
第VIII因子と低密度リポタンパク質レセプター関連タンパク質との相互作用のアンタゴニスト
【課題】インビトロおよびインビボにおいて第VIII因子の安定性おおよび半減期を増大させる、利用可能な第VIII因子調製物を提供すること。
【解決手段】本発明は、第VIII因子由来のペプチドおよび第VIII因子に対して作製された抗体の使用、ならびに第VIII因子のLRPとの相互作用の阻害に関する。さらに本発明は、第VIIIとのLRPの相互作用を阻害するための方法ならびに第VIII因子の分解を減少させる方法、生物学的流体における第VIII因子の半減期を延長させる方法、および/または血液凝固障害(特に血友病A)に罹患する患者を処置する方法に関する。本発明はまた、血液凝固障害(特に血友病A)の処置のための、生物学的流体における第VIII因子の分解の減少、LRPとの第VIII因子の相互作用の阻害、および/または生物学的流体における第VIII因子の半減期の延長に有用な薬学的組成物に関する。
【解決手段】本発明は、第VIII因子由来のペプチドおよび第VIII因子に対して作製された抗体の使用、ならびに第VIII因子のLRPとの相互作用の阻害に関する。さらに本発明は、第VIIIとのLRPの相互作用を阻害するための方法ならびに第VIII因子の分解を減少させる方法、生物学的流体における第VIII因子の半減期を延長させる方法、および/または血液凝固障害(特に血友病A)に罹患する患者を処置する方法に関する。本発明はまた、血液凝固障害(特に血友病A)の処置のための、生物学的流体における第VIII因子の分解の減少、LRPとの第VIII因子の相互作用の阻害、および/または生物学的流体における第VIII因子の半減期の延長に有用な薬学的組成物に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、第VIII因子由来のペプチドおよび第VIII因子に対して産生された抗体の使用および第VIII因子のLRPとの相互作用の阻害に関する。さらに、本発明は、LRPの第VIII因子との相互作用を阻害する方法、ならびに第VIII因子の分解を減少させる方法、および/または生物学的流体における第VIII因子の半減期を延長させる方法、および/または血液凝固障害(特に血友病A)に罹患する患者を処置する方法に関する。本発明はまた、血液凝固障害(特に血友病A)の処置のための、生物学的流体における第VIII因子の分解の減少、LRPとの第VIII因子の相互作用の阻害、および/または生物学的流体における第VIII因子の半減期の延長に有用な薬学的組成物に関する。
【背景技術】
【0002】
血液凝固には、異なる止血反応経路(最終的に血栓の形成につながる)の組合せが関係する。血栓は、脈管壁の表面上の血液構成成分の塊であり、そして、凝集した血小板および不溶性の架橋フィブリンから主として成る。フィブリンの形成は、凝固酵素トロンビンによるフィブリノーゲンの限定的なタンパク質分解によって誘導される。この酵素は、凝固カスケード(活性型血小板および白血球ならびに多くの脈管細胞の表面で起こる一連のチモーゲン活性化である)の最終生成物である(例えば、K.G.Mannら、Blood,1990,Vol.76,1−16頁を参照のこと;本明細書中に参考として援用される)。
【0003】
この凝固カスケードの重要な工程は、活性型第IX因子(第IXa因子)と活性型第VIII因子(第VIII因子a)の複合体による、第X因子の活性化である。
【0004】
第VIII凝固因子は、このように凝固カスケードの内因性の経路における重要なタンパク質である。FVIIIの機能は、酵素FIXaのための補因子として役立ち、そして、このプロテアーゼの触媒効率を5〜6倍の規模で増加させる(van Dieijeら、1981.The Journal of Biological Chemistry 256:3433−3442;本明細書中に参考として援用される)。FVIIIの重要性は、重度の出血障害である血友病A(活性型FVIIIの欠如によって特徴付けられる)によって示される(Kazazianら、1995.The Metabolic and Molecular Basis of Inherited Disease. In:Scriver,Beadet,SlyおよびValle,編 New York,McGraw−Hill Inc.III:3241−3267;本明細書中に参考として援用される)。
【0005】
血友病Aは、第VIII因子の機能不全によって特徴付けられる伴性の出血障害である。この疾患は、男性の人口の約0.01%で発症する。血友病Aは、健全なドナーから得た第VIII因子を含む血漿を投与することによって処置され得る。しかし、この処置には、いくつかの欠点がある。第VIII因子の供給は限られており、非常に高価である;血中の第VIII因子濃度は、わずか100ng/mlであり、そして、現在の血漿分別方法を用いると、収量が低い。第VIII因子の供給源は、プールされたドナーの血液なので、レシピエントは、種々の感染症(ドナーの血中に存在し得る、非A型肝炎ウイルス、非B型肝炎ウイルス、B型肝炎ウイルスまたはAIDSウイルスを含む)を得る高いリスクを負う。さらに、レシピエントは、外因性の第VIII因子に対して抗体を産生し得、そのうちのいくらかは第VIII因子の有効性を大きく減少させ得る。
【0006】
上記のように、第VIII凝固因子(FVIII)は、第X因子のタンパク質分解性の活性化における活性型第IX因子(FIXa)のための補因子として、内因性凝固経路において役割を果たす(総説については、Fay,P.J.(1999)Thromb.Haemostasis 82,193−200;Lenting,P.J.,van Mourik,J.A.,およびMertens,K.(1998)Blood 92,3983−3996(本明細書中に参考として援用される)を参照のこと)。第VIII因子は、分離性のドメイン構造(Al−al−A2−a2−B−a3−A3−C1−C2)を有する、300kDaの糖タンパク質である(Lenting,P.J.,van Mourik,J.A.,およびMertens,K.(1998)Blood 92,3983−3996;Vehar,G.A.,Keyt,B.,Eaton,D.,Rodriguez,H.,O’Brien,D.P.,Rotblat,F.,Opperman,H.,Keck,R.,Wood,W.I.,Harkins,R.N.,Tuddenham,E.G.D.,Lawn,R.M.,およびCapon,D.J.(1984)Nature 312,337−342;本明細書中に参考として援用される)。AドメインおよびCドメインは、構造上関連するタンパク質である第V因子のAドメインおよびCドメインと30〜40%の相応性を共有し、一方、Bドメインならびに短い酸性領域a1、a2およびa3は、FVIIIに固有である(Church,W.R.,Jernigan,R.L.,Toole,J.,Hewick,R.M.,Knopf,J.,Knutson,G.J.,Nesheim,M.E.,Mann,K.G.,およびFass,D.N.(1984)Proc.Natl.Acad.Sci.U.S.A.81,6934−6937;本明細書中に参考として援用される)。
【0007】
血漿において、FVIIIは、90〜220kDaの重鎖(A1−a1−A2−a2−B)および80kDaの軽鎖(a3−A3−C1−C2)から成る金属イオン連結ヘテロ二量体として循環する(Rotblat,F.,O’Brien,D.P.,O’Brien,F.J.,Goodall,A.H.,およびTuddenham,E.G.D.(1985)Biochemistry 24,4294−4300;Kaufman,R.J.,Wasly,L.C.,およびDomer,A.J.(1988)J.Biol.Chem.263,6352−6362;本明細書中に参考として援用される)。不活性タンパク質は、そのキャリアタンパク質であるフォン・ヴィレブランド因子(vWF)と強固に会合する(Lollar,P.,Hill−Eubanks,D.C.,およびParker,C.G.(1988)J.Biol.Chem.263,10451−10455;本明細書中に参考として援用される)。トロンビンまたは第Xa因子のいずれかによる限定的なタンパク質分解は、FVIII前駆体をその活性型誘導体に変換する(Lollar,P.,Knutson,G.J.,およびFass,D.N.(1985)Biochemistry 24,8056−8064;Eaton,D.,Rodriguez,H.,およびVehar G.A.(1986)Biochemistry 25,505−512;本明細書中に参考として援用される)。次いで、BドメインおよびA3ドメインに接している酸性の領域が、分子から取り除かれ(Fay,P.J.,Haidaris,P.J.,およびSmudzin,T.M.(1991)J.Biol.Chem.266,8957−8962;本明細書中に参考として援用される)、vWFに対する高い結合親和性の損失を生じる(Lollar,P.,ら、(1988)前出)。得られた活性型FVIII(FVIIIa)分子は、A2−a2ドメイン(金属イオン連結A1−a1/A3−C1−C2部分と非共有結合している)を含むヘテロ三量体から成る(Fay,P.J.,ら、(1991)前出)。
【0008】
FVIIIの重鎖および軽鎖内で、いくつかの領域がFIXa相互作用部位として同定されている(Fay P.J.,Beattie,T.,Huggins,C.F.,およびRegan,L.M.(1994)J.Biol.Chem.269,20522−20527;Bajaj,S.P.,Schmidt,A.E.,Mathur,A.,Padmanabhan,K.,Zhong,D.,Mastri,M.,およびFay,P.J.(2001)J.Biol.Chem.276.16302−16309;Lenting,P.J.,van de Loo,J.W.H.P.,Donath,M.J.,S.,H.,van Mourik,J.,A.,およびMertens,K.(1996)J.Biol.Chem.271,1935−1940;本明細書中に参考として援用される)。A2ドメイン残基Arg484−Phe5O9、Ser558−Gln565およびArg698−Asp712は、重鎖のFIXaへの結合に寄与する(Fay P.J.,(1994)前出;Bajaj,.(2001)前出;Fay,P.J.,およびScandella,D.(1999)J.Biol.Chem.274,29826−29830;本明細書中に参考として援用される)。FVIII軽鎖内では、A3ドメイン領域Glu1811−Lys1818が、FIXa相互作用部位として同定されている(Lenting,P.J.,(1996)前出)。さらに、FVIII領域Arg484−Phe509およびLys1804−Lys1818はまた、血友病患者において産生され得る抗体に対する標的エピトープとして同定されている。このような抗体は、FVIIIaと活性型FIXとの複合体アセンブリと干渉することによってFVIII活性化を阻害する(Haeley,J.F.,Lubin,I.M.,Nakai,H.,Saenko,E.L.,Hoyer,L.W.,Scandella,D.,およびLollar,P.(1995)J.Biol.Chem.270,14505−14509;Fijnvandraat,K.,Celie,P.H.N.,Turenhout,E.A.M.,van Mourik,J.A.,ten Cate,J.W.,Mertens,K.,Peters,M.,およびVoorberg,J.(1998)Blood 91,2347−2352;Zhong,D.,Saenko,E.L.,Shima,M.,Felch,M.,およびScandella,D.(1998)Blood 92,136−142;本明細書中に参考として援用される)。
【0009】
第VIII因子の半減期は、例えば、第VIII因子を関連したクリアランスのレセプターのためのその結合部位で改変することによって直接的にか、または、第VIII因子とそのレセプターとの間の結合に影響を及ぼす化合物を用いることによって間接的にのいずれかで、そのクリアランスにおいて役割を果たすレセプターに対する第VIII因子の親和性を減少することにより、第VIII因子分解(クリアランス)のメカニズムに影響を与えることによって延長され得る。前記のプロセスに関する細胞内レセプターおよび第VIII因子−レセプター相互作用に関する分子の部位が知られていないので、このような薬剤の産生は、困難であった。
【0010】
非活性型第VIII因子ヘテロ二量体の半減期は、第VIII因子に対して大きな親和性を有し(しかし、第VIIIa因子に対しては持たない)、キャリアタンパク質として役立つフォン・ヴィレブランド因子の存在に依存する(Sadler,J.E.およびDavie,E.W.:血友病A,Haernophilia B and von Willebrand’s Disease,in Starnatoyannopoulos,G.ら(編)、The Molecular basis of blood diseases.W.B.Saunders Co.,Philadelphia,1987,576−602頁)。3型フォン・ヴィレブランド病を有し、自身の循環系に検出可能なフォン・ヴィレブランド因子を示さない患者はまた、第VIII因子の二次性機能不全を有することが知られている。さらに、これらの患者における静脈内に投与された第VIII因子の半減期は、2〜4時間であり、それは、血友病A患者について報告される10〜40時間より明らかに短い。
【0011】
これらの調査結果は、第VIII因子が循環系から迅速に除去される傾向があり、このプロセスは、天然のキャリアであるフォン・ヴィレブランド因子との複合体の形成によってある程度阻害されることを意味する。
【0012】
最近、トロンビンによって活性化された第VIII因子は、低密度リポタンパク質レセプタータンパク質(本明細書中で以降、LRPと称する)との結合に関連付けられている(Yakhyaev,A.ら、Blood,vol.90(補遺1),1997,126−I,本明細書中に参考として援用される)。この文献の要約は、トロンビンで活性化された第VIII因子フラグメントの細胞取り込みおよび分解を記載し、そして、第VIIIa因子ヘテロ三量体の他の2つのサブユニットでなく、A2ドメインが細胞LRPと相互作用すると報告する。著者は、LRPに結合するA2ドメインが、さらに第VIIIa因子へテロ三量体のA2ドメイン間の相互作用を不安定化し、それによって第VIIIa因子活性がダウンレギュレートされることを提言する。
【0013】
非活性型FVIIIが多機能性のエンドサイトーシスレセプターである低密度リポタンパク質レセプター関連タンパク質(LRP)と相互作用することがまた示されている(Lenting,P.J.,Neels,J.G.,van den Berg,B.M.M.,Clijsters,P.P.F.M.,Meijerman,D.W.E.,Pannekoek,H.,van Mourik,J.A.,Mertens,K.,およびvan Zonneveld,A.,−J.(1999)J.Biol.Chem.274,23734−23739;WO 00/28021;Saenko,E.L.,Yakhyaev,A.V.,Mikhailenko,I.,Strickland,D.K.,およびSarafanov,A.G.(1999)J.Biol.Chem.274,37685−37692;本明細書中に参考として援用される)。このレセプターが、循環からのFVIIIのクリアランスにおいて役割を果たすということが示唆される(Saenko,E.L.ら、前出;Schwarz,H.P.,Lenting,P.J.,Binder,B.,Mihaly,J.,Denis,C.,Domer,F.,およびTurecek,P.L.(2000)Blood 95,1703−1708;本明細書中に参考として援用される)。
【0014】
LRPは、低密度リポタンパク質(LDL)レセプターファミリー(LDLレセプター、超低密度リポタンパク質レセプター、ポリリポタンパク質Eレセプター2およびメガリンをまた含む)のメンバーである(総説としては、Neels J.G.,Horn,I.R.,van den Berg,B.M.M.,Pannekoek,H.,およびvan Zonneveld,A.−J.(1998)Fibrinolysis Proteolysis 12,219−240;Herz,J.,およびStrickland,D.K.(2001)J.Clin.Invest.108,779−784(本明細書中に参考として援用される)を参照のこと)。LRPは、肝臓、肺、胎盤および脳を含む種々の組織において発現する(Moestrup,S.K.,Gliemann,J.,およびPallesen,G.(1992)Cell Tissue Res.269,375−382;本明細書中に参考として援用される)。このレセプターは、膜貫通型の85kDaのβ鎖に非共有結合性に連結される、細胞外の515kDaのα鎖から成る(Herz,J.,Kowal,R.C.,Goldstein,J.L.,およびBrown,M.S.(1990)EMBO J.9,1769−1776;本明細書中に参考として援用される)。α鎖は、多くの構造的かつ機能的に無関係のリガンドの結合を媒介する、可変数の相補型反復の4つのクラスターを含む(Moestrup,S.K.,Hotlet,T.L.,Etzerodt,M.,Thogersen,H.C.,Nykjaer,A.,Andreasen,P.A.,Rasmussen,H.H.,SottrupJensen,L.,およびGliemann,J.(1993)J.Biol.Chem.268,13691−13696;Willnow,T.E.,Orth,K.,およびHerz,J.(1994)J.Biol.Chem.269,15827−15832;Neels,J.G.,van den Berg,B.M.M.,Lookene,A.,Olivecrona,G.,Pannekoek,H.,およびvan Zonneveld,A.−J.(1999)J.Biol.Chem.274,31305−31311;本明細書中に参考として援用される)。
【0015】
β鎖は、膜貫通ドメインおよびエンドサイトーシスに不可欠である短い細胞質テールを含む。α鎖は、大きな外部ドメインとして機能し、そして、3つの型の反復を含む:表皮性増殖因子様ドメイン、Tyr−Trp−Thr−Asp配列およびLDLレレプタークラスAドメイン。これらのクラスAドメイン(リガンド結合に関連付けられている)は、クラスターI(2つのドメイン)、クラスターII(8つのドメイン)、クラスターIII(10のドメイン)および、クラスターIV(11のドメイン)と呼ばれる4つの別々のクラスターに存在する。
【0016】
LRPはまた、サル腎臓細胞(COS)またはチャイニーズハムスター卵巣細胞(CHO)のような細胞型において発現しており(FitzGerald,D.J.,ら、J.Cell Biol.Vol.129,1995,1533−1541頁;本明細書中に参考として援用される)、これらは、第VIII因子を含む補乳動物のタンパク質の発現のためにしばしば使用される(Kaufman,R.J.ら、Blood,Coag.Fibrinol,vol.8(補遺2),1997,3−14頁;本明細書中に参考として援用される)。
【0017】
LRPは、プロテアーゼ、Kunitz型インヒビター、プロテアーゼ−セルピン複合体、リパーゼおよびリポタンパク質を含む、多数のリガンドのクリアランスにおいて役割を果たし、これらは、いくらかの生理学的かつ病態生理学的なクリアランスのプロセスにおいてLRPが役割を果たすと言われている(Naritaら、Blood,vol.2,555−560頁,1998;Orthら、Proc.Natl.Acad.Sci.,vol,89,7422−7426頁,1992;Kounnasら、J.Biol.Chem.,vol.271,6523−6529頁,1996;本明細書中に参考として援用される)。
【0018】
LRPはまた、活性型の非酵素の補因子である第VIIIa因子に結合する(Yakhyaev,A.ら、Blood,vol.90,(補遺1),1997,126−I)。この文献の開示は、第VIIIa因子の調節においてLRPを関連付けているが、非活性型ヘテロ二量体である第VIII因子の調節におけるLRPの役割の示唆はない。
【0019】
FVIII軽鎖は、組換えLRPクラスターIIおよびIVと相互作用することが示されたが、LRPクラスターIおよびIIIへの結合は観察されなかった(Neels,J.G.,ら、(1999)前出)。
【0020】
第VIII因子の薬動力学的プロファイルを向上させるために、第VIII因子ポリペプチドのくつかの領域を改変するいくつかの試みがなされている:
WO 87/07144は、アルギニン残基およびリジン残基を含むタンパク分解性の切断部位のいくらかの改変を示し、これは、特異的なプロテアーゼが触媒する切断(例えば、Arg1721およびAla1722の間の第Xa因子の切断部位)について分子の不安定性を減少する。
【0021】
WO 95/18827、WO 95/18828およびWO 95/18829は、重鎖のA2領域に改変を有する第VIII因子の誘導体を記載する。
【0022】
WO 97/03193は、第VIII因子ポリペプチドアナログを開示し、これらの修飾が分子の金属結合特性を変更する。
【0023】
WO 97/03195において、第VIII:C因子ポリペプチドアナログが記載され、Arg残基に隣接して位置する1つ以上のアミノ酸残基の改変が提供される。
【0024】
EP 808 901は、第VIII因子の少なくとも1つの免疫優性領域に少なくとも1つの変異を有する第VIII因子改変体の構成、および、第VIII因子インヒビターを有する患者の処置のためのこの第VIII因子改変体の使用を記載する。これらの改変は、半減期インビボまたはインビトロにおいて、第VIII因子改変体の半減期を延長せず、また、第VIII因子改変体の安定性を増大もしない。
【0025】
さらに、WO 00/28021は、軽鎖のA3および/またはC1もしくはC2ドメインに改変を有し、そして、その改変が低密度リポ蛋白受容体タンパク質(LRP)との結合親和性に影響を与えるという点で、特徴付けられる、第VIII:C因子活性を有する第VIII因子ポリペプチドを記載する。
【0026】
ヒトのmRNAから得られた第VIII因子cDNAの分子クローニング、ならびに、その後の哺乳動物細胞、酵母細胞、および細菌細胞において第VIII因子活性を有するタンパク質の産生が、報告されている(WO 85/01961、EP 160 4577、EP 150 735およびEP 253 455を参照のこと;本明細書中に参考として援用される)。形質転換された微生物を用いて第VIII因子活性を有するタンパク質を産生する方法は、EP 253 455(本明細書中に参考として援用される)に開示される。欧州特許出願EP 150 735および同EP 123 945ならびにBrinkhousら、(1985)は、第VIII因子のタンパク分解性の分解産物における第VIII因子の活性を開示する(参照によってここに組み込まれる)。第VIII因子の2つのタンパク分解性の分解産物の複合体(92kDaおよび80kDaのポリペプチド)は、増強された第VIII因子活性を示す(Fayら、Biochem.Biophys.Acta(1986)871:268−278;Fatonら、Biochemistry(1986)25:505−512;本明細書中に参考として援用される)。
【発明の概要】
【発明が解決しようとする課題】
【0027】
従って、本発明者らは、インビトロおよびインビボにおいて第VIII因子の安定性おおよび半減期を増大させる、利用可能な第VIII因子調製物の作製を目指した。
【課題を解決するための手段】
【0028】
(発明の要旨)
驚くべきことに、本発明者らは、第VIII因子の部分的なアミノ酸配列、ならびに特定のペプチド内の特異的なエピトープに対する抗体を含む特定のペプチドが、インビトロおよびインビボにおいて第VIII因子の安定性を有意に改善し得ることを発見した。
【0029】
従って、本発明は一般に、第VIII因子由来のペプチドおよび第VIII因子に対して作製された抗体の使用、ならびに第VIII因子のLRPとの相互作用の阻害に関する。さらに、本発明は、LRPの第VIII因子との相互作用を阻害する方法、第VIII因子の分解を減少させる方法、および/または生物学的流体における第VIII因子の半減期を延長させる方法、および/または血液凝固障害(特に血友病A)に罹患する患者を処置する方法に関する。本発明はまた、血液凝固障害(特に血友病A)の処置のための、生物学的流体における第VIII因子の分解の減少、第VIII因子のLRPとの相互作用の阻害、および/または生物学的流体における第VIII因子の半減期の延長に有用な薬学的組成物に関する。
本発明はまた、以下の項目を提供する。
(項目1)
第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸配列を含むペプチド、または、該アミノ酸配列内の1つ以上のエピトープに特異的に結合し、第VIII因子とLRPとの相互作用を阻害する抗体の使用。
(項目2)
第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸配列を含むペプチド、または、該アミノ酸配列内の1つ以上のエピトープに特異的に結合し、生物学的流体における第VIII因子の分解を減少させる抗体の使用。
