
細胞の分泌活性化方法
【課題】本発明は、小胞体膜上でのポリソーム形成を促進すること、そしてその結果として目的とするタンパク質の生成を促進すること、並びに目的とするタンパク質の産生能が促進された細胞を提供することを課題とする。
【解決手段】本発明は、細胞において、哺乳動物p180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供することにより、上記課題を解決することができることを示した。本発明はまた、目的とするタンパク質をコードする核酸分子を形質転換した細胞または目的とするタンパク質の産生量が増加された細胞において、ヒトp180タンパク質の全長またはその部分を発現させ、細胞内の小胞体膜上でのポリソーム形成を促進することを含む、前記細胞中での目的とするタンパク質の生成を促進する方法を提供することにより、上記課題を解決することができることを示した。
【解決手段】本発明は、細胞において、哺乳動物p180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供することにより、上記課題を解決することができることを示した。本発明はまた、目的とするタンパク質をコードする核酸分子を形質転換した細胞または目的とするタンパク質の産生量が増加された細胞において、ヒトp180タンパク質の全長またはその部分を発現させ、細胞内の小胞体膜上でのポリソーム形成を促進することを含む、前記細胞中での目的とするタンパク質の生成を促進する方法を提供することにより、上記課題を解決することができることを示した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、細胞内におけるポリソーム形成を促進すること、また細胞内におけるポリソーム形成を促進することを通じて細胞内における目的のタンパク質の生成を促進することに関するものである。
【背景技術】
【0002】
mRNAの翻訳は、潜在的には複数のレベルで制御される一方、イニシエーションでの制御は、重要なステップであるとして、もっともよく研究されてきた(Sonenberg, N. and Hinnebusch, A.G. (2009) Cell, 136, 731-745;Gebauer, F. and Hentze, M.W. (2004) Nat Rev Mol Cell Biol, 5, 827-835)。mRNAの5’非翻訳領域とmRNAの3’非翻訳領域は、mRNAの翻訳効率、安定性および局在化を含む翻訳制御の様々な段階で重要な役割を果たしている(Abaza, I. and Gebauer, F. (2008) RNA, 14, 404-409)。ポリソームアッセンブリの程度は、細胞における翻訳効率に対する直接的なインパクトを介した翻訳調節の別の重要な側面であると想定できる。低温電子断層撮影における最近の進展が、細胞融解物および無傷細胞における翻訳ポリソームの組織化に関する重要な洞察をもたらした一方(Brandt, F., et al., (2010) Mol Cell, 39, 560-569;Brandt, F., et al., (2009) Cell, 136, 261-271)、mRNAのリボソーム占有が、mRNAの長さにのみ依存しているか、または何等か未知のメカニズムによって制御されているかについては、依然としてあいまいなままである(Arava, Y., et al., (2003) Proc Natl Acad Sci U S A, 100, 3889-3894;Kopeina, G.S., et al., (2008) Nucleic Acids Res, 36, 2476-2488;Nelson, E.M. and Winkler, M.M. (1987) J Biol Chem, 262, 11501-11506)。
【0003】
特に、膜タンパク質および分泌タンパク質に関して、その後の膜を通過した転位のために、状況はずっと複雑である。リボソーム装置およびトランスロコン装置が、構造的にそして機能的にどのように共役しているのかについては、限られた情報しか得られていない(Pool, M.R. (2009) J Cell Biol, 185, 889-902)。
【0004】
膜-結合型リボソーム遊離の細胞質リボソームから構造的に識別されるのかどうかは依然としてわかっていないが、遊離のリボソームと比較して、小胞体-結合型のリボソームは、より効率的な生合成を媒介することは知られている(Stephens, S.B. and Nicchitta, C.V. (2008) Mol Biol Cell, 19, 623-632)。さらに、単一のポリソーム上で、協調的な翻訳とその後の膜を通過する転位を達成するためには、リボソーム/トランスロコン複合体の各ユニット間で、より高次の協調が必要不可欠である。しかしながら、そのような協調についての分子的基礎については、基本的に全く分かっていない。
【0005】
コラーゲンは、皮膚、腱および骨などの結合組織における細胞外マトリクスの主要な構成成分の一つである。それらは、前駆体型(すなわち、プロコラーゲン)として小胞体膜上で合成され、そして繊維芽細胞、軟骨芽細胞および骨芽細胞を含む専門的分泌細胞により分泌される。コラーゲンの分泌のための専門化細胞は、膵臓細胞およびプラズマ細胞などのその他の分泌細胞と同様に、高速合成を提供するために粗面小胞体の高度に発達したネットワークを有する。しかし、専門的分泌細胞におけるタンパク質生合成の非常に効率的な活性を裏付けるメカニズムについては、ほとんどわかっていない。
【0006】
アスコルビン酸は、in vitro培養のあいだのプロコラーゲン分泌について、一般的にそして長く使用されてきた刺激物質である。コラーゲン-分泌性繊維芽細胞において、アスコルビン酸処理は、小胞体からゴルジ体への輸送の回復とそれに続くトラフィック需要の増大に応答して、de novo生合成を活性化するものと考えられている。しかしながら、de novo生合成がどのようにして専門的分泌細胞中で活性化されるのかについては、いまだわかっていない。
【0007】
p180タンパク質は、必須の小胞体膜タンパク質であり、そして分泌組織中で多く発現されている(Langley, R., et al., (1998) DNA Cell Biol, 17, 449-460)。p180タンパク質は、初めは、粗面小胞体膜上のリボソーム受容体候補の一つとして同定された(Savitz, A.J. and Meyer, D.I. (1990) Nature, 346, 540-544)。その独特なリピートドメインが、リボソームに対して結合能力を有し、そして非常に塩基性のpIを有するドデカペプチドの54回のタンデムリピートが含まれることが報告された(Wanker, E.E., et al., (1995) J Cell Biol, 130, 29-39)。哺乳動物において、p180タンパク質のアミノ酸配列、ヌクレオチド配列は、種を超えて高度に保存されており、ヒトとの関係で最も相同性が低いブタであっても、80%の相同性を有していることが知られている。しかしながら、p180タンパク質の機能に付いては十分に解析が進んでおらず、特にp180タンパク質が動物細胞中のリボソームに対して直接結合するかどうかは、いまだよくわかっていない。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Sonenberg, N. and Hinnebusch, A.G. (2009) Cell, 136, 731-745
【非特許文献2】Gebauer, F. and Hentze, M.W. (2004) Nat Rev Mol Cell Biol, 5, 827-835
【非特許文献3】Abaza, I. and Gebauer, F. (2008) RNA, 14, 404-409
【非特許文献4】Brandt, F., et al., (2010) Mol Cell, 39, 560-569
【非特許文献5】Brandt, F., et al., (2009) Cell, 136, 261-271
【非特許文献6】Arava, Y., et al., (2003) Proc Natl Acad Sci U S A, 100, 3889-3894
【非特許文献7】Kopeina, G.S., et al., (2008) Nucleic Acids Res, 36, 2476-2488
【非特許文献8】Nelson, E.M. and Winkler, M.M. (1987) J Biol Chem, 262, 11501-11506
【非特許文献9】Pool, M.R. (2009) J Cell Biol, 185, 889-902
【非特許文献10】Stephens, S.B. and Nicchitta, C.V. (2008) Mol Biol Cell, 19, 623-632
【非特許文献11】Langley, R., et al., (1998) DNA Cell Biol, 17, 449-460
【非特許文献12】Savitz, A.J. and Meyer, D.I. (1990) Nature, 346, 540-544
【非特許文献13】Wanker, E.E., et al., (1995) J Cell Biol, 130, 29-39
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、 小胞体膜上でのポリソーム形成を促進すること、そしてその結果として目的とするタンパク質の生成を促進すること、並びに目的とするタンパク質の産生能が促進された細胞を提供することを課題とする。
【課題を解決するための手段】
【0010】
本発明は、細胞において、p180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供することにより、上記課題を解決することができることを示した。本発明はまた、目的とするタンパク質をコードする核酸分子を形質転換した細胞または目的とするタンパク質の産生量が増加された細胞において、p180タンパク質の全長またはその部分を発現させ、細胞内の小胞体膜上でのポリソーム形成を促進することを含む、前記細胞中での目的とするタンパク質の生成を促進する方法を提供することにより、上記課題を解決することができることを示した。
【発明の効果】
【0011】
本発明により、発現系を構成する細胞内において、小胞体膜上のポリソーム形成を促進させることができることから、結果として、細胞中での目的とするタンパク質の生成を促進することができる。
【図面の簡単な説明】
【0012】
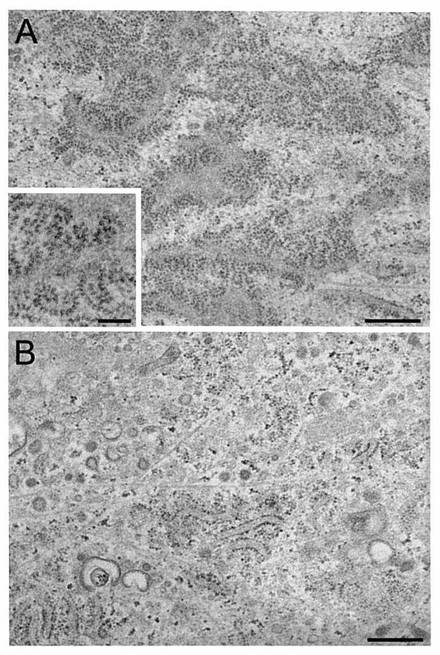
【図1】図1は、コラーゲン分泌性HEL線維芽細胞における粗面小胞体表面の透過電子顕微鏡観察の様子を示す。
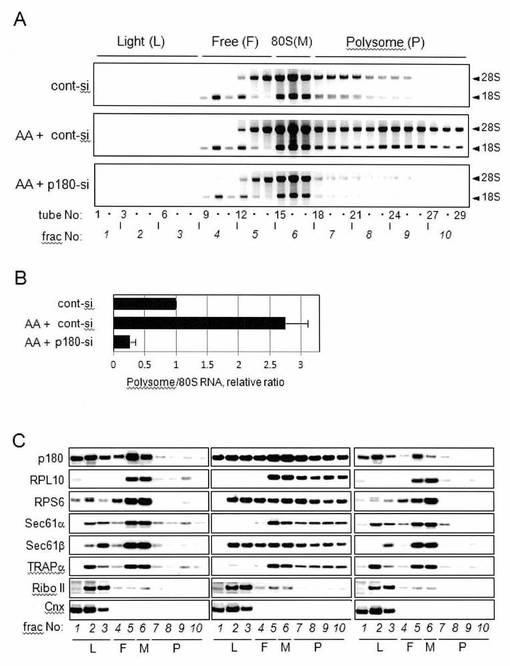
【図2】図2は、p180タンパク質が膜結合型ポリソームのアセンブリーを促進することを示す図である。
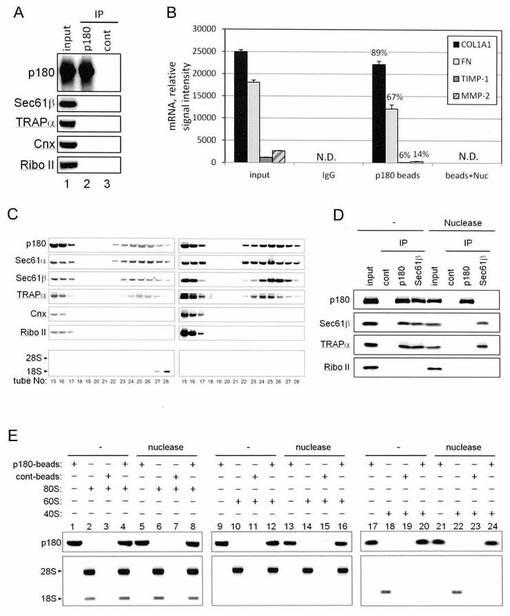
【図3】図3は、p180タンパク質がmRNAとの強い相互作用活性と、リボソームとの結合活性をそれぞれもつことを示した図である。
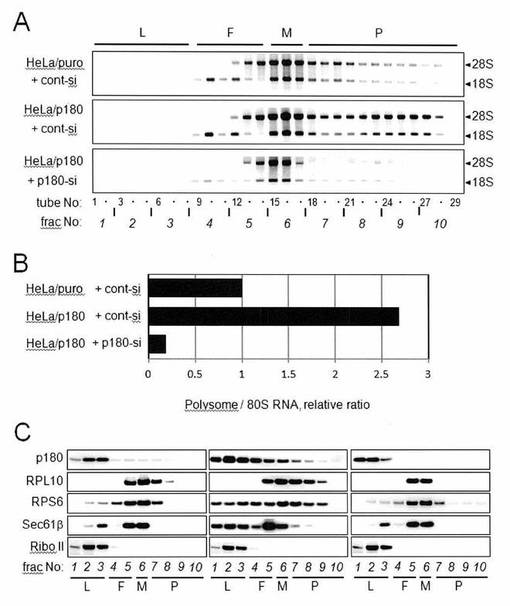
【図4】図4は、p180タンパク質の過剰発現は、膜結合型ポリソームの形成を亢進することを示した図である。
【図5】図5は、アスコルビン酸刺激下における小胞体膜結合型ポリソームの生合成活性は、p180タンパク質の発現抑制に伴って低下することを示した図である。
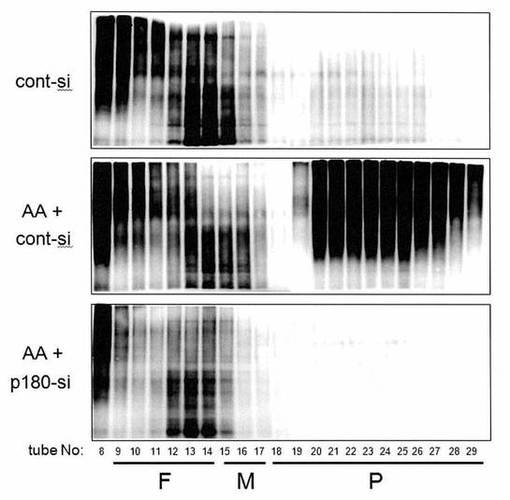
【図6】図6は、アスコルビン酸刺激下で起こる、p180タンパク質依存的な膜結合型ポリソームの形成促進はCOL,FNなどに対して優先的に作用することを示した図である。
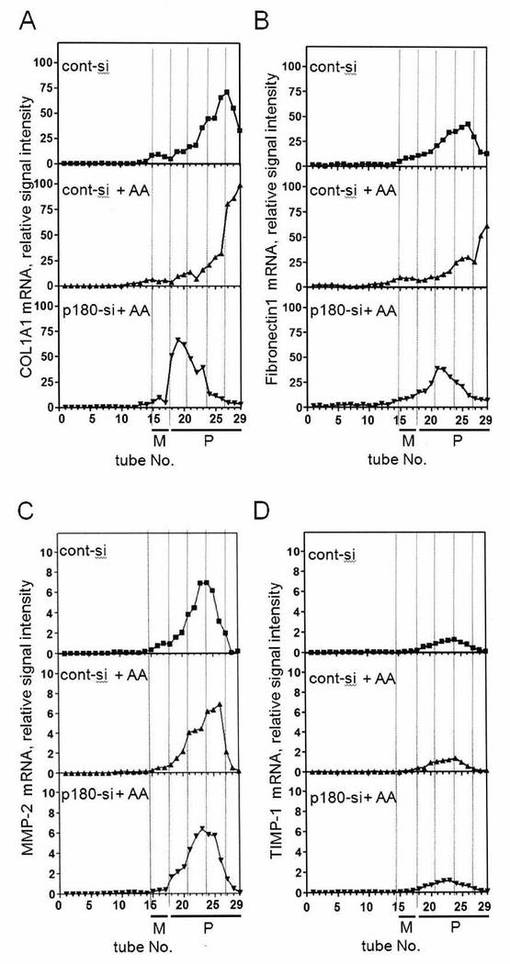
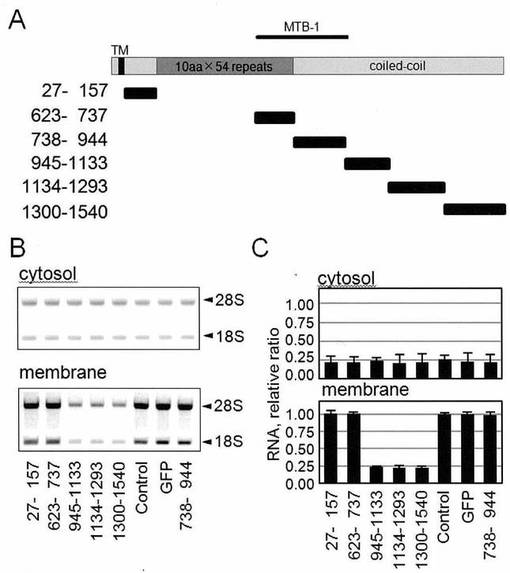
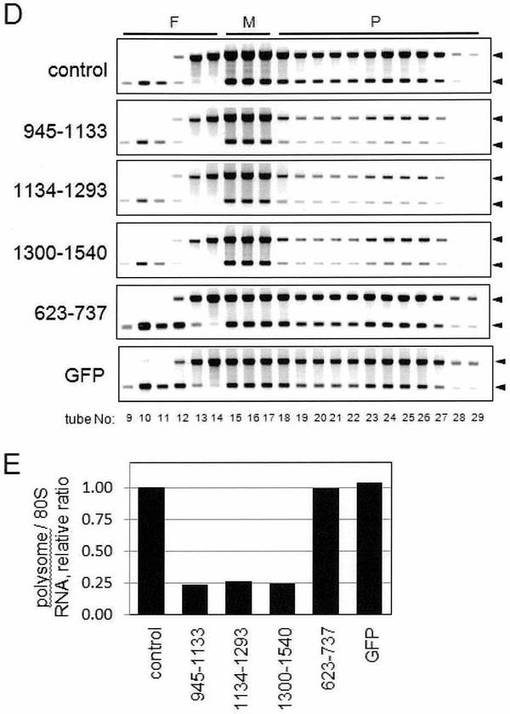
【図7−1】図7は、膜結合型ポリソームの形成に重要なp180タンパク質の責任領域はcoiled-coilドメインであることを示した図である。
【図7−2】図7は、膜結合型ポリソームの形成に重要なp180タンパク質の責任領域はcoiled-coilドメインであることを示した図である。
【図7−3】図7は、膜結合型ポリソームの形成に重要なp180タンパク質の責任領域はcoiled-coilドメインであることを示した図である。
【図7−4】図7は、膜結合型ポリソームの形成に重要なp180タンパク質の責任領域はcoiled-coilドメインであることを示した図である。
【図8】図8は、図7Aで示された様々な領域の完全長構造および短縮型構造の発現タンパク質が、正しいサイズで発現されたことを示す図である。
【発明を実施するための形態】
【0013】
本発明においては、細胞においてp180タンパク質の全長またはその部分を発現させることにより、細胞内の小胞体膜上でのポリソーム形成を促進することができることを見出し、本発明の第一の態様を完成するに至った。
【0014】
すなわち、本発明においては、細胞において、哺乳動物のp180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供する。ここで、哺乳動物のp180タンパク質のアミノ酸配列という場合、ヒトのp180タンパク質(GenBank Accession No. AB287347)のほか、マウスのp180タンパク質(GenBank Accession No.NP_077243)、ラット(GenBank Accession No. XP_230637)、イヌ(GenBank Accession No. NP_001003179)、ウマ(GenBank Accession No.XP_001915027)、サル(GenBank Accession No. XP_002798281)、チンパンジー(GenBank Accession No.XP_514527)、ブタ(GenBank Accession No.XP_001926148)などが含まれる。
【0015】
例えば、哺乳動物のp180タンパク質としてヒトのp180タンパク質を使用する場合、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域、623〜737番アミノ酸からなる領域、738〜944番アミノ酸からなる領域、のいずれかを含む部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供する。
【0016】
この態様において、p180タンパク質の全長または部分を細胞中で発現させることにより、その細胞内の小胞体膜上でのポリソーム形成を促進することができる。ここで、ポリソームは、細胞内の小胞体膜上に存在する複数のリボソームに対して、1分子のmRNAが結合したもののことをいう。
