
稲由来のオーキシン受容体タンパク質コード遺伝子及びその用途
稲由来の植物の細胞膜水素ポンプの活性化に関与するオーキシン受容体タンパク質、前記タンパク質をコードする遺伝子、前記遺伝子を含む組み換えベクター、前記組み換えベクターにより形質転換された宿主細胞、前記組み換え植物発現ベクターにより植物体を形質転換させて植物体の形質を改善する方法、前記組み換え植物発現ベクターにより形質転換されて形質の改善された植物体及びその種子、及び前記遺伝子を含む植物体の形質を改善するための組成物を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、多様な作物の農業特性に関与する、細胞膜水素ポンプの活性化に関与するオーキシン受容体タンパク質コード遺伝子及びその用途に関する。さらに詳細には、稲、トウモロコシなどの植物由来のオーキシン受容体ABP57タンパク質、遺伝子及び類似遺伝子と、これらを成長促進などの植物の水素ポンプの活性に関連する植物形質の改善のための活用に関する。
【背景技術】
【0002】
オーキシンは、多様な植物形質に関与する植物ホルモンである。ホルモンの生理調節活性は、受容体タンパク質とホルモンとの結合及びこれから派生する一連の信号伝達過程を通じて具現され、したがって、ホルモン受容体の発見は、ホルモンの調節メカニズムの理解のための最も基本的な要件である〔例えば、非特許文献1参照〕。植物の基本ホルモンの一つであるオーキシン受容体を発見するための研究は長期間行われてきており、その結果、ABP1、TIR1、ABP57などのタンパク質がオーキシン受容体として機能するということが報告されている〔例えば、非特許文献2参照〕。
【0003】
トウモロコシの二葉から最初に分離されたABP1の場合、22kDタンパク質であって、遺伝子分離、タンパク質のオーキシン結合、生理的機能などについて集中的な研究が蓄積されて、ABP1がオーキシンの生理的な効果に関連するということが明かになった。しかし、ABP1のIAA電離定数が比較的高く、オーキシン結合後にオーキシン信号伝逹のメカニズムを説明することができないため、ABP1が本当のオーキシン受容体であるか否かは未だに不明確である。オーキシンホルモンの処理時、発現が変化する遺伝子群を対象とする一連の研究を通じて明かになったTIR1は、タンパク質分解ターンオーバー(turn over)に関与するオーキシンホルモン受容体であることが明らかになった。しかし、TIR1は、オーキシンの遺伝子発現の調節以外には、オーキシンホルモンの多様な調節機能、例えば、オーキシンによる細胞壁の酸性化と種状のオーキシン用量反応曲線(dose−response curve)など、オーキシンホルモンの生理的な作用メカニズムをすべて説明できないという問題点がある。
【0004】
本発明者らは、以前の研究を通じて稲の幼苗及び根からオーキシンの細胞膜水素ポンプの活性化に直接関与する水溶性タンパク質を見い出した。〔例えば、非特許文献3、非特許文献4、非特許文献5、非特許文献6参照〕。本発明者らが見い出したタンパク質は、サイズが57kDであり、タンパク質とタンパク質との相互作用による水素ポンプの活性化、IAA結合定数、種状の水素ポンプの活性化に及ぼすIAA−dose responseを直接中継するという生化学的な特性を示して、オーキシン受容体としての特性をよく示しており、タンパク質のサイズを考慮してABP57と名付けた。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Badescu and Napier (2006) Trends Plant Sci 11, 217−223.
【非特許文献2】Hertel et al. (1972) Planta 107, 325−340.
【非特許文献3】Kim et al. (1997) FEBS Lett 409, 273−276.
【非特許文献4】Kim et al. (1998) FEBS Lett 438,241−244.
【非特許文献5】Kim et al. (2000) Plant Growth Regul 32, 143−150.
【非特許文献6】Kim et al. (2001) J Biol Chem 276, 10730−10736.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、上記のような要求に応じてなされたものであり、本発明者らは、植物の細胞膜水素ポンプの活性化に関与するオーキシン受容体ABP57の遺伝子を新たに分離し、それを植物の成長や発達など、農業的な有用形質改善に活用できるということを明らかにすることで本発明を完成するに至った。
【課題を解決するための手段】
【0007】
上記目的を達成するために、本発明は、稲由来の植物の細胞膜水素ポンプの活性化に関与するオーキシン受容体タンパク質を提供する。また、本発明は、前記タンパク質をコードする遺伝子を提供する。また、本発明は、前記遺伝子を含む組み換えベクターを提供する。また、本発明は、前記組み換えベクターにより形質転換された宿主細胞を提供する。また、本発明は、前記組み換え植物発現ベクターにより植物体を形質転換させて植物体の形質を改善する方法を提供する。
また、本発明は、前記組み換え植物発現ベクターにより形質転換されて形質の改善された植物体及びその種子を提供する。また、本発明は、前記遺伝子を含む植物体の形質を改善するための組成物を提供する。
【発明の効果】
【0008】
本発明によれば、オーキシン受容体ABP57遺伝子を新たに糾明し、試験管内の実験及び形質転換体を利用した植物体内の実験を通じて、ABP57遺伝子が植物の細胞膜水素ポンプの活性化やオーキシンホルモンIAAに結合する機能を有し、オーキシンの生理的機能に関連する植物の成長に関与するということを最初に提示した。植物の細胞膜水素ポンプが有する多様な生理及び生化学的機能を考慮すれば、本発明を通じて確保した新規な遺伝子を活用した優秀な形質の新作物の開発、ABP57と水素ポンプ及びIAAとの相互作用を利用した作物生長促進剤または作物保護剤の開発など、多様な産業上の利用可能性を期待することができる。
【図面の簡単な説明】
【0009】
【図1】稲の幼苗由来のオーキシン受容体ABP57を精製した後、SDS−PAGEを利用した電気泳動写真と、トリプシンで切断したペプチド断片のアミノ酸配列の分析結果を示す図である。
【図2】ペプチド断片のアミノ酸配列情報を利用してNCBIのタンパク質DBを活用したBLASTP分析を通じて取得したABP57遺伝子のORF(open reading frame)アミノ酸配列を示す図である。
【図3】稲の幼苗由来のtotal RNA(レーン1)及びgenomic DNA(レーン2)を増幅したABP57遺伝子の断片の電気泳動写真と、KOMEの稲完全長cDNA DBを活用したBLASTN分析を通じて取得したABP57の類似遺伝子群によってコードされた推定タンパク質の特性を示す図である。
【図4】ABP57の類似遺伝子群によってコードされたタンパク質のアミノ酸配列を比較した結果を示す図である。
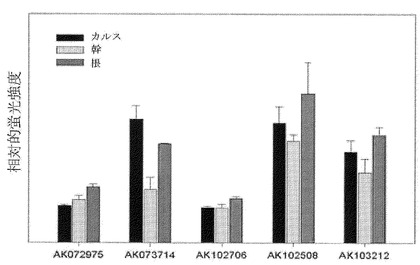
【図5】マイクロアレイ分析データを活用した稲カルス、幹及び根におけるABP57の類似遺伝子群の発現分析結果を示すグラフである。
【図6】大腸菌システムを利用したMBP(maltose binding protein)融合ABP57の生産と、カラムを利用したABP57の精製過程を示す図面代用写真である。
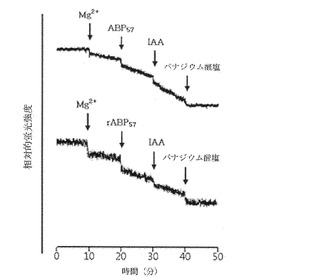
【図7】組み換えABP57による稲の細胞膜水素ポンプの活性化の検定結果を示すグラフである。
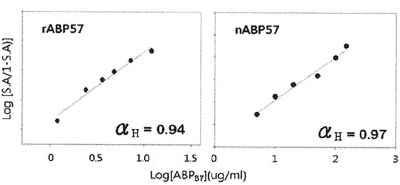
