
細胞によるタンパク質および抗体収量を改善するために適用される全ゲノム進化技術
全ゲノム進化技術は治療抗体の規模拡大可能な製造のための必要性を支持する広いツールとみなすことができる。ランダムな性質およびインビボでの作用様式が他の相補技術と本方法とを隔てるので、宿主細胞の製造性能を改善するための代替的解決が提供される。既存の生成株の速度が最適化できることによって、本方法は、前臨床、臨床または商品段階での高いタイターの抗体を示す速く増殖する細胞を生成するように、迅速な細胞株最適化のための現在の必要性を満足するために適切になる。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、2006年4月17日に出願された米国出願番号第60/792,937号の35 U.S.C §119(e)下の利益を主張し、本明細書にその全体を参照によって援用される。
【0002】
(発明の分野)
本発明は、親細胞と比べて改善された増殖特性(例えば、速い増殖率、高細胞密度での増大された生体分子生成、および/または高細胞密度での増大された細胞増殖または細胞生存能力)を有する細胞の生成に関する。さらに、本発明は、規模拡大可能な製造における使用のための改善された増殖特性について細胞のスクリーニングおよび細胞ベース製造に関する増大された増殖に関わる経路の発見に関する。
【背景技術】
【0003】
(発明の背景)
治療のタンパク質およびモノクローナル抗体(mAb)は、疾患に関連する特定の標的を特異的に置換または阻害する能力のために過去何十年も最も首尾よい種類の医薬の1つになっている(1)。Remicade(登録商標)、Rituxan(登録商標)、Herceptin(登録商標)、Humira(登録商標)、およびAvastin(登録商標)などの市販mAbの首尾よい市場性能によって、モノクローナル抗体は最も重要な薬物種類の1つとして現れ、現在全ての新しい薬物出現物の約半分を示す。現在まで出現した全てのmAbのうち、40%は大成功薬物であるかまたは大成功収益を潜在的にする。治療mAbの世界売り上げは、2004年に100億ドルを超えており、2008年までに300億ドルを超えるように計画されている(Needham & Company, 2005)。次の5年で、mAb市場は、キメラ(一部ヒトおよび一部マウス)からヒト化(CDR移植)から完全ヒト抗体までの技術的な進化によって、バイオテクノロジーおよび医薬産業の、最速で成長し、かつ最も有利な部門であり続けるようである。従来の低分子アプローチと比較して、mAbは高い特異性、低い毒性、および治療までのより迅速な開発経路を提供する。標的に結合する際に機能を発揮する低分子と異なり、mAbは、標的特異的な細胞傷害活性を引き出すように、体の免疫系を利用することによって大きな治療インパクトを与えることができる。
【0004】
mAbの大きな成功にも関わらず、効率的な開発は全く課題のままである。治療mAbを開発する会社は、究極的に市場までの時間、有効性、安全性および商品費用に影響を及ぼす、首尾よいGMP製造、精製および製品プロセスの開発初期段階で、適切なシステムを提供しなければならない。治療mAbの製造は、治療のための高用量を支持するのに必要な年収量と関連する高い製造コストおよび付加製造能力に関連する高い資本コストのために課題のままである。時節にあわせたやり方で大量の製品を作り出すことができる製造システムが、市場での余剰の発酵能力の維持により達成可能な短い開発スケジュールおよび低い全体的商品コストのために必要とされる。
【0005】
mAb製造を専門とする会社は、高いmAb収量を生み出すことができるいくつかの細胞ベースシステムを開発した。これらのシステムは、細菌、酵母、植物および哺乳動物細胞を含む(2)。微生物ベースプラットフォームは、ある最高の全タイター収量となった;しかし、活性タンパク質の量は、適切なフォールディングまたはプロセシングのために生じた全タンパク質のうち一部分だけを典型的に示した。また、微生物システムは、翻訳後修飾などのさらなるプロセシングを要する、抗体を含む複数鎖巨大分子などの複雑なタンパク質を効率的に作り出すように首尾よく適合されていなかった。これらの微生物ベース製造の制限の点から、哺乳動物細胞は、治療抗体および非抗体タンパク質のサブセットを製造する、大規模の優良製造規則(GMP)のための最も信頼可能で広く使用されるシステムの1つとなっている。
【0006】
臨床物質および最終的に市場供給のための有意な製造能力を要する現在進行中の300の抗体開発プログラムがある(3)。これらのプログラムの大半は、製造のための流加または還流バイオリアクターを用いた現在標準のGMP哺乳動物細胞培養発酵法の下でチャイニーズハムスター卵巣(CHO)およびマウスミエローマ(NS0、SP2)細胞株のような組み換え哺乳動物細胞株を使用する(4)。細胞ベース製造能力への負担は生成物の増大が要求されるにつれて増大し続け、これらのmAbプログラムのそれぞれは、多くの臨床試行から最終的に市場へと進む。このリスクの点から、十分な製造能力が市場に存在し続けること、および全体的商品コストが今日の市場の現在コストと類似のままか、または低いままであることを確実にするように代替的手順が要求される。
【0007】
様々なアプローチが、全体的生成および精製収量の改善を含む哺乳動物細胞からの抗体の大規模製造のための将来の必要性ならびにトランスジェニック動物ベース戦略のような代替的製造供給源の使用を支持するように実施されている(5、6)。代替的宿主システムは費用効果規模可能性および規制コンプライアンスについて確認されているが、当該技術分野の水準は哺乳動物細胞培養のままである。哺乳動物細胞における抗体生成の収率を改善する可能性が、1)培養条件および/または培地最適化によるバイオリアクター性能の改善;2)組み込んでいる非常に活性なプロモーターによる改善されたベクター発現または増幅による増大ベクターコピー数;および/または3)大規模バイオリアクターにおける良好なタイター収量および改善された細胞増殖を提供する宿主細胞内の内在性経路の増進による細胞宿主の最適化によって達成することができる。これらの改善またはこれらの組み合わせのいずれかが年間製品要求を作り出すのに必要な製造運転数を短くするプロセスとなり、それによって市場での全体製造制約を軽減することができる。
【0008】
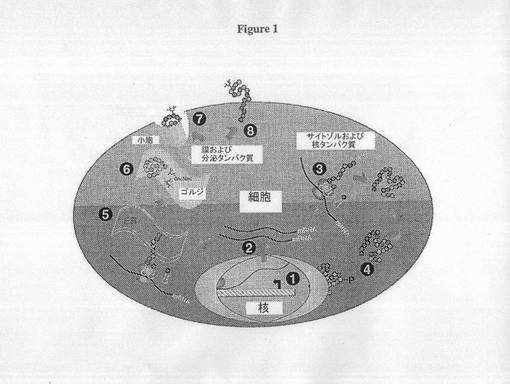
細胞宿主最適化は、a)mRNA転写および成熟、b)タンパク質合成および翻訳後修飾、c)タンパク質分泌および細胞副次局在(sub-localization)、d)細胞質と細胞小器官との間のタンパク質移動、ならびにe)細胞周期および生存調節を含む、内在性経路の操作によって達成することができる。図1に示すように、これらのプロセスの1つ以上の調節に関わるタンパク質の微妙な構造変化が生成細胞株の全体性能に直接または間接的に影響を及ぼすことがある。機能的な細胞表現型をスクリーニングできるランダム化ゲノムワイド突然変異生成アプローチは、増殖率、非常に高い細胞密度での生存、タンパク質合成および分泌速度を調節する複雑な細胞プロセスを増進するアプローチを提供する。ほとんどの細胞突然変異生成スキームでは不幸にも、突然変異薬剤の使用によって、GMP製造に適切でない不安定な細胞株を生じるゲノムワイド染色体不安定となる。
【0009】
以前の研究によって、ミスマッチ修復と呼ばれる複製後DNA修復機構の阻害はDNA複製中のDNAポリメラーゼによって組み込まれる点変異の増大のために安定な細胞内に遺伝的多様性をもたらし得ることが示された(7、8)。DNA複製は、親細胞が遺伝情報を子孫細胞に伝えるために全ての細胞が増殖中に受ける複雑なプロセスである。細胞がDNAを複製する時に、変異が、ポリメラーゼ忠実度を含む様々な機構から新しく合成された鋳型内に生じる。ミスマッチ修復(MMR)システム(11〜13)のような一連の複製後DNA修復プロセスは、自然で進化し、個体が遺伝型同一性を保持するために原核生物細胞および真核生物細胞にユビキタスに存在する。MMRは、親DNA鋳型と子孫DNA鋳型との間に存在する不一致な遺伝情報を訂正する二次プルーフリーディングプロセスを介した「天然」転移および転換変異の蓄積を妨げる。
【0010】
細胞集団内の遺伝的多様性を生じるミスマッチ修復を阻害するプロセスは、本明細書で「全ゲノム進化技術(whole genome evolution technology)」と呼ばれる。全ゲノム進化技術は、MMRの可逆阻害に基づいており、例えば新規治療抗体またはタンパク質を生じる細胞内でMMRを阻害することができるドミナントネガティブタンパク質インヒビターまたは化学インヒビターの活性によって媒介される。抑制されたMMRはDNA複製中に生じる変異のために子孫細胞のゲノムにおける点変異の遺伝となる(8、11、12)。細胞中のMMR抑制は、MMR熟練(proficient)細胞に典型的に観察されるものよりも高い頻度(1000倍の増大まで)で遺伝する天然変異を可能にする。全ゲノム進化技術に由来する子孫細胞の遺伝的に多様な集団は、新規な特徴を有するサブクローンを同定する自動化機能的ハイスループットスクリーニング(HTS)を介してスクリーニングすることができる細胞のライブラリーとなる。
【0011】
全ゲノム進化技術は、望ましい表現型を有する細胞の開発のための進化の力を利用する。この技術と他の進化ベース技術とを隔てる重要な区別は、ランダムなインビボ性質のプロセスである。全ての細胞が本来的に有する多くの遺伝子および経路が利用できることによって、機能的な細胞スクリーニングで同定される予測されない変異体の生成を可能にし、望ましい表現型を有する子孫細胞および遺伝子産物をもたらす。この技術は、全ゲノム進化を増大するようにインビボで既存生成株に適用され、それによってもはやインビトロ操作を要しないために、時間およびコストで効率的である。全ゲノム進化技術は、例えば、CHO、NSO、およびハイブリドーマ細胞において、増大されたタイター生成を有する進化した子孫細胞に由来する組み換え治療抗体を生成するいくつかの哺乳動物細胞株に首尾よく適用された(8〜10)。また、全ゲノム進化技術は改善された生物学的特性を有するタンパク質または抗体を生成する進化した子孫細胞に由来する組み換え治療抗体を生成する哺乳動物細胞株に適用されている(15〜17)。
【0012】
特に、哺乳動物細胞株による抗体生成のための生産性を増大するために細胞宿主性能を改善する方法が本明細書に提供される。ミスマッチ修復は、改善された抗体製造収量をもたらすことができる増殖特性に影響を与える細胞プロセスを改善するように、生成細胞株で阻害される。
【0013】
(発明の要旨)
本明細書に、細胞宿主性能を改善する方法、例えば、細胞株による生産性を増大する方法が提供される。本発明の方法は、(a)親細胞集団の親細胞のミスマッチ修復を阻害する工程;(b)MMR阻害親細胞をインキュベートまたは拡大して突然変異生成を可能にし、それによって超突然変異娘細胞を生成する工程;(c)改善された増殖特性を有する超突然変異娘細胞を検出する工程;および(d)改善された増殖特性を有する超突然変異娘細胞の遺伝的安定性を保存する工程を含む、親細胞集団と比べて少なくとも1つの改善された増殖特性を有する細胞を生成する方法を含む。
【0014】
いくつかの態様において、ミスマッチ修復を阻害する工程がミスマッチ修復の化学インヒビターまたはミスマッチ修復のタンパク質インヒビターに親細胞を曝露する工程を含む。
【0015】
本発明のいくつかの局面において、MMR阻害親細胞をインキュベートまたは拡大して突然変異生成を可能にし、それによって超突然変異娘細胞を生成する工程が好ましくは少なくとも20回の継代、より好ましくは少なくとも30回の継代でMMR阻害細胞を継代する工程を含む。
【0016】
本発明の方法において遺伝的安定性を保存する工程が好ましくは超突然変異娘細胞からミスマッチ修復の化学インヒビターを回収する工程、またはミスマッチ修復のタンパク質インヒビターを不活性化する工程を含む。遺伝的安定性の保存が改善された増殖特性を有する超突然変異娘細胞を検出する工程の前、後または同時に生じてもよい。
【0017】
本発明の方法において、改善された増殖特性を有する超突然変異娘細胞を検出する工程が好ましくは改善された増殖特性を有する超突然変異娘細胞のハイスループットスクリーニング(HTS)を含む。
【0018】
好ましい態様において、改善された増殖特性は、速い増殖率、高細胞密度での増大された細胞増殖または細胞生存能力、および/または高細胞密度での増大された生体分子生成である。いくつかの態様において、速い増殖率を有する超突然変異娘細胞の検出が、同じ培養条件下で同等の長さの培養時間を続けた後に、超突然変異娘細胞によって生じた細胞集団のサイズと、親細胞によって生じた細胞集団のサイズとを比較する工程を含み、大きな娘細胞集団が速い増殖率を示す。いくつかの態様において、速い増殖率を有する超突然変異娘細胞の検出が、親細胞の増殖率よりも大きな増殖率を有する娘細胞を同定する工程を含む。いくつかの態様において、高細胞密度での増大された細胞生存能力の検出は、同じ培養条件下で同等の長さの培養時間を続けた後に、超突然変異娘細胞によって生じた高密度での生存可能細胞集団のサイズと、親細胞によって生じた高密度での生存可能細胞集団のサイズとを比較する工程を含み、大きな生存可能娘細胞集団が高細胞密度での増大された細胞生存能力を示す。細胞集団(例えば、親細胞集団および/または娘細胞集団)のサイズが好ましくは光学画像システムを用いて決定される。いくつかの態様において、増大された生体分子生成を示す超突然変異娘細胞の検出は、同じ培養条件下で親細胞よりも娘細胞による生体分子の高い収量を決定する工程を含む。いくつかの態様において、親および娘細胞集団のサイズは、それぞれの各集団によって生成される生体分子の収量を決定する場合に同等である。
【0019】
本発明の方法における使用のための親細胞は、好ましくは抗体生成細胞である。いくつかの態様において、親細胞はハイブリドーマ細胞である。親細胞は真核生物細胞または原核生物細胞であってもよい。本発明における使用のための親細胞は、好ましくは哺乳動物細胞であり、より好ましくはヒトまたはげっ歯類(例えば、ハムスター、マウス)細胞であり、例えば、限定されないが、NS0、SP2、またはチャイニーズハムスター卵巣(CHO)細胞である。本発明の方法における使用のための親細胞は、細菌細胞、酵母細胞、植物細胞または両生類細胞であってもよい。
【0020】
また、本発明の方法によって生成した細胞、ならびに均一および不均一組成のかかる細胞が本発明の範囲に含まれる。
【0021】
本発明の細胞を培養し、細胞または細胞の培養培地から生体分子を単離することによって生体分子を製造する方法が本発明によってさらに提供される。生体分子は、化学薬剤および/または生物学的薬剤、例えば、限定されないが、タンパク質、例えば、抗体を含んでいてもよい。本発明の方法によって生成した生体分子およびその医薬組成物は同様に、本発明の範囲に含まれる。
【0022】
また、改善された増殖特性の原因遺伝子を同定する方法であって、本発明の方法によって生成した細胞のゲノムと、親細胞のゲノムとを比較して変異を同定する工程を含み、改善された増殖特性の原因遺伝子が少なくとも1つのかかる変異を含む方法が、本発明の範囲に含まれる。
【0023】
本発明のこれらおよび他の局面は以下に記載される1つ以上の態様によって提供される。
【0024】
(例示態様の詳細な説明)
本明細書で言及されるGenBankデータベース配列のアクセッション番号を含む、参考文献、特許、特許出願、および科学文献は、当業者の知識を確立し、あたかもそれぞれが特異的にかつ個別に参照によって援用されて示されるように同じ程度でその全体を参照によって本明細書に援用される。本明細書に引用される任意の参考文献と本明細書の具体的な教示との間に矛盾があれば、後者によって解決される。同様に、単語または句の当該技術で理解される定義と本明細書で具体的に教示される単語または句の定義との間に矛盾があれば、後者によって解決される。
【0025】
本明細書に引用されるそれぞれの範囲は、範囲の全ての組み合わせおよび下位の組み合わせ、ならびにそこに含まれる特定の数を含む。
【0026】
別に規定されない場合、本明細書に使用される全ての技術用語および科学用語は、発明の属する当業者によって一般に理解されるものと同じ意味を有する。本明細書に記載されるものと類似または同等の任意の方法および物質が本発明の試験の実施に使用できるが、好ましい物質および方法が本明細書に記載される。本発明を記載し、かつ特許請求する時に、以下の技術用語が使用される。
【0027】
本明細書および添付の特許請求の範囲に使用される場合、単数形の「a」、「an」および「the」は内容が明らかに他のものを示さない場合に複数の参照を含む。従って、例えば、「細胞(a cell)」への参照には、2つ以上の細胞の組み合わせが含まれる等である。
【0028】
量、時間継続期間等のような測定可能な値を参照する場合に本明細書で使用される用語「約」は、変化が開示された方法を行うために適切であるように、特定の値から、±20%または±10%、より好ましくは±5%、さらに好ましくは±1%、さらにより好ましくは±0.1%の変化を包含することを意味する。
【0029】
「保存的に改変(modified)されたバリアント」はアミノ酸配列および核酸配列の両方に適用する。特定の核酸配列に関して、保存的に改変されたバリアントは、同一のアミノ酸配列または本質的に同一のアミノ酸配列をコードする核酸、または核酸がアミノ酸配列をコードしない場合の本質的に同一の配列のことをいう。遺伝コードの縮重のために、多数の機能的に同一な核酸が任意の所定のタンパク質をコードする。例えば、コドンGCA、GCC、GCG、およびGCUの全てはアミノ酸アラニンをコードする。従って、アラニンがコドンで特定される全ての位置で、コドンは、コードされたポリペプチドを改変せずに、記載される対応のコドンのいずれかに改変することができる。かかる核酸変異は、「サイレント変異」であり、1種の保存的に改変された変異である。また、ポリペプチドをコードする本明細書の全ての核酸配列は、核酸の全て可能なサイレント変異を記載する。当業者は、核酸の各コドン(通常メチオニンの唯一のコドンであるAUG、および通常トリプトファンの唯一のコドンであるTGGを除く)が機能的に同一の分子を生じるように改変できることを認識する。従って、ポリペプチドをコードする核酸の各サイレント変異は、記載されるそれぞれの配列に示される。
【0030】
例えば、細胞、核酸、タンパク質、またはベクターを参照して使用される場合「組み換え」は、細胞、核酸、タンパク質またはベクターが異種の核酸もしくはタンパク質の導入またはネイティブの核酸もしくはタンパク質の改変によって改変されたこと、または細胞が上記改変された細胞に由来することを示す。従って、例えば、組み換え細胞は、ネイティブ(非組み換え)形式の細胞で見られない遺伝子を発現するか、または他の方法で異常発現し、過小発現(underexpress)するか、または全く発現しないネイティブ遺伝子を発現する。
【0031】