【0017】
細胞において発現させるタンパク質は、p180タンパク質の全長またはその部分である。ここで、ヒトp180タンパク質の全長は、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質であり(GenBank Accession No. AB287347)、このタンパク質は、SEQ ID NO: 2に示されるヌクレオチド配列によりコードされる(GenBank Accession No.AB287347)。このほか、上述したマウスのp180タンパク質は、GenBank Accession No. NP_077243により示されるヌクレオチド配列によりコードされ、ラットのp180タンパク質は、GenBank Accession No. XP_230637により示されるヌクレオチド配列によりコードされ、、イヌのp180タンパク質は、GenBank Accession No. NP_001003179により示されるヌクレオチド配列によりコードされ、ウマのp180タンパク質は、GenBank Accession No. XP_001915027により示されるヌクレオチド配列によりコードされ、サルのp180タンパク質は、GenBank Accession No. XP_002798281により示されるヌクレオチド配列によりコードされ、チンパンジーのp180タンパク質は、GenBank Accession No. XP_514527により示されるヌクレオチド配列によりコードされ、ブタのp180タンパク質は、GenBank Accession No. XP_001926148により示されるヌクレオチド配列によりコードされる。
【0018】
本発明において、例えば、p180タンパク質の部分という場合、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分などの部分のことを意味する。このような部分を含むものは、ポリソーム形成を促進する能力を示すことができる。このようにして規定されるヒトp180タンパク質の部分としては、上述したSEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域、623〜737番アミノ酸からなる領域、738〜944番アミノ酸からなる領域、などを含む部分そのものの他、ヒトp180タンパク質のN末端膜貫通ドメインのC末端側に隣接するMTB-2ドメイン、またはリボソーム-結合リピートドメインを含む高度に塩基性のN-末端領域、高度に塩基性のタンデムリピートドメイン、または微小管結合および束形成ドメイン(MTB-1ドメイン)、などが一例として含まれる(図7A)。
【0019】
他の哺乳動物のp180タンパク質の部分を使用する場合も、ヒトのp180タンパク質のアミノ酸配列とその他の哺乳動物のp180タンパク質のアミノ酸配列が、全体的に、高度に保存されていることから、他の哺乳動物のp180タンパク質の対応する部分または領域を含む断片を使用することができる。
【0020】
本発明において使用する細胞は、タンパク質発現に適した哺乳動物の細胞であればどのようなものであってもよく、例えば、CHO細胞、HEK293細胞、HeLa細胞などの哺乳動物由来の細胞を使用することができる。
【0021】
前述した哺乳動物p180タンパク質の全長またはその部分を、これらの細胞内において発現させるためには、当該技術分野において一般的に使用されている形質転換方法を使用することができる。その形質転換を行うためには、例えば、pcDNA、pEGFP、pCAGGSなどの発現ベクターを使用することができる。
【0022】
上述したように、哺乳動物p180タンパク質の全長または部分を細胞中で発現させることにより、ポリソーム形成を促進することができることから、この性質を利用することにより、細胞中で産生させたい目的とするタンパク質の生成を促進することができる。
【0023】
したがって、本発明においてはまた、哺乳動物のp180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供する方法を提供する。目的とするタンパク質をコードする核酸分子を形質転換した細胞または目的とするタンパク質の産生量が増加された細胞において、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、などの部分を発現させ、細胞内の小胞体膜上でのポリソーム形成を促進することを含む、前記細胞中での目的とするタンパク質の生成を促進する方法を提供する。
【0024】
この方法は、目的とするタンパク質を産生させるための細胞において、目的とするタンパク質の産生をさらに亢進させたい場合に使用することができる。例えば、目的とするタンパク質の産生のためにそのタンパク質をコードする遺伝子を細胞にトランスフェクトして形質転換細胞を作出したが、目的とするタンパク質の産生が十分でないか、またはその産生をより亢進させたい場合に、その細胞にさらに哺乳動物p180タンパク質の全長またはその部分を発現させることにより、目的とするタンパク質の産生を亢進させることができる。あるいは、細胞に対して特定の刺激を加えることにより目的とするタンパク質の産生量が増加されるが、目的とするタンパク質の産生が十分でないか、またはその産生をより亢進させたい場合に、その細胞にさらに哺乳動物p180タンパク質の全長またはその部分を発現させることにより、目的とするタンパク質の産生を亢進させることができる。
【0025】
前述したリポソーム形成を促進する方法の場合と同様に、この目的とするタンパク質の生成を促進する方法においてもまた、細胞において発現させるタンパク質は、哺乳動物p180タンパク質の全長またはその部分である。目的とするタンパク質の産生を亢進させたい細胞に対して、哺乳動物p180タンパク質の全長またはその部分を発現させるためには、前述したような当該技術分野において一般的に使用されている形質転換方法を使用することができる。
【0026】
細胞中で生成を促進したい目的とするタンパク質は、前述したように、細胞に対して目的とするタンパク質をコードする核酸分子を形質転換することにより、または細胞に対して特定の刺激を加えることにより目的とするタンパク質の産生量が増加されることにより、細胞内においてその生成を促進させることができるものである。目的とするタンパク質としては、例えば、コラーゲン、フィブロネクチン、ラミニンなどを挙げることができるが、これらには限定されない。
【0027】
目的とするタンパク質としてコラーゲンを使用する場合、例えば、コラーゲンをコードする遺伝子としてヒトI型コラーゲン、アルファ1(GenBank Accession No. NM_000088.3)を例えばpcDNAベクターを使用してHeLa細胞に形質転換してコラーゲン産生亢進細胞株を作成し、またはコラーゲンの産生が亢進されているHEL細胞に対してさらにコラーゲンの発現を促進するアスコルビン酸添加刺激処理を行うことによってコラーゲン高発現状態の細胞を調製し、その細胞株に対してヒトp180タンパク質の全長またはその部分を発現させることができる。その結果、細胞内の小胞体膜上におけるコラーゲン遺伝子のmRNAが取り込まれたポリソームの形成を促進し、結果的にコラーゲンの産生を亢進することができる。
【0028】
また、目的とするタンパク質としてフィブロネクチンを使用する場合、フィブロネクチンをコードする遺伝子としてヒトフィブロネクチン1(GenBank Accession No. NM_212482)をpcDNAベクターを使用してHeLa細胞に形質転換してフィブロネクチン産生亢進細胞株を作成し、またはフィブロネクチンの産生が亢進されているHEL細胞に対してヒトp180タンパク質の全長またはその部分を発現させる。その結果、細胞内の小胞体膜上におけるフィブロネクチン遺伝子のmRNAが取り込まれたポリソームの形成を促進し、結果的にフィブロネクチンの産生を亢進することができる。
【0029】
本発明の第一の態様において、細胞においてp180タンパク質の全長またはその部分を発現させることにより、細胞内の小胞体上でのポリソーム形成を促進することができることを見出したことに基づいて、第一の態様の方法を実現するための細胞を提供することができ、本発明の第二の態様を完成するに至った。
【0030】
すなわち、本発明においては、哺乳動物のp180タンパク質のアミノ酸の特定の部分を発現させた細胞、その具体的な好ましい態様としては、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、などの部分を発現させた細胞、を提供する。この様な構成上の特徴を有する細胞は、細胞内の小胞体膜上でのポリソーム形成能が促進されるという特徴を有するものであり、そのような特徴を有する結果、細胞内において転写されるmRNAからのタンパク質の生成を亢進することができる。
【0031】
本発明のこの態様における細胞は、タンパク質発現に適した哺乳動物の細胞であればどのようなものを起源として作出した細胞であってもよく、例えば、起源となる細胞としては、CHO細胞、HEK293細胞、HeLa細胞などの哺乳動物由来の細胞を使用することができる。これらの細胞に対して、前述した哺乳動物p180タンパク質の全長またはその部分を、当該技術分野において一般的に使用されている方法を使用してトランスフェクトして、これらの細胞内において哺乳動物p180タンパク質の全長またはその部分を発現させることができる。
【0032】
このような性質を有する細胞は、ポリソーム中に、細胞中で産生を亢進させたい目的とするタンパク質をコードする遺伝子から転写されたmRNAが含まれるものであってもよい。目的とするタンパク質としては、例えば、コラーゲン、フィブロネクチン、ラミニンなどを挙げることができるが、これらには限定されない。
【0033】
本明細書においては、ここまでに説明した本発明をさらに具体的に説明することを目的として、以下の実施例を記載するが、しかしながら、以下の実施例の記載は、本発明を限定することを目的としたものではない。
【実施例】
【0034】
実施例1:コラーゲン-分泌性HEL繊維芽細胞における粗面小胞体表面の透過電子顕微鏡観察
本発明者らは、アスコルビン酸処理により、mRNAレベルを変化させることなく、膜-結合型リボソームが増加することを報告した(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。これらの知見は、アスコルビン酸が、リボソーム-トランスロコン複合体の変化またはポリソーム組織化の亢進のいずれかにより、高いトラフィック負荷に応じて、新たな翻訳調節メカニズムを介して、生合成を上方制御していることを示唆していた。
【0035】
本実施例においては、p180タンパク質がポリソーム組織化に影響を与えるかどうかを決定するため、対照細胞およびp180-除去細胞の超微細構造を調べることを目的として、電子顕微鏡観察を行った。
【0036】
本実施例における電子顕微鏡解析のため、対照HEL細胞(A)およびp180 siRNA-形質転換HEL細胞(B)を、2%ウシ胎児血清(FBS)(Intergen)を含有するF12/DMEM(Nissui)中、アスコルビン酸の存在下にて増殖させた。特に示さない場合には、L-アスコルビン酸リン酸マグネシウム塩n-水和物(Wako)を、0.2 mMの最終濃度となるように添加した。細胞培養およびsiRNAトランスフェクションのプロトコルは、以前に記載したように行った(Ueno, T., et al., (2010) Exp Cell Res, 316, 329-340)。
【0037】
透過型電子顕微鏡解析用に、2%FBS/DMEM中で培養された細胞を、10μMタキソールを含有する培養液を用いて37℃にて3分間処理し、その後以前に記載したように(Ogawa-Goto, et al., (2002) J Virol, 76, 2350-2362)、透過型電子顕微鏡解析用に、従来から使用されている固定化手順を行った。すなわち、超薄切片を、基質と並行または垂直に切り出し、そしてModel 7650 EM(Hitachi)を用いて80 kVにて可視化した。
【0038】
電子顕微鏡像の調査の結果、アスコルビン酸-刺激HEL繊維芽細胞由来の水平方向に切断した切片の小胞体膜表面上に、多数の極めて長いポリソームが存在することが示された(図1A)。高倍率像では、約25〜30個のリボソームからなるリボソームのらせん状アレイまたは28個以上のリボソームから成るヘアピン型ポリソームが示され(図1A、挿入図)、ラット皮膚繊維芽細胞で報告されたものに非常に似ていた(Christensen, A.K. and Bourne, C.M. (1999) Anat Rec, 255, 116-129)。対照的に、p180タンパク質に対するsiRNAを用いたp180タンパク質の除去では、アスコルビン酸刺激にもかかわらず、小胞体膜上のポリソームでは劇的な減少が引き起こされた(図1B)。バー:図1Aおよび図1Bでは500 nm:図1A中の挿入図では200 nmを示す。
【0039】
実施例2:p180タンパク質による小胞体膜上のポリソームアッセンブリの促進
本実施例においては、小胞体膜上でのポリソーム形成におけるp180タンパク質の具体的な役割をさらに調べるため、高-塩濃度バッファーを用いたストリンジェントな条件下でのポリソーム勾配解析を行った。
【0040】
HEL細胞を、アスコルビン酸(AA)の存在下または非存在下にて培養し、そして対照siRNA またはp180-特異的siRNA(siRNA)により処置した。siRNAのトランスフェクションの4日後に、連続的ジギトニン抽出を行って、膜画分を得て、その後15〜50%のスクロース勾配を使用してポリソーム解析を行った。
【0041】
細胞質画分および膜画分の抽出は、ジギトニンを以前に記載した方法(Lerner, R.S., et al., (2003) RNA, 9, 1123-1137)から、わずかに修飾して使用して(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)、連続的界面活性剤抽出により行った。簡単に述べると、6ウェルプレート中でのHEL細胞の単層を、200μMのシクロヘキシミドとともに15分間事前培養し、そしてPBSで洗浄した。抽出は、手順を通じて200μMのシクロヘキシミドの存在下にて行った。
【0042】
細胞を0.2 mlの浸透バッファー(110 mM KOAc、25 mM HEPES、pH 7.5、2.5 mM Mg(OAc)2、1 mM EGTA、0.015%ジギトニン、プロテアーゼ阻害剤カクテル、200μMのシクロヘキシミド、80 U/ml RNase阻害剤(Toyobo)、1 mMジチオスレイトール)とともに5分間氷上でインキュベートさせた。上清(細胞質画分)の回収後、細胞を洗浄バッファー(110 mM KOAc、25 mM HEPES、pH 7.5、2.5 mM Mg(OAc)2、1 mM EGTA、0.004%ジギトニン、プロテアーゼ阻害剤カクテル)を用いて2回、注意深く洗浄した。
