
細胞内シグナル変換操作によるT細胞応答の調節方法
【課題】T細胞共刺激に関連する細胞内シグナルを操作することによってT細胞応答を調節すること。
【解決手段】D3ホスホイノシチドの産生は、T細胞をホスファチジルイノシトール3−キナーゼのインヒビターまたはアクチベーターと接触させることによって操作しうる。本発明方法にしたがったT細胞応答の阻害は、臓器あるいは骨髄移植および自己免疫疾患などの抗原に対する免疫応答を阻害することが望ましい状況において治療上有用である。別法として、本発明方法にしたがったT細胞応答の刺激は、被検体における抗腫瘍応答を刺激すること、病原体あるいはワクチン化に対する応答を刺激することなどの抗原に対する免疫応答を強化することにおいて治療上有用である。
【解決手段】D3ホスホイノシチドの産生は、T細胞をホスファチジルイノシトール3−キナーゼのインヒビターまたはアクチベーターと接触させることによって操作しうる。本発明方法にしたがったT細胞応答の阻害は、臓器あるいは骨髄移植および自己免疫疾患などの抗原に対する免疫応答を阻害することが望ましい状況において治療上有用である。別法として、本発明方法にしたがったT細胞応答の刺激は、被検体における抗腫瘍応答を刺激すること、病原体あるいはワクチン化に対する応答を刺激することなどの抗原に対する免疫応答を強化することにおいて治療上有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、細胞内シグナル変換操作によるT細胞応答の調節に関する。
【背景技術】
【0002】
抗原特異的T細胞応答は、T細胞上の細胞表面受容体と抗原付与細胞(APCs)上のリガンドとの間の多重の相互作用によって誘発される。第1の相互作用は、T細胞上のT細胞受容体(TCR)/CD3複合体と抗原付与細胞上の主要組織適合遺伝子複合体(MHC)分子/抗原ペプチド複合体のとの間の作用である。この相互作用は、T細胞における第1の、抗原特異的、活性化シグナルの引金となる。第1の活性化シグナルに加えて、T細胞応答を誘発するには、第2の、共刺激性(costimulatory)シグナルが必要である。適当な共刺激がないと、TCRシグナリングはT細胞にアネルギー状態を誘発する。アネルギーT細胞に対し、引き続いて抗原を適当に付与すると、特有の応答を起こさなくなる[シュワルツ,R.H.らの(1990)Science248:1349を参照]。
【0003】
共刺激シグナルは、T細胞内でCD28などのT細胞表面受容体を通って発信される。たとえば、T細胞のサブ最適ポリクローナル刺激[たとえば、抗CD3抗体またはホルボールエステル(これらは第1の活性化シグナルを提供しうる)による刺激]は、CD28と抗CD28抗体の架橋によって有効化されることが説明されている[リンスリー,P.S.らの(1991)J.Exp.Med.173:721;ギミ,C.D.らの(1991)Proc.Natl.Acad.Sci.U.S.A.88:6575]。さらに、CD28の刺激は、T細胞クローンにおけるアネルギー状態の誘発を妨げることができる[ハーデイング,F.A.の(1992)Nature356:607〜609]。CD28に対する天然のリガンドはAPCs上で同定されている。CD28リガンドには、B7−1(CD80)およびB7−2(CD70などのプロテインのB7ファミリーのメンバーが含まれる[フリードマン,A.S.らの(1987)J.Immunol.137:3260〜3267;フリーマン,G.J.らの(1989)J.Immunol.143:2714〜2722;フリーマン,G.J.らの(1991)J.Exp.Med.174:625〜631;フリーマン,G.J.らの(1993)Science262:909〜911;アズマ,M.らの(1993)Nature366:76〜79;フリーマン,G.J.らの(1993)J.Exp.Med.178:2185〜2192]。CD28に加えて、B7ファミリーのプロテインも、CTLA4と呼ばれるCD28に関連するT細胞上の他の表面受容体(これは、またT細胞共刺激においても役割を演じる)に結合することがわかっている[リンスリー,P.S.らの(1991)J.Exp.Med.174:561〜569;フリーマン,G.J.らの(1993)Science262:909〜911]。
