
稔性のカプシクム(Capsicum)植物
【課題】トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対して抵抗性を示す、カプシクム(Capsicum)属の植物およびその産生方法を提供する。
【解決手段】SNFD表現型に関与する遺伝情報が存在しない、または植物のゲノムにおいて少なくともSNFD表現型が発現されない程度まで抑制されている、植物のゲノムにおけるL4抵抗性対立遺伝子の存在によって、PMMoV病原型1.2.3に対して抵抗性を示す、カプシクム属の植物。
【解決手段】SNFD表現型に関与する遺伝情報が存在しない、または植物のゲノムにおいて少なくともSNFD表現型が発現されない程度まで抑制されている、植物のゲノムにおけるL4抵抗性対立遺伝子の存在によって、PMMoV病原型1.2.3に対して抵抗性を示す、カプシクム属の植物。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、耐病性植物に関する。より詳しく述べると、本発明は、トウガラシマイルドモットルウイルス(PMMoV)の特異的病原型に対して抵抗性であるトウガラシ植物およびそのような植物を産生する方法に関する。
【背景技術】
【0002】
発明の背景
カプシクム(ナス科またはイネホオズキ科)は、植物の属であり、その甘く芳香のある果実は、スパイス、野菜、および薬剤として用いられている。この属は約40種を含む。ほとんどの品種はカプサイシン(メチルバニリルノネナミド)を含み、これは口の中で強い焼けるような感覚を生じ、薬剤における循環刺激剤および疼痛軽減剤として用いることができる刺激性の化学物質である。植物は中央アメリカおよび南アメリカ原産であるが、それらはほぼあらゆる気候に耐えることから、果実は世界中で産生されている。市販のトウガラシは主に、カプシクム・アンヌーム(Capsicum annuum)(ピーマン、カイエンペッパー、ハラペーニョペッパー、アナハイムペッパー)、キダチトウガラシ(Capsicum frutescens)(タバスコペッパー)、およびカプシクム・キネンセ(Capsicum chinense)(ハバネロペッパー)種である。
【0003】
パプリカまたはピメントとしても知られるカプシクム・アンヌームは、栽培品種によって長さ、色、および辛みが多様である果実を有する草本の一年草である。種は、世界中で、例えば西ヨーロッパおよびアメリカ(USA)において栽培されている。キダチトウガラシおよびC.キネンセは小さい極めて辛みの強い果実を有し、タバスコおよび他のホットペッパー製品において用いられている。強い芳香のためにはかなり暖かい気候が必要であることから、これらの種は、主に熱帯地域およびアメリカのより暖かい地域で栽培されている。
【0004】
カプシクム種の野菜は、栽培のあいだ多くのトバモウイルスによって障害を受ける。トバモウイルス属には、タバコモザイクウイルス(TMV)のタイプ種、および血清学的に関連するトマトモザイクウイルス(ToMV)、トウガラシマイルドモットルウイルス(PMMoV)およびいくつかの他の植物ウイルスが含まれる。ウイルス感染症は植物の活力を低減させうるが、一般的に植物を殺すことはない。例えば、トウガラシ、トマト、およびナスを含む広範な植物に影響を及ぼすTMVは、トウガラシの果実に重度の壊死を引き起こして、果実のほとんどを市場に出せなくする。TMVは、何年ものあいだ土壌において生存しうることから、非常に持続的な病気である。
【0005】
PMMoVは、TMVおよびToMVに対して抵抗性である栽培品種を含む、全てのカプシクム種の植物全体に感染する。ピーマン植物における病気の症状には、若い植物の発育停止、葉の縮れおよび黄色の斑点が含まれる。果実は変形して(こぶだらけで斑点がある)、大きさがわずかに低減されている。
【0006】
1980年代に、その当時公知であるPMMoV抵抗性カプシクム系統に感染することができるTMV様のウイルス株(tm-3)が単離された(Boukema et al., 1980)(非特許文献1)。単離された株は、TMVのP14株と呼ばれた。P14株の発見以降、この新しいウイルスに対して抵抗性を示す植物種の系統C.カコエンス(C. chacoense)(系統PI 260429)が1982年に発見された(Boukema, 1982)(非特許文献2)。後に、この植物における抵抗性は、L-座のL4対立遺伝子によって付与されること(Boukema, 1984)(非特許文献3)、およびP14株がL3対立遺伝子によって付与された前者の抵抗性を克服することができるが、L4対立遺伝子によって付与された抵抗性を打破することができないことが決定された。このL3によって付与された抵抗性打破株は当初TMVとして分類されたが、後に特異的PMMoV病原型として再分類され、病原型は病原型1.2.3として割付された。
【0007】
育種業者において、PMMoV病原型1.2.3に対して抵抗性を示すC.カコエンス系統と共にそれに由来する商業的トウガラシ系統は、一般的にTm3-抵抗性系統と呼ばれる。以降、多くの種苗会社が、商業的に好ましい特徴を有する抵抗性トウガラシ植物を得るために、L4対立遺伝子を含むこのC.カコエンス系統の遺伝子材料をその育種系統に遺伝子移入してきた。
【0008】
L4対立遺伝子が、ToMV、TMVを含む多くのウイルス、ならびにPMMoV病原型1、1.2、および1.2.3に対して抵抗性を付与することは現在公知である。L4対立遺伝子によって付与された抵抗性は、本明細書において以降「PMMoV抵抗性」と呼ばれるが、厳密に言えば、これは上記のようにより広い抵抗性を付与する。
【0009】
トバモウイルスの命名は1980および2004年のあいだに数回の改訂を受けた。本明細書において、1994年4月11日にUnion for the Protection of New Varieties of Plants」(UPOV)によって発行された甘トウガラシ(C. annuum L.)に関する「明瞭性、均一性、および安定性に関する試験を行うためのガイドライン」(TG/76/7)の命名法を利用する。これらのガイドラインにおいて、ペッパートバモウイルス病原型に対する遺伝的抵抗性は、同じ座(L-座)に存在する対立遺伝子5個(L-、L1、L2、L3およびL4)によって制御されると見なされている。本明細書において、様々なウイルス病原型に対する抵抗性とトウガラシにおける抵抗性付与対立遺伝子組成との関係が示されている、先に参照したUPOVガイドラインTG/76/7の21〜22頁の表を明確に参照する。C.アンヌームのホモ接合L3L3遺伝子型は、タバコモザイクウイルス(TMV)、トマトモザイクウイルス(ToMV)、ピーマンモザイクウイルス(BePMV)、タバコマイルドグリーンモザイクウイルス(TMGMV)、ダルカマライエローフレックウイルス(DYFV)、およびトウガラシマイルドモットルウイルス(PMMoV)病原型1.2に対する抵抗性を付与するが、ホモ接合L4L4遺伝子型は、トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対するさらなる抵抗性を提供する。
【0010】
UPOVの命名法は、ウイルス学に関する科学論文において通常用いられる命名とはいくぶん異なる。一般的に科学文献において、命名は、単離体の種の名称および呼称に限定される。病原型の呼称はごくまれに記述されるに過ぎない。例えば、PMMoVの病原型1.2.3は、国際ウイルス分類委員会のユニバーサルウイルスデータベース(ICTV、The Universal Virus Database of the International Committee on Taxonomy of Viruses)には組み入れられていない。さらに、命名法およびその経時的な発展に関して、所定のトバモウイルスは、二つ以上の慣用名を得る可能性があり、以下により詳細に説明するようにさらに再分類される可能性があることに注意すべきである。ウイルス単離体の任意の新しい割付によらず、本発明はL4対立遺伝子によって付与される任意のウイルス株に対する植物の抵抗性に関すると理解される。
【0011】
先に述べたように、トバモウイルスに対するカプシクム種の抵抗性は、L-遺伝子の病原型特異的対立遺伝子(L1、L2、L3、L4)によって付与され、PMMoV病原型1.2.3に対する抵抗性はL4対立遺伝子によって付与される。L-座が存在する領域は第11染色体南のテロメアに位置する(Lefebvre et al, 2002)(非特許文献4)。抵抗性対立遺伝子は、過敏反応(HR)の誘発によって作用する。
【0012】
C.カコエンスから他のカプシクム種に遺伝子移入する場合の、L4対立遺伝子によって付与される抵抗性の顕著な望ましくない特徴は、新しい植物系統における抵抗性が非メンデル性で遺伝される点である。分離に関するメンデルの法則は、配偶子形成の際に対立遺伝子対が分離して、受精の際に無作為に合体すると述べている。それぞれの形質に関して、二倍体生物は、それぞれの親から一つずつの二つの対立遺伝子が遺伝する。二つの対立遺伝子が異なる場合、一つの、優性の対立遺伝子が生物の表現型において完全に発現される;もう一方の劣性の対立遺伝子は表現型に目立った効果を示さない。通常のメンデル遺伝に従って、ホモ接合優性対立遺伝子を有する植物と、ホモ接合劣性対立遺伝子を有する植物との交雑によって、遺伝的(全集団がヘテロ接合である)と共に表現型(全集団が優性形質を発現する)の上でも均一なF1、または雑種第一代が得られるであろう。そのようなF1集団は、その優性形質に関して非分離であると言われる(当然、F2集団では分離が起こる)。このように、F1集団は、親の少なくとも一つがその形質に関してホモ接合である場合には、優性形質に関して分離しない。同様に、非分離F1が存在すれば、親系統の一つにおいて優性形質がホモ接合性であることが確認される。しかし、これはL4対立遺伝子が遺伝子移入される商業的植物系統の場合には当てはまらない。
【0013】
植物育種業者にとって、育種の結果は好ましくは予測可能でなければならないことから、育種系統がホモ接合(真の育種)であることは重要である。例えば、商業的ハイブリッドは二つの(ホモ接合)近交系から産生され、得られたヘテロ接合F1植物は、雑種強勢またはヘテロシスの結果としてその近交系の親より適合である。植物育種業者は、より強健な後代を生成するためにそのようなヘテロシス交雑を意図的に利用して、ホモ接合近交系は経済的価値が高い産生系統として役立つ。したがって、育種の実践において、その植物系統を自家受粉させて、その子孫を非分離に関してスクリーニングすることによって、植物系統が、単一の遺伝子によって付与された優性の抵抗性形質(すなわち、一遺伝子優性形質)に関してホモ接合であるか否かを評価することは通例である。または、予想されるホモ接合優性植物系統をホモ接合劣性系統と交雑させて、それによって分離するF1によって、試験した「優性の」親がホモ接合ではないことが明らかとなる。
【0014】
L4抵抗性対立遺伝子は、通常の一遺伝子優性抵抗性対立遺伝子であると考えられる(Boukema, 1983(非特許文献5);Van Duin, 1998(非特許文献6))。しかし、L4抵抗性対立遺伝子によって付与されたPMMoV抵抗性の場合、育種業者は一般的に、ホモ接合系統からの交雑の予測可能性およびそれらの系統の安定性に関する問題を観察する。予測されるホモ接合(L4L4)抵抗性植物の自家受粉によって常に、均一な抵抗性表現型を有する子孫が得られるが(それによって親植物のホモ接合性を確認する)、一方、そのようなホモ接合抵抗性の親植物と、感受性のある親植物(すなわちL4対立遺伝子を欠損する個体)との交雑によって、しばしば抵抗性で感受性がある植物を含むF1、すなわち分離するF1が得られる。植物の感受性は、例えばPMMoV病原型1.2.3を接種後の全身性のモザイクの存在によって検出してもよい。この現象は、抵抗性がF1子孫植物のいくつかにおいて驚くほど失われており、先に概要した理由から、育種業者にとっては非常に不都合である。育種業者はしばしば、そのような予測不可能なホモ接合親植物を、「分離」植物と呼ぶが、厳密に言えば、それらのF1子孫が分離している。
【0015】
改善されたPMMoV抵抗性近交系を開発するため、ならびに通常の植物選択および試験交雑を用いることによって、育種業者は最近、正常な非分離F1(すなわち、メンデルの法則に従う)を産生するホモ接合植物を得ることができ、それによってその系統におけるL4対立遺伝子によって付与される抵抗性が固定されたことを示した。現在のところ、これがどのように行われるかは不明である。しかし、これらの「非分離」植物の主要な問題は、それらが稔性の低減と矮小生長を示す点である。稔性の低減の重要な結末は、そのような植物が限られた量の種子および/または限られた量の花粉を産生するということである。このように、そのような植物は抵抗性で「非分離である」が、それらは低い稔性および矮性の生長を特徴とする表現型を示し、これは商業的種子産生にとって望ましくない。矮性の生長は、例えば、種子の晩期成熟(すなわち、遅い成長)または植物の生長(速度)の低減によって示される可能性がある。この表現型は、本出願において分離非稔性矮性に関する「SNFD表現型」としてさらに命名されるであろう。現在のところ、生長、稔性、および不稔性に関する問題を含むこれらの負の形質と、C.カコエンスからのL4対立遺伝子のカプシクム系統への遺伝子移入との関連に関する説明はない(例えばVan Duin, 1998(非特許文献6)を参照されたい)。
【0016】
L-遺伝子のいずれのL1、L2、およびL3対立遺伝子をカプシクム種に遺伝子移入しても、ホモ接合抵抗性植物においてSNFD表現型を示すことなく、メンデル遺伝集団を産生することが常に見いだされている。C.カコエンスからのL4遺伝子移入は、今初めて上記の問題を生じる。
【0017】
現在のところ、L4対立遺伝子によって付与されたPMMoV病原型1.2.3に対して抵抗性であって、「非分離」であり、正常な生育(速度)を示し、および適当量の種子を産生するトウガラシ植物が必要である。したがって、本発明の目的は、L4対立遺伝子によって付与されたPMMoV病原型1.2.3に対する抵抗性を、作物栽培学または良好な生長および正常な種子産生のような商業的に良好な形質と組み合わせて、このように、SNFD表現型の望ましくない特徴を有しないカプシクム属の植物を提供することである。
【先行技術文献】
【非特許文献】
【0018】
【非特許文献1】Boukema et al., 1980
【非特許文献2】Boukema, 1982
【非特許文献3】Boukema, 1984
【非特許文献4】Lefebvre et al, 2002
【非特許文献5】Boukema, 1983
【非特許文献6】Van Duin, 1998
【発明の概要】
【0019】
発明の概要
本発明の目的は、本発明によって達成される。本発明に従って、L4対立遺伝子を含むゲノム遺伝子移入の大きさを低減させることによって、先に記述したホモ接合植物において観察されるPMMoV病原型1.2.3に対する抵抗性とSNFD表現型とのあいだの連鎖を断つことが可能であることが判明した。特に、これはL4対立遺伝子のPMMoV抵抗性付与部分の直接近位に存在するL4抵抗性対立遺伝子から遺伝子情報を除去することによって得られる。その結果は、PMMoV病原型1.2.3に対する抵抗性が、SNFD表現型付与部分を移入することなく、一つの植物からもう一つの植物に転移される可能性があり、それによってSNFD表現型を示さないPMMoV抵抗性および作物栽培学上貴重な植物を産生することが可能となったことである。
【0020】
理論に拘束されることなく、SNFD表現型の特徴である負の副作用は、L4対立遺伝子が存在するC.カコエンス染色体断片によって引き起こされ、C.カコエンス染色体断片を例えばC.アンヌームのゲノムに挿入すると、L4対立遺伝子自身と遺伝的に連鎖するL4対立遺伝子の抵抗性付与部分の近位における劣性遺伝子によって、SNFD表現型が引き起こされ、レシピエント植物ゲノムの染色体構築を破壊すると考えられる。したがって、SNFD表現型は、完全なL4対立遺伝子のC.カコエンス染色体遺伝子移入に関してホモ接合である植物に限って発現される。
【0021】
L4対立遺伝子によって付与された望ましい表現型と望ましくない表現型とのあいだの連鎖を断つ可能性に関するこの重要な洞察が得られたことから、SNFD表現型に関する遺伝情報が、L4対立遺伝子の抵抗性付与部分の近位から除去されている、またはホモ接合植物においてSNFD表現型がその組み合わせにおいてもはや発現されない程度まで変化している植物を選択することが可能となった。
【0022】
さて本発明は、第一の局面において、植物のゲノムにおけるL4抵抗性対立遺伝子の存在により、トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対する抵抗性を示し、該L4対立遺伝子が切断されているカプシクム属の植物を提供する。切断は、低減された長さの対立遺伝子を提供し、それによって稔性で正常な生長を示す、すなわちSNFD表現型が存在しない非分離植物が得られる。
【0023】
SNFD表現型に関与する遺伝情報の欠如は、それによってSNFD表現型が得られるL4対立遺伝子の近位における遺伝情報の除去または変化によって行ってもよく、好ましくはL4対立遺伝子の近位における組換え事象によってもたらされる。SNFD情報が対立遺伝子の一つの側にのみ存在する場合、この情報を除去するためには、1回の組換え事象で十分である。SNFD表現型に関する遺伝情報が抵抗性対立遺伝子の両側に存在する場合、この遺伝情報の双方の部分が好ましくは変化または除去される。本発明の好ましい態様に従って、少なくとも一つの組換え事象、好ましくは第11染色体の南側にあるL4対立遺伝子の北側(セントロメア側)で組換え事象が起こる植物が選択されるが、テロメア側での切断も同様に可能である。二重組換えの場合、組換え事象は、一世代で起こる必要はないが、連続世代からの組換え事象によって共に、抵抗性対立遺伝子の片側または両側での遺伝情報の最終的な除去または変化が起こってもよい。少なくとも一つの組換え事象は、抵抗性の付与に関与する遺伝情報が除去されず、それによってL4抵抗性対立遺伝子の抵抗性付与部分の存在を維持するように行わなければならない。
【0024】
本発明の植物の好ましい態様において、該L4抵抗性対立遺伝子は、C.カコエンスのゲノムに由来する。
【0025】
適切な遺伝情報が除去されたまたは変化したか否かを評価するために、当業者は、遺伝子マーカーを適宜利用してもよい。さて本発明は、例えば組換え植物のマーカー支援選抜において用いられる可能性があるいくつかの遺伝子マーカーを記述する。そのような用途は、当技術分野で周知であり、以下により詳細に記述されるであろう。
【0026】
好ましい態様において、本発明は、L4抵抗性対立遺伝子に連鎖した少なくとも一つのマーカーの不在によって示されるように、SNFD表現型に関与する遺伝情報が存在しない植物を提供し、このとき該マーカーは2群のマーカー(E35/M49-F-90、E39/M58-F-65、E39/M51-F-380、E58/M62-F-168、E66/M54-F-600、およびTm3-DRS)、3群のマーカー(E60/M54-F-447)、および1/3群のマーカー(E63/M61-F-501、E66/M43-F-387、E66/M49-F-387、E66/M61-F-99、E67/M50-F-150、E67/M62-F-214、E70/M54-F-133、E71/M47-F-550、E74/M61-F-385)からなる群より選択され、好ましくは3群および2群のマーカー、より好ましくは2群のマーカー(E35/M49-F-90、E39/M58-F-65、E39/M51-F-380、E58/M62-F-168、E66/M54-F-600、およびTm3-DRS)から選択される。
【0027】
もう一つの好ましい態様において、本発明は、L4抵抗性対立遺伝子の存在が、1群のマーカー(E58/M50-F-580、E39/M58-F-95、E58/M60-F-255、およびE54/M55-F-101)、および1/3群のマーカー(E63/M61-F-501、E66/M43-F-387、E66/M49-F-387、E66/M61-F-99、E67/M50-F-150、E67/M62-F-214、E70/M54-F-133、E71/M47-F-550、E74/M61-F-385)から選択され、好ましくは1群のマーカーから選択され、より好ましくはマーカーE58/M50-F-580およびE54/M55-F-101から選択されるL4抵抗性対立遺伝子に連鎖している少なくとも一つのマーカーの存在によって示される、植物を提供する。
【0028】
先に述べたマーカーは、制限酵素EcoRI(コードE)およびMseI(コードM)の制限部位にそれぞれ対応する「コアプライマー」

プラス、プライマーコードによって示されるさらなる選択ヌクレオチド3個からなるAFLPプライマーに基づく。プライマーコードは、http://www.keygene.com/publications/index.htmおよび具体的にはそこで開示される「KF Nomenclature Primer Enzyme Combinations.pdf」と題する文書において見いだされうる。増幅された断片の大きさ(塩基対)は、プライマー酵素の組み合わせ後に認められる。例えば、マーカーE58/M60-255は、プライマー酵素の組み合わせ[EcoRI+プライマーコード58/MseI+プライマーコード60]によってDNAを切断した後に得られた255塩基対の増幅された断片を示す。
【0029】
プライマーコードは以下のとおりである:35:ACA;39:AGA;43:ATA;47:CAA;49:CAG;50:CAT;51:CCA;54:CCT;55:CGA;58:CGT;60:CTC;61:CTG;62:CTT;63:GAA;66:GAT;67:GCA;70:GCT;71:GGA;74:GGT。プライマー酵素組み合わせコードE63/M61-F-501の例は、EcoRI+GAA(=E63)/MseI+CTG(=M61)である。
【0030】
マーカーTG036に由来するマーカーTm3-DRS(Lefebvre et al., 2002)は、PCRプライマー対P118
およびプライマーP119
によって定義されるマーカーであり、PCR産物をTru1I(制限部位TTAA)によって消化した後にプライマーP118およびプライマーP119によってPCRを行うことによって検出することができる。約350塩基対の増幅断片の存在は、L4対立遺伝子の存在を示している。
【0031】
SNFD表現型に関与する遺伝情報が存在しないまたは除去されている植物の供給はまた、以下により詳細に記述される切断型L4対立遺伝子を植物に提供することに関して記述されてもよい。そのような切断に関連する局面は、先に記述した遺伝情報の除去または変化に同様に適用される。
【0032】
本発明の植物のもう一つの好ましい態様において、植物はC.アンヌーム種の植物である。
【0033】
本発明の植物のなおもう一つの好ましい態様において、植物はL4抵抗性対立遺伝子に関してホモ接合であり、ホモ接合植物は好ましくは近交系植物である。
【0034】
もう一つの局面において、本発明は、L4対立遺伝子が切断されていて、切断がテロメアおよび/またはセントロメア側から約0.001〜10 cM、好ましくは対立遺伝子のセントロメア側から0.001〜10 cMの遺伝距離に及ぶヌクレオチド配列の除去を含む、植物のゲノムにおけるL4抵抗性対立遺伝子の存在により、トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対して抵抗性を示すカプシクム属の植物を提供する。先に記述した好ましい態様はまた、本発明のこの局面に従う植物にも当てはまる。
【0035】
コアTM-3抵抗性遺伝子を含む1群のマーカーと、3群のマーカーとのあいだで観察された組換え頻度は、距離約2 cMと同等であった。1群のマーカーと2群のマーカーとのあいだで観察された組換え頻度も同様に、距離約2 cMに等しかった。本明細書において同定されたマーカー群内での任意のマーカーのあいだの距離は、組換えが存在しなかったために決定することができなかった。このように、本明細書において記述された切断は、好ましくは、1群のマーカーの任意の位置からセントロメア(2群)またはテロメア(3群)側に存在する任意のマーカーから2 cM未満の距離に及ぶ遺伝的欠失を表す。
【0036】
もう一つの局面において、本発明は、本発明のホモ接合、好ましくは近交系植物を、商業的に望ましい特徴を示すもう一つのホモ接合、好ましくは近交系のトウガラシ植物と交雑させることによって得ることができる、PMMoV病原型1.2.3に対して抵抗性を示すハイブリッドトウガラシ植物を提供する。
【0037】
さらにもう一つの局面において、本発明は、切断型L4抵抗性対立遺伝子が、カプシクム属の植物において発現されうる遺伝情報を含み、それによって該植物にPMMoV病原型1.2.3に対する抵抗性を付与し、およびSNFD表現型を付与する遺伝情報が該対立遺伝子に存在しない、切断型L4抵抗性対立遺伝子を含む単離された核酸配列を提供する。
【0038】
そのような単離核酸配列の好ましい態様において、抵抗性付与遺伝情報は、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択される、好ましくは、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580からなる群より選択される、より好ましくはマーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される少なくとも一つのマーカーを含み、抵抗性付与遺伝情報はさらに、SNFD表現型を付与する遺伝情報をさらに含み、2群マーカーおよび3群マーカー、好ましくは2群マーカーからなる群より選択される少なくとも一つのマーカーがL4抵抗性対立遺伝子に存在しないことを含む。最も好ましい態様において、本発明の単離核酸配列に含まれるL4抵抗性対立遺伝子は、C.カコエンスのゲノムに由来する。
【0039】
もう一つの局面において、本発明は、a)PMMoV病原型1.2.3に対して感受性であるカプシクム属のレシピエント植物またはその一部を提供する段階、およびb)切断型L4抵抗性対立遺伝子を含むゲノム領域を該レシピエント植物、その一部、またはその後代植物のゲノムに導入する段階を含む、PMMoV病原型1.2.3に対する抵抗性を示すカプシクム属の植物を産生する方法であって、該対立遺伝子が、該植物、植物の一部、または後代植物において発現されうる遺伝情報を含み、それによって該植物、植物の一部、または後代植物にPMMoV病原型1.2.3に対する抵抗性を付与し、およびSNFD表現型を付与する遺伝情報が、少なくともSNFD表現型が発現されない程度まで該対立遺伝子に存在しない方法を提供する。段階b)の前に、例えば本明細書に記述のL4抵抗性対立遺伝子の切断型を有する組換え型ドナー植物の形で、またはインサートとして該ゲノム領域を含むベクターの形で、切断型L4抵抗性対立遺伝子を含むゲノム領域を提供する段階が存在してもよい。
【0040】
本発明の方法において用いられる切断型L4抵抗性対立遺伝子を含むゲノム領域は、SNFD表現型を有する植物が得られるL4対立遺伝子と比較してその大きさが低減されているL4抵抗性対立遺伝子を含む。L4抵抗性対立遺伝子の大きさは、大きさの低減がPMMoV病原型1.2.3に対する抵抗性の付与に関与する遺伝情報に影響を及ぼさないように低減される。
【0041】
切断型L4抵抗性対立遺伝子を含むゲノム領域の導入を実現するための非常に適した方法は、レシピエント植物と、L4抵抗性対立遺伝子を含むカプシクム属のPMMoV病原型1.2.3抵抗性ドナー植物との交雑に由来する後代植物におけるゲノム遺伝子移入によって行われる。遺伝子移入のプロセスは当業者に周知であり、一般的に、その親の一つとの種間ハイブリッドの反復戻し交雑によって、一つの種の一つまたは複数の遺伝子をもう一つの遺伝子プールに導入する段階を必然的に伴う。
【0042】
しかし当業者は、遺伝子移入が、L4抵抗性対立遺伝子またはL4抵抗性対立遺伝子を含むゲノム領域が植物のゲノムに導入されうる唯一の方法ではないことを容易に理解するであろう。例えば、他の方法は、半数体細胞融合、植物形質転換等のようなトランスジェニック技術を用いる段階を必然的に伴ってもよい。したがって、ゲノム領域の導入は、インビトロ培養技術、プロトプラスト融合、形質転換、または倍加半数体法によって行ってもよい。しばしば、そのような技術は、無傷の植物に関して行われることはなく、植物の一部、一般的に単細胞の組織培養物またはプロトプラストのような植物の繁殖材料について行われる。したがって、本発明の植物を得るために、そのような方法には、任意で該植物の一部をトウガラシ植物に生長させるさらなる段階が含まれてもよい。一般的に、これは、植物を再生する段階、または異種DNAによって形質転換された植物または植物の一部もしくは組織の双方の繁殖材料を再生する段階、および任意で該植物もしくは繁殖材料またはその双方を生物学的に複製する段階を必然的に伴うであろう。
【0043】
切断型L4抵抗性対立遺伝子が、遺伝子移入のプロセスによってカプシクム属のレシピエント植物に導入される本発明の方法によって、好ましくは、切断型L4抵抗性対立遺伝子を含むゲノム領域がそのゲノムに遺伝子移入された後代植物が得られる。切断型L4抵抗性対立遺伝子を有する後代植物を得るプロセスは、組換えを必然的に伴うことから、後代植物の表現型または遺伝子型のいずれかに基づく選択を行う前に分離集団を産生することが好ましい。したがって、産生されてそのゲノムに切断型L4抵抗性遺伝子を有する後代植物は好ましくは、例えば先に述べた交雑から得られたF1植物の自家受粉によって産生された、または該交雑から得られたF1植物ともう一つのトウガラシ植物との交雑によって得られた分離集団の植物である。
【0044】
本発明の方法において、ゲノム領域は、好ましくはC.カコエンスのゲノムから導入され、そのような方法において用いられるレシピエント植物は好ましくはC.アンヌーム植物である。
【0045】
本発明の植物を産生する方法の基本的な要素を記述してきたが、本発明の方法はさらなる段階を含んでもよい。好ましい態様において、本発明の方法は、c)PMMoV病原型1.2.3抵抗性植物またはそのPMMoV病原型1.2.3抵抗性後代植物を選択する段階、およびd)SNFD表現型を発現しない抵抗性植物または抵抗性後代植物を選択する段階をさらに含む。
【0046】
