFPを異なるいくつかの細胞タイプに同時トランスフェクトして、ポリペプチド凝集の内部抗体媒介阻害の統計分析を測定した。
【0203】
特に、インサイチューでの病的なHD凝集体の形成は、ポリグルタミンの長さが40残基またはそれ以上である場合に(すなわちQ72、またはQ104、しかしQ25では認めない;図14)顕著な斑点状の蛍光形態を示すことが認められた。さらに、これらのポリペプチド凝集体は洗浄剤不溶性であり、トランスフェクト細胞のSDS溶解後に残留蛍光体の改善された定量方法を行うことができる(カザンツェフ(Kazantsev)ら、PNAS 96:11404〜11409(1999))。巣状塊の平面は実験における変数として消去されるため、この改善されたアッセイによって、ポリペプチド凝集体をより正確に計数することができる。したがって、このアッセイを、ポリQ抗原凝集体単独またはC4 sFvもしくは対照内部抗体の存在下で定量のために利用した。COS-7、BHK、および293細胞にC4または対照内部抗体を発現するプラスミドと、Q25、Q72、またはQ104を含むHD-ポリQ-eGFP融合体とを同時トランスフェクトさせた。HD-ポリQ72-eGFPまたはHD-ポリQ104-eGFPを用いて、内部抗体対抗原プラスミドの比が3:1の場合に凝集体の程度の定量的な減少を認めたが、最も有意な定量的効果は5:1の比で認めた(図14)。C4 sFvについて認められた凝集体における顕著な減少と比較して、無関係な内部抗体構築物による対照トランスフェクタント(C8、またはML3-9 sFv)は、親の空のベクターと比較して凝集体形成に統計学的に有意な減少を示さなかった。Q25プラスミド単独をトランスフェクトした細胞は、SDS不溶性の凝集体を決して示さなかった。
【0204】
典型的な内部抗体構築物をC末端血液凝集素(HA)ペプチドタグに融合させて、蛋白質を細胞質に存在させながら蛋白質の免疫的検出を容易にした。二重標識免疫蛍光試験は、HD-ポリQ72-GFPまたはHD-Q104-GFP(緑色標識、図15)プラス抗HD C4 sFv(赤色標識、図15)を発現するBHK細胞が、実質的な散在するGFP標識を示すが、蛍光凝集体を示すことは時折に過ぎないこと(通常HAによっても標識する;図15の矢印を確認のこと)を確認した。内部抗体を発現しないように思われるそれらの細胞は、大きい凝集体の証拠を示した(図15、矢印の先、上のパネル)。散在する標識はまた、内部抗体との同時トランスフェクションによらず、正常な長さの(25Q)構築物の特徴であった(図15)。二重トランスフェクタントについて認められた凝集体形成の減少は、イムノブロット法によって異なるトランスフェクションの典型において区別できない産物レベルが示されたために、HD蛋白質発現の抑制に由来しないように思われた(図15、下のパネル)。
【0205】
したがって、これらの結果は、内部抗体が、神経疾患(例えば、ハンチントン病)に関連するポリペプチド凝集体に特異的に結合して形成を阻害することができることを確認する。さらに、これらの結果はまた、ターゲティングシグナルを有する内部抗体を用いて望ましくない細胞内ポリペプチドを再ターゲティングできることを示している。
【0206】
実施例4
哺乳類細胞内の内部抗体の細胞内特異性を分析する方法
本実施例において、標的抗原を発現する哺乳類細胞において内部抗体の特異性を分析する方法を示す。
【0207】
抗ハンチンチン内部抗体(α-Nt-HD-C4 sFv)の特異性を証明するために、哺乳類細胞(COS-7)をモデルHD標的ポリペプチドと、α-Nt-HD-C4 sFvまたは無関係なポリペプチドに対して特異的な内部抗体のいずれかと同時トランスフェクトした。モデルHD標的ポリペプチドはc-myc(EQKLISEEDL(配列番号:45))エピトープタグ(HD-Q104-myc)に融合した。結合した抗原が細胞核に再ターゲティングされるように、内部抗体配列をHAタグおよびSV40核局在シグナル(sFv-HA-NLS)と融合させた。トランスフェクションの48時間後、細胞を固定して透過性にした。HD-Q104-myc蛋白質は、抗mycエピトープ抗体(すなわちMAb 9E10 IgG)の後にFITC標識ヤギ抗マウスIgGを用いて検出可能であった。sFv-HA-NLS内部抗体は、ポリクローナルウサギ抗HA IgGの後にローダミン結合ヤギ抗ウサギIgG抗体によって検出した。
【0208】
図12A〜Fに示す実験結果は、双方の内部抗体が細胞核に存在するが(図12A〜B)、α-Nt-HD-C4 sFv-HA-NLSのみがモデル変化ハンチンチンポリペプチドHD-Q104-mycを細胞質から核に再配置できることを示している(それぞれ、図12D、12F対12C、12E)。さらに、対照内部抗体は、HD-Q104抗原の凝集を防止しなかった(パネルCおよびEの矢印の先で示す)。
【0209】
細胞内にターゲティングした抗原に対する内部抗体の特異性を証明するもう一つの方法は、ターゲティングした抗原または無関係な抗原のいずれかと共に特異的内部抗体を同時発現する細胞を分析することである。図13A〜Fは、グルタミン反復配列104個を有するモデル変化ハンチンチンポリペプチド(図13B、D、F)またはグルタミン反復配列81個を有するモデル変化DRPLAポリペプチド(図13A、C、E)のいずれかをコードするプラスミドによってさらにトランスフェクトした、α-Nt-HD-C4 sFv-HA-NLSを発現する安定にトランスフェクトしたCOS-7細胞を示している。双方のモデル変化ポリペプチドをGFPに融合した(それぞれ、GFP-HD-Q104およびGFP-DRPLA-Q81)。抗Nt-HD-C4 sFv-HA-NLS内部抗体は、GFP-HD-Q104のみが細胞核内部で内部抗体によって顕色されたために、明らかに、GFP-HD-Q104(図13D、F)に対する特異性を示すが、GFP-DRPLA-Q81(図13C、E)に対する特異性を示さない。
【0210】
これらの方法によって、関連する細胞内環境で内部抗体特異性を迅速に評価することができ、本明細書において考察した抗原に適用できるのみならず、他の内部抗体/ターゲティング抗原対を調べるために用いてもよい。
【0211】
実施例5
哺乳類の脳組織におけるポリペプチド凝集をアッセイする方法
本実施例では、哺乳類の脳における変異型ハンチンチンポリペプチドが細胞内凝集体を形成できることを検出する方法を示す。
【0212】
変化したハンチンチンポリペプチドが哺乳類の脳において細胞内凝集体を形成できることを証明するために、マウス(P12 Balb)からの小脳切片に、正常および変化したハンチンチンポリペプチドを表すポリペプチドをコードするプラスミドを遺伝子銃によってバイオリスティックにトランスフェクトして、24時間調べた。グルタミン残基104個を有するモデル変化ハンチンチンGFPポリペプチド(HD-104Q-GFP)をコードするプラスミドを24時間培養物にトランスフェクトすると、輝く蛍光点として示される大きい凝集体を脳組織全体に認めた。同様に、グルタミン残基72個を有する変化したハンチンチンGFPポリペプチド(HD-72Q-GFP)をコードするプラスミドを24時間培養物にトランスフェクトすると、輝く蛍光点によって示されるより小さく、より明確な凝集体が認められた。
【0213】
さらに、このアッセイはまた、モデルハンチンチンポリペプチドを発現する特定の細胞タイプの同定能を提供する。例えば、プルキンエ細胞は、カルビンジンに対する抗体を用いて同定することができ、ニューロンは神経細糸に対する抗体を用いて可視化することができる。したがって、これらの細胞特異的抗体を可視化するために赤色標識を用いることによって、HD-ポリQ-GFP融合蛋白質のGFP部分に対して陽性である細胞も同時に同定することができる。次に、切片をレーザー共焦点顕微鏡を用いて調べる。
【0214】
重要なことは、例えば、実施例2に示す方法を用いて、このアッセイは、内部抗体が細胞内ポリペプチド凝集(例えば、ハンチンチンポリペプチドの)の形成を阻害できるか否か、または多数の細胞タイプを有する脳組織においてそのようなポリペプチドを再ターゲティングできるか否かをアッセイするように適合させることができる。さらに、このアッセイは、ポリペプチド凝集体の形成を破壊することができる他の結合分子のスクリーニング能を提供する。最後に、このアッセイは、長期間培養して、本明細書に記載のようにアッセイするために特定の神経細胞型(例えば、線条体細胞)を単離するために用いてもよい。
【0215】
実施例6
神経ポリペプチドに対するsFvファージライブラリを作製する方法
本実施例において、DNAワクチン接種を用いて選択された抗原に対して内部抗体ファージライブラリを作成する方法を示す。
【0216】
広範囲の可能なポリペプチドに対して特異的な内部抗体を作製することができるように、DNAワクチン接種戦略を開発した。この方法は、「本来の」ライブラリにおいてまれな内部抗体を選択するより、特定のエピトープに対して「向けられた」ライブラリを作製する長所を有する。したがって、スクリーニングしなければならないクローン数は大きく減少する。得られたファージディスプレイライブラリが、蛋白質がインビトロで取りうる形のみに対するエピトープよりむしろ、その細胞内または細胞内構造のエピトープに対する抗体を発現するように活性化された脾臓から作製することができるように、蛋白質の代わりに、DNAを用いてマウスを免疫した。
【0217】
ハンチンチンの最初のアミノ酸17個、異なる長さのポリグルタミン反復配列、および改変された緑色蛍光蛋白質を含む融合蛋白質を発現するプラスミド構築物を用いて、雌性マウス8匹(BalbCBy/J遺伝的背景)においてインビボで抗原を発現させた。GFPに融合したモデルハンチンチンN末端をコードし、正常数のグルタミン残基またはハンチントン病に関連した変化した残基数のいずれかを有するプラスミド構築物を、正常生理食塩液媒体においてマウスに0.5 μg/μlの濃度で注入した。
【0218】
免疫前血清を得るために、免疫する前にマウスから血液を採取した。全てのマウスにプラスミド構築物の一つを1日目(プラスミドDNA 50 μgを筋肉内注射、プラスミドDNA 50 μgを皮内注射−合計100 μg)に注射した。注射は14日目に繰り返してマウスを21日目に採血した。次に、マウス血清を、血清の希釈を変化させて、pHD-25QトランスフェクトCOS7細胞溶解物のイムノブロットをプロービングすることによって陽性液性応答に関してスクリーニングした。陽性反応マウスの刺激された脾臓は、モノクローナル抗体、内部抗体、およびファージディスプレイを作製するための候補物質であると決定された。
【0219】
注射したマウス8匹中、50%(8匹中4匹)が、最小の血清希釈100倍でイムノブロットアッセイ法における融合蛋白質を検出することができる陽性抗体反応を示した(表5を参照)。これらの中で、75%(4匹中3匹)がグルタミン反復配列104個の構築物によって免疫された。
【0220】
【表5】
【0221】
このように、これらの結果は、本明細書に記載の方法を用いて抗原特異的内部抗体結合ドメインを、上記の原料、そして例えば特異的抗体結合分子を作製するためにファージディスプレイライブラリを用いて操作できることを示している。その上、これらの方法は、実質的に他のいかなる当該抗原にも容易に適用できる。
【0222】
実施例7
哺乳類における神経疾患の内部抗体阻害を検定する方法
本実施例において、内部抗体が生きている哺乳類において神経疾患を治療または治癒できるか否かを検定する方法を示す。
【0223】
内部抗体が生きている哺乳類における神経疾患に関連した変化した神経ポリペプチド凝集体の形成を阻害することができるか否かを決定するために、トランスジェニックマウスを用いる方法が開発されている。
【0224】
特に、ハンチントン病およびSCAの動物モデルが開発されている(例えば、表2を参照のこと)。
【0225】
例えば、変化したハンチンチンポリペプチドを発現するように遺伝子操作されたトランスジェニックマウスのコロニーが確立されており(ベイテス(Bates)ら、(3)(29)に記載のように)、これらの動物はハンチントン病のいくつかの神経学的特徴を示す。マウスは症候性であり、8週齢までに、皮質、線条体および小脳のプルキンエ細胞において神経ハンチンチンポリペプチド凝集体を示し始める。
【0226】
さらに、変化したアタキシン発現によって引き起こされるSCA神経疾患を表す第二の動物モデルが確立されている。これらの動物は、プルキンエ細胞特異的プロモーターの制御下で発現される拡大された反復配列アタキシン遺伝子を有する。これらの動物は、12週までに症状を示し、4週までにプルキンエ細胞において最初の組織学を示す。
【0227】
なお第3のアプローチにおいて、変化したアトロフィン1によって引き起こされるDRPLAを表す動物モデルを確立することができる(シリング(Schilling)ら、Neuron 24:275〜286(1999))。
【0228】
したがって、これらの動物は、内部抗体が神経ポリペプチド凝集体の形成を阻害して、このように疾患の発症を予防、治療、または遅らせることができるか否かを調べる3つのインビボアッセイ系となる。例えば、内部抗体はポリペプチドとして、または本明細書に記載のいかなる方法も用いて発現可能な核酸構築物として頭蓋内に輸送してもよい。
【0229】
さらに、内部抗体が神経疾患の発症を予防、治療、または遅らせるか否かを証明するさらにもう一つの方法において、これらの動物を条件によって発現することができる内部抗体を発現する動物と交配させることができる。このように、疾患の臨床経過のあいだの異なる時点で、内部抗体をそのインビボ有効性に関して調べてもよい。特に、ネスラー(Nestler)とその共同研究者らのTet調節系を用いることができ、それは、2つのトランスジェニックマウス系を含み、一方は神経組織における発現を指示するニューロン特異的エノラーゼ(NSE)プロモーターの制御下でTet調節トランス活性化因子(tTA)を含み、もう一方は当該遺伝子、例えば抗ハンチンチン内部抗体(すなわち、α-Nt-HD-C4 sFv)、またはTet調節プロモーターの下流にクローニングされた抗アタキシン内部抗体(TetOp)を有する構築物を含む(チェン(Chen)ら、Mol. Pharmacol. 54:495〜503(1998);アーノルド(Arnold)ら、PNAS 94:8842〜8847(1997);およびクラーク(Clark)ら、J. of Neurosci. 17:7325〜7395(1997)))。選択された内部抗体の発現を活性化するために、内部抗体遺伝子と変化した神経ポリペプチド(例えば、ハンチンチンまたはアタキシンのいずれか)を有する子孫の飲料水からドキシサイクリン(Tet誘導体)を除去する。
【0230】
交配スケジュールは以下の通りである:第一に、双方のトランスジーンの存在がゲノムDNAのPCRによって決定され、遺伝子の発現がイムノブロット法によって確認される、NSE-tTAおよびHDトランスジーンの双方を有する二遺伝子マウスを作製する。これらのマウスを、ニューロンにおけるTetOpイントラC4プラスミドの発現を容易にする脳切片培養源としてまず用いることができ、後にこれをTetOpトランスジェニックマウスと交配させる。
【0231】
NSE-tTA脳切片を用いて、HDポリQおよびTetOpイントラC4の双方をトランスフェクトさせて、発現された内部抗体が形成された凝集体の数を減少させるように機能するか否かをアッセイすることができる(例えば、凝集体をライソゾーム分解されるようにターゲティングする)。次に、成功したプラスミドを注入してトランスジェニック創始マウスを作製することができる。これらを二世代にわたって交配することができ、内部抗体の非誘導レベルに関して試験することができる。
【0232】
トランスジーン発現を完全に阻害するために、ドキシサイクリンを25 μg/mlの低用量で動物に持続的に投与することができる(チェン(Chen)ら、Mol. Pharmacol. 54:495〜503(1998))。この低用量では、ドキシサイクリンの除去による内部抗体の誘導は迅速となりうる。疾患進行のどの段階で内部抗体の治療が最も有効であるかを調べるために、ドキシサイクリンを様々な年齢群のHD-NSE-tTA×TetOpイントラC4マウスの飲料水から4週目、6週目、8週目、および10週目に除去することができる。これらの動物からの脳を摘出して、内部抗体治療の有効性の指標としてその野生型子孫と比較して凝集体の有無および数をアッセイすることができる。
【0233】
上記のように、これらの実験を用いて、HDトランスジェニックマウスによって抗HD内部抗体を調べ、SCA1マウスによって抗アタキシン内部抗体を調べ、双方の系統のマウスによって抗ポリグルタミン内部抗体を調べることができる。二遺伝子NSE-tTA/HDまたはアタキシンマウスを、特異的およびより一般的な内部抗体交配の双方のために用いることができる。
【0234】
したがって、本方法によって、インビトロアッセイにおいて同定された内部抗体の最も有効なものをインビボで決定することができる。さらに、これらの方法によって、HD(またはSCA1マウス)において神経症状の行動学的救出を認めるために必要なレベルまで凝集体の毒性を減少させるか否かを決定することができる。最後に、これらの方法によって、内部抗体発現の誘導が最も有効であるのは疾患進行のどの段階であるかも決定することができる。
【0235】
実施例8
望ましくない細胞内ポリペプチド複合体に対する治療的宿主抗体免疫応答を誘発するための核酸ワクチンのインビボ証明
本実施例において、望ましくない細胞内ポリペプチド凝集体に対して宿主によって内部抗体を誘発する方法および組成物を示す。
【0236】
免疫系によって作製された内部抗体が、外から投与される代わりに、神経疾患に関連した変化した神経ポリペプチド凝集体の形成を阻害するか否かを決定するために、病原性細胞内ポリペプチドを表すポリペプチド抗原、すなわちハンチンチンポリペプチドをコードするDNAワクチンによって試験動物を免疫した。対照と比較して、試験ポリペプチドをコードするDNAワクチンは、免疫した動物において治療的液性応答を誘発し、その結果内因性細胞内ハンチンチンポリペプチドと共にDNAコードハンチンチン免疫原に対する宿主内部抗体(すなわち、抗体)の形成が起こる。
【0237】
簡単に説明すると、実験アプローチは以下の通りであった。グルタミン残基約165個を含む(正常は35コピー未満)変異型ヒトハンチンチンの断片を発現する試験動物(HDR6/2トランスジェニックマウス)を得た。これらのマウスは、5週齢までに測定可能な微妙な行動学的および神経化学的変化を示し、重度の神経疾患は11〜12週で明らかになった。15週を超える生存はまれである。重要なことは、これらのマウスは同様に、絶食時の血清グルコースレベルの増加から証明されるように(10週齢で)、ヒトにおいて認められるものと類似のハンチンチン関連糖尿病症候群を発症する。ハンチンチン関連糖尿病は、膵臓の島細胞における変異型HD蛋白質の異常な蓄積に帰因している。
【0238】
この実験に関して、本明細書に示したこれまでの実験において内部抗体を選択するために用いたHD蛋白質断片の一部に対する抗体を産生するためにマウスを免疫した。特に、HDR6/2マウスを5週齢時に、緑色蛍光蛋白質(GFP)に結合したHDトランスジーン断片を発現するプラスミド100 μgを注射することによって免疫して、7週齢時に追加免疫した。2回の注射は同じ週齢で行い、一方は脚の筋肉に、そしてもう一方は尾の真下の皮内に行った。8週目にマウスから採血して、当初の注射プラスミドのトランスフェクションの24時間後にHSK-293細胞の溶解物から調製したイムノブロットを用いて反応性を決定した。
【0239】
次に、陽性免疫応答対反応しない免疫応答を示したHDマウスについて、血液中グルコースレベルを比較した。血中グルコースは、6時間の絶食後にコンフォートカーブ試験片を備えたアドバンテージグルコメータを用いて1試料あたり2本ずつ実施した。無処置トランスジェニックHDマウスの絶食時の血中グルコースレベルは、10週齢から始めた野生型マウスのレベルより有意に高かった。プラスミド免疫に反応しないトランスジェニックマウス、またはGFP配列のみを含むプラスミドによって免疫したマウスは、無処置マウスの範囲のレベルを示した。しかし、プラスミドを免疫してイムノブロット法によってアッセイして反応性であったマウスは、血中グルコースレベルが有意に低下し、13および14週齢では完全に野生型マウスの範囲内であった。このように、HD免疫は、膵臓の表現型の発生を防止しうる(図16参照)。
【0240】
グルコースレベルの他に、インスリンをコードするmRNAの膵臓レベルも同様に測定した。一連のマウスを14週目に屠殺して、それぞれの膵臓からRNAを抽出した。試料を遺伝子型、治療、および反応に基づいてプールした。インスリンmRNAレベルは、逆転写ポリメラーゼ連鎖反応(RT-PCR)法によって決定した。野生型マウスからの試料は、RT-PCR反応法において用いられるプライマーに基づいて予測される位置で強いバンドを示した。無処理および非反応マウスは、同じサイズのかすかなバンドのみを示し、インスリン生合成の欠損を示した。免疫して反応性であったマウスは、密度測定法によって確認すると、野生型の強度に近いインスリンmRNAを示すバンドを示した。このように、治療的改善は、インスリンmRNAレベルの回復であるように思われる。
【0241】
最後に、DNAワクチン接種に反応するマウスからの血清を、上記の治療的内部抗体を選択するために用いられるポリペプチド抗原に結合する抗体を動物が産生したか否かを決定するために調べた。したがって、ハンチンチンの最初のアミノ酸17個に対応するペプチドを合成して、試験皿に固定した。次に、陽性反応者に存在するエピトープのELISA試験を行った;イムノブロット法によって陽性であり、異常なグルコースレベルの低下とインスリンmRNAの増加の双方を誘発したマウス血清は同様に、アミノ酸17個のペプチドに対する結合を示した。
【0242】
要約すると、これらの知見は、例えば、凝集または蓄積による変異型ポリペプチド機能による病態を予防または逆転させるために細胞内で作用する能力を有する免疫によって産生された抗体と一致する。
【0243】
引用文献の詳述
【0244】
同義
当業者は、日常的な実験のみを用いて、本明細書に記載の本発明の特定の態様に対する多くの同等物を認識、または確認できるであろう。そのような同等物は添付の特許請求の範囲に含まれると解釈される。
【図面の簡単な説明】
【0245】
【図1】病的な長さのポリグルタミン(すなわち、ポリQ)に変異した場合の、ポリグルタミンに富み、神経疾患に関連するいくつかのポリペプチドの野生型N末端アミノ酸配列を示す。比較のために、神経疾患に関連しないポリグルタミン鎖を有するTBPポリペプチドを示す。ハンチンチン特異的C4 sFv内部抗体を作製するために用いたペプチドも同様に示す。
【図2】特定の細胞内位置に異種ポリペプチドをターゲティングすることができる多くのターゲティングシグナルのアミノ酸配列を示す。血液凝集素(HA)タグは、抗原とは個別に内部抗体を可視化するために用いられる免疫染色法のために、適当な抗体に強く結合するペプチドである。
【図3】蛍光顕微鏡を用いて、グルタミン残基の数が増加している(すなわち25、47、72、104個)モデルハンチンチンGFP融合ポリペプチド(すなわち、HD-25Q-GFP、HD-47Q-GFP、HD-72Q-GFP、HD-104Q-GFP)の長さ依存的凝集を示す。細胞において発現されると(この図においてCOS-7)、それぞれグルタミン残基72および104個を有するモデルハンチンチンGFP融合蛋白質(HD-72Q、HD-104Q)は、小さく輝く蛍光斑に容易に凝集する(図3C〜D)。対照的に、グルタミン残基が野生型の数(25個)である(HD-25Q)モデルハンチンチンGFPポリペプチドは、凝集しないままであり、散在した細胞質GFP蛍光を示すに過ぎないか、またはポリQの長さがわずかに病的なポリペプチド(HD-47Q)では、凝集体を形成する傾向を認めるが、多くの細胞が細胞全体に均一に分布する抗原を維持している(図3A〜B)。
【図4】蛍光顕微鏡を用いて、グルタミン残基72および104個を有するモデルハンチンチンGFP融合ポリペプチド(pHD-72Q、図3B;pHD-104Q、図3A)が、マウスに由来する小脳切片において発現され、遺伝子銃を用いてバイオリスティックにトランスフェクトすると、小さく強い染色領域に凝集できることを示す。
【図5A−D】内部抗体が、細胞においてモデルハンチンチンポリペプチドに特異的に結合してこれを再ターゲティングできることを示す。図5Aおよび5Bでは、位相差光学顕微鏡を用いた細胞の分布を示す。図5C、5E、および5Gは、蛍光顕微鏡を用いて、モデルハンチンチンGFP融合ポリペプチド(HD-25Q-GFP)と、モデルハンチンチンポリペプチドに特異的に結合する内部抗体とを同時発現して、核ターゲティングシグナルを含む細胞(αNt-HD-C4 sFv-NLS)(図5E)が、核における染色の集合によって証明されるように、モデルハンチンチンポリペプチドが核に分布するように再ターゲティングできることを示している(図5G)。対照的に、無関係なポリペプチドに結合する内部抗体を発現する細胞は、モデルハンチンチンポリペプチドの分布を再ターゲティングすることができない(図5D、5F、およびFH)。
【図5E−H】内部抗体が、細胞においてモデルハンチンチンポリペプチドに特異的に結合してこれを再ターゲティングできることを示す。図5Aおよび5Bでは、位相差光学顕微鏡を用いた細胞の分布を示す。図5C、5E、および5Gは、蛍光顕微鏡を用いて、モデルハンチンチンGFP融合ポリペプチド(HD-25Q-GFP)と、モデルハンチンチンポリペプチドに特異的に結合する内部抗体とを同時発現して、核ターゲティングシグナルを含む細胞(αNt-HD-C4 sFv-NLS)(図5E)が、核における染色の集合によって証明されるように、モデルハンチンチンポリペプチドが核に分布するように再ターゲティングできることを示している(図5G)。対照的に、無関係なポリペプチドに結合する内部抗体を発現する細胞は、モデルハンチンチンポリペプチドの分布を再ターゲティングすることができない(図5D、5F、およびFH)。
【図6A−C】内部抗体の特定のポリペプチド位置の再ターゲティング能が、内部抗体の結合特異性の関数であることを示す。例えば、図6C〜Gにおいて、グルタミンに富むDRPLAポリペプチド(GFP-DRPLA-35Q)と、モデルハンチンチンポリペプチド(H-HD-C4 sFv;図6D、6F)またはもう一つの無関係なポリペプチド(陰性対照sFv;図6C、6E、および6G)のいずれかに対する内部抗体とを同時発現する細胞(COS-7)は、DRPLAポリペプチドの細胞分布に変化を示さない。図6Aおよび6Bは、位相差光学顕微鏡を用いて細胞の全体的な分布を示す。
【図6D−G】内部抗体の特定のポリペプチド位置の再ターゲティング能が、内部抗体の結合特異性の関数であることを示す。例えば、図6C〜Gにおいて、グルタミンに富むDRPLAポリペプチド(GFP-DRPLA-35Q)と、モデルハンチンチンポリペプチド(H-HD-C4 sFv;図6D、6F)またはもう一つの無関係なポリペプチド(陰性対照sFv;図6C、6E、および6G)のいずれかに対する内部抗体とを同時発現する細胞(COS-7)は、DRPLAポリペプチドの細胞分布に変化を示さない。図6Aおよび6Bは、位相差光学顕微鏡を用いて細胞の全体的な分布を示す。
【図7A−D】蛍光顕微鏡を用いて、内部抗体が変異型モデルハンチンチンポリペプチド(すなわち、グルタミン反復配列104個を有する)の細胞内分布を核に特異的に再ターゲティングできるが(図7C、7E、および7F)、無関係な内部抗体は再ターゲティングできないことを示している(図7B、7D、および7G)。図7Aおよび7Bでは、位相差光学顕微鏡を用いた細胞の分布を示す。
【図7E−H】蛍光顕微鏡を用いて、内部抗体が変異型モデルハンチンチンポリペプチド(すなわち、グルタミン反復配列104個を有する)の細胞内分布を核に特異的に再ターゲティングできるが(図7C、7E、および7F)、無関係な内部抗体は再ターゲティングできないことを示している(図7B、7D、および7G)。図7Aおよび7Bでは、位相差光学顕微鏡を用いた細胞の分布を示す。
【図8】蛍光顕微鏡を用いて、細胞内ポリペプチド凝集体の形成が内部抗体の発現によって阻害されたことを示す。ヒトにおいて神経疾患に関連することが知られている変異を有するハンチンチンポリペプチドを表す異なる2つのポリペプチドは、細胞において発現されると、細胞内凝集体を形成することが示された(図8Aおよび8E)。これらの凝集体の形成は、ターゲティングシグナルを有しない(図8B、8Fを参照)、核ターゲティングシグナルを有する(図8C、8Gを参照)、またはライソゾームターゲティングシグナルを有する(図8D、8G)異なる3つの内部抗体によって阻害された。図8C〜D、8F〜Gはまた、ターゲティングシグナルを有する(核またはライソゾーム)内部抗体は、細胞内ポリペプチド凝集体を減少させたのみならずに、モデルハンチンチンポリペプチドの再ターゲティングを引き起こしたことを示している。
【図9】モデルハンチンチン特異的C4 sFv内部抗体の結合特異性を示すために用いられた様々なポリペプチドの存在および見かけの分子量を示すクーマシー染色電気泳動ゲルの写真を示す(図10のイムノブロットによってプロービングする)。
【図10】ハンチンチンポリペプチドエピトープ(レーン1〜3、6〜7)のみが、モデルハンチンチンポリペプチド特異的C4 sFv内部抗体をアフィニティ生成するために用いることができるが、無関係なDRPLAポリペプチド(レーン2および5)は、C4 sFv内部抗体と相互作用できないことを示すイムノブロットである。
【図11】AはELISA法によって分析した場合の、固定GST融合蛋白質に対する抗ハンチンチン内部抗体C4 sFvの直接結合試験の結果を示す。 BはBIAコアによって分析した場合の、抗ハンチンチン内部抗体C4 sFvの速度論的結合アフィニティ試験の結果を示す。抗ハンチンチン内部抗体C4 sFvの濃度範囲(60〜100 nM)を用いて、ストレプトアビジンセンサーチップに結合したビオチン結合HDペプチド50 RU上で結合定数(kon)を測定した。
