
細胞におけるタンパク質の発現を増大させる方法
本発明は、タンパク質の核酸配列のコードおよび/または非コード領域におけるRNase Lの切断部位数を減少させることによって、細胞における、好ましくは真核細胞における該タンパク質の発現を増大させる方法に関する。さらに、本発明は、減少したRNase Lの切断部位数を示す核酸配列に加えて、そのような配列から翻訳されたタンパク質に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、タンパク質の核酸配列のコードおよび/または非コード領域におけるRNase Lの切断部位数を減少させることによって、細胞における、好ましくは真核細胞における該タンパク質の発現を増大させる方法に関する。さらに、本発明は、減少したRNase L切断部位数を示す核酸配列に加えて、そのような配列から翻訳されたタンパク質に関する。
【0002】
一過性応答遺伝子は、細胞増殖、シグナル伝達事象、ならびに炎症性刺激、微生物、および放射線などの外因性因子に対する応答を含めた、重大な生物学的応答をレギュレーションする(Lai et al., 2006; Lal et al., 2004; Lopez de Silanes et al., 2005)。それらは、特異的配列エレメントなどのシス作用因子と、ある種のRNA結合タンパク質のようなトランス作用因子との両方によってコントロールされる。主として3’非翻訳領域(3’UTR)における配列エレメントおよびRNA結合タンパク質によるモデュレーションは、メッセンジャーRNA(mRNA)の安定性およびタンパク質の翻訳に影響を及ぼすことがある。
【0003】
かなりの数の遺伝子が、そのmRNAの3’UTRに不安定化配列エレメントを、主としてアデニル酸ウリジル酸(AU)リッチエレメント(ARE)を有する。これらのAREは、タンパク質とmRNAとの相互作用に影響を及ぼすおそれのある不均一な群の配列クラスを含むことから、mRNAの分解特性を左右する(Bakheet et al., 2006; Barreau et al., 2005)。細胞性mRNAの安定化は、HuRタンパク質などのmRNA安定化促進タンパク質の活性によって、またはジンクフィンガータンパク質であるトリステトラプロリン(TTP)などのRNA分解促進タンパク質の不活性化によって起こる場合がある。
【0004】
細胞性mRNAに影響を及ぼすおそれのある、別のクラスのトランス作用因子は、普遍的細胞内酵素である、エンドリボヌクレアーゼのリボヌクレアーゼL(RNase L)であり、それは、以前はウイルスmRNAに特異的であると考えられていた。しかし、最近の研究は、RNase Lがある種の生物学的過程の一過性応答に関与する可能性があることを示した(Khabar et al., 2003a; Li et al., 2000)。RNase Lは、インターフェロン(IFN)系の一部であると見なされている。IFNは、酵素オリゴアデニル酸シンターゼ(OAS)の遺伝子発現を誘導し、それはウイルス二本鎖RNA中間体に結合すると活性化し、短い2’−5’オリゴアデニル酸(2−5A)を合成する。これらは、今度はウイルスmRNAを強力に分解するRNase Lを活性化する。RNase Lは、ナノモル濃度以下のレベルの2−5Aによって活性化され、優先的にウイルスmRNAのUUおよびUAジヌクレオチドの後でウイルスRNAの1本鎖領域の切断を招く(Wrechester et al., 1981; Han et al., 2004)。さらに高いレベルでは、RNase Lは18Sおよび28SリボソームRNAの切断などのより広い効果を誘導することがある(Wreschner et al., 1981)。
【0005】
近年、選ばれた細胞性mRNAの分解にRNase Lが関与することが広く受け入れられるようになった(Bisbal et al., 2000; Chandrasekaran et al., 2004; Khabar et al., 2003b; Le Roy et al., 2001; Li et al., 2000)。具体的には、RNase Lは、PKR mRNAをダウンレギュレーションすることが示された(Khabar et al., 2003b)。正常細胞におけるIFN抗ウイルス応答中に、PKR mRNAの発現は一過性であるが、RNase L−ヌル細胞では、mRNAの安定性増大が原因で長期動態のPKR mRNA発現が観察される(Khabar et al., 2003b)。その効果は、ウイルスタンパク質の合成阻害を誘導する過程である真核生物翻訳開始因子2サブユニット(eIF2a)のPKR依存性リン酸化の延長を招く(Khabar et al., 2003b)。したがって、RNase Lは、PKRによって付与される翻訳停止が短時間であることを確実にするために、IFN応答が一過性であることを担っている。RNase LがIFN応答をネガティブレギュレーションする同様の役割は、また、ISG43と呼ばれる43kDユビキチン特異的プロテアーゼをコードする新規なIFN活性化遺伝子がRNase Lによってダウンレギュレーションされるという報告によって示唆されている(Li et al., 2000)。ISG43 mRNAの半減期がRNase L−ヌル細胞で増大することから、このレギュレーションは、mRNA安定性のレベルで起こる(Li et al., 2000)。RNase Lは、筋分化に不可欠な重要な転写因子をコードする、別の機能的に重要な細胞性mRNAであるmyoDをダウンレギュレーションすることができる。RNase Lおよびその抑制因子RLIは、C2細胞系が筋芽細胞から筋管に分化する間に連続的に誘導される(Bisbal et al., 2000)。RNase Lの阻害または過剰発現は、MyoD mRNAの半減期をそれぞれ延長または短縮する(Bisbal et al., 2000)。RNase L分子のプールがミトコンドリアに局在し、IFN−α処置後に増大することから、CYTB、ATPase 6(ATP6)、およびチトクロームオキシダーゼII(CO)のmRNAなどのミトコンドリアmRNAのダウンレギュレーションにRNase Lが果たす役割が、IFNの抗増殖作用のメカニズムとして提案されている(Le Roy et al., 2001)。これは、アンチセンス構築物の誘導を介してRNase L活性を低下させることによって、または2−5AによってRNase L活性を直接活性化することによって実証された(Le Roy et al., 2001)。RNase Lは、特に2−5Aのレベルが限定的な条件で、非ウイルス性mRNAに比べてウイルス、例えば脳心筋炎ウイルス(EMCV)のmRNAに対して選択性を示す(Li et al., 1998a)。細胞性mRNAに対するRNase Lの効果は、特定のmRNAに大いに限定されると考えられる。というのも、この主題を扱った研究で細胞性mRNAに対する全体的な効果が観察されないからである。さらに、上記研究のいずれによっても、細胞性ターゲットmRNAへのRNase Lの直接結合も、ウイルスmRNAに対して示されたような配列特異性も実証されていない。したがって、非ウイルス性細胞性mRNAに関連するRNase L活性のメカニズムは、大部分が不明なままである。
【0006】
以前にドミナントネガティブ型のRNase Lが、アミノ酸置換または完全長タンパク質の短縮化のいずれかによって生成された。例えばドミナントネガティブRNase LであるZB1(カルボキシ末端の89個のアミノ酸を欠如した短縮型マウスRNase L)は、野生型RNase Lの抗ウイルス作用および抗増殖作用を阻害する(Hassel et al., 1993)。Dongら(2001)は、RNase Lの他の短縮化および点変異、例えばヌクレアーゼドメインにおける変異(R667A)を記載している。
【0007】
米国特許第6,762,038号は、トランスフェクションされた遺伝子の発現強化のために、RNase L遺伝子にホモ接合性破壊を有するマウスから作出された変異型胚性線維芽細胞系(MEF)の使用を示唆している(Zhou et al., 1997)。しかし、発現強化の効果は、これらの特定の細胞系に限られる。さらに、新しいRNase L−ヌル細胞系の創出は、非常に骨の折れる工程であって、全ての哺乳動物および/または真核生物発現系に容易に適用できるわけではない。
【0008】
哺乳動物発現系は、治療用タンパク質および抗体の産生のための重要な手段となっており、原核生物発現系に比べて、例えばタンパク質の正しいフォールディングおよび解糖などの翻訳後修飾に関していくつかの利点を有する。しかし、多数の系においてタンパク質発現が低収率であることは、費用のかかる技術的障害となる。所望のタンパク質、例えばG−タンパク質結合レセプター(GPCR)などの膜タンパク質、大型タンパク質、抗体、融合タンパク質、タンパク質複合体、ワクチン、および血漿タンパク質を発現させることが本質的に困難ならば、これは特に問題である。この困難の理由は、タンパク質自体の本質的な不安定性、AUリッチエレメント含有mRNAの場合のようなmRNAの本質的な不安定性、または弱いプロモーター活性であろう。
【0009】
これらの問題に共通の解決は、CMVプロモーターなどの強力なプロモーター、およびCMVのイントロンAなどの特定種類のイントロンを含めた、そのプロモーターの上流または下流のエンハンサーエレメントを提供することによってタンパク質の発現を増大させることに焦点を当てていた。他の解決は、核および染色体の構築に重要な300〜3000塩基対長のDNAエレメントであるクロマチンマトリックス付着領域(MAR)を関与させた。これらのエレメントは、隣接クロマチンが導入遺伝子の発現に影響を及ぼすことを阻止し、そのことが、所望のレギュレーション性発現を示すクローンを単離する確率の増大を誘導すると提案された(米国特許第7,129,062号)。この特定のアプローチは、その効果を達成するためにいくつかの異なるベクター構築物を必要としうることから、潜在的に問題がある。これらのアプローチが利用可能であるにもかかわらず、上記のタンパク質のように、発現が困難なタンパク質のタンパク質発現をさらに増大させる必要性は、依然として存在する。
【0010】
コドン最適化(またはコドン使用頻度最適化)は、タンパク質産生を押し上げるための、当技術分野において公知の別法である。それは、発現されるべきタンパク質をコードする核酸配列がホストによって稀に使用されるコドン(トリプレット)を有するならば、その発現レベルは最大ではないという観察に基づく。コドン最適化は、基本的にターゲット核酸配列における稀なコドンを変化させることにより、それらがホストのコドン使用頻度をより厳密に反映させることを含む。したがって、普通、最適化工程に使用される情報は、最適化されるDNAまたはタンパク質配列、およびそれぞれのホストのコドン使用頻度表である(例えばヒトゲノムについては表1参照)。コドン使用頻度表は、所与の生物における特定のアミノ酸についてそれぞれの可能なコドンの相対頻度を一覧している。全ての生物におけるコドン使用頻度のバイアスの全一覧は、次のウェブサイトから見出すことができる:http://www.kazusa.or.jp/codon/。所与のホスト生物に対するコドン使用頻度のバイアスに基づいてコドンを最適化するために、いくつかのウェブベースのプログラムも利用可能である。コドン最適化は、非ヒトまたは非哺乳動物起源の遺伝子がヒトまたは他の哺乳動物ホスト細胞において発現される(またはその逆)一部の状況で成功することがある。しかし、コドン使用頻度は、タンパク質の発現レベルを左右する多数の要因のわずか一つであって、コドン最適化の効果は多くの場合に限定的である。
【0011】
任意の生物の細胞において、好ましくは真核細胞において発現される、本質的に発現が困難なタンパク質を含めた内因性および外因性(リコンビナント)タンパク質の収率を顕著に改善する方法を提供することが、本発明の一目的であった。この方法は、時間および費用効率的であるべきであり、原核細胞および真核細胞を含めた任意の種類の生物の細胞におけるタンパク質の大規模産生を可能にするべきである。治療的に使用されるタンパク質の産生を可能にするために、使用される細胞系、リコンビナントタンパク質の特性、または外因性材料の使用などの、リコンビナントタンパク質を産生するために規制当局に認可された特徴を変更せずに、発現収率におけるこの改善を達成することが、本発明の別の目的であった。
【0012】
本発明の目的は、タンパク質の核酸配列におけるRNase L切断部位を減少させる工程を含む、細胞における、好ましくは真核細胞における該タンパク質の発現を増大させる方法によって解決される。一態様では、該細胞は原核細胞であり、別の態様では、該細胞は真核細胞である。
【0013】
タンパク質の核酸配列は、コード領域および非コード領域、すなわちアミノ酸配列に翻訳される領域(エクソンとも呼ばれる)およびアミノ酸配列に翻訳されない領域の両方を含む。核酸配列の非コード領域は、例えば5’非翻訳領域(5’UTR)、3’非翻訳領域(3’UTR)、およびイントロンである。これらのエレメントの全て(5’UTR、3’UTR、イントロン、およびコード領域)は、遺伝子およびタンパク質の発現をコントロールすることができることから、上記方法に関するターゲットである。本発明により、RNase Lの切断部位数を減少させる該工程は、コード領域または非コード領域のいずれかまたは両方における該数を減少させる。
【0014】
一態様ではRNase Lの該切断部位数は、(野生型核酸配列におけるRNase Lの切断部位数に比べて)少なくとも10%、好ましくは少なくとも25%、さらに好ましくは少なくとも50%減少する。
【0015】
好ましくは、該切断部位は、UUおよび/またはUAジヌクレオチドである。
【0016】
