
窒素同化の増加を示すトランスジェニック植物
【課題】増大した窒素同化/利用能、より速くより活発な成長、より多い栄養体および生殖体の収穫量、そして栄養および生殖部分の富化されたまたは改変された窒素含量が改良された農業および栄養特性を有する植物の作出方法を提供する。
【解決手段】窒素同化/利用経路で鍵となる重要な酵素の発現を改変すべく修飾される植物の遺伝子工学的操作を行い、天然のまたは修飾された窒素同化酵素の異所性過剰発現のための植物により達成される。
【解決手段】窒素同化/利用経路で鍵となる重要な酵素の発現を改変すべく修飾される植物の遺伝子工学的操作を行い、天然のまたは修飾された窒素同化酵素の異所性過剰発現のための植物により達成される。
【発明の詳細な説明】
【技術分野】
【0001】
1.序文
本発明は、一般に、増強された窒素同化および利用能を示し、より大きく、より効率的に、より速く成長し、かつ/また、栄養および/または生殖に関係した植物部分の窒素含量および/または生物量(バイオマス)の増加を示す植物体を遺伝子工学的に作出することに関するものである。より詳細には、本発明は、遺伝子工学的手法により窒素同化/利用経路で鍵となる酵素の発現を改変させたトランスジェニック植物を作出することに関するものである。このように遺伝子工学的に作出された植物は、低い窒素肥料投入量の条件下でまたは窒素の乏しい土壌で生産的に栽培することができる。また、遺伝子工学的に作出された植物を用いると、理想的な栽培条件下でより速く成長または成熟する作物、より高い作物収穫量および/またはより栄養分の多い生産物を得ることが可能となる。
【背景技術】
【0002】
2.発明の背景
窒素はしばしば植物の成長の律速要因になり、基本的に野外作物はすべて無機窒素肥料に依存している。肥料は多くの種類の土壌から急速に枯渇してしまうので、生育期に2,3回、成長しつつある作物に肥料を施さなければならない。窒素肥料は、通常は硝酸アンモニウム、硝酸カリウムまたは尿素として供給され、一般にトウモロコシやコムギのような農作物に関連したコストの40%を占めている。北アメリカと西ヨーロッパの両地域では毎年約1100万トンの窒素肥料が使われており、そのために農業従事者は毎年22億ドルを出費していると概算された (Sheldrick, 1987, World Nitrogen Survey, Technical Paper no. 59, Washington, D.C.) 。さらに、世界銀行の試算では、1年間の窒素肥料の需要量が世界中で今後10年間に約9000万トンから1億3000万トン以上に増加するであろうと推論された。植物による窒素の使用効率を上げれば、より低い肥料投入量でまたは比較的やせた土地で作物を栽培することが可能となり、それゆえ、開発されたまたは開発されつつある農業システムに相当の経済的効果を及ぼすことができるだろう。
【0003】
通常の選別技術を用いて、植物の育種家らは、トウモロコシ、コムギ、イネ、その他の作物種の天然集団から得られる変種を利用することによって窒素使用効率を改良しようと試みた。しかしながら、野外条件下で評価することが困難な諸特性に関する従来の育種計画には、広範な集団のスクリーニングに伴う困難が立ちはだかり、こうした選別法はほとんど成功しなかった。
【0004】
2.1. 植物の窒素同化経路
植物は窒素をその環境から無機化合物の形で、すなわち根から吸収される硝酸とアンモニア、そして窒素固定根粒でアンモニアに還元される大気中のN2の形で得ている。若干の硝酸とアンモニアが輸送道管(木部および篩部)において検出され得るが、大部分の窒素はまず有機体(例:アミノ酸)に同化されてから、植物体内に輸送される。
【0005】
無機窒素を有機体に同化する第一段階は、主にグルタミンを形成するためのグルタミン酸によるアンモニアの取り込みを含み、これはグルタミンシンテターゼ酵素 (GS;EC 6.3.1.2)により触媒される。次いで、斯く形成されたグルタミンは、アスパラギンシンテターゼ酵素 (AS;E.C. 6.3.5.4)により触媒されるアスパラギンの形成においてそのアミド基を供与する。この経路でのアンモニアからアスパラギンへの窒素の定常流はグルタミン酸とα−ケトグルタル酸とアスパラギン酸の循環に依存しており、それぞれグルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ(GOGAT;E.C.)とアスパラギン酸アミノトランスフェラーゼ(AspAT;E.C.)により触媒される(図1参照)。かくして、GS、AS、AspATおよびGOGATは高等植物の主要な窒素同化経路の不可欠な酵素といえる。
【0006】
アンモニアの取り込みがGSにより触媒される経路以外の代替経路により進行しうることを示す証拠が存在する(図1)。Knight and Langston-Unkefer, 1988, Science 241:951-954(非特許文献1)を参照されたい。一つの経路はグルタミン酸を形成するためのα−ケトグルタル酸によるアンモニアの取り込みを含み、これはグルタミン酸デヒドロゲナーゼ(GDH)により触媒される。もう一つの経路はアスパラギンを形成するためのアスパラギン酸によるアンモニアの取り込みを含み、これはアスパラギンシンテターゼにより触媒される (Oaks and Ross, 1984, Can. J. Bot. 62:68-73(非特許文献2); Stulen and Oaks, 1977, Plant Physiol. 60:680-683(非特許文献3)) 。これらの酵素(GDHおよびAS)は両方ともアンモニアに対して高いKmを有し、正常な生育条件下(例えば、低濃度の内部アンモニア)でのこの代替窒素同化経路の役割は依然として不明のままである。ある研究により、これらまたは他の代替窒素同化経路は細胞内のアンモニウム濃度が正常レベルを越えて上昇したときの植物の窒素同化能に対して大いに寄与していることが示唆された (Knight and Langston-Unkefer,前掲)。
【0007】
2.2. 窒素の輸送および利用
グルタミンおよびアスパラギンは主要な長距離窒素輸送化合物を代表するもので、篩部の液汁中に豊富に存在している。窒素担体としてのそれらの共通した役割のほかに、これら2つのアミノ酸は植物窒素代謝においてやや異なる役割を担っている。グルタミンは2つのアミノ酸のうちで代謝的により活性で、さまざまな同化反応で多数の基質にそのアミド窒素を直接供与することができる。その反応性ゆえに、一般的に植物は窒素の貯蔵のためにグルタミンを利用することはない。
【0008】
対照的に、アスパラギンはその高いN:C比のためにグルタミンと比べてより効率のよい窒素輸送用の化合物となる。さらに、アスパラギンはグルタミンよりも安定しており、液胞中に高レベルで蓄積され得る。実際、窒素同化能の高い植物では、アスパラギンが窒素の輸送および代謝において支配的な役割を果たしているようである。The Biochemistry of Plants: A Comprehensive Treatise, vol 5, Amino acid and derivatives, Miflin 編, Academic Press, New York (1980) pp. 569-607(非特許文献4) 中の Lea and Miflin による「植物におけるアスパラギンと他の窒素化合物の輸送および代謝」;および Sieciechowiczら, 1988, Phytochemistry 27:663-671(非特許文献5) を参照されたい。その相対的安定性のために、アスパラギンは窒素代謝に直接関与することはないが、アスパラギナーゼ酵素(ANS;E.C. 3.5.1.1)によってまず加水分解されてアスパラギン酸とアンモニアを生成し、次いでこれらはアミノ酸とタンパク質の合成に利用されるにちがいない(図1参照)。
【0009】
2.3. 窒素の同化および利用に関与する植物遺伝子
植物の窒素同化/利用に係わる酵素をコードしている遺伝子の多くがクローン化され、研究されてきた。植物のグルタミンシンテターゼ (GS)遺伝子およびアスパラギンシンテターゼ(AS)遺伝子の論議については、Transgenic Plants, Vol. 1, Kung and Wu編, Academic Press, San Diego, CA, (1993) pp. 181-194(非特許文献6) 中の Tsai and Coruzzi による「アミノ酸生合成酵素をコードする遺伝子を研究するためのトランスジェニック植物」ならびにその中で引用された文献;アルファルファのアスパラギン酸アミノトランスフェラーゼ遺伝子の論議については、Udvardi and Kahn, 1991, Mol. Gen. Genet. 231:97-105(非特許文献7) ;タバコのグルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ(GOGAT、グルタミン酸シンテターゼとも言う)遺伝子の論議については、Zehnacker ら, 1992, Planta 187:266-274(非特許文献8);ハウチワマメ (lupin)のアスパラギナーゼ遺伝子の論議については、Lough ら, 1992, Plant Mol. Biol. 19:391-399(非特許文献9) および Dicksonら, 1992, Plant Mol. Biol. 20:333-336(非特許文献10) を参照されたい。
【0010】
植物の窒素同化/利用遺伝子の中で、最も広く研究されているものはグルタミンシンテターゼ遺伝子とアスパラギンシンテターゼ遺伝子である。GSとASには複数の遺伝子が存在し、これら遺伝子の分子特性付けにより、それらが異なる発現パターンを有することが示された。
【0011】
2.3.1. グルタミンシンテターゼ遺伝子
GSは植物の生育中に多くの器官で活発に機能している (McNally ら, 1983, Plant Physiol. 72:22-25(非特許文献11))。根において、それは土の中の水から得られたアンモニアを同化し (Oaks and Hirel, 1985, Ann. Rev. Plant Physiol. 36:345-365(非特許文献12)、また、マメ科植物の根粒ではGSが根粒菌により固定されたアンモニアを同化する (Cullimore ら, 1983, Planta 157:245-253(非特許文献13)) 。子葉にあっては、GSが発芽中に集められた窒素貯蔵物を再同化し (Recent Advances in Phythochemistry: Mobilization of Reserves in Germination, Nozolilloら編, Plenum Press, (1983) p. 77-109(非特許文献14)中の Lea and Joyによる「発芽種子におけるアミノ酸相互変換」)、そして葉では葉緑体GS2が光呼吸で放出されたアンモニアを同化する (Givan ら, 1988, TIBS 13:433-437(非特許文献15))。GSの多種多様な役割は、差次的に発現される異なる遺伝子から誘導されたさまざまのGSイソ型が担っている (Gebhardtら, 1986, EMBO J. 5:1429-1435(非特許文献16); Tingey ら, 1987, EMBO J. 6:1-9(非特許文献17))。
【0012】
エンドウのPhaseolusおよび Arabidopsisでは、葉緑体GS2が単一の核遺伝子によってコードされているが、これらの種のそれぞれには細胞質ゾルGSの複数の遺伝子が存在している (Bennett ら, 1989, Plant Mol. Biol. 12:553-565(非特許文献18); Tingey ら, 1988, J. Biol. Chem. 263:9651-9657(非特許文献19); Peterman and Goodman, 1991, Mol. Gen. Genet. 230:145-154(非特許文献20)) 。in vivo およびトランスジェニック宿主植物におけるこれらGS遺伝子の発現の分析は、植物の窒素代謝での各種GSイソ型の役割を解明する上で役に立った。
【0013】
エンドウのGS遺伝子ファミリーは4つの別個のしかし相同の核遺伝子を含んでいる。3つは細胞質ゾルのGSイソ型をコードし、1つは葉緑体のGS2イソ型をコードする (Tingeyら, 1987, EMBO J. 6:1-9(非特許文献21); Tingey ら, 1988, J. Biol. Chem. 263:9651-9657(非特許文献22)) 。ノーザンブロット分析により、葉緑体GS2の遺伝子は、一部にはフィトクロムにより、また、一部には光呼吸作用により光依存的に葉で発現されることが実証された (Edwards and Coruzzi, 1989, Plant Cell 1:241-248(非特許文献23))。また、細胞質ゾルGSの3つの遺伝子(GS1、GS3AおよびGS3B)も異なる役割を果たしているようである。根では、細胞質ゾルGS1が主要なイソ型であるが、それは結節でも発現される。細胞質ゾルGS3AおよびGS3Bは結節で高度に発現され、さらに発芽中の種子の子葉でも発現される (Tingeyら, 1987, EMBO J. 6:1-9(非特許文献24); Walker and Coruzzi, 1989, Plant Physiol. 91:702-708(非特許文献25))。GS3A遺伝子とGS3B遺伝子は配列が同一に近いものの、遺伝子特異的S1−ヌクレアーゼ分析により、GS3Aの発現がGS3Bの発現よりも常に高いことが明らかになった (Walker and Coruzzi, 1989, Plant Physiol. 91:702-708(非特許文献25))。プロモーター−GUS融合体およびトランスジェニック植物の分析により、葉緑体GS2は光合成細胞型でのみ発現され、また、細胞質ゾルGS3Aは大部分の器官の脈管構造の篩部細胞でもっぱら発現されることが分かった。さらに、GS3Aは根や結節の分裂組織でも強く発現される (Edwards ら, 1990, Proc. Natl. Acad. Sci. USA, 87:3459-3463(非特許文献26); Brearsら, 1991, The Plant Journal, vol. 1, pp. 235-244(非特許文献27)) 。細胞型および器官レベルでの厳密に制御された調節からすると、GSの各種遺伝子はアンモニア同化において重複することのない別個の役割を果たしていると考えられる。
【0014】
2.3.2. アスパラギンシンテターゼ遺伝子
2つのAS遺伝子がエンドウからクローン化され(AS1およびAS2)、両方とも根粒と子葉で最高レベルに発現される。AS1およびAS2はともに根において発現される。AS2は構成的に根で発現され、一方、AS1は暗黒で育てた植物の根にのみ発現される (Tsai and Coruzzi, 1990, EMBO J 9:323-332(非特許文献28)) 。さらに、AS1とAS2は暗順応植物の成熟葉で発現されるが、その発現は光によって阻害される。暗黒でのこの高レベルのAS遺伝子発現は、光合成炭素の利用能が低下した条件下で合成される長距離窒素輸送化合物としてのアスパラギンの使用と合致している(アスパラギンはグルタミンより高いN:C比を有する)。トランスジェニック植物におけるAS1プロモーター−GUS融合体の研究から、AS1遺伝子もGS3A遺伝子と同様に篩部細胞でもっぱら発現されることが示された。細胞型および器官レベルでの厳密に制御された調節からすると、各種AS遺伝子も植物窒素代謝において異なる役割を果たしていると思われる。
【0015】
2.4. 植物の窒素同化/利用過程の遺伝子工学的操作
植物では、窒素同化過程の遺伝子工学的操作がさまざまな結果をもたらした。一つの事例を挙げると、タバコにおいて原核生物のアンモニウム依存性アスパラギンシンテターゼ(ASN−A)遺伝子を発現させることが、種々のグルタミンシンテターゼ(GS)阻害剤に対する耐性を付与した (Duditsら, Transgenic Plants expressing a prokaryotic ammonium dependent asparagine synthetase, WO 9111524, Aug. 8, 1991(特許文献1))。また、これらの同じ植物が成長速度の増加、植物発育の促進、早期開花、植物の新鮮重量および乾燥重量の増加を含めて多くの成長変化を示した。GS阻害剤処理は遺伝子工学的に作出した植物の成長を低下させるどころかむしろ高めたので、ASN−A発現の成長効果は理屈に合わないことになる。
【0016】
対照的に、グルタミンシンテターゼ(GS)の過剰発現を試験する多くの研究は、その過剰発現が植物の成長に及ぼすポジティブな効果を報告することができなかった。Lea and Forde, 1994, Plant Molec. Biol. 17:541-558(非特許文献29); Eckes ら,1989, Molec. Gen. Genet. 217:263-268(非特許文献30) (アルファルファGSを過剰発現するトランスジェニックタバコ植物); Hemonら, 1990, Plant Mol. Biol. 15:895-904(非特許文献31)(細胞質またはミトコンドリアでマメGSを過剰発現するトランスジェニックタバコ植物); Hirelら, 1992, Plant Mol. Biol. 20:207-218(非特許文献32) (タバコ植物内でダイズGSを過剰発現するトランスジェニックタバコ植物)を参照されたい。ある研究が、アルファルファGS1遺伝子を過剰発現するトランスジェニックタバコ植物における総可溶性タンパク質含量の増加を認めたと報告した。しかしながら、この同じ研究がGS1遺伝子に対するアンチセンスRNAを発現するトランスジェニックタバコ植物における総可溶性タンパク質含量の同様の増加をも報告したことから、GS1発現と可溶性タンパク質の増加との関係ははっきりしていないようである (Templeら, 1993, Mol. Gen. Genet. 236:315-325(非特許文献33)) 。植物におけるGS過剰発現の明確に確立された効果は、GS阻害性除草剤であるホスフィノトリシンに対する耐性である (Eckes ら, 前掲; Donnら, 1984, J. Molec. Appl. Genet. 2:621-635(非特許文献34) (ホスフィノトリシン耐性アルファルファ細胞系はGS遺伝子の増幅を含んでいた))。また、アルファルファGS遺伝子を過剰発現するように遺伝子操作された植物は遺伝子操作されていない植物よりも速く成長するという報告があった (Eckes ら, 1988, オーストラリア特許庁文書番号:AU-A-17321/88(特許文献2))。しかし、報じられたような速い成長は低窒素生育条件下でのみ起こり、普通または高窒素生育条件下では認められなかった。その上、より速い成長がより多い生物量または生産量でもって成熟植物を生み出すのかどうか不明である。Eckes らの前掲を Eckesら, 1989, Molec. Gen. Genet. 217:263-268(非特許文献35)と比較されたい。
【先行技術文献】
【特許文献】
【0017】
【特許文献1】WO 9111524
【特許文献2】AU-A-17321/88
【非特許文献】
【0018】
【非特許文献1】Knight and Langston-Unkefer, 1988, Science 241:951-954
【非特許文献2】Oaks and Ross, 1984, Can. J. Bot. 62:68-73
【非特許文献3】Stulen and Oaks, 1977, Plant Physiol. 60:680-683
【非特許文献4】The Biochemistry of Plants: A Comprehensive Treatise, vol 5, Amino acid and derivatives, Miflin 編, Academic Press, New York (1980) pp. 569-607
【非特許文献5】Sieciechowiczら, 1988, Phytochemistry 27:663-671
【非特許文献6】Transgenic Plants, Vol. 1, Kung and Wu編, Academic Press, San Diego, CA, (1993) pp. 181-194
【非特許文献7】Udvardi and Kahn, 1991, Mol. Gen. Genet. 231:97-105
【非特許文献8】Zehnacker ら, 1992, Planta 187:266-274
【非特許文献9】Lough ら, 1992, Plant Mol. Biol. 19:391-399
【非特許文献10】Dicksonら, 1992, Plant Mol. Biol. 20:333-336
【非特許文献11】McNally ら, 1983, Plant Physiol. 72:22-25
【非特許文献12】Oaks and Hirel, 1985, Ann. Rev. Plant Physiol. 36:345-365
【非特許文献13】Cullimore ら, 1983, Planta 157:245-253
【非特許文献14】Recent Advances in Phythochemistry: Mobilization of Reserves in Germination, Nozolilloら編, Plenum Press, (1983) p. 77-109
【非特許文献15】Givan ら, 1988, TIBS 13:433-437
【非特許文献16】Gebhardtら, 1986, EMBO J. 5:1429-1435
【非特許文献17】Tingey ら, 1987, EMBO J. 6:1-9
【非特許文献18】Bennett ら, 1989, Plant Mol. Biol. 12:553-565
【非特許文献19】Tingey ら, 1988, J. Biol. Chem. 263:9651-9657
【非特許文献20】Peterman and Goodman, 1991, Mol. Gen. Genet. 230:145-154
【非特許文献21】Tingeyら, 1987, EMBO J. 6:1-9
【非特許文献22】Tingey ら, 1988, J. Biol. Chem. 263:9651-9657
【非特許文献23】Edwards and Coruzzi, 1989, Plant Cell 1:241-248
【非特許文献24】Tingeyら, 1987, EMBO J. 6:1-9
【非特許文献25】Walker and Coruzzi, 1989, Plant Physiol. 91:702-708
【非特許文献26】Edwards ら, 1990, Proc. Natl. Acad. Sci. USA, 87:3459-3463
【非特許文献27】Brearsら, 1991, The Plant Journal, vol. 1, pp. 235-244
【非特許文献28】Tsai and Coruzzi, 1990, EMBO J 9:323-332
【非特許文献29】Lea and Forde, 1994, Plant Molec. Biol. 17:541-558
【非特許文献30】Eckes ら,1989, Molec. Gen. Genet. 217:263-268
【非特許文献31】Hemonら, 1990, Plant Mol. Biol. 15:895-904
【非特許文献32】Hirelら, 1992, Plant Mol. Biol. 20:207-218
【非特許文献33】Templeら, 1993, Mol. Gen. Genet. 236:315-325
【非特許文献34】Donnら, 1984, J. Molec. Appl. Genet. 2:621-635
【非特許文献35】Eckesら, 1989, Molec. Gen. Genet. 217:263-268
【発明の概要】
【0019】
3.発明の概要
本発明は、窒素の同化および利用に係わる主要な酵素(これら酵素のそれぞれの役割は図1に示される)の発現レベルおよび/または細胞特異的な発現パターンを変化させたトランスジェニック植物の作出に関するものであり、かくして、得られる植物は増強された窒素同化および/または利用能ならびに改良された農業特性を有するものである。本発明は、特に、グルタミンシンテターゼ、アスパラギンシンテターゼ、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ(グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼはグルタミン酸シンテターゼとしても知られている)、アスパラギン酸アミノトランスフェラーゼ、グルタミン酸デヒドロゲナーゼおよびアスパラギナーゼの発現を変えることに関するものである(図1参照)。
【0020】
本発明は作物の重要な農業特性を改良することに用いられる。改良点の一つは、より低い窒素肥料投入量でまたは窒素の乏しい土壌で生産的に栽培される遺伝子操作植物の能力であるだろう。更なる改良点としては、より活発な(すなわち、より速い)成長ならびに普通の栽培条件(すなわち、非限定栄養条件)下でのより多い栄養体および/または生殖体の収穫量が含まれる。これらの改良点を達成するには、伝統的な作物育種法は大きな分離集団のスクリーニングを必要とするだろう。本発明は、大半の植物ではないにしても、その多くが所望の特性を備えている植物を作出することによって大規模スクリーニングの必要性を回避するものである。
【0021】
本発明によると、所望の植物改良を達成するには、いくつかの場合に、窒素同化または利用酵素をコードする1個または複数個の遺伝子の異所性過剰発現が必要となる。かかる発現の改変は次の遺伝子:すなわち、a)酵素のコーディング配列が強力な構成プロモーターに機能しうる状態で結合されているトランスジーン;b)所望の酵素をコードする、追加のコピー数の天然遺伝子;c)窒素同化または利用のための目的遺伝子(類)の発現を活性化する調節遺伝子(類);d)発現を高めるように修飾されたその調節領域を有する、1コピーの天然遺伝子;およびe)変異型、改変型またはキメラ型の窒素同化または利用酵素を発現するトランスジーン;のいずれか1個または数個を用いて植物を遺伝子操作することを包含する。
【0022】
他の場合には、所望の植物改良を達成するのに、窒素同化または利用酵素の発現パターンを変えることが必要となる。かかる改変発現パターンは次の遺伝子:すなわち、a)酵素のコーディング配列が所望の発現パターンを有するプロモーター(該プロモーターは組織−または発生−特異的発現パターンをもつと思われるものを含む)に機能しうる状態で結合されているトランスジーン;b)好ましいパターンでの酵素コード化遺伝子の発現を活性化する修飾調節遺伝子;c)好ましいパターンで発現するように修飾されたその調節領域を有する、天然コピー数の酵素コード化遺伝子;のいずれか1個または数個を用いて植物を遺伝子操作することを包含する。
【0023】
さらに他の場合には、所望の植物改良を達成するために、窒素同化または利用酵素の発現レベルおよび/またはパターンを抑制することが必要となる。かかる発現の抑制はアンチセンスRNA、リボザイム、共抑制構築物または「優性ネガティブ (dominant negative)」変異体をコードする遺伝子を用いて植物を遺伝子操作することを包含する(優性ネガティブ変異体による遺伝子抑制の作用機序の説明については、Herskowitz, 1967, Nature 329:219-222を参照されたい)。さらに、遺伝子の抑制は、天然遺伝子を1コピーの欠陥遺伝子または所望の発現レベルおよび/またはパターンを有するプロモーターの制御下にある酵素コード化配列と置き換えた相同的組換え構築物を用いて植物を遺伝子操作することによっても達成される。
【0024】
さらに他の場合には、所望の植物改良を達成するために、窒素同化または利用経路において改変された酵素または異なる型の酵素を発現させることが必要となる。かかる目標達成の作業は、宿主植物の窒素同化または利用酵素の触媒作用とは異なる触媒作用を有する対応酵素をコードする植物発現可能遺伝子を開発し、この遺伝子構築物で植物を遺伝子操作することを包含する。この種の酵素をコードする遺伝子配列は細菌、酵母、藻類、動物および植物を含むがこれらに限らないさまざまな供給源から得ることができる。ある場合には、かかるコーディング配列が該配列を所望の植物活性プロモーターと機能的に連結させることによる植物発現可能遺伝子融合体の構築に直接用いられる。また、他の場合には、遺伝子融合体での該コーディング配列の利用は、宿主植物におけるそれらの翻訳能を高めるために、あるいはそこにコード化された酵素の触媒作用を変えるために、in vitro突然変異誘発またはde novo 合成による先行修飾を必要とする。有用な改変には基質結合および/または触媒作用に関与する残基の修飾が含まれるが、これらに限らない。好ましい改変にはハイブリッド酵素の構築も含まれる。例えば、同一のまたは異なる生物から得られた関連酵素の異なるドメインを組み合わせて新規な性質を有する酵素を作ることができる。
【0025】
あらゆる場合に、望まれる改良点を有する植物は、窒素同化または利用酵素の改変された発現パターンまたはレベル、対応するmRNAまたはタンパク質の改変された発現パターンまたはレベル、改変された窒素同化または利用能、増加した成長速度、増加した栄養体収穫高、または改良された生殖体収穫高(例えば、より多いまたはより大きい種子または果実)について遺伝子操作植物をスクリーニングすることによって単離することができる。遺伝子操作植物のスクリーニングは、酵素/タンパク質レベルを測定するためのエンザイムアッセイおよびイムノアッセイ;mRNAレベルを測定するためのノーザン分析、RNアーゼ保護、プライマー伸長、逆転写/PCRなど;各種植物組織のアミノ酸組成、遊離アミノ酸プールまたは総窒素含量の測定;経時的新鮮重量増加に基づく成長速度の測定;または総乾燥重量および/または総種子重量に基づく植物収穫高の測定を包含する。
【0026】
本発明は、一部には、植物の窒素同化または利用酵素の発現を高めることが、成長特性の向上または栄養体や生殖体の収穫高の増加をもたらしたという驚くべき発見に基づいている。本発明は、ここでは、エンドウ・グルタミンシンターゼ(GS)遺伝子またはエンドウ・アスパラギンシンターゼ(AS)遺伝子をコードする配列に機能的に連結された強力な構成植物プロモーターのカリフラワーモザイクウイルス(CaMV)35Sプロモーターをコードする組換え構築物を用いてタバコ植物を遺伝子工学的に操作した実施例により説明される。RNAおよびタンパク質の分析により、大多数の遺伝子操作植物がGSまたはASの異所性過剰発現を示すことがわかった。GSまたはASを過剰発現する系統は、対照の野生型植物よりも高い窒素含量、活発な成長特性、増加した栄養体収穫高または良好な種子収穫高および品質を示した。
【0027】
3.1. 定義
本明細書に使用される、以下にリストされる用語は、示される意味を有する。
35S =35S転写のためのカリフラワーモザイクウイルスプロモーター
AS =アスパラギンシンテターゼ
AspAT =アスパラギン酸アミノトランスフェラーゼ(またAAT として知られている)
CaMV =カリフラワーモザイクウイルス
cDNA =相補DNA
DNA =デオキシリボ核酸
GDH =グルタミン酸デヒドロゲナーゼ
遺伝子融合体=異種遺伝子に操作により連結されたプロモーターを含む遺伝子構築(前記プロモーターは異種遺伝子の転写を調節する)
GOGAT =グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ
(また、グルタミン酸シンテターゼとして知られている)
Fd-GOGAT =フェレドキシン依存性グルタミン酸シンターゼ
NADH-GOGAT=NADH依存性グルタミン酸シンターゼ
GS =グルタミンシンテターゼ
異種遺伝子=遺伝子構築の状況において、異種遺伝子は、その遺伝子が前記遺伝子が自然に連結されていないプロモーターに連結されることを意味する。異種遺伝子は前記プロモーターを寄与する生物からのものであってもよく、またそうでなくてもよい。異種遺伝子はメッセンジャーRNA(mRNA)、アンチセンスRNAまたはリボザイムをコードし得る。
【0028】
窒素非制限生育条件=窒素非制限生育条件は、土壌または培地が健全な植物成長を持続するのに充分な量の窒素栄養を含み、または受容する条件である。窒素非制限生育条件の例は第5.2.3.節に示される。更に、当業者は、重要な穀物および観賞植物の殆どの種および変種につきこのような土壌、培地及び肥料を構成するものを認識するであろう。
PCR =ポリメラーゼ連鎖反応
前駆植物 =形質転換されていない、野生型植物
RNA =リボ核酸
【図面の簡単な説明】
【0029】
【図1】植物における窒素同化/代謝の経路。窒素同化の主要な経路はグルタミンシンテターゼ(GS)およびグルタミン酸シンターゼ(GOGAT) によるものである。グルタミン酸デヒドロゲナーゼ(GDH) は生合成の役割においてアンモニア毒性の条件下で機能するものと考えられ、または触媒量のグルタミン酸を与えてGS/GOGATサイクルを支持し得る。GDH はおそらくグルタミン酸からアンモニアを放出するその触媒的役割において更に活性である(例えば、発芽中)。アスパラギン酸アミノトランスフェラーゼ(AspAT) は可逆反応を触媒作用する。アスパラギンシンテターゼ(AS)は二つの活性;グルタミン依存性活性およびアンモニア依存性活性を有する。アスパラギン代謝はアスパラギナーゼ(ANS) により起こってアスパラギン酸およびアンモニアを放出する。
【図2】キメラFd/NADH GOGAT 酵素の操作。植物フェレドキシン-GOGAT(Fd-GOGAT)の大きいサブユニットはFd結合ドメイン(斜めのクロスバー)を含む。植物およびE.coli NADH-GOGAT:大きいサブユニット(オープンバー)、小さいサブユニットはNADH結合ドメイン(垂直ハッチ)を含む。キメラFd/NADH GOGAT はFd-GOGATの大きいサブユニット(Fd 結合ドメイン)+植物またはE.coliのNADH-GOGATの小さいサブユニットを含むように操作される。その操作は植物Fd-GOGATをコードする配列およびNADH結合ドメインを含む、植物またはE.coli NADH-GOGAT の小さいサブユニットをコードする配列のイン−フレーム転写融合をつくることにより行われる。そのキメラタンパク質は、還元剤としてFdまたはNADHを使用し得る二特異性または二機能性のGOGAT 酵素をコードする。
【図3】2成分植物発現ベクターの地図。2成分発現ベクターpTEV4 、pTEV5 、pTEV8 およびpTEV9 はトランスジェニックタバコ中のcDNAの高レベルの発現のために構築されたpBIN19(Bevan, 1984, Nucleic Acids Res. 12:8711-8721)の誘導体である。構築の詳細につき、第6.1.1.節を参照のこと。
【図4】トランスジェニックタバコに移入されたキメラ35S CaMV-GS cDNA構築物。エンドウ豆GScDNAをストラスボルグ株CaMV 35Sプロモーター(35S) の後の発現のためにpTEV発現ベクター(図3、および第6.1.1.節を参照のこと)にクローン化した。GS3AおよびGS2 につき、イントロンをゲノム配列からcDNAにとり込む“修飾" クローンを構築した(第6.1.2.節を参照のこと)。GScDNAクローンの起源は、GS2(また(aka)GS185として知られている); GS1(aka GS299); GS3A (aka GS341)(Tingeyら, 1988, J.Biol.Chem. 263:9651-9657; Tingeyら,1987, EBBO J. 6:1-9)であった。
【図5】GSトランスジーンを含む一次(T1)形質転換体中のGSタンパク質の分析。上のパネル:一次形質転換体中のGSポリペプチドのウェスタン分析。レーン1および2:細胞質ゾルGS3A遺伝子を有する一次形質転換体Z17-6 およびZ17-12はそれぞれ過剰発現および同時抑制表現型を示す。レーン3−6:葉緑体GS2 遺伝子を有する一次形質転換体Z41-20、Z54-2 、Z54-7 、およびZ54-8 は葉緑体GS2(GSを比較のこと)につき全て同時抑制される。対照は、TL- タバコの葉、PL- エンドウ豆の葉、およびPR- エンドウ豆の根。全GS活性がウェスタンパネルの下に示される(対照=(100%)に対する%として)。下のパネル:サンプルのほぼ等しい負荷を実証するルビスコの大きいサブユニットタンパク質のクーマシー染色。ctGS- 葉緑体GS2(約45 kD); cyGS- 細胞質ゾルGS( 約38 kD)。
【図6A】エンドウ豆トランスジーンを含むT2子孫トランスジェニック植物からのGSタンパク質、RNAおよびホロ酵素の分析。典型的に分析されたそれぞれの一次形質転換体からの4種のT2植物のうちの単一の代表的な植物がこの図に含まれた。Z17-9 の場合、T2子孫は二つの異なるプロフィールを示し、その両方が示される(Z17-9A およびZ17-9B) 。対照:TL/T- タバコの葉、P- エンドウ豆の葉。パネルA(上):トランスジェニック植物中のGSポリペプチドのウェスタン分析。パネルA(下):サンプルのほぼ等しい負荷を示すためのルビスコの大きいサブユニットタンパク質のクーマシー染色。
【図6B】エンドウ豆トランスジーンを含むT2子孫トランスジェニック植物からのGSタンパク質、RNAおよびホロ酵素の分析。典型的に分析されたそれぞれの一次形質転換体からの4種のT2植物のうちの単一の代表的な植物がこの図に含まれた。Z17-9 の場合、T2子孫は二つの異なるプロフィールを示し、その両方が示される(Z17-9A およびZ17-9B) 。対照:TL/T- タバコの葉、P- エンドウ豆の葉。パネルB(上):GS1(左)、GS3A(中央)およびGS2(右)の推定cDNAプローブとハイブリッドを形成されたノーザンブロット。パネルB(下):エンドウ豆rRNA遺伝子プローブによる対照ハイブリダイゼーション。
【図6C】エンドウ豆トランスジーンを含むT2子孫トランスジェニック植物からのGSタンパク質、RNAおよびホロ酵素の分析。典型的に分析されたそれぞれの一次形質転換体からの4種のT2植物のうちの単一の代表的な植物がこの図に含まれた。Z17-9 の場合、T2子孫は二つの異なるプロフィールを示し、その両方が示される(Z17-9A およびZ17-9B) 。対照:TL/T- タバコの葉、P- エンドウ豆の葉。パネルC:トランスジェニック植物中のGSホロ酵素A* 、B、およびCを示す非変性ゲルおよびGS活性分析。GS活性は対照と比較して%として表される(対照=100 %の活性)。
【図7A】GSホロ酵素の活性ゲル分析。植物を過剰発現するGS1 およびGS3Aのホロ酵素の移動に対する葉緑体および細胞質ゾル濃縮GSタンパク質サンプルの移動を実証する、エンドウ豆葉緑体(PC)、エンドウ豆の根(PR)、タバコ葉緑体(TC)およびタバコの根(TR)からのタンパク質抽出物。レーン1:エンドウ豆葉緑体タンパク質(PC)はGSホロ酵素Bのみを有する;レーン2:エンドウ豆の根タンパク質(PR)はGSホロ酵素Cのみを有する;レーン3:タバコ葉緑体タンパク質(TC)はGSホロ酵素Bのみを有する;レーン4:タバコの根タンパク質はGSホロ酵素Cのみを有する。レーン5:植物Z17-7(35S-GS3A構築物を有する)からのタンパク質はGSホロ酵素A* およびBを有する;レーン5:植物Z3-1(35S-GS1構築物を有する)からのタンパク質はGSホロ酵素BおよびCを有する。
【図7B】GSホロ酵素A* 、B、およびCから単離されたGSタンパク質のウェスタン分析。GS3AおよびGS1 を過剰発現するトランスジェニックタバコ中で観察されたホロ酵素A* およびCを非変性ゲルから切除し、タンパク質単離緩衝液中で再度抽出し、そしてGS抗体を使用してウェスタン分析のための変性条件下で電気泳動にかけた。レーン1:対照としてのタバコの葉のタンパク質;レーン2:Z17-7 からのGSホロ酵素A* ; レーン3:対照としての単離された葉緑体GS2(ホロ酵素B);レーン4:Z3-1からのGSホロ酵素C。
【図8】細胞質ゾルGS1 またはGS3Aを異所性発現する成長分析に選択されたトランスジェニック植物中のGSタンパク質およびRNAのウェスタンおよびノーザン分析。上のパネル:GSタンパク質に関するウェスタンブロット。下のパネル:GS mRNAに関するノーザンブロット。P1およびT1はエンドウ豆およびタバコの葉の対照である。レーン1および2、並びに5および6はGS1 を過剰発現する植物であり、そしてレーン3および4、並びに7および8はGS3Aを過剰発現する植物である。破線の左のトランスジェニック植物を生育実験Aにおいて分析し、そして右のトランスジェニック植物を生育実験Bにおいて分析した。相当するプローブをノーザンブロットにおいて使用した。左のエンドウ豆対照をGS1 にハイブリッドを形成し、右手のエンドウ豆対照をGS3Aにハイブリッドを形成した。
【図9A】細胞質ゾルGS1 (Z3)または細胞質ゾルGS3A (Z17)を過剰発現するトランスジェニック系統の新鮮重量の増加。パネルA:トランスジェニック系統Z3-1、Z3- 2 、Z17-6 、Z17-7 、および形質転換されなかった対照(C) を用いる実験Aの結果。これは表2に示され、そして表3に統計的に分析されたデータのグラフ表示である。