【図12A−D】抗ハンチンチン内部抗体Nt-HD C4 sFv-NLSによるGFP融合HD断片の核への再ターゲティングを示す。パネルB、D、およびFは、COS-7細胞において48時間同時トランスフェクションした後の、HD-Q104-Myc-HIS6およびC4 sFv-NLSまたは陰性sFv-NLS内部抗体(パネルA、C、およびE)の免疫蛍光を示す。HD-Q104-Myc-HIS6は、抗myc 9E10 mAbによる免疫染色の後、FITC標識ヤギ抗マウスIgG抗体による免疫染色によって可視化した(パネルA、B)。内部抗体は、ポリクローナル抗HA抗体の後、ローダミン標識ヤギ抗ウサギIgG抗体によって免疫染色した(パネルC、D;パネルEおよびFは同じ領域の二重染色を表す)。
【図12E−F】抗ハンチンチン内部抗体Nt-HD C4 sFv-NLSによるGFP融合HD断片の核への再ターゲティングを示す。パネルB、D、およびFは、COS-7細胞において48時間同時トランスフェクションした後の、HD-Q104-Myc-HIS6およびC4 sFv-NLSまたは陰性sFv-NLS内部抗体(パネルA、C、およびE)の免疫蛍光を示す。HD-Q104-Myc-HIS6は、抗myc 9E10 mAbによる免疫染色の後、FITC標識ヤギ抗マウスIgG抗体による免疫染色によって可視化した(パネルA、B)。内部抗体は、ポリクローナル抗HA抗体の後、ローダミン標識ヤギ抗ウサギIgG抗体によって免疫染色した(パネルC、D;パネルEおよびFは同じ領域の二重染色を表す)。
【図13A−D】抗ハンチンチン内部抗体C4 sFv-NLSを安定に発現するCOS-7細胞におけるGFP-HD-Q104(b、d、f;40倍)またはGFP-DRPLA-Q81(a、c、e;陰性対照として;20倍)の再配置を示す。パネルAおよびBは、GFP発現を示し、パネルCおよびDはC4 sFv-NLS内部抗体のローダミン染色を示し、そしてパネルEおよびFは、同じ領域の二重染色を示す。
【図13E−F】抗ハンチンチン内部抗体C4 sFv-NLSを安定に発現するCOS-7細胞におけるGFP-HD-Q104(b、d、f;40倍)またはGFP-DRPLA-Q81(a、c、e;陰性対照として;20倍)の再配置を示す。パネルAおよびBは、GFP発現を示し、パネルCおよびDはC4 sFv-NLS内部抗体のローダミン染色を示し、そしてパネルEおよびFは、同じ領域の二重染色を示す。
【図14】病原性の長さのHD-ポリQ-GFP凝集体の抗HD-C4 sFv阻害を示す。上のパネルは、プラスミドトランスフェクション比5:1(sFv:HD)でHD-ポリQ-GFP(Q=25、72、および104)単独またはC4または対照sFv(ML3〜9)をトランスフェクトしたBHK-21細胞のデジタル蛍光顕微鏡画像を示す。リン酸カルシウム法によるトランスフェクションの48時間後に生存して撮影した細胞を示す;凝集体は、GFPシグナルの強い斑として見える。類似の結果がHD-72Q-GFPの同時トランスフェクションについても認められた。下のパネルは、COS-7、BHK-21、およびHEK293細胞におけるsFv(C4またはML3-9)または親ベクター(pcDNA)とHD-72Q-GFPとの5:1同時トランスフェクション後の凝集体の定量を表す。同時トランスフェクションの48時間後に、細胞を2%SDS/2%トライトンX100/50 mMトリスによって溶解した。無作為な視野6または8個における不溶性の蛍光凝集体の数を計数した。バーは、同時トランスフェクション6個の平均値±SEMを表す。C4は、対照ML3-9またはpcDNAと比較すると、凝集体の数を有意に減少させた(p<0.0005)。類似の結果はHD-104Q-GFPについても得られた。
【図15】病原性の長さのHD-ポリQ-GFP凝集体の抗HD C4 sFv阻害を示す。上のパネルは、C4:HD-ポリQ-GFP HD-ポリQ-GFP(緑)、C4 sFv(赤、抗HA抗体およびアレキサ568結合二次抗体によって検出される)を5:1で同時トランスフェクトした二重標識細胞のデジタル蛍光顕微鏡画像、および合併画像を示す。104Q同時トランスフェクト体にはいくつかの凝集体を認め(25Qでは認めない)、C4を発現していない細胞ではしばしば認めるが(矢印の先)、いくつかの二重トランスフェクト細胞においても認めた(矢印)。下のパネルは、同時トランスフェクト体におけるHD-72Q-GFP蛋白質発現のイムノブロット分析のデジタル画像を示す。細胞溶解物を24時間目に(顕著な凝集体形成と、不溶性区画にHD-72Q-GFPが失われる前)5:1同時トランスフェクト体(sFv:HD-72Q-GFP)から1試料あたり2本ずつ採取してSDS-PAGE法を行った。1000倍希釈した抗AFP一次抗体(クアンタム社)をプローブとしてメンブレンを調べ、剥離して、500倍希釈抗アクチン抗体(シグマ社)によって再プロービングした。HD-72Q-GFPバンドは、HRP結合二次抗体と化学発光による検出後の強度と類似であった。
【図16】ハンチンチンエピトープをコードするDNAワクチンによって処理した、および処理していないマウス(野生型およびトランスジェニックハンチンチン系(HD))におけるグルコースレベルを示す。
【技術分野】
【0001】
関連情報
本出願は、参照として本明細書に組み入れられる、1999年7月27日に提出された「神経障害に関連したポリペプチド蓄積を阻害する方法と組成物」と題する米国特許仮出願番号第60/146,047号の優先権を主張する。本明細書において引用した特許、特許出願、および参考文献は全て、その全文が参照として本明細書に組み入れられる。
【0002】
政府助成研究
本発明は、その一部を米国保健省と遺伝疾患基金の助成金(第NS38002号)の補助を受けた。
【背景技術】
【0003】
発明の背景
神経変性障害は、人類を襲う最も怖れられている疾患に属する。例えば、年齢65歳以上の10人に1人がアルツハイマー病に罹患すると推定されているが、この疾患は、記憶の喪失と、最終的に死に至る徐々に衰弱する疾患である。実際、現在のところ、北米において400万人以上の人々がこの疾患に罹患しており、現在の治療は本質的に待期的である。同様に、パーキンソン病は、北米においてほぼ100万人に罹患しており、現在、疾患の発病を遅らせる、または実質的に疾患の進行を遅らせるために利用できる治療はない。もう一つの潜伏性の神経障害はハンチントン病(HD)であり、これは現在、北米において30,000人以上が罹患している。あまり一般的ではないが、等しく衰弱するのが、新しい変異型クロイツフェルトヤコブ病のようなプリオン疾患であり、これも同様に精神の痴呆と最終的に死に至る。前頭側頭痴呆(FTD)、筋萎縮性側索硬化症(ALS)、脊髄延髄筋萎縮(SBMA、またはケネディ病)、歯状赤核淡蒼球萎縮症(DRPLA)、および脊髄小脳運動失調(例えば、SCA1〜SCA7)のような他の多くの神経変性疾患も同様に、類似の経過を示す。これらの疾患の多くの病因はなおも完全には理解されていないが、これらの疾患のほとんどが神経系の細胞において変化したポリペプチドが産生されたことが原因である可能性があるように思われる(例えば、非特許文献1、非特許文献2、非特許文献3、および非特許文献4、非特許文献5を参照のこと)。
【0004】
例えば、ハンチントン病は、主に皮質と線条体に起こる選択的な神経細胞死に関連する(非特許文献6、非特許文献7)。障害は、ハンチンチンポリペプチドにおいてポリグルタミン伸長部をコードするht遺伝子の最初のエキソンにおける伸長CAG反復配列によって引き起こされるように思われる;これは、ヒト細胞において広汎に認められる機能不明の大きい〜350 kDaのポリペプチドであるが、皮質と線条体細胞においてその最高濃度が存在する(非特許文献8)。CAG反復配列は非常に多形性であり、非罹患者の染色体では6〜39反復まで変化し、HD患者の染色体では36〜180反復まで変化する(非特許文献9、非特許文献10)。成人発症例の大多数は、40〜55単位の伸長部を有するが、伸長部が70およびそれ以上になると、疾患の若年性型を必ず引き起こす。脳において、ハンチンチンポリペプチドは、主にニューロンに認められて、主として細胞質ポリペプチドであり、その分画は、小胞および/または微小管に関連しており、このことは、これが細胞骨格接着または小胞の輸送において機能的役割を有する可能性があることを示唆している(非特許文献11、非特許文献12、非特許文献13)。ハンチンチンはまた、核にも検出され(非特許文献14、非特許文献15)、このことは、転写の調節がこのポリペプチドの可能性のある機能として除外できないことを示唆している。ハンチンチンはまた、膵臓の細胞にも認められ、この場合、ハンチンチンにおける病的に伸長した反復配列の存在は、ハンチントン病に関連した糖尿病の発症に関連している(非特許文献16、非特許文献17)。
【0005】
HDの他に、CAG/ポリグルタミン伸長部は、脊髄延髄筋萎縮症、歯状赤核淡蒼球萎縮症、ならびに1、2、3および6型脊髄小脳運動失調を含む他の少なくとも6つの遺伝性神経変性障害において認められている(非特許文献18、非特許文献19)。正常で伸長した大きさの範囲はSCA6を例外として同等であり、SCA6の場合、伸長した対立遺伝子がより小さく、変異がおそらく異なる経路によって作用する可能性がある。しかし、全ての場合において、CAG反復配列は、コード領域内に存在して、ポリグルタミン残基の鎖に翻訳される。ポリグルタミン配列を有するポリペプチドは無関係で、ほとんどの機能が不明であるが、変異は、類似のメカニズムを通じて作用する可能性がある(非特許文献20)。これらのポリペプチドは通常、広く発現されて、一般的に核および細胞質の双方に存在する。さらに、アルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆、筋萎縮性側索硬化症、脊髄延髄筋萎縮症、歯状赤核淡蒼球萎縮症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、4型脊髄小脳運動失調、5型脊髄小脳運動失調、6型脊髄小脳運動失調、および7型脊髄小脳運動失調を含む神経変性疾患はまた、神経系の細胞における変化したポリペプチドの産生にも関連している(非特許文献3)。
【0006】
したがって、神経系の細胞における変化したポリペプチドの蓄積が、細胞のサブセットの神経変性および疾患の発症の前兆となる可能性があると推測されている。
【非特許文献1】ハートレー(Hurtley, S.)、Science 282:1071(1998)
【非特許文献2】ショールソン(Shoulson, I.)、Science 282:1072〜1074(1998)
【非特許文献3】ハーディ(Hardy)ら、Science 282:1075〜1079(1998)
【非特許文献4】プライス(Price)ら、Science 282:1079〜1083(1998)
【非特許文献5】クレメント(Klement)ら、Cell 95:41〜53(1998)
【非特許文献6】ハーパー(Harper)、「ハンチントン病(Huntington's Disease)」、第22版、ソーンダース社(1991)
【非特許文献7】フォンサッテル(Vonsattel)ら、J. Neuropath. Exp. Neurol. 44:559〜577、(1985)
【非特許文献8】ハンチントン病共同研究グループ、Cell 72:971〜983(1983)
【非特許文献9】ルビンスタイン(Rubinsztein)ら、Am. J. Hum. Genet. 59:16〜22(1996)
【非特許文献10】サタシバム(Sathasivam)ら、Hum. Genet. 99:692〜695(1997)
【非特許文献11】ジフィグリア(DiFiglia)ら、Neuron 14:1075〜1081(1995)
【非特許文献12】グテクンスト(Gutekunst)ら、P.N.A.S. 92:8710〜8714(1995)
【非特許文献13】シャープ(Sharp)ら、Neuron 14:1065〜1074(1995)
【非特許文献14】フーゲベーン(Hoogeveen)ら、Hum. Mol. Genet. 2:2069〜2073(1993)
【非特許文献15】デルーイ(de Rooij)ら、Hum. Mel. Genet. 5:1093〜1098(1996)
【非特許文献16】ハールバート(Hurlbert)ら、Diabetes 48:649〜651(1999)
【非特許文献17】ファラー(Farrer, L.)、Clin. Genet. 27:62〜67(1985)
【非特許文献18】ベイテス(Bates)ら、Human Mol. Genet. 6:1633〜1637(1997)
【非特許文献19】トロッティア(Trottier)ら、Nature 378:403〜406(1995)
【非特許文献20】シソディア(Sisodia, S.)、Cell 95:1〜4(1998)
【発明の開示】
【0007】
発明の概要
細胞内ポリペプチドの蓄積を異常な多量体または凝集体として含む神経障害の衰弱する作用を治療する方法は、病態が発生する前にこれらの複合体または凝集体の形成を防止することである。
【0008】
この目的のため、これらの神経変性ポリペプチドおよびその複合体または凝集体の形成を阻害する新規組成物および方法が開発されている。
【0009】
本発明は、細胞内の病的ハンチンチンポリペプチドを内部抗体(intrabody)に接触させると、その異常な病態の特徴であるポリペプチド凝集体の形成を予防できる、という発見に基づいている。その上、そのような内部抗体は、細胞内で正常なポリペプチドを切断させるようにして凝集体の形成を阻害することができ、ターゲティングシグナルと結合すると、望ましくないポリペプチドが、例えば、ライソゾームまたはプロテアソームにおいて破壊されるように、再度ターゲティングすることができる。
【0010】
したがって、一つの局面において、本発明は、複合体を形成することができるポリペプチドをポリペプチド結合分子、例えば凝集を最小限にして、それによって細胞内凝集体の形成を減少させるようにポリペプチドに特異的に結合する内部抗体、に接触させる段階を含む、選択されたポリペプチドの細胞内凝集体の形成を阻害する方法を提供する。
【0011】
第二の局面において、本発明は、細胞内凝集体を有する危険にさらされている対象に、ポリペプチド結合分子、例えば凝集を最小限にして、それによって細胞内凝集体の形成を減少させるようにポリペプチドに特異的に結合する内部抗体、を投与することを含む、対象において選択されたポリペプチドの細胞内凝集体の形成を阻害する方法を提供する。一つの態様において、対象は神経障害のリスクを有し、好ましくはヒト対象である。もう一つの態様において、対象は、実験動物、好ましくはハンチントン病の動物モデルである。
【0012】
第三の局面において、本発明は、ポリペプチド結合分子、例えば神経障害に関連したポリペプチド凝集体または複合体を形成することができ、それによって凝集体または複合体が形成されることを阻害するポリペプチドに特異的に結合する内部抗体、を対象に投与する段階を含む、神経障害を有する、または有する可能性がある対象を治療する方法を提供する。
【0013】
第四の局面において、本発明は、細胞内ポリペプチド凝集体を形成することができるポリペプチドを提供する段階;ポリペプチドを、試験ポリペプチド結合分子、例えば、内部抗体またはその機能的断片、に接触させる段階;および試験ポリペプチド結合分子、例えば内部抗体またはその機能的断片がポリペプチドを特異的に認識する能力を決定し、それによってポリペプチド結合分子、例えば、細胞内ポリペプチド凝集体を形成することができるポリペプチドを特異的に認識する内部抗体またはその機能的断片であることが同定される段階を含む、ポリペプチド結合分子、例えば細胞内ポリペプチド凝集体または複合体を形成することができるポリペプチドを特異的に認識する内部抗体、またはその機能的断片を同定する方法を提供する。
【0014】
上記の局面の一つの態様において、本発明の内部抗体は、多価であってもよく、スペーサー領域を有してもよく、ファージディスプレイライブラリのような発現ライブラリから選択してもよく、例えば、Fv配列が含まれる。
【0015】
好ましい態様において、内部抗体は、所定のポリペプチドに対して少なくとも105 M-1、より好ましくは少なくとも106 M-1の結合親和性を有するが(解離定数、Ka)、内部抗体が多価である場合、それぞれの原子価は、ポリペプチドに対して少なくとも102 M-1、好ましくは103 M-1の結合親和性を有してもよい。一つの態様において、選択された内部抗体は、野生型ポリペプチドと比較して変異型ポリペプチドに対する親和性が高い。もう一つの態様において、選択された内部抗体は、野生型ポリペプチド、例えば野生型ポリペプチドと変異型ポリペプチドの双方に共通するポリペプチド領域に対して特異的親和性を有する。
【0016】
第五の局面において、本発明は、細胞内凝集体または複合体を形成することができるポリペプチドを提供する段階;ポリペプチドに結合する試験ポリペプチド結合分子、例えば内部抗体またはその機能的断片を提供する段階;ポリペプチドおよび内部抗体断片またはその断片を結合分子と共にインキュベートする段階;ならびに試験化合物が、ポリペプチド結合分子、例えば内部抗体またはその機能的断片の結合を変化させるか否かを決定して、内部抗体に結合するそれらの結合分子が消失すれば、それによって試験化合物が、細胞内ポリペプチド凝集体を形成することができるポリペプチドと相互作用することができると同定される段階を含む、望まない細胞内ポリペプチド凝集体または複合体を形成することができるポリペプチドを特異的に認識する化合物を同定する方法を提供する。好ましい態様において、方法は、異なる結合分子少なくとも102、103、104、105、106、107、108個および好ましくは109個またはそれ以上の多様なライブラリに適用する、および再適用することができる。
【0017】
関連する態様において、試験化合物は小さい有機分子、ペプチド、または天然物抽出物である。
【0018】
第六の局面において、本発明は、神経障害に関連した細胞内ポリペプチド凝集体を形成することができる選択されたポリペプチドに結合する、内部抗体またはその機能的断片をコードする単離核酸分子を提供する。
【0019】
一つの態様において、本発明は、それぞれ配列番号:2、配列番号:4および配列番号:6に記載したアミノ酸配列をコードする、配列番号:1または配列番号:3に記載した核酸配列、および好ましくは配列番号:5に記載した核酸配列を提供し、これらの配列は、好ましい内部抗体、またはその断片に対応する。
【0020】
もう一つの局面において、本発明の内部抗体は、配列番号:1、配列番号:3または配列番号:5の核酸分子と、ストリンジェントな条件下でハイブリダイズする核酸分子によってコードされる。好ましい態様において、核酸は哺乳類に由来する。そのような核酸はまた、ベクター、好ましくは例えばアデノウイルス、アデノ随伴ウイルス、レトロウイルス、または単純ヘルペスウイルスに由来する発現ベクターのような発現ベクター、によってコードされてもよい。その上、内部抗体またはポリペプチド内部抗体をコードするベクターは、薬学的に許容される担体を含む薬学的組成物の形であってもよい。
【0021】
さらにもう一つの態様において、本発明は、宿主細胞が好ましくは、神経起源である、または神経起源の組織の一部である、上記のベクターのいかなるものも含む宿主細胞を提供する。
【0022】
第七の局面において、本発明は、病態、例えば神経障害、に関連した細胞内ポリペプチド凝集体を形成することができる選択されたポリペプチドに結合するポリペプチド骨格のポリペプチド結合分子、好ましくは内部抗体またはその機能的断片を提供する。
【0023】
第八の局面において、本発明は、免疫が凝集体、例えば選択されたポリペプチドの細胞内凝集体の形成、を阻害するために十分な宿主抗体免疫応答を誘発する、動物、例えばヒト患者、を選択されたポリペプチドと共通のエピトープを有する免疫原によって免疫することによって、動物において、選択されたポリペプチドの細胞内凝集体の形成を阻害する方法を提供する。好ましい態様において、免疫原は、例えば、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、SCA7、および好ましくはハンチンチンのようなポリペプチドと共有するエピトープを含む、ポリペプチドをコードする発現可能な核酸ワクチン、例えばDNAワクチンである。
【0024】
関連する態様において、動物は、ハンチントン病またはハンチントン関連糖尿病(例えば、高い絶食時血糖レベルおよび/または低いインスリンレベル)を有する、または有する危険にさらされている。
【0025】
さらにもう一つの態様において、本発明は、好ましくは内部抗体が、神経障害、例えば、ハンチンチンポリペプチド、例えば、対応する野生型ハンチンチンポリペプチドと比較してさらなるグルタミン残基を有するハンチンチンポリペプチド、によって引き起こされるハンチントン病、に関連する凝集体を形成することができるポリペプチドに選択的に結合しうる、内部抗体をコードする核酸を発現するように遺伝子操作されたトランスジェニック動物を提供する。
【0026】
上記の局面のそれぞれにおいて、本発明は、好ましい態様において、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、またはSCA7、および好ましくはハンチンチン、タウ、または野生型ポリペプチドもしくは変異型ポリペプチドのいかなる断片も含むその変異型ポリペプチドのようなポリペプチドを含む細胞内凝集体の形成を阻害する方法を提供する。
【0027】
上記の局面のもう一つの態様において、ポリペプチドには、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドが含まれ、この場合、好ましくは、ポリペプチドはハンチンチンであり、より好ましくは対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有するハンチンチンポリペプチドである。
【0028】
上記の局面のなおもう一つの態様において、本発明の内部抗体は多価であってもよく、または同じもしくは異なる特異性を有する結合部位のあいだにスペーサー領域を有してもよい。
【0029】
上記の局面のさらにもう一つの態様において、本発明の内部抗体は、低分子ペプチド、ペプチド模倣抗体、抗体断片、または好ましくは内部抗体、例えばターゲティングシグナルに対応するアミノ酸配列、例えばそれによって内部抗体が標的ポリペプチドをプロテアソームに再ターゲティングさせることができるユビキチン、を含む内部抗体を含んでもよい。
【0030】
上記の局面のさらにもう一つの態様において、本発明のポリペプチド結合分子は、ターゲティングシグナルに対応し、それによって内部抗体が、特定の細胞位置に標的ポリペプチドを再ターゲティングさせることができるアミノ酸配列を含んでもよい。好ましい態様において、ターゲティングシグナルは、細胞質、核、ライソゾーム、細胞膜関連、小胞体関連、ペルオキシソーム、またはプロテオソームシグナルであり、好ましくは核またはライソゾームシグナルである。
【0031】
上記の局面のさらにもう一つの態様において、内部抗体には、配列番号:2または配列番号:4に提供されたハンチンチンに対して特異的な最小のsFv結合領域が含まれ、好ましくは、配列番号:6に提供される内部抗体α-Nt-HD-C4 sFvのアミノ酸配列を含む。もう一つの態様において、ポリペプチドをインビボまたはインビトロで内部抗体と接触させてもよい。
【0032】
したがって、本発明は、多くの長所を含むと認識される。例えば、内部抗体を、細胞内で作用して、細胞内レベルで細胞に保護を与える発現可能な遺伝子の形で投与してもよい。さらに、本発明の組成物および方法は、それらが免疫系とは独立して機能し、いかなる望ましくない免疫応答も誘発する可能性が低いように設計される。
【0033】
本発明のその他の特徴および長所は、以下の詳細な説明および請求の範囲から明らかとなるであろう。
【0034】
発明の詳細な説明
本発明の完全な範囲を明らかに理解するために、以下の定義を提供する。
【0035】
I.定義
本明細書において用いられるように、「形成の阻害」という用語には、化合物もしくは内部抗体の減少もしくは消失能、例えば、凝集体形成を阻害して、HD類似体ポリペプチドのポリペプチド切断を可能にする可溶性状態に抗体抗原複合体を維持することによって、細胞において望ましくないポリペプチドの蓄積速度を減少させる、蓄積を防止するために利用できるポリペプチドを減少させる、またはそうでなければ蓄積もしくは形成を相殺もしくは消失させる消失させることができる能力が含まれると解釈される。
【0036】
「ポリペプチド凝集体」という用語には、ポリペプチドのいかなる望ましくない、逸脱した、もしくは異常な蓄積、ポリペプチド・ポリペプチド相互作用、ポリペプチド複合体、またはポリペプチド凝集体が含まれると解釈される。この用語にはまた、特定の細胞内空間に攻撃ポリペプチドを局在させることによって、細胞に対する害を最小限にしようとして細胞が形成する可能性があるポリペプチド凝集体が含まれると解釈される。好ましい態様において、ポリペプチドは、疾患、例えば神経疾患に関連した変化したポリペプチドである。この用語には、細胞、例えば神経起源の細胞において認められる、例えば、ハンチンチン凝集体、アミロイド斑、神経原線維濃縮体、レーヴィ小体、プリオン斑、レーヴィ様小体、および望ましくないいかなる複合体または異常な蓄積または封入体も含まれると解釈される。さらになお、この用語はまた、細胞外ポリペプチド凝集体を意味してもよい。「ポリペプチド」または「複数のポリペプチド」、および「蛋白質」という用語は、明細書全体において互換的に用いられる。
【0037】
「選択されたポリペプチド」または「標的ポリペプチド」という用語は、望ましくない蓄積または凝集、例えばポリペプチド凝集体を形成するいかなるポリペプチドも含まれると解釈される。典型的に、選択または標的ポリペプチドは、対応する野生型ポリペプチドと比較して変化したポリペプチドである。
【0038】
「ポリペプチド結合分子」という用語には、例えば、望ましくない蓄積、立体構造、または凝集体を形成することができるポリペプチドに特異的に結合することができるいかなる分子も含まれる。したがって、この用語には、低分子、ペプチド、ペプチド模倣体、抗体(例えば、内因性抗体、すなわち、宿主動物の免疫系によって産生される;外来の抗体、すなわち、発現可能な核酸(複数)、外来核酸(複数)を含む細胞(複数)の形で、またはポリペプチド治療として投与される抗体、を含む)、抗体断片(例えば、Fab断片)ならびに好ましくは内部抗体、または細胞において細胞内で抗原に結合するように作用する免疫グロブリンスーパーファミリーからの類似の遺伝子操作蛋白質が含まれる。
【0039】
「内部抗体(intrabody)」という用語には、標的ポリペプチドに細胞内で特異的に結合することができるいかなる一本鎖ポリペプチド結合物質も含まれると解釈される。典型的に、内部抗体は、重鎖可変領域に結合した最小の軽鎖可変領域を含む一本鎖Fv(sFv)である。内部抗体は典型的に、ポリペプチド治療の形で投与されるが、発現可能な核酸の形、またはそのような核酸を発現する細胞の形で投与してもよい。
【0040】
「多価」という用語には、例えば、分子内において異なるsFv成分によって決定される一つまたはそれ以上の結合特異性を有する、一つ以上の結合部位を含むいかなる物質または内部抗体も含まれると解釈される。
【0041】
「スペーサー領域」という用語には、一つまたはそれ以上の機能的ドメインに、例えば一つのsFvをもう一つのsFvに、またはsFv結合部位をターゲティングシグナルに、結合させるように設計されたペプチド骨格の結合物質または内部抗体のいかなる領域も含まれると解釈される。
【0042】
「リンカー領域」という用語は、リンカーが遺伝子レベルでコードされる、VHリンカーVL、またはVLリンカーVHとして軽鎖可変領域と重鎖領域とを結合させるsFvのペプチド配列を含むと解釈される。
【0043】
「再ターゲティングする」という用語は、標的ポリペプチドに結合した場合に、結合物質または内部抗体がポリペプチドの細胞内位置を再び向け直す能力が含まれると解釈される。
【0044】
「ターゲティングシグナル」という用語は、異種ポリペプチドと結合すると、細胞内の特定の位置にポリペプチドの細胞内輸送を向けることができる、当技術分野で認識されたいかなるアミノ酸配列も含まれると解釈される。この用語には、例えば、核、細胞質、細胞膜関連、小胞体関連、ライソゾーム、ペルオキシソーム、またはプロテアソームのターゲティングシグナルが含まれると解釈される。
【0045】
「野生型ハンチンチンポリペプチド」という用語は、疾患に関連しないグルタミン残基の正常数のみを有するハンチンチンポリペプチドが含まれると解釈される。典型的に、グルタミン残基が38個未満であるいかなるハンチンチンポリペプチドも疾患に関連しておらず、これらは正常または野生型ポリペプチドであると見なしてもよい。
【0046】
「神経障害」という用語には、中枢神経系および/または末梢神経系の細胞を含むいかなる疾患または病態も含まれると解釈される。この用語には、例えば、アルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆(FTD)、筋萎縮性側索硬化症(ALS)、脊髄延髄筋萎縮症(SBMAまたはケネディ病)、歯状赤核淡蒼球萎縮症(DRPLA)、および脊髄小脳運動失調(例えば、SCA-1〜SCA-7)が含まれると解釈される。
【0047】
「発現ライブラリ」という用語には、発現可能な核酸を含むいかなるライブラリも含まれると解釈される。
【0048】
「ファージディスプレイライブラリ」という用語には、当技術分野で認識された意味が含まれると解釈され、典型的に、その発現および/または増殖がファージをパッケージングした核酸配列によって促進される、発現可能な核酸のいかなる集合体も意味する。一つの典型的な態様において、sFvは、sFv抗体結合部位(その特異的結合が表現型である)が、ファージ表面上で抗原結合に利用できるように、そしてその遺伝子型(sFvをコードするDNA)がファージ抗体遺伝子材料内に含まれるように、ファージ外膜蛋白質に融合させる。
【0049】
「Fv」という用語は、例えば、Fab、Fv、Fd、VH、またはVLのような抗体のいかなる抗原結合断片も含まれると解釈される。「sFv」という用語は一本鎖Fvであり、エピトープと相互作用することができる結合ポリペプチドを形成するために必要な最小の重鎖可変領域に結合した最小の軽鎖可変領域が含まれると解釈される。典型的に、内部抗体はFab、Fd、Fv、またはsFv領域を少なくとも一つ含む。
【0050】
