
細胞に分子を送達する方法及び担体複合体
【課題】本発明は担体複合体及び細胞に分子を送達する方法およびそのための担体複合体を提供する。
【解決手段】本発明の担体複合体は分子及び本発明の芳香族陽イオンペプチドを含む。ある実施態様では、本発明は細胞を担体複合体と接触させることを含む。また別の実施態様では、本発明は細胞を分子及び芳香族陽イオンペプチドと接触させることを含む。
【解決手段】本発明の担体複合体は分子及び本発明の芳香族陽イオンペプチドを含む。ある実施態様では、本発明は細胞を担体複合体と接触させることを含む。また別の実施態様では、本発明は細胞を分子及び芳香族陽イオンペプチドと接触させることを含む。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、米国仮特許出願60/467,516号(2003年5月1日出願)の優先権を主張する。米国仮特許出願60/467,516号の明細は引用によりその全体が本明細書に含まれる。
本発明は、グラント番号P01-DA-08924によりNational Institute on Drug Abuseから政府の援助を受けて達成された。合衆国政府は本発明に関して一定の権利を有する。
【背景技術】
【0002】
生物学的細胞は、一般的に細胞膜を通過する分子について高度に選択性を有する。したがって、化合物(例えば小分子および生物学的分子)の細胞内への送達は通常は化合物の物理的特性によって制限される。前記小分子及び生物学的分子は例えば医薬的に活性を有する化合物であろう。
in vivoでのそのような分子(巨大分子、たとえばタンパク質及び核酸を含む)が細胞内へ送達されないことは、潜在的に有効な多数の化合物の治療的、予防的及び/又は診断的使用の妨げであった。さらにまた、in vitroでは有望とされる多くの化合物が、in vivoでは細胞内に効率的に前記化合物を送達する能力の欠如のために潜在的薬剤として廃棄されてきた。
いくつかの報告は、“タンパク質導入ドメイン(protein transduction domain)”(PTD)と前記化合物を共有結合することによって、細胞への化合物の送達の問題を扱ってきた。Schwarzeら(Trends Pharmacol. Sci. 2000, 21:45-8)及び米国特許6,221,355号(Dowdy)は、濃度依存態様で細胞の脂質二重層を通過することができるいくつかのPTDを開示した。前記開示されたPTDにはHIV-1 tatタンパク質由来PTD、アンテナペディア(ANTPと略される)遺伝子によってコードされるショウジョウバエのホメオティック転写因子由来のPTD、及び単純ヘルペスウイルスVP22転写因子由来のPTDが含まれる。HIV-1 tat PTDは長さが11個のアミノ酸であり、ANTP PTDは16個のアミノ酸であり、VP22 PTDは34個のアミノ酸である。
しかしながら、最近の文献では、これらPTDはエネルギー依存エンドサイトーシスを介して細胞内に入ることが示されている。したがって、“PTD-カーゴ”複合体は細胞のエンドソーム小胞内に収納され、例えば細胞の細胞質には利用できない。したがって、“PTD-カーゴ”複合体は、生物活性を示すためにはエンドソーム小胞から放出される必要がある(Richard et al., J. Biol. Chem. 2003, 278:585-590;Drin et al., J. Biol. Chem. 2003, 278:381192-31201)。さらにまた、最近の報告ではこれらPTDは細胞に対して有毒であることが示されている。
したがって、エネルギー非依存的非エンドサイトーシス態様で細胞の脂質膜を通過することができるペプチドが希求されている。さらにまた、大きなペプチドについて一般的に知られている免疫反応を回避するために、より小さな、ペプチダーゼ耐性ペプチドが求められている。最後に、前記ペプチド担体は細胞に対して毒性がないことが重要である。
【発明の概要】
【発明が解決しようとする課題】
【0003】
エネルギー非依存性メカニズムによって細胞膜を通過し、分子を細胞内に送達する方法および、そのための担体複合体を提供する。
【課題を解決するための手段】
【0004】
前述の要求は、分子を細胞に送達する方法を提供する本発明によって充足される。本方法は、細胞を担体複合体と接触させることを含み、前記担体複合体は、送達する分子及び芳香族陽イオンペプチドを含み、さらに前記芳香族陽イオンペプチドは以下を含む:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmはr+1以下である最大の数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下である最大の数であるが、ただしaが1であるときはptもまた1であることができる関係。
また別の実施態様では、本発明は、分子及び芳香族陽イオンペプチドを含む担体複合体を提供し、前記芳香族陽イオンペプチドは以下を含む:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmはr+1以下である最大の数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下である最大の数であるが、ただしaが1であるときはptもまた1であることができる関係。
さらに別の実施態様では、本発明は分子を細胞に送達する方法を提供する。本方法は、細胞を分子及び芳香族陽イオンペプチドと接触させることを含み、前記芳香族陽イオンペプチドは以下を含む:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmはr+1以下である最大の数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下である最大の数であるが、ただしaが1であるときはptもまた1であることができる関係。
【図面の簡単な説明】
【0005】
【図1】Caco-2細胞のペプチドの取り込み。[3H][Dmt1]DALDA(A)及び[14C]Gly-Sar(B)の経時的取り込み。Caco-2細胞を[3H][Dmt1]DALDA(250nM, 47Ci/mmol)又は[14C]Gly-Sar(50μM, 56.7mCi/mmol)と37℃又は4℃で1時間インキュベートした。その後、放射能を溶解細胞で測定した。(C)[3H][Dmt1]DALDAの蓄積に対する酸洗浄の影響。Caco-2細胞を[3H][Dmt1]DALDAとともに37℃で1時間インキュベートした。細胞の溶解前に、細胞を酸洗浄に付して細胞表面結合放射能を除去した。(D)[Dmt1]DALDA取り込みに対する[Dmt1]DALDAの濃度の影響。細胞をある範囲の濃度(1μMから3mM)の[Dmt1]DALDAとともに1時間37℃でインキュベートした。全てのデータは、3つのそれぞれ別個の単層細胞の平均±S.E.として示されている。誤差バーが示されていない場合は、誤差バーは記号サイズより小さい。
【図2】Caco-2細胞における[3H][Dmt1]DALDA取り込み(A及びC)及び[14C]Gly-Sar取り込み(B及びD)に対するpH及びDEPCの影響。Caco-2細胞を[3H][Dmt1]DALDA(250nM, 47Ci/mmol)又は[14C]Gly-Sar(50μM, 56.7mCi/mmol)とともに種々のpH条件下で37℃で1時間インキュベートした(A及びB)。[3H][Dmt1]DALDA(250nM, 47Ci/mmol)又は[14C]Gly-Sar(50μM, 56.7mCi/mmol)と37℃で1時間インキュベートする前に、細胞を0.2mMのDEPCと10分間25℃でプレインキュベートした(C及びD)。全てのデータは、3つのそれぞれ別個の単層細胞の平均±S.E.として示されている。
【図3】(A)種々の細胞株における[3H][Dmt1]DALDAの取り込み。細胞を[3H][Dmt1]DALDA(250nM, 47Ci/mmol)とともに37℃で1時間インキュベートした。細胞の溶解前に、細胞を酸洗浄に付して細胞表面結合放射能を除去した。提示データは酸耐性放射能を示し、3つのそれぞれ別個の単層細胞の平均±S.E.として表示されている。(B) [3H][Dmt1]DALDAの細胞膜への特異的結合。SH-SY5Y細胞及びCaco-2細胞から調製した膜を[3H][Dmt1]DALDA(15から960pM)とともに25℃で1時間インキュベートした。非特異的結合は、1μMの非標識[Dmt1]DALDAを含めることによって評価した。遊離放射性リガンドは迅速ろ過によって結合放射性リガンドから分離した。特異的結合はCaco-2細胞では観察されなかった。SH-SY5Y細胞については、Kd値は118pM(87から149の範囲)で、Bmax値は96 fmol/mgタンパク質であった。
【図4】(A)[3H][Dmt1]DALDA(黒棒線)及び[14C]Gly-Sar(白棒線)の流出。Caco-2細胞に[3H][Dmt1]DALDA(250nM, 47Ci/mmol)又は[14C]Gly-Sar(50μM, 56.7mCi/mmol)を37℃又は4℃で1時間予備装荷した。続いて細胞を洗浄し、さらに培養液とともに37℃又は4℃で1時間インキュベートした。培養液及び細胞溶解物の両方で放射能を測定した。データは培養液中に流出したペプチドの百分率として表されている。(B)[3H][Dmt1]DALDA流出に対するDEPCの影響。[3H][Dmt1]DALDAを装荷する前に、細胞を25℃で10分間、0.2mMのDEPCとともにインキュベートした。(C)[3H][Dmt1]DALDAの流出に対するベラパミル(p-糖タンパク質の阻害剤)の影響、及び(D)[3H][Dmt1]DALDAの取り込みに対するベラパミルの影響。
【図5】単層Caco-2細胞を通過する[3H][Dmt1]DALDA及び[14C]Gly-Sarの輸送。Caco-2細胞(2x105)をトランスウェル細胞培養チャンバー内の微小孔を有するメンブレンフィルター上に播種した。先端部から外側基底部へのペプチド輸送は、 [3H] [Dmt1]DALDA又は[14C]Gly-Sarを先端部区画に添加することによって決定した。放射能の計測のために、ペプチド添加後種々の時間に20μLのアリコットを先端部区画及び外側基底部区画の両方から取り出した。
【図6】[Dmt1,dnsDap4]DALDA及び[Dmt1,atnDap4]DALDAの細胞内取り込み。Caco-2細胞を0.1μMの[Dmt1,dnsDap4]DALDAとともに37℃で15分間インキュベートした。続いて細胞を洗浄し、PBSで被覆した。顕微鏡検査は室温で10分以内に実施した。励起は340nmで実施し、放射は520nmで測定した。蛍光は細胞質全体に拡散しているように見えたが、核からは完全に排除されていた。37℃で小胞体内濃縮が認められないことから、非エンドサイトーシス性取り込みが示唆される。
【図7】3つのペプチドと架橋剤SMCCとのカップリングの質量分析による確認。SMCC(1μg)及びペプチド(5μg)を一緒に2mLのPBSに溶解し、室温で30分間インキュベートし、4℃で保存した。前記サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。サンプルはMALDI-TOF MS(Matrix Assisted Laser Desorption Ionization Time-of-Flight Mass Spectrometry)によって分析した。ペプチド及びそれらの対応するSMCC共役物の分子量はスペクトル上に表示されている。
【図8】β-ガラクトシダーゼ(β-Gal)のN2A神経芽腫細胞内への取り込みを強化するペプチドの能力。細胞(N2A神経芽腫細胞又はCaco-2細胞)を96ウェルプレートにプレーティングし(2x104細胞/ウェル)、β-Gal又は(SMCCを介して)ペプチドと共役させたβ-Galとともに37℃で1時間インキュベートした。続いて細胞をリン酸緩衝液で4回洗浄した。次に、細胞をβ-gal染色セット(Roche)で37℃で少なくとも2時間染色し、顕微鏡下で調べた。(A)CaCo-2細胞をβ-Galとともにインキュベートしたときにはβ-Galの取り込みは観察されなかった。(B)青い細胞の存在は[Dmt1]DALDAに共役したβ-GalのCaco-2細胞内取り込みを示している。(C)[D-Arg-Dmt-Lys-Phe-NH2]に共役したβ-GalのCaco-2細胞内取り込み強化。(D)[Phe1]DALDAに共役したβ-GalのCaco-2細胞内取り込みの強化。SMCCのみと共役したβ-Galでは取り込みは強化されなかった。
【図9】[Dmt 1]DALDA−SMCC共役物との同時インキュベーションは緑色蛍光タンパク質(GFP)のHuh7細胞内への取り込みを強化する。Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて3μgの緑色蛍光タンパク質(GFP)単独(A)、3μgのGFP及び40μLの[Dmt1]DALDA(B)、又は3μgのGFP及びSMCCと共役させた40μL[Dmt1]DALDA(C)を含む0.5mLのMEMとともに37℃で60分間インキュベートした。続いて2mLの細胞培養液を細胞に添加し、これを細胞培養インキュベーターでさらに24時間インキュベートした。インキュベーション後に細胞を細胞培養液で4回洗浄し生細胞中に保持されているGFPを共焦点レーザー走査型顕微鏡で可視化した。励起は340nmで実施し、放射は520nmで測定した。上段パネルは、Huh7細胞の0.8μm厚の中央部水平光学切片を通したGFPの画像を示す。下段パネルは、同じ顕微鏡野の微分干渉コントラスト像を示す。
【図10】[Dmt1]DALDAとRNAオリゴとの共役。γ-32P-ATP及びポリヌクレオチドキナーゼを用いて合成RNAオリゴ(長さは40ヌクレオチド)の5'端をリン酸化した。生成物をゲル電気泳動で精製した。ゲル精製RNAオリゴの500,000cpmを1mgのEDC(N-[3-ジメチルアミノプロピル-N'-エチルカルボジイミド])の存在下で[Dmt1]DALDAと結合させた。前記結合反応の生成物([Dmt1]DALDA−RNAオリゴ)及び単独RNAオリゴは15%ポリアクリルアミド尿素ゲルで分析した。
【図11】[Dmt1]DALDA−[32P]RNAオリゴ共役物のCaco-2細胞内への取り込み。Caco-2細胞(1x106)をDMEM培養液で3回洗浄し、DMEMで5分間プレインキュベートした。続いて細胞を[Dmt1]DALDA−[32P]RNAオリゴ共役物又は対照RNA(約20,000cpm)と37℃で60分間インキュベートした。インキュベーション後に、細胞を洗浄して溶解し、細胞溶解物中の放射能を測定した。[Dmt1]DALDA−[32P]RNAオリゴ共役物の取り込みは、RNA単独と比較して3倍を超えて強化された。
【図12】RNAオリゴのHuh7細胞内への取り込みを強化するペプチド-SMCC共役物の効果。(A)RNAオリゴの細胞取り込みに対する時間の影響。Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて[32P]RNAオリゴ(一本鎖、11塩基;約100,000cpm)単独又は40mLの[Dmt1]DALDA−SMCC共役物を含む1.0mLのDMEMと37℃で15分間又は60分間インキュベートした。続いて、細胞をDMEM中で4回、さらに酢酸ナトリウム溶液で1回洗浄して非特異的結合を取り除いた後、溶解緩衝液で30分間インキュベートし、保持されている放射能を測定した。RNAオリゴと[Dmt1]DALDA−SMCCと37℃で同時インキュベートすることによって、RNAオリゴの取り込みは15分間インキュベーション後に10倍、60分間インキュベーション後に20倍増加した。(B)RNAオリゴの細胞取り込みに対する温度の影響。[Dmt1]DALDA−SMCCのRNA取り込みを強化する能力は4℃では低かったが、それでもなお前記は取り込みを10倍増加させた。(C)種々のペプチド−SMCC共役物によるRNAの細胞内取り込みの強化。Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて[32P]RNAオリゴ単独又は40mLのペプチド−SMCC共役物を含む1.0mLのDMEMと37℃で15分間インキュベートした。3つのペプチド−SMCC共役物はいずれもRNA取り込みを増加させた。
【図13】[Dmt1]DALDA−SMCC共役物との同時インキュベーションは異なる長さの2つのRNAの取り込みを強化した。[Dmt1]DALDAをSMCCと共役させ、質量分析によって確認した。11-mer RNAオリゴ及び1350-mer RNAを[Dmt1]DALDA−SMCC共役物と室温で15分間混合した。Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いてRNA(約100,000cpm)単独又は[Dmt1]DALDA−SMCC共役物と混合したRNAを含む1.0mLのDMEMと37℃及び5%CO2で60分間インキュベートした。続いて、洗浄した細胞をDMEM中で4回、さらに酢酸ナトリウム溶液で1回洗浄して非特異的結合を取り除いた。続いて細胞を溶解緩衝液で30分間インキュベートし、保持されている放射能を測定した。RNA単独とのインキュベーションと比較して、[Dmt1]DALDA−SMCC共役物とのインキュベーションによって11-mer RNAは22倍、1350-mer RNAは3倍、取り込みが増加した。
【図14】DNAオリゴの[Dmt1]DALDAへの共役。SMCC(1μg)及び[Dmt1]DALDA(5μg)を一緒に2mLのPBSに溶解し、室温で30分間インキュベートし、脱保護した3'-チオールDNAオリゴと4℃で24時間混合した。インキュベーション後に、サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。サンプルはMALDI-TOF MSで分析した(A)。3'-チオールDNAオリゴ及び[Dmt1]DALDA共有結合複合体の分子量はそれぞれ6392及び7171であることが判明した。γ-32P-ATP及びポリヌクレオチドキナーゼを用いて、共役オリゴ及び非共役オリゴの両者の5'端をリン酸化し、キナーゼ反応の生成物を15%ポリアクリルアミド尿素ゲルで分析し、細胞取り込み実験のためにゲル精製した(B)。
【図15】[Dmt1]DALDAと共役したDNAオリゴの細胞内取り込み。SMCCを用いて3'-チオール改変20-mer DNAを[Dmt1]DALDAと共役させ、共役物の生成は質量分析によって確認した。共役及び非共役DNAオリゴの5'末端を32Pで放射能標識し、さらにゲル精製した。神経細胞系N2A細胞(1x106細胞/ウェル)をDMEMで洗浄し、[Dmt1]DALDA共役又は非共役DNAオリゴ(約100,000cpm)のどちらかを含む1mLのDMEMとともに37℃及び5%CO2で2時間又は19時間インキュベートした。続いて細胞を4回DMEMで、さらに1回酢酸ナトリウム溶液中で洗浄して非特異的結合を除去した。続いて細胞を溶解緩衝液中で30分間インキュベートし、保持されている放射能を測定した。Y軸は、総放射能の百部率で表したDNAの取り込みを示している。
【図16】[Dmt1]DALDAは培養細胞に毒性を示さない。神経細胞系N2A細胞を [Dmt1]DALDA(1nMから10μM)とともに24時間インキュベートし、細胞生存度をMTTアッセイによって決定した。
【図17】[Dmt1]DALDA−SMCC共役物はHuh7細胞にアポトーシスを誘導しない。Huh7細胞(1x106細胞/ウェル)をDMEM中で3回洗浄し、1mLの新しい培養液を適用した。続いて50μL のPBS中の[Dmt1]DALDA−SMCC共役物(1mM)又はPBSのみ(対照)を前記細胞培養液に添加し、5%CO2で37℃24時間インキュベートした。インキュベーション後に、アポトーシス核染色用ヘキスト色素(Molecular Probes, Eugene, OR)の1mLを細胞に添加し、さらに15分間インキュベートした。過剰なヘキスト色素を細胞培養液(pHインジケーターを含まない)で細胞を洗浄することによって除去し、蛍光顕微鏡検査(励起350nm、放射461nm)を用いて[Dmt1]DALDA−SMCC共役物で処理した細胞を対照細胞と比較した。
【発明を実施するための形態】
【0006】
発明の詳細な説明
本発明は、少なくとも1つの分子及び芳香族陽イオンペプチドを含むある種の担体複合体は、エネルギー非依存性メカニズムによって細胞膜を通過し、前記分子を細胞内に送達することができるという本発明者らの驚くべき発見によるものである。
芳香族陽イオンペプチド
本発明で有用な芳香族陽イオンペプチドは、以下に説明するように正味の正電荷を有し、水溶性であり強い極性を示す。前記ペプチドは最小限3つのアミノ酸を含み、好ましくは最小限4つのアミノ酸を含み、前記はペプチド結合によって共有結合されている。
前記芳香族陽イオンペプチドに存在するアミノ酸の最大数は10、好ましくは約8、もっとも好ましくは約6である。最適には、前記ペプチドに存在するアミノ酸の数は約4である。アミノ酸の最大数の定義で用いられる“約”という用語は、+1又は−1個のアミノ酸を意味する。
本発明で有用な芳香族陽イオンペプチドのアミノ酸は任意のアミノ酸であってよい。本明細書において、“アミノ酸”という用語は、少なくとも1つのアミノ基及び少なくとも1つのカルボキシル基を含む任意の有機分子を指すために用いられる。好ましくは、少なくとも1つのアミノ基はカルボキシル基に対してα位に存在する。
前記アミノ酸は天然に存在するであり得る。天然に存在するアミノ酸には、例えば、タンパク質に通常見出される最も一般的な20のアミノ酸、すなわちアラニン(Ala)、アルギニン(Arg)、アスパラギン(Asn)、アスパラギン酸(Asp)、システイン(Cys)、グルタミン(Glu)、グルタミン酸(Glu)、グリシン(Gly)、ヒスチジン(His)、イソロイシン(Ileu)、ロイシン(Leu)、リジン(Lys)、メチオニン(Met)、フェニルアラニン(Phe)、プロリン(Pro)、セリン(Ser)、スレオニン(Thr)、トリプトファン(Trp)、チロシン(Tyr)、及びバリン(Val)が含まれる。
【0007】
天然に存在する他のアミノ酸には、例えばタンパク質合成に関連しない代謝過程で合成されるアミノ酸が含まれる。例えば、アミノ酸であるオルニチンは哺乳動物の代謝で尿素生成時に合成される。
本発明で有用な芳香族陽イオンペプチドは、場合によって天然には存在しない1つ又は2つ以上のアミノ酸を含む。ある実施態様では、前記ペプチドは天然に存在するアミノ酸を含まない。
天然に存在しないアミノ酸は、典型的には生きている生物の通常の代謝過程では合成されず、タンパク質中に通常出現しないアミノ酸である。
さらに、本発明で有用な天然に存在しないアミノ酸は好ましくは一般的なプロテアーゼによって認識されない。したがって、前記天然に存在しないアミノ酸は、一般的なプロテアーゼに対して好ましくは耐性であり、より好ましくは非感受性である。
天然に存在しないアミノ酸は前記ペプチドのいずれの位置にあってもよい。例えば、天然に存在しないアミノ酸はN-末端、C-末端及び/又はN-末端とC-末端の間の1つ又は2つ以上の任意の位置に存在することができる。
非天然アミノ酸は、例えばアルキル基、アリール基又はアルキルアリール基を含むことができる。アルキルアミノ酸のいくつかの例にはα-アミノ酪酸、β-アミノ酪酸、γ-アミノ酪酸、δ-アミノ吉草酸及びε-アミノカプロン酸が含まれる。アリールアミノ酸のいくつかの例には、オルト-、メタ-及びパラ-アミノ安息香酸が含まれる。アルキルアリールアミノ酸のいくつかの例には、オルト-、メタ-、及びパラ-アミノフェニル酢酸並びにγ-フェニル-β-アミノ酪酸が含まれる。
天然に存在しないアミノ酸にはまた天然に存在するアミノ酸の誘導体が含まれる。天然に存在するアミノ酸の誘導体は、例えば天然に存在するアミノ酸に対し1つ又は2つ以上の化学基の付加を含む。
【0008】
例えば、1つ又は2つ以上の化学基は、フェニルアラニン又はチロシン残基の芳香環の2'、3'、4'、5'若しくは6'位の1つ又は2つ以上に、又はトリプトファン残基のベンゾ環の4'、5'、6'若しくは7'位の1つ又は2つ以上に付加することができる。この基は芳香環に付加することができる任意の化学基であり得る。そのような基のいくつかの例には、分枝又は非分枝C1‐C4アルキル、例えばメチル、エチル、n-プロピル、イソプロピル、ブチル、イソブチル若しくはt-ブチル、C1‐C4アルコキシ(すなわちアルコキシ)、アミノ、C1‐C4アルキルアミノ(例えばメチルアミノ)及びC1‐C4ジアルキルアミノ(例えばジメチルアミノ)、ニトロ、ヒドロキシル、ハロ(すなわちフルオロ、クロロ、ブロモ又はヨード)が含まれる。天然に存在するアミノ酸の天然には存在しない誘導体のいくつかの具体例には、ノルバリン(Nva)、ノルロイシン(Nle)及びヒドロキシプロリン(Hyp)が含まれる。
本発明で有用なペプチドのアミノ酸改変のまた別の例は、前記ペプチドのアスパラギン酸又はグルタミン酸のカルボキシル基の誘導体化である。誘導体化の一例は、アンモニア又は第一若しくは第二アミン(例えばメチルアミン、エチルアミン、ジメチルアミン又はジエチルアミン)によるアミド化である。誘導体化のまた別の例には例えばメチル又はエチルアルコールによるエステル化が含まれる。
また別の改変にはリジン、アルギニン又はヒスチジンのアミノ基の改変が含まれる。例えば、これらのアミノ基はアシル化することができる。いくつかの適切なアシル基には、例えばベンゾイル基又はアルカノイル基が含まれ、前記は上記で述べた任意のC1‐C4アルキル基、例えばアセチル又はプロピオニル基を含む。
天然に存在しないアミノ酸は、一般的には左旋性(L-)でも右旋性(D-)でも又はその混合物でもあり得る。天然に存在しない適切なアミノ酸の例にはまた、上記の天然に存在するL-アミノ酸のいずれかの右旋性型が、天然には存在しないアミノ酸のL-及び/又はD-型とともに含まれる。これに関して、D-アミノ酸は通常はタンパク質に存在しないことは留意されるべきである(ただしそれらは細胞の正常なリボソームによるタンパク質合成機構以外の手段によって合成されるある種のペプチド抗生物質で見出される)。本明細書において、そのようなD-アミノ酸は天然に存在しないアミノ酸と考えられる。
【0009】
プロテアーゼに対する感受性を最小限にするために、本発明で有用なペプチドは、天然に存在するか否かに関係なく、一般的なプロテアーゼによって認識される、5つ未満、好ましくは4つ未満、より好ましくは3つ未満、もっとも好ましくは2つ未満の連続するL-アミノ酸を有するべきである。ある実施態様では、前記ペプチドはD-アミノ酸のみであり、L-アミノ酸は存在しない。
ペプチドがプロテアーゼ感受性アミノ酸配列を含む場合は、少なくとも1つのアミノ酸は好ましくは天然に存在しないD-アミノ酸であり、それによってプロテアーゼ耐性を付与する。プロテアーゼ感受性配列の例には、一般的なプロテアーゼ(例えばエンドペプチダーゼ及びトリプシン)によって切断される2つ以上の連続する塩基性アミノ酸が含まれる。塩基性アミノ酸の例にはアルギニン、リジン、及びヒスチジンが含まれる。
芳香族陽イオンペプチドが、ペプチド中のアミノ酸の総数に対して生理学的pHで最少数の正味の正電荷を有するということが重要である。生理学的pHでの正味の正電荷の最小数は下記では(pm)と称される。ペプチド内のアミノ酸残基の総数は下記では(r)と称される。
下記で考察される正味の正電荷の最小数は全て生理学的pHでのものである。本明細書で用いられる“生理学的pH”という用語は、哺乳動物の体内の組織及び器官の細胞の正常なpHを指す。例えばヒトの生理学的pHは通常ほぼ7.4であるが、哺乳動物の正常な生理学的pHは約7.0から約7.8の間の任意のpHであり得る。
【0010】
本明細書で用いられる“正味の電荷”は、ペプチド内に存在するアミノ酸が保持する正電荷数と負電荷数の差し引きを指す。本明細書では、正味の電荷は生理学的pHで測定されると理解される。生理学的pHで正に荷電する天然に存在するアミノ酸にはL-リジン、L-アルギニンおよびL-ヒスチジンが含まれる。生理学的pHで負に荷電する天然に存在するアミノ酸にはL-アスパラギン酸及びL-グルタミン酸が含まれる。
典型的には、ペプチドは正に荷電したN-末端アミノ基及び負に荷電したC-末端カルボキシル基を有する。電荷は生理学的pHで互いに相殺される。正味の電荷を計算する例として、ペプチドTyr-Arg-Phe-Lys-Glu-His-Trp-Argは、1つの負に荷電したアミノ酸(すなわちGlu)及び4つの正に荷電したアミノ酸(すなわち2つのArg残基、1つのLys及び1つのHis)を有する。したがって、上記のペプチドは正味の正電荷3を有する。
本発明のある実施態様では、芳香族陽イオンペプチドは、生理学的pHでの正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間に、3pmがr+1以下である最大数であるという関係を有する。この実施態様では、正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間の関係は以下のとおりである:

別の実施態様では、芳香族陽イオンペプチドは、正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間に、2pmがr+1以下である最大数であるという関係を有する。この実施態様では、正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間の関係は以下のとおりである:
ある実施態様では、正味の正電荷の数(pm)とアミノ酸残基の数(r)は等しい。別の好ましい実施態様では、前記ペプチドは、3つから4つのアミノ酸残基、及び最小限1つの正味の正電荷、好ましくは最小限2つの正味の正電荷、より好ましくは最小限3つの正味の正電荷を有する。
芳香族陽イオンペプチドが正味の正電荷の総数(pt)に対し最小数の芳香族基を有することもまた重要である。芳香族基の最小数は以下では(a)と称する。
【0011】
芳香族基をもつ天然に存在するアミノ酸には、ヒスチジン、トリプトファン、チロシン及びフェニルアラニンが含まれる。例えば、ヘキサペプチドであるLys-Gln-Tyr-Arg-Phe-Trpは正味の正電荷が2であり(リジン及びアルギニン残基が寄与している)、3つの芳香族基(チロシン、フェニルアラニン及びトリプトファン残基によって付与される)を有する。
