
空キャプシドを実質的に含まない組換えAAVビリオン調製物を生成するための方法
【課題】AAVベクター粒子ストックから空キャプシドを除去するかまたは空キャプシド数を減少させて、製造能力を増強する新規な方法を提供すること。
【解決手段】本発明は、空キャプシドの量が減少したrAAVビリオンストックを調製するための、効率的かつ商業的に実現可能な方法を開発したことに基づく。本発明者らは、本明細書において、空キャプシドが、カラムクロマトグラフィー技術を使用することによって、遺伝物質を含むrAAVビリオン(「AAVベクター粒子」)から分離され得ることを見出した。AAVベクター粒子とAAVの空キャプシドとの混合物からAAVの空キャプシドを分離するための方法が、記載される。この方法は、カラムクロマトグラフィー技術を使用し、商業的に実現可能なレベルの組換えAAVビリオンを提供する。
【解決手段】本発明は、空キャプシドの量が減少したrAAVビリオンストックを調製するための、効率的かつ商業的に実現可能な方法を開発したことに基づく。本発明者らは、本明細書において、空キャプシドが、カラムクロマトグラフィー技術を使用することによって、遺伝物質を含むrAAVビリオン(「AAVベクター粒子」)から分離され得ることを見出した。AAVベクター粒子とAAVの空キャプシドとの混合物からAAVの空キャプシドを分離するための方法が、記載される。この方法は、カラムクロマトグラフィー技術を使用し、商業的に実現可能なレベルの組換えAAVビリオンを提供する。
【発明の詳細な説明】
【技術分野】
【0001】
(技術分野)
本発明は、アデノ随伴ウイルス(AAV)ビリオンを精製するための方法に関する。より具体的には、本発明は、パッケージングされたゲノムを含む組換えAAV(rAAV)ビリオンを、パッケージングされたrAAVビリオンとそのパッケージングされたゲノムを欠くAAVの空キャプシドとの両方を含むAAVビリオン混合物から精製するための方法に関する。
【背景技術】
【0002】
(背景)
治療上有効な量の遺伝子産物を安全かつ持続的に患者へと送達する遺伝子治療方法が、現在開発中である。これらの方法を使用して、核酸分子が、患者へと直接導入され得る(インビボ遺伝子治療)か、または患者もしくはドナーから単離された細胞中へと導入され得、その後、その細胞は上記患者へと戻される(エキソビボ遺伝子治療)。その後、導入された核酸は、その患者自身の細胞または移植細胞が望ましい治療産物を産生するように仕向ける。遺伝子治療はまた、臨床家が、治療のために特定の器官または細胞標的(例えば、筋肉、血球、脳細胞など)を選択するのを可能にする。
【0003】
核酸は、いくつかの方法で患者の細胞中へと導入され得る。その方法としては、ウイルス媒介性遺伝子送達、裸DNA送達、およびトランスフェクション法が挙げられる。ウイルス媒介性遺伝子送達は、大部分の遺伝子治療試験において使用されている(C.P.Hodgson,Biotechnology(1995)13:222〜225)。最も一般的に使用される組換えウイルスは、レトロウイルス、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびアデノ随伴ウイルス(AAV)を基にしている。
【0004】
組換えアデノ随伴ウイルスベクターは、ヒト遺伝子治療のための遺伝子送達ベクターとしての見込みを保持していた。しかし、そのようなベクターを薬物として使用するための1つの重要な障害は、商業的に実現可能なレベルでそのベクターを生成および精製するための、真に拡張可能なプロセスの開発である。商業的使用のためにAAVベクター生成を拡張することに関与する難問の概説について、QuおよびWright,Cur.Opin.Drug Disc.and Develop.(2000)3:750〜755を参照のこと。最近、rAAVビリオンを精製するための拡張可能性のあるいくつかのカラムクロマトグラフィー技術が、開発された。これらのカラムクロマトグラフィーベースの精製方法は、rAAVビリオンが、大規模精製され得ることを実証したが、カラムクロマトグラフィーを使用して精製ビリオンを調製すると、かなりの量のAAVの空キャプシドを含む。空キャプシドと、目的とする異種遺伝子を含むビリオン(「AAVベクター粒子」)との代表的な比率は、約10以上である。すなわち、回収されたベクターのうちの約90%は、空キャプシドである。
【0005】
大量の空キャプシドの存在は、例えば、そのキャプシドタンパク質に対する望ましくない免疫応答を惹起することによって、または標的細胞表面結合部位について競合することによって、臨床的適用を妨害し得る。結果として、rAAVビリオン調製物から上記空キャプシドを除去する技術が、開発されている。これらの技術は、代表的には、超遠心分離(例えば、塩化セシウムまたはイオジキサノール中で勾配遠心分離)に依存する。そのような遠心分離技術は、労働集約的であり、代表的には、低ベクター収率を生じ、かつ拡張可能ではない。Kaludovら(2002)Hum.Gene Ther.13:1235〜1243は、アニオン交換カラムを使用して、rAAV−2ベクター、rAAV−4ベクター、およびrAAV−5ベクターを精製する方法を記載する。しかし、その実験は、溶出物をプールして画分を濃縮した後でさえ、パッケージされたゲノムとしてそれぞれ、2%、0.6%、および6.3%しか回収可能ではなかった。
【発明の概要】
【発明が解決しようとする課題】
【0006】
従って、AAVベクター粒子ストックから空キャプシドを除去するかまたは空キャプシド数を減少させて、製造能力を増強する新規な方法についての必要性が、存在する。
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明は、空キャプシドの量が減少したrAAVビリオンストックを調製するための、効率的かつ商業的に実現可能な方法を開発したことに基づく。本発明者らは、本明細書において、空キャプシドが、カラムクロマトグラフィー技術を使用することによって、遺伝物質を含むrAAVビリオン(「AAVベクター粒子」)から分離され得ることを見出した。この結果は、驚くべきものである。なぜなら、空キャプシドと、パッケージングされたキャプシドとは、同一の表面特性を有すると、以前には考えられていたからである。本発明者らが知る限りでは、これは、ウイルス粒子の電荷および/または電荷密度が、空キャプシドと完全粒子との間では異なることについての最初の証明である。本明細書中に記載される技術は、AAVの空キャプシドをAAVベクター粒子から分離するための効率的かつ拡張可能な方法を提供する。
【0008】
従って、一実施形態において、本発明は、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法に関する。上記方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)上記宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)上記粗細胞溶解物を、第一のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(d)上記AAVベクター粒子および上記AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(e)上記工程(d)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(f)低塩緩衝液を、上記工程(e)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが上記カラムに結合したままである条件下で添加する工程;ならびに
(g)AAVベクター粒子を含む上記工程(f)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する。
【0009】
さらなる実施形態において、上記方法は、
(h)上記工程(g)からの画分を、アニオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが存在する場合には上記カラムに結合する条件下で、適用する工程;
(i)低塩緩衝液を、上記工程(h)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が上記カラムに結合したままである条件下で添加する工程;
(j)高塩緩衝液を、上記工程(i)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(k)AAVベクター粒子を含む上記工程(i)からの画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する。
【0010】
なおさらなる実施形態において、本発明は、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法に関する。上記方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)上記宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)上記粗細胞溶解物を清澄化して、清澄化した細胞溶解物を提供する工程;
(d)上記清澄化した細胞溶解物を、機能的リガンドR−SO3−を有するマトリックスを含む第一のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(e)上記AAVベクター粒子および上記AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(f)上記工程(e)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(g)低塩緩衝液を、上記工程(f)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが上記カラムに結合したままである条件下で添加する工程;
(h)AAVベクター粒子を含む工程(g)からの溶出画分を収集する工程;
(i)上記工程(h)からの画分を、アニオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが存在する場合には上記カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、上記工程(i)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が上記カラムに結合したままである条件下で添加する工程;
(k)高塩緩衝液を、上記工程(j)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;ならびに
(l)AAVベクター粒子を含む上記工程(k)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する。
【0011】
別の実施形態において、本発明は、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法に関する。上記方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)上記宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)上記粗細胞溶解物を、カチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(d)上記AAVベクター粒子および上記AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(e)上記工程(d)からのAAV調製物を、アニオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(f)低塩緩衝液を、上記工程(e)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が上記カラムに結合したままである条件下で添加する工程;
(g)高塩緩衝液を、上記工程(f)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(h)AAVベクター粒子を含む工程(g)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する。
【0012】
さらなる実施形態において、上記方法は、
(i)上記工程(h)からのAAV調製物を、第二のアニオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが存在する場合には上記カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、上記工程(i)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が上記カラムに結合したままである条件下で添加する工程;
(k)高塩緩衝液を、上記工程(j)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(l)AAVベクター粒子を含む上記工程(k)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する。
【0013】
代替的な実施形態において、上記方法は、
(i)上記工程(h)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、上記工程(i)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが上記カラムに結合したままである条件下で添加する工程;ならびに
(k)AAVベクター粒子を含む上記工程(j)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する。
【0014】
上記の方法すべての特定の実施形態において、上記第一のカチオン交換カラムおよび/または第二のカチオン交換カラムは、カルボキシメチル化マトリックスまたはスルホン化マトリックス(例えば、機能的リガンドR−SO3−を含むマトリックス)を含む。
【0015】
上記の方法すべてのさらなる実施形態において、上記AAVベクター粒子は、上記AAV生成物中に少なくとも50%の量(例えば少なくとも75%の量、例えば少なくとも85%または少なくとも90%の量)で存在する。
【0016】
上記の方法すべてのなおさらなる実施形態において、上記AAVベクター粒子は、AAV−2またはAAV−5に由来する。
【0017】
本発明はまた、以下の項目を提供する。
(項目1)
AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法であって、該方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)該宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)該粗細胞溶解物を、第一のカチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(d)該AAVベクター粒子および該AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(e)該工程(d)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(f)低塩緩衝液を、該工程(e)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが該カラムに結合したままである条件下で添加する工程;ならびに
(g)AAVベクター粒子を含む該工程(f)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する、方法。
(項目2)
項目1に記載の方法であって、前記第一のカチオン交換カラムは、カルボキシメチル化マトリックスまたはスルホン化マトリックスを含む、方法。
(項目3)
項目2に記載の方法であって、前記マトリックスは、機能的リガンドR−SO3−を含む、方法。
(項目4)
項目1に記載の方法であって、前記第二のカチオン交換カラムは、カルボキシメチル化マトリックスまたはスルホン化マトリックスを含む、方法。
(項目5)
項目4に記載の方法であって、前記マトリックスは、機能的リガンドR−SO3−を含む、方法。
(項目6)
項目1に記載の方法であって、
(h)前記工程(g)からの画分を、アニオン交換クロマトグラフィーカラムに対して、前記AAVベクター粒子および前記AAVの空キャプシドが存在する場合には該カラムに結合する条件下で、適用する工程;
(i)低塩緩衝液を、該工程(h)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が該カラムに結合したままである条件下で添加する工程;
(j)高塩緩衝液を、該工程(i)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(k)AAVベクター粒子を含む該工程(j)からの画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する、方法。
(項目7)
AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法であって、該方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)該宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)該粗細胞溶解物を清澄化して、清澄化した細胞溶解物を提供する工程;
(d)該清澄化した細胞溶解物を、機能的リガンドR−SO3−を有するマトリックスを含む第一のカチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(e)該AAVベクター粒子および該AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(f)該工程(e)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(g)低塩緩衝液を、該工程(f)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが該カラムに結合したままである条件下で添加する工程;
(h)AAVベクター粒子を含む工程(g)からの溶出画分を収集する工程;
(i)該工程(h)からの画分を、アニオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが存在する場合には該カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、該工程(i)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が該カラムに結合したままである条件下で添加する工程;
(k)高塩緩衝液を、該工程(j)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;ならびに
(l)AAVベクター粒子を含む該工程(k)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する、方法。
(項目8)
AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法であって、該方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)該宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)該粗細胞溶解物を、カチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(d)該AAVベクター粒子および該AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(e)該工程(d)からのAAV調製物を、アニオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(f)低塩緩衝液を、該工程(e)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が該カラムに結合したままである条件下で添加する工程;
(g)高塩緩衝液を、該工程(f)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(h)AAVベクター粒子を含む工程(g)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する、方法。
(項目9)
項目8に記載の方法であって、
(i)前記工程(h)からのAAV調製物を、第二のアニオン交換クロマトグラフィーカラムに対して、前記AAVベクター粒子および前記AAVの空キャプシドが存在する場合には該カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、該工程(i)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が該カラムに結合したままである条件下で添加する工程;
(k)高塩緩衝液を、該工程(j)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(l)AAVベクター粒子を含む該工程(k)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する、方法。
(項目10)
項目8に記載の方法であって、
(i)前記工程(h)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、前記AAVベクター粒子および前記AAVの空キャプシドが該カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、該工程(i)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが該カラムに結合したままである条件下で添加する工程;ならびに
(k)AAVベクターを含む該工程(j)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する、方法。
(項目11)
項目1〜10のうちのいずれか1項に記載の方法であって、AAVベクター粒子は、前記AAV生成物中に少なくとも75%の量で存在する、方法。
(項目12)
項目1〜10のうちのいずれか1項に記載の方法であって、AAVベクター粒子は、前記AAV生成物中に少なくとも85%の量で存在する、方法。
(項目13)
項目1〜10のうちのいずれか1項に記載の方法であって、AAVベクター粒子は、前記AAV生成物中に少なくとも90%の量で存在する、方法。
(項目14)
項目1〜10のうちのいずれか1項に記載の方法であって、前記AAVベクター粒子は、AAV−2に由来する、方法。
(項目15)
項目1〜10のうちのいずれか1項に記載の方法であって、前記AAVベクター粒子は、AAV−5に由来する、方法。
本発明のこれらの実施形態および他の実施形態は、本明細書中の開示を考慮すると当業者には容易に思い浮かぶ。
【図面の簡単な説明】
【0018】
【図1】図1Aおよび1Bは、AAVベクター粒子およびAAV空キャプシドの両方を含む粗溶解物の結合特徴を示す。図1Aにおいて、試験した樹脂は、以下の通りであった:レーン1:コントロール;レーン2および3:MACRO PREP Q(BioRad,Hercules,CAから入手可能な強アニオン交換体);レーン4および5:UNOSPHERE Q(BioRad,Hercules,CAから入手可能な強アニオン交換体);レーン6および7:POROS 50HQ(Applied Biosystems,Foster City,CAから入手可能な強アニオン交換体);レーン8および9:POROS 50D(Applied Biosystems,Foster City,CAから入手可能な弱アニオン交換体)。図1Bにおいて、試験した樹脂は、以下の通りであった:レーン1:コントロール;レーン2および3:POROS 50PI(Applied Biosystems,Foster City,CAから入手可能な弱アニオン交換体);レーン4および5:SOURCE 30Q(Amersham Biosciences,Piscataway,NJから入手可能な強アニオン交換体);レーン6および7:DEAE SEPHAROSE(Amersham Biosciences,Piscataway,NJから入手可能な弱アニオン交換体);レーン8および9:Q SEPHAROSE(Amersham Biosciences,Piscataway,NJから入手可能な強アニオン交換体)。図1Aおよび1Bの両方について、レーン2、4、6および8は、低塩(50mM NaCl)洗浄画分を使用し;レーン3、5、7および9は、高塩(1M NaCl)洗浄画分を使用した。
【図2】図2は、実施例において記載されるように、アニオン交換クロマトグラフィーを使用して分離する前および分離した後の、AAV空キャプシドおよびAAVベクター粒子(Vgs)の分析を示す。
【図3】図3は、実施例において記載されるように、アニオン交換カラムからの画分の銀染色したSDS−PAGEゲルを示す。レーン1:AAVベクター粒子;レーン2〜5:ベクター溶出画分;レーン6:タンパク質分子量標準物質。
【図4】図4は、実施例に記載されるように、カチオン交換カラムからの溶出画分(レーン12〜21)を示す銀染色したSDS−PAGEゲルを示す。
【図5】図5は、カチオン交換カラムクロマトグラフィーを用いて、AAVベクター粒子からAAV空粒子を分離することを示す、銀染色したSDS−PAGEゲルを示す。レーン1:出発物質;レーン2〜4:カチオン交換カラムから溶出したベクターの3つの独立したサンプル。
【発明を実施するための形態】
【0019】
(発明の詳細な説明)
