
細胞内でのCAG伸長遺伝子産物の発現および凝集を調節するための方法ならびに同様に調節するために有用である薬剤を特定するための方法
【課題】細胞内での第1遺伝子の発現を調節するための方法を提供する。
【解決手段】第1遺伝子は36を超えるCAGトリヌクレオチドリピートを含み、かつグルタミン媒介タンパク質凝集を形成するタンパク質をコードするものである。第1遺伝子の抑制は、SPT4遺伝子またはSUPT4H遺伝子の発現を減少させることによって達成される。また、それは、Spt4/Spt5複合体またはSupt4h/Supt5h複合体の形成を阻害することによっても達成可能である。また、CAG伸長遺伝子産物の発現および凝集の調節またはハンチントン病などのポリグルタミン病の治療のために、有用な薬剤を特定するための方法。
【解決手段】第1遺伝子は36を超えるCAGトリヌクレオチドリピートを含み、かつグルタミン媒介タンパク質凝集を形成するタンパク質をコードするものである。第1遺伝子の抑制は、SPT4遺伝子またはSUPT4H遺伝子の発現を減少させることによって達成される。また、それは、Spt4/Spt5複合体またはSupt4h/Supt5h複合体の形成を阻害することによっても達成可能である。また、CAG伸長遺伝子産物の発現および凝集の調節またはハンチントン病などのポリグルタミン病の治療のために、有用な薬剤を特定するための方法。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
この非仮出願は、米国特許法第119条第(a)項に基づき、2010年12月10日に、中華民国、台湾において出願された、特許出願第099143336号による優先権を主張し、その内容全体は本明細書において参照により援用される。
【0002】
配列表
本発明は配列表を含む。
【0003】
本発明は、細胞内での伸長CAGリピートを含む遺伝子の発現を調節するための方法に関する。また、本発明はポリグルタミン媒介性タンパク質凝集を調節または阻止するための方法、および前記調節方法の実行において有用である薬剤を特定するための方法を提供する。
【背景技術】
【0004】
ポリグルタミン(PolyQ)病は9つの遺伝学的に異なる疾患から成る部類の疾患である。それらはハンチントン病(HD)、歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、SBMAならびに脊髄小脳失調症1、2、3、6、7および17(SCA1/2/3/6/7/17)を含む。これらの疾患は、グルタミンをコードするCAGリピートの翻訳物の伸長によって引き起こされるため、それらはCAGリピート病としても知られている。
【0005】
これらの遺伝学的に異なる疾患の間で共有される1つの共通な生理学的特徴は、該疾患にかかる患者は全て、彼らの脳内にタンパク質性の沈着物を有することが発見されていることである。これらの各疾患において、タンパク質性沈着物は種々のタンパク質と関連しているが、全てのタンパク質はグルタミンの伸長ストレッチを含んでいる。現在まで、疾患関連タンパク質におけるこのpolyQ配列の伸長ストレッチは、全てのpolyQ病に関与する唯一知られている遺伝子変異である。
【0006】
一般に、遺伝子内のCAGリピート数は、36未満の良性の数字から37以上の病理学的な数字に及び得る。CAGリピートのより大きな数字は病理学的表現型と相関すると考えられている。なぜなら、長いストレッチのグルタミンを含むタンパク質およびポリペプチドは、生体内においてアミロイド様繊維(βシート構造を有するタンパク質凝集体の重合)を形成する特有の傾向を有しており(Scherzingerら、1997年)、かつ伸長polyQ鎖を有する変異タンパク質は、異なるタンパク質構造をもたらし、それは凝集および最終的には神経細胞死をもたらすと考えられているからである(ZoghbiおよびOrr、2000年)。
【0007】
ヒトにおいて、伸長polyQ変異タンパク質は、中枢神経系(CNS)の細胞において広範囲にわたって発現されている。しかしながら、それぞれの異なる疾患において、ニューロンの特定集団は他のものよりも脆弱である。そのため、脆弱性における差異によって、各9つの異なる疾患に対する神経変性の特徴的パターンおよび臨床的特徴がもたらされる。疾患の重症度はCAGリピート数に相関し得る。例えば、HDでは、28−35の間のCAGリピート数は中間と考えられ、35−40は不完全浸透と考えられ、かつ40を超える数は完全浸透と考えられている。
【0008】
表1は、伸長CAGリピート、疾患遺伝子、およびそれらを定義付ける病原リピート長を原因とする8つの疾患を記載している。SCA6はその他のpolyQ病と異なり、SCA6におけるCAGリピート長が症状が現れる年齢に対する決定的要因ではないため、本リストには含まれていない。また、SCA6における病理学的リピート長はその他のpolyQ病よりもはるかに短く、21−30の間の数は病理学的表現型をもたらすのに十分である。
【0009】
【表1】

【0010】
上記の8つの疾患のうちで、HDは患者に対するそれの破壊的な影響によって、おそらく世間で最もよく知られている疾患であろう。該疾患は主に皮質および線条体において発生する選択的神経細胞死と関連している。それは、患者から、体の動き、認識力、および人格を徐々に奪う、致死的かつ残酷な疾患であり、患者およびその家族に重大な経済的および感情的な負担を強いる。HDの頻度においては、西欧系の人々の間で特によく見られる(約20,000人に1人)。残念ながら、現在のところ、この恐ろしい疾患に対する治療法はない。
【0011】
現在、HDに対する可能な治療は、主に肉眼的症状に対処することに限定されている。例えば、FDAに承認された最新の化合物の1つであるテトラベナジンは、HD患者において運動過剰を減少させるための薬である。テトラベナジンは神経伝達物質の早期の分解を促進する小胞モノアミン輸送体(VMAT)阻害剤である。従って、この薬は単に症状を治療するのみであり、疾患の根源を治療するものではない。現在HDを治療するために使用されているその他の薬としては、神経遮断薬およびベンゾジアゼピンが挙げられる。疾患が進行するにつれて、異なる症状に対処するための、抗精神病薬、および運動機能低下症のための薬を含めた、より幅広い範囲の薬局方が必要とされている。現在知られているどの治療においても、HDの根本的原因に取り組む努力が行われていない。
【0012】
上述のように、HDの根本的原因は、CNS細胞内の遺伝子、特にハンチンチンタンパク質(Htt)をコードする遺伝子HttにおけるCAGリピートの異常な伸長である。健常人では、Htt遺伝子内に約8−25のCAGヌクレオチド配列の構成リピートがある。HD患者では、CAGリピートの数は36以上に伸長されている。このタイプの変異は優性であるため、人は1コピーの変異ハンチンチン遺伝子を受け継ぐだけでHDを発症する。
【0013】
最近の細胞および動物モデルの研究において、変異Httによって形成された凝集体はHDの進行において重要な役割を果たしていることが示されている。変異Httタンパク質は、タンパク質分解的切断にさらされるときに、polyQ伸長部分からのより短い断片を置き去りにし得ることが観察されている。グルタミンの過剰なコピーが変異Httにおいて存在すると、グルタミンの極性性質はその他のタンパク質との望ましくない相互作用をもたらす。特に、グルタミンの過剰なコピーを有する変異Httは互いに水素結合を形成し、機能性タンパク質に折り畳まれるよりも凝集する。時間とともに、蓄積されたタンパク質凝集体は、患者において神経細胞に損傷を与え、細胞死および神経障害をもたらす。タンパク質凝集体による損傷の影響は、マウスモデルにおいて、タンパク質凝集体の形成を阻害可能な化学試薬が、細胞の生存を高め、かつHDの病的状態を改善可能であることを示す実験によって裏付けられている(Sanchezら、2003年;Tanakaら、2004年)。
【0014】
タンパク質凝集を阻止するために阻害分子を使用することの他に、変異ハンチンチン遺伝子の発現を減少させることは、原理的に不溶性タンパク質凝集体の発生を阻害するための代替的手法となる。生体内研究において、polyQタンパク質凝集の程度はタンパク質の濃度に関連することが示されている(Scherzingerら、1999年)。従って、変異ハンチンチン遺伝子の発現レベルを下げることによって、より低いレベルにおいて伸長PolyQタンパク質が発現され、その結果、それはタンパク質凝集体の形成を減少させ、かつHDの発症を遅らせるであろう。
【0015】
これらの結果は、Htt内のCAGリピート変異から生じる不溶性タンパク質凝集体の形成を調節することで、HD病因と闘うための、潜在的に単純かつ強力な方針を提示している。例えば、polyQ変異遺伝子の発現またはpolyQ凝集体の形成を調節可能な治療薬は、それらの生理学的な症状だけでなく、polyQ病の根本的原因に対処することが可能である。残念ながら、変異polyQ遺伝子の発現を調節可能な細胞性因子および薬剤に関する知識の不足が、本治療方針の実用的な開発を妨げてきた。
【0016】
したがって、伸長CAGリピート変異を伴う遺伝子の発現を調節または減少させることが可能な方法および手段、ならびに変異遺伝子の発現の調節または減少に効果的な薬剤を特定および開発するための方法および手段が、今もなお緊急に必要とされている。
【発明の概要】
【課題を解決するための手段】
【0017】
上記のことを考慮して、本発明の目的は、伸長CAGリピートを含む遺伝子の発現を調節または減少させることが可能な方法および手段を提供することである。
【0018】
また、本発明の目的は、polyQ病を治療するための治療方法および治療薬を提供することである。
【0019】
さらに、本発明の目的は、伸長CAGリピートを含む遺伝子の発現の調節または減少に効果的な薬剤を、スクリーニングおよび開発するための方法および手段を提供することである。
【0020】
本発明の上記の目的は、微生物転写伸長因子Spt4およびそれの哺乳類オーソログであるSupt4hが、伸長CAGリピートを含む遺伝子の発現において調節の役割を担っているという予期せぬ発見によって充足された。
【0021】
特に、発明者は、伸長CAGリピートを含む遺伝子の発現およびpolyQ配列の伸長ストレッチを有するタンパク質の凝集は、両方ともSpt4−/Supt4h−欠損細胞において軽減されることを発見した。さらに、発明者は、Spt4/Supt4hは、CAGリピートが短いかまたはないものを含む遺伝子に対しては、ごくわずかしか影響しないことを発見した。これらの予期せぬ発見によって、Spt4/Supt4hはpolyQ病介入のための有用な標的として確立される。
【0022】
本発明の他の発見では、発明者は、Spt4−/Supt4h−欠損による軽減効果は、CAG伸長遺伝子における転写伸長障害に起因し得るものであり、それは対応するmRNAおよびタンパク質の産生の減少につながることも発見した。本発見は、Spt4およびSupt4hのいくつかの周知特性において一貫した見解を可能とするものであり、それらは本発明において治療手段計画を誘導する機構モデルの基礎を形成している。
【0023】
例えば、いくつかの最近の研究は、Spt4は酵母細胞における転写およびヘテロクロマチン形成の制御機構において、重要な役割を果たすことに関与するとしている(CrottiおよびBasrai、2004年;Rondonら、2003年)。転写制御へのそれの影響の分析によって、Spt4は伸長過程において正の制御因子であることが示されている。特に、Spt4は、酵母細胞内でGCを高含有率で有するかまたは3Kbを超える大きさであるDNA鋳型からの、RNAポリメラーゼII媒介転写産物の合成を促進する(Rondonら、2003年)。さらに、最近の証明において、Spt4は、転写の間、RNAポリメラーゼIIの処理能力に影響を及ぼすことが明らかとなり、それはSpt4をクロマチン鋳型に沿ったポリメラーゼIIの持続性に対する重要な要因として特定するものである(MasonおよびStruhl、2005年)。