本発明のある実施態様では、本発明の方法で有用な芳香族陽イオンペプチドは、芳香族基の最小数(a)と生理学的pHでの正味の正電荷の総数(pt)との間に、3aがpt+1以下である最大の数であるが、ただしptが1であるときはaもまた1であることができる関係を有する。この実施態様では、芳香族基の最小数(a)と正味の正電荷の数(pt)との間の関係は以下のとおりである:
別の実施態様では、芳香族陽イオンペプチドは、芳香族基の最小数(a)と正味の正電荷の総数(pt)との関係において、2aがpt+1以下である最大の数である。この実施態様では、芳香族アミノ酸残基の最小数(a)と正味の正電荷の総数(pt)との間の関係は以下のとおりである:
別の実施態様では、芳香族基の数(a)と正味の正電荷の総数(pt)は等しい。
【0012】
カルボキシル基、特にC-末端アミノ酸の末端カルボキシル基は好ましくは、例えばアンモニアでアミド化されて、C-末端アミドを形成する。あるいは、C-末端アミノ酸の末端カルボキシル基は、第一又は第二アミンでアミド化されてもよい。前記第一又は第二アミンは、例えばアルキル、特に分枝又は非分枝C1‐C4アルキル又はアリールアミンであり得る。したがって、本ペプチドのC-末端のアミノ酸は、アミド、N-メチルアミド、N-エチルアミド、N,N-ジメチルアミド、N,N-ジエチルアミド、N-メチル-N-エチルアミド、N-フェニルアミド又はN-フェニル-N-エチルアミド基に変換することができる。
さらに、2つ以上のカルボキシレート基(例えばアスパラギン、グルタミン、アスパラギン酸又はグルタミン酸残基)を有するアミノ酸残基の遊離カルボキシレート基もまた、それらがどこに存在していてもアミド化することができる。これらの位置におけるアミド化は、上記のようにアンモニアを用いても、又は第一若しくは第二アミンのいずれを用いてもよい。
ある実施態様では、本発明の方法で有用な芳香族陽イオンペプチドは、正味の正電荷が2で少なくとも1つの芳香族アミノ酸を有するトリペプチドである。具体的な実施態様では、本発明の方法で有用な芳香族陽イオンペプチドは、正味の正電荷が2で2つの芳香族アミノ酸を有するトリペプチドである。
本発明の方法で有用な芳香族陽イオンペプチドには以下のペプチド例が含まれる(ただしこれらに限定されない):
Lys-D-Arg-Tyr-NH2
Phe-D-Arg-His
D-Tyr-Trp-Lys- NH2
Trp-D-Lys-Tyr-Arg- NH2
Tyr-His-D-Gly-Met
Phe-Arg-D-His-Asp
Tyr-D-Arg-Phe-Lys-Glu- NH2
Met-Tyr-D-Lys-Phe-Arg
D-His-Glu-Lys-Tyr-D-Phe-Arg
Lys-D-Gln-Tyr-Arg-D-Phe-Trp-NH2
Phe-D-Arg-Lys-Trp-Tyr-D-Arg-His
Gly-D-Phe-Lys-Tyr-His-D-Arg-Tyr-NH2
Val-D-Lys-His-Tyr-D-Phe-Ser-Tyr-Arg-NH2
Trp-Lys-Phe-D-Asp-Arg-Tyr-D-His-Lys
Lys-Trp-D-Tyr-Arg-Asn-Phe-Tyr-D-His-NH2
Thr-Gly-Tyr-Arg-D-His-Phe-Trp-D-His-Lys
Asp-D-Trp-Lys-Tyr-D-His-Phe-Arg-D-Gly-Lys-NH2
D-His-Lys-Tyr-D-Phe-Glu-D-Asp-D-His-D-Lys-Arg-Trp-NH2 及び
Ala-D-Phe-D-Arg-Tyr-Lys-D-Trp-His-D-Tyr-Gly-Phe
【0013】
特に好ましい実施態様では、芳香族陽イオンペプチドは、式Tyr-D-Arg-Phe-Lys-NH2(簡便に頭文字でDALDAと表わす)を有する。DALDAは正味の正電荷が3で(前記はチロシン、アルギニン及びリジンによって付与される)、フェニルアラニン及びチロシンの寄与による2つの芳香族基を有する。DALDAのチロシンは、例えば2',6'-ジメチルチロシンのようにチロシンの改変誘導体であってよく、式2',6'-Dmt-D-Arg-Phe-Lys-NH2(すなわちDmt1-DALDA)を有する化合物を生成することができる。チロシンの他の改変誘導体には以下が含まれる:2'-メチルチロシン(Mmt);N,2',6'-トリメチルチロシン(Tmt);及び2'-ヒドロキシ-6'-メチルチロシン(Hmt)。
別の好ましい実施態様では、DALDAのN-末端のアミノ酸はフェニルアラニン又はその誘導体であり得る。N-末端にフェニルアラニンを有する芳香族陽イオンペプチドは式Phe-D-Arg-Phe-Lys-NH2(すなわちPhe1-DALDA)を有する。好ましいフェニルアラニンの誘導体には以下が含まれる:2'-メチルフェニルアラニン(Mmp);2',6'-ジメチルフェニルアラニン(Dmp);N,2',6'-トリメチルフェニルアラニン(Tmp);及び2'-ヒドロキシ-6'-メチルフェニルアラニン(Hmp)。
別の実施態様では、Dmt1-DALDAのアミノ酸配列は、DmtがN-末端に存在しないように再編成される。そのような芳香族陽イオンペプチドの例は式D-Arg-2',6'Dmt-Lys-Phe-NH2を有する。
【0014】
本明細書に開示した具体的なペプチドのいずれも、例えば上記で述べたペプチド及び下記(例えば表1)で述べるペプチド(Dmt1-DALDA、DALDA、Phe1-DALDA、D-Arg-2',6'Dmt-Lys-Phe-NH2及びそれらの誘導体を含む)、さらに機能的な類似体を含むことができる。ペプチドは、類似体がDmt1-DALDA、DALDA、Phe1-DALDA又はD-Arg-2',6'Dmt-Lys-Phe-NH2と同じ機能を有するならば、Dmt1-DALDA、DALDA、Phe1-DALDA、D-Arg-2',6'Dmt-Lys-Phe-NH2の機能的類似体と考えられる。例えば前記類似体は、1つ又は2つ以上のアミノ酸が別のアミノ酸によって置換された、Dmt1-DALDA、DALDA、Phe1-DALDA、D-Arg-2',6'Dmt-Lys-Phe-NH2の置換変種であり得る。
Dmt1-DALDA、DALDA、Phe1-DALDA、D-Arg-2',6'Dmt-Lys-Phe-NH2の適切な置換変種は保存的アミノ酸置換を含む。アミノ酸はそれらの物理化学的特性に従って以下のように分類できる:
(a)非極性アミノ酸:Ala(A)、Ser(S)、Thr(T)、Pro(P)、Gly(G);
(b)酸性アミノ酸:Asn(N)、Asp(D)、Glu(E)、Gln(Q);
(c)塩基性アミノ酸:His(H)、Arg(R)、Lys(K);
(d)疎水性アミノ酸:Met(M)、Leu(L)、Ile(I)、Val(V);及び
(e)芳香族アミノ酸:Phe(F)、Tyr(Y)、Trp(W)、His(H)。
ペプチド内のアミノ酸の同じグループ内のアミノ酸による置換は保存的置換と称される。保存的置換は元のペプチドの物理化学的特性を保存する傾向がある。対照的に、ペプチド内のアミノ酸の異なるグループ内の別のアミノ酸による置換は、一般的に元のペプチドの特性を変化させる傾向がある。
【0015】
本発明の実施で有用な類似体の例には表1及び2に示す芳香族陽イオンペプチドが含まれる(ただしこれらに限定されない)。
表1:
Dab=ジアミノ酪酸
Dap=ジアミノプロピオン酸
Dmt=ジメチルチロシン
Mmt=2'-メチルチロシン
Tmt=N,2',6'-トリメチルチロシン
Hmt=2'-ヒドロキシ,6'-メチルチロシン
dnsDap=β-ダンシル-L-α,β-ジアミノプロピオン酸
atnDap=β-アントラニロイル-L-α,β-ジアミノプロピオン酸
Bio=ビオチン
【0016】
表2:
Cha=シクロヘキシル
表1及び2に示されたペプチドのアミノ酸はL-又はD-立体配置のいずれでもよい。
さらに別の陽イオンペプチドは米国仮特許出願60/444,777号(2003年2月4日出願)で見出すことができる(前記出願は参照により本明細書に含まれる)。
【0017】
分子
分子は生物学的分子又は小分子であり得る。好ましくは、前記生物学的分子又は小分子は医薬的に活性な分子である。本明細書で用いられる医薬的に活性な分子は、in vivoで有益な作用を示す任意の分子である。
生物学的な分子は、核酸又はアミノ酸配列を含み、分子量が450を超える任意の分子である。そのような核酸及びアミノ酸配列は、本明細書ではそれぞれ“ポリヌクレオチド”及び“ポリアミノ酸”と称される。
生物学的分子にはポリヌクレオチド、ペプチド核酸及びポリアミノ酸、例えばペプチド、ポリペプチド及びタンパク質が含まれる。医薬的に活性を有する生物学的分子の例には、内因性ペプチド(例えばバゾプレッシン、グルタチオン)、タンパク質(例えばインターフェロン)、ホルモン(例えばヒト成長ホルモン)、酵素(例えばα-ガラクトシダーゼ)、抗体(例えばβ-アミロイドに対する抗体、前記はアルツハイマー病の治療に用いることができる)、神経栄養性増殖因子(例えば神経増殖因子NGF、脳由来神経栄養性因子BDNF)、サイトカイン(例えば血小板由来増殖因子PDGF、血管内皮細胞増殖因子VEGF)及びオリゴヌクレオチドが含まれる。
前記オリゴヌクレオチドは、任意のヌクレオチド配列(例えばDNA又はRNA)を含むことができる。前記DNA及びRNA配列は一本鎖でも二本鎖でもよい。例えば、ストレス時の細胞の生存の促進に有益なタンパク質をコードするDNAを本発明のペプチドに共役させることができる。そのようなタンパク質の例には熱ショックタンパク質(例えばhsp60、hsp70など)が含まれる。
一本鎖RNA分子の例にはリボザイム、RNAデコイ、リボザイムのための外側ガイド配列、アンチセンスRNA及びmRNAが含まれる。これら一本鎖RNA分子の総説については以下の文献を参照されたい:Sullenger et al. Nature 418:252-247(2002)。前記文献(Sullenger)で開示されている、これら一本鎖RNAのついての記述、並びにリボザイム、RNAデコイ、リボザイムのための外側ガイド配列、アンチセンスRNA及びmRNA分子により治療することができる病気及び疾患についての記述は、引用により本明細書に含まれる。
【0018】
二本鎖RNAの例はRNA干渉性分子(すなわちRNAi、例えばsiRNA(すなわち干渉性小RNA))である。前記siRNAは当技術分野で公知である任意のsiRNAであり得る。
siRNAは、例えば、疾患、症状又は病気で示唆されているタンパク質の翻訳を阻害することができるほど充分にmRNAに相補的であり得る。そのようなタンパク質の例には、例えばアルツハイマー病と関連しているβ-アミロイド及び癌と関連しているrasタンパク質が含まれる。
あるいは、siRNAは、例えばウイルスによって生成されるRNAと十分に相補的であり得る。ウイルスによって生成されるRNAは、一般的に宿主細胞の感染、ウイルスの生存及び/又はウイルスの増殖に必要ないずれのRNAでもあり得る。そのようなRNAの例には、内部リボソームエントリー部位、RNA依存ポリメラーゼ開始部位、並びにウイルスのエンベロープタンパク質、ウイルスヌクレアーゼ及びウイルスプロテアーゼをコードするRNAが含まれる。
ウイルスの例には、例えば肝炎ウイルス(例えばA、BおよびC型肝炎ウイルス)、ヒト免疫不全ウイルス、エプスタイン-バールウイルス、サイトメガロウイルス及びヒトパピローマウイルスが含まれる。
ウイルスRNAを標的とするsiRNAは当業者には公知である。例えば、C型肝炎ウイルスRNAを標的とするsiRNAは当業者に公知であり、下記の文献を参照されたい:Randall et al. PNAS 100:235-240(2003)。
この分子は小分子でもよい。小分子には有機化合物、有機金属化合物、有機及び有機金属化合物の塩、単糖類、アミノ酸及びヌクレオチドが含まれる。小分子にはさらに、分子量が450を越えない点を除けば生物学的分子と考えられる分子が含まれよう。したがって、小分子は脂質、オリゴ糖、オリゴペプチド及びオリゴヌクレオチド、並びにそれらの誘導体で、分子量が450以下を有するものであり得る。
小分子は任意の分子量を有することができることは強調されよう。それらは単に生物学的分子とみなされないために小分子と称されるだけであり、典型的には450未満の分子量を有する。小分子には、合成化合物とともに天然に見出される化合物が含まれる。医薬的に活性を有する小分子の例には抗生物質(例えばテトラサイクリン、ペニシリン、エリスロマイシン)、細胞毒性物質(例えばドキソルビシン、アドリアマイシン)、及び抗酸化剤(例えばビタミンE、ビタミンC、ベータカロチン)が含まれる。
【0019】
担体複合体
上記で述べた少なくとも1つの分子、及び上記で述べた少なくとも1つの芳香族陽イオンペプチドが会合して担体複合体が形成される。前記分子及び芳香族陽イオンペプチドは当業者に公知の任意の方法によって会合される。適切な会合の型には化学的結合及び物理的結合が含まれる。化学的結合には、例えば共有結合及び配位結合が含まれる。物理的結合には、例えば水素結合、二極性相互作用、ファンデルワールス力、静電気的相互作用、疎水性相互作用及び芳香族スタッキングが含まれる。
分子と芳香族陽イオンペプチドとの間の会合のタイプは、典型的には例えば分子上の利用可能な官能基、芳香族陽イオンペプチド上の利用可能な官能基に依存する。
化学的結合及び物理的結合では、分子上の官能基は典型的には芳香族陽イオンペプチド上の官能基と会合する。あるいは、芳香族陽イオンペプチド上の官能基が分子上の官能基と会合する。
分子上及び芳香族陽イオンペプチド上の官能基を直接結合させることもできる。例えば、分子上の官能基(例えばスルフヒドリル基)は、芳香族陽イオンペプチド上の官能基(例えばスルフヒドリル基と結合してジスルフィドを形成することができる。
あるいは、前記官能基は架橋剤(すなわちリンカー)を介して会合させることができる。架橋剤のいくつかの例は下記に記載されている。架橋剤は分子又は芳香族陽イオンペプチドのどちらに付加することもできる。
リンカーは芳香族陽イオンペプチドの正味の電荷数に影響を与えることも与えないこともある。典型的にはリンカーは芳香族陽イオンペプチドの正味の電荷に影響しないであろう。リンカー内に存在する各アミノ基(それが存在する場合)は、芳香族陽イオンペプチドの正味の正電荷に寄与するであろう。リンカー内に存在する各カルボキシル基(それが存在する場合)は、芳香族陽イオンペプチドの正味の負電荷に寄与するであろう。
【0020】
担体複合体の分子又は芳香族陽イオンペプチドの数は、多数の分子を収納することができるペプチドの能力、又は多数のペプチドを収納することができる分子の能力によって制限を受ける。例えば立体的障害は、特に大型の分子を収納するペプチドの能力を妨害するであろう。あるいは、立体的障害は比較的大型の(例えば長さが7、8、9又は10アミノ酸)芳香族陽イオンペプチドを収納する前記分子の能力を妨害するであろう。
担体複合体中の分子又は芳香族陽イオンペプチドの数はまた、他方に存在する官能基の数によって制限を受ける。例えば、ペプチドと会合できる分子の最大数は、ペプチドに存在する官能基の数に左右される。あるいは、分子と会合できるペプチドの最大数は、分子に存在する官能基の数に左右される。
ある実施態様では、担体複合体は、芳香族陽イオンペプチドと会合した少なくとも1つの分子、好ましくは少なくとも2つの分子を含む。いくつかの(例えば3、4、5又それより多い)官能基を含む比較的大きなペプチド(例えば長さが8、10アミノ酸)は、いくつかの(例えば3、4、5又それより多い)分子と会合することができる。
別の実施態様では、担体複合体は、分子と会合した少なくとも1つの芳香族陽イオンペプチド、好ましくは少なくとも2つの芳香族陽イオンペプチドを含む。例えば、いくつかの(例えば3、4、5又それより多い)官能基を含む分子は、いくつかの(例えば3、4、5又それより多い)ペプチドと会合することができる。
さらにまた別の実施態様では、担体複合体は1つの分子と会合した1つの芳香族陽イオンペプチドを含む。
ある実施態様では、担体複合体は、少なくとも1つの芳香族陽イオンペプチドと化学的に結合した(例えば共役した)少なくとも1つの分子を含む。前記分子は、当業者に公知の任意の方法によって芳香族陽イオンペプチドと化学的に結合させることができる。例えば、前記分子の官能基を芳香族陽イオンペプチドの官能基に直接付加することができる。適切な官能基のいくつかの例には、例えばアミノ、カルボキシル、スルフヒドリル、マレイミド、イソシアネート、イソチオシアネート及びヒドロキシルが含まれる。
【0021】
前記分子はまた、架橋剤(例えばジアルデヒド、カルボジイミド、ジマレイミドなど)の手段によって芳香族陽イオンペプチドと化学的に結合させることができる。架橋剤は、例えばピアースバイオテクノロジー社(Pierce Biotechnology, Inc., Rockford, IL)から入手できる。以下のURLのウェブサイト(http://www.piercenet.com/products/browse.cfm?fldID=26436A16-60A0-4A56-85F7-213A50830440)が資料を提供している。さらに別の架橋剤には白金架橋剤が含まれ、前記は、米国特許5,580,990号;5,985,566号及び6,133,038号(Kreatech Biotechnology, B.V., Amsterdam, The Netherlands)に記載されている。
分子上の官能基はペプチドの官能基と異なっていてもよい。例えばスルフヒドリル基が分子に(例えばβ-ガラクトシダーゼに、又は5'及び/又は3'末端チオール改変DNA及びRNAオリゴヌクレオチドに)存在する場合、前記分子は、ペプチド(例えば(Dmt1)DALDA)と、架橋剤SMCC(すなわちスクシンイミジル4-(N-マレイミドメチル)シクロヘキサン-1-カルボキシレート)(ピアースバイオテクノロジー(Pierce Biotechnology)から入手できる)を用いてリジンの4-アミノ基を介して架橋することができる(下記実施例10を参照されたい)。別の実施例では、DALDAのリジンの4-アミノ基は、架橋剤EDC(すなわちN-[3-ジメチルアミノプロピル-N'-エチルカルボイミド])(ピアースバイオテクノロジー(Pierce Biotechnology)から入手できる)を用いてRNA又はDNAの5'-末端のα-リン酸基と直接共役させることができる(下記実施例13を参照されたい)。
あるいは、分子上及びペプチド上の官能基は同じでもよい。ホモ二官能基性架橋剤は典型的には同一の官能基を架橋するために用いられる。ホモ二官能基性架橋剤の例にはEGS(すなわちエチレングリコールビス[スクシンイミジルスクシネート])、DSS(すなわちジスクシンイミジルスベレート)、DMA(すなわちジメチルアジピミデート・2HCl)、DTSSP(すなわち3,3'-ジチオビス[スルホスクシンイミジルプロピオネート])、DPDPB(すなわち1,4-ジ-[3'-(2'-ピリジルジチオ)-プロピオンアミド]ブタン)及びBMH(すなわちビス-マレイミドヘキサン)が含まれる。そのようなホモ二官能基性架橋剤もピアースバイオテクノロジー(Pierce Biotechnology)から入手できる。
【0022】
前記分子とペプチドを化学的に結合させるために、典型的には分子、ペプチド及び架橋剤を一緒に混合する。分子、ペプチド及び架橋剤の添加順序は重要ではない。例えば、ペプチドを架橋剤と混合し、続いて分子を添加してもよい。あるいは、分子を架橋剤と混合し、続いてペプチドを添加してもよい。最適には、分子及びペプチドを混合し、続いて架橋剤を添加する。
化学的に結合した担体複合体は分子を細胞に送達する。いくつかの事例では、分子は芳香族陽イオンペプチドから切り離されないで細胞内で機能する。例えば、芳香族陽イオンペプチドが分子の触媒部位を阻害しない場合は、芳香族陽イオンペプチドから分子の切り離しは不要である(下記の実施例11を参照されたい)。
他の事例では、分子を芳香族陽イオンペプチドから切断することは有益かもしれない。上記のピアースバイオテクノロジー社のウェブサイトはまた、細胞内で例えば酵素によって切断することができる適切な架橋剤を当業者が選択するための助けとなる。したがって、前記分子は芳香族陽イオンペプチドから分離することができる。切断可能なリンカーの例には、SMPT(すなわち4-スクシンイミジルオキシカルボニル-メチル-a-[2-ピリジルジチオ]トルエン)、スルホ-LC-SPDP(すなわちスルホスクシンイミジル6-(3-[2-ピリジルジチオ]-プロピオンアミド)ヘキサノエート)、LC-SPDP(すなわちスクシンイミジル6-(3-[2-ピリジルジチオ]-プロピオンアミド)ヘキサノエート)、スルホ-LC-SPDP(すなわちスルホスクシンイミジル6-(3-[2-ピリジルジチオ]-プロピオンアミド)ヘキサノエート)、SPDP(すなわちN-スクシンイミジル3-[2-ピリジルジチオ]-プロピオンアミドヘキサノエート)及びAEDP(すなわち3-[(2-アミノエチル)ジチオ]-プロピオン酸・HCl)が含まれる。
【0023】
また別の実施態様では、担体複合体は、少なくとも1つの芳香族陽イオンペプチドと物理的に結合した少なくとも1つの分子を含む。当業者に知られた任意の方法を用いて分子を芳香族陽イオンペプチドと物理的に結合させることができる。
例えば、芳香族陽イオンペプチド及び分子は当業者に公知の任意の方法によって一緒に混合することができる。混合の順序は重要ではない。例えば、分子は改変又は未改変芳香族陽イオンペプチドと当業者に公知の方法によって物理的に混合することができる。あるいは、改変又は未改変芳香族陽イオンペプチドは当業者に公知の任意の方法によって分子と物理的に混合することができる。
例えば、芳香族陽イオンペプチド及び分子を容器に入れ、例えば容器を振盪することによって攪拌し前記芳香族陽イオンペプチド及び分子を混合することができる。
芳香族陽イオンペプチドは当業者に公知の任意の方法によって改変することができる。例えば、芳香族陽イオンペプチドは上記で述べたように架橋剤又は官能基によって改変してもよい。リンカーは芳香族陽イオンペプチドの正味の電荷に影響を与ることも与えないこともある。典型的にはリンカーは芳香族陽イオンペプチドの正味の電荷に寄与しないであろう。リンカー内に存在する各アミノ基(それが存在する場合)は、芳香族陽イオンペプチドの正味の正電荷に寄与するであろう。リンカー内に存在する各カルボキシル基(それが存在する場合)は、芳香族陽イオンペプチドの正味の負電荷に寄与するであろう。
例えば、[Dmt1]DALDAを、架橋剤SMCC(すなわちスクシンイミジル4-(N-マレイミドメチル)シクロヘキサン-1-カルボキシレート)(ピアースバイオテクノロジー(Pierce Biotechnology)から入手できる)を用いてリジンの4-アミノ基を介して改変することができる(下記実施例10を参照されたい)。担体複合体を形成するために、改変芳香族陽イオンペプチドを通常先ず初めに形成させ、続いて分子と混合する。
物理的に結合させた担体複合体の1つの利点は、分子が芳香族陽イオンペプチドと化学的に結合した担体複合体のように芳香族陽イオンペプチドを除去する必要なく分子が細胞内で機能するということである。さらにまた、芳香族陽イオンペプチドが分子の触媒部位を阻害しない場合、複合体の解離も必要ではない(下記実施例12参照)。
【0024】
芳香族陽イオンペプチドの合成
本発明の方法で有用なペプチドは当業者に公知の任意の方法によって化学的に合成することができる。タンパク質を合成する適切な方法には、例えばStuart & Youngの文献に記載されたものが含まれる: “Solid Phase Peptide Synthesis”, Second Edition, Pierce Chemical Company (1984)及び”Solid Phase Peptide Synthesis”, Methods Enzymology, 289, Academic Press, Inc, New York (1977)。
投与の態様
ある実施態様では、本発明は分子を細胞に送達する方法に関する。前記方法は細胞を前記分子及び芳香族陽イオンペプチドと接触させることを含む。細胞は、当業者に公知の任意の方法によって前記分子及び芳香族陽イオンペプチドと接触させることができる。例えば、細胞をin vitroで前記分子及び芳香族陽イオンペプチドとインキュベートすることができる。ある特徴では、前記分子及び芳香族陽イオンペプチドは、化学的又は物理的に結合した前記分子及び芳香族陽イオンペプチドを含む担体複合体の形態(例えば上記に記載した担体複合体)として存在することができる。
また別の実施態様では、分子を細胞に送達する方法は細胞を担体複合体と接触させることを含む。分子は、前記分子及び芳香族陽イオンペプチドを含む担体複合体と細胞を接触させることによって前記細胞に送達される。細胞は、当業者に公知の任意の方法によって担体複合体と接触させることができる。
例えば、細胞をin vitroで担体複合体とインキュベートすることができる。この細胞は任意の細胞でよい。細胞は植物、動物又は細菌由来であり得る。植物細胞の例にはアラビドプシス(Arabidopsis)細胞が含まれる。細菌細胞の例にはサッカロミセス(Saccharomyces)及びラクトバシルス(Lactobacillus)が含まれる。動物細胞には哺乳動物細胞、例えば神経細胞、腎上皮細胞、腎臓細胞、血管内皮細胞。グリア細胞、腸上皮細胞及び肝細胞が含まれる。血管内皮細胞の例には血液脳関門内皮細胞が含まれる。
【0025】
あるいは、前記担体複合体は哺乳動物にin vivoで投与することができる。医薬化合物を投与するための多数の周知の方法のいずれかによって、好ましくは医薬組成物として有効量の担体複合体を投与の必要性がある哺乳動物に投与することができる。
前記担体複合体は全身的又は局所的に投与することができる。ある実施態様では、前記担体複合体は静脈内に投与される。例えば、担体複合体は瞬時静脈内ボーラス注射により投与することができる。しかしながら好ましくは、担体複合体は静脈内定常速度輸液として投与される。
担体複合体は、哺乳動物の組織に、例えば注射器が利用できる組織に注射によって局所的に投与することができる。例えば、前記担体複合体が腫瘍に送達されるべき細胞毒性物質を含んでいる場合は、その腫瘍は局所投与に利用できることが好ましい。そのような腫瘍には例えば皮膚癌及び乳癌が含まれる。
担体複合体はまた、経口的、局部的、鼻腔内、筋肉内、皮下又は経皮的に投与することができる。好ましい実施態様では、担体複合体の経皮的投与はイオン導入療法によって実施される。前記方法では、担体複合体は電流によって皮膚を通して送達される。
他の投与ルートには脳室内又はくも膜下腔内投与が含まれる。脳室内とは脳の脳室系内への投与を指す。くも膜下腔内投与は、脳又は脊髄のくも膜下腔内への投与を意味する。したがって、脳室内又はくも膜下腔内投与は、中枢神経系の器官又は組織に影響を与える疾患及び症状に対して好まれるであろう。
本発明の方法において有用な担体複合体は、当業界で知られたように持続性放出によって哺乳動物に投与することができる。持続性放出投与は、特定の期間にわたって一定レベルの薬剤を達成するドラッグデリバリーのための方法である。前記レベルは典型的には血清濃度によって測定される。
調剤分野で公知のいずれの製剤も担体複合体の投与に適切である。経口投与のためには液体又は固体製剤を用いることができる。いくつかの製剤例には錠剤、ゼラチンカプセル、ピル、トローチ、エリキシル、懸濁液、シロップ、ウェファース、チューインガムなどが含まれる。ペプチドは、当業者に公知の適切な医薬担体(ビヒクル)又は賦形剤とともに混合することができる。担体及び賦形剤の例にはデンプン、ミルク、糖、あるタイプの粘度、ゼラチン、ステアリン酸又はその塩(ステアリン酸マグネシウム又はカルシウムを含む)、植物脂肪又は油、ゴム及びグリコールが含まれる。
【0026】
全身的、脳室内、くも膜下腔内、局部、鼻腔内、皮下又は経皮投与のためには、前記担体複合体製剤は通常の希釈剤、担体又は賦形剤など(例えば当業界で公知のもの)を利用して前記担体複合体を送達することができる。例えば、前記製剤は以下の1つ又は2つ以上を含むことができる:安定化剤、界面活性剤(好ましくは非イオン性界面活性剤)及び場合によって塩及び/又は緩衝剤。前記担体複合体は水溶液の形態で又は凍結乾燥形態として輸送することができる。
安定化剤は、例えばアミノ酸(例えばグリシン)、又はオリゴ糖(例えばシュクロース、テトラオース、ラクトース又はデキストラン)であり得る。あるいは、安定剤は糖アルコール(例えばマンニトール)又は前記の組合せであってよい。好ましくは安定剤又は安定剤の組合せは担体複合体の重量の約0.1から約10重量%を構成する。
界面活性剤は好ましくは非イオン性界面活性剤、例えばポリソルベートである。適切な界面活性剤のいくつかの例には、トゥイーン20、トゥイーン80;ポリエチレングリコール又はポリオキシエチレンポリオキシプロピレングリコール、例えばプルロニックF-68(約0.001%(w/v)から約10%(w/v))が含まれる。
塩又は緩衝剤は任意の塩又は緩衝剤、例えばそれぞれ塩化ナトリウム又はリン酸ナトリウム/カリウムであり得る。好ましくは、緩衝剤は前記医薬組成物のpHを約5.5から約7.5の範囲で維持する。前記塩及び/又は緩衝剤はまた、浸透圧濃度をヒト又は動物への投与に適したレベルに維持するために有用である。好ましくは、前記塩又は緩衝剤は、約150mMから約300mMのほぼ等張濃度で存在する。
本発明の方法で有用な担体複合体製剤はさらにまた1つ又は2つ以上の一般的な添加物を含むことができる。そのような添加物のいくつかの例には、可溶化剤、例えばグリセロール;抗酸化剤、例えば塩化ベンザルコニウム(第四アンモニウム化合物の混合物、“クォーツ(quats)”として知られている)、ベンジルアルコール、クロレトン又はクロロブタノール;麻酔剤、例えばモルヒネ誘導体;又は等張剤(例えば上記に記載したもの)などが含まれる。酸化又は他の腐敗に対するまた別の予防として、前記医薬組成物は、非透過性の栓で密封されたバイアルで窒素ガス下で保存することができる。
前記哺乳動物は任意の哺乳動物でよい。それらには例えば家畜(例えばヒツジ、ブタ、乳牛及びウマ);愛玩動物(例えばイヌ及びネコ);実験動物(例えばラット、マウス及びウサギ)が含まれる。好ましい実施態様ではこの哺乳動物はヒトである。
【0027】
有用性
本担体複合体はエネルギー非依存機構で細胞膜を通過することができるので、多数のin vivo及びin vitroでの応用が可能である。
本担体複合体は、例えば研究用ツールとしてin vitroで用いることができる。例えば、本担体複合体は、分子(例えばタンパク質)を細胞に送達し、それによって前記分子の機能的役割を研究することができる。そのような分子には、例えば細胞シグナル伝達タンパク質(例えば核内因子NF-κB、キナーゼ、例えばJAK)が含まれる。
