
細胞における局在性が改変された免疫ポリペプチドに基づく抗腫瘍組成物
【課題】少なくとも1つがその天然の局在性とは異なる細胞局在性を有するよう改変された、1または数種の免疫ポリペプチドを治療薬として含んでなる抗腫瘍組成物を提供する。
【解決手段】免疫ポリペプチドを発現する組換えベクターに基づく組成物。それはさらに、局在性が改変されたパピローマウイルスの初期および/または後期領域に由来する免疫ポリペプチドをコードする少なくとも1つの配列を含んでなる組換えベクター、ならびにそのベクターを含んでなるウイルス粒子。この組成物、組換えベクターおよびウイルス粒子の治療上の使用。
【解決手段】免疫ポリペプチドを発現する組換えベクターに基づく組成物。それはさらに、局在性が改変されたパピローマウイルスの初期および/または後期領域に由来する免疫ポリペプチドをコードする少なくとも1つの配列を含んでなる組換えベクター、ならびにそのベクターを含んでなるウイルス粒子。この組成物、組換えベクターおよびウイルス粒子の治療上の使用。
【発明の詳細な説明】
【発明の背景】
【0001】
本発明の主題は、天然の局在性とは異なる細胞での局在性を有するよう改変された免疫ポリペプチドを治療薬または予防薬として含んでなる抗腫瘍組成物である。それはまた、そのポリペプチドを発現する組換えベクターに基づく組成物に関する。かかる組成物はさらに詳しくは、パピローマウイルスに関連した病巣の治療または予防を意図したものである。
【0002】
一般に、癌は細胞増殖の制御の欠如に起因する疾病であると解釈されている。その原因にはいろいろあろうが、特に細胞の遺伝子の機能不全(潜在的癌遺伝子の例えば体細胞変異による活性化;発現の脱制御;腫瘍抑制遺伝子の発現の阻害)またはウイルス遺伝子の望ましくない発現による可能性がある。
【0003】
ヒトにおいては、パピローマウイルス(HPV)は皮膚の良性感染から疣贅、および悪性腫瘍にわたる病院に関与する。これまでに確認された75タイプのHPVのうち、20の異なる分離株が生殖道に極めて特異的であり、そのうち5株(HPV−16および18、そしてより程度は低いがHPV−31、33および45)が子宮頸部および生殖道下方の癌に明確に関与している。一連の全体の研究は、これらのウイルスの形質転換に果たす役割、新生細胞のゲノムへのそれらの特異的な組み込み、癌性細胞におけるそれらの遺伝子の活性、および悪性の表現型のHPV陽性新生細胞を維持する際のあるウイルス遺伝子の発現の重要性を実証するものである(Monsenego, J. Impact Medecin, 11 March 1994)。
【0004】
一般に、パピローマウイルスは、キャプシドタンパク質に取り巻かれた約7900塩基対の環状ゲノムを有るDNAウイルスである。いくつかのタイプのパピローマウイルス、特にウシ・パピローマウイルス(BPV)およびヒト・パピローマウイルス(HPV)が確認されており(Pfister, 1987, in The papovaviridae: The Papillomaviruses, Salzman and Howley edition, Plenum Press, New York, p 1-38)。それらのゲノムは、リーディングフレームE1、E2、E4、E5、E6およびE7を包含する初期領域と、キャプシドタンパク質L1およびL2をコードする後期領域を含んでなる。
【0005】
初期タンパク質はDNAに結合する能力を有し、優先的に核で見られる。E1およびE2の発現産物はウイルスの複製とウイルス遺伝子の発現を調節し、一方、E5、E6およびE7領域の発現産物は感染細胞の発癌転換のプロセスに関与する。このことに関して、BPV−1 E5タンパク質がin vitroで細胞を形質転換可能なことが実験的に示されている(Schlegel et al., 1986, Science 233, 464-467)。E7の形質転換力はHPV−16およびHPV−18に関して実証されており(Kanda et al., 1988, J. Virol. 62, 610-613; Vousden et al., 1988, Oncogene Res. 3, 1-9; Bedell et al., 1987, J. Viorol. 61, 3635-3640)、網膜芽細胞種(Rb)遺伝子の産物と結合する能力と関連している(Munger et al., 1989, EMBO J. 8, 4099-4105; Heck et al., 1992, Proc. Natl. Acad. Sci. USA 89, 4442-4446)。さらに、Crook et al (1991, Cell 67, 547-556)はHPV−6および18のE6抗原がp53遺伝子の産物を合成することができることを示したが、このことは細胞の形質転換におけるその有力な役割を説明している。
【0006】
HPVウイルスに関連する病状はそれらの持続性および再発性のために治療上問題がある。通常のアプローチは依然として外科手術と化学療法であるが、これらの疾病の治療のため、今般、免疫療法が考案されている。理想的なワクチン候補は、予防目的(免疫予防)としては、感染が永続的に確立されないようにする、また隣接する組織に広がらないようにし、治療目的(免疫療法)としては、感染した患者における腫瘍発達を軽減するべきである。これまでには、キャプシド抗原を使用して、ウイルス粒子の表面に局在するエピトープに対する抗体の産生を誘導すること、および初期タンパク質を使用して、ウイルスDNAの組み込み後に感染した細胞に対して細胞性免疫を確立することが提案されている。
【0007】
このことに関して、欧州特許EP0,462,187では、パピローマウイルス初期遺伝子を発現するポックスウイルスの投与を含む治療的アプローチが記載されている。WO93/02184に記載されている予防接種的アプローチは、免疫物質としてのキャプシド抗原、特にin vitroで再構成されたDNAを含まないウイルス粒子(ウイルス様粒子のVLP)の使用に基づいている。仏国出願第96 09584号では、パピローマウイルス初期ペプチドによってもたらせれる予防効果と後期ペプチドによって与えられる治療効果を組み合わせた組成物が開示されている。しかしながらこれまでに、所望によりそれらの形質転換活性を無効にするよう変異させてはいるが、それらの細胞局在性という観点では天然型であるウイルスタンパク質が使用されてきた。
【発明の概要】
【0008】
本発明は、それが特異的であれ、非特異的であれ、また体液型(抗体の産生)であれ、細胞型(細胞傷害性応答CTL)であれ、治療される腫瘍または癌に対する免疫応答を増強するまたは刺激するために、宿主の免疫系に対するそれらの接近性を高める目的でその局在性が改変された免疫タンパク質を使用することを提案する。
【0009】
今般、HPV−16 E6およびE7核抗原が、適当な付着配列および分泌配列を導入して、そこでのトランスメンブラン型での提供を付与することにより改変された。局在性の変化は免疫応答に関して有益な効果を持ち、その結果、膜E6およびE7抗原およびヒトIL−2を同時発現するワクシニアウイルスで処理した動物は、核抗原を産生する同等のウイルスを受容させたものに比べて抗腫瘍活性が高くなる。本発明の目的は、腫瘍もしくは癌の確立または発達を少なくとも部分的に阻害することに関して先行技術の組成物よりも効果的な公知の抗腫瘍組成物へ利用できるようにすることである。特に有用な用途はHPV感染の、さらに詳しくは子宮頸部癌などの重篤な病状の治療である。
【0010】
従って、本発明の主題は、1以上の免疫ポリペプチドを治療薬または予防薬として含んでなる組成物であり、これらポリペプチドの少なくとも1種はその天然の局在性とは異なる局在性を持つよう改変されている。
【0011】
本発明の目的のため、「免疫ポリペプチド」とは、それと同じものが正常な細胞には存在しないポリペプチドを表す。好ましい例としては、腫瘍特異的抗原がある。例としては、その発現が胎胚期に起こり、誕生時にそれが消失するまで退縮する細胞抗原、通常は非常に低レベルで発現し、高レベルで発現すると腫瘍の特徴を持つようになる抗原、その構造またはコンホメーションが改変されている細胞抗原、あるいは非細胞抗原、特に発癌ウイルスに由来するウイルス抗原が挙げられる。それらは、例えば、遺伝子BRCA−1(Miki et al., 1994, Science 226, 66-71)、BRCA−2(Wooster et al., 1995, Nature 378, 789-792)、MUC−1(Hareuveni et al., 1990, Proc. Natl. Acad. Sci. USA 87, 9498-9502)、CEAなどの発現産物であり得、これらのある変異または過剰発現が癌の発達に関与している。ウイルス抗原については、より詳しくは、パピローマウイルス初期もしくは後期遺伝子、エプスタイン・バーウイルスEBNA−1の発現産物、HTLV(ヒトTリンパ球ウイルス)IおよびIIウイルスまたはB型およびC型肝炎ウイルスの抗原が挙げられる。腫瘍特異的抗原は当業者が入手できる文献に広く記載されている。
【発明の具体的説明】
【0012】
本発明で使用されるポリペプチドの不可欠な特徴としては、その天然の局在性とは異なる局在性を持つことである。輸送のメカニズムおよび関与するシグナルは細胞生物学の書物に記載されている(例えば、Molecular Biology of the Cell, Third Ed. garland Publishing Inc. NY & Londonを参照)。便宜には、大部分のポリペプチドがサイトソル中の遊離リボゾームで合成され、そこでそれらは活性を示す。しかしながら、いくつかのポリペプチドは異なる細胞内指向性を持っており(それは一般に適当なペプチドシグナルの存在によって確認される)、そこまで輸送されなければならない。このように、原形質膜への輸送されるよう、または細胞外へ分泌されるよう意図されたポリペプチドは、小胞体(ER)と会合したリボゾームによって、通常はそれらのアミノ末端に、ERへの送達を開始させる分泌配列(またはシグナルペプチド)を含んでなる前駆体の形態で合成される。次いでそれは特異的エンドペプチダーゼによって除去され、成熟ポリペプチドとなる。分泌配列は通常、15〜35疎水性の必須アミノ酸を含んでなる。非共通配列が存在し、これがエンドペプチダーゼによる認識を決定する分泌配列であるものと思われる。しかしながら、グリシン、セリンまたはアラニン残基の後でタンパク質分解的切断がしばしば起こらなければならない。
【0013】
膜タンパク質は一般に、原形質膜に依然として挿入されている、疎水性の高い付着配列を含んでなる。ポリペプチドはトランスメンブラン型であると考えられ、一方の末端は細胞外に露出し、付着配列は膜をわたり、もう一方の末端はサイトソル側にある。ほとんどの場合、ポリペプチド鎖は膜の脂質二重層に埋め込まれており、αらせんコンホメーションを有している(例えば、Branden and Tooze, 1991, in Introduvtion to Protein Structure p. 202-214, New Garlandを参照)。
【0014】
核局在性に関しては、主としてリジンおよびアルギニンなどの陽電荷残基からなった、短い、いわゆる核局在化配列(NLS)の存在によって与えられると考えられる。例としては、HPV L1およびL2ポリペプチドに存在する核転座シグナルKRKKRKおよびRKRRKRが挙げられる(Zhou et al., 1991, Virology 185, 625-632)。しかしながら、核内でそれらの機能を発揮するポリペプチドには、典型的なNLS配列を持っていないものがある。パピローマウイルスE6およびE7抗原の場合がそうである。
【0015】
本発明の組成物中に含有される免疫ポリペプチドは、天然のポリペプチド、その断片、異種起源の配列を含んでなるキメラ、または変異体(1以上のアミノ酸の欠失、挿入および/または置換)内の適当な局在化シグナルの導入および/または欠失から得てもよい。さらに詳しくは、そのアミノ酸配列は、それに由来する天然ポリペプチドの配列の総てまたは一部と70%を越える、有利には80%を越える、特には90%を越える同一性を示す。この同一性は適当なコンピュータープログラムを用いて、あるいは最高の相同性が得られるように配列を並べることにより、そして2つの配列のアミノ酸が全番号の位置と比べて同一であるとわかった位置の数を計数することにより容易に算出できる。
【0016】
当業者ならば、ポリペプチドの細胞への提供を変化させるシグナルを知っている。免疫ポリペプチドが分泌することが望ましい場合、そのアミノ末端へ分泌配列を付加すれば、ERを介してそれを宿主細胞の外へ輸送することが可能となる。この挿入は、翻訳の開始に関するコドンのすぐ下流で起こることが好ましい。本発明のでは、干渉を避けるために、天然の局在性を決定する残基の総てまたは一部を変異/欠失させることが有利であろう。例えば、天然のポリペプチドが膜指向性を持つならば、それは分泌配列をすでに含んでなり、疎水性付着配列を所望により変異/欠失させる。さらに、天然のシグナルの不活性化(変異/欠失による)により細胞質への局在化が可能となる。通常、細胞での局在性が異なる(例えば、核、膜、分泌など)免疫ペプチドを細胞質に供されると、in vivoにおいては主要組織適合性複合体のクラスI抗原とCTL応答によって媒介されるポリペプチドの提を促すことができる(Tobery and Siliciano, 1997, J. Exp. Med. 5, 909-920)。意図し得るもう1つの具体例としては、有効なCTR応答を刺激する目的でも、免疫ポリペプチドとユビキチンを融合させることである。
【0017】
有利には、本発明の組成物は、好ましくは膜付着配列を、および天然のポリペプチドが分泌配列を欠いている場合にはそれを挿入することにより、膜局在性を持つよう改変された免疫ポリペプチドを含んでなる。分泌配列の好ましい挿入部位は、前記されたようにN末端であり、膜付着配列のそれは、例えば停止コドンのすぐ上流のC末端である。本明細書ではまた、天然の局在化シグナル(例えば、NLS配列)の総てまたは一部の変異/欠失させて、新たな場所で干渉することがないようにすることが有利であり得る。
【0018】
本発明で使用できる局在化シグナルの選択肢は広い。それは、それが処置される細胞に認識される限り、それを含んでなる、真核生物起源またはそれ以外(ウイルス、寄生体、真菌類など)いずれのタンパク質に由来するものであってもよい。それは天然でも合成でもよく、後者に関して異種であっても同種であってもよい。それはまた、それが由来するシグナルに対する1以上の改変を含んなる(ただし、それ/それらはその機能に影響を及ぼさない)。指針としては、狂犬病糖タンパク質、HIVウイルスenv糖タンパク質、または麻疹ウイルスFタンパク質の膜付着配列および/または分泌配列を使用することが好ましいであろう。免疫ポリペプチドの改変がいくつかの局在化シグナルを含む場合には(例えば、膜付着配列および分泌配列)、これらは共通に起源であっても、異なる起源であってもよい。
【0019】
また、特定の細胞コンパートメントをターゲッティングする局在化シグナルを使用することもできる。特に、エンドサイトーシスに関する共通配列(例えば、配列IPNYRNMを有する免疫グロブリンIgG1重鎖のC末端領域に存在する;Kaisho et al., 1997, Science 276, 412-414)、またはゴルジ体膜へのターゲッティングを可能にする配列(Mochamer and Rose, 1987, J. Cell Biol. 105, 1205-1214; Mochamer, 1993, Curr. Opin. Cell Biol. 5, 606-612)が挙げられる。特に、P11222の受託番号でSwiss−Protデータバンクに開示されているコロナウイルスE1糖タンパク質由来配列の使用が意図される。免疫ポリペプチドのC末端にこの種の配列を組み込むことが好ましい。
【0020】
さらに、細胞局在性の改変は、いずれの常法により行ってもよく、特に位置指定突然変異誘発、異種シグナルの連結、またはPCRによって行ってもよい。
【0021】
好ましい具体例によれば、本発明の抗腫瘍組成物は、パピローマウイルス感染およびその結果起こる疾患、特に軽度子宮頸部形成異常および子宮頸部癌の治療または予防を意図したものである。この具体例によれば、それはパピローマウイルス、特にHPV−16、18、31、33または45などの危険性の高いウイルスの初期および/または後期領域に由来する少なくとも1種の免疫ポリペプチドを含んでなる。
【0022】
本発明によって追求される目的によれば、1種異常のいずれかの種の免疫パピローマウイルスポリペプチドが使用できる。先に想起されたように、それらのゲノムは8種のポリペプチド、すなわちウイルスキャプシドを含んでなる2種の後期ポリペプチドL1およびL2と、調節、ウイルスゲノムの維持および感染した細胞の形質転換に関与する6種の初期ポリペプチド(E1、E2、E4、E5、E6およびE7)をコードする。
【0023】
初期型の免疫ポリペプチドに関しては、特に膜局在性を有するよう改変されたE6またはE7由来のポリペプチドを使用することを選択するのが有利である。先に想起された観察を形質転換力に与えると、細胞の形質転換プロセス関与する領域で変異した非発癌性変異体を使用するのが好ましい。かかる変異体は文献に記載されている(Munger et al., 1989, EMBO J. 8, 4099-4105; Crook et al., 1991, Cell 67, 547-556; Heck et al., 1992, Proc. Natl. Acad. Sci. USA 89, 4442-4446; Phelps et al., 1992, J. Virol. 66, 2418-2427)。本発明の目的に特に好適な免疫ポリペプチドは、残基111〜115(+1、天然のウイルス抗原の1番目のアミノ酸を表す)が欠失し、かつ、麻疹ウイルスFタンパク質の分泌または付着シグナルと融合したHPV−16 E6抗原である(配列番号1)。また、残基22〜25が欠失し、狂犬病糖タンパク質の付着配列および分泌配列と融合したHPV−16 E6抗原も使用できる(配列番号2)。
【0024】
本発明の抗腫瘍組成物はまた、L1またはL2に由来の、パピローマウイルスの後期領域に由来する免疫ポリペプチドを含んでなってもよい。
【0025】
もちろん、本発明の抗腫瘍組成物は、そのうちの少なくとも1つがその天然の局在性とは異なる細胞局在性を有する、数種の免疫ポリペプチドを含んでなってもよい。パピローマウイルス由来の数種のポリペプチドを組み合わた組成物の例としては、共通の起源であっても異なる起源であってもよいもの(例えば、活性スペクトルを広くする目的では、HPV−16および18)が挙げられる。初期起源の数種のポリペプチドを組み合わせた組成物は治療効果を増強することができる。L1およびL2由来のポリペプチドの組合せは、組成物の予防特性に有益な効果を持ち得た。最後に、本発明により追求される目的に最も特に好適な組成物は、予防効果と治療効果を併せ持つよう、パピローマウイルスの少なくとも1種の初期ポリペプチドと少なくとも1種の後期ポリペプチドを含んでなる。好ましい具体例によれば、初期起源の少なくとも1種の免疫ポリペプチドが、先に挙げたもののような分泌または付着配列の付加によって膜局在性を有するよう改変される。
【0026】
これに関して、本発明の好ましい具体例は、
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド
を含んでなる。
【0027】
「相同」とは、その配列との70%を越える、有利には80%を越える、好ましくは90%を越える、最も好ましくは95%を越える同一性の程度をいう。
【0028】
有利には、本発明の抗腫瘍組成物は、免疫応答の強度を特異的に、または特異的にではなく増強する目的で、その抗腫瘍作用を増強する少なくとも1種の化合物さらに含んでなってもよい。アジュバントの他、免疫刺激剤は特に好ましい化合物を表す。「免疫刺激剤」とは、免疫ポリペプチドに向けられた抗体の産生を増幅するために体液性免疫応答を、あるいは、腫瘍または感染細胞に対する有意な細胞傷害性応答を誘発するために細胞性免疫応答を強める能力を有する化合物を意味するものと理解される。指針としては、免疫刺激剤は、癌の動物モデルにおいて、免疫ポリペプチドで処置した動物の拒否反応率を免疫刺激剤の存在下と不在下で比較することにより評価すればよい。より一般には、免疫刺激を証明する手段はRoitt(Immunology, 4th edition, Moby Ltd)に示されている。かかる組成物の利点の1つは、それが免疫ポリペプチドによって誘導される特異的免疫と免疫刺激分子によって誘導される非特異的免疫を併せ持つことである。
【0029】
本発明では、特にヒト起源の天然の免疫刺激剤、その一部、異なる起源の配列との融合から得られたキメラ、または変異体(しかしながら免疫刺激作用は保存されている条件で)を使用することができる。考えられ得るあらゆる分子のうち、インターロイキン−2、インターロイキン−7、インターロイキン−12および付着分子B7.1およびB7.2から選択される免疫刺激剤を使用することが好ましい。
【0030】
一般に、免疫ポリペプチドおよび免疫刺激ポリペプチドは通常の化学合成法によって、または組換えDNA技術によって作製され得る(例えば、Maniatis et al., 1989, Laboratory Manual, Cold Spring harbor, Laboratory Press, Cold Spring Harbor, NY)。さらに詳しくは、製造方法は、問題のポリペプチドをコードするDNA断片で形質転換した細胞を培養して産生細胞を作出する行為と、その培養物からポリペプチドを回収する行為を含んでなる。産生細胞はいずれの起源であってもよく、限定されるものではないが、考慮されているDNA断片がそのゲノムに組み込まれているか、または適当な発現ベクターに組み込まれているかのいずれかである限り、細菌、酵母菌、または哺乳類細胞であってもよい。もちろん、DNA断片は、産生細胞中でその発現を可能にする転写および翻訳シグナルの制御下に置かれる。発現ベクターおよび制御シグナルは当業者に公知である。
【0031】
本発明はまた、1以上の免疫ポリペプチドをコードする配列を含んでなる少なくとも1種の組換えベクター、その天然の局在性とは異なる局在性を有するよう改変されている少なくとも1種のポリペプチド、および所望により抗腫瘍作用を増強する化合物を、治療薬または予防薬として含んでなる抗腫瘍組成物に関する。この種の組成物は生産が高い費用がかからない、また、種々の環境条件下で安定性が高いという利点を持つ。特に、保存条件は制限が少ない。これらのポリペプチドは先に定義したような特徴を持つ。
【0032】
免疫ポリペプチドをコードする、または抗腫瘍作用を増強する配列は、一般の用法で、また文献のデータを用いて、クローン化によって、PCR(ポリメラーゼ連鎖反応)によって、または化学合成によって得られる。好ましい具体例に関しては、パピローマウイルスポリペプチドをコードする配列は、患者からまたはコレクションから得られるパピローマウイルス陽性細胞から選択すればよい。適当な局在化シグナルの挿入は、分子生物学的技術によって行ってよい。免疫刺激剤をコードする配列は、細胞DNAから、またはそれが発現している細胞のメッセンジャーRNAからクローン化すればよい。当業者ならば、公開されたデータから適当なプローブまたはプライマーを作製することができる。HPV−16および18ゲノムのヌクレオチド配列はそれぞれ受託番号K02718およびX05015でGenbankに開示されていることを記載しておくべきであろう。ヒトIL−2遺伝子の配列は、仏国特許第85 09480号およびTaniguchi et al. (1983, Nature 302, 305-311)に記載され、Freeman et al. (1989, J. of Immunology 143, 2714-2722)においてはB7.1をコードしている。
【0033】
本発明で使用できるベクターは、プラスミドまたは特にポックスウイルス、アデノウイルス、レトロウイルス、ヘルペスウイルスまたはアデノ随伴ウイルスに由来するウイルスベクターであってよい。病原性を弱めた非組み込みベクターであれば有利である。かかるベクターおよびそれらの調製法は当業者に公知である。
【0034】
アデノウイルスベクターが使用される場合には、複製のために不可欠な領域、特に宿主生物または環境にそれが伝播しないようにするにはE1領域の大部分を欠失させることにより、非反復因子を使用することが好ましい。欠陥のある必須機能がトランスで相補される限り、アデノウイルスゲノムの他の領域、特にE2、E4および/またはL1〜L5領域内を改変または欠失させてもよい。これらの具体例を例示するためには、E2A領域のDBP(DNA結合タンパク質に関するもの)遺伝子に作用する熱感受性変異体が挙げられる(Ensinger et al., 1972, J. Virol. 10, 328-339)。オープンリーディングフレーム(ORF)6および7をコードする配列を除く、E4領域の部分的欠失もまた意図され得る(Ketner et al., 1989, Nucleic Acids Res. 17, 3037-3048)。もう1つの可能性としては転写単位E4の全欠失がある。さらに、本発明のアデノウイルスベクターは、非必須領域E3の総てまたは一部を欠いていてもよい。この選択肢によれば、やはり、宿主の免疫系を逃れることが可能となるポリペプチド、特に糖タンパク質gp19kをコードするE3配列を保存することが有利であり得る(Gooding et al., 1990, Critical Review of Immunology 10, 53-71)。本発明の好ましいアデノウイルスベクターは包膜に不可欠な配列、すなわち5’および3’ITR(逆方向末端反復)および包膜領域の最小限の保持を示すであろう。それはヒトまたは動物のアデノウイルスに、またいずれかの抗原型に由来してもよい。C亜群ヒト・アデノウイルス、特にアデノウイルス2(Ad2)および5(Ad5)が、本発明を実施するのに特に最も適している。種々のアデノウイルスベクターならびにそれらの調製技術は通常のものであり、Graham and Prevect (1991, in Methods in Molecular Biology, vol 7, p 109-128; Ed: E.J. Murey, The Human Press Inc.)および国際出願WO94/28152に記載されている。例えば、それはin vitroにおいて大腸菌(E. coli)で、連結反応または相同組換え(例えば、国際出願WO96/17070を参照)によって、または相補ラインにおける組換えによって作出してもよい。