【図8】稲細胞膜活性化のABP57濃度依存度の測定によるHill Plot分析を通じて水素ポンプとABP57との相互作用の検定結果を示すグラフである。rABP57:recombinant ABP57 protein、nABP57:natural ABP57 protein。
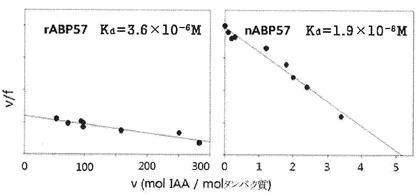
【図9】Scatchard plotを活用したABP57のIAA(indole−3−acetic acid)結合特性の検定結果を示すグラフである。rABP57: recombinant ABP57 protein、 nABP57: natural ABP57 protein。
【図10】ABP57遺伝子のシロイヌナズナ形質転換ベクターの模式図及び形質転換体遺伝子の発現分析結果を示す図である。
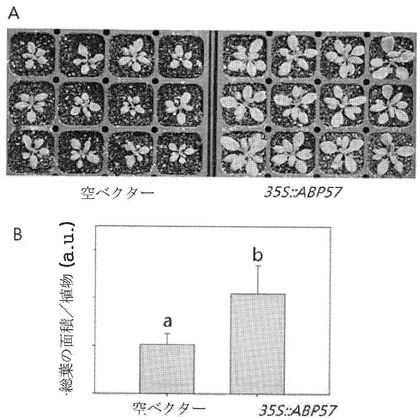
【図11】ABP57遺伝子の形質転換シロイヌナズナと対照区との成長特性を比較した図面代用写真(A)及びグラフ(B)である。
【図12】ABP57遺伝子により形質転換された稲(過発現系統)と対照区(Nakdong)との発芽特性を比較した図面代用写真である。
【発明を実施するための形態】
【0010】
本発明の目的を達成するために、本発明は、配列番号1乃至5のアミノ酸配列からなる群から選択される、稲由来の植物の細胞膜水素ポンプ(H+−ATPase)の活性化に関与するオーキシン受容体タンパク質を提供する。配列番号1乃至5のアミノ酸配列は、Protein IDがそれぞれBAC10824(ABP57)、BAC10825、BAC79737、BAC79739及びBAC83785である。好ましくは、上記オーキシン受容体タンパク質は、配列番号1のアミノ酸配列からなるABP57タンパク質である。
【0011】
本発明に係るオーキシン受容体タンパク質の範囲は、稲(Oryza sativa)から分離された配列番号1乃至5のアミノ酸配列を有するタンパク質及び上記タンパク質の機能的同等物を含む。“機能的同等物”とは、アミノ酸の付加、置換または欠失の結果、配列番号1乃至5のアミノ酸配列と少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上、さらに好ましくは95%以上の配列相同性を有するものであって、配列番号1乃至5のタンパク質と実質的に同質の生理活性を示すタンパク質を言う。
【0012】
また、本発明は、上述のようなオーキシン受容体タンパク質をコードする遺伝子を提供する。本発明の遺伝子は、オーキシン受容体タンパク質をコードするゲノムDNA及びcDNAをいずれも含む。好ましくは、本発明の遺伝子は、配列番号6乃至10の塩基配列を含むことができる。配列番号6乃至10の塩基配列は、cDNA IDがそれぞれAK072975、AK073714、AK102706、AK102508及びAK103212である。さらに好ましくは、本発明の遺伝子は、配列番号6の塩基配列からなるAK072975である。また、上述のような塩基配列の変異体が本発明の範囲内に含まれる。具体的に、上記遺伝子は、配列番号6乃至10の塩基配列とそれぞれ70%以上、好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上の配列相同性を有する塩基配列を含むことができる。ポリヌクレオチドについての“配列相同性の%”は、二つの最適に配列された配列と比較領域を比較することによって確認され、比較領域でのポリヌクレオチド配列の一部は、二つの配列の最適配列に対する参照配列(追加または削除を含まない)に比べて追加または削除(すなわち、ギャップ)を含むことができる。
【0013】
本発明者らは、稲の幼苗の幹からオーキシン受容体ABP57を精製し、精製されたタンパク質をトリプシンで切断した後、ESI−MS/MSで分析して4個のペプチドについてのアミノ酸情報を獲得した。NCBIのnon−redundant protein sequence DBを対象としてペプチド4種のアミノ酸配列情報を活用してBLAST検索を行って、現在までその機能が明らかになっていない遺伝子(AK072975)を確認した。登録された遺伝子の塩基配列に基づいてPCR(Polymerase Chain Reaction)プライマーをデザインして、稲の幼苗由来のtotal RNA及びgenomic DNAを増幅して遺伝子の断片を確保し、塩基配列の分析結果、AK072975と100%一致した。遺伝子のORFは1,539bpであり、512アミノ酸から構成されたpI 7.97、58.7kDaのタンパク質をコードした。
【0014】
上記のような遺伝子塩基配列情報を利用してKOME(Knowledge−based Oryza Molecular Biological Encyclopedia)の完全長cDNA DBとトウモロコシ染色体の塩基配列コンソーシアム(Maize Genome Sequence Consortium)のESTDB検索を行った結果、稲ではAK072975以外に機能が明らかになっていない4種の類似遺伝子(AK073714、AK102508、AK102706、AK103212)を確認し、各遺伝子のORFはそれぞれ53ないし73%のアミノ酸相同性を示し、トウモロコシにも相同性を示す機能未確認の遺伝子1種があることが確認された。稲60kのマイクロアレイ分析データを検討した結果、AK072975及び類似遺伝子は稲のカルス、幹及び根で発現された。
【0015】
上記のような遺伝子がコードするタンパク質が水素ポンプ活性化の機能を有するか否かを確認するために分離した遺伝子が含まれたタンパク質発現ベクターを作製し、大腸菌で発現させた後、組み換えタンパク質を純粋分離した。前記組み換えタンパク質を試料として稲の細胞膜水素ポンプ活性化の機能及びオーキシン結合機能を測定した結果、組み換えタンパク質がすべての特性を有することを観察し、ABP57をコードする遺伝子であるということを確認した。
【0016】
また、本発明は、本発明に係るオーキシン受容体タンパク質コード遺伝子を含む組み換えベクターを提供する。本発明の組み換えベクターは、好ましくは組み換え植物発現ベクターである。
【0017】
用語“組み換え”は、細胞が異種の核酸を複製するか、核酸を発現するか、またはペプチド、異種のペプチドまたは異種の核酸によって暗号化されたタンパク質を発現する細胞を指称する。組み換え細胞は、細胞の天然形態では発見されない遺伝子または遺伝子の断片を、センスまたはアンチセンスの形態のうち一つとして発現することができる。また、組み換え細胞は、天然状態の細胞で発見される遺伝子を発現することができるが、この遺伝子は変形されたものであって、人為的な手段によって細胞内に再導入されたものである。
【0018】
用語“ベクター”は、細胞内に伝達するDNA断片、核酸分子を指称するときに使用される。ベクターは、DNAを複製させ、宿主細胞から独立して再生産されることができる。用語“伝達体”は、よく“ベクター”と互換して使用される。用語“発現ベクター”は、目的としたコード配列と、特定の宿主生物で作動可能に連結されたコード配列の発現に必須な適正の核酸配列を含む組み換えDNA分子を意味する。真核細胞で利用可能なプロモーター、エンハンサー、終結信号及びポリアデニル化信号は公知のものである。
【0019】