【0024】
特定の理論になんら束縛されるものではないが、発明者は、Spt4がpolyQ含有タンパク質の発現に必要とされる理由は、ポリメラーゼIIが移動するのに長時間を要するそれらのDNA鋳型(例えば、CAG伸長領域を含む鋳型)から、転写機構が早期に解離することを阻止するその役割のためであると確信している。このモデルは、Spt4−/Supt4h−欠損細胞がCAG伸長遺伝子の発現の軽減を示す結果を説明する。従って、Spt4またはSupt4hを標的とすることで、治療方法を有利に考案することが可能である。
【0025】
さらに、本発明の他の発見では、発明者は、Spt4欠損による軽減効果は、Spt4との相互作用に不具合を有する特異的Spt5変異体によって再現可能であることを発見した。一般に、Spt4は細胞内でSpt5と複合体を形成するため、本発見は、Spt4/Spt5複合体もまた治療標的であることをさらに確立する。
【0026】
本発明の上記の様々な驚くべき発見に基づいて、発明者は、伸長CAGリピート配列を含む遺伝子の発現を調節するための様々な手段および方法を着想かつ具体化した。
【0027】
したがって、一態様において、本発明は36を超えるリピート数を有する伸長CAGリピートを含む遺伝子の発現を調節するための方法を提供する。本発明のこの態様による実施形態では、概して、伸長CAGリピートを含む遺伝子の発現に関与する転写伸長因子を標的とすることに努める。転写伸長因子を標的とすることは、直接的または間接的のどちらであってもよい。
【0028】
いくつかの好ましい実施形態において、本発明のこの態様による方法は、一般に、細胞内での転写伸長因子遺伝子の発現を抑制する段階を含む。転写伸長因子遺伝子は、好ましくは、酵母細胞に対してはSPT4遺伝子、または哺乳類細胞に対してはSUPT4H遺伝子である。伸長CAG遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子、またはそれらの組み合わせから成る群より選択されるものである。
【0029】
その他の好ましい実施形態において、本発明のこの態様による方法は、一般に、転写伸長因子複合体の形成を阻害する段階を含む。転写伸長因子複合体は、好ましくは、酵母細胞内ではSpt4/Spt5複合体、または哺乳類細胞内ではSupt4h/Supt5h複合体である。また、伸長遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子、またはそれらの組み合わせから成る群より選択されるものである。
【0030】
さらに、その他の好ましい実施形態において、本発明のこの態様による方法は、伸長CAGリピートを含む遺伝子の発現に関連する、転写伸長因子の抑制および転写因子複合体の形成の阻害における両段階を含み得る。転写伸長因子遺伝子は、好ましくは、酵母細胞に対してはSPT4、または哺乳類細胞に対してはSUPT4Hである。転写因子複合体は、好ましくは、酵母細胞内ではSpt4/Spt5、または哺乳類細胞内ではSupt4h/Supt5hである。また、伸長遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子、またはそれらの組み合わせから成る群より選択されるものである。
【0031】
他の態様では、本発明は、伸長CAGリピートを含む遺伝子の発現の調節またはポリグルタミン病の治療のために、有用な医薬剤を特定するための方法をさらに提供する。伸長遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子から成る群より選択される。CAGリピートの数は、好ましくは36を超える。
【0032】
本発明のこの態様による方法は、一般に、Spt4/Spt5複合体またはSupt4h/Supt5h複合体形成の破壊に対するその有効性を評価するために候補化合物を試験する段階、および所定の水準において効果的であるかどうか、前記化合物を先導化合物として特定する段階を含む。好ましくは、タンパク質間相互作用アッセイが候補化合物の有効性を評価するために使用される。
【0033】
本発明のその他の態様および利点は、以下の説明および添付の特許請求の範囲から明らかとなるであろう。
【図面の簡単な説明】
【0034】
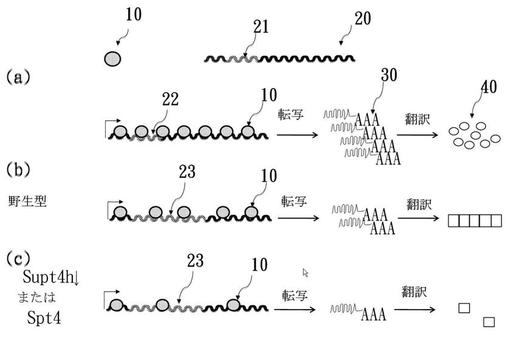
【図1】図1はSpt4−/Supt4h−欠損がCAG含有遺伝子の発現に与える下方制御効果を示している。(a)は、非病原数のCAGリピート(灰色)を含むDNA鋳型(波線で表示)に沿って移動するRNAポリメラーゼII(斑点球体で表示)で開始する遺伝子転写工程により、mRNAが産生され、その後タンパク質産物に翻訳されることを示している。(b)は、どのようにグルタミンの伸長数を含むタンパク質が凝集してタンパク質性沈着物が形成されるかを示している。(c)は、Spt4−およびSupt4h−が下方制御されたときに、Spt4−およびSupt4h−欠損がpolyQタンパク質の産生へ与える軽減効果を示している。
【図2A】図2Aは、RT−PCRによって決定された、線条体細胞内のSupt4h siRNAノックダウン後のHttmRNA発現レベルを示している。
【図2B】図2Bは、リアルタイムqRT−PCRによって評価された、Supt4h siRNAノックダウン後のHttmRNA発現レベルの変化を示している。
【図3】図3は、Supt4h siRNA形質移入を行う場合と行わない場合の細胞内タンパク質発現レベルを示している。
【図4】図4は、Supt4h下方制御が正常および変異Httそれぞれの発現に与える効果を示している。
【図5】図5は、Supt4h siRNAノックダウンを行う場合と行わない場合の、7Q−eGFPまたは81Q−eGFPを発現するST14A線条体細胞の生存率を示している。
【図6(a)】図6(a)および6(b)は、Spt4/Spt5複合体を阻害するSPT4欠損またはSPT5変異体が伸長CAGリピートを含む遺伝子の発現を抑制可能であり、それはコロニーカラーアッセイ(colony color assay)によって明らかとなるpolyQ凝集の減少につながることを示している。
【図6(b)】図6(a)および6(b)は、Spt4/Spt5複合体を阻害するSPT4欠損またはSPT5変異体が伸長CAGリピートを含む遺伝子の発現を抑制可能であり、それはコロニーカラーアッセイ(colony color assay)によって明らかとなるpolyQ凝集の減少につながることを示している。
【図7】図7は、Spt4/Spt5複合体形成を阻害することによって生じたpolyQ変異タンパク質の発現および凝集のパターン変化を示している。
【図8】図8は、Spt4/Spt5複合体形成を阻害することによって生じた伸長CAGリピートを含む転写産物の減少した発現を示している。
【発明を実施するための形態】
【0035】
本発明は、様々な結果の完全な理解を促進するために、例示的な実施形態および実施例によって次に示される。本発明は例示的な実施形態および実施例の観点から説明されるが、本明細書において開示される実施形態は例示を目的とするのみであり、様々な改良および変更が添付の特許請求の範囲に記載の本発明の精神および範囲から逸脱することなく当業者によって行われ得ることを理解されたい。
【0036】
定義
本開示を通して、遺伝子の名称はイタリック体の大文字で表示し、遺伝子と関連するタンパク質は最初の文字のみが大文字の非イタリック体で表示する。例えば、SPT4遺伝子の場合、用語「SPT4」は遺伝子を表示し、用語「Spt4」は遺伝子によって産生されたタンパク質を表示する。唯一の例外はハンチンチン遺伝子であり、遺伝子の名称は「Htt」で表示し、遺伝子産物(ハンチンチンタンパク質)は「Htt」で表示する。
【0037】
本明細書において、遺伝子SPT4は転写伸長タンパク質Spt4をコードする遺伝子を指す。遺伝子は(Maloneら、1993年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。タンパク質Spt4は(Malneら、1993年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。
【0038】
本明細書において、遺伝子SPT5は転写伸長タンパク質Spt5をコードする遺伝子を指す。遺伝子は(Swansonら、1991年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。タンパク質Spt5は(Swansonら、1991年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。
【0039】
本明細書において、遺伝子SUPT4Hは哺乳類転写伸長因子Supt4hをコードする遺伝子を指す。遺伝子は(Hartzogら、1996年;Chiangら、1996年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。タンパク質Supt4hは(Hartzogら、1996年;Chiangら、1996年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。
【0040】
本明細書において、遺伝子SUPT5Hは哺乳類転写伸長因子Supt5hをコードする遺伝子を指す。遺伝子は(Stachoraら、1997年;Chiangら、1998年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。タンパク質Supt5hは(Stachoraら、1997年;Chiangら、1998年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。
【0041】
本発明との関係においては、用語「polyQ病」は、表1に記載の8つの疾患を指す。
【0042】
本発明との関係においては、用語「polyQ変異タンパク質」は、36グルタミン残基より長いpolyQ鎖を有するタンパク質を指す。
【0043】
本発明との関係においては、用語「伸長CAGリピート」は、36よりも大きいCAGリピート数を指す。
【0044】
伸長CAGリピートを含む遺伝子の発現を抑制するための方法
上述の通り、本発明は細胞内での遺伝子の発現を調節するための方法を提供し、遺伝子は伸長CAGリピートを含む。好ましくは、CAGリピートの数は36コピーより多い。伸長CAGリピートを含む遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子から成る群より選択される。
【0045】
いくつかの好ましい実施形態において、本発明のこの態様による方法は、一般に、酵母細胞内でのSpt4または哺乳類細胞内でのSupt4hの発現を抑制する段階を含む。
【0046】
遺伝子産物の発現を抑制するための手段は特に限定されない。遺伝子産物(すなわち、Spt4およびSupt4hタンパク質)の発現を抑制可能である限り、それらは当技術分野におけるあらゆる周知の遺伝子サイレンシング法またはその他の将来開発される遺伝子抑制法であり得る。例示的な遺伝子サイレンシング法は、限定されるものではないが、SPT4またはSUPT4Hの遺伝子の発現を抑制可能である、遺伝子ノックダウン、遺伝子ノックアウト、化学試薬を用いて、またはそれらの混合を含み得る。