「変異型ポリペプチド」という用語は、少なくとも1つのアミノ酸置換または付加、例えばグルタミン付加を有することによって、その対応する野生型ポリペプチドとは異なるいかなるポリペプチドまたはその表示が含まれると解釈される。典型的に、変異型ポリペプチドは、アミノ酸置換およびフレームシフトもしくはスプライシング部位の変化、または一つもしくはそれ以上のグルタミンの伸長によるアミノ酸鎖の変化を有する。他の場合、「変異型ポリペプチド」は、野生型分子と比較して、その結果凝集したポリペプチド種の異常な形成が起こる異常な立体構造を意味する。
【0051】
「変化に富むライブラリ」という用語は、核酸または化合物、例えば有機分子または天然物抽出物のいかなる集合体も含まれると解釈される。
【0052】
「神経起源の細胞」という用語には、中枢神経系または末梢神経系の組織に由来するいかなる細胞も含まれると解釈される。この用語には、例えばニューロン(例えば、運動神経ニューロン、知覚神経ニューロンを含む)、シュワン細胞、プルキンエ細胞、星状細胞、小膠細胞、上衣細胞、および乏突起膠細胞が含まれると解釈される。この用語にはまた、例えば、胚幹細胞または他の全能性もしくは多能性細胞もしくは組織に由来する神経細胞特徴を有するいかなる細胞も含まれると解釈される。
【0053】
「神経起源の組織試料」という用語には、中枢神経系または末梢神経系の組織に由来する多細胞試料が含まれると解釈される。この用語には、例えば、典型的に一つまたはそれ以上の異なる細胞タイプを含む全脳切片および他の組織が含まれると解釈される。
【0054】
「プロテアソーム」という用語には、その通常の技術分野で認識される意味が含まれると解釈される。典型的にプロテアソームは、細胞蛋白質を分解して、多くの蛋白質の寿命の調節に関与して、同様にペプチド源を提供することができる細胞における多数のサブユニットポリペプチド部分を意味する。
【0055】
「ユビキチン」という用語には、その通常の技術分野で認識される意味が含まれると解釈される。この用語には、異種ポリペプチド、例えばプロテアソームと融合すると、異種ポリペプチドが破壊されるようターゲティングするために適している、ヒトを含むいかなる種からのユビキチンアミノ酸配列が含まれると解釈される。
【0056】
「特異的に認識する」という用語には、分子、例えば内部抗体または結合分子の、選択されたポリペプチドに対する特異的結合能が含まれると解釈される。
【0057】
「アミロイド前駆蛋白質」、「プレセニリン1」、「プレセニリン2」「α-2マクログロブリン」、「アポリポ蛋白質」、「αシヌクレイン」、「ハンチンチン」、「プリオン蛋白質」、「タウ」、「スーパーオキシドジスムターゼ(SOD)」、「アンドロゲン受容体(AR)」、「アトロフィン1」、「アタキシン1」、「アタキシン2」、「アタキシン3」、「CACNL1A4」、および「SCA7」という用語には、その通常の技術分野で認識される意味が含まれると解釈される。
【0058】
「アルツハイマー病」、「パーキンソン病」、「ハンチントン病」、「プリオン疾患」、「前頭側頭痴呆(FTD)」、「筋萎縮性側索硬化症(ALS)」、「脊髄延髄筋萎縮症(SBMAまたはケネディ病)」、「歯状赤核淡蒼球萎縮症(DRPLA)」、「1型脊髄小脳運動失調(SCA1)」、「2型脊髄小脳運動失調(SCA)」、「3型脊髄小脳運動失調(SCA3またはマチャドジョゼフ病(MJD)」、「4型脊髄小脳運動失調(SCA4)」、「5型脊髄小脳運動失調(SCA5)」、「6型脊髄小脳運動失調(SCA6)」、および「7型脊髄小脳運動失調(SCA7)」という用語は、当技術分野で認識されるその通常の意味が含まれると解釈される。
【0059】
「選択されたポリペプチド」という用語には、細胞における望ましくない蓄積またはポリペプチド凝集体を形成するとして同定されているいかなるポリペプチドも含まれると解釈される。典型的に、ポリペプチドは疾患に関連しており、変化している、例えば、対応する野生型ポリペプチドと比較してアミノ酸置換もしくはポリグルタミン伸長を含む、または野生型ポリペプチドの対応する三次構造と比較して異常な三次構造を呈する、またはその双方である。
【0060】
「宿主細胞」という用語は、例えば、トランスフェクション、リポフェクション、または感染によって異種ポリヌクレオチド配列を組み入れることができる遺伝子操作に適した細胞が含まれると解釈される。細胞は、微生物または高等真核細胞となりうる。この用語には、最初にトランスフェクトした細胞の子孫が含まれると解釈される。好ましい態様において、細胞は神経起源の細胞である。
【0061】
「異種ポリヌクレオチドセグメント」という用語には、一つまたはそれ以上のポリペプチド、またはポリペプチドの一部もしくは断片をコードするポリヌクレオチドセグメントが含まれると解釈される。異種ポリヌクレオチドセグメントはいかなる起源に由来してもよく、例えば、真核細胞、原核細胞、ウイルス、ファージ、または合成ポリヌクレオチド断片であってもよい。
【0062】
「由来する」という用語には、記載の起源からポリヌクレオチドセグメントを(全体または一部)の単離することが含まれると解釈される。この用語には、例えば、表示のポリヌクレオチド起源に関連した配列から、または配列に基づく直接クローニング、PCR増幅、または人工的な合成が含まれると解釈される。
【0063】
「トランスジェニック動物」という用語には、動物、例えば、動物の細胞の一つまたはそれ以上、好ましくは本質的に全てがトランスジーンを含む、ヒト以外の哺乳類、例えばブタ、サル、ヤギ、または齧歯類、例えばマウスが含まれると解釈される。トランスジーンは、例えば、マイクロインジェクション、トランスフェクション、または感染、例えば組み換え型ウイルスによる感染によって、細胞の前駆体に導入することによって、細胞に直接または間接的に導入される。遺伝子操作という用語は、組み換え型DNA分子の導入に向けられる。この分子は、染色体内に組み入れてもよく、または染色体外の複製DNAであってもよい。
【0064】
「投与する」という用語は、非蛍光または経口経路、筋肉内注射、皮下/皮内注射、静脈内注射、経頬投与、経皮輸送、頭蓋内輸送、のいずれかによる輸送を含む、動物またはヒトにおける所望の位置に治療物質を輸送するために適したいかなる経路によって、動物またはヒトに治療物質を投薬、輸送、または適用することを意味すると解釈される。「投与する」という用語はさらに、治療物質がその作用を細胞において発揮できるように、治療物質を細胞に近接させることを意味すると解釈される。
【0065】
「ベクター」という用語には、それに結合しているもう一つの核酸を輸送することができる核酸分子が含まれると解釈される。一つの態様において、ベクターに結合した核酸は、ペプチド結合分子または内部抗体をコードする。
【0066】
II.ポリペプチド結合分子
一般的に、本発明は、望ましくない凝集体、立体構造、または蓄積を形成することができ、この望ましくないポリペプチド活性を減少または阻害することができるポリペプチドに特異的に結合することができる分子に関する。したがって、本発明のポリペプチド結合分子には、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体が含まれる。本発明の抗体は典型的に、ポリペプチド骨格の治療物質として投与されるが、発現可能な核酸の形で投与してもよい。または、本発明のもう一つの態様において、本発明のポリペプチド結合分子は、ポリペプチドまたはペプチド骨格抗原またはそのような抗原をコードする核酸(すなわち核酸ワクチン)のいずれかを含むワクチンの投与に反応して宿主自身によって産生された内因性の抗体である。しかし、最も好ましい態様において、本発明のポリペプチド結合分子は内部抗体である。
【0067】
内部抗体
本発明の内部抗体は、望ましくない細胞内蓄積または凝集体を形成するポリペプチドに結合することができる。一つの態様において、内部抗体は神経疾患、例えばアルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆、筋萎縮性側索硬化症、脊髄延髄筋萎縮症、歯状赤核淡蒼球萎縮症、1型脊髄小脳運動失調、2型脊髄小脳運動失調、3型脊髄小脳運動失調、4型脊髄小脳運動失調、5型脊髄小脳運動失調、6型脊髄小脳運動失調、および7型脊髄小脳運動失調に関連したポリペプチドに結合する。
【0068】
本発明は、変化したポリペプチド、例えばアミロイド前駆蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、スーパーオキシドジスムターゼ(SOD)、アンドロゲン受容体(AR)、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7を含む神経疾患との関連を有するいかなるポリペプチド、野生型および/または異常な変異型にも結合することができる内部抗体のような分子を含むと解釈される。さらに、本発明はまた、伸長したポリグルタミン領域を有するいかなるポリペプチド、例えばAR、ハンチンチン、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7にも結合することができる内部抗体を含むと解釈される(例えば、ハートレー(Hurtley, S)、Science 282:1071(1998);ショールソン(Shoulson, I.)、Science 282:1072〜1074(1998);ハーディ(Hardy)ら、Science 282:1075〜1079(1998);およびプライス(Price)ら、Science 282:1079〜1083(1998)を参照のこと)。
【0069】
一つの態様において、本発明は、疾患に関連したポリペプチドを表す変化したポリペプチドを提供し、米国特許第5,132,405号;第5,091,513号;第5,084,398号;第5,525,491号;および第5,851,829号、または国際特許出願国際公開公報第99/14353号のいずれかに記載されている方法を用いること、ならびに対応する正常なポリペプチドと比較して疾患に関連する変化したポリペプチドに特異的なエピトープを認識することができる遺伝子操作された内部抗体を提供する(同様に、ザネッティ(Zanetti)ら、「抗体(The Antibodies)」、ハーウッド教育出版、4:1〜141(1997);チェン(Chen)ら、Human Gene Therapy 5:595〜601(1994);ジョーンズ(Jones)ら、Ad. Drug Delivery Reviews 31:153〜170(1998)も参照のこと)。好ましい態様において、内部抗体はハンチンチンポリペプチドに結合することができる。結合は例えば、神経細胞に蓄積して凝集してハンチントン病を引き起こす、正常および異常な全てのハンチンチンに対して特異的となりうる。
【0070】
典型的に、本発明のそのような操作された内部抗体は、本明細書において述べたポリペプチドのいずれかにおけるエピトープに対する可変領域断片(Fv)を含む最小のsFv抗体を含む。完全な抗原認識および結合部位を含むFv、sFv、または最小の抗体断片は、堅固に非共有結合している一つの重鎖と一つの軽鎖可変ドメインとのへテロダイマーからなる、この形状において、各可変ドメインの3つの相補性決定領域は相互作用して、VH-VLダイマーの表面上の抗原結合部位を定める。集合的に、6つの相補性決定領域(CDRs)が、内部抗体に対する抗原結合特異性を付与する。
【0071】
免疫グロブリンポリペプチドの構造に関するこの知識は、望ましくない細胞内蓄積または凝集体を形成するポリペプチドと会合する結合部位(複数)を有するFab、VH、FV、Fd、または一本鎖Fv(sFv)を開発するために利用されている。これらの抗体は、細胞内で機能することを意図しており、したがって内部抗体と呼ばれる。
【0072】
選択された親和性を有する内部抗体
本発明の態様の内部抗体の結合部位は、それらが合成DNA、すなわちポリメラーゼ連鎖反応によって得られた、および/または化学合成されたオリゴヌクレオチドであってもよい複数のDNAセグメントのライゲーションから作製された、またはハイブリドーマ、成熟B細胞クローン、cDNAライブラリ、もしくは哺乳類に由来する核酸を含むファージライブラリのゲノムに由来するDNA断片のライゲーションによって作製された、組み換え型DNAである合成DNAを発現するように作製された細胞宿主において合成されるという点において生合成的である、または生合成的であってもよい。好ましい態様において、ファージライブラリは、選択された抗原に対して免疫されている(免疫ライブラリ)、または所定の抗原に対して特に免疫されていない哺乳類(無傷のライブラリ)哺乳類に由来する。一つの態様において、哺乳類は、当技術分野で既知の方法を用いてDNAワクチン接種を用いて特定の抗原に対して免疫してもよい。または、抗原特異的ファージ抗体を無傷のファージディスプレイライブラリから選択してもよい。このように、本発明の内部抗体は、予め選択した抗原物質に対する特異性と親和性を有するように特に設計される。本発明の内部抗体は、その構造が抗原認識の原因であることが知られている無傷の抗体の領域後でパターン化されているという点において抗体に似ている。
【0073】
より詳しく述べると、ポリペプチドに結合特性を付与する領域におけるこれらの内部抗体の構造は、天然の抗体のFv領域と類似である。したがって、本発明の内部抗体は、ポリペプチドの特定の計画した有用性を念頭に置いて設計してもよく、既知の親和性および特異性を有する抗体、例えば、選択されたポリペプチドに対するマウス、ラット、またはヒトモノクローナル抗体の高度可変領域のCDRs、または類似のCDRsもしくはその一部を含む。
【0074】
もう一つの態様において、本発明は、抗原プロセシングおよび他の複雑な免疫応答を必要とせずに、エクスビボで適当な内部抗体結合を選択するファージディスプレイ方法を提供する。この方法は、溶媒条件制御能と抗原提示方法を与える(例えば、溶液中で遊離で、グリセロールまたは蔗糖のような共溶媒の存在下で、磁気ビーズに結合して、またはプラスチックマイクロタイタープレートに吸着させて)。
【0075】
一つの態様において、本発明の内部抗体は、HおよびL鎖可変遺伝子が遺伝子レベルで融合されている、一本鎖Fvの形でヒト抗体の最小結合部位を利用する。そのような抗体の構築は、大きい、例えばヒトsFv結合部位の7×109個のファージライブラリから結合部位を選択することによって促進される。これらのsFv遺伝子は典型的に、M13バクテリオファージの小外膜蛋白質(遺伝子3蛋白質)に典型的にライゲーションされており、sFv遺伝子3融合体が線維状ファージの先端で示される生存ファージに再構築されている。このように、表現型(すなわち、結合部位特異性および親和性)と遺伝子型はあらゆるファージ粒子に連結している。この方法は、免疫系が存在しない場合に結合部位の遺伝子選択を行う長所を提供する(例えば、実施例4および5を参照のこと)。
【0076】
その上、数ラウンド行った後(ファージ選択、感染、および感染細胞の再増殖)、本方法は、結合試験にとって十分量の対応するsFvを産生する条件で、細菌培養において増殖させることができる選択されたファージを増幅させる長所をさらに与えた。
【0077】
より大きい調製物が望ましい場合、選択の第3ラウンド後に、ファージを遊離のsFvを産生する大腸菌株の感染によって個々のプラークから増殖させ、これを例えばいかなるエピトープまたは融合免疫タグ(例えば、mycエピトープ(すなわち9E10)またはHis6タグ)に向けられたIMACクロマトグラフィーによってインビボで精製してもよく、PBS中でスーパーロース6カラム上でのFPLCによって存在する単量体または二量体を評価して、BIAコア2000バイオセンサー装置においてストレプトアビジンコーティングマイクロチップ上で固定された選択されたポリペプチド、例えば標的ポリグルタミンバイオポリマーに対して分析してもよい。このバイオセンサー装置は、結合反応に関する結合速度定数(Ka)および解離速度定数(Kd)、ならびにKa/Kdの商から計算した結合平衡定数Kaの測定を与える。したがって、そのような物理化学研究において示され、ファージ選択において用いられる溶媒条件は、関連するポリペプチド立体構造を、例えばBIAコア測定のあいだに保存するように設計することができる。
【0078】
二機能/多機能内部抗体
さらに、本発明の内部抗体は、生物活性領域を定義するさらなるポリペプチド領域、例えばターゲティングシグナル、酵素配列、毒素、またはそこに毒素もしくは治療薬、結合ポリペプチド、酵素もしくは酵素断片、または造影剤のような、遠隔操作で検出可能な物質を結合させることができる部位をさらに含んでもよい。
【0079】
好ましい態様において、内部抗体は、酵素、例えばポリペプチドの蛋白質溶解に至りうるプロテアーゼのようなもう一つの機能的ドメインを含む。もう一つの態様において、内部抗体は、その後破壊されるようにポリペプチドをプロテアソームにターゲティングすることができるターゲティングシグナル、例えば、ユビキチンを含む。さらにもう一つの態様において、本発明の内部抗体は、再ターゲティングすることができるターゲティングシグナルを含み、本発明の内部抗体はもう一つの細胞内位置に内部抗体結合ポリペプチドを再ターゲティングすることができるターゲティングシグナルを含む。例えば、そのような場所は、例えば細胞質、核、ライソゾーム、細胞質関連、小胞体関連、ペルオキシソーム、またはプロテアソームであってもよい。さらに、本発明の内部抗体または結合分子は、異種ポリペプチドの細胞位置を変化させるために当技術分野で認識されたいかなるターゲティングシグナルを含んでもよい。
【0080】
一つの態様において、内部抗体は、特定の細胞位置にポリペプチドをターゲティングするために適した当技術分野で認識された(例えば、国際公開公報第99/14353号に記載するように、または図2に示すように)いかなるターゲティングシグナルも含むように操作してもい。
【0081】
多価内部抗体
当然のこととして、本発明の内部抗体は、一つ以上の結合部位または単一の結合部位のコピー、および他の多くの機能的領域を含んでもよい。例えば、多価内部抗体は本発明の範囲に含まれ、そのような抗体は、選択されたポリペプチド内で認められる一つまたはそれ以上のエピトープに対して親和性を有してもよい。その上、好ましい態様において、多価内部抗体は、変化したポリペプチドにおいて認められるエピトープに対する一つまたはそれ以上の親和性の他に、正常なペプチド内に認められるエピトープに対して一つまたはそれ以上の親和性を有してもよい。もう一つの態様において、内部抗体は、変化したポリペプチド、例えば疾患に関連したポリペプチドにおいて認められる一つまたはそれ以上のエピトープに対する親和性を有する。もう一つの態様において、多価内部抗体は、変化したポリペプチドに対する選択的な結合能を有し、これは正常なポリペプチドとは異なる半減期を有してもよく、異なる速度でまたは異なる細胞内空間に蓄積してもよく(または分泌される)、変化した立体構造を呈してもよく、凝集してもよく(自身で、または他のポリペプチドと共に)、他のポリペプチドとの望ましくない相互作用を形成してもよく、細胞増殖の変化または細胞死を引き起こしてもよく、および/または障害または疾患を引き起こしてもよい。
【0082】
したがって、好ましい態様において、本発明の内部抗体は、例えば、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7のようないかなる一つまたはそれ以上のポリペプチドに対する親和性を有する一つまたはそれ以上の結合部位を有する。
【0083】
もう一つの好ましい態様において、本発明の内部抗体は、例えば、アルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆(FTD)、筋萎縮性側索硬化症(ALS)、脊髄延髄筋萎縮症(SBMAまたはケネディ病)、歯状赤核淡蒼球萎縮症(DRPLA)、1型脊髄小脳運動失調(SCA1)、2型脊髄小脳運動失調(SCA2)、3型脊髄小脳運動失調(SCA3またはマチャドジョゼフ病(MJD))、4型脊髄小脳運動失調(SCA4)、5型脊髄小脳運動失調(SCA5)、6型脊髄小脳運動失調(SCA6)、および7型脊髄小脳運動失調(SCA7)のような、本質的に神経学的な障害または疾患に関係するいかなるポリペプチドにも向けられる。
【0084】
その上、本発明の内部抗体は、短いポリグルタミンに富む領域を有する正常なポリペプチドと、疾患の状態に関連する可能性がある伸長したポリグルタミンに富む領域を有するポリペプチドとを区別するように操作することができる。長さが長いか短いかが異なるに過ぎないポリグルタミンを区別するために、一つの低親和性結合部位または多数の低親和性結合部位を有する内部抗体を、変化したポリペプチドを選択的に結合させるために用いてもよい(例えば、ハストン(Huston, J.S.)ら、(1992)、Biophysical Journal 62;87〜91;ハストン(Huston, J.S.)ら、(1996)、Adv. in Prot. Chem. 49:329〜450)。このように、本発明は、最小の可能性がある種において二つまたはそれ以上の結合部位を組み入れる内部抗体のsFv領域の二価または多価型を含む(ジョージおよびハストン(George, A.J., and Huston, J.S.)、(1997)、The Antibodies 4:99〜141;ハストン(Huston, J.S.)ら、(1996)、Quarterly Journal of Nuclear Medicine 40:320〜333)。さらに、他のドメイン、例えば四量体ストレプトアビジン融合体もまた、内部抗体の、選択されたポリペプチドの特異的ターゲティング能を促進するために用いてもよい(キプリヤノフ(Kipriyanov, S.M.)ら、(1996)、Protein Engineering 9:203〜211)。
【0085】
好ましい態様において、内部抗体、例えば単価内部抗体または結合分子のKaは、105 M-1であり、より好ましくは106 M-1である。もう一つの態様において、多価内部抗体または結合分子の所定の価数のKaは、102 M-1であり、より好ましくは103 M-1である。
【0086】
上記から明らかなように、本発明は、可変領域または天然の免疫グロブリン領域の後にパターン化される結合部位(複数)を含む内部抗体の大きいファミリーを提供する。本発明の本質が、単独で、または他のポリペプチドと様々なように結合して、選択されたポリペプチド、好ましくは、例えば不適当な細胞機能を呈する変化したポリペプチドに結合することができる限り、内部抗体に結合して、本発明の態様の試薬のために用いられるいかなるポリペプチド断片の性質も本質的に無限であることは明らかであろう。
【0087】
内部抗体をコードする核酸
本発明の内部抗体はDNAレベルで設計してもよい。したがって、本明細書に記載の内部抗体をコードする核酸もまた本発明の範囲に含まれる。
【0088】
ファージディスプレイライブラリは、その発現および/または増殖がファージをパッケージングした核酸配列によって促進される、発現可能な核酸の集合体を提供する。sFvファージディスプレイライブラリにおいて、sFvは、sFv抗体結合部位(その特異的結合が表現型である)がファージ表面上での抗原結合に利用でき、その遺伝子型(sFvをコードするDNA)がファージ抗体遺伝子材料内に含まれるように、ファージ外膜蛋白質に融合される。この遺伝子型DNAは、例えば、増幅とシークエンシングを行うためと共に、sFv遺伝子を内部抗体として用いるためのその後の操作を可能にするためにSfiIおよびNotI部位での制限的切断によって容易に単離される。遺伝子は、大腸菌において発現させるためにpHEN1プラスミドにスプライシングしてもよく、ポリペプチドを本来の折り畳まれたポリペプチドとして周辺質に発現および分泌してもよい。
【0089】
本発明の内部抗体を設計できるか否かは、当該モノクローナル抗体の可変領域においてアミノ酸配列またはそれらをコードするDNAを決定することができるかに依存する。ハイブリドーマ技術は、免疫応答を産生する本質的にいかなる望ましい物質に対する抗体を産生する細胞株の産生を可能にする。次に、免疫グロブリン軽鎖および重鎖をコードするRNAをハイブリドーマの細胞質から得ることができ、mRNAの5'末端部分を用いて、その後のシークエンシングのためにcDNAを調製し、または高度可変であって隣接するフレームワーク領域(FR)のアミノ酸配列を、HおよびL鎖ならびにそのV領域断片のアミノ酸シークエンシングによって決定することができる。そのような配列分析は現在、日常的に行われている。この知識によって、抗原に結合する可能性があるFRおよびCDR配列をコードする合成遺伝子の設計が可能となる。これらの合成遺伝子は既知の技術によって、または下記に開示の技術を用いて調製し、次にこれを適した宿主に挿入して発現させ、精製する。宿主細胞に応じて、適切な立体構造を得るために再生技術を必要としてもよい。次に、様々なポリペプチドの結合能を試験し、適当な親和性を有するポリペプチドを上記のタイプの試薬に組み入れるために選択する。必要であれば、従来のカセット変異誘発または他の蛋白質操作技術を用いて、最適な結合を得るためにDNAに一点置換を行ってもよい。
【0090】
当然のこととして、当該アミノ酸配列をコードするDNAを操作、増幅、および組み換えするプロセスは一般的に当技術分野で周知であり、したがって、本明細書において詳細に記述しない。当該内部抗体領域をコードする遺伝子を同定および単離する方法は十分に理解されており、本明細書ならびに例えば米国特許第5,132,405号;第5,091,513号;第5,084,398号;第5,525,491号;第5,851,829号、および国際特許出願国際公開公報第99/14353号に記載されており、これらの参考文献は参照として本明細書に組み入れられる。
【0091】
内部抗体抗原−標的ポリペプチド
単離された標的ポリペプチド、好ましくは望ましくない細胞内蓄積または凝集を形成することが知られているポリペプチド、より好ましくは、他の場所と比較して脳または神経起源の細胞において比較的高濃度で発現されるポリペプチド、または関連する非神経障害を引き起こすポリペプチド、またはその一部もしくは断片を、ファージライブラリ選択技術の標的として、またはポリクローナル抗体およびモノクローナル抗体調製のための標準的な技術を用いて標的ポリペプチドを結合する内部抗体を産生するための免疫原として用いることができる。完全長の標的ポリペプチドを用いることができ、または本発明は、免疫原もしくはファージディスプレイ選択における標的として用いられる標的ポリペプチドの抗原性ペプチド断片、例えばハンチンチンN末端配列(例えば、残基1〜17)を提供する。好ましくは、抗原性ペプチドは、ペプチドに対する内部抗体が天然の条件で標的ポリペプチドと特異的免疫複合体を形成するように、アミノ酸残基少なくとも5個、より好ましくはアミノ酸残基少なくとも15個によって形成される標的ポリペプチドのエピトープを含む。
【0092】
または、変化したポリペプチドのいかなる数にも共通する可能性があるポリグルタミンに富む領域を示すポリペプチドを、標的抗原として(例えばハンチンチンおよびアトロフィン)用いてもよい。そのようなポリペプチドによって、ポリグルタミンに富む領域を有するいかなる数のポリペプチドも結合することができる内部抗体を作製または選択することができる。したがって、そのような内部抗体は、それぞれが最小のポリグルタミンに富む領域を形成すれば、変化したポリペプチドのいくつかの型の蓄積、凝集体形成、または細胞内位置の再ターゲティングを阻害しうるであろう。しかし、さらなる特異性が非ポリグルタミン、ポリペプチド特異的エピトープに対して付与される、独自のクラスの当該ポリペプチド、例えばハンチンチンの特異性は、好ましくは双特異的もしくは多特異的sFv抗体もしくは二抗体、またはFvの関連型を必要とする。二抗体は二価または双特異的であり、リンカーがVドメインを結合する場合に形成する一本鎖sFvの時に三価および多特異的な型は、異常なほど短く、通常10残基またはそれより少なく、好ましくは5残基またはそれより少なく、その結果可変ドメイン融合蛋白質のあいだにFv結合部位を形成する。
【0093】
選択されたポリペプチド免疫原は典型的に、ファージライブラリから内部抗体を選択するために、または適した対象(例えば、ウサギ、ヤギ、マウス、または例えばヒト患者を含む他の哺乳類)を標的ポリペプチド免疫原によって免疫することによって抗体を誘発するために用いられる。適当な免疫原調製物は、例えば組み換え発現された標的ポリペプチドまたは化学合成されたポリペプチドを含みうる。ポリペプチドはさらに、検出または精製を促進するための部分と融合してもよく、そのような部分には、免疫反応性タグ、GFP、ビオチン等が含まれる。調製物はさらに、フロイントの完全もしくは不完全アジュバントのようなアジュバント、または類似の免疫刺激物質を含みうる。
【0094】
または、免疫原は、レシピエント動物の細胞によって核酸が取り込まれて発現されるように、宿主細胞免疫応答、好ましくは液性応答を誘発するために十分量のポリペプチドが得られるように、発現可能な核酸の形で、例えばDNAワクチンとして投与してもよい。適した対象を免疫原性標的ポリペプチド調製物によって免疫すると、抗標的ポリペプチド抗体反応を誘発する。
【0095】
したがって、内部抗体は、標準的な技術を用いて得られたモノクローナル抗体に由来してもよい。より好ましくは、内部抗体は、DNAワクチン接種を用いて標的ポリペプチドに対して免疫した動物に由来する。さらにより好ましくは、内部抗体は、標的ポリペプチドまたは標的ポリペプチドをコードする発現可能なDNAによって免疫した動物に由来するファージライブラリに由来する。特定の場合において、同じ標的ポリペプチドに由来する副疾患を有する神経細胞または他の細胞によらず、標的ポリペプチドに対する抗体のインサイチュー産生によって、無傷のIgGを生成してもよく、または疾患を有する細胞に入って内部抗体として作用し、それによって標的ポリペプチドの病的な凝集に拮抗する他のクラスの抗体を生成してもよい。これらの場合、治療的抗体は、ポリペプチド免疫治療として、またはそれらが内部抗体ポリペプチドを分泌する、エクスビボで改変された細胞が患者に再投与される養子免疫細胞移入による免疫治療の形として投与してもよい(例えば、カバッツァーナカルボ(Cavazzana-Calvo, M.)ら、Science 288:669〜672)。
【0096】