RNase Lは、エンドリボヌクレアーゼであることから、RNA(一次RNA、すなわちスプライシングされていないRNAおよびmRNA、すなわちスプライシングされたRNA)レベルでのみ活性である。UUおよびUAジヌクレオチドは、RNAレベルでのみ存在する。
【0017】
しかし、該タンパク質の核酸配列におけるRNase L切断部位数を減少させる該工程は、DNAレベルで行われることが好ましく、RNA配列におけるUUおよびUAジヌクレオチドはDNA配列におけるTTおよびTAヌクレオチドに対応する。所与のDNA配列を特異的に変化させる技法は、当技術分野において周知であり、それには、非限定的に遺伝子合成、部位特異的変異誘発、制限酵素消化による欠失変異、および組換えによる変異導入が含まれる。遺伝子合成の技法は、本発明により特に好ましい。
【0018】
本発明の一態様では、RNase L切断部位数を減少させる該工程は、該核酸配列のコード領域における該数を減少させる。
【0019】
好ましくは、該核酸配列におけるRNase L切断部位数を減少させる該工程は、該タンパク質のアミノ酸配列を変更せずに行われる。したがって、オープンリーディングフレーム(ORF)は、該工程によって変更されない。
【0020】
RNase L切断部位数を減少させる該工程における一態様では、UUおよび/またはUAジヌクレオチドを含むコドンは、UUおよび/またはUAジヌクレオチドを含まない、同じアミノ酸をコードする代替コドンに交換される。
【0021】
RNase L切断部位数を減少させる該工程における一態様では、UUおよび/またはUAジヌクレオチドを含む隣接コドン対がもはやUUおよび/またはUAジヌクレオチドを含まないように、該隣接コドン対の少なくとも一つのコドンが、同じアミノ酸をコードする代替コドンに交換される。
【0022】
好ましくは、UUおよび/またはUAジヌクレオチドを含む該隣接コドン対の第一のコドンが交換される。
【0023】
一態様では、該代替コドンは、該細胞において、好ましくは該真核細胞において、より頻繁に使用されるコドンである。
【0024】
本発明の一態様では、RNase L切断部位数を減少させる該工程は、該核酸配列の非コード領域における該数を減少させる。
【0025】
好ましくは、該非コード領域は5’UTR、3’UTR、またはイントロンである。
【0026】
イントロンの例には、非限定的にCMVイントロン、SV40イントロン、ウサギβグロビンイントロン(RBTG)、および合成イントロンが含まれる。
【0027】
一態様では、該核酸配列の非コード領域におけるRNase Lの切断部位数を減少させる該工程は、ヌクレオチドの変異、欠失、または挿入によって行われる。
【0028】
好ましくは、該核酸配列の非コード領域におけるRNase Lの切断部位数を減少させる該工程は、開始コドン(ATG)に近接した5’UTRにおける配列のように、非コード領域における機能的に重要なエレメントを変更しないが、それは、それらが、翻訳強化配列(例えばコザック)、3’UTRにおけるポリAシグナル(例えばAAUAAAまたはAUUAAA)またはポリアデニル化に使用される他の必要もしくは補助配列エレメント、イントロン−エクソン接合部/境界域、スプライシングブランチポイントおよびイントロンにおけるエクソンのスプライス供与/受容部位、およびスプライス受容部位からブランチポイント終点の間のCTリッチ領域を有する場合があるからである。例えば、強力なポリAシグナル下流にU−リッチのヌクレオチド50個の領域があるが、それは、可能ならば変更すべきでない(Legendre and Gautheret, 2003)。
【0029】
好ましくは、非コード領域におけるRNase Lの切断部位数を減少させる該工程は、(a)イントロンのGC含量を80%よりも大きく、かつその長さを80%未満に変化させること、
(b)5’UTRのGC含量を80%よりも大きく変化させること、および
(c)3’UTRのGC含量を80%よりも大きく、かつその長さを80%未満に変化させること
はない。
【0030】
一態様では、上記方法はさらに、RNase Lの切断部位数を減少させる該工程の前にコドン最適化の工程を含む。
【0031】
一態様では、上記方法はさらに、RNase Lの切断部位数を減少させる該工程の後に、発現活性PCR産物の形のまたは発現ベクターに含まれる、該タンパク質の該核酸配列を、該細胞に、好ましくは該真核細胞にトランスフェクションする工程を含む。
【0032】
本明細書に使用される「発現活性PCR産物」という用語は、cDNA、オープンリーディングフレーム、または発現ベクターに含まれる遺伝子などの関心が持たれるDNA配列の隣接配列に相補的な2個のプライマーを使用したPCR増幅によって生成されるPCR産物を表すことを意図し、ここで、結果として生じるPCR産物は、プロモーター、関心が持たれるDNA配列および終結配列を含み、ホスト細胞にトランスフェクションされた場合に、関心が持たれるDNAを発現させる(Al-Zoghaibi et al., 2007も参照されたい)。
【0033】
本発明によると、任意の発現ベクターを使用することができるけれども、真核生物/哺乳動物発現ベクターが好ましい。哺乳動物発現ベクターは、タンパク質の生物学的機能を研究するための一般に普及したツールであり、様々な種類(例えばプラスミドベクターまたはウイルスベクター)が当技術分野において公知である。適切な発現ベクターは、特定のプロモーター、5’UTR、3’UTR、またはイントロンに限定されない。それらは、構成的発現型、誘導型、または抑制型またはレギュレーション可能型でありうる。
【0034】
本発明によると、プロモーターが真核生物性であり、終結部位が、ポリアデニル化シグナルを有するポリA配列であることが好ましい。真核生物または哺乳動物プロモーターは、当技術分野において公知であり、それらには、非限定的にサイトメガロウイルス(CMV)前初期プロモーター、SV40プロモーター、伸長因子(EF)プロモーター、RSVプロモーター、およびニワトリβ−アクチンプロモーターが含まれる。好ましい真核生物ポリアデニル化シグナルは、ウシ成長因子(BGH)ポリ部位、成長ホルモンポリA、SV40ポリA、およびHSKポリAである(Foecking and Hofstetter, 1986; Kobayashi et al., 1997)。真核生物ターミネーターの例は、ウシ成長因子(BGH)ポリA部位、SV40ポリA、HSKポリA、および合成ポリAである。
【0035】
細胞にDNA/ベクターを一過性または安定的にトランスフェクションする方法は、当技術分野において周知である。これらには、非限定的にリン酸カルシウム共沈、エレクトロポレーション、陽イオンポリマートランスフェクション、およびリポソーム介在性トランスフェクション(例えばリポフェクション)が含まれる。リポソーム介在性トランスフェクション用の試薬は、市販されており、例えばリポフェクタミン(Invitrogen)およびポリエチレンイミン(Sigma)である。細胞は、また、ウイルストランスフェクションによって、またはウイルスコート粒子を介してトランスフェクションすることができる。本発明により細胞をトランスフェクションする別の好ましい方法は、該発現活性PCR産物または該発現ベクターをin vivoマイクロインジェクションすること、および該発現活性PCR産物または該発現ベクターを有する細胞を選択薬の助けを借りてまたは借りずに選択的に成長させることである。したがって、本発明による発現活性PCR産物または発現ベクターは、追加的に選択マーカーを含むことがある。安定な細胞系を産生させるために、クローンは、非限定的にネオマイシン、ブラストサイジン、ピューロマイシン、ゼオシン(zeocin)、ハイグロマイシン、およびジヒドロ葉酸レダクターゼ(dhfr)が含まれる様々な選択マーカーを使用して選択することができる。
【0036】
上記方法の全ての適切な真核生物/哺乳動物細胞(ホスト細胞)もまた、当技術分野において周知であって、それらには、非限定的にCHO、HEK293、HeLa、およびCOS−7細胞が含まれる。
【0037】
一態様では、上記方法は、さらに、該細胞における、好ましくは該真核細胞における該発現活性PCR産物または発現ベクターから該タンパク質を翻訳させる工程を含む。
【0038】
好ましくは、該タンパク質は、レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択される。
【0039】
本発明の目的は、また、RNase Lの切断部位数が(野生型核酸配列におけるRNase Lの切断部位数に比べて)少なくとも10%、好ましくは少なくとも25%、さらに好ましくは少なくとも50%減少している核酸配列によって解決される。
【0040】
好ましくは、該核酸配列は、上記方法によって産生される。
【0041】
好ましくは、該核酸配列は、レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択されるタンパク質の核酸配列である。
【0042】
一態様では、該核酸配列は、配列番号:2、3、5、7、9、11、13、15、17、19、23、24、26、28、30〜36、好ましくは配列番号:9、15、17、23、30〜36より選択される配列を有する。
【0043】
本発明の目的は、また、上記核酸を含む発現活性PCR産物または発現ベクターによって解決される。
【0044】
本発明の目的は、さらに、上記の発現活性PCR産物または発現ベクターを有するホスト細胞によって解決される。
【0045】
最終的に、本発明の目的は、また、上記方法によって産生されるタンパク質であって、レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択されるタンパク質によって解決される。
【0046】
本発明によると、「治療用タンパク質」という用語は、抗体フラグメント、免疫グロブリン鎖、重鎖および軽鎖、Fabフラグメント、酵素、成長因子、インターフェロン、サイトカイン、リンホカイン、接着分子、レセプターに加えて、その誘導体またはフラグメントなどの、薬学的に関連して使用されるタンパク質を包含することが意図される。
【0047】
本発明によると、レポータータンパク質(単独または別のタンパク質と融合したもの)は、上記方法のために特に好ましい「ターゲット」である。(非限定的に)緑色蛍光タンパク質(GFP)、赤色蛍光タンパク質(RFP)、黄色蛍光タンパク質(YFP)、青色シアニン蛍光タンパク質(CFP)、ルシフェラーゼ、分泌型アルカリホスファターゼ(SEAP)、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、分泌型ホルモン、分泌型サイトカイン、β−ガラクトシダーゼ、ならびに他の蛍光および生物発光タンパク質を含めた、多数の蛍光および非蛍光レポータータンパク質がある。レポータータンパク質の選択は、使用される細胞系(内因性活性)、実験の性質(例えば遺伝子発現の動力学およびトランスフェクション効率)、および選択された検出方法に対するアッセイの適応性に依存する(Naylor 1999)。迅速な応答および変化のなどのレポーター性能を改善するために、レポータータンパク質自体のいくつかの改変、例えば不安定化エレメントの使用が探究されている(Li et al., 1998b; Zhao et al., 1995)。緑色蛍光タンパク質(GFP)および他の蛍光タンパク質は、生組織、細胞および実験動物における遺伝子発現を非侵襲的にモニタリングするためにますます普及しつつある(Naylor 1999)。したがって、GFP(およびEGFPなどのその誘導体)、他の蛍光タンパク質(上記参照)に加えて、蛍光タンパク質を含む融合タンパク質は、本発明による方法のためになおさらに好ましい「ターゲット」タンパク質である。
【0048】
驚くことに本発明者らは、真核生物および/または哺乳動物発現系で発現されるべきタンパク質の核酸配列におけるRNase Lの切断部位、好ましくはUUおよびUAジヌクレオチドの数を減少させることが、該タンパク質の発現および収率の顕著な改善を招くことを見出した。一定の理論に縛られることを望むわけではないが、RNase L切断部位の減少(すなわちこれらの部位の頻度減少)が、ホスト細胞において、好ましくは真核生物ホスト細胞において内因性RNase Lの攻撃を受けにくい(すなわち抵抗性が高い)核酸配列、すなわち(一次RNAおよびmRNAの両方の)RNAを生じ、mRNAの安定性増大を、したがってタンパク質の発現増大を誘導すると考えられる。
【0049】
関係する普遍的な概念のせいで、本発明は、任意の配列に適用することができる。さらに具体的には、本発明による方法は、内因性および外因性(リコンビナント)タンパク質/遺伝子の両方に、安定的に組み込まれた遺伝子および一過性発現遺伝子の両方に、発現が困難なタンパク質/遺伝子に、ならびに少量に内因性発現されるタンパク質/遺伝子にも適用可能である。それは、原核生物および真核生物系の両方に適用することができる。本明細書に記載されたアプローチは、タンパク質産生に費やされる時間およびコストの顕著な減少を誘導することから、エリスロポエチン、成長因子、インターフェロン、インスリン、治療用および診断用抗体、ならびにタンパク質またはペプチド系ワクチンなどの生物学的製剤として使用されるタンパク質/遺伝子のさらに効率的な産生を可能にするであろう。それは、大量に発現/産生させることが非常に困難であることが証明された遺伝子/タンパク質の発現に特に有用である。例には、非限定的にGタンパク質結合レセプター(GPCR)などの膜タンパク質、大型タンパク質、抗体、融合タンパク質、タンパク質複合体、ワクチン、および血漿タンパク質が含まれる。それは、また、レポータータンパク質の、例えばGFPまたはルシフェラーゼなどの蛍光タンパク質の発現および性能を顕著に改善することに役立つことから、そのようなレポータータンパク質を使用する方法の感度(蛍光または発光顕微鏡法、蛍光ベースのマイクロアレイ、細胞選別など)を増大することができる。さらにこのアプローチは、(非限定的に)ハムスターCHO1およびHEK293を含めた細胞系などの、生物工学工業に使用される任意の細胞系に適用可能であることから、特に実践的であり、かつ簡単である。