本発明の実施は、別段記載されなければ、ウイルス学、微生物学、分子生物学および組換えDNA技術の従来の方法を使用し、これらの方法は、当該分野の技術範囲内である。このような技術は、文献中に十分に説明されている。例えば、Sambrookら.Molecular Cloning:A Laboratory Manual(最新版);DNA Cloning:A Practical Approach,Vol.I&II(D.Glover,編);Oligonucleotide Synthesis.(N.Gait,編,最新版);Nucleic Acid Hybridization(B.Hames&S.Higgins,編,最新版);Transcription and Translation(B.Hames&S.Higgins,編、最新版);CRC Handbook of Parvoviruses,vol.I&II(P.Tijssen,編);Fundamental Virology,第2版,vol.I&II(B.N.Fields and D.M.Knipe,編);Freshney Culture of Animal Cells,A Manual of Basic Technique(Wiley−Liss,第3版);ならびにAusubelら(1991)Current Protocols in Molecular Biology(Wiley Interscience,NY)を参照のこと。
(1.定義)
本発明を記載するにあたって、以下の用語が使用され、以下に示されるように定義されると解釈される。
【0020】
本明細書および添付の特許請求の範囲において使用される場合、単数形「a」、「an」、および「the」は、文脈が明らかに別のことを示さなければ、複数形の指示対象を包含することに注意しなければならない。従って、例えば、「(1つの)パッケージされたキャプシド」への言及は、2つ以上のこのようなキャプシドの混合物などを包含する。
【0021】
「ベクター」とは、任意の遺伝的エレメント(例えば、プラスミド、ファージ、トランスポゾン、コスミド、染色体、ウイルス、ビリオンなど)を意味し、これらは、適切な制御エレメントと会合した場合に複製し得、細胞間で遺伝子配列を運び得る。従って、この用語は、クローニングビヒクルおよび発現ビヒクル、ならびにウイルスベクターを包含する。
【0022】
「AAVベクター」とは、以下の(しかし、それらの限定されない)アデノ随伴ウイルス血清型に由来するベクターを意味する:AAV−1、AAV−2、AAV−3、AAV−4、AAV−5、AAV−6、AAV−7およびAAV−8。AAVベクターは、AAV野生型遺伝子のうちの1つ以上(好ましくはrep遺伝子および/またはcap遺伝子)が、全てまたは一部欠失されているが、機能的隣接ITR配列を保持している。機能的ITR配列は、AAVビリオンのレスキュー、複製およびパッケージングに必要である。従って、AAVベクターは、ウイルスの複製およびパッケージングに、シスで必要とされる少なくともそれらの配列(例えば、機能的ITR)を含むことが、本明細書で規定される。このITRは、野生型ヌクレオチド配列である必要はなく、その配列が、機能的レスキュー、複製およびパッケージングを提供する限りにおいて、例えば、ヌクレオチドの挿入、欠失もしくは置換によって改変されていてもよい。
【0023】
「AAVヘルパー機能」とは、AAV遺伝子生成物(これは、次に、生産的なAAV複製のためにトランスで機能する)を提供するように発現され得るAAV由来のコード配列をいう。従って、AAVヘルパー機能は、主要AAVオープンリーディングフレーム(ORF)、repおよびcapをともに含む。このRep発現生成物は、多くの機能(とりわけ、以下が挙げられる:AAV DNA複製起点の認識、結合およびニック形成;DNAヘリカーゼ活性;ならびにAAV(または他の異種)プロモーターからの転写の調節)を有することが示された。このCap発現生成物は、必要なパッケージング機能を供給する。AAVヘルパー機能は、AAVベクターから失われているトランスでのAAV機能を補完するために、本明細書で用いられる。
【0024】
用語「AAVヘルパー構築物」は、一般に、AAVベクター(これは、目的のヌクレオチド配列を送達するための導入ベクターを生成するために使用される)から欠失されたAAV機能を提供するヌクレオチド配列を含む核酸分子をいう。AAVヘルパー構築物は、AAVのrep遺伝子および/またはcap遺伝子の一過性の発現を提供して、失われている、AAV複製に必須のAAV機能を補完するために一般に使用される;しかし、ヘルパー構築物は、AAV ITRを欠いており、それら自体では、複製もパッケージングもすることができない。AAVヘルパー構築物は、プラスミド、ファージ、トランスポゾン、コスミド、ウイルスまたはビリオンの形態であり得る。多くのAAVヘルパー構築物(例えば、一般的に使用されるプラスミドpAAV/AdおよびpIM29+45)が記載されており、これらは、Rep発現生成物およびCap発現生成物の両方をコードする。例えば、Samulskiら(1989)J.Virol.63:3822−3828;およびMcCartyら(1991)J.Virol.65:2936−2945を参照のこと。多くの他のベクターが記載されており、これらは、Rep発現生成物および/またはCap発現生成物をコードする。例えば、米国特許第5,139,941号および同第6,376,237号を参照のこと。
【0025】
用語「補助機能」とは、AAVが、その複製に関して依存性である、非AAV由来のウイルス機能および/または細胞機能をいう。従って、この用語は、AAV複製に必要とされるタンパク質およびRNA(AAV遺伝子転写の活性化、段階特異的AAV mRNAスプライシング、AAV DNA複製、Cap発現生成物の合成ならびにAAVキャプシドアセンブリに関与する部分を含む)を表す。ウイルスベースの補助機能は、既知のヘルパーウイルス(例えば、アデノウイルス、ヘルペスウイルス(単純ヘルペスウイルス1型以外)およびワクシニアウイルス)のいずれかに由来し得る。
【0026】
用語「補助機能ベクター」とは、一般に、補助機能を提供するヌクレオチド配列を含む核酸分子をいう。補助機能ベクターは、適切な宿主細胞にトランスフェクトされ得る。ここで、このベクターは、次いで、この宿主細胞におけるAAVビリオン生成を支援し得る。この用語から明示的に除外されるのは、天然に存在するままの感染性ウイルス粒子(例えば、アデノウイルス、ヘルペスウイルスまたはワクシニアウイルスの粒子である。従って、補助機能ベクターは、プラスミド、ファージ、トランスポゾン、またはコスミドの形態であり得る。特に、アデノウイルス遺伝子の完全な補完が、補助ヘルパー機能に必ずしも必須でないことが実証された。例えば、DNA複製および後期遺伝子合成ができないアデノウイルス変異体は、AAV複製に許容されることが示された。Itoら,(1970)J.Gen.Virol.9:243;Ishibashiら(1971)Virology 45:317。同様に、E2B領域およびE3領域内の変異は、AAV複製を支援することが示され、このことは、このE2B領域およびE3領域が、補助機能を提供することにおそらく関与しないことを示す。Carterら(1983)Virology 126:505。しかし、E1領域を欠いたアデノウイルスまたはE4領域が欠失したアデノウイルスは、AAV複製を支援することができない。従って、E1A領域またはE4領域は、直接的にせよ間接的にせよ、AAV複製に必要とされるようである。Laughlimら(1982)J.Virol.41:868;Janikら(1981)Proc.Natl.Acad.Sci.USA 78;1925;Carterら(1983)Virology 126:505。他の特徴づけられたAd変異体は、以下を含む:E1B(Laughlimら(1982)前出;Janikら(1981)前出;Ostroveら(1980)Virology,104:502);E2A(Handaら(1975)J.Gen.Virol.29:239;Straussら(1976)J.Virol.17:140;Myersら(1980)J.Virol.35:665;Jayら(1981)Proc.Natl.Acad.Sci.USA 78:2927;Myersら(1981)J.Biol.Chem.256:567);E2B(Carter,Adeno−Associated Virus Helper Functions,I CRC Handbook of Parvoviruses(P.Tijssen編、1990));E3(Carterら(1983)前出);E4(Carterら(1983)前出;Carter(1995))。E1Bコード領域に変異を有するアデノウイルスによって提供される補助機能の研究により、矛盾する結果がもたらされたが、Samulskiら(1988)J.Virol.62:206−210は、近年、E1B55kが、AAVビリオン生成に必須である一方で、E1B19kは必須でないことを報告した。さらに、国際公開WO97/17458およびMatshushitaら(1998)Gene Therapy 5:938−945は、種々のAd遺伝子をコードする補助機能ベクターを記載する。
【0027】
特に好ましい補助機能ベクターは、アデノウイルスVA RNAコード領域、アデノウイルスE4 ORF6コード領域、アデノウイルスE2A 72kDコード領域、アデノウイルスE1Aコード領域、およびインタクトなE1B55kコード領域を欠くアデノウイルスE1B領域を含む。このようなベクターは、国際公開番号WO01/83797に記載される。
【0028】
「組換えウイルス」とは、例えば、異種核酸構築物を付加するかまたはこの構築物を粒子に挿入することによって遺伝的に改変されたウイルスを意味する。
【0029】
「AAVビリオン」とは、完全なウイルス粒子(例えば、野生型(wt)AAVウイルス粒子(AAVキャプシドタンパク質コードと会合した直鎖状の一本鎖AAV核酸ゲノムを含む))を意味する。この点に関して、いずれかの相補的センス(例えば、「センス」鎖または「アンチセンス」鎖)の一本鎖AAV核酸分子は、いずれか1つのAAVビリオンにパッケージングされ得、両方の鎖は、等しく感染性である。
【0030】
用語「組換えAAVビリオン」、「rAAVビリオン」、「AAVベクター粒子」、「完全キャプシド」、「完全体」または「完全粒子」とは、AAV ITRが両側に隣接している、目的の異種ヌクレオチド配列をキャプシドに包んでいる、AAVタンパク質殻を含む感染性の複製欠損ウイルスとして本明細書中に規定される。rAAVビリオンは、そこに導入されるAAVベクターを特定する配列、AAVヘルパー機能および補助機能を有している適切な宿主細胞において生成される。このようにして、この宿主細胞は、その後の遺伝子送達のための感染性組換えビリオン粒子へのAAVベクター(目的の組換えヌクレオチド配列を含む)のパッケージングに必要とされるAAVポリペプチドをコードできるようにされる。
【0031】
用語「空キャプシド」、「空粒子」、および「空」とは、AVVタンパク質の殻を含むが、AAV ITRが両側に隣接した目的の異種ヌクレオチド配列を含むポリヌクレオチド構築物の全体または一部を欠く、AAVビリオンをいう。従って、この空キャプシドは、目的の遺伝子を宿主細胞へと移入するためには機能しない。
【0032】
用語「宿主細胞」は、AAVヘルパー構築物、AAVベクタープラスミド、補助機能ベクター、または他の移入DNAのレシピエントとして用いられ得るかまたは用いられた、例えば、微生物、酵母細胞、昆虫細胞、および哺乳動物細胞を示す。この用語は、トランスフェクトされた元の細胞の子孫を包含する。従って、「宿主細胞」は、本明細書中で用いられる場合、一般に、外因性DNA配列でトランスフェクトされた細胞をいう。単一の親細胞の子孫が、自然変異、偶発変異または意図的変異に起因して、元の親と形態またはゲノムDNA相補体もしくはDNA相補体全体において必ずしも完全に同一というわけではないことが理解される。
【0033】
用語「トランスフェクション」は、細胞による外来DNAの取り込みを言及するために用いられ、そして細胞は、外因性DNAが細胞膜の内側に導入されている場合、「トランスフェクト」されている。多数のトランスフェクション技術が当該分野で一般に公知である。例えば、Grahamら(1973)Virology,52:456,Sambrookら(1989)Molecular Cloning,a laboratory manual,Clod Spring Harbor Laboratories,New York,Davisら(1986)Basic Methods in Molecular Biology,Elsevier,およびChuら(1981)Gene 13:197を参照のこと。このような技術を用いて、1以上の外因性DNA部分を適切な宿主細胞へと導入し得る。
【0034】
本明細書中で用いられる場合、用語「細胞株」とは、連続増殖または長期増殖および分裂をインビトロで行い得る細胞集団をいう。しばしば、細胞株は、単一の子孫細胞に由来するクローン集団である。このようなクローン集団の保存または継代の間の核型における自然変化または誘導された変化が生じ得ることがさらに当該分野で公知である。それゆえ、言及される細胞株に由来する細胞は、先祖細胞または先祖培養物と正確には同一ではないかもしれず、そして言及される細胞株は、このような改変体を包含する。
【0035】
AAVベクター粒子(パッケージングされたゲノム)を含むrAAVビリオンのストックまたは調製物は、ストック中に存在するビリオンのうちの少なくとも約50%〜99%以上が、パッケージングされたゲノムを含むrAAVビリオン(すなわち、AAVベクター粒子)である場合、AAVの空キャプシドを「実質的に含まない」。好ましくは、このAAVベクター粒子は、ストック中に存在するビリオンのうちの、少なくとも約75%〜85%、より好ましくは約90%を構成し、さらにより好ましくは少なくとも約95重量%、またはさらにはストック中に存在するビリオンのうちの99重量%以上、またはこれらの範囲の間の任意の整数を構成する。従って、ストックは、得られるストックの約40%〜約1%以下、好ましくは約25%〜約15%以下、さらに好ましくは約10%以下、さらにより好ましくは約5%〜約1%以下が空キャプシドを含む場合、AAVの空キャプシドを実質的に含まない。
【0036】
「核酸」配列とは、DNA配列またはRNA配列をいう。この用語は、DNAおよびRNAの公知の塩基アナログ(例えば、4−アセチルシトシン、8−ヒドロキシ−N6−メチルアデノシン、アジリジニルシトシン、プソイドイソシトシン、5−(カルボキシヒドロキシルメチル)ウラシル、5−フルオロウラシル、5−ブロモウラシル、5−カルボキシメチルアミノメチル−2−チオウラシル、5−カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、イノシン、N6−イソペンテニルアデニン、1−メチルアデニン、1−メチルプソイドウラシル、1−メチルグアニン、1−メチルイノシン、2,2−ジメチルグアニン、2−メチルアデニン、2−メチルグアニン、3−メチルシトシン、5−メチルシトシン、N6−メチルアデニン、7−メチルグアニン、5−メチルアミノメチルウラシル、5−メトキシアミノメチル−2−チオウラシル、β−D−マンノシルキューオシン、5’−メトキシカルボニルメチルウラシル、5−メトキシウラシル、2−メチルチオ−N6−イソペンテニルアデニン、ウラシル−5−オキシ酢酸メチルエステル、ウラシル−5−オキシ酢酸、オキシブトキソシン、プロイドウラシル、キューオシン、2−チオシトシン、5−メチル−2−チオウラシル、2−チオウラシル、4−チオウラシル、5−メチルウラシル、Bウラシル−5−オキシ酢酸メチルエステル、ウラシル−5−オキシ酢酸、プソイドウラシル、キューオシン、2−チオシトシン、および2,6−ジアミノプリンであるがこれらに限定されない)のいずれかを含む配列を含む。
【0037】
「コード配列」または選択されたポリペプチドを「コード」する配列は、適切な制御配列の制御下においた場合に、インビボで、(DNAの場合)転写される、および(mRNAの場合)ポリペプチドへと翻訳される、核酸分子である。コード配列の境界は、5’(アミノ)末端における開始コドンおよび3’(カルボキシ)末端における翻訳停止コドンによって決定される。転写終結配列は、コード配列の3’側に配置され得る。
【0038】
用語DNA「制御配列」とは、レシピエント細胞におけるコード配列の複製、転写および翻訳をまとまって提供する、プロモーター配列、ポリアデニル化シグナル、転写終結配列、上流調節ドメイン、複製起点、内部リボソーム進入部位(「IRES」)、エンハンサーなどをまとめていう。選択されたコード配列が、適切な宿主細胞において複製され得、転写され得、そして翻訳され得る限り、これらの制御配列の全てがいつも存在する必要があるわけではない。
【0039】
用語「プロモーター」は、本明細書中で、DNA調節配列を含むヌクレオチド領域を言及するために、その元々の意味で用いられ、ここで、この調節配列は、RNAポリメラーゼに結合し得かつ下流の(3’方向の)コード配列の転写を開始し得る遺伝子に由来する。転写プロモーターは、「誘導性プロモーター」(ここで、このプロモーターに作動可能に連結されたポリヌクレオチド配列の発現は、分析物、補因子、調節タンパク質などによって誘導される)、「抑制性プロモーター」(ここで、このプロモーターに作動可能に連結されたポリヌクレオチド配列の発現は、分析物、補因子、調節タンパク質などによって誘導される)および「構成性プロモーター」を包含し得る。
【0040】
「作動可能に連結される」とは、そのように記載される成分がそれらの通常の機能を果たすように構成された構成要素の再配置をいう。従って、コード配列に作動可能に連結された制御配列は、コード配列の発現をもたらしえる。この制御配列は、その発現を導くように機能する限り、コード配列と連続する必要はない。従って、例えば、介在する、転写されるが翻訳されない配列は、プロモーター配列とコード配列との間に存在し得、そしてプロモーター配列は、なおも、コード配列に「作動可能に連結され」ていると考えられる。
【0041】
本出願全体で特定の核酸分子中でのヌクレオチド配列の相対位置を記載する目的で、例えば、特定のヌクレオチド配列が別の配列に対して「上流」、「下流」、「3’」、または「5’」に位置することが記載されている場合、これは、当該分野で慣習的であるように言及されるDNA分子の「センス」鎖または「コーディング」鎖におけるこの配列の位置であると理解されるべきである。
【0042】
用語「異種」とは、核酸配列(例えば、コード配列および制御配列)に関する場合、通常は一緒に連結していない配列、および/または特定の細胞に通常は関連していない配列を示す。従って、核酸構築物またはベクターのうちの「異種」領域は、他の分子とは天然で関連して見出されない別の核酸分子内のまたは別の核酸分子に結合した核酸のセグメントである。例えば、核酸構築物のうちの異種領域は、コード配列と天然では関連して見出されない配列が隣接したコード配列を含み得る。異種コード配列の別の例は、コード配列自体が天然には見出されない構築物である(例えば、ネイティブ遺伝子とは異なるコドンを有する合成配列)。同様に、細胞中に通常は存在しない構築物で形質転換された細胞は、本発明の目的では異種であると考えられる。対立遺伝子改変体または天然に存在する変異事象は、本明細書中で用いられる場合、異種DNAを生じない。
【0043】
「単離された」によって、ヌクレオチド配列を言及する場合、同一の分子が同じ型の他の生物学的高分子が実質的に存在せずに、存在することを意味する。従って、「特定のポリペプチドをコードする単離された核酸分子」とは、対象のポリペプチドをコードしない他の核酸分子を実質的に含まない核酸分子をいう;しかし、この分子は、その組成物の基本的特徴に有害な影響を与えない、いくつかのさらなる塩基または部分を含み得る。
【0044】
(2.発明を実施する形態)
本発明を詳細に記載する前に、特定の処方または特定のプロセスのパラメーターももちろん変化し得るので、本発明が特定の処方にも特定のプロセスのパラメーターにも限定されないことが理解されるべきである。本明細書中で用いられる用語は、単に本発明の特定の実施形態を記載する目的のためのものであり、限定することを意図していないこともまた理解されるべきである。
【0045】
本明細書中に記載される方法および材料と類似するかまたは等価な多数の方法および材料が本発明の実施において用いられ得るが、好ましい材料および方法は、本明細書中に記載される。
【0046】
本発明は、AAVビリオンの精製ストック中に含まれるAAVの空キャプシドの数を、その中に含まれるAAVベクター粒子の損失を最少にしながらも、減らすかまたは除去することを包含する。本発明の方法は、rAAVビリオンが精製されるプロセスと関係なく用いられ得る。
【0047】
rAAVビリオンを生成するために当該分野で周知のいくつかの方法が存在する;例えば、AAVヘルパーウイルス(例えば、アデノウイルス、ヘルペスウイルスまたはワクシニアウイルス)のうちの1つとの同時感染を伴う、ベクターおよびAAVヘルパー配列を用いたトランスフェクション、または組換えAAVベクター、AAVヘルパーベクター、および補助機能ベクターを用いたトランスフェクション。rAAVビリオンを生成するための方法の詳細な記載については、米国特許第6,001,650号および同第6,004,797号を参照のこと。
【0048】
例えば、野生型AAVウイルスおよびヘルパーウイルスは、rAAVビリオンを生成するための必要な複製機能を提供するために用いられ得る(例えば、米国特許第5.139,941号を参照のこと)。あるいは、周知のヘルパーウイルスのうちの1つによる感染に伴ったヘルパー機能遺伝子を含むプラスミドは、複製機能の供給源として用いられ得る(例えば、米国特許第5,622,856号および同第5,139,941号を参照のこと)。同様に、補助機能遺伝子を含むプラスミドは、野生型AAVによる感染と組み合わせて用いられて必要な複製機能を提供し得る。これらの3つのアプローチは、rAAVベクターと関連して組み合わされた場合、各々、rAAVビリオンを生成するために充分である。他のアプローチは、当該分野で周知であり、そしてまた、rAAVビリオンを生成するために当業者に用いられ得る。
【0049】
本発明の好ましい実施形態では、三重トランスフェクション法(米国特許第6,001,650号に詳細に記載される)を用いて、rAAVビリオンが生成される。なぜなら、この方法は、感染性ヘルパーウイルスの使用を必要とせずに、rAAVビリオンが、検出可能なヘルパーウイルスが全く存在せずに生成されるのを可能にするからである。これは、rAAVビリオン生成のための3つのベクター(AAVヘルパー機能ベクター、補助機能ベクター、およびrAAV発現ベクター)の使用によって達成される。しかし、当業者は、これらのベクターによってコードされる核酸配列が種々の組合せの2以上のベクター上で提供され得ることを理解する。
【0050】
本明細書中に説明したように、AAVヘルパー機能ベクターは、生産的AAV複製およびキャプシド形成のためにトランスで機能する「AAVヘルパー機能」配列(すなわち、repおよびcap)をコードする。好ましくは、AAVヘルパー機能ベクターは、検出可能なwtAAVビリオンを全く生成することなく、効率的なAAVベクター(すなわち、機能的なrep遺伝子およびcap遺伝子を含むAAVビリオン)生成を支持する。このようなベクターの一例であるpHLP19は、米国特許第6,001,650号に記載される。AAVヘルパー機能ベクターのrep遺伝子およびcap遺伝子は、上記で説明したように、公知のAAV血清型のいずれかに由来し得る。例えば、AAVヘルパー機能ベクターは、AAV−2由来のrep遺伝子およびAAV−6由来のcap遺伝子を有し得る;当業者は、他のrep遺伝子およびcap遺伝子の組合せが可能であり、定義する特徴がrAAVビリオン生成を支持する能力であることを認識する。
【0051】
補助機能ベクターは、AAVが複製を依存する非AAV由来ウイルス機能および/または細胞機能(すなわち、補助機能)についてのヌクレオチド配列をコードする。補助機能は、AAV複製に必要とされる機能(AAV遺伝子の転写、段階特異的AAV mRNAスプライシング、AAV DNA複製、cap発現産物の合成、およびAAVキャプシドアセンブリの活性化に関与する部分が挙げられるがこれらに限定されない)を包含する。ウイルスベースの補助機能は、周知のヘルパーウイルス(例えば、アデノウイルス、ヘルペスウイルス(1型単純疱疹ウイルス以外)、およびワクシニアウイルス)のいずれかに由来し得る。好ましい実施形態では、補助機能プラスミドpLadeno5が用いられる(pLadeno5に関する詳細は、米国特許第6,004,797号に記載される)。このプラスミドは、AAVベクター生成についてのアデノウイルス補助機能の完全なセットを提供するが、複製能力があるアデノウイルスを形成するために必要な成分を欠いている。
【0052】
一旦AAVビリオンのストックが生成されると、多数の方法(以下に詳述される)を用いて、感染性力価が決定され得、そしてAAVの空キャプシドからAAVベクター粒子が精製され得る。
【0053】
本発明をさらに理解するために、より詳細な考察を、組換えAAV発現ベクター、AAVヘルパーおよび補助機能、AAVビリオンを含む組成物、ならびにビリオンの送達に関して以下に提供する。
【0054】
(組換えAAV発現ベクター)
組換えAAV(rAAV)発現ベクターは、制御エレメント(転写開始領域を含む)、目的のポリヌクレオチド、および転写末端領域を転写の方向に作動可能に連結した成分として提供するように、公知の技術を使用して構築される。この制御エレメントは、目的の宿主細胞において機能的であるように選択される。作動可能に連結した成分を含む、結果の構築物は、機能性AAV ITR配列と結合される(5’および3’)。
【0055】
AAV ITR領域のヌクレオチド配列は公知である。たとえば、AAV−2配列については、Kotin,R.M.(1994)Human Gene Therapy 5:793−801;Berns,K.I.「Parvoviridae and their Replication」Fundamental Virology,第2版,(B.N.FieldsおよびD.M.Knipe(編))を参照のこと。本発明のベクターで使用されるAAV ITRは、野生型ヌクレオチド配列を有する必要はなく、例えば、ヌクレオチドの挿入、欠失、または置換によって変化され得る。加えて、AAV ITRは、いくつかのAAV抗原型(AAV−1、AAV−2、AAV−3、AAV−4、AAV−5、AAV−6、AAV−7、およびAAV−8などが挙げられるが、これらに限定されない)のいずれかに由来し得る。さらに、5’および3’のITRは、AAV発現ベクター中の選択されたヌクレオチド配列に隣接し、それらが意図されるように機能する限り(すなわち、宿主細胞ゲノムまたはベクターからの所定の配列の切断およびレスキューを可能にし、そして、AAV Rep遺伝子産物が受容細胞に存在する場合、受容細胞ゲノム中にDNA分子を組込むことを可能にする)、必ずしも同一である必要はなく、同一のAAV抗原型または単離物に由来する必要もない。