【0004】
CD28/CTLA4とB7ファミリープロテインの受容体:リガンド相互関係の解明および共刺激におけるこの相互作用の役割の解明によって、共刺激分子を結合するというT細胞表面受容体の細胞外相互作用処置などの治療用途へのアプローチが導かれた。たとえば、B7−1とB7−2の両方に結合し、それらのCD28/CTLA4との相互作用を遮断するCTLA4Ig融合プロテインは、同種および異種移植における拒絶反応の阻害のために用いられている[ターカ,L.A.らの(1992)Proc.Natl.Acad.Sci.U.S.A.89:11102〜11105;レンスコー,D.J.らの(1992)Science257:789〜792などを参照]。同様に、B7−1および/またはB7−2と反応する抗体が、インビトロにおけるT細胞の増殖およびIL−2の産生の阻害およびインビトロにおける抗原に対する第1免疫応答の阻害に用いられている[ハスコック,K.S.らの(1993)Science262:905〜907;アズマ,M.らの(1993)Nature366:76〜79;パワーズ,G.D.らの(1994)Cell.Immunol.153:298〜311;チェン,C.らの(1994)J.Immunol.152:2105〜2114]。同時に、これらの研究では、B7−1およびB7−2などの共刺激性分子を結合するT細胞表面受容体が免疫応答を操作するための所望標的であることが示されている。
【0005】
CD28/CTLA4とそれらのリガンド間の細胞外相互作用は、ある程度詳細に解明されているけれども、これらの分子の連結に続いてT細胞内で起きる細胞内のイベントについては少ししかわかっていない。T細胞の共刺激経路はシクロスポリンAの阻害効果に対して耐性があるので、T細胞の共刺激は、TCRを経るシグナリングとは異なる細胞内シグナル変換経路をもつと考えられている[ジューン,C.H.らの(1990)Immunology Today11:211〜216を参照]。プロテインチロシンのリン酸化は、T細胞内でCD28との連結により起こることがわかっており、プロテインチロシンキナーゼインヒビターであるハービマイシンAがCD28誘発IL−2産生を阻害しうることが説明されている[ヴァンデンバーグ,P.らの(1992)J.Exp.Med.175:951〜960;ルー,Y.らの(1992)J.Immunol.149:24〜29]。
【0006】
CD28受容体は、抗原受容体に関係するシグナルならびに抗原受容体とは無関係のシグナルの引金をひくことができることが研究によってわかっている。さらに、CD28のオリゴマー形成の度合は、CD28媒介シグナル変換において重要な決定要素である[リンスリー,P.S.およびレドベター,J.A.の(1993)Annu.Rev.Immunol.11:191およびレドベター,J.A.らの(1990)Blood75:1531]。抗CD28および抗CD3モノクローナル抗体を用いる研究から、カルシウムシグナリング、ホスホリパーゼC(PLC)活性化、チロシンキナーゼ活性の増加およびp21ras活性化のすべてが、受容体結合後の早期に起こることがわかっている[ジューン,C.H.らの(1994)Immunology Today15:321;およびラッド,C.E.らの(1994)IImmunol.Today15:225]。プロテインチロシンキナーゼp72itkおよびp56lck[オーガスト,A.およびデュポン,B.の(1994)Biochem.Biophys.Res.Commun.199:1466およびオーガスト,A.らの(1994)Proc.Natl.Acad.Sci.U.S.A.91:9347]は、CD28とT細胞抗原受容体の両方によって活性化される、あるいはそれらと物理的に会合すると報告されている。