
粘膜ワクチン生成のためのキメラキャリア分子
本発明は、粘膜表面に、および粘膜表面を通して抗原を送達するためのタンパク質複合体、および植物のような宿主細胞での該複合体の生成に関する。少なくとも二つ、好ましくは同一のサブユニットを含むタンパク質複合体であって、少なくとも一つのサブユニットは未改変であり、そして少なくとも一つのサブユニットは対象となる第一分子と融合しており、さらにタンパク質複合体は該サブユニットを介して細胞表面レセプターと相互作用できるタンパク質複合体を提供する。また、本発明のタンパク質複合体を生成するための方法であって、a)未改変サブユニットをコードするヌクレオチド配列および対象となる分子をコードするヌクレオチド配列を持つ宿主細胞であって、少なくとも対象となる分子の一つがサブユニットに融合する宿主細胞を提供する工程;b)該宿主細胞を培養し、それによって該ヌクレオチド配列を発現させ、タンパク質複合体の組み立てを可能にする工程;c)複合体を単離する工程;d)細胞表面レセプターへの、または細胞表面レセプターを模擬する分子への複合体の結合を決定する工程を含む方法も提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は粘膜(例えば口腔または鼻)ワクチンの開発、組成および生成に関する。より具体的には、本発明は粘膜表面への、および表面を通し抗原を送達するためのタンパク質複合体、ならびに植物細胞のような宿主細胞内での該複合体の生成に関する。
【背景技術】
【0002】
経口ワクチン接種は、一般的に経口ワクチンは利用し易く、比較的安価および安全であることから、注射ワクチンに代る魅力的な代替法である。より重要なことは、それが粘膜レベル、即ち多くの病原菌の侵入部位に防御を誘導できることである。例えば経口または鼻内送達による粘膜部位へのワクチン投与は、現在ワクチンが全く利用できないある種の病原菌(例えば呼吸系発疹ウイルス、およびおそらくはHIVでさえ)に対するワクチンの生成にとっては必要条件でもあるだろう。これに加え、経口ワクチン接種は、食物または飲料水による大量ワクチン接種を可能にする。
【0003】
しかしながら、経口ワクチン接種はそれほど有効でない場合が多いと思われる。免疫応答は一時的であり、一般的には、望ましい効果を誘導するには、生きている微生物を使用する場合でさえ大用量の抗原を必要とする。これは、一部は、抗原の取り込みが非効率であることによるものである(Lavell et al. Adv Drug Delivery Rev 1995; 18:5-22)。その結果、粘膜経路から抗原を効率的に送達するための様々な方策が研究されてきた(例えばO'Hagan, J Pharm Pharmacol (1997) 49:1-10; Husband, Vaccine (1193) 11:107-112参照)。粘膜域を裏打ちしている上皮細胞のバリアを通過して送達できた抗原は、粘膜関連リンパ組織(MALT)下部にある誘導部位を刺激する。MALT内で感作した抗原特異的リンパ細胞は循環系を通り移動して、遠方の粘膜部位に定着する。したがって粘膜免疫は、局所と全身防御の両方を提供できる。
【0004】
粘膜ワクチンを開発する一つの方策は、コレラ菌(Vibrio cholera)のこれらトキシン(CT)および近縁な大腸菌(E. coli)易熱性エテロトキシン(LT)のような細菌エンテロトキシンを用いることであり、これらは粘膜送達された場合に極めて免疫源性であり、またキャリア分子およびアジュバントとして機能して、特に後者はホロトキシンを使用した時、無縁な抗原に対する応答を増強できる。
【0005】
毒素原性大腸菌/エシェリキア(Escherichia coli)(LT)の易熱性エンテロトキシンおよびコレラ菌由来の関連コレラトキシン(CT)は極めて強力な免疫原であり、経口投与による粘膜アジュバントである(概説についてはSpangler, 1992およびWilliams et al., 1999参照)。両毒素はAサブユニットと、同一Bサブユニットの五量体環を含む。Aサブユニットはキメラ分子の毒性部分であり、Gsα活性化アデニレートシクラーゼをADPリボシル化して、サイクリックAMPレベルを上昇させる。これが最終的に、水分を多く含む下痢を特徴とする胃腔内の水損失を引き起こす。五量体Bサブユニットを含む、強力な非共有結合性複合体の一次機能は、レセプター相互作用を媒介して毒性のAサブユニットの内在化および取込みを起こすことである。Bサブユニット環の一次レセプターは、小腸を含む哺乳動物細胞の細胞表面上に普遍的に見出されるグリコスフィンゴリピド(Holmgren, 1973; Holmgren et al., 1973, 1975)のモノシアロガングリオシドGM1[Gal(β1-3)GalNAcβ(1-4)(NeuAc(α2-3))Galβ(1-4)Glc(β1-1)セラミド]である。この固有の特徴は、Bサブユニットをアジュバント活性に関係する重要な免疫調節現象を始動する上で重要なものにしている。それはまたBサブユニットを、強力な、いわゆる粘膜キャリア分子にもしている。粘膜キャリア分子は、例えばレセプターを介して、小腸の腸上皮粘膜のような粘膜表面に局在する免疫活性細胞と相互作用する分子である。
【0006】
LT(LTB)およびCT(CTB)両方のBサブユニットは、様々な抗原との翻訳融合において、これらの抗原を、レセプター介在取込みにより胃粘膜上皮を通し行き来させる粘膜キャリア分子としてうまく用いられている(例えばDertzbaugh and Elson, 1993; Jagusztyn-Krynicka et al., 1993)。おそらくは、キャリア分子への抗原の融合が、MALT誘導部位に送達される抗原量と、それに続く抗原特異的B-およびT-リンパ球の刺激を増強するのだろう。抗原は、LTBに化学的に共役させることもできる(O'Dowd et al. Vaccine (1999) 17:1442-1453; Green et al. Vaccine (1996) 14:949-958)。これに加えて、LTBとCTBは共に、抗原と一緒に同時投与した時の経口取込みによる免疫応答を改善する(Bowen et al., 1994; Wilson et al., 1993)。
【0007】
Riganoら(2003, Vaccine 21:809-811)は、トランスジェニックシロイヌナズナでのLTB-ESAT6抗原融合複合体の生成を開示している。融合タンパク質の発現は、ホモ五量体(LTB-ESAT6)5複合体を生ずる。ESAT6は、非常に小型の抗原であり、それゆえに、LTBと融合してもLTB五量体形成を妨害しない。Kim etら(2003; Plant Cell Reports Vol. 21, no.9, pp.884-890)は、ジャガイモでのCTB-NSP175融合タンパク質の発現を報告しており、この発現はホモ五量体複合体を生じた。LTBまたはCTBへの抗原またはエピトープの遺伝的融合は、いくつかの例で成功しているが、Bサブユニットへの非相同性エピトープの翻訳融合は、例えばSandkvistら(J Bacteriol 1987 169:4570)、Schodelら(Gene 1991;99-255)およびDertzbaughら(Infect Immun 1993; 61:384)らが報告しているように、LTBまたはCTB融合タンパク質の構造、分泌、GM1-結合および免疫原性に干渉することがあると思われる。
【0008】
明らかに、五量体化およびGM1結合が保持されるようにLTBまたはCTBに付与できる抗原のサイズとタイプには制限がある。LTBまたはCTベースのワクチンの開発に関しては、最も機能的なワクチンは大型構造を持つタンパク質から構成されることから、この制限は特に実際的な意味を持つ。これに加えて、多くの感染防御抗原(ウイルス、細菌およびその他)は、ホモ多量体またはヘテロ多量体を問わず、多量体複合体から構成されており、そのような形で送達されたときにのみ防御免疫応答を誘導する。例えば、ブタコレラ(a classical swine fever)(CSFV)E2糖タンパク質ベースワクチンは、CSFV-E2ホモ二量体を含有する。様々な感染防御抗原のこの多量体特性は、これらタイプの抗原を通常に遺伝的に融合して用いる、LTBおよび/またはCTBのキャリア分子としての使用を大きく損ねることになるが、それはこの様にして五つの同一融合タンパク質の複合体を形成するからである。
【0009】
商品化および市場導入のためには、融合タンパク質の大規模生産が必要であり、製品は安全であり、またほぼ純粋である必要がある。細菌および酵母で、LTBおよびCTBの融合タンパク質、ならびに抗原をベースにしたこのようなワクチンを生産するには、十分な量の、経口送達のための組換えタンパク質を得るための大規模な発酵技術と厳密な精製プロトコールが必要である。トランスジェニック植物、特に食用植物部分は、経口送達用ワクチンのための安全な発現システムである(Langridge, 2000; Mason and Arntzen, 1995; Sala, 2003参照)。最近、タバコ、ジャガイモおよびトウモロコシを含む様々な植物種で、LTBおよびCTBの製造に成功している(Arakawa et al., 1997; Haq et al., 1995; Hein et al., 1995; Lauterslager et al., 2001; Streatfield et al., 2001)。驚くべき事に、植物もまた大腸菌やコレラ菌と同様にBサブユニットの五量体も生成できるらしい。
【0010】
いくつかのグループが、LTBまたはCTBのいずれかを蓄積したリョウゼンツラン、ジャガイモ塊茎またはトウモロコシを食べさせることによりマウスを経口免疫し、血清IgGおよび分泌性sIgA反応の両方を引き起こしたと報告している(Arakawa et al., 1998; Haq et al., 1995; Lauterslager et al., 2001; Mason et al., 1998; Streatfield et al., 2001)。WO-A-991825は、植物で生産したSEKDEL配列に融合させたCTBサブユニットを用いたマウスの経口免疫を記載している。
【発明の開示】
【0011】
LTBを蓄積したトランスジェニック塊茎50〜100グラムを摂食したヒトボランティアによる前臨床試験でも、特異的血清IgGおよび分泌性sIgA反応が起こった(Tacket et al., 1998)。したがって、トランスジェニック宿主細胞系は、LTBまたはCTB融合タンパク質のような粘膜キャリア分子の(大規模)生産に、本質的に、好都合に用いられる。しかしながら、融合タンパク質の発現レベルは、融合タンパク質のサイズおよび複雑さの増加に伴って大きく低下することが示されている。例えば、ジャガイモ塊茎での、CSFV-E2糖タンパク質(約35kDa、および約70kDaの二量体として防御)と融合したLTBをコードする遺伝子構築体の発現は、約15kDaのLTBサブユニットをコードする遺伝子の発現に比べ、100倍以上低い(Lauterslager, 2002; PhD thesis University of Utrecht)。おそらく、モノマーが、組み立てられた多量体構造に比べ(酵素)分解を受けやすいことが多く観察されていることから、大型のLTBまたはCTB融合タンパク質の低い発現レベルは、大型の五量体タンパク質複合体の不安定性を基本的に反映したものだろう。この見方は、ジャガイモへのCTB-GAD融合タンパク質の蓄積を報告したArakawaら(1999; Transgenics, Harwood Academic Publishers, Basel, Vol. 3, no.1, pp.51-60)と一致する。GAD(ヒトグルタミン酸デカルボキシラーゼ)は65kDaの自己抗原である。CTB-GADの定量化は、発現レベルが全可溶性タンパク質のわずか0.001%であることを明らかにし、このような高分子量複合体を相当レベル蓄積しようとするとき遭遇する可能性がある問題を明示した。しかしながらマウスに新鮮なトランスジェニックジャガイモ物質を長期間、かつ大量に摂食させた時には、少量の活性ホモ五量体複合体は免疫原性であると思われた。
【0012】
したがって、本発明の目的は、比較的大型の抗原を対象となる部位に、例えば組換え体宿主細胞内に、複合体の発現レベルを犠牲にすることなしに送達できる機能的キャリア複合体を提供することである。
【0013】
次に本発明は、全てではなく、幾つかのサブユニットだけが抗原またはその他の対象となる分子に融合した時に、多サブユニットキャリア分子の組み立て、機能性および安定性が高まることについて洞察する。少なくとも二つの、好ましくは同一のサブユニットを含み、そのうちの少なくとも一つのサブユニットは未改変であり、少なくとも一つのサブユニットは関心の第一分子と融合しているタンパク質複合体を提供し、このときタンパク質複合体は細胞表面レセプターに結合できる。
【0014】
好ましい態様では、該タンパク質複合体は、対象となる分子を運搬または移動させるのに用いることができるキャリア分子である。例えば、本発明のタンパク質複合体は、対象となる分子を対象となる部位、例えば粘膜表面まで、および/または横切り送達するためのキャリア複合体またはキャリア分子である。本発明によれば、対象となる分子は、複合体のキャリア特性を用いて生物内の特定部位に送達、その中に分配、移動、または保持することを望む、様々な分類のタンパク質またはポリペプチド、あるいはその領域を含むことができる。それは特に、キャリアの全サブユニットに付与しようとした場合に、例えば立体障害によって、キャリ分子の安定で機能的な多量体構造の形成を妨害するであろう、サブユニットに融合または付加された成分を指す。例えば、サブユニットに融合した該対象となる分子のサイズは、サブユニット単独のサイズの1/4、1/3、1/2、1倍、2倍、3倍または4倍のこともある。
【0015】
本明細書で使用する、用語「未改変」は対象となる分子に融合していないサブユニットまたはモノマーを指す。しかしながらそれは、未変性のサブユニットまたはモノマーに限定されない;例えば、それは、一または複数のアミノ酸が未変性サブユニットから取り除かれた、交換された、および/または追加された組換えタンパク質である。これは、(真核)宿主細胞での組換えタンパク質の安定性、および/または生成工程、例えば発現もしくは分泌を変えるために行うことができる。例えば、未改変サブユニットは、シグナルペプチド、または小胞体(ER)中の保持のためC末端に(SE)KDEL配列を付与したサブユニットを含んでもよい。用語「未改変サブユニット」は、実質的には、未変性サブユニットまたはその修飾が多量体化を妨害しない若干修飾されたものを指す。
【0016】
WO-A-9612801は、発現により一つになってCTホロ毒素タンパク質複合体を形成する、LTAサブユニットおよび、SEKDEL配列に融合したLTBサブユニットの協調発現を開示している。このようなタンパク質複合体は、それが本発明の対象となる分子に融合したサブユニットを含んでいないことから、本発明の複合体とは異なる。SEKDELヘキサペプチドは、対象となる分子とは見なさない。むしろ、上記のように、LTB-SEKDELサブユニットは「未改変」サブユニットと見なされる。したがって本発明の用語法によれば、WO-A-9612801の複合体は、それが未改変サブユニットだけを含むことから、本明細書に示すタンパク質複合体とは別のものである。
【0017】
US6,103,243は、例えばLTBのような粘膜キャリア複合体を用いることによって、免疫原の取込みを改良する経口ワクチンおよび方法を記載している。LTBサブユニットには対象となる抗原が、化学結合、または遺伝的融合のいずれかによって付与されている。これにより、全てのサブユニットが1タイプの対象となる分子に融合しているホモ五量体タンパク質複合体ができる。このように、本発明とは異なり、US6,103,243の複合体は、少なくとも一つの未改変サブユニットおよび対象となる分子に融合している少なくとも一つのサブユニットを含んでいない。
【0018】
本発明の好ましい態様では、タンパク質複合体は、少なくとも二つの同一サブユニット、例えばLTBサブユニットを含み、そのうちの少なくとも一つのサブユニットは変更されており、また少なくとも一つのサブユニットは対象となる分子に融合している。本発明の一つの局面では、タンパク質複合体は、少なくとも一つのサブユニットが未改変であること、そして少なくとも一つのサブユニットが対象となる第一分子に融合していることを特徴とする、少なくとも二つの、好ましくは同一のサブユニットを含み、このとき該対象となる第一分子は、対象となる第二分子と結合して対象となる多量体を形成することができ、またこのときタンパク質複合体は該サブユニットを介して細胞表面レセプターと相互作用できる。好ましくは、該第一分子は、分子間共有結合を介して、例えば一または複数のジスルフィド架橋結合を介して、該第二分子と結合または相互作用する。本発明の対象となる多量体は、例えば、ホモ二量体またはヘテロ二量体抗原のような多量体感染防御抗原である。公知の多量体感染防御抗原としては、CSFV-E2ホモ二量体、ウイルス性出血性敗血症ウイルスの三量体糖タンパク質G(VHSV-G)(Lorenzen, N., Lorenzen, E., Einer-Jensen, K., Heppell, J., Wu, T., Davis, H. (1998) Protective immunity to VHS in rainbow trout (Oncorhynchus mykiss, Walbaum) following DNA vaccination. Fish & Shellfish Immunology 8: 261-270; Lorenzen, N., Olesen, N.J. (1997) Immunisation with viral antigens: viral haemorrhagic septicaemia. In: Fish Vaccinology. Gudding, R., Lillehaug, A., Midlyng, P.J., Brown, F. (eds). Dev. Biol. Stand. Basel, Karger, vol. 90, p 201-209) および狂犬病ウイルス(RV G)および水疱性口内炎ウイルス(VSV G)の三量体糖タンパク質G;センダイ(Sendai)ウイルスのホモ四量体リンタンパク質P(SeV P)が挙げられる。
【0019】
本発明によれば、対象となる第二分子はサブユニットと融合してもよい。例えば、本発明の一つの態様では、タンパク質複合体は、三つのサブユニットを含み、そのうちの二つは同一(例えばCSFV-E2)モノマーに融合しており、そして一つは未改変サブユニットである。サブユニットは互いに近接しており、融合モノマーが相互作用でき、そしてホモ多量体、例えば抗原(例えばCSFV-E2)ホモ二量体を形成できることが望ましい。未改変サブユニットの存在は、タンパク質複合体の多サブユニット構造の安定性を高め、複合体のコンホメーションが保持され、それがサブユニットを介してそのレセプターと相互作用できることを保証する。融合モノマーがヘテロ多量体を形成できるように、少なくとも三つのサブユニットのタンパク質複合体が異なるモノマーと融合している二またはそれ以上のサブユニットと、少なくとも一つの未改変モノマーを含むことは、明らかに可能である。
【0020】
もちろん、対象となる第二分子は、対象となる第一分子と相互作用して対象となる多量体を形成するために、サブユニットと融合する必要はない。そのうえ、本発明によれば、例えば立体障害に関して、対象となる多量体を含むタンパク質複合体を、該対象となる多量体が、全てがサブユニットに融合しているわけではない対象となる多量体から構成されるようにデザインすることが有利なことも時にある。例えば、全てがあるサブユニットに融合している対象となる分子間の相互作用の強さは、それらが融合している個々のサブユニットの相対的な方向によりある程度決定される。したがって、互いに正常に(即ち、それらの未変性で、融合していないコンホメーションにある)相互作用し合える分子は、サブユニットと融合した結果分子間の相互作用を低下、ときに完全に失うことにより、立体的に制限されることもある。したがって、本発明によれば、タンパク質複合体は、例えばジスルフィド架橋結合を通して、多量体構造を形成することができる対象となる多量体分子から構成される対象となる多量体を含んでよい。好都合にも、少なくとも一つの未改変サブユニットおよび抗原と融合したサブユニットを少なくとも一つ含む本発明のタンパク質複合体は、一般的に、一つまたは複数(多量体)抗原の、一抗原成分へのより良い折りたたみ構造を示す。前述したように、対象となる一つ、幾つか、または全てがサブユニットと融合する。当業者は、サブユニットおよび対象となる多量体の四次構造および対称性に基づいて、最適なコンホメーション、例えば最適な抗原特性を持つ対象となる多量体を含むタンパク質複合体を得るために、融合型分子と非融合型分子の最適数を選ぶことができるだろう。
【0021】
一つの態様では、タンパク質複合体は、腸上皮上にある細胞表面レセプター、例えばGM1のようなガングリオシド分子に結合できる。対象となる分子を含む、このようなタンパク質複合体は、粘膜キャリア分子として有利に用いられる。
【0022】
好ましい態様では、本発明のタンパク質複合体は、大腸菌の易熱性エンテロトキシン(LT)またはコレラ菌のコレラトキシン(CT)、好ましくはそのBサブユニットに本質的に基づいている。本発明の複合体は、例えば五つのLTまたはCTのBサブユニットから構成される環構造体であり、この場合そのうち少なくとも一つのサブユニットは未改変であり、少なくとも一つのサブユニットは対象となる分子に融合している。このような本発明のキメラまたはヘテロ五量体環構造体は、一、二、三または四つの未改変LTB-またはCTB-モノマーと、四、三、二または一つのL/CTB-融合タンパク質をそれぞれ含む。好ましい態様では、対象となる分子は、サブユニットに、それがレセプター結合ポケットから見て分子の反対側に位置するように、または少なくとも該サブユニットのレセプター結合特性を妨害しないように融合している。例えば、五量体LTB-またはCTB-構造体に基づく本発明の複合体では、これは対象となる分子がBサブユニットのC末端に融合することが好ましいことを意味している(Sixma et al. 1991 Nature 351;371-377)。この本発明の方針に従って、それらのコンホメーションの完全性を保持したキメラ五量体を得ている。LTBサブユニットおよびCTBサブユニットは相同的であることから、それらを用いて、少なくとも一つのサブユニット(LTBまたはCTBのいずれでも)が対象となる分子と融合しており、そして少なくとも一つのサブユニットが未改変である、キメラLTB/CTB五量体から構成されたタンパク質複合体を形成できる。LTBおよび/またはCTBをベースとした本発明のキメラタンパク質複合体は、五つ全てのサブユニットが対象となる分子に融合しているホモ五量体構造と比べると、GM1レセプターへの結合の向上および発現レベルの上昇を示す。
【0023】
別の態様では、本発明のタンパク質複合体は、西洋ワサビペルオキシダーゼ(HRP)に基づいている。本発明の複合体は、例えば、少なくとも一つのサブユニットは未改変であり、そして少なくとも一つのサブユニットは対象となる分子と融合している、六つのHRPを含む構造体または組成物である。このような本発明のキメラまたはヘテロ六量構造体は、一、二、三、四または五つの未改変HRPサブユニット(モノマー)と五、四、三、二または一つのHRP融合タンパク質をそれぞれ含む。
【0024】
本発明の複合体は、多量体抗原の最適な折りたたみ構造と分子内相互作用(例えばジスルフィド架橋結合を介した)を可能にし、したがって最適な免疫原特性を有している。一つの態様では、対象となる分子は、生物、好ましくは哺乳動物、より好ましくはヒトの免疫系に変化をもたらす、または影響することができる。好ましい態様では、対象となる分子は抗原である。抗原は、免疫系を刺激する任意の物質である。抗原は、身体に侵入する細菌またはウイルスの様な外来性微生物、またはタンパク質またはタンパク質フラグメントのような該微生物の成分であることが多い。対象となる分子は、好ましくは、細菌抗原、ウイルス抗原、原虫抗原、線虫抗原および真菌抗原からなる群より選択される。本発明は、各種抗原、例えば線状B細胞エピトープCPV(イヌパルボウイルスエピトープ)およびT細胞特異エピトープHA(インフルエンザウイルス血球凝集素エピトープ)のようなTおよび/またはB細胞エピトープを送達するためのタンパク質複合体、ならびにそのようなタンパク質複合体を含む組成物を提供する。本発明の複合体は、例えばCSFV(ブタコレラウイルス)のE2タンパク質のような、ウイルス(糖)タンパク質のような大型の抗原の送達にも用いることができる。一つのサブユニット融合タンパク質が複数の対象となる分子と融合してもよい。例えば、少なくとも一つの未改変サブユニットと、二つのHAエピトープおよび二つのパルボエピトープと転写的に融合した少なくとも一つのサブユニットとを含むタンパク質複合体が提供される(図1参照)。本発明の一つの態様では、タンパク質複合体(例えばLTBをベースとした複合体)は、少なくとも一つの未改変のサブユニット、およびCSFV E2糖タンパク質に融合している少なくとも一つ、好ましくは二つのサブユニットを含む。別の態様では、本発明のタンパク質複合体は、少なくとも一つの未改変サブユニット、パルボエピトープに融合した少なくとも一つのサブユニット、および複数の対象となる分子に融合している少なくとも一つのサブユニットを含み、後者は二つのHAエピトープおよび二つのパルボエピトープに融合したサブユニットを含む。
【0025】
更なる態様では、本発明のタンパク質複合体は、未改変である少なくとも一つのサブユニット、および免疫調節分子(またはその一部分)、例えばサイトカインもしくは熱ショックタンパク質(HSP)に融合している少なくとも一つのサブユニットを含む。サイトカインは、免疫応答中に細胞間シグナル伝達に関係する分子の大きなグループを表す一般的用語である。サイトカインはタンパク質またはポリペプチドであり、一部には糖タンパク質が結合している(糖タンパク質)。サイトカインは幾つかの群:インターフェロン(IFNアルファ、ベータ、ガンマ);インターロイキン(IL-1〜IL-15);コロニー刺激因子(CSF)および、腫瘍壊死因子(TNF)アルファおよびベータまたは形質転換増殖因子(TGF)ベータのようなその他サイトカインに区別できる。HSPは、CD91と呼ばれるレセプターを介したそれらとマクロファージおよび樹状細胞との相互作用に由来する、顕著な免疫調節特性を有している。例えば、HSP70-2は、炎症刺激に対する反応で誘導される、重要な免疫調節タンパク質である。本発明のタンパク質複合体に含まれる対象となるHSPとしては、HSP-60、HSP-70、HSP-90およびGp-96が挙げられる。所望する免疫学的応答のタイプに応じて(例えばTh1対Th2、抗体応答、抗炎症応答)、一つまたは複数(異なる)免疫調節タンパク質またはその一部分を本発明のタンパク質複合体のサブユニットに融合することができる。例えば、本発明の多成分ワクチンは、抗原および、抗体応答(T、B細胞)を誘導する免疫調節分子、好ましくはサイトカインを含むタンパク質複合体を含む。一方、例えば自己免疫疾患(例えば糖尿病、多発性硬化症)の治療または予防については、自己抗原および、寛容を誘導する免疫調節タンパク質を含む多成分ワクチンまたはワクチンを使用することが有利だろう。
【0026】