組み換えベクターの好ましい例は、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)のような適当な宿主に存在するとき、そのものの一部、いわゆるT−領域を植物細胞に転移させ得るTi−プラスミドベクターである。他の類型のTi−プラスミドベクター(欧州特許第0116718号(B1)明細書参照)は、現在植物細胞、または雑種DNAを植物のゲノム内に適切に挿入させる新たな植物が生産され得る原形質体であって、雑種DNA配列を転移させるために利用されている。Ti−プラスミドベクターの特に好ましい形態は、欧州特許第0120516号(B1)明細書及び米国特許第4940838号明細書に請求されているような、いわゆるバイナリー(binary)ベクターである。本発明に係るDNAの植物宿主への導入に利用し得る他の適したベクターは、二重鎖植物ウイルス(例えば、CaMV)、一本鎖ウイルス、ゲミニウイルスなどに由来し得るウイルスベクター、例えば、非完全性植物ウイルスベクターから選択することができる。そのようなベクターの使用は、特に植物宿主を適切に形質転換することが難しいときに有利である。
【0020】
発現ベクターは、好ましくは、一つ以上の選択性マーカーを備えるであろう。このマーカーは、通常、化学的な方法によって選択され得る特性を有する核酸配列であって、形質転換された細胞を非形質転換細胞から区別できるすべての遺伝子がこれに該当する。その例としては、グリホサート(glyphosate)またはフォスフィノスリシン(phosphinothricin)のような除草剤抵抗性遺伝子、カナマイシン(Kanamycin)、G418、ブレオマイシン(Bleomycin)、ハイグロマイシン(hygromycin)、クロラムフェニコール(chloramphenicol)のような抗生剤耐性遺伝子があるが、これらに限定されるものではない。
【0021】
本発明の組み換えベクターにおいて、プロモーターは、CaMV 35S、アクチン、ユビキチン、pEMU、MASまたはヒストンプロモーターであり得るが、これらに制限されない。“プロモーター”とは、構造遺伝子からのDNAアップストリームの領域を意味し、転写を開始するためにRNAポリメラーゼが結合するDNA分子を言う。“植物プロモーター”とは、植物細胞で転写を開始することができるプロモーターである。“構成的(constitutive)プロモーター”は、大部分の環境条件及び発達状態または細胞分化下で活性のあるプロモーターである。形質転換体の選択が各ステップで各種の組織によって行われることができるため、構成的プロモーターが本発明で好ましい。したがって、構成的プロモーターは選択可能性を制限しない。
【0022】
本発明の組み換えベクターにおいて、通常のターミネータを使用可能であり、その例としては、ノパリンシンターゼ(NOS)、稲α−アミラーゼRAmy1 Aターミネータ、ファゼオリン(phaseoline)ターミネータ、アグロバクテリウム・ツメファシエンスのオクトピン(Octopine)遺伝子のターミネータなどがあるが、これらに限定されるものではない。ターミネータの必要性に関し、そのような領域が植物細胞における転写の確実性及び効率を向上させることが一般的に知られている。したがって、ターミネータの使用は本発明の内容で非常に好ましい。
【0023】
また、本発明は、本発明の組み換えベクターにより形質転換された宿主細胞を提供する。本発明のベクターを原核細胞に安定的かつ連続的にクローニング及び発現させることができる宿主細胞は、当業界に公知の如何なる宿主細胞も利用することが可能であり、例えば、E.coli JM109,E.coli BL21,E.coli RR1,E.coli LE392,E.coli B,E.coli X1776,E.coli W3110、枯草菌、バチルス・チューリンゲンシスのようなバチルス属菌株、そしてネズミチフス菌、セラチア・マルセッセンス及び多様なシュードモナス種のような腸内菌科菌株などがある。
【0024】
また、本発明のベクターを真核細胞に形質転換させる場合には、宿主細胞として、酵母(Saccharomyce cerevisiae)、昆虫細胞、ヒト細胞(例えば、CHO細胞株(Chinese hamster ovary)、W138、BHK、COS−7、293、HepG2、3T3、RIN及びMDCK細胞株)及び植物細胞などを利用することができる。宿主細胞は、好ましくは植物細胞である。
【0025】
本発明のベクターを宿主細胞内へ運搬する方法は、宿主細胞が原核細胞である場合、CaCl2法、ハナハン法〔Hanahan,D.,J.Mol.Biol.,166,557−580(1983)〕及び電気穿孔法などによって実施することができる。また、宿主細胞が真核細胞である場合には、微細注入法、リン酸カルシウム沈澱法、電気穿孔法、リポソーム−媒介形質感染法、DEAE−デキストラン処理法、及び遺伝子衝撃などによってベクターを宿主細胞内に注入することができる。
【0026】
また、本発明は、本発明の組み換え植物発現ベクターにより植物体を形質転換させるステップを含む植物体の形質を改善する方法を提供する。このような植物体の形質改善は、好ましくは、植物の成長促進でありうるが、これに制限されない。
【0027】
生体内のシステムを利用した遺伝子生理的機能の確認は非常に重要な過程である。本発明者らは、ABP57遺伝子をモデル植物であるシロイヌナズナで過発現させた後、成長特性を観察した結果、対照植物に比べて成長速度が2倍以上速くなったことを観察した。水素ポンプの活性化に関連する試験管内の実験結果を考慮すれば、シロイヌナズナで発現されたABP57遺伝子による成長促進は、シロイヌナズナの細胞膜にある水素ポンプの活性化と密接な関連があると推定される。
【0028】
また、本発明は、本発明の組み換え植物発現ベクターにより形質転換されて形質の改善された植物体を提供する。ここで植物体は、好ましくは成長の促進された植物体である。植物体としては、単子葉植物または双子葉植物が含まれる。単子葉植物の例としてはこれに限定されるものではないが、稲、小麦、麦、竹の子、トウモロコシ、サトイモ、アスパラガス、タマネギ、ニンニク、ネギ、ニラ、ヒメニラ、ナガイモ及びショウガがある。双子葉植物の例としてはこれに限定されるものではないが、シロイヌナズナ、ナス、タバコ、唐辛子、トマト、ゴボウ、シュンギク、チサ、キキョウ、ホウレン草、フダンソウ、サツマイモ、サラリ、ニンジン、セリ、パセリ、白菜、キャベツ、ガッム(大根の一種であって、葉はカラシナに、根は白菜に似ている)、スイカ、マクワウリ、キュウリ、カボチャ、フクベ、イチゴ、大豆、リョクトウ、隠元豆及びエンドウがある。このうち植物体は、好ましくはシロイヌナズナ(Arabidopsis thaliana)または稲である。
【0029】
また、本発明は、本発明の組み換え植物発現ベクターにより形質転換されて形質の改善された植物体の種子を提供する。
【0030】
また、本発明は、本発明の遺伝子を含む植物体の形質を改善するための組成物を提供する。本発明の組成物において、前記遺伝子は、好ましくは配列番号6乃至10の塩基配列からなり、さらに好ましくは、配列番号6の塩基配列からなることができる。前記植物体の形質改善は、好ましくは植物体の成長促進である。
【実施例】
【0031】
以下、本発明を実施例によって詳細に説明する。ただし、下記実施例は単に本発明を例示するためのものであり、本発明の内容が下記実施例に限定されるものではない。
【0032】
〔実施例1:RT−PCR法によるABP57遺伝子の増幅及び塩基配列の決定〕
Nakdong稲の硫化幼苗抽出液からDEAE及びCMイオン交換クロマトグラフィーとトリプトファン親和性クロマトグラフィー(tryptophan affinity chromatography)を用いてABP57を精製した後、トリプシンでタンパク質を切断し、切断されたタンパク質をESI−MS/MSを利用してアミノ酸配列を分析し、その結果、4種のペプチドについてのアミノ酸配列情報を得た(図1)。ペプチドのアミノ酸配列情報に基づいて、NCBIのタンパク質配列DBを検索した結果、4個のペプチドをすべて含む遺伝子1種(AK072975)を確保し、この遺伝子の塩基配列情報に基づいてPCR増幅用プライマーを作製した(図2)。