また別のin vitroでの利用には、例えば、細胞の子孫(系統)を決定するためのマーカー(例えばβ-ガラクトシダーゼ)の細胞(例えば幹細胞、造血細胞又は胚細胞)内へのデリバリーが含まれる。
他のin vitroでの応用には例えば検出可能な抗体の細胞へのデリバリーが含まれる(これによって特定のタンパク質の細胞内の存在が決定される)。
本担体複合体はまたin vivoで治療薬としての用途を有する。例えば前記芳香族陽イオンペプチドを用いてアンチセンスポリヌクレオチドを哺乳動物の細胞に送達してタンパク質の過剰発現をダウンレギュレートすることができる。さらにまた、RNA干渉(RNAi)のためのオリゴヌクレオチドの送達に芳香族陽イオンペプチドを用いることができる。
本明細書においてRNAiは、ウイルス若しくは細菌の遺伝子の発現又は複製を制御する細胞内メカニズムを指す。このメカニズムには、遺伝子生成物(典型的にはRNA)を標的として二本鎖RNA(例えばsiRNA)を導入することが含まれる。
脳血液関門は特に選択的である。したがって、また別のin vivoでの応用には脳血液関門を通過して分子を細胞に送達することが含まれる。そのような分子には、例えばアルツハイマー病患者の治療におけるβ-アミロイドに対する抗体が含まれる。
化学療法剤に付随する典型的な問題は、細胞内で適切なレベルを達成することである。例えば、化学療法剤は細胞膜を通過するには大きすぎるか、又は十分な芳香族性をもたないであろう。したがって、また別のin vivoでの応用には化学療法剤(例えば上記に記載したような細胞毒性物質)の細胞内への送達が含まれる。
【0028】
本発明のいくつかの態様を以下に列挙する。
(1) 分子を細胞に送達する方法であって、細胞を担体複合体と接触させることを含み、前記複合体は送達する分子及び芳香族陽イオンペプチドを含み、前記芳香族陽イオンペプチドが以下を含む、前記細胞に分子を送達する前記方法:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmがr+1以下の最大数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下の最大数であるが、ただしaが1であるときはptもまた1であることができる関係。
(2) 2aがpt+1以下の最大の数である、(1)に記載の方法。
(3) aがpに等しい、(1)に記載の方法。
(4) ペプチドが最小限2つの正電荷を有する、(1)に記載の方法。
(5) ペプチドが最小限3つの正電荷を有する、(1)に記載の方法。
(6) ペプチドが、2つの正に荷電したアミノ酸及び1つの芳香族アミノ酸を含むトリペプチドである、(1)に記載の方法。
(7) ペプチドが水溶性である、(1)に記載の方法。
(8) ペプチドが1つ又は2つ以上のD-アミノ酸を含む、(1)に記載の方法。
(9) C-末端アミノ酸のC-末端カルボキシル基がアミド化されている、(1)に記載の方法。
(10) ペプチドが1つ又は2つ以上の天然には存在しないアミノ酸を含む、(1)に記載の方法。
(11) ペプチドが最小限4つのアミノ酸を有する、(1)に記載の方法。
(12) ペプチドが最大限約8つのアミノ酸を有する、(1)に記載の方法。
(13) ペプチドが最大限約6つのアミノ酸を有する、(1)に記載の方法。
(14) ペプチドが4つのアミノ酸を有する、(1)に記載の方法。
(15) ペプチドが式Tyr-D-Arg-Phe-Lys-NH2(DALDA)を有する、(1)に記載の方法。
(16) ペプチドが式2’,6’-Dmt-D-Arg-Phe-Lys-NH2(Dmt1-DALDA)を有する、(1)に記載の方法。
(17) ペプチドが式Phe-D-Arg-Phe-Lys-NH2(Phe1-DALDA)を有する、(1)に記載の方法。
(18) ペプチドが式D-Arg-2’,6’ Dmt-Lys-Phe-NH2を有する、(1)に記載の方法。
(19) ペプチドが式2’,6’-Dmp-D-Arg-Phe-Lys-NH2(Dmp1-DALDA)を有する、(1)に記載の方法。
(20) 分子が小分子である、(1)に記載の方法。
(21) 小分子が医薬的に活性を有する分子である、(20)に記載の方法。
(22) 医薬的に活性を有する分子が抗生物質である、(21)に記載の方法。
(23) 医薬的に活性を有する分子が細胞毒性物質である、(21)に記載の方法。
(24) 細胞毒性物質がドキソルビシンである、(23)に記載の方法。
(25) 細胞毒性物質がアドリアマイシンである、(23)に記載の方法。
(26) 医薬的に活性を有する分子が抗酸化剤である、(21)に記載の方法。
(27) 抗酸化剤がビタミンEである、(26)に記載の方法。
(28) 抗酸化剤がビタミンCである、(26)に記載の方法。
(29) 抗酸化剤がベータカロチンである、(26)に記載の方法。
(30) 分子が生物学的分子である、(1)に記載の方法。
(31) 生物学的分子がポリアミノ酸である、(30)に記載の方法。
(32) 生物学的分子が医薬的に活性を有する分子である、(30)に記載の方法。
(33) 医薬的に活性を有する分子が内因性ペプチド又はタンパク質である、(32)に記載の方法。
(34) 医薬的に活性を有する分子が酵素である、(32)に記載の方法。
(35) 医薬的に活性を有する分子が抗体である、(32)に記載の方法。
(36) 医薬的に活性を有する分子が神経栄養性増殖因子である、(32)に記載の方法。
(37) 医薬的に活性を有する分子がサイトカインである、(32)に記載の方法。
(38) 医薬的に活性を有する分子がポリヌクレオチドである、(32)に記載の方法。
(39) 医薬的に活性を有する分子がオリゴヌクレオチドである、(32)に記載の方法。
(40) オリゴヌクレオチドがRNAである、(39)に記載の方法。
(41) RNAが二本鎖RNAである、(40)に記載の方法。
(42) 二本鎖RNAがsiRNAである、(41)に記載の方法。
(43) オリゴヌクレオチドがDNAである、(39)に記載の方法。
(44) オリゴヌクレオチドが一本鎖RNAである、(39)に記載の方法。
(45) 一本鎖RNAがメッセンジャーRNA(mRNA)である、(44)に記載の方法。
(46) オリゴヌクレオチドがリボザイムである、(39)に記載の方法。
(47) オリゴヌクレオチドがアンチセンスRNAである、(39)に記載の方法。
(48) オリゴヌクレオチドがリボザイムのための外側ガイド配列である、(39)に記載の方法。
(49) オリゴヌクレオチドがRNAデコイである、(39)に記載の方法。
(50) 生物学的分子がポリアミノ酸である、(30)に記載の方法。
(51) 細胞が細菌細胞である、(1)に記載の方法。
(52) 細胞が植物細胞である、(1)に記載の方法。
(53) 細胞が動物細胞である、(1)に記載の方法。
(54) 動物細胞が哺乳動物細胞である、(53)に記載の方法。
(55) 細胞が神経細胞である、(53)に記載の方法。
(56) 細胞が腎上皮細胞である、(53)に記載の方法。
(57) 細胞が腸上皮細胞である、(53)に記載の方法。
(58) 細胞が血管内皮細胞である、(53)に記載の方法。
(59) 内皮細胞が血管-脳関門内皮細胞である、(53)に記載の方法。
(60) 細胞がグリア細胞である、(53)に記載の方法。
(61) 細胞が肝細胞である、(53)に記載の方法。
(62) 芳香族-陽イオンペプチドがリンカーを含む、(1)に記載の方法。
(63) 分子がリンカーを含む、(1)に記載の方法。
(64) 分子と芳香族陽イオンペプチドが化学的に結合されている、(1)に記載の方法。
(65) 分子と芳香族陽イオンペプチドが物理的に結合されている、(1)に記載の方法。
(66) 分子及び芳香族陽イオンペプチドを含む担体複合体であって、前記芳香族陽イオンペプチドが以下を含む、前記担体複合体:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmがr+1以下の最大数である関係;さらに
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下の最大数であるが、ただしaが1であるときはptもまた1であることができる関係。
(67) 2aはpt+1以下の最大の数である、(66)に記載の担体複合体。
(68) aがptに等しい、(66)に記載の担体複合体。
(69) ペプチドが最小限2つの正電荷を有する、(66)に記載の担体複合体。
(70) ペプチドが最小限3つの正電荷を有する、(66)に記載の担体複合体。
(71) ペプチドが、2つの正に荷電したアミノ酸及び1つの芳香族アミノ酸を含むトリペプチドである、(66)に記載の担体複合体。
(72) ペプチドが水溶性である、(66)に記載の担体複合体。
(73) ペプチドが1つ又は2つ以上のD-アミノ酸を含む、(66)に記載の担体複合体。
(74) C-末端アミノ酸のC-末端カルボキシル基がアミド化されている、(66)に記載の担体複合体。
(75) ペプチドが1つ又は2つ以上の天然には存在しないアミノ酸を含む、(66)に記載の担体複合体。
(76) ペプチドが最小限4つのアミノ酸を有する、(66)に記載の担体複合体。
(77) ペプチドが最大限約8つのアミノ酸を有する、(66)に記載の担体複合体。
(78) ペプチドが最大限約6つのアミノ酸を有する、(66)に記載の担体複合体。
(79) ペプチドが4つのアミノ酸を有する、(66)に記載の担体複合体。
(80) ペプチドが式Tyr-D-Arg-Phe-Lys-NH2(DALDA)を有する、(66)に記載の担体複合体。
(81) ペプチドが式2’,6’-Dmt-D-Arg-Phe-Lys-NH2(Dmt1-DALDA)を有する、(66)に記載の担体複合体。
(82) ペプチドが式Phe-D-Arg-Phe-Lys-NH2(Phe1-DALDA)を有する、(66)に記載の担体複合体。
(83) ペプチドが式D-Arg-2’,6’ Dmt-Lys-Phe-NH2を有する、(66)に記載の担体複合体。
(84) ペプチドが式2’,6’-Dmp-D-Arg-Phe-Lys-NH2(Dmp1-DALDA)を有する、(66)に記載の担体複合体。
(85) 分子が小分子である、(66)に記載の担体複合体。
(86) 小分子が生物学的分子である、(66)に記載の担体複合体。
(87) 生物学的分子がポリアミノ酸である、(86)に記載の担体複合体。
(88) 小分子が医薬的に活性を有する分子である、(85)に記載の担体複合体。
(89) 医薬的に活性を有する分子が抗生物質である、(88)に記載の担体複合体。
(90) 医薬的に活性を有する分子が細胞毒性物質である、(88)に記載の担体複合体。
(91) 細胞毒性物質がドキソルビシンである、(90)に記載の担体複合体。
(92) 細胞毒性物質がアドリアマイシンである、(90)に記載の担体複合体。
(93) 医薬的に活性を有する分子が抗酸化剤である、(88)に記載の担体複合体。
(94) 抗酸化剤がビタミンEである、(93)に記載の担体複合体。
(95) 抗酸化剤がビタミンCである、(93)に記載の担体複合体。
(96) 抗酸化剤がベータカロチンである、(93)に記載の担体複合体。
(97) 分子が生物学的分子である、(66)に記載の担体複合体。
(98) 生物学的分子が医薬的に活性を有する分子である、(97)に記載の担体複合体。
(99) 医薬的に活性を有する分子が内因性ペプチド又はタンパク質である、(98)に記載の担体複合体。
(100) 医薬的に活性を有する分子が酵素である、(98)に記載の担体複合体。
(101) 医薬的に活性を有する分子が抗体である、(98)に記載の担体複合体。
(102) 医薬的に活性を有する分子が神経栄養性増殖因子である、(98)に記載の担体複合体。
(103) 医薬的に活性を有する分子がサイトカインである、(98)に記載の担体複合体。
(104) 医薬的に活性を有する分子がポリヌクレオチドである、(98)に記載の担体複合体。
(105) 医薬的に活性を有する分子がオリゴヌクレオチドである、(98)に記載の担体複合体。
(106) オリゴヌクレオチドがRNAである、(105)に記載の担体複合体。
(107) RNAが二本鎖RNAである、(106)に記載の担体複合体。
(108) 二本鎖RNAがsiRNAである、(107)に記載の担体複合体。
(109) オリゴヌクレオチドがDNAである、(106)に記載の担体複合体。
(110) オリゴヌクレオチドが一本鎖RNAである、(105)に記載の担体複合体。
(111) 一本鎖RNAがメッセンジャーRNA(mRNA)である、(110)に記載の担体複合体。
(112) オリゴヌクレオチドがリボザイムである、(105)に記載の担体複合体。
(113) オリゴヌクレオチドがアンチセンスRNAである、(105)に記載の担体複合体。
(114) オリゴヌクレオチドがリボザイムのための外側ガイド配列である、(105)に記載の担体複合体。
(115) オリゴヌクレオチドがRNAデコイである、(105)に記載の担体複合体。
(116) 生物学的分子がポリアミノ酸である、(97)に記載の担体複合体。
(117) 芳香族-陽イオンペプチドがリンカーを含む、(66)に記載の担体複合体。
(118) 分子がリンカーを含む、(66)に記載の担体複合体。
(119) 分子と芳香族陽イオンペプチドが化学的に結合されている、(66)に記載の担体複合体。
(120) 分子と芳香族陽イオンペプチドが物理的に結合されている、(66)に記載の担体複合体。
(121) 分子を細胞に送達する方法であって、前記方法が細胞を分子及び芳香族陽イオンペプチドと接触させることを含み、前記芳香族陽イオンペプチドが以下を含む、前記分子を細胞に送達する方法:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmがr+1以下の最大数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下の最大数であるが、ただしaが1であるときはptもまた1であることができる関係。
(122) 分子と芳香族陽イオンペプチドが化学的に結合されている、(121)に記載の方法。
(123) 分子と芳香族陽イオンペプチドが物理的に結合されている、(121)に記載の担体複合体。
【実施例1】
【0029】
実施例1:材料及び方法
薬剤及び化学物質:[Dmt1]DALDA及び[3H] [Dmt1]DALDA(47Ci/mmol)は以前に記載された方法(Schiller et al., Eur. J. Med. Chem. 2000, 35:895-901;Zhao et al., J. Pharmacol. Exp. Ther. 2002, 302:188-196)にしたがって合成した。[14C]Gly-Sar(56.7mCi/mmol)及び[3H][D-Ala2,N-Me-Phe4,Gly5-オール]-エンケファリン(50Ci/mmol)はAmersham Biosciences(Piscataway, NJ)から購入した。その他の全ての薬剤及び化学物質はSigma-Aldrich(St. Louis, MO)から入手した。
【0030】
細胞培養:全ての細胞株はアメリカンタイプカルチャーコレクション(American Type Culture Collection, Manassas, VA)から入手し、さらに細胞培養用品はInvitrogen(Carlsbad, CA)から得た。Caco-2細胞はMEMで増殖させ、一方SH-SY5Y、HEK293及びHuh7細胞はダルベッコー改変イーグル培養液で増殖させた。増殖培地には10%ウシ胎児血清、200μg/mLペニシリン及び100μg/mL硫酸ストレプトマイシンを補充した。CRFK細胞はMEM+10%ウマ血清、非必須アミノ酸及びペニシリン/ストレプトマイシンで増殖させた。全ての細胞株は95%空気及び5%CO2の加湿大気中で37℃で維持した。
【0031】
ペプチド取り込みアッセイ:ペプチドの内在化は最初にCaco-2細胞を用いて調べ、続いてSH-SY5Y、HEK293及びCRFK細胞で確認した。コラーゲンを被覆した12ウェルプレートで単層細胞を3日間増殖させた(5x105細胞/ウェル)。4日目に、予め温めたHBSSで細胞を洗浄し、続いて250nMの[3H] [Dmt1]DALDA又は50μMの[14C]Gly-Sarのどちらかを含む、0.2mLのHBSSとともに1時間まで37℃にて種々の時間インキュベートした。別の実験で、非標識 [Dmt1]DALDA(1μMから3mM)の存在下で、同じ濃度の[3H] [Dmt1]DALDAとともに37℃で1時間細胞をインキュベートした。4℃での取り込み実験のためには、 [3H] [Dmt1]DALDA又は[14C]Gly-Sarとのインキュベーションの前に細胞を20分氷上に静置した。インキュベーションの終わりに、細胞を4回HBSSで洗浄し、1%のSDSを含む0.1NのNaOHの0.2mLを添加した。続いて細胞の内容物をシンチレーションバイアルに移し、放射能を計測した。細胞溶解物のアリコットをブラッドフォード法(Bio-Rad, Hercules, CA)によるタンパク質含有量の決定に用いた。内在化放射能を表面結合放射能と区別するために、酸洗浄工程を含めた。細胞溶解の前に、0.2mLの0.2M酢酸/0.05MNaClとともに細胞を氷中でインキュベートした。
【0032】
Caco-2細胞からのペプチド流出アッセイ:単層CaCo-2細胞を12ウェルプレートで3日間増殖させた(5x105細胞/ウェル)。4日目に、細胞を[3H] [Dmt1]DALDA又は[14C]Gly-Sarで37℃で1時間予備装荷した。続いて細胞を氷冷インキュベーション溶液で4回洗浄して取り込みを停止させ、続いて37℃又は4℃で1時間0.5mLのMEMでインキュベートして細胞から前記インキュベーション溶液へのペプチドの流出を測定した。放射能の量は細胞溶解物及びインキュベーション媒体で測定した。ペプチド取り込み及び細胞からの流出に対するP-糖タンパク質の役割を調べるために、[Dmt1]DALDAの取り込み及び流出を100μMのベラパミル(P-糖タンパク質の阻害剤)の存在下でも測定した。
【0033】
Caco-2単層を通過するペプチド移動のアッセイ:単層Caco-2細胞は以前に記載されたたように調製した(Irie et al., J. Pharmacol. Exp. Ther. 2001, 298:711-717)。Caco-2細胞(2x105)をトランスウェル細胞培養チャンバー(Corning Glassworks, Corning, NY)内の微小孔を有するメンブレンフィルター(24mm、0.4μm)上に播種した。各トランスウェルチャンバーの先端部区画を1.5mLの培養液で、外側基底部区画を2.5mLの培養液で満たした。前記細胞単層に新しい培養液を1から2日毎に与え、28日目に輸送実験に使用した。先端部から外側基底部へのペプチド輸送は、0.2μMの[3H] [Dmt1]DALDA又は100μMの[14C]Gly-Sarを先端部区画に添加することによって決定した。放射能の計測のために、ペプチド添加後種々の時間に50μLのアリコットを先端部区画及び外側基底部区画の両方から取り出した。
見かけの透過率係数は以下の等式によって計算した:Papp=X/(t・A・Co)
式中、X/tはレシーバー区画における取り込み速度、Aは拡散面積(4.72cm2)、及びCoはドナー区画における最初の濃度である。
【0034】
共焦点レーザー走査型顕微鏡法:芳香族陽イオンペプチドの細胞内取り込みは、以下の2つの蛍光ペプチドを用いて共焦点レーザー走査型顕微鏡法(CLSM)によって確認した:[Dmt1,dnsDap4]DALDA(Dmt-D-Arg-Phe-dnsDap-NH2、ここでdnsDapはβ-ダンシル-1-α,β-ジアミノ-プロピオン酸である)及び[Dmt1,atnDap4]DALDA(Dmt-D-Arg-Phe-atnDap-NH2、ここでatnDapはβ-アントラニロイル-L-α,β-ジアミノ-プロピオン酸である)。Caco-2細胞又はSH-SY5Y細胞を上記のように増殖させ、(35mmの)ガラス底の培養皿(MatTek, Ashland, MA)で2日間平板培養した。続いて培養液を取り除き、0.1μMの蛍光ペプチドを含むHBSSの1mLとともに細胞を4℃又は37℃で15分インキュベートした。続いて細胞を3回氷冷HBSSで洗浄し、さらに200μLのPBSで被覆し、C-Apochromat63x1.2Wcorr対物レンズ(Nikon, Tokyo, Japan)を取り付けた共焦点レーザー走査型顕微鏡を用いて10分以内に室温で顕微鏡観察を実施した。励起/放射波長は、[Dmt1,dnsDap4]DALDA については340/520nm、[Dmt1,atnDap4]DALDAについては320/420nmにそれぞれ設定した。Z方向の光学切片形成のために、2.0μmの5から10枠を作製した。
【0035】
細胞膜を用いる放射性リガンド結合アッセイ:Caco-2細胞及びSH-SY5Y細胞から調製した膜を用いて、[3H] [Dmt1]DALDAの細胞表面レセプターへの特異的結合を測定した。培養4日後に、細胞をPBS緩衝液で2回洗浄してから掻き取った。細胞を500gで5分遠心し、ペレットを-80℃で保存した。氷冷の50mMトリス塩酸緩衝液(5μg/mLロイペプチン、2μg/mLキモスタチン、10μg/mL及び1mMのEGTA、pH7.4)中で細胞をホモジナイズした。前記ホモジネートを36000gで20分遠心した。ペレットを50mMトリス塩酸緩衝液で再懸濁させた。膜ホモジネート(約140μgのタンパク質)のアリコットを[3H][Dmt1]DALDA(15から960pM)とともに25℃で60分インキュベートした。非特異的結合は1μMの非標識[3H] [Dmt1]DALDAを添加して評価した。遊離放射性リガンドは、細胞採集装置(Brandel Inc., Gaithersburg, MD)を用いGF/Bフィルター(Whatman, Maidstone, UK)で迅速ろ過して結合放射能リガンドから分離した。フィルターを3回10mLのトリス緩衝液で洗浄し、放射能を液体シンチレーション計測によって決定した。結合親和性(Kd)及びレセプター数(Bmax)は非直線回帰(GraphPad Software, San Diego, CA)を用いて決定した。
【0036】
タンパク質と[Dmt1]DALDAとの共役:[Dmt1]DALDAをβ-ガラクトシダーゼ(組換え大腸菌、Sigma-Aldrich)と架橋剤SMCC(スクシンイミジル4-(N-マレイミドメチル)シクロヘキサン-1-カルボキシレート)(Pierce)を用いて架橋した。SMCCはアミン含有分子([Dmt1]DALDAのLys4)と反応して安定なアミド結合を生成する。続いて前記のマレイミド端をスルフヒドリル含有化合物と結合させてチオエーテル結合を形成する(Greg T. Hermanson, “Bioconjugate Techniques”, Academic Press, page 234-237)。β-Galはその天然の状態で豊富な遊離スルフヒドリル基を含んでいる。β-Galの取り込みは、X-Galの使用による簡便な読み出しを提供する。簡単に記せば、1mLの[Dmt1]DALDA(5x10-3M)をリン酸緩衝液中の1mgのSMCCと室温で1時間混合した。前記操作によって“活性化ペプチド”が生成される。前記“活性化ペプチド”をリン酸緩衝液で1:10に希釈した。1mgのβ-Galを前記1:10の“活性化ペプチド”1mLに添加し、4℃で2時間又は一晩混合した。
[Dmt1]DALDAと架橋剤SMCCとのカップリング及び質量分析による確認:SMCC(1μg)及び[Dmt1]DALDA(5μg)を一緒に2mLのPBSに溶解し、室温で30分インキュベートし、4℃で保存した。前記サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールのターゲットプレートにスポットした。サンプルはMALDI-TOF MS(Matrix Assisted Laser Desorption Ionization−Time-of-Flight Mass Spectrometry, Applied Biosystems(Voyager DE Pro))によってポジティブリフレクトロンモードで分析した。
【0037】
RNAと[Dmt1]DALDAとの共役及びゲル電気泳動による確認:γ-32P-ATP及びポリヌクレオチドキナーゼを用いて合成RNAオリゴ(長さは40ヌクレオチド)の5'端をリン酸化した。生成物をゲル電気泳動で精製した。ゲル精製RNAオリゴの500,000cpmを1mgのEDC(N-[3-ジメチルアミノプロピル-N'-エチルカルボジイミド])の存在下で[Dmt1]DALDAと結合させた。前記結合反応の生成物([Dmt1]DALDA−RNAオリゴ)及び単独RNAオリゴを15%ポリアクリルアミド尿素ゲルで分析した。
DNAと[Dmt1]DALDAとの共役及び質量分析による確認:SMCC(1μg)及び[Dmt1]DALDA(5μg)を一緒に2mLのPBSに溶解し、室温で30分インキュベートし、さらに脱保護3'-チオールDNAオリゴと4℃で24時間混合した。インキュベーション後、サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。サンプルはMALDI-TOF MSによって分析した。
【0038】
RNA及び[Dmt1]DALDA−SMCC共役物の物理的混合による担体複合体の形成:[Dmt1]DALDA−SMCC共役物は上記のように調製した。RNA分子は、細胞取り込み実験で使用する前に、[Dmt1]DALDA−SMCC共役物と室温で15分間PBS中で混合した。
タンパク質及び[Dmt1]DALDA−SMCC共役物の物理的混合による担体複合体の形成:[Dmt1]DALDA−SMCC共役物は上記のように調製した。タンパク質分子(すなわち緑色蛍光タンパク質、GFP)は、細胞取り込み実験で使用する前に、[Dmt1]DALDA−SMCC結合物と室温で15分混合した。
[Dmt1]DALDA−RNA共役物の細胞内取り込みアッセイ:γ-32P-ATP及びポリヌクレオチドキナーゼを用いて合成RNAオリゴの5'端をリン酸化し、生成物をゲル電気泳動で精製した。ゲル精製RNAオリゴの500,000cpmを1mgのEDC(N-[3-ジメチルアミノプロピル-N'-エチルカルボジイミド])の存在下で[Dmt1]DALDAと結合させた。Caco-2細胞(1x106)をDMEM培養液で3回洗浄し、DMEMで5分間プレインキュベートした。続いて細胞を[Dmt1]DALDA−[32P]RNAオリゴ共役物又は非共役RNA(約20,000cpm)と37℃で60分インキュベートした。インキュベーション後に、細胞を3回DMEMで洗浄し、溶解緩衝液でインキュベートし、細胞溶解物中の放射能を測定した。
【0039】
[Dmt1]DALDA−架橋剤共役物と混合した場合の細胞内へのRNA取り込みアッセイ
Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、[32P]RNAオリゴのみまたは40μlの[Dmt1]DALDA−SMCC共役物を含む1.0mlのDMEMとともに37℃または4℃にて60分間インキュベートした。次に細胞をDMEMで4回洗浄し、酢酸ナトリウム溶液で1回洗浄して非特異的結合を低減させて、溶解緩衝液中で30分間インキュベートし、細胞溶解物中の放射能を測定した。
[Dmt1]DALDA−タンパク質共役物の細胞内取り込みアッセイ:細胞(N2A神経芽腫細胞又はCaco-2細胞)を96ウェルプレートでにプレーティングし(2x104細胞/ウェル)、[Dmt1]DALDA架橋β-Gal又はβ-Galのみとともに37℃で1時間インキュベートした。続いて細胞をPBSで4回洗浄した。次に細胞をβ-Gal染色セット(Roche)で37℃で少なくとも2時間染色し、顕微鏡下で調べた。
[Dmt1]DALDA−SMCC共役物と同時インキュベートした場合の細胞内タンパク質取り込みアッセイ:Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて3μgの緑色蛍光タンパク質(GFP)単独(A)、3μgのGFP及び40μL[Dmt1]DALDA(B)、又は3μgのGFP及びSMCCに共役させた40μLの[Dmt1]DALDA(C)を含む0.5mLのMEMとともに37℃で60分インキュベートした。続いて2mLの細胞培養液を細胞に添加し、これを細胞培養インキュベーターでさらに24時間インキュベートした。インキュベーション後に細胞を細胞培養液で4回洗浄し生細胞中に保持されているGFPを共焦点レーザー走査型顕微鏡で可視化した。励起は340nmで実施し、放射は520nmで測定した。
アポトーシスアッセイ:アポトーシスは、アポトーシス核染色用ヘキスト色素(Molecular Probes, Eugene, OR)を使用して決定した。ヘキスト色素を細胞培養に添加し15分間インキュベートした。過剰のヘキスト色素は細胞培養液(pH指示薬を含まない)で細胞を洗浄して除去し、蛍光顕微鏡を用いて細胞を調べた(励起350nm、放射461nm)。
【実施例2】
【0040】
実施例2:[Dmt1]DALDA及びGly-SarのCaco-2細胞への取り込みの経時的変化
37℃でCaco-2細胞とインキュベートしたときに、[3H][Dmt1]DALDAは細胞溶解物中に5分という早期に観察され、30分までに定常レベルに達した(図1A)。1時間のインキュベーション後に細胞溶解物中で回収された[3H][Dmt1]DALDAの総量は総薬剤の約1%を占めた。対照的に、同じ実験条件下で、[14C]Gly-Sarは5分間から45分間増加し続けた(図1B)。測定された放射能は[Dmt1]DALDAの濃度を反映していると考えられる。なぜならば、本発明者らは[Dmt1]DALDAがペプチダーゼによる分解に対して耐性を有することを以前に示しているからである(Szeto et al., J. Pharmacol. Exp. Ther., 2001, 298:57-61)。測定された放射能が細胞膜と結合しているか否かを決定するために、細胞を酸洗浄に付して表面結合を除去した。図1Cは、[3H][Dmt1]DALDAの80.8%が酸洗浄に耐性であり、したがって細胞内に存在すると考えられることを示している。[Dmt1]DALDAの取り込みは広範囲の濃度にわたって濃度依存性であることが判明した(図1D)。
【実施例3】
【0041】
実施例3:[Dmt1]DALDA及びGly-Sarの取り込みに対する温度依存性及びpHの影響