(項目3)
第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸配列を含むペプチド、または、該アミノ酸配列内の1つ以上のエピトープに特異的に結合し、血中における第VIII因子の半減期を延長させる抗体の使用。
(項目4)
第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸を含むペプチド、または、該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の、血液凝固障害および/または血栓溶解系もしくは線維素溶解系の一時的な機能障害の予防または処置のため医薬の調製のための使用。
(項目5)
前記血液凝固障害が血友病Aまたはフォン・ヴィレブランド病であり、かつ、前記一時的な機能障害が外科的手術の間または外科的手術後に起こる、項目4に記載の使用。
(項目6)
前記抗体が、AF 234247に記載のアミノ酸配列(VHKM33)、またはAF 234258に記載のアミノ酸配列(VLKM41)から本質的になる、項目1〜5のいずれか1項に記載の使用。
(項目7)
生物学的流体における第VIII因子の分解を減少させる方法であって、第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるペプチド、または該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体が、該生物学的流体に添加される、方法。
(項目8)
少なくとも10%まで第VIII因子の分解を減少させるために十分な量の前記ペプチドまたは前記抗体の量が添加される、項目7に記載の方法。
(項目9)
前記減少が少なくとも20%である、項目8に記載の方法。
(項目10)
前記減少が少なくとも50%である、項目9に記載の方法。
(項目11)
前記減少が本質的に100%である、項目10に記載の方法。
(項目12)
生物学的流体における第VIII因子の半減期を延長させる方法であって、第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるペプチド、または該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体が、該生物学的流体に添加される、方法。
(項目13)
前記生物学的流体における前記第VIII因子の半減期が少なくとも2倍延長される、項目12に記載の方法。
(項目14)
前記生物学的流体における前記第VIII因子の半減期が少なくとも5倍延長される、項目13に記載の方法。
(項目15)
前記生物学的流体における前記第VIII因子の半減期が少なくとも10倍に延長される、項目14に記載の方法。
(項目16)
生物学的流体において、第VIII因子とLRPとの相互作用を阻害する方法であって、第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるペプチド、または該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体が、該生物学的流体に添加される、方法。
(項目17)
第VIII因子とLRPとの相互作用を少なくとも20%の阻害程度を達成する量で、前記ペプチドまたは前記抗体が添加される、項目16に記載の方法。
(項目18)
相互作用の少なくとも50%の程度の阻害程度が達成される、項目17に記載の方法。
(項目19)
前記抗体が、配列番号20および配列番号22に記載のアミノ酸配列(KM33)、または配列番号16および配列番号18に記載のアミノ酸配列(KM41)を含む、項目7〜18のいずれか1項に記載の方法。
(項目20)
前記抗体が、配列番号26に記載のアミノ酸配列(KM33)または配列番号25に記載のアミノ酸配列(KM41)から本質的になる、項目19に記載の方法。
(項目21)
少なくとも1つのペプチドを含む薬学的組成物であって、該ペプチドは、第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸配列を含むか、または該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体を含み、さらに、生理学的に受容可能な賦形剤、キャリア、希釈剤および/または安定化剤を含む、薬学的組成物。
(項目22)
前記調製物が凍結乾燥されている、項目20に記載の薬学的組成物。
(項目23)
生物学的流体における第VIII因子の分解を減少し、そして/あるいは第VIII因子の半減期を延長し、そして/あるいは、LRPへの第VIII因子の結合を阻害し、そして/あるいは血液凝固障害および/または血栓溶解系もしくは線維素溶解系の一時的な機能障害の予防または処置ための項目21または22に記載の薬学的組成物。
(項目24)
前記ペプチドまたは前記抗体が項目8〜11のいずれか1項で定義される第VIII因子の分解の減少を達成するか、項目17もしくは18に記載のいずれか1項で定義されるLRPと第VIII因子との相互作用の阻害を達成するか、そして/または、項目13〜15のいずれか1項で定義される生物学的流体における第VIII因子の半減期の延長を達成するのに十分な量で含まれる、項目21または22のいずれか1項に記載の薬学的組成物。
(項目25)
前記血液凝固障害が血友病Aまたはフォン・ヴィレブランド病であって、かつ一時的な機能障害が外科的手術の間、または外科的手術後に起こる、項目23に記載の調製物。
(項目26)
さらに第VIII因子を含む、項目21〜25のいずれか1項に記載の薬学的組成物。
(項目27)
前記抗体が配列番号20および配列番号22に記載のアミノ酸配列(KM33)、または配列番号16および配列番号18に記載のアミノ酸配列(KM41)を含む、項目21〜26のいずれか記載の薬学的組成物。
(項目28)
前記抗体が、配列番号26に記載のアミノ酸配列(KM33)または配列番号25に記載のアミノ酸配列(KM41)から本質的になる、項目27に記載の薬学的組成物。
(項目29)
血液凝固障害に罹患する患者を処置する方法であって、前記方法が前記患者に項目21〜28のいずれか1項に記載の薬学的組成物を投与する工程を包含する、方法。
(項目30)
前記血液凝固障害が血友病Aまたはフォン・ヴィレブランド病であり、かつ前記一時的な機能障害が外科的手術の間、または外科的手術後に起こる、項目29に記載の方法。
【図面の簡単な説明】
【0030】
【図1】FVIII軽鎖フラグメントの固定されたLRPへの結合。16fmol/mm2でCM5センサ−チップに固定されたLRPを、以下と共にインキュベートした:A、FVIII軽鎖(150nM)(実線)および第Xa因子切断軽鎖(150nM)(点線)。B、a3−A3−C1フラグメント(150nM)(実線)および単離したC2ドメイン(750nM)(点線)。インキュベーションを、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH7.4)中にて25℃で2分間、20μl/分の流量で行った。分離を、緩衝液でリガンド溶液を置換して開始した。反応は、共鳴単位(RU)として示し、非特異的な結合(LRPコーティングしたチャネルと比較して5%未満)について補正する。
【図2】FVIII軽鎖の組換えLRPフラグメントへの結合。FVIII軽鎖(LCh)(25nM)を、種々の濃度の組換えLRPクラスターII(0〜600nM)の存在下または非存在下で、150mM NaCl、5mM CaCl2、1%(w/v) HSA、0.1%Tween(登録商標)20および50mM Tris(pH7.4)の50μl容積中にて、37℃で2時間、固定されたLRPクラスターIVと共にインキュベートした。同じ緩衝液で洗浄した後、結合したFVIII軽鎖を、37℃で15分間、ペルオキシダーゼ結合体化抗FVIII抗体CLB−CAg 12と共にインキュベートすることによって定量した。残留結合は、競合物質の非存在下での結合の割合として表し、非特異的な結合(LRPクラスターIIを固定化したウェルへの結合と比較して5%未満)について補正する。挿入図、FVIII軽鎖の連続希釈物を固定されたLRPクラスターII(1pmol/ウェル)と共に、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%Tween(登録商標)20および50mM Tris(pH7.4)の50μl容積中にて、37℃で2時間インキュベートした。同じ緩衝液で洗浄した後、結合したFVIII軽鎖を、上記のようにして定量した。データは、3つの実験の平均±S.D.を表す。
【図3】LRPクラスターIIの固定化されたFVa軽鎖への結合。LRPクラスターII(100nM)を、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH7.4)中にて、固定化されたFVIII軽鎖(71fmol/mm2)(I)または固定化されたFVa軽鎖(76fmol/mm2)(II)のいずれかと共に、20μ/分の流量にて25℃で2分間インキュベートした。反応を、共鳴単位(RU)として示し、非特異的結合(コーティングされたチャネルと比較して5%未満)について補正する。
【図4】FVIII軽鎖とLRPとの間の相互作用に及ぼすscFv抗体フラグメントの影響。A、FVIII軽鎖(LCh)(25nM)を、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%Tween(登録商標)20および50mM Tris(pH7.4)の50μl容積中にて、種々の濃度(0〜100nM)のscFv KM41(黒丸)またはscFv KM36(白丸)の存在下で、固定されたLRPクラスターII(1pmol/ウェル)と共に37℃で2時間インキュベートした。同じ緩衝液で洗浄した後、結合したFVIII軽鎖を、ペルオキシダーゼ結合体化FVIII抗体CLB−CAg 12と共に37℃で15分間インキュベートすることによって定量した。残留結合は、競合物質の非存在下における結合の割合を表し、非特異的な結合(LRPクラスターIIを固定化したウェルへの結合と比べて5%未満)について補正する。データは、3つの実験の平均±S.D.を表す。B,FVIII軽鎖(50nM)を、図1の説明文に記載されたように、固定されたLRP(16fmol/mm2)と共にインキュベートした。結合を、scFv KM41の非存在下(曲線1)または増加した濃度のscFv KM41(20nM、60nM、300nM、500nM(それぞれ曲線2〜5))において評価した。複合体は、SPR分析の前に、30分間にわたって形成させた。
【図5】FVIII軽鎖とLRPとの間の相互作用に及ぼすscFv KM33の影響。FVIII軽鎖(LCh)(25nM)を、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%Tween(登録商標)20および50mM Tris(pH7.4)の50μl容積中にて、種々の濃度(0〜30nM)のscFv KM36(白丸)またはscFv KM33(黒丸)の存在下、固定化したLRPクラスターII(1pmol/ウェル)と共に37℃で2時間インキュベートした。同じ緩衝液で洗浄した後、結合したFVIII軽鎖を、ペルオキシダーゼ結合体化抗FVIII抗体CLB−CAg 12と共に37℃で15分間インキュベートすることによって定量した。残留結合は、競合物質の非存在下の結合の割合として表し、非特異的結合(クラスターでコーティングしたウェルへの結合と比較して5%未満)について補正する。データは、3つの実験の平均±S.D.を表す。
【図6】FVIII/FV1811−1818軽鎖キメラへのLRPクラスターIIへの結合。CM5センサチップ上のScFv EL14(67fmol/mm2)を、150mM NaCl、50mM Tris(pH 7.4)中20fmol/mm2の密度までの組換え野生型FVIII軽鎖または組換えFVIII/FV1811−1818キメラのいずれかと共にインキュベートした。LRPクラスターII(25〜125nM)を、FVIII/FV1811−1818キメラ(白丸)またはインタクトなFVIII軽鎖(黒丸)をそれぞれ固定化した2つの別々のチャネル、および1つのコントロール(scFv EL14でコーティングした)チャネルを、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH7.4)中、25℃、20μl/molの流速で2分間通過させた。結合したLRPクラスターIIを2分後の会合量として表す。
【図7】FVIII/FV1811−1818軽鎖キメラへのscFv KM41の結合。CM5センサチップのscFv EL14(67fmol/mm2)を、150mM NaCl、50mM Tris(pH7.4)中、20fmol/mm2の密度までの組換え野生型FVIII軽鎖または組換えFVIII/FV1811−1818キメラのいずれかと共にインキュベートした。ScFv KM41(40nM)を、それぞれインタクトなFVIII軽鎖(I)またはFVIII/FV1811−1818キメラ(II)を固定化した2つの別々のチャネルを通過させた。反応を、共鳴単位(RU)として示し、非特異的結合について補正する。
【図8】実施例IIに従って、マウスに、ヒトFVIII単体、コントロールのscFvフラグメント(KM38)の存在下でのヒトFVIII、および、推定FVIII−LRP相互作用と特異的に干渉するscFvフラグメント(KM33)の存在下でのヒトFVIIIのいずれかで注射した。
【発明を実施するための形態】
【0031】
(発明の詳細な説明)
本発明の1つの実施形態は、第VIII因子由来であるが第VIII因子のLRPとの相互作用を阻害する任意の実質的な第VIII因子活性を有さない配列番号1、2、3または4のいずれかで定義されるアミノ酸配列を含むペプチドの使用に関する。
用語「ペプチド」は、ペプチド結合を介して結合された一連のアミノ酸から成る分子に関する。好ましくは本発明に有用なペプチドは、タンパク質において一般に見出される20のアミノ酸からなる(例えば、周知の教科書である「Biochemistry」(A.Lehninger,第2版、Worth Publicshers,NY,NY(1975))を参照のこと)。
【0032】
本発明に有用なペプチドはまた、第VIII因子由来であるが、本質的に第VIII因子活性を有さず、改変され、その結果、そのアミノ酸配列が、上記ペプチドの第VIII因子とLRPタンパク質との間の相互作用を減少する能力を有意に変更しない、1つ以上の欠失、付加、置換および/または変換を含む、配列番号1、2、3または4のいずれかで定義されるアミノ酸配列を含むペプチドの誘導体であり得る。
【0033】
本発明に有用なペプチドが1つまたはアミノ酸置換を含む場合、前記1つ以上のアミノ酸置換は保存的アミノ酸置換であることが、好ましい。例えば、置換がアラニン、ロイシン、イソロイシン、バリン、プロリン、フェニルアラニン、トリプトファンおよびメチオニンのような非極性の疎水性アミノ酸において発生する場合、置換されるアミノ酸がアラニン、ロイシン、イソロイシン、バリン、プロリン、フェニルアラニン、トリプトファンおよびメチオニンのような非極性の疎水性アミノ酸の中から選択されることが好ましい。同様に、置換がセリン、トレオニン、チロシン、アスパラギン、グルタミン、システインまたはグリシンのような非荷電極性アミノ酸において発生する場合、置換されるアミノ酸がセリン、トレオニン、チロシン、アスパラギン、グルタミン、システインまたはグリシンのような非荷電極性アミノ酸の中から選択されることが好ましい。置換がリジン、アルギニンまたはヒスチジンのような正に荷電した(塩基性)アミノ酸において発生する場合、置換されるアミノ酸がリジン、アルギニンまたはヒスチジンのような正に荷電した(塩基性)アミノ酸の中から選択されることが好ましい。同様に、置換がアスパラギン酸またはグルタミン酸のような負に荷電した(酸性)アミノ酸において発生する場合、置換されるアミノ酸がアスパラギン酸またはグルタミン酸のような負に荷電した(酸性)アミノ酸の中から選択されることが好ましい。本発明に有用なペプチドは、D−アミノ酸、L−アミノ酸またはD−およびL−アミノ酸の混合物からなり得る。
【0034】
本発明に有用なペプチドはまた、例えば、これらのペプチドの1つ以上のタンデム反復または交互反復のような、配列番号1、2、3または4で定義されるペプチドの任意の重合形態を包含する。
【0035】
前記のペプチドの特定のアミノ酸の化学改変(特に、N末端および/またはC末端アミノ酸残基の化学改変)がまた包含される。この化学改変は、末端アミノ基および/またはカルボキシ基を遮蔽し、インビボもしくはインビトロでの分解に対する上記ペプチドの安定性を増加し得、または、キャリア機能(アルブミンまたは他の血漿タンパク質を含む)、標的化機能もしくはペプチドの溶解性を改変するような機能を有する分子もしくは部分をペプチドに添加する。用語「改変」は、グリコシル残基の付加または除去をさらに包含する。
【0036】
本発明に有用なペプチドは、標準のペプチド合成技術によって、または、組換えDNA技術によって、全体または一部が作製され得る。
【0037】
本発明に有用な1つの特定のペプチドは、配列番号1(Glu1811−Lys1818)のアミノ酸配列を含む。本発明に有用なさらなる特定のペプチドは、配列番号2(Lys1804−Lys1818)または配列番号3(Tyr1815−Ala1834)由来のアミノ酸配列を含む。このように、本発明に有用なさらなるペプチドは、アミノ酸配列Lys1804−Ala1834(配列番号4)を含む。
【0038】
好ましくは、本発明に有用なペプチドは、上記の第VIII因子活性についてのアッセイの1つで測定された、本発明の前記のペプチドが由来する天然に存在する第VIII因子分子の対応する第VIII因子活性と比較して、5.0%未満の第VIII因子活性を有し、より好ましくは、1.0%未満の第VIII因子活性を有し、そして最も好ましくは、本質的に活性を有さない。
【0039】
第VIII因子活性の評価は、適切なアッセイ手段(特に、サンプルの第VIII因子活性を決定するために典型的に実施される任意のアッセイ(例えば、MikaelssonおよびOswaldson,Scan.J.Hematol.Suppl.33,79−86,1984に記載されるような1ステップクロットアッセイ、または、第VIII因子 IMMUNOCHROM(Baxter)のような色素発色アッセイ))によって実行され得る。
【0040】
第VIII因子活性はまた、第X因子の第Xa因子への変換において第IXa因子のための補因子として作用する第VIII因子の能力を測定することによって実施され得、色素発色基質が第X因子について使用される(Coatest第VIII因子、Chomogenix、Moeindal、Sweden)。
【0041】
本発明のさらなる実施形態は、第VIII因子由来であるが、本質的に第VIII因子活性を有さず、第VIII因子とLRPとの相互作用を阻害する、配列番号1、2、3または4のいずれかで定義されたアミノ酸配列を含むペプチド内の1つ以上のエピトープに特異的に結合する抗体の使用に関する。
【0042】
本明細書中で使用される場合、用語「抗体」は、ポリクローナル抗体またはモノクローナル抗体(好ましくはモノクローナル抗体)およびそれらのフラグメントまたは領域、ならびに第VIII因子もしくは本発明に有用なペプチド内の1つ以上のエピトープに特異的に結合することおよび第VIII因子分子と低密度リポ蛋白受容体関連タンパク質(LRP)との間の相互作用を干渉することが可能であるそれらの誘導体を含むことを意味する。
【0043】
本明細書中で使用される場合、用語「エピトープ」は、本発明に有用なペプチドの全てまたは部分をいうことを意味し、このペプチドは、第VIII因子において見出される全てまたは対応するアミノ酸の一部の1°、2°および/または3°構造を模倣し、第VIII因子とLRPとの間の相互作用を阻害する抗体によって特異的に認識され得る。エピトープは、ペプチド配列自体(すなわち、ペプチドの1°構造)、種々の三次元の形態におけるペプチド(すなわち、ペプチド配列の2°または3°構造)および/またはこのペプチドのアミノ酸への改変(例えば、糖分子の付加(例えば、グリコシル化ペプチド))を含み得る。作用の任意の特定の論理または生物学的メカニズムに束縛されずに、本発明を実施するのに有用なペプチドが少なくとも2つの異なるエピトープを含むことが可能である。1つのエピトープは、配列番号1(Gly1811−Lys1818)内に位置する主要なアミノ酸配列のようである。もう一方のエピトープは、前述の主要なエピトープに隣接して位置するアミノ酸配列の組合せから成り得る;すなわち、もう一方のエピトープは、配列番号2(Lys1804−Lys1818)および/または配列番号3(Tyr1815−Ala1834)から形成され得る。このエピトープはまた、二次構造または三次構造エピトープであり得、そして、アミノ酸配列Lys1804−Lys1818およびTyr1815−Ala1834の両方を含み得る。
【0044】
抗体のフラグメントは、例えば、Fab、Fab’、F(ab’)2、FvおよびscFvフラグメントを含む。これらのフラグメントは、当該分野で周知の方法(たとえば、パパインのような酵素でタンパク質分解性に切断してFabフラグメントを生成するか、または、ペプシンのような酵素によりF(ab’)2フラグメントを生成するか)を用いて、インタクトな抗体から産生され得る。
【0045】
1つの好ましい実施形態において、本発明の実施に有用な抗体は、scFv抗体フラグメントである。好ましくは、抗体は、実施例において、KM33として記載される抗体のアミノ酸配列(KM33の重鎖のアミノ酸配列については配列番号20を、KM33の軽鎖のアミノ酸配列については配列番号22を、そして、ペプチドリンカーによって連結されるKM33の軽鎖および重鎖を含む融合タンパク質については配列番号26を参照のこと)、および/または、GenBank 登録番号AF234247(VHKM33)によって提供された抗体のアミノ酸配列、あるいは実施例においてKM41として記載される抗体のアミノ酸配列(KM41の重鎖のアミノ酸配列については配列番号16、KM41の軽鎖のアミノ酸配列については配列番号18、そして、ペプチドリンカーで結合されるKM41の軽鎖および重鎖を含む融合タンパク質については配列番号25を参照のこと)、および/またはGenBank 登録番号AF234258(VLKM41)によって提供された抗体のアミノ酸配列を含む。
【0046】
これらおよび他のscFvは、van der Brink,E.N.ら、Blood 97(4),966−972(2001)(本明細書中に参考として援用される)に記載されたようにして産生され得る。
【0047】
好ましくは本発明に有用な抗体は、第VIII因子と低密度リポタンパク質レセプター関連タンパク質(LRP)との間の相互作用を阻害することが可能であり、その結果、この相互作用は、たとえば、所定の供給源由来の第VIII因子とLRPとの間の相互作用と比較すると、少なくとも20%、より好ましくは少なくとも50%、なおより好ましくは少なくとも90%または95%かそれ以上、減少される。
【0048】
第VIII因子とLRPとの間の相互作用は、本明細書中に記載されたような表面プラズモン共鳴または固相結合アッセイを用いて決定され得る。
【0049】
本明細書中で使用される場合、用語「低密度リポタンパク質レセプター関連タンパク質」は、「LRP」として省略され、これらの用語は、タンパク質の低密度リポタンパク質(LDL)レセプターファミリーのメンバーである膜糖タンパク質をいうために交換可能に使用される(Gliemann,J.in Biol.Chem.379,951−964(1998)(参考として援用される)を参照のこと)。ヒトLRPは、31のクラスAのシステインリッチな反復(LDLRAとして知られている)ドメイン(クラスターI、II、IIIおよびIVとして知られている4つのクラスターにおけるLRP分子において分配される)を含む。クラスターIIは、CL−II−1/2(アミノ末端基の側面にある表皮性の成長因子の反復E4およびLDLRAドメインC3−C7にわたる)として知られているドメインを含む。このCL−II−1/2ドメインは、第VIII因子と相互作用する(Neels、J.G.ら、前出を参照のこと)。
【0050】
本発明はまた、前期ペプチドが第VIII因子由来であるが、本質的に第VIII因子活性を有さない配列番号1、2、3または4に定義されるアミノ酸配列を含む、少なくとも1つのペプチド、または前記アミノ酸配列の中の1つ以上のエピトープに特異的に結合する抗体、ならびに生理学的に受容可能な賦形剤、キャリア、希釈剤および/または安定化剤を含む薬学的組成物に関する。
【0051】
本発明に従う薬学的組成物は、本発明に有用な1つ以上のペプチド、または本発明に有用な1つ以上の抗体またはそれらの誘導体を含み得る。本発明に従う薬学的組成物はまた、さらに、治療活性成分または予防活性成分のうちの1つ以上を含むことができる。1つの実施形態において、薬学的組成物は、本発明に有用な1つ以上のペプチドを含む。別の実施形態において、薬学的組成物は、前記ペプチドに対する本発明に有用な1つ以上の抗体を含む。薬学的組成物が発明に有用な1つ以上の抗体を含む場合、前記薬学的組成物はさらに、第VIII因子を含み得る。そのような薬学的組成物における第VIII因子は、あらゆる自然に存在する、合成の、または、組換えの供給源から得られ得る。
【0052】
