
移植細胞により産生されるホルモンを用いた魚の卵及び/又は魚の精子の発生及び成熟を改善するための手段及び方法
本発明は、ホルモン投与を用いた魚の卵及び/又は魚の精子の発生及び/又は成熟を改善するための手段及び方法であって、前記魚に前記ホルモンを産生する細胞を提供することを含む、前記方法に関する。好ましいホルモンは、黄体形成ホルモン(LH)、卵胞刺激ホルモン(FSH)又は絨毛性ゴナドトロピン(GC)又はそれらの機能的一部、派生物及び/若しくはアナログのような排卵誘発ホルモンである。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、魚の養殖の分野に関する。本発明はとりわけ、魚の卵及び/又は魚の精子の発生及び成熟のホルモン促進型改善の分野に関する。
【背景技術】
【0002】
多くの魚類種は、環境因子に応答して成熟する。光周期、温度、季節、圧力及びエネルギー備蓄のようなこれらの因子は、当該動物によって感知され、下垂体の視床下部中枢の抑制作用を制御する。このようにして、下垂体によるゴナドトロピンの産生は通常は、特定の環境条件下でのみ低下され、活性化される。活性化される際、下垂体はオス及びメスの両方の生殖腺の発育及び発生を刺激するゴナドトロピンを放出する。
【発明の開示】
【発明が解決しようとする課題】
【0003】
水産業にとって、成熟を概して、たいてい未精製の下垂体抽出物からなる同一のホルモンの定期的な注入により人工的に刺激することができることは重要である。定期的な注入は、ある程度までオスとメスの両方の生殖腺の発生に関する環境誘因を凌駕する。しかしながら、現行方式の卵及び/又は精子の発生及び/又は成熟の人工的な改善は、完全に満足できるものではない。例として、まだ未成熟尾ウナギにおいては、それらが産卵回遊を開始する際に、卵母細胞が排卵にとって十分に成熟する前の3ヶ月間−5ヶ月間の間、メスを毎週処理しなければならない。これは時間のかかる処理であり、ウナギにとってもストレスが多い。
【課題を解決するための手段】
【0004】
本発明は、ホルモン投与を用いた魚の卵及び/又は魚の精子の発生及び/又は成熟を改善するための方法であって、前記魚に前記ホルモンを産生する細胞を提供することを含む、前記方法を提供する。魚に移植される当該細胞は1つ以上のホルモンを放出し、それによって少なくとも当該ホルモン自身の定期的な注入に関する必要性を低減させる。分泌ホルモンが循環系へ放出され、全ての組織、とりわけ生殖器に十分量で到達する限りは、当該細胞は種々の方法で移植されてもよい。当該細胞は、体のどこに移植されてもよい。ホルモン産生細胞で構成される移植は概して、黄体形成ホルモン(LH)、卵胞刺激ホルモン(FSH)又は絨毛性ゴナドトロピン(CG)のようなホルモンを産生するために脳下垂体をバイパスするために用いられる。ホルモン産生細胞の移植は好ましくは、血流にアクセス可能な場所に挿入される。好ましい挿入方法は、腹腔内注入及び皮下注入である。血流にアクセス可能な場所は、LH、FSH及びCGのような二次的なホルモンを産生する細胞に極めて適している。
【0005】
細胞の移植は哺乳動物においてよく用いられ、細胞を移植し少なくともしばらくの間維持する方法に関する多くの経験が得られてきた。魚が概して、成熟卵又は成熟精子の採卵又は収穫後に始末されることを考慮すると、移植細胞の制御に関する大きな必要性は存在しない。重要なのは、当該細胞が生殖腺の完全な発生及び成熟を可能にするのに十分な量で存在したままの状態であることである。従って、細胞の数は、発生及び成熟に関して必要とされる数未満であってはならないかもしれないし、生殖腺の発生及び成熟又は当該動物の身体全体の健康を妨げるほど多数であってはならないかもしれない(又は、多数になってはならないかもしれない)。当該細胞によるホルモンの産生レベルは、あまり重要ではない。注入される細胞の数は、成熟に関して要求される総量によって決まる。ホルモン産生は、バイオアッセイにより定量化される。
【0006】
注入される細胞のホルモン放出は、卵及び/又は精子の発生及び/又は成熟を刺激するのに十分に高い必要がある。排卵誘発ホルモン(fertility hormone)の過剰発現はそれ自体有毒ではなく、卵及び/又は精子の成熟及び/又は発生の刺激にマイナスに作用しないので、当該ホルモンの発現に関する上限境界は重要ではない。哺乳類界において、種々の移植補助物が、当該細胞の長期の残留を可能にし又は当該細胞の分化を可能にするために開発されてきた。これらの補助物もまた当然ながら、本発明において用いることができる。当該補助物としては、細胞の移植及び接着のためのコラーゲン又は合成マトリクスが挙げられるが、これらに限定されない。
【0007】
当該細胞は、多くの場合、極めて長い時間存在する必要がないので、多くの様々な種由来の魚細胞をレシピエント魚へ移植することが可能である。移植細胞と当該レシピエントとの間の進化的な差異が大きい場合、当該レシピエント魚は、当該移植細胞(移植片)への免疫応答を備え付けるように思われる。しかしながら、当該細胞はたいてい限られた時間の間の存在だけが必要とされるので、そのような免疫応答は概して容認することができる。当該処理の丈夫さ及び予見可能性の増大のために、当該移植片は、レシピエント魚種と同じ属又は科に由来することが好ましい。好ましくは、2つとも同じ種に由来する。魚は概して非近交系集団であるので、同一種の魚間の免疫学的相違が存在する。これは概して、我々の実施例の1つのウナギの実験に示されるとおり問題とはならないが、共通の免疫学的マーカーに関して当該移植片及びレシピエントをさらに調和させることが可能である。代表的なマーカーは、主要組織適合性抗原及び非主要組織適合性抗原である。当該移植細胞の移植は、当該レシピエント魚にシクロスポリンのような免疫抑制剤を提供することによってさらに促進されてもよい。
【0008】
本発明のホルモン産生細胞の移植は、卵及び/又は精子の成熟、卵及び/又は精子の生殖能、卵の授精、結果として生じる胚の質、受精卵及び未受精卵の生存並びに胚の生存を改善するために用いることができる。これらの改善は全て、魚の卵及び/又は精子の改善された発生及び成熟をもたらす。魚の卵及び/又は精子の発生及び成熟なる用語は従って、自然過程の産卵に限定されず、卵の授精に関する人工的な方法にもまた関連する。従って当該用語は、本発明の方法で処理される魚の未受精卵及び/又は精子の収穫にも関連する。当該未受精卵はまた、子孫を創る以外の目的で用いられてもよい。その限定されない例は、キャビアのような人間の食用卵の製造である。
【0009】
魚の卵及び/又は精子の発生及び成熟は、種々の手段で刺激することができる。本発明のある実施態様において、魚の卵及び/又は精子の発生及び成熟は、卵、精子又は受精卵から生じた胚の絶対数又は質が上昇される際に、刺激される。
【0010】
生殖は、高度に制御された生物学的過程である。生殖の種々の側面は、種々のホルモンによって制御される。しかしながら、いくつかのホルモンは、魚に移植される細胞によって発現される際に、おおよそ類似した効果をもたらすことができる。これらのホルモンとしては、成長ホルモン、コルチコリベリン(Corticoliberin)(副腎皮質刺激ホルモン)、甲状腺刺激ホルモン、FSH、LH、プロラクチン、CG(絨毛性ゴナドトロピン)、MG(閉経期ゴナドトロピン)、ソマトロピン又はそれらの組み合わせが挙げられる。上記で述べたホルモンはまた、違う名前でも知られる。基礎を成すアミノ酸配列が同一であるので、同義語で呼ばれるホルモンもまた、本発明の範囲内に包含される。例えば、成長ホルモンは時々また、ソマトトロピン、ソマトトロピックホルモン(somatotropic hormone)、脳下垂体成長ホルモン、ソマトトロピックホルモン(somatotropic hormone)又はSTHとも称される。コルチコリベリン(Corticoliberin)はまた、放出性副腎皮質刺激ホルモン(releasing corticotropin hormone)とも称される。副腎皮質刺激ホルモンはまた、コルチコトロピン、アドレノコルチコトロピン(adrenocorticotropin)、アドレノトロピン(adrenotropin)、コルチコトロピン、ACTH又はアドレノコルチコトロピック(adrenocorticotropic)ホルモンとも称される。甲状腺刺激ホルモンはまた、TSH、チロトロピン又はチロトロピック(thyrotropic)ホルモンとも称される。FSHはまた、卵胞刺激ホルモン、フォリトロピン又はガメトチネティック(gametocinetic)ホルモンとも称される。LHはまた、黄体形成ホルモン、ルテオトロピン又は間質細胞刺激ホルモン(ICSH)とも称される。プロラクチンはまた、PRL、乳腺刺激ホルモン(lactogenic hormone)、乳腺刺激ホルモン(mammotropic hormone)、ガラクトポイエティック(galactopoietic)ホルモン又はラクトトロピン(lactotropin)とも称される。CG(絨毛性ゴナドトロピン)はまた、絨毛性性腺刺激ホルモン、絨毛性性腺刺激ホルモン、コリオゴナドトロピン(choriogonadotropin)又は絨毛性ゴナドトロピンとも称され、さらに、閉経期ゴナドトロピン(MG)はまた、ウロゴナドトロピン(urogonadotropin)、メノトロピン又は閉経期性腺刺激ホルモンとも称される。
【発明を実施するための最良の形態】
【0011】
本発明の好ましい実施態様において、前記ホルモンは卵の発生及び成熟に直接的に関与するホルモンである。そのような排卵誘発ホルモン(fertility hormone)は概して、下垂体又は生殖器によって産生される。好ましい実施態様において、当該排卵誘発ホルモン(fertility hormone)は、黄体形成ホルモン(LH)、卵胞刺激ホルモン(FSH)又は絨毛性ゴナドトロピン(GC)又はそのようなホルモンの機能的部分、派生物及び/若しくはアナログを含む。これらのホルモンは、魚の卵及び/又は精子の発生及び成熟の非常に強力な刺激要因である。これらのホルモンは、本質的に極めて保存されており、種の壁をほとんど有していない。例えば、尿中のヒト排卵誘発ホルモン(fertility hormone)の存在は、それらをカエルの卵と共にインキュベートすることによって検出することができる。同様に、ヒト絨毛性ゴナドトロピン(hGC)はまた、ウナギにも作用し、さらに、コイ及びサケの下垂体抽出物は、ウナギ、タイ及びマスのような多くの様々な魚種に作用する。
【0012】
当該レシピエントは、異種ホルモンに対しての免疫応答を発生することが可能である。この免疫応答は概して、あまりにも遅すぎて卵及び/又は精子の成熟及び/又は発生の刺激に作用することができないが、当該ホルモンは、魚ホルモン又はその機能的部分、誘導体及び/又はアナログであることが好ましい。これは、提供されるホルモンと内生的なホルモンとの間の相違を制限し、その結果、当該レシピエントにおいて、提供されるホルモンに対する免疫応答の発生を少なくともある程度制限する。好ましくは、当該ホルモンは、当該レシピエントと同一の属に属する種に由来する。このようにして、免疫応答が発生する可能性はさらに低減される。とりわけ好ましい実施態様において、提供されるホルモンは、当該レシピエントにおけるホルモン等価物と免疫学的に同一である。これは、提供されるホルモンに対するどんな有害な免疫応答の発生も完全に回避する。
【0013】
当該細胞は、処理なしで好ましいホルモンを発現することができるか又は好ましいホルモンを発現するように操作することができるかのどちらかである。当該細胞が既にホルモンを発現しないか又は十分なホルモンを発現しない際には、ホルモンを発現するための遺伝子情報を当該細胞に提供することができる。
【0014】
好ましい実施態様において、当該細胞はホルモンを発現するための遺伝子情報を提供される。これは、当該細胞にホルモンに関するコード配列を含む発現カセットを提供することによって行うことができる。しかしながら、活性制御配列をそれぞれのホルモンに関するコード配列付近に挿入することによって内生的な遺伝子を活性化することもまた可能である。これは例えば、相同組換えを通じて行うことができる。
【0015】
LHタンパク及びFSHタンパクは、関連タンパクのファミリーに属する。双方は、共通のα−サブユニット及び異なるβ−サブユニットからなるヘテロダイマーとして機能しうる特性を共有する。1つよりも多いタンパク鎖からなるホルモンの場合は、細胞がホルモンを生成するために必要とされる全ての鎖を発現しないことも起こり得る。これらの場合には、発現カセットだけが、欠けている鎖に必要である。従って、ホルモンのサブユニットの全てのサブユニットではないが1つ以上のサブユニットが当該細胞で十分に発現される場合には、当該細胞において残りのサブユニットを発現する必要があるだけである。
【0016】
当該サブユニットが1つも発現されない場合には、当該細胞を当該サブユニットの全てが十分なレベルで発現されるように処理しなければならない。好ましい実施態様において、当該細胞は、ホルモンのサブユニットに関する発現カセットを提供される。好ましい実施態様において、当該細胞は、LH及びFSHを作り出す3つのタンパク鎖に関する(すなわち、それぞれのホルモンに関する共通のα−サブユニット及び独特なβ−サブユニットに関する)発現カセットを提供されるか又は別個の株化細胞において、βLH + α 及び βFSH + α の組み合わせが発現される。従って好ましい実施態様において、前記細胞は、前記ホルモンを発現するために遺伝学的に改変される。好ましくは、当該細胞は前記ホルモンをコードする1つ以上の遺伝子を提供される。
【0017】