【0056】
AAVベクターに使用するために適切なポリヌクレオチド分子は、約5キロベース(kb)未満の大きさである。選択されるポリヌクレオチド配列は、インビボで被験体におけるそれらの転写または発現を方向付ける制御エレメントに作動可能に連結される。このような制御エレメントは、通常、選択遺伝子に関連する制御配列を含み得る。あるいは、異種制御配列が、使用され得る。一般的に、有用な異種制御配列は、哺乳動物遺伝子またはウイルス遺伝子をコードする配列に由来する制御配列を含む。例としては、ニューロン特異的エノラーゼプロモーター、GFAPプロモーター、SV40初期プロモーター、マウス乳腺癌ウイルスLTRプロモーター;アデノウイルス主要後期プロモーター(Ad MLP);単純ヘルペスウイルス(HSV)プロモーター、サイトメガロウイルス(CMV)プロモーター(例えば、CMV前初期プロモーター領域(CMVIE))、ラウス肉腫ウイルス(RSV)プロモーター、CAGプロモーター、合成プロモーター、ハイブリッドプロモーターなどが挙げられるが、これらに限定されない。加えて、非ウイルス遺伝子由来の配列(例えば、マウスメタロチオネイン遺伝子)もまた、本明細書中で使用を見出す。このようなプロモーター配列は、例えば、Stratagene(San Diego,CA)から市販されている。
【0057】
上記AAV発現ベクターは、AAV ITRによって結合される目的のポリヌクレオチド分子を含み、AAVゲノムから切断される主要なAAVオープンリーディングフレーム(「ORF」)を有するAAVゲノム中に選択配列を直接挿入することによって、構築され得る。AAVゲノムの他の部分はまた、複製およびパッケージング機能を可能にするようにITRの十分な部分が残る場合に限り、削除され得る。このような構築物は、当該分野で周知の技術を使用して設計され得る。例えば、米国特許第5,173,414号および同第5,139,941号;国際公開番号WO 92/01070(1992年1月23日公開)およびWO 93/03769(1993年3月4日公開);Lebkowskiら(1988)Molec.Cell.Biol.8:3988−3996;Vincentら(1990)Vaccines 90(Cold Spring Harbor Laboratory Press);Carter(1992)Current Opinion in Biotechnology 3:533−539;Muzyczka(1992)Current Topics in Microbiol.and Immunol.158:97−129;Kotin(1994)Human Gene Therapy 5:793−801;ShellingおよびSmith(1994)Gene Therapy 1:165−169;ならびにZhouら(1994)J.Exp.Med.179:1867−1875を参照のこと。
【0058】
あるいは、AAV ITRは、ウイルスゲノムからか、または別のベクター中に存在する、選択核酸構築物の同一で融合された5’および3’を含むAAVベクターから、標準的なライゲーション技術(例えば、Sambrookら,前出に記載される)を使用して切断され得る。例えば、ライゲーションは、20mM Tris−Cl(pH7.5)、10mM MgCl2、10mM DTT、33μg/ml BSA、10mM〜50mM NaCl、および0℃における40μM ATP、0.01〜0.02(Weiss)単位 T4 DNAリガーゼ(「付着末端」ライゲーション用)か、または14℃における1mM ATP、0.3〜0.6(Weiss)単位 T4 DNAリガーゼ(「平滑末端」ライゲーション用)において達成され得る。分子内の「付着末端」ライゲーションは、通常、30〜100μg/mlの総DNA濃度(総最終濃度5〜100nM)において実行される。ITRを含むAAVベクターは、例えば、米国特許第5,139,941号に記載されている。特に、いくつかのAAVベクターが上記特許明細書中に記載され、アメリカン タイプ カルチャー コレクション(「ATCC」)から入手可能である(受託番号53222、53223、53224、53225および53226)。
【0059】
本発明の目的のために、AAV発現ベクターからrAAVビリオンを生成するために適切な宿主細胞としては、微生物、酵母細胞、昆虫細胞、および哺乳動物細胞が挙げられる。これらの細胞は、異種DNA分子のレシピエントとして使用され得るかもしくは使用されており、例えば、浮遊培養物、バイオリアクターなどで増殖し得る。この用語は、トランスフェクトされたもとの細胞の子孫を含む。従って、本明細書中で使用される場合、「宿主細胞」は、一般的に、外因性DNA配列によってトランスフェクトされた細胞をいう。安定なヒト細胞株である293(例えば、アメリカン タイプ カルチャー コレクション(受託番号ATCC CRL1573)を通じて容易に入手可能)由来の細胞が、本発明の実施において好ましい。特に、ヒト細胞株293は、アデノウイルス5型のDNAフラグメントでトランスフェクトされたヒト胚性腎細胞株であり(Grahamら(1977)J.Gen.Virol.36:59)、アデノウイルスのE1a遺伝子およびE1b遺伝子を発現する(Aielloら(1979)Virology 94:460)。上記293細胞株は、容易にトランスフェフェクトされ、rAAVビリオンを生成する特に都合のよいプラットフォームを提供する。
【0060】
(AAVヘルパー機能)
上記AAV発現ベクターを含む宿主細胞は、AAV ITRに隣接して、rAAVビリオンを生成するためのヌクレオチド配列を複製し、そしてキャプシド形成するために、AAVヘルパー機能を提供し得なければならない。AAVヘルパー機能は、一般に、AAVに由来するコード配列である。この配列は、順生産的なAAV複製のためにトランスで機能するAAV遺伝子産物を提供するように発現され得る。AAVヘルパー機能は、本明細書中で、AAV発現ベクターから失われる必須のAAV機能を補完するために使用される。従って、AAVヘルパー機能は、主要なAAV ORF(すなわち、repコード領域およびcapコード領域)の1つもしくは両方、またはそれらの機能的ホモログを含む。
【0061】
「AAV repコード領域」によって、複製タンパク質のRep78、Rep68、Rep52およびRep40をコードするAAVゲノムのart認識(art−recognized)領域が意味される。これらのRep発現産物は、多くの機能(DNA複製のAAV起点の認識、結合、およびニッキング、DNAヘリカーゼ活性、ならびにAAV(または他の異種)プロモーターからの転写の調節を含む)を有することが示されている。このRep発現産物は、AAVゲノムを複製するために集合的に必要とされる。AAVのrepコード領域の記載については、例えば、Muzyczka,N.(1992)Current Topics in Microbiol.and Immunol.158:97−129;およびKotin,R.M.(1994)Human Gene Therapy 5:793−801を参照のこと。AAVのrepコード領域の適切なホモログとしては、AAV−2のDNA複製を媒介することもまた公知であるヒトヘルペスウイルス6(HHV−6)rep遺伝子が挙げられる(Thomsonら(1994)Virology 204:304−311)。
【0062】
「AAV capコード領域」によって、キャプシドタンパク質のVP1、VP2、およびVP3をコードするAAVゲノムのart認識領域、またはそれらの機能的ホモログが意味される。これらのCap発現産物は、ウイルスゲノムのパッケージングのために集合的に必要とされるパッケージング機能を与える。AAV capコード領域の記載については、例えば、Muzyczka,N.およびKotin,R.M.(前出)を参照のこと。
【0063】
AAVヘルパー機能は、AAV発現ベクターのトランスフェクションの前か、または同時に、AAVヘルパー構築物を用いて宿主細胞をトランスフェクトすることによって、宿主細胞中に導入される。従って、AAVヘルパー構築物が使用され、AAV rep遺伝子および/またはcap遺伝子の、少なくとも一過性の発現を提供し、生産的なAAV感染のために必須の失われたAAV機能を補完する。AAVヘルパー構築物は、AAV ITRを欠き、それ自体を複製もパッケージングもし得ない。
【0064】
これらの構築物は、プラスミド、ファージ、トランスポゾン、コスミド、ウイルス、またはビリオンの形態であり得る。多くのAAVヘルパー構築物(例えば、共通に使用されるプラスミドpAAV/AdおよびRepとCapとの両方の発現産物をコードするpIM29+45)が記載されている。例えば、Samulskiら(1989)J.Virol.63:3822−3828;およびMcCartyら(1991)J.Virol.65:2936−2945を参照のこと。Repおよび/またはCapの発現産物をコードする多くの他のベクターが、記載されている。例えば、米国特許第5,139,941号を参照のこと。
【0065】
(AAV補助機能)
宿主細胞(またはパッケージング細胞)はまた、rAAVビリオンを生成するために、非AAVの機能、または「補助機能(accessory function)」を提供され得なければならない。補助機能は、AAVがその複製のために依存する、非AAV由来のウイルス機能および/または細胞機能である。従って、補助機能としては、AAV複製(AAV遺伝子転写の活性化、段階特異的なAAV mRNAスプライシング、AAV DNA複製、Cap発現産物の合成およびAAVキャプシド集合に関係する機能を含む)の際に必要とされる、少なくともこれらの非AAVタンパク質およびRNAが挙げられる。ウイルスベースの補助機能は、任意の公知のヘルパーウイルスに由来し得る。特に、補助機能は、当業者に公知の方法を使用して宿主細胞に導入され得、次いで宿主細胞中で発現される。代表的には、補助機能は、宿主細胞を関連のないヘルパーウイルスに感染させることによって提供される。多くの適切なヘルパーウイルスが公知である。これらとしては、アデノウイルス;ヘルペスウイルス(例えば、単純ヘルペスウイルス1型および2型);およびワクシニアウイルスが挙げられる。非ウイルス性の補助機能(例えば、種々の公知の薬剤のいずれかを使用する細胞の同期化(cell synchronization)によって提供される機能)もまた、本明細書中で使用を見出される。例えば、Bullerら(1981)J.Virol.40:241−247;McPhersonら(1985)Virology 147:217−222;Schlehoferら(1986)Virology 152:110−117を参照のこと。あるいは、補助機能は、上に記載されるような補助機能ベクターを使用して提供され得る。例えば、米国特許第6,004,797号および国際公開番号WO 01/83797を参照のこと。
【0066】
補助機能を提供する核酸配列は、天然の供給源(例えば、アデノウイルス粒子のゲノムから)から得られ得か、または当該分野で公知の組換え方法もしくは合成方法を使用して構築され得る。上に説明したように、アデノウイルス遺伝子の完全な相補体が、補助ヘルパー機能のために必要とされるわけではないことが示されている。特に、DNA複製および後期遺伝子の合成が不可能なアデノウイルス変異体が、AAV複製に許容性であることが示されている。Itoら,(1970)J.Gen.Virol.9:243;Ishibashiら,(1971)Virology 45:317。同様に、E2B領域およびE3領域内の変異体は、AAV複製を支持することが示されており、このことは、上記E2B領域およびE3領域は、おそらく補助機能の提供に関与していないことを示している。Carterら,(1983)Virology 126:505。しかしながら、E1領域に欠陥のあるアデノウイルス、またはE4領域を欠くアデノウイルスは、AAV複製を支持し得ない。従って、E1A領域およびE4領域は、直接的または間接的にAAV複製に必要とされる可能性が高い。Laughlinら,(1982)J.Virol.41:868;Janikら,(1981)Proc.Natl.Acad.Sci.USA 78:1925;Carterら,(1983)Virology 126:505。他の特徴付けられたAd変異体としては、以下が挙げられる:E1B(Laughlinら(1982),前出;Janikら(1981),前出;Ostroveら,(1980)Virology 104:502);E2A(Handaら,(1975)J.Gen.Virol.29:239;Straussら,(1976)J Virol.17:140;Myersら,(1980)J.Virol.35:665;Jayら,(1981)Proc.Natl.Acad.Sci.USA 78:2927;Myersら,(1981)J.Biol.Chem.256:567);E2B(Carter,Adeno−Associated Virus Helper Functions,I CRC Handbook of Parvoviruses(P.Tijssen(編),1990));E3(Carterら(1983),前出);およびE4(Carterら(1983),前出;Carter(1995))。E1Bコード領域に変異を有するアデノウイルスによって提供された補助機能の研究が相反する結果を示しているにもかかわらず、E1B55kが、AAVビリオン生成に必要とされる一方、E1B19kは必要とされないことが、最近報告された(Samulskiら,(1988)J.Virol.62:206−210)。加えて、国際公開WO 97/17458およびMatshushitaら,(1998)Gene Therapy 5:938−945は、種々のAd遺伝子をコードする補助機能ベクターを記載する。特に好ましい補助機能ベクターは、アデノウイルスVA RNAコード領域、アデノウイルスE4 ORF6コード領域、アデノウイルスE2A 72kDコード領域、アデノウイルスE1Aコード領域、およびインタクトなE1B55kコード領域を欠くアデノウイルスE1B領域を含む。このようなベクターは、国際公開番号WO 01/83797に記載される。
【0067】
宿主細胞をヘルパーウイルスに感染させるか、または宿主細胞を補助機能ベクターを用いてトランスフェクションすることの結果として、AAVヘルパー構築物をトランス活性化する補助機能が発現され、AAV Repタンパク質および/またはCapタンパク質を産生する。このRep発現産物は、AAV発現ベクターから組換えDNA(目的のDNAを含む)を切断する。Repタンパク質はまた、AAVゲノムを二重にするために役割を果たす。この発現されたCapタンパク質は、キャプシド中に集合し、組換えAAVゲノムはキャプシド中にパッケージングされる。従って、生産性のAAV複製が後に続き、DNAは、rAAVビリオン中にパッケージングされる。
【0068】
(rAAVビリオンの精製)
組換えAAV複製に続いて、rAAVビリオンは、種々の慣用的な精製方法(例えば、カラムクロマトグラフィー、CsCl勾配など)を使用して、宿主細胞から精製され得る。例えば、多くのカラム精製の工程(例えば、アニオン交換カラム、アフィニティーカラムおよび/またはカチオン交換カラムによる精製)が、使用され得る。例えば、国際公開番号WO 02/12455を参照のこと。さらに、感染が適用されて補助機能が発現される場合、残りのヘルパーウイルスは、公知の方法を使用して不活化され得る。例えば、アデノウイルスは、約60℃の温度まで、例えば20分間以上加熱することによって不活化され得る。この処理は、ヘルパーウイルスのみを効果的に不活化する。なぜならば、AAVが非常に熱安定性である一方、ヘルパーアデノウイルスは熱不安定性だからである。
【0069】
任意の数のレポーター遺伝子を含む組換えAAVベクターが、感染性力価を決定するために使用され得る。例えば、アルカリホスファターゼ、β−ガラクトシダーゼ(LacZ)、緑色蛍光タンパク質、またはルシフェラーゼが、使用され得る。トランスフェクトされた宿主細胞を収集した後、大きなスケールの生成物のために適切な技術を使用して(例えば、微流動化(microfluidization))、トランスフェクトされた宿主細胞を破壊することによって、溶解物が形成される。次いで、この溶解物はフィルター処理され(例えば、0.45μmフィルターを通して)、そして、本明細書中に記載されるようなカラムクロマトグラフィー方法を使用して精製される。任意のAAVベクターの感染性力価を決定するための他の技術もまた、報告されている。例えば、Zhenら,「An Infectious Titer Assay for Adeno−associated Virus(AAV)Vectors with Sensitivity Sufficient to Detect Single Infectious Events.」Hum.Gene Ther.(2004)In,Pressを参照のこと。
【0070】
次いで、精製されたAAVストックが処理され、カラムクロマトグラフィー技術を使用して空キャプシドを除去する。本発明の特に好ましい方法において、rAAV調製物は、トランスフェクトされた細胞を溶解し、粗細胞溶解物を得ることによって得られる。次いで、この粗細胞溶解物は清澄化され、当該分野で周知の技術(例えば、フィルター処理、遠心分離など)によって細胞の破片を除去して清澄化細胞溶解物にされ得る。粗細胞溶解物または清澄化細胞溶解物は、AAVベクター粒子とAAVの空キャプシドとの両方を含み、次いで、非分離条件下で第一のカチオン交換カラムに対して適用される。上記第一のカチオン交換カラムは、細胞溶解物の調製物中に存在する細胞成分および他の成分から、AAVベクター粒子およびAAVの空キャプシドをさらに分離するために機能する。細胞溶解物の最初の精製を行うための方法は、公知である。1つの代表的な方法が、米国特許第6,593,123号に記載される。第一のカチオン交換カラムから回収された画分は、次いで、AAVの空キャプシドをAAVベクター粒子から分離する異なる溶出条件を使用して、第二のイオン交換体(すなわち、第二のカチオン交換カラムおよび/またはアニオン交換カラム)に対して、適用される。
【0071】
第一のカチオン交換カラムと第二のカチオン交換カラムとの両方に適切なカチオン交換体が使用される場合、当該分野で公知の多種多様な材料を含む。特に好ましいのは、広範なpH範囲にわたってrAAVビリオンを結合し得る強力なカチオン交換体である。例えば、カルボキシメチル化カチオン交換マトリックスおよびスルホン化カチオン交換マトリックスが、本明細書中の用途のために特に有用である。有用なマトリックス材料としては、セルロースマトリックス(例えば、繊維性マトリックス、微粒子性マトリックス、およびビーズマトリックス);アガロースマトリックス、デキストランマトリックス、ポリアクリレートマトリックス、ポリビニルマトリックス、ポリスチレンマトリックス、シリカマトリックス、およびポリエーテルマトリックス;ならびに合成物が挙げられるが、これらに限定されない。本明細書中で特に好ましいのは、機能的リガンドR−SO3−(好ましくは、スルホプロピル樹脂またはスルホエチル樹脂)を含むマトリックスである。代表的なマトリックスとしては、POROS HS、POROS SP、POROS S(すべてApplied Biosystems,Foster City,CAから入手可能な強力なカチオン交換体)、POROS CM(Applied Biosystems,Foster City,CAから入手可能な弱いカチオン交換体)、TOSOHAAS TOYOPEARL SP550CおよびMERCK FRACTOGEL EMD SO3−−650(m)、ならびにSOURCE 15S、SOURCE 30S、SEPHAROSE SP FF、SEPHAROSE SP XL(すべてAmersham Bioscience,Piscataway,NJから入手可能)が挙げられるが、これらに限定されない。
【0072】
下に与えられるすべてのカラムクロマトグラフィープロトコルについて、カラムは、当該分野で公知の標準的プロトコルを使用して、適切な緩衝液溶液で調製され得る。次いで、サンプルがロードされる。使用される第一のカチオン交換カラムについて、条件は、空キャプシドとAAVベクター粒子との両方がカラム樹脂に結合し、続いて一緒に溶出されるが、細胞溶解物中に存在する他の細胞成分および細胞の破片から分離されるような条件である。例えば、空キャプシドおよび完全キャプシドは、適切なイオン強度の緩衝液を使用して溶出される。適切な緩衝液としては、例えば、10〜50mMリン酸ナトリウム、好ましくは、15〜40(例えば、15、20、25、30、35、40など)mMリン酸ナトリウムが挙げられ、このリン酸ナトリウムは、塩(例えば、NaClまたはKCl)を例えば、100〜700mMの濃度(例えば、200〜400mM(例えば、200、300、325、350、370、380、400など、またはこれらの範囲内の任意の濃度))で含む。緩衝液のpHは、約3〜約9.5であり得、例えば、4〜8であり、例えば、pH4、4.5、5、5.5、6など、またはこれらの範囲内の任意のpHであり得る。これらの画分は回収され、次いで、分離条件化でアニオン交換カラムおよび/または第二のカチオン交換カラムのどちらかに適用され得る。
【0073】
続いて空のAAVキャプシドをAAVベクター粒子から分離するための工程において、第二のカチオン交換カラムが使用される場合、2種の溶出緩衝液が使用される。1つは低塩緩衝液であり、1つは高塩緩衝液である。特に、空キャプシドは、AAVベクター粒子から、約pH6〜pH12のpHにおける適切な緩衝液を使用して分離される。このpHは、好ましくは、pH7〜pH10であり、さらにより好ましくはpH7.5〜pH9.5であり、例えば、pH7.5、8.0、8.1、8.2、8.3、8.4、8.5、9.0、9.1、9.2、9.3、9.4、9.5、または示された範囲の間の任意のpHである。適切な緩衝液は、当該分野で周知であり、以下の緩衝液イオンを含む緩衝液が挙げられるが、これらに限定されない:酢酸;マロン酸;MES:リン酸;HEPES、BICINEなど。サンプルを溶出するために、開始緩衝液のイオン強度は、塩(例えば、NaCl、KCl、硫酸アンモニアまたは硫酸塩、ギ酸塩、酢酸塩、クエン酸塩および/もしくはリン酸塩を含む任意の他の塩)を使用することによって増加される。本発明の一実施形態において、カラムはまず低塩濃度で処理される。この濃度は、例えば、10〜200mMの酢酸アンモニウムであり、例えば、20、25、30、35、40、45、50、55、60、65、100mMまたはこれらの範囲内の任意の濃度である。この処理は、AAVベクター粒子のカラム樹脂からの溶出を生じる。続いて、このカラムは、AAVの空キャプシドを溶出するために、より高塩濃度で処理される。第二の緩衝液としての使用についての1つの例は、100〜800mMの濃度、好ましくは、500〜700mMであり、例えば、500、550、600、650、700、800mM、またはこれらの示された範囲内の任意の濃度の酢酸アンモニウムである。これら条件を使用して、AAVベクター粒子を早い画分に溶出し、空粒子を遅い画分に溶出する。
【0074】
上に説明されるように、本発明の代替的方法において、第一のカチオン交換カラムからの調製物は、第二のカチオン交換カラムの代わりにか、または第二のカチオン交換カラムに加えてのいずれかで、アニオン交換カラムに対して適用される。アニオン交換カラムが第二のカチオン交換カラムに加えて使用される場合、アニオン交換カラムは、第二のカチオン交換カラムの前か、または第二のカチオン交換カラムに続いて使用され得る。さらに、第二のアニオン交換カラムは、第一のアニオン交換カラムの後に使用され得る。本発明での使用についての多くの適切なアニオン交換体が公知であり、以下が挙げられるが、これらに限定されない:MACRO PREP Q(BioRad,Hercules,CAから入手可能な強力なアニオン交換体);UNOSPHERE Q(BioRad,Hercules,CAから入手可能な強力なアニオン交換体);POROS 50HQ(Applied Biosystems,Foster City,CAから入手可能な強力なアニオン交換体);POROS 50D(Applied Biosystems,Foster City,CAから入手可能な弱いアニオン交換体);POROS 50PI(Applied Biosystems,Foster City,CAから入手可能な弱いアニオン交換体);SOURCE 30Q(Amersham Biosciences,Piscataway,NJから入手可能な強力なアニオン交換体);DEAE SEPHAROSE(Amersham Biosciences,Piscataway,NJから入手可能な弱いアニオン交換体);Q SEPHAROSE(Amersham Biosciences,Piscataway,NJから入手可能な強力なアニオン交換体)。
【0075】
上記アニオン交換カラムはまず、標準的な緩衝液を使用して、製造者の仕様書に従って平衡化される。例えば、カラムは、例えば、5〜50mM(好ましくは、7〜20mM、例えば、10mM)のリン酸ナトリウム緩衝液で平衡化され得る。次いでサンプルがロードされ、そして2種の溶出緩衝液が使用される。1つは低塩緩衝液であり、1つは高塩緩衝液である。低塩洗浄および高塩洗浄のそれぞれに続いて、画分が回収され、そして画分中のタンパク質が、標準的技術(例えば、260nmおよび280nmにおけるUV吸光度のモニタリング)を使用して検出される。アニオン交換体を使用して、低塩溶出からのタンパク質ピークは、AAVの空キャプシドを含み、そして高塩画分は、AAVベクター粒子を含む。
【0076】
特に、アニオン交換カラムに関して、空キャプシドは、約pH5〜pH12のpHにおける適切な緩衝液を使用してAAVベクター粒子から分離され得る。このpHは、好ましくは、pH6〜pH10であり、さらにより好ましくはpH7〜pH9.5であり、例えば、pH7.1、7.2、7.3、7.4、8.0、8.1、8.2、8.3、8.4、8.5、9.0、9.1、9.2、9.3、9.4、9.5、または示された範囲の間の任意のpHである。アニオン交換カラムでの使用に適切な緩衝液は、当該分野で周知であり、一般的に本質的に陽イオン性であるか、または両性イオン性である。このような緩衝液としては、以下の緩衝液イオンを含む緩衝液が挙げられるが、これらに限定されない:N−メチルピペラジン;ピペラジン;ビス−Tris;ビス−Trisプロパン;トリエタノールアミン;Tris;N−メチルジエタノールアミン;1,3−ジアミノプロパン;エタノールアミン;酢酸など。サンプルを溶出するために、開始緩衝液のイオン強度は、塩(例えば、NaCl塩、KCl塩、硫酸塩、フマル酸塩または酢酸塩)を使用して、適切なpHにおいて増加される。
