
粒子の誘導アッセンブリーをベースとするバイオセンサー
エフェクター又は補助因子を検出するセンサーシステムは、(a)核酸酵素;(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;(c)前記基質と少なくとも部分的に相補的である第二のポリヌクレオチドを含む第一の粒子セット(前記ポリヌクレオチドはその3'末端で前記粒子に付着している);及び(d)前記基質と少なくとも部分的に相補的である第三のポリヌクレオチドを含む第二の粒子セット(前記ポリヌクレオチドはその5'末端で前記粒子に付着している)を含む。

【発明の詳細な説明】
【技術分野】
【0001】
本発明はバイオセンサーの分野に関する。より具体的には、本発明は、触媒的に活性な核酸による、加熱を要しないバイオセンサーに関する。
【背景技術】
【0002】
タンパク質の他に、核酸もまた触媒活性を有することが近年見出された。触媒的に活性な核酸は触媒性DNA/RNAであり、DNAザイム/RNAザイム、デオキシリボザイム/リボザイム及びDNA酵素/RNA酵素としても知られている。触媒的に活性な核酸はまた改変核酸を含むことができる。核酸系酵素の触媒活性は、常に一定の補助因子(例えば金属イオン)の存在に左右される。したがって、これら補助因子のための核酸酵素をベースとするバイオセンサー(例えば金属イオンのためのバイオセンサー)を、対応する核酸酵素の活性を基準にして設計することができる。他方、広範囲の分析物と高い親和性及び特異性で結合するように、核酸を選択することができる。これらの結合核酸はアプタマーとして知られている。
アプタマーは、高い親和性及び特異性で標的を認識する核酸(例えばDNA又はRNA)である(Ellington and Szostak 1990;Jayasena 1999)。アプタザイム(アロステリックDNA/RNAザイム又はアロステリック(デオキシ)リボザイムとも称される)は、エフェクター(標的分子)によって調節されるDNA/RNAザイムである。前記は典型的には、エフェクターを認識するアプタマードメイン及び触媒ドメインを含む(Hesselberth et al. 2000;Soukup and Breaker 2000;Tang and Breaker 1997)。エフェクターは、アプタマードメインと触媒ドメインとの間の特異的な相互作用を介して、アプタザイムの触媒活性を低下又は増加させることができる。したがって、アプタザイムの活性を用いて前記エフェクターの有無及び量をモニターすることができる。この方法は、診断及び探知の目的でアプタザイムセンサーを選別及び設計するために用いられた(Breaker 2002;Robertson and Ellington 1999;Seetharaman et al. 2001)。DNAザイム及びDNAアプタザイムはセンサーの開発にもっとも魅力的な候補物質である。なぜならば、DNAはRNAよりも合成がはるかに廉価であり、さらにRNAよりも安定であるからである。さらにまた、DNAザイムの触媒コア近くにアプタマーモチーフを導入することによるDNAアプタザイムの設計の一般的方法が利用可能である(Wang et al. 2002)。高い切断活性には、アプタマーモチーフとの結合時に、前記アプタザイムの触媒コア部分の活性をアロステリックに調節することができるエフェクター分子の存在が必要とされる。
【0003】
in vitroセレクション方法を用いて、極めて高い親和性(ピコモル範囲の高い解離定数を有する)をもつ、広範囲の標的分子のためのアプタマーを得ることができる(Brody and Gold 2000;Jayasena 1999;Wilson and Szostak 1999)。例えば、アプタマーは、金属イオン(例えばZN(II);Ciesiolka et al.(1995)及びNi(II);Hofmann et al. 1997)、ヌクレオチド(例えばアデノシン三燐酸(ATP);Huizenga and Szostak(1995)及びグアニン;Kiga et al. 1998)、補助因子(例えばNAD;Kiga et al.(1998)及びフラビン;Lauhon and Szostak 1995)、抗生物質(例えばヴィオマイシン;Wallis et al.(1997)及びストレプトマイシン;Wallace and Schroeder 1998)、タンパク質(例えばHIV逆転写酵素;Chaloin et al.(2002)及びC型肝炎ウイルスRNA依存RNAポリメラーゼ;Biroccio et al. 2002)、毒素(例えばコレラ完全毒素及びスタフィロコッカス内毒素B;Bruno and Kiel 2002)及び細菌芽胞(例えば炭疽;Bruno and Kiel 1999)を認識するために開発された。抗体と比べて、DNA/RNA系アプタマーは、in vitroで短時間に(数日に対して数週)、及び限られた費用で製造することができるために入手が容易で、より安価に製造される。さらにまた、DNA/RNAアプタマーは、その生物関連物質の認識能力を失うことなく何度も変性及び再生させることができる。これらの固有の特性のゆえに、アプタマーは、高度な感受性及び選択性を有するバイオセンサーの設計のための理想的な土台となる(Hesselberth et al. 2000)。
【0004】
DNA/RNAザイム又はアプタザイム活性をアッセイするために、検出可能な標識が用いられる。しかしながら、これらの多くは顕著な欠点を有する。放射性同位元素は安全性と廃棄に関する懸念があり、一方、蛍光発色団は光退色を受け、アッセイしようとしている生物学的活性を阻害する可能性がある(すなわち、有機色素(例えばコカイン検出アプタマーシステム(Stojanovic and Landry, 2002)で用いられるもの)は視覚的検出に高い濃度を必要とし、高価な試行錯誤の後でやっと特異的なアプタマーとの組合せが得られる。
金属粒子はこれらの問題の全てを解決する。前記粒子は、有機色素のいずれの欠点も伴わずに、検出剤として少量(ナノモル濃度)でアプタマーとともに用いることができる。発色検出のために金属粒子を用いるアプタマー系センサーでは、(エフェクター結合時の)アプタザイムによる核酸基質の切断は、色の変化によって検出することができる。
典型的には、DNA/RNAザイム又はアプタザイムをベースとするセンサーは以下の3つの部分を有する:
(1)核酸酵素(以下の記述では、核酸酵素はDNA/RNAザイム及びアプタザイムと称されるであろう)及び補助因子(例えば基質の切断を触媒する金属イオン);
(2)核酸酵素のための核酸基質(この場合基質配列の内部部分は前記酵素配列の部分と相補的である);及び
(3)基質の3'-及び5'-末端と相補的なポリヌクレオチドと結合した粒子。
【0005】
標的補助因子又はエフェクターを検出するために、ポリヌクレオチドの相補性部分(基質鎖の3'-及び5'-末端と相補的な、粒子に結合したポリヌクレオチド、並びに基質鎖内部配列と相補的な核酸酵素の5'-及び3'-末端)を、標的の補助因子又はエフェクターが含まれていると思われるサンプルの存在下でアニールさせる。補助因子又はエフェクターが存在しない場合、アプタザイムは不活性であるか、又は実質的に低い活性を示し、基質切断は全く又はほとんど生じず、したがって粒子の凝集をもたらす。補助因子又はエフェクターが存在する場合、酵素は活性化し基質を切断して凝集形成は妨げられる(なぜならば、粒子間の連結が酵素性切断によって破壊されるからである)。表1はそのような系の例示である。
*金粒子が検出に用いられた場合。
【0006】
金粒子の事例では、凝集状態では青色が表示されるが、分散状態(又は非凝集状態)では赤色である。補助因子又はエフェクターのような標的分析物の存在は、センサーシステムの発色を基準にして検出することができる。
切断の程度は色の変化の度合いに反映されるので、標的補助因子又はエフェクターの濃度を定量することができる。単純な分光法を鋭敏な検出に用いることができる。検出及び定量に色の変化を用いるだけでなく、他の切断の成果、例えば沈殿を利用してもよい。アプタマーを予め選択したエフェクターを認識するアプタマーに置き換えることによって、任意の所望のエフェクターのための比色センサーを容易に製造し用いることができる。
これまでの研究を基にして、DNAザイムに誘導される(directed)金ナノ粒子アッセンブリーを基にしたPb(II)のための比色バイオセンサー、及びアプタザイムによって誘導される金ナノ粒子アッセンブリーを基にしたアデノシンのための比色バイオセンサーが設計された(例えば以下を参照されたい:米国特許出願09/605,558;10/144,094;10/144,679;10/384,497)。高感度及び高選択性ではあるが、このタイプの分析センサーは、検出のために50℃を超える数分間の加熱及び2時間に及ぶ室温への緩徐な冷却を必要とする。
【発明の開示】
【0007】
(発明の要旨)
第一の特徴では、本発明は、エフェクター又は補助因子を検出することを目的とするセンサーシステムであり、前記センサーシステムは次のものを含む:(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;(c)第二のポリヌクレオチドを含む第一の粒子セット、ここで前記第二のポリヌクレオチドはその3'末端で前記粒子に付着している;及び(d)第三のポリヌクレオチドを含む第二の粒子セット、ここで前記第三のポリヌクレオチドはその5'末端で前記粒子に付着している。前記第一のポリヌクレオチドは、前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的である。
第二の特徴では、本発明は、エフェクター又は補助因子を検出するためのセンサーシステムであり、前記センサーシステムは次のものを含む:(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;(c)第二のポリヌクレオチドを含む第一の粒子セット;及び(d)第三のポリヌクレオチドを含む第二の粒子セット。前記第一のポリヌクレオチドは、前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的である。前記第二の粒子セットは少なくとも20nmの直径を有する。
第三の特徴では、本発明はサンプル中のエフェクター又は補助因子の検出方法であり、前記方法は、少なくとも
(a)核酸酵素(補助因子結合部位と、任意にエフェクター結合部位とを含む);
(b)前記核酸酵素の基質(第一のポリヌクレオチドを含む);
(c)第二のポリヌクレオチドを含む第一の粒子セット;
(d)第三のポリヌクレオチドを含む第二の粒子セット;及び
(e)サンプル;
を一緒に混合して、混合物を生成することを含む。前記第一のポリヌクレオチドは、前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的である。前記混合物は、加熱することなく前記混合物の生成から10分以内に、エフェクター又は補助因子のサンプル中の存在を示す色の変化を生じる。
【0008】
(詳細な説明)
本発明は、以前のDNAザイム-及びアプタザイム-金属粒子センサーシステムで要求される加熱を回避し、実験室だけでなく消費者が家庭で又は検査技師が野外でこれらセンサーを使用することを可能にする。本発明は、次のことが粒子をベースとする比色バイオセンサーの性能に影響を与えるという発見を利用している:(1)粒子の整列様式を頭部-尾部整列様式から尾部-尾部整列様式に変更すること;(2)より大きな粒子を使用すること;及び(3)イオン強度、アプタマー濃度及びpHを制御すること。
粒子の凝集は、粒子の互いに対する整列様式に影響される。粒子は2種の態様、“頭部-尾部”整列様式又は“尾部-尾部”整列様式で整列化することができる(図3)。例えば、初期の、DNAザイムをベースとするPb2+センサーでは、粒子は“頭部-尾部”様式で整列化される。粒子が“頭部-尾部”様式で整列化される場合、ただ1種のチオール改変DNAが、粒子にDNAを付着させるために必要である。この立体配置では、おそらく立体的影響のため、粒子が凝集することは困難である。したがって、加熱及び冷却が粒子のアッセンブリーを促進するために必要である。しかしながら、“尾部-尾部”整列様式を用いる場合、粒子は周囲温度で凝集できることを発見した(図5)。“尾部-尾部”整列化する粒子には、3'-チオール改変ポリヌクレオチド及び5'-チオール改変ポリヌクレオチドの両方が必要である。
粒子凝集体の色は、主としてその凝集体のサイズによって支配される。したがって、凝集速度が等しい場合、色の変化の速度は、より大きな粒子を用いたときに増す。下記の実施例で示すように、直径が13nm及び35nmの粒子の混合物を用いた場合、色の変化の速度は直径13nmの粒子のみを用いた場合よりも速い。色の変化の速度は、直径35nmの粒子のみを用いた場合にさらに速くなる。
同様に、溶液のイオン強度は、システムの性能に影響する。高い塩濃度は凝集に有利に働き、これは、おそらく、塩が負の電荷を有するポリヌクレオチド鎖間の静電気的反発を低下させるためである。2つの粒子を連結させるのに基質鎖のみが必要とされるが、酵素鎖の非存在下では、基質は“ふらふらして(floppy)”いて、粒子を集合させることができない。したがって、酵素の濃度は重要なパラメーターである。
pHもまた凝集に影響する。pHの影響は、おそらくポリヌクレオチドの塩基のプロトン化のためであり、塩基対形成に影響し、したがって凝集速度に影響を及ぼす。
【0009】
[定義]
“補助因子”は、核酸酵素によって触媒される反応の触媒過程に関与するイオン又は分子であり、触媒活性に必要とされるものである。
“エフェクター”は、エフェクター結合部位を有する酵素と結合した場合に、酵素の触媒作用を強化又は阻害することができる分子である。“エフェクター結合部位”は“特異的”、すなわち、他のエフェクター分子の存在下でただ1つのエフェクター分子とのみ結合するものであってもよい。エフェクター結合部位の特異性の例は、多くの他の類似分子(例えばシチジン、グアノシン及びウリジン)の存在下で、アデノシン分子のみが結合するといった場合である。あるいは、エフェクター結合部位は、“部分的に”特異的(1種類の分子とだけ結合する)であってもよく、又は“非特異的” (分子に対する無差別性を有する)であってもよい。エフェクターの例には、環境汚染物質(例えば窒素肥料、農薬、ダイオキシン、フェノール又は2,4-ジクロロフェノキシ酢酸)、重金属イオン(例えばPb(II)、Hg(II)、As(III)、UO2(II)、Fe(III)、Zn(II)、Cu(II)又はCo(II))、生物学的分子(例えばグルコース、インシュリン、hCGホルモン、HIV、HIVタンパク質)、化学的及び生物学的テロ関連物質(例えば炭疽、天然痘又は神経ガス)、爆発物(例えばTNT又はDNT)、薬剤(例えばコカイン又は抗生物質)が含まれる。
“核酸酵素”は、主として核酸を含む酵素、例えばリボザイム(RNAザイム)、デオキシリボザイム(DNAザイム)及びアプタザイムである。核酸は、天然の核酸でも非天然の核酸でも又は改変された核酸であってもよい。ペプチド核酸(PNA)もまた含まれる。核酸酵素は、効率的な基質切断及び/又は特異的なエフェクター結合のために“補助因子”を必要とする。一般的な補助因子には、Mg(II)、Ca(II)、Zn(II)、Mn(II)、Co(II)及びPb(II)が含まれる。
【0010】
“ポリヌクレオチド”は、少なくとも2つ以上のヌクレオチドを有する核酸配列をさす。ポリヌクレオチドは、天然に存在するヌクレオチド及び合成ヌクレオチドを含み得る。PNA分子も、この用語に包含される。
“感度”は、センサーによって検出することができる、補助因子又はエフェクターの濃度のもっとも小さな増加を指す。
“検出限界”は、分析装置の検出限界を指す。本発明のDNAザイムをベースとするセンサー及びアプタザイムをベースとするセンサーとの関連において、検出限界は、センサーがバックグラウンドから区別することができる補助因子又はエフェクターの最低濃度を指す。
“塩基対形成”は、ポリヌクレオチドが、低ストリンジェンシー条件下でヌクレオチドと少なくとも1つの水素結合を形成する能力を指す。前記ヌクレオチドは、第二のヌクレオチドの一部であっても、第一のポリヌクレオチド内で見出されるヌクレオチドであってもよい。第一のポリヌクレオチドが第二のポリヌクレオチドと少なくとも1つの水素結合を形成することができるとき、ポリヌクレオチドは第二のポリヌクレオチドと部分的に相補的である。部分的に相補的であるためには、ポリヌクレオチドは、塩基対を形成する領域に取り囲まれている、塩基対が形成されない領域を有することができ、ループ、ステム-ループ及び他の二次構造を形成する。
“アプタマー”はエフェクター結合部位を含むポリヌクレオチドを指す。“エフェクター結合部位”は“特異的”、すなわち、他のエフェクター分子の存在下でただ1つのエフェクター分子とのみ結合するものであってもよい。エフェクター結合部位の特異性の例は、多くの他の類似分子(例えばシチジン、グアノシン及びウリジン)の存在下で、アデノシン分子のみが結合するといった場合である。あるいは、エフェクター結合部位は、“部分的に”特異的(1種類の分子とだけ結合する)であってもよく、又は“非特異的” (分子に対して無差別性を有する)であってもよい。
“アプタザイム”は、エフェクターと結合するアプタマー領域を含む核酸酵素を指す。エフェクターの結合は、触媒作用を強化又は阻害することができる。
【0011】
[1のエフェクター(又は複数のエフェクター)の結合部位をもつ核酸酵素]
多様な触媒活性を有する、多数の核酸酵素が発見又は開発されてきた(表1及び2)。触媒機能について、酵素は通常1又は2以上の補助因子に依存する。in vitroセレクションは、特定のイオンについての選択性及び感度を“強化する”ために用いることができる。分子の会合(連結、リン酸化及びアミド結合形成)又は解離(切断又は転移)を触媒する核酸酵素は、本発明で特に有用である。
エフェクターの存在下で核酸の切断を触媒する核酸酵素が用いられる。前記核酸酵素は、RNA(リボザイム)、DNA(デオキシリボザイム)、DNA/RNAハイブリッド酵素、又はペプチド核酸(PNA)酵素であってもよい。PNAはポリアミド骨格及びヌクレオシド塩基を含む(例えばBiosearch, Inc.(Bedford, MA)から入手できる)。使用することができるリボザイムには、グループI及びグループIIのイントロン、細菌リボヌクレアーゼPのRNA成分、ハンマーヘッドリボザイム、ヘアピンリボザイム、肝炎デルタウイルスリボザイム及びニューロスポーラVSリボザイムが含まれる。さらに、例えば以前に単離されたもの(Tang and Breaker 2000))もin vitroセレクションされたリボザイムに含まれる。リボザイムは、デオキシリボザイムより不安定な傾向があり、したがってデオキシリボザイムが好ましい。拡張された化学的官能性をもつデオキシリボザイムもまた所望される(Santoro et al. 2000)。
【0012】
表1:in vitroセレクション実験から単離されたリボザイムによって触媒される反応
a:in vitroセレクション実験から単離されたリボザイムによって触媒される反応。Kcat/Kuncatは、未触媒反応を超える速度増大である。
参考文献:1(Vaish et al. 1998);2(Tsang and Joyce 1996);3(Ekland et al. 1995);4(Lorsch and Szostak 1994);5(Ekland and Bartel 1996);6(Illangasekare and Yarus 1997);7(Piccirilli et al. 1992);8(Lohse and Szostak 1996);9(Wilson and Szostak 1995);10(Wecker et al. 1996);11(Dai et al. 1995);12(Wiegand et al. 1997);13(Zhang and Cech 1997);14(Tarasow et al. 1997);15(Prudent et al. 1994);16(Conn et al. 1996)。
【0013】
表2:in vitroセレクションにより単離されたデオキシリボザイム
b:Kmaxは至適条件下で得られた最大速度定数である。
参考文献:1(Breaker and Joyce 1994);2(Breaker et al. 1995);3(Faulhammer and Famulok 1997);4(Santoro and Joyce 1997);5Geyer and Sen 1997);6(Roth and Breaker 1998);7(Li et al. 2000);8(Carmi et al. 1996);9(Cuenoud and Szostak 1995);10(Li and Breaker 1999);11(Li et al. 2000);12(Li and Sen 1996)。
【0014】
リボザイム及びデオキシリボザイムを製造する方法には、オリゴヌクレオチドの化学的合成、ポリメラーゼ連鎖反応(PCR)、DNAクローニング及び複製が含まれる。好ましくは、核酸酵素はDNA/RNAハイブリッド及びPNAである。改変した塩基、ホスフェート又は糖を含むヌクレオチドも用いることができる。いくつかの事例では、これらの改変したヌクレオチドは、安定性の利点を有し、又はエフェクター特異性を付与する。改変した塩基の例には、イノシン、ネブラリン、2-アミノプリンリボシド、N7-デアザアデノシン及びO6-メチルグアノシンが含まれる(Earnshaw and Gait 1998)。改変した糖及びホスフェートには、2'-デオキシヌクレオシド、脱塩基(abasic)、プロピル、ホスホロチオエート及び2'-O-アリルヌクレオシドが含まれる(Earnshaw and Gait 1998)。
酵素を含む鎖から離れた場所の核酸の鎖を切断する核酸酵素はトランス作動性酵素である。トランス作動性酵素の使用は、酵素生成物が取り除かれるために何回もの基質切断を可能にする。そのような核酸酵素の例は17E(配列番号1)である。対応する基質は17DS(配列番号2:rは、1個のリボヌクレオチドを表す)である。両者は表3A及び図1に示されている。“8-17”DNAザイムシステムの二次構造(酵素鎖(17E)及び基質鎖(17DS)を含む)は図1aに示されている。切断部位は矢印で示されている。切断部位のリボヌクレオシドアデノシン(rA)を除いて、他の全てのヌクレオシドがデオキシリボヌクレオシドである。Pb(II)の存在下で、酵素鎖は基質鎖を切断する(図1b)。したがって、図1cに示されているように、酵素鎖及び基質鎖を、金粒子のDNAザイム誘導アッセンブリーで用いることができ、前記アッセンブリーの金属イオン(例えばPb(II))についてのバイオセンサーとしての用途で用いることができる。このシステムでは、17DSはその3' 末端及び5'末端の両方で12塩基伸長されており、前記塩基は13nmの金粒子に付着させた12-merのDNA(DNAAU)と相補的である。他の例もまた表3Bに提示されている。
【0015】
表3A:DNA酵素及び基質
c"r"は1個のリボヌクレオチドを表す。
【0016】
表3B:RNA/DNAをベースとするアプタマー及びRNA/DNAザイム
【0017】
定方向変異導入を用いて、核酸酵素又はその基質の1又は2以上の特性を変化させることができる。一例として17E及び17DSを用い、ハイブリダイズした酵素と基質の2本のアームのアビディティーを変化させるようとすることもできる。前記“アーム”は、図1でワトソン-クリックの塩基対形成を示す領域である。アビディティーを変化させるために、アームの長さを変化させる。アームの長さを増加させることによって、ワトソン-クリックの塩基対形成の数が増加し、したがってアビディティーが増加する;アームの長さを減少させることによってアビディティーは低下する。アームのアビディティーを低下させることによって酵素から基質が除去されやすくなり、したがってより迅速な酵素のターンオーバーが可能になる。
アビディティーを低下させる別の方法には、酵素と基質との間にミスマッチを作り出すことが含まれる。あるいは、アームのG-C含有量を変化させてもよい。いずれの誘導変化の影響もモニターし、酵素が所望の活性(イオンに対する感度及び選択性を含む)を維持していることを確実にすべきである。例えば、変異酵素がアデノシンに対する感度及び選択性を維持していることを確実にするために、変異酵素がアデノシンの存在下で反応性を維持しているか否か(感度)及び他のエフェクターの存在下でより低レベルの活性を維持しているか否か(選択性)を決定する試験を実施できよう。
【0018】
[アプタマーのin vitroセレクション]
所望のエフェクターと結合するアプタマー及びアプタザイムは、in vitroセレクションによって単離することができる。in vitroセレクションは、一定の機能をもつRNA分子又はDNA分子を選別と増幅との複数回ものサイクルにより多数の配列変種から単離する技術である(Chapman and Szostak 1994;Joyce 1994)。最大の活性又は新規な触媒能力をもつDNAザイム及びRNAザイムが、アプタマーと同様に、例えば指数関数的濃縮によるリガンドの系統的進化(SELEX, systematic evolution of ligands by exponential enrichment)技術を用いて入手することができる(Tuerk and Gold 1990)。
in vitroセレクションは、典型的には、ランダム化された配列の大きな集合(プール)(通常は1013〜1016の配列変種を含む)を用いて開始される。そのようなランダム化されたプールの作製には、標準的なホスホアミダイト化学反応を用いた縮退ポリヌクレオチドのセットの化学合成を用いることができる。4種のヌクレオシド(アデノシン、シトシン、グアニン、チミジン)の3'-ホスホアミダイト化合物を予め混合し、これをポリヌクレオチドの合成に使用する;ランダム化は4種のホスホアミダイトの比率を制御することによって得られる。ホスホアミダイト定数を特定の位置で保持するだけでなく、バイアスを達成することもできる。ランダム化DNAライブラリーを作製する他のストラテジーには、変異導入ポリメラーゼ連鎖反応(PCR)及び鋳型特異的(template-directed)変異導入が含まれる(Cadwell and Joyce 1992;Cadwell and Joyce 1994;Tsang and Joyce 1996)。RNA分子のin vitroセレクションを所望する場合は、先ず初めにin vitro転写によってランダム化DNAライブラリーをRNAライブラリーに変換する。
【0019】