PMMoV病原型1.2.3抵抗性植物または後代植物を選択する段階は、抵抗性バイオアッセイを用いるような、例えばPMMoV病原型1.2.3の浮遊液を植物の葉に接種する段階および感染の発生を評価する段階によるような、表現型の決定に基づく方法によって行ってもよい。しかし、好ましい態様において、PMMoV病原型1.2.3抵抗性植物を選択する段階は、該植物におけるL4対立遺伝子の抵抗性付与部分の存在を示す1群または1/3群のマーカーを含む群から選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーの存在に関して、植物または後代植物のゲノムをスクリーニングするプロセスを含む。1群の適したマーカーには、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580が含まれ、好ましくは、マーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される少なくとも一つのマーカーの存在が決定される。抵抗性形質の存在を評価するために適した1/3群のマーカーには、抵抗性植物に存在することを当業者が確立したマーカーが含まれる。1/3群のマーカーはいずれも、L4抵抗性対立遺伝子を示すマーカーと共に、その対立遺伝子におけるSNFD表現型付与部分を示すマーカーを含む。このように、いくつかの1/3群のマーカーは、L4抵抗性対立遺伝子におけるSNFD表現型付与部分を除去し、3群のマーカーと類似である適した切断を示すが、他の1/3群のマーカーは、コア抵抗性遺伝子または調節配列の存在を示し、したがって1群のマーカーと類似である。
【0047】
PMMoV病原型1.2.3抵抗性植物の中から、SNFD表現型(先に記述した段階d)を発現しない植物または後代植物を選択する段階は、本発明によって開示されるマーカー支援育種プロセスの中心となる。SNFD表現型を発現しない抵抗性植物または後代植物の選択は、2群のマーカーおよび3群のマーカーからなる群(任意で1/3群のマーカーからも選択される)、好ましくは2群のマーカーより選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーに関して、抵抗性植物または後代植物のゲノムにおける切断型L4抵抗性対立遺伝子をスクリーニングする段階、ならびに該マーカーの少なくとも一つが存在せず、それによって切断を示す、植物または後代植物を選択する段階を含む。
【0048】
もう一つの局面において、本発明は、先に記述したPMMoV病原型1.2.3に対する抵抗性を示すカプシクム属の植物を産生する方法を行う段階を含み、さらに以下の段階:e)選択した植物を自家受粉する段階;f)該自家受粉から得られた種子を植えて、該種子を植物に生長させる段階;g)PMMoV病原型1.2.3抵抗性を示し、商業的に望ましい特徴を保有する段階g)からの植物を同定する段階、およびh)PMMoV病原型1.2.3抵抗性を示し、商業的に望ましい特徴を保有する近交系トウガラシ植物が産生されるまでe)〜g)の段階を繰り返す段階を含む、PMMoV病原型1.2.3に対する抵抗性を示すカプシクム属の近交系植物を産生するための方法を提供する。本発明の状況における商業的に望ましい特徴は、外観の改善、より高い種子収率、稔性の改善、より高い果実収率、より大きいもしくはより小さい果実、または改善された果実の色もしくは味のような果実特徴のような任意の特徴に関連してもよく;または例えば、幹の強度の改善、根のシステムの改善、ストレス抵抗性の改善、耐病性の改善等のような特徴に関連してもよい。実践において、商業的に望ましい特徴は、野生型植物に対して、植物をより商業的に貴重にするであろう如何なる特徴であってもよい。
【0049】
もう一つの局面において、本発明は、先に記述した本発明の方法の任意の一つによって得ることが可能なPMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物または近交系植物を提供する。
【0050】
他の局面において、本発明は、本発明に従う植物の後代、葉、茎、根、根の先端、台木、シュート、果実等のような部分、または本発明に従う植物の細胞、プロトプラスト、カルス、細胞塊、(体細胞)胚、葯、葉柄、花粉、胚珠、花、培養細胞、種子等のような部分に関する。そのような部分は、好ましくは(器官)組織培養による繁殖に適していてもよく、または果実のような、消費に適してもよい。
【0051】
もう一つの局面において、本発明は、本発明のトウガラシ植物を生長させることによって産生されたトウガラシ種子を提供する。
【0052】
もう一つの局面において、本発明は、本発明の方法によって得ることができる本発明の近交系トウガラシ植物を、好ましくは商業的に望ましい特徴を示す近交系トウガラシ植物と交雑することによって得ることができるPMMoV病原型1.2.3に対して抵抗性を示す、ハイブリッドトウガラシ植物またはその一部を提供する。
【図面の簡単な説明】
【0053】
【図1】この座に連鎖したマーカーの異なる群(1、2、3、1/3群)を示すL4座のマイクロマップを示す。これらの群のそれぞれに存在するマーカーを、実施例1において示すように表1に記載する。
【図2】個々のマーカーが示された、同じマイクロマップを示す。
【図3】本明細書において用いられるマーカー、および特にヌクレオチド配列情報およびマーカーの長さに関する詳細な情報を提供する。マーカーの長さは、C.カコエンスアクセッション番号PI 260429から単離されたゲノムDNAを鋳型DNAとして用いて、表記のプライマーとのPCR反応を行うことによって増幅されたDNA断片の塩基対におけるおおよその長さである。
【発明を実施するための形態】
【0054】
発明の詳細な説明
定義
本明細書において提供された見出しは、全体として明細書を参照することによって得られうる本発明の様々な局面または態様の制限ではない。したがって、下記に定義される用語は、明細書を全体として参照することによってより十分に定義される。
【0055】
「L4抵抗性対立遺伝子」または「L4対立遺伝子」という用語は、本明細書において、遺伝子がL-座の一部であり、Boukema(1984)によって最初に記述された、トマトモザイクウイルス(ToMV)ならびにトウガラシマイルドモットルウイルス(PMMoV)病原型1、1.2、および1.2.3に対する抵抗性を植物に提供する、カプシクム種のL遺伝子の病原型特異的抵抗性対立遺伝子を指すために用いられる。L座が存在する領域は第11染色体南のテロメア上に位置する。非切断型のL4対立遺伝子は、マーカーTm3-DRSと会合する。したがって、L4対立遺伝子は、例えば、本明細書において定義される一つまたは複数の2群マーカー(E35/M49-F-90、E39/M58-F-65、E39/M51-F-380、E58/M62-F-168、E66/M54-F-600、およびTm3-DRS)と一つまたは複数の3群または1/3群マーカー(E60/M54-F-447、E63/M61-F-501、E66/M43-F-387、E66/M49-F-387、E66/M61-F-99、E67/M50-F-150、E67/M62-F-214、E70/M54-F-133、E71/M47-F-550、E74/M61-F-385)とによって境界を定められるC.カコエンスアクセッション番号PI 260429号に由来し、そのヌクレオチド配列がPMMoV病原型1.2.3に対する抵抗性を付与するヌクレオチド配列を表す。
【0056】
本明細書において用いられるように、「トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3」という用語は、L3対立遺伝子によって付与される抵抗性を克服することができる、カプシクムに感染するマイルドモットルウイルスの株を示す。文献において、そのような株は一般的に、PMMoVの抵抗性を打破する株という用語で示される。ICTVが承認した頭字語PMMoVおよびPMMVはいずれも、トウガラシマイルドモットルウイルスを示すために用いられる可能性があると理解されるべきである。文献において認められる可能性があるPMMoVの同義語は:タバコモザイクウイルスのサムスン潜在株(SL-TMV)、ペッパーモザイクウイルス、およびカプシクムモザイクウイルスである。ウイルス分類における急速な発達により、いくつかの株、特に当初トウガラシマイルドモットルウイルス(PMMoV;ICTV10進コード71.0.1.0.007)として分類された最初のダッチ株(Tobias et al., 1982)は、現在では、パプリカマイルドモットルウイルス(PaMMV;ICTV 10進コード71.0.1.0.006)として分類されている。したがって、本明細書において用いられる「トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3」という用語にはまた、トウガラシ植物におけるL3対立遺伝子によって付与される抵抗性を克服することができるPaMMVが含まれる。一般的に、本明細書において用いられるように、「トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3」という用語によって示されるウイルスの例には、以下の株が含まれるがそれらに限定されるわけではない:TMVの単離体P14(Boukema, 1982;Tobias et al., 1982;Rast, 1988);PMMoV-Iによって示されるイタリア株(Wetter et al, 1984;Rodriguez-Cerezo et al., 1989;Velasco et al, 2002;Berzal-Herranz et al., 1995;Garcia-Luque et al., 1993);PMMoV-Sによって示されるスペイン株(Tenllado et al., 1997;Velasco et al., 2002;Tenllado et al., 1996;Berzal-Herranz et al., 1995;Garcia-Luque et al., 1993;Alonso et al., 1991;Avila-Rincon et al., 1989;Rodriguez-Cerezo et al., 1989);PMMoV-JおよびPMMoV-Ijによって示される日本株(Tsuda et al., 1998;Hagiwara et al., 2002;Kirita et al., 1997);PMMV-kによって示される韓国株(Lim et al., 1997);およびPaMMV(Garcia-Luque et al., 1993;Ruiz del Pino et al., 2003;Gilardi et al., 2004)。トウガラシマイルドモットルウイルスおよびパプリカマイルドモットルウイルスの記述に関する典拠の確かな参考文献は、i.a. Brunt, A.A., Crabtree, K., Dallwitz, M.J., Gibbs, A.J., Watson, L. and Zurcher, E.J.(eds.)(1996以降)「Plant Viruses Online : Descriptions and Lists from the VIDE DatabaseVersion : 16 th January 1997」、およびThe Universal Virus Database of the International Committee on Taxonomy of Viruses(http://www.ncbi.nlm.nih.gov/ICTVdb/index.htm)である。
【0057】
「切断型対立遺伝子」は、本明細書において、その遺伝距離に基づいて完全な対立遺伝子より大きさが小さい対立遺伝子として定義される。例えば大きさの低減は、少なくとも一つの連結したマーカーの不在および/または区別可能な表現型、本発明の場合ではSNFD表現型の不在によって示されてもよい。
【0058】
本明細書において用いられるように、「対立遺伝子」という用語は、その全てが少なくとも一つの形質または特徴に関連する遺伝子の一つまたは複数の代替形を意味する。二倍体細胞において、所定の遺伝子の二つの対立遺伝子は、相同な染色体の対において対応する座を占める。本発明は、QTL、すなわち一つまたは複数の遺伝子または調節配列を含んでもよいゲノム領域に関連することから、いくつかの場合において「対立遺伝子」の代わりに「ハプロタイプ」(すなわち、染色体セグメントの対立遺伝子)を指すことがより正確であるが、それらの場合、「対立遺伝子」という用語は、「ハプロタイプ」という用語を含むと理解すべきである。対立遺伝子は、それらが類似の表現型を発現する場合、同一であると見なされる。配列における差は起こりうるが、それらが表現型に影響を及ぼさない限り重要ではない。
【0059】
「ゲノム領域」は、本明細書において、遺伝子および調節要素領域のような様々なゲノム機能を有する配列を含んでもよいヌクレオチド配列として、好ましくはDNA配列として定義される。ゲノム領域は、ヌクレオチド構築物であってもよく、ベクターに含まれていてもよい。または、ゲノム領域は、該植物の交雑後の染色体組換えによって一つの植物からもう一つの植物に転移されてもよい。ゲノム領域は、原則的に一つまたは複数の種に由来する遺伝子材料を含んでもよい。
【0060】
「遺伝子」は、本明細書において、染色体上の特異的な位置を占有し、生物における特定の特徴または形質に関する遺伝的インストラクションを含むDNAの配列からなる遺伝単位として定義される。
【0061】
「座」は、本明細書において、所定の遺伝子が所定の種の染色体上で占有する遺伝子マップ上の位置として定義される。
【0062】
本明細書において用いられるように、「ヘテロ接合」という用語は、異なる対立遺伝子が相同染色体上の対応する座に存在する場合に存在する遺伝子状態を意味する。
【0063】
本明細書において用いられるように、「ホモ接合」という用語は、同一の対立遺伝子が相同染色体上の対応する座に存在する場合に存在する遺伝子状態を意味する。ホモ接合性は、個々の植物の自家受粉後の分離の不在、または感受性がある植物と交雑した場合にF1における分離の不在として定義される。
【0064】
本明細書において用いられるように、「ハイブリッド」という用語は、二つの遺伝的に異なる個体間の交雑の任意の子孫を意味する(Rieger et al., 1968)。
【0065】
本明細書において用いられるように、「近交系」という用語は、実質的にホモ接合の個体または系統を意味する。
【0066】
本出願において、「組換え事象」は、(減数分裂)染色体交差を意味すると理解される。
【0067】
本明細書において用いられるように、「遺伝子移入」、「遺伝子移入された」、および「遺伝子移入する」という用語は、種を交雑することによって、一つの種、品種または栽培品種の遺伝子がもう一つの種、品種または栽培品種のゲノムに移動するプロセスを指す。交雑は、天然または人工的であってもよい。プロセスは任意で、戻し交雑親に戻し交雑することによって完了してもよく、この場合遺伝子移入は、種間ハイブリッドとその親の一つとの反復戻し交雑によって、一つの種の遺伝子をもう一つの種の遺伝子プールに浸潤させることを指す。遺伝子移入はまた、レシピエント植物のゲノムにおいて安定に組み入れられたヘテロ接合遺伝子材料として記述されてもよい。
【0068】
「遺伝子操作」、「形質転換」および「遺伝子改変」は全て、本明細書において標的植物のDNAへの、通常、もう一つの生物の染色体DNAまたはゲノムの、しかしこれらに限定されない任意の種の遺伝情報の移入に関する同義語として用いられる。遺伝子操作は、レシピエント植物のゲノムに異種遺伝子材料を安定に組み入れる一つの方法であり、これには遺伝子またはその対立遺伝子バリアントをコードする外来ヌクレオチド配列を含む異種DNAと共に、植物細胞または組織における異種DNAの安定な組み込みを引き起こすことができる要素、および植物細胞または植物組織において外来ヌクレオチド配列の発現を可能にする要素から選択される調節要素を含むDNA組換え体によって、植物の細胞または組織を形質転換する段階を含むプロセスが含まれてもよい。
【0069】
本明細書において用いられるように、「分子マーカー」という用語は、制限断片長多型(RFLP)マーカー、増幅断片長多型(AFLP)マーカー、一塩基多型(SNP)マーカー、マイクロサテライトマーカー、配列特徴増幅反復配列(SCAR)マーカー、もしくはイソ酵素マーカー、または特異的遺伝子および染色体位置を定義して、二つの対立遺伝子間の多型を検出する本明細書に記述のマーカーの組み合わせのような任意の技術を用いることによって得ることができるマーカーを意味する。本明細書において提供された1、2、3および1/3群のマーカーは、カプシクムゲノムDNAを鋳型として用い、図3において示したプライマー配列を増幅プライマーの対の第一および第二のプライマーとして用いる核酸増幅反応を行うことによって得られた二本鎖または一本鎖ヌクレオチド配列からなるマーカー(特にAFLPマーカー)を表し、該プライマー配列および/またはその相補体がいずれかの側に隣接するカプシクム特異的ヌクレオチド配列および/またはその相補体からなる二本鎖または一本鎖ヌクレオチド配列を提供する。カプシクム特異的ヌクレオチド配列という用語は、本明細書において鋳型DNAとしてC.カコエンスアクセッション番号PI 260429号のゲノムDNAを用いて、およびそれと配列類似性少なくとも90%、好ましくは95%、より好ましくは97%、さらにより好ましくは98%より高い配列類似性を有する配列を用いた場合に得られた配列を示すために用いられる。
【0070】
本明細書において用いられるように、「制限断片長多型」または「RFLP」という用語は、特異的な制限酵素によって切断されたDNA断片の大きさにおける個体間の変動を意味する。RFLPをもたらす多型配列は、遺伝子連鎖マップ上のマーカーとして用いられる。
【0071】
本明細書において用いられるように、「集団」という用語は、共通の遺伝的起源を共有する遺伝的に均一または不均一な植物コレクションを意味する。
【0072】
本明細書において用いられるように、「品種」または「栽培品種」という用語は、構造的特徴および性能によって同じ種における他の品種から同定されうる類似の植物群を意味する。本明細書において用いられるように「品種」という用語は、1972年11月10日、1978年10月28日、および1991年3月19日にジュネーブで改正された、1961年12月2日の植物の新品種の保護に関する国際会議(UPOV条約)での対応する定義と同一の意味を有する。このように、「品種」は、公知の最低のランクの単一の植物学的分類内での植物のグループ分けを意味し、育成者権の付与に関する条件が十分に満たされているか否かによらず、そのグループ分けはi)所定の遺伝子型または遺伝子型の組み合わせに起因する特徴の発現によって定義されうる、ii)該特徴の少なくとも一つの発現によって任意の他の植物のグループ分けと区別されうる、およびiii)繁殖に関するその適切性に関する単位が不変であると見なされうる。
【0073】
本明細書において用いられるように、「トウガラシ」または「カプシクム」という用語は、カプシクム属の任意の種、品種、栽培品種、または集団を意味する。
【0074】
一つの局面において本発明は、トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対して抵抗性を示すカプシクム属の植物を提供する。本発明に従うカプシクム属の植物は、C.アンヌーム、黄色トウガラシ(C. baccatum)、C.カルデナセイ(C. cardenasei)、C.カコエンス、C.キネンセ、C.エキシミウム(C. eximium)、キダチトウガラシ、C.ミクロカルプム(C. microcarpum)、C.ミニマム(C. minimum)、C.ペンデュラム(C. pendulum)、C.プラテルミッスム(C. praetermissum)、およびC.プベスケンス(C. pubescens)が含まれるがそれらに限定されるわけではないカプシクム属の任意の種であってもよい。好ましくは、本発明の植物は、白い花の咲くカプシクム種、好ましくは、C.アンヌーム、キダチトウガラシ、C.キネンセ、またはC.カコエンス植物であり、より好ましくは、C.アンヌーム、キダチトウガラシ、またはC.キネンセ植物であり、さらにより好ましくはC.アンヌームまたはキダチトウガラシ植物であり、最も好ましくはC.アンヌームである。さらにより好ましい態様において、本発明は、作物栽培学上または商業的に望ましい特徴を有するC.アンヌーム植物を提供する。
【0075】
本明細書において用いられるように、「C.アンヌーム」という用語は、その植物のゲノムが遺伝的バックグラウンドとしてC.アンヌームゲノムを含む、カプシクム属のトウガラシ植物を意味する。そのような植物は本質的に、組換え、形質転換または任意の他のプロセスの結果として、他の種のDNAが組み入れられているC.アンヌームゲノムを有するであろう。当業者は、二つの植物種のあいだの育種によって、双方の親の特徴および遺伝子材料を有し、このように種間の分界線を曖昧にする植物が得られるであろうことを理解するであろう。そのような植物は全て本明細書においてC.アンヌーム植物と呼ばれる。
【0076】
トバモウイルス、トウガラシマイルドモットルウイルス(PMMoV)は、1952年にアメリカの南カリフォルニアにおいてC.アンヌームにおいて初めて報告された(McKinney, 1952)。症状には、軽度の萎黄病および特に植物が若いあいだに感染した場合には生長停止が含まれる。果実は小さく、変形して斑点を有することがあり、壊死性の陥没を有する可能性があり、このようにそれらを市場に出せなくする。PMMoVは、植物間の密接な接触によって、種子表面上のビリオンによって、および移植によって伝搬する。ウイルスは、多くの国において報告されている(アルゼンチン、オーストラリア、カナダ、デンマーク、フランス、ハンガリー、アイスランド、イタリア、日本、韓国、オランダ、スペイン、イギリス、およびアメリカ)。病気の制御は、抵抗性栽培品種を用いることによって最善に達成される。L4対立遺伝子によって付与された抵抗性を克服することができると報告されている新しい株が報告されている(Antignus et al., 2000)。
【0077】
本発明の植物におけるトウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対する抵抗性は、植物のゲノムにおけるL4抵抗性対立遺伝子の存在による。PMMoV病原型1.2.3に対して抵抗性を付与する本発明の植物のゲノムに存在するL4抵抗性対立遺伝子は、如何なる起源に由来してもよい。本発明の好ましい態様に従って、L4抵抗性対立遺伝子は、C.カコエンスに由来し、より好ましくはアクセッション番号PI 260429号およびSA 185号に由来する(Boukema, 1983)。
【0078】
本発明の植物は、後代にPMMoV抵抗性を伝搬することができ、L4抵抗性対立遺伝子に関してヘテロ接合であってもよいが、好ましくはL4抵抗性対立遺伝子に関してホモ接合である。L4抵抗性対立遺伝子に関してホモ接合である本発明の植物は、稔性の低減を示さず、および/または矮性の生長を示さない。
【0079】
植物が、稔性の低減および/または矮性生長を示さない場合、SNFD表現型を引き起こす遺伝情報の十分量が紛失している、またはこの遺伝情報が十分に不活化されていると推測される。SNFD表現型に関与する遺伝情報の不在は、少なくとも、SNFD表現型が発現されない程度までである。
【0080】
本発明者らは、AFLPによってL4対立遺伝子の近傍における様々な遺伝子マーカーの存在を決定して、マーカー間、またはマーカーとL4対立遺伝子とのあいだの組換えに基づいて、これらのマーカーを異なる群に分けることができることを発見した(実施例における表1を参照されたい)。L4座に連結した異なる群の位置を図1に示す。1群のマーカーは、PMMoV病原型1.2.3バイオアッセイにおいてマーカーと抵抗性スコアとのあいだに組換えが観察されない限り、L4対立遺伝子自身に対して最も近位に存在すると考えられ、L4-対立遺伝子の存在を示している。2群および3群のマーカーは、いずれかの側で1群マーカーに隣接している。第4の群は、1群と3群の双方に及ぶ領域に存在するマーカーからなる。この群のマーカーを、本明細書において1/3群のマーカーと呼ぶ。
【0081】
1/3群に存在するマーカーは、所定のマーカーと1群マーカーとのあいだ、さらに所定のマーカーと3群のマーカーとのあいだにも組換えが観察されなかったため、今のところこれらのマーカーのそれぞれが1群または3群内に存在することができないという点において異なる。組換えは1/3群のマーカーと2群のマーカーとのあいだに観察されたことから、これら二つの群からのマーカーは、明らかにその遺伝子の位置が異なり、これらのマーカーの実際の位置は1群または3群のいずれかである。
【0082】
このように、群のそれぞれにおいて組換えは観察されなかった。
【0083】
染色体の残りに向かっての図1のマップの方向は、2群のマーカーが、第11染色体南のセントロメア側に存在して、3群のマーカーがテロメア側(末端に向かって)に存在するような方向である。すなわち、対立遺伝子の主な部分は、それに対して図が明らかな参照を行うLefebvre et al., 2002の図1において公表されたように、トウガラシの種間コンセンサス連鎖マップにおいて示されるように、分離集団PYに関して第11染色体(Chr. P11(brun))のマーカーTG036より下に存在する。
【0084】
抵抗性であって、L4対立遺伝子に関してホモ接合であり、2群、3群、または1/3群のいずれかのマーカー、特に2群のマーカーを紛失した植物は、SNFD表現型を示さないことが見いだされた。このように、L4対立遺伝子に通常連鎖している特定のゲノム領域を他の遺伝子型の相同なDNAに置換してもよく、それでも抵抗性に影響を及ぼすことはないが、生長および稔性と共に抵抗性形質の子孫植物への遺伝挙動に強く影響を及ぼす。
【0085】
SNFD表現型を有しない植物は、例えば通常の雑種形成(植物交雑)技術によって得ることができる。しかし、その条件は、正確な選択基準を用いることである。本発明に従って、現在では適した選択基準、すなわちL4抵抗性対立遺伝子の近傍からの遺伝情報の小片の、完全または部分的欠如または不活性が定義されている。本発明に従って、SNFD表現型が、植物およびその後代に存在しない限り、抵抗性対立遺伝子周囲の遺伝情報のどれほど多くが組換え体に存在しないもしくは抑制されているか、またはどのようにしてSNFD表現型を付与する遺伝子が不活化されているかは無関係である。
【0086】
組換えがL4対立遺伝子およびSNFD表現型を引き起こす遺伝子のあいだで起こっている植物を発見するために、分離集団またはそのような集団の後代を、組換え型表現型、すなわち非SNFD表現型と組み合わせてPMMoV P 1.2.3に対するホモ接合抵抗性を有する植物に関して実際にスクリーニングする。
【0087】
このようにして適した組換え体を発見することは可能であるが、作物栽培学上貴重な組換え体を発見することが複雑になりうる多くの理由がある。先に述べたように、ホモ接合体はSNFD表現型を示す。しかし、そのようなホモ接合体を、感受性がある(すなわち、L4対立遺伝子を含まない)植物と交雑すれば、F1は抵抗性を示すがSNFDを示さないことが見いだされた。これは、L4抵抗性対立遺伝子は優性であるが、SNFD表現型に関与する遺伝子が劣性であるという事実によると思われた。
【0088】
C.カコエンス染色体断片1コピーを有する植物の後代は、正常な(非SNFD)表現型と抵抗性を併せ持つであろう。しかし、そのF1子孫は、抵抗性表現型に関して分離し、商業的カプシクム品種は、流通させるために遺伝子の均一性に関する厳密な要件に従わなければならないことから、そのような植物は、商業的に用いることができない。
【0089】
上記の困難が起こる可能性があるにもかかわらず、SNFD表現型に関する遺伝情報が存在しない所望の植物の選択が、通常の方法で非常に可能であることは明らかであろう。しかし、このような選択は、同定のために、比較的大きい分離集団または多数の雑種形成を必要とする。
【0090】
したがって、適した植物の選択において分子生物学的ツールを利用することはより効率的である可能性がある。有用な技術は、Vos et al., 1995によって記述されるように、AFLP技術である。
【0091】
本発明に応用した場合、この技術は、L4対立遺伝子に遺伝的に連鎖しているDNAマーカーのマッピングに基づき、その後、雑種形成実験の子孫において組換え事象がL4対立遺伝子の近位で起こったか否かを比較的単純な方法で決定することができる。L4対立遺伝子に関するホモ接合性とSNFD表現型の欠如とを併せ持つ植物を選択する効率は、分子マーカーを用いることによって有意に増加させることができる。L4連鎖マーカーが存在しない場合に、(したがって)L4対立遺伝子とそのマーカーとの交差が起こった。対立遺伝子から様々な遺伝的距離でマーカーを選択することによって、組換えの位置を定義して、それによって対立遺伝子の近傍から多数または少数が消失したか否かを決定することができる。したがって、連鎖したマーカーの一つまたは複数が存在しない後代は、対立遺伝子の望ましくない近位の少なくとも小片を欠損しており、したがってSNFD表現型をもたらす遺伝情報が存在しないL4対立遺伝子を有する染色体が存在する機会を平均より多く有する。
【0092】
対立遺伝子の近位での遺伝子マーカーのマッピングは、例えばLefebvre and Chevre, 1995;Michelmore, 1995;Winter and Kahl, 1995において記述される、分子生物学技術の当業者によってかなり容易に行うことができる技法である。AFLP技術に関する一般的な情報はVos et al.(前記)において認められうる。
【0093】
L4抵抗性対立遺伝子がホモ接合条件で存在し、SNFD表現型を発現しない態様における本発明に従う植物は、他の作物栽培学上貴重なカプシクムタイプにPMMoV P1.2.3抵抗性を転移させるために適切に用いることができる。このように、ハイブリッドトウガラシ植物は、ホモ接合植物、好ましくは近交系植物をもう一つのトウガラシ植物と交雑することによって作出されうる。もう一つのトウガラシ植物は、PMMoV P1.2.3に対して抵抗性であってもなくてもよいが、好ましくはホモ接合であり、さらにより好ましくは近交系である。
【0094】