順方向プライマー(ABP57−5’):5’−ATGGCAGAGATTGTTAGTTC−3’(配列番号11)。
逆方向プライマー(RAPK1−2):5’−CTAAAATTTCAGGCGCAGTA−3’(配列番号12)。
遺伝子を分離するために、4週間成長させたNakdong稲の幼苗の幹からRNAとgenomic DNAとを分離してRT−PCRを行った。増幅された1.5kbのcDNA切片をpGEMT−easy vector(Promega)にクローニングし、全体の塩基配列を決定した。
増幅されたABP57遺伝子は、1,539bpと512アミノ酸から構成された58.7kDaのタンパク質をコードし(図3)、NCBIに登録されたAK072975と100%の相同性を示した。
【0033】
〔実施例2:ABP57類似遺伝子の塩基配列情報の確保及び遺伝子発現の分析〕
上述の遺伝子の塩基配列情報を利用してKOMEの完全長cDNA DB及びトウモロコシ染色体の塩基配列コンソーシアムのEST DBの検索を行った結果、稲ではAK072975遺伝子以外に4種の類似遺伝子(AK073714、AK102508、AK102706、AK103212)が確認され、各遺伝子のORFはそれぞれ53ないし73%のアミノ酸相同性を示し、トウモロコシにも相同性を示す遺伝子1種が確認された(図3及び図4)。稲60kのマイクロアレイ分析データを検討した結果、AK072975及び類似遺伝子は稲のカルス、幹及び根で発現された(図5)。
【0034】
〔実施例3:ABP57組み換えタンパク質による細胞膜水素ポンプの活性化〕
ABP57組み換えタンパク質を大腸菌で生産するために、pMAL−c2x(New England Biolabs)のBamH1及びHindIIIサイトにABP57 cDNAのORF全長をクローニングした。IPTGをpMAL−c2x/ABP57ベクターを有する菌(E. coli Rosetta(DE3)/pLysS)培養液に添加してABP57の生産を誘導し、菌抽出液中のMBP−fused ABP57をamylase resinを利用して精製した。融合タンパク質にTEV(Tobacco Etch Virus)protease 1%を添加して4℃で12時間反応させて、ABP57に結合されたMBPをとり除き、最終的にHiPrepTM Phenyl FF 16/10 columnを利用したFPLCを通じて精製した(図6)。
【0035】
精製されたタンパク質と稲から分離した細胞膜水素ポンプとを混合した後、蛍光分析器を利用して水素ポンプの活性を測定した結果、水素ポンプの活性が増大することが確認された(図7)。組み換えタンパク質の濃度による水素ポンプの活性化程度の変化を測定してHill Plot分析を行った結果、Hill定数は0.94とタンパク質とリガンドのnon−cooperative interactionである定数1に近い値を示し、以前研究の結果から確認された水素ポンプとnatural ABP57との相互作用に対するHill定数0.97と非常に類似していた(図8)。
組み換えタンパク質とオーキシンホルモンIAA結合定数の測定のためのScatchard分析を行った結果、組み換えタンパク質のIAA電離定数は3.6×10−6M、結合桁数は1個であり、natural ABP57のlow affinity binding siteに該当することが分かった(図9)。
【0036】
〔実施例4:形質転換植物を利用したABP57機能の糾明〕
ABP57遺伝子をPCRで増幅して植物形質転換ベクターを構築するために、一対のPCRプライマーを作製した。
ABP57−5’−1:5’−AGTCGGATCCATGGCAGAGA−3’(配列番号13)。
ABP57−3’−1:5’−AGTCCTCGAGCTAAAATTTC−3’(配列番号14)。
増幅されたABP57のORFを含む遺伝子の断片をpBI111LベクターのBamHI及びXhoIサイトに挿入し(図10)、それをアグロバクテリウム・ストレインC58C1に導入した。シロイヌナズナCol−0生態型宿主をflower dipping法で形質転換した後、収穫した種子をカナマイシンの含まれた培地で発芽させて形質転換体を選抜した後、ポットに移して成長させ、葉で導入遺伝子の発現をRT−PCRで確認し(図10)、形質転換体の成長をpBI111Lベクターのみを形質転換した対照区と比較した結果、大部分の形質転換体の成長速度が対照区に比べて約2倍速いことが確認された(図11)。一方、ABP57遺伝子により形質転換された稲の場合、対照区に比べて発芽及び初期成長が促進され(図12)、種子のサイズも対照区に比べて約4ないし8%大きくなった(表1:ABP57遺伝子により形質転換された稲種子と対照区である玄米100粒中との比較)。
【0037】
【表1】
【技術分野】
【0001】
本発明は、多様な作物の農業特性に関与する、細胞膜水素ポンプの活性化に関与するオーキシン受容体タンパク質コード遺伝子及びその用途に関する。さらに詳細には、稲、トウモロコシなどの植物由来のオーキシン受容体ABP57タンパク質、遺伝子及び類似遺伝子と、これらを成長促進などの植物の水素ポンプの活性に関連する植物形質の改善のための活用に関する。
【背景技術】
【0002】
オーキシンは、多様な植物形質に関与する植物ホルモンである。ホルモンの生理調節活性は、受容体タンパク質とホルモンとの結合及びこれから派生する一連の信号伝達過程を通じて具現され、したがって、ホルモン受容体の発見は、ホルモンの調節メカニズムの理解のための最も基本的な要件である〔例えば、非特許文献1参照〕。植物の基本ホルモンの一つであるオーキシン受容体を発見するための研究は長期間行われてきており、その結果、ABP1、TIR1、ABP57などのタンパク質がオーキシン受容体として機能するということが報告されている〔例えば、非特許文献2参照〕。
【0003】
トウモロコシの二葉から最初に分離されたABP1の場合、22kDタンパク質であって、遺伝子分離、タンパク質のオーキシン結合、生理的機能などについて集中的な研究が蓄積されて、ABP1がオーキシンの生理的な効果に関連するということが明かになった。しかし、ABP1のIAA電離定数が比較的高く、オーキシン結合後にオーキシン信号伝逹のメカニズムを説明することができないため、ABP1が本当のオーキシン受容体であるか否かは未だに不明確である。オーキシンホルモンの処理時、発現が変化する遺伝子群を対象とする一連の研究を通じて明かになったTIR1は、タンパク質分解ターンオーバー(turn over)に関与するオーキシンホルモン受容体であることが明らかになった。しかし、TIR1は、オーキシンの遺伝子発現の調節以外には、オーキシンホルモンの多様な調節機能、例えば、オーキシンによる細胞壁の酸性化と種状のオーキシン用量反応曲線(dose−response curve)など、オーキシンホルモンの生理的な作用メカニズムをすべて説明できないという問題点がある。
【0004】
本発明者らは、以前の研究を通じて稲の幼苗及び根からオーキシンの細胞膜水素ポンプの活性化に直接関与する水溶性タンパク質を見い出した。〔例えば、非特許文献3、非特許文献4、非特許文献5、非特許文献6参照〕。本発明者らが見い出したタンパク質は、サイズが57kDであり、タンパク質とタンパク質との相互作用による水素ポンプの活性化、IAA結合定数、種状の水素ポンプの活性化に及ぼすIAA−dose responseを直接中継するという生化学的な特性を示して、オーキシン受容体としての特性をよく示しており、タンパク質のサイズを考慮してABP57と名付けた。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Badescu and Napier (2006) Trends Plant Sci 11, 217−223.