当該細胞は、初代細胞又はインビトロで長期間培養される株化細胞であってもよい。好ましい実施態様において、当該細胞はクローン集団の細胞に由来する。このようにして、当該細胞を使用に先立ってきめ細かい品質管理にさらすことができる。これはまた、同一の性質を有する細胞バンクの生成も可能にする。さらに、クローン集団をさらなる処理にさらすことができる。例えば、当該レシピエントにおける免疫応答を低減したい場合には、主要組織適合性抗原及び/又は非主要組織適合性抗原の発現をノックアウトすることが可能である。従って好ましい実施態様において、当該細胞は、当該レシピエントにおける低減された免疫原性に関して選択される。本発明の方法で用いるための又は本発明の使用のための細胞は、初代細胞又は培養細胞であってもよい。好ましくは、前記細胞は培養細胞であり、より好ましくは、不死化した培養細胞である。培養細胞は概して、株化細胞である。株化細胞は、前記細胞の表現型の実質的な変化なしに少なくとも5継代の間増殖することができる。不死化した株化細胞は、そのような表現型の変化を受けることなく少なくとも50回継代することができる。不死化した細胞は、種々の方法で初代細胞から取得することができる。好ましくは、前記の不死化した細胞は、培養物中で初代細胞に関する一般的な難局を経験してきた初代細胞の培養物から獲得される。別の好ましい実施態様において、前記細胞は、初代細胞への1つ以上の遺伝子の導入により不死化されている。
【0018】
本発明の方法は、あらゆるタイプの魚に用いられてもよい。好ましい魚は、ウナギ、スズキ(シーバス)、タイ、ハリバ、サケ、マス、タラ、コイ、ナマズ及びチョウザメである。しかしながら、本発明は、とりわけ回遊性魚、好ましくは1回生殖の魚に関して好都合である。これらの魚は、産卵までに時間がかかり、概して外部のシグナルへ全て同じように応答するわけではない。本発明の方法で、少なくともある程度環境刺激と関係なく魚の発生及び成熟を刺激することが可能である。これは、当該魚養殖の出発点に関する大量の予測可能性をもたらす。回遊性魚において、当該生産工程において作業をよりうまく予定することができるように卵の発生及び成熟を同調させることがさらに可能である。
【0019】
本発明は、排卵誘発ホルモン(fertility hormone)を産生する単離魚細胞及び/又は組み換え型魚細胞をさらに提供する。好ましい実施態様において、前記細胞は、前記ホルモンを発現するために遺伝学的に改変される。本発明は、排卵誘発ホルモン(fertility hormone)を発現する能力を提供される魚細胞をさらに提供する。好ましくは、当該魚細胞は、前記排卵誘発ホルモン(fertility hormone)をコードする組み換え型塩基配列及び/又は単離塩基配列を提供される。当該ホルモンが1つ以上のサブユニットからなる場合には、当該魚細胞は好ましくは、前記ホルモンの少なくとも1つのサブユニットをコードする単離塩基配列及び/又は組み換え型塩基配列を提供される。好ましくは、当該魚細胞は、前記ホルモンの全てのサブユニットをコードする塩基配列を提供される。好ましくは、前記細胞は、一般消費者向けの魚である。好ましい実施態様において、前記細胞は、ウナギ、スズキ(シーバス)、タイ、ハリバ、サケ、マス、タラ、コイ、ナマズ及びチョウザメに由来する。好ましくは、前記細胞は、上記で記載されたとおりの細胞、すなわち、初代細胞又はインビトロで長期間培養される株化細胞に由来する細胞である。好ましい実施態様において、当該細胞は、クローン集団の細胞に由来する。好ましくは、前記細胞は、培養細胞、より好ましくは、不死化した培養細胞である。
【0020】
好ましい実施態様において、使用が培養することのできる細胞の移植片で構成される。さらに、これらの細胞は好ましくはクローンであって、好ましくは特性に関して選択可能である。また一方、初代培養物、組織、受精卵又は胚性材料(embryonic material)を活用することも可能である。遺伝子導入した魚の卵の調製方法が公表されている(Morita et al 2004, Transgenic Research 13, 551)。しかしながら、記載された遺伝子導入した魚の卵は、さらに増殖させることができない。さらに、導入される遺伝子の発現は、Moritaらによって記載されたとおり、安定ではない。卵中の導入遺伝子の選択は困難であり、一つ一つの卵へのマイクロインジェクションを用いなければならないので、バイオリアクターとしての魚の卵の使用は培養可能な細胞に対してかなり時間がかかる一方で、細胞培養物では、我々は標準的なトランスフェクション技術を活用することができる。培養可能な細胞は、容易に保存され、いつでも利用可能にされるという利点を有することができる。さらに、遺伝学的に改変される卵と比較して、培養可能な細胞での操作に伴う倫理的な問題は全くない。培養細胞の懸濁液の注入は、卵の移植及び/又は胚の移植と比較して、かなり容易である。
【0021】
本発明は、本発明に従う細胞を含む魚をさらに提供する。好ましくは、前記魚は、一般消費者向けの魚である。好ましくは、前記魚は、一般消費者向けの幼魚又は成魚である。生殖までの時間は、好ましくは短い。初代胚性材料(primary embryonic material)は、利用可能でないか又は十分な量で利用可能でないという不都合を有する。好ましい実施態様において、前記魚は、ウナギ、スズキ(シーバス)、タイ、ハリバ、サケ、マス、タラ、コイ、ナマズ及びチョウザメである。好ましい実施態様において、前記魚は、遺伝学的に改変されない精子細胞、卵母細胞及び/又はそれらの前駆細胞を含む。
【実施例】
【0022】
ゼブラフィッシュ遺伝子LHβ、FSHβ及びαのクローニング
ゼブラフィッシュの遺伝子LHβ(AY424304)、FSHβ(AY424303)及びα(AY424306)は既に公表され、またウナギの遺伝子も Anguilla japonica のLHβ(AB175835)、Anguilla anguilla のFSHβ(AY169722)及び Anguilla japonica のα(AB175834)に関して報告される。LHβ、FSHβ及びαをクローニングするために、プライマーを各自のcDNA配列に基づいて設計した。
【0023】
LHβのプライマー設計に関して用いた配列は下記のとおりである。
atatataaat ctggacacgc agagacactt acaacagcct gctgagcaac cgcaacgcct gtcaag[atgt
tattggctgg aaatggtgtc ttctttctct tctctttgtt tttcctgctg gcggctgctc agagcttggt
ttttccacgc tgtgagctag taaatgagac ggtatcggtg gaaaaagagg gctgtccaaa atgcctggtg
tttcagacca ccatctgcag cggccactgc gtaacaaggg atcccgttta caagagcccg ttttccaccg
tccaccagac agtgtgcatg taccgggacg tccgctatga gaccattaac ctgcccgact gttccgccgg
cgtggacccg cagatcacat acccggtggc gctgagctgc gactgcagtc tgtgcaccat aaacacttcc
gactgcacca tccagagcct gcagcccgac ttctgcatgt cccagagaga ggatttcccc gcatactag]a
cctcgggcaa ctcacgtcaa cctacgcaca tagtcgagct cagcattatt agccctcctg tatgtttttt
ccattaatat atatactttc aagacactag tattcagctt aaagtgacat ttaaagacta aactaggtta
attaggggga aaagtagagt aagtcattgt ataatagtgg tttgttctgg agacaatcca aaactaatat
tgcttaaggg ggctaataaa attgacctta aaatgaattt aaataattta aaaactgcat ttattctagt
cgaaataaaa gaaataagac tttctttaga agaaaaaaca ttataggaaa tactgcaaaa aaattcctga
atctgttcaa catcattcgg gaaatcaaag gagggctaat aactgtgact tcagctgtac atcaataaag
aggctggttc ttaaattcaa aaaaaaaaaa aaaaaaaaaa aaaaaaaa
【0024】
FSHβのプライマー設計に関して用いた配列は下記のとおりである。
ggtctccacg aaactcccgc agatgaggat gcgtgtgctt gttctggcgc tgctgttgcc ggtgttaatg
agcgcagaat cagaatgcag gtgcagctgt cgactcacca acatctccat cactgtggag agcgaagaat
gtgggagctg cgtcacaatc gacaccacag cctgtgcagg act[atgctgg acaatggatc gagtttaccc
tagttccatg gcacagcaca cccagaaggt ctgtaacttc aagaacttga tgtacaagag ctacgagttt
aaaggctgtc ctgcaggggt tgattcagtc ttcgtgtacc ccgtggctct gagctgtgag tgcaaccagg
ttaactcaga cacaacagac tggggagcta tcagcccgca gaccaccagc tgcagcatac actag]agcac
tgtatcatga ccttaacaac atgtacgttg cagaatcaaa ttaagtaagg agtacaatta gaccatttaa
ggatatcaat tatttacaaa acctttagtt tttcatgcat cccacacaca tggtaatttg gttacttgaa
ttaatctgtt gtgttaattc tatagttggt actatggtaa ctagagtact agagtatcca atgctatact
agttttaatt acagttaatt atagaaaagt atgctacagt atttattaca gtttttctgt tttcaatatt
tagtactaca gtatgctagt gcattcatta acaataagct gtaaatacta taataaatac aggttaatac
actttactat agtatgcttg atcaacacta ttatttaatg tgagttacta tagtactttt caattgggat
ttgtcatttt ggatattgtg ggcttttttg gctattcata aagttttttt tatttttttt ttatttaatt
ttcagtcaaa tggaaacaag tccaccataa tacacttgtg tttcttttgt caaacttatc aatttgtgtc
tgtagatttc aattacaata catattttaa aggccaaaaa aaaaaaaaaa aaaaaaaa
【0025】
αサブユニットのプライマー設計に関して用いた配列は下記のとおりである。
gaagacactc atcacgctcc gccggaagtc gaggacaaag ccatc[atgtt ttggacaaga tacgctgaag
caagcatttt cttgttgtta atgattcttc atgtcggaca actgtattca agaaacgatg tgtctaacta
tggatgtgaa gagtgcaaac tcaagatgaa cgaacgtttc tccaaacccg gggctccggt ctatcagtgc
gtgggctgct gcttttcgag agcttacccc acacccctga ggtccaagaa aaccatgctt gtcccaaaaa
acatcacatc agaagccact tgctgtgtag caaaagaatc taaaatggtt gccacgaata tcccactata
caaccacaca gactgccact gcagcacctg ttactatcat aagtcttaa]a acacactctc ttcacatttc
tcaaatgctc atttcctgtt cttaaatcac agtgactcat gaaatatgat ttttatgtag ctttccatat
ttcaactgtg gccatttcca attcgtttct aaaatggttg gcataagtat tgtaaactgc atattctgtc
actatccctt taagagcgta atatgccatc ctttactatc attaaatcgc ttatttattt tgttgccttt
actgtgacat tcttcaaatc tataaatgaa ataaaagatt gctgaaggca aaaaaaaaaa aaaaaaaaaa
aaaaaaaaaa
【0026】
いずれの場合にも、括弧でくくられる部分はコード配列を表し、予想される増幅領域には下線が引かれる。
プライマーは下記のとおり設計された。
上流 - LHβ/EcoRI 5'-CAA CCG AAT TCA ACG CCT TCA AGA TGT-3'
下流 - LHβ/EcoRV 5'-CCG ATA TCT AGT ATG CGG GGA AAT-3'
上流 - FSHβ3 5'-AGG ATG CGT GTG CTT GTT CT-3'
下流 - FSHβ2/3 5'-TGT TGT TAA GGT CAT GAT ACA GTG C-3'
上流 - α1 5'-GTC GAG GAC AAA GCC ATC AT-3'
下流 - α1 5'-TGC CAA CCA TTT TAG AAA CGA-3'
さらなるクローニングステップを円滑にするために、LHβオリゴは、上流オリゴ及び下流オリゴ中にEcoRI及びEcoRVに関する制限酵素サイトを含む。
【0027】