続いて、ランダム化ライブラリーを、所望の機能(例えば標的化エフェクターとの結合)をもつ分子についてスクリーニングして、単離する。分離は、アフィニティーカラムクロマトグラフィー(例えば標的化エフェクターを用いる)、ゲル電気泳動、又はタグ付加反応中間体の選択的増幅を用いて達成することができる。選別した分子を、例えばDNAのためのPCR、又はRNAのための等温増幅反応を用いて増幅する。続いて、これらの選別増幅分子に、例えば変異導入PCRを用いて変異を導入し(多様性の再導入)、さらに高い活性をもつ分子の選別を試みる。所望の活性を持つ配列がプール中で優位を占めるまで、これら3つの工程、選別、増幅及び変異導入を、しばしば選別のストリンジェンシーを高めながら、繰り返す。
ランダム配列からin vitroで単離された新規な核酸酵素は、RNA及びDNAの触媒レパートリーを拡大させた(表1)。デオキシリボザイムは、リボザイムと比較してより少ない反応タイプを触媒する(表2)。ほとんどのデオキシリボザイムの触媒速度(Kcat)は、同じ反応を触媒するリボザイムの触媒速度に匹敵する。ある種の事例では、核酸酵素の触媒効率(Kcat/Km)は、タンパク質酵素の触媒効率を超えさえする(Santoro and Joyce 1997)。
in vitroセレクションを用いて、現存の核酸酵素のイオン特異性又は結合親和性を変化させることができ、又は所望の基質に特異的な核酸酵素を得ることができる。例えば、in vitroセレクションを用いて、最適なハンマーヘッドリボザイム活性に必要とされるMg2+濃度を低下させ、生理的条件下での酵素の性能が改善された(Conaty et al. 1999;Zillmann et al. 1997)。
【0020】
in vitroセレクションによって特定の補助因子のために開発された核酸酵素は、しばしば、他の分子の存在下で活性を有するであろう。例えば、17Eデオキシリボザイムは、Zn2+の存在下での活性のためにin vitroセレクションによって開発された。しかしながら、この酵素は、Zn2+よりもPb2+の存在下でより強い活性を示した。Zn2+関連活性を捜し求める過程で製造されたが、17EはPb2+についての鋭敏で選択的なセンサーとして用いることができる。より強い選択性をもつ核酸酵素を製造するために、負の選別工程を導入することができる(Peter J. Bruesehoff, Jing Li, Anthony J. Augustine III, and Yi Lu, "Immproving Metal Ion Specificity During In Vitro Selection of Catalytic DNA", Combinatorial Chemistry and High Throughput Screening 5, 327-355 (2002))。
他のヌクレオチド配列も有用であり、例えば米国特許出願第09/605,558号(2000年6月27日出願)に記載されたものが挙げられる(前記文献は参照により本明細書に含まれるものとする)。
【0021】
[アデノシンセンサー]
図2Aに示したアプタザイムはアデノシンに特異的である。アプタザイムの触媒コアは“8-17”DNAザイムから適合化され、Pb2+の存在下での高活性について最適化された。前記DNAザイムの3'-末端はアデノシンアプタマーモチーフに連結されている。したがって、アデノシンの存在は、DNAザイムの活性な三次元構造の形成を促進することができる。アプタザイムをベースとするセンサーでは、この複合体が、ただ1つのリボアデノシン部位での基質鎖の切断を促進する。アデノシンがなければ、3成分がワトソン-クリック塩基対形成によって合体することができたとしても、切断活性は劇的に低下する。また、アプタザイムの両端にハイブリダイズしている直径13nmの2つのDNA官能化金粒子も示されている。
図2Bに示されているように、基質鎖(遊離基質及び酵素と調節因子鎖とにハイブリダイズしている基質の両方)は、DNA官能化金粒子のためのリンカーとして機能し、青色をもつ凝集体を形成する(反応A)。アデノシン及び金属イオンの存在下では、基質は切断されることができる(反応B)。切断された基質はもはや粒子のためのリンカーとして機能することができず、色は赤色のままである(反応C)。
このタイプのセンサーでは、基質鎖、酵素鎖及び調節因子鎖の存在は、室温での凝集体形成のためには不充分である。加熱無しに、3つの鎖全てがハイブリダイズして、リンカーとして機能することは困難である。この問題を解決するために、別のDNA鎖(“Ade-リンカー”)を導入する(図8)。Ade-リンカーの目的は、高効率で基質鎖にハイブリダイズし、室温での粒子の凝集を促進させることである。ハイブリダイゼーションの後、Ade-リンカーは、10ヌクレオチドを有する、ふくらんだ(bulged)二次構造を形成する。このふくらみ(bulge)は、Ade-リンカーが切断された基質を用いて粒子を集合させるのを妨げる。このふくらみにより、たとえいくつかのAde-リンカーが切断基質とハイブリダイズしたとしても、全体構造は依然としてふらふらしていて、粒子を集合させることはできない。
アデノシンが存在しない場合、アプタザイムの基質はAde-リンカーとハイブリダイズし、ハイブリダイズした生成物は粒子を集合させて青色の凝集体を形成することができる(反応A)。アデノシンの存在下では、基質鎖は切断され(反応B)、切断された基質は金粒子を集合させることができず、赤色の分離している粒子を生じる(反応C)。
【0022】
[核酸酵素基質の3'及び5'末端と相補的なポリヌクレオチドタグを付した粒子]
センサーが酵素活性を表示するためには、ポリヌクレオチドを付着させた粒子の凝集の変化に際して検出可能な変化が生じなければならない。さらにまた、粒子の組成は、基質の切断を妨害しないようなものでなければならない。粒子は、例えば金属、半導体及び磁性物質から製造することができる;ZnS、ZnO、TiO2、AgI、AgBr、HgI2、PbS、PbSe、ZnTe、CdTe、In2S3、In2Se3、Cd3P2、Cd3As2、InAs及びGaAsで(例えば以下を参照されたい:Mirkin et al. 2002)、好ましくは金粒子である(商業的に入手可能である。例えば:Amersham Biosciences, Piscataway, NJ;Nanoprobes, Inc., Yaphank, NY)。非金属粒子もまた用いることができ、セラミック及びポリマー、例えばポリスチレンラテックス粒子又は色素含有ラテックス粒子が含まれる。好ましくは、粒子は、少なくとも20nm、より好ましくは少なくとも30nm、さらに好ましくは少なくとも35nmの平均粒子直径(15〜500nm、20〜200nm及び35〜100nmを含む)を有する。
少なくとも35nmの直径をもつ金コロイド粒子が好ましい。金コロイド粒子は、濃い色を生じるバンドに対して高い吸光係数を有する。これらの色は、粒子サイズ、濃度、粒子間距離、凝集の度合い及び凝集体の形状とともに変動する。例えば、粒子に付着させたポリヌクレオチドのハイブリダイゼーションによってもたらされる金粒子の凝集は、裸眼で観察することができる迅速な色の変化を生じる(例えば以下を参照されたい:Mirkin et al. 2002)。
粒子、ポリヌクレオチド又はその両方は、付着のために誘導体化される。例えば、3'-又は5'-末端がアルカンチオールで誘導体化されたポリヌクレオチドは、金粒子に容易に付着する(Whitesides 1995)。3'チオールDNAを平坦な金表面に付着させる方法も、ポリヌクレオチドの粒子への付着のために用いることができる(Mucic et al. 1996)。アルカンチオール誘導体化粒子をポリヌクレオチドに付着させるために用いてもよい。ポリヌクレオチドを固体表面に付着させる他の官能基には、ホスホロチオエート(ポリヌクレオチドを金表面に付着させる:Beebe and Rabke-Clemmer 1995)、置換アルキルシロキサン(シリカ及びガラス表面にポリヌクレオチドを結合させる)、アミノアルキルシロキサン及びメルカプトアルキルシロキサン(Grabar et al. 1995)が含まれる。5'-チオヌクレオシド又は3'-チオヌクレオシドで終わるポリヌクレオチドもまた、固体表面にポリヌクレオチドを付着させるために用いることができる。ポリヌクレオチドを付着させるいくつかの方法は、表4に提示されている。
【0023】
表4:ポリヌクレオチドを粒子に付着させるための系
【0024】
[粒子整列様式]
粒子が“尾部-尾部”整列様式で並ぶように、以下の様式で粒子を官能化する。基質の5'部位で基質と結合する粒子は、その3'末端で前記粒子と連結されている相補性ポリヌクレオチド鎖で官能化され、一方、基質の3'側で基質と結合する粒子は、その5'末端で前記粒子と連結される相補性ポリヌクレオチド鎖で官能化されている。基質-粒子複合体は、図3Aに示された構造ではなく、図3Bに示された構造を有する。
好ましくは、基質は、例えば“粘着末端”として機能する数塩基による3'-末端及び5'-末端の伸長によって、改変されて、粒子に付着させた相補的ポリヌクレオチド鎖とのアニーリングが促進される。基質の改変は、核酸酵素/基質の相互作用を阻害することなく基質に連結された粒子を含む複合体の形成を可能にする。しかしながら、基質が核酸酵素との相互作用に重要ではない領域を含む場合、改変は必ずしも必要ではない。
粒子を基質に直接付着させることも可能であり、あるいは基質の一方の端を粒子に付着させ、他方の端は粒子に付着したポリヌクレオチドと相補的にすることも可能である。したがって、相補的ポリヌクレオチドは任意である。
標的補助因子又はエフェクターを検出するために、核酸酵素、基質及び標識粒子を、前記酵素が感受性を有する標的の補助因子又はエフェクター(例えばアデノシン)を含んでいると疑われるサンプルの存在下で混合する(図3)。補助因子又はエフェクターの存在下では、酵素は基質を切断して、凝集体の形成を妨げる。
【0025】
粒子の種々の凝集状態によって種々の色が生じる。例えば、金粒子の大規模な凝集は青色を呈し、粒子の小規模の凝集は赤色を呈する。さらにまた、基質の切断量、よって凝集の度合いは、補助因子又はエフェクターの濃度に依存する。低い補助因子又はエフェクター濃度は、単独の粒子と凝集体との混合物を生じる部分的な基質の切断のみをもたらし、半定量的又は定量的アッセイを可能にする。色の相違を増幅して感度を改善することができる。定量的測定のために、アッセイ混合物の光学スペクトルが決定される。色の変化の他に、粒子の凝集体の形成又は塊状粒子(aggregated particles)の沈殿をモニターすることもできる。色の変化は、裸眼又は分光光度計によって観察することができる。凝集体の形成は、電子顕微鏡又はネフェロメーターで観察することができる。塊状粒子の沈殿は、裸眼又は顕微鏡で観察することができる。
色の変化の観察を容易にするために、対照的な色の背景で色を観察することができる。金粒子を用いるときは、白色の固体表面(例えばシリカ又はアルミナのTLCプレート、ろ紙、硝酸セルロース膜、及びナイロン膜)上にハイブリダイゼーション溶液のサンプルをスポットし、このスポットを乾燥させることによって、色の変化の観察が容易にされる。先ず初めに、スポット(点)は、ハイブリダイゼーション溶液の色を維持している(凝集の非存在下での桃色/赤から、金粒子の凝集が存在する場合の紫がかった赤色/紫の範囲にある)。乾燥時に、スポットを滴下する前に凝集が存在する場合は、青色のスポットが生じる;分散が発生した場合は、桃色のスポットが生じる。青色と桃色のスポットは安定で、その後の冷却若しくは加熱でも、又は時間が経過しても変化しない。これらは簡便で永久的な検査記録を提供する。色の変化を観察するために他の工程は必要とされない。
あるいは、アッセイ結果は、35nmの金粒子とともに使用して、サンプルをガラス繊維フィルター上にスポットすることによって可視化してもよい。水でリンスした後で、凝集体を含むスポットを観察する。アッセイ結果を可視化するため、また別の方法も利用可能である(Mirkin et al. 2002)。
標的とする補助因子又はエフェクターは、生物学的サンプルを含む多様なサンプルで検出することができる。既知量の補助因子又はエフェクターを含む標準試料を未知サンプルと並べてアッセイし、色の変化を比較することができる。あるいは、基準カラーチャート(pH試験紙とともに用いられるものと類似するもの)を用意してもよい。
【0026】
[キット]
本発明はまた、補助因子又はエフェクターのような分析物を検出するキットを提供する。ある実施態様では、前記キットは少なくとも1つの容器を含み、前記容器は、ポリヌクレオチドが付着している少なくとも2つのタイプの粒子と、少なくとも3つの部分をもつ基質(第一の部分は第二の部分に対して5'側であり、第二の部分は分析物の存在下で核酸酵素によって切断され、第三の部分は第二の部分に対して3' 側である)と、核酸酵素とを保持する。第一の粒子に付着しているポリヌクレオチドは、前記基質の少なくとも第一の部分の配列と相補的な配列を有し、且つその5'末端で前記粒子に付着している。第二の粒子に付着しているポリヌクレオチドは、前記基質の少なくとも第三の部分の配列と相補的な配列を有し、且つその3'末端で前記粒子に付着している。
キットが供給される場合、組成物の種々の成分は別々の容器に包装されてあって、使用直前に混合することができる。そのように成分を別々に包装することによって、活性成分の長期保存が可能になる。例えば、ポリヌクレオチドが付着している粒子の1又は2以上、基質、及び核酸酵素は、別々の容器で供給される。
キットに含まれる試薬は、種々の成分の耐用期間が維持され、且つ容器の材料によって吸着されたり変化させられたりしない任意の種類の容器で供給することができる。例えば、密封ガラスアンプルには、中性又は非反応性ガス(例えば窒素ガス)下で封入された、1若しくは2以上の試薬又は緩衝液を含むことができる。アンプルは、任意の適切な物質、例えばガラス、有機ポリマー(例えばポリカーボネート、ポリスチレンなど)、セラミック、金属、又は同様の試薬を保持するために典型的に用いられる他の任意の材料から成るであろう。その他の適切な容器の例には、アンプルと類似の材料から製造することができる単純なビン、金属箔(例えばアルミニウム又は合金)で裏打ちされた内部を含み得るエンベロープが含まれる。その他の容器には、試験管、バイアル、フラスコ、ビン、注射筒などが含まれる。容器は無菌的な出入口を有することができる。前記容器は、例えば、皮下注射針で突き刺すことができる栓を有するビンである。他の容器は、2つの区画を有していてもよい。これら区画は容易に取り外し可能な膜で分離されてあり、取り外しに際して成分の混合が可能にされる。取り外し可能な膜は、ガラス、プラスチック、ゴムなどであってもよい。
キットはまた、標的とする補助因子又はエフェクターの検出に有用な他の試薬及び物品を含むことができる。前記試薬には、既知量の補助因子又はエフェクターを含む標準溶液、希釈液及び他の緩衝液、予備処理試薬などが含まれる。部品として提供することができる他の物品には、支持体(凝集体の分解を可視化するため、例えばTLCシリカプレート);多孔性材料、注射筒、ピペット、キュベット及び容器が含まれる。被検対象の様々な量の補助因子又はエフェクターの存在に対応して、多様な凝集状態の粒子の外観を表示する基準チャートを提供することができる。
キットは、使用説明資料とともに提供してもよい。使用説明は、紙又は他の物体に印刷されていてもよく、及び/又は読み出し可能な電子媒体(例えばフロッピーディスク、CD-ROM、DVD-ROM、Zipディスク、ビデオテープ、オーディオテープなど)として供給することができる。詳細な使用説明はキットに物理的に付随していなくてもよく、それに代わって、ユーザーは、キットの製造元又は販売元によって特定されるインターネットウェッブサイトで指示を得ることもでき、又は電子メールとして供給されることもできる。
【実施例1】
【0027】
(実施例1:Pb2+比色バイオセンサー)
[ポリヌクレオチド及び試薬]
全てのポリヌクレオチドは、インテグレーテッドDNAテクノロジー社(Integrated DNA Technology Inc., Coralville, IA)から購入した。アデノシン及び他のヌクレオシドはシグマ・アルドリッチ(Sigma Aldrich, St. Louis, MO)から購入した。直径35nmの金粒子を調製し、3'-及び5'-チオール-改変した12-merのDNAを付着させた(Storhoff et al. 1998)。基質及び酵素はHPLCによって精製した。
[金粒子の調製及び官能化]
直径35nmの金粒子は、HAuCl4をクエン酸ナトリウムで還元して調製した。調製に使用したガラス製品は王水に浸漬し、ミリポア水で充分にリンスした。250mLの2首フラスコで、200mLのHAuCl4(0.3mM)を攪拌しながら加熱還流した。続いて、2mLのクエン酸ナトリウム(38.8mM)を添加した。数分以内に、色が淡黄色から深紅に変化した。色が変化した後、この系をさらに半時間還流して、還元を完了させた。続いて、この系をゆっくりと室温まで冷却し、粒子をパイレックスロートでろ過した。粒子をJEOL2010電子顕微鏡による透過電子顕微鏡法(TEM)を用いて性状を調べ、サイズを35±6nmと決定した。粒子の大半は球形で、ほんの一部の粒子は縦横比が2未満の棍棒状であった。金コロイドは532nmで〜1の吸光ピークを有し、濃度は、全ての粒子が35nmの半径を有する球形であると仮定して、およそ0.23nMであると計算した。全てのHAuCl4が反応し、粒子の密度はバルクの金と同じであった。この溶液は532nmで4.4x109M-1cm-1の吸光係数を示した。
【0028】
[3'-及び5'-チオール改変したDNAによる粒子の官能化]
チオール改変したDNAを50mMのジチオスレイトール(DTT)とともにインキュベートして活性化した。典型的には、60μLの1mMのDNAを60μLの100mMのDTTと室温(20〜22℃)で2時間インキュベートした。続いて、混合物をSep-Pak C18カラムにのせてDTTを除去した。カラムは、10mLの95%CH3CN、CH3CNとメタノールと水との混合物(容積比1:1:1)10mL、20mLの水、及び10mLの2MのNH4OAcで処理した。続いて、DNAとDTT混合物をカラムにロードした。20mLの脱イオン水でDTTを洗い流し、1mLの95%CH3CN、CH3CNとメタノールと水との混合物(容積比1:1:1)10mLによってDNAを溶出させた。溶出したDNAを12mLの金粒子(0.23nM)に添加した。1日インキュベートした後、1MのNaCl及び0.1Mのトリス-酢酸(TA)(pH7.2)を含む1.4mLの緩衝液を添加した。さらに2日間インキュベーションした後、8000rpm、10分間の遠心分離によって、粒子を収集した。上清を除去し、DNA官能化した粒子を25mLのTA緩衝液(pH7.2)、100mMのNaClに再分散させた。このプロセスを3回繰り返して、遊離DNAの除去を確実にした。最後に粒子を同じ緩衝液に分散させて、532nmでの吸光係数を〜2.2に調整した(〜0.5nMの濃度に対応する)。
【0029】
[DNAザイムによる粒子の凝集]
典型的な実験では、20μLの3'-チオール改変DNA官能化金粒子と20μLの5'-チオール改変DNA官能化金粒子と(両者とも〜0.5nMの濃度)を混合した。NaCl、酵素(17E)及びPb2+を添加し、最終容積を基質の添加時に100μLに調整した。凝集を基質鎖の添加によって開始させ、吸光をヒューレットパッカード(Hewlett-Packard)8453分光光度計でモニターした。緩衝液は、pH依存実験を除く全ての実験でpH7.2のTAとした。基質添加から最初のスペクトルのモニターまでの間の時間は、全てのサンプルについて15秒となるように制御した。
図4に示した実験では、比較を容易にするために吸光比を正規化した。これらの実験は2μMの酵素で実施し、基質濃度は、13nm粒子に対しては160nMで、13nM〜35nM混合粒子に対しては120nMで、35nm-35nm粒子に対しては3nMであった。緩衝液は25mMのTA(pH7.2)であった。
【0030】
[新規Pb2+センサーシステムの最適化]
(f)凝集10分後の吸光比
透明な赤色から青色への色の変化が35nmの金粒子の凝集に際して観察することができ、したがって、凝集過程はUV可視化分光光度計によって簡便にモニターすることができる。基質鎖の添加後の種々の時点における、連結基質(3nM)、NaCl(250mM)及び17E酵素(1μM)を用いた新規なセンサーシステムのいくつかのスペクトルが図6Aに示されている。基質鎖を添加する前、このシステムは、532nmで吸光ピークを有する赤色を示し、13nmの直径の粒子と比較して10nm赤色にシフトしていた。最初の3分では、色の変化は非常に小さかった。この3分の間に、基質鎖が酵素鎖と塩基対を形成し、ハイブリダイズしたDNAザイムが金粒子に連結されたと考えられる。次の数分では、色の変化の速度が非常に速く、粒子が架橋されて大きな凝集体を形成したことを示している。凝集体の形成とともに、532nmでの吸収ピークは減少し、一方、700nm領域で吸光が増加して赤色から青色への色の遷移が生じた。10分後、凝集体は沈殿し始め、全ての波長で観察される吸光の減少が生じた。サンプル調製物及び粒子濃度における相違の影響を最小限にするために、532nmと700nmの吸光比を用いて色の変化の速度をモニターした(より高い比は赤色に関連し、より低い比は青色に関連する)。
種々の基質濃度の存在下における色の変化の速度が図6Bに示されている。より高い基質濃度は、より速い速度の色の変化を生じた。したがって、基質濃度の低下は所定の時間で“青色の少ない”または“赤色の強い”色を生じた。異なる濃度のPb2+は、異なる速度で基質を切断した。したがって、センサーによって表示される色を精査することによって、Pb2+を検出し定量した。この実験は3nMの基質、250mMのNaCl及び1μMの17Eを用いて実施した。
【0031】
[凝集に影響を与える付加的な因子]
上記に概略したように、粒子の凝集速度に影響する他の因子が見出された。前記因子は、例えば酵素濃度、塩濃度及びpH値である。図6Cに示した実験は、全て3nMの基質濃度及び1μMの17E酵素濃度を用いて実施し、NaCl濃度を変化させた。より高いNaCl濃度は凝集を促進し、おそらくは、NaClが負の電荷をもつDNAの負電荷間の静電気的反発を低下させたためであろう。
酵素濃度もまた粒子の凝集に顕著な影響を有した。図6Dに示した実験は全て0.3nMの基質濃度及び250mMのNaCl濃度を用いて実施し、17E酵素濃度を変化させた。より高い酵素鎖濃度は、より速い凝集速度をもたらした。酵素の非存在下又は酵素濃度が非常に低いとき(図6D、最上部の曲線)、凝集は阻害された。ただし物理的には基質鎖のみがリンカーとして作用して粒子を集合させるので、酵素鎖の存在は任意であるようにみえる。これらの観察から、凝集の最初の過程は基質鎖と酵素鎖とのハイブリダイゼーションであるとの仮説を立てた。続いて、ハイブリダイズしたDNAザイムは、金粒子を集合させることができる。酵素鎖の非存在下では、基質は単独ではおそらくふらふらしていて、金粒子を集合させられないだろう。
種々のpH値もまた凝集に影響を与える。図6Eに示したように、pH5.2からpH10.2(pH5.2、酢酸緩衝液;pH6.2、MES緩衝液;pH7.2〜10.2、TA緩衝液)では、全てのサンプルで、室温(20〜22℃)での金粒子のDNAザイム誘導アッセンブリーに特徴的であったS字状曲線が得られた。pH5.2のサンプルでは、粒子の凝集は阻害されているようであった。10分の凝集の後の吸光比をプロットし図6Fに提示した。pH7.2から9.2では、類似する吸光特性が観察され、同様な凝集速度が示唆されているが、それより高いpH及びそれより低いpHではともに凝集速度は低下する。したがって、センサーは約6.2から約10.2の間のpH範囲で用いることができるが、最適な性能には約7.2から約9.2のpHで用いることができる。粒子の凝集速度の変化は、おそらくは、低いpH又は高いpHでのDNA塩基のプロトン化及び脱プロトン化が塩基対形成に影響を与え、したがって凝集速度の変化を生じるためであろう。
【0032】
[室温での凝集]
本発明の新規なセンサーの種々のPb2+濃度による室温(20〜22℃)での凝集の反応速度(kinetics)をUV可視化分光光度計によってモニターした。Pb2+の存在下では、基質と酵素鎖のハイブリダイゼーション後に、基質鎖は酵素鎖によって切断されるか、又は粒子を集合させるリンカーでありえると仮説を立てた。2つの過程の相対速度は、Pb2+濃度に左右される。図7Aに示したように、Pb2+濃度が増加するにつれ、凝集速度は抑制された。Pb2+の添加がないサンプルの場合、凝集は10分後にほぼ完了した。10分での吸光比を用いてPb2+濃度を定量した(図7B)。
【実施例2】
【0033】
(実施例2:アデノシン比色バイオセンサー)
[アプタザイムによる粒子の凝集]
アプタザイムをベースとするセンサーについて基質鎖の最適濃度を決定するために、各粒子溶液(E532=2.2)の20μLを混合し、Ade-リンカーを最終濃度5μMとなるように、NaClを最終濃度250mMとなるように添加し、基質添加後に容積を100μLに調整した。その溶液は25mMのTA緩衝液(pH7.2)で調整した。
この溶液を用いて、最終濃度0.15μMの基質鎖、3μMの酵素及び調節鎖、5mMのアデノシン、250mMのNaCl及び25mMのTA緩衝液(pH7.2)を含む第二の溶液を調製した。この第二の溶液の98μLを反応チューブに移し、このときを0時点と設定した。金属イオンのストック溶液2μLを前記反応チューブに添加して切断反応を開始させ、種々の時点で2μLアリコートを検出チューブに移した。反応チューブから一度検出チューブに各アリコートを移したら、EDTAを前記検出チューブに添加して切断反応を停止させた。反応チューブに由来する2μLアリコートは0.3pmolの基質を含み、切断が発生しない場合は、検出チューブ中3nMの基質濃度を与えるであろう。反応チューブの各サンプルの吸光スペクトルを、反応チューブからアリコートを移した後20分してモニターした。
センサーの感度及び選択性を調べるために、種々の濃度のアデノシン又は他のヌクレオシドを含む種々の反応チューブを調製した。30分のインキュベーション後に、2μLアリコートを検出チューブに移し、検出チューブの吸光スペクトルを20分のインキュベーション後に測定した。
図9Aは、凝集20分後の吸光スペクトルによって測定された、粒子の基質濃度依存性の凝集を示している。図9Bは、アプタザイムによる基質の切断の反応速度を示している。図9Cは、新規に設計した、アデノシンのためのアプタザイムをベースとするセンサーの感度及び選択性を示している。桃色、赤色及び青色の点は、それぞれ5mMのシチジン、グアノシン及びウリジンの吸光比である。
【0034】
(参考文献:表3Bに示すものは、別にして下に示す)
Allara D, Nuzzo R. (1985) Spontaneously organized molecular assemblies. 1. Formation, dynamics and physical properties of n-alkanoic acids adsorbed from solution on an oxidized aluminum surface. Langmuir 1: 45-52.