本発明に有用なペプチドおよび/または抗体は、薬学的に受容可能なビヒクル(たとえば、1つ以上の賦形剤、キャリア、希釈剤および安定化剤)と組み合わされて、溶液、懸濁液、エマルジョンまたは凍結乾燥粉末として処方され得る。適切なキャリアとしては以下が挙げられるがこれらに限定されない:希釈剤またはフィラー、無菌水媒体および種々の無毒性の有機溶剤。このようなビヒクルの例は、水、生理食塩水、リンガー溶液、デキストロース溶液およびヒト血清アルブミン溶液である。リポソームおよび不揮発性油のような非水性ビヒクルがまた使用され得る。ビヒクルまたは凍結乾燥物は等張性(例えば、NaCl、スクロース、マンニトール)および安定性(例えば、緩衝液および/または防腐剤)を維持する添加物を含み得る。適切な薬学的に受容可能なビヒクルは、A.OsolによるRemington’s Pharmaceutical Sciences(この分野における標準教科書)(参考として援用される)に記載されている。これらの組成物は、錠剤、カプセル、粉末、水性懸濁液または溶液、注射剤等の形で処方され得る。
【0053】
好ましくは本発明に従う薬学的組成物は、凍結乾燥されている。
【0054】
本発明の方法に基づいた薬学的組成物の投与は、好ましくは、改善が達成されるまで、最大の治療応答を保証する投薬レジメンを用いて実行されるべきである。概して、用量は、使われるペプチドの分子の大きさに依存する。約15アミノ酸のペプチドについて、静脈内投与のための投薬量は、約0.1〜1000mg/kgの間、好ましくは1〜500mg/kgの間、そして、最も好ましくは10〜100mg/kgの間で変化し得る。さらに大きなペプチドについては、その用量は、漸増し得る。経口投与のために、用量は、もちろん、適切な投薬がまた、薬物に対する応答に影響を与え得る、患者の身体全体の健康、年齢および他の因子に依存し得ることに留意して、大幅に高いかもしれない。薬物は、所望のレベルで治療効果を維持するために、連続的注入によって、または、約4〜50時間の定期的な間隔で投与され得る。
【0055】
本発明に従う薬学的組成物は、1つの成分調製物の形式であり得るか、または、キットにおいて1つ以上の他の成分と組み合わせて存在し得る。
【0056】
本発明に従う薬学的組成物は、哺乳動物(好ましくはヒト)への投与が意図される。本発明に有用なペプチドが特定の哺乳動物(例えばヒト)への投与が意図される場合、前記ペプチドは、特定の哺乳動物由来であることが好ましい。本発明に有用な抗体が特定の哺乳動物(例えばヒト)への投与が意図される場合、前記抗体は、特定の哺乳動物由来のエピトープを用いて生成されることが好ましい。さらに、ヒトへの投与が意図される本発明に有用な抗体は、ヒト抗体またはヒト化抗体であることが好ましい。
【0057】
本発明に有用なペプチド、本発明に有用な抗体およびこれらの誘導体ならびに前記ペプチド、抗体および誘導体を含む薬学的組成物は、前記ペプチド、抗体おまたはこれらの誘導体が体内の作用部位に到達し得る任意の手段によって被験者に投与され得る。好ましくは発明に従うペプチド、抗体およびこれらの誘導体、ならびに、前記のペプチド、抗体および誘導体を含む薬学的組成物は、非経口(すなわち、静脈内、皮下、または筋肉内)で投与される。
【0058】
本発明に従う調製物は、特に血液凝固障害を持つ患者の予防または処置のために、第VIII因子とLRPとの相互作用を阻害し、第VIII因子の分解を減少させ、または、第VIII因子の半減期を延長させる薬剤の投与が必要な全ての被験者を処置するために使われ得る。このような血液凝固障害は、少なくとも部分的に、第VIII因子の機能障害(例えば遺伝性または先天性の機能障害あるいは後天性の機能障害)によって引き起こされ得る。このような血液凝固障害はまた、第IX因子、第V因子、第X因子またはヴォン・ヴィレブランド因子を含む、凝固経路に関連している他のタンパク質における機能障害(例えば、遺伝性または先天性の機能障害あるいは後天性の機能障害)によって少なくとも部分的に引き起こされ得る。好ましくは、前記血液凝固障害は血友病Aまたはフォン・ヴィレブランド病である。
【0059】
このように、本発明はさらに、血液凝固障害の処置のための医薬の調製のための、第VIII因子由来であるが、本質的に第VIII因子活性を有さない配列番号1、2、3もしくは4のいずれかに定義されるアミノ酸を含むペプチドの使用、または、上記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の使用に関する。
【0060】
好ましい実施形態において、血液凝固障害は、血友病Aまたはフォン・ヴィレブランド病である。
【0061】
本発明に従う薬学的調製物はまた、例えば、手術もしくは外科的手術の直前、間、または後に、血栓溶解系または繊維素溶解系の一時的な障害を有する患者の予防または処置のために使われ得る。
【0062】
本発明はまた、生物学的流体における第VIII因子の分解を減少させるための、第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるアミノ酸配列を含むペプチドの使用または前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の使用に関する。
【0063】
さらに関連の実施形態において、本発明は、生物学的流体における第VIII因子の分解を減少させる方法を提供し、ここで、第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるペプチドまたは前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体が前記生物学的流体に添加される。
【0064】
本明細書中で使用される場合、「生物学的な流体」は、血液のような哺乳動物から単離された流体、または血漿もしくは血清のようなその画分、ならびに、真核細胞または原核細胞、細胞株または組織の培養物から得られた媒体またはそれらの画分を含む。
【0065】
好ましくは前記ペプチドまたは前記抗体は、第VIII因子の分解を少なくとも10%、好ましくは少なくとも20%、より好ましくは少なくとも50%ならびに最も好ましくは本質的に100%減少させるのに十分な量で添加される。
【0066】
本発明はさらに、生物学的流体における第VIII因子の半減期を延長させるための、第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるアミノ酸配列を含むペプチドまたは前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の使用および関連する方法に関する。
【0067】
好ましくは前記ペプチドまたは前記抗体は、生物学的流体における第VIII因子の半減期を少なくとも2倍、より好ましくは少なくとも5倍、そして最も好ましくは少なくとも10倍延長するのに十分な量で添加される。
【0068】
本発明はさらに、(生物学的流体における第VIII因子とLRPとの相互作用を阻害するための、)第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるペプチドまたは前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の使用に関する。好ましくは、少なくとも10%、好ましくは少なくとも20%、最も好ましくは少なくとも50%の第VIII因子のLRPとの阻害程度を達成するのに十分な量の前記ペプチドまたは前記抗体が添加される。
【0069】
本発明は、さらに血液凝固障害に罹患した患者を処置する方法であって、該方法は、第VIII因子のLRPとの相互作用を阻害するための、第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるペプチドまたは前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の有効量を前記患者に投与する工程を包含する、方法、または、前記ペプチドの1つ以上もしくは前記抗体の1つ以上を含む薬学的組成物に関する。
【0070】
本発明は、引き続いて記載される実施例に例示されるが、本発明はこれに制限されることを意図しない。
【実施例】
【0071】
(実施例I)
材料−CNBr−セファロース4Bは、Amersham Pharmacia Biotech製であった。マイクロタイタープレート(Maxisorp)、細胞培養フラスコ、Optimem I培地、ウシ胎仔血清(FCS)、ペニシリンおよびストレプトマイシンは、Life Technologies(Life Technologies Inc.,Breda,The Netherlands)製であった。グレースのInsect培地(TNM−FH)およびInsect−XPRESS培地は、BioWhiftaker (Alkmaar、The Netherlands)から購入した。
【0072】
タンパク質−血漿由来のFVIII軽鎖、およびその第Xa因子切断誘導体を、以前に示されたように調製した(Lenting,P.J.,Donath,M.J.S.H.,van Mourik,J.A.,およびMertens,K.(1994)J.Biol.Chem.269,7150−7155;Donath,M.S.J.H.,Lenting,P.J.,van Mourik,J.A.,およびMertens,K.(1995)J.Biol.Chem.270.3648−3655;本明細書中に参考として援用される)。抗FVIIIモノクローナル抗体CLB−CAg A、CLB−CAg 117およびCLB−CAg 12が、以前に記載されている(Lenting,P.J.ら、(1994),前出;Leyte,A.,Mertens,K.,Distel,B.,Evers,R.F.,de Keyzer−Nellen,M.J.,Groenen−van Dooren,M.M.,de Bruin,J.,Pannekoek,H.,van Mourik,J.A.,およびVerbeet,M.P.(1989)Biochem.J.263,187−194;本明細書中に参考として援用される)。抗FVa軽鎖モノクローナル抗体CLB−FV 5は、標準のハイブリドーマ技術によって得た。
【0073】
FVIIIの軽鎖に対するドメイン可変単鎖抗体フラグメント(scFvs)を、Escherichia.coli株TG 1に発現させ、そして、金属器レートクロマトグラフィー(Qiagen,Hilden,Germany)によって、scFvs KM36、KM41およびKM33を150mM NaCl、5mM CaCl2、100mM イミダゾールおよび20mM Hepes(pH 7.4)中で溶出したことを除き、以前に記載されるように(32〜34)精製した。
【0074】
ヒトFVIII領域Trp1707−Arg1721(WDYGMSSPHVLRNR)(配列番号5)、Lys1804−Lys1818(KNFVKPNETKTYFWK)(配列番号2)、Tyr1815−Ala1834(YFWKVQHHMAPTKDEFDCKA)(配列番号3)、His1822−Ala1834(HMAPTKDEFDCKA)(配列番号6)、Thr1892−Ala1901(TENMERNCRA)(配列番号7)、Glu1908−His1919(EDPTFKENYRFH)(配列番号8)、Thr1964−Lys1972(TVRKKEEYK)(配列番号9)、Lys2049−Gly2057(KLARLHYSG)(配列番号10)およびAsp2108−Gly2117(DGKKWQTYRG)(配列番号11)を包含する合成ペプチドを、手動の「T−bag」法(Houghton,R.A.(1985)PNAS U.S.A.82,5131−5135;WO96/41816;本明細書中に参考として援用される)によってFmoc(N−(9−フルオレニル)メトキシカルボニル)化学によって、または430A Applied Biosystems装置(Pharmacia LKB Biotechnology,Roosendaal,the Netherlands;Medprobe AS,Oslo,Norway)を使用して合成した。
【0075】
ペプチドは、95%以上純粋(高圧液体クロマトグラフィー(HPLC)分析によって測定)であり、これらの同一性を質量分析によって裏付けた。精製した胎盤由来のLRPを、記載されたように得た(Moestrup,S.K.,およびGliemann,J.(1991)J.Biol.Chem.266,14011−14017;本明細書中に参考として援用される)。グルタチオンS−トラスフェラーゼ融合レセプター関連タンパク質(GST−RAP)をEscherichia coli株DH5αに発現させ、記載されたとおりにグルタチオン−セファロースを用いて精製した(Herz,J.,Goldstein,J.L.,Strickland,D.K.,Ho,Y.K.,およびBrown,M.S.(1991)J.Biol.Chem.266,21232−21238;本明細書中に参考として援用される)。組換えLRPリガンド結合クラスターIIおよびIVを発現するBaby Hamster Kidney(BHK)細胞が以前に記載されている(Neels、J.G.ら、(1999)、前出)。ヒト血清アルブミン(HSA)は、CLB(Amsterdam、The Netherlands)の製品部門製であった。タンパク質を、標準としてHASを用い、Bradfordの方法(Bradford,M.M.(1976)Anal.Biolchem.72,248−254;本明細書中に参考として援用される)によって定量した。
【0076】
組換えタンパク質−FVIII Bドメイン欠失変異体であるFVIII−del(868−1562)をコードするプラスミドpCLB−BPVdB695は以前に記載されており(Mertens,K.,Donath,M.J.S.H.,van Leen,R.W.,de Keyzer−Nellen,M.J.M.,Verbeet,M.P.,Klaasse Bos,J.M.,Leyte,A.,およびvan Mourik,J.A.(1993)Br.J.Haematol.85,133−142;本明細書中に参考として援用される)、そして、FVIII/FV1811−1818キメラをコードするプラスミドを構築するためのテンプレートとして用いた。FVIII/FVコドン置換を含むFVIII軽鎖配列由来のオリゴヌクレオチドプライマー(以下の表II)を用い、Overlap Extension PCR変異誘発法を用いてプラスミドを構築した(Tao,B.Y.,およびLee K.C.P.(1994)PCR Technology Current Innovations(Griffin,H.G.,およびGriffin,A.M.編),71−72,CRC Press,Boca Raton,FL;本明細書中に参考として援用される)。配列決定分析を、プラスミドにおける変異の存在を証明するために、実施した。FVIIIをコードするプラスミドのマウス線維芽(C127)細胞へのトランスフェクションを、以前に記載されたとおりに実施した(Mertens、K.ら、(1993)、前出)。野生型FVIIIまたはFVIII/FV1811−1818キメラを発現する安定な細胞株を、5% FCS、100U/ml ペニシリン、100μg/mlストレプトマイシン、1μg/ml アンホテリシンBおよび0.8μg/ml デスオキシコール酸を補充したRPMI 1640培地中の1−I細胞工場(cell factory)に維持した。FVIII1を含む培地を1週間に3回回収した。引き続いて、細胞片を除去するためにこの培地を濾過し、中空糸カートリッジ(Hemofiow F5,Fresenius,Bad Homburg,Germany)を用いておおよそ10倍に濃縮した。ベンズアミジンを、10mMの最終濃度で加え、濃縮物を−20℃で保管した。FVIIIを、確立した手順(Mertens、K.ら、(1993)、前出)に従って、抗体CLB−CAg 117およびQ−セファロースクロマトグラフィーを用いる、免疫親和性クロマトグラフィーによって濃縮した培地から精製した。FVIII軽鎖を、40mM EDTA、100mM NaClおよび50mM Tris(pH 7.4)を含む緩衝液中、FVIII/FV1811−1818キメラおよび野生型FVIIIを25℃で4時間インキュベートすることによって調製した。続いて、FVIII/FV1811−1818軽鎖キメラおよび野生型FVIII軽鎖を、Q−セファロースクロマトグラフィーを用いて精製した。組換タンパク質を、1M NaClおよび50mM Tris(pH 7.4)を含む緩衝液中で溶出し、150mM NaClおよび50mM Tris(pH 7.4)に対して透析し、そして、4℃で保管した。組換えC2ドメイン(すなわち、残基Ser2173−Tyr2332)をコードするプラスミドの構築は、以前に記載されている(Fijnvandraat、K.、ら、(1998)、前出)。FVIIIa3−A3−C1フラグメント(すなわち、残基Glu1649−Asn2172)をコードするプラスミドpACgp67b−His−a3−A3−C1を、pCLB−BPVdB695を鋳型として用い、オリゴヌクレオチドプライマー、5’−TTACTCGAGGAAATAACTCGTACTACTC−3’(センス)(配列番号13)、および5’−AATGCGGCCGCTTCAATTTAAATCACAGCCCAT−3’(アンチセンス)(配列番号14)を用いてポリメラーゼ連鎖反応によって構築した(Mertens、K.ら、(1993)、前出)。増幅されたDNAフラグメントを精製し、XhoIおよびNotIで消化し、pBluescriptに連結した。得られた構築物は、配列決定によって証明した。続いて、pBluescript−a3−A3−Clを、EspIおよびNotlで消化し、そして、得られたフラグメントを精製し、EspI/NotIで消化したpACgp67b−80Kプラスミドに連結した(Fijnvandraat,K.,Turenhout,E.A.M.,van denBrink,E.N.,ten Cate,J.W.,van Mourik,J.A.,Peters,M.,およびVoorberg,J.(1997)Blood 89,4371−4377;本明細書中に参考として援用される)。His−タグ(5’−ATTGGATCCGGCCATCATCATCATCATCATGGCGGCAGCCCCCGCAGCTTTCAAAAGCCCGGGGCCATGGGA−3’)(配列番号15)をコードするDNAフラグメントを、BamHIおよびNcoIで消化し、そして、BamHI/NcoIで消化したpACgp67b−a3−A3−C1プラスミド中にクローニングした。バキュロ・ウイルス表現系を使って、組換えa3−A3−C1およびC2フラグメントを、記載されたとおりに昆虫細胞の感染によって得た(Fijnvandraat、K.ら(1998)、前出)。a3−A3−C1フラグメントを免疫親和性クロマトグラフィーを用いてInsect−XPRESS培地から精製した(親和性マトリクスとしてCNBr−セファロース4Bと連結された抗A3ドメイン抗体CLB−CAg Aを用いる)。CLB−CAg A−セファロースを、a3−A3−C1フラグメントを含む培地と共に4℃で16時間インキュベートした。結合の後、免疫親和性マトリクスを回収して、1M NaClおよび50mM Tris(pH 7.4)を含む緩衝液で洗浄し、そして、150mM NaCl、55%(v/v)エチレングリコールおよび50mMリジン(pH 11)で溶出した。溶離画分をすぐに、1M イミダゾール(pH 6.0)で中和し、150mM NaCl、50%(v/v)グリセロールおよび50mM Tris(pH 7.4)に対して透析し、そして、−20℃で保管した。同様の免疫親和性クロマトグラフィー技術を用いて組換えC2ドメインを精製した。但し、抗C2ドメイン抗体CLB−CAg 117をCLB−CAg Aの代わりに用いた。
【0077】
第Va因子軽鎖の精製−ヒト第V因子(FV)を、本発明者らの組織(CLB、オランダ)によって供給されたヒト血漿から得た。全長FVを、免疫親和性クロマトグラフィーを用いて精製した。FVa軽鎖を、100mM NaCl、5mM CaCl2および50mM Tris(pH 7.4)を含む緩衝液中、FV(10μM)をトロンビン(2μM)と共に37℃で2時間インキュベートすることによって調製した。トロンビンを、ヒルジン(Sigma−Aldrich、St.Louis、MO)によって不活化し、そして、抗第V因子軽鎖モノクローナル抗体CLB−FV 5(5mg/ml)に結合したCLBr−セファロース4Bを用いる免疫親和性クロマトグラフィーを用いて、FVa軽鎖を精製した。免疫親和性マトリックスを、100mM NaCl、25mM EDTAおよび50mM Tris(pH 7.4)で洗浄し、そして、100mM NaCl、5mM CaCl2、55%(v/v)エチレングリコールおよび50mM Tris(pH 7.4)で溶出した。精製したFVa軽鎖を、150mM NaCl、5mM CaCl2、50%(v/v)グリセロールおよび50mM Tris(pH 7.4)に対して透析し、そして、−20℃で保管した。
【0078】
組換え型LRPフラグメントの発現および精製−組換えLRPクラスターIIおよびIVを、1単位/ml ペニシリンおよび100μg/ml ストレプトマイシン100を補充したOptimem I培地を用いてBHK細胞中に発現させた(Neels、J.G.ら、(1999)、前出)。培地を回収した後、CaCl2を、10mMの最終濃度まで添加した。馴化培地からのLRPクラスターIIおよびIVの精製は、1つの精製工程によって行った(親和性マトリクスとしてCNBr−セファロース4Bと連結されたGST−RAPを使って)。このマトリックスを、カラムに回収し、150mM NaCl、5mM CaCl2および50mM Hepes(pH 7.4)を含む緩衝液で洗浄し、そして、150mM NaCl、20mM EDTAおよび50mM Hepes(pH 7.4)で溶出した。続いて、精製したLRPクラスター調製物を、Centricon 10コンセントレーター(Millipore,Bedford,−MA)を用いて、4℃、4000×gで1時間の連続回転の遠心分離により濃縮した。最終的に、それらの調製物を、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH 7.4)に対して透析し、そして、−4℃で保管した。
【0079】
固相結合アッセイ−組換えLRPクラスターIIまたはIV(1pmol/ウェル)を、50μlの容量の50mM NaHCO3(pH 9.8)中のマイクロタイターウェルに、4℃で16時間吸着させた。ウェルを、200μlの容量の2%(w/v)HSA、150mM NaCl、5mM CaCl2および50mM Tris(pH 7.4)で、37℃で1時間ブロッキングした。150mM NaCl、5mM CaCl2、0.1%(V/V)Tween(登録商標)20および50mM Tris(pH 7.4)で3回急速に洗浄した(各5秒未満)後、FVIII軽鎖またはその誘導体を、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%(v/v)Tween(登録商標)20および50mM Tris(pH 7.4)を含む50μlの容量の緩衝液中にて、37℃で2時間、種々の濃度でインキュベートした。結合したリガンドを、同じ緩衝液中でペルオキシダーゼ結合体化モノクローナル抗体CLB−CAg 12と共に37℃で15分間インキュベートすることによって検出した。抗体CLB−CAg 12は、FVIIIフラグメントのLRPまたはLRPクラスターのフラグメントへの結合をに干渉しなかった(データ示さず)。競合実験において、FVIII軽鎖(25nM)を、50μlの容量の競合物質の限界希釈の存在下または非存在下のいずれかにおいて、固定されたLRPクラスターを含むウェルにて、37℃で2時間インキュベートした。上で記載されたように、残留FVIII結合を検出した。データを、空のマイクロタイターウェル(固定されたLRPクラスターを含むマイクロタイターウェルへの結合と比較して5%未満であった)について補正した。
【0080】
表面プラズモン共鳴−BIAcoreTM2000バイオセンサーシステム(Biacore AB、Uppsala、Sweden)を使って、タンパク質相互作用の動力学を、表面プラズモン共鳴(SPR)分析によって決定した。LRP(16fmol/mm2)、FVIII軽鎖(71fmol/mm2)、a3−A3−Clフラグメント(67fmol/mm2)、FVa軽鎖(76fmol/mm2)またはscFv EL14(67fmol/mm2)は、製造業者によって指定されたようなアミン結合キットを用いて、第1級アミノ基を介しての活性化されたCM5−センサチップのデキストラン表面に共有結合した。1つのコントロールフローチャネルを、タンパク質の非存在下で、慣例的に活性化し、ブロックした。分析物の会合を、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH 7.4)中、25℃で20μl/分の流速で2分間評価した。同じ緩衝液流中で2分間解離させた。センサチップを、100mM H3PO4または20mM EDTA、1M NaClおよび50mM Hepes(pH 7.4)のいずれかの中にて、20μl/分の流速で数回のパルスを用いて再生した。会合速度定数(kon)および解離速度定数(koff)を、BlAevaluationソフトウェア3.1(Biacore AB、Uppsala、Sweden)を使うことによって決定した。データを、バルク屈折率変化について較正し、2部位結合モデルに従う非線形回帰分析によって適合した。平衡解離定数(kd)を、比率koff/konから計算した。低親和性の相互作用についてのkd値は、BlAevaluationソフトウェアを使うことによって定常状態の親和性の分析を用いて見積った。競合実験において、FVIII軽鎖(50nM)を、競合物質の限界希釈の存在下もしくは非存在下のいずれかにおいて、20μl/分の流速で2分間、固定化されたLRP(16fmol/mm2)と共にインキュベートした。