【図9B】細胞質ゾルGS1 (Z3)または細胞質ゾルGS3A (Z17)を過剰発現するトランスジェニック系統の新鮮重量の増加。パネルB:トランスジェニック系統Z3-3、Z3-4、Z17-3 、Z17-11、および2種の形質転換されなかった対照(C1 およびC2) を用いる実験Bの結果。これは表2に示され、そして表3に統計的に分析されたデータのグラフ表示である。
【図10A】変化されたGS発現パターンを有する植物の定性的成長パターン。それぞれのパネル中の植物を同時に種をまき、そして約3週間にわたって土壌中で生育した。対照パネル:SR1 未形質転換タバコ(100%のGS活性)。
【図10B】変化されたGS発現パターンを有する植物の定性的成長パターン。それぞれのパネル中の植物を同時に種をまき、そして約3週間にわたって土壌中で生育した。Z3-A1 パネル:過剰発現GS1 を有するトランスジェニック植物(123%のGS活性)。
【図10C】変化されたGS発現パターンを有する植物の定性的成長パターン。それぞれのパネル中の植物を同時に種をまき、そして約3週間にわたって土壌中で生育した。Z54-A2パネル:GS2 につき同時抑制されたトランスジェニック植物(28 %のGS活性)。
【図10D】変化されたGS発現パターンを有する植物の定性的成長パターン。それぞれのパネル中の植物を同時に種をまき、そして約3週間にわたって土壌中で生育した。Z17-B7パネル:GS3 を過剰発現するトランスジェニック植物(107%のGS活性)。
【図11A】GS活性と植物新鮮重量または全葉タンパク質の線形関係。トランスジーンと関連するKan R 表現型の分離を示さなかった一次形質転換体のT2子孫を成長分析につき選択した。Kan R T2植物をMSK 培地 (R.B.Horschら,Science 227:1229 (1985))で選択し、18日で砂に移した。植物を下灌漑し(subirrigated)、2日毎に10mMのKNO3を含む1Xホアグランド液(D.R.Hoagland ら, Circ. Calif.Agric.Exp.Stn. 347:461 (1938))50mlで表面で栄養補給した。それぞれの系統につき、8種のT2子孫を合計植物新鮮重量(グラム)、トランスフェラーゼアッセイ(B.M.Shapiroら, Methods Enzymol. 17A:910 (1970))により測定されるような合計の葉GSの比活性およびタンパク質/新鮮重量(グラム)につき個々に分析した。分析した植物は、対照、SR1 未形質転換タバコ;GS2 により同時抑制されたZ54-4; GS3A を過剰発現するZ17-7; GS1を過剰発現するZ3-1であった。図11A;植物新鮮重量vs.GS 活性。
【図11B】GS活性と植物新鮮重量または全葉タンパク質の線形関係。トランスジーンと関連するKan R 表現型の分離を示さなかった一次形質転換体のT2子孫を成長分析につき選択した。Kan R T2植物をMSK 培地 (R.B.Horschら,Science 227:1229 (1985))で選択し、18日で砂に移した。植物を下灌漑し(subirrigated)、2日毎に10mMのKNO3を含む1Xホアグランド液(D.R.Hoagland ら, Circ. Calif.Agric.Exp.Stn. 347:461 (1938))50mlで表面で栄養補給した。それぞれの系統につき、8種のT2子孫を合計植物新鮮重量(グラム)、トランスフェラーゼアッセイ(B.M.Shapiroら, Methods Enzymol. 17A:910 (1970))により測定されるような合計の葉GSの比活性およびタンパク質/新鮮重量(グラム)につき個々に分析した。分析した植物は、対照、SR1 未形質転換タバコ;GS2 により同時抑制されたZ54-4; GS3A を過剰発現するZ17-7; GS1を過剰発現するZ3-1であった。図11B; タンパク質/新鮮重量(グラム)vs.GS活性。
【図12】トランスジェニックタバコに移入されたキメラ35 S CaMV-AS構築物。AS1 遺伝子のcDNAおよびgln ΔAS1 遺伝子を、2成分発現ベクターpTEV5 を使用してタバコへの移入のために35S プロモーターおよびノパリンシンターゼ転写ターミネーターに融合した。
【図13】AS1 またはgln ΔAS1 を発現するトランスジェニック植物のノーザン分析。個々の形質転換体の葉から単離された全RNA10μg を浸出レーン中に負荷した。ブロットをエンドウ豆からのAS1 cDNAで探査した。陽性対照は暗所で生育されたエンドウ豆の葉(PL)中のASmRNAを含む。陰性対照は明るい所で生育されたタバコの葉(TL)中のASmRNAを含む。
【図14】AS1 およびgln ΔAS1 を過剰発現するトランスジェニック系統の新鮮重量の増加が発芽後の3週から6週までグラフで表される。これは表(5)中に示され、そして表(6)中で統計的に分析されたデータのグラフ表示である。
【発明を実施するための形態】
【0030】
5. 発明の詳細な説明
本発明は植物における窒素代謝の遺伝子操作に関する。特に、本発明は、良好な成長特性、強化された栄養特性、改良された栄養および収穫量および/または増進された種子の収穫量または品質を有する植物を操作するために、窒素同化に関係する酵素および/またはそれらの発現を変化することに関する。
【0031】
それ故、特別な機構に制限されることを目的としないで、操作の標的はアミノ酸、グルタミン、アスパラギン酸、アスパラギンまたはグルタミン酸へのアンモニアの同化、または生合成反応におけるこれらの同じアミノ酸の利用に関係する酵素をコードする遺伝子である。標的遺伝子として、グルタミンシンテターゼ
(GS)、アスパラギンシンテターゼ(AS)、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ(GOGAT) 、アスパラギン酸アミノトランスフェラーゼ(AspAT) 、グルタミン酸デヒドロゲナーゼ(GDH) およびアスパラギナーゼ(ANS) をコードする遺伝子が挙げられる。窒素同化および利用においてこれらの酵素により果たされる役割の線図につき図1を参照のこと。
【0032】
これらの酵素は変性され、またはそれらの発現が増進、抑制またはそれ以外に修飾(例えば、異所性発現)されて所望の性質を有する植物を操作し得る。操作は植物を本明細書に記載された核酸構築物で形質転換することにより行われる。形質転換された植物またはそれらの子孫は、所望の変性された酵素を発現し、または窒素同化または利用酵素の所望の変化された発現、相当するmRNAの変化された発現、変化された窒素同化または利用能、増大された成長速度、増進された栄養収穫量、および/または改良された再生産収穫量を示す植物につきスクリーニングされる。
【0033】
所望の生理学的変化および/または作物栽培学的変化を示す操作された植物は植物育種または直接に農業生産に使用し得る。また、一つの変性された酵素を有するこれらの植物はその他の窒素同化または利用酵素の変化でもって操作されたその他の変性植物と交雑されて(例えば、GS過剰発現植物をAS過剰発現植物に交雑する)、親と較べて更に増強された生理学的性質および/または作物栽培学的性質を有する系統を生産し得る。
【0034】
本発明が、GSまたはASの異所性過剰発現につき操作された植物の実施例により説明される。全ての場合、GSまたはASの異所性過剰発現を示す操作された植物はまた対照の野生型植物よりも良好な成長特性、強化された栄養特性、改良された栄養収穫量および/または強化された種子の性質または収穫量を示す。
【0035】
5.1. 窒素同化および利用経路の変化
本発明の一つの局面によれば、望ましい植物は、アミノ酸グルタミン、アスパラギンまたはグルタミン酸へのアンモニアの初期同化そして更にアスパラギン酸への変換に関係する酵素の異所性過剰発現を操作することにより得られる。異所性という用語は、特別な遺伝子または当該酵素に関する異常な細胞下の(例えば、オルガネラ局在化と細胞質ゾル局在化の間のスイッチ)、細胞型、組織型および/または発育的もしくは一時的な発現(例えば、明/暗所)パターンを意味するために本明細書で使用される。このような異所性発現は前記酵素の正常な組織または発育段階における発現を必ずしも排除するのではないが、前記酵素の正常ではない組織または発育段階における発現を伴う。過剰発現という用語は前記酵素につき特別な組織、全部および/または発育または一時的な段階における正常な発現レベルを上回ることを意味するために本明細書で使用される。
【0036】
グルタミンへのアンモニアの同化そしてグルタミン酸、アスパラギン酸、およびアスパラギンへのその更なる代謝に関係する主要な酵素は、グルタミンシンテターゼ、アスパラギンシンテターゼ、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ、アスパラギン酸アミノトランスフェラーゼ、グルタミン酸デヒドロゲナーゼおよびアスパラギナーゼである。本発明は、これらの酵素の一種以上の異所性過剰発現の操作が所望の生理学的性質および作物栽培学的性質を有する植物を生産することを提供する。好ましい実施態様において、植物はグルタミンシンテターゼまたはアスパラギンシンテターゼの異所性発現につき操作される。酵素の細胞質ゾル形態または葉緑体形態が存在するGSに関して、細胞質ゾル形態の増進された発現の操作が好ましい。GSの細胞質ゾル形態として、小節−特異的酵素(例えば、エンドウ豆GS3A&B)および根−特異的酵素(例えば、エンドウ豆GS1)の両方が挙げられる。“根−特異的" 細胞質ゾルGS(例えば、エンドウ豆GS1)の増進された発現の操作が特に好ましい。また、本発明は前記酵素の細胞下の局在化を変化する操作を提供する。例えば、ASの如き細胞質ゾル酵素への葉緑体標的配列の操作は、植物中の窒素同化を改良し得る。これは光呼吸性アンモニアを再同化するために葉肉細胞中で特に有益であろう。
【0037】
本発明の別の局面によれば、望ましい植物は別の窒素同化経路により増進されたアンモニアとり込みを操作することにより得られる。特に、操作はグルタミンシンテターゼによる窒素同化の正常な主要経路を抑制することにより行われる。多重GSアイソザイムをコードする植物種において、これは内在性GS遺伝子の抑制を必要とし得る。好ましい実施態様において、抑制されたGS発現でもって操作された植物は、アスパラギンシンテターゼ(AS)および/またはグルタミンデヒドロゲナーゼ(GDH) の如きオルタナチブN同化性酵素の異所性過剰発現につき更に操作される。最も好ましい実施態様において、GSおよび/またはAS/GDH操作された植物は窒素同化または利用プロセスに関係するその他の酵素の一種以上の増進された発現につき更に操作される(図1を参照のこと)。
【0038】
本発明の第三の局面によれば、望ましい植物は同化された窒素の利用に関係する酵素の異所性過剰発現を操作することにより得られる。本発明のこの局面の実施態様は代謝反応においてグルタミン、グルタミン酸およびアスパラギンの使用を触媒作用する酵素の異所性過剰発現でもって植物を操作することを伴い得る。好ましい実施態様において、植物はアスパラギナーゼの異所性過剰発現につき操作される。
【0039】
本発明の第四の局面によれば、望ましい植物は窒素の同化または利用に関係する酵素の変化され、突然変異されたキメラ形態または異種形態の発現を操作することにより得られる。本発明のこの局面の実施態様は、異種起源からの窒素同化酵素または利用酵素(即ち、異なる植物または動物もしくは微生物を含む、生物からの酵素)を発現するように植物を操作することを伴い得る。付加的な実施態様は、例えば、基質結合、触媒作用、および/または生産物放出において増大された効率を有する窒素同化酵素または利用酵素を開発し、そしてこのような新規な酵素を発現するように植物を操作することを伴い得る。これらの新規な酵素は前記プロセスに影響する主要アミノ酸残基のin vitro突然変異誘発により開発し得る。また、このような新規な酵素は関連酵素からのドメインを組換えることにより開発し得る。例えば、キメラ二機能性GOGAT 酵素はNADH-GOGATのNADH結合ドメインをFd-GOGAT遺伝子にスプライシングすることによりフェレドキシン-GOGAT活性およびNADH-GOGAT活性の両方を含むように操作し得る(図2を参照のこと)。このようなキメラGOGAT 酵素はGOGAT 反応においてNADHまたはフェレドキシンを還元剤として利用できるという利点を有するであろう。この新規な酵素の異所性発現はグルタミン酸の更に有効な合成をもたらし得る。本明細書に示された酵素修飾の別の例(第7.0 節を参照のこと)は、その基質特異性を変化するためにドメインを欠失されたAS酵素の操作である。
【0040】
本発明によれば、窒素同化酵素または利用酵素の組織および発育の発現パターンの調節は所望の植物改良を得るのに重要であり得る。植物が正常または別のアンモニア同化経路に関係する酵素の異所性過剰発現につき操作される場合、本発明の好ましい実施態様は植物の多くまたは全部において変化された発現を行うことを伴う。植物が同化窒素の使用を触媒作用する酵素の異所性過剰発現につき操作される場合、本発明の好ましい実施態様はこのような発現を窒素“シンク" 組織および構造、例えば、葉および種子に制限する。
【0041】
5.2. トランスジェニック植物の発生
5.2.1. 核酸構築物
核酸配列の性質は、種々の潜在的な宿主植物細胞の遺伝子構造であるように変化される。本発明の好ましい実施態様は、当業者が絶対に必須ではないが、明らかに有利であると認識し得る幾つかの特徴を記載するであろう。これらは遺伝子構造の単離、合成または構築の方法、植物細胞に導入される遺伝子構築物の操作、遺伝子構築物の或る特徴、および遺伝子構築物と関連するベクターの或る特徴を含む。
【0042】
更に、本発明の遺伝子構築物はDNA分子またはRNA分子につきコードし得る。本発明によれば、標的植物の所望の安定な遺伝子型の変化は外因的に導入された一種以上の核酸構築物、特に組換えDNA構築物のゲノム組込みにより行われることが好ましい。それにもかかわらず、本発明によれば、このような遺伝子型の変化はまた自律的に複製でき、しかも染色体上かつ生殖上安定であるエピソーム(DNAまたはRNA)の導入により影響し得る。導入された核酸構築物がRNAを含む場合、このような構築物からの植物形質転換または遺伝子発現は逆転写により生成されたDNA中間体により進行し得る。
【0043】
本明細書に記載された核酸構築物は、当業者に公知の方法を使用して生産し得る。当業者は構築物の成分を単離、特性決定、そして操作するだけでなく、構築物それ自体をつくるのに使用し得る組換えDNA方法の教示につきSambrookら,1989, 分子クローニング:実験マニュアル, コールドスプリング ハーバー ラボラトリー プレス, プレインビュー, N.Y.のような出典を参考にし得る。所望の成分の核酸配列が知られている、幾つかの場合には、生物起源からそれを単離するのではなく、それを合成することが有利であり得る。このような場合、当業者はCaruthers ら, 1980, Nuc.Acids.Res.Symp.Ser. 7:215-233 、およびChowおよびKempe, 1981, Nuc.Acids.Res. 9:2807-2817 のような教示を参考にし得る。その他の場合、所望の成分はポリメラーゼ連鎖反応(PCR) 増幅により有利に生産し得る。PCR 教示につき、当業者はGelfand, 1989, PCR 技術、DNA増幅に関する原理および応用, H.A.Erlich編集, ストックトン プレス, N.Y.、分子生物学における現行のプロトコル, 2巻, 15章, Ausubel ら編集, ジョン ウィリィ&サンズ, 1988を参考にし得る。
【0044】
5.2.1.1. 発現構築物
本発明によれば、窒素同化または利用酵素の異所性過剰発現を有する植物は、所望の酵素をコードする配列と操作により関連された植物プロモーターを含む遺伝子構築物で植物細胞を形質転換することにより操作し得る。(操作により関連されたは“関連”プロモーターにより調節された転写が機能性メッセンジャーRNA(その翻訳が酵素を生産するであろう)を生産することを意味するために本明細書で使用される)。本発明の好ましい実施態様において、関連プロモーターは強力かつ非組織特異性または非発育特異性の植物プロモーター(例えば、多くまたは全ての組織型中で強く発現するプロモーター)である。このような強力な“構成的”プロモーターとして、CaMV 35Sプロモーター、T-DNAマンノピンシンテターゼプロモーター、およびそれらの種々の誘導体が挙げられるが、これらに限定されない。
【0045】
本発明の別の実施態様において、組織特異性または発育特異性のプロモーターを所望の酵素をコードする配列で操作により関連させる遺伝子構築物で植物を操作することが有利であり得る。例えば、光合成組織および器官中の発現が所望される場合、リブロースビホスフェートカルボキシラーゼ(ルビスコ)遺伝子または葉緑体a/b 結合タンパク質(CAB) 遺伝子のプロモーターの如きプロモーターが使用し得る。種子中の発現が所望される場合、種々の種子貯蔵タンパク質遺伝子のプロモーターの如きプロモーターが使用し得る。窒素固定小節中の発現が所望される場合、レグヘモグロビン遺伝子またはノジュリン遺伝子のプロモーターの如きプロモーターが使用し得る。根特異性発現が所望される場合、根特異性グルタミンシンテターゼ遺伝子をコードするプロモーターの如きプロモーターが使用し得る(Tingeyら, 1987, EMBO J. 6:1-9; Edwardsら, 1990, Proc.Nat.Acad.Sci.USA 87:3459-3463を参照のこと)。
【0046】
本発明の更に別の実施態様において、誘導プロモーターを所望の酵素をコードする配列と操作により関連させる遺伝子構築物で植物を形質転換することが有利であり得る。このようなプロモーターの例は多く、そして多岐にわたる。それらとして、二三の名前を挙げると、熱ショック遺伝子、防御応答遺伝子(例えば、フェニルアラニンアンモニアリアーゼ遺伝子)、傷誘発遺伝子(例えば、ヒドロキシプロリンに富む細胞壁タンパク質遺伝子)、化学誘導遺伝子(例えば、ニトレート還元酵素遺伝子、グルコナーゼ遺伝子、キチナーゼ遺伝子、等)、暗所誘導遺伝子(例えば、アスパラギンシンテターゼ遺伝子(CoruzziおよびTsai, 米国特許第5,256,558 号, 1993年10月26日, 植物アスパラギンシンテターゼをコードする遺伝子)が挙げられるが、これらに限定されない。
【0047】
本発明の更に別の実施態様において、修飾プロモーターまたは人工プロモーターを所望の酵素をコードする配列に操作により連結する遺伝子構築物で植物を形質転換することが有利であり得る。典型的には、異なるプロモーターの構造要素を組換えることにより構築された、このプロモーターは天然プロモーターには見られない特異な発現パターンおよび/またはレベルを有する。例えば、シス調節要素をプロモーターコアーと組み合わせることにより構築された人工プロモーターの例につき、Salinaら, 1992, Plant Cell 4:1485-1493を参照のこと。
【0048】
本発明の更に別の実施態様において、窒素同化または利用酵素の異所性過剰発現は所望の酵素をコードする遺伝子のコピー数を増加することにより操作し得る。所望の遺伝子の増加されたコピーを有する植物細胞を生産するための一つのアプローチは、遺伝子の多重コピーを含む核酸構築で形質転換することである。また、所望の酵素をコードする遺伝子は、増幅選択可能なマーカー(ASM) 遺伝子、例えば、グルタミンシンテターゼまたはジヒドロ葉酸還元酵素遺伝子を含む核酸構築物中に入れられる。このような構築物で形質転換された細胞は、ASM 遺伝子の増加されたコピーを有する細胞系を選択する培養レジメにかけられる。GS遺伝子の増幅されたコピーを含む植物細胞系の単離に使用される選択プロトコルにつき、Donnら, 1984, J.Mol.Appl.Genet. 2:549-562 を参照のこと。所望の遺伝子がASM遺伝子の近くに連結されるので、ASM 遺伝子を増幅した細胞系はまたおそらく所望の酵素をコードする遺伝子を増幅させるであろう。
【0049】
本発明のもう一つの実施態様において、窒素同化または利用酵素の異所性過剰発現は所望の酵素をコードする内在性遺伝子またはトランスジーンの発現を調節する調節遺伝子をコードする核酸構築物で植物細胞を形質転換することにより操作されてもよく、この場合、導入された調節遺伝子は所望の組織および/または発育段階、シンテターゼプロモーター、およびそれらの種々の誘導体中の酵素の強い発現を可能にするように修飾される。
【0050】
5.2.1.2. 抑制構築物
本発明によれば、所望の植物は窒素同化/代謝においてGS活性またはその他の酵素の活性を抑制することにより操作し得る(図1)。実施態様において、抑制は成熟標的mRNAを含む、宿主標的RNA転写産物のセグメントまたは全部に相補性のアンチセンスRNAをコードする遺伝子構築物で植物細胞を形質転換することにより操作し得る。別の実施態様において、標的遺伝子(例えば、GSmRNA)抑制は宿主標的RNA転写産物(例えば、成熟GSmRNAを含む、GSRNA転写産物)を開裂するリボザイムをコードする遺伝子構築物で植物細胞を形質転換することにより操作し得る。
【0051】
更に別の実施態様において、標的遺伝子抑制は“優性の負の”突然変異を含む標的酵素をコードする遺伝子構築物で植物細胞を形質転換することにより操作し得る。好ましい突然変異は触媒作用、基質結合(例えば、GSにつき、グルタミン酸またはアンモニウムイオンの結合部位)、または生産物放出に影響する突然変異である。有益な突然変異は上記のプロセスに関係する重要な一つ以上の残基の欠失または点突然変異であり得る。当業者は優性の負の突然変異を構築するためのアプローチおよび戦略につき本明細書中の教示およびHerskowitz (Nature, 329:219-222, 1987)の教示を参考にし得る。
【0052】
上記の抑制構築物の全てにつき、このような遺伝子構築物は標的遺伝子と同じ組織および発育特異性で発現することが好ましい。こうして、これらの抑制構築物は標的遺伝子のプロモーターと操作により関連されることが好ましい。また、抑制構築物を構成的に発現させることが好ましい場合がある。こうして、CaMV 35S プロモーターの如き強力な構成的プロモーターがまた抑制構築物を発現するのに使用し得る。これらの抑制構築物に最も好ましいプロモーターは標的遺伝子の修飾プロモーターであり、この場合、その修飾は組織特異性または発育特異性の変化なしに標的遺伝子プロモーターの増進された発現をもたらす。
【0053】
本発明によれば、標的遺伝子発現の抑制された所望の植物はまた同時抑制構築物で植物細胞を形質転換することにより操作し得る。同時抑制構築物は標的遺伝子の完全または部分コード配列と操作により関連された機能性プロモーターを含む。操作により関連されたプロモーターは強力な構成的プロモーター、例えば、CaMV 35Sプロモーターであることが好ましい。また、同時抑制構築物プロモーターは、標的遺伝子と同じ組織および発育特異性で発現するプロモーターであってもよい。このような別のプロモーターは標的遺伝子それ自体のプロモーター(例えば、GS同時抑制構築物の発現を誘導するためのGSプロモーター)を含み得る。 本発明によれば、同時抑制構築物は不完全標的mRNAまたは欠損標的酵素をコードすることが好ましいが、完全に機能性の標的mRNAまたは酵素をコードする構築物がまた同時抑制を行うのに有益であり得る。
【0054】
実施態様において、GSアイソザイムの全てではないとしても殆どの抑制が所望される場合、同時抑制構築物は葉緑体GSmRNA(例えば、エンドウ豆GS2 mRNA)の完全または部分コピーをコードすることが好ましい。本明細書(第6.2.2.節)に開示されるように、このような構築物は標的遺伝子の発現を抑制するのに特に有効である。
【0055】
本発明によれば、標的遺伝子発現が抑制された所望の植物はまた内在性標的遺伝子の部位誘導突然変異誘発を行い得る構築物で植物細胞を形質転換することにより操作し得る(植物中の標的遺伝子の部位誘導突然変異誘発を行うための核酸構築物の説明につき、Offringaら, 1990, EMBO J. 9:3077-84;およびKanevskiiら, 1990, Dokl.Akad.Nauk.SSSR 312:1505-1507 を参照のこと)。このような構築物は不活性コード配列による相同組換えにより、またはそれによらない相同組換えにより内在性標的遺伝子配列を置換することにより標的遺伝子の抑制を行うことが好ましい。
【0056】
5.2.1.3. 組換え核酸構築物のその他の特徴
本発明の組換え構築物は、その構築物の伝播のための選択可能なマーカーを含んでもよい。例えば、細菌中で伝播される構築物は抗生物質耐性遺伝子、例えば、カナマイシン、テトラサイクリン、ストレプトマイシン、またはクロラムフェニコールに対する耐性を与える遺伝子を含むことが好ましい。構築物を伝播するのに適したベクターとして、二三名を挙げると、プラスミド、コスミド、バクテリオファージまたはウイルスが挙げられる。
【0057】
加えて、組換え構築物はこれらの構築物により形質転換された植物細胞の単離、同定またはトラッキングのための植物発現性の選択可能なマーカー遺伝子またはスクリーニング可能なマーカー遺伝子を含んでもよい。選択可能なマーカーとして、抗生物質耐性(例えば、カナマイシンまたはハイグロマイシンに対する耐性)または除草剤耐性(例えば、スルホニル尿素、ホスフィノトリシン、またはグリフォセートに対する耐性)を与える遺伝子が挙げられるが、これらに限定されない。スクリーニング可能なマーカーとして、β−グルクロニダーゼをコードする遺伝子(Jefferson, 1987, Plant Molec Biol.Rep 5:387-405) 、ルシフェラーゼをコードする遺伝子(Ow ら, 1986, Science 234:856-859)、アントシアニン色素産生を調節するBおよびC1遺伝子産物(Goff ら, 1990, EMBO J 9:2517-2522) が挙げられるが、これらに限定されない。
【0058】
植物を形質転換するためにアグロバクテリウム系を利用する本発明の実施態様(以下を参照のこと)において、組換えDNA構築物は更に植物細胞に形質転換されるDNA配列に隣接する右のT-DNAボーダー配列を少なくとも含む。好ましい実施態様において、移入される配列は左右のT-DNAボーダー配列により隣接される。このようなT-DNAをベースとする形質転換ベクターの適切な設計および構築は当業者に公知である。
【0059】
5.2.2.植物および植物細胞の形質転換
本発明によれば、望ましい植物は本明細書に記載された核酸構築物で植物細胞を形質転換することにより得られてもよい。幾つかの場合、植物または植物細胞を幾つかの異なる遺伝子構築物で操作することが望ましい場合がある。このような操作は植物または植物細胞を所望の構築物の全部で同時に形質転換することにより行い得る。また、操作は連続的に行われてもよい。それは、一つの遺伝子で形質転換し、選択およびスクリーニング後に所望の形質転換体を得、形質転換体を第二遺伝子構築物で形質転換すること、等である。好ましい実施態様において、それぞれの遺伝子構築物は、多重遺伝子インサートを含む植物形質転換体の同定を促進するように異なる選択可能なマーカー遺伝子またはスクリーニング可能なマーカー遺伝子に連結されるであろう。別の実施態様において、幾つかの異なる遺伝子がそれぞれの遺伝子につき操作された親系統を交雑することにより一種の植物にとり込まれ得る。
【0060】
本発明の実施態様において、アグロバクテリウムが遺伝子構築物を植物に導入するのに使用される。このような形質転換は2成分アグロバクテリウムT-DNAベクター(Bevan, 1984, Nuc.Acid Res. 12:8711-8721) 、および同時培養操作(Horsch ら, 1985, Science 227:1229-1231)を使用することが好ましい。一般に、アグロバクテリウム形質転換系が双子葉植物を操作するのに使用される(Bevanら,1982, Ann.Rev.Genet 16:357-384; Rogersら, 1986, Methods Enzymol. 118:627-641)。アグロバクテリウム形質転換系がまた単子葉植物および植物細胞を形質転換するだけでなく、DNAをそれらに移入するのに使用し得る(Hernalsteenら,1984, EMBO J 3:3039-3041; Hooykass-Van Slogterenら, 1984, Nature 311:763-764; Grimsley ら, 1987, Nature 325:1677-179; Boultonら, 1989, Plant Mol.Biol. 12:31-40; Gould ら, 1991, Plant Physiol. 95:426-434 を参照のこと)。
【0061】
別の実施態様において、組換え核酸構築物を植物および植物細胞に導入するための種々の別法がまた使用し得る。これらのその他の方法は、標的が単子葉植物または植物細胞である場合に特に有益である。別の遺伝子移入方法および形質転換方法として、裸のDNAのカルシウム、ポリエチレングリコール(PEG) またはエレクトロポレーション介在性とり込みによるプロトプラスト形質転換(Paszko-wskiら, 1984, EMBO J 3:2717-2722, Potrykusら, 1985, Molec.Gen.Genet. 199:169-177; Frommら, 1985, Proc.Nat.Acad.Sci.USA 82:5824-5828; Shimamoto,1989, Nature 338:274-276を参照のこと) および植物組織のエレクトロポレーション(D'Halluinら, 1992, Plant Cell 4:1495-1505) が挙げられるが、これらに限定されない。植物細胞形質転換の更に別の方法として、マイクロインジェクション、炭化ケイ素介在性DNAとり込み(Kaeppler ら, 1990, Plant Cell Reporter 9:415-418) 、およびマイクロプロジェクティル衝撃(Kleinら, 1988, Proc.Nat.Acad.Sci.USA 85:4305-4309; Gordon-Kammら, 1990, Plant Cell 2:603-618を参照のこと)が挙げられる。
【0062】
本発明によれば、多種の植物および植物細胞系が、本発明の核酸構築物および上記の種々の形質転換方法を使用して本明細書に記載された所望の生理学的特徴および作物栽培学的特徴につき操作し得る。好ましい実施態様において、操作のための標的植物および植物細胞として、トウモロコシ、小麦、米、大豆、トマト、タバコ、ニンジン、ジャガイモ、てんさい、ヒマワリ、ヤムイモ、アラビドプシス、ナタネ、およびペチュニアが挙げられるが、これらに限定されない。
【0063】
5.2.3. 形質転換された植物および植物細胞の選択および同定
本発明によれば、所望の植物は、開示された遺伝子構築物をプロトプラスト、組織培養細胞、組織および器官外植体、花粉、胚、並びに植物全体を含むが、これらに限定されない種々の植物細胞型に操作することにより得られる。本発明の実施態様において、操作された植物体は下記のアプローチおよび方法に従って形質転換体(導入された一つ以上の遺伝子構築物をとり込み、または組込んだ形質転換体)につき選択またはスクリーニングされる。次に単離された形質転換体が植物中で再生し得る。また、操作された植物体は、誘導された植物または幼植物体をマーカー遺伝子形質につき選択またはスクリーニングにかける前に植物または幼植物体に再生し得る。一つ以上のマーカー遺伝子を選択またはスクリーニングする前または後に、植物を植物細胞、組織または器官から再生するための方法は当業者に公知である。
【0064】
形質転換された植物細胞、カルス、組織または植物は、操作された植物体を形質転換DNAに存在するマーカー遺伝子によりコードされた形質につき選択またはスクリーニングすることにより同定され、単離し得る。例えば、選択は形質転換遺伝子構築物が耐性を与える抑制量の抗生物質または除草剤を含む培地で操作された植物体を生育することにより行い得る。更に、形質転換された植物および植物細胞はまた本発明の組換え核酸構築物に存在し得る可視のマーカー遺伝子(例えば、β−グルクロニダーゼ遺伝子、ルシフェラーゼ遺伝子、B遺伝子またはC1遺伝子)の活性につきスクリーニングすることにより同定し得る。このような選択方法およびスクリーニング方法が当業者に公知である。
【0065】
また、物理的方法および生化学的方法が本発明の遺伝子構築物を含む植物または植物細胞形質転換体を同定するのに使用し得る。これらの方法として、1)組換
えDNAインサートの構造を検出し、測定するためのサザン分析またはPCR 増幅;
2)遺伝子構築物のRNA転写産物を検出し、試験するためのノーザンブロット、S1 RNase保護、プライマー伸長PCR 増幅または逆転写酵素PCR 増幅;3)このような遺伝子産物が遺伝子構築物によりコードされる場合、酵素またはリボザイム活性を検出するための酵素アッセイ;4)遺伝子構築産物がタンパク質である場合、タンパク質ゲル電気泳動、ウェスタンブロット技術、免疫沈殿、またはエンザイムリンクドイムノアッセイが挙げられるが、これらに限定されない。更に別の技術、例えば、in situ ハイブリダイゼーション、酵素染色、および免疫染色がまた特定の植物器官および組織中の組換え構築物の存在または発現を検出するのに使用し得る。全てのこれらのアッセイを行う方法が当業者に公知である。
【0066】
5.2.4. 改良された作物栽培学的形質を有する植物に関する形質転換された植物のスクリーニング
本発明によれば、改良された作物栽培学的特徴を有する植物を得るために、形質転換された植物が所望の生理学的変化を示す植物につきスクリーニングし得る。例えば、植物がGS酵素の異所性過剰発現につき操作された場合、形質転換された植物が所望のレベルで所望の組織および発育段階中にGS酵素を発現する植物につき試験される。植物が標的遺伝子の抑制につき操作された場合、形質転換された植物が種々の組織中で減少されたレベルで標的遺伝子産物(例えば、RNAまたはタンパク質)を発現する植物につき試験される。次に、所望の生理学的変化、例えば、異所性GS過剰発現またはGS抑制を示す植物が所望の作物栽培学的変化を有する植物につき続いてスクリーニングし得る。
【0067】
また、形質転換された植物は所望の作物栽培学的変化を示す植物につき直接にスクリーニングし得る。一つの実施態様において、このようなスクリーニングは窒素栄養欠乏条件下の形質転換された植物の生産的生育に関するものであってもよい。それは、野生型植物の成長を停止し、または野生型植物のサイズまたは品質をかなり低下するように低減させる、利用可能な窒素栄養に関する、条件下の形質転換された植物の成長に関するスクリーンである。タバコおよび同様の窒素栄養要件を有する植物に関する窒素栄養欠乏条件の例は、土壌または合成培地中の唯一の窒素栄養が(a)0.5mM以下の濃度で供給または周期的に適用される硝酸塩、または(b)0.5mM以下の硝酸塩に生理学的に均等である濃度で供給または周期的に適用される硝酸塩の生理学的に均等物(例えば、アンモニウムまたは硝酸塩とアンモニウムの混合物)(Eckes ら, 1988, オーストラリア特許庁書類番号AU-A-17321/88 を参照のこと)である条件である。窒素栄養欠乏条件の別の例は、土壌または合成培地中の利用可能な窒素栄養の定常状態レベルが約0.02mM未満の硝酸塩またはその生理学的均等物である条件である。本明細書に使用される硝酸塩という用語は植物窒素肥料として普通に使用される硝酸塩、例えば、硝酸カリウム、硝酸カルシウム、硝酸ナトリウム、硝酸アンモニウム、等のいずれか一種またはあらゆる混合物を意味する。本明細書に使用されるアンモニウムという用語は植物窒素肥料として普通に使用されるアンモニウム塩、例えば、硝酸アンモニウム、塩化アンモニウム、硫酸アンモニウム、等のいずれか一種またはあらゆる混合物を意味する。
【0068】
その他の実施態様において、形質転換された植物のスクリーニングは、窒素非限定生育条件下で培養される(即ち、健全な植物成長を持続するのに充分な量の窒素栄養を含み、または受容する土壌または培地を使用して培養される)時に、未操作の前駆植物と較べて、改良された作物栽培学的特徴(例えば、早い成長、大きな栄養収穫量または再生産収穫量、または改良されたタンパク質含量、等)に関するものであってもよい。タバコおよび同様の窒素栄養要件を有する植物に関する窒素非限定条件の例は、土壌または合成培地中の唯一の窒素栄養が(a) 10mM以上の濃度で供給または周期的に適用される硝酸塩、または(b)10 mM以上の硝酸塩に生理学的に均等である濃度で供給または周期的に適用される硝酸塩の生理学的に均等物である条件である。窒素非限定条件の別の例は、土壌または合成培地中の利用可能な窒素栄養の定常状態レベルが少なくとも約1.0 mMの硝酸カリウムまたはその生理学的均等物である条件である。植物生育に関する窒素栄養欠乏条件または“非限定" 条件であることに関する更に別のガイダンスが当業界に見られる。例えば、Hewitt, E.J., 植物栄養の研究に使用される砂培養方法および水培養方法, 第2編, Farnham Royal (Bucks), Commonwealth Agricultural Bu-reaux, 1966;およびHewitt, E.J., 植物無機栄養, ロンドン, 英国大学プレス,1975を参照のこと。
【0069】
形質転換された植物がマメ科牧草類である実施態様において、所望の作物栽培学的変化および改良による形質転換された植物の直接のスクリーニングは、小節形成または窒素固定が抑制される条件下である以外は上記のようにして行われてもよい。
【0070】
本発明によれば、窒素同化プロセスまたは利用プロセスの変化で操作された植物は改良された窒素含量、変化されたアミノ酸またはタンパク質組成、活発な成長特性、増大された栄養収穫量または良好な種子収穫量および品質を示し得る。このような改良された作物栽培学的特徴を有する操作された植物および植物系統は下記のパラメーター:1)新鮮重量または乾燥重量の増加の速度に関して測定される成長の速度;2)新鮮重量または乾燥重量に関する、成熟植物の栄養収穫量;3)種子または果実収穫量;4)種子または果実重量;5)植物の合計窒素含量;6)果実または種子の全窒素含量;7)植物の遊離アミノ酸含量;8)果実または種子の遊離アミノ酸含量;9)植物の合計タンパク質含量;および10) 果実または種子の合計タンパク質含量のいずれかを試験することにより同定し得る。これらのパラメーターを試験するための操作および方法が当業者に公知である。
【0071】
本発明によれば、所望の植物は、前記パラメーターの一つ以上において対照植物(即ち、前駆植物)に対し改良を示す植物である。実施態様において、所望の植物は、少なくとも一つのパラメーターにおいて対照植物に対し少なくとも5%の増加を示す植物である。好ましい実施態様において、所望の植物は、少なくとも一つのパラメーターにおいて対照植物に対し少なくとも20%の増加を示す植物である。少なくとも一つのパラメーターにおいて少なくとも50%の増加を示す植物が最も好ましい。
【0072】
5.3. 発明の有用性