【0047】
好ましい実施形態では、siRNAおよび遺伝子ノックアウトは、哺乳類細胞内のSUPT4H遺伝子および酵母細胞内のSPT4遺伝子それぞれを制御するために使用される。遺伝子ノックダウンを生じさせるために使用されるRNA干渉配列は、好ましくはSUPT4Hの相補的遺伝子配列であり、より好ましくは80%以上の配列相同性を有するものである。
【0048】
特定の理論になんら束縛されるものではないが、SPT4およびSUPT4Hを抑制することによって、長いCAGリピートを有する遺伝子転写産物を生成する細胞の能力が減少するため、伸長polyQ鎖を有するタンパク質の発現が軽減すると考えられている。
【0049】
図1はHtt遺伝子が転写されてHttタンパク質に翻訳される工程を図示している。
【0050】
図1(a)を参照すると、正常Htt遺伝子20上に(斜線部分で示される)CAGリピート21を有するDNA鋳型に沿って移動するRNAポリメラーゼII(灰色球体)10が示されている。転写の間、RNAポリメラーゼII10は、初めにHtt遺伝子20のDNA鋳型と関連し、次いで鋳型に沿って移動することで、CAGリピートを含む遺伝子のmRNA(つまり、Htt遺伝子mRNA)30が生産される。その後、Htt遺伝子mRNA30を翻訳することで、Httタンパク質40が産生される。つまり、図1(a)は正常CAGリピート、すなわち「短いCAGリピート」22を含む遺伝子の場合において起こることを示している。
【0051】
図1(b)は、伸長CAGリピート、すなわち「長いCAGリピート」23を含む変異Htt遺伝子20の場合に起こることを示している。上述の通り、伸長polyQリピートのストレッチを有するHttタンパク質40が産生されると、臓器または組織内で凝集および蓄積する傾向があり、それはHD表現型の原因となる。
【0052】
図1(c)では、SUPT4H遺伝子は下方制御されている。その結果、ポリメラーゼII10は、転写伸長の間、変異Htt遺伝子20と持続的に関連することが不可能となり、Httタンパク質産生の減少をもたらし、polyQ病表現型を阻止または改善する。
【0053】
転写伸長因子遺伝子SPT4およびSUPT4Hの発現を抑制することに加えて、Spt4およびSupt4hタンパク質を直接標的にするかまたはそれらの相互作用パートナーを間接的に標的にすることによって、同様のpolyQ変異タンパク質軽減効果も達成可能である。例えば、Spt4タンパク質はSpt5タンパク質と関連することで、酵母細胞内で複合体を形成することが知られている。発明者は、Spt4/Spt5複合体の形成を破壊することによっても、変異polyQタンパク質の発現が軽減することを発見した。同様に、Supt4hおよびSupt5hも哺乳類細胞(例えばヒト細胞)内で複合体を形成する。したがって、2つの複合体を抑制することは、ヒトまたは酵母の細胞内でのCAG伸長遺伝子の発現を抑制、およびpolyQ凝集を阻止するための、代替的かつ実用的な方法である。
【0054】
したがって、いくつかの好ましい実施形態において、本発明のこの態様による方法は、一般に、転写伸長タンパク質複合体の形成を破壊する段階を含む。好ましくは、転写伸長タンパク質複合体は、酵母細胞内ではSpt4/Spt5、または哺乳類細胞内ではSupt4h/Supt5hである。
【0055】
タンパク質複合体の形成を破壊するための手段は、特に限定されない。それらは当技術分野においてあらゆる周知の方法によって達成可能であり、例えば、酵母細胞の場合にはSpt4またはSpt5、および哺乳類細胞の場合にはSupt4hまたはSupt5hに対する、周知または後に開発される阻害剤を用いることを含む。また、siRNA、RNAi、または当技術分野におけるあらゆるその他の周知の遺伝子サイレンシング法などの遺伝子サイレンシング法は、対応する複合体において少なくとも1つの発現因子を抑制するために有利に使用可能である。例えば、Spt4/Spt5の場合、SPT4、SPT5、または両方の遺伝子抑制は、複合体の形成を未然に防ぐために有利に使用可能である。あるいは、変異のSpt4またはSpt5が、野生型Spt4/Spt5複合体の形成を破壊するための破壊剤として導入されてもよい。
【0056】
その他のさらなる実施形態では、遺伝子抑制段階もまた、タンパク質複合体破壊段階とともに有利に使用可能である。例えば、例示的な実施形態では、細胞内で変異polyQタンパク質の発現を調節するための方法は、酵母細胞に対してはSPT4遺伝子、または哺乳類細胞に対してはSUPT4H遺伝子の発現を抑制する段階、および酵母細胞内ではSpt4/Spt5複合体、または哺乳類細胞内ではSupt4h/Supt5h複合体の形成を阻害する段階を含み得る。
【0057】
polyQ病の治療方法
さらに他の態様では、本発明はpolyQ病を治療するための方法も提供する。polyQ病は、脊髄小脳失調症1、2、3、7、17型、歯状核赤核淡蒼球ルイ体萎縮症、球脊髄性筋萎縮症、およびハンチントン病から成る群より選択されるものである。
【0058】
いくつかの好ましい実施形態において、本発明のこの態様による方法は、SUPT4H遺伝子、SUPT5H遺伝子、または両方の発現を抑制するための、遺伝子サイレンシング剤の効果的な量を投与する一般的な段階を含み得る。遺伝子サイレンシング剤の種類は特に限定されない。薬剤がSUPT4HまたはSUPT5Hの遺伝子発現を抑制可能である限り、当技術分野において周知のあらゆる遺伝子サイレンシング剤、または将来開発される遺伝子サイレンシング剤を使用可能である。当業者は、種々の遺伝子サイレンシング剤には適切に選択された投与経路が必要であることを理解するであろう。例示的な実施形態では、遺伝子サイレンシング剤は、siRNA、RNAi、またはD−RNAiである。
【0059】
いくつかのその他の好ましい実施形態において、本発明のこの態様による方法は、Supt4h/Supt5h複合体の形成を破壊可能な医薬剤を投与する一般的な段階を含み得る。また、生体内でSupt4h/Supt5h複合体の形成を破壊可能である限り、医薬剤は特に限定されない。適した医薬剤はSupt4hまたはSupt5hの特異的阻害剤であってもよい。例示的な薬剤は、限定されるものではないが、小分子、ペプチド、または抗体を含み得る。それはまた、Supt4hまたはSupt5hの工学的変異体であってもよい。
【0060】
先導化合物として有用である薬剤を特定するための方法
さらに、他の態様では、本発明はpolyQ病を治療するための先導化合物をスクリーニングおよび特定するための方法をさらに提供する。本発明のこの態様による方法は、一般に、タンパク質複合体の形成の破壊に対する候補化合物の有効性を試験する段階、および既定の限界量において有効であるかどうか、化合物を先導化合物として特定する段階を含む。
【0061】
試験段階は、Spt4/Spt5相互作用、またはSupt4h/Supt5h相互作用に対して、タンパク質間相互作用アッセイを用いて実施可能である。タンパク質間相互作用アッセイの設計は、特に限定されない。例えば、Spt4/Spt5複合体またはSupt4h/Supt5h複合体から生じるタンパク質間相互作用は、これらのタンパク質の全長または断片を用いて、生体内または生体外のどちらかにおいて確立可能である。アッセイが生体内アッセイである場合、Spt4/Spt5相互作用に対しては酵母細胞をベースにしたアッセイ、およびSupt4h/Supt5h相互作用に対しては哺乳類細胞をベースにしたアッセイが好ましい。
【0062】
タンパク質間相互作用の読み取りは、例えば、蛍光シグナル、FRETシグナル、または酵素反応であり得る。
【0063】
このタンパク質間相互作用を妨げる、小分子、ペプチド、または抗体が、候補化合物として選択される。
【0064】
先導化合物のタンパク質間相互作用に基づいた特定は、先導化合物を特定するための効果的で一般的な手法として示されていることに特に留意されたい。例えば、MDM2は、腫瘍形成に関して、p53に結合して否定的に制御することが知られている。癌治療に対して有効性を示す小分子、Nutlin−3は、MDM2とp53タンパク質との間の相互作用を阻止するそれの能力によって特定された(Vassilevら、Science,303:844−848、その内容全体は本明細書において参照により援用される)。この例示的な事例によって、その他のいくつかの研究とともに、本明細書において開示されるタンパク質間相互作用に基づいた薬物の発見方法の妥当性が支持される。
【0065】
好ましい実施形態では、使用されるアッセイは、発明者によって以前に開発されたカラーコロニーアッセイである。本アッセイの詳細は、米国特許第7,375,190号B2に記載されており、その内容全体は本明細書において参照により援用される。
【0066】
本発明をさらに説明するために、以下の具体的な実施例を提供する。
実施例
【実施例1】
【0067】
変異HttmRNA発現はSupt4h siRNAノックダウンによって阻害される。
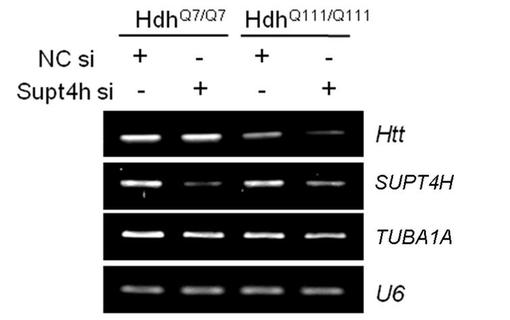
マウス線条体神経細胞株ST14A(ラット)、HdhQ7/Q7(マウス)、HdhQ111/Q111(マウス)、およびHdhQ7/Q111(マウス)を、33℃で5%CO2とともに10%ウシ胎仔血清(FBS)が追加されたダルベッコ変法イーグル培地(SH30022、HyClone)中で培養した。ST14Aは、pTet−Offプラスミド(BD Biosciences)で形質移入されることで安定な細胞株ST14Atetを確立し、それはテトラサイクリンの非存在下でpTRE2−7Q−eGFPまたはpTRE2−81Q−eGFPを発現した。DNAおよびsiRNAの形質移入は、LipofectAMINE 2000(Invitrogen)を使用して行った。100nMのSupt4h siRNA(5’−CUAUAGACCAGUUCGAAUA−3’(SEQ ID 1)、5’−UCAAAUACCAAUAAAGCGA−3’(SEQ ID 2)、5’−GGGAGUGUCUGGGCGGAUU−3’(SEQ ID 3)、5’−CCCAAGGAAUCGUGCGGGA−3’(SEQ ID 4)を含む、DHARMACON,ON−TARGET plus SMART pool,L−048866−01)、および(DHARMACON,J−086342−10,5’−UGGCCUACAAAUCGAGAGAUU−3’(SEQ ID 5))を使用して、マウスおよびラットの細胞それぞれにおいてSupt4hの発現を阻害した。いかなる遺伝子も標的としない等量のアニールされた二本鎖オリゴヌクレオチド(5’−UUCUCCGAACGUGUCACGUTT−3’(SEQ ID 6)、および5’−ACGUGACACGUUCGGAGAATT−3’(SEQ ID 7))の形質移入は、コントロールとしての役割を果たした。
【0068】
図2Aでは、ホモ接合の野生型(HdhQ7/Q7)または変異(HdhQ111/Q111)のハンチンチン対立遺伝子を所有する線条体細胞において、Supt4h siRNAノックダウン後に、RT−PCRによってHttmRNAレベルを調査した。RNAポリメラーゼIIIによって転写されたU6は、ローディングコントロールとしての役割を果たした。TUBA1Aは、Supt4hがハウスキーピング遺伝子のポリメラーゼII依存性の転写に及ぼす影響を調査するために含めた。変異HttmRNAレベルは、HdhQ111/Q111細胞において、SUPT4H遺伝子ノックダウンに応答して減少することが示されている。しかしながら、同程度のSUPT4H下方制御を有しながらも、HdhQ7/Q7における正常HttmRNAの発現において検出可能な変化はない。