【非特許文献2】Hertel et al. (1972) Planta 107, 325−340.
【非特許文献3】Kim et al. (1997) FEBS Lett 409, 273−276.
【非特許文献4】Kim et al. (1998) FEBS Lett 438,241−244.
【非特許文献5】Kim et al. (2000) Plant Growth Regul 32, 143−150.
【非特許文献6】Kim et al. (2001) J Biol Chem 276, 10730−10736.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、上記のような要求に応じてなされたものであり、本発明者らは、植物の細胞膜水素ポンプの活性化に関与するオーキシン受容体ABP57の遺伝子を新たに分離し、それを植物の成長や発達など、農業的な有用形質改善に活用できるということを明らかにすることで本発明を完成するに至った。
【課題を解決するための手段】
【0007】
上記目的を達成するために、本発明は、稲由来の植物の細胞膜水素ポンプの活性化に関与するオーキシン受容体タンパク質を提供する。また、本発明は、前記タンパク質をコードする遺伝子を提供する。また、本発明は、前記遺伝子を含む組み換えベクターを提供する。また、本発明は、前記組み換えベクターにより形質転換された宿主細胞を提供する。また、本発明は、前記組み換え植物発現ベクターにより植物体を形質転換させて植物体の形質を改善する方法を提供する。
また、本発明は、前記組み換え植物発現ベクターにより形質転換されて形質の改善された植物体及びその種子を提供する。また、本発明は、前記遺伝子を含む植物体の形質を改善するための組成物を提供する。
【発明の効果】
【0008】
本発明によれば、オーキシン受容体ABP57遺伝子を新たに糾明し、試験管内の実験及び形質転換体を利用した植物体内の実験を通じて、ABP57遺伝子が植物の細胞膜水素ポンプの活性化やオーキシンホルモンIAAに結合する機能を有し、オーキシンの生理的機能に関連する植物の成長に関与するということを最初に提示した。植物の細胞膜水素ポンプが有する多様な生理及び生化学的機能を考慮すれば、本発明を通じて確保した新規な遺伝子を活用した優秀な形質の新作物の開発、ABP57と水素ポンプ及びIAAとの相互作用を利用した作物生長促進剤または作物保護剤の開発など、多様な産業上の利用可能性を期待することができる。
【図面の簡単な説明】
【0009】
【図1】稲の幼苗由来のオーキシン受容体ABP57を精製した後、SDS−PAGEを利用した電気泳動写真と、トリプシンで切断したペプチド断片のアミノ酸配列の分析結果を示す図である。
【図2】ペプチド断片のアミノ酸配列情報を利用してNCBIのタンパク質DBを活用したBLASTP分析を通じて取得したABP57遺伝子のORF(open reading frame)アミノ酸配列を示す図である。
【図3】稲の幼苗由来のtotal RNA(レーン1)及びgenomic DNA(レーン2)を増幅したABP57遺伝子の断片の電気泳動写真と、KOMEの稲完全長cDNA DBを活用したBLASTN分析を通じて取得したABP57の類似遺伝子群によってコードされた推定タンパク質の特性を示す図である。
【図4】ABP57の類似遺伝子群によってコードされたタンパク質のアミノ酸配列を比較した結果を示す図である。
【図5】マイクロアレイ分析データを活用した稲カルス、幹及び根におけるABP57の類似遺伝子群の発現分析結果を示すグラフである。
【図6】大腸菌システムを利用したMBP(maltose binding protein)融合ABP57の生産と、カラムを利用したABP57の精製過程を示す図面代用写真である。
【図7】組み換えABP57による稲の細胞膜水素ポンプの活性化の検定結果を示すグラフである。
【図8】稲細胞膜活性化のABP57濃度依存度の測定によるHill Plot分析を通じて水素ポンプとABP57との相互作用の検定結果を示すグラフである。rABP57:recombinant ABP57 protein、nABP57:natural ABP57 protein。
【図9】Scatchard plotを活用したABP57のIAA(indole−3−acetic acid)結合特性の検定結果を示すグラフである。rABP57: recombinant ABP57 protein、 nABP57: natural ABP57 protein。
【図10】ABP57遺伝子のシロイヌナズナ形質転換ベクターの模式図及び形質転換体遺伝子の発現分析結果を示す図である。
【図11】ABP57遺伝子の形質転換シロイヌナズナと対照区との成長特性を比較した図面代用写真(A)及びグラフ(B)である。
【図12】ABP57遺伝子により形質転換された稲(過発現系統)と対照区(Nakdong)との発芽特性を比較した図面代用写真である。
【発明を実施するための形態】
【0010】
本発明の目的を達成するために、本発明は、配列番号1乃至5のアミノ酸配列からなる群から選択される、稲由来の植物の細胞膜水素ポンプ(H+−ATPase)の活性化に関与するオーキシン受容体タンパク質を提供する。配列番号1乃至5のアミノ酸配列は、Protein IDがそれぞれBAC10824(ABP57)、BAC10825、BAC79737、BAC79739及びBAC83785である。好ましくは、上記オーキシン受容体タンパク質は、配列番号1のアミノ酸配列からなるABP57タンパク質である。
【0011】
本発明に係るオーキシン受容体タンパク質の範囲は、稲(Oryza sativa)から分離された配列番号1乃至5のアミノ酸配列を有するタンパク質及び上記タンパク質の機能的同等物を含む。“機能的同等物”とは、アミノ酸の付加、置換または欠失の結果、配列番号1乃至5のアミノ酸配列と少なくとも70%以上、好ましくは80%以上、さらに好ましくは90%以上、さらに好ましくは95%以上の配列相同性を有するものであって、配列番号1乃至5のタンパク質と実質的に同質の生理活性を示すタンパク質を言う。
【0012】
また、本発明は、上述のようなオーキシン受容体タンパク質をコードする遺伝子を提供する。本発明の遺伝子は、オーキシン受容体タンパク質をコードするゲノムDNA及びcDNAをいずれも含む。好ましくは、本発明の遺伝子は、配列番号6乃至10の塩基配列を含むことができる。配列番号6乃至10の塩基配列は、cDNA IDがそれぞれAK072975、AK073714、AK102706、AK102508及びAK103212である。さらに好ましくは、本発明の遺伝子は、配列番号6の塩基配列からなるAK072975である。また、上述のような塩基配列の変異体が本発明の範囲内に含まれる。具体的に、上記遺伝子は、配列番号6乃至10の塩基配列とそれぞれ70%以上、好ましくは80%以上、さらに好ましくは90%以上、最も好ましくは95%以上の配列相同性を有する塩基配列を含むことができる。ポリヌクレオチドについての“配列相同性の%”は、二つの最適に配列された配列と比較領域を比較することによって確認され、比較領域でのポリヌクレオチド配列の一部は、二つの配列の最適配列に対する参照配列(追加または削除を含まない)に比べて追加または削除(すなわち、ギャップ)を含むことができる。
