
等比体積の形態変換
【解決手段】N−末端アミノ基または置換アミノ基、C−末端カルボキシ基またはカルボキシ誘導体、骨格およびL−アミノ酸側鎖を有するL−配置の前駆体の三次元座標および配列情報から、少なくとも部分的にD−アミノ酸を含有するポリペプチドの等比体積構造を生成する方法であって、
−少なくとも部分的に、骨格のCO基をNH基に、およびその逆に、置換する工程、
−同時に、前駆体のL−アミノ酸側鎖、N−末端アミノ基または置換アミノ基およびC−末端カルボキシ基またはカルボキシ誘導体の三次元座標を一定に保持する工程
を含む方法。

−少なくとも部分的に、骨格のCO基をNH基に、およびその逆に、置換する工程、
−同時に、前駆体のL−アミノ酸側鎖、N−末端アミノ基または置換アミノ基およびC−末端カルボキシ基またはカルボキシ誘導体の三次元座標を一定に保持する工程
を含む方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、retro-inverso型ペプチドの設計および合成分野に存する。本発明の主題は、慣用技術より容易かつ迅速に実施される、改良された等比体積の形態変換法である。また、本発明は、新規な方法によって得られるポリペプチドに関する。
【背景技術】
【0002】
多数の新規な生物工学的薬剤はタンパク質であり、体内で受容体と相互作用する。そのような成功した薬剤開発の例には、インターロイキン2およびGM−CSFまたはG−CSFのようなサイトカインがある。タンパク質薬剤の別の重要な種類は、ファージ提示法(Phage Display)等の全組換え系由来またはヒト化マウス抗体由来の抗体である。成功した薬剤ではあるが、薬剤としてのタンパク質は、多数の不利点を有することから、長期にわたりこれらの生物工学的薬剤に代わる、いわゆる「スモール・ドラッグ(small drugs)」の研究が継続されている。最も突出した不利点は、タンパク質薬剤がタンパク分解的に切断されるので、インビボでの半減期が短いこと、および、タンパク質薬剤に対する免疫反応の可能性が高いことである。誤って折りたたまれたタンパク質がわずかに寄与してタンパク質製剤が汚染されるか、またはTAGが精製または製造目的に使用されると、抗体またはアレルギー性免疫反応の不活化さえも起こり得る。抗体のヒト型化−通常、時間およびコスト集約的方法−は、これらのリスクをかなり縮小するが、廃することはできない。
【0003】
しかしながら、古典的薬剤の「スモール・ドラッグ」即ち有機小分子に至る取り組みは、リスクが高く、これまでのところあまり成功したアプローチではないことが判明した。スモール・ドラッグが複雑で大型のタンパク質受容体を標的にすることに成功した例はほとんど知られておらず、確立されていないであろう。
【0004】
有効な代替物の中で、ペプチドは際立った役割を演じる。基本的に、ペプチドは完全タンパク質に由来し、より大きなタンパク質の必須の結合ドメインを含むことができる。それらは、所与の標的に対し、天然タンパク質の作動薬または拮抗薬の両者として設計することができる。それらの大きさにより、通常、組換え産物の代替として化学的に合成し得る。さらに、小さなペプチドは、通常、大きなタンパク質より免疫原性が少ない。小さなペプチドは、経口適用の特別の生薬製剤においても使用されることがある。所望の薬剤適用に好適なペプチドを発見することは、同等のスモール・ドラッグを発見するよりもはるかに容易かつ迅速であることがわかっている。
【0005】
それでもやはり、免疫原性は完全には廃することはできず、−より重要なことには−、L−ペプチドは体液中で多数のプロテアーゼによって容易に分解され、完全タンパク質より半減期がさらに短い傾向がある。しかしながら、タンパク質の自然発生結合ドメイン由来のL−ペプチドを、最終的薬剤候補としてのL−ペプチドの非生分解性模倣剤を設計する出発点として使用することは可能である。このような目的に用いるという概念のうちで、非天然の、しばしばD−アミノ酸によって完全に構築されるペプチド等比体積物の設計は非常に魅力的である。このことは、合成および生薬製剤の同一の原理が、L−アミノ酸を基にした前駆体に関して、そのようなD−アミノ酸を基にしたペプチドに適用することができるという事実によるものである。これらのL−ペプチドの模倣剤は、なおもペプチドである。さらに、さらなる予期されない毒性が増加する危険性は、慣用の有機スモール・ドラッグよりも、L−ペプチドのペプチド性模倣剤のほうがはるかに小さい。
【0006】
D−アミノ酸を基にしたL−ペプチド模倣薬に至る1つの重要なアプローチは、retro-inverso型ペプチドの概念である。これらのペプチドは、アミノ酸配列の反転、続く、骨格のα−炭素原子での立体化学の反転(L−アミノ酸からD−アミノ酸へ)によって得られる。生物活性の相当の損失なく、所与のL−ペプチドに対応するretro-inverso型ペプチドを合成することは時に可能であるが、そのような「試行錯誤」のアプローチはリスクが高く、普通失敗に終わる。所与のペプチドおよび意図された立体化学の転回に関する基本的な構造的問題を明らかにするために、分子のモデリングを基にした合成のような、失敗のリスクを最小限にする戦略的計画が必要である。残念なことに、ペプチドのそのような分子モデリングは簡単ではなく、多くの時間、努力および熟練したスタッフを要する。
【0007】
ペプチドのretro-inverso型類似体の成功と失敗の両方について、多数の例が、引例[1、2]に概説されている。「鏡像ファージ提示法(mirror-image phage display)」のような組換え技術も、所与のL−ペプチドの、D−ペプチドを基にした等比体積体を得るために用いられるかもしれないが、実際に使用できるコンピュータに基づくアプローチと同様に、リスクがあり面倒である。
【0008】
今までのところ、新規なD−アミノ酸を基にした等比体積のペプチドを設計する標準的手法は、多大な時間を要し、天然ペプチドの受容体結合活性立体構造を模倣した立体構造に自動的にはならない。一般に、所与のL−ペプチドの立体構造的類似体としてretro-inverso型ペプチドを産生する、当業界のコンピュータに基づく手法は、以下の3工程を含む[3]。
【0009】
最初に、原型のペプチドの配列を反転させなければならない。
第2工程において、α−炭素原子における側鎖の立体化学をLからDに変えなければならない。
原型のペプチドとretro-inverso型ペプチドとの間で立体構造の類似性を達成するため、骨格のねじれ角度phiおよびpsiを、それぞれ対応する残基について入れ替えなければならない。
【0010】
これらの基本的な3工程では中間体構造となるだけであり、この構造を構造最適化および分子動力学に対応させる前に、さらなる形態変換を実施しなければならない。
【0011】
この全方法には、主な不利点が3点ある:
第一に、この方法は、複雑で時間がかかり、高価である。これは、工程1および工程2が原型のL−ペプチドの所与の構造を完全に破壊するという事実によるものである。配列の反転およびD−アミノ酸への変化により、アミノ酸の側鎖の配置が再構成される。これら側鎖は、工程3および工程4において「L−ペプチドに似た」立体配置へ再構成されなければならない。実際に、このアプローチは、出発点としてL−ペプチドの配列を使用しているだけであり、構造的情報(側鎖の位置および空間的定位)はまったく使用せず、L−ペプチドがあれば使用できる。このように、全体の方法は非常に時間がかかる「最初から」のモデリング方法である。
【0012】
第二に、この方法は、分子モデリングソフトにおいて、ほとんど自動化または実行できない。
【0013】
第三に、天然のペプチドと、側鎖の完全な重ねあわせはなお達成できない。この問題は、retro-inverso型ペプチドが、ペプチド−受容体複合体から設計して、受容体−結合立体構造を有する、D−アミノ酸を基にした等比体積のペプチドを得なければならない場合、より難しくなるであろう。
【0014】
全ての古典的なretro-inversoアプローチの主要な問題は、構造的情報が全く保存されず、失敗のハイ・リスクが、L−ペプチドから構造的に類似するD−ペプチドへ、「試行錯誤」で合成されること、および/または、「破壊的/再建的」に形態変換されることの、両者に関係することである。
【0015】
本発明は、簡単な方法を説明することによって、当業界のこれらの不利点を克服し、所与のL−ペプチドに関する構造的情報を保つ。さらに、本発明は、所与のL−ペプチド前駆体の意図された立体化学的転回の間に発生する、典型的な構造および配列に関連する問題の解決策を提示する。
【発明の開示】
【0016】
概説
本発明は、等比体積の形態変換、即ち、対応する天然L−ペプチド前駆体の等比体積体として働くD−アミノ酸を基にしたペプチドの、構造に基づく合理的設計を可能にする、新規かつ迅速な方法に関する。上述した慣用法と対照的に、本発明は、所与のL−ペプチド配列に関する構造的情報(例えば、結晶学的データ、NMRデータ)を使用することができ、通常、このL−ペプチドがその標的結合部位にドッキングされることを示す。
【0017】
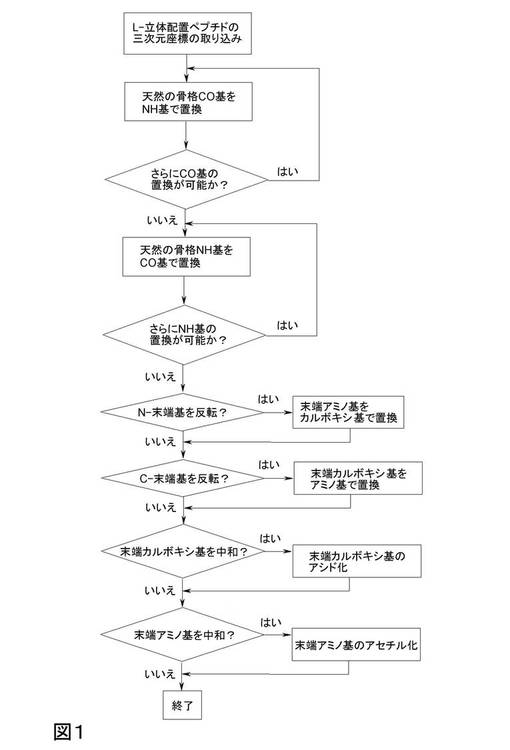
本発明の主題は、請求項1〜29のいずれかの、方法、ポリペプチド、化合物、使用および医薬製剤である。本発明による合理的設計の中心思想は、全方法手順にわたり、アミノ酸の側鎖の空間的定位を保持することである。従って、phiおよびpsi2面角の交換、および意図された分子の完全なデノボ構造の必要がない。本発明による方法手順は、前駆体ペプチドの骨格構造を変えるだけでこれを達成し、一方、完全なアミノ酸部分の交換を回避する(L−アミノ酸のD−アミノ酸による正式の置換なし)。骨格のペプチド結合が完全な平面構造であるので、このアプローチは、ジスルフィド架橋のような共有結合側鎖修飾、例えばらせんまたはβ−面のような、らせんの制約または他の構造的特性を含む、側鎖の定位または構造的特性を変化させない。これらの操作工程のワークフローは、標準的モデリング・ソフトウェアで容易に自動化することができる。また本発明はあらゆるソフトウェアまたはアルゴリズムにも及び、それらにより本発明の工程がコンピュータで自動化される。特に、本発明の方法は、図1に示される工程に従って実施される。
【0018】
等比体積の形態変換により、L−ペプチド前駆体の意図されたD−類似体の構造的データへ高速でアクセスされる。このことにより、二つの主要な問題の即時の評価が可能になり、その前駆体と立体化学的に適合しない、新たに設計された分子を提供することができる:
即ち、前駆体の配列中のL−プロリンの存在、および、ペプチドの末端の帯電である。
【0019】
要すれば、ペプチド末端の帯電の修飾のための合成的解決策が入手可能であるが、ここで開示される本発明は、プロリンの問題のための多数の合成的解決策を提供し、それは等比体積の形態変換の間、プロリンの構造的に正確な置換のために使用することができる。
【0020】
従って、等比体積の形態変換は、L−ペプチド前駆体−ペプチド含有プロリンであっても−をそれぞれのD−ペプチド類似体へ、立体化学的に映すための、高速で魅力的な解決策を提供する。
【0021】
また、本発明は、これらの新たに設計された化合物の合成に関し、かつ、本発明によるビルディング・ブロックおよびD−ペプチドにおける末端の修飾を含有するペプチドに関する。一般に、本発明は、出発点となるL−ペプチド構造に関する充分な情報が入手可能ならば、殆ど全てのペプチド構造に適応できる解決策を提供する。