【0081】
FVIII/FV1811−1818軽鎖キメラのLRPクラスターIIへの結合−150mM NaClおよび50mM Tris(pH 7.4)を含む緩衝液中で、組換えFVIII/FV1811−1818軽鎖キメラまたは組換え野生型FVIII軽鎖を、20fmol/mm2の密度まで、固定されたscFv EL14に結合した。LRPクラスターII(25〜125nM)を、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH 7.4)中、20μl/分の流速にて、25℃で2分間、それぞれ、FVIII/FV1811−1818軽鎖キメラまたは野生型組換えFVIII軽鎖を固定化した別々のチャネル、および、1つのコントロール(scFv EL14でコーティングされている)チャネルを通した。
【0082】
(結果:)
LRPとFVIII軽鎖フラグメントとの間の相互作用−単離されたFVIII C2ドメイン(すなわち、残基Ser2173−Tyr2332)は、インタクトなFVIII軽鎖よりも効果的にLRPに会合しない(Lenting、P.J.、ら、(1999)、前出;WO00/28021、前出)。本研究において、FVIII軽鎖の別の部位がLRP結合に寄与する可能性を探究した。この目的のために、SPR分析を用いて、固定化されたLRPとの4つのFVIII誘導体の相互作用をモニタリングした。これらの誘導体としては、FVIII軽鎖、a3−A3−C1部分(すなわち、残基Glu1649−Asn2172)、C末端C2ドメインおよびFVIII軽鎖フラグメント(第X因子による位置Arg1721での切断を用いて、N末端の酸性の領域を欠失している(すなわち、残基Ala1722−Tyr2332))が挙げられる。
【0083】
図1において示されたように、全てのFVIIIフラグメントは、固定されたLRPとの時間依存的な会合を示し、その後分離した(最も高い反応が最高LRP密度で観察されたので、用量依存的であるように思われる(データ示さず))。FVIII軽鎖、第X因子切断軽鎖およびa3−A3−C1フラグメントの得られたデータを適切に示すのに、2部位結合モデルを必要とした。このモデルに従う計算された会合速度定数(kon)および解離速度定数(koff)は、これらのフラグメントについて同じオーダーにあった(以下の表I)。これは、固定されたLRPとの高い親和性および僅かに低い親和性の相互作用を示す、同等のKd値(すなわちFVIII軽鎖について18nMおよび59nM、第X因子切断軽鎖について22nMおよび60nM、ならびにa3−A3−C1誘導体について26nMおよび74nMを生じる(以下の表I)。対照的に、単離したC2ドメインは、任意の結合モデルによって正確に記載されるにはあまりにも速い解離速度を示した。従って、定常状態の親和性の計算を用いて、このフラグメント(3.4μM)の親和性(Kd)を推測によって見積った。まとめると、これらの結果は、A3−C1領域(すなわち、残基A1722−Asn2172)に高い親和性LRP結合部位が存在し、そしてC2ドメインに低い親和性の部位が存在することを示す。
【0084】
【表1】

【0085】
(表1に対する注意事項:FVIII軽鎖およびその誘導体の固定化されたLRPへの結合についての動力学的パラメーター。固定されたLRP(16fmol/mm2)への、種々の濃度のFVIII軽鎖(10〜250nM)、67kDaフラグメント(10〜250nM)、a3−A3−C1フラグメント(10〜250nM)またはC2ドメイン(500〜2000nM)の会合および解離を「実験手順」で示されたように評価した。得られたデータを2部位結合モデルを用いて分析して、会合速度定数(kon)および解離速度定数(koff)を計算した。各クラスの結合部位を、それぞれ1および2と称する。親和定数(Kd)は、比率 koff/konから計算した。C2ドメインとLRPとの間の相互作用を、定常親和性分析を用いて評価した。データを、各測定について少なくとも5つの異なる濃度を用いた、3〜6回の測定に基づく。データは、平均±S.D.を表す。)
固定されたLRPクラスターIIおよびIVへのFVIII軽鎖の結合−以前の研究において、LRPリガンド結合クラスターIIおよびIVが、VIII軽鎖との相互作用を媒介することが示された(Neels、J.G.ら(1999)、前出)。本研究において、LRPクラスターIIおよびIVがFVIII軽鎖への結合に競合し得るか否かという問題に取り組むために固相結合アッセイを用いた。図2の挿入図において示したように、FVIII軽鎖は、固定されたLRPクラスターIIに用量依存性の様式で結合し得た。この観察は、以前の調査結果(SPIR分析を用いて、LRPクラスターIIと固定されたFVIII軽鎖との間の相互作用をモニタリングした)(Neels、J.G.ら(1999)、前出)と一致している。競合研究は、LRPクラスターIIおよびIVが、FVIII軽鎖への結合に競合することを明らかにした(図2)。LRPクラスターIIは、固定されたLRPに結合するFVIII軽鎖クラスターIVの用量依存性の阻害を示した。これらのデータは、LRPクラスターIIおよびIVが、FVIII軽鎖内の同様の結合領域を共有することを意味する。
【0086】
LRPクラスターIIの固定化されたFVa軽鎖への結合−FVIIIとFVとの間の公知の相同性(Church,W.R.ら(1984)、前出)を考慮すると、FVIIIおよび活性化されたFVの軽鎖がLRPクラスターIIへの結合特性を共有するか否かという問題が生じ得る。この目的のために、LRPクラスターIIの連続希釈を、固定化されたFVIII軽鎖およびFVa軽鎖と共にインキュベートした。図3に示すように、FVaおよびFVIIIの軽鎖は、FVIIIのみがLRPクラスターIIに対して高親和性の結合を示すという点で異なることを証明した。これらの観察は、FVIII軽鎖のa3−A3−Clドメインは、FVa軽鎖には保存されてない高親和性のLRP相互作用領域を含むことを示す。
【0087】
合成のペプチドの、FVIII軽鎖のLRPクラスターIIへの結合に及ぼす影響−a3−A3−Clドメインの表面ループを模倣する合成ペプチドのパネルを構築した(WO 96/41816、前出)。FVa軽鎖がLRPクラスターIIと効率的に会合しないという観察を、FVIIIに固有の合成のペプチドの構築のための選択基準として使用した。これらのループの溶媒接触能を、ヒドロパシー分析(Lenting、P.J.ら(1996)、前出)を用いて、および、インタクトなFVIIIヘテロ二量体の三次元モデル(Stoilova−McPhie,S.,Villoutreix,B.O.,Mertens,K.,Kemball−Cook,G.,およびHolzenburg,A.(2002)Blood 99,1215−1223;本明細書中に参考として援用される)を研究することによって確定した。合成のペプチドは、以下の残基を含んだ:Trp1707−Arg1721、Lys1804−Lys1818、Tyr1815−Ala1834、His1822−Ala1834、Thr1892−Ala1901、Glu1908−His1919、Thr964−Lys1972、Lys2049−Gly2057およびAsp2108−Gly2117(表II)。
【0088】
【表2】
【0089】
(表IIに対する注意事項:合成のペプチド由来のFVIII a3−A3−C1フラグメントの、FVIII軽鎖とLRPクラスターIIとの間の相互作用への影響。FVIII軽鎖(25nM)を、種々の濃度の合成ペプチド(0〜1mM)の存在下または非存在下において、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%Tween(登録商標)20および50mM Tris(pH 7.4)50μlの容量中にて、固定されたLRPクラスターII(1pmol/ウェル)と共に37℃で2時間インキュベートした。同じ緩衝液で洗浄した後、FVIII軽鎖を、ペルオキシダーゼ結合体化抗−FVIII抗体CLB−CAg 12と共に37℃で15分間インキュベートする事によって定量した。最大半減阻害定数(IC50)は、3回の実験の平均値±S.D.を表す。)
続いて、これらのペプチドを、FVIII軽鎖と固定されたLRPクラスターIIとの間の相互作用を阻害するそれらの能力について試験した。表IIに示したように、合成ペプチドLys1804−Lys1818(配列番号2)およびTyr1815−Ala1834(配列番号3)は、FVIII軽鎖と固定されたLRPクラスターIIとの間の相互作用を効率的に阻害した。最大半減阻害(IC50)は、各々、約1.9μMおよび16.8μMのペプチド濃度に達した。他の合成ペプチドは、このような阻害効果を示さなかった。これらの観察は、FVIIIのA3ドメイン内の配列Lys1804−Ala1834が、LRPとの相互作用に関する重要な残基を含むことを示唆する。
【0090】
ScFv抗体フラグメントがLRPクラスターIIへのFVIII軽鎖の結合に及ぼす影響−以前に、領域Gln1778−Asp1840内の残基に対する阻害性抗体を持つ患者由来の組換えscFv抗体フラグメントを分離するために、ファージディスプレイを用いた(van den Brink,E.N.,Turenhout,E.A.M.,Bovenschen,N.,Heijnen,B.G.,Mertens,K.,Peters,M.およびVoorberg,J(2001),Blood 97,966−972;Voorberg,J.,ら、Method for diagnosis and treatment of haemophilia A patients with an inhibitor;国際特許出願番号WO 99/58680;これらは本明細書中に参考として援用される)。これらのscFvsを、FVIII軽鎖とLRPまたはクラスターIIとの間の相互作用を阻害するそれらの能力について評価した。第1のscFv(scFv KM36称される)は、Gln1778−Asp1840内の領域に指向するが、FVIIIへの結合のために残基Arg1803−Lys1818を必要としない(van den Brinkら(2001)、前出;WO 99/58680、前出)。第2のscFv(scFv KM41として命名された)は、領域Arg1803−Lys1818に指向し、かつFVIIIの凝固促進性活性を阻害する(van den Brinkら(2001)、前出;WO 99/58680、前出)。scFv KM41の重鎖のアミノ酸配列ならびに対応するDNA塩基配列を配列番号16に示す。scFv KM41の軽鎖のアミノ酸配列ならびに対応するDNA塩基配列を配列番号18に示す。KM41の重鎖および軽鎖は、ペプチドリンカー(配列番号24)で結合され得、かつ、C末端にヒスチジンタグを有し得る。発現されたscFv KM41タンパク質のアミノ酸配列を、配列番号25に示す。第3のscFv(scFv KM33と称される)は、scFv KM41と類似している。但し、そのFVIII軽鎖との相互作用のためにA3ドメイン領域1778−1818を必要としない(van den Brinkら(2001)、前出;WO 99/58680、前出)。scFv KM33の重鎖のアミノ酸配列ならびに対応するDNA塩基配列を配列番号20に示す。scFv KM33の軽鎖のアミノ酸配列ならびに対応するDNA塩基配列を配列番号22に示す。重鎖および軽鎖は、ペプチドリンカー(配列番号24)で連結され得、かつ、C末端にヒスチジンタグを有し得る。発現されたscFv KM33タンパク質のアミノ酸配列を、配列番号26に示す。図4Aにおいて、scFv KM36がFVIII軽鎖と固定されたLRPクラスターIIとの間の相互作用に影響を及ぼさなかったことを示した。対照的に、scFv KM41の存在は、FVIII軽鎖のLRPクラスターIIへの結合を阻害した(図4A)。scFv KM41がFVIII軽鎖とLRPとの間の相互作用に及ぼす影響を、SPIR分析を用いてさらに研究した。図4Bに示したように、FVIII軽鎖の固定されたLRPへの会合は、scFv KM41の存在下で効率的に阻害された。図5に示したように、scFv KM33は、FVIII軽鎖の固定されたLRPクラスターIIへの結合を用量依存性の様式で効率的に阻害した。見かけ上の阻害定数は、約5nMであったが、これは、同じscFvへのFVIII軽鎖の結合についてのKd値と類似している(van den Brinkら(2001)、前出;WO 99/58680、前出)。これらのデータは、scFv KM33、scFv KM41およびLRPがFVIIIの軽鎖内の重複する結合部位を共有することを強力に示唆する。
【0091】
FVIII軽鎖配列Glu1811−Lys1818は、LRPのための結合部位を含む−合成のペプチドLys1804−Lys1818およびTyr1815−Ala1834は、FVIIIa−FIXa複合体のアセンブリを妨害することによる、FVIII凝固促進活性の効果的なインヒビターである(Lenting、P.J.、ら、(1996)、前出;WO 96/41816、前出)が、scFvs KM33およびKM41は、FVIIIa−FIXa複合体のアセンブリを妨害することによる、FVIII凝固促進活性の効果的なインヒビターである(Lenting、P.J.、ら、(1996)、前出;van der Brinkら、(2001)、前出;WO 99/58680、前出)。FVIII A3ドメイン残基Glu1811−Lys1818は、FIXaとの相互作用に関与する(Lenting、P.J.、ら、(1996)、前出;WO 96/41816、前出)ので、この特定のFVIII軽鎖領域を、LRPとの相互作用におけるその役割に関して調査した。FVa軽鎖がLRPクラスターIIと相互作用しなかったので、FVの対応する残基(すなわち、残基1704SSYTYVWH1711;(配列番号12))によって残基Glu1811−Lys1818を交換してFVIII軽鎖キメラを、構築した。単離したキメラ(FVIII/FV1811−1818)を、SPR分析を用いて野生型組換えFVIII軽鎖とLRPクラスターIIとの会合に関して比較した。図6に示したように。LRPクラスターIIは、野生型のFVIII軽鎖と比較して固定化したFVIII/FV1811−1818との2〜3倍減少した会合を示した。
【0092】
次いで、単離したキメラ(FVIII/FV1811−1818)を、SPR分析を用いて、scFv KM41と相互作用する能力について評価した。図7に示すように、scFv KM41は、固定化したFVIII/FV1811−1818キメラを認識せず、一方、固定化した野生型の組換えFVIII軽鎖と容易に反応した。これらの観察は、FVIII A3ドメイン領域Glu1811−Lys1818が、scFv KM41に結合するために重要な残基を含むことを示す。これらのデータは、FVIII軽鎖領域Glu1811−Lys1818が、LRP−FVIII軽鎖複合体のアセンブリに重要な役割を果たすことを示す。
【0093】
本発明の研究において、FVIII軽鎖のA3ドメイン領域Glu1811−Lys1818は、LRPとの高い親和性の相互作用に寄与することが示される。いくつかの系統の証拠が、この結論を支持する。第1に、合成ペプチドLys1804−Lys1818およびTyr1815−Ala1834由来のA3ドメインは、FVIII軽鎖とLRPクラスターIIとの間の相互作用に影響を及ぼした(表II)。第2に、FVIII軽鎖キメラ(領域Glu1811−Lys1818がFVの対応する残基と交換される)は、野生型FVIII軽鎖と比較してLRPクラスターIIとの会合の減少を示した(図6)。第3に、領域Glu1811−Lys1818に対する組換えscFv抗体フラグメントが、FVIII軽鎖のLRPまたはLRPクラスターIIフラグメントへの結合を阻害した(図4)。
【0094】
多くのLRPリガンド(RAP、リポタンパク質リパーゼ、およびα2−マクログロブリンが挙げられる)について、リガンド表面の正に荷電した残基がLRPとの相互作用に関与することが確立されている(Meiman,L.,Cao,Z.F.,Rennke,S.,Paz Marzolo,M.,Wardell,M.R.およびBu,G.(2001)J.Biol.Chem.276.29338−29346;Chappell,D.A.,Fry,G.L.,Waknitz,M.A.,Muhonen,L.E.,Pladet,M.W.,Iverius,P.H.およびStrickland,D.K.(1993)J.Biol.Chem.268,14168−14175;Howard,G.C.,Yamaguchi,Y.,Misra,U.K.,Gawdi,G.,Nelsen,A.,DeCamp,D.L.およびPizzo,S.V.(1996)J.Biol.Chem.271,14105−14111;Nielsen,K.L.,Holtet,T.L.,Etzerodt,M.,Moestrup,S.K.,Gliemann,J.,Sottrup−Jensen,L.およびThorgersen,H.C.(1996)J.Biol.Chem.271,12909−12912;本明細書中に参考として援用される)。興味深いことに、FVIII A3ドメイン領域Glu1811−Lys1818(すなわち、残基1811ETKTYFWK1818(配列番号1))もまた、1813位および1818位に2つの露出された正に荷電したリジン残基を含む。FVのA3ドメイン内の相同体部分(すなわち、残基1704SSYTYVWH1711)と比較すると、これらのリジン残基は、FVIII A3ドメインに対して固有であるようである(Church,W.R.ら、(1984)、前出)。FVIII残基Glu1811−Lys1818のFVの対応する残基への置換は、LRPクラスターIIへの低下した結合を生じた(図6)。これらの結果は、領域Glu1811−Lys1818内の正に荷電した残基がLRPとの静電的相互作用を媒介することを示唆する。
【0095】
今日までに、LRP−FVIII軽鎖複合体のアセンブリに寄与する、FVIII軽鎖内の2つのアミノ酸領域が確認されている。本発明の研究において発見されたA3ドメイン領域Glu1811−Lys1818についての役割に加えて、カルボキシ末端のC2ドメインもまた、LRPとの相互作用に寄与することが知られている(Lenting,P.J.、Neels,J.G.、van den Berg,B.M.M.、Clijsters,P.P.F.M.、Meijerman,D.W.E.、Pannekoek,H.、van Mourik,J.A.、Mertens,K.およびvan Zonneveld,A.,−J.(1999)J.Biol.Chem.274,23734−23739;Lenting,P.J.ら A factor VIII polypeptide with factor VIILC−activity.国際特許出願WO 00/28021;本明細書中に参考として援用される)。単離したC2ドメインはLRPとの低い親和性の相互作用(Kd 約3.4μM)を示すので、A3ドメインのLRP相互作用部位は、C2ドメインのうちの1つよりも優性であるようにみえる(表I)。これは、単離したC2ドメインがLRPとわずかな会合のみを示した過去の研究と一致する(Lenting,P.J.ら(1999)、前出;WO 00/28021、前出)。さらに、FVIII軽鎖のLRPへの結合親和性は、C2ドメインの欠失に影響されない(表I)。しかし、抗C2ドメインモノクローナル抗体(ESHH4)は、FVIII軽鎖とLRPとの間の相互作用を完全に阻害することが示された(Lenting、P.J.ら(1999)、前出;WO 00/28021、前出)。抗体ESH4がLRP結合を阻害するメカニズムは、まだ解明されていない。抗C2抗体は、そのFVIII軽鎖との相互作用のために領域Glu1811−Lys1818を必要としない(Scandella,D.,Gilbert,G.E.,Shima,M.,Nakai,H.,Ealeson,C.,Felch,M.,Prescott,R.,Rajalaksmi,K.J.,Hoyer,L.W.およびSaenko,E.(1995)Blood 86,1811−1819;本明細書中に参考として援用される)ので、ESH4がA3ドメインにおける同様の部位への結合に関してLRPと競合することはなさそうである。従って、阻害に寄与し得るメカニズムのうちの1つは、立体障害を含む。
【0096】
対照的に、scFv KM41は、ほんの部分的にFVIII軽鎖とLRPとの間の相互作用を阻害する(図4)。これは、完全抗体(約150kDa)と比較して、比較的小さいサイズのscFv抗体フラグメント(約30kDa)に起因し得る。これらの観察は、領域Glu1811−Lys1818に加えて、A3−C1ドメイン内の他の表面の露出された構造エレメント(すなわち、残基Ala1722−Asn2172)がLRP−FVIII軽鎖複合体のアセンブリに寄与することを示唆する。これは、FVIII/FV1811−1818軽鎖キメラが、LRPクラスターIIに対する残基結合を示したという観察の系統にある(図6)。この文脈において、FVIII軽鎖のA3ドメイン内のGlu1811−Lys1818領域が、タンパク質表面に露出されるより大きなセグメントの一部(すなわち、残基Glu1804−Lys1818)であることを言及すべきである(Lenting、P.J.ら(1996)、前出)。Lys1813位およびLys1818位のリジン残基に加えて、この領域は、Lys1804位およびLys1808位に2つのさらなるFVIII固有のリジン残基を含み、これは、LRPとの相互作用において役割を果たし得る。
【0097】
(実施例II)
実験手順:vWFノックアウトマウス(健全な動物の第VIII因子と比較して、内因的な第VIII因子が20%減少している)を、ヒトの組換え型第VIII因子(RecombinateTM;200U/kg)および過剰量の組換えscFv(KM33)(3OnM 対 3nM 血漿中の第VIII因子)で処理した。対照実験において、第VIII因子および第VIII因子分子上の提案した追加のLRP結合部位として異なるドメインに結合する対照scFvフラグメント(KM38)を同じ濃度で使用した。任意に付加することなく組換えFVIIIの注入を用いて、このモデル系における第VIII因子のクリアランスをモニタリングした。
【0098】
注射後の確定時点(15分、30分、60分、120分および240分)で、血液を心臓穿刺によって回収し、引き続いて血漿を単離し、ショック凍結し、そして、第VIII因子活性について分析した。各時点にて10匹のマウスを用い、血漿を3つの濃度(1:20、1:60および1:100)に希薄し、そして、2連で分析した。第VIII因子レベルは、定量的ELISAを実施することによって定量した。
【0099】
結果:第VIII因子対照群からのデータは、15分で0.35U/mLの第VIII因子活性を示し、これは、14%の回復に対応する。30分で第VIII因子活性は約0.2U/mLに減少し、これは、これらのマウスの第VIII因子活性バックグラウンド(8%の回復に対応する)である。FVIII濃度のそれ以上の変化は、次の時点で検出されなかった。データは、このマウス系において、組換え型第VIII因子が最初の30分以内に動物循環から除去されることを示す。
【0100】
FVIIIおよびLRPでブロッキングしたscFvフラグメント(KM33)で処置したマウス由来の血漿サンプルの分析は、適用の15分後に、約0.8U/mLの第VIII因子が検出され得ることを示した。これは、30.4%の回復に変換された。第VIII因子活性レベルは、次の15分間で0.5U/mL(20%の回復)まで減少し、そして、60分後に0.35U/mL(14%の回復)に達した。注射の4時間後、FVIII活性レベルは、マウスの第VIII因子のバックグラウンド活性に達した。FVIII対照群と比較して、15分後および30分後の循環におけるこの高用量のFVIII活性は、FVIIIのクリアランスからの非常に良好な保護を示す。
【0101】
第2群の動物に、第VIII因子にLRPとは異なる部位で結合するscFvフラグメント(KM38)とヒトFVIIIとの同時注射を与えた。これらの動物に関して、第VIII因子の対照群と同様の結果が得られた(図8)。
【0102】
FVIII−LRP相互作用と干渉する抗体フラグメントを用いると、vWFノックアウトマウスにおけるRecombinateTMの半減期が有意に増加することが可能であった。さらに、マウスのvWF−KO系におけるヒトFVIIIの回復は、倍以上に増加し得ることが明白に示され得る。
【0103】
当業者の範囲内の変更が、本発明の範囲内であると考えられるべきである。特に定義されない限り、本明細書中で使用される全ての技術科学用語は、本発明が属する分野の当業者によって一般と理解されるものと同じ意味を有する。本明細書中に記載されるものと類似または相当する任意の方法および材料が、本発明の実施または試験に使用され得るが、好ましい方法および材料をここに記載する。
【技術分野】
【0001】