CD28媒介カルシウム、p21rasおよびp70s6キナーゼシグナルは休止期のT細胞よりも幼若化T細胞において顕著であり、抗原受容体シグナルと比較して一時的に遅い[レドベター,J.A.らの(1987)Proc.Natl.Acad.Sci.U.S.A.84:1384;ナンズ,J.S.らの(1993)Biochem.J.293:835;シーゲル,J.N.らの(1993)J.Immunol.151:4116;およびパイ,S.V.らの(1994)J.Immunol.24:2364]。さらに、CD8+T細胞およびCD4+T細胞はその両方がCD28シグナルに応答するけれども、抗CD28誘発カルシウムシグナルはCD4+T細胞に対して第1に制限される[アベ,R.P.らの(1995)J.Immunol.印刷中]。これらの研究は共に、CD28シグナル変換が数個のシグナル変換カスケードに連結し、シグナルの性質がT細胞発達中に規定されることを示している。
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明は、細胞内シグナル変換操作によるT細胞応答の調節に関する。さらに詳しくは本発明は、T細胞の共刺激において起こる細胞内シグナリングイベントを操作することに関する。
【課題を解決するための手段】
【0008】
本発明は、共刺激性分子を結合するT細胞の表面受容体の連結から得られる1種あるいはそれ以上の細胞内シグナルを阻害または刺激することによるT細胞応答の調節方法を含む。現在、CD28受容体刺激によって、T細胞内にD3−ホスホイノシチドが産生されることが発見されている。さらに、T細胞内のホスファチジルイノシトール3−キナーゼの活性が、リンホカイン産生および細胞増殖などのT細胞応答を阻害しうることが発見されている。これらの発見は、共刺激性シグナル変換経路におけるD3−ホスホイノシチドの機能的役割を示し、T細胞応答を調節するための細胞内標的としてのホスファチジルイノシトール3−キナーゼを提供する。したがって、D3−ホスホイノシチドを含む細胞内シグナリングイベントは、共刺激シグナルの阻害およびそれによるT細胞不応答の誘発、あるいは共刺激シグナルの発信およびそれによるT細胞応答の産生のいずれかによって調節される。さらに、T細胞応答を阻害または刺激するのに用いうる、ホスファチジルイノシトール3−キナーゼのインヒビターまたはアクチベーターを同定するための新規なスクリーニングアッセイも本発明の範囲に含まれる。
【0009】
さらに本発明は、共刺激性分子を結合する表面受容体を発現するというT細胞による応答の阻害方法に関する。このような方法には、T細胞中のD3−ホスホイノシチドの産生を阻害する作用剤にT細胞を接触させることが包含される。ひとつの具体例においては、該作用剤は、真菌代謝物であるウォルトマンニンまたはバイオフラボノイドであるクエルセチンあるいはそれらの類縁体など(たとえばLY294002)のホスファチジルイノシトール3−キナーゼのインヒビターである。本発明方法の別の具体例においては、プロテインチロシンリン酸化などの共刺激に関連する異なる細胞内シグナルを阻害する作用剤の少なくとも1種にT細胞を接触させる。たとえば、ホスファチジルイノシトール3−キナーゼおよびプロテインチロシンキナーゼのインヒビターにT細胞を接触させることができる。好ましいプロテインチロシンキナーゼのインヒビターはハービマイシンAである。別法として、チロシンホスファターゼまたはチロシンホスファターゼのアクチベーターによってT細胞内でのプロテインチロシンリン酸化を阻害することができる。この具体例においては、ホスファチジルイノシトール3−キナーゼおよび細胞チロシンホスファターゼに結合して活性化させる抗体などの分子(たとえばCD45またはHcph)にT細胞を接触させることができる。本発明方法の別の好ましい具体例においては、プロテインセリンキナーゼのインヒビターまたはセリンホスファターゼのアクチベーターにT細胞を接触させる。