全RNAを液体窒素中でホモジナイズし、TRIZOL試薬を用いて製造業者の使用説明書に従い抽出したゼブラフィッシュの頭部から単離した。微量のDNAをDNaseIと共にインキュベーションすることによって除去し、その後、フェノール/クロロホルム抽出及びエタノール沈殿を行った。RT−PCRを、プラチナを有するSuperscript IIワンステップRT−PCRシステムを用いて行った。Taq反応を、25 pmol の上流プライマー及び下流プライマーを用いて 100 ng の全RNAで行った。逆転写を 50 ℃ で30分間行った。PCR条件は、94 ℃ で20秒間の変性、LHβに関して 55 ℃ で30秒間及びFSHβに関して 50 ℃ で30秒間のアニーリング、さらに 72 ℃ で1分間の伸張の40サイクルに引き続き、72 ℃ で10分間の最終伸張ステップであった。PCR産物をアガロースの 1 % ゲル中での電気泳動によって分離し、エチジウムブロマイドで染色した。FSHβのPCR産物はアガロースゲル中で観察した際にかすかなバンドを形成したので、最適化するために、我々はこのPCR産物をテンプレートとして用いてPCRを行い、反応はPCR産物の1/20で、10 μM の上流プライマー及び下流プライマーを用いて行った。PCR条件は、94 ℃ で20秒間の変性、アニーリングを 50 ℃ − 60 ℃ のグラジエントで30秒間行い、72 ℃ で1分間の伸張の40サイクル、引き続き、72 ℃ で10分間の最終伸張ステップであった。はっきりしたバンドが、その結果観察された。増幅配列の同一性を確認するために、PCR産物を pCRII-TOPO ベクターにクローニングし、正しい方向を確認するために制限酵素で消化し、配列決定した。配列解析は、我々がゼブラフィッシュのLHβサブユニット、αサブユニット及びFSHβサブユニットをクローニングしたことを示した。これらのコンストラクトにより、常時発現プロモーター下での当該遺伝子のサブクローニングが可能である。
【0028】
常時発現プロモーター(CMV)制御下のLHβ、FSHβ及びαのクローニング
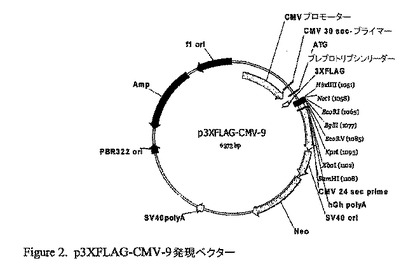
p3XFLAG-CMV-9発現ベクターを用いて、一過性の融合タンパク又は安定的な融合タンパクを確立した。当該ベクターは、マルチクローニング領域の上流の3つの隣接するFLAGエピトープをコードする。これは結果として、抗−FLAG抗体を用いた増大した検出をもたらす。当該CMVのプロモーター制御領域は、FLAG融合コンストラクトの転写を促進する。プレプロトリプシンリーダー配列はFLAG配列の前に位置し、タンパクの分泌を促進する。アミノグリコシドホスホトランスフェラーゼ遺伝子(Neo)は、ジェネテシン(G418)のようなアミノグリコシドへの耐性を与え、安定発現株の選択を可能にする。
【0029】
我々は当該ベクターを用いたが、その理由は、合成されるタンパクがプレプロトリプシンリーダーによって促進されて分泌され、我々が抗−FLAG抗体でのウェスタンブロッティングによってタンパクの発現を検出することができ、さらに我々がジェネテシンで選択して安定な株化細胞を作ることができるという利点を有しているからである。
【0030】
LHβのクローニング
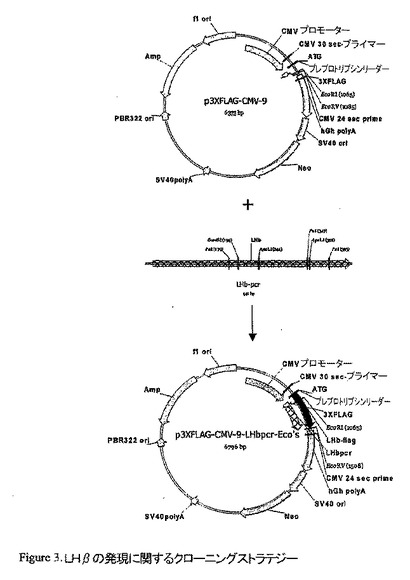
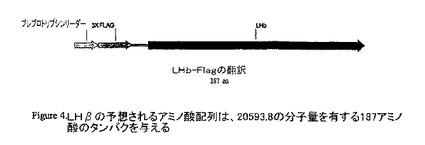
PCR産物を精製し、p3XFLAG-CMV-9 発現ベクターと同様にEcoRI及びEcoRVで消化した。当該DNA断片を、4 ℃ で一晩連結した。次に連結混合物を用いて、ケミカルコンピテント細胞をトランスフォーメーションした。酵素消化は、陽性クローンを選び出した。正しい様式の消化を生じさせるそれらのクローンを配列決定した。当面、配列決定されたクローンはLHβの間違った挿入を示したので、当該クローニングステップを繰り返さなければならなかった。予想されるアミノ酸配列は、20593.8 の分子量を有する187アミノ酸のタンパクを与える。
【0031】
FSHβのクローニング
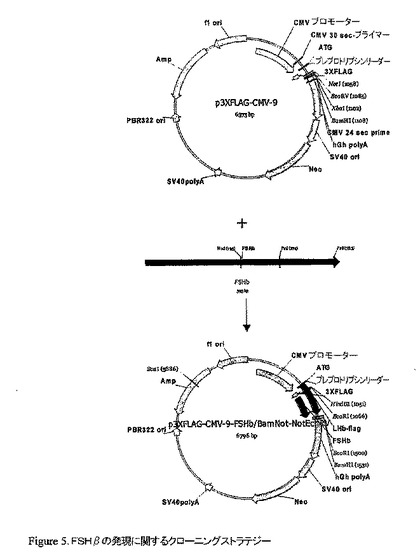
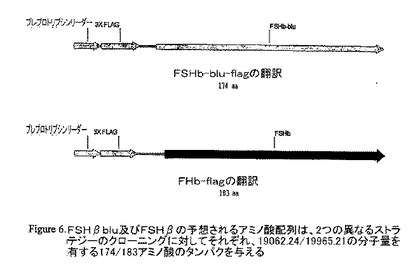
pCRII-TOPO FSHβをBamHI/NotIで消化し、FSHβ cDNAに相当するバンドを精製し、p3XFLAG-CMV-9 にサブクローニングした。陽性クローンを次に、Not/EcoRV及びマグビーンヌクレアーゼ(mug bean nuclease)で消化し、再連結してプレプロトリプシンリーダーを有するフレーム中のFSHβを得て、これらのコンストラクトをCMV−FSHβと命名した。もう1つの方法として、pCRII-TOPO FSHβをXba/BamHIで消化し、結果として生じたバンドを p3XFLAG-CMV-9 にサブクローニングした。陽性クローンを次にEcoRVで消化し、再連結してプレプロトリプシンリーダーを有するフレーム中のFSHβを得て、このコンストラクトをCMV−FSHβと命名した。酵素消化は、陽性クローンを選び出した。正しい様式の消化を生じさせるそれらのクローンを配列決定した。異なるストラテジーのクローニングに相当する2つの異なるコロニーの配列は正しかった。これらのコンストラクトを、ZF4株化細胞のトランスフォーメーションに用いた。予想されるアミノ酸配列は、2つの異なるストラテジーのクローニングに対してそれぞれ、19062.24/19965.21の分子量を有する174/183アミノ酸のタンパクを与える。
【0032】
αのクローニング
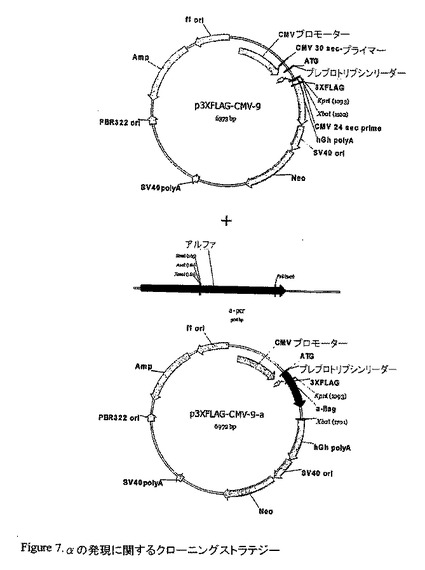
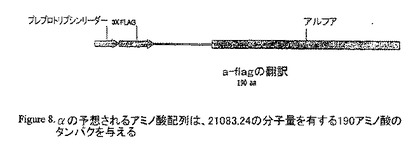
pCRII-TOPO αをKpnI/XbaIで消化し、結果として得られたバンドを p3XFLAG-CMV-9 にサブクローニングした。酵素消化は、陽性クローンを選び出した。正しい様式の消化を生じさせるそれらのクローンを配列決定した。配列解析は、我々が正しい配向で、FLAG及びプレプロトリプシンリーダー配列を有するフレーム中にaをクローニングしたことを示し、このコンストラクトを、CMV−αと命名した。予想されるアミノ酸配列は、21083.24の分子量を有する190アミノ酸のタンパクを与える。
【0033】
CMV−FSHβプラスミド及びCMV−αプラスミドを、QIAプレップスピンミニプレップキット(QIAprep spin Miniprep kit)を用いるSDSを含むアルカリ溶菌法によって精製した。精製されたプラスミドを、ScaIで直線化した。直線化された生成物を、アガロースの 1 % ゲル中の電気泳動によって分離し、エチジウムブロマイドで染色した。直線化されたプラスミドを精製し、定量化した。
【0034】
ゼブラフィッシュ繊維芽細胞株化細胞(ZF4)におけるFSHβのトランスフェクション
ZF4(ATCC番号:CRL-2050)細胞は、生まれて1日のゼブラフィッシュの胚由来の繊維芽細胞である。凍結した一定分量のZF4細胞を、液体窒素から移し、直ちに氷上に10分間置いた。解凍された細胞懸濁液をバイアルから移し、10 mL の完全培養液(1.2 g/L の重炭酸ナトリウム、2.5 mM のL−グルタミン、15 mM のHEPES、0.5 mM のピルビン酸ナトリウム、10 % のウシ胎仔血清及び 10 % のペニシリン/ストレプトマイシンを含むダルベッコ変法イーグル培養液とハムF12培養液の1:1の混合物)中に室温で希釈した。上清を 1200 rpm で8分間の遠心分離によって廃棄した。細胞を 8 mL の完全培養液中に再懸濁し、組織フラスコ(T25)に移し、28 ℃ で培養した。細胞密度を調べるために、倒立顕微鏡下で細胞を検査した。80 % 集密の培養で、培養液を除去し、3 mL のPBSで洗浄して細胞残屑及び血清を除去した。次に、0.5 mL のトリプシン溶液(0.25 %)を加え、単層の細胞がフラスコから剥離するまで室温でインキュベートし、単一細胞懸濁液が得られたら、5 mL の完全培養液を加えてトリプシン処理を停止した。生存細胞及び生育不能な細胞を、Fuchs-Rosenthal血球計算板を用いてカウントした。新たなフラスコ中に 100 cells/mm2 − 150 cells/mm2(各々のT25フラスコ中に 5 × 105)の密度で細胞を播種した。当該細胞を、次の継代前に 28 ℃ で最大4日間インキュベートした。トランスフェクションは、ゼブラフィッシュ繊維芽細胞が 50 % − 60 % コンフルエントになった際に、Fugene 6 を用いて製造業者の使用説明書に従い、ゼブラフィッシュ繊維芽細胞で行った。Fugene 6/DNA複合体は、無血清培養液中に Fugene 6 :DNA=6:1の割合で調製した。培養液を細胞から除去し、無血清培養液に交換した。Fugene 6/DNA複合体を思慮深く滴下して加え、混合した。当該細胞を 28 ℃ で5時間インキュベートした。次に、培養液を除去し、完全培養液に交換した。コントロールとして、Zf4細胞をpEYFP-N1プラスミドでコトンラスフェクトし、トランスフェクションの後、16時間及び24時間において、ライカ共焦点下で陽性細胞を解析した。種々の濃度のDNAを用いて、当該プラスミドに関する至適濃度を確立し、結果として、21 cm2 の表面中に 1 μg のDNAが最善の条件であり、およそ 30 % のトランスフォーメーションが実現された。
【0035】
LHβ、FSHβ及びαの発現の検出
(免疫組織化学)
トランスフェクトした細胞をまた免疫組織化学によって解析し、細胞中のタンパク発現を検出した。細胞を、2 % パラホルムアルデヒド及び 0.1 % グルタルアルデヒドのPBSで10分間固定し、固定液を除去するために、細胞をPBSで2回洗浄した。次に、細胞を 0.2 % Triton-X-100 のPBSで10分間透過処理した。固定液によって起こる自己蛍光を低減するために、2 mg/mL NaBH4 のPBSを10分間加えた。ブロッキングを 0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチンで30分間行った。抗−FLAG抗体(1:250)とのインキュベーションを、4 ℃ において一晩、0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチン中で行った。抗体を除去し、0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチンで 各々10分間、2回洗浄した。二次抗体(抗−ラビット Alexa 488)を、0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチン中に 1:1000 希釈で加え、室温で60分間インキュベートした。次に、0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチンで3回洗浄し、DABCO/Gelvatolに搭載した。ライカ共焦点での細胞の解析は、トランスフェクトした細胞の小胞中のタンパク発現を示した。この局在は、分泌されるタンパクに関して予想される部位と一致する。
【0036】
(ウェスタンブロット)
種々のコンストラクトをT25フラスコ中でFugene 6を用いて2回トランスフェクトし、コントロールとして、細胞をDNAでトランスフェクトせず、空のベクター及びポジティブコントロールのCMV−BAPでトランスフェクトした。タンパクを、トランスフェクション後3日目及び5日目に上清から獲得した。上清を、アミコンカラム(amicon column)を用いて、製造業者の使用説明書に従い濃縮した。FLAG融合タンパクを、抗−FLAG M2アフィニティーゲルを用いて製造業者の使用説明書に従い免疫沈降した。タンパクサンプルを、サンプルバッファーで1:4に希釈し、ボイルして、-80 ℃ で維持した。ウェスタンブロットの条件を決めるために、ポジティブコントロール(CMV−BAP)及びネガティブコントロール(DNAなし)のみ解析した。サンプルを 12 % アクリルアミドゲル中にロードし、50 mA で60分間ランした。ゲルをニトロセルロースメンブランにブロットした。メンブランを 4 ℃ において一晩、5 % ミルクでブロックし、免疫検出を室温において1時間、5 % ミルクで 1:250 希釈した抗−FLAG抗体に対して行った。免疫検出を、ECLで製造業者の使用説明書に従い明らかにした。当該抗体は、予想されるサイズのバンドを示す発現されたBAPタンパクを認識した。これらの実験で我々は、一過性にトランスフェクトした細胞が所定のタンパクを産生することを明らかにした。
【0037】
(安定的な株化細胞の作製)
細胞に6ウェルチャンバー中で、直線化し精製したDNAをFugene 6を用いてトランスフェクトした。コントロールとして、細胞にDNAなしでトランスフェクトした。トランスフェクションの3日後又は5日後に、完全培養液を、G418を加えた完全培養液に交換した。コンストラクトを発現していない細胞を死滅させるG418の量は、株化細胞間で異なる。ZF4株化細胞に関しては、製造業者の推奨する濃度は、0.8 mg/mL − 1 mg/mL である。細胞を4つ組で、0.8 mg/mL 及び 1 mg/mL で処理し、培養液を毎日交換して死細胞を洗い流した。これを、DNAを有さない細胞が死滅し当該細胞を含むプレート中に少しも細胞を観察することができなくなるまで、15日間行った。次にトランスフェクトした細胞を、同一の濃度のG418と共に5日間インキュベートした。コンフルエントの単層が得られたら、細胞をT25フラスコ中に継代培養し、G418の濃度を、完全培養液中 0.5 mg/mL まで下げた。安定的な株化細胞を、抗−FLAG抗体を用いた免疫組織化学解析及びウェスタンブロット解析で試験してタンパク発現を検出した。
【0038】
(バイオアッセイ)
トランスフェクトした細胞によって発現されるホルモンがアクティブであるかどうかを検証するために、簡便なバイオアッセイを行った。ゼブラフィッシュの濾胞細胞培養物は、種々の遺伝子の発現を上方制御するか又は下方制御することによって下垂体抽出物及び/又はヒト絨毛性ゴナドトロピン(hCG)に応答する。hCG(15 IU/mL)は、アクチビンβAの発現レベルを時間依存的に上昇させる。この効果は、処理40分で明らかであり、2時間で最大レベルに到達し、より長い処理(4時間)により当該効果は消失する。その一方、アクチビンβBは、同一の条件で抑制される。種々の濃度のhCGを用いた実験が行われる際、濃度依存的応答が観察される。キンギョ下垂体抽出物もまた、濃度依存的にアクチビンβAの発現を刺激しアクチビンβBを抑制する。我々は、培養物中の濾胞細胞のこの特性を用いてFSHβ及びLHβがアクティブであるか否かを検証することを計画した。
【0039】
(インビトロ濾胞細胞培養)
(濾胞細胞の単離)