【0022】
ここで開示される本発明の原理が、例えば配列情報単独に基づく詳細な構造的データを利用しなくとも使用できることは、当業者には明白である。しかしながら、このことは、合成研究所における「試行錯誤」のアプローチが、活性があり構造的に有効なD−アミノ酸を基にした物質を同定する、好適な、例えばインビトロの試験システムと関連して使用されなければならないことを暗示する。本発明の原理の使用を通じて、このアプローチは、以下に開示する通りの本発明の完全な構造を基にした実行と、全ての優位点−特に高速で先頭を検出する好機−を共有しない。しかしながら、発明の原理のこのような部分的実行もまた、本発明の方法手順の範囲に包含されることは明白である。
【0023】
詳細な説明
本発明は、工程a〜cを含む、1つ以上(全部まで)のペプチド結合の反転による、天然のペプチドの構造の座標を基にした、ペプチド等比体積体またはペプチド様物質を設計する方法を開示する:
所与のL−ペプチドの構造データを、所与のペプチドの立体構造の構造データを処理することが可能なソフトウェアをロードした、好適なコンピュータ装置に取り込む工程、
天然のカルボニル(CO)骨格基の原子をアミド(NH)基で置換する工程、
天然のアミド(NH)骨格基の原子をカルボニル(CO)基で置換する工程。
【0024】
工程a〜cは、所与のソフトウェア・ユーティリティを使用して熟練したスタッフによって手作業で実施することができ、または、コンピュータ/ソフトウェアユニットの適宜のプログラミングによって自動化することができる。複雑なペプチド構造でも、工程a〜cは数時間以内で終わることができる。これらの工程は、中間体構造において終了するであろうが、この構造は、全ての側鎖を正確な空間的定位に含有し、D−ペプチド構造に関して既に反転した骨格を示す。ペプチドの末端部は、まだ反転されない。従って、初期の生成物の構造は、末端未反転骨格を有する(以下の例を参照、2次元の略図に変形する)。
【0025】
【化1】
【0026】
本発明のさらなる様相において、C−末端および/またはN−末端基は、非末端骨格の反転に加えて、修飾することができる。C−末端カルボキシル基は、アミノ基と入れ換えることができ、および/または、N−末端アミノ基はカルボキシル基と入れ換えることができる:
【0027】
【化2】
【0028】
これらの付加的工程により、3つのさらなる分子となり、それらは、初期の生成物の構造と共に、本発明による全体の設計の最適化のための出発点となる(以下参照)。これらの場合のいくつかにおいて、gem−ジアミノアルキルおよびC−2−置換マロニル残基は、末端基として取り込まれる。このようなビルディング・ブロックの合成は、細部にわたり精査されている。[4]
【0029】
これまでのところ、この方法は、等比体積体の設計の古典的な方法と比較して、3つの主要な優位点を既に有している:
容易かつ迅速に実施される。
分子モデリング・プログラムにおいて、容易に自動化または実行することができる。
【0030】
得られる構造の側鎖の配置および立体構造は、さらなる最適化および動力学の出発点であるが、天然のペプチドと比較して変化しない。このことは、この方法が天然のペプチドと同一の側鎖の立体構造を維持する等比体積体を生成するため、特に受容体結合ペプチドの設計に非常に有益である。従って、得られる初期の生成物の構造は等比体積体であり、側鎖の受容体タンパク質との相互作用は元のL−ペプチドと同等である。
【0031】
以下の2次元の図は、等比体積の形態変換法のこれらの優位点を示す:殆ど操作を要せず、側鎖の配置は決して変わらない。
【0032】
【化3】
【0033】
また本発明は、上述の方法によって得られるD−アミノ酸を基にしたポリペプチドを提供する。この方法がL−ペプチドの等比体積構造を生成する一般的な方法であるという事実によって、本発明はまた、プロリン含有ペプチドの等比体積化合物も提供する。
【0034】
従って、本発明はまた、天然のペプチドが1つ以上のプロリン残基を含有する場合に重要なビルディング・ブロックを提供する。プロリンは、プロリン含有retro-inverso型ペプチドの例が生物学的活性を保持していたとしても[6]、しばしば、慣用のretro-inversoアプローチ[5]と適合性がないと見なされる。プロリンは、その側鎖がα−アミノ基に拘束され、従って骨格に戻る、唯一の天然の環状アミノ酸である。この特性は、等比体積の形態変換にとって特別なことを明確するが、これは骨格の反転に基づくものであり、従ってプロリンの場合−アミノ酸の中での例外として−等比体積の形態変換の間、立体化学的ひずみをもたらす。さらに、この制約は、プロリンを含まないペプチドに見られるものと異なる、制限された骨格の2面角を決定する。標準的なretro-inverso型ペプチドにおいて、プロリンの方向の逆転が、天然のペプチドと比較して、ピロリジン環のトポケミカルな置換をもたらす。またこのことは、ここで提示される等比体積の形態変換法についても当てはまる。しかしながら、上述した通りの等比体積の形態変換によって、非末端的に骨格が反転したペプチドを、例えば、元のL−ペプチド構造で規定される構造的要求との見込まれる非適合性について分子動力学によって試験することができる。従って、合成およびインビトロ試験以前に、プロリンの問題が実際に所与の構造の重要な様相であろうとなかろうと、理論的解釈を展開することができる。骨格の反転が受け入れ難いほどにプロリン位置での構造をゆがめる場合、等比体積の形態変換法によって、この特有であるが頻発する問題:β−回転またはβ−ヘアピンのような構造において、プロリンは、等比体積の形態変換後、グリシンによってしばしば置換され得る:について、多数の解決策が提供される:
【0035】
【化4】
【0036】
本発明のさらなる様相において、等比体積の形態変換に加えて、二つのアミノ酸ユニット(プロリンおよび直接隣接する残基)は、5−アミノバレリン酸またはその誘導体のような、1個のビルディング・ブロックで置換することができる:
【0037】
【化5】
【0038】
一般に、等比体積の形態変換後、2つのアミノ酸(プロリンおよび直接隣接する残基)は、一般式
...-(CO)-X1-X2-X3-X4-NH-…
で表されるビルディング・ブロックで置換することができる:
式中、X1、X2、X3、およびX4は独立してCH2、(C=O)、NH、NR、O、(CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基である。以下の図は、この一般式によって表されるビルディング・ブロックの幾つかの使用例を示す:
【0039】
【化6】
【0040】
また本発明は、等比体積の形態変換の後、プロリン残基の立体構造を模倣し、かつ、プロリンおよびその近接するアミノ酸残基を置換する、環状のビルディング・ブロックを提供する:
【0041】
【化7】
【0042】
このようなビルディング・ブロックの使用は、グリシンまたは5−アミノバレリン酸誘導体のような延長したグリシン等価構造が所与のペプチドの立体特性を安定化するのに充分でない場合に、有用であり、かつ、必要であろう。標的構造への「誘導適合」が、プロリン特異的立体構造の適切な再構築がないペプチドに問題になる場合に、このことは特に当てはまる。プロリンおよび近接するグリシンの置換のためにここで示したもののような、プロリン置換ビルディング・ブロックは、以下の一般式に従って構築することができる:
【0043】
本発明のさらなる様相において、等比体積の形態変換後、2つの近接するアミノ酸(ひとつはプロリンであった)は、一般式
【化8】
で表されるビルディング・ブロックによって置換することができる。
【0044】
式中、X1、X2およびX3は独立してCH2、(C=O)、O、S、NH、NR、 (CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基である。以下の図は、この一般式で表されるビルディング・ブロックのいくつかの例を示す:
【0045】
【化9】
【0046】
このようなビルディング・ブロックの使用は、グリシンまたは5−アミノバレリン酸誘導体のような延長したグリシン等価構造が所与のペプチドの立体特性を安定化するのに充分でない場合に、有用であり、かつ、必要であろう。標的構造への「誘導適合」が、プロリン特異的立体構造の適切な再構築がないペプチドに問題になる場合に、このことは特に当てはまる。
【0047】
本発明のさらなる様相において、等比体積の形態変換後、2つのアミノ酸は一般式
【化10】
で表されるビルディング・ブロックによって置換することができる。
【0048】
式中、X1、X2、X3およびX4は独立してCH2、(C=O)、O、S、NH、NR、 (CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基である。以下の図は、この一般式で表されるビルディング・ブロックのいくつかの例を示す:
【0049】
【化11】
【0050】
このようなビルディング・ブロックの使用は、グリシンまたは5−アミノバレリン酸誘導体のような延長したグリシン等価構造が所与のペプチドの立体特性を安定化するのに充分でない場合に、有用であり、かつ、必要であろう。標的構造への「誘導適合」が、プロリン特異的立体構造の適切な再構築がないペプチドに問題になる場合に、このことは特に当てはまる。ここで示される一般式を基にした分子は、以前に示したピロリジンを基にした構造の代わりに使用することができる。
【0051】
上記で概説した通りのビルディング・ブロックおよびビルディング・ブロックの使用は、本発明によるプロリンを含むポリペプチドの等比体積の形態変換において有用である。しかしながら、それらビルディング・ブロックは、D−ペプチドおよびretro-inverso型ペプチドの設計および製造のための慣用技術においても有用である。
【0052】
本発明において記載されるアミノ酸は、自然発生のL−立体異性体のものであってもよく、エナンチオマーのD体であってもよい。一文字の記号は、容認されている標準的なポリペプチド命名法を参照するが、D−またはL−アミノ酸を択一的に意味することができる。小文字はDアミノ酸に言及するものである。
【0053】
記号 アミノ酸
A L−アラニンまたはD−アラニン
V L−バリンまたはD−バリン
L L−ロイシンまたはD−ロイシン
I L−イソロイシンまたはD−イソロイシンまたはD−アロ−イソロイシン
M L−メチオニンまたはD−メチオニン
F L−フェニルアラニンまたはD−フェニルアラニン
Y L−チロシンまたはD−チロシン
W L−トリプトファンまたはD−トリプトファン
H L−ヒスチジンまたはD−ヒスチジン
S L−セリンまたはD−セリン
T L−トレオニンまたはD−トレオニンまたはD−アロ−トレオニン
C L−システインまたはD−システイン
N L−アスパラギンまたはD−アスパラギン
Q L−グルタミンまたはD−グルタミン
D L−アスパラギン酸またはD−アスパラギン酸
E L−グルタミン酸またはD−グルタミン酸
K L−リジンまたはD−リジン
R L−アルギニンまたはD−アルギニン
P L−プロリンまたはD−プロリン
G グリシン
a D−アラニン
v D−バリン
l D−ロイシン
i D−イソロイシンまたはD−アロ−イソロイシン
m D−メチオニン
f D−フェニルアラニン
y D−チロシン
w D−トリプトファン
h D−ヒスチジン
s D−セリン
t D−トレオニンまたはD−アロ−トレオニン
c D−システイン
n D−アスパラギン
q D−グルタミン