本発明は、第VIII因子由来のペプチドおよび第VIII因子に対して産生された抗体の使用および第VIII因子のLRPとの相互作用の阻害に関する。さらに、本発明は、LRPの第VIII因子との相互作用を阻害する方法、ならびに第VIII因子の分解を減少させる方法、および/または生物学的流体における第VIII因子の半減期を延長させる方法、および/または血液凝固障害(特に血友病A)に罹患する患者を処置する方法に関する。本発明はまた、血液凝固障害(特に血友病A)の処置のための、生物学的流体における第VIII因子の分解の減少、LRPとの第VIII因子の相互作用の阻害、および/または生物学的流体における第VIII因子の半減期の延長に有用な薬学的組成物に関する。
【背景技術】
【0002】
血液凝固には、異なる止血反応経路(最終的に血栓の形成につながる)の組合せが関係する。血栓は、脈管壁の表面上の血液構成成分の塊であり、そして、凝集した血小板および不溶性の架橋フィブリンから主として成る。フィブリンの形成は、凝固酵素トロンビンによるフィブリノーゲンの限定的なタンパク質分解によって誘導される。この酵素は、凝固カスケード(活性型血小板および白血球ならびに多くの脈管細胞の表面で起こる一連のチモーゲン活性化である)の最終生成物である(例えば、K.G.Mannら、Blood,1990,Vol.76,1−16頁を参照のこと;本明細書中に参考として援用される)。
【0003】
この凝固カスケードの重要な工程は、活性型第IX因子(第IXa因子)と活性型第VIII因子(第VIII因子a)の複合体による、第X因子の活性化である。
【0004】
第VIII凝固因子は、このように凝固カスケードの内因性の経路における重要なタンパク質である。FVIIIの機能は、酵素FIXaのための補因子として役立ち、そして、このプロテアーゼの触媒効率を5〜6倍の規模で増加させる(van Dieijeら、1981.The Journal of Biological Chemistry 256:3433−3442;本明細書中に参考として援用される)。FVIIIの重要性は、重度の出血障害である血友病A(活性型FVIIIの欠如によって特徴付けられる)によって示される(Kazazianら、1995.The Metabolic and Molecular Basis of Inherited Disease. In:Scriver,Beadet,SlyおよびValle,編 New York,McGraw−Hill Inc.III:3241−3267;本明細書中に参考として援用される)。
【0005】
血友病Aは、第VIII因子の機能不全によって特徴付けられる伴性の出血障害である。この疾患は、男性の人口の約0.01%で発症する。血友病Aは、健全なドナーから得た第VIII因子を含む血漿を投与することによって処置され得る。しかし、この処置には、いくつかの欠点がある。第VIII因子の供給は限られており、非常に高価である;血中の第VIII因子濃度は、わずか100ng/mlであり、そして、現在の血漿分別方法を用いると、収量が低い。第VIII因子の供給源は、プールされたドナーの血液なので、レシピエントは、種々の感染症(ドナーの血中に存在し得る、非A型肝炎ウイルス、非B型肝炎ウイルス、B型肝炎ウイルスまたはAIDSウイルスを含む)を得る高いリスクを負う。さらに、レシピエントは、外因性の第VIII因子に対して抗体を産生し得、そのうちのいくらかは第VIII因子の有効性を大きく減少させ得る。
【0006】
上記のように、第VIII凝固因子(FVIII)は、第X因子のタンパク質分解性の活性化における活性型第IX因子(FIXa)のための補因子として、内因性凝固経路において役割を果たす(総説については、Fay,P.J.(1999)Thromb.Haemostasis 82,193−200;Lenting,P.J.,van Mourik,J.A.,およびMertens,K.(1998)Blood 92,3983−3996(本明細書中に参考として援用される)を参照のこと)。第VIII因子は、分離性のドメイン構造(Al−al−A2−a2−B−a3−A3−C1−C2)を有する、300kDaの糖タンパク質である(Lenting,P.J.,van Mourik,J.A.,およびMertens,K.(1998)Blood 92,3983−3996;Vehar,G.A.,Keyt,B.,Eaton,D.,Rodriguez,H.,O’Brien,D.P.,Rotblat,F.,Opperman,H.,Keck,R.,Wood,W.I.,Harkins,R.N.,Tuddenham,E.G.D.,Lawn,R.M.,およびCapon,D.J.(1984)Nature 312,337−342;本明細書中に参考として援用される)。AドメインおよびCドメインは、構造上関連するタンパク質である第V因子のAドメインおよびCドメインと30〜40%の相応性を共有し、一方、Bドメインならびに短い酸性領域a1、a2およびa3は、FVIIIに固有である(Church,W.R.,Jernigan,R.L.,Toole,J.,Hewick,R.M.,Knopf,J.,Knutson,G.J.,Nesheim,M.E.,Mann,K.G.,およびFass,D.N.(1984)Proc.Natl.Acad.Sci.U.S.A.81,6934−6937;本明細書中に参考として援用される)。
【0007】
血漿において、FVIIIは、90〜220kDaの重鎖(A1−a1−A2−a2−B)および80kDaの軽鎖(a3−A3−C1−C2)から成る金属イオン連結ヘテロ二量体として循環する(Rotblat,F.,O’Brien,D.P.,O’Brien,F.J.,Goodall,A.H.,およびTuddenham,E.G.D.(1985)Biochemistry 24,4294−4300;Kaufman,R.J.,Wasly,L.C.,およびDomer,A.J.(1988)J.Biol.Chem.263,6352−6362;本明細書中に参考として援用される)。不活性タンパク質は、そのキャリアタンパク質であるフォン・ヴィレブランド因子(vWF)と強固に会合する(Lollar,P.,Hill−Eubanks,D.C.,およびParker,C.G.(1988)J.Biol.Chem.263,10451−10455;本明細書中に参考として援用される)。トロンビンまたは第Xa因子のいずれかによる限定的なタンパク質分解は、FVIII前駆体をその活性型誘導体に変換する(Lollar,P.,Knutson,G.J.,およびFass,D.N.(1985)Biochemistry 24,8056−8064;Eaton,D.,Rodriguez,H.,およびVehar G.A.(1986)Biochemistry 25,505−512;本明細書中に参考として援用される)。次いで、BドメインおよびA3ドメインに接している酸性の領域が、分子から取り除かれ(Fay,P.J.,Haidaris,P.J.,およびSmudzin,T.M.(1991)J.Biol.Chem.266,8957−8962;本明細書中に参考として援用される)、vWFに対する高い結合親和性の損失を生じる(Lollar,P.,ら、(1988)前出)。得られた活性型FVIII(FVIIIa)分子は、A2−a2ドメイン(金属イオン連結A1−a1/A3−C1−C2部分と非共有結合している)を含むヘテロ三量体から成る(Fay,P.J.,ら、(1991)前出)。
【0008】
FVIIIの重鎖および軽鎖内で、いくつかの領域がFIXa相互作用部位として同定されている(Fay P.J.,Beattie,T.,Huggins,C.F.,およびRegan,L.M.(1994)J.Biol.Chem.269,20522−20527;Bajaj,S.P.,Schmidt,A.E.,Mathur,A.,Padmanabhan,K.,Zhong,D.,Mastri,M.,およびFay,P.J.(2001)J.Biol.Chem.276.16302−16309;Lenting,P.J.,van de Loo,J.W.H.P.,Donath,M.J.,S.,H.,van Mourik,J.,A.,およびMertens,K.(1996)J.Biol.Chem.271,1935−1940;本明細書中に参考として援用される)。A2ドメイン残基Arg484−Phe5O9、Ser558−Gln565およびArg698−Asp712は、重鎖のFIXaへの結合に寄与する(Fay P.J.,(1994)前出;Bajaj,.(2001)前出;Fay,P.J.,およびScandella,D.(1999)J.Biol.Chem.274,29826−29830;本明細書中に参考として援用される)。FVIII軽鎖内では、A3ドメイン領域Glu1811−Lys1818が、FIXa相互作用部位として同定されている(Lenting,P.J.,(1996)前出)。さらに、FVIII領域Arg484−Phe509およびLys1804−Lys1818はまた、血友病患者において産生され得る抗体に対する標的エピトープとして同定されている。このような抗体は、FVIIIaと活性型FIXとの複合体アセンブリと干渉することによってFVIII活性化を阻害する(Haeley,J.F.,Lubin,I.M.,Nakai,H.,Saenko,E.L.,Hoyer,L.W.,Scandella,D.,およびLollar,P.(1995)J.Biol.Chem.270,14505−14509;Fijnvandraat,K.,Celie,P.H.N.,Turenhout,E.A.M.,van Mourik,J.A.,ten Cate,J.W.,Mertens,K.,Peters,M.,およびVoorberg,J.(1998)Blood 91,2347−2352;Zhong,D.,Saenko,E.L.,Shima,M.,Felch,M.,およびScandella,D.(1998)Blood 92,136−142;本明細書中に参考として援用される)。
【0009】
第VIII因子の半減期は、例えば、第VIII因子を関連したクリアランスのレセプターのためのその結合部位で改変することによって直接的にか、または、第VIII因子とそのレセプターとの間の結合に影響を及ぼす化合物を用いることによって間接的にのいずれかで、そのクリアランスにおいて役割を果たすレセプターに対する第VIII因子の親和性を減少することにより、第VIII因子分解(クリアランス)のメカニズムに影響を与えることによって延長され得る。前記のプロセスに関する細胞内レセプターおよび第VIII因子−レセプター相互作用に関する分子の部位が知られていないので、このような薬剤の産生は、困難であった。
【0010】
非活性型第VIII因子ヘテロ二量体の半減期は、第VIII因子に対して大きな親和性を有し(しかし、第VIIIa因子に対しては持たない)、キャリアタンパク質として役立つフォン・ヴィレブランド因子の存在に依存する(Sadler,J.E.およびDavie,E.W.:血友病A,Haernophilia B and von Willebrand’s Disease,in Starnatoyannopoulos,G.ら(編)、The Molecular basis of blood diseases.W.B.Saunders Co.,Philadelphia,1987,576−602頁)。3型フォン・ヴィレブランド病を有し、自身の循環系に検出可能なフォン・ヴィレブランド因子を示さない患者はまた、第VIII因子の二次性機能不全を有することが知られている。さらに、これらの患者における静脈内に投与された第VIII因子の半減期は、2〜4時間であり、それは、血友病A患者について報告される10〜40時間より明らかに短い。
【0011】
これらの調査結果は、第VIII因子が循環系から迅速に除去される傾向があり、このプロセスは、天然のキャリアであるフォン・ヴィレブランド因子との複合体の形成によってある程度阻害されることを意味する。
【0012】
最近、トロンビンによって活性化された第VIII因子は、低密度リポタンパク質レセプタータンパク質(本明細書中で以降、LRPと称する)との結合に関連付けられている(Yakhyaev,A.ら、Blood,vol.90(補遺1),1997,126−I,本明細書中に参考として援用される)。この文献の要約は、トロンビンで活性化された第VIII因子フラグメントの細胞取り込みおよび分解を記載し、そして、第VIIIa因子ヘテロ三量体の他の2つのサブユニットでなく、A2ドメインが細胞LRPと相互作用すると報告する。著者は、LRPに結合するA2ドメインが、さらに第VIIIa因子へテロ三量体のA2ドメイン間の相互作用を不安定化し、それによって第VIIIa因子活性がダウンレギュレートされることを提言する。
【0013】
非活性型FVIIIが多機能性のエンドサイトーシスレセプターである低密度リポタンパク質レセプター関連タンパク質(LRP)と相互作用することがまた示されている(Lenting,P.J.,Neels,J.G.,van den Berg,B.M.M.,Clijsters,P.P.F.M.,Meijerman,D.W.E.,Pannekoek,H.,van Mourik,J.A.,Mertens,K.,およびvan Zonneveld,A.,−J.(1999)J.Biol.Chem.274,23734−23739;WO 00/28021;Saenko,E.L.,Yakhyaev,A.V.,Mikhailenko,I.,Strickland,D.K.,およびSarafanov,A.G.(1999)J.Biol.Chem.274,37685−37692;本明細書中に参考として援用される)。このレセプターが、循環からのFVIIIのクリアランスにおいて役割を果たすということが示唆される(Saenko,E.L.ら、前出;Schwarz,H.P.,Lenting,P.J.,Binder,B.,Mihaly,J.,Denis,C.,Domer,F.,およびTurecek,P.L.(2000)Blood 95,1703−1708;本明細書中に参考として援用される)。
【0014】
LRPは、低密度リポタンパク質(LDL)レセプターファミリー(LDLレセプター、超低密度リポタンパク質レセプター、ポリリポタンパク質Eレセプター2およびメガリンをまた含む)のメンバーである(総説としては、Neels J.G.,Horn,I.R.,van den Berg,B.M.M.,Pannekoek,H.,およびvan Zonneveld,A.−J.(1998)Fibrinolysis Proteolysis 12,219−240;Herz,J.,およびStrickland,D.K.(2001)J.Clin.Invest.108,779−784(本明細書中に参考として援用される)を参照のこと)。LRPは、肝臓、肺、胎盤および脳を含む種々の組織において発現する(Moestrup,S.K.,Gliemann,J.,およびPallesen,G.(1992)Cell Tissue Res.269,375−382;本明細書中に参考として援用される)。このレセプターは、膜貫通型の85kDaのβ鎖に非共有結合性に連結される、細胞外の515kDaのα鎖から成る(Herz,J.,Kowal,R.C.,Goldstein,J.L.,およびBrown,M.S.(1990)EMBO J.9,1769−1776;本明細書中に参考として援用される)。α鎖は、多くの構造的かつ機能的に無関係のリガンドの結合を媒介する、可変数の相補型反復の4つのクラスターを含む(Moestrup,S.K.,Hotlet,T.L.,Etzerodt,M.,Thogersen,H.C.,Nykjaer,A.,Andreasen,P.A.,Rasmussen,H.H.,SottrupJensen,L.,およびGliemann,J.(1993)J.Biol.Chem.268,13691−13696;Willnow,T.E.,Orth,K.,およびHerz,J.(1994)J.Biol.Chem.269,15827−15832;Neels,J.G.,van den Berg,B.M.M.,Lookene,A.,Olivecrona,G.,Pannekoek,H.,およびvan Zonneveld,A.−J.(1999)J.Biol.Chem.274,31305−31311;本明細書中に参考として援用される)。
【0015】
β鎖は、膜貫通ドメインおよびエンドサイトーシスに不可欠である短い細胞質テールを含む。α鎖は、大きな外部ドメインとして機能し、そして、3つの型の反復を含む:表皮性増殖因子様ドメイン、Tyr−Trp−Thr−Asp配列およびLDLレレプタークラスAドメイン。これらのクラスAドメイン(リガンド結合に関連付けられている)は、クラスターI(2つのドメイン)、クラスターII(8つのドメイン)、クラスターIII(10のドメイン)および、クラスターIV(11のドメイン)と呼ばれる4つの別々のクラスターに存在する。
【0016】
LRPはまた、サル腎臓細胞(COS)またはチャイニーズハムスター卵巣細胞(CHO)のような細胞型において発現しており(FitzGerald,D.J.,ら、J.Cell Biol.Vol.129,1995,1533−1541頁;本明細書中に参考として援用される)、これらは、第VIII因子を含む補乳動物のタンパク質の発現のためにしばしば使用される(Kaufman,R.J.ら、Blood,Coag.Fibrinol,vol.8(補遺2),1997,3−14頁;本明細書中に参考として援用される)。
【0017】
LRPは、プロテアーゼ、Kunitz型インヒビター、プロテアーゼ−セルピン複合体、リパーゼおよびリポタンパク質を含む、多数のリガンドのクリアランスにおいて役割を果たし、これらは、いくらかの生理学的かつ病態生理学的なクリアランスのプロセスにおいてLRPが役割を果たすと言われている(Naritaら、Blood,vol.2,555−560頁,1998;Orthら、Proc.Natl.Acad.Sci.,vol,89,7422−7426頁,1992;Kounnasら、J.Biol.Chem.,vol.271,6523−6529頁,1996;本明細書中に参考として援用される)。
【0018】
LRPはまた、活性型の非酵素の補因子である第VIIIa因子に結合する(Yakhyaev,A.ら、Blood,vol.90,(補遺1),1997,126−I)。この文献の開示は、第VIIIa因子の調節においてLRPを関連付けているが、非活性型ヘテロ二量体である第VIII因子の調節におけるLRPの役割の示唆はない。
【0019】
FVIII軽鎖は、組換えLRPクラスターIIおよびIVと相互作用することが示されたが、LRPクラスターIおよびIIIへの結合は観察されなかった(Neels,J.G.,ら、(1999)前出)。
【0020】
第VIII因子の薬動力学的プロファイルを向上させるために、第VIII因子ポリペプチドのくつかの領域を改変するいくつかの試みがなされている:
WO 87/07144は、アルギニン残基およびリジン残基を含むタンパク分解性の切断部位のいくらかの改変を示し、これは、特異的なプロテアーゼが触媒する切断(例えば、Arg1721およびAla1722の間の第Xa因子の切断部位)について分子の不安定性を減少する。
【0021】
WO 95/18827、WO 95/18828およびWO 95/18829は、重鎖のA2領域に改変を有する第VIII因子の誘導体を記載する。
【0022】
WO 97/03193は、第VIII因子ポリペプチドアナログを開示し、これらの修飾が分子の金属結合特性を変更する。
【0023】
WO 97/03195において、第VIII:C因子ポリペプチドアナログが記載され、Arg残基に隣接して位置する1つ以上のアミノ酸残基の改変が提供される。
【0024】
EP 808 901は、第VIII因子の少なくとも1つの免疫優性領域に少なくとも1つの変異を有する第VIII因子改変体の構成、および、第VIII因子インヒビターを有する患者の処置のためのこの第VIII因子改変体の使用を記載する。これらの改変は、半減期インビボまたはインビトロにおいて、第VIII因子改変体の半減期を延長せず、また、第VIII因子改変体の安定性を増大もしない。
【0025】
さらに、WO 00/28021は、軽鎖のA3および/またはC1もしくはC2ドメインに改変を有し、そして、その改変が低密度リポ蛋白受容体タンパク質(LRP)との結合親和性に影響を与えるという点で、特徴付けられる、第VIII:C因子活性を有する第VIII因子ポリペプチドを記載する。
【0026】
ヒトのmRNAから得られた第VIII因子cDNAの分子クローニング、ならびに、その後の哺乳動物細胞、酵母細胞、および細菌細胞において第VIII因子活性を有するタンパク質の産生が、報告されている(WO 85/01961、EP 160 4577、EP 150 735およびEP 253 455を参照のこと;本明細書中に参考として援用される)。形質転換された微生物を用いて第VIII因子活性を有するタンパク質を産生する方法は、EP 253 455(本明細書中に参考として援用される)に開示される。欧州特許出願EP 150 735および同EP 123 945ならびにBrinkhousら、(1985)は、第VIII因子のタンパク分解性の分解産物における第VIII因子の活性を開示する(参照によってここに組み込まれる)。第VIII因子の2つのタンパク分解性の分解産物の複合体(92kDaおよび80kDaのポリペプチド)は、増強された第VIII因子活性を示す(Fayら、Biochem.Biophys.Acta(1986)871:268−278;Fatonら、Biochemistry(1986)25:505−512;本明細書中に参考として援用される)。
【発明の概要】
【発明が解決しようとする課題】
【0027】
従って、本発明者らは、インビトロおよびインビボにおいて第VIII因子の安定性おおよび半減期を増大させる、利用可能な第VIII因子調製物の作製を目指した。
【課題を解決するための手段】
【0028】
(発明の要旨)
驚くべきことに、本発明者らは、第VIII因子の部分的なアミノ酸配列、ならびに特定のペプチド内の特異的なエピトープに対する抗体を含む特定のペプチドが、インビトロおよびインビボにおいて第VIII因子の安定性を有意に改善し得ることを発見した。
【0029】
従って、本発明は一般に、第VIII因子由来のペプチドおよび第VIII因子に対して作製された抗体の使用、ならびに第VIII因子のLRPとの相互作用の阻害に関する。さらに、本発明は、LRPの第VIII因子との相互作用を阻害する方法、第VIII因子の分解を減少させる方法、および/または生物学的流体における第VIII因子の半減期を延長させる方法、および/または血液凝固障害(特に血友病A)に罹患する患者を処置する方法に関する。本発明はまた、血液凝固障害(特に血友病A)の処置のための、生物学的流体における第VIII因子の分解の減少、第VIII因子のLRPとの相互作用の阻害、および/または生物学的流体における第VIII因子の半減期の延長に有用な薬学的組成物に関する。
本発明はまた、以下の項目を提供する。
(項目1)
第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸配列を含むペプチド、または、該アミノ酸配列内の1つ以上のエピトープに特異的に結合し、第VIII因子とLRPとの相互作用を阻害する抗体の使用。
(項目2)
第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸配列を含むペプチド、または、該アミノ酸配列内の1つ以上のエピトープに特異的に結合し、生物学的流体における第VIII因子の分解を減少させる抗体の使用。
(項目3)
第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸配列を含むペプチド、または、該アミノ酸配列内の1つ以上のエピトープに特異的に結合し、血中における第VIII因子の半減期を延長させる抗体の使用。
(項目4)
第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸を含むペプチド、または、該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の、血液凝固障害および/または血栓溶解系もしくは線維素溶解系の一時的な機能障害の予防または処置のため医薬の調製のための使用。
(項目5)
前記血液凝固障害が血友病Aまたはフォン・ヴィレブランド病であり、かつ、前記一時的な機能障害が外科的手術の間または外科的手術後に起こる、項目4に記載の使用。
(項目6)
前記抗体が、AF 234247に記載のアミノ酸配列(VHKM33)、またはAF 234258に記載のアミノ酸配列(VLKM41)から本質的になる、項目1〜5のいずれか1項に記載の使用。
(項目7)
生物学的流体における第VIII因子の分解を減少させる方法であって、第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるペプチド、または該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体が、該生物学的流体に添加される、方法。
(項目8)
少なくとも10%まで第VIII因子の分解を減少させるために十分な量の前記ペプチドまたは前記抗体の量が添加される、項目7に記載の方法。
(項目9)
前記減少が少なくとも20%である、項目8に記載の方法。
(項目10)
前記減少が少なくとも50%である、項目9に記載の方法。