【0010】
さらに本発明は、T細胞内の共刺激に関連する細胞内シグナルを妨害しながらT細胞における第1の、抗原特異的シグナルの引金をひくことによってT細胞内に抗原に対する不応答を誘発する方法を提供する。共刺激性シグナル変換の妨害の結果として、抗原の存在下でT細胞が適当な共刺激性シグナルを受け取ることができず、抗原特異的不応答がT細胞内に誘発される。T細胞の不応答を誘発するために、T細胞における第1の活性化シグナルの刺激に適した形体の抗原およびT細胞内におけるD3−ホスホイノシチドの産生を阻害する作用剤に抗原特異的T細胞を接触させる。たとえば、APCによって供与される抗原およびウォルトマンニンまたはクエルセチンあるいはそれらの誘導体または類縁体など(たとえばLY294002)のホスファチジルイノシトール3−キナーゼのインヒビターにT細胞を接触させることができる。さらに、プロテインチロシンリン酸化および/またはプロテインセリンリン酸化などの共刺激に関連する他の細胞内シグナルをT細胞内で阻害することができる。
【0011】
T細胞応答の阻害方法およびT細胞不応答の誘発方法は、たとえば、移植を受ける被験体(臓器移植または骨髄移植など)または自己免疫疾患あるいは異常免疫応答に関連する他の疾病に罹患している被験体などの免疫応答を下降方向に調節することが望ましい状況において有用である。共刺激に関連するシグナル変換を阻害する作用剤(たとえばイノシトールリン酸3−キナーゼのインヒビターなど)を被験体に投与することができ、また別法として被験体からT細胞を採取し、本明細書に記載したようにインビトロで処理し、それを被験体に投与することもできる。
【0012】
さらに本発明は、第1の活性化シグナルを受け、共刺激分子を結合する表面受容体を発現したT細胞によって応答を刺激する方法に関する。このような方法には、ホスファチジルイノシトール3−キナーゼなどのT細胞中のD−3ホスホイノシチドの産生を刺激する作用剤にT細胞を接触させることが包含される。別の具体例においては、D3−ホスホイノシチドの産生を刺激する作用剤およびプロテインチロシンリン酸化などの共刺激に関連する異なる細胞内シグナルを刺激する作用剤の少なくとも1種にT細胞を接触させる。たとえば、ホスファチジルイノシトール3−キナーゼおよび過バナジウム酸塩などのプロテインチロシンキナーゼのアクチベーターにT細胞を接触させることができる。本発明の別の具体例においては、さらに、プロテインセリンキナーゼを活性化する作用剤にT細胞を接触させる。別法として、CD45またはHcphなどの細胞ホスファターゼのインヒビターをPI3Kアクチベーターとともに用いることができる。さらなる本発明の他の具体例においては、抗原特異的T細胞を抗原およびT細胞内でD−3ホスホイノシチドの産生を刺激する作用剤に接触させることによって抗原特異的T細胞応答を刺激し、それによってT細胞内における第1の活性化シグナルおよび共刺激性シグナルの両方を刺激する。本発明の別の具体例においては、1種またはそれ以上のプロテインチロシンおよび/またはプロテインセリンキナーゼを活性化する作用剤に抗原特異的T細胞を接触させることによって、抗原特異的T細胞応答をさらに刺激する。
【0013】
T細胞応答を刺激する方法は、免疫応答を上昇方向に調節することが望ましい状況において有用である。たとえば、腫瘍ができている被験体における腫瘍に対する応答、あるいは被験体の病原体(たとえばバクテリア、HIVなどのウイルス、真菌、寄生虫など)に対する応答を刺激することができる。さらに、本発明方法は、ワクチン接種の効能を増強するのに用いることができる。共刺激に関連する細胞内シグナルを刺激する作用剤(イノシトールホスファターゼ3−キナーゼのアクチベーターなど)を被験体に投与するかまたは別法としてインビトロにてT細胞を刺激し、次いでそれを被験体に投与することができる。
【0014】