L4抵抗性対立遺伝子がホモ接合状態で存在し、SNFD表現型を発現しない態様における本発明に従う植物はまた、他の作物栽培学上貴重なカプシクムタイプにPMMoV P1.2.3抵抗性を固定するために用いることができる。これは、例えば標準的な戻し交雑技法(例えば、Briggs and Knowles, 1967)の後に植物を少なくとも2世代自家受粉させて、抵抗性対立遺伝子に関してホモ接合であって、SNFD表現型を発現しない株を選択することによって起こってもよい。
【0095】
選択プログラムの特定の段階において、他家受粉を用いる。しかし、自家受粉植物の他家受粉は、雌性の親として用いられる植物において自家受精が防止されることを必要とする。これは、生殖器官の雄性部分を手で除去することによって行うことができる。これは、その物理的除去によって、化学物質によって、および/または花にやる水を用いることによって行うことができる。生殖器官の雄性部分を除去または機能障害にするこれらの方法は全て、当技術分野で周知である。雑種形成の後代は、雑種形成の雌性の親に種子を産生させて、F1または戻し交雑の種子を採取して、それを撒いて新しい植物を得ることによって得ることができる。F1植物を自家受粉させて、F2世代を産生する、または戻し交雑スキームの戻し交雑親と戻し交雑することができる。戻し交雑植物をさらに、(戻し交雑)親と交雑させて、その後の世代における植物の作物栽培学上の価値を改善することができる、または自家受粉させて、L4対立遺伝子に関してホモ接合であって、SNFD表現型を発現しない植物を産生することができる。
【0096】
本発明は、本明細書においてカプシクム・アンヌームを参照して説明する。本発明の原理が同様にカプシクム属の他の種、およびより一般的にPMMoV抵抗性を付与するためにL4対立遺伝子を導入することができるナス科の植物に適応可能であることは、当業者には明らかであろう。基本的に、本発明の様々な局面は、PMMoVによって影響を受けうる全ての植物に関する可能性があり、本発明の原理はPMMoVの宿主範囲における如何なる植物種にも応用可能である。この文脈において、Brunt, A.A., Crabtree, K., Dallwitz, M.J., Gibbs, A.J., Watson, L. and Zurcher, E.J.(eds.)(1996以降)「Plant Viruses Online : Descriptions and Lists from the VIDE Database」Version : 20 th August 1996、URL: http://biology.anu.edu.au/Groups/MES/vide/として公式に引用される、しかしhttp://image.fs.uidaho.edu/vide/refs.htmにおいて認められる可能性があるウェブサイト刊行物において記述されているように、PMMoVに関する宿主範囲を特に参照されたい。この文脈において、Dallwitz, 1980およびDallwitz et al., 1993も同様に引用されなければならない。したがって、C.アンヌームに対する参照は本発明を制限すると解釈してはならない。
【0097】
カプシクム植物自身および果実のような消費に適したその一部の他に、本発明は繁殖に適した植物の部分を含む。先に述べたように、本発明は、カプシクム植物の植物の部分に限定されず、PMMoVの宿主範囲における任意の植物の部分に関する。繁殖に適した部分の例は、種子、葉、幹、根、シュート等、プロトプラスト、体細胞胚、葯、葉柄、培養細胞等のような器官組織である。本発明に従う植物は、通常の方法で、または植物の部分からの組織培養技術によっても栽培または繁殖させることができる。
【0098】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法は、PMMoV病原型1.2.3に対して感受性があるカプシクム属の適したレシピエント植物を提供する段階を含む。
【0099】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法はさらに、L4抵抗性対立遺伝子がレシピエント植物に対してPMMoV病原型1.2.3に対する抵抗性を付与する、例えばドナー植物からのL4抵抗性対立遺伝子をレシピエント植物に導入する段階を含む。
【0100】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法はさらに、抵抗性植物から分離集団を産生する段階を含んでもよい。
【0101】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法はさらに、分離集団の実質的にそれぞれの植物の後代を産生する段階を含んでもよい。例えば植物を交雑または自家受粉させることによって植物に種子を産生させる段階、および該種子を新しい植物に生長させる段階によって、後代または子孫を産生してもよい。または、後代は他の適した技術を用いて得てもよい。そのような代用法は、植物の育種領域において全く一般的である。
【0102】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法は、そのゲノムにL4抵抗性対立遺伝子が存在し、SNFD表現型に関与する遺伝情報が、少なくともSNFD表現型が発現されない程度まで存在しない植物を、該後代の少なくとも一つから選択する段階を含む。
【0103】
本発明の植物のゲノムにおけるL4抵抗性対立遺伝子の存在は、天然または人工の如何なる原因の結果であってもよく、例えば遺伝子移入、遺伝子操作(形質転換)、またはプロトプラスト融合の結果であってもよい。
【0104】
ドナー植物から、PMMoVに対して非抵抗性であるまたは低い抵抗性レベルを保有するレシピエント植物への、L4抵抗性をコードする一つまたは複数の遺伝子の遺伝子移入は、当技術分野で公知の技術を用いて行うことができる。例えば、PMMoV抵抗性をコードする一つまたは複数の遺伝子、すなわちL4対立遺伝子を、従来の育種技術を用いて、任意で例えばマーカーを用いる分子スクリーニング法と併用して、PMMoVに対して非抵抗性であるまたは低い抵抗性レベルを保有するレシピエントトウガラシ植物に遺伝子移入することができる。
【0105】
ドナー植物から、PMMoVに対して非抵抗性であるまたは低い抵抗性レベルを保有するレシピエント植物にPMMoV抵抗性をコードする一つまたは複数の遺伝子を遺伝子移入するために、従来の育種技術を適切に用いることができる。これらの方法は、そのような種、品種、または栽培品種を互いに交雑することができる限り、種、品種、または栽培品種の従来の交雑を必然的に伴う。計画されたドナーおよびレシピエント種を限られた程度に互いに交雑することができる場合に限り、ブリッジ(bridge)種を用いてもよい。
【0106】
系統図育種と呼ばれる一つの従来の育種法において、PMMoVに対して抵抗性を示し、PMMoV抵抗性をコードするL4対立遺伝子を含む第一のトウガラシ植物を、PMMoVに対して非抵抗性であるか、またはPMMoVに対して低レベルの抵抗性を保有し、および耐病性、害虫抵抗性、貴重な果実特徴等のような、しかしこれらに限定されない作物栽培学上または商業的に望ましい特徴を示す第二のトウガラシ植物と交雑させる。次に、得られた植物集団(すなわち、F1)を自家受粉させて種子(F2種子)を産生させる。
【0107】
F2種子から生長させたF2植物を、PMMoVに対する抵抗性に関してスクリーニングする。集団を多数の異なる方法でスクリーニングすることができる。第一に、集団を、本明細書において抵抗性バイオアッセイとも呼ばれる従来の病理疾患スクリーニングを用いてスクリーニングすることができる。PMMoVに関するそのようなバイオアッセイは当技術分野で公知である。具体的には、インキュベーターまたは温室において、個々の植物またはその一部をPMMoVに曝露して各植物で得られた抵抗性または感受性表現型を採点する。限定されない例として、植物を、以下のように温室においてスクリーニングすることができる:トウガラシ植物をその栽培に関して一般的に適用され、当業者に公知である生長条件で、例えば昼間の温度22℃、夜間温度20℃で通常の夏の光の条件で温室においてF2種子から成長させることができる。F2植物を、例えば同様に、一つまたは複数の抵抗性植物および一つまたは複数の感受性植物のような、対照植物を含むトレイにおいて生長させてもよい。接種物として感染した葉を用いてもよく、これを凍結保存してもよい。試験ウイルスを含む接種物は、感染した葉を水ですりつぶして(例えば、水100 mlにおいて凍結した葉1 g)、カーボランダム(シリコンカーバイド)を加えることによって調製してもよい。試験植物の接種は、例えばスポンジを用いて植物の第一の真の葉の表面領域(例えば最小で3.5×2 cm)に接種物をこすりつけることによって行ってもよい。初回の観察後、肉眼的に見て感受性がある(肉眼的症状は全身性のモザイク)である植物を除去してもよい。1週間後の2回目の観察の際に、感受性のある抵抗性植物の数を計数してもよい。葉の試験に関して、葉(植物の上部から4〜5 cm)を、湿った濾紙を含むトレイに置いてもよく、葉を濾紙の上に載せる。感受性および抵抗性対照をトレイに加えて、トレイをプラスチックで包んでもよく、光をあてる時間16時間/日で20℃でインキュベートしてもよい。接種は先に記述したようにスポンジによって起こってもよく、評価は6〜8日後に行う。
【0108】
または、従来の育種法を分子スクリーニング法と組み合わせて行ってもよい。そのような方法は一般的に、マーカー支援選抜またはマーカー支援育種と呼ばれ、所望の形質をコードする一つまたは複数の遺伝子(すなわち、L4対立遺伝子、またはその切断型)を含む子孫植物を同定するために、一つまたは複数の分子マーカーを用いることを必然的に伴う。ゲノムの遺伝子組成は、植物の表現型を予測して、冗長なバイオアッセイは不要となる。マーカー支援選抜を用いて、バイオアッセイスクリーニングから得た結果を確認してもよい。PMMoV抵抗性表現型を示すF2ハイブリッド植物は、PMMoV抵抗性をコードする所望の遺伝子を含み、次に商業的に望ましい特徴をさらに保有する植物を選択して、トウガラシ植物をますます近交系にするために多数の世代にわたって自家受粉させる。そのような育種および選択の結果は、PMMoV抵抗性に関連した遺伝子と共に商業的に重要な形質に関連した他の遺伝子に関して遺伝的にホモ接合である系統の産生である。
【0109】
次に本発明は、以下の段階を含む、PMMoV病原型1.2.3に対する抵抗性を示すカプシクム属の植物を産生する方法を提供する:
a)PMMoV病原型1.2.3に対して感受性があるカプシクム属の第一の植物またはその一部を提供する段階;
b)該植物のゲノムにおけるL4抵抗性対立遺伝子の存在により、PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の第二の植物を提供する段階;
c)PMMoV病原型1.2.3に対して抵抗性を示す後代植物を産生するために該第一および第二の植物を交雑させる段階;ならびに
d)対立遺伝子が、PMMoV病原型1.2.3に対する抵抗性を該後代植物に付与する遺伝情報を含み、SNFD表現型を付与する遺伝情報が該対立遺伝子に、少なくともSNFD表現型が発現されない程度まで存在しない、切断型L4抵抗性対立遺伝子の存在に関して該抵抗性後代植物のゲノムをさらにスクリーニングする段階。
【0110】
本方法の好ましい態様において、段階d)におけるスクリーニングは、図3に示されるように2群マーカー、1/3群マーカー、および3群マーカーからなる群より選択される、好ましくは図3に示される2群マーカーから選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーの不在を評価することによって行われる。
【0111】
本発明の方法に従って、選択される商業的に望ましい特徴には、SNFD表現型の不在が含まれ、このことは、PMMoV病原型1.2.3に対して抵抗性ではない商業的カプシクム品種と比較して、植物が非分離で、稔性で、正常な生長を示すことを意味する。これらの特徴は、任意の適した方法によって植物において同定されてもよい。例えば、正常な生長特徴は、矮性生長の不在が正常な生長を示している生長成績の肉眼的検分によって同定してもよい。稔性は、種子および/または花粉の量の肉眼的検分によって評価してもよい。非分離表現型は、先に記述したように交雑(抵抗性×感受性)のF1種子に関する抵抗性スクリーニングの試験結果の評価によって評価してもよい。
【0112】
または、SNFD表現型の不在は、特異的マーカーの不在に関して植物をスクリーニングすることによって評価してもよい。本発明の植物および方法の一つの特に好ましい態様において、本明細書において記述される2群のマーカーの一つまたは複数が存在しない。このように、一つの好ましい態様において、PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法は、抵抗性植物からの分離集団の後代の少なくとも一つから、そのゲノムにおいて本明細書に記述される1群の一つまたは複数、好ましくは全てのマーカーが存在し、およびそのゲノムにおいて本明細書に記述される2群の一つまたは複数のマーカーが存在しない植物を選択する段階を含む。
【0113】
本発明の植物のもう一つの態様において、本明細書において記述される3群のマーカーの一つまたは複数が存在しない。本発明の植物のさらにもう一つの態様において、本明細書に記述される1/3群のマーカーの一つまたは複数が存在しない。この文脈において、1/3群のマーカーの一つまたは複数が存在しないことは、そのような情報が存在しないことによって、その植物のゲノムにおけるL4抵抗性対立遺伝子の存在によりPMMoV病原型1.2.3に対して抵抗性を示す植物が得られないであろうことから、PMMoV病原型1.2.3に対する抵抗性に関与するゲノム領域に存在しないマーカーのみに関連すると理解すべきである。したがって、1/3群の一つまたは複数のマーカーの不在は、好ましくは、3群のマーカーの領域に対応する領域に存在するマーカーのみに関連し、好ましくは1群のマーカーの領域には関連しない。最も好ましい態様において、L4対立遺伝子は、一つまたは複数の2群マーカーが存在しないように切断される。この場合、切断はセントロメア側であるが、3群のマーカーの切断の場合、切断はテロメア側である。もう一つの最も好ましい態様において、切断は、TG036(Lefebvre et al., 2002;Ben-Chaim et al., 2001)マーカー(2群マーカーのメンバー)が存在しないように起こる。
【0114】
このように、L4対立遺伝子を提供された植物におけるSNFD表現型の不在は、本明細書において切断型L4対立遺伝子とも呼ばれるL4対立遺伝子を含むC.カコエンス遺伝子移入の切断型を植物に導入することによって実現してもよい。切断の程度は、当業者によって容易に決定することができる。例えば、1/3群のマーカーを含む領域が除去された場合に、PMMoV病原型1.2.3に対する抵抗性に関与する情報を有しないそれらの部分のみが除去される限り、トランスジェニック技法または組換えおよび選択のプロセスのいずれかによる、2群、3群、および/または1/3群のマーカーを含むそれらの遺伝子移入領域の任意の一つの部分的または完全な除去によって、適したレベルの切断を行うことができる。好ましくは、該切断は、2群のマーカーを含むL4対立遺伝子を含むC.カコエンス遺伝子移入領域の部分的または完全な除去を必然的に伴う。このように、トランスジェニック技法または組換えおよび選択のプロセスのいずれかによって、一つまたは複数の2群マーカーを含む領域を除去すると、もはやSNFD表現型を示さない植物が得られるであろう。最も好ましくは、この切断によって、TG036マーカーの除去が得られる(Lefebvre et al., 2002;Ben-Chaim et al., 2001)。
【0115】
切断レベルはまた、遺伝距離として定義してもよい。遺伝距離は、同じ染色体上の座のあいだの交差頻度によって測定される。二つの座が離れているほど、それらのあいだで交差が起こる可能性がより高くなる。逆に、二つの座が互いに近ければ、それらのあいだで交差が起こる可能性はより低い。座は、本明細書において定義される様々なマーカー群のマーカーによって示されてもよい。規則として、1センチモルガン(cM)は、座(マーカー)のあいだの1%組換えに等しい。それによって2群、3群および/または1/3群の一つまたは複数のマーカーの不在をもたらす切断レベルは、適切に0.001〜10 cM、より好ましくは0.01〜10 cMである可能性がある。または、遺伝子移入の存在を示すマーカー(例えば、1群の一つまたは複数のマーカー)と、切断されているマーカーとの遺伝距離は、適切に0.001〜10 cM、より好ましくは0.01〜10 cMであってもよい。
【0116】
新規かつ優れたPMMoV抵抗性近交系トウガラシ植物系統はまた、反復選択技術および戻し交雑技術を用いて開発することができる。この方法において、戻し交雑親を第一のドナー植物(戻し交雑親とは異なり、本明細書において「非戻し交雑親」と呼ばれる)と交雑させることによって、L4抵抗性を標的レシピエント植物(戻し交雑親と呼ばれる)に遺伝子移入することができる。戻し交雑親は、PMMoVに対して非抵抗性であるか、または低レベルの抵抗性を保有し、かつ耐病性、害虫抵抗性、貴重な果実特徴等のような、しかしこれらに限定されない商業的に望ましい特徴を保有し、SNFD表現型が存在しない植物である。戻し交雑技法は、「ドナー親」からの遺伝子を作物栽培学上の価値が高い遺伝的バックグラウンドに交雑するために広く応用されている。一般的に、優性遺伝子の栽培学上許容される表現型への遺伝子移入は、戻し交雑3〜5回の後に、例えば2〜3回の自家受粉によって得ることができ、全ての段階で栽培学上の価値に対する選択が行われる。非戻し交雑親は、PMMoV抵抗性を示し、L4抵抗性対立遺伝子を含む。非戻し交雑親またはドナー植物は、戻し交雑親または標的レシピエント植物と他家受精される任意の植物品種または近交系となりうる。戻し交雑親と非戻し交雑親との交雑によって得られた後代を、戻し交雑親と戻し交雑する。次に、得られた植物集団をスクリーニングする。集団は、多数の異なる方法でスクリーニングすることができる。第一に、集団は、本明細書において先に記述したように従来の病理スクリーニングを用いてスクリーニングすることができる。
【0117】
第二に、L4抵抗性対立遺伝子を含み、SNFD表現型に関連するマーカーの一つまたは複数が存在しない後代を同定するために、本明細書において先に記述した分子マーカーの一つまたは複数を用いて、マーカー支援選抜を行うことができる。または、マーカー支援選抜を用いて、抵抗性バイオアッセイスクリーニングから得られた結果を確認することができる。
【0118】
適切な選択を行った後、プロセスを繰り返す。戻し交雑親に対する戻し交雑、ならびにPMMoV抵抗性に関しておよびSNFD表現型(またはそれに関連したマーカー)の不在に関して選択するプロセスは、約5世代以上繰り返される。このプロセスから得られる後代は、L4対立遺伝子に関してヘテロ接合である。次に、L4対立遺伝子付与PMMoV抵抗性を含み、SNFD表現型を欠損するホモ接合の純粋な育種後代を提供するために、最後の戻し交雑世代を自家受粉させる。
【0119】
L4対立遺伝子に関してホモ接合であって、SNFD表現型を発現しない本明細書において記述されるPMMoV抵抗性近交系トウガラシ系統は、PMMoV抵抗性ハイブリッド植物を作製するためにさらなる交雑に用いることができる。例えば、第一のPMMoV抵抗性近交系トウガラシ植物を、耐病性、害虫抵抗性、望ましい果実特徴等のような、しかしこれに限定されない商業的に望ましい形質を保有する第二の近交系トウガラシ植物と交雑させることができる。この第二の近交系トウガラシ系統はPMMoVに対して抵抗性であってもなくてもよい。
【0120】
本発明の局面において用いられる選択技法は、植物からの核酸試料、好ましくは当技術分野において周知の核酸単離技法を用いて得られる可能性があるような、DNAの供給、およびマーカーの有無に関して該核酸の配列を試験することを必然的に伴う可能性がある。本明細書において定義されるマーカーは、その定義に至るAFLP反応を行うことによって、またはプライマーがその特異的鋳型にアニールすることができ、特異的増幅産物を産生することができる条件で、マーカー特異的プライマーの一つまたは複数の組によるPCRのような増幅反応を行うことによって、検出される可能性があるAFLPマーカーである。特異的増幅産物の存在は、鋳型配列におけるマーカーの存在を示す一方、増幅産物の不在はマーカーの不在を示す。L4遺伝子移入(L4対立遺伝子)の存在を確認するために、1群の一つまたは複数、好ましくは全てのマーカー、最も好ましくは本明細書において記述されるマーカーE58/M50-F-580および/またはマーカーE54/M55-F-101を核酸試料において検出する。適切に切断された対立遺伝子の存在を確認するために、好ましくは本明細書において記述される2群の一つまたは複数のマーカーが存在しない。AFLPマーカーのいくつかはまた、配列タグ部位マーカーに変換されてもよく、これによって交雑の後代の迅速かつ簡便なスクリーニングが可能となる(Werner et al., 2001)。
【0121】
または、植物試料のゲノムにおけるL4マーカーの有無に関してスクリーニングする方法は、プローブが、標的ポリヌクレオチド配列と選択的に結合して、安定なハイブリダイゼーション複合体を形成する条件で試料に接触される、一つまたは複数のマーカー配列を含む染色体領域上の標的ポリヌクレオチド配列に選択的に結合するプローブに、植物からの核酸試料を接触させる段階、およびハイブリダイゼーション複合体の有無を検出して、それによって植物における該マーカーの有無に関してスクリーニングする段階を含んでもよい。
【0122】
本明細書において先に記述した方法において用いられるマーカー支援選抜は、例えば、段階的に行うことができ、それによってL4対立遺伝子の存在およびSNFD表現型の不在が1代より多い世代において選択される。L4対立遺伝子付与抵抗性に関するマーカー支援選抜は、耐病性、害虫抵抗性、望ましい果実特徴等のような他の商業的に望ましい形質に関して試験および選択する前、同時、または後に行ってもよい。同様に、PMMoV抵抗性またはSNFD表現型の不在が評価される順序は、特に制限的ではない。SNFD表現型を付与する遺伝情報を含む植物の(好ましくはマーカー支援)排除は、PMMoV抵抗性植物の(好ましくはマーカー支援)選択前に、または同時に行ってもよいと理解すべきである。
【0123】
L4対立遺伝子が天然において周囲の環境から単離され、L4抵抗性を付与する遺伝情報のみが得られ、SNFD表現型を付与する遺伝情報を得ない材料源は、例えばC.カコエンスアクセッション番号PI 260429またはSA 185(Boukema, 1983)植物に由来する。そのようなアクセッション番号は、例えばCentre for Genetic Resources, the Netherlands(CGN), Wageningen, The NetherlandsのPlant Genetic Resources(PGR)クラスタから入手可能である。
【0124】
本発明に従う切断型L4抵抗性対立遺伝子を含む単離された核酸配列は、例えば、PMMoV病原型1.2.3に対して抵抗性である植物からDNAを単離すること、および該DNAからL4抵抗性対立遺伝子を単離することによって産生されてもよい。そのような単離は、例えば該L4対立遺伝子のPMMoV抵抗性付与部分を含む遺伝子領域を、例えば適したフォワードおよびリバースプライマーを用いて増幅することによって行ってもよい。適したプライマーは、例えば、3群のマーカーの定義のために用いられるマーカーと共に、2群のマーカーの定義のために用いられたプライマーである。増幅断片の大きさは、あまり離れていない、例えば1群のマーカーの位置により近い、C.カコエンスゲノムの位置においてアニールする他のプライマーを用いるまたは開発することによって低減させることができる。このようにして、SNFD表現型を付与する遺伝情報がL4対立遺伝子には存在しないことを達成することができる。当業者は、カプシクム属の植物において発現されるためにそのような核酸において存在しなければならず、それによって該植物にPMMoV病原型1.2.3に対する抵抗性を付与する様々な調節要素を承知している。
【0125】
L4対立遺伝子またはL4遺伝子は、例えば当技術分野で周知の標準的なDNA単離法を用いて、カプシクム植物からのゲノムDNAの試料を提供することによって単離してもよい。大きいDNA断片を検索するためには、単離体の質が比較的高いこと、すなわちゲノムDNAの重度の分解が本質的に回避されることが必要である。一般的に、高分子量(HMW)DNA調製物を得るための単離技術を用いる場合には、十分な大きさのDNAを得てもよい。HMW DNAは、調製の際に物理的剪断から保護されて得られるDNAである。カプシクム植物組織からのHMW DNAの調製は、例えば、細胞壁加水分解を用いてプロトプラストを単離する段階、およびアガロースにプロトプラストを抱埋する段階を必然的に伴ってもよい。もう一つの方法は、植物の核からHMW DNAを調製することである。メガベースDNAの単離は、多数の植物種に関して首尾よく行われ、そのような方法は、細菌の人工染色体(BAC)および酵母人工染色体(YAC)クローニングベクターにおいて大きいインサートDNAライブラリを構築するために適したメガベースの大きさのDNAを提供するために一般的に適用される。ゲノムDNAを単離した後、例えばPCR増幅反応において用いられる制限酵素を用いることによって、処置してもよく、または本明細書において提供されたマーカー配列のいずれかによってクローニングもしくはプロービングしてもよい。
【0126】
本発明はさらに、PMMoV感染症に対する抵抗性を示すトランスジェニック植物を提供するために、当技術分野において公知の技術を用いて、そのような単離および精製された遺伝子(または対立遺伝子)のトウガラシまたは他の植物への挿入を企図する。植物の形質転換は、植物細胞において機能するであろう発現ベクターの構築を必然的に伴う。本発明において、そのようなベクターは、プロモーターのような調節要素の制御下である、または機能的に連結したPMMoV抵抗性をコードする遺伝子を含むDNAを含む。発現ベクターは、組み合わせに含まれる遺伝子の少なくとも一つがPMMoV抵抗性をコードする限り、一つまたは複数のそのような機能的に連結した遺伝子/調節要素の組み合わせを含んでもよい。ベクターは、プラスミドの形であってもよく、アグロバクテリウム(Agrobacterium)形質転換系のような当技術分野において公知の形質転換法を用いて、PMMoVに対して抵抗性であるトランスジェニック植物を提供するために、単独で、または他のプラスミドと併用して用いることができる。
【0127】
発現ベクターには、マーカーを含む形質転換細胞を負の選択(選択マーカー遺伝子を含まない細胞の増殖を阻害することによって)、正の選択(遺伝子マーカーによってコードされる産物に関してスクリーニングすることによって)によって回収させる調節要素(プロモーターのような)に機能的に連結した少なくとも一つの遺伝子マーカーが含まれうる。植物の形質転換に関して多くの一般的に用いられる選択マーカー遺伝子は、当技術分野において公知であり、これには例えば抗生物質もしくは除草剤であってもよい選択的化学物質を代謝的に解毒する酵素をコードする遺伝子、または阻害剤に対して非感受性である変更された標的をコードする遺伝子が含まれる。マンノース選択のようないくつかの陽性選択法が当技術分野において公知である。または、マーカーを含まない形質転換を用いて先に述べたマーカー遺伝子を有しない植物を得ることができ、その技術は当技術分野において公知である。
【0128】
発現ベクターを植物に導入するための一つの方法は、アグロバクテリウムの天然形質転換系に基づく(例えば、Horsch et al., 1985を参照されたい)。A.ツメファシエンス(A. tumefaciens)およびA.リゾゲネス(A. rhizogenes)は、植物細胞を遺伝的に形質転換する植物病原性土壌細菌である。A.ツメファシエンスおよびA.リゾゲネスのTiおよびRiプラスミドはそれぞれ、植物の遺伝子形質転換に関与する遺伝子を有する(例えば、Kado, 1991を参照されたい)。植物組織に発現ベクターを導入する方法には、アグロバクテリウム・ツメファシエンスによる植物細胞の直接感染または同時培養が含まれる(Horsch et al., 1985)。アグロバクテリウムベクター系およびアグロバクテリウム媒介遺伝子移入のための方法の記述は、Gruber and Crosby, 1993およびMoloney et al., 1989によって提供される。同様に、米国特許第5,591,616号を参照されたい。植物発現ベクター、レポーター遺伝子、および形質転換プロトコールに関する一般的な記述、アグロバクテリウムベクター系に関する記述、ならびにアグロバクテリウム媒介遺伝子移入のための方法は、Gruber and Crosby, 1993において見いだされうる。植物組織を培養する一般的な方法は、例えば、Miki et al., 1993およびPhillips et al., 1988によって提供される。分子クローニング技術および適した発現ベクターに関する適切な参照ハンドブックは、Sambrook, J., E.F. Fritsch, and T. Maniatis,「Molecular Cloning : a Laboratory Manual」(3rd edition), Cold Spring Harbor Laboratory Press, Plainview, N.Y., 2000である。
【0129】
発現ベクターを植物に導入するためのもう一つの方法は、DNAがマイクロプロジェクタイルの表面において運ばれるマイクロプロジェクタイル媒介形質転換に基づく。発現ベクターは、植物細胞壁および膜を貫通するために十分である速度300から600 m/sでマイクロプロジェクタイルを加速する微粒子銃装置によって植物組織に導入される(Sanford et al., 1987, 1993;Sanford, 1988, 1990;Klein et al., 1988, 1992を参照されたい)。植物にDNAを導入するためのもう一つの方法は、標的細胞の超音波処理による(Zhang et al., 1991を参照されたい)。または、リポソームまたはスフェロプラスト融合が、発現ベクターを植物に導入するために用いられている(例えば、Deshayes et al., 1985およびChristou et al., 1987を参照されたい)。CaCl2沈殿、ポリビニルアルコール、またはポリ-L-オルニチンを用いるプロトプラストへのDNAの直接取り込みも同様に報告されている(例えば、Hain et al., 1985およびDraper et al., 1982を参照されたい)。プロトプラストならびに細胞および組織全体の電気穿孔も同様に記述されている(D'Halluin et al., 1992およびLaursen et al., 1994)。