d D−アスパラギン酸
e D−グルタミン酸
k D−リジン
r D−アルギニン
p D−プロリン
【実施例】
【0054】
1.ペプチドの等比体積の形態変換
1.1 インターロイキン2受容体のβ鎖に結合する、プロリンを含まない、らせんペプチドの等比体積の形態変換
以下の実施例は、インターロイキン2受容体のβ鎖に結合する、らせんペプチドの形態変換を実証する。
【0055】
このペプチド−インターロイキン2(IL−2)に類似−は、高い親和性で、IL−2特異的受容体に結合する能力を有する。IL−2は、腫瘍治療において使用されるサイトカインである。IL−2は特異的受容体(IL−2R)に結合し、それによってIL−2特異的細胞内シグナルの引き金を引く。
【0056】
IL−2は、TおよびBリンパ球の成長に対する刺激効果を有し、細胞傷害性および細胞溶解性のNK細胞を活性化する。従って、IL−2は、免疫応答の調節において、中心的意義を有する。このように、IL−2は、腫瘍および炎症反応に対する免疫応答において、基本的に重要である。IL−2による腫瘍防御のために重要なメカニズムの1つは、LAK(「リンフォカイン活性化キラー細胞」)の誘導であると思われる。これらの細胞は、腫瘍細胞を破壊する能力がある。
【0057】
天然ペプチドの組み合わせを有する構造ファイルは、式STKKTQLQLEHLLLDLQMILNGINNYを有するらせんペプチドと、インターロイキン2受容体のβおよびγ鎖との複合体構造を含有する。このらせんペプチドは、等比体積の形態変換の実施の、理想的な例を提供する:
【0058】
受容体に結合した立体構造中のホモ二量体ペプチドの組み合わせを、分子モデリング・ソフトウェアに取り込む。
天然のカルボニル(CO)骨格基の原子は、アミド(NH)基で置換される。
天然のアミド(NH)骨格基の原子は、カルボニル(CO)基で置換される。
【0059】
等比体積の形態変換の方法手順では、修飾は骨格にのみ起こるので、側鎖原子の立体配置は変わらない。従って、この方法は、D−アミノ酸を基にした受容体結合ペプチドの設計および最適化のための出発立体構造を産生する、良好な方法手順である。
【0060】
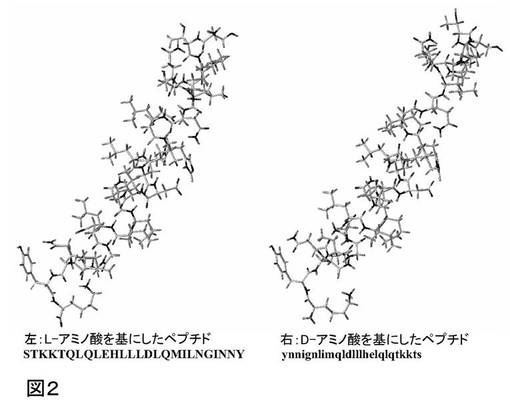
天然のらせんペプチドの構造を、等比体積の形態変換および位置最適化の後に得られる構造と比較することによって、図2に示す通り、天然のペプチドと形態変換された分子との構造的等価性が実証される。
【0061】
骨格反転の効果の1つには、アミノ酸配列の反転だけではなく、アミノ酸のα炭素原子の立体配置の反転もあり、これはLからDへ変化する。このことは、本発明では、アミノ酸配列において小文字で示される。トレオニンは2つのキラル中心を含有し、従って、側鎖のキラリティーを維持し、しかるべき等比体積体はD−アロ−Thrであり、ペプチド配列式中、小文字「t」で示される。
【0062】
概して、式STKKTQLQLEHLLLDLQMILNGINNYのL−ペプチドは、等比体積の形態変換によって、式ynnignlimqldlllhelqlqtkktsで表すことができるDペプチドに形態変換される。本発明は、このペプチドの等比体積の、受容体にドッキングされた構造の組み合わせを産生する、迅速な方法を開示する。
【0063】
得られるペプチドは、完全にD−アミノ酸より成る。このことから、天然のL−ペプチドと比較して、生物学的半減期がより長くなる。D−アミノ酸を基にしたペプチドは、タンパク質分解酵素にはより安定である。従って、等比体積的に形態変換したペプチド模倣体は、医薬品として、L−ペプチドより良好な特性を有するように設計される。
【0064】
D−ペプチドは、免疫系疾患、例えば、炎症および関節炎の進行、または全てのタイプおよび起源の免疫不全症候群;細胞の進展する増殖に関係する疾患、例えば、癌腫、肉腫、リンパ腫および白血病等の形態の癌腫症;または感染過程の治療に好適である。
【0065】
1.2 エリスロポエチン受容体に結合するペプチドの等比体積の形態変換
以下の実施例は、エリスロポエチン受容体に結合するペプチドの形態変換を実証する。
【0066】
天然の構造を代表する組み合わせは、プロテイン・データ・バンク(Protein Data Bank)で公的に入手可能である(PDBコード:1EBP)。このファイルは、式TYSCHFGPLTWVCKPQを有する二量体のアゴニスト・ペプチドと、二量体のエリスロポエチン受容体との複合体の構造を含む[7]。この構造的に充分立証されたペプチドは、等比体積の形態変換の実施の理想的な例となる:
【0067】
受容体に結合した立体構造におけるホモ二量体ペプチドの組み合わせは、シビル(Sybyl)モデリング・ソフトウェアの適宜の分子に組み込まれる。
天然のカルボニル(CO)骨格基の原子は、アミド(NH)基で置換される。
天然のアミド(NH)骨格基の原子は、カルボニル(CO)基で置換される。
【0068】
ペプチド配列が2つのプロリン残基を包含するので、プロリン位置での構造的ひずみが、等比体積の形態変換によって得られる初期の構造において発生する。しかしながら、「誘導適合」位置のペプチドの第二の構造が多数の他の分子の相互作用によって安定化され、この構造はプロリン特異的特性によって引き起こされる特異的な立体構造を場合によっては必要としないことも明らかである。従って、プロリンは−誘導適合立体構造を支持するが−、そのために必須ではなく、最も単純な場合、非常にたわみやすいグリシンで置換することができる。
【0069】
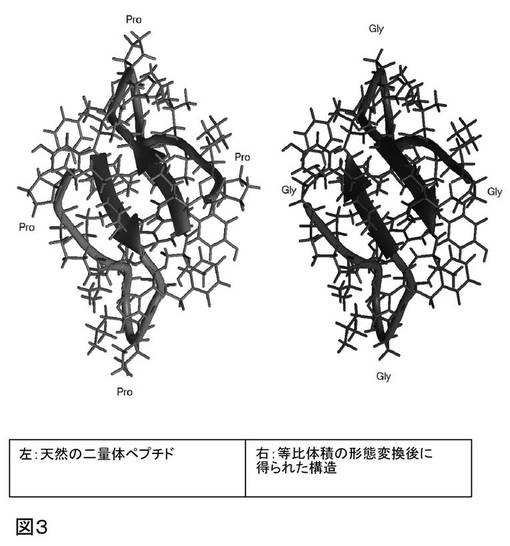
等比体積の形態変換の方法手順では、修飾が骨格にのみ起こるので、側鎖原子の立体構造は変化しない(天然のプロリン残基の例外はある)。天然の二量体ペプチドの構造を、等比体積の形態変換およびプロリンをグリシンで置換した後に得られる構造と比較することにより、図3に示される通り、天然ペプチドと形態変換した分子との構造的等価性が実証される。
【0070】
図3中、矢印の方向は、骨格のN−末端からC−末端への方向を指す。等比体積の形態変換後、構造は、さらなる位置の最適化および分子の動力学刺激を受けられる状態である。
【0071】
骨格反転の効果の1つには、アミノ酸配列の反転だけではなく、アミノ酸のα炭素原子の立体配置の反転もあり、これはLからDへ変化する。このことは、本発明では、アミノ酸配列において小文字で示される。トレオニンは2つのキラル中心を含有し、従って、側鎖のキラリティーを維持し、しかるべき等比体積体はD−アロ−Thrであり、ペプチド配列式中、小文字「t」で示される。
【0072】
得られる分子の構造は、以下の式:
【化12】
で表すことができる。
【0073】
上記の工程に加えて、C−末端カルボキシル基はアミノ基と入れ替え、N−末端アミノ基はカルボキシル基と入れ替える。
【0074】
概して、式TYSCHFGPLTWVCKPQのL−ペプチドは、等比体積の形態変換によって、式qGkcvwtlGGfhcsytで表すことができるD−ペプチドに形態変換される。
【0075】
この式は、例えば、グリシンまたはアラニン等のさらなる非結合アミノ酸の添加、並びに、N−およびC−末端のアミド化および/またはアセチル化による、ペプチドのC−およびN−末端の可能な修飾を含む。
【0076】
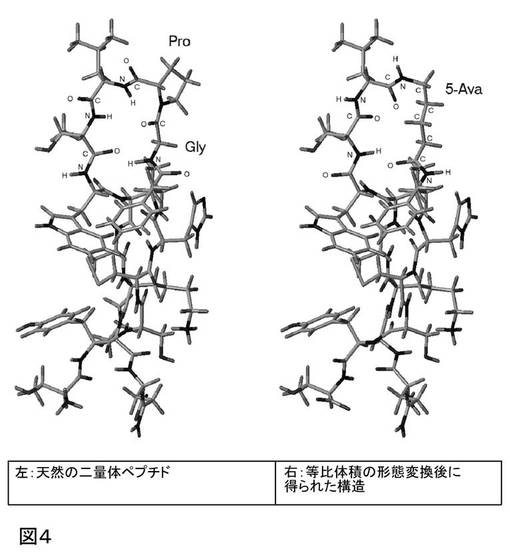
1.3 5−アミノバレリン酸による2個のアミノ酸の置換
1.2において示される、天然のペプチドの等比体積の形態変換のさらなる例として、1.2において述べた操作に加えて、2個のアミノ酸を、5−アミノバレリン酸(5−Ava)で置換する。概して、式TYSCHFGPLTWVCKPQのL−ペプチドは、等比体積の形態変換によって、式qGkcvwtl-(5-Ava)-fhcsytで表すことができるD−ペプチドに形態変換される。
このビルディング・ブロックの使用は、図4に示される構造で図示される。
【0077】
2.ビルディング・ブロックの合成
3−(2S−アリルオキシカルボニルアミノ−ピロリジン−1−イル)−3−オキソ−プロピオン酸:
グリシンのような構造的に不活性であるかまたは単にたわみやすい残基によるプロリンの置換に加えて、等比体積の形態変化によって得られたペプチド中の、レトロ−プロリンを模倣する適宜の合成ビルディング・ブロックによる前駆体ペプチドにおいて、L−プロリンによって誘導された特性を再構成することは必要であるか有利であろう。以下に、分子の合成を記載するが、この分子は、例えば、プロリンおよびその近接するグリシン残基:
【化13】
を置換した上述のエリスロポエチン模倣ペプチドにおいて、この種類のビルディング・ブロック機能を果たすことができる。
【0078】
前駆体1−(2−メトキシカルボニル−アセチル)−ピロリジン−2S−カルボン酸:
L−プロリン(3.45g, 30mmol)を30mlの1N NaOHに溶解し、30mlの水で希釈した。激しく攪拌しながら、3−クロロ−3−オキソプロピオン酸メチル(4.10g, 30mmol)および30mlの1N NaOHを0℃で同時に滴下した。溶液を0℃で1時間攪拌し、室温でさらに1時間攪拌した。混合物を飽和NaHSO4溶液でpH=1〜2に酸性化し、酢酸エチルで数回抽出した。合わせた有機層を乾燥し(Na2SO4)、溶媒を真空除去し、得られた粗油状物質をシリカカラムクロマトグラフィーで精製した。
【0079】
前駆体:3−(2S−アリルオキシカルボニルアミノ−ピロリジン−1−イル)−3−オキソ−3−プロピオン酸メチルエステル:
ジイソプロピルエチルアミン(0.97g, 7.5mmol)を、1−(2−メトキシカルボニル−アセチル)−ピロリジン−2S−カルボン酸(1.08g, 5.0mmol)のアセトン(15ml)および水(0.5ml)の溶液に0℃で滴下した。クロロ蟻酸エチル(0.76g, 7.0mmol)を滴下し、溶液を0℃で30分間攪拌した。4N NaN3水溶液(2.0ml, 8.0mmol)を添加し、溶液を0℃でさらに3時間攪拌した。溶液を水(20ml)およびEt2O(30ml)に分配し、水層をEt2O(4x30ml)で抽出した。合わせた有機層を乾燥し(MgSO4)、トルエン(20ml)を添加した。Et2Oを減圧下で除去し、アリルアルコール(1.37ml, 20mmol)を添加した。反応混合物を4時間、還流温度に加熱した。溶媒を減圧下で除去し、得られた油状物質をシリカカラムクロマトグラフィーで精製した。
【0080】
3−(2S−アリルオキシカルボニルアミノ−ピロリジン−1−イル)−3−オキソ−プロピオン酸:
3−(2S−アリルオキシカルボニルアミノ−ピロリジン−1−イル)−3−オキソ−プロピオン酸メチルエステル(1.35g, 5.0mmol)を2−ヘキサノン(6ml)に溶解した。乾燥臭化リチウム(0.48g, 5.5mmol)を添加し、反応混合物を1時間、還流温度に加熱した。冷却した混合物を、水(20ml)およびEt2O(30ml)に分配した。水層をEt2O(3x30ml)で抽出し、合わせた有機層を乾燥し(MgSO4)、溶媒を真空除去した。得られた固体を、酢酸エチル/ヘキサンで再結晶した。
引用:
1. ADDIN EN.REFLIST MHVvレゲンモーテル(Regenmortel)、Sミュラー(Muller):免疫原および診断薬としてのD−ペプチド(D-peptides as immunogens and diagnostic reagents):カレント・オピニオン・イン・バイオテクノロジー(Current Opinion in Biotechnology)、1998年、9巻:377-382頁。