【0077】
本発明の一実施形態において、アニオン交換カラムは、まず、低塩濃度で処理される。その濃度は例えば、10〜100mMのNaClであり、例えば、10、20、25、30、35、40、45、50、55、60、65、100mM、またはこれらの範囲内の任意の濃度である。最初の処理に続いて、次いでカラムは、空キャプシドを溶出するために、高塩濃度(たとえば、より高いNaCl濃度、またはより強力なイオン強度を有する別の緩衝液で)で処理される。第二の緩衝液としての使用についての1つの例は、酢酸ナトリウム緩衝液、または100〜300mMの濃度を有するTrisベースの緩衝液である。この濃度は、好ましくは、125〜200mMであり、例えば、125、130、140、150、160、170、180、190、200mM、またはこれらの示された範囲内の任意の濃度である。空キャプシドがカラムから溶出された後、AAVベクター粒子は、高濃度の塩を使用して回収され得る。溶出緩衝液としての使用についての1つの例は、酢酸ナトリウムを含む10mM Tris緩衝液である。酢酸ナトリウムの濃度は100〜500mMの範囲であり、好ましくは、130〜300mMであり、例えば、100、130、150、200、250、300、350、400、450、500mM、またはこれら示された範囲内の任意の濃度である。
【0078】
上に記載される技術を使用して、90%以上のAAVの空キャプシドが、AAVベクター粒子から分離され得る。さらに、AAVベクター粒子の高い回収は、容易に達成される。すなわち、10%以上、好ましくは25%以上、より好ましくは50%以上、例えば、60%以上のAAVベクター粒子が回収され得る。
【0079】
空キャプシドおよびパッケージされたゲノムを有するAAVベクター粒子をアッセイするための方法は、当該分野において公知である。例えば、Grimmら、Gene Therapy(1999)6:1322−1330;Sommerら、Molec.Ther.(2003)7:122−128を参照のこと。変性キャプシドの試験のために、その方法は、処理されたAAVストックを上記3つのキャプシドタンパク質を分離可能な任意のゲル(例えば、緩衝液中に3〜8%トリス−酢酸を含む勾配ゲル)からなるSDS−ポリアクリルアミドゲル電気泳動に供する工程、次いでサンプル物質が分離されるまでそのゲルを泳動する工程、およびそのゲルをナイロンまたはニトロセルロース膜(好ましくはナイロン)にブロットする工程を包含する。抗AAVキャプシド抗体(好ましくは抗AAVキャプシドモノクローナル抗体、最も好ましくは上記B1抗AAV−2モノクローナル抗体)が次いで、変性キャプシドタンパク質に結合する一次抗体として使用される(Wobusら、J.Virol.(2000)74:9281−9293)。次いで二次抗体が使用される。これは、その一次抗体に結合しその一次抗体との結合を検出するための手段を含有するもの、より好ましくは、自身に共有結合された検出分子を含有する抗IgG抗体であり、最も好ましくは、西洋ワサビペルオキシダーゼに共有結合されたヒツジ抗マウスIgG抗体である。結合を検出するための方法は、その一次抗体とその二次抗体との間の結合を半定量的に決定するために使用され、好ましくは検出方法が放射性同位元素放射、電磁放射線または比色変化を検出可能であり、最も好ましくは化学発光検出キットである。
【0080】
感染力価の試験のための、その方法は、約100,000の宿主細胞(好ましくはヒト由来、最も好ましくはHeLa細胞)を、組織培養処理プレート(好ましくは24ウェル組織培養処理プレート)に播種し、そしてアデノウイルス(好ましくはアデノウイルス−2血清型)および処理されたrAAVストックをその宿主細胞に加えてから約24時間インキュベートする工程を包含する。その宿主細胞、アデノウイルスおよびrAAVストックは、24時間インキュベートされ、その後その宿主細胞は好ましくはホルムアルデヒドおよびグルタルアルデヒドで固定され、そしてrAAV発現導入遺伝子を検出する適切な薬剤で染色される(例えば、rAAV−LacZ、X−galが染色剤として企図される)。他のレポーター遺伝子に対する他の薬剤は、当該分野において周知である。任意の導入遺伝子を含有するベクターの感染力価を決定するためのより一般的な方法もまた、当該分野において公知である。例えば、Zhenら、「An Infectious Titer Assay for Adeno−associated Virus(AAV) Vectors with Sensitivity Sufficient to Detect Single Infectious Events」Hum.Gene Ther.(2004)(印刷中)を参照のこと。
【実施例】
【0081】
(3.実験)
以下は、本発明を実施するための特定の実施形態の例である。この例は、例示目的のみのために提供され、任意の方法において本発明の範囲を制限することを意図されない。
【0082】
使用された数字(例えば量、温度など)に関して精度を保証するための努力はなされたが、それに対するいくつかの実験誤差または偏差は、当然許容されるべきである。
【0083】
(材料および方法)
(rAAV産生および精製)
ヒトIX因子(rAAV−hFIX)をコードする遺伝子を含有する組み換えAAVビリオンを、米国特許第6,001,650号および同第6,004,797号に記載されるトリプルトランスフェクション(triple−transfection)法により産生した。使用したプラスミドは、補助機能プラスミド「pladeno5」、AAVヘルパー機能プラスミド「pHLP19」、および組み換えAAVプラスミド「phFIX−16」であった。ヒト胎児腎臓(HEK)293細胞を、rAAVビリオンの産生のために宿主細胞として使用した。
【0084】
トランスフェクションされた293細胞を、トランスフェクション後約72時間で回収し、MicrofluidizerTM(Microfluidics International Corp.,Newton,MA)を使用した微細流動化(microfluidization)により破砕し、そしてその粗製溶解物を収集し、連絡的フィルタを通して細胞細片を除去して、清澄な細胞溶解物を作製した。パッケージされたキャプシドおよび空キャプシドを含有するその清澄な細胞溶解物を、次いで、他の細胞成分からAAVベクター粒子およびAAVの空キャプシドを精製するために、POROS 50HSカラムにロードした。このAAVベクター粒子および空キャプシドを、このPOROS 50HSカラムから、20mMリン酸ナトリウム、370mM NaCl(pH5.5)を含有する緩衝液を使用して溶出した。AAVベクター粒子および空キャプシドの両方を含有するこのAAV調製物を、その塩濃度を減少させるために2倍希釈し、Q SEPHAROSEカラムを通してさらに精製し、流れて通過したAAVベクター粒子および空キャプシドを収集し、そして濃度および緩衝液の交換を限外濾過および透析濾過(dialfiltration)技術を使用して行った。
【0085】
最終的なPOROS 50HSカラム精製産物は、AAV空キャプシドおよび組み換えベクターゲノムを含有するAAVベクター粒子の両方を含有した。各々に対するコントロールサンプルを生成するために、塩化セシウムを攪拌しながら1.41g/mlの密度までPOROS 50HSカラム精製物質に加え、次いでTi70ローターを使用して45,000rpmで23時間超遠心分離へ供した。この可視の空キャプシドおよびAAVベクター粒子のバンドを、シリンジを使用して別々に汲み出した。この精製された空キャプシドおよびAAVベクター粒子を、10mMリン酸ナトリウム(pH7.4)、140mM NaClおよび5%ソルビタール(sorbital)からなる緩衝液に対して、透析カセット(Pierce)を使用し、4℃での、3回の緩衝液交換によって、別々に透析した。
【0086】
(樹脂およびカラムクロマトグラフィー)
アニオン交換樹脂、カチオン交換樹脂、等電点樹脂および疎水的相互作用(HIC)樹脂を含む種々の型のクロマトグラフィー樹脂を、AAV空キャプシドおよびAAVベクター粒子を分離するための、その能力について試験した。ハイスループットの多岐管を、樹脂のスクリーニングのために使用した。樹脂を、多岐管に設置された層高3cmの使い捨ての空のカラム(Bio−Rad Laboratories)に充填した。このカラムを、10mMのリン酸ナトリウムを含有する緩衝液を使用して平衡化した。次いで0.2μmで濾過された清澄なHEK293細胞溶解物のAAVベクターおよび空キャプシドを、50mMの濃度のNaCl(塩)のカラム平衡緩衝液中に希釈し、次いでそのカラムにロードした。このカラムを、最初に平衡緩衝液で洗浄し、次いで10mMリン酸ナトリウムおよび40mM NaCl(pH7.4)を含有する緩衝液でさらに洗浄し(低塩洗浄)、非結合物質を洗い出した。カラムを、次いで、10mMリン酸ナトリウムおよび1M NaClを含有する溶出緩衝液で処理した(高塩洗浄)。低塩洗浄を使用して洗い出された物質を1つの画分として収集し、高塩洗浄を使用して洗い出された物質を別の画分として収集した。
【0087】
使い捨てカラムを使用した初期スクリーニングに引き続き、より大きなスケールのカラムクロマトグラフィー(BIOCAD700(Applied Biosystems)上で行われる)を使用し、有望な樹脂および条件をさらに調べた。XK16ガラスカラム
(Amersham Pharmacia)を、線速度150cm/時間の定流量パッキング法を使用して、種々の樹脂で充填した。クロマトグラフィーパラメーターを、種々の方法(詳細は各実験結果を参照のこと)を使用してプログラムした。一般的に、そのプログラムは、カラム平衡化工程、サンプルローディング工程、低塩洗浄工程、線形勾配工程または段階的勾配工程、そして高塩洗浄工程の機能的なブロックを包含する。種々の画分において溶出したAAVベクター粒子および空キャプシドをモニターし、以下に記載された技術を使用して分析した。
【0088】
(SDS−PAGEおよびウエスタンブロッティング)
カラム画分からのサンプルを取り、還元剤(DTT)を含有するSDS−PAGEローディング緩衝液中で加熱し、そしてキャプシドタンパク質をプレキャスト勾配ポリアクリルアミドゲル(Novex)上で分解した。銀染色を、SilverXpress(Invitrogen,CA)を使用して、製造者の説明書に従って実施した。同様に、ウエスタンブロット分析を、SDS−PAGEにより分離されたタンパク質のニトロセルロース膜への移送に引き続いて行った。モノクローナル抗体(B1、American Research Products,MA)を、ウエスタンブロットによりAAVキャプシドタンパク質(VP1、VP2およびVP3)を検出するための一次抗体として使用した。ヒツジ抗マウスIgG HRP結合体(Promega,WI)を二次抗体として使用し、ECLキット(Amersham,UK)により検出した。
【0089】
(定量リアルタイムPCR)
カラム画分中のAAVベクターゲノム(vg)濃度を、定量リアルタイムPCR(Q−PCR)により測定した。サンプルを希釈し、DNase Iで消化して、外来性DNAを除去した。DNaseの不活化後、サンプルをさらに希釈し、プライマーおよびそのプライマー間のDNA配列に対して特異的なTaqManTM蛍光発生プローブを使用して増幅した。規定されたレベルの蛍光に達するのに必要なサイクル数(閾値サイクル、Ct)を、各サンプルに対して、Applied Biosystems Prism 7700 Sequence Detection Systemで測定した。AAVベクターに含まれる配列と同一な配列を含有するプラスミドDNAを、そのQ−PCR反応における標準曲線を作成するために利用した。上記サンプルから得られるCt値を使用して、そのプラスミドの標準曲線のCt値に対して正規化することによりベクターゲノムの力価を決定した。
【0090】
(実施例1)
(空キャプシドおよびパッケージされたゲノムを有するAAVベクター粒子の結合特性)
AAVベクター粒子がアニオン交換カラムへ結合するか否か試験するために、複数の樹脂を、ハイスループット多岐管技術を使用してスクリーニングした。AAVベクター粒子および空キャプシドを含有する293細胞培養液の細胞溶解物を、NaCl濃度を約50mMまで減少させるために20mMリン酸ナトリウム緩衝液を使用して希釈し、そして各カラムにロードした。このカラムを次いで洗浄し、上記された手順に従って溶出した。その収集されたサンプルをウエスタンブロットに供し、画分中の上記AAV粒子の分布を検出した。図1Aおよび図1Bに示されるように、そのAAVキャプシドタンパク質特異的モノクローナル抗体は、高塩(1M NaCl)溶出画分のみ(図1、3レーン、5レーン、7レーンおよび9レーン)においてタンパク質を検出した。このデータは明らかに、AAVベクター粒子および空キャプシドの両方が、スクリーニングされた全てのアニオン交換樹脂(弱交換体および強交換体の両方)に結合することを示す。
【0091】
(実施例2)
(空キャプシドとAAVベクター粒子との間の電荷の差異)
AAV空キャプシドおよびAAVベクター粒子を、CsCl勾配遠心分離を使用して分離した。上記空キャプシドは、より低塩密度(約1.3gm/cc)で可視バンドとして生じ、上記AAVベクター粒子は、より高塩密度(約1.38〜1.41gm/cc)で可視バンドとして分布した。この空キャプシドおよびAAVベクター粒子を、上に記載されたように調製し、そしてカラムに別々にロードした。Q SEPHAROSE、POROS 50HQおよびUNOSPHERE Qの3つの樹脂を、空粒子と完全(full)粒子との間の溶出プロファイルを比較するために、同じプログラムを使用して行ったカラムクロマトグラフィー泳動のために使用した。AAV空キャプシドは、AAVベクター粒子(より後の画分に溶出する)と比較して、より低塩濃度で溶出(より先の画分に溶出)し;この現象は、全ての3つの試験されたアニオン樹脂に対して観察された。UNOSPHERE Q樹脂の場合、空キャプシドは、0〜500mM NaClの塩勾配における画分10に溶出するが、AAVベクター粒子は、画分11に溶出し;Q SEPHAROSEカラムの場合、空キャプシドは画分11、画分12および画分13に溶出するが、AAVベクター粒子は画分13、画分14および画分15に溶出した。酢酸ナトリウムのより浅い勾配を使用した場合、同様の現象がPOROS 50HQカラムで観察され;空キャプシドは画分30〜画分36で溶出したが、AAVベクター粒子は画分36〜画分40で溶出した。これらのデータは、明らかに、AAV空キャプシドとAAVベクター粒子との間に基礎的な電荷の差異が存在すること、そしてこの2つの集団がカラムクロマトグラフィー技術により、これらの特性を使用して分離され得ることを示す。これらの樹脂による空キャプシドおよびAAVベクター粒子の分離を確認したところ、260/280nmでのUV吸収の比が溶出画分において次第に増加し、これは差次的な溶出と一致する(DNAを欠くより低い260/280比を有する空キャプシドが先に溶出する)。
【0092】
(実施例3)
(空キャプシドとAAVベクター粒子との間の溶出効率および分解能における樹脂および塩の効果)
このプロセスにおいてスクリーニングされる樹脂は、全てアニオン交換樹脂であるが、差異(電荷密度、ビーズサイズおよび組成など)を示した。空キャプシドおよびAAVベクター粒子の結合および分離の効率は、樹脂により変動した。この空キャプシドは、Q SEPHAROSEカラム内で110mMのNaClを使用して溶出した。対照的に、約65mMのNaClが、UNOSPHEREカラムを使用して空キャプシドを溶出するために、十分であった。そしてPOROS HQカラムの場合は、約130mMのNaClが、この空キャプシドを溶出するために十分であった。
【0093】
複数の異なる塩もまた、溶出プロファイルの研究のために使用した。塩化ナトリウム、酢酸ナトリウム、酢酸カリウム、酢酸アンモニウムおよびクエン酸ナトリウムを、並行比較(side−by−side comparison)に供した。上記のカラムクロマトグラフィー泳動を行うための同じプログラムを使用すると、塩化ナトリウムと比較して酢酸ナトリウムまたは酢酸アンモニウムを使用した場合、空キャプシド溶出は複数の画分にわたって遅延した。これは、異なる塩はアニオン性カラムクロマトグラフィーにおいて異なる特性を有し、そしてこれらの特性は上記AAVベクター粒子からの空キャプシドの分離における分解能を強化するために使用され得ることを示唆する。
【0094】
(実施例4)
(空キャプシドに対する溶出条件を定義する)
上で議論された実験データに基づいて、POROS 50HQ樹脂を使用して、AAVベクター粒子から空キャプシドを分離するための、カラムクロマトグラフィーベースの技術を開発した。上で記載されたようなCsCl遠心分離を使用して調製された空キャプシドを、POROS 50HQ樹脂を詰めたカラムに充填した。低塩(50mM NaCl)の初期洗浄工程に引き続く分離泳動において、このカラムを、150mM酢酸ナトリウム、160mM酢酸ナトリウムまたは170mM酢酸ナトリウムのいずれかで洗浄した。この空キャプシドは、170mM NaAcを使用した場合に最も効率的に溶出した。この同じ塩の溶出プロフィールを使用した分離クロマトグラフィー泳動において、精製されたAAVベクター粒子はカラム内に残り、次いで高い濃度の塩化ナトリウムを使用した線形の塩濃度を加えたとき、溶出した。
【0095】
(実施例5)
(アニオン交換クロマトグラフィーを使用したAAVベクター粒子からの空キャプシドの分離)
空キャプシドおよびAAVベクター粒子がカラムクロマトグラフィー技術を使用して分離され得ることを実証するために、精製された空キャプシドおよびAAVベクター粒子を一緒に混合し、空キャプシドおよびAAVベクター粒子の両方を含有するサンプルを生成した。9E+13粒子/mlの濃度の3mlの空キャプシドを、7E+12ベクターゲノム(vg)/mlの濃度の2mlの完全粒子と混合した。その物質を、20mMのリン酸ナトリウム緩衝液(pH7.4)を使用して、さらに3倍に希釈した。14mlのこの希釈物質を、20mLのベッド容量(bed volume)で、POROS 50HQカラムに充填した。上に記載されたカラムクロマトグラフィー溶出プロファイルを使用し、そしてそのデータは、その空キャプシドが170mM酢酸ナトリウムの塩条件で溶出するがそのAAVベクター粒子はカラム内に残り、次いでNaCl塩の線形勾配を使用して溶出することを示した。その分離の過程にわたってそのカラムから溶出される物質のUV吸収パターン(260:280nm比)は、そのAAVベクター粒子からのその空キャプシドの効率的な分離と完全に一致し、そしてさらにそれを実証した。そのカラムから溶出される種々の画分を、キャプシドELISA、Q−PCR、SDS−PAGEおよびOD分析を使用してさらに分析した。得られた全てのデータは、AAVベクター粒子からの空キャプシドの分離が完全であり、そのAAVベクター粒子中のベクターゲノムに対する粒子の比は1:1であることを示した。このようにして、(そのカラムに充填された)出発物質がベクター粒子より約16倍多い空キャプシドを含んだとしても、このAAVベクター粒子はその空キャプシドから完全に分離された(図2)。
【0096】
(実施例6)
(アニオン交換クロマトグラフィーを使用したAAVベクター粒子からの空キャプシドの分離におけるpHの効果)
空キャプシドを除去するための完全なカラムベースの精製プロセスを開発するために、空キャプシドおよびAAVベクター粒子の両方を含有するカチオン交換カラム(POROS HS)から回収したビリオンを、そのPOROS 50HQカラムに加えた。CsCl精製サンプルから空キャプシドおよびAAVベクター粒子を分離するための類似の方法を使用した場合、CsCl勾配の前精製および混合したサンプルに対して観察されたような明確な分離は確認されなかったが、その2つの粒子の型は識別可能なままであった。すなわち、空キャプシドおよびAAVベクター粒子の、部分的には分離するが不完全な分離を観察した。カラムクロマトグラフィーの分解能に影響し得る複数のパラメーターを、その分離効率を最適化するために試験した。試験されたパラメーターの中で、pHがその分離分解能を有意に強化することが見出された。pH7.4、pH8.0、pH8.5およびpH9.0の緩衝液を試験した。空AAVキャプシドは、高いpH(例えばpH9.0)がそのプロセスにおいて使用された場合、そのカラムから早期の画分中に選択的に取り出されたが、AAVベクター粒子は後期の画分中に、より高い塩濃度において溶出した。pH9.0を使用して、その空キャプシドを、より高いUV280シグナルおよびより低いUV260シグナルのUV吸収シグネチャーパターンの単一ピークとして取り出した。対照的に、第二のピークはAAVベクター粒子を含み、DNAを含有する完全AAVベクター粒子に対して予想されたものと反対のUV吸収パターン(UV260がUV280より優位を占める)を示した。この知見は、Q−PCRおよびSDS−PAGEアッセイによりさらに確認され、その第二のピークがAAVベクター粒子を含み、第一のピークが空キャプシドを含むことを示した。
【0097】
(実施例7)
(アニオン交換カラムクロマトグラフィーを使用したAAVベクター粒子からのAAV空キャプシド分離の手順)
上に記載された知見に基づいて、XK−16ガラスカラム(Amersham Biosciences,Piscataway,NJ)において、トリスベースの緩衝液(pH8.5)およびPOROS 50HQカラム樹脂を使用して、上記AAVベクター粒子からAAV空キャプシドを分離するために、手順を設計した。UV吸収パターンにより示されたように、早期のセグメントに溶出された物質は空キャプシドを含有し、後期のセグメントはAAVベクター粒子を含有した。これはQ−PCR分析によりさらに確認された。より高い塩濃度で溶出したベクターを、純度を決定するために、SDS−PAGEで分析した。ベクターゲノムはQ−PCRにより決定し、複製されたサンプルをSDS−PAGEに充填し、そして銀染色により染色した。コントロールとして利用するために、AAVベクター粒子のみを含有するrAAVビリオンを、CsCl遠心分離技術を使用して精製し、そして同じベクターゲノムをSDS−PAGEゲルで泳動した。
【0098】
これらの技術を使用して、ベクターの約60%を回収した。SDS−PAGE銀染色分析に基づくと、回収されたベクターには、基本的に空キャプシドが存在しなかった(10未満の粒子が空キャプシドであった)(図3)。図3に示されるように、全てのレーンに等しい数のベクターゲノムを充填した場合、空キャプシドを含まないベクターのコントロール(レーン1)とカラムから溶出された画分に含まれるベクター(レーン2〜5)との間には、バンドの濃さ(総タンパク質に対応する)において有意な差はなかった。
【0099】
(実施例8)
(カチオン交換カラムクロマトグラフィーを使用したAAVベクター粒子からの空キャプシドの分離)
カチオン交換カラムクロマトグラフィーについてもまた、AAVベクター粒子から空キャプシドを分離するその能力を調査した。空キャプシドおよびAAVベクター粒子の両方を含む第一のカチオン交換カラムからのAAV調製物(材料および方法の節に記載される)を、POROS HS樹脂に加えた。そのカラムを最初に、200mM NaClを含む20mMリン酸ナトリウム緩衝液(pH7.4)で平衡化した。第一のカチオン交換カラムから得られたAAVベクター粒子および空キャプシドの両方を含有するAAV調製物を、20mMのリン酸ナトリウム(pH7.4)を使用して希釈し、塩濃度を約200mM NaClまで減少させた。この物質を次いで、POROS HSカラムに充填した。次いでpH8.5の10mMトリスを含む500mM〜700mM酢酸アンモニウム勾配を、そのカラムに適用した。この勾配溶出プロトコールを使用して、上記AAVベクター粒子を最初に溶出し、空キャプシドから効率的に分離し、この空キャプシドを続いてより高い酢酸アンモニウム濃度で溶出した。この分離に対する対照性は、空キャプシドがAAVベクターと比較してより低い塩濃度で溶出したアニオン交換樹脂(上記の実施例7)を使用しても観察された。
【0100】
そのカチオン交換カラムにおける空キャプシドおよびAAVベクター粒子の相対的な結合パターンおよび溶出パターンに関するさらなる情報は、このカラム溶出液のUV吸収により提供された。例えば、カチオン交換カラムをpH8.5で泳動した場合、そのクロマトグラフは、早期の画分において260nm吸収が優位なUV吸収パターン(AAVベクター粒子に対応する)、および後期の溶出画分において280nm吸収が優位なUV吸収パターン(空キャプシドに対応する)を示した。図4に示されるように、同じベクターゲノムをSDS−PAGE銀染色分析に供する場合、タンパク質の量は早期の画分から後期の画分に向かって有意に増加し、これは早期の画分がほとんど空キャプシドを含まないことを示す。
【0101】
これらの知見に基づいて、カチオン交換クロマトグラフィーを使用したAAVベクター(DNA含有)粒子からの空AAVキャプシドの分離は達成され得る。図5は、SDS−PAGEゲルが実証する分離を示す。カチオン交換カラムから溶出された空キャプシドおよびAAVベクター粒子の両方を含むAAVベクター調製物(分離を達成するために必要とされる条件は使用されていなかった)を、次いでカチオン交換カラムに充填し、そして酢酸アンモニウムを使用した段階勾配で溶出した。ベクターゲノムを含む画分をQ−PCRを使用して同定し、これらの画分のアリコートを、SDS−PAGE銀染色分析に供した。溶出された画分におけるタンパク質のシグナルが、その第二のカラムに充填された物質に対して、vgインプット当たりのタンパク質シグナルをほとんど含有しなかったことが明らかである。これは空キャプシドがそのAAVベクター粒子から除去されたことを示している。レーン2〜レーン4で示されるのは、3つの独立した実験から溶出されたベクターであり、この知見の一貫性および再現性を示している。
【0102】
このように、AAVベクター粒子からの空キャプシドの分離のための方法が記載される。本発明の好ましい実施形態がいくらか詳細に記載されたが、添付された特許請求の範囲により定義される本発明の精神および範囲から逸脱することなく、明らかな改変がなされ得ることが理解される。
【技術分野】
【0001】
(技術分野)