【0130】
トウガラシ標的組織の形質転換後、上記の選択マーカー遺伝子の発現によって、当技術分野において現在は周知である再生および選択法を用いて、形質転換された細胞、組織、および/または植物の選択的選択が可能となる。
【0131】
前述の形質転換法は、ナス科の種のような、しかしこれに限定されないトランスジェニックトウガラシ植物または他の植物種を産生するために用いることができるであろう。先に述べたように、本発明の様々な局面は、原則的に、PMMoVによって影響を受けうる全ての植物に関し、このように、本発明に従うトランスジェニック植物はPMMoVの宿主範囲内の任意の植物種において産生される可能性がある。
【0132】
次に、PMMoV感染に対して抵抗性であるトウガラシまたは他の植物種のトランスジェニックハイブリッドを産生するために、そのようなトランスジェニック植物をもう一つの(非形質転換または形質転換)植物と交雑させることができる。または、本明細書に記述の形質転換技術を用いてPMMoV抵抗性をコードする外来(ヘテロ接合またはホモ接合)遺伝子を含むように操作されているトランスジェニックトウガラシまたは他の植物種におけるPMMoV抵抗性に関する外来の(異種)遺伝子を、当技術分野で周知である従来の育種技術(戻し交雑のような)を用いてもう一つの植物に移入することができるであろう。例えば、本明細書において先に考察したように、戻し交雑は、PMMoV抵抗性をコードするが、SNFD表現型に関与する遺伝情報を含まない外来(異種)遺伝子を含むトランスジェニックPMMoV抵抗性近交系トウガラシまたは他の植物系統から、その遺伝子を含まない非抵抗性トウガラシ植物または他の穀物にPMMoV抵抗性を遺伝子移入するために、またはPMMoV抵抗性をコードする外来遺伝子を含むトランスジェニックハイブリッドPMMoV抵抗性トウガラシ植物または他の植物から、その遺伝子を含まない系統に遺伝子移入するために用いることができるであろう。
【0133】
もう一つの態様において、プロトプラスト融合は、優れた新しいPMMoV抵抗性植物を作製するために用いることができる。より具体的には、第一のプロトプラストを、PMMoVによる感染に対して抵抗性を示すが、SNFD表現型に関与する遺伝情報を含まないトウガラシ植物または他の植物から得ることができる。第二のプロトプラストは、耐病性、害虫抵抗性、望ましい果実特徴等のような、しかしこれらに限定されない商業的に望ましい特徴を含む第二のトウガラシまたは他の植物品種から得ることができる。次に、プロトプラストを当技術分野において公知の従来のプロトプラスト融合技法を用いて融合させる。例えば、プロトプラスト融合は、膜の融合を促進するためにポリエチレングリコール(PEG)溶液を用いることによって行うことができる。そのような体細胞ハイブリダイゼーションは、種間ハイブリッドまたはその改変を産生するために、Sundberg et al., 1986によって開示された条件下で行ってもよい。しかし、当業者は、ポリエチレングリコール(PEG)を用いる以外の他の方法においてプロトプラスト融合を行うことができると認識するであろう。例えば、プロトプラストは、Koop and Spangenberg, 1989によって記述されるように電場誘導融合技術を用いて融合させることができる。さらに、プロトプラスト融合はデキストランおよびポリビニルアルコールによって行うことができる。プロトプラスト融合のためのさらなる方法は、Gleba et al., 1984およびDodds and Roberts, 1995において見いだされる可能性がある。
【0134】
例として、限定的ではなく、本発明の例を以下に示す。
【実施例】
【0135】
実施例
Sylvia(S & G品種「Cuby」の近交系)の雄性の親と、Manito(S & G品種「Tasty」の近交系)の雄性の親と呼ばれる二つの近交系親系統を産生した。いずれの品種SylviaおよびManitoも、PMMoV抵抗性に関して最初に分離した。植物の選択および試験交雑によって、双方の品種において抵抗性を固定することが可能であった。しかし、不安定な雄性不稔性/稔性不良および矮性生長のような形質が、双方の品種の雄性の親に存在することが認められた。
【0136】
これらの実施例において用いられるPMMoV病原型1.2.3は、Boukema, 1982によって言及された単離体P14と類似の特徴を有するウイルス単離体である。以下の実施例において、様々な実験において用いられたウイルス単離体は、PTG(Proefstation voor Tuinbouw onder Glas)、現在はPPO、Naaldwijk, The Netherlandsから得られ、これを機械的に繁殖させた。
【0137】
実施例1.同じ品種内の抵抗性および感受性トウガラシ植物におけるL4抵抗性マーカー
L4座の連鎖マップは、Keygene(Wageningen, The Netherlands)によって産生された。L座がマッピングされている領域は第11染色体南のテロメアに存在した(Lefebvre et al., 2002)。
【0138】
この座の周囲のマーカーは、四つの群に分けることができるであろう。L4座に連鎖した異なる群の位置を図1に示す。各群は、多くのマーカーを含み、各群の中に組換えは見いだされなかった。連鎖マップの染色体の残りに向けての方向は不明である。各群に存在するマーカーを表1に示す。
【0139】
(表1)それぞれの群におけるマーカーの概要
【0140】
分離品種Manito およびSylviaから、F1植物と共に抵抗性親株を、先に記述した方法と全く同様にバイオアッセイにおいて試験した。全体として、トレイは、感受性対照1個および抵抗性対照1個を含む試験植物35個からなった。幹および葉の試験のバイオアッセイは、本質的に先に記述したとおりに行った。バイオアッセイの結果を表2に示す。
【0141】
(表2)品種SylviaおよびManitoならびにその抵抗性親系統におけるバイオアッセイの結果
【0142】
表2に示すバイオアッセイから、抵抗性植物3個および感受性植物3個を、分離品種ManitoおよびSylviaの双方のF1植物から無作為に選択すると共に、その対応する抵抗性の雄性の親から植物5個を無作為に選択した。選択した植物を、L4座に連鎖した、表1に記載する4個のマーカーによってさらに分析した。試験が行われた当時、必ずしも表1に記載された全てのマーカーが同定されていたわけではない。それぞれの品種の中で、抵抗性植物3個および感受性植物3個を、様々なマーカーの存在に関して試験することによって分析した。対応する抵抗性の親から、植物5個を分析した。マーカー分析の結果を表3に記載する。
【0143】
(表3)バイオアッセイから選択された植物のマーカースコア(優性、本文を参照されたい)
番号はマーカー試験における試験番号に対応する;R、抵抗性;S、感受性;+、存在;−、不在。
【0144】
表3から認められうるように、L4対立遺伝子に連鎖したマーカーは全て、バイオアッセイ(R)において表現型の抵抗性を示すF1植物に存在した。しかし、感受性植物(S)では、L4マーカーはいずれも存在しなかった。Manito(番号29〜33)およびSylvia(番号40〜44)の抵抗性の雄性の親は、バイオアッセイにおいて分離を示さず、L4対立遺伝子に連鎖した全てのマーカーがこれらの植物において存在した。マーカーは、単に存在または不在(優性)として採点されたことから、遺伝子移入セグメントがホモ接合で存在するか、ヘテロ接合で存在するかを結論付けることはできなかった。F1における感受性はL4遺伝子移入の不在によって引き起こされると結論付けられた。さらなる試験から、L4抵抗性の遺伝における母株の効果が存在しないことが判明した。
【0145】
実施例2.非分離Sylvia品種の開発
非分離品種Sylviaを開発するために、抵抗性親系統(系統4578)の個々の植物を感受性の親植物と交雑した。L4抵抗性の分離の発生を、得られた個々のF1集団において評価した。結果を表4に記載する。
【0146】
植物番号4578-8を自家受粉させることによって、系統6636を得た。系統6636の個々の植物は、これらの植物が異なる(しかし関連する)感受性の雌性系統における雄性系統として用いられたことから1回より多く記載される。
【0147】
系統4578の植物はいずれも完全に抵抗性のF1を生じなかった。しかし、系統4578(vide系統6636)を自家受粉させることによって、抵抗性遺伝子を100%転移した株を発見することが可能であった。10例中4例において、これによって完全に抵抗性のF1が得られた。植物番号6636-4に由来するF1の分離は、他の系統とは異なるパターンを示す。表4に示すように、分離するF1集団(2R/3S−3R/2S)における抵抗性植物の数には変動がある。分離は、分離するF1集団における抵抗性植物と感受性植物の数を合わせると、より信頼できると期待される。全てのF1集団において同じ抵抗性起源が用いられていることから、非安定(分離)抵抗性の原因は同じである。表8は、表4において言及した三個の交雑(4578、6636交雑1、6636交雑2;6636の2回の交雑は、異なる二つの感受性の親を同一の抵抗性植物と交雑した結果である)のそれぞれの編集した結果を示す。
【0148】
(表4)個々の植物に由来するSylviaのF1分離
表4におけるF1集団を産生するために用いられる系統6636の個々の植物の自家受粉を、PMMoV抵抗性に関しても試験した。自家受粉した系統は全て完全な抵抗性を示した。結果を表5に記載する。
【0149】
(表5)親系統からの自家受粉の結果
【0150】
この実験から、L4対立遺伝子の望ましくない特徴が明確になり、非分離子孫をもたらす個々の植物の自家受粉ではホモ接合性が確認できるが、個々の抵抗性植物と感受性植物との交雑は分離F1子孫をもたらし、ヘテロ接合性が確認できると結論付けることができる。
【0151】
実施例3:非分離Manito品種の開発
実施例2における記述と類似のアプローチに従って非分離Manito株を開発した。抵抗性親系統の植物を感受性の親植物と個々に交雑させて、個々のF1集団における分離を評価した。結果を表6に記載する。低い稔性(SNFD表現型)を示す植物は全て交雑の前に除去し、これらの植物を表6において「試験していない」として記載する。低い稔性は当初、植物番号0025-5では観察されなかったが、後に存在するように思われた。
【0152】
(表6)個々の植物に由来するManitoのF1分離(初回スクリーニング)
【0153】
植物0025-5のみが非分離ハイブリッドを生じた。全体で60株中14株が稔性が低い(SNFD表現型)と印された。この表におけるF1集団の分離も同様に表8において編集する。
【0154】
非分離F1が得られる抵抗性植物の数を増加させるために、2回目の試験を行った。稔性が低いとして印をつけられた植物を交雑前に除去しなかった。結果を表7に記載して、分離を同様に、表8において編集する。全体で50株中12株(25%)が完全に抵抗性のF1集団を生じた。
【0155】
(表7)個々の植物に由来するManitoのF1分離(2回目のスクリーニング)
【0156】
(表8)実施例2および3における分離F1の編集結果
【0157】
全ての場合において同様に、感受性植物より抵抗性植物の数が多い。4578および6636-交雑1による分離は、1:1に近く、抵抗性の親がヘテロ接合である一遺伝子モデルに適合する。6636-交雑2および2825-2834において、分離は9:7により近い。9:7の分離は、F2集団に由来する。二つのホモ接合系統(系統は抵抗性に関して分離しない)を交雑することによって、F1において9:7の分離を得ることは可能ではない。ここでも、1:1分離が存在する可能性が高い。0021-0026の分離は、他とは異なり、3:1分離に最善に適合する。おそらくこの差は、植物が稔性に関して選択され、稔性の低い植物は交雑を行う前に除去されたという事実によって引き起こされる。
【0158】
実施例4.L4対立遺伝子の観察された非メンデル遺伝の説明としての連鎖ドラグ(drag)
緒言
表8において交雑のために用いた植物の中で、稔性および矮性の生長(SNFD表現型)を採点した。非分離F1をもたらす植物は全て、低い稔性を示した。これは、低い稔性と非分離との連鎖を示している。表7に示す結果は、植物の約25%が非分離ハイブリッドを生じることを示した。表6において、植物の25%は、稔性が低いと印され、F1の評価の前に除去された。稔性の低い植物が良好な抵抗性移入親植物であると見なした場合、表6および表7の結果は同等である。本明細書において言及されるように、これらの非分離植物に由来する近交系は実際に矮性生長を示した。このことから、負の形質(不安定な抵抗性、低い稔性、矮性生長)は、L4対立遺伝子に密接に連鎖している、または多面発現性であると結論付けることができる。
【0159】
F1における分離の性質に基づいて、実施例1〜3における観察に関して可能性がある説明を系統立てて、試験した。致死遺伝子が感受性対立遺伝子に連鎖する可能性、または致死遺伝子が抵抗性対立遺伝子に連鎖する可能性を試験したが、否定された。遺伝子サイレンシングの可能性、L4対立遺伝子の発現に関する第二の遺伝子の必要性、またはL4対立遺伝子の移入に関する第二の遺伝子の必要性も同様に否定された。さらに、トランスポゾンまたはインプリンティングの存在に関する証拠は見いだされなかった(遺伝子は、種子発達の際の特異的条件により沈黙化される)。さらに、抵抗性形質が一遺伝子性ではない可能性は、試験スキームの作製後に除外された。C.カコエンスの遺伝子移入セグメントによってC.アンヌーム染色体の染色体フォールディングの問題が起こるか否かをその後試験した。不規則なフォールディングは、染色体不均衡をもたらし、ホモ接合(L4L4)植物の減数分裂において問題を引き起こすであろう。次に、染色体対形成の際に、染色体の一つが損傷を受けて、それによって「-」対立遺伝子と呼ばれる欠損対立遺伝子が起こりうる。次に、これによって、抵抗性の喪失をもたらすであろう。分離F1において感受性植物ではマーカーが見いだされないことを考慮すると、染色体が切断されている可能性が高い。欠損対立遺伝子のホモ接合の存在が致死的である場合、L4遺伝子型の自家受粉によって、実際にバイオアッセイにおいて完全な抵抗性系統が得られるであろう。同様に、非分離F1は、このように誘導することができ、L4遺伝子型を感受性系統と交雑すると、F1は1:1の比で分離するであろう。ホモ接合L4遺伝子型(L4L4)を有する植物は、L4遺伝子型を自家受粉した後に見いだされうるが、これらの植物は減数分裂において問題に遭遇するであろうと予測される。次に、これは、バイオアッセイにおいて試験した場合に系統が完全に抵抗性である(すなわち、試験した全ての植物が抵抗性である)理由、および感受性の親と交雑した場合に分離F1集団が得られることがなおも見いだされる理由を説明するであろう。同様に、マーカーが、感受性があるF1集団の植物において見いだされない理由を説明するであろう。
【0160】
この説明は、非分離F1をもたらすL4L4遺伝子型が、染色体フォールディングの再配列の結果であることを予測する。この再配列フォールディングの状況において、染色体の対形成によって、染色体のアームの切断は起こらないであろう。しかし、植物内の他のプロセスが破壊される可能性はより高く、それによって低い稔性および低減された生長(矮性植物)をもたらす。これらの植物に由来する花粉の量はごく限られていることは公知である。同様に、そのような系統に由来する種子の量は少ないことが報告されている。
【0161】
したがって上記の実験から、L4対立遺伝子を含むC.カコエンスのゲノムの断片が大きすぎて、分離F1集団を引き起こすと結論された。したがって、このセグメントの大きさを減少させれば、問題を解決することができるであろう。
【0162】
実験のセットアップ
L4座の様々なマーカーの存在を、多数のトウガラシ系統において評価した。第一に、二つの抵抗性C.アンヌーム系統M-873およびM-3751を、遺伝子移入セグメントの大きさに関して評価した。これらの系統は、それによって非分離F1および正常な稔性をもたらすことが公知であった。セグメントの大きさを、L4対立遺伝子に隣接するマーカーに関してスクリーニングすることによって、S & G近交系(Manitoの父株)において存在する遺伝子移入と比較した。マーカーの位置に関しては、図2を参照されたい。結果を表9に記載する。
【0163】
系統を、L4座に連鎖したマーカー4個において試験し、L4対立遺伝子に対するマーカーの位置を表1に示す。表9に記載したマーカーは、ホモ接合が存在する、ヘテロ接合が存在する、または存在しないと採点される。同様に、抵抗性親系統の稔性およびF1の抵抗性を記載する。稔性の評価は花粉の量を「限られた量の花粉」(-:稔性不良)、「中間量の花粉」(+/-:中間の稔性)、および「非抵抗性の稔性対照植物と類似の花粉量」(+:良好な稔性)のように採点することによって、肉眼で行った。
【0164】
系統M-873およびM-3751における抵抗性は、De Ruiter Seeds, Bergschenhoek, The Netherlandsのトウガラシ育種系統に由来した。他の起源は全て、L4ドナーとしてS & G近交系を共有した。系統M-873およびM-3751のうち、良好なおよび中間の稔性を有する植物を、マーカーの存在または不在に関してスクリーニングした。中間の稔性は全ての場合において、Manitoの父系統における稔性より良好であった。他の系統は、稔性に基づいて選択された育種系統であった、またはそれらによって非分離ハイブリッドが起こったためであった。
【0165】
(表9)いくつかのトウガラシ系統におけるC.カコエンス遺伝子移入の評価
++:ホモ接合マーカーが存在する、抵抗性;-:ホモ接合マーカーが存在しない、感受性;+/-:ヘテロ接合マーカーが存在する、抵抗性;+:マーカーが存在する、対立遺伝子の差は試験していない;?:試験していない。
【0166】
結果は、系統M-873およびM-3751において、より小さい遺伝子移入セグメントが存在することを示している。2群のマーカー(E39/M58-F-65)は存在しない。これらの系統の系図を評価することによって、いずれも、vp-nr 91Pa0424、F3T13 7310を共有した。
【0167】
系統8972、8609、および8613の植物は、バイオアッセイにおいて完全な抵抗性スコアを示したにもかかわらず、ヘテロ接合スコアを生じた。株8972に関して、交雑を行った。結果を表10に記載する。
【0168】
(表10)表9のヘテロ接合スコア系統8972に由来するF1集団の結果
【0169】
系統8972から得られたF1集団の結果は、期待通りであった。さらなる組換え体を同定するために、分離系統を、L4-座マーカーの存在に関して分析した。3:1に最も近いF3分離に基づいて株を選択した。F1からF2への自家受粉の際に組換えが起こるはずであることから、それぞれのF3集団の植物をプールした。組換えが起こった場合、F3集団は組換えに関してホモ接合でなければならない。選択された株を、表11に記載する。
【0170】
(表11)組換え体同定のために選択された系統
これらの植物に関するマーカー分析の結果を表12に記載する。L4座のマーカーを表1に示すように示す。マーカースコア「+」は、マーカーの存在を示し、マーカースコア「-」は不在を示す。
【0171】
(表12)選択されたL4抵抗性および非SNFD表現型系統のマーカースコア
【0172】
L4抵抗性対立遺伝子のマーカー(1群マーカー)を含むこれらの選択系統において新しい組換え体は見いだされなかったが、2群のマーカーの位置を含む遺伝子材料を含まなかったと結論付けることができる。これらの系統は抵抗性であるが、SNFD表現型は観察されないことが見いだされた。したがって、2群のマーカー、または2群および3群のマーカーの位置を含む遺伝子材料の喪失に至る組換えによって、SNFD表現型の喪失をもたらすと結論付けられた。
【0173】
参照文献
【技術分野】
【0001】
発明の分野
本発明は、耐病性植物に関する。より詳しく述べると、本発明は、トウガラシマイルドモットルウイルス(PMMoV)の特異的病原型に対して抵抗性であるトウガラシ植物およびそのような植物を産生する方法に関する。
【背景技術】
【0002】
発明の背景
カプシクム(ナス科またはイネホオズキ科)は、植物の属であり、その甘く芳香のある果実は、スパイス、野菜、および薬剤として用いられている。この属は約40種を含む。ほとんどの品種はカプサイシン(メチルバニリルノネナミド)を含み、これは口の中で強い焼けるような感覚を生じ、薬剤における循環刺激剤および疼痛軽減剤として用いることができる刺激性の化学物質である。植物は中央アメリカおよび南アメリカ原産であるが、それらはほぼあらゆる気候に耐えることから、果実は世界中で産生されている。市販のトウガラシは主に、カプシクム・アンヌーム(Capsicum annuum)(ピーマン、カイエンペッパー、ハラペーニョペッパー、アナハイムペッパー)、キダチトウガラシ(Capsicum frutescens)(タバスコペッパー)、およびカプシクム・キネンセ(Capsicum chinense)(ハバネロペッパー)種である。
【0003】
パプリカまたはピメントとしても知られるカプシクム・アンヌームは、栽培品種によって長さ、色、および辛みが多様である果実を有する草本の一年草である。種は、世界中で、例えば西ヨーロッパおよびアメリカ(USA)において栽培されている。キダチトウガラシおよびC.キネンセは小さい極めて辛みの強い果実を有し、タバスコおよび他のホットペッパー製品において用いられている。強い芳香のためにはかなり暖かい気候が必要であることから、これらの種は、主に熱帯地域およびアメリカのより暖かい地域で栽培されている。
【0004】
カプシクム種の野菜は、栽培のあいだ多くのトバモウイルスによって障害を受ける。トバモウイルス属には、タバコモザイクウイルス(TMV)のタイプ種、および血清学的に関連するトマトモザイクウイルス(ToMV)、トウガラシマイルドモットルウイルス(PMMoV)およびいくつかの他の植物ウイルスが含まれる。ウイルス感染症は植物の活力を低減させうるが、一般的に植物を殺すことはない。例えば、トウガラシ、トマト、およびナスを含む広範な植物に影響を及ぼすTMVは、トウガラシの果実に重度の壊死を引き起こして、果実のほとんどを市場に出せなくする。TMVは、何年ものあいだ土壌において生存しうることから、非常に持続的な病気である。
【0005】
PMMoVは、TMVおよびToMVに対して抵抗性である栽培品種を含む、全てのカプシクム種の植物全体に感染する。ピーマン植物における病気の症状には、若い植物の発育停止、葉の縮れおよび黄色の斑点が含まれる。果実は変形して(こぶだらけで斑点がある)、大きさがわずかに低減されている。
【0006】
1980年代に、その当時公知であるPMMoV抵抗性カプシクム系統に感染することができるTMV様のウイルス株(tm-3)が単離された(Boukema et al., 1980)(非特許文献1)。単離された株は、TMVのP14株と呼ばれた。P14株の発見以降、この新しいウイルスに対して抵抗性を示す植物種の系統C.カコエンス(C. chacoense)(系統PI 260429)が1982年に発見された(Boukema, 1982)(非特許文献2)。後に、この植物における抵抗性は、L-座のL4対立遺伝子によって付与されること(Boukema, 1984)(非特許文献3)、およびP14株がL3対立遺伝子によって付与された前者の抵抗性を克服することができるが、L4対立遺伝子によって付与された抵抗性を打破することができないことが決定された。このL3によって付与された抵抗性打破株は当初TMVとして分類されたが、後に特異的PMMoV病原型として再分類され、病原型は病原型1.2.3として割付された。
【0007】
育種業者において、PMMoV病原型1.2.3に対して抵抗性を示すC.カコエンス系統と共にそれに由来する商業的トウガラシ系統は、一般的にTm3-抵抗性系統と呼ばれる。以降、多くの種苗会社が、商業的に好ましい特徴を有する抵抗性トウガラシ植物を得るために、L4対立遺伝子を含むこのC.カコエンス系統の遺伝子材料をその育種系統に遺伝子移入してきた。
【0008】
L4対立遺伝子が、ToMV、TMVを含む多くのウイルス、ならびにPMMoV病原型1、1.2、および1.2.3に対して抵抗性を付与することは現在公知である。L4対立遺伝子によって付与された抵抗性は、本明細書において以降「PMMoV抵抗性」と呼ばれるが、厳密に言えば、これは上記のようにより広い抵抗性を付与する。
【0009】
トバモウイルスの命名は1980および2004年のあいだに数回の改訂を受けた。本明細書において、1994年4月11日にUnion for the Protection of New Varieties of Plants」(UPOV)によって発行された甘トウガラシ(C. annuum L.)に関する「明瞭性、均一性、および安定性に関する試験を行うためのガイドライン」(TG/76/7)の命名法を利用する。これらのガイドラインにおいて、ペッパートバモウイルス病原型に対する遺伝的抵抗性は、同じ座(L-座)に存在する対立遺伝子5個(L-、L1、L2、L3およびL4)によって制御されると見なされている。本明細書において、様々なウイルス病原型に対する抵抗性とトウガラシにおける抵抗性付与対立遺伝子組成との関係が示されている、先に参照したUPOVガイドラインTG/76/7の21〜22頁の表を明確に参照する。C.アンヌームのホモ接合L3L3遺伝子型は、タバコモザイクウイルス(TMV)、トマトモザイクウイルス(ToMV)、ピーマンモザイクウイルス(BePMV)、タバコマイルドグリーンモザイクウイルス(TMGMV)、ダルカマライエローフレックウイルス(DYFV)、およびトウガラシマイルドモットルウイルス(PMMoV)病原型1.2に対する抵抗性を付与するが、ホモ接合L4L4遺伝子型は、トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対するさらなる抵抗性を提供する。
【0010】
UPOVの命名法は、ウイルス学に関する科学論文において通常用いられる命名とはいくぶん異なる。一般的に科学文献において、命名は、単離体の種の名称および呼称に限定される。病原型の呼称はごくまれに記述されるに過ぎない。例えば、PMMoVの病原型1.2.3は、国際ウイルス分類委員会のユニバーサルウイルスデータベース(ICTV、The Universal Virus Database of the International Committee on Taxonomy of Viruses)には組み入れられていない。さらに、命名法およびその経時的な発展に関して、所定のトバモウイルスは、二つ以上の慣用名を得る可能性があり、以下により詳細に説明するようにさらに再分類される可能性があることに注意すべきである。ウイルス単離体の任意の新しい割付によらず、本発明はL4対立遺伝子によって付与される任意のウイルス株に対する植物の抵抗性に関すると理解される。
【0011】
先に述べたように、トバモウイルスに対するカプシクム種の抵抗性は、L-遺伝子の病原型特異的対立遺伝子(L1、L2、L3、L4)によって付与され、PMMoV病原型1.2.3に対する抵抗性はL4対立遺伝子によって付与される。L-座が存在する領域は第11染色体南のテロメアに位置する(Lefebvre et al, 2002)(非特許文献4)。抵抗性対立遺伝子は、過敏反応(HR)の誘発によって作用する。
【0012】
C.カコエンスから他のカプシクム種に遺伝子移入する場合の、L4対立遺伝子によって付与される抵抗性の顕著な望ましくない特徴は、新しい植物系統における抵抗性が非メンデル性で遺伝される点である。分離に関するメンデルの法則は、配偶子形成の際に対立遺伝子対が分離して、受精の際に無作為に合体すると述べている。それぞれの形質に関して、二倍体生物は、それぞれの親から一つずつの二つの対立遺伝子が遺伝する。二つの対立遺伝子が異なる場合、一つの、優性の対立遺伝子が生物の表現型において完全に発現される;もう一方の劣性の対立遺伝子は表現型に目立った効果を示さない。通常のメンデル遺伝に従って、ホモ接合優性対立遺伝子を有する植物と、ホモ接合劣性対立遺伝子を有する植物との交雑によって、遺伝的(全集団がヘテロ接合である)と共に表現型(全集団が優性形質を発現する)の上でも均一なF1、または雑種第一代が得られるであろう。そのようなF1集団は、その優性形質に関して非分離であると言われる(当然、F2集団では分離が起こる)。このように、F1集団は、親の少なくとも一つがその形質に関してホモ接合である場合には、優性形質に関して分離しない。同様に、非分離F1が存在すれば、親系統の一つにおいて優性形質がホモ接合性であることが確認される。しかし、これはL4対立遺伝子が遺伝子移入される商業的植物系統の場合には当てはまらない。
【0013】
植物育種業者にとって、育種の結果は好ましくは予測可能でなければならないことから、育種系統がホモ接合(真の育種)であることは重要である。例えば、商業的ハイブリッドは二つの(ホモ接合)近交系から産生され、得られたヘテロ接合F1植物は、雑種強勢またはヘテロシスの結果としてその近交系の親より適合である。植物育種業者は、より強健な後代を生成するためにそのようなヘテロシス交雑を意図的に利用して、ホモ接合近交系は経済的価値が高い産生系統として役立つ。したがって、育種の実践において、その植物系統を自家受粉させて、その子孫を非分離に関してスクリーニングすることによって、植物系統が、単一の遺伝子によって付与された優性の抵抗性形質(すなわち、一遺伝子優性形質)に関してホモ接合であるか否かを評価することは通例である。または、予想されるホモ接合優性植物系統をホモ接合劣性系統と交雑させて、それによって分離するF1によって、試験した「優性の」親がホモ接合ではないことが明らかとなる。
【0014】