【0050】
以下に、本発明にしたがってRNase Lの切断部位数を減少させる工程の一般的原理を説明する。
【0051】
コード領域におけるUUおよび/またはUAジヌクレオチド数の減少
表2および表3は、引き起こすことのできる変化を示す。これらの改変は、所与の生物におけるそれらのコドンバイアス(コドン使用頻度)に基づきタンパク質発現のためにコドンを最適化するために使用されるコドン使用頻度表(比較のために表1参照)と全く別であることに留意することが重要である。本方法は、コドン使用頻度に基づいてではなく、RNase Lの切断部位/ターゲット(UUおよびUAジヌクレオチド)数を減少させるためにコドンを変化させることを対象とする。さらに、本発明による方法は、「生物依存的」ではなく、UUおよび/またはUAジヌクレオチドの数/頻度は、対応するアミノ酸配列が変更されないままである限り(表2および3参照)、任意の生物由来の任意の遺伝子(核酸配列)において減少させることができる。さらに、UUおよび/またはUAジヌクレオチド数の減少は、また、所望の生物についての古典的コドン使用頻度最適化(例えば表1参照)と組み合わせることができ、可能性があることには、いっそうさらなる発現増大を招く。
【0052】
表2は、UUおよび/またはUAジヌクレオチドを有するコドンに加えて、それらの非UU/UA含有代替物を示す。フェニルアラニン(Phe)についてUUまたはUAを完全に欠如した他のコドンがないことから、フェニルアラニンをコードするUUUは、UUCだけに変化させることができる。これは、UACだけに変化させることができるチロシン(UAU)の場合も同様である。いったん表2により変化を加えたら、表3により、UUまたはUAを形成するジ−トリプレット、すなわちNNU UNNまたはNNU ANN(Nは任意のヌクレオチドである)を変化させる。代替物が1個を超えて存在する場合は、それぞれの生物または高発現される遺伝子において最も高頻度に使用されるコドンを優先する。
【0053】
コード領域におけるUUおよび/またはUAジヌクレオチドを減少させるための工程:
1. 表2によりコドンを変化させる。
2. 代替コドンが1個を超えて存在する場合は、所望の生物においてより高頻度に使用されるコドンを使用する(随意)。
3. 表3により、一緒になってUUまたはUAジヌクレオチドを形成するジ−トリプレット(NNU UNN、NNU ANN)の最初のコドンを変化させる。
4. 1個を超える代替コドンが存在する場合は、所望の生物においてより高頻度に使用されるコドンまたは発現が最強のコドンを使用する(随意、表4および5参照)。
5. 古典的コドン使用頻度最適化(ホスト細胞生物、または高発現遺伝子において最も高頻度に使用されるコドンの一覧に基づく)を、所望により、そして好ましくはUUおよびUA減少の前に行うことができる。
【0054】
イントロンにおけるUUおよび/またはUAジヌクレオチド数の減少
イントロンにおけるUUおよび/またはUAジヌクレオチドを減少させるための工程:
1. UU/UAジヌクレオチドにおける2個のヌクレオチドのうち1個または2個を変異または欠失させる:UUまたはUAをUC、UG、GA、CA、またはTAにする。または1個のヌクレオチドを挿入する。
2. イントロンの全GC含量は、80%を超えるべきではなく、かつ長さはその本来の長さの80%未満に変化させるべきではない。
3. エクソンのスプライス供与部位および受容部位ならびにブランチポイントを含めたエクソン−イントロン境界部を変化させてはならない。スプライスアクセプター部位からブランチポイント終末にわたるCT−リッチ領域を破壊することを避ける。
【0055】
5’UTRにおけるUUおよび/またはUAジヌクレオチド数の減少
5’UTRにおけるUUおよび/またはUAジヌクレオチドを減少させる工程:
1. UUまたはUAをUC、UG、GA、CA、またはTAに変異させる。
2. 5’UTRの全GC含量は、80%を超えるべきではない。
3. コンテクスト配列はコザックなどの翻訳強化配列を有することがあるため、開始コドンATG近くのコンテクスト配列を避ける。
【0056】
3’UTRにおけるUUおよび/またはUAジヌクレオチド数の減少
3’UTRにおけるUUおよび/またはUAジヌクレオチドを減少させる工程:
1. UU/UAジヌクレオチドにおける2個のヌクレオチドのうち1個または2個を変異または欠失させる:すなわちUUまたはUAからUC、UG、GA、CA、またはTAにする。または、1個のヌクレオチドを挿入する。
2. 3’UTRの全GC含量は80%を超えるべきではなく、かつ長さはその本来の長さの80%未満に変化させるべきではない。
3. AAUAAAまたはAUUAAAなどのポリAシグナルを変化させてはならず、ポリアデニル化に使用される任意の必要または補助配列エレメントが認められる場合は、それを変更することを避ける。
【0057】
非コード領域における可能性のある変化
1. 変異:NUUNまたはNUANからNUSNへ(SはGまたはCであり、Nは任意のヌクレオチドである)
2. 挿入:NUUNまたはNUANからNUSUNまたはNUSAへ(SはGまたはCであり、Nは任意のヌクレオチドである)
3. 欠失:NUUNからNUSへ(SはGまたはCであり、Nは任意のヌクレオチドである)
4. 欠失:NUANからNANまたはNUSへ(SはGまたはCであり、Nは任意のヌクレオチドである)
【0058】
表
【0059】
【表1】

【0060】
【表2】
【0061】
【表3】
【0062】
【表4】
【0063】
【表5】
【0064】
これから以下の図を参照する。
【図面の簡単な説明】
【0065】
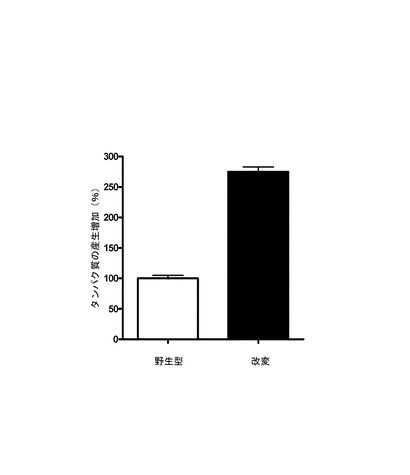
【図1】哺乳動物発現系におけるEGFPの発現に及ぼすUUおよびUAジヌクレオチドの減少の効果を野生型配列と比べて示すグラフである。
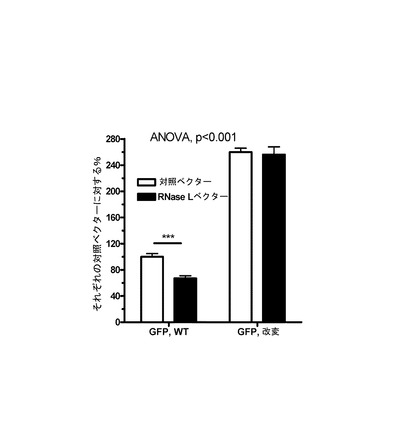
【図2】RNase Lの同時過剰発現を伴う哺乳動物発現系におけるEGFPの発現に及ぼすUUおよびUAジヌクレオチドの減少の効果を、野生型配列と比べて示すグラフである。
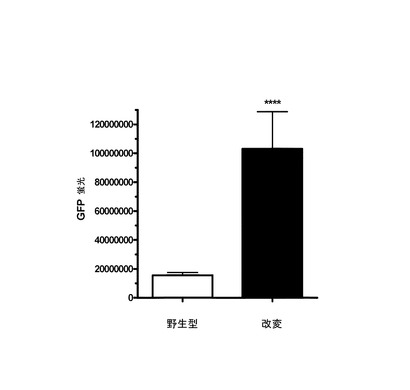
【図3】UU/UA減少型改変EGFP配列または野生型EGFP配列を有するPCR産物をトランスフェクションされた哺乳動物細胞におけるGFP蛍光を示すグラフである。
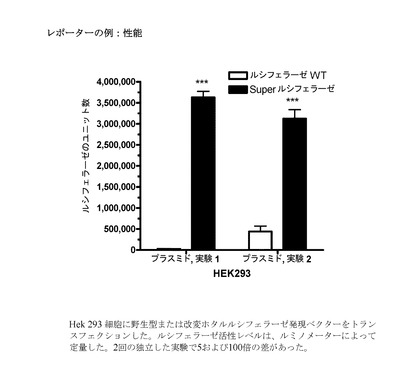
【図4】野生型または改変(すなわちUU/AGを減少させた)ホタルルシフェラーゼ発現ベクターのいずれかをトランスフェクションされたHek293細胞におけるルシフェラーゼ活性を示すグラフである。
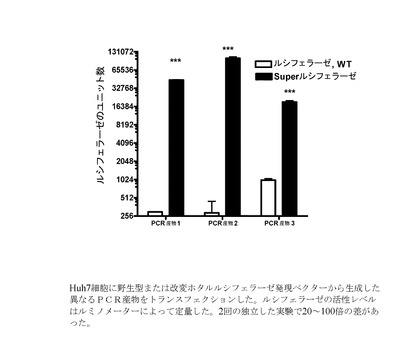
【図5】野生型またはUU/UA減少型ホタルルシフェラーゼ発現ベクターから生成した異なるPCR産物を、トランスフェクションされたHuh7細胞におけるルシフェラーゼ活性を示すグラフである。
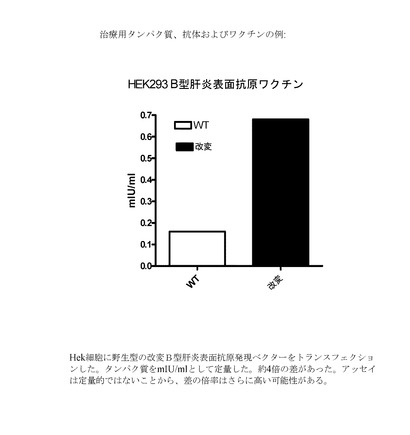
【図6】野生型またはUU/UA減少型B型肝炎表面抗原発現ベクターをトランスフェクションされたHek293細胞におけるB型肝炎表面抗原の発現を示すグラフである。
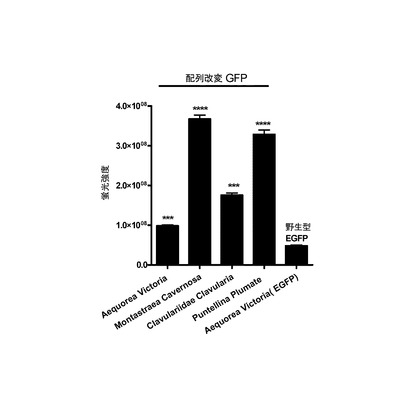
【図7】MODCで不安定化された野生型およびMODCで不安定化されたUU/UA減少型緑色蛍光タンパク質をトランスフェクションされたHek293細胞における様々な緑色蛍光タンパク質の発現を示すグラフである。
【0066】
これから以下の実施例を参照して本発明をさらに説明するが、それらの実施例は、例示であって、本発明の範囲を限定するつもりはない。
【0067】
実施例1:イントロンにおけるUUおよび/またはUAジヌクレオチドの減少
/=スプライシング部位
下線=コンセンサス機能的部位
太字の下線付き斜体=ブランチポイント
【0068】
配列番号:1: 野生型ウサギβグロビン−イントロン1(RBTG1):
【0069】
【化1】
【0070】
配列番号:2: UU/UA減少型RBTG1(CTリッチ領域にUU/UA減少を有さず):
【0071】
【化2】
【0072】
配列番号:3: UU/UA減少型RBTG1(CTリッチ領域に最低限のUU/UA減少を有する):
【0073】
【化3】
【0074】
実施例2: 3’UTRにおけるUUおよび/またはUAジヌクレオチドの減少
.=欠失
下線=突然変異
太字の下線付き=ポリAシグナル
【0075】
配列番号:4: 野生型SV40 3’UTR
【0076】
【化4】
【0077】
配列番号:5: UU/UA減少型SV40 3’UTR
【0078】
【化5】
【0079】
配列番号:6: 野生型ウシ成長ホルモン(BGH)3’UTR
【0080】
【化6】
【0081】
配列番号:7: UU/UA減少型BGH 3’UTR
【0082】
【化7】
【0083】
実施例3: コード領域におけるUUおよび/またはUAジヌクレオチドの減少
配列番号:8: 野生型の強化緑色蛍光タンパク質(EGFP)
【0084】
【化8】
【0085】
配列番号:9: UU/UA減少型EGFP(「スーパーGFP」)
【0086】
【化9】
【0087】
配列番号:10
MONTASTREA CAVERNOSA由来野生型GFP配列
【0088】
【化10】
【0089】
配列番号:11
MONSTER−OM:UU/UAジヌクレオチド数を減少させることによる改変型の配列番号:10
【0090】
【化11】
【0091】
配列番号:12
CLAVULARIA種 − 野生型配列
【0092】
【化12】
【0093】
配列番号:13
CLAVULARIA種−OM:改変(UU/UA減少)
【0094】
【化13】
【0095】
配列番号:14
ホタルルシフェラーゼ+:野生型配列:
【0096】
【化14】
【0097】
配列番号:15
LUC+DU(SuperLuciferase):(UU/UA減少型)
【0098】
【化15】
【0099】
配列番号:16
ホタルLUC2ルシフェラーゼ野生型:
【0100】
【化16】
【0101】
配列番号:17
LUC2OM:LUC2改変SuperLuciferase2(UU/UA減少型)
【0102】
【化17】
【0103】
配列番号:18
Puntellina Plumate(GFP)野生型:
【0104】
【化18】
【0105】
配列番号:19
Puntellina Plumate(GFP):改変配列(UU/UA減少型)
【0106】
【化19】
【0107】
配列番号:20
Discosoma野生型配列由来赤色蛍光タンパク質
【0108】
【化20】
【0109】
配列番号:21
赤色蛍光タンパク質の改変配列
【0110】
【化21】
【0111】
配列番号:22
B型肝炎表面抗原野生型、adrB型肝炎ウイルス株:
【0112】
【化22】
【0113】
配列番号:23
HBSAGOM:B型肝炎表面抗原改変配列:
【0114】
【化23】
【0115】
配列番号:24
HBSAGM:HBSAGOM: B型肝炎表面抗原改変配列2
【0116】
【化24】
【0117】
この配列は、UU/UA減少およびヒト化の両方を含む。
【0118】
配列番号:25
IFN−α、ヒト
【0119】
【化25】
【0120】
配列番号:26
IFN−αOM:改変配列
【0121】
【化26】
【0122】
配列番号:27
CSF3Mコロニー刺激因子野生型配列、ヒト:
【0123】
【化27】
【0124】
配列番号:28
CSF3Mコロニー刺激因子改変配列:
【0125】
【化28】
【0126】
配列番号:29
MODC野生型配列(マウスオルニチンデカルボキシラーゼ)
【0127】
【化29】
【0128】
配列番号:30
MODC改変配列:
【0129】
【化30】
【0130】
不安定化および配列改変されたレポーター:
不安定化および改変されたレポーター配列の例:
配列番号:31
MONTASTRAEA CAVERNOSA
【0131】
【化31】
MODC配列は太字
【0132】
配列番号:32
Clavulariidae Clavularia−OM:改変
【0133】
【化32】
【0134】
配列番号:33
ホタルLUC+DU(SuperLuciferase):
【0135】
【化33】
【0136】
配列番号:34
ホタルLUC2OM:LUC2改変SuperLuciferase2
【0137】
【化34】
【0138】
配列番号:35
Puntellina Plumate GFP:改変配列
【0139】
【化35】
【0140】
配列番号:36
赤色蛍光タンパク質の改変配列
【0141】
【化36】
【0142】
実施例4:UU/UA減少型EGFPのin vivo分析