インキュベーションを4℃で実施したとき、[3H][Dmt1]DALDAの取り込みは37℃よりも遅かったが、45分までに76.5%(図1A)に、さらに1時間までに86.3%(図1A)に達した。対照的に、[14C]Gly-Sarの取り込みは4℃では完全に停止した(図1B)。PEPT1によるGly-Sarの取り込みはpH依存性であり、最適取り込みはpH6.0で生じることが知られている(Terada et al., 1999, Am. J. Physiol. 276:G1435-G1441)。これは我々の実験で確認された(図2B)。対照的に、[3H][Dmt1]DALDAの取り込みは、pHを4.0から7.4の間で変化させたときには変化がなかった(図2A)。温度及びpHに対する依存性の欠如は、Caco-2細胞の[Dmt1]DALDAの取り込みはPEPT1(ペプチドトランスポーター1)を介したものではないことを示唆している。
【実施例4】
【0042】
実施例4:[Dmt1]DALDA及びGly-Sarの取り込みに対するDEPCの影響
PEPT1が[Dmt1]DALDAの輸送に関与していないことをさらに明瞭に示すために、[3H][Dmt1]DALDA 及び[14C]Gly-Sarの取り込みに対する DEPC(ジエチルピロカルボネート;0.2mM)の影響を調べた。DEPCは、Caco-2細胞でPEPT1を阻害することが示されているヒスチジン残基修飾試薬である(Terada et al., FEBS. Lett., 1996, 394:196-200)。インキュベーション培養液へのDEPCの添加は[14C]Gly-Sar取り込みを顕著に阻害した(図2D)。驚いたことには、DEPCは[3H][Dmt1]DALDAの取り込みを阻害しなかっただけでなく、実際には[3H][Dmt1]DALDAの取り込みを34倍増加させた(図2C)。
【実施例5】
【0043】
実施例5:種々の細胞タイプにおける[Dmt1]DALDAの内在化
[Dmt1]DALDAの内在化はCaco-2細胞に限定されないことを示すために、異なるいくつかの細胞株で[Dmt1]DALDAの内在化を比較した。内在化放射能(酸耐性)を表面結合放射能(酸感受性)と区別するために酸洗浄工程を含めた。図3Aでは、Caco-2、SH-SY5Y、HEK293及びCRFK細胞における酸耐性放射能レベルが比較されている。結果は、[3H][Dmt1]DALDAは全ての細胞株で大量に取り込まれることを示している。
【実施例6】
【0044】
実施例6:[3H] [Dmt1]DALDAによる放射性リガンド能結合アッセイ
[Dmt1]DALDAがレセプター仲介機構によって内在化されるか否かを決定するために、Caco-2細胞及びSH-SY5Y細胞から調製した膜を用いて放射能リガンド([3H][Dmt1]DALDA)結合アッセイを実施した。図3Bは、[3H][Dmt1]DALDAのSH-SY5Y膜との特異的結合を示している。算出されたKd値は118pM(87から149の範囲)であり、Bmax値は96 fmol/mgタンパク質(88から104)であると概算された。これは、チャイニーズハムスター卵巣細胞で発現されたリコンビナントヒトμ-オピオイドレセプターを用いて得られた値に匹敵する(G.-M. Zhao and H.H. Szeto, 未発表データ)。高い親和性を示す特異的結合はCaco-2膜では観察されなかった(図3B)。HEK293細胞はオピオイドレセプターを持たないことが知られている(Blake et al., J. Biol. Chem. 1997, 272:782-790)。
【実施例7】
【0045】
実施例7:Caco-2細胞からの[Dmt1]DALDA及びGly-Sarの流出
30分未満のインキュベーション後にCaco-2細胞で定常状態の[3H][Dmt1]DALDAレベルが達成されるということは、前記細胞のペプチド流出速度はその時点での取り込み速度と等しいことを示唆している。細胞からのGly-Sar及び[Dmt1]DALDAの流出を調べるために、Caco-2細胞に[14C]Gly-Sar又は[3H][Dmt1]DALDAを予備装荷し、続いて前記ペプチドを含まない新しい培養液と交換した。図4Aは、[14C]Gly-Sarの39%は37℃で1時間後に培養液中で見出されることを示している。[14C]Gly-Sarの流出は4℃で顕著に低下した。Caco-2細胞からの[3H][Dmt1]DALDAの流出ははるかに速く、80%のペプチドが1時間までに培養液中に流出した。 [3H][Dmt1]DALDAの内在化(図1A)とは対照的に、細胞からの[3H][Dmt1]DALDA流出に対して温度は顕著な影響を示した(図4A)。[Dmt1]DALDAの流出はDEPCで処理された細胞で減少した(図4B)。DEPCによる[3H][Dmt1]DALDA流出の低下は、DEPCの存在下で[3H][Dmt1]DALDAの取り込みが大きく増加することと一致する(図2C)。他方、[3H][Dmt1]DALDAの流出はベラパミル(P-糖タンパク質の阻害剤)の影響を受けなかった(図4C)。ベラパミルはまた[3H][Dmt1]DALDAの細胞内取り込みに影響を与えなかった(図4D)。
細胞からの[Dmt1]DALDAの流出は、[3H][Dmt1]DALDA−タンパク質共役物が細胞内に取り込まれた後に酵素的に切断されて[Dmt1]DALDAが細胞から流出し、一方、運ばれたタンパク質が細胞内に留まるならば有益なことであろう。
【実施例8】
【0046】
実施例8:[Dmt1]DALDA及びGly-Sarの細胞間輸送
トランスウェルで増殖させたCaco-2単層細胞を用いて、[Dmt1,dnsDap4]DALDA及び[14C]Gly-Sarの先端部から外側基底部への輸送を調べた。図5は、ペプチドをトランスウェルの先端部位に添加した後種々の時間における [14C]Gly-Sar及び[3H][Dmt1]DALDAの外側基底部位への輸送を示している。60分で先端部から外側基底部位へ移動した[3H][Dmt1]DALDAの百分率(10.4%)は輸送された[14C]Gly-Sarの百分率(11.9%)に匹敵した。見かけの透過率係数は、[3H][Dmt1]DALDAについては1.24x10-5cm/s、Gly-Sarについては1.26x10-5cm/sと概算された。
【実施例9】
【0047】
実施例9:CLSMを用いた芳香族陽イオンペプチドの細胞内取り込みの可視化
芳香族陽イオンペプチドの取り込み及び細胞内内在化態様を可視化するために、2つの蛍光ペプチド([Dmt1,dnsDap4]DALDA及び[Dmt1,atnDap4]DALDA)をCLSMによって調べた。図6は、0.1μMの[Dmt1,dnsDap4]DALDAとともに37℃で15分間インキュベートした後のCaco-2細胞への蛍光ペプチドの内在化を示している。蛍光は細胞質全体に拡散して出現し、明白な小胞体内分布は認められなかった。このことは、ペプチドの取り込みはエンドサイトーシスを必要とせず、ペプチドはエンドソームに封入されないことが示唆される。さらにまた、ペプチドは完全に核から排除されていたことは留意されたい。4℃にて30分間0.1μMの[Dmt1,antDap4]DALDAとともに SH-SY5Y細胞をインキュベートした後の前記細胞内への[Dmt1,atnDap4]DALDAの内在化は、エネルギー非依存性の、非エンドサイトーシス性取り込みメカニズムを支持する。なぜならば、エンドサイトーシスはエネルギー依存性の過程だからである。
【実施例10】
【0048】
実施例10:ペプチドと架橋剤SMCCとのカップリング及び質量分析による確認
SMCC(1μg)及び5μgの[Dmt1]DALDA、[Phe1]DALDA又は[d-Arg-Dmt-Lys-Phe-NH2]を一緒に2mLのPBSに溶解し、室温で30分間インキュベートし、4℃で保存した。前記サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。サンプルはMALDI-TOF MS(Matrix Assisted Laser Desorption Ionization−Time-of-Flight Mass Spectrometry, Applied Biosystems(Voyager DE Pro))によってポジティブリフレクトロンモードで分析した。
ペプチド及びそれらの対応するペプチド-SMCC結合物の分子量は質量スペクトルに示されている(図7)。
【実施例11】
【0049】
実施例11:タンパク質カーゴ(protein cargo)に共役させたペプチドは前記タンパク質カーゴを細胞内に輸送する
種々のペプチドをβ-ガラクトシダーゼ(組換え大腸菌、Sigma-Aldrich)と架橋剤SMCC(Pierce)を用いて架橋した。SMCCはアミン含有分子([Dmt1]DALDAのLys4)と反応して安定なアミド結合を生成する。ペプチド−SMCC共役物の形成は質量分析によって確認する(図7)。続いて前記のマレイミド端をスルフヒドリル含有化合物と結合させてチオエーテル結合を形成する(Greg T. Hermanson, “Bioconjugate Techniques”, Academic Press, page 234-237)。β-Galはその天然の状態で豊富な遊離スルフヒドリル基を含んでいる。β-Galの取り込みは、X-Galの使用による簡便な読み出しを提供する。
簡単に記せば、1mLの5x10-3M の[Dmt1]DALDA、[Phe1]DALDA又は[d-Arg-Dmt-Lys-Phe-NH2]をリン酸緩衝液中の1mgのSMCCと室温で1時間混合した。前記操作によって“活性化ペプチド”が生成される。前記“活性化ペプチド”をリン酸緩衝液で1:10に希釈した。1mgのβ-Galを前記1:10の“活性化ペプチド”1mLに添加し、4℃で2時間又は一晩混合した。
細胞(N2A神経芽腫細胞又はCaco-2細胞)を96ウェルプレートにプレーティングし(2x104細胞/ウェル)、β-Gal若しくは[Dmt1]DALDA、[Phe1]DALDA又は[d-Arg-Dmt-Lys-Phe-NH2]と架橋したβ-Galとともに37℃で1時間インキュベートした。続いて細胞をリン酸緩衝液で4回洗浄した。次に細胞をβ-gal染色セット(Roche)で37℃で少なくとも2時間染色し、顕微鏡下で調べた。
β-Galとともにインキュベートしたときにはβ-Galの取り込みは観察されなかった(図8A)。青い細胞の存在は[Dmt1]DALDAと共役したβ-GalのCaco-2細胞内取り込みを示している(図8B)。β-Galの取り込み強化はまた、β-Galを[d-Arg-Dmt-Lys-Phe-NH2](図8C)又は[Phe1]DALDA(図8D)と共役させたときにも認められた。β-GalとSMCCのみとの共役では取り込みは強化されなかった。
同様な結果が神経細胞系N2A細胞又はCHO細胞(チャイニーズハムスター卵巣細胞)を用いたときにも得られた。
【実施例12】
【0050】
実施例12:[Dmt1]DALDA−SMCC共役物との同時インキュベーションは緑色蛍光タンパク質(GFP)のHuh7細胞への取り込みを強化する
Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて3μgの緑色蛍光タンパク質(GFP)単独、3μgのGFP及び40μlの[Dmt1]DALDA、又は3μgのGFP及び40μlのSMCCと共役させた[Dmt1]DALDAを含む0.5mLのMEMとともに37℃で60分間インキュベートした。続いて2mLの細胞培養液を細胞に添加し、これを細胞培養インキュベーターでさらに24時間インキュベートした。インキュベーション後に細胞を細胞培養液で4回洗浄し生細胞中に保持されているGFPを共焦点レーザー走査型顕微鏡で可視化した。励起は340nmで実施し、放射は520nmで測定した。
図9(上段パネル)は、Huh7細胞の0.8μm厚の中央水平光学切片を通したGFPの画像を示す。図9(下段パネル)は、同じ顕微鏡野の微分干渉コントラスト像を示す。
GFPと[Dmt1]DALDAとの同時インキュベーションは、GFPだけでインキュベートした場合(図9A)と比較して、前記細胞の細胞質内の緑色蛍光を中程度に増加させることを示した(図9B)。核内には緑色蛍光は観察されなかった。GFPと[Dmt1]DALDA−SMCC共役物との同時インキュベーションはさらに強いGFPの取り込みを示した(図9C)。これらのデータは、[Dmt1]DALDAは、この改変ペプチドをタンパク質と単に物理的に混合することによって細胞内へのそのタンパク質の取り込みを促進し、ペプチドとタンパク質との間の化学的結合は不要であることを示している。
【実施例13】
【0051】
実施例13:[Dmt1]DALDAとRNAオリゴとの共役
γ-32P-ATPを用いたポリヌクレオチドキナーゼ反応により合成RNAオリゴ(長さは40ヌクレオチド)の5'端をリン酸化した。生成物を反応のためにゲル精製した。ゲル精製RNAオリゴの500,000cpmを1mgのEDC(N-[3-ジメチルアミノプロピル-N'-エチルカルボジイミド])の存在下で10mMの[Dmt1]DALDAとの反応で結合させた。前記結合反応生成物([Dmt1]DALDA−RNAオリゴ)及び対照であるRNAオリゴ単独を15%ポリアクリルアミド尿素ゲルで分析した。ゲル上の2つの別個のバンドは単独のRNAオリゴ及び[Dmt1]DALDA−RNAオリゴ共役物であることを示す(図10)。
【実施例14】
【0052】
実施例14:Caco-2細胞への[Dmt1]DALDA−RNAオリゴ共役物の取り込み
オリゴの添加前に、Caco-2細胞(1x106)をDMEM培養液で3回洗浄し、DMEMで5分間プレインキュベートした。続いて [Dmt1]DALDA−RNAオリゴ共役物又は非共役RNA(約20,000cpm)を細胞培養液に添加し、37℃で60分間インキュベートした。
インキュベーション後に、反応培養液を除去し、細胞を4回DMEMで、さらに1回酢酸ナトリウム溶液中で洗浄して非特異的結合を減少させた。最後に細胞を溶解緩衝液中で30分間インキュベートし、細胞溶解物中の放射能を測定した。
Caco-2細胞は、非共役RNAオリゴと比較して、3倍を超える強い[Dmt1]DALDA−RNAオリゴ共役物の取り込みを示した(図11)。したがって[Dmt1]DALDAは、細胞膜を横断するRNAオリゴの通過を促進する。
【実施例15】
【0053】
実施例15:RNAと[Dmt1]DALDA−SMCCリンカーとの混合はRNAの細胞内取り込みを増加させる
RNA及び[Dmt1]DALDA−SMCC共役物を物理的に混合することによって担体複合体を生成した。[Dmt1]DALDA-SMCC共役物は[Dmt1]DALDAを方法の項で述べたように架橋剤SMCCと混合することによって調製した。細胞取り込み実験で使用する前に、一本鎖の11mer[32P]RNAオリゴを[Dmt1]DALDA−SMCC共役物と15分間室温で混合した。
Huh7細胞(1x106細胞/ウェル)をDMEM培養液で洗浄し、続いて[32P]RNAオリゴ(約100,000cpm)のみを含む1.0mLのDMEM、又は40mLの[Dmt1]DALDA−SMCC共役物と37℃又は4℃にてインキュベートした。続いて細胞を4回DMEMで、さらに1回酢酸ナトリウム溶液中で洗浄して非特異的結合を除去し、その後、細胞を溶解緩衝液中で30分間インキュベートし、保持されている放射能を測定した。
RNAオリゴと[Dmt1]DALDA−SMCCとの37℃での同時インキュベーションはRNAオリゴの取り込みを時間の関数として増加させた(図12A)。RNAだけでインキュベートした場合と比較して、[Dmt1]DALDA−SMCCの存在下におけるRNAオリゴの取り込みは、1時間で約20倍に増加した。RNAの取り込みは、4℃でさえも[Dmt1]DALDA−SMCCによって顕著に強化された(図12B)。これらのデータは、[Dmt1]DALDAとの化学的共役がなくてもRNAの取り込みを強化することができることを示している。4℃における取り込みは、エネルギー非依存性非エンドサイトーシス的過程による取り込みを示唆しており、[Dmt1]DALDAの受動拡散による細胞膜透過能力と一致する。
[Dmt1]DALDA−SMCCの他に、[Phe1]DALDA−SMCC又は[d-Arg-Dmt-Lys-Phe-NH2]−SMCCとの同時インキュベーションもまた11mer RNAオリゴの取り込みを強化した。図12Cは、3つの異なるペプチド−SMCC共役物と37℃で15分間インキュベートしたときRNAの取り込みが増加したことを示している。
[Dmt1]DALDA−SMCC共役物との同時インキュベーションはまた、図13に示すようにはるかに大きいRNA分子(1350-mer)の細胞内取り込みを促進することができるが、ただしより小さなオリゴの場合ほど強くはない。
【実施例16】
【0054】
実施例16:[Dmt1]DALDAとDNAオリゴとの共役
SMCC(1μg)及び[Dmt1]DALDA(SS002;5μg)を一緒に2mLのPBSに溶解し、室温で30分間インキュベートし、さらに脱保護3'-チオールDNAオリゴと4℃で24時間混合した。インキュベーション後、サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。
DNA−[Dmt1]DALDA共役物の生成はMALDI-TOF MSによって確認した。3'-チオールDNAオリゴ及び[Dmt1]DALDA−DNA共有結合複合体の分子量は、それぞれ6392及び7171であることが判明した(図14A)。
【実施例17】
【0055】
実施例17:[Dmt1]DALDA−DNAオリゴ共役物のCaco-2細胞への取り込み
3'-チオール修飾20mer DNAをSMCCを用いて[Dmt1]DALDAに結合させ、共役物の生成は質量分析によって確認した。共役及び非共役DNAオリゴの5'末端を32Pで放射能標識し、さらにゲル精製した(図14B)。
神経細胞系N2A細胞(1x106細胞/ウェル)をDMEMで洗浄し、[Dmt1]DALDA共役又は非共役DNAオリゴ(約100,000cpm)のどちらかを含む1 mLのDMEMと37℃及び5%CO2で2時間又は19時間インキュベートした。続いて細胞を4回DMEMで、さらに1回酢酸ナトリウム溶液中で洗浄して非特異的結合を減少させた。続いて細胞を溶解緩衝液中で30分間インキュベートし、保持されている放射能を測定した。
19時間のインキュベーション後、[Dmt1]DALDAと共役したDNAの取り込みは、非共役DNAと比較して増加し、DNA取り込みは[Dmt1]DALDAとの共役によって強化できることが示された。
【実施例18】
【0056】
実施例18:ペプチド及びペプチド−SMCC共役物は細胞に対して毒性を示さない
ペプチドもペプチド−SMCC共役物も培養細胞に毒性を示さない。[Dmt1]DALDA(1nMから10μM)による24時間の処理は、N2A細胞(図16)、SH-SY5Y細胞又はCaco-2細胞においてMTTアッセイ(MTSアッセイ、Promega, Madison, WI)によって測定したところ、細胞の生存度へ影響を与えなかった。
培養細胞とペプチド−SMCC共役物とのインキュベーションもまた、トリパンブルーの取り込み測定によれば細胞の生存度に影響を与えなかった。トリパンブルーは膜透過性が増加した細胞によってのみ取り込まれる。Huh7細胞(1x106)をDMEM中で3回洗浄し、1mLの新しい培養液又は1mMの[Dmt1]DALDA−SMCC共役物、[D-Arg-Dmt-Lys-Phe-NH2]−SMCC共役物、若しくは [Phe1]DALDA−SMCC共役物の50μLを含む培養液を添加し、37℃で24時間、5%CO2でインキュベートした。続いて細胞をDMEMで3回洗浄し、0.4%トリパンブルーの1mLを前記細胞に2分間添加した。過剰な色素を細胞培養液中で細胞を洗浄することによって除去し、光学顕微鏡で細胞を調べた。
光学顕微鏡による細胞の検査で、培養液のみでインキュベートされた細胞は最小限のトリパンブルーの取り込みを示すことが判明した。トリパンブルーの取り込み増加は、[Dmt1]DALDA−SMCC、[D-Arg-Dmt-Lys-Phe-NH2]−SMCC又は[Phe1]DALDAとインキュベートした細胞で観察されなかった。対照的に、DEPC(ジエチルピロカルボネート)と細胞をインキュベートすることによって顕著なトリパンブルーの取り込みが生じた。
培養細胞と[Dmt1]DALDA−SMCC共役物とのインキュベーションは、Huh7培養細胞においてアポトーシスを誘導しなかった。Huh7細胞(1x106細胞/ウェル)をDMEM中で3回洗浄し、1mLの新しい培養液を適用した。続いてPBS中の改変[Dmt1]DALDA(1mM)又はPBSのみ(対照)50μLを前記細胞培養液に添加し、37℃で24時間、5%CO2でインキュベートした。インキュベーション後に、アポトーシス核の染色のために1mLのヘキスト色素(Molecular Probes, Eugene, OR)を細胞に添加し、さらに15分間インキュベートした。過剰な色素を細胞培養液(pH指示薬を含まない)で細胞を洗浄することによって除去し、蛍光顕微鏡検査(励起350nm、放射461nm)を用いて[Dmt1]DALDA−SMCC共役物で処理した細胞を対照細胞と比較した。アポトーシスは核内の蛍光の濃度によって示される。図17は、[Dmt1]DALDA−SMCCで処理したHuh7細胞のアポトーシスのレベルは対照細胞のそれと同じであることを示している。
【技術分野】
【0001】
本出願は、米国仮特許出願60/467,516号(2003年5月1日出願)の優先権を主張する。米国仮特許出願60/467,516号の明細は引用によりその全体が本明細書に含まれる。
本発明は、グラント番号P01-DA-08924によりNational Institute on Drug Abuseから政府の援助を受けて達成された。合衆国政府は本発明に関して一定の権利を有する。
【背景技術】
【0002】
生物学的細胞は、一般的に細胞膜を通過する分子について高度に選択性を有する。したがって、化合物(例えば小分子および生物学的分子)の細胞内への送達は通常は化合物の物理的特性によって制限される。前記小分子及び生物学的分子は例えば医薬的に活性を有する化合物であろう。
in vivoでのそのような分子(巨大分子、たとえばタンパク質及び核酸を含む)が細胞内へ送達されないことは、潜在的に有効な多数の化合物の治療的、予防的及び/又は診断的使用の妨げであった。さらにまた、in vitroでは有望とされる多くの化合物が、in vivoでは細胞内に効率的に前記化合物を送達する能力の欠如のために潜在的薬剤として廃棄されてきた。
いくつかの報告は、“タンパク質導入ドメイン(protein transduction domain)”(PTD)と前記化合物を共有結合することによって、細胞への化合物の送達の問題を扱ってきた。Schwarzeら(Trends Pharmacol. Sci. 2000, 21:45-8)及び米国特許6,221,355号(Dowdy)は、濃度依存態様で細胞の脂質二重層を通過することができるいくつかのPTDを開示した。前記開示されたPTDにはHIV-1 tatタンパク質由来PTD、アンテナペディア(ANTPと略される)遺伝子によってコードされるショウジョウバエのホメオティック転写因子由来のPTD、及び単純ヘルペスウイルスVP22転写因子由来のPTDが含まれる。HIV-1 tat PTDは長さが11個のアミノ酸であり、ANTP PTDは16個のアミノ酸であり、VP22 PTDは34個のアミノ酸である。
しかしながら、最近の文献では、これらPTDはエネルギー依存エンドサイトーシスを介して細胞内に入ることが示されている。したがって、“PTD-カーゴ”複合体は細胞のエンドソーム小胞内に収納され、例えば細胞の細胞質には利用できない。したがって、“PTD-カーゴ”複合体は、生物活性を示すためにはエンドソーム小胞から放出される必要がある(Richard et al., J. Biol. Chem. 2003, 278:585-590;Drin et al., J. Biol. Chem. 2003, 278:381192-31201)。さらにまた、最近の報告ではこれらPTDは細胞に対して有毒であることが示されている。
したがって、エネルギー非依存的非エンドサイトーシス態様で細胞の脂質膜を通過することができるペプチドが希求されている。さらにまた、大きなペプチドについて一般的に知られている免疫反応を回避するために、より小さな、ペプチダーゼ耐性ペプチドが求められている。最後に、前記ペプチド担体は細胞に対して毒性がないことが重要である。
【発明の概要】
【発明が解決しようとする課題】
【0003】
エネルギー非依存性メカニズムによって細胞膜を通過し、分子を細胞内に送達する方法および、そのための担体複合体を提供する。
【課題を解決するための手段】
【0004】
前述の要求は、分子を細胞に送達する方法を提供する本発明によって充足される。本方法は、細胞を担体複合体と接触させることを含み、前記担体複合体は、送達する分子及び芳香族陽イオンペプチドを含み、さらに前記芳香族陽イオンペプチドは以下を含む:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmはr+1以下である最大の数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下である最大の数であるが、ただしaが1であるときはptもまた1であることができる関係。
また別の実施態様では、本発明は、分子及び芳香族陽イオンペプチドを含む担体複合体を提供し、前記芳香族陽イオンペプチドは以下を含む:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmはr+1以下である最大の数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下である最大の数であるが、ただしaが1であるときはptもまた1であることができる関係。
さらに別の実施態様では、本発明は分子を細胞に送達する方法を提供する。本方法は、細胞を分子及び芳香族陽イオンペプチドと接触させることを含み、前記芳香族陽イオンペプチドは以下を含む:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmはr+1以下である最大の数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下である最大の数であるが、ただしaが1であるときはptもまた1であることができる関係。
【図面の簡単な説明】
【0005】
【図1】Caco-2細胞のペプチドの取り込み。[3H][Dmt1]DALDA(A)及び[14C]Gly-Sar(B)の経時的取り込み。Caco-2細胞を[3H][Dmt1]DALDA(250nM, 47Ci/mmol)又は[14C]Gly-Sar(50μM, 56.7mCi/mmol)と37℃又は4℃で1時間インキュベートした。その後、放射能を溶解細胞で測定した。(C)[3H][Dmt1]DALDAの蓄積に対する酸洗浄の影響。Caco-2細胞を[3H][Dmt1]DALDAとともに37℃で1時間インキュベートした。細胞の溶解前に、細胞を酸洗浄に付して細胞表面結合放射能を除去した。(D)[Dmt1]DALDA取り込みに対する[Dmt1]DALDAの濃度の影響。細胞をある範囲の濃度(1μMから3mM)の[Dmt1]DALDAとともに1時間37℃でインキュベートした。全てのデータは、3つのそれぞれ別個の単層細胞の平均±S.E.として示されている。誤差バーが示されていない場合は、誤差バーは記号サイズより小さい。
【図2】Caco-2細胞における[3H][Dmt1]DALDA取り込み(A及びC)及び[14C]Gly-Sar取り込み(B及びD)に対するpH及びDEPCの影響。Caco-2細胞を[3H][Dmt1]DALDA(250nM, 47Ci/mmol)又は[14C]Gly-Sar(50μM, 56.7mCi/mmol)とともに種々のpH条件下で37℃で1時間インキュベートした(A及びB)。[3H][Dmt1]DALDA(250nM, 47Ci/mmol)又は[14C]Gly-Sar(50μM, 56.7mCi/mmol)と37℃で1時間インキュベートする前に、細胞を0.2mMのDEPCと10分間25℃でプレインキュベートした(C及びD)。全てのデータは、3つのそれぞれ別個の単層細胞の平均±S.E.として示されている。
【図3】(A)種々の細胞株における[3H][Dmt1]DALDAの取り込み。細胞を[3H][Dmt1]DALDA(250nM, 47Ci/mmol)とともに37℃で1時間インキュベートした。細胞の溶解前に、細胞を酸洗浄に付して細胞表面結合放射能を除去した。提示データは酸耐性放射能を示し、3つのそれぞれ別個の単層細胞の平均±S.E.として表示されている。(B) [3H][Dmt1]DALDAの細胞膜への特異的結合。SH-SY5Y細胞及びCaco-2細胞から調製した膜を[3H][Dmt1]DALDA(15から960pM)とともに25℃で1時間インキュベートした。非特異的結合は、1μMの非標識[Dmt1]DALDAを含めることによって評価した。遊離放射性リガンドは迅速ろ過によって結合放射性リガンドから分離した。特異的結合はCaco-2細胞では観察されなかった。SH-SY5Y細胞については、Kd値は118pM(87から149の範囲)で、Bmax値は96 fmol/mgタンパク質であった。
【図4】(A)[3H][Dmt1]DALDA(黒棒線)及び[14C]Gly-Sar(白棒線)の流出。Caco-2細胞に[3H][Dmt1]DALDA(250nM, 47Ci/mmol)又は[14C]Gly-Sar(50μM, 56.7mCi/mmol)を37℃又は4℃で1時間予備装荷した。続いて細胞を洗浄し、さらに培養液とともに37℃又は4℃で1時間インキュベートした。培養液及び細胞溶解物の両方で放射能を測定した。データは培養液中に流出したペプチドの百分率として表されている。(B)[3H][Dmt1]DALDA流出に対するDEPCの影響。[3H][Dmt1]DALDAを装荷する前に、細胞を25℃で10分間、0.2mMのDEPCとともにインキュベートした。(C)[3H][Dmt1]DALDAの流出に対するベラパミル(p-糖タンパク質の阻害剤)の影響、及び(D)[3H][Dmt1]DALDAの取り込みに対するベラパミルの影響。
【図5】単層Caco-2細胞を通過する[3H][Dmt1]DALDA及び[14C]Gly-Sarの輸送。Caco-2細胞(2x105)をトランスウェル細胞培養チャンバー内の微小孔を有するメンブレンフィルター上に播種した。先端部から外側基底部へのペプチド輸送は、 [3H] [Dmt1]DALDA又は[14C]Gly-Sarを先端部区画に添加することによって決定した。放射能の計測のために、ペプチド添加後種々の時間に20μLのアリコットを先端部区画及び外側基底部区画の両方から取り出した。