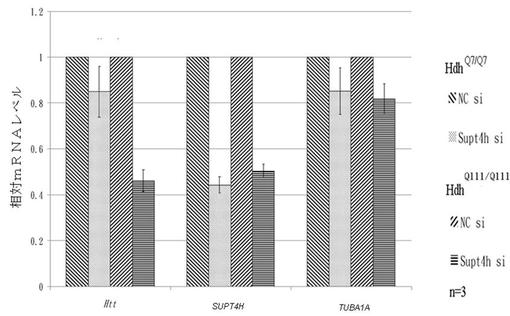
【0069】
図2Bでは、Supt4h siRNAノックダウン後のmRNAの発現レベルにおける変化を、リアルタイムqRT−PCRによって評価した。各mRNAをU6を用いて正常化し、コントロールsiRNA(NC siRNA)で形質移入された細胞内の転写レベルを1に設定した。本図表において、横軸はHtt遺伝子、SUPT4H遺伝子、TUBA1A遺伝子を表し、縦軸は相対mRNAレベルを表している。SUPT4H遺伝子ノックダウンによって、HdhQ111/Q111内の変異HttmRNAレベルは大幅に減少する一方で、HdhQ7/Q7内の正常HttmRNAはわずかに減少するだけであることが示されている。
【0070】
図2Aおよび2Bの両方は、変異Htt遺伝子の発現がSupt4hによって制御されることを示している。
【実施例2】
【0071】
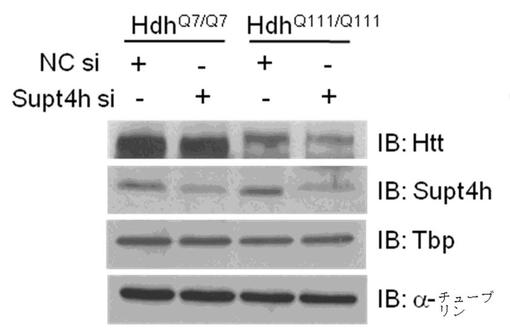
Supt4h siRNAノックダウンを行う場合と行わない場合の細胞内でのHttタンパク質発現の分析。
図3では、実施例1から収集された細胞可溶化物を免疫ブロット法によって分析し、Supt4h siRNAノックダウンに応答したタンパク質発現の変化を決定した。全タンパク質抽出物の等量を、Htt、Supt4h、Tbp、およびα−Tubulinに対して免疫ブロットした。TATA−box結合タンパク質であるTbpを、短いストレッチのCAGリピートを有するコード遺伝子のタンパク質を代表するものとして含めた。
【0072】
変異Httタンパク質のレベルは、NC siRNAで処理したものと比較して、Supt4h siRNAで処理したHdhQ111/Q111細胞内においてより低いことが示されている。変異Httタンパク質の減少は、実施例1において観察されたように、HdhQ111/Q111細胞内での対応するmRNAの変化と一致する。さらに、HttQ7およびTpbタンパク質などの正常CAGリピート(CAGトリヌクレオチドリピート数が36以下)を有するコード遺伝子のタンパク質レベルは、Supt4h siRNAノックダウンによって影響されない。
【実施例3】
【0073】
SUPT4H遺伝子の下方制御は変異Htt発現を阻害するが、正常Htt発現は阻害しない。
図4では、ヘテロ接合Htt対立遺伝子(HdhQ7/Q111)を有する線条体細胞にSupt4h siRNAが形質移入され、タンパク質発現を実施例2に記載されるように分析した。HttQ7およびHttQ111の位置を矢印で示す。SUPT4Hノックダウンに応答して、HttQ111のタンパク質レベルは減少したが、HttQ7のタンパク質レベルは不変である。それは伸長CAGリピートを含む遺伝子のみがSupt4hによって影響を受けることをさらに確認するものである。
【実施例4】
【0074】
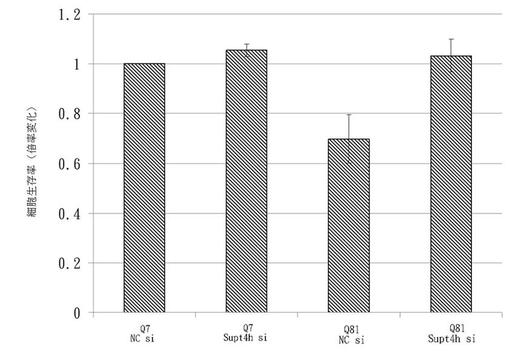
81Q−eGFPを発現するST14A細胞の生存率はSupt4h siRNAノックダウンによって向上した。
7Q−eGFPまたは81Q−eGFPを発現するST14A細胞の生存率を、Supt4h siRNAノックダウンを行う場合と行わない場合で測定した。示されたpolyQ−eGFPおよびsiRNAで形質移入された後、最小量(0.5%)の血清を含み39℃に維持された増殖培地において細胞を培養し、神経細胞分化を誘導した。コントロールsiRNA存在下において7Q−eGFPを発現する生細胞の数を1に設定し、その他の試料の相対細胞生存率を図5に示す。7Q−eGFPで形質移入された細胞と比較すると、81Q−eGFPで形質移入されたST14Aは細胞生存率において減少を示したが、それはsiRNA介在SUPT4Hノックダウンによって反転した。一方で、私たちはSupt4h siRNAは7Q−eGFPを発現する細胞の生存に影響を及ぼさないことを観察した。これは伸長polyQリピートを含む凝集傾向にあるタンパク質の発現によって引き起こされる細胞生存率の減少は、SUPT4H遺伝子の下方制御によって改善可能であることを実証している。
【実施例5】
【0075】
酵母細胞内でのSpt4/Spt5複合体の形成を阻止することによるpolyQ凝集の阻害
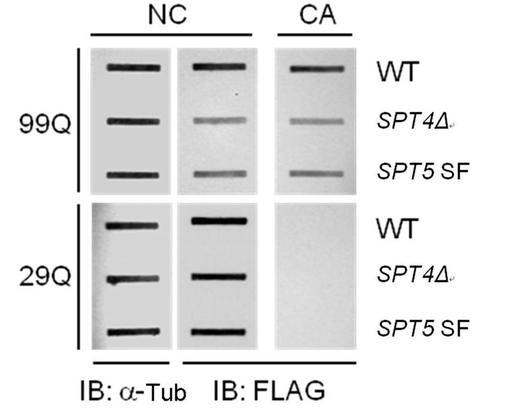
この目的のために、私達の研究室によって開発されたコロニーカラーアッセイ(米国特許第7,375,190号B2)を使用して、polyQ含有タンパク質の凝集を決定した。本アッセイシステムにおいて、レポータータンパク質であるAde2は、29Qまたは99Qのストレッチのどちらかに融合される。融合タンパク質が溶解したときに、それは機能し、Ade2酵素活性によって細胞の色は白くなる。対照的に、タンパク質が凝集すると、Ade2は十分機能せず、細胞の色は赤くなる。29Q−Ade2はそれの短いpolyQリピート(36Q未満)のために凝集不可能であり、正常Htt遺伝子などの短いCAGトリヌクレオチドリピートを有する遺伝子を代表するものとして含める。図6では、29Q−ADE2および99Q−ADE2を発現する野生型(WT)細胞は、白色および赤色をそれぞれ示すことが明らかである。しかしながら、SPT4遺伝子欠損(SPT4Δ)またはSPT5 SF変異体がこれらの細胞に導入されると、99Q−ADE2発現細胞において赤色の細胞色は白色に変化する。SPT5 SF細胞は、Spt4/Spt5複合体の形成を阻害する特異的SPT5 S324F点突然変異を保有している。これらの結果は、polyQ凝集はSpt4/Spt5複合体の形成を妨げることによって影響を受けることを示唆している。
【実施例6】
【0076】
SPT4遺伝子欠損またはSPT5 SF変異を有する細胞内のpolyQ−Ade2タンパク質の発現および凝集の分析
polyQ−Ade2タンパク質の発現および凝集は、スロットブロット法およびフィルタートラップ法それぞれによって調査した。フィルタートラップ法は酢酸セルロース(CA)膜を使用して凝集タンパク質のみを補足し、一方で、スロットブロット法はニトロセルロース(NC)膜を使用し、それは全てのローディングされたタンパク質を保持する。膜上でのpolyQ−Ade2の保持は、抗−FLAG抗体を用いて免疫ブロット法によって検出した。α−チューブリン(α−Tub)を、各試料において均等なタンパク質ローディングを確保するために含めた。図7では、99Q−Ade2タンパク質の凝集(CA、上方パネル)は、WT細胞と比較してSPT4ΔおよびSPT5 SF変異細胞において著しく減少することを示している。この発見は、実施例5において観察されるように、これらの変異細胞ではpolyQ凝集がより少ないという概念と一致している。凝集の減少に加えて、私たちは99Q−Ade2タンパク質の発現(NC、上方パネル)が、SPT4ΔおよびSPT5 SF細胞において減少していることも発見した。しかしながら、これらの変異細胞は29Q−Ade2の発現(NC、下方パネル)には影響しなかった。これらの結果は、Spt4/Spt5複合体の形成を妨げることによるpolyQ凝集の阻害が、現実的であることを確認するものである。さらに、短いものはそうではないが、長いpolyQの伸長ストレッチ(>36Q)を有するタンパク質の発現は、Spt4/Spt5複合体形成を阻止する変異に影響されやすい。
【実施例7】
【0077】
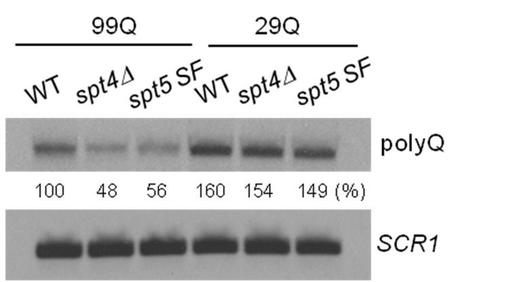
SPT4遺伝子欠損またはSPT5 SF変異を有する細胞内のノーザンブロット法によるpolyQ−ADE2 mRNAレベルの分析
99Q−Ade2および29Q−Ade2をコードする転写産物の発現は、実施例5において説明したように、細胞内のノーザンブロット法によって分析した。polyQ−ADE2 mRNAsをpolyQプローブによって検出し、SCR1をローディングコントロールとして含めた。正常化の後、99Q−ADE2を発現するWT細胞内のmRNAレベルを100%に設定した。示された細胞における相対的99Q−ADE2および29Q−ADE2のmRNAレベルを図8に示す。WTと比較すると、SPT4ΔおよびSPT5 SFの細胞の両方は、99Q−ADE2のmRNA発現において減少を示しているが、29Q−ADE2のmRNA発現においては減少を示していない。この観察結果は、実施例6でのSPT4ΔおよびSPT5 SFの変異細胞内における99Q−Ade2タンパク質発現の変化と一致している。
【0078】
本発見はまた、Spt4/Spt5複合体の形成を妨げることによって、短いpolyQリピートをコードする遺伝子に影響することなく、伸長CAGリピートを有する遺伝子の発現を阻害することが現実的であることも立証する。
【技術分野】
【0001】
関連出願の相互参照
この非仮出願は、米国特許法第119条第(a)項に基づき、2010年12月10日に、中華民国、台湾において出願された、特許出願第099143336号による優先権を主張し、その内容全体は本明細書において参照により援用される。
【0002】
配列表
本発明は配列表を含む。
【0003】
本発明は、細胞内での伸長CAGリピートを含む遺伝子の発現を調節するための方法に関する。また、本発明はポリグルタミン媒介性タンパク質凝集を調節または阻止するための方法、および前記調節方法の実行において有用である薬剤を特定するための方法を提供する。
【背景技術】
【0004】
ポリグルタミン(PolyQ)病は9つの遺伝学的に異なる疾患から成る部類の疾患である。それらはハンチントン病(HD)、歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)、SBMAならびに脊髄小脳失調症1、2、3、6、7および17(SCA1/2/3/6/7/17)を含む。これらの疾患は、グルタミンをコードするCAGリピートの翻訳物の伸長によって引き起こされるため、それらはCAGリピート病としても知られている。
【0005】
これらの遺伝学的に異なる疾患の間で共有される1つの共通な生理学的特徴は、該疾患にかかる患者は全て、彼らの脳内にタンパク質性の沈着物を有することが発見されていることである。これらの各疾患において、タンパク質性沈着物は種々のタンパク質と関連しているが、全てのタンパク質はグルタミンの伸長ストレッチを含んでいる。現在まで、疾患関連タンパク質におけるこのpolyQ配列の伸長ストレッチは、全てのpolyQ病に関与する唯一知られている遺伝子変異である。
【0006】
一般に、遺伝子内のCAGリピート数は、36未満の良性の数字から37以上の病理学的な数字に及び得る。CAGリピートのより大きな数字は病理学的表現型と相関すると考えられている。なぜなら、長いストレッチのグルタミンを含むタンパク質およびポリペプチドは、生体内においてアミロイド様繊維(βシート構造を有するタンパク質凝集体の重合)を形成する特有の傾向を有しており(Scherzingerら、1997年)、かつ伸長polyQ鎖を有する変異タンパク質は、異なるタンパク質構造をもたらし、それは凝集および最終的には神経細胞死をもたらすと考えられているからである(ZoghbiおよびOrr、2000年)。
【0007】
ヒトにおいて、伸長polyQ変異タンパク質は、中枢神経系(CNS)の細胞において広範囲にわたって発現されている。しかしながら、それぞれの異なる疾患において、ニューロンの特定集団は他のものよりも脆弱である。そのため、脆弱性における差異によって、各9つの異なる疾患に対する神経変性の特徴的パターンおよび臨床的特徴がもたらされる。疾患の重症度はCAGリピート数に相関し得る。例えば、HDでは、28−35の間のCAGリピート数は中間と考えられ、35−40は不完全浸透と考えられ、かつ40を超える数は完全浸透と考えられている。