さらに、内部抗体は、組み換え型組み合わせ免疫グロブリンライブラリ(例えば、内部抗体ファージディスプレイライブラリ)を標的ポリペプチドによってスクリーニングして、それによって、標的ポリペプチドに結合する免疫グロブリンライブラリメンバーを単離することによって同定および単離することができる。ファージディスプレイライブラリを作製およびスクリーニングするキットは市販されている(例えば、ファルマシア社の組み換え型ファージ抗体システム、カタログ番号27-9400-01;およびストラタジーン社のSurfZAP(商標)ファージディスプレイキット、カタログ番号240612)。さらに、特定の親和性を有する内部抗体を作製、スクリーニング、および選択するために特に用いやすい方法および試薬の例は、例えば、米国特許第5,132,405号;第5,091,513号;第5,084,398号;第5,525,491号;第5,851,829号、国際特許出願国際公開公報第99/14353号、およびシーツ(Sheets)ら、P.N.A.S. 95:6157〜6162(1998)に見ることができる。
【0097】
このように、本発明は、以下の少なくとも内部抗体の概念を(下記の実施例はさらなる証拠となるであろう)示し、それらは:1)内部抗体は遺伝子治療として輸送することができること;2)内部抗体は標的細胞に入る(免疫毒素のように)蛋白質免疫治療として輸送することができ、標的抗原との結合を通して、細胞内で機能することができること;3)内部抗体は、静脈内投与する代わりに、例えば宿主液性応答の形で免疫治療のための内部抗体蛋白質が免疫系によって作製されるように、適当な免疫によって誘発することができること;および4)異常なポリペプチド蓄積は、非常に低レベルで存在しうること、そして蛋白質・蛋白質相互作用を通じて細胞機能を有意に破壊でき、このように、内部抗体の中等度の細胞内レベルでも治療的となることである。
【0098】
III. 発現ベクターと宿主細胞
本発明のもう一つの局面は、ベクター、好ましくは内部抗体(またはその一部)をコードする核酸を含む発現ベクターに関する。一つのタイプのベクターは「プラスミド」であり、これは、その中にさらなるDNAセグメントがライゲーションされうる環状二本鎖DNAループを意味する。もう一つのタイプのベクターは、さらなるDNAセグメントをウイルスゲノムの中にライゲーションすることができるウイルスベクターである。特定のベクターは、それらが導入された宿主細胞において自律的に複製することができる(例えば、細菌の複製開始点を有する細菌ベクターおよびエピソーム哺乳類ベクター)。他のベクター(例えば、非エピソーム哺乳類ベクター)は、宿主細胞に導入されると宿主細胞のゲノムに組み入れられ、それによって宿主ゲノムと共に複製される。その上、特定のベクターは、それらが機能的に結合する遺伝子の発現を指示することができる。そのようなベクターは、本明細書において「発現ベクター」と呼ぶ。一般的に、組み換えDNA技術において用いられる発現ベクターはしばしば、プラスミドの形である。プラスミドは、最も一般的に用いられるベクターの型であるため、本明細書において、「プラスミド」および「ベクター」は互換的に用いることができる。しかし、本発明には、同等の機能を提供するウイルスベクター(例えば、複製欠損レトロウイルス、レンチウイルス、アデノウイルスおよびアデノ随伴ウイルス)のような発現ベクターのそのような他の形も含まれると解釈される。
【0099】
本発明の組み換え発現ベクターは、宿主細胞における発現に適した形で本発明の内部抗体をコードする核酸を含み、このことは、組み換え型発現ベクターが、発現される核酸配列に機能的に結合している、発現のために用いられる宿主細胞に基づいて選択される一つまたはそれ以上の調節配列を含むことを意味する。組み換え発現ベクターにおいて、「機能的に結合」とは、当該ヌクレオチド配列が調節配列(複数)に対して、ヌクレオチド配列を発現させるように結合していることを意味すると解釈される(例えば、インビトロ転写/翻訳系において、またはベクターが宿主細胞に導入される場合は宿主細胞において)。「調節配列」という用語には、プロモーター、誘導型プロモーター、エンハンサー、および他の発現制御エレメント(例えば、ポリアデニル配列)が含まれると解釈される。そのような調節配列は、例えば、ゴッデル(Goeddel)(「遺伝子発現技術(Gene Expression Technology)」Methods in Enzymology 185、アカデミック出版、サンディエゴ、カリフォルニア州(1990))に記載されている。調節配列には、多くのタイプの宿主細胞においてヌクレオチド配列の構成的発現を指示する配列、および特定の宿主細胞に限ってヌクレオチド配列の発現を指示する配列(例えば、組織特異的調節配列)が含まれる。発現ベクターの設計は、形質転換すべき宿主細胞の選択、望ましい蛋白質の発現レベル等のような要因に依存しうることは当業者によって認識されるであろう。本発明の発現ベクターは、宿主細胞に導入されて、それによって内部抗体を産生することができ、その後これをポリペプチド製剤として投与してもよい。または、内部抗体をコードする核酸を、好ましくは哺乳類、より好ましくはヒト、より好ましくは哺乳類の神経起源の細胞において発現することができるベクターに導入する。
【0100】
多くのプロモーターおよび発現ベクターが哺乳類細胞において高い発現を与えるとして当技術分野で既知である。例えば、本発明の核酸分子は哺乳類の発現ベクターを用いて発現させることができる。哺乳類発現ベクターの例には、pCDM8(シード(Seed, B.)(1987)Nature 329:840)およびpMT2PC(カウフマン(Kaufman)ら(1987)、EMBO J 6:187〜195)が含まれる。哺乳類細胞において用いる場合、発現ベクターの制御機能はしばしば、ウイルス調節エレメントによって提供される。例えば、一般的に用いられるプロモーターは、ポリオーマ、アデノウイルス2、サイトメガロウイルス、およびシミアンウイルス40に由来する。原核細胞と真核細胞の双方に適した他の発現系に関しては、サムブルック、フリッチュおよびマニアティス(Sambrook, J., Fritsh, E.F., and Maniatis, T.)、「分子のクローニング:実験マニュアル(Molecular Cloning:A Laboratry Manual)」第二版、コールドスプリングハーバー出版、コールドスプリングハーバー、(1989);アウスユベール(Ausubel)ら、(1998)、「分子生物学の現行プロトコール(Current Protocols in Molecular Biology)」、ジョンウィリー&サンズインクを参照のこと。
【0101】
もう一つの態様において、組み換え型哺乳類発現ベクターは、選択的に特定の細胞タイプにおいて核酸の発現を指示することができる(例えば、組織特異的調節エレメントを用いて核酸を発現させる)。組織特異的調節エレメントは当技術分野で既知である。適した組織特異的プロモーターの非制限的な例には、アルブミンプロモーター(肝臓特異的;ピンカート(Pinkert)ら、(1987)、Genes Dev. 1:268〜277)、リンパ様特異的プロモーター(カラムおよびイートン(Calame and Eaton)(1988)、Adv. Immunol. 43:235〜275)、特にT細胞受容体(ウィノトおよびバルチモア(Winoto and Baltimore)(1989)EMBO J. 8:729〜733)および免疫グロブリン(バネリ(Banerji)ら(1983)、Cell 33:729〜740;クイーンおよびバルチモア(Queen and Baltimore)(1983)Cell 33:741〜748)のプロモーター、ニューロン特異的プロモーター(例えば、神経細糸プロモーター;ビルネおよびラドル(Byrne and Ruddle)(1989)PNAS 86:5473〜5477)、膵臓特異的プロモーター(エドルンド(Edlund)ら、(1983)Science 230:912〜916)、および乳腺特異的プロモーター(例えば、乳清プロモーター;米国特許第4,873,316号および欧州特許第246,166号)が含まれる。発達的に調節されるプロモーター、例えばマウスホックスプロモーター(ケッセルおよびグルス(Kessel and Gruss)(1990)、Science 249:374〜379)およびαフェトプロテインプロモーター(カンペスおよびチルフマン(Campes and Tilghman)(1989)Genes Dev.3:537〜546)も同様に含まれる。
【0102】
本発明のもう一つの局面は、本発明の組み換え型発現ベクターが導入される宿主細胞に関する。「宿主細胞」および「組み換え型宿主細胞」という用語は本明細書において互換的に用いられる。そのような用語は、特定の被験細胞を意味するのみならず、そのような細胞の子孫または可能性がある子孫も意味すると理解される。変異または環境的影響によりその後の世代に特定の改変が起こる可能性があるため、そのような子孫は、実際に親細胞と同一でない可能性があるが、なおも本明細書において用いられる用語の範囲に含まれる。
【0103】
宿主細胞は、いかなる原核または真核細胞となりうる。例えば、内部抗体蛋白質は、大腸菌のような細菌細胞、昆虫細胞、酵母または哺乳類細胞(例えば、チャイニーズハムスター卵巣細胞(CHO)、哺乳類腎細胞(COS)、ハムスター腎細胞(BHK-21)、ヒト上皮細胞(293T)のような)において発現されうる。その他の適した宿主細胞は当業者に既知である。
【0104】
ベクターDNAは、従来の形質転換またはトランスフェクション技術によって原核細胞または真核細胞に導入することができる。本明細書において用いられるように、「形質転換」および「トランスフェクション」という用語は、リン酸カルシウムもしくは塩化カルシウム共沈殿、DEAEデキストラン媒介トランスフェクション、リポフェクション、または電気穿孔を含む、外来核酸(例えば、DNA)を宿主細胞に導入する当技術分野で認識された多様な技術を意味すると解釈される。好ましい態様において、内部抗体をコードする核酸は、プロタミン骨格の非ウイルスベクターを用いてトランスフェクションしてもよい(リチャードソン(Richardson, J.H.)(1995)、Trends in Biotechnology, 13:306〜310)。宿主細胞を形質転換またはトランスフェクトする適した方法は、上記のサムブルック(Sambrook)ら、ならびに例えば、本明細書において言及した他の実験マニュアルおよび参考文献に見ることができる。
【0105】
哺乳類細胞の安定なトランスフェクションに関して、用いる発現ベクターとトランスフェクション技術に応じて、外来DNAをそのゲノムに組み入れるのはごく小さい分画の細胞に過ぎない場合があることが知られている。これらの組み込み体を同定および選択するために、選択マーカー(例えば、抗生物質に対する抵抗性)をコードする遺伝子を一般的に当該遺伝子と共に宿主細胞に導入する。好ましい選択マーカーには、G418、ヒグロマイシン、およびメソトレキセートのような薬剤に対する抵抗性を付与するマーカーが含まれる。選択マーカーをコードする核酸は、内部抗体をコードするベクターと同じベクター上で宿主細胞に導入することができ、または異なるベクターに導入することができる。導入した核酸によって安定にトランスフェクトされた細胞は、薬剤選択によって同定することができる(例えば、選択マーカー遺伝子を組み入れた細胞は生存するが、他の細胞は死滅する)。
【0106】
原核または真核宿主培養細胞のような本発明の宿主細胞は、内部抗体を産生する(すなわち発現する)ために用いることができる。したがって、本発明はさらに、本発明の宿主細胞を用いて内部抗体ポリペプチドを産生する方法を提供する。一つの態様において、方法は、本発明の宿主細胞(その中に内部抗体をコードする組み換え型発現ベクターが導入される)を適した培地において、内部抗体が産生されるように培養する段階を含む。もう一つの態様において、方法は、培地または宿主細胞から内部抗体を単離することをさらに含む。
【0107】
本発明の宿主細胞はまた、ヒト以外のトランスジェニック動物を作製するために用いることができる。例えば、一つの態様において、本発明の宿主細胞は、その中に内部抗体コード配列が導入される受精卵または胚肝細胞である。次に、そのような宿主細胞を用いてその中に外来内部抗体配列がそのゲノムに導入されているヒト以外のトランスジェニック動物、または内因性Fv配列が変化している相同組み換え動物を作製することができる。そのような動物は、内部抗体の機能および/または活性を調べるために、および内部抗体/標的ポリペプチド活性の調節物質を同定および/または評価するために有用である。本明細書において用いられるように、「トランスジェニック動物」は、ヒト以外の動物、好ましくは哺乳類、より好ましくは動物の一つまたはそれ以上の細胞がトランスジーンを含む、ラットまたはマウスのような齧歯類である。トランスジェニック動物の他の例には、ヒト以外の霊長類、ヒツジ、イヌ、ウシ、ヤギ、ニワトリ、両生類等が含まれる。トランスジーンはそこからトランスジェニック動物が発生する細胞のゲノムに組み入れられ、成熟動物のゲノムに留まり、それによってトランスジェニック動物の一つまたはそれ以上の細胞タイプまたは組織においてコードされた遺伝子産物の発現を指向する外来DNAである。本明細書において用いられるように、「相同的組み換え型動物」とは、ヒト以外の動物、好ましくは哺乳類、より好ましくは内因性の遺伝子と、動物が発達する前に動物の細胞、例えば動物の胚細胞に導入された外来DNA分子とのあいだの相同的組み換えによって内因性標的ポリペプチド遺伝子が変化しているマウスである。この動物は、内部抗体を発現するようにさらに改変してもよく、または内部抗体を発現する動物と交配させることができる。より好ましくは、トランスジェニック動物は、当該ポリペプチドの一つまたはそれ以上、すなわち標的ポリペプチド、またはより好ましくは内部抗体が例えば、実施例6に記載のように条件的制御下にあるように操作される。さらに、当技術分野で認識されるいかなる技術も、本発明のトランスジェニック動物および/または相同的組み換え動物を産生するために用いてもよい。
【0108】
IV.スクリーニングアッセイ
本発明は、調節物質、すなわち標的ポリペプチド、例えばハンチンチンポリペプチドに結合する候補結合分子(例えば、ペプチド、ペプチド模倣体、低分子または他の薬剤)を同定する方法(本明細書において「スクリーニングアッセイ」と呼ぶ)を提供する。方法は、所定の疾患のモデルを表す異常を示す細胞内でインサイチューで行う。このように、標的ポリペプチドの病的な凝集をインサイチューで相殺する能力は、さらなる治療物質開発のための必要条件となる。
【0109】
特に、本発明の一つまたはそれ以上の方法は、選択されたポリペプチドの望ましくない蓄積、複合体形成、または凝集を変化させる化合物をスクリーニングするためにヒト以外の対象の細胞において内部抗体を用いることを含む。好ましい態様において、選択されるポリペプチドは、例えば、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、スーパーオキシドジスムターゼ(SOD)、アンドロゲン受容体(AR)、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、またはSCA7である。もう一つの好ましい態様において、ポリペプチドはアルツハイマー病に関連する。さらにもう一つの好ましい態様において、ポリペプチドは、ハンチンチンであり、ハンチンチンの異常型はハンチントン病に関係している。したがって、特定の疾患に関係しているポリペプチドに対して特異的な内部抗体を、細胞に基づくアッセイにおいて、インビトロで、または動物全体においてポリペプチドに接触させ、そして一つまたはそれ以上の化合物をほぼ同じ時期に投与し、内部抗体/ポリペプチド相互作用による凝集の変化をモニターする。
【0110】
インビトロにおいて、そのような変化は、結合の変化または他のいくつかの生化学パラメータの変化として測定してもよい。細胞に基づくアッセイにおいて、内部抗体/ポリペプチド相互作用の変化は、標的ポリペプチドの蓄積、複合体形成、または凝集における変化、または宿主細胞の細胞生物学における変化(例えば、細胞死レベルの変化)として測定してもよい。生物全体の意味において、内部抗体と標的ポリペプチドとのインビボ相互作用のそのような変化は、疾患の症状の変化として測定してもよい。
【0111】
一つまたは全てのアッセイを用いて、内部抗体/ポリペプチド相互作用に関して凝集の蓄積を変化させる化合物の可能性がある治療的有効性を確認してもよい。そのような化合物は、さらなる分析のために選択してもよい。理想的には、好ましい態様において、化合物は標的ポリペプチドに対する特異的結合として同定される。もう一つの好ましい態様において、化合物は正常および変異型ポリペプチドの双方を認識することができ、それらを適当なレベルで複合体を形成させることによって、正常な細胞機能と病理プロセスの排除の双方を促進することができる。なおもう一つの好ましい態様において、化合物は、野生型ポリペプチドより高い特異性を有する変異型ポリペプチドを認識することができ、またはこのように機能するように変化することができる。なおもう一つの好ましい態様において、化合物は、例えば、毒性、血液脳関門の通過能、半減期、もう一つの機能的部分との結合能、またはさらに改変を受ける能力のような特定の薬理学的特性または長所に関して選択してもよい。
【0112】
当該結合分子はまた、組織切片培養、例えば動物、例えば、神経疾患(例えば、AD、HD、SCA疾患等)のトランスジェニック動物モデルからの臓器型切片培養を用いてアッセイしてもよい。好ましい態様において、HDまたはSCA1を有するマウスを用いる。さらに、前述のいかなる実験動物も、興奮毒のような化合物または酸化的障害を引き起こすことが知られている化合物に対する感受性に関して、野生型対照と平行して調べることができる。神経変性、例えば、線条体のHD様変性、または他の神経毒性作用を誘導するために適している当技術分野で記載の多くの化合物を、インビボで、またはそのような動物に由来する臓器型切片培養を用いて試験することができる(例えば表1、および例えば、ボウリング(Bowling)ら、Life Sci. 56:1151〜1171(1995);ジャンセル(Jansel)ら、Brain Res. 532:351〜354(1990);ロススタイン(Rothstein)ら、P.N.A.S. 90:6591〜6595(1993);およびシュワルツ(Schwarcz)ら、Life Sci. 35:19〜32(1963)を参照のこと)。短期間の暴露の後に野生型細胞または組織に対して明白な毒性を示さない用量を用いることによって、通常誘発される初期の臨床下病態を中和することによる疾患を有する細胞の異なる脆弱性を確認することができる。
【0113】
好ましい態様において、表1に記載した化合物を用いて、変異型ニューロン、野生型ニューロン、または内部抗体もしくは結合分子を有するニューロンのいずれかにおいて異なる作用を誘発してもよい。
【0114】
【表1】
【0115】
一つの態様において、培養は、本明細書に記述した方法を用いて調製することができ、2週間成熟させた。例えばHDまたはSCA1トランスジーンのいずれかの存在による認識可能な病態は、この時点で最小であるはずであるが、切片におけるニューロンは、なおもリスクが増加している可能性がある。次に、上記に示した化合物を加えて、結果を形態学的にモニターすることができる。
【0116】
したがって、このアッセイ系は、低分子治療物質をスクリーニングするために理想的である。一つの態様において、特異的結合に対する構造的基礎を発見するために、ポリグルタミン領域の伸長した反復配列に特異的なsFv結合部位(例えば、相補性決定領域、CDRs)におけるアミノ酸配列の差の比較を行う。特に、ハンチンチンポリペプチドに対して特異的なsFvのH3 CDRループは、内部抗体/ポリペプチド結合相互作用のほとんどの原因であり、したがってH3ペプチド模倣体をこの系を用いてスクリーニングして、無傷の内部抗体の代わりに用いてもよい。次に、この情報を用いて、立体構造エピトープのような異常なポリグルタミン領域と相互作用する他の結合分子を同定することができ、これらを例えばペプチドファージディスプレイライブラリ(ニューイングランドバイオラブス社)を用いて適当な類似体を発見するためにスクリーニングしてもよい。
【0117】
好ましい態様において、上記のアッセイを用いてポリペプチドの蓄積または凝集に対して阻害作用を有する候補結合分子を同定する。もう一つの態様において、候補化合物は、内部抗体の生物活性を模倣する、例えば、凝集を防止してポリペプチドを再ターゲティングしてもよい。
【0118】
本発明の試験化合物は、以下を含む当技術分野で既知の組み合わせライブラリ法を含む様々なアプローチのいずれかを用いて得ることができる:生物ライブラリ;空間的に位置特定可能な平行な固相または液相ライブラリ;脱重畳積分を必要とする合成ライブラリ法;「1ビーズ1化合物」ライブラリ法;およびアフィニティクロマトグラフィー選択を用いる合成ライブラリ法。生物ライブラリアプローチはペプチドライブラリに限定されるが、他の4つのアプローチはペプチド、非ペプチドオリゴマーまたは低分子化合物ライブラリに適用できる(ラム(Lam, K.S.)(1997)Anticancer Drug Des. 12:145)。
【0119】
分子ライブラリを合成するための方法の例は、例えば、デウィット(DeWitt)ら(1993)Proc. Natl. Acad. Sci. USA 90:6909;エルブ(Erb)ら(1994、Proc. Natl. Acad. Sci. USA91:11422);ズッカーマン(Zuckermann)ら(1994、J. Med. Chem. 37:2678);チョー(Cho)ら(1993)Science 261:1303;カレル(Carell)ら(1994)Angew. Chem. Int. Ed. Engl. 33:2059;カレル(Carell)ら(1994)Angrew. Chem. Int. Ed. Engl. 33:2061;およびギャロップ(Gallop)ら(1994)J. Med. Chem. 37:1233に見ることができる。
【0120】
化合物のライブラリは、溶液(例えば、ホーテン(Houghten)、(1992)、Biotechniques 13:412〜421)、またはビーズ(ラム(Lam)、(1991)、Nature 354:82〜84)、チップ(フォドー(Fodor)(1993)、Nature 364:555〜556)、細菌(ラドナー(Ladner)、米国特許第5,223,409号)、胞子(ラドナー(Ladner)、米国特許第5,223,409号)、プラスミド(カル(Cull)ら、(1992)、Proc. Natl. Acad. Sci. USA 89:1865〜1869)、またはファージ(スコットおよびスミス(Scott and Smith)(1990)Science 249:386〜390;デブリン(Devlin)、(1990)、Science 249:404〜406;チャーラ(Cwirla)ら(1990)Proc. Natl. Acad. Sci. USA 87:6378〜6382;フェリチ(Felici)(1991)、J. Mol. Biol. 222:301〜310;およびラドナー(Ladner)、上記)に存在してもよい。
【0121】
一つの態様において、アッセイは、その中で標的ポリペプチド、例えばハンチンチンと内部抗体とを発現する細胞を試験化合物に接触させて、試験化合物がポリペプチド/内部抗体相互作用を変化させるか否かを決定する、細胞に基づくアッセイである。例えば、細胞は、哺乳類起源または酵母細胞となりうる。試験化合物がポリペプチドに結合するか否かを決定することは、例えば試験化合物とポリペプチドとの結合が複合体における標識化合物の検出によって決定されうるように、放射性同位元素または酵素標識に化合物をカップリングさせることによって行うことができる。例えば、試験化合物は、125I、35S、14C、3H、または32Pによって直接または間接的に標識することができ、放射性同位元素は、放射線放出の直接計数またはシンチレーション計数によって検出することができる。または、試験化合物は、例えば、ホースラディッシュペルオキシダーゼ、アルカリホスファターゼ、またはルシフェラーゼによって酵素標識することができ、酵素標識は、適当な基質の産物への変換を測定することによって検出される。
【0122】
いかなる相互作用体も標識せずに、試験化合物が標的ポリペプチドと相互作用できるか否かを決定することも本発明の範囲内である。例えば、マイクロフィジオメーターを用いて、試験化合物または標的ポリペプチドのいずれも標識せずに、標的ポリペプチドと試験化合物との相互作用を検出することができる(マッコネル(McConnell, H.M.)ら、(1992)、Science 257:1906〜1912)。さらにもう一つの態様において、本発明のアッセイは、標的ポリペプチドと内部抗体を試験化合物に接触させ、試験化合物がポリペプチド/内部抗体相互作用を変化させるか否かを決定する細胞不含アッセイである。試験化合物と標的ポリペプチドとの結合は、直接または間接的に決定することができる。候補化合物が標的ポリペプチドに結合するか否かを決定することは、リアルタイム生体分子相互作用分析(BIA)のような技術を用いて行うことができる(シェランダーおよびウルバニクスキー(Sjolander, S. and Urbaniczky, C.)、(1991)、Anal. Chem. 63:2338〜2345およびスザボ(Szabo)ら(1995)、Curr. Opin. Struct. Biol. 5:699〜705)。本明細書において用いられるように、「BIA」は、いかなる相互作用も標識せずにリアルタイムで生体特異的相互作用を調べる技術である(例えば、BIAcoa(商標))。光学現象表面プラズモン共鳴(SPR)の変化は、生体分子間のリアルタイム反応の指標として用いることができる。
【0123】
化合物および天然抽出物の試験ライブラリの多くの薬剤スクリーニングプログラムにおいて、所定の期間において調べる化合物数を最大限にするために高処理量アッセイが望ましい。精製、または半精製蛋白質を用いて行ってもよいアッセイのような細胞不含系において行われるアッセイは、しばしばその中で試験化合物によって媒介される分子標的の変化を迅速に開始させて比較的容易に検出するために作製することができる「一次」スクリーニングと呼ばれる。その上、試験化合物の細胞毒性および/または生物学的利用性の影響は、一般的にインビトロ系では無視することができ、アッセイはその代わりに上流または下流のエレメントによる結合親和性の変化として現れる可能性がある分子標的上の薬剤の影響に主に重点が置かれる。したがって、本発明の例としてのスクリーニングアッセイにおいて、当該化合物を標的ポリペプチドに接触させ、標的ポリペプチドの蓄積または凝集の定量は望ましくない複合体または凝集体の形成を阻害する化合物の有効性を評価することによって決定される。化合物の有効性は、試験化合物の様々な濃度を用いて得られるデータから用量反応曲線を作製することによって評価することができる。その上、対照アッセイも同様に、比較の基準値を提供するために行うことができる。
【0124】
本発明の上記のアッセイ方法の一つ以上の態様において、複合体型と非複合体との分離を促進するため、またはアッセイの自動化に適合させるために標的ポリペプチドを固定することが望ましいかもしれない。試験化合物の標的ポリペプチドへの結合は、反応物質を含むために適したいかなる容器においても行うことができる。そのような容器の例には、マイクロタイタープレート、試験管、および微量遠心管が含まれる。一つの態様において、一つまたはそれ以上の蛋白質をマトリクスに結合させるドメインを付加する融合蛋白質を提供することができる。例えば、グルタチオンSトランスフェラーゼ/標的ポリペプチド融合蛋白質をグルタチオンセファロースビーズ(シグマケミカル社、セントルイス、ミズーリ州)またはグルタチオン誘導体化マイクロタイタープレートに吸着させることができ、次にこれを試験化合物または試験化合物および内部抗体と組み合わせて、複合体形成を行うための条件(例えば、イオン強度およびpHの生理的条件)でインキュベートする。インキュベーション後、ビーズまたはマイクロタイタープレートウェルを洗浄して、いかなる未結合成分も除去して、ビーズの場合にはマトリクスを固定して、複合体を例えば上記のように直接または間接的に測定する。または、複合体は、マトリクスから解離させることができ、標的ポリペプチド結合または活性のレベルを標準的な技術を用いて決定することができる。
【0125】
そのようなスクリーニング方法によって選択された化合物の試験は、モデルの階層性を用いて、トランスフェクト細胞モデルから開始して、適当な正常または変異体マウスからの脳切片について行い、次にウイルスを形質導入したマウスまたは疾患のトランスジェニックマウスモデルにおいて試験を行って、ハンチントンのような所定の神経疾患に対する関連を調べてもよい。これらの平均によって、試験化合物を臨床試験で利益が得られる可能性がある少数の化合物まで狭めてもよい。
【0126】
マトリクス上に蛋白質を固定する他の技術はまた、本発明のスクリーニングアッセイにおいて用いることができる。例えば、いずれかの標的ポリペプチドをビオチンおよびストレプトアビジンの結合を利用して固定することができる。ビオチン結合標的ポリペプチドまたは内部抗体は、当技術分野で既知の技術を用いて(例えば、ビオチン結合キット、ピアスケミカル社、ロックフォード、イリノイ州)ビオチンNHS(Nヒドロキシスクシニミド)から調製して、ストレプトアビジンをコーティングした96ウェルプレート(ピアスケミカル社)のウェルに固定することができる。
【0127】
本発明のさらにもう一つの局面において、標的ポリペプチドまたは内部抗体結合領域は、二ハイブリッドアッセイまたは三ハイブリッドアッセイにおいて、標的ポリペプチドおよび/または内部抗体に結合または相互作用する他の蛋白質また化合物を同定するための「おとり蛋白質」として用いることができる(例えば、米国特許第5,283,317号;ゼルボス(Zervos)ら(1993)、Cell 72:223〜232;マデュラ(Madura)ら、(1993)、J. Biol. Chem. 268:12046〜12054;バーテル(Bartel)ら(1993)、Biotechniques 14:920〜924;イワブチ(Iwabuchi)ら、(1993)、Oncogene 8:1693〜1696;およびブレント(Brent)、国際公開公報第94/10300号を参照のこと)。