Beebe T, Rabke-Clemmer C, (1995) Thiol labeling of DNA for attachment to gold surfaces. Patent No. 5,472,881 USA.
Biroccio A, Hamm J, Incitti I, De Francesco R, Tomei L. (2002) Selection of RNA aptamers that are specific and high-affinity ligands of the hepatitis C virus RNA-dependent RNA polymerase. J Virol 76: 3688-3696.
Breaker RR, Joyce GF, Breaker RR, Joyce GF. (1995) A DNA enzyme with Mg(2+)-dependent RNA phosphoesterase activity.; A DNA enzyme that cleaves RNA. Chem Biol; Chem Biol 2; 1: 223-229.
Breaker RR, Joyce GF. (1994) A DNA enzyme that cleaves RNA. Chem Biol 1: 223-229.
Breaker RR. (2002) Engineered allosteric ribozymes as biosensor components. Curr Opin Biotechnol 13: 31-39.
Brody EN, Gold L. (2000) Aptamers as therapeutic and diagnostic agents. J Biotechnol 74: 5-13.
Brown A, Pavot C, Li J, Lu Y. A lead-dependent DNAzyme with a two-step mechanism. submitted.
Bruno JG, Kiel JL. (1999) In vitro selection of DNA aptamers to anthrax spores with electrochemiluminescence detection. Biosens Bioelectron 14: 457-464.
Bruno JG, Kiel JL. (2002) Use of magnetic beads in selection and detection of biotoxin aptamers by electrochemiluminescence and enzymatic methods. Biotechniques 32: 178-80, 182-3.
【0035】
Cadwell RC, Joyce GF. (1992) Randomization of genes by PCR mutagenesis. PCR Methods Appl 2: 28-33.
Cadwell RC, Joyce GF. (1994) Mutagenic PCR. PCR Methods Appl 3: S136-40.
Cao Y, Jin R, Mirkin CA. (2001) DNA-modified core-shell Ag/Au particles. J Am Chem Soc 123: 7961-7962.
Carmi N, Shultz LA, Breaker RR. (1996) In vitro selection of self-cleaving DNAs. Chem Biol 3: 1039-1046.
Chaloin L, Lehmann MJ, Sczakiel G, Restle T. (2002) Endogenous expression of a high-affinity pseudoknot RNA aptamer suppresses replication of HIV-1. Nucleic Acids Res 30: 4001-4008.
Chapman KB, Szostak JW. (1994) In vitro selection of catalytic RNAs. Curr Opin Struct Biol 4: 618-622.
Ciesiolka J, Gorski J, Yarus M. (1995) Selection of an RNA domain that binds Zn2+. RNA 1: 538-550.
Conaty J, Hendry P, Lockett T. (1999) Selected classes of minimised hammerhead ribozyme have very high cleavage rates at low Mg2+ concentration. Nucleic Acids Res 27: 2400-2407.
Conn M, Prudent J, Schultz P. (1996) Porphyrin Metallation Catalyzed by a Small RNA Molecule. J Am Chem Soc 118: 7012-7013.
Cuenoud B, Szostak JW. (1995) A DNA metalloenzyme with DNA ligase activity. Nature 375: 611-614.
【0036】
Dai X, De Mesmaeker A, Joyce GF. (1995) Cleavage of an amide bond by a ribozyme. Science 267: 237-240.
Earnshaw, Gait. (1998) Modified oligoribonucleotides as site-specific probes of RNA structure and function. Biopolymers 48: 39-55.
Ekland EH, Bartel DP. (1996) RNA-catalysed RNA polymerization using nucleoside triphosphates. Nature 382: 373-376.
Ekland EH, Szostak JW, Bartel DP. (1995) Structurally complex and highly active RNA ligases derived from random RNA sequences. Science 269: 364-370.
Ellington AD, Szostak JW. (1990) In vitro selection of RNA molecules that bind specific ligands. Nature 346: 818-822.
Faulhammer D, Famulok M. (1997) Angew Chem Int Ed Engl 35: 2837-2841.
Geyer CR, Sen D. (1997) Evidence for the metal-cofactor independence of an RNA phosphodiester-cleaving DNA enzyme. Chem Biol 4: 579-593.
Grabar K, Freeman R, Hommer M, Natan M. (1995) Preparation and characterization of Au colloid Monolayers. Anal. Chem. 67: 735-743.
Hesselberth J, Robertson MP, Jhaveri S, Ellington AD. (2000) In vitro selection of nucleic acids for diagnostic applications. J Biotechnol 74: 15-25.
Hofmann HP, Limmer S, Hornung V, Sprinzl M. (1997) Ni2+-binding RNA motifs with an asymmetric purine-rich internal loop and a G-A base pair. RNA 3: 1289-1300.
Huizenga DE, Szostak JW. (1995) A DNA aptamer that binds adenosine and ATP. Biochemistry 34: 656-665.
Iler R. (1979) Chapter 6. In: anonymous (eds) The chemistry of silica. Wiley, New York.
【0037】
Illangasekare M, Yarus M. (1997) Small-molecule-substrate interactions with a self-aminoacylating ribozyme. J Mol Biol 268: 631-639.
Jayasena SD. (1999) Aptamers: an emerging class of molecules that rival antibodies in diagnostics. Clin Chem 45: 1628-1650.
Jhaveri S, Kirby R, Conrad R, Maglott E, Bowser M, Kennedy R, Glick G, Ellington A. (2000) Designed signaling aptamers that transduce molecular recognition to changes in fluorescence intensity. J. Am. Chem. Soc. 122: 2469-2473.
Jhaveri S, Rajendran M, Ellington AD. (2000) In vitro selection of signaling aptamers. Nat Biotechnol 18: 1293-1297.
Joyce GF. (1994) In vitro evolution of nucleic acids. Curr Opin Struct Biol 4: 331-336.
Kiga D, Futamura Y, Sakamoto K, Yokoyama S. (1998) An RNA aptamer to the xanthine/guanine base with a distinctive mode of purine recognition. Nucleic Acids Res 26: 1755-1760.
Lauhon CT, Szostak JW. (1995) RNA aptamers that bind flavin and nicotinamide redox cofactors. J Am Chem Soc 117: 1246-1257.
Li J, Lu Y. (2000) A highly sensitive and selective catalytic DNA biosensor for lead ions. J Am Chem Soc 122: 10466-10467.
Li J, Zheng W, Kwon AH, Lu Y. (2000) In vitro selection and characterization of a highly efficient Zn(II)-dependent RNA-cleaving deoxyribozyme. Nucleic Acids Res 28: 481-488.
Li Y, Breaker RR. (1999) Phosphorylating DNA with DNA. Proc Natl Acad Sci U S A 96: 2746-2751.
Li Y, Sen D. (1996) A catalytic DNA for porphyrin metallation. Nat Struct Biol 3: 743-7.
【0038】
Link S, Wang Z, El-Sayed M. (1999) Alloy formation of gold-silver particles and the dependence of the plasmon absorption on their compositions. J Phys Chem B 103: 3529-3533.
Lohse PA, Szostak JW. (1996) Ribozyme-catalysed amino-acid transfer reactions. Nature 381: 442-444.
Lorsch JR, Szostak JW. (1994) In vitro evolution of new ribozymes with polynucleotide kinase activity. Nature 371: 31-36.
Lu Y. (2002) New transition-metal-dependent DNAzymes as efficient endonucleases and as selective metal biosensors. Chemistry 8: 4589-4596
Lu Y, Liu J, SIMPLE CATALYTIC DNA BIOSENSORS FOR IONS BASED ON COLOR CHANGES, Application Serial No. 09/605,558, USA.
Maoz R, Sagiv J. (1987) Penetration-controlled reactions in organized monolayer assemblies. 1. Aqueous permanganate interaction with monolayer and multilayer films of long-chain surfactants. Langmuir 3: 1034-1044.
Mirkin CA, Letsinger RL, Mucic RC, Storhoff JJ. (1996) A DNA-based method for rationally assembling particles into macroscopic materials. Nature 382: 607-609.
Mirkin, C.A., Letsinger, L. R, Mucic, C. R, Storhoff, J. J, Elghanian R, (2002) Particles having polynucleotides attached thereto and uses therefor. Patent No. 6,361,944 USA.
Mucic R, Herrlein M, Mirkin, C.A., Letsinger R. (1996) Synthesis and characterization of DNA with ferrocenyl groups attached to their 5’-termini: Electrochemical characterization of a redox-active nucleotide monolayer. Chem. Commun.555.
Nuzzo R, Fusco F, Allara D. (1987) Spontaneously organized molecular assemblies, 3. Preparation and properties of solution adsorbed monolayers of organic disulfides on gold surfaces. J Am Chem Soc 109: 2358.
【0039】
Piccirilli JA, McConnell TS, Zaug AJ, Noller HF, Cech TR. (1992) Aminoacyl esterase activity of the Tetrahymena ribozyme. Science 256: 1420-1424.
Prudent JR, Uno T, Schultz PG. (1994) Expanding the scope of RNA catalysis. Science 264: 1924-1927.
Rakow NA, Suslick KS. (2000) A colorimetric sensor array for odour visualization. Nature 406: 710-713.
Robertson MP, Ellington AD. (1999) In vitro selection of an allosteric ribozyme that transduces analytes to amplicons. Nat Biotechnol 17: 62-66.
Roth A, Breaker RR. (1998) An amino acid as a cofactor for a catalytic polynucleotide. Proc Natl Acad Sci U S A 95: 6027-6031.
Rusconi CP, Scardino E, Layzer J, Pitoc GA, Ortel TL, Monroe D, Sullenger BA. (2002) RNA aptamers as reversible antagonists of coagulation factor IXa. Nature 419: 90-94.
Santoro SW, Joyce GF. (1997) A general purpose RNA-cleaving DNA enzyme. Proc Natl Acad Sci U S A 94: 4262-4266.
Seetharaman S, Zivarts M, Sudarsan N, Breaker RR. (2001) Immobilized RNA switches for the analysis of complex chemical and biological mixtures. Nat Biotechnol 19: 336-341.
Shaiu WL, Larson DD, Vesenka J, Henderson E. (1993) Atomic force microscopy of oriented linear DNA molecules labeled with 5nm gold spheres. Nucleic Acids Res 21: 99-103.
Sillen LG, (1964) Stability constants of metal-ion complexes. Edition: 2d ed.
Smith J, Olson D, Armitage B. (1999) Molecular recognition of PNA-containing hybrids: Spontaneous assembly of helical cyanine dye aggregates on PNA templates. J. Am. Chem. Soc. 121: 2686-2695.
【0040】
Soriaga M, Hubbard A. (1982) Determination of the orientation of aromatic molecules adsorbed on platinum electrodes: The influence of solute concentration. J Am Chem Soc 104.
Soukup GA, Breaker RR. (2000) Allosteric nucleic acid catalysts. Curr Opin Struct Biol 10: 318-325.
Stojanovic MN, Landry DW. (2002) Aptamer-based colorimetric probe for cocaine. J Am Chem Soc 124: 9678-9679.
Storhoff J, Elghanian R, Mucic R, Mirkin C, Letsinger RL. (1998) One-pot colorimetric differentiation of polynucleotides with single base imperfections using gold particle probes. J Am Chem Soc 120: 1959-1964.
Tang J, Breaker RR. (1997) Rational design of allosteric ribozymes. Chem Biol 4: 453-459.
Tang J, Breaker RR. (2000) Structural diversity of self-cleaving ribozymes. Proc Natl Acad Sci U S A 97: 5784-5789.
Tarasow TM, Tarasow SL, Eaton BE. (1997) RNA-catalysed carbon-carbon bond formation. Nature 389: 54-57.
Timmons, Zisman. (1965) J. Phys. Chem. 69: 984-990.
Tompkins H, Allara D. (1974) The study of the gas-solid interaction of acetic acid with a cuprous oxide surface using reflection-absorption spectroscopy. J. Colloid and Interface Sci. 49410.
Tsang J, Joyce GF. (1996) In vitro evolution of randomized ribozymes. Methods Enzymol 267: 410-426.
Tuerk C, Gold L. (1990) Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 249: 505-510.
Vaish NK, Heaton PA, Fedorova O, Eckstein F. (1998) In vitro selection of a purine nucleotide-specific hammerheadlike ribozyme. Proc Natl Acad Sci U S A 95: 2158-2162.
【0041】
Wallace ST, Schroeder R. (1998) In vitro selection and characterization of streptomycin-binding RNAs: recognition discrimination between antibiotics. RNA 4: 112-123.
Wallis MG, Streicher B, Wank H, von Ahsen U, Clodi E, Wallace ST, Famulok M, Schroeder R. (1997) In vitro selection of a viomycin-binding RNA pseudoknot. Chem Biol 4: 357-366.
Wang DY, Lai BH, Sen D. (2002) A general strategy for effector-mediated control of RNA-cleaving ribozymes and DNA enzymes. J Mol Biol 318: 33-43.
Wecker M, Smith D, Gold L. (1996) In vitro selection of a novel catalytic RNA: characterization of a sulfur alkylation reaction and interaction with a small peptide. RNA 2: 982-994.
Whitesides, (1995) Proceedings of the Robert A. Welch Foundation 39th Conference On Chemical Research Nanophase Chemistry., Houston, TX.
Wiegand TW, Janssen RC, Eaton BE. (1997) Selection of RNA amide synthases. Chem Biol 4: 675-683.
Wilson C, Szostak JW. (1995) In vitro evolution of a self-alkylating ribozyme. Nature 374: 777-782.
Wilson DS, Szostak JW. (1999) In vitro selection of functional nucleic acids. Annu Rev Biochem 68: 611-647.
Zhang B, Cech TR. (1997) Peptide bond formation by in vitro selected ribozymes. Nature 390: 96-100.
Zillmann M, Limauro SE, Goodchild J. (1997) In vitro optimization of truncated stem-loop II variants of the hammerhead ribozyme for cleavage in low concentrations of magnesium under non-turnover conditions. RNA 3: 734-747.
【0042】
(表3Bに右上添字として示す参考文献)
REFERENCES for Table 3B
1. Ellington, A.D. & Conrad, R. (1995). Aptamers as potential nucleic acid pharmaceuticals. Biotechnol. Annu. Rev. 1: 185-214.
2. Jayasena, S.D. (1999). Aptamers: an emerging class of molecules that rival antibodies in diagnostics. Clin. Chem. (Washington, D. C.) 45: 1628-50.
3. Sun, L.Q., Cairns, M.J., Saravolac, E.G., Baker, A. & Gerlach, W.L. (2000). Catalytic nucleic acids: From lab to applications. Pharmacol. Rev. 52: 325-47.
4. Hesselberth, J., Robertson, M.P., Jhaveri, S. & Ellington, A.D. (2000). In vitro selection of nucleic acids for diagnostic applications. Rev. Mol. Biotechnol. 74: 15-25.
5. Joyce, G.F. (1999). Reactions Catalyzed by RNA and DNA Enzymes. In The RNA World, vol. 37 (Gesteland, R.F., Cech, T.R. & Atkins, J.F., ed.), pp. 687-9, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
6. Breaker, R.R. (1997). DNA enzymes. Nat. Biotechnol. 15: 427-31.
7. Sen, D. & Geyer, C.R. (1998). DNA enzymes. Curr. Opin. Chem. Biol. 2: 680-7.
8. Breaker, R.R. (1999). Catalytic DNA: in training and seeking employment. Nat. Biotechnol. 17: 422-3.
9. Breaker, R.R. (2000). Making catalytic DNAs. Science (Washington, D. C.) 290: 2095-6.
【0043】
10. Derose, V.J. (2002). Two Decades of RNA Catalysis. Chemistry & Biology 9: 961-9.
11. Ellington, A.D. & Szostak, J.W. (1990). In vitro selection of RNA molecules that bind specific ligands. Nature (London) 346: 818-22.
12. Ellington, A.D. & Szostak, J.W. (1992). Selection in vitro of single-stranded DNA molecules that fold into specific ligand-binding structures. Nature (London) 355: 850-2.
13. Santoro, S.W. & Joyce, G.F. (1997). A general purpose RNA-cleaving DNA enzyme. Proc. Natl. Acad. Sci. U. S. A. 94: 4262-6.
14. Faulhammer, D. & Famulok, M. (1996). The Ca2+ ion as a cofactor for a novel RNA-cleaving deoxyribozyme. Angew. Chem., Int. Ed. Engl. 35: 2837-41.
15. Li, J., Zheng, W., Kwon, A.H. & Lu, Y. (2000). In vitro selection and characterization of a highly efficient Zn(II)-dependent RNA-cleaving deoxyribozyme. Nucleic Acids Res. 28: 481-8.
16. Jenison, R.D., Gill, S.C., Pardi, A. & Polisky, B. (1994). High-resolution molecular discrimination by RNA. Science (Washington, DC, United States) 263: 1425-9.
17. Santoro, S.W. & Joyce, G.F. (1998). Mechanism and utility of an RNA-cleaving DNA enzyme. Biochemistry 37: 13330-42.
18. Mannironi, C., Di Nardo, A., Fruscoloni, P. & Tocchini-Valentini, G.P. (1997). In vitro selection of dopamine RNA ligands. Biochemistry 36: 9726-34.
19. Sigurdsson, S.T., Thomson, J.B. & Eckstein, F. (1998). Small ribozymes. Cold Spring Harbor Monogr. Ser. 35: 339-76.
【0044】
20. Stage-Zimmermann, T.K. & Uhlenbeck, O.C. (1998). Hammerhead ribozyme kinetics. RNA 4: 875-89.
21. Werstuck, G. & Green, M.R. (1998). Controlling gene expression in living cells through small molecule-RNA interactions. Science (Washington, D. C.) 282: 296-8.
22. Walter, N.G. & Burke, J.M. (1998). The hairpin ribozyme: structure, assembly and catalysis. Curr. Opin. Chem. Biol. 2: 24-30.
23. Holeman, L.A., Robinson, S.L., Szostak, J.W. & Wilson, C. (1998). Isolation and characterization of fluorophore-binding RNA aptamers. Folding Des. 3: 423-31.
24. Wilson, C. & Szostak, J.W. (1998). Isolation of a fluorophore-specific DNA aptamer with weak redox activity. Chem. Biol. 5: 609-17.
25. Pan, T. & Uhlenbeck, O.C. (1992). A small metalloribozyme with a two-step mechanism. Nature 358: 560-3.
26. Yang, Q., Goldstein, I.J., Mei, H.-Y. & Engelke, D.R. (1998). DNA ligands that bind tightly and selectively to cellobiose. Proc. Natl. Acad. Sci. U. S. A. 95: 5462-7.
27. Been, M.D. & Wickham, G.S. (1997). Self-cleaving ribozymes of hepatitis delta virus RNA. Eur. J. Biochem. 247: 741-53.
28. Tanner, N.K. (1998). Biochemistry of hepatitis delta virus catalytic RNAs. Ribozymes Gene Ther. Cancer: 23-38.
29. Famulok, M. & Szostak, J.W. (1992). Stereospecific recognition of tryptophan agarose by in vitro selected RNA. J. Am. Chem. Soc. 114: 3990-1.
【0045】
30. Cech, T.R. (1993). Structure and mechanism of the large catalytic RNAs: group I and group II introns and ribonuclease P. In The RNA World (Gesteland, R.F. & Atkins, J.F., ed.), pp. 239-70, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
31. Cech, T.R. & Herschlag, D. (1996). Group I ribozymes: substrate recognition, catalytic strategies, and comparative mechanistic analysis. Nucleic Acids Mol. Biol. 10: 1-17.
32. Famulok, M. (1994). Molecular Recognition of Amino Acids by RNA-Aptamers: An L-Citrulline Binding RNA Motif and Its Evolution into an L-Arginine Binder. J. Am. Chem. Soc. 116: 1698-706.