対象となる別の分子は、本発明の複合体の一部である場合に、「レポーター」分子として用いて体内のキャリア複合体の局在を知らせることができるような分子を含む。これは、レセプター分子への複合体のインビボ結合のモニタリング、および粘膜上皮を通しての複合体の(レセプター介在)輸送にとりわけ役立つ。対象となるレポーター分子は、例えば、酵素(クロラムフェニコールトランスアセチラーゼ(CAT)、ネオマイシンホスホトランスフェラーゼ(neo)、ベータ−グルコシダーゼ(GUS)またはホタルルシフェラーゼ;等)またはグリーンフルオレセントタンパク質(GFP)またはそのスペクトル変種のような蛍光タンパク質である。
【0027】
本発明の一つの局面では、タンパク質複合体は、そのうちの少なくとも二つのサブユニットに対象となる分子が付与されている少なくとも三つのサブユニットを含む。該少なくとも二つのサブユニットは、対象となる同一の分子または対象となる異なる分子に翻訳的に融合できる。少なくとも二種類の対象となる分子を持つ本発明のタンパク質複合体は、少なくとも二種類の抗原のため、例えば多成分ワクチンの生成のためのキャリア分子として有利に用いられる。別のタイプの対象となる分子のその他組合せ、例えば一または複数の抗原と一または複数の免疫調節(促進的または阻害的)タンパク質との組合せも、もちろん可能である。
【0028】
コレラトキシンをベースとする多成分ワクチンがYuとLangridge(Nature, 2001, vol.19:548)により記載されており、かれらはネズミロタウイルスエンテロトキシンNSP4の22アミノ酸免疫優性エピトープに融合したコレラトキシンBサブユニット、およびCTA2サブユニットに融合したETECフィムブリエ定着因子CFA/Iの発現を研究した。本発明の複合体とは異なり、報告されたコレラトキシン複合体の全てのサブユニット(BおよびA2の両方)がある抗原に融合していた。この具体例では、CTB発現または機能的CTB/A2複合体の形成に問題がなかったという事実は、おそらくはBサブユニットに融合したNSP4エピトープのサイズが極めて小さい(約5kDa)ことに関係しているだろう。既に述べたように、機能的五量体の形成の問題は、立体障害に大きく帰すべき問題であり、サブユニットに融合したこのような小さいエピトープは五量体形成を妨害することはないだろう。本発明によれば、抗原が5kDaよりはるかに大きい(多成分)ワクチンも生産できるようになる。例えば、対象となる分子に融合した少なくとも一つの(CT/LT)Bサブユニット、少なくとも一つの未改変Bサブユニット、および対象となる異なる分子に融合したA2サブユニットを含むタンパク質複合体を提供する。好ましくは、Bサブユニットに融合した該分子は、7kDaより大きく、より好ましくは10kDaより大きく、よりさらに好ましくは15もしくは25kDaよりさらに大きい。しかしながら、本発明の別の態様では、同一キャリア複合体の(LT/CT)Bサブユニットおよび(LT/CT)A2サブユニットは、同一抗原に融合しており、このとき該抗原は免疫防御特性を持つ多量体(二量体、三量体、四量体)タンパク質複合体の一部分である。
【0029】
本発明はさらに、本発明のタンパク質複合体を生成するための方法であって、a)未改変サブユニットをコードする第一ヌクレオチド配列および対象となる分子と融合したサブユニットをコードする第二ヌクレオチド配列を持つ宿主細胞を提供する工程;b)該宿主細胞を培養し、それによって該第一および第二ヌクレオチド配列を発現させ、タンパク質複合体を組み立てさせる工程;c)複合体を分離する工程;およびd)複合体を細胞表面レセプターに結合する工程を含む方法を提供する。
【0030】
用語「宿主細胞」は、異質遺伝子を複製および/または転写および/または翻訳できる任意の細胞を指す。一つの態様では、宿主細胞は植物細胞、ファージまたは細菌である。好ましい態様では、本発明の宿主細胞は食用宿主細胞であり、これは消費時に有害な影響を生じない。好ましい例はジャガイモ、トマト、タバコ、トウモロコシおよび乳酸桿菌である。したがって、宿主細胞は、インビトロに在ろうとインビボに在ろうと、任意の真核生物または前核生物細胞(例えば大腸菌のような細菌細胞、メタノール資化酵母(Pichia pastoris)のような酵母細胞、チャイニーズハムスター卵巣細胞のような哺乳動物細胞、トリ細胞、両生類細胞、植物細胞、魚類細胞、マッシュルーム(Agaricus bisporis)のような真菌細胞、およびヨウガ(Spodoptera frugiperda)のような昆虫細胞)を指す。例えば、宿主細胞はトランスジェニック植物内に在ってもよい。
【0031】
上記のように、特定態様において、植物を本発明の方法に用いることができる。該植物宿主は、トウモロコシ(Zea mays)、コムギ(Triticum aestivum)、イネ(Oryza sativa)またはアオウキグサ種(Lemna spp)のような単子葉植物、またはタバコ種(N. tabacum)のようなタバコ(Nicotinana)属、トマトのようなトマト(Lycopersicon)属、ウマゴヤシ(Medicago)を含むマメ(Leguminosae)科、またはヒメツリガネゴケ(Physcomitrella patens)のような蘚類でよい。好ましい態様では、該宿主植物はジャガイモ(Solanum tuberosum)である。
【0032】
本発明によれば、宿主細胞には、少なくとも二種類の核酸配列:未改変(対象となる抗原と融合していない)サブユニットをコードする核酸配列、および対象となる分子と翻訳的に融合したサブユニットをコードする核酸配列が与えられる。例えば、(植物)宿主細胞に、未改変LTBサブユニットをコードする第一ヌクレオチド構築体および対象となる抗原に融合したLTBサブユニットをコードする第二構築体をトランスフェクションする。対象となる分子に融合したサブユニットをコードするヌクレオチド配列は、生ずる融合タンパク質の可撓性を増すために、サブユニットと対象となる分子の間の領域にリンカーまたはヒンジを含んでもよい。適合条件の下でトランスフェクションおよび培養した後、該細胞は二つのポリペプチドを発現し、抗原が「負荷された」、機能的なLTBをベースとしたキメラタンパク質複合体を組み立てるだろう。少なくとも二種類の対象となる分子を持つ本発明のタンパク質複合体を生成するためには、宿主細胞には、当然少なくとも三種類のヌクレオチドを付与する。これら異なる核酸配列は、それぞれが異なるヌクレオチド配列を担持している、異なるベクターを用いた(例えばT-DNA)該宿主細胞のコトランスフェクションによって、該宿主細胞内に導入できる。または、二もしくはそれ以上の種類の遺伝子構築体を、交差によって、または一つのバイナリーベクター上に二もしくはそれ以上の発現カセットを用いることによって、一つの宿主細胞に導入できる。より更には、既に本発明のタンパク質複合体の一(または複数の)成分を含んでいる樹立宿主細胞株に、再形質転換を用いて、追加の核酸配列を付与できる。さらに別の態様では、少なくとも二種類の核酸配列を発現している宿主細胞は、もう一方の配列を安定的に発現している宿主細胞に一方の配列を一過性(ウイルス媒介性)に発現させることで得られる。
【0033】
異なるベクターを使用することの利点は、それによって、宿主細胞に様々な割合で核酸構築体を与えることができるという事実に見いだせる。これにより、未改変の「フリー」サブユニットの量を、宿主細胞が発現した融合サブユニットの量に対して滴定でき、生じた多量体タンパク質複合体の組成を最適化できる。例えば、融合サブユニットを多く含む複合体を所望するのであれば、宿主細胞を構築体A(未改変サブユニット)および構築体B(融合サブユニット)と、構築体Bが構築体Aより過剰であるようにして(例えばA:B=1:3または1:5または、さらに1:10)コトランスフェクションまたはコトランスフォーメーションする。これと逆に、組み立てたタンパク質複合体が融合サブユニットを比較的少なく含むように変えるには、構築体Aを構築体Bより過剰にするのが好ましい。後者はもちろん、本発明の機能的タンパク質複合体の組み立てに対する、融合サブユニットの負の立体的影響または妨害を最小にしたい場合に好ましい。
【0034】
さらに別の態様では、宿主細胞は対象となる分子、例えばLTB-CSFV E2に融合したサブユニットをコードする核酸構築体を用いて形質転換され、このとき該構築体は、サブユニットをコードする核酸配列と対象となる分子をコードする核酸配列との間にタンパク質分解性切断部位を含んでいる。この融合タンパク質がプロテアーゼ(宿主細胞内で異種内因的に発現された)によって部分的インビボ切断されると、宿主細胞は未改変サブユニットと対象となる分子に融合したサブユニットの両方を含むことになり、これらは本発明のキメラタンパク質複合体を形成できる。
【0035】
当技術分野で公知の様々な方法を用いて、宿主細胞に組換えまたは単離された核酸(DNAまたはRNA)を提供することができる。これらには、形質転換、トランスフェクション(例えばリン酸カルシウム沈殿またはカチオン性リポソーム試薬を用いた)、エレクトロポレーション、バーティクルガンおよびアグロバクテリウム(Agrobacterium)介在T-DNA転移が挙げられる。当業者は、宿主細胞のタイプに応じて、選択する方法を認めるだろう。
【0036】
宿主細胞に該外来ヌクレオチドを与えた後、宿主細胞を培養して該第一および第二ヌクレオチド配列を(同時)発現させ、生じたポリペプチドを本発明のタンパク質複合体に組み立てる。いくつかの例、特に形質転換またはトランスフェクションの方法が比較的非効率な例では、該ヌクレオチドを実際に受け取っている宿主細胞を選択し、このように選択された宿主細胞だけを培養することが有利である。形質転換またはトランスフェクション(または宿主細胞に単離核酸を提供するその他の方法)後の宿主細胞選択は、標準の方法に従って実施できる。例えば、対象となるDNA配列を宿主に送達するのに用いられる最も一般的なベクターは、効率的な宿主細胞への送達および宿主細胞による発現により、該宿主細胞に選択薬剤に対する耐性を付与するタンパク質(酵素のような)をコードする核酸配列も含んでいる。頻繁に用いられる選択薬剤は、ネオマシン、カナマイシン、アンピシリン、カルベニシリン等のような抗生物質である。
【0037】
本発明の方法では、少なくとも二種類の核酸(バイナリーベクターまたは異なるベクターのいずれかを用いても)が付与された(選択された)宿主細胞は、少なくとも二種類のポリペプチド、例えば未改変(非融合)サブユニットXおよび融合サブユニットYを発現するだろう。
【0038】
本発明の別の態様では、第一宿主細胞、例えば微生物宿主細胞は、未改変サブユニットXを提供し、第二宿主細胞は対象となる分子、例えば抗原を含む融合サブユニットYを提供する。場合によっては、第三宿主細胞は、第二対象となる分子、例えば第二抗原またはサイトカインのような免疫調節分子を含む融合サブユニットZを産生できる。別々に産生されたXおよびY(および場合によりZ)サブユニットを単離した後、それらを、少なくとも一つのXサブユニットおよび少なくとも一つのYサブユニットを含むタンパク質複合体の形成にとって好ましい条件の下で、互いに接触させることができる。本発明のキメラタンパク質複合体をインビトロ再構成するこの方法は、それらをある割合で互いに接触させることによって、一つの複合体の中にある未改変サブユニットと融合サブユニット数の滴定を可能にする。未改変および融合サブユニットの両方を産生する単一宿主細胞を用いる場合と対照的に、上記再構築法によるキメラタンパク質複合体の作製は、サブユニットの相対発現レベルには依存しない。むしろ、互いに接触している未改変と融合サブユニットとの割合は、制御することができる(実施例9も参照)。それにより本発明は、本発明のタンパク質複合体を生成する方法であって:a)未改変サブユニットをコードするヌクレオチド配列を第一宿主細胞に提供し、また対象となる分子をコードするヌクレオチド配列を第二宿主細胞に提供し、このとき対象となる少なくとも一分子がサブユニットに融合している工程;b)該宿主細胞を培養し、それにより該ヌクレオチド配列を発現させる工程;c)該ヌクレオチドがコードするタンパク質を単離する工程;d)タンパク質複合体の組み立てを可能にする条件の下に単離されたタンパク質を接触させる工程;e)複合体を単離する工程;f)細胞表面レセプターまたは細胞表面レセプターを模した分子への複合体の結合を決定する工程を含む方法を提供する。上記のうち工程d)では、特定サブユニット組成を持ったタンパク質複合体が形成されるように、タンパク質を特定の割合で混合することができる。
【0039】
該少なくとも二つのポリペプチドの多量体複合体への組み立ては、それぞれが異なるサブユニット組成を持った多様な複合体になることができる。理論的には、2タイプの複合体:未改変Xサブユニットのみ、または融合Yサブユニットのみを含むホモメリック複合体、およびXおよびYサブユニットの混合体を含むヘテロメリック、またはキメラ複合体が形成できる。複合体中に存在するXおよびYサブユニットの数に応じて、各種タイプのキメラ複合体が可能である。例えば、三量体は、二つのXサブユニットと一つのYサブユニット、またはその逆を含むことができる;四量体複合体はXサブユニットを三、二または一のみ、そしてYサブユニットを一、二または三つ、それぞれ含むことができる;といった具合である。推察のとおり、本発明のタンパク質複合体は、後者(キメラ)のタンパク質複合体に関係する;ホモメリックX複合体は対象となる分子を欠いており、ホモメリックY複合体は、仮に組み立てたとして、融合した(嵩高の)対象となる分子数が多いことによる立体障害の存在により、おそらくはレセプターへ結合することができないだろう。そのうえ、全てのサブユニットが抗原と融合しているホモメリックタンパク質複合体は、個々の抗原モノマーの配向/コンホメーションが免疫特性に関して最適でないことから、多量体抗原にとって有用なキャリア分子ではないだろう。それゆえに、本発明の一つの態様では、本明細書で提供するキメラタンパク質複合体が、キメラおよびホモメリック複合体の混合物から単離される。しかしながら、キメラタンパク質複合体をホモメリック複合体から分離することは常に必要とされるものではないことを理解しなければならない。一つの態様では、キメラおよびホモメリック複合体を含む組成物は、ワクチンとしての好適に用いられる。しかしながら、キメラ複合体がホモメリック複合体より大量に存在することが好ましい。例えば、キメラタンパク質複合体が、タンパク質複合体総数の少なくとも50%、好ましくは少なくとも60%、より好ましくは少なくとも70%を構成することである。
【0040】
キメラおよびホモメリックタンパク質複合体は、それらのサイズに基づいて互いを分離できる。融合サブユニットは、未改変サブユニットより明らかに大きいことから、サブユニット組成が異なる複合体はサイズが異なるだろう。この差により、複合体のサイズに従って、所望のキメラ複合体を、ホモメリックとキメラタンパク質複合体との混合物から
単離できる。好ましい態様では、本発明のタンパク質複合体は、ゲル濾過クロマトグラフィーを用いて単離される。ゲル濾過クロマトグラフィー(サイズ排除クロマトグラフィーまたは分子篩クロマトグラフィーとしても知られる)を用いて、タンパク質をそれらのサイズに従って分離することができる。タンパク質クロマトグラフィーに関する一般的情報は、RK Scopes(Springer-Verlag3rd edition, January1994; ISBN 0387940723)による "Protein Purification: Principles and Practice" のような本技術分野で用いられている手引き書から得ることができる。
【0041】
簡単に述べると、ゲル濾過では、タンパク質(の混合物)溶液を半透性多孔性樹脂を詰めたカラムに通す。半透性樹脂はある範囲の孔径を有しており、これがカラムで分離できるタンパク質のサイズを決定する。これは樹脂の画分域または排除域と呼ばれる。樹脂の排除域より大きなタンパク質は孔の中に入ることはできず、樹脂の間の空間を通り、カラムを素早く通過する。これはカラムの空隙容量として知られる。樹脂の排除域以下の小さいタンパク質、およびその他の低分子物質は全て樹脂の孔に侵入し、またそれらはカラムの全容積を通過しなければならないため、それらのカラムを通過する動きは遅くなる。カラムの排除域内に納まるサイズのタンパク質は、孔の一部のみに侵入する。このようなタンパク質の動きは、それらのサイズに従って遅くなるだろう;小さいタンパク質ほど、より大きな容積を通過しなければならないため、カラムをゆっくりと通過することになる。ゲル濾過クロマトグラフィーによってタンパク質サンプルを分離するためには、カラムをまず所望の緩衝液で平衡化しなければならない。これは、カラムに、カラム容積の数倍量の緩衝液を通すだけで達成できる。平衡緩衝液はタンパク質サンプルが溶出していく緩衝液であることから、平衡化は、重要な工程である。次にサンプルをカラムに負荷し、樹脂に浸透させる。次により一層多い量の平衡緩衝液をカラムに通して、サンプルを分離し、カラムからそれを溶出する。画分はカラムからのサンプル溶出物として集められる。より大きなタンパク質は早い画分に溶出し、より小さなタンパク質はそれに続く画分に溶出する。
【0042】
ゲル濾過は、理想的に言えば、タンパク質複合体の分解を減らすことに加えて、濾過中のサンプルの拡散を小さくするのにも役立ち、これが解像度を向上することから、低温で行うべきである。タンパク質の分離は、より長いカラムを使用することで向上するが、分離時間が長くなるほどタンパク質の分解は増加する。
【0043】
クロマトグラフィー媒体の選択は、ゲル濾過クロマトグラフィーでは重要な考慮事項である。幾つかの、一般的なゲル濾過クロマトグラフィー媒体は次の通りである;Sephadex G-50(1〜30kDaの範囲にあるタンパク質の画分化に適している);Sephadex G-75; Sephadex G-100 (4-150kD); Sephadex G-200 (5-600kD); Bio-Gel P-10 (1.5-20kD); Bio-Gel P-30 (2.4-40kD); Bio-Gel P-100 (5-100kD)およびBio-Gel P-300 (60-400kD)。SephadexはPharmaciaの登録商標である。Bio-GelはBio-Radの登録商標である。カラム容積の約0.6倍のところに溶出される分子が最も良く分離するが、溶出が遅いものほど一般的にピークは広がる。一つの態様では、最善の分離のために、画分域が広い材料を用いて最初に分断を行い、極めて大きなタンパク質と極めて小さなタンパク質を取り除き、続いてより画分域が狭い材料を用いる。ゲル濾過カラムは、分子量が分かっている標準物質(タンパク質)を用いてキャリブレーションできる。キャリブレーションカーブは、log MW(分子量の対数)の関数として、保持時間を表すように作製できる。本発明の方法では、ホモメリックタンパク質複合体は、カラムから溶出するヘテロメリック複合体の範囲を示すのに都合良く用いられる。これについては、上記サンプルの一つについて行われた以下実施例で詳しく説明する予定である。サブユニットXおよびYを発現し、両サブユニットともそれ自身および/または互いに四量体を形成できる宿主細胞(「XY」と称す)が与えられる。さらに、サブユニットX(未改変サブユニット)のみ発現する宿主細胞「X」およびサブユニットY(対象となる分子と融合したサブユニット)のみ発現する宿主細胞「Y」が与えられる。ホモメリックXおよびY複合体の推定サイズ(サブユニットXおよびYそれぞれのサイズの四倍)に従って、好適なゲル濾過媒体を選択する。定義により、本発明のキメラタンパク質複合体は、Xホモマーより大きく、かつYホモマーより小さなサイズを有する。それゆえに、全てのキメラタンパク質複合体(XYYY;XXYY;およびXXXY)の溶出容積は、Yホモ四量体(最大の複合体)より大きいが、しかしXホモ四量体(最小の複合体)よりは小さくなるだろう。Yホモ四量体のみ含有する宿主細胞「Y」のタンパク質サンプルおよびXホモ四量体のみ含有する宿主細胞「X」のタンパク質サンプルをゲル濾過カラムに装填することで、宿主細胞「XY」が産生するキメラタンパク質の溶出容積の「境界」を容易に知ることができる。例えば、カラム画分を集め、SDS-PAGEに続き、特異的試薬、例えば抗体を用いたウエスタンブロットを行い、XまたはYサブユニットの存在について分析する。所望であれば、条件(カラムサイズ、カラム直径、流速等)を調整して、Y四量体からのX四量体の分離を最適化できる。もちろん、XとYとのサイズ差が大きいほど(即ち対象となる分子が大きいほど)、異なる四量体複合体の相互の分離はより容易になるだろう。しかしながら、本発明は特に、大きなサイズ差(例えば大きな抗原を有する複合体)に起因する問題を解決することに関係する。それ故に、本発明の複合体の分離は、大きな問題にはならないだろう。キメラ複合体の溶出容積が一度決定されれば、ホモ四量体によるキャリブレーションに用いたものと同一の条件下で、宿主細胞「XY」のタンパク質サンプルをカラムに装填し、溶出することができる。カラム画分を集め、上記のようにしてXおよび/またはYサブユニットの存在を分析する。煮沸していないタンパク質画分のSDS-PAGEを行い、インタクトタンパク質複合体のサイズを分析することができる。ホモメリック複合体を欠いている本発明のキメラ複合体を含有する画分を、更なる使用のために保存しておく。それとも画分を一つにまとめてプールし、本発明のタンパク質複合体の混合体を得てもよい。画分を個別のままにしてもよく、例えば別々に単離された複合体として、複合体XYYY、XXYYおよびXXXYを得ることもできる。
【0044】
通常のゲル濾過クロマトグラフィーは時間のかかる工程である。作業は、クロマトグラフィー媒体の粒子サイズまたは樹脂を小さくし、カラムをより小型にすることで著しく迅速化できる。これには、液体を、高圧を利用してカラムの中を流すための特別な装置が必要である。好ましい態様では、本発明のタンパク質複合体は高圧液体クロマトグラフィー(HPLC)または高性能液体クロマトグラフィー(FPLC)を用いて単離する。
【0045】
本発明の方法では、単離されたタンパク質複合体はさらに特徴付けされ、細胞表面レセプターまたはレセプター結合成分を模した分子に対する、その結合能力を決定し、例えばGM1レセプターを模擬するD-ガラクトースは、本発明のLTBまたはCTBをベースとするタンパク質複合体の特徴付け(または精製)に用いることができる。好ましい態様では、一過性結合よりも容易に検出できることから、結合は、永久結合を含む。単離または精製されたレセプターを用いることができるが、レセプターを発現している細胞、またはそれら細胞に由来する膜材料を用いてもよい。本発明のタンパク質複合体を、複合体がそのレセプターへの結合に適した条件の下で、レセプターに接触させる。レセプター結合に影響する可能性のある条件としては、イオン強度(pH;塩)および温度が挙げられる。その後、結合した複合体の量を決定する。好ましい態様では、本発明の、単離されたタンパク質複合体の細胞表面レセプターへの結合は、酵素結合免疫吸着アッセイ(ELISA)またはそれに実質基づく方法により行われる。ELISAの基本原理は、酵素を用いて、抗体(Ab)への抗原(Ag)の結合を検出することである。酵素は無色の基質(発色団)を着色された生成物に変換し、Ag:Ab結合の存在を示す。本発明の特定の態様では、タンパク質複合体は大腸菌の易熱性エンテロトキシン(LT)のBサブユニットに基づいている。Bサブユニットのインビボ五量体化およびその本来のレセプターGM1への五量体の結合は、トキシンの取り込みを先導し、最終的に粘膜免疫に伴う免疫調節現象を誘導する重要な現象である。本明細書に例示するように、LTBサブユニットに基づく本発明のタンパク質複合体の結合は、De Haanら(Vaccine 1996; 1:777-783)記載のGM1-ELISAを用いて容易に決定される。このようなタイプのアッセイを用いることで、少なくとも一の未改変LTBサブユニットと少なくとも一つのLTB融合タンパク質を含む、本発明のLTBベースのタンパク質複合体が、その本来の五量体の構造を保持していることが明らかになるだろう。
【0046】
本発明のさらなる局面では、本発明のタンパク質複合体を含む宿主細胞が提供される。宿主細胞は、例えば、本発明の複合体の構成成分をコードする一または複数の核酸構築体が付与された微生物細胞である。一つの態様では、該宿主細胞は細菌細胞、例えば形質転換した大腸菌細胞である。上記したように、細胞は、好ましくは、粘膜表面に、および粘膜表面を横切り、一または複数の抗原を送達することができるタンパク質複合体を含む食用細胞である。食用細胞の例は、食用植物、例えばジャガイモである。このような複合体は、粘膜キャリア分子として有利に用いられる。従って、本発明の粘膜キャリア分子を含む食用細胞、またはその抽出物は、例えば食物もしくは飲料水を用いた経口ワクチン接種に用いることができる。
【0047】
前に述べたように、キャリ分子の通常の使用は、比較的小さな抗原の送達に実質的に限定されている。しかしながら本発明のキメラ複合体は、融合タンパク質のサイズとは実質的に無関係に、細胞表面レセプターに結合する能力を保持していることから、ここでは比較的大きな対象となる分子(抗原等)を送達するためにキャリア分子を用いることができる。抗原が体内の適当な部位に一度送達すると、免疫応答を引き起こせることは周知である。たとえば、粘膜域を裏打ちする上皮細胞のバリア横切り送達できた抗原は、粘膜関連リンパ組織(MALT)の下にある誘導部位を刺激する。本発明のタンパク質複合体には、一または複数の対象となる抗原性分子を、実質的に対象となる分子のサイズおよび複雑さに関係なく、そして、重要なことであるが、上皮バリアを通して複合体を送達し、MALTを刺激するキャリア特性を失わずに、与えることができる。それゆえに、本発明の複合体は、被験体、好ましくはマウスもしくはヒトのような哺乳動物被験体に免疫応答を誘導するのに、都合良く用いられる。一つの態様では、対象となる抗原を含む本発明のキメラタンパク質複合体は、ワクチンに用いられる。本発明の複合体を、特定抗原(または各種抗原の組合せ)と一緒に含むワクチンは、被験体に投与されると、被験体を、例えばその抗原を原因とする疾患から保護する免疫応答を誘導できる。ワクチンは感染後に投与でき、疾患の進行を遅らせるか、もしくは停止するための治療(処置)ワクチンでもよい。好ましくは、それは被験体の初期感染を防止できる、予防性(予防)ワクチンである。
【0048】
さらには、本発明のタンパク質複合体または該複合体を含む細胞を含む(植物ベースの)ワクチン、ならびに有効量の該ワクチンを含む薬学的組成物を提供する。本発明のワクチンは、少なくとも二種類の抗原を含む本発明のタンパク質複合体を含む多成分ワクチンでよい。
【0049】
本発明のさらに別の局面では、本発明のタンパク質複合体の有効量の少なくとも一回投与量を被験体に投与すること、好ましくは経口投与することを含み、ここで対象となる分子が抗原である、特定病原菌に対する被験体の免疫応答を高めるための方法が提供される。この方法はまた、大型(構造)抗原を含む保護抗原を、宿主(植物)または宿主細胞組成物を摂食することにより、複合体を経口的に送り込み、細胞腸管内に在る免疫系に送達する道を開く。本明細書には、好ましい免疫化の経路を通し、本発明のワクチンを被験体に投与することを含む、粘膜(鼻、直腸または膣)免疫化のための方法も記載される。このように本発明は、キャリア分子をベースとするワクチンに取り込むことができる抗原性化合物のサイズおよび多様性を拡大する。
【0050】
実施例
実施例1:LTBサブユニットワクチン発現カセットの構築
バイナリー型植物発現ベクターpBINPLUS(Van Engelen et al., 1995)のT-DNA部分および本明細書で報告した全てのpLANTIGENワクチン構築体の遺伝子挿入物の図式的概観を図1に示す。全ての遺伝子は、塊茎だけに発現するためにクラスIパタチンプロモーター(Ppat)の制御下に置かれ、さらにERに保持されるためにそれぞれの融合タンパク質のC末端にKDEL(Lys-Asn-Gln-Leu)配列をコードするDNA配列を保有する(Munro and Pelham, 1987)。