【0128】
本発明はさらに、上記のスクリーニングアッセイによって同定される新規物質およびこれらのアッセイを用いることによってそのような物質を産生するためのプロセスに関する。したがって、一つの態様において、本発明は、上記のスクリーニングアッセイ(例えば、細胞に基づくアッセイまたは細胞不含アッセイ)のいずれか一つの段階を含む方法によって得ることができる化合物または物質を含む。例えば、一つの態様において、本発明には、本明細書に記載の方法のいずれかによって得ることができる化合物または物質が含まれる。
【0129】
したがって、物質、例えば適当な動物モデルにおいて本明細書に記載のように同定された化合物または内部抗体をさらに用いることも本発明の範囲内である。例えば、本明細書において記述されたように同定された物質を動物モデルにおいて用いて、そのような物質による治療の有効性、毒性、または副作用を決定することができる。または、本明細書において同定された物質を動物モデルにおいて用いて、そのような物質の作用機序を決定することができる。さらに、適当であると思われる場合には、そのような物質をヒト対象、好ましくは神経障害の危険にさらされている対象に投与してもよい。
【0130】
本発明はまた、本明細書に記載したいかなる神経障害も診断、予後を決定、および治療するために、上記のスクリーニングアッセイによって同定された新規物質を用いることに関する。したがって、本明細書に記載のいかなる神経障害の診断、予後、または治療において用いられる薬剤または薬学的組成物の設計、処方、合成、製造、および/または産生においてそのような物質を用いることは本発明の範囲内である。
【0131】
V.使用方法
予防的方法
一つの局面において、本発明は、ポリペプチドの蓄積またはそのような複合体もしくは凝集体の形成を阻害する物質を対象に投与することによって、対象において、神経ポリペプチドの異常または望ましくない細胞内蓄積または凝集に関連した疾患または病態を予防する方法を提供する。好ましい態様において、疾患または病態は神経疾患(例えば、ハーディ(Hardy)ら、Science 282:1075〜1079(1998)から抜粋した表2を参照のこと)である。そのような異常または望ましくないポリペプチド活性によって引き起こされる、またはそれらに関係する神経疾患の危険にさらされている対象は、例えば、当技術分野で認識された診断または予後的技法によって同定することができる。したがって、陽性診断が示されると、本明細書に記載した適当な化合物または内部抗体のいかなる投与も、神経疾患または障害が予防される、その重症度が減少する、またはその進行もしくは発症が遅れるように、異常に特徴的な症状が発現する前に予防的物質として投与してもよい。
【0132】
常染色体優性原発性神経変性疾患
【表2】
Ch、染色体;PrP、プリオン蛋白質;T、タングル;LB、レーヴィ小体;+、存在する;AD、アルツハイマー病;PD、パーキンソン病;HD、ハンチントン病;SOD;スーパーオキシドジスムターゼ。
* SBMAは技術的には常染色体優性ではないが、その細胞での作用機序からみておそらく優性である。
【0133】
治療的方法
本発明のもう一つの局面は、標的ポリペプチド、好ましくは神経細胞において発現されたポリペプチドの異常な発現または活性を特徴とする疾患または障害に罹患した人を治療する方法に関する。
【0134】
したがって、本発明は、中枢または末梢神経系の細胞に関連する疾患または病態を治療する方法を提供する。例えば、本発明は、以下の細胞のいずれかの異常な細胞増殖、分化、または細胞死に至る可能性がある異常なポリペプチド蓄積または凝集に関連した神経系の病変を治療する方法を提供する:ニューロン(例えば、運動神経ニューロン、知覚神経ニューロン)、シュワン細胞、プルキンエ細胞、星状細胞、小膠細胞、上衣細胞、乏突起膠細胞または他のいかなるタイプの神経細胞。神経系の障害には、アルツハイマー病、パーキンソン病、ハンチントン病、プリオン疾患、前頭側頭痴呆(FTD)、筋萎縮性側索硬化症(ALS)、脊髄延髄筋萎縮症(SBMAまたはケネディ病)、歯状赤核淡蒼球萎縮症(DRPLA)、1型脊髄小脳運動失調(SCA1)、2型脊髄小脳運動失調(SCA2)、3型脊髄小脳運動失調(SCA3またはマチャドジョゼフ病(MJD))、4型脊髄小脳運動失調(SCA4)、5型脊髄小脳運動失調(SCA5)、6型脊髄小脳運動失調(SCA6)、および7型脊髄小脳運動失調(SCA7)が含まれるが、これらに限定しない。
【0135】
望ましくない細胞内ポリペプチド複合体/蓄積の形成の阻害
一つの態様において、本発明は、望ましくない細胞内ポリペプチド凝集または蓄積の形成を阻害する方法を提供する。特に、本発明の内部抗体は、ポリペプチドに特異的に結合することによって望ましくないポリペプチドが複合体または凝集体を蓄積または形成することを阻害するように設計する。いくつかの状況において、ポリペプチドに対して内部抗体が単に特異的結合しただけで、ポリペプチドを含む複合体または凝集体の形成を阻害するために十分であるかもしれない。そのような内部抗体は、標的ポリペプチドに対して親和性を有する一つまたはそれ以上の原子価、すなわち結合部位を有し、例えば、立体的な妨害によって、または蛋白質をより不安定にする変化した立体構造を誘導することによって、および/またはポリペプチド複合体がポリペプチドの蓄積または凝集に至る相互作用を形成できないようにする、例えばその溶解性を増加させる、または凝集体を形成する臨界濃度を増加させることによって、ポリペプチドの蓄積または凝集を阻害してもよい。
【0136】
したがって、本発明は、本明細書に記載の内部抗体のいずれかを、単独、またはもう一つの内部抗体、または望ましくない細胞内ポリペプチド蓄積もしくは凝集を形成することが知られているポリペプチドをターゲティングするために適した他の治療物質と組み合わせて、対象に投与する方法を提供する。一つの態様において、対象は実験動物であってもよい。好ましい態様において、本発明の方法は、ヒト患者を治療するために適している。もう一つの好ましい態様において、内部抗体または結合分子の投与は、アルツハイマー病の治療または治癒のためである。なおもう一つの好ましい態様において、内部抗体または結合分子の投与は、ハンチントン病の治療のためである。本発明の一つの態様において、内部抗体を投与する方法は、ポリペプチドとしてである。本発明の好ましい態様において、内部抗体は、下記の章に記載するように、遺伝子治療の形で発現可能な核酸として投与される。
【0137】
遺伝子治療
本発明の内部抗体またはペプチド骨格の結合分子は、遺伝子治療として投与した場合に、本明細書に記載のいかなる神経疾患の治療においても特に有用である。一般的アプローチは、導入された遺伝子材料によってコードされる一つまたはそれ以上の遺伝子産物が、望ましくないポリペプチド蓄積または凝集を阻害するために細胞において産生されるように、内部抗体またはペプチド骨格の結合分子をコードする核酸を細胞に導入することを含む。
【0138】
内部抗体またはペプチド骨格の結合分子をコードする本発明の核酸分子は、核酸によってコードされ、ベクターに挿入され、遺伝子治療ベクターとして用いることができる。遺伝子治療ベクターは、例えば静脈内注射、局所投与(例えば、米国特許第5,328,470号を参照のこと)または定位注入(例えば、チェン(Chen)ら、(1994)、PNAS 91:3054〜3057)によって対象に輸送することができる。遺伝子治療ベクターの薬学的調製物は、許容される希釈剤において遺伝子治療ベクターを含みうる、またはその中で遺伝子輸送媒体が埋め込まれている徐放性マトリクスを含みうる。または、組み換え型細胞から完全な遺伝子輸送ベクター、例えばレトロウイルスベクターを無傷で産生することができる場合、薬学的調製物は、遺伝子輸送系を産生する一つまたはそれ以上の細胞を含みうる。薬学的組成物は容器に含めることができる。遺伝子治療ベクターは、典型的に、内因性の転写因子に反応する構成的な調節エレメントを利用する。好ましい態様において、内部抗体またはペプチド結合分子をコードする遺伝子治療ベクターは、アデノウイルス、アデノ随伴ウイルス、レトロウイルス、または単純ヘルペスウイルスであるウイルスに由来する発現ベクターである(例えば、デュリング(During)ら、Mol. Med. Today 4:485〜93(1998);デュリング(During)ら、Nat. Med. 4:1131〜5(1998);デュリング(During)ら、Gene Ther. 5:820〜7(1998);カプリット(Kaplitt)ら、Pediatr. Neurosurg. 28:3〜14(1998);フリース(Freese)ら、Epilepsia 38:759〜66(1997);オコナー(O'Connor)ら、Exp. Neurol. 148:167〜78(1997);デュリング(During)ら、Exp. Neurol. 144:74〜81(1997);フリース(Freese)ら、Mov. Disord. 11:469〜88(1996);デュリング(During)、Lancet 348:618(1996);フリース(Freese)ら、J. Clin. Endocrinol. Metab. 81:2404〜4(1996);デュリング(During)ら、Clin. Neurosci. 3:292〜300(1995〜6);デュリング(During)ら、Science 266:1399〜403(1994);およびカプリット(Kaplitt)ら、Nat. Genet. 8:148〜54(1994)を参照のこと)。
【0139】
非ウイルス遺伝子輸送媒体はまた、本発明の治療的プラスミドの細胞特異的輸送を行うための手段である。これらは、DNA複合体形成剤とカップリング、または融合した従来の抗体または一本鎖Fv抗体である(ウエレク(Uherek)ら、J. Biol. Chem. 273:8835〜8841(1998);フォスター(Foster)ら、Human Gene Therapy 8:719〜727(1997);チェン(Chen)ら、Gene Therapy 2:116〜123(1995)を参照のこと)。このクラスの遺伝子輸送媒体はまた、リポソームにカップリングした抗体またはその断片が含まれる(ヒュアン(Huang)ら、米国特許第4,925,661号/第4,957,735号/第6,008,202号)。
【0140】
遺伝子治療のアプローチに関する検討に関しては、アンダーソン(Anderson, W.F.)(1992)Science 256:808〜813;ミラー(Miller, A.D.)(1992)Nature 357:455〜460;フリードマン(Friedmann, T.)(1989)Science 244:1275〜1281;およびクーノヤー(Cournoyer, D.)ら(1990)Curr. Opin. Biotech. 1:196〜208を参照のこと。
【0141】
細胞タイプ、遺伝子、および遺伝子治療の方法に関する詳しい記述は、例えば、ウィルソン(Wilson, J.M.)ら(1988)Proc. Natl. Acad. Sci. USA 85:3014〜3018;アーメンターノ(Armentano, D.)ら(1990)Proc. Natl. Acad. Sci. USA 87:6141〜6145;ウルフ(Wolff, J.A.)ら(1990)Science 247:1465〜1468;チョウデュリー(Chowdhury, J.R.)ら、(1991)Science 254:1802〜1805;フェリー(Ferry, N.)ら(1991)Proc. Natl. Acad. Sci. USA 88:8377〜8381;ウィルソン(Wilson, J.M.)ら(1992)J. Biol. Chem. 267:963〜967;クアンチン(Quantin, B.)ら(1992)Proc. Natl. Acad. Sci. USA 89:2581〜2584;ダイ(Dai, Y.)ら(1992)Proc. Natl. Acad. Sci. USA 89:10892〜10895;バン・ビューセケム(van Beusechem, V.W.)ら(1992)Proc. Natl. Acad. Sci. USA 89:7640〜7644;ローゼンフェルド(Rosenfeld, M.A.)ら(1992)Cell 68:143〜155;ケイ(Kay, M.A.)ら、(1992)Human Gene Therapy 3:641〜647;クリスチャーノ(Christiano, R.J.)ら(1993)Proc. Natl. Acad. Sci. USA 90:2122〜2126;ヒュ(Hwu, P.)ら(1993)J. Immunol. 150:4104〜4115;およびヘルツおよびジェラルド(Herz, J. and Gerard, R.D.)(1993)Proc. Natl. Acad. Sci. USA 90:2812〜2816;バコード・レーヴィ(Bachoud-Levi)ら(1998)Progress in Brain Research, 117:511〜524;ローウェンスタイン(Lowenstein)ら(1998)Progress in Brain Research 117:485〜501;ウェイヤーブロック(Weyerbrock)ら(1999)Current Opinion in Oncology 11:168〜173;カルパチ(Karpati)ら(1996)Trends Neurosci. 19:49〜54;スケイパー(Skaper)ら(1998)Mol. and Cell Neurosci. 12:179〜193;スール(Suhr)ら(1999)Arch. Neurol. 56:287〜292;ラビノヴィッツ(Rabinowitz)ら(1998)Current Opinion in Biotechnology 9:470〜475;タイラー(Tyler)ら(1999)P.N.A.S. 96:7053〜7058を参照のこと。
【0142】
VI.薬学的組成物
本発明の内部抗体は、投与に適した薬学的組成物に組み入れることができる。そのような組成物は典型的に、内部抗体と薬学的に許容される担体とを含む。本明細書において用いられるように、「薬学的に許容される担体」という用語には、例えばポリペプチド、核酸、ペプチド骨格の結合分子、または低分子の形での内部抗体または結合分子の薬学的な投与に適合する、いかなるおよび全ての溶媒、分散媒体、コーティング、抗菌および抗真菌剤、等張および吸収遅延剤等も含まれると解釈される。そのような媒体および物質を薬学的に活性な物質に用いることは、当技術分野で周知である。従来の培地または物質が内部抗体と不適合性である場合を除き、組成物におけるその使用が意図される。補助活性化合物もまた、組成物に組み入れることができる。
【0143】
本発明の薬学的組成物は、その意図する投与経路と適合するように製剤化される。投与経路の例には、非経口、例えば静脈内、皮内、くも膜下、皮下、経口(例えば吸入)、経皮(局所)、経粘膜、および直腸投与が含まれる。非経口、皮内、または皮下投与に用いられる溶液または懸濁液は、以下の成分を含みうる:注射用水、生理食塩液、固定油、ポリエチレングリコール、グリセリン、プロピレングリコール、または他の合成溶媒のような滅菌希釈剤;ベンジルアルコールまたはメチルパラベンのような抗菌剤;アスコルビン酸または重炭酸ナトリウムのような抗酸化剤;エチレンジアミン四酢酸のようなキレート化剤;酢酸塩、クエン酸塩またはリン酸塩のような緩衝剤および塩化ナトリウムまたはデキストロースのような等張性調節剤。pHは、塩酸または水酸化ナトリウムのような酸または塩基によって調節することができる。非経口調製物は、アンプル、使い捨てシリンジ、またはガラスもしくはプラスチック製の多用量バイアルに封入することができる。
【0144】
注射可能な用途に適した薬学的組成物には、滅菌水溶液(水溶性の場合)または分散剤および滅菌注射溶液または分散液の即時調製用の滅菌粉末が含まれる。静脈内投与の場合、適した担体には、生理食塩液、静菌水、CremophorEL(商標)(BASF、パルシッパニー、ニュージャージー州)またはリン酸緩衝生理食塩液(PBS)が含まれる。全ての場合において、組成物は滅菌でなければならず、容易なシリンジ操作性が存在する程度に流動性でなければならない。これは製造および保存条件で安定でなければならず、細菌および真菌のような微生物の混入作用に対して保護されなければならない。担体は、例えば、水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコール、液体ポリエチレングリコール等)、およびその適した混合物を含む溶媒または分散媒体となりうる。適切な流動性は、例えば、レシチンのようなコーティングを用いることによって、分散剤の場合には必要な粒子径を維持することによって、そして界面活性剤を利用することによって維持することができる。微生物の作用の予防は、様々な抗菌剤および抗真菌剤、例えばパラベン、クロロブタノール、フェノール、アスコルビン酸、チメロサル等によって行うことができる。多くの場合において、等張剤、例えば糖、マンニトール、ソルビトールのような多価アルコール、塩化ナトリウムを組成物に含めることが好ましいであろう。注射可能組成物の持続的な吸収は、吸収を遅らせる物質、例えばモノステアリン酸アルミニウムおよびゼラチンを組成物に含めることによって得ることができる。
【0145】
滅菌注射溶液は、先に列挙した成分の一つまたは組み合わせと共に適当な溶媒に内部抗体の必要量を組み入れて、必要に応じてその後濾過滅菌することによって調製することができる。一般的に、分散剤は、基礎となる分散媒体と先に列挙した必要な他の成分とを含む滅菌媒体に活性化合物を組み入れることによって調製される。滅菌注射溶液を調製するための滅菌粉末の場合、調製の好ましい方法は、先に濾過滅菌したその溶液から活性成分プラスさらに望ましい成分の粉末を生じる真空乾燥および凍結乾燥である。
【0146】
経口組成物は一般的に、不活性希釈剤または食用担体を含む。それらは、ゼラチンカプセルに封入することができ、または錠剤に圧縮することができる。経口治療的投与の目的の場合、活性化合物を賦形剤と共に組み入れて、錠剤、トローチ剤、またはカプセル剤の形で用いることができる。経口組成物はまた、液体担体中の化合物が経口適用され、ヒューッと音を立てて喀出されるか、または飲み込まれる、口腔洗浄剤として用いるための液体担体を用いて調製することができる。薬学的に適合性の結合物質、および/またはアジュバント材料は組成物の一部として含めることができる。錠剤、丸剤、カプセル剤、トローチ剤等は、以下の成分のいかなるものも、または類似の性質の以下の化合物を含みうる:微結晶セルロース、トラガカントガム、またはゼラチンのような結合剤;デンプンまたは乳糖のような賦形剤;アルギン酸、プリモゲル、またはコーンスターチのような崩壊剤;ステアリン酸マグネシウムまたはステローツのような潤滑剤;コロイド二酸化珪素のような滑り剤;蔗糖またはサッカリンのような甘味料;またはペパーミント、サリチル酸メチル、またはオレンジ香料のような着香料。
【0147】
吸入によって投与するために、内部抗体は、適した噴射剤、例えば二酸化炭素のような気体を含む圧縮容器もしくはディスペンサー、またはネブライザーからエアロゾルスプレーの形で輸送することができる。
【0148】
全身投与はまた、経粘膜または経皮手段によって行うことができる。経粘膜または経皮投与の場合、浸透すべき障壁に対して適当な浸透剤を製剤に用いる。そのような浸透剤は、一般的に当技術分野で既知であり、例えば、経粘膜投与の場合、洗浄剤、胆汁酸塩、およびフシジン酸誘導体が含まれる。経粘膜投与は、点鼻スプレーまたは坐剤を利用することによって行うことができる。経皮投与の場合、活性化合物を、当技術分野で一般的に既知の軟膏、軟膏、ゲル、またはクリームに製剤化する。
【0149】
内部抗体はまた、坐剤(例えば、カカオバターおよび他のグリセリドのような従来の坐剤基剤と共に)の形で、または直腸輸送のための固定浣腸の形で調製することができる。
【0150】
一つの態様において、内部抗体は、インプラントおよび微量封入輸送系を含む徐放製剤のような体内からの急速な排泄に対して内部抗体を保護する担体と共に調製される。エチレン酢酸ビニル、多価無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、およびポリ乳酸のような生体分解性の生体適合性のポリマーを用いることができる。そのような製剤を調製する方法は、当業者に明らかであろう。材料はまた、アルザコーポレーションおよびノバファーマシューティカルズインクから購入することができる。リポソーム懸濁液(ウイルス抗原に対するモノクローナル抗体によって感染細胞にターゲティングされるリポソームを含む)もまた、薬学的に許容される担体として用いることができる。これらは、例えば、米国特許第4,522,811号に記載されるように、当業者に既知の方法に従って調製することができる。
【0151】
投与を容易にするため、そして用量を均一にするために、経口または非経口組成物を単位投与剤形に処方することは特に都合がよい。本明細書において用いられる単位投与剤形は、治療すべき対象の単位用量として適している物理的に個別の単位を意味する;各単位は、必要な薬学的担体に関連して望ましい治療効果を生じるように計算された内部抗体の既定量を含む。本発明の単位投与剤形の仕様は、内部抗体の独自の特徴、得られる特定の治療効果、および対象を治療するためにそのような内部抗体を合成する技術分野に固有の制限によって影響を受け、直接左右される。
【0152】
そのような内部抗体の毒性および治療効果は、例えば、LD50(集団の50%に対して致死的な用量)およびED50(集団の50%において治療的に有効な用量)を決定するために、細胞培養または実験動物において標準的な薬学的技法によって決定することができる。毒性と治療効果の用量比が治療指数であり、これはLD50/ED50の比として表すことができる。大きい治療指数を示す内部抗体が好ましい。毒性副作用を示す内部抗体を用いてもよいが、非感染細胞に対する起こりうる障害を最小限にして、それによって副作用を減少させるために、罹患組織部位にそのような内部抗体をターゲティングする輸送システムを設計する場合には注意しなければならない。
【0153】
細胞培養アッセイおよび動物実験から得られたデータは、ヒトにおいて用いられる用量範囲を決定するために用いることができる。そのような化合物の用量は、ほとんど、または全く毒性を示さないED50を含む循環中の濃度範囲内に存在する。用量は、用いる投与剤形および利用する投与経路に応じてこの用量範囲内で変化してもよい。本発明の方法において用いられる内部抗体に関して、治療的有効量は、細胞培養アッセイからまず推定することができる。用量は、細胞培養において決定したIC50(すなわち、症状の半最大阻害が得られる試験内部抗体の濃度)を含む循環中の血漿濃度範囲が得られるように動物モデルにおいて処方してもよい。そのような情報を用いて、ヒトにおいて有用な用量をより正確に決定することができる。血漿レベルは、例えば、高速液体クロマトグラフィーによって測定してもよい。
【0154】
本発明の内部抗体をコードする遺伝子をベクターに挿入して、遺伝子治療ベクターとして用いることができる。遺伝子治療ベクターは、上記の章において記載したように、対象に輸送することができる。
【0155】
薬学的組成物は、投与説明書と共に、容器、パック、またはディスペンサーに含めることができる。
【0156】
本発明は、制限的に解釈してはならない以下の実施例によってさらに説明する。
【0157】
実施例
実施例全体を通じて、特に明記していない限り、以下の材料と方法を用いた。
【0158】
材料と方法
一般的に、本発明の実践は、特に明記していない限り、化学、分子生物学、組み換え型DNA技術、PCR技術、免疫学(特に、例えば抗体技術)および必要な細胞培養または動物飼育技術に関する従来の技術を用い、これらは当業者の範囲内であり、文献に説明されている。例えばサムブルック、フリッチュおよびマニアティス(Sambrook, J., Fritsh, E.F., and Maniatis, T.)、「分子のクローニング(Molecular Cloning)」、コールドスプリングハーバー研究所出版(1989);「DNAクローニング(DNA Cloning)」、第1巻および2巻(グロバー(D.N. Glover)編、1985);「PCRハンドブック:核酸化学の現行プロトコール(PCR Handbook Current Protocols in Mucleic Acid Chemistry)」、ビューケージ(Beaucage)編、ジョンウィリー&サンズ(1999);「抗体操作プロトコール(分子生物学の方法)(Antibody Engineering Protocols)(Methods in Molecular Biology)」、510、ポール(Paul, S.)、ヒュマナ出版(1996);「抗体の操作:実践アプローチ(Antibody Engineering:A Practical Approach(実践アプローチシリーズ、169)」、マッカファーティ(McCafferty)編、アール出版(1996);「抗体:実験マニュアル(Antibodies:A Laboratory Manual)」、ハーロウ(Harlow)ら、C.S.H.J.出版、(1999);「分子生物学の現行プロトコール(Current Protocols in Molecular Biology)」、アウスユベール(Ausubel)ら、編、ジョンウィリー&サンズ(1992);「哺乳類細胞大量培養技術(Large-Scale Mammalian Cell Culture Technology)」、ルビニエッキ(Lubiniecki, A.)編、マーセルデッカー出版(1990);ならびに「マウス胚の操作(Manipulating the Mouse Embryo)」、ホーガン(Hogan)ら、C.S.H.L.出版、(1994)を参照のこと。
【0159】
GST融合蛋白質およびGFP融合蛋白質の構築
全てのプラスミド構築に関して標準的な技術を用いた(例えば、アウスユベール(Ausubel)ら、およびサムブルック(Sambrook)ら、上記を参照のこと)。
【0160】
DRPLA構築物は、オノデラ(Onodera)らから得るか、またはオノデラ(Onodera)ら(FEBS letters(1996)399:135〜139)が記述したように誘導する。
【0161】
HD構築物、例えばHD-Q25およびHD-Q104 cDNAは、pcDNA-HD17およびpcDNA-HD14(カザンツェフ(Kazantsev)らからの寄贈、Proc. Natl. Acad. Sci. USA 96:11404〜11409(1999))から、BamHI部位を含むHDセンスプライマー
およびc-mycアンチセンスプライマー
を用いるPCR(95℃で1分間、50℃で1分間、および72℃で1分間を25サイクル)によって増幅した。PCR産物は、プロテナーゼK(最終濃度1mg/ml)によって37℃で30分間処理してから、QIAEX IIキット(キアゲン社)を用いてゲル精製を行った。次に、精製したHD-Q25およびQ104 PCR産物をXmaIおよびBamHI制限酵素によって消化してpGEXベクターに個別にライゲーションした。HD-Q25およびQ104 PCR産物はまた、BamHIおよびSmaI制限酵素によって消化して、BamHIおよびNruIによって消化したpQBi25c3ベクター(QUANTUMバイオテクノロジー社、モントリオール、ケベック州、カナダ)に個別にライゲーションした。
【0162】
プラスミドpQBi-25c3-HD-Q(-)は、pQBi-25c3-HD-Q104をEcoRIおよびHindIIIによって消化することによって調製した。酵素を75℃で10分不活化した後、消化した産物をdNTPの存在下でクレノウポリメラーゼによって30℃で30分処理して、平滑末端DNAを生じた。次に、消化したベクターをゲル精製して、自己ライゲーションした。
【0163】
プラスミドpGEX-HD-42QおよびHD-65Qは以下のように調製した:42Qおよび65QをコードするcDNAセグメントを、pBSK-42QおよびpBSK-65QからHindIIIおよびXmaIによる消化によって単離した。ゲル精製を行った後、cDNAセグメントを、同じ制限酵素によって消化してHD-25Q cDNAコード配列を置換したpGEX-HD 25Qにクローニングした。本発明の発現された実験ポリペプチドの関連するアミノ酸配列を下記の表3に要約する。
【0164】
実験ポリペプチドのアミノ酸配列
【表3】
略語:DRPLA、歯状赤核淡蒼球萎縮症(アトロフィン1);HD、ハンチントン病(ハンチンチン);GST、グルタチオントランスフェラーゼ;GFP、緑色蛍光蛋白質。アトロフィン1およびハンチンチン蛋白質配列には下線を示す。隣接する配列は発現ベクターの一部である。GSTとGFPのポリペプチド配列は当技術分野において認識されており、例えばGFP:プラスミドpQB125-fC3(カタログ番号AFP2133)、カンタムバイオテクノロジーズ社(1801 de Maissonneuve Bvld. West Montreal、Quebec, h3H 159、カナダ);GST:プラスミドpGEX-4T-3(カタログ番号27-4583-01)、ファルマシア社(ゲンバンクアクセッション番号U13855号);およびオノデラ(Onodera)ら(1996)「伸長したポリグルタミンドメイン蛋白質の大腸菌における毒性(Toxicity of expanded polyglutamine-domain proteins in Escherichia coli)」)に記載のGST-DRPLA構築物を参照のこと。
【0165】
蛋白質発現と精製