33. Geiger, A., Burgstaller, P., Von Der Eltz, H., Roeder, A. & Famulok, M. (1996). RNA aptamers that bind L-arginine with sub-micromolar dissociation constants and high enantioselectivity. Nucleic Acids Res. 24: 1029-36.
34. Tao, J. & Frankel, A.D. (1996). Arginine-Binding RNAs Resembling TAR Identified by in Vitro Selection. Biochemistry 35: 2229-38.
35. Connell, G.J., Illangesekare, M. & Yarus, M. (1993). Three small ribooligonucleotides with specific arginine sites. Biochemistry 32: 5497-502.
36. Burgstaller, P., Kochoyan, M. & Famulok, M. (1995). Structural probing and damage selection of citrulline- and arginine-specific RNA aptamers identify base positions required for binding. Nucleic Acids Res. 23: 4769-76.
37. Nolte, A., Klussmann, S., Bald, R., Erdmann, V.A. & Fuerste, J.P. (1996). Mirror-design of L-oligonucleotide ligands binding to L-arginine. Nature Biotechnology 14: 1116-9.
38. Valadkhan, S. & Manley, J.L. (2001). Splicing-related catalysis by protein-free snRNAs. Nature (London, United Kingdom) 413: 701-7.
39. Nissen, P., Hansen, J., Ban, N., Moore, P.B. & Steitz, T.A. (2000). The structural basis of ribosome activity in peptide bond synthesis. Science (Washington, D. C.) 289: 920-30.
【0046】
40. Harada, K. & Frankel, A.D. (1995). Identification of two novel arginine binding DNAs. EMBO J. 14: 5798-811.
41. Carmi, N., Balkhi, H.R. & Breaker, R.R. (1998). Cleaving DNA with DNA. Proc. Natl. Acad. Sci. U. S. A. 95: 2233-7.
42. Majerfeld, I. & Yarus, M. (1994). An RNA pocket for an aliphatic hydrophobe. Nat. Struct. Biol. 1: 287-92.
43. Cuenoud, B. & Szostak, J.W. (1995). A DNA metalloenzyme with DNA ligase activity. Nature 375: 611-4.
44. Majerfeld, I. & Yarus, M. (1998). Isoleucine:RNA sites with associated coding sequences. Rna 4: 471-8.
45. Li, Y. & Breaker, R.R. (1999). Phosphorylating DNA with DNA. Proc. Natl. Acad. Sci. U. S. A. 96: 2746-51.
46. Sassanfar, M. & Szostak, J.W. (1993). An RNA motif that binds ATP. Nature (London) 364: 550-3.
47. Burgstaller, P. & Famulok, M. (1994). Isolation of RNA aptamers for biological cofactors by in vitro selection. Angew. Chem. 106: 1163-6 (See also Angew. Chem., Int. Ed. Engl., 994, 33(10), 084-7).
48. Burke, D.H. & Gold, L. (1997). RNA aptamers to the adenosine moiety of S-adenosyl methionine: structural inferences from variations on a theme and the reproducibility of SELEX. Nucleic Acids Res. 25: 2020-4.
49. Huizenga, D.E. & Szostak, J.W. (1995). A DNA Aptamer That Binds Adenosine and ATP. Biochemistry 34: 656-65.
【0047】
50. Klussmann, S., Nolte, A., Bald, R., Erdmann, V.A. & Fuerste, J.P. (1996). Mirror-image RNA that binds D-adenosine. Nat. Biotechnol. 14: 1112-5.
51. Burmeister, J., Von Kiedrowski, G. & Ellington, A.D. (1997). Cofactor-assisted self-cleavage in DNA libraries with a 3’-’5’-phosphoramidate bond. Angew. Chem., Int. Ed. Engl. 36: 1321-4.
52. Connell, G.J. & Yarus, M. (1994). RNAs with dual specificity and dual RNAs with similar specificity. Science (Washington, D. C.) 264: 1137-41.
53. Li, Y. & Sen, D. (1996). A catalytic DNA for porphyrin metallation. Nat. Struct. Biol. 3: 743-7.
54. Lauhon, C.T. & Szostak, J.W. (1995). RNA aptamers that bind flavin and nicotinamide redox cofactors. J. Am. Chem. Soc. 117: 1246-57.
55. Travascio, P., Bennet, A.J., Wang, D.Y. & Sen, D. (1999). A ribozyme and a catalytic DNA with peroxidase activity: active sites versus cofactor-binding sites. Chemistry & Biology 6: 779-87.
56. Lorsch, J.R. & Szostak, J.W. (1994). In vitro selection of RNA aptamers specific for cyanocobalamin. Biochemistry 33: 973-82.
57. Rink, S.M., Shen, J.-C. & Loeb, L.A. (1998). Creation of RNA molecules that recognize the oxidative lesion 7,8-dihydro-8-hydroxy-2’-deoxyguanosine (8-oxodG) in DNA. Proc. Natl. Acad. Sci. U. S. A. 95: 11619-24.
58. Haller, A.A. & Sarnow, P. (1997). In vitro selection of a 7-methyl-guanosine binding RNA that inhibits translation of capped mRNA molecules. Proc. Natl. Acad. Sci. U. S. A. 94: 8521-6.
59. Kiga, D., Futamura, Y., Sakamoto, K. & Yokoyama, S. (1998). An RNA aptamer to the xanthine/guanine base with a distinctive mode of purine recognition. Nucleic Acids Res. 26: 1755-60.
【0048】
60. Lato, S.M., Boles, A.R. & Ellington, A.D. (1995). In vitro selection of RNA lectins: Using combinatorial chemistry to interpret ribozyme evolution. Chem. Biol. 2: 291-303.
61. Wang, Y., Killian, J., Hamasaki, K. & Rando, R.R. (1996). RNA Molecules That Specifically and Stoichiometrically Bind Aminoglycoside Antibiotics with High Affinities. Biochemistry 35: 12338-46.
62. Wallis, M.G., Von Ahsen, U., Schroeder, R. & Famulok, M. (1995). A novel RNA motif for neomycin recognition. Chem. Biol. 2: 543-52.
63. Famulok, M. & Huettenhofer, A. (1996). In Vitro Selection Analysis of Neomycin Binding RNAs with a Mutagenized Pool of Variants of the 16S rRNA Decoding Region. Biochemistry 35: 4265-70.
64. Wallis, M.G., Streicher, B., Wank, H., Von Ahsen, U., Clodi, E., Wallace, S.T., Famulok, M. & Schroeder, R. (1997). In vitro selection of a viomycin-binding RNA pseudoknot. Chem. Biol. 4: 357-66.
65. Burke, D.H., Hoffman, D.C., Brown, A., Hansen, M., Pardi, A. & Gold, L. (1997). RNA aptamers to the peptidyl transferase inhibitor chloramphenicol. Chem. Biol. 4: 833-43.
66. Wallace, S.T. & Schroeder, R. (1998). In vitro selection and characterization of streptomycin-binding RNAs: recognition discrimination between antibiotics. Rna 4: 112-23.
67. Giver, L., Bartel, D.P., Zapp, M.L., Green, M.R. & Ellington, A.D. (1993). Selection and design of high-affinity RNA ligands for HIV-1 Rev. Gene 137: 19-24.
68. Giver, L., Bartel, D., Zapp, M., Pawul, A., Green, M. & Ellington, A.D. (1993). Selective optimization of the Rev-binding element of HIV-1. Nucleic Acids Res. 21: 5509-16.
69. Williams, K.P., Liu, X.-H., Schumacher, T.N.M., Lin, H.Y., Ausiello, D.A., Kim, P.S. & Bartel, D.P. (1997). Bioactive and nuclease-resistant L-DNA ligand of vasopressin. Proc. Natl. Acad. Sci. U. S. A. 94: 11285-90.
【0049】
70. Zimmerman, J.M. & Maher, L.J., Iii (2002). In vivo selection of spectinomycin-binding RNAs. Nucleic Acids Res. 30: 5425-35.
71. Vianini, E., Palumbo, M. & Gatto, B. (2001). In vitro selection of DNA aptamers that bind L-tyrosinamide. Bioorganic & Medicinal Chemistry 9: 2543-8.
72. Andreola, M.-L., Pileur, F., Calmels, C., Ventura, M., Tarrago-Litvak, L., Toulme, J.-J. & Litvak, S. (2001). DNA aptamers selected against the HIV-1 RNase H display in vitro antiviral activity. Biochemistry 40: 10087-94.
73. Fukusaki, E.-I., Kato, T., Maeda, H., Kawazoe, N., Ito, Y., Okazawa, A., Kajiyama, S.-I. & Kobayashi, A. (2000). DNA aptamers that bind to chitin. Bioorg. Med. Chem. Lett. 10: 423-5.
74. Bock, L.C., Griffin, L.C., Latham, J.A., Vermaas, E.H. & Toole, J.J. (1992). Selection of single-stranded DNA molecules that bind and inhibit human thrombin. Nature (London) 355: 564-6.
75. Koizumi, M. & Breaker, R.R. (2000). Molecular Recognition of cAMP by an RNA Aptamer. Biochemistry 39: 8983-92.
76. Kato, T., Takemura, T., Yano, K., Ikebukuro, K. & Karube, I. (2000). In vitro selection of DNA aptamers which bind to cholic acid. Biochim. Biophys. Acta 1493: 12-8.
77. Okazawa, A., Maeda, H., Fukusaki, E., Katakura, Y. & Kobayashi, A. (2000). In vitro selection of hematoporphyrin binding DNA aptamers. Bioorg. Med. Chem. Lett. 10: 2653-6.
78. Kawakami, J., Imanaka, H., Yokota, Y. & Sugimoto, N. (2000). In vitro selection of aptamers that act with Zn2+. J. Inorg. Biochem. 82: 197-206.
79. Bruno, J.G. & Kiel, J.L. (1999). In vitro selection of DNA aptamers to anthrax spores with electrochemiluminescence detection. Biosensors & Bioelectronics 14: 457-64.
【図面の簡単な説明】
【0050】
【図1】図1Aは酵素鎖(17E)及び基質鎖(17DS)から成る“8-17”DNAザイムシステムの二次構造を示し、図1BはPb(II)の存在下での17Eによる17DSの切断を示す。図1Cは、DNAザイムによって誘導される金粒子のアッセンブリーと、その金属イオン(例えばPb(II))のバイオセンサーとしての利用を表す模式図を示す。
【図2】図2Aは、“8-17”DNAザイム土台上に構築されたアデノシンアプタザイムの一次構造及び提案する二次構造を示す。図2Bは、アデノシンの比色検出の模式図を占め得s。
【図3】DNA連結金粒子の2つの整列様式を示す。図3Aは、“頭部-尾部”整列様式の粒子を示し、この場合ただ1種類のチオール改変DNAが必要である。図3Bは、“尾部-尾部”整列様式の粒子を示す。
【図4】色の変化の速度に対する粒子の整列様式及びサイズの影響を示す。
【図5】Pb2+の比色バイオセンサーを示す。前記センサーでは、粒子の直径は〜35nmであり、粒子は“尾部-尾部”整列様式の態様で並べられている。
【図6】新規なPb2+センサーシステムを示す。図6Aは、35nmの金粒子の種々の時間における吸光スペクトルを示す。図6Bは、凝集速度に対する基質濃度の影響を示す。図6Cは、凝集速度に対するNaCl濃度の影響を示す。図6Dは、凝集速度に対する17E濃度の影響を示す。図6Eは、種々のpHの緩衝液中での粒子の凝集を示す。図6Fは、凝集10分後の吸光比を示す。
【図7】新規なPb2+センサーの感度及び選択性を示す。図7Aは、種々のPb2+濃度による金粒子の凝集の反応速度を示す。図7Bは、開始後10分の吸光比を示す。
【図8】アプタザイムをベースとするアデノシンセンサーの新規な設計を示す。反応A:アデノシンの非存在下で青色凝集体が形成される。反応B:基質はアデノシンの存在下で切断される。反応C:切断された基質は金粒子を集させることができず、赤色の分離している粒子が生じる。
【図9】図9Aは粒子の基質濃度依存性の凝集を示す。図9Bはアプタザイムによる基質の切断の反応速度を示す。図9Cはアプタザイムをベースとするセンサーのアデノシンに対する感度及び選択性を示す。
【図1A】
【図1B】
【図1C】
【図2A】
【図2B】
【図3A】
【図3B】
【技術分野】
【0001】
本発明はバイオセンサーの分野に関する。より具体的には、本発明は、触媒的に活性な核酸による、加熱を要しないバイオセンサーに関する。
【背景技術】
【0002】
タンパク質の他に、核酸もまた触媒活性を有することが近年見出された。触媒的に活性な核酸は触媒性DNA/RNAであり、DNAザイム/RNAザイム、デオキシリボザイム/リボザイム及びDNA酵素/RNA酵素としても知られている。触媒的に活性な核酸はまた改変核酸を含むことができる。核酸系酵素の触媒活性は、常に一定の補助因子(例えば金属イオン)の存在に左右される。したがって、これら補助因子のための核酸酵素をベースとするバイオセンサー(例えば金属イオンのためのバイオセンサー)を、対応する核酸酵素の活性を基準にして設計することができる。他方、広範囲の分析物と高い親和性及び特異性で結合するように、核酸を選択することができる。これらの結合核酸はアプタマーとして知られている。
アプタマーは、高い親和性及び特異性で標的を認識する核酸(例えばDNA又はRNA)である(Ellington and Szostak 1990;Jayasena 1999)。アプタザイム(アロステリックDNA/RNAザイム又はアロステリック(デオキシ)リボザイムとも称される)は、エフェクター(標的分子)によって調節されるDNA/RNAザイムである。前記は典型的には、エフェクターを認識するアプタマードメイン及び触媒ドメインを含む(Hesselberth et al. 2000;Soukup and Breaker 2000;Tang and Breaker 1997)。エフェクターは、アプタマードメインと触媒ドメインとの間の特異的な相互作用を介して、アプタザイムの触媒活性を低下又は増加させることができる。したがって、アプタザイムの活性を用いて前記エフェクターの有無及び量をモニターすることができる。この方法は、診断及び探知の目的でアプタザイムセンサーを選別及び設計するために用いられた(Breaker 2002;Robertson and Ellington 1999;Seetharaman et al. 2001)。DNAザイム及びDNAアプタザイムはセンサーの開発にもっとも魅力的な候補物質である。なぜならば、DNAはRNAよりも合成がはるかに廉価であり、さらにRNAよりも安定であるからである。さらにまた、DNAザイムの触媒コア近くにアプタマーモチーフを導入することによるDNAアプタザイムの設計の一般的方法が利用可能である(Wang et al. 2002)。高い切断活性には、アプタマーモチーフとの結合時に、前記アプタザイムの触媒コア部分の活性をアロステリックに調節することができるエフェクター分子の存在が必要とされる。
【0003】
in vitroセレクション方法を用いて、極めて高い親和性(ピコモル範囲の高い解離定数を有する)をもつ、広範囲の標的分子のためのアプタマーを得ることができる(Brody and Gold 2000;Jayasena 1999;Wilson and Szostak 1999)。例えば、アプタマーは、金属イオン(例えばZN(II);Ciesiolka et al.(1995)及びNi(II);Hofmann et al. 1997)、ヌクレオチド(例えばアデノシン三燐酸(ATP);Huizenga and Szostak(1995)及びグアニン;Kiga et al. 1998)、補助因子(例えばNAD;Kiga et al.(1998)及びフラビン;Lauhon and Szostak 1995)、抗生物質(例えばヴィオマイシン;Wallis et al.(1997)及びストレプトマイシン;Wallace and Schroeder 1998)、タンパク質(例えばHIV逆転写酵素;Chaloin et al.(2002)及びC型肝炎ウイルスRNA依存RNAポリメラーゼ;Biroccio et al. 2002)、毒素(例えばコレラ完全毒素及びスタフィロコッカス内毒素B;Bruno and Kiel 2002)及び細菌芽胞(例えば炭疽;Bruno and Kiel 1999)を認識するために開発された。抗体と比べて、DNA/RNA系アプタマーは、in vitroで短時間に(数日に対して数週)、及び限られた費用で製造することができるために入手が容易で、より安価に製造される。さらにまた、DNA/RNAアプタマーは、その生物関連物質の認識能力を失うことなく何度も変性及び再生させることができる。これらの固有の特性のゆえに、アプタマーは、高度な感受性及び選択性を有するバイオセンサーの設計のための理想的な土台となる(Hesselberth et al. 2000)。
【0004】
DNA/RNAザイム又はアプタザイム活性をアッセイするために、検出可能な標識が用いられる。しかしながら、これらの多くは顕著な欠点を有する。放射性同位元素は安全性と廃棄に関する懸念があり、一方、蛍光発色団は光退色を受け、アッセイしようとしている生物学的活性を阻害する可能性がある(すなわち、有機色素(例えばコカイン検出アプタマーシステム(Stojanovic and Landry, 2002)で用いられるもの)は視覚的検出に高い濃度を必要とし、高価な試行錯誤の後でやっと特異的なアプタマーとの組合せが得られる。
金属粒子はこれらの問題の全てを解決する。前記粒子は、有機色素のいずれの欠点も伴わずに、検出剤として少量(ナノモル濃度)でアプタマーとともに用いることができる。発色検出のために金属粒子を用いるアプタマー系センサーでは、(エフェクター結合時の)アプタザイムによる核酸基質の切断は、色の変化によって検出することができる。
典型的には、DNA/RNAザイム又はアプタザイムをベースとするセンサーは以下の3つの部分を有する:
(1)核酸酵素(以下の記述では、核酸酵素はDNA/RNAザイム及びアプタザイムと称されるであろう)及び補助因子(例えば基質の切断を触媒する金属イオン);
(2)核酸酵素のための核酸基質(この場合基質配列の内部部分は前記酵素配列の部分と相補的である);及び
(3)基質の3'-及び5'-末端と相補的なポリヌクレオチドと結合した粒子。
【0005】
標的補助因子又はエフェクターを検出するために、ポリヌクレオチドの相補性部分(基質鎖の3'-及び5'-末端と相補的な、粒子に結合したポリヌクレオチド、並びに基質鎖内部配列と相補的な核酸酵素の5'-及び3'-末端)を、標的の補助因子又はエフェクターが含まれていると思われるサンプルの存在下でアニールさせる。補助因子又はエフェクターが存在しない場合、アプタザイムは不活性であるか、又は実質的に低い活性を示し、基質切断は全く又はほとんど生じず、したがって粒子の凝集をもたらす。補助因子又はエフェクターが存在する場合、酵素は活性化し基質を切断して凝集形成は妨げられる(なぜならば、粒子間の連結が酵素性切断によって破壊されるからである)。表1はそのような系の例示である。
*金粒子が検出に用いられた場合。
【0006】
金粒子の事例では、凝集状態では青色が表示されるが、分散状態(又は非凝集状態)では赤色である。補助因子又はエフェクターのような標的分析物の存在は、センサーシステムの発色を基準にして検出することができる。
切断の程度は色の変化の度合いに反映されるので、標的補助因子又はエフェクターの濃度を定量することができる。単純な分光法を鋭敏な検出に用いることができる。検出及び定量に色の変化を用いるだけでなく、他の切断の成果、例えば沈殿を利用してもよい。アプタマーを予め選択したエフェクターを認識するアプタマーに置き換えることによって、任意の所望のエフェクターのための比色センサーを容易に製造し用いることができる。
これまでの研究を基にして、DNAザイムに誘導される(directed)金ナノ粒子アッセンブリーを基にしたPb(II)のための比色バイオセンサー、及びアプタザイムによって誘導される金ナノ粒子アッセンブリーを基にしたアデノシンのための比色バイオセンサーが設計された(例えば以下を参照されたい:米国特許出願09/605,558;10/144,094;10/144,679;10/384,497)。高感度及び高選択性ではあるが、このタイプの分析センサーは、検出のために50℃を超える数分間の加熱及び2時間に及ぶ室温への緩徐な冷却を必要とする。
【発明の開示】
【0007】
(発明の要旨)
第一の特徴では、本発明は、エフェクター又は補助因子を検出することを目的とするセンサーシステムであり、前記センサーシステムは次のものを含む:(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;(c)第二のポリヌクレオチドを含む第一の粒子セット、ここで前記第二のポリヌクレオチドはその3'末端で前記粒子に付着している;及び(d)第三のポリヌクレオチドを含む第二の粒子セット、ここで前記第三のポリヌクレオチドはその5'末端で前記粒子に付着している。前記第一のポリヌクレオチドは、前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的である。
第二の特徴では、本発明は、エフェクター又は補助因子を検出するためのセンサーシステムであり、前記センサーシステムは次のものを含む:(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;(c)第二のポリヌクレオチドを含む第一の粒子セット;及び(d)第三のポリヌクレオチドを含む第二の粒子セット。前記第一のポリヌクレオチドは、前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的である。前記第二の粒子セットは少なくとも20nmの直径を有する。