(1)改変EGFP配列は、遺伝子合成会社によってカスタム合成され、隣接するSalIおよびBamHI部位を有するようにpUC19ベクター中に供給した。0.1μg/ml BSAを含有する緩衝液中に10ユニットのSalIで、10μgのベクターを37℃で1時間消化させ、続いてBamHI緩衝液中のBamHIで37℃でさらに1時間消化させた。消化されたDNAをフェノール−クロロホルム法を用いて抽出し、続いてエタノール沈殿を行った。CMVプロモーターおよびBHG 3’UTRを有する発現ベクターを同じ制限酵素(SalIおよびXbaI)で消化しておいたものにに合成EGFP−コード領域をライゲーションし、フェノール−クロロホルム抽出によって精製し、続いてエタノール沈澱を行った。発現ベクターへのEGFP−DNAのクローニングは、以下のライゲーション反応を用いて行った:消化されたベクターDNA 30μgを消化されたEGFP−DNA 90μgと共に、T4 DNAリガーゼを含有する反応液10μl中で混合した。ライゲーション産物を使用して、DH5αコンピテントE. coli細胞をトランスフォーメーションし、続いて結果として得られたコロニーを細菌培養培地中でエクスパンションさせた。Qiagenプラスミド精製キット(Qiagen, Germany)を使用してリコンビナントDNAを抽出した。挿入物を有するベクターのサイズは、ゲル電気泳動を用いて検証した。野生型DNAを有するベクターに加えて、UU/UA減少型改変コード領域を有する、結果として得られた発現ベクターを機能的実験に使用して、コードされたタンパク質の発現を確認した。10% FBSおよび抗生物質(Invitrogen)を補充したDMEM培地中で標準培養条件(37℃、5% CO2)でHEK293細胞を成長させた。96ウェルプレートのウェル1個あたり2.5×104個の細胞に、EGFPのUU/UA減少型改変コード領域を有するベクターまたは野生型EGFP−DNAを有するベクター100ngをトランスフェクションした。Lipofectamine 2000(Invitrogen)を使用して無血清培地中でトランスフェクションを5時間行い、続いて培地を血清補充培地に置換した。約24または48時間後に、プレートをイメージングし、BDハイコンテントイメージング装置を使用して定量した。定量は、Proxcellイメージングアルゴリズムを用いて行った。
【0143】
データは、EGFPのUU/UA減少型コード領域の使用が、野生型EGFP−DNAに比べて真核細胞におけるEGFPの有意に高い発現(2.5〜3倍)を可能にすることを明らかに示している(図1)。
【0144】
(2)10% FBSおよび抗生物質(Invitrogen)を補充したDMEM培地中で標準培養条件(37℃、5% CO2)でHEK293細胞を成長させた。96ウェルプレートのウェル1個あたり2.5×104個の細胞に、EGFPのUU/UA減少型改変コード領域を有するベクターまたは野生型EGFP−DNAを有するベクター100ngをトランスフェクションした。また細胞に、空の対照ベクター(pcDNA3.1)またはRNase Lベクター(pcDNA3.1)のいずれかを同時トランスフェクションした。トランスフェクションは、Lipofectamine 2000(Invitrogen)を用いて無血清培地中で5時間行い、続いて培地を血清補充培地と置換した。約24または48時間後に、プレートをイメージングし、BDハイコンテントイメージング装置を使用して定量した。定量は、Proxcellイメージングアルゴリズムを用いて行った。
【0145】
データは、野生型EGFP配列に比べて/対照的に、改変EGFP配列の使用がより高い発現を招き、それがRNase Lの同時発現によって制約されなかったことを示している(図2)。
【0146】
(3)フォワード(5’)プライマーがCMVプロモーター領域の開始部またはCMVプロモーター上流の配列に相補性であって、リバース(3’)プライマーがBGHポリA部位またはこの部位の下流の配列に相補的な、プライマーを使用することによって、発現活性PCR産物を生成させた。100μlの反応液中でTaqおよびPfuポリメラーゼ混合物を使用して以下のサイクル条件でPCRを実施した:95℃、12分(ホットスタートポリメラーゼを活性化する)、94℃、1分;52℃、1分;72℃、4分を32サイクル、および72℃で7分の最終伸長。プライマー、小型PCR産物、緩衝剤、および酵素を除去するためにQiagen PCR精製カラムを使用してPCR産物を精製し、滅菌水に溶出した。10% FBSおよび抗生物質(Invitrogen)を補充したDMEM培地中で標準培養条件(37℃、5% CO2)でHEK293細胞を成長させた。96ウェルプレートのウェル1個あたり2.5×104個の細胞に、野生型または改変配列を有するEGFP発現ベクターから生成した精製PCR産物100ngをトランスフェクションした。トランスフェクションは、Lipofectamine 2000(Invitrogen)を使用して無血清培地中で5時間行い、続いて培地を血清補充培地と置換した。約24または48時間後に、プレートをイメージングし、BDハイコンテントイメージング装置を使用して定量した。定量は、Proxcellイメージングアルゴリズムを用いて行った。
【0147】
データは、EGFPのUU/UA減少型コード領域を有するPCR産物が、野生型配列を有するPCR産物よりも高いEGFP発現(5〜10倍の増大)を誘導したことを示している(図3)。
【0148】
(4)(2)および(3)に記載したものと同じ方法を用いて、野生型またはUU/UAを減少させたホタルルシフェラーゼ発現ベクター(「SuperLuciferase」、配列番号:15)をHek293細胞にトランスフェクションした。ルシフェラーゼの活性レベルは、ルミノメーターによって定量した。データは、2回の独立した実験の中で約5倍および約100倍の差があったことを示している(図4)。
【0149】
同様に、上記(3)に概略した方法によって野生型または改変ホタルルシフェラーゼ発現ベクター(「SuperLuciferase」、配列番号:15)から生成した異なるPCR産物をHuh7細胞にトランスフェクションした。ルシフェラーゼ活性レベルは、今回もルミノメーターで定量した。そのデータは、SuperLuciferaseのUU/UA減少型コード領域を有するPCR産物が野生型配列を有するPCR産物よりも実質的に高いルシフェラーゼの発現(20〜100倍の増大)を誘導したことを示している(図5)。これは、本発明による方法が、真に多様なレポータータンパク質で有効であることを実証している。
【0150】
同様に、上記(2)と同じ方法を用いて野生型またはUU/UA減少型B型肝炎表面(配列番号:23)抗原発現ベクターをHek293細胞にトランスフェクションした。発現されたタンパク質をmIU/mlとして定量した。約4倍の差があった(図6)が、これは、おそらく独立した実験ではいっそう高い場合があるであろう。
【0151】
これは、本発明による方法が、改変された、すなわちコード配列がUU/UA減少型である治療用タンパク質、抗体およびワクチンでもまた有効であり、このことが実質的な発現増大を誘導することの一例である。
【0152】
(5)転写変化およびその後のレポートされたタンパク質レベルに対する効果を反映させるために、PEST配列(=プロリン、グルタミン酸、セリンおよびトレオニンに富むペプチド配列)を含むタンパク質不安定化アミノ酸領域を使用して、様々なレポータータンパク質の半減期を短縮した。PEST配列は、短い細胞内半減期を有するタンパク質と関連する。Liら(J. Biol. Chem., 1998, 273, pp. 34970-34975)は、EGFPを不安定化するためのMODCのPEST配列の使用を記載し、Leclercら(Biotechniques, 2000, 29, pp. 590-591, pp. 594-596は、ホタルルシフェラーゼのタンパク質半減期を短縮するためにPEST配列を使用した。
【0153】
そのようなMODCドメイン(マウスオルニチンデカルボキシラーゼ)、さらに具体的には高度不安定性MODCの分解ドメインのアミノ酸422〜461を使用して、本発明者らは、いくつかのレポーター遺伝子を前記MODCドメインと融合させることによって不安定にした。さらに、本発明者らは、Aequorea Victoria、Montastrea Cavernosa、ClavulariaおよびPuntelina Plumate由来EGFPに関して、本発明にしたがってレポーター遺伝子およびMODCドメインの両方におけるUU/UAジヌクレオチド数を減少させることによって、そのような融合体を改変した。UU/UAジヌクレオチド数は、融合体のEGFP部およびMODC部の両方で減少させた。
【0154】
それぞれフォワードおよびリバースプライマーにEcoRIおよびBamHI部位を有する特異的プライマーを使用して、マウス線維芽細胞のゲノムDNAからMODCドメインを増幅させた。同じ制限部位を使用して、増幅したcDNAを、GFPコード領域を有するフレーム内にクローニングした。図7に示すように、Hek293細胞に不安定化されたGFPをトランスフェクションした。イメージング装置およびソフトウェアによって蛍光強度を定量した。Aequorea Victoria由来の不安定化野生型EGFPに比べて(すなわち評価の基準は、野生型MODC(配列番号:29)と融合した野生型EGFP(配列番号:8)であった)、改変Aequorea Victoria由来蛍光からは2倍の増大、改変Montastrea Cavernosa緑色蛍光タンパク質からは8倍の増大、改変Clavularia緑色蛍光タンパク質からは4倍の増大、および改変Puntelina Plumate緑色蛍光タンパク質からは7倍の増大があった(図7参照)。本明細書における「改変」という用語は、「UU/UA減少型であって、MODC(それ自体もUU/UA減少型)と融合された」ことを意味する。
【0155】
結果として、このことは、通常発現シグナルがより弱い状況でも本発明が有効であり、そのような状況で本発明が発現の増大倍率を改善することを示している。
【0156】
【表6】
【技術分野】
【0001】
本発明は、タンパク質の核酸配列のコードおよび/または非コード領域におけるRNase Lの切断部位数を減少させることによって、細胞における、好ましくは真核細胞における該タンパク質の発現を増大させる方法に関する。さらに、本発明は、減少したRNase L切断部位数を示す核酸配列に加えて、そのような配列から翻訳されたタンパク質に関する。
【0002】
一過性応答遺伝子は、細胞増殖、シグナル伝達事象、ならびに炎症性刺激、微生物、および放射線などの外因性因子に対する応答を含めた、重大な生物学的応答をレギュレーションする(Lai et al., 2006; Lal et al., 2004; Lopez de Silanes et al., 2005)。それらは、特異的配列エレメントなどのシス作用因子と、ある種のRNA結合タンパク質のようなトランス作用因子との両方によってコントロールされる。主として3’非翻訳領域(3’UTR)における配列エレメントおよびRNA結合タンパク質によるモデュレーションは、メッセンジャーRNA(mRNA)の安定性およびタンパク質の翻訳に影響を及ぼすことがある。
【0003】
かなりの数の遺伝子が、そのmRNAの3’UTRに不安定化配列エレメントを、主としてアデニル酸ウリジル酸(AU)リッチエレメント(ARE)を有する。これらのAREは、タンパク質とmRNAとの相互作用に影響を及ぼすおそれのある不均一な群の配列クラスを含むことから、mRNAの分解特性を左右する(Bakheet et al., 2006; Barreau et al., 2005)。細胞性mRNAの安定化は、HuRタンパク質などのmRNA安定化促進タンパク質の活性によって、またはジンクフィンガータンパク質であるトリステトラプロリン(TTP)などのRNA分解促進タンパク質の不活性化によって起こる場合がある。
【0004】
細胞性mRNAに影響を及ぼすおそれのある、別のクラスのトランス作用因子は、普遍的細胞内酵素である、エンドリボヌクレアーゼのリボヌクレアーゼL(RNase L)であり、それは、以前はウイルスmRNAに特異的であると考えられていた。しかし、最近の研究は、RNase Lがある種の生物学的過程の一過性応答に関与する可能性があることを示した(Khabar et al., 2003a; Li et al., 2000)。RNase Lは、インターフェロン(IFN)系の一部であると見なされている。IFNは、酵素オリゴアデニル酸シンターゼ(OAS)の遺伝子発現を誘導し、それはウイルス二本鎖RNA中間体に結合すると活性化し、短い2’−5’オリゴアデニル酸(2−5A)を合成する。これらは、今度はウイルスmRNAを強力に分解するRNase Lを活性化する。RNase Lは、ナノモル濃度以下のレベルの2−5Aによって活性化され、優先的にウイルスmRNAのUUおよびUAジヌクレオチドの後でウイルスRNAの1本鎖領域の切断を招く(Wrechester et al., 1981; Han et al., 2004)。さらに高いレベルでは、RNase Lは18Sおよび28SリボソームRNAの切断などのより広い効果を誘導することがある(Wreschner et al., 1981)。
【0005】