本発明の遺伝子操作された植物は、野生型植物の成長を停止させたり、成長を小さくして野生型植物を実際に無用のものにする、窒素栄養素が欠乏した条件下(すなわち、土壌に窒素が少なく、また、窒素肥料の投入量が少ない)で生産的に栽培することができる。遺伝子操作された植物はまた、より早い成熟、より速い成長および/またはより高い収穫量の達成、および/または窒素無制限成長条件(すなわち、健康的な植物の成長を維持するのに充分な量の窒素栄養素を含むか受け取る土壌または培地)を使用して栽培するときのより栄養に富む食品および動物飼料の生産に有利に使用することができる。窒素無制限成長条件は、種同士の間および一つの種の亜種に対して変わる。しかし、当業者であれば、全部でなくても、ほとんどの重要な作物および観賞植物の栽培に対して、何が窒素無制限成長条件を構成するかを知っている。例えば、小麦の栽培に対しては、Alcozら、Agronomy Journal 85:1198-1203 (1993)、Rao および Dao, J. Am. Soc. Agronomy 84:1028-1032 (1992)、Howardおよび Lessman, Agronomy Jornal 83:208-211 (1991);トウモロコシの栽培に対しては、Tollenear ら、Agronomy Journal 85:251-255 (1993)、Straw ら、Tennessee Farm and Home Science: Progress Report, 166:20-24 (Spring 1993) 、Miles, S.R., J. Am. Soc. Agronomy 26:129-137 (1934) 、Daraら、J. Am. Soc. Agronomy 84:1006-1010 (1992)、Binford ら、Agronomy Journal 84:53-39 (1992); 大豆の栽培に対しては、Chenら、Canadian Journal of Plant Science 72:1049-1056 (1992) 、Wallace ら、Journal of Plant Nutrition 13:1523-1537 (1990); 米の栽培に対しては、Oritani および Yoshida, Japanese Journal of Crop Science 53:204-212 (1984); 亜麻仁の栽培に対しては、Diepenbrock および Porksen, Industrial Crops and Products 1:165-173 (1992); トマトの栽培に対しては、Grubinger ら、Journal of the American Society for Horticultural Science 118:212-216 (1993)、Cerne, M., Acta Horticulture 277:179-182 (1990); パイナップルの栽培に対しては、Magistadら、J. Am. Soc. Agronomy 24:610-622 (1992)、Asoegwu, S.N., Fertilizer Research 15:203-210 (1988)、Asoegwu, S.N., Fruits 42:505-509 (1987); レタスの栽培に対しては、Richardson および Hardgrave, Journal of the Science of Food and Agriculture 59:345-349 (1992); ミントの栽培に対しては、Munsi, P.S., Acta Horticulturae 306:436-443 (1992); カモミールの栽培に対しては、Letchamo, W., Acta Horticulturae 306:375-384 (1992); タバコの栽培に対しては、Sissonら、Crop Science 31:1615-1620 (1991); 馬鈴薯の栽培に対しては、Porterおよび Sisson, American Patato Journal, 68:493-505 (1991);アブラナ科の栽培に対しては、Rahnら、Conference "Proceedings, second congress of the European Society for Agronomy" Warwick Univ., p.424-425 (August 23-28 1992); バナナの栽培に対しては、Hegde および Srinivas, Tropical Agriculture 68:331-334 (1991) 、Langenegger および Smith, Fruits 43:639-643 (1988); イチゴの栽培に対しては、Human および Kotze, Communications in Soil Science and Plant Analysis 21:771-782 (1990);モロコシの栽培に対しては、Mahalle および Seth, Indian Journal of Agricultural Sciences 59:395-397 (1989); オオバコの栽培に対しては、Anjorin および Obigbesan, Conference "International Cooperation for Effective Plantain and Banana Research" Proceedings of the third meeting. Abidjan, Ivory Coast, p.115-117 (May 27-31, 1985); サトウキビの栽培に対しては、Yadav, R.L., Fertiliser News 31:17-22 (1986)、Yadav および Sharma, Indian Journal of Agricultural Sciences 53:38-43 (1983); テンサイの栽培に対しては、Draycottら、Conference "Symposium Nitrogen and Sugar Beet" International Institute for Sugar Beet Research - Brussels Belgium, p.293-303 (1983)参照。また、Goh および Haynes, "Nitrogen and Agronomic Practice" in Meneral Nitrogen in the Plant-Soil System, Academic Press, Inc., Orlando, Florida, p.379-468 (1986), Engelstad, O.P., Fertilizer Technology and Use, Third Edition, Soil Science Society of America, p.633 (1985), Yadav and Sharmna,
Indian Journal of Agricultural Sciences, 53:3-43 (1983) も参照。
【0073】
GS抑制は、いくつかのGS抑制された植物、特に豆科植物が、非抑制植物よりもより速く成長する、あるいはより高い窒素含量を有するという点で有用である(Knightおよび Langston-Unkefer, Science, 241:951-954 参照)。GS抑制された植物はまた、アミノ酸またはタンパク質含量を変えて、特定のダイエット食品の製造に有用であり得る。さらに、本明細書に開示する遺伝子操作された全植物は、農業的に有用なプラントラインを展開するための育種ストックとして役立つと考えられる。
【実施例】
【0074】
6.実施例:グルタミンシンテターゼの植物における異所性過剰発現は、植物成長表現型の増加を引き起こす
ここでは、トランスジェニック植物において窒素使用効率を操作するための分子遺伝学法を記載する。その方法は、通常はGS発現が認められない細胞型および/またはレベルでGSを発現する、グルタミンシンテターゼの異所性発現に頼るものである。トランスジェニック植物における細胞特異的GS発現のパターンは、細胞質ゾルGS(通常は、篩部で発現するだけである。)を全細胞型で構成的に過剰発現することにより変えられる。GSのそのような異所性発現は、窒素同化酵素の区画化および細胞型特異性から生じる生理的制限を回避することができる。葉肉細胞での細胞質ゾルGSの異所性高レベル発現は、光呼吸によって失われたアンモニアの再同化に対して別のルートを提供すると考えられる。このことは、光呼吸によって失われるアンモニアの量が最初の窒素同化の10倍を超えるとき、成長の上での利点を与えると考えられる(Wallsgroveら、1983, Plant Cell Environ. 6:301-309; Keys ら、1978, Nature, 275:741-743 )。ここに開示する研究は、細胞質ゾルGSに対する異種GSサブユニットの構成的過剰発現がGS mRNA、GSタンパク質、全GS活性、天然のGSホロ酵素の増加ならびに一例においては新規GSホロ酵素の産生をもたらすことを示す。細胞質ゾルGSを過剰発現する形質転換された植物は、野生型と比較して、統計的に重要な成長上の利点を有する。形質転換された植物は、栄養成長期の際に、形質転換されていない前駆植物よりも速く成長し、最終の新鮮重量をより高くし、溶解性のより高いタンパク質を有する。しかし、場合によっては、細胞質ゾルGSおよび/または葉緑体GSの過剰発現が、内在遺伝子発現または共同抑制の下方調整を引き起こす。細胞質ゾルGS過剰発現構成体を含むいくつかの形質転換された植物および葉緑体GS2構成体を含む全ての形質転換された植物はGSを過剰発現しないで、むしろGS発現を抑制し、内在GS遺伝子の抑制も含む(すなわち、共同抑制)。そのようなGSが共同抑制された植物は、小さい成長特性を示す可能性があるが、窒素が他の窒素同化/代謝経路にそれて入ることによりアミノ酸およびタンパク質含量が変わると考えられる。
【0075】
6.1. 材料および方法
6.1.1. 植物発現ベクターの構築
植物発現ベクター pTEV 4 、5 、7 および 8を次のように構築した。カリフラワーモザイクウイルス(CaMV)の Strasbourg 株の 35Sプロモーターを含む、転写開始に対して -941 〜 +26まで伸びる HindIII−EcoRI 断片を、pBluescript KS II-(pT109) に挿入した(Hohnら、1982, Curr. Topics Microbiol. Immunol. 96:194-236 )。次いで、HindIII および XhoI 部位の間のポリリンカー配列を、XbaI、SstIおよび StuI 部位を含むように修飾した(pT145 )。こうして、pBIN19 (Clontech) 由来の、ノパリンシンターゼ転写ターミネーターを含むT4ポリメラーゼ処理したSstI−EcoRI 断片を StuI 部位に挿入することができ、pT161 を作った。このように構築された発現カセットは、隣がEcoRI 部位であり、修飾ポリリンカーを含む、pBIN19 (Bevan, 1984, Nucleic Acids Res. 12:8711-8721) 由来のプラスミドである pW3に移入された。プロモーターの5’末端がpW3の左端に隣接するように配向したクローンを選択し (pW63) 、プロモーターとターミネーターとの間に多数のクローニング部位を挿入した。こうして、列挙したユニークなクローニング部位を有する次のバイナリーベクターを作った(図3):pTEV4 (HindIII-XbaI-BamHI-XhoI) 、pTEV5 (HindIII-StuI-SstI-KpnI)、pTEV8 (HindIII-XhoI-BamHI-XbaI) 、pTEV9 (HindIII-KpnI-SstI-StuI)。
【0076】
6.1.2. GS cDNAのバイナリー発現ベクターへの移入
細胞質ゾルGS1およびGS3Aならびに葉緑体GS2のエンドウ遺伝子に対応するcDNAを pBluescriptから上記のバイナリー発現ベクターへ移入した(図4参照)。これらのcDNAは、以前は、各々GS299、GS341およびGS185として記載された(Tingeyら、1987, EMBO J. 6:1-9; Tingey ら、1988, J. Biol. Chem. 263:9651-9657)。葉緑体GS2に対しては、ゲノム配列の第一イントロンをcDNAの適切な位置に挿入して修飾cDNAを構築した(Z54)。これは、ポリメラーゼ連鎖反応法を使用してcDNAの5’末端からエキソン2内に位置する BsmI 部位(アミノ酸 43 )まで伸びる断片を増幅することにより行い、これは次いで、pBluescript におけるcDNAにクローン化することができた。細胞質ゾルGS3Aに対しては、ゲノムGS3Aクローンの BgIII-KpnI 断片を、pBluescript cDNAクローンに交換クローン化することにより修飾DNA(Z17)を構築し、全てのゲノムイントロン(アミノ酸6より前方)が挿入されたcDNA配列を作った。イントロンを入れてcDNAを構築することの目的は、単子葉植物で示されたように(Sinibaldi および Mettler, 1991)、トランスジェニック植物での発現を高めるために試みることであった。そのcDNAをpBluescript から下記のバイナリー発現ベクターに移入した。すなわち、GS1−pTEV4 を XbaI-XhoI部位に入れてpZ3(NRRL受理番号 B-21330)を作製し、GS3Aおよび修飾GS3A−pTEV4 を XbaI-XhoI部位に入れて、各々、pZ9(NRRL受理番号 B-21331)およびpZ17(NRRL受理番号 B-21332)を作製し、GS2および修飾GS2−pTEV5 を StuI-KpnI部位に入れて、各々、pZ41(NRRL受理番号 B-21333)およびpZ54(NRRL受理番号 B-21334)を作製した。
【0077】
6.1.3. 植物の形質転換
バイナリーベクター構築体を、以前に記載された方法を使用して三親交配することにより、無力のアグロバクテリウム (Agrobacterium)株 LBA4404に移入した (Bevan, 1984, Nucleic Acids Res. 12:8711-8721)。Nicotiana tabacum 系 SR1を、葉接種法により形質転換し (Horschら、1985, Science 227-1299-1231)、再生した苗条(shoot) を、 200μg/mlのカナマイシンを含む培地上で選択した。一次形質転換体を滅菌培地で保持し、次いで、成熟するまで土壌で成長させた。トランスジェニック種子を 10%次亜塩素酸ナトリウムで滅菌し、100 μg/mlカナマイシンを含む培地上で発芽させた。
【0078】
6.1.4. GSタンパク質および酵素活性分析
可溶タンパク質を、以前に記載されたように、タバコおよびエンドウの葉の組織から抽出した(Tingeyおよび Coruzzi, 1987, Plant Physiol. 84:366-373)。タンパク質を変性し、12% アクリルアミドのSDS−PAGEにより分離し、ニトロセルロース上に電気ブロットした。ウェスタン分析法を、Promega 製の ProtoBlotキットならびにタバコ葉緑体GS2および Phaseolus細胞質ゾルGSに対して高められた抗体の混合物を使用して行った (Hirel ら、1984, Plant Physiol. 74:448-450; Lara ら、1984, Plant Physiol. 76:1019-1023)。形質転換体における全GS活性を、以前に記載されたADP−依存トランスフェラーゼアッセイを使用して測定した (Shapiro および Stadtman, 1970, Methods Enzymol. 17A:910-922)。非変性ゲル電気泳動は、GSアイソザイム検出のためのADP−依存トランスフェラーゼアッセイと共に公知のプロトコール (Davis, 1964, Annals New York Acad. Sci. 121:404-427)に従った。
【0079】
6.1.5. RNA分析
RNAを、Bio101製の「RNAマトリックス」を使用し、製造者の示唆する以下のプロトコールに従って単離した。全RNAを、40 mM のトリエタノールアミン、2 mMのEDTAおよび 3.2% のホルムアルデヒド/ 1.2% アガロースで電気泳動した (Thomas, 1983, Methods Enzymol. 100:255-266) 。ゲルを 10 mMのリン酸ナトリウムに浸漬し、Hybond-Nナイロン膜 (Amersham) 上にキャピラリーブロットした。cDNAを、NEN 製のランダムプライマーおよび拡張試薬標識システムを使用して標識化し、鎖特異的リボプローブを、StratageneRNA転写キットを使用して作った。水性ハイブリダイーションを、膜の製造者のプロトコールに従って行い、ブロットを 0.1xSSRE、0.1%xSDSで洗浄した。
【0080】
6.1.6. 植物の成長条件
先に高いレベルでGS1またはGS3Aを発現すると解析された一次形質転換体の子孫を、100 μg/mlのカナマイシンを含む Murashige-Skoog (MS) 培地で発芽させた。14日後、カナマイシン耐性苗を、白砂を入れた4インチのポットに移し、Saran Wrap (登録商標) で約1週間覆って、過剰な蒸散を防ぎ、苗が定着できるようにした。ポットを、唯一の窒素源として 10 mMの硝酸カリウムを含む過剰の 1X Hoagland溶液で定期的に湿らした。次いで、毎週、3〜7本の植物を切って新鮮重量を測定し、隣り合った植物の陰になるのが顕著になるまで4週間続けた。植物を、明−暗のサイクルを 16-8 時間とし、温度のサイクルを 24-18℃として成長させた。昼間の光の強さは 1000 lux であった。
【0081】
6.2. 結果
6.2.1. トランスジェニック植物に導入したGS構築体
葉緑体GS2 (aka GS185 (Tingey ら、1988, J. Biol. Chem. 263:9651-9657))、細胞質ゾルGS1 (aka GS299 (Tingey ら、1988, J. Biol. Chem. 263:9651-9657))およびGS3A (aka GS341 (Tingey ら、1987, EMBO J. 6:1-9)) に対する Pisum sativum cDNAを pTEV バイナリー発現ベクターに挿入して(図3および4参照)、CaMV 35Sプロモーターの後ろを発現させ、トランスジェニックタバコに移入した。GS2(図4、構築体Z54)およびGS3A(図4、構築体Z17)に対しては、1個以上のイントロンを挿入したcDNAを構築し、CaMV 35Sプロモーターの後ろを発現させた。イントロンを挿入したcDNAを構成することの目的は、単子葉植物で示されたように(Sinibaldi および Mettler, 1991, Progress in Nucleic Acid Research and Molecular Biology 42:1991)、トランスジェニック植物での発現を高めるために試みることであった。さらに、未修飾の全長GS cDNAも、GS2(Z41)、G3A(Z9)およびGS1(Z3)に対して、35S-CaMVプロモーター下で発現させた(図4参照)。図4に詳述した 35S-CaMV-GS構築体の各々に対して、少なくとも8個の一次(T1)形質転換体を分析し、代表的サンプルを図5に示す。選択した一次形質転換体に対しては、4個のカナマイシン耐性T2子孫植物も分析した(図6)。以下に示すT1およびT2植物の分析は、ウェスタン分析法(図5および図6、パネルA);ノーザンブロット分析法(図6、パネルB)、GSホロ酵素分析(図6、パネルC)およびGS酵素活性分析(図6、パネルCならびに表1Aおよび1B)を含み、分析した全トランスジェニック系を代表している。
【表1A】

【表1B】
【0082】
6.2.2. 35S−葉緑体GS2遺伝子融合体を有するトランスジェニック植物の分析
35S−GS2構築体(図4、Z41またはZ54)のいずれかを含むトランスジェニック植物を分析した。35S−GS2(Z41)および修飾した(イントロンを含む)35S−GS2構築体(Z54)は共に、一次T1形質転換体およびT2子孫植物の両方に対して同様の結果を与えた。全一次形質転換体のウェスタンブロット分析は、野生型タバコ(図5、レーンTL)と比較して、葉緑体GS2ポリペプチド(ctGS)の量がかなり減少したことを示した(図5、レーン3〜6)。ポリクローナルGS2抗体は、エンドウおよびタバコの両方のGS2を認識することが分かっているので(Tingeyおよび Coruzzi, 1987, Plant Physiol. 84:366-373; Tingey ら、1988, J. Biol. Chem. 263:9651-9657) 、この減少は、宿主のタバコGS2遺伝子およびエンドウGS2トランス遺伝子の両方の下方調整を表す。細胞質ゾルGSポリペプチド(cyGS)の量は、形質転換していない対照の野生型タバコ(図5、レーンTL)と比較して変わらないことがこれらの形質転換体(図5、レーン3〜6)で認められた。Z41の場合は、全部で14個の独立した一次形質転換体の全GS活性が下方調整され、高くて野生型の活性の85%、低い方は野生型GS活性の22%であった(表1Aおよび1B)。Z54構築体の場合は、再生された全部で9個の独立した一次形質転換体が野生型GS活性の50%以下に下方調整され、その範囲は49%〜11%であった(表1Aおよび1B)。これらのデータから、イントロンを含むZ54構築体がかなり共同抑制されたことは明らかである。対照的に、Z41構築体は、内在性タバコ葉緑体GS2の下方調整の点であまり有効でなく、これらの植物はより広い範囲の共同抑制表現型を示した(表1Aおよび1BのZ41の個体間のGS活性の変化を参照)。典型的には、GS2が共同抑制された植物は、野生型よりもゆっくり成長し、光呼吸中のアンモニアの蓄積と関連する毒性またはグルタミンの欠乏のいずれかにより、介在する黄白化を発生した(図10参照)。従って、これらの形質転換体は、以前に記載されたオオムギのGS2突然変異体に類似していた(Wallsgroveら、1987, Plant Physiol. 83:155-158)。高められた (1.2%) CO2 の雰囲気下(光呼吸の抑制のため)で成長させた、またはグルタミンを補足したZ41またはZ54型のいずれかの共同抑制された植物は、あまり重い症状を示さなかった。このことも、これらの植物がGS2欠乏であるという結果を支持するものである。
【0083】
一次Z41およびZ54形質転換体由来の4個のカナマイシン耐性T2子孫植物も分析した(図6)。子孫のウェスタン分析により得られた結果および全GS活性に対して得られた結果は、一次形質転換体に対して認められた結果と同様であった(図6、パネルA、および表1B)。図6は、いくつかのZ54またはZ41一次形質転換体の代表的な一つのT2子孫のデータを示す。これらの植物のウェスタンブロット分析から、葉緑体GS2タンパク質の量が少ないことが確認され(図6、パネルA)、非変性GS活性ゲル分析により、GS2ホロ酵素の量が野生型のタバコ(図6、パネルC、レーンTL)と比較して少ないことが確認された(図6、パネルC、レーン9〜14)。ノーザン分析は、GS2トランス遺伝子からの転写体が、対照のエンドウRNA(図6、パネルB、レーンP)に存在するものと比較して検出できないことを示した(図6、パネルB、レーン9〜14)。これらの結果は、エンドウGS2トランス遺伝子の挿入によるタバコ葉緑体GS2の特異的共同抑制を示唆する。さらに、エンドウGS2トランス遺伝子も抑えられた。細胞質ゾルGSmRNAおよびタンパク質のレベルは、これらのGS2共同抑制植物において影響を受けなかった。
【0084】
6.2.3. 35−S細胞質ゾルGS3A遺伝子融合体を有するトランスジェニック植物の分析
35S−GS3A構築体(Z17またはZ9;図4)のいずれかを含むトランスジェニック植物を分析した。Z17(イントロンを含む系)の場合、GS活性を分析した13個の独立した一次形質転換体のうち、6個はGS活性の過剰発現を示した (119 〜 145%)が、7個は、形質転換しなかった対照(100%)と比較して、共同抑制(52〜28%)を示した(表1Aおよび1B)。図5および6は、Z17の代表的な過剰発現体および共同抑制された系のデータを含む。形質転換体Z17−12は、GS酵素活性が共同抑制されており(野生型の27%)、葉緑体GS2および細胞質ゾルGSタンパク質の両方とも、野生型のタバコ(図5、レーンTL)と比較して低い(図5、レーン2)。対照的に、形質転換体Z17−6は、全GS活性レベルが高められ(127%)、細胞質ゾルGSタンパク質レベルが野生型のタバコ(図5、レーンTL)と比較して増加した(図5、レーン1)。他の独立形質転換体のT2子孫の分析では、細胞質ゾルGSタンパク質が下方調整された形質転換体がさらに示されたが(Z17−9BおよびZ17−10;図6、パネルA、レーン6および7)、他は、細胞質ゾルGSのレベルが高められた(Z17−7およびZ17−9A;図6、パネルA、レーン4および5)。Z17植物に対して認められた共同抑制現象(Z17−9B、Z17−10およびZ17−12)は、GS2形質転換体(Z54およびZ41)に対して認められた現象とは、葉緑体GS2および細胞質ゾルGSの両方がGS3A共同抑制植物において下方調整されるという点で明らかに異なる(図6、パネルA、レーン6〜8、レーン9〜14参照)。図6は、35S−GS3A(Z17−9B、Z17−10、Z17−12)によって引き起こされる共同抑制がGSの量の減少(ウェスタンおよびGS活性ゲル分析;図6、パネルAおよびC、レーン6〜8)およびGS3Aトランス遺伝子の本質的に検出できない転写(ノーザン分析法;図6、パネルB、レーン6〜8)を伴うことを示す。GS3A構築体を過剰発現する形質転換体(Z17−6、Z17−7およびZ17−9A)では、GS3A転写体が非常に多く(図6、パネルB、レーン3〜5)、このことは、ウェスタンブロット分析(図6、パネルA、レーン3〜5)およびGS活性分析(表1)によって検出可能な細胞質ゾルGSの量がより多いことを表す。細胞質ゾルGS3Aを過剰発現するこれらのZ17形質転換体の可溶タンパク質の非変性GS活性ゲル分析は、野生型タバコの葉において支配的な葉緑体GS2ホロ酵素(バンドB、図6、パネルC、レーンT)よりもゆっくり移動する新規GSホロ酵素の存在を示す(バンドA* 、図6、パネルC、レーン3〜5)。同じGS3Aトランス遺伝子構築体を有する個々のZ17形質転換体が二つの異なる表現型である共同抑制型(図6、レーン6〜8)および過剰発現型(図6、レーン3〜5)を与えるというのは興味深いことである。
【0085】
分析するトランスジェニック植物集団の大きさを拡大するために、形質転換の第二ラウンドを行い、上記と同様の結果を得た。分析した全部で23個の独立した一次Z17形質転換体のうち、5個はGSが共同抑制され、8個はGSを過剰発現した。さらに、修飾していない(イントロンを含まない)GS3AcDNAを含む一次形質転換体を分析した(Z9、図4)。分析した4個のZ9一次形質転換体のうち、1個はGSが共同抑制され、2個は細胞質ゾルGSを過剰発現した。このことは、Z17(イントロンを含む35S−GS3A)およびZ9(35S−GS3A cDNA)構築体の間に定性的な相違はないことを示唆した。特に興味深いのは、Z17−9AおよびZ17−9B(図6、レーン5および6)のこれらの二つのT2植物が、単一の一次形質転換体からの自家受粉によって誘導されたけれども、種々の表現型を有するはずであるという観察である。Z17−9一次形質転換体の全GS活性を分析し、活性が減少し、従って共同抑制されたことを見出した(表1参照)。Z17−9の他の二つのT2子孫植物を分析し(Z17−9CおよびZ17−9D)、これらが共に共同抑制されて、この集団における共同抑制の割合が3:1と優勢であることが分かった。
【0086】
6.2.4. 35S−細胞質ゾルGS1遺伝子融合体を有するトランスジェニック植物の分析
35S−GS1構築体(Z3;図4参照)を含むトランスジェニック植物も分析した。独立した8個のZ3一次形質転換体のうち、5個は、ウェスタンおよびノーザンブロット分析から、明らかに過剰発現の表現型であり、共抑制型はなかった。これらのZ3形質転換体のうち二つのT2子孫を図6に示す。Z3−1およびA3−2のどちらも、細胞質ゾルGSタンパク質の量の増加を示し(図6、パネルA、レーン1および2)、これは、GS mRNAのレベルの増加によって表される(図6、パネルB、レーン1および2)。非変性活性ゲル分析は、タバコの葉の葉緑体GS2ホロ酵素(図6、パネルC、レーンT)よりも速く移動するGSホロ酵素(バンドC)(図6、パネルC、レーン1および2)を示した。このZ3植物中のより速く移動するGSホロ酵素(バンドC)は、天然のエンドウ細胞質ゾルGSと大きさが対応する。
【0087】
6.2.5. トランスジェニック植物における天然および新規の細胞質ゾルGSホロ酵素の分析
細胞質ゾルGS3A(Z17)およびGS1(Z3)の異所性発現は、さらに、しかし、野生型タバコの葉に見られる葉緑体GS2(バンドB)と比較して異なるGSホロ酵素活性バンド(例えば、バンドA* およびC)を与えた。これらのトランスジェニック植物からの抽出物の電気泳動を非変性活性ゲルで繰り返した。該ゲルには、比較のために、細胞質ゾルGSホロ酵素(バンドC)に富むエンドウの根(PR)およびタバコの根(TR)のタンパク質のレーン(図7A、レーン2および4)、ならびに葉緑体GS3ホロ酵素(バンドB)に富む精製したエンドウの葉緑体(PC)およびタバコの葉緑体(TC)から得た抽出物(図7A、レーン1および3)を含めた。トランスジェニックタバコZ3−1の抽出物(図7A、レーン6)に見られる別のGS1ホロ酵素活性(バンドC)は、天然のエンドウの細胞質ゾルGSバンド(バンドC、図7A、レーン2および4)と共に移動する。対照してみると、Z17−7トランスジェニック植物の葉(図7A、レーン5)に見られる新規GS3A活性(バンドA* )は、細胞質ゾルGS(バンドC)とも、葉緑体GS2バンド(バンドB)とも一緒に移動しない。そして、大きさはより大きく、電荷はより酸性側である。GS活性バンドA* 、BおよびCのサブユニット組成物を決定するために、これらのバンドを調製ゲルから切り取り、変性SDSゲルに再び載せ、次いで、GSサブニットのウェスタンブロット分析を行った(図7B)。この分析は、GS活性バンドA* およびCが共に、もっぱら細胞質ゾルGSポリペプチドを含むことを示した(図7B、レーン2および4)。この知見では、より大きいGS3A活性バンドA* が、異種GS3A細胞質ゾルサブユニットと内在性タバコ葉緑体前駆体GS2サブユニットとの会合の結果であるという可能性は少なかった。GS活性バンドA* は、トランスジェニックGS3Aサブユニットとシャペロニン型タンパク質との集合を表すと考えられるが、該複合体をATPで解離するという試みは成功しなかった。その結果、新規GSホロ酵素の性質は不明のままである。
【0088】
6.2.6. 細胞質ゾルGS1またはGS3Aを異所的に過剰発現する形質転換体の成長分析のための選択
細胞質ゾルGS3A(Z17)または細胞質ゾルGS1(Z3)を異所的に過剰発現する2組の植物を選択して成長分析を行った。第一ラウンドの形質転換から(下記実験A参照)、植物Z3−1およびZ3−2をGS1高レベル発現体として選択し(図6、レーン1および2;図8、レーン1および2)、植物Z17−6およびZ17−7をGS3A高レベル発現体として選択した(図5、レーン1;図6、レーン3および4;図8、レーン3および4)。これらの形質転換体のカナマイシン耐性T2子孫を以下に記載する実験Aでの成長分析のために選択した。第二ラウンドの形質転換から、二つのさらに独立して形質転換したGS1−過剰発現植物(Z3−3およびZ3−4)(図8、レーン5および6)および二つのさらに独立して形質転換したGS3A−過剰発現植物(Z17−3およびZ17−11)(図8、レーン7および8)を選択して分析した。これらの植物のカナマイシン耐性T2子孫を、第二成長実験で分析した(下記実験B)。
【0089】
6.2.7. 植物成長実験の設計
植物成長分析を、図8でGSタンパク質およびRNAの分析を行ったT2子孫植物に対して行った。個々のT2植物を白砂中で成長させ、その成長を、タイムポイントごとに4〜7個の植物の新鮮重量を測定することにより評価した。新鮮重量の測定は、植物が急速に成長し、供給した窒素にもっぱら依存し、抽薹および開花中に生じる可能性のある窒素の大きな内部源を流動させることのない栄養成長期中のみ行った。植物は、窒素が無制限であり(すなわち、10 mM の硝酸塩の施肥が定期的に行われる。)、隣接する植物の光合成妨害が少ない条件下で成長させ、そのような妨害が現れたときに成長分析を終わらせた。分析した全ての植物は同じ週齢であり、0.1 〜0.3 g の新鮮重量で分析を開始し、植物が約6週齢で、隣接する植物の妨害が抽薹の開始時に現れるまで続けた。
【0090】
6.2.8. 植物成長実験A
表2に、Z3−1およびZ3−2の系(過剰発現GS1)ならびにZ17−6およびZ17−7の系(過剰発現GS3A)の全新鮮重量測定値の平均の結果を示す。これらの結果を、図9、パネルAでグラフにより示し、表3で統計的に分析した。トランスジェニック系の全4個の過剰発現するエンドウの細胞質ゾルGSは、コントロールよりも35% 〜 114% だけ大きくなった。これは、3個の系、すなわち、Z3−2(P=0.08)、Z17−6(P=0.0015)およびZ17−7(P=0.013 )に対して統計的に重要であった(表3)。
【0091】
6.2.9. 植物成長実験B
成長実験を、同じGS1(Z3)およびGS3A(Z17)構築物を有する種々のトランスジェニック系で繰り返して、上記で得られた結果を確認した。この実験は、統計的分析のために、より大きい植物集団を含む。表2は、トランスジェニック系Z3−3、Z3−4、Z17−3およびZ17−11ならびに2個のコントロール系(C1、C2)の4回のタイムポイントに対する平均データを示す。Z3−4を除く全ての系は、コントロールよりも 40%〜44% だけ大きくなり、6週目の新鮮重量の違いは、統計的に有意であった(表3)。これらの結果はまた、図9、パネルBにグラフにより示す。第二の成長実験は第一の実験結果を確証するものであることは明らかであり、これは、エンドウの細胞質ゾルGS1またはGS3Aのいずれかの異所性過剰発現がタバコの成長速度を高めたことを示唆するものである。テストした全ての系において、GS3A過剰発現は、非形質転換コントロールと比較して成長速度を増加させ、これは、トランスジェニックタバコに対して統計的に有意な成長速度の増加であった。
【表2】
【表3】
【0092】
6.2.10. 植物の成長に対するGS過剰発現の定性的影響
図10は、GSを過剰発現する植物の成長表現型(Z3−A1およびZ17−B7)のコントロール植物およびGSが共同抑制される植物(Z54−A2)の表現型に対する定性的比較を示す。その結果は、GS過剰発現のレベルがたとえ低くても、容易に認められる成長の改善が得られることを示す(図10、Z17−B7およびZ3−A1の成長をコントロール植物の成長と比較)。さらに、これらの結果は、成長の改善がGS過剰発現によるものであり、CaMV−35S GS構成体による植物の単なる遺伝子操作によるのではないことを示す。例えば、CaMV35S−GS2により遺伝子操作され、GS発現が共同抑制されたZ54−A1は、かなり小さい成長を示した。さらに、これらの結果は、GS活性が植物成長の律速段階であることを示す。というのは、この酵素の阻害により成長に対する重大な表現型の影響が引き起こされるからである。
【0093】
6.2.11. GS活性と最終の新鮮重量および全タンパク質との間の相関性
GS遺伝子の異所性過剰発現または共同抑制と関連するGS活性の変化が、栄養成長期の終わりの「最終」の新鮮重量に対して影響があるかどうかを調べるための実験を行った。成長分析は、GS2によって共同抑制される系(Z54−4)、GS1を過剰発現する系(Z3−1)、GS3Aを過剰発現する系(Z17−7)および形質転換されていないタバコのコントロール(SR1)に対するT2世代植物において行った。植物を砂土で成長させ、10 mM のKNO3 を含む Hoagland 溶液で定期的に湿らした。指定のタイムポイントで、各系の8個のT2植物を秤量し、葉のGS活性を各々について測定した。このデータを分析すると、32日および43日の両日に測定した個々の全ての「最終」の新鮮重量とGS特異的活性との間には直線的関係があることが分かる(図11A)。例えば、GS活性が共同抑制されるZ54−4植物(野生型GS活性の 27%)はコントロールの半分の重量であり、GS3A(136 % GS活性)またはGS1(284 % GS活性)を過剰発現する植物は、各々、コントロールよりも1.5 倍および2倍重い。これらの同じT2植物の場合、葉の全タンパク質(μg タンパク質/ gm 新鮮重量)と葉のGS活性との間にも直線的関係が存在する。最も高いレベルのGS活性を発現する植物(284 % )は、可溶タンパク質レベル/ gm 新鮮重量が、コントロールと比較して 1.5倍高かった(図11B)。このデータの無対T−テスト分析は、GS過剰発現系(Z3−1、Z17−7)が、p<0.0001のときにかなり大きいGS活性、新鮮重量、および葉の可溶タンパク質を有することを示した。ただし、p値が 0.0007 であるZ17−7に対する新鮮重量は除く。同様に、GS2によって共同抑制される系(Z54−4)は、p<0.001 のときに、コントロールSR1よりもかなり小さいGS活性、新鮮重量、および葉の可溶タンパク質を有した。成長実験で使用されるGS過剰発現T2系(Z3、Z17)のGS活性プロフィルは、T2世代ではGS活性が一貫して高かったことを除き、親のT0系およびT1子孫に匹敵する。このことは、恐らく、GSトランス遺伝子と会合した KanR 表現型の分離が認められなかったので、トランス遺伝子のいくつかまたは全部が、T2世代でホモ接合体になったという事実によると考えられる。成長実験の終わりには、GSを過剰発現するトランスジェニック系は、コントロールよりも視覚的に緑が強くなり、劇的に大きくなった。
【0094】
6.3. 考察
遺伝子工学が収穫のある植物の改善において重要であると考えられ始めるにつれて、選択した遺伝子の過剰発現において決定的なパラメータを理解することがますます重要になってくる。宿主植物相同体が存在する遺伝子の過剰発現が、ウイルスコートタンパク質およびBT毒性遺伝子などの相同体のない遺伝子の過剰発現より複雑であることは明らかである(Powell-Abel ら、1986, Science 232:738-743; Vacckら、1987, Nature 328:33-37)。これは、トランスジェニック植物が検出でき、宿主相同体が存在するトランス遺伝子を、恐らくフィードバック阻害またはいくつかの他の機構により抑えることができる共同抑制現象による(van der Krolら、1990, Plant Cell 2:291-299; Napoliら、1990, Plant Cell 2:279-289)。本発明では、同じ構成プロモーター(35S−CaMV)の後ろの葉緑体または細胞質ゾルGSに対する3種類のエンドウGS遺伝子をトランスジェニックタバコで異所的に過剰発現する試みを呈示する。その試みの結果、各GS遺伝子に対して異なる過剰発現および/または共同抑制が得られた。さらに、過剰発現に成功した細胞質ゾルGSに対する2種類の遺伝子(GS1およびGS3A)の場合、その過剰発現により、GS RNA、タンパク質および酵素の過剰産生だけでなく、窒素使用効率が改善された表現型が得られた。
【0095】
細胞質ゾルGS1に対するエンドウ遺伝子のタバコでの過剰発現は、GS活性および細胞質ゾルGSタンパク質が増加し、トランス遺伝子mRNAのレベルが高い純粋な表現型を与える。さらに、GS1タンパク質は、会合して、大きさおよび電荷が天然のエンドウの細胞質ゾルGSと同じであるGSホロ酵素になる。細胞質ゾルGS3Aを過剰発現するトランスジェニック植物においては、状況がいくらか異なる。高レベルのGS3Aトランス遺伝子mRNAは、ウェスタンブロットで可視化される細胞質ゾルGSのレベルの増加を引き起こす。しかし、GS3Aの過剰発現は、エンドウまたはタバコの天然の葉緑体または細胞質ゾルGSホロ酵素より大きい新規GSホロ酵素の出現を引き起こす。これらのトランスジェニック植物において、細胞質ゾルGS遺伝子は、通常は見られない細胞の型(例えば、葉肉細胞)で発現され、GS3AトランスジェニックリーフのGSホロ酵素がより大きいのは、細胞質ゾルGSサブユニットが天然の葉緑体前駆体GS2と共会合することによると考えられた。しかし、この新規GS3Aホロ酵素は、もっぱら細胞質ゾルGSサブユニットから構成されることが分かっており、従って、トランスジェニックGS3Aサブユニットと内在性のタバコ葉緑体前駆体GS2との共会合によるのではない。二つの他の可能性が存在する。より大きいGS3Aホロ酵素は、トランスジェニックGS3Aサブユニットが集合して、通常の八面体構造以外の立体配置になった結果であると考えられる。あるいは、新規のGS3Aホロ酵素は、過剰発現した細胞質ゾルサブユニットが会合したシャペロン型から解離し損なった結果であるかもしれない。実際、GSと groEL様タンパク質との密接な会合が、以前、エンドウで確認された (Tsuprun ら、1992, Biochim. Biophys. Acta 1099:67-73)。しかし、新規GS3A活性バンドを潜在的なシャペロン型からATPによって分離する試みは失敗に終わった。新規GS3Aホロ酵素は、(GS活性ゲル分析での検出から)明らかにGS活性を有するにちがいないが、この新規GSアイソザイムが、天然の細胞質ゾルGSまたは葉緑体GS2ホロ酵素と等モルベースで同様の活性を有するかどうかを模索するのは興味深い。もし有するとすれば、35S−GS3Aを過剰発現し、従って新規GSホロ酵素を有する植物は、高められた全GS活性を有すると予想することができる。実際はそうではなく、4個のZ17−6のT2子孫植物(GS3Aを発現)の全GS活性の平均(野生型との比較)は100%であることが分かり、4個のZ17−7の子孫植物の平均は、野生型と比較すると 107% であった。対照すると、Z3−1およびZ3−2(GS1天然ホロ酵素を過剰発現)のT2子孫に対して得られるGS活性値は、野生型と比較して、各々、123%および120%であった。これは、Z3の過剰発現形質転換体でGS1サブユニットが集合して天然の大きさのGSホロ酵素になることは全GS活性に対して有利であることを示唆する。
【0096】
ここで、窒素の使用効率を、細胞質ゾルGS1または細胞質ゾルGS3Aの過剰発現に成功したトランスジェニックタバコの栄養成長期に評価した。栄養成長期中には、栄養の急速な吸収および光合成能の最大限の活用を特徴とする急速な葉の発達がある。窒素は、最もしばしば制限する微量栄養であり、植物内でのその吸収および使用の生理機能は、栄養期と生殖期とでは異なる。第一に、土壌からの窒素の混入、伸び広がる細胞へのその混入および光呼吸による損失の制限があり、次いで、抽薹の開始とともに、生殖期中には、種子の収穫に変換するための窒素保存物の流動がある。恐らく、窒素使用効率は、栄養成長期中はあまり複雑ではなく、本発明者らのトランスジェニック植物成長分析の焦点はこの成長期にある。