2.PMフィッシャー(Fischer):立体化学および方向性ペプチド異性体の設計、合成および適用(The Design, Synthesis and Application of Stereochemical and Directional Peptide Isomers):批判的批評。カレント・プロテイン・アンド・ペプチド・サイエンス(Current Protein and Peptide Science)、2003年、4巻:339-356頁。
3.DTナイヤー(Nair)、KJカウル(Kaur)、Kシン(Singh)、Pムクヘリー(Mukherjee)、Dラジャゴパ(Rajagopa)、Aジョージ(George)、Vバル(Bal)、Sラス(Rath)、KVSラオ(Rao)、DMサルンケ(Salunke):天然のペプチド抗原の、対応するRetro-Inverso類似体による模倣は、それらの固有の構造および相互作用傾向に依存する(Mimicry of Native Peptide Antigens by the Corresponding Retro-Inverso Analogs Is Dependent on Their Intrinsic Structure and Interaction Propensities)1。ザ・ジャーナル・オブ・イムノロジー(The Journal of Immunology)、2003年、170巻:1362-1373頁。
4.MDフレッチャー(Fletcher)、MMキャンベル(Campbell):部分的に修飾されたRetro-inverso型ペプチド:開発、合成、および立体構造的性質(Partially Modified Retro-Inverso Peptides: Development, Synthesis, and Conformational Behavior)。ケミカル・レビューズ(Chemical Reviews)、1998年、98巻:763-796頁。
5.Kヴォグラー(Vogler)、Pランズ(Lanz)、Wレーギアー(Lergier)、Wハエフェリー(Haefely):[D−アミノ酸を用いたブラジキニン類似体の合成(全−D−ブラジキニンおよび全−D−レトロ−ブラジキニン)]([Synthesis of bradykinin analogs with D-amino acids (all-D-bradykinin and all-D-retro-bradykinin)])。Helv Chim Acta 1966年、49巻:390-403頁。
6.Tウィーランド(Wieland)、Bペンケ(Penke)、Cビル(Birr):[アンタマニド。XVI。全−D−レトロ−アンタマニドおよびD−tyr 6−全−D−レトロ−アンタマニド]([Antamanide. XVI. All-D-retro-antamanide and D-tyr 6 -all-D-retro-antamanide].)。Justus Liebigs Ann Chem 1972年、759巻:71-5頁.
7.Oリブナー(Livnah)、EAスツラ(Stura)、DLジョンソン(Johnson)、SAミドルトン(Middleton)、LSムルカヒー(Mulcahy)、NCライトン(Wrighton)、WJドウアー(Dower)、 LKジョリフィー(Jolliffe)、IAウィルソン(Wilson):ペプチド作動薬によるタンパク質ホルモンの機能的模倣(Functional mimicry of a protein hormone by a peptide agonist):EPO受容体複合体2.8A(the EPO receptor complex at 2.8 A)。サイエンス(Science)、1996年、273巻:464-471頁。
【図面の簡単な説明】
【0081】
【図1】本発明の方法の工程の説明図。
【図2】天然のペプチドと形態変換された分子との構造的等価性を示す。
【図3】天然の二量体ペプチドの構造について、天然ペプチドと形態変換した分子との構造的等価性を示す。
【図4】天然のペプチドの等比体積の形態変換のさらなる例を示す。
【技術分野】
【0001】
本発明は、retro-inverso型ペプチドの設計および合成分野に存する。本発明の主題は、慣用技術より容易かつ迅速に実施される、改良された等比体積の形態変換法である。また、本発明は、新規な方法によって得られるポリペプチドに関する。
【背景技術】
【0002】
多数の新規な生物工学的薬剤はタンパク質であり、体内で受容体と相互作用する。そのような成功した薬剤開発の例には、インターロイキン2およびGM−CSFまたはG−CSFのようなサイトカインがある。タンパク質薬剤の別の重要な種類は、ファージ提示法(Phage Display)等の全組換え系由来またはヒト化マウス抗体由来の抗体である。成功した薬剤ではあるが、薬剤としてのタンパク質は、多数の不利点を有することから、長期にわたりこれらの生物工学的薬剤に代わる、いわゆる「スモール・ドラッグ(small drugs)」の研究が継続されている。最も突出した不利点は、タンパク質薬剤がタンパク分解的に切断されるので、インビボでの半減期が短いこと、および、タンパク質薬剤に対する免疫反応の可能性が高いことである。誤って折りたたまれたタンパク質がわずかに寄与してタンパク質製剤が汚染されるか、またはTAGが精製または製造目的に使用されると、抗体またはアレルギー性免疫反応の不活化さえも起こり得る。抗体のヒト型化−通常、時間およびコスト集約的方法−は、これらのリスクをかなり縮小するが、廃することはできない。
【0003】
しかしながら、古典的薬剤の「スモール・ドラッグ」即ち有機小分子に至る取り組みは、リスクが高く、これまでのところあまり成功したアプローチではないことが判明した。スモール・ドラッグが複雑で大型のタンパク質受容体を標的にすることに成功した例はほとんど知られておらず、確立されていないであろう。
【0004】
有効な代替物の中で、ペプチドは際立った役割を演じる。基本的に、ペプチドは完全タンパク質に由来し、より大きなタンパク質の必須の結合ドメインを含むことができる。それらは、所与の標的に対し、天然タンパク質の作動薬または拮抗薬の両者として設計することができる。それらの大きさにより、通常、組換え産物の代替として化学的に合成し得る。さらに、小さなペプチドは、通常、大きなタンパク質より免疫原性が少ない。小さなペプチドは、経口適用の特別の生薬製剤においても使用されることがある。所望の薬剤適用に好適なペプチドを発見することは、同等のスモール・ドラッグを発見するよりもはるかに容易かつ迅速であることがわかっている。
【0005】
それでもやはり、免疫原性は完全には廃することはできず、−より重要なことには−、L−ペプチドは体液中で多数のプロテアーゼによって容易に分解され、完全タンパク質より半減期がさらに短い傾向がある。しかしながら、タンパク質の自然発生結合ドメイン由来のL−ペプチドを、最終的薬剤候補としてのL−ペプチドの非生分解性模倣剤を設計する出発点として使用することは可能である。このような目的に用いるという概念のうちで、非天然の、しばしばD−アミノ酸によって完全に構築されるペプチド等比体積物の設計は非常に魅力的である。このことは、合成および生薬製剤の同一の原理が、L−アミノ酸を基にした前駆体に関して、そのようなD−アミノ酸を基にしたペプチドに適用することができるという事実によるものである。これらのL−ペプチドの模倣剤は、なおもペプチドである。さらに、さらなる予期されない毒性が増加する危険性は、慣用の有機スモール・ドラッグよりも、L−ペプチドのペプチド性模倣剤のほうがはるかに小さい。
【0006】
D−アミノ酸を基にしたL−ペプチド模倣薬に至る1つの重要なアプローチは、retro-inverso型ペプチドの概念である。これらのペプチドは、アミノ酸配列の反転、続く、骨格のα−炭素原子での立体化学の反転(L−アミノ酸からD−アミノ酸へ)によって得られる。生物活性の相当の損失なく、所与のL−ペプチドに対応するretro-inverso型ペプチドを合成することは時に可能であるが、そのような「試行錯誤」のアプローチはリスクが高く、普通失敗に終わる。所与のペプチドおよび意図された立体化学の転回に関する基本的な構造的問題を明らかにするために、分子のモデリングを基にした合成のような、失敗のリスクを最小限にする戦略的計画が必要である。残念なことに、ペプチドのそのような分子モデリングは簡単ではなく、多くの時間、努力および熟練したスタッフを要する。
【0007】
ペプチドのretro-inverso型類似体の成功と失敗の両方について、多数の例が、引例[1、2]に概説されている。「鏡像ファージ提示法(mirror-image phage display)」のような組換え技術も、所与のL−ペプチドの、D−ペプチドを基にした等比体積体を得るために用いられるかもしれないが、実際に使用できるコンピュータに基づくアプローチと同様に、リスクがあり面倒である。
【0008】
今までのところ、新規なD−アミノ酸を基にした等比体積のペプチドを設計する標準的手法は、多大な時間を要し、天然ペプチドの受容体結合活性立体構造を模倣した立体構造に自動的にはならない。一般に、所与のL−ペプチドの立体構造的類似体としてretro-inverso型ペプチドを産生する、当業界のコンピュータに基づく手法は、以下の3工程を含む[3]。
【0009】
最初に、原型のペプチドの配列を反転させなければならない。
第2工程において、α−炭素原子における側鎖の立体化学をLからDに変えなければならない。
原型のペプチドとretro-inverso型ペプチドとの間で立体構造の類似性を達成するため、骨格のねじれ角度phiおよびpsiを、それぞれ対応する残基について入れ替えなければならない。
【0010】
これらの基本的な3工程では中間体構造となるだけであり、この構造を構造最適化および分子動力学に対応させる前に、さらなる形態変換を実施しなければならない。
【0011】
この全方法には、主な不利点が3点ある:
第一に、この方法は、複雑で時間がかかり、高価である。これは、工程1および工程2が原型のL−ペプチドの所与の構造を完全に破壊するという事実によるものである。配列の反転およびD−アミノ酸への変化により、アミノ酸の側鎖の配置が再構成される。これら側鎖は、工程3および工程4において「L−ペプチドに似た」立体配置へ再構成されなければならない。実際に、このアプローチは、出発点としてL−ペプチドの配列を使用しているだけであり、構造的情報(側鎖の位置および空間的定位)はまったく使用せず、L−ペプチドがあれば使用できる。このように、全体の方法は非常に時間がかかる「最初から」のモデリング方法である。
【0012】
第二に、この方法は、分子モデリングソフトにおいて、ほとんど自動化または実行できない。
【0013】
第三に、天然のペプチドと、側鎖の完全な重ねあわせはなお達成できない。この問題は、retro-inverso型ペプチドが、ペプチド−受容体複合体から設計して、受容体−結合立体構造を有する、D−アミノ酸を基にした等比体積のペプチドを得なければならない場合、より難しくなるであろう。
【0014】
全ての古典的なretro-inversoアプローチの主要な問題は、構造的情報が全く保存されず、失敗のハイ・リスクが、L−ペプチドから構造的に類似するD−ペプチドへ、「試行錯誤」で合成されること、および/または、「破壊的/再建的」に形態変換されることの、両者に関係することである。
【0015】