L4抵抗性対立遺伝子は、通常の一遺伝子優性抵抗性対立遺伝子であると考えられる(Boukema, 1983(非特許文献5);Van Duin, 1998(非特許文献6))。しかし、L4抵抗性対立遺伝子によって付与されたPMMoV抵抗性の場合、育種業者は一般的に、ホモ接合系統からの交雑の予測可能性およびそれらの系統の安定性に関する問題を観察する。予測されるホモ接合(L4L4)抵抗性植物の自家受粉によって常に、均一な抵抗性表現型を有する子孫が得られるが(それによって親植物のホモ接合性を確認する)、一方、そのようなホモ接合抵抗性の親植物と、感受性のある親植物(すなわちL4対立遺伝子を欠損する個体)との交雑によって、しばしば抵抗性で感受性がある植物を含むF1、すなわち分離するF1が得られる。植物の感受性は、例えばPMMoV病原型1.2.3を接種後の全身性のモザイクの存在によって検出してもよい。この現象は、抵抗性がF1子孫植物のいくつかにおいて驚くほど失われており、先に概要した理由から、育種業者にとっては非常に不都合である。育種業者はしばしば、そのような予測不可能なホモ接合親植物を、「分離」植物と呼ぶが、厳密に言えば、それらのF1子孫が分離している。
【0015】
改善されたPMMoV抵抗性近交系を開発するため、ならびに通常の植物選択および試験交雑を用いることによって、育種業者は最近、正常な非分離F1(すなわち、メンデルの法則に従う)を産生するホモ接合植物を得ることができ、それによってその系統におけるL4対立遺伝子によって付与される抵抗性が固定されたことを示した。現在のところ、これがどのように行われるかは不明である。しかし、これらの「非分離」植物の主要な問題は、それらが稔性の低減と矮小生長を示す点である。稔性の低減の重要な結末は、そのような植物が限られた量の種子および/または限られた量の花粉を産生するということである。このように、そのような植物は抵抗性で「非分離である」が、それらは低い稔性および矮性の生長を特徴とする表現型を示し、これは商業的種子産生にとって望ましくない。矮性の生長は、例えば、種子の晩期成熟(すなわち、遅い成長)または植物の生長(速度)の低減によって示される可能性がある。この表現型は、本出願において分離非稔性矮性に関する「SNFD表現型」としてさらに命名されるであろう。現在のところ、生長、稔性、および不稔性に関する問題を含むこれらの負の形質と、C.カコエンスからのL4対立遺伝子のカプシクム系統への遺伝子移入との関連に関する説明はない(例えばVan Duin, 1998(非特許文献6)を参照されたい)。
【0016】
L-遺伝子のいずれのL1、L2、およびL3対立遺伝子をカプシクム種に遺伝子移入しても、ホモ接合抵抗性植物においてSNFD表現型を示すことなく、メンデル遺伝集団を産生することが常に見いだされている。C.カコエンスからのL4遺伝子移入は、今初めて上記の問題を生じる。
【0017】
現在のところ、L4対立遺伝子によって付与されたPMMoV病原型1.2.3に対して抵抗性であって、「非分離」であり、正常な生育(速度)を示し、および適当量の種子を産生するトウガラシ植物が必要である。したがって、本発明の目的は、L4対立遺伝子によって付与されたPMMoV病原型1.2.3に対する抵抗性を、作物栽培学または良好な生長および正常な種子産生のような商業的に良好な形質と組み合わせて、このように、SNFD表現型の望ましくない特徴を有しないカプシクム属の植物を提供することである。
【先行技術文献】
【非特許文献】
【0018】
【非特許文献1】Boukema et al., 1980
【非特許文献2】Boukema, 1982
【非特許文献3】Boukema, 1984
【非特許文献4】Lefebvre et al, 2002
【非特許文献5】Boukema, 1983
【非特許文献6】Van Duin, 1998
【発明の概要】
【0019】
発明の概要
本発明の目的は、本発明によって達成される。本発明に従って、L4対立遺伝子を含むゲノム遺伝子移入の大きさを低減させることによって、先に記述したホモ接合植物において観察されるPMMoV病原型1.2.3に対する抵抗性とSNFD表現型とのあいだの連鎖を断つことが可能であることが判明した。特に、これはL4対立遺伝子のPMMoV抵抗性付与部分の直接近位に存在するL4抵抗性対立遺伝子から遺伝子情報を除去することによって得られる。その結果は、PMMoV病原型1.2.3に対する抵抗性が、SNFD表現型付与部分を移入することなく、一つの植物からもう一つの植物に転移される可能性があり、それによってSNFD表現型を示さないPMMoV抵抗性および作物栽培学上貴重な植物を産生することが可能となったことである。
【0020】
理論に拘束されることなく、SNFD表現型の特徴である負の副作用は、L4対立遺伝子が存在するC.カコエンス染色体断片によって引き起こされ、C.カコエンス染色体断片を例えばC.アンヌームのゲノムに挿入すると、L4対立遺伝子自身と遺伝的に連鎖するL4対立遺伝子の抵抗性付与部分の近位における劣性遺伝子によって、SNFD表現型が引き起こされ、レシピエント植物ゲノムの染色体構築を破壊すると考えられる。したがって、SNFD表現型は、完全なL4対立遺伝子のC.カコエンス染色体遺伝子移入に関してホモ接合である植物に限って発現される。
【0021】
L4対立遺伝子によって付与された望ましい表現型と望ましくない表現型とのあいだの連鎖を断つ可能性に関するこの重要な洞察が得られたことから、SNFD表現型に関する遺伝情報が、L4対立遺伝子の抵抗性付与部分の近位から除去されている、またはホモ接合植物においてSNFD表現型がその組み合わせにおいてもはや発現されない程度まで変化している植物を選択することが可能となった。
【0022】
さて本発明は、第一の局面において、植物のゲノムにおけるL4抵抗性対立遺伝子の存在により、トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対する抵抗性を示し、該L4対立遺伝子が切断されているカプシクム属の植物を提供する。切断は、低減された長さの対立遺伝子を提供し、それによって稔性で正常な生長を示す、すなわちSNFD表現型が存在しない非分離植物が得られる。
【0023】
SNFD表現型に関与する遺伝情報の欠如は、それによってSNFD表現型が得られるL4対立遺伝子の近位における遺伝情報の除去または変化によって行ってもよく、好ましくはL4対立遺伝子の近位における組換え事象によってもたらされる。SNFD情報が対立遺伝子の一つの側にのみ存在する場合、この情報を除去するためには、1回の組換え事象で十分である。SNFD表現型に関する遺伝情報が抵抗性対立遺伝子の両側に存在する場合、この遺伝情報の双方の部分が好ましくは変化または除去される。本発明の好ましい態様に従って、少なくとも一つの組換え事象、好ましくは第11染色体の南側にあるL4対立遺伝子の北側(セントロメア側)で組換え事象が起こる植物が選択されるが、テロメア側での切断も同様に可能である。二重組換えの場合、組換え事象は、一世代で起こる必要はないが、連続世代からの組換え事象によって共に、抵抗性対立遺伝子の片側または両側での遺伝情報の最終的な除去または変化が起こってもよい。少なくとも一つの組換え事象は、抵抗性の付与に関与する遺伝情報が除去されず、それによってL4抵抗性対立遺伝子の抵抗性付与部分の存在を維持するように行わなければならない。
【0024】
本発明の植物の好ましい態様において、該L4抵抗性対立遺伝子は、C.カコエンスのゲノムに由来する。
【0025】
適切な遺伝情報が除去されたまたは変化したか否かを評価するために、当業者は、遺伝子マーカーを適宜利用してもよい。さて本発明は、例えば組換え植物のマーカー支援選抜において用いられる可能性があるいくつかの遺伝子マーカーを記述する。そのような用途は、当技術分野で周知であり、以下により詳細に記述されるであろう。
【0026】
好ましい態様において、本発明は、L4抵抗性対立遺伝子に連鎖した少なくとも一つのマーカーの不在によって示されるように、SNFD表現型に関与する遺伝情報が存在しない植物を提供し、このとき該マーカーは2群のマーカー(E35/M49-F-90、E39/M58-F-65、E39/M51-F-380、E58/M62-F-168、E66/M54-F-600、およびTm3-DRS)、3群のマーカー(E60/M54-F-447)、および1/3群のマーカー(E63/M61-F-501、E66/M43-F-387、E66/M49-F-387、E66/M61-F-99、E67/M50-F-150、E67/M62-F-214、E70/M54-F-133、E71/M47-F-550、E74/M61-F-385)からなる群より選択され、好ましくは3群および2群のマーカー、より好ましくは2群のマーカー(E35/M49-F-90、E39/M58-F-65、E39/M51-F-380、E58/M62-F-168、E66/M54-F-600、およびTm3-DRS)から選択される。
【0027】
もう一つの好ましい態様において、本発明は、L4抵抗性対立遺伝子の存在が、1群のマーカー(E58/M50-F-580、E39/M58-F-95、E58/M60-F-255、およびE54/M55-F-101)、および1/3群のマーカー(E63/M61-F-501、E66/M43-F-387、E66/M49-F-387、E66/M61-F-99、E67/M50-F-150、E67/M62-F-214、E70/M54-F-133、E71/M47-F-550、E74/M61-F-385)から選択され、好ましくは1群のマーカーから選択され、より好ましくはマーカーE58/M50-F-580およびE54/M55-F-101から選択されるL4抵抗性対立遺伝子に連鎖している少なくとも一つのマーカーの存在によって示される、植物を提供する。
【0028】
先に述べたマーカーは、制限酵素EcoRI(コードE)およびMseI(コードM)の制限部位にそれぞれ対応する「コアプライマー」
プラス、プライマーコードによって示されるさらなる選択ヌクレオチド3個からなるAFLPプライマーに基づく。プライマーコードは、http://www.keygene.com/publications/index.htmおよび具体的にはそこで開示される「KF Nomenclature Primer Enzyme Combinations.pdf」と題する文書において見いだされうる。増幅された断片の大きさ(塩基対)は、プライマー酵素の組み合わせ後に認められる。例えば、マーカーE58/M60-255は、プライマー酵素の組み合わせ[EcoRI+プライマーコード58/MseI+プライマーコード60]によってDNAを切断した後に得られた255塩基対の増幅された断片を示す。
【0029】
プライマーコードは以下のとおりである:35:ACA;39:AGA;43:ATA;47:CAA;49:CAG;50:CAT;51:CCA;54:CCT;55:CGA;58:CGT;60:CTC;61:CTG;62:CTT;63:GAA;66:GAT;67:GCA;70:GCT;71:GGA;74:GGT。プライマー酵素組み合わせコードE63/M61-F-501の例は、EcoRI+GAA(=E63)/MseI+CTG(=M61)である。
【0030】
マーカーTG036に由来するマーカーTm3-DRS(Lefebvre et al., 2002)は、PCRプライマー対P118
およびプライマーP119
によって定義されるマーカーであり、PCR産物をTru1I(制限部位TTAA)によって消化した後にプライマーP118およびプライマーP119によってPCRを行うことによって検出することができる。約350塩基対の増幅断片の存在は、L4対立遺伝子の存在を示している。
【0031】
SNFD表現型に関与する遺伝情報が存在しないまたは除去されている植物の供給はまた、以下により詳細に記述される切断型L4対立遺伝子を植物に提供することに関して記述されてもよい。そのような切断に関連する局面は、先に記述した遺伝情報の除去または変化に同様に適用される。
【0032】
本発明の植物のもう一つの好ましい態様において、植物はC.アンヌーム種の植物である。
【0033】
本発明の植物のなおもう一つの好ましい態様において、植物はL4抵抗性対立遺伝子に関してホモ接合であり、ホモ接合植物は好ましくは近交系植物である。
【0034】
もう一つの局面において、本発明は、L4対立遺伝子が切断されていて、切断がテロメアおよび/またはセントロメア側から約0.001〜10 cM、好ましくは対立遺伝子のセントロメア側から0.001〜10 cMの遺伝距離に及ぶヌクレオチド配列の除去を含む、植物のゲノムにおけるL4抵抗性対立遺伝子の存在により、トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対して抵抗性を示すカプシクム属の植物を提供する。先に記述した好ましい態様はまた、本発明のこの局面に従う植物にも当てはまる。
【0035】
コアTM-3抵抗性遺伝子を含む1群のマーカーと、3群のマーカーとのあいだで観察された組換え頻度は、距離約2 cMと同等であった。1群のマーカーと2群のマーカーとのあいだで観察された組換え頻度も同様に、距離約2 cMに等しかった。本明細書において同定されたマーカー群内での任意のマーカーのあいだの距離は、組換えが存在しなかったために決定することができなかった。このように、本明細書において記述された切断は、好ましくは、1群のマーカーの任意の位置からセントロメア(2群)またはテロメア(3群)側に存在する任意のマーカーから2 cM未満の距離に及ぶ遺伝的欠失を表す。
【0036】
もう一つの局面において、本発明は、本発明のホモ接合、好ましくは近交系植物を、商業的に望ましい特徴を示すもう一つのホモ接合、好ましくは近交系のトウガラシ植物と交雑させることによって得ることができる、PMMoV病原型1.2.3に対して抵抗性を示すハイブリッドトウガラシ植物を提供する。
【0037】
さらにもう一つの局面において、本発明は、切断型L4抵抗性対立遺伝子が、カプシクム属の植物において発現されうる遺伝情報を含み、それによって該植物にPMMoV病原型1.2.3に対する抵抗性を付与し、およびSNFD表現型を付与する遺伝情報が該対立遺伝子に存在しない、切断型L4抵抗性対立遺伝子を含む単離された核酸配列を提供する。
【0038】
そのような単離核酸配列の好ましい態様において、抵抗性付与遺伝情報は、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択される、好ましくは、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580からなる群より選択される、より好ましくはマーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される少なくとも一つのマーカーを含み、抵抗性付与遺伝情報はさらに、SNFD表現型を付与する遺伝情報をさらに含み、2群マーカーおよび3群マーカー、好ましくは2群マーカーからなる群より選択される少なくとも一つのマーカーがL4抵抗性対立遺伝子に存在しないことを含む。最も好ましい態様において、本発明の単離核酸配列に含まれるL4抵抗性対立遺伝子は、C.カコエンスのゲノムに由来する。
【0039】
もう一つの局面において、本発明は、a)PMMoV病原型1.2.3に対して感受性であるカプシクム属のレシピエント植物またはその一部を提供する段階、およびb)切断型L4抵抗性対立遺伝子を含むゲノム領域を該レシピエント植物、その一部、またはその後代植物のゲノムに導入する段階を含む、PMMoV病原型1.2.3に対する抵抗性を示すカプシクム属の植物を産生する方法であって、該対立遺伝子が、該植物、植物の一部、または後代植物において発現されうる遺伝情報を含み、それによって該植物、植物の一部、または後代植物にPMMoV病原型1.2.3に対する抵抗性を付与し、およびSNFD表現型を付与する遺伝情報が、少なくともSNFD表現型が発現されない程度まで該対立遺伝子に存在しない方法を提供する。段階b)の前に、例えば本明細書に記述のL4抵抗性対立遺伝子の切断型を有する組換え型ドナー植物の形で、またはインサートとして該ゲノム領域を含むベクターの形で、切断型L4抵抗性対立遺伝子を含むゲノム領域を提供する段階が存在してもよい。
【0040】
本発明の方法において用いられる切断型L4抵抗性対立遺伝子を含むゲノム領域は、SNFD表現型を有する植物が得られるL4対立遺伝子と比較してその大きさが低減されているL4抵抗性対立遺伝子を含む。L4抵抗性対立遺伝子の大きさは、大きさの低減がPMMoV病原型1.2.3に対する抵抗性の付与に関与する遺伝情報に影響を及ぼさないように低減される。
【0041】
切断型L4抵抗性対立遺伝子を含むゲノム領域の導入を実現するための非常に適した方法は、レシピエント植物と、L4抵抗性対立遺伝子を含むカプシクム属のPMMoV病原型1.2.3抵抗性ドナー植物との交雑に由来する後代植物におけるゲノム遺伝子移入によって行われる。遺伝子移入のプロセスは当業者に周知であり、一般的に、その親の一つとの種間ハイブリッドの反復戻し交雑によって、一つの種の一つまたは複数の遺伝子をもう一つの遺伝子プールに導入する段階を必然的に伴う。
【0042】
しかし当業者は、遺伝子移入が、L4抵抗性対立遺伝子またはL4抵抗性対立遺伝子を含むゲノム領域が植物のゲノムに導入されうる唯一の方法ではないことを容易に理解するであろう。例えば、他の方法は、半数体細胞融合、植物形質転換等のようなトランスジェニック技術を用いる段階を必然的に伴ってもよい。したがって、ゲノム領域の導入は、インビトロ培養技術、プロトプラスト融合、形質転換、または倍加半数体法によって行ってもよい。しばしば、そのような技術は、無傷の植物に関して行われることはなく、植物の一部、一般的に単細胞の組織培養物またはプロトプラストのような植物の繁殖材料について行われる。したがって、本発明の植物を得るために、そのような方法には、任意で該植物の一部をトウガラシ植物に生長させるさらなる段階が含まれてもよい。一般的に、これは、植物を再生する段階、または異種DNAによって形質転換された植物または植物の一部もしくは組織の双方の繁殖材料を再生する段階、および任意で該植物もしくは繁殖材料またはその双方を生物学的に複製する段階を必然的に伴うであろう。
【0043】
切断型L4抵抗性対立遺伝子が、遺伝子移入のプロセスによってカプシクム属のレシピエント植物に導入される本発明の方法によって、好ましくは、切断型L4抵抗性対立遺伝子を含むゲノム領域がそのゲノムに遺伝子移入された後代植物が得られる。切断型L4抵抗性対立遺伝子を有する後代植物を得るプロセスは、組換えを必然的に伴うことから、後代植物の表現型または遺伝子型のいずれかに基づく選択を行う前に分離集団を産生することが好ましい。したがって、産生されてそのゲノムに切断型L4抵抗性遺伝子を有する後代植物は好ましくは、例えば先に述べた交雑から得られたF1植物の自家受粉によって産生された、または該交雑から得られたF1植物ともう一つのトウガラシ植物との交雑によって得られた分離集団の植物である。
【0044】
本発明の方法において、ゲノム領域は、好ましくはC.カコエンスのゲノムから導入され、そのような方法において用いられるレシピエント植物は好ましくはC.アンヌーム植物である。
【0045】
本発明の植物を産生する方法の基本的な要素を記述してきたが、本発明の方法はさらなる段階を含んでもよい。好ましい態様において、本発明の方法は、c)PMMoV病原型1.2.3抵抗性植物またはそのPMMoV病原型1.2.3抵抗性後代植物を選択する段階、およびd)SNFD表現型を発現しない抵抗性植物または抵抗性後代植物を選択する段階をさらに含む。
【0046】
PMMoV病原型1.2.3抵抗性植物または後代植物を選択する段階は、抵抗性バイオアッセイを用いるような、例えばPMMoV病原型1.2.3の浮遊液を植物の葉に接種する段階および感染の発生を評価する段階によるような、表現型の決定に基づく方法によって行ってもよい。しかし、好ましい態様において、PMMoV病原型1.2.3抵抗性植物を選択する段階は、該植物におけるL4対立遺伝子の抵抗性付与部分の存在を示す1群または1/3群のマーカーを含む群から選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーの存在に関して、植物または後代植物のゲノムをスクリーニングするプロセスを含む。1群の適したマーカーには、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580が含まれ、好ましくは、マーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される少なくとも一つのマーカーの存在が決定される。抵抗性形質の存在を評価するために適した1/3群のマーカーには、抵抗性植物に存在することを当業者が確立したマーカーが含まれる。1/3群のマーカーはいずれも、L4抵抗性対立遺伝子を示すマーカーと共に、その対立遺伝子におけるSNFD表現型付与部分を示すマーカーを含む。このように、いくつかの1/3群のマーカーは、L4抵抗性対立遺伝子におけるSNFD表現型付与部分を除去し、3群のマーカーと類似である適した切断を示すが、他の1/3群のマーカーは、コア抵抗性遺伝子または調節配列の存在を示し、したがって1群のマーカーと類似である。
【0047】
PMMoV病原型1.2.3抵抗性植物の中から、SNFD表現型(先に記述した段階d)を発現しない植物または後代植物を選択する段階は、本発明によって開示されるマーカー支援育種プロセスの中心となる。SNFD表現型を発現しない抵抗性植物または後代植物の選択は、2群のマーカーおよび3群のマーカーからなる群(任意で1/3群のマーカーからも選択される)、好ましくは2群のマーカーより選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーに関して、抵抗性植物または後代植物のゲノムにおける切断型L4抵抗性対立遺伝子をスクリーニングする段階、ならびに該マーカーの少なくとも一つが存在せず、それによって切断を示す、植物または後代植物を選択する段階を含む。
【0048】
もう一つの局面において、本発明は、先に記述したPMMoV病原型1.2.3に対する抵抗性を示すカプシクム属の植物を産生する方法を行う段階を含み、さらに以下の段階:e)選択した植物を自家受粉する段階;f)該自家受粉から得られた種子を植えて、該種子を植物に生長させる段階;g)PMMoV病原型1.2.3抵抗性を示し、商業的に望ましい特徴を保有する段階g)からの植物を同定する段階、およびh)PMMoV病原型1.2.3抵抗性を示し、商業的に望ましい特徴を保有する近交系トウガラシ植物が産生されるまでe)〜g)の段階を繰り返す段階を含む、PMMoV病原型1.2.3に対する抵抗性を示すカプシクム属の近交系植物を産生するための方法を提供する。本発明の状況における商業的に望ましい特徴は、外観の改善、より高い種子収率、稔性の改善、より高い果実収率、より大きいもしくはより小さい果実、または改善された果実の色もしくは味のような果実特徴のような任意の特徴に関連してもよく;または例えば、幹の強度の改善、根のシステムの改善、ストレス抵抗性の改善、耐病性の改善等のような特徴に関連してもよい。実践において、商業的に望ましい特徴は、野生型植物に対して、植物をより商業的に貴重にするであろう如何なる特徴であってもよい。
【0049】
もう一つの局面において、本発明は、先に記述した本発明の方法の任意の一つによって得ることが可能なPMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物または近交系植物を提供する。
【0050】
他の局面において、本発明は、本発明に従う植物の後代、葉、茎、根、根の先端、台木、シュート、果実等のような部分、または本発明に従う植物の細胞、プロトプラスト、カルス、細胞塊、(体細胞)胚、葯、葉柄、花粉、胚珠、花、培養細胞、種子等のような部分に関する。そのような部分は、好ましくは(器官)組織培養による繁殖に適していてもよく、または果実のような、消費に適してもよい。
【0051】
もう一つの局面において、本発明は、本発明のトウガラシ植物を生長させることによって産生されたトウガラシ種子を提供する。
【0052】
もう一つの局面において、本発明は、本発明の方法によって得ることができる本発明の近交系トウガラシ植物を、好ましくは商業的に望ましい特徴を示す近交系トウガラシ植物と交雑することによって得ることができるPMMoV病原型1.2.3に対して抵抗性を示す、ハイブリッドトウガラシ植物またはその一部を提供する。
【図面の簡単な説明】
【0053】
【図1】この座に連鎖したマーカーの異なる群(1、2、3、1/3群)を示すL4座のマイクロマップを示す。これらの群のそれぞれに存在するマーカーを、実施例1において示すように表1に記載する。
【図2】個々のマーカーが示された、同じマイクロマップを示す。
【図3】本明細書において用いられるマーカー、および特にヌクレオチド配列情報およびマーカーの長さに関する詳細な情報を提供する。マーカーの長さは、C.カコエンスアクセッション番号PI 260429から単離されたゲノムDNAを鋳型DNAとして用いて、表記のプライマーとのPCR反応を行うことによって増幅されたDNA断片の塩基対におけるおおよその長さである。
【発明を実施するための形態】
【0054】
発明の詳細な説明
定義
本明細書において提供された見出しは、全体として明細書を参照することによって得られうる本発明の様々な局面または態様の制限ではない。したがって、下記に定義される用語は、明細書を全体として参照することによってより十分に定義される。
【0055】
「L4抵抗性対立遺伝子」または「L4対立遺伝子」という用語は、本明細書において、遺伝子がL-座の一部であり、Boukema(1984)によって最初に記述された、トマトモザイクウイルス(ToMV)ならびにトウガラシマイルドモットルウイルス(PMMoV)病原型1、1.2、および1.2.3に対する抵抗性を植物に提供する、カプシクム種のL遺伝子の病原型特異的抵抗性対立遺伝子を指すために用いられる。L座が存在する領域は第11染色体南のテロメア上に位置する。非切断型のL4対立遺伝子は、マーカーTm3-DRSと会合する。したがって、L4対立遺伝子は、例えば、本明細書において定義される一つまたは複数の2群マーカー(E35/M49-F-90、E39/M58-F-65、E39/M51-F-380、E58/M62-F-168、E66/M54-F-600、およびTm3-DRS)と一つまたは複数の3群または1/3群マーカー(E60/M54-F-447、E63/M61-F-501、E66/M43-F-387、E66/M49-F-387、E66/M61-F-99、E67/M50-F-150、E67/M62-F-214、E70/M54-F-133、E71/M47-F-550、E74/M61-F-385)とによって境界を定められるC.カコエンスアクセッション番号PI 260429号に由来し、そのヌクレオチド配列がPMMoV病原型1.2.3に対する抵抗性を付与するヌクレオチド配列を表す。
【0056】
本明細書において用いられるように、「トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3」という用語は、L3対立遺伝子によって付与される抵抗性を克服することができる、カプシクムに感染するマイルドモットルウイルスの株を示す。文献において、そのような株は一般的に、PMMoVの抵抗性を打破する株という用語で示される。ICTVが承認した頭字語PMMoVおよびPMMVはいずれも、トウガラシマイルドモットルウイルスを示すために用いられる可能性があると理解されるべきである。文献において認められる可能性があるPMMoVの同義語は:タバコモザイクウイルスのサムスン潜在株(SL-TMV)、ペッパーモザイクウイルス、およびカプシクムモザイクウイルスである。ウイルス分類における急速な発達により、いくつかの株、特に当初トウガラシマイルドモットルウイルス(PMMoV;ICTV10進コード71.0.1.0.007)として分類された最初のダッチ株(Tobias et al., 1982)は、現在では、パプリカマイルドモットルウイルス(PaMMV;ICTV 10進コード71.0.1.0.006)として分類されている。したがって、本明細書において用いられる「トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3」という用語にはまた、トウガラシ植物におけるL3対立遺伝子によって付与される抵抗性を克服することができるPaMMVが含まれる。一般的に、本明細書において用いられるように、「トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3」という用語によって示されるウイルスの例には、以下の株が含まれるがそれらに限定されるわけではない:TMVの単離体P14(Boukema, 1982;Tobias et al., 1982;Rast, 1988);PMMoV-Iによって示されるイタリア株(Wetter et al, 1984;Rodriguez-Cerezo et al., 1989;Velasco et al, 2002;Berzal-Herranz et al., 1995;Garcia-Luque et al., 1993);PMMoV-Sによって示されるスペイン株(Tenllado et al., 1997;Velasco et al., 2002;Tenllado et al., 1996;Berzal-Herranz et al., 1995;Garcia-Luque et al., 1993;Alonso et al., 1991;Avila-Rincon et al., 1989;Rodriguez-Cerezo et al., 1989);PMMoV-JおよびPMMoV-Ijによって示される日本株(Tsuda et al., 1998;Hagiwara et al., 2002;Kirita et al., 1997);PMMV-kによって示される韓国株(Lim et al., 1997);およびPaMMV(Garcia-Luque et al., 1993;Ruiz del Pino et al., 2003;Gilardi et al., 2004)。トウガラシマイルドモットルウイルスおよびパプリカマイルドモットルウイルスの記述に関する典拠の確かな参考文献は、i.a. Brunt, A.A., Crabtree, K., Dallwitz, M.J., Gibbs, A.J., Watson, L. and Zurcher, E.J.(eds.)(1996以降)「Plant Viruses Online : Descriptions and Lists from the VIDE DatabaseVersion : 16 th January 1997」、およびThe Universal Virus Database of the International Committee on Taxonomy of Viruses(http://www.ncbi.nlm.nih.gov/ICTVdb/index.htm)である。