【0097】
本発明の知見は、タバコで異所的に発現されるエンドウ細胞質ゾルグルタミンシンテターゼがトランスジェニックタバコの栄養成長期にかなりの利点を付与することを示す。細胞質ゾルGS1またはGS3Aを異所的に(すなわち、全細胞型で)過剰発現する植物は、全新鮮重量がコントロールよりも高い。特に顕著なのは、GS3Aを発現する全て系(Z17)の全新鮮重量が6週目のコントロールより高く、これらが統計的に常に重要であることである。各ケースで、コントロールとトランスジェニック系との間の相違がサンプルの差異による確率は 5%未満であった。分析したGS1発現系(Z3)の場合、4個のうち3個はコントロールより大きくなり、3個のうち2個は、その相違が 10%のレベルで統計的に重要であった。この増加した窒素使用効率は、作物を同様に遺伝子操作して、通常の窒素量で良好に成長させたり、肥料の少ない投入で栽培したり、あるいは品質の悪い土壌で栽培することができ、従って、開発された、または開発中の農業システムにおいて、経済的にかなり影響がある。
【0098】
GS過剰発現は、以前にトランスジェニックタバコで試みられたが(Eckes ら、Mol. Gen. Genet, 217: 263-268; Hemonら、1990 Plant Mol. Biol. 15: 895-904; Hirel ら、1992, Plant Mol. Biol. 20:207-218; Temple ら、1993, Mol. Gen. Genet. 236:315-325) 、GSの過剰発現をGS活性のかなりの増加ならびに植物の成長および栄養特性における改善と相関させた最初の報告は本発明である。Templeらは、GS mRNAおよびタンパク質の増加を報告したが、トランスジェニック植物でのGS活性の対応する増加は報告しなかった(Templeら、同書)。Hemon らは、GS発現構築体により遺伝子操作したトランスジェニック植物でのGS mRNAレベルの増加を報告したが、GSタンパク質または酵素活性の対応する増加の報告はなかった(Hemon ら、同書)。他の二つの報告では、トランスジェニック植物でのGS遺伝子の過剰発現の結果、GS酵素のレベルが増加したが、GS過剰産生の表現型による影響についての報告はなかった(Eckes ら、同書; Hirelら、同書)。植物成長速度を約 20%改善するアルファルファGS遺伝子の過剰発現の報告が一つある(Eckes ら、1988, オーストラリア特許出願 No. AU-A-17321/88)。しかし、ここで報告された改善は、低窒素条件下での成長のみに限られていると思われる。同様に遺伝子操作した植物について、窒素無制限媒体で行った続く分析では、コントロール植物と比較して、表現型の変化を示さないことが報告された(Eckes ら、1989, Mol. Gen. Genet. 217:263-268)。さらに、より速い成長速度により、遺伝子操作した植物とコントロール植物との間で最終の新鮮重量に相違が生じるという報告はない。これらの以前の研究に対して、本発明は、窒素条件とは無関係に、GS過剰発現が植物の成長、収率および/または栄養特性を改善することができることをはっきり例証するものである。
【0099】
本発明の農業上の有用性は、栄養器官が採取される作物の種類に直接関係し、これらには、全ての飼料穀物、馬鈴薯、テンサイおよびサトウキビならびにタバコが含まれる。最終の新鮮重量の記録の1週間以内に、植物は、節間の伸長を受け始め、各集団に対する次の新鮮重量の測定の標準偏差は、植物の異なる生理学的段階の結果、増加した。増加した栄養成長速度も種子の収率をかなり有利にするかどうかは重要な問題であり、まだ回答は出されていない。種子の収率および種子の窒素含量に関係する生理学的パラメーターは、窒素吸収効率だけでなく、抽薹開始時の保存物の再流動および畑の集団密度の成りゆきも含む。そのような研究は、種子の収穫に対して選択され、収穫生理学に関していくらか知識があるトランスジェニック種において行うとより良好である。
【0100】
細胞質ゾルGS1によるのではなく、エンドウの葉緑体GS2および細胞質ゾルGS3Aをコードする遺伝子による内在性タバコの共同抑制も興味深い。これは、エンドウGS2がタバコ葉緑体GS2型のみを抑制し、一方、GS3Aがタバコ葉緑体GS2および細胞質ゾルGSの両方を抑制するときに、特にそうである。以前、ペチュニアカルコンシンテターゼおよびジヒドロフラバノール−4−リダクターゼが、トランスジェニックペチュニアで、内在物およびトランス遺伝子の両方を抑制することが示された(van der Krolら、1990, Plant Cell 2:291-299; Napoliら、1990, Plant Cell 2:279-289)。さらに最近、カルコンシンテターゼ遺伝子の5’または3’端は、共同抑制を引き起こすのに十分であるが、プロモーターの小さい遺伝子はそうではないことが報告された(Jorgensen, 1992, Agbiotech News and Information Sept: 1992)。これは、転写開始剤の必要性を示唆するものである。一時的な異所性配列対合が、遺伝子抑制の可能な機構として引き合いに出され、これは、たぶん、転写の開始と関係するDNAの巻き戻しに依存すると考えられる(Jorgensen, 1990, Trends in Biotechnology 8:340-344; Jorgensen, 1991, Trends in Biotechnology 9:255-267; Jorgensen, 1992, Agbiotech News and Information Sept: 1992) 。エンドウGS遺伝子発現に対する本発明の知見から、共同抑制現象は、ヌクレオチドレベルでの配列の完全な相同性には依存しないことは明らかである。
【0101】
窒素同化酵素の発現を加減することによる窒素利用の増加はまた、トウモロコシなどのトランスジェニック植物での収率を高めるための実行可能な方法であると考えられる。作物における窒素使用効率は、高められた収率として測定され、従って、農業的尺度となる。この種の適応または限定は、生存のために、環境条件に対する種々の反応に依存して、より高い収率には依存しない野生型植物に対しては、真に有利であるとはいえない(Sechley ら、1992, Int. Rev. Cyt. 134:85-163 )。従って、作物の収率の増加は、従来の品種改良法よりもむしろ、本明細書に記載したような遺伝子工学法によってより容易に実現することができる。
【0102】
7.0. 実施例:植物中のアスパラギンシンテターゼの異所性過剰発現が植物増殖表現型の増加を引き起こす
以下の研究は、アスパラギン製造の増加及び植物成長に対する影響を試験するとを目的とする、植物中でのAS遺伝子発現の操作に係わる。窒素運搬/貯蔵化合物としてアスパラギンがグルタミンよりも好ましいいくつかの特徴があり、従ってアスパラギンへの窒素の同化の増加はin vivo で価値があり得る。アスパラギンは、グルタミンよりも高いN:C 比率を有する長距離窒素輸送化合物である。従って、それは窒素輸送のためのより経済的な化合物である。それに加えて、アスパラギンはグルタミンより安定であり、液胞中で高いレベルに蓄積することができる(Sieciechowicz et al., 1988, Phytochemistry 27:663-671; Lea and Fowden, 1975, Proc. R. Soc. Lond. 192:13-26)。エンドウの葉の分化の間にはアスパラギンが活発に代謝されるが、窒素を成長のためにもはや必要としない成熟した葉ではアスパラギンは容易には代謝されず、葉から活発な成長領域、例えば分化中の葉や種子に再輸送(師部中)される(Sieciechowicz et al., 1988, Phytochemistry 27:663-671; Ta et al., 1984, Plant Physiol 74:822-826)。ASは通常暗所でのみ発現され(Tsai and Coruzzi, 1990, EMBO J. 9:323-332) 、従って35-AS1は構成的に発現され、細胞型に関して異所的に発現されるだけでなく一時的発現についてもそうである。即ち、本明細書に示した研究は、あらゆる細胞型中での光非依存性形態のASの異所性過剰発現がアスパラギン製造を増加させるかどうかを調べるものである。増加されたアスパラギン製造がトランスジェニック植物の窒素使用効率と成長表現型において利点を与えるかどうかについても本明細書で試験する。
【0103】
過剰発現野性型ASに加えて、本研究ではグルタミン結合ドメインを欠いているAS酵素の改変形態(glnΔAs1)の異所性過剰発現を調べた。この研究により調べられた問題は、その酵素の glnΔAs1形態の異所性過剰発現が増強されたアンモニア依存性AS活性を有する新規な植物AS酵素を生成するか、あるいはそのような突然変異が、内因性野性型ASサブユニットとともに集合することにより優勢ネガティブ効果を有し(Herskowitz, 1987, Nature 329:219-222)、ヘテロダイマーを形成するかどうか(Rognes, 1975, Phytochemistry 14:1975-1982; Hongo and Sato, 1983, Biochim et Biophys Acta 742:484-489)ということである。エンドウASを異所的に発現するトランスジェニック植物の分析により、アスパラギンの増加した蓄積と改善された成長表現型(35S-AS1の場合)、そして成長表現型に対する有害な作用を伴うアスパラギンの増加した蓄積が示された(35S-glnΔAS1の場合)。これらの結果は、窒素代謝と成長表現型をAS遺伝子の異所性過剰発現によって操作することが可能なことを示している。窒素が植物成長の律速元素であることが多く、典型的にはその成長期に作物に数回施用されるので、作物植物における窒素利用効率を改善する分子設計技術は農業にとって大きな利益をもたらすものである。
【0104】
7.1. 方法及び材料
7.1.1. AS遺伝子構築物
以前にエンドウからクローン化されているAS1 cDNA(Tsai and Coruzzi, 1990, EMBO J 9:323-332)を、pTZ18UからpBluescript KS- (Stratagene)のEcoRI部位に移入した。 glnΔAS1欠失突然変異体を、「内部−外部」PCRを使用して構築した(Innis et al., 1990, PCR Protocols: A guide to Methods and Applications. New York, Academic Press pp.1-461) 。アミノ酸2-4(CGI)に対応するコード配列をAS1 cDNAのアミノ末端から削除し、開始メチオニンと非翻訳リーダーをそのまま残した。この欠失が、動物ASについて定義されたアミノ酸MCGIを含んでいるAS酵素の推定グルタミン結合ドメインに対応した(Pfeiffer et al., 1986,J. Biol. Chem. 261:1914-1919; Pfeiffer et al., 1987, J. Biol. Chem. 252:11565-11570)。野性型AS1及び glnΔAS1に対応するcDNAを次にpBluescript からバイナリ発現ベクターpTEV5へ移入した。このベクターは、CaMV 35Sプロモーター(-941から+26まで)、多回クローニングサイト、及びノパリンシンターゼターミネーターを含む。図12は、AS1 cDNA pZ127 (NRRL受託番号 B-21335) 及び glnΔAS1 cDNA pZ167 (NRRL受託番号 B-21336)を含むバイナリーベクター構築物の詳細を示し、これをタバコに形質転換した。
【0105】
7.1.2. 植物形質転換体
バイナリーベクター構築物を、安全化Agrobacterium 株LBA4404 に、その後Nicotiana tabacum SR1 に、他の文献に記載された標準的手順を使用して移入した(Bevan, 1984, Nucleic Acids Res. 12:8711-8721; Horsch et al., 1985, Science 227:1229-1231)。
【0106】
7.1.3. 形質転換体のRNA分析
Bio101からの「RNAマトリックス」を使用してRNAを単離し、全RNAをこれま
でに記載されたように電気泳動にかけた(Thomas, 1983, Methods Enzymol. 100:255-266)。ゲルを毛細管でHybond-Nナイロン膜(Amersham)上へブロットした。cDNAを、NEN により供給されるランダムプライマーと延長試薬標識システムを使用して標識した。ハイブリダイゼーションを水溶液中で行い、ブロットを0.1X SSPE, 0.1% SDS中で洗浄した。ノーザンブロットをエンドウAS1 cDNA, pAS1でプロービングした(Tsai and Coruzzi, 1990, EMBO J 9:323-332)。
【0107】
7.1.4. 遊離アミノ酸の抽出
タバコ葉組織試料を液体窒素中で凍結し、メタノール:クロロホルム:水(12:5:3、v/v/v)からなる抽出培地の10のml中で抽出した。ホモジネートを、12,000 X Gで15分間遠心分離した。ペレットを再び抽出し、上清を合わせた。上清へ2.5 mlのクロロホルム及び3.8 mlの蒸留水を添加すると分離した。メタノール:水相を集め、減圧下に乾燥し、1 mlの蒸留水中に再溶解した。その溶液を12,000 gで2分間遠心分離して0.45μm ナイロンフィルターマイクロ遠心分離チューブフィルターシステムを通しろ過した。
【0108】
7.1.5. アミノ酸プールのHPLC測定
アミノ酸は、Microsorb Type OAA Analysis カラム(Rainin)上でDuPontのHPLC機器を使用してo-フタルジアルデヒド(OPA) 誘導体として測定した。試料(100μL)をOPA 作用試薬の100μl で誘導化した。2分の誘導化の後、誘導化された試料の50μlを注入した。この勾配は 2種の溶離液;A. 4.5% メタノール及び0.5%テトラヒドロフルオランを含む95% 0.1M酢酸ナトリウム(pH 7.2); B. 100 %メタノールを使用して生成した。溶離液をろ過し、使用の前にHeで脱気した。OPA誘導化アミノ酸の検出は、UV分光光度計で340 nmにおいて行った。各測定は二回行い、値はその平均値を表す。
【0109】
7.1.6. 植物成長条件
高レベルのAS1 mRNAまたは変異 glnΔAS1 mRNAを発現するものとして特徴付けられる一次形質転換体の後代を、100 μg/mlのカナマイシンを含むMS培地上で生育させた。14日後、カナマイシン耐性苗木を白砂で満たした4インチの鉢に移し、約1週間サランラップで包んで過剰な蒸発を防ぎ、苗木が根付くようにした。鉢に唯一の窒素源として10mM硝酸カリウムを含む1X Hoagland溶液を定期的に与えた。その後、3〜7の植物を各週に新鮮重量を測定するために採取し、隣り合った植物の陰になるのが顕著になるまで4週間継続した。植物を16-8時間の光-暗所サイクルの下に24-18℃の温度サイクルで成長させた。昼間光度は1000ルクスであった。
【0110】
7.2. 結果
7.2.1. エンドウAS1と glnΔAS1 を発現するトランスジェニック植物の構築
35S-CaMVプロモーターから発現されたエンドウAS1 cDNA (Tsai and Coruzzi,1990, EMBO J 9:323-332) をトランスジェニックタバコに移し(図12及び7.1 材料及び方法の項を参照) 、5つの独立した一次形質転換体(Z127; 1-5)が高いレベルのAS1 mRNAを発現することが示された(下記参照)。グルタミン結合ドメインに欠失を有するAS1 cDNA(glnΔAS1)を含む3つの独立したトランスジェニック系(Z167;1-3)もトランスジーンRNA の高いレベルを含むことが示された (下記参照)。
【0111】
7.2.2. AS1と glnΔAS1を発現する形質転換体のノーザン分析
トランスジェニック植物から抽出されたRNA のノーザンブロット分析を行い、35S-AS1 トランスジーンが高いレベルで発現した植物を同定した(図13)。陽性対照として、ASのRNAを暗所で生育したエンドウ植物の葉中に検出した(図13、レーンPL)。これに対して、光中で生育した野性型タバコの葉にはAS mRNA は検出されなかった(図13、TL)。以前の研究により、タバコAS mRNA は暗所で生育した植物の組織中だけに発現されることが示されている(Tsai and Coruzzi, 1991, Mol Cell Biol 11:4966-4972)。AS1 を過剰発現する形質転換体(Z127-1、-3、-4及び-5)は全て、それらが光中で生育されたにもかかわらず、AS1 mRNAの高いレベルを含んでいた(図13)。このように、35S CaMVプロモーターは、エンドウAS1の構成的な発現を生成するのに対して、内因性AS mRNAは、光中ではタバコ葉中に発現されない。 glnΔAS1形質転換体も、タバコ対照と比較して高水準のmRNAの構成的な発現を示した(Z167-2、-3及び-4) (図13)。AS酵素のよく知られた不安定性のために、AS酵素を均質物にまで精製できず、植物ASの抗体をASタンパク分析のために使用できなかった。それに加えて、in vitroアッセイでは、酵素の不安定さのためにAS活性が検出されなかった。
【0112】
7.2.3. AS1及び glnΔAS1 を発現するトランスジェニック系のアミノ酸分析
ノーザン分析の結果に基づいて、AS1 mRNAの高いレベルを示した2つの独立のトランスジェニック系(Z127-1及びZ127-4)をさらに分析するために選択した。同様に、Z167-2及びZ167-4系を glnΔAS1 構築物の高発現体の代表として選択した。これらの植物のT2後代について、以下に記載するアミノ酸分析及び成長分析を行った。
【0113】
7.2.4. AS1-過剰発現系
選択した両方のZ127系(Z127-1及びZ127-4)は、アスパラギンの増加されたレベル(野性型対照より10〜100 倍高い)を示した(表4)。個々のT2植物の間で見られるばらつきは、個体のホモ接合性またはヘテロ接合性を反映している可能性が最も高く、中間レベル:高いアスパラギンレベルの約2:1 の比は、この考え方の証拠となるものである。しかし、すべての場合においてアスパラギンのかなりの増加は対照濃度のほぼ100 倍まで及んでいることが判る。面白いことには、これらの植物においてグルタミン濃度の対応する減少が存在し(Z127-4のデータは唯一高い値ではずれていたが)、これはAS反応における基質としてのグルタミンの使用を反映しており、同じく予測できるのは他の基質アスパルテートの濃度の減少である。しかし、Z127系中のグルタメートの低下した濃度はいくらか予想外である。生化学的予測とAS反応に関与する他の3つのアミノ酸について集めたデータから、グルタメートの増加が予測された。グルタメートの見かけ上の減少は、アミノ基転移のようないくつかの関連するプロセスにおける基質として使用されたために起こった高い代謝回転の結果であり得る。
【0114】
7.2.5. Gln ΔAS1 過剰発現系
glnΔAS1 を過剰発現する選択された二つの系において調べた問題は、ASのグルタミン結合ドメインの欠失がアスパラギン生合成に対して優勢−ネガティブ効果を有しているかどうかということである。これらの系(Z167-2及びZ167-4)について集めたデータはデータ値のばらつきのために解釈するのがやや困難である(表4)。しかし、ほとんど全ての場合において、野性型非トランスジェニックタバコと比較して3から19倍の範囲のアスパラギン濃度の実質的な増加が存在する。これらの結果は、トランスジェニック系がアスパルテート、グルタメートまたはグルタミンプールに対して殆ど影響を及ぼさずにアスパラギンを蓄積する能力を有することを示唆している。一つの可能性は、 glnΔAS1酵素がアンモニアとアスパルテートから直接アスパラギンを合成できるということである。
【0115】
7.2.6. AS1 及び glnΔAS1を発現する形質転換体についての植物成長実験
白砂中で生育された個体トランスジェニックT2植物を使用して成長分析を行った。これらの研究は、近隣の植物からの干渉を最小にした条件下に成長速度を調べることを目的とする。このため、生育の栄養成長段階(発芽後6週間まで)の間にのみ新鮮重量測定を行った。この間、植物は急速に成長し、供給された窒素にのみ依存し、薹立や顕花の間におこるように内部窒素源を移動させない。植物は、窒素について非制限的(即ち10 mM 硝酸塩の規則的な施肥)で隣接植物からの光合成干渉を減らすような条件下に生育された。その成長分析は、そのような干渉が顕著になったときに終了した。分析したすべての植物は各時点で同じ加齢を有し、 0.1〜0.3 gの新鮮重量/植物の間に分析を開始し、近隣の植物の干渉が顕著になり薹立が間近の約6週齢になるまで継続した。
【表4】
【表5】
【0116】
表5及び6は、系Z127-1及びZ127-4(野性型AS1を過剰発現する)とZ167-2及びZ167-4(glnΔAS1を過剰発現する)についての平均全新鮮重量測定の結果を示し、これらの結果を図13にグラフとして示す。野性型ASを過剰発現しているトランスジェニック系は、対照(100%)と比較して133%及び110%成長したが (表5) 、無対t検定により分析した場合、いずれの場合も統計学的に有意でなかった(表6)。 glnΔAS1 構築物(Z167)を過剰発現するトランスジェニック系は同等に成長しなかった。6週まで生存したZ167-4植物は成長において対照から区別できず、生存したZ167-2植物は対照よりも非常に小さかった(P-0.041;5%レベルにおいて有意)(表5及び6、図14も参照)。実験における3つの異なる系を比較すると、カナマイシン耐性Z167植物の大部分が死滅したことは興味深い。典型的に、Z167植物は鉢で生育されると発芽するのが遅く、不健康に見えた。これは、Z167-2について集められた新鮮重量のデータに明確に反映されていたが、Z167-4データについてはそれほど明確ではなく、glnΔAS1 遺伝子生成物が植物成長に対し実際に優勢−ネガティブ効果を有していたことを示唆している。
【表6】
【0117】
7.3. 考察
本明細書に開示した研究では、ASをトランスジェニック植物中に異所的に過剰発現させ、この操作の一次窒素同化と植物成長に対する影響を試験するものであった。特に、ASの細胞特異的発現パターンを変更し、光に関してASの制御も改変した。野性型植物においては、ASは通常その師部中に発現されるだけであり(Tsai, 1991, Molecular Biology Studies of the Light-Repressed and Organ-Specific Expression of Plant Asparagine Synthetase Genes. Ph.D. Thesis, The Rockefeller University, New York, NY)、その発現は光合成及び非光合成組織の両方において光によって劇的に抑制される(Tsai and Coruzzi, 1990, EMBO J 9:323-332; Tsai and Coruzzi, 1991, Mol Cell Biol 11:4966-4972)。ここで、エンドウの野性型AS1及びAS1 の変異形態(glnΔAS1)は、構成的プロモーター(35S-CaMV)の制御下にトランスジェニックタバコ中に発現され、AS1 がすべての細胞型中に光から独立して発現される。それが通常は発現されない細胞において構成的に発現されるAS1の生理的な意義は、植物窒素代謝に対して重大な影響を有し得る。例えば、アスパラギンは光呼吸窒素リサイクルに関与しており(Givan et al., 1988, TIBS 13:433-437; Ta et al., 1984, Plant Physiol 74:822-826)、従って光合成細胞中のASの異所性発現は光呼吸に劇的な影響を与え得る。さらに、この点でアンモニア依存性AS酵素の発現は光呼吸アンモニアの再同化を助け得る。
【0118】
35S-AS1 を発現する4つの独立したトランスジェニックタバコ系が野生型エンドウAS1トランスジーンを構成的に発現することが示された。2つの系をさらに分析し(Z127-1及びZ127-4)、遊離アスパラギンが高いレベルにトランスジェニック葉組織中に蓄積し、典型的にはトランスジェニック系Z127-1及びZ121-4が対照非形質転換タバコ系の10〜100 倍のアスパラギンを蓄積したことから、発現されたAS1遺伝子が機能的であったことが示された。これらの増加したアスパラギンレベルは、予測されるようにAS基質、グルタミン及びアスパルテートの減少を伴った。しかし、ASの基質であるグルタミンのより高い内因性レベルを与えることによって、より多くの無機窒素を窒素運搬化合物のアスパラギンに導入することはなお可能であり得る。
【0119】
Z127トランスジェニック植物の植物成長実験は、AS1 過剰発現植物中のアスパラギンの蓄積が植物分化の栄養成長期の間の成長に対して、ポジティブな効果を持つかどうかということを調べることを目的とするものであった。栄養成長の間に起こる急速な葉の分化は、栄養の有効性を強く要求し、この時点では組織を広げて大きくすることにおける新しいタンパク質の合成のために窒素が典型的に最も臨界的な栄養素である。このとき同化され蓄積された窒素はその後その植物中でリサイクルされて種子貯蔵物中に蓄積され、また主要な長距離輸送アミノ酸であるアスパラギンも種子貯蔵物の形成に重要な役割を果たす(Dilworth and Dure, 1978, Plant Physiol 61:698-702; Sieciechowicz et al., 1988, Phytochemistry 27:663-671)。2つのZ127系が、栄養成長の終了までの6週間にわたって非形質転換対照より大きくなり、10% 及び33% の成長の優位性が得られたことが判明した。しかしこれらの値は、t検定を行うと統計学的には有意でなかった。即ち、植物は10〜100 倍より高いレベルのアスパラギンを製造するが、グルタミンレベルが成長中の増加に対して制限的であることがあり得る。さらに本明細書に示したのは、トランスジェニックタバコ中でGSを過剰発現することは統計学的に有意な(上出)より大きい成長優位性を与えることができるという知見である。グルタミンがアスパラギン生合成のための基質であるので、両者は無機窒素の一次的同化において中枢となるアミノ酸である。従って、GSとASを高いレベルで発現するトランスジェニック系を形成する(AS及びGS過剰発現体を交雑することによって)ことは、いずれの親よりも有利な成長形質も持つことがあり得ると予測できる。特に本明細書で記載した方法は、トランスジェニック系中の同化が2、3の細胞型に制限されず、すべての植物細胞中で使用可能な窒素が利用されることを可能にするという長所を有する。一つの植物中でのGS及びASの両方の異所性過剰発現はグルタミン蓄積を回避する利点を有し得、これはグルタミンの高い濃度の存在下ではグルタミンが活性な代謝産物であって細胞の代謝を破壊し得るからである。これに対し、アスパラギンは比較的不活性な化合物であり、経済的に窒素を蓄えることができる。さらに、アスパラギンは、グルタメートの分子を遊離する反応において形成され、アンモニア単位をさらに受容するのに利用され得る(Lea and Fowden, 1975, Proc. R. Soc. Lond. 192:13-26).
野性型ASの異所性過剰発現に加えて、1つの試みにおいて植物グルタミン依存性ASを、そのアンモニア依存活性を増強するように改変した。特に、ASのグルタミン結合ドメインに対する抗体が同じASポリペプチドに存在しているグルタミン依存AS活性を抑制するが、アンモニア依存活性は増強することが動物中で示されている(Pfeiffer et al., 1986, J. Biol. Chem. 261:1914-1919; Pfeiffer et al., 1987, J. Biol. Chem. 252:11565-11570)。類推として、突然変異によりグルタミン結合のために必要とされる3つのアミノ酸を特異的に削除された部位特異的変異体(glnΔAS1)がエンドウAS1 cDNAに形成されている(Tsai and Coruzzi, 1990, EMBO J 9:323-332)。この glnΔAS1をトランスジェニック植物に導入することによって、アンモニア依存AS活性を増強すること及び/またはサブユニット活性低下及び野性型と突然変異サブユニットのヘテロダイマーの形成を通して内因性グルタミン依存AS活性を抑制することが可能であり得る。 glnΔAS1トランスジーンを過剰発現する2つの独立のトランスジェニック系、Z167-2及びZ167-4が非形質転換タバコ対照よりも約3〜19倍高いアスパラギンレベルを蓄積し得ることが見出された。アスパラギンの同化における glnΔAS1遺伝子の活性は、グルタミン以外の窒素基質(例えばアンモニア)を利用する能力を持っている改変酵素を示唆するものである。E.coli AsnA遺伝子と哺乳類ASの既知のアンモニア依存AS活性に対する類推から、突然変異植物 glnΔAS1酵素を発現するトランスジェニック植物におけるアスパラギンの高いレベルは、 glnΔAS1 酵素が直接アンモニアをアスパラギンに同化でき、従って一次窒素同化においてGSを迂回し得ることを示唆している。この提案が正しいならば、これらのトランスジェニック植物中のアスパラギンの相対レベル(Z167 対Z127) に基づいて、gln ΔAS1 遺伝子がアスパラギンの合成において過剰発現された野性型AS1ほどは効率的でないことは明らかである。
【0120】
glnΔAS1 を発現するトランスジェニック系(Z167-2 及びZ167-4) は非形質転換対照より大きくならず、実際にはZ167-2の成果と実験の終了前におけるZ167植物の高い死滅割合によって示されるように、それらの成長は非形質転換植物より劣っているのが典型的であった。これらの植物がその葉中に3〜19倍高いレベルのアスパラギンを蓄積するのに成長が劣ることは奇妙である。植物ASはホモダイマーとして集合すると思われる(Rognes, 1975, Phytochemistry, 14:1975-1982)。野性型ASが通常発現されない葉の葉肉組織においては、 glnΔAS1 形態はホモダイマーへ自己集合することができ、アスパラギンを生成することができる酵素を形成する。しかし師部細胞においては、 glnΔAS1 サブユニットは野性型ASサブユニットと共に集合し得、これにより野性型ASを優勢−ネガティブ突然変異として不活性化する(Herkowitz, 1987, Nature 329:219-222) 。 glnΔAS1植物中では、葉の葉肉細胞中に合成されたアスパラギンは、師部に輸送され、ロードされることができないことがあり得、これがこれらのトランスジェニック系の劣った成長表現型の要因となっている可能性がある。これらの観察は、細胞型機能、窒素代謝遺伝子の細胞特異的遺伝子発現及び植物窒素代謝に対するその影響の特性化を明らかにするものである。
【0121】
8. 微生物の寄託
以下の微生物は、Agricultural Research Culture Collection, Northern Regional Research Center (NRRL), Peoria, Illinois寄託されており、以下の受託番号が付与されている。
【0122】
株 プラスミド NRRL受託番号
Escherichia coli, Z3 pZ3 B-21330
Escherichia coli, Z9 pZ9 B-21331
Escherichia coli, Z17 PZ17 B-21332
Escherichia coli, Z41 pZ41 B-21333
Escherichia coli, Z54 pZ54 B-21334
Escherichia coli, Z127 pZ127 B-21335
Escherichia coli, Z167 pZ167 B-21336
本発明をその特異的態様を参照して詳細に記載したが、機能的に等価な改変は本発明の範囲の範囲内であることが理解されるであろう。実際、本明細書に示し記載したものに加えて本発明の種々の改変は、これまでの記載及び添付の図面から当業者に明らかであり、そのような改変は添付の請求の範囲に包含されることを意図するものである。
【0123】
種々の刊行物を本明細書中で引用したが、その全体を引用により本明細書の一部とするものである。
【0124】
【0125】
Form PCT/RO/134 (続き)
アグリカルチュラル リサーチ カルチャー コレクション (NRRL)
インターナショナル ディポジタリー オーソリティー
アメリカ合衆国 61604 イリノイ州
ペオリア,エヌ.ユニバーシティー ストリート 1815
受託番号 寄託日
B−21331 1994年 9月28日
B−21332 1994年 9月28日
B−21333 1994年 9月28日
B−21334 1994年 9月28日
B−21335 1994年 9月28日
B−21336 1994年 9月28日
【受託番号】
【0126】
NRRL B-21330
NRRL B-21331
NRRL B-21332
NRRL B-21333
NRRL B-21334
NRRL B-21335
NRRL B-21336
【技術分野】
【0001】
1.序文
本発明は、一般に、増強された窒素同化および利用能を示し、より大きく、より効率的に、より速く成長し、かつ/また、栄養および/または生殖に関係した植物部分の窒素含量および/または生物量(バイオマス)の増加を示す植物体を遺伝子工学的に作出することに関するものである。より詳細には、本発明は、遺伝子工学的手法により窒素同化/利用経路で鍵となる酵素の発現を改変させたトランスジェニック植物を作出することに関するものである。このように遺伝子工学的に作出された植物は、低い窒素肥料投入量の条件下でまたは窒素の乏しい土壌で生産的に栽培することができる。また、遺伝子工学的に作出された植物を用いると、理想的な栽培条件下でより速く成長または成熟する作物、より高い作物収穫量および/またはより栄養分の多い生産物を得ることが可能となる。
【背景技術】
【0002】
2.発明の背景
窒素はしばしば植物の成長の律速要因になり、基本的に野外作物はすべて無機窒素肥料に依存している。肥料は多くの種類の土壌から急速に枯渇してしまうので、生育期に2,3回、成長しつつある作物に肥料を施さなければならない。窒素肥料は、通常は硝酸アンモニウム、硝酸カリウムまたは尿素として供給され、一般にトウモロコシやコムギのような農作物に関連したコストの40%を占めている。北アメリカと西ヨーロッパの両地域では毎年約1100万トンの窒素肥料が使われており、そのために農業従事者は毎年22億ドルを出費していると概算された (Sheldrick, 1987, World Nitrogen Survey, Technical Paper no. 59, Washington, D.C.) 。さらに、世界銀行の試算では、1年間の窒素肥料の需要量が世界中で今後10年間に約9000万トンから1億3000万トン以上に増加するであろうと推論された。植物による窒素の使用効率を上げれば、より低い肥料投入量でまたは比較的やせた土地で作物を栽培することが可能となり、それゆえ、開発されたまたは開発されつつある農業システムに相当の経済的効果を及ぼすことができるだろう。
【0003】
通常の選別技術を用いて、植物の育種家らは、トウモロコシ、コムギ、イネ、その他の作物種の天然集団から得られる変種を利用することによって窒素使用効率を改良しようと試みた。しかしながら、野外条件下で評価することが困難な諸特性に関する従来の育種計画には、広範な集団のスクリーニングに伴う困難が立ちはだかり、こうした選別法はほとんど成功しなかった。
【0004】
2.1. 植物の窒素同化経路
植物は窒素をその環境から無機化合物の形で、すなわち根から吸収される硝酸とアンモニア、そして窒素固定根粒でアンモニアに還元される大気中のN2の形で得ている。若干の硝酸とアンモニアが輸送道管(木部および篩部)において検出され得るが、大部分の窒素はまず有機体(例:アミノ酸)に同化されてから、植物体内に輸送される。
【0005】
無機窒素を有機体に同化する第一段階は、主にグルタミンを形成するためのグルタミン酸によるアンモニアの取り込みを含み、これはグルタミンシンテターゼ酵素 (GS;EC 6.3.1.2)により触媒される。次いで、斯く形成されたグルタミンは、アスパラギンシンテターゼ酵素 (AS;E.C. 6.3.5.4)により触媒されるアスパラギンの形成においてそのアミド基を供与する。この経路でのアンモニアからアスパラギンへの窒素の定常流はグルタミン酸とα−ケトグルタル酸とアスパラギン酸の循環に依存しており、それぞれグルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ(GOGAT;E.C.)とアスパラギン酸アミノトランスフェラーゼ(AspAT;E.C.)により触媒される(図1参照)。かくして、GS、AS、AspATおよびGOGATは高等植物の主要な窒素同化経路の不可欠な酵素といえる。
【0006】
アンモニアの取り込みがGSにより触媒される経路以外の代替経路により進行しうることを示す証拠が存在する(図1)。Knight and Langston-Unkefer, 1988, Science 241:951-954(非特許文献1)を参照されたい。一つの経路はグルタミン酸を形成するためのα−ケトグルタル酸によるアンモニアの取り込みを含み、これはグルタミン酸デヒドロゲナーゼ(GDH)により触媒される。もう一つの経路はアスパラギンを形成するためのアスパラギン酸によるアンモニアの取り込みを含み、これはアスパラギンシンテターゼにより触媒される (Oaks and Ross, 1984, Can. J. Bot. 62:68-73(非特許文献2); Stulen and Oaks, 1977, Plant Physiol. 60:680-683(非特許文献3)) 。これらの酵素(GDHおよびAS)は両方ともアンモニアに対して高いKmを有し、正常な生育条件下(例えば、低濃度の内部アンモニア)でのこの代替窒素同化経路の役割は依然として不明のままである。ある研究により、これらまたは他の代替窒素同化経路は細胞内のアンモニウム濃度が正常レベルを越えて上昇したときの植物の窒素同化能に対して大いに寄与していることが示唆された (Knight and Langston-Unkefer,前掲)。
【0007】
2.2. 窒素の輸送および利用
グルタミンおよびアスパラギンは主要な長距離窒素輸送化合物を代表するもので、篩部の液汁中に豊富に存在している。窒素担体としてのそれらの共通した役割のほかに、これら2つのアミノ酸は植物窒素代謝においてやや異なる役割を担っている。グルタミンは2つのアミノ酸のうちで代謝的により活性で、さまざまな同化反応で多数の基質にそのアミド窒素を直接供与することができる。その反応性ゆえに、一般的に植物は窒素の貯蔵のためにグルタミンを利用することはない。
【0008】
対照的に、アスパラギンはその高いN:C比のためにグルタミンと比べてより効率のよい窒素輸送用の化合物となる。さらに、アスパラギンはグルタミンよりも安定しており、液胞中に高レベルで蓄積され得る。実際、窒素同化能の高い植物では、アスパラギンが窒素の輸送および代謝において支配的な役割を果たしているようである。The Biochemistry of Plants: A Comprehensive Treatise, vol 5, Amino acid and derivatives, Miflin 編, Academic Press, New York (1980) pp. 569-607(非特許文献4) 中の Lea and Miflin による「植物におけるアスパラギンと他の窒素化合物の輸送および代謝」;および Sieciechowiczら, 1988, Phytochemistry 27:663-671(非特許文献5) を参照されたい。その相対的安定性のために、アスパラギンは窒素代謝に直接関与することはないが、アスパラギナーゼ酵素(ANS;E.C. 3.5.1.1)によってまず加水分解されてアスパラギン酸とアンモニアを生成し、次いでこれらはアミノ酸とタンパク質の合成に利用されるにちがいない(図1参照)。
【0009】
2.3. 窒素の同化および利用に関与する植物遺伝子