(項目11)
前記減少が本質的に100%である、項目10に記載の方法。
(項目12)
生物学的流体における第VIII因子の半減期を延長させる方法であって、第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるペプチド、または該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体が、該生物学的流体に添加される、方法。
(項目13)
前記生物学的流体における前記第VIII因子の半減期が少なくとも2倍延長される、項目12に記載の方法。
(項目14)
前記生物学的流体における前記第VIII因子の半減期が少なくとも5倍延長される、項目13に記載の方法。
(項目15)
前記生物学的流体における前記第VIII因子の半減期が少なくとも10倍に延長される、項目14に記載の方法。
(項目16)
生物学的流体において、第VIII因子とLRPとの相互作用を阻害する方法であって、第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるペプチド、または該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体が、該生物学的流体に添加される、方法。
(項目17)
第VIII因子とLRPとの相互作用を少なくとも20%の阻害程度を達成する量で、前記ペプチドまたは前記抗体が添加される、項目16に記載の方法。
(項目18)
相互作用の少なくとも50%の程度の阻害程度が達成される、項目17に記載の方法。
(項目19)
前記抗体が、配列番号20および配列番号22に記載のアミノ酸配列(KM33)、または配列番号16および配列番号18に記載のアミノ酸配列(KM41)を含む、項目7〜18のいずれか1項に記載の方法。
(項目20)
前記抗体が、配列番号26に記載のアミノ酸配列(KM33)または配列番号25に記載のアミノ酸配列(KM41)から本質的になる、項目19に記載の方法。
(項目21)
少なくとも1つのペプチドを含む薬学的組成物であって、該ペプチドは、第VIII因子由来であるが、本質的な第VIII因子活性を持たないものとして配列番号1、2、3または4のいずれかに定義されるアミノ酸配列を含むか、または該アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体を含み、さらに、生理学的に受容可能な賦形剤、キャリア、希釈剤および/または安定化剤を含む、薬学的組成物。
(項目22)
前記調製物が凍結乾燥されている、項目20に記載の薬学的組成物。
(項目23)
生物学的流体における第VIII因子の分解を減少し、そして/あるいは第VIII因子の半減期を延長し、そして/あるいは、LRPへの第VIII因子の結合を阻害し、そして/あるいは血液凝固障害および/または血栓溶解系もしくは線維素溶解系の一時的な機能障害の予防または処置ための項目21または22に記載の薬学的組成物。
(項目24)
前記ペプチドまたは前記抗体が項目8〜11のいずれか1項で定義される第VIII因子の分解の減少を達成するか、項目17もしくは18に記載のいずれか1項で定義されるLRPと第VIII因子との相互作用の阻害を達成するか、そして/または、項目13〜15のいずれか1項で定義される生物学的流体における第VIII因子の半減期の延長を達成するのに十分な量で含まれる、項目21または22のいずれか1項に記載の薬学的組成物。
(項目25)
前記血液凝固障害が血友病Aまたはフォン・ヴィレブランド病であって、かつ一時的な機能障害が外科的手術の間、または外科的手術後に起こる、項目23に記載の調製物。
(項目26)
さらに第VIII因子を含む、項目21〜25のいずれか1項に記載の薬学的組成物。
(項目27)
前記抗体が配列番号20および配列番号22に記載のアミノ酸配列(KM33)、または配列番号16および配列番号18に記載のアミノ酸配列(KM41)を含む、項目21〜26のいずれか記載の薬学的組成物。
(項目28)
前記抗体が、配列番号26に記載のアミノ酸配列(KM33)または配列番号25に記載のアミノ酸配列(KM41)から本質的になる、項目27に記載の薬学的組成物。
(項目29)
血液凝固障害に罹患する患者を処置する方法であって、前記方法が前記患者に項目21〜28のいずれか1項に記載の薬学的組成物を投与する工程を包含する、方法。
(項目30)
前記血液凝固障害が血友病Aまたはフォン・ヴィレブランド病であり、かつ前記一時的な機能障害が外科的手術の間、または外科的手術後に起こる、項目29に記載の方法。
【図面の簡単な説明】
【0030】
【図1】FVIII軽鎖フラグメントの固定されたLRPへの結合。16fmol/mm2でCM5センサ−チップに固定されたLRPを、以下と共にインキュベートした:A、FVIII軽鎖(150nM)(実線)および第Xa因子切断軽鎖(150nM)(点線)。B、a3−A3−C1フラグメント(150nM)(実線)および単離したC2ドメイン(750nM)(点線)。インキュベーションを、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH7.4)中にて25℃で2分間、20μl/分の流量で行った。分離を、緩衝液でリガンド溶液を置換して開始した。反応は、共鳴単位(RU)として示し、非特異的な結合(LRPコーティングしたチャネルと比較して5%未満)について補正する。
【図2】FVIII軽鎖の組換えLRPフラグメントへの結合。FVIII軽鎖(LCh)(25nM)を、種々の濃度の組換えLRPクラスターII(0〜600nM)の存在下または非存在下で、150mM NaCl、5mM CaCl2、1%(w/v) HSA、0.1%Tween(登録商標)20および50mM Tris(pH7.4)の50μl容積中にて、37℃で2時間、固定されたLRPクラスターIVと共にインキュベートした。同じ緩衝液で洗浄した後、結合したFVIII軽鎖を、37℃で15分間、ペルオキシダーゼ結合体化抗FVIII抗体CLB−CAg 12と共にインキュベートすることによって定量した。残留結合は、競合物質の非存在下での結合の割合として表し、非特異的な結合(LRPクラスターIIを固定化したウェルへの結合と比較して5%未満)について補正する。挿入図、FVIII軽鎖の連続希釈物を固定されたLRPクラスターII(1pmol/ウェル)と共に、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%Tween(登録商標)20および50mM Tris(pH7.4)の50μl容積中にて、37℃で2時間インキュベートした。同じ緩衝液で洗浄した後、結合したFVIII軽鎖を、上記のようにして定量した。データは、3つの実験の平均±S.D.を表す。
【図3】LRPクラスターIIの固定化されたFVa軽鎖への結合。LRPクラスターII(100nM)を、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH7.4)中にて、固定化されたFVIII軽鎖(71fmol/mm2)(I)または固定化されたFVa軽鎖(76fmol/mm2)(II)のいずれかと共に、20μ/分の流量にて25℃で2分間インキュベートした。反応を、共鳴単位(RU)として示し、非特異的結合(コーティングされたチャネルと比較して5%未満)について補正する。
【図4】FVIII軽鎖とLRPとの間の相互作用に及ぼすscFv抗体フラグメントの影響。A、FVIII軽鎖(LCh)(25nM)を、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%Tween(登録商標)20および50mM Tris(pH7.4)の50μl容積中にて、種々の濃度(0〜100nM)のscFv KM41(黒丸)またはscFv KM36(白丸)の存在下で、固定されたLRPクラスターII(1pmol/ウェル)と共に37℃で2時間インキュベートした。同じ緩衝液で洗浄した後、結合したFVIII軽鎖を、ペルオキシダーゼ結合体化FVIII抗体CLB−CAg 12と共に37℃で15分間インキュベートすることによって定量した。残留結合は、競合物質の非存在下における結合の割合を表し、非特異的な結合(LRPクラスターIIを固定化したウェルへの結合と比べて5%未満)について補正する。データは、3つの実験の平均±S.D.を表す。B,FVIII軽鎖(50nM)を、図1の説明文に記載されたように、固定されたLRP(16fmol/mm2)と共にインキュベートした。結合を、scFv KM41の非存在下(曲線1)または増加した濃度のscFv KM41(20nM、60nM、300nM、500nM(それぞれ曲線2〜5))において評価した。複合体は、SPR分析の前に、30分間にわたって形成させた。
【図5】FVIII軽鎖とLRPとの間の相互作用に及ぼすscFv KM33の影響。FVIII軽鎖(LCh)(25nM)を、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%Tween(登録商標)20および50mM Tris(pH7.4)の50μl容積中にて、種々の濃度(0〜30nM)のscFv KM36(白丸)またはscFv KM33(黒丸)の存在下、固定化したLRPクラスターII(1pmol/ウェル)と共に37℃で2時間インキュベートした。同じ緩衝液で洗浄した後、結合したFVIII軽鎖を、ペルオキシダーゼ結合体化抗FVIII抗体CLB−CAg 12と共に37℃で15分間インキュベートすることによって定量した。残留結合は、競合物質の非存在下の結合の割合として表し、非特異的結合(クラスターでコーティングしたウェルへの結合と比較して5%未満)について補正する。データは、3つの実験の平均±S.D.を表す。
【図6】FVIII/FV1811−1818軽鎖キメラへのLRPクラスターIIへの結合。CM5センサチップ上のScFv EL14(67fmol/mm2)を、150mM NaCl、50mM Tris(pH 7.4)中20fmol/mm2の密度までの組換え野生型FVIII軽鎖または組換えFVIII/FV1811−1818キメラのいずれかと共にインキュベートした。LRPクラスターII(25〜125nM)を、FVIII/FV1811−1818キメラ(白丸)またはインタクトなFVIII軽鎖(黒丸)をそれぞれ固定化した2つの別々のチャネル、および1つのコントロール(scFv EL14でコーティングした)チャネルを、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH7.4)中、25℃、20μl/molの流速で2分間通過させた。結合したLRPクラスターIIを2分後の会合量として表す。
【図7】FVIII/FV1811−1818軽鎖キメラへのscFv KM41の結合。CM5センサチップのscFv EL14(67fmol/mm2)を、150mM NaCl、50mM Tris(pH7.4)中、20fmol/mm2の密度までの組換え野生型FVIII軽鎖または組換えFVIII/FV1811−1818キメラのいずれかと共にインキュベートした。ScFv KM41(40nM)を、それぞれインタクトなFVIII軽鎖(I)またはFVIII/FV1811−1818キメラ(II)を固定化した2つの別々のチャネルを通過させた。反応を、共鳴単位(RU)として示し、非特異的結合について補正する。
【図8】実施例IIに従って、マウスに、ヒトFVIII単体、コントロールのscFvフラグメント(KM38)の存在下でのヒトFVIII、および、推定FVIII−LRP相互作用と特異的に干渉するscFvフラグメント(KM33)の存在下でのヒトFVIIIのいずれかで注射した。
【発明を実施するための形態】
【0031】
(発明の詳細な説明)
本発明の1つの実施形態は、第VIII因子由来であるが第VIII因子のLRPとの相互作用を阻害する任意の実質的な第VIII因子活性を有さない配列番号1、2、3または4のいずれかで定義されるアミノ酸配列を含むペプチドの使用に関する。
用語「ペプチド」は、ペプチド結合を介して結合された一連のアミノ酸から成る分子に関する。好ましくは本発明に有用なペプチドは、タンパク質において一般に見出される20のアミノ酸からなる(例えば、周知の教科書である「Biochemistry」(A.Lehninger,第2版、Worth Publicshers,NY,NY(1975))を参照のこと)。
【0032】
本発明に有用なペプチドはまた、第VIII因子由来であるが、本質的に第VIII因子活性を有さず、改変され、その結果、そのアミノ酸配列が、上記ペプチドの第VIII因子とLRPタンパク質との間の相互作用を減少する能力を有意に変更しない、1つ以上の欠失、付加、置換および/または変換を含む、配列番号1、2、3または4のいずれかで定義されるアミノ酸配列を含むペプチドの誘導体であり得る。
【0033】
本発明に有用なペプチドが1つまたはアミノ酸置換を含む場合、前記1つ以上のアミノ酸置換は保存的アミノ酸置換であることが、好ましい。例えば、置換がアラニン、ロイシン、イソロイシン、バリン、プロリン、フェニルアラニン、トリプトファンおよびメチオニンのような非極性の疎水性アミノ酸において発生する場合、置換されるアミノ酸がアラニン、ロイシン、イソロイシン、バリン、プロリン、フェニルアラニン、トリプトファンおよびメチオニンのような非極性の疎水性アミノ酸の中から選択されることが好ましい。同様に、置換がセリン、トレオニン、チロシン、アスパラギン、グルタミン、システインまたはグリシンのような非荷電極性アミノ酸において発生する場合、置換されるアミノ酸がセリン、トレオニン、チロシン、アスパラギン、グルタミン、システインまたはグリシンのような非荷電極性アミノ酸の中から選択されることが好ましい。置換がリジン、アルギニンまたはヒスチジンのような正に荷電した(塩基性)アミノ酸において発生する場合、置換されるアミノ酸がリジン、アルギニンまたはヒスチジンのような正に荷電した(塩基性)アミノ酸の中から選択されることが好ましい。同様に、置換がアスパラギン酸またはグルタミン酸のような負に荷電した(酸性)アミノ酸において発生する場合、置換されるアミノ酸がアスパラギン酸またはグルタミン酸のような負に荷電した(酸性)アミノ酸の中から選択されることが好ましい。本発明に有用なペプチドは、D−アミノ酸、L−アミノ酸またはD−およびL−アミノ酸の混合物からなり得る。
【0034】
本発明に有用なペプチドはまた、例えば、これらのペプチドの1つ以上のタンデム反復または交互反復のような、配列番号1、2、3または4で定義されるペプチドの任意の重合形態を包含する。
【0035】
前記のペプチドの特定のアミノ酸の化学改変(特に、N末端および/またはC末端アミノ酸残基の化学改変)がまた包含される。この化学改変は、末端アミノ基および/またはカルボキシ基を遮蔽し、インビボもしくはインビトロでの分解に対する上記ペプチドの安定性を増加し得、または、キャリア機能(アルブミンまたは他の血漿タンパク質を含む)、標的化機能もしくはペプチドの溶解性を改変するような機能を有する分子もしくは部分をペプチドに添加する。用語「改変」は、グリコシル残基の付加または除去をさらに包含する。
【0036】
本発明に有用なペプチドは、標準のペプチド合成技術によって、または、組換えDNA技術によって、全体または一部が作製され得る。
【0037】
本発明に有用な1つの特定のペプチドは、配列番号1(Glu1811−Lys1818)のアミノ酸配列を含む。本発明に有用なさらなる特定のペプチドは、配列番号2(Lys1804−Lys1818)または配列番号3(Tyr1815−Ala1834)由来のアミノ酸配列を含む。このように、本発明に有用なさらなるペプチドは、アミノ酸配列Lys1804−Ala1834(配列番号4)を含む。
【0038】
好ましくは、本発明に有用なペプチドは、上記の第VIII因子活性についてのアッセイの1つで測定された、本発明の前記のペプチドが由来する天然に存在する第VIII因子分子の対応する第VIII因子活性と比較して、5.0%未満の第VIII因子活性を有し、より好ましくは、1.0%未満の第VIII因子活性を有し、そして最も好ましくは、本質的に活性を有さない。
【0039】
第VIII因子活性の評価は、適切なアッセイ手段(特に、サンプルの第VIII因子活性を決定するために典型的に実施される任意のアッセイ(例えば、MikaelssonおよびOswaldson,Scan.J.Hematol.Suppl.33,79−86,1984に記載されるような1ステップクロットアッセイ、または、第VIII因子 IMMUNOCHROM(Baxter)のような色素発色アッセイ))によって実行され得る。
【0040】
第VIII因子活性はまた、第X因子の第Xa因子への変換において第IXa因子のための補因子として作用する第VIII因子の能力を測定することによって実施され得、色素発色基質が第X因子について使用される(Coatest第VIII因子、Chomogenix、Moeindal、Sweden)。
【0041】
本発明のさらなる実施形態は、第VIII因子由来であるが、本質的に第VIII因子活性を有さず、第VIII因子とLRPとの相互作用を阻害する、配列番号1、2、3または4のいずれかで定義されたアミノ酸配列を含むペプチド内の1つ以上のエピトープに特異的に結合する抗体の使用に関する。
【0042】
本明細書中で使用される場合、用語「抗体」は、ポリクローナル抗体またはモノクローナル抗体(好ましくはモノクローナル抗体)およびそれらのフラグメントまたは領域、ならびに第VIII因子もしくは本発明に有用なペプチド内の1つ以上のエピトープに特異的に結合することおよび第VIII因子分子と低密度リポ蛋白受容体関連タンパク質(LRP)との間の相互作用を干渉することが可能であるそれらの誘導体を含むことを意味する。
【0043】
本明細書中で使用される場合、用語「エピトープ」は、本発明に有用なペプチドの全てまたは部分をいうことを意味し、このペプチドは、第VIII因子において見出される全てまたは対応するアミノ酸の一部の1°、2°および/または3°構造を模倣し、第VIII因子とLRPとの間の相互作用を阻害する抗体によって特異的に認識され得る。エピトープは、ペプチド配列自体(すなわち、ペプチドの1°構造)、種々の三次元の形態におけるペプチド(すなわち、ペプチド配列の2°または3°構造)および/またはこのペプチドのアミノ酸への改変(例えば、糖分子の付加(例えば、グリコシル化ペプチド))を含み得る。作用の任意の特定の論理または生物学的メカニズムに束縛されずに、本発明を実施するのに有用なペプチドが少なくとも2つの異なるエピトープを含むことが可能である。1つのエピトープは、配列番号1(Gly1811−Lys1818)内に位置する主要なアミノ酸配列のようである。もう一方のエピトープは、前述の主要なエピトープに隣接して位置するアミノ酸配列の組合せから成り得る;すなわち、もう一方のエピトープは、配列番号2(Lys1804−Lys1818)および/または配列番号3(Tyr1815−Ala1834)から形成され得る。このエピトープはまた、二次構造または三次構造エピトープであり得、そして、アミノ酸配列Lys1804−Lys1818およびTyr1815−Ala1834の両方を含み得る。
【0044】
抗体のフラグメントは、例えば、Fab、Fab’、F(ab’)2、FvおよびscFvフラグメントを含む。これらのフラグメントは、当該分野で周知の方法(たとえば、パパインのような酵素でタンパク質分解性に切断してFabフラグメントを生成するか、または、ペプシンのような酵素によりF(ab’)2フラグメントを生成するか)を用いて、インタクトな抗体から産生され得る。
【0045】
1つの好ましい実施形態において、本発明の実施に有用な抗体は、scFv抗体フラグメントである。好ましくは、抗体は、実施例において、KM33として記載される抗体のアミノ酸配列(KM33の重鎖のアミノ酸配列については配列番号20を、KM33の軽鎖のアミノ酸配列については配列番号22を、そして、ペプチドリンカーによって連結されるKM33の軽鎖および重鎖を含む融合タンパク質については配列番号26を参照のこと)、および/または、GenBank 登録番号AF234247(VHKM33)によって提供された抗体のアミノ酸配列、あるいは実施例においてKM41として記載される抗体のアミノ酸配列(KM41の重鎖のアミノ酸配列については配列番号16、KM41の軽鎖のアミノ酸配列については配列番号18、そして、ペプチドリンカーで結合されるKM41の軽鎖および重鎖を含む融合タンパク質については配列番号25を参照のこと)、および/またはGenBank 登録番号AF234258(VLKM41)によって提供された抗体のアミノ酸配列を含む。
【0046】
これらおよび他のscFvは、van der Brink,E.N.ら、Blood 97(4),966−972(2001)(本明細書中に参考として援用される)に記載されたようにして産生され得る。
【0047】
好ましくは本発明に有用な抗体は、第VIII因子と低密度リポタンパク質レセプター関連タンパク質(LRP)との間の相互作用を阻害することが可能であり、その結果、この相互作用は、たとえば、所定の供給源由来の第VIII因子とLRPとの間の相互作用と比較すると、少なくとも20%、より好ましくは少なくとも50%、なおより好ましくは少なくとも90%または95%かそれ以上、減少される。
【0048】
第VIII因子とLRPとの間の相互作用は、本明細書中に記載されたような表面プラズモン共鳴または固相結合アッセイを用いて決定され得る。
【0049】
本明細書中で使用される場合、用語「低密度リポタンパク質レセプター関連タンパク質」は、「LRP」として省略され、これらの用語は、タンパク質の低密度リポタンパク質(LDL)レセプターファミリーのメンバーである膜糖タンパク質をいうために交換可能に使用される(Gliemann,J.in Biol.Chem.379,951−964(1998)(参考として援用される)を参照のこと)。ヒトLRPは、31のクラスAのシステインリッチな反復(LDLRAとして知られている)ドメイン(クラスターI、II、IIIおよびIVとして知られている4つのクラスターにおけるLRP分子において分配される)を含む。クラスターIIは、CL−II−1/2(アミノ末端基の側面にある表皮性の成長因子の反復E4およびLDLRAドメインC3−C7にわたる)として知られているドメインを含む。このCL−II−1/2ドメインは、第VIII因子と相互作用する(Neels、J.G.ら、前出を参照のこと)。
【0050】
本発明はまた、前期ペプチドが第VIII因子由来であるが、本質的に第VIII因子活性を有さない配列番号1、2、3または4に定義されるアミノ酸配列を含む、少なくとも1つのペプチド、または前記アミノ酸配列の中の1つ以上のエピトープに特異的に結合する抗体、ならびに生理学的に受容可能な賦形剤、キャリア、希釈剤および/または安定化剤を含む薬学的組成物に関する。
【0051】
本発明に従う薬学的組成物は、本発明に有用な1つ以上のペプチド、または本発明に有用な1つ以上の抗体またはそれらの誘導体を含み得る。本発明に従う薬学的組成物はまた、さらに、治療活性成分または予防活性成分のうちの1つ以上を含むことができる。1つの実施形態において、薬学的組成物は、本発明に有用な1つ以上のペプチドを含む。別の実施形態において、薬学的組成物は、前記ペプチドに対する本発明に有用な1つ以上の抗体を含む。薬学的組成物が発明に有用な1つ以上の抗体を含む場合、前記薬学的組成物はさらに、第VIII因子を含み得る。そのような薬学的組成物における第VIII因子は、あらゆる自然に存在する、合成の、または、組換えの供給源から得られ得る。
【0052】
本発明に有用なペプチドおよび/または抗体は、薬学的に受容可能なビヒクル(たとえば、1つ以上の賦形剤、キャリア、希釈剤および安定化剤)と組み合わされて、溶液、懸濁液、エマルジョンまたは凍結乾燥粉末として処方され得る。適切なキャリアとしては以下が挙げられるがこれらに限定されない:希釈剤またはフィラー、無菌水媒体および種々の無毒性の有機溶剤。このようなビヒクルの例は、水、生理食塩水、リンガー溶液、デキストロース溶液およびヒト血清アルブミン溶液である。リポソームおよび不揮発性油のような非水性ビヒクルがまた使用され得る。ビヒクルまたは凍結乾燥物は等張性(例えば、NaCl、スクロース、マンニトール)および安定性(例えば、緩衝液および/または防腐剤)を維持する添加物を含み得る。適切な薬学的に受容可能なビヒクルは、A.OsolによるRemington’s Pharmaceutical Sciences(この分野における標準教科書)(参考として援用される)に記載されている。これらの組成物は、錠剤、カプセル、粉末、水性懸濁液または溶液、注射剤等の形で処方され得る。
【0053】
好ましくは本発明に従う薬学的組成物は、凍結乾燥されている。
【0054】
本発明の方法に基づいた薬学的組成物の投与は、好ましくは、改善が達成されるまで、最大の治療応答を保証する投薬レジメンを用いて実行されるべきである。概して、用量は、使われるペプチドの分子の大きさに依存する。約15アミノ酸のペプチドについて、静脈内投与のための投薬量は、約0.1〜1000mg/kgの間、好ましくは1〜500mg/kgの間、そして、最も好ましくは10〜100mg/kgの間で変化し得る。さらに大きなペプチドについては、その用量は、漸増し得る。経口投与のために、用量は、もちろん、適切な投薬がまた、薬物に対する応答に影響を与え得る、患者の身体全体の健康、年齢および他の因子に依存し得ることに留意して、大幅に高いかもしれない。薬物は、所望のレベルで治療効果を維持するために、連続的注入によって、または、約4〜50時間の定期的な間隔で投与され得る。