本発明は、アデノ随伴ウイルス(AAV)ビリオンを精製するための方法に関する。より具体的には、本発明は、パッケージングされたゲノムを含む組換えAAV(rAAV)ビリオンを、パッケージングされたrAAVビリオンとそのパッケージングされたゲノムを欠くAAVの空キャプシドとの両方を含むAAVビリオン混合物から精製するための方法に関する。
【背景技術】
【0002】
(背景)
治療上有効な量の遺伝子産物を安全かつ持続的に患者へと送達する遺伝子治療方法が、現在開発中である。これらの方法を使用して、核酸分子が、患者へと直接導入され得る(インビボ遺伝子治療)か、または患者もしくはドナーから単離された細胞中へと導入され得、その後、その細胞は上記患者へと戻される(エキソビボ遺伝子治療)。その後、導入された核酸は、その患者自身の細胞または移植細胞が望ましい治療産物を産生するように仕向ける。遺伝子治療はまた、臨床家が、治療のために特定の器官または細胞標的(例えば、筋肉、血球、脳細胞など)を選択するのを可能にする。
【0003】
核酸は、いくつかの方法で患者の細胞中へと導入され得る。その方法としては、ウイルス媒介性遺伝子送達、裸DNA送達、およびトランスフェクション法が挙げられる。ウイルス媒介性遺伝子送達は、大部分の遺伝子治療試験において使用されている(C.P.Hodgson,Biotechnology(1995)13:222〜225)。最も一般的に使用される組換えウイルスは、レトロウイルス、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびアデノ随伴ウイルス(AAV)を基にしている。
【0004】
組換えアデノ随伴ウイルスベクターは、ヒト遺伝子治療のための遺伝子送達ベクターとしての見込みを保持していた。しかし、そのようなベクターを薬物として使用するための1つの重要な障害は、商業的に実現可能なレベルでそのベクターを生成および精製するための、真に拡張可能なプロセスの開発である。商業的使用のためにAAVベクター生成を拡張することに関与する難問の概説について、QuおよびWright,Cur.Opin.Drug Disc.and Develop.(2000)3:750〜755を参照のこと。最近、rAAVビリオンを精製するための拡張可能性のあるいくつかのカラムクロマトグラフィー技術が、開発された。これらのカラムクロマトグラフィーベースの精製方法は、rAAVビリオンが、大規模精製され得ることを実証したが、カラムクロマトグラフィーを使用して精製ビリオンを調製すると、かなりの量のAAVの空キャプシドを含む。空キャプシドと、目的とする異種遺伝子を含むビリオン(「AAVベクター粒子」)との代表的な比率は、約10以上である。すなわち、回収されたベクターのうちの約90%は、空キャプシドである。
【0005】
大量の空キャプシドの存在は、例えば、そのキャプシドタンパク質に対する望ましくない免疫応答を惹起することによって、または標的細胞表面結合部位について競合することによって、臨床的適用を妨害し得る。結果として、rAAVビリオン調製物から上記空キャプシドを除去する技術が、開発されている。これらの技術は、代表的には、超遠心分離(例えば、塩化セシウムまたはイオジキサノール中で勾配遠心分離)に依存する。そのような遠心分離技術は、労働集約的であり、代表的には、低ベクター収率を生じ、かつ拡張可能ではない。Kaludovら(2002)Hum.Gene Ther.13:1235〜1243は、アニオン交換カラムを使用して、rAAV−2ベクター、rAAV−4ベクター、およびrAAV−5ベクターを精製する方法を記載する。しかし、その実験は、溶出物をプールして画分を濃縮した後でさえ、パッケージされたゲノムとしてそれぞれ、2%、0.6%、および6.3%しか回収可能ではなかった。
【発明の概要】
【発明が解決しようとする課題】
【0006】
従って、AAVベクター粒子ストックから空キャプシドを除去するかまたは空キャプシド数を減少させて、製造能力を増強する新規な方法についての必要性が、存在する。
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明は、空キャプシドの量が減少したrAAVビリオンストックを調製するための、効率的かつ商業的に実現可能な方法を開発したことに基づく。本発明者らは、本明細書において、空キャプシドが、カラムクロマトグラフィー技術を使用することによって、遺伝物質を含むrAAVビリオン(「AAVベクター粒子」)から分離され得ることを見出した。この結果は、驚くべきものである。なぜなら、空キャプシドと、パッケージングされたキャプシドとは、同一の表面特性を有すると、以前には考えられていたからである。本発明者らが知る限りでは、これは、ウイルス粒子の電荷および/または電荷密度が、空キャプシドと完全粒子との間では異なることについての最初の証明である。本明細書中に記載される技術は、AAVの空キャプシドをAAVベクター粒子から分離するための効率的かつ拡張可能な方法を提供する。
【0008】
従って、一実施形態において、本発明は、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法に関する。上記方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)上記宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)上記粗細胞溶解物を、第一のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(d)上記AAVベクター粒子および上記AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(e)上記工程(d)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(f)低塩緩衝液を、上記工程(e)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが上記カラムに結合したままである条件下で添加する工程;ならびに
(g)AAVベクター粒子を含む上記工程(f)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する。
【0009】
さらなる実施形態において、上記方法は、
(h)上記工程(g)からの画分を、アニオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが存在する場合には上記カラムに結合する条件下で、適用する工程;
(i)低塩緩衝液を、上記工程(h)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が上記カラムに結合したままである条件下で添加する工程;
(j)高塩緩衝液を、上記工程(i)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(k)AAVベクター粒子を含む上記工程(i)からの画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する。
【0010】
なおさらなる実施形態において、本発明は、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法に関する。上記方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)上記宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)上記粗細胞溶解物を清澄化して、清澄化した細胞溶解物を提供する工程;
(d)上記清澄化した細胞溶解物を、機能的リガンドR−SO3−を有するマトリックスを含む第一のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(e)上記AAVベクター粒子および上記AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(f)上記工程(e)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(g)低塩緩衝液を、上記工程(f)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが上記カラムに結合したままである条件下で添加する工程;
(h)AAVベクター粒子を含む工程(g)からの溶出画分を収集する工程;
(i)上記工程(h)からの画分を、アニオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが存在する場合には上記カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、上記工程(i)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が上記カラムに結合したままである条件下で添加する工程;
(k)高塩緩衝液を、上記工程(j)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;ならびに
(l)AAVベクター粒子を含む上記工程(k)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する。
【0011】
別の実施形態において、本発明は、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法に関する。上記方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)上記宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)上記粗細胞溶解物を、カチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(d)上記AAVベクター粒子および上記AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(e)上記工程(d)からのAAV調製物を、アニオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で適用する工程;
(f)低塩緩衝液を、上記工程(e)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が上記カラムに結合したままである条件下で添加する工程;
(g)高塩緩衝液を、上記工程(f)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(h)AAVベクター粒子を含む工程(g)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する。
【0012】
さらなる実施形態において、上記方法は、
(i)上記工程(h)からのAAV調製物を、第二のアニオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが存在する場合には上記カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、上記工程(i)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が上記カラムに結合したままである条件下で添加する工程;
(k)高塩緩衝液を、上記工程(j)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(l)AAVベクター粒子を含む上記工程(k)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する。
【0013】
代替的な実施形態において、上記方法は、
(i)上記工程(h)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、上記AAVベクター粒子および上記AAVの空キャプシドが上記カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、上記工程(i)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが上記カラムに結合したままである条件下で添加する工程;ならびに
(k)AAVベクター粒子を含む上記工程(j)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する。
【0014】
上記の方法すべての特定の実施形態において、上記第一のカチオン交換カラムおよび/または第二のカチオン交換カラムは、カルボキシメチル化マトリックスまたはスルホン化マトリックス(例えば、機能的リガンドR−SO3−を含むマトリックス)を含む。
【0015】
上記の方法すべてのさらなる実施形態において、上記AAVベクター粒子は、上記AAV生成物中に少なくとも50%の量(例えば少なくとも75%の量、例えば少なくとも85%または少なくとも90%の量)で存在する。
【0016】
上記の方法すべてのなおさらなる実施形態において、上記AAVベクター粒子は、AAV−2またはAAV−5に由来する。
【0017】
本発明はまた、以下の項目を提供する。
(項目1)
AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法であって、該方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)該宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)該粗細胞溶解物を、第一のカチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(d)該AAVベクター粒子および該AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(e)該工程(d)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(f)低塩緩衝液を、該工程(e)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが該カラムに結合したままである条件下で添加する工程;ならびに
(g)AAVベクター粒子を含む該工程(f)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する、方法。
(項目2)
項目1に記載の方法であって、前記第一のカチオン交換カラムは、カルボキシメチル化マトリックスまたはスルホン化マトリックスを含む、方法。
(項目3)
項目2に記載の方法であって、前記マトリックスは、機能的リガンドR−SO3−を含む、方法。
(項目4)
項目1に記載の方法であって、前記第二のカチオン交換カラムは、カルボキシメチル化マトリックスまたはスルホン化マトリックスを含む、方法。
(項目5)
項目4に記載の方法であって、前記マトリックスは、機能的リガンドR−SO3−を含む、方法。
(項目6)
項目1に記載の方法であって、
(h)前記工程(g)からの画分を、アニオン交換クロマトグラフィーカラムに対して、前記AAVベクター粒子および前記AAVの空キャプシドが存在する場合には該カラムに結合する条件下で、適用する工程;
(i)低塩緩衝液を、該工程(h)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が該カラムに結合したままである条件下で添加する工程;
(j)高塩緩衝液を、該工程(i)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(k)AAVベクター粒子を含む該工程(j)からの画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する、方法。
(項目7)
AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法であって、該方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)該宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)該粗細胞溶解物を清澄化して、清澄化した細胞溶解物を提供する工程;
(d)該清澄化した細胞溶解物を、機能的リガンドR−SO3−を有するマトリックスを含む第一のカチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(e)該AAVベクター粒子および該AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(f)該工程(e)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(g)低塩緩衝液を、該工程(f)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが該カラムに結合したままである条件下で添加する工程;
(h)AAVベクター粒子を含む工程(g)からの溶出画分を収集する工程;
(i)該工程(h)からの画分を、アニオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが存在する場合には該カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、該工程(i)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が該カラムに結合したままである条件下で添加する工程;
(k)高塩緩衝液を、該工程(j)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;ならびに
(l)AAVベクター粒子を含む該工程(k)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する、方法。
(項目8)
AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物からAAVベクター粒子を精製して、AAVの空キャプシドを実質的に含まないAAV生成物を提供するための方法であって、該方法は、
(a)AAVベクター粒子を含む宿主細胞を提供する工程;
(b)該宿主細胞を溶解して、AAVベクター粒子とAAVの空キャプシドとを含む粗細胞溶解物を得る工程;
(c)該粗細胞溶解物を、カチオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(d)該AAVベクター粒子および該AAVの空キャプシドを、非分離条件下で溶出して、AAVベクター粒子とAAVの空キャプシドとを含むAAV調製物を提供する工程;
(e)該工程(d)からのAAV調製物を、アニオン交換クロマトグラフィーカラムに対して、該AAVベクター粒子および該AAVの空キャプシドが該カラムに結合する条件下で適用する工程;
(f)低塩緩衝液を、該工程(e)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が該カラムに結合したままである条件下で添加する工程;
(g)高塩緩衝液を、該工程(f)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(h)AAVベクター粒子を含む工程(g)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
を包含する、方法。
(項目9)
項目8に記載の方法であって、
(i)前記工程(h)からのAAV調製物を、第二のアニオン交換クロマトグラフィーカラムに対して、前記AAVベクター粒子および前記AAVの空キャプシドが存在する場合には該カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、該工程(i)からのカラムに対して、AAVの空キャプシドが溶出されAAVベクター粒子が該カラムに結合したままである条件下で添加する工程;
(k)高塩緩衝液を、該工程(j)からのカラムに対して、AAVベクター粒子が溶出される条件下で添加する工程;
(l)AAVベクター粒子を含む該工程(k)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する、方法。
(項目10)
項目8に記載の方法であって、
(i)前記工程(h)からのAAV調製物を、第二のカチオン交換クロマトグラフィーカラムに対して、前記AAVベクター粒子および前記AAVの空キャプシドが該カラムに結合する条件下で、適用する工程;
(j)低塩緩衝液を、該工程(i)からのカラムに対して、AAVベクター粒子が溶出されAAVの空キャプシドが該カラムに結合したままである条件下で添加する工程;ならびに
(k)AAVベクターを含む該工程(j)からの溶出画分を収集して、AAVの空キャプシドを実質的に含まないAAV生成物を提供する工程、
をさらに包含する、方法。
(項目11)
項目1〜10のうちのいずれか1項に記載の方法であって、AAVベクター粒子は、前記AAV生成物中に少なくとも75%の量で存在する、方法。
(項目12)
項目1〜10のうちのいずれか1項に記載の方法であって、AAVベクター粒子は、前記AAV生成物中に少なくとも85%の量で存在する、方法。
(項目13)
項目1〜10のうちのいずれか1項に記載の方法であって、AAVベクター粒子は、前記AAV生成物中に少なくとも90%の量で存在する、方法。
(項目14)
項目1〜10のうちのいずれか1項に記載の方法であって、前記AAVベクター粒子は、AAV−2に由来する、方法。
(項目15)
項目1〜10のうちのいずれか1項に記載の方法であって、前記AAVベクター粒子は、AAV−5に由来する、方法。
本発明のこれらの実施形態および他の実施形態は、本明細書中の開示を考慮すると当業者には容易に思い浮かぶ。
【図面の簡単な説明】
【0018】
【図1】図1Aおよび1Bは、AAVベクター粒子およびAAV空キャプシドの両方を含む粗溶解物の結合特徴を示す。図1Aにおいて、試験した樹脂は、以下の通りであった:レーン1:コントロール;レーン2および3:MACRO PREP Q(BioRad,Hercules,CAから入手可能な強アニオン交換体);レーン4および5:UNOSPHERE Q(BioRad,Hercules,CAから入手可能な強アニオン交換体);レーン6および7:POROS 50HQ(Applied Biosystems,Foster City,CAから入手可能な強アニオン交換体);レーン8および9:POROS 50D(Applied Biosystems,Foster City,CAから入手可能な弱アニオン交換体)。図1Bにおいて、試験した樹脂は、以下の通りであった:レーン1:コントロール;レーン2および3:POROS 50PI(Applied Biosystems,Foster City,CAから入手可能な弱アニオン交換体);レーン4および5:SOURCE 30Q(Amersham Biosciences,Piscataway,NJから入手可能な強アニオン交換体);レーン6および7:DEAE SEPHAROSE(Amersham Biosciences,Piscataway,NJから入手可能な弱アニオン交換体);レーン8および9:Q SEPHAROSE(Amersham Biosciences,Piscataway,NJから入手可能な強アニオン交換体)。図1Aおよび1Bの両方について、レーン2、4、6および8は、低塩(50mM NaCl)洗浄画分を使用し;レーン3、5、7および9は、高塩(1M NaCl)洗浄画分を使用した。
【図2】図2は、実施例において記載されるように、アニオン交換クロマトグラフィーを使用して分離する前および分離した後の、AAV空キャプシドおよびAAVベクター粒子(Vgs)の分析を示す。
【図3】図3は、実施例において記載されるように、アニオン交換カラムからの画分の銀染色したSDS−PAGEゲルを示す。レーン1:AAVベクター粒子;レーン2〜5:ベクター溶出画分;レーン6:タンパク質分子量標準物質。
【図4】図4は、実施例に記載されるように、カチオン交換カラムからの溶出画分(レーン12〜21)を示す銀染色したSDS−PAGEゲルを示す。
【図5】図5は、カチオン交換カラムクロマトグラフィーを用いて、AAVベクター粒子からAAV空粒子を分離することを示す、銀染色したSDS−PAGEゲルを示す。レーン1:出発物質;レーン2〜4:カチオン交換カラムから溶出したベクターの3つの独立したサンプル。
【発明を実施するための形態】
【0019】
(発明の詳細な説明)
本発明の実施は、別段記載されなければ、ウイルス学、微生物学、分子生物学および組換えDNA技術の従来の方法を使用し、これらの方法は、当該分野の技術範囲内である。このような技術は、文献中に十分に説明されている。例えば、Sambrookら.Molecular Cloning:A Laboratory Manual(最新版);DNA Cloning:A Practical Approach,Vol.I&II(D.Glover,編);Oligonucleotide Synthesis.(N.Gait,編,最新版);Nucleic Acid Hybridization(B.Hames&S.Higgins,編,最新版);Transcription and Translation(B.Hames&S.Higgins,編、最新版);CRC Handbook of Parvoviruses,vol.I&II(P.Tijssen,編);Fundamental Virology,第2版,vol.I&II(B.N.Fields and D.M.Knipe,編);Freshney Culture of Animal Cells,A Manual of Basic Technique(Wiley−Liss,第3版);ならびにAusubelら(1991)Current Protocols in Molecular Biology(Wiley Interscience,NY)を参照のこと。
(1.定義)
本発明を記載するにあたって、以下の用語が使用され、以下に示されるように定義されると解釈される。
【0020】
本明細書および添付の特許請求の範囲において使用される場合、単数形「a」、「an」、および「the」は、文脈が明らかに別のことを示さなければ、複数形の指示対象を包含することに注意しなければならない。従って、例えば、「(1つの)パッケージされたキャプシド」への言及は、2つ以上のこのようなキャプシドの混合物などを包含する。
【0021】