句「核酸」または「ポリヌクレオチド配列」とは、5’から3’末端に読まれるデオキシリボヌクレオチドまたはリボヌクレオチドの単鎖または二重鎖ポリマーのことをいう。また、核酸はポリメラーゼによる正確な読み通しを許容する改変ヌクレオチドを含むことができるが、例えば、保存的に改変されたバリアントを含む、該核酸によってコードされるポリペプチド発現を改変しない。
【0032】
「ポリペプチド」、「ペプチド」および「タンパク質」は、アミノ酸残基のポリマーを示すために本明細書で交換可能に使用される。該用語は、1つ以上のアミノ酸残基が対応する天然アミノ酸の人工化学模倣物であるアミノ酸ポリマー、ならびに天然アミノ酸ポリマーおよび非天然アミノ酸ポリマーに適用する。ポリペプチドは、保存的に改変されたバリアントを含む。当業者は、コードされる配列における単一アミノ酸または小さい割合のアミノ酸を改変、付加、または欠失する核酸配列、ペプチド配列、ポリペプチド配列、またはタンパク質配列に対する置換、欠失または付加が、改変がアミノ酸の化学的に類似するアミノ酸での置換となる場合の「保存的に改変されたバリアント」であることを認識する。機能的に類似するアミノ酸を提供する保存的置換表は、当該技術分野で周知である。かかる保存的に改変されたバリアントはまた、本発明の多形バリアント、種間ホモログ、および対立遺伝子であり、これらを排除しない。以下の8つのグループはそれぞれ、別の保存的置換であるアミノ酸を含む:1)アラニン(A)、グリシン(G);2)アスパラギン酸(D)、グルタミン酸(E);3)アスパラギン(N)、グルタミン(Q);4)アルギニン(R)、リジン(K);5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);7)セリン(S)、トレオニン(T);および8)システイン(C)、メチオニン(M)(33)。また、用語「保存的置換」は、かかるポリペプチドがまた必須の結合活性を示すことを条件として、非置換親アミノ酸の代わりに置換アミノ酸の使用を含む。
【0033】
「アミノ酸」とは、天然アミノ酸および合成アミノ酸、ならびに天然アミノ酸に類似する様式で機能するアミノ酸アナログおよびアミノ酸模倣物のことをいう。天然アミノ酸は、遺伝コードによってコードされるもの、および後に改変されるアミノ酸、例えば、ヒドロキシプロリン、γ-カルボキシグルタミン酸およびO-ホスホセリンである。「アミノ酸アナログ」とは、天然アミノ酸と同じ基本化学構造、即ち、水素、カルボキシル基、アミノ基、およびR基に結合するα炭素、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムを有する化合物のことをいう。かかるアナログは改変R基(例えば、ノルロイシン)または改変ペプチド骨格を有するが、天然アミノ酸と同じ基本化学構造を保持する。「アミノ酸模倣物」とは、アミノ酸の一般化学構造と異なるが天然アミノ酸と類似する様式で機能する構造を有する化学化合物のことをいう。
【0034】
アミノ酸は一般的に既知である3文字記号またはIUPAC-IUB 生化学命名委員会(Biochemical Nomenclature Commission)によって推奨された1文字記号のいずれかによって、本明細書に示すことができる(以下の表1を参照)。ヌクレオチドは、同様に、一般的に受け入れられている1文字コードによって参照できる。
【0035】
全てのアミノ酸配列は、左から右への向きが従来のアミノ末端からカルボキシ末端への方向である方式によって本明細書に表されるということに注意されたい。
【0036】
本明細書で使用する場合、用語「インビトロ」または「エキソビボ」とは、人工的環境、ならびに例えば、限定されないが、試験管および細胞培養の人工的環境で生じるプロセスまたは反応のことをいう。用語「インビボ」とは、天然環境(例えば、動物または細胞)および天然環境で生じるプロセスまたは反応のことをいう。
【0037】
「薬学的に許容できる」、「生理学的に寛容できる」およびそれらの文法的バリエーションは、組成物、担体、希釈剤および試薬を示す場合、交換可能に使用され、物質が組成物の投与を禁止する程度までの望ましくない生理学的効果を生成せずにヒトにまたはヒトで投与できることを表す。
【0038】
用語「薬学的に許容できる担体」とは、過剰な毒性、炎症、アレルギー反応、または他の合併症が無く、良さそうな医学判断の範囲内で、ヒトおよび動物の組織と接触して使用するために適切であり、合理的な利益/リスク比と釣り合う試薬、賦形剤、細胞、化合物、物質、組成物、および/または用量形態のことをいう。本明細書でより詳細に記載されるように、本発明における使用に適切な薬学的に許容できる担体としては、気体、液体、ならびに半固体および固体物質が挙げられる。
【0039】
「免疫グロブリン」または「抗体」は、抗体分子および様々な抗体由来分子の両方のことをいうために広く使用され、免疫系の主要成分である高等な哺乳動物で生じる糖タンパク質のグループの任意の一員を含む。用語「抗体」は最も広い意味で使用され、所望の生物学的活性(例えば、標的抗原への特異的結合)を示す限り、モノクローナル抗体、ポリエピトープ特異性を有する抗体組成物、二重特異的抗体、ダイアボディ(diabody)、および単鎖分子、ならびに抗体断片(例えば、Fab、F(ab')2、およびFv)を特異的に含む。免疫グロブリン分子は、それぞれが軽鎖および重鎖の末端部分を含む抗原結合ドメインを含み、補体結合のような様々な機能に必要なFc領域を含む。5種類の免疫グロブリンがあり、Fc領域における重鎖の一次構造が免疫グロブリンの種類を決定する。具体的に、α鎖、δ鎖、ε鎖、γ鎖、およびμ鎖は、それぞれIgA、IgD、IgE、IgG、およびIgMに対応する。本明細書で使用される場合、「免疫グロブリン」または「抗体」は全てのサブクラスのα、δ、ε、γ、およびμを含み、また4つの鎖の免疫グロブリン構造の任意の天然多量体(例えば、IgAおよびIgM)または合成多量体のことをいう。抗体は、非共有的に、特異的に、かつ可逆的に抗原と結合する。本明細書で使用される場合、用語「モノクローナル抗体」とは、実質的に均一な抗体の集団から得られた抗体のことをいい、即ち、該集団を構成する個々の抗体は少量で存在し得る可能な天然変異を除いて同一である。例えば、モノクローナル抗体は、単一クローンの抗体生成細胞によって生成されてもよい。ポリクローナル抗体と異なり、モノクローナル抗体は単一特異的である(例えば、単一抗原の単一エピトープに特異的)。修飾句「モノクローナル」は実質的に均一な集団の抗体から得られる抗体の特性を示し、任意の特定の方法による抗体生成が必要であると解釈されるべきでない。例えば、本発明によって使用されるモノクローナル抗体はKohler et al.(18)に最初に記載されたハイブリドーマ法によって作製することができるか、または組み換えDNA法によって作製することができる。また、「モノクローナル抗体」は例えばMarks et al.(19)に記載される技術を用いてファージ抗体ライブラリーから単離することができる。
【0040】
抗体由来分子は、抗原結合特異性を保持するインタクト抗体の部分を含み、例えば、少なくとも1つの可変領域(重鎖または軽鎖可変領域のいずれか)を含む。例えば、抗体由来分子としては、Fab断片、Fab'断片、F(ab')2断片、Fd断片、F(v)断片、Fabc 断片、Fd断片、Fabc断片、Sc抗体(単鎖抗体)、ダイアボディ、個々の抗体軽鎖、個々の抗体重鎖、抗体鎖と他の分子との間のキメラ融合物、重鎖モノマーまたはダイマー、軽鎖モノマーまたはダイマー、1つの重鎖および1つの軽鎖からなるダイマー等の分子が挙げられる。免疫グロブリンの全てのクラス(例えば、IgA、IgD、IgE、IgG、およびIgM)およびそのサブクラスが含まれる。
【0041】
抗体は、毒素部分または非毒素部分を標識/コンジュゲートすることができる。毒素部分としては、例えば、細菌毒素、ウイルス毒素、放射性アイソトープ等が挙げられる。抗体は、抗体の検出を補助するための生物学的アッセイ(例えば、放射性アイソトープ標識、蛍光標識)における使用のために標識できる。また、抗体は、例えば、放射性免疫治療(20)、画像技術および放射性免疫指示(guided)手術のような適用のために所望の部位に直接的に放射線を送達する放射活性アイソトープ、または特異的抗体/抗原複合体のインビボ画像化または検出を可能にする標識を、診断目的または治療目的で、標識/コンジュゲートすることができる。また、抗体は毒素とコンジュゲートして免疫毒素を提供してもよい(21)。
【0042】
抗体に関して、用語「免疫学的に特異的」とは、標的分子の1つ以上のエピトープに結合するが、抗原性生物学的分子の混合集団を含む試料中の他の分子を実質的に認識せず、かつ結合しない抗体のことをいう。
【0043】
「キメラ」または「キメラ化」抗体(免疫グロブリン)とは、所望の生物学的活性を示す限り、重鎖および/または軽鎖の一部が特定のクラスに由来する抗体の対応配列に同一または相同であるか、あるいは特定の抗体クラスまたはサブクラスに属するが、残りの鎖は別のクラスに由来する抗体の対応配列に同一または相同であるか、あるいは別の抗体クラスまたはサブクラスに属する抗体、ならびにかかる抗体の断片のことをいう(22)。
【0044】
「ヒト化」形態の非ヒト(例えば、マウス)抗体は、非ヒト免疫グロブリンに由来する最小配列を含むキメラ免疫グロブリン、免疫グロブリン鎖またはその断片(例えば、Fv、Fab、Fab'、F(ab’)2または抗体の他の抗原結合部分配列)である。ほとんどの部分について、ヒト化抗体は、レシピエントの相補決定領域(CDR)からの残基が所望の特異性、親和性、および能力を有するマウス、ラットまたはウサギ等の非ヒト種(ドナー抗体)のCDRからの残基によって置換されるヒト免疫グロブリン(レシピエント抗体)である。いくつかの例において、ヒト免疫グロブリンのFvフレームワーク領域(FR)残基は対応する非ヒト残基によって置換される。さらに、ヒト化抗体は、レシピエント抗体にも移入CDRまたはフレームワーク配列にも見られない残基を含み得る。これらの改変は、抗体性能をさらに改良し、かつ最適化するために行われる。一般的に、ヒト化抗体は、CDR領域の全てまたは実質的に全てが非ヒト免疫グロブリンのものに対応し、FR領域の全てまたは実質的に全てがヒト免疫グロブリン配列のものであるように、実質的に全てのうち少なくとも1つ、典型的に2つの可変領域を含む。また、ヒト化抗体は最適に、免疫グロブリン定常領域(Fc)の少なくとも一部、典型的にヒト免疫グロブリンのものを含む。更なる詳細について、Jones et al.(23); Reichmann et al.,(24);Presta(25)を参照。
【0045】
「完全にヒト」とは、分子全体がヒト起源であるか、またはヒト形態の抗体に同一のアミノ酸配列からなる抗体等の免疫グロブリンのことをいう。
【0046】
「エピトープ」とは、抗体結合部位として働く抗原の免疫学的決定基のことをいう。本明細書で使用される場合、用語「コンフォメーションエピトープ」とは、連続する一連のアミノ酸以外の抗原のアミノ酸間の空間関係によって形成される不連続エピトープのことをいう。
【0047】
「ハイブリドーマ」とは、親細胞の特異的な免疫潜在性を発現する培養新生物形成リンパ球と刺激されたBリンパ球またはTリンパ球との間での細胞融合の生成物のことをいう。
【0048】
本明細書で使用される場合、「高いタイター」とは、親細胞株よりも少なくとも約1.5倍高いタイターのことをいう。いくつかの態様において、タイターは、親細胞株よりも、少なくとも約1.5〜3倍高く、3〜5倍高く、5〜7倍高く、7〜9倍高く、または9〜10倍高い。
【0049】
本明細書で使用される場合、「高い親和性」とは、式Ka=8/3(It−Tt)(式中、「It」は50%トレーサーでのインヒビター取り込みの全モル濃度であり、「Tt」はトレーサーの全モル濃度である)による標準方法で計算できる高い抗体結合親和性のことをいう(26)。結合親和性はまた、式B/T=n・NAb・W108[(V-Vm)K+Q・W]を用いて計算できる(27)。本明細書で使用される場合、「高い親和性」は、約1×107M-1未満である。いくつかの態様において、抗体は約1×108M-1未満の親和性を有する。他の態様において、抗体は約1×109M-1未満の親和性を有する。他の態様において、抗体は約1×1010M-1未満の親和性を有する。他の態様において、抗体は約1×1011M-1未満の親和性を有する。他の態様において、抗体は約1×1012M-1未満の親和性を有する。他の態様において、抗体は約1×1013M-1未満の親和性を有する。他の態様において、抗体は約1×1014M-1未満の親和性を有する。
【0050】
本明細書で使用される場合、用語「速い増殖率」とは、所定の集団の細胞、例えば超突然変異娘細胞集団などの娘集団が、参照細胞集団、例えば、ミスマッチ修復が阻害されていない親細胞集団などの親細胞集団よりも大きな増殖率を示す状態、または所定の集団が同じ培養条件(例えば、温度、培養培地、培養期間の長さ、湿度、CO2、O2等)下で参照細胞集団よりも速い倍加時間を示す状態のことをいう。「増殖率」は一般的に、第一時点で決定された細胞コロニーの面積を第二時点での細胞コロニーの面積で割り、得られた数を2つのそれぞれの時点の間の時間差で割ることによって算出できる。例えば、増殖率は14日目の代表的なクローンの面積に対する17日目のコロニーの面積を3日で割ることによって決定されてもよい(増殖率=17日目面積/14日目面積÷3日)。
【0051】
本明細書で使用される場合、用語「増大された高密度生成」または「高細胞密度での増大された生体分子生成」とは、所定の細胞集団、例えば、超突然変異娘細胞集団などの娘集団が、同じ培養条件(例えば、温度、培養培地、培養期間の長さ、湿度、CO2、O2等)下で、参照細胞集団、例えばミスマッチ修復が阻害されていない親細胞集団などの親細胞集団よりも大きな生体分子(例えば、抗体)生成を示す状態のことをいう。
【0052】
本明細書で使用される場合、「高細胞密度での増大された細胞増殖」とは、所定の細胞集団、例えば超突然変異娘細胞集団などの娘細胞集団が、同じ培養条件(例えば、温度、培養培地、培養期間の長さ、湿度、CO2、O2等)下で、参照細胞集団、例えばミスマッチ修復が阻害されていない親細胞集団などの親細胞集団と比べて細胞数を測定した場合に増大したレベルの増殖を示す状態のことをいう。
【0053】
本明細書で使用される場合、「高細胞密度での増大された細胞生存能力」とは、所定の細胞集団、例えば超突然変異娘細胞集団などの娘細胞集団が、同じ培養条件(例えば、温度、培養培地、培養期間の長さ、湿度、CO2、O2等)下で、参照集団、例えばミスマッチ修復が阻害されていない親細胞集団などの親細胞集団と比べて増大した数の生存細胞を示す状態のことをいう。
【0054】
本明細書で使用される場合、「高細胞密度」とは、約1.0×106細胞/mLよりも大きい細胞濃度のことをいう。「低細胞密度」とは、約1.0×106細胞/mL未満の細胞濃度のことをいう。
【0055】
本明細書で使用される場合、「取り除かれる(cured)」とは、ミスマッチ修復のタンパク質インヒビターが細胞から排除されているか、またはミスマッチ修復のタンパク質インヒビターの発現が消失されるまたはノックアウトされる細胞の状態のことをいい、安定化されたゲノムがもたらされ、免疫グロブリンなどの安定な生物学的生成物が生成される。同様に、ミスマッチ修復のインヒビターの「不活性化」とは、細胞からのインヒビターの排除もしくは除去、またはミスマッチ修復のタンパク質インヒビターの不連続な発現(例えば、インヒビターの発現を消失もしくはノックアウトする)のことをいい、安定化されたゲノムがもたらされ、免疫グロブリンなどの安定な生体分子が生成される。
【0056】
本明細書で使用される場合、「スクリーニング」とは、限定されないが、核酸配列、タンパク質配列、タンパク質機能(例えば、結合、酵素活性、阻害活性、交差阻害活性、中和活性など)を含む細胞の遺伝型もしくは表現型または細胞生成物を評価するアッセイのことをいう。該アッセイとしては、ELISA系アッセイ、Biacore分析などが挙げられる。
【0057】
本明細書で使用される場合、「単離される」とは、天然環境の成分から分離および/または回収された核酸またはタンパク質のことをいう。天然環境の夾雑成分は、ポリペプチドの診断使用または治療使用に干渉する物質であり、酵素、ホルモン、および他のタンパク質溶質または非タンパク質溶質を挙げることができる。いくつかの態様において、核酸またはタンパク質は、タンパク質の95重量%よりも高く精製される。他の態様において、核酸またはタンパク質は、タンパク質の99重量%よりも高く精製される。タンパク質精製の測定は、Lowry法などの当該技術分野で公知の任意の手段、クーマシーブルーまたは銀染色などの染色を用いた還元または非還元条件下でのSDS-PAGEによるものであってもよい。核酸の精製は限定されないが、分光法、例えばメチレンブルーのような蛍光染色または化学染色を用いたアガロースまたはポリアクリルアミド分離を含む、任意の公知の方法によって評価されてもよい。
【0058】
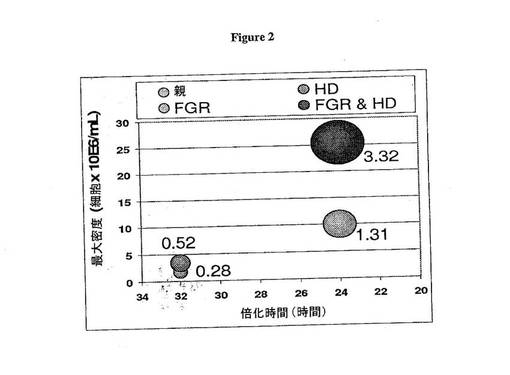
哺乳動物細胞株による抗体生成の生産性を増進するために細胞宿主性能を改善する方法が本明細書に提供される。ミスマッチ修復が生成細胞株で阻害され、増殖特性に影響を及ぼす細胞性プロセスを改善して、改善された製造収量をもたらすことができる。例えば、細胞は、速い増殖、高細胞密度での増殖および/または細胞生存能力、および/または長い発酵運転のための高密度での生産性を維持する能力を遺伝的に高めることができる。いかなる理論にも拘束されることを意図しないが、これらの増大は、増殖および生存を阻害する代謝副産物の蓄積を検出する増殖因子または生化学レセプターの構造変化を介して生じることが考えられる。図2は、速い増殖率(FGR)の細胞および/または規模拡大可能な製造のための全体的生産性を高密度(HD)で増殖および生成する細胞による、増大された増殖の劇的な効果のモデルを提供する。
【0059】
増殖率、高細胞密度での細胞増殖および細胞生存能力、および/または生成細胞株、好ましくは哺乳動物生成細胞株の高細胞密度での生産性を含む増殖特性を改善するために本明細書に提供される方法は、適時でかつ費用が効果的である様式で、モノクローナル抗体(mAb)の高い収量を生み出す能力を細胞に付与する。スケールでの全体的生成を改善するための増大された細胞増殖パラメーターを有する細胞が提供される。本明細書に記載される方法は、組み換え哺乳動物細胞の増殖率を改善するが、特異的な生産性にはほとんど影響を与えない。
【0060】