近年、選ばれた細胞性mRNAの分解にRNase Lが関与することが広く受け入れられるようになった(Bisbal et al., 2000; Chandrasekaran et al., 2004; Khabar et al., 2003b; Le Roy et al., 2001; Li et al., 2000)。具体的には、RNase Lは、PKR mRNAをダウンレギュレーションすることが示された(Khabar et al., 2003b)。正常細胞におけるIFN抗ウイルス応答中に、PKR mRNAの発現は一過性であるが、RNase L−ヌル細胞では、mRNAの安定性増大が原因で長期動態のPKR mRNA発現が観察される(Khabar et al., 2003b)。その効果は、ウイルスタンパク質の合成阻害を誘導する過程である真核生物翻訳開始因子2サブユニット(eIF2a)のPKR依存性リン酸化の延長を招く(Khabar et al., 2003b)。したがって、RNase Lは、PKRによって付与される翻訳停止が短時間であることを確実にするために、IFN応答が一過性であることを担っている。RNase LがIFN応答をネガティブレギュレーションする同様の役割は、また、ISG43と呼ばれる43kDユビキチン特異的プロテアーゼをコードする新規なIFN活性化遺伝子がRNase Lによってダウンレギュレーションされるという報告によって示唆されている(Li et al., 2000)。ISG43 mRNAの半減期がRNase L−ヌル細胞で増大することから、このレギュレーションは、mRNA安定性のレベルで起こる(Li et al., 2000)。RNase Lは、筋分化に不可欠な重要な転写因子をコードする、別の機能的に重要な細胞性mRNAであるmyoDをダウンレギュレーションすることができる。RNase Lおよびその抑制因子RLIは、C2細胞系が筋芽細胞から筋管に分化する間に連続的に誘導される(Bisbal et al., 2000)。RNase Lの阻害または過剰発現は、MyoD mRNAの半減期をそれぞれ延長または短縮する(Bisbal et al., 2000)。RNase L分子のプールがミトコンドリアに局在し、IFN−α処置後に増大することから、CYTB、ATPase 6(ATP6)、およびチトクロームオキシダーゼII(CO)のmRNAなどのミトコンドリアmRNAのダウンレギュレーションにRNase Lが果たす役割が、IFNの抗増殖作用のメカニズムとして提案されている(Le Roy et al., 2001)。これは、アンチセンス構築物の誘導を介してRNase L活性を低下させることによって、または2−5AによってRNase L活性を直接活性化することによって実証された(Le Roy et al., 2001)。RNase Lは、特に2−5Aのレベルが限定的な条件で、非ウイルス性mRNAに比べてウイルス、例えば脳心筋炎ウイルス(EMCV)のmRNAに対して選択性を示す(Li et al., 1998a)。細胞性mRNAに対するRNase Lの効果は、特定のmRNAに大いに限定されると考えられる。というのも、この主題を扱った研究で細胞性mRNAに対する全体的な効果が観察されないからである。さらに、上記研究のいずれによっても、細胞性ターゲットmRNAへのRNase Lの直接結合も、ウイルスmRNAに対して示されたような配列特異性も実証されていない。したがって、非ウイルス性細胞性mRNAに関連するRNase L活性のメカニズムは、大部分が不明なままである。
【0006】
以前にドミナントネガティブ型のRNase Lが、アミノ酸置換または完全長タンパク質の短縮化のいずれかによって生成された。例えばドミナントネガティブRNase LであるZB1(カルボキシ末端の89個のアミノ酸を欠如した短縮型マウスRNase L)は、野生型RNase Lの抗ウイルス作用および抗増殖作用を阻害する(Hassel et al., 1993)。Dongら(2001)は、RNase Lの他の短縮化および点変異、例えばヌクレアーゼドメインにおける変異(R667A)を記載している。
【0007】
米国特許第6,762,038号は、トランスフェクションされた遺伝子の発現強化のために、RNase L遺伝子にホモ接合性破壊を有するマウスから作出された変異型胚性線維芽細胞系(MEF)の使用を示唆している(Zhou et al., 1997)。しかし、発現強化の効果は、これらの特定の細胞系に限られる。さらに、新しいRNase L−ヌル細胞系の創出は、非常に骨の折れる工程であって、全ての哺乳動物および/または真核生物発現系に容易に適用できるわけではない。
【0008】
哺乳動物発現系は、治療用タンパク質および抗体の産生のための重要な手段となっており、原核生物発現系に比べて、例えばタンパク質の正しいフォールディングおよび解糖などの翻訳後修飾に関していくつかの利点を有する。しかし、多数の系においてタンパク質発現が低収率であることは、費用のかかる技術的障害となる。所望のタンパク質、例えばG−タンパク質結合レセプター(GPCR)などの膜タンパク質、大型タンパク質、抗体、融合タンパク質、タンパク質複合体、ワクチン、および血漿タンパク質を発現させることが本質的に困難ならば、これは特に問題である。この困難の理由は、タンパク質自体の本質的な不安定性、AUリッチエレメント含有mRNAの場合のようなmRNAの本質的な不安定性、または弱いプロモーター活性であろう。
【0009】
これらの問題に共通の解決は、CMVプロモーターなどの強力なプロモーター、およびCMVのイントロンAなどの特定種類のイントロンを含めた、そのプロモーターの上流または下流のエンハンサーエレメントを提供することによってタンパク質の発現を増大させることに焦点を当てていた。他の解決は、核および染色体の構築に重要な300〜3000塩基対長のDNAエレメントであるクロマチンマトリックス付着領域(MAR)を関与させた。これらのエレメントは、隣接クロマチンが導入遺伝子の発現に影響を及ぼすことを阻止し、そのことが、所望のレギュレーション性発現を示すクローンを単離する確率の増大を誘導すると提案された(米国特許第7,129,062号)。この特定のアプローチは、その効果を達成するためにいくつかの異なるベクター構築物を必要としうることから、潜在的に問題がある。これらのアプローチが利用可能であるにもかかわらず、上記のタンパク質のように、発現が困難なタンパク質のタンパク質発現をさらに増大させる必要性は、依然として存在する。
【0010】
コドン最適化(またはコドン使用頻度最適化)は、タンパク質産生を押し上げるための、当技術分野において公知の別法である。それは、発現されるべきタンパク質をコードする核酸配列がホストによって稀に使用されるコドン(トリプレット)を有するならば、その発現レベルは最大ではないという観察に基づく。コドン最適化は、基本的にターゲット核酸配列における稀なコドンを変化させることにより、それらがホストのコドン使用頻度をより厳密に反映させることを含む。したがって、普通、最適化工程に使用される情報は、最適化されるDNAまたはタンパク質配列、およびそれぞれのホストのコドン使用頻度表である(例えばヒトゲノムについては表1参照)。コドン使用頻度表は、所与の生物における特定のアミノ酸についてそれぞれの可能なコドンの相対頻度を一覧している。全ての生物におけるコドン使用頻度のバイアスの全一覧は、次のウェブサイトから見出すことができる:http://www.kazusa.or.jp/codon/。所与のホスト生物に対するコドン使用頻度のバイアスに基づいてコドンを最適化するために、いくつかのウェブベースのプログラムも利用可能である。コドン最適化は、非ヒトまたは非哺乳動物起源の遺伝子がヒトまたは他の哺乳動物ホスト細胞において発現される(またはその逆)一部の状況で成功することがある。しかし、コドン使用頻度は、タンパク質の発現レベルを左右する多数の要因のわずか一つであって、コドン最適化の効果は多くの場合に限定的である。
【0011】
任意の生物の細胞において、好ましくは真核細胞において発現される、本質的に発現が困難なタンパク質を含めた内因性および外因性(リコンビナント)タンパク質の収率を顕著に改善する方法を提供することが、本発明の一目的であった。この方法は、時間および費用効率的であるべきであり、原核細胞および真核細胞を含めた任意の種類の生物の細胞におけるタンパク質の大規模産生を可能にするべきである。治療的に使用されるタンパク質の産生を可能にするために、使用される細胞系、リコンビナントタンパク質の特性、または外因性材料の使用などの、リコンビナントタンパク質を産生するために規制当局に認可された特徴を変更せずに、発現収率におけるこの改善を達成することが、本発明の別の目的であった。
【0012】
本発明の目的は、タンパク質の核酸配列におけるRNase L切断部位を減少させる工程を含む、細胞における、好ましくは真核細胞における該タンパク質の発現を増大させる方法によって解決される。一態様では、該細胞は原核細胞であり、別の態様では、該細胞は真核細胞である。
【0013】
タンパク質の核酸配列は、コード領域および非コード領域、すなわちアミノ酸配列に翻訳される領域(エクソンとも呼ばれる)およびアミノ酸配列に翻訳されない領域の両方を含む。核酸配列の非コード領域は、例えば5’非翻訳領域(5’UTR)、3’非翻訳領域(3’UTR)、およびイントロンである。これらのエレメントの全て(5’UTR、3’UTR、イントロン、およびコード領域)は、遺伝子およびタンパク質の発現をコントロールすることができることから、上記方法に関するターゲットである。本発明により、RNase Lの切断部位数を減少させる該工程は、コード領域または非コード領域のいずれかまたは両方における該数を減少させる。
【0014】
一態様ではRNase Lの該切断部位数は、(野生型核酸配列におけるRNase Lの切断部位数に比べて)少なくとも10%、好ましくは少なくとも25%、さらに好ましくは少なくとも50%減少する。
【0015】
好ましくは、該切断部位は、UUおよび/またはUAジヌクレオチドである。
【0016】
RNase Lは、エンドリボヌクレアーゼであることから、RNA(一次RNA、すなわちスプライシングされていないRNAおよびmRNA、すなわちスプライシングされたRNA)レベルでのみ活性である。UUおよびUAジヌクレオチドは、RNAレベルでのみ存在する。
【0017】
しかし、該タンパク質の核酸配列におけるRNase L切断部位数を減少させる該工程は、DNAレベルで行われることが好ましく、RNA配列におけるUUおよびUAジヌクレオチドはDNA配列におけるTTおよびTAヌクレオチドに対応する。所与のDNA配列を特異的に変化させる技法は、当技術分野において周知であり、それには、非限定的に遺伝子合成、部位特異的変異誘発、制限酵素消化による欠失変異、および組換えによる変異導入が含まれる。遺伝子合成の技法は、本発明により特に好ましい。
【0018】
本発明の一態様では、RNase L切断部位数を減少させる該工程は、該核酸配列のコード領域における該数を減少させる。
【0019】
好ましくは、該核酸配列におけるRNase L切断部位数を減少させる該工程は、該タンパク質のアミノ酸配列を変更せずに行われる。したがって、オープンリーディングフレーム(ORF)は、該工程によって変更されない。
【0020】
RNase L切断部位数を減少させる該工程における一態様では、UUおよび/またはUAジヌクレオチドを含むコドンは、UUおよび/またはUAジヌクレオチドを含まない、同じアミノ酸をコードする代替コドンに交換される。
【0021】
RNase L切断部位数を減少させる該工程における一態様では、UUおよび/またはUAジヌクレオチドを含む隣接コドン対がもはやUUおよび/またはUAジヌクレオチドを含まないように、該隣接コドン対の少なくとも一つのコドンが、同じアミノ酸をコードする代替コドンに交換される。
【0022】
好ましくは、UUおよび/またはUAジヌクレオチドを含む該隣接コドン対の第一のコドンが交換される。
【0023】
一態様では、該代替コドンは、該細胞において、好ましくは該真核細胞において、より頻繁に使用されるコドンである。
【0024】
本発明の一態様では、RNase L切断部位数を減少させる該工程は、該核酸配列の非コード領域における該数を減少させる。
【0025】
好ましくは、該非コード領域は5’UTR、3’UTR、またはイントロンである。
【0026】
イントロンの例には、非限定的にCMVイントロン、SV40イントロン、ウサギβグロビンイントロン(RBTG)、および合成イントロンが含まれる。
【0027】
一態様では、該核酸配列の非コード領域におけるRNase Lの切断部位数を減少させる該工程は、ヌクレオチドの変異、欠失、または挿入によって行われる。
【0028】
好ましくは、該核酸配列の非コード領域におけるRNase Lの切断部位数を減少させる該工程は、開始コドン(ATG)に近接した5’UTRにおける配列のように、非コード領域における機能的に重要なエレメントを変更しないが、それは、それらが、翻訳強化配列(例えばコザック)、3’UTRにおけるポリAシグナル(例えばAAUAAAまたはAUUAAA)またはポリアデニル化に使用される他の必要もしくは補助配列エレメント、イントロン−エクソン接合部/境界域、スプライシングブランチポイントおよびイントロンにおけるエクソンのスプライス供与/受容部位、およびスプライス受容部位からブランチポイント終点の間のCTリッチ領域を有する場合があるからである。例えば、強力なポリAシグナル下流にU−リッチのヌクレオチド50個の領域があるが、それは、可能ならば変更すべきでない(Legendre and Gautheret, 2003)。
【0029】
好ましくは、非コード領域におけるRNase Lの切断部位数を減少させる該工程は、(a)イントロンのGC含量を80%よりも大きく、かつその長さを80%未満に変化させること、
(b)5’UTRのGC含量を80%よりも大きく変化させること、および
(c)3’UTRのGC含量を80%よりも大きく、かつその長さを80%未満に変化させること
はない。
【0030】
一態様では、上記方法はさらに、RNase Lの切断部位数を減少させる該工程の前にコドン最適化の工程を含む。
【0031】
一態様では、上記方法はさらに、RNase Lの切断部位数を減少させる該工程の後に、発現活性PCR産物の形のまたは発現ベクターに含まれる、該タンパク質の該核酸配列を、該細胞に、好ましくは該真核細胞にトランスフェクションする工程を含む。
【0032】