【0055】
本発明に従う薬学的組成物は、1つの成分調製物の形式であり得るか、または、キットにおいて1つ以上の他の成分と組み合わせて存在し得る。
【0056】
本発明に従う薬学的組成物は、哺乳動物(好ましくはヒト)への投与が意図される。本発明に有用なペプチドが特定の哺乳動物(例えばヒト)への投与が意図される場合、前記ペプチドは、特定の哺乳動物由来であることが好ましい。本発明に有用な抗体が特定の哺乳動物(例えばヒト)への投与が意図される場合、前記抗体は、特定の哺乳動物由来のエピトープを用いて生成されることが好ましい。さらに、ヒトへの投与が意図される本発明に有用な抗体は、ヒト抗体またはヒト化抗体であることが好ましい。
【0057】
本発明に有用なペプチド、本発明に有用な抗体およびこれらの誘導体ならびに前記ペプチド、抗体および誘導体を含む薬学的組成物は、前記ペプチド、抗体おまたはこれらの誘導体が体内の作用部位に到達し得る任意の手段によって被験者に投与され得る。好ましくは発明に従うペプチド、抗体およびこれらの誘導体、ならびに、前記のペプチド、抗体および誘導体を含む薬学的組成物は、非経口(すなわち、静脈内、皮下、または筋肉内)で投与される。
【0058】
本発明に従う調製物は、特に血液凝固障害を持つ患者の予防または処置のために、第VIII因子とLRPとの相互作用を阻害し、第VIII因子の分解を減少させ、または、第VIII因子の半減期を延長させる薬剤の投与が必要な全ての被験者を処置するために使われ得る。このような血液凝固障害は、少なくとも部分的に、第VIII因子の機能障害(例えば遺伝性または先天性の機能障害あるいは後天性の機能障害)によって引き起こされ得る。このような血液凝固障害はまた、第IX因子、第V因子、第X因子またはヴォン・ヴィレブランド因子を含む、凝固経路に関連している他のタンパク質における機能障害(例えば、遺伝性または先天性の機能障害あるいは後天性の機能障害)によって少なくとも部分的に引き起こされ得る。好ましくは、前記血液凝固障害は血友病Aまたはフォン・ヴィレブランド病である。
【0059】
このように、本発明はさらに、血液凝固障害の処置のための医薬の調製のための、第VIII因子由来であるが、本質的に第VIII因子活性を有さない配列番号1、2、3もしくは4のいずれかに定義されるアミノ酸を含むペプチドの使用、または、上記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の使用に関する。
【0060】
好ましい実施形態において、血液凝固障害は、血友病Aまたはフォン・ヴィレブランド病である。
【0061】
本発明に従う薬学的調製物はまた、例えば、手術もしくは外科的手術の直前、間、または後に、血栓溶解系または繊維素溶解系の一時的な障害を有する患者の予防または処置のために使われ得る。
【0062】
本発明はまた、生物学的流体における第VIII因子の分解を減少させるための、第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるアミノ酸配列を含むペプチドの使用または前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の使用に関する。
【0063】
さらに関連の実施形態において、本発明は、生物学的流体における第VIII因子の分解を減少させる方法を提供し、ここで、第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるペプチドまたは前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体が前記生物学的流体に添加される。
【0064】
本明細書中で使用される場合、「生物学的な流体」は、血液のような哺乳動物から単離された流体、または血漿もしくは血清のようなその画分、ならびに、真核細胞または原核細胞、細胞株または組織の培養物から得られた媒体またはそれらの画分を含む。
【0065】
好ましくは前記ペプチドまたは前記抗体は、第VIII因子の分解を少なくとも10%、好ましくは少なくとも20%、より好ましくは少なくとも50%ならびに最も好ましくは本質的に100%減少させるのに十分な量で添加される。
【0066】
本発明はさらに、生物学的流体における第VIII因子の半減期を延長させるための、第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるアミノ酸配列を含むペプチドまたは前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の使用および関連する方法に関する。
【0067】
好ましくは前記ペプチドまたは前記抗体は、生物学的流体における第VIII因子の半減期を少なくとも2倍、より好ましくは少なくとも5倍、そして最も好ましくは少なくとも10倍延長するのに十分な量で添加される。
【0068】
本発明はさらに、(生物学的流体における第VIII因子とLRPとの相互作用を阻害するための、)第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるペプチドまたは前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の使用に関する。好ましくは、少なくとも10%、好ましくは少なくとも20%、最も好ましくは少なくとも50%の第VIII因子のLRPとの阻害程度を達成するのに十分な量の前記ペプチドまたは前記抗体が添加される。
【0069】
本発明は、さらに血液凝固障害に罹患した患者を処置する方法であって、該方法は、第VIII因子のLRPとの相互作用を阻害するための、第VIII因子由来であるが、本質的に第VIII因子活性を持たない配列番号1、2、3または4のいずれかで定義されるペプチドまたは前記アミノ酸配列内の1つ以上のエピトープに特異的に結合する抗体の有効量を前記患者に投与する工程を包含する、方法、または、前記ペプチドの1つ以上もしくは前記抗体の1つ以上を含む薬学的組成物に関する。
【0070】
本発明は、引き続いて記載される実施例に例示されるが、本発明はこれに制限されることを意図しない。
【実施例】
【0071】
(実施例I)
材料−CNBr−セファロース4Bは、Amersham Pharmacia Biotech製であった。マイクロタイタープレート(Maxisorp)、細胞培養フラスコ、Optimem I培地、ウシ胎仔血清(FCS)、ペニシリンおよびストレプトマイシンは、Life Technologies(Life Technologies Inc.,Breda,The Netherlands)製であった。グレースのInsect培地(TNM−FH)およびInsect−XPRESS培地は、BioWhiftaker (Alkmaar、The Netherlands)から購入した。
【0072】
タンパク質−血漿由来のFVIII軽鎖、およびその第Xa因子切断誘導体を、以前に示されたように調製した(Lenting,P.J.,Donath,M.J.S.H.,van Mourik,J.A.,およびMertens,K.(1994)J.Biol.Chem.269,7150−7155;Donath,M.S.J.H.,Lenting,P.J.,van Mourik,J.A.,およびMertens,K.(1995)J.Biol.Chem.270.3648−3655;本明細書中に参考として援用される)。抗FVIIIモノクローナル抗体CLB−CAg A、CLB−CAg 117およびCLB−CAg 12が、以前に記載されている(Lenting,P.J.ら、(1994),前出;Leyte,A.,Mertens,K.,Distel,B.,Evers,R.F.,de Keyzer−Nellen,M.J.,Groenen−van Dooren,M.M.,de Bruin,J.,Pannekoek,H.,van Mourik,J.A.,およびVerbeet,M.P.(1989)Biochem.J.263,187−194;本明細書中に参考として援用される)。抗FVa軽鎖モノクローナル抗体CLB−FV 5は、標準のハイブリドーマ技術によって得た。
【0073】
FVIIIの軽鎖に対するドメイン可変単鎖抗体フラグメント(scFvs)を、Escherichia.coli株TG 1に発現させ、そして、金属器レートクロマトグラフィー(Qiagen,Hilden,Germany)によって、scFvs KM36、KM41およびKM33を150mM NaCl、5mM CaCl2、100mM イミダゾールおよび20mM Hepes(pH 7.4)中で溶出したことを除き、以前に記載されるように(32〜34)精製した。
【0074】
ヒトFVIII領域Trp1707−Arg1721(WDYGMSSPHVLRNR)(配列番号5)、Lys1804−Lys1818(KNFVKPNETKTYFWK)(配列番号2)、Tyr1815−Ala1834(YFWKVQHHMAPTKDEFDCKA)(配列番号3)、His1822−Ala1834(HMAPTKDEFDCKA)(配列番号6)、Thr1892−Ala1901(TENMERNCRA)(配列番号7)、Glu1908−His1919(EDPTFKENYRFH)(配列番号8)、Thr1964−Lys1972(TVRKKEEYK)(配列番号9)、Lys2049−Gly2057(KLARLHYSG)(配列番号10)およびAsp2108−Gly2117(DGKKWQTYRG)(配列番号11)を包含する合成ペプチドを、手動の「T−bag」法(Houghton,R.A.(1985)PNAS U.S.A.82,5131−5135;WO96/41816;本明細書中に参考として援用される)によってFmoc(N−(9−フルオレニル)メトキシカルボニル)化学によって、または430A Applied Biosystems装置(Pharmacia LKB Biotechnology,Roosendaal,the Netherlands;Medprobe AS,Oslo,Norway)を使用して合成した。
【0075】
ペプチドは、95%以上純粋(高圧液体クロマトグラフィー(HPLC)分析によって測定)であり、これらの同一性を質量分析によって裏付けた。精製した胎盤由来のLRPを、記載されたように得た(Moestrup,S.K.,およびGliemann,J.(1991)J.Biol.Chem.266,14011−14017;本明細書中に参考として援用される)。グルタチオンS−トラスフェラーゼ融合レセプター関連タンパク質(GST−RAP)をEscherichia coli株DH5αに発現させ、記載されたとおりにグルタチオン−セファロースを用いて精製した(Herz,J.,Goldstein,J.L.,Strickland,D.K.,Ho,Y.K.,およびBrown,M.S.(1991)J.Biol.Chem.266,21232−21238;本明細書中に参考として援用される)。組換えLRPリガンド結合クラスターIIおよびIVを発現するBaby Hamster Kidney(BHK)細胞が以前に記載されている(Neels、J.G.ら、(1999)、前出)。ヒト血清アルブミン(HSA)は、CLB(Amsterdam、The Netherlands)の製品部門製であった。タンパク質を、標準としてHASを用い、Bradfordの方法(Bradford,M.M.(1976)Anal.Biolchem.72,248−254;本明細書中に参考として援用される)によって定量した。
【0076】
組換えタンパク質−FVIII Bドメイン欠失変異体であるFVIII−del(868−1562)をコードするプラスミドpCLB−BPVdB695は以前に記載されており(Mertens,K.,Donath,M.J.S.H.,van Leen,R.W.,de Keyzer−Nellen,M.J.M.,Verbeet,M.P.,Klaasse Bos,J.M.,Leyte,A.,およびvan Mourik,J.A.(1993)Br.J.Haematol.85,133−142;本明細書中に参考として援用される)、そして、FVIII/FV1811−1818キメラをコードするプラスミドを構築するためのテンプレートとして用いた。FVIII/FVコドン置換を含むFVIII軽鎖配列由来のオリゴヌクレオチドプライマー(以下の表II)を用い、Overlap Extension PCR変異誘発法を用いてプラスミドを構築した(Tao,B.Y.,およびLee K.C.P.(1994)PCR Technology Current Innovations(Griffin,H.G.,およびGriffin,A.M.編),71−72,CRC Press,Boca Raton,FL;本明細書中に参考として援用される)。配列決定分析を、プラスミドにおける変異の存在を証明するために、実施した。FVIIIをコードするプラスミドのマウス線維芽(C127)細胞へのトランスフェクションを、以前に記載されたとおりに実施した(Mertens、K.ら、(1993)、前出)。野生型FVIIIまたはFVIII/FV1811−1818キメラを発現する安定な細胞株を、5% FCS、100U/ml ペニシリン、100μg/mlストレプトマイシン、1μg/ml アンホテリシンBおよび0.8μg/ml デスオキシコール酸を補充したRPMI 1640培地中の1−I細胞工場(cell factory)に維持した。FVIII1を含む培地を1週間に3回回収した。引き続いて、細胞片を除去するためにこの培地を濾過し、中空糸カートリッジ(Hemofiow F5,Fresenius,Bad Homburg,Germany)を用いておおよそ10倍に濃縮した。ベンズアミジンを、10mMの最終濃度で加え、濃縮物を−20℃で保管した。FVIIIを、確立した手順(Mertens、K.ら、(1993)、前出)に従って、抗体CLB−CAg 117およびQ−セファロースクロマトグラフィーを用いる、免疫親和性クロマトグラフィーによって濃縮した培地から精製した。FVIII軽鎖を、40mM EDTA、100mM NaClおよび50mM Tris(pH 7.4)を含む緩衝液中、FVIII/FV1811−1818キメラおよび野生型FVIIIを25℃で4時間インキュベートすることによって調製した。続いて、FVIII/FV1811−1818軽鎖キメラおよび野生型FVIII軽鎖を、Q−セファロースクロマトグラフィーを用いて精製した。組換タンパク質を、1M NaClおよび50mM Tris(pH 7.4)を含む緩衝液中で溶出し、150mM NaClおよび50mM Tris(pH 7.4)に対して透析し、そして、4℃で保管した。組換えC2ドメイン(すなわち、残基Ser2173−Tyr2332)をコードするプラスミドの構築は、以前に記載されている(Fijnvandraat、K.、ら、(1998)、前出)。FVIIIa3−A3−C1フラグメント(すなわち、残基Glu1649−Asn2172)をコードするプラスミドpACgp67b−His−a3−A3−C1を、pCLB−BPVdB695を鋳型として用い、オリゴヌクレオチドプライマー、5’−TTACTCGAGGAAATAACTCGTACTACTC−3’(センス)(配列番号13)、および5’−AATGCGGCCGCTTCAATTTAAATCACAGCCCAT−3’(アンチセンス)(配列番号14)を用いてポリメラーゼ連鎖反応によって構築した(Mertens、K.ら、(1993)、前出)。増幅されたDNAフラグメントを精製し、XhoIおよびNotIで消化し、pBluescriptに連結した。得られた構築物は、配列決定によって証明した。続いて、pBluescript−a3−A3−Clを、EspIおよびNotlで消化し、そして、得られたフラグメントを精製し、EspI/NotIで消化したpACgp67b−80Kプラスミドに連結した(Fijnvandraat,K.,Turenhout,E.A.M.,van denBrink,E.N.,ten Cate,J.W.,van Mourik,J.A.,Peters,M.,およびVoorberg,J.(1997)Blood 89,4371−4377;本明細書中に参考として援用される)。His−タグ(5’−ATTGGATCCGGCCATCATCATCATCATCATGGCGGCAGCCCCCGCAGCTTTCAAAAGCCCGGGGCCATGGGA−3’)(配列番号15)をコードするDNAフラグメントを、BamHIおよびNcoIで消化し、そして、BamHI/NcoIで消化したpACgp67b−a3−A3−C1プラスミド中にクローニングした。バキュロ・ウイルス表現系を使って、組換えa3−A3−C1およびC2フラグメントを、記載されたとおりに昆虫細胞の感染によって得た(Fijnvandraat、K.ら(1998)、前出)。a3−A3−C1フラグメントを免疫親和性クロマトグラフィーを用いてInsect−XPRESS培地から精製した(親和性マトリクスとしてCNBr−セファロース4Bと連結された抗A3ドメイン抗体CLB−CAg Aを用いる)。CLB−CAg A−セファロースを、a3−A3−C1フラグメントを含む培地と共に4℃で16時間インキュベートした。結合の後、免疫親和性マトリクスを回収して、1M NaClおよび50mM Tris(pH 7.4)を含む緩衝液で洗浄し、そして、150mM NaCl、55%(v/v)エチレングリコールおよび50mMリジン(pH 11)で溶出した。溶離画分をすぐに、1M イミダゾール(pH 6.0)で中和し、150mM NaCl、50%(v/v)グリセロールおよび50mM Tris(pH 7.4)に対して透析し、そして、−20℃で保管した。同様の免疫親和性クロマトグラフィー技術を用いて組換えC2ドメインを精製した。但し、抗C2ドメイン抗体CLB−CAg 117をCLB−CAg Aの代わりに用いた。
【0077】
第Va因子軽鎖の精製−ヒト第V因子(FV)を、本発明者らの組織(CLB、オランダ)によって供給されたヒト血漿から得た。全長FVを、免疫親和性クロマトグラフィーを用いて精製した。FVa軽鎖を、100mM NaCl、5mM CaCl2および50mM Tris(pH 7.4)を含む緩衝液中、FV(10μM)をトロンビン(2μM)と共に37℃で2時間インキュベートすることによって調製した。トロンビンを、ヒルジン(Sigma−Aldrich、St.Louis、MO)によって不活化し、そして、抗第V因子軽鎖モノクローナル抗体CLB−FV 5(5mg/ml)に結合したCLBr−セファロース4Bを用いる免疫親和性クロマトグラフィーを用いて、FVa軽鎖を精製した。免疫親和性マトリックスを、100mM NaCl、25mM EDTAおよび50mM Tris(pH 7.4)で洗浄し、そして、100mM NaCl、5mM CaCl2、55%(v/v)エチレングリコールおよび50mM Tris(pH 7.4)で溶出した。精製したFVa軽鎖を、150mM NaCl、5mM CaCl2、50%(v/v)グリセロールおよび50mM Tris(pH 7.4)に対して透析し、そして、−20℃で保管した。
【0078】
組換え型LRPフラグメントの発現および精製−組換えLRPクラスターIIおよびIVを、1単位/ml ペニシリンおよび100μg/ml ストレプトマイシン100を補充したOptimem I培地を用いてBHK細胞中に発現させた(Neels、J.G.ら、(1999)、前出)。培地を回収した後、CaCl2を、10mMの最終濃度まで添加した。馴化培地からのLRPクラスターIIおよびIVの精製は、1つの精製工程によって行った(親和性マトリクスとしてCNBr−セファロース4Bと連結されたGST−RAPを使って)。このマトリックスを、カラムに回収し、150mM NaCl、5mM CaCl2および50mM Hepes(pH 7.4)を含む緩衝液で洗浄し、そして、150mM NaCl、20mM EDTAおよび50mM Hepes(pH 7.4)で溶出した。続いて、精製したLRPクラスター調製物を、Centricon 10コンセントレーター(Millipore,Bedford,−MA)を用いて、4℃、4000×gで1時間の連続回転の遠心分離により濃縮した。最終的に、それらの調製物を、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH 7.4)に対して透析し、そして、−4℃で保管した。
【0079】
固相結合アッセイ−組換えLRPクラスターIIまたはIV(1pmol/ウェル)を、50μlの容量の50mM NaHCO3(pH 9.8)中のマイクロタイターウェルに、4℃で16時間吸着させた。ウェルを、200μlの容量の2%(w/v)HSA、150mM NaCl、5mM CaCl2および50mM Tris(pH 7.4)で、37℃で1時間ブロッキングした。150mM NaCl、5mM CaCl2、0.1%(V/V)Tween(登録商標)20および50mM Tris(pH 7.4)で3回急速に洗浄した(各5秒未満)後、FVIII軽鎖またはその誘導体を、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%(v/v)Tween(登録商標)20および50mM Tris(pH 7.4)を含む50μlの容量の緩衝液中にて、37℃で2時間、種々の濃度でインキュベートした。結合したリガンドを、同じ緩衝液中でペルオキシダーゼ結合体化モノクローナル抗体CLB−CAg 12と共に37℃で15分間インキュベートすることによって検出した。抗体CLB−CAg 12は、FVIIIフラグメントのLRPまたはLRPクラスターのフラグメントへの結合をに干渉しなかった(データ示さず)。競合実験において、FVIII軽鎖(25nM)を、50μlの容量の競合物質の限界希釈の存在下または非存在下のいずれかにおいて、固定されたLRPクラスターを含むウェルにて、37℃で2時間インキュベートした。上で記載されたように、残留FVIII結合を検出した。データを、空のマイクロタイターウェル(固定されたLRPクラスターを含むマイクロタイターウェルへの結合と比較して5%未満であった)について補正した。
【0080】
表面プラズモン共鳴−BIAcoreTM2000バイオセンサーシステム(Biacore AB、Uppsala、Sweden)を使って、タンパク質相互作用の動力学を、表面プラズモン共鳴(SPR)分析によって決定した。LRP(16fmol/mm2)、FVIII軽鎖(71fmol/mm2)、a3−A3−Clフラグメント(67fmol/mm2)、FVa軽鎖(76fmol/mm2)またはscFv EL14(67fmol/mm2)は、製造業者によって指定されたようなアミン結合キットを用いて、第1級アミノ基を介しての活性化されたCM5−センサチップのデキストラン表面に共有結合した。1つのコントロールフローチャネルを、タンパク質の非存在下で、慣例的に活性化し、ブロックした。分析物の会合を、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH 7.4)中、25℃で20μl/分の流速で2分間評価した。同じ緩衝液流中で2分間解離させた。センサチップを、100mM H3PO4または20mM EDTA、1M NaClおよび50mM Hepes(pH 7.4)のいずれかの中にて、20μl/分の流速で数回のパルスを用いて再生した。会合速度定数(kon)および解離速度定数(koff)を、BlAevaluationソフトウェア3.1(Biacore AB、Uppsala、Sweden)を使うことによって決定した。データを、バルク屈折率変化について較正し、2部位結合モデルに従う非線形回帰分析によって適合した。平衡解離定数(kd)を、比率koff/konから計算した。低親和性の相互作用についてのkd値は、BlAevaluationソフトウェアを使うことによって定常状態の親和性の分析を用いて見積った。競合実験において、FVIII軽鎖(50nM)を、競合物質の限界希釈の存在下もしくは非存在下のいずれかにおいて、20μl/分の流速で2分間、固定化されたLRP(16fmol/mm2)と共にインキュベートした。
【0081】
FVIII/FV1811−1818軽鎖キメラのLRPクラスターIIへの結合−150mM NaClおよび50mM Tris(pH 7.4)を含む緩衝液中で、組換えFVIII/FV1811−1818軽鎖キメラまたは組換え野生型FVIII軽鎖を、20fmol/mm2の密度まで、固定されたscFv EL14に結合した。LRPクラスターII(25〜125nM)を、150mM NaCl、2mM CaCl2、0.005%(v/v)Tween(登録商標)20および20mM Hepes(pH 7.4)中、20μl/分の流速にて、25℃で2分間、それぞれ、FVIII/FV1811−1818軽鎖キメラまたは野生型組換えFVIII軽鎖を固定化した別々のチャネル、および、1つのコントロール(scFv EL14でコーティングされている)チャネルを通した。
【0082】
(結果:)