第三の特徴では、本発明はサンプル中のエフェクター又は補助因子の検出方法であり、前記方法は、少なくとも
(a)核酸酵素(補助因子結合部位と、任意にエフェクター結合部位とを含む);
(b)前記核酸酵素の基質(第一のポリヌクレオチドを含む);
(c)第二のポリヌクレオチドを含む第一の粒子セット;
(d)第三のポリヌクレオチドを含む第二の粒子セット;及び
(e)サンプル;
を一緒に混合して、混合物を生成することを含む。前記第一のポリヌクレオチドは、前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的である。前記混合物は、加熱することなく前記混合物の生成から10分以内に、エフェクター又は補助因子のサンプル中の存在を示す色の変化を生じる。
【0008】
(詳細な説明)
本発明は、以前のDNAザイム-及びアプタザイム-金属粒子センサーシステムで要求される加熱を回避し、実験室だけでなく消費者が家庭で又は検査技師が野外でこれらセンサーを使用することを可能にする。本発明は、次のことが粒子をベースとする比色バイオセンサーの性能に影響を与えるという発見を利用している:(1)粒子の整列様式を頭部-尾部整列様式から尾部-尾部整列様式に変更すること;(2)より大きな粒子を使用すること;及び(3)イオン強度、アプタマー濃度及びpHを制御すること。
粒子の凝集は、粒子の互いに対する整列様式に影響される。粒子は2種の態様、“頭部-尾部”整列様式又は“尾部-尾部”整列様式で整列化することができる(図3)。例えば、初期の、DNAザイムをベースとするPb2+センサーでは、粒子は“頭部-尾部”様式で整列化される。粒子が“頭部-尾部”様式で整列化される場合、ただ1種のチオール改変DNAが、粒子にDNAを付着させるために必要である。この立体配置では、おそらく立体的影響のため、粒子が凝集することは困難である。したがって、加熱及び冷却が粒子のアッセンブリーを促進するために必要である。しかしながら、“尾部-尾部”整列様式を用いる場合、粒子は周囲温度で凝集できることを発見した(図5)。“尾部-尾部”整列化する粒子には、3'-チオール改変ポリヌクレオチド及び5'-チオール改変ポリヌクレオチドの両方が必要である。
粒子凝集体の色は、主としてその凝集体のサイズによって支配される。したがって、凝集速度が等しい場合、色の変化の速度は、より大きな粒子を用いたときに増す。下記の実施例で示すように、直径が13nm及び35nmの粒子の混合物を用いた場合、色の変化の速度は直径13nmの粒子のみを用いた場合よりも速い。色の変化の速度は、直径35nmの粒子のみを用いた場合にさらに速くなる。
同様に、溶液のイオン強度は、システムの性能に影響する。高い塩濃度は凝集に有利に働き、これは、おそらく、塩が負の電荷を有するポリヌクレオチド鎖間の静電気的反発を低下させるためである。2つの粒子を連結させるのに基質鎖のみが必要とされるが、酵素鎖の非存在下では、基質は“ふらふらして(floppy)”いて、粒子を集合させることができない。したがって、酵素の濃度は重要なパラメーターである。
pHもまた凝集に影響する。pHの影響は、おそらくポリヌクレオチドの塩基のプロトン化のためであり、塩基対形成に影響し、したがって凝集速度に影響を及ぼす。
【0009】
[定義]
“補助因子”は、核酸酵素によって触媒される反応の触媒過程に関与するイオン又は分子であり、触媒活性に必要とされるものである。
“エフェクター”は、エフェクター結合部位を有する酵素と結合した場合に、酵素の触媒作用を強化又は阻害することができる分子である。“エフェクター結合部位”は“特異的”、すなわち、他のエフェクター分子の存在下でただ1つのエフェクター分子とのみ結合するものであってもよい。エフェクター結合部位の特異性の例は、多くの他の類似分子(例えばシチジン、グアノシン及びウリジン)の存在下で、アデノシン分子のみが結合するといった場合である。あるいは、エフェクター結合部位は、“部分的に”特異的(1種類の分子とだけ結合する)であってもよく、又は“非特異的” (分子に対する無差別性を有する)であってもよい。エフェクターの例には、環境汚染物質(例えば窒素肥料、農薬、ダイオキシン、フェノール又は2,4-ジクロロフェノキシ酢酸)、重金属イオン(例えばPb(II)、Hg(II)、As(III)、UO2(II)、Fe(III)、Zn(II)、Cu(II)又はCo(II))、生物学的分子(例えばグルコース、インシュリン、hCGホルモン、HIV、HIVタンパク質)、化学的及び生物学的テロ関連物質(例えば炭疽、天然痘又は神経ガス)、爆発物(例えばTNT又はDNT)、薬剤(例えばコカイン又は抗生物質)が含まれる。
“核酸酵素”は、主として核酸を含む酵素、例えばリボザイム(RNAザイム)、デオキシリボザイム(DNAザイム)及びアプタザイムである。核酸は、天然の核酸でも非天然の核酸でも又は改変された核酸であってもよい。ペプチド核酸(PNA)もまた含まれる。核酸酵素は、効率的な基質切断及び/又は特異的なエフェクター結合のために“補助因子”を必要とする。一般的な補助因子には、Mg(II)、Ca(II)、Zn(II)、Mn(II)、Co(II)及びPb(II)が含まれる。
【0010】
“ポリヌクレオチド”は、少なくとも2つ以上のヌクレオチドを有する核酸配列をさす。ポリヌクレオチドは、天然に存在するヌクレオチド及び合成ヌクレオチドを含み得る。PNA分子も、この用語に包含される。
“感度”は、センサーによって検出することができる、補助因子又はエフェクターの濃度のもっとも小さな増加を指す。
“検出限界”は、分析装置の検出限界を指す。本発明のDNAザイムをベースとするセンサー及びアプタザイムをベースとするセンサーとの関連において、検出限界は、センサーがバックグラウンドから区別することができる補助因子又はエフェクターの最低濃度を指す。
“塩基対形成”は、ポリヌクレオチドが、低ストリンジェンシー条件下でヌクレオチドと少なくとも1つの水素結合を形成する能力を指す。前記ヌクレオチドは、第二のヌクレオチドの一部であっても、第一のポリヌクレオチド内で見出されるヌクレオチドであってもよい。第一のポリヌクレオチドが第二のポリヌクレオチドと少なくとも1つの水素結合を形成することができるとき、ポリヌクレオチドは第二のポリヌクレオチドと部分的に相補的である。部分的に相補的であるためには、ポリヌクレオチドは、塩基対を形成する領域に取り囲まれている、塩基対が形成されない領域を有することができ、ループ、ステム-ループ及び他の二次構造を形成する。
“アプタマー”はエフェクター結合部位を含むポリヌクレオチドを指す。“エフェクター結合部位”は“特異的”、すなわち、他のエフェクター分子の存在下でただ1つのエフェクター分子とのみ結合するものであってもよい。エフェクター結合部位の特異性の例は、多くの他の類似分子(例えばシチジン、グアノシン及びウリジン)の存在下で、アデノシン分子のみが結合するといった場合である。あるいは、エフェクター結合部位は、“部分的に”特異的(1種類の分子とだけ結合する)であってもよく、又は“非特異的” (分子に対して無差別性を有する)であってもよい。
“アプタザイム”は、エフェクターと結合するアプタマー領域を含む核酸酵素を指す。エフェクターの結合は、触媒作用を強化又は阻害することができる。
【0011】
[1のエフェクター(又は複数のエフェクター)の結合部位をもつ核酸酵素]
多様な触媒活性を有する、多数の核酸酵素が発見又は開発されてきた(表1及び2)。触媒機能について、酵素は通常1又は2以上の補助因子に依存する。in vitroセレクションは、特定のイオンについての選択性及び感度を“強化する”ために用いることができる。分子の会合(連結、リン酸化及びアミド結合形成)又は解離(切断又は転移)を触媒する核酸酵素は、本発明で特に有用である。
エフェクターの存在下で核酸の切断を触媒する核酸酵素が用いられる。前記核酸酵素は、RNA(リボザイム)、DNA(デオキシリボザイム)、DNA/RNAハイブリッド酵素、又はペプチド核酸(PNA)酵素であってもよい。PNAはポリアミド骨格及びヌクレオシド塩基を含む(例えばBiosearch, Inc.(Bedford, MA)から入手できる)。使用することができるリボザイムには、グループI及びグループIIのイントロン、細菌リボヌクレアーゼPのRNA成分、ハンマーヘッドリボザイム、ヘアピンリボザイム、肝炎デルタウイルスリボザイム及びニューロスポーラVSリボザイムが含まれる。さらに、例えば以前に単離されたもの(Tang and Breaker 2000))もin vitroセレクションされたリボザイムに含まれる。リボザイムは、デオキシリボザイムより不安定な傾向があり、したがってデオキシリボザイムが好ましい。拡張された化学的官能性をもつデオキシリボザイムもまた所望される(Santoro et al. 2000)。
【0012】
表1:in vitroセレクション実験から単離されたリボザイムによって触媒される反応
a:in vitroセレクション実験から単離されたリボザイムによって触媒される反応。Kcat/Kuncatは、未触媒反応を超える速度増大である。
参考文献:1(Vaish et al. 1998);2(Tsang and Joyce 1996);3(Ekland et al. 1995);4(Lorsch and Szostak 1994);5(Ekland and Bartel 1996);6(Illangasekare and Yarus 1997);7(Piccirilli et al. 1992);8(Lohse and Szostak 1996);9(Wilson and Szostak 1995);10(Wecker et al. 1996);11(Dai et al. 1995);12(Wiegand et al. 1997);13(Zhang and Cech 1997);14(Tarasow et al. 1997);15(Prudent et al. 1994);16(Conn et al. 1996)。
【0013】
表2:in vitroセレクションにより単離されたデオキシリボザイム
b:Kmaxは至適条件下で得られた最大速度定数である。
参考文献:1(Breaker and Joyce 1994);2(Breaker et al. 1995);3(Faulhammer and Famulok 1997);4(Santoro and Joyce 1997);5Geyer and Sen 1997);6(Roth and Breaker 1998);7(Li et al. 2000);8(Carmi et al. 1996);9(Cuenoud and Szostak 1995);10(Li and Breaker 1999);11(Li et al. 2000);12(Li and Sen 1996)。
【0014】
リボザイム及びデオキシリボザイムを製造する方法には、オリゴヌクレオチドの化学的合成、ポリメラーゼ連鎖反応(PCR)、DNAクローニング及び複製が含まれる。好ましくは、核酸酵素はDNA/RNAハイブリッド及びPNAである。改変した塩基、ホスフェート又は糖を含むヌクレオチドも用いることができる。いくつかの事例では、これらの改変したヌクレオチドは、安定性の利点を有し、又はエフェクター特異性を付与する。改変した塩基の例には、イノシン、ネブラリン、2-アミノプリンリボシド、N7-デアザアデノシン及びO6-メチルグアノシンが含まれる(Earnshaw and Gait 1998)。改変した糖及びホスフェートには、2'-デオキシヌクレオシド、脱塩基(abasic)、プロピル、ホスホロチオエート及び2'-O-アリルヌクレオシドが含まれる(Earnshaw and Gait 1998)。
酵素を含む鎖から離れた場所の核酸の鎖を切断する核酸酵素はトランス作動性酵素である。トランス作動性酵素の使用は、酵素生成物が取り除かれるために何回もの基質切断を可能にする。そのような核酸酵素の例は17E(配列番号1)である。対応する基質は17DS(配列番号2:rは、1個のリボヌクレオチドを表す)である。両者は表3A及び図1に示されている。“8-17”DNAザイムシステムの二次構造(酵素鎖(17E)及び基質鎖(17DS)を含む)は図1aに示されている。切断部位は矢印で示されている。切断部位のリボヌクレオシドアデノシン(rA)を除いて、他の全てのヌクレオシドがデオキシリボヌクレオシドである。Pb(II)の存在下で、酵素鎖は基質鎖を切断する(図1b)。したがって、図1cに示されているように、酵素鎖及び基質鎖を、金粒子のDNAザイム誘導アッセンブリーで用いることができ、前記アッセンブリーの金属イオン(例えばPb(II))についてのバイオセンサーとしての用途で用いることができる。このシステムでは、17DSはその3' 末端及び5'末端の両方で12塩基伸長されており、前記塩基は13nmの金粒子に付着させた12-merのDNA(DNAAU)と相補的である。他の例もまた表3Bに提示されている。
【0015】
表3A:DNA酵素及び基質
c"r"は1個のリボヌクレオチドを表す。
【0016】
表3B:RNA/DNAをベースとするアプタマー及びRNA/DNAザイム
【0017】
定方向変異導入を用いて、核酸酵素又はその基質の1又は2以上の特性を変化させることができる。一例として17E及び17DSを用い、ハイブリダイズした酵素と基質の2本のアームのアビディティーを変化させるようとすることもできる。前記“アーム”は、図1でワトソン-クリックの塩基対形成を示す領域である。アビディティーを変化させるために、アームの長さを変化させる。アームの長さを増加させることによって、ワトソン-クリックの塩基対形成の数が増加し、したがってアビディティーが増加する;アームの長さを減少させることによってアビディティーは低下する。アームのアビディティーを低下させることによって酵素から基質が除去されやすくなり、したがってより迅速な酵素のターンオーバーが可能になる。
アビディティーを低下させる別の方法には、酵素と基質との間にミスマッチを作り出すことが含まれる。あるいは、アームのG-C含有量を変化させてもよい。いずれの誘導変化の影響もモニターし、酵素が所望の活性(イオンに対する感度及び選択性を含む)を維持していることを確実にすべきである。例えば、変異酵素がアデノシンに対する感度及び選択性を維持していることを確実にするために、変異酵素がアデノシンの存在下で反応性を維持しているか否か(感度)及び他のエフェクターの存在下でより低レベルの活性を維持しているか否か(選択性)を決定する試験を実施できよう。
【0018】
[アプタマーのin vitroセレクション]
所望のエフェクターと結合するアプタマー及びアプタザイムは、in vitroセレクションによって単離することができる。in vitroセレクションは、一定の機能をもつRNA分子又はDNA分子を選別と増幅との複数回ものサイクルにより多数の配列変種から単離する技術である(Chapman and Szostak 1994;Joyce 1994)。最大の活性又は新規な触媒能力をもつDNAザイム及びRNAザイムが、アプタマーと同様に、例えば指数関数的濃縮によるリガンドの系統的進化(SELEX, systematic evolution of ligands by exponential enrichment)技術を用いて入手することができる(Tuerk and Gold 1990)。
in vitroセレクションは、典型的には、ランダム化された配列の大きな集合(プール)(通常は1013〜1016の配列変種を含む)を用いて開始される。そのようなランダム化されたプールの作製には、標準的なホスホアミダイト化学反応を用いた縮退ポリヌクレオチドのセットの化学合成を用いることができる。4種のヌクレオシド(アデノシン、シトシン、グアニン、チミジン)の3'-ホスホアミダイト化合物を予め混合し、これをポリヌクレオチドの合成に使用する;ランダム化は4種のホスホアミダイトの比率を制御することによって得られる。ホスホアミダイト定数を特定の位置で保持するだけでなく、バイアスを達成することもできる。ランダム化DNAライブラリーを作製する他のストラテジーには、変異導入ポリメラーゼ連鎖反応(PCR)及び鋳型特異的(template-directed)変異導入が含まれる(Cadwell and Joyce 1992;Cadwell and Joyce 1994;Tsang and Joyce 1996)。RNA分子のin vitroセレクションを所望する場合は、先ず初めにin vitro転写によってランダム化DNAライブラリーをRNAライブラリーに変換する。
【0019】
続いて、ランダム化ライブラリーを、所望の機能(例えば標的化エフェクターとの結合)をもつ分子についてスクリーニングして、単離する。分離は、アフィニティーカラムクロマトグラフィー(例えば標的化エフェクターを用いる)、ゲル電気泳動、又はタグ付加反応中間体の選択的増幅を用いて達成することができる。選別した分子を、例えばDNAのためのPCR、又はRNAのための等温増幅反応を用いて増幅する。続いて、これらの選別増幅分子に、例えば変異導入PCRを用いて変異を導入し(多様性の再導入)、さらに高い活性をもつ分子の選別を試みる。所望の活性を持つ配列がプール中で優位を占めるまで、これら3つの工程、選別、増幅及び変異導入を、しばしば選別のストリンジェンシーを高めながら、繰り返す。
ランダム配列からin vitroで単離された新規な核酸酵素は、RNA及びDNAの触媒レパートリーを拡大させた(表1)。デオキシリボザイムは、リボザイムと比較してより少ない反応タイプを触媒する(表2)。ほとんどのデオキシリボザイムの触媒速度(Kcat)は、同じ反応を触媒するリボザイムの触媒速度に匹敵する。ある種の事例では、核酸酵素の触媒効率(Kcat/Km)は、タンパク質酵素の触媒効率を超えさえする(Santoro and Joyce 1997)。
in vitroセレクションを用いて、現存の核酸酵素のイオン特異性又は結合親和性を変化させることができ、又は所望の基質に特異的な核酸酵素を得ることができる。例えば、in vitroセレクションを用いて、最適なハンマーヘッドリボザイム活性に必要とされるMg2+濃度を低下させ、生理的条件下での酵素の性能が改善された(Conaty et al. 1999;Zillmann et al. 1997)。
【0020】
in vitroセレクションによって特定の補助因子のために開発された核酸酵素は、しばしば、他の分子の存在下で活性を有するであろう。例えば、17Eデオキシリボザイムは、Zn2+の存在下での活性のためにin vitroセレクションによって開発された。しかしながら、この酵素は、Zn2+よりもPb2+の存在下でより強い活性を示した。Zn2+関連活性を捜し求める過程で製造されたが、17EはPb2+についての鋭敏で選択的なセンサーとして用いることができる。より強い選択性をもつ核酸酵素を製造するために、負の選別工程を導入することができる(Peter J. Bruesehoff, Jing Li, Anthony J. Augustine III, and Yi Lu, "Immproving Metal Ion Specificity During In Vitro Selection of Catalytic DNA", Combinatorial Chemistry and High Throughput Screening 5, 327-355 (2002))。
他のヌクレオチド配列も有用であり、例えば米国特許出願第09/605,558号(2000年6月27日出願)に記載されたものが挙げられる(前記文献は参照により本明細書に含まれるものとする)。
【0021】
[アデノシンセンサー]
図2Aに示したアプタザイムはアデノシンに特異的である。アプタザイムの触媒コアは“8-17”DNAザイムから適合化され、Pb2+の存在下での高活性について最適化された。前記DNAザイムの3'-末端はアデノシンアプタマーモチーフに連結されている。したがって、アデノシンの存在は、DNAザイムの活性な三次元構造の形成を促進することができる。アプタザイムをベースとするセンサーでは、この複合体が、ただ1つのリボアデノシン部位での基質鎖の切断を促進する。アデノシンがなければ、3成分がワトソン-クリック塩基対形成によって合体することができたとしても、切断活性は劇的に低下する。また、アプタザイムの両端にハイブリダイズしている直径13nmの2つのDNA官能化金粒子も示されている。
図2Bに示されているように、基質鎖(遊離基質及び酵素と調節因子鎖とにハイブリダイズしている基質の両方)は、DNA官能化金粒子のためのリンカーとして機能し、青色をもつ凝集体を形成する(反応A)。アデノシン及び金属イオンの存在下では、基質は切断されることができる(反応B)。切断された基質はもはや粒子のためのリンカーとして機能することができず、色は赤色のままである(反応C)。
このタイプのセンサーでは、基質鎖、酵素鎖及び調節因子鎖の存在は、室温での凝集体形成のためには不充分である。加熱無しに、3つの鎖全てがハイブリダイズして、リンカーとして機能することは困難である。この問題を解決するために、別のDNA鎖(“Ade-リンカー”)を導入する(図8)。Ade-リンカーの目的は、高効率で基質鎖にハイブリダイズし、室温での粒子の凝集を促進させることである。ハイブリダイゼーションの後、Ade-リンカーは、10ヌクレオチドを有する、ふくらんだ(bulged)二次構造を形成する。このふくらみ(bulge)は、Ade-リンカーが切断された基質を用いて粒子を集合させるのを妨げる。このふくらみにより、たとえいくつかのAde-リンカーが切断基質とハイブリダイズしたとしても、全体構造は依然としてふらふらしていて、粒子を集合させることはできない。
アデノシンが存在しない場合、アプタザイムの基質はAde-リンカーとハイブリダイズし、ハイブリダイズした生成物は粒子を集合させて青色の凝集体を形成することができる(反応A)。アデノシンの存在下では、基質鎖は切断され(反応B)、切断された基質は金粒子を集合させることができず、赤色の分離している粒子を生じる(反応C)。
【0022】
[核酸酵素基質の3'及び5'末端と相補的なポリヌクレオチドタグを付した粒子]
センサーが酵素活性を表示するためには、ポリヌクレオチドを付着させた粒子の凝集の変化に際して検出可能な変化が生じなければならない。さらにまた、粒子の組成は、基質の切断を妨害しないようなものでなければならない。粒子は、例えば金属、半導体及び磁性物質から製造することができる;ZnS、ZnO、TiO2、AgI、AgBr、HgI2、PbS、PbSe、ZnTe、CdTe、In2S3、In2Se3、Cd3P2、Cd3As2、InAs及びGaAsで(例えば以下を参照されたい:Mirkin et al. 2002)、好ましくは金粒子である(商業的に入手可能である。例えば:Amersham Biosciences, Piscataway, NJ;Nanoprobes, Inc., Yaphank, NY)。非金属粒子もまた用いることができ、セラミック及びポリマー、例えばポリスチレンラテックス粒子又は色素含有ラテックス粒子が含まれる。好ましくは、粒子は、少なくとも20nm、より好ましくは少なくとも30nm、さらに好ましくは少なくとも35nmの平均粒子直径(15〜500nm、20〜200nm及び35〜100nmを含む)を有する。
少なくとも35nmの直径をもつ金コロイド粒子が好ましい。金コロイド粒子は、濃い色を生じるバンドに対して高い吸光係数を有する。これらの色は、粒子サイズ、濃度、粒子間距離、凝集の度合い及び凝集体の形状とともに変動する。例えば、粒子に付着させたポリヌクレオチドのハイブリダイゼーションによってもたらされる金粒子の凝集は、裸眼で観察することができる迅速な色の変化を生じる(例えば以下を参照されたい:Mirkin et al. 2002)。
粒子、ポリヌクレオチド又はその両方は、付着のために誘導体化される。例えば、3'-又は5'-末端がアルカンチオールで誘導体化されたポリヌクレオチドは、金粒子に容易に付着する(Whitesides 1995)。3'チオールDNAを平坦な金表面に付着させる方法も、ポリヌクレオチドの粒子への付着のために用いることができる(Mucic et al. 1996)。アルカンチオール誘導体化粒子をポリヌクレオチドに付着させるために用いてもよい。ポリヌクレオチドを固体表面に付着させる他の官能基には、ホスホロチオエート(ポリヌクレオチドを金表面に付着させる:Beebe and Rabke-Clemmer 1995)、置換アルキルシロキサン(シリカ及びガラス表面にポリヌクレオチドを結合させる)、アミノアルキルシロキサン及びメルカプトアルキルシロキサン(Grabar et al. 1995)が含まれる。5'-チオヌクレオシド又は3'-チオヌクレオシドで終わるポリヌクレオチドもまた、固体表面にポリヌクレオチドを付着させるために用いることができる。ポリヌクレオチドを付着させるいくつかの方法は、表4に提示されている。
【0023】
表4:ポリヌクレオチドを粒子に付着させるための系
【0024】
[粒子整列様式]
粒子が“尾部-尾部”整列様式で並ぶように、以下の様式で粒子を官能化する。基質の5'部位で基質と結合する粒子は、その3'末端で前記粒子と連結されている相補性ポリヌクレオチド鎖で官能化され、一方、基質の3'側で基質と結合する粒子は、その5'末端で前記粒子と連結される相補性ポリヌクレオチド鎖で官能化されている。基質-粒子複合体は、図3Aに示された構造ではなく、図3Bに示された構造を有する。
好ましくは、基質は、例えば“粘着末端”として機能する数塩基による3'-末端及び5'-末端の伸長によって、改変されて、粒子に付着させた相補的ポリヌクレオチド鎖とのアニーリングが促進される。基質の改変は、核酸酵素/基質の相互作用を阻害することなく基質に連結された粒子を含む複合体の形成を可能にする。しかしながら、基質が核酸酵素との相互作用に重要ではない領域を含む場合、改変は必ずしも必要ではない。
粒子を基質に直接付着させることも可能であり、あるいは基質の一方の端を粒子に付着させ、他方の端は粒子に付着したポリヌクレオチドと相補的にすることも可能である。したがって、相補的ポリヌクレオチドは任意である。
標的補助因子又はエフェクターを検出するために、核酸酵素、基質及び標識粒子を、前記酵素が感受性を有する標的の補助因子又はエフェクター(例えばアデノシン)を含んでいると疑われるサンプルの存在下で混合する(図3)。補助因子又はエフェクターの存在下では、酵素は基質を切断して、凝集体の形成を妨げる。
【0025】
粒子の種々の凝集状態によって種々の色が生じる。例えば、金粒子の大規模な凝集は青色を呈し、粒子の小規模の凝集は赤色を呈する。さらにまた、基質の切断量、よって凝集の度合いは、補助因子又はエフェクターの濃度に依存する。低い補助因子又はエフェクター濃度は、単独の粒子と凝集体との混合物を生じる部分的な基質の切断のみをもたらし、半定量的又は定量的アッセイを可能にする。色の相違を増幅して感度を改善することができる。定量的測定のために、アッセイ混合物の光学スペクトルが決定される。色の変化の他に、粒子の凝集体の形成又は塊状粒子(aggregated particles)の沈殿をモニターすることもできる。色の変化は、裸眼又は分光光度計によって観察することができる。凝集体の形成は、電子顕微鏡又はネフェロメーターで観察することができる。塊状粒子の沈殿は、裸眼又は顕微鏡で観察することができる。
色の変化の観察を容易にするために、対照的な色の背景で色を観察することができる。金粒子を用いるときは、白色の固体表面(例えばシリカ又はアルミナのTLCプレート、ろ紙、硝酸セルロース膜、及びナイロン膜)上にハイブリダイゼーション溶液のサンプルをスポットし、このスポットを乾燥させることによって、色の変化の観察が容易にされる。先ず初めに、スポット(点)は、ハイブリダイゼーション溶液の色を維持している(凝集の非存在下での桃色/赤から、金粒子の凝集が存在する場合の紫がかった赤色/紫の範囲にある)。乾燥時に、スポットを滴下する前に凝集が存在する場合は、青色のスポットが生じる;分散が発生した場合は、桃色のスポットが生じる。青色と桃色のスポットは安定で、その後の冷却若しくは加熱でも、又は時間が経過しても変化しない。これらは簡便で永久的な検査記録を提供する。色の変化を観察するために他の工程は必要とされない。
あるいは、アッセイ結果は、35nmの金粒子とともに使用して、サンプルをガラス繊維フィルター上にスポットすることによって可視化してもよい。水でリンスした後で、凝集体を含むスポットを観察する。アッセイ結果を可視化するため、また別の方法も利用可能である(Mirkin et al. 2002)。
標的とする補助因子又はエフェクターは、生物学的サンプルを含む多様なサンプルで検出することができる。既知量の補助因子又はエフェクターを含む標準試料を未知サンプルと並べてアッセイし、色の変化を比較することができる。あるいは、基準カラーチャート(pH試験紙とともに用いられるものと類似するもの)を用意してもよい。
【0026】
[キット]