「ベクター」とは、任意の遺伝的エレメント(例えば、プラスミド、ファージ、トランスポゾン、コスミド、染色体、ウイルス、ビリオンなど)を意味し、これらは、適切な制御エレメントと会合した場合に複製し得、細胞間で遺伝子配列を運び得る。従って、この用語は、クローニングビヒクルおよび発現ビヒクル、ならびにウイルスベクターを包含する。
【0022】
「AAVベクター」とは、以下の(しかし、それらの限定されない)アデノ随伴ウイルス血清型に由来するベクターを意味する:AAV−1、AAV−2、AAV−3、AAV−4、AAV−5、AAV−6、AAV−7およびAAV−8。AAVベクターは、AAV野生型遺伝子のうちの1つ以上(好ましくはrep遺伝子および/またはcap遺伝子)が、全てまたは一部欠失されているが、機能的隣接ITR配列を保持している。機能的ITR配列は、AAVビリオンのレスキュー、複製およびパッケージングに必要である。従って、AAVベクターは、ウイルスの複製およびパッケージングに、シスで必要とされる少なくともそれらの配列(例えば、機能的ITR)を含むことが、本明細書で規定される。このITRは、野生型ヌクレオチド配列である必要はなく、その配列が、機能的レスキュー、複製およびパッケージングを提供する限りにおいて、例えば、ヌクレオチドの挿入、欠失もしくは置換によって改変されていてもよい。
【0023】
「AAVヘルパー機能」とは、AAV遺伝子生成物(これは、次に、生産的なAAV複製のためにトランスで機能する)を提供するように発現され得るAAV由来のコード配列をいう。従って、AAVヘルパー機能は、主要AAVオープンリーディングフレーム(ORF)、repおよびcapをともに含む。このRep発現生成物は、多くの機能(とりわけ、以下が挙げられる:AAV DNA複製起点の認識、結合およびニック形成;DNAヘリカーゼ活性;ならびにAAV(または他の異種)プロモーターからの転写の調節)を有することが示された。このCap発現生成物は、必要なパッケージング機能を供給する。AAVヘルパー機能は、AAVベクターから失われているトランスでのAAV機能を補完するために、本明細書で用いられる。
【0024】
用語「AAVヘルパー構築物」は、一般に、AAVベクター(これは、目的のヌクレオチド配列を送達するための導入ベクターを生成するために使用される)から欠失されたAAV機能を提供するヌクレオチド配列を含む核酸分子をいう。AAVヘルパー構築物は、AAVのrep遺伝子および/またはcap遺伝子の一過性の発現を提供して、失われている、AAV複製に必須のAAV機能を補完するために一般に使用される;しかし、ヘルパー構築物は、AAV ITRを欠いており、それら自体では、複製もパッケージングもすることができない。AAVヘルパー構築物は、プラスミド、ファージ、トランスポゾン、コスミド、ウイルスまたはビリオンの形態であり得る。多くのAAVヘルパー構築物(例えば、一般的に使用されるプラスミドpAAV/AdおよびpIM29+45)が記載されており、これらは、Rep発現生成物およびCap発現生成物の両方をコードする。例えば、Samulskiら(1989)J.Virol.63:3822−3828;およびMcCartyら(1991)J.Virol.65:2936−2945を参照のこと。多くの他のベクターが記載されており、これらは、Rep発現生成物および/またはCap発現生成物をコードする。例えば、米国特許第5,139,941号および同第6,376,237号を参照のこと。
【0025】
用語「補助機能」とは、AAVが、その複製に関して依存性である、非AAV由来のウイルス機能および/または細胞機能をいう。従って、この用語は、AAV複製に必要とされるタンパク質およびRNA(AAV遺伝子転写の活性化、段階特異的AAV mRNAスプライシング、AAV DNA複製、Cap発現生成物の合成ならびにAAVキャプシドアセンブリに関与する部分を含む)を表す。ウイルスベースの補助機能は、既知のヘルパーウイルス(例えば、アデノウイルス、ヘルペスウイルス(単純ヘルペスウイルス1型以外)およびワクシニアウイルス)のいずれかに由来し得る。
【0026】
用語「補助機能ベクター」とは、一般に、補助機能を提供するヌクレオチド配列を含む核酸分子をいう。補助機能ベクターは、適切な宿主細胞にトランスフェクトされ得る。ここで、このベクターは、次いで、この宿主細胞におけるAAVビリオン生成を支援し得る。この用語から明示的に除外されるのは、天然に存在するままの感染性ウイルス粒子(例えば、アデノウイルス、ヘルペスウイルスまたはワクシニアウイルスの粒子である。従って、補助機能ベクターは、プラスミド、ファージ、トランスポゾン、またはコスミドの形態であり得る。特に、アデノウイルス遺伝子の完全な補完が、補助ヘルパー機能に必ずしも必須でないことが実証された。例えば、DNA複製および後期遺伝子合成ができないアデノウイルス変異体は、AAV複製に許容されることが示された。Itoら,(1970)J.Gen.Virol.9:243;Ishibashiら(1971)Virology 45:317。同様に、E2B領域およびE3領域内の変異は、AAV複製を支援することが示され、このことは、このE2B領域およびE3領域が、補助機能を提供することにおそらく関与しないことを示す。Carterら(1983)Virology 126:505。しかし、E1領域を欠いたアデノウイルスまたはE4領域が欠失したアデノウイルスは、AAV複製を支援することができない。従って、E1A領域またはE4領域は、直接的にせよ間接的にせよ、AAV複製に必要とされるようである。Laughlimら(1982)J.Virol.41:868;Janikら(1981)Proc.Natl.Acad.Sci.USA 78;1925;Carterら(1983)Virology 126:505。他の特徴づけられたAd変異体は、以下を含む:E1B(Laughlimら(1982)前出;Janikら(1981)前出;Ostroveら(1980)Virology,104:502);E2A(Handaら(1975)J.Gen.Virol.29:239;Straussら(1976)J.Virol.17:140;Myersら(1980)J.Virol.35:665;Jayら(1981)Proc.Natl.Acad.Sci.USA 78:2927;Myersら(1981)J.Biol.Chem.256:567);E2B(Carter,Adeno−Associated Virus Helper Functions,I CRC Handbook of Parvoviruses(P.Tijssen編、1990));E3(Carterら(1983)前出);E4(Carterら(1983)前出;Carter(1995))。E1Bコード領域に変異を有するアデノウイルスによって提供される補助機能の研究により、矛盾する結果がもたらされたが、Samulskiら(1988)J.Virol.62:206−210は、近年、E1B55kが、AAVビリオン生成に必須である一方で、E1B19kは必須でないことを報告した。さらに、国際公開WO97/17458およびMatshushitaら(1998)Gene Therapy 5:938−945は、種々のAd遺伝子をコードする補助機能ベクターを記載する。
【0027】
特に好ましい補助機能ベクターは、アデノウイルスVA RNAコード領域、アデノウイルスE4 ORF6コード領域、アデノウイルスE2A 72kDコード領域、アデノウイルスE1Aコード領域、およびインタクトなE1B55kコード領域を欠くアデノウイルスE1B領域を含む。このようなベクターは、国際公開番号WO01/83797に記載される。
【0028】
「組換えウイルス」とは、例えば、異種核酸構築物を付加するかまたはこの構築物を粒子に挿入することによって遺伝的に改変されたウイルスを意味する。
【0029】
「AAVビリオン」とは、完全なウイルス粒子(例えば、野生型(wt)AAVウイルス粒子(AAVキャプシドタンパク質コードと会合した直鎖状の一本鎖AAV核酸ゲノムを含む))を意味する。この点に関して、いずれかの相補的センス(例えば、「センス」鎖または「アンチセンス」鎖)の一本鎖AAV核酸分子は、いずれか1つのAAVビリオンにパッケージングされ得、両方の鎖は、等しく感染性である。
【0030】
用語「組換えAAVビリオン」、「rAAVビリオン」、「AAVベクター粒子」、「完全キャプシド」、「完全体」または「完全粒子」とは、AAV ITRが両側に隣接している、目的の異種ヌクレオチド配列をキャプシドに包んでいる、AAVタンパク質殻を含む感染性の複製欠損ウイルスとして本明細書中に規定される。rAAVビリオンは、そこに導入されるAAVベクターを特定する配列、AAVヘルパー機能および補助機能を有している適切な宿主細胞において生成される。このようにして、この宿主細胞は、その後の遺伝子送達のための感染性組換えビリオン粒子へのAAVベクター(目的の組換えヌクレオチド配列を含む)のパッケージングに必要とされるAAVポリペプチドをコードできるようにされる。
【0031】
用語「空キャプシド」、「空粒子」、および「空」とは、AVVタンパク質の殻を含むが、AAV ITRが両側に隣接した目的の異種ヌクレオチド配列を含むポリヌクレオチド構築物の全体または一部を欠く、AAVビリオンをいう。従って、この空キャプシドは、目的の遺伝子を宿主細胞へと移入するためには機能しない。
【0032】
用語「宿主細胞」は、AAVヘルパー構築物、AAVベクタープラスミド、補助機能ベクター、または他の移入DNAのレシピエントとして用いられ得るかまたは用いられた、例えば、微生物、酵母細胞、昆虫細胞、および哺乳動物細胞を示す。この用語は、トランスフェクトされた元の細胞の子孫を包含する。従って、「宿主細胞」は、本明細書中で用いられる場合、一般に、外因性DNA配列でトランスフェクトされた細胞をいう。単一の親細胞の子孫が、自然変異、偶発変異または意図的変異に起因して、元の親と形態またはゲノムDNA相補体もしくはDNA相補体全体において必ずしも完全に同一というわけではないことが理解される。
【0033】
用語「トランスフェクション」は、細胞による外来DNAの取り込みを言及するために用いられ、そして細胞は、外因性DNAが細胞膜の内側に導入されている場合、「トランスフェクト」されている。多数のトランスフェクション技術が当該分野で一般に公知である。例えば、Grahamら(1973)Virology,52:456,Sambrookら(1989)Molecular Cloning,a laboratory manual,Clod Spring Harbor Laboratories,New York,Davisら(1986)Basic Methods in Molecular Biology,Elsevier,およびChuら(1981)Gene 13:197を参照のこと。このような技術を用いて、1以上の外因性DNA部分を適切な宿主細胞へと導入し得る。
【0034】
本明細書中で用いられる場合、用語「細胞株」とは、連続増殖または長期増殖および分裂をインビトロで行い得る細胞集団をいう。しばしば、細胞株は、単一の子孫細胞に由来するクローン集団である。このようなクローン集団の保存または継代の間の核型における自然変化または誘導された変化が生じ得ることがさらに当該分野で公知である。それゆえ、言及される細胞株に由来する細胞は、先祖細胞または先祖培養物と正確には同一ではないかもしれず、そして言及される細胞株は、このような改変体を包含する。
【0035】
AAVベクター粒子(パッケージングされたゲノム)を含むrAAVビリオンのストックまたは調製物は、ストック中に存在するビリオンのうちの少なくとも約50%〜99%以上が、パッケージングされたゲノムを含むrAAVビリオン(すなわち、AAVベクター粒子)である場合、AAVの空キャプシドを「実質的に含まない」。好ましくは、このAAVベクター粒子は、ストック中に存在するビリオンのうちの、少なくとも約75%〜85%、より好ましくは約90%を構成し、さらにより好ましくは少なくとも約95重量%、またはさらにはストック中に存在するビリオンのうちの99重量%以上、またはこれらの範囲の間の任意の整数を構成する。従って、ストックは、得られるストックの約40%〜約1%以下、好ましくは約25%〜約15%以下、さらに好ましくは約10%以下、さらにより好ましくは約5%〜約1%以下が空キャプシドを含む場合、AAVの空キャプシドを実質的に含まない。
【0036】
「核酸」配列とは、DNA配列またはRNA配列をいう。この用語は、DNAおよびRNAの公知の塩基アナログ(例えば、4−アセチルシトシン、8−ヒドロキシ−N6−メチルアデノシン、アジリジニルシトシン、プソイドイソシトシン、5−(カルボキシヒドロキシルメチル)ウラシル、5−フルオロウラシル、5−ブロモウラシル、5−カルボキシメチルアミノメチル−2−チオウラシル、5−カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、イノシン、N6−イソペンテニルアデニン、1−メチルアデニン、1−メチルプソイドウラシル、1−メチルグアニン、1−メチルイノシン、2,2−ジメチルグアニン、2−メチルアデニン、2−メチルグアニン、3−メチルシトシン、5−メチルシトシン、N6−メチルアデニン、7−メチルグアニン、5−メチルアミノメチルウラシル、5−メトキシアミノメチル−2−チオウラシル、β−D−マンノシルキューオシン、5’−メトキシカルボニルメチルウラシル、5−メトキシウラシル、2−メチルチオ−N6−イソペンテニルアデニン、ウラシル−5−オキシ酢酸メチルエステル、ウラシル−5−オキシ酢酸、オキシブトキソシン、プロイドウラシル、キューオシン、2−チオシトシン、5−メチル−2−チオウラシル、2−チオウラシル、4−チオウラシル、5−メチルウラシル、Bウラシル−5−オキシ酢酸メチルエステル、ウラシル−5−オキシ酢酸、プソイドウラシル、キューオシン、2−チオシトシン、および2,6−ジアミノプリンであるがこれらに限定されない)のいずれかを含む配列を含む。
【0037】
「コード配列」または選択されたポリペプチドを「コード」する配列は、適切な制御配列の制御下においた場合に、インビボで、(DNAの場合)転写される、および(mRNAの場合)ポリペプチドへと翻訳される、核酸分子である。コード配列の境界は、5’(アミノ)末端における開始コドンおよび3’(カルボキシ)末端における翻訳停止コドンによって決定される。転写終結配列は、コード配列の3’側に配置され得る。
【0038】
用語DNA「制御配列」とは、レシピエント細胞におけるコード配列の複製、転写および翻訳をまとまって提供する、プロモーター配列、ポリアデニル化シグナル、転写終結配列、上流調節ドメイン、複製起点、内部リボソーム進入部位(「IRES」)、エンハンサーなどをまとめていう。選択されたコード配列が、適切な宿主細胞において複製され得、転写され得、そして翻訳され得る限り、これらの制御配列の全てがいつも存在する必要があるわけではない。
【0039】
用語「プロモーター」は、本明細書中で、DNA調節配列を含むヌクレオチド領域を言及するために、その元々の意味で用いられ、ここで、この調節配列は、RNAポリメラーゼに結合し得かつ下流の(3’方向の)コード配列の転写を開始し得る遺伝子に由来する。転写プロモーターは、「誘導性プロモーター」(ここで、このプロモーターに作動可能に連結されたポリヌクレオチド配列の発現は、分析物、補因子、調節タンパク質などによって誘導される)、「抑制性プロモーター」(ここで、このプロモーターに作動可能に連結されたポリヌクレオチド配列の発現は、分析物、補因子、調節タンパク質などによって誘導される)および「構成性プロモーター」を包含し得る。
【0040】
「作動可能に連結される」とは、そのように記載される成分がそれらの通常の機能を果たすように構成された構成要素の再配置をいう。従って、コード配列に作動可能に連結された制御配列は、コード配列の発現をもたらしえる。この制御配列は、その発現を導くように機能する限り、コード配列と連続する必要はない。従って、例えば、介在する、転写されるが翻訳されない配列は、プロモーター配列とコード配列との間に存在し得、そしてプロモーター配列は、なおも、コード配列に「作動可能に連結され」ていると考えられる。
【0041】
本出願全体で特定の核酸分子中でのヌクレオチド配列の相対位置を記載する目的で、例えば、特定のヌクレオチド配列が別の配列に対して「上流」、「下流」、「3’」、または「5’」に位置することが記載されている場合、これは、当該分野で慣習的であるように言及されるDNA分子の「センス」鎖または「コーディング」鎖におけるこの配列の位置であると理解されるべきである。
【0042】
用語「異種」とは、核酸配列(例えば、コード配列および制御配列)に関する場合、通常は一緒に連結していない配列、および/または特定の細胞に通常は関連していない配列を示す。従って、核酸構築物またはベクターのうちの「異種」領域は、他の分子とは天然で関連して見出されない別の核酸分子内のまたは別の核酸分子に結合した核酸のセグメントである。例えば、核酸構築物のうちの異種領域は、コード配列と天然では関連して見出されない配列が隣接したコード配列を含み得る。異種コード配列の別の例は、コード配列自体が天然には見出されない構築物である(例えば、ネイティブ遺伝子とは異なるコドンを有する合成配列)。同様に、細胞中に通常は存在しない構築物で形質転換された細胞は、本発明の目的では異種であると考えられる。対立遺伝子改変体または天然に存在する変異事象は、本明細書中で用いられる場合、異種DNAを生じない。
【0043】
「単離された」によって、ヌクレオチド配列を言及する場合、同一の分子が同じ型の他の生物学的高分子が実質的に存在せずに、存在することを意味する。従って、「特定のポリペプチドをコードする単離された核酸分子」とは、対象のポリペプチドをコードしない他の核酸分子を実質的に含まない核酸分子をいう;しかし、この分子は、その組成物の基本的特徴に有害な影響を与えない、いくつかのさらなる塩基または部分を含み得る。
【0044】
(2.発明を実施する形態)
本発明を詳細に記載する前に、特定の処方または特定のプロセスのパラメーターももちろん変化し得るので、本発明が特定の処方にも特定のプロセスのパラメーターにも限定されないことが理解されるべきである。本明細書中で用いられる用語は、単に本発明の特定の実施形態を記載する目的のためのものであり、限定することを意図していないこともまた理解されるべきである。
【0045】
本明細書中に記載される方法および材料と類似するかまたは等価な多数の方法および材料が本発明の実施において用いられ得るが、好ましい材料および方法は、本明細書中に記載される。
【0046】
本発明は、AAVビリオンの精製ストック中に含まれるAAVの空キャプシドの数を、その中に含まれるAAVベクター粒子の損失を最少にしながらも、減らすかまたは除去することを包含する。本発明の方法は、rAAVビリオンが精製されるプロセスと関係なく用いられ得る。
【0047】
rAAVビリオンを生成するために当該分野で周知のいくつかの方法が存在する;例えば、AAVヘルパーウイルス(例えば、アデノウイルス、ヘルペスウイルスまたはワクシニアウイルス)のうちの1つとの同時感染を伴う、ベクターおよびAAVヘルパー配列を用いたトランスフェクション、または組換えAAVベクター、AAVヘルパーベクター、および補助機能ベクターを用いたトランスフェクション。rAAVビリオンを生成するための方法の詳細な記載については、米国特許第6,001,650号および同第6,004,797号を参照のこと。
【0048】
例えば、野生型AAVウイルスおよびヘルパーウイルスは、rAAVビリオンを生成するための必要な複製機能を提供するために用いられ得る(例えば、米国特許第5.139,941号を参照のこと)。あるいは、周知のヘルパーウイルスのうちの1つによる感染に伴ったヘルパー機能遺伝子を含むプラスミドは、複製機能の供給源として用いられ得る(例えば、米国特許第5,622,856号および同第5,139,941号を参照のこと)。同様に、補助機能遺伝子を含むプラスミドは、野生型AAVによる感染と組み合わせて用いられて必要な複製機能を提供し得る。これらの3つのアプローチは、rAAVベクターと関連して組み合わされた場合、各々、rAAVビリオンを生成するために充分である。他のアプローチは、当該分野で周知であり、そしてまた、rAAVビリオンを生成するために当業者に用いられ得る。
【0049】
本発明の好ましい実施形態では、三重トランスフェクション法(米国特許第6,001,650号に詳細に記載される)を用いて、rAAVビリオンが生成される。なぜなら、この方法は、感染性ヘルパーウイルスの使用を必要とせずに、rAAVビリオンが、検出可能なヘルパーウイルスが全く存在せずに生成されるのを可能にするからである。これは、rAAVビリオン生成のための3つのベクター(AAVヘルパー機能ベクター、補助機能ベクター、およびrAAV発現ベクター)の使用によって達成される。しかし、当業者は、これらのベクターによってコードされる核酸配列が種々の組合せの2以上のベクター上で提供され得ることを理解する。
【0050】
本明細書中に説明したように、AAVヘルパー機能ベクターは、生産的AAV複製およびキャプシド形成のためにトランスで機能する「AAVヘルパー機能」配列(すなわち、repおよびcap)をコードする。好ましくは、AAVヘルパー機能ベクターは、検出可能なwtAAVビリオンを全く生成することなく、効率的なAAVベクター(すなわち、機能的なrep遺伝子およびcap遺伝子を含むAAVビリオン)生成を支持する。このようなベクターの一例であるpHLP19は、米国特許第6,001,650号に記載される。AAVヘルパー機能ベクターのrep遺伝子およびcap遺伝子は、上記で説明したように、公知のAAV血清型のいずれかに由来し得る。例えば、AAVヘルパー機能ベクターは、AAV−2由来のrep遺伝子およびAAV−6由来のcap遺伝子を有し得る;当業者は、他のrep遺伝子およびcap遺伝子の組合せが可能であり、定義する特徴がrAAVビリオン生成を支持する能力であることを認識する。
【0051】
補助機能ベクターは、AAVが複製を依存する非AAV由来ウイルス機能および/または細胞機能(すなわち、補助機能)についてのヌクレオチド配列をコードする。補助機能は、AAV複製に必要とされる機能(AAV遺伝子の転写、段階特異的AAV mRNAスプライシング、AAV DNA複製、cap発現産物の合成、およびAAVキャプシドアセンブリの活性化に関与する部分が挙げられるがこれらに限定されない)を包含する。ウイルスベースの補助機能は、周知のヘルパーウイルス(例えば、アデノウイルス、ヘルペスウイルス(1型単純疱疹ウイルス以外)、およびワクシニアウイルス)のいずれかに由来し得る。好ましい実施形態では、補助機能プラスミドpLadeno5が用いられる(pLadeno5に関する詳細は、米国特許第6,004,797号に記載される)。このプラスミドは、AAVベクター生成についてのアデノウイルス補助機能の完全なセットを提供するが、複製能力があるアデノウイルスを形成するために必要な成分を欠いている。
【0052】
一旦AAVビリオンのストックが生成されると、多数の方法(以下に詳述される)を用いて、感染性力価が決定され得、そしてAAVの空キャプシドからAAVベクター粒子が精製され得る。
【0053】
本発明をさらに理解するために、より詳細な考察を、組換えAAV発現ベクター、AAVヘルパーおよび補助機能、AAVビリオンを含む組成物、ならびにビリオンの送達に関して以下に提供する。
【0054】
(組換えAAV発現ベクター)
組換えAAV(rAAV)発現ベクターは、制御エレメント(転写開始領域を含む)、目的のポリヌクレオチド、および転写末端領域を転写の方向に作動可能に連結した成分として提供するように、公知の技術を使用して構築される。この制御エレメントは、目的の宿主細胞において機能的であるように選択される。作動可能に連結した成分を含む、結果の構築物は、機能性AAV ITR配列と結合される(5’および3’)。
【0055】
AAV ITR領域のヌクレオチド配列は公知である。たとえば、AAV−2配列については、Kotin,R.M.(1994)Human Gene Therapy 5:793−801;Berns,K.I.「Parvoviridae and their Replication」Fundamental Virology,第2版,(B.N.FieldsおよびD.M.Knipe(編))を参照のこと。本発明のベクターで使用されるAAV ITRは、野生型ヌクレオチド配列を有する必要はなく、例えば、ヌクレオチドの挿入、欠失、または置換によって変化され得る。加えて、AAV ITRは、いくつかのAAV抗原型(AAV−1、AAV−2、AAV−3、AAV−4、AAV−5、AAV−6、AAV−7、およびAAV−8などが挙げられるが、これらに限定されない)のいずれかに由来し得る。さらに、5’および3’のITRは、AAV発現ベクター中の選択されたヌクレオチド配列に隣接し、それらが意図されるように機能する限り(すなわち、宿主細胞ゲノムまたはベクターからの所定の配列の切断およびレスキューを可能にし、そして、AAV Rep遺伝子産物が受容細胞に存在する場合、受容細胞ゲノム中にDNA分子を組込むことを可能にする)、必ずしも同一である必要はなく、同一のAAV抗原型または単離物に由来する必要もない。
【0056】
AAVベクターに使用するために適切なポリヌクレオチド分子は、約5キロベース(kb)未満の大きさである。選択されるポリヌクレオチド配列は、インビボで被験体におけるそれらの転写または発現を方向付ける制御エレメントに作動可能に連結される。このような制御エレメントは、通常、選択遺伝子に関連する制御配列を含み得る。あるいは、異種制御配列が、使用され得る。一般的に、有用な異種制御配列は、哺乳動物遺伝子またはウイルス遺伝子をコードする配列に由来する制御配列を含む。例としては、ニューロン特異的エノラーゼプロモーター、GFAPプロモーター、SV40初期プロモーター、マウス乳腺癌ウイルスLTRプロモーター;アデノウイルス主要後期プロモーター(Ad MLP);単純ヘルペスウイルス(HSV)プロモーター、サイトメガロウイルス(CMV)プロモーター(例えば、CMV前初期プロモーター領域(CMVIE))、ラウス肉腫ウイルス(RSV)プロモーター、CAGプロモーター、合成プロモーター、ハイブリッドプロモーターなどが挙げられるが、これらに限定されない。加えて、非ウイルス遺伝子由来の配列(例えば、マウスメタロチオネイン遺伝子)もまた、本明細書中で使用を見出す。このようなプロモーター配列は、例えば、Stratagene(San Diego,CA)から市販されている。
【0057】