【0043】
次いで、膜画分を0.2 mlの溶解バッファー(400 mM KOAc、25 mM HEPES、pH 7.5、15 mM Mg(OAc)2、1 mM EGTA、1%NP-40、0.5% DOC、プロテアーゼ阻害剤カクテル)を用いて30分間、氷上で抽出した。膜画分を含有する溶液を注意深く回収し、そして7500×gで10分間、4℃にて遠心分離により清澄化して、不溶性の残渣を除去した。得られた上清を、その後の解析に使用した。
HeLa細胞の界面活性剤抽出を、0.05%ジギトニンを含有する浸透バッファーを用いて、10分間、氷上で行い、その後上記と同一の手順を行った。
【0044】
膜画分を、400 mM KOAcおよび25 mM K-HEPES(pH 7.5)バッファー(15 mM Mg(OAc)2、1 mM EGTA、プロテアーゼ阻害剤カクテル、200μMシクロヘキシミド、1 mMジチオスレイトール、および80 U/ml RNase阻害剤を含有するもの)中で、直線的な15〜50%スクロース勾配上に充填した。勾配は、150,000×gにて70分間、TLS-55ローター(Beckman)を用いて、4℃で遠心分離した。画分を手作業にて回収した。勾配中のrRNA、mRNA、およびタンパク質を解析した。
【0045】
全DNA含有量により標準化した等量のサンプルを、ポリソーム解析に供した。(A)各画分におけるリボソームRNAを、アガロースゲル電気泳動により解析した。40Sサブユニットよりも軽い画分(Light(L))、40S、60Sサブユニットの位置(Free(F))、80Sモノソームの位置(80S(M))、そしてポリソームの位置(Polysome(P))を、上部に示す。上段パネル:非-刺激対照細胞(対照siRNA処置細胞);中段パネル:アスコルビン酸-刺激した対照細胞(アスコルビン酸刺激+対照siRNA処置細胞);下段パネル:アスコルビン酸-刺激p180タンパク質特異的siRNA-処理細胞。結果を図2Aに示す。
【0046】
非-刺激細胞(図2A、上段、“cont-si”)におけるリボソーム形成と比較して、アスコルビン酸刺激は、膜画分におけるポリソーム形成を大幅に亢進した(図2A、中段、“AA+cont-si”)。実施例1の透過電子顕微鏡像(図1)と同様に、重い画分が、アスコルビン酸刺激後に、顕著に集められた。p180タンパク質の喪失は、膜-結合型ポリソームの劇的な減少を引き起こした(図2A、下図、“AA+p180-si”)。
【0047】
次いで、ポリソーム画分の全RNA含有量を、ゲルの密度スキャニングにより推定し、相対量を示す(平均±SD、n=3)。この結果を、図2Bに示す。アスコルビン酸-処理細胞(AA+cont-si)は、非-処理細胞(cont-si)と比較して、2.7倍高いレベルのポリソームRNAを含有したことが示された(図2B)。p180タンパク質の喪失(AA+p180-si)は、ポリソームのRNAレベルを、対照細胞の10%未満にまで減少させた(図2B)。これらのデータから、p180タンパク質がアスコルビン酸刺激に際して高いトラフィック負荷に応じて、小胞体膜上でのポリソーム形成を亢進するために必要とされることが示唆される。
【0048】
実施例3:アスコルビン酸-処理細胞および/またはp180-除去細胞由来の全膜画分中でのトランスロコン-関連タンパク質の発現レベル
高レベルのp180タンパク質が小胞体膜上で発現される場合、アスコルビン酸刺激は、生合成活性を、特に特に膜画分における生合成活性を、亢進させることが知られている(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。このことは、リボソーム-トランスロコン複合体の組織化が、p180-依存的に変化する可能性があることを示唆している。
【0049】
本実施例においては、ポリソームの組織化をより詳細に特徴づけることを目的として、p180タンパク質およびトランスロコンタンパク質の分布を、回収されたスクロース勾配の画分を使用して、特異的抗体を使用した免疫ブロッティング解析により解析した。
【0050】
回収された画分におけるp180タンパク質およびその他のトランスロコン-関連タンパク質の分布を、ウェスタンブロットにより解析した結果を、図2Cに示す。p180タンパク質のウェスタンブロット解析は、以前に記載したように、強力なブロッティング条件のもと行った(Ogawa-Goto, et al., (2002) J Virol, 76, 2350-2362)。ウェスタンブロット解析においては、以下の抗体を使用した:GFPに対するウサギ抗体(Clontech)、タンパク質ジスルフィドイソメラーゼおよびカルネキシンに対するウサギ抗体(Stressgen)、リボソームタンパク質L10(RPL10)に対するウサギ抗体(Santa Cruz Biotechnology)、リボソームタンパク質S6(RPS6 )に対するウサギ抗体(Cell Signaling)、ビオチンに対するウサギ抗体(Bethyl Laboratories)、Sec61βに対するウサギ抗体(Upstate)、Sec61αおよびTRAPαに対するウサギ抗体(C.V.Nicchitta, Duke University Medical Centerより供与を受けた)、ヒトp180タンパク質に対するウサギ抗体(N1)(Ogawa-Goto, K., et al., (2007) Mol Biol Cell, 18, 3741-3751);リボフォリンII(RiboII)に対するヤギ抗体(Santa Cruz Biotechnology);およびCRIMP-63に対するマウスモノクローナル抗体(Klopfenstein, D.R., et al., (1998) EMBO J, 17, 6168-6177)(H.P. Hauri, University of Baselより供与を受けた)、(His)5 tagに対するマウスモノクローナル抗体(Qiagen)。
【0051】
ウェスタンブロット解析用のサンプルを、特に指示しない限り、DNA量により正規化した。プロテアーゼ阻害剤として、EDTA-不含のコンプリートインヒビターミクスチャー(Complete Inhibitor Mixture)(Roche Applied Science)を使用した。ミクロコッカスS1ヌクレアーゼは、New England Biolabsから購入した。
【0052】
非-刺激対照細胞(図2C左)、アスコルビン酸-刺激対照細胞(図2C中)、およびアスコルビン酸-刺激p180タンパク質特異的siRNA-処理細胞(図2C右)が示される。
相対的タンパク質レベルを密度スキャニングによる定量し、そして対応するバンドのそばに示した。データは、3回の異なる実験の平均±SDを示し、疑似トランスフェクト細胞におけるレベルの%として示される。
【0053】
非-刺激細胞において、ポリソーム画分(各図の“P”、画分7〜10)は、少量のp180タンパク質、SEC61α、SEC61β、およびTRAPαを、リボソームマーカーRPL10およびRPS6とともに含有した(図2C、左)。アスコルビン酸処理に際して、6種のタンパク質全てのレベルが、特にポリソーム画分において上昇したが、リボホリンII、カルネキシンのレベルは、変化しなかった(図2C中、画分7〜10)。
【0054】
アスコルビン酸刺激細胞において、トランスロコンサブユニットSec61βの特異的増加が見出され(図2C中、“P”、画分7〜10)、これはリボソームマーカータンパク質(たとえば、RPL10およびRPS6)や以前に報告されたp180タンパク質と一致していた(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。全膜画分におけるSec61αおよびTRAPαの含量は、アスコルビン酸刺激後も変化しなかったが(図2C中)、それらのシフトは、ポリソーム画分中で、Sec61βおよびリボソームマーカーと付随して生じた。p180タンパク質除去に際して、リボソームと同時にSec61β、Sec61αおよびTRAPαのレベルの減少が見出され(図2C、右、“P”、画分7〜10)、これも全膜画分中での含有量の減少と付随していた(図2C、右)。画分9中で検出された微量の残留p180タンパク質は、少量のポリソームの存在(図2A、下段)と一致していた様である。
【0055】
対照的に、その他のトランスロコン-関連タンパク質であるリボホリンIIまたは別の微小管-結合小胞体タンパク質CLIMP-63(Klopfenstein, D.R., et al., (1998) EMBO J, 17, 6168-6177)は、アスコルビン酸処理および/またはp180タンパク質ノックダウンにより影響を受けなかった。このことから、アッセンブリが進んだポリソームが、軽いポリソームと同様の構成成分を有することが示唆される。
実施例4:p180タンパク質のmRNAとの強い相互作用活性と、ribosomeとの結合活性
p180タンパク質がポリソーム/トランスロコンとどのように複合体を形成しているのか、明らかにすることを目的として、まずp180タンパク質がポリソームの構成成分であるmRNA, トランスロコン複合体、リボソームとそれぞれ結合しうるか調べた。
【0056】
まず、p180タンパク質とmRNAとの結合について調べるため、アスコルビン酸刺激下でHEL細胞を一定期間培養した。細胞質画分を除いたHEL細胞に対して、EDTA処理を行い、膜に結合しているリボソームを剥離させ、洗い流した。その後、1%NP-40を含むバッファーを用いて膜画分を抽出し、超遠心により残存するリボソームを沈殿除去した。得られた膜画分ライセートに対し、0.5%デオキシコール酸、0.1%SDSを追加し、0.4 M KOAc存在下、4℃で1時間インキュベーションした。
【0057】
そして超遠心によりデブリを除いたライセートに対し、抗p180抗体で免疫沈降を行い、p180タンパク質結合ビーズを調製した。このp180タンパク質ビーズにはトランスロコン複合体の構成成分や、小胞体のマーカータンパク質、リボソームが混入していないことをウェスタンブロット法とrRNAの電気泳動で確認した(図3A)。
【0058】
このビーズをqPCR法で解析したところ、COL1A1、FN、TIMP-1、MMP-2のmRNAが検出され、高い塩濃度存在下でもp180タンパク質とmRNAが共沈する結果となった。このときCOL1A1、FNのインプットに対する結合量はTIMP-1、MMP-2よりも著しく高かった(図3B)。
【0059】
以上の結果から、p180タンパク質はmRNAと高塩濃度による影響を受けないほど、強く相互作用しており、さらに結合しうるmRNAには選択性がある可能性が示された。
次にp180タンパク質が、トランスロコンと結合しうるか調べるため、前述のリボソームを除去した膜画分ライセートを5〜20%のショ糖密度勾配遠心分離にかけ解析したところ、トランスロコンの構成タンパク質であるSec61α、Sec61β、TRAPαとp180タンパク質は22〜28フラクションに共に検出された(図3C)。さらにこのライセートを用いてp180タンパク質、sec61βに対する抗体で免疫沈降を行った。その結果、p180タンパク質とトランスロコン複合体の構成成分であるsec61β、TRAPαが共沈した(図3D)。しかし、膜画分ライセートをヌクレアーゼ処理し、mRNAを分解すると共沈されなかった。このことから、p180タンパク質とトランスロコンは直接的に相互作用しておらず、mRNAのようにヌクレアーゼ感受性の核酸を介して間接的に相互作用していることが考えられた。
【0060】
次にp180タンパク質とリボソームの結合を調べるため、前述のp180タンパク質結合ビーズと精製した80Sリボソームおよび、60S、40Sリボソームサブユニットをそれぞれ混合し、ビーズとの共沈を調べたところ、80S、60Sとp180タンパク質結合ビーズとの共沈は認められたが、40Sとは共沈しなかった(図3E、1〜4、9〜12、17〜20)。さらにヌクレアーゼ処理した試料を用いた場合も同様の結果となったことから(図3E、5〜8、13〜16、21〜24)、p180タンパク質結合ビーズと80S、60Sとの結合はmRNAの影響によるものではないと考えられた。以上のことから、p180タンパク質は60Sと結合する能力があることが示された。
【0061】
実施例5:p180タンパク質の過剰発現による膜-結合型ポリソームの形成の亢進
本実施例においては、膜-結合型ポリソームの形成におけるp180タンパク質の具体的な役割をさらに明らかにするため、p180タンパク質を過剰発現するHeLa形質転換細胞(HeLa/p180 cells)においてポリソーム組織化を調べた。
【0062】
小胞体-結合型ポリソームの量を、p180タンパク質を過剰発現する安定的HeLa形質転換細胞(HeLa/p180)および対照細胞株(HeLa/puro)を使用して解析した。膜画分を、HeLa/puro細胞、非処理HeLa/p180細胞、およびsiRNA-処理HeLa/p180細胞からの連続的ジギトニン抽出により得て、そして15〜50%スクロース勾配を使用したポリソーム解析に供した。
【0063】
膜画分のスクロース勾配解析により、p180タンパク質の過剰発現が、p180-依存的に膜-結合型ポリソームの劇的な増加が引き起こされることが示された(図4A)。
ポリソーム画分のRNA含有量を、ゲルの密度スキャニングにより推定し、そして相対量を示す(図4B)。HeLa/puro細胞と比較した相対的RNAレベルでは、約2.5倍増加した(図4B)。対照的に、細胞質画分におけるポリソームパターンは、p180タンパク質の操作により顕著には影響を受けなかった(データは示さず)。
【0064】
回収された画分におけるp180タンパク質およびその他のトランスロコン-関連タンパク質の分布を、ウェスタンブロットにより解析した。ウェスタンブロット解析に使用された条件(使用された抗体を含む実験条件)は、実施例2に記載した通りである。HeLa/puro細胞(図4C左)、非処理HeLa/p180細胞(図4C中)およびsiRNA-処理HeLa/p180細胞(図4C右)が示される。その結果、図3Cのデータと同様に、p180タンパク質のポリソームとの同時移動が観察された(図4C)。さらに、Sec61βの発現レベルの上昇がポリソーム画分において見られたが、リボホリンIIでは見られなかったことは、注目された。
【0065】
実施例6:小胞体-結合型ポリソームにおける生合成活性の亢進
十分なp180タンパク質を含む膜画分は、p180-依存的に高い生合成活性を示す(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。そこで、本実施例においては、活性化された生合成が、ポリソーム組織化の変化に伴っているかどうかを調べるため、メチオニンについての代わりとして新たに合成されタンパク質中に特異的に取り込まれるAHAを含む非-RIラベリングシステムを用いて、de novo生合成の活性を推定した(Dieterich, D.C., et al., (2006) Proc Natl Acad Sci U S A, 103, 9482-9487)。