N-HD-C4 sFvからのVHリンカーVL cDNAコード配列は、NcoIおよびNotI制限酵素による消化によってpHEN-1ベクターから切除した。ゲル精製を行った後、cDNAを、同じ制限酵素によって先に消化されたpSYN1ベクターにライゲーションした。次に、ライゲーションしたプラスミドを電気穿孔によってDH5αコンピテント細菌に導入して、増幅されたプラスミドDNAをQIAプレップスピンミニプレップキット(キアゲンインク、バレンシア、カリフォルニア州)によって精製した。N-HD-C4 sFvは、以下のように大腸菌株TG1において発現させた:形質転換体を100 μg/mlアンピシリンと1%グルコースを含む2×TY培地10 mlに接種した。37℃で一晩インキュベートした後、培養を100 μg/mlアンピシリンと0.1%グルコースを含む2XTY培地1Lに接種して、Abs600 nm=1が得られるまで37℃で強く攪拌した。次に、IPTGを加えて最終濃度0.1 nMとして、培養をRTで200 rpmで一晩インキュベートした。遠心によって細胞を回収して、pH 8.0のPBS 30 mlに浮遊させた。ライソザイムを最終濃度1mg/mlとなるように加えて、混合物をRTで5分インキュベートした。次に、細菌溶解物を氷中で超音波処理して、染色体DNAを切断して、25,000×gで4℃で30分間遠心して細菌の破片および他の不溶性材料を除去した。上清を、予め洗浄したNi-NTA樹脂(キアゲン社)の20 mMイミダゾールを含むPBS溶液1mlと共に4℃で2時間インキュベートした。次に、樹脂をPBS 30 mlによって洗浄した後、35 mMイミダゾールを含むPBS 10 mlおよび40 mMイミダゾールを含むPBS 10 mlによって洗浄した。次に、sFv分子を、250 mMイミダゾールを含むPBSによって1ml分画に溶出した。当該分画をプールしてPBS中で透析した。
【0166】
GST融合蛋白質は、以下のように大腸菌株BL21において発現させた:新鮮な形質転換体を、100 μg/mlアンピシリンと0.1%グルコースを含むLB培地1Lに接種して、Abs600 nm=1が得られるまで37℃で強く攪拌した。次に、IPTGを最終濃度0.1 nMとなるように加えて、培養を200 rpmで3時間インキュベートした。遠心によって細胞を回収して、pH 8.0のPBS 30 mlに浮遊させた。ライソザイムを最終濃度1mg/mlとなるように加えて、混合物をRTで5分インキュベートした。次に、細菌溶解物を氷中で超音波処理して、25,000×gで4℃で30分間遠心した。上清を、予め洗浄したグルタチオンセファロース4Bビーズ(ファルマシア社)のPBS溶液1mlと共に4℃で2時間インキュベートした。次に、樹脂をPBS 50 mlによって洗浄した後、蛋白質を10 mM還元グルタチオンを含む50 mMトリス、pH 8.0によって1ml分画に溶出した。当該分画をプールしてPBS中で透析した。
【0167】
免疫精製実験
N-HD-C4 sFv約3μgをGST-DRPLA-Q35、GST-HD-Q25、またはGST-HD-Q42のPBS溶液のいずれか4μg(最終容量200 μl)と共に混合して、RTで1時間揺り動かした。グルタチオンセファロース4Bビーズ(50%スラリー、PBSで洗浄済み)20 μlを加えて、RTでさらに30分インキュベートして、sFv/GST融合蛋白質複合体を捕獲した。ビーズを12,000×gで10分間遠心することによって回収して、PBS1mlで2回洗浄した。複合体を、10 mM還元グルタチオンを含むpH 8.0の50 mMトリス10 μlと共に室温で10分間溶出した。次に、試験管を12,000×gで1分間遠心して、可溶性複合体を含む上清を新しい試験管に移し、2×SDS試料緩衝液の等量と混合して、95℃に加熱してからSDS-PAGEにローディングした。
【0168】
COS7細胞の同時トランスフェクションと免疫蛍光
COS7細胞を6ウェルプレートのカバーガラス(細胞105個/ウェル)上で増殖させて、スーパーフェクトトランスフェクション試薬(キアゲン社)5μlを用いて、N-HD-C4 sFvまたは陰性対照sFv(HAタグおよび核配置配列(NLS)を含む)のいずれかを含むプラスミドと、GFP標的化融合蛋白質を含むプラスミドを3:1の比率(標的に対するsFv)で同時トランスフェクトした。トランスフェクションの24時間後、細胞を以下のように免疫蛍光のために調製した:細胞を4%パラホルムアルデヒドによって固定して、0.2%トライトンX100によって透過させた。二次抗体の非特異的結合を防止するために、細胞を10%正常ヤギ血清の3%BSA溶液によってブロッキングした。次に、予め吸収させたポリクローナルウサギ抗HA IgGを加えてsFv抗体を検出し、結合したIgGをローダミン結合ヤギ抗ウサギIgG抗体(ピアス社)によって顕色させた。
【0169】
内部抗体
再ターゲティングする内部抗体は、以下のように構築した。簡単に説明すると、プラスミドpcDNA3.1(+)(インビトロゲン社、カールスバッド、カリフォルニア州)を、HA免疫タグを表すインフルエンザ血液凝集素エピトープ(YPYDVPDYA(配列番号:43))と、SV40核ターゲティング配列(TPPLLLRLV(配列番号:44))、ライソゾームターゲティングシグナル、またはターゲティングシグナルなし(ターゲティングシグナルが存在しなければsFvは細胞質に残される)のいずれか、およびハンチンチンに特異的なC4 sFv(抗HD-C4)、または無関係なsFv対照をコードするように改変した。sFv内部抗体構築物の発現は、サイトメガロウイルス(CMV)プロモーターの制御下であった。
【0170】
速度論的結合分析
精製scFvクローンC4のKdは、BIAコア2000(ビアコアAG社、アップサラ、スウェーデン)において表面プラズモン共鳴を用いて決定した。BIAコアフローセルにおいて、ビオチン結合HDペプチド(1M NaClおよび50 mM NaOH中で250 nM)の約50共鳴単位(RU)を、1M NaClの50 mM NaOH溶液を連続的に1分間3回注入して予め条件を設定したストレプトアビジンセンサーチップにカップリングさせた。カップリングしたペプチドのこの量によって、結合した内部抗体の最大量40〜80 RUが得られた。内部抗体が結合した後の表面を再生するために、1M NaClを含む50 mM NaOH5μlを注入して、基準値に回復させた。濃度範囲60〜100 nMで5μl/分の連続的な流れの下で結合を測定した。konはln(dR/dt)/t対濃度のプロットから決定し、式中Rは反応であり、tは時間である(カールッソン(Karlsson)ら、J. Immunol. Methods 145:229〜240(1991))。koffは、流速20 μl/分を用いることによって分析した内部抗体の最高濃度でのセンサーグラムの解離部分から決定した。Kdはkoff/konとして計算した。
【0171】
脳組織培養
臓器型切片培養は、公表された方法に軽微な改変を加えて(1)(14)、P12〜P14小脳、大脳皮質、または線条体から作製した。脳をマウスから摘出して、高マグネシウム(1.8 mM CaCl2、および2.4 mM MgCl2)HEPES緩衝ハンクス生理食塩液(HBHS)の4℃のビーカーに入れた。次に脳をブロックにして、プレキシグラス(厚さ0.3 cm)の1cm2片上で2%アガロースブロックの隣にSuperGlue(登録商標)を用いて載せる。歯科用ワックスを切片チャンバー組織ホルダー上のプラスチックに載せる。載せた後、組織をビブラトームの切片チャンバーに置く。ビブラトームの水槽は、冷却した4℃の高マグネシウムHBHSを含む。温度は切片チャンバーに接続したペルティエ熱電流冷蔵システム(FHC、ブランズウィック、メイン州)によって維持する。厚さ300、または400 μmの切片を当該組織から切断する。新しい各切片を作製すると、それを切片の回収が終了するまで4℃の増殖培地に入れた。一つの調製物からの切片を封入したフィルター単位に入れて、高マグネシウム人工脳脊髄液(12 mM NaCl、0.33 mM KCl、0.12 mM NaH2PO4、2.5 mM NaHCO3、1mMデキストロース、2.4 mM MgCl2、1.8 mM CaCl2;全ての物質をSigma(登録商標)、セントルイス、ミズーリ州から得た)中で37℃で1時間インキュベートした。溶液には95%O2と5%CO2を絶えず通気した。空気培地界面で増殖した切片を0.4 μmのTranswell(登録商標)フィルター(Corning Costar(登録商標)、ケンブリッジ、マサチューセッツ州)で維持した。増殖培地は、25%熱不活化ウシ血清、25%HBHS、重炭酸塩を含まない50%MEM、4mM Lグルタミン、30 mM Dグルコース、50 mM重炭酸ナトリウム、および12.5 mM HEPES(全ての物質をシグマ社、セントルイス、ミズーリ州から得た)を含む。温度は33℃で5%CO2で維持した。培地は、週に3回交換した。
【0172】
脳組織培養のトランスフェクション
プラスミドを、バイオラド社からのバイオリスティックPDS-1000/He粒子輸送システム、「遺伝子銃」を用いて神経組織に導入した。プラスミドは、金またはタングステンからなるミクロ担体上にコーティングした。次に、ミクロ担体をマクロ担体にローディングした。マクロ担体は、高圧ヘリウムと、停止スクリーン方向の部分的真空とによって加速された。停止スクリーンは、マクロ担体を停止させるが、微小担体を標的細胞に向けさせ、浸透させる。臓器型培養を衝突のために1%寒天を含むペトリ皿に入れ、その後培養を増殖チャンバーに戻した。典型的に、厚さ300〜400 μmの切片を用いた。
【0173】
顕微鏡
標準的な倒立および直立蛍光顕微鏡を用いた。しかしさらに、これまでに記載されているようにレーザー共焦点顕微鏡を適用してもよい(例えば、ベッカー(Becker)ら、IEEE Transactions on Biomed. Eng. 45:105〜118(1998);ターナー(Turner)ら、Neuron 4:833〜845(1990);およびターナー(Turner)ら、International Rev. Exp. Path. 36:53〜72(1996))を参照のこと。
【0174】
一本鎖Fvファージライブラリ構築およびアミノ末端HDペプチドに対して特異的なヒトsFv抗体の選択
グルタミン残基の数が変化しており、GFPと融合したN末端ハンチンチンペプチドに対して免疫したマウスの脾臓からの一本鎖Fvライブラリ構築は、以下のように実施してもよい。簡単に説明するとVHおよびVL遺伝子に対応するcDNAは、スーパースクリプトII RNアーゼH逆転写酵素(ギブコBRL社)とプライマーを用いて、ガンマ重鎖とカッパ軽鎖のそれぞれ、可変および定常領域のあいだの接合部に対して免疫したマウスの総脾臓RNAから作製することができる。VHおよびVL遺伝子は、例えば、PCR 25サイクル(94℃を1分間、55℃を1分間、72℃を2分間)によって、cDNAからPfuポリメラーゼ(ストラタジーン社)およびVHプライマー[VH1 BACK(34)およびVH1 FOR2(49)、およびVLプライマーVK2BACKおよびVK4FOR(10)]によってそれぞれ増幅する。リンカーDNAは、反応をふさぐためにクレノウポリメラーゼおよびデオキシヌクレオチド三リン酸(dNTPs)を用いて、以下のプライマーの等モル比を混合することによって調製してもよい。
【0175】
次に、VHおよびVL PCR産物と二本鎖リンカーDNAをアガロースゲル上で精製し、VHおよびVL産物各1μgとDNAリンカー300 ngとをプライマーを加えずにPCR反応混合物50 μl中で混合した。7サイクル(94℃で2分、72℃で4分)後、VH1 BACK-SfiI(SfiI制限部位を有するVH1 BACK)およびVK4FOR-NotI(NotI制限部位を有するVK4FOR)プライマー各25 pmolを反応混合物に加えて、20サイクル(94℃で1.5分、および72℃で2.5分)増幅する。ゲル精製後、集合したVHリンカーVL PCR産物をSfiIおよびNotIによって十分に消化して、pHEN1ベクターにクローニングする。ライゲーション混合物の少量をTG1コンピテント細胞に電気穿孔する。ライブラリの多様性はVH1 BACKとVH4FORプライマーによる組み換え型コロニーのPCRスクリーニングの後、BstNI制限酵素による消化によって分析してもよい(グッソウ(Gussow)ら、Nucleic Acid Res. 17:4000(1989))。次に、ファージライブラリを、100 mg/mlアンピシリンおよび1%グルコースを含む2×TY培地50 mlにコロニーを採取して、37℃で45分振とうさせ、ヘルパーファージを加えて、ファージを救出することによって増幅させることができる。
【0176】
ビオチン標識抗原は、例えばeアミノ基またはリジン残基と非特異的に反応するNHSビオチン(ピアスケミカル社)を用いて調製することができる。所定のライブラリに関して、少量をその対応する抗原で平衡にしてもよい。次に、ストレプトアビジンコーティング磁気ビーズを加えて、ストレプトアビジンを抗原ファージ抗体複合体に結合させ、これをライブラリ溶液からのビーズの磁石による分離によって単離する;次にビーズを洗浄する。結合したsFvファージをpH 2の酸性溶液によってビーズから溶出し、直ちに中和して、感染細菌によって救出する。救出されたファージは、増幅して別のラウンドの選択のために用いることができる。
【0177】
選択は、例えばCOS-7細胞に発現された融合蛋白質を用いて、HDに特異的な隣接エピトープとポリグルタミン反復配列の長さとの組み合わせを有する当初の構築物から行ってもよい。後者は、異種ポリグルタミン蛋白質(例えば、DRPLAを用いて)に対する二回目の選択によって同定することができ、前者はグルタミン(Gln)の25コピー以上を含まない蛋白質のイソ型との交叉反応性によって同定することができる。
【0178】
実施例2に記載するように、選択の4ラウンドは、各ラウンドで抗原濃度を減少させることによって(100 nM、50 nM、10 nM、1nM)実施した。第3ラウンドの後、細菌クローンをスクリーニングして、N末端HD配列に対して特異的なsFv配列を分泌するクローンを単離する。スクリーニングはELISA法によって行った。分泌されたsFvを含む細菌の上清を、陰性対照として2%ウシ血清アルブミン(BSA)またはビオチン結合抗原を含む2%BSA(0.1 μg/ウェル、最終濃度)のいずれかと共に、予めブロッキングしたマイクロタイタープレートにおいて4℃で一晩インキュベートした。次に、混合物をストレプトアビジンコーティング(0.1μg/ウェル、最終濃度)したマイクロタイタープレートに移した。一晩インキュベートした後、プレートを洗浄して、マウス抗myc IgG(9E10、0.1 μg/ウェル最終濃度)を加えた。結合したIgGを、ヤギ抗マウスIgGに結合したアルカリホスファターゼによって検出した。
【0179】
さらなる特徴付けのために、可溶性sFv蛋白質を精製する必要があった。したがって、選択されたクローンのVHリンカーVLをコードするcDNAをpHENからpSAYN-1ベクターに移した。pSYN-1ベクターは、6×His尾部を有する蛋白質を発現させ、これによってNi-NTAカラムでの迅速な精製が容易となる。これらのsFv分子の結合特異性は、以下のようなアフィニティ精製によって調べることができる。
【0180】
各クローン約3μgを、GST-DRPLA-Q35、GST-HD-Q25、またはGST-HD-Q42のいずれか4μgと共に室温で1時間溶液中でインキュベートした。次に、グルタチオンセファロース4B(ファルマシア社)の底面容積10 μlを各試験管に加えて、GST融合蛋白質/sFv複合体を沈殿させた。30分インキュベートした後、試験管を遠心して、沈殿物をPBS1mlによって3回洗浄した。10 mM還元グルタチオンを沈殿物に加えることによって、複合体をグルタチオンセファロースビーズから溶出した。
【0181】
遠心後、溶出した複合体を含む上清を新しい試験管に移して、等量の2×SDSローディング緩衝液と共に混合した。試料を沸騰させて、蛋白質を12%SDS-PAGE上で分離した。蛋白質のバンドは、クーマシーブルー染色によって可視化して、および/またはsFv分子をイムノブロット法によって検出するためにニトロセルロースメンブレンに転写した。
【0182】
実施例1
細胞内ポリペプチド凝集の効果をアッセイする方法
本実施例において、細胞内ポリペプチド凝集をアッセイする方法および細胞内ポリペプチド凝集が細胞死に至ることを証明する方法を提供する。
【0183】
細胞内ポリペプチド凝集の影響を調べるために、ハンチントン病患者において認められるポリペプチドを表すハンチンチンポリペプチドを発現する細胞と比較して、正常なハンチンチンポリペプチドを表すポリペプチドを発現する細胞を調べた。霊長類細胞(COS-7)に正常なポリペプチドまたはグルタミン残基47、72、もしくは104個を有する疾患に関連した変化したハンチンチンポリペプチドを表すモデルハンチンチンGFP融合ポリペプチドをコードするプラスミド構築物をトランスフェクトさせた。24時間のトランスフェクション後、試験蛋白質のそれぞれに融合させたGFPタグによって示されるように、大量の細胞がハンチンチンポリペプチドを発現する。観察は3回の時点で記録した(表4を参照のこと)。
【0184】
【表4】
【0185】
48時間の時点での例を図3に示す。蛍光顕微鏡を用いて、グルタミン残基104個を有するハンチンチン-GFPポリペプチドを発現する細胞(図3D)は、大きい輝く凝集体を有することが認められた。最も多くの細胞死は、これらの細胞に認められた。グルタミン残基がより少ない(72Q)モデルハンチンチンポリペプチドを発現する細胞では、極めて明るくより多様な凝集体を認め、細胞死は多少少なかった。グルタミン残基47個を有するモデルハンチンチンポリペプチドを発現する細胞では、蛍光強度は多少散在していた。正常範囲のグルタミン残基(25)を有するモデルハンチンチンポリペプチドを発現する細胞は、ポリペプチド凝集の証拠を認めず散在した蛍光を示した。
【0186】
このように、この実験は、変化したハンチンチンポリペプチド、細胞内凝集および細胞死が一致することを証明する。したがって、このアッセイ系は、ハンチントン病の病態の範囲をインビトロで再現し、細胞内ポリペプチド凝集およびポリペプチド凝集体媒介細胞死を変化させる治療をスクリーニングするために用いることができる(実施例2を参照のこと)。
【0187】
実施例2
神経ポリペプチドに対して結合特異性を有する内部抗体を操作および選択する方法
本実施例において、選択したポリペプチドに対して親和性を有する内部抗体を同定および選択する方法を示す。
【0188】
ハンチンチンポリペプチドを含む細胞内ポリペプチド凝集体の形成を阻害することができる内部抗体を作製するために、ハンチンチンのN末端アミノ鎖残基17個(Nt-HD)に対応するビオチン結合ペプチドを、この配列に対して特異的なsFv分子を示すファージを捕獲するための抗原として作製した。簡単に説明すると、異なるクローン109個を含むヒトsFvファージライブラリを、タフツ大学(ボストン、マサチューセッツ州)の蛋白質中央施設で合成されたペプチドを用いてビオチン結合Nt-HDと共にインキュベートした。次に、ストレプトアビジンコーティング磁気ビーズを加えて、関連するsFvファージ抗体と複合体を形成したストレプトアビジンを単離して洗浄した。結合したsFvファージを酸によって溶出して、中和し、感染細菌によって救出した。次に、救出したファージを増幅して、もう一つのラウンドの選択に用いた。
【0189】
選択の4ラウンドは、各ラウンドで抗原の濃度を減少させて(すなわち、100 nM、50 nM、10 nM、1nM)行った。第3ラウンドの後、細菌クローンをスクリーニングして、Nt-HD配列に対して特異的なsFv分子を分泌するクローンを単離した。スクリーニングはELISA法を用いて実施した。クローン90個をスクリーニングしたところ、クローン20個がビオチン結合Nt-HDペプチドに対して高い結合(陰性対照の結合より少なくとも10倍)を示した。sFv種の結合特異性を調べるために、ELISA法においてOD 0.2を示したクローン8個を、大規模sFv発現のためにクローン20個から選択した。sFv分子を含む細菌上清はストレプトアビジン、GST-HD-42Q、GST-HD-65Q、およびGST-DRPLA-35Q(陰性対照として、オノデラ(Onodera)ら(33))をコーティングしたマイクロタイタープレート上でELISA法によってアッセイした。陽性対照に関しては、細菌上清をビオチン結合N末端HDペプチドと共に溶液中でインキュベートしてから、ストレプトアビジンをコーティングしたマイクロタイタープレートに移した。結合したsFvを、α-9E10 IgGの後、アルカリホスファターゼ結合ヤギ抗マウスIgGによって検出した。この実験から、Nt-HD-隣接ペプチド配列を含む抗原と選択的に反応するクローン1個(α-Nt-HD-C4 sFvと命名する)を同定した。
【0190】
さらに特徴を調べるために、可溶性sFv蛋白質を精製する必要があった。したがって、選択されたクローンのVHリンカーVLをコードするcDNAをpHEN-1ベクターからpSYN-1ベクターに移入した。pSYN-1ベクターは、蛋白質の発現を可能にして、ヘキサHis尾部に融合したsFvの周辺質への分泌を可能にし、これによって迅速な精製が促進される。したがって、α-Nt-HD-C4 sFvの2つのサブクローンを細菌において発現させ、得られたポリペプチドをNi-NTAカラムにおいて精製した(Qiagen(登録商標))。
【0191】
選択したsFvクローンの結合特異性は、以下のように、完全な範囲の抗原に対するアフィニティ結合によって調べた:(a)図9および10の実験結果に対応する試料数を用いて、α-Nt-HD-C4 sFvクローン1(レーン2〜4)、またはα-Nt-HD-C4 sFvクローン2(レーン5〜7)約3μgを以下の抗原4μgと共に混合して、溶液中で室温で1時間インキュベートした:(レーン2および5)GST-DRPLA-35Q(レーン3および6)GST-HD-25Q、(レーン4および7)GST-HD-42Q;(b)グルタチオンセファロース4B(ファルマシア社)の底面容量10 μlを各試験管に加えて全てのGST含有蛋白質複合体を結合させた;ビーズを10 mMグルタチオン溶液によって洗浄し、これによってGST抗原またはsFv抗原複合体を溶出して、これらの溶出物をSDS-PAGE法によって分析した。蛋白質のバンドはクーマシーブルー染色(図9)によって可視化し、同様にイムノブロット法によってsFv分子を検出するためにニトロセルロースメンブレンに転写した(図10)。いずれのサブクローンもHD隣接ペプチドのみに結合する所望の特異性を示した。
【0192】
例えば、図9は、Nt-HD配列を含む試料に限って(すなわちGST-HD-25QおよびGST-HD-42Q)、α-Nt-HD-C4 sFvに対応するバンドが他の蛋白質とほぼ同じ強度で存在することを示している。イムノブロット分析はまた、双方の抗C4 sFvクローンがN末端HD配列を含む試料中に存在するが、GST-DRPLA-Q35を含む試料では検出されないことを確認した(図10)。
【0193】
本発明の内部抗体の正確な結合特性の特徴を調べるために、固定した抗原に結合するsFvのさらに定性的および定量的試験をインビトロで実施した。C4 sFv抗原結合特性はELISAアッセイによってモニターし、同様にC4 sFvがN末端HD配列に特異的に結合することを確認した(図11A)。定量的結合実験に関して、大量輸送の人為的結果が無視できる条件で速度論の測定を行うために、C末端ビオチンを有するHDペプチド(1〜17位)を非常に低濃度でストレプトアビジンコーティングBIAコアセンサーチップ上にローディングした。結合と解離の速度論は、BIAコア2000においてC4 sFv抗体について決定した(図11B)。Kdは、単純な生体分子相互作用に関して1:1ラングミュアモデルを用いて測定したKaおよびKd速度定数に基づいて7.9 nMであると計算された(BIA評価3.0ソフトウェア、バイアコアインターナショナルAB、アップサラ、スウェーデン)(図11B、右上の挿入図)。対応する結合親和性(Ka)1.3×108 M-1は、この大きいファージディスプレイライブラリから厳密な抗原に対して選択された抗体に典型的で、これは特に短いペプチド配列に対してかなり高い親和性sFvであることを示している(シーツ(Sheets)ら、Proc. Natl. Acad. Sci. USA 95:6157〜6162)。これらの結果は、HD(1〜17位)ペプチドが、見かけの結合定数を有意に減少させる広範囲の非構造類似体よりむしろ、C4 sFv複合部位によって認識される比較的堅固なエピトープを形成する可能性があることを示唆している。
【0194】
これらの結果は、ハンチンチンのN末端配列のような特定のエピトープに対して特異的であるα-Nt-HD-C4 sFv内部抗体分子を同定および選択できることを示している。したがって、選択されたポリペプチドポリペプチドに対して独自の特異的N末端またはC末端エピトープを検出するいかなる内部抗体を選択してもよい。その上、上記の方法を用いて、ポリグルタミンに富むドメインに対して特異的な内部抗体を作製することができる。そのような内部抗体は、ポリグルタミンに富む領域を有するいかなる当該ポリペプチド、例えば、ハンチンチン、アタキシン、アトロフィン等にも結合しうるであろう。最後に、これらの方法は、選択されたポリペプチドに対して二重または多重の特異性を有する多価内部抗体の操作を提供する。多価内部抗体は、例えば標的ポリペプチドの標的ポリグルタミンに富む領域と共に、ポリペプチドの他のN末端および/またはC末端隣接エピトープに結合することができるであろう。
【0195】
実施例3
病的ハンチンチンポリペプチドを発現する哺乳類細胞におけるポリペプチド凝集を阻害する方法
本実施例において、哺乳類細胞において凝集体の形成を阻害して、細胞内ポリペプチドを再ターゲティングする方法を示す。
【0196】
内部抗体が細胞内ポリペプチドに特異的に結合して、ポリペプチド凝集を阻害できることを証明するために、哺乳類細胞(COS-7)に、GFPに融合したモデルハンチンチンポリペプチドをコードする第一のプラスミドと、モデルハンチンチンポリペプチドに特異的に結合する内部抗体をコードする第二のプラスミドとを同時トランスフェクトさせた。アッセイは、双方の標的抗原、すなわちモデルハンチンチンポリペプチドと内部抗体を可視化できるように設計する。特に、ハンチンチンポリペプチドは、ハンチンチンGFP融合ポリペプチドのGFPドメインが放出する蛍光を検出することによって可視化され、調べる内部抗体は、内部抗体に特異的に結合することができるローダミン結合抗体を用いて可視化される。
【0197】
したがって、図5A〜Hに示す実験結果は、蛍光顕微鏡を用いて、内部抗体が細胞においてハンチンチンポリペプチドに特異的に結合してこれを再ターゲティングできることを示している。図5Aおよび5Bにおいて、細胞の分布をまず、位相差光学顕微鏡を用いて示す。図5C、5E、および5Gにおいて、モデルハンチンチンGFP融合ポリペプチド(HD-25Q-GFP)(図5E)と、核ターゲティングシグナルをさらに含むモデルハンチンチンポリペプチドに特異的に結合する内部抗体(α-Nt-HD-C4 sFv-NLS)とを同時発現する細胞は、核におけるローダミンおよびGFP染色の集合によって示されるように、ハンチンチンポリペプチドを核に分布するよう再ターゲティングできることが認められた(図5G)。対照的に、無関係な蛋白質に結合する内部抗体を発現する細胞は、ハンチンチンポリペプチドの分布を再ターゲティングすることができない(図5D、5F、およびFH)。
【0198】
これらの実験を拡大して、上記のように試験した内部抗体が同様に、ハンチントン病に関連するポリグルタミン反復配列を有する変化したハンチンチンポリペプチドに対しても同じ有効性を有すること、そして細胞内凝集体を形成する傾向が最も高いことを示した。このように、哺乳類細胞に、ポリグルタミン104個を有するモデル変化ハンチンチンポリペプチド(pHD-104Q-GFP)をコードするプラスミドと、ハンチンチン特異的内部抗体(α-Nt-HD-C4 sFv)または無関係なポリペプチドに対して結合特異性を有する内部抗体のいずれかをコードする第二のプラスミドとをトランスフェクトさせた。図7A〜Hは、蛍光顕微鏡を用いて、ハンチンチン特異的内部抗体のみがモデル変化ハンチンチンポリペプチド(すなわち、グルタミン反復配列104個を有する)の細胞分布を核に特異的に再ターゲティングできるが(図7C、7E、および7G)、無関係な内部抗体は作用を示さないことを示している(図7D、7F、および7H)。
【0199】
ハンチンチン特異的内部抗体(α-Nt-HD-C4 sFv)の特異性をさらに証明するために、哺乳類細胞に、ポリグルタミンに富み、凝集して神経疾患(DRPLA)を引き起こすことができるもう一つの標的ポリペプチドを同時トランスフェクトさせた。モデルDRPLA標的ポリペプチドをGFPドメインと融合させて、トランスフェクトした細胞における標的ポリペプチドの発現および細胞内位置を追跡した。さらに、一つはハンチンチンに特異的であり、もう一つは無関係なポリペプチドに特異的な内部抗体である異なる2つの内部抗体を、個々にDRPLA標的ポリペプチドと共に同時トランスフェクトした。グルタミンに富むDRPLAポリペプチド(GFP-DRPLA-35Q)(図6C〜G)と、ハンチンチンポリペプチド(α-Nt-HD-C4 sFv;図6D、6F)またはもう一つの無関係なポリペプチド(陰性対照sFv;図6C、6E、および6G)のいずれかに対する内部抗体とを同時発現する細胞(COS-7)は、DRPLAポリペプチドの細胞分布に変化を示さない。
【0200】
さらに、ハンチンチン特異的内部抗体が細胞内ポリペプチド凝集体の形成を防止できるのみならず、異なる細胞の位置に望ましくないポリペプチドを再ターゲティングしうることを確立する実験を実施した。哺乳類細胞に、ポリグルタミン含有量が最も高く(グルタミン残基72または104個)、凝集して疾患を誘発する傾向が大きい、変化したハンチンチンポリペプチドをコードするプラスミドを、ターゲティングシグナルなし、核ターゲティングシグナル、またはライソゾームターゲティングシグナルのいずれかを有するハンチンチン特異的内部抗体の一つと共にトランスフェクトした。
【0201】
蛍光顕微鏡を用いて調べた3つの内部抗体のそれぞれは、対照(図8A、8E)と比較して細胞内ポリペプチド凝集体の形成を阻害することが示された(8B〜8D、8F〜7H)。これは、ポリペプチド凝集体によって放出された明るいGFPシグナルの減少によって証明される。さらに、同様に、核ターゲティングシグナル(図8C、8Gを参照)またはライソゾームターゲティングシグナル(図8D、8G)のいずれかを有する内部抗体と接触させると、モデルハンチンチンポリペプチドの再ターゲティングが得られることも証明された。これは、核ターゲティングシグナルを有する内部抗体を発現するそれらの細胞の核にGFPシグナルが蓄積することによって示された(図8C、8G)。ライソゾームターゲティングシグナルを有する内部抗体をトランスフェクトした細胞では、細胞質全体により散在するシグナルを認めた(図8D、8H)。