【図6】[Dmt1,dnsDap4]DALDA及び[Dmt1,atnDap4]DALDAの細胞内取り込み。Caco-2細胞を0.1μMの[Dmt1,dnsDap4]DALDAとともに37℃で15分間インキュベートした。続いて細胞を洗浄し、PBSで被覆した。顕微鏡検査は室温で10分以内に実施した。励起は340nmで実施し、放射は520nmで測定した。蛍光は細胞質全体に拡散しているように見えたが、核からは完全に排除されていた。37℃で小胞体内濃縮が認められないことから、非エンドサイトーシス性取り込みが示唆される。
【図7】3つのペプチドと架橋剤SMCCとのカップリングの質量分析による確認。SMCC(1μg)及びペプチド(5μg)を一緒に2mLのPBSに溶解し、室温で30分間インキュベートし、4℃で保存した。前記サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。サンプルはMALDI-TOF MS(Matrix Assisted Laser Desorption Ionization Time-of-Flight Mass Spectrometry)によって分析した。ペプチド及びそれらの対応するSMCC共役物の分子量はスペクトル上に表示されている。
【図8】β-ガラクトシダーゼ(β-Gal)のN2A神経芽腫細胞内への取り込みを強化するペプチドの能力。細胞(N2A神経芽腫細胞又はCaco-2細胞)を96ウェルプレートにプレーティングし(2x104細胞/ウェル)、β-Gal又は(SMCCを介して)ペプチドと共役させたβ-Galとともに37℃で1時間インキュベートした。続いて細胞をリン酸緩衝液で4回洗浄した。次に、細胞をβ-gal染色セット(Roche)で37℃で少なくとも2時間染色し、顕微鏡下で調べた。(A)CaCo-2細胞をβ-Galとともにインキュベートしたときにはβ-Galの取り込みは観察されなかった。(B)青い細胞の存在は[Dmt1]DALDAに共役したβ-GalのCaco-2細胞内取り込みを示している。(C)[D-Arg-Dmt-Lys-Phe-NH2]に共役したβ-GalのCaco-2細胞内取り込み強化。(D)[Phe1]DALDAに共役したβ-GalのCaco-2細胞内取り込みの強化。SMCCのみと共役したβ-Galでは取り込みは強化されなかった。
【図9】[Dmt 1]DALDA−SMCC共役物との同時インキュベーションは緑色蛍光タンパク質(GFP)のHuh7細胞内への取り込みを強化する。Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて3μgの緑色蛍光タンパク質(GFP)単独(A)、3μgのGFP及び40μLの[Dmt1]DALDA(B)、又は3μgのGFP及びSMCCと共役させた40μL[Dmt1]DALDA(C)を含む0.5mLのMEMとともに37℃で60分間インキュベートした。続いて2mLの細胞培養液を細胞に添加し、これを細胞培養インキュベーターでさらに24時間インキュベートした。インキュベーション後に細胞を細胞培養液で4回洗浄し生細胞中に保持されているGFPを共焦点レーザー走査型顕微鏡で可視化した。励起は340nmで実施し、放射は520nmで測定した。上段パネルは、Huh7細胞の0.8μm厚の中央部水平光学切片を通したGFPの画像を示す。下段パネルは、同じ顕微鏡野の微分干渉コントラスト像を示す。
【図10】[Dmt1]DALDAとRNAオリゴとの共役。γ-32P-ATP及びポリヌクレオチドキナーゼを用いて合成RNAオリゴ(長さは40ヌクレオチド)の5'端をリン酸化した。生成物をゲル電気泳動で精製した。ゲル精製RNAオリゴの500,000cpmを1mgのEDC(N-[3-ジメチルアミノプロピル-N'-エチルカルボジイミド])の存在下で[Dmt1]DALDAと結合させた。前記結合反応の生成物([Dmt1]DALDA−RNAオリゴ)及び単独RNAオリゴは15%ポリアクリルアミド尿素ゲルで分析した。
【図11】[Dmt1]DALDA−[32P]RNAオリゴ共役物のCaco-2細胞内への取り込み。Caco-2細胞(1x106)をDMEM培養液で3回洗浄し、DMEMで5分間プレインキュベートした。続いて細胞を[Dmt1]DALDA−[32P]RNAオリゴ共役物又は対照RNA(約20,000cpm)と37℃で60分間インキュベートした。インキュベーション後に、細胞を洗浄して溶解し、細胞溶解物中の放射能を測定した。[Dmt1]DALDA−[32P]RNAオリゴ共役物の取り込みは、RNA単独と比較して3倍を超えて強化された。
【図12】RNAオリゴのHuh7細胞内への取り込みを強化するペプチド-SMCC共役物の効果。(A)RNAオリゴの細胞取り込みに対する時間の影響。Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて[32P]RNAオリゴ(一本鎖、11塩基;約100,000cpm)単独又は40mLの[Dmt1]DALDA−SMCC共役物を含む1.0mLのDMEMと37℃で15分間又は60分間インキュベートした。続いて、細胞をDMEM中で4回、さらに酢酸ナトリウム溶液で1回洗浄して非特異的結合を取り除いた後、溶解緩衝液で30分間インキュベートし、保持されている放射能を測定した。RNAオリゴと[Dmt1]DALDA−SMCCと37℃で同時インキュベートすることによって、RNAオリゴの取り込みは15分間インキュベーション後に10倍、60分間インキュベーション後に20倍増加した。(B)RNAオリゴの細胞取り込みに対する温度の影響。[Dmt1]DALDA−SMCCのRNA取り込みを強化する能力は4℃では低かったが、それでもなお前記は取り込みを10倍増加させた。(C)種々のペプチド−SMCC共役物によるRNAの細胞内取り込みの強化。Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて[32P]RNAオリゴ単独又は40mLのペプチド−SMCC共役物を含む1.0mLのDMEMと37℃で15分間インキュベートした。3つのペプチド−SMCC共役物はいずれもRNA取り込みを増加させた。
【図13】[Dmt1]DALDA−SMCC共役物との同時インキュベーションは異なる長さの2つのRNAの取り込みを強化した。[Dmt1]DALDAをSMCCと共役させ、質量分析によって確認した。11-mer RNAオリゴ及び1350-mer RNAを[Dmt1]DALDA−SMCC共役物と室温で15分間混合した。Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いてRNA(約100,000cpm)単独又は[Dmt1]DALDA−SMCC共役物と混合したRNAを含む1.0mLのDMEMと37℃及び5%CO2で60分間インキュベートした。続いて、洗浄した細胞をDMEM中で4回、さらに酢酸ナトリウム溶液で1回洗浄して非特異的結合を取り除いた。続いて細胞を溶解緩衝液で30分間インキュベートし、保持されている放射能を測定した。RNA単独とのインキュベーションと比較して、[Dmt1]DALDA−SMCC共役物とのインキュベーションによって11-mer RNAは22倍、1350-mer RNAは3倍、取り込みが増加した。
【図14】DNAオリゴの[Dmt1]DALDAへの共役。SMCC(1μg)及び[Dmt1]DALDA(5μg)を一緒に2mLのPBSに溶解し、室温で30分間インキュベートし、脱保護した3'-チオールDNAオリゴと4℃で24時間混合した。インキュベーション後に、サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。サンプルはMALDI-TOF MSで分析した(A)。3'-チオールDNAオリゴ及び[Dmt1]DALDA共有結合複合体の分子量はそれぞれ6392及び7171であることが判明した。γ-32P-ATP及びポリヌクレオチドキナーゼを用いて、共役オリゴ及び非共役オリゴの両者の5'端をリン酸化し、キナーゼ反応の生成物を15%ポリアクリルアミド尿素ゲルで分析し、細胞取り込み実験のためにゲル精製した(B)。
【図15】[Dmt1]DALDAと共役したDNAオリゴの細胞内取り込み。SMCCを用いて3'-チオール改変20-mer DNAを[Dmt1]DALDAと共役させ、共役物の生成は質量分析によって確認した。共役及び非共役DNAオリゴの5'末端を32Pで放射能標識し、さらにゲル精製した。神経細胞系N2A細胞(1x106細胞/ウェル)をDMEMで洗浄し、[Dmt1]DALDA共役又は非共役DNAオリゴ(約100,000cpm)のどちらかを含む1mLのDMEMとともに37℃及び5%CO2で2時間又は19時間インキュベートした。続いて細胞を4回DMEMで、さらに1回酢酸ナトリウム溶液中で洗浄して非特異的結合を除去した。続いて細胞を溶解緩衝液中で30分間インキュベートし、保持されている放射能を測定した。Y軸は、総放射能の百部率で表したDNAの取り込みを示している。
【図16】[Dmt1]DALDAは培養細胞に毒性を示さない。神経細胞系N2A細胞を [Dmt1]DALDA(1nMから10μM)とともに24時間インキュベートし、細胞生存度をMTTアッセイによって決定した。
【図17】[Dmt1]DALDA−SMCC共役物はHuh7細胞にアポトーシスを誘導しない。Huh7細胞(1x106細胞/ウェル)をDMEM中で3回洗浄し、1mLの新しい培養液を適用した。続いて50μL のPBS中の[Dmt1]DALDA−SMCC共役物(1mM)又はPBSのみ(対照)を前記細胞培養液に添加し、5%CO2で37℃24時間インキュベートした。インキュベーション後に、アポトーシス核染色用ヘキスト色素(Molecular Probes, Eugene, OR)の1mLを細胞に添加し、さらに15分間インキュベートした。過剰なヘキスト色素を細胞培養液(pHインジケーターを含まない)で細胞を洗浄することによって除去し、蛍光顕微鏡検査(励起350nm、放射461nm)を用いて[Dmt1]DALDA−SMCC共役物で処理した細胞を対照細胞と比較した。
【発明を実施するための形態】
【0006】
発明の詳細な説明
本発明は、少なくとも1つの分子及び芳香族陽イオンペプチドを含むある種の担体複合体は、エネルギー非依存性メカニズムによって細胞膜を通過し、前記分子を細胞内に送達することができるという本発明者らの驚くべき発見によるものである。
芳香族陽イオンペプチド
本発明で有用な芳香族陽イオンペプチドは、以下に説明するように正味の正電荷を有し、水溶性であり強い極性を示す。前記ペプチドは最小限3つのアミノ酸を含み、好ましくは最小限4つのアミノ酸を含み、前記はペプチド結合によって共有結合されている。
前記芳香族陽イオンペプチドに存在するアミノ酸の最大数は10、好ましくは約8、もっとも好ましくは約6である。最適には、前記ペプチドに存在するアミノ酸の数は約4である。アミノ酸の最大数の定義で用いられる“約”という用語は、+1又は−1個のアミノ酸を意味する。
本発明で有用な芳香族陽イオンペプチドのアミノ酸は任意のアミノ酸であってよい。本明細書において、“アミノ酸”という用語は、少なくとも1つのアミノ基及び少なくとも1つのカルボキシル基を含む任意の有機分子を指すために用いられる。好ましくは、少なくとも1つのアミノ基はカルボキシル基に対してα位に存在する。
前記アミノ酸は天然に存在するであり得る。天然に存在するアミノ酸には、例えば、タンパク質に通常見出される最も一般的な20のアミノ酸、すなわちアラニン(Ala)、アルギニン(Arg)、アスパラギン(Asn)、アスパラギン酸(Asp)、システイン(Cys)、グルタミン(Glu)、グルタミン酸(Glu)、グリシン(Gly)、ヒスチジン(His)、イソロイシン(Ileu)、ロイシン(Leu)、リジン(Lys)、メチオニン(Met)、フェニルアラニン(Phe)、プロリン(Pro)、セリン(Ser)、スレオニン(Thr)、トリプトファン(Trp)、チロシン(Tyr)、及びバリン(Val)が含まれる。
【0007】
天然に存在する他のアミノ酸には、例えばタンパク質合成に関連しない代謝過程で合成されるアミノ酸が含まれる。例えば、アミノ酸であるオルニチンは哺乳動物の代謝で尿素生成時に合成される。
本発明で有用な芳香族陽イオンペプチドは、場合によって天然には存在しない1つ又は2つ以上のアミノ酸を含む。ある実施態様では、前記ペプチドは天然に存在するアミノ酸を含まない。
天然に存在しないアミノ酸は、典型的には生きている生物の通常の代謝過程では合成されず、タンパク質中に通常出現しないアミノ酸である。
さらに、本発明で有用な天然に存在しないアミノ酸は好ましくは一般的なプロテアーゼによって認識されない。したがって、前記天然に存在しないアミノ酸は、一般的なプロテアーゼに対して好ましくは耐性であり、より好ましくは非感受性である。
天然に存在しないアミノ酸は前記ペプチドのいずれの位置にあってもよい。例えば、天然に存在しないアミノ酸はN-末端、C-末端及び/又はN-末端とC-末端の間の1つ又は2つ以上の任意の位置に存在することができる。
非天然アミノ酸は、例えばアルキル基、アリール基又はアルキルアリール基を含むことができる。アルキルアミノ酸のいくつかの例にはα-アミノ酪酸、β-アミノ酪酸、γ-アミノ酪酸、δ-アミノ吉草酸及びε-アミノカプロン酸が含まれる。アリールアミノ酸のいくつかの例には、オルト-、メタ-及びパラ-アミノ安息香酸が含まれる。アルキルアリールアミノ酸のいくつかの例には、オルト-、メタ-、及びパラ-アミノフェニル酢酸並びにγ-フェニル-β-アミノ酪酸が含まれる。
天然に存在しないアミノ酸にはまた天然に存在するアミノ酸の誘導体が含まれる。天然に存在するアミノ酸の誘導体は、例えば天然に存在するアミノ酸に対し1つ又は2つ以上の化学基の付加を含む。
【0008】
例えば、1つ又は2つ以上の化学基は、フェニルアラニン又はチロシン残基の芳香環の2'、3'、4'、5'若しくは6'位の1つ又は2つ以上に、又はトリプトファン残基のベンゾ環の4'、5'、6'若しくは7'位の1つ又は2つ以上に付加することができる。この基は芳香環に付加することができる任意の化学基であり得る。そのような基のいくつかの例には、分枝又は非分枝C1‐C4アルキル、例えばメチル、エチル、n-プロピル、イソプロピル、ブチル、イソブチル若しくはt-ブチル、C1‐C4アルコキシ(すなわちアルコキシ)、アミノ、C1‐C4アルキルアミノ(例えばメチルアミノ)及びC1‐C4ジアルキルアミノ(例えばジメチルアミノ)、ニトロ、ヒドロキシル、ハロ(すなわちフルオロ、クロロ、ブロモ又はヨード)が含まれる。天然に存在するアミノ酸の天然には存在しない誘導体のいくつかの具体例には、ノルバリン(Nva)、ノルロイシン(Nle)及びヒドロキシプロリン(Hyp)が含まれる。
本発明で有用なペプチドのアミノ酸改変のまた別の例は、前記ペプチドのアスパラギン酸又はグルタミン酸のカルボキシル基の誘導体化である。誘導体化の一例は、アンモニア又は第一若しくは第二アミン(例えばメチルアミン、エチルアミン、ジメチルアミン又はジエチルアミン)によるアミド化である。誘導体化のまた別の例には例えばメチル又はエチルアルコールによるエステル化が含まれる。
また別の改変にはリジン、アルギニン又はヒスチジンのアミノ基の改変が含まれる。例えば、これらのアミノ基はアシル化することができる。いくつかの適切なアシル基には、例えばベンゾイル基又はアルカノイル基が含まれ、前記は上記で述べた任意のC1‐C4アルキル基、例えばアセチル又はプロピオニル基を含む。
天然に存在しないアミノ酸は、一般的には左旋性(L-)でも右旋性(D-)でも又はその混合物でもあり得る。天然に存在しない適切なアミノ酸の例にはまた、上記の天然に存在するL-アミノ酸のいずれかの右旋性型が、天然には存在しないアミノ酸のL-及び/又はD-型とともに含まれる。これに関して、D-アミノ酸は通常はタンパク質に存在しないことは留意されるべきである(ただしそれらは細胞の正常なリボソームによるタンパク質合成機構以外の手段によって合成されるある種のペプチド抗生物質で見出される)。本明細書において、そのようなD-アミノ酸は天然に存在しないアミノ酸と考えられる。
【0009】
プロテアーゼに対する感受性を最小限にするために、本発明で有用なペプチドは、天然に存在するか否かに関係なく、一般的なプロテアーゼによって認識される、5つ未満、好ましくは4つ未満、より好ましくは3つ未満、もっとも好ましくは2つ未満の連続するL-アミノ酸を有するべきである。ある実施態様では、前記ペプチドはD-アミノ酸のみであり、L-アミノ酸は存在しない。
ペプチドがプロテアーゼ感受性アミノ酸配列を含む場合は、少なくとも1つのアミノ酸は好ましくは天然に存在しないD-アミノ酸であり、それによってプロテアーゼ耐性を付与する。プロテアーゼ感受性配列の例には、一般的なプロテアーゼ(例えばエンドペプチダーゼ及びトリプシン)によって切断される2つ以上の連続する塩基性アミノ酸が含まれる。塩基性アミノ酸の例にはアルギニン、リジン、及びヒスチジンが含まれる。
芳香族陽イオンペプチドが、ペプチド中のアミノ酸の総数に対して生理学的pHで最少数の正味の正電荷を有するということが重要である。生理学的pHでの正味の正電荷の最小数は下記では(pm)と称される。ペプチド内のアミノ酸残基の総数は下記では(r)と称される。
下記で考察される正味の正電荷の最小数は全て生理学的pHでのものである。本明細書で用いられる“生理学的pH”という用語は、哺乳動物の体内の組織及び器官の細胞の正常なpHを指す。例えばヒトの生理学的pHは通常ほぼ7.4であるが、哺乳動物の正常な生理学的pHは約7.0から約7.8の間の任意のpHであり得る。
【0010】
本明細書で用いられる“正味の電荷”は、ペプチド内に存在するアミノ酸が保持する正電荷数と負電荷数の差し引きを指す。本明細書では、正味の電荷は生理学的pHで測定されると理解される。生理学的pHで正に荷電する天然に存在するアミノ酸にはL-リジン、L-アルギニンおよびL-ヒスチジンが含まれる。生理学的pHで負に荷電する天然に存在するアミノ酸にはL-アスパラギン酸及びL-グルタミン酸が含まれる。
典型的には、ペプチドは正に荷電したN-末端アミノ基及び負に荷電したC-末端カルボキシル基を有する。電荷は生理学的pHで互いに相殺される。正味の電荷を計算する例として、ペプチドTyr-Arg-Phe-Lys-Glu-His-Trp-Argは、1つの負に荷電したアミノ酸(すなわちGlu)及び4つの正に荷電したアミノ酸(すなわち2つのArg残基、1つのLys及び1つのHis)を有する。したがって、上記のペプチドは正味の正電荷3を有する。
本発明のある実施態様では、芳香族陽イオンペプチドは、生理学的pHでの正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間に、3pmがr+1以下である最大数であるという関係を有する。この実施態様では、正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間の関係は以下のとおりである:
別の実施態様では、芳香族陽イオンペプチドは、正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間に、2pmがr+1以下である最大数であるという関係を有する。この実施態様では、正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間の関係は以下のとおりである:
ある実施態様では、正味の正電荷の数(pm)とアミノ酸残基の数(r)は等しい。別の好ましい実施態様では、前記ペプチドは、3つから4つのアミノ酸残基、及び最小限1つの正味の正電荷、好ましくは最小限2つの正味の正電荷、より好ましくは最小限3つの正味の正電荷を有する。
芳香族陽イオンペプチドが正味の正電荷の総数(pt)に対し最小数の芳香族基を有することもまた重要である。芳香族基の最小数は以下では(a)と称する。
【0011】
芳香族基をもつ天然に存在するアミノ酸には、ヒスチジン、トリプトファン、チロシン及びフェニルアラニンが含まれる。例えば、ヘキサペプチドであるLys-Gln-Tyr-Arg-Phe-Trpは正味の正電荷が2であり(リジン及びアルギニン残基が寄与している)、3つの芳香族基(チロシン、フェニルアラニン及びトリプトファン残基によって付与される)を有する。
本発明のある実施態様では、本発明の方法で有用な芳香族陽イオンペプチドは、芳香族基の最小数(a)と生理学的pHでの正味の正電荷の総数(pt)との間に、3aがpt+1以下である最大の数であるが、ただしptが1であるときはaもまた1であることができる関係を有する。この実施態様では、芳香族基の最小数(a)と正味の正電荷の数(pt)との間の関係は以下のとおりである:
別の実施態様では、芳香族陽イオンペプチドは、芳香族基の最小数(a)と正味の正電荷の総数(pt)との関係において、2aがpt+1以下である最大の数である。この実施態様では、芳香族アミノ酸残基の最小数(a)と正味の正電荷の総数(pt)との間の関係は以下のとおりである:
別の実施態様では、芳香族基の数(a)と正味の正電荷の総数(pt)は等しい。
【0012】
カルボキシル基、特にC-末端アミノ酸の末端カルボキシル基は好ましくは、例えばアンモニアでアミド化されて、C-末端アミドを形成する。あるいは、C-末端アミノ酸の末端カルボキシル基は、第一又は第二アミンでアミド化されてもよい。前記第一又は第二アミンは、例えばアルキル、特に分枝又は非分枝C1‐C4アルキル又はアリールアミンであり得る。したがって、本ペプチドのC-末端のアミノ酸は、アミド、N-メチルアミド、N-エチルアミド、N,N-ジメチルアミド、N,N-ジエチルアミド、N-メチル-N-エチルアミド、N-フェニルアミド又はN-フェニル-N-エチルアミド基に変換することができる。
さらに、2つ以上のカルボキシレート基(例えばアスパラギン、グルタミン、アスパラギン酸又はグルタミン酸残基)を有するアミノ酸残基の遊離カルボキシレート基もまた、それらがどこに存在していてもアミド化することができる。これらの位置におけるアミド化は、上記のようにアンモニアを用いても、又は第一若しくは第二アミンのいずれを用いてもよい。
ある実施態様では、本発明の方法で有用な芳香族陽イオンペプチドは、正味の正電荷が2で少なくとも1つの芳香族アミノ酸を有するトリペプチドである。具体的な実施態様では、本発明の方法で有用な芳香族陽イオンペプチドは、正味の正電荷が2で2つの芳香族アミノ酸を有するトリペプチドである。
本発明の方法で有用な芳香族陽イオンペプチドには以下のペプチド例が含まれる(ただしこれらに限定されない):
Lys-D-Arg-Tyr-NH2
Phe-D-Arg-His
D-Tyr-Trp-Lys- NH2
Trp-D-Lys-Tyr-Arg- NH2
Tyr-His-D-Gly-Met
Phe-Arg-D-His-Asp
Tyr-D-Arg-Phe-Lys-Glu- NH2
Met-Tyr-D-Lys-Phe-Arg
D-His-Glu-Lys-Tyr-D-Phe-Arg
Lys-D-Gln-Tyr-Arg-D-Phe-Trp-NH2
Phe-D-Arg-Lys-Trp-Tyr-D-Arg-His
Gly-D-Phe-Lys-Tyr-His-D-Arg-Tyr-NH2
Val-D-Lys-His-Tyr-D-Phe-Ser-Tyr-Arg-NH2
Trp-Lys-Phe-D-Asp-Arg-Tyr-D-His-Lys
Lys-Trp-D-Tyr-Arg-Asn-Phe-Tyr-D-His-NH2
Thr-Gly-Tyr-Arg-D-His-Phe-Trp-D-His-Lys
Asp-D-Trp-Lys-Tyr-D-His-Phe-Arg-D-Gly-Lys-NH2
D-His-Lys-Tyr-D-Phe-Glu-D-Asp-D-His-D-Lys-Arg-Trp-NH2 及び
Ala-D-Phe-D-Arg-Tyr-Lys-D-Trp-His-D-Tyr-Gly-Phe
【0013】
特に好ましい実施態様では、芳香族陽イオンペプチドは、式Tyr-D-Arg-Phe-Lys-NH2(簡便に頭文字でDALDAと表わす)を有する。DALDAは正味の正電荷が3で(前記はチロシン、アルギニン及びリジンによって付与される)、フェニルアラニン及びチロシンの寄与による2つの芳香族基を有する。DALDAのチロシンは、例えば2',6'-ジメチルチロシンのようにチロシンの改変誘導体であってよく、式2',6'-Dmt-D-Arg-Phe-Lys-NH2(すなわちDmt1-DALDA)を有する化合物を生成することができる。チロシンの他の改変誘導体には以下が含まれる:2'-メチルチロシン(Mmt);N,2',6'-トリメチルチロシン(Tmt);及び2'-ヒドロキシ-6'-メチルチロシン(Hmt)。
別の好ましい実施態様では、DALDAのN-末端のアミノ酸はフェニルアラニン又はその誘導体であり得る。N-末端にフェニルアラニンを有する芳香族陽イオンペプチドは式Phe-D-Arg-Phe-Lys-NH2(すなわちPhe1-DALDA)を有する。好ましいフェニルアラニンの誘導体には以下が含まれる:2'-メチルフェニルアラニン(Mmp);2',6'-ジメチルフェニルアラニン(Dmp);N,2',6'-トリメチルフェニルアラニン(Tmp);及び2'-ヒドロキシ-6'-メチルフェニルアラニン(Hmp)。
別の実施態様では、Dmt1-DALDAのアミノ酸配列は、DmtがN-末端に存在しないように再編成される。そのような芳香族陽イオンペプチドの例は式D-Arg-2',6'Dmt-Lys-Phe-NH2を有する。
【0014】
本明細書に開示した具体的なペプチドのいずれも、例えば上記で述べたペプチド及び下記(例えば表1)で述べるペプチド(Dmt1-DALDA、DALDA、Phe1-DALDA、D-Arg-2',6'Dmt-Lys-Phe-NH2及びそれらの誘導体を含む)、さらに機能的な類似体を含むことができる。ペプチドは、類似体がDmt1-DALDA、DALDA、Phe1-DALDA又はD-Arg-2',6'Dmt-Lys-Phe-NH2と同じ機能を有するならば、Dmt1-DALDA、DALDA、Phe1-DALDA、D-Arg-2',6'Dmt-Lys-Phe-NH2の機能的類似体と考えられる。例えば前記類似体は、1つ又は2つ以上のアミノ酸が別のアミノ酸によって置換された、Dmt1-DALDA、DALDA、Phe1-DALDA、D-Arg-2',6'Dmt-Lys-Phe-NH2の置換変種であり得る。
Dmt1-DALDA、DALDA、Phe1-DALDA、D-Arg-2',6'Dmt-Lys-Phe-NH2の適切な置換変種は保存的アミノ酸置換を含む。アミノ酸はそれらの物理化学的特性に従って以下のように分類できる:
(a)非極性アミノ酸:Ala(A)、Ser(S)、Thr(T)、Pro(P)、Gly(G);
(b)酸性アミノ酸:Asn(N)、Asp(D)、Glu(E)、Gln(Q);
(c)塩基性アミノ酸:His(H)、Arg(R)、Lys(K);
(d)疎水性アミノ酸:Met(M)、Leu(L)、Ile(I)、Val(V);及び
(e)芳香族アミノ酸:Phe(F)、Tyr(Y)、Trp(W)、His(H)。
ペプチド内のアミノ酸の同じグループ内のアミノ酸による置換は保存的置換と称される。保存的置換は元のペプチドの物理化学的特性を保存する傾向がある。対照的に、ペプチド内のアミノ酸の異なるグループ内の別のアミノ酸による置換は、一般的に元のペプチドの特性を変化させる傾向がある。
【0015】
本発明の実施で有用な類似体の例には表1及び2に示す芳香族陽イオンペプチドが含まれる(ただしこれらに限定されない)。
表1:
Dab=ジアミノ酪酸
Dap=ジアミノプロピオン酸
Dmt=ジメチルチロシン
Mmt=2'-メチルチロシン
Tmt=N,2',6'-トリメチルチロシン
Hmt=2'-ヒドロキシ,6'-メチルチロシン
dnsDap=β-ダンシル-L-α,β-ジアミノプロピオン酸
atnDap=β-アントラニロイル-L-α,β-ジアミノプロピオン酸
Bio=ビオチン
【0016】
表2:
Cha=シクロヘキシル
表1及び2に示されたペプチドのアミノ酸はL-又はD-立体配置のいずれでもよい。
さらに別の陽イオンペプチドは米国仮特許出願60/444,777号(2003年2月4日出願)で見出すことができる(前記出願は参照により本明細書に含まれる)。
【0017】
分子
分子は生物学的分子又は小分子であり得る。好ましくは、前記生物学的分子又は小分子は医薬的に活性な分子である。本明細書で用いられる医薬的に活性な分子は、in vivoで有益な作用を示す任意の分子である。
生物学的な分子は、核酸又はアミノ酸配列を含み、分子量が450を超える任意の分子である。そのような核酸及びアミノ酸配列は、本明細書ではそれぞれ“ポリヌクレオチド”及び“ポリアミノ酸”と称される。
生物学的分子にはポリヌクレオチド、ペプチド核酸及びポリアミノ酸、例えばペプチド、ポリペプチド及びタンパク質が含まれる。医薬的に活性を有する生物学的分子の例には、内因性ペプチド(例えばバゾプレッシン、グルタチオン)、タンパク質(例えばインターフェロン)、ホルモン(例えばヒト成長ホルモン)、酵素(例えばα-ガラクトシダーゼ)、抗体(例えばβ-アミロイドに対する抗体、前記はアルツハイマー病の治療に用いることができる)、神経栄養性増殖因子(例えば神経増殖因子NGF、脳由来神経栄養性因子BDNF)、サイトカイン(例えば血小板由来増殖因子PDGF、血管内皮細胞増殖因子VEGF)及びオリゴヌクレオチドが含まれる。
前記オリゴヌクレオチドは、任意のヌクレオチド配列(例えばDNA又はRNA)を含むことができる。前記DNA及びRNA配列は一本鎖でも二本鎖でもよい。例えば、ストレス時の細胞の生存の促進に有益なタンパク質をコードするDNAを本発明のペプチドに共役させることができる。そのようなタンパク質の例には熱ショックタンパク質(例えばhsp60、hsp70など)が含まれる。
一本鎖RNA分子の例にはリボザイム、RNAデコイ、リボザイムのための外側ガイド配列、アンチセンスRNA及びmRNAが含まれる。これら一本鎖RNA分子の総説については以下の文献を参照されたい:Sullenger et al. Nature 418:252-247(2002)。前記文献(Sullenger)で開示されている、これら一本鎖RNAのついての記述、並びにリボザイム、RNAデコイ、リボザイムのための外側ガイド配列、アンチセンスRNA及びmRNA分子により治療することができる病気及び疾患についての記述は、引用により本明細書に含まれる。
【0018】
二本鎖RNAの例はRNA干渉性分子(すなわちRNAi、例えばsiRNA(すなわち干渉性小RNA))である。前記siRNAは当技術分野で公知である任意のsiRNAであり得る。