【0066】
de novo生合成活性は、HEL細胞においてAHAを用いた非-RI標識システムを使用して推定した。細胞を、アスコルビン酸の存在下または非存在下にて培養し、そして対照siRNA またはp180-特異的siRNAにより処置した。AHAとともにインキュベーションしたのち、膜画分を、15〜50%スクロース勾配上で分画した。AHA-取り込みタンパク質を、クリック化学によりビオチン-アルキンを用いて標識し、そして抗-ビオチン抗体を用いてイムノブロットした。結果を図5に示す。この図において、それぞれのパネルは以下のものを示す。上段パネル:非-刺激対照細胞;中段パネル:アスコルビン酸-刺激対照細胞;下段パネル:アスコルビン酸-刺激siRNA-処理細胞。
【0067】
膜画分の密度勾配解析により、アスコルビン酸刺激ののち、ポリソーム画分中でタンパク質生合成のきわめて高レベルの活性化が示された(図5、上段および中段)。さらに、p180タンパク質の喪失により、対応する画分における生合成のほぼ完全な阻害が引き起こされた(図5、下段)。これらの知見は、翻訳活性がポリソームの占有レベルと相関しており、そしてこの上方制御が主として小胞体膜上のp180/トランスロコン複合体の増大により引き起こされるようであることを、直接的に示している。
【0068】
実施例7:アスコルビン酸刺激に際した、特異的タンパク質についてのmRNAのリボソーム占有による180-依存的な活性化
以前の研究において、本発明者らは、p180タンパク質のノックダウンが、プロコラーゲンおよびフィブロネクチンを含む大型の細胞外マトリクスタンパク質の分泌の選択的な阻害を生じることを見出した(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。そこで、これらの知見の分子的なバックグラウンドをさらに明らかにするため、本実施例においては、スクロース勾配中のプロコラーゲンmRNAおよびフィブロネクチンmRNAのパターンを、非-反応性タンパク質のmRNAパターンと比較した。
【0069】
HEL細胞を、アスコルビン酸の存在下または非存在下にて培養し、そして対照siRNAまたはp180-特異的siRNA(siRNA)により処理した。連続的ジギトニン抽出により得られた膜画分を、15〜50%スクロース勾配を使用したポリソーム解析に供した。cDNAを、個々のチューブから抽出したRNAサンプルから合成した。対照細胞(対照)、アスコルビン酸-処理細胞(AA)、およびアスコルビン酸およびp180-特異的siRNA-処理細胞(AA+siRNA)から得られたmRNAの分布をプロットした。結果を図6に示し、この図において1型プロコラーゲンα鎖(図6A)、フィブロネクチン(図6B)、MMP-2(図6C)およびTIMP-1(図6D)が示される。
【0070】
対照細胞においてはアスコルビン酸添加の前に、プロコラーゲンmRNAおよびフィブロネクチンmRNAは、チューブ26〜27でピークに達した(図6A、図6B、上段)。アスコルビン酸刺激は、きわめて重いポリソームへのこれらのmRNAの顕著なシフトを引き起こした(チューブ28および29、中段)。対照的に、p180タンパク質の喪失は、ポリソーム画分に存在するものの、チューブ19および20にピークを持つより軽い画分への顕著なシフトを引きおこした(下段)。一方、MMP-2およびTIMP-1のmRNAパターンは、p180タンパク質操作の有無にかかわらず、チューブ23および24付近で変化は見られなかった(図6Cおよび図6D)。このことは、それらの分泌レベルが一定であることと一致していた(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。
【0071】
実施例8:野生型p180タンパク質または変異型p180タンパク質の発現の、膜結合型RPL10のレベルへの影響
推定一次配列に基づいて、ヒトp180タンパク質は、リボソーム-結合リピートドメインを含む高度に塩基性のN-末端領域、MTB-1微小管-束形成ドメイン、および推定コイルドコイルドメインからなる酸性C-末端領域を有することが予想される(Wanker, E.E., et al., (1995) J Cell Biol, 130, 29-39;Ogawa-Goto, K., et al., (2007) Mol Biol Cell, 18, 3741-3751)。そこで、本実施例においては、膜-結合型ポリソーム形成についての責任領域を調べるため、ヒトp180タンパク質の異なる領域を含有するGFP-タグ付加ポリペプチドのシリーズを、HEL細胞において発現させ、そして膜におけるリボソーム/ポリソームに対するそれらの作用を調べた。
【0072】
野生型ヒトp180タンパク質および変異型ヒトp180タンパク質に対する発現プラスミドを、以前に記載したように使用した(Ogawa-Goto, K., et al., (2007) Mol Biol Cell, 18, 3741-3751)。トランスフェクトのために使用した、p180タンパク質の様々な領域の完全長構造および短縮型構造の概略図を、図7Aおよび図7Fに示す。ヒトp180タンパク質は、N-末端に近い予想膜貫通ドメイン(TM)、高度に塩基性のタンデムリピートドメイン(濃灰色ボックス)、微小管結合および束形成ドメイン(MTB-1)およびC-末端酸性コイルドコイルドメイン(灰色ボックス)を有する。それぞれの短縮型変異体の左側の番号は、ヒトp180タンパク質のアミノ酸残基を意味する(DDBJ accession number: AB287347)。HEL細胞には、Amaxa Nucleofectorトランスフェクションシステム(Lonza)を、製造者のプロトコルにしたがって使用して、5μgのプラスミドを用いてトランスフェクトした。トランスフェクションの2日後、細胞を、細胞質画分および膜画分の連続的界面活性剤抽出に供した。
【0073】
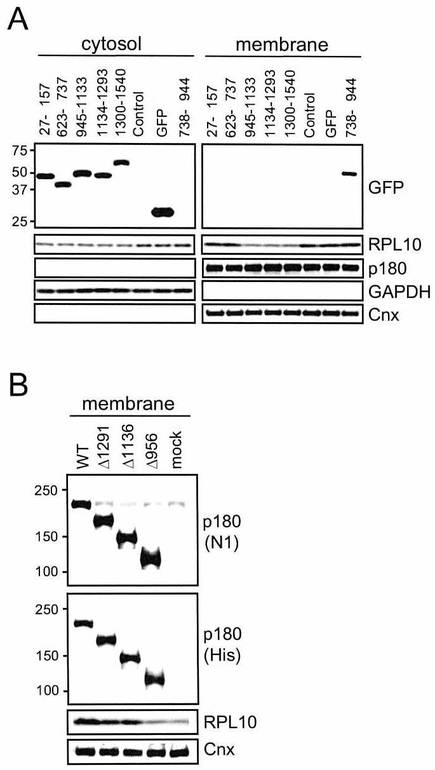
この様にして、6種類のヒトp180タンパク質の異なる領域を含有するGFP-タグ付加ポリペプチドのシリーズ(図7A)を、HEL細胞中で発現させ、そして膜および細胞質画分を連続的ジギトニン抽出により得た。抗-GFP抗体を用いたキメラタンパク質のウェスタンブロット解析により、ほとんどのタンパク質が、細胞質画分において正しいサイズで発現されたことが確認された(図8A)。
【0074】
ほとんどの短縮型ペプチドは、細胞質画分において同程度検出され、例外として、アミノ酸738〜944(MTB-1のC-末端側)を含む配列は、膜画分においてのみ限定的に検出される。これはおそらくは微小管束形成を介してのものであった(Ogawa-Goto, K., et al., (2007) Mol Biol Cell, 18, 3741-3751)。ここでは、膜画分中のリボソームRNA(それぞれ28Sおよび18Sを示される)が示される(図7B)。膜-結合型画分中のRNA含有量は、密度スキャニングにより推定され、そして相対量が示される(図7C)。データは、平均±SD(n=3)を示す。
【0075】
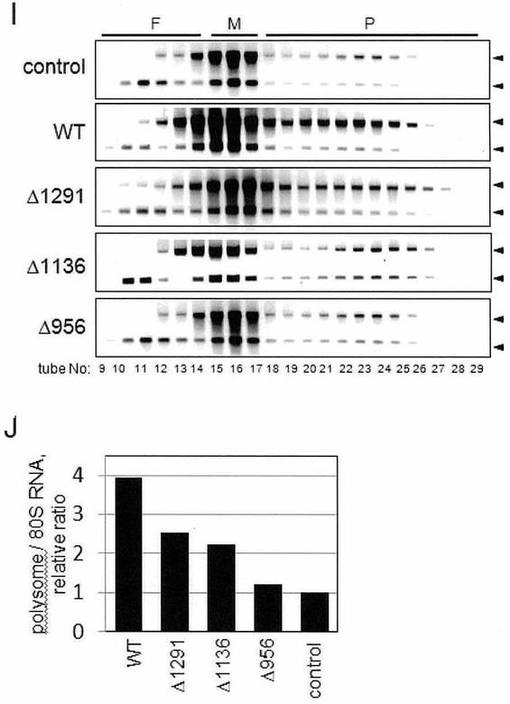
ポリソーム解析を、対照細胞の膜画分と比較して、アミノ酸残基945〜1133、1134〜1293、および1300〜1540を含有するペプチドを過剰発現する細胞の膜画分を使用して行った。すべての細胞において、p180タンパク質レベルは変化がなく、一方、解析された6種のキメラタンパク質のうち、コイルドコイルドメインにおける3種のC-末端ポリペプチド(アミノ酸945〜1133、アミノ酸1134〜1293およびアミノ酸1300〜1540)が、膜-結合型リボソームにおける顕著な減少を予期せず引き起こした(図7B)。それらの阻害作用は、75%よりも高く(図7C)、そして膜上でのポリソームアッセンブリを阻害することにより引き起こされ(図7D、図7E)、このことから細胞質中で発現されるこれらのコイルドコイルポリペプチドが、ドミナントネガティブ作用を示すことを示唆している。
【0076】
膜(カルネキシン、cnx)および細胞質(GAPDH)マーカーは、予想された抽出を確認することが示される。
リピートドメインおよびMTB-1を含むその他のキメラタンパク質は、顕著な作用は何も有さなかった(図7B、図7C)。同様の結果が、HeLa/p180形質転換細胞の解析においても見出された(データは示さず)。
【0077】
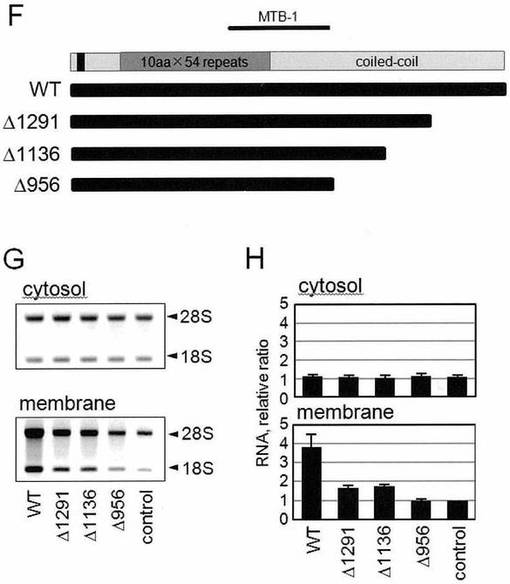
次に、コイルドコイルポリペプチドの機能を、非常に低レベルの内在性p180タンパク質しか有さない親HeLa細胞においてさらに調べた。安定な形質転換細胞と同様に(図4)、野生型p180タンパク質の一過性の過剰発現により、膜上でのリボソームおよびポリソームアッセンブリを増加させたが、一方C-末端ドメインを欠失する短縮型変異体(△956、△1136および△1291)はアッセンブリを亢進させることができなかった(図7F〜図7J、図8B)。本実施例において使用した野生型p180タンパク質およびp180タンパク質の様々なC-末端欠失変異体の概略図を、図7Fに示す。そして、野生型p180タンパク質およびp180タンパク質変異体をコードする発現プラスミドを、HeLa細胞中にトランスフェクトし、膜画分中のリボソームRNAを解析した結果を図7Gに、そして膜-結合型画分中の相対的RNA含有量を図7Hに、それぞれ示す。データは、平均±SD(n=3)を示す。したがって、外来性で発現させたp180タンパク質が、C-末端ドメイン依存的に、小胞体-結合型ポリソームのアッセンブリに対して要求に叶うものである。
【0078】
図7Fに示されるヒトp180タンパク質の短縮型変異体のシリーズが、HeLa細胞において一過性に発現された。抗-p180抗体または抗-(His)6抗体を用いた免疫ブロッティングにより、正しいサイズのタンパク質が膜画分中で発現されていることが確認された。そして、疑似トランスフェクト細胞と比較して、野生型p180タンパク質の一過性過剰発現は、膜画分においてRPL10のレベルの上昇を引き起こし(約3.8倍)、一方そのレベルは、その他の変異体についてはより低いものである(2倍未満)(図8B))。
【産業上の利用可能性】
【0079】
本発明により、発現系を構成する細胞内において、小胞体膜上のポリソーム形成を促進させることができることから、結果として、細胞中での目的とするタンパク質の生成を促進することができる。
【配列表フリーテキスト】
【0080】
SEQ ID NO: 1:ヒトp180タンパク質のアミノ酸配列
SEQ ID NO: 2:ヒトp180タンパク質をコードする核酸のヌクレオチド配列
SEQ ID NO: 3:ヒトコラーゲンタンパク質のアミノ酸配列
SEQ ID NO: 4:ヒトコラーゲンタンパク質をコードする核酸のヌクレオチド配列
【技術分野】
【0001】
本発明は、細胞内におけるポリソーム形成を促進すること、また細胞内におけるポリソーム形成を促進することを通じて細胞内における目的のタンパク質の生成を促進することに関するものである。
【背景技術】
【0002】
mRNAの翻訳は、潜在的には複数のレベルで制御される一方、イニシエーションでの制御は、重要なステップであるとして、もっともよく研究されてきた(Sonenberg, N. and Hinnebusch, A.G. (2009) Cell, 136, 731-745;Gebauer, F. and Hentze, M.W. (2004) Nat Rev Mol Cell Biol, 5, 827-835)。mRNAの5’非翻訳領域とmRNAの3’非翻訳領域は、mRNAの翻訳効率、安定性および局在化を含む翻訳制御の様々な段階で重要な役割を果たしている(Abaza, I. and Gebauer, F. (2008) RNA, 14, 404-409)。ポリソームアッセンブリの程度は、細胞における翻訳効率に対する直接的なインパクトを介した翻訳調節の別の重要な側面であると想定できる。低温電子断層撮影における最近の進展が、細胞融解物および無傷細胞における翻訳ポリソームの組織化に関する重要な洞察をもたらした一方(Brandt, F., et al., (2010) Mol Cell, 39, 560-569;Brandt, F., et al., (2009) Cell, 136, 261-271)、mRNAのリボソーム占有が、mRNAの長さにのみ依存しているか、または何等か未知のメカニズムによって制御されているかについては、依然としてあいまいなままである(Arava, Y., et al., (2003) Proc Natl Acad Sci U S A, 100, 3889-3894;Kopeina, G.S., et al., (2008) Nucleic Acids Res, 36, 2476-2488;Nelson, E.M. and Winkler, M.M. (1987) J Biol Chem, 262, 11501-11506)。
【0003】
特に、膜タンパク質および分泌タンパク質に関して、その後の膜を通過した転位のために、状況はずっと複雑である。リボソーム装置およびトランスロコン装置が、構造的にそして機能的にどのように共役しているのかについては、限られた情報しか得られていない(Pool, M.R. (2009) J Cell Biol, 185, 889-902)。
【0004】
膜-結合型リボソーム遊離の細胞質リボソームから構造的に識別されるのかどうかは依然としてわかっていないが、遊離のリボソームと比較して、小胞体-結合型のリボソームは、より効率的な生合成を媒介することは知られている(Stephens, S.B. and Nicchitta, C.V. (2008) Mol Biol Cell, 19, 623-632)。さらに、単一のポリソーム上で、協調的な翻訳とその後の膜を通過する転位を達成するためには、リボソーム/トランスロコン複合体の各ユニット間で、より高次の協調が必要不可欠である。しかしながら、そのような協調についての分子的基礎については、基本的に全く分かっていない。
【0005】
コラーゲンは、皮膚、腱および骨などの結合組織における細胞外マトリクスの主要な構成成分の一つである。それらは、前駆体型(すなわち、プロコラーゲン)として小胞体膜上で合成され、そして繊維芽細胞、軟骨芽細胞および骨芽細胞を含む専門的分泌細胞により分泌される。コラーゲンの分泌のための専門化細胞は、膵臓細胞およびプラズマ細胞などのその他の分泌細胞と同様に、高速合成を提供するために粗面小胞体の高度に発達したネットワークを有する。しかし、専門的分泌細胞におけるタンパク質生合成の非常に効率的な活性を裏付けるメカニズムについては、ほとんどわかっていない。
【0006】
アスコルビン酸は、in vitro培養のあいだのプロコラーゲン分泌について、一般的にそして長く使用されてきた刺激物質である。コラーゲン-分泌性繊維芽細胞において、アスコルビン酸処理は、小胞体からゴルジ体への輸送の回復とそれに続くトラフィック需要の増大に応答して、de novo生合成を活性化するものと考えられている。しかしながら、de novo生合成がどのようにして専門的分泌細胞中で活性化されるのかについては、いまだわかっていない。