【0008】
表1は、伸長CAGリピート、疾患遺伝子、およびそれらを定義付ける病原リピート長を原因とする8つの疾患を記載している。SCA6はその他のpolyQ病と異なり、SCA6におけるCAGリピート長が症状が現れる年齢に対する決定的要因ではないため、本リストには含まれていない。また、SCA6における病理学的リピート長はその他のpolyQ病よりもはるかに短く、21−30の間の数は病理学的表現型をもたらすのに十分である。
【0009】
【表1】
【0010】
上記の8つの疾患のうちで、HDは患者に対するそれの破壊的な影響によって、おそらく世間で最もよく知られている疾患であろう。該疾患は主に皮質および線条体において発生する選択的神経細胞死と関連している。それは、患者から、体の動き、認識力、および人格を徐々に奪う、致死的かつ残酷な疾患であり、患者およびその家族に重大な経済的および感情的な負担を強いる。HDの頻度においては、西欧系の人々の間で特によく見られる(約20,000人に1人)。残念ながら、現在のところ、この恐ろしい疾患に対する治療法はない。
【0011】
現在、HDに対する可能な治療は、主に肉眼的症状に対処することに限定されている。例えば、FDAに承認された最新の化合物の1つであるテトラベナジンは、HD患者において運動過剰を減少させるための薬である。テトラベナジンは神経伝達物質の早期の分解を促進する小胞モノアミン輸送体(VMAT)阻害剤である。従って、この薬は単に症状を治療するのみであり、疾患の根源を治療するものではない。現在HDを治療するために使用されているその他の薬としては、神経遮断薬およびベンゾジアゼピンが挙げられる。疾患が進行するにつれて、異なる症状に対処するための、抗精神病薬、および運動機能低下症のための薬を含めた、より幅広い範囲の薬局方が必要とされている。現在知られているどの治療においても、HDの根本的原因に取り組む努力が行われていない。
【0012】
上述のように、HDの根本的原因は、CNS細胞内の遺伝子、特にハンチンチンタンパク質(Htt)をコードする遺伝子HttにおけるCAGリピートの異常な伸長である。健常人では、Htt遺伝子内に約8−25のCAGヌクレオチド配列の構成リピートがある。HD患者では、CAGリピートの数は36以上に伸長されている。このタイプの変異は優性であるため、人は1コピーの変異ハンチンチン遺伝子を受け継ぐだけでHDを発症する。
【0013】
最近の細胞および動物モデルの研究において、変異Httによって形成された凝集体はHDの進行において重要な役割を果たしていることが示されている。変異Httタンパク質は、タンパク質分解的切断にさらされるときに、polyQ伸長部分からのより短い断片を置き去りにし得ることが観察されている。グルタミンの過剰なコピーが変異Httにおいて存在すると、グルタミンの極性性質はその他のタンパク質との望ましくない相互作用をもたらす。特に、グルタミンの過剰なコピーを有する変異Httは互いに水素結合を形成し、機能性タンパク質に折り畳まれるよりも凝集する。時間とともに、蓄積されたタンパク質凝集体は、患者において神経細胞に損傷を与え、細胞死および神経障害をもたらす。タンパク質凝集体による損傷の影響は、マウスモデルにおいて、タンパク質凝集体の形成を阻害可能な化学試薬が、細胞の生存を高め、かつHDの病的状態を改善可能であることを示す実験によって裏付けられている(Sanchezら、2003年;Tanakaら、2004年)。
【0014】
タンパク質凝集を阻止するために阻害分子を使用することの他に、変異ハンチンチン遺伝子の発現を減少させることは、原理的に不溶性タンパク質凝集体の発生を阻害するための代替的手法となる。生体内研究において、polyQタンパク質凝集の程度はタンパク質の濃度に関連することが示されている(Scherzingerら、1999年)。従って、変異ハンチンチン遺伝子の発現レベルを下げることによって、より低いレベルにおいて伸長PolyQタンパク質が発現され、その結果、それはタンパク質凝集体の形成を減少させ、かつHDの発症を遅らせるであろう。
【0015】
これらの結果は、Htt内のCAGリピート変異から生じる不溶性タンパク質凝集体の形成を調節することで、HD病因と闘うための、潜在的に単純かつ強力な方針を提示している。例えば、polyQ変異遺伝子の発現またはpolyQ凝集体の形成を調節可能な治療薬は、それらの生理学的な症状だけでなく、polyQ病の根本的原因に対処することが可能である。残念ながら、変異polyQ遺伝子の発現を調節可能な細胞性因子および薬剤に関する知識の不足が、本治療方針の実用的な開発を妨げてきた。
【0016】
したがって、伸長CAGリピート変異を伴う遺伝子の発現を調節または減少させることが可能な方法および手段、ならびに変異遺伝子の発現の調節または減少に効果的な薬剤を特定および開発するための方法および手段が、今もなお緊急に必要とされている。
【発明の概要】
【課題を解決するための手段】
【0017】
上記のことを考慮して、本発明の目的は、伸長CAGリピートを含む遺伝子の発現を調節または減少させることが可能な方法および手段を提供することである。
【0018】
また、本発明の目的は、polyQ病を治療するための治療方法および治療薬を提供することである。
【0019】
さらに、本発明の目的は、伸長CAGリピートを含む遺伝子の発現の調節または減少に効果的な薬剤を、スクリーニングおよび開発するための方法および手段を提供することである。
【0020】
本発明の上記の目的は、微生物転写伸長因子Spt4およびそれの哺乳類オーソログであるSupt4hが、伸長CAGリピートを含む遺伝子の発現において調節の役割を担っているという予期せぬ発見によって充足された。
【0021】
特に、発明者は、伸長CAGリピートを含む遺伝子の発現およびpolyQ配列の伸長ストレッチを有するタンパク質の凝集は、両方ともSpt4−/Supt4h−欠損細胞において軽減されることを発見した。さらに、発明者は、Spt4/Supt4hは、CAGリピートが短いかまたはないものを含む遺伝子に対しては、ごくわずかしか影響しないことを発見した。これらの予期せぬ発見によって、Spt4/Supt4hはpolyQ病介入のための有用な標的として確立される。
【0022】
本発明の他の発見では、発明者は、Spt4−/Supt4h−欠損による軽減効果は、CAG伸長遺伝子における転写伸長障害に起因し得るものであり、それは対応するmRNAおよびタンパク質の産生の減少につながることも発見した。本発見は、Spt4およびSupt4hのいくつかの周知特性において一貫した見解を可能とするものであり、それらは本発明において治療手段計画を誘導する機構モデルの基礎を形成している。
【0023】
例えば、いくつかの最近の研究は、Spt4は酵母細胞における転写およびヘテロクロマチン形成の制御機構において、重要な役割を果たすことに関与するとしている(CrottiおよびBasrai、2004年;Rondonら、2003年)。転写制御へのそれの影響の分析によって、Spt4は伸長過程において正の制御因子であることが示されている。特に、Spt4は、酵母細胞内でGCを高含有率で有するかまたは3Kbを超える大きさであるDNA鋳型からの、RNAポリメラーゼII媒介転写産物の合成を促進する(Rondonら、2003年)。さらに、最近の証明において、Spt4は、転写の間、RNAポリメラーゼIIの処理能力に影響を及ぼすことが明らかとなり、それはSpt4をクロマチン鋳型に沿ったポリメラーゼIIの持続性に対する重要な要因として特定するものである(MasonおよびStruhl、2005年)。
【0024】
特定の理論になんら束縛されるものではないが、発明者は、Spt4がpolyQ含有タンパク質の発現に必要とされる理由は、ポリメラーゼIIが移動するのに長時間を要するそれらのDNA鋳型(例えば、CAG伸長領域を含む鋳型)から、転写機構が早期に解離することを阻止するその役割のためであると確信している。このモデルは、Spt4−/Supt4h−欠損細胞がCAG伸長遺伝子の発現の軽減を示す結果を説明する。従って、Spt4またはSupt4hを標的とすることで、治療方法を有利に考案することが可能である。
【0025】
さらに、本発明の他の発見では、発明者は、Spt4欠損による軽減効果は、Spt4との相互作用に不具合を有する特異的Spt5変異体によって再現可能であることを発見した。一般に、Spt4は細胞内でSpt5と複合体を形成するため、本発見は、Spt4/Spt5複合体もまた治療標的であることをさらに確立する。
【0026】
本発明の上記の様々な驚くべき発見に基づいて、発明者は、伸長CAGリピート配列を含む遺伝子の発現を調節するための様々な手段および方法を着想かつ具体化した。
【0027】
したがって、一態様において、本発明は36を超えるリピート数を有する伸長CAGリピートを含む遺伝子の発現を調節するための方法を提供する。本発明のこの態様による実施形態では、概して、伸長CAGリピートを含む遺伝子の発現に関与する転写伸長因子を標的とすることに努める。転写伸長因子を標的とすることは、直接的または間接的のどちらであってもよい。
【0028】
いくつかの好ましい実施形態において、本発明のこの態様による方法は、一般に、細胞内での転写伸長因子遺伝子の発現を抑制する段階を含む。転写伸長因子遺伝子は、好ましくは、酵母細胞に対してはSPT4遺伝子、または哺乳類細胞に対してはSUPT4H遺伝子である。伸長CAG遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子、またはそれらの組み合わせから成る群より選択されるものである。
【0029】
その他の好ましい実施形態において、本発明のこの態様による方法は、一般に、転写伸長因子複合体の形成を阻害する段階を含む。転写伸長因子複合体は、好ましくは、酵母細胞内ではSpt4/Spt5複合体、または哺乳類細胞内ではSupt4h/Supt5h複合体である。また、伸長遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子、またはそれらの組み合わせから成る群より選択されるものである。
【0030】
さらに、その他の好ましい実施形態において、本発明のこの態様による方法は、伸長CAGリピートを含む遺伝子の発現に関連する、転写伸長因子の抑制および転写因子複合体の形成の阻害における両段階を含み得る。転写伸長因子遺伝子は、好ましくは、酵母細胞に対してはSPT4、または哺乳類細胞に対してはSUPT4Hである。転写因子複合体は、好ましくは、酵母細胞内ではSpt4/Spt5、または哺乳類細胞内ではSupt4h/Supt5hである。また、伸長遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子、またはそれらの組み合わせから成る群より選択されるものである。
【0031】
他の態様では、本発明は、伸長CAGリピートを含む遺伝子の発現の調節またはポリグルタミン病の治療のために、有用な医薬剤を特定するための方法をさらに提供する。伸長遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子から成る群より選択される。CAGリピートの数は、好ましくは36を超える。
【0032】
本発明のこの態様による方法は、一般に、Spt4/Spt5複合体またはSupt4h/Supt5h複合体形成の破壊に対するその有効性を評価するために候補化合物を試験する段階、および所定の水準において効果的であるかどうか、前記化合物を先導化合物として特定する段階を含む。好ましくは、タンパク質間相互作用アッセイが候補化合物の有効性を評価するために使用される。