siRNAは、例えば、疾患、症状又は病気で示唆されているタンパク質の翻訳を阻害することができるほど充分にmRNAに相補的であり得る。そのようなタンパク質の例には、例えばアルツハイマー病と関連しているβ-アミロイド及び癌と関連しているrasタンパク質が含まれる。
あるいは、siRNAは、例えばウイルスによって生成されるRNAと十分に相補的であり得る。ウイルスによって生成されるRNAは、一般的に宿主細胞の感染、ウイルスの生存及び/又はウイルスの増殖に必要ないずれのRNAでもあり得る。そのようなRNAの例には、内部リボソームエントリー部位、RNA依存ポリメラーゼ開始部位、並びにウイルスのエンベロープタンパク質、ウイルスヌクレアーゼ及びウイルスプロテアーゼをコードするRNAが含まれる。
ウイルスの例には、例えば肝炎ウイルス(例えばA、BおよびC型肝炎ウイルス)、ヒト免疫不全ウイルス、エプスタイン-バールウイルス、サイトメガロウイルス及びヒトパピローマウイルスが含まれる。
ウイルスRNAを標的とするsiRNAは当業者には公知である。例えば、C型肝炎ウイルスRNAを標的とするsiRNAは当業者に公知であり、下記の文献を参照されたい:Randall et al. PNAS 100:235-240(2003)。
この分子は小分子でもよい。小分子には有機化合物、有機金属化合物、有機及び有機金属化合物の塩、単糖類、アミノ酸及びヌクレオチドが含まれる。小分子にはさらに、分子量が450を越えない点を除けば生物学的分子と考えられる分子が含まれよう。したがって、小分子は脂質、オリゴ糖、オリゴペプチド及びオリゴヌクレオチド、並びにそれらの誘導体で、分子量が450以下を有するものであり得る。
小分子は任意の分子量を有することができることは強調されよう。それらは単に生物学的分子とみなされないために小分子と称されるだけであり、典型的には450未満の分子量を有する。小分子には、合成化合物とともに天然に見出される化合物が含まれる。医薬的に活性を有する小分子の例には抗生物質(例えばテトラサイクリン、ペニシリン、エリスロマイシン)、細胞毒性物質(例えばドキソルビシン、アドリアマイシン)、及び抗酸化剤(例えばビタミンE、ビタミンC、ベータカロチン)が含まれる。
【0019】
担体複合体
上記で述べた少なくとも1つの分子、及び上記で述べた少なくとも1つの芳香族陽イオンペプチドが会合して担体複合体が形成される。前記分子及び芳香族陽イオンペプチドは当業者に公知の任意の方法によって会合される。適切な会合の型には化学的結合及び物理的結合が含まれる。化学的結合には、例えば共有結合及び配位結合が含まれる。物理的結合には、例えば水素結合、二極性相互作用、ファンデルワールス力、静電気的相互作用、疎水性相互作用及び芳香族スタッキングが含まれる。
分子と芳香族陽イオンペプチドとの間の会合のタイプは、典型的には例えば分子上の利用可能な官能基、芳香族陽イオンペプチド上の利用可能な官能基に依存する。
化学的結合及び物理的結合では、分子上の官能基は典型的には芳香族陽イオンペプチド上の官能基と会合する。あるいは、芳香族陽イオンペプチド上の官能基が分子上の官能基と会合する。
分子上及び芳香族陽イオンペプチド上の官能基を直接結合させることもできる。例えば、分子上の官能基(例えばスルフヒドリル基)は、芳香族陽イオンペプチド上の官能基(例えばスルフヒドリル基と結合してジスルフィドを形成することができる。
あるいは、前記官能基は架橋剤(すなわちリンカー)を介して会合させることができる。架橋剤のいくつかの例は下記に記載されている。架橋剤は分子又は芳香族陽イオンペプチドのどちらに付加することもできる。
リンカーは芳香族陽イオンペプチドの正味の電荷数に影響を与えることも与えないこともある。典型的にはリンカーは芳香族陽イオンペプチドの正味の電荷に影響しないであろう。リンカー内に存在する各アミノ基(それが存在する場合)は、芳香族陽イオンペプチドの正味の正電荷に寄与するであろう。リンカー内に存在する各カルボキシル基(それが存在する場合)は、芳香族陽イオンペプチドの正味の負電荷に寄与するであろう。
【0020】
担体複合体の分子又は芳香族陽イオンペプチドの数は、多数の分子を収納することができるペプチドの能力、又は多数のペプチドを収納することができる分子の能力によって制限を受ける。例えば立体的障害は、特に大型の分子を収納するペプチドの能力を妨害するであろう。あるいは、立体的障害は比較的大型の(例えば長さが7、8、9又は10アミノ酸)芳香族陽イオンペプチドを収納する前記分子の能力を妨害するであろう。
担体複合体中の分子又は芳香族陽イオンペプチドの数はまた、他方に存在する官能基の数によって制限を受ける。例えば、ペプチドと会合できる分子の最大数は、ペプチドに存在する官能基の数に左右される。あるいは、分子と会合できるペプチドの最大数は、分子に存在する官能基の数に左右される。
ある実施態様では、担体複合体は、芳香族陽イオンペプチドと会合した少なくとも1つの分子、好ましくは少なくとも2つの分子を含む。いくつかの(例えば3、4、5又それより多い)官能基を含む比較的大きなペプチド(例えば長さが8、10アミノ酸)は、いくつかの(例えば3、4、5又それより多い)分子と会合することができる。
別の実施態様では、担体複合体は、分子と会合した少なくとも1つの芳香族陽イオンペプチド、好ましくは少なくとも2つの芳香族陽イオンペプチドを含む。例えば、いくつかの(例えば3、4、5又それより多い)官能基を含む分子は、いくつかの(例えば3、4、5又それより多い)ペプチドと会合することができる。
さらにまた別の実施態様では、担体複合体は1つの分子と会合した1つの芳香族陽イオンペプチドを含む。
ある実施態様では、担体複合体は、少なくとも1つの芳香族陽イオンペプチドと化学的に結合した(例えば共役した)少なくとも1つの分子を含む。前記分子は、当業者に公知の任意の方法によって芳香族陽イオンペプチドと化学的に結合させることができる。例えば、前記分子の官能基を芳香族陽イオンペプチドの官能基に直接付加することができる。適切な官能基のいくつかの例には、例えばアミノ、カルボキシル、スルフヒドリル、マレイミド、イソシアネート、イソチオシアネート及びヒドロキシルが含まれる。
【0021】
前記分子はまた、架橋剤(例えばジアルデヒド、カルボジイミド、ジマレイミドなど)の手段によって芳香族陽イオンペプチドと化学的に結合させることができる。架橋剤は、例えばピアースバイオテクノロジー社(Pierce Biotechnology, Inc., Rockford, IL)から入手できる。以下のURLのウェブサイト(http://www.piercenet.com/products/browse.cfm?fldID=26436A16-60A0-4A56-85F7-213A50830440)が資料を提供している。さらに別の架橋剤には白金架橋剤が含まれ、前記は、米国特許5,580,990号;5,985,566号及び6,133,038号(Kreatech Biotechnology, B.V., Amsterdam, The Netherlands)に記載されている。
分子上の官能基はペプチドの官能基と異なっていてもよい。例えばスルフヒドリル基が分子に(例えばβ-ガラクトシダーゼに、又は5'及び/又は3'末端チオール改変DNA及びRNAオリゴヌクレオチドに)存在する場合、前記分子は、ペプチド(例えば(Dmt1)DALDA)と、架橋剤SMCC(すなわちスクシンイミジル4-(N-マレイミドメチル)シクロヘキサン-1-カルボキシレート)(ピアースバイオテクノロジー(Pierce Biotechnology)から入手できる)を用いてリジンの4-アミノ基を介して架橋することができる(下記実施例10を参照されたい)。別の実施例では、DALDAのリジンの4-アミノ基は、架橋剤EDC(すなわちN-[3-ジメチルアミノプロピル-N'-エチルカルボイミド])(ピアースバイオテクノロジー(Pierce Biotechnology)から入手できる)を用いてRNA又はDNAの5'-末端のα-リン酸基と直接共役させることができる(下記実施例13を参照されたい)。
あるいは、分子上及びペプチド上の官能基は同じでもよい。ホモ二官能基性架橋剤は典型的には同一の官能基を架橋するために用いられる。ホモ二官能基性架橋剤の例にはEGS(すなわちエチレングリコールビス[スクシンイミジルスクシネート])、DSS(すなわちジスクシンイミジルスベレート)、DMA(すなわちジメチルアジピミデート・2HCl)、DTSSP(すなわち3,3'-ジチオビス[スルホスクシンイミジルプロピオネート])、DPDPB(すなわち1,4-ジ-[3'-(2'-ピリジルジチオ)-プロピオンアミド]ブタン)及びBMH(すなわちビス-マレイミドヘキサン)が含まれる。そのようなホモ二官能基性架橋剤もピアースバイオテクノロジー(Pierce Biotechnology)から入手できる。
【0022】
前記分子とペプチドを化学的に結合させるために、典型的には分子、ペプチド及び架橋剤を一緒に混合する。分子、ペプチド及び架橋剤の添加順序は重要ではない。例えば、ペプチドを架橋剤と混合し、続いて分子を添加してもよい。あるいは、分子を架橋剤と混合し、続いてペプチドを添加してもよい。最適には、分子及びペプチドを混合し、続いて架橋剤を添加する。
化学的に結合した担体複合体は分子を細胞に送達する。いくつかの事例では、分子は芳香族陽イオンペプチドから切り離されないで細胞内で機能する。例えば、芳香族陽イオンペプチドが分子の触媒部位を阻害しない場合は、芳香族陽イオンペプチドから分子の切り離しは不要である(下記の実施例11を参照されたい)。
他の事例では、分子を芳香族陽イオンペプチドから切断することは有益かもしれない。上記のピアースバイオテクノロジー社のウェブサイトはまた、細胞内で例えば酵素によって切断することができる適切な架橋剤を当業者が選択するための助けとなる。したがって、前記分子は芳香族陽イオンペプチドから分離することができる。切断可能なリンカーの例には、SMPT(すなわち4-スクシンイミジルオキシカルボニル-メチル-a-[2-ピリジルジチオ]トルエン)、スルホ-LC-SPDP(すなわちスルホスクシンイミジル6-(3-[2-ピリジルジチオ]-プロピオンアミド)ヘキサノエート)、LC-SPDP(すなわちスクシンイミジル6-(3-[2-ピリジルジチオ]-プロピオンアミド)ヘキサノエート)、スルホ-LC-SPDP(すなわちスルホスクシンイミジル6-(3-[2-ピリジルジチオ]-プロピオンアミド)ヘキサノエート)、SPDP(すなわちN-スクシンイミジル3-[2-ピリジルジチオ]-プロピオンアミドヘキサノエート)及びAEDP(すなわち3-[(2-アミノエチル)ジチオ]-プロピオン酸・HCl)が含まれる。
【0023】
また別の実施態様では、担体複合体は、少なくとも1つの芳香族陽イオンペプチドと物理的に結合した少なくとも1つの分子を含む。当業者に知られた任意の方法を用いて分子を芳香族陽イオンペプチドと物理的に結合させることができる。
例えば、芳香族陽イオンペプチド及び分子は当業者に公知の任意の方法によって一緒に混合することができる。混合の順序は重要ではない。例えば、分子は改変又は未改変芳香族陽イオンペプチドと当業者に公知の方法によって物理的に混合することができる。あるいは、改変又は未改変芳香族陽イオンペプチドは当業者に公知の任意の方法によって分子と物理的に混合することができる。
例えば、芳香族陽イオンペプチド及び分子を容器に入れ、例えば容器を振盪することによって攪拌し前記芳香族陽イオンペプチド及び分子を混合することができる。
芳香族陽イオンペプチドは当業者に公知の任意の方法によって改変することができる。例えば、芳香族陽イオンペプチドは上記で述べたように架橋剤又は官能基によって改変してもよい。リンカーは芳香族陽イオンペプチドの正味の電荷に影響を与ることも与えないこともある。典型的にはリンカーは芳香族陽イオンペプチドの正味の電荷に寄与しないであろう。リンカー内に存在する各アミノ基(それが存在する場合)は、芳香族陽イオンペプチドの正味の正電荷に寄与するであろう。リンカー内に存在する各カルボキシル基(それが存在する場合)は、芳香族陽イオンペプチドの正味の負電荷に寄与するであろう。
例えば、[Dmt1]DALDAを、架橋剤SMCC(すなわちスクシンイミジル4-(N-マレイミドメチル)シクロヘキサン-1-カルボキシレート)(ピアースバイオテクノロジー(Pierce Biotechnology)から入手できる)を用いてリジンの4-アミノ基を介して改変することができる(下記実施例10を参照されたい)。担体複合体を形成するために、改変芳香族陽イオンペプチドを通常先ず初めに形成させ、続いて分子と混合する。
物理的に結合させた担体複合体の1つの利点は、分子が芳香族陽イオンペプチドと化学的に結合した担体複合体のように芳香族陽イオンペプチドを除去する必要なく分子が細胞内で機能するということである。さらにまた、芳香族陽イオンペプチドが分子の触媒部位を阻害しない場合、複合体の解離も必要ではない(下記実施例12参照)。
【0024】
芳香族陽イオンペプチドの合成
本発明の方法で有用なペプチドは当業者に公知の任意の方法によって化学的に合成することができる。タンパク質を合成する適切な方法には、例えばStuart & Youngの文献に記載されたものが含まれる: “Solid Phase Peptide Synthesis”, Second Edition, Pierce Chemical Company (1984)及び”Solid Phase Peptide Synthesis”, Methods Enzymology, 289, Academic Press, Inc, New York (1977)。
投与の態様
ある実施態様では、本発明は分子を細胞に送達する方法に関する。前記方法は細胞を前記分子及び芳香族陽イオンペプチドと接触させることを含む。細胞は、当業者に公知の任意の方法によって前記分子及び芳香族陽イオンペプチドと接触させることができる。例えば、細胞をin vitroで前記分子及び芳香族陽イオンペプチドとインキュベートすることができる。ある特徴では、前記分子及び芳香族陽イオンペプチドは、化学的又は物理的に結合した前記分子及び芳香族陽イオンペプチドを含む担体複合体の形態(例えば上記に記載した担体複合体)として存在することができる。
また別の実施態様では、分子を細胞に送達する方法は細胞を担体複合体と接触させることを含む。分子は、前記分子及び芳香族陽イオンペプチドを含む担体複合体と細胞を接触させることによって前記細胞に送達される。細胞は、当業者に公知の任意の方法によって担体複合体と接触させることができる。
例えば、細胞をin vitroで担体複合体とインキュベートすることができる。この細胞は任意の細胞でよい。細胞は植物、動物又は細菌由来であり得る。植物細胞の例にはアラビドプシス(Arabidopsis)細胞が含まれる。細菌細胞の例にはサッカロミセス(Saccharomyces)及びラクトバシルス(Lactobacillus)が含まれる。動物細胞には哺乳動物細胞、例えば神経細胞、腎上皮細胞、腎臓細胞、血管内皮細胞。グリア細胞、腸上皮細胞及び肝細胞が含まれる。血管内皮細胞の例には血液脳関門内皮細胞が含まれる。
【0025】
あるいは、前記担体複合体は哺乳動物にin vivoで投与することができる。医薬化合物を投与するための多数の周知の方法のいずれかによって、好ましくは医薬組成物として有効量の担体複合体を投与の必要性がある哺乳動物に投与することができる。
前記担体複合体は全身的又は局所的に投与することができる。ある実施態様では、前記担体複合体は静脈内に投与される。例えば、担体複合体は瞬時静脈内ボーラス注射により投与することができる。しかしながら好ましくは、担体複合体は静脈内定常速度輸液として投与される。
担体複合体は、哺乳動物の組織に、例えば注射器が利用できる組織に注射によって局所的に投与することができる。例えば、前記担体複合体が腫瘍に送達されるべき細胞毒性物質を含んでいる場合は、その腫瘍は局所投与に利用できることが好ましい。そのような腫瘍には例えば皮膚癌及び乳癌が含まれる。
担体複合体はまた、経口的、局部的、鼻腔内、筋肉内、皮下又は経皮的に投与することができる。好ましい実施態様では、担体複合体の経皮的投与はイオン導入療法によって実施される。前記方法では、担体複合体は電流によって皮膚を通して送達される。
他の投与ルートには脳室内又はくも膜下腔内投与が含まれる。脳室内とは脳の脳室系内への投与を指す。くも膜下腔内投与は、脳又は脊髄のくも膜下腔内への投与を意味する。したがって、脳室内又はくも膜下腔内投与は、中枢神経系の器官又は組織に影響を与える疾患及び症状に対して好まれるであろう。
本発明の方法において有用な担体複合体は、当業界で知られたように持続性放出によって哺乳動物に投与することができる。持続性放出投与は、特定の期間にわたって一定レベルの薬剤を達成するドラッグデリバリーのための方法である。前記レベルは典型的には血清濃度によって測定される。
調剤分野で公知のいずれの製剤も担体複合体の投与に適切である。経口投与のためには液体又は固体製剤を用いることができる。いくつかの製剤例には錠剤、ゼラチンカプセル、ピル、トローチ、エリキシル、懸濁液、シロップ、ウェファース、チューインガムなどが含まれる。ペプチドは、当業者に公知の適切な医薬担体(ビヒクル)又は賦形剤とともに混合することができる。担体及び賦形剤の例にはデンプン、ミルク、糖、あるタイプの粘度、ゼラチン、ステアリン酸又はその塩(ステアリン酸マグネシウム又はカルシウムを含む)、植物脂肪又は油、ゴム及びグリコールが含まれる。
【0026】
全身的、脳室内、くも膜下腔内、局部、鼻腔内、皮下又は経皮投与のためには、前記担体複合体製剤は通常の希釈剤、担体又は賦形剤など(例えば当業界で公知のもの)を利用して前記担体複合体を送達することができる。例えば、前記製剤は以下の1つ又は2つ以上を含むことができる:安定化剤、界面活性剤(好ましくは非イオン性界面活性剤)及び場合によって塩及び/又は緩衝剤。前記担体複合体は水溶液の形態で又は凍結乾燥形態として輸送することができる。
安定化剤は、例えばアミノ酸(例えばグリシン)、又はオリゴ糖(例えばシュクロース、テトラオース、ラクトース又はデキストラン)であり得る。あるいは、安定剤は糖アルコール(例えばマンニトール)又は前記の組合せであってよい。好ましくは安定剤又は安定剤の組合せは担体複合体の重量の約0.1から約10重量%を構成する。
界面活性剤は好ましくは非イオン性界面活性剤、例えばポリソルベートである。適切な界面活性剤のいくつかの例には、トゥイーン20、トゥイーン80;ポリエチレングリコール又はポリオキシエチレンポリオキシプロピレングリコール、例えばプルロニックF-68(約0.001%(w/v)から約10%(w/v))が含まれる。
塩又は緩衝剤は任意の塩又は緩衝剤、例えばそれぞれ塩化ナトリウム又はリン酸ナトリウム/カリウムであり得る。好ましくは、緩衝剤は前記医薬組成物のpHを約5.5から約7.5の範囲で維持する。前記塩及び/又は緩衝剤はまた、浸透圧濃度をヒト又は動物への投与に適したレベルに維持するために有用である。好ましくは、前記塩又は緩衝剤は、約150mMから約300mMのほぼ等張濃度で存在する。
本発明の方法で有用な担体複合体製剤はさらにまた1つ又は2つ以上の一般的な添加物を含むことができる。そのような添加物のいくつかの例には、可溶化剤、例えばグリセロール;抗酸化剤、例えば塩化ベンザルコニウム(第四アンモニウム化合物の混合物、“クォーツ(quats)”として知られている)、ベンジルアルコール、クロレトン又はクロロブタノール;麻酔剤、例えばモルヒネ誘導体;又は等張剤(例えば上記に記載したもの)などが含まれる。酸化又は他の腐敗に対するまた別の予防として、前記医薬組成物は、非透過性の栓で密封されたバイアルで窒素ガス下で保存することができる。
前記哺乳動物は任意の哺乳動物でよい。それらには例えば家畜(例えばヒツジ、ブタ、乳牛及びウマ);愛玩動物(例えばイヌ及びネコ);実験動物(例えばラット、マウス及びウサギ)が含まれる。好ましい実施態様ではこの哺乳動物はヒトである。
【0027】
有用性
本担体複合体はエネルギー非依存機構で細胞膜を通過することができるので、多数のin vivo及びin vitroでの応用が可能である。
本担体複合体は、例えば研究用ツールとしてin vitroで用いることができる。例えば、本担体複合体は、分子(例えばタンパク質)を細胞に送達し、それによって前記分子の機能的役割を研究することができる。そのような分子には、例えば細胞シグナル伝達タンパク質(例えば核内因子NF-κB、キナーゼ、例えばJAK)が含まれる。
また別のin vitroでの利用には、例えば、細胞の子孫(系統)を決定するためのマーカー(例えばβ-ガラクトシダーゼ)の細胞(例えば幹細胞、造血細胞又は胚細胞)内へのデリバリーが含まれる。
他のin vitroでの応用には例えば検出可能な抗体の細胞へのデリバリーが含まれる(これによって特定のタンパク質の細胞内の存在が決定される)。
本担体複合体はまたin vivoで治療薬としての用途を有する。例えば前記芳香族陽イオンペプチドを用いてアンチセンスポリヌクレオチドを哺乳動物の細胞に送達してタンパク質の過剰発現をダウンレギュレートすることができる。さらにまた、RNA干渉(RNAi)のためのオリゴヌクレオチドの送達に芳香族陽イオンペプチドを用いることができる。
本明細書においてRNAiは、ウイルス若しくは細菌の遺伝子の発現又は複製を制御する細胞内メカニズムを指す。このメカニズムには、遺伝子生成物(典型的にはRNA)を標的として二本鎖RNA(例えばsiRNA)を導入することが含まれる。
脳血液関門は特に選択的である。したがって、また別のin vivoでの応用には脳血液関門を通過して分子を細胞に送達することが含まれる。そのような分子には、例えばアルツハイマー病患者の治療におけるβ-アミロイドに対する抗体が含まれる。
化学療法剤に付随する典型的な問題は、細胞内で適切なレベルを達成することである。例えば、化学療法剤は細胞膜を通過するには大きすぎるか、又は十分な芳香族性をもたないであろう。したがって、また別のin vivoでの応用には化学療法剤(例えば上記に記載したような細胞毒性物質)の細胞内への送達が含まれる。
【0028】
本発明のいくつかの態様を以下に列挙する。
(1) 分子を細胞に送達する方法であって、細胞を担体複合体と接触させることを含み、前記複合体は送達する分子及び芳香族陽イオンペプチドを含み、前記芳香族陽イオンペプチドが以下を含む、前記細胞に分子を送達する前記方法:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmがr+1以下の最大数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下の最大数であるが、ただしaが1であるときはptもまた1であることができる関係。
(2) 2aがpt+1以下の最大の数である、(1)に記載の方法。
(3) aがpに等しい、(1)に記載の方法。
(4) ペプチドが最小限2つの正電荷を有する、(1)に記載の方法。
(5) ペプチドが最小限3つの正電荷を有する、(1)に記載の方法。
(6) ペプチドが、2つの正に荷電したアミノ酸及び1つの芳香族アミノ酸を含むトリペプチドである、(1)に記載の方法。
(7) ペプチドが水溶性である、(1)に記載の方法。
(8) ペプチドが1つ又は2つ以上のD-アミノ酸を含む、(1)に記載の方法。
(9) C-末端アミノ酸のC-末端カルボキシル基がアミド化されている、(1)に記載の方法。
(10) ペプチドが1つ又は2つ以上の天然には存在しないアミノ酸を含む、(1)に記載の方法。
(11) ペプチドが最小限4つのアミノ酸を有する、(1)に記載の方法。
(12) ペプチドが最大限約8つのアミノ酸を有する、(1)に記載の方法。
(13) ペプチドが最大限約6つのアミノ酸を有する、(1)に記載の方法。
(14) ペプチドが4つのアミノ酸を有する、(1)に記載の方法。
(15) ペプチドが式Tyr-D-Arg-Phe-Lys-NH2(DALDA)を有する、(1)に記載の方法。
(16) ペプチドが式2’,6’-Dmt-D-Arg-Phe-Lys-NH2(Dmt1-DALDA)を有する、(1)に記載の方法。
(17) ペプチドが式Phe-D-Arg-Phe-Lys-NH2(Phe1-DALDA)を有する、(1)に記載の方法。
(18) ペプチドが式D-Arg-2’,6’ Dmt-Lys-Phe-NH2を有する、(1)に記載の方法。
(19) ペプチドが式2’,6’-Dmp-D-Arg-Phe-Lys-NH2(Dmp1-DALDA)を有する、(1)に記載の方法。
(20) 分子が小分子である、(1)に記載の方法。
(21) 小分子が医薬的に活性を有する分子である、(20)に記載の方法。
(22) 医薬的に活性を有する分子が抗生物質である、(21)に記載の方法。
(23) 医薬的に活性を有する分子が細胞毒性物質である、(21)に記載の方法。
(24) 細胞毒性物質がドキソルビシンである、(23)に記載の方法。
(25) 細胞毒性物質がアドリアマイシンである、(23)に記載の方法。
(26) 医薬的に活性を有する分子が抗酸化剤である、(21)に記載の方法。
(27) 抗酸化剤がビタミンEである、(26)に記載の方法。
(28) 抗酸化剤がビタミンCである、(26)に記載の方法。
(29) 抗酸化剤がベータカロチンである、(26)に記載の方法。
(30) 分子が生物学的分子である、(1)に記載の方法。
(31) 生物学的分子がポリアミノ酸である、(30)に記載の方法。
(32) 生物学的分子が医薬的に活性を有する分子である、(30)に記載の方法。
(33) 医薬的に活性を有する分子が内因性ペプチド又はタンパク質である、(32)に記載の方法。
(34) 医薬的に活性を有する分子が酵素である、(32)に記載の方法。
(35) 医薬的に活性を有する分子が抗体である、(32)に記載の方法。
(36) 医薬的に活性を有する分子が神経栄養性増殖因子である、(32)に記載の方法。
(37) 医薬的に活性を有する分子がサイトカインである、(32)に記載の方法。
(38) 医薬的に活性を有する分子がポリヌクレオチドである、(32)に記載の方法。
(39) 医薬的に活性を有する分子がオリゴヌクレオチドである、(32)に記載の方法。
(40) オリゴヌクレオチドがRNAである、(39)に記載の方法。
(41) RNAが二本鎖RNAである、(40)に記載の方法。
(42) 二本鎖RNAがsiRNAである、(41)に記載の方法。
(43) オリゴヌクレオチドがDNAである、(39)に記載の方法。
(44) オリゴヌクレオチドが一本鎖RNAである、(39)に記載の方法。
(45) 一本鎖RNAがメッセンジャーRNA(mRNA)である、(44)に記載の方法。
(46) オリゴヌクレオチドがリボザイムである、(39)に記載の方法。
(47) オリゴヌクレオチドがアンチセンスRNAである、(39)に記載の方法。
(48) オリゴヌクレオチドがリボザイムのための外側ガイド配列である、(39)に記載の方法。
(49) オリゴヌクレオチドがRNAデコイである、(39)に記載の方法。
(50) 生物学的分子がポリアミノ酸である、(30)に記載の方法。
(51) 細胞が細菌細胞である、(1)に記載の方法。
(52) 細胞が植物細胞である、(1)に記載の方法。
(53) 細胞が動物細胞である、(1)に記載の方法。
(54) 動物細胞が哺乳動物細胞である、(53)に記載の方法。
(55) 細胞が神経細胞である、(53)に記載の方法。
(56) 細胞が腎上皮細胞である、(53)に記載の方法。
(57) 細胞が腸上皮細胞である、(53)に記載の方法。
(58) 細胞が血管内皮細胞である、(53)に記載の方法。
(59) 内皮細胞が血管-脳関門内皮細胞である、(53)に記載の方法。
(60) 細胞がグリア細胞である、(53)に記載の方法。
(61) 細胞が肝細胞である、(53)に記載の方法。
(62) 芳香族-陽イオンペプチドがリンカーを含む、(1)に記載の方法。
(63) 分子がリンカーを含む、(1)に記載の方法。
(64) 分子と芳香族陽イオンペプチドが化学的に結合されている、(1)に記載の方法。
(65) 分子と芳香族陽イオンペプチドが物理的に結合されている、(1)に記載の方法。
(66) 分子及び芳香族陽イオンペプチドを含む担体複合体であって、前記芳香族陽イオンペプチドが以下を含む、前記担体複合体:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmがr+1以下の最大数である関係;さらに
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下の最大数であるが、ただしaが1であるときはptもまた1であることができる関係。
(67) 2aはpt+1以下の最大の数である、(66)に記載の担体複合体。
(68) aがptに等しい、(66)に記載の担体複合体。
(69) ペプチドが最小限2つの正電荷を有する、(66)に記載の担体複合体。
(70) ペプチドが最小限3つの正電荷を有する、(66)に記載の担体複合体。
(71) ペプチドが、2つの正に荷電したアミノ酸及び1つの芳香族アミノ酸を含むトリペプチドである、(66)に記載の担体複合体。
(72) ペプチドが水溶性である、(66)に記載の担体複合体。
(73) ペプチドが1つ又は2つ以上のD-アミノ酸を含む、(66)に記載の担体複合体。
(74) C-末端アミノ酸のC-末端カルボキシル基がアミド化されている、(66)に記載の担体複合体。
(75) ペプチドが1つ又は2つ以上の天然には存在しないアミノ酸を含む、(66)に記載の担体複合体。
(76) ペプチドが最小限4つのアミノ酸を有する、(66)に記載の担体複合体。
(77) ペプチドが最大限約8つのアミノ酸を有する、(66)に記載の担体複合体。
(78) ペプチドが最大限約6つのアミノ酸を有する、(66)に記載の担体複合体。
(79) ペプチドが4つのアミノ酸を有する、(66)に記載の担体複合体。
(80) ペプチドが式Tyr-D-Arg-Phe-Lys-NH2(DALDA)を有する、(66)に記載の担体複合体。
(81) ペプチドが式2’,6’-Dmt-D-Arg-Phe-Lys-NH2(Dmt1-DALDA)を有する、(66)に記載の担体複合体。
(82) ペプチドが式Phe-D-Arg-Phe-Lys-NH2(Phe1-DALDA)を有する、(66)に記載の担体複合体。
(83) ペプチドが式D-Arg-2’,6’ Dmt-Lys-Phe-NH2を有する、(66)に記載の担体複合体。
(84) ペプチドが式2’,6’-Dmp-D-Arg-Phe-Lys-NH2(Dmp1-DALDA)を有する、(66)に記載の担体複合体。
(85) 分子が小分子である、(66)に記載の担体複合体。
(86) 小分子が生物学的分子である、(66)に記載の担体複合体。