【0035】
レトロウイルスが使用されるならば、LTR(長い末端反復)および包膜配列(例えばNaviaux and Verma, 1992, Current Opinion in Biotechnology 3, 540-547を参照)が保存される。免疫ポリペプチドをコードする配列はレトロウイルスLTR、または下記に記載されるもののような内部プロモーターの制御下に置いてもよい。それはいずれの起源(ネズミ、霊長類、ネコ、ヒトなど)のレトロウイルスに由来してもよく、特にMoMuLV(モロニーネズミ白血病ウイルス)、MVS(ネズミ肉腫ウイルス)またはフレンドネズミレトロウイルス(Fb29)に由来していてもよい。それはまた、特にLTR(真核生物プロモーターによるプロモーター領域の置換)のレベル、または包膜領域(異種の包膜領域、例えばVL30型による置換)のレベルでの改変を含んでなる(仏国出願第94 08300号および第97 05203を参照)。
【0036】
有利な具体例によれば、本発明の組換えベクターはポックスウイルス、特にカナリア痘ウイルスのような鳥類痘ウイルス、鶏痘ウイルス、またはワクシニアウイルスに由来し、後者が好ましい。本発明において意図され得るあらゆるワクシニアウイルスのうち、コペンハーゲン、ワイエスおよび改変型アンカラ(MVAは、改変型ワクシニアウイルスアンカラ)株が選択されることが好ましい。
【0037】
一般に、挿入部位は、組換えウイルスの複製能および増殖能が損なわれないように、複製に不可欠でない領域中で選択される。指針としては、コペンハーゲン株のウイルスを使用する場合は、好ましい挿入部位はTK遺伝子座であり、それは後者を後者を不活性化する作用を有し、それにより組換え体の選択が容易になる。また、K1L遺伝子座を使用することもできる。MVAウイルスに関しては、組換え体(免疫型および免疫刺激型)配列の挿入は、切除部位I〜VIの少なくとも1つ、特にIIまたはIII内で行われてよい(Mayer et al., 1991, J. Gen. Virol. 72, 1031-1038; Sutter et al., 1994, Vaccine 12, 1032-1040)挿入はまた、D4R領域などのウイルスの必須領域で起こってもよく、欠損のある機能は、例えば相補ラインによりトランスで提供することができる。
【0038】
もちろん本発明では、免疫ポリペプチドをコードする、または抗腫瘍作用を増強する配列は、宿主細胞または宿主生物での発現のために必要なエレメントの制御下に置かれる。それらは転写を調節するエレメント、ならびに翻訳の開始および終結のシグナルを含む。それらの中で、プロモーターが特に重要である。一般に、宿主生物または宿主細胞中で機能し、治療に望ましく、かつ、使用されるベクターに好適なプロモーターが使用されよう。さらに、それは調節配列、例えば転写を活性化するエレメントまたは特定の細胞シグナルに応答する配列を含むよう改変してもよい。このことに関して、パピローマウイルスに関連する病巣は生殖道のレベルに位置するので組織特異的プロモーターを、あるいは腫瘍細胞のみに対して過剰発現を制限するよう、腫瘍に特異的なシグナルに応答するプロモーター(例えば、一般に腫瘍細胞によって過剰発現する増殖因子の存在下で活性化される)を使用することが有利であろう。
【0039】
本発明において意図し得るプロモーターの中で、SV40(シミアンウイルス40)プロモーター、HMG(ヒドロキシ−メチル−グルタリル−補酵素A)プロモーター、TK(チミジンキナーゼ)プロモーター、CMV(サイトメガロウイルス)プロモーター、RSV(ラウス肉腫ウイルス)プロモーター、アデノウイルスベクターに好適なMLPプロモーター(主要後期プロモーター)およびレトロウイルスベクターにより特異的なMo−MLV(モロニーネズミ白血病ウイルス)のLTRが挙げられる。サイトメガロウイルス(CMV)初期プロモーターが最も特に好ましい。それはまた腫瘍または癌細胞中での発現を刺激するプロモーターであってもよい。特に、乳癌および前立腺癌で過剰発現するMUC−1遺伝子のプロモーター(Chen et al., 1995, J. Clin. Invest. 96, 2775-2782)、黒色腫で過剰発現するチロシノース遺伝子(Vile et al., 1993, Cancer Res. 53, 3860-3864)および乳癌および膵臓癌で過剰発現するERS−2遺伝子(Harris et al., 1994, Gene Thetapy 1, 170-175)が挙げられる。問題のプロモーターが文献に記載されており、常法によって細胞ゲノムまたはウイルスゲノムからクローン化され得る。
【0040】
ポックスウイルスベクターに関しては、ポックスプロモーター、例えば7.5K、H5R、TK、p.28、p.11またはワクシニアウイルスのK1Lを使用してもよい。合成プロモーターもまた本発明の実施に適している(例えば、Chakrabarti et al., 1997, Biotechniques 23, 1094-1097; Hammond et al., 1997, J. Virological Methods 66, 135-138およびKumar and Boyle, 1990, Virology 179, 151-158を参照)。これに関しては、後期プロモーターと初期プロモーターとの間のキメラプロモーターであることが有利である。
【0041】
さらに、発現に必要なエレメントはまた、宿主細胞中での発現または維持を増大させる配列(イントロン、転写を終結させる配列、翻訳の開始部位など)を含んでももよい。しかしながら、ポックスウイルスベクターの場合には、イントロンの使用は避けられる。
【0042】
本発明の組成物は、独立または共通のエレメントの制御下に置かれた選ばれたポリペプチドに相当する配列を発現する1以上の組換えベクターを含んでもよい。後者の選択によれば、内部に翻訳を開始させる配列(IRES)または異なる遺伝子相における融合物を使用してもよい。
【0043】
本発明で使用される組換えベクターを産生する一般条件は、当技術分野の現状において広く記載されている。ポックスウイルスベクターに関しては、欧州特許EP 83 286に言及されており、その内容は引用することにより本明細書の開示の一部とされる。これらの条件は、発現単位が組み込まれるゲノム領域を有するベクターとして許容されるその他のウイルスにも適用できる。もちろん、それらを同一の遺伝子座または異なる遺伝子座に挿入してもよい。
【0044】
本発明によって追求される目的に従って、組換えウイルスの単離および精製工程を容易にするため、組換えベクターは選択可能なマーカー遺伝子を発現するための単位をさらに含んでなってもよい。特に、抗生物質G418耐性を与えるneo遺伝子、プロマイシン耐性を与えるpac遺伝子、ガンシクロビルまたはアシクロビルのようなあるヌクレオシド類似体に対する感受性を与える1型単純ヘルペスウイルス(HSV−1)のTK遺伝子、gpt(キサンチングアニンホスホリボシルトランスフェラーゼ)遺伝子、β−ガラクトシダーゼをコードする細菌遺伝子LacZおよびβ−グルクロニダーゼをコードするgus Aが挙げられる。後者2つの酵素マーカーによって、それぞれ基質X−Gal(5−ブロモ−4−クロロ−3−インドリル−β−D−ガラクトピラノシド)およびXglcA(5−ブロモ−6−クロロ−3−インドリル−β−D−グルコロニド)の存在下での染色により組換えウイルスを同定することが可能になる。
【0045】
好ましい抗腫瘍組成物はパピローマウイルス感染または腫瘍の治療および予防を意図し、
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド
が挿入された少なくとも1種のワクシニアウイルスのコペンハーゲン株またはMVA株を含んでなる。
【0046】
この組成物はまた、好ましくはIL−2またはB7.1から選択される免疫刺激剤をコードする配列を含んでなってもよい。免疫遺伝子を発現させる組換えベクターの1つによって、または独立のベクターによってそれを行ってもよい。
【0047】
本発明の組成物はワクチンの分野で公知の方法に従って調製してもよく、利用可能用量は広い範囲で変化させ得る。それらは特に、用いられるポリペプチドおよびベクター、治療される病状、患者の状態および臨床医が評価し得るその他のパラメーターに依存する。しかしながら、一般に、治療薬がウイルスベクターである場合にはウイルスの用量は104〜1013、有利には105〜1012、好ましくは105〜109プラーク形成単位(pfu)であり、治療薬がポリペプチド由来のものである場合には0.05〜500mg、有利には0.5〜200mg、好ましくは1〜100mgである。
【0048】
本発明の組成物は通常の投与経路のいずれかによって、好ましくは全身的経路によって、特には筋肉内、静脈内、肺内、皮下もしくは上皮下経路によって、または乱刺法によって投与してもよい。接触可能な腫瘍の場合には、腫瘍の部位もしくはその付近への直接注射、または局所塗布を用いることもできる。ワクチンとしては、本発明の組成物を当技術分野で一般的な実施に従い、例えば1回用量として、またはある一定の時間間隔の後に1回もしくは数回繰り返す用量として投与してもよい。他方、治療処置に関しては、治療が効果的であるに十分な周期で頻繁にそれを投与することができる。治療薬がウイルスベクターである場合には、ウイルスは生きている形態であることが好ましい。ポックルウイルスベクターに関しては、MVA株またはチミジンキナーゼ陰性コペンハーゲン株のような弱毒株を使用するのが好ましい。最後に、組換えウイルスベクターは、当業者にとって公知である適当な化学処理によって弱毒化してもよい。しかしながら、死滅した組換えベクターの注射を意図することもできる。
【0049】
好ましい具体例によれば、本発明の抗腫瘍組成物は、治療上有効量の治療薬を医薬上許容される担体と組み合わせて含んでなる。担体は、注射によってヒトまたは動物へ投与できるように選択する。それはまたビヒクル、賦形剤および/またはアジュバントを含んでもよく、液体または凍結乾燥形態で提供されてもよい。これに関して、ベクターのトランスフェクション効率および/または安定性を増大させることができる1以上の物質の組み合わせが意図され得る。これらの物質は当業者が利用できる文献に広く記載されている(例えば、Felgner et al., 1989, Proc. West. Pharmacol. Soc. 32, 115-121; Hodgson and Solaiman, 1996, Nature Biotechnology 14, 339-342, Remy et al., 1994, Bioconjugte chemistry 5, 647-654を参照)。限定されるものではないが例として、それらはポリマー、脂質、特に陽イオン脂質、リポソーム、核タンパク質および中性脂質であってもよい。意図し得る組み合わせは、陽イオン脂質(DC−Chol、DOGSなど)、および中性脂質(DOPE)と組み合わせたプラスミドベクターがある。この物はまた、その他の物質、特に抗癌物質と組み合わせてもよく、後者は個々にまたは同時に投与することが可能である。リガンドFlt3はその他の一例である(Lynch et al., 1997, Nature Medicine 3, 625; Brasel et al., 1996, Blood 88, 2001-2012; Marakovsky et al., 1996, J. Exp. Med. 184, 1953-1962)。この組成物はポリペプチドL1および/またはL2を含んでなる場合には模倣粒子の形態であってよいことを記載すべきであろう。
【0050】
本発明の主題はまた、パピローマウイルスの初期および/または後期領域に由来して先に定義された特徴を有する免疫ポリペプチドをコードする、少なくとも1つの配列を含んでなる組換えベクターである。それは注目されるその他の配列、例えば先に定義されたように、1以上の免疫ポリペプチドおよび/または免疫刺激ポリペプチドをコードする配列をさらに有してもよい。
【0051】
本発明のベクターの選択肢は広い。それは先に挙げたもののようなプラスミドまたはウイルスベクターであってもよい。好ましい具体例はポックスウイルスベクターからなり、最も特別にはワクシニアウイルスのコペンハーゲンまたはMVA株からなる。発現される注目の配列の挿入部位および発現に必要な要素は前記のものから選択され得る。
【0052】
好ましいベクターは、
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列
が挿入されたワクシニアウイルスのコペンハーゲンまたはMVA株である。
【0053】
それは所望によりIL2またはポリペプチドB7.1をコードする配列を含んでもよい。
【0054】
本発明はまた、本発明の組換えウイルスベクターから調製したウイルス粒子に関する。アデノウイルスベクターに関しては、このウイルス粒子はその欠損に好適な相補細胞にベクターをトランスフェクトすることによって作出してもよい。例えば、E1機能を効果的に相補する、ヒト胎児腎臓細胞から確立された293ライン(Graham et al.,1977, J. Gen. Virol. 36, 59-72)、A549−E1ライン(Imler et al., 1996, Gene Therapy 3, 75-84)または二重相補を可能にするライン(Yeh et al., 1996, J. Virol. 70, 559-565; Krougliak and Graham, 1995, Human Gene Therapy 6,1575-1586; Wang et al., 1995 Gene Therapy 2, 775-783;国際出願WO97/14119)を使用してもよい。相補細胞とは、ウイルス粒子を作出するために欠損ベクターをトランスで相補することができる細胞を意味するものと理解される。この細胞はベクターの欠損機能の総てをそれ自身で相補するわけではなく、この場合には部分的な相補としてヘルパーウイルスを使用することができる。ウイルス粒子は培養上清からだけでなく細胞からも回収してもよい。一般に用いられる方法の1つとしては、連続凍結/解凍サイクルによって細胞を溶解させて細胞溶解の上清中のビリオンを回収することである。これらを増幅して先行技術(クロマトグラフィー法、特に塩化セリウム勾配による超遠心分離など)に従い精製してもよい。
【0055】
レトロウイルス粒子は、適切なライン、例えば、それらの配列がベクター中において欠損している/機能しないウイルスポリペプチドgag、polおよび/またはenvをトランスで提供することができるラインにレトロウイルスベクターをトランスフェクトすることによって得ることができる。かかるラインは文献に記載されている(PA317、Psi CRIPGP + Am−12など)。
【0056】
ポックスウイルスベクターに関しては、異種遺伝子を発現することができるワクシニアウイルスを産生するための一般条件は、欧州特許EP 83 286および出願EP 206 920に教示されている。MVAウイルスに関しては、Mayr et al (1975, Infection 3, 6-14)およびSutter and Moss (1992, Proc. Natl. Acad. Sci. USA 89, 10847-10851)にさらに詳しく記載されている。便宜には、in vivoにおけるその(それらの)発現に適当なエレメントの制御下に置かれた導入される遺伝子を、挿入部位のいずれかの側にウイルス配列を含む導入ベクター中に挿入する。それを感染性ワクシニアウイルスに感染させた細胞中に導入する。組換え遺伝子は、この感染性ウイルスと導入ベクターの相同配列の間の相同組換えによってウイルスのゲノムへ組み込まれる。
【0057】
組換えベクターまたはウイルス粒子は、所望により、ベクターのトランスフェクション効率および/または安定性を増大させる1以上の前記物質と組み合わせてもよい。
【0058】
本発明では、この免疫ポリペプチドはウイルス粒子を取り巻くタンパク質構造(キャプシド、包膜など)に付着されていてもよい。
【0059】
ポックスウイルスは多重膜に取り巻かれた複雑な構造である。2種のウイルス粒子が産生され、それは細胞内成熟ビリオン(IMV)と細胞外包膜ビリオン(EEV)である。IMVの表面は二重膜からなり、EEVの包膜は原形質膜をはじめとする4つの膜からなる。本発明により追求される目的に従い、免疫ポリペプチドをコードする配列はまた、免疫応答を増強する目的とする、ポックスウイルス膜の一方または他方(IMVまたはEEV)への挿入によって改変されていてもよい。有利な具体例によれば、免疫ポリペプチドはEEV粒子の包膜の外膜に付着している(Katz et al. 1997, AIDS Res. Hum. Retrov. 13, 1497-1500)。これを行うために、その外膜のタンパク質成分であるB5Rタンパク質のC末端部分をコードする配列が、停止コドンのすぐ上流の免疫ポリペプチドのコード領域の3’に挿入される。全く好ましい例としては、ポリペプチドE7またはその非発癌性変異体とB5Rタンパク質の42個のC末端残基との間の融合物がある。免疫応答に対する有益な効果は、種々の組成(天然の局在性、細胞膜における付着、ウイルス膜における付着)を比較する以下の実施例に記載されたプロトコールに従う免疫予防および免疫治療研究によって評価され得る。
【0060】
本発明はまた、癌または腫瘍の、特に子宮頸部癌、軽度子宮頸部形成異常またはパピローマウイルス感染の治療または予防用医薬を製造するための、本発明の抗腫瘍組成物、組換えベクターまたはウイルス粒子の使用に関する。好ましい使用は筋肉内経路により注射可能な医薬の製造のためもののである。
【0061】
最後に、本発明はまた、医薬上有効量の本発明の抗腫瘍組成物、組換えベクターまたはウイルス粒子をかかる治療を必要とする患者に投与することにより、先に挙げた病状の治療または予防方法に関する。
【実施例】
【0062】
以下、本発明を実施例によって詳しく説明するが、結果として限定されるものではない。
【0063】
以下に記載した構築体は、Maniatis et al., (1989, 前記)に詳述されている一般的な遺伝子工学および分子クローニング技術に従い、または市販のキットを用いる場合には製造業者の推奨に従って作製する。in vitroでの合成オリゴヌクレオチド指定突然変異誘発は、Amershamにより供給されるキットによって実施する。PCR増幅技術は当業者にとっては公知である(例えば、Innis, Gelfand, Sninsky and White, Academic Press Incにより出版されているPCR Protocols - A guide to methods and applications, 1990を参照)。制限部位の修復に関しては、用いられる技術は大腸菌(Klenow)のDNAポリメラーゼIの大断片で、突出している5’末端を埋めることからなる。クローニング工程に関しては、組換えM13バクテリオファージを寒天最小培地(寒天7.5%)または豊富な液体LBM培地中の大腸菌NM522株(Stratagene)上で増殖させる。100μg/mlの抗性物質を補った寒天または液体培地で大腸菌C600株(Stratagene)、BJ5183(Hanahan, 1983, J. Mol. Biol. 166, 557-580)およびNM522においてアンピシリン耐性遺伝子を運ぶ組換えプラスミドを反復させる。クローニングが相同組換えによって行われる場合には、BJ5183株を使用することが好ましい(Bubeck et al., 1993, Nucleic Acid Res. 21, 3601-3602)。
【0064】
組換えワクシニアウイルス構築体は、すでに挙げた文献ならびにMackett et al., (1982, Proc. Natl. Acad. Sci. USA 79, 7415-7419)およびMackett et al. (1984, J. Virol. 49, 857-864)に開示されている当技術分野の標準技術に従って行う。
【0065】
in vivoでの試験(免疫予防または免疫治療)は、6〜8週齢のメスC57B16マウス(C. Rivers Rouen, France)またはヌードマウス(Janvier, Le Genest St. Isle, France)について行う。一般に、試験されるウイルス、用量または投与経路によって動物を20の群に分ける(指定されている場合は除く)。腫瘍細胞TC1(Wu, John Hopkins University, Baltimore, USA)は、2種のレトロウイルス、すなわちHPV−16天然E6およびE7遺伝子を発現するものと、ras癌遺伝子を発現するもので形質転換したC57B16マウスから採取した肺細胞から得る。この細胞は、G418(0.5mg/ml)の存在下、DMEM培地(ダルベッコ改変イーグル培地)で培養する。その細胞をトリプシンで処理して等張液で3回洗浄した後に動物の抗原投与に使用する。
【0066】
実施例1: トランスメンブラン局在化シグナルを供給したHPV−16 E6およびE7の非発癌性変異体をコードする配列を運ぶベクターの構築
E6およびE7遺伝子を欧州特許EP 0,462,187の実施例2および3に記載されているCaski細胞系から単離する。それ以後のクローニング工程を容易にするように、2つの構築体をHPV−16 E6およびE7遺伝子を含有するクローンM13E7/E6から誘導した。1つめのM13TG8188と示されるものは指定突然変異誘発によってE7遺伝子のそれぞれ上流および下流のPstIおよびBamHI部位の導入の結果得られ、2つめのM13TG8189はE6遺伝子の上流のPstI部位を含んでなる。開始暗号ATGの上流および停止コドンの下流における点突然変異の導入は、当業者の能力の範囲内である。
【0067】
HPV−16 E7タンパク質と網膜芽細胞腫遺伝子の産物との組合せが、種々の著者によって実証され(例えば、Munger et al., 1989, EMBO J. 8, 4099-4105を参照)、その形質転換力と相関している。明白な安全上の理由で、形質転換作用に関与している天然のE7タンパク質のコドン21〜26を欠失させた非発癌性変異体は、オリゴヌクレオチドoTG5118(配列番号3)によるベクターM13TG8188の指定突然変異誘発によって作出する。以下E7*と示される変異型E7タンパク質を有するM13TG9104が得られる。
【0068】
狂犬病糖タンパク質に対する分泌シグナルおよび付着シグナルは、Bg1IIの形態でpBR327(仏国特許83 15716に記載されているpTG150)に挿入された狂犬病糖タンパク質遺伝子から単離し、ベクターM13(M13TG177)にサブクローン化する。BamHI部位はオリゴヌクレオチドoTG5745(配列番号4)による指定突然変異誘発によって、それらと同じ位相にあるこれらシグナルの間に導入する。得られた構築体はM13TG9128と呼ばれる。次いで分泌配列および付着配列をPstI消化によってM13TG9128から単離し、PstIで線状化したベクターpTG5003にワクシニアウイルスp7.5の天然のプロモーターの3’に挿入すると、pTG5016が得られる。指針としては、pTG5003をSalI消化によってpTG186ポリ(仏国特許2,583,429に記載)から誘導し、Klenow断片で処理し、次いでSmaIで消化して再連結すると、結果としてそれはここでそのポリリンカー中に他のもの総てを除きPstIおよびEcoRIのみを含むようになる。位相中にE7*配列および狂犬病糖タンパク質の分泌シグナルおよび付着シグナル(本明細書では以下E7*TMRと示す)をもたらすように、ベクターM13TG9104をオリゴヌクレオチドoTG6390およびoTG6880(配列番号5および6)による指定突然変異誘発によって改変する。得られた構築体はM13TG9150と呼ばれる。
【0069】
同様に、HPV−16 E6タンパク質は腫瘍抑制遺伝子p53の発現産物と相互作用することができるということが実証されている(Crook et al., 1991, Cell 67, 547-556)。この相互作用に関与するドメインは明確に定義されており、天然のタンパク質の残基111と115との間に位置している。ベクターM13TG9125は、オリゴヌクレオチドoTG5377(配列番号7)によるM13TG8189の突然変異誘発によって作出される。E6遺伝子λ111−115を以下E6*と呼ぶ。
【0070】