本発明は、簡単な方法を説明することによって、当業界のこれらの不利点を克服し、所与のL−ペプチドに関する構造的情報を保つ。さらに、本発明は、所与のL−ペプチド前駆体の意図された立体化学的転回の間に発生する、典型的な構造および配列に関連する問題の解決策を提示する。
【発明の開示】
【0016】
概説
本発明は、等比体積の形態変換、即ち、対応する天然L−ペプチド前駆体の等比体積体として働くD−アミノ酸を基にしたペプチドの、構造に基づく合理的設計を可能にする、新規かつ迅速な方法に関する。上述した慣用法と対照的に、本発明は、所与のL−ペプチド配列に関する構造的情報(例えば、結晶学的データ、NMRデータ)を使用することができ、通常、このL−ペプチドがその標的結合部位にドッキングされることを示す。
【0017】
本発明の主題は、請求項1〜29のいずれかの、方法、ポリペプチド、化合物、使用および医薬製剤である。本発明による合理的設計の中心思想は、全方法手順にわたり、アミノ酸の側鎖の空間的定位を保持することである。従って、phiおよびpsi2面角の交換、および意図された分子の完全なデノボ構造の必要がない。本発明による方法手順は、前駆体ペプチドの骨格構造を変えるだけでこれを達成し、一方、完全なアミノ酸部分の交換を回避する(L−アミノ酸のD−アミノ酸による正式の置換なし)。骨格のペプチド結合が完全な平面構造であるので、このアプローチは、ジスルフィド架橋のような共有結合側鎖修飾、例えばらせんまたはβ−面のような、らせんの制約または他の構造的特性を含む、側鎖の定位または構造的特性を変化させない。これらの操作工程のワークフローは、標準的モデリング・ソフトウェアで容易に自動化することができる。また本発明はあらゆるソフトウェアまたはアルゴリズムにも及び、それらにより本発明の工程がコンピュータで自動化される。特に、本発明の方法は、図1に示される工程に従って実施される。
【0018】
等比体積の形態変換により、L−ペプチド前駆体の意図されたD−類似体の構造的データへ高速でアクセスされる。このことにより、二つの主要な問題の即時の評価が可能になり、その前駆体と立体化学的に適合しない、新たに設計された分子を提供することができる:
即ち、前駆体の配列中のL−プロリンの存在、および、ペプチドの末端の帯電である。
【0019】
要すれば、ペプチド末端の帯電の修飾のための合成的解決策が入手可能であるが、ここで開示される本発明は、プロリンの問題のための多数の合成的解決策を提供し、それは等比体積の形態変換の間、プロリンの構造的に正確な置換のために使用することができる。
【0020】
従って、等比体積の形態変換は、L−ペプチド前駆体−ペプチド含有プロリンであっても−をそれぞれのD−ペプチド類似体へ、立体化学的に映すための、高速で魅力的な解決策を提供する。
【0021】
また、本発明は、これらの新たに設計された化合物の合成に関し、かつ、本発明によるビルディング・ブロックおよびD−ペプチドにおける末端の修飾を含有するペプチドに関する。一般に、本発明は、出発点となるL−ペプチド構造に関する充分な情報が入手可能ならば、殆ど全てのペプチド構造に適応できる解決策を提供する。
【0022】
ここで開示される本発明の原理が、例えば配列情報単独に基づく詳細な構造的データを利用しなくとも使用できることは、当業者には明白である。しかしながら、このことは、合成研究所における「試行錯誤」のアプローチが、活性があり構造的に有効なD−アミノ酸を基にした物質を同定する、好適な、例えばインビトロの試験システムと関連して使用されなければならないことを暗示する。本発明の原理の使用を通じて、このアプローチは、以下に開示する通りの本発明の完全な構造を基にした実行と、全ての優位点−特に高速で先頭を検出する好機−を共有しない。しかしながら、発明の原理のこのような部分的実行もまた、本発明の方法手順の範囲に包含されることは明白である。
【0023】
詳細な説明
本発明は、工程a〜cを含む、1つ以上(全部まで)のペプチド結合の反転による、天然のペプチドの構造の座標を基にした、ペプチド等比体積体またはペプチド様物質を設計する方法を開示する:
所与のL−ペプチドの構造データを、所与のペプチドの立体構造の構造データを処理することが可能なソフトウェアをロードした、好適なコンピュータ装置に取り込む工程、
天然のカルボニル(CO)骨格基の原子をアミド(NH)基で置換する工程、
天然のアミド(NH)骨格基の原子をカルボニル(CO)基で置換する工程。
【0024】
工程a〜cは、所与のソフトウェア・ユーティリティを使用して熟練したスタッフによって手作業で実施することができ、または、コンピュータ/ソフトウェアユニットの適宜のプログラミングによって自動化することができる。複雑なペプチド構造でも、工程a〜cは数時間以内で終わることができる。これらの工程は、中間体構造において終了するであろうが、この構造は、全ての側鎖を正確な空間的定位に含有し、D−ペプチド構造に関して既に反転した骨格を示す。ペプチドの末端部は、まだ反転されない。従って、初期の生成物の構造は、末端未反転骨格を有する(以下の例を参照、2次元の略図に変形する)。
【0025】
【化1】
【0026】
本発明のさらなる様相において、C−末端および/またはN−末端基は、非末端骨格の反転に加えて、修飾することができる。C−末端カルボキシル基は、アミノ基と入れ換えることができ、および/または、N−末端アミノ基はカルボキシル基と入れ換えることができる:
【0027】
【化2】
【0028】
これらの付加的工程により、3つのさらなる分子となり、それらは、初期の生成物の構造と共に、本発明による全体の設計の最適化のための出発点となる(以下参照)。これらの場合のいくつかにおいて、gem−ジアミノアルキルおよびC−2−置換マロニル残基は、末端基として取り込まれる。このようなビルディング・ブロックの合成は、細部にわたり精査されている。[4]
【0029】
これまでのところ、この方法は、等比体積体の設計の古典的な方法と比較して、3つの主要な優位点を既に有している:
容易かつ迅速に実施される。
分子モデリング・プログラムにおいて、容易に自動化または実行することができる。
【0030】
得られる構造の側鎖の配置および立体構造は、さらなる最適化および動力学の出発点であるが、天然のペプチドと比較して変化しない。このことは、この方法が天然のペプチドと同一の側鎖の立体構造を維持する等比体積体を生成するため、特に受容体結合ペプチドの設計に非常に有益である。従って、得られる初期の生成物の構造は等比体積体であり、側鎖の受容体タンパク質との相互作用は元のL−ペプチドと同等である。
【0031】
以下の2次元の図は、等比体積の形態変換法のこれらの優位点を示す:殆ど操作を要せず、側鎖の配置は決して変わらない。
【0032】
【化3】
【0033】
また本発明は、上述の方法によって得られるD−アミノ酸を基にしたポリペプチドを提供する。この方法がL−ペプチドの等比体積構造を生成する一般的な方法であるという事実によって、本発明はまた、プロリン含有ペプチドの等比体積化合物も提供する。
【0034】
従って、本発明はまた、天然のペプチドが1つ以上のプロリン残基を含有する場合に重要なビルディング・ブロックを提供する。プロリンは、プロリン含有retro-inverso型ペプチドの例が生物学的活性を保持していたとしても[6]、しばしば、慣用のretro-inversoアプローチ[5]と適合性がないと見なされる。プロリンは、その側鎖がα−アミノ基に拘束され、従って骨格に戻る、唯一の天然の環状アミノ酸である。この特性は、等比体積の形態変換にとって特別なことを明確するが、これは骨格の反転に基づくものであり、従ってプロリンの場合−アミノ酸の中での例外として−等比体積の形態変換の間、立体化学的ひずみをもたらす。さらに、この制約は、プロリンを含まないペプチドに見られるものと異なる、制限された骨格の2面角を決定する。標準的なretro-inverso型ペプチドにおいて、プロリンの方向の逆転が、天然のペプチドと比較して、ピロリジン環のトポケミカルな置換をもたらす。またこのことは、ここで提示される等比体積の形態変換法についても当てはまる。しかしながら、上述した通りの等比体積の形態変換によって、非末端的に骨格が反転したペプチドを、例えば、元のL−ペプチド構造で規定される構造的要求との見込まれる非適合性について分子動力学によって試験することができる。従って、合成およびインビトロ試験以前に、プロリンの問題が実際に所与の構造の重要な様相であろうとなかろうと、理論的解釈を展開することができる。骨格の反転が受け入れ難いほどにプロリン位置での構造をゆがめる場合、等比体積の形態変換法によって、この特有であるが頻発する問題:β−回転またはβ−ヘアピンのような構造において、プロリンは、等比体積の形態変換後、グリシンによってしばしば置換され得る:について、多数の解決策が提供される:
【0035】
【化4】
【0036】
本発明のさらなる様相において、等比体積の形態変換に加えて、二つのアミノ酸ユニット(プロリンおよび直接隣接する残基)は、5−アミノバレリン酸またはその誘導体のような、1個のビルディング・ブロックで置換することができる:
【0037】
【化5】
【0038】
一般に、等比体積の形態変換後、2つのアミノ酸(プロリンおよび直接隣接する残基)は、一般式
...-(CO)-X1-X2-X3-X4-NH-…
で表されるビルディング・ブロックで置換することができる:
式中、X1、X2、X3、およびX4は独立してCH2、(C=O)、NH、NR、O、(CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基である。以下の図は、この一般式によって表されるビルディング・ブロックの幾つかの使用例を示す:
【0039】
【化6】
【0040】
また本発明は、等比体積の形態変換の後、プロリン残基の立体構造を模倣し、かつ、プロリンおよびその近接するアミノ酸残基を置換する、環状のビルディング・ブロックを提供する:
【0041】
【化7】
【0042】
このようなビルディング・ブロックの使用は、グリシンまたは5−アミノバレリン酸誘導体のような延長したグリシン等価構造が所与のペプチドの立体特性を安定化するのに充分でない場合に、有用であり、かつ、必要であろう。標的構造への「誘導適合」が、プロリン特異的立体構造の適切な再構築がないペプチドに問題になる場合に、このことは特に当てはまる。プロリンおよび近接するグリシンの置換のためにここで示したもののような、プロリン置換ビルディング・ブロックは、以下の一般式に従って構築することができる:
【0043】
本発明のさらなる様相において、等比体積の形態変換後、2つの近接するアミノ酸(ひとつはプロリンであった)は、一般式
【化8】
で表されるビルディング・ブロックによって置換することができる。
【0044】
式中、X1、X2およびX3は独立してCH2、(C=O)、O、S、NH、NR、 (CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基である。以下の図は、この一般式で表されるビルディング・ブロックのいくつかの例を示す:
【0045】
【化9】
【0046】
このようなビルディング・ブロックの使用は、グリシンまたは5−アミノバレリン酸誘導体のような延長したグリシン等価構造が所与のペプチドの立体特性を安定化するのに充分でない場合に、有用であり、かつ、必要であろう。標的構造への「誘導適合」が、プロリン特異的立体構造の適切な再構築がないペプチドに問題になる場合に、このことは特に当てはまる。