【0051】
pL4:LTBに対する合成遺伝子(synLT-B)の設計および構築ならびにバイナリー型植物発現ベクターpLANTIGEN4(pL4)の発生は、以前に記載された(Lauterslager et al., 2001)。pL4は、LTBに対する合成遺伝子(synLT-B)を保有し、成熟LTBタンパク質をコードする配列の直後およびKDELをコードする配列の前にユニークなBamHI制限部位を有する。全てがLTBおよびカルボキシ末端のKDEL配列と同フレーム内に入るように全ての合成配列を作製し、このユニークな部位にクローニングした。
【0052】
pL12:pLANTIGEN12(pL12)にクローニングされたイヌパルボウイルス(CPV)エピトープをコードするフラグメントのコアは、アミノ酸配列

をコードする(Langeveld et al., 1994)。イヌパルボウイルス(CPV)のウイルスタンパク質VP2のアミノ末端領域をコードする合成BamHI/BglIIフラグメントを、pL4のユニークなBamHI部位にクローニングすることによって、pLANTIGEN12を作製した。以前に記載されたようなオリゴから得られたフラグメントの連結によって合成フラグメントを作製した(Florack et al., 1994)。オリゴは、Eurogentec(ベルギー)からのものであった。
【0053】
pL13:pLANTIGEN13(pL13)では、C末端膜貫通(TM)領域を欠くCSFV E2糖タンパク質をコードするフラグメントを連結した。野生型CSFVでは、E2糖タンパク質は膜貫通的に結合している。ブタコレラウイルス(CSFV)のE2成熟タンパク質をコードするBamHIフラグメントをpL4のユニークなBamHI部位にクローニングすることによって、pL13を構築した。CSFV E2をコードするフラグメントは、オリゴ
を使用したpPRb2(Hulst et al., 1993)のPCRによって得られた。
【0054】
pL15:pLANTIGEN15(pL15)では、二つのHAエピトープ配列と共にCPV配列を2回、間隔を空けるための2個のアラニン残基でそれぞれ分離してクローニングした。HAエピトープは、デカペプチドFERFEIFPKEをコードし、PR8 HA-1のアミノ酸111〜120に相当する(Hackett et al., 1985)。CPVは、線状B細胞エピトープであるが、HAはT細胞特異的である。pL12にクローニングしたものと類似した二つのCPVエピトープと共にインフルエンザウイルスヘマグルチニン(HA)H鎖の二つのデカペプチドからなる四量体配列をコードする合成フラグメントを、pL4のユニークなBamHI部位にクローニングすることによって、pL15を構築した。四つのエピトープ配列全てをそれぞれ2個のアラニン残基で分離するようにクローニングした。
【0055】
実施例2:宿主細胞の形質転換、成長およびタンパク質抽出
以前に我々は、16ロットの塊茎を生じたpL4遺伝子構築体を含有する22個の独立したトランスジェニックジャガイモ植物の発生について記載した(Lauterslager et al., 2001)。それ以来さらに形質転換実験を行い、それによってpL4遺伝子構築体を保有する追加の31ロットの独立した塊茎が産出した。
【0056】
本発明では、実施例1に記載されたバイナリー型発現ベクターおよびその組み合わせをエレクトロポレーションによりアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)AglO(Lazo et al., 1991)に導入し、ジャガイモデジレ(Desiree)カルティバー(De Z.P.C., Leeuwarden, The Netherland)の形質転換に使用した。トランスジェニックシュートの形質転換、成長、選択および塊茎の産生は以前に記載されている(Lauterslager et al., 2001)。pL12を用いたジャガイモデジレカルティバーの茎節間の形質転換によって、27個の独立したトランスジェニック植物が発生し、そのうち22個が温室内で塊茎を産生した。pL13を用いた形質転換によって23個の植物が発生し、そのうち20個が塊茎を形成した。pL15を用いた形質転換は31個の植物を産出し、そのうち20個が塊茎を産生した。
【0057】
典型的には、2〜4か月後に温室で成長した植物から200から300グラムの塊茎を収穫した。貯蔵および塊茎の齢が引き起こす効果を減らすために、ほぼ同サイズの新鮮収穫された塊茎材料から抽出液を作製した。ジャガイモ宿主細胞から生成したタンパク質複合体を単離するために、新鮮収穫した直径約5cmの塊茎を使用した。皮を除いた塊茎を薄切りしたものを25mMリン酸ナトリウム(pH6.6)、100mM NaCl、1mMエチレンジアミン四酢酸(EDTA)、50mMアスコルビン酸ナトリウム、1% TritonX-100および20mMメタ重亜硫酸ナトリウム中で抽出した。組織ホモジネートを4℃、12000rpmで5分間遠心分離し、上清を集め、新しい試験管に移した。標準としてウシ血清アルブミン(BSA)を使用したBradford法(Bradford, 1976)で全可溶性タンパク質を推定した。LTB五量体複合体の発現または蓄積をGM1-ELISAによって確認した(実施例3)。
【0058】
実施例3:レセプターとタンパク質複合体との結合の決定
修正ガングリオシドGM1酵素結合イムノソルベントアッセイ(GM1-ELISA)によって、トランスジェニックジャガイモ塊茎から単離した機能的キメラタンパク質複合体の量を推定した。マイクロタイタープレート(PolySorp Immunoplates, Nunc)に、リン酸緩衝生理食塩水(PBS)に溶かした5μg/mLウシ脳由来モノシアロガングリオシドGM1(Sigma Aldrich, St. Louis, USA)をコーティングした。コーティング後のプレートを脱イオン水で3回洗浄して前処置し、一定量、典型的には全可溶性抽出可能塊茎タンパク質5μgを負荷して、PBSに溶かした2%スキムミルク、0.1% ウシ血清アルブミンおよび0.1% Tween20を用いて室温にて1時間、100rpmで連続的に振盪しながらブロッキングした。大腸菌発現ベクターによって生成した組換えLTBを連続2倍希釈したものを、pBINPLUSPAT塊茎からの等量の全タンパク質を入れた各プレートに添加した。結合を4℃で16時間行った。脱イオン水でプレートを3回洗浄後に、LTB五量体に特異的なモノクローナル抗体VD12と共に1:1000希釈で室温にて1時間プレートをインキュベートした。脱イオン水で3回すすいだ後に、AP標識ヒツジ抗マウス抗体と共にプレートをさらにインキュベートし、結合した標識を4-ニトロフェニルリン酸(二ナトリウム塩六水和物、Janssen Chimica)で検出した。標準曲線の分析、濃度および標準偏差の計算のためにMicroplate Manager/PCバージョン4.0ソフトウェアを使用してBio-Rad Benchmark Microplate Reader(Bio Rad, Veenendaal, The Netherlands)を用いて405nmで検出を行った。単離した五量体を含むサンプルの読み取り値を、標準曲線からの結果と比較することによって五量体LTBの量を推定した。独立した実験で少なくとも2回サンプルを分析した。モル比を計算するために、塊茎の新鮮重量1gあたりの全タンパク質の量を7mg/gに調整した。融合タンパク質のモル重量を、それぞれのpLANTIGEN融合構築体によってコードされる成熟タンパク質の公知のアミノ酸配列から推定した。トランスジェニック塊茎に生成した五量体タンパク質複合体の平均発現レベルを表1に示す。これらのデータは、対象となる分子のサイズが大きいほど、機能的(すなわちGM1結合性)ホモ五量体タンパク質複合体の発現レベルが低くなることを示す。
【0059】
実施例4:LTBおよびLTB-CSFV E2を有するキメラタンパク質複合体
未改変サブユニットをコードするバイナリー型発現ベクターpL4(Lauterslager et al., 2001)およびサブユニットに融合した対象となる分子をコードするpLl3(LTB-CSFV E2融合体、概要については実施例1を参照されたい)を、本質的に以前に記載し(Lauterslager et al., 2001)、上記(実施例1)に概略を述べたように、アグロバクテリウム・ツメファシエンス介在性のジャガイモcv.デジレ(De Z.P.C., Leeuwarden, The Netherland)の形質転換を使用した共形質転換によりジャガイモに導入した。共形質転換を可能にするために、茎節間の感染前に、pL4およびpL13を保有する両方の組換えアグロバクテリウムを、OD595=1で1:1の比に混合した。混合後の細菌懸濁液を形質転換実験および形質転換細胞の再生に使用し、トランスジェニックシュートの選択は該の通りに行った。独立したトランスジェニックシュート71個を得て、53個を選択し、かつpL4および/またはpL13遺伝子構築体の存在について分析して、共形質転換の発生を明らかにし、それらをさらなる分析のために選択した。このために、尿素抽出緩衝液(62% 尿素、0.5M NaCl、70mMトリスHCl(pH8.0)、30mM EDTA(pH8.0)、1.5% サルコシル)50μl中で直径約5mmの円盤形の葉を摩砕することによって、個別の植物から集めた葉材料からゲノムDNAを単離した。等容のフェノール/クロロホルム(1:1)を添加して、サンプルを混合し、室温で15分間放置した。混合後に、サンプルを3000rpmで10分間遠心分離し、上清を新しい試験管に移し、4.4M酢酸アンモニウム(pH5.2)10μlを加えた。ゲノムDNAを沈殿させるためにイソプロパノール120μlを添加し、混合した。サンプルを1000rpmで3分間遠心分離して上清を除去した。残ったペレットを乾燥して、水または10mMトリスHCl(pH8.0)および1mM EDTAを含有する緩衝液に懸濁した。プライマー
を例とするがそれに限定されるわけではないpL4およびpL13遺伝子構築体を識別できる特異的プライマーを使用したPCRにマイクロリットル量のゲノムDNAサンプルを供した。増幅条件は、製造業者による最適条件でAccuTaqポリメラーゼ(Sigma-Aldrich)を使用し、それぞれ変性を可能にする94℃30秒間、56℃45秒間でのアニーリングに続く68℃2分間の伸長の40サイクルであった。PCR反応混合物を0.5×TBEでの1.2%アガロースゲル電気泳動に供し、予測サイズが周知である増幅された遺伝子構築体pL4および/またはpL13に対応するフラグメントの存在についてゲルをスキャンし、それらの公知の遺伝子配列ならびにプライマーLTB11およびTnosminus50の使用から推定された。植物22個は両遺伝子構築体を含んでいたが、28個はpL4のみを含み、顕著にはわずか3個がpL13遺伝子構築体を有した。本明細書においてさらにpL(4+13)8と名付けた第8番の植物または植物pL(4+13)16、pL(4+13)31、pL(4+13)39、もしくはpL(4+13)46のようにpL4およびpL13遺伝子構築体の両方について陽性であった22個の植物を、温室に移し、キメラタンパク質複合体の蓄積をさらに分析する目的で塊茎材料を産生させるために成熟するまで成長させた。
【0060】
塊茎を収穫し、GM1結合性LTB五量体の量を実施例2および3に記載したように評価した。数個の植物、例えば植物pL(4+13)15、pL(4+13)16、pL(4+13)39、pL(4+13)46、pL(4+13)60、pL(4+13)64およびpL(4+13)67は、GM1結合性LTB五量体のかなりの蓄積を示した(表2参照)。これらの植物をさらにキャラクタリゼーションした。pL(4+13)16およびpL(4+13)46のような選択された共形質転換されたジャガイモ植物の塊茎材料は、それぞれ塊茎の新鮮重量(FW)1グラムあたり約8および20マイクログラムでガングリオシドGM1と結合することから推定されるようにLTB5五量体を蓄積した(表2)。
【0061】
ウエスタンブロットによりキメラ植物pL(4+13)16をさらに評価した。このために、植物の塊茎抽出物の全タンパク質25マイクログラムを10% SDS-PAGEゲルに負荷し、半未変性(semi-native)の非還元条件下で運転してタンパク質複合体の分離を可能にした。半未変性SDS-PAGEの後に、抗LTB5コンホメーションのモノクローナル抗体であるVD12(図2A)または抗CSFV E2コンホメーションのモノクローナル抗体であるV3(図2B)のいずれかである特異的抗体を1:1000希釈で使用したその後のウエスタン分析のために、CAPS(10% エタノールに溶かした0.22% 3-[シクロヘキシルアミノ]-1-プロパン-スルホン酸(pH11))緩衝液中で標準法を使用してニトロセルロース上で、分離した後のタンパク質を転写した。ホースラディッシュペルオキシダーゼ標識二次抗体およびLumiLight基質を使用して特異的抗体の結合/認識を可視化して、LumiImager(Roche、Boehringer, Germany)で可視化した。対照として両方ともpL13遺伝子構築体を保有するpL1317および1331を使用した。pL1317は、20個の独立したpL13トランスジェニック植物の間でLTB-CSFV E2融合タンパク質についてのGM1結合性五量体の最高の発現体として以前に同定された。さらに、pL4遺伝子構築体を保有し、約17マイクログラム/gのFW GM1結合性LTB五量体を蓄積しているpL417およびGM1結合性五量体についての陰性対照を使用した。
【0062】
図2Aは、コンホメーションの抗LTB5モノクローナル抗体VD12を使用してこれらの植物の全タンパク質抽出物25マイクログラム量を分析した結果を、図2Bは、コンホメーションのCSFV E2に特異的なV3を用いた分析の際の結果を描いたものである。共形質転換された植物pL(4+13)16(レーン1;pL4およびpL13で共形質転換)、タンパク質分子量マーカー(レーン2)、PAT4を保有する対照抽出物、空の発現カセットベクター(レーン3)、両方ともLTB-CSFV E2遺伝子融合体のみを保有するpL1331(レーン4)およびpL1317(レーン5)の抽出物、ならびにpL4で形質転換され未改変rec-LTBサブユニットのみを蓄積するpL(4)17(レーン6)を負荷した。
【0063】
図2から、pL(4+13)16はホモ五量体LTB5よりもサイズが有意に大きく(レーン1と6とを比較)、VD12およびV3の両方のコンホメーション特異的モノクローナル抗体によって認識される(AおよびBのレーン1を比較)かなりの量のタンパク質複合体を蓄積することが明白である。これは、VD12によって認識される五量体LTB5の構造およびV3によって認識される抗原性CSFV E2二量体エピトープを保有するキメラタンパク質複合体の存在を実証している。重要には、後者の抗原性CSFV E2二量体エピトープは、pL13にほとんど存在せず(レーン4および5)、pL13はLTB-CSFV E2融合タンパク質のみを有し、未改変LTBサブユニットを有さない。さらに、約300kDaのホモ五量体LTB-CSFV E2複合体は、VD12およびV3を用いた分析後にほとんど目視不可能である。考えられるところでは、これは以前に観察されたようにその非常に低い発現レベルが原因である。図2およびGM1 ELISAから、植物pL(4+13)16に存在するようにLTB(pL4)およびLTB-CSFV E2(pL13)の両方を含むキメラ複合体だけが、GM1結合性質および抗原性CSFV E2エピトープの両方を保有するかなりの量(FW塊茎1グラムあたり8マイクログラム)のタンパク質複合体の蓄積を促進することも明白である(図2AおよびB、レーン1)。対照的に、例えば植物pL1317のようにLTB-CSFV E2遺伝子融合体のみを発現する植物は、GM1細胞表面レセプターと結合可能な抗原性タンパク質複合体を形成しない(図2AおよびBのレーン1および4を比較)。
【0064】
GM1のELISA(表2)によると全てがかなりの量の機能的LTB5を蓄積するキメラ植物pL(4+13)16、pL(4+13)31、pL(4+13)39、pL(4+13)46、pL(4+13)60、pL(4+13)64およびpL(4+13)67を、すでに記載したようなウエスタンブロットによってさらに評価した。図3Aは、VD12と共にインキュベーションした後の結果を示し、LTB5のみを含有するpL4(21)(レーン2)と(LTB-CSFV E2)5を含有するpL13(17)(レーン3)との間の位置に泳動することから明白なように、pL(4+13)16からpL(4+13)67の抽出物中にLTB5を有する多数のキメラ複合体が存在することを表す。予想通りこれらのキメラ複合体は、CSFV E2に特異的なV3 mAbとも反応する(パネルB、図3)。図3Bから、pL13(17)(レーン2)に類似した量の全タンパク質を負荷したが、キメラ植物からの抽出物(レーン6〜10)がV3 mAbとより強く反応して、GM1結合性LTB5複合体への機能的CSFV E2二量体の蓄積をさらに強調していることもはっきりしている。
【0065】
実施例5:LTBおよびLTB-パルボを有するキメラタンパク質複合体
バイナリー型発現ベクターpL4(LTB)およびpL12(LTB-CPVパルボ;概観については図1および実施例1参照)を、実施例4に本質的に記載したようにジャガイモcv.デジレのアグロバクテリウム・ツメファシエンス介在性形質転換を使用した共形質転換によって、両方ともジャガイモに導入した。茎節間の感染前に、pL4またはpL12のいずれかを保有する組換えアグロバクテリウムをOD595=1で1:1の比で混合し、混合した細菌懸濁液を形質転換実験に使用して共形質転換を可能にした。45個を超える独立したトランスジェニックシュートを選択し、pL4および/またはpL12遺伝子構築体の存在について分析して、共形質転換の発生を明らかにし、それらをさらなる分析のために選択した。20個のトランスジェニック植物は、両遺伝子構築体を含有すると思われたが、19個の植物はpL4のみを有し、6個の植物はpL12のみを有した。塊茎を形成させるために20個の二重形質転換体全てを温室内で成長させ、塊茎を収穫し、GM1結合性LTB五量体の量を立証した。選択されたpL(4+12)植物の一覧を表3に示す。
【0066】
VD12およびイヌパルボウイルスに特異的なmAbである3C9を使用したウエスタンブロット(図4)は、pL(4+12)19、pL(4+12)20、pL(4+12)23、pL(4+12)31、pL(4+12)41、pL(4+12)46、pL(4+12)51、pL(4+12)52、pL(4+12)57、pL(4+12)62およびpL(4+12)65のようないくつかの植物がLTB5(pL4、レーン2)およびLTB-CPV5(pL12、レーン3)の間にSDS-PAGEゲル上を泳動するキメラタンパク質複合体を含有することを示した。図4AおよびBから、pL(4+12)31、pL(4+12)57およびpL(4+12)62のようないくつかの植物のタンパク質抽出物がVD12(抗LTB5)および3C9(抗パルボ)の両方によって認識された多数のバンドを現したことが明白であり、これは、例えば未改変LTBサブユニット4個およびLTB-CPV融合サブユニット1個またはLTB3個およびLTB-CPV2個、またはLTB2個およびLTB-CPV3個またはLTB1個およびLTB-CPV4個のいずれかを含有するキメラ複合体が存在することを示す(図4AおよびB)。pL(4+12)57の抽出物では特にホモ五量体LTB5およびホモ五量体(LTB-パルボ)5以外に4個のキメラ複合体が存在することがはっきりしている。
【0067】
実施例6:LTB、LTB-パルボおよびLTB-iippを有するキメラタンパク質複合体
バイナリー型発現ベクターpL4(LTB)、pL12(LTB-CPV パルボ)およびpL15(LTB-iipp:iipp=インフルエンザ-インフルエンザ-パルボ-パルボ、二つのパルボエピトープと結合した二つのインフルエンザエピトープ、概要については実施例1参照)を、本質的に上記のようにジャガイモcv.デジレのアグロバクテリウム・ツメファシエンス介在性形質転換を使用した共形質転換によって、ジャガイモに導入した。独立したトランスジェニックシュート29個を選択し、pL4、pL12およびpL15遺伝子構築体の存在について分析して、共形質転換の発生を明らかにした。これらの植物をさらなる分析にために選択した。トランスジェニック植物5個は三つの遺伝子構築体全てを含有すると思われたが、10個は、pL(4+12)、pL(4+15)またはpL(12+15)のいずれかである二つの遺伝子構築体を含有した。残りの14個の植物はわずか一つの遺伝子構築体を含有した。二つ以上の遺伝子構築体を保有する全ての植物を温室に移し、成熟するまで成長させた。5個の三重形質転換体pL(4+12+15)7、pL(4+12+15)9、pL(4+12+15)11、pL(4+12+15)16およびpL(4+12+15)19を、GM1 ELISAならびにVD12および3C9 mAbを使用したウエスタンブロットにより分析した(図5)。
【0068】
図5A左欄から、三つの遺伝子構築体全てを有するこれら5つの植物(レーン6〜10)が、pL(4)21(レーン3)およびpL(12)01(レーン4)の間またはpL(12)01(レーン4)およびpL(15)16(レーン5)の間に泳動する複合体を含有することが明らかであり、これはLTB、LTB-パルボおよび/またはLTB-iippの多様な組み合わせであることを示唆している。図5A右欄から、これらの複合体がmAb 3C9と正の反応をすることが明白であり、これは少なくとも一つのパルボエピトープが存在することを表している。図5Bから、特にpL(4+12+15)7(レーン6)およびpL(4+12+15)11(レーン8)がLTB-パルボおよびLTB-iippの両方を含有することがはっきりとしている。よって、これらの植物ではLTB、LTB-パルボおよびLTB-iippを含有するキメラGM1結合タンパク質複合体が一緒に産生する。
【0069】
実施例7:大腸菌LTBサブユニット発現カセットの構築
本発明に記載のタンパク質複合体を、様々な種類の組換え宿主細胞に産生させることができる。実施例7および8は、微生物である大腸菌における少なくとも一つの未改変サブユニット、およびこの場合はレポーター分子GFPである対象となる分子に融合した少なくとも一つのLTBサブユニットを有するキメラタンパク質複合体の産生を記載する。
【0070】
大腸菌および他の原核生物での発現のために、LTBについて本来の野生型大腸菌の配列(EtxB)を使用した。ブタ大腸菌株(Dallas and Falkow, 1980; Leong et al., 1985)からもともと単離されたヌクレオチド配列ECELTBP(SWISS-PROT P32890)に大きく類似するLTBコード配列を、pYA3047(Jagusztyn-Krynicka et al., 1993)からクローニングした。プライマー
およびテンプレートとして野生型EtxB配列を有するベクターを使用したPCRによってフラグメントを増幅させた。生じたBpiI/EcoRIを、NcoI/EcoRIで消化したpET21dベクター(Novagen)にクローニングしてpET-wiLTB1を発生させた。ヌクレオチド配列の確認後に、生じたクローンを、発現の研究のために形質転換によりTOP10F'細胞(Invitrogen)に導入した。プライマー
を使用したPCRによりGFP配列末端のBamHI/BpiI部位に導入することによってLTB-GFP融合タンパク質を作製して、BamHIで消化したpET-wiLTBにクローニングした。ヌクレオチド配列の確認後に、発現の研究のために生じたクローンpET-wiLTB-GFP2を形質転換によってTOP10F'細胞に導入した(実施例8)。
【0071】
実施例8:大腸菌細胞の形質転換、成長およびタンパク質抽出
二つの選択されたクローン、pET-wiLTB1およびpET-wiLTB-GFP2を形質転換により大腸菌Rosetta株(Novagen)に導入して、50mg/Lアンピシリンおよび34mg/Lクロラムフェニコールを補充したLB培地中で37℃で一晩成長させた。o/n培養物2mLを新鮮培地50mLに入れて希釈し、OD595が0.2に達するまで連続的に振盪しながら20℃で成長させた。IPTGを終濃度1mMになるまで添加して発現を誘導し、培養物をOD595=0.5になるまでさらに成長させた。4℃で14000rpmで10分間遠心分離することによって細胞を集め、ペレットをBugBuster(Novagen)抽出試薬に再懸濁し、終濃度1μg/μlになるまでリゾチームを添加した。サンプルを室温で5分間インキュべートして、Benzonaseを添加して室温でさらに15分間インキュベートした。サンプルをその後4℃で14000rpmで5分間遠心分離した。LTB5を精製するためにpET-wiLTBの上清を固定化D-ガラクトースカラムに負荷した(実施例10も参照)。溶出プロフィルを図6に示す。図6Aから、溶出緩衝液の適用直後にそれぞれの画分中のタンパク質を測定できることが明白である。図6Bから、GM1 ELISAから明白なようにこれらの画分がGM1結合性LTB5複合体を含有していることがはっきりしている。RecLTB-GFPは、封入体に蓄積すると思われた。封入体を含有する生じたペレットを、本質的にSambrookら(1989)が記載したものをKhouryおよびMeinersmann(A genetic hybrid of the Campylobacter jejuni flaA gene with Escherichia coli and assessment of the efficacy of the hybrid as an oral vaccine. Avian disease 39 (1995) 812-820)に従ってやや修正してさらに処理した。封入体を標準法を使用して単離し、8M尿素で抽出した。尿素可溶性画分を0.1Mトリス(pH7.4)で透析し、沈殿した画分を10000gで10分間遠心分離することによって除去した。GM1結合性複合体を含有する上清をD-ガラクトースを用いたアフィニティクロマトグラフィーによってさらに精製して、精製recLTB-GFPを得た。
【0072】
実施例9:LTBおよびLTB-GFPキメラ複合体の産生
少なくとも一つの未改変LTBサブユニットおよび少なくとも一つのLTB-GFP融合サブユニットを有する、本発明のキメラ複合体を産生させる代替法は、未改変サブユニットを有する第一組成物および融合サブユニットを有する第二組成物を提供すること、その後に両組成物を1:4、2:3、3:2または4:1モル比で混合することを伴う。次に、両種の(すなわち未改変および改変)サブユニットの混合物を、五量体誘導条件で変性および再生する。例えば、実施例7および8に記載されたように組換え大腸菌2株に産生され、D-ガラクトースを用いたアフィニティクロマトグラフィーで精製されたrecLTBおよびrecLTB-GFPを8M尿素に溶かした。または、精製recLTBをrecLTB-GFPを有する封入体に添加して、上記のようにさらに処理することもできる。尿素可溶性画分を0.1Mトリス(pH7.4)で透析して、不溶性材料を4℃で10000gで10分間遠心分離することによって除去できる。または、0.3M D-ガラクトースをトリス緩衝液に添加して五量体化を促進できる。キメラLTBおよびLTB-GFP分子を含む五量体複合体を含有する上清を、例えば固定化D-ガラクトースを用いたアフィニティクロマトグラフィーによってさらに精製できる。タンパク質複合体のキメラサブユニット組成物を上記のような半未変性SDS-PAGEおよびウエスタンブロットによって検証できる。GM1との結合を上記のようにGM1-ELISAを使用して決定できる。