幼魚のゼブラフィッシュを鳥獣店から購入し、オス及びメスを分離せずに維持した。メスを 0.01 % トリカインメタンスルホナート(tricaine methansulfonate)溶液で2分間又はまだ耐えているまで麻酔し、解剖前に斬首した。卵巣を次に取り外し、10 mm 培養ディッシュL-15(Gibco)に設置した。5匹のメスの濾胞を、注意深くインスリン針の補助で切り離した。切り離した濾胞を解剖顕微鏡の接眼マイクロメーターで測定し、およそ 0.45 mm の正常な卵黄形成濾胞を選択し、プールし、10 % ウシ胎仔血清を追加したM199培養液中、6日間T25フラスコで、28 ℃ 及び 5 % CO2 において培養した。培養液は、インキュベーションの3日目に交換した。6日間のインキュベーションの間に、濾胞細胞は著しく増殖し、当該実験に関する細胞の産生量は上昇した。細胞を洗浄し、28 ℃ で15分間トリプシン処理した。その後細胞を 1000 rpm で2分間の遠心分離を介してM199培養液で3回洗浄し、次にホルモン処理の前に、完全M199中で24時間、1ウェルあたり 1 × 105 cells/mL の密度で24ウェルプレートで継代培養した。細胞の量が実験に十分ではなかったので、我々は十分な材料を得るために20匹のメスで着手することにした。
【0040】
(ホルモン処理)
種々の濃度の上清を用い、ポジティブコントロールとしてhCGを 15 IU/mL で用い、さらにコイ下垂体抽出物もまた含む。当該下垂体抽出物条件を用いて、我々はメスウナギの生殖能力における効果を観察するために用いるべき細胞の量を比較することができる。
【0041】
(インビボ注入)
hCGを、20 IU/mL の濃度で 0.9 % NaCl溶液に溶解した。各々の魚は、ネガティブコントロールとしての 50 μL の生理食塩水、ポジティブコントロールとしてのhCG及び種々の濃度の上清又は精製されたFSHβ及び/又はLHβを受けた。注入してから1時間後、2時間後、4時間後、6時間後及び12時間後で、魚を殺生し、RNA抽出のために卵巣を取り外した。
【0042】
(RNA抽出)
ホルモン処理の終わりに、全RNAをゼブラフィッシュの卵巣又は濾胞細胞から単離し、液体窒素中でホモジナイズし、TRIZOL試薬を用いて製造業者の使用説明書に従い抽出した。微量のDNAをDNaseIとのインキュベーションによって除去し、続いてフェノール/クロロホルム抽出及びエタノール沈殿を行った。プラチナTaqを有するSuperscript II ワンステップRT−PCRシステムを用いて、RT−PCRを行った。反応を 25 pmol の上流プライマー及び下流プライマーを用いて 100 ng の全RNAで行った。逆転写を 50 ℃ で30分間行った。PCR条件は、94 ℃ で20秒間の変性、56 ℃ で30秒間のアニーリング及び 72 ℃ で1分間の伸張の40サイクルに引き続き 72 ℃ で10分間の最終伸張であった。PCR産物を、アガロースの 1 % ゲル中の電気泳動によって分離し、エチジウムブロマイドで染色した。
【0043】
(アクチビンβA,アクチビンβB及びβアクチンを増幅するために設計されたプライマー)
上流−アクチビンA 5'- TGC TGC AAG CGA CAA TTT TA -3'
下流−アクチビンA 5'- CAT TCG TTT CGG ACT CAA G -3'
上流−アクチビンB 5'- CAA CTT AGA TGG ACA CGC TG -3'
下流−アクチビンB 5'- GTG GAT GTC GAG GTC TTG TC -3'
上流−βアクチン 5'- CCC CTT GTT CAC AAT AAC CT -3'
下流−βアクチン 5'- TCT GTG GCT TTG GGA TTC A -3'
【0044】
(ウナギにおける安定的な株化細胞の移植)
安定的な株化細胞を、ウナギの腹腔内に移植した。ウナギを麻酔し、腹部に小さな切込みを入れた。一定量のホルモンを産生する正確な量の細胞を移植した。ネガティブコントロールとして、ホルモンを発現せず、空のベクターのみを有する細胞を移植し、さらにポジティブコントロールとして、ウナギの1つの群に下垂体抽出物を注入した。
【0045】
(成熟過程及び卵の成熟の測定)
メスウナギの最終成熟を定量化するために、成熟過程に従って変化する種々の特性が存在するが、それらは著しい体重の増加、眼の直径の増加及び卵母細胞の発生の間の形態変化である。最終卵母細胞成熟の7つの形態学的ステージが、下垂体抽出物処理の間に観察される。不透明な卵母細胞(ステージ0)はまだ小さく、全体的に脂肪小滴で満たされる。ステージ1への最終水和反応オンセット発生は、増加する透明度及び中心核を有する卵母細胞を示す。ステージ2において、卵母細胞は完全に透明であり、脂肪小滴は群れを成し、集まっている。ステージ3において、卵核胞(GV)移行が起こる。ステージ4において、卵核胞は表面(periphery)に発見され、脂肪小滴は反対側に位置づけられる。ステージ5において、脂肪小滴は脂肪融合に起因して減少し、結果としてより大きな脂肪小滴を生じさせる。ステージ6では、減数分裂IIが卵核胞の消失と共に完了し、少数の大きな脂肪小滴が観察される。ステージ7において、卵母細胞は、ただ1つの脂肪小滴を有する。初めの2つのステージの間に卵母細胞の直径が変化するが、これは水和反応の過程によるものである。卵母細胞成熟におけるこれらの形態変化により、我々は細胞移植の効果を判断することができる。細胞の移植後、ウナギは体重測定され、眼の直径が定期的に測定される。処理の間に、卵母細胞サンプルは、卵母細胞が当該処理に応答するかどうかを判断するために取得される。
【0046】
ホルモン産生細胞の移植
序論
CPE(コイ下垂体抽出物)注入の標準的技術を用いたホルモン産生ZF4細胞の移植の効果を比較するための試験を行った。性成熟は、最近Palstraらによって記載された(2005年)とおり、CPEの繰り返し注入によって獲得することができる。繰り返し注入は、結果として高品質の卵及び生存能力のある子孫をもたらさないが、これは誘導されるストレスによるものであろう。従って、CPE注入手順を、本実施例中に記載されるようなホルモン産生細胞の移植のようなよりストレスの少ない手順で置き換えなければならない。ウナギにおけるCPEの効果は、形態学的変化、組織学的変化及び内分泌学的変化によって数週間後に観察することができるので、メスの銀ウナギを用いた4週間の試験を行った。注入される細胞の量は、実験用ウナギ下垂体の推定の重さに関連した(1 kg のウナギにつき、およそ 6 μLの組織)。
【0047】
材料及び方法
(ホルモン産生細胞)
3つの異なる安定的な株化細胞を独立して成長させ、FSHβ、LHβ及びFSH/LHαに関する遺伝子をCMV−プロモーターに挿入し、ZF4細胞に安定的にトランスフェクトした。上記に記載されたとおりの手順に従った(Fugene 6 でトランスフェクトし、G418で選択した)。
【0048】
(GFP−β−ガラクトシダーゼ安定的株化細胞)
pMP2838プラスミドを用いて、緑色蛍光タンパク(GFP)とβ−ガラクトシダーゼタンパクの融合タンパクを発現するZF4の安定的株化細胞を作製した。当該プラスミドはBakkersによって記載された(2000年)。手短に言えば、pUAS-gfpN_LacZプラスミドのgfpN-LacZ遺伝子を取り出し、pEGFP-C3プラスミド中のgfp遺伝子に関する代替品として用いた。従って、β−ガラクトシダーゼ活性(gfpN-LacZ)を有する緑色蛍光融合タンパクは、CMVプロモーターの制御下で発現された。pMP2838プラスミドはまた、安定的株化細胞を作製するための上記に記載されたとおりゲンタマイシン(G418)での陽性クローンの選択を可能にするネオマイシン耐性(Neor)遺伝子も含む。同様に、ZF4安定的株化細胞を、上記に記載されたのと同じ手順で生成した(Fugene 6 でトランスフェクトし、G418で選択した)。
【0049】
注入の日に、細胞を回収し、各々の株化細胞に関して 108 cells の総数に定量化した。4つの株化細胞を均等な細胞濃度に混合し、1 mL 中に 2 × 107 cells の最終濃度まで血清なしのDMEM−F12培養液に希釈した。
【0050】
(β−ガラクトシダーゼ組織染色)
組織を簡単にPBS(リン酸緩衝生理食塩水)中でリンスし、2 mM MgCl2、5 mM EDTA 及び 0.02 % NP−40を含む 1 % パラホルムアルデヒド− 0.1 % グルタルアルデヒドのPBSで室温において30分間、速やかに固定した。次に、室温において洗浄溶液(2 mM MgCl2、5 mM EDTA、0.02 % NP−40 及び 0.01 % デオキシコール酸ナトリウムを含むPBS)で5分間、2回洗浄した。その後組織を、37 ℃ において12時間、染色溶液(2 mM MgCl2、0.02 % NP−40、0.01 % デオキシコール酸ナトリウム、5 mM K3Fe(CN)6、5 mM K4Fe(CN)6 及び 1 mg/mL X−galを含むPBS)で染色した。染色した組織を次にPBSで洗浄し、パラフィン包埋した。
【0051】
(動物及び手順)
60匹のメスの銀ウナギ(900 g −1600 g)をグレベリンゲン(Grevelingen)湖(オランダ)で降海回遊の期間に野生で捕獲した。全てのウナギに到着次第すぐに識別のためのマイクロチップ(Trovan)を備え付け、外部パラメーターを測定した。10匹のウナギを、コントロール動物として速やかに屠殺した。残りのメスウナギを、18 ℃ の温度において 3000 L の人工海水(3.5 %)の再循環システム中で維持した。処理日における細菌感染を予防するために、当該動物を3時間、大きな個々の水槽中で抗生物質フルメキン(50 mg/mL)に曝した。24匹のウナギの一つの群に、前に記載された方法(Palstraら、2005年)に従い、毎週、コイ下垂体抽出物(CPE:20 mg/kg)を注入し、もう一つの群には、1 mL の4つのタイプの細胞(β−gal、LHβ、FSHβ、LH/FSHα)の混合物を実験の開始時に一度だけ注入した。細胞は、背ビレの始まりよりも下かつ側線よりも上の皮下に懸濁液として注入した。
【0052】
1週間毎に、各々の群から6匹のウナギを屠殺して処理効果を解析した。これらは総重量、アイインデックス(eye index)、生殖腺重量及び胸びれの長さを含んでいた。さらに、組織サンプル(血液、下垂体、肝臓、生殖腺)を後の解析のために採取した。さらに、注入箇所のサンプルを獲得してβ−ガラクトシダーゼ陽性細胞の存在を検証した。同一のウナギから、サンプルを、細胞が注入されなかった部位から獲得した。
【0053】
結果及び考察
(細胞移植)
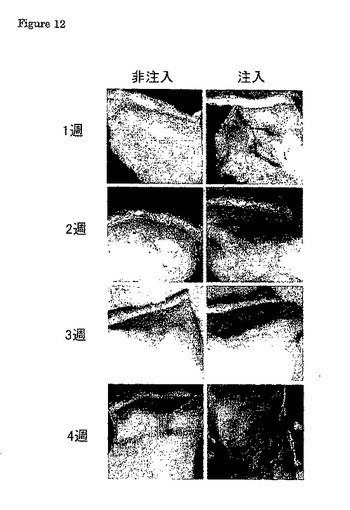
皮下移植の組織学的解析を、β−ガラクトシダーゼ染色に関して確認した。全てのサンプルは、当該細胞の存在を示した(図12)。発色は初めに、皮下層に関して特有な脂肪/コラーゲン組織で生じたが、また多くの場合、筋層の周りの浸潤も観察することができた。1週と4週のサンプルを比較すると、多くの場合、β−ガラクトシダーゼ染色細胞の全体集団の増加すら存在し、それらの細胞が宿主組織に浸潤する能力があることをはっきりと示唆した。拒絶反応に関する兆候は全くなく、ZF4細胞が進化的にウナギから極めて遠いゼブラフィッシュに由来するので、これは驚くべき知見である。
【0054】
(形態変化)
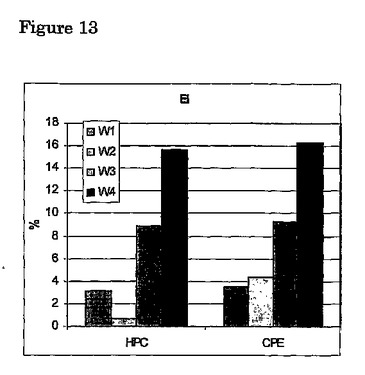
成熟のオンセットを示唆する最も顕著かつ直接の(proximate)変化は、銀ウナギの眼のサイズの増大である(Durifら、2004年)。図13から、細胞移植によって誘導されるアイインデックス(eye index)(長さに対する表面積、Pankhurst 1982年)において観察される変化が、CPE注入によって誘導されるものとほとんど等しいことは明らかである。当該結果は、細胞移植の効果をはっきりと示す。さらなる証拠を、3−4ヶ月刺激の銀ウナギから得ることができ、これは結果として最終成熟をもたらすはずである。
【0055】
結論
上記の実験から少なくとも2つの結論を出すことができる。1)ゼブラフィッシュZF4細胞(繊維芽細胞株化細胞)は、4週間を越えて銀ウナギによって拒絶されず、増殖することすら観察された。2)移植された細胞混合物は、アイインデックス(eye index)において、CPEを用いて標準的な手法を行ったのと同様の効果を有し、従ってホルモン刺激に関する証拠を示した。
【0056】
上記に記載された手法及びコンストラクトの使用
ウナギ株化細胞を生成し、ウナギに移植した。さらに両方のユニット(α+β)を発現する魚細胞を生成した。
【0057】
参考文献
Palstra, A.P., Cohen, E.G.H., Niemantsverdriet, P.R.W., van Ginneken, V.J.T., van den Thillart, G.E.E.J.M. (2005) Artificial maturation and reproduction of European silver eel: Development of oocytes during final maturation. Aquaculture, 2005.
Durif, C., Dufour, S., Elie, P. (2005) The silvering process of the eel : a new classification from the yellow resident stage to the silver migrating stage. J. Fish Biol. 66, 1-19
Pankhurst, W.N. (1982) The relation of visual changes to the onset of sexual maturation in European eel, Anguilla anguilla L. J. Fish Biol. 21, 179-196
Bakker J. (2000) Chitin oligosacharides in Zebrafish development. Investigations at the molecular and cellular level. (Thesis University of Leiden).