【0007】
p180タンパク質は、必須の小胞体膜タンパク質であり、そして分泌組織中で多く発現されている(Langley, R., et al., (1998) DNA Cell Biol, 17, 449-460)。p180タンパク質は、初めは、粗面小胞体膜上のリボソーム受容体候補の一つとして同定された(Savitz, A.J. and Meyer, D.I. (1990) Nature, 346, 540-544)。その独特なリピートドメインが、リボソームに対して結合能力を有し、そして非常に塩基性のpIを有するドデカペプチドの54回のタンデムリピートが含まれることが報告された(Wanker, E.E., et al., (1995) J Cell Biol, 130, 29-39)。哺乳動物において、p180タンパク質のアミノ酸配列、ヌクレオチド配列は、種を超えて高度に保存されており、ヒトとの関係で最も相同性が低いブタであっても、80%の相同性を有していることが知られている。しかしながら、p180タンパク質の機能に付いては十分に解析が進んでおらず、特にp180タンパク質が動物細胞中のリボソームに対して直接結合するかどうかは、いまだよくわかっていない。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Sonenberg, N. and Hinnebusch, A.G. (2009) Cell, 136, 731-745
【非特許文献2】Gebauer, F. and Hentze, M.W. (2004) Nat Rev Mol Cell Biol, 5, 827-835
【非特許文献3】Abaza, I. and Gebauer, F. (2008) RNA, 14, 404-409
【非特許文献4】Brandt, F., et al., (2010) Mol Cell, 39, 560-569
【非特許文献5】Brandt, F., et al., (2009) Cell, 136, 261-271
【非特許文献6】Arava, Y., et al., (2003) Proc Natl Acad Sci U S A, 100, 3889-3894
【非特許文献7】Kopeina, G.S., et al., (2008) Nucleic Acids Res, 36, 2476-2488
【非特許文献8】Nelson, E.M. and Winkler, M.M. (1987) J Biol Chem, 262, 11501-11506
【非特許文献9】Pool, M.R. (2009) J Cell Biol, 185, 889-902
【非特許文献10】Stephens, S.B. and Nicchitta, C.V. (2008) Mol Biol Cell, 19, 623-632
【非特許文献11】Langley, R., et al., (1998) DNA Cell Biol, 17, 449-460
【非特許文献12】Savitz, A.J. and Meyer, D.I. (1990) Nature, 346, 540-544
【非特許文献13】Wanker, E.E., et al., (1995) J Cell Biol, 130, 29-39
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、 小胞体膜上でのポリソーム形成を促進すること、そしてその結果として目的とするタンパク質の生成を促進すること、並びに目的とするタンパク質の産生能が促進された細胞を提供することを課題とする。
【課題を解決するための手段】
【0010】
本発明は、細胞において、p180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供することにより、上記課題を解決することができることを示した。本発明はまた、目的とするタンパク質をコードする核酸分子を形質転換した細胞または目的とするタンパク質の産生量が増加された細胞において、p180タンパク質の全長またはその部分を発現させ、細胞内の小胞体膜上でのポリソーム形成を促進することを含む、前記細胞中での目的とするタンパク質の生成を促進する方法を提供することにより、上記課題を解決することができることを示した。
【発明の効果】
【0011】
本発明により、発現系を構成する細胞内において、小胞体膜上のポリソーム形成を促進させることができることから、結果として、細胞中での目的とするタンパク質の生成を促進することができる。
【図面の簡単な説明】
【0012】
【図1】図1は、コラーゲン分泌性HEL線維芽細胞における粗面小胞体表面の透過電子顕微鏡観察の様子を示す。
【図2】図2は、p180タンパク質が膜結合型ポリソームのアセンブリーを促進することを示す図である。
【図3】図3は、p180タンパク質がmRNAとの強い相互作用活性と、リボソームとの結合活性をそれぞれもつことを示した図である。
【図4】図4は、p180タンパク質の過剰発現は、膜結合型ポリソームの形成を亢進することを示した図である。
【図5】図5は、アスコルビン酸刺激下における小胞体膜結合型ポリソームの生合成活性は、p180タンパク質の発現抑制に伴って低下することを示した図である。
【図6】図6は、アスコルビン酸刺激下で起こる、p180タンパク質依存的な膜結合型ポリソームの形成促進はCOL,FNなどに対して優先的に作用することを示した図である。
【図7−1】図7は、膜結合型ポリソームの形成に重要なp180タンパク質の責任領域はcoiled-coilドメインであることを示した図である。
【図7−2】図7は、膜結合型ポリソームの形成に重要なp180タンパク質の責任領域はcoiled-coilドメインであることを示した図である。
【図7−3】図7は、膜結合型ポリソームの形成に重要なp180タンパク質の責任領域はcoiled-coilドメインであることを示した図である。
【図7−4】図7は、膜結合型ポリソームの形成に重要なp180タンパク質の責任領域はcoiled-coilドメインであることを示した図である。
【図8】図8は、図7Aで示された様々な領域の完全長構造および短縮型構造の発現タンパク質が、正しいサイズで発現されたことを示す図である。
【発明を実施するための形態】
【0013】
本発明においては、細胞においてp180タンパク質の全長またはその部分を発現させることにより、細胞内の小胞体膜上でのポリソーム形成を促進することができることを見出し、本発明の第一の態様を完成するに至った。
【0014】
すなわち、本発明においては、細胞において、哺乳動物のp180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供する。ここで、哺乳動物のp180タンパク質のアミノ酸配列という場合、ヒトのp180タンパク質(GenBank Accession No. AB287347)のほか、マウスのp180タンパク質(GenBank Accession No.NP_077243)、ラット(GenBank Accession No. XP_230637)、イヌ(GenBank Accession No. NP_001003179)、ウマ(GenBank Accession No.XP_001915027)、サル(GenBank Accession No. XP_002798281)、チンパンジー(GenBank Accession No.XP_514527)、ブタ(GenBank Accession No.XP_001926148)などが含まれる。
【0015】
例えば、哺乳動物のp180タンパク質としてヒトのp180タンパク質を使用する場合、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域、623〜737番アミノ酸からなる領域、738〜944番アミノ酸からなる領域、のいずれかを含む部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供する。
【0016】
この態様において、p180タンパク質の全長または部分を細胞中で発現させることにより、その細胞内の小胞体膜上でのポリソーム形成を促進することができる。ここで、ポリソームは、細胞内の小胞体膜上に存在する複数のリボソームに対して、1分子のmRNAが結合したもののことをいう。
【0017】
細胞において発現させるタンパク質は、p180タンパク質の全長またはその部分である。ここで、ヒトp180タンパク質の全長は、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質であり(GenBank Accession No. AB287347)、このタンパク質は、SEQ ID NO: 2に示されるヌクレオチド配列によりコードされる(GenBank Accession No.AB287347)。このほか、上述したマウスのp180タンパク質は、GenBank Accession No. NP_077243により示されるヌクレオチド配列によりコードされ、ラットのp180タンパク質は、GenBank Accession No. XP_230637により示されるヌクレオチド配列によりコードされ、、イヌのp180タンパク質は、GenBank Accession No. NP_001003179により示されるヌクレオチド配列によりコードされ、ウマのp180タンパク質は、GenBank Accession No. XP_001915027により示されるヌクレオチド配列によりコードされ、サルのp180タンパク質は、GenBank Accession No. XP_002798281により示されるヌクレオチド配列によりコードされ、チンパンジーのp180タンパク質は、GenBank Accession No. XP_514527により示されるヌクレオチド配列によりコードされ、ブタのp180タンパク質は、GenBank Accession No. XP_001926148により示されるヌクレオチド配列によりコードされる。
【0018】
本発明において、例えば、p180タンパク質の部分という場合、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分などの部分のことを意味する。このような部分を含むものは、ポリソーム形成を促進する能力を示すことができる。このようにして規定されるヒトp180タンパク質の部分としては、上述したSEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域、623〜737番アミノ酸からなる領域、738〜944番アミノ酸からなる領域、などを含む部分そのものの他、ヒトp180タンパク質のN末端膜貫通ドメインのC末端側に隣接するMTB-2ドメイン、またはリボソーム-結合リピートドメインを含む高度に塩基性のN-末端領域、高度に塩基性のタンデムリピートドメイン、または微小管結合および束形成ドメイン(MTB-1ドメイン)、などが一例として含まれる(図7A)。
【0019】
他の哺乳動物のp180タンパク質の部分を使用する場合も、ヒトのp180タンパク質のアミノ酸配列とその他の哺乳動物のp180タンパク質のアミノ酸配列が、全体的に、高度に保存されていることから、他の哺乳動物のp180タンパク質の対応する部分または領域を含む断片を使用することができる。
【0020】
本発明において使用する細胞は、タンパク質発現に適した哺乳動物の細胞であればどのようなものであってもよく、例えば、CHO細胞、HEK293細胞、HeLa細胞などの哺乳動物由来の細胞を使用することができる。
【0021】
前述した哺乳動物p180タンパク質の全長またはその部分を、これらの細胞内において発現させるためには、当該技術分野において一般的に使用されている形質転換方法を使用することができる。その形質転換を行うためには、例えば、pcDNA、pEGFP、pCAGGSなどの発現ベクターを使用することができる。
【0022】
上述したように、哺乳動物p180タンパク質の全長または部分を細胞中で発現させることにより、ポリソーム形成を促進することができることから、この性質を利用することにより、細胞中で産生させたい目的とするタンパク質の生成を促進することができる。
【0023】
したがって、本発明においてはまた、哺乳動物のp180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法を提供する方法を提供する。目的とするタンパク質をコードする核酸分子を形質転換した細胞または目的とするタンパク質の産生量が増加された細胞において、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、などの部分を発現させ、細胞内の小胞体膜上でのポリソーム形成を促進することを含む、前記細胞中での目的とするタンパク質の生成を促進する方法を提供する。
【0024】
この方法は、目的とするタンパク質を産生させるための細胞において、目的とするタンパク質の産生をさらに亢進させたい場合に使用することができる。例えば、目的とするタンパク質の産生のためにそのタンパク質をコードする遺伝子を細胞にトランスフェクトして形質転換細胞を作出したが、目的とするタンパク質の産生が十分でないか、またはその産生をより亢進させたい場合に、その細胞にさらに哺乳動物p180タンパク質の全長またはその部分を発現させることにより、目的とするタンパク質の産生を亢進させることができる。あるいは、細胞に対して特定の刺激を加えることにより目的とするタンパク質の産生量が増加されるが、目的とするタンパク質の産生が十分でないか、またはその産生をより亢進させたい場合に、その細胞にさらに哺乳動物p180タンパク質の全長またはその部分を発現させることにより、目的とするタンパク質の産生を亢進させることができる。
【0025】
前述したリポソーム形成を促進する方法の場合と同様に、この目的とするタンパク質の生成を促進する方法においてもまた、細胞において発現させるタンパク質は、哺乳動物p180タンパク質の全長またはその部分である。目的とするタンパク質の産生を亢進させたい細胞に対して、哺乳動物p180タンパク質の全長またはその部分を発現させるためには、前述したような当該技術分野において一般的に使用されている形質転換方法を使用することができる。
【0026】
細胞中で生成を促進したい目的とするタンパク質は、前述したように、細胞に対して目的とするタンパク質をコードする核酸分子を形質転換することにより、または細胞に対して特定の刺激を加えることにより目的とするタンパク質の産生量が増加されることにより、細胞内においてその生成を促進させることができるものである。目的とするタンパク質としては、例えば、コラーゲン、フィブロネクチン、ラミニンなどを挙げることができるが、これらには限定されない。
【0027】