【0033】
本発明のその他の態様および利点は、以下の説明および添付の特許請求の範囲から明らかとなるであろう。
【図面の簡単な説明】
【0034】
【図1】図1はSpt4−/Supt4h−欠損がCAG含有遺伝子の発現に与える下方制御効果を示している。(a)は、非病原数のCAGリピート(灰色)を含むDNA鋳型(波線で表示)に沿って移動するRNAポリメラーゼII(斑点球体で表示)で開始する遺伝子転写工程により、mRNAが産生され、その後タンパク質産物に翻訳されることを示している。(b)は、どのようにグルタミンの伸長数を含むタンパク質が凝集してタンパク質性沈着物が形成されるかを示している。(c)は、Spt4−およびSupt4h−が下方制御されたときに、Spt4−およびSupt4h−欠損がpolyQタンパク質の産生へ与える軽減効果を示している。
【図2A】図2Aは、RT−PCRによって決定された、線条体細胞内のSupt4h siRNAノックダウン後のHttmRNA発現レベルを示している。
【図2B】図2Bは、リアルタイムqRT−PCRによって評価された、Supt4h siRNAノックダウン後のHttmRNA発現レベルの変化を示している。
【図3】図3は、Supt4h siRNA形質移入を行う場合と行わない場合の細胞内タンパク質発現レベルを示している。
【図4】図4は、Supt4h下方制御が正常および変異Httそれぞれの発現に与える効果を示している。
【図5】図5は、Supt4h siRNAノックダウンを行う場合と行わない場合の、7Q−eGFPまたは81Q−eGFPを発現するST14A線条体細胞の生存率を示している。
【図6(a)】図6(a)および6(b)は、Spt4/Spt5複合体を阻害するSPT4欠損またはSPT5変異体が伸長CAGリピートを含む遺伝子の発現を抑制可能であり、それはコロニーカラーアッセイ(colony color assay)によって明らかとなるpolyQ凝集の減少につながることを示している。
【図6(b)】図6(a)および6(b)は、Spt4/Spt5複合体を阻害するSPT4欠損またはSPT5変異体が伸長CAGリピートを含む遺伝子の発現を抑制可能であり、それはコロニーカラーアッセイ(colony color assay)によって明らかとなるpolyQ凝集の減少につながることを示している。
【図7】図7は、Spt4/Spt5複合体形成を阻害することによって生じたpolyQ変異タンパク質の発現および凝集のパターン変化を示している。
【図8】図8は、Spt4/Spt5複合体形成を阻害することによって生じた伸長CAGリピートを含む転写産物の減少した発現を示している。
【発明を実施するための形態】
【0035】
本発明は、様々な結果の完全な理解を促進するために、例示的な実施形態および実施例によって次に示される。本発明は例示的な実施形態および実施例の観点から説明されるが、本明細書において開示される実施形態は例示を目的とするのみであり、様々な改良および変更が添付の特許請求の範囲に記載の本発明の精神および範囲から逸脱することなく当業者によって行われ得ることを理解されたい。
【0036】
定義
本開示を通して、遺伝子の名称はイタリック体の大文字で表示し、遺伝子と関連するタンパク質は最初の文字のみが大文字の非イタリック体で表示する。例えば、SPT4遺伝子の場合、用語「SPT4」は遺伝子を表示し、用語「Spt4」は遺伝子によって産生されたタンパク質を表示する。唯一の例外はハンチンチン遺伝子であり、遺伝子の名称は「Htt」で表示し、遺伝子産物(ハンチンチンタンパク質)は「Htt」で表示する。
【0037】
本明細書において、遺伝子SPT4は転写伸長タンパク質Spt4をコードする遺伝子を指す。遺伝子は(Maloneら、1993年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。タンパク質Spt4は(Malneら、1993年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。
【0038】
本明細書において、遺伝子SPT5は転写伸長タンパク質Spt5をコードする遺伝子を指す。遺伝子は(Swansonら、1991年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。タンパク質Spt5は(Swansonら、1991年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。
【0039】
本明細書において、遺伝子SUPT4Hは哺乳類転写伸長因子Supt4hをコードする遺伝子を指す。遺伝子は(Hartzogら、1996年;Chiangら、1996年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。タンパク質Supt4hは(Hartzogら、1996年;Chiangら、1996年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。
【0040】
本明細書において、遺伝子SUPT5Hは哺乳類転写伸長因子Supt5hをコードする遺伝子を指す。遺伝子は(Stachoraら、1997年;Chiangら、1998年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。タンパク質Supt5hは(Stachoraら、1997年;Chiangら、1998年)によって特徴付けられ、その内容全体が本明細書において参照により援用される。
【0041】
本発明との関係においては、用語「polyQ病」は、表1に記載の8つの疾患を指す。
【0042】
本発明との関係においては、用語「polyQ変異タンパク質」は、36グルタミン残基より長いpolyQ鎖を有するタンパク質を指す。
【0043】
本発明との関係においては、用語「伸長CAGリピート」は、36よりも大きいCAGリピート数を指す。
【0044】
伸長CAGリピートを含む遺伝子の発現を抑制するための方法
上述の通り、本発明は細胞内での遺伝子の発現を調節するための方法を提供し、遺伝子は伸長CAGリピートを含む。好ましくは、CAGリピートの数は36コピーより多い。伸長CAGリピートを含む遺伝子は、好ましくは、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR,およびHttの遺伝子から成る群より選択される。
【0045】
いくつかの好ましい実施形態において、本発明のこの態様による方法は、一般に、酵母細胞内でのSpt4または哺乳類細胞内でのSupt4hの発現を抑制する段階を含む。
【0046】
遺伝子産物の発現を抑制するための手段は特に限定されない。遺伝子産物(すなわち、Spt4およびSupt4hタンパク質)の発現を抑制可能である限り、それらは当技術分野におけるあらゆる周知の遺伝子サイレンシング法またはその他の将来開発される遺伝子抑制法であり得る。例示的な遺伝子サイレンシング法は、限定されるものではないが、SPT4またはSUPT4Hの遺伝子の発現を抑制可能である、遺伝子ノックダウン、遺伝子ノックアウト、化学試薬を用いて、またはそれらの混合を含み得る。
【0047】
好ましい実施形態では、siRNAおよび遺伝子ノックアウトは、哺乳類細胞内のSUPT4H遺伝子および酵母細胞内のSPT4遺伝子それぞれを制御するために使用される。遺伝子ノックダウンを生じさせるために使用されるRNA干渉配列は、好ましくはSUPT4Hの相補的遺伝子配列であり、より好ましくは80%以上の配列相同性を有するものである。
【0048】
特定の理論になんら束縛されるものではないが、SPT4およびSUPT4Hを抑制することによって、長いCAGリピートを有する遺伝子転写産物を生成する細胞の能力が減少するため、伸長polyQ鎖を有するタンパク質の発現が軽減すると考えられている。
【0049】
図1はHtt遺伝子が転写されてHttタンパク質に翻訳される工程を図示している。
【0050】
図1(a)を参照すると、正常Htt遺伝子20上に(斜線部分で示される)CAGリピート21を有するDNA鋳型に沿って移動するRNAポリメラーゼII(灰色球体)10が示されている。転写の間、RNAポリメラーゼII10は、初めにHtt遺伝子20のDNA鋳型と関連し、次いで鋳型に沿って移動することで、CAGリピートを含む遺伝子のmRNA(つまり、Htt遺伝子mRNA)30が生産される。その後、Htt遺伝子mRNA30を翻訳することで、Httタンパク質40が産生される。つまり、図1(a)は正常CAGリピート、すなわち「短いCAGリピート」22を含む遺伝子の場合において起こることを示している。
【0051】
図1(b)は、伸長CAGリピート、すなわち「長いCAGリピート」23を含む変異Htt遺伝子20の場合に起こることを示している。上述の通り、伸長polyQリピートのストレッチを有するHttタンパク質40が産生されると、臓器または組織内で凝集および蓄積する傾向があり、それはHD表現型の原因となる。
【0052】
図1(c)では、SUPT4H遺伝子は下方制御されている。その結果、ポリメラーゼII10は、転写伸長の間、変異Htt遺伝子20と持続的に関連することが不可能となり、Httタンパク質産生の減少をもたらし、polyQ病表現型を阻止または改善する。
【0053】
転写伸長因子遺伝子SPT4およびSUPT4Hの発現を抑制することに加えて、Spt4およびSupt4hタンパク質を直接標的にするかまたはそれらの相互作用パートナーを間接的に標的にすることによって、同様のpolyQ変異タンパク質軽減効果も達成可能である。例えば、Spt4タンパク質はSpt5タンパク質と関連することで、酵母細胞内で複合体を形成することが知られている。発明者は、Spt4/Spt5複合体の形成を破壊することによっても、変異polyQタンパク質の発現が軽減することを発見した。同様に、Supt4hおよびSupt5hも哺乳類細胞(例えばヒト細胞)内で複合体を形成する。したがって、2つの複合体を抑制することは、ヒトまたは酵母の細胞内でのCAG伸長遺伝子の発現を抑制、およびpolyQ凝集を阻止するための、代替的かつ実用的な方法である。
【0054】
したがって、いくつかの好ましい実施形態において、本発明のこの態様による方法は、一般に、転写伸長タンパク質複合体の形成を破壊する段階を含む。好ましくは、転写伸長タンパク質複合体は、酵母細胞内ではSpt4/Spt5、または哺乳類細胞内ではSupt4h/Supt5hである。
【0055】
タンパク質複合体の形成を破壊するための手段は、特に限定されない。それらは当技術分野においてあらゆる周知の方法によって達成可能であり、例えば、酵母細胞の場合にはSpt4またはSpt5、および哺乳類細胞の場合にはSupt4hまたはSupt5hに対する、周知または後に開発される阻害剤を用いることを含む。また、siRNA、RNAi、または当技術分野におけるあらゆるその他の周知の遺伝子サイレンシング法などの遺伝子サイレンシング法は、対応する複合体において少なくとも1つの発現因子を抑制するために有利に使用可能である。例えば、Spt4/Spt5の場合、SPT4、SPT5、または両方の遺伝子抑制は、複合体の形成を未然に防ぐために有利に使用可能である。あるいは、変異のSpt4またはSpt5が、野生型Spt4/Spt5複合体の形成を破壊するための破壊剤として導入されてもよい。
【0056】