速い増殖率、高細胞密度での増大された細胞増殖および細胞生存能力、および高細胞密度での増大された生体分子生成の少なくとも1つの増殖特性を含む、改善された増殖特性を有する細胞を生成する方法が本明細書に提供される。いくつかの好ましい態様において、改善された増殖特性としては、速い増殖率および高細胞密度での増大された生体分子生成;速い増殖率および高細胞密度での増大された細胞増殖および細胞生存能力;高細胞密度での増大された細胞増殖および細胞生存能力ならびに高細胞密度での増大された生体分子生成;または速い増殖率、高細胞密度での増大された細胞増殖および細胞生存能力、ならびに高細胞密度での増大された生体分子生成が挙げられる。いくつかの態様において、細胞は、高アフィニティー抗体の生成および高いタイター抗体の生成の少なくとも1つの特性をさらに示す。改善された増殖特性を有する細胞を生成する方法は、本明細書に教示される全ゲノム進化技術の適用を含む。全ゲノム進化技術の利点は、増殖特性、規模拡大可能な製造のためのタイター収量またはその両方をさらに最適化する必要がある以下のもの、細胞株、例えば、抗体を発現する細胞株(即ち、生成細胞株)、好ましくは哺乳動物細胞株、より好ましくは抗体を発現する哺乳動物細胞株(即ち、哺乳動物生成細胞株または哺乳動物製造細胞株)に、該技術を直接適用できるということである。本発明は、製造のための増大された増殖の抗体生成細胞株の生成を容易にする。全ゲノム進化技術は、適時で規模拡大可能なGMP製造のための十分な抗体量を生成する細胞株に適用されてもよい。全ゲノム進化技術は、微生物、植物および哺乳動物細胞系システムでの高い程度のMMRの保存および機能のために、任意の種の細胞の宿主増殖パラメーターを最適化するために利用することができる。
【0061】
改善された増殖特性を有する遺伝的に改変された宿主細胞を生成する方法は、生成物開発のための細胞宿主を作製するための価値のある方法を提供し、改変が増大された細胞増殖特性となる場合に構造または発現レベルを改変する下流遺伝子の発見に有用な試薬の生成を可能にする。本明細書に記載される本発明は、MMRの阻害を介して増大された増殖特性を有する遺伝的に改変された細胞宿主の作製に関し、次に増大された増殖株の指向された改変および生成について改変された遺伝子座をスクリーニングおよび同定するために使用することができる。
【0062】
哺乳動物細胞における遺伝的進化を増大するために、MMRが宿主細胞で抑制される。MMRは、好ましくはMMRのタンパク質インヒビターの導入(例えば、MMR阻害タンパク質をコードする発現ベクターの導入)、または化学MMRインヒビターへの宿主細胞の曝露(例えば、MMRの化学インヒビターの存在下で細胞のインキュベート)によって抑制される(8、9)。両方の方法は、MMRを阻害する点および遺伝的に進化した子孫細胞となる点で有効であることが示され、交換可能に使用されている(L. Grasso, 個人的意見)。
【0063】
宿主細胞(即ち、親細胞)は真核生物または原核生物であってもよい。好ましくは、宿主細胞は、哺乳動物細胞、より好ましくはヒト細胞またはげっ歯類(例えば、マウス、ハムスター)細胞である。本発明の方法の特定の態様における使用に適切な哺乳動物細胞としては、限定されないが、sp20細胞、NS0細胞、チャイニーズハムスター卵巣細胞(CHO細胞(28))、胎児ハムスター腎臓(BHK細胞)、ヒト胚腎臓株293(HeLa細胞(29))、正常イヌ腎臓細胞株(例えば、MDCK、ATCC CCL 34)、正常ネコ腎臓細胞株(CRFK細胞)、サル腎臓細胞(CV1 ATCC CCL 70);アフリカミドリサル腎臓細胞(VERO-76、ATCC CRL-1587)、COS(例えば、COS-7)細胞、および非腫瘍性マウス筋芽G8細胞(例えば、ATCC CRL 1246)、線維芽細胞株(例えば、ヒト、マウスまたはニワトリ胚性線維芽細胞株)、ミエローマ細胞株、マウスNIH/3T3細胞、LMTK31細胞、マウスセルトリ細胞(TM4、(30));ヒト頚部癌細胞(HELA、ATCC CCL2);バッファローラット肝臓細胞(BRL 3A、ATCC CRL 1442);ヒト肺細胞(W138、ATCC CCL 75);ヒト肝臓細胞(Hep G2、HB8065);マウス乳腺癌細胞(MMT 060562、ATCC CCL51)、TRI細胞(31);MRC5細胞;FS4細胞;ならびにヒト肝癌株(Hep G2)が挙げられる。
【0064】
哺乳動物発現細胞に代わるものとして、他の非哺乳動物細胞が本発明の方法に使用されてもよい。かかる非哺乳動物細胞としては、限定されないが、細菌細胞、酵母細胞、植物細胞、両生類細胞、および昆虫細胞(例えば、Spodoptera frugiperda細胞等)を含む真核生物細胞および原核生物細胞が挙げられる。ベクターおよび非哺乳動物宿主細胞が当該分野で周知であり、継続して最適化され、開発されている。抗体を発現できる任意の宿主細胞系は、本発明の方法に使用されてもよい。
【0065】
宿主細胞は、好ましくは抗体生成細胞である。例えば、宿主細胞はハイブリドーマ細胞であってもよい。
【0066】
細胞はミスマッチ修復(MMR)のタンパク質インヒビターの導入によって超突然変異可能にされてもよい。例えば、タンパク質インヒビターは、MMR遺伝子のドミナントネガティブ対立遺伝子を含むポリヌクレオチドによってコードされてもよい。MMRのタンパク質インヒビターをコードする核酸は、細胞、例えば、発現細胞、ハイブリドーマ細胞(即ち、免疫グロブリン生成細胞のミエローマ細胞との融合後)、またはミエローマ細胞(即ち、融合前に細胞に導入されてもよい)に導入されてもよい。
【0067】
MMRのタンパク質インヒビターをコードする核酸は、ゲノムDNA、cDNA、RNA、または化学合成ポリヌクレオチドであってもよい。ポリヌクレオチドは、構成的に活性なプロモーターセグメント(例えば、限定されないが、CMV、SV40、EF-1 DまたはLTR配列)またはテトラサイクリン、もしくはエクダイソン/グルココルチコイド誘導ベクターに由来するものなどの誘導プロモーター配列を含む発現ベクターにクローニングすることができ、インヒビターの発現を調節することができる。ポリヌクレオチドは、トランスフェクションによって細胞に導入することができる。
【0068】
トランスフェクションは、ポリヌクレオチドが細胞に導入される任意の方法である。トランスフェクションの方法は、例えば、培養において1つ以上の単離細胞の懸濁液を用いてインビトロで行うことができる。細胞はモノクローナル抗体の生成のためのハイブリドーマを作製するために使用される任意の不死細胞であってもよく、または細胞はハイブリドーマ自体であってもよい。ハイブリドーマは、ヘテロハイブリドーマ細胞(例えば、ヒト-マウス細胞融合物)またはホモハイブリドーマ細胞(例えば、ヒト-ヒトハイブリドーマ細胞およびマウス-マウスハイブリドーマ細胞)であってもよい。
【0069】
一般的に、トランスフェクションは、複数の細胞または単一の細胞の懸濁液を用いて行うが、他の方法も、トランスフェクトされた細胞が増殖および利用できるように処理された細胞または組織の十分な部分がポリヌクレオチドを取り込む限り、利用することができる。ポリヌクレオチドのタンパク質生成物は、細胞で一過的または安定的に発現されてもよい。トランスフェクションの技術は周知である。ポリヌクレオチドを導入するために利用可能な技術は、限定されないが、エレクトロポレーション、形質転換、細胞融合、塩化カルシウムの使用、目的の細胞との融合のための脂質と共にポリヌクレオチドのパッケージングを含む。細胞がミスマッチ修復のタンパク質インヒビターをコードする核酸でトランスフェクトされると、細胞は培養中に増殖でき、再生産できる。ミスマッチ修復のタンパク質インヒビターをコードする核酸が多くの細胞世代について一定レベルで発現されるように、トランスフェクションが安定である場合に、細胞株が得られる。
【0070】
ミスマッチ修復のドミナントネガティブタンパク質インヒビターをコードする核酸は、限定されないが、それぞれが参照によって本明細書に援用される米国特許第6,146,894号および第6,808,894号および米国出願公開第20040115695号および第20050048621号に記載されるように、PMS2、PMS2-134、PMS1、PMSR3、PMSR2、PMSR6、MLH1、GTBP、MSH3、MSH2、MLH3、またはMSH1、およびPMSR遺伝子のホモログを含む任意の公知のミスマッチ修復遺伝子に由来してもよい;Nicolaides et al.(32)およびHorii et al(33)。かかる効果を生じる任意の対立遺伝子が本発明に使用できる。ミスマッチ修復のドミナントネガティブタンパク質インヒビターは、ヒト、動物、酵母、細菌、または他の生物の細胞から得ることができる。ミスマッチ修復のタンパク質インヒビターの非限定例は、PMS2の最初の133アミノ酸を有するPMS2-134である。本明細書に使用されるPMS2およびPMS2-134としては、ヒトPMS2およびPMS2-134およびそれらの種(例えば、マウス;シロイヌナズナ等の植物;等)の同等物が挙げられる。PMS2タンパク質のC末端の欠失は、PMS2のMLH1との結合に干渉すると考えられている。ミスマッチ修復を阻害するmutLホモログのアミノ酸のさらなる記述によって、共通のアミノ酸配列
またはその断片が明らかにされる。従って、ミスマッチ修復のタンパク質インヒビターとしては、配列番号1または2のアミノ酸配列を含むポリペプチドおよびその断片が挙げられる。
【0071】
細胞をミスマッチ修復活性の欠損についてスクリーニングすることによって、ミスマッチ修復のさらなるタンパク質インヒビターを同定することができる。癌を有する動物またはヒト由来の細胞を欠損ミスマッチ修復についてスクリーニングすることができる。結腸癌患者由来の細胞が、特に有用であってもよい。ミスマッチ修復のタンパク質インヒビターをコードする任意の細胞由来のゲノムDNA、cDNA、またはmRNAは、野生型配列からの変異について分析することができる。ミスマッチ修復遺伝子のドミナントネガティブ対立遺伝子はまた、例えば、hPMS2-134対立遺伝子のバリアントまたは他のミスマッチ修復遺伝子を生成することによって人工的に作製することができる。部位特異的突然変異生成の様々な技術を使用することができる。天然または人工的であろうと、超突然変異可能な細胞または動物の生成における使用のためのかかる対立遺伝子の適切さが、1つ以上の野生型対立遺伝子の存在下での対立遺伝子によって生じたミスマッチ修復活性の試験によって評価され、ドミナントネガティブ対立遺伝子であるかどうかを決定することができる。
【0072】
ミスマッチ修復のドミナントネガティブタンパク質インヒビターによって、DNA修復の有効性の低減による自発的変異の割合が増大され、それによって細胞または動物が超突然変異可能になる。これは、かかる細胞または動物の自発性変異の割合がかかる対立遺伝子がない細胞または動物と比較して上昇することを意味する。自発的変異の割合の上昇程度は、正常細胞または正常動物の割合よりも少なくとも2倍、5倍、10倍、20倍、50倍、100倍、200倍、500倍または1000倍であってもよい。超突然変異可能な細胞によって、遺伝子に新しい変異が蓄積され、新しい出力特性が生じる。細胞は所望の特性についてスクリーニングされてもよく、これらの特性を生じる細胞株が拡大されてもよい。さらに、細胞はミスマッチ修復欠陥について修復されてもよく、遺伝的に安定な細胞がもたらされる。好ましい態様において、ミスマッチ修復のタンパク質インヒビターが不活性化される。例えば、ミスマッチ修復のタンパク質インヒビターは所望の増殖特性を有する細胞の同定の前または後に不活性化されてもよい。ミスマッチ修復のタンパク質インヒビターの不活性化は、当該分野で公知の任意の手段、例えば、誘導剤の除去または細胞からのミスマッチ修復のタンパク質インヒビターの除去(即ち、ミスマッチ修復のタンパク質インヒビターから細胞を取り除くこと)によるものであってもよい。ミスマッチ修復のインヒビター不活性化は超突然変異細胞のゲノムを安定にする。
【0073】
本発明の別の局面は、MMRを欠く細胞(内在性ミスマッチ修復遺伝子の欠失、またはMMRのドミナントネガティブタンパク質インヒビターの導入のいずれかによる)および宿主ゲノムにおける増大した変異割合を生じる化学突然変異原の使用である。MMR活性の欠失によって、細胞はDNA損傷剤の毒性効果に耐性になることが分かっている。本発明は、MMRのドミナントネガティブタンパク質インヒビターの発現を介して熟練MMR細胞をミスマッチ修復欠陥にすること、DNA突然変異薬剤の使用によってゲノム超突然変異性を増大することを含む。化学突然変異剤、例えば、アルキル化剤、架橋剤等は化学特性によって分類可能である。本発明によれば、本明細書に他のものを記載しない場合に、以下の化学突然変異剤が有用であり、本発明の方法の態様のいずれかにおける変異割合をさらに増大するために使用されてもよい:N-エチル-N-ニトロソ尿素(ENU)、N-メチル-N-ニトロソ尿素(MNU)、プロカルバジン塩酸、クロラムブシル、シクロホスファミド、メチルメタンスルホネート(MMS)、エチルメタンスルホネート(EMS)、ジエチルスルフェート、アクリルアミドモノマー、トリエチレンメラミン(TEM)、メルファラン、窒素イペリット、ビンクリスチン、ジメチルニトロソアミン、N-メチル-N'-ニトロ-ニトロソグアニジン(MNNG)、7,12ジメチルベンズアントラセン(DMBA)、エチレンオキシド、ヘキサメチルホスホルアミド、ビスルファン。本発明の好ましい局面において、100遺伝子ごとに1つの変異;1000遺伝子あたり1つの変異の範囲における変異割合を付与する突然変異生成技術が使用される。かかる組み合わせ(MMR欠乏および化学突然変異剤)の使用は、各特定の薬剤によって優先的に誘導される多様なゲノム改変(限定されないが、遺伝子コーディング領域、遺伝子イントロン領域、または5'もしくは3'近位および/または遠位領域の背景でのDNAセグメントの拡張または欠失、点変異、改変された繰り返し配列)の生成を可能にする。
【0074】
変異は、細胞または動物の遺伝型における改変を分析することによって、例えば、目的の遺伝子と関連するゲノムDNA、cDNA、メッセンジャーRNA、またはアミノ酸の配列を調べることによって、検出することができる。また、変異は遺伝子の表現型をスクリーニングすることによって検出することができる。改変された表現型は、電気泳動移動度、分光学的特性、または変異遺伝子によってコードされるタンパク質の他の物理的または構造的特性における改変を同定することによって検出できる。また、インサイチュで、単離形態で、またはモデル系でタンパク質の改変された機能をスクリーニングすることができる。限定されないが、タンパク質分泌、化学耐性、病原耐性等を測定すること等の、目的の遺伝子の機能と関連する細胞または動物の任意の特性の改変をスクリーニングすることができる。
【0075】
本発明の改善された増殖特性を有する細胞を生成する方法のいくつかの態様において、細胞がミスマッチ修復の化学インヒビターに曝露される。本発明の方法の特定態様に使用されるミスマッチ修復の化学インヒビターとしては、限定されないが、アントラセン、ATPaseインヒビター、ヌクレアーゼインヒビター、RNA干渉分子、ポリメラーゼインヒビターおよびミスマッチ修復タンパク質をコードするヌクレオチドに特異的にハイブリダイズするアンチセンスオリゴヌクレオチドの少なくとも1つが挙げられる(WO2004/046330)。好ましい態様において、化学インヒビターは、以下の式:
(式中、R1〜R10は独立して、水素、ヒドロキシル、アミノ、アルキル、置換アルキル、アルケニル、置換アルケニル、アルキニル、置換アルキニル、O-アルキル、S-アルキル、N−アルキル、O-アルケニル、S-アルケニル、N-アルケニル、O-アルキニル、S-アルキニル、N-アルキニル、アリール、置換アリール、アリールオキシ、置換アリールオキシ、ヘテロアリール、置換へテロアリール、アラルキルオキシ、アリールアルキル、アルキルアリール、アルキルアリールオキシ、アリールスルホニル、アルキルスルホニル、アルコキシカルボニル、アリールオキシカルボニル、グアニジノ、カルボキシ、アルコール、アミノ酸、スルホネート、アルキルスルホネート、CN、NO2、アルデヒド基、エステル、エーテル、環状エーテル、ケトン、オルガノ硫黄化合物、オルガノ金属基、カルボン酸、オルガノケイ素、または1つ以上のアルキル化ヒドロキシル基を任意に含む炭水化物である;へテロアルキル、ヘテロアリール、および置換へテロアリールは、酸素、硫黄、金属原子、リン、ケイ素または窒素である少なくとも1つのヘテロ原子を含む;置換アルキル、置換アルケニル、置換アルキニル、置換アリール、および置換へテロアリールの置換基は、ハロゲン、CN、NO2、低級アルキル、アリール、ヘテロアリール、アラルキル、アラルコキシ、グアニジノ、アルコキシカルボニル、アルコキシ、ヒドロキシ、カルボキシおよびアミノである;アミノ基はアシル基、または1〜3のアリールまたは低級アルキル基で任意に置換される)
を有するアントラセン化合物である。特定の態様において、R5およびR6は水素である。他の態様において、R1〜R10は独立して、水素、ヒドロキシル、メチル、エチル、プロピル、イソプロピル、ブチル、イソブチル、フェニル、トリル、ヒドロキシメチル、ヒドロキシプロピル、またはヒドロキシブチルである。アントラセンの非限定例としては、1,2-ジメチルアントラセン、9,10-ジメチルアントラセン、7,8-ジメチルアントラセン、9,10-ジフェニルアントラセン、9,10-ジヒドロキシメチルアントラセン、9-ヒドロキシメチル-10-メチルアントラセン、ジメチルアントラセン-1,2-ジオール、9-ヒドロキシメチル-10-メチルアントラセン-1,2-ジオール、9-ヒドロキシメチル-10-メチルアントラセン-3,4-ジオール、および9,10-ジ-m-トリルアントラセンが挙げられる。
【0076】
化学インヒビターは、細胞の増殖培地に導入されてもよい。いくつかの態様において、化学インヒビターは、超突然変異細胞から回収されるか、または細胞が、細胞のゲノムを再安定化するために化学インヒビターから除去される。あるいは、該方法は、ミスマッチ修復の化学インヒビターの不活性化を含み、それによって超突然変異細胞のゲノムを安定化してもよい。
【0077】
MMR抑制細胞はインキュベートされて突然変異生成を可能にする。これによって、超突然変異娘細胞が生じる。好ましくは、MMR抑制細胞は、少なくとも約20世代、より好ましくは少なくとも約30世代継代され、その時にゲノムワイド変異が娘細胞に蓄積する。
【0078】
細胞の遺伝的に多様なプールは、好ましくは当該技術分野の公知の任意の方法によってサブクローニングされる。例えば、超突然変異娘細胞は、限界希釈でサブクローニングされた単一細胞であってもよく、拡大されてもよい。クローンは好ましくは、2週間まで拡大される。
【0079】
親細胞株と比べて1つ以上の改善された増殖特性(例えば、親細胞集団よりも速い増殖率(FGR)または高密度(HD)での生成)を示す娘細胞が次に同定される。いくつかの好ましい態様において、改善された増殖特性としては、速い増殖率と高密度での生成の両方が挙げられる。いくつかの態様において、細胞は、高い親和性の抗体生成および高いタイターの抗体生成の少なくとも1つの特性をさらに示す。例えば、いくつかの態様において、速い増殖(FGR)および高い抗体タイターを示す細胞が検出される。高い特異的な生産性を有する速い増殖率の細胞を生じる能力によって、多くの細胞種類の抗体製造の全体的生成における劇的な増大をもたらすことができる(図2)。