【図面の簡単な説明】
【0058】
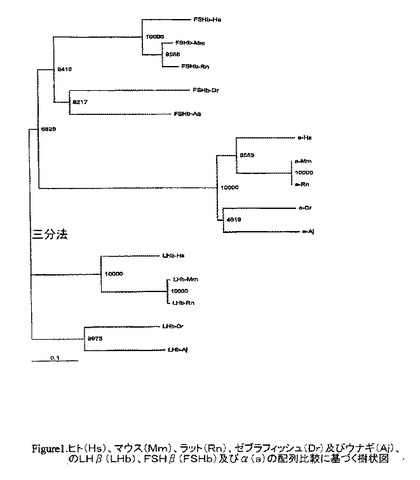
【図1】ヒト(Hs)、マウス(Mm)、ラット(Rn)、ゼブラフィッシュ(Dr)及びウナギ(Aj)のa(a)に由来するLHβ(LHβ)、FSHβ(FSHβ)の配列比較に基づく樹上図
【図2】p3XFLAG-CMV-9発現ベクター
【図3】LHβの発現に関するクローニングストラテジー
【図4】LHβの予想されるアミノ酸配列は、20593.8の分子量を有する187アミノ酸のタンパクを与える。
【図5】FSHβの発現に関するクローニングストラテジー
【図6】FSHbblu及びFSHbの予想されるアミノ酸配列は、2つの異なるストラテジーのクローニングに対してそれぞれ、19062.24/19965.21の分子量を有する174/183アミノ酸のタンパクを与える。
【図7】αの発現に関するクローニングストラテジー
【図8】aの予想されるアミノ酸配列は、21083.24の分子量を有する190アミノ酸のタンパクを与える。
【図9】ZF4細胞のpEYFP-N1でのトランスフェクション。トランスフェクション後、24時間に観察した。
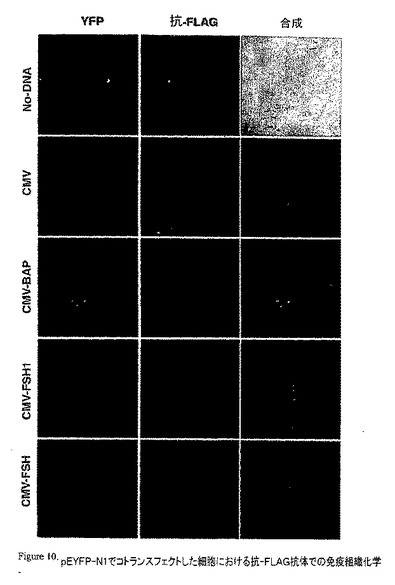
【図10】pEYFP-N1でコトランスフェクトした細胞における抗−FLAG抗体での免疫組織化学
【図11】抗−FLAGでのウェスタンブロット。抗体は、FLAG−BAPに関して予想されるサイズのタンパクを認識した。
【図12】β−ガラクトシダーゼに関する染色は、銀ウナギに皮下的に注入されたβ−gal−ZF4細胞の存在を示唆した。同一のウナギから、皮膚/筋肉組織を、注入した領域及びさらに尾に向かう領域から解剖した。皮下の脂肪組織は、真皮及び筋層がそうであるように図中で容易に認識できる。
【図13】銀ウナギの初期ステージに関するホルモン産生細胞(HPC)又はコイ下垂体抽出物(CPE)の4週間処理(W1−W4)の間のアイインデックス(EI)の変化(%)。初期ステージ及び各々の週に関して、6匹の十分成長したメスの銀ウナギが取得された(体重 1300 g ± 300 g)。
【技術分野】
【0001】
本発明は、魚の養殖の分野に関する。本発明はとりわけ、魚の卵及び/又は魚の精子の発生及び成熟のホルモン促進型改善の分野に関する。
【背景技術】
【0002】
多くの魚類種は、環境因子に応答して成熟する。光周期、温度、季節、圧力及びエネルギー備蓄のようなこれらの因子は、当該動物によって感知され、下垂体の視床下部中枢の抑制作用を制御する。このようにして、下垂体によるゴナドトロピンの産生は通常は、特定の環境条件下でのみ低下され、活性化される。活性化される際、下垂体はオス及びメスの両方の生殖腺の発育及び発生を刺激するゴナドトロピンを放出する。
【発明の開示】
【発明が解決しようとする課題】
【0003】
水産業にとって、成熟を概して、たいてい未精製の下垂体抽出物からなる同一のホルモンの定期的な注入により人工的に刺激することができることは重要である。定期的な注入は、ある程度までオスとメスの両方の生殖腺の発生に関する環境誘因を凌駕する。しかしながら、現行方式の卵及び/又は精子の発生及び/又は成熟の人工的な改善は、完全に満足できるものではない。例として、まだ未成熟尾ウナギにおいては、それらが産卵回遊を開始する際に、卵母細胞が排卵にとって十分に成熟する前の3ヶ月間−5ヶ月間の間、メスを毎週処理しなければならない。これは時間のかかる処理であり、ウナギにとってもストレスが多い。
【課題を解決するための手段】
【0004】
本発明は、ホルモン投与を用いた魚の卵及び/又は魚の精子の発生及び/又は成熟を改善するための方法であって、前記魚に前記ホルモンを産生する細胞を提供することを含む、前記方法を提供する。魚に移植される当該細胞は1つ以上のホルモンを放出し、それによって少なくとも当該ホルモン自身の定期的な注入に関する必要性を低減させる。分泌ホルモンが循環系へ放出され、全ての組織、とりわけ生殖器に十分量で到達する限りは、当該細胞は種々の方法で移植されてもよい。当該細胞は、体のどこに移植されてもよい。ホルモン産生細胞で構成される移植は概して、黄体形成ホルモン(LH)、卵胞刺激ホルモン(FSH)又は絨毛性ゴナドトロピン(CG)のようなホルモンを産生するために脳下垂体をバイパスするために用いられる。ホルモン産生細胞の移植は好ましくは、血流にアクセス可能な場所に挿入される。好ましい挿入方法は、腹腔内注入及び皮下注入である。血流にアクセス可能な場所は、LH、FSH及びCGのような二次的なホルモンを産生する細胞に極めて適している。
【0005】
細胞の移植は哺乳動物においてよく用いられ、細胞を移植し少なくともしばらくの間維持する方法に関する多くの経験が得られてきた。魚が概して、成熟卵又は成熟精子の採卵又は収穫後に始末されることを考慮すると、移植細胞の制御に関する大きな必要性は存在しない。重要なのは、当該細胞が生殖腺の完全な発生及び成熟を可能にするのに十分な量で存在したままの状態であることである。従って、細胞の数は、発生及び成熟に関して必要とされる数未満であってはならないかもしれないし、生殖腺の発生及び成熟又は当該動物の身体全体の健康を妨げるほど多数であってはならないかもしれない(又は、多数になってはならないかもしれない)。当該細胞によるホルモンの産生レベルは、あまり重要ではない。注入される細胞の数は、成熟に関して要求される総量によって決まる。ホルモン産生は、バイオアッセイにより定量化される。
【0006】
注入される細胞のホルモン放出は、卵及び/又は精子の発生及び/又は成熟を刺激するのに十分に高い必要がある。排卵誘発ホルモン(fertility hormone)の過剰発現はそれ自体有毒ではなく、卵及び/又は精子の成熟及び/又は発生の刺激にマイナスに作用しないので、当該ホルモンの発現に関する上限境界は重要ではない。哺乳類界において、種々の移植補助物が、当該細胞の長期の残留を可能にし又は当該細胞の分化を可能にするために開発されてきた。これらの補助物もまた当然ながら、本発明において用いることができる。当該補助物としては、細胞の移植及び接着のためのコラーゲン又は合成マトリクスが挙げられるが、これらに限定されない。
【0007】
当該細胞は、多くの場合、極めて長い時間存在する必要がないので、多くの様々な種由来の魚細胞をレシピエント魚へ移植することが可能である。移植細胞と当該レシピエントとの間の進化的な差異が大きい場合、当該レシピエント魚は、当該移植細胞(移植片)への免疫応答を備え付けるように思われる。しかしながら、当該細胞はたいてい限られた時間の間の存在だけが必要とされるので、そのような免疫応答は概して容認することができる。当該処理の丈夫さ及び予見可能性の増大のために、当該移植片は、レシピエント魚種と同じ属又は科に由来することが好ましい。好ましくは、2つとも同じ種に由来する。魚は概して非近交系集団であるので、同一種の魚間の免疫学的相違が存在する。これは概して、我々の実施例の1つのウナギの実験に示されるとおり問題とはならないが、共通の免疫学的マーカーに関して当該移植片及びレシピエントをさらに調和させることが可能である。代表的なマーカーは、主要組織適合性抗原及び非主要組織適合性抗原である。当該移植細胞の移植は、当該レシピエント魚にシクロスポリンのような免疫抑制剤を提供することによってさらに促進されてもよい。
【0008】
本発明のホルモン産生細胞の移植は、卵及び/又は精子の成熟、卵及び/又は精子の生殖能、卵の授精、結果として生じる胚の質、受精卵及び未受精卵の生存並びに胚の生存を改善するために用いることができる。これらの改善は全て、魚の卵及び/又は精子の改善された発生及び成熟をもたらす。魚の卵及び/又は精子の発生及び成熟なる用語は従って、自然過程の産卵に限定されず、卵の授精に関する人工的な方法にもまた関連する。従って当該用語は、本発明の方法で処理される魚の未受精卵及び/又は精子の収穫にも関連する。当該未受精卵はまた、子孫を創る以外の目的で用いられてもよい。その限定されない例は、キャビアのような人間の食用卵の製造である。
【0009】
魚の卵及び/又は精子の発生及び成熟は、種々の手段で刺激することができる。本発明のある実施態様において、魚の卵及び/又は精子の発生及び成熟は、卵、精子又は受精卵から生じた胚の絶対数又は質が上昇される際に、刺激される。
【0010】
生殖は、高度に制御された生物学的過程である。生殖の種々の側面は、種々のホルモンによって制御される。しかしながら、いくつかのホルモンは、魚に移植される細胞によって発現される際に、おおよそ類似した効果をもたらすことができる。これらのホルモンとしては、成長ホルモン、コルチコリベリン(Corticoliberin)(副腎皮質刺激ホルモン)、甲状腺刺激ホルモン、FSH、LH、プロラクチン、CG(絨毛性ゴナドトロピン)、MG(閉経期ゴナドトロピン)、ソマトロピン又はそれらの組み合わせが挙げられる。上記で述べたホルモンはまた、違う名前でも知られる。基礎を成すアミノ酸配列が同一であるので、同義語で呼ばれるホルモンもまた、本発明の範囲内に包含される。例えば、成長ホルモンは時々また、ソマトトロピン、ソマトトロピックホルモン(somatotropic hormone)、脳下垂体成長ホルモン、ソマトトロピックホルモン(somatotropic hormone)又はSTHとも称される。コルチコリベリン(Corticoliberin)はまた、放出性副腎皮質刺激ホルモン(releasing corticotropin hormone)とも称される。副腎皮質刺激ホルモンはまた、コルチコトロピン、アドレノコルチコトロピン(adrenocorticotropin)、アドレノトロピン(adrenotropin)、コルチコトロピン、ACTH又はアドレノコルチコトロピック(adrenocorticotropic)ホルモンとも称される。甲状腺刺激ホルモンはまた、TSH、チロトロピン又はチロトロピック(thyrotropic)ホルモンとも称される。FSHはまた、卵胞刺激ホルモン、フォリトロピン又はガメトチネティック(gametocinetic)ホルモンとも称される。LHはまた、黄体形成ホルモン、ルテオトロピン又は間質細胞刺激ホルモン(ICSH)とも称される。プロラクチンはまた、PRL、乳腺刺激ホルモン(lactogenic hormone)、乳腺刺激ホルモン(mammotropic hormone)、ガラクトポイエティック(galactopoietic)ホルモン又はラクトトロピン(lactotropin)とも称される。CG(絨毛性ゴナドトロピン)はまた、絨毛性性腺刺激ホルモン、絨毛性性腺刺激ホルモン、コリオゴナドトロピン(choriogonadotropin)又は絨毛性ゴナドトロピンとも称され、さらに、閉経期ゴナドトロピン(MG)はまた、ウロゴナドトロピン(urogonadotropin)、メノトロピン又は閉経期性腺刺激ホルモンとも称される。
【発明を実施するための最良の形態】
【0011】
本発明の好ましい実施態様において、前記ホルモンは卵の発生及び成熟に直接的に関与するホルモンである。そのような排卵誘発ホルモン(fertility hormone)は概して、下垂体又は生殖器によって産生される。好ましい実施態様において、当該排卵誘発ホルモン(fertility hormone)は、黄体形成ホルモン(LH)、卵胞刺激ホルモン(FSH)又は絨毛性ゴナドトロピン(GC)又はそのようなホルモンの機能的部分、派生物及び/若しくはアナログを含む。これらのホルモンは、魚の卵及び/又は精子の発生及び成熟の非常に強力な刺激要因である。これらのホルモンは、本質的に極めて保存されており、種の壁をほとんど有していない。例えば、尿中のヒト排卵誘発ホルモン(fertility hormone)の存在は、それらをカエルの卵と共にインキュベートすることによって検出することができる。同様に、ヒト絨毛性ゴナドトロピン(hGC)はまた、ウナギにも作用し、さらに、コイ及びサケの下垂体抽出物は、ウナギ、タイ及びマスのような多くの様々な魚種に作用する。
【0012】
当該レシピエントは、異種ホルモンに対しての免疫応答を発生することが可能である。この免疫応答は概して、あまりにも遅すぎて卵及び/又は精子の成熟及び/又は発生の刺激に作用することができないが、当該ホルモンは、魚ホルモン又はその機能的部分、誘導体及び/又はアナログであることが好ましい。これは、提供されるホルモンと内生的なホルモンとの間の相違を制限し、その結果、当該レシピエントにおいて、提供されるホルモンに対する免疫応答の発生を少なくともある程度制限する。好ましくは、当該ホルモンは、当該レシピエントと同一の属に属する種に由来する。このようにして、免疫応答が発生する可能性はさらに低減される。とりわけ好ましい実施態様において、提供されるホルモンは、当該レシピエントにおけるホルモン等価物と免疫学的に同一である。これは、提供されるホルモンに対するどんな有害な免疫応答の発生も完全に回避する。
【0013】
当該細胞は、処理なしで好ましいホルモンを発現することができるか又は好ましいホルモンを発現するように操作することができるかのどちらかである。当該細胞が既にホルモンを発現しないか又は十分なホルモンを発現しない際には、ホルモンを発現するための遺伝子情報を当該細胞に提供することができる。
【0014】
好ましい実施態様において、当該細胞はホルモンを発現するための遺伝子情報を提供される。これは、当該細胞にホルモンに関するコード配列を含む発現カセットを提供することによって行うことができる。しかしながら、活性制御配列をそれぞれのホルモンに関するコード配列付近に挿入することによって内生的な遺伝子を活性化することもまた可能である。これは例えば、相同組換えを通じて行うことができる。
【0015】
LHタンパク及びFSHタンパクは、関連タンパクのファミリーに属する。双方は、共通のα−サブユニット及び異なるβ−サブユニットからなるヘテロダイマーとして機能しうる特性を共有する。1つよりも多いタンパク鎖からなるホルモンの場合は、細胞がホルモンを生成するために必要とされる全ての鎖を発現しないことも起こり得る。これらの場合には、発現カセットだけが、欠けている鎖に必要である。従って、ホルモンのサブユニットの全てのサブユニットではないが1つ以上のサブユニットが当該細胞で十分に発現される場合には、当該細胞において残りのサブユニットを発現する必要があるだけである。
【0016】
当該サブユニットが1つも発現されない場合には、当該細胞を当該サブユニットの全てが十分なレベルで発現されるように処理しなければならない。好ましい実施態様において、当該細胞は、ホルモンのサブユニットに関する発現カセットを提供される。好ましい実施態様において、当該細胞は、LH及びFSHを作り出す3つのタンパク鎖に関する(すなわち、それぞれのホルモンに関する共通のα−サブユニット及び独特なβ−サブユニットに関する)発現カセットを提供されるか又は別個の株化細胞において、βLH + α 及び βFSH + α の組み合わせが発現される。従って好ましい実施態様において、前記細胞は、前記ホルモンを発現するために遺伝学的に改変される。好ましくは、当該細胞は前記ホルモンをコードする1つ以上の遺伝子を提供される。
【0017】
当該細胞は、初代細胞又はインビトロで長期間培養される株化細胞であってもよい。好ましい実施態様において、当該細胞はクローン集団の細胞に由来する。このようにして、当該細胞を使用に先立ってきめ細かい品質管理にさらすことができる。これはまた、同一の性質を有する細胞バンクの生成も可能にする。さらに、クローン集団をさらなる処理にさらすことができる。例えば、当該レシピエントにおける免疫応答を低減したい場合には、主要組織適合性抗原及び/又は非主要組織適合性抗原の発現をノックアウトすることが可能である。従って好ましい実施態様において、当該細胞は、当該レシピエントにおける低減された免疫原性に関して選択される。本発明の方法で用いるための又は本発明の使用のための細胞は、初代細胞又は培養細胞であってもよい。好ましくは、前記細胞は培養細胞であり、より好ましくは、不死化した培養細胞である。培養細胞は概して、株化細胞である。株化細胞は、前記細胞の表現型の実質的な変化なしに少なくとも5継代の間増殖することができる。不死化した株化細胞は、そのような表現型の変化を受けることなく少なくとも50回継代することができる。不死化した細胞は、種々の方法で初代細胞から取得することができる。好ましくは、前記の不死化した細胞は、培養物中で初代細胞に関する一般的な難局を経験してきた初代細胞の培養物から獲得される。別の好ましい実施態様において、前記細胞は、初代細胞への1つ以上の遺伝子の導入により不死化されている。