本発明はまた、補助因子又はエフェクターのような分析物を検出するキットを提供する。ある実施態様では、前記キットは少なくとも1つの容器を含み、前記容器は、ポリヌクレオチドが付着している少なくとも2つのタイプの粒子と、少なくとも3つの部分をもつ基質(第一の部分は第二の部分に対して5'側であり、第二の部分は分析物の存在下で核酸酵素によって切断され、第三の部分は第二の部分に対して3' 側である)と、核酸酵素とを保持する。第一の粒子に付着しているポリヌクレオチドは、前記基質の少なくとも第一の部分の配列と相補的な配列を有し、且つその5'末端で前記粒子に付着している。第二の粒子に付着しているポリヌクレオチドは、前記基質の少なくとも第三の部分の配列と相補的な配列を有し、且つその3'末端で前記粒子に付着している。
キットが供給される場合、組成物の種々の成分は別々の容器に包装されてあって、使用直前に混合することができる。そのように成分を別々に包装することによって、活性成分の長期保存が可能になる。例えば、ポリヌクレオチドが付着している粒子の1又は2以上、基質、及び核酸酵素は、別々の容器で供給される。
キットに含まれる試薬は、種々の成分の耐用期間が維持され、且つ容器の材料によって吸着されたり変化させられたりしない任意の種類の容器で供給することができる。例えば、密封ガラスアンプルには、中性又は非反応性ガス(例えば窒素ガス)下で封入された、1若しくは2以上の試薬又は緩衝液を含むことができる。アンプルは、任意の適切な物質、例えばガラス、有機ポリマー(例えばポリカーボネート、ポリスチレンなど)、セラミック、金属、又は同様の試薬を保持するために典型的に用いられる他の任意の材料から成るであろう。その他の適切な容器の例には、アンプルと類似の材料から製造することができる単純なビン、金属箔(例えばアルミニウム又は合金)で裏打ちされた内部を含み得るエンベロープが含まれる。その他の容器には、試験管、バイアル、フラスコ、ビン、注射筒などが含まれる。容器は無菌的な出入口を有することができる。前記容器は、例えば、皮下注射針で突き刺すことができる栓を有するビンである。他の容器は、2つの区画を有していてもよい。これら区画は容易に取り外し可能な膜で分離されてあり、取り外しに際して成分の混合が可能にされる。取り外し可能な膜は、ガラス、プラスチック、ゴムなどであってもよい。
キットはまた、標的とする補助因子又はエフェクターの検出に有用な他の試薬及び物品を含むことができる。前記試薬には、既知量の補助因子又はエフェクターを含む標準溶液、希釈液及び他の緩衝液、予備処理試薬などが含まれる。部品として提供することができる他の物品には、支持体(凝集体の分解を可視化するため、例えばTLCシリカプレート);多孔性材料、注射筒、ピペット、キュベット及び容器が含まれる。被検対象の様々な量の補助因子又はエフェクターの存在に対応して、多様な凝集状態の粒子の外観を表示する基準チャートを提供することができる。
キットは、使用説明資料とともに提供してもよい。使用説明は、紙又は他の物体に印刷されていてもよく、及び/又は読み出し可能な電子媒体(例えばフロッピーディスク、CD-ROM、DVD-ROM、Zipディスク、ビデオテープ、オーディオテープなど)として供給することができる。詳細な使用説明はキットに物理的に付随していなくてもよく、それに代わって、ユーザーは、キットの製造元又は販売元によって特定されるインターネットウェッブサイトで指示を得ることもでき、又は電子メールとして供給されることもできる。
【実施例1】
【0027】
(実施例1:Pb2+比色バイオセンサー)
[ポリヌクレオチド及び試薬]
全てのポリヌクレオチドは、インテグレーテッドDNAテクノロジー社(Integrated DNA Technology Inc., Coralville, IA)から購入した。アデノシン及び他のヌクレオシドはシグマ・アルドリッチ(Sigma Aldrich, St. Louis, MO)から購入した。直径35nmの金粒子を調製し、3'-及び5'-チオール-改変した12-merのDNAを付着させた(Storhoff et al. 1998)。基質及び酵素はHPLCによって精製した。
[金粒子の調製及び官能化]
直径35nmの金粒子は、HAuCl4をクエン酸ナトリウムで還元して調製した。調製に使用したガラス製品は王水に浸漬し、ミリポア水で充分にリンスした。250mLの2首フラスコで、200mLのHAuCl4(0.3mM)を攪拌しながら加熱還流した。続いて、2mLのクエン酸ナトリウム(38.8mM)を添加した。数分以内に、色が淡黄色から深紅に変化した。色が変化した後、この系をさらに半時間還流して、還元を完了させた。続いて、この系をゆっくりと室温まで冷却し、粒子をパイレックスロートでろ過した。粒子をJEOL2010電子顕微鏡による透過電子顕微鏡法(TEM)を用いて性状を調べ、サイズを35±6nmと決定した。粒子の大半は球形で、ほんの一部の粒子は縦横比が2未満の棍棒状であった。金コロイドは532nmで〜1の吸光ピークを有し、濃度は、全ての粒子が35nmの半径を有する球形であると仮定して、およそ0.23nMであると計算した。全てのHAuCl4が反応し、粒子の密度はバルクの金と同じであった。この溶液は532nmで4.4x109M-1cm-1の吸光係数を示した。
【0028】
[3'-及び5'-チオール改変したDNAによる粒子の官能化]
チオール改変したDNAを50mMのジチオスレイトール(DTT)とともにインキュベートして活性化した。典型的には、60μLの1mMのDNAを60μLの100mMのDTTと室温(20〜22℃)で2時間インキュベートした。続いて、混合物をSep-Pak C18カラムにのせてDTTを除去した。カラムは、10mLの95%CH3CN、CH3CNとメタノールと水との混合物(容積比1:1:1)10mL、20mLの水、及び10mLの2MのNH4OAcで処理した。続いて、DNAとDTT混合物をカラムにロードした。20mLの脱イオン水でDTTを洗い流し、1mLの95%CH3CN、CH3CNとメタノールと水との混合物(容積比1:1:1)10mLによってDNAを溶出させた。溶出したDNAを12mLの金粒子(0.23nM)に添加した。1日インキュベートした後、1MのNaCl及び0.1Mのトリス-酢酸(TA)(pH7.2)を含む1.4mLの緩衝液を添加した。さらに2日間インキュベーションした後、8000rpm、10分間の遠心分離によって、粒子を収集した。上清を除去し、DNA官能化した粒子を25mLのTA緩衝液(pH7.2)、100mMのNaClに再分散させた。このプロセスを3回繰り返して、遊離DNAの除去を確実にした。最後に粒子を同じ緩衝液に分散させて、532nmでの吸光係数を〜2.2に調整した(〜0.5nMの濃度に対応する)。
【0029】
[DNAザイムによる粒子の凝集]
典型的な実験では、20μLの3'-チオール改変DNA官能化金粒子と20μLの5'-チオール改変DNA官能化金粒子と(両者とも〜0.5nMの濃度)を混合した。NaCl、酵素(17E)及びPb2+を添加し、最終容積を基質の添加時に100μLに調整した。凝集を基質鎖の添加によって開始させ、吸光をヒューレットパッカード(Hewlett-Packard)8453分光光度計でモニターした。緩衝液は、pH依存実験を除く全ての実験でpH7.2のTAとした。基質添加から最初のスペクトルのモニターまでの間の時間は、全てのサンプルについて15秒となるように制御した。
図4に示した実験では、比較を容易にするために吸光比を正規化した。これらの実験は2μMの酵素で実施し、基質濃度は、13nm粒子に対しては160nMで、13nM〜35nM混合粒子に対しては120nMで、35nm-35nm粒子に対しては3nMであった。緩衝液は25mMのTA(pH7.2)であった。
【0030】
[新規Pb2+センサーシステムの最適化]
(f)凝集10分後の吸光比
透明な赤色から青色への色の変化が35nmの金粒子の凝集に際して観察することができ、したがって、凝集過程はUV可視化分光光度計によって簡便にモニターすることができる。基質鎖の添加後の種々の時点における、連結基質(3nM)、NaCl(250mM)及び17E酵素(1μM)を用いた新規なセンサーシステムのいくつかのスペクトルが図6Aに示されている。基質鎖を添加する前、このシステムは、532nmで吸光ピークを有する赤色を示し、13nmの直径の粒子と比較して10nm赤色にシフトしていた。最初の3分では、色の変化は非常に小さかった。この3分の間に、基質鎖が酵素鎖と塩基対を形成し、ハイブリダイズしたDNAザイムが金粒子に連結されたと考えられる。次の数分では、色の変化の速度が非常に速く、粒子が架橋されて大きな凝集体を形成したことを示している。凝集体の形成とともに、532nmでの吸収ピークは減少し、一方、700nm領域で吸光が増加して赤色から青色への色の遷移が生じた。10分後、凝集体は沈殿し始め、全ての波長で観察される吸光の減少が生じた。サンプル調製物及び粒子濃度における相違の影響を最小限にするために、532nmと700nmの吸光比を用いて色の変化の速度をモニターした(より高い比は赤色に関連し、より低い比は青色に関連する)。
種々の基質濃度の存在下における色の変化の速度が図6Bに示されている。より高い基質濃度は、より速い速度の色の変化を生じた。したがって、基質濃度の低下は所定の時間で“青色の少ない”または“赤色の強い”色を生じた。異なる濃度のPb2+は、異なる速度で基質を切断した。したがって、センサーによって表示される色を精査することによって、Pb2+を検出し定量した。この実験は3nMの基質、250mMのNaCl及び1μMの17Eを用いて実施した。
【0031】
[凝集に影響を与える付加的な因子]
上記に概略したように、粒子の凝集速度に影響する他の因子が見出された。前記因子は、例えば酵素濃度、塩濃度及びpH値である。図6Cに示した実験は、全て3nMの基質濃度及び1μMの17E酵素濃度を用いて実施し、NaCl濃度を変化させた。より高いNaCl濃度は凝集を促進し、おそらくは、NaClが負の電荷をもつDNAの負電荷間の静電気的反発を低下させたためであろう。
酵素濃度もまた粒子の凝集に顕著な影響を有した。図6Dに示した実験は全て0.3nMの基質濃度及び250mMのNaCl濃度を用いて実施し、17E酵素濃度を変化させた。より高い酵素鎖濃度は、より速い凝集速度をもたらした。酵素の非存在下又は酵素濃度が非常に低いとき(図6D、最上部の曲線)、凝集は阻害された。ただし物理的には基質鎖のみがリンカーとして作用して粒子を集合させるので、酵素鎖の存在は任意であるようにみえる。これらの観察から、凝集の最初の過程は基質鎖と酵素鎖とのハイブリダイゼーションであるとの仮説を立てた。続いて、ハイブリダイズしたDNAザイムは、金粒子を集合させることができる。酵素鎖の非存在下では、基質は単独ではおそらくふらふらしていて、金粒子を集合させられないだろう。
種々のpH値もまた凝集に影響を与える。図6Eに示したように、pH5.2からpH10.2(pH5.2、酢酸緩衝液;pH6.2、MES緩衝液;pH7.2〜10.2、TA緩衝液)では、全てのサンプルで、室温(20〜22℃)での金粒子のDNAザイム誘導アッセンブリーに特徴的であったS字状曲線が得られた。pH5.2のサンプルでは、粒子の凝集は阻害されているようであった。10分の凝集の後の吸光比をプロットし図6Fに提示した。pH7.2から9.2では、類似する吸光特性が観察され、同様な凝集速度が示唆されているが、それより高いpH及びそれより低いpHではともに凝集速度は低下する。したがって、センサーは約6.2から約10.2の間のpH範囲で用いることができるが、最適な性能には約7.2から約9.2のpHで用いることができる。粒子の凝集速度の変化は、おそらくは、低いpH又は高いpHでのDNA塩基のプロトン化及び脱プロトン化が塩基対形成に影響を与え、したがって凝集速度の変化を生じるためであろう。
【0032】
[室温での凝集]
本発明の新規なセンサーの種々のPb2+濃度による室温(20〜22℃)での凝集の反応速度(kinetics)をUV可視化分光光度計によってモニターした。Pb2+の存在下では、基質と酵素鎖のハイブリダイゼーション後に、基質鎖は酵素鎖によって切断されるか、又は粒子を集合させるリンカーでありえると仮説を立てた。2つの過程の相対速度は、Pb2+濃度に左右される。図7Aに示したように、Pb2+濃度が増加するにつれ、凝集速度は抑制された。Pb2+の添加がないサンプルの場合、凝集は10分後にほぼ完了した。10分での吸光比を用いてPb2+濃度を定量した(図7B)。
【実施例2】
【0033】
(実施例2:アデノシン比色バイオセンサー)
[アプタザイムによる粒子の凝集]
アプタザイムをベースとするセンサーについて基質鎖の最適濃度を決定するために、各粒子溶液(E532=2.2)の20μLを混合し、Ade-リンカーを最終濃度5μMとなるように、NaClを最終濃度250mMとなるように添加し、基質添加後に容積を100μLに調整した。その溶液は25mMのTA緩衝液(pH7.2)で調整した。
この溶液を用いて、最終濃度0.15μMの基質鎖、3μMの酵素及び調節鎖、5mMのアデノシン、250mMのNaCl及び25mMのTA緩衝液(pH7.2)を含む第二の溶液を調製した。この第二の溶液の98μLを反応チューブに移し、このときを0時点と設定した。金属イオンのストック溶液2μLを前記反応チューブに添加して切断反応を開始させ、種々の時点で2μLアリコートを検出チューブに移した。反応チューブから一度検出チューブに各アリコートを移したら、EDTAを前記検出チューブに添加して切断反応を停止させた。反応チューブに由来する2μLアリコートは0.3pmolの基質を含み、切断が発生しない場合は、検出チューブ中3nMの基質濃度を与えるであろう。反応チューブの各サンプルの吸光スペクトルを、反応チューブからアリコートを移した後20分してモニターした。
センサーの感度及び選択性を調べるために、種々の濃度のアデノシン又は他のヌクレオシドを含む種々の反応チューブを調製した。30分のインキュベーション後に、2μLアリコートを検出チューブに移し、検出チューブの吸光スペクトルを20分のインキュベーション後に測定した。
図9Aは、凝集20分後の吸光スペクトルによって測定された、粒子の基質濃度依存性の凝集を示している。図9Bは、アプタザイムによる基質の切断の反応速度を示している。図9Cは、新規に設計した、アデノシンのためのアプタザイムをベースとするセンサーの感度及び選択性を示している。桃色、赤色及び青色の点は、それぞれ5mMのシチジン、グアノシン及びウリジンの吸光比である。
【0034】
(参考文献:表3Bに示すものは、別にして下に示す)
Allara D, Nuzzo R. (1985) Spontaneously organized molecular assemblies. 1. Formation, dynamics and physical properties of n-alkanoic acids adsorbed from solution on an oxidized aluminum surface. Langmuir 1: 45-52.
Beebe T, Rabke-Clemmer C, (1995) Thiol labeling of DNA for attachment to gold surfaces. Patent No. 5,472,881 USA.
Biroccio A, Hamm J, Incitti I, De Francesco R, Tomei L. (2002) Selection of RNA aptamers that are specific and high-affinity ligands of the hepatitis C virus RNA-dependent RNA polymerase. J Virol 76: 3688-3696.
Breaker RR, Joyce GF, Breaker RR, Joyce GF. (1995) A DNA enzyme with Mg(2+)-dependent RNA phosphoesterase activity.; A DNA enzyme that cleaves RNA. Chem Biol; Chem Biol 2; 1: 223-229.
Breaker RR, Joyce GF. (1994) A DNA enzyme that cleaves RNA. Chem Biol 1: 223-229.
Breaker RR. (2002) Engineered allosteric ribozymes as biosensor components. Curr Opin Biotechnol 13: 31-39.
Brody EN, Gold L. (2000) Aptamers as therapeutic and diagnostic agents. J Biotechnol 74: 5-13.
Brown A, Pavot C, Li J, Lu Y. A lead-dependent DNAzyme with a two-step mechanism. submitted.
Bruno JG, Kiel JL. (1999) In vitro selection of DNA aptamers to anthrax spores with electrochemiluminescence detection. Biosens Bioelectron 14: 457-464.
Bruno JG, Kiel JL. (2002) Use of magnetic beads in selection and detection of biotoxin aptamers by electrochemiluminescence and enzymatic methods. Biotechniques 32: 178-80, 182-3.
【0035】
Cadwell RC, Joyce GF. (1992) Randomization of genes by PCR mutagenesis. PCR Methods Appl 2: 28-33.
Cadwell RC, Joyce GF. (1994) Mutagenic PCR. PCR Methods Appl 3: S136-40.
Cao Y, Jin R, Mirkin CA. (2001) DNA-modified core-shell Ag/Au particles. J Am Chem Soc 123: 7961-7962.
Carmi N, Shultz LA, Breaker RR. (1996) In vitro selection of self-cleaving DNAs. Chem Biol 3: 1039-1046.
Chaloin L, Lehmann MJ, Sczakiel G, Restle T. (2002) Endogenous expression of a high-affinity pseudoknot RNA aptamer suppresses replication of HIV-1. Nucleic Acids Res 30: 4001-4008.
Chapman KB, Szostak JW. (1994) In vitro selection of catalytic RNAs. Curr Opin Struct Biol 4: 618-622.
Ciesiolka J, Gorski J, Yarus M. (1995) Selection of an RNA domain that binds Zn2+. RNA 1: 538-550.
Conaty J, Hendry P, Lockett T. (1999) Selected classes of minimised hammerhead ribozyme have very high cleavage rates at low Mg2+ concentration. Nucleic Acids Res 27: 2400-2407.
Conn M, Prudent J, Schultz P. (1996) Porphyrin Metallation Catalyzed by a Small RNA Molecule. J Am Chem Soc 118: 7012-7013.
Cuenoud B, Szostak JW. (1995) A DNA metalloenzyme with DNA ligase activity. Nature 375: 611-614.
【0036】
Dai X, De Mesmaeker A, Joyce GF. (1995) Cleavage of an amide bond by a ribozyme. Science 267: 237-240.
Earnshaw, Gait. (1998) Modified oligoribonucleotides as site-specific probes of RNA structure and function. Biopolymers 48: 39-55.
Ekland EH, Bartel DP. (1996) RNA-catalysed RNA polymerization using nucleoside triphosphates. Nature 382: 373-376.
Ekland EH, Szostak JW, Bartel DP. (1995) Structurally complex and highly active RNA ligases derived from random RNA sequences. Science 269: 364-370.
Ellington AD, Szostak JW. (1990) In vitro selection of RNA molecules that bind specific ligands. Nature 346: 818-822.
Faulhammer D, Famulok M. (1997) Angew Chem Int Ed Engl 35: 2837-2841.
Geyer CR, Sen D. (1997) Evidence for the metal-cofactor independence of an RNA phosphodiester-cleaving DNA enzyme. Chem Biol 4: 579-593.
Grabar K, Freeman R, Hommer M, Natan M. (1995) Preparation and characterization of Au colloid Monolayers. Anal. Chem. 67: 735-743.
Hesselberth J, Robertson MP, Jhaveri S, Ellington AD. (2000) In vitro selection of nucleic acids for diagnostic applications. J Biotechnol 74: 15-25.
Hofmann HP, Limmer S, Hornung V, Sprinzl M. (1997) Ni2+-binding RNA motifs with an asymmetric purine-rich internal loop and a G-A base pair. RNA 3: 1289-1300.
Huizenga DE, Szostak JW. (1995) A DNA aptamer that binds adenosine and ATP. Biochemistry 34: 656-665.
Iler R. (1979) Chapter 6. In: anonymous (eds) The chemistry of silica. Wiley, New York.
【0037】
Illangasekare M, Yarus M. (1997) Small-molecule-substrate interactions with a self-aminoacylating ribozyme. J Mol Biol 268: 631-639.
Jayasena SD. (1999) Aptamers: an emerging class of molecules that rival antibodies in diagnostics. Clin Chem 45: 1628-1650.
Jhaveri S, Kirby R, Conrad R, Maglott E, Bowser M, Kennedy R, Glick G, Ellington A. (2000) Designed signaling aptamers that transduce molecular recognition to changes in fluorescence intensity. J. Am. Chem. Soc. 122: 2469-2473.
Jhaveri S, Rajendran M, Ellington AD. (2000) In vitro selection of signaling aptamers. Nat Biotechnol 18: 1293-1297.
Joyce GF. (1994) In vitro evolution of nucleic acids. Curr Opin Struct Biol 4: 331-336.
Kiga D, Futamura Y, Sakamoto K, Yokoyama S. (1998) An RNA aptamer to the xanthine/guanine base with a distinctive mode of purine recognition. Nucleic Acids Res 26: 1755-1760.
Lauhon CT, Szostak JW. (1995) RNA aptamers that bind flavin and nicotinamide redox cofactors. J Am Chem Soc 117: 1246-1257.
Li J, Lu Y. (2000) A highly sensitive and selective catalytic DNA biosensor for lead ions. J Am Chem Soc 122: 10466-10467.
Li J, Zheng W, Kwon AH, Lu Y. (2000) In vitro selection and characterization of a highly efficient Zn(II)-dependent RNA-cleaving deoxyribozyme. Nucleic Acids Res 28: 481-488.
Li Y, Breaker RR. (1999) Phosphorylating DNA with DNA. Proc Natl Acad Sci U S A 96: 2746-2751.
Li Y, Sen D. (1996) A catalytic DNA for porphyrin metallation. Nat Struct Biol 3: 743-7.
【0038】
Link S, Wang Z, El-Sayed M. (1999) Alloy formation of gold-silver particles and the dependence of the plasmon absorption on their compositions. J Phys Chem B 103: 3529-3533.
Lohse PA, Szostak JW. (1996) Ribozyme-catalysed amino-acid transfer reactions. Nature 381: 442-444.
Lorsch JR, Szostak JW. (1994) In vitro evolution of new ribozymes with polynucleotide kinase activity. Nature 371: 31-36.
Lu Y. (2002) New transition-metal-dependent DNAzymes as efficient endonucleases and as selective metal biosensors. Chemistry 8: 4589-4596
Lu Y, Liu J, SIMPLE CATALYTIC DNA BIOSENSORS FOR IONS BASED ON COLOR CHANGES, Application Serial No. 09/605,558, USA.
Maoz R, Sagiv J. (1987) Penetration-controlled reactions in organized monolayer assemblies. 1. Aqueous permanganate interaction with monolayer and multilayer films of long-chain surfactants. Langmuir 3: 1034-1044.
Mirkin CA, Letsinger RL, Mucic RC, Storhoff JJ. (1996) A DNA-based method for rationally assembling particles into macroscopic materials. Nature 382: 607-609.