【0073】
実施例10:LTBサブユニット担体複合体の精製
ホモ五量体性RecLTB5およびLTB-サブユニット担体の複合体を、Uesakaら(Uesaka et al. (1994) Simple method of purification of Escherichia coli heat-labile enterotoxin and cholera toxin using immobilized galactose. Microbial Pathogenesis 16 : 71-76)に従って固定化D-ガラクトースアガロース(Pierce、カタログ番号20372)を用いたアフィニティクロマトグラフィーによって精製した。
【0074】
実施例7に記載したように微生物LTB発現カセットを備える大腸菌細胞を、実施例8に記載したように成長させ、4℃で6500gで30分間遠心分離することによって集め、超音波処理して、記載されたようにさらに処理した(Uesaka et al., 1994)。超音波処理した大腸菌の上清を、50mMトリスHCl(pH7.4)、0.2M NaCl、1mM EDTAおよび20mMメタ重亜硫酸ナトリウム(TEAN)に調整し、FPLC(BioRad, Veenendaal, The Netherlands)に取り付けたD-ガラクトースカラムに負荷した。カラムをTEAN緩衝液で洗浄し、当業者に公知の標準法により0.3M D-ガラクトースまたは0.5M D-ガラクトースのいずれかを含有するTEAN緩衝液で溶出させた。図6は、大腸菌からのrecLTBの精製結果を示す。画分を集め、GM1結合性LTBの存在を実施例3に記載するようにGM1-ELISAによって決定した。図6Aに示すように大腸菌から精製したrecLTBについての結果を図6Bに示す。GM1-ELISAから明白なように、タンパク質について陽性の全ての画分はGM1結合性複合体を含有した。固定化D-ガラクトースを用いた大腸菌からのrecLTBの精製についての溶出プロフィルおよび対応する画分のGM1 ELISAによるGM1結合活性の存在の分析を図6に表示する。
【0075】
次に、D-ガラクトースクロマトグラフィーによってpL4(21)由来塊茎抽出物の上清からタンパク質複合体を精製した(図7)。図7Aおよび7Bから、そのような複合体を塊茎からも精製できることがはっきりしている。
【0076】
次に、キメラタンパク質複合体をpL(4+13)植物から精製した。実施例2に記載したように抽出緩衝液中で塊茎材料を摩砕することによってpL(4+13)46の塊茎材料から抽出物を調製し、抽出物を4℃で6500gで30分間遠心分離して不溶性材料およびデンプン粒を除去した(図8)。上清をTEAN緩衝液条件に調整し、D-ガラクトースカラムに負荷し、記載したようにさらに処理した。図8から、画分5でタンパク質がカラムから溶出し始め、画分10〜11に最大吸光度となることがはっきりしている。GM1-ELISAから、VD12 mAbで検出されたように大部分のLTBは画分5〜15に存在し、画分10で最大であることも明白である(図8B)。さらに、第二抗体がコンホメーションの抗CSFV E2 mAb V3である修正GM1 ELISAから明白なように、これらの画分はCSFV E2エピトープについても陽性であること(図8C)は、そのような複合体にLTBおよびLTB-CSFV E2の両方が存在することを示し、これはウエスタンブロットから得られた以前の結果と一致している(図3)。
【0077】
実施例11:LTBおよびLTB-VHSV Gのキメラ複合体
LTBと、配列X66134(EMBL)に類似しLorenzenら(Molecular cloning and expression in Escherichia coli of the glycoprotein gene of VHS virus, and immunization of rainbow trout with the recombinant protein. J. Gen. Virol. 74 (1993) 623-630)によって発表されたウイルス性出血性敗血症ウイルス由来スパイク糖タンパク質Gとの遺伝子融合体を以下のように作製する:VHSV G配列およびプライマー
のPCR増幅によってユニークなBamHI部位を導入する。PCR条件は、Pwoポリメラーゼを使用して96℃30秒間、60℃30秒間および72℃45秒間を30サイクルである。得られたフラグメントをBamHIおよびSmaIで切り出し、LTBに対する合成遺伝子を保有するpLANTIGEN4のユニークなBamHI部位にフレーム内にクローニングする。パタチンプロモーターおよびノパリンシンターゼターミネーター配列の制御下の生じた遺伝子配列をpLANTIGEN24(pL24)と名付ける。pL4およびpL24でジャガイモの共形質転換を行い多数のpL(4+24)植物を発生させる。pL4および/またはpL24遺伝子構築体の存在を以前に記載したようなPCRによって確認する。両遺伝子構築体について陽性の植物に塊茎を形成させる。塊茎を収穫し、GM1 ELISAを使用してGM1結合複合体に関して分析する。複合体にVHSV Gタンパク質が存在することを、モノクローナル抗体IP1H3、3F1H10および3F1A2(Lorenzen et al., 2000. Three monoclonal antibodies to the VHS virus glycoprotein: comparison of reactivity in relation to differences in immunoglobulin variable domain gene sequences. Fish & Shellfish immunology 10: 129-142)と共にインキュベートすることによって確認する。半未変性条件下で、抗LTB5 mAbであるVD12および1P1H3、3F1H10および3F1A2を使用して行ったSDS-PAGEで塊茎抽出物のウエスタンブロットによって、キメラ複合体をさらにキャラクタリゼーションできる。
【0078】
実施例12:LTBおよびLTB-SVCV Gのキメラ複合体
コイ春ウイルス血症ウイルスの完全SVCV G遺伝子(Genbankアクセッション番号NC002803)を有しLTBとの遺伝子融合体を作製するためのBamHI/BglIIフラグメントを、オリゴヌクレオチド
を使用して増幅させた。生じたフラグメントをBamHIおよびXhoIで切り出し、LTB;pをコードするpL4のBamHI/XhoI部位にクローニングすることによってpLANTIGEN27(pL27)を発生させた。塊茎特異的パタチンプロモーター(pLANTIGEN27)の制御下に置かれた生じた遺伝子を、ジャガイモの形質転換のためにA.ツメファシエンスに導入した。pL4およびpL27の共形質転換を行い、記載したようなPCRによって両遺伝子構築体の存在についてトランスジェニック植物を評価した。pL(4+27)遺伝子構築体を保有するトランスジェニック植物を選択し、温室内で成熟するまで成長させた。GM1 ELISAによりGM1結合性複合体の蓄積について、および特異的mAbを使用してSVCV Gタンパク質の存在について塊茎を分析した。記載したような半未変性SDS-PAGE後のウエスタンブロットによって、タンパク質複合体のサブユニット組成を可視化した。
【0079】
実施例13:LTBおよびLTB-ClyIIAのキメラ複合体
アクチノバチルス・プリューロニューモニエ(Actinobacillus pleuropneumoniae)血清型9参考株CVI13261を、0.1% V因子(NAD)を含有するハート・インフュージョン寒天培地(Difco)上で成長させる。プロテイナーゼK/SDS溶解によって高分子量DNAを単離した後に、フェノール/クロロホルム抽出を行い、得られたゲノムDNAを沈殿させる。アクチノバチルス・プリューロニューモニエ血清型9(GenBank-EMBLアクセッション番号X61111)由来ClyIIA遺伝子を、オリゴヌクレオチド
を使用したPCRによって血清型9株から単離したゲノムDNAからクローニングする。次に、PCRによってBamHI部位もClyIIA遺伝子のアミノ末端に導入し、BamHIで切り出して生じたフラグメントを、大腸菌での発現について記載したようにpET-wiLTBにクローニングし、pET-wiLTB-ClyIIAを発生させた。IPTGで誘導した際にpET-wiLTBおよびpET-wiLTB-ClyIIAの共発現によってキメラ複合体を得ることができる。または、大腸菌でのpET-wiLTB-ClyIIAの過剰発現の際に得られた封入体および精製recLTBを混合して8M尿素で可溶化し、記載したようにトリス緩衝液で透析して五量体複合体を再生させる。
【0080】
実施例14:LTB、LTB-H5およびLTB N1を有する多価タンパク質複合体
共に植物での発現に関して最適化されたA型インフルエンザウイルスH5N1亜型ヘマグルチニン(Genbankアクセッション番号AF028709)およびノイラミニダーゼ(AF028708; Claas et al. (1998) Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet 351 (9101), 472-477)に対する合成遺伝子を作製する。H5およびN1配列のアミノ末端およびカルボキシ末端にBamHI部位を導入し、pLANTIGEN4遺伝子構築体のユニークなBamHI部位に合成H5遺伝子またはN1遺伝子を有するそれぞれのBamHIフラグメントをクローニングすることによって遺伝子融合体を作製する。LTB-H5およびLTB-N1融合タンパク質をコードする生じた遺伝子を実施例1に記載したようにパタチンプロモーターの後にクローニングする。LTB、LTB-H5およびLTB-N1の共形質転換を行い、前に記載したようにPCRによってそれぞれの遺伝子の存在についてジャガイモ植物を分析する。三つ全ての発現カセットについてトランスジェニックである植物を単離して、成熟するまで温室内でさらに成長させることができる。GM1結合複合体の存在についてGM1-ELISAならびにVD12とH5およびN1特異的抗体とを使用したウエスタンブロットによって塊茎を分析する。別の態様では、LTB-H5またはLTB-N1のいずれかを保有する植物を発生させ、それぞれの複合体の蓄積について分析する。塊茎からrecLTB-H5およびrecLTB-N1を精製でき、精製recLTBとそれぞれ1:1:3、1:2:2、1:3:1、2:1:2、2:2:1または3:1:1の比で混合して、溶解し、8M尿素を使用して変性させ、実施例9に記載したようにさらに処理してキメラ複合体を発生させることができる。
【0081】
(表1)GM1 ELISAから得られたように、構築体pLANTIGEN4、12、13および15のいずれか一つを保有するトランスジェニックジャガイモ塊茎で産生したホモ五量体タンパク質複合体のモル発現データの平均とそれぞれの標準偏差および共分散との比較。構築体間の対の差を、分散を安定化するためにデータを対数変換した後でANOVAを使用して解析した。発現レベルをmMで表す。最終列は標準偏差(Sd)を示す。
【0082】
(表2)GM1 ELISAから得られたようにpL4およびpL13を有するキメラタンパク質複合体を保有するトランスジェニックジャガイモ塊茎の発現データの比較。トランスジェニック性質ならびにpL4および/またはpL13遺伝子構築体の存在(第二列および第三列)は、実施例4に記載するように個別の植物から単離したゲノムDNAのPCRによった。記載したようにVD12およびV3 mAbを使用した半未変性ウエスタンブロット分析の結果を比較すること、ならびに図2(第四列)に示すように高分子量複合体の存在をスクリーニングすることにより、キメラの存在を立証した。GM1 ELISAによって決定したように塊茎新鮮重量1グラムあたりのGM1結合性LTB5部分のマイクログラムで発現データを表現する(第五列)。比較のために、pL4遺伝子構築体のみを有する塊茎は平均10〜15μg/g FW LTB5を含有したが、pL13遺伝子構築体の最高の発現体であるpL13(17)は1μg/g FW塊茎未満を有した。
【0083】
(表3)GM1 ELISAから得られたようにpL4およびpL12からなるキメラタンパク質複合体を保有するトランスジェニックジャガイモ塊茎の発現データの比較。トランスジェニック性質ならびにpL4および/またはpL12遺伝子構築体の存在(第二列および第三列)は、実施例5に記載するように個別の植物から単離されたゲノムDNAのPCRによった。記載したようにVD12および3C9 mAbを使用した半未変性ウエスタンブロット分析の結果を比較すること、ならびに図2に示すように高分子量複合体の存在をスクリーニングすることにより、キメラの存在を立証した。GM1 ELISAによって決定したように塊茎新鮮重量1グラムあたりのGM1結合性部分のマイクログラムで発現データを表現する。
【0084】
参考文献
【図面の簡単な説明】
【0085】
【図1】異なるLTBおよびLTBサブユニットワクチン遺伝子構築体を含有する、バイナリーベクターpLANTIGEN4、12、13および15のT-DNAを示した図。LB、左側T-DNA境界配列;PNOSノパリン合成酵素プロモータ;NPTII、ネオマイシンホスホトランスフェラーゼII遺伝子、選択可能カナマイシン耐性マーカー;TNOS、ノパリン合成酵素ターミネータ配列;PPAT、クラスIパタチンプロモータ;遺伝子、PPATプロモータ制御下での発現に関するクローニング部位;RB、右側T-DNA境界配列;SP、シグナルペプチド;LT-B、植物での発現に最適化されたLTBの合成遺伝子;KDEL、小胞体保持シグナル;パルボ、イヌパルボウイルス(CPV)エピトープ;Ala、アラニン;インフルエンザ、HAインフルエンザウイルス血球凝集素エピトープ;CSFV E2、膜貫通ドメインを欠くブタコレラウイルスE2糖タンパク質。
【図2】LTB5コンホメーションのモノクローナル抗体VD12(A)およびCSFV E2コンホメーションのmAb V3(B)を用いた、塊茎抽出物(各総タンパク質25マイクログラム)のウエスタンブロット分析。レーン1、pL(4+13)16;レーン2、タンパク質サイズマーカー;レーン3、コントロール抽出物;レーン4、pL13(31);レーン5、pL13(17)およびレーン6、pL4(17)。Aの左側矢印は、五量体複合体を含有するLTB5を示し、Bの矢印は複合体を含有するCSFV E2コンホメーションのエピトープを示す。
【図3】条件。Aでは、左側のブロットをVD12(抗LTB5)とインキュベーションし、またBでは右側ブロットをV3(抗CSFV E2)とインキュベーションした。サンプルpL(4+13)16(レーン4)およびpL(4+13)46(レーン7)について25マイクログラムを負荷した以外は、それぞれに50マイクログラムの全塊茎タンパク質を負荷した。レーン1、野生型塊茎抽出物;レーン2、pL(4)21抽出物;レーン3、pL(13)17抽出物;レーン4、pL(4+13)16抽出物;レーン5、pL(4+13)31抽出物;レーン6、pL(4+13)39抽出物;レーン7、pL(4+13)46抽出物;レーン8、pL(4+13)60抽出物;レーン9、pL(4+13)64抽出物およびレーン10、pL(4+13)67抽出物。レーン2の左側の矢印は、下側矢印がホモ五量体(LTB)5を示し、上側矢印がホモ五量体(LTB-CSFV E2)5を示している。右側矢印は、本発明のキメラ複合体を示している。分子サイズマーカーを左側に示した。
【図4】半未変性条件の下、12%SDS-PAGEゲルによるpL(4+12)植物塊茎抽出物(各25マイクログラム)のウエスタンブロットの図。A、左側ブロットはVD12(抗LTB5)とインキュベーションし、B、右側ブロットは3C9(抗パルボ)とインキュベーションした。レーン1、M、フルレンジレインボウ(full range rainbow)分子量マーカー;レーン2、pL4(pL417)抽出物;レーン3、pL12(pL(12)01)抽出物;レーン4、pL4抽出物とpL12抽出物の1:1混合物(pL(4)17とpL(12)02);レーン5、V、PAT4ベクター陰性コントロール;レーン6〜10、それぞれpL(4+12)16、pL(4+12)23、pL(4+12)51、pL(4+12)52およびpL(4+12)57の抽出物。左側の矢印は、LTBおよびLTB-パルボ(LTB-P)を表し、右側の矢印は本発明のキメラ複合体を示す。左側の実線ダッシュはタンパク質マーカーを示す。レーン3の、LTBより若干高い位置に移動した下側のバンドは、負荷緩衝液中でサンプルを加熱した際に生じたLTB-Pの分解産物である。
【図5】半未変性条件下で10% SDS-PAGEゲル(A)、または還元条件下で15% SDS-PAGE(B)によるpL(4+12+15)植物塊茎抽出物のウエスタンブロット。A、左ブロットは、VD12(抗LTB5)とインキュベーションし、右ブロットは3C9(抗パルボ)とインキュベーションした。B、ブロットは3C9(抗パルボ)とインキュベーションした。それぞれレーンあたり25マイクログラムの全塊茎タンパク質抽出物を負荷した。レーン1、分子量標準物質;レーン2、野生型塊茎抽出物;レーン3、pL421抽出物;レーン4、pL1201抽出物;レーン5、pL1516抽出物;レーン6〜10、pL(4+12+15)塊茎抽出物;レーン6、pL(4+12+15)7;レーン7、pL(4+12+15)9;レーン8、pL(4+12+15)11;レーン9、pL(4+12+15)16およびレーン10、pL(4+12+15)19抽出物。A、左側矢印は、下矢印がLTB5を;中矢印が(LTB-パルボ)5を、上矢印が(LTB-iipp)5を示す。右の矢印は、キメラ複合体を示す。B、左、下矢印はLTB-パルボを、上矢印はLTB-iippを示す。
【図6】固定化Dガラクトース(Pierce)を用いたアフィニティークロマトグラフィーによる大腸菌recLTBの精製を示す図。カラムを6容積量のTEAN緩衝液で徹底的に洗った後、遠心分離によって集めた、超音波処理大腸菌細胞の上清を、FPLC装置に装着した5cmのDガラクトースカラムに負荷した。プロテアーゼインヒビターカクテル(Roche)を加えた47.5mLのTEAN緩衝液に溶解した大腸菌からの粗タンパク質抽出物を、0.5mL/分の流速でカラムに負荷した。3倍容積量(142.5mL)のTEAN緩衝液(50mM トリス-HCl (pH7.4);0.2M NaCl;1mM EDTA)を0.5mL/分の流速で洗浄した。溶出液を、0.5M Dガラクトースを追加した洗浄緩衝液で、0.5mL/分の流速で洗浄した。1mLの画分を集め、収集後すぐに氷上に置いた。検出器は0.1 A.U.に設定し、記録計の速度は0.25cm/mLであった。A. 溶出プロフィール。実線矢印は、0.5M Dガラクトース添加TEAN緩衝液の溶出開始点を示す。記録下の実線は、画分番号16〜22(主ピーク)を表す。B. GM1-ELISAプール画分1〜32(各1mL)。LTB5の量(ng/μl画分)は、GM1 ELISAによって決定した。C. クマジー染色されたゲル画分17〜22。プレキャスト10%SDS-PAGEゲル(Bio-Rad)を、還元条件下で行なった。レーンM、分子サイズマーカー;レーンE、10μl粗抽出物(カラム負荷前);レーン17〜22、各画分10μl。サンプルは、負荷前に3分間煮沸し、泳動は標準的な条件で行われた。ゲルをクマジーブリリアントブルーで一晩染色し、0.3% Tween-20で脱色した。左側矢印は、LTBモノマーを表しており、右側にサイズを表す(kDa)分子量マーカーが示してある。
【図7】固定化Dガラクトース(Pierce)を用いたアフィニティークロマトグラフィーによる、38グラム新鮮重量pL421植物塊茎からのrecLTBの精製を示す図。塊茎の皮を剥き、小片に細断してから70mLの冷抽出緩衝液を加え、ステンレス鋼製ブレンダーで粉砕および抽出を実施した。粗タンパク抽出物およびrecLTBを含有する上清を5分間、15300rpm、4℃で遠心分離して粒子およびデンプン顆粒を取り除いた。上清が完全に清澄になるまで遠心分離工程を繰り返し、残った47mLの粗抽出物をカラムに負荷した。負荷は0.5mL/分で行い、カラムを4℃に冷却した。負荷後、カラムを42mLのTEAN緩衝液で、0.75mL/分の速度で洗い、溶出液を0.3MのDガラクトースを添加した(0.5Mの代わりに;合計26mL)TEAN緩衝液で洗浄した。画分を集めた。画分サイズは0.75mLで、画分は収集後すぐに氷上に置いた。検出器は0.2 A.U.に設定し、記録計の速度は0.25cm/mLであった。A. 溶出プロフィール。実線矢印は、0.3M Dガラクトース添加TEAN緩衝液の溶出開始点を示す。記録下の実線は、0.75mLの画分番号1〜15を示す。主ピークは画分10〜11にあった。B. GM1-ELISA画分1〜21(各0.75mL)。LTB5の量(ng/画分)は、GM1 ELISAによって決定した。
【図8】6.5グラムの凍結乾燥したpL(4+13)46植物の塊茎からの、固定化Dガラクトース(Pierce)を用いたアフィニティークロマトグラフィーによるキメラタンパク質複合体pL(4+13)の精製を示す図。抽出の前に、凍結乾燥した塊茎材料に、水を26mL、続いてプロテアーゼインヒビターカクテル(Roche)を添加した抽出緩衝液を65mL加えた。抽出は、氷上で22分間連続振盪しながら行い、抽出物は80μMのメッシュ布を使って濾過し、4℃、10分間、15300rpmの遠心分離を2回行って、残ったデンプン顆粒を除去した。残った57mLの、清澄な粗抽出液をカラムに負荷した。負荷は0.5mL/分で行い、カラムを4℃に冷却した。負荷後、カラムをTEAN緩衝液で、0.75mL/分の速度で61分間洗浄し、溶出液を0.3MのDガラクトースを添加したTEAN緩衝液で洗浄した。画分を集めた。画分サイズは0.75mLで、画分は収集後すぐに氷上に置いた。検出器は0.2 A.U.に設定し、記録計の速度は0.25cm/mLであった。A. 溶出プロフィール。実線矢印は、0.3M Dガラクトース添加TEAN緩衝液の溶出開始点を示す。記録下の実線は、0.75mLの画分番号1〜15を表す。主ピークは画分10〜11にあった。B. 標準GM1-ELISA画分5〜19(各0.75mL)。LTB5の量(ng/画分)は、GM1 ELISAによって決定した。C.修飾GM1-ELISA画分5〜19。結合の検出は、CSFV E2に特異的なV3 mAbおよびアルカリホスファターゼ標識ヒツジ抗マウスIgG(VD12、抗LTB5の代わりに)を用いて行った。
【技術分野】
【0001】
本発明は粘膜(例えば口腔または鼻)ワクチンの開発、組成および生成に関する。より具体的には、本発明は粘膜表面への、および表面を通し抗原を送達するためのタンパク質複合体、ならびに植物細胞のような宿主細胞内での該複合体の生成に関する。
【背景技術】
【0002】
経口ワクチン接種は、一般的に経口ワクチンは利用し易く、比較的安価および安全であることから、注射ワクチンに代る魅力的な代替法である。より重要なことは、それが粘膜レベル、即ち多くの病原菌の侵入部位に防御を誘導できることである。例えば経口または鼻内送達による粘膜部位へのワクチン投与は、現在ワクチンが全く利用できないある種の病原菌(例えば呼吸系発疹ウイルス、およびおそらくはHIVでさえ)に対するワクチンの生成にとっては必要条件でもあるだろう。これに加え、経口ワクチン接種は、食物または飲料水による大量ワクチン接種を可能にする。
【0003】
しかしながら、経口ワクチン接種はそれほど有効でない場合が多いと思われる。免疫応答は一時的であり、一般的には、望ましい効果を誘導するには、生きている微生物を使用する場合でさえ大用量の抗原を必要とする。これは、一部は、抗原の取り込みが非効率であることによるものである(Lavell et al. Adv Drug Delivery Rev 1995; 18:5-22)。その結果、粘膜経路から抗原を効率的に送達するための様々な方策が研究されてきた(例えばO'Hagan, J Pharm Pharmacol (1997) 49:1-10; Husband, Vaccine (1193) 11:107-112参照)。粘膜域を裏打ちしている上皮細胞のバリアを通過して送達できた抗原は、粘膜関連リンパ組織(MALT)下部にある誘導部位を刺激する。MALT内で感作した抗原特異的リンパ細胞は循環系を通り移動して、遠方の粘膜部位に定着する。したがって粘膜免疫は、局所と全身防御の両方を提供できる。
【0004】
粘膜ワクチンを開発する一つの方策は、コレラ菌(Vibrio cholera)のこれらトキシン(CT)および近縁な大腸菌(E. coli)易熱性エテロトキシン(LT)のような細菌エンテロトキシンを用いることであり、これらは粘膜送達された場合に極めて免疫源性であり、またキャリア分子およびアジュバントとして機能して、特に後者はホロトキシンを使用した時、無縁な抗原に対する応答を増強できる。
【0005】
毒素原性大腸菌/エシェリキア(Escherichia coli)(LT)の易熱性エンテロトキシンおよびコレラ菌由来の関連コレラトキシン(CT)は極めて強力な免疫原であり、経口投与による粘膜アジュバントである(概説についてはSpangler, 1992およびWilliams et al., 1999参照)。両毒素はAサブユニットと、同一Bサブユニットの五量体環を含む。Aサブユニットはキメラ分子の毒性部分であり、Gsα活性化アデニレートシクラーゼをADPリボシル化して、サイクリックAMPレベルを上昇させる。これが最終的に、水分を多く含む下痢を特徴とする胃腔内の水損失を引き起こす。五量体Bサブユニットを含む、強力な非共有結合性複合体の一次機能は、レセプター相互作用を媒介して毒性のAサブユニットの内在化および取込みを起こすことである。Bサブユニット環の一次レセプターは、小腸を含む哺乳動物細胞の細胞表面上に普遍的に見出されるグリコスフィンゴリピド(Holmgren, 1973; Holmgren et al., 1973, 1975)のモノシアロガングリオシドGM1[Gal(β1-3)GalNAcβ(1-4)(NeuAc(α2-3))Galβ(1-4)Glc(β1-1)セラミド]である。この固有の特徴は、Bサブユニットをアジュバント活性に関係する重要な免疫調節現象を始動する上で重要なものにしている。それはまたBサブユニットを、強力な、いわゆる粘膜キャリア分子にもしている。粘膜キャリア分子は、例えばレセプターを介して、小腸の腸上皮粘膜のような粘膜表面に局在する免疫活性細胞と相互作用する分子である。
【0006】
LT(LTB)およびCT(CTB)両方のBサブユニットは、様々な抗原との翻訳融合において、これらの抗原を、レセプター介在取込みにより胃粘膜上皮を通し行き来させる粘膜キャリア分子としてうまく用いられている(例えばDertzbaugh and Elson, 1993; Jagusztyn-Krynicka et al., 1993)。おそらくは、キャリア分子への抗原の融合が、MALT誘導部位に送達される抗原量と、それに続く抗原特異的B-およびT-リンパ球の刺激を増強するのだろう。抗原は、LTBに化学的に共役させることもできる(O'Dowd et al. Vaccine (1999) 17:1442-1453; Green et al. Vaccine (1996) 14:949-958)。これに加えて、LTBとCTBは共に、抗原と一緒に同時投与した時の経口取込みによる免疫応答を改善する(Bowen et al., 1994; Wilson et al., 1993)。
【0007】