麻疹ウイルスFタンパク質に対する分泌シグナルおよび付着シグナルは、麻疹ウイルスFタンパク質をコードするDNAを含有するプラスミド構築体pTG2148(欧州特許EP 0,305,229に記載)からPCRによって単離する。これらの配列とE6*をコードするものとの間の融合は、直接PCRによって行われ、すなわち、分泌配列を5’にXbaI部位を有するオリゴヌクレオチドoTG10829(配列番号8)およびE6の5’末端を覆うoTG10830(配列番号9)により増幅し、変異型E6*をコードする配列をFタンパク質の分泌シグナルの3’末端との融合を可能にするオリゴヌクレオチドoTG10835(配列番号10)およびFタンパク質の付着配列の5’末端との融合を可能にするoTG10836(配列番号11)により、M13TG9125から増幅される。付着配列に関しては、E6*の3’と付着配列の5’との間における融合を可能にするプライマーoTG10833(配列番号12)ならびに3’にKpnIおよびSphI部位を作り出すoTG10834(配列番号13)が使用される。それぞれNおよびC末端でFタンパク質の膜分泌配列および付着配列と融合したE6*配列を有する増幅断片(本明細書では以下E6*TMFと示す)をXbaIおよびSphIで消化し、次いで同一部位にベクターM13TG131に挿入する(Kieny et al., 1983, Gene 26, 91-99)。このようにして得られた構築体はM13TG9199と呼ばれる。
【0071】
実施例2: トランスメンブラン局在性のE6およびE7抗原ならびにヒトIL−2を発現する組換えウイルスMVATG8042の構築
MVAウイルスは、ワクシニアウイルスのアンカラ株に由来する。それは哺乳類細胞では感染性粒子を生ずることはできないが、胎芽鶏繊維芽細胞では正常に発達する。これらの細胞への適応は、この種の細胞でのその発達およびその感染サイクルに不可欠でない6領域の除去を引き起こした(ウイルスゲノムの約15%の消失;Meyer et al., 1991, J. Gen. Virol. 72, 1031-1038)。外来の遺伝物質の組み込みは、これらの除去領域のいずれのレベルで達成されてもよい。本発明では、HindIII制限断片NおよびA各々のレベルに位置する除去IIおよびIIIが用いられる(Altenburger et al., 1989, Arch. Virol. 105, 15-27)。
【0072】
第一に、MVAウイルスの除去III領域への挿入を可能にするベクターpTG6019を構築する。除去領域IIIのいずれかの側の相同組換えアームは、PCRによりウイルスゲノム、ならびに左のアームについてはプライマーoTG7637およびoTG7638(配列番号14および15)、ならびに右のアームについてはoTG7635およびoTG7636(配列番号16および17)から単離する(米国特許第5,185,146号を参照)。増幅した断片をベクターpTG1EのEcoRI部位にクローン化し、pTG6039を得る。導入される遺伝物質は2個の組換えアームの間に挿入される。ベクターpTG1Eは多重クローニング部位の代わりにEcoRIアダプターが存在することを除けばpTG1H(仏国特許第2,583,429号)と同様である。
【0073】
第一に、マーカー遺伝子gusAを発現するカセットを挿入する。まず7.5KプロモーターをpTG6019のBamHI部位へクローン化する。そのBamHI部位へBglII−BamHI断片の形で作出されたgusA遺伝子が挿入されているpTG6021が得られる。後者は文献に開示されている配列から得てもよい。得られた構築物はoTG6022と呼ばれる。マーカーの存在により、XglcA基質を用いてGUS酵素活性を検出することによって野生型ウイルスを組換えウイルスと識別することが可能となろう。赤色はβ−ガラクトシダーゼ活性を示す。しかしながら臨床適用の点では、組換えウイルスを選択した後の最終産物から、この細菌性マーカーを除去できることが有用であろう。このために2つの相同部位間の配列を欠失させるワクシニアの能力が利用される。従って、第2のp7.5Kプロモーターは、後者の発現を命令するのに適切なセンス方向でgusA遺伝子の下流に挿入される。ベクターpTG6025は、付着端を提供したp7.5断片のpTG6022のBamHIとSacI部位間への挿入の結果生ずる。
【0074】
さらに、ヒトインターロイキン−2をコードするcDNAを、PstI消化によってプラスミドpTG36(仏国特許第2,583,770号)から単離し、次いでプラスミドpTG186(仏国特許第2,583,429号)のPstI部位へ挿入すると、pTG188が得られる。相同組換えによって得られるウイルスはVVTG188と呼ばれる。BglII/EcoRI消化後、M13TG9132のBamHI/EcoRI部位でIL−2配列を天然のワクシニアプロモーターpH5Rの3’に挿入し、M13TG9185を得る。指針として、ベクターM13TG9132は、PCRによってウイルスゲノムから単離されたH5R遺伝子のプロモーターの、多重クローニング部位の外に位置する内部BglII部位の突然変異によってM13TG131(Kieny et al., 1983, 前記) から誘導されるファージM13TG6131への挿入から得られる。
【0075】
次いで、HPV−16およびIL−2遺伝子をMVAゲノムの除去III領域へクローン化する。E7*TMR配列はBglII/HindIII消化によって単離し、次いでワクシニアプロモーターp7.5の3'においてpTG6025のBamHIとHindIII部位の間に挿入する。得られる構築体はpTG6050と呼ばれる。ヒトIL−2遺伝子の発現カセットの挿入部位は、相同組換えにより、オリゴヌクレオチドoTG10502およびoTG10503(配列番号18および19)の挿入によってpTG6050のHindIIIとKpnI部位の間に作り出される。次いで作出したベクターpTG6074をHindII/KpnI消化により線状化し、さらにBglII/EcoRI消化によってM13TG9185から単離された発現カセットpH5R−IL−2を用いて相同組換えを行う。最後に、得られた構築体pTG6088をKpnIで線状化し、次いでKpnI/XbaI消化によってM13TG9199から単離されたE6*TMF遺伝子、およびXbaI/KpaI消化によってM13TG9136から単離されたプロモーターp7.5と連結する。得られた構築体はpTG8042と呼ばれる(図1)。ベクターM13TG9136は、oTG5925を用いる指定突然変異誘発(p7.5プロモーターの3'におけるPstI部位の作出;配列番号20)ならびにoTG5924を用いる指定突然変異誘発(p7.5プロモーターの5’におけるBamHIおよびKpnI部位の作出;配列番号21)によって改変されたM13TG5107から得られる。
【0076】
ウイルスMVATG8042は、先行技術の規定に従ってMVAゲノムを用いる相同組換えによって作出する。組換え体の単離はgusAマーカー遺伝子の存在によって容易になる。
【0077】
HPV初期抗原の細胞局在性の改変がその発現を阻害しないことを確かめる。抗E7抗体を用いてMVATG8042に感染させた細胞抽出物のウェスタンブロット解析は、20〜35kDaの間の分子量を有する3個のバンドの検出を可能にする。この不均一性は狂犬病糖タンパク質の膜付着領域における可能性のあるO−糖分解部位の存在によって説明され得る。ウェスタンブロット解析がフェニル−Gal−Nac(フェニルN−アセチル−α−D−ガラクトピラノシド、Sigma、p4023)の存在下で繰り返す場合には、20kDaの1形態のみが検出され、これはE7*TMR遺伝子の発現産物のO−糖分解を確実にする。
【0078】
E6*TMF遺伝子の発現産物の抗E6抗体によるウェスタンブロット検出は、予測された20kDaの分子量に移動する1個のバンドのみを示し、これは翻訳後の修飾がないことを確実にする。指針として、 前記抗E6およびE7抗体は精製された抗原の投与によって得られたウサギ抗血清である。しかしながら、モノクローナルでもポリクローナルでも、いずれの他の特異的抗体も好適であり得る。
【0079】
hIL−2遺伝子の発現は、ELISA(Quantikine R&D系)およびIL−2依存性細胞増殖試験により評価する。培養条件にもよるが、産生されるIL−2の量は0.1pfu/細胞を用いて感染させた106細胞あたり200〜800ng/ml/24時間まで様々である。
【0080】
実施例3: ウイルスMVATG8042のin vivoにおける効率(免疫療法)
C57BL6マウスに皮下経路によって植え付けられた103個のBMK−16myc細胞を接種する。指針として、 細胞系統はHPV−16ゲノムおよびネズミc myc遺伝子でトランスフェクトした新生マウスの腎臓細胞に由来する。次いで、MVATG8042ウイルスの107pfu(プラーク形成単位)もまた、D3、D6およびD9に皮下経路により投与し、腫瘍の発達を定期的にモニターする。処理されたマウスは非組換えMVAウイルスを受容した動物からなる対照と比べてD15までの腫瘍成長において遅延を示す。さらに、処理はD30〜35での腫瘍の部分的な抑制を伴い、これは同等のMVAを用いる場合には観察されず、またこれは天然の核局在性を有するE6*およびE7*変異体を発現する。
【0081】
実施例4: トランスメンブラン局在性のE7*抗原を発現する組換えウイルスVVTG5095の構築
E7*TMR配列をBglII/BamHI消化によってM13TG9150から単離し、pTG5016のBamHI部位に挿入する。得られた構築体はpTG5095と呼ばれ、p7.5プロモーターの制御下に狂犬病糖タンパク質の分泌シグナルおよび付着シグナルと融合したE7*配列を含む。ウイルスVVTG5095はワクシニアゲノムを用いた相同組換えによって作出する。
【0082】
実施例5: トランスメンブラン局在性のE7*抗原および免疫刺激剤B7.1を発現する組換えウイルスVVTG6002の構築
ヒトB7.1遺伝子をプライマーoTG6353およびoTG6352(配列番号22および23)を用いた逆PCR(RT−PCR)によってダウジ(Daudi)ヒト細胞系統から単離し、M13TG6131のBglIIとEcoRI部位の間に挿入する。M13TG9149は、p7.5プロモーターの3’においてM13TG5107の同じ部位の間にサブクローン化されている、BglII−EcoRI断片を単離したものから得られる(M13TG5107はM13TG130のBamHI部位へクローン化されたp7.5プロモーター配列を運ぶ)。p7.5−B7.1カセットは、このようにして得たM13TG9152と呼ばれる構築体からEcoRI/PstI消化によって単離し、oTG1086(5’AATTGCA3’)を用いてpTG5095のEcoRI部位へ導入する。得られた構築体はpTG6002と呼ばれ、次いでワクシニアゲノムを用いる相同組換えによって組換えウイルスVVTG6002を作出する。
同等の構築体は、実施例2で開発されたような同様の技術に従う、E7*およびB7.1遺伝子を発現するカセットのMVAN33遺伝子の除去領域IIIへの挿入によって調製される。
【0083】
実施例6: ウイルスVVTG5095およびVVTG6002のin vivoにおける効率(免疫予防)
C57BL6マウスに107pfuのVVTG5095またはVVTG6002を用いて皮下経路によって3回ワクチン接種した。最後の免疫化の3日後、皮下に植え付けた103個のE7W1細胞を用いて、これらの動物に抗原投与する。指針として、E7W1細胞はHPV−16の発癌性E7遺伝子を発現するベクターでトランスフェクトされたネズミリンパ腫系統から得られる。時間の関数としての動物の生存率(%)を107pfuの非組換えワクシニアウイルスVVTG186(前記に記載のベクターTG186に由来する)で処置した対照マウスで得られたものと比較する。死亡率のモニタリングは3群の間の差異を示す。D36で対照群では100%の動物が死亡したのに対し、VVTG5095でワクチン接種した動物では約4分の1の生存が観察される。防御はB7.1抗原をコードする配列を含む構築体VVTG6002により実質的に増強される。
【0084】
実施例7: HPV16の改変された初期遺伝子および後期遺伝子を発現するMVATG9936の構築
HPV16L1およびL2タンパク質をコードする断片をPCRによって一般的な先行技術に従ってカスキ(Caski)細胞のゲノムDNA(ATCC1550)から単離する。L1配列を保持する増幅断片をM13TG130(Kieny et al., 1983, Gene 26, 91-99)へサブクローン化し、構築体M13TG8171を得る。クローン化されたL1遺伝子の配列はGenebankに含まれる配列(受託番号K02718)と比較して数箇所の突然変異、すなわち、248番におけるAの代わりのC、253番におけるAの代わりのC、537番におけるAの代わりのG、682番におけるCの代わりのG、874番におけるAの代わりのG、1393番におけるトリプレットACTの挿入、1390番におけるトリプレットGATの欠損を示す。この配列をZhou et alによって発表されたもの(1991, Virology 185, 251-257)と適合させるよう、かつ、ワクシニアの初期段階の発達を妨げることのできる初期転写終結に関して可能性ある部位を構成するTTTTTNT配列のレベルでサイレント変異を導入するよう点突然変異によって補正する。部位指定突然変異誘発の技術は当業者の能力の範囲内である。補整された配列を運ぶベクターはM13TG4041と呼ばれる。
【0085】
L2配列を運ぶPCR断片のベクターM13TG6131への挿入により、M13TG9126が得られる。Genebankに開示された配列と比較して5個の点突然変異、すなわち、378番におけるTの代わりのC、691番におけるGの代わりのA、702番におけるGの代わりのA、990番におけるAの代わりのGおよび1092番におけるAの代わりのCが確認される。指針として、ベクターM13TG6131は、多重クローニング部位の外に位置する内部BglII部位の突然変異によってM13TG131(Kieny et al., 1983, Gene 26, 91-99)から誘導される。
【0086】
ベクターM13TG4041をXbaI−SacIを用いて消化し、次いでL1配列を有する断片をL2遺伝子の下流でかつ、反対の方向に、M13TG9126のXbaI−SacI部位の間に挿入する。このようにして作出された構築体はM13TG4042と呼ばれる。次いで、L1およびL2配列をBglII消化によって単離し、後期p11Kと初期p7.5プロモーターの融合の結果生じる合成プロモーターp11K7.5を含むベクターM13TG4052のBglIIとBamHI部位の間にサブクローン化する。2つの方向のうち、M13TG4055と呼ばれるプロモーターのL1遺伝子の下流に位置するものを選択する。次いで、pH5RプロモーターおよびhIL−2遺伝子を有する発現カセットを、L1とL2遺伝子の間に位置するM13TG4055のHindIII部位へ導入するのに先立ち、HindIII消化によってpTG8042から単離する。最後に、得られた構築体M13TG4057をBglIIで消化し、次いでHPV16後期配列を有する断片を、合成プロモーターp4BK1Lのクローン化の結果生じるM13TG4060のBamHI部位へ挿入する。後者は初期プロモーターp4B(Davidson and Moss, 1989, J. Mol. Biol. 210, 749-769)と後期プロモーターpK1L(Davidson and Moss, 1989, J. Mol. Biol. 210, 771-784)間のハイブリッドプロモーターである。p11K7.5の制御下にあるL1遺伝子、L1配列に対して反対の方向でp4BK1Lの制御下にカセットpH5R−IL2およびL2遺伝子を含んでなるM13TG4062が得られる。
【0087】
E7*TMRおよびE6*TMFをコードする配列を含む多シストロン性カセットを構築する。第一に、EMCウイルス(脳心筋炎ウイルス;Genebank受託番号M22458)のIRES配列を常法によってpTG6002のEcoRIとNcoI部位の間にE7*TMR遺伝子の下流が導入されたEcoRI−NcoI断片の形で単離して(実施例5)pTG8084を得る。次いでE6*TMF遺伝子を各5’末端にNcoI部位を作り出す適当なプライマーを用いてPCRによって増幅する。E7*TMR遺伝子およびpTG8084から得られたIRES配列を有するBglII−NcoI断片ならびにNcoIおよびSacIによって消化されたE6*TMF遺伝子を有するPCR配列を、予めBglIIおよびSacIで消化されたベクターM13TG6131(実施例2)中で再構築する。M13TG4059が得られる。次いで、後者からE7*TMR−IRES−E6*TMF単位をBglII断片の形で単離し、p7.5KプロモーターのpTG8093下流のBamHI部位へ挿入する(pTG8093はp7.5KプロモーターのベクターpTG6025へのクローン化の結果生じる)。このようにして得られた構築体はpTG9901と呼ばれる。
【0088】
M13TG4062のSacI断片をpTG9901のSacI部位へクローン化し、pTG9902を得る。以下の方法でベクターpポリII(Lathe et al., 1987, Gene 57, 193-201)において、選抜可能なgpt遺伝子(Falkner and Moss, 1988, J. Virol. 62, 1849-1854)を後期およびIL2遺伝子とともに構築する。最初のものはBglII−NcoI消化によってM13TG4076(このベクターはその2つの末端にp4BK1Lプロモーター配列によってフランクされたgpt遺伝子を含む)から、2番目のものはNcoI−SacI消化によってpTG9902から単離する。精製された断片をpポリIIのBglII−SacI部位の間にクローン化する。pTG9933は、SacIによって切断されたベクターpTG9901へ導入されているSacI断片が得られるものから得られる。このようにして得られた構築体pTG9936は図2に示されている。
MVATG9936ウイルスは、MVAN33ゲノムを用いる相同組換えによって作出する。クローンの単離は先行技術の規定に従って行ってよい。
【0089】
実施例8: 免疫予防におけるウイルスの効率
A.ウイルスの処方の重要性(実験N121)
この前臨床試験の目的はウイルス株(MVAに対するコペンハーゲンワクシニアウイルス)および抗腫瘍防御に関する処方(天然の核局在性に対するトランスメンブラン型での提供)を比較することである。
【0090】
C57B16マウスを107pfuのウイルスで3回ワクチン接種する。注射は腹膜内経路によって10日毎に(D1、D11、D21)行う。最後の免疫化の7日後に、それらの右手側面への5×104個のTC1細胞の皮下投与によって動物に抗原投与する。ウイルスおよび投与される溶液、各々:
1 MVATG8042(実施例2、E6*TMF、E7*TMR、IL−2)、
2 MVATG6037(実施例5、E7*TMR、B7.1)、
3 VVTG5095(実施例4、E7*TMR)、
4 MVATG6090(E6*、E7*、IL−2)、
5 VVTG5061x188(E6*、E7*、IL−2)、
6 VVTG186(非組換えワクシニアウイルス)、
7 MVAN33(非組換えMVAウイルス)、および
8 塩溶液(PBS)
に従って20個体の動物の8群を形成する。
【0091】
1〜3群は少なくとも1種のトランスメンブランHPV抗原を発現する本発明のウイルスでワクチン接種するが、4および5群は天然の核局在性のHPV16初期抗原を発現するウイルスを、また6〜8群は非組換え対照ウイルスまたは塩溶液を施す。ウイルスMVATG6090およびVVTG5061x188は国際出願WO98/04705に記載されているということを記載すべきであろう。種々の群の動物の生存率(%)を腫瘍の抗原投与に続いて12週間モニターする。結果は以下のように要約できる。
【0092】
一般に、3つの対照群における死亡率は高く、95%(ウイルスMVAN33およびVV186を用いて)および81%(PBSを用いて)に達する。
HPV核抗原を発現するウイルスで免疫化した動物の生存においては有意な増加が観察される。例えば、ウイルスMVATG6090を受容したマウスの55%は腫瘍を持たないが、VVTG5061x188の投与は75%の腫瘍排除レベルを誘導する。
【0093】
他方、HPV16トランスメンブラン抗原を発現するウイルスでワクチン接種された動物の大部分では腫瘍が排除されたか、または腫瘍成長において実質的な遅延を示す。さらに正確には、MVATG8042によれば100%の排除が、MVATG6037によれば95%の排除が、またVVTG5095によれば90%の排除が観察される。
【0094】
図3は対照(MVAN33およびPBS)と比較して、MVATG8042およびMVATG6090でワクチン接種された動物を用いて得られた生存曲線を示す。
【0095】
全体として、これらのデータはコペンハーゲンワクシニアウイルスとMVAに由来するウイルス間に有意な差異がないこと、ならびに膜型での提供によって与えられたより優れた免疫原性を実証するものである。試験した総てのウイルスのうち、この免疫予防の動物モデルにおいて100%の腫瘍排除が起こるので、ウイルスMVATG8042が最も効果的である。
【0096】
B.記憶反応の研究(実験N122)
C57B16マウスに、D82にそれらの右手側面への5×104個のTC1細胞の皮下投与によって抗原投与するに先立ち、D1、D11およびD21に107pfuのウイルスを用いて腹膜内経路によって免疫化する。従前の実験(N121)と同一の8群を形成する。腫瘍の発達を時間の関数としてモニターする。腫瘍の抗原投与の50日後に得られたデータは腫瘍の排除に関して最も効果的なウイルスはウイルスMVATG8042であるということを確実とする。その投与は100%の動物を防御し、長期免疫を誘導する能力を示す。
【0097】
C.用量効果(実験N127)
C57B16マウスに、D28に5×104個のTC1細胞のそれらの右手側面への皮下投与によって抗原投与するに先立ち、D1、D11およびD21に105、106または107pfuのMVATG8042ウイルスを用いて腹膜内経路によって免疫化する。対照動物には同量のMVAN33を施す。腫瘍の発達を1週間に2回、12週間モニターする。結果(図4)は、投与されたMVATG8042の用量にかかわらず100%の防御を示すが、対照動物の大部分は腫瘍を発達させる。これらのデータは、低用量であってもMVATG8042ウイルスの効力を確実とする。
【0098】
D.投与経路の効果(実験N134)
C57B16マウスに、D44に5×104個のTC1細胞のそれらの右手側面への皮下投与によって抗原投与するに先立ち、D1、D11およびD21に107pfuのウイルスを用いて種々の投与経路(乱刺、皮下、腹膜内または筋肉内)によって免疫化する。腫瘍の発達を週に2回、12週間モニターする。図5に示されるように、腹膜内または筋肉内経路は、ウイルスMVATG8042による最高レベルの防御を与える。
【0099】
実施例9: 免疫治療におけるウイルスの効力
A.免疫治療の観点におけるウイルスの効力(実験N125)
この研究の目的は予め確立された腫瘍に対する、膜または核形態でHPV抗原を示すウイルスの治療能力を比較することである。このために、5×104個のTC1細胞を皮下経路によってC57B16マウスの右手側面へ投与する(D0)。次いで107pfuのウイルスをD7、D14およびD21に腹膜内経路によって注射する。4つの動物群を、各々MVAN33(負の対照)、トリス−HCl/NaClの塩溶液(負の対照)、MVATG8042(トランスメンブラン型)およびMVATG6090(核型)という投与されるウイルスに従って形成する。腫瘍の発達を週に2回、12週間評価する。結果は治療上の観点での抗腫瘍防御に関して、膜型での提供のより優れた効力を示す。MVATG8042を施したマウスは100%が腫瘍細胞の植え込み140日後に生存したが、MVATG6090を接種した動物に関しては生存率は約60%であり、対照動物に関しては極めて低い。
【0100】
B.毒性研究−用量の効果
ヒト腫瘍へのそれらの適用を意図するに先立ち、ウイルスの毒性がないことを確かめることは重要である。106または107pfuのMVATG8042またはMVAN33ウイルスまたは野生型コペンハーゲンワクシニアウイルス(VVwt)を頭蓋内経路によってヌードマウス(5個体/群)へ投与する。マウスの生存率を20日間モニターし、得られたものを以下の表1に示す。
【0101】
表1
ウイルス 生存しているヌードマウスの数
106pfu 107pfu
MVAN33 5/5 5/5
MVATG8042 5/5 5/5
VVwt − 0/5
【0102】
107pfuのMVATG8042ウイルスの頭蓋内投与後の副作用は検出されないが、野生型ワクシニアウイルスで処理したマウスの総てが注射後3日で死亡した。対照MVAウイルスはどちらの動物に対しても有毒でなく、これは、すでに文献に記載されているワクシニアウイルスに比べ、その弱毒性を確実とするものである。
【0103】
[配列表]
(1)一般情報
(i)特許出願人:
(A)名称: Transgene SA
(B)通り: 11 rue de Molsheim
(C)市: Strasbourg
(D)国:フランス
(F)郵便番号:67082
(G)電話:(33)03 88 27 91 00
(H)テレファックス:(33) 03 88 27 91 11
(ii)発明の名称:細胞における局在性が改変された免疫ポリペプチドに基づく抗腫瘍組成物
(iii)配列数:23
(iv)読み取り可能なコンピューター:
(A)媒体形態:テープ