その他のさらなる実施形態では、遺伝子抑制段階もまた、タンパク質複合体破壊段階とともに有利に使用可能である。例えば、例示的な実施形態では、細胞内で変異polyQタンパク質の発現を調節するための方法は、酵母細胞に対してはSPT4遺伝子、または哺乳類細胞に対してはSUPT4H遺伝子の発現を抑制する段階、および酵母細胞内ではSpt4/Spt5複合体、または哺乳類細胞内ではSupt4h/Supt5h複合体の形成を阻害する段階を含み得る。
【0057】
polyQ病の治療方法
さらに他の態様では、本発明はpolyQ病を治療するための方法も提供する。polyQ病は、脊髄小脳失調症1、2、3、7、17型、歯状核赤核淡蒼球ルイ体萎縮症、球脊髄性筋萎縮症、およびハンチントン病から成る群より選択されるものである。
【0058】
いくつかの好ましい実施形態において、本発明のこの態様による方法は、SUPT4H遺伝子、SUPT5H遺伝子、または両方の発現を抑制するための、遺伝子サイレンシング剤の効果的な量を投与する一般的な段階を含み得る。遺伝子サイレンシング剤の種類は特に限定されない。薬剤がSUPT4HまたはSUPT5Hの遺伝子発現を抑制可能である限り、当技術分野において周知のあらゆる遺伝子サイレンシング剤、または将来開発される遺伝子サイレンシング剤を使用可能である。当業者は、種々の遺伝子サイレンシング剤には適切に選択された投与経路が必要であることを理解するであろう。例示的な実施形態では、遺伝子サイレンシング剤は、siRNA、RNAi、またはD−RNAiである。
【0059】
いくつかのその他の好ましい実施形態において、本発明のこの態様による方法は、Supt4h/Supt5h複合体の形成を破壊可能な医薬剤を投与する一般的な段階を含み得る。また、生体内でSupt4h/Supt5h複合体の形成を破壊可能である限り、医薬剤は特に限定されない。適した医薬剤はSupt4hまたはSupt5hの特異的阻害剤であってもよい。例示的な薬剤は、限定されるものではないが、小分子、ペプチド、または抗体を含み得る。それはまた、Supt4hまたはSupt5hの工学的変異体であってもよい。
【0060】
先導化合物として有用である薬剤を特定するための方法
さらに、他の態様では、本発明はpolyQ病を治療するための先導化合物をスクリーニングおよび特定するための方法をさらに提供する。本発明のこの態様による方法は、一般に、タンパク質複合体の形成の破壊に対する候補化合物の有効性を試験する段階、および既定の限界量において有効であるかどうか、化合物を先導化合物として特定する段階を含む。
【0061】
試験段階は、Spt4/Spt5相互作用、またはSupt4h/Supt5h相互作用に対して、タンパク質間相互作用アッセイを用いて実施可能である。タンパク質間相互作用アッセイの設計は、特に限定されない。例えば、Spt4/Spt5複合体またはSupt4h/Supt5h複合体から生じるタンパク質間相互作用は、これらのタンパク質の全長または断片を用いて、生体内または生体外のどちらかにおいて確立可能である。アッセイが生体内アッセイである場合、Spt4/Spt5相互作用に対しては酵母細胞をベースにしたアッセイ、およびSupt4h/Supt5h相互作用に対しては哺乳類細胞をベースにしたアッセイが好ましい。
【0062】
タンパク質間相互作用の読み取りは、例えば、蛍光シグナル、FRETシグナル、または酵素反応であり得る。
【0063】
このタンパク質間相互作用を妨げる、小分子、ペプチド、または抗体が、候補化合物として選択される。
【0064】
先導化合物のタンパク質間相互作用に基づいた特定は、先導化合物を特定するための効果的で一般的な手法として示されていることに特に留意されたい。例えば、MDM2は、腫瘍形成に関して、p53に結合して否定的に制御することが知られている。癌治療に対して有効性を示す小分子、Nutlin−3は、MDM2とp53タンパク質との間の相互作用を阻止するそれの能力によって特定された(Vassilevら、Science,303:844−848、その内容全体は本明細書において参照により援用される)。この例示的な事例によって、その他のいくつかの研究とともに、本明細書において開示されるタンパク質間相互作用に基づいた薬物の発見方法の妥当性が支持される。
【0065】
好ましい実施形態では、使用されるアッセイは、発明者によって以前に開発されたカラーコロニーアッセイである。本アッセイの詳細は、米国特許第7,375,190号B2に記載されており、その内容全体は本明細書において参照により援用される。
【0066】
本発明をさらに説明するために、以下の具体的な実施例を提供する。
実施例
【実施例1】
【0067】
変異HttmRNA発現はSupt4h siRNAノックダウンによって阻害される。
マウス線条体神経細胞株ST14A(ラット)、HdhQ7/Q7(マウス)、HdhQ111/Q111(マウス)、およびHdhQ7/Q111(マウス)を、33℃で5%CO2とともに10%ウシ胎仔血清(FBS)が追加されたダルベッコ変法イーグル培地(SH30022、HyClone)中で培養した。ST14Aは、pTet−Offプラスミド(BD Biosciences)で形質移入されることで安定な細胞株ST14Atetを確立し、それはテトラサイクリンの非存在下でpTRE2−7Q−eGFPまたはpTRE2−81Q−eGFPを発現した。DNAおよびsiRNAの形質移入は、LipofectAMINE 2000(Invitrogen)を使用して行った。100nMのSupt4h siRNA(5’−CUAUAGACCAGUUCGAAUA−3’(SEQ ID 1)、5’−UCAAAUACCAAUAAAGCGA−3’(SEQ ID 2)、5’−GGGAGUGUCUGGGCGGAUU−3’(SEQ ID 3)、5’−CCCAAGGAAUCGUGCGGGA−3’(SEQ ID 4)を含む、DHARMACON,ON−TARGET plus SMART pool,L−048866−01)、および(DHARMACON,J−086342−10,5’−UGGCCUACAAAUCGAGAGAUU−3’(SEQ ID 5))を使用して、マウスおよびラットの細胞それぞれにおいてSupt4hの発現を阻害した。いかなる遺伝子も標的としない等量のアニールされた二本鎖オリゴヌクレオチド(5’−UUCUCCGAACGUGUCACGUTT−3’(SEQ ID 6)、および5’−ACGUGACACGUUCGGAGAATT−3’(SEQ ID 7))の形質移入は、コントロールとしての役割を果たした。
【0068】
図2Aでは、ホモ接合の野生型(HdhQ7/Q7)または変異(HdhQ111/Q111)のハンチンチン対立遺伝子を所有する線条体細胞において、Supt4h siRNAノックダウン後に、RT−PCRによってHttmRNAレベルを調査した。RNAポリメラーゼIIIによって転写されたU6は、ローディングコントロールとしての役割を果たした。TUBA1Aは、Supt4hがハウスキーピング遺伝子のポリメラーゼII依存性の転写に及ぼす影響を調査するために含めた。変異HttmRNAレベルは、HdhQ111/Q111細胞において、SUPT4H遺伝子ノックダウンに応答して減少することが示されている。しかしながら、同程度のSUPT4H下方制御を有しながらも、HdhQ7/Q7における正常HttmRNAの発現において検出可能な変化はない。
【0069】
図2Bでは、Supt4h siRNAノックダウン後のmRNAの発現レベルにおける変化を、リアルタイムqRT−PCRによって評価した。各mRNAをU6を用いて正常化し、コントロールsiRNA(NC siRNA)で形質移入された細胞内の転写レベルを1に設定した。本図表において、横軸はHtt遺伝子、SUPT4H遺伝子、TUBA1A遺伝子を表し、縦軸は相対mRNAレベルを表している。SUPT4H遺伝子ノックダウンによって、HdhQ111/Q111内の変異HttmRNAレベルは大幅に減少する一方で、HdhQ7/Q7内の正常HttmRNAはわずかに減少するだけであることが示されている。
【0070】
図2Aおよび2Bの両方は、変異Htt遺伝子の発現がSupt4hによって制御されることを示している。
【実施例2】
【0071】
Supt4h siRNAノックダウンを行う場合と行わない場合の細胞内でのHttタンパク質発現の分析。
図3では、実施例1から収集された細胞可溶化物を免疫ブロット法によって分析し、Supt4h siRNAノックダウンに応答したタンパク質発現の変化を決定した。全タンパク質抽出物の等量を、Htt、Supt4h、Tbp、およびα−Tubulinに対して免疫ブロットした。TATA−box結合タンパク質であるTbpを、短いストレッチのCAGリピートを有するコード遺伝子のタンパク質を代表するものとして含めた。
【0072】
変異Httタンパク質のレベルは、NC siRNAで処理したものと比較して、Supt4h siRNAで処理したHdhQ111/Q111細胞内においてより低いことが示されている。変異Httタンパク質の減少は、実施例1において観察されたように、HdhQ111/Q111細胞内での対応するmRNAの変化と一致する。さらに、HttQ7およびTpbタンパク質などの正常CAGリピート(CAGトリヌクレオチドリピート数が36以下)を有するコード遺伝子のタンパク質レベルは、Supt4h siRNAノックダウンによって影響されない。
【実施例3】
【0073】
SUPT4H遺伝子の下方制御は変異Htt発現を阻害するが、正常Htt発現は阻害しない。
図4では、ヘテロ接合Htt対立遺伝子(HdhQ7/Q111)を有する線条体細胞にSupt4h siRNAが形質移入され、タンパク質発現を実施例2に記載されるように分析した。HttQ7およびHttQ111の位置を矢印で示す。SUPT4Hノックダウンに応答して、HttQ111のタンパク質レベルは減少したが、HttQ7のタンパク質レベルは不変である。それは伸長CAGリピートを含む遺伝子のみがSupt4hによって影響を受けることをさらに確認するものである。
【実施例4】
【0074】
81Q−eGFPを発現するST14A細胞の生存率はSupt4h siRNAノックダウンによって向上した。
7Q−eGFPまたは81Q−eGFPを発現するST14A細胞の生存率を、Supt4h siRNAノックダウンを行う場合と行わない場合で測定した。示されたpolyQ−eGFPおよびsiRNAで形質移入された後、最小量(0.5%)の血清を含み39℃に維持された増殖培地において細胞を培養し、神経細胞分化を誘導した。コントロールsiRNA存在下において7Q−eGFPを発現する生細胞の数を1に設定し、その他の試料の相対細胞生存率を図5に示す。7Q−eGFPで形質移入された細胞と比較すると、81Q−eGFPで形質移入されたST14Aは細胞生存率において減少を示したが、それはsiRNA介在SUPT4Hノックダウンによって反転した。一方で、私たちはSupt4h siRNAは7Q−eGFPを発現する細胞の生存に影響を及ぼさないことを観察した。これは伸長polyQリピートを含む凝集傾向にあるタンパク質の発現によって引き起こされる細胞生存率の減少は、SUPT4H遺伝子の下方制御によって改善可能であることを実証している。
【実施例5】
【0075】
酵母細胞内でのSpt4/Spt5複合体の形成を阻止することによるpolyQ凝集の阻害