【0080】
好ましくは、改善された増殖特性を有する細胞の同定には、ハイスループットスクリーニングが含まれる。いくつかの態様において、様々な機能的で自動化ハイスループットスクリーニング(HTS)アッセイが実施され、少なくとも1つの改善された増殖特性(例えば、増大した薬理学的活性でタンパク質を生じる細胞;増大されたタイター収量を有する細胞;速い増殖率の細胞;高細胞密度での増大された増殖および生存能力を有する細胞;高細胞密度での増大された生体分子生成を有する細胞)を有するサブクローンを同定する。かかる細胞は改善された規模拡大可能な製造に適切であってもよい。
【0081】
速い増殖率の娘細胞の検出は、当該技術分野で公知の任意の方法、例えば、限定されないが、細胞カウント、および親細胞の集団に対する娘細胞の集団の比較によるものであってもよい。いくつかの態様において、親細胞よりも速い増殖率を有する速い増殖率の娘細胞の検出は、同じ培養条件下で同等の長さの培養時間を続けた後に、超突然変異娘細胞によって生じた細胞集団のサイズと、親細胞によって生じた細胞集団のサイズとを比較することを含み、親細胞集団に対する大きな娘細胞集団は、速い増殖率の娘細胞を示す。いくつかの態様において、速い増殖率の娘細胞の検出は、親細胞の増殖率よりも大きい増殖率を有する娘細胞の同定を含む。また、高細胞密度での増大された生体分子生成を示す細胞の検出は、当該技術分野で公知の任意の手段、例えば、限定されないが、同じ培養条件下で同等の長さの培養時間を続けた後に、娘細胞集団の細胞密度と、親細胞集団の細胞密度とを比較することであってもよく、高い細胞密度の娘細胞集団がHD細胞株を示す。
【0082】
細胞集団のサイズは、当該技術分野で公知の任意の方法によって決定されてもよい。細胞集団のサイズは好ましくは、光学画像化システム、例えば、MetaMorph(登録商標)システム(Molecular Devices Corp.)を用いて決定される。
【0083】
MMRが所望の表現型を示すサブクローンに保存される。遺伝的安定性は、以前に記載されたようにMMRインヒビターを除去することによって保存されてもよい(8、9)。例えば、細胞は、MMRの化学インヒビターを含む培養培地から除去されてもよい。あるいは、化学インヒビターは、細胞から回収されてもよい。いくつかの態様において、MMRのタンパク質インヒビターが不活性化されるか、または細胞が取り除かれる。遺伝的安定性は、所望の表現型を示す娘細胞の検出の前、後、または同時に細胞に保存されてもよい。
【0084】
所望の表現型を示すMMR熟練サブクローンは、i)細胞または抗体の増大した特性の保存および安定;ii)野生型MMR活性の保存および宿主ゲノムの安定化;および/またはiii)タンパク質構造の完全性および機能を確認するために分析される。
【0085】
本発明の方法によって生成した細胞は、本発明の範囲に含まれる。
【0086】
本発明の方法によって生成した細胞を培養する工程、および該細胞または該細胞の培養培地から生体分子を単離する工程を含む生体分子を製造する方法がまた、本発明の範囲に含まれる。本明細書で使用される場合、用語「生体分子」とは、細胞によって生成される分子のことをいい、化学薬剤(例えば、フコシルまたはグリコシル部分)および/または生物学的薬剤(例えば、限定されないが抗体を含むタンパク質)を含んでいてもよい。本発明の方法によって生成した生体分子、ならびに該生体分子および薬学的に許容できる担体を含む医薬組成物は、同様に本発明の範囲に含まれる。
【0087】
全ゲノム進化技術法によって、最適化された細胞増殖またはタイター生成に関わる未知の遺伝子および経路に対する様々なゲノムツールおよびタンパク質ツールによって分析できる同遺伝子型細胞の開発が可能とされる。染色体不安定の結果として異数性を誘導する標準の化学突然変異薬剤(7)に比べて、全ゲノム進化技術法は、宿主細胞の染色体安定および長期生存能力をインタクトのままにすると同時にわずかな点変異となるために、同等な遺伝的アプローチが保証されることを可能にする。この特徴は、遺伝的不安定ゲノムおよび再発現表現型の結果の両方となる高い変異バックグラウンドおよび化学突然変異細胞で見られる変異「ホットスポット」を回避する。このアプローチはより強固な結果を生ずるだけでなく、増大した増殖および生成と関連する経路に関わる標的物を明確に同定することによって種々の遺伝子/タンパク質発見も容易にする。全ゲノム進化技術法が適用されて、改善された細胞の増殖特性に重要な遺伝子を同定する方法が、本発明の範囲に含まれる。該方法は、所望の表現型を有する本発明の方法によって生じた娘細胞のゲノムと、親細胞のゲノムとを比較する工程、および2つのゲノム間の遺伝的差異を検出する工程を含み、かかる遺伝的差異または変異の少なくとも1つを含む遺伝子は所望の表現型に重要な遺伝子である。例えば、全ゲノム進化技術由来mAb生成細胞は、同遺伝子型の親および高タイター全ゲノム進化技術由来子孫セット間での遺伝子発現のRNAマイクロアレイ分析を行うことによって高いタイター生産性に関わる経路を同定するために使用された(9)。
【0088】
高いタイターまたは速い増殖などの改善された増殖特性のための進化した経路の同定によって、多くの生成物の高い生成のための高性能細胞の直接操作が可能になる。これは、増大した特性を有する全ゲノム進化技術由来細胞における改変経路の発見、およびこれらの経路を親細胞骨格で細胞操作することによる変異表現型の繰り返しによって達成できる。例えば、本明細書に記載される方法によって同定された改善された増殖特性に重要な遺伝子が、当該技術分野で公知の方法によって、細胞、好ましくは生成細胞で組み換え発現することができる。親細胞株の改善された増殖特性に重要な遺伝子の組み換え発現(全ゲノム進化技術または全ゲノム進化技術からもたらされる指向された経路改変のいずれかによる)は、次に安定な生成細胞株を生成するために必要な時間を低減することによって治療への速度を加速できる。
【0089】
以下の実施例は本発明のいくつかの局面をより詳細に記載する。実施例は、限定されないが本明細書に記載される本発明の局面をさらに示すために提供される。
【実施例】
【0090】
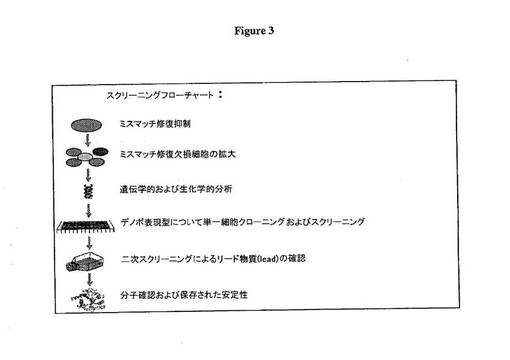
全ゲノム進化技術法は、図3に概略したように適用された。MMRはMMR化学インヒビター9,10-ジメチルアントラセンを用いて親細胞で抑制された。MMR抑制細胞株が増殖され、次にサブクローン化され、およそ10,000個の子孫細胞を生じ、低密度で細胞増殖をモニターできるカスタム化視覚プラットフォームおよびソフトウェアを用いて速い増殖率の表現型をスクリーニングした。
【0091】
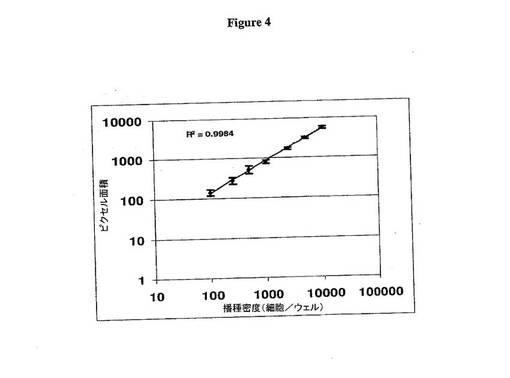
全ゲノム進化技術由来CHO-MAb細胞プールにおける速い増殖率(FGR)クローンをスクリーニングするために、新規画像ベースHTS法が開発された。この方法は、従来の時間消費細胞カウント法の要件を回避する。代わりに、これは、Meta 画像システム(Molecular Devices, Downingtown, PA)を使用し、該システムは、播種し、生育し、無菌条件下で全ゲノム進化技術を介して生じた子孫細胞クローンのコロニーサイズを分析するように設計されたORCA自動化プラットフォーム(Beckman Coulter, Fullerton, CA)に接続されたデジタルカメラを介して細胞コロニーサイズを画像化する。Metamorphソフトウェアパッケージ(Ver.6.3r0)を用いて、コロニー面積画像がピクセルで定量され、14日目と17日目のサイズを比較して、3日増殖期間中の細胞コロニーサイズの割合を計算するスプレッドシートに出力される。画像システムは、画像ピクセル面積と細胞数との間の線形関係を生じるように改良された(図4)。
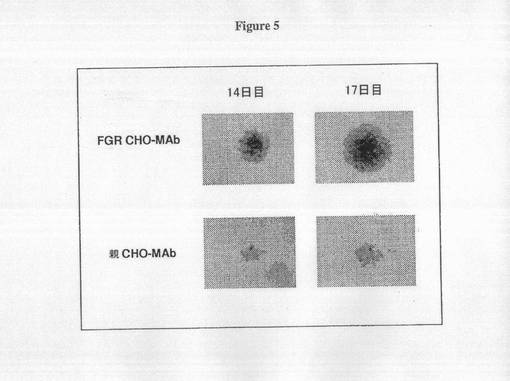
【0092】
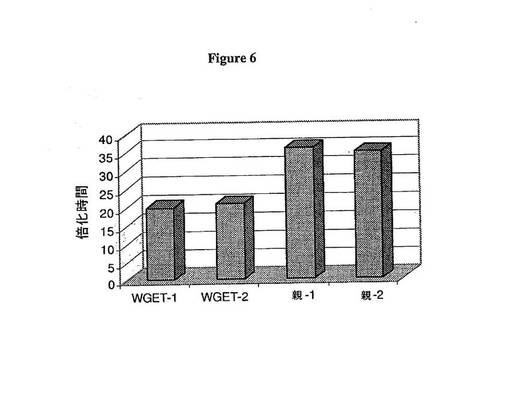
全ゲノム進化技術由来CHO-MAb細胞における速い増殖率の子孫を同定するために、細胞は、Biomek FX ロボットシステム(Beckman Coulter, Fullerton CA)を用いて100〜200バーコード丸底96ウェルプレートに0.8細胞/ウェルで播種され、ウェルあたりの単一細胞クローンを確実にする。画像化方法は、定量のために細胞を犠牲にする必要はなく、従って、レプリカプレートはこの手順を用いて必要とされない。クローナル播種の開始2週間後に、プレートは顕微鏡画像によって決定されるような、クローン増殖の高出力分析のためのORCAシステムによってスクリーニングされる。プレートは、重層インキュベーター内で生育され、コロニーの如何なる阻害もせずに画像解析後にインキュベーターに戻される。この工程は、3日間繰り返された後にコロニーの拡大サイズを決定して、次に14日目と17日目の時点間の細胞の増殖率を決定するために使用される。細胞増殖が17日目と14日目に測定されたコロニーサイズ値の間の率として、17日目のコロニー面積を14日目のコロニー面積で割って、これを3日で割ることによって算出される。図5は親またはFGR CHO-MAbクローンが14日目、次に再度17日目に画像化される典型的な分析を示す。予測されるように、FGR細胞は親CHO-MAb由来のクローンと比較して17日目と14日目との間の大きな率を示す。この細胞株を平均して、親クローンは、およそ20,000個の独立した親CHO-MAb由来サブクローンの一次スクリーニングから決定された約0.5の増殖率を示すが、FGRクローンは>0.8の増殖率を示す。標準として、各プレートは比較物として親CHO-MAb細胞由来のクローンを含む。図5は親株および全ゲノム進化技術由来細胞に由来するサブクローンの典型的なプロファイルを示す。全ゲノム進化技術由来CHO-MAb細胞において、スクリーニングされたウェルの約5%が0.8以上の増殖率を示す。これらのもののうち、このアッセイを用いた他の細胞株のスクリーニングと一致して拡大される場合、親株のものと比較して50%が改善された増殖率を有することが確認された。確認されたクローンは48時間定量増殖アッセイ中に3mL静止培養で増殖され、それによって細胞は、Cedex自動化細胞カウンターを用いて0日目、1日目および2日目に物理的にカウントされる。図5は代表的な48時間定量増殖アッセイ結果を示す。速い増殖率クローンを行う一番のものが20mL振とうフラスコアッセイでさらに拡大され、評価される。このレベルに達するクローンの大半は、典型的に速い増殖率を維持すると同時に高い抗体特異的生産性を保持する(図6)。
【0093】
速い増殖率サブクローンに由来する抗体の構造解析によって、これらの細胞によって生成される抗体は親抗体のものと類似する遺伝的特性および生化学特性を保持することが確認される。また、速い増殖率の細胞の拡大培養によって、全ゲノム進化技術によって媒介される増大した増殖率が安定であったこと、および3ヶ月間の細胞の全体タイターは親CHOおよびより速い増殖率由来細胞の両方で一定のままであったことが示される(図6)。
【0094】
【図面の簡単な説明】
【0095】
【図1】図1は哺乳動物細胞における抗体生成の経路を示す。発酵中にタンパク質生成および細胞増殖を調節する多くの経路が、哺乳動物細胞に存在する。これらの経路の1つ以上の改変は、宿主細胞株の生成性能および増殖性能を改善する遺伝子操作によって達成することができる。これらの経路は:(1)遺伝子転写;(2)mRNA安定化およびトランスロケーション;(3)タンパク質合成;(4)タンパク質翻訳後修飾および輸送;(5)小胞体(ER)でのフォールディング;(6)ゴルジでのタンパク質成熟およびグリコシル化;(7)分泌経路;および(8)細胞膜発現を含む。(GlcNAc=N-アセチルグルコサミン;P=リン酸化残基)。
【図2】図2は増大された増殖率および高細胞密度での安定性を有する細胞の生成効果を示す。10日間発酵運転は、拡大された期間で増大された増殖率または高密度での増殖能力を有する細胞をシミュレートした。モデルは、i)細胞密度が増大するとき、増殖率が減少する;ii)抗体の蓄積は細胞数およびpcdのみに依存する;およびiii)0日目での播種は500,000/mLであることを仮定する。親株の倍化時間は32時間であり、最大密度の1.7×106/mLは5日目で達成され、その特異的な生産性は25pg/細胞/日(pcd)である。これらの条件下で、親細胞株は発酵運転の完了時におよそ0.28g/Lを生じる。ここに示すように、球状サイズおよび1Lあたりの発現gに近い数によって表されるこのグラフでは、7日目で高細胞密度(HD)の3.4×106/mLに達する親株由来の細胞株は高い抗体タイターを有する。24時間対32時間の、速い増殖率(FGR)または速い倍化時間を有する別の株は、高い密度で増殖できる細胞によって達成されるものよりさらに高い収量を生じる。高密度増殖および増殖率(それぞれHDおよびFGR)における組み合わされた改善を示す細胞株はこのモデルで最良に実施され、3.32g/Lに達する。
【図3】図3は全ゲノム進化技術プロセスフローチャートを示す。
【図4】図4は顕微鏡画像解析を介した細胞計算を示す。CHO-MAb細胞は96ウェルU底面プレートにウェルあたり100〜10,000細胞の密度で播種され、画像プログラムの線形性を決定した。ウェルを、Meta画像システムを用いて20倍で画像化した。コロニーサイズを、Metamorphソフトウェアパッケージ(Ver.6.3r0)を用いて細胞によってカバーされたウェルのピクセル面積を統合することによって決定した。データ点は12ウェルの平均±標準偏差である。
【図5】図5は親CHO-MAbまたは全ゲノム進化技術由来クローンのコロニーサイズを比較する。親CHO-MAb細胞または全ゲノム進化技術CHO-MAb細胞は、96ウェルバーコードプレートにサブクローニングして37℃のCO2インキュベーターで12日間増殖させる。代表的なCHO-MAbコロニーは、コロニーのピクセル面積を計算するMetamorph画像ソフトウェアを用いて14日目に画像化され、次に17日目に再画像化される。増殖率は、14日目の代表的クローンのコロニー面積に対する17日目のコロニーの面蹟を3日で割ることで決定される(増殖率=17日目面蹟/14日目面蹟÷3日)。CHO-MAb株の親細胞は約0.5の増殖率を有する。>0.8の増殖率を示す細胞を拡大し、標準化増殖アッセイで分析する。固有の緩やかな増殖率の親細胞は典型的に、予測される速い増殖を有するように進化したクローンに比べて、14日目の小さなコロニーとなる。
【図6】図6は、速く増殖するサブクローンを確認する代表的な増殖データを示す。速い増殖率の親CHO-MAb細胞を7日間の振とうフラスコアッセイで増殖させ、1、4および7日目に細胞を計算して増殖率を分析した。速い増殖率のサブセット(WGET-1およびWGET-2)および親細胞由来(親-1および親-2)CHO-MAbサブクローンに対する倍化時間の関数として増殖率を示す。
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、2006年4月17日に出願された米国出願番号第60/792,937号の35 U.S.C §119(e)下の利益を主張し、本明細書にその全体を参照によって援用される。
【0002】
(発明の分野)
本発明は、親細胞と比べて改善された増殖特性(例えば、速い増殖率、高細胞密度での増大された生体分子生成、および/または高細胞密度での増大された細胞増殖または細胞生存能力)を有する細胞の生成に関する。さらに、本発明は、規模拡大可能な製造における使用のための改善された増殖特性について細胞のスクリーニングおよび細胞ベース製造に関する増大された増殖に関わる経路の発見に関する。
【背景技術】
【0003】
(発明の背景)
治療のタンパク質およびモノクローナル抗体(mAb)は、疾患に関連する特定の標的を特異的に置換または阻害する能力のために過去何十年も最も首尾よい種類の医薬の1つになっている(1)。Remicade(登録商標)、Rituxan(登録商標)、Herceptin(登録商標)、Humira(登録商標)、およびAvastin(登録商標)などの市販mAbの首尾よい市場性能によって、モノクローナル抗体は最も重要な薬物種類の1つとして現れ、現在全ての新しい薬物出現物の約半分を示す。現在まで出現した全てのmAbのうち、40%は大成功薬物であるかまたは大成功収益を潜在的にする。治療mAbの世界売り上げは、2004年に100億ドルを超えており、2008年までに300億ドルを超えるように計画されている(Needham & Company, 2005)。次の5年で、mAb市場は、キメラ(一部ヒトおよび一部マウス)からヒト化(CDR移植)から完全ヒト抗体までの技術的な進化によって、バイオテクノロジーおよび医薬産業の、最速で成長し、かつ最も有利な部門であり続けるようである。従来の低分子アプローチと比較して、mAbは高い特異性、低い毒性、および治療までのより迅速な開発経路を提供する。標的に結合する際に機能を発揮する低分子と異なり、mAbは、標的特異的な細胞傷害活性を引き出すように、体の免疫系を利用することによって大きな治療インパクトを与えることができる。
【0004】
mAbの大きな成功にも関わらず、効率的な開発は全く課題のままである。治療mAbを開発する会社は、究極的に市場までの時間、有効性、安全性および商品費用に影響を及ぼす、首尾よいGMP製造、精製および製品プロセスの開発初期段階で、適切なシステムを提供しなければならない。治療mAbの製造は、治療のための高用量を支持するのに必要な年収量と関連する高い製造コストおよび付加製造能力に関連する高い資本コストのために課題のままである。時節にあわせたやり方で大量の製品を作り出すことができる製造システムが、市場での余剰の発酵能力の維持により達成可能な短い開発スケジュールおよび低い全体的商品コストのために必要とされる。
【0005】
mAb製造を専門とする会社は、高いmAb収量を生み出すことができるいくつかの細胞ベースシステムを開発した。これらのシステムは、細菌、酵母、植物および哺乳動物細胞を含む(2)。微生物ベースプラットフォームは、ある最高の全タイター収量となった;しかし、活性タンパク質の量は、適切なフォールディングまたはプロセシングのために生じた全タンパク質のうち一部分だけを典型的に示した。また、微生物システムは、翻訳後修飾などのさらなるプロセシングを要する、抗体を含む複数鎖巨大分子などの複雑なタンパク質を効率的に作り出すように首尾よく適合されていなかった。これらの微生物ベース製造の制限の点から、哺乳動物細胞は、治療抗体および非抗体タンパク質のサブセットを製造する、大規模の優良製造規則(GMP)のための最も信頼可能で広く使用されるシステムの1つとなっている。
【0006】