LRPとFVIII軽鎖フラグメントとの間の相互作用−単離されたFVIII C2ドメイン(すなわち、残基Ser2173−Tyr2332)は、インタクトなFVIII軽鎖よりも効果的にLRPに会合しない(Lenting、P.J.、ら、(1999)、前出;WO00/28021、前出)。本研究において、FVIII軽鎖の別の部位がLRP結合に寄与する可能性を探究した。この目的のために、SPR分析を用いて、固定化されたLRPとの4つのFVIII誘導体の相互作用をモニタリングした。これらの誘導体としては、FVIII軽鎖、a3−A3−C1部分(すなわち、残基Glu1649−Asn2172)、C末端C2ドメインおよびFVIII軽鎖フラグメント(第X因子による位置Arg1721での切断を用いて、N末端の酸性の領域を欠失している(すなわち、残基Ala1722−Tyr2332))が挙げられる。
【0083】
図1において示されたように、全てのFVIIIフラグメントは、固定されたLRPとの時間依存的な会合を示し、その後分離した(最も高い反応が最高LRP密度で観察されたので、用量依存的であるように思われる(データ示さず))。FVIII軽鎖、第X因子切断軽鎖およびa3−A3−C1フラグメントの得られたデータを適切に示すのに、2部位結合モデルを必要とした。このモデルに従う計算された会合速度定数(kon)および解離速度定数(koff)は、これらのフラグメントについて同じオーダーにあった(以下の表I)。これは、固定されたLRPとの高い親和性および僅かに低い親和性の相互作用を示す、同等のKd値(すなわちFVIII軽鎖について18nMおよび59nM、第X因子切断軽鎖について22nMおよび60nM、ならびにa3−A3−C1誘導体について26nMおよび74nMを生じる(以下の表I)。対照的に、単離したC2ドメインは、任意の結合モデルによって正確に記載されるにはあまりにも速い解離速度を示した。従って、定常状態の親和性の計算を用いて、このフラグメント(3.4μM)の親和性(Kd)を推測によって見積った。まとめると、これらの結果は、A3−C1領域(すなわち、残基A1722−Asn2172)に高い親和性LRP結合部位が存在し、そしてC2ドメインに低い親和性の部位が存在することを示す。
【0084】
【表1】
【0085】
(表1に対する注意事項:FVIII軽鎖およびその誘導体の固定化されたLRPへの結合についての動力学的パラメーター。固定されたLRP(16fmol/mm2)への、種々の濃度のFVIII軽鎖(10〜250nM)、67kDaフラグメント(10〜250nM)、a3−A3−C1フラグメント(10〜250nM)またはC2ドメイン(500〜2000nM)の会合および解離を「実験手順」で示されたように評価した。得られたデータを2部位結合モデルを用いて分析して、会合速度定数(kon)および解離速度定数(koff)を計算した。各クラスの結合部位を、それぞれ1および2と称する。親和定数(Kd)は、比率 koff/konから計算した。C2ドメインとLRPとの間の相互作用を、定常親和性分析を用いて評価した。データを、各測定について少なくとも5つの異なる濃度を用いた、3〜6回の測定に基づく。データは、平均±S.D.を表す。)
固定されたLRPクラスターIIおよびIVへのFVIII軽鎖の結合−以前の研究において、LRPリガンド結合クラスターIIおよびIVが、VIII軽鎖との相互作用を媒介することが示された(Neels、J.G.ら(1999)、前出)。本研究において、LRPクラスターIIおよびIVがFVIII軽鎖への結合に競合し得るか否かという問題に取り組むために固相結合アッセイを用いた。図2の挿入図において示したように、FVIII軽鎖は、固定されたLRPクラスターIIに用量依存性の様式で結合し得た。この観察は、以前の調査結果(SPIR分析を用いて、LRPクラスターIIと固定されたFVIII軽鎖との間の相互作用をモニタリングした)(Neels、J.G.ら(1999)、前出)と一致している。競合研究は、LRPクラスターIIおよびIVが、FVIII軽鎖への結合に競合することを明らかにした(図2)。LRPクラスターIIは、固定されたLRPに結合するFVIII軽鎖クラスターIVの用量依存性の阻害を示した。これらのデータは、LRPクラスターIIおよびIVが、FVIII軽鎖内の同様の結合領域を共有することを意味する。
【0086】
LRPクラスターIIの固定化されたFVa軽鎖への結合−FVIIIとFVとの間の公知の相同性(Church,W.R.ら(1984)、前出)を考慮すると、FVIIIおよび活性化されたFVの軽鎖がLRPクラスターIIへの結合特性を共有するか否かという問題が生じ得る。この目的のために、LRPクラスターIIの連続希釈を、固定化されたFVIII軽鎖およびFVa軽鎖と共にインキュベートした。図3に示すように、FVaおよびFVIIIの軽鎖は、FVIIIのみがLRPクラスターIIに対して高親和性の結合を示すという点で異なることを証明した。これらの観察は、FVIII軽鎖のa3−A3−Clドメインは、FVa軽鎖には保存されてない高親和性のLRP相互作用領域を含むことを示す。
【0087】
合成のペプチドの、FVIII軽鎖のLRPクラスターIIへの結合に及ぼす影響−a3−A3−Clドメインの表面ループを模倣する合成ペプチドのパネルを構築した(WO 96/41816、前出)。FVa軽鎖がLRPクラスターIIと効率的に会合しないという観察を、FVIIIに固有の合成のペプチドの構築のための選択基準として使用した。これらのループの溶媒接触能を、ヒドロパシー分析(Lenting、P.J.ら(1996)、前出)を用いて、および、インタクトなFVIIIヘテロ二量体の三次元モデル(Stoilova−McPhie,S.,Villoutreix,B.O.,Mertens,K.,Kemball−Cook,G.,およびHolzenburg,A.(2002)Blood 99,1215−1223;本明細書中に参考として援用される)を研究することによって確定した。合成のペプチドは、以下の残基を含んだ:Trp1707−Arg1721、Lys1804−Lys1818、Tyr1815−Ala1834、His1822−Ala1834、Thr1892−Ala1901、Glu1908−His1919、Thr964−Lys1972、Lys2049−Gly2057およびAsp2108−Gly2117(表II)。
【0088】
【表2】
【0089】
(表IIに対する注意事項:合成のペプチド由来のFVIII a3−A3−C1フラグメントの、FVIII軽鎖とLRPクラスターIIとの間の相互作用への影響。FVIII軽鎖(25nM)を、種々の濃度の合成ペプチド(0〜1mM)の存在下または非存在下において、150mM NaCl、5mM CaCl2、1%(w/v)HSA、0.1%Tween(登録商標)20および50mM Tris(pH 7.4)50μlの容量中にて、固定されたLRPクラスターII(1pmol/ウェル)と共に37℃で2時間インキュベートした。同じ緩衝液で洗浄した後、FVIII軽鎖を、ペルオキシダーゼ結合体化抗−FVIII抗体CLB−CAg 12と共に37℃で15分間インキュベートする事によって定量した。最大半減阻害定数(IC50)は、3回の実験の平均値±S.D.を表す。)
続いて、これらのペプチドを、FVIII軽鎖と固定されたLRPクラスターIIとの間の相互作用を阻害するそれらの能力について試験した。表IIに示したように、合成ペプチドLys1804−Lys1818(配列番号2)およびTyr1815−Ala1834(配列番号3)は、FVIII軽鎖と固定されたLRPクラスターIIとの間の相互作用を効率的に阻害した。最大半減阻害(IC50)は、各々、約1.9μMおよび16.8μMのペプチド濃度に達した。他の合成ペプチドは、このような阻害効果を示さなかった。これらの観察は、FVIIIのA3ドメイン内の配列Lys1804−Ala1834が、LRPとの相互作用に関する重要な残基を含むことを示唆する。
【0090】
ScFv抗体フラグメントがLRPクラスターIIへのFVIII軽鎖の結合に及ぼす影響−以前に、領域Gln1778−Asp1840内の残基に対する阻害性抗体を持つ患者由来の組換えscFv抗体フラグメントを分離するために、ファージディスプレイを用いた(van den Brink,E.N.,Turenhout,E.A.M.,Bovenschen,N.,Heijnen,B.G.,Mertens,K.,Peters,M.およびVoorberg,J(2001),Blood 97,966−972;Voorberg,J.,ら、Method for diagnosis and treatment of haemophilia A patients with an inhibitor;国際特許出願番号WO 99/58680;これらは本明細書中に参考として援用される)。これらのscFvsを、FVIII軽鎖とLRPまたはクラスターIIとの間の相互作用を阻害するそれらの能力について評価した。第1のscFv(scFv KM36称される)は、Gln1778−Asp1840内の領域に指向するが、FVIIIへの結合のために残基Arg1803−Lys1818を必要としない(van den Brinkら(2001)、前出;WO 99/58680、前出)。第2のscFv(scFv KM41として命名された)は、領域Arg1803−Lys1818に指向し、かつFVIIIの凝固促進性活性を阻害する(van den Brinkら(2001)、前出;WO 99/58680、前出)。scFv KM41の重鎖のアミノ酸配列ならびに対応するDNA塩基配列を配列番号16に示す。scFv KM41の軽鎖のアミノ酸配列ならびに対応するDNA塩基配列を配列番号18に示す。KM41の重鎖および軽鎖は、ペプチドリンカー(配列番号24)で結合され得、かつ、C末端にヒスチジンタグを有し得る。発現されたscFv KM41タンパク質のアミノ酸配列を、配列番号25に示す。第3のscFv(scFv KM33と称される)は、scFv KM41と類似している。但し、そのFVIII軽鎖との相互作用のためにA3ドメイン領域1778−1818を必要としない(van den Brinkら(2001)、前出;WO 99/58680、前出)。scFv KM33の重鎖のアミノ酸配列ならびに対応するDNA塩基配列を配列番号20に示す。scFv KM33の軽鎖のアミノ酸配列ならびに対応するDNA塩基配列を配列番号22に示す。重鎖および軽鎖は、ペプチドリンカー(配列番号24)で連結され得、かつ、C末端にヒスチジンタグを有し得る。発現されたscFv KM33タンパク質のアミノ酸配列を、配列番号26に示す。図4Aにおいて、scFv KM36がFVIII軽鎖と固定されたLRPクラスターIIとの間の相互作用に影響を及ぼさなかったことを示した。対照的に、scFv KM41の存在は、FVIII軽鎖のLRPクラスターIIへの結合を阻害した(図4A)。scFv KM41がFVIII軽鎖とLRPとの間の相互作用に及ぼす影響を、SPIR分析を用いてさらに研究した。図4Bに示したように、FVIII軽鎖の固定されたLRPへの会合は、scFv KM41の存在下で効率的に阻害された。図5に示したように、scFv KM33は、FVIII軽鎖の固定されたLRPクラスターIIへの結合を用量依存性の様式で効率的に阻害した。見かけ上の阻害定数は、約5nMであったが、これは、同じscFvへのFVIII軽鎖の結合についてのKd値と類似している(van den Brinkら(2001)、前出;WO 99/58680、前出)。これらのデータは、scFv KM33、scFv KM41およびLRPがFVIIIの軽鎖内の重複する結合部位を共有することを強力に示唆する。
【0091】
FVIII軽鎖配列Glu1811−Lys1818は、LRPのための結合部位を含む−合成のペプチドLys1804−Lys1818およびTyr1815−Ala1834は、FVIIIa−FIXa複合体のアセンブリを妨害することによる、FVIII凝固促進活性の効果的なインヒビターである(Lenting、P.J.、ら、(1996)、前出;WO 96/41816、前出)が、scFvs KM33およびKM41は、FVIIIa−FIXa複合体のアセンブリを妨害することによる、FVIII凝固促進活性の効果的なインヒビターである(Lenting、P.J.、ら、(1996)、前出;van der Brinkら、(2001)、前出;WO 99/58680、前出)。FVIII A3ドメイン残基Glu1811−Lys1818は、FIXaとの相互作用に関与する(Lenting、P.J.、ら、(1996)、前出;WO 96/41816、前出)ので、この特定のFVIII軽鎖領域を、LRPとの相互作用におけるその役割に関して調査した。FVa軽鎖がLRPクラスターIIと相互作用しなかったので、FVの対応する残基(すなわち、残基1704SSYTYVWH1711;(配列番号12))によって残基Glu1811−Lys1818を交換してFVIII軽鎖キメラを、構築した。単離したキメラ(FVIII/FV1811−1818)を、SPR分析を用いて野生型組換えFVIII軽鎖とLRPクラスターIIとの会合に関して比較した。図6に示したように。LRPクラスターIIは、野生型のFVIII軽鎖と比較して固定化したFVIII/FV1811−1818との2〜3倍減少した会合を示した。
【0092】
次いで、単離したキメラ(FVIII/FV1811−1818)を、SPR分析を用いて、scFv KM41と相互作用する能力について評価した。図7に示すように、scFv KM41は、固定化したFVIII/FV1811−1818キメラを認識せず、一方、固定化した野生型の組換えFVIII軽鎖と容易に反応した。これらの観察は、FVIII A3ドメイン領域Glu1811−Lys1818が、scFv KM41に結合するために重要な残基を含むことを示す。これらのデータは、FVIII軽鎖領域Glu1811−Lys1818が、LRP−FVIII軽鎖複合体のアセンブリに重要な役割を果たすことを示す。
【0093】
本発明の研究において、FVIII軽鎖のA3ドメイン領域Glu1811−Lys1818は、LRPとの高い親和性の相互作用に寄与することが示される。いくつかの系統の証拠が、この結論を支持する。第1に、合成ペプチドLys1804−Lys1818およびTyr1815−Ala1834由来のA3ドメインは、FVIII軽鎖とLRPクラスターIIとの間の相互作用に影響を及ぼした(表II)。第2に、FVIII軽鎖キメラ(領域Glu1811−Lys1818がFVの対応する残基と交換される)は、野生型FVIII軽鎖と比較してLRPクラスターIIとの会合の減少を示した(図6)。第3に、領域Glu1811−Lys1818に対する組換えscFv抗体フラグメントが、FVIII軽鎖のLRPまたはLRPクラスターIIフラグメントへの結合を阻害した(図4)。
【0094】
多くのLRPリガンド(RAP、リポタンパク質リパーゼ、およびα2−マクログロブリンが挙げられる)について、リガンド表面の正に荷電した残基がLRPとの相互作用に関与することが確立されている(Meiman,L.,Cao,Z.F.,Rennke,S.,Paz Marzolo,M.,Wardell,M.R.およびBu,G.(2001)J.Biol.Chem.276.29338−29346;Chappell,D.A.,Fry,G.L.,Waknitz,M.A.,Muhonen,L.E.,Pladet,M.W.,Iverius,P.H.およびStrickland,D.K.(1993)J.Biol.Chem.268,14168−14175;Howard,G.C.,Yamaguchi,Y.,Misra,U.K.,Gawdi,G.,Nelsen,A.,DeCamp,D.L.およびPizzo,S.V.(1996)J.Biol.Chem.271,14105−14111;Nielsen,K.L.,Holtet,T.L.,Etzerodt,M.,Moestrup,S.K.,Gliemann,J.,Sottrup−Jensen,L.およびThorgersen,H.C.(1996)J.Biol.Chem.271,12909−12912;本明細書中に参考として援用される)。興味深いことに、FVIII A3ドメイン領域Glu1811−Lys1818(すなわち、残基1811ETKTYFWK1818(配列番号1))もまた、1813位および1818位に2つの露出された正に荷電したリジン残基を含む。FVのA3ドメイン内の相同体部分(すなわち、残基1704SSYTYVWH1711)と比較すると、これらのリジン残基は、FVIII A3ドメインに対して固有であるようである(Church,W.R.ら、(1984)、前出)。FVIII残基Glu1811−Lys1818のFVの対応する残基への置換は、LRPクラスターIIへの低下した結合を生じた(図6)。これらの結果は、領域Glu1811−Lys1818内の正に荷電した残基がLRPとの静電的相互作用を媒介することを示唆する。
【0095】
今日までに、LRP−FVIII軽鎖複合体のアセンブリに寄与する、FVIII軽鎖内の2つのアミノ酸領域が確認されている。本発明の研究において発見されたA3ドメイン領域Glu1811−Lys1818についての役割に加えて、カルボキシ末端のC2ドメインもまた、LRPとの相互作用に寄与することが知られている(Lenting,P.J.、Neels,J.G.、van den Berg,B.M.M.、Clijsters,P.P.F.M.、Meijerman,D.W.E.、Pannekoek,H.、van Mourik,J.A.、Mertens,K.およびvan Zonneveld,A.,−J.(1999)J.Biol.Chem.274,23734−23739;Lenting,P.J.ら A factor VIII polypeptide with factor VIILC−activity.国際特許出願WO 00/28021;本明細書中に参考として援用される)。単離したC2ドメインはLRPとの低い親和性の相互作用(Kd 約3.4μM)を示すので、A3ドメインのLRP相互作用部位は、C2ドメインのうちの1つよりも優性であるようにみえる(表I)。これは、単離したC2ドメインがLRPとわずかな会合のみを示した過去の研究と一致する(Lenting,P.J.ら(1999)、前出;WO 00/28021、前出)。さらに、FVIII軽鎖のLRPへの結合親和性は、C2ドメインの欠失に影響されない(表I)。しかし、抗C2ドメインモノクローナル抗体(ESHH4)は、FVIII軽鎖とLRPとの間の相互作用を完全に阻害することが示された(Lenting、P.J.ら(1999)、前出;WO 00/28021、前出)。抗体ESH4がLRP結合を阻害するメカニズムは、まだ解明されていない。抗C2抗体は、そのFVIII軽鎖との相互作用のために領域Glu1811−Lys1818を必要としない(Scandella,D.,Gilbert,G.E.,Shima,M.,Nakai,H.,Ealeson,C.,Felch,M.,Prescott,R.,Rajalaksmi,K.J.,Hoyer,L.W.およびSaenko,E.(1995)Blood 86,1811−1819;本明細書中に参考として援用される)ので、ESH4がA3ドメインにおける同様の部位への結合に関してLRPと競合することはなさそうである。従って、阻害に寄与し得るメカニズムのうちの1つは、立体障害を含む。
【0096】
対照的に、scFv KM41は、ほんの部分的にFVIII軽鎖とLRPとの間の相互作用を阻害する(図4)。これは、完全抗体(約150kDa)と比較して、比較的小さいサイズのscFv抗体フラグメント(約30kDa)に起因し得る。これらの観察は、領域Glu1811−Lys1818に加えて、A3−C1ドメイン内の他の表面の露出された構造エレメント(すなわち、残基Ala1722−Asn2172)がLRP−FVIII軽鎖複合体のアセンブリに寄与することを示唆する。これは、FVIII/FV1811−1818軽鎖キメラが、LRPクラスターIIに対する残基結合を示したという観察の系統にある(図6)。この文脈において、FVIII軽鎖のA3ドメイン内のGlu1811−Lys1818領域が、タンパク質表面に露出されるより大きなセグメントの一部(すなわち、残基Glu1804−Lys1818)であることを言及すべきである(Lenting、P.J.ら(1996)、前出)。Lys1813位およびLys1818位のリジン残基に加えて、この領域は、Lys1804位およびLys1808位に2つのさらなるFVIII固有のリジン残基を含み、これは、LRPとの相互作用において役割を果たし得る。
【0097】
(実施例II)
実験手順:vWFノックアウトマウス(健全な動物の第VIII因子と比較して、内因的な第VIII因子が20%減少している)を、ヒトの組換え型第VIII因子(RecombinateTM;200U/kg)および過剰量の組換えscFv(KM33)(3OnM 対 3nM 血漿中の第VIII因子)で処理した。対照実験において、第VIII因子および第VIII因子分子上の提案した追加のLRP結合部位として異なるドメインに結合する対照scFvフラグメント(KM38)を同じ濃度で使用した。任意に付加することなく組換えFVIIIの注入を用いて、このモデル系における第VIII因子のクリアランスをモニタリングした。
【0098】
注射後の確定時点(15分、30分、60分、120分および240分)で、血液を心臓穿刺によって回収し、引き続いて血漿を単離し、ショック凍結し、そして、第VIII因子活性について分析した。各時点にて10匹のマウスを用い、血漿を3つの濃度(1:20、1:60および1:100)に希薄し、そして、2連で分析した。第VIII因子レベルは、定量的ELISAを実施することによって定量した。
【0099】
結果:第VIII因子対照群からのデータは、15分で0.35U/mLの第VIII因子活性を示し、これは、14%の回復に対応する。30分で第VIII因子活性は約0.2U/mLに減少し、これは、これらのマウスの第VIII因子活性バックグラウンド(8%の回復に対応する)である。FVIII濃度のそれ以上の変化は、次の時点で検出されなかった。データは、このマウス系において、組換え型第VIII因子が最初の30分以内に動物循環から除去されることを示す。
【0100】
FVIIIおよびLRPでブロッキングしたscFvフラグメント(KM33)で処置したマウス由来の血漿サンプルの分析は、適用の15分後に、約0.8U/mLの第VIII因子が検出され得ることを示した。これは、30.4%の回復に変換された。第VIII因子活性レベルは、次の15分間で0.5U/mL(20%の回復)まで減少し、そして、60分後に0.35U/mL(14%の回復)に達した。注射の4時間後、FVIII活性レベルは、マウスの第VIII因子のバックグラウンド活性に達した。FVIII対照群と比較して、15分後および30分後の循環におけるこの高用量のFVIII活性は、FVIIIのクリアランスからの非常に良好な保護を示す。
【0101】
第2群の動物に、第VIII因子にLRPとは異なる部位で結合するscFvフラグメント(KM38)とヒトFVIIIとの同時注射を与えた。これらの動物に関して、第VIII因子の対照群と同様の結果が得られた(図8)。
【0102】
FVIII−LRP相互作用と干渉する抗体フラグメントを用いると、vWFノックアウトマウスにおけるRecombinateTMの半減期が有意に増加することが可能であった。さらに、マウスのvWF−KO系におけるヒトFVIIIの回復は、倍以上に増加し得ることが明白に示され得る。
【0103】
当業者の範囲内の変更が、本発明の範囲内であると考えられるべきである。特に定義されない限り、本明細書中で使用される全ての技術科学用語は、本発明が属する分野の当業者によって一般と理解されるものと同じ意味を有する。本明細書中に記載されるものと類似または相当する任意の方法および材料が、本発明の実施または試験に使用され得るが、好ましい方法および材料をここに記載する。
【特許請求の範囲】
【請求項1】
本明細書中に記載の発明。
【請求項1】
本明細書中に記載の発明。
【図1】

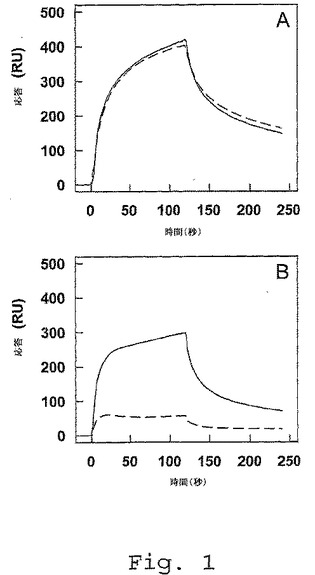
【図2】
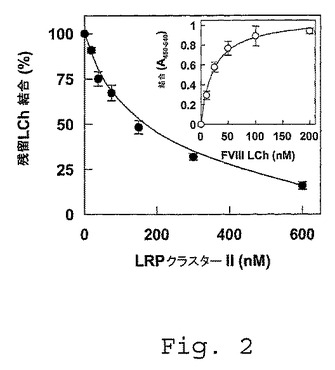
【図3】
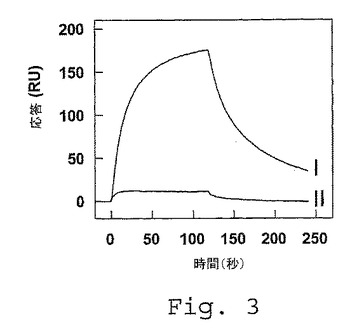
【図4】
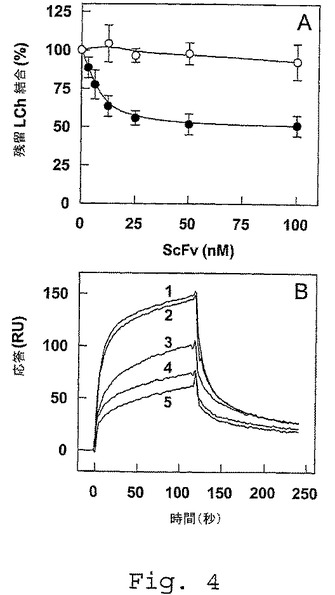
【図5】
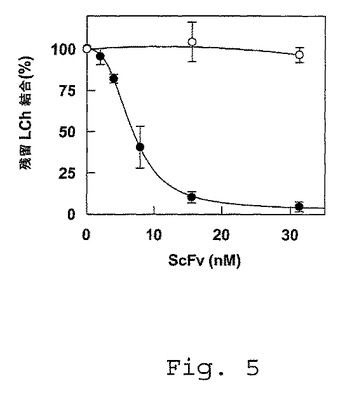
【図6】
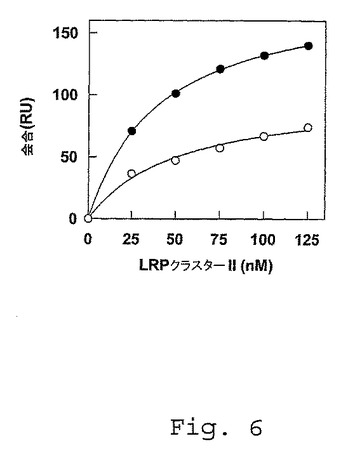
【図7】
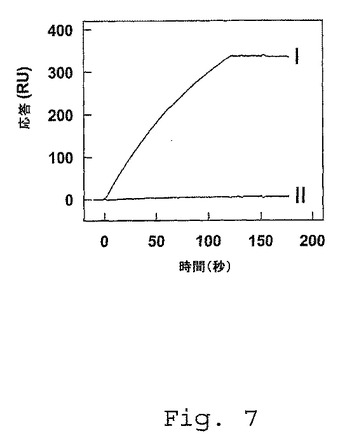
【図8】
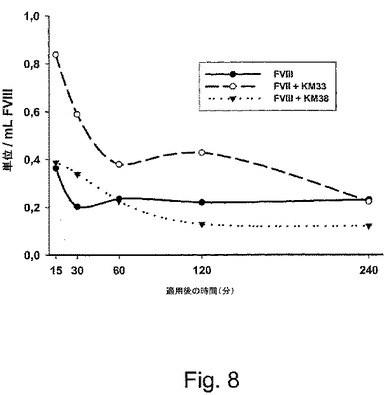
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2010−202664(P2010−202664A)
【公開日】平成22年9月16日(2010.9.16)
【国際特許分類】
【出願番号】特願2010−130425(P2010−130425)
【出願日】平成22年6月7日(2010.6.7)
【分割の表示】特願2004−501452(P2004−501452)の分割
【原出願日】平成15年4月28日(2003.4.28)
【出願人】(504403840)
【Fターム(参考)】
【公開日】平成22年9月16日(2010.9.16)
【国際特許分類】
【出願日】平成22年6月7日(2010.6.7)
【分割の表示】特願2004−501452(P2004−501452)の分割
【原出願日】平成15年4月28日(2003.4.28)
【出願人】(504403840)
【Fターム(参考)】
[ Back to top ]