【0047】
本発明のさらなる様相において、等比体積の形態変換後、2つのアミノ酸は一般式
【化10】
で表されるビルディング・ブロックによって置換することができる。
【0048】
式中、X1、X2、X3およびX4は独立してCH2、(C=O)、O、S、NH、NR、 (CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基である。以下の図は、この一般式で表されるビルディング・ブロックのいくつかの例を示す:
【0049】
【化11】
【0050】
このようなビルディング・ブロックの使用は、グリシンまたは5−アミノバレリン酸誘導体のような延長したグリシン等価構造が所与のペプチドの立体特性を安定化するのに充分でない場合に、有用であり、かつ、必要であろう。標的構造への「誘導適合」が、プロリン特異的立体構造の適切な再構築がないペプチドに問題になる場合に、このことは特に当てはまる。ここで示される一般式を基にした分子は、以前に示したピロリジンを基にした構造の代わりに使用することができる。
【0051】
上記で概説した通りのビルディング・ブロックおよびビルディング・ブロックの使用は、本発明によるプロリンを含むポリペプチドの等比体積の形態変換において有用である。しかしながら、それらビルディング・ブロックは、D−ペプチドおよびretro-inverso型ペプチドの設計および製造のための慣用技術においても有用である。
【0052】
本発明において記載されるアミノ酸は、自然発生のL−立体異性体のものであってもよく、エナンチオマーのD体であってもよい。一文字の記号は、容認されている標準的なポリペプチド命名法を参照するが、D−またはL−アミノ酸を択一的に意味することができる。小文字はDアミノ酸に言及するものである。
【0053】
記号 アミノ酸
A L−アラニンまたはD−アラニン
V L−バリンまたはD−バリン
L L−ロイシンまたはD−ロイシン
I L−イソロイシンまたはD−イソロイシンまたはD−アロ−イソロイシン
M L−メチオニンまたはD−メチオニン
F L−フェニルアラニンまたはD−フェニルアラニン
Y L−チロシンまたはD−チロシン
W L−トリプトファンまたはD−トリプトファン
H L−ヒスチジンまたはD−ヒスチジン
S L−セリンまたはD−セリン
T L−トレオニンまたはD−トレオニンまたはD−アロ−トレオニン
C L−システインまたはD−システイン
N L−アスパラギンまたはD−アスパラギン
Q L−グルタミンまたはD−グルタミン
D L−アスパラギン酸またはD−アスパラギン酸
E L−グルタミン酸またはD−グルタミン酸
K L−リジンまたはD−リジン
R L−アルギニンまたはD−アルギニン
P L−プロリンまたはD−プロリン
G グリシン
a D−アラニン
v D−バリン
l D−ロイシン
i D−イソロイシンまたはD−アロ−イソロイシン
m D−メチオニン
f D−フェニルアラニン
y D−チロシン
w D−トリプトファン
h D−ヒスチジン
s D−セリン
t D−トレオニンまたはD−アロ−トレオニン
c D−システイン
n D−アスパラギン
q D−グルタミン
d D−アスパラギン酸
e D−グルタミン酸
k D−リジン
r D−アルギニン
p D−プロリン
【実施例】
【0054】
1.ペプチドの等比体積の形態変換
1.1 インターロイキン2受容体のβ鎖に結合する、プロリンを含まない、らせんペプチドの等比体積の形態変換
以下の実施例は、インターロイキン2受容体のβ鎖に結合する、らせんペプチドの形態変換を実証する。
【0055】
このペプチド−インターロイキン2(IL−2)に類似−は、高い親和性で、IL−2特異的受容体に結合する能力を有する。IL−2は、腫瘍治療において使用されるサイトカインである。IL−2は特異的受容体(IL−2R)に結合し、それによってIL−2特異的細胞内シグナルの引き金を引く。
【0056】
IL−2は、TおよびBリンパ球の成長に対する刺激効果を有し、細胞傷害性および細胞溶解性のNK細胞を活性化する。従って、IL−2は、免疫応答の調節において、中心的意義を有する。このように、IL−2は、腫瘍および炎症反応に対する免疫応答において、基本的に重要である。IL−2による腫瘍防御のために重要なメカニズムの1つは、LAK(「リンフォカイン活性化キラー細胞」)の誘導であると思われる。これらの細胞は、腫瘍細胞を破壊する能力がある。
【0057】
天然ペプチドの組み合わせを有する構造ファイルは、式STKKTQLQLEHLLLDLQMILNGINNYを有するらせんペプチドと、インターロイキン2受容体のβおよびγ鎖との複合体構造を含有する。このらせんペプチドは、等比体積の形態変換の実施の、理想的な例を提供する:
【0058】
受容体に結合した立体構造中のホモ二量体ペプチドの組み合わせを、分子モデリング・ソフトウェアに取り込む。
天然のカルボニル(CO)骨格基の原子は、アミド(NH)基で置換される。
天然のアミド(NH)骨格基の原子は、カルボニル(CO)基で置換される。
【0059】
等比体積の形態変換の方法手順では、修飾は骨格にのみ起こるので、側鎖原子の立体配置は変わらない。従って、この方法は、D−アミノ酸を基にした受容体結合ペプチドの設計および最適化のための出発立体構造を産生する、良好な方法手順である。
【0060】
天然のらせんペプチドの構造を、等比体積の形態変換および位置最適化の後に得られる構造と比較することによって、図2に示す通り、天然のペプチドと形態変換された分子との構造的等価性が実証される。
【0061】
骨格反転の効果の1つには、アミノ酸配列の反転だけではなく、アミノ酸のα炭素原子の立体配置の反転もあり、これはLからDへ変化する。このことは、本発明では、アミノ酸配列において小文字で示される。トレオニンは2つのキラル中心を含有し、従って、側鎖のキラリティーを維持し、しかるべき等比体積体はD−アロ−Thrであり、ペプチド配列式中、小文字「t」で示される。
【0062】
概して、式STKKTQLQLEHLLLDLQMILNGINNYのL−ペプチドは、等比体積の形態変換によって、式ynnignlimqldlllhelqlqtkktsで表すことができるDペプチドに形態変換される。本発明は、このペプチドの等比体積の、受容体にドッキングされた構造の組み合わせを産生する、迅速な方法を開示する。
【0063】
得られるペプチドは、完全にD−アミノ酸より成る。このことから、天然のL−ペプチドと比較して、生物学的半減期がより長くなる。D−アミノ酸を基にしたペプチドは、タンパク質分解酵素にはより安定である。従って、等比体積的に形態変換したペプチド模倣体は、医薬品として、L−ペプチドより良好な特性を有するように設計される。
【0064】
D−ペプチドは、免疫系疾患、例えば、炎症および関節炎の進行、または全てのタイプおよび起源の免疫不全症候群;細胞の進展する増殖に関係する疾患、例えば、癌腫、肉腫、リンパ腫および白血病等の形態の癌腫症;または感染過程の治療に好適である。
【0065】
1.2 エリスロポエチン受容体に結合するペプチドの等比体積の形態変換
以下の実施例は、エリスロポエチン受容体に結合するペプチドの形態変換を実証する。
【0066】
天然の構造を代表する組み合わせは、プロテイン・データ・バンク(Protein Data Bank)で公的に入手可能である(PDBコード:1EBP)。このファイルは、式TYSCHFGPLTWVCKPQを有する二量体のアゴニスト・ペプチドと、二量体のエリスロポエチン受容体との複合体の構造を含む[7]。この構造的に充分立証されたペプチドは、等比体積の形態変換の実施の理想的な例となる:
【0067】
受容体に結合した立体構造におけるホモ二量体ペプチドの組み合わせは、シビル(Sybyl)モデリング・ソフトウェアの適宜の分子に組み込まれる。
天然のカルボニル(CO)骨格基の原子は、アミド(NH)基で置換される。
天然のアミド(NH)骨格基の原子は、カルボニル(CO)基で置換される。
【0068】
ペプチド配列が2つのプロリン残基を包含するので、プロリン位置での構造的ひずみが、等比体積の形態変換によって得られる初期の構造において発生する。しかしながら、「誘導適合」位置のペプチドの第二の構造が多数の他の分子の相互作用によって安定化され、この構造はプロリン特異的特性によって引き起こされる特異的な立体構造を場合によっては必要としないことも明らかである。従って、プロリンは−誘導適合立体構造を支持するが−、そのために必須ではなく、最も単純な場合、非常にたわみやすいグリシンで置換することができる。
【0069】
等比体積の形態変換の方法手順では、修飾が骨格にのみ起こるので、側鎖原子の立体構造は変化しない(天然のプロリン残基の例外はある)。天然の二量体ペプチドの構造を、等比体積の形態変換およびプロリンをグリシンで置換した後に得られる構造と比較することにより、図3に示される通り、天然ペプチドと形態変換した分子との構造的等価性が実証される。
【0070】
図3中、矢印の方向は、骨格のN−末端からC−末端への方向を指す。等比体積の形態変換後、構造は、さらなる位置の最適化および分子の動力学刺激を受けられる状態である。
【0071】
骨格反転の効果の1つには、アミノ酸配列の反転だけではなく、アミノ酸のα炭素原子の立体配置の反転もあり、これはLからDへ変化する。このことは、本発明では、アミノ酸配列において小文字で示される。トレオニンは2つのキラル中心を含有し、従って、側鎖のキラリティーを維持し、しかるべき等比体積体はD−アロ−Thrであり、ペプチド配列式中、小文字「t」で示される。
【0072】
得られる分子の構造は、以下の式:
【化12】
で表すことができる。
【0073】
上記の工程に加えて、C−末端カルボキシル基はアミノ基と入れ替え、N−末端アミノ基はカルボキシル基と入れ替える。
【0074】
概して、式TYSCHFGPLTWVCKPQのL−ペプチドは、等比体積の形態変換によって、式qGkcvwtlGGfhcsytで表すことができるD−ペプチドに形態変換される。
【0075】
この式は、例えば、グリシンまたはアラニン等のさらなる非結合アミノ酸の添加、並びに、N−およびC−末端のアミド化および/またはアセチル化による、ペプチドのC−およびN−末端の可能な修飾を含む。
【0076】
1.3 5−アミノバレリン酸による2個のアミノ酸の置換
1.2において示される、天然のペプチドの等比体積の形態変換のさらなる例として、1.2において述べた操作に加えて、2個のアミノ酸を、5−アミノバレリン酸(5−Ava)で置換する。概して、式TYSCHFGPLTWVCKPQのL−ペプチドは、等比体積の形態変換によって、式qGkcvwtl-(5-Ava)-fhcsytで表すことができるD−ペプチドに形態変換される。
このビルディング・ブロックの使用は、図4に示される構造で図示される。
【0077】
2.ビルディング・ブロックの合成
3−(2S−アリルオキシカルボニルアミノ−ピロリジン−1−イル)−3−オキソ−プロピオン酸:
グリシンのような構造的に不活性であるかまたは単にたわみやすい残基によるプロリンの置換に加えて、等比体積の形態変化によって得られたペプチド中の、レトロ−プロリンを模倣する適宜の合成ビルディング・ブロックによる前駆体ペプチドにおいて、L−プロリンによって誘導された特性を再構成することは必要であるか有利であろう。以下に、分子の合成を記載するが、この分子は、例えば、プロリンおよびその近接するグリシン残基:
【化13】
を置換した上述のエリスロポエチン模倣ペプチドにおいて、この種類のビルディング・ブロック機能を果たすことができる。