【0057】
「切断型対立遺伝子」は、本明細書において、その遺伝距離に基づいて完全な対立遺伝子より大きさが小さい対立遺伝子として定義される。例えば大きさの低減は、少なくとも一つの連結したマーカーの不在および/または区別可能な表現型、本発明の場合ではSNFD表現型の不在によって示されてもよい。
【0058】
本明細書において用いられるように、「対立遺伝子」という用語は、その全てが少なくとも一つの形質または特徴に関連する遺伝子の一つまたは複数の代替形を意味する。二倍体細胞において、所定の遺伝子の二つの対立遺伝子は、相同な染色体の対において対応する座を占める。本発明は、QTL、すなわち一つまたは複数の遺伝子または調節配列を含んでもよいゲノム領域に関連することから、いくつかの場合において「対立遺伝子」の代わりに「ハプロタイプ」(すなわち、染色体セグメントの対立遺伝子)を指すことがより正確であるが、それらの場合、「対立遺伝子」という用語は、「ハプロタイプ」という用語を含むと理解すべきである。対立遺伝子は、それらが類似の表現型を発現する場合、同一であると見なされる。配列における差は起こりうるが、それらが表現型に影響を及ぼさない限り重要ではない。
【0059】
「ゲノム領域」は、本明細書において、遺伝子および調節要素領域のような様々なゲノム機能を有する配列を含んでもよいヌクレオチド配列として、好ましくはDNA配列として定義される。ゲノム領域は、ヌクレオチド構築物であってもよく、ベクターに含まれていてもよい。または、ゲノム領域は、該植物の交雑後の染色体組換えによって一つの植物からもう一つの植物に転移されてもよい。ゲノム領域は、原則的に一つまたは複数の種に由来する遺伝子材料を含んでもよい。
【0060】
「遺伝子」は、本明細書において、染色体上の特異的な位置を占有し、生物における特定の特徴または形質に関する遺伝的インストラクションを含むDNAの配列からなる遺伝単位として定義される。
【0061】
「座」は、本明細書において、所定の遺伝子が所定の種の染色体上で占有する遺伝子マップ上の位置として定義される。
【0062】
本明細書において用いられるように、「ヘテロ接合」という用語は、異なる対立遺伝子が相同染色体上の対応する座に存在する場合に存在する遺伝子状態を意味する。
【0063】
本明細書において用いられるように、「ホモ接合」という用語は、同一の対立遺伝子が相同染色体上の対応する座に存在する場合に存在する遺伝子状態を意味する。ホモ接合性は、個々の植物の自家受粉後の分離の不在、または感受性がある植物と交雑した場合にF1における分離の不在として定義される。
【0064】
本明細書において用いられるように、「ハイブリッド」という用語は、二つの遺伝的に異なる個体間の交雑の任意の子孫を意味する(Rieger et al., 1968)。
【0065】
本明細書において用いられるように、「近交系」という用語は、実質的にホモ接合の個体または系統を意味する。
【0066】
本出願において、「組換え事象」は、(減数分裂)染色体交差を意味すると理解される。
【0067】
本明細書において用いられるように、「遺伝子移入」、「遺伝子移入された」、および「遺伝子移入する」という用語は、種を交雑することによって、一つの種、品種または栽培品種の遺伝子がもう一つの種、品種または栽培品種のゲノムに移動するプロセスを指す。交雑は、天然または人工的であってもよい。プロセスは任意で、戻し交雑親に戻し交雑することによって完了してもよく、この場合遺伝子移入は、種間ハイブリッドとその親の一つとの反復戻し交雑によって、一つの種の遺伝子をもう一つの種の遺伝子プールに浸潤させることを指す。遺伝子移入はまた、レシピエント植物のゲノムにおいて安定に組み入れられたヘテロ接合遺伝子材料として記述されてもよい。
【0068】
「遺伝子操作」、「形質転換」および「遺伝子改変」は全て、本明細書において標的植物のDNAへの、通常、もう一つの生物の染色体DNAまたはゲノムの、しかしこれらに限定されない任意の種の遺伝情報の移入に関する同義語として用いられる。遺伝子操作は、レシピエント植物のゲノムに異種遺伝子材料を安定に組み入れる一つの方法であり、これには遺伝子またはその対立遺伝子バリアントをコードする外来ヌクレオチド配列を含む異種DNAと共に、植物細胞または組織における異種DNAの安定な組み込みを引き起こすことができる要素、および植物細胞または植物組織において外来ヌクレオチド配列の発現を可能にする要素から選択される調節要素を含むDNA組換え体によって、植物の細胞または組織を形質転換する段階を含むプロセスが含まれてもよい。
【0069】
本明細書において用いられるように、「分子マーカー」という用語は、制限断片長多型(RFLP)マーカー、増幅断片長多型(AFLP)マーカー、一塩基多型(SNP)マーカー、マイクロサテライトマーカー、配列特徴増幅反復配列(SCAR)マーカー、もしくはイソ酵素マーカー、または特異的遺伝子および染色体位置を定義して、二つの対立遺伝子間の多型を検出する本明細書に記述のマーカーの組み合わせのような任意の技術を用いることによって得ることができるマーカーを意味する。本明細書において提供された1、2、3および1/3群のマーカーは、カプシクムゲノムDNAを鋳型として用い、図3において示したプライマー配列を増幅プライマーの対の第一および第二のプライマーとして用いる核酸増幅反応を行うことによって得られた二本鎖または一本鎖ヌクレオチド配列からなるマーカー(特にAFLPマーカー)を表し、該プライマー配列および/またはその相補体がいずれかの側に隣接するカプシクム特異的ヌクレオチド配列および/またはその相補体からなる二本鎖または一本鎖ヌクレオチド配列を提供する。カプシクム特異的ヌクレオチド配列という用語は、本明細書において鋳型DNAとしてC.カコエンスアクセッション番号PI 260429号のゲノムDNAを用いて、およびそれと配列類似性少なくとも90%、好ましくは95%、より好ましくは97%、さらにより好ましくは98%より高い配列類似性を有する配列を用いた場合に得られた配列を示すために用いられる。
【0070】
本明細書において用いられるように、「制限断片長多型」または「RFLP」という用語は、特異的な制限酵素によって切断されたDNA断片の大きさにおける個体間の変動を意味する。RFLPをもたらす多型配列は、遺伝子連鎖マップ上のマーカーとして用いられる。
【0071】
本明細書において用いられるように、「集団」という用語は、共通の遺伝的起源を共有する遺伝的に均一または不均一な植物コレクションを意味する。
【0072】
本明細書において用いられるように、「品種」または「栽培品種」という用語は、構造的特徴および性能によって同じ種における他の品種から同定されうる類似の植物群を意味する。本明細書において用いられるように「品種」という用語は、1972年11月10日、1978年10月28日、および1991年3月19日にジュネーブで改正された、1961年12月2日の植物の新品種の保護に関する国際会議(UPOV条約)での対応する定義と同一の意味を有する。このように、「品種」は、公知の最低のランクの単一の植物学的分類内での植物のグループ分けを意味し、育成者権の付与に関する条件が十分に満たされているか否かによらず、そのグループ分けはi)所定の遺伝子型または遺伝子型の組み合わせに起因する特徴の発現によって定義されうる、ii)該特徴の少なくとも一つの発現によって任意の他の植物のグループ分けと区別されうる、およびiii)繁殖に関するその適切性に関する単位が不変であると見なされうる。
【0073】
本明細書において用いられるように、「トウガラシ」または「カプシクム」という用語は、カプシクム属の任意の種、品種、栽培品種、または集団を意味する。
【0074】
一つの局面において本発明は、トウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対して抵抗性を示すカプシクム属の植物を提供する。本発明に従うカプシクム属の植物は、C.アンヌーム、黄色トウガラシ(C. baccatum)、C.カルデナセイ(C. cardenasei)、C.カコエンス、C.キネンセ、C.エキシミウム(C. eximium)、キダチトウガラシ、C.ミクロカルプム(C. microcarpum)、C.ミニマム(C. minimum)、C.ペンデュラム(C. pendulum)、C.プラテルミッスム(C. praetermissum)、およびC.プベスケンス(C. pubescens)が含まれるがそれらに限定されるわけではないカプシクム属の任意の種であってもよい。好ましくは、本発明の植物は、白い花の咲くカプシクム種、好ましくは、C.アンヌーム、キダチトウガラシ、C.キネンセ、またはC.カコエンス植物であり、より好ましくは、C.アンヌーム、キダチトウガラシ、またはC.キネンセ植物であり、さらにより好ましくはC.アンヌームまたはキダチトウガラシ植物であり、最も好ましくはC.アンヌームである。さらにより好ましい態様において、本発明は、作物栽培学上または商業的に望ましい特徴を有するC.アンヌーム植物を提供する。
【0075】
本明細書において用いられるように、「C.アンヌーム」という用語は、その植物のゲノムが遺伝的バックグラウンドとしてC.アンヌームゲノムを含む、カプシクム属のトウガラシ植物を意味する。そのような植物は本質的に、組換え、形質転換または任意の他のプロセスの結果として、他の種のDNAが組み入れられているC.アンヌームゲノムを有するであろう。当業者は、二つの植物種のあいだの育種によって、双方の親の特徴および遺伝子材料を有し、このように種間の分界線を曖昧にする植物が得られるであろうことを理解するであろう。そのような植物は全て本明細書においてC.アンヌーム植物と呼ばれる。
【0076】
トバモウイルス、トウガラシマイルドモットルウイルス(PMMoV)は、1952年にアメリカの南カリフォルニアにおいてC.アンヌームにおいて初めて報告された(McKinney, 1952)。症状には、軽度の萎黄病および特に植物が若いあいだに感染した場合には生長停止が含まれる。果実は小さく、変形して斑点を有することがあり、壊死性の陥没を有する可能性があり、このようにそれらを市場に出せなくする。PMMoVは、植物間の密接な接触によって、種子表面上のビリオンによって、および移植によって伝搬する。ウイルスは、多くの国において報告されている(アルゼンチン、オーストラリア、カナダ、デンマーク、フランス、ハンガリー、アイスランド、イタリア、日本、韓国、オランダ、スペイン、イギリス、およびアメリカ)。病気の制御は、抵抗性栽培品種を用いることによって最善に達成される。L4対立遺伝子によって付与された抵抗性を克服することができると報告されている新しい株が報告されている(Antignus et al., 2000)。
【0077】
本発明の植物におけるトウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対する抵抗性は、植物のゲノムにおけるL4抵抗性対立遺伝子の存在による。PMMoV病原型1.2.3に対して抵抗性を付与する本発明の植物のゲノムに存在するL4抵抗性対立遺伝子は、如何なる起源に由来してもよい。本発明の好ましい態様に従って、L4抵抗性対立遺伝子は、C.カコエンスに由来し、より好ましくはアクセッション番号PI 260429号およびSA 185号に由来する(Boukema, 1983)。
【0078】
本発明の植物は、後代にPMMoV抵抗性を伝搬することができ、L4抵抗性対立遺伝子に関してヘテロ接合であってもよいが、好ましくはL4抵抗性対立遺伝子に関してホモ接合である。L4抵抗性対立遺伝子に関してホモ接合である本発明の植物は、稔性の低減を示さず、および/または矮性の生長を示さない。
【0079】
植物が、稔性の低減および/または矮性生長を示さない場合、SNFD表現型を引き起こす遺伝情報の十分量が紛失している、またはこの遺伝情報が十分に不活化されていると推測される。SNFD表現型に関与する遺伝情報の不在は、少なくとも、SNFD表現型が発現されない程度までである。
【0080】
本発明者らは、AFLPによってL4対立遺伝子の近傍における様々な遺伝子マーカーの存在を決定して、マーカー間、またはマーカーとL4対立遺伝子とのあいだの組換えに基づいて、これらのマーカーを異なる群に分けることができることを発見した(実施例における表1を参照されたい)。L4座に連結した異なる群の位置を図1に示す。1群のマーカーは、PMMoV病原型1.2.3バイオアッセイにおいてマーカーと抵抗性スコアとのあいだに組換えが観察されない限り、L4対立遺伝子自身に対して最も近位に存在すると考えられ、L4-対立遺伝子の存在を示している。2群および3群のマーカーは、いずれかの側で1群マーカーに隣接している。第4の群は、1群と3群の双方に及ぶ領域に存在するマーカーからなる。この群のマーカーを、本明細書において1/3群のマーカーと呼ぶ。
【0081】
1/3群に存在するマーカーは、所定のマーカーと1群マーカーとのあいだ、さらに所定のマーカーと3群のマーカーとのあいだにも組換えが観察されなかったため、今のところこれらのマーカーのそれぞれが1群または3群内に存在することができないという点において異なる。組換えは1/3群のマーカーと2群のマーカーとのあいだに観察されたことから、これら二つの群からのマーカーは、明らかにその遺伝子の位置が異なり、これらのマーカーの実際の位置は1群または3群のいずれかである。
【0082】
このように、群のそれぞれにおいて組換えは観察されなかった。
【0083】
染色体の残りに向かっての図1のマップの方向は、2群のマーカーが、第11染色体南のセントロメア側に存在して、3群のマーカーがテロメア側(末端に向かって)に存在するような方向である。すなわち、対立遺伝子の主な部分は、それに対して図が明らかな参照を行うLefebvre et al., 2002の図1において公表されたように、トウガラシの種間コンセンサス連鎖マップにおいて示されるように、分離集団PYに関して第11染色体(Chr. P11(brun))のマーカーTG036より下に存在する。
【0084】
抵抗性であって、L4対立遺伝子に関してホモ接合であり、2群、3群、または1/3群のいずれかのマーカー、特に2群のマーカーを紛失した植物は、SNFD表現型を示さないことが見いだされた。このように、L4対立遺伝子に通常連鎖している特定のゲノム領域を他の遺伝子型の相同なDNAに置換してもよく、それでも抵抗性に影響を及ぼすことはないが、生長および稔性と共に抵抗性形質の子孫植物への遺伝挙動に強く影響を及ぼす。
【0085】
SNFD表現型を有しない植物は、例えば通常の雑種形成(植物交雑)技術によって得ることができる。しかし、その条件は、正確な選択基準を用いることである。本発明に従って、現在では適した選択基準、すなわちL4抵抗性対立遺伝子の近傍からの遺伝情報の小片の、完全または部分的欠如または不活性が定義されている。本発明に従って、SNFD表現型が、植物およびその後代に存在しない限り、抵抗性対立遺伝子周囲の遺伝情報のどれほど多くが組換え体に存在しないもしくは抑制されているか、またはどのようにしてSNFD表現型を付与する遺伝子が不活化されているかは無関係である。
【0086】
組換えがL4対立遺伝子およびSNFD表現型を引き起こす遺伝子のあいだで起こっている植物を発見するために、分離集団またはそのような集団の後代を、組換え型表現型、すなわち非SNFD表現型と組み合わせてPMMoV P 1.2.3に対するホモ接合抵抗性を有する植物に関して実際にスクリーニングする。
【0087】
このようにして適した組換え体を発見することは可能であるが、作物栽培学上貴重な組換え体を発見することが複雑になりうる多くの理由がある。先に述べたように、ホモ接合体はSNFD表現型を示す。しかし、そのようなホモ接合体を、感受性がある(すなわち、L4対立遺伝子を含まない)植物と交雑すれば、F1は抵抗性を示すがSNFDを示さないことが見いだされた。これは、L4抵抗性対立遺伝子は優性であるが、SNFD表現型に関与する遺伝子が劣性であるという事実によると思われた。
【0088】
C.カコエンス染色体断片1コピーを有する植物の後代は、正常な(非SNFD)表現型と抵抗性を併せ持つであろう。しかし、そのF1子孫は、抵抗性表現型に関して分離し、商業的カプシクム品種は、流通させるために遺伝子の均一性に関する厳密な要件に従わなければならないことから、そのような植物は、商業的に用いることができない。
【0089】
上記の困難が起こる可能性があるにもかかわらず、SNFD表現型に関する遺伝情報が存在しない所望の植物の選択が、通常の方法で非常に可能であることは明らかであろう。しかし、このような選択は、同定のために、比較的大きい分離集団または多数の雑種形成を必要とする。
【0090】
したがって、適した植物の選択において分子生物学的ツールを利用することはより効率的である可能性がある。有用な技術は、Vos et al., 1995によって記述されるように、AFLP技術である。
【0091】
本発明に応用した場合、この技術は、L4対立遺伝子に遺伝的に連鎖しているDNAマーカーのマッピングに基づき、その後、雑種形成実験の子孫において組換え事象がL4対立遺伝子の近位で起こったか否かを比較的単純な方法で決定することができる。L4対立遺伝子に関するホモ接合性とSNFD表現型の欠如とを併せ持つ植物を選択する効率は、分子マーカーを用いることによって有意に増加させることができる。L4連鎖マーカーが存在しない場合に、(したがって)L4対立遺伝子とそのマーカーとの交差が起こった。対立遺伝子から様々な遺伝的距離でマーカーを選択することによって、組換えの位置を定義して、それによって対立遺伝子の近傍から多数または少数が消失したか否かを決定することができる。したがって、連鎖したマーカーの一つまたは複数が存在しない後代は、対立遺伝子の望ましくない近位の少なくとも小片を欠損しており、したがってSNFD表現型をもたらす遺伝情報が存在しないL4対立遺伝子を有する染色体が存在する機会を平均より多く有する。
【0092】
対立遺伝子の近位での遺伝子マーカーのマッピングは、例えばLefebvre and Chevre, 1995;Michelmore, 1995;Winter and Kahl, 1995において記述される、分子生物学技術の当業者によってかなり容易に行うことができる技法である。AFLP技術に関する一般的な情報はVos et al.(前記)において認められうる。
【0093】
L4抵抗性対立遺伝子がホモ接合条件で存在し、SNFD表現型を発現しない態様における本発明に従う植物は、他の作物栽培学上貴重なカプシクムタイプにPMMoV P1.2.3抵抗性を転移させるために適切に用いることができる。このように、ハイブリッドトウガラシ植物は、ホモ接合植物、好ましくは近交系植物をもう一つのトウガラシ植物と交雑することによって作出されうる。もう一つのトウガラシ植物は、PMMoV P1.2.3に対して抵抗性であってもなくてもよいが、好ましくはホモ接合であり、さらにより好ましくは近交系である。
【0094】
L4抵抗性対立遺伝子がホモ接合状態で存在し、SNFD表現型を発現しない態様における本発明に従う植物はまた、他の作物栽培学上貴重なカプシクムタイプにPMMoV P1.2.3抵抗性を固定するために用いることができる。これは、例えば標準的な戻し交雑技法(例えば、Briggs and Knowles, 1967)の後に植物を少なくとも2世代自家受粉させて、抵抗性対立遺伝子に関してホモ接合であって、SNFD表現型を発現しない株を選択することによって起こってもよい。
【0095】
選択プログラムの特定の段階において、他家受粉を用いる。しかし、自家受粉植物の他家受粉は、雌性の親として用いられる植物において自家受精が防止されることを必要とする。これは、生殖器官の雄性部分を手で除去することによって行うことができる。これは、その物理的除去によって、化学物質によって、および/または花にやる水を用いることによって行うことができる。生殖器官の雄性部分を除去または機能障害にするこれらの方法は全て、当技術分野で周知である。雑種形成の後代は、雑種形成の雌性の親に種子を産生させて、F1または戻し交雑の種子を採取して、それを撒いて新しい植物を得ることによって得ることができる。F1植物を自家受粉させて、F2世代を産生する、または戻し交雑スキームの戻し交雑親と戻し交雑することができる。戻し交雑植物をさらに、(戻し交雑)親と交雑させて、その後の世代における植物の作物栽培学上の価値を改善することができる、または自家受粉させて、L4対立遺伝子に関してホモ接合であって、SNFD表現型を発現しない植物を産生することができる。
【0096】
本発明は、本明細書においてカプシクム・アンヌームを参照して説明する。本発明の原理が同様にカプシクム属の他の種、およびより一般的にPMMoV抵抗性を付与するためにL4対立遺伝子を導入することができるナス科の植物に適応可能であることは、当業者には明らかであろう。基本的に、本発明の様々な局面は、PMMoVによって影響を受けうる全ての植物に関する可能性があり、本発明の原理はPMMoVの宿主範囲における如何なる植物種にも応用可能である。この文脈において、Brunt, A.A., Crabtree, K., Dallwitz, M.J., Gibbs, A.J., Watson, L. and Zurcher, E.J.(eds.)(1996以降)「Plant Viruses Online : Descriptions and Lists from the VIDE Database」Version : 20 th August 1996、URL: http://biology.anu.edu.au/Groups/MES/vide/として公式に引用される、しかしhttp://image.fs.uidaho.edu/vide/refs.htmにおいて認められる可能性があるウェブサイト刊行物において記述されているように、PMMoVに関する宿主範囲を特に参照されたい。この文脈において、Dallwitz, 1980およびDallwitz et al., 1993も同様に引用されなければならない。したがって、C.アンヌームに対する参照は本発明を制限すると解釈してはならない。
【0097】
カプシクム植物自身および果実のような消費に適したその一部の他に、本発明は繁殖に適した植物の部分を含む。先に述べたように、本発明は、カプシクム植物の植物の部分に限定されず、PMMoVの宿主範囲における任意の植物の部分に関する。繁殖に適した部分の例は、種子、葉、幹、根、シュート等、プロトプラスト、体細胞胚、葯、葉柄、培養細胞等のような器官組織である。本発明に従う植物は、通常の方法で、または植物の部分からの組織培養技術によっても栽培または繁殖させることができる。
【0098】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法は、PMMoV病原型1.2.3に対して感受性があるカプシクム属の適したレシピエント植物を提供する段階を含む。
【0099】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法はさらに、L4抵抗性対立遺伝子がレシピエント植物に対してPMMoV病原型1.2.3に対する抵抗性を付与する、例えばドナー植物からのL4抵抗性対立遺伝子をレシピエント植物に導入する段階を含む。
【0100】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法はさらに、抵抗性植物から分離集団を産生する段階を含んでもよい。
【0101】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法はさらに、分離集団の実質的にそれぞれの植物の後代を産生する段階を含んでもよい。例えば植物を交雑または自家受粉させることによって植物に種子を産生させる段階、および該種子を新しい植物に生長させる段階によって、後代または子孫を産生してもよい。または、後代は他の適した技術を用いて得てもよい。そのような代用法は、植物の育種領域において全く一般的である。
【0102】
PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法は、そのゲノムにL4抵抗性対立遺伝子が存在し、SNFD表現型に関与する遺伝情報が、少なくともSNFD表現型が発現されない程度まで存在しない植物を、該後代の少なくとも一つから選択する段階を含む。
【0103】
本発明の植物のゲノムにおけるL4抵抗性対立遺伝子の存在は、天然または人工の如何なる原因の結果であってもよく、例えば遺伝子移入、遺伝子操作(形質転換)、またはプロトプラスト融合の結果であってもよい。
【0104】
ドナー植物から、PMMoVに対して非抵抗性であるまたは低い抵抗性レベルを保有するレシピエント植物への、L4抵抗性をコードする一つまたは複数の遺伝子の遺伝子移入は、当技術分野で公知の技術を用いて行うことができる。例えば、PMMoV抵抗性をコードする一つまたは複数の遺伝子、すなわちL4対立遺伝子を、従来の育種技術を用いて、任意で例えばマーカーを用いる分子スクリーニング法と併用して、PMMoVに対して非抵抗性であるまたは低い抵抗性レベルを保有するレシピエントトウガラシ植物に遺伝子移入することができる。
【0105】
ドナー植物から、PMMoVに対して非抵抗性であるまたは低い抵抗性レベルを保有するレシピエント植物にPMMoV抵抗性をコードする一つまたは複数の遺伝子を遺伝子移入するために、従来の育種技術を適切に用いることができる。これらの方法は、そのような種、品種、または栽培品種を互いに交雑することができる限り、種、品種、または栽培品種の従来の交雑を必然的に伴う。計画されたドナーおよびレシピエント種を限られた程度に互いに交雑することができる場合に限り、ブリッジ(bridge)種を用いてもよい。
【0106】
系統図育種と呼ばれる一つの従来の育種法において、PMMoVに対して抵抗性を示し、PMMoV抵抗性をコードするL4対立遺伝子を含む第一のトウガラシ植物を、PMMoVに対して非抵抗性であるか、またはPMMoVに対して低レベルの抵抗性を保有し、および耐病性、害虫抵抗性、貴重な果実特徴等のような、しかしこれらに限定されない作物栽培学上または商業的に望ましい特徴を示す第二のトウガラシ植物と交雑させる。次に、得られた植物集団(すなわち、F1)を自家受粉させて種子(F2種子)を産生させる。
【0107】
F2種子から生長させたF2植物を、PMMoVに対する抵抗性に関してスクリーニングする。集団を多数の異なる方法でスクリーニングすることができる。第一に、集団を、本明細書において抵抗性バイオアッセイとも呼ばれる従来の病理疾患スクリーニングを用いてスクリーニングすることができる。PMMoVに関するそのようなバイオアッセイは当技術分野で公知である。具体的には、インキュベーターまたは温室において、個々の植物またはその一部をPMMoVに曝露して各植物で得られた抵抗性または感受性表現型を採点する。限定されない例として、植物を、以下のように温室においてスクリーニングすることができる:トウガラシ植物をその栽培に関して一般的に適用され、当業者に公知である生長条件で、例えば昼間の温度22℃、夜間温度20℃で通常の夏の光の条件で温室においてF2種子から成長させることができる。F2植物を、例えば同様に、一つまたは複数の抵抗性植物および一つまたは複数の感受性植物のような、対照植物を含むトレイにおいて生長させてもよい。接種物として感染した葉を用いてもよく、これを凍結保存してもよい。試験ウイルスを含む接種物は、感染した葉を水ですりつぶして(例えば、水100 mlにおいて凍結した葉1 g)、カーボランダム(シリコンカーバイド)を加えることによって調製してもよい。試験植物の接種は、例えばスポンジを用いて植物の第一の真の葉の表面領域(例えば最小で3.5×2 cm)に接種物をこすりつけることによって行ってもよい。初回の観察後、肉眼的に見て感受性がある(肉眼的症状は全身性のモザイク)である植物を除去してもよい。1週間後の2回目の観察の際に、感受性のある抵抗性植物の数を計数してもよい。葉の試験に関して、葉(植物の上部から4〜5 cm)を、湿った濾紙を含むトレイに置いてもよく、葉を濾紙の上に載せる。感受性および抵抗性対照をトレイに加えて、トレイをプラスチックで包んでもよく、光をあてる時間16時間/日で20℃でインキュベートしてもよい。接種は先に記述したようにスポンジによって起こってもよく、評価は6〜8日後に行う。
【0108】
または、従来の育種法を分子スクリーニング法と組み合わせて行ってもよい。そのような方法は一般的に、マーカー支援選抜またはマーカー支援育種と呼ばれ、所望の形質をコードする一つまたは複数の遺伝子(すなわち、L4対立遺伝子、またはその切断型)を含む子孫植物を同定するために、一つまたは複数の分子マーカーを用いることを必然的に伴う。ゲノムの遺伝子組成は、植物の表現型を予測して、冗長なバイオアッセイは不要となる。マーカー支援選抜を用いて、バイオアッセイスクリーニングから得た結果を確認してもよい。PMMoV抵抗性表現型を示すF2ハイブリッド植物は、PMMoV抵抗性をコードする所望の遺伝子を含み、次に商業的に望ましい特徴をさらに保有する植物を選択して、トウガラシ植物をますます近交系にするために多数の世代にわたって自家受粉させる。そのような育種および選択の結果は、PMMoV抵抗性に関連した遺伝子と共に商業的に重要な形質に関連した他の遺伝子に関して遺伝的にホモ接合である系統の産生である。
【0109】
次に本発明は、以下の段階を含む、PMMoV病原型1.2.3に対する抵抗性を示すカプシクム属の植物を産生する方法を提供する:
a)PMMoV病原型1.2.3に対して感受性があるカプシクム属の第一の植物またはその一部を提供する段階;
b)該植物のゲノムにおけるL4抵抗性対立遺伝子の存在により、PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の第二の植物を提供する段階;
c)PMMoV病原型1.2.3に対して抵抗性を示す後代植物を産生するために該第一および第二の植物を交雑させる段階;ならびに
d)対立遺伝子が、PMMoV病原型1.2.3に対する抵抗性を該後代植物に付与する遺伝情報を含み、SNFD表現型を付与する遺伝情報が該対立遺伝子に、少なくともSNFD表現型が発現されない程度まで存在しない、切断型L4抵抗性対立遺伝子の存在に関して該抵抗性後代植物のゲノムをさらにスクリーニングする段階。
【0110】
本方法の好ましい態様において、段階d)におけるスクリーニングは、図3に示されるように2群マーカー、1/3群マーカー、および3群マーカーからなる群より選択される、好ましくは図3に示される2群マーカーから選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーの不在を評価することによって行われる。