本発明はさらに、ホスファチジルイノシトール3−キナーゼのインヒビターまたはアクチベーターを同定するためのスクリーニングアッセイに関する。ひとつの具体例においては、共刺激性分子を結合する細胞表面受容体を発現するT細胞(CD28など)が用いられる。インヒビターを同定するには、T細胞における受容体に関連する細胞内シグナル変換経路を、被検作用剤の存在下に刺激し、T細胞におけるD−3ホスホイノシチドの少なくとも1種の産生を阻害する能力に基づいてインヒビターを同定する。アクチベーターを同定するには、被検作用剤にT細胞を接触させ、T細胞におけるD−3ホスホイノシチドの少なくとも1種の産生を刺激する能力に基づいてアクチベーターを同定する。
本発明はさらに、CD28を介して刺激されたT細胞におけるCD28に関連するおよそ67kDaの新規なチロシン−リン酸化プロテインに関する。該プロテインの活性化あるいは該プロテインのCD28との相互作用を遮断する方法、または別法として、該67Kプロテインの活性化あるいは該プロテインのCD28との相互作用を刺激する方法もまた本発明に包含される。
【0015】
本発明は、T細胞上の表面受容体の共刺激分子への結合などの共刺激によってT細胞において産生される細胞内シグナルを調節することによるT細胞応答の調節方法に関するものである。さらに詳しくは本発明は、共刺激性シグナルを阻害または刺激し、それによってT細胞応答を阻害または刺激するためのT細胞におけるD−3ホスホイノイシチド産生の調節に関する。共刺激に関連するシグナル変換の妨害による共刺激性シグナルの阻害を利用してさらに、T細胞不応答を誘発することができる。本発明は、CD28表面受容体を介するT細胞の刺激によってT細胞にD−3ホスホイノシチドが産生され、CD28が連結したT細胞においてホスファチジルイノシトール3−キナーゼ(本明細書においてPI3Kとも称する)のインヒビターがD−3ホスホイノシチドの産生を阻害するという発見に少なくとも部分的には基づいている。さらに本発明は、T細胞におけるPI3K活性を阻害すると、サイトカイン産生および細胞増殖などのT細胞応答が阻害されるという発見に少なくとも部分的には基づいている。
【0016】
したがって、さらに本発明は、シグナル変換に関連する細胞内シグナル変換を妨害することによるT細胞の応答を阻害する方法に関する。ひとつの具体例においては、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤に、共刺激性分子を結合する細胞表面受容体を発現しているT細胞を接触させることによって細胞内シグナルを阻害する。語句「共刺激性分子に結合する細胞表面受容体を発現しているT細胞」とは、CD28および/またはCRLA4、あるいはB7−1、B7−2または他のB7ファミリーのメンバーなどの共刺激性分子を結合する能力をもつ他の受容体を発現しているT細胞を意味する。
【0017】
T細胞による「応答」とは、T細胞における第1の活性化シグナルおよび共刺激性シグナルの引金をひくことによって生じるT細胞応答を意味し、リンホカインの産生(IL−2産生など)およびT細胞の増殖が含まれる。T細胞応答の阻害には、応答の完全遮断(無応答など)または応答の大きさの程度の減少(応答の部分的阻害など)も含まれる。
「D−3ホスホイノシチド」とは、イノシトール環のD−3位でリン酸化されるホスファチジルイノシトールの誘導体を意味し、ホスファチジルイノシトール(3)−モノリン酸塩(PtdIns(3)P)、ホスファチジルイノシトール(3,4)−ビスリン酸塩(PtdIns(3,4)P2)およびホスファチジルイノシトール(3,4,5)−トリリン酸塩(PtdIns(3,4,5)P3)などの化合物が含まれる。
D−3ホスホイノシチドはホスファチジルイノシトール3−キナーゼ(PI3K)の活性によって細胞内的に産生される。PI3Kは、そのSH2ドメインを介してチロシル−リン酸化プロテインを結合する85kDaのサブユニットおよび110kDaの触媒サブユニットからなるヘテロ二量体である。PI3Kは最初、ホスファチジルイノシトール、PtdIns(4)PおよびPtdIns(4,5)P2のイノシトール環のD−3位をリン酸化する脂質キナーゼとして同定された。最近の2つの研究では、PI3Kが、事実、脂質およびセリンキナーゼ活性の両方を有する2重特異的キナーゼであることが説明されている[ジャンド,R.らの(1991)EMBO J.13:522およびカーペンター,C.L.らの(1993)Mol.Crll Biol.13:1657]。