(B)コンピューター:IBM PC互換機
(C)オペレーティングシステム:PC−DOS/MS−DOS
(D)ソフトウエア:PatentIn リリース#1.0、バーション1.25(EPO)
(2)配列番号1の情報:
(i)配列の特徴:
(A)長さ:243個のアミノ酸
(B)型:アミノ酸
(D)トポロジー:直鎖
(ii)分子型:タンパク質
(iii)ハイポセティカル:NO
(vi)起源:
(A)生物名:ヒト・パピローマウイルス
(B)株:HPV−16
(C)個体/単離物:E6タンパク質とFタンパク質の融合シグナル
(vii)直接の起源:
(B)クローン:E6*TMF
(xi)配列の記載:配列番号1
Met Gly Leu Lys Val Asn Val Ser Ala Ile Phe Met Ala Val Leu Leu
1 5 10 15
Thr Leu Gln Thr Pro Thr Gly Gln Ile His Trp Gly Met His Gln Lys
20 25 30
Arg Thr Ala Met Phe Gln Asp Pro Gln Glu Arg Pro Arg Lys Leu Pro
35 40 45
Gln Leu Cys Thr Glu Leu Gln Thr Thr Ile His Asp Ile Ile Leu Glu
50 55 60
Cys Val Tyr Cys Lys Gln Gln Leu Leu Arg Arg Glu Val Tyr Asp Phe
65 70 75 80
Ala Phe Arg Asp Leu Cys Ile Val Tyr Arg Asp Gly Asn Pro Tyr Ala
85 90 95
Val Cys Asp Lys Cys Leu Lys Phe Tyr Ser Lys Ile Ser Glu Tyr Arg
100 105 110
His Tyr Cys Tyr Ser Leu Tyr Gly Thr Thr Leu Glu Gln Gln Tyr Asn
115 120 125
Lys Pro Leu Cys Asp Leu Leu Ile Arg Cys Ile Asn Cys Gln Lys Pro
130 135 140
Leu Gln Arg His Leu Asp Lys Lys Gln Arg Phe His Asn Ile Arg Gly
145 150 155 160
Arg Trp Thr Gly Arg Cys Met Ser Cys Cys Arg Ser Ser Arg Thr Arg
165 170 175
Arg Glu Thr Gln Leu Gly Leu Ser Ser Thr Ser Ile Val Tyr Ile Leu
180 185 190
Ile Ala Val Cys Leu Gly Gly Leu Ile Gly Ile Pro Ala Leu Ile Cys
195 200 205
Cys Cys Arg Gly Arg Cys Asn Lys Lys Gly Glu Gln Val Gly Met Ser
210 215 220
Arg Pro Gly Leu Lys Pro Asp Leu Thr Gly Thr Ser Lys Ser Tyr Val
225 230 235 240
Arg Ser Leu
(2)配列番号2の情報:
(i)配列の特徴:
(A)長さ:185個のアミノ酸
(B)型:アミノ酸
(D)トポロジー:直鎖
(ii)分子型:タンパク質
(iii)ハイポセティカル:NO
(vi)起源:
(A)生物名:ヒト・パピローマウイルス
(B)株:HPV−16
(C)個体/単離物:狂犬病糖タンパク質のE7融合シグナル
(vii)直接の起源:
(B)クローン:E7*TMR
(xi)配列の記載:配列番号2
Met Val Pro Gln Ala Leu Leu Phe Val Pro Leu Leu Val Phe Pro Leu
1 5 10 15
Cys Phe Gly Lys Phe Pro Ile Gly Ser Met His Gly Asp Thr Pro Thr
20 25 30
Leu His Glu Tyr Met Leu Asp Leu Gln Pro Glu Thr Thr Gln Leu Asn
35 40 45
Asp Ser Ser Glu Glu Glu Asp Glu Ile Asp Gly Pro Ala Gly Gln Ala
50 55 60
Glu Pro Asp Arg Ala His Tyr Asn Ile Val Thr Phe Cys Cys Lys Cys
65 70 75 80
Asp Ser Thr Leu Arg Leu Cys Val Gln Ser Thr His Val Asp Ile Arg
85 90 95
Thr Leu Glu Asp Leu Leu Met Gly Thr Leu Gly Ile Val Cys Pro Ile
100 105 110
Cys Ser Gln Lys Pro Arg Ser Tyr Val Leu Leu Ser Ala Gly Ala Leu
115 120 125
Thr Ala Leu Met Leu Ile Ile Phe Leu Met Thr Cys Cys Arg Arg Val
130 135 140
Asn Arg Ser Glu Pro Thr Gln His Asn Leu Arg Gly Thr Gly Arg Glu
145 150 155 160
Val Ser Val Thr Pro Gln Ser Gly Lys Ile Ile Ser Ser Trp Glu Ser
165 170 175
His Lys Ser Gly Gly Glu Thr Arg Leu
180 185
(2)配列番号3の情報:
(i)配列の特徴:
(A)長さ:36個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(A)生物名:ヒト・パピローマウイルス
(B)株:HPV−16
(C)個体/単離物:合成オリゴヌクレオチドoTG5118(E7欠損2126)
(xi)配列の記載:配列番号3
TCTGAGCTGT CATTTAATTG AGTTGTCTCT GGTTGC 36
(2)配列番号4の情報:
(i)配列の特徴:
(A)長さ:42個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:狂犬病ウイルス
(B)株:HPV−16
(C)個体/単離物:突然変異誘発オリゴヌクレオチドoTG5745
(xi)配列の記載:配列番号4
TGCACTCAGT AATACATAGG ATCCAATAGG GAATTTCCCA AA 42
(2)配列番号5の情報:
(i)配列の特徴:
(A)長さ:38個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG6390
(xi)配列の記載:配列番号5
GTATCTCCAT GCATGGATCC TGCAGGGTTT CTCTACGT 38
(2)配列番号6の情報:
(i)配列の特徴:
(A)長さ:36個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG6880
(xi)配列の記載:配列番号6
GGATCCGCCA TGGTAGATCT TGGTTTCTGA GAACAG 36
(2)配列番号7の情報:
(i)配列の特徴:
(A)長さ:32個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(A)生物名:狂犬病ウイルス
(B)株:HPV−16
(C)個体/単離物:合成オリゴヌクレオチドoTG5377(E6欠損111ないし115)
(xi)配列の記載:配列番号7
TGTCCAGATG TCTTTGCAGT GGCTTTTGAC AG 32
(2)配列番号8の情報:
(i)配列の特徴:
(A)長さ:34個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10829
(xi)配列の記載:配列番号8
GCGCGCTCTA GAATTATGGG TCTCAAGGTG AACG 34
(2)配列番号9の情報:
(i)配列の特徴:
(A)長さ:35個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10830
(xi)配列の記載:配列番号9
CAGTTCTCTT TTGGTGCATG CCCCAATGGA TTTGA 35
(2)配列番号10の情報:
(i)配列の特徴:
(A)長さ:38個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10835
(xi)配列の記載:配列番号10
ATGCTAGTGC TCGATAAACC CAGCTGGGTT TCTCTACG 38
(2)配列番号11の情報:
(i)配列の特徴:
(A)長さ:35個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10836
(xi)配列の記載:配列番号11
TCAAATCCAT TGGGGCATGC ACCAAAAGAG AACTG 35
(2)配列番号12の情報:
(i)配列の特徴:
(A)長さ:38個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG10833
(xi)配列の記載:配列番号12
CGTAGAGAAA CCCAGCTGGG TTTATCGAGC ACTAGCAT 38
(2)配列番号13の情報:
(i)配列の特徴:
(A)長さ:36個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG10834
(xi)配列の記載:配列番号13
GCGGGCATGC GGTACCTCAG AGCGACCTTA CATAGG 36
(2)配列番号14の情報:
(i)配列の特徴:
(A)長さ:32個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:ワクシニアウイルス
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG7637(PCR領域III)
(xi)配列の記載:配列番号14
GGGGGGGAAT TCAGTAAACT TGACTAAATC TT 32
(2)配列番号15の情報:
(i)配列の特徴:
(A)長さ:39個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(A)生物名:ワクシニアウイルス
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG7638(PCR領域III)
(xi)配列の記載:配列番号15
GGGGGGGGAT CCGAGCTCAC CAGCCACCGA AAGAGCAAT 39
(2)配列番号16の情報:
(i)配列の特徴:
(A)長さ:32個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:ワクシニアウイルス
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG7635(PCR領域III)
(xi)配列の記載:配列番号16
GGGGGGGGAT CCGGAAAGTT TTATAGGTAG TT 32
(2)配列番号17の情報:
(i)配列の特徴:
(A)長さ:30個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(A)生物名:ワクシニアウイルス
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG7636(PCR領域III)
(xi)配列の記載:配列番号17
GGGGGGGAAT TCTTTGTATT TACGTGAACG 30
(2)配列番号18の情報:
(i)配列の特徴:
(A)長さ:77個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10502
(xi)配列の記載:配列番号18
AGCTTTTTAT TCTATACTTA AAAAATGAAA ATAAACTCGA GTTGTCAAAG CATCATCTCA 60
ACACTGACTT GAGGTAC 77
(2)配列番号19の情報:
(i)配列の特徴:
(A)長さ:69個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10503
(xi)配列の記載:配列番号19
CTCAAGTCAG TGTTGAGATG ATGCTTTGAC AACTCGAGTT TATTTTCATT TTTTAAGTAT 60
AGAATAAAA 69
(2)配列番号20の情報:
(i)配列の特徴:
(A)長さ:39個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG5925
(xi)配列の記載:配列番号20
TCAGATCTGT CGAGGGATCT GCAGCTTCTT CTAGAGGTA 39
(2)配列番号21の情報:
(i)配列の特徴:
(A)長さ:44個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG5924
(xi)配列の記載:配列番号21
AGTGAATTGC TGCAGGTACC CGGATCCGCA TCGACTATCG ACAT 44
(2)配列番号22の情報:
(i)配列の特徴:
(A)長さ:35個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:ホモサピエンス
(B)株:ダウジ細胞系統
(C)個体/単離物:PCRプライマーoTG6353(クローンニングB7.1)
(xi)配列の記載:配列番号22
TCAGCCCCTG AATTCTGCGG ACACTGTTAT ACAGG 35
(2)配列番号23の情報:
(i)配列の特徴:
(A)長さ:33個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:ホモサピエンス
(B)株:ダウジ細胞系統
(C)個体/単離物:PCRプライマーoTG6352(クローンニングB7.1)
(xi)配列の記載:配列番号23
TTGACCCTAA AGATCTGAAG CCATGGGCCA CAC 33
【図面の簡単な説明】
【0104】
【図1】pTG8042の模式図である。BRG3およびBRD3は、MVAウイルスの除去領域IIIのレベルでの挿入が可能な左および右の組換えアームを表す。pTG8042は、7.5kプロモーターによって命令される、麻疹ウイルスFタンパク質のシグナル配列(SF)およびトランスメンブラン領域(TMF)と融合したHPV16 E6遺伝子を発現させるためのカセット;プロモーターpH5Rの制御下に置かれたIL−2遺伝子を発現させるためのカセット;p7.5プロモーターの制御下に置かれた狂犬病糖タンパク質のシグナル配列(SF)およびトランスメンブラン領域(TMF)と融合したHPV16 E7遺伝子を発現させるためのカセットならびにp7.5プロモーターの制御下に置かれたgusA遺伝子(uidA)を発現させるためのカセットを含んでなる。
【図2】ベクターpTG9936の模式図である。BRG3およびBRD3は、MVAゲノムの除去領域IIIのレベルでの挿入が可能な左および右の組換えアームを表す。pTG9936は、gptマーカー遺伝子の発現を命令するプロモーターp4BK1L、HPV16 L2遺伝子の発現を命令するプロモーターp4BK1L、IL−2の発現を命令するプロモーターpH5R、HPV16 L1遺伝子の発現を命令するプロモーターp11k7.5、狂犬病糖タンパク質のシグナル配列(SF)およびトランスメンブラン領域(TMF)、続いてIRES配列と融合したHPV16 E7遺伝子、ならびに麻疹ウイルスFタンパク質のシグナル配列(SF)およびトランスメンブラン領域(TMF)と融合したHPV16 E6遺伝子の発現を命令するプロモーターp7.5を有する。
【図3】ウイルスMVATG8042、MVATG6090、MVAN33で、またはPBSによる腫瘍の抗原投与前にワクチン接種した動物の生存率(%)を時間(日)の関数として表すことによって実験N121を図示している。
【図4】107、106および105pfuのウイルスMVAN33またはMVATG8042の投与後の腫瘍を持たない動物の割合(%)を時間(日)の関数として表す(実験N127)。
【図5】乱刺法、皮下(SC)、腹膜内(IP)または筋肉内(IM)により、ウイルスMVATG8042でまたは対照MVAN33で処置した動物の生存率(%)を時間(日)の関数として表すことによって実験N134を図示している。
【発明の背景】
【0001】
本発明の主題は、天然の局在性とは異なる細胞での局在性を有するよう改変された免疫ポリペプチドを治療薬または予防薬として含んでなる抗腫瘍組成物である。それはまた、そのポリペプチドを発現する組換えベクターに基づく組成物に関する。かかる組成物はさらに詳しくは、パピローマウイルスに関連した病巣の治療または予防を意図したものである。
【0002】
一般に、癌は細胞増殖の制御の欠如に起因する疾病であると解釈されている。その原因にはいろいろあろうが、特に細胞の遺伝子の機能不全(潜在的癌遺伝子の例えば体細胞変異による活性化;発現の脱制御;腫瘍抑制遺伝子の発現の阻害)またはウイルス遺伝子の望ましくない発現による可能性がある。
【0003】
ヒトにおいては、パピローマウイルス(HPV)は皮膚の良性感染から疣贅、および悪性腫瘍にわたる病院に関与する。これまでに確認された75タイプのHPVのうち、20の異なる分離株が生殖道に極めて特異的であり、そのうち5株(HPV−16および18、そしてより程度は低いがHPV−31、33および45)が子宮頸部および生殖道下方の癌に明確に関与している。一連の全体の研究は、これらのウイルスの形質転換に果たす役割、新生細胞のゲノムへのそれらの特異的な組み込み、癌性細胞におけるそれらの遺伝子の活性、および悪性の表現型のHPV陽性新生細胞を維持する際のあるウイルス遺伝子の発現の重要性を実証するものである(Monsenego, J. Impact Medecin, 11 March 1994)。
【0004】
一般に、パピローマウイルスは、キャプシドタンパク質に取り巻かれた約7900塩基対の環状ゲノムを有るDNAウイルスである。いくつかのタイプのパピローマウイルス、特にウシ・パピローマウイルス(BPV)およびヒト・パピローマウイルス(HPV)が確認されており(Pfister, 1987, in The papovaviridae: The Papillomaviruses, Salzman and Howley edition, Plenum Press, New York, p 1-38)。それらのゲノムは、リーディングフレームE1、E2、E4、E5、E6およびE7を包含する初期領域と、キャプシドタンパク質L1およびL2をコードする後期領域を含んでなる。
【0005】
初期タンパク質はDNAに結合する能力を有し、優先的に核で見られる。E1およびE2の発現産物はウイルスの複製とウイルス遺伝子の発現を調節し、一方、E5、E6およびE7領域の発現産物は感染細胞の発癌転換のプロセスに関与する。このことに関して、BPV−1 E5タンパク質がin vitroで細胞を形質転換可能なことが実験的に示されている(Schlegel et al., 1986, Science 233, 464-467)。E7の形質転換力はHPV−16およびHPV−18に関して実証されており(Kanda et al., 1988, J. Virol. 62, 610-613; Vousden et al., 1988, Oncogene Res. 3, 1-9; Bedell et al., 1987, J. Viorol. 61, 3635-3640)、網膜芽細胞種(Rb)遺伝子の産物と結合する能力と関連している(Munger et al., 1989, EMBO J. 8, 4099-4105; Heck et al., 1992, Proc. Natl. Acad. Sci. USA 89, 4442-4446)。さらに、Crook et al (1991, Cell 67, 547-556)はHPV−6および18のE6抗原がp53遺伝子の産物を合成することができることを示したが、このことは細胞の形質転換におけるその有力な役割を説明している。
【0006】
HPVウイルスに関連する病状はそれらの持続性および再発性のために治療上問題がある。通常のアプローチは依然として外科手術と化学療法であるが、これらの疾病の治療のため、今般、免疫療法が考案されている。理想的なワクチン候補は、予防目的(免疫予防)としては、感染が永続的に確立されないようにする、また隣接する組織に広がらないようにし、治療目的(免疫療法)としては、感染した患者における腫瘍発達を軽減するべきである。これまでには、キャプシド抗原を使用して、ウイルス粒子の表面に局在するエピトープに対する抗体の産生を誘導すること、および初期タンパク質を使用して、ウイルスDNAの組み込み後に感染した細胞に対して細胞性免疫を確立することが提案されている。
【0007】
このことに関して、欧州特許EP0,462,187では、パピローマウイルス初期遺伝子を発現するポックスウイルスの投与を含む治療的アプローチが記載されている。WO93/02184に記載されている予防接種的アプローチは、免疫物質としてのキャプシド抗原、特にin vitroで再構成されたDNAを含まないウイルス粒子(ウイルス様粒子のVLP)の使用に基づいている。仏国出願第96 09584号では、パピローマウイルス初期ペプチドによってもたらせれる予防効果と後期ペプチドによって与えられる治療効果を組み合わせた組成物が開示されている。しかしながらこれまでに、所望によりそれらの形質転換活性を無効にするよう変異させてはいるが、それらの細胞局在性という観点では天然型であるウイルスタンパク質が使用されてきた。
【発明の概要】
【0008】
本発明は、それが特異的であれ、非特異的であれ、また体液型(抗体の産生)であれ、細胞型(細胞傷害性応答CTL)であれ、治療される腫瘍または癌に対する免疫応答を増強するまたは刺激するために、宿主の免疫系に対するそれらの接近性を高める目的でその局在性が改変された免疫タンパク質を使用することを提案する。
【0009】
今般、HPV−16 E6およびE7核抗原が、適当な付着配列および分泌配列を導入して、そこでのトランスメンブラン型での提供を付与することにより改変された。局在性の変化は免疫応答に関して有益な効果を持ち、その結果、膜E6およびE7抗原およびヒトIL−2を同時発現するワクシニアウイルスで処理した動物は、核抗原を産生する同等のウイルスを受容させたものに比べて抗腫瘍活性が高くなる。本発明の目的は、腫瘍もしくは癌の確立または発達を少なくとも部分的に阻害することに関して先行技術の組成物よりも効果的な公知の抗腫瘍組成物へ利用できるようにすることである。特に有用な用途はHPV感染の、さらに詳しくは子宮頸部癌などの重篤な病状の治療である。
【0010】
従って、本発明の主題は、1以上の免疫ポリペプチドを治療薬または予防薬として含んでなる組成物であり、これらポリペプチドの少なくとも1種はその天然の局在性とは異なる局在性を持つよう改変されている。
【0011】
本発明の目的のため、「免疫ポリペプチド」とは、それと同じものが正常な細胞には存在しないポリペプチドを表す。好ましい例としては、腫瘍特異的抗原がある。例としては、その発現が胎胚期に起こり、誕生時にそれが消失するまで退縮する細胞抗原、通常は非常に低レベルで発現し、高レベルで発現すると腫瘍の特徴を持つようになる抗原、その構造またはコンホメーションが改変されている細胞抗原、あるいは非細胞抗原、特に発癌ウイルスに由来するウイルス抗原が挙げられる。それらは、例えば、遺伝子BRCA−1(Miki et al., 1994, Science 226, 66-71)、BRCA−2(Wooster et al., 1995, Nature 378, 789-792)、MUC−1(Hareuveni et al., 1990, Proc. Natl. Acad. Sci. USA 87, 9498-9502)、CEAなどの発現産物であり得、これらのある変異または過剰発現が癌の発達に関与している。ウイルス抗原については、より詳しくは、パピローマウイルス初期もしくは後期遺伝子、エプスタイン・バーウイルスEBNA−1の発現産物、HTLV(ヒトTリンパ球ウイルス)IおよびIIウイルスまたはB型およびC型肝炎ウイルスの抗原が挙げられる。腫瘍特異的抗原は当業者が入手できる文献に広く記載されている。
【発明の具体的説明】
【0012】
本発明で使用されるポリペプチドの不可欠な特徴としては、その天然の局在性とは異なる局在性を持つことである。輸送のメカニズムおよび関与するシグナルは細胞生物学の書物に記載されている(例えば、Molecular Biology of the Cell, Third Ed. garland Publishing Inc. NY & Londonを参照)。便宜には、大部分のポリペプチドがサイトソル中の遊離リボゾームで合成され、そこでそれらは活性を示す。しかしながら、いくつかのポリペプチドは異なる細胞内指向性を持っており(それは一般に適当なペプチドシグナルの存在によって確認される)、そこまで輸送されなければならない。このように、原形質膜への輸送されるよう、または細胞外へ分泌されるよう意図されたポリペプチドは、小胞体(ER)と会合したリボゾームによって、通常はそれらのアミノ末端に、ERへの送達を開始させる分泌配列(またはシグナルペプチド)を含んでなる前駆体の形態で合成される。次いでそれは特異的エンドペプチダーゼによって除去され、成熟ポリペプチドとなる。分泌配列は通常、15〜35疎水性の必須アミノ酸を含んでなる。非共通配列が存在し、これがエンドペプチダーゼによる認識を決定する分泌配列であるものと思われる。しかしながら、グリシン、セリンまたはアラニン残基の後でタンパク質分解的切断がしばしば起こらなければならない。