Riganoら(2003, Vaccine 21:809-811)は、トランスジェニックシロイヌナズナでのLTB-ESAT6抗原融合複合体の生成を開示している。融合タンパク質の発現は、ホモ五量体(LTB-ESAT6)5複合体を生ずる。ESAT6は、非常に小型の抗原であり、それゆえに、LTBと融合してもLTB五量体形成を妨害しない。Kim etら(2003; Plant Cell Reports Vol. 21, no.9, pp.884-890)は、ジャガイモでのCTB-NSP175融合タンパク質の発現を報告しており、この発現はホモ五量体複合体を生じた。LTBまたはCTBへの抗原またはエピトープの遺伝的融合は、いくつかの例で成功しているが、Bサブユニットへの非相同性エピトープの翻訳融合は、例えばSandkvistら(J Bacteriol 1987 169:4570)、Schodelら(Gene 1991;99-255)およびDertzbaughら(Infect Immun 1993; 61:384)らが報告しているように、LTBまたはCTB融合タンパク質の構造、分泌、GM1-結合および免疫原性に干渉することがあると思われる。
【0008】
明らかに、五量体化およびGM1結合が保持されるようにLTBまたはCTBに付与できる抗原のサイズとタイプには制限がある。LTBまたはCTベースのワクチンの開発に関しては、最も機能的なワクチンは大型構造を持つタンパク質から構成されることから、この制限は特に実際的な意味を持つ。これに加えて、多くの感染防御抗原(ウイルス、細菌およびその他)は、ホモ多量体またはヘテロ多量体を問わず、多量体複合体から構成されており、そのような形で送達されたときにのみ防御免疫応答を誘導する。例えば、ブタコレラ(a classical swine fever)(CSFV)E2糖タンパク質ベースワクチンは、CSFV-E2ホモ二量体を含有する。様々な感染防御抗原のこの多量体特性は、これらタイプの抗原を通常に遺伝的に融合して用いる、LTBおよび/またはCTBのキャリア分子としての使用を大きく損ねることになるが、それはこの様にして五つの同一融合タンパク質の複合体を形成するからである。
【0009】
商品化および市場導入のためには、融合タンパク質の大規模生産が必要であり、製品は安全であり、またほぼ純粋である必要がある。細菌および酵母で、LTBおよびCTBの融合タンパク質、ならびに抗原をベースにしたこのようなワクチンを生産するには、十分な量の、経口送達のための組換えタンパク質を得るための大規模な発酵技術と厳密な精製プロトコールが必要である。トランスジェニック植物、特に食用植物部分は、経口送達用ワクチンのための安全な発現システムである(Langridge, 2000; Mason and Arntzen, 1995; Sala, 2003参照)。最近、タバコ、ジャガイモおよびトウモロコシを含む様々な植物種で、LTBおよびCTBの製造に成功している(Arakawa et al., 1997; Haq et al., 1995; Hein et al., 1995; Lauterslager et al., 2001; Streatfield et al., 2001)。驚くべき事に、植物もまた大腸菌やコレラ菌と同様にBサブユニットの五量体も生成できるらしい。
【0010】
いくつかのグループが、LTBまたはCTBのいずれかを蓄積したリョウゼンツラン、ジャガイモ塊茎またはトウモロコシを食べさせることによりマウスを経口免疫し、血清IgGおよび分泌性sIgA反応の両方を引き起こしたと報告している(Arakawa et al., 1998; Haq et al., 1995; Lauterslager et al., 2001; Mason et al., 1998; Streatfield et al., 2001)。WO-A-991825は、植物で生産したSEKDEL配列に融合させたCTBサブユニットを用いたマウスの経口免疫を記載している。
【発明の開示】
【0011】
LTBを蓄積したトランスジェニック塊茎50〜100グラムを摂食したヒトボランティアによる前臨床試験でも、特異的血清IgGおよび分泌性sIgA反応が起こった(Tacket et al., 1998)。したがって、トランスジェニック宿主細胞系は、LTBまたはCTB融合タンパク質のような粘膜キャリア分子の(大規模)生産に、本質的に、好都合に用いられる。しかしながら、融合タンパク質の発現レベルは、融合タンパク質のサイズおよび複雑さの増加に伴って大きく低下することが示されている。例えば、ジャガイモ塊茎での、CSFV-E2糖タンパク質(約35kDa、および約70kDaの二量体として防御)と融合したLTBをコードする遺伝子構築体の発現は、約15kDaのLTBサブユニットをコードする遺伝子の発現に比べ、100倍以上低い(Lauterslager, 2002; PhD thesis University of Utrecht)。おそらく、モノマーが、組み立てられた多量体構造に比べ(酵素)分解を受けやすいことが多く観察されていることから、大型のLTBまたはCTB融合タンパク質の低い発現レベルは、大型の五量体タンパク質複合体の不安定性を基本的に反映したものだろう。この見方は、ジャガイモへのCTB-GAD融合タンパク質の蓄積を報告したArakawaら(1999; Transgenics, Harwood Academic Publishers, Basel, Vol. 3, no.1, pp.51-60)と一致する。GAD(ヒトグルタミン酸デカルボキシラーゼ)は65kDaの自己抗原である。CTB-GADの定量化は、発現レベルが全可溶性タンパク質のわずか0.001%であることを明らかにし、このような高分子量複合体を相当レベル蓄積しようとするとき遭遇する可能性がある問題を明示した。しかしながらマウスに新鮮なトランスジェニックジャガイモ物質を長期間、かつ大量に摂食させた時には、少量の活性ホモ五量体複合体は免疫原性であると思われた。
【0012】
したがって、本発明の目的は、比較的大型の抗原を対象となる部位に、例えば組換え体宿主細胞内に、複合体の発現レベルを犠牲にすることなしに送達できる機能的キャリア複合体を提供することである。
【0013】
次に本発明は、全てではなく、幾つかのサブユニットだけが抗原またはその他の対象となる分子に融合した時に、多サブユニットキャリア分子の組み立て、機能性および安定性が高まることについて洞察する。少なくとも二つの、好ましくは同一のサブユニットを含み、そのうちの少なくとも一つのサブユニットは未改変であり、少なくとも一つのサブユニットは関心の第一分子と融合しているタンパク質複合体を提供し、このときタンパク質複合体は細胞表面レセプターに結合できる。
【0014】
好ましい態様では、該タンパク質複合体は、対象となる分子を運搬または移動させるのに用いることができるキャリア分子である。例えば、本発明のタンパク質複合体は、対象となる分子を対象となる部位、例えば粘膜表面まで、および/または横切り送達するためのキャリア複合体またはキャリア分子である。本発明によれば、対象となる分子は、複合体のキャリア特性を用いて生物内の特定部位に送達、その中に分配、移動、または保持することを望む、様々な分類のタンパク質またはポリペプチド、あるいはその領域を含むことができる。それは特に、キャリアの全サブユニットに付与しようとした場合に、例えば立体障害によって、キャリ分子の安定で機能的な多量体構造の形成を妨害するであろう、サブユニットに融合または付加された成分を指す。例えば、サブユニットに融合した該対象となる分子のサイズは、サブユニット単独のサイズの1/4、1/3、1/2、1倍、2倍、3倍または4倍のこともある。
【0015】
本明細書で使用する、用語「未改変」は対象となる分子に融合していないサブユニットまたはモノマーを指す。しかしながらそれは、未変性のサブユニットまたはモノマーに限定されない;例えば、それは、一または複数のアミノ酸が未変性サブユニットから取り除かれた、交換された、および/または追加された組換えタンパク質である。これは、(真核)宿主細胞での組換えタンパク質の安定性、および/または生成工程、例えば発現もしくは分泌を変えるために行うことができる。例えば、未改変サブユニットは、シグナルペプチド、または小胞体(ER)中の保持のためC末端に(SE)KDEL配列を付与したサブユニットを含んでもよい。用語「未改変サブユニット」は、実質的には、未変性サブユニットまたはその修飾が多量体化を妨害しない若干修飾されたものを指す。
【0016】
WO-A-9612801は、発現により一つになってCTホロ毒素タンパク質複合体を形成する、LTAサブユニットおよび、SEKDEL配列に融合したLTBサブユニットの協調発現を開示している。このようなタンパク質複合体は、それが本発明の対象となる分子に融合したサブユニットを含んでいないことから、本発明の複合体とは異なる。SEKDELヘキサペプチドは、対象となる分子とは見なさない。むしろ、上記のように、LTB-SEKDELサブユニットは「未改変」サブユニットと見なされる。したがって本発明の用語法によれば、WO-A-9612801の複合体は、それが未改変サブユニットだけを含むことから、本明細書に示すタンパク質複合体とは別のものである。
【0017】
US6,103,243は、例えばLTBのような粘膜キャリア複合体を用いることによって、免疫原の取込みを改良する経口ワクチンおよび方法を記載している。LTBサブユニットには対象となる抗原が、化学結合、または遺伝的融合のいずれかによって付与されている。これにより、全てのサブユニットが1タイプの対象となる分子に融合しているホモ五量体タンパク質複合体ができる。このように、本発明とは異なり、US6,103,243の複合体は、少なくとも一つの未改変サブユニットおよび対象となる分子に融合している少なくとも一つのサブユニットを含んでいない。
【0018】
本発明の好ましい態様では、タンパク質複合体は、少なくとも二つの同一サブユニット、例えばLTBサブユニットを含み、そのうちの少なくとも一つのサブユニットは変更されており、また少なくとも一つのサブユニットは対象となる分子に融合している。本発明の一つの局面では、タンパク質複合体は、少なくとも一つのサブユニットが未改変であること、そして少なくとも一つのサブユニットが対象となる第一分子に融合していることを特徴とする、少なくとも二つの、好ましくは同一のサブユニットを含み、このとき該対象となる第一分子は、対象となる第二分子と結合して対象となる多量体を形成することができ、またこのときタンパク質複合体は該サブユニットを介して細胞表面レセプターと相互作用できる。好ましくは、該第一分子は、分子間共有結合を介して、例えば一または複数のジスルフィド架橋結合を介して、該第二分子と結合または相互作用する。本発明の対象となる多量体は、例えば、ホモ二量体またはヘテロ二量体抗原のような多量体感染防御抗原である。公知の多量体感染防御抗原としては、CSFV-E2ホモ二量体、ウイルス性出血性敗血症ウイルスの三量体糖タンパク質G(VHSV-G)(Lorenzen, N., Lorenzen, E., Einer-Jensen, K., Heppell, J., Wu, T., Davis, H. (1998) Protective immunity to VHS in rainbow trout (Oncorhynchus mykiss, Walbaum) following DNA vaccination. Fish & Shellfish Immunology 8: 261-270; Lorenzen, N., Olesen, N.J. (1997) Immunisation with viral antigens: viral haemorrhagic septicaemia. In: Fish Vaccinology. Gudding, R., Lillehaug, A., Midlyng, P.J., Brown, F. (eds). Dev. Biol. Stand. Basel, Karger, vol. 90, p 201-209) および狂犬病ウイルス(RV G)および水疱性口内炎ウイルス(VSV G)の三量体糖タンパク質G;センダイ(Sendai)ウイルスのホモ四量体リンタンパク質P(SeV P)が挙げられる。
【0019】
本発明によれば、対象となる第二分子はサブユニットと融合してもよい。例えば、本発明の一つの態様では、タンパク質複合体は、三つのサブユニットを含み、そのうちの二つは同一(例えばCSFV-E2)モノマーに融合しており、そして一つは未改変サブユニットである。サブユニットは互いに近接しており、融合モノマーが相互作用でき、そしてホモ多量体、例えば抗原(例えばCSFV-E2)ホモ二量体を形成できることが望ましい。未改変サブユニットの存在は、タンパク質複合体の多サブユニット構造の安定性を高め、複合体のコンホメーションが保持され、それがサブユニットを介してそのレセプターと相互作用できることを保証する。融合モノマーがヘテロ多量体を形成できるように、少なくとも三つのサブユニットのタンパク質複合体が異なるモノマーと融合している二またはそれ以上のサブユニットと、少なくとも一つの未改変モノマーを含むことは、明らかに可能である。
【0020】
もちろん、対象となる第二分子は、対象となる第一分子と相互作用して対象となる多量体を形成するために、サブユニットと融合する必要はない。そのうえ、本発明によれば、例えば立体障害に関して、対象となる多量体を含むタンパク質複合体を、該対象となる多量体が、全てがサブユニットに融合しているわけではない対象となる多量体から構成されるようにデザインすることが有利なことも時にある。例えば、全てがあるサブユニットに融合している対象となる分子間の相互作用の強さは、それらが融合している個々のサブユニットの相対的な方向によりある程度決定される。したがって、互いに正常に(即ち、それらの未変性で、融合していないコンホメーションにある)相互作用し合える分子は、サブユニットと融合した結果分子間の相互作用を低下、ときに完全に失うことにより、立体的に制限されることもある。したがって、本発明によれば、タンパク質複合体は、例えばジスルフィド架橋結合を通して、多量体構造を形成することができる対象となる多量体分子から構成される対象となる多量体を含んでよい。好都合にも、少なくとも一つの未改変サブユニットおよび抗原と融合したサブユニットを少なくとも一つ含む本発明のタンパク質複合体は、一般的に、一つまたは複数(多量体)抗原の、一抗原成分へのより良い折りたたみ構造を示す。前述したように、対象となる一つ、幾つか、または全てがサブユニットと融合する。当業者は、サブユニットおよび対象となる多量体の四次構造および対称性に基づいて、最適なコンホメーション、例えば最適な抗原特性を持つ対象となる多量体を含むタンパク質複合体を得るために、融合型分子と非融合型分子の最適数を選ぶことができるだろう。
【0021】
一つの態様では、タンパク質複合体は、腸上皮上にある細胞表面レセプター、例えばGM1のようなガングリオシド分子に結合できる。対象となる分子を含む、このようなタンパク質複合体は、粘膜キャリア分子として有利に用いられる。
【0022】
好ましい態様では、本発明のタンパク質複合体は、大腸菌の易熱性エンテロトキシン(LT)またはコレラ菌のコレラトキシン(CT)、好ましくはそのBサブユニットに本質的に基づいている。本発明の複合体は、例えば五つのLTまたはCTのBサブユニットから構成される環構造体であり、この場合そのうち少なくとも一つのサブユニットは未改変であり、少なくとも一つのサブユニットは対象となる分子に融合している。このような本発明のキメラまたはヘテロ五量体環構造体は、一、二、三または四つの未改変LTB-またはCTB-モノマーと、四、三、二または一つのL/CTB-融合タンパク質をそれぞれ含む。好ましい態様では、対象となる分子は、サブユニットに、それがレセプター結合ポケットから見て分子の反対側に位置するように、または少なくとも該サブユニットのレセプター結合特性を妨害しないように融合している。例えば、五量体LTB-またはCTB-構造体に基づく本発明の複合体では、これは対象となる分子がBサブユニットのC末端に融合することが好ましいことを意味している(Sixma et al. 1991 Nature 351;371-377)。この本発明の方針に従って、それらのコンホメーションの完全性を保持したキメラ五量体を得ている。LTBサブユニットおよびCTBサブユニットは相同的であることから、それらを用いて、少なくとも一つのサブユニット(LTBまたはCTBのいずれでも)が対象となる分子と融合しており、そして少なくとも一つのサブユニットが未改変である、キメラLTB/CTB五量体から構成されたタンパク質複合体を形成できる。LTBおよび/またはCTBをベースとした本発明のキメラタンパク質複合体は、五つ全てのサブユニットが対象となる分子に融合しているホモ五量体構造と比べると、GM1レセプターへの結合の向上および発現レベルの上昇を示す。
【0023】
別の態様では、本発明のタンパク質複合体は、西洋ワサビペルオキシダーゼ(HRP)に基づいている。本発明の複合体は、例えば、少なくとも一つのサブユニットは未改変であり、そして少なくとも一つのサブユニットは対象となる分子と融合している、六つのHRPを含む構造体または組成物である。このような本発明のキメラまたはヘテロ六量構造体は、一、二、三、四または五つの未改変HRPサブユニット(モノマー)と五、四、三、二または一つのHRP融合タンパク質をそれぞれ含む。
【0024】
本発明の複合体は、多量体抗原の最適な折りたたみ構造と分子内相互作用(例えばジスルフィド架橋結合を介した)を可能にし、したがって最適な免疫原特性を有している。一つの態様では、対象となる分子は、生物、好ましくは哺乳動物、より好ましくはヒトの免疫系に変化をもたらす、または影響することができる。好ましい態様では、対象となる分子は抗原である。抗原は、免疫系を刺激する任意の物質である。抗原は、身体に侵入する細菌またはウイルスの様な外来性微生物、またはタンパク質またはタンパク質フラグメントのような該微生物の成分であることが多い。対象となる分子は、好ましくは、細菌抗原、ウイルス抗原、原虫抗原、線虫抗原および真菌抗原からなる群より選択される。本発明は、各種抗原、例えば線状B細胞エピトープCPV(イヌパルボウイルスエピトープ)およびT細胞特異エピトープHA(インフルエンザウイルス血球凝集素エピトープ)のようなTおよび/またはB細胞エピトープを送達するためのタンパク質複合体、ならびにそのようなタンパク質複合体を含む組成物を提供する。本発明の複合体は、例えばCSFV(ブタコレラウイルス)のE2タンパク質のような、ウイルス(糖)タンパク質のような大型の抗原の送達にも用いることができる。一つのサブユニット融合タンパク質が複数の対象となる分子と融合してもよい。例えば、少なくとも一つの未改変サブユニットと、二つのHAエピトープおよび二つのパルボエピトープと転写的に融合した少なくとも一つのサブユニットとを含むタンパク質複合体が提供される(図1参照)。本発明の一つの態様では、タンパク質複合体(例えばLTBをベースとした複合体)は、少なくとも一つの未改変のサブユニット、およびCSFV E2糖タンパク質に融合している少なくとも一つ、好ましくは二つのサブユニットを含む。別の態様では、本発明のタンパク質複合体は、少なくとも一つの未改変サブユニット、パルボエピトープに融合した少なくとも一つのサブユニット、および複数の対象となる分子に融合している少なくとも一つのサブユニットを含み、後者は二つのHAエピトープおよび二つのパルボエピトープに融合したサブユニットを含む。
【0025】
更なる態様では、本発明のタンパク質複合体は、未改変である少なくとも一つのサブユニット、および免疫調節分子(またはその一部分)、例えばサイトカインもしくは熱ショックタンパク質(HSP)に融合している少なくとも一つのサブユニットを含む。サイトカインは、免疫応答中に細胞間シグナル伝達に関係する分子の大きなグループを表す一般的用語である。サイトカインはタンパク質またはポリペプチドであり、一部には糖タンパク質が結合している(糖タンパク質)。サイトカインは幾つかの群:インターフェロン(IFNアルファ、ベータ、ガンマ);インターロイキン(IL-1〜IL-15);コロニー刺激因子(CSF)および、腫瘍壊死因子(TNF)アルファおよびベータまたは形質転換増殖因子(TGF)ベータのようなその他サイトカインに区別できる。HSPは、CD91と呼ばれるレセプターを介したそれらとマクロファージおよび樹状細胞との相互作用に由来する、顕著な免疫調節特性を有している。例えば、HSP70-2は、炎症刺激に対する反応で誘導される、重要な免疫調節タンパク質である。本発明のタンパク質複合体に含まれる対象となるHSPとしては、HSP-60、HSP-70、HSP-90およびGp-96が挙げられる。所望する免疫学的応答のタイプに応じて(例えばTh1対Th2、抗体応答、抗炎症応答)、一つまたは複数(異なる)免疫調節タンパク質またはその一部分を本発明のタンパク質複合体のサブユニットに融合することができる。例えば、本発明の多成分ワクチンは、抗原および、抗体応答(T、B細胞)を誘導する免疫調節分子、好ましくはサイトカインを含むタンパク質複合体を含む。一方、例えば自己免疫疾患(例えば糖尿病、多発性硬化症)の治療または予防については、自己抗原および、寛容を誘導する免疫調節タンパク質を含む多成分ワクチンまたはワクチンを使用することが有利だろう。
【0026】
対象となる別の分子は、本発明の複合体の一部である場合に、「レポーター」分子として用いて体内のキャリア複合体の局在を知らせることができるような分子を含む。これは、レセプター分子への複合体のインビボ結合のモニタリング、および粘膜上皮を通しての複合体の(レセプター介在)輸送にとりわけ役立つ。対象となるレポーター分子は、例えば、酵素(クロラムフェニコールトランスアセチラーゼ(CAT)、ネオマイシンホスホトランスフェラーゼ(neo)、ベータ−グルコシダーゼ(GUS)またはホタルルシフェラーゼ;等)またはグリーンフルオレセントタンパク質(GFP)またはそのスペクトル変種のような蛍光タンパク質である。
【0027】
本発明の一つの局面では、タンパク質複合体は、そのうちの少なくとも二つのサブユニットに対象となる分子が付与されている少なくとも三つのサブユニットを含む。該少なくとも二つのサブユニットは、対象となる同一の分子または対象となる異なる分子に翻訳的に融合できる。少なくとも二種類の対象となる分子を持つ本発明のタンパク質複合体は、少なくとも二種類の抗原のため、例えば多成分ワクチンの生成のためのキャリア分子として有利に用いられる。別のタイプの対象となる分子のその他組合せ、例えば一または複数の抗原と一または複数の免疫調節(促進的または阻害的)タンパク質との組合せも、もちろん可能である。
【0028】
コレラトキシンをベースとする多成分ワクチンがYuとLangridge(Nature, 2001, vol.19:548)により記載されており、かれらはネズミロタウイルスエンテロトキシンNSP4の22アミノ酸免疫優性エピトープに融合したコレラトキシンBサブユニット、およびCTA2サブユニットに融合したETECフィムブリエ定着因子CFA/Iの発現を研究した。本発明の複合体とは異なり、報告されたコレラトキシン複合体の全てのサブユニット(BおよびA2の両方)がある抗原に融合していた。この具体例では、CTB発現または機能的CTB/A2複合体の形成に問題がなかったという事実は、おそらくはBサブユニットに融合したNSP4エピトープのサイズが極めて小さい(約5kDa)ことに関係しているだろう。既に述べたように、機能的五量体の形成の問題は、立体障害に大きく帰すべき問題であり、サブユニットに融合したこのような小さいエピトープは五量体形成を妨害することはないだろう。本発明によれば、抗原が5kDaよりはるかに大きい(多成分)ワクチンも生産できるようになる。例えば、対象となる分子に融合した少なくとも一つの(CT/LT)Bサブユニット、少なくとも一つの未改変Bサブユニット、および対象となる異なる分子に融合したA2サブユニットを含むタンパク質複合体を提供する。好ましくは、Bサブユニットに融合した該分子は、7kDaより大きく、より好ましくは10kDaより大きく、よりさらに好ましくは15もしくは25kDaよりさらに大きい。しかしながら、本発明の別の態様では、同一キャリア複合体の(LT/CT)Bサブユニットおよび(LT/CT)A2サブユニットは、同一抗原に融合しており、このとき該抗原は免疫防御特性を持つ多量体(二量体、三量体、四量体)タンパク質複合体の一部分である。
【0029】
本発明はさらに、本発明のタンパク質複合体を生成するための方法であって、a)未改変サブユニットをコードする第一ヌクレオチド配列および対象となる分子と融合したサブユニットをコードする第二ヌクレオチド配列を持つ宿主細胞を提供する工程;b)該宿主細胞を培養し、それによって該第一および第二ヌクレオチド配列を発現させ、タンパク質複合体を組み立てさせる工程;c)複合体を分離する工程;およびd)複合体を細胞表面レセプターに結合する工程を含む方法を提供する。
【0030】
用語「宿主細胞」は、異質遺伝子を複製および/または転写および/または翻訳できる任意の細胞を指す。一つの態様では、宿主細胞は植物細胞、ファージまたは細菌である。好ましい態様では、本発明の宿主細胞は食用宿主細胞であり、これは消費時に有害な影響を生じない。好ましい例はジャガイモ、トマト、タバコ、トウモロコシおよび乳酸桿菌である。したがって、宿主細胞は、インビトロに在ろうとインビボに在ろうと、任意の真核生物または前核生物細胞(例えば大腸菌のような細菌細胞、メタノール資化酵母(Pichia pastoris)のような酵母細胞、チャイニーズハムスター卵巣細胞のような哺乳動物細胞、トリ細胞、両生類細胞、植物細胞、魚類細胞、マッシュルーム(Agaricus bisporis)のような真菌細胞、およびヨウガ(Spodoptera frugiperda)のような昆虫細胞)を指す。例えば、宿主細胞はトランスジェニック植物内に在ってもよい。
【0031】
上記のように、特定態様において、植物を本発明の方法に用いることができる。該植物宿主は、トウモロコシ(Zea mays)、コムギ(Triticum aestivum)、イネ(Oryza sativa)またはアオウキグサ種(Lemna spp)のような単子葉植物、またはタバコ種(N. tabacum)のようなタバコ(Nicotinana)属、トマトのようなトマト(Lycopersicon)属、ウマゴヤシ(Medicago)を含むマメ(Leguminosae)科、またはヒメツリガネゴケ(Physcomitrella patens)のような蘚類でよい。好ましい態様では、該宿主植物はジャガイモ(Solanum tuberosum)である。
【0032】
本発明によれば、宿主細胞には、少なくとも二種類の核酸配列:未改変(対象となる抗原と融合していない)サブユニットをコードする核酸配列、および対象となる分子と翻訳的に融合したサブユニットをコードする核酸配列が与えられる。例えば、(植物)宿主細胞に、未改変LTBサブユニットをコードする第一ヌクレオチド構築体および対象となる抗原に融合したLTBサブユニットをコードする第二構築体をトランスフェクションする。対象となる分子に融合したサブユニットをコードするヌクレオチド配列は、生ずる融合タンパク質の可撓性を増すために、サブユニットと対象となる分子の間の領域にリンカーまたはヒンジを含んでもよい。適合条件の下でトランスフェクションおよび培養した後、該細胞は二つのポリペプチドを発現し、抗原が「負荷された」、機能的なLTBをベースとしたキメラタンパク質複合体を組み立てるだろう。少なくとも二種類の対象となる分子を持つ本発明のタンパク質複合体を生成するためには、宿主細胞には、当然少なくとも三種類のヌクレオチドを付与する。これら異なる核酸配列は、それぞれが異なるヌクレオチド配列を担持している、異なるベクターを用いた(例えばT-DNA)該宿主細胞のコトランスフェクションによって、該宿主細胞内に導入できる。または、二もしくはそれ以上の種類の遺伝子構築体を、交差によって、または一つのバイナリーベクター上に二もしくはそれ以上の発現カセットを用いることによって、一つの宿主細胞に導入できる。より更には、既に本発明のタンパク質複合体の一(または複数の)成分を含んでいる樹立宿主細胞株に、再形質転換を用いて、追加の核酸配列を付与できる。さらに別の態様では、少なくとも二種類の核酸配列を発現している宿主細胞は、もう一方の配列を安定的に発現している宿主細胞に一方の配列を一過性(ウイルス媒介性)に発現させることで得られる。