臨床物質および最終的に市場供給のための有意な製造能力を要する現在進行中の300の抗体開発プログラムがある(3)。これらのプログラムの大半は、製造のための流加または還流バイオリアクターを用いた現在標準のGMP哺乳動物細胞培養発酵法の下でチャイニーズハムスター卵巣(CHO)およびマウスミエローマ(NS0、SP2)細胞株のような組み換え哺乳動物細胞株を使用する(4)。細胞ベース製造能力への負担は生成物の増大が要求されるにつれて増大し続け、これらのmAbプログラムのそれぞれは、多くの臨床試行から最終的に市場へと進む。このリスクの点から、十分な製造能力が市場に存在し続けること、および全体的商品コストが今日の市場の現在コストと類似のままか、または低いままであることを確実にするように代替的手順が要求される。
【0007】
様々なアプローチが、全体的生成および精製収量の改善を含む哺乳動物細胞からの抗体の大規模製造のための将来の必要性ならびにトランスジェニック動物ベース戦略のような代替的製造供給源の使用を支持するように実施されている(5、6)。代替的宿主システムは費用効果規模可能性および規制コンプライアンスについて確認されているが、当該技術分野の水準は哺乳動物細胞培養のままである。哺乳動物細胞における抗体生成の収率を改善する可能性が、1)培養条件および/または培地最適化によるバイオリアクター性能の改善;2)組み込んでいる非常に活性なプロモーターによる改善されたベクター発現または増幅による増大ベクターコピー数;および/または3)大規模バイオリアクターにおける良好なタイター収量および改善された細胞増殖を提供する宿主細胞内の内在性経路の増進による細胞宿主の最適化によって達成することができる。これらの改善またはこれらの組み合わせのいずれかが年間製品要求を作り出すのに必要な製造運転数を短くするプロセスとなり、それによって市場での全体製造制約を軽減することができる。
【0008】
細胞宿主最適化は、a)mRNA転写および成熟、b)タンパク質合成および翻訳後修飾、c)タンパク質分泌および細胞副次局在(sub-localization)、d)細胞質と細胞小器官との間のタンパク質移動、ならびにe)細胞周期および生存調節を含む、内在性経路の操作によって達成することができる。図1に示すように、これらのプロセスの1つ以上の調節に関わるタンパク質の微妙な構造変化が生成細胞株の全体性能に直接または間接的に影響を及ぼすことがある。機能的な細胞表現型をスクリーニングできるランダム化ゲノムワイド突然変異生成アプローチは、増殖率、非常に高い細胞密度での生存、タンパク質合成および分泌速度を調節する複雑な細胞プロセスを増進するアプローチを提供する。ほとんどの細胞突然変異生成スキームでは不幸にも、突然変異薬剤の使用によって、GMP製造に適切でない不安定な細胞株を生じるゲノムワイド染色体不安定となる。
【0009】
以前の研究によって、ミスマッチ修復と呼ばれる複製後DNA修復機構の阻害はDNA複製中のDNAポリメラーゼによって組み込まれる点変異の増大のために安定な細胞内に遺伝的多様性をもたらし得ることが示された(7、8)。DNA複製は、親細胞が遺伝情報を子孫細胞に伝えるために全ての細胞が増殖中に受ける複雑なプロセスである。細胞がDNAを複製する時に、変異が、ポリメラーゼ忠実度を含む様々な機構から新しく合成された鋳型内に生じる。ミスマッチ修復(MMR)システム(11〜13)のような一連の複製後DNA修復プロセスは、自然で進化し、個体が遺伝型同一性を保持するために原核生物細胞および真核生物細胞にユビキタスに存在する。MMRは、親DNA鋳型と子孫DNA鋳型との間に存在する不一致な遺伝情報を訂正する二次プルーフリーディングプロセスを介した「天然」転移および転換変異の蓄積を妨げる。
【0010】
細胞集団内の遺伝的多様性を生じるミスマッチ修復を阻害するプロセスは、本明細書で「全ゲノム進化技術(whole genome evolution technology)」と呼ばれる。全ゲノム進化技術は、MMRの可逆阻害に基づいており、例えば新規治療抗体またはタンパク質を生じる細胞内でMMRを阻害することができるドミナントネガティブタンパク質インヒビターまたは化学インヒビターの活性によって媒介される。抑制されたMMRはDNA複製中に生じる変異のために子孫細胞のゲノムにおける点変異の遺伝となる(8、11、12)。細胞中のMMR抑制は、MMR熟練(proficient)細胞に典型的に観察されるものよりも高い頻度(1000倍の増大まで)で遺伝する天然変異を可能にする。全ゲノム進化技術に由来する子孫細胞の遺伝的に多様な集団は、新規な特徴を有するサブクローンを同定する自動化機能的ハイスループットスクリーニング(HTS)を介してスクリーニングすることができる細胞のライブラリーとなる。
【0011】
全ゲノム進化技術は、望ましい表現型を有する細胞の開発のための進化の力を利用する。この技術と他の進化ベース技術とを隔てる重要な区別は、ランダムなインビボ性質のプロセスである。全ての細胞が本来的に有する多くの遺伝子および経路が利用できることによって、機能的な細胞スクリーニングで同定される予測されない変異体の生成を可能にし、望ましい表現型を有する子孫細胞および遺伝子産物をもたらす。この技術は、全ゲノム進化を増大するようにインビボで既存生成株に適用され、それによってもはやインビトロ操作を要しないために、時間およびコストで効率的である。全ゲノム進化技術は、例えば、CHO、NSO、およびハイブリドーマ細胞において、増大されたタイター生成を有する進化した子孫細胞に由来する組み換え治療抗体を生成するいくつかの哺乳動物細胞株に首尾よく適用された(8〜10)。また、全ゲノム進化技術は改善された生物学的特性を有するタンパク質または抗体を生成する進化した子孫細胞に由来する組み換え治療抗体を生成する哺乳動物細胞株に適用されている(15〜17)。
【0012】
特に、哺乳動物細胞株による抗体生成のための生産性を増大するために細胞宿主性能を改善する方法が本明細書に提供される。ミスマッチ修復は、改善された抗体製造収量をもたらすことができる増殖特性に影響を与える細胞プロセスを改善するように、生成細胞株で阻害される。
【0013】
(発明の要旨)
本明細書に、細胞宿主性能を改善する方法、例えば、細胞株による生産性を増大する方法が提供される。本発明の方法は、(a)親細胞集団の親細胞のミスマッチ修復を阻害する工程;(b)MMR阻害親細胞をインキュベートまたは拡大して突然変異生成を可能にし、それによって超突然変異娘細胞を生成する工程;(c)改善された増殖特性を有する超突然変異娘細胞を検出する工程;および(d)改善された増殖特性を有する超突然変異娘細胞の遺伝的安定性を保存する工程を含む、親細胞集団と比べて少なくとも1つの改善された増殖特性を有する細胞を生成する方法を含む。
【0014】
いくつかの態様において、ミスマッチ修復を阻害する工程がミスマッチ修復の化学インヒビターまたはミスマッチ修復のタンパク質インヒビターに親細胞を曝露する工程を含む。
【0015】
本発明のいくつかの局面において、MMR阻害親細胞をインキュベートまたは拡大して突然変異生成を可能にし、それによって超突然変異娘細胞を生成する工程が好ましくは少なくとも20回の継代、より好ましくは少なくとも30回の継代でMMR阻害細胞を継代する工程を含む。
【0016】
本発明の方法において遺伝的安定性を保存する工程が好ましくは超突然変異娘細胞からミスマッチ修復の化学インヒビターを回収する工程、またはミスマッチ修復のタンパク質インヒビターを不活性化する工程を含む。遺伝的安定性の保存が改善された増殖特性を有する超突然変異娘細胞を検出する工程の前、後または同時に生じてもよい。
【0017】
本発明の方法において、改善された増殖特性を有する超突然変異娘細胞を検出する工程が好ましくは改善された増殖特性を有する超突然変異娘細胞のハイスループットスクリーニング(HTS)を含む。
【0018】
好ましい態様において、改善された増殖特性は、速い増殖率、高細胞密度での増大された細胞増殖または細胞生存能力、および/または高細胞密度での増大された生体分子生成である。いくつかの態様において、速い増殖率を有する超突然変異娘細胞の検出が、同じ培養条件下で同等の長さの培養時間を続けた後に、超突然変異娘細胞によって生じた細胞集団のサイズと、親細胞によって生じた細胞集団のサイズとを比較する工程を含み、大きな娘細胞集団が速い増殖率を示す。いくつかの態様において、速い増殖率を有する超突然変異娘細胞の検出が、親細胞の増殖率よりも大きな増殖率を有する娘細胞を同定する工程を含む。いくつかの態様において、高細胞密度での増大された細胞生存能力の検出は、同じ培養条件下で同等の長さの培養時間を続けた後に、超突然変異娘細胞によって生じた高密度での生存可能細胞集団のサイズと、親細胞によって生じた高密度での生存可能細胞集団のサイズとを比較する工程を含み、大きな生存可能娘細胞集団が高細胞密度での増大された細胞生存能力を示す。細胞集団(例えば、親細胞集団および/または娘細胞集団)のサイズが好ましくは光学画像システムを用いて決定される。いくつかの態様において、増大された生体分子生成を示す超突然変異娘細胞の検出は、同じ培養条件下で親細胞よりも娘細胞による生体分子の高い収量を決定する工程を含む。いくつかの態様において、親および娘細胞集団のサイズは、それぞれの各集団によって生成される生体分子の収量を決定する場合に同等である。
【0019】
本発明の方法における使用のための親細胞は、好ましくは抗体生成細胞である。いくつかの態様において、親細胞はハイブリドーマ細胞である。親細胞は真核生物細胞または原核生物細胞であってもよい。本発明における使用のための親細胞は、好ましくは哺乳動物細胞であり、より好ましくはヒトまたはげっ歯類(例えば、ハムスター、マウス)細胞であり、例えば、限定されないが、NS0、SP2、またはチャイニーズハムスター卵巣(CHO)細胞である。本発明の方法における使用のための親細胞は、細菌細胞、酵母細胞、植物細胞または両生類細胞であってもよい。
【0020】
また、本発明の方法によって生成した細胞、ならびに均一および不均一組成のかかる細胞が本発明の範囲に含まれる。
【0021】
本発明の細胞を培養し、細胞または細胞の培養培地から生体分子を単離することによって生体分子を製造する方法が本発明によってさらに提供される。生体分子は、化学薬剤および/または生物学的薬剤、例えば、限定されないが、タンパク質、例えば、抗体を含んでいてもよい。本発明の方法によって生成した生体分子およびその医薬組成物は同様に、本発明の範囲に含まれる。
【0022】
また、改善された増殖特性の原因遺伝子を同定する方法であって、本発明の方法によって生成した細胞のゲノムと、親細胞のゲノムとを比較して変異を同定する工程を含み、改善された増殖特性の原因遺伝子が少なくとも1つのかかる変異を含む方法が、本発明の範囲に含まれる。
【0023】
本発明のこれらおよび他の局面は以下に記載される1つ以上の態様によって提供される。
【0024】
(例示態様の詳細な説明)
本明細書で言及されるGenBankデータベース配列のアクセッション番号を含む、参考文献、特許、特許出願、および科学文献は、当業者の知識を確立し、あたかもそれぞれが特異的にかつ個別に参照によって援用されて示されるように同じ程度でその全体を参照によって本明細書に援用される。本明細書に引用される任意の参考文献と本明細書の具体的な教示との間に矛盾があれば、後者によって解決される。同様に、単語または句の当該技術で理解される定義と本明細書で具体的に教示される単語または句の定義との間に矛盾があれば、後者によって解決される。
【0025】
本明細書に引用されるそれぞれの範囲は、範囲の全ての組み合わせおよび下位の組み合わせ、ならびにそこに含まれる特定の数を含む。
【0026】
別に規定されない場合、本明細書に使用される全ての技術用語および科学用語は、発明の属する当業者によって一般に理解されるものと同じ意味を有する。本明細書に記載されるものと類似または同等の任意の方法および物質が本発明の試験の実施に使用できるが、好ましい物質および方法が本明細書に記載される。本発明を記載し、かつ特許請求する時に、以下の技術用語が使用される。
【0027】
本明細書および添付の特許請求の範囲に使用される場合、単数形の「a」、「an」および「the」は内容が明らかに他のものを示さない場合に複数の参照を含む。従って、例えば、「細胞(a cell)」への参照には、2つ以上の細胞の組み合わせが含まれる等である。
【0028】
量、時間継続期間等のような測定可能な値を参照する場合に本明細書で使用される用語「約」は、変化が開示された方法を行うために適切であるように、特定の値から、±20%または±10%、より好ましくは±5%、さらに好ましくは±1%、さらにより好ましくは±0.1%の変化を包含することを意味する。
【0029】
「保存的に改変(modified)されたバリアント」はアミノ酸配列および核酸配列の両方に適用する。特定の核酸配列に関して、保存的に改変されたバリアントは、同一のアミノ酸配列または本質的に同一のアミノ酸配列をコードする核酸、または核酸がアミノ酸配列をコードしない場合の本質的に同一の配列のことをいう。遺伝コードの縮重のために、多数の機能的に同一な核酸が任意の所定のタンパク質をコードする。例えば、コドンGCA、GCC、GCG、およびGCUの全てはアミノ酸アラニンをコードする。従って、アラニンがコドンで特定される全ての位置で、コドンは、コードされたポリペプチドを改変せずに、記載される対応のコドンのいずれかに改変することができる。かかる核酸変異は、「サイレント変異」であり、1種の保存的に改変された変異である。また、ポリペプチドをコードする本明細書の全ての核酸配列は、核酸の全て可能なサイレント変異を記載する。当業者は、核酸の各コドン(通常メチオニンの唯一のコドンであるAUG、および通常トリプトファンの唯一のコドンであるTGGを除く)が機能的に同一の分子を生じるように改変できることを認識する。従って、ポリペプチドをコードする核酸の各サイレント変異は、記載されるそれぞれの配列に示される。
【0030】
例えば、細胞、核酸、タンパク質、またはベクターを参照して使用される場合「組み換え」は、細胞、核酸、タンパク質またはベクターが異種の核酸もしくはタンパク質の導入またはネイティブの核酸もしくはタンパク質の改変によって改変されたこと、または細胞が上記改変された細胞に由来することを示す。従って、例えば、組み換え細胞は、ネイティブ(非組み換え)形式の細胞で見られない遺伝子を発現するか、または他の方法で異常発現し、過小発現(underexpress)するか、または全く発現しないネイティブ遺伝子を発現する。
【0031】
句「核酸」または「ポリヌクレオチド配列」とは、5’から3’末端に読まれるデオキシリボヌクレオチドまたはリボヌクレオチドの単鎖または二重鎖ポリマーのことをいう。また、核酸はポリメラーゼによる正確な読み通しを許容する改変ヌクレオチドを含むことができるが、例えば、保存的に改変されたバリアントを含む、該核酸によってコードされるポリペプチド発現を改変しない。
【0032】
「ポリペプチド」、「ペプチド」および「タンパク質」は、アミノ酸残基のポリマーを示すために本明細書で交換可能に使用される。該用語は、1つ以上のアミノ酸残基が対応する天然アミノ酸の人工化学模倣物であるアミノ酸ポリマー、ならびに天然アミノ酸ポリマーおよび非天然アミノ酸ポリマーに適用する。ポリペプチドは、保存的に改変されたバリアントを含む。当業者は、コードされる配列における単一アミノ酸または小さい割合のアミノ酸を改変、付加、または欠失する核酸配列、ペプチド配列、ポリペプチド配列、またはタンパク質配列に対する置換、欠失または付加が、改変がアミノ酸の化学的に類似するアミノ酸での置換となる場合の「保存的に改変されたバリアント」であることを認識する。機能的に類似するアミノ酸を提供する保存的置換表は、当該技術分野で周知である。かかる保存的に改変されたバリアントはまた、本発明の多形バリアント、種間ホモログ、および対立遺伝子であり、これらを排除しない。以下の8つのグループはそれぞれ、別の保存的置換であるアミノ酸を含む:1)アラニン(A)、グリシン(G);2)アスパラギン酸(D)、グルタミン酸(E);3)アスパラギン(N)、グルタミン(Q);4)アルギニン(R)、リジン(K);5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);7)セリン(S)、トレオニン(T);および8)システイン(C)、メチオニン(M)(33)。また、用語「保存的置換」は、かかるポリペプチドがまた必須の結合活性を示すことを条件として、非置換親アミノ酸の代わりに置換アミノ酸の使用を含む。
【0033】
「アミノ酸」とは、天然アミノ酸および合成アミノ酸、ならびに天然アミノ酸に類似する様式で機能するアミノ酸アナログおよびアミノ酸模倣物のことをいう。天然アミノ酸は、遺伝コードによってコードされるもの、および後に改変されるアミノ酸、例えば、ヒドロキシプロリン、γ-カルボキシグルタミン酸およびO-ホスホセリンである。「アミノ酸アナログ」とは、天然アミノ酸と同じ基本化学構造、即ち、水素、カルボキシル基、アミノ基、およびR基に結合するα炭素、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムを有する化合物のことをいう。かかるアナログは改変R基(例えば、ノルロイシン)または改変ペプチド骨格を有するが、天然アミノ酸と同じ基本化学構造を保持する。「アミノ酸模倣物」とは、アミノ酸の一般化学構造と異なるが天然アミノ酸と類似する様式で機能する構造を有する化学化合物のことをいう。
【0034】
アミノ酸は一般的に既知である3文字記号またはIUPAC-IUB 生化学命名委員会(Biochemical Nomenclature Commission)によって推奨された1文字記号のいずれかによって、本明細書に示すことができる(以下の表1を参照)。ヌクレオチドは、同様に、一般的に受け入れられている1文字コードによって参照できる。
【0035】
全てのアミノ酸配列は、左から右への向きが従来のアミノ末端からカルボキシ末端への方向である方式によって本明細書に表されるということに注意されたい。
【0036】
本明細書で使用する場合、用語「インビトロ」または「エキソビボ」とは、人工的環境、ならびに例えば、限定されないが、試験管および細胞培養の人工的環境で生じるプロセスまたは反応のことをいう。用語「インビボ」とは、天然環境(例えば、動物または細胞)および天然環境で生じるプロセスまたは反応のことをいう。
【0037】
「薬学的に許容できる」、「生理学的に寛容できる」およびそれらの文法的バリエーションは、組成物、担体、希釈剤および試薬を示す場合、交換可能に使用され、物質が組成物の投与を禁止する程度までの望ましくない生理学的効果を生成せずにヒトにまたはヒトで投与できることを表す。
【0038】
用語「薬学的に許容できる担体」とは、過剰な毒性、炎症、アレルギー反応、または他の合併症が無く、良さそうな医学判断の範囲内で、ヒトおよび動物の組織と接触して使用するために適切であり、合理的な利益/リスク比と釣り合う試薬、賦形剤、細胞、化合物、物質、組成物、および/または用量形態のことをいう。本明細書でより詳細に記載されるように、本発明における使用に適切な薬学的に許容できる担体としては、気体、液体、ならびに半固体および固体物質が挙げられる。
【0039】