Mirkin, C.A., Letsinger, L. R, Mucic, C. R, Storhoff, J. J, Elghanian R, (2002) Particles having polynucleotides attached thereto and uses therefor. Patent No. 6,361,944 USA.
Mucic R, Herrlein M, Mirkin, C.A., Letsinger R. (1996) Synthesis and characterization of DNA with ferrocenyl groups attached to their 5’-termini: Electrochemical characterization of a redox-active nucleotide monolayer. Chem. Commun.555.
Nuzzo R, Fusco F, Allara D. (1987) Spontaneously organized molecular assemblies, 3. Preparation and properties of solution adsorbed monolayers of organic disulfides on gold surfaces. J Am Chem Soc 109: 2358.
【0039】
Piccirilli JA, McConnell TS, Zaug AJ, Noller HF, Cech TR. (1992) Aminoacyl esterase activity of the Tetrahymena ribozyme. Science 256: 1420-1424.
Prudent JR, Uno T, Schultz PG. (1994) Expanding the scope of RNA catalysis. Science 264: 1924-1927.
Rakow NA, Suslick KS. (2000) A colorimetric sensor array for odour visualization. Nature 406: 710-713.
Robertson MP, Ellington AD. (1999) In vitro selection of an allosteric ribozyme that transduces analytes to amplicons. Nat Biotechnol 17: 62-66.
Roth A, Breaker RR. (1998) An amino acid as a cofactor for a catalytic polynucleotide. Proc Natl Acad Sci U S A 95: 6027-6031.
Rusconi CP, Scardino E, Layzer J, Pitoc GA, Ortel TL, Monroe D, Sullenger BA. (2002) RNA aptamers as reversible antagonists of coagulation factor IXa. Nature 419: 90-94.
Santoro SW, Joyce GF. (1997) A general purpose RNA-cleaving DNA enzyme. Proc Natl Acad Sci U S A 94: 4262-4266.
Seetharaman S, Zivarts M, Sudarsan N, Breaker RR. (2001) Immobilized RNA switches for the analysis of complex chemical and biological mixtures. Nat Biotechnol 19: 336-341.
Shaiu WL, Larson DD, Vesenka J, Henderson E. (1993) Atomic force microscopy of oriented linear DNA molecules labeled with 5nm gold spheres. Nucleic Acids Res 21: 99-103.
Sillen LG, (1964) Stability constants of metal-ion complexes. Edition: 2d ed.
Smith J, Olson D, Armitage B. (1999) Molecular recognition of PNA-containing hybrids: Spontaneous assembly of helical cyanine dye aggregates on PNA templates. J. Am. Chem. Soc. 121: 2686-2695.
【0040】
Soriaga M, Hubbard A. (1982) Determination of the orientation of aromatic molecules adsorbed on platinum electrodes: The influence of solute concentration. J Am Chem Soc 104.
Soukup GA, Breaker RR. (2000) Allosteric nucleic acid catalysts. Curr Opin Struct Biol 10: 318-325.
Stojanovic MN, Landry DW. (2002) Aptamer-based colorimetric probe for cocaine. J Am Chem Soc 124: 9678-9679.
Storhoff J, Elghanian R, Mucic R, Mirkin C, Letsinger RL. (1998) One-pot colorimetric differentiation of polynucleotides with single base imperfections using gold particle probes. J Am Chem Soc 120: 1959-1964.
Tang J, Breaker RR. (1997) Rational design of allosteric ribozymes. Chem Biol 4: 453-459.
Tang J, Breaker RR. (2000) Structural diversity of self-cleaving ribozymes. Proc Natl Acad Sci U S A 97: 5784-5789.
Tarasow TM, Tarasow SL, Eaton BE. (1997) RNA-catalysed carbon-carbon bond formation. Nature 389: 54-57.
Timmons, Zisman. (1965) J. Phys. Chem. 69: 984-990.
Tompkins H, Allara D. (1974) The study of the gas-solid interaction of acetic acid with a cuprous oxide surface using reflection-absorption spectroscopy. J. Colloid and Interface Sci. 49410.
Tsang J, Joyce GF. (1996) In vitro evolution of randomized ribozymes. Methods Enzymol 267: 410-426.
Tuerk C, Gold L. (1990) Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 249: 505-510.
Vaish NK, Heaton PA, Fedorova O, Eckstein F. (1998) In vitro selection of a purine nucleotide-specific hammerheadlike ribozyme. Proc Natl Acad Sci U S A 95: 2158-2162.
【0041】
Wallace ST, Schroeder R. (1998) In vitro selection and characterization of streptomycin-binding RNAs: recognition discrimination between antibiotics. RNA 4: 112-123.
Wallis MG, Streicher B, Wank H, von Ahsen U, Clodi E, Wallace ST, Famulok M, Schroeder R. (1997) In vitro selection of a viomycin-binding RNA pseudoknot. Chem Biol 4: 357-366.
Wang DY, Lai BH, Sen D. (2002) A general strategy for effector-mediated control of RNA-cleaving ribozymes and DNA enzymes. J Mol Biol 318: 33-43.
Wecker M, Smith D, Gold L. (1996) In vitro selection of a novel catalytic RNA: characterization of a sulfur alkylation reaction and interaction with a small peptide. RNA 2: 982-994.
Whitesides, (1995) Proceedings of the Robert A. Welch Foundation 39th Conference On Chemical Research Nanophase Chemistry., Houston, TX.
Wiegand TW, Janssen RC, Eaton BE. (1997) Selection of RNA amide synthases. Chem Biol 4: 675-683.
Wilson C, Szostak JW. (1995) In vitro evolution of a self-alkylating ribozyme. Nature 374: 777-782.
Wilson DS, Szostak JW. (1999) In vitro selection of functional nucleic acids. Annu Rev Biochem 68: 611-647.
Zhang B, Cech TR. (1997) Peptide bond formation by in vitro selected ribozymes. Nature 390: 96-100.
Zillmann M, Limauro SE, Goodchild J. (1997) In vitro optimization of truncated stem-loop II variants of the hammerhead ribozyme for cleavage in low concentrations of magnesium under non-turnover conditions. RNA 3: 734-747.
【0042】
(表3Bに右上添字として示す参考文献)
REFERENCES for Table 3B
1. Ellington, A.D. & Conrad, R. (1995). Aptamers as potential nucleic acid pharmaceuticals. Biotechnol. Annu. Rev. 1: 185-214.
2. Jayasena, S.D. (1999). Aptamers: an emerging class of molecules that rival antibodies in diagnostics. Clin. Chem. (Washington, D. C.) 45: 1628-50.
3. Sun, L.Q., Cairns, M.J., Saravolac, E.G., Baker, A. & Gerlach, W.L. (2000). Catalytic nucleic acids: From lab to applications. Pharmacol. Rev. 52: 325-47.
4. Hesselberth, J., Robertson, M.P., Jhaveri, S. & Ellington, A.D. (2000). In vitro selection of nucleic acids for diagnostic applications. Rev. Mol. Biotechnol. 74: 15-25.
5. Joyce, G.F. (1999). Reactions Catalyzed by RNA and DNA Enzymes. In The RNA World, vol. 37 (Gesteland, R.F., Cech, T.R. & Atkins, J.F., ed.), pp. 687-9, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
6. Breaker, R.R. (1997). DNA enzymes. Nat. Biotechnol. 15: 427-31.
7. Sen, D. & Geyer, C.R. (1998). DNA enzymes. Curr. Opin. Chem. Biol. 2: 680-7.
8. Breaker, R.R. (1999). Catalytic DNA: in training and seeking employment. Nat. Biotechnol. 17: 422-3.
9. Breaker, R.R. (2000). Making catalytic DNAs. Science (Washington, D. C.) 290: 2095-6.
【0043】
10. Derose, V.J. (2002). Two Decades of RNA Catalysis. Chemistry & Biology 9: 961-9.
11. Ellington, A.D. & Szostak, J.W. (1990). In vitro selection of RNA molecules that bind specific ligands. Nature (London) 346: 818-22.
12. Ellington, A.D. & Szostak, J.W. (1992). Selection in vitro of single-stranded DNA molecules that fold into specific ligand-binding structures. Nature (London) 355: 850-2.
13. Santoro, S.W. & Joyce, G.F. (1997). A general purpose RNA-cleaving DNA enzyme. Proc. Natl. Acad. Sci. U. S. A. 94: 4262-6.
14. Faulhammer, D. & Famulok, M. (1996). The Ca2+ ion as a cofactor for a novel RNA-cleaving deoxyribozyme. Angew. Chem., Int. Ed. Engl. 35: 2837-41.
15. Li, J., Zheng, W., Kwon, A.H. & Lu, Y. (2000). In vitro selection and characterization of a highly efficient Zn(II)-dependent RNA-cleaving deoxyribozyme. Nucleic Acids Res. 28: 481-8.
16. Jenison, R.D., Gill, S.C., Pardi, A. & Polisky, B. (1994). High-resolution molecular discrimination by RNA. Science (Washington, DC, United States) 263: 1425-9.
17. Santoro, S.W. & Joyce, G.F. (1998). Mechanism and utility of an RNA-cleaving DNA enzyme. Biochemistry 37: 13330-42.
18. Mannironi, C., Di Nardo, A., Fruscoloni, P. & Tocchini-Valentini, G.P. (1997). In vitro selection of dopamine RNA ligands. Biochemistry 36: 9726-34.
19. Sigurdsson, S.T., Thomson, J.B. & Eckstein, F. (1998). Small ribozymes. Cold Spring Harbor Monogr. Ser. 35: 339-76.
【0044】
20. Stage-Zimmermann, T.K. & Uhlenbeck, O.C. (1998). Hammerhead ribozyme kinetics. RNA 4: 875-89.
21. Werstuck, G. & Green, M.R. (1998). Controlling gene expression in living cells through small molecule-RNA interactions. Science (Washington, D. C.) 282: 296-8.
22. Walter, N.G. & Burke, J.M. (1998). The hairpin ribozyme: structure, assembly and catalysis. Curr. Opin. Chem. Biol. 2: 24-30.
23. Holeman, L.A., Robinson, S.L., Szostak, J.W. & Wilson, C. (1998). Isolation and characterization of fluorophore-binding RNA aptamers. Folding Des. 3: 423-31.
24. Wilson, C. & Szostak, J.W. (1998). Isolation of a fluorophore-specific DNA aptamer with weak redox activity. Chem. Biol. 5: 609-17.
25. Pan, T. & Uhlenbeck, O.C. (1992). A small metalloribozyme with a two-step mechanism. Nature 358: 560-3.
26. Yang, Q., Goldstein, I.J., Mei, H.-Y. & Engelke, D.R. (1998). DNA ligands that bind tightly and selectively to cellobiose. Proc. Natl. Acad. Sci. U. S. A. 95: 5462-7.
27. Been, M.D. & Wickham, G.S. (1997). Self-cleaving ribozymes of hepatitis delta virus RNA. Eur. J. Biochem. 247: 741-53.
28. Tanner, N.K. (1998). Biochemistry of hepatitis delta virus catalytic RNAs. Ribozymes Gene Ther. Cancer: 23-38.
29. Famulok, M. & Szostak, J.W. (1992). Stereospecific recognition of tryptophan agarose by in vitro selected RNA. J. Am. Chem. Soc. 114: 3990-1.
【0045】
30. Cech, T.R. (1993). Structure and mechanism of the large catalytic RNAs: group I and group II introns and ribonuclease P. In The RNA World (Gesteland, R.F. & Atkins, J.F., ed.), pp. 239-70, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
31. Cech, T.R. & Herschlag, D. (1996). Group I ribozymes: substrate recognition, catalytic strategies, and comparative mechanistic analysis. Nucleic Acids Mol. Biol. 10: 1-17.
32. Famulok, M. (1994). Molecular Recognition of Amino Acids by RNA-Aptamers: An L-Citrulline Binding RNA Motif and Its Evolution into an L-Arginine Binder. J. Am. Chem. Soc. 116: 1698-706.
33. Geiger, A., Burgstaller, P., Von Der Eltz, H., Roeder, A. & Famulok, M. (1996). RNA aptamers that bind L-arginine with sub-micromolar dissociation constants and high enantioselectivity. Nucleic Acids Res. 24: 1029-36.
34. Tao, J. & Frankel, A.D. (1996). Arginine-Binding RNAs Resembling TAR Identified by in Vitro Selection. Biochemistry 35: 2229-38.
35. Connell, G.J., Illangesekare, M. & Yarus, M. (1993). Three small ribooligonucleotides with specific arginine sites. Biochemistry 32: 5497-502.
36. Burgstaller, P., Kochoyan, M. & Famulok, M. (1995). Structural probing and damage selection of citrulline- and arginine-specific RNA aptamers identify base positions required for binding. Nucleic Acids Res. 23: 4769-76.
37. Nolte, A., Klussmann, S., Bald, R., Erdmann, V.A. & Fuerste, J.P. (1996). Mirror-design of L-oligonucleotide ligands binding to L-arginine. Nature Biotechnology 14: 1116-9.
38. Valadkhan, S. & Manley, J.L. (2001). Splicing-related catalysis by protein-free snRNAs. Nature (London, United Kingdom) 413: 701-7.
39. Nissen, P., Hansen, J., Ban, N., Moore, P.B. & Steitz, T.A. (2000). The structural basis of ribosome activity in peptide bond synthesis. Science (Washington, D. C.) 289: 920-30.
【0046】
40. Harada, K. & Frankel, A.D. (1995). Identification of two novel arginine binding DNAs. EMBO J. 14: 5798-811.
41. Carmi, N., Balkhi, H.R. & Breaker, R.R. (1998). Cleaving DNA with DNA. Proc. Natl. Acad. Sci. U. S. A. 95: 2233-7.
42. Majerfeld, I. & Yarus, M. (1994). An RNA pocket for an aliphatic hydrophobe. Nat. Struct. Biol. 1: 287-92.
43. Cuenoud, B. & Szostak, J.W. (1995). A DNA metalloenzyme with DNA ligase activity. Nature 375: 611-4.
44. Majerfeld, I. & Yarus, M. (1998). Isoleucine:RNA sites with associated coding sequences. Rna 4: 471-8.
45. Li, Y. & Breaker, R.R. (1999). Phosphorylating DNA with DNA. Proc. Natl. Acad. Sci. U. S. A. 96: 2746-51.
46. Sassanfar, M. & Szostak, J.W. (1993). An RNA motif that binds ATP. Nature (London) 364: 550-3.
47. Burgstaller, P. & Famulok, M. (1994). Isolation of RNA aptamers for biological cofactors by in vitro selection. Angew. Chem. 106: 1163-6 (See also Angew. Chem., Int. Ed. Engl., 994, 33(10), 084-7).
48. Burke, D.H. & Gold, L. (1997). RNA aptamers to the adenosine moiety of S-adenosyl methionine: structural inferences from variations on a theme and the reproducibility of SELEX. Nucleic Acids Res. 25: 2020-4.
49. Huizenga, D.E. & Szostak, J.W. (1995). A DNA Aptamer That Binds Adenosine and ATP. Biochemistry 34: 656-65.
【0047】
50. Klussmann, S., Nolte, A., Bald, R., Erdmann, V.A. & Fuerste, J.P. (1996). Mirror-image RNA that binds D-adenosine. Nat. Biotechnol. 14: 1112-5.
51. Burmeister, J., Von Kiedrowski, G. & Ellington, A.D. (1997). Cofactor-assisted self-cleavage in DNA libraries with a 3’-’5’-phosphoramidate bond. Angew. Chem., Int. Ed. Engl. 36: 1321-4.
52. Connell, G.J. & Yarus, M. (1994). RNAs with dual specificity and dual RNAs with similar specificity. Science (Washington, D. C.) 264: 1137-41.
53. Li, Y. & Sen, D. (1996). A catalytic DNA for porphyrin metallation. Nat. Struct. Biol. 3: 743-7.
54. Lauhon, C.T. & Szostak, J.W. (1995). RNA aptamers that bind flavin and nicotinamide redox cofactors. J. Am. Chem. Soc. 117: 1246-57.
55. Travascio, P., Bennet, A.J., Wang, D.Y. & Sen, D. (1999). A ribozyme and a catalytic DNA with peroxidase activity: active sites versus cofactor-binding sites. Chemistry & Biology 6: 779-87.
56. Lorsch, J.R. & Szostak, J.W. (1994). In vitro selection of RNA aptamers specific for cyanocobalamin. Biochemistry 33: 973-82.
57. Rink, S.M., Shen, J.-C. & Loeb, L.A. (1998). Creation of RNA molecules that recognize the oxidative lesion 7,8-dihydro-8-hydroxy-2’-deoxyguanosine (8-oxodG) in DNA. Proc. Natl. Acad. Sci. U. S. A. 95: 11619-24.
58. Haller, A.A. & Sarnow, P. (1997). In vitro selection of a 7-methyl-guanosine binding RNA that inhibits translation of capped mRNA molecules. Proc. Natl. Acad. Sci. U. S. A. 94: 8521-6.
59. Kiga, D., Futamura, Y., Sakamoto, K. & Yokoyama, S. (1998). An RNA aptamer to the xanthine/guanine base with a distinctive mode of purine recognition. Nucleic Acids Res. 26: 1755-60.
【0048】
60. Lato, S.M., Boles, A.R. & Ellington, A.D. (1995). In vitro selection of RNA lectins: Using combinatorial chemistry to interpret ribozyme evolution. Chem. Biol. 2: 291-303.
61. Wang, Y., Killian, J., Hamasaki, K. & Rando, R.R. (1996). RNA Molecules That Specifically and Stoichiometrically Bind Aminoglycoside Antibiotics with High Affinities. Biochemistry 35: 12338-46.
62. Wallis, M.G., Von Ahsen, U., Schroeder, R. & Famulok, M. (1995). A novel RNA motif for neomycin recognition. Chem. Biol. 2: 543-52.
63. Famulok, M. & Huettenhofer, A. (1996). In Vitro Selection Analysis of Neomycin Binding RNAs with a Mutagenized Pool of Variants of the 16S rRNA Decoding Region. Biochemistry 35: 4265-70.
64. Wallis, M.G., Streicher, B., Wank, H., Von Ahsen, U., Clodi, E., Wallace, S.T., Famulok, M. & Schroeder, R. (1997). In vitro selection of a viomycin-binding RNA pseudoknot. Chem. Biol. 4: 357-66.
65. Burke, D.H., Hoffman, D.C., Brown, A., Hansen, M., Pardi, A. & Gold, L. (1997). RNA aptamers to the peptidyl transferase inhibitor chloramphenicol. Chem. Biol. 4: 833-43.
66. Wallace, S.T. & Schroeder, R. (1998). In vitro selection and characterization of streptomycin-binding RNAs: recognition discrimination between antibiotics. Rna 4: 112-23.
67. Giver, L., Bartel, D.P., Zapp, M.L., Green, M.R. & Ellington, A.D. (1993). Selection and design of high-affinity RNA ligands for HIV-1 Rev. Gene 137: 19-24.
68. Giver, L., Bartel, D., Zapp, M., Pawul, A., Green, M. & Ellington, A.D. (1993). Selective optimization of the Rev-binding element of HIV-1. Nucleic Acids Res. 21: 5509-16.
69. Williams, K.P., Liu, X.-H., Schumacher, T.N.M., Lin, H.Y., Ausiello, D.A., Kim, P.S. & Bartel, D.P. (1997). Bioactive and nuclease-resistant L-DNA ligand of vasopressin. Proc. Natl. Acad. Sci. U. S. A. 94: 11285-90.
【0049】
70. Zimmerman, J.M. & Maher, L.J., Iii (2002). In vivo selection of spectinomycin-binding RNAs. Nucleic Acids Res. 30: 5425-35.
71. Vianini, E., Palumbo, M. & Gatto, B. (2001). In vitro selection of DNA aptamers that bind L-tyrosinamide. Bioorganic & Medicinal Chemistry 9: 2543-8.
72. Andreola, M.-L., Pileur, F., Calmels, C., Ventura, M., Tarrago-Litvak, L., Toulme, J.-J. & Litvak, S. (2001). DNA aptamers selected against the HIV-1 RNase H display in vitro antiviral activity. Biochemistry 40: 10087-94.
73. Fukusaki, E.-I., Kato, T., Maeda, H., Kawazoe, N., Ito, Y., Okazawa, A., Kajiyama, S.-I. & Kobayashi, A. (2000). DNA aptamers that bind to chitin. Bioorg. Med. Chem. Lett. 10: 423-5.
74. Bock, L.C., Griffin, L.C., Latham, J.A., Vermaas, E.H. & Toole, J.J. (1992). Selection of single-stranded DNA molecules that bind and inhibit human thrombin. Nature (London) 355: 564-6.
75. Koizumi, M. & Breaker, R.R. (2000). Molecular Recognition of cAMP by an RNA Aptamer. Biochemistry 39: 8983-92.
76. Kato, T., Takemura, T., Yano, K., Ikebukuro, K. & Karube, I. (2000). In vitro selection of DNA aptamers which bind to cholic acid. Biochim. Biophys. Acta 1493: 12-8.
77. Okazawa, A., Maeda, H., Fukusaki, E., Katakura, Y. & Kobayashi, A. (2000). In vitro selection of hematoporphyrin binding DNA aptamers. Bioorg. Med. Chem. Lett. 10: 2653-6.
78. Kawakami, J., Imanaka, H., Yokota, Y. & Sugimoto, N. (2000). In vitro selection of aptamers that act with Zn2+. J. Inorg. Biochem. 82: 197-206.
79. Bruno, J.G. & Kiel, J.L. (1999). In vitro selection of DNA aptamers to anthrax spores with electrochemiluminescence detection. Biosensors & Bioelectronics 14: 457-64.