上記AAV発現ベクターは、AAV ITRによって結合される目的のポリヌクレオチド分子を含み、AAVゲノムから切断される主要なAAVオープンリーディングフレーム(「ORF」)を有するAAVゲノム中に選択配列を直接挿入することによって、構築され得る。AAVゲノムの他の部分はまた、複製およびパッケージング機能を可能にするようにITRの十分な部分が残る場合に限り、削除され得る。このような構築物は、当該分野で周知の技術を使用して設計され得る。例えば、米国特許第5,173,414号および同第5,139,941号;国際公開番号WO 92/01070(1992年1月23日公開)およびWO 93/03769(1993年3月4日公開);Lebkowskiら(1988)Molec.Cell.Biol.8:3988−3996;Vincentら(1990)Vaccines 90(Cold Spring Harbor Laboratory Press);Carter(1992)Current Opinion in Biotechnology 3:533−539;Muzyczka(1992)Current Topics in Microbiol.and Immunol.158:97−129;Kotin(1994)Human Gene Therapy 5:793−801;ShellingおよびSmith(1994)Gene Therapy 1:165−169;ならびにZhouら(1994)J.Exp.Med.179:1867−1875を参照のこと。
【0058】
あるいは、AAV ITRは、ウイルスゲノムからか、または別のベクター中に存在する、選択核酸構築物の同一で融合された5’および3’を含むAAVベクターから、標準的なライゲーション技術(例えば、Sambrookら,前出に記載される)を使用して切断され得る。例えば、ライゲーションは、20mM Tris−Cl(pH7.5)、10mM MgCl2、10mM DTT、33μg/ml BSA、10mM〜50mM NaCl、および0℃における40μM ATP、0.01〜0.02(Weiss)単位 T4 DNAリガーゼ(「付着末端」ライゲーション用)か、または14℃における1mM ATP、0.3〜0.6(Weiss)単位 T4 DNAリガーゼ(「平滑末端」ライゲーション用)において達成され得る。分子内の「付着末端」ライゲーションは、通常、30〜100μg/mlの総DNA濃度(総最終濃度5〜100nM)において実行される。ITRを含むAAVベクターは、例えば、米国特許第5,139,941号に記載されている。特に、いくつかのAAVベクターが上記特許明細書中に記載され、アメリカン タイプ カルチャー コレクション(「ATCC」)から入手可能である(受託番号53222、53223、53224、53225および53226)。
【0059】
本発明の目的のために、AAV発現ベクターからrAAVビリオンを生成するために適切な宿主細胞としては、微生物、酵母細胞、昆虫細胞、および哺乳動物細胞が挙げられる。これらの細胞は、異種DNA分子のレシピエントとして使用され得るかもしくは使用されており、例えば、浮遊培養物、バイオリアクターなどで増殖し得る。この用語は、トランスフェクトされたもとの細胞の子孫を含む。従って、本明細書中で使用される場合、「宿主細胞」は、一般的に、外因性DNA配列によってトランスフェクトされた細胞をいう。安定なヒト細胞株である293(例えば、アメリカン タイプ カルチャー コレクション(受託番号ATCC CRL1573)を通じて容易に入手可能)由来の細胞が、本発明の実施において好ましい。特に、ヒト細胞株293は、アデノウイルス5型のDNAフラグメントでトランスフェクトされたヒト胚性腎細胞株であり(Grahamら(1977)J.Gen.Virol.36:59)、アデノウイルスのE1a遺伝子およびE1b遺伝子を発現する(Aielloら(1979)Virology 94:460)。上記293細胞株は、容易にトランスフェフェクトされ、rAAVビリオンを生成する特に都合のよいプラットフォームを提供する。
【0060】
(AAVヘルパー機能)
上記AAV発現ベクターを含む宿主細胞は、AAV ITRに隣接して、rAAVビリオンを生成するためのヌクレオチド配列を複製し、そしてキャプシド形成するために、AAVヘルパー機能を提供し得なければならない。AAVヘルパー機能は、一般に、AAVに由来するコード配列である。この配列は、順生産的なAAV複製のためにトランスで機能するAAV遺伝子産物を提供するように発現され得る。AAVヘルパー機能は、本明細書中で、AAV発現ベクターから失われる必須のAAV機能を補完するために使用される。従って、AAVヘルパー機能は、主要なAAV ORF(すなわち、repコード領域およびcapコード領域)の1つもしくは両方、またはそれらの機能的ホモログを含む。
【0061】
「AAV repコード領域」によって、複製タンパク質のRep78、Rep68、Rep52およびRep40をコードするAAVゲノムのart認識(art−recognized)領域が意味される。これらのRep発現産物は、多くの機能(DNA複製のAAV起点の認識、結合、およびニッキング、DNAヘリカーゼ活性、ならびにAAV(または他の異種)プロモーターからの転写の調節を含む)を有することが示されている。このRep発現産物は、AAVゲノムを複製するために集合的に必要とされる。AAVのrepコード領域の記載については、例えば、Muzyczka,N.(1992)Current Topics in Microbiol.and Immunol.158:97−129;およびKotin,R.M.(1994)Human Gene Therapy 5:793−801を参照のこと。AAVのrepコード領域の適切なホモログとしては、AAV−2のDNA複製を媒介することもまた公知であるヒトヘルペスウイルス6(HHV−6)rep遺伝子が挙げられる(Thomsonら(1994)Virology 204:304−311)。
【0062】
「AAV capコード領域」によって、キャプシドタンパク質のVP1、VP2、およびVP3をコードするAAVゲノムのart認識領域、またはそれらの機能的ホモログが意味される。これらのCap発現産物は、ウイルスゲノムのパッケージングのために集合的に必要とされるパッケージング機能を与える。AAV capコード領域の記載については、例えば、Muzyczka,N.およびKotin,R.M.(前出)を参照のこと。
【0063】
AAVヘルパー機能は、AAV発現ベクターのトランスフェクションの前か、または同時に、AAVヘルパー構築物を用いて宿主細胞をトランスフェクトすることによって、宿主細胞中に導入される。従って、AAVヘルパー構築物が使用され、AAV rep遺伝子および/またはcap遺伝子の、少なくとも一過性の発現を提供し、生産的なAAV感染のために必須の失われたAAV機能を補完する。AAVヘルパー構築物は、AAV ITRを欠き、それ自体を複製もパッケージングもし得ない。
【0064】
これらの構築物は、プラスミド、ファージ、トランスポゾン、コスミド、ウイルス、またはビリオンの形態であり得る。多くのAAVヘルパー構築物(例えば、共通に使用されるプラスミドpAAV/AdおよびRepとCapとの両方の発現産物をコードするpIM29+45)が記載されている。例えば、Samulskiら(1989)J.Virol.63:3822−3828;およびMcCartyら(1991)J.Virol.65:2936−2945を参照のこと。Repおよび/またはCapの発現産物をコードする多くの他のベクターが、記載されている。例えば、米国特許第5,139,941号を参照のこと。
【0065】
(AAV補助機能)
宿主細胞(またはパッケージング細胞)はまた、rAAVビリオンを生成するために、非AAVの機能、または「補助機能(accessory function)」を提供され得なければならない。補助機能は、AAVがその複製のために依存する、非AAV由来のウイルス機能および/または細胞機能である。従って、補助機能としては、AAV複製(AAV遺伝子転写の活性化、段階特異的なAAV mRNAスプライシング、AAV DNA複製、Cap発現産物の合成およびAAVキャプシド集合に関係する機能を含む)の際に必要とされる、少なくともこれらの非AAVタンパク質およびRNAが挙げられる。ウイルスベースの補助機能は、任意の公知のヘルパーウイルスに由来し得る。特に、補助機能は、当業者に公知の方法を使用して宿主細胞に導入され得、次いで宿主細胞中で発現される。代表的には、補助機能は、宿主細胞を関連のないヘルパーウイルスに感染させることによって提供される。多くの適切なヘルパーウイルスが公知である。これらとしては、アデノウイルス;ヘルペスウイルス(例えば、単純ヘルペスウイルス1型および2型);およびワクシニアウイルスが挙げられる。非ウイルス性の補助機能(例えば、種々の公知の薬剤のいずれかを使用する細胞の同期化(cell synchronization)によって提供される機能)もまた、本明細書中で使用を見出される。例えば、Bullerら(1981)J.Virol.40:241−247;McPhersonら(1985)Virology 147:217−222;Schlehoferら(1986)Virology 152:110−117を参照のこと。あるいは、補助機能は、上に記載されるような補助機能ベクターを使用して提供され得る。例えば、米国特許第6,004,797号および国際公開番号WO 01/83797を参照のこと。
【0066】
補助機能を提供する核酸配列は、天然の供給源(例えば、アデノウイルス粒子のゲノムから)から得られ得か、または当該分野で公知の組換え方法もしくは合成方法を使用して構築され得る。上に説明したように、アデノウイルス遺伝子の完全な相補体が、補助ヘルパー機能のために必要とされるわけではないことが示されている。特に、DNA複製および後期遺伝子の合成が不可能なアデノウイルス変異体が、AAV複製に許容性であることが示されている。Itoら,(1970)J.Gen.Virol.9:243;Ishibashiら,(1971)Virology 45:317。同様に、E2B領域およびE3領域内の変異体は、AAV複製を支持することが示されており、このことは、上記E2B領域およびE3領域は、おそらく補助機能の提供に関与していないことを示している。Carterら,(1983)Virology 126:505。しかしながら、E1領域に欠陥のあるアデノウイルス、またはE4領域を欠くアデノウイルスは、AAV複製を支持し得ない。従って、E1A領域およびE4領域は、直接的または間接的にAAV複製に必要とされる可能性が高い。Laughlinら,(1982)J.Virol.41:868;Janikら,(1981)Proc.Natl.Acad.Sci.USA 78:1925;Carterら,(1983)Virology 126:505。他の特徴付けられたAd変異体としては、以下が挙げられる:E1B(Laughlinら(1982),前出;Janikら(1981),前出;Ostroveら,(1980)Virology 104:502);E2A(Handaら,(1975)J.Gen.Virol.29:239;Straussら,(1976)J Virol.17:140;Myersら,(1980)J.Virol.35:665;Jayら,(1981)Proc.Natl.Acad.Sci.USA 78:2927;Myersら,(1981)J.Biol.Chem.256:567);E2B(Carter,Adeno−Associated Virus Helper Functions,I CRC Handbook of Parvoviruses(P.Tijssen(編),1990));E3(Carterら(1983),前出);およびE4(Carterら(1983),前出;Carter(1995))。E1Bコード領域に変異を有するアデノウイルスによって提供された補助機能の研究が相反する結果を示しているにもかかわらず、E1B55kが、AAVビリオン生成に必要とされる一方、E1B19kは必要とされないことが、最近報告された(Samulskiら,(1988)J.Virol.62:206−210)。加えて、国際公開WO 97/17458およびMatshushitaら,(1998)Gene Therapy 5:938−945は、種々のAd遺伝子をコードする補助機能ベクターを記載する。特に好ましい補助機能ベクターは、アデノウイルスVA RNAコード領域、アデノウイルスE4 ORF6コード領域、アデノウイルスE2A 72kDコード領域、アデノウイルスE1Aコード領域、およびインタクトなE1B55kコード領域を欠くアデノウイルスE1B領域を含む。このようなベクターは、国際公開番号WO 01/83797に記載される。
【0067】
宿主細胞をヘルパーウイルスに感染させるか、または宿主細胞を補助機能ベクターを用いてトランスフェクションすることの結果として、AAVヘルパー構築物をトランス活性化する補助機能が発現され、AAV Repタンパク質および/またはCapタンパク質を産生する。このRep発現産物は、AAV発現ベクターから組換えDNA(目的のDNAを含む)を切断する。Repタンパク質はまた、AAVゲノムを二重にするために役割を果たす。この発現されたCapタンパク質は、キャプシド中に集合し、組換えAAVゲノムはキャプシド中にパッケージングされる。従って、生産性のAAV複製が後に続き、DNAは、rAAVビリオン中にパッケージングされる。
【0068】
(rAAVビリオンの精製)
組換えAAV複製に続いて、rAAVビリオンは、種々の慣用的な精製方法(例えば、カラムクロマトグラフィー、CsCl勾配など)を使用して、宿主細胞から精製され得る。例えば、多くのカラム精製の工程(例えば、アニオン交換カラム、アフィニティーカラムおよび/またはカチオン交換カラムによる精製)が、使用され得る。例えば、国際公開番号WO 02/12455を参照のこと。さらに、感染が適用されて補助機能が発現される場合、残りのヘルパーウイルスは、公知の方法を使用して不活化され得る。例えば、アデノウイルスは、約60℃の温度まで、例えば20分間以上加熱することによって不活化され得る。この処理は、ヘルパーウイルスのみを効果的に不活化する。なぜならば、AAVが非常に熱安定性である一方、ヘルパーアデノウイルスは熱不安定性だからである。
【0069】
任意の数のレポーター遺伝子を含む組換えAAVベクターが、感染性力価を決定するために使用され得る。例えば、アルカリホスファターゼ、β−ガラクトシダーゼ(LacZ)、緑色蛍光タンパク質、またはルシフェラーゼが、使用され得る。トランスフェクトされた宿主細胞を収集した後、大きなスケールの生成物のために適切な技術を使用して(例えば、微流動化(microfluidization))、トランスフェクトされた宿主細胞を破壊することによって、溶解物が形成される。次いで、この溶解物はフィルター処理され(例えば、0.45μmフィルターを通して)、そして、本明細書中に記載されるようなカラムクロマトグラフィー方法を使用して精製される。任意のAAVベクターの感染性力価を決定するための他の技術もまた、報告されている。例えば、Zhenら,「An Infectious Titer Assay for Adeno−associated Virus(AAV)Vectors with Sensitivity Sufficient to Detect Single Infectious Events.」Hum.Gene Ther.(2004)In,Pressを参照のこと。
【0070】
次いで、精製されたAAVストックが処理され、カラムクロマトグラフィー技術を使用して空キャプシドを除去する。本発明の特に好ましい方法において、rAAV調製物は、トランスフェクトされた細胞を溶解し、粗細胞溶解物を得ることによって得られる。次いで、この粗細胞溶解物は清澄化され、当該分野で周知の技術(例えば、フィルター処理、遠心分離など)によって細胞の破片を除去して清澄化細胞溶解物にされ得る。粗細胞溶解物または清澄化細胞溶解物は、AAVベクター粒子とAAVの空キャプシドとの両方を含み、次いで、非分離条件下で第一のカチオン交換カラムに対して適用される。上記第一のカチオン交換カラムは、細胞溶解物の調製物中に存在する細胞成分および他の成分から、AAVベクター粒子およびAAVの空キャプシドをさらに分離するために機能する。細胞溶解物の最初の精製を行うための方法は、公知である。1つの代表的な方法が、米国特許第6,593,123号に記載される。第一のカチオン交換カラムから回収された画分は、次いで、AAVの空キャプシドをAAVベクター粒子から分離する異なる溶出条件を使用して、第二のイオン交換体(すなわち、第二のカチオン交換カラムおよび/またはアニオン交換カラム)に対して、適用される。
【0071】
第一のカチオン交換カラムと第二のカチオン交換カラムとの両方に適切なカチオン交換体が使用される場合、当該分野で公知の多種多様な材料を含む。特に好ましいのは、広範なpH範囲にわたってrAAVビリオンを結合し得る強力なカチオン交換体である。例えば、カルボキシメチル化カチオン交換マトリックスおよびスルホン化カチオン交換マトリックスが、本明細書中の用途のために特に有用である。有用なマトリックス材料としては、セルロースマトリックス(例えば、繊維性マトリックス、微粒子性マトリックス、およびビーズマトリックス);アガロースマトリックス、デキストランマトリックス、ポリアクリレートマトリックス、ポリビニルマトリックス、ポリスチレンマトリックス、シリカマトリックス、およびポリエーテルマトリックス;ならびに合成物が挙げられるが、これらに限定されない。本明細書中で特に好ましいのは、機能的リガンドR−SO3−(好ましくは、スルホプロピル樹脂またはスルホエチル樹脂)を含むマトリックスである。代表的なマトリックスとしては、POROS HS、POROS SP、POROS S(すべてApplied Biosystems,Foster City,CAから入手可能な強力なカチオン交換体)、POROS CM(Applied Biosystems,Foster City,CAから入手可能な弱いカチオン交換体)、TOSOHAAS TOYOPEARL SP550CおよびMERCK FRACTOGEL EMD SO3−−650(m)、ならびにSOURCE 15S、SOURCE 30S、SEPHAROSE SP FF、SEPHAROSE SP XL(すべてAmersham Bioscience,Piscataway,NJから入手可能)が挙げられるが、これらに限定されない。
【0072】
下に与えられるすべてのカラムクロマトグラフィープロトコルについて、カラムは、当該分野で公知の標準的プロトコルを使用して、適切な緩衝液溶液で調製され得る。次いで、サンプルがロードされる。使用される第一のカチオン交換カラムについて、条件は、空キャプシドとAAVベクター粒子との両方がカラム樹脂に結合し、続いて一緒に溶出されるが、細胞溶解物中に存在する他の細胞成分および細胞の破片から分離されるような条件である。例えば、空キャプシドおよび完全キャプシドは、適切なイオン強度の緩衝液を使用して溶出される。適切な緩衝液としては、例えば、10〜50mMリン酸ナトリウム、好ましくは、15〜40(例えば、15、20、25、30、35、40など)mMリン酸ナトリウムが挙げられ、このリン酸ナトリウムは、塩(例えば、NaClまたはKCl)を例えば、100〜700mMの濃度(例えば、200〜400mM(例えば、200、300、325、350、370、380、400など、またはこれらの範囲内の任意の濃度))で含む。緩衝液のpHは、約3〜約9.5であり得、例えば、4〜8であり、例えば、pH4、4.5、5、5.5、6など、またはこれらの範囲内の任意のpHであり得る。これらの画分は回収され、次いで、分離条件化でアニオン交換カラムおよび/または第二のカチオン交換カラムのどちらかに適用され得る。
【0073】
続いて空のAAVキャプシドをAAVベクター粒子から分離するための工程において、第二のカチオン交換カラムが使用される場合、2種の溶出緩衝液が使用される。1つは低塩緩衝液であり、1つは高塩緩衝液である。特に、空キャプシドは、AAVベクター粒子から、約pH6〜pH12のpHにおける適切な緩衝液を使用して分離される。このpHは、好ましくは、pH7〜pH10であり、さらにより好ましくはpH7.5〜pH9.5であり、例えば、pH7.5、8.0、8.1、8.2、8.3、8.4、8.5、9.0、9.1、9.2、9.3、9.4、9.5、または示された範囲の間の任意のpHである。適切な緩衝液は、当該分野で周知であり、以下の緩衝液イオンを含む緩衝液が挙げられるが、これらに限定されない:酢酸;マロン酸;MES:リン酸;HEPES、BICINEなど。サンプルを溶出するために、開始緩衝液のイオン強度は、塩(例えば、NaCl、KCl、硫酸アンモニアまたは硫酸塩、ギ酸塩、酢酸塩、クエン酸塩および/もしくはリン酸塩を含む任意の他の塩)を使用することによって増加される。本発明の一実施形態において、カラムはまず低塩濃度で処理される。この濃度は、例えば、10〜200mMの酢酸アンモニウムであり、例えば、20、25、30、35、40、45、50、55、60、65、100mMまたはこれらの範囲内の任意の濃度である。この処理は、AAVベクター粒子のカラム樹脂からの溶出を生じる。続いて、このカラムは、AAVの空キャプシドを溶出するために、より高塩濃度で処理される。第二の緩衝液としての使用についての1つの例は、100〜800mMの濃度、好ましくは、500〜700mMであり、例えば、500、550、600、650、700、800mM、またはこれらの示された範囲内の任意の濃度の酢酸アンモニウムである。これら条件を使用して、AAVベクター粒子を早い画分に溶出し、空粒子を遅い画分に溶出する。
【0074】
上に説明されるように、本発明の代替的方法において、第一のカチオン交換カラムからの調製物は、第二のカチオン交換カラムの代わりにか、または第二のカチオン交換カラムに加えてのいずれかで、アニオン交換カラムに対して適用される。アニオン交換カラムが第二のカチオン交換カラムに加えて使用される場合、アニオン交換カラムは、第二のカチオン交換カラムの前か、または第二のカチオン交換カラムに続いて使用され得る。さらに、第二のアニオン交換カラムは、第一のアニオン交換カラムの後に使用され得る。本発明での使用についての多くの適切なアニオン交換体が公知であり、以下が挙げられるが、これらに限定されない:MACRO PREP Q(BioRad,Hercules,CAから入手可能な強力なアニオン交換体);UNOSPHERE Q(BioRad,Hercules,CAから入手可能な強力なアニオン交換体);POROS 50HQ(Applied Biosystems,Foster City,CAから入手可能な強力なアニオン交換体);POROS 50D(Applied Biosystems,Foster City,CAから入手可能な弱いアニオン交換体);POROS 50PI(Applied Biosystems,Foster City,CAから入手可能な弱いアニオン交換体);SOURCE 30Q(Amersham Biosciences,Piscataway,NJから入手可能な強力なアニオン交換体);DEAE SEPHAROSE(Amersham Biosciences,Piscataway,NJから入手可能な弱いアニオン交換体);Q SEPHAROSE(Amersham Biosciences,Piscataway,NJから入手可能な強力なアニオン交換体)。
【0075】
上記アニオン交換カラムはまず、標準的な緩衝液を使用して、製造者の仕様書に従って平衡化される。例えば、カラムは、例えば、5〜50mM(好ましくは、7〜20mM、例えば、10mM)のリン酸ナトリウム緩衝液で平衡化され得る。次いでサンプルがロードされ、そして2種の溶出緩衝液が使用される。1つは低塩緩衝液であり、1つは高塩緩衝液である。低塩洗浄および高塩洗浄のそれぞれに続いて、画分が回収され、そして画分中のタンパク質が、標準的技術(例えば、260nmおよび280nmにおけるUV吸光度のモニタリング)を使用して検出される。アニオン交換体を使用して、低塩溶出からのタンパク質ピークは、AAVの空キャプシドを含み、そして高塩画分は、AAVベクター粒子を含む。
【0076】
特に、アニオン交換カラムに関して、空キャプシドは、約pH5〜pH12のpHにおける適切な緩衝液を使用してAAVベクター粒子から分離され得る。このpHは、好ましくは、pH6〜pH10であり、さらにより好ましくはpH7〜pH9.5であり、例えば、pH7.1、7.2、7.3、7.4、8.0、8.1、8.2、8.3、8.4、8.5、9.0、9.1、9.2、9.3、9.4、9.5、または示された範囲の間の任意のpHである。アニオン交換カラムでの使用に適切な緩衝液は、当該分野で周知であり、一般的に本質的に陽イオン性であるか、または両性イオン性である。このような緩衝液としては、以下の緩衝液イオンを含む緩衝液が挙げられるが、これらに限定されない:N−メチルピペラジン;ピペラジン;ビス−Tris;ビス−Trisプロパン;トリエタノールアミン;Tris;N−メチルジエタノールアミン;1,3−ジアミノプロパン;エタノールアミン;酢酸など。サンプルを溶出するために、開始緩衝液のイオン強度は、塩(例えば、NaCl塩、KCl塩、硫酸塩、フマル酸塩または酢酸塩)を使用して、適切なpHにおいて増加される。
【0077】
本発明の一実施形態において、アニオン交換カラムは、まず、低塩濃度で処理される。その濃度は例えば、10〜100mMのNaClであり、例えば、10、20、25、30、35、40、45、50、55、60、65、100mM、またはこれらの範囲内の任意の濃度である。最初の処理に続いて、次いでカラムは、空キャプシドを溶出するために、高塩濃度(たとえば、より高いNaCl濃度、またはより強力なイオン強度を有する別の緩衝液で)で処理される。第二の緩衝液としての使用についての1つの例は、酢酸ナトリウム緩衝液、または100〜300mMの濃度を有するTrisベースの緩衝液である。この濃度は、好ましくは、125〜200mMであり、例えば、125、130、140、150、160、170、180、190、200mM、またはこれらの示された範囲内の任意の濃度である。空キャプシドがカラムから溶出された後、AAVベクター粒子は、高濃度の塩を使用して回収され得る。溶出緩衝液としての使用についての1つの例は、酢酸ナトリウムを含む10mM Tris緩衝液である。酢酸ナトリウムの濃度は100〜500mMの範囲であり、好ましくは、130〜300mMであり、例えば、100、130、150、200、250、300、350、400、450、500mM、またはこれら示された範囲内の任意の濃度である。