この目的のために、私達の研究室によって開発されたコロニーカラーアッセイ(米国特許第7,375,190号B2)を使用して、polyQ含有タンパク質の凝集を決定した。本アッセイシステムにおいて、レポータータンパク質であるAde2は、29Qまたは99Qのストレッチのどちらかに融合される。融合タンパク質が溶解したときに、それは機能し、Ade2酵素活性によって細胞の色は白くなる。対照的に、タンパク質が凝集すると、Ade2は十分機能せず、細胞の色は赤くなる。29Q−Ade2はそれの短いpolyQリピート(36Q未満)のために凝集不可能であり、正常Htt遺伝子などの短いCAGトリヌクレオチドリピートを有する遺伝子を代表するものとして含める。図6では、29Q−ADE2および99Q−ADE2を発現する野生型(WT)細胞は、白色および赤色をそれぞれ示すことが明らかである。しかしながら、SPT4遺伝子欠損(SPT4Δ)またはSPT5 SF変異体がこれらの細胞に導入されると、99Q−ADE2発現細胞において赤色の細胞色は白色に変化する。SPT5 SF細胞は、Spt4/Spt5複合体の形成を阻害する特異的SPT5 S324F点突然変異を保有している。これらの結果は、polyQ凝集はSpt4/Spt5複合体の形成を妨げることによって影響を受けることを示唆している。
【実施例6】
【0076】
SPT4遺伝子欠損またはSPT5 SF変異を有する細胞内のpolyQ−Ade2タンパク質の発現および凝集の分析
polyQ−Ade2タンパク質の発現および凝集は、スロットブロット法およびフィルタートラップ法それぞれによって調査した。フィルタートラップ法は酢酸セルロース(CA)膜を使用して凝集タンパク質のみを補足し、一方で、スロットブロット法はニトロセルロース(NC)膜を使用し、それは全てのローディングされたタンパク質を保持する。膜上でのpolyQ−Ade2の保持は、抗−FLAG抗体を用いて免疫ブロット法によって検出した。α−チューブリン(α−Tub)を、各試料において均等なタンパク質ローディングを確保するために含めた。図7では、99Q−Ade2タンパク質の凝集(CA、上方パネル)は、WT細胞と比較してSPT4ΔおよびSPT5 SF変異細胞において著しく減少することを示している。この発見は、実施例5において観察されるように、これらの変異細胞ではpolyQ凝集がより少ないという概念と一致している。凝集の減少に加えて、私たちは99Q−Ade2タンパク質の発現(NC、上方パネル)が、SPT4ΔおよびSPT5 SF細胞において減少していることも発見した。しかしながら、これらの変異細胞は29Q−Ade2の発現(NC、下方パネル)には影響しなかった。これらの結果は、Spt4/Spt5複合体の形成を妨げることによるpolyQ凝集の阻害が、現実的であることを確認するものである。さらに、短いものはそうではないが、長いpolyQの伸長ストレッチ(>36Q)を有するタンパク質の発現は、Spt4/Spt5複合体形成を阻止する変異に影響されやすい。
【実施例7】
【0077】
SPT4遺伝子欠損またはSPT5 SF変異を有する細胞内のノーザンブロット法によるpolyQ−ADE2 mRNAレベルの分析
99Q−Ade2および29Q−Ade2をコードする転写産物の発現は、実施例5において説明したように、細胞内のノーザンブロット法によって分析した。polyQ−ADE2 mRNAsをpolyQプローブによって検出し、SCR1をローディングコントロールとして含めた。正常化の後、99Q−ADE2を発現するWT細胞内のmRNAレベルを100%に設定した。示された細胞における相対的99Q−ADE2および29Q−ADE2のmRNAレベルを図8に示す。WTと比較すると、SPT4ΔおよびSPT5 SFの細胞の両方は、99Q−ADE2のmRNA発現において減少を示しているが、29Q−ADE2のmRNA発現においては減少を示していない。この観察結果は、実施例6でのSPT4ΔおよびSPT5 SFの変異細胞内における99Q−Ade2タンパク質発現の変化と一致している。
【0078】
本発見はまた、Spt4/Spt5複合体の形成を妨げることによって、短いpolyQリピートをコードする遺伝子に影響することなく、伸長CAGリピートを有する遺伝子の発現を阻害することが現実的であることも立証する。
【特許請求の範囲】
【請求項1】
細胞内での第1遺伝子の発現を調節するための方法であって、前記第1遺伝子は36を超えるリピート数を有する伸長CAGリピートを含み、前記方法は第2遺伝子の発現を抑制する段階を含み、前記第2遺伝子はSPT4、SPT5、SUPT4HおよびSUPT5Hから成る群より選択されるものである方法。
【請求項2】
請求項1の方法であって、前記第1遺伝子は、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR、およびHttの遺伝子から成る群より選択される方法。
【請求項3】
請求項1の方法であって、前記第1遺伝子は、36を超えるグルタミン残基を有する伸長ポリグルタミンストレッチを含み、かつ細胞内で凝集体を形成するタンパク質をコードする方法。
【請求項4】
請求項1の方法であって、前記抑制段階は、遺伝子ノックダウン、遺伝子ノックアウト、化学阻害剤、またはそれらの組み合わせから選択される遺伝子抑制法によって実施される方法。
【請求項5】
請求項1の方法であって、前記細胞は動物細胞または酵母細胞である方法。
【請求項6】
請求項5の方法であって、前記細胞は哺乳類細胞である方法。
【請求項7】
細胞内での第1遺伝子の発現を調節するための方法であって、該方法は、細胞内のSpt4/Spt5またはSupt4h/Supt5hの複合体形成を阻害する段階を含み、前記第1遺伝子は36を超えるリピートを有する伸長CAGリピートを含むものである方法。
【請求項8】
請求項7の方法であって、前記第1遺伝子は、36を超えるグルタミン残基を有する伸長ポリグルタミンストレッチを含み、かつ細胞内で凝集体を形成するタンパク質をコードする方法。
【請求項9】
請求項7の方法であって、前記第1遺伝子は、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR、およびHttの遺伝子から成る群より選択される方法。
【請求項10】
請求項7の方法であって、前記阻害段階は、抗体、小試薬、またはペプチドから選択される阻害剤を細胞に投与することによって実施される方法。
【請求項11】
請求項7の方法であって、前記細胞は動物細胞または酵母細胞である方法。
【請求項12】
請求項11の方法であって、前記細胞は哺乳類細胞である方法。
【請求項13】
ポリグルタミン病を治療するための方法であって、請求項1の方法を前記ポリグルタミン病を患う対象者に適用する段階を含む方法。
【請求項14】
請求項13の方法であって、前記ポリグルタミン病は、脊髄小脳失調症1、2、3、7、17型、歯状核赤核淡蒼球ルイ体萎縮症、球脊髄性筋萎縮症、およびハンチントン病から成る群より選択される方法。
【請求項15】
ポリグルタミン病を治療するための方法であって、請求項7の方法を前記ポリグルタミン病を患う対象者に適用する段階を含む方法。
【請求項16】
請求項15の方法であって、前記ポリグルタミン病は、脊髄小脳失調症1、2、3、7、17型、歯状核赤核淡蒼球ルイ体萎縮症、球脊髄性筋萎縮症、およびハンチントン病から成る群より選択される方法。
【請求項17】
第1遺伝子の調節またはポリグルタミン病の治療に有用な化合物を特定するための方法であって、前記方法は、Spt4/Spt5複合体またはSupt4h/Supt5h複合体の形成を破壊可能な阻害活性を有するものを特定するために、複数の試験化合物をスクリーニングする段階を含み、前記第1遺伝子は伸長CAGリピートを含む方法。
【請求項1】
細胞内での第1遺伝子の発現を調節するための方法であって、前記第1遺伝子は36を超えるリピート数を有する伸長CAGリピートを含み、前記方法は第2遺伝子の発現を抑制する段階を含み、前記第2遺伝子はSPT4、SPT5、SUPT4HおよびSUPT5Hから成る群より選択されるものである方法。
【請求項2】
請求項1の方法であって、前記第1遺伝子は、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR、およびHttの遺伝子から成る群より選択される方法。
【請求項3】
請求項1の方法であって、前記第1遺伝子は、36を超えるグルタミン残基を有する伸長ポリグルタミンストレッチを含み、かつ細胞内で凝集体を形成するタンパク質をコードする方法。
【請求項4】
請求項1の方法であって、前記抑制段階は、遺伝子ノックダウン、遺伝子ノックアウト、化学阻害剤、またはそれらの組み合わせから選択される遺伝子抑制法によって実施される方法。
【請求項5】
請求項1の方法であって、前記細胞は動物細胞または酵母細胞である方法。
【請求項6】
請求項5の方法であって、前記細胞は哺乳類細胞である方法。
【請求項7】
細胞内での第1遺伝子の発現を調節するための方法であって、該方法は、細胞内のSpt4/Spt5またはSupt4h/Supt5hの複合体形成を阻害する段階を含み、前記第1遺伝子は36を超えるリピートを有する伸長CAGリピートを含むものである方法。
【請求項8】
請求項7の方法であって、前記第1遺伝子は、36を超えるグルタミン残基を有する伸長ポリグルタミンストレッチを含み、かつ細胞内で凝集体を形成するタンパク質をコードする方法。
【請求項9】
請求項7の方法であって、前記第1遺伝子は、SCA1,SCA2,SCA3,SCA7,SCA17,DRPLA,AR、およびHttの遺伝子から成る群より選択される方法。
【請求項10】
請求項7の方法であって、前記阻害段階は、抗体、小試薬、またはペプチドから選択される阻害剤を細胞に投与することによって実施される方法。
【請求項11】
請求項7の方法であって、前記細胞は動物細胞または酵母細胞である方法。
【請求項12】
請求項11の方法であって、前記細胞は哺乳類細胞である方法。
【請求項13】
ポリグルタミン病を治療するための方法であって、請求項1の方法を前記ポリグルタミン病を患う対象者に適用する段階を含む方法。
【請求項14】
請求項13の方法であって、前記ポリグルタミン病は、脊髄小脳失調症1、2、3、7、17型、歯状核赤核淡蒼球ルイ体萎縮症、球脊髄性筋萎縮症、およびハンチントン病から成る群より選択される方法。
【請求項15】
ポリグルタミン病を治療するための方法であって、請求項7の方法を前記ポリグルタミン病を患う対象者に適用する段階を含む方法。
【請求項16】
請求項15の方法であって、前記ポリグルタミン病は、脊髄小脳失調症1、2、3、7、17型、歯状核赤核淡蒼球ルイ体萎縮症、球脊髄性筋萎縮症、およびハンチントン病から成る群より選択される方法。
【請求項17】
第1遺伝子の調節またはポリグルタミン病の治療に有用な化合物を特定するための方法であって、前記方法は、Spt4/Spt5複合体またはSupt4h/Supt5h複合体の形成を破壊可能な阻害活性を有するものを特定するために、複数の試験化合物をスクリーニングする段階を含み、前記第1遺伝子は伸長CAGリピートを含む方法。
【図1】

【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6(a)】
【図6(b)】
【図7】
【図8】
【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6(a)】
【図6(b)】
【図7】
【図8】
【公開番号】特開2012−125240(P2012−125240A)
【公開日】平成24年7月5日(2012.7.5)
【国際特許分類】
【出願番号】特願2011−265291(P2011−265291)
【出願日】平成23年12月2日(2011.12.2)
【出願人】(509069917)ナショナル ヤン−ミン ユニバーシティ (2)
【Fターム(参考)】
【公開日】平成24年7月5日(2012.7.5)
【国際特許分類】
【出願日】平成23年12月2日(2011.12.2)
【出願人】(509069917)ナショナル ヤン−ミン ユニバーシティ (2)
【Fターム(参考)】
[ Back to top ]