【0013】
膜タンパク質は一般に、原形質膜に依然として挿入されている、疎水性の高い付着配列を含んでなる。ポリペプチドはトランスメンブラン型であると考えられ、一方の末端は細胞外に露出し、付着配列は膜をわたり、もう一方の末端はサイトソル側にある。ほとんどの場合、ポリペプチド鎖は膜の脂質二重層に埋め込まれており、αらせんコンホメーションを有している(例えば、Branden and Tooze, 1991, in Introduvtion to Protein Structure p. 202-214, New Garlandを参照)。
【0014】
核局在性に関しては、主としてリジンおよびアルギニンなどの陽電荷残基からなった、短い、いわゆる核局在化配列(NLS)の存在によって与えられると考えられる。例としては、HPV L1およびL2ポリペプチドに存在する核転座シグナルKRKKRKおよびRKRRKRが挙げられる(Zhou et al., 1991, Virology 185, 625-632)。しかしながら、核内でそれらの機能を発揮するポリペプチドには、典型的なNLS配列を持っていないものがある。パピローマウイルスE6およびE7抗原の場合がそうである。
【0015】
本発明の組成物中に含有される免疫ポリペプチドは、天然のポリペプチド、その断片、異種起源の配列を含んでなるキメラ、または変異体(1以上のアミノ酸の欠失、挿入および/または置換)内の適当な局在化シグナルの導入および/または欠失から得てもよい。さらに詳しくは、そのアミノ酸配列は、それに由来する天然ポリペプチドの配列の総てまたは一部と70%を越える、有利には80%を越える、特には90%を越える同一性を示す。この同一性は適当なコンピュータープログラムを用いて、あるいは最高の相同性が得られるように配列を並べることにより、そして2つの配列のアミノ酸が全番号の位置と比べて同一であるとわかった位置の数を計数することにより容易に算出できる。
【0016】
当業者ならば、ポリペプチドの細胞への提供を変化させるシグナルを知っている。免疫ポリペプチドが分泌することが望ましい場合、そのアミノ末端へ分泌配列を付加すれば、ERを介してそれを宿主細胞の外へ輸送することが可能となる。この挿入は、翻訳の開始に関するコドンのすぐ下流で起こることが好ましい。本発明のでは、干渉を避けるために、天然の局在性を決定する残基の総てまたは一部を変異/欠失させることが有利であろう。例えば、天然のポリペプチドが膜指向性を持つならば、それは分泌配列をすでに含んでなり、疎水性付着配列を所望により変異/欠失させる。さらに、天然のシグナルの不活性化(変異/欠失による)により細胞質への局在化が可能となる。通常、細胞での局在性が異なる(例えば、核、膜、分泌など)免疫ペプチドを細胞質に供されると、in vivoにおいては主要組織適合性複合体のクラスI抗原とCTL応答によって媒介されるポリペプチドの提を促すことができる(Tobery and Siliciano, 1997, J. Exp. Med. 5, 909-920)。意図し得るもう1つの具体例としては、有効なCTR応答を刺激する目的でも、免疫ポリペプチドとユビキチンを融合させることである。
【0017】
有利には、本発明の組成物は、好ましくは膜付着配列を、および天然のポリペプチドが分泌配列を欠いている場合にはそれを挿入することにより、膜局在性を持つよう改変された免疫ポリペプチドを含んでなる。分泌配列の好ましい挿入部位は、前記されたようにN末端であり、膜付着配列のそれは、例えば停止コドンのすぐ上流のC末端である。本明細書ではまた、天然の局在化シグナル(例えば、NLS配列)の総てまたは一部の変異/欠失させて、新たな場所で干渉することがないようにすることが有利であり得る。
【0018】
本発明で使用できる局在化シグナルの選択肢は広い。それは、それが処置される細胞に認識される限り、それを含んでなる、真核生物起源またはそれ以外(ウイルス、寄生体、真菌類など)いずれのタンパク質に由来するものであってもよい。それは天然でも合成でもよく、後者に関して異種であっても同種であってもよい。それはまた、それが由来するシグナルに対する1以上の改変を含んなる(ただし、それ/それらはその機能に影響を及ぼさない)。指針としては、狂犬病糖タンパク質、HIVウイルスenv糖タンパク質、または麻疹ウイルスFタンパク質の膜付着配列および/または分泌配列を使用することが好ましいであろう。免疫ポリペプチドの改変がいくつかの局在化シグナルを含む場合には(例えば、膜付着配列および分泌配列)、これらは共通に起源であっても、異なる起源であってもよい。
【0019】
また、特定の細胞コンパートメントをターゲッティングする局在化シグナルを使用することもできる。特に、エンドサイトーシスに関する共通配列(例えば、配列IPNYRNMを有する免疫グロブリンIgG1重鎖のC末端領域に存在する;Kaisho et al., 1997, Science 276, 412-414)、またはゴルジ体膜へのターゲッティングを可能にする配列(Mochamer and Rose, 1987, J. Cell Biol. 105, 1205-1214; Mochamer, 1993, Curr. Opin. Cell Biol. 5, 606-612)が挙げられる。特に、P11222の受託番号でSwiss−Protデータバンクに開示されているコロナウイルスE1糖タンパク質由来配列の使用が意図される。免疫ポリペプチドのC末端にこの種の配列を組み込むことが好ましい。
【0020】
さらに、細胞局在性の改変は、いずれの常法により行ってもよく、特に位置指定突然変異誘発、異種シグナルの連結、またはPCRによって行ってもよい。
【0021】
好ましい具体例によれば、本発明の抗腫瘍組成物は、パピローマウイルス感染およびその結果起こる疾患、特に軽度子宮頸部形成異常および子宮頸部癌の治療または予防を意図したものである。この具体例によれば、それはパピローマウイルス、特にHPV−16、18、31、33または45などの危険性の高いウイルスの初期および/または後期領域に由来する少なくとも1種の免疫ポリペプチドを含んでなる。
【0022】
本発明によって追求される目的によれば、1種異常のいずれかの種の免疫パピローマウイルスポリペプチドが使用できる。先に想起されたように、それらのゲノムは8種のポリペプチド、すなわちウイルスキャプシドを含んでなる2種の後期ポリペプチドL1およびL2と、調節、ウイルスゲノムの維持および感染した細胞の形質転換に関与する6種の初期ポリペプチド(E1、E2、E4、E5、E6およびE7)をコードする。
【0023】
初期型の免疫ポリペプチドに関しては、特に膜局在性を有するよう改変されたE6またはE7由来のポリペプチドを使用することを選択するのが有利である。先に想起された観察を形質転換力に与えると、細胞の形質転換プロセス関与する領域で変異した非発癌性変異体を使用するのが好ましい。かかる変異体は文献に記載されている(Munger et al., 1989, EMBO J. 8, 4099-4105; Crook et al., 1991, Cell 67, 547-556; Heck et al., 1992, Proc. Natl. Acad. Sci. USA 89, 4442-4446; Phelps et al., 1992, J. Virol. 66, 2418-2427)。本発明の目的に特に好適な免疫ポリペプチドは、残基111〜115(+1、天然のウイルス抗原の1番目のアミノ酸を表す)が欠失し、かつ、麻疹ウイルスFタンパク質の分泌または付着シグナルと融合したHPV−16 E6抗原である(配列番号1)。また、残基22〜25が欠失し、狂犬病糖タンパク質の付着配列および分泌配列と融合したHPV−16 E6抗原も使用できる(配列番号2)。
【0024】
本発明の抗腫瘍組成物はまた、L1またはL2に由来の、パピローマウイルスの後期領域に由来する免疫ポリペプチドを含んでなってもよい。
【0025】
もちろん、本発明の抗腫瘍組成物は、そのうちの少なくとも1つがその天然の局在性とは異なる細胞局在性を有する、数種の免疫ポリペプチドを含んでなってもよい。パピローマウイルス由来の数種のポリペプチドを組み合わた組成物の例としては、共通の起源であっても異なる起源であってもよいもの(例えば、活性スペクトルを広くする目的では、HPV−16および18)が挙げられる。初期起源の数種のポリペプチドを組み合わせた組成物は治療効果を増強することができる。L1およびL2由来のポリペプチドの組合せは、組成物の予防特性に有益な効果を持ち得た。最後に、本発明により追求される目的に最も特に好適な組成物は、予防効果と治療効果を併せ持つよう、パピローマウイルスの少なくとも1種の初期ポリペプチドと少なくとも1種の後期ポリペプチドを含んでなる。好ましい具体例によれば、初期起源の少なくとも1種の免疫ポリペプチドが、先に挙げたもののような分泌または付着配列の付加によって膜局在性を有するよう改変される。
【0026】
これに関して、本発明の好ましい具体例は、
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド
を含んでなる。
【0027】
「相同」とは、その配列との70%を越える、有利には80%を越える、好ましくは90%を越える、最も好ましくは95%を越える同一性の程度をいう。
【0028】
有利には、本発明の抗腫瘍組成物は、免疫応答の強度を特異的に、または特異的にではなく増強する目的で、その抗腫瘍作用を増強する少なくとも1種の化合物さらに含んでなってもよい。アジュバントの他、免疫刺激剤は特に好ましい化合物を表す。「免疫刺激剤」とは、免疫ポリペプチドに向けられた抗体の産生を増幅するために体液性免疫応答を、あるいは、腫瘍または感染細胞に対する有意な細胞傷害性応答を誘発するために細胞性免疫応答を強める能力を有する化合物を意味するものと理解される。指針としては、免疫刺激剤は、癌の動物モデルにおいて、免疫ポリペプチドで処置した動物の拒否反応率を免疫刺激剤の存在下と不在下で比較することにより評価すればよい。より一般には、免疫刺激を証明する手段はRoitt(Immunology, 4th edition, Moby Ltd)に示されている。かかる組成物の利点の1つは、それが免疫ポリペプチドによって誘導される特異的免疫と免疫刺激分子によって誘導される非特異的免疫を併せ持つことである。
【0029】
本発明では、特にヒト起源の天然の免疫刺激剤、その一部、異なる起源の配列との融合から得られたキメラ、または変異体(しかしながら免疫刺激作用は保存されている条件で)を使用することができる。考えられ得るあらゆる分子のうち、インターロイキン−2、インターロイキン−7、インターロイキン−12および付着分子B7.1およびB7.2から選択される免疫刺激剤を使用することが好ましい。
【0030】
一般に、免疫ポリペプチドおよび免疫刺激ポリペプチドは通常の化学合成法によって、または組換えDNA技術によって作製され得る(例えば、Maniatis et al., 1989, Laboratory Manual, Cold Spring harbor, Laboratory Press, Cold Spring Harbor, NY)。さらに詳しくは、製造方法は、問題のポリペプチドをコードするDNA断片で形質転換した細胞を培養して産生細胞を作出する行為と、その培養物からポリペプチドを回収する行為を含んでなる。産生細胞はいずれの起源であってもよく、限定されるものではないが、考慮されているDNA断片がそのゲノムに組み込まれているか、または適当な発現ベクターに組み込まれているかのいずれかである限り、細菌、酵母菌、または哺乳類細胞であってもよい。もちろん、DNA断片は、産生細胞中でその発現を可能にする転写および翻訳シグナルの制御下に置かれる。発現ベクターおよび制御シグナルは当業者に公知である。
【0031】
本発明はまた、1以上の免疫ポリペプチドをコードする配列を含んでなる少なくとも1種の組換えベクター、その天然の局在性とは異なる局在性を有するよう改変されている少なくとも1種のポリペプチド、および所望により抗腫瘍作用を増強する化合物を、治療薬または予防薬として含んでなる抗腫瘍組成物に関する。この種の組成物は生産が高い費用がかからない、また、種々の環境条件下で安定性が高いという利点を持つ。特に、保存条件は制限が少ない。これらのポリペプチドは先に定義したような特徴を持つ。
【0032】
免疫ポリペプチドをコードする、または抗腫瘍作用を増強する配列は、一般の用法で、また文献のデータを用いて、クローン化によって、PCR(ポリメラーゼ連鎖反応)によって、または化学合成によって得られる。好ましい具体例に関しては、パピローマウイルスポリペプチドをコードする配列は、患者からまたはコレクションから得られるパピローマウイルス陽性細胞から選択すればよい。適当な局在化シグナルの挿入は、分子生物学的技術によって行ってよい。免疫刺激剤をコードする配列は、細胞DNAから、またはそれが発現している細胞のメッセンジャーRNAからクローン化すればよい。当業者ならば、公開されたデータから適当なプローブまたはプライマーを作製することができる。HPV−16および18ゲノムのヌクレオチド配列はそれぞれ受託番号K02718およびX05015でGenbankに開示されていることを記載しておくべきであろう。ヒトIL−2遺伝子の配列は、仏国特許第85 09480号およびTaniguchi et al. (1983, Nature 302, 305-311)に記載され、Freeman et al. (1989, J. of Immunology 143, 2714-2722)においてはB7.1をコードしている。
【0033】
本発明で使用できるベクターは、プラスミドまたは特にポックスウイルス、アデノウイルス、レトロウイルス、ヘルペスウイルスまたはアデノ随伴ウイルスに由来するウイルスベクターであってよい。病原性を弱めた非組み込みベクターであれば有利である。かかるベクターおよびそれらの調製法は当業者に公知である。
【0034】
アデノウイルスベクターが使用される場合には、複製のために不可欠な領域、特に宿主生物または環境にそれが伝播しないようにするにはE1領域の大部分を欠失させることにより、非反復因子を使用することが好ましい。欠陥のある必須機能がトランスで相補される限り、アデノウイルスゲノムの他の領域、特にE2、E4および/またはL1〜L5領域内を改変または欠失させてもよい。これらの具体例を例示するためには、E2A領域のDBP(DNA結合タンパク質に関するもの)遺伝子に作用する熱感受性変異体が挙げられる(Ensinger et al., 1972, J. Virol. 10, 328-339)。オープンリーディングフレーム(ORF)6および7をコードする配列を除く、E4領域の部分的欠失もまた意図され得る(Ketner et al., 1989, Nucleic Acids Res. 17, 3037-3048)。もう1つの可能性としては転写単位E4の全欠失がある。さらに、本発明のアデノウイルスベクターは、非必須領域E3の総てまたは一部を欠いていてもよい。この選択肢によれば、やはり、宿主の免疫系を逃れることが可能となるポリペプチド、特に糖タンパク質gp19kをコードするE3配列を保存することが有利であり得る(Gooding et al., 1990, Critical Review of Immunology 10, 53-71)。本発明の好ましいアデノウイルスベクターは包膜に不可欠な配列、すなわち5’および3’ITR(逆方向末端反復)および包膜領域の最小限の保持を示すであろう。それはヒトまたは動物のアデノウイルスに、またいずれかの抗原型に由来してもよい。C亜群ヒト・アデノウイルス、特にアデノウイルス2(Ad2)および5(Ad5)が、本発明を実施するのに特に最も適している。種々のアデノウイルスベクターならびにそれらの調製技術は通常のものであり、Graham and Prevect (1991, in Methods in Molecular Biology, vol 7, p 109-128; Ed: E.J. Murey, The Human Press Inc.)および国際出願WO94/28152に記載されている。例えば、それはin vitroにおいて大腸菌(E. coli)で、連結反応または相同組換え(例えば、国際出願WO96/17070を参照)によって、または相補ラインにおける組換えによって作出してもよい。
【0035】
レトロウイルスが使用されるならば、LTR(長い末端反復)および包膜配列(例えばNaviaux and Verma, 1992, Current Opinion in Biotechnology 3, 540-547を参照)が保存される。免疫ポリペプチドをコードする配列はレトロウイルスLTR、または下記に記載されるもののような内部プロモーターの制御下に置いてもよい。それはいずれの起源(ネズミ、霊長類、ネコ、ヒトなど)のレトロウイルスに由来してもよく、特にMoMuLV(モロニーネズミ白血病ウイルス)、MVS(ネズミ肉腫ウイルス)またはフレンドネズミレトロウイルス(Fb29)に由来していてもよい。それはまた、特にLTR(真核生物プロモーターによるプロモーター領域の置換)のレベル、または包膜領域(異種の包膜領域、例えばVL30型による置換)のレベルでの改変を含んでなる(仏国出願第94 08300号および第97 05203を参照)。
【0036】
有利な具体例によれば、本発明の組換えベクターはポックスウイルス、特にカナリア痘ウイルスのような鳥類痘ウイルス、鶏痘ウイルス、またはワクシニアウイルスに由来し、後者が好ましい。本発明において意図され得るあらゆるワクシニアウイルスのうち、コペンハーゲン、ワイエスおよび改変型アンカラ(MVAは、改変型ワクシニアウイルスアンカラ)株が選択されることが好ましい。
【0037】
一般に、挿入部位は、組換えウイルスの複製能および増殖能が損なわれないように、複製に不可欠でない領域中で選択される。指針としては、コペンハーゲン株のウイルスを使用する場合は、好ましい挿入部位はTK遺伝子座であり、それは後者を後者を不活性化する作用を有し、それにより組換え体の選択が容易になる。また、K1L遺伝子座を使用することもできる。MVAウイルスに関しては、組換え体(免疫型および免疫刺激型)配列の挿入は、切除部位I〜VIの少なくとも1つ、特にIIまたはIII内で行われてよい(Mayer et al., 1991, J. Gen. Virol. 72, 1031-1038; Sutter et al., 1994, Vaccine 12, 1032-1040)挿入はまた、D4R領域などのウイルスの必須領域で起こってもよく、欠損のある機能は、例えば相補ラインによりトランスで提供することができる。
【0038】
もちろん本発明では、免疫ポリペプチドをコードする、または抗腫瘍作用を増強する配列は、宿主細胞または宿主生物での発現のために必要なエレメントの制御下に置かれる。それらは転写を調節するエレメント、ならびに翻訳の開始および終結のシグナルを含む。それらの中で、プロモーターが特に重要である。一般に、宿主生物または宿主細胞中で機能し、治療に望ましく、かつ、使用されるベクターに好適なプロモーターが使用されよう。さらに、それは調節配列、例えば転写を活性化するエレメントまたは特定の細胞シグナルに応答する配列を含むよう改変してもよい。このことに関して、パピローマウイルスに関連する病巣は生殖道のレベルに位置するので組織特異的プロモーターを、あるいは腫瘍細胞のみに対して過剰発現を制限するよう、腫瘍に特異的なシグナルに応答するプロモーター(例えば、一般に腫瘍細胞によって過剰発現する増殖因子の存在下で活性化される)を使用することが有利であろう。
【0039】
本発明において意図し得るプロモーターの中で、SV40(シミアンウイルス40)プロモーター、HMG(ヒドロキシ−メチル−グルタリル−補酵素A)プロモーター、TK(チミジンキナーゼ)プロモーター、CMV(サイトメガロウイルス)プロモーター、RSV(ラウス肉腫ウイルス)プロモーター、アデノウイルスベクターに好適なMLPプロモーター(主要後期プロモーター)およびレトロウイルスベクターにより特異的なMo−MLV(モロニーネズミ白血病ウイルス)のLTRが挙げられる。サイトメガロウイルス(CMV)初期プロモーターが最も特に好ましい。それはまた腫瘍または癌細胞中での発現を刺激するプロモーターであってもよい。特に、乳癌および前立腺癌で過剰発現するMUC−1遺伝子のプロモーター(Chen et al., 1995, J. Clin. Invest. 96, 2775-2782)、黒色腫で過剰発現するチロシノース遺伝子(Vile et al., 1993, Cancer Res. 53, 3860-3864)および乳癌および膵臓癌で過剰発現するERS−2遺伝子(Harris et al., 1994, Gene Thetapy 1, 170-175)が挙げられる。問題のプロモーターが文献に記載されており、常法によって細胞ゲノムまたはウイルスゲノムからクローン化され得る。
【0040】
ポックスウイルスベクターに関しては、ポックスプロモーター、例えば7.5K、H5R、TK、p.28、p.11またはワクシニアウイルスのK1Lを使用してもよい。合成プロモーターもまた本発明の実施に適している(例えば、Chakrabarti et al., 1997, Biotechniques 23, 1094-1097; Hammond et al., 1997, J. Virological Methods 66, 135-138およびKumar and Boyle, 1990, Virology 179, 151-158を参照)。これに関しては、後期プロモーターと初期プロモーターとの間のキメラプロモーターであることが有利である。
【0041】
さらに、発現に必要なエレメントはまた、宿主細胞中での発現または維持を増大させる配列(イントロン、転写を終結させる配列、翻訳の開始部位など)を含んでももよい。しかしながら、ポックスウイルスベクターの場合には、イントロンの使用は避けられる。
【0042】
本発明の組成物は、独立または共通のエレメントの制御下に置かれた選ばれたポリペプチドに相当する配列を発現する1以上の組換えベクターを含んでもよい。後者の選択によれば、内部に翻訳を開始させる配列(IRES)または異なる遺伝子相における融合物を使用してもよい。
【0043】
本発明で使用される組換えベクターを産生する一般条件は、当技術分野の現状において広く記載されている。ポックスウイルスベクターに関しては、欧州特許EP 83 286に言及されており、その内容は引用することにより本明細書の開示の一部とされる。これらの条件は、発現単位が組み込まれるゲノム領域を有するベクターとして許容されるその他のウイルスにも適用できる。もちろん、それらを同一の遺伝子座または異なる遺伝子座に挿入してもよい。
【0044】
本発明によって追求される目的に従って、組換えウイルスの単離および精製工程を容易にするため、組換えベクターは選択可能なマーカー遺伝子を発現するための単位をさらに含んでなってもよい。特に、抗生物質G418耐性を与えるneo遺伝子、プロマイシン耐性を与えるpac遺伝子、ガンシクロビルまたはアシクロビルのようなあるヌクレオシド類似体に対する感受性を与える1型単純ヘルペスウイルス(HSV−1)のTK遺伝子、gpt(キサンチングアニンホスホリボシルトランスフェラーゼ)遺伝子、β−ガラクトシダーゼをコードする細菌遺伝子LacZおよびβ−グルクロニダーゼをコードするgus Aが挙げられる。後者2つの酵素マーカーによって、それぞれ基質X−Gal(5−ブロモ−4−クロロ−3−インドリル−β−D−ガラクトピラノシド)およびXglcA(5−ブロモ−6−クロロ−3−インドリル−β−D−グルコロニド)の存在下での染色により組換えウイルスを同定することが可能になる。
【0045】
好ましい抗腫瘍組成物はパピローマウイルス感染または腫瘍の治療および予防を意図し、
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド
が挿入された少なくとも1種のワクシニアウイルスのコペンハーゲン株またはMVA株を含んでなる。
【0046】
この組成物はまた、好ましくはIL−2またはB7.1から選択される免疫刺激剤をコードする配列を含んでなってもよい。免疫遺伝子を発現させる組換えベクターの1つによって、または独立のベクターによってそれを行ってもよい。
【0047】
本発明の組成物はワクチンの分野で公知の方法に従って調製してもよく、利用可能用量は広い範囲で変化させ得る。それらは特に、用いられるポリペプチドおよびベクター、治療される病状、患者の状態および臨床医が評価し得るその他のパラメーターに依存する。しかしながら、一般に、治療薬がウイルスベクターである場合にはウイルスの用量は104〜1013、有利には105〜1012、好ましくは105〜109プラーク形成単位(pfu)であり、治療薬がポリペプチド由来のものである場合には0.05〜500mg、有利には0.5〜200mg、好ましくは1〜100mgである。
【0048】