【0018】
本発明の方法は、あらゆるタイプの魚に用いられてもよい。好ましい魚は、ウナギ、スズキ(シーバス)、タイ、ハリバ、サケ、マス、タラ、コイ、ナマズ及びチョウザメである。しかしながら、本発明は、とりわけ回遊性魚、好ましくは1回生殖の魚に関して好都合である。これらの魚は、産卵までに時間がかかり、概して外部のシグナルへ全て同じように応答するわけではない。本発明の方法で、少なくともある程度環境刺激と関係なく魚の発生及び成熟を刺激することが可能である。これは、当該魚養殖の出発点に関する大量の予測可能性をもたらす。回遊性魚において、当該生産工程において作業をよりうまく予定することができるように卵の発生及び成熟を同調させることがさらに可能である。
【0019】
本発明は、排卵誘発ホルモン(fertility hormone)を産生する単離魚細胞及び/又は組み換え型魚細胞をさらに提供する。好ましい実施態様において、前記細胞は、前記ホルモンを発現するために遺伝学的に改変される。本発明は、排卵誘発ホルモン(fertility hormone)を発現する能力を提供される魚細胞をさらに提供する。好ましくは、当該魚細胞は、前記排卵誘発ホルモン(fertility hormone)をコードする組み換え型塩基配列及び/又は単離塩基配列を提供される。当該ホルモンが1つ以上のサブユニットからなる場合には、当該魚細胞は好ましくは、前記ホルモンの少なくとも1つのサブユニットをコードする単離塩基配列及び/又は組み換え型塩基配列を提供される。好ましくは、当該魚細胞は、前記ホルモンの全てのサブユニットをコードする塩基配列を提供される。好ましくは、前記細胞は、一般消費者向けの魚である。好ましい実施態様において、前記細胞は、ウナギ、スズキ(シーバス)、タイ、ハリバ、サケ、マス、タラ、コイ、ナマズ及びチョウザメに由来する。好ましくは、前記細胞は、上記で記載されたとおりの細胞、すなわち、初代細胞又はインビトロで長期間培養される株化細胞に由来する細胞である。好ましい実施態様において、当該細胞は、クローン集団の細胞に由来する。好ましくは、前記細胞は、培養細胞、より好ましくは、不死化した培養細胞である。
【0020】
好ましい実施態様において、使用が培養することのできる細胞の移植片で構成される。さらに、これらの細胞は好ましくはクローンであって、好ましくは特性に関して選択可能である。また一方、初代培養物、組織、受精卵又は胚性材料(embryonic material)を活用することも可能である。遺伝子導入した魚の卵の調製方法が公表されている(Morita et al 2004, Transgenic Research 13, 551)。しかしながら、記載された遺伝子導入した魚の卵は、さらに増殖させることができない。さらに、導入される遺伝子の発現は、Moritaらによって記載されたとおり、安定ではない。卵中の導入遺伝子の選択は困難であり、一つ一つの卵へのマイクロインジェクションを用いなければならないので、バイオリアクターとしての魚の卵の使用は培養可能な細胞に対してかなり時間がかかる一方で、細胞培養物では、我々は標準的なトランスフェクション技術を活用することができる。培養可能な細胞は、容易に保存され、いつでも利用可能にされるという利点を有することができる。さらに、遺伝学的に改変される卵と比較して、培養可能な細胞での操作に伴う倫理的な問題は全くない。培養細胞の懸濁液の注入は、卵の移植及び/又は胚の移植と比較して、かなり容易である。
【0021】
本発明は、本発明に従う細胞を含む魚をさらに提供する。好ましくは、前記魚は、一般消費者向けの魚である。好ましくは、前記魚は、一般消費者向けの幼魚又は成魚である。生殖までの時間は、好ましくは短い。初代胚性材料(primary embryonic material)は、利用可能でないか又は十分な量で利用可能でないという不都合を有する。好ましい実施態様において、前記魚は、ウナギ、スズキ(シーバス)、タイ、ハリバ、サケ、マス、タラ、コイ、ナマズ及びチョウザメである。好ましい実施態様において、前記魚は、遺伝学的に改変されない精子細胞、卵母細胞及び/又はそれらの前駆細胞を含む。
【実施例】
【0022】
ゼブラフィッシュ遺伝子LHβ、FSHβ及びαのクローニング
ゼブラフィッシュの遺伝子LHβ(AY424304)、FSHβ(AY424303)及びα(AY424306)は既に公表され、またウナギの遺伝子も Anguilla japonica のLHβ(AB175835)、Anguilla anguilla のFSHβ(AY169722)及び Anguilla japonica のα(AB175834)に関して報告される。LHβ、FSHβ及びαをクローニングするために、プライマーを各自のcDNA配列に基づいて設計した。
【0023】
LHβのプライマー設計に関して用いた配列は下記のとおりである。
atatataaat ctggacacgc agagacactt acaacagcct gctgagcaac cgcaacgcct gtcaag[atgt
tattggctgg aaatggtgtc ttctttctct tctctttgtt tttcctgctg gcggctgctc agagcttggt
ttttccacgc tgtgagctag taaatgagac ggtatcggtg gaaaaagagg gctgtccaaa atgcctggtg
tttcagacca ccatctgcag cggccactgc gtaacaaggg atcccgttta caagagcccg ttttccaccg
tccaccagac agtgtgcatg taccgggacg tccgctatga gaccattaac ctgcccgact gttccgccgg
cgtggacccg cagatcacat acccggtggc gctgagctgc gactgcagtc tgtgcaccat aaacacttcc
gactgcacca tccagagcct gcagcccgac ttctgcatgt cccagagaga ggatttcccc gcatactag]a
cctcgggcaa ctcacgtcaa cctacgcaca tagtcgagct cagcattatt agccctcctg tatgtttttt
ccattaatat atatactttc aagacactag tattcagctt aaagtgacat ttaaagacta aactaggtta
attaggggga aaagtagagt aagtcattgt ataatagtgg tttgttctgg agacaatcca aaactaatat
tgcttaaggg ggctaataaa attgacctta aaatgaattt aaataattta aaaactgcat ttattctagt
cgaaataaaa gaaataagac tttctttaga agaaaaaaca ttataggaaa tactgcaaaa aaattcctga
atctgttcaa catcattcgg gaaatcaaag gagggctaat aactgtgact tcagctgtac atcaataaag
aggctggttc ttaaattcaa aaaaaaaaaa aaaaaaaaaa aaaaaaaa
【0024】
FSHβのプライマー設計に関して用いた配列は下記のとおりである。
ggtctccacg aaactcccgc agatgaggat gcgtgtgctt gttctggcgc tgctgttgcc ggtgttaatg
agcgcagaat cagaatgcag gtgcagctgt cgactcacca acatctccat cactgtggag agcgaagaat
gtgggagctg cgtcacaatc gacaccacag cctgtgcagg act[atgctgg acaatggatc gagtttaccc
tagttccatg gcacagcaca cccagaaggt ctgtaacttc aagaacttga tgtacaagag ctacgagttt
aaaggctgtc ctgcaggggt tgattcagtc ttcgtgtacc ccgtggctct gagctgtgag tgcaaccagg
ttaactcaga cacaacagac tggggagcta tcagcccgca gaccaccagc tgcagcatac actag]agcac
tgtatcatga ccttaacaac atgtacgttg cagaatcaaa ttaagtaagg agtacaatta gaccatttaa
ggatatcaat tatttacaaa acctttagtt tttcatgcat cccacacaca tggtaatttg gttacttgaa
ttaatctgtt gtgttaattc tatagttggt actatggtaa ctagagtact agagtatcca atgctatact
agttttaatt acagttaatt atagaaaagt atgctacagt atttattaca gtttttctgt tttcaatatt
tagtactaca gtatgctagt gcattcatta acaataagct gtaaatacta taataaatac aggttaatac
actttactat agtatgcttg atcaacacta ttatttaatg tgagttacta tagtactttt caattgggat
ttgtcatttt ggatattgtg ggcttttttg gctattcata aagttttttt tatttttttt ttatttaatt
ttcagtcaaa tggaaacaag tccaccataa tacacttgtg tttcttttgt caaacttatc aatttgtgtc
tgtagatttc aattacaata catattttaa aggccaaaaa aaaaaaaaaa aaaaaaaa
【0025】
αサブユニットのプライマー設計に関して用いた配列は下記のとおりである。
gaagacactc atcacgctcc gccggaagtc gaggacaaag ccatc[atgtt ttggacaaga tacgctgaag
caagcatttt cttgttgtta atgattcttc atgtcggaca actgtattca agaaacgatg tgtctaacta
tggatgtgaa gagtgcaaac tcaagatgaa cgaacgtttc tccaaacccg gggctccggt ctatcagtgc
gtgggctgct gcttttcgag agcttacccc acacccctga ggtccaagaa aaccatgctt gtcccaaaaa
acatcacatc agaagccact tgctgtgtag caaaagaatc taaaatggtt gccacgaata tcccactata
caaccacaca gactgccact gcagcacctg ttactatcat aagtcttaa]a acacactctc ttcacatttc
tcaaatgctc atttcctgtt cttaaatcac agtgactcat gaaatatgat ttttatgtag ctttccatat
ttcaactgtg gccatttcca attcgtttct aaaatggttg gcataagtat tgtaaactgc atattctgtc
actatccctt taagagcgta atatgccatc ctttactatc attaaatcgc ttatttattt tgttgccttt
actgtgacat tcttcaaatc tataaatgaa ataaaagatt gctgaaggca aaaaaaaaaa aaaaaaaaaa
aaaaaaaaaa
【0026】
いずれの場合にも、括弧でくくられる部分はコード配列を表し、予想される増幅領域には下線が引かれる。
プライマーは下記のとおり設計された。
上流 - LHβ/EcoRI 5'-CAA CCG AAT TCA ACG CCT TCA AGA TGT-3'
下流 - LHβ/EcoRV 5'-CCG ATA TCT AGT ATG CGG GGA AAT-3'
上流 - FSHβ3 5'-AGG ATG CGT GTG CTT GTT CT-3'
下流 - FSHβ2/3 5'-TGT TGT TAA GGT CAT GAT ACA GTG C-3'
上流 - α1 5'-GTC GAG GAC AAA GCC ATC AT-3'
下流 - α1 5'-TGC CAA CCA TTT TAG AAA CGA-3'
さらなるクローニングステップを円滑にするために、LHβオリゴは、上流オリゴ及び下流オリゴ中にEcoRI及びEcoRVに関する制限酵素サイトを含む。
【0027】
全RNAを液体窒素中でホモジナイズし、TRIZOL試薬を用いて製造業者の使用説明書に従い抽出したゼブラフィッシュの頭部から単離した。微量のDNAをDNaseIと共にインキュベーションすることによって除去し、その後、フェノール/クロロホルム抽出及びエタノール沈殿を行った。RT−PCRを、プラチナを有するSuperscript IIワンステップRT−PCRシステムを用いて行った。Taq反応を、25 pmol の上流プライマー及び下流プライマーを用いて 100 ng の全RNAで行った。逆転写を 50 ℃ で30分間行った。PCR条件は、94 ℃ で20秒間の変性、LHβに関して 55 ℃ で30秒間及びFSHβに関して 50 ℃ で30秒間のアニーリング、さらに 72 ℃ で1分間の伸張の40サイクルに引き続き、72 ℃ で10分間の最終伸張ステップであった。PCR産物をアガロースの 1 % ゲル中での電気泳動によって分離し、エチジウムブロマイドで染色した。FSHβのPCR産物はアガロースゲル中で観察した際にかすかなバンドを形成したので、最適化するために、我々はこのPCR産物をテンプレートとして用いてPCRを行い、反応はPCR産物の1/20で、10 μM の上流プライマー及び下流プライマーを用いて行った。PCR条件は、94 ℃ で20秒間の変性、アニーリングを 50 ℃ − 60 ℃ のグラジエントで30秒間行い、72 ℃ で1分間の伸張の40サイクル、引き続き、72 ℃ で10分間の最終伸張ステップであった。はっきりしたバンドが、その結果観察された。増幅配列の同一性を確認するために、PCR産物を pCRII-TOPO ベクターにクローニングし、正しい方向を確認するために制限酵素で消化し、配列決定した。配列解析は、我々がゼブラフィッシュのLHβサブユニット、αサブユニット及びFSHβサブユニットをクローニングしたことを示した。これらのコンストラクトにより、常時発現プロモーター下での当該遺伝子のサブクローニングが可能である。
【0028】
常時発現プロモーター(CMV)制御下のLHβ、FSHβ及びαのクローニング
p3XFLAG-CMV-9発現ベクターを用いて、一過性の融合タンパク又は安定的な融合タンパクを確立した。当該ベクターは、マルチクローニング領域の上流の3つの隣接するFLAGエピトープをコードする。これは結果として、抗−FLAG抗体を用いた増大した検出をもたらす。当該CMVのプロモーター制御領域は、FLAG融合コンストラクトの転写を促進する。プレプロトリプシンリーダー配列はFLAG配列の前に位置し、タンパクの分泌を促進する。アミノグリコシドホスホトランスフェラーゼ遺伝子(Neo)は、ジェネテシン(G418)のようなアミノグリコシドへの耐性を与え、安定発現株の選択を可能にする。
【0029】
我々は当該ベクターを用いたが、その理由は、合成されるタンパクがプレプロトリプシンリーダーによって促進されて分泌され、我々が抗−FLAG抗体でのウェスタンブロッティングによってタンパクの発現を検出することができ、さらに我々がジェネテシンで選択して安定な株化細胞を作ることができるという利点を有しているからである。