【0078】
前駆体1−(2−メトキシカルボニル−アセチル)−ピロリジン−2S−カルボン酸:
L−プロリン(3.45g, 30mmol)を30mlの1N NaOHに溶解し、30mlの水で希釈した。激しく攪拌しながら、3−クロロ−3−オキソプロピオン酸メチル(4.10g, 30mmol)および30mlの1N NaOHを0℃で同時に滴下した。溶液を0℃で1時間攪拌し、室温でさらに1時間攪拌した。混合物を飽和NaHSO4溶液でpH=1〜2に酸性化し、酢酸エチルで数回抽出した。合わせた有機層を乾燥し(Na2SO4)、溶媒を真空除去し、得られた粗油状物質をシリカカラムクロマトグラフィーで精製した。
【0079】
前駆体:3−(2S−アリルオキシカルボニルアミノ−ピロリジン−1−イル)−3−オキソ−3−プロピオン酸メチルエステル:
ジイソプロピルエチルアミン(0.97g, 7.5mmol)を、1−(2−メトキシカルボニル−アセチル)−ピロリジン−2S−カルボン酸(1.08g, 5.0mmol)のアセトン(15ml)および水(0.5ml)の溶液に0℃で滴下した。クロロ蟻酸エチル(0.76g, 7.0mmol)を滴下し、溶液を0℃で30分間攪拌した。4N NaN3水溶液(2.0ml, 8.0mmol)を添加し、溶液を0℃でさらに3時間攪拌した。溶液を水(20ml)およびEt2O(30ml)に分配し、水層をEt2O(4x30ml)で抽出した。合わせた有機層を乾燥し(MgSO4)、トルエン(20ml)を添加した。Et2Oを減圧下で除去し、アリルアルコール(1.37ml, 20mmol)を添加した。反応混合物を4時間、還流温度に加熱した。溶媒を減圧下で除去し、得られた油状物質をシリカカラムクロマトグラフィーで精製した。
【0080】
3−(2S−アリルオキシカルボニルアミノ−ピロリジン−1−イル)−3−オキソ−プロピオン酸:
3−(2S−アリルオキシカルボニルアミノ−ピロリジン−1−イル)−3−オキソ−プロピオン酸メチルエステル(1.35g, 5.0mmol)を2−ヘキサノン(6ml)に溶解した。乾燥臭化リチウム(0.48g, 5.5mmol)を添加し、反応混合物を1時間、還流温度に加熱した。冷却した混合物を、水(20ml)およびEt2O(30ml)に分配した。水層をEt2O(3x30ml)で抽出し、合わせた有機層を乾燥し(MgSO4)、溶媒を真空除去した。得られた固体を、酢酸エチル/ヘキサンで再結晶した。
引用:
1. ADDIN EN.REFLIST MHVvレゲンモーテル(Regenmortel)、Sミュラー(Muller):免疫原および診断薬としてのD−ペプチド(D-peptides as immunogens and diagnostic reagents):カレント・オピニオン・イン・バイオテクノロジー(Current Opinion in Biotechnology)、1998年、9巻:377-382頁。
2.PMフィッシャー(Fischer):立体化学および方向性ペプチド異性体の設計、合成および適用(The Design, Synthesis and Application of Stereochemical and Directional Peptide Isomers):批判的批評。カレント・プロテイン・アンド・ペプチド・サイエンス(Current Protein and Peptide Science)、2003年、4巻:339-356頁。
3.DTナイヤー(Nair)、KJカウル(Kaur)、Kシン(Singh)、Pムクヘリー(Mukherjee)、Dラジャゴパ(Rajagopa)、Aジョージ(George)、Vバル(Bal)、Sラス(Rath)、KVSラオ(Rao)、DMサルンケ(Salunke):天然のペプチド抗原の、対応するRetro-Inverso類似体による模倣は、それらの固有の構造および相互作用傾向に依存する(Mimicry of Native Peptide Antigens by the Corresponding Retro-Inverso Analogs Is Dependent on Their Intrinsic Structure and Interaction Propensities)1。ザ・ジャーナル・オブ・イムノロジー(The Journal of Immunology)、2003年、170巻:1362-1373頁。
4.MDフレッチャー(Fletcher)、MMキャンベル(Campbell):部分的に修飾されたRetro-inverso型ペプチド:開発、合成、および立体構造的性質(Partially Modified Retro-Inverso Peptides: Development, Synthesis, and Conformational Behavior)。ケミカル・レビューズ(Chemical Reviews)、1998年、98巻:763-796頁。
5.Kヴォグラー(Vogler)、Pランズ(Lanz)、Wレーギアー(Lergier)、Wハエフェリー(Haefely):[D−アミノ酸を用いたブラジキニン類似体の合成(全−D−ブラジキニンおよび全−D−レトロ−ブラジキニン)]([Synthesis of bradykinin analogs with D-amino acids (all-D-bradykinin and all-D-retro-bradykinin)])。Helv Chim Acta 1966年、49巻:390-403頁。
6.Tウィーランド(Wieland)、Bペンケ(Penke)、Cビル(Birr):[アンタマニド。XVI。全−D−レトロ−アンタマニドおよびD−tyr 6−全−D−レトロ−アンタマニド]([Antamanide. XVI. All-D-retro-antamanide and D-tyr 6 -all-D-retro-antamanide].)。Justus Liebigs Ann Chem 1972年、759巻:71-5頁.
7.Oリブナー(Livnah)、EAスツラ(Stura)、DLジョンソン(Johnson)、SAミドルトン(Middleton)、LSムルカヒー(Mulcahy)、NCライトン(Wrighton)、WJドウアー(Dower)、 LKジョリフィー(Jolliffe)、IAウィルソン(Wilson):ペプチド作動薬によるタンパク質ホルモンの機能的模倣(Functional mimicry of a protein hormone by a peptide agonist):EPO受容体複合体2.8A(the EPO receptor complex at 2.8 A)。サイエンス(Science)、1996年、273巻:464-471頁。
【図面の簡単な説明】
【0081】
【図1】本発明の方法の工程の説明図。
【図2】天然のペプチドと形態変換された分子との構造的等価性を示す。
【図3】天然の二量体ペプチドの構造について、天然ペプチドと形態変換した分子との構造的等価性を示す。
【図4】天然のペプチドの等比体積の形態変換のさらなる例を示す。
【特許請求の範囲】
【請求項1】
N−末端アミノ基または置換アミノ基、C−末端カルボキシ基またはカルボキシ誘導体、骨格およびL−アミノ酸側鎖を有するL−配置の前駆体の三次元座標および配列情報から、少なくとも部分的にD−アミノ酸を含有するポリペプチドの等比体積構造を生成する方法であって、
−少なくとも部分的に、骨格のCO基をNH基に、およびその逆に、置換する工程、
−同時に、前駆体のL−アミノ酸側鎖、N−末端アミノ基または置換アミノ基およびC−末端カルボキシ基またはカルボキシ誘導体の三次元座標を一定に保持する工程
を含む方法。
【請求項2】
請求項1に記載の方法であって、
−少なくとも部分的に、骨格のCO基をNH基に、およびその逆に、置換する工程、
−同時に、前駆体のL−アミノ酸側鎖の三次元座標を一定に保持し、かつ、N−末端アミノ基または置換アミノ基をカルボキシ基またはカルボキシ誘導体で置換する工程および/またはC−末端カルボキシ基またはカルボキシ誘導体をアミノ基または置換アミノ基で置換する工程
を含む方法。
【請求項3】
請求項1〜2に記載の方法であって、前駆体の骨格の全てのCO基をNH基に、およびその逆に置換する方法。
【請求項4】
請求項1〜3に記載の方法であって、
−前駆体の構造および配列中のプロリン残基またはプロリン残基およびその隣接する残基を、新たに構成された骨格中のプロリンまたはプロリンおよびその直接隣接する残基の立体構造の特性を模倣したビルディング・ブロックとしての有機分子で、少なくとも部分的に置換する
ことを特徴とする方法。
【請求項5】
図1に記載の工程を含む、請求項1〜4に記載の方法。
【請求項6】
コンピュータ装置で実施される、請求項1〜5に記載の方法。
【請求項7】
少なくとも1つのD−アミノ酸および/または合成アミノ酸を含むポリペプチドを生成する方法であって、請求項1〜6のいずれかの方法によって等比体積構造を得る工程、および該等比体積構造のポリペプチドを合成する工程を含む方法。
【請求項8】
請求項7または8の方法であって、ポリペプチドがD−アミノ酸および/または合成アミノ酸よりなる方法。
【請求項9】
請求項1〜8に記載の方法によって得られるポリペプチド。
【請求項10】
請求項9に記載のポリペプチドであって、100残基未満、特に60以下、または40以下の残基で、少なくとも7残基を有する、ポリペプチド。
【請求項11】
請求項9〜10に記載のポリペプチドであって、式1:
【化1】
に図示され非排他的に例示される通り、骨格のCOがNH基で、およびその逆に置換され、同時にC−末端カルボキシおよびN−末端アミノ基が変化していないことを特徴とする、ポリペプチド。
【請求項12】
請求項9〜11に記載のポリペプチドであって、式2:
【化2】
に図示され非排他的に例示される通り、末端アミノ基がカルボキシ基で置換され、および/または、末端カルボキシ基がアミノ基で置換されたか、あるいは、N−およびC−末端が互いに交換された、ポリペプチド。
【請求項13】
請求項9〜12に記載のポリペプチドであって、前駆体の少なくとも1つのプロリン残基がグリシンで置換された、ポリペプチド。
【請求項14】
請求項9〜13に記載のポリペプチドであって、5−アミノバレリン酸のビルディング・ブロックとしての使用を示す式4、およびビルディング・ブロックとしての5−アミノバレリン酸の、例としての非排他的誘導体の使用を示す式5によって、非排他的に図示される通り、5−アミノバレリン酸および一般式:…-(CO)-X1-X2-X3-X4-NH-…で表されるその誘導体を用いて、前駆体配列中のプロリン残基およびその隣接する残基を置換した、ポリペプチド:式中、X1、X2、X3、およびX4は独立してCH2、(C=O)、NH、NR、O、(CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基である。
式4:
【化3】
式5:
【化4】
【請求項15】
式7を有する化合物、特に、3−(2S−アリルオキシカルボニル−アミノ−ピロリジン−1−イル)−3−オキソ−プロピオン酸(式6):式中、X1、X2およびX3は独立してCH2、(C=O)、O、S、NH、NR、(CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基であり;一般式で記載される分子は、式8において非排他的に図示される。
式6:
【化5】
式7:
【化6】
式8:
【化7】
【請求項16】
請求項9〜15に記載のポリペプチドであって、式9:
【化8】