本発明の組成物は通常の投与経路のいずれかによって、好ましくは全身的経路によって、特には筋肉内、静脈内、肺内、皮下もしくは上皮下経路によって、または乱刺法によって投与してもよい。接触可能な腫瘍の場合には、腫瘍の部位もしくはその付近への直接注射、または局所塗布を用いることもできる。ワクチンとしては、本発明の組成物を当技術分野で一般的な実施に従い、例えば1回用量として、またはある一定の時間間隔の後に1回もしくは数回繰り返す用量として投与してもよい。他方、治療処置に関しては、治療が効果的であるに十分な周期で頻繁にそれを投与することができる。治療薬がウイルスベクターである場合には、ウイルスは生きている形態であることが好ましい。ポックルウイルスベクターに関しては、MVA株またはチミジンキナーゼ陰性コペンハーゲン株のような弱毒株を使用するのが好ましい。最後に、組換えウイルスベクターは、当業者にとって公知である適当な化学処理によって弱毒化してもよい。しかしながら、死滅した組換えベクターの注射を意図することもできる。
【0049】
好ましい具体例によれば、本発明の抗腫瘍組成物は、治療上有効量の治療薬を医薬上許容される担体と組み合わせて含んでなる。担体は、注射によってヒトまたは動物へ投与できるように選択する。それはまたビヒクル、賦形剤および/またはアジュバントを含んでもよく、液体または凍結乾燥形態で提供されてもよい。これに関して、ベクターのトランスフェクション効率および/または安定性を増大させることができる1以上の物質の組み合わせが意図され得る。これらの物質は当業者が利用できる文献に広く記載されている(例えば、Felgner et al., 1989, Proc. West. Pharmacol. Soc. 32, 115-121; Hodgson and Solaiman, 1996, Nature Biotechnology 14, 339-342, Remy et al., 1994, Bioconjugte chemistry 5, 647-654を参照)。限定されるものではないが例として、それらはポリマー、脂質、特に陽イオン脂質、リポソーム、核タンパク質および中性脂質であってもよい。意図し得る組み合わせは、陽イオン脂質(DC−Chol、DOGSなど)、および中性脂質(DOPE)と組み合わせたプラスミドベクターがある。この物はまた、その他の物質、特に抗癌物質と組み合わせてもよく、後者は個々にまたは同時に投与することが可能である。リガンドFlt3はその他の一例である(Lynch et al., 1997, Nature Medicine 3, 625; Brasel et al., 1996, Blood 88, 2001-2012; Marakovsky et al., 1996, J. Exp. Med. 184, 1953-1962)。この組成物はポリペプチドL1および/またはL2を含んでなる場合には模倣粒子の形態であってよいことを記載すべきであろう。
【0050】
本発明の主題はまた、パピローマウイルスの初期および/または後期領域に由来して先に定義された特徴を有する免疫ポリペプチドをコードする、少なくとも1つの配列を含んでなる組換えベクターである。それは注目されるその他の配列、例えば先に定義されたように、1以上の免疫ポリペプチドおよび/または免疫刺激ポリペプチドをコードする配列をさらに有してもよい。
【0051】
本発明のベクターの選択肢は広い。それは先に挙げたもののようなプラスミドまたはウイルスベクターであってもよい。好ましい具体例はポックスウイルスベクターからなり、最も特別にはワクシニアウイルスのコペンハーゲンまたはMVA株からなる。発現される注目の配列の挿入部位および発現に必要な要素は前記のものから選択され得る。
【0052】
好ましいベクターは、
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列
が挿入されたワクシニアウイルスのコペンハーゲンまたはMVA株である。
【0053】
それは所望によりIL2またはポリペプチドB7.1をコードする配列を含んでもよい。
【0054】
本発明はまた、本発明の組換えウイルスベクターから調製したウイルス粒子に関する。アデノウイルスベクターに関しては、このウイルス粒子はその欠損に好適な相補細胞にベクターをトランスフェクトすることによって作出してもよい。例えば、E1機能を効果的に相補する、ヒト胎児腎臓細胞から確立された293ライン(Graham et al.,1977, J. Gen. Virol. 36, 59-72)、A549−E1ライン(Imler et al., 1996, Gene Therapy 3, 75-84)または二重相補を可能にするライン(Yeh et al., 1996, J. Virol. 70, 559-565; Krougliak and Graham, 1995, Human Gene Therapy 6,1575-1586; Wang et al., 1995 Gene Therapy 2, 775-783;国際出願WO97/14119)を使用してもよい。相補細胞とは、ウイルス粒子を作出するために欠損ベクターをトランスで相補することができる細胞を意味するものと理解される。この細胞はベクターの欠損機能の総てをそれ自身で相補するわけではなく、この場合には部分的な相補としてヘルパーウイルスを使用することができる。ウイルス粒子は培養上清からだけでなく細胞からも回収してもよい。一般に用いられる方法の1つとしては、連続凍結/解凍サイクルによって細胞を溶解させて細胞溶解の上清中のビリオンを回収することである。これらを増幅して先行技術(クロマトグラフィー法、特に塩化セリウム勾配による超遠心分離など)に従い精製してもよい。
【0055】
レトロウイルス粒子は、適切なライン、例えば、それらの配列がベクター中において欠損している/機能しないウイルスポリペプチドgag、polおよび/またはenvをトランスで提供することができるラインにレトロウイルスベクターをトランスフェクトすることによって得ることができる。かかるラインは文献に記載されている(PA317、Psi CRIPGP + Am−12など)。
【0056】
ポックスウイルスベクターに関しては、異種遺伝子を発現することができるワクシニアウイルスを産生するための一般条件は、欧州特許EP 83 286および出願EP 206 920に教示されている。MVAウイルスに関しては、Mayr et al (1975, Infection 3, 6-14)およびSutter and Moss (1992, Proc. Natl. Acad. Sci. USA 89, 10847-10851)にさらに詳しく記載されている。便宜には、in vivoにおけるその(それらの)発現に適当なエレメントの制御下に置かれた導入される遺伝子を、挿入部位のいずれかの側にウイルス配列を含む導入ベクター中に挿入する。それを感染性ワクシニアウイルスに感染させた細胞中に導入する。組換え遺伝子は、この感染性ウイルスと導入ベクターの相同配列の間の相同組換えによってウイルスのゲノムへ組み込まれる。
【0057】
組換えベクターまたはウイルス粒子は、所望により、ベクターのトランスフェクション効率および/または安定性を増大させる1以上の前記物質と組み合わせてもよい。
【0058】
本発明では、この免疫ポリペプチドはウイルス粒子を取り巻くタンパク質構造(キャプシド、包膜など)に付着されていてもよい。
【0059】
ポックスウイルスは多重膜に取り巻かれた複雑な構造である。2種のウイルス粒子が産生され、それは細胞内成熟ビリオン(IMV)と細胞外包膜ビリオン(EEV)である。IMVの表面は二重膜からなり、EEVの包膜は原形質膜をはじめとする4つの膜からなる。本発明により追求される目的に従い、免疫ポリペプチドをコードする配列はまた、免疫応答を増強する目的とする、ポックスウイルス膜の一方または他方(IMVまたはEEV)への挿入によって改変されていてもよい。有利な具体例によれば、免疫ポリペプチドはEEV粒子の包膜の外膜に付着している(Katz et al. 1997, AIDS Res. Hum. Retrov. 13, 1497-1500)。これを行うために、その外膜のタンパク質成分であるB5Rタンパク質のC末端部分をコードする配列が、停止コドンのすぐ上流の免疫ポリペプチドのコード領域の3’に挿入される。全く好ましい例としては、ポリペプチドE7またはその非発癌性変異体とB5Rタンパク質の42個のC末端残基との間の融合物がある。免疫応答に対する有益な効果は、種々の組成(天然の局在性、細胞膜における付着、ウイルス膜における付着)を比較する以下の実施例に記載されたプロトコールに従う免疫予防および免疫治療研究によって評価され得る。
【0060】
本発明はまた、癌または腫瘍の、特に子宮頸部癌、軽度子宮頸部形成異常またはパピローマウイルス感染の治療または予防用医薬を製造するための、本発明の抗腫瘍組成物、組換えベクターまたはウイルス粒子の使用に関する。好ましい使用は筋肉内経路により注射可能な医薬の製造のためもののである。
【0061】
最後に、本発明はまた、医薬上有効量の本発明の抗腫瘍組成物、組換えベクターまたはウイルス粒子をかかる治療を必要とする患者に投与することにより、先に挙げた病状の治療または予防方法に関する。
【実施例】
【0062】
以下、本発明を実施例によって詳しく説明するが、結果として限定されるものではない。
【0063】
以下に記載した構築体は、Maniatis et al., (1989, 前記)に詳述されている一般的な遺伝子工学および分子クローニング技術に従い、または市販のキットを用いる場合には製造業者の推奨に従って作製する。in vitroでの合成オリゴヌクレオチド指定突然変異誘発は、Amershamにより供給されるキットによって実施する。PCR増幅技術は当業者にとっては公知である(例えば、Innis, Gelfand, Sninsky and White, Academic Press Incにより出版されているPCR Protocols - A guide to methods and applications, 1990を参照)。制限部位の修復に関しては、用いられる技術は大腸菌(Klenow)のDNAポリメラーゼIの大断片で、突出している5’末端を埋めることからなる。クローニング工程に関しては、組換えM13バクテリオファージを寒天最小培地(寒天7.5%)または豊富な液体LBM培地中の大腸菌NM522株(Stratagene)上で増殖させる。100μg/mlの抗性物質を補った寒天または液体培地で大腸菌C600株(Stratagene)、BJ5183(Hanahan, 1983, J. Mol. Biol. 166, 557-580)およびNM522においてアンピシリン耐性遺伝子を運ぶ組換えプラスミドを反復させる。クローニングが相同組換えによって行われる場合には、BJ5183株を使用することが好ましい(Bubeck et al., 1993, Nucleic Acid Res. 21, 3601-3602)。
【0064】
組換えワクシニアウイルス構築体は、すでに挙げた文献ならびにMackett et al., (1982, Proc. Natl. Acad. Sci. USA 79, 7415-7419)およびMackett et al. (1984, J. Virol. 49, 857-864)に開示されている当技術分野の標準技術に従って行う。
【0065】
in vivoでの試験(免疫予防または免疫治療)は、6〜8週齢のメスC57B16マウス(C. Rivers Rouen, France)またはヌードマウス(Janvier, Le Genest St. Isle, France)について行う。一般に、試験されるウイルス、用量または投与経路によって動物を20の群に分ける(指定されている場合は除く)。腫瘍細胞TC1(Wu, John Hopkins University, Baltimore, USA)は、2種のレトロウイルス、すなわちHPV−16天然E6およびE7遺伝子を発現するものと、ras癌遺伝子を発現するもので形質転換したC57B16マウスから採取した肺細胞から得る。この細胞は、G418(0.5mg/ml)の存在下、DMEM培地(ダルベッコ改変イーグル培地)で培養する。その細胞をトリプシンで処理して等張液で3回洗浄した後に動物の抗原投与に使用する。
【0066】
実施例1: トランスメンブラン局在化シグナルを供給したHPV−16 E6およびE7の非発癌性変異体をコードする配列を運ぶベクターの構築
E6およびE7遺伝子を欧州特許EP 0,462,187の実施例2および3に記載されているCaski細胞系から単離する。それ以後のクローニング工程を容易にするように、2つの構築体をHPV−16 E6およびE7遺伝子を含有するクローンM13E7/E6から誘導した。1つめのM13TG8188と示されるものは指定突然変異誘発によってE7遺伝子のそれぞれ上流および下流のPstIおよびBamHI部位の導入の結果得られ、2つめのM13TG8189はE6遺伝子の上流のPstI部位を含んでなる。開始暗号ATGの上流および停止コドンの下流における点突然変異の導入は、当業者の能力の範囲内である。
【0067】
HPV−16 E7タンパク質と網膜芽細胞腫遺伝子の産物との組合せが、種々の著者によって実証され(例えば、Munger et al., 1989, EMBO J. 8, 4099-4105を参照)、その形質転換力と相関している。明白な安全上の理由で、形質転換作用に関与している天然のE7タンパク質のコドン21〜26を欠失させた非発癌性変異体は、オリゴヌクレオチドoTG5118(配列番号3)によるベクターM13TG8188の指定突然変異誘発によって作出する。以下E7*と示される変異型E7タンパク質を有するM13TG9104が得られる。
【0068】
狂犬病糖タンパク質に対する分泌シグナルおよび付着シグナルは、Bg1IIの形態でpBR327(仏国特許83 15716に記載されているpTG150)に挿入された狂犬病糖タンパク質遺伝子から単離し、ベクターM13(M13TG177)にサブクローン化する。BamHI部位はオリゴヌクレオチドoTG5745(配列番号4)による指定突然変異誘発によって、それらと同じ位相にあるこれらシグナルの間に導入する。得られた構築体はM13TG9128と呼ばれる。次いで分泌配列および付着配列をPstI消化によってM13TG9128から単離し、PstIで線状化したベクターpTG5003にワクシニアウイルスp7.5の天然のプロモーターの3’に挿入すると、pTG5016が得られる。指針としては、pTG5003をSalI消化によってpTG186ポリ(仏国特許2,583,429に記載)から誘導し、Klenow断片で処理し、次いでSmaIで消化して再連結すると、結果としてそれはここでそのポリリンカー中に他のもの総てを除きPstIおよびEcoRIのみを含むようになる。位相中にE7*配列および狂犬病糖タンパク質の分泌シグナルおよび付着シグナル(本明細書では以下E7*TMRと示す)をもたらすように、ベクターM13TG9104をオリゴヌクレオチドoTG6390およびoTG6880(配列番号5および6)による指定突然変異誘発によって改変する。得られた構築体はM13TG9150と呼ばれる。
【0069】
同様に、HPV−16 E6タンパク質は腫瘍抑制遺伝子p53の発現産物と相互作用することができるということが実証されている(Crook et al., 1991, Cell 67, 547-556)。この相互作用に関与するドメインは明確に定義されており、天然のタンパク質の残基111と115との間に位置している。ベクターM13TG9125は、オリゴヌクレオチドoTG5377(配列番号7)によるM13TG8189の突然変異誘発によって作出される。E6遺伝子λ111−115を以下E6*と呼ぶ。
【0070】
麻疹ウイルスFタンパク質に対する分泌シグナルおよび付着シグナルは、麻疹ウイルスFタンパク質をコードするDNAを含有するプラスミド構築体pTG2148(欧州特許EP 0,305,229に記載)からPCRによって単離する。これらの配列とE6*をコードするものとの間の融合は、直接PCRによって行われ、すなわち、分泌配列を5’にXbaI部位を有するオリゴヌクレオチドoTG10829(配列番号8)およびE6の5’末端を覆うoTG10830(配列番号9)により増幅し、変異型E6*をコードする配列をFタンパク質の分泌シグナルの3’末端との融合を可能にするオリゴヌクレオチドoTG10835(配列番号10)およびFタンパク質の付着配列の5’末端との融合を可能にするoTG10836(配列番号11)により、M13TG9125から増幅される。付着配列に関しては、E6*の3’と付着配列の5’との間における融合を可能にするプライマーoTG10833(配列番号12)ならびに3’にKpnIおよびSphI部位を作り出すoTG10834(配列番号13)が使用される。それぞれNおよびC末端でFタンパク質の膜分泌配列および付着配列と融合したE6*配列を有する増幅断片(本明細書では以下E6*TMFと示す)をXbaIおよびSphIで消化し、次いで同一部位にベクターM13TG131に挿入する(Kieny et al., 1983, Gene 26, 91-99)。このようにして得られた構築体はM13TG9199と呼ばれる。
【0071】
実施例2: トランスメンブラン局在性のE6およびE7抗原ならびにヒトIL−2を発現する組換えウイルスMVATG8042の構築
MVAウイルスは、ワクシニアウイルスのアンカラ株に由来する。それは哺乳類細胞では感染性粒子を生ずることはできないが、胎芽鶏繊維芽細胞では正常に発達する。これらの細胞への適応は、この種の細胞でのその発達およびその感染サイクルに不可欠でない6領域の除去を引き起こした(ウイルスゲノムの約15%の消失;Meyer et al., 1991, J. Gen. Virol. 72, 1031-1038)。外来の遺伝物質の組み込みは、これらの除去領域のいずれのレベルで達成されてもよい。本発明では、HindIII制限断片NおよびA各々のレベルに位置する除去IIおよびIIIが用いられる(Altenburger et al., 1989, Arch. Virol. 105, 15-27)。
【0072】
第一に、MVAウイルスの除去III領域への挿入を可能にするベクターpTG6019を構築する。除去領域IIIのいずれかの側の相同組換えアームは、PCRによりウイルスゲノム、ならびに左のアームについてはプライマーoTG7637およびoTG7638(配列番号14および15)、ならびに右のアームについてはoTG7635およびoTG7636(配列番号16および17)から単離する(米国特許第5,185,146号を参照)。増幅した断片をベクターpTG1EのEcoRI部位にクローン化し、pTG6039を得る。導入される遺伝物質は2個の組換えアームの間に挿入される。ベクターpTG1Eは多重クローニング部位の代わりにEcoRIアダプターが存在することを除けばpTG1H(仏国特許第2,583,429号)と同様である。
【0073】
第一に、マーカー遺伝子gusAを発現するカセットを挿入する。まず7.5KプロモーターをpTG6019のBamHI部位へクローン化する。そのBamHI部位へBglII−BamHI断片の形で作出されたgusA遺伝子が挿入されているpTG6021が得られる。後者は文献に開示されている配列から得てもよい。得られた構築物はoTG6022と呼ばれる。マーカーの存在により、XglcA基質を用いてGUS酵素活性を検出することによって野生型ウイルスを組換えウイルスと識別することが可能となろう。赤色はβ−ガラクトシダーゼ活性を示す。しかしながら臨床適用の点では、組換えウイルスを選択した後の最終産物から、この細菌性マーカーを除去できることが有用であろう。このために2つの相同部位間の配列を欠失させるワクシニアの能力が利用される。従って、第2のp7.5Kプロモーターは、後者の発現を命令するのに適切なセンス方向でgusA遺伝子の下流に挿入される。ベクターpTG6025は、付着端を提供したp7.5断片のpTG6022のBamHIとSacI部位間への挿入の結果生ずる。
【0074】
さらに、ヒトインターロイキン−2をコードするcDNAを、PstI消化によってプラスミドpTG36(仏国特許第2,583,770号)から単離し、次いでプラスミドpTG186(仏国特許第2,583,429号)のPstI部位へ挿入すると、pTG188が得られる。相同組換えによって得られるウイルスはVVTG188と呼ばれる。BglII/EcoRI消化後、M13TG9132のBamHI/EcoRI部位でIL−2配列を天然のワクシニアプロモーターpH5Rの3’に挿入し、M13TG9185を得る。指針として、ベクターM13TG9132は、PCRによってウイルスゲノムから単離されたH5R遺伝子のプロモーターの、多重クローニング部位の外に位置する内部BglII部位の突然変異によってM13TG131(Kieny et al., 1983, 前記) から誘導されるファージM13TG6131への挿入から得られる。
【0075】
次いで、HPV−16およびIL−2遺伝子をMVAゲノムの除去III領域へクローン化する。E7*TMR配列はBglII/HindIII消化によって単離し、次いでワクシニアプロモーターp7.5の3'においてpTG6025のBamHIとHindIII部位の間に挿入する。得られる構築体はpTG6050と呼ばれる。ヒトIL−2遺伝子の発現カセットの挿入部位は、相同組換えにより、オリゴヌクレオチドoTG10502およびoTG10503(配列番号18および19)の挿入によってpTG6050のHindIIIとKpnI部位の間に作り出される。次いで作出したベクターpTG6074をHindII/KpnI消化により線状化し、さらにBglII/EcoRI消化によってM13TG9185から単離された発現カセットpH5R−IL−2を用いて相同組換えを行う。最後に、得られた構築体pTG6088をKpnIで線状化し、次いでKpnI/XbaI消化によってM13TG9199から単離されたE6*TMF遺伝子、およびXbaI/KpaI消化によってM13TG9136から単離されたプロモーターp7.5と連結する。得られた構築体はpTG8042と呼ばれる(図1)。ベクターM13TG9136は、oTG5925を用いる指定突然変異誘発(p7.5プロモーターの3'におけるPstI部位の作出;配列番号20)ならびにoTG5924を用いる指定突然変異誘発(p7.5プロモーターの5’におけるBamHIおよびKpnI部位の作出;配列番号21)によって改変されたM13TG5107から得られる。
【0076】
ウイルスMVATG8042は、先行技術の規定に従ってMVAゲノムを用いる相同組換えによって作出する。組換え体の単離はgusAマーカー遺伝子の存在によって容易になる。
【0077】
HPV初期抗原の細胞局在性の改変がその発現を阻害しないことを確かめる。抗E7抗体を用いてMVATG8042に感染させた細胞抽出物のウェスタンブロット解析は、20〜35kDaの間の分子量を有する3個のバンドの検出を可能にする。この不均一性は狂犬病糖タンパク質の膜付着領域における可能性のあるO−糖分解部位の存在によって説明され得る。ウェスタンブロット解析がフェニル−Gal−Nac(フェニルN−アセチル−α−D−ガラクトピラノシド、Sigma、p4023)の存在下で繰り返す場合には、20kDaの1形態のみが検出され、これはE7*TMR遺伝子の発現産物のO−糖分解を確実にする。
【0078】
E6*TMF遺伝子の発現産物の抗E6抗体によるウェスタンブロット検出は、予測された20kDaの分子量に移動する1個のバンドのみを示し、これは翻訳後の修飾がないことを確実にする。指針として、 前記抗E6およびE7抗体は精製された抗原の投与によって得られたウサギ抗血清である。しかしながら、モノクローナルでもポリクローナルでも、いずれの他の特異的抗体も好適であり得る。
【0079】
hIL−2遺伝子の発現は、ELISA(Quantikine R&D系)およびIL−2依存性細胞増殖試験により評価する。培養条件にもよるが、産生されるIL−2の量は0.1pfu/細胞を用いて感染させた106細胞あたり200〜800ng/ml/24時間まで様々である。
【0080】
実施例3: ウイルスMVATG8042のin vivoにおける効率(免疫療法)
C57BL6マウスに皮下経路によって植え付けられた103個のBMK−16myc細胞を接種する。指針として、 細胞系統はHPV−16ゲノムおよびネズミc myc遺伝子でトランスフェクトした新生マウスの腎臓細胞に由来する。次いで、MVATG8042ウイルスの107pfu(プラーク形成単位)もまた、D3、D6およびD9に皮下経路により投与し、腫瘍の発達を定期的にモニターする。処理されたマウスは非組換えMVAウイルスを受容した動物からなる対照と比べてD15までの腫瘍成長において遅延を示す。さらに、処理はD30〜35での腫瘍の部分的な抑制を伴い、これは同等のMVAを用いる場合には観察されず、またこれは天然の核局在性を有するE6*およびE7*変異体を発現する。
【0081】
実施例4: トランスメンブラン局在性のE7*抗原を発現する組換えウイルスVVTG5095の構築
E7*TMR配列をBglII/BamHI消化によってM13TG9150から単離し、pTG5016のBamHI部位に挿入する。得られた構築体はpTG5095と呼ばれ、p7.5プロモーターの制御下に狂犬病糖タンパク質の分泌シグナルおよび付着シグナルと融合したE7*配列を含む。ウイルスVVTG5095はワクシニアゲノムを用いた相同組換えによって作出する。
【0082】
実施例5: トランスメンブラン局在性のE7*抗原および免疫刺激剤B7.1を発現する組換えウイルスVVTG6002の構築