本明細書に使用される「発現活性PCR産物」という用語は、cDNA、オープンリーディングフレーム、または発現ベクターに含まれる遺伝子などの関心が持たれるDNA配列の隣接配列に相補的な2個のプライマーを使用したPCR増幅によって生成されるPCR産物を表すことを意図し、ここで、結果として生じるPCR産物は、プロモーター、関心が持たれるDNA配列および終結配列を含み、ホスト細胞にトランスフェクションされた場合に、関心が持たれるDNAを発現させる(Al-Zoghaibi et al., 2007も参照されたい)。
【0033】
本発明によると、任意の発現ベクターを使用することができるけれども、真核生物/哺乳動物発現ベクターが好ましい。哺乳動物発現ベクターは、タンパク質の生物学的機能を研究するための一般に普及したツールであり、様々な種類(例えばプラスミドベクターまたはウイルスベクター)が当技術分野において公知である。適切な発現ベクターは、特定のプロモーター、5’UTR、3’UTR、またはイントロンに限定されない。それらは、構成的発現型、誘導型、または抑制型またはレギュレーション可能型でありうる。
【0034】
本発明によると、プロモーターが真核生物性であり、終結部位が、ポリアデニル化シグナルを有するポリA配列であることが好ましい。真核生物または哺乳動物プロモーターは、当技術分野において公知であり、それらには、非限定的にサイトメガロウイルス(CMV)前初期プロモーター、SV40プロモーター、伸長因子(EF)プロモーター、RSVプロモーター、およびニワトリβ−アクチンプロモーターが含まれる。好ましい真核生物ポリアデニル化シグナルは、ウシ成長因子(BGH)ポリ部位、成長ホルモンポリA、SV40ポリA、およびHSKポリAである(Foecking and Hofstetter, 1986; Kobayashi et al., 1997)。真核生物ターミネーターの例は、ウシ成長因子(BGH)ポリA部位、SV40ポリA、HSKポリA、および合成ポリAである。
【0035】
細胞にDNA/ベクターを一過性または安定的にトランスフェクションする方法は、当技術分野において周知である。これらには、非限定的にリン酸カルシウム共沈、エレクトロポレーション、陽イオンポリマートランスフェクション、およびリポソーム介在性トランスフェクション(例えばリポフェクション)が含まれる。リポソーム介在性トランスフェクション用の試薬は、市販されており、例えばリポフェクタミン(Invitrogen)およびポリエチレンイミン(Sigma)である。細胞は、また、ウイルストランスフェクションによって、またはウイルスコート粒子を介してトランスフェクションすることができる。本発明により細胞をトランスフェクションする別の好ましい方法は、該発現活性PCR産物または該発現ベクターをin vivoマイクロインジェクションすること、および該発現活性PCR産物または該発現ベクターを有する細胞を選択薬の助けを借りてまたは借りずに選択的に成長させることである。したがって、本発明による発現活性PCR産物または発現ベクターは、追加的に選択マーカーを含むことがある。安定な細胞系を産生させるために、クローンは、非限定的にネオマイシン、ブラストサイジン、ピューロマイシン、ゼオシン(zeocin)、ハイグロマイシン、およびジヒドロ葉酸レダクターゼ(dhfr)が含まれる様々な選択マーカーを使用して選択することができる。
【0036】
上記方法の全ての適切な真核生物/哺乳動物細胞(ホスト細胞)もまた、当技術分野において周知であって、それらには、非限定的にCHO、HEK293、HeLa、およびCOS−7細胞が含まれる。
【0037】
一態様では、上記方法は、さらに、該細胞における、好ましくは該真核細胞における該発現活性PCR産物または発現ベクターから該タンパク質を翻訳させる工程を含む。
【0038】
好ましくは、該タンパク質は、レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択される。
【0039】
本発明の目的は、また、RNase Lの切断部位数が(野生型核酸配列におけるRNase Lの切断部位数に比べて)少なくとも10%、好ましくは少なくとも25%、さらに好ましくは少なくとも50%減少している核酸配列によって解決される。
【0040】
好ましくは、該核酸配列は、上記方法によって産生される。
【0041】
好ましくは、該核酸配列は、レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択されるタンパク質の核酸配列である。
【0042】
一態様では、該核酸配列は、配列番号:2、3、5、7、9、11、13、15、17、19、23、24、26、28、30〜36、好ましくは配列番号:9、15、17、23、30〜36より選択される配列を有する。
【0043】
本発明の目的は、また、上記核酸を含む発現活性PCR産物または発現ベクターによって解決される。
【0044】
本発明の目的は、さらに、上記の発現活性PCR産物または発現ベクターを有するホスト細胞によって解決される。
【0045】
最終的に、本発明の目的は、また、上記方法によって産生されるタンパク質であって、レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択されるタンパク質によって解決される。
【0046】
本発明によると、「治療用タンパク質」という用語は、抗体フラグメント、免疫グロブリン鎖、重鎖および軽鎖、Fabフラグメント、酵素、成長因子、インターフェロン、サイトカイン、リンホカイン、接着分子、レセプターに加えて、その誘導体またはフラグメントなどの、薬学的に関連して使用されるタンパク質を包含することが意図される。
【0047】
本発明によると、レポータータンパク質(単独または別のタンパク質と融合したもの)は、上記方法のために特に好ましい「ターゲット」である。(非限定的に)緑色蛍光タンパク質(GFP)、赤色蛍光タンパク質(RFP)、黄色蛍光タンパク質(YFP)、青色シアニン蛍光タンパク質(CFP)、ルシフェラーゼ、分泌型アルカリホスファターゼ(SEAP)、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、分泌型ホルモン、分泌型サイトカイン、β−ガラクトシダーゼ、ならびに他の蛍光および生物発光タンパク質を含めた、多数の蛍光および非蛍光レポータータンパク質がある。レポータータンパク質の選択は、使用される細胞系(内因性活性)、実験の性質(例えば遺伝子発現の動力学およびトランスフェクション効率)、および選択された検出方法に対するアッセイの適応性に依存する(Naylor 1999)。迅速な応答および変化のなどのレポーター性能を改善するために、レポータータンパク質自体のいくつかの改変、例えば不安定化エレメントの使用が探究されている(Li et al., 1998b; Zhao et al., 1995)。緑色蛍光タンパク質(GFP)および他の蛍光タンパク質は、生組織、細胞および実験動物における遺伝子発現を非侵襲的にモニタリングするためにますます普及しつつある(Naylor 1999)。したがって、GFP(およびEGFPなどのその誘導体)、他の蛍光タンパク質(上記参照)に加えて、蛍光タンパク質を含む融合タンパク質は、本発明による方法のためになおさらに好ましい「ターゲット」タンパク質である。
【0048】
驚くことに本発明者らは、真核生物および/または哺乳動物発現系で発現されるべきタンパク質の核酸配列におけるRNase Lの切断部位、好ましくはUUおよびUAジヌクレオチドの数を減少させることが、該タンパク質の発現および収率の顕著な改善を招くことを見出した。一定の理論に縛られることを望むわけではないが、RNase L切断部位の減少(すなわちこれらの部位の頻度減少)が、ホスト細胞において、好ましくは真核生物ホスト細胞において内因性RNase Lの攻撃を受けにくい(すなわち抵抗性が高い)核酸配列、すなわち(一次RNAおよびmRNAの両方の)RNAを生じ、mRNAの安定性増大を、したがってタンパク質の発現増大を誘導すると考えられる。
【0049】
関係する普遍的な概念のせいで、本発明は、任意の配列に適用することができる。さらに具体的には、本発明による方法は、内因性および外因性(リコンビナント)タンパク質/遺伝子の両方に、安定的に組み込まれた遺伝子および一過性発現遺伝子の両方に、発現が困難なタンパク質/遺伝子に、ならびに少量に内因性発現されるタンパク質/遺伝子にも適用可能である。それは、原核生物および真核生物系の両方に適用することができる。本明細書に記載されたアプローチは、タンパク質産生に費やされる時間およびコストの顕著な減少を誘導することから、エリスロポエチン、成長因子、インターフェロン、インスリン、治療用および診断用抗体、ならびにタンパク質またはペプチド系ワクチンなどの生物学的製剤として使用されるタンパク質/遺伝子のさらに効率的な産生を可能にするであろう。それは、大量に発現/産生させることが非常に困難であることが証明された遺伝子/タンパク質の発現に特に有用である。例には、非限定的にGタンパク質結合レセプター(GPCR)などの膜タンパク質、大型タンパク質、抗体、融合タンパク質、タンパク質複合体、ワクチン、および血漿タンパク質が含まれる。それは、また、レポータータンパク質の、例えばGFPまたはルシフェラーゼなどの蛍光タンパク質の発現および性能を顕著に改善することに役立つことから、そのようなレポータータンパク質を使用する方法の感度(蛍光または発光顕微鏡法、蛍光ベースのマイクロアレイ、細胞選別など)を増大することができる。さらにこのアプローチは、(非限定的に)ハムスターCHO1およびHEK293を含めた細胞系などの、生物工学工業に使用される任意の細胞系に適用可能であることから、特に実践的であり、かつ簡単である。
【0050】
以下に、本発明にしたがってRNase Lの切断部位数を減少させる工程の一般的原理を説明する。
【0051】
コード領域におけるUUおよび/またはUAジヌクレオチド数の減少
表2および表3は、引き起こすことのできる変化を示す。これらの改変は、所与の生物におけるそれらのコドンバイアス(コドン使用頻度)に基づきタンパク質発現のためにコドンを最適化するために使用されるコドン使用頻度表(比較のために表1参照)と全く別であることに留意することが重要である。本方法は、コドン使用頻度に基づいてではなく、RNase Lの切断部位/ターゲット(UUおよびUAジヌクレオチド)数を減少させるためにコドンを変化させることを対象とする。さらに、本発明による方法は、「生物依存的」ではなく、UUおよび/またはUAジヌクレオチドの数/頻度は、対応するアミノ酸配列が変更されないままである限り(表2および3参照)、任意の生物由来の任意の遺伝子(核酸配列)において減少させることができる。さらに、UUおよび/またはUAジヌクレオチド数の減少は、また、所望の生物についての古典的コドン使用頻度最適化(例えば表1参照)と組み合わせることができ、可能性があることには、いっそうさらなる発現増大を招く。
【0052】
表2は、UUおよび/またはUAジヌクレオチドを有するコドンに加えて、それらの非UU/UA含有代替物を示す。フェニルアラニン(Phe)についてUUまたはUAを完全に欠如した他のコドンがないことから、フェニルアラニンをコードするUUUは、UUCだけに変化させることができる。これは、UACだけに変化させることができるチロシン(UAU)の場合も同様である。いったん表2により変化を加えたら、表3により、UUまたはUAを形成するジ−トリプレット、すなわちNNU UNNまたはNNU ANN(Nは任意のヌクレオチドである)を変化させる。代替物が1個を超えて存在する場合は、それぞれの生物または高発現される遺伝子において最も高頻度に使用されるコドンを優先する。
【0053】
コード領域におけるUUおよび/またはUAジヌクレオチドを減少させるための工程:
1. 表2によりコドンを変化させる。
2. 代替コドンが1個を超えて存在する場合は、所望の生物においてより高頻度に使用されるコドンを使用する(随意)。
3. 表3により、一緒になってUUまたはUAジヌクレオチドを形成するジ−トリプレット(NNU UNN、NNU ANN)の最初のコドンを変化させる。
4. 1個を超える代替コドンが存在する場合は、所望の生物においてより高頻度に使用されるコドンまたは発現が最強のコドンを使用する(随意、表4および5参照)。
5. 古典的コドン使用頻度最適化(ホスト細胞生物、または高発現遺伝子において最も高頻度に使用されるコドンの一覧に基づく)を、所望により、そして好ましくはUUおよびUA減少の前に行うことができる。
【0054】
イントロンにおけるUUおよび/またはUAジヌクレオチド数の減少
イントロンにおけるUUおよび/またはUAジヌクレオチドを減少させるための工程:
1. UU/UAジヌクレオチドにおける2個のヌクレオチドのうち1個または2個を変異または欠失させる:UUまたはUAをUC、UG、GA、CA、またはTAにする。または1個のヌクレオチドを挿入する。
2. イントロンの全GC含量は、80%を超えるべきではなく、かつ長さはその本来の長さの80%未満に変化させるべきではない。
3. エクソンのスプライス供与部位および受容部位ならびにブランチポイントを含めたエクソン−イントロン境界部を変化させてはならない。スプライスアクセプター部位からブランチポイント終末にわたるCT−リッチ領域を破壊することを避ける。
【0055】
5’UTRにおけるUUおよび/またはUAジヌクレオチド数の減少
5’UTRにおけるUUおよび/またはUAジヌクレオチドを減少させる工程:
1. UUまたはUAをUC、UG、GA、CA、またはTAに変異させる。
2. 5’UTRの全GC含量は、80%を超えるべきではない。
3. コンテクスト配列はコザックなどの翻訳強化配列を有することがあるため、開始コドンATG近くのコンテクスト配列を避ける。
【0056】
3’UTRにおけるUUおよび/またはUAジヌクレオチド数の減少
3’UTRにおけるUUおよび/またはUAジヌクレオチドを減少させる工程:
1. UU/UAジヌクレオチドにおける2個のヌクレオチドのうち1個または2個を変異または欠失させる:すなわちUUまたはUAからUC、UG、GA、CA、またはTAにする。または、1個のヌクレオチドを挿入する。
2. 3’UTRの全GC含量は80%を超えるべきではなく、かつ長さはその本来の長さの80%未満に変化させるべきではない。
3. AAUAAAまたはAUUAAAなどのポリAシグナルを変化させてはならず、ポリアデニル化に使用される任意の必要または補助配列エレメントが認められる場合は、それを変更することを避ける。
【0057】
非コード領域における可能性のある変化
1. 変異:NUUNまたはNUANからNUSNへ(SはGまたはCであり、Nは任意のヌクレオチドである)
2. 挿入:NUUNまたはNUANからNUSUNまたはNUSAへ(SはGまたはCであり、Nは任意のヌクレオチドである)
3. 欠失:NUUNからNUSへ(SはGまたはCであり、Nは任意のヌクレオチドである)
4. 欠失:NUANからNANまたはNUSへ(SはGまたはCであり、Nは任意のヌクレオチドである)
【0058】
表
【0059】
【表1】
【0060】
【表2】
【0061】
【表3】
【0062】
【表4】
【0063】
【表5】
【0064】
これから以下の図を参照する。
【図面の簡単な説明】
【0065】
【図1】哺乳動物発現系におけるEGFPの発現に及ぼすUUおよびUAジヌクレオチドの減少の効果を野生型配列と比べて示すグラフである。
【図2】RNase Lの同時過剰発現を伴う哺乳動物発現系におけるEGFPの発現に及ぼすUUおよびUAジヌクレオチドの減少の効果を、野生型配列と比べて示すグラフである。
【図3】UU/UA減少型改変EGFP配列または野生型EGFP配列を有するPCR産物をトランスフェクションされた哺乳動物細胞におけるGFP蛍光を示すグラフである。
【図4】野生型または改変(すなわちUU/AGを減少させた)ホタルルシフェラーゼ発現ベクターのいずれかをトランスフェクションされたHek293細胞におけるルシフェラーゼ活性を示すグラフである。
【図5】野生型またはUU/UA減少型ホタルルシフェラーゼ発現ベクターから生成した異なるPCR産物を、トランスフェクションされたHuh7細胞におけるルシフェラーゼ活性を示すグラフである。
【図6】野生型またはUU/UA減少型B型肝炎表面抗原発現ベクターをトランスフェクションされたHek293細胞におけるB型肝炎表面抗原の発現を示すグラフである。
【図7】MODCで不安定化された野生型およびMODCで不安定化されたUU/UA減少型緑色蛍光タンパク質をトランスフェクションされたHek293細胞における様々な緑色蛍光タンパク質の発現を示すグラフである。
【0066】
これから以下の実施例を参照して本発明をさらに説明するが、それらの実施例は、例示であって、本発明の範囲を限定するつもりはない。
【0067】
実施例1:イントロンにおけるUUおよび/またはUAジヌクレオチドの減少
/=スプライシング部位
下線=コンセンサス機能的部位
太字の下線付き斜体=ブランチポイント
【0068】
配列番号:1: 野生型ウサギβグロビン−イントロン1(RBTG1):
【0069】
【化1】
【0070】
配列番号:2: UU/UA減少型RBTG1(CTリッチ領域にUU/UA減少を有さず):
【0071】
【化2】
【0072】
配列番号:3: UU/UA減少型RBTG1(CTリッチ領域に最低限のUU/UA減少を有する):
【0073】
【化3】
【0074】
実施例2: 3’UTRにおけるUUおよび/またはUAジヌクレオチドの減少
.=欠失
下線=突然変異
太字の下線付き=ポリAシグナル
【0075】
配列番号:4: 野生型SV40 3’UTR
【0076】
【化4】
【0077】
配列番号:5: UU/UA減少型SV40 3’UTR
【0078】
【化5】
【0079】
配列番号:6: 野生型ウシ成長ホルモン(BGH)3’UTR
【0080】
【化6】
【0081】
配列番号:7: UU/UA減少型BGH 3’UTR
【0082】
【化7】
【0083】
実施例3: コード領域におけるUUおよび/またはUAジヌクレオチドの減少
配列番号:8: 野生型の強化緑色蛍光タンパク質(EGFP)
【0084】
【化8】
【0085】
配列番号:9: UU/UA減少型EGFP(「スーパーGFP」)
【0086】
【化9】
【0087】
配列番号:10
MONTASTREA CAVERNOSA由来野生型GFP配列
【0088】
【化10】
【0089】
配列番号:11
MONSTER−OM:UU/UAジヌクレオチド数を減少させることによる改変型の配列番号:10
【0090】
【化11】
【0091】
配列番号:12
CLAVULARIA種 − 野生型配列
【0092】
【化12】
【0093】
配列番号:13
CLAVULARIA種−OM:改変(UU/UA減少)
【0094】
【化13】
【0095】
配列番号:14
ホタルルシフェラーゼ+:野生型配列:
【0096】
【化14】
【0097】
配列番号:15
LUC+DU(SuperLuciferase):(UU/UA減少型)
【0098】
【化15】
【0099】
配列番号:16
ホタルLUC2ルシフェラーゼ野生型:
【0100】
【化16】
【0101】
配列番号:17
LUC2OM:LUC2改変SuperLuciferase2(UU/UA減少型)
【0102】
【化17】
【0103】
配列番号:18
Puntellina Plumate(GFP)野生型:
【0104】
【化18】
【0105】
配列番号:19
Puntellina Plumate(GFP):改変配列(UU/UA減少型)
【0106】
【化19】
【0107】
配列番号:20
Discosoma野生型配列由来赤色蛍光タンパク質
【0108】
【化20】
【0109】
配列番号:21
赤色蛍光タンパク質の改変配列
【0110】
【化21】
【0111】
配列番号:22
B型肝炎表面抗原野生型、adrB型肝炎ウイルス株:
【0112】
【化22】
【0113】
配列番号:23
HBSAGOM:B型肝炎表面抗原改変配列:
【0114】
【化23】
【0115】
配列番号:24
HBSAGM:HBSAGOM: B型肝炎表面抗原改変配列2
【0116】
【化24】
【0117】
この配列は、UU/UA減少およびヒト化の両方を含む。
【0118】
配列番号:25
IFN−α、ヒト
【0119】
【化25】
【0120】
配列番号:26
IFN−αOM:改変配列
【0121】
【化26】
【0122】
配列番号:27
CSF3Mコロニー刺激因子野生型配列、ヒト:
【0123】
【化27】
【0124】
配列番号:28
CSF3Mコロニー刺激因子改変配列:
【0125】
【化28】
【0126】
配列番号:29
MODC野生型配列(マウスオルニチンデカルボキシラーゼ)
【0127】
【化29】
【0128】
配列番号:30
MODC改変配列:
【0129】
【化30】
【0130】
不安定化および配列改変されたレポーター:
不安定化および改変されたレポーター配列の例:
配列番号:31
MONTASTRAEA CAVERNOSA
【0131】
【化31】
MODC配列は太字
【0132】
配列番号:32
Clavulariidae Clavularia−OM:改変
【0133】
【化32】
【0134】
配列番号:33
ホタルLUC+DU(SuperLuciferase):
【0135】
【化33】
【0136】
配列番号:34
ホタルLUC2OM:LUC2改変SuperLuciferase2
【0137】
【化34】
【0138】
配列番号:35
Puntellina Plumate GFP:改変配列
【0139】
【化35】
【0140】
配列番号:36
赤色蛍光タンパク質の改変配列
【0141】
【化36】
【0142】
実施例4:UU/UA減少型EGFPのin vivo分析
(1)改変EGFP配列は、遺伝子合成会社によってカスタム合成され、隣接するSalIおよびBamHI部位を有するようにpUC19ベクター中に供給した。0.1μg/ml BSAを含有する緩衝液中に10ユニットのSalIで、10μgのベクターを37℃で1時間消化させ、続いてBamHI緩衝液中のBamHIで37℃でさらに1時間消化させた。消化されたDNAをフェノール−クロロホルム法を用いて抽出し、続いてエタノール沈殿を行った。CMVプロモーターおよびBHG 3’UTRを有する発現ベクターを同じ制限酵素(SalIおよびXbaI)で消化しておいたものにに合成EGFP−コード領域をライゲーションし、フェノール−クロロホルム抽出によって精製し、続いてエタノール沈澱を行った。発現ベクターへのEGFP−DNAのクローニングは、以下のライゲーション反応を用いて行った:消化されたベクターDNA 30μgを消化されたEGFP−DNA 90μgと共に、T4 DNAリガーゼを含有する反応液10μl中で混合した。ライゲーション産物を使用して、DH5αコンピテントE. coli細胞をトランスフォーメーションし、続いて結果として得られたコロニーを細菌培養培地中でエクスパンションさせた。Qiagenプラスミド精製キット(Qiagen, Germany)を使用してリコンビナントDNAを抽出した。挿入物を有するベクターのサイズは、ゲル電気泳動を用いて検証した。野生型DNAを有するベクターに加えて、UU/UA減少型改変コード領域を有する、結果として得られた発現ベクターを機能的実験に使用して、コードされたタンパク質の発現を確認した。10% FBSおよび抗生物質(Invitrogen)を補充したDMEM培地中で標準培養条件(37℃、5% CO2)でHEK293細胞を成長させた。96ウェルプレートのウェル1個あたり2.5×104個の細胞に、EGFPのUU/UA減少型改変コード領域を有するベクターまたは野生型EGFP−DNAを有するベクター100ngをトランスフェクションした。Lipofectamine 2000(Invitrogen)を使用して無血清培地中でトランスフェクションを5時間行い、続いて培地を血清補充培地に置換した。約24または48時間後に、プレートをイメージングし、BDハイコンテントイメージング装置を使用して定量した。定量は、Proxcellイメージングアルゴリズムを用いて行った。
【0143】
データは、EGFPのUU/UA減少型コード領域の使用が、野生型EGFP−DNAに比べて真核細胞におけるEGFPの有意に高い発現(2.5〜3倍)を可能にすることを明らかに示している(図1)。
【0144】
(2)10% FBSおよび抗生物質(Invitrogen)を補充したDMEM培地中で標準培養条件(37℃、5% CO2)でHEK293細胞を成長させた。96ウェルプレートのウェル1個あたり2.5×104個の細胞に、EGFPのUU/UA減少型改変コード領域を有するベクターまたは野生型EGFP−DNAを有するベクター100ngをトランスフェクションした。また細胞に、空の対照ベクター(pcDNA3.1)またはRNase Lベクター(pcDNA3.1)のいずれかを同時トランスフェクションした。トランスフェクションは、Lipofectamine 2000(Invitrogen)を用いて無血清培地中で5時間行い、続いて培地を血清補充培地と置換した。約24または48時間後に、プレートをイメージングし、BDハイコンテントイメージング装置を使用して定量した。定量は、Proxcellイメージングアルゴリズムを用いて行った。
【0145】
データは、野生型EGFP配列に比べて/対照的に、改変EGFP配列の使用がより高い発現を招き、それがRNase Lの同時発現によって制約されなかったことを示している(図2)。
【0146】
(3)フォワード(5’)プライマーがCMVプロモーター領域の開始部またはCMVプロモーター上流の配列に相補性であって、リバース(3’)プライマーがBGHポリA部位またはこの部位の下流の配列に相補的な、プライマーを使用することによって、発現活性PCR産物を生成させた。100μlの反応液中でTaqおよびPfuポリメラーゼ混合物を使用して以下のサイクル条件でPCRを実施した:95℃、12分(ホットスタートポリメラーゼを活性化する)、94℃、1分;52℃、1分;72℃、4分を32サイクル、および72℃で7分の最終伸長。プライマー、小型PCR産物、緩衝剤、および酵素を除去するためにQiagen PCR精製カラムを使用してPCR産物を精製し、滅菌水に溶出した。10% FBSおよび抗生物質(Invitrogen)を補充したDMEM培地中で標準培養条件(37℃、5% CO2)でHEK293細胞を成長させた。96ウェルプレートのウェル1個あたり2.5×104個の細胞に、野生型または改変配列を有するEGFP発現ベクターから生成した精製PCR産物100ngをトランスフェクションした。トランスフェクションは、Lipofectamine 2000(Invitrogen)を使用して無血清培地中で5時間行い、続いて培地を血清補充培地と置換した。約24または48時間後に、プレートをイメージングし、BDハイコンテントイメージング装置を使用して定量した。定量は、Proxcellイメージングアルゴリズムを用いて行った。
【0147】
データは、EGFPのUU/UA減少型コード領域を有するPCR産物が、野生型配列を有するPCR産物よりも高いEGFP発現(5〜10倍の増大)を誘導したことを示している(図3)。
【0148】
(4)(2)および(3)に記載したものと同じ方法を用いて、野生型またはUU/UAを減少させたホタルルシフェラーゼ発現ベクター(「SuperLuciferase」、配列番号:15)をHek293細胞にトランスフェクションした。ルシフェラーゼの活性レベルは、ルミノメーターによって定量した。データは、2回の独立した実験の中で約5倍および約100倍の差があったことを示している(図4)。
【0149】