において非排他的に図示される通り、請求項15に記載のビルディング・ブロックを使用して、少なくとも1つのプロリン残基およびその直接隣接する残基を置換した、ポリペプチド。
【請求項17】
式10の化合物:式中、X1、X2、X3およびX4はは独立してCH2、(C=O)、O、S、NH、NR、(CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基であり、それらにより各分子の例は非排他的に式11に示される。
式10
【化9】
式11
【化10】
【請求項18】
請求項9〜14、16に記載のポリペプチドであって、請求項15に記載のビルディング・ブロックを使用して、少なくとも1つのプロリン残基およびその直接隣接する残基を置換した、ポリペプチド。
【請求項19】
請求項9によって得られ、かつ、プロリン、またはプロリンおよびその直接隣接する残基の置換基として請求項12〜16に特定される少なくとも1つのまたは自由な組み合わせのビルディング・ブロックを用いた、ポリペプチド。
【請求項20】
請求項9〜19に記載のポリペプチドであって、N−末端のアセチル化またはC−末端のアミド化によって、または、N−末端のアセチル化およびC−末端のアミド化によって修飾された、ポリペプチド。
【請求項21】
請求項9〜20に記載のポリペプチドであって、C−末端またはN−末端のいずれか、または両末端での、非結合アミノ酸による前駆体配列の延長によって修飾され、それによって付加された残基の数が全部で15以下、好ましい場合は6以下である、ポリペプチド。
【請求項22】
請求項9〜20に記載のポリペプチドであって、プロリン以外の1つ以上のアミノ酸残基が、物理化学的に関連する天然または非天然のアミノ酸残基を使用する保存的交換によって置換され、一方、結合様式および結合に要される構造が維持される、ポリペプチド。
【請求項23】
少なくとも1つのD−アミノ酸および/または合成アミノ酸および5−アミノバレリン酸を含む、ポリペプチド。
【請求項24】
請求項23のポリペプチドであって、D−アミノ酸に5−アミノバレリン酸が続き、次にD−アミノ酸が続く配列を含む、ポリペプチド。
【請求項25】
請求項23または24のポリペプチドであって、D−アミノ酸および/または合成アミノ酸および少なくとも1つの5−アミノバレリン酸よりなる、ポリペプチド。
【請求項26】
請求項23〜25のいずれかのポリペプチドであって、5−アミノバレリン酸が、請求項14、15および/または16のいずれか1項のビルディング・ブロックで置換される、ポリペプチド。
【請求項27】
アミノ酸配列:
ynnignlimqldlllhelqlqtkkts
のポリペプチド。
【請求項28】
ワクチン接種のため、または診断的、医薬的または化粧的目的のための、請求項9〜22に記載の化合物の使用。
【請求項29】
請求項9〜22に記載の化合物を含む、医薬製剤。
【請求項1】
N−末端アミノ基または置換アミノ基、C−末端カルボキシ基またはカルボキシ誘導体、骨格およびL−アミノ酸側鎖を有するL−配置の前駆体の三次元座標および配列情報から、少なくとも部分的にD−アミノ酸を含有するポリペプチドの等比体積構造を生成する方法であって、
−少なくとも部分的に、骨格のCO基をNH基に、およびその逆に、置換する工程、
−同時に、前駆体のL−アミノ酸側鎖、N−末端アミノ基または置換アミノ基およびC−末端カルボキシ基またはカルボキシ誘導体の三次元座標を一定に保持する工程
を含む方法。
【請求項2】
請求項1に記載の方法であって、
−少なくとも部分的に、骨格のCO基をNH基に、およびその逆に、置換する工程、
−同時に、前駆体のL−アミノ酸側鎖の三次元座標を一定に保持し、かつ、N−末端アミノ基または置換アミノ基をカルボキシ基またはカルボキシ誘導体で置換する工程および/またはC−末端カルボキシ基またはカルボキシ誘導体をアミノ基または置換アミノ基で置換する工程
を含む方法。
【請求項3】
請求項1〜2に記載の方法であって、前駆体の骨格の全てのCO基をNH基に、およびその逆に置換する方法。
【請求項4】
請求項1〜3に記載の方法であって、
−前駆体の構造および配列中のプロリン残基またはプロリン残基およびその隣接する残基を、新たに構成された骨格中のプロリンまたはプロリンおよびその直接隣接する残基の立体構造の特性を模倣したビルディング・ブロックとしての有機分子で、少なくとも部分的に置換する
ことを特徴とする方法。
【請求項5】
図1に記載の工程を含む、請求項1〜4に記載の方法。
【請求項6】
コンピュータ装置で実施される、請求項1〜5に記載の方法。
【請求項7】
少なくとも1つのD−アミノ酸および/または合成アミノ酸を含むポリペプチドを生成する方法であって、請求項1〜6のいずれかの方法によって等比体積構造を得る工程、および該等比体積構造のポリペプチドを合成する工程を含む方法。
【請求項8】
請求項7または8の方法であって、ポリペプチドがD−アミノ酸および/または合成アミノ酸よりなる方法。
【請求項9】
請求項1〜8に記載の方法によって得られるポリペプチド。
【請求項10】
請求項9に記載のポリペプチドであって、100残基未満、特に60以下、または40以下の残基で、少なくとも7残基を有する、ポリペプチド。
【請求項11】
請求項9〜10に記載のポリペプチドであって、式1:
【化1】
に図示され非排他的に例示される通り、骨格のCOがNH基で、およびその逆に置換され、同時にC−末端カルボキシおよびN−末端アミノ基が変化していないことを特徴とする、ポリペプチド。
【請求項12】
請求項9〜11に記載のポリペプチドであって、式2:
【化2】
に図示され非排他的に例示される通り、末端アミノ基がカルボキシ基で置換され、および/または、末端カルボキシ基がアミノ基で置換されたか、あるいは、N−およびC−末端が互いに交換された、ポリペプチド。
【請求項13】
請求項9〜12に記載のポリペプチドであって、前駆体の少なくとも1つのプロリン残基がグリシンで置換された、ポリペプチド。
【請求項14】
請求項9〜13に記載のポリペプチドであって、5−アミノバレリン酸のビルディング・ブロックとしての使用を示す式4、およびビルディング・ブロックとしての5−アミノバレリン酸の、例としての非排他的誘導体の使用を示す式5によって、非排他的に図示される通り、5−アミノバレリン酸および一般式:…-(CO)-X1-X2-X3-X4-NH-…で表されるその誘導体を用いて、前駆体配列中のプロリン残基およびその隣接する残基を置換した、ポリペプチド:式中、X1、X2、X3、およびX4は独立してCH2、(C=O)、NH、NR、O、(CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基である。
式4:
【化3】
式5:
【化4】
【請求項15】
式7を有する化合物、特に、3−(2S−アリルオキシカルボニル−アミノ−ピロリジン−1−イル)−3−オキソ−プロピオン酸(式6):式中、X1、X2およびX3は独立してCH2、(C=O)、O、S、NH、NR、(CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基であり;一般式で記載される分子は、式8において非排他的に図示される。
式6:
【化5】
式7:
【化6】
式8:
【化7】
【請求項16】
請求項9〜15に記載のポリペプチドであって、式9:
【化8】
において非排他的に図示される通り、請求項15に記載のビルディング・ブロックを使用して、少なくとも1つのプロリン残基およびその直接隣接する残基を置換した、ポリペプチド。
【請求項17】
式10の化合物:式中、X1、X2、X3およびX4はは独立してCH2、(C=O)、O、S、NH、NR、(CHR)、または(CR2)より選択され、Rはアミノ基、アルコール、ハロゲンまたはいずれかの有機残基であり、それらにより各分子の例は非排他的に式11に示される。
式10
【化9】
式11
【化10】
【請求項18】
請求項9〜14、16に記載のポリペプチドであって、請求項15に記載のビルディング・ブロックを使用して、少なくとも1つのプロリン残基およびその直接隣接する残基を置換した、ポリペプチド。
【請求項19】
請求項9によって得られ、かつ、プロリン、またはプロリンおよびその直接隣接する残基の置換基として請求項12〜16に特定される少なくとも1つのまたは自由な組み合わせのビルディング・ブロックを用いた、ポリペプチド。
【請求項20】
請求項9〜19に記載のポリペプチドであって、N−末端のアセチル化またはC−末端のアミド化によって、または、N−末端のアセチル化およびC−末端のアミド化によって修飾された、ポリペプチド。
【請求項21】
請求項9〜20に記載のポリペプチドであって、C−末端またはN−末端のいずれか、または両末端での、非結合アミノ酸による前駆体配列の延長によって修飾され、それによって付加された残基の数が全部で15以下、好ましい場合は6以下である、ポリペプチド。
【請求項22】
請求項9〜20に記載のポリペプチドであって、プロリン以外の1つ以上のアミノ酸残基が、物理化学的に関連する天然または非天然のアミノ酸残基を使用する保存的交換によって置換され、一方、結合様式および結合に要される構造が維持される、ポリペプチド。
【請求項23】
少なくとも1つのD−アミノ酸および/または合成アミノ酸および5−アミノバレリン酸を含む、ポリペプチド。
【請求項24】
請求項23のポリペプチドであって、D−アミノ酸に5−アミノバレリン酸が続き、次にD−アミノ酸が続く配列を含む、ポリペプチド。
【請求項25】
請求項23または24のポリペプチドであって、D−アミノ酸および/または合成アミノ酸および少なくとも1つの5−アミノバレリン酸よりなる、ポリペプチド。
【請求項26】
請求項23〜25のいずれかのポリペプチドであって、5−アミノバレリン酸が、請求項14、15および/または16のいずれか1項のビルディング・ブロックで置換される、ポリペプチド。
【請求項27】
アミノ酸配列:
ynnignlimqldlllhelqlqtkkts
のポリペプチド。
【請求項28】
ワクチン接種のため、または診断的、医薬的または化粧的目的のための、請求項9〜22に記載の化合物の使用。
【請求項29】
請求項9〜22に記載の化合物を含む、医薬製剤。
【図1】

【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公表番号】特表2007−529475(P2007−529475A)
【公表日】平成19年10月25日(2007.10.25)
【国際特許分類】
【出願番号】特願2007−503353(P2007−503353)
【出願日】平成17年3月17日(2005.3.17)
【国際出願番号】PCT/EP2005/051256
【国際公開番号】WO2005/090389
【国際公開日】平成17年9月29日(2005.9.29)
【出願人】(503470263)アプラーゲン ゲゼルシャフト ミット ベシュレンクテル ハフツング (5)
【Fターム(参考)】
【公表日】平成19年10月25日(2007.10.25)
【国際特許分類】
【出願日】平成17年3月17日(2005.3.17)
【国際出願番号】PCT/EP2005/051256
【国際公開番号】WO2005/090389
【国際公開日】平成17年9月29日(2005.9.29)
【出願人】(503470263)アプラーゲン ゲゼルシャフト ミット ベシュレンクテル ハフツング (5)
【Fターム(参考)】
[ Back to top ]