植物の窒素同化/利用に係わる酵素をコードしている遺伝子の多くがクローン化され、研究されてきた。植物のグルタミンシンテターゼ (GS)遺伝子およびアスパラギンシンテターゼ(AS)遺伝子の論議については、Transgenic Plants, Vol. 1, Kung and Wu編, Academic Press, San Diego, CA, (1993) pp. 181-194(非特許文献6) 中の Tsai and Coruzzi による「アミノ酸生合成酵素をコードする遺伝子を研究するためのトランスジェニック植物」ならびにその中で引用された文献;アルファルファのアスパラギン酸アミノトランスフェラーゼ遺伝子の論議については、Udvardi and Kahn, 1991, Mol. Gen. Genet. 231:97-105(非特許文献7) ;タバコのグルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ(GOGAT、グルタミン酸シンテターゼとも言う)遺伝子の論議については、Zehnacker ら, 1992, Planta 187:266-274(非特許文献8);ハウチワマメ (lupin)のアスパラギナーゼ遺伝子の論議については、Lough ら, 1992, Plant Mol. Biol. 19:391-399(非特許文献9) および Dicksonら, 1992, Plant Mol. Biol. 20:333-336(非特許文献10) を参照されたい。
【0010】
植物の窒素同化/利用遺伝子の中で、最も広く研究されているものはグルタミンシンテターゼ遺伝子とアスパラギンシンテターゼ遺伝子である。GSとASには複数の遺伝子が存在し、これら遺伝子の分子特性付けにより、それらが異なる発現パターンを有することが示された。
【0011】
2.3.1. グルタミンシンテターゼ遺伝子
GSは植物の生育中に多くの器官で活発に機能している (McNally ら, 1983, Plant Physiol. 72:22-25(非特許文献11))。根において、それは土の中の水から得られたアンモニアを同化し (Oaks and Hirel, 1985, Ann. Rev. Plant Physiol. 36:345-365(非特許文献12)、また、マメ科植物の根粒ではGSが根粒菌により固定されたアンモニアを同化する (Cullimore ら, 1983, Planta 157:245-253(非特許文献13)) 。子葉にあっては、GSが発芽中に集められた窒素貯蔵物を再同化し (Recent Advances in Phythochemistry: Mobilization of Reserves in Germination, Nozolilloら編, Plenum Press, (1983) p. 77-109(非特許文献14)中の Lea and Joyによる「発芽種子におけるアミノ酸相互変換」)、そして葉では葉緑体GS2が光呼吸で放出されたアンモニアを同化する (Givan ら, 1988, TIBS 13:433-437(非特許文献15))。GSの多種多様な役割は、差次的に発現される異なる遺伝子から誘導されたさまざまのGSイソ型が担っている (Gebhardtら, 1986, EMBO J. 5:1429-1435(非特許文献16); Tingey ら, 1987, EMBO J. 6:1-9(非特許文献17))。
【0012】
エンドウのPhaseolusおよび Arabidopsisでは、葉緑体GS2が単一の核遺伝子によってコードされているが、これらの種のそれぞれには細胞質ゾルGSの複数の遺伝子が存在している (Bennett ら, 1989, Plant Mol. Biol. 12:553-565(非特許文献18); Tingey ら, 1988, J. Biol. Chem. 263:9651-9657(非特許文献19); Peterman and Goodman, 1991, Mol. Gen. Genet. 230:145-154(非特許文献20)) 。in vivo およびトランスジェニック宿主植物におけるこれらGS遺伝子の発現の分析は、植物の窒素代謝での各種GSイソ型の役割を解明する上で役に立った。
【0013】
エンドウのGS遺伝子ファミリーは4つの別個のしかし相同の核遺伝子を含んでいる。3つは細胞質ゾルのGSイソ型をコードし、1つは葉緑体のGS2イソ型をコードする (Tingeyら, 1987, EMBO J. 6:1-9(非特許文献21); Tingey ら, 1988, J. Biol. Chem. 263:9651-9657(非特許文献22)) 。ノーザンブロット分析により、葉緑体GS2の遺伝子は、一部にはフィトクロムにより、また、一部には光呼吸作用により光依存的に葉で発現されることが実証された (Edwards and Coruzzi, 1989, Plant Cell 1:241-248(非特許文献23))。また、細胞質ゾルGSの3つの遺伝子(GS1、GS3AおよびGS3B)も異なる役割を果たしているようである。根では、細胞質ゾルGS1が主要なイソ型であるが、それは結節でも発現される。細胞質ゾルGS3AおよびGS3Bは結節で高度に発現され、さらに発芽中の種子の子葉でも発現される (Tingeyら, 1987, EMBO J. 6:1-9(非特許文献24); Walker and Coruzzi, 1989, Plant Physiol. 91:702-708(非特許文献25))。GS3A遺伝子とGS3B遺伝子は配列が同一に近いものの、遺伝子特異的S1−ヌクレアーゼ分析により、GS3Aの発現がGS3Bの発現よりも常に高いことが明らかになった (Walker and Coruzzi, 1989, Plant Physiol. 91:702-708(非特許文献25))。プロモーター−GUS融合体およびトランスジェニック植物の分析により、葉緑体GS2は光合成細胞型でのみ発現され、また、細胞質ゾルGS3Aは大部分の器官の脈管構造の篩部細胞でもっぱら発現されることが分かった。さらに、GS3Aは根や結節の分裂組織でも強く発現される (Edwards ら, 1990, Proc. Natl. Acad. Sci. USA, 87:3459-3463(非特許文献26); Brearsら, 1991, The Plant Journal, vol. 1, pp. 235-244(非特許文献27)) 。細胞型および器官レベルでの厳密に制御された調節からすると、GSの各種遺伝子はアンモニア同化において重複することのない別個の役割を果たしていると考えられる。
【0014】
2.3.2. アスパラギンシンテターゼ遺伝子
2つのAS遺伝子がエンドウからクローン化され(AS1およびAS2)、両方とも根粒と子葉で最高レベルに発現される。AS1およびAS2はともに根において発現される。AS2は構成的に根で発現され、一方、AS1は暗黒で育てた植物の根にのみ発現される (Tsai and Coruzzi, 1990, EMBO J 9:323-332(非特許文献28)) 。さらに、AS1とAS2は暗順応植物の成熟葉で発現されるが、その発現は光によって阻害される。暗黒でのこの高レベルのAS遺伝子発現は、光合成炭素の利用能が低下した条件下で合成される長距離窒素輸送化合物としてのアスパラギンの使用と合致している(アスパラギンはグルタミンより高いN:C比を有する)。トランスジェニック植物におけるAS1プロモーター−GUS融合体の研究から、AS1遺伝子もGS3A遺伝子と同様に篩部細胞でもっぱら発現されることが示された。細胞型および器官レベルでの厳密に制御された調節からすると、各種AS遺伝子も植物窒素代謝において異なる役割を果たしていると思われる。
【0015】
2.4. 植物の窒素同化/利用過程の遺伝子工学的操作
植物では、窒素同化過程の遺伝子工学的操作がさまざまな結果をもたらした。一つの事例を挙げると、タバコにおいて原核生物のアンモニウム依存性アスパラギンシンテターゼ(ASN−A)遺伝子を発現させることが、種々のグルタミンシンテターゼ(GS)阻害剤に対する耐性を付与した (Duditsら, Transgenic Plants expressing a prokaryotic ammonium dependent asparagine synthetase, WO 9111524, Aug. 8, 1991(特許文献1))。また、これらの同じ植物が成長速度の増加、植物発育の促進、早期開花、植物の新鮮重量および乾燥重量の増加を含めて多くの成長変化を示した。GS阻害剤処理は遺伝子工学的に作出した植物の成長を低下させるどころかむしろ高めたので、ASN−A発現の成長効果は理屈に合わないことになる。
【0016】
対照的に、グルタミンシンテターゼ(GS)の過剰発現を試験する多くの研究は、その過剰発現が植物の成長に及ぼすポジティブな効果を報告することができなかった。Lea and Forde, 1994, Plant Molec. Biol. 17:541-558(非特許文献29); Eckes ら,1989, Molec. Gen. Genet. 217:263-268(非特許文献30) (アルファルファGSを過剰発現するトランスジェニックタバコ植物); Hemonら, 1990, Plant Mol. Biol. 15:895-904(非特許文献31)(細胞質またはミトコンドリアでマメGSを過剰発現するトランスジェニックタバコ植物); Hirelら, 1992, Plant Mol. Biol. 20:207-218(非特許文献32) (タバコ植物内でダイズGSを過剰発現するトランスジェニックタバコ植物)を参照されたい。ある研究が、アルファルファGS1遺伝子を過剰発現するトランスジェニックタバコ植物における総可溶性タンパク質含量の増加を認めたと報告した。しかしながら、この同じ研究がGS1遺伝子に対するアンチセンスRNAを発現するトランスジェニックタバコ植物における総可溶性タンパク質含量の同様の増加をも報告したことから、GS1発現と可溶性タンパク質の増加との関係ははっきりしていないようである (Templeら, 1993, Mol. Gen. Genet. 236:315-325(非特許文献33)) 。植物におけるGS過剰発現の明確に確立された効果は、GS阻害性除草剤であるホスフィノトリシンに対する耐性である (Eckes ら, 前掲; Donnら, 1984, J. Molec. Appl. Genet. 2:621-635(非特許文献34) (ホスフィノトリシン耐性アルファルファ細胞系はGS遺伝子の増幅を含んでいた))。また、アルファルファGS遺伝子を過剰発現するように遺伝子操作された植物は遺伝子操作されていない植物よりも速く成長するという報告があった (Eckes ら, 1988, オーストラリア特許庁文書番号:AU-A-17321/88(特許文献2))。しかし、報じられたような速い成長は低窒素生育条件下でのみ起こり、普通または高窒素生育条件下では認められなかった。その上、より速い成長がより多い生物量または生産量でもって成熟植物を生み出すのかどうか不明である。Eckes らの前掲を Eckesら, 1989, Molec. Gen. Genet. 217:263-268(非特許文献35)と比較されたい。
【先行技術文献】
【特許文献】
【0017】
【特許文献1】WO 9111524
【特許文献2】AU-A-17321/88
【非特許文献】
【0018】
【非特許文献1】Knight and Langston-Unkefer, 1988, Science 241:951-954
【非特許文献2】Oaks and Ross, 1984, Can. J. Bot. 62:68-73
【非特許文献3】Stulen and Oaks, 1977, Plant Physiol. 60:680-683
【非特許文献4】The Biochemistry of Plants: A Comprehensive Treatise, vol 5, Amino acid and derivatives, Miflin 編, Academic Press, New York (1980) pp. 569-607
【非特許文献5】Sieciechowiczら, 1988, Phytochemistry 27:663-671
【非特許文献6】Transgenic Plants, Vol. 1, Kung and Wu編, Academic Press, San Diego, CA, (1993) pp. 181-194
【非特許文献7】Udvardi and Kahn, 1991, Mol. Gen. Genet. 231:97-105
【非特許文献8】Zehnacker ら, 1992, Planta 187:266-274
【非特許文献9】Lough ら, 1992, Plant Mol. Biol. 19:391-399
【非特許文献10】Dicksonら, 1992, Plant Mol. Biol. 20:333-336
【非特許文献11】McNally ら, 1983, Plant Physiol. 72:22-25
【非特許文献12】Oaks and Hirel, 1985, Ann. Rev. Plant Physiol. 36:345-365
【非特許文献13】Cullimore ら, 1983, Planta 157:245-253
【非特許文献14】Recent Advances in Phythochemistry: Mobilization of Reserves in Germination, Nozolilloら編, Plenum Press, (1983) p. 77-109
【非特許文献15】Givan ら, 1988, TIBS 13:433-437
【非特許文献16】Gebhardtら, 1986, EMBO J. 5:1429-1435
【非特許文献17】Tingey ら, 1987, EMBO J. 6:1-9
【非特許文献18】Bennett ら, 1989, Plant Mol. Biol. 12:553-565
【非特許文献19】Tingey ら, 1988, J. Biol. Chem. 263:9651-9657
【非特許文献20】Peterman and Goodman, 1991, Mol. Gen. Genet. 230:145-154
【非特許文献21】Tingeyら, 1987, EMBO J. 6:1-9
【非特許文献22】Tingey ら, 1988, J. Biol. Chem. 263:9651-9657
【非特許文献23】Edwards and Coruzzi, 1989, Plant Cell 1:241-248
【非特許文献24】Tingeyら, 1987, EMBO J. 6:1-9
【非特許文献25】Walker and Coruzzi, 1989, Plant Physiol. 91:702-708
【非特許文献26】Edwards ら, 1990, Proc. Natl. Acad. Sci. USA, 87:3459-3463
【非特許文献27】Brearsら, 1991, The Plant Journal, vol. 1, pp. 235-244
【非特許文献28】Tsai and Coruzzi, 1990, EMBO J 9:323-332
【非特許文献29】Lea and Forde, 1994, Plant Molec. Biol. 17:541-558
【非特許文献30】Eckes ら,1989, Molec. Gen. Genet. 217:263-268
【非特許文献31】Hemonら, 1990, Plant Mol. Biol. 15:895-904
【非特許文献32】Hirelら, 1992, Plant Mol. Biol. 20:207-218
【非特許文献33】Templeら, 1993, Mol. Gen. Genet. 236:315-325
【非特許文献34】Donnら, 1984, J. Molec. Appl. Genet. 2:621-635
【非特許文献35】Eckesら, 1989, Molec. Gen. Genet. 217:263-268
【発明の概要】
【0019】
3.発明の概要
本発明は、窒素の同化および利用に係わる主要な酵素(これら酵素のそれぞれの役割は図1に示される)の発現レベルおよび/または細胞特異的な発現パターンを変化させたトランスジェニック植物の作出に関するものであり、かくして、得られる植物は増強された窒素同化および/または利用能ならびに改良された農業特性を有するものである。本発明は、特に、グルタミンシンテターゼ、アスパラギンシンテターゼ、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ(グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼはグルタミン酸シンテターゼとしても知られている)、アスパラギン酸アミノトランスフェラーゼ、グルタミン酸デヒドロゲナーゼおよびアスパラギナーゼの発現を変えることに関するものである(図1参照)。
【0020】
本発明は作物の重要な農業特性を改良することに用いられる。改良点の一つは、より低い窒素肥料投入量でまたは窒素の乏しい土壌で生産的に栽培される遺伝子操作植物の能力であるだろう。更なる改良点としては、より活発な(すなわち、より速い)成長ならびに普通の栽培条件(すなわち、非限定栄養条件)下でのより多い栄養体および/または生殖体の収穫量が含まれる。これらの改良点を達成するには、伝統的な作物育種法は大きな分離集団のスクリーニングを必要とするだろう。本発明は、大半の植物ではないにしても、その多くが所望の特性を備えている植物を作出することによって大規模スクリーニングの必要性を回避するものである。
【0021】
本発明によると、所望の植物改良を達成するには、いくつかの場合に、窒素同化または利用酵素をコードする1個または複数個の遺伝子の異所性過剰発現が必要となる。かかる発現の改変は次の遺伝子:すなわち、a)酵素のコーディング配列が強力な構成プロモーターに機能しうる状態で結合されているトランスジーン;b)所望の酵素をコードする、追加のコピー数の天然遺伝子;c)窒素同化または利用のための目的遺伝子(類)の発現を活性化する調節遺伝子(類);d)発現を高めるように修飾されたその調節領域を有する、1コピーの天然遺伝子;およびe)変異型、改変型またはキメラ型の窒素同化または利用酵素を発現するトランスジーン;のいずれか1個または数個を用いて植物を遺伝子操作することを包含する。
【0022】
他の場合には、所望の植物改良を達成するのに、窒素同化または利用酵素の発現パターンを変えることが必要となる。かかる改変発現パターンは次の遺伝子:すなわち、a)酵素のコーディング配列が所望の発現パターンを有するプロモーター(該プロモーターは組織−または発生−特異的発現パターンをもつと思われるものを含む)に機能しうる状態で結合されているトランスジーン;b)好ましいパターンでの酵素コード化遺伝子の発現を活性化する修飾調節遺伝子;c)好ましいパターンで発現するように修飾されたその調節領域を有する、天然コピー数の酵素コード化遺伝子;のいずれか1個または数個を用いて植物を遺伝子操作することを包含する。
【0023】
さらに他の場合には、所望の植物改良を達成するために、窒素同化または利用酵素の発現レベルおよび/またはパターンを抑制することが必要となる。かかる発現の抑制はアンチセンスRNA、リボザイム、共抑制構築物または「優性ネガティブ (dominant negative)」変異体をコードする遺伝子を用いて植物を遺伝子操作することを包含する(優性ネガティブ変異体による遺伝子抑制の作用機序の説明については、Herskowitz, 1967, Nature 329:219-222を参照されたい)。さらに、遺伝子の抑制は、天然遺伝子を1コピーの欠陥遺伝子または所望の発現レベルおよび/またはパターンを有するプロモーターの制御下にある酵素コード化配列と置き換えた相同的組換え構築物を用いて植物を遺伝子操作することによっても達成される。
【0024】
さらに他の場合には、所望の植物改良を達成するために、窒素同化または利用経路において改変された酵素または異なる型の酵素を発現させることが必要となる。かかる目標達成の作業は、宿主植物の窒素同化または利用酵素の触媒作用とは異なる触媒作用を有する対応酵素をコードする植物発現可能遺伝子を開発し、この遺伝子構築物で植物を遺伝子操作することを包含する。この種の酵素をコードする遺伝子配列は細菌、酵母、藻類、動物および植物を含むがこれらに限らないさまざまな供給源から得ることができる。ある場合には、かかるコーディング配列が該配列を所望の植物活性プロモーターと機能的に連結させることによる植物発現可能遺伝子融合体の構築に直接用いられる。また、他の場合には、遺伝子融合体での該コーディング配列の利用は、宿主植物におけるそれらの翻訳能を高めるために、あるいはそこにコード化された酵素の触媒作用を変えるために、in vitro突然変異誘発またはde novo 合成による先行修飾を必要とする。有用な改変には基質結合および/または触媒作用に関与する残基の修飾が含まれるが、これらに限らない。好ましい改変にはハイブリッド酵素の構築も含まれる。例えば、同一のまたは異なる生物から得られた関連酵素の異なるドメインを組み合わせて新規な性質を有する酵素を作ることができる。
【0025】
あらゆる場合に、望まれる改良点を有する植物は、窒素同化または利用酵素の改変された発現パターンまたはレベル、対応するmRNAまたはタンパク質の改変された発現パターンまたはレベル、改変された窒素同化または利用能、増加した成長速度、増加した栄養体収穫高、または改良された生殖体収穫高(例えば、より多いまたはより大きい種子または果実)について遺伝子操作植物をスクリーニングすることによって単離することができる。遺伝子操作植物のスクリーニングは、酵素/タンパク質レベルを測定するためのエンザイムアッセイおよびイムノアッセイ;mRNAレベルを測定するためのノーザン分析、RNアーゼ保護、プライマー伸長、逆転写/PCRなど;各種植物組織のアミノ酸組成、遊離アミノ酸プールまたは総窒素含量の測定;経時的新鮮重量増加に基づく成長速度の測定;または総乾燥重量および/または総種子重量に基づく植物収穫高の測定を包含する。
【0026】
本発明は、一部には、植物の窒素同化または利用酵素の発現を高めることが、成長特性の向上または栄養体や生殖体の収穫高の増加をもたらしたという驚くべき発見に基づいている。本発明は、ここでは、エンドウ・グルタミンシンターゼ(GS)遺伝子またはエンドウ・アスパラギンシンターゼ(AS)遺伝子をコードする配列に機能的に連結された強力な構成植物プロモーターのカリフラワーモザイクウイルス(CaMV)35Sプロモーターをコードする組換え構築物を用いてタバコ植物を遺伝子工学的に操作した実施例により説明される。RNAおよびタンパク質の分析により、大多数の遺伝子操作植物がGSまたはASの異所性過剰発現を示すことがわかった。GSまたはASを過剰発現する系統は、対照の野生型植物よりも高い窒素含量、活発な成長特性、増加した栄養体収穫高または良好な種子収穫高および品質を示した。
【0027】
3.1. 定義
本明細書に使用される、以下にリストされる用語は、示される意味を有する。
35S =35S転写のためのカリフラワーモザイクウイルスプロモーター
AS =アスパラギンシンテターゼ
AspAT =アスパラギン酸アミノトランスフェラーゼ(またAAT として知られている)
CaMV =カリフラワーモザイクウイルス
cDNA =相補DNA
DNA =デオキシリボ核酸
GDH =グルタミン酸デヒドロゲナーゼ
遺伝子融合体=異種遺伝子に操作により連結されたプロモーターを含む遺伝子構築(前記プロモーターは異種遺伝子の転写を調節する)
GOGAT =グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ
(また、グルタミン酸シンテターゼとして知られている)
Fd-GOGAT =フェレドキシン依存性グルタミン酸シンターゼ
NADH-GOGAT=NADH依存性グルタミン酸シンターゼ
GS =グルタミンシンテターゼ
異種遺伝子=遺伝子構築の状況において、異種遺伝子は、その遺伝子が前記遺伝子が自然に連結されていないプロモーターに連結されることを意味する。異種遺伝子は前記プロモーターを寄与する生物からのものであってもよく、またそうでなくてもよい。異種遺伝子はメッセンジャーRNA(mRNA)、アンチセンスRNAまたはリボザイムをコードし得る。
【0028】
窒素非制限生育条件=窒素非制限生育条件は、土壌または培地が健全な植物成長を持続するのに充分な量の窒素栄養を含み、または受容する条件である。窒素非制限生育条件の例は第5.2.3.節に示される。更に、当業者は、重要な穀物および観賞植物の殆どの種および変種につきこのような土壌、培地及び肥料を構成するものを認識するであろう。
PCR =ポリメラーゼ連鎖反応
前駆植物 =形質転換されていない、野生型植物
RNA =リボ核酸
【図面の簡単な説明】
【0029】
【図1】植物における窒素同化/代謝の経路。窒素同化の主要な経路はグルタミンシンテターゼ(GS)およびグルタミン酸シンターゼ(GOGAT) によるものである。グルタミン酸デヒドロゲナーゼ(GDH) は生合成の役割においてアンモニア毒性の条件下で機能するものと考えられ、または触媒量のグルタミン酸を与えてGS/GOGATサイクルを支持し得る。GDH はおそらくグルタミン酸からアンモニアを放出するその触媒的役割において更に活性である(例えば、発芽中)。アスパラギン酸アミノトランスフェラーゼ(AspAT) は可逆反応を触媒作用する。アスパラギンシンテターゼ(AS)は二つの活性;グルタミン依存性活性およびアンモニア依存性活性を有する。アスパラギン代謝はアスパラギナーゼ(ANS) により起こってアスパラギン酸およびアンモニアを放出する。
【図2】キメラFd/NADH GOGAT 酵素の操作。植物フェレドキシン-GOGAT(Fd-GOGAT)の大きいサブユニットはFd結合ドメイン(斜めのクロスバー)を含む。植物およびE.coli NADH-GOGAT:大きいサブユニット(オープンバー)、小さいサブユニットはNADH結合ドメイン(垂直ハッチ)を含む。キメラFd/NADH GOGAT はFd-GOGATの大きいサブユニット(Fd 結合ドメイン)+植物またはE.coliのNADH-GOGATの小さいサブユニットを含むように操作される。その操作は植物Fd-GOGATをコードする配列およびNADH結合ドメインを含む、植物またはE.coli NADH-GOGAT の小さいサブユニットをコードする配列のイン−フレーム転写融合をつくることにより行われる。そのキメラタンパク質は、還元剤としてFdまたはNADHを使用し得る二特異性または二機能性のGOGAT 酵素をコードする。
【図3】2成分植物発現ベクターの地図。2成分発現ベクターpTEV4 、pTEV5 、pTEV8 およびpTEV9 はトランスジェニックタバコ中のcDNAの高レベルの発現のために構築されたpBIN19(Bevan, 1984, Nucleic Acids Res. 12:8711-8721)の誘導体である。構築の詳細につき、第6.1.1.節を参照のこと。
【図4】トランスジェニックタバコに移入されたキメラ35S CaMV-GS cDNA構築物。エンドウ豆GScDNAをストラスボルグ株CaMV 35Sプロモーター(35S) の後の発現のためにpTEV発現ベクター(図3、および第6.1.1.節を参照のこと)にクローン化した。GS3AおよびGS2 につき、イントロンをゲノム配列からcDNAにとり込む“修飾" クローンを構築した(第6.1.2.節を参照のこと)。GScDNAクローンの起源は、GS2(また(aka)GS185として知られている); GS1(aka GS299); GS3A (aka GS341)(Tingeyら, 1988, J.Biol.Chem. 263:9651-9657; Tingeyら,1987, EBBO J. 6:1-9)であった。
【図5】GSトランスジーンを含む一次(T1)形質転換体中のGSタンパク質の分析。上のパネル:一次形質転換体中のGSポリペプチドのウェスタン分析。レーン1および2:細胞質ゾルGS3A遺伝子を有する一次形質転換体Z17-6 およびZ17-12はそれぞれ過剰発現および同時抑制表現型を示す。レーン3−6:葉緑体GS2 遺伝子を有する一次形質転換体Z41-20、Z54-2 、Z54-7 、およびZ54-8 は葉緑体GS2(GSを比較のこと)につき全て同時抑制される。対照は、TL- タバコの葉、PL- エンドウ豆の葉、およびPR- エンドウ豆の根。全GS活性がウェスタンパネルの下に示される(対照=(100%)に対する%として)。下のパネル:サンプルのほぼ等しい負荷を実証するルビスコの大きいサブユニットタンパク質のクーマシー染色。ctGS- 葉緑体GS2(約45 kD); cyGS- 細胞質ゾルGS( 約38 kD)。
【図6A】エンドウ豆トランスジーンを含むT2子孫トランスジェニック植物からのGSタンパク質、RNAおよびホロ酵素の分析。典型的に分析されたそれぞれの一次形質転換体からの4種のT2植物のうちの単一の代表的な植物がこの図に含まれた。Z17-9 の場合、T2子孫は二つの異なるプロフィールを示し、その両方が示される(Z17-9A およびZ17-9B) 。対照:TL/T- タバコの葉、P- エンドウ豆の葉。パネルA(上):トランスジェニック植物中のGSポリペプチドのウェスタン分析。パネルA(下):サンプルのほぼ等しい負荷を示すためのルビスコの大きいサブユニットタンパク質のクーマシー染色。
【図6B】エンドウ豆トランスジーンを含むT2子孫トランスジェニック植物からのGSタンパク質、RNAおよびホロ酵素の分析。典型的に分析されたそれぞれの一次形質転換体からの4種のT2植物のうちの単一の代表的な植物がこの図に含まれた。Z17-9 の場合、T2子孫は二つの異なるプロフィールを示し、その両方が示される(Z17-9A およびZ17-9B) 。対照:TL/T- タバコの葉、P- エンドウ豆の葉。パネルB(上):GS1(左)、GS3A(中央)およびGS2(右)の推定cDNAプローブとハイブリッドを形成されたノーザンブロット。パネルB(下):エンドウ豆rRNA遺伝子プローブによる対照ハイブリダイゼーション。
【図6C】エンドウ豆トランスジーンを含むT2子孫トランスジェニック植物からのGSタンパク質、RNAおよびホロ酵素の分析。典型的に分析されたそれぞれの一次形質転換体からの4種のT2植物のうちの単一の代表的な植物がこの図に含まれた。Z17-9 の場合、T2子孫は二つの異なるプロフィールを示し、その両方が示される(Z17-9A およびZ17-9B) 。対照:TL/T- タバコの葉、P- エンドウ豆の葉。パネルC:トランスジェニック植物中のGSホロ酵素A* 、B、およびCを示す非変性ゲルおよびGS活性分析。GS活性は対照と比較して%として表される(対照=100 %の活性)。
【図7A】GSホロ酵素の活性ゲル分析。植物を過剰発現するGS1 およびGS3Aのホロ酵素の移動に対する葉緑体および細胞質ゾル濃縮GSタンパク質サンプルの移動を実証する、エンドウ豆葉緑体(PC)、エンドウ豆の根(PR)、タバコ葉緑体(TC)およびタバコの根(TR)からのタンパク質抽出物。レーン1:エンドウ豆葉緑体タンパク質(PC)はGSホロ酵素Bのみを有する;レーン2:エンドウ豆の根タンパク質(PR)はGSホロ酵素Cのみを有する;レーン3:タバコ葉緑体タンパク質(TC)はGSホロ酵素Bのみを有する;レーン4:タバコの根タンパク質はGSホロ酵素Cのみを有する。レーン5:植物Z17-7(35S-GS3A構築物を有する)からのタンパク質はGSホロ酵素A* およびBを有する;レーン5:植物Z3-1(35S-GS1構築物を有する)からのタンパク質はGSホロ酵素BおよびCを有する。
【図7B】GSホロ酵素A* 、B、およびCから単離されたGSタンパク質のウェスタン分析。GS3AおよびGS1 を過剰発現するトランスジェニックタバコ中で観察されたホロ酵素A* およびCを非変性ゲルから切除し、タンパク質単離緩衝液中で再度抽出し、そしてGS抗体を使用してウェスタン分析のための変性条件下で電気泳動にかけた。レーン1:対照としてのタバコの葉のタンパク質;レーン2:Z17-7 からのGSホロ酵素A* ; レーン3:対照としての単離された葉緑体GS2(ホロ酵素B);レーン4:Z3-1からのGSホロ酵素C。
【図8】細胞質ゾルGS1 またはGS3Aを異所性発現する成長分析に選択されたトランスジェニック植物中のGSタンパク質およびRNAのウェスタンおよびノーザン分析。上のパネル:GSタンパク質に関するウェスタンブロット。下のパネル:GS mRNAに関するノーザンブロット。P1およびT1はエンドウ豆およびタバコの葉の対照である。レーン1および2、並びに5および6はGS1 を過剰発現する植物であり、そしてレーン3および4、並びに7および8はGS3Aを過剰発現する植物である。破線の左のトランスジェニック植物を生育実験Aにおいて分析し、そして右のトランスジェニック植物を生育実験Bにおいて分析した。相当するプローブをノーザンブロットにおいて使用した。左のエンドウ豆対照をGS1 にハイブリッドを形成し、右手のエンドウ豆対照をGS3Aにハイブリッドを形成した。
【図9A】細胞質ゾルGS1 (Z3)または細胞質ゾルGS3A (Z17)を過剰発現するトランスジェニック系統の新鮮重量の増加。パネルA:トランスジェニック系統Z3-1、Z3- 2 、Z17-6 、Z17-7 、および形質転換されなかった対照(C) を用いる実験Aの結果。これは表2に示され、そして表3に統計的に分析されたデータのグラフ表示である。
【図9B】細胞質ゾルGS1 (Z3)または細胞質ゾルGS3A (Z17)を過剰発現するトランスジェニック系統の新鮮重量の増加。パネルB:トランスジェニック系統Z3-3、Z3-4、Z17-3 、Z17-11、および2種の形質転換されなかった対照(C1 およびC2) を用いる実験Bの結果。これは表2に示され、そして表3に統計的に分析されたデータのグラフ表示である。
【図10A】変化されたGS発現パターンを有する植物の定性的成長パターン。それぞれのパネル中の植物を同時に種をまき、そして約3週間にわたって土壌中で生育した。対照パネル:SR1 未形質転換タバコ(100%のGS活性)。
【図10B】変化されたGS発現パターンを有する植物の定性的成長パターン。それぞれのパネル中の植物を同時に種をまき、そして約3週間にわたって土壌中で生育した。Z3-A1 パネル:過剰発現GS1 を有するトランスジェニック植物(123%のGS活性)。
【図10C】変化されたGS発現パターンを有する植物の定性的成長パターン。それぞれのパネル中の植物を同時に種をまき、そして約3週間にわたって土壌中で生育した。Z54-A2パネル:GS2 につき同時抑制されたトランスジェニック植物(28 %のGS活性)。
【図10D】変化されたGS発現パターンを有する植物の定性的成長パターン。それぞれのパネル中の植物を同時に種をまき、そして約3週間にわたって土壌中で生育した。Z17-B7パネル:GS3 を過剰発現するトランスジェニック植物(107%のGS活性)。
【図11A】GS活性と植物新鮮重量または全葉タンパク質の線形関係。トランスジーンと関連するKan R 表現型の分離を示さなかった一次形質転換体のT2子孫を成長分析につき選択した。Kan R T2植物をMSK 培地 (R.B.Horschら,Science 227:1229 (1985))で選択し、18日で砂に移した。植物を下灌漑し(subirrigated)、2日毎に10mMのKNO3を含む1Xホアグランド液(D.R.Hoagland ら, Circ. Calif.Agric.Exp.Stn. 347:461 (1938))50mlで表面で栄養補給した。それぞれの系統につき、8種のT2子孫を合計植物新鮮重量(グラム)、トランスフェラーゼアッセイ(B.M.Shapiroら, Methods Enzymol. 17A:910 (1970))により測定されるような合計の葉GSの比活性およびタンパク質/新鮮重量(グラム)につき個々に分析した。分析した植物は、対照、SR1 未形質転換タバコ;GS2 により同時抑制されたZ54-4; GS3A を過剰発現するZ17-7; GS1を過剰発現するZ3-1であった。図11A;植物新鮮重量vs.GS 活性。
【図11B】GS活性と植物新鮮重量または全葉タンパク質の線形関係。トランスジーンと関連するKan R 表現型の分離を示さなかった一次形質転換体のT2子孫を成長分析につき選択した。Kan R T2植物をMSK 培地 (R.B.Horschら,Science 227:1229 (1985))で選択し、18日で砂に移した。植物を下灌漑し(subirrigated)、2日毎に10mMのKNO3を含む1Xホアグランド液(D.R.Hoagland ら, Circ. Calif.Agric.Exp.Stn. 347:461 (1938))50mlで表面で栄養補給した。それぞれの系統につき、8種のT2子孫を合計植物新鮮重量(グラム)、トランスフェラーゼアッセイ(B.M.Shapiroら, Methods Enzymol. 17A:910 (1970))により測定されるような合計の葉GSの比活性およびタンパク質/新鮮重量(グラム)につき個々に分析した。分析した植物は、対照、SR1 未形質転換タバコ;GS2 により同時抑制されたZ54-4; GS3A を過剰発現するZ17-7; GS1を過剰発現するZ3-1であった。図11B; タンパク質/新鮮重量(グラム)vs.GS活性。
【図12】トランスジェニックタバコに移入されたキメラ35 S CaMV-AS構築物。AS1 遺伝子のcDNAおよびgln ΔAS1 遺伝子を、2成分発現ベクターpTEV5 を使用してタバコへの移入のために35S プロモーターおよびノパリンシンターゼ転写ターミネーターに融合した。
【図13】AS1 またはgln ΔAS1 を発現するトランスジェニック植物のノーザン分析。個々の形質転換体の葉から単離された全RNA10μg を浸出レーン中に負荷した。ブロットをエンドウ豆からのAS1 cDNAで探査した。陽性対照は暗所で生育されたエンドウ豆の葉(PL)中のASmRNAを含む。陰性対照は明るい所で生育されたタバコの葉(TL)中のASmRNAを含む。
【図14】AS1 およびgln ΔAS1 を過剰発現するトランスジェニック系統の新鮮重量の増加が発芽後の3週から6週までグラフで表される。これは表(5)中に示され、そして表(6)中で統計的に分析されたデータのグラフ表示である。
【発明を実施するための形態】
【0030】
5. 発明の詳細な説明
本発明は植物における窒素代謝の遺伝子操作に関する。特に、本発明は、良好な成長特性、強化された栄養特性、改良された栄養および収穫量および/または増進された種子の収穫量または品質を有する植物を操作するために、窒素同化に関係する酵素および/またはそれらの発現を変化することに関する。
【0031】
それ故、特別な機構に制限されることを目的としないで、操作の標的はアミノ酸、グルタミン、アスパラギン酸、アスパラギンまたはグルタミン酸へのアンモニアの同化、または生合成反応におけるこれらの同じアミノ酸の利用に関係する酵素をコードする遺伝子である。標的遺伝子として、グルタミンシンテターゼ
(GS)、アスパラギンシンテターゼ(AS)、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ(GOGAT) 、アスパラギン酸アミノトランスフェラーゼ(AspAT) 、グルタミン酸デヒドロゲナーゼ(GDH) およびアスパラギナーゼ(ANS) をコードする遺伝子が挙げられる。窒素同化および利用においてこれらの酵素により果たされる役割の線図につき図1を参照のこと。
【0032】
これらの酵素は変性され、またはそれらの発現が増進、抑制またはそれ以外に修飾(例えば、異所性発現)されて所望の性質を有する植物を操作し得る。操作は植物を本明細書に記載された核酸構築物で形質転換することにより行われる。形質転換された植物またはそれらの子孫は、所望の変性された酵素を発現し、または窒素同化または利用酵素の所望の変化された発現、相当するmRNAの変化された発現、変化された窒素同化または利用能、増大された成長速度、増進された栄養収穫量、および/または改良された再生産収穫量を示す植物につきスクリーニングされる。
【0033】
所望の生理学的変化および/または作物栽培学的変化を示す操作された植物は植物育種または直接に農業生産に使用し得る。また、一つの変性された酵素を有するこれらの植物はその他の窒素同化または利用酵素の変化でもって操作されたその他の変性植物と交雑されて(例えば、GS過剰発現植物をAS過剰発現植物に交雑する)、親と較べて更に増強された生理学的性質および/または作物栽培学的性質を有する系統を生産し得る。
【0034】
本発明が、GSまたはASの異所性過剰発現につき操作された植物の実施例により説明される。全ての場合、GSまたはASの異所性過剰発現を示す操作された植物はまた対照の野生型植物よりも良好な成長特性、強化された栄養特性、改良された栄養収穫量および/または強化された種子の性質または収穫量を示す。
【0035】
5.1. 窒素同化および利用経路の変化
本発明の一つの局面によれば、望ましい植物は、アミノ酸グルタミン、アスパラギンまたはグルタミン酸へのアンモニアの初期同化そして更にアスパラギン酸への変換に関係する酵素の異所性過剰発現を操作することにより得られる。異所性という用語は、特別な遺伝子または当該酵素に関する異常な細胞下の(例えば、オルガネラ局在化と細胞質ゾル局在化の間のスイッチ)、細胞型、組織型および/または発育的もしくは一時的な発現(例えば、明/暗所)パターンを意味するために本明細書で使用される。このような異所性発現は前記酵素の正常な組織または発育段階における発現を必ずしも排除するのではないが、前記酵素の正常ではない組織または発育段階における発現を伴う。過剰発現という用語は前記酵素につき特別な組織、全部および/または発育または一時的な段階における正常な発現レベルを上回ることを意味するために本明細書で使用される。