【図面の簡単な説明】
【0050】
【図1】図1Aは酵素鎖(17E)及び基質鎖(17DS)から成る“8-17”DNAザイムシステムの二次構造を示し、図1BはPb(II)の存在下での17Eによる17DSの切断を示す。図1Cは、DNAザイムによって誘導される金粒子のアッセンブリーと、その金属イオン(例えばPb(II))のバイオセンサーとしての利用を表す模式図を示す。
【図2】図2Aは、“8-17”DNAザイム土台上に構築されたアデノシンアプタザイムの一次構造及び提案する二次構造を示す。図2Bは、アデノシンの比色検出の模式図を占め得s。
【図3】DNA連結金粒子の2つの整列様式を示す。図3Aは、“頭部-尾部”整列様式の粒子を示し、この場合ただ1種類のチオール改変DNAが必要である。図3Bは、“尾部-尾部”整列様式の粒子を示す。
【図4】色の変化の速度に対する粒子の整列様式及びサイズの影響を示す。
【図5】Pb2+の比色バイオセンサーを示す。前記センサーでは、粒子の直径は〜35nmであり、粒子は“尾部-尾部”整列様式の態様で並べられている。
【図6】新規なPb2+センサーシステムを示す。図6Aは、35nmの金粒子の種々の時間における吸光スペクトルを示す。図6Bは、凝集速度に対する基質濃度の影響を示す。図6Cは、凝集速度に対するNaCl濃度の影響を示す。図6Dは、凝集速度に対する17E濃度の影響を示す。図6Eは、種々のpHの緩衝液中での粒子の凝集を示す。図6Fは、凝集10分後の吸光比を示す。
【図7】新規なPb2+センサーの感度及び選択性を示す。図7Aは、種々のPb2+濃度による金粒子の凝集の反応速度を示す。図7Bは、開始後10分の吸光比を示す。
【図8】アプタザイムをベースとするアデノシンセンサーの新規な設計を示す。反応A:アデノシンの非存在下で青色凝集体が形成される。反応B:基質はアデノシンの存在下で切断される。反応C:切断された基質は金粒子を集させることができず、赤色の分離している粒子が生じる。
【図9】図9Aは粒子の基質濃度依存性の凝集を示す。図9Bはアプタザイムによる基質の切断の反応速度を示す。図9Cはアプタザイムをベースとするセンサーのアデノシンに対する感度及び選択性を示す。
【図1A】
【図1B】
【図1C】
【図2A】
【図2B】
【図3A】
【図3B】
【特許請求の範囲】
【請求項1】
エフェクター又は補助因子を検出するためのセンサーシステムであって、前記センサーシステムが、
(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;
(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;
(c)第二のポリヌクレオチドを含む第一の粒子セットであって、前記ポリヌクレオチドがその3'末端で前記粒子に付着している、前記第一の粒子セット;及び
(d)第三のポリヌクレオチドを含む第二の粒子セットであって、前記ポリヌクレオチドがその5'末端で前記粒子に付着している、前記第二の粒子セット;
を含み、ここで、前記第一のポリヌクレオチドは、前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的である、前記センサーシステム。
【請求項2】
核酸酵素がDNAを含む、請求項1のセンサーシステム。
【請求項3】
第一の粒子セット及び第二の粒子セットが金を含む、請求項2のセンサーシステム。
【請求項4】
第一の粒子セット及び第二の粒子セットが、金属、半導体及びラテックスから成る群から選択される物質を含む、請求項2のセンサー。
【請求項5】
エフェクター又は補助因子が、窒素肥料、農薬、ダイオキシン、フェノール、2,4-ジクロロフェノキシ酢酸、Pb(II)、Hg(II)、As(III)、UO2(II)、Fe(III)、Zn(II)、Cu(II)、Co(II)、グルコース、インシュリン、hCGホルモン、HIV、HIVタンパク質、炭疽、天然痘、神経ガス、TNT、DNT、コカイン及び抗生物質からなる群から選択される、請求項2のセンサー。
【請求項6】
核酸酵素が配列番号1の配列を有するポリヌクレオチドを含み、且つ基質が配列番号2の配列を有するポリヌクレオチドを含む、請求項2のセンサー。
【請求項7】
核酸酵素が配列番号5の配列を有するポリヌクレオチドと配列番号6の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号4の配列のポリヌクレオチドを含む、請求項2のセンサー。
【請求項8】
核酸酵素が配列番号8の配列を有するポリヌクレオチドと配列番号9の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号7の配列を有するポリヌクレオチドを含む、請求項2のセンサー。
【請求項9】
請求項1のセンサーをサンプルと混合する工程を含む、エフェクター又は補助因子を検出する方法。
【請求項10】
サンプルを色の変化について分析する工程をさらに含む、請求項9の方法。
【請求項11】
色の変化が混合後10分で少なくとも95%は完了する、請求項10の方法。
【請求項12】
混合が10〜45℃の温度で実施される、請求項9の方法。
【請求項13】
サンプル中の分析物の量が塊状粒子の形成又は沈殿に反比例している、請求項9の方法。
【請求項14】
塊状粒子の形成又は沈殿が混合後10分で少なくとも95%は完了する、請求項13の方法。
【請求項15】
Mg(II)、Ca(II)、Zn(II)、Mn(II)、Co(II)及びPb(II)から成る群から選択されるイオンを添加する工程をさらに含む、請求項9の方法。
【請求項16】
サンプルが生物学的サンプルを含む、請求項9の方法。
【請求項17】
エフェクター又は補助因子を検出するためのセンサーシステムであって、前記センサーシステムが、
(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;
(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;
(c)第二のポリヌクレオチドを含む、第一の粒子セット;及び
(d)第三のポリヌクレオチドを含む、第二の粒子セット;
を含み、ここで、前記第一のポリヌクレオチドは前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的であり、前記第二の粒子セットは少なくとも20nmの直径を有する、前記センサーシステム。
【請求項18】
第二の粒子セットが少なくとも30nmの直径を有する、請求項17のセンサーシステム。
【請求項19】
核酸酵素がDNAを含む、請求項18のセンサーシステム。
【請求項20】
第一の粒子セット及び第二の粒子セットが金を含む、請求項18のセンサーシステム。
【請求項21】
第一の粒子セット及び第二の粒子セットが、金属、半導体及びラテックスから成る群から選択される物質を含む、請求項18のセンサー。
【請求項22】
エフェクター又は補助因子が、窒素肥料、農薬、ダイオキシン、フェノール、2,4-ジクロロフェノキシ酢酸、Pb(II)、Hg(II)、As(III)、UO2(II)、Fe(III)、Zn(II)、Cu(II)、Co(II)、グルコース、インシュリン、hCGホルモン、HIV、HIVタンパク質、炭疽、天然痘、神経ガス、TNT、DNT、コカイン及び抗生物質からなる群から選択される、請求項18のセンサー。
【請求項23】
核酸酵素が配列番号1の配列を有するポリヌクレオチドを含み、且つ基質が配列番号2の配列を有するポリヌクレオチドを含む、請求項18のセンサー。
【請求項24】
核酸酵素が配列番号5の配列を有するポリヌクレオチドと配列番号6の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号4の配列のポリヌクレオチドを含む、請求項18のセンサー。
【請求項25】
核酸酵素が配列番号8の配列を有するポリヌクレオチドと配列番号9の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号7の配列を有するポリヌクレオチドを含む、請求項18のセンサー。
【請求項26】
請求項17のセンサーをサンプルと混合する工程を含む、エフェクター又は補助因子を検出する方法。
【請求項27】
サンプルを色の変化について分析する工程をさらに含む、請求項26の方法。
【請求項28】
色の変化が混合後10分で少なくとも95%完了する、請求項27の方法。
【請求項29】
混合が10〜45℃の温度で実施される、請求項26の方法。
【請求項30】
サンプル中の分析物の量が塊状粒子の形成又は沈殿に反比例している、請求項26の方法。
【請求項31】
塊状粒子の形成又は沈殿が混合後10分で95%完了する、請求項26の方法。
【請求項32】
Mg(II)、Ca(II)、Zn(II)、Mn(II)、Co(II)及びPb(II)から成る群から選択されるイオンを添加する工程をさらに含む、請求項30の方法。
【請求項33】
サンプルが生物学的サンプルを含む、請求項26の方法。
【請求項34】
サンプル中のエフェクター又は補助因子の検出方法であって、前記方法が、少なくとも(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;
(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;
(c)第二のポリヌクレオチドを含む、第一の粒子セット;
(d)第三のポリヌクレオチドを含む、第二の粒子セット;及び
(e)サンプル;
を一緒に混合して混合物を生成する工程を含み、ここで、前記第一のポリヌクレオチドは前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的であり、また、前記混合物は、加熱することなく前記混合物の生成から10分以内に、エフェクター又は補助因子のサンプル中の存在を示す色の変化を生じる、前記検出方法。
【請求項35】
核酸酵素がDNAを含む、請求項34の方法。
【請求項36】
第一の粒子セット及び第二の粒子セットが金を含む、請求項34の方法。
【請求項37】
第一の粒子セット及び第二の粒子セットが、金属、半導体及びラテックスから成る群から選択される物質を含む、請求項34の方法。
【請求項38】
エフェクター又は補助因子が、窒素肥料、農薬、ダイオキシン、フェノール、2,4-ジクロロフェノキシ酢酸、Pb(II)、Hg(II)、As(III)、UO2(II)、Fe(III)、Zn(II)、Cu(II)、Co(II)、グルコース、インシュリン、hCG-ホルモン、HIV、HIVタンパク質、炭疽、天然痘、神経ガス、TNT、DNT、コカイン及び抗生物質からなる群から選択される、請求項34の方法。
【請求項39】
核酸酵素が配列番号1の配列を有するポリヌクレオチドを含み、且つ基質が配列番号2の配列を有するポリヌクレオチドを含む、請求項34の方法。
【請求項40】
核酸酵素が配列番号5の配列を有するポリヌクレオチドと配列番号6の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号4の配列のポリヌクレオチドを含む、請求項34の方法。
【請求項41】
核酸酵素が配列番号8の配列を有するポリヌクレオチドと配列番号9の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号7の配列を有するポリヌクレオチドを含む、請求項34の方法。
【請求項1】
エフェクター又は補助因子を検出するためのセンサーシステムであって、前記センサーシステムが、
(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;
(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;
(c)第二のポリヌクレオチドを含む第一の粒子セットであって、前記ポリヌクレオチドがその3'末端で前記粒子に付着している、前記第一の粒子セット;及び
(d)第三のポリヌクレオチドを含む第二の粒子セットであって、前記ポリヌクレオチドがその5'末端で前記粒子に付着している、前記第二の粒子セット;
を含み、ここで、前記第一のポリヌクレオチドは、前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的である、前記センサーシステム。
【請求項2】
核酸酵素がDNAを含む、請求項1のセンサーシステム。
【請求項3】
第一の粒子セット及び第二の粒子セットが金を含む、請求項2のセンサーシステム。
【請求項4】
第一の粒子セット及び第二の粒子セットが、金属、半導体及びラテックスから成る群から選択される物質を含む、請求項2のセンサー。
【請求項5】
エフェクター又は補助因子が、窒素肥料、農薬、ダイオキシン、フェノール、2,4-ジクロロフェノキシ酢酸、Pb(II)、Hg(II)、As(III)、UO2(II)、Fe(III)、Zn(II)、Cu(II)、Co(II)、グルコース、インシュリン、hCGホルモン、HIV、HIVタンパク質、炭疽、天然痘、神経ガス、TNT、DNT、コカイン及び抗生物質からなる群から選択される、請求項2のセンサー。
【請求項6】
核酸酵素が配列番号1の配列を有するポリヌクレオチドを含み、且つ基質が配列番号2の配列を有するポリヌクレオチドを含む、請求項2のセンサー。
【請求項7】
核酸酵素が配列番号5の配列を有するポリヌクレオチドと配列番号6の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号4の配列のポリヌクレオチドを含む、請求項2のセンサー。
【請求項8】
核酸酵素が配列番号8の配列を有するポリヌクレオチドと配列番号9の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号7の配列を有するポリヌクレオチドを含む、請求項2のセンサー。
【請求項9】
請求項1のセンサーをサンプルと混合する工程を含む、エフェクター又は補助因子を検出する方法。
【請求項10】
サンプルを色の変化について分析する工程をさらに含む、請求項9の方法。
【請求項11】
色の変化が混合後10分で少なくとも95%は完了する、請求項10の方法。
【請求項12】
混合が10〜45℃の温度で実施される、請求項9の方法。
【請求項13】
サンプル中の分析物の量が塊状粒子の形成又は沈殿に反比例している、請求項9の方法。
【請求項14】
塊状粒子の形成又は沈殿が混合後10分で少なくとも95%は完了する、請求項13の方法。
【請求項15】
Mg(II)、Ca(II)、Zn(II)、Mn(II)、Co(II)及びPb(II)から成る群から選択されるイオンを添加する工程をさらに含む、請求項9の方法。
【請求項16】
サンプルが生物学的サンプルを含む、請求項9の方法。
【請求項17】
エフェクター又は補助因子を検出するためのセンサーシステムであって、前記センサーシステムが、
(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;
(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;
(c)第二のポリヌクレオチドを含む、第一の粒子セット;及び
(d)第三のポリヌクレオチドを含む、第二の粒子セット;
を含み、ここで、前記第一のポリヌクレオチドは前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的であり、前記第二の粒子セットは少なくとも20nmの直径を有する、前記センサーシステム。
【請求項18】
第二の粒子セットが少なくとも30nmの直径を有する、請求項17のセンサーシステム。
【請求項19】
核酸酵素がDNAを含む、請求項18のセンサーシステム。
【請求項20】
第一の粒子セット及び第二の粒子セットが金を含む、請求項18のセンサーシステム。
【請求項21】
第一の粒子セット及び第二の粒子セットが、金属、半導体及びラテックスから成る群から選択される物質を含む、請求項18のセンサー。
【請求項22】
エフェクター又は補助因子が、窒素肥料、農薬、ダイオキシン、フェノール、2,4-ジクロロフェノキシ酢酸、Pb(II)、Hg(II)、As(III)、UO2(II)、Fe(III)、Zn(II)、Cu(II)、Co(II)、グルコース、インシュリン、hCGホルモン、HIV、HIVタンパク質、炭疽、天然痘、神経ガス、TNT、DNT、コカイン及び抗生物質からなる群から選択される、請求項18のセンサー。
【請求項23】
核酸酵素が配列番号1の配列を有するポリヌクレオチドを含み、且つ基質が配列番号2の配列を有するポリヌクレオチドを含む、請求項18のセンサー。
【請求項24】
核酸酵素が配列番号5の配列を有するポリヌクレオチドと配列番号6の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号4の配列のポリヌクレオチドを含む、請求項18のセンサー。
【請求項25】
核酸酵素が配列番号8の配列を有するポリヌクレオチドと配列番号9の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号7の配列を有するポリヌクレオチドを含む、請求項18のセンサー。
【請求項26】
請求項17のセンサーをサンプルと混合する工程を含む、エフェクター又は補助因子を検出する方法。
【請求項27】
サンプルを色の変化について分析する工程をさらに含む、請求項26の方法。
【請求項28】
色の変化が混合後10分で少なくとも95%完了する、請求項27の方法。
【請求項29】
混合が10〜45℃の温度で実施される、請求項26の方法。
【請求項30】
サンプル中の分析物の量が塊状粒子の形成又は沈殿に反比例している、請求項26の方法。
【請求項31】
塊状粒子の形成又は沈殿が混合後10分で95%完了する、請求項26の方法。
【請求項32】
Mg(II)、Ca(II)、Zn(II)、Mn(II)、Co(II)及びPb(II)から成る群から選択されるイオンを添加する工程をさらに含む、請求項30の方法。
【請求項33】
サンプルが生物学的サンプルを含む、請求項26の方法。
【請求項34】
サンプル中のエフェクター又は補助因子の検出方法であって、前記方法が、少なくとも(a)補助因子結合部位と、任意にエフェクター結合部位とを含む、核酸酵素;
(b)第一のポリヌクレオチドを含む、前記核酸酵素の基質;
(c)第二のポリヌクレオチドを含む、第一の粒子セット;
(d)第三のポリヌクレオチドを含む、第二の粒子セット;及び
(e)サンプル;
を一緒に混合して混合物を生成する工程を含み、ここで、前記第一のポリヌクレオチドは前記第二のポリヌクレオチドを含むか又は前記第二のポリヌクレオチドと少なくとも部分的に相補的であり、且つ前記第一のポリヌクレオチドは、前記第三のポリヌクレオチドを含むか又は前記第三のポリヌクレオチドと少なくとも部分的に相補的であり、また、前記混合物は、加熱することなく前記混合物の生成から10分以内に、エフェクター又は補助因子のサンプル中の存在を示す色の変化を生じる、前記検出方法。
【請求項35】
核酸酵素がDNAを含む、請求項34の方法。
【請求項36】
第一の粒子セット及び第二の粒子セットが金を含む、請求項34の方法。
【請求項37】
第一の粒子セット及び第二の粒子セットが、金属、半導体及びラテックスから成る群から選択される物質を含む、請求項34の方法。
【請求項38】
エフェクター又は補助因子が、窒素肥料、農薬、ダイオキシン、フェノール、2,4-ジクロロフェノキシ酢酸、Pb(II)、Hg(II)、As(III)、UO2(II)、Fe(III)、Zn(II)、Cu(II)、Co(II)、グルコース、インシュリン、hCG-ホルモン、HIV、HIVタンパク質、炭疽、天然痘、神経ガス、TNT、DNT、コカイン及び抗生物質からなる群から選択される、請求項34の方法。
【請求項39】
核酸酵素が配列番号1の配列を有するポリヌクレオチドを含み、且つ基質が配列番号2の配列を有するポリヌクレオチドを含む、請求項34の方法。
【請求項40】
核酸酵素が配列番号5の配列を有するポリヌクレオチドと配列番号6の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号4の配列のポリヌクレオチドを含む、請求項34の方法。
【請求項41】
核酸酵素が配列番号8の配列を有するポリヌクレオチドと配列番号9の配列を有するポリヌクレオチドとを含み、且つ基質が配列番号7の配列を有するポリヌクレオチドを含む、請求項34の方法。
【図4】

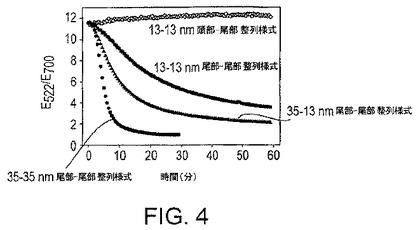
【図5】
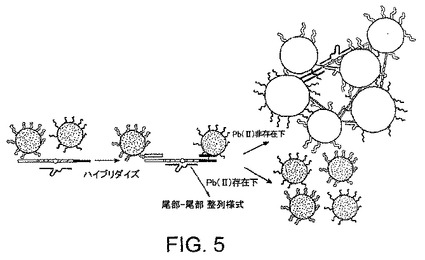
【図6A】
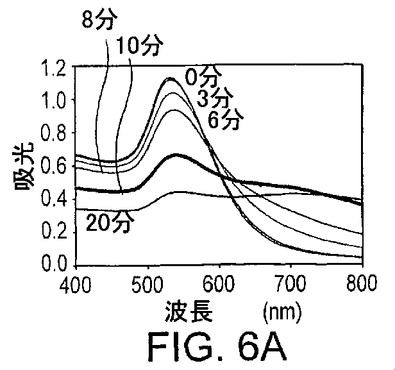
【図6B】
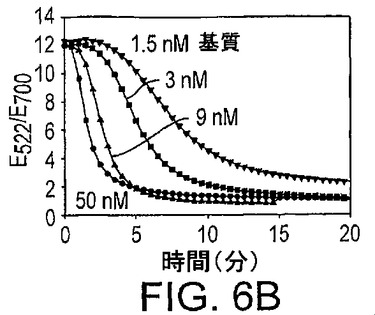
【図6C】
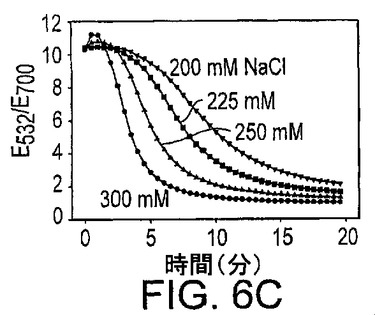
【図6D】
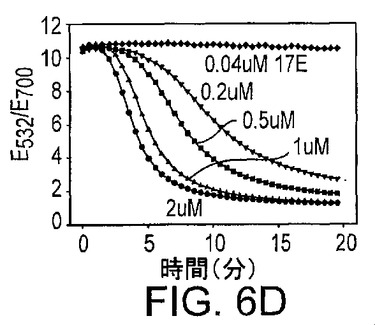
【図6E】
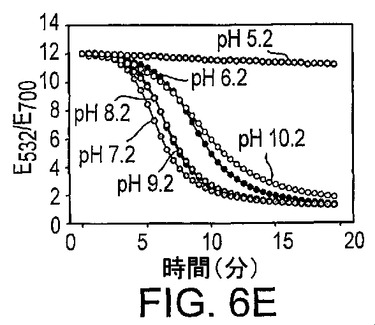
【図6F】
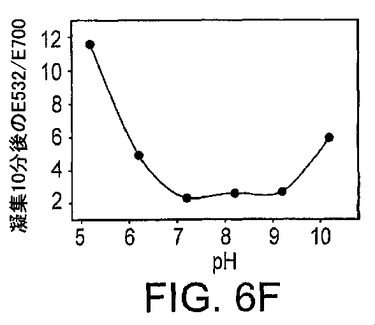
【図7A】
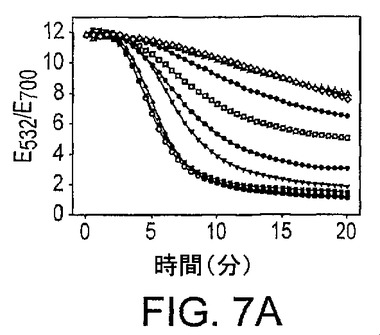
【図7B】
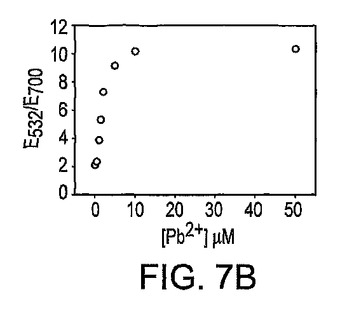
【図8】
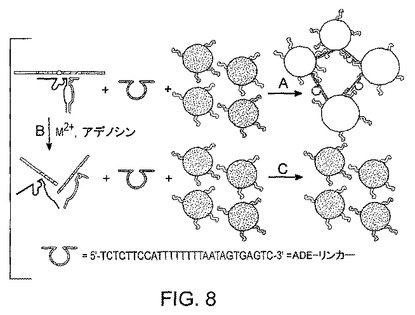
【図9A】
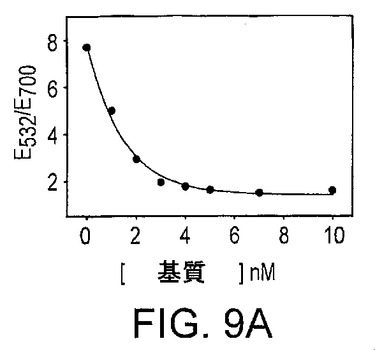
【図9B】
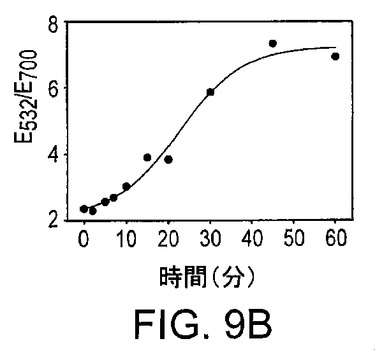
【図9C】
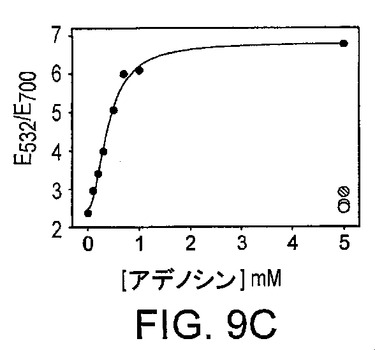
【図5】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【図7A】
【図7B】
【図8】
【図9A】
【図9B】
【図9C】
【公表番号】特表2007−522802(P2007−522802A)
【公表日】平成19年8月16日(2007.8.16)
【国際特許分類】
【出願番号】特願2006−549584(P2006−549584)
【出願日】平成17年1月12日(2005.1.12)
【国際出願番号】PCT/US2005/001060
【国際公開番号】WO2005/100602
【国際公開日】平成17年10月27日(2005.10.27)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
2.パイレックス
【出願人】(500033634)ザ・ボード・オブ・トラスティーズ・オブ・ザ・ユニバーシティ・オブ・イリノイ (21)
【氏名又は名称原語表記】THE BOARD OF TRUSTEES OF THE UNIVERSITY OF ILLINOIS
【住所又は居所原語表記】506 South Wright Street, Urbana, IL 61801
【Fターム(参考)】
【公表日】平成19年8月16日(2007.8.16)
【国際特許分類】
【出願日】平成17年1月12日(2005.1.12)
【国際出願番号】PCT/US2005/001060
【国際公開番号】WO2005/100602
【国際公開日】平成17年10月27日(2005.10.27)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
2.パイレックス
【出願人】(500033634)ザ・ボード・オブ・トラスティーズ・オブ・ザ・ユニバーシティ・オブ・イリノイ (21)
【氏名又は名称原語表記】THE BOARD OF TRUSTEES OF THE UNIVERSITY OF ILLINOIS
【住所又は居所原語表記】506 South Wright Street, Urbana, IL 61801
【Fターム(参考)】
[ Back to top ]