【0202】
さらになお、本発明の内部抗体がHDの細胞内凝集の形成を阻害できるか否かを調べるさらにもう一つの試験において、抗HD C4 sFv内部抗体およびHD-ポリQ-GFPを異なるいくつかの細胞タイプに同時トランスフェクトして、ポリペプチド凝集の内部抗体媒介阻害の統計分析を測定した。
【0203】
特に、インサイチューでの病的なHD凝集体の形成は、ポリグルタミンの長さが40残基またはそれ以上である場合に(すなわちQ72、またはQ104、しかしQ25では認めない;図14)顕著な斑点状の蛍光形態を示すことが認められた。さらに、これらのポリペプチド凝集体は洗浄剤不溶性であり、トランスフェクト細胞のSDS溶解後に残留蛍光体の改善された定量方法を行うことができる(カザンツェフ(Kazantsev)ら、PNAS 96:11404〜11409(1999))。巣状塊の平面は実験における変数として消去されるため、この改善されたアッセイによって、ポリペプチド凝集体をより正確に計数することができる。したがって、このアッセイを、ポリQ抗原凝集体単独またはC4 sFvもしくは対照内部抗体の存在下で定量のために利用した。COS-7、BHK、および293細胞にC4または対照内部抗体を発現するプラスミドと、Q25、Q72、またはQ104を含むHD-ポリQ-eGFP融合体とを同時トランスフェクトさせた。HD-ポリQ72-eGFPまたはHD-ポリQ104-eGFPを用いて、内部抗体対抗原プラスミドの比が3:1の場合に凝集体の程度の定量的な減少を認めたが、最も有意な定量的効果は5:1の比で認めた(図14)。C4 sFvについて認められた凝集体における顕著な減少と比較して、無関係な内部抗体構築物による対照トランスフェクタント(C8、またはML3-9 sFv)は、親の空のベクターと比較して凝集体形成に統計学的に有意な減少を示さなかった。Q25プラスミド単独をトランスフェクトした細胞は、SDS不溶性の凝集体を決して示さなかった。
【0204】
典型的な内部抗体構築物をC末端血液凝集素(HA)ペプチドタグに融合させて、蛋白質を細胞質に存在させながら蛋白質の免疫的検出を容易にした。二重標識免疫蛍光試験は、HD-ポリQ72-GFPまたはHD-Q104-GFP(緑色標識、図15)プラス抗HD C4 sFv(赤色標識、図15)を発現するBHK細胞が、実質的な散在するGFP標識を示すが、蛍光凝集体を示すことは時折に過ぎないこと(通常HAによっても標識する;図15の矢印を確認のこと)を確認した。内部抗体を発現しないように思われるそれらの細胞は、大きい凝集体の証拠を示した(図15、矢印の先、上のパネル)。散在する標識はまた、内部抗体との同時トランスフェクションによらず、正常な長さの(25Q)構築物の特徴であった(図15)。二重トランスフェクタントについて認められた凝集体形成の減少は、イムノブロット法によって異なるトランスフェクションの典型において区別できない産物レベルが示されたために、HD蛋白質発現の抑制に由来しないように思われた(図15、下のパネル)。
【0205】
したがって、これらの結果は、内部抗体が、神経疾患(例えば、ハンチントン病)に関連するポリペプチド凝集体に特異的に結合して形成を阻害することができることを確認する。さらに、これらの結果はまた、ターゲティングシグナルを有する内部抗体を用いて望ましくない細胞内ポリペプチドを再ターゲティングできることを示している。
【0206】
実施例4
哺乳類細胞内の内部抗体の細胞内特異性を分析する方法
本実施例において、標的抗原を発現する哺乳類細胞において内部抗体の特異性を分析する方法を示す。
【0207】
抗ハンチンチン内部抗体(α-Nt-HD-C4 sFv)の特異性を証明するために、哺乳類細胞(COS-7)をモデルHD標的ポリペプチドと、α-Nt-HD-C4 sFvまたは無関係なポリペプチドに対して特異的な内部抗体のいずれかと同時トランスフェクトした。モデルHD標的ポリペプチドはc-myc(EQKLISEEDL(配列番号:45))エピトープタグ(HD-Q104-myc)に融合した。結合した抗原が細胞核に再ターゲティングされるように、内部抗体配列をHAタグおよびSV40核局在シグナル(sFv-HA-NLS)と融合させた。トランスフェクションの48時間後、細胞を固定して透過性にした。HD-Q104-myc蛋白質は、抗mycエピトープ抗体(すなわちMAb 9E10 IgG)の後にFITC標識ヤギ抗マウスIgGを用いて検出可能であった。sFv-HA-NLS内部抗体は、ポリクローナルウサギ抗HA IgGの後にローダミン結合ヤギ抗ウサギIgG抗体によって検出した。
【0208】
図12A〜Fに示す実験結果は、双方の内部抗体が細胞核に存在するが(図12A〜B)、α-Nt-HD-C4 sFv-HA-NLSのみがモデル変化ハンチンチンポリペプチドHD-Q104-mycを細胞質から核に再配置できることを示している(それぞれ、図12D、12F対12C、12E)。さらに、対照内部抗体は、HD-Q104抗原の凝集を防止しなかった(パネルCおよびEの矢印の先で示す)。
【0209】
細胞内にターゲティングした抗原に対する内部抗体の特異性を証明するもう一つの方法は、ターゲティングした抗原または無関係な抗原のいずれかと共に特異的内部抗体を同時発現する細胞を分析することである。図13A〜Fは、グルタミン反復配列104個を有するモデル変化ハンチンチンポリペプチド(図13B、D、F)またはグルタミン反復配列81個を有するモデル変化DRPLAポリペプチド(図13A、C、E)のいずれかをコードするプラスミドによってさらにトランスフェクトした、α-Nt-HD-C4 sFv-HA-NLSを発現する安定にトランスフェクトしたCOS-7細胞を示している。双方のモデル変化ポリペプチドをGFPに融合した(それぞれ、GFP-HD-Q104およびGFP-DRPLA-Q81)。抗Nt-HD-C4 sFv-HA-NLS内部抗体は、GFP-HD-Q104のみが細胞核内部で内部抗体によって顕色されたために、明らかに、GFP-HD-Q104(図13D、F)に対する特異性を示すが、GFP-DRPLA-Q81(図13C、E)に対する特異性を示さない。
【0210】
これらの方法によって、関連する細胞内環境で内部抗体特異性を迅速に評価することができ、本明細書において考察した抗原に適用できるのみならず、他の内部抗体/ターゲティング抗原対を調べるために用いてもよい。
【0211】
実施例5
哺乳類の脳組織におけるポリペプチド凝集をアッセイする方法
本実施例では、哺乳類の脳における変異型ハンチンチンポリペプチドが細胞内凝集体を形成できることを検出する方法を示す。
【0212】
変化したハンチンチンポリペプチドが哺乳類の脳において細胞内凝集体を形成できることを証明するために、マウス(P12 Balb)からの小脳切片に、正常および変化したハンチンチンポリペプチドを表すポリペプチドをコードするプラスミドを遺伝子銃によってバイオリスティックにトランスフェクトして、24時間調べた。グルタミン残基104個を有するモデル変化ハンチンチンGFPポリペプチド(HD-104Q-GFP)をコードするプラスミドを24時間培養物にトランスフェクトすると、輝く蛍光点として示される大きい凝集体を脳組織全体に認めた。同様に、グルタミン残基72個を有する変化したハンチンチンGFPポリペプチド(HD-72Q-GFP)をコードするプラスミドを24時間培養物にトランスフェクトすると、輝く蛍光点によって示されるより小さく、より明確な凝集体が認められた。
【0213】
さらに、このアッセイはまた、モデルハンチンチンポリペプチドを発現する特定の細胞タイプの同定能を提供する。例えば、プルキンエ細胞は、カルビンジンに対する抗体を用いて同定することができ、ニューロンは神経細糸に対する抗体を用いて可視化することができる。したがって、これらの細胞特異的抗体を可視化するために赤色標識を用いることによって、HD-ポリQ-GFP融合蛋白質のGFP部分に対して陽性である細胞も同時に同定することができる。次に、切片をレーザー共焦点顕微鏡を用いて調べる。
【0214】
重要なことは、例えば、実施例2に示す方法を用いて、このアッセイは、内部抗体が細胞内ポリペプチド凝集(例えば、ハンチンチンポリペプチドの)の形成を阻害できるか否か、または多数の細胞タイプを有する脳組織においてそのようなポリペプチドを再ターゲティングできるか否かをアッセイするように適合させることができる。さらに、このアッセイは、ポリペプチド凝集体の形成を破壊することができる他の結合分子のスクリーニング能を提供する。最後に、このアッセイは、長期間培養して、本明細書に記載のようにアッセイするために特定の神経細胞型(例えば、線条体細胞)を単離するために用いてもよい。
【0215】
実施例6
神経ポリペプチドに対するsFvファージライブラリを作製する方法
本実施例において、DNAワクチン接種を用いて選択された抗原に対して内部抗体ファージライブラリを作成する方法を示す。
【0216】
広範囲の可能なポリペプチドに対して特異的な内部抗体を作製することができるように、DNAワクチン接種戦略を開発した。この方法は、「本来の」ライブラリにおいてまれな内部抗体を選択するより、特定のエピトープに対して「向けられた」ライブラリを作製する長所を有する。したがって、スクリーニングしなければならないクローン数は大きく減少する。得られたファージディスプレイライブラリが、蛋白質がインビトロで取りうる形のみに対するエピトープよりむしろ、その細胞内または細胞内構造のエピトープに対する抗体を発現するように活性化された脾臓から作製することができるように、蛋白質の代わりに、DNAを用いてマウスを免疫した。
【0217】
ハンチンチンの最初のアミノ酸17個、異なる長さのポリグルタミン反復配列、および改変された緑色蛍光蛋白質を含む融合蛋白質を発現するプラスミド構築物を用いて、雌性マウス8匹(BalbCBy/J遺伝的背景)においてインビボで抗原を発現させた。GFPに融合したモデルハンチンチンN末端をコードし、正常数のグルタミン残基またはハンチントン病に関連した変化した残基数のいずれかを有するプラスミド構築物を、正常生理食塩液媒体においてマウスに0.5 μg/μlの濃度で注入した。
【0218】
免疫前血清を得るために、免疫する前にマウスから血液を採取した。全てのマウスにプラスミド構築物の一つを1日目(プラスミドDNA 50 μgを筋肉内注射、プラスミドDNA 50 μgを皮内注射−合計100 μg)に注射した。注射は14日目に繰り返してマウスを21日目に採血した。次に、マウス血清を、血清の希釈を変化させて、pHD-25QトランスフェクトCOS7細胞溶解物のイムノブロットをプロービングすることによって陽性液性応答に関してスクリーニングした。陽性反応マウスの刺激された脾臓は、モノクローナル抗体、内部抗体、およびファージディスプレイを作製するための候補物質であると決定された。
【0219】
注射したマウス8匹中、50%(8匹中4匹)が、最小の血清希釈100倍でイムノブロットアッセイ法における融合蛋白質を検出することができる陽性抗体反応を示した(表5を参照)。これらの中で、75%(4匹中3匹)がグルタミン反復配列104個の構築物によって免疫された。
【0220】
【表5】
【0221】
このように、これらの結果は、本明細書に記載の方法を用いて抗原特異的内部抗体結合ドメインを、上記の原料、そして例えば特異的抗体結合分子を作製するためにファージディスプレイライブラリを用いて操作できることを示している。その上、これらの方法は、実質的に他のいかなる当該抗原にも容易に適用できる。
【0222】
実施例7
哺乳類における神経疾患の内部抗体阻害を検定する方法
本実施例において、内部抗体が生きている哺乳類において神経疾患を治療または治癒できるか否かを検定する方法を示す。
【0223】
内部抗体が生きている哺乳類における神経疾患に関連した変化した神経ポリペプチド凝集体の形成を阻害することができるか否かを決定するために、トランスジェニックマウスを用いる方法が開発されている。
【0224】
特に、ハンチントン病およびSCAの動物モデルが開発されている(例えば、表2を参照のこと)。
【0225】
例えば、変化したハンチンチンポリペプチドを発現するように遺伝子操作されたトランスジェニックマウスのコロニーが確立されており(ベイテス(Bates)ら、(3)(29)に記載のように)、これらの動物はハンチントン病のいくつかの神経学的特徴を示す。マウスは症候性であり、8週齢までに、皮質、線条体および小脳のプルキンエ細胞において神経ハンチンチンポリペプチド凝集体を示し始める。
【0226】
さらに、変化したアタキシン発現によって引き起こされるSCA神経疾患を表す第二の動物モデルが確立されている。これらの動物は、プルキンエ細胞特異的プロモーターの制御下で発現される拡大された反復配列アタキシン遺伝子を有する。これらの動物は、12週までに症状を示し、4週までにプルキンエ細胞において最初の組織学を示す。
【0227】
なお第3のアプローチにおいて、変化したアトロフィン1によって引き起こされるDRPLAを表す動物モデルを確立することができる(シリング(Schilling)ら、Neuron 24:275〜286(1999))。
【0228】
したがって、これらの動物は、内部抗体が神経ポリペプチド凝集体の形成を阻害して、このように疾患の発症を予防、治療、または遅らせることができるか否かを調べる3つのインビボアッセイ系となる。例えば、内部抗体はポリペプチドとして、または本明細書に記載のいかなる方法も用いて発現可能な核酸構築物として頭蓋内に輸送してもよい。
【0229】
さらに、内部抗体が神経疾患の発症を予防、治療、または遅らせるか否かを証明するさらにもう一つの方法において、これらの動物を条件によって発現することができる内部抗体を発現する動物と交配させることができる。このように、疾患の臨床経過のあいだの異なる時点で、内部抗体をそのインビボ有効性に関して調べてもよい。特に、ネスラー(Nestler)とその共同研究者らのTet調節系を用いることができ、それは、2つのトランスジェニックマウス系を含み、一方は神経組織における発現を指示するニューロン特異的エノラーゼ(NSE)プロモーターの制御下でTet調節トランス活性化因子(tTA)を含み、もう一方は当該遺伝子、例えば抗ハンチンチン内部抗体(すなわち、α-Nt-HD-C4 sFv)、またはTet調節プロモーターの下流にクローニングされた抗アタキシン内部抗体(TetOp)を有する構築物を含む(チェン(Chen)ら、Mol. Pharmacol. 54:495〜503(1998);アーノルド(Arnold)ら、PNAS 94:8842〜8847(1997);およびクラーク(Clark)ら、J. of Neurosci. 17:7325〜7395(1997)))。選択された内部抗体の発現を活性化するために、内部抗体遺伝子と変化した神経ポリペプチド(例えば、ハンチンチンまたはアタキシンのいずれか)を有する子孫の飲料水からドキシサイクリン(Tet誘導体)を除去する。
【0230】
交配スケジュールは以下の通りである:第一に、双方のトランスジーンの存在がゲノムDNAのPCRによって決定され、遺伝子の発現がイムノブロット法によって確認される、NSE-tTAおよびHDトランスジーンの双方を有する二遺伝子マウスを作製する。これらのマウスを、ニューロンにおけるTetOpイントラC4プラスミドの発現を容易にする脳切片培養源としてまず用いることができ、後にこれをTetOpトランスジェニックマウスと交配させる。
【0231】
NSE-tTA脳切片を用いて、HDポリQおよびTetOpイントラC4の双方をトランスフェクトさせて、発現された内部抗体が形成された凝集体の数を減少させるように機能するか否かをアッセイすることができる(例えば、凝集体をライソゾーム分解されるようにターゲティングする)。次に、成功したプラスミドを注入してトランスジェニック創始マウスを作製することができる。これらを二世代にわたって交配することができ、内部抗体の非誘導レベルに関して試験することができる。
【0232】
トランスジーン発現を完全に阻害するために、ドキシサイクリンを25 μg/mlの低用量で動物に持続的に投与することができる(チェン(Chen)ら、Mol. Pharmacol. 54:495〜503(1998))。この低用量では、ドキシサイクリンの除去による内部抗体の誘導は迅速となりうる。疾患進行のどの段階で内部抗体の治療が最も有効であるかを調べるために、ドキシサイクリンを様々な年齢群のHD-NSE-tTA×TetOpイントラC4マウスの飲料水から4週目、6週目、8週目、および10週目に除去することができる。これらの動物からの脳を摘出して、内部抗体治療の有効性の指標としてその野生型子孫と比較して凝集体の有無および数をアッセイすることができる。
【0233】
上記のように、これらの実験を用いて、HDトランスジェニックマウスによって抗HD内部抗体を調べ、SCA1マウスによって抗アタキシン内部抗体を調べ、双方の系統のマウスによって抗ポリグルタミン内部抗体を調べることができる。二遺伝子NSE-tTA/HDまたはアタキシンマウスを、特異的およびより一般的な内部抗体交配の双方のために用いることができる。
【0234】
したがって、本方法によって、インビトロアッセイにおいて同定された内部抗体の最も有効なものをインビボで決定することができる。さらに、これらの方法によって、HD(またはSCA1マウス)において神経症状の行動学的救出を認めるために必要なレベルまで凝集体の毒性を減少させるか否かを決定することができる。最後に、これらの方法によって、内部抗体発現の誘導が最も有効であるのは疾患進行のどの段階であるかも決定することができる。
【0235】
実施例8
望ましくない細胞内ポリペプチド複合体に対する治療的宿主抗体免疫応答を誘発するための核酸ワクチンのインビボ証明
本実施例において、望ましくない細胞内ポリペプチド凝集体に対して宿主によって内部抗体を誘発する方法および組成物を示す。
【0236】
免疫系によって作製された内部抗体が、外から投与される代わりに、神経疾患に関連した変化した神経ポリペプチド凝集体の形成を阻害するか否かを決定するために、病原性細胞内ポリペプチドを表すポリペプチド抗原、すなわちハンチンチンポリペプチドをコードするDNAワクチンによって試験動物を免疫した。対照と比較して、試験ポリペプチドをコードするDNAワクチンは、免疫した動物において治療的液性応答を誘発し、その結果内因性細胞内ハンチンチンポリペプチドと共にDNAコードハンチンチン免疫原に対する宿主内部抗体(すなわち、抗体)の形成が起こる。
【0237】
簡単に説明すると、実験アプローチは以下の通りであった。グルタミン残基約165個を含む(正常は35コピー未満)変異型ヒトハンチンチンの断片を発現する試験動物(HDR6/2トランスジェニックマウス)を得た。これらのマウスは、5週齢までに測定可能な微妙な行動学的および神経化学的変化を示し、重度の神経疾患は11〜12週で明らかになった。15週を超える生存はまれである。重要なことは、これらのマウスは同様に、絶食時の血清グルコースレベルの増加から証明されるように(10週齢で)、ヒトにおいて認められるものと類似のハンチンチン関連糖尿病症候群を発症する。ハンチンチン関連糖尿病は、膵臓の島細胞における変異型HD蛋白質の異常な蓄積に帰因している。
【0238】
この実験に関して、本明細書に示したこれまでの実験において内部抗体を選択するために用いたHD蛋白質断片の一部に対する抗体を産生するためにマウスを免疫した。特に、HDR6/2マウスを5週齢時に、緑色蛍光蛋白質(GFP)に結合したHDトランスジーン断片を発現するプラスミド100 μgを注射することによって免疫して、7週齢時に追加免疫した。2回の注射は同じ週齢で行い、一方は脚の筋肉に、そしてもう一方は尾の真下の皮内に行った。8週目にマウスから採血して、当初の注射プラスミドのトランスフェクションの24時間後にHSK-293細胞の溶解物から調製したイムノブロットを用いて反応性を決定した。
【0239】
次に、陽性免疫応答対反応しない免疫応答を示したHDマウスについて、血液中グルコースレベルを比較した。血中グルコースは、6時間の絶食後にコンフォートカーブ試験片を備えたアドバンテージグルコメータを用いて1試料あたり2本ずつ実施した。無処置トランスジェニックHDマウスの絶食時の血中グルコースレベルは、10週齢から始めた野生型マウスのレベルより有意に高かった。プラスミド免疫に反応しないトランスジェニックマウス、またはGFP配列のみを含むプラスミドによって免疫したマウスは、無処置マウスの範囲のレベルを示した。しかし、プラスミドを免疫してイムノブロット法によってアッセイして反応性であったマウスは、血中グルコースレベルが有意に低下し、13および14週齢では完全に野生型マウスの範囲内であった。このように、HD免疫は、膵臓の表現型の発生を防止しうる(図16参照)。
【0240】
グルコースレベルの他に、インスリンをコードするmRNAの膵臓レベルも同様に測定した。一連のマウスを14週目に屠殺して、それぞれの膵臓からRNAを抽出した。試料を遺伝子型、治療、および反応に基づいてプールした。インスリンmRNAレベルは、逆転写ポリメラーゼ連鎖反応(RT-PCR)法によって決定した。野生型マウスからの試料は、RT-PCR反応法において用いられるプライマーに基づいて予測される位置で強いバンドを示した。無処理および非反応マウスは、同じサイズのかすかなバンドのみを示し、インスリン生合成の欠損を示した。免疫して反応性であったマウスは、密度測定法によって確認すると、野生型の強度に近いインスリンmRNAを示すバンドを示した。このように、治療的改善は、インスリンmRNAレベルの回復であるように思われる。
【0241】
最後に、DNAワクチン接種に反応するマウスからの血清を、上記の治療的内部抗体を選択するために用いられるポリペプチド抗原に結合する抗体を動物が産生したか否かを決定するために調べた。したがって、ハンチンチンの最初のアミノ酸17個に対応するペプチドを合成して、試験皿に固定した。次に、陽性反応者に存在するエピトープのELISA試験を行った;イムノブロット法によって陽性であり、異常なグルコースレベルの低下とインスリンmRNAの増加の双方を誘発したマウス血清は同様に、アミノ酸17個のペプチドに対する結合を示した。
【0242】
要約すると、これらの知見は、例えば、凝集または蓄積による変異型ポリペプチド機能による病態を予防または逆転させるために細胞内で作用する能力を有する免疫によって産生された抗体と一致する。
【0243】
引用文献の詳述
【0244】
同義
当業者は、日常的な実験のみを用いて、本明細書に記載の本発明の特定の態様に対する多くの同等物を認識、または確認できるであろう。そのような同等物は添付の特許請求の範囲に含まれると解釈される。
【図面の簡単な説明】
【0245】
【図1】病的な長さのポリグルタミン(すなわち、ポリQ)に変異した場合の、ポリグルタミンに富み、神経疾患に関連するいくつかのポリペプチドの野生型N末端アミノ酸配列を示す。比較のために、神経疾患に関連しないポリグルタミン鎖を有するTBPポリペプチドを示す。ハンチンチン特異的C4 sFv内部抗体を作製するために用いたペプチドも同様に示す。
【図2】特定の細胞内位置に異種ポリペプチドをターゲティングすることができる多くのターゲティングシグナルのアミノ酸配列を示す。血液凝集素(HA)タグは、抗原とは個別に内部抗体を可視化するために用いられる免疫染色法のために、適当な抗体に強く結合するペプチドである。
【図3】蛍光顕微鏡を用いて、グルタミン残基の数が増加している(すなわち25、47、72、104個)モデルハンチンチンGFP融合ポリペプチド(すなわち、HD-25Q-GFP、HD-47Q-GFP、HD-72Q-GFP、HD-104Q-GFP)の長さ依存的凝集を示す。細胞において発現されると(この図においてCOS-7)、それぞれグルタミン残基72および104個を有するモデルハンチンチンGFP融合蛋白質(HD-72Q、HD-104Q)は、小さく輝く蛍光斑に容易に凝集する(図3C〜D)。対照的に、グルタミン残基が野生型の数(25個)である(HD-25Q)モデルハンチンチンGFPポリペプチドは、凝集しないままであり、散在した細胞質GFP蛍光を示すに過ぎないか、またはポリQの長さがわずかに病的なポリペプチド(HD-47Q)では、凝集体を形成する傾向を認めるが、多くの細胞が細胞全体に均一に分布する抗原を維持している(図3A〜B)。
【図4】蛍光顕微鏡を用いて、グルタミン残基72および104個を有するモデルハンチンチンGFP融合ポリペプチド(pHD-72Q、図3B;pHD-104Q、図3A)が、マウスに由来する小脳切片において発現され、遺伝子銃を用いてバイオリスティックにトランスフェクトすると、小さく強い染色領域に凝集できることを示す。
【図5A−D】内部抗体が、細胞においてモデルハンチンチンポリペプチドに特異的に結合してこれを再ターゲティングできることを示す。図5Aおよび5Bでは、位相差光学顕微鏡を用いた細胞の分布を示す。図5C、5E、および5Gは、蛍光顕微鏡を用いて、モデルハンチンチンGFP融合ポリペプチド(HD-25Q-GFP)と、モデルハンチンチンポリペプチドに特異的に結合する内部抗体とを同時発現して、核ターゲティングシグナルを含む細胞(αNt-HD-C4 sFv-NLS)(図5E)が、核における染色の集合によって証明されるように、モデルハンチンチンポリペプチドが核に分布するように再ターゲティングできることを示している(図5G)。対照的に、無関係なポリペプチドに結合する内部抗体を発現する細胞は、モデルハンチンチンポリペプチドの分布を再ターゲティングすることができない(図5D、5F、およびFH)。
【図5E−H】内部抗体が、細胞においてモデルハンチンチンポリペプチドに特異的に結合してこれを再ターゲティングできることを示す。図5Aおよび5Bでは、位相差光学顕微鏡を用いた細胞の分布を示す。図5C、5E、および5Gは、蛍光顕微鏡を用いて、モデルハンチンチンGFP融合ポリペプチド(HD-25Q-GFP)と、モデルハンチンチンポリペプチドに特異的に結合する内部抗体とを同時発現して、核ターゲティングシグナルを含む細胞(αNt-HD-C4 sFv-NLS)(図5E)が、核における染色の集合によって証明されるように、モデルハンチンチンポリペプチドが核に分布するように再ターゲティングできることを示している(図5G)。対照的に、無関係なポリペプチドに結合する内部抗体を発現する細胞は、モデルハンチンチンポリペプチドの分布を再ターゲティングすることができない(図5D、5F、およびFH)。
【図6A−C】内部抗体の特定のポリペプチド位置の再ターゲティング能が、内部抗体の結合特異性の関数であることを示す。例えば、図6C〜Gにおいて、グルタミンに富むDRPLAポリペプチド(GFP-DRPLA-35Q)と、モデルハンチンチンポリペプチド(H-HD-C4 sFv;図6D、6F)またはもう一つの無関係なポリペプチド(陰性対照sFv;図6C、6E、および6G)のいずれかに対する内部抗体とを同時発現する細胞(COS-7)は、DRPLAポリペプチドの細胞分布に変化を示さない。図6Aおよび6Bは、位相差光学顕微鏡を用いて細胞の全体的な分布を示す。
【図6D−G】内部抗体の特定のポリペプチド位置の再ターゲティング能が、内部抗体の結合特異性の関数であることを示す。例えば、図6C〜Gにおいて、グルタミンに富むDRPLAポリペプチド(GFP-DRPLA-35Q)と、モデルハンチンチンポリペプチド(H-HD-C4 sFv;図6D、6F)またはもう一つの無関係なポリペプチド(陰性対照sFv;図6C、6E、および6G)のいずれかに対する内部抗体とを同時発現する細胞(COS-7)は、DRPLAポリペプチドの細胞分布に変化を示さない。図6Aおよび6Bは、位相差光学顕微鏡を用いて細胞の全体的な分布を示す。
【図7A−D】蛍光顕微鏡を用いて、内部抗体が変異型モデルハンチンチンポリペプチド(すなわち、グルタミン反復配列104個を有する)の細胞内分布を核に特異的に再ターゲティングできるが(図7C、7E、および7F)、無関係な内部抗体は再ターゲティングできないことを示している(図7B、7D、および7G)。図7Aおよび7Bでは、位相差光学顕微鏡を用いた細胞の分布を示す。
【図7E−H】蛍光顕微鏡を用いて、内部抗体が変異型モデルハンチンチンポリペプチド(すなわち、グルタミン反復配列104個を有する)の細胞内分布を核に特異的に再ターゲティングできるが(図7C、7E、および7F)、無関係な内部抗体は再ターゲティングできないことを示している(図7B、7D、および7G)。図7Aおよび7Bでは、位相差光学顕微鏡を用いた細胞の分布を示す。
【図8】蛍光顕微鏡を用いて、細胞内ポリペプチド凝集体の形成が内部抗体の発現によって阻害されたことを示す。ヒトにおいて神経疾患に関連することが知られている変異を有するハンチンチンポリペプチドを表す異なる2つのポリペプチドは、細胞において発現されると、細胞内凝集体を形成することが示された(図8Aおよび8E)。これらの凝集体の形成は、ターゲティングシグナルを有しない(図8B、8Fを参照)、核ターゲティングシグナルを有する(図8C、8Gを参照)、またはライソゾームターゲティングシグナルを有する(図8D、8G)異なる3つの内部抗体によって阻害された。図8C〜D、8F〜Gはまた、ターゲティングシグナルを有する(核またはライソゾーム)内部抗体は、細胞内ポリペプチド凝集体を減少させたのみならずに、モデルハンチンチンポリペプチドの再ターゲティングを引き起こしたことを示している。
【図9】モデルハンチンチン特異的C4 sFv内部抗体の結合特異性を示すために用いられた様々なポリペプチドの存在および見かけの分子量を示すクーマシー染色電気泳動ゲルの写真を示す(図10のイムノブロットによってプロービングする)。
【図10】ハンチンチンポリペプチドエピトープ(レーン1〜3、6〜7)のみが、モデルハンチンチンポリペプチド特異的C4 sFv内部抗体をアフィニティ生成するために用いることができるが、無関係なDRPLAポリペプチド(レーン2および5)は、C4 sFv内部抗体と相互作用できないことを示すイムノブロットである。
【図11】AはELISA法によって分析した場合の、固定GST融合蛋白質に対する抗ハンチンチン内部抗体C4 sFvの直接結合試験の結果を示す。 BはBIAコアによって分析した場合の、抗ハンチンチン内部抗体C4 sFvの速度論的結合アフィニティ試験の結果を示す。抗ハンチンチン内部抗体C4 sFvの濃度範囲(60〜100 nM)を用いて、ストレプトアビジンセンサーチップに結合したビオチン結合HDペプチド50 RU上で結合定数(kon)を測定した。