【0013】
本発明者らは、稲の幼苗の幹からオーキシン受容体ABP57を精製し、精製されたタンパク質をトリプシンで切断した後、ESI−MS/MSで分析して4個のペプチドについてのアミノ酸情報を獲得した。NCBIのnon−redundant protein sequence DBを対象としてペプチド4種のアミノ酸配列情報を活用してBLAST検索を行って、現在までその機能が明らかになっていない遺伝子(AK072975)を確認した。登録された遺伝子の塩基配列に基づいてPCR(Polymerase Chain Reaction)プライマーをデザインして、稲の幼苗由来のtotal RNA及びgenomic DNAを増幅して遺伝子の断片を確保し、塩基配列の分析結果、AK072975と100%一致した。遺伝子のORFは1,539bpであり、512アミノ酸から構成されたpI 7.97、58.7kDaのタンパク質をコードした。
【0014】
上記のような遺伝子塩基配列情報を利用してKOME(Knowledge−based Oryza Molecular Biological Encyclopedia)の完全長cDNA DBとトウモロコシ染色体の塩基配列コンソーシアム(Maize Genome Sequence Consortium)のESTDB検索を行った結果、稲ではAK072975以外に機能が明らかになっていない4種の類似遺伝子(AK073714、AK102508、AK102706、AK103212)を確認し、各遺伝子のORFはそれぞれ53ないし73%のアミノ酸相同性を示し、トウモロコシにも相同性を示す機能未確認の遺伝子1種があることが確認された。稲60kのマイクロアレイ分析データを検討した結果、AK072975及び類似遺伝子は稲のカルス、幹及び根で発現された。
【0015】
上記のような遺伝子がコードするタンパク質が水素ポンプ活性化の機能を有するか否かを確認するために分離した遺伝子が含まれたタンパク質発現ベクターを作製し、大腸菌で発現させた後、組み換えタンパク質を純粋分離した。前記組み換えタンパク質を試料として稲の細胞膜水素ポンプ活性化の機能及びオーキシン結合機能を測定した結果、組み換えタンパク質がすべての特性を有することを観察し、ABP57をコードする遺伝子であるということを確認した。
【0016】
また、本発明は、本発明に係るオーキシン受容体タンパク質コード遺伝子を含む組み換えベクターを提供する。本発明の組み換えベクターは、好ましくは組み換え植物発現ベクターである。
【0017】
用語“組み換え”は、細胞が異種の核酸を複製するか、核酸を発現するか、またはペプチド、異種のペプチドまたは異種の核酸によって暗号化されたタンパク質を発現する細胞を指称する。組み換え細胞は、細胞の天然形態では発見されない遺伝子または遺伝子の断片を、センスまたはアンチセンスの形態のうち一つとして発現することができる。また、組み換え細胞は、天然状態の細胞で発見される遺伝子を発現することができるが、この遺伝子は変形されたものであって、人為的な手段によって細胞内に再導入されたものである。
【0018】
用語“ベクター”は、細胞内に伝達するDNA断片、核酸分子を指称するときに使用される。ベクターは、DNAを複製させ、宿主細胞から独立して再生産されることができる。用語“伝達体”は、よく“ベクター”と互換して使用される。用語“発現ベクター”は、目的としたコード配列と、特定の宿主生物で作動可能に連結されたコード配列の発現に必須な適正の核酸配列を含む組み換えDNA分子を意味する。真核細胞で利用可能なプロモーター、エンハンサー、終結信号及びポリアデニル化信号は公知のものである。
【0019】
組み換えベクターの好ましい例は、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)のような適当な宿主に存在するとき、そのものの一部、いわゆるT−領域を植物細胞に転移させ得るTi−プラスミドベクターである。他の類型のTi−プラスミドベクター(欧州特許第0116718号(B1)明細書参照)は、現在植物細胞、または雑種DNAを植物のゲノム内に適切に挿入させる新たな植物が生産され得る原形質体であって、雑種DNA配列を転移させるために利用されている。Ti−プラスミドベクターの特に好ましい形態は、欧州特許第0120516号(B1)明細書及び米国特許第4940838号明細書に請求されているような、いわゆるバイナリー(binary)ベクターである。本発明に係るDNAの植物宿主への導入に利用し得る他の適したベクターは、二重鎖植物ウイルス(例えば、CaMV)、一本鎖ウイルス、ゲミニウイルスなどに由来し得るウイルスベクター、例えば、非完全性植物ウイルスベクターから選択することができる。そのようなベクターの使用は、特に植物宿主を適切に形質転換することが難しいときに有利である。
【0020】
発現ベクターは、好ましくは、一つ以上の選択性マーカーを備えるであろう。このマーカーは、通常、化学的な方法によって選択され得る特性を有する核酸配列であって、形質転換された細胞を非形質転換細胞から区別できるすべての遺伝子がこれに該当する。その例としては、グリホサート(glyphosate)またはフォスフィノスリシン(phosphinothricin)のような除草剤抵抗性遺伝子、カナマイシン(Kanamycin)、G418、ブレオマイシン(Bleomycin)、ハイグロマイシン(hygromycin)、クロラムフェニコール(chloramphenicol)のような抗生剤耐性遺伝子があるが、これらに限定されるものではない。
【0021】
本発明の組み換えベクターにおいて、プロモーターは、CaMV 35S、アクチン、ユビキチン、pEMU、MASまたはヒストンプロモーターであり得るが、これらに制限されない。“プロモーター”とは、構造遺伝子からのDNAアップストリームの領域を意味し、転写を開始するためにRNAポリメラーゼが結合するDNA分子を言う。“植物プロモーター”とは、植物細胞で転写を開始することができるプロモーターである。“構成的(constitutive)プロモーター”は、大部分の環境条件及び発達状態または細胞分化下で活性のあるプロモーターである。形質転換体の選択が各ステップで各種の組織によって行われることができるため、構成的プロモーターが本発明で好ましい。したがって、構成的プロモーターは選択可能性を制限しない。
【0022】
本発明の組み換えベクターにおいて、通常のターミネータを使用可能であり、その例としては、ノパリンシンターゼ(NOS)、稲α−アミラーゼRAmy1 Aターミネータ、ファゼオリン(phaseoline)ターミネータ、アグロバクテリウム・ツメファシエンスのオクトピン(Octopine)遺伝子のターミネータなどがあるが、これらに限定されるものではない。ターミネータの必要性に関し、そのような領域が植物細胞における転写の確実性及び効率を向上させることが一般的に知られている。したがって、ターミネータの使用は本発明の内容で非常に好ましい。
【0023】