【0030】
LHβのクローニング
PCR産物を精製し、p3XFLAG-CMV-9 発現ベクターと同様にEcoRI及びEcoRVで消化した。当該DNA断片を、4 ℃ で一晩連結した。次に連結混合物を用いて、ケミカルコンピテント細胞をトランスフォーメーションした。酵素消化は、陽性クローンを選び出した。正しい様式の消化を生じさせるそれらのクローンを配列決定した。当面、配列決定されたクローンはLHβの間違った挿入を示したので、当該クローニングステップを繰り返さなければならなかった。予想されるアミノ酸配列は、20593.8 の分子量を有する187アミノ酸のタンパクを与える。
【0031】
FSHβのクローニング
pCRII-TOPO FSHβをBamHI/NotIで消化し、FSHβ cDNAに相当するバンドを精製し、p3XFLAG-CMV-9 にサブクローニングした。陽性クローンを次に、Not/EcoRV及びマグビーンヌクレアーゼ(mug bean nuclease)で消化し、再連結してプレプロトリプシンリーダーを有するフレーム中のFSHβを得て、これらのコンストラクトをCMV−FSHβと命名した。もう1つの方法として、pCRII-TOPO FSHβをXba/BamHIで消化し、結果として生じたバンドを p3XFLAG-CMV-9 にサブクローニングした。陽性クローンを次にEcoRVで消化し、再連結してプレプロトリプシンリーダーを有するフレーム中のFSHβを得て、このコンストラクトをCMV−FSHβと命名した。酵素消化は、陽性クローンを選び出した。正しい様式の消化を生じさせるそれらのクローンを配列決定した。異なるストラテジーのクローニングに相当する2つの異なるコロニーの配列は正しかった。これらのコンストラクトを、ZF4株化細胞のトランスフォーメーションに用いた。予想されるアミノ酸配列は、2つの異なるストラテジーのクローニングに対してそれぞれ、19062.24/19965.21の分子量を有する174/183アミノ酸のタンパクを与える。
【0032】
αのクローニング
pCRII-TOPO αをKpnI/XbaIで消化し、結果として得られたバンドを p3XFLAG-CMV-9 にサブクローニングした。酵素消化は、陽性クローンを選び出した。正しい様式の消化を生じさせるそれらのクローンを配列決定した。配列解析は、我々が正しい配向で、FLAG及びプレプロトリプシンリーダー配列を有するフレーム中にaをクローニングしたことを示し、このコンストラクトを、CMV−αと命名した。予想されるアミノ酸配列は、21083.24の分子量を有する190アミノ酸のタンパクを与える。
【0033】
CMV−FSHβプラスミド及びCMV−αプラスミドを、QIAプレップスピンミニプレップキット(QIAprep spin Miniprep kit)を用いるSDSを含むアルカリ溶菌法によって精製した。精製されたプラスミドを、ScaIで直線化した。直線化された生成物を、アガロースの 1 % ゲル中の電気泳動によって分離し、エチジウムブロマイドで染色した。直線化されたプラスミドを精製し、定量化した。
【0034】
ゼブラフィッシュ繊維芽細胞株化細胞(ZF4)におけるFSHβのトランスフェクション
ZF4(ATCC番号:CRL-2050)細胞は、生まれて1日のゼブラフィッシュの胚由来の繊維芽細胞である。凍結した一定分量のZF4細胞を、液体窒素から移し、直ちに氷上に10分間置いた。解凍された細胞懸濁液をバイアルから移し、10 mL の完全培養液(1.2 g/L の重炭酸ナトリウム、2.5 mM のL−グルタミン、15 mM のHEPES、0.5 mM のピルビン酸ナトリウム、10 % のウシ胎仔血清及び 10 % のペニシリン/ストレプトマイシンを含むダルベッコ変法イーグル培養液とハムF12培養液の1:1の混合物)中に室温で希釈した。上清を 1200 rpm で8分間の遠心分離によって廃棄した。細胞を 8 mL の完全培養液中に再懸濁し、組織フラスコ(T25)に移し、28 ℃ で培養した。細胞密度を調べるために、倒立顕微鏡下で細胞を検査した。80 % 集密の培養で、培養液を除去し、3 mL のPBSで洗浄して細胞残屑及び血清を除去した。次に、0.5 mL のトリプシン溶液(0.25 %)を加え、単層の細胞がフラスコから剥離するまで室温でインキュベートし、単一細胞懸濁液が得られたら、5 mL の完全培養液を加えてトリプシン処理を停止した。生存細胞及び生育不能な細胞を、Fuchs-Rosenthal血球計算板を用いてカウントした。新たなフラスコ中に 100 cells/mm2 − 150 cells/mm2(各々のT25フラスコ中に 5 × 105)の密度で細胞を播種した。当該細胞を、次の継代前に 28 ℃ で最大4日間インキュベートした。トランスフェクションは、ゼブラフィッシュ繊維芽細胞が 50 % − 60 % コンフルエントになった際に、Fugene 6 を用いて製造業者の使用説明書に従い、ゼブラフィッシュ繊維芽細胞で行った。Fugene 6/DNA複合体は、無血清培養液中に Fugene 6 :DNA=6:1の割合で調製した。培養液を細胞から除去し、無血清培養液に交換した。Fugene 6/DNA複合体を思慮深く滴下して加え、混合した。当該細胞を 28 ℃ で5時間インキュベートした。次に、培養液を除去し、完全培養液に交換した。コントロールとして、Zf4細胞をpEYFP-N1プラスミドでコトンラスフェクトし、トランスフェクションの後、16時間及び24時間において、ライカ共焦点下で陽性細胞を解析した。種々の濃度のDNAを用いて、当該プラスミドに関する至適濃度を確立し、結果として、21 cm2 の表面中に 1 μg のDNAが最善の条件であり、およそ 30 % のトランスフォーメーションが実現された。
【0035】
LHβ、FSHβ及びαの発現の検出
(免疫組織化学)
トランスフェクトした細胞をまた免疫組織化学によって解析し、細胞中のタンパク発現を検出した。細胞を、2 % パラホルムアルデヒド及び 0.1 % グルタルアルデヒドのPBSで10分間固定し、固定液を除去するために、細胞をPBSで2回洗浄した。次に、細胞を 0.2 % Triton-X-100 のPBSで10分間透過処理した。固定液によって起こる自己蛍光を低減するために、2 mg/mL NaBH4 のPBSを10分間加えた。ブロッキングを 0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチンで30分間行った。抗−FLAG抗体(1:250)とのインキュベーションを、4 ℃ において一晩、0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチン中で行った。抗体を除去し、0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチンで 各々10分間、2回洗浄した。二次抗体(抗−ラビット Alexa 488)を、0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチン中に 1:1000 希釈で加え、室温で60分間インキュベートした。次に、0.1 % BSA−c、0.02 % 冷水魚(cold water fish)皮膚ゼラチンで3回洗浄し、DABCO/Gelvatolに搭載した。ライカ共焦点での細胞の解析は、トランスフェクトした細胞の小胞中のタンパク発現を示した。この局在は、分泌されるタンパクに関して予想される部位と一致する。
【0036】
(ウェスタンブロット)
種々のコンストラクトをT25フラスコ中でFugene 6を用いて2回トランスフェクトし、コントロールとして、細胞をDNAでトランスフェクトせず、空のベクター及びポジティブコントロールのCMV−BAPでトランスフェクトした。タンパクを、トランスフェクション後3日目及び5日目に上清から獲得した。上清を、アミコンカラム(amicon column)を用いて、製造業者の使用説明書に従い濃縮した。FLAG融合タンパクを、抗−FLAG M2アフィニティーゲルを用いて製造業者の使用説明書に従い免疫沈降した。タンパクサンプルを、サンプルバッファーで1:4に希釈し、ボイルして、-80 ℃ で維持した。ウェスタンブロットの条件を決めるために、ポジティブコントロール(CMV−BAP)及びネガティブコントロール(DNAなし)のみ解析した。サンプルを 12 % アクリルアミドゲル中にロードし、50 mA で60分間ランした。ゲルをニトロセルロースメンブランにブロットした。メンブランを 4 ℃ において一晩、5 % ミルクでブロックし、免疫検出を室温において1時間、5 % ミルクで 1:250 希釈した抗−FLAG抗体に対して行った。免疫検出を、ECLで製造業者の使用説明書に従い明らかにした。当該抗体は、予想されるサイズのバンドを示す発現されたBAPタンパクを認識した。これらの実験で我々は、一過性にトランスフェクトした細胞が所定のタンパクを産生することを明らかにした。
【0037】
(安定的な株化細胞の作製)
細胞に6ウェルチャンバー中で、直線化し精製したDNAをFugene 6を用いてトランスフェクトした。コントロールとして、細胞にDNAなしでトランスフェクトした。トランスフェクションの3日後又は5日後に、完全培養液を、G418を加えた完全培養液に交換した。コンストラクトを発現していない細胞を死滅させるG418の量は、株化細胞間で異なる。ZF4株化細胞に関しては、製造業者の推奨する濃度は、0.8 mg/mL − 1 mg/mL である。細胞を4つ組で、0.8 mg/mL 及び 1 mg/mL で処理し、培養液を毎日交換して死細胞を洗い流した。これを、DNAを有さない細胞が死滅し当該細胞を含むプレート中に少しも細胞を観察することができなくなるまで、15日間行った。次にトランスフェクトした細胞を、同一の濃度のG418と共に5日間インキュベートした。コンフルエントの単層が得られたら、細胞をT25フラスコ中に継代培養し、G418の濃度を、完全培養液中 0.5 mg/mL まで下げた。安定的な株化細胞を、抗−FLAG抗体を用いた免疫組織化学解析及びウェスタンブロット解析で試験してタンパク発現を検出した。
【0038】
(バイオアッセイ)
トランスフェクトした細胞によって発現されるホルモンがアクティブであるかどうかを検証するために、簡便なバイオアッセイを行った。ゼブラフィッシュの濾胞細胞培養物は、種々の遺伝子の発現を上方制御するか又は下方制御することによって下垂体抽出物及び/又はヒト絨毛性ゴナドトロピン(hCG)に応答する。hCG(15 IU/mL)は、アクチビンβAの発現レベルを時間依存的に上昇させる。この効果は、処理40分で明らかであり、2時間で最大レベルに到達し、より長い処理(4時間)により当該効果は消失する。その一方、アクチビンβBは、同一の条件で抑制される。種々の濃度のhCGを用いた実験が行われる際、濃度依存的応答が観察される。キンギョ下垂体抽出物もまた、濃度依存的にアクチビンβAの発現を刺激しアクチビンβBを抑制する。我々は、培養物中の濾胞細胞のこの特性を用いてFSHβ及びLHβがアクティブであるか否かを検証することを計画した。
【0039】
(インビトロ濾胞細胞培養)
(濾胞細胞の単離)
幼魚のゼブラフィッシュを鳥獣店から購入し、オス及びメスを分離せずに維持した。メスを 0.01 % トリカインメタンスルホナート(tricaine methansulfonate)溶液で2分間又はまだ耐えているまで麻酔し、解剖前に斬首した。卵巣を次に取り外し、10 mm 培養ディッシュL-15(Gibco)に設置した。5匹のメスの濾胞を、注意深くインスリン針の補助で切り離した。切り離した濾胞を解剖顕微鏡の接眼マイクロメーターで測定し、およそ 0.45 mm の正常な卵黄形成濾胞を選択し、プールし、10 % ウシ胎仔血清を追加したM199培養液中、6日間T25フラスコで、28 ℃ 及び 5 % CO2 において培養した。培養液は、インキュベーションの3日目に交換した。6日間のインキュベーションの間に、濾胞細胞は著しく増殖し、当該実験に関する細胞の産生量は上昇した。細胞を洗浄し、28 ℃ で15分間トリプシン処理した。その後細胞を 1000 rpm で2分間の遠心分離を介してM199培養液で3回洗浄し、次にホルモン処理の前に、完全M199中で24時間、1ウェルあたり 1 × 105 cells/mL の密度で24ウェルプレートで継代培養した。細胞の量が実験に十分ではなかったので、我々は十分な材料を得るために20匹のメスで着手することにした。
【0040】
(ホルモン処理)
種々の濃度の上清を用い、ポジティブコントロールとしてhCGを 15 IU/mL で用い、さらにコイ下垂体抽出物もまた含む。当該下垂体抽出物条件を用いて、我々はメスウナギの生殖能力における効果を観察するために用いるべき細胞の量を比較することができる。
【0041】
(インビボ注入)
hCGを、20 IU/mL の濃度で 0.9 % NaCl溶液に溶解した。各々の魚は、ネガティブコントロールとしての 50 μL の生理食塩水、ポジティブコントロールとしてのhCG及び種々の濃度の上清又は精製されたFSHβ及び/又はLHβを受けた。注入してから1時間後、2時間後、4時間後、6時間後及び12時間後で、魚を殺生し、RNA抽出のために卵巣を取り外した。
【0042】
(RNA抽出)
ホルモン処理の終わりに、全RNAをゼブラフィッシュの卵巣又は濾胞細胞から単離し、液体窒素中でホモジナイズし、TRIZOL試薬を用いて製造業者の使用説明書に従い抽出した。微量のDNAをDNaseIとのインキュベーションによって除去し、続いてフェノール/クロロホルム抽出及びエタノール沈殿を行った。プラチナTaqを有するSuperscript II ワンステップRT−PCRシステムを用いて、RT−PCRを行った。反応を 25 pmol の上流プライマー及び下流プライマーを用いて 100 ng の全RNAで行った。逆転写を 50 ℃ で30分間行った。PCR条件は、94 ℃ で20秒間の変性、56 ℃ で30秒間のアニーリング及び 72 ℃ で1分間の伸張の40サイクルに引き続き 72 ℃ で10分間の最終伸張であった。PCR産物を、アガロースの 1 % ゲル中の電気泳動によって分離し、エチジウムブロマイドで染色した。
【0043】
(アクチビンβA,アクチビンβB及びβアクチンを増幅するために設計されたプライマー)
上流−アクチビンA 5'- TGC TGC AAG CGA CAA TTT TA -3'
下流−アクチビンA 5'- CAT TCG TTT CGG ACT CAA G -3'
上流−アクチビンB 5'- CAA CTT AGA TGG ACA CGC TG -3'
下流−アクチビンB 5'- GTG GAT GTC GAG GTC TTG TC -3'
上流−βアクチン 5'- CCC CTT GTT CAC AAT AAC CT -3'
下流−βアクチン 5'- TCT GTG GCT TTG GGA TTC A -3'
【0044】
(ウナギにおける安定的な株化細胞の移植)
安定的な株化細胞を、ウナギの腹腔内に移植した。ウナギを麻酔し、腹部に小さな切込みを入れた。一定量のホルモンを産生する正確な量の細胞を移植した。ネガティブコントロールとして、ホルモンを発現せず、空のベクターのみを有する細胞を移植し、さらにポジティブコントロールとして、ウナギの1つの群に下垂体抽出物を注入した。
【0045】
(成熟過程及び卵の成熟の測定)