【図12A−D】抗ハンチンチン内部抗体Nt-HD C4 sFv-NLSによるGFP融合HD断片の核への再ターゲティングを示す。パネルB、D、およびFは、COS-7細胞において48時間同時トランスフェクションした後の、HD-Q104-Myc-HIS6およびC4 sFv-NLSまたは陰性sFv-NLS内部抗体(パネルA、C、およびE)の免疫蛍光を示す。HD-Q104-Myc-HIS6は、抗myc 9E10 mAbによる免疫染色の後、FITC標識ヤギ抗マウスIgG抗体による免疫染色によって可視化した(パネルA、B)。内部抗体は、ポリクローナル抗HA抗体の後、ローダミン標識ヤギ抗ウサギIgG抗体によって免疫染色した(パネルC、D;パネルEおよびFは同じ領域の二重染色を表す)。
【図12E−F】抗ハンチンチン内部抗体Nt-HD C4 sFv-NLSによるGFP融合HD断片の核への再ターゲティングを示す。パネルB、D、およびFは、COS-7細胞において48時間同時トランスフェクションした後の、HD-Q104-Myc-HIS6およびC4 sFv-NLSまたは陰性sFv-NLS内部抗体(パネルA、C、およびE)の免疫蛍光を示す。HD-Q104-Myc-HIS6は、抗myc 9E10 mAbによる免疫染色の後、FITC標識ヤギ抗マウスIgG抗体による免疫染色によって可視化した(パネルA、B)。内部抗体は、ポリクローナル抗HA抗体の後、ローダミン標識ヤギ抗ウサギIgG抗体によって免疫染色した(パネルC、D;パネルEおよびFは同じ領域の二重染色を表す)。
【図13A−D】抗ハンチンチン内部抗体C4 sFv-NLSを安定に発現するCOS-7細胞におけるGFP-HD-Q104(b、d、f;40倍)またはGFP-DRPLA-Q81(a、c、e;陰性対照として;20倍)の再配置を示す。パネルAおよびBは、GFP発現を示し、パネルCおよびDはC4 sFv-NLS内部抗体のローダミン染色を示し、そしてパネルEおよびFは、同じ領域の二重染色を示す。
【図13E−F】抗ハンチンチン内部抗体C4 sFv-NLSを安定に発現するCOS-7細胞におけるGFP-HD-Q104(b、d、f;40倍)またはGFP-DRPLA-Q81(a、c、e;陰性対照として;20倍)の再配置を示す。パネルAおよびBは、GFP発現を示し、パネルCおよびDはC4 sFv-NLS内部抗体のローダミン染色を示し、そしてパネルEおよびFは、同じ領域の二重染色を示す。
【図14】病原性の長さのHD-ポリQ-GFP凝集体の抗HD-C4 sFv阻害を示す。上のパネルは、プラスミドトランスフェクション比5:1(sFv:HD)でHD-ポリQ-GFP(Q=25、72、および104)単独またはC4または対照sFv(ML3〜9)をトランスフェクトしたBHK-21細胞のデジタル蛍光顕微鏡画像を示す。リン酸カルシウム法によるトランスフェクションの48時間後に生存して撮影した細胞を示す;凝集体は、GFPシグナルの強い斑として見える。類似の結果がHD-72Q-GFPの同時トランスフェクションについても認められた。下のパネルは、COS-7、BHK-21、およびHEK293細胞におけるsFv(C4またはML3-9)または親ベクター(pcDNA)とHD-72Q-GFPとの5:1同時トランスフェクション後の凝集体の定量を表す。同時トランスフェクションの48時間後に、細胞を2%SDS/2%トライトンX100/50 mMトリスによって溶解した。無作為な視野6または8個における不溶性の蛍光凝集体の数を計数した。バーは、同時トランスフェクション6個の平均値±SEMを表す。C4は、対照ML3-9またはpcDNAと比較すると、凝集体の数を有意に減少させた(p<0.0005)。類似の結果はHD-104Q-GFPについても得られた。
【図15】病原性の長さのHD-ポリQ-GFP凝集体の抗HD C4 sFv阻害を示す。上のパネルは、C4:HD-ポリQ-GFP HD-ポリQ-GFP(緑)、C4 sFv(赤、抗HA抗体およびアレキサ568結合二次抗体によって検出される)を5:1で同時トランスフェクトした二重標識細胞のデジタル蛍光顕微鏡画像、および合併画像を示す。104Q同時トランスフェクト体にはいくつかの凝集体を認め(25Qでは認めない)、C4を発現していない細胞ではしばしば認めるが(矢印の先)、いくつかの二重トランスフェクト細胞においても認めた(矢印)。下のパネルは、同時トランスフェクト体におけるHD-72Q-GFP蛋白質発現のイムノブロット分析のデジタル画像を示す。細胞溶解物を24時間目に(顕著な凝集体形成と、不溶性区画にHD-72Q-GFPが失われる前)5:1同時トランスフェクト体(sFv:HD-72Q-GFP)から1試料あたり2本ずつ採取してSDS-PAGE法を行った。1000倍希釈した抗AFP一次抗体(クアンタム社)をプローブとしてメンブレンを調べ、剥離して、500倍希釈抗アクチン抗体(シグマ社)によって再プロービングした。HD-72Q-GFPバンドは、HRP結合二次抗体と化学発光による検出後の強度と類似であった。
【図16】ハンチンチンエピトープをコードするDNAワクチンによって処理した、および処理していないマウス(野生型およびトランスジェニックハンチンチン系(HD))におけるグルコースレベルを示す。
【特許請求の範囲】
【請求項1】
凝集体を形成することができるポリペプチドを、凝集を最小限にするように該ポリペプチドに特異的に結合し、それによって細胞内凝集体の形成を阻害する、ポリペプチド結合分子またはその機能的断片に接触させる段階を含む、選択されたポリペプチドの細胞内凝集体の形成を阻害する方法。
【請求項2】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項1記載の方法。
【請求項3】
ポリペプチドが対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項1または2記載の方法。
【請求項4】
ポリペプチド結合分子が、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体(intrabody)からなる群より選択される、請求項1〜3のいずれか一項記載の方法。
【請求項5】
ポリペプチド結合分子が内部抗体である、請求項1〜3のいずれか一項記載の方法。
【請求項6】
内部抗体が、以下の(a)〜(g)からなる群より選択される1つまたはそれ以上の特性を有する、請求項5記載の方法:
(a)一価のポリペプチドである;
(b)ハンチンチンポリペプチドのアミノ末端残基1〜17位に特異的に結合する;
(c)ポリグルタミンを除外してエピトープを特異的に結合する;
(d)多価である;
(e)スペーサー領域を含む;
(f)ユビキチンに対応するアミノ酸配列を含む;および
(g)ポリペプチドをプロテアソームに再ターゲティングする。
【請求項7】
内部抗体がターゲティングシグナルに対応するアミノ酸配列を含む、請求項5記載の方法。
【請求項8】
内部抗体がポリペプチドの細胞内位置を再ターゲティングする、請求項7記載の方法。
【請求項9】
ターゲティングシグナルが細胞質、核、ライソゾーム、細胞膜関連、小胞体関連、ペルオキシソーム、またはプロテオソームのターゲティングシグナルである、請求項7記載の方法。
【請求項10】
内部抗体が以下の(a)〜(c)からなる群より選択されるアミノ酸配列を含む、請求項5記載の方法:
(a)配列番号:2;
(b)配列番号:4;および
(c)配列番号:6に提供される抗Nt-HD C4 sFvのアミノ酸配列。
【請求項11】
ポリペプチドがインビボで内部抗体と接触される、請求項5記載の方法。
【請求項12】
細胞内凝集体を有する危険にさらされている対象に、凝集体の形成を最小限にして、それによって細胞内凝集体の形成を阻害するようにポリペプチドに特異的に結合するポリペプチド結合分子またはその機能的断片を投与する段階を含む、対象において選択されたポリペプチドの細胞内凝集体の形成を阻害する方法。
【請求項13】
対象が神経疾患または神経疾患関連糖尿病の危険にさらされている、請求項12記載の方法。
【請求項14】
対象が(a)ヒト、(b)実験動物、または(c)ハンチントン病動物モデルである、請求項12記載の方法。
【請求項15】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項12記載の方法。
【請求項16】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項12記載の方法。
【請求項17】
ポリペプチド結合分子が、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体からなる群より選択される、請求項12記載の方法。
【請求項18】
ポリペプチド結合分子が内部抗体である、請求項12記載の方法。
【請求項19】
内部抗体が対象において発現可能な核酸として投与される、請求項18記載の方法。
【請求項20】
内部抗体が配列番号:6に提供された抗Nt-HD C4 sFvのアミノ酸配列を含む、請求項18記載の方法。
【請求項21】
神経障害に関連したポリペプチド凝集体を形成することができるポリペプチドを特異的に結合し、それによって凝集体の形成を阻害するポリペプチド結合分子またはその機能的断片を対象に投与することを含む、神経障害を有する、または有する可能性がある対象を治療する方法。
【請求項22】
神経障害が、アルツハイマー病、パーキンソン病、ハンチントン病、ハンチントン病関連糖尿病、プリオン疾患、FTD、ALS、SBMA、DRPLA、SCA1、SCA2、SCA3/MJD、SCA4、SCA5、SCA6、およびSCA7からなる群より選択される、請求項21記載の方法。
【請求項23】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項21記載の方法。
【請求項24】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項21記載の方法。
【請求項25】
ポリペプチド結合分子が、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体からなる群より選択される、請求項21記載の方法。
【請求項26】
ポリペプチド結合分子が内部抗体である、請求項21記載の方法。
【請求項27】
内部抗体が対象において発現可能な核酸として投与される、請求項26記載の方法。
【請求項28】
神経障害がハンチントン病であり、かつ内部抗体が配列番号:6に提供される抗Nt-HD C4 sFvのアミノ酸配列を含む、請求項26または27記載の方法。
【請求項29】
以下の段階を含む、細胞内ポリペプチド凝集体を形成することができるポリペプチドを特異的に認識するポリペプチド結合分子またはその機能的断片を同定する方法:
細胞内ポリペプチド凝集体を形成することができるポリペプチドを提供する段階;
該ポリペプチドを試験ポリペプチド結合分子またはその機能的断片に接触させる段階;および
該試験ポリペプチド結合分子またはその機能的断片が、該ポリペプチドを特異的に認識する能力を決定し、
それによって、細胞内ポリペプチド凝集体を形成することができるポリペプチドを特異的に認識するポリペプチド結合分子を同定する段階。
【請求項30】
ポリペプチド結合分子が、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体からなる群より選択される、請求項29記載の方法。
【請求項31】
ポリペプチド結合分子が内部抗体である、請求項29記載の方法。
【請求項32】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項29記載の方法。
【請求項33】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項29記載の方法。
【請求項34】
以下の(a)〜(f)の段階を含む、望ましくない細胞内ポリペプチド凝集体を形成することができるポリペプチドを特異的に認識する化合物を同定する方法:
(a)細胞内ポリペプチド凝集体を形成することができる該ポリペプチドを提供する段階;
(b)該ポリペプチドを特異的に結合する内部抗体またはその機能的断片を提供する段階;
(c)1つまたはそれ以上の試験結合分子を提供する段階;
(d)該ポリペプチドおよび該内部抗体またはその断片を、該試験結合分子と共にインキュベートする段階;
(e)該ポリペプチドに対する該内部抗体またはその機能的断片の結合を変化させる、該試験結合分子の能力を決定する段階;ならびに
(f)段階(e)の結果、内部抗体を結合する分子が、試験結合分子から消去される段階、
それによって段階(f)の結果、該試験分子が、細胞内ポリペプチド凝集体を形成することができる該ポリペプチドと特異的に相互作用することができる化合物として同定される。
【請求項35】
方法の段階が、異なる結合分子102、103、104、105、106、107、108、および109個からなる群より選択される複雑性を有する多様なライブラリに関して繰り返される、請求項34記載の方法。
【請求項36】
試験結合分子が、有機低分子、ペプチド、および天然物抽出物からなる群より選択される、請求項34記載の方法。
【請求項37】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項34記載の方法。
【請求項38】
ポリペプチドが、野生型ポリペプチドと比較してさらなるグルタミン残基を含む、請求項34記載の方法。
【請求項39】
神経障害に関連した細胞内ポリペプチド凝集体を形成することができる選択されたポリペプチドに結合する、内部抗体またはその機能的断片をコードする単離核酸分子。
【請求項40】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項39記載の分子。
【請求項41】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項39または40記載の分子。
【請求項42】
神経障害に関連した細胞内ポリペプチド凝集体を形成することができる選択されたポリペプチドに結合する内部抗体またはその機能的断片。
【請求項43】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項42記載の分子。
【請求項44】
ポリペプチドが、対応する野生型ポリペプチドと比較して、さらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項42または43記載の分子。
【請求項45】
以下の段階を含む、動物において選択されたポリペプチドの細胞内凝集体の形成を阻害する方法:
免疫が、選択されたポリペプチドの細胞内凝集体の形成阻害に十分な宿主抗体免疫応答を誘発する、選択されたポリペプチドと共通のエピトープを有する免疫原によって動物を免疫する段階。
【請求項46】
動物がヒトである、請求項45記載の方法。
【請求項47】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項45記載の方法。
【請求項48】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項45〜47のいずれか一項記載の方法。
【請求項49】
免疫原が、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択されるポリペプチドと共通のエピトープを含むポリペプチドである、請求項45〜48のいずれか一項記載の方法。
【請求項50】
免疫原が、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択されるポリペプチドと共通のエピトープを含むポリペプチドをコードする発現可能な核酸ワクチンである、請求項45〜48のいずれか一項記載の方法。
【請求項51】
動物が以下の(a)〜(d)の1つまたはそれ以上の状態を有する、または有する危険にさらされている、請求項45〜50のいずれか一項記載の方法:
(a)ハンチントン病;
(b)ハンチントン関連糖尿病;
(c)高い絶食時血糖レベル;または
(d)低いインスリンレベル。
【請求項1】
凝集体を形成することができるポリペプチドを、凝集を最小限にするように該ポリペプチドに特異的に結合し、それによって細胞内凝集体の形成を阻害する、ポリペプチド結合分子またはその機能的断片に接触させる段階を含む、選択されたポリペプチドの細胞内凝集体の形成を阻害する方法。
【請求項2】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項1記載の方法。
【請求項3】
ポリペプチドが対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項1または2記載の方法。
【請求項4】
ポリペプチド結合分子が、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体(intrabody)からなる群より選択される、請求項1〜3のいずれか一項記載の方法。
【請求項5】
ポリペプチド結合分子が内部抗体である、請求項1〜3のいずれか一項記載の方法。
【請求項6】
内部抗体が、以下の(a)〜(g)からなる群より選択される1つまたはそれ以上の特性を有する、請求項5記載の方法:
(a)一価のポリペプチドである;
(b)ハンチンチンポリペプチドのアミノ末端残基1〜17位に特異的に結合する;
(c)ポリグルタミンを除外してエピトープを特異的に結合する;
(d)多価である;
(e)スペーサー領域を含む;
(f)ユビキチンに対応するアミノ酸配列を含む;および
(g)ポリペプチドをプロテアソームに再ターゲティングする。
【請求項7】
内部抗体がターゲティングシグナルに対応するアミノ酸配列を含む、請求項5記載の方法。
【請求項8】
内部抗体がポリペプチドの細胞内位置を再ターゲティングする、請求項7記載の方法。
【請求項9】
ターゲティングシグナルが細胞質、核、ライソゾーム、細胞膜関連、小胞体関連、ペルオキシソーム、またはプロテオソームのターゲティングシグナルである、請求項7記載の方法。
【請求項10】
内部抗体が以下の(a)〜(c)からなる群より選択されるアミノ酸配列を含む、請求項5記載の方法:
(a)配列番号:2;
(b)配列番号:4;および
(c)配列番号:6に提供される抗Nt-HD C4 sFvのアミノ酸配列。
【請求項11】
ポリペプチドがインビボで内部抗体と接触される、請求項5記載の方法。
【請求項12】
細胞内凝集体を有する危険にさらされている対象に、凝集体の形成を最小限にして、それによって細胞内凝集体の形成を阻害するようにポリペプチドに特異的に結合するポリペプチド結合分子またはその機能的断片を投与する段階を含む、対象において選択されたポリペプチドの細胞内凝集体の形成を阻害する方法。
【請求項13】
対象が神経疾患または神経疾患関連糖尿病の危険にさらされている、請求項12記載の方法。
【請求項14】
対象が(a)ヒト、(b)実験動物、または(c)ハンチントン病動物モデルである、請求項12記載の方法。
【請求項15】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項12記載の方法。
【請求項16】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項12記載の方法。
【請求項17】
ポリペプチド結合分子が、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体からなる群より選択される、請求項12記載の方法。
【請求項18】
ポリペプチド結合分子が内部抗体である、請求項12記載の方法。
【請求項19】
内部抗体が対象において発現可能な核酸として投与される、請求項18記載の方法。
【請求項20】
内部抗体が配列番号:6に提供された抗Nt-HD C4 sFvのアミノ酸配列を含む、請求項18記載の方法。
【請求項21】
神経障害に関連したポリペプチド凝集体を形成することができるポリペプチドを特異的に結合し、それによって凝集体の形成を阻害するポリペプチド結合分子またはその機能的断片を対象に投与することを含む、神経障害を有する、または有する可能性がある対象を治療する方法。
【請求項22】
神経障害が、アルツハイマー病、パーキンソン病、ハンチントン病、ハンチントン病関連糖尿病、プリオン疾患、FTD、ALS、SBMA、DRPLA、SCA1、SCA2、SCA3/MJD、SCA4、SCA5、SCA6、およびSCA7からなる群より選択される、請求項21記載の方法。
【請求項23】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項21記載の方法。
【請求項24】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項21記載の方法。
【請求項25】
ポリペプチド結合分子が、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体からなる群より選択される、請求項21記載の方法。
【請求項26】
ポリペプチド結合分子が内部抗体である、請求項21記載の方法。
【請求項27】
内部抗体が対象において発現可能な核酸として投与される、請求項26記載の方法。
【請求項28】
神経障害がハンチントン病であり、かつ内部抗体が配列番号:6に提供される抗Nt-HD C4 sFvのアミノ酸配列を含む、請求項26または27記載の方法。
【請求項29】
以下の段階を含む、細胞内ポリペプチド凝集体を形成することができるポリペプチドを特異的に認識するポリペプチド結合分子またはその機能的断片を同定する方法:
細胞内ポリペプチド凝集体を形成することができるポリペプチドを提供する段階;
該ポリペプチドを試験ポリペプチド結合分子またはその機能的断片に接触させる段階;および
該試験ポリペプチド結合分子またはその機能的断片が、該ポリペプチドを特異的に認識する能力を決定し、
それによって、細胞内ポリペプチド凝集体を形成することができるポリペプチドを特異的に認識するポリペプチド結合分子を同定する段階。
【請求項30】
ポリペプチド結合分子が、低分子、ペプチド、ペプチド模倣体、抗体、抗体断片、および内部抗体からなる群より選択される、請求項29記載の方法。
【請求項31】
ポリペプチド結合分子が内部抗体である、請求項29記載の方法。
【請求項32】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項29記載の方法。
【請求項33】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項29記載の方法。
【請求項34】
以下の(a)〜(f)の段階を含む、望ましくない細胞内ポリペプチド凝集体を形成することができるポリペプチドを特異的に認識する化合物を同定する方法:
(a)細胞内ポリペプチド凝集体を形成することができる該ポリペプチドを提供する段階;
(b)該ポリペプチドを特異的に結合する内部抗体またはその機能的断片を提供する段階;
(c)1つまたはそれ以上の試験結合分子を提供する段階;
(d)該ポリペプチドおよび該内部抗体またはその断片を、該試験結合分子と共にインキュベートする段階;
(e)該ポリペプチドに対する該内部抗体またはその機能的断片の結合を変化させる、該試験結合分子の能力を決定する段階;ならびに
(f)段階(e)の結果、内部抗体を結合する分子が、試験結合分子から消去される段階、
それによって段階(f)の結果、該試験分子が、細胞内ポリペプチド凝集体を形成することができる該ポリペプチドと特異的に相互作用することができる化合物として同定される。
【請求項35】
方法の段階が、異なる結合分子102、103、104、105、106、107、108、および109個からなる群より選択される複雑性を有する多様なライブラリに関して繰り返される、請求項34記載の方法。
【請求項36】
試験結合分子が、有機低分子、ペプチド、および天然物抽出物からなる群より選択される、請求項34記載の方法。
【請求項37】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項34記載の方法。
【請求項38】
ポリペプチドが、野生型ポリペプチドと比較してさらなるグルタミン残基を含む、請求項34記載の方法。
【請求項39】
神経障害に関連した細胞内ポリペプチド凝集体を形成することができる選択されたポリペプチドに結合する、内部抗体またはその機能的断片をコードする単離核酸分子。
【請求項40】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項39記載の分子。
【請求項41】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項39または40記載の分子。
【請求項42】
神経障害に関連した細胞内ポリペプチド凝集体を形成することができる選択されたポリペプチドに結合する内部抗体またはその機能的断片。
【請求項43】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項42記載の分子。
【請求項44】
ポリペプチドが、対応する野生型ポリペプチドと比較して、さらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項42または43記載の分子。
【請求項45】
以下の段階を含む、動物において選択されたポリペプチドの細胞内凝集体の形成を阻害する方法:
免疫が、選択されたポリペプチドの細胞内凝集体の形成阻害に十分な宿主抗体免疫応答を誘発する、選択されたポリペプチドと共通のエピトープを有する免疫原によって動物を免疫する段階。
【請求項46】
動物がヒトである、請求項45記載の方法。
【請求項47】
ポリペプチドが、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択される、請求項45記載の方法。
【請求項48】
ポリペプチドが、対応する野生型ポリペプチドと比較してさらなるグルタミン残基を有する自然界に存在するポリペプチドを含む、請求項45〜47のいずれか一項記載の方法。
【請求項49】
免疫原が、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択されるポリペプチドと共通のエピトープを含むポリペプチドである、請求項45〜48のいずれか一項記載の方法。
【請求項50】
免疫原が、アミロイド前駆体蛋白質、プレセニリン1、プレセニリン2、α-2マクログロブリン、アポリポ蛋白質、αシヌクレイン、ハンチンチン、プリオン蛋白質、タウ、SOD、AR、アトロフィン1、アタキシン1、アタキシン2、アタキシン3、CACNL1A4、およびSCA7からなる群より選択されるポリペプチドと共通のエピトープを含むポリペプチドをコードする発現可能な核酸ワクチンである、請求項45〜48のいずれか一項記載の方法。
【請求項51】
動物が以下の(a)〜(d)の1つまたはそれ以上の状態を有する、または有する危険にさらされている、請求項45〜50のいずれか一項記載の方法:
(a)ハンチントン病;
(b)ハンチントン関連糖尿病;
(c)高い絶食時血糖レベル;または
(d)低いインスリンレベル。
【図1】


【図2】

【図3】
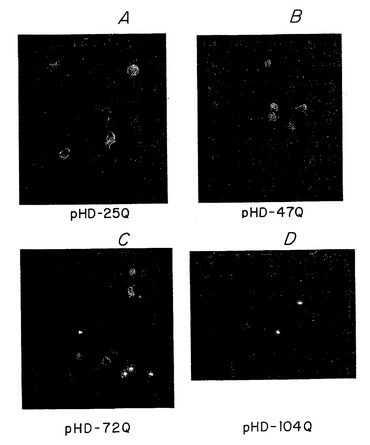
【図4】

【図5A−D】

【図5E−H】
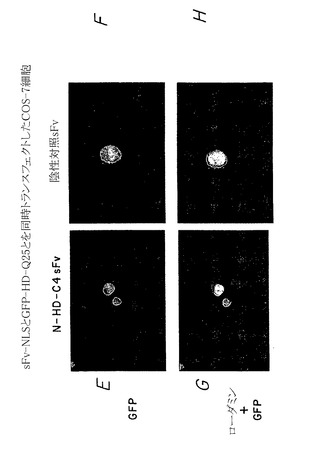
【図6A−C】

【図6D−G】
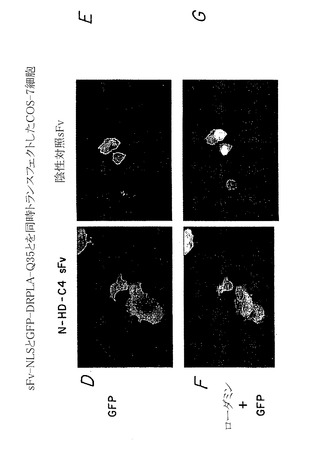
【図7A−D】

【図7E−H】
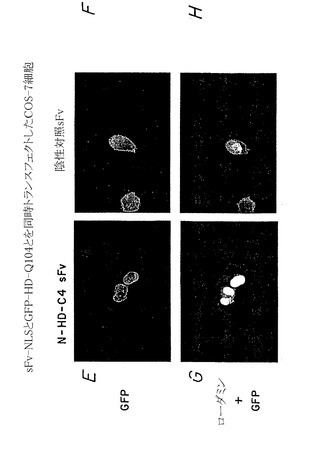
【図8】

【図9】
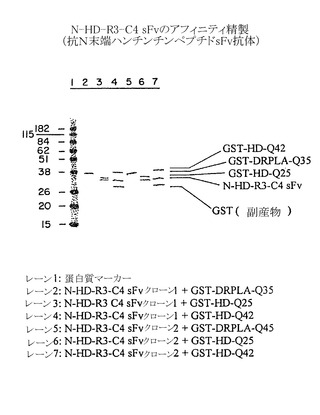
【図10】
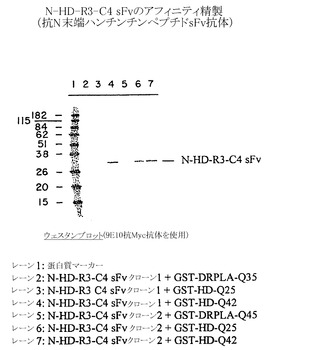
【図11】
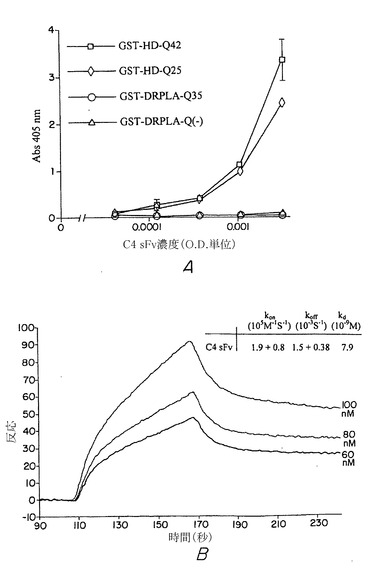
【図12A−D】
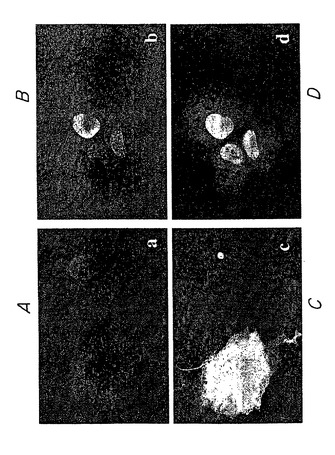
【図12E−F】
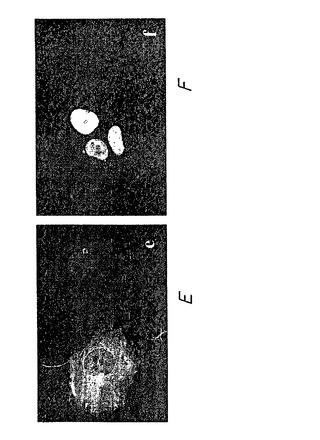
【図13A−D】
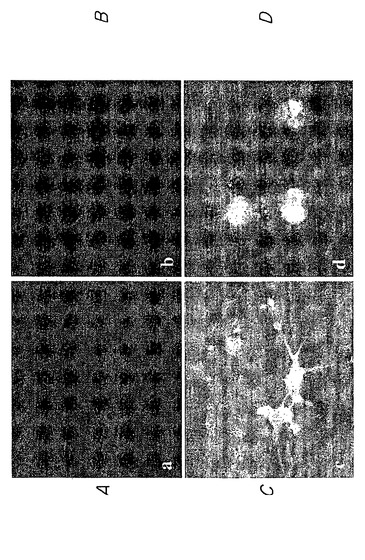
【図13E−F】
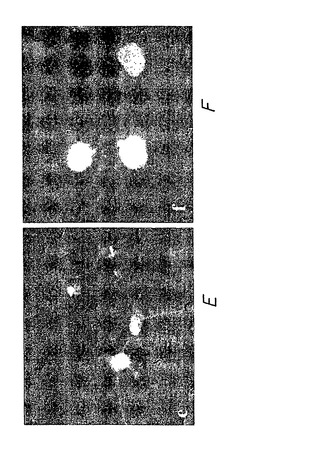
【図14】
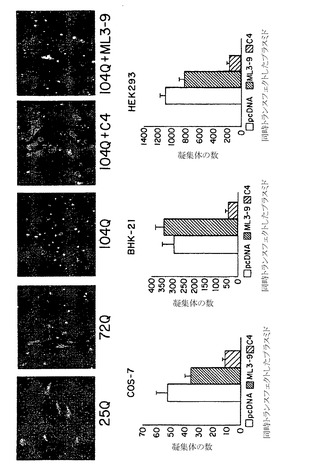
【図15】
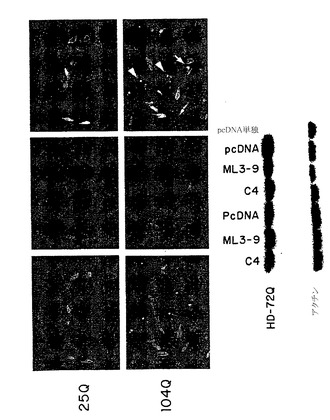
【図16】
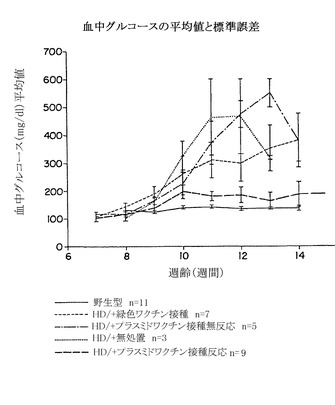
【図2】
【図3】
【図4】
【図5A−D】
【図5E−H】
【図6A−C】
【図6D−G】
【図7A−D】
【図7E−H】
【図8】
【図9】
【図10】
【図11】
【図12A−D】
【図12E−F】
【図13A−D】
【図13E−F】
【図14】
【図15】
【図16】
【公開番号】特開2007−6894(P2007−6894A)
【公開日】平成19年1月18日(2007.1.18)
【国際特許分類】
【出願番号】特願2006−189911(P2006−189911)
【出願日】平成18年7月11日(2006.7.11)
【分割の表示】特願2001−511881(P2001−511881)の分割
【原出願日】平成12年7月24日(2000.7.24)
【出願人】(398005777)アブジェニックス インク. (40)
【出願人】(502032323)ニューヨーク ステイト デパートメント オブ ヘルス (1)
【Fターム(参考)】
【公開日】平成19年1月18日(2007.1.18)
【国際特許分類】
【出願日】平成18年7月11日(2006.7.11)
【分割の表示】特願2001−511881(P2001−511881)の分割
【原出願日】平成12年7月24日(2000.7.24)
【出願人】(398005777)アブジェニックス インク. (40)
【出願人】(502032323)ニューヨーク ステイト デパートメント オブ ヘルス (1)
【Fターム(参考)】
[ Back to top ]