また、本発明は、本発明の組み換えベクターにより形質転換された宿主細胞を提供する。本発明のベクターを原核細胞に安定的かつ連続的にクローニング及び発現させることができる宿主細胞は、当業界に公知の如何なる宿主細胞も利用することが可能であり、例えば、E.coli JM109,E.coli BL21,E.coli RR1,E.coli LE392,E.coli B,E.coli X1776,E.coli W3110、枯草菌、バチルス・チューリンゲンシスのようなバチルス属菌株、そしてネズミチフス菌、セラチア・マルセッセンス及び多様なシュードモナス種のような腸内菌科菌株などがある。
【0024】
また、本発明のベクターを真核細胞に形質転換させる場合には、宿主細胞として、酵母(Saccharomyce cerevisiae)、昆虫細胞、ヒト細胞(例えば、CHO細胞株(Chinese hamster ovary)、W138、BHK、COS−7、293、HepG2、3T3、RIN及びMDCK細胞株)及び植物細胞などを利用することができる。宿主細胞は、好ましくは植物細胞である。
【0025】
本発明のベクターを宿主細胞内へ運搬する方法は、宿主細胞が原核細胞である場合、CaCl2法、ハナハン法〔Hanahan,D.,J.Mol.Biol.,166,557−580(1983)〕及び電気穿孔法などによって実施することができる。また、宿主細胞が真核細胞である場合には、微細注入法、リン酸カルシウム沈澱法、電気穿孔法、リポソーム−媒介形質感染法、DEAE−デキストラン処理法、及び遺伝子衝撃などによってベクターを宿主細胞内に注入することができる。
【0026】
また、本発明は、本発明の組み換え植物発現ベクターにより植物体を形質転換させるステップを含む植物体の形質を改善する方法を提供する。このような植物体の形質改善は、好ましくは、植物の成長促進でありうるが、これに制限されない。
【0027】
生体内のシステムを利用した遺伝子生理的機能の確認は非常に重要な過程である。本発明者らは、ABP57遺伝子をモデル植物であるシロイヌナズナで過発現させた後、成長特性を観察した結果、対照植物に比べて成長速度が2倍以上速くなったことを観察した。水素ポンプの活性化に関連する試験管内の実験結果を考慮すれば、シロイヌナズナで発現されたABP57遺伝子による成長促進は、シロイヌナズナの細胞膜にある水素ポンプの活性化と密接な関連があると推定される。
【0028】
また、本発明は、本発明の組み換え植物発現ベクターにより形質転換されて形質の改善された植物体を提供する。ここで植物体は、好ましくは成長の促進された植物体である。植物体としては、単子葉植物または双子葉植物が含まれる。単子葉植物の例としてはこれに限定されるものではないが、稲、小麦、麦、竹の子、トウモロコシ、サトイモ、アスパラガス、タマネギ、ニンニク、ネギ、ニラ、ヒメニラ、ナガイモ及びショウガがある。双子葉植物の例としてはこれに限定されるものではないが、シロイヌナズナ、ナス、タバコ、唐辛子、トマト、ゴボウ、シュンギク、チサ、キキョウ、ホウレン草、フダンソウ、サツマイモ、サラリ、ニンジン、セリ、パセリ、白菜、キャベツ、ガッム(大根の一種であって、葉はカラシナに、根は白菜に似ている)、スイカ、マクワウリ、キュウリ、カボチャ、フクベ、イチゴ、大豆、リョクトウ、隠元豆及びエンドウがある。このうち植物体は、好ましくはシロイヌナズナ(Arabidopsis thaliana)または稲である。
【0029】
また、本発明は、本発明の組み換え植物発現ベクターにより形質転換されて形質の改善された植物体の種子を提供する。
【0030】
また、本発明は、本発明の遺伝子を含む植物体の形質を改善するための組成物を提供する。本発明の組成物において、前記遺伝子は、好ましくは配列番号6乃至10の塩基配列からなり、さらに好ましくは、配列番号6の塩基配列からなることができる。前記植物体の形質改善は、好ましくは植物体の成長促進である。
【実施例】
【0031】
以下、本発明を実施例によって詳細に説明する。ただし、下記実施例は単に本発明を例示するためのものであり、本発明の内容が下記実施例に限定されるものではない。
【0032】
〔実施例1:RT−PCR法によるABP57遺伝子の増幅及び塩基配列の決定〕
Nakdong稲の硫化幼苗抽出液からDEAE及びCMイオン交換クロマトグラフィーとトリプトファン親和性クロマトグラフィー(tryptophan affinity chromatography)を用いてABP57を精製した後、トリプシンでタンパク質を切断し、切断されたタンパク質をESI−MS/MSを利用してアミノ酸配列を分析し、その結果、4種のペプチドについてのアミノ酸配列情報を得た(図1)。ペプチドのアミノ酸配列情報に基づいて、NCBIのタンパク質配列DBを検索した結果、4個のペプチドをすべて含む遺伝子1種(AK072975)を確保し、この遺伝子の塩基配列情報に基づいてPCR増幅用プライマーを作製した(図2)。
順方向プライマー(ABP57−5’):5’−ATGGCAGAGATTGTTAGTTC−3’(配列番号11)。
逆方向プライマー(RAPK1−2):5’−CTAAAATTTCAGGCGCAGTA−3’(配列番号12)。
遺伝子を分離するために、4週間成長させたNakdong稲の幼苗の幹からRNAとgenomic DNAとを分離してRT−PCRを行った。増幅された1.5kbのcDNA切片をpGEMT−easy vector(Promega)にクローニングし、全体の塩基配列を決定した。
増幅されたABP57遺伝子は、1,539bpと512アミノ酸から構成された58.7kDaのタンパク質をコードし(図3)、NCBIに登録されたAK072975と100%の相同性を示した。
【0033】
〔実施例2:ABP57類似遺伝子の塩基配列情報の確保及び遺伝子発現の分析〕
上述の遺伝子の塩基配列情報を利用してKOMEの完全長cDNA DB及びトウモロコシ染色体の塩基配列コンソーシアムのEST DBの検索を行った結果、稲ではAK072975遺伝子以外に4種の類似遺伝子(AK073714、AK102508、AK102706、AK103212)が確認され、各遺伝子のORFはそれぞれ53ないし73%のアミノ酸相同性を示し、トウモロコシにも相同性を示す遺伝子1種が確認された(図3及び図4)。稲60kのマイクロアレイ分析データを検討した結果、AK072975及び類似遺伝子は稲のカルス、幹及び根で発現された(図5)。
【0034】
〔実施例3:ABP57組み換えタンパク質による細胞膜水素ポンプの活性化〕
ABP57組み換えタンパク質を大腸菌で生産するために、pMAL−c2x(New England Biolabs)のBamH1及びHindIIIサイトにABP57 cDNAのORF全長をクローニングした。IPTGをpMAL−c2x/ABP57ベクターを有する菌(E. coli Rosetta(DE3)/pLysS)培養液に添加してABP57の生産を誘導し、菌抽出液中のMBP−fused ABP57をamylase resinを利用して精製した。融合タンパク質にTEV(Tobacco Etch Virus)protease 1%を添加して4℃で12時間反応させて、ABP57に結合されたMBPをとり除き、最終的にHiPrepTM Phenyl FF 16/10 columnを利用したFPLCを通じて精製した(図6)。