【0033】
異なるベクターを使用することの利点は、それによって、宿主細胞に様々な割合で核酸構築体を与えることができるという事実に見いだせる。これにより、未改変の「フリー」サブユニットの量を、宿主細胞が発現した融合サブユニットの量に対して滴定でき、生じた多量体タンパク質複合体の組成を最適化できる。例えば、融合サブユニットを多く含む複合体を所望するのであれば、宿主細胞を構築体A(未改変サブユニット)および構築体B(融合サブユニット)と、構築体Bが構築体Aより過剰であるようにして(例えばA:B=1:3または1:5または、さらに1:10)コトランスフェクションまたはコトランスフォーメーションする。これと逆に、組み立てたタンパク質複合体が融合サブユニットを比較的少なく含むように変えるには、構築体Aを構築体Bより過剰にするのが好ましい。後者はもちろん、本発明の機能的タンパク質複合体の組み立てに対する、融合サブユニットの負の立体的影響または妨害を最小にしたい場合に好ましい。
【0034】
さらに別の態様では、宿主細胞は対象となる分子、例えばLTB-CSFV E2に融合したサブユニットをコードする核酸構築体を用いて形質転換され、このとき該構築体は、サブユニットをコードする核酸配列と対象となる分子をコードする核酸配列との間にタンパク質分解性切断部位を含んでいる。この融合タンパク質がプロテアーゼ(宿主細胞内で異種内因的に発現された)によって部分的インビボ切断されると、宿主細胞は未改変サブユニットと対象となる分子に融合したサブユニットの両方を含むことになり、これらは本発明のキメラタンパク質複合体を形成できる。
【0035】
当技術分野で公知の様々な方法を用いて、宿主細胞に組換えまたは単離された核酸(DNAまたはRNA)を提供することができる。これらには、形質転換、トランスフェクション(例えばリン酸カルシウム沈殿またはカチオン性リポソーム試薬を用いた)、エレクトロポレーション、バーティクルガンおよびアグロバクテリウム(Agrobacterium)介在T-DNA転移が挙げられる。当業者は、宿主細胞のタイプに応じて、選択する方法を認めるだろう。
【0036】
宿主細胞に該外来ヌクレオチドを与えた後、宿主細胞を培養して該第一および第二ヌクレオチド配列を(同時)発現させ、生じたポリペプチドを本発明のタンパク質複合体に組み立てる。いくつかの例、特に形質転換またはトランスフェクションの方法が比較的非効率な例では、該ヌクレオチドを実際に受け取っている宿主細胞を選択し、このように選択された宿主細胞だけを培養することが有利である。形質転換またはトランスフェクション(または宿主細胞に単離核酸を提供するその他の方法)後の宿主細胞選択は、標準の方法に従って実施できる。例えば、対象となるDNA配列を宿主に送達するのに用いられる最も一般的なベクターは、効率的な宿主細胞への送達および宿主細胞による発現により、該宿主細胞に選択薬剤に対する耐性を付与するタンパク質(酵素のような)をコードする核酸配列も含んでいる。頻繁に用いられる選択薬剤は、ネオマシン、カナマイシン、アンピシリン、カルベニシリン等のような抗生物質である。
【0037】
本発明の方法では、少なくとも二種類の核酸(バイナリーベクターまたは異なるベクターのいずれかを用いても)が付与された(選択された)宿主細胞は、少なくとも二種類のポリペプチド、例えば未改変(非融合)サブユニットXおよび融合サブユニットYを発現するだろう。
【0038】
本発明の別の態様では、第一宿主細胞、例えば微生物宿主細胞は、未改変サブユニットXを提供し、第二宿主細胞は対象となる分子、例えば抗原を含む融合サブユニットYを提供する。場合によっては、第三宿主細胞は、第二対象となる分子、例えば第二抗原またはサイトカインのような免疫調節分子を含む融合サブユニットZを産生できる。別々に産生されたXおよびY(および場合によりZ)サブユニットを単離した後、それらを、少なくとも一つのXサブユニットおよび少なくとも一つのYサブユニットを含むタンパク質複合体の形成にとって好ましい条件の下で、互いに接触させることができる。本発明のキメラタンパク質複合体をインビトロ再構成するこの方法は、それらをある割合で互いに接触させることによって、一つの複合体の中にある未改変サブユニットと融合サブユニット数の滴定を可能にする。未改変および融合サブユニットの両方を産生する単一宿主細胞を用いる場合と対照的に、上記再構築法によるキメラタンパク質複合体の作製は、サブユニットの相対発現レベルには依存しない。むしろ、互いに接触している未改変と融合サブユニットとの割合は、制御することができる(実施例9も参照)。それにより本発明は、本発明のタンパク質複合体を生成する方法であって:a)未改変サブユニットをコードするヌクレオチド配列を第一宿主細胞に提供し、また対象となる分子をコードするヌクレオチド配列を第二宿主細胞に提供し、このとき対象となる少なくとも一分子がサブユニットに融合している工程;b)該宿主細胞を培養し、それにより該ヌクレオチド配列を発現させる工程;c)該ヌクレオチドがコードするタンパク質を単離する工程;d)タンパク質複合体の組み立てを可能にする条件の下に単離されたタンパク質を接触させる工程;e)複合体を単離する工程;f)細胞表面レセプターまたは細胞表面レセプターを模した分子への複合体の結合を決定する工程を含む方法を提供する。上記のうち工程d)では、特定サブユニット組成を持ったタンパク質複合体が形成されるように、タンパク質を特定の割合で混合することができる。
【0039】
該少なくとも二つのポリペプチドの多量体複合体への組み立ては、それぞれが異なるサブユニット組成を持った多様な複合体になることができる。理論的には、2タイプの複合体:未改変Xサブユニットのみ、または融合Yサブユニットのみを含むホモメリック複合体、およびXおよびYサブユニットの混合体を含むヘテロメリック、またはキメラ複合体が形成できる。複合体中に存在するXおよびYサブユニットの数に応じて、各種タイプのキメラ複合体が可能である。例えば、三量体は、二つのXサブユニットと一つのYサブユニット、またはその逆を含むことができる;四量体複合体はXサブユニットを三、二または一のみ、そしてYサブユニットを一、二または三つ、それぞれ含むことができる;といった具合である。推察のとおり、本発明のタンパク質複合体は、後者(キメラ)のタンパク質複合体に関係する;ホモメリックX複合体は対象となる分子を欠いており、ホモメリックY複合体は、仮に組み立てたとして、融合した(嵩高の)対象となる分子数が多いことによる立体障害の存在により、おそらくはレセプターへ結合することができないだろう。そのうえ、全てのサブユニットが抗原と融合しているホモメリックタンパク質複合体は、個々の抗原モノマーの配向/コンホメーションが免疫特性に関して最適でないことから、多量体抗原にとって有用なキャリア分子ではないだろう。それゆえに、本発明の一つの態様では、本明細書で提供するキメラタンパク質複合体が、キメラおよびホモメリック複合体の混合物から単離される。しかしながら、キメラタンパク質複合体をホモメリック複合体から分離することは常に必要とされるものではないことを理解しなければならない。一つの態様では、キメラおよびホモメリック複合体を含む組成物は、ワクチンとしての好適に用いられる。しかしながら、キメラ複合体がホモメリック複合体より大量に存在することが好ましい。例えば、キメラタンパク質複合体が、タンパク質複合体総数の少なくとも50%、好ましくは少なくとも60%、より好ましくは少なくとも70%を構成することである。
【0040】
キメラおよびホモメリックタンパク質複合体は、それらのサイズに基づいて互いを分離できる。融合サブユニットは、未改変サブユニットより明らかに大きいことから、サブユニット組成が異なる複合体はサイズが異なるだろう。この差により、複合体のサイズに従って、所望のキメラ複合体を、ホモメリックとキメラタンパク質複合体との混合物から
単離できる。好ましい態様では、本発明のタンパク質複合体は、ゲル濾過クロマトグラフィーを用いて単離される。ゲル濾過クロマトグラフィー(サイズ排除クロマトグラフィーまたは分子篩クロマトグラフィーとしても知られる)を用いて、タンパク質をそれらのサイズに従って分離することができる。タンパク質クロマトグラフィーに関する一般的情報は、RK Scopes(Springer-Verlag3rd edition, January1994; ISBN 0387940723)による "Protein Purification: Principles and Practice" のような本技術分野で用いられている手引き書から得ることができる。
【0041】
簡単に述べると、ゲル濾過では、タンパク質(の混合物)溶液を半透性多孔性樹脂を詰めたカラムに通す。半透性樹脂はある範囲の孔径を有しており、これがカラムで分離できるタンパク質のサイズを決定する。これは樹脂の画分域または排除域と呼ばれる。樹脂の排除域より大きなタンパク質は孔の中に入ることはできず、樹脂の間の空間を通り、カラムを素早く通過する。これはカラムの空隙容量として知られる。樹脂の排除域以下の小さいタンパク質、およびその他の低分子物質は全て樹脂の孔に侵入し、またそれらはカラムの全容積を通過しなければならないため、それらのカラムを通過する動きは遅くなる。カラムの排除域内に納まるサイズのタンパク質は、孔の一部のみに侵入する。このようなタンパク質の動きは、それらのサイズに従って遅くなるだろう;小さいタンパク質ほど、より大きな容積を通過しなければならないため、カラムをゆっくりと通過することになる。ゲル濾過クロマトグラフィーによってタンパク質サンプルを分離するためには、カラムをまず所望の緩衝液で平衡化しなければならない。これは、カラムに、カラム容積の数倍量の緩衝液を通すだけで達成できる。平衡緩衝液はタンパク質サンプルが溶出していく緩衝液であることから、平衡化は、重要な工程である。次にサンプルをカラムに負荷し、樹脂に浸透させる。次により一層多い量の平衡緩衝液をカラムに通して、サンプルを分離し、カラムからそれを溶出する。画分はカラムからのサンプル溶出物として集められる。より大きなタンパク質は早い画分に溶出し、より小さなタンパク質はそれに続く画分に溶出する。
【0042】
ゲル濾過は、理想的に言えば、タンパク質複合体の分解を減らすことに加えて、濾過中のサンプルの拡散を小さくするのにも役立ち、これが解像度を向上することから、低温で行うべきである。タンパク質の分離は、より長いカラムを使用することで向上するが、分離時間が長くなるほどタンパク質の分解は増加する。
【0043】
クロマトグラフィー媒体の選択は、ゲル濾過クロマトグラフィーでは重要な考慮事項である。幾つかの、一般的なゲル濾過クロマトグラフィー媒体は次の通りである;Sephadex G-50(1〜30kDaの範囲にあるタンパク質の画分化に適している);Sephadex G-75; Sephadex G-100 (4-150kD); Sephadex G-200 (5-600kD); Bio-Gel P-10 (1.5-20kD); Bio-Gel P-30 (2.4-40kD); Bio-Gel P-100 (5-100kD)およびBio-Gel P-300 (60-400kD)。SephadexはPharmaciaの登録商標である。Bio-GelはBio-Radの登録商標である。カラム容積の約0.6倍のところに溶出される分子が最も良く分離するが、溶出が遅いものほど一般的にピークは広がる。一つの態様では、最善の分離のために、画分域が広い材料を用いて最初に分断を行い、極めて大きなタンパク質と極めて小さなタンパク質を取り除き、続いてより画分域が狭い材料を用いる。ゲル濾過カラムは、分子量が分かっている標準物質(タンパク質)を用いてキャリブレーションできる。キャリブレーションカーブは、log MW(分子量の対数)の関数として、保持時間を表すように作製できる。本発明の方法では、ホモメリックタンパク質複合体は、カラムから溶出するヘテロメリック複合体の範囲を示すのに都合良く用いられる。これについては、上記サンプルの一つについて行われた以下実施例で詳しく説明する予定である。サブユニットXおよびYを発現し、両サブユニットともそれ自身および/または互いに四量体を形成できる宿主細胞(「XY」と称す)が与えられる。さらに、サブユニットX(未改変サブユニット)のみ発現する宿主細胞「X」およびサブユニットY(対象となる分子と融合したサブユニット)のみ発現する宿主細胞「Y」が与えられる。ホモメリックXおよびY複合体の推定サイズ(サブユニットXおよびYそれぞれのサイズの四倍)に従って、好適なゲル濾過媒体を選択する。定義により、本発明のキメラタンパク質複合体は、Xホモマーより大きく、かつYホモマーより小さなサイズを有する。それゆえに、全てのキメラタンパク質複合体(XYYY;XXYY;およびXXXY)の溶出容積は、Yホモ四量体(最大の複合体)より大きいが、しかしXホモ四量体(最小の複合体)よりは小さくなるだろう。Yホモ四量体のみ含有する宿主細胞「Y」のタンパク質サンプルおよびXホモ四量体のみ含有する宿主細胞「X」のタンパク質サンプルをゲル濾過カラムに装填することで、宿主細胞「XY」が産生するキメラタンパク質の溶出容積の「境界」を容易に知ることができる。例えば、カラム画分を集め、SDS-PAGEに続き、特異的試薬、例えば抗体を用いたウエスタンブロットを行い、XまたはYサブユニットの存在について分析する。所望であれば、条件(カラムサイズ、カラム直径、流速等)を調整して、Y四量体からのX四量体の分離を最適化できる。もちろん、XとYとのサイズ差が大きいほど(即ち対象となる分子が大きいほど)、異なる四量体複合体の相互の分離はより容易になるだろう。しかしながら、本発明は特に、大きなサイズ差(例えば大きな抗原を有する複合体)に起因する問題を解決することに関係する。それ故に、本発明の複合体の分離は、大きな問題にはならないだろう。キメラ複合体の溶出容積が一度決定されれば、ホモ四量体によるキャリブレーションに用いたものと同一の条件下で、宿主細胞「XY」のタンパク質サンプルをカラムに装填し、溶出することができる。カラム画分を集め、上記のようにしてXおよび/またはYサブユニットの存在を分析する。煮沸していないタンパク質画分のSDS-PAGEを行い、インタクトタンパク質複合体のサイズを分析することができる。ホモメリック複合体を欠いている本発明のキメラ複合体を含有する画分を、更なる使用のために保存しておく。それとも画分を一つにまとめてプールし、本発明のタンパク質複合体の混合体を得てもよい。画分を個別のままにしてもよく、例えば別々に単離された複合体として、複合体XYYY、XXYYおよびXXXYを得ることもできる。
【0044】
通常のゲル濾過クロマトグラフィーは時間のかかる工程である。作業は、クロマトグラフィー媒体の粒子サイズまたは樹脂を小さくし、カラムをより小型にすることで著しく迅速化できる。これには、液体を、高圧を利用してカラムの中を流すための特別な装置が必要である。好ましい態様では、本発明のタンパク質複合体は高圧液体クロマトグラフィー(HPLC)または高性能液体クロマトグラフィー(FPLC)を用いて単離する。
【0045】
本発明の方法では、単離されたタンパク質複合体はさらに特徴付けされ、細胞表面レセプターまたはレセプター結合成分を模した分子に対する、その結合能力を決定し、例えばGM1レセプターを模擬するD-ガラクトースは、本発明のLTBまたはCTBをベースとするタンパク質複合体の特徴付け(または精製)に用いることができる。好ましい態様では、一過性結合よりも容易に検出できることから、結合は、永久結合を含む。単離または精製されたレセプターを用いることができるが、レセプターを発現している細胞、またはそれら細胞に由来する膜材料を用いてもよい。本発明のタンパク質複合体を、複合体がそのレセプターへの結合に適した条件の下で、レセプターに接触させる。レセプター結合に影響する可能性のある条件としては、イオン強度(pH;塩)および温度が挙げられる。その後、結合した複合体の量を決定する。好ましい態様では、本発明の、単離されたタンパク質複合体の細胞表面レセプターへの結合は、酵素結合免疫吸着アッセイ(ELISA)またはそれに実質基づく方法により行われる。ELISAの基本原理は、酵素を用いて、抗体(Ab)への抗原(Ag)の結合を検出することである。酵素は無色の基質(発色団)を着色された生成物に変換し、Ag:Ab結合の存在を示す。本発明の特定の態様では、タンパク質複合体は大腸菌の易熱性エンテロトキシン(LT)のBサブユニットに基づいている。Bサブユニットのインビボ五量体化およびその本来のレセプターGM1への五量体の結合は、トキシンの取り込みを先導し、最終的に粘膜免疫に伴う免疫調節現象を誘導する重要な現象である。本明細書に例示するように、LTBサブユニットに基づく本発明のタンパク質複合体の結合は、De Haanら(Vaccine 1996; 1:777-783)記載のGM1-ELISAを用いて容易に決定される。このようなタイプのアッセイを用いることで、少なくとも一の未改変LTBサブユニットと少なくとも一つのLTB融合タンパク質を含む、本発明のLTBベースのタンパク質複合体が、その本来の五量体の構造を保持していることが明らかになるだろう。
【0046】
本発明のさらなる局面では、本発明のタンパク質複合体を含む宿主細胞が提供される。宿主細胞は、例えば、本発明の複合体の構成成分をコードする一または複数の核酸構築体が付与された微生物細胞である。一つの態様では、該宿主細胞は細菌細胞、例えば形質転換した大腸菌細胞である。上記したように、細胞は、好ましくは、粘膜表面に、および粘膜表面を横切り、一または複数の抗原を送達することができるタンパク質複合体を含む食用細胞である。食用細胞の例は、食用植物、例えばジャガイモである。このような複合体は、粘膜キャリア分子として有利に用いられる。従って、本発明の粘膜キャリア分子を含む食用細胞、またはその抽出物は、例えば食物もしくは飲料水を用いた経口ワクチン接種に用いることができる。
【0047】
前に述べたように、キャリ分子の通常の使用は、比較的小さな抗原の送達に実質的に限定されている。しかしながら本発明のキメラ複合体は、融合タンパク質のサイズとは実質的に無関係に、細胞表面レセプターに結合する能力を保持していることから、ここでは比較的大きな対象となる分子(抗原等)を送達するためにキャリア分子を用いることができる。抗原が体内の適当な部位に一度送達すると、免疫応答を引き起こせることは周知である。たとえば、粘膜域を裏打ちする上皮細胞のバリア横切り送達できた抗原は、粘膜関連リンパ組織(MALT)の下にある誘導部位を刺激する。本発明のタンパク質複合体には、一または複数の対象となる抗原性分子を、実質的に対象となる分子のサイズおよび複雑さに関係なく、そして、重要なことであるが、上皮バリアを通して複合体を送達し、MALTを刺激するキャリア特性を失わずに、与えることができる。それゆえに、本発明の複合体は、被験体、好ましくはマウスもしくはヒトのような哺乳動物被験体に免疫応答を誘導するのに、都合良く用いられる。一つの態様では、対象となる抗原を含む本発明のキメラタンパク質複合体は、ワクチンに用いられる。本発明の複合体を、特定抗原(または各種抗原の組合せ)と一緒に含むワクチンは、被験体に投与されると、被験体を、例えばその抗原を原因とする疾患から保護する免疫応答を誘導できる。ワクチンは感染後に投与でき、疾患の進行を遅らせるか、もしくは停止するための治療(処置)ワクチンでもよい。好ましくは、それは被験体の初期感染を防止できる、予防性(予防)ワクチンである。
【0048】
さらには、本発明のタンパク質複合体または該複合体を含む細胞を含む(植物ベースの)ワクチン、ならびに有効量の該ワクチンを含む薬学的組成物を提供する。本発明のワクチンは、少なくとも二種類の抗原を含む本発明のタンパク質複合体を含む多成分ワクチンでよい。
【0049】
本発明のさらに別の局面では、本発明のタンパク質複合体の有効量の少なくとも一回投与量を被験体に投与すること、好ましくは経口投与することを含み、ここで対象となる分子が抗原である、特定病原菌に対する被験体の免疫応答を高めるための方法が提供される。この方法はまた、大型(構造)抗原を含む保護抗原を、宿主(植物)または宿主細胞組成物を摂食することにより、複合体を経口的に送り込み、細胞腸管内に在る免疫系に送達する道を開く。本明細書には、好ましい免疫化の経路を通し、本発明のワクチンを被験体に投与することを含む、粘膜(鼻、直腸または膣)免疫化のための方法も記載される。このように本発明は、キャリア分子をベースとするワクチンに取り込むことができる抗原性化合物のサイズおよび多様性を拡大する。
【0050】
実施例
実施例1:LTBサブユニットワクチン発現カセットの構築
バイナリー型植物発現ベクターpBINPLUS(Van Engelen et al., 1995)のT-DNA部分および本明細書で報告した全てのpLANTIGENワクチン構築体の遺伝子挿入物の図式的概観を図1に示す。全ての遺伝子は、塊茎だけに発現するためにクラスIパタチンプロモーター(Ppat)の制御下に置かれ、さらにERに保持されるためにそれぞれの融合タンパク質のC末端にKDEL(Lys-Asn-Gln-Leu)配列をコードするDNA配列を保有する(Munro and Pelham, 1987)。
【0051】
pL4:LTBに対する合成遺伝子(synLT-B)の設計および構築ならびにバイナリー型植物発現ベクターpLANTIGEN4(pL4)の発生は、以前に記載された(Lauterslager et al., 2001)。pL4は、LTBに対する合成遺伝子(synLT-B)を保有し、成熟LTBタンパク質をコードする配列の直後およびKDELをコードする配列の前にユニークなBamHI制限部位を有する。全てがLTBおよびカルボキシ末端のKDEL配列と同フレーム内に入るように全ての合成配列を作製し、このユニークな部位にクローニングした。
【0052】
pL12:pLANTIGEN12(pL12)にクローニングされたイヌパルボウイルス(CPV)エピトープをコードするフラグメントのコアは、アミノ酸配列
をコードする(Langeveld et al., 1994)。イヌパルボウイルス(CPV)のウイルスタンパク質VP2のアミノ末端領域をコードする合成BamHI/BglIIフラグメントを、pL4のユニークなBamHI部位にクローニングすることによって、pLANTIGEN12を作製した。以前に記載されたようなオリゴから得られたフラグメントの連結によって合成フラグメントを作製した(Florack et al., 1994)。オリゴは、Eurogentec(ベルギー)からのものであった。
【0053】
pL13:pLANTIGEN13(pL13)では、C末端膜貫通(TM)領域を欠くCSFV E2糖タンパク質をコードするフラグメントを連結した。野生型CSFVでは、E2糖タンパク質は膜貫通的に結合している。ブタコレラウイルス(CSFV)のE2成熟タンパク質をコードするBamHIフラグメントをpL4のユニークなBamHI部位にクローニングすることによって、pL13を構築した。CSFV E2をコードするフラグメントは、オリゴ
を使用したpPRb2(Hulst et al., 1993)のPCRによって得られた。
【0054】
pL15:pLANTIGEN15(pL15)では、二つのHAエピトープ配列と共にCPV配列を2回、間隔を空けるための2個のアラニン残基でそれぞれ分離してクローニングした。HAエピトープは、デカペプチドFERFEIFPKEをコードし、PR8 HA-1のアミノ酸111〜120に相当する(Hackett et al., 1985)。CPVは、線状B細胞エピトープであるが、HAはT細胞特異的である。pL12にクローニングしたものと類似した二つのCPVエピトープと共にインフルエンザウイルスヘマグルチニン(HA)H鎖の二つのデカペプチドからなる四量体配列をコードする合成フラグメントを、pL4のユニークなBamHI部位にクローニングすることによって、pL15を構築した。四つのエピトープ配列全てをそれぞれ2個のアラニン残基で分離するようにクローニングした。
【0055】
実施例2:宿主細胞の形質転換、成長およびタンパク質抽出
以前に我々は、16ロットの塊茎を生じたpL4遺伝子構築体を含有する22個の独立したトランスジェニックジャガイモ植物の発生について記載した(Lauterslager et al., 2001)。それ以来さらに形質転換実験を行い、それによってpL4遺伝子構築体を保有する追加の31ロットの独立した塊茎が産出した。
【0056】
本発明では、実施例1に記載されたバイナリー型発現ベクターおよびその組み合わせをエレクトロポレーションによりアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)AglO(Lazo et al., 1991)に導入し、ジャガイモデジレ(Desiree)カルティバー(De Z.P.C., Leeuwarden, The Netherland)の形質転換に使用した。トランスジェニックシュートの形質転換、成長、選択および塊茎の産生は以前に記載されている(Lauterslager et al., 2001)。pL12を用いたジャガイモデジレカルティバーの茎節間の形質転換によって、27個の独立したトランスジェニック植物が発生し、そのうち22個が温室内で塊茎を産生した。pL13を用いた形質転換によって23個の植物が発生し、そのうち20個が塊茎を形成した。pL15を用いた形質転換は31個の植物を産出し、そのうち20個が塊茎を産生した。
【0057】
典型的には、2〜4か月後に温室で成長した植物から200から300グラムの塊茎を収穫した。貯蔵および塊茎の齢が引き起こす効果を減らすために、ほぼ同サイズの新鮮収穫された塊茎材料から抽出液を作製した。ジャガイモ宿主細胞から生成したタンパク質複合体を単離するために、新鮮収穫した直径約5cmの塊茎を使用した。皮を除いた塊茎を薄切りしたものを25mMリン酸ナトリウム(pH6.6)、100mM NaCl、1mMエチレンジアミン四酢酸(EDTA)、50mMアスコルビン酸ナトリウム、1% TritonX-100および20mMメタ重亜硫酸ナトリウム中で抽出した。組織ホモジネートを4℃、12000rpmで5分間遠心分離し、上清を集め、新しい試験管に移した。標準としてウシ血清アルブミン(BSA)を使用したBradford法(Bradford, 1976)で全可溶性タンパク質を推定した。LTB五量体複合体の発現または蓄積をGM1-ELISAによって確認した(実施例3)。
【0058】
実施例3:レセプターとタンパク質複合体との結合の決定