(87) 生物学的分子がポリアミノ酸である、(86)に記載の担体複合体。
(88) 小分子が医薬的に活性を有する分子である、(85)に記載の担体複合体。
(89) 医薬的に活性を有する分子が抗生物質である、(88)に記載の担体複合体。
(90) 医薬的に活性を有する分子が細胞毒性物質である、(88)に記載の担体複合体。
(91) 細胞毒性物質がドキソルビシンである、(90)に記載の担体複合体。
(92) 細胞毒性物質がアドリアマイシンである、(90)に記載の担体複合体。
(93) 医薬的に活性を有する分子が抗酸化剤である、(88)に記載の担体複合体。
(94) 抗酸化剤がビタミンEである、(93)に記載の担体複合体。
(95) 抗酸化剤がビタミンCである、(93)に記載の担体複合体。
(96) 抗酸化剤がベータカロチンである、(93)に記載の担体複合体。
(97) 分子が生物学的分子である、(66)に記載の担体複合体。
(98) 生物学的分子が医薬的に活性を有する分子である、(97)に記載の担体複合体。
(99) 医薬的に活性を有する分子が内因性ペプチド又はタンパク質である、(98)に記載の担体複合体。
(100) 医薬的に活性を有する分子が酵素である、(98)に記載の担体複合体。
(101) 医薬的に活性を有する分子が抗体である、(98)に記載の担体複合体。
(102) 医薬的に活性を有する分子が神経栄養性増殖因子である、(98)に記載の担体複合体。
(103) 医薬的に活性を有する分子がサイトカインである、(98)に記載の担体複合体。
(104) 医薬的に活性を有する分子がポリヌクレオチドである、(98)に記載の担体複合体。
(105) 医薬的に活性を有する分子がオリゴヌクレオチドである、(98)に記載の担体複合体。
(106) オリゴヌクレオチドがRNAである、(105)に記載の担体複合体。
(107) RNAが二本鎖RNAである、(106)に記載の担体複合体。
(108) 二本鎖RNAがsiRNAである、(107)に記載の担体複合体。
(109) オリゴヌクレオチドがDNAである、(106)に記載の担体複合体。
(110) オリゴヌクレオチドが一本鎖RNAである、(105)に記載の担体複合体。
(111) 一本鎖RNAがメッセンジャーRNA(mRNA)である、(110)に記載の担体複合体。
(112) オリゴヌクレオチドがリボザイムである、(105)に記載の担体複合体。
(113) オリゴヌクレオチドがアンチセンスRNAである、(105)に記載の担体複合体。
(114) オリゴヌクレオチドがリボザイムのための外側ガイド配列である、(105)に記載の担体複合体。
(115) オリゴヌクレオチドがRNAデコイである、(105)に記載の担体複合体。
(116) 生物学的分子がポリアミノ酸である、(97)に記載の担体複合体。
(117) 芳香族-陽イオンペプチドがリンカーを含む、(66)に記載の担体複合体。
(118) 分子がリンカーを含む、(66)に記載の担体複合体。
(119) 分子と芳香族陽イオンペプチドが化学的に結合されている、(66)に記載の担体複合体。
(120) 分子と芳香族陽イオンペプチドが物理的に結合されている、(66)に記載の担体複合体。
(121) 分子を細胞に送達する方法であって、前記方法が細胞を分子及び芳香族陽イオンペプチドと接触させることを含み、前記芳香族陽イオンペプチドが以下を含む、前記分子を細胞に送達する方法:
(a)少なくとも1の正味の正電荷;
(b)最小限3つのアミノ酸;
(c)最大限10のアミノ酸;
(d)正味の正電荷の最小数(pm)とアミノ酸残基の総数(r)との間において、3pmがr+1以下の最大数である関係;及び
(e)芳香族基の最小数(a)と正味の正電荷の総数(pt)との間において、3aはpt+1以下の最大数であるが、ただしaが1であるときはptもまた1であることができる関係。
(122) 分子と芳香族陽イオンペプチドが化学的に結合されている、(121)に記載の方法。
(123) 分子と芳香族陽イオンペプチドが物理的に結合されている、(121)に記載の担体複合体。
【実施例1】
【0029】
実施例1:材料及び方法
薬剤及び化学物質:[Dmt1]DALDA及び[3H] [Dmt1]DALDA(47Ci/mmol)は以前に記載された方法(Schiller et al., Eur. J. Med. Chem. 2000, 35:895-901;Zhao et al., J. Pharmacol. Exp. Ther. 2002, 302:188-196)にしたがって合成した。[14C]Gly-Sar(56.7mCi/mmol)及び[3H][D-Ala2,N-Me-Phe4,Gly5-オール]-エンケファリン(50Ci/mmol)はAmersham Biosciences(Piscataway, NJ)から購入した。その他の全ての薬剤及び化学物質はSigma-Aldrich(St. Louis, MO)から入手した。
【0030】
細胞培養:全ての細胞株はアメリカンタイプカルチャーコレクション(American Type Culture Collection, Manassas, VA)から入手し、さらに細胞培養用品はInvitrogen(Carlsbad, CA)から得た。Caco-2細胞はMEMで増殖させ、一方SH-SY5Y、HEK293及びHuh7細胞はダルベッコー改変イーグル培養液で増殖させた。増殖培地には10%ウシ胎児血清、200μg/mLペニシリン及び100μg/mL硫酸ストレプトマイシンを補充した。CRFK細胞はMEM+10%ウマ血清、非必須アミノ酸及びペニシリン/ストレプトマイシンで増殖させた。全ての細胞株は95%空気及び5%CO2の加湿大気中で37℃で維持した。
【0031】
ペプチド取り込みアッセイ:ペプチドの内在化は最初にCaco-2細胞を用いて調べ、続いてSH-SY5Y、HEK293及びCRFK細胞で確認した。コラーゲンを被覆した12ウェルプレートで単層細胞を3日間増殖させた(5x105細胞/ウェル)。4日目に、予め温めたHBSSで細胞を洗浄し、続いて250nMの[3H] [Dmt1]DALDA又は50μMの[14C]Gly-Sarのどちらかを含む、0.2mLのHBSSとともに1時間まで37℃にて種々の時間インキュベートした。別の実験で、非標識 [Dmt1]DALDA(1μMから3mM)の存在下で、同じ濃度の[3H] [Dmt1]DALDAとともに37℃で1時間細胞をインキュベートした。4℃での取り込み実験のためには、 [3H] [Dmt1]DALDA又は[14C]Gly-Sarとのインキュベーションの前に細胞を20分氷上に静置した。インキュベーションの終わりに、細胞を4回HBSSで洗浄し、1%のSDSを含む0.1NのNaOHの0.2mLを添加した。続いて細胞の内容物をシンチレーションバイアルに移し、放射能を計測した。細胞溶解物のアリコットをブラッドフォード法(Bio-Rad, Hercules, CA)によるタンパク質含有量の決定に用いた。内在化放射能を表面結合放射能と区別するために、酸洗浄工程を含めた。細胞溶解の前に、0.2mLの0.2M酢酸/0.05MNaClとともに細胞を氷中でインキュベートした。
【0032】
Caco-2細胞からのペプチド流出アッセイ:単層CaCo-2細胞を12ウェルプレートで3日間増殖させた(5x105細胞/ウェル)。4日目に、細胞を[3H] [Dmt1]DALDA又は[14C]Gly-Sarで37℃で1時間予備装荷した。続いて細胞を氷冷インキュベーション溶液で4回洗浄して取り込みを停止させ、続いて37℃又は4℃で1時間0.5mLのMEMでインキュベートして細胞から前記インキュベーション溶液へのペプチドの流出を測定した。放射能の量は細胞溶解物及びインキュベーション媒体で測定した。ペプチド取り込み及び細胞からの流出に対するP-糖タンパク質の役割を調べるために、[Dmt1]DALDAの取り込み及び流出を100μMのベラパミル(P-糖タンパク質の阻害剤)の存在下でも測定した。
【0033】
Caco-2単層を通過するペプチド移動のアッセイ:単層Caco-2細胞は以前に記載されたたように調製した(Irie et al., J. Pharmacol. Exp. Ther. 2001, 298:711-717)。Caco-2細胞(2x105)をトランスウェル細胞培養チャンバー(Corning Glassworks, Corning, NY)内の微小孔を有するメンブレンフィルター(24mm、0.4μm)上に播種した。各トランスウェルチャンバーの先端部区画を1.5mLの培養液で、外側基底部区画を2.5mLの培養液で満たした。前記細胞単層に新しい培養液を1から2日毎に与え、28日目に輸送実験に使用した。先端部から外側基底部へのペプチド輸送は、0.2μMの[3H] [Dmt1]DALDA又は100μMの[14C]Gly-Sarを先端部区画に添加することによって決定した。放射能の計測のために、ペプチド添加後種々の時間に50μLのアリコットを先端部区画及び外側基底部区画の両方から取り出した。
見かけの透過率係数は以下の等式によって計算した:Papp=X/(t・A・Co)
式中、X/tはレシーバー区画における取り込み速度、Aは拡散面積(4.72cm2)、及びCoはドナー区画における最初の濃度である。
【0034】
共焦点レーザー走査型顕微鏡法:芳香族陽イオンペプチドの細胞内取り込みは、以下の2つの蛍光ペプチドを用いて共焦点レーザー走査型顕微鏡法(CLSM)によって確認した:[Dmt1,dnsDap4]DALDA(Dmt-D-Arg-Phe-dnsDap-NH2、ここでdnsDapはβ-ダンシル-1-α,β-ジアミノ-プロピオン酸である)及び[Dmt1,atnDap4]DALDA(Dmt-D-Arg-Phe-atnDap-NH2、ここでatnDapはβ-アントラニロイル-L-α,β-ジアミノ-プロピオン酸である)。Caco-2細胞又はSH-SY5Y細胞を上記のように増殖させ、(35mmの)ガラス底の培養皿(MatTek, Ashland, MA)で2日間平板培養した。続いて培養液を取り除き、0.1μMの蛍光ペプチドを含むHBSSの1mLとともに細胞を4℃又は37℃で15分インキュベートした。続いて細胞を3回氷冷HBSSで洗浄し、さらに200μLのPBSで被覆し、C-Apochromat63x1.2Wcorr対物レンズ(Nikon, Tokyo, Japan)を取り付けた共焦点レーザー走査型顕微鏡を用いて10分以内に室温で顕微鏡観察を実施した。励起/放射波長は、[Dmt1,dnsDap4]DALDA については340/520nm、[Dmt1,atnDap4]DALDAについては320/420nmにそれぞれ設定した。Z方向の光学切片形成のために、2.0μmの5から10枠を作製した。
【0035】
細胞膜を用いる放射性リガンド結合アッセイ:Caco-2細胞及びSH-SY5Y細胞から調製した膜を用いて、[3H] [Dmt1]DALDAの細胞表面レセプターへの特異的結合を測定した。培養4日後に、細胞をPBS緩衝液で2回洗浄してから掻き取った。細胞を500gで5分遠心し、ペレットを-80℃で保存した。氷冷の50mMトリス塩酸緩衝液(5μg/mLロイペプチン、2μg/mLキモスタチン、10μg/mL及び1mMのEGTA、pH7.4)中で細胞をホモジナイズした。前記ホモジネートを36000gで20分遠心した。ペレットを50mMトリス塩酸緩衝液で再懸濁させた。膜ホモジネート(約140μgのタンパク質)のアリコットを[3H][Dmt1]DALDA(15から960pM)とともに25℃で60分インキュベートした。非特異的結合は1μMの非標識[3H] [Dmt1]DALDAを添加して評価した。遊離放射性リガンドは、細胞採集装置(Brandel Inc., Gaithersburg, MD)を用いGF/Bフィルター(Whatman, Maidstone, UK)で迅速ろ過して結合放射能リガンドから分離した。フィルターを3回10mLのトリス緩衝液で洗浄し、放射能を液体シンチレーション計測によって決定した。結合親和性(Kd)及びレセプター数(Bmax)は非直線回帰(GraphPad Software, San Diego, CA)を用いて決定した。
【0036】
タンパク質と[Dmt1]DALDAとの共役:[Dmt1]DALDAをβ-ガラクトシダーゼ(組換え大腸菌、Sigma-Aldrich)と架橋剤SMCC(スクシンイミジル4-(N-マレイミドメチル)シクロヘキサン-1-カルボキシレート)(Pierce)を用いて架橋した。SMCCはアミン含有分子([Dmt1]DALDAのLys4)と反応して安定なアミド結合を生成する。続いて前記のマレイミド端をスルフヒドリル含有化合物と結合させてチオエーテル結合を形成する(Greg T. Hermanson, “Bioconjugate Techniques”, Academic Press, page 234-237)。β-Galはその天然の状態で豊富な遊離スルフヒドリル基を含んでいる。β-Galの取り込みは、X-Galの使用による簡便な読み出しを提供する。簡単に記せば、1mLの[Dmt1]DALDA(5x10-3M)をリン酸緩衝液中の1mgのSMCCと室温で1時間混合した。前記操作によって“活性化ペプチド”が生成される。前記“活性化ペプチド”をリン酸緩衝液で1:10に希釈した。1mgのβ-Galを前記1:10の“活性化ペプチド”1mLに添加し、4℃で2時間又は一晩混合した。
[Dmt1]DALDAと架橋剤SMCCとのカップリング及び質量分析による確認:SMCC(1μg)及び[Dmt1]DALDA(5μg)を一緒に2mLのPBSに溶解し、室温で30分インキュベートし、4℃で保存した。前記サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールのターゲットプレートにスポットした。サンプルはMALDI-TOF MS(Matrix Assisted Laser Desorption Ionization−Time-of-Flight Mass Spectrometry, Applied Biosystems(Voyager DE Pro))によってポジティブリフレクトロンモードで分析した。
【0037】
RNAと[Dmt1]DALDAとの共役及びゲル電気泳動による確認:γ-32P-ATP及びポリヌクレオチドキナーゼを用いて合成RNAオリゴ(長さは40ヌクレオチド)の5'端をリン酸化した。生成物をゲル電気泳動で精製した。ゲル精製RNAオリゴの500,000cpmを1mgのEDC(N-[3-ジメチルアミノプロピル-N'-エチルカルボジイミド])の存在下で[Dmt1]DALDAと結合させた。前記結合反応の生成物([Dmt1]DALDA−RNAオリゴ)及び単独RNAオリゴを15%ポリアクリルアミド尿素ゲルで分析した。
DNAと[Dmt1]DALDAとの共役及び質量分析による確認:SMCC(1μg)及び[Dmt1]DALDA(5μg)を一緒に2mLのPBSに溶解し、室温で30分インキュベートし、さらに脱保護3'-チオールDNAオリゴと4℃で24時間混合した。インキュベーション後、サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。サンプルはMALDI-TOF MSによって分析した。
【0038】
RNA及び[Dmt1]DALDA−SMCC共役物の物理的混合による担体複合体の形成:[Dmt1]DALDA−SMCC共役物は上記のように調製した。RNA分子は、細胞取り込み実験で使用する前に、[Dmt1]DALDA−SMCC共役物と室温で15分間PBS中で混合した。
タンパク質及び[Dmt1]DALDA−SMCC共役物の物理的混合による担体複合体の形成:[Dmt1]DALDA−SMCC共役物は上記のように調製した。タンパク質分子(すなわち緑色蛍光タンパク質、GFP)は、細胞取り込み実験で使用する前に、[Dmt1]DALDA−SMCC結合物と室温で15分混合した。
[Dmt1]DALDA−RNA共役物の細胞内取り込みアッセイ:γ-32P-ATP及びポリヌクレオチドキナーゼを用いて合成RNAオリゴの5'端をリン酸化し、生成物をゲル電気泳動で精製した。ゲル精製RNAオリゴの500,000cpmを1mgのEDC(N-[3-ジメチルアミノプロピル-N'-エチルカルボジイミド])の存在下で[Dmt1]DALDAと結合させた。Caco-2細胞(1x106)をDMEM培養液で3回洗浄し、DMEMで5分間プレインキュベートした。続いて細胞を[Dmt1]DALDA−[32P]RNAオリゴ共役物又は非共役RNA(約20,000cpm)と37℃で60分インキュベートした。インキュベーション後に、細胞を3回DMEMで洗浄し、溶解緩衝液でインキュベートし、細胞溶解物中の放射能を測定した。
【0039】
[Dmt1]DALDA−架橋剤共役物と混合した場合の細胞内へのRNA取り込みアッセイ
Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、[32P]RNAオリゴのみまたは40μlの[Dmt1]DALDA−SMCC共役物を含む1.0mlのDMEMとともに37℃または4℃にて60分間インキュベートした。次に細胞をDMEMで4回洗浄し、酢酸ナトリウム溶液で1回洗浄して非特異的結合を低減させて、溶解緩衝液中で30分間インキュベートし、細胞溶解物中の放射能を測定した。
[Dmt1]DALDA−タンパク質共役物の細胞内取り込みアッセイ:細胞(N2A神経芽腫細胞又はCaco-2細胞)を96ウェルプレートでにプレーティングし(2x104細胞/ウェル)、[Dmt1]DALDA架橋β-Gal又はβ-Galのみとともに37℃で1時間インキュベートした。続いて細胞をPBSで4回洗浄した。次に細胞をβ-Gal染色セット(Roche)で37℃で少なくとも2時間染色し、顕微鏡下で調べた。
[Dmt1]DALDA−SMCC共役物と同時インキュベートした場合の細胞内タンパク質取り込みアッセイ:Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて3μgの緑色蛍光タンパク質(GFP)単独(A)、3μgのGFP及び40μL[Dmt1]DALDA(B)、又は3μgのGFP及びSMCCに共役させた40μLの[Dmt1]DALDA(C)を含む0.5mLのMEMとともに37℃で60分インキュベートした。続いて2mLの細胞培養液を細胞に添加し、これを細胞培養インキュベーターでさらに24時間インキュベートした。インキュベーション後に細胞を細胞培養液で4回洗浄し生細胞中に保持されているGFPを共焦点レーザー走査型顕微鏡で可視化した。励起は340nmで実施し、放射は520nmで測定した。
アポトーシスアッセイ:アポトーシスは、アポトーシス核染色用ヘキスト色素(Molecular Probes, Eugene, OR)を使用して決定した。ヘキスト色素を細胞培養に添加し15分間インキュベートした。過剰のヘキスト色素は細胞培養液(pH指示薬を含まない)で細胞を洗浄して除去し、蛍光顕微鏡を用いて細胞を調べた(励起350nm、放射461nm)。
【実施例2】
【0040】
実施例2:[Dmt1]DALDA及びGly-SarのCaco-2細胞への取り込みの経時的変化
37℃でCaco-2細胞とインキュベートしたときに、[3H][Dmt1]DALDAは細胞溶解物中に5分という早期に観察され、30分までに定常レベルに達した(図1A)。1時間のインキュベーション後に細胞溶解物中で回収された[3H][Dmt1]DALDAの総量は総薬剤の約1%を占めた。対照的に、同じ実験条件下で、[14C]Gly-Sarは5分間から45分間増加し続けた(図1B)。測定された放射能は[Dmt1]DALDAの濃度を反映していると考えられる。なぜならば、本発明者らは[Dmt1]DALDAがペプチダーゼによる分解に対して耐性を有することを以前に示しているからである(Szeto et al., J. Pharmacol. Exp. Ther., 2001, 298:57-61)。測定された放射能が細胞膜と結合しているか否かを決定するために、細胞を酸洗浄に付して表面結合を除去した。図1Cは、[3H][Dmt1]DALDAの80.8%が酸洗浄に耐性であり、したがって細胞内に存在すると考えられることを示している。[Dmt1]DALDAの取り込みは広範囲の濃度にわたって濃度依存性であることが判明した(図1D)。
【実施例3】
【0041】
実施例3:[Dmt1]DALDA及びGly-Sarの取り込みに対する温度依存性及びpHの影響
インキュベーションを4℃で実施したとき、[3H][Dmt1]DALDAの取り込みは37℃よりも遅かったが、45分までに76.5%(図1A)に、さらに1時間までに86.3%(図1A)に達した。対照的に、[14C]Gly-Sarの取り込みは4℃では完全に停止した(図1B)。PEPT1によるGly-Sarの取り込みはpH依存性であり、最適取り込みはpH6.0で生じることが知られている(Terada et al., 1999, Am. J. Physiol. 276:G1435-G1441)。これは我々の実験で確認された(図2B)。対照的に、[3H][Dmt1]DALDAの取り込みは、pHを4.0から7.4の間で変化させたときには変化がなかった(図2A)。温度及びpHに対する依存性の欠如は、Caco-2細胞の[Dmt1]DALDAの取り込みはPEPT1(ペプチドトランスポーター1)を介したものではないことを示唆している。
【実施例4】
【0042】
実施例4:[Dmt1]DALDA及びGly-Sarの取り込みに対するDEPCの影響
PEPT1が[Dmt1]DALDAの輸送に関与していないことをさらに明瞭に示すために、[3H][Dmt1]DALDA 及び[14C]Gly-Sarの取り込みに対する DEPC(ジエチルピロカルボネート;0.2mM)の影響を調べた。DEPCは、Caco-2細胞でPEPT1を阻害することが示されているヒスチジン残基修飾試薬である(Terada et al., FEBS. Lett., 1996, 394:196-200)。インキュベーション培養液へのDEPCの添加は[14C]Gly-Sar取り込みを顕著に阻害した(図2D)。驚いたことには、DEPCは[3H][Dmt1]DALDAの取り込みを阻害しなかっただけでなく、実際には[3H][Dmt1]DALDAの取り込みを34倍増加させた(図2C)。
【実施例5】
【0043】
実施例5:種々の細胞タイプにおける[Dmt1]DALDAの内在化
[Dmt1]DALDAの内在化はCaco-2細胞に限定されないことを示すために、異なるいくつかの細胞株で[Dmt1]DALDAの内在化を比較した。内在化放射能(酸耐性)を表面結合放射能(酸感受性)と区別するために酸洗浄工程を含めた。図3Aでは、Caco-2、SH-SY5Y、HEK293及びCRFK細胞における酸耐性放射能レベルが比較されている。結果は、[3H][Dmt1]DALDAは全ての細胞株で大量に取り込まれることを示している。
【実施例6】
【0044】
実施例6:[3H] [Dmt1]DALDAによる放射性リガンド能結合アッセイ
[Dmt1]DALDAがレセプター仲介機構によって内在化されるか否かを決定するために、Caco-2細胞及びSH-SY5Y細胞から調製した膜を用いて放射能リガンド([3H][Dmt1]DALDA)結合アッセイを実施した。図3Bは、[3H][Dmt1]DALDAのSH-SY5Y膜との特異的結合を示している。算出されたKd値は118pM(87から149の範囲)であり、Bmax値は96 fmol/mgタンパク質(88から104)であると概算された。これは、チャイニーズハムスター卵巣細胞で発現されたリコンビナントヒトμ-オピオイドレセプターを用いて得られた値に匹敵する(G.-M. Zhao and H.H. Szeto, 未発表データ)。高い親和性を示す特異的結合はCaco-2膜では観察されなかった(図3B)。HEK293細胞はオピオイドレセプターを持たないことが知られている(Blake et al., J. Biol. Chem. 1997, 272:782-790)。
【実施例7】
【0045】
実施例7:Caco-2細胞からの[Dmt1]DALDA及びGly-Sarの流出
30分未満のインキュベーション後にCaco-2細胞で定常状態の[3H][Dmt1]DALDAレベルが達成されるということは、前記細胞のペプチド流出速度はその時点での取り込み速度と等しいことを示唆している。細胞からのGly-Sar及び[Dmt1]DALDAの流出を調べるために、Caco-2細胞に[14C]Gly-Sar又は[3H][Dmt1]DALDAを予備装荷し、続いて前記ペプチドを含まない新しい培養液と交換した。図4Aは、[14C]Gly-Sarの39%は37℃で1時間後に培養液中で見出されることを示している。[14C]Gly-Sarの流出は4℃で顕著に低下した。Caco-2細胞からの[3H][Dmt1]DALDAの流出ははるかに速く、80%のペプチドが1時間までに培養液中に流出した。 [3H][Dmt1]DALDAの内在化(図1A)とは対照的に、細胞からの[3H][Dmt1]DALDA流出に対して温度は顕著な影響を示した(図4A)。[Dmt1]DALDAの流出はDEPCで処理された細胞で減少した(図4B)。DEPCによる[3H][Dmt1]DALDA流出の低下は、DEPCの存在下で[3H][Dmt1]DALDAの取り込みが大きく増加することと一致する(図2C)。他方、[3H][Dmt1]DALDAの流出はベラパミル(P-糖タンパク質の阻害剤)の影響を受けなかった(図4C)。ベラパミルはまた[3H][Dmt1]DALDAの細胞内取り込みに影響を与えなかった(図4D)。
細胞からの[Dmt1]DALDAの流出は、[3H][Dmt1]DALDA−タンパク質共役物が細胞内に取り込まれた後に酵素的に切断されて[Dmt1]DALDAが細胞から流出し、一方、運ばれたタンパク質が細胞内に留まるならば有益なことであろう。
【実施例8】
【0046】
実施例8:[Dmt1]DALDA及びGly-Sarの細胞間輸送
トランスウェルで増殖させたCaco-2単層細胞を用いて、[Dmt1,dnsDap4]DALDA及び[14C]Gly-Sarの先端部から外側基底部への輸送を調べた。図5は、ペプチドをトランスウェルの先端部位に添加した後種々の時間における [14C]Gly-Sar及び[3H][Dmt1]DALDAの外側基底部位への輸送を示している。60分で先端部から外側基底部位へ移動した[3H][Dmt1]DALDAの百分率(10.4%)は輸送された[14C]Gly-Sarの百分率(11.9%)に匹敵した。見かけの透過率係数は、[3H][Dmt1]DALDAについては1.24x10-5cm/s、Gly-Sarについては1.26x10-5cm/sと概算された。
【実施例9】
【0047】
実施例9:CLSMを用いた芳香族陽イオンペプチドの細胞内取り込みの可視化
芳香族陽イオンペプチドの取り込み及び細胞内内在化態様を可視化するために、2つの蛍光ペプチド([Dmt1,dnsDap4]DALDA及び[Dmt1,atnDap4]DALDA)をCLSMによって調べた。図6は、0.1μMの[Dmt1,dnsDap4]DALDAとともに37℃で15分間インキュベートした後のCaco-2細胞への蛍光ペプチドの内在化を示している。蛍光は細胞質全体に拡散して出現し、明白な小胞体内分布は認められなかった。このことは、ペプチドの取り込みはエンドサイトーシスを必要とせず、ペプチドはエンドソームに封入されないことが示唆される。さらにまた、ペプチドは完全に核から排除されていたことは留意されたい。4℃にて30分間0.1μMの[Dmt1,antDap4]DALDAとともに SH-SY5Y細胞をインキュベートした後の前記細胞内への[Dmt1,atnDap4]DALDAの内在化は、エネルギー非依存性の、非エンドサイトーシス性取り込みメカニズムを支持する。なぜならば、エンドサイトーシスはエネルギー依存性の過程だからである。
【実施例10】
【0048】
実施例10:ペプチドと架橋剤SMCCとのカップリング及び質量分析による確認
SMCC(1μg)及び5μgの[Dmt1]DALDA、[Phe1]DALDA又は[d-Arg-Dmt-Lys-Phe-NH2]を一緒に2mLのPBSに溶解し、室温で30分間インキュベートし、4℃で保存した。前記サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。サンプルはMALDI-TOF MS(Matrix Assisted Laser Desorption Ionization−Time-of-Flight Mass Spectrometry, Applied Biosystems(Voyager DE Pro))によってポジティブリフレクトロンモードで分析した。
ペプチド及びそれらの対応するペプチド-SMCC結合物の分子量は質量スペクトルに示されている(図7)。
【実施例11】
【0049】
実施例11:タンパク質カーゴ(protein cargo)に共役させたペプチドは前記タンパク質カーゴを細胞内に輸送する
種々のペプチドをβ-ガラクトシダーゼ(組換え大腸菌、Sigma-Aldrich)と架橋剤SMCC(Pierce)を用いて架橋した。SMCCはアミン含有分子([Dmt1]DALDAのLys4)と反応して安定なアミド結合を生成する。ペプチド−SMCC共役物の形成は質量分析によって確認する(図7)。続いて前記のマレイミド端をスルフヒドリル含有化合物と結合させてチオエーテル結合を形成する(Greg T. Hermanson, “Bioconjugate Techniques”, Academic Press, page 234-237)。β-Galはその天然の状態で豊富な遊離スルフヒドリル基を含んでいる。β-Galの取り込みは、X-Galの使用による簡便な読み出しを提供する。
簡単に記せば、1mLの5x10-3M の[Dmt1]DALDA、[Phe1]DALDA又は[d-Arg-Dmt-Lys-Phe-NH2]をリン酸緩衝液中の1mgのSMCCと室温で1時間混合した。前記操作によって“活性化ペプチド”が生成される。前記“活性化ペプチド”をリン酸緩衝液で1:10に希釈した。1mgのβ-Galを前記1:10の“活性化ペプチド”1mLに添加し、4℃で2時間又は一晩混合した。
細胞(N2A神経芽腫細胞又はCaco-2細胞)を96ウェルプレートにプレーティングし(2x104細胞/ウェル)、β-Gal若しくは[Dmt1]DALDA、[Phe1]DALDA又は[d-Arg-Dmt-Lys-Phe-NH2]と架橋したβ-Galとともに37℃で1時間インキュベートした。続いて細胞をリン酸緩衝液で4回洗浄した。次に細胞をβ-gal染色セット(Roche)で37℃で少なくとも2時間染色し、顕微鏡下で調べた。