【0035】
精製されたタンパク質と稲から分離した細胞膜水素ポンプとを混合した後、蛍光分析器を利用して水素ポンプの活性を測定した結果、水素ポンプの活性が増大することが確認された(図7)。組み換えタンパク質の濃度による水素ポンプの活性化程度の変化を測定してHill Plot分析を行った結果、Hill定数は0.94とタンパク質とリガンドのnon−cooperative interactionである定数1に近い値を示し、以前研究の結果から確認された水素ポンプとnatural ABP57との相互作用に対するHill定数0.97と非常に類似していた(図8)。
組み換えタンパク質とオーキシンホルモンIAA結合定数の測定のためのScatchard分析を行った結果、組み換えタンパク質のIAA電離定数は3.6×10−6M、結合桁数は1個であり、natural ABP57のlow affinity binding siteに該当することが分かった(図9)。
【0036】
〔実施例4:形質転換植物を利用したABP57機能の糾明〕
ABP57遺伝子をPCRで増幅して植物形質転換ベクターを構築するために、一対のPCRプライマーを作製した。
ABP57−5’−1:5’−AGTCGGATCCATGGCAGAGA−3’(配列番号13)。
ABP57−3’−1:5’−AGTCCTCGAGCTAAAATTTC−3’(配列番号14)。
増幅されたABP57のORFを含む遺伝子の断片をpBI111LベクターのBamHI及びXhoIサイトに挿入し(図10)、それをアグロバクテリウム・ストレインC58C1に導入した。シロイヌナズナCol−0生態型宿主をflower dipping法で形質転換した後、収穫した種子をカナマイシンの含まれた培地で発芽させて形質転換体を選抜した後、ポットに移して成長させ、葉で導入遺伝子の発現をRT−PCRで確認し(図10)、形質転換体の成長をpBI111Lベクターのみを形質転換した対照区と比較した結果、大部分の形質転換体の成長速度が対照区に比べて約2倍速いことが確認された(図11)。一方、ABP57遺伝子により形質転換された稲の場合、対照区に比べて発芽及び初期成長が促進され(図12)、種子のサイズも対照区に比べて約4ないし8%大きくなった(表1:ABP57遺伝子により形質転換された稲種子と対照区である玄米100粒中との比較)。
【0037】
【表1】
【特許請求の範囲】
【請求項1】
配列番号1乃至5のアミノ酸配列からなる群から選択される稲由来の植物の細胞膜水素ポンプの活性化に関与するオーキシン受容体タンパク質。
【請求項2】
請求項1に記載のタンパク質をコードする遺伝子。
【請求項3】
前記遺伝子は、配列番号6乃至10の塩基配列からなる群から選択されることを特徴とする請求項2に記載の遺伝子。
【請求項4】
請求項2に記載の遺伝子を含む組み換えベクター。
【請求項5】
組み換え植物発現ベクターであることを特徴とする請求項4に記載の組み換えベクター。
【請求項6】
請求項4に記載の組み換えベクターにより形質転換された宿主細胞。
【請求項7】
請求項5に記載の組み換え植物発現ベクターにより植物体を形質転換させるステップを含む植物体の形質を改善する方法。
【請求項8】
前記植物体の形質改善は植物の成長促進であることを特徴とする請求項7に記載の方法。
【請求項9】
請求項5に記載の組み換え植物発現ベクターにより形質転換されて形質の改善された植物体。
【請求項10】
前記植物体の形質改善は植物の成長促進であることを特徴とする請求項9に記載の植物体。
【請求項11】
請求項9に記載の植物体の種子。
【請求項12】
請求項2に記載の遺伝子を含む植物体の形質を改善するための組成物。
【請求項1】
配列番号1乃至5のアミノ酸配列からなる群から選択される稲由来の植物の細胞膜水素ポンプの活性化に関与するオーキシン受容体タンパク質。
【請求項2】
請求項1に記載のタンパク質をコードする遺伝子。
【請求項3】
前記遺伝子は、配列番号6乃至10の塩基配列からなる群から選択されることを特徴とする請求項2に記載の遺伝子。
【請求項4】
請求項2に記載の遺伝子を含む組み換えベクター。
【請求項5】
組み換え植物発現ベクターであることを特徴とする請求項4に記載の組み換えベクター。
【請求項6】
請求項4に記載の組み換えベクターにより形質転換された宿主細胞。
【請求項7】
請求項5に記載の組み換え植物発現ベクターにより植物体を形質転換させるステップを含む植物体の形質を改善する方法。
【請求項8】
前記植物体の形質改善は植物の成長促進であることを特徴とする請求項7に記載の方法。
【請求項9】
請求項5に記載の組み換え植物発現ベクターにより形質転換されて形質の改善された植物体。
【請求項10】
前記植物体の形質改善は植物の成長促進であることを特徴とする請求項9に記載の植物体。
【請求項11】
請求項9に記載の植物体の種子。
【請求項12】
請求項2に記載の遺伝子を含む植物体の形質を改善するための組成物。
【図1】


【図2】

【図3】

【図4】

【図5】
【図6】

【図7】
【図8】
【図9】
【図10】
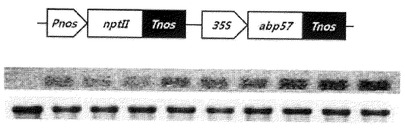
【図11】
【図12】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2011−526627(P2011−526627A)
【公表日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願番号】特願2011−525999(P2011−525999)
【出願日】平成22年7月21日(2010.7.21)
【国際出願番号】PCT/KR2010/004754
【国際公開番号】WO2011/016632
【国際公開日】平成23年2月10日(2011.2.10)
【出願人】(599151282)大韓民国農村振興庁 (16)
【氏名又は名称原語表記】REPUBLIC OF KOREA(MANAGEMENT RURAL DEVELOPMENT ADMINISTRATION)
【住所又は居所原語表記】250 Seodundong,Kwonseongu,Suwon,Gyeongido 441−707,Republic of Korea
【Fターム(参考)】
【公表日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願日】平成22年7月21日(2010.7.21)
【国際出願番号】PCT/KR2010/004754
【国際公開番号】WO2011/016632
【国際公開日】平成23年2月10日(2011.2.10)
【出願人】(599151282)大韓民国農村振興庁 (16)
【氏名又は名称原語表記】REPUBLIC OF KOREA(MANAGEMENT RURAL DEVELOPMENT ADMINISTRATION)
【住所又は居所原語表記】250 Seodundong,Kwonseongu,Suwon,Gyeongido 441−707,Republic of Korea
【Fターム(参考)】
[ Back to top ]