同様に、上記(3)に概略した方法によって野生型または改変ホタルルシフェラーゼ発現ベクター(「SuperLuciferase」、配列番号:15)から生成した異なるPCR産物をHuh7細胞にトランスフェクションした。ルシフェラーゼ活性レベルは、今回もルミノメーターで定量した。そのデータは、SuperLuciferaseのUU/UA減少型コード領域を有するPCR産物が野生型配列を有するPCR産物よりも実質的に高いルシフェラーゼの発現(20〜100倍の増大)を誘導したことを示している(図5)。これは、本発明による方法が、真に多様なレポータータンパク質で有効であることを実証している。
【0150】
同様に、上記(2)と同じ方法を用いて野生型またはUU/UA減少型B型肝炎表面(配列番号:23)抗原発現ベクターをHek293細胞にトランスフェクションした。発現されたタンパク質をmIU/mlとして定量した。約4倍の差があった(図6)が、これは、おそらく独立した実験ではいっそう高い場合があるであろう。
【0151】
これは、本発明による方法が、改変された、すなわちコード配列がUU/UA減少型である治療用タンパク質、抗体およびワクチンでもまた有効であり、このことが実質的な発現増大を誘導することの一例である。
【0152】
(5)転写変化およびその後のレポートされたタンパク質レベルに対する効果を反映させるために、PEST配列(=プロリン、グルタミン酸、セリンおよびトレオニンに富むペプチド配列)を含むタンパク質不安定化アミノ酸領域を使用して、様々なレポータータンパク質の半減期を短縮した。PEST配列は、短い細胞内半減期を有するタンパク質と関連する。Liら(J. Biol. Chem., 1998, 273, pp. 34970-34975)は、EGFPを不安定化するためのMODCのPEST配列の使用を記載し、Leclercら(Biotechniques, 2000, 29, pp. 590-591, pp. 594-596は、ホタルルシフェラーゼのタンパク質半減期を短縮するためにPEST配列を使用した。
【0153】
そのようなMODCドメイン(マウスオルニチンデカルボキシラーゼ)、さらに具体的には高度不安定性MODCの分解ドメインのアミノ酸422〜461を使用して、本発明者らは、いくつかのレポーター遺伝子を前記MODCドメインと融合させることによって不安定にした。さらに、本発明者らは、Aequorea Victoria、Montastrea Cavernosa、ClavulariaおよびPuntelina Plumate由来EGFPに関して、本発明にしたがってレポーター遺伝子およびMODCドメインの両方におけるUU/UAジヌクレオチド数を減少させることによって、そのような融合体を改変した。UU/UAジヌクレオチド数は、融合体のEGFP部およびMODC部の両方で減少させた。
【0154】
それぞれフォワードおよびリバースプライマーにEcoRIおよびBamHI部位を有する特異的プライマーを使用して、マウス線維芽細胞のゲノムDNAからMODCドメインを増幅させた。同じ制限部位を使用して、増幅したcDNAを、GFPコード領域を有するフレーム内にクローニングした。図7に示すように、Hek293細胞に不安定化されたGFPをトランスフェクションした。イメージング装置およびソフトウェアによって蛍光強度を定量した。Aequorea Victoria由来の不安定化野生型EGFPに比べて(すなわち評価の基準は、野生型MODC(配列番号:29)と融合した野生型EGFP(配列番号:8)であった)、改変Aequorea Victoria由来蛍光からは2倍の増大、改変Montastrea Cavernosa緑色蛍光タンパク質からは8倍の増大、改変Clavularia緑色蛍光タンパク質からは4倍の増大、および改変Puntelina Plumate緑色蛍光タンパク質からは7倍の増大があった(図7参照)。本明細書における「改変」という用語は、「UU/UA減少型であって、MODC(それ自体もUU/UA減少型)と融合された」ことを意味する。
【0155】
結果として、このことは、通常発現シグナルがより弱い状況でも本発明が有効であり、そのような状況で本発明が発現の増大倍率を改善することを示している。
【0156】
【表6】
【特許請求の範囲】
【請求項1】
タンパク質の核酸配列におけるRNase Lの切断部位数を減少させる工程を含む、細胞における、好ましくは真核細胞における該タンパク質の発現を増大させる方法。
【請求項2】
RNase Lの切断部位数が、少なくとも10%、好ましくは少なくとも25%、さらに好ましくは少なくとも50%減少される、請求項1記載の方法。
【請求項3】
切断部位が、UUおよび/またはUAジヌクレオチドである、請求項1または2記載の方法。
【請求項4】
RNase Lの切断部位数を減少させる工程が、核酸配列のコード領域における該数を減少させる、請求項1〜3のいずれか一項記載の方法。
【請求項5】
核酸配列におけるRNase Lの切断部位数を減少させる工程が、タンパク質のアミノ酸配列を変更せずに行われる、請求項4記載の方法。
【請求項6】
RNase Lの切断部位数を減少させる工程において、UUおよび/またはUAジヌクレオチドを含むコドンが、UUおよび/またはUAジヌクレオチドを含まず、同じアミノ酸をコードする代替コドンに交換される、請求項5記載の方法。
【請求項7】
RNase Lの切断部位数を減少させる工程において、該隣接コドン対の少なくとも一つのコドンが、同じアミノ酸をコードする代替コドンに交換され、UUおよび/またはUAジヌクレオチドを含む隣接コドン対がもはやUUおよび/またはUAジヌクレオチドを含まない、請求項5記載の方法。
【請求項8】
代替コドンが、細胞において、好ましくは真核細胞において、より頻繁に使用されるコドンである、請求項6または7記載の方法。
【請求項9】
RNase Lの切断部位数を減少させる工程が、核酸配列の非コード領域における該数を減少させる、請求項1〜3のいずれか記載の方法。
【請求項10】
非コード領域が、5’UTR、3’UTR、またはイントロンである、請求項9記載の方法。
【請求項11】
RNase Lの切断部位数を減少させる工程が、ヌクレオチドの突然変異、欠失、または挿入によって行われる、請求項9または10記載の方法。
【請求項12】
RNase Lの切断部位数を減少させる工程の前に、コドン最適化の工程をさらに含む、請求項1〜11のいずれか一項記載の方法。
【請求項13】
RNase Lの切断部位数を減少させる工程の後に、細胞内に、好ましくは真核細胞内に、発現活性PCR産物の形のまたは発現ベクターに含まれる、タンパク質の核酸配列をトランスフェクションする工程をさらに含む、請求項1〜12のいずれか一項記載の方法。
【請求項14】
細胞において、好ましくは真核細胞において発現活性PCR産物または発現ベクターからタンパク質を翻訳させる工程をさらに含む、請求項13記載の方法。
【請求項15】
タンパク質が、レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択される、請求項1〜14のいずれか一項記載の方法。
【請求項16】
RNase Lの切断部位数が、少なくとも10%、好ましくは少なくとも25%、さらに好ましくは少なくとも50%減少している、核酸配列。
【請求項17】
レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択されるタンパク質の核酸配列である、請求項16記載の核酸配列。
【請求項18】
配列番号:2、3、5、7、9、11、13、15、17、19、23、24、26、28、30〜36より選択される配列を有する、請求項16記載の核酸配列。
【請求項19】
請求項16〜18のいずれか記載の核酸配列を含む、発現活性PCR産物または発現ベクター。
【請求項20】
請求項19記載の発現活性PCR産物または発現ベクターを有する任意の生物のホスト細胞。
【請求項21】
レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択される、請求項14記載の方法によって産生されたタンパク質。
【請求項1】
タンパク質の核酸配列におけるRNase Lの切断部位数を減少させる工程を含む、細胞における、好ましくは真核細胞における該タンパク質の発現を増大させる方法。
【請求項2】
RNase Lの切断部位数が、少なくとも10%、好ましくは少なくとも25%、さらに好ましくは少なくとも50%減少される、請求項1記載の方法。
【請求項3】
切断部位が、UUおよび/またはUAジヌクレオチドである、請求項1または2記載の方法。
【請求項4】
RNase Lの切断部位数を減少させる工程が、核酸配列のコード領域における該数を減少させる、請求項1〜3のいずれか一項記載の方法。
【請求項5】
核酸配列におけるRNase Lの切断部位数を減少させる工程が、タンパク質のアミノ酸配列を変更せずに行われる、請求項4記載の方法。
【請求項6】
RNase Lの切断部位数を減少させる工程において、UUおよび/またはUAジヌクレオチドを含むコドンが、UUおよび/またはUAジヌクレオチドを含まず、同じアミノ酸をコードする代替コドンに交換される、請求項5記載の方法。
【請求項7】
RNase Lの切断部位数を減少させる工程において、該隣接コドン対の少なくとも一つのコドンが、同じアミノ酸をコードする代替コドンに交換され、UUおよび/またはUAジヌクレオチドを含む隣接コドン対がもはやUUおよび/またはUAジヌクレオチドを含まない、請求項5記載の方法。
【請求項8】
代替コドンが、細胞において、好ましくは真核細胞において、より頻繁に使用されるコドンである、請求項6または7記載の方法。
【請求項9】
RNase Lの切断部位数を減少させる工程が、核酸配列の非コード領域における該数を減少させる、請求項1〜3のいずれか記載の方法。
【請求項10】
非コード領域が、5’UTR、3’UTR、またはイントロンである、請求項9記載の方法。
【請求項11】
RNase Lの切断部位数を減少させる工程が、ヌクレオチドの突然変異、欠失、または挿入によって行われる、請求項9または10記載の方法。
【請求項12】
RNase Lの切断部位数を減少させる工程の前に、コドン最適化の工程をさらに含む、請求項1〜11のいずれか一項記載の方法。
【請求項13】
RNase Lの切断部位数を減少させる工程の後に、細胞内に、好ましくは真核細胞内に、発現活性PCR産物の形のまたは発現ベクターに含まれる、タンパク質の核酸配列をトランスフェクションする工程をさらに含む、請求項1〜12のいずれか一項記載の方法。
【請求項14】
細胞において、好ましくは真核細胞において発現活性PCR産物または発現ベクターからタンパク質を翻訳させる工程をさらに含む、請求項13記載の方法。
【請求項15】
タンパク質が、レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択される、請求項1〜14のいずれか一項記載の方法。
【請求項16】
RNase Lの切断部位数が、少なくとも10%、好ましくは少なくとも25%、さらに好ましくは少なくとも50%減少している、核酸配列。
【請求項17】
レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択されるタンパク質の核酸配列である、請求項16記載の核酸配列。
【請求項18】
配列番号:2、3、5、7、9、11、13、15、17、19、23、24、26、28、30〜36より選択される配列を有する、請求項16記載の核酸配列。
【請求項19】
請求項16〜18のいずれか記載の核酸配列を含む、発現活性PCR産物または発現ベクター。
【請求項20】
請求項19記載の発現活性PCR産物または発現ベクターを有する任意の生物のホスト細胞。
【請求項21】
レポータータンパク質、治療用タンパク質、抗体、ワクチン、膜タンパク質、融合タンパク質、血漿タンパク質、サイトカイン、インターフェロン、成長因子、ケモカイン、およびGPCRを含む群より選択される、請求項14記載の方法によって産生されたタンパク質。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2012−514986(P2012−514986A)
【公表日】平成24年7月5日(2012.7.5)
【国際特許分類】
【出願番号】特願2011−545697(P2011−545697)
【出願日】平成22年1月19日(2010.1.19)
【国際出願番号】PCT/EP2010/000271
【国際公開番号】WO2010/081741
【国際公開日】平成22年7月22日(2010.7.22)
【出願人】(511174465)
【氏名又は名称原語表記】KING FAISAL SPECIALIST HOSPITAL AND RESEARCH CENTRE
【出願人】(511174476)テラマーク・マーケンクリエーション・ゲーエムベーハー (1)
【氏名又は名称原語表記】TERRAMARK MARKENCREATION GMBH
【Fターム(参考)】
【公表日】平成24年7月5日(2012.7.5)
【国際特許分類】
【出願日】平成22年1月19日(2010.1.19)
【国際出願番号】PCT/EP2010/000271
【国際公開番号】WO2010/081741
【国際公開日】平成22年7月22日(2010.7.22)
【出願人】(511174465)
【氏名又は名称原語表記】KING FAISAL SPECIALIST HOSPITAL AND RESEARCH CENTRE
【出願人】(511174476)テラマーク・マーケンクリエーション・ゲーエムベーハー (1)
【氏名又は名称原語表記】TERRAMARK MARKENCREATION GMBH
【Fターム(参考)】
[ Back to top ]