β-Galとともにインキュベートしたときにはβ-Galの取り込みは観察されなかった(図8A)。青い細胞の存在は[Dmt1]DALDAと共役したβ-GalのCaco-2細胞内取り込みを示している(図8B)。β-Galの取り込み強化はまた、β-Galを[d-Arg-Dmt-Lys-Phe-NH2](図8C)又は[Phe1]DALDA(図8D)と共役させたときにも認められた。β-GalとSMCCのみとの共役では取り込みは強化されなかった。
同様な結果が神経細胞系N2A細胞又はCHO細胞(チャイニーズハムスター卵巣細胞)を用いたときにも得られた。
【実施例12】
【0050】
実施例12:[Dmt1]DALDA−SMCC共役物との同時インキュベーションは緑色蛍光タンパク質(GFP)のHuh7細胞への取り込みを強化する
Huh7細胞(1x106細胞/ウェル)をDMEMで洗浄し、続いて3μgの緑色蛍光タンパク質(GFP)単独、3μgのGFP及び40μlの[Dmt1]DALDA、又は3μgのGFP及び40μlのSMCCと共役させた[Dmt1]DALDAを含む0.5mLのMEMとともに37℃で60分間インキュベートした。続いて2mLの細胞培養液を細胞に添加し、これを細胞培養インキュベーターでさらに24時間インキュベートした。インキュベーション後に細胞を細胞培養液で4回洗浄し生細胞中に保持されているGFPを共焦点レーザー走査型顕微鏡で可視化した。励起は340nmで実施し、放射は520nmで測定した。
図9(上段パネル)は、Huh7細胞の0.8μm厚の中央水平光学切片を通したGFPの画像を示す。図9(下段パネル)は、同じ顕微鏡野の微分干渉コントラスト像を示す。
GFPと[Dmt1]DALDAとの同時インキュベーションは、GFPだけでインキュベートした場合(図9A)と比較して、前記細胞の細胞質内の緑色蛍光を中程度に増加させることを示した(図9B)。核内には緑色蛍光は観察されなかった。GFPと[Dmt1]DALDA−SMCC共役物との同時インキュベーションはさらに強いGFPの取り込みを示した(図9C)。これらのデータは、[Dmt1]DALDAは、この改変ペプチドをタンパク質と単に物理的に混合することによって細胞内へのそのタンパク質の取り込みを促進し、ペプチドとタンパク質との間の化学的結合は不要であることを示している。
【実施例13】
【0051】
実施例13:[Dmt1]DALDAとRNAオリゴとの共役
γ-32P-ATPを用いたポリヌクレオチドキナーゼ反応により合成RNAオリゴ(長さは40ヌクレオチド)の5'端をリン酸化した。生成物を反応のためにゲル精製した。ゲル精製RNAオリゴの500,000cpmを1mgのEDC(N-[3-ジメチルアミノプロピル-N'-エチルカルボジイミド])の存在下で10mMの[Dmt1]DALDAとの反応で結合させた。前記結合反応生成物([Dmt1]DALDA−RNAオリゴ)及び対照であるRNAオリゴ単独を15%ポリアクリルアミド尿素ゲルで分析した。ゲル上の2つの別個のバンドは単独のRNAオリゴ及び[Dmt1]DALDA−RNAオリゴ共役物であることを示す(図10)。
【実施例14】
【0052】
実施例14:Caco-2細胞への[Dmt1]DALDA−RNAオリゴ共役物の取り込み
オリゴの添加前に、Caco-2細胞(1x106)をDMEM培養液で3回洗浄し、DMEMで5分間プレインキュベートした。続いて [Dmt1]DALDA−RNAオリゴ共役物又は非共役RNA(約20,000cpm)を細胞培養液に添加し、37℃で60分間インキュベートした。
インキュベーション後に、反応培養液を除去し、細胞を4回DMEMで、さらに1回酢酸ナトリウム溶液中で洗浄して非特異的結合を減少させた。最後に細胞を溶解緩衝液中で30分間インキュベートし、細胞溶解物中の放射能を測定した。
Caco-2細胞は、非共役RNAオリゴと比較して、3倍を超える強い[Dmt1]DALDA−RNAオリゴ共役物の取り込みを示した(図11)。したがって[Dmt1]DALDAは、細胞膜を横断するRNAオリゴの通過を促進する。
【実施例15】
【0053】
実施例15:RNAと[Dmt1]DALDA−SMCCリンカーとの混合はRNAの細胞内取り込みを増加させる
RNA及び[Dmt1]DALDA−SMCC共役物を物理的に混合することによって担体複合体を生成した。[Dmt1]DALDA-SMCC共役物は[Dmt1]DALDAを方法の項で述べたように架橋剤SMCCと混合することによって調製した。細胞取り込み実験で使用する前に、一本鎖の11mer[32P]RNAオリゴを[Dmt1]DALDA−SMCC共役物と15分間室温で混合した。
Huh7細胞(1x106細胞/ウェル)をDMEM培養液で洗浄し、続いて[32P]RNAオリゴ(約100,000cpm)のみを含む1.0mLのDMEM、又は40mLの[Dmt1]DALDA−SMCC共役物と37℃又は4℃にてインキュベートした。続いて細胞を4回DMEMで、さらに1回酢酸ナトリウム溶液中で洗浄して非特異的結合を除去し、その後、細胞を溶解緩衝液中で30分間インキュベートし、保持されている放射能を測定した。
RNAオリゴと[Dmt1]DALDA−SMCCとの37℃での同時インキュベーションはRNAオリゴの取り込みを時間の関数として増加させた(図12A)。RNAだけでインキュベートした場合と比較して、[Dmt1]DALDA−SMCCの存在下におけるRNAオリゴの取り込みは、1時間で約20倍に増加した。RNAの取り込みは、4℃でさえも[Dmt1]DALDA−SMCCによって顕著に強化された(図12B)。これらのデータは、[Dmt1]DALDAとの化学的共役がなくてもRNAの取り込みを強化することができることを示している。4℃における取り込みは、エネルギー非依存性非エンドサイトーシス的過程による取り込みを示唆しており、[Dmt1]DALDAの受動拡散による細胞膜透過能力と一致する。
[Dmt1]DALDA−SMCCの他に、[Phe1]DALDA−SMCC又は[d-Arg-Dmt-Lys-Phe-NH2]−SMCCとの同時インキュベーションもまた11mer RNAオリゴの取り込みを強化した。図12Cは、3つの異なるペプチド−SMCC共役物と37℃で15分間インキュベートしたときRNAの取り込みが増加したことを示している。
[Dmt1]DALDA−SMCC共役物との同時インキュベーションはまた、図13に示すようにはるかに大きいRNA分子(1350-mer)の細胞内取り込みを促進することができるが、ただしより小さなオリゴの場合ほど強くはない。
【実施例16】
【0054】
実施例16:[Dmt1]DALDAとDNAオリゴとの共役
SMCC(1μg)及び[Dmt1]DALDA(SS002;5μg)を一緒に2mLのPBSに溶解し、室温で30分間インキュベートし、さらに脱保護3'-チオールDNAオリゴと4℃で24時間混合した。インキュベーション後、サンプルのアリコットをマトリックス(50%アセトニトリル中の飽和3-ヒドロキシピコリン酸(HPA)、10mg/mLクエン酸アンモニウム)と1:10の割合で混合し、ステンレススチールの標的プレートにスポットした。
DNA−[Dmt1]DALDA共役物の生成はMALDI-TOF MSによって確認した。3'-チオールDNAオリゴ及び[Dmt1]DALDA−DNA共有結合複合体の分子量は、それぞれ6392及び7171であることが判明した(図14A)。
【実施例17】
【0055】
実施例17:[Dmt1]DALDA−DNAオリゴ共役物のCaco-2細胞への取り込み
3'-チオール修飾20mer DNAをSMCCを用いて[Dmt1]DALDAに結合させ、共役物の生成は質量分析によって確認した。共役及び非共役DNAオリゴの5'末端を32Pで放射能標識し、さらにゲル精製した(図14B)。
神経細胞系N2A細胞(1x106細胞/ウェル)をDMEMで洗浄し、[Dmt1]DALDA共役又は非共役DNAオリゴ(約100,000cpm)のどちらかを含む1 mLのDMEMと37℃及び5%CO2で2時間又は19時間インキュベートした。続いて細胞を4回DMEMで、さらに1回酢酸ナトリウム溶液中で洗浄して非特異的結合を減少させた。続いて細胞を溶解緩衝液中で30分間インキュベートし、保持されている放射能を測定した。
19時間のインキュベーション後、[Dmt1]DALDAと共役したDNAの取り込みは、非共役DNAと比較して増加し、DNA取り込みは[Dmt1]DALDAとの共役によって強化できることが示された。
【実施例18】
【0056】
実施例18:ペプチド及びペプチド−SMCC共役物は細胞に対して毒性を示さない
ペプチドもペプチド−SMCC共役物も培養細胞に毒性を示さない。[Dmt1]DALDA(1nMから10μM)による24時間の処理は、N2A細胞(図16)、SH-SY5Y細胞又はCaco-2細胞においてMTTアッセイ(MTSアッセイ、Promega, Madison, WI)によって測定したところ、細胞の生存度へ影響を与えなかった。
培養細胞とペプチド−SMCC共役物とのインキュベーションもまた、トリパンブルーの取り込み測定によれば細胞の生存度に影響を与えなかった。トリパンブルーは膜透過性が増加した細胞によってのみ取り込まれる。Huh7細胞(1x106)をDMEM中で3回洗浄し、1mLの新しい培養液又は1mMの[Dmt1]DALDA−SMCC共役物、[D-Arg-Dmt-Lys-Phe-NH2]−SMCC共役物、若しくは [Phe1]DALDA−SMCC共役物の50μLを含む培養液を添加し、37℃で24時間、5%CO2でインキュベートした。続いて細胞をDMEMで3回洗浄し、0.4%トリパンブルーの1mLを前記細胞に2分間添加した。過剰な色素を細胞培養液中で細胞を洗浄することによって除去し、光学顕微鏡で細胞を調べた。
光学顕微鏡による細胞の検査で、培養液のみでインキュベートされた細胞は最小限のトリパンブルーの取り込みを示すことが判明した。トリパンブルーの取り込み増加は、[Dmt1]DALDA−SMCC、[D-Arg-Dmt-Lys-Phe-NH2]−SMCC又は[Phe1]DALDAとインキュベートした細胞で観察されなかった。対照的に、DEPC(ジエチルピロカルボネート)と細胞をインキュベートすることによって顕著なトリパンブルーの取り込みが生じた。
培養細胞と[Dmt1]DALDA−SMCC共役物とのインキュベーションは、Huh7培養細胞においてアポトーシスを誘導しなかった。Huh7細胞(1x106細胞/ウェル)をDMEM中で3回洗浄し、1mLの新しい培養液を適用した。続いてPBS中の改変[Dmt1]DALDA(1mM)又はPBSのみ(対照)50μLを前記細胞培養液に添加し、37℃で24時間、5%CO2でインキュベートした。インキュベーション後に、アポトーシス核の染色のために1mLのヘキスト色素(Molecular Probes, Eugene, OR)を細胞に添加し、さらに15分間インキュベートした。過剰な色素を細胞培養液(pH指示薬を含まない)で細胞を洗浄することによって除去し、蛍光顕微鏡検査(励起350nm、放射461nm)を用いて[Dmt1]DALDA−SMCC共役物で処理した細胞を対照細胞と比較した。アポトーシスは核内の蛍光の濃度によって示される。図17は、[Dmt1]DALDA−SMCCで処理したHuh7細胞のアポトーシスのレベルは対照細胞のそれと同じであることを示している。
【特許請求の範囲】
【請求項1】
担体複合体の、分子を細胞に送達するための医薬の製造における使用であって、前記担体複合体は芳香族陽イオンペプチドに結合した分子を含み、前記芳香族陽イオンペプチドが以下から選ばれ、前記分子が、ポリヌクレオチド、ポリアミノ酸、細胞毒性物質、抗生物質、抗酸化剤、増殖因子およびサイトカインからなる群から選ばれる、前記使用:
Tyr-D-Arg-Phe-Lys-NH2 (DALDA),
2',6'-Dmt-D-Arg-Phe-Lys-NH2 (Dmt1-DALDA),
Phe-D-Arg-Phe-Lys-NH2 (Phe1-DALDA),
D-Arg-2',6'Dmt-Lys-Phe-NH2, および、
2',6'-Dmp-D-Arg-Phe-Lys-NH2 (Dmp1-DALDA)。
【請求項2】
分子が抗生物質である、請求項1に記載の使用。
【請求項3】
分子が細胞毒性物質である、請求項1に記載の使用。
【請求項4】
細胞毒性物質がドキソルビシンである、請求項3に記載の使用。
【請求項5】
細胞毒性物質がアドリアマイシンである、請求項3に記載の使用。
【請求項6】
分子が抗酸化剤である、請求項1に記載の使用。
【請求項7】
抗酸化剤がビタミンEである、請求項6に記載の使用。
【請求項8】
抗酸化剤がビタミンCである、請求項6に記載の使用。
【請求項9】
抗酸化剤がベータカロチンである、請求項6に記載の使用。
【請求項10】
分子がポリアミノ酸である、請求項1に記載の使用。
【請求項11】
ポリアミノ酸が内因性ペプチド又はタンパク質である、請求項10に記載の使用。
【請求項12】
ポリアミノ酸が酵素である、請求項10に記載の使用。
【請求項13】
ポリアミノ酸が抗体である、請求項10に記載の使用。
【請求項14】
増殖因子が神経栄養性増殖因子である、請求項1に記載の使用。
【請求項15】
分子がサイトカインである、請求項1に記載の使用。
【請求項16】
分子がポリヌクレオチドである、請求項1に記載の使用。
【請求項17】
分子がオリゴヌクレオチドである、請求項1に記載の使用。
【請求項18】
オリゴヌクレオチドがRNAである、請求項17に記載の使用。
【請求項19】
RNAが二本鎖RNAである、請求項18に記載の使用。
【請求項20】
二本鎖RNAがsiRNAである、請求項19に記載の使用。
【請求項21】
オリゴヌクレオチドがDNAである、請求項17に記載の使用。
【請求項22】
オリゴヌクレオチドが一本鎖RNAである、請求項17に記載の使用。
【請求項23】
一本鎖RNAがメッセンジャーRNA(mRNA)である、請求項22に記載の使用。
【請求項24】
オリゴヌクレオチドがリボザイムである、請求項17に記載の使用。
【請求項25】
オリゴヌクレオチドがアンチセンスRNAである、請求項17に記載の使用。
【請求項26】
オリゴヌクレオチドがリボザイムのための外側ガイド配列である、請求項17に記載の使用。
【請求項27】
オリゴヌクレオチドがRNAデコイである、請求項17に記載の使用。
【請求項28】
細胞が細菌細胞である、請求項1に記載の使用。
【請求項29】
細胞が植物細胞である、請求項1に記載の使用。
【請求項30】
細胞が動物細胞である、請求項1に記載の使用。
【請求項31】
動物細胞が哺乳動物細胞である、請求項30に記載の使用。
【請求項32】
細胞が神経細胞である、請求項30に記載の使用。
【請求項33】
細胞が腎上皮細胞である、請求項30に記載の使用。
【請求項34】
細胞が腸上皮細胞である、請求項30に記載の使用。
【請求項35】
細胞が血管内皮細胞である、請求項30に記載の使用。
【請求項36】
内皮細胞が血管-脳関門内皮細胞である、請求項30に記載の使用。
【請求項37】
細胞がグリア細胞である、請求項30に記載の使用。
【請求項38】
細胞が肝細胞である、請求項30に記載の使用。
【請求項39】
芳香族-陽イオンペプチドがリンカーを含む、請求項1に記載の使用。
【請求項40】
分子がリンカーを含む、請求項1に記載の使用。
【請求項41】
分子と芳香族陽イオンペプチドが化学結合している、請求項1に記載の使用。
【請求項42】
芳香族陽イオンペプチドに結合した分子を含む担体複合体であって、前記芳香族陽イオンペプチドが以下を含み、前記分子が、ポリヌクレオチド、ポリアミノ酸、細胞毒性物質、抗生物質、抗酸化剤、増殖因子およびサイトカインからなる群から選ばれる、前記担体複合体:
Tyr-D-Arg-Phe-Lys-NH2 (DALDA),
2',6'-Dmt-D-Arg-Phe-Lys-NH2 (Dmt1-DALDA),
Phe-D-Arg-Phe-Lys-NH2 (Phe1-DALDA),
D-Arg-2',6'Dmt-Lys-Phe-NH2, および、
2',6'-Dmp-D-Arg-Phe-Lys-NH2 (Dmp1-DALDA)。
【請求項43】
分子が抗生物質である、請求項42に記載の担体複合体。
【請求項44】
分子が細胞毒性物質である、請求項42に記載の担体複合体。
【請求項45】
細胞毒性物質がドキソルビシンである、請求項44に記載の担体複合体。
【請求項46】
細胞毒性物質がアドリアマイシンである、請求項44に記載の担体複合体。
【請求項47】
分子が抗酸化剤である、請求項42に記載の担体複合体。
【請求項48】
抗酸化剤がビタミンEである、請求項47に記載の担体複合体。
【請求項49】
抗酸化剤がビタミンCである、請求項47に記載の担体複合体。
【請求項50】
抗酸化剤がベータカロチンである、請求項47に記載の担体複合体。
【請求項51】
ポリアミノ酸が内因性ペプチド又はタンパク質である、請求項42に記載の担体複合体。
【請求項52】
ポリアミノ酸が酵素である、請求項42に記載の担体複合体。
【請求項53】
分子が抗体である、請求項42に記載の担体複合体。
【請求項54】
増殖因子が神経栄養性増殖因子である、請求項42に記載の担体複合体。
【請求項55】
分子がサイトカインである、請求項42に記載の担体複合体。
【請求項56】
分子がポリヌクレオチドである、請求項42に記載の担体複合体。
【請求項57】
分子がオリゴヌクレオチドである、請求項56に記載の担体複合体。
【請求項58】
オリゴヌクレオチドがRNAである、請求項57に記載の担体複合体。
【請求項59】
RNAが二本鎖RNAである、請求項58に記載の担体複合体。
【請求項60】
二本鎖RNAがsiRNAである、請求項59に記載の担体複合体。
【請求項61】
オリゴヌクレオチドがDNAである、請求項57に記載の担体複合体。
【請求項62】
オリゴヌクレオチドが一本鎖RNAである、請求項57に記載の担体複合体。
【請求項63】
一本鎖RNAがメッセンジャーRNA(mRNA)である、請求項62に記載の担体複合体。
【請求項64】
オリゴヌクレオチドがリボザイムである、請求項57に記載の担体複合体。
【請求項65】
オリゴヌクレオチドがアンチセンスRNAである、請求項57に記載の担体複合体。
【請求項66】
オリゴヌクレオチドがリボザイムのための外側ガイド配列である、請求項57に記載の担体複合体。
【請求項67】
オリゴヌクレオチドがRNAデコイである、請求項57に記載の担体複合体。
【請求項68】
生物学的分子がポリアミノ酸である、請求項42に記載の担体複合体。
【請求項69】
芳香族-陽イオンペプチドがリンカーを含む、請求項42に記載の担体複合体。
【請求項70】
分子がリンカーを含む、請求項42に記載の担体複合体。
【請求項71】
分子と芳香族陽イオンペプチドが化学結合している、請求項42に記載の担体複合体。
【請求項72】
分子を細胞にin vitroで送達する方法であって、前記方法が細胞をin vitroで芳香族陽イオンペプチドに結合した分子と接触させることを含み、前記芳香族陽イオンペプチドが以下を含み、前記分子が、ポリヌクレオチド、ポリアミノ酸、細胞毒性物質、抗生物質、抗酸化剤、増殖因子およびサイトカインからなる群から選ばれる、前記方法:
Tyr-D-Arg-Phe-Lys-NH2 (DALDA),
2',6'-Dmt-D-Arg-Phe-Lys-NH2 (Dmt1-DALDA),
Phe-D-Arg-Phe-Lys-NH2 (Phe1-DALDA),
D-Arg-2',6'Dmt-Lys-Phe-NH2,および、
2',6'-Dmp-D-Arg-Phe-Lys-NH2 (Dmp1-DALDA)。
【請求項73】
分子と芳香族陽イオンペプチドが化学結合している、請求項72に記載の方法。
【請求項1】
担体複合体の、分子を細胞に送達するための医薬の製造における使用であって、前記担体複合体は芳香族陽イオンペプチドに結合した分子を含み、前記芳香族陽イオンペプチドが以下から選ばれ、前記分子が、ポリヌクレオチド、ポリアミノ酸、細胞毒性物質、抗生物質、抗酸化剤、増殖因子およびサイトカインからなる群から選ばれる、前記使用:
Tyr-D-Arg-Phe-Lys-NH2 (DALDA),
2',6'-Dmt-D-Arg-Phe-Lys-NH2 (Dmt1-DALDA),
Phe-D-Arg-Phe-Lys-NH2 (Phe1-DALDA),
D-Arg-2',6'Dmt-Lys-Phe-NH2, および、
2',6'-Dmp-D-Arg-Phe-Lys-NH2 (Dmp1-DALDA)。
【請求項2】
分子が抗生物質である、請求項1に記載の使用。
【請求項3】
分子が細胞毒性物質である、請求項1に記載の使用。
【請求項4】
細胞毒性物質がドキソルビシンである、請求項3に記載の使用。
【請求項5】
細胞毒性物質がアドリアマイシンである、請求項3に記載の使用。
【請求項6】
分子が抗酸化剤である、請求項1に記載の使用。
【請求項7】
抗酸化剤がビタミンEである、請求項6に記載の使用。
【請求項8】
抗酸化剤がビタミンCである、請求項6に記載の使用。
【請求項9】
抗酸化剤がベータカロチンである、請求項6に記載の使用。
【請求項10】
分子がポリアミノ酸である、請求項1に記載の使用。
【請求項11】
ポリアミノ酸が内因性ペプチド又はタンパク質である、請求項10に記載の使用。
【請求項12】
ポリアミノ酸が酵素である、請求項10に記載の使用。
【請求項13】
ポリアミノ酸が抗体である、請求項10に記載の使用。
【請求項14】
増殖因子が神経栄養性増殖因子である、請求項1に記載の使用。
【請求項15】
分子がサイトカインである、請求項1に記載の使用。
【請求項16】
分子がポリヌクレオチドである、請求項1に記載の使用。
【請求項17】
分子がオリゴヌクレオチドである、請求項1に記載の使用。
【請求項18】
オリゴヌクレオチドがRNAである、請求項17に記載の使用。
【請求項19】
RNAが二本鎖RNAである、請求項18に記載の使用。
【請求項20】
二本鎖RNAがsiRNAである、請求項19に記載の使用。
【請求項21】
オリゴヌクレオチドがDNAである、請求項17に記載の使用。
【請求項22】
オリゴヌクレオチドが一本鎖RNAである、請求項17に記載の使用。
【請求項23】
一本鎖RNAがメッセンジャーRNA(mRNA)である、請求項22に記載の使用。
【請求項24】
オリゴヌクレオチドがリボザイムである、請求項17に記載の使用。
【請求項25】
オリゴヌクレオチドがアンチセンスRNAである、請求項17に記載の使用。
【請求項26】
オリゴヌクレオチドがリボザイムのための外側ガイド配列である、請求項17に記載の使用。
【請求項27】
オリゴヌクレオチドがRNAデコイである、請求項17に記載の使用。
【請求項28】
細胞が細菌細胞である、請求項1に記載の使用。
【請求項29】
細胞が植物細胞である、請求項1に記載の使用。
【請求項30】
細胞が動物細胞である、請求項1に記載の使用。
【請求項31】
動物細胞が哺乳動物細胞である、請求項30に記載の使用。
【請求項32】
細胞が神経細胞である、請求項30に記載の使用。
【請求項33】
細胞が腎上皮細胞である、請求項30に記載の使用。
【請求項34】
細胞が腸上皮細胞である、請求項30に記載の使用。
【請求項35】
細胞が血管内皮細胞である、請求項30に記載の使用。
【請求項36】
内皮細胞が血管-脳関門内皮細胞である、請求項30に記載の使用。
【請求項37】
細胞がグリア細胞である、請求項30に記載の使用。
【請求項38】
細胞が肝細胞である、請求項30に記載の使用。
【請求項39】
芳香族-陽イオンペプチドがリンカーを含む、請求項1に記載の使用。
【請求項40】
分子がリンカーを含む、請求項1に記載の使用。
【請求項41】
分子と芳香族陽イオンペプチドが化学結合している、請求項1に記載の使用。
【請求項42】
芳香族陽イオンペプチドに結合した分子を含む担体複合体であって、前記芳香族陽イオンペプチドが以下を含み、前記分子が、ポリヌクレオチド、ポリアミノ酸、細胞毒性物質、抗生物質、抗酸化剤、増殖因子およびサイトカインからなる群から選ばれる、前記担体複合体:
Tyr-D-Arg-Phe-Lys-NH2 (DALDA),
2',6'-Dmt-D-Arg-Phe-Lys-NH2 (Dmt1-DALDA),
Phe-D-Arg-Phe-Lys-NH2 (Phe1-DALDA),
D-Arg-2',6'Dmt-Lys-Phe-NH2, および、
2',6'-Dmp-D-Arg-Phe-Lys-NH2 (Dmp1-DALDA)。
【請求項43】
分子が抗生物質である、請求項42に記載の担体複合体。
【請求項44】
分子が細胞毒性物質である、請求項42に記載の担体複合体。
【請求項45】
細胞毒性物質がドキソルビシンである、請求項44に記載の担体複合体。
【請求項46】
細胞毒性物質がアドリアマイシンである、請求項44に記載の担体複合体。
【請求項47】
分子が抗酸化剤である、請求項42に記載の担体複合体。
【請求項48】
抗酸化剤がビタミンEである、請求項47に記載の担体複合体。
【請求項49】
抗酸化剤がビタミンCである、請求項47に記載の担体複合体。
【請求項50】
抗酸化剤がベータカロチンである、請求項47に記載の担体複合体。
【請求項51】
ポリアミノ酸が内因性ペプチド又はタンパク質である、請求項42に記載の担体複合体。
【請求項52】
ポリアミノ酸が酵素である、請求項42に記載の担体複合体。
【請求項53】
分子が抗体である、請求項42に記載の担体複合体。
【請求項54】
増殖因子が神経栄養性増殖因子である、請求項42に記載の担体複合体。
【請求項55】
分子がサイトカインである、請求項42に記載の担体複合体。
【請求項56】
分子がポリヌクレオチドである、請求項42に記載の担体複合体。
【請求項57】
分子がオリゴヌクレオチドである、請求項56に記載の担体複合体。
【請求項58】
オリゴヌクレオチドがRNAである、請求項57に記載の担体複合体。
【請求項59】
RNAが二本鎖RNAである、請求項58に記載の担体複合体。
【請求項60】
二本鎖RNAがsiRNAである、請求項59に記載の担体複合体。
【請求項61】
オリゴヌクレオチドがDNAである、請求項57に記載の担体複合体。
【請求項62】
オリゴヌクレオチドが一本鎖RNAである、請求項57に記載の担体複合体。
【請求項63】
一本鎖RNAがメッセンジャーRNA(mRNA)である、請求項62に記載の担体複合体。
【請求項64】
オリゴヌクレオチドがリボザイムである、請求項57に記載の担体複合体。
【請求項65】
オリゴヌクレオチドがアンチセンスRNAである、請求項57に記載の担体複合体。
【請求項66】
オリゴヌクレオチドがリボザイムのための外側ガイド配列である、請求項57に記載の担体複合体。
【請求項67】
オリゴヌクレオチドがRNAデコイである、請求項57に記載の担体複合体。
【請求項68】
生物学的分子がポリアミノ酸である、請求項42に記載の担体複合体。
【請求項69】
芳香族-陽イオンペプチドがリンカーを含む、請求項42に記載の担体複合体。
【請求項70】
分子がリンカーを含む、請求項42に記載の担体複合体。
【請求項71】
分子と芳香族陽イオンペプチドが化学結合している、請求項42に記載の担体複合体。
【請求項72】
分子を細胞にin vitroで送達する方法であって、前記方法が細胞をin vitroで芳香族陽イオンペプチドに結合した分子と接触させることを含み、前記芳香族陽イオンペプチドが以下を含み、前記分子が、ポリヌクレオチド、ポリアミノ酸、細胞毒性物質、抗生物質、抗酸化剤、増殖因子およびサイトカインからなる群から選ばれる、前記方法:
Tyr-D-Arg-Phe-Lys-NH2 (DALDA),
2',6'-Dmt-D-Arg-Phe-Lys-NH2 (Dmt1-DALDA),
Phe-D-Arg-Phe-Lys-NH2 (Phe1-DALDA),
D-Arg-2',6'Dmt-Lys-Phe-NH2,および、
2',6'-Dmp-D-Arg-Phe-Lys-NH2 (Dmp1-DALDA)。
【請求項73】
分子と芳香族陽イオンペプチドが化学結合している、請求項72に記載の方法。
【図1】

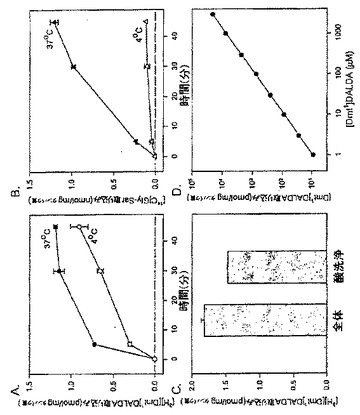
【図2】
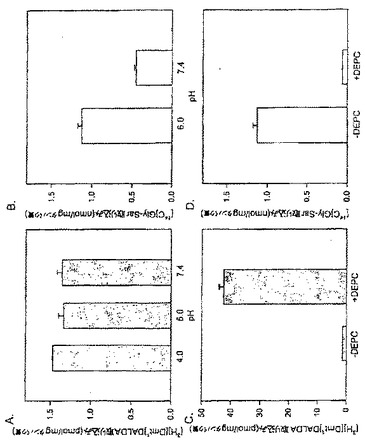
【図3】
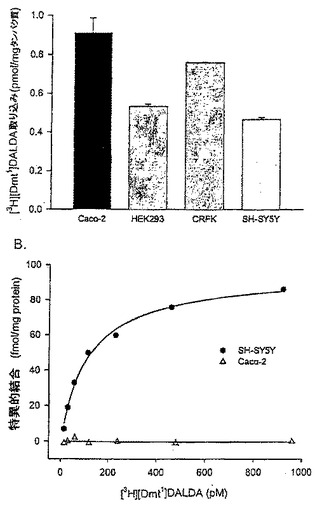
【図4】

【図5】
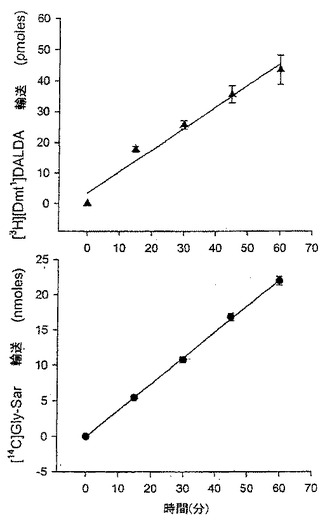
【図6】
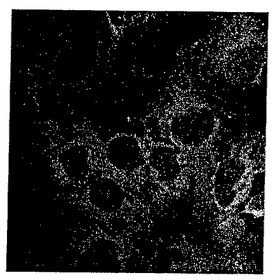
【図7】
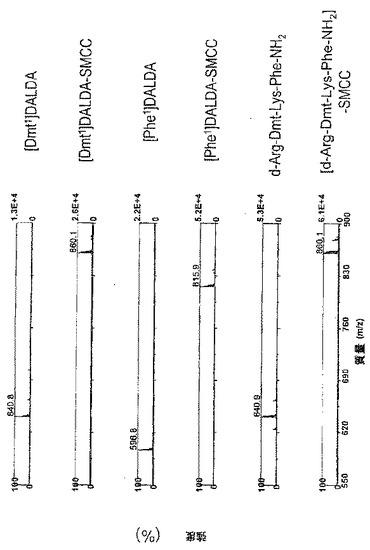
【図8】
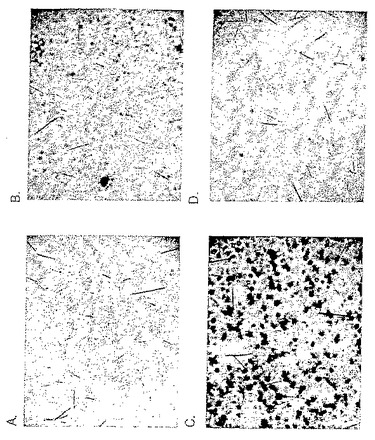
【図9】
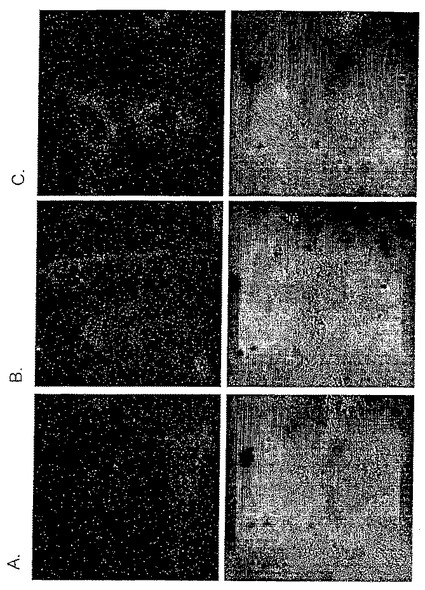
【図10】
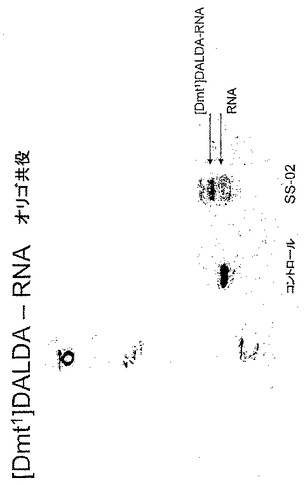
【図11】
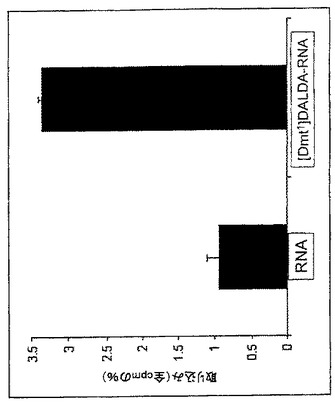
【図12】
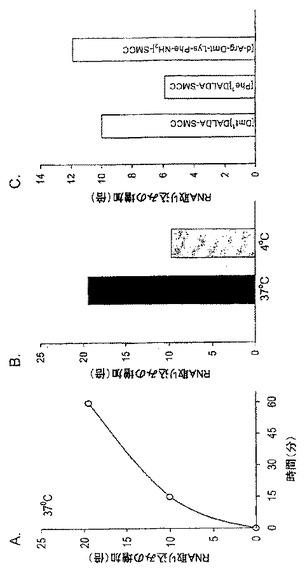
【図13】
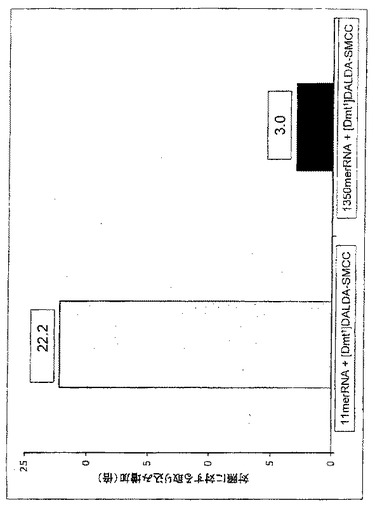
【図14】
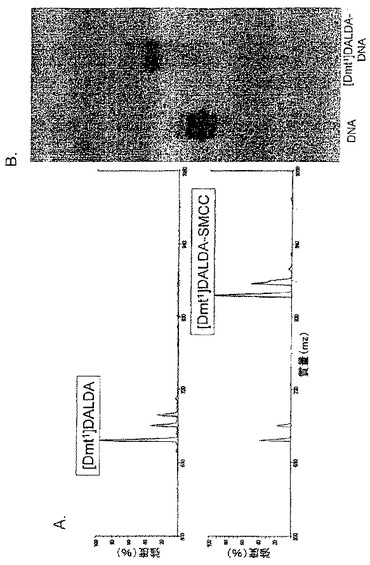
【図15】
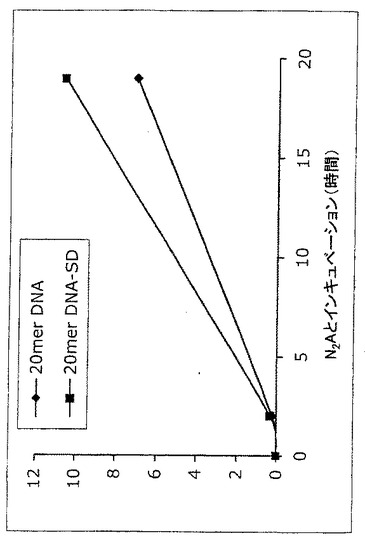
【図16】
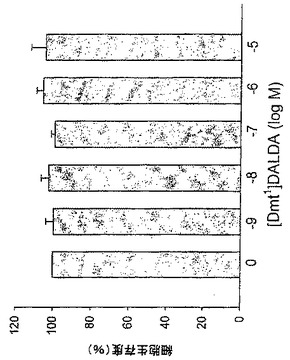
【図17】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公開番号】特開2011−168603(P2011−168603A)
【公開日】平成23年9月1日(2011.9.1)
【国際特許分類】
【出願番号】特願2011−87496(P2011−87496)
【出願日】平成23年4月11日(2011.4.11)
【分割の表示】特願2006−532552(P2006−532552)の分割
【原出願日】平成16年5月3日(2004.5.3)
【出願人】(505294425)コーネル リサーチ ファウンデイション インコーポレイテッド (11)
【Fターム(参考)】
【公開日】平成23年9月1日(2011.9.1)
【国際特許分類】
【出願日】平成23年4月11日(2011.4.11)
【分割の表示】特願2006−532552(P2006−532552)の分割
【原出願日】平成16年5月3日(2004.5.3)
【出願人】(505294425)コーネル リサーチ ファウンデイション インコーポレイテッド (11)
【Fターム(参考)】
[ Back to top ]