【0111】
本発明の方法に従って、選択される商業的に望ましい特徴には、SNFD表現型の不在が含まれ、このことは、PMMoV病原型1.2.3に対して抵抗性ではない商業的カプシクム品種と比較して、植物が非分離で、稔性で、正常な生長を示すことを意味する。これらの特徴は、任意の適した方法によって植物において同定されてもよい。例えば、正常な生長特徴は、矮性生長の不在が正常な生長を示している生長成績の肉眼的検分によって同定してもよい。稔性は、種子および/または花粉の量の肉眼的検分によって評価してもよい。非分離表現型は、先に記述したように交雑(抵抗性×感受性)のF1種子に関する抵抗性スクリーニングの試験結果の評価によって評価してもよい。
【0112】
または、SNFD表現型の不在は、特異的マーカーの不在に関して植物をスクリーニングすることによって評価してもよい。本発明の植物および方法の一つの特に好ましい態様において、本明細書において記述される2群のマーカーの一つまたは複数が存在しない。このように、一つの好ましい態様において、PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生するための本発明の方法は、抵抗性植物からの分離集団の後代の少なくとも一つから、そのゲノムにおいて本明細書に記述される1群の一つまたは複数、好ましくは全てのマーカーが存在し、およびそのゲノムにおいて本明細書に記述される2群の一つまたは複数のマーカーが存在しない植物を選択する段階を含む。
【0113】
本発明の植物のもう一つの態様において、本明細書において記述される3群のマーカーの一つまたは複数が存在しない。本発明の植物のさらにもう一つの態様において、本明細書に記述される1/3群のマーカーの一つまたは複数が存在しない。この文脈において、1/3群のマーカーの一つまたは複数が存在しないことは、そのような情報が存在しないことによって、その植物のゲノムにおけるL4抵抗性対立遺伝子の存在によりPMMoV病原型1.2.3に対して抵抗性を示す植物が得られないであろうことから、PMMoV病原型1.2.3に対する抵抗性に関与するゲノム領域に存在しないマーカーのみに関連すると理解すべきである。したがって、1/3群の一つまたは複数のマーカーの不在は、好ましくは、3群のマーカーの領域に対応する領域に存在するマーカーのみに関連し、好ましくは1群のマーカーの領域には関連しない。最も好ましい態様において、L4対立遺伝子は、一つまたは複数の2群マーカーが存在しないように切断される。この場合、切断はセントロメア側であるが、3群のマーカーの切断の場合、切断はテロメア側である。もう一つの最も好ましい態様において、切断は、TG036(Lefebvre et al., 2002;Ben-Chaim et al., 2001)マーカー(2群マーカーのメンバー)が存在しないように起こる。
【0114】
このように、L4対立遺伝子を提供された植物におけるSNFD表現型の不在は、本明細書において切断型L4対立遺伝子とも呼ばれるL4対立遺伝子を含むC.カコエンス遺伝子移入の切断型を植物に導入することによって実現してもよい。切断の程度は、当業者によって容易に決定することができる。例えば、1/3群のマーカーを含む領域が除去された場合に、PMMoV病原型1.2.3に対する抵抗性に関与する情報を有しないそれらの部分のみが除去される限り、トランスジェニック技法または組換えおよび選択のプロセスのいずれかによる、2群、3群、および/または1/3群のマーカーを含むそれらの遺伝子移入領域の任意の一つの部分的または完全な除去によって、適したレベルの切断を行うことができる。好ましくは、該切断は、2群のマーカーを含むL4対立遺伝子を含むC.カコエンス遺伝子移入領域の部分的または完全な除去を必然的に伴う。このように、トランスジェニック技法または組換えおよび選択のプロセスのいずれかによって、一つまたは複数の2群マーカーを含む領域を除去すると、もはやSNFD表現型を示さない植物が得られるであろう。最も好ましくは、この切断によって、TG036マーカーの除去が得られる(Lefebvre et al., 2002;Ben-Chaim et al., 2001)。
【0115】
切断レベルはまた、遺伝距離として定義してもよい。遺伝距離は、同じ染色体上の座のあいだの交差頻度によって測定される。二つの座が離れているほど、それらのあいだで交差が起こる可能性がより高くなる。逆に、二つの座が互いに近ければ、それらのあいだで交差が起こる可能性はより低い。座は、本明細書において定義される様々なマーカー群のマーカーによって示されてもよい。規則として、1センチモルガン(cM)は、座(マーカー)のあいだの1%組換えに等しい。それによって2群、3群および/または1/3群の一つまたは複数のマーカーの不在をもたらす切断レベルは、適切に0.001〜10 cM、より好ましくは0.01〜10 cMである可能性がある。または、遺伝子移入の存在を示すマーカー(例えば、1群の一つまたは複数のマーカー)と、切断されているマーカーとの遺伝距離は、適切に0.001〜10 cM、より好ましくは0.01〜10 cMであってもよい。
【0116】
新規かつ優れたPMMoV抵抗性近交系トウガラシ植物系統はまた、反復選択技術および戻し交雑技術を用いて開発することができる。この方法において、戻し交雑親を第一のドナー植物(戻し交雑親とは異なり、本明細書において「非戻し交雑親」と呼ばれる)と交雑させることによって、L4抵抗性を標的レシピエント植物(戻し交雑親と呼ばれる)に遺伝子移入することができる。戻し交雑親は、PMMoVに対して非抵抗性であるか、または低レベルの抵抗性を保有し、かつ耐病性、害虫抵抗性、貴重な果実特徴等のような、しかしこれらに限定されない商業的に望ましい特徴を保有し、SNFD表現型が存在しない植物である。戻し交雑技法は、「ドナー親」からの遺伝子を作物栽培学上の価値が高い遺伝的バックグラウンドに交雑するために広く応用されている。一般的に、優性遺伝子の栽培学上許容される表現型への遺伝子移入は、戻し交雑3〜5回の後に、例えば2〜3回の自家受粉によって得ることができ、全ての段階で栽培学上の価値に対する選択が行われる。非戻し交雑親は、PMMoV抵抗性を示し、L4抵抗性対立遺伝子を含む。非戻し交雑親またはドナー植物は、戻し交雑親または標的レシピエント植物と他家受精される任意の植物品種または近交系となりうる。戻し交雑親と非戻し交雑親との交雑によって得られた後代を、戻し交雑親と戻し交雑する。次に、得られた植物集団をスクリーニングする。集団は、多数の異なる方法でスクリーニングすることができる。第一に、集団は、本明細書において先に記述したように従来の病理スクリーニングを用いてスクリーニングすることができる。
【0117】
第二に、L4抵抗性対立遺伝子を含み、SNFD表現型に関連するマーカーの一つまたは複数が存在しない後代を同定するために、本明細書において先に記述した分子マーカーの一つまたは複数を用いて、マーカー支援選抜を行うことができる。または、マーカー支援選抜を用いて、抵抗性バイオアッセイスクリーニングから得られた結果を確認することができる。
【0118】
適切な選択を行った後、プロセスを繰り返す。戻し交雑親に対する戻し交雑、ならびにPMMoV抵抗性に関しておよびSNFD表現型(またはそれに関連したマーカー)の不在に関して選択するプロセスは、約5世代以上繰り返される。このプロセスから得られる後代は、L4対立遺伝子に関してヘテロ接合である。次に、L4対立遺伝子付与PMMoV抵抗性を含み、SNFD表現型を欠損するホモ接合の純粋な育種後代を提供するために、最後の戻し交雑世代を自家受粉させる。
【0119】
L4対立遺伝子に関してホモ接合であって、SNFD表現型を発現しない本明細書において記述されるPMMoV抵抗性近交系トウガラシ系統は、PMMoV抵抗性ハイブリッド植物を作製するためにさらなる交雑に用いることができる。例えば、第一のPMMoV抵抗性近交系トウガラシ植物を、耐病性、害虫抵抗性、望ましい果実特徴等のような、しかしこれに限定されない商業的に望ましい形質を保有する第二の近交系トウガラシ植物と交雑させることができる。この第二の近交系トウガラシ系統はPMMoVに対して抵抗性であってもなくてもよい。
【0120】
本発明の局面において用いられる選択技法は、植物からの核酸試料、好ましくは当技術分野において周知の核酸単離技法を用いて得られる可能性があるような、DNAの供給、およびマーカーの有無に関して該核酸の配列を試験することを必然的に伴う可能性がある。本明細書において定義されるマーカーは、その定義に至るAFLP反応を行うことによって、またはプライマーがその特異的鋳型にアニールすることができ、特異的増幅産物を産生することができる条件で、マーカー特異的プライマーの一つまたは複数の組によるPCRのような増幅反応を行うことによって、検出される可能性があるAFLPマーカーである。特異的増幅産物の存在は、鋳型配列におけるマーカーの存在を示す一方、増幅産物の不在はマーカーの不在を示す。L4遺伝子移入(L4対立遺伝子)の存在を確認するために、1群の一つまたは複数、好ましくは全てのマーカー、最も好ましくは本明細書において記述されるマーカーE58/M50-F-580および/またはマーカーE54/M55-F-101を核酸試料において検出する。適切に切断された対立遺伝子の存在を確認するために、好ましくは本明細書において記述される2群の一つまたは複数のマーカーが存在しない。AFLPマーカーのいくつかはまた、配列タグ部位マーカーに変換されてもよく、これによって交雑の後代の迅速かつ簡便なスクリーニングが可能となる(Werner et al., 2001)。
【0121】
または、植物試料のゲノムにおけるL4マーカーの有無に関してスクリーニングする方法は、プローブが、標的ポリヌクレオチド配列と選択的に結合して、安定なハイブリダイゼーション複合体を形成する条件で試料に接触される、一つまたは複数のマーカー配列を含む染色体領域上の標的ポリヌクレオチド配列に選択的に結合するプローブに、植物からの核酸試料を接触させる段階、およびハイブリダイゼーション複合体の有無を検出して、それによって植物における該マーカーの有無に関してスクリーニングする段階を含んでもよい。
【0122】
本明細書において先に記述した方法において用いられるマーカー支援選抜は、例えば、段階的に行うことができ、それによってL4対立遺伝子の存在およびSNFD表現型の不在が1代より多い世代において選択される。L4対立遺伝子付与抵抗性に関するマーカー支援選抜は、耐病性、害虫抵抗性、望ましい果実特徴等のような他の商業的に望ましい形質に関して試験および選択する前、同時、または後に行ってもよい。同様に、PMMoV抵抗性またはSNFD表現型の不在が評価される順序は、特に制限的ではない。SNFD表現型を付与する遺伝情報を含む植物の(好ましくはマーカー支援)排除は、PMMoV抵抗性植物の(好ましくはマーカー支援)選択前に、または同時に行ってもよいと理解すべきである。
【0123】
L4対立遺伝子が天然において周囲の環境から単離され、L4抵抗性を付与する遺伝情報のみが得られ、SNFD表現型を付与する遺伝情報を得ない材料源は、例えばC.カコエンスアクセッション番号PI 260429またはSA 185(Boukema, 1983)植物に由来する。そのようなアクセッション番号は、例えばCentre for Genetic Resources, the Netherlands(CGN), Wageningen, The NetherlandsのPlant Genetic Resources(PGR)クラスタから入手可能である。
【0124】
本発明に従う切断型L4抵抗性対立遺伝子を含む単離された核酸配列は、例えば、PMMoV病原型1.2.3に対して抵抗性である植物からDNAを単離すること、および該DNAからL4抵抗性対立遺伝子を単離することによって産生されてもよい。そのような単離は、例えば該L4対立遺伝子のPMMoV抵抗性付与部分を含む遺伝子領域を、例えば適したフォワードおよびリバースプライマーを用いて増幅することによって行ってもよい。適したプライマーは、例えば、3群のマーカーの定義のために用いられるマーカーと共に、2群のマーカーの定義のために用いられたプライマーである。増幅断片の大きさは、あまり離れていない、例えば1群のマーカーの位置により近い、C.カコエンスゲノムの位置においてアニールする他のプライマーを用いるまたは開発することによって低減させることができる。このようにして、SNFD表現型を付与する遺伝情報がL4対立遺伝子には存在しないことを達成することができる。当業者は、カプシクム属の植物において発現されるためにそのような核酸において存在しなければならず、それによって該植物にPMMoV病原型1.2.3に対する抵抗性を付与する様々な調節要素を承知している。
【0125】
L4対立遺伝子またはL4遺伝子は、例えば当技術分野で周知の標準的なDNA単離法を用いて、カプシクム植物からのゲノムDNAの試料を提供することによって単離してもよい。大きいDNA断片を検索するためには、単離体の質が比較的高いこと、すなわちゲノムDNAの重度の分解が本質的に回避されることが必要である。一般的に、高分子量(HMW)DNA調製物を得るための単離技術を用いる場合には、十分な大きさのDNAを得てもよい。HMW DNAは、調製の際に物理的剪断から保護されて得られるDNAである。カプシクム植物組織からのHMW DNAの調製は、例えば、細胞壁加水分解を用いてプロトプラストを単離する段階、およびアガロースにプロトプラストを抱埋する段階を必然的に伴ってもよい。もう一つの方法は、植物の核からHMW DNAを調製することである。メガベースDNAの単離は、多数の植物種に関して首尾よく行われ、そのような方法は、細菌の人工染色体(BAC)および酵母人工染色体(YAC)クローニングベクターにおいて大きいインサートDNAライブラリを構築するために適したメガベースの大きさのDNAを提供するために一般的に適用される。ゲノムDNAを単離した後、例えばPCR増幅反応において用いられる制限酵素を用いることによって、処置してもよく、または本明細書において提供されたマーカー配列のいずれかによってクローニングもしくはプロービングしてもよい。
【0126】
本発明はさらに、PMMoV感染症に対する抵抗性を示すトランスジェニック植物を提供するために、当技術分野において公知の技術を用いて、そのような単離および精製された遺伝子(または対立遺伝子)のトウガラシまたは他の植物への挿入を企図する。植物の形質転換は、植物細胞において機能するであろう発現ベクターの構築を必然的に伴う。本発明において、そのようなベクターは、プロモーターのような調節要素の制御下である、または機能的に連結したPMMoV抵抗性をコードする遺伝子を含むDNAを含む。発現ベクターは、組み合わせに含まれる遺伝子の少なくとも一つがPMMoV抵抗性をコードする限り、一つまたは複数のそのような機能的に連結した遺伝子/調節要素の組み合わせを含んでもよい。ベクターは、プラスミドの形であってもよく、アグロバクテリウム(Agrobacterium)形質転換系のような当技術分野において公知の形質転換法を用いて、PMMoVに対して抵抗性であるトランスジェニック植物を提供するために、単独で、または他のプラスミドと併用して用いることができる。
【0127】
発現ベクターには、マーカーを含む形質転換細胞を負の選択(選択マーカー遺伝子を含まない細胞の増殖を阻害することによって)、正の選択(遺伝子マーカーによってコードされる産物に関してスクリーニングすることによって)によって回収させる調節要素(プロモーターのような)に機能的に連結した少なくとも一つの遺伝子マーカーが含まれうる。植物の形質転換に関して多くの一般的に用いられる選択マーカー遺伝子は、当技術分野において公知であり、これには例えば抗生物質もしくは除草剤であってもよい選択的化学物質を代謝的に解毒する酵素をコードする遺伝子、または阻害剤に対して非感受性である変更された標的をコードする遺伝子が含まれる。マンノース選択のようないくつかの陽性選択法が当技術分野において公知である。または、マーカーを含まない形質転換を用いて先に述べたマーカー遺伝子を有しない植物を得ることができ、その技術は当技術分野において公知である。
【0128】
発現ベクターを植物に導入するための一つの方法は、アグロバクテリウムの天然形質転換系に基づく(例えば、Horsch et al., 1985を参照されたい)。A.ツメファシエンス(A. tumefaciens)およびA.リゾゲネス(A. rhizogenes)は、植物細胞を遺伝的に形質転換する植物病原性土壌細菌である。A.ツメファシエンスおよびA.リゾゲネスのTiおよびRiプラスミドはそれぞれ、植物の遺伝子形質転換に関与する遺伝子を有する(例えば、Kado, 1991を参照されたい)。植物組織に発現ベクターを導入する方法には、アグロバクテリウム・ツメファシエンスによる植物細胞の直接感染または同時培養が含まれる(Horsch et al., 1985)。アグロバクテリウムベクター系およびアグロバクテリウム媒介遺伝子移入のための方法の記述は、Gruber and Crosby, 1993およびMoloney et al., 1989によって提供される。同様に、米国特許第5,591,616号を参照されたい。植物発現ベクター、レポーター遺伝子、および形質転換プロトコールに関する一般的な記述、アグロバクテリウムベクター系に関する記述、ならびにアグロバクテリウム媒介遺伝子移入のための方法は、Gruber and Crosby, 1993において見いだされうる。植物組織を培養する一般的な方法は、例えば、Miki et al., 1993およびPhillips et al., 1988によって提供される。分子クローニング技術および適した発現ベクターに関する適切な参照ハンドブックは、Sambrook, J., E.F. Fritsch, and T. Maniatis,「Molecular Cloning : a Laboratory Manual」(3rd edition), Cold Spring Harbor Laboratory Press, Plainview, N.Y., 2000である。
【0129】
発現ベクターを植物に導入するためのもう一つの方法は、DNAがマイクロプロジェクタイルの表面において運ばれるマイクロプロジェクタイル媒介形質転換に基づく。発現ベクターは、植物細胞壁および膜を貫通するために十分である速度300から600 m/sでマイクロプロジェクタイルを加速する微粒子銃装置によって植物組織に導入される(Sanford et al., 1987, 1993;Sanford, 1988, 1990;Klein et al., 1988, 1992を参照されたい)。植物にDNAを導入するためのもう一つの方法は、標的細胞の超音波処理による(Zhang et al., 1991を参照されたい)。または、リポソームまたはスフェロプラスト融合が、発現ベクターを植物に導入するために用いられている(例えば、Deshayes et al., 1985およびChristou et al., 1987を参照されたい)。CaCl2沈殿、ポリビニルアルコール、またはポリ-L-オルニチンを用いるプロトプラストへのDNAの直接取り込みも同様に報告されている(例えば、Hain et al., 1985およびDraper et al., 1982を参照されたい)。プロトプラストならびに細胞および組織全体の電気穿孔も同様に記述されている(D'Halluin et al., 1992およびLaursen et al., 1994)。
【0130】
トウガラシ標的組織の形質転換後、上記の選択マーカー遺伝子の発現によって、当技術分野において現在は周知である再生および選択法を用いて、形質転換された細胞、組織、および/または植物の選択的選択が可能となる。
【0131】
前述の形質転換法は、ナス科の種のような、しかしこれに限定されないトランスジェニックトウガラシ植物または他の植物種を産生するために用いることができるであろう。先に述べたように、本発明の様々な局面は、原則的に、PMMoVによって影響を受けうる全ての植物に関し、このように、本発明に従うトランスジェニック植物はPMMoVの宿主範囲内の任意の植物種において産生される可能性がある。
【0132】
次に、PMMoV感染に対して抵抗性であるトウガラシまたは他の植物種のトランスジェニックハイブリッドを産生するために、そのようなトランスジェニック植物をもう一つの(非形質転換または形質転換)植物と交雑させることができる。または、本明細書に記述の形質転換技術を用いてPMMoV抵抗性をコードする外来(ヘテロ接合またはホモ接合)遺伝子を含むように操作されているトランスジェニックトウガラシまたは他の植物種におけるPMMoV抵抗性に関する外来の(異種)遺伝子を、当技術分野で周知である従来の育種技術(戻し交雑のような)を用いてもう一つの植物に移入することができるであろう。例えば、本明細書において先に考察したように、戻し交雑は、PMMoV抵抗性をコードするが、SNFD表現型に関与する遺伝情報を含まない外来(異種)遺伝子を含むトランスジェニックPMMoV抵抗性近交系トウガラシまたは他の植物系統から、その遺伝子を含まない非抵抗性トウガラシ植物または他の穀物にPMMoV抵抗性を遺伝子移入するために、またはPMMoV抵抗性をコードする外来遺伝子を含むトランスジェニックハイブリッドPMMoV抵抗性トウガラシ植物または他の植物から、その遺伝子を含まない系統に遺伝子移入するために用いることができるであろう。
【0133】
もう一つの態様において、プロトプラスト融合は、優れた新しいPMMoV抵抗性植物を作製するために用いることができる。より具体的には、第一のプロトプラストを、PMMoVによる感染に対して抵抗性を示すが、SNFD表現型に関与する遺伝情報を含まないトウガラシ植物または他の植物から得ることができる。第二のプロトプラストは、耐病性、害虫抵抗性、望ましい果実特徴等のような、しかしこれらに限定されない商業的に望ましい特徴を含む第二のトウガラシまたは他の植物品種から得ることができる。次に、プロトプラストを当技術分野において公知の従来のプロトプラスト融合技法を用いて融合させる。例えば、プロトプラスト融合は、膜の融合を促進するためにポリエチレングリコール(PEG)溶液を用いることによって行うことができる。そのような体細胞ハイブリダイゼーションは、種間ハイブリッドまたはその改変を産生するために、Sundberg et al., 1986によって開示された条件下で行ってもよい。しかし、当業者は、ポリエチレングリコール(PEG)を用いる以外の他の方法においてプロトプラスト融合を行うことができると認識するであろう。例えば、プロトプラストは、Koop and Spangenberg, 1989によって記述されるように電場誘導融合技術を用いて融合させることができる。さらに、プロトプラスト融合はデキストランおよびポリビニルアルコールによって行うことができる。プロトプラスト融合のためのさらなる方法は、Gleba et al., 1984およびDodds and Roberts, 1995において見いだされる可能性がある。
【0134】
例として、限定的ではなく、本発明の例を以下に示す。
【実施例】
【0135】
実施例
Sylvia(S & G品種「Cuby」の近交系)の雄性の親と、Manito(S & G品種「Tasty」の近交系)の雄性の親と呼ばれる二つの近交系親系統を産生した。いずれの品種SylviaおよびManitoも、PMMoV抵抗性に関して最初に分離した。植物の選択および試験交雑によって、双方の品種において抵抗性を固定することが可能であった。しかし、不安定な雄性不稔性/稔性不良および矮性生長のような形質が、双方の品種の雄性の親に存在することが認められた。
【0136】
これらの実施例において用いられるPMMoV病原型1.2.3は、Boukema, 1982によって言及された単離体P14と類似の特徴を有するウイルス単離体である。以下の実施例において、様々な実験において用いられたウイルス単離体は、PTG(Proefstation voor Tuinbouw onder Glas)、現在はPPO、Naaldwijk, The Netherlandsから得られ、これを機械的に繁殖させた。
【0137】
実施例1.同じ品種内の抵抗性および感受性トウガラシ植物におけるL4抵抗性マーカー
L4座の連鎖マップは、Keygene(Wageningen, The Netherlands)によって産生された。L座がマッピングされている領域は第11染色体南のテロメアに存在した(Lefebvre et al., 2002)。
【0138】
この座の周囲のマーカーは、四つの群に分けることができるであろう。L4座に連鎖した異なる群の位置を図1に示す。各群は、多くのマーカーを含み、各群の中に組換えは見いだされなかった。連鎖マップの染色体の残りに向けての方向は不明である。各群に存在するマーカーを表1に示す。
【0139】
(表1)それぞれの群におけるマーカーの概要
【0140】
分離品種Manito およびSylviaから、F1植物と共に抵抗性親株を、先に記述した方法と全く同様にバイオアッセイにおいて試験した。全体として、トレイは、感受性対照1個および抵抗性対照1個を含む試験植物35個からなった。幹および葉の試験のバイオアッセイは、本質的に先に記述したとおりに行った。バイオアッセイの結果を表2に示す。
【0141】
(表2)品種SylviaおよびManitoならびにその抵抗性親系統におけるバイオアッセイの結果
【0142】
表2に示すバイオアッセイから、抵抗性植物3個および感受性植物3個を、分離品種ManitoおよびSylviaの双方のF1植物から無作為に選択すると共に、その対応する抵抗性の雄性の親から植物5個を無作為に選択した。選択した植物を、L4座に連鎖した、表1に記載する4個のマーカーによってさらに分析した。試験が行われた当時、必ずしも表1に記載された全てのマーカーが同定されていたわけではない。それぞれの品種の中で、抵抗性植物3個および感受性植物3個を、様々なマーカーの存在に関して試験することによって分析した。対応する抵抗性の親から、植物5個を分析した。マーカー分析の結果を表3に記載する。
【0143】
(表3)バイオアッセイから選択された植物のマーカースコア(優性、本文を参照されたい)
番号はマーカー試験における試験番号に対応する;R、抵抗性;S、感受性;+、存在;−、不在。
【0144】
表3から認められうるように、L4対立遺伝子に連鎖したマーカーは全て、バイオアッセイ(R)において表現型の抵抗性を示すF1植物に存在した。しかし、感受性植物(S)では、L4マーカーはいずれも存在しなかった。Manito(番号29〜33)およびSylvia(番号40〜44)の抵抗性の雄性の親は、バイオアッセイにおいて分離を示さず、L4対立遺伝子に連鎖した全てのマーカーがこれらの植物において存在した。マーカーは、単に存在または不在(優性)として採点されたことから、遺伝子移入セグメントがホモ接合で存在するか、ヘテロ接合で存在するかを結論付けることはできなかった。F1における感受性はL4遺伝子移入の不在によって引き起こされると結論付けられた。さらなる試験から、L4抵抗性の遺伝における母株の効果が存在しないことが判明した。
【0145】
実施例2.非分離Sylvia品種の開発
非分離品種Sylviaを開発するために、抵抗性親系統(系統4578)の個々の植物を感受性の親植物と交雑した。L4抵抗性の分離の発生を、得られた個々のF1集団において評価した。結果を表4に記載する。
【0146】
植物番号4578-8を自家受粉させることによって、系統6636を得た。系統6636の個々の植物は、これらの植物が異なる(しかし関連する)感受性の雌性系統における雄性系統として用いられたことから1回より多く記載される。
【0147】
系統4578の植物はいずれも完全に抵抗性のF1を生じなかった。しかし、系統4578(vide系統6636)を自家受粉させることによって、抵抗性遺伝子を100%転移した株を発見することが可能であった。10例中4例において、これによって完全に抵抗性のF1が得られた。植物番号6636-4に由来するF1の分離は、他の系統とは異なるパターンを示す。表4に示すように、分離するF1集団(2R/3S−3R/2S)における抵抗性植物の数には変動がある。分離は、分離するF1集団における抵抗性植物と感受性植物の数を合わせると、より信頼できると期待される。全てのF1集団において同じ抵抗性起源が用いられていることから、非安定(分離)抵抗性の原因は同じである。表8は、表4において言及した三個の交雑(4578、6636交雑1、6636交雑2;6636の2回の交雑は、異なる二つの感受性の親を同一の抵抗性植物と交雑した結果である)のそれぞれの編集した結果を示す。
【0148】
(表4)個々の植物に由来するSylviaのF1分離
表4におけるF1集団を産生するために用いられる系統6636の個々の植物の自家受粉を、PMMoV抵抗性に関しても試験した。自家受粉した系統は全て完全な抵抗性を示した。結果を表5に記載する。
【0149】
(表5)親系統からの自家受粉の結果
【0150】
この実験から、L4対立遺伝子の望ましくない特徴が明確になり、非分離子孫をもたらす個々の植物の自家受粉ではホモ接合性が確認できるが、個々の抵抗性植物と感受性植物との交雑は分離F1子孫をもたらし、ヘテロ接合性が確認できると結論付けることができる。
【0151】
実施例3:非分離Manito品種の開発
実施例2における記述と類似のアプローチに従って非分離Manito株を開発した。抵抗性親系統の植物を感受性の親植物と個々に交雑させて、個々のF1集団における分離を評価した。結果を表6に記載する。低い稔性(SNFD表現型)を示す植物は全て交雑の前に除去し、これらの植物を表6において「試験していない」として記載する。低い稔性は当初、植物番号0025-5では観察されなかったが、後に存在するように思われた。
【0152】
(表6)個々の植物に由来するManitoのF1分離(初回スクリーニング)
【0153】
植物0025-5のみが非分離ハイブリッドを生じた。全体で60株中14株が稔性が低い(SNFD表現型)と印された。この表におけるF1集団の分離も同様に表8において編集する。
【0154】
非分離F1が得られる抵抗性植物の数を増加させるために、2回目の試験を行った。稔性が低いとして印をつけられた植物を交雑前に除去しなかった。結果を表7に記載して、分離を同様に、表8において編集する。全体で50株中12株(25%)が完全に抵抗性のF1集団を生じた。
【0155】
(表7)個々の植物に由来するManitoのF1分離(2回目のスクリーニング)