修正ガングリオシドGM1酵素結合イムノソルベントアッセイ(GM1-ELISA)によって、トランスジェニックジャガイモ塊茎から単離した機能的キメラタンパク質複合体の量を推定した。マイクロタイタープレート(PolySorp Immunoplates, Nunc)に、リン酸緩衝生理食塩水(PBS)に溶かした5μg/mLウシ脳由来モノシアロガングリオシドGM1(Sigma Aldrich, St. Louis, USA)をコーティングした。コーティング後のプレートを脱イオン水で3回洗浄して前処置し、一定量、典型的には全可溶性抽出可能塊茎タンパク質5μgを負荷して、PBSに溶かした2%スキムミルク、0.1% ウシ血清アルブミンおよび0.1% Tween20を用いて室温にて1時間、100rpmで連続的に振盪しながらブロッキングした。大腸菌発現ベクターによって生成した組換えLTBを連続2倍希釈したものを、pBINPLUSPAT塊茎からの等量の全タンパク質を入れた各プレートに添加した。結合を4℃で16時間行った。脱イオン水でプレートを3回洗浄後に、LTB五量体に特異的なモノクローナル抗体VD12と共に1:1000希釈で室温にて1時間プレートをインキュベートした。脱イオン水で3回すすいだ後に、AP標識ヒツジ抗マウス抗体と共にプレートをさらにインキュベートし、結合した標識を4-ニトロフェニルリン酸(二ナトリウム塩六水和物、Janssen Chimica)で検出した。標準曲線の分析、濃度および標準偏差の計算のためにMicroplate Manager/PCバージョン4.0ソフトウェアを使用してBio-Rad Benchmark Microplate Reader(Bio Rad, Veenendaal, The Netherlands)を用いて405nmで検出を行った。単離した五量体を含むサンプルの読み取り値を、標準曲線からの結果と比較することによって五量体LTBの量を推定した。独立した実験で少なくとも2回サンプルを分析した。モル比を計算するために、塊茎の新鮮重量1gあたりの全タンパク質の量を7mg/gに調整した。融合タンパク質のモル重量を、それぞれのpLANTIGEN融合構築体によってコードされる成熟タンパク質の公知のアミノ酸配列から推定した。トランスジェニック塊茎に生成した五量体タンパク質複合体の平均発現レベルを表1に示す。これらのデータは、対象となる分子のサイズが大きいほど、機能的(すなわちGM1結合性)ホモ五量体タンパク質複合体の発現レベルが低くなることを示す。
【0059】
実施例4:LTBおよびLTB-CSFV E2を有するキメラタンパク質複合体
未改変サブユニットをコードするバイナリー型発現ベクターpL4(Lauterslager et al., 2001)およびサブユニットに融合した対象となる分子をコードするpLl3(LTB-CSFV E2融合体、概要については実施例1を参照されたい)を、本質的に以前に記載し(Lauterslager et al., 2001)、上記(実施例1)に概略を述べたように、アグロバクテリウム・ツメファシエンス介在性のジャガイモcv.デジレ(De Z.P.C., Leeuwarden, The Netherland)の形質転換を使用した共形質転換によりジャガイモに導入した。共形質転換を可能にするために、茎節間の感染前に、pL4およびpL13を保有する両方の組換えアグロバクテリウムを、OD595=1で1:1の比に混合した。混合後の細菌懸濁液を形質転換実験および形質転換細胞の再生に使用し、トランスジェニックシュートの選択は該の通りに行った。独立したトランスジェニックシュート71個を得て、53個を選択し、かつpL4および/またはpL13遺伝子構築体の存在について分析して、共形質転換の発生を明らかにし、それらをさらなる分析のために選択した。このために、尿素抽出緩衝液(62% 尿素、0.5M NaCl、70mMトリスHCl(pH8.0)、30mM EDTA(pH8.0)、1.5% サルコシル)50μl中で直径約5mmの円盤形の葉を摩砕することによって、個別の植物から集めた葉材料からゲノムDNAを単離した。等容のフェノール/クロロホルム(1:1)を添加して、サンプルを混合し、室温で15分間放置した。混合後に、サンプルを3000rpmで10分間遠心分離し、上清を新しい試験管に移し、4.4M酢酸アンモニウム(pH5.2)10μlを加えた。ゲノムDNAを沈殿させるためにイソプロパノール120μlを添加し、混合した。サンプルを1000rpmで3分間遠心分離して上清を除去した。残ったペレットを乾燥して、水または10mMトリスHCl(pH8.0)および1mM EDTAを含有する緩衝液に懸濁した。プライマー
を例とするがそれに限定されるわけではないpL4およびpL13遺伝子構築体を識別できる特異的プライマーを使用したPCRにマイクロリットル量のゲノムDNAサンプルを供した。増幅条件は、製造業者による最適条件でAccuTaqポリメラーゼ(Sigma-Aldrich)を使用し、それぞれ変性を可能にする94℃30秒間、56℃45秒間でのアニーリングに続く68℃2分間の伸長の40サイクルであった。PCR反応混合物を0.5×TBEでの1.2%アガロースゲル電気泳動に供し、予測サイズが周知である増幅された遺伝子構築体pL4および/またはpL13に対応するフラグメントの存在についてゲルをスキャンし、それらの公知の遺伝子配列ならびにプライマーLTB11およびTnosminus50の使用から推定された。植物22個は両遺伝子構築体を含んでいたが、28個はpL4のみを含み、顕著にはわずか3個がpL13遺伝子構築体を有した。本明細書においてさらにpL(4+13)8と名付けた第8番の植物または植物pL(4+13)16、pL(4+13)31、pL(4+13)39、もしくはpL(4+13)46のようにpL4およびpL13遺伝子構築体の両方について陽性であった22個の植物を、温室に移し、キメラタンパク質複合体の蓄積をさらに分析する目的で塊茎材料を産生させるために成熟するまで成長させた。
【0060】
塊茎を収穫し、GM1結合性LTB五量体の量を実施例2および3に記載したように評価した。数個の植物、例えば植物pL(4+13)15、pL(4+13)16、pL(4+13)39、pL(4+13)46、pL(4+13)60、pL(4+13)64およびpL(4+13)67は、GM1結合性LTB五量体のかなりの蓄積を示した(表2参照)。これらの植物をさらにキャラクタリゼーションした。pL(4+13)16およびpL(4+13)46のような選択された共形質転換されたジャガイモ植物の塊茎材料は、それぞれ塊茎の新鮮重量(FW)1グラムあたり約8および20マイクログラムでガングリオシドGM1と結合することから推定されるようにLTB5五量体を蓄積した(表2)。
【0061】
ウエスタンブロットによりキメラ植物pL(4+13)16をさらに評価した。このために、植物の塊茎抽出物の全タンパク質25マイクログラムを10% SDS-PAGEゲルに負荷し、半未変性(semi-native)の非還元条件下で運転してタンパク質複合体の分離を可能にした。半未変性SDS-PAGEの後に、抗LTB5コンホメーションのモノクローナル抗体であるVD12(図2A)または抗CSFV E2コンホメーションのモノクローナル抗体であるV3(図2B)のいずれかである特異的抗体を1:1000希釈で使用したその後のウエスタン分析のために、CAPS(10% エタノールに溶かした0.22% 3-[シクロヘキシルアミノ]-1-プロパン-スルホン酸(pH11))緩衝液中で標準法を使用してニトロセルロース上で、分離した後のタンパク質を転写した。ホースラディッシュペルオキシダーゼ標識二次抗体およびLumiLight基質を使用して特異的抗体の結合/認識を可視化して、LumiImager(Roche、Boehringer, Germany)で可視化した。対照として両方ともpL13遺伝子構築体を保有するpL1317および1331を使用した。pL1317は、20個の独立したpL13トランスジェニック植物の間でLTB-CSFV E2融合タンパク質についてのGM1結合性五量体の最高の発現体として以前に同定された。さらに、pL4遺伝子構築体を保有し、約17マイクログラム/gのFW GM1結合性LTB五量体を蓄積しているpL417およびGM1結合性五量体についての陰性対照を使用した。
【0062】
図2Aは、コンホメーションの抗LTB5モノクローナル抗体VD12を使用してこれらの植物の全タンパク質抽出物25マイクログラム量を分析した結果を、図2Bは、コンホメーションのCSFV E2に特異的なV3を用いた分析の際の結果を描いたものである。共形質転換された植物pL(4+13)16(レーン1;pL4およびpL13で共形質転換)、タンパク質分子量マーカー(レーン2)、PAT4を保有する対照抽出物、空の発現カセットベクター(レーン3)、両方ともLTB-CSFV E2遺伝子融合体のみを保有するpL1331(レーン4)およびpL1317(レーン5)の抽出物、ならびにpL4で形質転換され未改変rec-LTBサブユニットのみを蓄積するpL(4)17(レーン6)を負荷した。
【0063】
図2から、pL(4+13)16はホモ五量体LTB5よりもサイズが有意に大きく(レーン1と6とを比較)、VD12およびV3の両方のコンホメーション特異的モノクローナル抗体によって認識される(AおよびBのレーン1を比較)かなりの量のタンパク質複合体を蓄積することが明白である。これは、VD12によって認識される五量体LTB5の構造およびV3によって認識される抗原性CSFV E2二量体エピトープを保有するキメラタンパク質複合体の存在を実証している。重要には、後者の抗原性CSFV E2二量体エピトープは、pL13にほとんど存在せず(レーン4および5)、pL13はLTB-CSFV E2融合タンパク質のみを有し、未改変LTBサブユニットを有さない。さらに、約300kDaのホモ五量体LTB-CSFV E2複合体は、VD12およびV3を用いた分析後にほとんど目視不可能である。考えられるところでは、これは以前に観察されたようにその非常に低い発現レベルが原因である。図2およびGM1 ELISAから、植物pL(4+13)16に存在するようにLTB(pL4)およびLTB-CSFV E2(pL13)の両方を含むキメラ複合体だけが、GM1結合性質および抗原性CSFV E2エピトープの両方を保有するかなりの量(FW塊茎1グラムあたり8マイクログラム)のタンパク質複合体の蓄積を促進することも明白である(図2AおよびB、レーン1)。対照的に、例えば植物pL1317のようにLTB-CSFV E2遺伝子融合体のみを発現する植物は、GM1細胞表面レセプターと結合可能な抗原性タンパク質複合体を形成しない(図2AおよびBのレーン1および4を比較)。
【0064】
GM1のELISA(表2)によると全てがかなりの量の機能的LTB5を蓄積するキメラ植物pL(4+13)16、pL(4+13)31、pL(4+13)39、pL(4+13)46、pL(4+13)60、pL(4+13)64およびpL(4+13)67を、すでに記載したようなウエスタンブロットによってさらに評価した。図3Aは、VD12と共にインキュベーションした後の結果を示し、LTB5のみを含有するpL4(21)(レーン2)と(LTB-CSFV E2)5を含有するpL13(17)(レーン3)との間の位置に泳動することから明白なように、pL(4+13)16からpL(4+13)67の抽出物中にLTB5を有する多数のキメラ複合体が存在することを表す。予想通りこれらのキメラ複合体は、CSFV E2に特異的なV3 mAbとも反応する(パネルB、図3)。図3Bから、pL13(17)(レーン2)に類似した量の全タンパク質を負荷したが、キメラ植物からの抽出物(レーン6〜10)がV3 mAbとより強く反応して、GM1結合性LTB5複合体への機能的CSFV E2二量体の蓄積をさらに強調していることもはっきりしている。
【0065】
実施例5:LTBおよびLTB-パルボを有するキメラタンパク質複合体
バイナリー型発現ベクターpL4(LTB)およびpL12(LTB-CPVパルボ;概観については図1および実施例1参照)を、実施例4に本質的に記載したようにジャガイモcv.デジレのアグロバクテリウム・ツメファシエンス介在性形質転換を使用した共形質転換によって、両方ともジャガイモに導入した。茎節間の感染前に、pL4またはpL12のいずれかを保有する組換えアグロバクテリウムをOD595=1で1:1の比で混合し、混合した細菌懸濁液を形質転換実験に使用して共形質転換を可能にした。45個を超える独立したトランスジェニックシュートを選択し、pL4および/またはpL12遺伝子構築体の存在について分析して、共形質転換の発生を明らかにし、それらをさらなる分析のために選択した。20個のトランスジェニック植物は、両遺伝子構築体を含有すると思われたが、19個の植物はpL4のみを有し、6個の植物はpL12のみを有した。塊茎を形成させるために20個の二重形質転換体全てを温室内で成長させ、塊茎を収穫し、GM1結合性LTB五量体の量を立証した。選択されたpL(4+12)植物の一覧を表3に示す。
【0066】
VD12およびイヌパルボウイルスに特異的なmAbである3C9を使用したウエスタンブロット(図4)は、pL(4+12)19、pL(4+12)20、pL(4+12)23、pL(4+12)31、pL(4+12)41、pL(4+12)46、pL(4+12)51、pL(4+12)52、pL(4+12)57、pL(4+12)62およびpL(4+12)65のようないくつかの植物がLTB5(pL4、レーン2)およびLTB-CPV5(pL12、レーン3)の間にSDS-PAGEゲル上を泳動するキメラタンパク質複合体を含有することを示した。図4AおよびBから、pL(4+12)31、pL(4+12)57およびpL(4+12)62のようないくつかの植物のタンパク質抽出物がVD12(抗LTB5)および3C9(抗パルボ)の両方によって認識された多数のバンドを現したことが明白であり、これは、例えば未改変LTBサブユニット4個およびLTB-CPV融合サブユニット1個またはLTB3個およびLTB-CPV2個、またはLTB2個およびLTB-CPV3個またはLTB1個およびLTB-CPV4個のいずれかを含有するキメラ複合体が存在することを示す(図4AおよびB)。pL(4+12)57の抽出物では特にホモ五量体LTB5およびホモ五量体(LTB-パルボ)5以外に4個のキメラ複合体が存在することがはっきりしている。
【0067】
実施例6:LTB、LTB-パルボおよびLTB-iippを有するキメラタンパク質複合体
バイナリー型発現ベクターpL4(LTB)、pL12(LTB-CPV パルボ)およびpL15(LTB-iipp:iipp=インフルエンザ-インフルエンザ-パルボ-パルボ、二つのパルボエピトープと結合した二つのインフルエンザエピトープ、概要については実施例1参照)を、本質的に上記のようにジャガイモcv.デジレのアグロバクテリウム・ツメファシエンス介在性形質転換を使用した共形質転換によって、ジャガイモに導入した。独立したトランスジェニックシュート29個を選択し、pL4、pL12およびpL15遺伝子構築体の存在について分析して、共形質転換の発生を明らかにした。これらの植物をさらなる分析にために選択した。トランスジェニック植物5個は三つの遺伝子構築体全てを含有すると思われたが、10個は、pL(4+12)、pL(4+15)またはpL(12+15)のいずれかである二つの遺伝子構築体を含有した。残りの14個の植物はわずか一つの遺伝子構築体を含有した。二つ以上の遺伝子構築体を保有する全ての植物を温室に移し、成熟するまで成長させた。5個の三重形質転換体pL(4+12+15)7、pL(4+12+15)9、pL(4+12+15)11、pL(4+12+15)16およびpL(4+12+15)19を、GM1 ELISAならびにVD12および3C9 mAbを使用したウエスタンブロットにより分析した(図5)。
【0068】
図5A左欄から、三つの遺伝子構築体全てを有するこれら5つの植物(レーン6〜10)が、pL(4)21(レーン3)およびpL(12)01(レーン4)の間またはpL(12)01(レーン4)およびpL(15)16(レーン5)の間に泳動する複合体を含有することが明らかであり、これはLTB、LTB-パルボおよび/またはLTB-iippの多様な組み合わせであることを示唆している。図5A右欄から、これらの複合体がmAb 3C9と正の反応をすることが明白であり、これは少なくとも一つのパルボエピトープが存在することを表している。図5Bから、特にpL(4+12+15)7(レーン6)およびpL(4+12+15)11(レーン8)がLTB-パルボおよびLTB-iippの両方を含有することがはっきりとしている。よって、これらの植物ではLTB、LTB-パルボおよびLTB-iippを含有するキメラGM1結合タンパク質複合体が一緒に産生する。
【0069】
実施例7:大腸菌LTBサブユニット発現カセットの構築
本発明に記載のタンパク質複合体を、様々な種類の組換え宿主細胞に産生させることができる。実施例7および8は、微生物である大腸菌における少なくとも一つの未改変サブユニット、およびこの場合はレポーター分子GFPである対象となる分子に融合した少なくとも一つのLTBサブユニットを有するキメラタンパク質複合体の産生を記載する。
【0070】
大腸菌および他の原核生物での発現のために、LTBについて本来の野生型大腸菌の配列(EtxB)を使用した。ブタ大腸菌株(Dallas and Falkow, 1980; Leong et al., 1985)からもともと単離されたヌクレオチド配列ECELTBP(SWISS-PROT P32890)に大きく類似するLTBコード配列を、pYA3047(Jagusztyn-Krynicka et al., 1993)からクローニングした。プライマー
およびテンプレートとして野生型EtxB配列を有するベクターを使用したPCRによってフラグメントを増幅させた。生じたBpiI/EcoRIを、NcoI/EcoRIで消化したpET21dベクター(Novagen)にクローニングしてpET-wiLTB1を発生させた。ヌクレオチド配列の確認後に、生じたクローンを、発現の研究のために形質転換によりTOP10F'細胞(Invitrogen)に導入した。プライマー
を使用したPCRによりGFP配列末端のBamHI/BpiI部位に導入することによってLTB-GFP融合タンパク質を作製して、BamHIで消化したpET-wiLTBにクローニングした。ヌクレオチド配列の確認後に、発現の研究のために生じたクローンpET-wiLTB-GFP2を形質転換によってTOP10F'細胞に導入した(実施例8)。
【0071】
実施例8:大腸菌細胞の形質転換、成長およびタンパク質抽出
二つの選択されたクローン、pET-wiLTB1およびpET-wiLTB-GFP2を形質転換により大腸菌Rosetta株(Novagen)に導入して、50mg/Lアンピシリンおよび34mg/Lクロラムフェニコールを補充したLB培地中で37℃で一晩成長させた。o/n培養物2mLを新鮮培地50mLに入れて希釈し、OD595が0.2に達するまで連続的に振盪しながら20℃で成長させた。IPTGを終濃度1mMになるまで添加して発現を誘導し、培養物をOD595=0.5になるまでさらに成長させた。4℃で14000rpmで10分間遠心分離することによって細胞を集め、ペレットをBugBuster(Novagen)抽出試薬に再懸濁し、終濃度1μg/μlになるまでリゾチームを添加した。サンプルを室温で5分間インキュべートして、Benzonaseを添加して室温でさらに15分間インキュベートした。サンプルをその後4℃で14000rpmで5分間遠心分離した。LTB5を精製するためにpET-wiLTBの上清を固定化D-ガラクトースカラムに負荷した(実施例10も参照)。溶出プロフィルを図6に示す。図6Aから、溶出緩衝液の適用直後にそれぞれの画分中のタンパク質を測定できることが明白である。図6Bから、GM1 ELISAから明白なようにこれらの画分がGM1結合性LTB5複合体を含有していることがはっきりしている。RecLTB-GFPは、封入体に蓄積すると思われた。封入体を含有する生じたペレットを、本質的にSambrookら(1989)が記載したものをKhouryおよびMeinersmann(A genetic hybrid of the Campylobacter jejuni flaA gene with Escherichia coli and assessment of the efficacy of the hybrid as an oral vaccine. Avian disease 39 (1995) 812-820)に従ってやや修正してさらに処理した。封入体を標準法を使用して単離し、8M尿素で抽出した。尿素可溶性画分を0.1Mトリス(pH7.4)で透析し、沈殿した画分を10000gで10分間遠心分離することによって除去した。GM1結合性複合体を含有する上清をD-ガラクトースを用いたアフィニティクロマトグラフィーによってさらに精製して、精製recLTB-GFPを得た。
【0072】
実施例9:LTBおよびLTB-GFPキメラ複合体の産生
少なくとも一つの未改変LTBサブユニットおよび少なくとも一つのLTB-GFP融合サブユニットを有する、本発明のキメラ複合体を産生させる代替法は、未改変サブユニットを有する第一組成物および融合サブユニットを有する第二組成物を提供すること、その後に両組成物を1:4、2:3、3:2または4:1モル比で混合することを伴う。次に、両種の(すなわち未改変および改変)サブユニットの混合物を、五量体誘導条件で変性および再生する。例えば、実施例7および8に記載されたように組換え大腸菌2株に産生され、D-ガラクトースを用いたアフィニティクロマトグラフィーで精製されたrecLTBおよびrecLTB-GFPを8M尿素に溶かした。または、精製recLTBをrecLTB-GFPを有する封入体に添加して、上記のようにさらに処理することもできる。尿素可溶性画分を0.1Mトリス(pH7.4)で透析して、不溶性材料を4℃で10000gで10分間遠心分離することによって除去できる。または、0.3M D-ガラクトースをトリス緩衝液に添加して五量体化を促進できる。キメラLTBおよびLTB-GFP分子を含む五量体複合体を含有する上清を、例えば固定化D-ガラクトースを用いたアフィニティクロマトグラフィーによってさらに精製できる。タンパク質複合体のキメラサブユニット組成物を上記のような半未変性SDS-PAGEおよびウエスタンブロットによって検証できる。GM1との結合を上記のようにGM1-ELISAを使用して決定できる。
【0073】
実施例10:LTBサブユニット担体複合体の精製
ホモ五量体性RecLTB5およびLTB-サブユニット担体の複合体を、Uesakaら(Uesaka et al. (1994) Simple method of purification of Escherichia coli heat-labile enterotoxin and cholera toxin using immobilized galactose. Microbial Pathogenesis 16 : 71-76)に従って固定化D-ガラクトースアガロース(Pierce、カタログ番号20372)を用いたアフィニティクロマトグラフィーによって精製した。
【0074】
実施例7に記載したように微生物LTB発現カセットを備える大腸菌細胞を、実施例8に記載したように成長させ、4℃で6500gで30分間遠心分離することによって集め、超音波処理して、記載されたようにさらに処理した(Uesaka et al., 1994)。超音波処理した大腸菌の上清を、50mMトリスHCl(pH7.4)、0.2M NaCl、1mM EDTAおよび20mMメタ重亜硫酸ナトリウム(TEAN)に調整し、FPLC(BioRad, Veenendaal, The Netherlands)に取り付けたD-ガラクトースカラムに負荷した。カラムをTEAN緩衝液で洗浄し、当業者に公知の標準法により0.3M D-ガラクトースまたは0.5M D-ガラクトースのいずれかを含有するTEAN緩衝液で溶出させた。図6は、大腸菌からのrecLTBの精製結果を示す。画分を集め、GM1結合性LTBの存在を実施例3に記載するようにGM1-ELISAによって決定した。図6Aに示すように大腸菌から精製したrecLTBについての結果を図6Bに示す。GM1-ELISAから明白なように、タンパク質について陽性の全ての画分はGM1結合性複合体を含有した。固定化D-ガラクトースを用いた大腸菌からのrecLTBの精製についての溶出プロフィルおよび対応する画分のGM1 ELISAによるGM1結合活性の存在の分析を図6に表示する。
【0075】
次に、D-ガラクトースクロマトグラフィーによってpL4(21)由来塊茎抽出物の上清からタンパク質複合体を精製した(図7)。図7Aおよび7Bから、そのような複合体を塊茎からも精製できることがはっきりしている。
【0076】
次に、キメラタンパク質複合体をpL(4+13)植物から精製した。実施例2に記載したように抽出緩衝液中で塊茎材料を摩砕することによってpL(4+13)46の塊茎材料から抽出物を調製し、抽出物を4℃で6500gで30分間遠心分離して不溶性材料およびデンプン粒を除去した(図8)。上清をTEAN緩衝液条件に調整し、D-ガラクトースカラムに負荷し、記載したようにさらに処理した。図8から、画分5でタンパク質がカラムから溶出し始め、画分10〜11に最大吸光度となることがはっきりしている。GM1-ELISAから、VD12 mAbで検出されたように大部分のLTBは画分5〜15に存在し、画分10で最大であることも明白である(図8B)。さらに、第二抗体がコンホメーションの抗CSFV E2 mAb V3である修正GM1 ELISAから明白なように、これらの画分はCSFV E2エピトープについても陽性であること(図8C)は、そのような複合体にLTBおよびLTB-CSFV E2の両方が存在することを示し、これはウエスタンブロットから得られた以前の結果と一致している(図3)。
【0077】
実施例11:LTBおよびLTB-VHSV Gのキメラ複合体
LTBと、配列X66134(EMBL)に類似しLorenzenら(Molecular cloning and expression in Escherichia coli of the glycoprotein gene of VHS virus, and immunization of rainbow trout with the recombinant protein. J. Gen. Virol. 74 (1993) 623-630)によって発表されたウイルス性出血性敗血症ウイルス由来スパイク糖タンパク質Gとの遺伝子融合体を以下のように作製する:VHSV G配列およびプライマー
のPCR増幅によってユニークなBamHI部位を導入する。PCR条件は、Pwoポリメラーゼを使用して96℃30秒間、60℃30秒間および72℃45秒間を30サイクルである。得られたフラグメントをBamHIおよびSmaIで切り出し、LTBに対する合成遺伝子を保有するpLANTIGEN4のユニークなBamHI部位にフレーム内にクローニングする。パタチンプロモーターおよびノパリンシンターゼターミネーター配列の制御下の生じた遺伝子配列をpLANTIGEN24(pL24)と名付ける。pL4およびpL24でジャガイモの共形質転換を行い多数のpL(4+24)植物を発生させる。pL4および/またはpL24遺伝子構築体の存在を以前に記載したようなPCRによって確認する。両遺伝子構築体について陽性の植物に塊茎を形成させる。塊茎を収穫し、GM1 ELISAを使用してGM1結合複合体に関して分析する。複合体にVHSV Gタンパク質が存在することを、モノクローナル抗体IP1H3、3F1H10および3F1A2(Lorenzen et al., 2000. Three monoclonal antibodies to the VHS virus glycoprotein: comparison of reactivity in relation to differences in immunoglobulin variable domain gene sequences. Fish & Shellfish immunology 10: 129-142)と共にインキュベートすることによって確認する。半未変性条件下で、抗LTB5 mAbであるVD12および1P1H3、3F1H10および3F1A2を使用して行ったSDS-PAGEで塊茎抽出物のウエスタンブロットによって、キメラ複合体をさらにキャラクタリゼーションできる。
【0078】
実施例12:LTBおよびLTB-SVCV Gのキメラ複合体
コイ春ウイルス血症ウイルスの完全SVCV G遺伝子(Genbankアクセッション番号NC002803)を有しLTBとの遺伝子融合体を作製するためのBamHI/BglIIフラグメントを、オリゴヌクレオチド