【0078】
上に記載される技術を使用して、90%以上のAAVの空キャプシドが、AAVベクター粒子から分離され得る。さらに、AAVベクター粒子の高い回収は、容易に達成される。すなわち、10%以上、好ましくは25%以上、より好ましくは50%以上、例えば、60%以上のAAVベクター粒子が回収され得る。
【0079】
空キャプシドおよびパッケージされたゲノムを有するAAVベクター粒子をアッセイするための方法は、当該分野において公知である。例えば、Grimmら、Gene Therapy(1999)6:1322−1330;Sommerら、Molec.Ther.(2003)7:122−128を参照のこと。変性キャプシドの試験のために、その方法は、処理されたAAVストックを上記3つのキャプシドタンパク質を分離可能な任意のゲル(例えば、緩衝液中に3〜8%トリス−酢酸を含む勾配ゲル)からなるSDS−ポリアクリルアミドゲル電気泳動に供する工程、次いでサンプル物質が分離されるまでそのゲルを泳動する工程、およびそのゲルをナイロンまたはニトロセルロース膜(好ましくはナイロン)にブロットする工程を包含する。抗AAVキャプシド抗体(好ましくは抗AAVキャプシドモノクローナル抗体、最も好ましくは上記B1抗AAV−2モノクローナル抗体)が次いで、変性キャプシドタンパク質に結合する一次抗体として使用される(Wobusら、J.Virol.(2000)74:9281−9293)。次いで二次抗体が使用される。これは、その一次抗体に結合しその一次抗体との結合を検出するための手段を含有するもの、より好ましくは、自身に共有結合された検出分子を含有する抗IgG抗体であり、最も好ましくは、西洋ワサビペルオキシダーゼに共有結合されたヒツジ抗マウスIgG抗体である。結合を検出するための方法は、その一次抗体とその二次抗体との間の結合を半定量的に決定するために使用され、好ましくは検出方法が放射性同位元素放射、電磁放射線または比色変化を検出可能であり、最も好ましくは化学発光検出キットである。
【0080】
感染力価の試験のための、その方法は、約100,000の宿主細胞(好ましくはヒト由来、最も好ましくはHeLa細胞)を、組織培養処理プレート(好ましくは24ウェル組織培養処理プレート)に播種し、そしてアデノウイルス(好ましくはアデノウイルス−2血清型)および処理されたrAAVストックをその宿主細胞に加えてから約24時間インキュベートする工程を包含する。その宿主細胞、アデノウイルスおよびrAAVストックは、24時間インキュベートされ、その後その宿主細胞は好ましくはホルムアルデヒドおよびグルタルアルデヒドで固定され、そしてrAAV発現導入遺伝子を検出する適切な薬剤で染色される(例えば、rAAV−LacZ、X−galが染色剤として企図される)。他のレポーター遺伝子に対する他の薬剤は、当該分野において周知である。任意の導入遺伝子を含有するベクターの感染力価を決定するためのより一般的な方法もまた、当該分野において公知である。例えば、Zhenら、「An Infectious Titer Assay for Adeno−associated Virus(AAV) Vectors with Sensitivity Sufficient to Detect Single Infectious Events」Hum.Gene Ther.(2004)(印刷中)を参照のこと。
【実施例】
【0081】
(3.実験)
以下は、本発明を実施するための特定の実施形態の例である。この例は、例示目的のみのために提供され、任意の方法において本発明の範囲を制限することを意図されない。
【0082】
使用された数字(例えば量、温度など)に関して精度を保証するための努力はなされたが、それに対するいくつかの実験誤差または偏差は、当然許容されるべきである。
【0083】
(材料および方法)
(rAAV産生および精製)
ヒトIX因子(rAAV−hFIX)をコードする遺伝子を含有する組み換えAAVビリオンを、米国特許第6,001,650号および同第6,004,797号に記載されるトリプルトランスフェクション(triple−transfection)法により産生した。使用したプラスミドは、補助機能プラスミド「pladeno5」、AAVヘルパー機能プラスミド「pHLP19」、および組み換えAAVプラスミド「phFIX−16」であった。ヒト胎児腎臓(HEK)293細胞を、rAAVビリオンの産生のために宿主細胞として使用した。
【0084】
トランスフェクションされた293細胞を、トランスフェクション後約72時間で回収し、MicrofluidizerTM(Microfluidics International Corp.,Newton,MA)を使用した微細流動化(microfluidization)により破砕し、そしてその粗製溶解物を収集し、連絡的フィルタを通して細胞細片を除去して、清澄な細胞溶解物を作製した。パッケージされたキャプシドおよび空キャプシドを含有するその清澄な細胞溶解物を、次いで、他の細胞成分からAAVベクター粒子およびAAVの空キャプシドを精製するために、POROS 50HSカラムにロードした。このAAVベクター粒子および空キャプシドを、このPOROS 50HSカラムから、20mMリン酸ナトリウム、370mM NaCl(pH5.5)を含有する緩衝液を使用して溶出した。AAVベクター粒子および空キャプシドの両方を含有するこのAAV調製物を、その塩濃度を減少させるために2倍希釈し、Q SEPHAROSEカラムを通してさらに精製し、流れて通過したAAVベクター粒子および空キャプシドを収集し、そして濃度および緩衝液の交換を限外濾過および透析濾過(dialfiltration)技術を使用して行った。
【0085】
最終的なPOROS 50HSカラム精製産物は、AAV空キャプシドおよび組み換えベクターゲノムを含有するAAVベクター粒子の両方を含有した。各々に対するコントロールサンプルを生成するために、塩化セシウムを攪拌しながら1.41g/mlの密度までPOROS 50HSカラム精製物質に加え、次いでTi70ローターを使用して45,000rpmで23時間超遠心分離へ供した。この可視の空キャプシドおよびAAVベクター粒子のバンドを、シリンジを使用して別々に汲み出した。この精製された空キャプシドおよびAAVベクター粒子を、10mMリン酸ナトリウム(pH7.4)、140mM NaClおよび5%ソルビタール(sorbital)からなる緩衝液に対して、透析カセット(Pierce)を使用し、4℃での、3回の緩衝液交換によって、別々に透析した。
【0086】
(樹脂およびカラムクロマトグラフィー)
アニオン交換樹脂、カチオン交換樹脂、等電点樹脂および疎水的相互作用(HIC)樹脂を含む種々の型のクロマトグラフィー樹脂を、AAV空キャプシドおよびAAVベクター粒子を分離するための、その能力について試験した。ハイスループットの多岐管を、樹脂のスクリーニングのために使用した。樹脂を、多岐管に設置された層高3cmの使い捨ての空のカラム(Bio−Rad Laboratories)に充填した。このカラムを、10mMのリン酸ナトリウムを含有する緩衝液を使用して平衡化した。次いで0.2μmで濾過された清澄なHEK293細胞溶解物のAAVベクターおよび空キャプシドを、50mMの濃度のNaCl(塩)のカラム平衡緩衝液中に希釈し、次いでそのカラムにロードした。このカラムを、最初に平衡緩衝液で洗浄し、次いで10mMリン酸ナトリウムおよび40mM NaCl(pH7.4)を含有する緩衝液でさらに洗浄し(低塩洗浄)、非結合物質を洗い出した。カラムを、次いで、10mMリン酸ナトリウムおよび1M NaClを含有する溶出緩衝液で処理した(高塩洗浄)。低塩洗浄を使用して洗い出された物質を1つの画分として収集し、高塩洗浄を使用して洗い出された物質を別の画分として収集した。
【0087】
使い捨てカラムを使用した初期スクリーニングに引き続き、より大きなスケールのカラムクロマトグラフィー(BIOCAD700(Applied Biosystems)上で行われる)を使用し、有望な樹脂および条件をさらに調べた。XK16ガラスカラム
(Amersham Pharmacia)を、線速度150cm/時間の定流量パッキング法を使用して、種々の樹脂で充填した。クロマトグラフィーパラメーターを、種々の方法(詳細は各実験結果を参照のこと)を使用してプログラムした。一般的に、そのプログラムは、カラム平衡化工程、サンプルローディング工程、低塩洗浄工程、線形勾配工程または段階的勾配工程、そして高塩洗浄工程の機能的なブロックを包含する。種々の画分において溶出したAAVベクター粒子および空キャプシドをモニターし、以下に記載された技術を使用して分析した。
【0088】
(SDS−PAGEおよびウエスタンブロッティング)
カラム画分からのサンプルを取り、還元剤(DTT)を含有するSDS−PAGEローディング緩衝液中で加熱し、そしてキャプシドタンパク質をプレキャスト勾配ポリアクリルアミドゲル(Novex)上で分解した。銀染色を、SilverXpress(Invitrogen,CA)を使用して、製造者の説明書に従って実施した。同様に、ウエスタンブロット分析を、SDS−PAGEにより分離されたタンパク質のニトロセルロース膜への移送に引き続いて行った。モノクローナル抗体(B1、American Research Products,MA)を、ウエスタンブロットによりAAVキャプシドタンパク質(VP1、VP2およびVP3)を検出するための一次抗体として使用した。ヒツジ抗マウスIgG HRP結合体(Promega,WI)を二次抗体として使用し、ECLキット(Amersham,UK)により検出した。
【0089】
(定量リアルタイムPCR)
カラム画分中のAAVベクターゲノム(vg)濃度を、定量リアルタイムPCR(Q−PCR)により測定した。サンプルを希釈し、DNase Iで消化して、外来性DNAを除去した。DNaseの不活化後、サンプルをさらに希釈し、プライマーおよびそのプライマー間のDNA配列に対して特異的なTaqManTM蛍光発生プローブを使用して増幅した。規定されたレベルの蛍光に達するのに必要なサイクル数(閾値サイクル、Ct)を、各サンプルに対して、Applied Biosystems Prism 7700 Sequence Detection Systemで測定した。AAVベクターに含まれる配列と同一な配列を含有するプラスミドDNAを、そのQ−PCR反応における標準曲線を作成するために利用した。上記サンプルから得られるCt値を使用して、そのプラスミドの標準曲線のCt値に対して正規化することによりベクターゲノムの力価を決定した。
【0090】
(実施例1)
(空キャプシドおよびパッケージされたゲノムを有するAAVベクター粒子の結合特性)
AAVベクター粒子がアニオン交換カラムへ結合するか否か試験するために、複数の樹脂を、ハイスループット多岐管技術を使用してスクリーニングした。AAVベクター粒子および空キャプシドを含有する293細胞培養液の細胞溶解物を、NaCl濃度を約50mMまで減少させるために20mMリン酸ナトリウム緩衝液を使用して希釈し、そして各カラムにロードした。このカラムを次いで洗浄し、上記された手順に従って溶出した。その収集されたサンプルをウエスタンブロットに供し、画分中の上記AAV粒子の分布を検出した。図1Aおよび図1Bに示されるように、そのAAVキャプシドタンパク質特異的モノクローナル抗体は、高塩(1M NaCl)溶出画分のみ(図1、3レーン、5レーン、7レーンおよび9レーン)においてタンパク質を検出した。このデータは明らかに、AAVベクター粒子および空キャプシドの両方が、スクリーニングされた全てのアニオン交換樹脂(弱交換体および強交換体の両方)に結合することを示す。
【0091】
(実施例2)
(空キャプシドとAAVベクター粒子との間の電荷の差異)
AAV空キャプシドおよびAAVベクター粒子を、CsCl勾配遠心分離を使用して分離した。上記空キャプシドは、より低塩密度(約1.3gm/cc)で可視バンドとして生じ、上記AAVベクター粒子は、より高塩密度(約1.38〜1.41gm/cc)で可視バンドとして分布した。この空キャプシドおよびAAVベクター粒子を、上に記載されたように調製し、そしてカラムに別々にロードした。Q SEPHAROSE、POROS 50HQおよびUNOSPHERE Qの3つの樹脂を、空粒子と完全(full)粒子との間の溶出プロファイルを比較するために、同じプログラムを使用して行ったカラムクロマトグラフィー泳動のために使用した。AAV空キャプシドは、AAVベクター粒子(より後の画分に溶出する)と比較して、より低塩濃度で溶出(より先の画分に溶出)し;この現象は、全ての3つの試験されたアニオン樹脂に対して観察された。UNOSPHERE Q樹脂の場合、空キャプシドは、0〜500mM NaClの塩勾配における画分10に溶出するが、AAVベクター粒子は、画分11に溶出し;Q SEPHAROSEカラムの場合、空キャプシドは画分11、画分12および画分13に溶出するが、AAVベクター粒子は画分13、画分14および画分15に溶出した。酢酸ナトリウムのより浅い勾配を使用した場合、同様の現象がPOROS 50HQカラムで観察され;空キャプシドは画分30〜画分36で溶出したが、AAVベクター粒子は画分36〜画分40で溶出した。これらのデータは、明らかに、AAV空キャプシドとAAVベクター粒子との間に基礎的な電荷の差異が存在すること、そしてこの2つの集団がカラムクロマトグラフィー技術により、これらの特性を使用して分離され得ることを示す。これらの樹脂による空キャプシドおよびAAVベクター粒子の分離を確認したところ、260/280nmでのUV吸収の比が溶出画分において次第に増加し、これは差次的な溶出と一致する(DNAを欠くより低い260/280比を有する空キャプシドが先に溶出する)。
【0092】
(実施例3)
(空キャプシドとAAVベクター粒子との間の溶出効率および分解能における樹脂および塩の効果)
このプロセスにおいてスクリーニングされる樹脂は、全てアニオン交換樹脂であるが、差異(電荷密度、ビーズサイズおよび組成など)を示した。空キャプシドおよびAAVベクター粒子の結合および分離の効率は、樹脂により変動した。この空キャプシドは、Q SEPHAROSEカラム内で110mMのNaClを使用して溶出した。対照的に、約65mMのNaClが、UNOSPHEREカラムを使用して空キャプシドを溶出するために、十分であった。そしてPOROS HQカラムの場合は、約130mMのNaClが、この空キャプシドを溶出するために十分であった。
【0093】
複数の異なる塩もまた、溶出プロファイルの研究のために使用した。塩化ナトリウム、酢酸ナトリウム、酢酸カリウム、酢酸アンモニウムおよびクエン酸ナトリウムを、並行比較(side−by−side comparison)に供した。上記のカラムクロマトグラフィー泳動を行うための同じプログラムを使用すると、塩化ナトリウムと比較して酢酸ナトリウムまたは酢酸アンモニウムを使用した場合、空キャプシド溶出は複数の画分にわたって遅延した。これは、異なる塩はアニオン性カラムクロマトグラフィーにおいて異なる特性を有し、そしてこれらの特性は上記AAVベクター粒子からの空キャプシドの分離における分解能を強化するために使用され得ることを示唆する。
【0094】
(実施例4)
(空キャプシドに対する溶出条件を定義する)
上で議論された実験データに基づいて、POROS 50HQ樹脂を使用して、AAVベクター粒子から空キャプシドを分離するための、カラムクロマトグラフィーベースの技術を開発した。上で記載されたようなCsCl遠心分離を使用して調製された空キャプシドを、POROS 50HQ樹脂を詰めたカラムに充填した。低塩(50mM NaCl)の初期洗浄工程に引き続く分離泳動において、このカラムを、150mM酢酸ナトリウム、160mM酢酸ナトリウムまたは170mM酢酸ナトリウムのいずれかで洗浄した。この空キャプシドは、170mM NaAcを使用した場合に最も効率的に溶出した。この同じ塩の溶出プロフィールを使用した分離クロマトグラフィー泳動において、精製されたAAVベクター粒子はカラム内に残り、次いで高い濃度の塩化ナトリウムを使用した線形の塩濃度を加えたとき、溶出した。
【0095】
(実施例5)
(アニオン交換クロマトグラフィーを使用したAAVベクター粒子からの空キャプシドの分離)
空キャプシドおよびAAVベクター粒子がカラムクロマトグラフィー技術を使用して分離され得ることを実証するために、精製された空キャプシドおよびAAVベクター粒子を一緒に混合し、空キャプシドおよびAAVベクター粒子の両方を含有するサンプルを生成した。9E+13粒子/mlの濃度の3mlの空キャプシドを、7E+12ベクターゲノム(vg)/mlの濃度の2mlの完全粒子と混合した。その物質を、20mMのリン酸ナトリウム緩衝液(pH7.4)を使用して、さらに3倍に希釈した。14mlのこの希釈物質を、20mLのベッド容量(bed volume)で、POROS 50HQカラムに充填した。上に記載されたカラムクロマトグラフィー溶出プロファイルを使用し、そしてそのデータは、その空キャプシドが170mM酢酸ナトリウムの塩条件で溶出するがそのAAVベクター粒子はカラム内に残り、次いでNaCl塩の線形勾配を使用して溶出することを示した。その分離の過程にわたってそのカラムから溶出される物質のUV吸収パターン(260:280nm比)は、そのAAVベクター粒子からのその空キャプシドの効率的な分離と完全に一致し、そしてさらにそれを実証した。そのカラムから溶出される種々の画分を、キャプシドELISA、Q−PCR、SDS−PAGEおよびOD分析を使用してさらに分析した。得られた全てのデータは、AAVベクター粒子からの空キャプシドの分離が完全であり、そのAAVベクター粒子中のベクターゲノムに対する粒子の比は1:1であることを示した。このようにして、(そのカラムに充填された)出発物質がベクター粒子より約16倍多い空キャプシドを含んだとしても、このAAVベクター粒子はその空キャプシドから完全に分離された(図2)。
【0096】
(実施例6)
(アニオン交換クロマトグラフィーを使用したAAVベクター粒子からの空キャプシドの分離におけるpHの効果)
空キャプシドを除去するための完全なカラムベースの精製プロセスを開発するために、空キャプシドおよびAAVベクター粒子の両方を含有するカチオン交換カラム(POROS HS)から回収したビリオンを、そのPOROS 50HQカラムに加えた。CsCl精製サンプルから空キャプシドおよびAAVベクター粒子を分離するための類似の方法を使用した場合、CsCl勾配の前精製および混合したサンプルに対して観察されたような明確な分離は確認されなかったが、その2つの粒子の型は識別可能なままであった。すなわち、空キャプシドおよびAAVベクター粒子の、部分的には分離するが不完全な分離を観察した。カラムクロマトグラフィーの分解能に影響し得る複数のパラメーターを、その分離効率を最適化するために試験した。試験されたパラメーターの中で、pHがその分離分解能を有意に強化することが見出された。pH7.4、pH8.0、pH8.5およびpH9.0の緩衝液を試験した。空AAVキャプシドは、高いpH(例えばpH9.0)がそのプロセスにおいて使用された場合、そのカラムから早期の画分中に選択的に取り出されたが、AAVベクター粒子は後期の画分中に、より高い塩濃度において溶出した。pH9.0を使用して、その空キャプシドを、より高いUV280シグナルおよびより低いUV260シグナルのUV吸収シグネチャーパターンの単一ピークとして取り出した。対照的に、第二のピークはAAVベクター粒子を含み、DNAを含有する完全AAVベクター粒子に対して予想されたものと反対のUV吸収パターン(UV260がUV280より優位を占める)を示した。この知見は、Q−PCRおよびSDS−PAGEアッセイによりさらに確認され、その第二のピークがAAVベクター粒子を含み、第一のピークが空キャプシドを含むことを示した。
【0097】
(実施例7)
(アニオン交換カラムクロマトグラフィーを使用したAAVベクター粒子からのAAV空キャプシド分離の手順)
上に記載された知見に基づいて、XK−16ガラスカラム(Amersham Biosciences,Piscataway,NJ)において、トリスベースの緩衝液(pH8.5)およびPOROS 50HQカラム樹脂を使用して、上記AAVベクター粒子からAAV空キャプシドを分離するために、手順を設計した。UV吸収パターンにより示されたように、早期のセグメントに溶出された物質は空キャプシドを含有し、後期のセグメントはAAVベクター粒子を含有した。これはQ−PCR分析によりさらに確認された。より高い塩濃度で溶出したベクターを、純度を決定するために、SDS−PAGEで分析した。ベクターゲノムはQ−PCRにより決定し、複製されたサンプルをSDS−PAGEに充填し、そして銀染色により染色した。コントロールとして利用するために、AAVベクター粒子のみを含有するrAAVビリオンを、CsCl遠心分離技術を使用して精製し、そして同じベクターゲノムをSDS−PAGEゲルで泳動した。
【0098】
これらの技術を使用して、ベクターの約60%を回収した。SDS−PAGE銀染色分析に基づくと、回収されたベクターには、基本的に空キャプシドが存在しなかった(10未満の粒子が空キャプシドであった)(図3)。図3に示されるように、全てのレーンに等しい数のベクターゲノムを充填した場合、空キャプシドを含まないベクターのコントロール(レーン1)とカラムから溶出された画分に含まれるベクター(レーン2〜5)との間には、バンドの濃さ(総タンパク質に対応する)において有意な差はなかった。
【0099】
(実施例8)
(カチオン交換カラムクロマトグラフィーを使用したAAVベクター粒子からの空キャプシドの分離)
カチオン交換カラムクロマトグラフィーについてもまた、AAVベクター粒子から空キャプシドを分離するその能力を調査した。空キャプシドおよびAAVベクター粒子の両方を含む第一のカチオン交換カラムからのAAV調製物(材料および方法の節に記載される)を、POROS HS樹脂に加えた。そのカラムを最初に、200mM NaClを含む20mMリン酸ナトリウム緩衝液(pH7.4)で平衡化した。第一のカチオン交換カラムから得られたAAVベクター粒子および空キャプシドの両方を含有するAAV調製物を、20mMのリン酸ナトリウム(pH7.4)を使用して希釈し、塩濃度を約200mM NaClまで減少させた。この物質を次いで、POROS HSカラムに充填した。次いでpH8.5の10mMトリスを含む500mM〜700mM酢酸アンモニウム勾配を、そのカラムに適用した。この勾配溶出プロトコールを使用して、上記AAVベクター粒子を最初に溶出し、空キャプシドから効率的に分離し、この空キャプシドを続いてより高い酢酸アンモニウム濃度で溶出した。この分離に対する対照性は、空キャプシドがAAVベクターと比較してより低い塩濃度で溶出したアニオン交換樹脂(上記の実施例7)を使用しても観察された。
【0100】
そのカチオン交換カラムにおける空キャプシドおよびAAVベクター粒子の相対的な結合パターンおよび溶出パターンに関するさらなる情報は、このカラム溶出液のUV吸収により提供された。例えば、カチオン交換カラムをpH8.5で泳動した場合、そのクロマトグラフは、早期の画分において260nm吸収が優位なUV吸収パターン(AAVベクター粒子に対応する)、および後期の溶出画分において280nm吸収が優位なUV吸収パターン(空キャプシドに対応する)を示した。図4に示されるように、同じベクターゲノムをSDS−PAGE銀染色分析に供する場合、タンパク質の量は早期の画分から後期の画分に向かって有意に増加し、これは早期の画分がほとんど空キャプシドを含まないことを示す。
【0101】
これらの知見に基づいて、カチオン交換クロマトグラフィーを使用したAAVベクター(DNA含有)粒子からの空AAVキャプシドの分離は達成され得る。図5は、SDS−PAGEゲルが実証する分離を示す。カチオン交換カラムから溶出された空キャプシドおよびAAVベクター粒子の両方を含むAAVベクター調製物(分離を達成するために必要とされる条件は使用されていなかった)を、次いでカチオン交換カラムに充填し、そして酢酸アンモニウムを使用した段階勾配で溶出した。ベクターゲノムを含む画分をQ−PCRを使用して同定し、これらの画分のアリコートを、SDS−PAGE銀染色分析に供した。溶出された画分におけるタンパク質のシグナルが、その第二のカラムに充填された物質に対して、vgインプット当たりのタンパク質シグナルをほとんど含有しなかったことが明らかである。これは空キャプシドがそのAAVベクター粒子から除去されたことを示している。レーン2〜レーン4で示されるのは、3つの独立した実験から溶出されたベクターであり、この知見の一貫性および再現性を示している。
【0102】
このように、AAVベクター粒子からの空キャプシドの分離のための方法が記載される。本発明の好ましい実施形態がいくらか詳細に記載されたが、添付された特許請求の範囲により定義される本発明の精神および範囲から逸脱することなく、明らかな改変がなされ得ることが理解される。
【特許請求の範囲】
【請求項1】
本明細書中に記載の発明。
【請求項1】
本明細書中に記載の発明。
【図1】


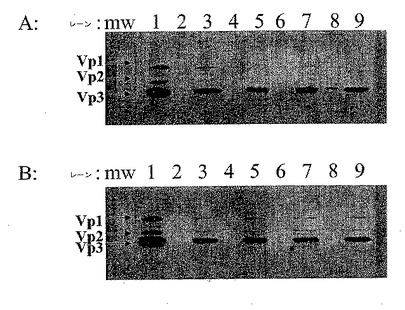
【図2】
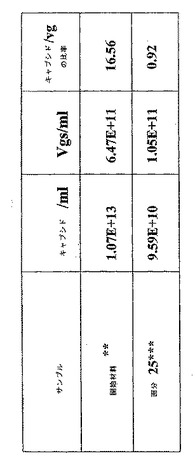
【図3】

【図4】
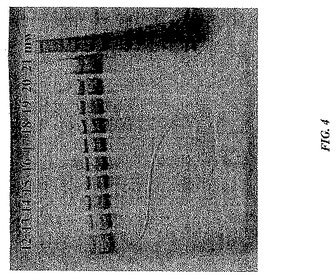
【図5】
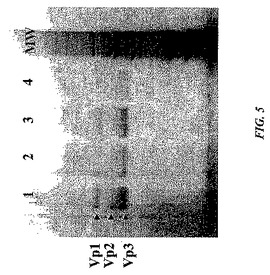
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2010−155868(P2010−155868A)
【公開日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願番号】特願2010−89074(P2010−89074)
【出願日】平成22年4月7日(2010.4.7)
【分割の表示】特願2006−533274(P2006−533274)の分割
【原出願日】平成16年5月21日(2004.5.21)
【出願人】(500579888)ジェンザイム・コーポレーション (34)
【Fターム(参考)】
【公開日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願日】平成22年4月7日(2010.4.7)
【分割の表示】特願2006−533274(P2006−533274)の分割
【原出願日】平成16年5月21日(2004.5.21)
【出願人】(500579888)ジェンザイム・コーポレーション (34)
【Fターム(参考)】
[ Back to top ]