【0036】
グルタミンへのアンモニアの同化そしてグルタミン酸、アスパラギン酸、およびアスパラギンへのその更なる代謝に関係する主要な酵素は、グルタミンシンテターゼ、アスパラギンシンテターゼ、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ、アスパラギン酸アミノトランスフェラーゼ、グルタミン酸デヒドロゲナーゼおよびアスパラギナーゼである。本発明は、これらの酵素の一種以上の異所性過剰発現の操作が所望の生理学的性質および作物栽培学的性質を有する植物を生産することを提供する。好ましい実施態様において、植物はグルタミンシンテターゼまたはアスパラギンシンテターゼの異所性発現につき操作される。酵素の細胞質ゾル形態または葉緑体形態が存在するGSに関して、細胞質ゾル形態の増進された発現の操作が好ましい。GSの細胞質ゾル形態として、小節−特異的酵素(例えば、エンドウ豆GS3A&B)および根−特異的酵素(例えば、エンドウ豆GS1)の両方が挙げられる。“根−特異的" 細胞質ゾルGS(例えば、エンドウ豆GS1)の増進された発現の操作が特に好ましい。また、本発明は前記酵素の細胞下の局在化を変化する操作を提供する。例えば、ASの如き細胞質ゾル酵素への葉緑体標的配列の操作は、植物中の窒素同化を改良し得る。これは光呼吸性アンモニアを再同化するために葉肉細胞中で特に有益であろう。
【0037】
本発明の別の局面によれば、望ましい植物は別の窒素同化経路により増進されたアンモニアとり込みを操作することにより得られる。特に、操作はグルタミンシンテターゼによる窒素同化の正常な主要経路を抑制することにより行われる。多重GSアイソザイムをコードする植物種において、これは内在性GS遺伝子の抑制を必要とし得る。好ましい実施態様において、抑制されたGS発現でもって操作された植物は、アスパラギンシンテターゼ(AS)および/またはグルタミンデヒドロゲナーゼ(GDH) の如きオルタナチブN同化性酵素の異所性過剰発現につき更に操作される。最も好ましい実施態様において、GSおよび/またはAS/GDH操作された植物は窒素同化または利用プロセスに関係するその他の酵素の一種以上の増進された発現につき更に操作される(図1を参照のこと)。
【0038】
本発明の第三の局面によれば、望ましい植物は同化された窒素の利用に関係する酵素の異所性過剰発現を操作することにより得られる。本発明のこの局面の実施態様は代謝反応においてグルタミン、グルタミン酸およびアスパラギンの使用を触媒作用する酵素の異所性過剰発現でもって植物を操作することを伴い得る。好ましい実施態様において、植物はアスパラギナーゼの異所性過剰発現につき操作される。
【0039】
本発明の第四の局面によれば、望ましい植物は窒素の同化または利用に関係する酵素の変化され、突然変異されたキメラ形態または異種形態の発現を操作することにより得られる。本発明のこの局面の実施態様は、異種起源からの窒素同化酵素または利用酵素(即ち、異なる植物または動物もしくは微生物を含む、生物からの酵素)を発現するように植物を操作することを伴い得る。付加的な実施態様は、例えば、基質結合、触媒作用、および/または生産物放出において増大された効率を有する窒素同化酵素または利用酵素を開発し、そしてこのような新規な酵素を発現するように植物を操作することを伴い得る。これらの新規な酵素は前記プロセスに影響する主要アミノ酸残基のin vitro突然変異誘発により開発し得る。また、このような新規な酵素は関連酵素からのドメインを組換えることにより開発し得る。例えば、キメラ二機能性GOGAT 酵素はNADH-GOGATのNADH結合ドメインをFd-GOGAT遺伝子にスプライシングすることによりフェレドキシン-GOGAT活性およびNADH-GOGAT活性の両方を含むように操作し得る(図2を参照のこと)。このようなキメラGOGAT 酵素はGOGAT 反応においてNADHまたはフェレドキシンを還元剤として利用できるという利点を有するであろう。この新規な酵素の異所性発現はグルタミン酸の更に有効な合成をもたらし得る。本明細書に示された酵素修飾の別の例(第7.0 節を参照のこと)は、その基質特異性を変化するためにドメインを欠失されたAS酵素の操作である。
【0040】
本発明によれば、窒素同化酵素または利用酵素の組織および発育の発現パターンの調節は所望の植物改良を得るのに重要であり得る。植物が正常または別のアンモニア同化経路に関係する酵素の異所性過剰発現につき操作される場合、本発明の好ましい実施態様は植物の多くまたは全部において変化された発現を行うことを伴う。植物が同化窒素の使用を触媒作用する酵素の異所性過剰発現につき操作される場合、本発明の好ましい実施態様はこのような発現を窒素“シンク" 組織および構造、例えば、葉および種子に制限する。
【0041】
5.2. トランスジェニック植物の発生
5.2.1. 核酸構築物
核酸配列の性質は、種々の潜在的な宿主植物細胞の遺伝子構造であるように変化される。本発明の好ましい実施態様は、当業者が絶対に必須ではないが、明らかに有利であると認識し得る幾つかの特徴を記載するであろう。これらは遺伝子構造の単離、合成または構築の方法、植物細胞に導入される遺伝子構築物の操作、遺伝子構築物の或る特徴、および遺伝子構築物と関連するベクターの或る特徴を含む。
【0042】
更に、本発明の遺伝子構築物はDNA分子またはRNA分子につきコードし得る。本発明によれば、標的植物の所望の安定な遺伝子型の変化は外因的に導入された一種以上の核酸構築物、特に組換えDNA構築物のゲノム組込みにより行われることが好ましい。それにもかかわらず、本発明によれば、このような遺伝子型の変化はまた自律的に複製でき、しかも染色体上かつ生殖上安定であるエピソーム(DNAまたはRNA)の導入により影響し得る。導入された核酸構築物がRNAを含む場合、このような構築物からの植物形質転換または遺伝子発現は逆転写により生成されたDNA中間体により進行し得る。
【0043】
本明細書に記載された核酸構築物は、当業者に公知の方法を使用して生産し得る。当業者は構築物の成分を単離、特性決定、そして操作するだけでなく、構築物それ自体をつくるのに使用し得る組換えDNA方法の教示につきSambrookら,1989, 分子クローニング:実験マニュアル, コールドスプリング ハーバー ラボラトリー プレス, プレインビュー, N.Y.のような出典を参考にし得る。所望の成分の核酸配列が知られている、幾つかの場合には、生物起源からそれを単離するのではなく、それを合成することが有利であり得る。このような場合、当業者はCaruthers ら, 1980, Nuc.Acids.Res.Symp.Ser. 7:215-233 、およびChowおよびKempe, 1981, Nuc.Acids.Res. 9:2807-2817 のような教示を参考にし得る。その他の場合、所望の成分はポリメラーゼ連鎖反応(PCR) 増幅により有利に生産し得る。PCR 教示につき、当業者はGelfand, 1989, PCR 技術、DNA増幅に関する原理および応用, H.A.Erlich編集, ストックトン プレス, N.Y.、分子生物学における現行のプロトコル, 2巻, 15章, Ausubel ら編集, ジョン ウィリィ&サンズ, 1988を参考にし得る。
【0044】
5.2.1.1. 発現構築物
本発明によれば、窒素同化または利用酵素の異所性過剰発現を有する植物は、所望の酵素をコードする配列と操作により関連された植物プロモーターを含む遺伝子構築物で植物細胞を形質転換することにより操作し得る。(操作により関連されたは“関連”プロモーターにより調節された転写が機能性メッセンジャーRNA(その翻訳が酵素を生産するであろう)を生産することを意味するために本明細書で使用される)。本発明の好ましい実施態様において、関連プロモーターは強力かつ非組織特異性または非発育特異性の植物プロモーター(例えば、多くまたは全ての組織型中で強く発現するプロモーター)である。このような強力な“構成的”プロモーターとして、CaMV 35Sプロモーター、T-DNAマンノピンシンテターゼプロモーター、およびそれらの種々の誘導体が挙げられるが、これらに限定されない。
【0045】
本発明の別の実施態様において、組織特異性または発育特異性のプロモーターを所望の酵素をコードする配列で操作により関連させる遺伝子構築物で植物を操作することが有利であり得る。例えば、光合成組織および器官中の発現が所望される場合、リブロースビホスフェートカルボキシラーゼ(ルビスコ)遺伝子または葉緑体a/b 結合タンパク質(CAB) 遺伝子のプロモーターの如きプロモーターが使用し得る。種子中の発現が所望される場合、種々の種子貯蔵タンパク質遺伝子のプロモーターの如きプロモーターが使用し得る。窒素固定小節中の発現が所望される場合、レグヘモグロビン遺伝子またはノジュリン遺伝子のプロモーターの如きプロモーターが使用し得る。根特異性発現が所望される場合、根特異性グルタミンシンテターゼ遺伝子をコードするプロモーターの如きプロモーターが使用し得る(Tingeyら, 1987, EMBO J. 6:1-9; Edwardsら, 1990, Proc.Nat.Acad.Sci.USA 87:3459-3463を参照のこと)。
【0046】
本発明の更に別の実施態様において、誘導プロモーターを所望の酵素をコードする配列と操作により関連させる遺伝子構築物で植物を形質転換することが有利であり得る。このようなプロモーターの例は多く、そして多岐にわたる。それらとして、二三の名前を挙げると、熱ショック遺伝子、防御応答遺伝子(例えば、フェニルアラニンアンモニアリアーゼ遺伝子)、傷誘発遺伝子(例えば、ヒドロキシプロリンに富む細胞壁タンパク質遺伝子)、化学誘導遺伝子(例えば、ニトレート還元酵素遺伝子、グルコナーゼ遺伝子、キチナーゼ遺伝子、等)、暗所誘導遺伝子(例えば、アスパラギンシンテターゼ遺伝子(CoruzziおよびTsai, 米国特許第5,256,558 号, 1993年10月26日, 植物アスパラギンシンテターゼをコードする遺伝子)が挙げられるが、これらに限定されない。
【0047】
本発明の更に別の実施態様において、修飾プロモーターまたは人工プロモーターを所望の酵素をコードする配列に操作により連結する遺伝子構築物で植物を形質転換することが有利であり得る。典型的には、異なるプロモーターの構造要素を組換えることにより構築された、このプロモーターは天然プロモーターには見られない特異な発現パターンおよび/またはレベルを有する。例えば、シス調節要素をプロモーターコアーと組み合わせることにより構築された人工プロモーターの例につき、Salinaら, 1992, Plant Cell 4:1485-1493を参照のこと。
【0048】
本発明の更に別の実施態様において、窒素同化または利用酵素の異所性過剰発現は所望の酵素をコードする遺伝子のコピー数を増加することにより操作し得る。所望の遺伝子の増加されたコピーを有する植物細胞を生産するための一つのアプローチは、遺伝子の多重コピーを含む核酸構築で形質転換することである。また、所望の酵素をコードする遺伝子は、増幅選択可能なマーカー(ASM) 遺伝子、例えば、グルタミンシンテターゼまたはジヒドロ葉酸還元酵素遺伝子を含む核酸構築物中に入れられる。このような構築物で形質転換された細胞は、ASM 遺伝子の増加されたコピーを有する細胞系を選択する培養レジメにかけられる。GS遺伝子の増幅されたコピーを含む植物細胞系の単離に使用される選択プロトコルにつき、Donnら, 1984, J.Mol.Appl.Genet. 2:549-562 を参照のこと。所望の遺伝子がASM遺伝子の近くに連結されるので、ASM 遺伝子を増幅した細胞系はまたおそらく所望の酵素をコードする遺伝子を増幅させるであろう。
【0049】
本発明のもう一つの実施態様において、窒素同化または利用酵素の異所性過剰発現は所望の酵素をコードする内在性遺伝子またはトランスジーンの発現を調節する調節遺伝子をコードする核酸構築物で植物細胞を形質転換することにより操作されてもよく、この場合、導入された調節遺伝子は所望の組織および/または発育段階、シンテターゼプロモーター、およびそれらの種々の誘導体中の酵素の強い発現を可能にするように修飾される。
【0050】
5.2.1.2. 抑制構築物
本発明によれば、所望の植物は窒素同化/代謝においてGS活性またはその他の酵素の活性を抑制することにより操作し得る(図1)。実施態様において、抑制は成熟標的mRNAを含む、宿主標的RNA転写産物のセグメントまたは全部に相補性のアンチセンスRNAをコードする遺伝子構築物で植物細胞を形質転換することにより操作し得る。別の実施態様において、標的遺伝子(例えば、GSmRNA)抑制は宿主標的RNA転写産物(例えば、成熟GSmRNAを含む、GSRNA転写産物)を開裂するリボザイムをコードする遺伝子構築物で植物細胞を形質転換することにより操作し得る。
【0051】
更に別の実施態様において、標的遺伝子抑制は“優性の負の”突然変異を含む標的酵素をコードする遺伝子構築物で植物細胞を形質転換することにより操作し得る。好ましい突然変異は触媒作用、基質結合(例えば、GSにつき、グルタミン酸またはアンモニウムイオンの結合部位)、または生産物放出に影響する突然変異である。有益な突然変異は上記のプロセスに関係する重要な一つ以上の残基の欠失または点突然変異であり得る。当業者は優性の負の突然変異を構築するためのアプローチおよび戦略につき本明細書中の教示およびHerskowitz (Nature, 329:219-222, 1987)の教示を参考にし得る。
【0052】
上記の抑制構築物の全てにつき、このような遺伝子構築物は標的遺伝子と同じ組織および発育特異性で発現することが好ましい。こうして、これらの抑制構築物は標的遺伝子のプロモーターと操作により関連されることが好ましい。また、抑制構築物を構成的に発現させることが好ましい場合がある。こうして、CaMV 35S プロモーターの如き強力な構成的プロモーターがまた抑制構築物を発現するのに使用し得る。これらの抑制構築物に最も好ましいプロモーターは標的遺伝子の修飾プロモーターであり、この場合、その修飾は組織特異性または発育特異性の変化なしに標的遺伝子プロモーターの増進された発現をもたらす。
【0053】
本発明によれば、標的遺伝子発現の抑制された所望の植物はまた同時抑制構築物で植物細胞を形質転換することにより操作し得る。同時抑制構築物は標的遺伝子の完全または部分コード配列と操作により関連された機能性プロモーターを含む。操作により関連されたプロモーターは強力な構成的プロモーター、例えば、CaMV 35Sプロモーターであることが好ましい。また、同時抑制構築物プロモーターは、標的遺伝子と同じ組織および発育特異性で発現するプロモーターであってもよい。このような別のプロモーターは標的遺伝子それ自体のプロモーター(例えば、GS同時抑制構築物の発現を誘導するためのGSプロモーター)を含み得る。 本発明によれば、同時抑制構築物は不完全標的mRNAまたは欠損標的酵素をコードすることが好ましいが、完全に機能性の標的mRNAまたは酵素をコードする構築物がまた同時抑制を行うのに有益であり得る。
【0054】
実施態様において、GSアイソザイムの全てではないとしても殆どの抑制が所望される場合、同時抑制構築物は葉緑体GSmRNA(例えば、エンドウ豆GS2 mRNA)の完全または部分コピーをコードすることが好ましい。本明細書(第6.2.2.節)に開示されるように、このような構築物は標的遺伝子の発現を抑制するのに特に有効である。
【0055】
本発明によれば、標的遺伝子発現が抑制された所望の植物はまた内在性標的遺伝子の部位誘導突然変異誘発を行い得る構築物で植物細胞を形質転換することにより操作し得る(植物中の標的遺伝子の部位誘導突然変異誘発を行うための核酸構築物の説明につき、Offringaら, 1990, EMBO J. 9:3077-84;およびKanevskiiら, 1990, Dokl.Akad.Nauk.SSSR 312:1505-1507 を参照のこと)。このような構築物は不活性コード配列による相同組換えにより、またはそれによらない相同組換えにより内在性標的遺伝子配列を置換することにより標的遺伝子の抑制を行うことが好ましい。
【0056】
5.2.1.3. 組換え核酸構築物のその他の特徴
本発明の組換え構築物は、その構築物の伝播のための選択可能なマーカーを含んでもよい。例えば、細菌中で伝播される構築物は抗生物質耐性遺伝子、例えば、カナマイシン、テトラサイクリン、ストレプトマイシン、またはクロラムフェニコールに対する耐性を与える遺伝子を含むことが好ましい。構築物を伝播するのに適したベクターとして、二三名を挙げると、プラスミド、コスミド、バクテリオファージまたはウイルスが挙げられる。
【0057】
加えて、組換え構築物はこれらの構築物により形質転換された植物細胞の単離、同定またはトラッキングのための植物発現性の選択可能なマーカー遺伝子またはスクリーニング可能なマーカー遺伝子を含んでもよい。選択可能なマーカーとして、抗生物質耐性(例えば、カナマイシンまたはハイグロマイシンに対する耐性)または除草剤耐性(例えば、スルホニル尿素、ホスフィノトリシン、またはグリフォセートに対する耐性)を与える遺伝子が挙げられるが、これらに限定されない。スクリーニング可能なマーカーとして、β−グルクロニダーゼをコードする遺伝子(Jefferson, 1987, Plant Molec Biol.Rep 5:387-405) 、ルシフェラーゼをコードする遺伝子(Ow ら, 1986, Science 234:856-859)、アントシアニン色素産生を調節するBおよびC1遺伝子産物(Goff ら, 1990, EMBO J 9:2517-2522) が挙げられるが、これらに限定されない。
【0058】
植物を形質転換するためにアグロバクテリウム系を利用する本発明の実施態様(以下を参照のこと)において、組換えDNA構築物は更に植物細胞に形質転換されるDNA配列に隣接する右のT-DNAボーダー配列を少なくとも含む。好ましい実施態様において、移入される配列は左右のT-DNAボーダー配列により隣接される。このようなT-DNAをベースとする形質転換ベクターの適切な設計および構築は当業者に公知である。
【0059】
5.2.2.植物および植物細胞の形質転換
本発明によれば、望ましい植物は本明細書に記載された核酸構築物で植物細胞を形質転換することにより得られてもよい。幾つかの場合、植物または植物細胞を幾つかの異なる遺伝子構築物で操作することが望ましい場合がある。このような操作は植物または植物細胞を所望の構築物の全部で同時に形質転換することにより行い得る。また、操作は連続的に行われてもよい。それは、一つの遺伝子で形質転換し、選択およびスクリーニング後に所望の形質転換体を得、形質転換体を第二遺伝子構築物で形質転換すること、等である。好ましい実施態様において、それぞれの遺伝子構築物は、多重遺伝子インサートを含む植物形質転換体の同定を促進するように異なる選択可能なマーカー遺伝子またはスクリーニング可能なマーカー遺伝子に連結されるであろう。別の実施態様において、幾つかの異なる遺伝子がそれぞれの遺伝子につき操作された親系統を交雑することにより一種の植物にとり込まれ得る。
【0060】
本発明の実施態様において、アグロバクテリウムが遺伝子構築物を植物に導入するのに使用される。このような形質転換は2成分アグロバクテリウムT-DNAベクター(Bevan, 1984, Nuc.Acid Res. 12:8711-8721) 、および同時培養操作(Horsch ら, 1985, Science 227:1229-1231)を使用することが好ましい。一般に、アグロバクテリウム形質転換系が双子葉植物を操作するのに使用される(Bevanら,1982, Ann.Rev.Genet 16:357-384; Rogersら, 1986, Methods Enzymol. 118:627-641)。アグロバクテリウム形質転換系がまた単子葉植物および植物細胞を形質転換するだけでなく、DNAをそれらに移入するのに使用し得る(Hernalsteenら,1984, EMBO J 3:3039-3041; Hooykass-Van Slogterenら, 1984, Nature 311:763-764; Grimsley ら, 1987, Nature 325:1677-179; Boultonら, 1989, Plant Mol.Biol. 12:31-40; Gould ら, 1991, Plant Physiol. 95:426-434 を参照のこと)。
【0061】
別の実施態様において、組換え核酸構築物を植物および植物細胞に導入するための種々の別法がまた使用し得る。これらのその他の方法は、標的が単子葉植物または植物細胞である場合に特に有益である。別の遺伝子移入方法および形質転換方法として、裸のDNAのカルシウム、ポリエチレングリコール(PEG) またはエレクトロポレーション介在性とり込みによるプロトプラスト形質転換(Paszko-wskiら, 1984, EMBO J 3:2717-2722, Potrykusら, 1985, Molec.Gen.Genet. 199:169-177; Frommら, 1985, Proc.Nat.Acad.Sci.USA 82:5824-5828; Shimamoto,1989, Nature 338:274-276を参照のこと) および植物組織のエレクトロポレーション(D'Halluinら, 1992, Plant Cell 4:1495-1505) が挙げられるが、これらに限定されない。植物細胞形質転換の更に別の方法として、マイクロインジェクション、炭化ケイ素介在性DNAとり込み(Kaeppler ら, 1990, Plant Cell Reporter 9:415-418) 、およびマイクロプロジェクティル衝撃(Kleinら, 1988, Proc.Nat.Acad.Sci.USA 85:4305-4309; Gordon-Kammら, 1990, Plant Cell 2:603-618を参照のこと)が挙げられる。
【0062】
本発明によれば、多種の植物および植物細胞系が、本発明の核酸構築物および上記の種々の形質転換方法を使用して本明細書に記載された所望の生理学的特徴および作物栽培学的特徴につき操作し得る。好ましい実施態様において、操作のための標的植物および植物細胞として、トウモロコシ、小麦、米、大豆、トマト、タバコ、ニンジン、ジャガイモ、てんさい、ヒマワリ、ヤムイモ、アラビドプシス、ナタネ、およびペチュニアが挙げられるが、これらに限定されない。
【0063】
5.2.3. 形質転換された植物および植物細胞の選択および同定
本発明によれば、所望の植物は、開示された遺伝子構築物をプロトプラスト、組織培養細胞、組織および器官外植体、花粉、胚、並びに植物全体を含むが、これらに限定されない種々の植物細胞型に操作することにより得られる。本発明の実施態様において、操作された植物体は下記のアプローチおよび方法に従って形質転換体(導入された一つ以上の遺伝子構築物をとり込み、または組込んだ形質転換体)につき選択またはスクリーニングされる。次に単離された形質転換体が植物中で再生し得る。また、操作された植物体は、誘導された植物または幼植物体をマーカー遺伝子形質につき選択またはスクリーニングにかける前に植物または幼植物体に再生し得る。一つ以上のマーカー遺伝子を選択またはスクリーニングする前または後に、植物を植物細胞、組織または器官から再生するための方法は当業者に公知である。
【0064】
形質転換された植物細胞、カルス、組織または植物は、操作された植物体を形質転換DNAに存在するマーカー遺伝子によりコードされた形質につき選択またはスクリーニングすることにより同定され、単離し得る。例えば、選択は形質転換遺伝子構築物が耐性を与える抑制量の抗生物質または除草剤を含む培地で操作された植物体を生育することにより行い得る。更に、形質転換された植物および植物細胞はまた本発明の組換え核酸構築物に存在し得る可視のマーカー遺伝子(例えば、β−グルクロニダーゼ遺伝子、ルシフェラーゼ遺伝子、B遺伝子またはC1遺伝子)の活性につきスクリーニングすることにより同定し得る。このような選択方法およびスクリーニング方法が当業者に公知である。
【0065】
また、物理的方法および生化学的方法が本発明の遺伝子構築物を含む植物または植物細胞形質転換体を同定するのに使用し得る。これらの方法として、1)組換
えDNAインサートの構造を検出し、測定するためのサザン分析またはPCR 増幅;
2)遺伝子構築物のRNA転写産物を検出し、試験するためのノーザンブロット、S1 RNase保護、プライマー伸長PCR 増幅または逆転写酵素PCR 増幅;3)このような遺伝子産物が遺伝子構築物によりコードされる場合、酵素またはリボザイム活性を検出するための酵素アッセイ;4)遺伝子構築産物がタンパク質である場合、タンパク質ゲル電気泳動、ウェスタンブロット技術、免疫沈殿、またはエンザイムリンクドイムノアッセイが挙げられるが、これらに限定されない。更に別の技術、例えば、in situ ハイブリダイゼーション、酵素染色、および免疫染色がまた特定の植物器官および組織中の組換え構築物の存在または発現を検出するのに使用し得る。全てのこれらのアッセイを行う方法が当業者に公知である。
【0066】
5.2.4. 改良された作物栽培学的形質を有する植物に関する形質転換された植物のスクリーニング
本発明によれば、改良された作物栽培学的特徴を有する植物を得るために、形質転換された植物が所望の生理学的変化を示す植物につきスクリーニングし得る。例えば、植物がGS酵素の異所性過剰発現につき操作された場合、形質転換された植物が所望のレベルで所望の組織および発育段階中にGS酵素を発現する植物につき試験される。植物が標的遺伝子の抑制につき操作された場合、形質転換された植物が種々の組織中で減少されたレベルで標的遺伝子産物(例えば、RNAまたはタンパク質)を発現する植物につき試験される。次に、所望の生理学的変化、例えば、異所性GS過剰発現またはGS抑制を示す植物が所望の作物栽培学的変化を有する植物につき続いてスクリーニングし得る。
【0067】
また、形質転換された植物は所望の作物栽培学的変化を示す植物につき直接にスクリーニングし得る。一つの実施態様において、このようなスクリーニングは窒素栄養欠乏条件下の形質転換された植物の生産的生育に関するものであってもよい。それは、野生型植物の成長を停止し、または野生型植物のサイズまたは品質をかなり低下するように低減させる、利用可能な窒素栄養に関する、条件下の形質転換された植物の成長に関するスクリーンである。タバコおよび同様の窒素栄養要件を有する植物に関する窒素栄養欠乏条件の例は、土壌または合成培地中の唯一の窒素栄養が(a)0.5mM以下の濃度で供給または周期的に適用される硝酸塩、または(b)0.5mM以下の硝酸塩に生理学的に均等である濃度で供給または周期的に適用される硝酸塩の生理学的に均等物(例えば、アンモニウムまたは硝酸塩とアンモニウムの混合物)(Eckes ら, 1988, オーストラリア特許庁書類番号AU-A-17321/88 を参照のこと)である条件である。窒素栄養欠乏条件の別の例は、土壌または合成培地中の利用可能な窒素栄養の定常状態レベルが約0.02mM未満の硝酸塩またはその生理学的均等物である条件である。本明細書に使用される硝酸塩という用語は植物窒素肥料として普通に使用される硝酸塩、例えば、硝酸カリウム、硝酸カルシウム、硝酸ナトリウム、硝酸アンモニウム、等のいずれか一種またはあらゆる混合物を意味する。本明細書に使用されるアンモニウムという用語は植物窒素肥料として普通に使用されるアンモニウム塩、例えば、硝酸アンモニウム、塩化アンモニウム、硫酸アンモニウム、等のいずれか一種またはあらゆる混合物を意味する。
【0068】
その他の実施態様において、形質転換された植物のスクリーニングは、窒素非限定生育条件下で培養される(即ち、健全な植物成長を持続するのに充分な量の窒素栄養を含み、または受容する土壌または培地を使用して培養される)時に、未操作の前駆植物と較べて、改良された作物栽培学的特徴(例えば、早い成長、大きな栄養収穫量または再生産収穫量、または改良されたタンパク質含量、等)に関するものであってもよい。タバコおよび同様の窒素栄養要件を有する植物に関する窒素非限定条件の例は、土壌または合成培地中の唯一の窒素栄養が(a) 10mM以上の濃度で供給または周期的に適用される硝酸塩、または(b)10 mM以上の硝酸塩に生理学的に均等である濃度で供給または周期的に適用される硝酸塩の生理学的に均等物である条件である。窒素非限定条件の別の例は、土壌または合成培地中の利用可能な窒素栄養の定常状態レベルが少なくとも約1.0 mMの硝酸カリウムまたはその生理学的均等物である条件である。植物生育に関する窒素栄養欠乏条件または“非限定" 条件であることに関する更に別のガイダンスが当業界に見られる。例えば、Hewitt, E.J., 植物栄養の研究に使用される砂培養方法および水培養方法, 第2編, Farnham Royal (Bucks), Commonwealth Agricultural Bu-reaux, 1966;およびHewitt, E.J., 植物無機栄養, ロンドン, 英国大学プレス,1975を参照のこと。
【0069】
形質転換された植物がマメ科牧草類である実施態様において、所望の作物栽培学的変化および改良による形質転換された植物の直接のスクリーニングは、小節形成または窒素固定が抑制される条件下である以外は上記のようにして行われてもよい。
【0070】
本発明によれば、窒素同化プロセスまたは利用プロセスの変化で操作された植物は改良された窒素含量、変化されたアミノ酸またはタンパク質組成、活発な成長特性、増大された栄養収穫量または良好な種子収穫量および品質を示し得る。このような改良された作物栽培学的特徴を有する操作された植物および植物系統は下記のパラメーター:1)新鮮重量または乾燥重量の増加の速度に関して測定される成長の速度;2)新鮮重量または乾燥重量に関する、成熟植物の栄養収穫量;3)種子または果実収穫量;4)種子または果実重量;5)植物の合計窒素含量;6)果実または種子の全窒素含量;7)植物の遊離アミノ酸含量;8)果実または種子の遊離アミノ酸含量;9)植物の合計タンパク質含量;および10) 果実または種子の合計タンパク質含量のいずれかを試験することにより同定し得る。これらのパラメーターを試験するための操作および方法が当業者に公知である。
【0071】
本発明によれば、所望の植物は、前記パラメーターの一つ以上において対照植物(即ち、前駆植物)に対し改良を示す植物である。実施態様において、所望の植物は、少なくとも一つのパラメーターにおいて対照植物に対し少なくとも5%の増加を示す植物である。好ましい実施態様において、所望の植物は、少なくとも一つのパラメーターにおいて対照植物に対し少なくとも20%の増加を示す植物である。少なくとも一つのパラメーターにおいて少なくとも50%の増加を示す植物が最も好ましい。
【0072】
5.3. 発明の有用性
本発明の遺伝子操作された植物は、野生型植物の成長を停止させたり、成長を小さくして野生型植物を実際に無用のものにする、窒素栄養素が欠乏した条件下(すなわち、土壌に窒素が少なく、また、窒素肥料の投入量が少ない)で生産的に栽培することができる。遺伝子操作された植物はまた、より早い成熟、より速い成長および/またはより高い収穫量の達成、および/または窒素無制限成長条件(すなわち、健康的な植物の成長を維持するのに充分な量の窒素栄養素を含むか受け取る土壌または培地)を使用して栽培するときのより栄養に富む食品および動物飼料の生産に有利に使用することができる。窒素無制限成長条件は、種同士の間および一つの種の亜種に対して変わる。しかし、当業者であれば、全部でなくても、ほとんどの重要な作物および観賞植物の栽培に対して、何が窒素無制限成長条件を構成するかを知っている。例えば、小麦の栽培に対しては、Alcozら、Agronomy Journal 85:1198-1203 (1993)、Rao および Dao, J. Am. Soc. Agronomy 84:1028-1032 (1992)、Howardおよび Lessman, Agronomy Jornal 83:208-211 (1991);トウモロコシの栽培に対しては、Tollenear ら、Agronomy Journal 85:251-255 (1993)、Straw ら、Tennessee Farm and Home Science: Progress Report, 166:20-24 (Spring 1993) 、Miles, S.R., J. Am. Soc. Agronomy 26:129-137 (1934) 、Daraら、J. Am. Soc. Agronomy 84:1006-1010 (1992)、Binford ら、Agronomy Journal 84:53-39 (1992); 大豆の栽培に対しては、Chenら、Canadian Journal of Plant Science 72:1049-1056 (1992) 、Wallace ら、Journal of Plant Nutrition 13:1523-1537 (1990); 米の栽培に対しては、Oritani および Yoshida, Japanese Journal of Crop Science 53:204-212 (1984); 亜麻仁の栽培に対しては、Diepenbrock および Porksen, Industrial Crops and Products 1:165-173 (1992); トマトの栽培に対しては、Grubinger ら、Journal of the American Society for Horticultural Science 118:212-216 (1993)、Cerne, M., Acta Horticulture 277:179-182 (1990); パイナップルの栽培に対しては、Magistadら、J. Am. Soc. Agronomy 24:610-622 (1992)、Asoegwu, S.N., Fertilizer Research 15:203-210 (1988)、Asoegwu, S.N., Fruits 42:505-509 (1987); レタスの栽培に対しては、Richardson および Hardgrave, Journal of the Science of Food and Agriculture 59:345-349 (1992); ミントの栽培に対しては、Munsi, P.S., Acta Horticulturae 306:436-443 (1992); カモミールの栽培に対しては、Letchamo, W., Acta Horticulturae 306:375-384 (1992); タバコの栽培に対しては、Sissonら、Crop Science 31:1615-1620 (1991); 馬鈴薯の栽培に対しては、Porterおよび Sisson, American Patato Journal, 68:493-505 (1991);アブラナ科の栽培に対しては、Rahnら、Conference "Proceedings, second congress of the European Society for Agronomy" Warwick Univ., p.424-425 (August 23-28 1992); バナナの栽培に対しては、Hegde および Srinivas, Tropical Agriculture 68:331-334 (1991) 、Langenegger および Smith, Fruits 43:639-643 (1988); イチゴの栽培に対しては、Human および Kotze, Communications in Soil Science and Plant Analysis 21:771-782 (1990);モロコシの栽培に対しては、Mahalle および Seth, Indian Journal of Agricultural Sciences 59:395-397 (1989); オオバコの栽培に対しては、Anjorin および Obigbesan, Conference "International Cooperation for Effective Plantain and Banana Research" Proceedings of the third meeting. Abidjan, Ivory Coast, p.115-117 (May 27-31, 1985); サトウキビの栽培に対しては、Yadav, R.L., Fertiliser News 31:17-22 (1986)、Yadav および Sharma, Indian Journal of Agricultural Sciences 53:38-43 (1983); テンサイの栽培に対しては、Draycottら、Conference "Symposium Nitrogen and Sugar Beet" International Institute for Sugar Beet Research - Brussels Belgium, p.293-303 (1983)参照。また、Goh および Haynes, "Nitrogen and Agronomic Practice" in Meneral Nitrogen in the Plant-Soil System, Academic Press, Inc., Orlando, Florida, p.379-468 (1986), Engelstad, O.P., Fertilizer Technology and Use, Third Edition, Soil Science Society of America, p.633 (1985), Yadav and Sharmna,
Indian Journal of Agricultural Sciences, 53:3-43 (1983) も参照。
【0073】
GS抑制は、いくつかのGS抑制された植物、特に豆科植物が、非抑制植物よりもより速く成長する、あるいはより高い窒素含量を有するという点で有用である(Knightおよび Langston-Unkefer, Science, 241:951-954 参照)。GS抑制された植物はまた、アミノ酸またはタンパク質含量を変えて、特定のダイエット食品の製造に有用であり得る。さらに、本明細書に開示する遺伝子操作された全植物は、農業的に有用なプラントラインを展開するための育種ストックとして役立つと考えられる。
【実施例】
【0074】
6.実施例:グルタミンシンテターゼの植物における異所性過剰発現は、植物成長表現型の増加を引き起こす
ここでは、トランスジェニック植物において窒素使用効率を操作するための分子遺伝学法を記載する。その方法は、通常はGS発現が認められない細胞型および/またはレベルでGSを発現する、グルタミンシンテターゼの異所性発現に頼るものである。トランスジェニック植物における細胞特異的GS発現のパターンは、細胞質ゾルGS(通常は、篩部で発現するだけである。)を全細胞型で構成的に過剰発現することにより変えられる。GSのそのような異所性発現は、窒素同化酵素の区画化および細胞型特異性から生じる生理的制限を回避することができる。葉肉細胞での細胞質ゾルGSの異所性高レベル発現は、光呼吸によって失われたアンモニアの再同化に対して別のルートを提供すると考えられる。このことは、光呼吸によって失われるアンモニアの量が最初の窒素同化の10倍を超えるとき、成長の上での利点を与えると考えられる(Wallsgroveら、1983, Plant Cell Environ. 6:301-309; Keys ら、1978, Nature, 275:741-743 )。ここに開示する研究は、細胞質ゾルGSに対する異種GSサブユニットの構成的過剰発現がGS mRNA、GSタンパク質、全GS活性、天然のGSホロ酵素の増加ならびに一例においては新規GSホロ酵素の産生をもたらすことを示す。細胞質ゾルGSを過剰発現する形質転換された植物は、野生型と比較して、統計的に重要な成長上の利点を有する。形質転換された植物は、栄養成長期の際に、形質転換されていない前駆植物よりも速く成長し、最終の新鮮重量をより高くし、溶解性のより高いタンパク質を有する。しかし、場合によっては、細胞質ゾルGSおよび/または葉緑体GSの過剰発現が、内在遺伝子発現または共同抑制の下方調整を引き起こす。細胞質ゾルGS過剰発現構成体を含むいくつかの形質転換された植物および葉緑体GS2構成体を含む全ての形質転換された植物はGSを過剰発現しないで、むしろGS発現を抑制し、内在GS遺伝子の抑制も含む(すなわち、共同抑制)。そのようなGSが共同抑制された植物は、小さい成長特性を示す可能性があるが、窒素が他の窒素同化/代謝経路にそれて入ることによりアミノ酸およびタンパク質含量が変わると考えられる。
【0075】
6.1. 材料および方法
6.1.1. 植物発現ベクターの構築