を使用して増幅させた。生じたフラグメントをBamHIおよびXhoIで切り出し、LTB;pをコードするpL4のBamHI/XhoI部位にクローニングすることによってpLANTIGEN27(pL27)を発生させた。塊茎特異的パタチンプロモーター(pLANTIGEN27)の制御下に置かれた生じた遺伝子を、ジャガイモの形質転換のためにA.ツメファシエンスに導入した。pL4およびpL27の共形質転換を行い、記載したようなPCRによって両遺伝子構築体の存在についてトランスジェニック植物を評価した。pL(4+27)遺伝子構築体を保有するトランスジェニック植物を選択し、温室内で成熟するまで成長させた。GM1 ELISAによりGM1結合性複合体の蓄積について、および特異的mAbを使用してSVCV Gタンパク質の存在について塊茎を分析した。記載したような半未変性SDS-PAGE後のウエスタンブロットによって、タンパク質複合体のサブユニット組成を可視化した。
【0079】
実施例13:LTBおよびLTB-ClyIIAのキメラ複合体
アクチノバチルス・プリューロニューモニエ(Actinobacillus pleuropneumoniae)血清型9参考株CVI13261を、0.1% V因子(NAD)を含有するハート・インフュージョン寒天培地(Difco)上で成長させる。プロテイナーゼK/SDS溶解によって高分子量DNAを単離した後に、フェノール/クロロホルム抽出を行い、得られたゲノムDNAを沈殿させる。アクチノバチルス・プリューロニューモニエ血清型9(GenBank-EMBLアクセッション番号X61111)由来ClyIIA遺伝子を、オリゴヌクレオチド
を使用したPCRによって血清型9株から単離したゲノムDNAからクローニングする。次に、PCRによってBamHI部位もClyIIA遺伝子のアミノ末端に導入し、BamHIで切り出して生じたフラグメントを、大腸菌での発現について記載したようにpET-wiLTBにクローニングし、pET-wiLTB-ClyIIAを発生させた。IPTGで誘導した際にpET-wiLTBおよびpET-wiLTB-ClyIIAの共発現によってキメラ複合体を得ることができる。または、大腸菌でのpET-wiLTB-ClyIIAの過剰発現の際に得られた封入体および精製recLTBを混合して8M尿素で可溶化し、記載したようにトリス緩衝液で透析して五量体複合体を再生させる。
【0080】
実施例14:LTB、LTB-H5およびLTB N1を有する多価タンパク質複合体
共に植物での発現に関して最適化されたA型インフルエンザウイルスH5N1亜型ヘマグルチニン(Genbankアクセッション番号AF028709)およびノイラミニダーゼ(AF028708; Claas et al. (1998) Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet 351 (9101), 472-477)に対する合成遺伝子を作製する。H5およびN1配列のアミノ末端およびカルボキシ末端にBamHI部位を導入し、pLANTIGEN4遺伝子構築体のユニークなBamHI部位に合成H5遺伝子またはN1遺伝子を有するそれぞれのBamHIフラグメントをクローニングすることによって遺伝子融合体を作製する。LTB-H5およびLTB-N1融合タンパク質をコードする生じた遺伝子を実施例1に記載したようにパタチンプロモーターの後にクローニングする。LTB、LTB-H5およびLTB-N1の共形質転換を行い、前に記載したようにPCRによってそれぞれの遺伝子の存在についてジャガイモ植物を分析する。三つ全ての発現カセットについてトランスジェニックである植物を単離して、成熟するまで温室内でさらに成長させることができる。GM1結合複合体の存在についてGM1-ELISAならびにVD12とH5およびN1特異的抗体とを使用したウエスタンブロットによって塊茎を分析する。別の態様では、LTB-H5またはLTB-N1のいずれかを保有する植物を発生させ、それぞれの複合体の蓄積について分析する。塊茎からrecLTB-H5およびrecLTB-N1を精製でき、精製recLTBとそれぞれ1:1:3、1:2:2、1:3:1、2:1:2、2:2:1または3:1:1の比で混合して、溶解し、8M尿素を使用して変性させ、実施例9に記載したようにさらに処理してキメラ複合体を発生させることができる。
【0081】
(表1)GM1 ELISAから得られたように、構築体pLANTIGEN4、12、13および15のいずれか一つを保有するトランスジェニックジャガイモ塊茎で産生したホモ五量体タンパク質複合体のモル発現データの平均とそれぞれの標準偏差および共分散との比較。構築体間の対の差を、分散を安定化するためにデータを対数変換した後でANOVAを使用して解析した。発現レベルをmMで表す。最終列は標準偏差(Sd)を示す。
【0082】
(表2)GM1 ELISAから得られたようにpL4およびpL13を有するキメラタンパク質複合体を保有するトランスジェニックジャガイモ塊茎の発現データの比較。トランスジェニック性質ならびにpL4および/またはpL13遺伝子構築体の存在(第二列および第三列)は、実施例4に記載するように個別の植物から単離したゲノムDNAのPCRによった。記載したようにVD12およびV3 mAbを使用した半未変性ウエスタンブロット分析の結果を比較すること、ならびに図2(第四列)に示すように高分子量複合体の存在をスクリーニングすることにより、キメラの存在を立証した。GM1 ELISAによって決定したように塊茎新鮮重量1グラムあたりのGM1結合性LTB5部分のマイクログラムで発現データを表現する(第五列)。比較のために、pL4遺伝子構築体のみを有する塊茎は平均10〜15μg/g FW LTB5を含有したが、pL13遺伝子構築体の最高の発現体であるpL13(17)は1μg/g FW塊茎未満を有した。
【0083】
(表3)GM1 ELISAから得られたようにpL4およびpL12からなるキメラタンパク質複合体を保有するトランスジェニックジャガイモ塊茎の発現データの比較。トランスジェニック性質ならびにpL4および/またはpL12遺伝子構築体の存在(第二列および第三列)は、実施例5に記載するように個別の植物から単離されたゲノムDNAのPCRによった。記載したようにVD12および3C9 mAbを使用した半未変性ウエスタンブロット分析の結果を比較すること、ならびに図2に示すように高分子量複合体の存在をスクリーニングすることにより、キメラの存在を立証した。GM1 ELISAによって決定したように塊茎新鮮重量1グラムあたりのGM1結合性部分のマイクログラムで発現データを表現する。
【0084】
参考文献
【図面の簡単な説明】
【0085】
【図1】異なるLTBおよびLTBサブユニットワクチン遺伝子構築体を含有する、バイナリーベクターpLANTIGEN4、12、13および15のT-DNAを示した図。LB、左側T-DNA境界配列;PNOSノパリン合成酵素プロモータ;NPTII、ネオマイシンホスホトランスフェラーゼII遺伝子、選択可能カナマイシン耐性マーカー;TNOS、ノパリン合成酵素ターミネータ配列;PPAT、クラスIパタチンプロモータ;遺伝子、PPATプロモータ制御下での発現に関するクローニング部位;RB、右側T-DNA境界配列;SP、シグナルペプチド;LT-B、植物での発現に最適化されたLTBの合成遺伝子;KDEL、小胞体保持シグナル;パルボ、イヌパルボウイルス(CPV)エピトープ;Ala、アラニン;インフルエンザ、HAインフルエンザウイルス血球凝集素エピトープ;CSFV E2、膜貫通ドメインを欠くブタコレラウイルスE2糖タンパク質。
【図2】LTB5コンホメーションのモノクローナル抗体VD12(A)およびCSFV E2コンホメーションのmAb V3(B)を用いた、塊茎抽出物(各総タンパク質25マイクログラム)のウエスタンブロット分析。レーン1、pL(4+13)16;レーン2、タンパク質サイズマーカー;レーン3、コントロール抽出物;レーン4、pL13(31);レーン5、pL13(17)およびレーン6、pL4(17)。Aの左側矢印は、五量体複合体を含有するLTB5を示し、Bの矢印は複合体を含有するCSFV E2コンホメーションのエピトープを示す。
【図3】条件。Aでは、左側のブロットをVD12(抗LTB5)とインキュベーションし、またBでは右側ブロットをV3(抗CSFV E2)とインキュベーションした。サンプルpL(4+13)16(レーン4)およびpL(4+13)46(レーン7)について25マイクログラムを負荷した以外は、それぞれに50マイクログラムの全塊茎タンパク質を負荷した。レーン1、野生型塊茎抽出物;レーン2、pL(4)21抽出物;レーン3、pL(13)17抽出物;レーン4、pL(4+13)16抽出物;レーン5、pL(4+13)31抽出物;レーン6、pL(4+13)39抽出物;レーン7、pL(4+13)46抽出物;レーン8、pL(4+13)60抽出物;レーン9、pL(4+13)64抽出物およびレーン10、pL(4+13)67抽出物。レーン2の左側の矢印は、下側矢印がホモ五量体(LTB)5を示し、上側矢印がホモ五量体(LTB-CSFV E2)5を示している。右側矢印は、本発明のキメラ複合体を示している。分子サイズマーカーを左側に示した。
【図4】半未変性条件の下、12%SDS-PAGEゲルによるpL(4+12)植物塊茎抽出物(各25マイクログラム)のウエスタンブロットの図。A、左側ブロットはVD12(抗LTB5)とインキュベーションし、B、右側ブロットは3C9(抗パルボ)とインキュベーションした。レーン1、M、フルレンジレインボウ(full range rainbow)分子量マーカー;レーン2、pL4(pL417)抽出物;レーン3、pL12(pL(12)01)抽出物;レーン4、pL4抽出物とpL12抽出物の1:1混合物(pL(4)17とpL(12)02);レーン5、V、PAT4ベクター陰性コントロール;レーン6〜10、それぞれpL(4+12)16、pL(4+12)23、pL(4+12)51、pL(4+12)52およびpL(4+12)57の抽出物。左側の矢印は、LTBおよびLTB-パルボ(LTB-P)を表し、右側の矢印は本発明のキメラ複合体を示す。左側の実線ダッシュはタンパク質マーカーを示す。レーン3の、LTBより若干高い位置に移動した下側のバンドは、負荷緩衝液中でサンプルを加熱した際に生じたLTB-Pの分解産物である。
【図5】半未変性条件下で10% SDS-PAGEゲル(A)、または還元条件下で15% SDS-PAGE(B)によるpL(4+12+15)植物塊茎抽出物のウエスタンブロット。A、左ブロットは、VD12(抗LTB5)とインキュベーションし、右ブロットは3C9(抗パルボ)とインキュベーションした。B、ブロットは3C9(抗パルボ)とインキュベーションした。それぞれレーンあたり25マイクログラムの全塊茎タンパク質抽出物を負荷した。レーン1、分子量標準物質;レーン2、野生型塊茎抽出物;レーン3、pL421抽出物;レーン4、pL1201抽出物;レーン5、pL1516抽出物;レーン6〜10、pL(4+12+15)塊茎抽出物;レーン6、pL(4+12+15)7;レーン7、pL(4+12+15)9;レーン8、pL(4+12+15)11;レーン9、pL(4+12+15)16およびレーン10、pL(4+12+15)19抽出物。A、左側矢印は、下矢印がLTB5を;中矢印が(LTB-パルボ)5を、上矢印が(LTB-iipp)5を示す。右の矢印は、キメラ複合体を示す。B、左、下矢印はLTB-パルボを、上矢印はLTB-iippを示す。
【図6】固定化Dガラクトース(Pierce)を用いたアフィニティークロマトグラフィーによる大腸菌recLTBの精製を示す図。カラムを6容積量のTEAN緩衝液で徹底的に洗った後、遠心分離によって集めた、超音波処理大腸菌細胞の上清を、FPLC装置に装着した5cmのDガラクトースカラムに負荷した。プロテアーゼインヒビターカクテル(Roche)を加えた47.5mLのTEAN緩衝液に溶解した大腸菌からの粗タンパク質抽出物を、0.5mL/分の流速でカラムに負荷した。3倍容積量(142.5mL)のTEAN緩衝液(50mM トリス-HCl (pH7.4);0.2M NaCl;1mM EDTA)を0.5mL/分の流速で洗浄した。溶出液を、0.5M Dガラクトースを追加した洗浄緩衝液で、0.5mL/分の流速で洗浄した。1mLの画分を集め、収集後すぐに氷上に置いた。検出器は0.1 A.U.に設定し、記録計の速度は0.25cm/mLであった。A. 溶出プロフィール。実線矢印は、0.5M Dガラクトース添加TEAN緩衝液の溶出開始点を示す。記録下の実線は、画分番号16〜22(主ピーク)を表す。B. GM1-ELISAプール画分1〜32(各1mL)。LTB5の量(ng/μl画分)は、GM1 ELISAによって決定した。C. クマジー染色されたゲル画分17〜22。プレキャスト10%SDS-PAGEゲル(Bio-Rad)を、還元条件下で行なった。レーンM、分子サイズマーカー;レーンE、10μl粗抽出物(カラム負荷前);レーン17〜22、各画分10μl。サンプルは、負荷前に3分間煮沸し、泳動は標準的な条件で行われた。ゲルをクマジーブリリアントブルーで一晩染色し、0.3% Tween-20で脱色した。左側矢印は、LTBモノマーを表しており、右側にサイズを表す(kDa)分子量マーカーが示してある。
【図7】固定化Dガラクトース(Pierce)を用いたアフィニティークロマトグラフィーによる、38グラム新鮮重量pL421植物塊茎からのrecLTBの精製を示す図。塊茎の皮を剥き、小片に細断してから70mLの冷抽出緩衝液を加え、ステンレス鋼製ブレンダーで粉砕および抽出を実施した。粗タンパク抽出物およびrecLTBを含有する上清を5分間、15300rpm、4℃で遠心分離して粒子およびデンプン顆粒を取り除いた。上清が完全に清澄になるまで遠心分離工程を繰り返し、残った47mLの粗抽出物をカラムに負荷した。負荷は0.5mL/分で行い、カラムを4℃に冷却した。負荷後、カラムを42mLのTEAN緩衝液で、0.75mL/分の速度で洗い、溶出液を0.3MのDガラクトースを添加した(0.5Mの代わりに;合計26mL)TEAN緩衝液で洗浄した。画分を集めた。画分サイズは0.75mLで、画分は収集後すぐに氷上に置いた。検出器は0.2 A.U.に設定し、記録計の速度は0.25cm/mLであった。A. 溶出プロフィール。実線矢印は、0.3M Dガラクトース添加TEAN緩衝液の溶出開始点を示す。記録下の実線は、0.75mLの画分番号1〜15を示す。主ピークは画分10〜11にあった。B. GM1-ELISA画分1〜21(各0.75mL)。LTB5の量(ng/画分)は、GM1 ELISAによって決定した。
【図8】6.5グラムの凍結乾燥したpL(4+13)46植物の塊茎からの、固定化Dガラクトース(Pierce)を用いたアフィニティークロマトグラフィーによるキメラタンパク質複合体pL(4+13)の精製を示す図。抽出の前に、凍結乾燥した塊茎材料に、水を26mL、続いてプロテアーゼインヒビターカクテル(Roche)を添加した抽出緩衝液を65mL加えた。抽出は、氷上で22分間連続振盪しながら行い、抽出物は80μMのメッシュ布を使って濾過し、4℃、10分間、15300rpmの遠心分離を2回行って、残ったデンプン顆粒を除去した。残った57mLの、清澄な粗抽出液をカラムに負荷した。負荷は0.5mL/分で行い、カラムを4℃に冷却した。負荷後、カラムをTEAN緩衝液で、0.75mL/分の速度で61分間洗浄し、溶出液を0.3MのDガラクトースを添加したTEAN緩衝液で洗浄した。画分を集めた。画分サイズは0.75mLで、画分は収集後すぐに氷上に置いた。検出器は0.2 A.U.に設定し、記録計の速度は0.25cm/mLであった。A. 溶出プロフィール。実線矢印は、0.3M Dガラクトース添加TEAN緩衝液の溶出開始点を示す。記録下の実線は、0.75mLの画分番号1〜15を表す。主ピークは画分10〜11にあった。B. 標準GM1-ELISA画分5〜19(各0.75mL)。LTB5の量(ng/画分)は、GM1 ELISAによって決定した。C.修飾GM1-ELISA画分5〜19。結合の検出は、CSFV E2に特異的なV3 mAbおよびアルカリホスファターゼ標識ヒツジ抗マウスIgG(VD12、抗LTB5の代わりに)を用いて行った。
【特許請求の範囲】
【請求項1】
少なくとも二つの、好ましくは同一のサブユニットを含むタンパク質複合体であって、そのうちの少なくとも一つのサブユニットは未改変であり、また少なくとも一つのサブユニットは対象となるの第一分子に融合しており、該サブユニットを介して細胞表面レセプターと相互作用できる該タンパク質複合体。
【請求項2】
対象となる第一分子が、好ましくは共有結合により、対象となる第二分子と結合して対象となる多量体を形成できる、請求項1記載のタンパク質複合体。
【請求項3】
複合体が、本質的に大腸菌(E. coli)の易熱性エンテロトキシン(LT)またはコレラ菌(Vibrio cholerae)のコレラトキシン(CT)、好ましくはそのBサブユニットに基づいている、請求項1または2記載のタンパク質複合体。
【請求項4】
複合体が、対象となる分子が供給されている少なくとも二つのサブユニットを含む、請求項1〜3のいずれか一項記載のタンパク質複合体。
【請求項5】
少なくとも二つのサブユニットに異なる対象となる分子が供給されている、請求項4記載のタンパク質複合体。
【請求項6】
細胞表面レセプターが、腸上皮に存在する、請求項1〜5のいずれか一項記載のタンパク質複合体。
【請求項7】
少なくとも一つの対象となる分子が、抗原である、請求項1〜6のいずれか一項記載のタンパク質複合体。
【請求項8】
抗原が、細菌抗原、ウイルス抗原、原虫抗原および真菌抗原からなる群より選択される、請求項7記載のタンパク質複合体。
【請求項9】
少なくとも一つの対象となる分子が、免疫調節タンパク質、好ましくはサイトカインまたは熱ショックタンパク質である、請求項1〜6のいずれか一項記載のタンパク質複合体。
【請求項10】
複合体が、大腸菌の易熱性エンテロトキシン(LT)またはコレラ菌のコレラトキシン(CT)の五つのBサブユニットを含み、そのうち一つのサブユニットが未改変である、請求項1〜9のいずれか一項記載のタンパク質複合体。
【請求項11】
細胞表面レセプターがガングリオシド分子、好ましくはGM1、またはその模擬体を含む、請求項1〜10のいずれか一項記載のタンパク質複合体。
【請求項12】
以下の工程を含む、請求項1〜11のいずれか一項記載のタンパク質複合体を生成するための方法:
a)宿主細胞に、未改変サブユニットをコードするヌクレオチド配列、および少なくとも一つの対象となる分子がサブユニットと融合している対象となる分子をコードするヌクレオチド配列を提供する工程;
b)該宿主細胞を培養し、それにより該ヌクレオチド配列を発現させてタンパク質複合体を組み立てさせる工程;
c)複合体を単離する工程;ならびに
d)複合体の細胞表面レセプターへの、または細胞表面レセプターを模擬する分子への結合を決定する工程。
【請求項13】
以下の工程を含む、請求項1〜11のいずれか一項記載のタンパク質複合体を生成するための方法:
a)第一宿主細胞に未改変サブユニットをコードするヌクレオチド配列、および第二宿主細胞に対象となる分子をコードするヌクレオチド配列を提供する工程であって、少なくとも一つの対象となる分子がサブユニットと融合している工程;
b)該宿主細胞を培養し、それにより該ヌクレオチド配列を発現させる工程;
c)該ヌクレオチドがコードするタンパク質を単離する工程;
d)単離したタンパク質をタンパク質複合体の組み立てが可能な条件下で接触する工程;
e)複合体を単離する工程;ならびに
f)複合体の細胞表面レセプターへの、または細胞表面レセプターを模擬する分子への結合を決定する工程。
【請求項14】
宿主細胞に、形質転換、共形質転換、交差、再形質転換または一過性トランスフェクションを用いて該ヌクレオチド配列を提供する、請求項13記載の方法。
【請求項15】
請求項1〜11のいずれか一項記載のタンパク質複合体を含む細胞。
【請求項16】
細胞が、植物細胞である、請求項15記載の細胞。
【請求項17】
細胞が、食用細胞である、請求項15または16記載の細胞。
【請求項18】
請求項1〜11のいずれか一項記載のタンパク質複合体、または請求項15〜17記載のいずれか一項記載の細胞を含む組成物。
【請求項19】
請求項1〜10のいずれか一項記載のタンパク質複合体、または請求項15〜17のいずれか一項記載の細胞、および薬学的に許容されるキャリアを含む、ワクチン。
【請求項20】
有効量の請求項19記載のワクチンを含む、医薬組成物。
【請求項21】
粘膜キャリア分子としての請求項1〜11のいずれか一項記載のタンパク質複合体の使用。
【請求項22】
有効量の請求項1〜11のいずれか一項記載のタンパク質複合体の少なくとも一投与量を被験体に投与する工程を含む、被験体の免疫応答を調節するための方法であって、対象となる分子が抗原である方法。
【請求項23】
請求項19記載のワクチンの被験体への投与を含む、粘膜免疫のための方法。
【請求項1】
少なくとも二つの、好ましくは同一のサブユニットを含むタンパク質複合体であって、そのうちの少なくとも一つのサブユニットは未改変であり、また少なくとも一つのサブユニットは対象となるの第一分子に融合しており、該サブユニットを介して細胞表面レセプターと相互作用できる該タンパク質複合体。
【請求項2】
対象となる第一分子が、好ましくは共有結合により、対象となる第二分子と結合して対象となる多量体を形成できる、請求項1記載のタンパク質複合体。
【請求項3】
複合体が、本質的に大腸菌(E. coli)の易熱性エンテロトキシン(LT)またはコレラ菌(Vibrio cholerae)のコレラトキシン(CT)、好ましくはそのBサブユニットに基づいている、請求項1または2記載のタンパク質複合体。
【請求項4】
複合体が、対象となる分子が供給されている少なくとも二つのサブユニットを含む、請求項1〜3のいずれか一項記載のタンパク質複合体。
【請求項5】
少なくとも二つのサブユニットに異なる対象となる分子が供給されている、請求項4記載のタンパク質複合体。
【請求項6】
細胞表面レセプターが、腸上皮に存在する、請求項1〜5のいずれか一項記載のタンパク質複合体。
【請求項7】
少なくとも一つの対象となる分子が、抗原である、請求項1〜6のいずれか一項記載のタンパク質複合体。
【請求項8】
抗原が、細菌抗原、ウイルス抗原、原虫抗原および真菌抗原からなる群より選択される、請求項7記載のタンパク質複合体。
【請求項9】
少なくとも一つの対象となる分子が、免疫調節タンパク質、好ましくはサイトカインまたは熱ショックタンパク質である、請求項1〜6のいずれか一項記載のタンパク質複合体。
【請求項10】
複合体が、大腸菌の易熱性エンテロトキシン(LT)またはコレラ菌のコレラトキシン(CT)の五つのBサブユニットを含み、そのうち一つのサブユニットが未改変である、請求項1〜9のいずれか一項記載のタンパク質複合体。
【請求項11】
細胞表面レセプターがガングリオシド分子、好ましくはGM1、またはその模擬体を含む、請求項1〜10のいずれか一項記載のタンパク質複合体。
【請求項12】
以下の工程を含む、請求項1〜11のいずれか一項記載のタンパク質複合体を生成するための方法:
a)宿主細胞に、未改変サブユニットをコードするヌクレオチド配列、および少なくとも一つの対象となる分子がサブユニットと融合している対象となる分子をコードするヌクレオチド配列を提供する工程;
b)該宿主細胞を培養し、それにより該ヌクレオチド配列を発現させてタンパク質複合体を組み立てさせる工程;
c)複合体を単離する工程;ならびに
d)複合体の細胞表面レセプターへの、または細胞表面レセプターを模擬する分子への結合を決定する工程。
【請求項13】
以下の工程を含む、請求項1〜11のいずれか一項記載のタンパク質複合体を生成するための方法:
a)第一宿主細胞に未改変サブユニットをコードするヌクレオチド配列、および第二宿主細胞に対象となる分子をコードするヌクレオチド配列を提供する工程であって、少なくとも一つの対象となる分子がサブユニットと融合している工程;
b)該宿主細胞を培養し、それにより該ヌクレオチド配列を発現させる工程;
c)該ヌクレオチドがコードするタンパク質を単離する工程;
d)単離したタンパク質をタンパク質複合体の組み立てが可能な条件下で接触する工程;
e)複合体を単離する工程;ならびに
f)複合体の細胞表面レセプターへの、または細胞表面レセプターを模擬する分子への結合を決定する工程。
【請求項14】
宿主細胞に、形質転換、共形質転換、交差、再形質転換または一過性トランスフェクションを用いて該ヌクレオチド配列を提供する、請求項13記載の方法。
【請求項15】
請求項1〜11のいずれか一項記載のタンパク質複合体を含む細胞。
【請求項16】
細胞が、植物細胞である、請求項15記載の細胞。
【請求項17】
細胞が、食用細胞である、請求項15または16記載の細胞。
【請求項18】
請求項1〜11のいずれか一項記載のタンパク質複合体、または請求項15〜17記載のいずれか一項記載の細胞を含む組成物。
【請求項19】
請求項1〜10のいずれか一項記載のタンパク質複合体、または請求項15〜17のいずれか一項記載の細胞、および薬学的に許容されるキャリアを含む、ワクチン。
【請求項20】
有効量の請求項19記載のワクチンを含む、医薬組成物。
【請求項21】
粘膜キャリア分子としての請求項1〜11のいずれか一項記載のタンパク質複合体の使用。
【請求項22】
有効量の請求項1〜11のいずれか一項記載のタンパク質複合体の少なくとも一投与量を被験体に投与する工程を含む、被験体の免疫応答を調節するための方法であって、対象となる分子が抗原である方法。
【請求項23】
請求項19記載のワクチンの被験体への投与を含む、粘膜免疫のための方法。
【図1】

【図2】

【図3】
【図4】

【図5】
【図6A】
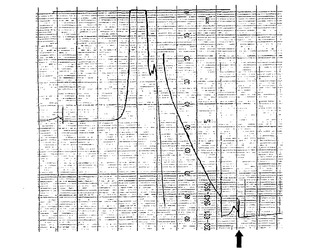
【図6B】
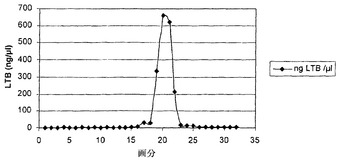
【図6C】

【図7】
【図8A】
【図8B】
【図8C】
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図6C】
【図7】
【図8A】
【図8B】
【図8C】
【公表番号】特表2008−500008(P2008−500008A)
【公表日】平成20年1月10日(2008.1.10)
【国際特許分類】
【出願番号】特願2006−532148(P2006−532148)
【出願日】平成16年10月8日(2004.10.8)
【国際出願番号】PCT/NL2004/000708
【国際公開番号】WO2005/033317
【国際公開日】平成17年4月14日(2005.4.14)
【出願人】(502152632)プラント リサーチ インターナショナル ベー.フェー. (1)
【氏名又は名称原語表記】Plant Research International B.V.
【住所又は居所原語表記】Droevendaalsesteeg 1,Wageningen,Netherlands
【Fターム(参考)】
【公表日】平成20年1月10日(2008.1.10)
【国際特許分類】
【出願日】平成16年10月8日(2004.10.8)
【国際出願番号】PCT/NL2004/000708
【国際公開番号】WO2005/033317
【国際公開日】平成17年4月14日(2005.4.14)
【出願人】(502152632)プラント リサーチ インターナショナル ベー.フェー. (1)
【氏名又は名称原語表記】Plant Research International B.V.
【住所又は居所原語表記】Droevendaalsesteeg 1,Wageningen,Netherlands
【Fターム(参考)】
[ Back to top ]