目的とするタンパク質としてコラーゲンを使用する場合、例えば、コラーゲンをコードする遺伝子としてヒトI型コラーゲン、アルファ1(GenBank Accession No. NM_000088.3)を例えばpcDNAベクターを使用してHeLa細胞に形質転換してコラーゲン産生亢進細胞株を作成し、またはコラーゲンの産生が亢進されているHEL細胞に対してさらにコラーゲンの発現を促進するアスコルビン酸添加刺激処理を行うことによってコラーゲン高発現状態の細胞を調製し、その細胞株に対してヒトp180タンパク質の全長またはその部分を発現させることができる。その結果、細胞内の小胞体膜上におけるコラーゲン遺伝子のmRNAが取り込まれたポリソームの形成を促進し、結果的にコラーゲンの産生を亢進することができる。
【0028】
また、目的とするタンパク質としてフィブロネクチンを使用する場合、フィブロネクチンをコードする遺伝子としてヒトフィブロネクチン1(GenBank Accession No. NM_212482)をpcDNAベクターを使用してHeLa細胞に形質転換してフィブロネクチン産生亢進細胞株を作成し、またはフィブロネクチンの産生が亢進されているHEL細胞に対してヒトp180タンパク質の全長またはその部分を発現させる。その結果、細胞内の小胞体膜上におけるフィブロネクチン遺伝子のmRNAが取り込まれたポリソームの形成を促進し、結果的にフィブロネクチンの産生を亢進することができる。
【0029】
本発明の第一の態様において、細胞においてp180タンパク質の全長またはその部分を発現させることにより、細胞内の小胞体上でのポリソーム形成を促進することができることを見出したことに基づいて、第一の態様の方法を実現するための細胞を提供することができ、本発明の第二の態様を完成するに至った。
【0030】
すなわち、本発明においては、哺乳動物のp180タンパク質のアミノ酸の特定の部分を発現させた細胞、その具体的な好ましい態様としては、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、などの部分を発現させた細胞、を提供する。この様な構成上の特徴を有する細胞は、細胞内の小胞体膜上でのポリソーム形成能が促進されるという特徴を有するものであり、そのような特徴を有する結果、細胞内において転写されるmRNAからのタンパク質の生成を亢進することができる。
【0031】
本発明のこの態様における細胞は、タンパク質発現に適した哺乳動物の細胞であればどのようなものを起源として作出した細胞であってもよく、例えば、起源となる細胞としては、CHO細胞、HEK293細胞、HeLa細胞などの哺乳動物由来の細胞を使用することができる。これらの細胞に対して、前述した哺乳動物p180タンパク質の全長またはその部分を、当該技術分野において一般的に使用されている方法を使用してトランスフェクトして、これらの細胞内において哺乳動物p180タンパク質の全長またはその部分を発現させることができる。
【0032】
このような性質を有する細胞は、ポリソーム中に、細胞中で産生を亢進させたい目的とするタンパク質をコードする遺伝子から転写されたmRNAが含まれるものであってもよい。目的とするタンパク質としては、例えば、コラーゲン、フィブロネクチン、ラミニンなどを挙げることができるが、これらには限定されない。
【0033】
本明細書においては、ここまでに説明した本発明をさらに具体的に説明することを目的として、以下の実施例を記載するが、しかしながら、以下の実施例の記載は、本発明を限定することを目的としたものではない。
【実施例】
【0034】
実施例1:コラーゲン-分泌性HEL繊維芽細胞における粗面小胞体表面の透過電子顕微鏡観察
本発明者らは、アスコルビン酸処理により、mRNAレベルを変化させることなく、膜-結合型リボソームが増加することを報告した(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。これらの知見は、アスコルビン酸が、リボソーム-トランスロコン複合体の変化またはポリソーム組織化の亢進のいずれかにより、高いトラフィック負荷に応じて、新たな翻訳調節メカニズムを介して、生合成を上方制御していることを示唆していた。
【0035】
本実施例においては、p180タンパク質がポリソーム組織化に影響を与えるかどうかを決定するため、対照細胞およびp180-除去細胞の超微細構造を調べることを目的として、電子顕微鏡観察を行った。
【0036】
本実施例における電子顕微鏡解析のため、対照HEL細胞(A)およびp180 siRNA-形質転換HEL細胞(B)を、2%ウシ胎児血清(FBS)(Intergen)を含有するF12/DMEM(Nissui)中、アスコルビン酸の存在下にて増殖させた。特に示さない場合には、L-アスコルビン酸リン酸マグネシウム塩n-水和物(Wako)を、0.2 mMの最終濃度となるように添加した。細胞培養およびsiRNAトランスフェクションのプロトコルは、以前に記載したように行った(Ueno, T., et al., (2010) Exp Cell Res, 316, 329-340)。
【0037】
透過型電子顕微鏡解析用に、2%FBS/DMEM中で培養された細胞を、10μMタキソールを含有する培養液を用いて37℃にて3分間処理し、その後以前に記載したように(Ogawa-Goto, et al., (2002) J Virol, 76, 2350-2362)、透過型電子顕微鏡解析用に、従来から使用されている固定化手順を行った。すなわち、超薄切片を、基質と並行または垂直に切り出し、そしてModel 7650 EM(Hitachi)を用いて80 kVにて可視化した。
【0038】
電子顕微鏡像の調査の結果、アスコルビン酸-刺激HEL繊維芽細胞由来の水平方向に切断した切片の小胞体膜表面上に、多数の極めて長いポリソームが存在することが示された(図1A)。高倍率像では、約25〜30個のリボソームからなるリボソームのらせん状アレイまたは28個以上のリボソームから成るヘアピン型ポリソームが示され(図1A、挿入図)、ラット皮膚繊維芽細胞で報告されたものに非常に似ていた(Christensen, A.K. and Bourne, C.M. (1999) Anat Rec, 255, 116-129)。対照的に、p180タンパク質に対するsiRNAを用いたp180タンパク質の除去では、アスコルビン酸刺激にもかかわらず、小胞体膜上のポリソームでは劇的な減少が引き起こされた(図1B)。バー:図1Aおよび図1Bでは500 nm:図1A中の挿入図では200 nmを示す。
【0039】
実施例2:p180タンパク質による小胞体膜上のポリソームアッセンブリの促進
本実施例においては、小胞体膜上でのポリソーム形成におけるp180タンパク質の具体的な役割をさらに調べるため、高-塩濃度バッファーを用いたストリンジェントな条件下でのポリソーム勾配解析を行った。
【0040】
HEL細胞を、アスコルビン酸(AA)の存在下または非存在下にて培養し、そして対照siRNA またはp180-特異的siRNA(siRNA)により処置した。siRNAのトランスフェクションの4日後に、連続的ジギトニン抽出を行って、膜画分を得て、その後15〜50%のスクロース勾配を使用してポリソーム解析を行った。
【0041】
細胞質画分および膜画分の抽出は、ジギトニンを以前に記載した方法(Lerner, R.S., et al., (2003) RNA, 9, 1123-1137)から、わずかに修飾して使用して(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)、連続的界面活性剤抽出により行った。簡単に述べると、6ウェルプレート中でのHEL細胞の単層を、200μMのシクロヘキシミドとともに15分間事前培養し、そしてPBSで洗浄した。抽出は、手順を通じて200μMのシクロヘキシミドの存在下にて行った。
【0042】
細胞を0.2 mlの浸透バッファー(110 mM KOAc、25 mM HEPES、pH 7.5、2.5 mM Mg(OAc)2、1 mM EGTA、0.015%ジギトニン、プロテアーゼ阻害剤カクテル、200μMのシクロヘキシミド、80 U/ml RNase阻害剤(Toyobo)、1 mMジチオスレイトール)とともに5分間氷上でインキュベートさせた。上清(細胞質画分)の回収後、細胞を洗浄バッファー(110 mM KOAc、25 mM HEPES、pH 7.5、2.5 mM Mg(OAc)2、1 mM EGTA、0.004%ジギトニン、プロテアーゼ阻害剤カクテル)を用いて2回、注意深く洗浄した。
【0043】
次いで、膜画分を0.2 mlの溶解バッファー(400 mM KOAc、25 mM HEPES、pH 7.5、15 mM Mg(OAc)2、1 mM EGTA、1%NP-40、0.5% DOC、プロテアーゼ阻害剤カクテル)を用いて30分間、氷上で抽出した。膜画分を含有する溶液を注意深く回収し、そして7500×gで10分間、4℃にて遠心分離により清澄化して、不溶性の残渣を除去した。得られた上清を、その後の解析に使用した。
HeLa細胞の界面活性剤抽出を、0.05%ジギトニンを含有する浸透バッファーを用いて、10分間、氷上で行い、その後上記と同一の手順を行った。
【0044】
膜画分を、400 mM KOAcおよび25 mM K-HEPES(pH 7.5)バッファー(15 mM Mg(OAc)2、1 mM EGTA、プロテアーゼ阻害剤カクテル、200μMシクロヘキシミド、1 mMジチオスレイトール、および80 U/ml RNase阻害剤を含有するもの)中で、直線的な15〜50%スクロース勾配上に充填した。勾配は、150,000×gにて70分間、TLS-55ローター(Beckman)を用いて、4℃で遠心分離した。画分を手作業にて回収した。勾配中のrRNA、mRNA、およびタンパク質を解析した。
【0045】
全DNA含有量により標準化した等量のサンプルを、ポリソーム解析に供した。(A)各画分におけるリボソームRNAを、アガロースゲル電気泳動により解析した。40Sサブユニットよりも軽い画分(Light(L))、40S、60Sサブユニットの位置(Free(F))、80Sモノソームの位置(80S(M))、そしてポリソームの位置(Polysome(P))を、上部に示す。上段パネル:非-刺激対照細胞(対照siRNA処置細胞);中段パネル:アスコルビン酸-刺激した対照細胞(アスコルビン酸刺激+対照siRNA処置細胞);下段パネル:アスコルビン酸-刺激p180タンパク質特異的siRNA-処理細胞。結果を図2Aに示す。
【0046】
非-刺激細胞(図2A、上段、“cont-si”)におけるリボソーム形成と比較して、アスコルビン酸刺激は、膜画分におけるポリソーム形成を大幅に亢進した(図2A、中段、“AA+cont-si”)。実施例1の透過電子顕微鏡像(図1)と同様に、重い画分が、アスコルビン酸刺激後に、顕著に集められた。p180タンパク質の喪失は、膜-結合型ポリソームの劇的な減少を引き起こした(図2A、下図、“AA+p180-si”)。
【0047】
次いで、ポリソーム画分の全RNA含有量を、ゲルの密度スキャニングにより推定し、相対量を示す(平均±SD、n=3)。この結果を、図2Bに示す。アスコルビン酸-処理細胞(AA+cont-si)は、非-処理細胞(cont-si)と比較して、2.7倍高いレベルのポリソームRNAを含有したことが示された(図2B)。p180タンパク質の喪失(AA+p180-si)は、ポリソームのRNAレベルを、対照細胞の10%未満にまで減少させた(図2B)。これらのデータから、p180タンパク質がアスコルビン酸刺激に際して高いトラフィック負荷に応じて、小胞体膜上でのポリソーム形成を亢進するために必要とされることが示唆される。
【0048】
実施例3:アスコルビン酸-処理細胞および/またはp180-除去細胞由来の全膜画分中でのトランスロコン-関連タンパク質の発現レベル
高レベルのp180タンパク質が小胞体膜上で発現される場合、アスコルビン酸刺激は、生合成活性を、特に特に膜画分における生合成活性を、亢進させることが知られている(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。このことは、リボソーム-トランスロコン複合体の組織化が、p180-依存的に変化する可能性があることを示唆している。
【0049】
本実施例においては、ポリソームの組織化をより詳細に特徴づけることを目的として、p180タンパク質およびトランスロコンタンパク質の分布を、回収されたスクロース勾配の画分を使用して、特異的抗体を使用した免疫ブロッティング解析により解析した。
【0050】
回収された画分におけるp180タンパク質およびその他のトランスロコン-関連タンパク質の分布を、ウェスタンブロットにより解析した結果を、図2Cに示す。p180タンパク質のウェスタンブロット解析は、以前に記載したように、強力なブロッティング条件のもと行った(Ogawa-Goto, et al., (2002) J Virol, 76, 2350-2362)。ウェスタンブロット解析においては、以下の抗体を使用した:GFPに対するウサギ抗体(Clontech)、タンパク質ジスルフィドイソメラーゼおよびカルネキシンに対するウサギ抗体(Stressgen)、リボソームタンパク質L10(RPL10)に対するウサギ抗体(Santa Cruz Biotechnology)、リボソームタンパク質S6(RPS6 )に対するウサギ抗体(Cell Signaling)、ビオチンに対するウサギ抗体(Bethyl Laboratories)、Sec61βに対するウサギ抗体(Upstate)、Sec61αおよびTRAPαに対するウサギ抗体(C.V.Nicchitta, Duke University Medical Centerより供与を受けた)、ヒトp180タンパク質に対するウサギ抗体(N1)(Ogawa-Goto, K., et al., (2007) Mol Biol Cell, 18, 3741-3751);リボフォリンII(RiboII)に対するヤギ抗体(Santa Cruz Biotechnology);およびCRIMP-63に対するマウスモノクローナル抗体(Klopfenstein, D.R., et al., (1998) EMBO J, 17, 6168-6177)(H.P. Hauri, University of Baselより供与を受けた)、(His)5 tagに対するマウスモノクローナル抗体(Qiagen)。
【0051】
ウェスタンブロット解析用のサンプルを、特に指示しない限り、DNA量により正規化した。プロテアーゼ阻害剤として、EDTA-不含のコンプリートインヒビターミクスチャー(Complete Inhibitor Mixture)(Roche Applied Science)を使用した。ミクロコッカスS1ヌクレアーゼは、New England Biolabsから購入した。
【0052】
非-刺激対照細胞(図2C左)、アスコルビン酸-刺激対照細胞(図2C中)、およびアスコルビン酸-刺激p180タンパク質特異的siRNA-処理細胞(図2C右)が示される。