植物発現ベクター pTEV 4 、5 、7 および 8を次のように構築した。カリフラワーモザイクウイルス(CaMV)の Strasbourg 株の 35Sプロモーターを含む、転写開始に対して -941 〜 +26まで伸びる HindIII−EcoRI 断片を、pBluescript KS II-(pT109) に挿入した(Hohnら、1982, Curr. Topics Microbiol. Immunol. 96:194-236 )。次いで、HindIII および XhoI 部位の間のポリリンカー配列を、XbaI、SstIおよび StuI 部位を含むように修飾した(pT145 )。こうして、pBIN19 (Clontech) 由来の、ノパリンシンターゼ転写ターミネーターを含むT4ポリメラーゼ処理したSstI−EcoRI 断片を StuI 部位に挿入することができ、pT161 を作った。このように構築された発現カセットは、隣がEcoRI 部位であり、修飾ポリリンカーを含む、pBIN19 (Bevan, 1984, Nucleic Acids Res. 12:8711-8721) 由来のプラスミドである pW3に移入された。プロモーターの5’末端がpW3の左端に隣接するように配向したクローンを選択し (pW63) 、プロモーターとターミネーターとの間に多数のクローニング部位を挿入した。こうして、列挙したユニークなクローニング部位を有する次のバイナリーベクターを作った(図3):pTEV4 (HindIII-XbaI-BamHI-XhoI) 、pTEV5 (HindIII-StuI-SstI-KpnI)、pTEV8 (HindIII-XhoI-BamHI-XbaI) 、pTEV9 (HindIII-KpnI-SstI-StuI)。
【0076】
6.1.2. GS cDNAのバイナリー発現ベクターへの移入
細胞質ゾルGS1およびGS3Aならびに葉緑体GS2のエンドウ遺伝子に対応するcDNAを pBluescriptから上記のバイナリー発現ベクターへ移入した(図4参照)。これらのcDNAは、以前は、各々GS299、GS341およびGS185として記載された(Tingeyら、1987, EMBO J. 6:1-9; Tingey ら、1988, J. Biol. Chem. 263:9651-9657)。葉緑体GS2に対しては、ゲノム配列の第一イントロンをcDNAの適切な位置に挿入して修飾cDNAを構築した(Z54)。これは、ポリメラーゼ連鎖反応法を使用してcDNAの5’末端からエキソン2内に位置する BsmI 部位(アミノ酸 43 )まで伸びる断片を増幅することにより行い、これは次いで、pBluescript におけるcDNAにクローン化することができた。細胞質ゾルGS3Aに対しては、ゲノムGS3Aクローンの BgIII-KpnI 断片を、pBluescript cDNAクローンに交換クローン化することにより修飾DNA(Z17)を構築し、全てのゲノムイントロン(アミノ酸6より前方)が挿入されたcDNA配列を作った。イントロンを入れてcDNAを構築することの目的は、単子葉植物で示されたように(Sinibaldi および Mettler, 1991)、トランスジェニック植物での発現を高めるために試みることであった。そのcDNAをpBluescript から下記のバイナリー発現ベクターに移入した。すなわち、GS1−pTEV4 を XbaI-XhoI部位に入れてpZ3(NRRL受理番号 B-21330)を作製し、GS3Aおよび修飾GS3A−pTEV4 を XbaI-XhoI部位に入れて、各々、pZ9(NRRL受理番号 B-21331)およびpZ17(NRRL受理番号 B-21332)を作製し、GS2および修飾GS2−pTEV5 を StuI-KpnI部位に入れて、各々、pZ41(NRRL受理番号 B-21333)およびpZ54(NRRL受理番号 B-21334)を作製した。
【0077】
6.1.3. 植物の形質転換
バイナリーベクター構築体を、以前に記載された方法を使用して三親交配することにより、無力のアグロバクテリウム (Agrobacterium)株 LBA4404に移入した (Bevan, 1984, Nucleic Acids Res. 12:8711-8721)。Nicotiana tabacum 系 SR1を、葉接種法により形質転換し (Horschら、1985, Science 227-1299-1231)、再生した苗条(shoot) を、 200μg/mlのカナマイシンを含む培地上で選択した。一次形質転換体を滅菌培地で保持し、次いで、成熟するまで土壌で成長させた。トランスジェニック種子を 10%次亜塩素酸ナトリウムで滅菌し、100 μg/mlカナマイシンを含む培地上で発芽させた。
【0078】
6.1.4. GSタンパク質および酵素活性分析
可溶タンパク質を、以前に記載されたように、タバコおよびエンドウの葉の組織から抽出した(Tingeyおよび Coruzzi, 1987, Plant Physiol. 84:366-373)。タンパク質を変性し、12% アクリルアミドのSDS−PAGEにより分離し、ニトロセルロース上に電気ブロットした。ウェスタン分析法を、Promega 製の ProtoBlotキットならびにタバコ葉緑体GS2および Phaseolus細胞質ゾルGSに対して高められた抗体の混合物を使用して行った (Hirel ら、1984, Plant Physiol. 74:448-450; Lara ら、1984, Plant Physiol. 76:1019-1023)。形質転換体における全GS活性を、以前に記載されたADP−依存トランスフェラーゼアッセイを使用して測定した (Shapiro および Stadtman, 1970, Methods Enzymol. 17A:910-922)。非変性ゲル電気泳動は、GSアイソザイム検出のためのADP−依存トランスフェラーゼアッセイと共に公知のプロトコール (Davis, 1964, Annals New York Acad. Sci. 121:404-427)に従った。
【0079】
6.1.5. RNA分析
RNAを、Bio101製の「RNAマトリックス」を使用し、製造者の示唆する以下のプロトコールに従って単離した。全RNAを、40 mM のトリエタノールアミン、2 mMのEDTAおよび 3.2% のホルムアルデヒド/ 1.2% アガロースで電気泳動した (Thomas, 1983, Methods Enzymol. 100:255-266) 。ゲルを 10 mMのリン酸ナトリウムに浸漬し、Hybond-Nナイロン膜 (Amersham) 上にキャピラリーブロットした。cDNAを、NEN 製のランダムプライマーおよび拡張試薬標識システムを使用して標識化し、鎖特異的リボプローブを、StratageneRNA転写キットを使用して作った。水性ハイブリダイーションを、膜の製造者のプロトコールに従って行い、ブロットを 0.1xSSRE、0.1%xSDSで洗浄した。
【0080】
6.1.6. 植物の成長条件
先に高いレベルでGS1またはGS3Aを発現すると解析された一次形質転換体の子孫を、100 μg/mlのカナマイシンを含む Murashige-Skoog (MS) 培地で発芽させた。14日後、カナマイシン耐性苗を、白砂を入れた4インチのポットに移し、Saran Wrap (登録商標) で約1週間覆って、過剰な蒸散を防ぎ、苗が定着できるようにした。ポットを、唯一の窒素源として 10 mMの硝酸カリウムを含む過剰の 1X Hoagland溶液で定期的に湿らした。次いで、毎週、3〜7本の植物を切って新鮮重量を測定し、隣り合った植物の陰になるのが顕著になるまで4週間続けた。植物を、明−暗のサイクルを 16-8 時間とし、温度のサイクルを 24-18℃として成長させた。昼間の光の強さは 1000 lux であった。
【0081】
6.2. 結果
6.2.1. トランスジェニック植物に導入したGS構築体
葉緑体GS2 (aka GS185 (Tingey ら、1988, J. Biol. Chem. 263:9651-9657))、細胞質ゾルGS1 (aka GS299 (Tingey ら、1988, J. Biol. Chem. 263:9651-9657))およびGS3A (aka GS341 (Tingey ら、1987, EMBO J. 6:1-9)) に対する Pisum sativum cDNAを pTEV バイナリー発現ベクターに挿入して(図3および4参照)、CaMV 35Sプロモーターの後ろを発現させ、トランスジェニックタバコに移入した。GS2(図4、構築体Z54)およびGS3A(図4、構築体Z17)に対しては、1個以上のイントロンを挿入したcDNAを構築し、CaMV 35Sプロモーターの後ろを発現させた。イントロンを挿入したcDNAを構成することの目的は、単子葉植物で示されたように(Sinibaldi および Mettler, 1991, Progress in Nucleic Acid Research and Molecular Biology 42:1991)、トランスジェニック植物での発現を高めるために試みることであった。さらに、未修飾の全長GS cDNAも、GS2(Z41)、G3A(Z9)およびGS1(Z3)に対して、35S-CaMVプロモーター下で発現させた(図4参照)。図4に詳述した 35S-CaMV-GS構築体の各々に対して、少なくとも8個の一次(T1)形質転換体を分析し、代表的サンプルを図5に示す。選択した一次形質転換体に対しては、4個のカナマイシン耐性T2子孫植物も分析した(図6)。以下に示すT1およびT2植物の分析は、ウェスタン分析法(図5および図6、パネルA);ノーザンブロット分析法(図6、パネルB)、GSホロ酵素分析(図6、パネルC)およびGS酵素活性分析(図6、パネルCならびに表1Aおよび1B)を含み、分析した全トランスジェニック系を代表している。
【表1A】
【表1B】
【0082】
6.2.2. 35S−葉緑体GS2遺伝子融合体を有するトランスジェニック植物の分析
35S−GS2構築体(図4、Z41またはZ54)のいずれかを含むトランスジェニック植物を分析した。35S−GS2(Z41)および修飾した(イントロンを含む)35S−GS2構築体(Z54)は共に、一次T1形質転換体およびT2子孫植物の両方に対して同様の結果を与えた。全一次形質転換体のウェスタンブロット分析は、野生型タバコ(図5、レーンTL)と比較して、葉緑体GS2ポリペプチド(ctGS)の量がかなり減少したことを示した(図5、レーン3〜6)。ポリクローナルGS2抗体は、エンドウおよびタバコの両方のGS2を認識することが分かっているので(Tingeyおよび Coruzzi, 1987, Plant Physiol. 84:366-373; Tingey ら、1988, J. Biol. Chem. 263:9651-9657) 、この減少は、宿主のタバコGS2遺伝子およびエンドウGS2トランス遺伝子の両方の下方調整を表す。細胞質ゾルGSポリペプチド(cyGS)の量は、形質転換していない対照の野生型タバコ(図5、レーンTL)と比較して変わらないことがこれらの形質転換体(図5、レーン3〜6)で認められた。Z41の場合は、全部で14個の独立した一次形質転換体の全GS活性が下方調整され、高くて野生型の活性の85%、低い方は野生型GS活性の22%であった(表1Aおよび1B)。Z54構築体の場合は、再生された全部で9個の独立した一次形質転換体が野生型GS活性の50%以下に下方調整され、その範囲は49%〜11%であった(表1Aおよび1B)。これらのデータから、イントロンを含むZ54構築体がかなり共同抑制されたことは明らかである。対照的に、Z41構築体は、内在性タバコ葉緑体GS2の下方調整の点であまり有効でなく、これらの植物はより広い範囲の共同抑制表現型を示した(表1Aおよび1BのZ41の個体間のGS活性の変化を参照)。典型的には、GS2が共同抑制された植物は、野生型よりもゆっくり成長し、光呼吸中のアンモニアの蓄積と関連する毒性またはグルタミンの欠乏のいずれかにより、介在する黄白化を発生した(図10参照)。従って、これらの形質転換体は、以前に記載されたオオムギのGS2突然変異体に類似していた(Wallsgroveら、1987, Plant Physiol. 83:155-158)。高められた (1.2%) CO2 の雰囲気下(光呼吸の抑制のため)で成長させた、またはグルタミンを補足したZ41またはZ54型のいずれかの共同抑制された植物は、あまり重い症状を示さなかった。このことも、これらの植物がGS2欠乏であるという結果を支持するものである。
【0083】
一次Z41およびZ54形質転換体由来の4個のカナマイシン耐性T2子孫植物も分析した(図6)。子孫のウェスタン分析により得られた結果および全GS活性に対して得られた結果は、一次形質転換体に対して認められた結果と同様であった(図6、パネルA、および表1B)。図6は、いくつかのZ54またはZ41一次形質転換体の代表的な一つのT2子孫のデータを示す。これらの植物のウェスタンブロット分析から、葉緑体GS2タンパク質の量が少ないことが確認され(図6、パネルA)、非変性GS活性ゲル分析により、GS2ホロ酵素の量が野生型のタバコ(図6、パネルC、レーンTL)と比較して少ないことが確認された(図6、パネルC、レーン9〜14)。ノーザン分析は、GS2トランス遺伝子からの転写体が、対照のエンドウRNA(図6、パネルB、レーンP)に存在するものと比較して検出できないことを示した(図6、パネルB、レーン9〜14)。これらの結果は、エンドウGS2トランス遺伝子の挿入によるタバコ葉緑体GS2の特異的共同抑制を示唆する。さらに、エンドウGS2トランス遺伝子も抑えられた。細胞質ゾルGSmRNAおよびタンパク質のレベルは、これらのGS2共同抑制植物において影響を受けなかった。
【0084】
6.2.3. 35−S細胞質ゾルGS3A遺伝子融合体を有するトランスジェニック植物の分析
35S−GS3A構築体(Z17またはZ9;図4)のいずれかを含むトランスジェニック植物を分析した。Z17(イントロンを含む系)の場合、GS活性を分析した13個の独立した一次形質転換体のうち、6個はGS活性の過剰発現を示した (119 〜 145%)が、7個は、形質転換しなかった対照(100%)と比較して、共同抑制(52〜28%)を示した(表1Aおよび1B)。図5および6は、Z17の代表的な過剰発現体および共同抑制された系のデータを含む。形質転換体Z17−12は、GS酵素活性が共同抑制されており(野生型の27%)、葉緑体GS2および細胞質ゾルGSタンパク質の両方とも、野生型のタバコ(図5、レーンTL)と比較して低い(図5、レーン2)。対照的に、形質転換体Z17−6は、全GS活性レベルが高められ(127%)、細胞質ゾルGSタンパク質レベルが野生型のタバコ(図5、レーンTL)と比較して増加した(図5、レーン1)。他の独立形質転換体のT2子孫の分析では、細胞質ゾルGSタンパク質が下方調整された形質転換体がさらに示されたが(Z17−9BおよびZ17−10;図6、パネルA、レーン6および7)、他は、細胞質ゾルGSのレベルが高められた(Z17−7およびZ17−9A;図6、パネルA、レーン4および5)。Z17植物に対して認められた共同抑制現象(Z17−9B、Z17−10およびZ17−12)は、GS2形質転換体(Z54およびZ41)に対して認められた現象とは、葉緑体GS2および細胞質ゾルGSの両方がGS3A共同抑制植物において下方調整されるという点で明らかに異なる(図6、パネルA、レーン6〜8、レーン9〜14参照)。図6は、35S−GS3A(Z17−9B、Z17−10、Z17−12)によって引き起こされる共同抑制がGSの量の減少(ウェスタンおよびGS活性ゲル分析;図6、パネルAおよびC、レーン6〜8)およびGS3Aトランス遺伝子の本質的に検出できない転写(ノーザン分析法;図6、パネルB、レーン6〜8)を伴うことを示す。GS3A構築体を過剰発現する形質転換体(Z17−6、Z17−7およびZ17−9A)では、GS3A転写体が非常に多く(図6、パネルB、レーン3〜5)、このことは、ウェスタンブロット分析(図6、パネルA、レーン3〜5)およびGS活性分析(表1)によって検出可能な細胞質ゾルGSの量がより多いことを表す。細胞質ゾルGS3Aを過剰発現するこれらのZ17形質転換体の可溶タンパク質の非変性GS活性ゲル分析は、野生型タバコの葉において支配的な葉緑体GS2ホロ酵素(バンドB、図6、パネルC、レーンT)よりもゆっくり移動する新規GSホロ酵素の存在を示す(バンドA* 、図6、パネルC、レーン3〜5)。同じGS3Aトランス遺伝子構築体を有する個々のZ17形質転換体が二つの異なる表現型である共同抑制型(図6、レーン6〜8)および過剰発現型(図6、レーン3〜5)を与えるというのは興味深いことである。
【0085】
分析するトランスジェニック植物集団の大きさを拡大するために、形質転換の第二ラウンドを行い、上記と同様の結果を得た。分析した全部で23個の独立した一次Z17形質転換体のうち、5個はGSが共同抑制され、8個はGSを過剰発現した。さらに、修飾していない(イントロンを含まない)GS3AcDNAを含む一次形質転換体を分析した(Z9、図4)。分析した4個のZ9一次形質転換体のうち、1個はGSが共同抑制され、2個は細胞質ゾルGSを過剰発現した。このことは、Z17(イントロンを含む35S−GS3A)およびZ9(35S−GS3A cDNA)構築体の間に定性的な相違はないことを示唆した。特に興味深いのは、Z17−9AおよびZ17−9B(図6、レーン5および6)のこれらの二つのT2植物が、単一の一次形質転換体からの自家受粉によって誘導されたけれども、種々の表現型を有するはずであるという観察である。Z17−9一次形質転換体の全GS活性を分析し、活性が減少し、従って共同抑制されたことを見出した(表1参照)。Z17−9の他の二つのT2子孫植物を分析し(Z17−9CおよびZ17−9D)、これらが共に共同抑制されて、この集団における共同抑制の割合が3:1と優勢であることが分かった。
【0086】
6.2.4. 35S−細胞質ゾルGS1遺伝子融合体を有するトランスジェニック植物の分析
35S−GS1構築体(Z3;図4参照)を含むトランスジェニック植物も分析した。独立した8個のZ3一次形質転換体のうち、5個は、ウェスタンおよびノーザンブロット分析から、明らかに過剰発現の表現型であり、共抑制型はなかった。これらのZ3形質転換体のうち二つのT2子孫を図6に示す。Z3−1およびA3−2のどちらも、細胞質ゾルGSタンパク質の量の増加を示し(図6、パネルA、レーン1および2)、これは、GS mRNAのレベルの増加によって表される(図6、パネルB、レーン1および2)。非変性活性ゲル分析は、タバコの葉の葉緑体GS2ホロ酵素(図6、パネルC、レーンT)よりも速く移動するGSホロ酵素(バンドC)(図6、パネルC、レーン1および2)を示した。このZ3植物中のより速く移動するGSホロ酵素(バンドC)は、天然のエンドウ細胞質ゾルGSと大きさが対応する。
【0087】
6.2.5. トランスジェニック植物における天然および新規の細胞質ゾルGSホロ酵素の分析
細胞質ゾルGS3A(Z17)およびGS1(Z3)の異所性発現は、さらに、しかし、野生型タバコの葉に見られる葉緑体GS2(バンドB)と比較して異なるGSホロ酵素活性バンド(例えば、バンドA* およびC)を与えた。これらのトランスジェニック植物からの抽出物の電気泳動を非変性活性ゲルで繰り返した。該ゲルには、比較のために、細胞質ゾルGSホロ酵素(バンドC)に富むエンドウの根(PR)およびタバコの根(TR)のタンパク質のレーン(図7A、レーン2および4)、ならびに葉緑体GS3ホロ酵素(バンドB)に富む精製したエンドウの葉緑体(PC)およびタバコの葉緑体(TC)から得た抽出物(図7A、レーン1および3)を含めた。トランスジェニックタバコZ3−1の抽出物(図7A、レーン6)に見られる別のGS1ホロ酵素活性(バンドC)は、天然のエンドウの細胞質ゾルGSバンド(バンドC、図7A、レーン2および4)と共に移動する。対照してみると、Z17−7トランスジェニック植物の葉(図7A、レーン5)に見られる新規GS3A活性(バンドA* )は、細胞質ゾルGS(バンドC)とも、葉緑体GS2バンド(バンドB)とも一緒に移動しない。そして、大きさはより大きく、電荷はより酸性側である。GS活性バンドA* 、BおよびCのサブユニット組成物を決定するために、これらのバンドを調製ゲルから切り取り、変性SDSゲルに再び載せ、次いで、GSサブニットのウェスタンブロット分析を行った(図7B)。この分析は、GS活性バンドA* およびCが共に、もっぱら細胞質ゾルGSポリペプチドを含むことを示した(図7B、レーン2および4)。この知見では、より大きいGS3A活性バンドA* が、異種GS3A細胞質ゾルサブユニットと内在性タバコ葉緑体前駆体GS2サブユニットとの会合の結果であるという可能性は少なかった。GS活性バンドA* は、トランスジェニックGS3Aサブユニットとシャペロニン型タンパク質との集合を表すと考えられるが、該複合体をATPで解離するという試みは成功しなかった。その結果、新規GSホロ酵素の性質は不明のままである。
【0088】
6.2.6. 細胞質ゾルGS1またはGS3Aを異所的に過剰発現する形質転換体の成長分析のための選択
細胞質ゾルGS3A(Z17)または細胞質ゾルGS1(Z3)を異所的に過剰発現する2組の植物を選択して成長分析を行った。第一ラウンドの形質転換から(下記実験A参照)、植物Z3−1およびZ3−2をGS1高レベル発現体として選択し(図6、レーン1および2;図8、レーン1および2)、植物Z17−6およびZ17−7をGS3A高レベル発現体として選択した(図5、レーン1;図6、レーン3および4;図8、レーン3および4)。これらの形質転換体のカナマイシン耐性T2子孫を以下に記載する実験Aでの成長分析のために選択した。第二ラウンドの形質転換から、二つのさらに独立して形質転換したGS1−過剰発現植物(Z3−3およびZ3−4)(図8、レーン5および6)および二つのさらに独立して形質転換したGS3A−過剰発現植物(Z17−3およびZ17−11)(図8、レーン7および8)を選択して分析した。これらの植物のカナマイシン耐性T2子孫を、第二成長実験で分析した(下記実験B)。
【0089】
6.2.7. 植物成長実験の設計
植物成長分析を、図8でGSタンパク質およびRNAの分析を行ったT2子孫植物に対して行った。個々のT2植物を白砂中で成長させ、その成長を、タイムポイントごとに4〜7個の植物の新鮮重量を測定することにより評価した。新鮮重量の測定は、植物が急速に成長し、供給した窒素にもっぱら依存し、抽薹および開花中に生じる可能性のある窒素の大きな内部源を流動させることのない栄養成長期中のみ行った。植物は、窒素が無制限であり(すなわち、10 mM の硝酸塩の施肥が定期的に行われる。)、隣接する植物の光合成妨害が少ない条件下で成長させ、そのような妨害が現れたときに成長分析を終わらせた。分析した全ての植物は同じ週齢であり、0.1 〜0.3 g の新鮮重量で分析を開始し、植物が約6週齢で、隣接する植物の妨害が抽薹の開始時に現れるまで続けた。
【0090】
6.2.8. 植物成長実験A
表2に、Z3−1およびZ3−2の系(過剰発現GS1)ならびにZ17−6およびZ17−7の系(過剰発現GS3A)の全新鮮重量測定値の平均の結果を示す。これらの結果を、図9、パネルAでグラフにより示し、表3で統計的に分析した。トランスジェニック系の全4個の過剰発現するエンドウの細胞質ゾルGSは、コントロールよりも35% 〜 114% だけ大きくなった。これは、3個の系、すなわち、Z3−2(P=0.08)、Z17−6(P=0.0015)およびZ17−7(P=0.013 )に対して統計的に重要であった(表3)。
【0091】
6.2.9. 植物成長実験B
成長実験を、同じGS1(Z3)およびGS3A(Z17)構築物を有する種々のトランスジェニック系で繰り返して、上記で得られた結果を確認した。この実験は、統計的分析のために、より大きい植物集団を含む。表2は、トランスジェニック系Z3−3、Z3−4、Z17−3およびZ17−11ならびに2個のコントロール系(C1、C2)の4回のタイムポイントに対する平均データを示す。Z3−4を除く全ての系は、コントロールよりも 40%〜44% だけ大きくなり、6週目の新鮮重量の違いは、統計的に有意であった(表3)。これらの結果はまた、図9、パネルBにグラフにより示す。第二の成長実験は第一の実験結果を確証するものであることは明らかであり、これは、エンドウの細胞質ゾルGS1またはGS3Aのいずれかの異所性過剰発現がタバコの成長速度を高めたことを示唆するものである。テストした全ての系において、GS3A過剰発現は、非形質転換コントロールと比較して成長速度を増加させ、これは、トランスジェニックタバコに対して統計的に有意な成長速度の増加であった。
【表2】
【表3】
【0092】
6.2.10. 植物の成長に対するGS過剰発現の定性的影響
図10は、GSを過剰発現する植物の成長表現型(Z3−A1およびZ17−B7)のコントロール植物およびGSが共同抑制される植物(Z54−A2)の表現型に対する定性的比較を示す。その結果は、GS過剰発現のレベルがたとえ低くても、容易に認められる成長の改善が得られることを示す(図10、Z17−B7およびZ3−A1の成長をコントロール植物の成長と比較)。さらに、これらの結果は、成長の改善がGS過剰発現によるものであり、CaMV−35S GS構成体による植物の単なる遺伝子操作によるのではないことを示す。例えば、CaMV35S−GS2により遺伝子操作され、GS発現が共同抑制されたZ54−A1は、かなり小さい成長を示した。さらに、これらの結果は、GS活性が植物成長の律速段階であることを示す。というのは、この酵素の阻害により成長に対する重大な表現型の影響が引き起こされるからである。
【0093】
6.2.11. GS活性と最終の新鮮重量および全タンパク質との間の相関性
GS遺伝子の異所性過剰発現または共同抑制と関連するGS活性の変化が、栄養成長期の終わりの「最終」の新鮮重量に対して影響があるかどうかを調べるための実験を行った。成長分析は、GS2によって共同抑制される系(Z54−4)、GS1を過剰発現する系(Z3−1)、GS3Aを過剰発現する系(Z17−7)および形質転換されていないタバコのコントロール(SR1)に対するT2世代植物において行った。植物を砂土で成長させ、10 mM のKNO3 を含む Hoagland 溶液で定期的に湿らした。指定のタイムポイントで、各系の8個のT2植物を秤量し、葉のGS活性を各々について測定した。このデータを分析すると、32日および43日の両日に測定した個々の全ての「最終」の新鮮重量とGS特異的活性との間には直線的関係があることが分かる(図11A)。例えば、GS活性が共同抑制されるZ54−4植物(野生型GS活性の 27%)はコントロールの半分の重量であり、GS3A(136 % GS活性)またはGS1(284 % GS活性)を過剰発現する植物は、各々、コントロールよりも1.5 倍および2倍重い。これらの同じT2植物の場合、葉の全タンパク質(μg タンパク質/ gm 新鮮重量)と葉のGS活性との間にも直線的関係が存在する。最も高いレベルのGS活性を発現する植物(284 % )は、可溶タンパク質レベル/ gm 新鮮重量が、コントロールと比較して 1.5倍高かった(図11B)。このデータの無対T−テスト分析は、GS過剰発現系(Z3−1、Z17−7)が、p<0.0001のときにかなり大きいGS活性、新鮮重量、および葉の可溶タンパク質を有することを示した。ただし、p値が 0.0007 であるZ17−7に対する新鮮重量は除く。同様に、GS2によって共同抑制される系(Z54−4)は、p<0.001 のときに、コントロールSR1よりもかなり小さいGS活性、新鮮重量、および葉の可溶タンパク質を有した。成長実験で使用されるGS過剰発現T2系(Z3、Z17)のGS活性プロフィルは、T2世代ではGS活性が一貫して高かったことを除き、親のT0系およびT1子孫に匹敵する。このことは、恐らく、GSトランス遺伝子と会合した KanR 表現型の分離が認められなかったので、トランス遺伝子のいくつかまたは全部が、T2世代でホモ接合体になったという事実によると考えられる。成長実験の終わりには、GSを過剰発現するトランスジェニック系は、コントロールよりも視覚的に緑が強くなり、劇的に大きくなった。
【0094】
6.3. 考察
遺伝子工学が収穫のある植物の改善において重要であると考えられ始めるにつれて、選択した遺伝子の過剰発現において決定的なパラメータを理解することがますます重要になってくる。宿主植物相同体が存在する遺伝子の過剰発現が、ウイルスコートタンパク質およびBT毒性遺伝子などの相同体のない遺伝子の過剰発現より複雑であることは明らかである(Powell-Abel ら、1986, Science 232:738-743; Vacckら、1987, Nature 328:33-37)。これは、トランスジェニック植物が検出でき、宿主相同体が存在するトランス遺伝子を、恐らくフィードバック阻害またはいくつかの他の機構により抑えることができる共同抑制現象による(van der Krolら、1990, Plant Cell 2:291-299; Napoliら、1990, Plant Cell 2:279-289)。本発明では、同じ構成プロモーター(35S−CaMV)の後ろの葉緑体または細胞質ゾルGSに対する3種類のエンドウGS遺伝子をトランスジェニックタバコで異所的に過剰発現する試みを呈示する。その試みの結果、各GS遺伝子に対して異なる過剰発現および/または共同抑制が得られた。さらに、過剰発現に成功した細胞質ゾルGSに対する2種類の遺伝子(GS1およびGS3A)の場合、その過剰発現により、GS RNA、タンパク質および酵素の過剰産生だけでなく、窒素使用効率が改善された表現型が得られた。
【0095】
細胞質ゾルGS1に対するエンドウ遺伝子のタバコでの過剰発現は、GS活性および細胞質ゾルGSタンパク質が増加し、トランス遺伝子mRNAのレベルが高い純粋な表現型を与える。さらに、GS1タンパク質は、会合して、大きさおよび電荷が天然のエンドウの細胞質ゾルGSと同じであるGSホロ酵素になる。細胞質ゾルGS3Aを過剰発現するトランスジェニック植物においては、状況がいくらか異なる。高レベルのGS3Aトランス遺伝子mRNAは、ウェスタンブロットで可視化される細胞質ゾルGSのレベルの増加を引き起こす。しかし、GS3Aの過剰発現は、エンドウまたはタバコの天然の葉緑体または細胞質ゾルGSホロ酵素より大きい新規GSホロ酵素の出現を引き起こす。これらのトランスジェニック植物において、細胞質ゾルGS遺伝子は、通常は見られない細胞の型(例えば、葉肉細胞)で発現され、GS3AトランスジェニックリーフのGSホロ酵素がより大きいのは、細胞質ゾルGSサブユニットが天然の葉緑体前駆体GS2と共会合することによると考えられた。しかし、この新規GS3Aホロ酵素は、もっぱら細胞質ゾルGSサブユニットから構成されることが分かっており、従って、トランスジェニックGS3Aサブユニットと内在性のタバコ葉緑体前駆体GS2との共会合によるのではない。二つの他の可能性が存在する。より大きいGS3Aホロ酵素は、トランスジェニックGS3Aサブユニットが集合して、通常の八面体構造以外の立体配置になった結果であると考えられる。あるいは、新規のGS3Aホロ酵素は、過剰発現した細胞質ゾルサブユニットが会合したシャペロン型から解離し損なった結果であるかもしれない。実際、GSと groEL様タンパク質との密接な会合が、以前、エンドウで確認された (Tsuprun ら、1992, Biochim. Biophys. Acta 1099:67-73)。しかし、新規GS3A活性バンドを潜在的なシャペロン型からATPによって分離する試みは失敗に終わった。新規GS3Aホロ酵素は、(GS活性ゲル分析での検出から)明らかにGS活性を有するにちがいないが、この新規GSアイソザイムが、天然の細胞質ゾルGSまたは葉緑体GS2ホロ酵素と等モルベースで同様の活性を有するかどうかを模索するのは興味深い。もし有するとすれば、35S−GS3Aを過剰発現し、従って新規GSホロ酵素を有する植物は、高められた全GS活性を有すると予想することができる。実際はそうではなく、4個のZ17−6のT2子孫植物(GS3Aを発現)の全GS活性の平均(野生型との比較)は100%であることが分かり、4個のZ17−7の子孫植物の平均は、野生型と比較すると 107% であった。対照すると、Z3−1およびZ3−2(GS1天然ホロ酵素を過剰発現)のT2子孫に対して得られるGS活性値は、野生型と比較して、各々、123%および120%であった。これは、Z3の過剰発現形質転換体でGS1サブユニットが集合して天然の大きさのGSホロ酵素になることは全GS活性に対して有利であることを示唆する。
【0096】
ここで、窒素の使用効率を、細胞質ゾルGS1または細胞質ゾルGS3Aの過剰発現に成功したトランスジェニックタバコの栄養成長期に評価した。栄養成長期中には、栄養の急速な吸収および光合成能の最大限の活用を特徴とする急速な葉の発達がある。窒素は、最もしばしば制限する微量栄養であり、植物内でのその吸収および使用の生理機能は、栄養期と生殖期とでは異なる。第一に、土壌からの窒素の混入、伸び広がる細胞へのその混入および光呼吸による損失の制限があり、次いで、抽薹の開始とともに、生殖期中には、種子の収穫に変換するための窒素保存物の流動がある。恐らく、窒素使用効率は、栄養成長期中はあまり複雑ではなく、本発明者らのトランスジェニック植物成長分析の焦点はこの成長期にある。
【0097】
本発明の知見は、タバコで異所的に発現されるエンドウ細胞質ゾルグルタミンシンテターゼがトランスジェニックタバコの栄養成長期にかなりの利点を付与することを示す。細胞質ゾルGS1またはGS3Aを異所的に(すなわち、全細胞型で)過剰発現する植物は、全新鮮重量がコントロールよりも高い。特に顕著なのは、GS3Aを発現する全て系(Z17)の全新鮮重量が6週目のコントロールより高く、これらが統計的に常に重要であることである。各ケースで、コントロールとトランスジェニック系との間の相違がサンプルの差異による確率は 5%未満であった。分析したGS1発現系(Z3)の場合、4個のうち3個はコントロールより大きくなり、3個のうち2個は、その相違が 10%のレベルで統計的に重要であった。この増加した窒素使用効率は、作物を同様に遺伝子操作して、通常の窒素量で良好に成長させたり、肥料の少ない投入で栽培したり、あるいは品質の悪い土壌で栽培することができ、従って、開発された、または開発中の農業システムにおいて、経済的にかなり影響がある。
【0098】
GS過剰発現は、以前にトランスジェニックタバコで試みられたが(Eckes ら、Mol. Gen. Genet, 217: 263-268; Hemonら、1990 Plant Mol. Biol. 15: 895-904; Hirel ら、1992, Plant Mol. Biol. 20:207-218; Temple ら、1993, Mol. Gen. Genet. 236:315-325) 、GSの過剰発現をGS活性のかなりの増加ならびに植物の成長および栄養特性における改善と相関させた最初の報告は本発明である。Templeらは、GS mRNAおよびタンパク質の増加を報告したが、トランスジェニック植物でのGS活性の対応する増加は報告しなかった(Templeら、同書)。Hemon らは、GS発現構築体により遺伝子操作したトランスジェニック植物でのGS mRNAレベルの増加を報告したが、GSタンパク質または酵素活性の対応する増加の報告はなかった(Hemon ら、同書)。他の二つの報告では、トランスジェニック植物でのGS遺伝子の過剰発現の結果、GS酵素のレベルが増加したが、GS過剰産生の表現型による影響についての報告はなかった(Eckes ら、同書; Hirelら、同書)。植物成長速度を約 20%改善するアルファルファGS遺伝子の過剰発現の報告が一つある(Eckes ら、1988, オーストラリア特許出願 No. AU-A-17321/88)。しかし、ここで報告された改善は、低窒素条件下での成長のみに限られていると思われる。同様に遺伝子操作した植物について、窒素無制限媒体で行った続く分析では、コントロール植物と比較して、表現型の変化を示さないことが報告された(Eckes ら、1989, Mol. Gen. Genet. 217:263-268)。さらに、より速い成長速度により、遺伝子操作した植物とコントロール植物との間で最終の新鮮重量に相違が生じるという報告はない。これらの以前の研究に対して、本発明は、窒素条件とは無関係に、GS過剰発現が植物の成長、収率および/または栄養特性を改善することができることをはっきり例証するものである。
【0099】
本発明の農業上の有用性は、栄養器官が採取される作物の種類に直接関係し、これらには、全ての飼料穀物、馬鈴薯、テンサイおよびサトウキビならびにタバコが含まれる。最終の新鮮重量の記録の1週間以内に、植物は、節間の伸長を受け始め、各集団に対する次の新鮮重量の測定の標準偏差は、植物の異なる生理学的段階の結果、増加した。増加した栄養成長速度も種子の収率をかなり有利にするかどうかは重要な問題であり、まだ回答は出されていない。種子の収率および種子の窒素含量に関係する生理学的パラメーターは、窒素吸収効率だけでなく、抽薹開始時の保存物の再流動および畑の集団密度の成りゆきも含む。そのような研究は、種子の収穫に対して選択され、収穫生理学に関していくらか知識があるトランスジェニック種において行うとより良好である。
【0100】
細胞質ゾルGS1によるのではなく、エンドウの葉緑体GS2および細胞質ゾルGS3Aをコードする遺伝子による内在性タバコの共同抑制も興味深い。これは、エンドウGS2がタバコ葉緑体GS2型のみを抑制し、一方、GS3Aがタバコ葉緑体GS2および細胞質ゾルGSの両方を抑制するときに、特にそうである。以前、ペチュニアカルコンシンテターゼおよびジヒドロフラバノール−4−リダクターゼが、トランスジェニックペチュニアで、内在物およびトランス遺伝子の両方を抑制することが示された(van der Krolら、1990, Plant Cell 2:291-299; Napoliら、1990, Plant Cell 2:279-289)。さらに最近、カルコンシンテターゼ遺伝子の5’または3’端は、共同抑制を引き起こすのに十分であるが、プロモーターの小さい遺伝子はそうではないことが報告された(Jorgensen, 1992, Agbiotech News and Information Sept: 1992)。これは、転写開始剤の必要性を示唆するものである。一時的な異所性配列対合が、遺伝子抑制の可能な機構として引き合いに出され、これは、たぶん、転写の開始と関係するDNAの巻き戻しに依存すると考えられる(Jorgensen, 1990, Trends in Biotechnology 8:340-344; Jorgensen, 1991, Trends in Biotechnology 9:255-267; Jorgensen, 1992, Agbiotech News and Information Sept: 1992) 。エンドウGS遺伝子発現に対する本発明の知見から、共同抑制現象は、ヌクレオチドレベルでの配列の完全な相同性には依存しないことは明らかである。
【0101】
窒素同化酵素の発現を加減することによる窒素利用の増加はまた、トウモロコシなどのトランスジェニック植物での収率を高めるための実行可能な方法であると考えられる。作物における窒素使用効率は、高められた収率として測定され、従って、農業的尺度となる。この種の適応または限定は、生存のために、環境条件に対する種々の反応に依存して、より高い収率には依存しない野生型植物に対しては、真に有利であるとはいえない(Sechley ら、1992, Int. Rev. Cyt. 134:85-163 )。従って、作物の収率の増加は、従来の品種改良法よりもむしろ、本明細書に記載したような遺伝子工学法によってより容易に実現することができる。
【0102】
7.0. 実施例:植物中のアスパラギンシンテターゼの異所性過剰発現が植物増殖表現型の増加を引き起こす
以下の研究は、アスパラギン製造の増加及び植物成長に対する影響を試験するとを目的とする、植物中でのAS遺伝子発現の操作に係わる。窒素運搬/貯蔵化合物としてアスパラギンがグルタミンよりも好ましいいくつかの特徴があり、従ってアスパラギンへの窒素の同化の増加はin vivo で価値があり得る。アスパラギンは、グルタミンよりも高いN:C 比率を有する長距離窒素輸送化合物である。従って、それは窒素輸送のためのより経済的な化合物である。それに加えて、アスパラギンはグルタミンより安定であり、液胞中で高いレベルに蓄積することができる(Sieciechowicz et al., 1988, Phytochemistry 27:663-671; Lea and Fowden, 1975, Proc. R. Soc. Lond. 192:13-26)。エンドウの葉の分化の間にはアスパラギンが活発に代謝されるが、窒素を成長のためにもはや必要としない成熟した葉ではアスパラギンは容易には代謝されず、葉から活発な成長領域、例えば分化中の葉や種子に再輸送(師部中)される(Sieciechowicz et al., 1988, Phytochemistry 27:663-671; Ta et al., 1984, Plant Physiol 74:822-826)。ASは通常暗所でのみ発現され(Tsai and Coruzzi, 1990, EMBO J. 9:323-332) 、従って35-AS1は構成的に発現され、細胞型に関して異所的に発現されるだけでなく一時的発現についてもそうである。即ち、本明細書に示した研究は、あらゆる細胞型中での光非依存性形態のASの異所性過剰発現がアスパラギン製造を増加させるかどうかを調べるものである。増加されたアスパラギン製造がトランスジェニック植物の窒素使用効率と成長表現型において利点を与えるかどうかについても本明細書で試験する。