【0156】
(表8)実施例2および3における分離F1の編集結果
【0157】
全ての場合において同様に、感受性植物より抵抗性植物の数が多い。4578および6636-交雑1による分離は、1:1に近く、抵抗性の親がヘテロ接合である一遺伝子モデルに適合する。6636-交雑2および2825-2834において、分離は9:7により近い。9:7の分離は、F2集団に由来する。二つのホモ接合系統(系統は抵抗性に関して分離しない)を交雑することによって、F1において9:7の分離を得ることは可能ではない。ここでも、1:1分離が存在する可能性が高い。0021-0026の分離は、他とは異なり、3:1分離に最善に適合する。おそらくこの差は、植物が稔性に関して選択され、稔性の低い植物は交雑を行う前に除去されたという事実によって引き起こされる。
【0158】
実施例4.L4対立遺伝子の観察された非メンデル遺伝の説明としての連鎖ドラグ(drag)
緒言
表8において交雑のために用いた植物の中で、稔性および矮性の生長(SNFD表現型)を採点した。非分離F1をもたらす植物は全て、低い稔性を示した。これは、低い稔性と非分離との連鎖を示している。表7に示す結果は、植物の約25%が非分離ハイブリッドを生じることを示した。表6において、植物の25%は、稔性が低いと印され、F1の評価の前に除去された。稔性の低い植物が良好な抵抗性移入親植物であると見なした場合、表6および表7の結果は同等である。本明細書において言及されるように、これらの非分離植物に由来する近交系は実際に矮性生長を示した。このことから、負の形質(不安定な抵抗性、低い稔性、矮性生長)は、L4対立遺伝子に密接に連鎖している、または多面発現性であると結論付けることができる。
【0159】
F1における分離の性質に基づいて、実施例1〜3における観察に関して可能性がある説明を系統立てて、試験した。致死遺伝子が感受性対立遺伝子に連鎖する可能性、または致死遺伝子が抵抗性対立遺伝子に連鎖する可能性を試験したが、否定された。遺伝子サイレンシングの可能性、L4対立遺伝子の発現に関する第二の遺伝子の必要性、またはL4対立遺伝子の移入に関する第二の遺伝子の必要性も同様に否定された。さらに、トランスポゾンまたはインプリンティングの存在に関する証拠は見いだされなかった(遺伝子は、種子発達の際の特異的条件により沈黙化される)。さらに、抵抗性形質が一遺伝子性ではない可能性は、試験スキームの作製後に除外された。C.カコエンスの遺伝子移入セグメントによってC.アンヌーム染色体の染色体フォールディングの問題が起こるか否かをその後試験した。不規則なフォールディングは、染色体不均衡をもたらし、ホモ接合(L4L4)植物の減数分裂において問題を引き起こすであろう。次に、染色体対形成の際に、染色体の一つが損傷を受けて、それによって「-」対立遺伝子と呼ばれる欠損対立遺伝子が起こりうる。次に、これによって、抵抗性の喪失をもたらすであろう。分離F1において感受性植物ではマーカーが見いだされないことを考慮すると、染色体が切断されている可能性が高い。欠損対立遺伝子のホモ接合の存在が致死的である場合、L4遺伝子型の自家受粉によって、実際にバイオアッセイにおいて完全な抵抗性系統が得られるであろう。同様に、非分離F1は、このように誘導することができ、L4遺伝子型を感受性系統と交雑すると、F1は1:1の比で分離するであろう。ホモ接合L4遺伝子型(L4L4)を有する植物は、L4遺伝子型を自家受粉した後に見いだされうるが、これらの植物は減数分裂において問題に遭遇するであろうと予測される。次に、これは、バイオアッセイにおいて試験した場合に系統が完全に抵抗性である(すなわち、試験した全ての植物が抵抗性である)理由、および感受性の親と交雑した場合に分離F1集団が得られることがなおも見いだされる理由を説明するであろう。同様に、マーカーが、感受性があるF1集団の植物において見いだされない理由を説明するであろう。
【0160】
この説明は、非分離F1をもたらすL4L4遺伝子型が、染色体フォールディングの再配列の結果であることを予測する。この再配列フォールディングの状況において、染色体の対形成によって、染色体のアームの切断は起こらないであろう。しかし、植物内の他のプロセスが破壊される可能性はより高く、それによって低い稔性および低減された生長(矮性植物)をもたらす。これらの植物に由来する花粉の量はごく限られていることは公知である。同様に、そのような系統に由来する種子の量は少ないことが報告されている。
【0161】
したがって上記の実験から、L4対立遺伝子を含むC.カコエンスのゲノムの断片が大きすぎて、分離F1集団を引き起こすと結論された。したがって、このセグメントの大きさを減少させれば、問題を解決することができるであろう。
【0162】
実験のセットアップ
L4座の様々なマーカーの存在を、多数のトウガラシ系統において評価した。第一に、二つの抵抗性C.アンヌーム系統M-873およびM-3751を、遺伝子移入セグメントの大きさに関して評価した。これらの系統は、それによって非分離F1および正常な稔性をもたらすことが公知であった。セグメントの大きさを、L4対立遺伝子に隣接するマーカーに関してスクリーニングすることによって、S & G近交系(Manitoの父株)において存在する遺伝子移入と比較した。マーカーの位置に関しては、図2を参照されたい。結果を表9に記載する。
【0163】
系統を、L4座に連鎖したマーカー4個において試験し、L4対立遺伝子に対するマーカーの位置を表1に示す。表9に記載したマーカーは、ホモ接合が存在する、ヘテロ接合が存在する、または存在しないと採点される。同様に、抵抗性親系統の稔性およびF1の抵抗性を記載する。稔性の評価は花粉の量を「限られた量の花粉」(-:稔性不良)、「中間量の花粉」(+/-:中間の稔性)、および「非抵抗性の稔性対照植物と類似の花粉量」(+:良好な稔性)のように採点することによって、肉眼で行った。
【0164】
系統M-873およびM-3751における抵抗性は、De Ruiter Seeds, Bergschenhoek, The Netherlandsのトウガラシ育種系統に由来した。他の起源は全て、L4ドナーとしてS & G近交系を共有した。系統M-873およびM-3751のうち、良好なおよび中間の稔性を有する植物を、マーカーの存在または不在に関してスクリーニングした。中間の稔性は全ての場合において、Manitoの父系統における稔性より良好であった。他の系統は、稔性に基づいて選択された育種系統であった、またはそれらによって非分離ハイブリッドが起こったためであった。
【0165】
(表9)いくつかのトウガラシ系統におけるC.カコエンス遺伝子移入の評価
++:ホモ接合マーカーが存在する、抵抗性;-:ホモ接合マーカーが存在しない、感受性;+/-:ヘテロ接合マーカーが存在する、抵抗性;+:マーカーが存在する、対立遺伝子の差は試験していない;?:試験していない。
【0166】
結果は、系統M-873およびM-3751において、より小さい遺伝子移入セグメントが存在することを示している。2群のマーカー(E39/M58-F-65)は存在しない。これらの系統の系図を評価することによって、いずれも、vp-nr 91Pa0424、F3T13 7310を共有した。
【0167】
系統8972、8609、および8613の植物は、バイオアッセイにおいて完全な抵抗性スコアを示したにもかかわらず、ヘテロ接合スコアを生じた。株8972に関して、交雑を行った。結果を表10に記載する。
【0168】
(表10)表9のヘテロ接合スコア系統8972に由来するF1集団の結果
【0169】
系統8972から得られたF1集団の結果は、期待通りであった。さらなる組換え体を同定するために、分離系統を、L4-座マーカーの存在に関して分析した。3:1に最も近いF3分離に基づいて株を選択した。F1からF2への自家受粉の際に組換えが起こるはずであることから、それぞれのF3集団の植物をプールした。組換えが起こった場合、F3集団は組換えに関してホモ接合でなければならない。選択された株を、表11に記載する。
【0170】
(表11)組換え体同定のために選択された系統
これらの植物に関するマーカー分析の結果を表12に記載する。L4座のマーカーを表1に示すように示す。マーカースコア「+」は、マーカーの存在を示し、マーカースコア「-」は不在を示す。
【0171】
(表12)選択されたL4抵抗性および非SNFD表現型系統のマーカースコア
【0172】
L4抵抗性対立遺伝子のマーカー(1群マーカー)を含むこれらの選択系統において新しい組換え体は見いだされなかったが、2群のマーカーの位置を含む遺伝子材料を含まなかったと結論付けることができる。これらの系統は抵抗性であるが、SNFD表現型は観察されないことが見いだされた。したがって、2群のマーカー、または2群および3群のマーカーの位置を含む遺伝子材料の喪失に至る組換えによって、SNFD表現型の喪失をもたらすと結論付けられた。
【0173】
参照文献
【特許請求の範囲】
【請求項1】
L4対立遺伝子が切断されている、植物のゲノムにおけるL4抵抗性対立遺伝子の存在によってトウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対して抵抗性を示すカプシクム(Capsicum)属の植物。
【請求項2】
L4抵抗性対立遺伝子が、C.カコエンス(C. chacoense)のゲノムに由来する、請求項1記載の植物。
【請求項3】
白い花が咲くカプシクムである、請求項1または2記載の植物。
【請求項4】
白い花が咲くカプシクム植物が、C.アンヌーム(C. annuum)植物である、請求項3記載の植物。
【請求項5】
切断が、図3において示される2群マーカー、1/3群マーカー、および3群マーカーからなる群より選択される、好ましくは図3において示される2群のマーカーから選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーの不在を必然的に伴う、前記請求項のいずれか一項記載の植物。
【請求項6】
切断が、対立遺伝子のセントロメア側および/またはテロメア側から、好ましくは対立遺伝子のセントロメア側から約0.001〜10 cMの遺伝距離に及ぶヌクレオチド配列の除去を含む、前記請求項のいずれか一項記載の植物。
【請求項7】
L4抵抗性対立遺伝子が、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択される、好ましくは、マーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される、少なくとも一つのマーカーを含む、前記請求項のいずれか一項記載の植物。
【請求項8】
L4抵抗性対立遺伝子に関してホモ接合である、前記請求項のいずれか一項記載の植物。
【請求項9】
近交系植物である、請求項8記載の植物。
【請求項10】
請求項1〜9のいずれか一項記載の植物を、商業的に望ましい特徴を示すホモ接合、好ましくは近交系のトウガラシ植物と交雑させることによって得ることができる、PMMoV病原型1.2.3に対して抵抗性を示すハイブリッドトウガラシ植物。
【請求項11】
切断型L4抵抗性対立遺伝子が、カプシクム属の植物において発現されうる遺伝情報を含み、それによってPMMoV病原型1.2.3に対する抵抗性を前記植物に付与し、かつSNFD表現型を付与する遺伝情報が該対立遺伝子に存在しない、切断型L4抵抗性対立遺伝子を含む単離された核酸配列。
【請求項12】
抵抗性付与遺伝情報が、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択される少なくとも一つのマーカー、好ましくはマーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される、少なくとも一つのマーカーを含み、かつSNFD表現型を付与する遺伝情報が2群マーカーおよび3群マーカー、好ましくは図3に示される2群マーカーからなる群より選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーを含む、請求項11記載の単離核酸配列。
【請求項13】
L4抵抗性対立遺伝子がC.カコエンスのゲノムに由来する、請求項11または12記載の単離核酸配列。
【請求項14】
以下の段階を含む、PMMoV病原型1.2.3に対して抵抗性を示す、カプシクム属の植物を産生する方法:
a)PMMoV病原型1.2.3に対して感受性があるカプシクム属のレシピエント植物またはその一部を提供する段階;
b)前記植物のゲノムにおけるL4抵抗性対立遺伝子の存在により、PMMoV病原型1.2.3に対する抵抗性を示す、カプシクム属のドナー植物を提供する段階;
c)PMMoV病原型1.2.3に対して抵抗性を示す後代植物を産生するために、該レシピエント植物とドナー植物とを交雑させる段階;
d)対立遺伝子がPMMoV病原型1.2.3に対する抵抗性を後代植物に付与する遺伝情報を含み、SNFD表現型を付与する遺伝情報が少なくともSNFD表現型が発現されない程度まで該対立遺伝子に存在しない、切断型L4抵抗性対立遺伝子の存在に関して抵抗性後代植物のゲノムをスクリーニングする段階、および切断型L4抵抗性対立遺伝子を有する後代植物を選択する段階。
【請求項15】
段階d)におけるスクリーニングが、図3において示される2群マーカー、1/3群マーカー、および3群マーカー、好ましくは図3において示される2群マーカーからなる群より選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーの不在に関して、後代植物のゲノムをスクリーニングすることによって行われる、請求項14記載の方法。
【請求項16】
段階d)におけるスクリーニングが、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカー、好ましくは、マーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される少なくとも一つのマーカーの存在に関して、後代植物のゲノムをスクリーニングすることによって行われる、請求項14または15記載の方法。
【請求項17】
段階c)が抵抗性バイオアッセイを行う段階を含む、請求項14〜16のいずれか一項記載の方法。
【請求項18】
後代植物が、段階c)における交雑から得られたF1植物の自家受粉によって、または該交雑から得られたF1植物ともう一つのトウガラシ植物とを交雑させることによって、産生された分離集団の植物である、請求項14〜17のいずれか一項記載の方法。
【請求項19】
以下の段階を含む、PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生する方法:
a)PMMoV病原型1.2.3に対して感受性を示すカプシクム属のレシピエント植物またはその一部を提供する段階、および
b)切断型L4抵抗性対立遺伝子を含むゲノム領域を該レシピエント植物、その一部、またはその後代植物のゲノムに導入する段階であって、ここで、該対立遺伝子が、該植物、植物の一部、または後代植物において発現されうる遺伝情報を含み、それによって該植物、植物の一部、または後代植物にPMMoV病原型1.2.3に対する抵抗性を付与し、かつSNFD表現型を付与する遺伝情報が、少なくともSNFD表現型が発現されない程度まで該対立遺伝子に存在せず、かつ該ゲノム領域の導入が、インビトロ培養技術、プロトプラスト融合、形質転換、または倍加半数体法によって行われる、段階。
【請求項20】
前記植物の一部をトウガラシ植物に生長させる段階をさらに含む、請求項19記載の方法。
【請求項21】
以下の段階をさらに含む、請求項19または20記載の方法。
c)PMMoV病原型1.2.3抵抗性植物または後代植物を選択する段階、および
d)SNFD表現型を発現しない抵抗性植物または後代植物を選択する段階。
【請求項22】
段階c)が、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカー、好ましくは、マーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される少なくとも一つのマーカーの存在に関して、前記植物または後代植物のゲノムをスクリーニングする段階を含む、請求項21記載の方法。
【請求項23】
段階c)が抵抗性バイオアッセイを行う段階を含む、請求項21記載の方法。
【請求項24】
段階d)が、2群マーカーおよび3群マーカー、好ましくは2群マーカーからなる群より選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーに関して抵抗性植物または後代植物のゲノムにおける切断型L4抵抗性対立遺伝子をスクリーニングする段階、ならびに該マーカーの少なくとも一つが存在しない植物または後代植物を選択する段階を含む、請求項21〜23のいずれか一項記載の方法。
【請求項25】
請求項14〜24のいずれか一項記載の方法を行う段階および以下の段階をさらに行う段階を含む、PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の近交系植物を産生する方法:
e)選択した植物を自家受粉する段階;
f)該自家受粉から得られた種子を播種して、該種子を植物に生長させる段階;
g)PMMoV病原型1.2.3抵抗性を示し、商業的に望ましい特徴を保有する、段階g)からの植物を同定する段階;および
h)PMMoV病原型1.2.3抵抗性を示し、商業的に望ましい特徴を保有する近交系トウガラシ植物が産生されるまで、段階e)〜g)を繰り返す段階。
【請求項26】
ドナー植物がC.カコエンス植物であって、切断型L4抵抗性対立遺伝子がC.カコエンスのゲノムに由来する、請求項14〜25のいずれか一項記載の方法。
【請求項27】
レシピエント植物がC.アンヌーム植物である、請求項14〜26のいずれか一項記載の方法。
【請求項28】
請求項14〜27のいずれか一項記載の方法によって得ることができるPMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物。
【請求項29】
近交系植物である、請求項28記載の植物。
【請求項30】
請求項1〜10、28、または29のいずれか一項記載の植物の後代。
【請求項31】
葉、茎、根、シュート、果実等、プロトプラスト、体細胞胚、葯、葉柄、培養細胞、種子等からなる群より選択される、好ましくは器官組織による繁殖に適した、請求項1〜10、28、または29のいずれか一項記載の植物の一部または誘導体。
【請求項32】
果実のような、消費に適した、請求項1〜10、28、または29のいずれか一項記載の植物の一部または誘導体。
【請求項33】
請求項1〜10、28、または29のいずれか一項記載のトウガラシ植物を生長させることによって産生されたトウガラシの種子。
【請求項34】
請求項29記載の植物を、商業的に望ましい特徴を示すホモ接合の、好ましくは近交系であるトウガラシ植物と交雑させることによって得ることができる、PMMoV病原型1.2.3に対して抵抗性を示すハイブリッドトウガラシ植物またはその一部。
【請求項1】
L4対立遺伝子が切断されている、植物のゲノムにおけるL4抵抗性対立遺伝子の存在によってトウガラシマイルドモットルウイルス(PMMoV)病原型1.2.3に対して抵抗性を示すカプシクム(Capsicum)属の植物。
【請求項2】
L4抵抗性対立遺伝子が、C.カコエンス(C. chacoense)のゲノムに由来する、請求項1記載の植物。
【請求項3】
白い花が咲くカプシクムである、請求項1または2記載の植物。
【請求項4】
白い花が咲くカプシクム植物が、C.アンヌーム(C. annuum)植物である、請求項3記載の植物。
【請求項5】
切断が、図3において示される2群マーカー、1/3群マーカー、および3群マーカーからなる群より選択される、好ましくは図3において示される2群のマーカーから選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーの不在を必然的に伴う、前記請求項のいずれか一項記載の植物。
【請求項6】
切断が、対立遺伝子のセントロメア側および/またはテロメア側から、好ましくは対立遺伝子のセントロメア側から約0.001〜10 cMの遺伝距離に及ぶヌクレオチド配列の除去を含む、前記請求項のいずれか一項記載の植物。
【請求項7】
L4抵抗性対立遺伝子が、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択される、好ましくは、マーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される、少なくとも一つのマーカーを含む、前記請求項のいずれか一項記載の植物。
【請求項8】
L4抵抗性対立遺伝子に関してホモ接合である、前記請求項のいずれか一項記載の植物。
【請求項9】
近交系植物である、請求項8記載の植物。
【請求項10】
請求項1〜9のいずれか一項記載の植物を、商業的に望ましい特徴を示すホモ接合、好ましくは近交系のトウガラシ植物と交雑させることによって得ることができる、PMMoV病原型1.2.3に対して抵抗性を示すハイブリッドトウガラシ植物。
【請求項11】
切断型L4抵抗性対立遺伝子が、カプシクム属の植物において発現されうる遺伝情報を含み、それによってPMMoV病原型1.2.3に対する抵抗性を前記植物に付与し、かつSNFD表現型を付与する遺伝情報が該対立遺伝子に存在しない、切断型L4抵抗性対立遺伝子を含む単離された核酸配列。
【請求項12】
抵抗性付与遺伝情報が、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択される少なくとも一つのマーカー、好ましくはマーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される、少なくとも一つのマーカーを含み、かつSNFD表現型を付与する遺伝情報が2群マーカーおよび3群マーカー、好ましくは図3に示される2群マーカーからなる群より選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーを含む、請求項11記載の単離核酸配列。
【請求項13】
L4抵抗性対立遺伝子がC.カコエンスのゲノムに由来する、請求項11または12記載の単離核酸配列。
【請求項14】
以下の段階を含む、PMMoV病原型1.2.3に対して抵抗性を示す、カプシクム属の植物を産生する方法:
a)PMMoV病原型1.2.3に対して感受性があるカプシクム属のレシピエント植物またはその一部を提供する段階;
b)前記植物のゲノムにおけるL4抵抗性対立遺伝子の存在により、PMMoV病原型1.2.3に対する抵抗性を示す、カプシクム属のドナー植物を提供する段階;
c)PMMoV病原型1.2.3に対して抵抗性を示す後代植物を産生するために、該レシピエント植物とドナー植物とを交雑させる段階;
d)対立遺伝子がPMMoV病原型1.2.3に対する抵抗性を後代植物に付与する遺伝情報を含み、SNFD表現型を付与する遺伝情報が少なくともSNFD表現型が発現されない程度まで該対立遺伝子に存在しない、切断型L4抵抗性対立遺伝子の存在に関して抵抗性後代植物のゲノムをスクリーニングする段階、および切断型L4抵抗性対立遺伝子を有する後代植物を選択する段階。
【請求項15】
段階d)におけるスクリーニングが、図3において示される2群マーカー、1/3群マーカー、および3群マーカー、好ましくは図3において示される2群マーカーからなる群より選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーの不在に関して、後代植物のゲノムをスクリーニングすることによって行われる、請求項14記載の方法。
【請求項16】
段階d)におけるスクリーニングが、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカー、好ましくは、マーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される少なくとも一つのマーカーの存在に関して、後代植物のゲノムをスクリーニングすることによって行われる、請求項14または15記載の方法。
【請求項17】
段階c)が抵抗性バイオアッセイを行う段階を含む、請求項14〜16のいずれか一項記載の方法。
【請求項18】
後代植物が、段階c)における交雑から得られたF1植物の自家受粉によって、または該交雑から得られたF1植物ともう一つのトウガラシ植物とを交雑させることによって、産生された分離集団の植物である、請求項14〜17のいずれか一項記載の方法。
【請求項19】
以下の段階を含む、PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物を産生する方法:
a)PMMoV病原型1.2.3に対して感受性を示すカプシクム属のレシピエント植物またはその一部を提供する段階、および
b)切断型L4抵抗性対立遺伝子を含むゲノム領域を該レシピエント植物、その一部、またはその後代植物のゲノムに導入する段階であって、ここで、該対立遺伝子が、該植物、植物の一部、または後代植物において発現されうる遺伝情報を含み、それによって該植物、植物の一部、または後代植物にPMMoV病原型1.2.3に対する抵抗性を付与し、かつSNFD表現型を付与する遺伝情報が、少なくともSNFD表現型が発現されない程度まで該対立遺伝子に存在せず、かつ該ゲノム領域の導入が、インビトロ培養技術、プロトプラスト融合、形質転換、または倍加半数体法によって行われる、段階。
【請求項20】
前記植物の一部をトウガラシ植物に生長させる段階をさらに含む、請求項19記載の方法。
【請求項21】
以下の段階をさらに含む、請求項19または20記載の方法。
c)PMMoV病原型1.2.3抵抗性植物または後代植物を選択する段階、および
d)SNFD表現型を発現しない抵抗性植物または後代植物を選択する段階。
【請求項22】
段階c)が、マーカーE39/M58-F-95、E54/M55-F-101、E58/M60-F-255、およびE58/M50-F-580を含む群から選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカー、好ましくは、マーカーE54/M55-F-101およびE58/M50-F-580からなる群より選択される少なくとも一つのマーカーの存在に関して、前記植物または後代植物のゲノムをスクリーニングする段階を含む、請求項21記載の方法。
【請求項23】
段階c)が抵抗性バイオアッセイを行う段階を含む、請求項21記載の方法。
【請求項24】
段階d)が、2群マーカーおよび3群マーカー、好ましくは2群マーカーからなる群より選択されるL4抵抗性対立遺伝子の少なくとも一つのマーカーに関して抵抗性植物または後代植物のゲノムにおける切断型L4抵抗性対立遺伝子をスクリーニングする段階、ならびに該マーカーの少なくとも一つが存在しない植物または後代植物を選択する段階を含む、請求項21〜23のいずれか一項記載の方法。
【請求項25】
請求項14〜24のいずれか一項記載の方法を行う段階および以下の段階をさらに行う段階を含む、PMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の近交系植物を産生する方法:
e)選択した植物を自家受粉する段階;
f)該自家受粉から得られた種子を播種して、該種子を植物に生長させる段階;
g)PMMoV病原型1.2.3抵抗性を示し、商業的に望ましい特徴を保有する、段階g)からの植物を同定する段階;および
h)PMMoV病原型1.2.3抵抗性を示し、商業的に望ましい特徴を保有する近交系トウガラシ植物が産生されるまで、段階e)〜g)を繰り返す段階。
【請求項26】
ドナー植物がC.カコエンス植物であって、切断型L4抵抗性対立遺伝子がC.カコエンスのゲノムに由来する、請求項14〜25のいずれか一項記載の方法。
【請求項27】
レシピエント植物がC.アンヌーム植物である、請求項14〜26のいずれか一項記載の方法。
【請求項28】
請求項14〜27のいずれか一項記載の方法によって得ることができるPMMoV病原型1.2.3に対して抵抗性を示すカプシクム属の植物。
【請求項29】
近交系植物である、請求項28記載の植物。
【請求項30】
請求項1〜10、28、または29のいずれか一項記載の植物の後代。
【請求項31】
葉、茎、根、シュート、果実等、プロトプラスト、体細胞胚、葯、葉柄、培養細胞、種子等からなる群より選択される、好ましくは器官組織による繁殖に適した、請求項1〜10、28、または29のいずれか一項記載の植物の一部または誘導体。
【請求項32】
果実のような、消費に適した、請求項1〜10、28、または29のいずれか一項記載の植物の一部または誘導体。
【請求項33】
請求項1〜10、28、または29のいずれか一項記載のトウガラシ植物を生長させることによって産生されたトウガラシの種子。
【請求項34】
請求項29記載の植物を、商業的に望ましい特徴を示すホモ接合の、好ましくは近交系であるトウガラシ植物と交雑させることによって得ることができる、PMMoV病原型1.2.3に対して抵抗性を示すハイブリッドトウガラシ植物またはその一部。
【図1】

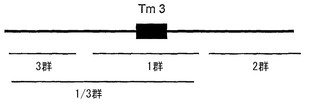
【図2】
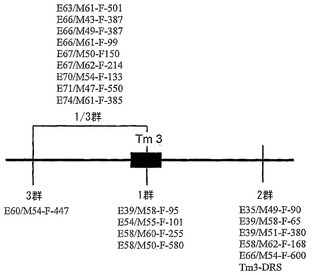
【図3】
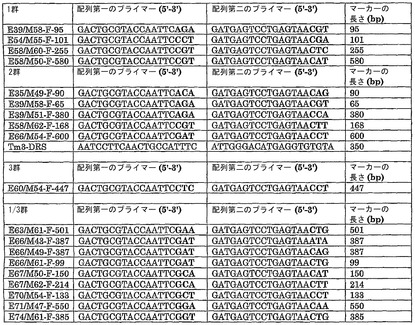
【図2】
【図3】
【公開番号】特開2013−78335(P2013−78335A)
【公開日】平成25年5月2日(2013.5.2)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−282235(P2012−282235)
【出願日】平成24年12月26日(2012.12.26)
【分割の表示】特願2007−534528(P2007−534528)の分割
【原出願日】平成17年9月30日(2005.9.30)
【出願人】(509316567)モンサント インベスト ビー.ブイ. (3)
【Fターム(参考)】
【公開日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願番号】特願2012−282235(P2012−282235)
【出願日】平成24年12月26日(2012.12.26)
【分割の表示】特願2007−534528(P2007−534528)の分割
【原出願日】平成17年9月30日(2005.9.30)
【出願人】(509316567)モンサント インベスト ビー.ブイ. (3)
【Fターム(参考)】
[ Back to top ]