相対的タンパク質レベルを密度スキャニングによる定量し、そして対応するバンドのそばに示した。データは、3回の異なる実験の平均±SDを示し、疑似トランスフェクト細胞におけるレベルの%として示される。
【0053】
非-刺激細胞において、ポリソーム画分(各図の“P”、画分7〜10)は、少量のp180タンパク質、SEC61α、SEC61β、およびTRAPαを、リボソームマーカーRPL10およびRPS6とともに含有した(図2C、左)。アスコルビン酸処理に際して、6種のタンパク質全てのレベルが、特にポリソーム画分において上昇したが、リボホリンII、カルネキシンのレベルは、変化しなかった(図2C中、画分7〜10)。
【0054】
アスコルビン酸刺激細胞において、トランスロコンサブユニットSec61βの特異的増加が見出され(図2C中、“P”、画分7〜10)、これはリボソームマーカータンパク質(たとえば、RPL10およびRPS6)や以前に報告されたp180タンパク質と一致していた(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。全膜画分におけるSec61αおよびTRAPαの含量は、アスコルビン酸刺激後も変化しなかったが(図2C中)、それらのシフトは、ポリソーム画分中で、Sec61βおよびリボソームマーカーと付随して生じた。p180タンパク質除去に際して、リボソームと同時にSec61β、Sec61αおよびTRAPαのレベルの減少が見出され(図2C、右、“P”、画分7〜10)、これも全膜画分中での含有量の減少と付随していた(図2C、右)。画分9中で検出された微量の残留p180タンパク質は、少量のポリソームの存在(図2A、下段)と一致していた様である。
【0055】
対照的に、その他のトランスロコン-関連タンパク質であるリボホリンIIまたは別の微小管-結合小胞体タンパク質CLIMP-63(Klopfenstein, D.R., et al., (1998) EMBO J, 17, 6168-6177)は、アスコルビン酸処理および/またはp180タンパク質ノックダウンにより影響を受けなかった。このことから、アッセンブリが進んだポリソームが、軽いポリソームと同様の構成成分を有することが示唆される。
実施例4:p180タンパク質のmRNAとの強い相互作用活性と、ribosomeとの結合活性
p180タンパク質がポリソーム/トランスロコンとどのように複合体を形成しているのか、明らかにすることを目的として、まずp180タンパク質がポリソームの構成成分であるmRNA, トランスロコン複合体、リボソームとそれぞれ結合しうるか調べた。
【0056】
まず、p180タンパク質とmRNAとの結合について調べるため、アスコルビン酸刺激下でHEL細胞を一定期間培養した。細胞質画分を除いたHEL細胞に対して、EDTA処理を行い、膜に結合しているリボソームを剥離させ、洗い流した。その後、1%NP-40を含むバッファーを用いて膜画分を抽出し、超遠心により残存するリボソームを沈殿除去した。得られた膜画分ライセートに対し、0.5%デオキシコール酸、0.1%SDSを追加し、0.4 M KOAc存在下、4℃で1時間インキュベーションした。
【0057】
そして超遠心によりデブリを除いたライセートに対し、抗p180抗体で免疫沈降を行い、p180タンパク質結合ビーズを調製した。このp180タンパク質ビーズにはトランスロコン複合体の構成成分や、小胞体のマーカータンパク質、リボソームが混入していないことをウェスタンブロット法とrRNAの電気泳動で確認した(図3A)。
【0058】
このビーズをqPCR法で解析したところ、COL1A1、FN、TIMP-1、MMP-2のmRNAが検出され、高い塩濃度存在下でもp180タンパク質とmRNAが共沈する結果となった。このときCOL1A1、FNのインプットに対する結合量はTIMP-1、MMP-2よりも著しく高かった(図3B)。
【0059】
以上の結果から、p180タンパク質はmRNAと高塩濃度による影響を受けないほど、強く相互作用しており、さらに結合しうるmRNAには選択性がある可能性が示された。
次にp180タンパク質が、トランスロコンと結合しうるか調べるため、前述のリボソームを除去した膜画分ライセートを5〜20%のショ糖密度勾配遠心分離にかけ解析したところ、トランスロコンの構成タンパク質であるSec61α、Sec61β、TRAPαとp180タンパク質は22〜28フラクションに共に検出された(図3C)。さらにこのライセートを用いてp180タンパク質、sec61βに対する抗体で免疫沈降を行った。その結果、p180タンパク質とトランスロコン複合体の構成成分であるsec61β、TRAPαが共沈した(図3D)。しかし、膜画分ライセートをヌクレアーゼ処理し、mRNAを分解すると共沈されなかった。このことから、p180タンパク質とトランスロコンは直接的に相互作用しておらず、mRNAのようにヌクレアーゼ感受性の核酸を介して間接的に相互作用していることが考えられた。
【0060】
次にp180タンパク質とリボソームの結合を調べるため、前述のp180タンパク質結合ビーズと精製した80Sリボソームおよび、60S、40Sリボソームサブユニットをそれぞれ混合し、ビーズとの共沈を調べたところ、80S、60Sとp180タンパク質結合ビーズとの共沈は認められたが、40Sとは共沈しなかった(図3E、1〜4、9〜12、17〜20)。さらにヌクレアーゼ処理した試料を用いた場合も同様の結果となったことから(図3E、5〜8、13〜16、21〜24)、p180タンパク質結合ビーズと80S、60Sとの結合はmRNAの影響によるものではないと考えられた。以上のことから、p180タンパク質は60Sと結合する能力があることが示された。
【0061】
実施例5:p180タンパク質の過剰発現による膜-結合型ポリソームの形成の亢進
本実施例においては、膜-結合型ポリソームの形成におけるp180タンパク質の具体的な役割をさらに明らかにするため、p180タンパク質を過剰発現するHeLa形質転換細胞(HeLa/p180 cells)においてポリソーム組織化を調べた。
【0062】
小胞体-結合型ポリソームの量を、p180タンパク質を過剰発現する安定的HeLa形質転換細胞(HeLa/p180)および対照細胞株(HeLa/puro)を使用して解析した。膜画分を、HeLa/puro細胞、非処理HeLa/p180細胞、およびsiRNA-処理HeLa/p180細胞からの連続的ジギトニン抽出により得て、そして15〜50%スクロース勾配を使用したポリソーム解析に供した。
【0063】
膜画分のスクロース勾配解析により、p180タンパク質の過剰発現が、p180-依存的に膜-結合型ポリソームの劇的な増加が引き起こされることが示された(図4A)。
ポリソーム画分のRNA含有量を、ゲルの密度スキャニングにより推定し、そして相対量を示す(図4B)。HeLa/puro細胞と比較した相対的RNAレベルでは、約2.5倍増加した(図4B)。対照的に、細胞質画分におけるポリソームパターンは、p180タンパク質の操作により顕著には影響を受けなかった(データは示さず)。
【0064】
回収された画分におけるp180タンパク質およびその他のトランスロコン-関連タンパク質の分布を、ウェスタンブロットにより解析した。ウェスタンブロット解析に使用された条件(使用された抗体を含む実験条件)は、実施例2に記載した通りである。HeLa/puro細胞(図4C左)、非処理HeLa/p180細胞(図4C中)およびsiRNA-処理HeLa/p180細胞(図4C右)が示される。その結果、図3Cのデータと同様に、p180タンパク質のポリソームとの同時移動が観察された(図4C)。さらに、Sec61βの発現レベルの上昇がポリソーム画分において見られたが、リボホリンIIでは見られなかったことは、注目された。
【0065】
実施例6:小胞体-結合型ポリソームにおける生合成活性の亢進
十分なp180タンパク質を含む膜画分は、p180-依存的に高い生合成活性を示す(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。そこで、本実施例においては、活性化された生合成が、ポリソーム組織化の変化に伴っているかどうかを調べるため、メチオニンについての代わりとして新たに合成されタンパク質中に特異的に取り込まれるAHAを含む非-RIラベリングシステムを用いて、de novo生合成の活性を推定した(Dieterich, D.C., et al., (2006) Proc Natl Acad Sci U S A, 103, 9482-9487)。
【0066】
de novo生合成活性は、HEL細胞においてAHAを用いた非-RI標識システムを使用して推定した。細胞を、アスコルビン酸の存在下または非存在下にて培養し、そして対照siRNA またはp180-特異的siRNAにより処置した。AHAとともにインキュベーションしたのち、膜画分を、15〜50%スクロース勾配上で分画した。AHA-取り込みタンパク質を、クリック化学によりビオチン-アルキンを用いて標識し、そして抗-ビオチン抗体を用いてイムノブロットした。結果を図5に示す。この図において、それぞれのパネルは以下のものを示す。上段パネル:非-刺激対照細胞;中段パネル:アスコルビン酸-刺激対照細胞;下段パネル:アスコルビン酸-刺激siRNA-処理細胞。
【0067】
膜画分の密度勾配解析により、アスコルビン酸刺激ののち、ポリソーム画分中でタンパク質生合成のきわめて高レベルの活性化が示された(図5、上段および中段)。さらに、p180タンパク質の喪失により、対応する画分における生合成のほぼ完全な阻害が引き起こされた(図5、下段)。これらの知見は、翻訳活性がポリソームの占有レベルと相関しており、そしてこの上方制御が主として小胞体膜上のp180/トランスロコン複合体の増大により引き起こされるようであることを、直接的に示している。
【0068】
実施例7:アスコルビン酸刺激に際した、特異的タンパク質についてのmRNAのリボソーム占有による180-依存的な活性化
以前の研究において、本発明者らは、p180タンパク質のノックダウンが、プロコラーゲンおよびフィブロネクチンを含む大型の細胞外マトリクスタンパク質の分泌の選択的な阻害を生じることを見出した(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。そこで、これらの知見の分子的なバックグラウンドをさらに明らかにするため、本実施例においては、スクロース勾配中のプロコラーゲンmRNAおよびフィブロネクチンmRNAのパターンを、非-反応性タンパク質のmRNAパターンと比較した。
【0069】
HEL細胞を、アスコルビン酸の存在下または非存在下にて培養し、そして対照siRNAまたはp180-特異的siRNA(siRNA)により処理した。連続的ジギトニン抽出により得られた膜画分を、15〜50%スクロース勾配を使用したポリソーム解析に供した。cDNAを、個々のチューブから抽出したRNAサンプルから合成した。対照細胞(対照)、アスコルビン酸-処理細胞(AA)、およびアスコルビン酸およびp180-特異的siRNA-処理細胞(AA+siRNA)から得られたmRNAの分布をプロットした。結果を図6に示し、この図において1型プロコラーゲンα鎖(図6A)、フィブロネクチン(図6B)、MMP-2(図6C)およびTIMP-1(図6D)が示される。
【0070】
対照細胞においてはアスコルビン酸添加の前に、プロコラーゲンmRNAおよびフィブロネクチンmRNAは、チューブ26〜27でピークに達した(図6A、図6B、上段)。アスコルビン酸刺激は、きわめて重いポリソームへのこれらのmRNAの顕著なシフトを引き起こした(チューブ28および29、中段)。対照的に、p180タンパク質の喪失は、ポリソーム画分に存在するものの、チューブ19および20にピークを持つより軽い画分への顕著なシフトを引きおこした(下段)。一方、MMP-2およびTIMP-1のmRNAパターンは、p180タンパク質操作の有無にかかわらず、チューブ23および24付近で変化は見られなかった(図6Cおよび図6D)。このことは、それらの分泌レベルが一定であることと一致していた(Ueno, T., et al., (2010) J Biol Chem, 285, 29941-29950)。
【0071】
実施例8:野生型p180タンパク質または変異型p180タンパク質の発現の、膜結合型RPL10のレベルへの影響
推定一次配列に基づいて、ヒトp180タンパク質は、リボソーム-結合リピートドメインを含む高度に塩基性のN-末端領域、MTB-1微小管-束形成ドメイン、および推定コイルドコイルドメインからなる酸性C-末端領域を有することが予想される(Wanker, E.E., et al., (1995) J Cell Biol, 130, 29-39;Ogawa-Goto, K., et al., (2007) Mol Biol Cell, 18, 3741-3751)。そこで、本実施例においては、膜-結合型ポリソーム形成についての責任領域を調べるため、ヒトp180タンパク質の異なる領域を含有するGFP-タグ付加ポリペプチドのシリーズを、HEL細胞において発現させ、そして膜におけるリボソーム/ポリソームに対するそれらの作用を調べた。
【0072】
野生型ヒトp180タンパク質および変異型ヒトp180タンパク質に対する発現プラスミドを、以前に記載したように使用した(Ogawa-Goto, K., et al., (2007) Mol Biol Cell, 18, 3741-3751)。トランスフェクトのために使用した、p180タンパク質の様々な領域の完全長構造および短縮型構造の概略図を、図7Aおよび図7Fに示す。ヒトp180タンパク質は、N-末端に近い予想膜貫通ドメイン(TM)、高度に塩基性のタンデムリピートドメイン(濃灰色ボックス)、微小管結合および束形成ドメイン(MTB-1)およびC-末端酸性コイルドコイルドメイン(灰色ボックス)を有する。それぞれの短縮型変異体の左側の番号は、ヒトp180タンパク質のアミノ酸残基を意味する(DDBJ accession number: AB287347)。HEL細胞には、Amaxa Nucleofectorトランスフェクションシステム(Lonza)を、製造者のプロトコルにしたがって使用して、5μgのプラスミドを用いてトランスフェクトした。トランスフェクションの2日後、細胞を、細胞質画分および膜画分の連続的界面活性剤抽出に供した。
【0073】
この様にして、6種類のヒトp180タンパク質の異なる領域を含有するGFP-タグ付加ポリペプチドのシリーズ(図7A)を、HEL細胞中で発現させ、そして膜および細胞質画分を連続的ジギトニン抽出により得た。抗-GFP抗体を用いたキメラタンパク質のウェスタンブロット解析により、ほとんどのタンパク質が、細胞質画分において正しいサイズで発現されたことが確認された(図8A)。
【0074】