「免疫グロブリン」または「抗体」は、抗体分子および様々な抗体由来分子の両方のことをいうために広く使用され、免疫系の主要成分である高等な哺乳動物で生じる糖タンパク質のグループの任意の一員を含む。用語「抗体」は最も広い意味で使用され、所望の生物学的活性(例えば、標的抗原への特異的結合)を示す限り、モノクローナル抗体、ポリエピトープ特異性を有する抗体組成物、二重特異的抗体、ダイアボディ(diabody)、および単鎖分子、ならびに抗体断片(例えば、Fab、F(ab')2、およびFv)を特異的に含む。免疫グロブリン分子は、それぞれが軽鎖および重鎖の末端部分を含む抗原結合ドメインを含み、補体結合のような様々な機能に必要なFc領域を含む。5種類の免疫グロブリンがあり、Fc領域における重鎖の一次構造が免疫グロブリンの種類を決定する。具体的に、α鎖、δ鎖、ε鎖、γ鎖、およびμ鎖は、それぞれIgA、IgD、IgE、IgG、およびIgMに対応する。本明細書で使用される場合、「免疫グロブリン」または「抗体」は全てのサブクラスのα、δ、ε、γ、およびμを含み、また4つの鎖の免疫グロブリン構造の任意の天然多量体(例えば、IgAおよびIgM)または合成多量体のことをいう。抗体は、非共有的に、特異的に、かつ可逆的に抗原と結合する。本明細書で使用される場合、用語「モノクローナル抗体」とは、実質的に均一な抗体の集団から得られた抗体のことをいい、即ち、該集団を構成する個々の抗体は少量で存在し得る可能な天然変異を除いて同一である。例えば、モノクローナル抗体は、単一クローンの抗体生成細胞によって生成されてもよい。ポリクローナル抗体と異なり、モノクローナル抗体は単一特異的である(例えば、単一抗原の単一エピトープに特異的)。修飾句「モノクローナル」は実質的に均一な集団の抗体から得られる抗体の特性を示し、任意の特定の方法による抗体生成が必要であると解釈されるべきでない。例えば、本発明によって使用されるモノクローナル抗体はKohler et al.(18)に最初に記載されたハイブリドーマ法によって作製することができるか、または組み換えDNA法によって作製することができる。また、「モノクローナル抗体」は例えばMarks et al.(19)に記載される技術を用いてファージ抗体ライブラリーから単離することができる。
【0040】
抗体由来分子は、抗原結合特異性を保持するインタクト抗体の部分を含み、例えば、少なくとも1つの可変領域(重鎖または軽鎖可変領域のいずれか)を含む。例えば、抗体由来分子としては、Fab断片、Fab'断片、F(ab')2断片、Fd断片、F(v)断片、Fabc 断片、Fd断片、Fabc断片、Sc抗体(単鎖抗体)、ダイアボディ、個々の抗体軽鎖、個々の抗体重鎖、抗体鎖と他の分子との間のキメラ融合物、重鎖モノマーまたはダイマー、軽鎖モノマーまたはダイマー、1つの重鎖および1つの軽鎖からなるダイマー等の分子が挙げられる。免疫グロブリンの全てのクラス(例えば、IgA、IgD、IgE、IgG、およびIgM)およびそのサブクラスが含まれる。
【0041】
抗体は、毒素部分または非毒素部分を標識/コンジュゲートすることができる。毒素部分としては、例えば、細菌毒素、ウイルス毒素、放射性アイソトープ等が挙げられる。抗体は、抗体の検出を補助するための生物学的アッセイ(例えば、放射性アイソトープ標識、蛍光標識)における使用のために標識できる。また、抗体は、例えば、放射性免疫治療(20)、画像技術および放射性免疫指示(guided)手術のような適用のために所望の部位に直接的に放射線を送達する放射活性アイソトープ、または特異的抗体/抗原複合体のインビボ画像化または検出を可能にする標識を、診断目的または治療目的で、標識/コンジュゲートすることができる。また、抗体は毒素とコンジュゲートして免疫毒素を提供してもよい(21)。
【0042】
抗体に関して、用語「免疫学的に特異的」とは、標的分子の1つ以上のエピトープに結合するが、抗原性生物学的分子の混合集団を含む試料中の他の分子を実質的に認識せず、かつ結合しない抗体のことをいう。
【0043】
「キメラ」または「キメラ化」抗体(免疫グロブリン)とは、所望の生物学的活性を示す限り、重鎖および/または軽鎖の一部が特定のクラスに由来する抗体の対応配列に同一または相同であるか、あるいは特定の抗体クラスまたはサブクラスに属するが、残りの鎖は別のクラスに由来する抗体の対応配列に同一または相同であるか、あるいは別の抗体クラスまたはサブクラスに属する抗体、ならびにかかる抗体の断片のことをいう(22)。
【0044】
「ヒト化」形態の非ヒト(例えば、マウス)抗体は、非ヒト免疫グロブリンに由来する最小配列を含むキメラ免疫グロブリン、免疫グロブリン鎖またはその断片(例えば、Fv、Fab、Fab'、F(ab’)2または抗体の他の抗原結合部分配列)である。ほとんどの部分について、ヒト化抗体は、レシピエントの相補決定領域(CDR)からの残基が所望の特異性、親和性、および能力を有するマウス、ラットまたはウサギ等の非ヒト種(ドナー抗体)のCDRからの残基によって置換されるヒト免疫グロブリン(レシピエント抗体)である。いくつかの例において、ヒト免疫グロブリンのFvフレームワーク領域(FR)残基は対応する非ヒト残基によって置換される。さらに、ヒト化抗体は、レシピエント抗体にも移入CDRまたはフレームワーク配列にも見られない残基を含み得る。これらの改変は、抗体性能をさらに改良し、かつ最適化するために行われる。一般的に、ヒト化抗体は、CDR領域の全てまたは実質的に全てが非ヒト免疫グロブリンのものに対応し、FR領域の全てまたは実質的に全てがヒト免疫グロブリン配列のものであるように、実質的に全てのうち少なくとも1つ、典型的に2つの可変領域を含む。また、ヒト化抗体は最適に、免疫グロブリン定常領域(Fc)の少なくとも一部、典型的にヒト免疫グロブリンのものを含む。更なる詳細について、Jones et al.(23); Reichmann et al.,(24);Presta(25)を参照。
【0045】
「完全にヒト」とは、分子全体がヒト起源であるか、またはヒト形態の抗体に同一のアミノ酸配列からなる抗体等の免疫グロブリンのことをいう。
【0046】
「エピトープ」とは、抗体結合部位として働く抗原の免疫学的決定基のことをいう。本明細書で使用される場合、用語「コンフォメーションエピトープ」とは、連続する一連のアミノ酸以外の抗原のアミノ酸間の空間関係によって形成される不連続エピトープのことをいう。
【0047】
「ハイブリドーマ」とは、親細胞の特異的な免疫潜在性を発現する培養新生物形成リンパ球と刺激されたBリンパ球またはTリンパ球との間での細胞融合の生成物のことをいう。
【0048】
本明細書で使用される場合、「高いタイター」とは、親細胞株よりも少なくとも約1.5倍高いタイターのことをいう。いくつかの態様において、タイターは、親細胞株よりも、少なくとも約1.5〜3倍高く、3〜5倍高く、5〜7倍高く、7〜9倍高く、または9〜10倍高い。
【0049】
本明細書で使用される場合、「高い親和性」とは、式Ka=8/3(It−Tt)(式中、「It」は50%トレーサーでのインヒビター取り込みの全モル濃度であり、「Tt」はトレーサーの全モル濃度である)による標準方法で計算できる高い抗体結合親和性のことをいう(26)。結合親和性はまた、式B/T=n・NAb・W108[(V-Vm)K+Q・W]を用いて計算できる(27)。本明細書で使用される場合、「高い親和性」は、約1×107M-1未満である。いくつかの態様において、抗体は約1×108M-1未満の親和性を有する。他の態様において、抗体は約1×109M-1未満の親和性を有する。他の態様において、抗体は約1×1010M-1未満の親和性を有する。他の態様において、抗体は約1×1011M-1未満の親和性を有する。他の態様において、抗体は約1×1012M-1未満の親和性を有する。他の態様において、抗体は約1×1013M-1未満の親和性を有する。他の態様において、抗体は約1×1014M-1未満の親和性を有する。
【0050】
本明細書で使用される場合、用語「速い増殖率」とは、所定の集団の細胞、例えば超突然変異娘細胞集団などの娘集団が、参照細胞集団、例えば、ミスマッチ修復が阻害されていない親細胞集団などの親細胞集団よりも大きな増殖率を示す状態、または所定の集団が同じ培養条件(例えば、温度、培養培地、培養期間の長さ、湿度、CO2、O2等)下で参照細胞集団よりも速い倍加時間を示す状態のことをいう。「増殖率」は一般的に、第一時点で決定された細胞コロニーの面積を第二時点での細胞コロニーの面積で割り、得られた数を2つのそれぞれの時点の間の時間差で割ることによって算出できる。例えば、増殖率は14日目の代表的なクローンの面積に対する17日目のコロニーの面積を3日で割ることによって決定されてもよい(増殖率=17日目面積/14日目面積÷3日)。
【0051】
本明細書で使用される場合、用語「増大された高密度生成」または「高細胞密度での増大された生体分子生成」とは、所定の細胞集団、例えば、超突然変異娘細胞集団などの娘集団が、同じ培養条件(例えば、温度、培養培地、培養期間の長さ、湿度、CO2、O2等)下で、参照細胞集団、例えばミスマッチ修復が阻害されていない親細胞集団などの親細胞集団よりも大きな生体分子(例えば、抗体)生成を示す状態のことをいう。
【0052】
本明細書で使用される場合、「高細胞密度での増大された細胞増殖」とは、所定の細胞集団、例えば超突然変異娘細胞集団などの娘細胞集団が、同じ培養条件(例えば、温度、培養培地、培養期間の長さ、湿度、CO2、O2等)下で、参照細胞集団、例えばミスマッチ修復が阻害されていない親細胞集団などの親細胞集団と比べて細胞数を測定した場合に増大したレベルの増殖を示す状態のことをいう。
【0053】
本明細書で使用される場合、「高細胞密度での増大された細胞生存能力」とは、所定の細胞集団、例えば超突然変異娘細胞集団などの娘細胞集団が、同じ培養条件(例えば、温度、培養培地、培養期間の長さ、湿度、CO2、O2等)下で、参照集団、例えばミスマッチ修復が阻害されていない親細胞集団などの親細胞集団と比べて増大した数の生存細胞を示す状態のことをいう。
【0054】
本明細書で使用される場合、「高細胞密度」とは、約1.0×106細胞/mLよりも大きい細胞濃度のことをいう。「低細胞密度」とは、約1.0×106細胞/mL未満の細胞濃度のことをいう。
【0055】
本明細書で使用される場合、「取り除かれる(cured)」とは、ミスマッチ修復のタンパク質インヒビターが細胞から排除されているか、またはミスマッチ修復のタンパク質インヒビターの発現が消失されるまたはノックアウトされる細胞の状態のことをいい、安定化されたゲノムがもたらされ、免疫グロブリンなどの安定な生物学的生成物が生成される。同様に、ミスマッチ修復のインヒビターの「不活性化」とは、細胞からのインヒビターの排除もしくは除去、またはミスマッチ修復のタンパク質インヒビターの不連続な発現(例えば、インヒビターの発現を消失もしくはノックアウトする)のことをいい、安定化されたゲノムがもたらされ、免疫グロブリンなどの安定な生体分子が生成される。
【0056】
本明細書で使用される場合、「スクリーニング」とは、限定されないが、核酸配列、タンパク質配列、タンパク質機能(例えば、結合、酵素活性、阻害活性、交差阻害活性、中和活性など)を含む細胞の遺伝型もしくは表現型または細胞生成物を評価するアッセイのことをいう。該アッセイとしては、ELISA系アッセイ、Biacore分析などが挙げられる。
【0057】
本明細書で使用される場合、「単離される」とは、天然環境の成分から分離および/または回収された核酸またはタンパク質のことをいう。天然環境の夾雑成分は、ポリペプチドの診断使用または治療使用に干渉する物質であり、酵素、ホルモン、および他のタンパク質溶質または非タンパク質溶質を挙げることができる。いくつかの態様において、核酸またはタンパク質は、タンパク質の95重量%よりも高く精製される。他の態様において、核酸またはタンパク質は、タンパク質の99重量%よりも高く精製される。タンパク質精製の測定は、Lowry法などの当該技術分野で公知の任意の手段、クーマシーブルーまたは銀染色などの染色を用いた還元または非還元条件下でのSDS-PAGEによるものであってもよい。核酸の精製は限定されないが、分光法、例えばメチレンブルーのような蛍光染色または化学染色を用いたアガロースまたはポリアクリルアミド分離を含む、任意の公知の方法によって評価されてもよい。
【0058】
哺乳動物細胞株による抗体生成の生産性を増進するために細胞宿主性能を改善する方法が本明細書に提供される。ミスマッチ修復が生成細胞株で阻害され、増殖特性に影響を及ぼす細胞性プロセスを改善して、改善された製造収量をもたらすことができる。例えば、細胞は、速い増殖、高細胞密度での増殖および/または細胞生存能力、および/または長い発酵運転のための高密度での生産性を維持する能力を遺伝的に高めることができる。いかなる理論にも拘束されることを意図しないが、これらの増大は、増殖および生存を阻害する代謝副産物の蓄積を検出する増殖因子または生化学レセプターの構造変化を介して生じることが考えられる。図2は、速い増殖率(FGR)の細胞および/または規模拡大可能な製造のための全体的生産性を高密度(HD)で増殖および生成する細胞による、増大された増殖の劇的な効果のモデルを提供する。
【0059】
増殖率、高細胞密度での細胞増殖および細胞生存能力、および/または生成細胞株、好ましくは哺乳動物生成細胞株の高細胞密度での生産性を含む増殖特性を改善するために本明細書に提供される方法は、適時でかつ費用が効果的である様式で、モノクローナル抗体(mAb)の高い収量を生み出す能力を細胞に付与する。スケールでの全体的生成を改善するための増大された細胞増殖パラメーターを有する細胞が提供される。本明細書に記載される方法は、組み換え哺乳動物細胞の増殖率を改善するが、特異的な生産性にはほとんど影響を与えない。
【0060】
速い増殖率、高細胞密度での増大された細胞増殖および細胞生存能力、および高細胞密度での増大された生体分子生成の少なくとも1つの増殖特性を含む、改善された増殖特性を有する細胞を生成する方法が本明細書に提供される。いくつかの好ましい態様において、改善された増殖特性としては、速い増殖率および高細胞密度での増大された生体分子生成;速い増殖率および高細胞密度での増大された細胞増殖および細胞生存能力;高細胞密度での増大された細胞増殖および細胞生存能力ならびに高細胞密度での増大された生体分子生成;または速い増殖率、高細胞密度での増大された細胞増殖および細胞生存能力、ならびに高細胞密度での増大された生体分子生成が挙げられる。いくつかの態様において、細胞は、高アフィニティー抗体の生成および高いタイター抗体の生成の少なくとも1つの特性をさらに示す。改善された増殖特性を有する細胞を生成する方法は、本明細書に教示される全ゲノム進化技術の適用を含む。全ゲノム進化技術の利点は、増殖特性、規模拡大可能な製造のためのタイター収量またはその両方をさらに最適化する必要がある以下のもの、細胞株、例えば、抗体を発現する細胞株(即ち、生成細胞株)、好ましくは哺乳動物細胞株、より好ましくは抗体を発現する哺乳動物細胞株(即ち、哺乳動物生成細胞株または哺乳動物製造細胞株)に、該技術を直接適用できるということである。本発明は、製造のための増大された増殖の抗体生成細胞株の生成を容易にする。全ゲノム進化技術は、適時で規模拡大可能なGMP製造のための十分な抗体量を生成する細胞株に適用されてもよい。全ゲノム進化技術は、微生物、植物および哺乳動物細胞系システムでの高い程度のMMRの保存および機能のために、任意の種の細胞の宿主増殖パラメーターを最適化するために利用することができる。
【0061】
改善された増殖特性を有する遺伝的に改変された宿主細胞を生成する方法は、生成物開発のための細胞宿主を作製するための価値のある方法を提供し、改変が増大された細胞増殖特性となる場合に構造または発現レベルを改変する下流遺伝子の発見に有用な試薬の生成を可能にする。本明細書に記載される本発明は、MMRの阻害を介して増大された増殖特性を有する遺伝的に改変された細胞宿主の作製に関し、次に増大された増殖株の指向された改変および生成について改変された遺伝子座をスクリーニングおよび同定するために使用することができる。
【0062】
哺乳動物細胞における遺伝的進化を増大するために、MMRが宿主細胞で抑制される。MMRは、好ましくはMMRのタンパク質インヒビターの導入(例えば、MMR阻害タンパク質をコードする発現ベクターの導入)、または化学MMRインヒビターへの宿主細胞の曝露(例えば、MMRの化学インヒビターの存在下で細胞のインキュベート)によって抑制される(8、9)。両方の方法は、MMRを阻害する点および遺伝的に進化した子孫細胞となる点で有効であることが示され、交換可能に使用されている(L. Grasso, 個人的意見)。
【0063】
宿主細胞(即ち、親細胞)は真核生物または原核生物であってもよい。好ましくは、宿主細胞は、哺乳動物細胞、より好ましくはヒト細胞またはげっ歯類(例えば、マウス、ハムスター)細胞である。本発明の方法の特定の態様における使用に適切な哺乳動物細胞としては、限定されないが、sp20細胞、NS0細胞、チャイニーズハムスター卵巣細胞(CHO細胞(28))、胎児ハムスター腎臓(BHK細胞)、ヒト胚腎臓株293(HeLa細胞(29))、正常イヌ腎臓細胞株(例えば、MDCK、ATCC CCL 34)、正常ネコ腎臓細胞株(CRFK細胞)、サル腎臓細胞(CV1 ATCC CCL 70);アフリカミドリサル腎臓細胞(VERO-76、ATCC CRL-1587)、COS(例えば、COS-7)細胞、および非腫瘍性マウス筋芽G8細胞(例えば、ATCC CRL 1246)、線維芽細胞株(例えば、ヒト、マウスまたはニワトリ胚性線維芽細胞株)、ミエローマ細胞株、マウスNIH/3T3細胞、LMTK31細胞、マウスセルトリ細胞(TM4、(30));ヒト頚部癌細胞(HELA、ATCC CCL2);バッファローラット肝臓細胞(BRL 3A、ATCC CRL 1442);ヒト肺細胞(W138、ATCC CCL 75);ヒト肝臓細胞(Hep G2、HB8065);マウス乳腺癌細胞(MMT 060562、ATCC CCL51)、TRI細胞(31);MRC5細胞;FS4細胞;ならびにヒト肝癌株(Hep G2)が挙げられる。
【0064】
哺乳動物発現細胞に代わるものとして、他の非哺乳動物細胞が本発明の方法に使用されてもよい。かかる非哺乳動物細胞としては、限定されないが、細菌細胞、酵母細胞、植物細胞、両生類細胞、および昆虫細胞(例えば、Spodoptera frugiperda細胞等)を含む真核生物細胞および原核生物細胞が挙げられる。ベクターおよび非哺乳動物宿主細胞が当該分野で周知であり、継続して最適化され、開発されている。抗体を発現できる任意の宿主細胞系は、本発明の方法に使用されてもよい。
【0065】
宿主細胞は、好ましくは抗体生成細胞である。例えば、宿主細胞はハイブリドーマ細胞であってもよい。
【0066】
細胞はミスマッチ修復(MMR)のタンパク質インヒビターの導入によって超突然変異可能にされてもよい。例えば、タンパク質インヒビターは、MMR遺伝子のドミナントネガティブ対立遺伝子を含むポリヌクレオチドによってコードされてもよい。MMRのタンパク質インヒビターをコードする核酸は、細胞、例えば、発現細胞、ハイブリドーマ細胞(即ち、免疫グロブリン生成細胞のミエローマ細胞との融合後)、またはミエローマ細胞(即ち、融合前に細胞に導入されてもよい)に導入されてもよい。