ヒトB7.1遺伝子をプライマーoTG6353およびoTG6352(配列番号22および23)を用いた逆PCR(RT−PCR)によってダウジ(Daudi)ヒト細胞系統から単離し、M13TG6131のBglIIとEcoRI部位の間に挿入する。M13TG9149は、p7.5プロモーターの3’においてM13TG5107の同じ部位の間にサブクローン化されている、BglII−EcoRI断片を単離したものから得られる(M13TG5107はM13TG130のBamHI部位へクローン化されたp7.5プロモーター配列を運ぶ)。p7.5−B7.1カセットは、このようにして得たM13TG9152と呼ばれる構築体からEcoRI/PstI消化によって単離し、oTG1086(5’AATTGCA3’)を用いてpTG5095のEcoRI部位へ導入する。得られた構築体はpTG6002と呼ばれ、次いでワクシニアゲノムを用いる相同組換えによって組換えウイルスVVTG6002を作出する。
同等の構築体は、実施例2で開発されたような同様の技術に従う、E7*およびB7.1遺伝子を発現するカセットのMVAN33遺伝子の除去領域IIIへの挿入によって調製される。
【0083】
実施例6: ウイルスVVTG5095およびVVTG6002のin vivoにおける効率(免疫予防)
C57BL6マウスに107pfuのVVTG5095またはVVTG6002を用いて皮下経路によって3回ワクチン接種した。最後の免疫化の3日後、皮下に植え付けた103個のE7W1細胞を用いて、これらの動物に抗原投与する。指針として、E7W1細胞はHPV−16の発癌性E7遺伝子を発現するベクターでトランスフェクトされたネズミリンパ腫系統から得られる。時間の関数としての動物の生存率(%)を107pfuの非組換えワクシニアウイルスVVTG186(前記に記載のベクターTG186に由来する)で処置した対照マウスで得られたものと比較する。死亡率のモニタリングは3群の間の差異を示す。D36で対照群では100%の動物が死亡したのに対し、VVTG5095でワクチン接種した動物では約4分の1の生存が観察される。防御はB7.1抗原をコードする配列を含む構築体VVTG6002により実質的に増強される。
【0084】
実施例7: HPV16の改変された初期遺伝子および後期遺伝子を発現するMVATG9936の構築
HPV16L1およびL2タンパク質をコードする断片をPCRによって一般的な先行技術に従ってカスキ(Caski)細胞のゲノムDNA(ATCC1550)から単離する。L1配列を保持する増幅断片をM13TG130(Kieny et al., 1983, Gene 26, 91-99)へサブクローン化し、構築体M13TG8171を得る。クローン化されたL1遺伝子の配列はGenebankに含まれる配列(受託番号K02718)と比較して数箇所の突然変異、すなわち、248番におけるAの代わりのC、253番におけるAの代わりのC、537番におけるAの代わりのG、682番におけるCの代わりのG、874番におけるAの代わりのG、1393番におけるトリプレットACTの挿入、1390番におけるトリプレットGATの欠損を示す。この配列をZhou et alによって発表されたもの(1991, Virology 185, 251-257)と適合させるよう、かつ、ワクシニアの初期段階の発達を妨げることのできる初期転写終結に関して可能性ある部位を構成するTTTTTNT配列のレベルでサイレント変異を導入するよう点突然変異によって補正する。部位指定突然変異誘発の技術は当業者の能力の範囲内である。補整された配列を運ぶベクターはM13TG4041と呼ばれる。
【0085】
L2配列を運ぶPCR断片のベクターM13TG6131への挿入により、M13TG9126が得られる。Genebankに開示された配列と比較して5個の点突然変異、すなわち、378番におけるTの代わりのC、691番におけるGの代わりのA、702番におけるGの代わりのA、990番におけるAの代わりのGおよび1092番におけるAの代わりのCが確認される。指針として、ベクターM13TG6131は、多重クローニング部位の外に位置する内部BglII部位の突然変異によってM13TG131(Kieny et al., 1983, Gene 26, 91-99)から誘導される。
【0086】
ベクターM13TG4041をXbaI−SacIを用いて消化し、次いでL1配列を有する断片をL2遺伝子の下流でかつ、反対の方向に、M13TG9126のXbaI−SacI部位の間に挿入する。このようにして作出された構築体はM13TG4042と呼ばれる。次いで、L1およびL2配列をBglII消化によって単離し、後期p11Kと初期p7.5プロモーターの融合の結果生じる合成プロモーターp11K7.5を含むベクターM13TG4052のBglIIとBamHI部位の間にサブクローン化する。2つの方向のうち、M13TG4055と呼ばれるプロモーターのL1遺伝子の下流に位置するものを選択する。次いで、pH5RプロモーターおよびhIL−2遺伝子を有する発現カセットを、L1とL2遺伝子の間に位置するM13TG4055のHindIII部位へ導入するのに先立ち、HindIII消化によってpTG8042から単離する。最後に、得られた構築体M13TG4057をBglIIで消化し、次いでHPV16後期配列を有する断片を、合成プロモーターp4BK1Lのクローン化の結果生じるM13TG4060のBamHI部位へ挿入する。後者は初期プロモーターp4B(Davidson and Moss, 1989, J. Mol. Biol. 210, 749-769)と後期プロモーターpK1L(Davidson and Moss, 1989, J. Mol. Biol. 210, 771-784)間のハイブリッドプロモーターである。p11K7.5の制御下にあるL1遺伝子、L1配列に対して反対の方向でp4BK1Lの制御下にカセットpH5R−IL2およびL2遺伝子を含んでなるM13TG4062が得られる。
【0087】
E7*TMRおよびE6*TMFをコードする配列を含む多シストロン性カセットを構築する。第一に、EMCウイルス(脳心筋炎ウイルス;Genebank受託番号M22458)のIRES配列を常法によってpTG6002のEcoRIとNcoI部位の間にE7*TMR遺伝子の下流が導入されたEcoRI−NcoI断片の形で単離して(実施例5)pTG8084を得る。次いでE6*TMF遺伝子を各5’末端にNcoI部位を作り出す適当なプライマーを用いてPCRによって増幅する。E7*TMR遺伝子およびpTG8084から得られたIRES配列を有するBglII−NcoI断片ならびにNcoIおよびSacIによって消化されたE6*TMF遺伝子を有するPCR配列を、予めBglIIおよびSacIで消化されたベクターM13TG6131(実施例2)中で再構築する。M13TG4059が得られる。次いで、後者からE7*TMR−IRES−E6*TMF単位をBglII断片の形で単離し、p7.5KプロモーターのpTG8093下流のBamHI部位へ挿入する(pTG8093はp7.5KプロモーターのベクターpTG6025へのクローン化の結果生じる)。このようにして得られた構築体はpTG9901と呼ばれる。
【0088】
M13TG4062のSacI断片をpTG9901のSacI部位へクローン化し、pTG9902を得る。以下の方法でベクターpポリII(Lathe et al., 1987, Gene 57, 193-201)において、選抜可能なgpt遺伝子(Falkner and Moss, 1988, J. Virol. 62, 1849-1854)を後期およびIL2遺伝子とともに構築する。最初のものはBglII−NcoI消化によってM13TG4076(このベクターはその2つの末端にp4BK1Lプロモーター配列によってフランクされたgpt遺伝子を含む)から、2番目のものはNcoI−SacI消化によってpTG9902から単離する。精製された断片をpポリIIのBglII−SacI部位の間にクローン化する。pTG9933は、SacIによって切断されたベクターpTG9901へ導入されているSacI断片が得られるものから得られる。このようにして得られた構築体pTG9936は図2に示されている。
MVATG9936ウイルスは、MVAN33ゲノムを用いる相同組換えによって作出する。クローンの単離は先行技術の規定に従って行ってよい。
【0089】
実施例8: 免疫予防におけるウイルスの効率
A.ウイルスの処方の重要性(実験N121)
この前臨床試験の目的はウイルス株(MVAに対するコペンハーゲンワクシニアウイルス)および抗腫瘍防御に関する処方(天然の核局在性に対するトランスメンブラン型での提供)を比較することである。
【0090】
C57B16マウスを107pfuのウイルスで3回ワクチン接種する。注射は腹膜内経路によって10日毎に(D1、D11、D21)行う。最後の免疫化の7日後に、それらの右手側面への5×104個のTC1細胞の皮下投与によって動物に抗原投与する。ウイルスおよび投与される溶液、各々:
1 MVATG8042(実施例2、E6*TMF、E7*TMR、IL−2)、
2 MVATG6037(実施例5、E7*TMR、B7.1)、
3 VVTG5095(実施例4、E7*TMR)、
4 MVATG6090(E6*、E7*、IL−2)、
5 VVTG5061x188(E6*、E7*、IL−2)、
6 VVTG186(非組換えワクシニアウイルス)、
7 MVAN33(非組換えMVAウイルス)、および
8 塩溶液(PBS)
に従って20個体の動物の8群を形成する。
【0091】
1〜3群は少なくとも1種のトランスメンブランHPV抗原を発現する本発明のウイルスでワクチン接種するが、4および5群は天然の核局在性のHPV16初期抗原を発現するウイルスを、また6〜8群は非組換え対照ウイルスまたは塩溶液を施す。ウイルスMVATG6090およびVVTG5061x188は国際出願WO98/04705に記載されているということを記載すべきであろう。種々の群の動物の生存率(%)を腫瘍の抗原投与に続いて12週間モニターする。結果は以下のように要約できる。
【0092】
一般に、3つの対照群における死亡率は高く、95%(ウイルスMVAN33およびVV186を用いて)および81%(PBSを用いて)に達する。
HPV核抗原を発現するウイルスで免疫化した動物の生存においては有意な増加が観察される。例えば、ウイルスMVATG6090を受容したマウスの55%は腫瘍を持たないが、VVTG5061x188の投与は75%の腫瘍排除レベルを誘導する。
【0093】
他方、HPV16トランスメンブラン抗原を発現するウイルスでワクチン接種された動物の大部分では腫瘍が排除されたか、または腫瘍成長において実質的な遅延を示す。さらに正確には、MVATG8042によれば100%の排除が、MVATG6037によれば95%の排除が、またVVTG5095によれば90%の排除が観察される。
【0094】
図3は対照(MVAN33およびPBS)と比較して、MVATG8042およびMVATG6090でワクチン接種された動物を用いて得られた生存曲線を示す。
【0095】
全体として、これらのデータはコペンハーゲンワクシニアウイルスとMVAに由来するウイルス間に有意な差異がないこと、ならびに膜型での提供によって与えられたより優れた免疫原性を実証するものである。試験した総てのウイルスのうち、この免疫予防の動物モデルにおいて100%の腫瘍排除が起こるので、ウイルスMVATG8042が最も効果的である。
【0096】
B.記憶反応の研究(実験N122)
C57B16マウスに、D82にそれらの右手側面への5×104個のTC1細胞の皮下投与によって抗原投与するに先立ち、D1、D11およびD21に107pfuのウイルスを用いて腹膜内経路によって免疫化する。従前の実験(N121)と同一の8群を形成する。腫瘍の発達を時間の関数としてモニターする。腫瘍の抗原投与の50日後に得られたデータは腫瘍の排除に関して最も効果的なウイルスはウイルスMVATG8042であるということを確実とする。その投与は100%の動物を防御し、長期免疫を誘導する能力を示す。
【0097】
C.用量効果(実験N127)
C57B16マウスに、D28に5×104個のTC1細胞のそれらの右手側面への皮下投与によって抗原投与するに先立ち、D1、D11およびD21に105、106または107pfuのMVATG8042ウイルスを用いて腹膜内経路によって免疫化する。対照動物には同量のMVAN33を施す。腫瘍の発達を1週間に2回、12週間モニターする。結果(図4)は、投与されたMVATG8042の用量にかかわらず100%の防御を示すが、対照動物の大部分は腫瘍を発達させる。これらのデータは、低用量であってもMVATG8042ウイルスの効力を確実とする。
【0098】
D.投与経路の効果(実験N134)
C57B16マウスに、D44に5×104個のTC1細胞のそれらの右手側面への皮下投与によって抗原投与するに先立ち、D1、D11およびD21に107pfuのウイルスを用いて種々の投与経路(乱刺、皮下、腹膜内または筋肉内)によって免疫化する。腫瘍の発達を週に2回、12週間モニターする。図5に示されるように、腹膜内または筋肉内経路は、ウイルスMVATG8042による最高レベルの防御を与える。
【0099】
実施例9: 免疫治療におけるウイルスの効力
A.免疫治療の観点におけるウイルスの効力(実験N125)
この研究の目的は予め確立された腫瘍に対する、膜または核形態でHPV抗原を示すウイルスの治療能力を比較することである。このために、5×104個のTC1細胞を皮下経路によってC57B16マウスの右手側面へ投与する(D0)。次いで107pfuのウイルスをD7、D14およびD21に腹膜内経路によって注射する。4つの動物群を、各々MVAN33(負の対照)、トリス−HCl/NaClの塩溶液(負の対照)、MVATG8042(トランスメンブラン型)およびMVATG6090(核型)という投与されるウイルスに従って形成する。腫瘍の発達を週に2回、12週間評価する。結果は治療上の観点での抗腫瘍防御に関して、膜型での提供のより優れた効力を示す。MVATG8042を施したマウスは100%が腫瘍細胞の植え込み140日後に生存したが、MVATG6090を接種した動物に関しては生存率は約60%であり、対照動物に関しては極めて低い。
【0100】
B.毒性研究−用量の効果
ヒト腫瘍へのそれらの適用を意図するに先立ち、ウイルスの毒性がないことを確かめることは重要である。106または107pfuのMVATG8042またはMVAN33ウイルスまたは野生型コペンハーゲンワクシニアウイルス(VVwt)を頭蓋内経路によってヌードマウス(5個体/群)へ投与する。マウスの生存率を20日間モニターし、得られたものを以下の表1に示す。
【0101】
表1
ウイルス 生存しているヌードマウスの数
106pfu 107pfu
MVAN33 5/5 5/5
MVATG8042 5/5 5/5
VVwt − 0/5
【0102】
107pfuのMVATG8042ウイルスの頭蓋内投与後の副作用は検出されないが、野生型ワクシニアウイルスで処理したマウスの総てが注射後3日で死亡した。対照MVAウイルスはどちらの動物に対しても有毒でなく、これは、すでに文献に記載されているワクシニアウイルスに比べ、その弱毒性を確実とするものである。
【0103】
[配列表]
(1)一般情報
(i)特許出願人:
(A)名称: Transgene SA
(B)通り: 11 rue de Molsheim
(C)市: Strasbourg
(D)国:フランス
(F)郵便番号:67082
(G)電話:(33)03 88 27 91 00
(H)テレファックス:(33) 03 88 27 91 11
(ii)発明の名称:細胞における局在性が改変された免疫ポリペプチドに基づく抗腫瘍組成物
(iii)配列数:23
(iv)読み取り可能なコンピューター:
(A)媒体形態:テープ
(B)コンピューター:IBM PC互換機
(C)オペレーティングシステム:PC−DOS/MS−DOS
(D)ソフトウエア:PatentIn リリース#1.0、バーション1.25(EPO)
(2)配列番号1の情報:
(i)配列の特徴:
(A)長さ:243個のアミノ酸
(B)型:アミノ酸
(D)トポロジー:直鎖
(ii)分子型:タンパク質
(iii)ハイポセティカル:NO
(vi)起源:
(A)生物名:ヒト・パピローマウイルス
(B)株:HPV−16
(C)個体/単離物:E6タンパク質とFタンパク質の融合シグナル
(vii)直接の起源:
(B)クローン:E6*TMF
(xi)配列の記載:配列番号1
Met Gly Leu Lys Val Asn Val Ser Ala Ile Phe Met Ala Val Leu Leu
1 5 10 15
Thr Leu Gln Thr Pro Thr Gly Gln Ile His Trp Gly Met His Gln Lys
20 25 30
Arg Thr Ala Met Phe Gln Asp Pro Gln Glu Arg Pro Arg Lys Leu Pro
35 40 45
Gln Leu Cys Thr Glu Leu Gln Thr Thr Ile His Asp Ile Ile Leu Glu
50 55 60
Cys Val Tyr Cys Lys Gln Gln Leu Leu Arg Arg Glu Val Tyr Asp Phe
65 70 75 80
Ala Phe Arg Asp Leu Cys Ile Val Tyr Arg Asp Gly Asn Pro Tyr Ala
85 90 95
Val Cys Asp Lys Cys Leu Lys Phe Tyr Ser Lys Ile Ser Glu Tyr Arg
100 105 110
His Tyr Cys Tyr Ser Leu Tyr Gly Thr Thr Leu Glu Gln Gln Tyr Asn
115 120 125
Lys Pro Leu Cys Asp Leu Leu Ile Arg Cys Ile Asn Cys Gln Lys Pro
130 135 140
Leu Gln Arg His Leu Asp Lys Lys Gln Arg Phe His Asn Ile Arg Gly
145 150 155 160
Arg Trp Thr Gly Arg Cys Met Ser Cys Cys Arg Ser Ser Arg Thr Arg
165 170 175
Arg Glu Thr Gln Leu Gly Leu Ser Ser Thr Ser Ile Val Tyr Ile Leu
180 185 190
Ile Ala Val Cys Leu Gly Gly Leu Ile Gly Ile Pro Ala Leu Ile Cys
195 200 205
Cys Cys Arg Gly Arg Cys Asn Lys Lys Gly Glu Gln Val Gly Met Ser
210 215 220
Arg Pro Gly Leu Lys Pro Asp Leu Thr Gly Thr Ser Lys Ser Tyr Val
225 230 235 240
Arg Ser Leu
(2)配列番号2の情報:
(i)配列の特徴:
(A)長さ:185個のアミノ酸
(B)型:アミノ酸
(D)トポロジー:直鎖
(ii)分子型:タンパク質
(iii)ハイポセティカル:NO
(vi)起源:
(A)生物名:ヒト・パピローマウイルス
(B)株:HPV−16
(C)個体/単離物:狂犬病糖タンパク質のE7融合シグナル
(vii)直接の起源:
(B)クローン:E7*TMR
(xi)配列の記載:配列番号2
Met Val Pro Gln Ala Leu Leu Phe Val Pro Leu Leu Val Phe Pro Leu
1 5 10 15
Cys Phe Gly Lys Phe Pro Ile Gly Ser Met His Gly Asp Thr Pro Thr
20 25 30
Leu His Glu Tyr Met Leu Asp Leu Gln Pro Glu Thr Thr Gln Leu Asn
35 40 45
Asp Ser Ser Glu Glu Glu Asp Glu Ile Asp Gly Pro Ala Gly Gln Ala
50 55 60
Glu Pro Asp Arg Ala His Tyr Asn Ile Val Thr Phe Cys Cys Lys Cys
65 70 75 80
Asp Ser Thr Leu Arg Leu Cys Val Gln Ser Thr His Val Asp Ile Arg
85 90 95
Thr Leu Glu Asp Leu Leu Met Gly Thr Leu Gly Ile Val Cys Pro Ile
100 105 110
Cys Ser Gln Lys Pro Arg Ser Tyr Val Leu Leu Ser Ala Gly Ala Leu
115 120 125
Thr Ala Leu Met Leu Ile Ile Phe Leu Met Thr Cys Cys Arg Arg Val
130 135 140
Asn Arg Ser Glu Pro Thr Gln His Asn Leu Arg Gly Thr Gly Arg Glu
145 150 155 160
Val Ser Val Thr Pro Gln Ser Gly Lys Ile Ile Ser Ser Trp Glu Ser
165 170 175
His Lys Ser Gly Gly Glu Thr Arg Leu
180 185
(2)配列番号3の情報:
(i)配列の特徴:
(A)長さ:36個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(A)生物名:ヒト・パピローマウイルス
(B)株:HPV−16
(C)個体/単離物:合成オリゴヌクレオチドoTG5118(E7欠損2126)
(xi)配列の記載:配列番号3
TCTGAGCTGT CATTTAATTG AGTTGTCTCT GGTTGC 36
(2)配列番号4の情報:
(i)配列の特徴:
(A)長さ:42個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:狂犬病ウイルス
(B)株:HPV−16
(C)個体/単離物:突然変異誘発オリゴヌクレオチドoTG5745
(xi)配列の記載:配列番号4
TGCACTCAGT AATACATAGG ATCCAATAGG GAATTTCCCA AA 42
(2)配列番号5の情報:
(i)配列の特徴:
(A)長さ:38個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG6390
(xi)配列の記載:配列番号5
GTATCTCCAT GCATGGATCC TGCAGGGTTT CTCTACGT 38
(2)配列番号6の情報:
(i)配列の特徴:
(A)長さ:36個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG6880
(xi)配列の記載:配列番号6
GGATCCGCCA TGGTAGATCT TGGTTTCTGA GAACAG 36
(2)配列番号7の情報:
(i)配列の特徴:
(A)長さ:32個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(A)生物名:狂犬病ウイルス
(B)株:HPV−16
(C)個体/単離物:合成オリゴヌクレオチドoTG5377(E6欠損111ないし115)
(xi)配列の記載:配列番号7
TGTCCAGATG TCTTTGCAGT GGCTTTTGAC AG 32
(2)配列番号8の情報:
(i)配列の特徴:
(A)長さ:34個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10829
(xi)配列の記載:配列番号8
GCGCGCTCTA GAATTATGGG TCTCAAGGTG AACG 34
(2)配列番号9の情報:
(i)配列の特徴:
(A)長さ:35個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10830
(xi)配列の記載:配列番号9
CAGTTCTCTT TTGGTGCATG CCCCAATGGA TTTGA 35
(2)配列番号10の情報:
(i)配列の特徴:
(A)長さ:38個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10835
(xi)配列の記載:配列番号10
ATGCTAGTGC TCGATAAACC CAGCTGGGTT TCTCTACG 38
(2)配列番号11の情報:
(i)配列の特徴:
(A)長さ:35個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10836
(xi)配列の記載:配列番号11
TCAAATCCAT TGGGGCATGC ACCAAAAGAG AACTG 35
(2)配列番号12の情報:
(i)配列の特徴:
(A)長さ:38個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG10833
(xi)配列の記載:配列番号12
CGTAGAGAAA CCCAGCTGGG TTTATCGAGC ACTAGCAT 38
(2)配列番号13の情報:
(i)配列の特徴:
(A)長さ:36個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG10834
(xi)配列の記載:配列番号13
GCGGGCATGC GGTACCTCAG AGCGACCTTA CATAGG 36
(2)配列番号14の情報:
(i)配列の特徴:
(A)長さ:32個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:ワクシニアウイルス
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG7637(PCR領域III)
(xi)配列の記載:配列番号14
GGGGGGGAAT TCAGTAAACT TGACTAAATC TT 32
(2)配列番号15の情報:
(i)配列の特徴:
(A)長さ:39個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(A)生物名:ワクシニアウイルス
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG7638(PCR領域III)
(xi)配列の記載:配列番号15
GGGGGGGGAT CCGAGCTCAC CAGCCACCGA AAGAGCAAT 39
(2)配列番号16の情報:
(i)配列の特徴:
(A)長さ:32個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:ワクシニアウイルス
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG7635(PCR領域III)
(xi)配列の記載:配列番号16
GGGGGGGGAT CCGGAAAGTT TTATAGGTAG TT 32
(2)配列番号17の情報:
(i)配列の特徴:
(A)長さ:30個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(A)生物名:ワクシニアウイルス
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG7636(PCR領域III)
(xi)配列の記載:配列番号17
GGGGGGGAAT TCTTTGTATT TACGTGAACG 30
(2)配列番号18の情報:
(i)配列の特徴:
(A)長さ:77個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10502
(xi)配列の記載:配列番号18
AGCTTTTTAT TCTATACTTA AAAAATGAAA ATAAACTCGA GTTGTCAAAG CATCATCTCA 60
ACACTGACTT GAGGTAC 77
(2)配列番号19の情報:
(i)配列の特徴:
(A)長さ:69個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(B)株:合成オリゴヌクレオチドoTG10503
(xi)配列の記載:配列番号19
CTCAAGTCAG TGTTGAGATG ATGCTTTGAC AACTCGAGTT TATTTTCATT TTTTAAGTAT 60
AGAATAAAA 69
(2)配列番号20の情報:
(i)配列の特徴:
(A)長さ:39個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:YES
(vi)起源:
(C)個体/単離物:合成オリゴヌクレオチドoTG5925
(xi)配列の記載:配列番号20
TCAGATCTGT CGAGGGATCT GCAGCTTCTT CTAGAGGTA 39
(2)配列番号21の情報:
(i)配列の特徴:
(A)長さ:44個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(B)株:改変型アンカラ
(C)個体/単離物:合成オリゴヌクレオチドoTG5924
(xi)配列の記載:配列番号21
AGTGAATTGC TGCAGGTACC CGGATCCGCA TCGACTATCG ACAT 44
(2)配列番号22の情報:
(i)配列の特徴:
(A)長さ:35個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:ホモサピエンス
(B)株:ダウジ細胞系統
(C)個体/単離物:PCRプライマーoTG6353(クローンニングB7.1)
(xi)配列の記載:配列番号22
TCAGCCCCTG AATTCTGCGG ACACTGTTAT ACAGG 35
(2)配列番号23の情報:
(i)配列の特徴:
(A)長さ:33個の塩基対
(B)型:核酸
(C)鎖の数:一本鎖
(D)トポロジー:直鎖
(ii)分子型:DNA(ゲノム)
(iii)ハイポセティカル:NO
(iii)アンチセンス:NO
(vi)起源:
(A)生物名:ホモサピエンス
(B)株:ダウジ細胞系統
(C)個体/単離物:PCRプライマーoTG6352(クローンニングB7.1)
(xi)配列の記載:配列番号23
TTGACCCTAA AGATCTGAAG CCATGGGCCA CAC 33
【図面の簡単な説明】
【0104】
【図1】pTG8042の模式図である。BRG3およびBRD3は、MVAウイルスの除去領域IIIのレベルでの挿入が可能な左および右の組換えアームを表す。pTG8042は、7.5kプロモーターによって命令される、麻疹ウイルスFタンパク質のシグナル配列(SF)およびトランスメンブラン領域(TMF)と融合したHPV16 E6遺伝子を発現させるためのカセット;プロモーターpH5Rの制御下に置かれたIL−2遺伝子を発現させるためのカセット;p7.5プロモーターの制御下に置かれた狂犬病糖タンパク質のシグナル配列(SF)およびトランスメンブラン領域(TMF)と融合したHPV16 E7遺伝子を発現させるためのカセットならびにp7.5プロモーターの制御下に置かれたgusA遺伝子(uidA)を発現させるためのカセットを含んでなる。
【図2】ベクターpTG9936の模式図である。BRG3およびBRD3は、MVAゲノムの除去領域IIIのレベルでの挿入が可能な左および右の組換えアームを表す。pTG9936は、gptマーカー遺伝子の発現を命令するプロモーターp4BK1L、HPV16 L2遺伝子の発現を命令するプロモーターp4BK1L、IL−2の発現を命令するプロモーターpH5R、HPV16 L1遺伝子の発現を命令するプロモーターp11k7.5、狂犬病糖タンパク質のシグナル配列(SF)およびトランスメンブラン領域(TMF)、続いてIRES配列と融合したHPV16 E7遺伝子、ならびに麻疹ウイルスFタンパク質のシグナル配列(SF)およびトランスメンブラン領域(TMF)と融合したHPV16 E6遺伝子の発現を命令するプロモーターp7.5を有する。
【図3】ウイルスMVATG8042、MVATG6090、MVAN33で、またはPBSによる腫瘍の抗原投与前にワクチン接種した動物の生存率(%)を時間(日)の関数として表すことによって実験N121を図示している。
【図4】107、106および105pfuのウイルスMVAN33またはMVATG8042の投与後の腫瘍を持たない動物の割合(%)を時間(日)の関数として表す(実験N127)。
【図5】乱刺法、皮下(SC)、腹膜内(IP)または筋肉内(IM)により、ウイルスMVATG8042でまたは対照MVAN33で処置した動物の生存率(%)を時間(日)の関数として表すことによって実験N134を図示している。
【特許請求の範囲】
【請求項1】
1以上の免疫ポリペプチドを治療薬または予防薬として含んでなる抗腫瘍組成物であって、少なくとも1種のポリペプチドが、その天然の局在性とは異なる局在性を有するよう改変されていることを特徴とする組成物。
【請求項2】
ポリペプチドの改変が適当な局在化シグナルの誘導および/または天然の局在化シグナルの欠損または不活性化によって得られる、請求項1記載の抗腫瘍組成物。
【請求項3】
天然には膜以外、特に核局在性を有する免疫ポリペプチドが膜局在性を有するよう改変されている、請求項1および2のいずれか一項に記載の抗腫瘍組成物。
【請求項4】
免疫ポリペプチドが膜付着配列および適当であれば分泌配列の挿入によって改変されている、請求項3記載の抗腫瘍組成物。
【請求項5】
ポリペプチドがその核局在化配列に関して欠失している、請求項1〜4のいずれか一項に記載の組成物。
【請求項6】
分泌および/または膜付着配列が、狂犬病糖タンパク質、HIVウイルスenv糖タンパク質および麻疹ウイルスFタンパク質のものからなる群から選択される、請求項1〜5のいずれか一項に記載の抗腫瘍組成物。
【請求項7】
免疫ポリペプチドが、特にその天然の局在化シグナルの変異/欠損によって細胞質局在性を有するよう改変されている、請求項1、2または5のいずれか一項に記載の抗腫瘍組成物。
【請求項8】
免疫ポリペプチドが、特定の細胞コンパートメントに局在させることを可能にする配列の、特にそのC末端での挿入によって改変されている、請求項1、2または5のいずれか一項に記載の抗腫瘍組成物。
【請求項9】
免疫ポリペプチドが、特にエンドサイトーシス配列、特に配列IPNYRNMの、特にそのC末端での挿入によって改変されている、請求項8記載の抗腫瘍組成物。
【請求項10】
免疫ポリペプチドが、ゴルジ体の膜における付着を可能にする配列の、特にそのC末端での挿入によって改変されている、請求項8記載の抗腫瘍組成物。
【請求項11】
免疫ポリペプチドがパピローマウイルス、特にヒトパピローマウイルス(HPV)16、18、31、33または45型の初期および/または後期領域に由来する、請求項1〜10のいずれか一項に記載の抗腫瘍組成物。
【請求項12】
免疫ポリペプチドがパピローマウイルスの初期領域のポリペプチド、特にE6またはE7に由来する、請求項11記載の抗腫瘍組成物。
【請求項13】
免疫ポリペプチドがパピローマウイルスのE6またはE7ポリペプチドの非発癌性変異体に由来する、請求項12記載の抗腫瘍組成物。
【請求項14】
免疫ポリペプチドがパピローマウイルスのL1またはL2ポリペプチドに由来する、請求項11記載の抗腫瘍組成物。
【請求項15】
パピローマウイルスの初期ポリペプチド由来の少なくとも1種の免疫ポリペプチドと後期ポリペプチド由来の少なくとも1種の免疫ポリペプチドを含んでなる、請求項11〜14のいずれか一項に記載の抗腫瘍組成物。
【請求項16】
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド
を含んでなる、請求項11〜15のいずれか一項に記載の抗腫瘍組成物。
【請求項17】
組成物の抗腫瘍作用を増強する少なくとも1種の化合物をさらに含んでなる、請求項1〜16のいずれか一項に記載の抗腫瘍組成物。
【請求項18】
化合物が免疫刺激剤である、請求項17記載の抗腫瘍組成物。
【請求項19】
免疫刺激化合物が、インターロイキン−2、インターロイキン−7、インターロイキン−12および付着分子B7.1およびB7.2からなる群から選択される、請求項18記載の抗腫瘍組成物。
【請求項20】
1以上の免疫ポリペプチドをコードする配列を含んでなる少なくとも1種の組換えベクター、その天然の局在性とは異なる局在性を有するよう改変されている少なくとも1種のポリペプチド、および所望によりその組成物の抗腫瘍作用を増強する化合物を、治療薬または予防薬として含んでなる抗腫瘍組成物。
【請求項21】
抗腫瘍作用を増強する免疫ポリペプチドが請求項1〜19のいずれか一項に記載の特徴を有する、請求項20記載の抗腫瘍組成物。
【請求項22】
組換えベクターがポックスウイルス、特にワクシニアウイルス、カナリア痘ウイルスおよび鶏痘ウイルスに由来する、請求項20記載の抗腫瘍組成物。
【請求項23】
組換えウイルスが、コペンハーゲン、ワイエスおよび改変型アンカラ(MVA)株から選択されるワクシニアウイルスに由来する、請求項22記載の抗腫瘍組成物。
【請求項24】
組換えウイルスがワクシニアウイルスのコペンハーゲン株に由来し、かつ、そのポリペプチドをコードする配列がそのワクシニアウイルスのTKおよび/またはK1Lのレベルで挿入されている、請求項23記載の抗腫瘍組成物。
【請求項25】
組換えウイルスがワクシニアウイルスのMVA株に由来し、かつ、そのポリペプチドをコードする配列がそのワクシニアウイルスの除去領域I〜VI、特にIIおよび/またはIIIのうち少なくとも1つのレベルで挿入されている、請求項23記載の抗腫瘍組成物。
【請求項26】
そのポリペプチドをコードする配列が、ワクシニアウイルス遺伝子のプロモーター、特にチミジンキナーゼ(TK)、7.5K、H5R p28、p11およびK1L遺伝子のプロモーターから選択されるプロモーターの制御下に置かれている、請求項22〜25のいずれか一項に記載の抗腫瘍組成物。
【請求項27】
パピローマウイルス感染または腫瘍の治療または予防を意図した、請求項22〜26のいずれか一項に記載の抗腫瘍組成物であって、
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列
が挿入されたワクシニアウイルスのコペンハーゲンまたはMVA株に由来する少なくとも1種の組換えベクターを含んでなる組成物。
【請求項28】
好ましくはIL−2またはB7.1から選択される免疫刺激化合物をコードする配列をさらに含んでなる、請求項27記載の抗腫瘍組成物。
【請求項29】
組換えベクターが生きている、または殺されている、請求項20〜28のいずれか一項に記載の抗腫瘍組成物。
【請求項30】
ヒトまたは動物への注射によりその投与が可能となる医薬上許容される担体を含んでなる、請求項1〜29のいずれか一項に記載の抗腫瘍組成物。
【請求項31】
パピローマウイルスの初期および/または後期領域に由来して請求項11〜16で定義された特徴を有する免疫ポリペプチドをコードする、少なくとも1つの配列を含んでなる組換えベクター。
【請求項32】
注目されるポリペプチド、特に免疫ポリペプチドおよび/または免疫刺激ポリペプチドをコードする1以上の配列をさらに含んでなる、請求項31記載の組換えベクター。
【請求項33】
プラスミドまたはウイルスベクター、特にポックスウイルス、特にワクシニアウイルスのコペンハーゲンまたはMVA株に由来する、請求項31または32記載の組換えベクター。
【請求項34】
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列
が挿入されたワクシニアウイルスのコペンハーゲンまたはMVA株に由来する、請求項33記載の組換えベクター。
【請求項35】
請求項31〜34のいずれか一項に記載の組換えベクターを含んでなるウイルス粒子。
【請求項36】
免疫ポリペプチドがそのウイルス粒子を取り巻くタンパク質構造に付着されている、請求項35記載のウイルス粒子。
【請求項37】
癌もしくは腫瘍の治療もしくは予防を意図した医薬を製造するための、請求項1〜30のいずれか一項に記載の抗腫瘍組成物、請求項31〜34のいずれか一項に記載の組換えベクター、または請求項35もしくは36記載のウイルス粒子の使用。
【請求項38】
子宮頸部癌、軽度子宮頸部形成異常およびパピローマウイルス感染の治療または予防を意図した医薬を製造するための、請求項37記載の使用。
【請求項39】
筋肉内経路により注射可能な医薬を製造するための、請求項37または38記載の使用。
【請求項1】
1以上の免疫ポリペプチドを治療薬または予防薬として含んでなる抗腫瘍組成物であって、少なくとも1種のポリペプチドが、その天然の局在性とは異なる局在性を有するよう改変されていることを特徴とする組成物。
【請求項2】
ポリペプチドの改変が適当な局在化シグナルの誘導および/または天然の局在化シグナルの欠損または不活性化によって得られる、請求項1記載の抗腫瘍組成物。
【請求項3】
天然には膜以外、特に核局在性を有する免疫ポリペプチドが膜局在性を有するよう改変されている、請求項1および2のいずれか一項に記載の抗腫瘍組成物。
【請求項4】
免疫ポリペプチドが膜付着配列および適当であれば分泌配列の挿入によって改変されている、請求項3記載の抗腫瘍組成物。
【請求項5】
ポリペプチドがその核局在化配列に関して欠失している、請求項1〜4のいずれか一項に記載の組成物。
【請求項6】
分泌および/または膜付着配列が、狂犬病糖タンパク質、HIVウイルスenv糖タンパク質および麻疹ウイルスFタンパク質のものからなる群から選択される、請求項1〜5のいずれか一項に記載の抗腫瘍組成物。
【請求項7】
免疫ポリペプチドが、特にその天然の局在化シグナルの変異/欠損によって細胞質局在性を有するよう改変されている、請求項1、2または5のいずれか一項に記載の抗腫瘍組成物。
【請求項8】
免疫ポリペプチドが、特定の細胞コンパートメントに局在させることを可能にする配列の、特にそのC末端での挿入によって改変されている、請求項1、2または5のいずれか一項に記載の抗腫瘍組成物。
【請求項9】
免疫ポリペプチドが、特にエンドサイトーシス配列、特に配列IPNYRNMの、特にそのC末端での挿入によって改変されている、請求項8記載の抗腫瘍組成物。
【請求項10】
免疫ポリペプチドが、ゴルジ体の膜における付着を可能にする配列の、特にそのC末端での挿入によって改変されている、請求項8記載の抗腫瘍組成物。
【請求項11】
免疫ポリペプチドがパピローマウイルス、特にヒトパピローマウイルス(HPV)16、18、31、33または45型の初期および/または後期領域に由来する、請求項1〜10のいずれか一項に記載の抗腫瘍組成物。
【請求項12】
免疫ポリペプチドがパピローマウイルスの初期領域のポリペプチド、特にE6またはE7に由来する、請求項11記載の抗腫瘍組成物。
【請求項13】
免疫ポリペプチドがパピローマウイルスのE6またはE7ポリペプチドの非発癌性変異体に由来する、請求項12記載の抗腫瘍組成物。
【請求項14】
免疫ポリペプチドがパピローマウイルスのL1またはL2ポリペプチドに由来する、請求項11記載の抗腫瘍組成物。
【請求項15】
パピローマウイルスの初期ポリペプチド由来の少なくとも1種の免疫ポリペプチドと後期ポリペプチド由来の少なくとも1種の免疫ポリペプチドを含んでなる、請求項11〜14のいずれか一項に記載の抗腫瘍組成物。
【請求項16】
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチド
を含んでなる、請求項11〜15のいずれか一項に記載の抗腫瘍組成物。
【請求項17】
組成物の抗腫瘍作用を増強する少なくとも1種の化合物をさらに含んでなる、請求項1〜16のいずれか一項に記載の抗腫瘍組成物。
【請求項18】
化合物が免疫刺激剤である、請求項17記載の抗腫瘍組成物。
【請求項19】
免疫刺激化合物が、インターロイキン−2、インターロイキン−7、インターロイキン−12および付着分子B7.1およびB7.2からなる群から選択される、請求項18記載の抗腫瘍組成物。
【請求項20】
1以上の免疫ポリペプチドをコードする配列を含んでなる少なくとも1種の組換えベクター、その天然の局在性とは異なる局在性を有するよう改変されている少なくとも1種のポリペプチド、および所望によりその組成物の抗腫瘍作用を増強する化合物を、治療薬または予防薬として含んでなる抗腫瘍組成物。
【請求項21】
抗腫瘍作用を増強する免疫ポリペプチドが請求項1〜19のいずれか一項に記載の特徴を有する、請求項20記載の抗腫瘍組成物。
【請求項22】
組換えベクターがポックスウイルス、特にワクシニアウイルス、カナリア痘ウイルスおよび鶏痘ウイルスに由来する、請求項20記載の抗腫瘍組成物。
【請求項23】
組換えウイルスが、コペンハーゲン、ワイエスおよび改変型アンカラ(MVA)株から選択されるワクシニアウイルスに由来する、請求項22記載の抗腫瘍組成物。
【請求項24】
組換えウイルスがワクシニアウイルスのコペンハーゲン株に由来し、かつ、そのポリペプチドをコードする配列がそのワクシニアウイルスのTKおよび/またはK1Lのレベルで挿入されている、請求項23記載の抗腫瘍組成物。
【請求項25】
組換えウイルスがワクシニアウイルスのMVA株に由来し、かつ、そのポリペプチドをコードする配列がそのワクシニアウイルスの除去領域I〜VI、特にIIおよび/またはIIIのうち少なくとも1つのレベルで挿入されている、請求項23記載の抗腫瘍組成物。
【請求項26】
そのポリペプチドをコードする配列が、ワクシニアウイルス遺伝子のプロモーター、特にチミジンキナーゼ(TK)、7.5K、H5R p28、p11およびK1L遺伝子のプロモーターから選択されるプロモーターの制御下に置かれている、請求項22〜25のいずれか一項に記載の抗腫瘍組成物。
【請求項27】
パピローマウイルス感染または腫瘍の治療または予防を意図した、請求項22〜26のいずれか一項に記載の抗腫瘍組成物であって、
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列
が挿入されたワクシニアウイルスのコペンハーゲンまたはMVA株に由来する少なくとも1種の組換えベクターを含んでなる組成物。
【請求項28】
好ましくはIL−2またはB7.1から選択される免疫刺激化合物をコードする配列をさらに含んでなる、請求項27記載の抗腫瘍組成物。
【請求項29】
組換えベクターが生きている、または殺されている、請求項20〜28のいずれか一項に記載の抗腫瘍組成物。
【請求項30】
ヒトまたは動物への注射によりその投与が可能となる医薬上許容される担体を含んでなる、請求項1〜29のいずれか一項に記載の抗腫瘍組成物。
【請求項31】
パピローマウイルスの初期および/または後期領域に由来して請求項11〜16で定義された特徴を有する免疫ポリペプチドをコードする、少なくとも1つの配列を含んでなる組換えベクター。
【請求項32】
注目されるポリペプチド、特に免疫ポリペプチドおよび/または免疫刺激ポリペプチドをコードする1以上の配列をさらに含んでなる、請求項31記載の組換えベクター。
【請求項33】
プラスミドまたはウイルスベクター、特にポックスウイルス、特にワクシニアウイルスのコペンハーゲンまたはMVA株に由来する、請求項31または32記載の組換えベクター。
【請求項34】
(1)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(2)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(3)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、および配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチドをコードする配列、
(4)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(5)配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列、
(6)配列番号1で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、配列番号2で示されるものの総てまたは一部と相同または同一の配列を有する免疫ポリペプチド、パピローマウイルスのタンパク質L1由来の免疫ポリペプチド、および/またはパピローマウイルスのタンパク質L2由来の免疫ポリペプチドをコードする配列
が挿入されたワクシニアウイルスのコペンハーゲンまたはMVA株に由来する、請求項33記載の組換えベクター。
【請求項35】
請求項31〜34のいずれか一項に記載の組換えベクターを含んでなるウイルス粒子。
【請求項36】
免疫ポリペプチドがそのウイルス粒子を取り巻くタンパク質構造に付着されている、請求項35記載のウイルス粒子。
【請求項37】
癌もしくは腫瘍の治療もしくは予防を意図した医薬を製造するための、請求項1〜30のいずれか一項に記載の抗腫瘍組成物、請求項31〜34のいずれか一項に記載の組換えベクター、または請求項35もしくは36記載のウイルス粒子の使用。
【請求項38】
子宮頸部癌、軽度子宮頸部形成異常およびパピローマウイルス感染の治療または予防を意図した医薬を製造するための、請求項37記載の使用。
【請求項39】
筋肉内経路により注射可能な医薬を製造するための、請求項37または38記載の使用。
【図1】


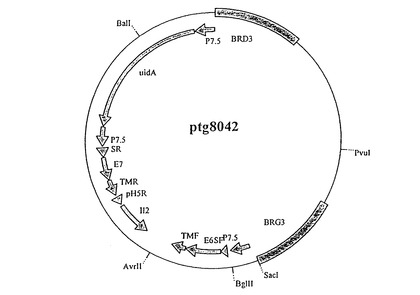
【図2】
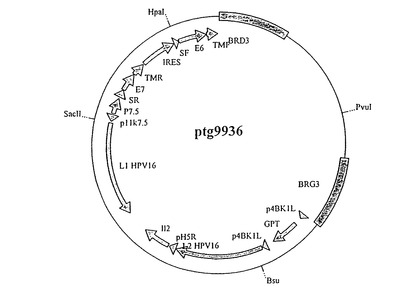
【図3】
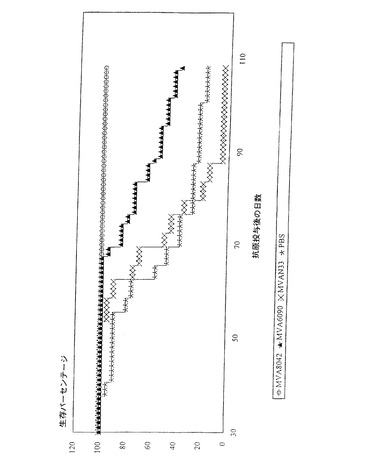
【図4】
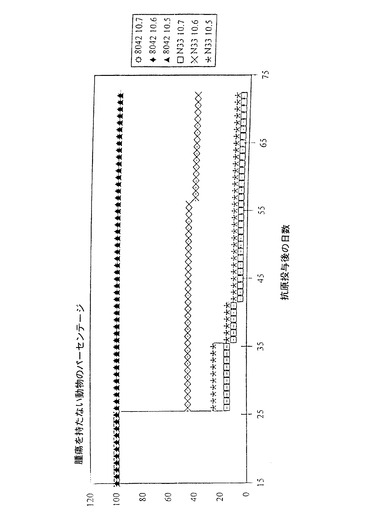
【図5】
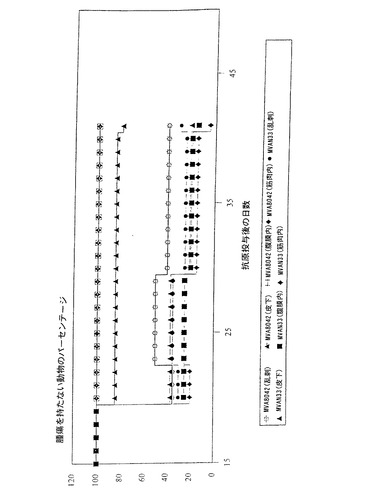
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2010−57500(P2010−57500A)
【公開日】平成22年3月18日(2010.3.18)
【国際特許分類】
【出願番号】特願2009−253035(P2009−253035)
【出願日】平成21年11月4日(2009.11.4)
【分割の表示】特願2000−503107(P2000−503107)の分割
【原出願日】平成10年7月17日(1998.7.17)
【出願人】(599082883)トランジェーヌ、ソシエテ、アノニム (32)
【氏名又は名称原語表記】TRANSGENE S.A.
【Fターム(参考)】
【公開日】平成22年3月18日(2010.3.18)
【国際特許分類】
【出願日】平成21年11月4日(2009.11.4)
【分割の表示】特願2000−503107(P2000−503107)の分割
【原出願日】平成10年7月17日(1998.7.17)
【出願人】(599082883)トランジェーヌ、ソシエテ、アノニム (32)
【氏名又は名称原語表記】TRANSGENE S.A.
【Fターム(参考)】
[ Back to top ]