ほとんどの短縮型ペプチドは、細胞質画分において同程度検出され、例外として、アミノ酸738〜944(MTB-1のC-末端側)を含む配列は、膜画分においてのみ限定的に検出される。これはおそらくは微小管束形成を介してのものであった(Ogawa-Goto, K., et al., (2007) Mol Biol Cell, 18, 3741-3751)。ここでは、膜画分中のリボソームRNA(それぞれ28Sおよび18Sを示される)が示される(図7B)。膜-結合型画分中のRNA含有量は、密度スキャニングにより推定され、そして相対量が示される(図7C)。データは、平均±SD(n=3)を示す。
【0075】
ポリソーム解析を、対照細胞の膜画分と比較して、アミノ酸残基945〜1133、1134〜1293、および1300〜1540を含有するペプチドを過剰発現する細胞の膜画分を使用して行った。すべての細胞において、p180タンパク質レベルは変化がなく、一方、解析された6種のキメラタンパク質のうち、コイルドコイルドメインにおける3種のC-末端ポリペプチド(アミノ酸945〜1133、アミノ酸1134〜1293およびアミノ酸1300〜1540)が、膜-結合型リボソームにおける顕著な減少を予期せず引き起こした(図7B)。それらの阻害作用は、75%よりも高く(図7C)、そして膜上でのポリソームアッセンブリを阻害することにより引き起こされ(図7D、図7E)、このことから細胞質中で発現されるこれらのコイルドコイルポリペプチドが、ドミナントネガティブ作用を示すことを示唆している。
【0076】
膜(カルネキシン、cnx)および細胞質(GAPDH)マーカーは、予想された抽出を確認することが示される。
リピートドメインおよびMTB-1を含むその他のキメラタンパク質は、顕著な作用は何も有さなかった(図7B、図7C)。同様の結果が、HeLa/p180形質転換細胞の解析においても見出された(データは示さず)。
【0077】
次に、コイルドコイルポリペプチドの機能を、非常に低レベルの内在性p180タンパク質しか有さない親HeLa細胞においてさらに調べた。安定な形質転換細胞と同様に(図4)、野生型p180タンパク質の一過性の過剰発現により、膜上でのリボソームおよびポリソームアッセンブリを増加させたが、一方C-末端ドメインを欠失する短縮型変異体(△956、△1136および△1291)はアッセンブリを亢進させることができなかった(図7F〜図7J、図8B)。本実施例において使用した野生型p180タンパク質およびp180タンパク質の様々なC-末端欠失変異体の概略図を、図7Fに示す。そして、野生型p180タンパク質およびp180タンパク質変異体をコードする発現プラスミドを、HeLa細胞中にトランスフェクトし、膜画分中のリボソームRNAを解析した結果を図7Gに、そして膜-結合型画分中の相対的RNA含有量を図7Hに、それぞれ示す。データは、平均±SD(n=3)を示す。したがって、外来性で発現させたp180タンパク質が、C-末端ドメイン依存的に、小胞体-結合型ポリソームのアッセンブリに対して要求に叶うものである。
【0078】
図7Fに示されるヒトp180タンパク質の短縮型変異体のシリーズが、HeLa細胞において一過性に発現された。抗-p180抗体または抗-(His)6抗体を用いた免疫ブロッティングにより、正しいサイズのタンパク質が膜画分中で発現されていることが確認された。そして、疑似トランスフェクト細胞と比較して、野生型p180タンパク質の一過性過剰発現は、膜画分においてRPL10のレベルの上昇を引き起こし(約3.8倍)、一方そのレベルは、その他の変異体についてはより低いものである(2倍未満)(図8B))。
【産業上の利用可能性】
【0079】
本発明により、発現系を構成する細胞内において、小胞体膜上のポリソーム形成を促進させることができることから、結果として、細胞中での目的とするタンパク質の生成を促進することができる。
【配列表フリーテキスト】
【0080】
SEQ ID NO: 1:ヒトp180タンパク質のアミノ酸配列
SEQ ID NO: 2:ヒトp180タンパク質をコードする核酸のヌクレオチド配列
SEQ ID NO: 3:ヒトコラーゲンタンパク質のアミノ酸配列
SEQ ID NO: 4:ヒトコラーゲンタンパク質をコードする核酸のヌクレオチド配列
【特許請求の範囲】
【請求項1】
細胞において、哺乳動物のp180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法。
【請求項2】
哺乳動物のp180タンパク質の全長またはその部分が、ヒトp180タンパク質(SEQ ID NO: 2)、マウスp180タンパク質(GenBank Accession No. NP_077243)、ラットp180タンパク質(GenBank Accession No. XP_230637)、イヌp180タンパク質(GenBank Accession No. NP_001003179)、ウマp180タンパク質(GenBank Accession No. XP_001915027)、サルp180タンパク質(GenBank Accession No. XP_002798281)、チンパンジーp180タンパク質(GenBank Accession No. XP_514527)、ブタp180タンパク質(GenBank Accession No. XP_001926148)、またはそれらの部分である、請求項1に記載の細胞内の小胞体膜上でのポリソーム形成を促進する方法。
【請求項3】
哺乳動物のp180タンパク質の部分が、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、から選択される、請求項1または2に記載の細胞内の小胞体膜上でのポリソーム形成を促進する方法。
【請求項4】
細胞内において発現されるタンパク質が、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の、リボソーム-結合リピートドメインを含む高度に塩基性のN-末端領域、高度に塩基性のタンデムリピートドメイン(濃灰色ボックス)、または微小管結合および束形成ドメイン(MTB-1)、またはSEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の全長である、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
目的とするタンパク質をコードする核酸分子を形質転換した細胞または目的とするタンパク質の産生量が増加された細胞において、哺乳動物のp180タンパク質の全長またはその部分を発現させ、細胞内の小胞体膜上でのポリソーム形成を促進することを含む、前記細胞中での目的とするタンパク質の生成を促進する方法。
【請求項6】
哺乳動物のp180タンパク質の全長またはその部分が、ヒトp180タンパク質(SEQ ID NO: 2)、マウスp180タンパク質(GenBank Accession No. NP_077243)、ラットp180タンパク質(GenBank Accession No. XP_230637)、イヌp180タンパク質(GenBank Accession No. NP_001003179)、ウマp180タンパク質(GenBank Accession No. XP_001915027)、サルp180タンパク質(GenBank Accession No. XP_002798281)、チンパンジーp180タンパク質(GenBank Accession No. XP_514527)、ブタp180タンパク質(GenBank Accession No. XP_001926148)、またはそれらの部分である、請求項5に記載の細胞中での目的とするタンパク質の生成を促進する方法。
【請求項7】
哺乳動物のp180タンパク質のアミノ酸の特定の部分が、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、からなる群から選択される、請求項5または6に記載の細胞中での目的とするタンパク質の生成を促進する方法。
【請求項8】
細胞内において発現されるタンパク質が、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の、リボソーム-結合リピートドメインを含む高度に塩基性のN-末端領域、高度に塩基性のタンデムリピートドメイン(濃灰色ボックス)、または微小管結合および束形成ドメイン(MTB-1)、またはSEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の全長である、請求項5〜7のいずれか1項に記載の方法。
【請求項9】
目的とするタンパク質が、コラーゲンまたはフィブロネクチンである、請求項5〜8のいずれか1項に記載の方法。
【請求項10】
SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、などの部分を発現させた細胞。
【請求項11】
ポリソームが、目的とするタンパク質をコードする遺伝子から転写されたmRNAが含まれるものである、請求項10に記載の細胞。
【請求項12】
目的とするタンパク質が、コラーゲンまたはフィブロネクチンである、請求項10または11に記載の方法。
【請求項1】
細胞において、哺乳動物のp180タンパク質の全長またはその部分を発現させることを含む、細胞内の小胞体膜上でのポリソーム形成を促進する方法。
【請求項2】
哺乳動物のp180タンパク質の全長またはその部分が、ヒトp180タンパク質(SEQ ID NO: 2)、マウスp180タンパク質(GenBank Accession No. NP_077243)、ラットp180タンパク質(GenBank Accession No. XP_230637)、イヌp180タンパク質(GenBank Accession No. NP_001003179)、ウマp180タンパク質(GenBank Accession No. XP_001915027)、サルp180タンパク質(GenBank Accession No. XP_002798281)、チンパンジーp180タンパク質(GenBank Accession No. XP_514527)、ブタp180タンパク質(GenBank Accession No. XP_001926148)、またはそれらの部分である、請求項1に記載の細胞内の小胞体膜上でのポリソーム形成を促進する方法。
【請求項3】
哺乳動物のp180タンパク質の部分が、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、から選択される、請求項1または2に記載の細胞内の小胞体膜上でのポリソーム形成を促進する方法。
【請求項4】
細胞内において発現されるタンパク質が、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の、リボソーム-結合リピートドメインを含む高度に塩基性のN-末端領域、高度に塩基性のタンデムリピートドメイン(濃灰色ボックス)、または微小管結合および束形成ドメイン(MTB-1)、またはSEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の全長である、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
目的とするタンパク質をコードする核酸分子を形質転換した細胞または目的とするタンパク質の産生量が増加された細胞において、哺乳動物のp180タンパク質の全長またはその部分を発現させ、細胞内の小胞体膜上でのポリソーム形成を促進することを含む、前記細胞中での目的とするタンパク質の生成を促進する方法。
【請求項6】
哺乳動物のp180タンパク質の全長またはその部分が、ヒトp180タンパク質(SEQ ID NO: 2)、マウスp180タンパク質(GenBank Accession No. NP_077243)、ラットp180タンパク質(GenBank Accession No. XP_230637)、イヌp180タンパク質(GenBank Accession No. NP_001003179)、ウマp180タンパク質(GenBank Accession No. XP_001915027)、サルp180タンパク質(GenBank Accession No. XP_002798281)、チンパンジーp180タンパク質(GenBank Accession No. XP_514527)、ブタp180タンパク質(GenBank Accession No. XP_001926148)、またはそれらの部分である、請求項5に記載の細胞中での目的とするタンパク質の生成を促進する方法。
【請求項7】
哺乳動物のp180タンパク質のアミノ酸の特定の部分が、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、からなる群から選択される、請求項5または6に記載の細胞中での目的とするタンパク質の生成を促進する方法。
【請求項8】
細胞内において発現されるタンパク質が、SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の、リボソーム-結合リピートドメインを含む高度に塩基性のN-末端領域、高度に塩基性のタンデムリピートドメイン(濃灰色ボックス)、または微小管結合および束形成ドメイン(MTB-1)、またはSEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の全長である、請求項5〜7のいずれか1項に記載の方法。
【請求項9】
目的とするタンパク質が、コラーゲンまたはフィブロネクチンである、請求項5〜8のいずれか1項に記載の方法。
【請求項10】
SEQ ID NO: 1に示されるアミノ酸配列を有するタンパク質(ヒトp180タンパク質)の27〜157番アミノ酸からなる領域を含む部分、623〜737番アミノ酸からなる領域を含む部分、738〜944番アミノ酸からなる領域を含む部分、などの部分を発現させた細胞。
【請求項11】
ポリソームが、目的とするタンパク質をコードする遺伝子から転写されたmRNAが含まれるものである、請求項10に記載の細胞。
【請求項12】
目的とするタンパク質が、コラーゲンまたはフィブロネクチンである、請求項10または11に記載の方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7−1】
【図7−2】
【図7−3】
【図7−4】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7−1】
【図7−2】
【図7−3】
【図7−4】
【図8】
【公開番号】特開2013−85498(P2013−85498A)
【公開日】平成25年5月13日(2013.5.13)
【国際特許分類】
【出願番号】特願2011−227462(P2011−227462)
【出願日】平成23年10月14日(2011.10.14)
【出願人】(000135151)株式会社ニッピ (18)
【Fターム(参考)】
【公開日】平成25年5月13日(2013.5.13)
【国際特許分類】
【出願日】平成23年10月14日(2011.10.14)
【出願人】(000135151)株式会社ニッピ (18)
【Fターム(参考)】
[ Back to top ]