【0067】
MMRのタンパク質インヒビターをコードする核酸は、ゲノムDNA、cDNA、RNA、または化学合成ポリヌクレオチドであってもよい。ポリヌクレオチドは、構成的に活性なプロモーターセグメント(例えば、限定されないが、CMV、SV40、EF-1 DまたはLTR配列)またはテトラサイクリン、もしくはエクダイソン/グルココルチコイド誘導ベクターに由来するものなどの誘導プロモーター配列を含む発現ベクターにクローニングすることができ、インヒビターの発現を調節することができる。ポリヌクレオチドは、トランスフェクションによって細胞に導入することができる。
【0068】
トランスフェクションは、ポリヌクレオチドが細胞に導入される任意の方法である。トランスフェクションの方法は、例えば、培養において1つ以上の単離細胞の懸濁液を用いてインビトロで行うことができる。細胞はモノクローナル抗体の生成のためのハイブリドーマを作製するために使用される任意の不死細胞であってもよく、または細胞はハイブリドーマ自体であってもよい。ハイブリドーマは、ヘテロハイブリドーマ細胞(例えば、ヒト-マウス細胞融合物)またはホモハイブリドーマ細胞(例えば、ヒト-ヒトハイブリドーマ細胞およびマウス-マウスハイブリドーマ細胞)であってもよい。
【0069】
一般的に、トランスフェクションは、複数の細胞または単一の細胞の懸濁液を用いて行うが、他の方法も、トランスフェクトされた細胞が増殖および利用できるように処理された細胞または組織の十分な部分がポリヌクレオチドを取り込む限り、利用することができる。ポリヌクレオチドのタンパク質生成物は、細胞で一過的または安定的に発現されてもよい。トランスフェクションの技術は周知である。ポリヌクレオチドを導入するために利用可能な技術は、限定されないが、エレクトロポレーション、形質転換、細胞融合、塩化カルシウムの使用、目的の細胞との融合のための脂質と共にポリヌクレオチドのパッケージングを含む。細胞がミスマッチ修復のタンパク質インヒビターをコードする核酸でトランスフェクトされると、細胞は培養中に増殖でき、再生産できる。ミスマッチ修復のタンパク質インヒビターをコードする核酸が多くの細胞世代について一定レベルで発現されるように、トランスフェクションが安定である場合に、細胞株が得られる。
【0070】
ミスマッチ修復のドミナントネガティブタンパク質インヒビターをコードする核酸は、限定されないが、それぞれが参照によって本明細書に援用される米国特許第6,146,894号および第6,808,894号および米国出願公開第20040115695号および第20050048621号に記載されるように、PMS2、PMS2-134、PMS1、PMSR3、PMSR2、PMSR6、MLH1、GTBP、MSH3、MSH2、MLH3、またはMSH1、およびPMSR遺伝子のホモログを含む任意の公知のミスマッチ修復遺伝子に由来してもよい;Nicolaides et al.(32)およびHorii et al(33)。かかる効果を生じる任意の対立遺伝子が本発明に使用できる。ミスマッチ修復のドミナントネガティブタンパク質インヒビターは、ヒト、動物、酵母、細菌、または他の生物の細胞から得ることができる。ミスマッチ修復のタンパク質インヒビターの非限定例は、PMS2の最初の133アミノ酸を有するPMS2-134である。本明細書に使用されるPMS2およびPMS2-134としては、ヒトPMS2およびPMS2-134およびそれらの種(例えば、マウス;シロイヌナズナ等の植物;等)の同等物が挙げられる。PMS2タンパク質のC末端の欠失は、PMS2のMLH1との結合に干渉すると考えられている。ミスマッチ修復を阻害するmutLホモログのアミノ酸のさらなる記述によって、共通のアミノ酸配列
またはその断片が明らかにされる。従って、ミスマッチ修復のタンパク質インヒビターとしては、配列番号1または2のアミノ酸配列を含むポリペプチドおよびその断片が挙げられる。
【0071】
細胞をミスマッチ修復活性の欠損についてスクリーニングすることによって、ミスマッチ修復のさらなるタンパク質インヒビターを同定することができる。癌を有する動物またはヒト由来の細胞を欠損ミスマッチ修復についてスクリーニングすることができる。結腸癌患者由来の細胞が、特に有用であってもよい。ミスマッチ修復のタンパク質インヒビターをコードする任意の細胞由来のゲノムDNA、cDNA、またはmRNAは、野生型配列からの変異について分析することができる。ミスマッチ修復遺伝子のドミナントネガティブ対立遺伝子はまた、例えば、hPMS2-134対立遺伝子のバリアントまたは他のミスマッチ修復遺伝子を生成することによって人工的に作製することができる。部位特異的突然変異生成の様々な技術を使用することができる。天然または人工的であろうと、超突然変異可能な細胞または動物の生成における使用のためのかかる対立遺伝子の適切さが、1つ以上の野生型対立遺伝子の存在下での対立遺伝子によって生じたミスマッチ修復活性の試験によって評価され、ドミナントネガティブ対立遺伝子であるかどうかを決定することができる。
【0072】
ミスマッチ修復のドミナントネガティブタンパク質インヒビターによって、DNA修復の有効性の低減による自発的変異の割合が増大され、それによって細胞または動物が超突然変異可能になる。これは、かかる細胞または動物の自発性変異の割合がかかる対立遺伝子がない細胞または動物と比較して上昇することを意味する。自発的変異の割合の上昇程度は、正常細胞または正常動物の割合よりも少なくとも2倍、5倍、10倍、20倍、50倍、100倍、200倍、500倍または1000倍であってもよい。超突然変異可能な細胞によって、遺伝子に新しい変異が蓄積され、新しい出力特性が生じる。細胞は所望の特性についてスクリーニングされてもよく、これらの特性を生じる細胞株が拡大されてもよい。さらに、細胞はミスマッチ修復欠陥について修復されてもよく、遺伝的に安定な細胞がもたらされる。好ましい態様において、ミスマッチ修復のタンパク質インヒビターが不活性化される。例えば、ミスマッチ修復のタンパク質インヒビターは所望の増殖特性を有する細胞の同定の前または後に不活性化されてもよい。ミスマッチ修復のタンパク質インヒビターの不活性化は、当該分野で公知の任意の手段、例えば、誘導剤の除去または細胞からのミスマッチ修復のタンパク質インヒビターの除去(即ち、ミスマッチ修復のタンパク質インヒビターから細胞を取り除くこと)によるものであってもよい。ミスマッチ修復のインヒビター不活性化は超突然変異細胞のゲノムを安定にする。
【0073】
本発明の別の局面は、MMRを欠く細胞(内在性ミスマッチ修復遺伝子の欠失、またはMMRのドミナントネガティブタンパク質インヒビターの導入のいずれかによる)および宿主ゲノムにおける増大した変異割合を生じる化学突然変異原の使用である。MMR活性の欠失によって、細胞はDNA損傷剤の毒性効果に耐性になることが分かっている。本発明は、MMRのドミナントネガティブタンパク質インヒビターの発現を介して熟練MMR細胞をミスマッチ修復欠陥にすること、DNA突然変異薬剤の使用によってゲノム超突然変異性を増大することを含む。化学突然変異剤、例えば、アルキル化剤、架橋剤等は化学特性によって分類可能である。本発明によれば、本明細書に他のものを記載しない場合に、以下の化学突然変異剤が有用であり、本発明の方法の態様のいずれかにおける変異割合をさらに増大するために使用されてもよい:N-エチル-N-ニトロソ尿素(ENU)、N-メチル-N-ニトロソ尿素(MNU)、プロカルバジン塩酸、クロラムブシル、シクロホスファミド、メチルメタンスルホネート(MMS)、エチルメタンスルホネート(EMS)、ジエチルスルフェート、アクリルアミドモノマー、トリエチレンメラミン(TEM)、メルファラン、窒素イペリット、ビンクリスチン、ジメチルニトロソアミン、N-メチル-N'-ニトロ-ニトロソグアニジン(MNNG)、7,12ジメチルベンズアントラセン(DMBA)、エチレンオキシド、ヘキサメチルホスホルアミド、ビスルファン。本発明の好ましい局面において、100遺伝子ごとに1つの変異;1000遺伝子あたり1つの変異の範囲における変異割合を付与する突然変異生成技術が使用される。かかる組み合わせ(MMR欠乏および化学突然変異剤)の使用は、各特定の薬剤によって優先的に誘導される多様なゲノム改変(限定されないが、遺伝子コーディング領域、遺伝子イントロン領域、または5'もしくは3'近位および/または遠位領域の背景でのDNAセグメントの拡張または欠失、点変異、改変された繰り返し配列)の生成を可能にする。
【0074】
変異は、細胞または動物の遺伝型における改変を分析することによって、例えば、目的の遺伝子と関連するゲノムDNA、cDNA、メッセンジャーRNA、またはアミノ酸の配列を調べることによって、検出することができる。また、変異は遺伝子の表現型をスクリーニングすることによって検出することができる。改変された表現型は、電気泳動移動度、分光学的特性、または変異遺伝子によってコードされるタンパク質の他の物理的または構造的特性における改変を同定することによって検出できる。また、インサイチュで、単離形態で、またはモデル系でタンパク質の改変された機能をスクリーニングすることができる。限定されないが、タンパク質分泌、化学耐性、病原耐性等を測定すること等の、目的の遺伝子の機能と関連する細胞または動物の任意の特性の改変をスクリーニングすることができる。
【0075】
本発明の改善された増殖特性を有する細胞を生成する方法のいくつかの態様において、細胞がミスマッチ修復の化学インヒビターに曝露される。本発明の方法の特定態様に使用されるミスマッチ修復の化学インヒビターとしては、限定されないが、アントラセン、ATPaseインヒビター、ヌクレアーゼインヒビター、RNA干渉分子、ポリメラーゼインヒビターおよびミスマッチ修復タンパク質をコードするヌクレオチドに特異的にハイブリダイズするアンチセンスオリゴヌクレオチドの少なくとも1つが挙げられる(WO2004/046330)。好ましい態様において、化学インヒビターは、以下の式:
(式中、R1〜R10は独立して、水素、ヒドロキシル、アミノ、アルキル、置換アルキル、アルケニル、置換アルケニル、アルキニル、置換アルキニル、O-アルキル、S-アルキル、N−アルキル、O-アルケニル、S-アルケニル、N-アルケニル、O-アルキニル、S-アルキニル、N-アルキニル、アリール、置換アリール、アリールオキシ、置換アリールオキシ、ヘテロアリール、置換へテロアリール、アラルキルオキシ、アリールアルキル、アルキルアリール、アルキルアリールオキシ、アリールスルホニル、アルキルスルホニル、アルコキシカルボニル、アリールオキシカルボニル、グアニジノ、カルボキシ、アルコール、アミノ酸、スルホネート、アルキルスルホネート、CN、NO2、アルデヒド基、エステル、エーテル、環状エーテル、ケトン、オルガノ硫黄化合物、オルガノ金属基、カルボン酸、オルガノケイ素、または1つ以上のアルキル化ヒドロキシル基を任意に含む炭水化物である;へテロアルキル、ヘテロアリール、および置換へテロアリールは、酸素、硫黄、金属原子、リン、ケイ素または窒素である少なくとも1つのヘテロ原子を含む;置換アルキル、置換アルケニル、置換アルキニル、置換アリール、および置換へテロアリールの置換基は、ハロゲン、CN、NO2、低級アルキル、アリール、ヘテロアリール、アラルキル、アラルコキシ、グアニジノ、アルコキシカルボニル、アルコキシ、ヒドロキシ、カルボキシおよびアミノである;アミノ基はアシル基、または1〜3のアリールまたは低級アルキル基で任意に置換される)
を有するアントラセン化合物である。特定の態様において、R5およびR6は水素である。他の態様において、R1〜R10は独立して、水素、ヒドロキシル、メチル、エチル、プロピル、イソプロピル、ブチル、イソブチル、フェニル、トリル、ヒドロキシメチル、ヒドロキシプロピル、またはヒドロキシブチルである。アントラセンの非限定例としては、1,2-ジメチルアントラセン、9,10-ジメチルアントラセン、7,8-ジメチルアントラセン、9,10-ジフェニルアントラセン、9,10-ジヒドロキシメチルアントラセン、9-ヒドロキシメチル-10-メチルアントラセン、ジメチルアントラセン-1,2-ジオール、9-ヒドロキシメチル-10-メチルアントラセン-1,2-ジオール、9-ヒドロキシメチル-10-メチルアントラセン-3,4-ジオール、および9,10-ジ-m-トリルアントラセンが挙げられる。
【0076】
化学インヒビターは、細胞の増殖培地に導入されてもよい。いくつかの態様において、化学インヒビターは、超突然変異細胞から回収されるか、または細胞が、細胞のゲノムを再安定化するために化学インヒビターから除去される。あるいは、該方法は、ミスマッチ修復の化学インヒビターの不活性化を含み、それによって超突然変異細胞のゲノムを安定化してもよい。
【0077】
MMR抑制細胞はインキュベートされて突然変異生成を可能にする。これによって、超突然変異娘細胞が生じる。好ましくは、MMR抑制細胞は、少なくとも約20世代、より好ましくは少なくとも約30世代継代され、その時にゲノムワイド変異が娘細胞に蓄積する。
【0078】
細胞の遺伝的に多様なプールは、好ましくは当該技術分野の公知の任意の方法によってサブクローニングされる。例えば、超突然変異娘細胞は、限界希釈でサブクローニングされた単一細胞であってもよく、拡大されてもよい。クローンは好ましくは、2週間まで拡大される。
【0079】
親細胞株と比べて1つ以上の改善された増殖特性(例えば、親細胞集団よりも速い増殖率(FGR)または高密度(HD)での生成)を示す娘細胞が次に同定される。いくつかの好ましい態様において、改善された増殖特性としては、速い増殖率と高密度での生成の両方が挙げられる。いくつかの態様において、細胞は、高い親和性の抗体生成および高いタイターの抗体生成の少なくとも1つの特性をさらに示す。例えば、いくつかの態様において、速い増殖(FGR)および高い抗体タイターを示す細胞が検出される。高い特異的な生産性を有する速い増殖率の細胞を生じる能力によって、多くの細胞種類の抗体製造の全体的生成における劇的な増大をもたらすことができる(図2)。
【0080】
好ましくは、改善された増殖特性を有する細胞の同定には、ハイスループットスクリーニングが含まれる。いくつかの態様において、様々な機能的で自動化ハイスループットスクリーニング(HTS)アッセイが実施され、少なくとも1つの改善された増殖特性(例えば、増大した薬理学的活性でタンパク質を生じる細胞;増大されたタイター収量を有する細胞;速い増殖率の細胞;高細胞密度での増大された増殖および生存能力を有する細胞;高細胞密度での増大された生体分子生成を有する細胞)を有するサブクローンを同定する。かかる細胞は改善された規模拡大可能な製造に適切であってもよい。
【0081】
速い増殖率の娘細胞の検出は、当該技術分野で公知の任意の方法、例えば、限定されないが、細胞カウント、および親細胞の集団に対する娘細胞の集団の比較によるものであってもよい。いくつかの態様において、親細胞よりも速い増殖率を有する速い増殖率の娘細胞の検出は、同じ培養条件下で同等の長さの培養時間を続けた後に、超突然変異娘細胞によって生じた細胞集団のサイズと、親細胞によって生じた細胞集団のサイズとを比較することを含み、親細胞集団に対する大きな娘細胞集団は、速い増殖率の娘細胞を示す。いくつかの態様において、速い増殖率の娘細胞の検出は、親細胞の増殖率よりも大きい増殖率を有する娘細胞の同定を含む。また、高細胞密度での増大された生体分子生成を示す細胞の検出は、当該技術分野で公知の任意の手段、例えば、限定されないが、同じ培養条件下で同等の長さの培養時間を続けた後に、娘細胞集団の細胞密度と、親細胞集団の細胞密度とを比較することであってもよく、高い細胞密度の娘細胞集団がHD細胞株を示す。
【0082】
細胞集団のサイズは、当該技術分野で公知の任意の方法によって決定されてもよい。細胞集団のサイズは好ましくは、光学画像化システム、例えば、MetaMorph(登録商標)システム(Molecular Devices Corp.)を用いて決定される。
【0083】
MMRが所望の表現型を示すサブクローンに保存される。遺伝的安定性は、以前に記載されたようにMMRインヒビターを除去することによって保存されてもよい(8、9)。例えば、細胞は、MMRの化学インヒビターを含む培養培地から除去されてもよい。あるいは、化学インヒビターは、細胞から回収されてもよい。いくつかの態様において、MMRのタンパク質インヒビターが不活性化されるか、または細胞が取り除かれる。遺伝的安定性は、所望の表現型を示す娘細胞の検出の前、後、または同時に細胞に保存されてもよい。
【0084】
所望の表現型を示すMMR熟練サブクローンは、i)細胞または抗体の増大した特性の保存および安定;ii)野生型MMR活性の保存および宿主ゲノムの安定化;および/またはiii)タンパク質構造の完全性および機能を確認するために分析される。
【0085】
本発明の方法によって生成した細胞は、本発明の範囲に含まれる。
【0086】
本発明の方法によって生成した細胞を培養する工程、および該細胞または該細胞の培養培地から生体分子を単離する工程を含む生体分子を製造する方法がまた、本発明の範囲に含まれる。本明細書で使用される場合、用語「生体分子」とは、細胞によって生成される分子のことをいい、化学薬剤(例えば、フコシルまたはグリコシル部分)および/または生物学的薬剤(例えば、限定されないが抗体を含むタンパク質)を含んでいてもよい。本発明の方法によって生成した生体分子、ならびに該生体分子および薬学的に許容できる担体を含む医薬組成物は、同様に本発明の範囲に含まれる。
【0087】
全ゲノム進化技術法によって、最適化された細胞増殖またはタイター生成に関わる未知の遺伝子および経路に対する様々なゲノムツールおよびタンパク質ツールによって分析できる同遺伝子型細胞の開発が可能とされる。染色体不安定の結果として異数性を誘導する標準の化学突然変異薬剤(7)に比べて、全ゲノム進化技術法は、宿主細胞の染色体安定および長期生存能力をインタクトのままにすると同時にわずかな点変異となるために、同等な遺伝的アプローチが保証されることを可能にする。この特徴は、遺伝的不安定ゲノムおよび再発現表現型の結果の両方となる高い変異バックグラウンドおよび化学突然変異細胞で見られる変異「ホットスポット」を回避する。このアプローチはより強固な結果を生ずるだけでなく、増大した増殖および生成と関連する経路に関わる標的物を明確に同定することによって種々の遺伝子/タンパク質発見も容易にする。全ゲノム進化技術法が適用されて、改善された細胞の増殖特性に重要な遺伝子を同定する方法が、本発明の範囲に含まれる。該方法は、所望の表現型を有する本発明の方法によって生じた娘細胞のゲノムと、親細胞のゲノムとを比較する工程、および2つのゲノム間の遺伝的差異を検出する工程を含み、かかる遺伝的差異または変異の少なくとも1つを含む遺伝子は所望の表現型に重要な遺伝子である。例えば、全ゲノム進化技術由来mAb生成細胞は、同遺伝子型の親および高タイター全ゲノム進化技術由来子孫セット間での遺伝子発現のRNAマイクロアレイ分析を行うことによって高いタイター生産性に関わる経路を同定するために使用された(9)。