【0103】
過剰発現野性型ASに加えて、本研究ではグルタミン結合ドメインを欠いているAS酵素の改変形態(glnΔAs1)の異所性過剰発現を調べた。この研究により調べられた問題は、その酵素の glnΔAs1形態の異所性過剰発現が増強されたアンモニア依存性AS活性を有する新規な植物AS酵素を生成するか、あるいはそのような突然変異が、内因性野性型ASサブユニットとともに集合することにより優勢ネガティブ効果を有し(Herskowitz, 1987, Nature 329:219-222)、ヘテロダイマーを形成するかどうか(Rognes, 1975, Phytochemistry 14:1975-1982; Hongo and Sato, 1983, Biochim et Biophys Acta 742:484-489)ということである。エンドウASを異所的に発現するトランスジェニック植物の分析により、アスパラギンの増加した蓄積と改善された成長表現型(35S-AS1の場合)、そして成長表現型に対する有害な作用を伴うアスパラギンの増加した蓄積が示された(35S-glnΔAS1の場合)。これらの結果は、窒素代謝と成長表現型をAS遺伝子の異所性過剰発現によって操作することが可能なことを示している。窒素が植物成長の律速元素であることが多く、典型的にはその成長期に作物に数回施用されるので、作物植物における窒素利用効率を改善する分子設計技術は農業にとって大きな利益をもたらすものである。
【0104】
7.1. 方法及び材料
7.1.1. AS遺伝子構築物
以前にエンドウからクローン化されているAS1 cDNA(Tsai and Coruzzi, 1990, EMBO J 9:323-332)を、pTZ18UからpBluescript KS- (Stratagene)のEcoRI部位に移入した。 glnΔAS1欠失突然変異体を、「内部−外部」PCRを使用して構築した(Innis et al., 1990, PCR Protocols: A guide to Methods and Applications. New York, Academic Press pp.1-461) 。アミノ酸2-4(CGI)に対応するコード配列をAS1 cDNAのアミノ末端から削除し、開始メチオニンと非翻訳リーダーをそのまま残した。この欠失が、動物ASについて定義されたアミノ酸MCGIを含んでいるAS酵素の推定グルタミン結合ドメインに対応した(Pfeiffer et al., 1986,J. Biol. Chem. 261:1914-1919; Pfeiffer et al., 1987, J. Biol. Chem. 252:11565-11570)。野性型AS1及び glnΔAS1に対応するcDNAを次にpBluescript からバイナリ発現ベクターpTEV5へ移入した。このベクターは、CaMV 35Sプロモーター(-941から+26まで)、多回クローニングサイト、及びノパリンシンターゼターミネーターを含む。図12は、AS1 cDNA pZ127 (NRRL受託番号 B-21335) 及び glnΔAS1 cDNA pZ167 (NRRL受託番号 B-21336)を含むバイナリーベクター構築物の詳細を示し、これをタバコに形質転換した。
【0105】
7.1.2. 植物形質転換体
バイナリーベクター構築物を、安全化Agrobacterium 株LBA4404 に、その後Nicotiana tabacum SR1 に、他の文献に記載された標準的手順を使用して移入した(Bevan, 1984, Nucleic Acids Res. 12:8711-8721; Horsch et al., 1985, Science 227:1229-1231)。
【0106】
7.1.3. 形質転換体のRNA分析
Bio101からの「RNAマトリックス」を使用してRNAを単離し、全RNAをこれま
でに記載されたように電気泳動にかけた(Thomas, 1983, Methods Enzymol. 100:255-266)。ゲルを毛細管でHybond-Nナイロン膜(Amersham)上へブロットした。cDNAを、NEN により供給されるランダムプライマーと延長試薬標識システムを使用して標識した。ハイブリダイゼーションを水溶液中で行い、ブロットを0.1X SSPE, 0.1% SDS中で洗浄した。ノーザンブロットをエンドウAS1 cDNA, pAS1でプロービングした(Tsai and Coruzzi, 1990, EMBO J 9:323-332)。
【0107】
7.1.4. 遊離アミノ酸の抽出
タバコ葉組織試料を液体窒素中で凍結し、メタノール:クロロホルム:水(12:5:3、v/v/v)からなる抽出培地の10のml中で抽出した。ホモジネートを、12,000 X Gで15分間遠心分離した。ペレットを再び抽出し、上清を合わせた。上清へ2.5 mlのクロロホルム及び3.8 mlの蒸留水を添加すると分離した。メタノール:水相を集め、減圧下に乾燥し、1 mlの蒸留水中に再溶解した。その溶液を12,000 gで2分間遠心分離して0.45μm ナイロンフィルターマイクロ遠心分離チューブフィルターシステムを通しろ過した。
【0108】
7.1.5. アミノ酸プールのHPLC測定
アミノ酸は、Microsorb Type OAA Analysis カラム(Rainin)上でDuPontのHPLC機器を使用してo-フタルジアルデヒド(OPA) 誘導体として測定した。試料(100μL)をOPA 作用試薬の100μl で誘導化した。2分の誘導化の後、誘導化された試料の50μlを注入した。この勾配は 2種の溶離液;A. 4.5% メタノール及び0.5%テトラヒドロフルオランを含む95% 0.1M酢酸ナトリウム(pH 7.2); B. 100 %メタノールを使用して生成した。溶離液をろ過し、使用の前にHeで脱気した。OPA誘導化アミノ酸の検出は、UV分光光度計で340 nmにおいて行った。各測定は二回行い、値はその平均値を表す。
【0109】
7.1.6. 植物成長条件
高レベルのAS1 mRNAまたは変異 glnΔAS1 mRNAを発現するものとして特徴付けられる一次形質転換体の後代を、100 μg/mlのカナマイシンを含むMS培地上で生育させた。14日後、カナマイシン耐性苗木を白砂で満たした4インチの鉢に移し、約1週間サランラップで包んで過剰な蒸発を防ぎ、苗木が根付くようにした。鉢に唯一の窒素源として10mM硝酸カリウムを含む1X Hoagland溶液を定期的に与えた。その後、3〜7の植物を各週に新鮮重量を測定するために採取し、隣り合った植物の陰になるのが顕著になるまで4週間継続した。植物を16-8時間の光-暗所サイクルの下に24-18℃の温度サイクルで成長させた。昼間光度は1000ルクスであった。
【0110】
7.2. 結果
7.2.1. エンドウAS1と glnΔAS1 を発現するトランスジェニック植物の構築
35S-CaMVプロモーターから発現されたエンドウAS1 cDNA (Tsai and Coruzzi,1990, EMBO J 9:323-332) をトランスジェニックタバコに移し(図12及び7.1 材料及び方法の項を参照) 、5つの独立した一次形質転換体(Z127; 1-5)が高いレベルのAS1 mRNAを発現することが示された(下記参照)。グルタミン結合ドメインに欠失を有するAS1 cDNA(glnΔAS1)を含む3つの独立したトランスジェニック系(Z167;1-3)もトランスジーンRNA の高いレベルを含むことが示された (下記参照)。
【0111】
7.2.2. AS1と glnΔAS1を発現する形質転換体のノーザン分析
トランスジェニック植物から抽出されたRNA のノーザンブロット分析を行い、35S-AS1 トランスジーンが高いレベルで発現した植物を同定した(図13)。陽性対照として、ASのRNAを暗所で生育したエンドウ植物の葉中に検出した(図13、レーンPL)。これに対して、光中で生育した野性型タバコの葉にはAS mRNA は検出されなかった(図13、TL)。以前の研究により、タバコAS mRNA は暗所で生育した植物の組織中だけに発現されることが示されている(Tsai and Coruzzi, 1991, Mol Cell Biol 11:4966-4972)。AS1 を過剰発現する形質転換体(Z127-1、-3、-4及び-5)は全て、それらが光中で生育されたにもかかわらず、AS1 mRNAの高いレベルを含んでいた(図13)。このように、35S CaMVプロモーターは、エンドウAS1の構成的な発現を生成するのに対して、内因性AS mRNAは、光中ではタバコ葉中に発現されない。 glnΔAS1形質転換体も、タバコ対照と比較して高水準のmRNAの構成的な発現を示した(Z167-2、-3及び-4) (図13)。AS酵素のよく知られた不安定性のために、AS酵素を均質物にまで精製できず、植物ASの抗体をASタンパク分析のために使用できなかった。それに加えて、in vitroアッセイでは、酵素の不安定さのためにAS活性が検出されなかった。
【0112】
7.2.3. AS1及び glnΔAS1 を発現するトランスジェニック系のアミノ酸分析
ノーザン分析の結果に基づいて、AS1 mRNAの高いレベルを示した2つの独立のトランスジェニック系(Z127-1及びZ127-4)をさらに分析するために選択した。同様に、Z167-2及びZ167-4系を glnΔAS1 構築物の高発現体の代表として選択した。これらの植物のT2後代について、以下に記載するアミノ酸分析及び成長分析を行った。
【0113】
7.2.4. AS1-過剰発現系
選択した両方のZ127系(Z127-1及びZ127-4)は、アスパラギンの増加されたレベル(野性型対照より10〜100 倍高い)を示した(表4)。個々のT2植物の間で見られるばらつきは、個体のホモ接合性またはヘテロ接合性を反映している可能性が最も高く、中間レベル:高いアスパラギンレベルの約2:1 の比は、この考え方の証拠となるものである。しかし、すべての場合においてアスパラギンのかなりの増加は対照濃度のほぼ100 倍まで及んでいることが判る。面白いことには、これらの植物においてグルタミン濃度の対応する減少が存在し(Z127-4のデータは唯一高い値ではずれていたが)、これはAS反応における基質としてのグルタミンの使用を反映しており、同じく予測できるのは他の基質アスパルテートの濃度の減少である。しかし、Z127系中のグルタメートの低下した濃度はいくらか予想外である。生化学的予測とAS反応に関与する他の3つのアミノ酸について集めたデータから、グルタメートの増加が予測された。グルタメートの見かけ上の減少は、アミノ基転移のようないくつかの関連するプロセスにおける基質として使用されたために起こった高い代謝回転の結果であり得る。
【0114】
7.2.5. Gln ΔAS1 過剰発現系
glnΔAS1 を過剰発現する選択された二つの系において調べた問題は、ASのグルタミン結合ドメインの欠失がアスパラギン生合成に対して優勢−ネガティブ効果を有しているかどうかということである。これらの系(Z167-2及びZ167-4)について集めたデータはデータ値のばらつきのために解釈するのがやや困難である(表4)。しかし、ほとんど全ての場合において、野性型非トランスジェニックタバコと比較して3から19倍の範囲のアスパラギン濃度の実質的な増加が存在する。これらの結果は、トランスジェニック系がアスパルテート、グルタメートまたはグルタミンプールに対して殆ど影響を及ぼさずにアスパラギンを蓄積する能力を有することを示唆している。一つの可能性は、 glnΔAS1酵素がアンモニアとアスパルテートから直接アスパラギンを合成できるということである。
【0115】
7.2.6. AS1 及び glnΔAS1を発現する形質転換体についての植物成長実験
白砂中で生育された個体トランスジェニックT2植物を使用して成長分析を行った。これらの研究は、近隣の植物からの干渉を最小にした条件下に成長速度を調べることを目的とする。このため、生育の栄養成長段階(発芽後6週間まで)の間にのみ新鮮重量測定を行った。この間、植物は急速に成長し、供給された窒素にのみ依存し、薹立や顕花の間におこるように内部窒素源を移動させない。植物は、窒素について非制限的(即ち10 mM 硝酸塩の規則的な施肥)で隣接植物からの光合成干渉を減らすような条件下に生育された。その成長分析は、そのような干渉が顕著になったときに終了した。分析したすべての植物は各時点で同じ加齢を有し、 0.1〜0.3 gの新鮮重量/植物の間に分析を開始し、近隣の植物の干渉が顕著になり薹立が間近の約6週齢になるまで継続した。
【表4】
【表5】
【0116】
表5及び6は、系Z127-1及びZ127-4(野性型AS1を過剰発現する)とZ167-2及びZ167-4(glnΔAS1を過剰発現する)についての平均全新鮮重量測定の結果を示し、これらの結果を図13にグラフとして示す。野性型ASを過剰発現しているトランスジェニック系は、対照(100%)と比較して133%及び110%成長したが (表5) 、無対t検定により分析した場合、いずれの場合も統計学的に有意でなかった(表6)。 glnΔAS1 構築物(Z167)を過剰発現するトランスジェニック系は同等に成長しなかった。6週まで生存したZ167-4植物は成長において対照から区別できず、生存したZ167-2植物は対照よりも非常に小さかった(P-0.041;5%レベルにおいて有意)(表5及び6、図14も参照)。実験における3つの異なる系を比較すると、カナマイシン耐性Z167植物の大部分が死滅したことは興味深い。典型的に、Z167植物は鉢で生育されると発芽するのが遅く、不健康に見えた。これは、Z167-2について集められた新鮮重量のデータに明確に反映されていたが、Z167-4データについてはそれほど明確ではなく、glnΔAS1 遺伝子生成物が植物成長に対し実際に優勢−ネガティブ効果を有していたことを示唆している。
【表6】
【0117】
7.3. 考察
本明細書に開示した研究では、ASをトランスジェニック植物中に異所的に過剰発現させ、この操作の一次窒素同化と植物成長に対する影響を試験するものであった。特に、ASの細胞特異的発現パターンを変更し、光に関してASの制御も改変した。野性型植物においては、ASは通常その師部中に発現されるだけであり(Tsai, 1991, Molecular Biology Studies of the Light-Repressed and Organ-Specific Expression of Plant Asparagine Synthetase Genes. Ph.D. Thesis, The Rockefeller University, New York, NY)、その発現は光合成及び非光合成組織の両方において光によって劇的に抑制される(Tsai and Coruzzi, 1990, EMBO J 9:323-332; Tsai and Coruzzi, 1991, Mol Cell Biol 11:4966-4972)。ここで、エンドウの野性型AS1及びAS1 の変異形態(glnΔAS1)は、構成的プロモーター(35S-CaMV)の制御下にトランスジェニックタバコ中に発現され、AS1 がすべての細胞型中に光から独立して発現される。それが通常は発現されない細胞において構成的に発現されるAS1の生理的な意義は、植物窒素代謝に対して重大な影響を有し得る。例えば、アスパラギンは光呼吸窒素リサイクルに関与しており(Givan et al., 1988, TIBS 13:433-437; Ta et al., 1984, Plant Physiol 74:822-826)、従って光合成細胞中のASの異所性発現は光呼吸に劇的な影響を与え得る。さらに、この点でアンモニア依存性AS酵素の発現は光呼吸アンモニアの再同化を助け得る。
【0118】
35S-AS1 を発現する4つの独立したトランスジェニックタバコ系が野生型エンドウAS1トランスジーンを構成的に発現することが示された。2つの系をさらに分析し(Z127-1及びZ127-4)、遊離アスパラギンが高いレベルにトランスジェニック葉組織中に蓄積し、典型的にはトランスジェニック系Z127-1及びZ121-4が対照非形質転換タバコ系の10〜100 倍のアスパラギンを蓄積したことから、発現されたAS1遺伝子が機能的であったことが示された。これらの増加したアスパラギンレベルは、予測されるようにAS基質、グルタミン及びアスパルテートの減少を伴った。しかし、ASの基質であるグルタミンのより高い内因性レベルを与えることによって、より多くの無機窒素を窒素運搬化合物のアスパラギンに導入することはなお可能であり得る。
【0119】
Z127トランスジェニック植物の植物成長実験は、AS1 過剰発現植物中のアスパラギンの蓄積が植物分化の栄養成長期の間の成長に対して、ポジティブな効果を持つかどうかということを調べることを目的とするものであった。栄養成長の間に起こる急速な葉の分化は、栄養の有効性を強く要求し、この時点では組織を広げて大きくすることにおける新しいタンパク質の合成のために窒素が典型的に最も臨界的な栄養素である。このとき同化され蓄積された窒素はその後その植物中でリサイクルされて種子貯蔵物中に蓄積され、また主要な長距離輸送アミノ酸であるアスパラギンも種子貯蔵物の形成に重要な役割を果たす(Dilworth and Dure, 1978, Plant Physiol 61:698-702; Sieciechowicz et al., 1988, Phytochemistry 27:663-671)。2つのZ127系が、栄養成長の終了までの6週間にわたって非形質転換対照より大きくなり、10% 及び33% の成長の優位性が得られたことが判明した。しかしこれらの値は、t検定を行うと統計学的には有意でなかった。即ち、植物は10〜100 倍より高いレベルのアスパラギンを製造するが、グルタミンレベルが成長中の増加に対して制限的であることがあり得る。さらに本明細書に示したのは、トランスジェニックタバコ中でGSを過剰発現することは統計学的に有意な(上出)より大きい成長優位性を与えることができるという知見である。グルタミンがアスパラギン生合成のための基質であるので、両者は無機窒素の一次的同化において中枢となるアミノ酸である。従って、GSとASを高いレベルで発現するトランスジェニック系を形成する(AS及びGS過剰発現体を交雑することによって)ことは、いずれの親よりも有利な成長形質も持つことがあり得ると予測できる。特に本明細書で記載した方法は、トランスジェニック系中の同化が2、3の細胞型に制限されず、すべての植物細胞中で使用可能な窒素が利用されることを可能にするという長所を有する。一つの植物中でのGS及びASの両方の異所性過剰発現はグルタミン蓄積を回避する利点を有し得、これはグルタミンの高い濃度の存在下ではグルタミンが活性な代謝産物であって細胞の代謝を破壊し得るからである。これに対し、アスパラギンは比較的不活性な化合物であり、経済的に窒素を蓄えることができる。さらに、アスパラギンは、グルタメートの分子を遊離する反応において形成され、アンモニア単位をさらに受容するのに利用され得る(Lea and Fowden, 1975, Proc. R. Soc. Lond. 192:13-26).
野性型ASの異所性過剰発現に加えて、1つの試みにおいて植物グルタミン依存性ASを、そのアンモニア依存活性を増強するように改変した。特に、ASのグルタミン結合ドメインに対する抗体が同じASポリペプチドに存在しているグルタミン依存AS活性を抑制するが、アンモニア依存活性は増強することが動物中で示されている(Pfeiffer et al., 1986, J. Biol. Chem. 261:1914-1919; Pfeiffer et al., 1987, J. Biol. Chem. 252:11565-11570)。類推として、突然変異によりグルタミン結合のために必要とされる3つのアミノ酸を特異的に削除された部位特異的変異体(glnΔAS1)がエンドウAS1 cDNAに形成されている(Tsai and Coruzzi, 1990, EMBO J 9:323-332)。この glnΔAS1をトランスジェニック植物に導入することによって、アンモニア依存AS活性を増強すること及び/またはサブユニット活性低下及び野性型と突然変異サブユニットのヘテロダイマーの形成を通して内因性グルタミン依存AS活性を抑制することが可能であり得る。 glnΔAS1トランスジーンを過剰発現する2つの独立のトランスジェニック系、Z167-2及びZ167-4が非形質転換タバコ対照よりも約3〜19倍高いアスパラギンレベルを蓄積し得ることが見出された。アスパラギンの同化における glnΔAS1遺伝子の活性は、グルタミン以外の窒素基質(例えばアンモニア)を利用する能力を持っている改変酵素を示唆するものである。E.coli AsnA遺伝子と哺乳類ASの既知のアンモニア依存AS活性に対する類推から、突然変異植物 glnΔAS1酵素を発現するトランスジェニック植物におけるアスパラギンの高いレベルは、 glnΔAS1 酵素が直接アンモニアをアスパラギンに同化でき、従って一次窒素同化においてGSを迂回し得ることを示唆している。この提案が正しいならば、これらのトランスジェニック植物中のアスパラギンの相対レベル(Z167 対Z127) に基づいて、gln ΔAS1 遺伝子がアスパラギンの合成において過剰発現された野性型AS1ほどは効率的でないことは明らかである。
【0120】
glnΔAS1 を発現するトランスジェニック系(Z167-2 及びZ167-4) は非形質転換対照より大きくならず、実際にはZ167-2の成果と実験の終了前におけるZ167植物の高い死滅割合によって示されるように、それらの成長は非形質転換植物より劣っているのが典型的であった。これらの植物がその葉中に3〜19倍高いレベルのアスパラギンを蓄積するのに成長が劣ることは奇妙である。植物ASはホモダイマーとして集合すると思われる(Rognes, 1975, Phytochemistry, 14:1975-1982)。野性型ASが通常発現されない葉の葉肉組織においては、 glnΔAS1 形態はホモダイマーへ自己集合することができ、アスパラギンを生成することができる酵素を形成する。しかし師部細胞においては、 glnΔAS1 サブユニットは野性型ASサブユニットと共に集合し得、これにより野性型ASを優勢−ネガティブ突然変異として不活性化する(Herkowitz, 1987, Nature 329:219-222) 。 glnΔAS1植物中では、葉の葉肉細胞中に合成されたアスパラギンは、師部に輸送され、ロードされることができないことがあり得、これがこれらのトランスジェニック系の劣った成長表現型の要因となっている可能性がある。これらの観察は、細胞型機能、窒素代謝遺伝子の細胞特異的遺伝子発現及び植物窒素代謝に対するその影響の特性化を明らかにするものである。
【0121】
8. 微生物の寄託
以下の微生物は、Agricultural Research Culture Collection, Northern Regional Research Center (NRRL), Peoria, Illinois寄託されており、以下の受託番号が付与されている。
【0122】
株 プラスミド NRRL受託番号
Escherichia coli, Z3 pZ3 B-21330
Escherichia coli, Z9 pZ9 B-21331
Escherichia coli, Z17 PZ17 B-21332
Escherichia coli, Z41 pZ41 B-21333
Escherichia coli, Z54 pZ54 B-21334
Escherichia coli, Z127 pZ127 B-21335
Escherichia coli, Z167 pZ167 B-21336
本発明をその特異的態様を参照して詳細に記載したが、機能的に等価な改変は本発明の範囲の範囲内であることが理解されるであろう。実際、本明細書に示し記載したものに加えて本発明の種々の改変は、これまでの記載及び添付の図面から当業者に明らかであり、そのような改変は添付の請求の範囲に包含されることを意図するものである。
【0123】
種々の刊行物を本明細書中で引用したが、その全体を引用により本明細書の一部とするものである。
【0124】
【0125】
Form PCT/RO/134 (続き)
アグリカルチュラル リサーチ カルチャー コレクション (NRRL)
インターナショナル ディポジタリー オーソリティー
アメリカ合衆国 61604 イリノイ州
ペオリア,エヌ.ユニバーシティー ストリート 1815
受託番号 寄託日
B−21331 1994年 9月28日
B−21332 1994年 9月28日
B−21333 1994年 9月28日
B−21334 1994年 9月28日
B−21335 1994年 9月28日
B−21336 1994年 9月28日
【受託番号】
【0126】
NRRL B-21330
NRRL B-21331
NRRL B-21332
NRRL B-21333
NRRL B-21334
NRRL B-21335
NRRL B-21336
【特許請求の範囲】
【請求項1】
1種または数種の窒素同化/代謝酵素の異所性過剰発現のための植物の遺伝子工学的操作により改良された農業または栄養特性を有する植物を作出する方法であって、上記の改良された農業または栄養特性を有する植物が、同様に栽培した遺伝子操作されていない前駆植物と比べて、該植物と該前駆植物を窒素非限定生育条件下で栽培するときに次の特性:
i)より速い成長速度、
ii)より多い成熟時の新鮮または乾燥重量、
iii)より多い果実または種子の収穫量、
iv)より高い総植物窒素含量、
v)より高い果実または種子の窒素含量、
vi)より高い全植物の遊離アミノ酸含量、
vii)より高い果実または種子の遊離アミノ酸含量、
viii)より高い果実または種子のタンパク質含量、または
ix)より高い栄養組織のタンパク質含量、
を示すものであり;過剰発現される窒素同化/代謝酵素がアスパラギン酸アミノトランスフェラーゼ、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ、グルタミン酸デヒドロゲナーゼ、アスパラギナーゼ、真核生物アスパラギンシンテターゼまたは細胞質ゾルグルタミンシンテターゼであり;そして1種または数種の窒素同化/代謝酵素の異所性過剰発現のための遺伝子工学的操作が、
i)1種または数種の窒素同化/代謝酵素の異所性過剰発現を与える1個または数個の遺伝子融合体で植物を形質転換し、
ii)該遺伝子融合体に連結されたマーカー遺伝子により付与された形質に基づいて上記の形質転換植物を選択または同定し、
iii)上記の形質転換植物を窒素非限定生育条件下で栽培して、上記の改良された農業または栄養特性の1以上について該形質転換植物をスクリーニングし、そして
iv)1以上の改良された農業または栄養特性を有する形質転換植物を選別する、
ことを含んでなる、上記の方法。
【請求項2】
遺伝子融合体が構成的に発現される強力な植物プロモーターに機能しうる状態で連結された窒素同化/代謝酵素をコードする遺伝子を含むものである、請求項1に記載の方法。
【請求項3】
構成的に発現される強力な植物プロモーターがCaMV 35Sプロモーターである、請求項2に記載の方法。
【請求項4】
窒素同化/代謝酵素が真核生物アスパラギンシンテターゼまたは細胞質ゾルグルタミンシンテターゼである、請求項3に記載の方法。
【請求項5】
窒素同化/代謝酵素が根−特異的グルタミンシンテターゼである、請求項4に記載の方法。
【請求項6】
遺伝子融合体がpZ3、pZ9またはpZ17の35S−GS遺伝子融合体である、請求項1に記載の方法。
【請求項7】
遺伝子融合体がpZ127の35S−AS遺伝子融合体である、請求項1に記載の方法。
【請求項8】
請求項1〜7のいずれか1項に記載の方法により作出された植物。
【請求項9】
1種または数種の窒素同化/代謝酵素の異所性過剰発現を与える1個または数個の上記遺伝子融合体を含有する、請求項1〜7のいずれか1項に記載の方法により作出された植物の種子。
【請求項10】
請求項9に記載の種子の植物。
【請求項11】
グルタミンシンテターゼ遺伝子の異所性過剰発現のための植物の遺伝子工学的操作により抑制されたグルタミンシンテターゼレベルを有する植物を作出する方法であって、上記の抑制されたグルタミンシンテターゼレベルが同様に栽培した遺伝子操作されていない前駆植物と比較してのことであり;そして植物の遺伝子工学的操作が、
i)グルタミンシンテターゼ遺伝子の異所性過剰発現を与えるようにデザインされた遺伝子融合体で植物を形質転換し、
ii)該遺伝子融合体に連結されたマーカー遺伝子により付与された形質に基づいて上記の形質転換植物を選択または同定し、
iii)異常に低いグルタミンシンテターゼレベルについて上記の形質転換植物をスクリーニングし、そして
iv)異常に低いグルタミンシンテターゼレベルを有する形質転換植物を選別する、
ことを含んでなる、上記の方法。
【請求項12】
グルタミンシンテターゼ遺伝子が葉緑体グルタミンシンテターゼをコードする遺伝子である、請求項11に記載の方法。
【請求項13】
遺伝子融合体がpZ41またはpZ54の35S−GSである、請求項12に記載の方法。
【請求項14】
不活性のアスパラギンシンテターゼの異所性過剰発現のための植物の遺伝子工学的操作により抑制されたアスパラギンシンテターゼレベルを有する植物を作出する方法であって、上記の抑制されたアスパラギンシンテターゼレベルが同様に栽培した遺伝子操作されていない前駆植物と比較してのことであり;そして植物の遺伝子工学的操作が、
i)不活性のアスパラギンシンテターゼの異所性過剰発現を与える遺伝子融合体で植物を形質転換し、
ii)該遺伝子融合体に連結されたマーカー遺伝子により付与された形質に基づいて上記の形質転換植物を選択または同定し、
iii)異常に低いアスパラギンシンテターゼレベルについて上記の形質転換植物をスクリーニングし、そして
iv)異常に低いアスパラギンシンテターゼレベルを有する形質転換植物を選別する、
ことを含んでなる、上記の方法。
【請求項15】
遺伝子融合体がpZ167の35S−ASである、請求項14に記載の方法。
【請求項16】
1種または数種の窒素同化/代謝酵素の異所性過剰発現を与える1個または数個の上記遺伝子融合体を含有する、請求項1〜7のいずれか1項に記載の方法により作出された植物の子孫、クローン、細胞系または細胞。
【請求項17】
(a)アスパラギン酸アミノトランスフェラーゼ、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ、グルタミン酸デヒドロゲナーゼ、アスパラギナーゼ、真核生物アスパラギンシンテターゼまたは細胞質ゾルグルタミンシンテターゼをコードする遺伝子を異所的に過剰発現し、そして(b)同様に栽培した遺伝子操作されていない前駆植物と比べて、該植物と該前駆植物を窒素非限定生育条件下で栽培するときに、次の改良された農業または栄養特性:
i)より速い成長速度、
ii)より多い成熟時の新鮮または乾燥重量、
iii)より多い果実または種子の収穫量、
iv)より高い総植物窒素含量、
v)より高い果実または種子の窒素含量、
vi)より高い全植物の遊離アミノ酸含量、
vii)より高い果実または種子の遊離アミノ酸含量、
viii)より高い果実または種子のタンパク質含量、または
ix)より高い栄養組織のタンパク質含量、
の1以上を示すものである、遺伝子工学的に作出された植物。
【請求項18】
細胞質ゾルグルタミンシンテターゼが根−特異的グルタミンシンテターゼである、請求項17に記載の遺伝子工学的に作出された植物。
【請求項1】
1種または数種の窒素同化/代謝酵素の異所性過剰発現のための植物の遺伝子工学的操作により改良された農業または栄養特性を有する植物を作出する方法であって、上記の改良された農業または栄養特性を有する植物が、同様に栽培した遺伝子操作されていない前駆植物と比べて、該植物と該前駆植物を窒素非限定生育条件下で栽培するときに次の特性:
i)より速い成長速度、
ii)より多い成熟時の新鮮または乾燥重量、
iii)より多い果実または種子の収穫量、
iv)より高い総植物窒素含量、
v)より高い果実または種子の窒素含量、
vi)より高い全植物の遊離アミノ酸含量、
vii)より高い果実または種子の遊離アミノ酸含量、
viii)より高い果実または種子のタンパク質含量、または
ix)より高い栄養組織のタンパク質含量、
を示すものであり;過剰発現される窒素同化/代謝酵素がアスパラギン酸アミノトランスフェラーゼ、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ、グルタミン酸デヒドロゲナーゼ、アスパラギナーゼ、真核生物アスパラギンシンテターゼまたは細胞質ゾルグルタミンシンテターゼであり;そして1種または数種の窒素同化/代謝酵素の異所性過剰発現のための遺伝子工学的操作が、
i)1種または数種の窒素同化/代謝酵素の異所性過剰発現を与える1個または数個の遺伝子融合体で植物を形質転換し、
ii)該遺伝子融合体に連結されたマーカー遺伝子により付与された形質に基づいて上記の形質転換植物を選択または同定し、
iii)上記の形質転換植物を窒素非限定生育条件下で栽培して、上記の改良された農業または栄養特性の1以上について該形質転換植物をスクリーニングし、そして
iv)1以上の改良された農業または栄養特性を有する形質転換植物を選別する、
ことを含んでなる、上記の方法。
【請求項2】
遺伝子融合体が構成的に発現される強力な植物プロモーターに機能しうる状態で連結された窒素同化/代謝酵素をコードする遺伝子を含むものである、請求項1に記載の方法。
【請求項3】
構成的に発現される強力な植物プロモーターがCaMV 35Sプロモーターである、請求項2に記載の方法。
【請求項4】
窒素同化/代謝酵素が真核生物アスパラギンシンテターゼまたは細胞質ゾルグルタミンシンテターゼである、請求項3に記載の方法。
【請求項5】
窒素同化/代謝酵素が根−特異的グルタミンシンテターゼである、請求項4に記載の方法。
【請求項6】
遺伝子融合体がpZ3、pZ9またはpZ17の35S−GS遺伝子融合体である、請求項1に記載の方法。
【請求項7】
遺伝子融合体がpZ127の35S−AS遺伝子融合体である、請求項1に記載の方法。
【請求項8】
請求項1〜7のいずれか1項に記載の方法により作出された植物。
【請求項9】
1種または数種の窒素同化/代謝酵素の異所性過剰発現を与える1個または数個の上記遺伝子融合体を含有する、請求項1〜7のいずれか1項に記載の方法により作出された植物の種子。
【請求項10】
請求項9に記載の種子の植物。
【請求項11】
グルタミンシンテターゼ遺伝子の異所性過剰発現のための植物の遺伝子工学的操作により抑制されたグルタミンシンテターゼレベルを有する植物を作出する方法であって、上記の抑制されたグルタミンシンテターゼレベルが同様に栽培した遺伝子操作されていない前駆植物と比較してのことであり;そして植物の遺伝子工学的操作が、
i)グルタミンシンテターゼ遺伝子の異所性過剰発現を与えるようにデザインされた遺伝子融合体で植物を形質転換し、
ii)該遺伝子融合体に連結されたマーカー遺伝子により付与された形質に基づいて上記の形質転換植物を選択または同定し、
iii)異常に低いグルタミンシンテターゼレベルについて上記の形質転換植物をスクリーニングし、そして
iv)異常に低いグルタミンシンテターゼレベルを有する形質転換植物を選別する、
ことを含んでなる、上記の方法。
【請求項12】
グルタミンシンテターゼ遺伝子が葉緑体グルタミンシンテターゼをコードする遺伝子である、請求項11に記載の方法。
【請求項13】
遺伝子融合体がpZ41またはpZ54の35S−GSである、請求項12に記載の方法。
【請求項14】
不活性のアスパラギンシンテターゼの異所性過剰発現のための植物の遺伝子工学的操作により抑制されたアスパラギンシンテターゼレベルを有する植物を作出する方法であって、上記の抑制されたアスパラギンシンテターゼレベルが同様に栽培した遺伝子操作されていない前駆植物と比較してのことであり;そして植物の遺伝子工学的操作が、
i)不活性のアスパラギンシンテターゼの異所性過剰発現を与える遺伝子融合体で植物を形質転換し、
ii)該遺伝子融合体に連結されたマーカー遺伝子により付与された形質に基づいて上記の形質転換植物を選択または同定し、
iii)異常に低いアスパラギンシンテターゼレベルについて上記の形質転換植物をスクリーニングし、そして
iv)異常に低いアスパラギンシンテターゼレベルを有する形質転換植物を選別する、
ことを含んでなる、上記の方法。
【請求項15】
遺伝子融合体がpZ167の35S−ASである、請求項14に記載の方法。
【請求項16】
1種または数種の窒素同化/代謝酵素の異所性過剰発現を与える1個または数個の上記遺伝子融合体を含有する、請求項1〜7のいずれか1項に記載の方法により作出された植物の子孫、クローン、細胞系または細胞。
【請求項17】
(a)アスパラギン酸アミノトランスフェラーゼ、グルタミン酸2−オキソグルタル酸アミノトランスフェラーゼ、グルタミン酸デヒドロゲナーゼ、アスパラギナーゼ、真核生物アスパラギンシンテターゼまたは細胞質ゾルグルタミンシンテターゼをコードする遺伝子を異所的に過剰発現し、そして(b)同様に栽培した遺伝子操作されていない前駆植物と比べて、該植物と該前駆植物を窒素非限定生育条件下で栽培するときに、次の改良された農業または栄養特性:
i)より速い成長速度、
ii)より多い成熟時の新鮮または乾燥重量、
iii)より多い果実または種子の収穫量、
iv)より高い総植物窒素含量、
v)より高い果実または種子の窒素含量、
vi)より高い全植物の遊離アミノ酸含量、
vii)より高い果実または種子の遊離アミノ酸含量、
viii)より高い果実または種子のタンパク質含量、または
ix)より高い栄養組織のタンパク質含量、
の1以上を示すものである、遺伝子工学的に作出された植物。
【請求項18】
細胞質ゾルグルタミンシンテターゼが根−特異的グルタミンシンテターゼである、請求項17に記載の遺伝子工学的に作出された植物。
【図1】

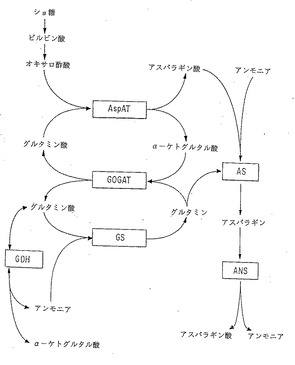
【図2】
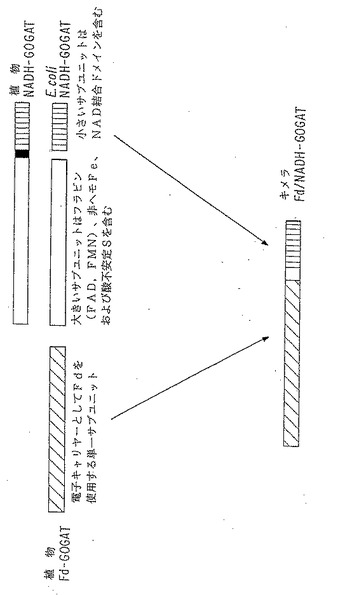
【図3】
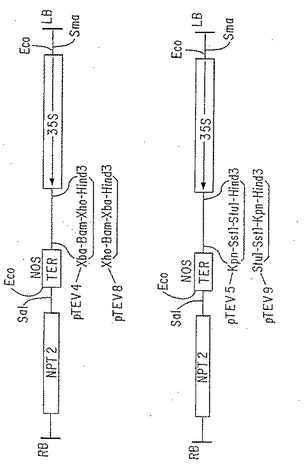
【図4】
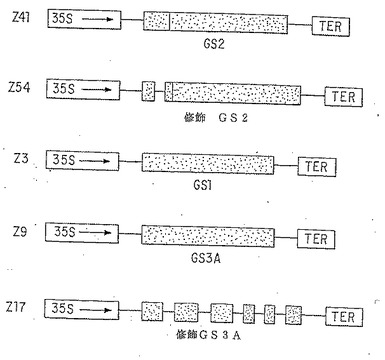
【図5】

【図6A】

【図6B】
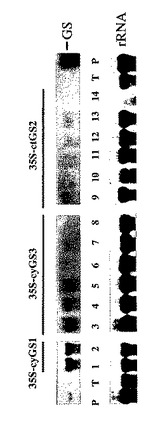
【図6C】
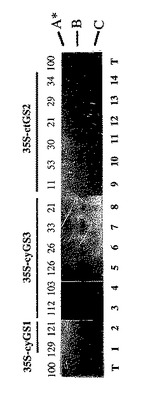
【図7A】
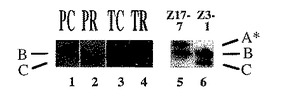
【図7B】
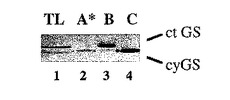
【図8】
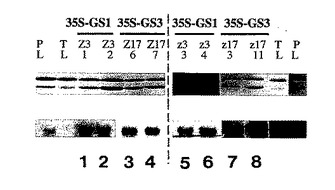
【図9A】
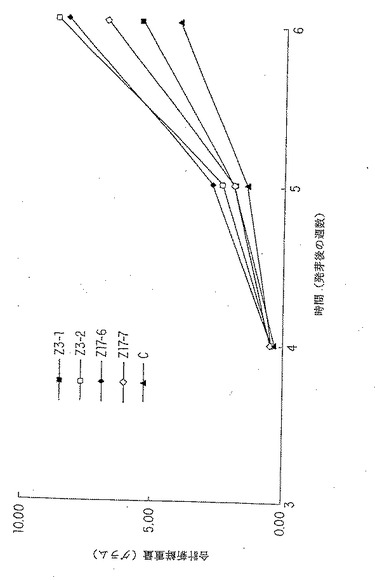
【図9B】
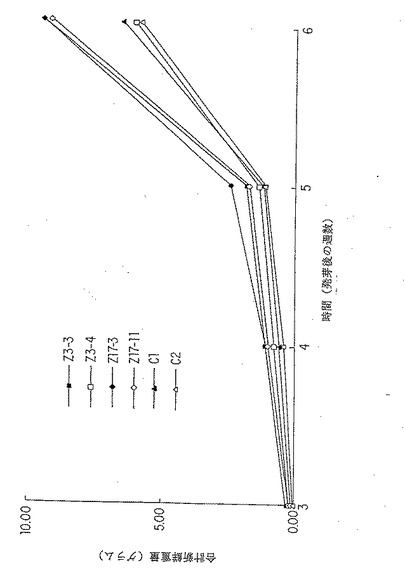
【図10A】

【図10B】

【図10C】

【図10D】

【図11A】
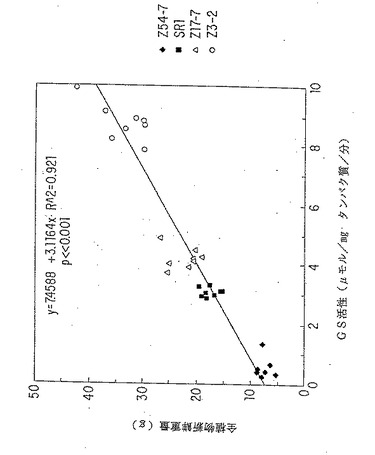
【図11B】
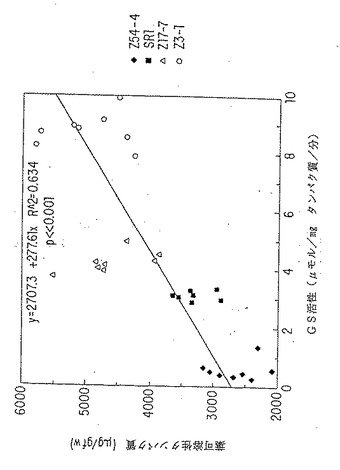
【図12】
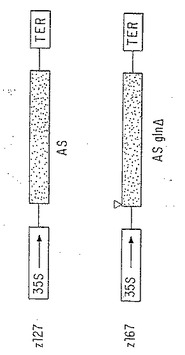
【図13】
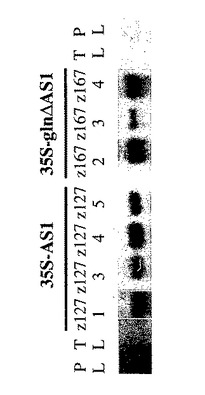
【図14】
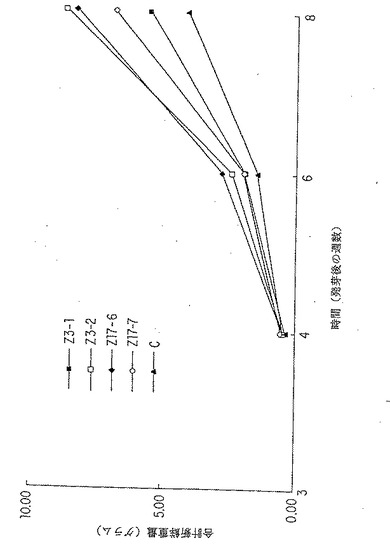
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【図8】
【図9A】
【図9B】
【図10A】
【図10B】
【図10C】
【図10D】
【図11A】
【図11B】
【図12】
【図13】
【図14】
【公開番号】特開2010−68805(P2010−68805A)
【公開日】平成22年4月2日(2010.4.2)
【国際特許分類】
【出願番号】特願2009−248278(P2009−248278)
【出願日】平成21年10月28日(2009.10.28)
【分割の表示】特願平7−510999の分割
【原出願日】平成6年10月6日(1994.10.6)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
【出願人】(502074002)ニューヨーク・ユニバーシティ (3)
【Fターム(参考)】
【公開日】平成22年4月2日(2010.4.2)
【国際特許分類】
【出願日】平成21年10月28日(2009.10.28)
【分割の表示】特願平7−510999の分割
【原出願日】平成6年10月6日(1994.10.6)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
【出願人】(502074002)ニューヨーク・ユニバーシティ (3)
【Fターム(参考)】
[ Back to top ]