メスウナギの最終成熟を定量化するために、成熟過程に従って変化する種々の特性が存在するが、それらは著しい体重の増加、眼の直径の増加及び卵母細胞の発生の間の形態変化である。最終卵母細胞成熟の7つの形態学的ステージが、下垂体抽出物処理の間に観察される。不透明な卵母細胞(ステージ0)はまだ小さく、全体的に脂肪小滴で満たされる。ステージ1への最終水和反応オンセット発生は、増加する透明度及び中心核を有する卵母細胞を示す。ステージ2において、卵母細胞は完全に透明であり、脂肪小滴は群れを成し、集まっている。ステージ3において、卵核胞(GV)移行が起こる。ステージ4において、卵核胞は表面(periphery)に発見され、脂肪小滴は反対側に位置づけられる。ステージ5において、脂肪小滴は脂肪融合に起因して減少し、結果としてより大きな脂肪小滴を生じさせる。ステージ6では、減数分裂IIが卵核胞の消失と共に完了し、少数の大きな脂肪小滴が観察される。ステージ7において、卵母細胞は、ただ1つの脂肪小滴を有する。初めの2つのステージの間に卵母細胞の直径が変化するが、これは水和反応の過程によるものである。卵母細胞成熟におけるこれらの形態変化により、我々は細胞移植の効果を判断することができる。細胞の移植後、ウナギは体重測定され、眼の直径が定期的に測定される。処理の間に、卵母細胞サンプルは、卵母細胞が当該処理に応答するかどうかを判断するために取得される。
【0046】
ホルモン産生細胞の移植
序論
CPE(コイ下垂体抽出物)注入の標準的技術を用いたホルモン産生ZF4細胞の移植の効果を比較するための試験を行った。性成熟は、最近Palstraらによって記載された(2005年)とおり、CPEの繰り返し注入によって獲得することができる。繰り返し注入は、結果として高品質の卵及び生存能力のある子孫をもたらさないが、これは誘導されるストレスによるものであろう。従って、CPE注入手順を、本実施例中に記載されるようなホルモン産生細胞の移植のようなよりストレスの少ない手順で置き換えなければならない。ウナギにおけるCPEの効果は、形態学的変化、組織学的変化及び内分泌学的変化によって数週間後に観察することができるので、メスの銀ウナギを用いた4週間の試験を行った。注入される細胞の量は、実験用ウナギ下垂体の推定の重さに関連した(1 kg のウナギにつき、およそ 6 μLの組織)。
【0047】
材料及び方法
(ホルモン産生細胞)
3つの異なる安定的な株化細胞を独立して成長させ、FSHβ、LHβ及びFSH/LHαに関する遺伝子をCMV−プロモーターに挿入し、ZF4細胞に安定的にトランスフェクトした。上記に記載されたとおりの手順に従った(Fugene 6 でトランスフェクトし、G418で選択した)。
【0048】
(GFP−β−ガラクトシダーゼ安定的株化細胞)
pMP2838プラスミドを用いて、緑色蛍光タンパク(GFP)とβ−ガラクトシダーゼタンパクの融合タンパクを発現するZF4の安定的株化細胞を作製した。当該プラスミドはBakkersによって記載された(2000年)。手短に言えば、pUAS-gfpN_LacZプラスミドのgfpN-LacZ遺伝子を取り出し、pEGFP-C3プラスミド中のgfp遺伝子に関する代替品として用いた。従って、β−ガラクトシダーゼ活性(gfpN-LacZ)を有する緑色蛍光融合タンパクは、CMVプロモーターの制御下で発現された。pMP2838プラスミドはまた、安定的株化細胞を作製するための上記に記載されたとおりゲンタマイシン(G418)での陽性クローンの選択を可能にするネオマイシン耐性(Neor)遺伝子も含む。同様に、ZF4安定的株化細胞を、上記に記載されたのと同じ手順で生成した(Fugene 6 でトランスフェクトし、G418で選択した)。
【0049】
注入の日に、細胞を回収し、各々の株化細胞に関して 108 cells の総数に定量化した。4つの株化細胞を均等な細胞濃度に混合し、1 mL 中に 2 × 107 cells の最終濃度まで血清なしのDMEM−F12培養液に希釈した。
【0050】
(β−ガラクトシダーゼ組織染色)
組織を簡単にPBS(リン酸緩衝生理食塩水)中でリンスし、2 mM MgCl2、5 mM EDTA 及び 0.02 % NP−40を含む 1 % パラホルムアルデヒド− 0.1 % グルタルアルデヒドのPBSで室温において30分間、速やかに固定した。次に、室温において洗浄溶液(2 mM MgCl2、5 mM EDTA、0.02 % NP−40 及び 0.01 % デオキシコール酸ナトリウムを含むPBS)で5分間、2回洗浄した。その後組織を、37 ℃ において12時間、染色溶液(2 mM MgCl2、0.02 % NP−40、0.01 % デオキシコール酸ナトリウム、5 mM K3Fe(CN)6、5 mM K4Fe(CN)6 及び 1 mg/mL X−galを含むPBS)で染色した。染色した組織を次にPBSで洗浄し、パラフィン包埋した。
【0051】
(動物及び手順)
60匹のメスの銀ウナギ(900 g −1600 g)をグレベリンゲン(Grevelingen)湖(オランダ)で降海回遊の期間に野生で捕獲した。全てのウナギに到着次第すぐに識別のためのマイクロチップ(Trovan)を備え付け、外部パラメーターを測定した。10匹のウナギを、コントロール動物として速やかに屠殺した。残りのメスウナギを、18 ℃ の温度において 3000 L の人工海水(3.5 %)の再循環システム中で維持した。処理日における細菌感染を予防するために、当該動物を3時間、大きな個々の水槽中で抗生物質フルメキン(50 mg/mL)に曝した。24匹のウナギの一つの群に、前に記載された方法(Palstraら、2005年)に従い、毎週、コイ下垂体抽出物(CPE:20 mg/kg)を注入し、もう一つの群には、1 mL の4つのタイプの細胞(β−gal、LHβ、FSHβ、LH/FSHα)の混合物を実験の開始時に一度だけ注入した。細胞は、背ビレの始まりよりも下かつ側線よりも上の皮下に懸濁液として注入した。
【0052】
1週間毎に、各々の群から6匹のウナギを屠殺して処理効果を解析した。これらは総重量、アイインデックス(eye index)、生殖腺重量及び胸びれの長さを含んでいた。さらに、組織サンプル(血液、下垂体、肝臓、生殖腺)を後の解析のために採取した。さらに、注入箇所のサンプルを獲得してβ−ガラクトシダーゼ陽性細胞の存在を検証した。同一のウナギから、サンプルを、細胞が注入されなかった部位から獲得した。
【0053】
結果及び考察
(細胞移植)
皮下移植の組織学的解析を、β−ガラクトシダーゼ染色に関して確認した。全てのサンプルは、当該細胞の存在を示した(図12)。発色は初めに、皮下層に関して特有な脂肪/コラーゲン組織で生じたが、また多くの場合、筋層の周りの浸潤も観察することができた。1週と4週のサンプルを比較すると、多くの場合、β−ガラクトシダーゼ染色細胞の全体集団の増加すら存在し、それらの細胞が宿主組織に浸潤する能力があることをはっきりと示唆した。拒絶反応に関する兆候は全くなく、ZF4細胞が進化的にウナギから極めて遠いゼブラフィッシュに由来するので、これは驚くべき知見である。
【0054】
(形態変化)
成熟のオンセットを示唆する最も顕著かつ直接の(proximate)変化は、銀ウナギの眼のサイズの増大である(Durifら、2004年)。図13から、細胞移植によって誘導されるアイインデックス(eye index)(長さに対する表面積、Pankhurst 1982年)において観察される変化が、CPE注入によって誘導されるものとほとんど等しいことは明らかである。当該結果は、細胞移植の効果をはっきりと示す。さらなる証拠を、3−4ヶ月刺激の銀ウナギから得ることができ、これは結果として最終成熟をもたらすはずである。
【0055】
結論
上記の実験から少なくとも2つの結論を出すことができる。1)ゼブラフィッシュZF4細胞(繊維芽細胞株化細胞)は、4週間を越えて銀ウナギによって拒絶されず、増殖することすら観察された。2)移植された細胞混合物は、アイインデックス(eye index)において、CPEを用いて標準的な手法を行ったのと同様の効果を有し、従ってホルモン刺激に関する証拠を示した。
【0056】
上記に記載された手法及びコンストラクトの使用
ウナギ株化細胞を生成し、ウナギに移植した。さらに両方のユニット(α+β)を発現する魚細胞を生成した。
【0057】
参考文献
Palstra, A.P., Cohen, E.G.H., Niemantsverdriet, P.R.W., van Ginneken, V.J.T., van den Thillart, G.E.E.J.M. (2005) Artificial maturation and reproduction of European silver eel: Development of oocytes during final maturation. Aquaculture, 2005.
Durif, C., Dufour, S., Elie, P. (2005) The silvering process of the eel : a new classification from the yellow resident stage to the silver migrating stage. J. Fish Biol. 66, 1-19
Pankhurst, W.N. (1982) The relation of visual changes to the onset of sexual maturation in European eel, Anguilla anguilla L. J. Fish Biol. 21, 179-196
Bakker J. (2000) Chitin oligosacharides in Zebrafish development. Investigations at the molecular and cellular level. (Thesis University of Leiden).
【図面の簡単な説明】
【0058】
【図1】ヒト(Hs)、マウス(Mm)、ラット(Rn)、ゼブラフィッシュ(Dr)及びウナギ(Aj)のa(a)に由来するLHβ(LHβ)、FSHβ(FSHβ)の配列比較に基づく樹上図
【図2】p3XFLAG-CMV-9発現ベクター
【図3】LHβの発現に関するクローニングストラテジー
【図4】LHβの予想されるアミノ酸配列は、20593.8の分子量を有する187アミノ酸のタンパクを与える。
【図5】FSHβの発現に関するクローニングストラテジー
【図6】FSHbblu及びFSHbの予想されるアミノ酸配列は、2つの異なるストラテジーのクローニングに対してそれぞれ、19062.24/19965.21の分子量を有する174/183アミノ酸のタンパクを与える。
【図7】αの発現に関するクローニングストラテジー
【図8】aの予想されるアミノ酸配列は、21083.24の分子量を有する190アミノ酸のタンパクを与える。
【図9】ZF4細胞のpEYFP-N1でのトランスフェクション。トランスフェクション後、24時間に観察した。
【図10】pEYFP-N1でコトランスフェクトした細胞における抗−FLAG抗体での免疫組織化学
【図11】抗−FLAGでのウェスタンブロット。抗体は、FLAG−BAPに関して予想されるサイズのタンパクを認識した。
【図12】β−ガラクトシダーゼに関する染色は、銀ウナギに皮下的に注入されたβ−gal−ZF4細胞の存在を示唆した。同一のウナギから、皮膚/筋肉組織を、注入した領域及びさらに尾に向かう領域から解剖した。皮下の脂肪組織は、真皮及び筋層がそうであるように図中で容易に認識できる。
【図13】銀ウナギの初期ステージに関するホルモン産生細胞(HPC)又はコイ下垂体抽出物(CPE)の4週間処理(W1−W4)の間のアイインデックス(EI)の変化(%)。初期ステージ及び各々の週に関して、6匹の十分成長したメスの銀ウナギが取得された(体重 1300 g ± 300 g)。
【特許請求の範囲】
【請求項1】
ホルモン投与を用いた魚の卵及び/又は魚の精子の発生及び/又は成熟を改善するための方法であって、前記魚に前記ホルモンを産生する細胞を提供することを含む、前記方法。
【請求項2】
前記ホルモンが黄体形成ホルモン(LH)、卵胞刺激ホルモン(FSH)又は絨毛性ゴナドトロピン(GC)又はそれらの機能的部分、派生物及び/若しくはアナログを含む、請求項1に記載の方法。
【請求項3】
前記細胞が前記ホルモンを発現するために遺伝子操作される、請求項1又は請求項2に記載の方法。
【請求項4】
前記ホルモンが前記魚と同じ属に由来する、請求項1〜請求項3のいずれか1項に記載の方法。
【請求項5】
前記細胞が前記ホルモンをコードする一つ以上の遺伝子を提供される、請求項1〜請求項4のいずれか1項に記載の方法。
【請求項6】
前記細胞がクローン集団である、請求項1〜請求項5のいずれか1項に記載の方法。
【請求項7】
前記細胞が前記魚における低減した免疫原性に関して選択される、請求項1〜請求項6のいずれか1項に記載の方法。
【請求項8】
前記魚が回遊性魚類である、請求項1〜請求項7のいずれか1項に記載の方法。
【請求項9】
排卵誘発ホルモンを産生する単離型魚細胞及び/又は組み換え型魚細胞。
【請求項10】
前記細胞が前記ホルモンを発現するために遺伝子操作される、請求項9に記載の魚細胞。
【請求項11】
前記細胞がウナギ細胞である、請求項9又は請求項10に記載の魚細胞。
【請求項12】
請求項9〜請求項11のいずれか1項に記載の細胞を含む、魚。
【請求項13】
前記魚が一般消費者向けの魚である、請求項12に記載の魚。
【請求項14】
前記魚がウナギである、請求項13に記載の魚。
【請求項15】
前記ウナギがAnguilla属に属する、請求項14に記載の魚。
【請求項1】
ホルモン投与を用いた魚の卵及び/又は魚の精子の発生及び/又は成熟を改善するための方法であって、前記魚に前記ホルモンを産生する細胞を提供することを含む、前記方法。
【請求項2】
前記ホルモンが黄体形成ホルモン(LH)、卵胞刺激ホルモン(FSH)又は絨毛性ゴナドトロピン(GC)又はそれらの機能的部分、派生物及び/若しくはアナログを含む、請求項1に記載の方法。
【請求項3】
前記細胞が前記ホルモンを発現するために遺伝子操作される、請求項1又は請求項2に記載の方法。
【請求項4】
前記ホルモンが前記魚と同じ属に由来する、請求項1〜請求項3のいずれか1項に記載の方法。
【請求項5】
前記細胞が前記ホルモンをコードする一つ以上の遺伝子を提供される、請求項1〜請求項4のいずれか1項に記載の方法。
【請求項6】
前記細胞がクローン集団である、請求項1〜請求項5のいずれか1項に記載の方法。
【請求項7】
前記細胞が前記魚における低減した免疫原性に関して選択される、請求項1〜請求項6のいずれか1項に記載の方法。
【請求項8】
前記魚が回遊性魚類である、請求項1〜請求項7のいずれか1項に記載の方法。
【請求項9】
排卵誘発ホルモンを産生する単離型魚細胞及び/又は組み換え型魚細胞。
【請求項10】
前記細胞が前記ホルモンを発現するために遺伝子操作される、請求項9に記載の魚細胞。
【請求項11】
前記細胞がウナギ細胞である、請求項9又は請求項10に記載の魚細胞。
【請求項12】
請求項9〜請求項11のいずれか1項に記載の細胞を含む、魚。
【請求項13】
前記魚が一般消費者向けの魚である、請求項12に記載の魚。
【請求項14】
前記魚がウナギである、請求項13に記載の魚。
【請求項15】
前記ウナギがAnguilla属に属する、請求項14に記載の魚。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】

【図12】
【図13】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2008−528027(P2008−528027A)
【公表日】平成20年7月31日(2008.7.31)
【国際特許分類】
【出願番号】特願2007−553056(P2007−553056)
【出願日】平成18年1月26日(2006.1.26)
【国際出願番号】PCT/NL2006/000043
【国際公開番号】WO2006/080841
【国際公開日】平成18年8月3日(2006.8.3)
【出願人】(502293566)ユニヴェルシテイト レイデン (1)
【Fターム(参考)】
【公表日】平成20年7月31日(2008.7.31)
【国際特許分類】
【出願日】平成18年1月26日(2006.1.26)
【国際出願番号】PCT/NL2006/000043
【国際公開番号】WO2006/080841
【国際公開日】平成18年8月3日(2006.8.3)
【出願人】(502293566)ユニヴェルシテイト レイデン (1)
【Fターム(参考)】
[ Back to top ]