【0088】
高いタイターまたは速い増殖などの改善された増殖特性のための進化した経路の同定によって、多くの生成物の高い生成のための高性能細胞の直接操作が可能になる。これは、増大した特性を有する全ゲノム進化技術由来細胞における改変経路の発見、およびこれらの経路を親細胞骨格で細胞操作することによる変異表現型の繰り返しによって達成できる。例えば、本明細書に記載される方法によって同定された改善された増殖特性に重要な遺伝子が、当該技術分野で公知の方法によって、細胞、好ましくは生成細胞で組み換え発現することができる。親細胞株の改善された増殖特性に重要な遺伝子の組み換え発現(全ゲノム進化技術または全ゲノム進化技術からもたらされる指向された経路改変のいずれかによる)は、次に安定な生成細胞株を生成するために必要な時間を低減することによって治療への速度を加速できる。
【0089】
以下の実施例は本発明のいくつかの局面をより詳細に記載する。実施例は、限定されないが本明細書に記載される本発明の局面をさらに示すために提供される。
【実施例】
【0090】
全ゲノム進化技術法は、図3に概略したように適用された。MMRはMMR化学インヒビター9,10-ジメチルアントラセンを用いて親細胞で抑制された。MMR抑制細胞株が増殖され、次にサブクローン化され、およそ10,000個の子孫細胞を生じ、低密度で細胞増殖をモニターできるカスタム化視覚プラットフォームおよびソフトウェアを用いて速い増殖率の表現型をスクリーニングした。
【0091】
全ゲノム進化技術由来CHO-MAb細胞プールにおける速い増殖率(FGR)クローンをスクリーニングするために、新規画像ベースHTS法が開発された。この方法は、従来の時間消費細胞カウント法の要件を回避する。代わりに、これは、Meta 画像システム(Molecular Devices, Downingtown, PA)を使用し、該システムは、播種し、生育し、無菌条件下で全ゲノム進化技術を介して生じた子孫細胞クローンのコロニーサイズを分析するように設計されたORCA自動化プラットフォーム(Beckman Coulter, Fullerton, CA)に接続されたデジタルカメラを介して細胞コロニーサイズを画像化する。Metamorphソフトウェアパッケージ(Ver.6.3r0)を用いて、コロニー面積画像がピクセルで定量され、14日目と17日目のサイズを比較して、3日増殖期間中の細胞コロニーサイズの割合を計算するスプレッドシートに出力される。画像システムは、画像ピクセル面積と細胞数との間の線形関係を生じるように改良された(図4)。
【0092】
全ゲノム進化技術由来CHO-MAb細胞における速い増殖率の子孫を同定するために、細胞は、Biomek FX ロボットシステム(Beckman Coulter, Fullerton CA)を用いて100〜200バーコード丸底96ウェルプレートに0.8細胞/ウェルで播種され、ウェルあたりの単一細胞クローンを確実にする。画像化方法は、定量のために細胞を犠牲にする必要はなく、従って、レプリカプレートはこの手順を用いて必要とされない。クローナル播種の開始2週間後に、プレートは顕微鏡画像によって決定されるような、クローン増殖の高出力分析のためのORCAシステムによってスクリーニングされる。プレートは、重層インキュベーター内で生育され、コロニーの如何なる阻害もせずに画像解析後にインキュベーターに戻される。この工程は、3日間繰り返された後にコロニーの拡大サイズを決定して、次に14日目と17日目の時点間の細胞の増殖率を決定するために使用される。細胞増殖が17日目と14日目に測定されたコロニーサイズ値の間の率として、17日目のコロニー面積を14日目のコロニー面積で割って、これを3日で割ることによって算出される。図5は親またはFGR CHO-MAbクローンが14日目、次に再度17日目に画像化される典型的な分析を示す。予測されるように、FGR細胞は親CHO-MAb由来のクローンと比較して17日目と14日目との間の大きな率を示す。この細胞株を平均して、親クローンは、およそ20,000個の独立した親CHO-MAb由来サブクローンの一次スクリーニングから決定された約0.5の増殖率を示すが、FGRクローンは>0.8の増殖率を示す。標準として、各プレートは比較物として親CHO-MAb細胞由来のクローンを含む。図5は親株および全ゲノム進化技術由来細胞に由来するサブクローンの典型的なプロファイルを示す。全ゲノム進化技術由来CHO-MAb細胞において、スクリーニングされたウェルの約5%が0.8以上の増殖率を示す。これらのもののうち、このアッセイを用いた他の細胞株のスクリーニングと一致して拡大される場合、親株のものと比較して50%が改善された増殖率を有することが確認された。確認されたクローンは48時間定量増殖アッセイ中に3mL静止培養で増殖され、それによって細胞は、Cedex自動化細胞カウンターを用いて0日目、1日目および2日目に物理的にカウントされる。図5は代表的な48時間定量増殖アッセイ結果を示す。速い増殖率クローンを行う一番のものが20mL振とうフラスコアッセイでさらに拡大され、評価される。このレベルに達するクローンの大半は、典型的に速い増殖率を維持すると同時に高い抗体特異的生産性を保持する(図6)。
【0093】
速い増殖率サブクローンに由来する抗体の構造解析によって、これらの細胞によって生成される抗体は親抗体のものと類似する遺伝的特性および生化学特性を保持することが確認される。また、速い増殖率の細胞の拡大培養によって、全ゲノム進化技術によって媒介される増大した増殖率が安定であったこと、および3ヶ月間の細胞の全体タイターは親CHOおよびより速い増殖率由来細胞の両方で一定のままであったことが示される(図6)。
【0094】
【図面の簡単な説明】
【0095】
【図1】図1は哺乳動物細胞における抗体生成の経路を示す。発酵中にタンパク質生成および細胞増殖を調節する多くの経路が、哺乳動物細胞に存在する。これらの経路の1つ以上の改変は、宿主細胞株の生成性能および増殖性能を改善する遺伝子操作によって達成することができる。これらの経路は:(1)遺伝子転写;(2)mRNA安定化およびトランスロケーション;(3)タンパク質合成;(4)タンパク質翻訳後修飾および輸送;(5)小胞体(ER)でのフォールディング;(6)ゴルジでのタンパク質成熟およびグリコシル化;(7)分泌経路;および(8)細胞膜発現を含む。(GlcNAc=N-アセチルグルコサミン;P=リン酸化残基)。
【図2】図2は増大された増殖率および高細胞密度での安定性を有する細胞の生成効果を示す。10日間発酵運転は、拡大された期間で増大された増殖率または高密度での増殖能力を有する細胞をシミュレートした。モデルは、i)細胞密度が増大するとき、増殖率が減少する;ii)抗体の蓄積は細胞数およびpcdのみに依存する;およびiii)0日目での播種は500,000/mLであることを仮定する。親株の倍化時間は32時間であり、最大密度の1.7×106/mLは5日目で達成され、その特異的な生産性は25pg/細胞/日(pcd)である。これらの条件下で、親細胞株は発酵運転の完了時におよそ0.28g/Lを生じる。ここに示すように、球状サイズおよび1Lあたりの発現gに近い数によって表されるこのグラフでは、7日目で高細胞密度(HD)の3.4×106/mLに達する親株由来の細胞株は高い抗体タイターを有する。24時間対32時間の、速い増殖率(FGR)または速い倍化時間を有する別の株は、高い密度で増殖できる細胞によって達成されるものよりさらに高い収量を生じる。高密度増殖および増殖率(それぞれHDおよびFGR)における組み合わされた改善を示す細胞株はこのモデルで最良に実施され、3.32g/Lに達する。
【図3】図3は全ゲノム進化技術プロセスフローチャートを示す。
【図4】図4は顕微鏡画像解析を介した細胞計算を示す。CHO-MAb細胞は96ウェルU底面プレートにウェルあたり100〜10,000細胞の密度で播種され、画像プログラムの線形性を決定した。ウェルを、Meta画像システムを用いて20倍で画像化した。コロニーサイズを、Metamorphソフトウェアパッケージ(Ver.6.3r0)を用いて細胞によってカバーされたウェルのピクセル面積を統合することによって決定した。データ点は12ウェルの平均±標準偏差である。
【図5】図5は親CHO-MAbまたは全ゲノム進化技術由来クローンのコロニーサイズを比較する。親CHO-MAb細胞または全ゲノム進化技術CHO-MAb細胞は、96ウェルバーコードプレートにサブクローニングして37℃のCO2インキュベーターで12日間増殖させる。代表的なCHO-MAbコロニーは、コロニーのピクセル面積を計算するMetamorph画像ソフトウェアを用いて14日目に画像化され、次に17日目に再画像化される。増殖率は、14日目の代表的クローンのコロニー面積に対する17日目のコロニーの面蹟を3日で割ることで決定される(増殖率=17日目面蹟/14日目面蹟÷3日)。CHO-MAb株の親細胞は約0.5の増殖率を有する。>0.8の増殖率を示す細胞を拡大し、標準化増殖アッセイで分析する。固有の緩やかな増殖率の親細胞は典型的に、予測される速い増殖を有するように進化したクローンに比べて、14日目の小さなコロニーとなる。
【図6】図6は、速く増殖するサブクローンを確認する代表的な増殖データを示す。速い増殖率の親CHO-MAb細胞を7日間の振とうフラスコアッセイで増殖させ、1、4および7日目に細胞を計算して増殖率を分析した。速い増殖率のサブセット(WGET-1およびWGET-2)および親細胞由来(親-1および親-2)CHO-MAbサブクローンに対する倍化時間の関数として増殖率を示す。
【特許請求の範囲】
【請求項1】
a.親細胞集団の親細胞のミスマッチ修復を阻害する工程、
b.親細胞をインキュベートして突然変異生成を可能にし、それによって超突然変異娘細胞を生成する工程、
c.改善された増殖特性を有する超突然変異娘細胞を検出する工程;および
d.改善された増殖特性を有する超突然変異娘細胞の遺伝的安定性を保存する工程
を含む、親細胞集団と比べて改善された増殖特性を有する細胞を生成する方法。
【請求項2】
前記改善された増殖特性が速い増殖率である、請求項1記載の方法。
【請求項3】
前記改善された増殖特性が高細胞密度での増大された生体分子生成である、請求項1記載の方法。
【請求項4】
前記改善された増殖特性が高細胞密度での増大された細胞生存能力である、請求項1記載の方法。
【請求項5】
前記親細胞が抗体生成細胞である、請求項1記載の方法。
【請求項6】
前記親細胞のミスマッチ修復を阻害する工程がミスマッチ修復の化学インヒビターに前記親細胞を曝露する工程を含む、請求項1記載の方法。
【請求項7】
前記親細胞のミスマッチ修復を阻害する工程がミスマッチ修復のタンパク質インヒビターに前記親細胞を曝露する工程を含む、請求項1記載の方法。
【請求項8】
前記インキュベートする工程が少なくとも20回の継代で前記超突然変異細胞を継代する工程を含む、請求項1記載の方法。
【請求項9】
前記遺伝的安定性を保存する工程が前記超突然変異娘細胞から前記化学インヒビターを回収する工程を含む、請求項6記載の方法。
【請求項10】
前記遺伝的安定性を保存する工程が前記タンパク質インヒビターを不活性化する工程を含む、請求項7記載の方法。
【請求項11】
前記遺伝的安定性を保存する工程が前記改善された増殖特性を有する超突然変異娘細胞を検出する工程の前に生じる、請求項1記載の方法。
【請求項12】
前記遺伝的安定性を保存する工程が前記改善された増殖特性を有する超突然変異娘細胞を検出する工程の後に生じる、請求項1記載の方法。
【請求項13】
前記改善された増殖特性を有する超突然変異娘細胞を検出する工程が改善された増殖特性を有する前記超突然変異娘細胞のハイスループットスクリーニングを含む、請求項1記載の方法。
【請求項14】
前記速い増殖率を有する超突然変異娘細胞を検出する工程が、同じ培養条件下で同等の長さの培養時間を続けた後に、超突然変異娘細胞によって生じた細胞集団のサイズと、親細胞によって生じた細胞集団のサイズとを比較する工程を含み、高い密度の娘細胞集団が前記速い増殖率を示す、請求項2記載の方法。
【請求項15】
前記速い増殖率を有する超突然変異娘細胞を検出する工程が親細胞の増殖率よりも大きな増殖率を有する前記娘細胞を同定する工程を含む、請求項2記載の方法。
【請求項16】
前記娘細胞集団のサイズが光学画像システムを用いて決定される、請求項14記載の方法。
【請求項17】
前記親細胞集団のサイズが光学画像システムを用いて決定される、請求項14記載の方法。
【請求項18】
前記親細胞が哺乳動物細胞である、請求項1記載の方法。
【請求項19】
前記親細胞がハイブリドーマ細胞である、請求項1記載の方法。
【請求項20】
請求項1記載の方法によって生成した細胞。
【請求項21】
請求項20記載の細胞を培養する工程、および該細胞または該細胞の培養培地から生体分子を単離する工程を含む、生体分子を製造する方法。
【請求項22】
前記生体分子が化学薬剤を含む、請求項21記載の方法。
【請求項23】
前記生体分子が生物学的薬剤を含む、請求項22記載の方法。
【請求項24】
前記生体分子が生物学的薬剤を含む、請求項21記載の方法。
【請求項25】
前記生物学的薬剤がタンパク質を含む、請求項24記載の方法。
【請求項26】
前記生物学的薬剤が抗体を含む、請求項24記載の方法。
【請求項27】
請求項21記載の方法によって生成した生体分子。
【請求項28】
請求項27記載の生体分子および薬学的に許容できる担体を含む医薬組成物。
【請求項29】
請求項20記載の細胞のゲノムと、親細胞のゲノムとを比較して変異を同定する工程を含み、改善された増殖特性の原因遺伝子が少なくとも1つの該変異を含む、改善された増殖特性の原因遺伝子を同定する方法。
【請求項1】
a.親細胞集団の親細胞のミスマッチ修復を阻害する工程、
b.親細胞をインキュベートして突然変異生成を可能にし、それによって超突然変異娘細胞を生成する工程、
c.改善された増殖特性を有する超突然変異娘細胞を検出する工程;および
d.改善された増殖特性を有する超突然変異娘細胞の遺伝的安定性を保存する工程
を含む、親細胞集団と比べて改善された増殖特性を有する細胞を生成する方法。
【請求項2】
前記改善された増殖特性が速い増殖率である、請求項1記載の方法。
【請求項3】
前記改善された増殖特性が高細胞密度での増大された生体分子生成である、請求項1記載の方法。
【請求項4】
前記改善された増殖特性が高細胞密度での増大された細胞生存能力である、請求項1記載の方法。
【請求項5】
前記親細胞が抗体生成細胞である、請求項1記載の方法。
【請求項6】
前記親細胞のミスマッチ修復を阻害する工程がミスマッチ修復の化学インヒビターに前記親細胞を曝露する工程を含む、請求項1記載の方法。
【請求項7】
前記親細胞のミスマッチ修復を阻害する工程がミスマッチ修復のタンパク質インヒビターに前記親細胞を曝露する工程を含む、請求項1記載の方法。
【請求項8】
前記インキュベートする工程が少なくとも20回の継代で前記超突然変異細胞を継代する工程を含む、請求項1記載の方法。
【請求項9】
前記遺伝的安定性を保存する工程が前記超突然変異娘細胞から前記化学インヒビターを回収する工程を含む、請求項6記載の方法。
【請求項10】
前記遺伝的安定性を保存する工程が前記タンパク質インヒビターを不活性化する工程を含む、請求項7記載の方法。
【請求項11】
前記遺伝的安定性を保存する工程が前記改善された増殖特性を有する超突然変異娘細胞を検出する工程の前に生じる、請求項1記載の方法。
【請求項12】
前記遺伝的安定性を保存する工程が前記改善された増殖特性を有する超突然変異娘細胞を検出する工程の後に生じる、請求項1記載の方法。
【請求項13】
前記改善された増殖特性を有する超突然変異娘細胞を検出する工程が改善された増殖特性を有する前記超突然変異娘細胞のハイスループットスクリーニングを含む、請求項1記載の方法。
【請求項14】
前記速い増殖率を有する超突然変異娘細胞を検出する工程が、同じ培養条件下で同等の長さの培養時間を続けた後に、超突然変異娘細胞によって生じた細胞集団のサイズと、親細胞によって生じた細胞集団のサイズとを比較する工程を含み、高い密度の娘細胞集団が前記速い増殖率を示す、請求項2記載の方法。
【請求項15】
前記速い増殖率を有する超突然変異娘細胞を検出する工程が親細胞の増殖率よりも大きな増殖率を有する前記娘細胞を同定する工程を含む、請求項2記載の方法。
【請求項16】
前記娘細胞集団のサイズが光学画像システムを用いて決定される、請求項14記載の方法。
【請求項17】
前記親細胞集団のサイズが光学画像システムを用いて決定される、請求項14記載の方法。
【請求項18】
前記親細胞が哺乳動物細胞である、請求項1記載の方法。
【請求項19】
前記親細胞がハイブリドーマ細胞である、請求項1記載の方法。
【請求項20】
請求項1記載の方法によって生成した細胞。
【請求項21】
請求項20記載の細胞を培養する工程、および該細胞または該細胞の培養培地から生体分子を単離する工程を含む、生体分子を製造する方法。
【請求項22】
前記生体分子が化学薬剤を含む、請求項21記載の方法。
【請求項23】
前記生体分子が生物学的薬剤を含む、請求項22記載の方法。
【請求項24】
前記生体分子が生物学的薬剤を含む、請求項21記載の方法。
【請求項25】
前記生物学的薬剤がタンパク質を含む、請求項24記載の方法。
【請求項26】
前記生物学的薬剤が抗体を含む、請求項24記載の方法。
【請求項27】
請求項21記載の方法によって生成した生体分子。
【請求項28】
請求項27記載の生体分子および薬学的に許容できる担体を含む医薬組成物。
【請求項29】
請求項20記載の細胞のゲノムと、親細胞のゲノムとを比較して変異を同定する工程を含み、改善された増殖特性の原因遺伝子が少なくとも1つの該変異を含む、改善された増殖特性の原因遺伝子を同定する方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2009−533072(P2009−533072A)
【公表日】平成21年9月17日(2009.9.17)
【国際特許分類】
【出願番号】特願2009−506703(P2009−506703)
【出願日】平成19年4月16日(2007.4.16)
【国際出願番号】PCT/US2007/066704
【国際公開番号】WO2007/121396
【国際公開日】平成19年10月25日(2007.10.25)
【出願人】(506031948)モルフォテック、インク. (16)
【Fターム(参考)】
【公表日】平成21年9月17日(2009.9.17)
【国際特許分類】
【出願日】平成19年4月16日(2007.4.16)
【国際出願番号】PCT/US2007/066704
【国際公開番号】WO2007/121396
【国際公開日】平成19年10月25日(2007.10.25)
【出願人】(506031948)モルフォテック、インク. (16)
【Fターム(参考)】
[ Back to top ]