【0018】
したがって、ひとつの具体例においては、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤は、PI3Kの活性を阻害する作用剤である。T細胞におけるPI3K活性を阻害する好ましい作用剤は、真菌代謝物であるウォルトマンニンまたはその誘導体あるいは類縁体である。ウォルトマンニンはT・ウォルトマンニイ(T.wortmannii)(協和発酵株式会社)またはP.フミクロスム(P.fumiculsum)(シグマ)から誘導されるインヒビターである。ウォルトマンニン誘導体または類縁体には、PI3KおよびT細胞応答を阻害する能力をもつウォルトマンニンに構造的に関連する化合物が含まれる。ウォルトマンニン誘導体よび類縁体の例は、ワイシンガー,D.らの(1974)Experientia30:135〜136;クロッス,A.らの(1981)J.Med.Chem.24:1465〜1471;およびバッジョリーニ,M.らの(1987)Exp.Cell Res.169:408〜418に開示されている。使用しうるその他のPI3K活性のインヒビターは、バイオフラボノイドであるクエルセチンまたはその誘導体あるいは類縁体である。クエルセチン誘導体または類縁体には、PI3KおよびT細胞応答を阻害する能力をもつクエルセチンに構造的に関連する化合物が含まれる。クエルセチン誘導体および類縁体の例は、ブラホス,C.J.らの(1994)J.Biol.Chem.269:5241〜5284に開示されている。PI3K活性を阻害する好ましいクエルセチン誘導体は、LY29400[2−(4−モルホリニル)−8−フェニル−4H−1−ベンゾピラン−4−オン](リリー、インデイアナポリス、IN)(上記ブラホスらに記載)。別法として、後記方法で同定される他のPI3Kインヒビターを用いることもできる。
【0019】
本発明の別の態様としては、共刺激に関連する2種またはそれ以上の細胞内シグナリングイベントの妨害によるT細胞応答の阻害が含まれる。たとえば、CD28の刺激によってT細胞におけるプロテインチロシンリン酸化が起こることが示されている[ヴァンデンバーグ,P.らの(1992)J.Exp.Med.175:951〜960;ルー,Y.らの(1992)J.Immunol.149:24〜29などを参照]。したがって、ひとつの具体例においては、T細胞応答は、T細胞における少なくとも1種のD−3ホスホイノシチドの産生を阻害する第1作用剤およびT細胞におけるチロシンリン酸化を阻害する第2作用剤にT細胞を接触させることによって阻害される。たとえば、T細胞をPI3K活性を阻害する作用剤およびプロテインチロシンキナーゼ活性を阻害する作用剤の両方に接触させることができる。好ましいプロテインチロシンキナーゼインヒビターは、srcプロテインチロシンキナーゼを阻害するものである。ひとつの具体例においては、該srcプロテインチロシンキナーゼインヒビターは、ハービマイシンAまたはその誘導体もしくは類縁体である。ハービマイシンAの誘導体もしくは類縁体には、構造的にハービマイシンAに関連し、プロテインチロシンキナーゼを阻害する能力をもつ化合物が含まれる。別の具体例においては、プロテインチロシンリン酸化を阻害する作用剤は、プロテインチロシンホスファターゼまたはプロテインチロシンホスファターゼのアクチベーターである。T細胞におけるチロシンホスファターゼ活性の増加によって、プロテインチロシンリン酸化の正味量が減少する。プロテインチロシンホスファターゼは、T細胞内においてはCD45またはHcphなどの細胞プロテインチロシンホスファターゼとなりうる。T細胞上の細胞表面チロシンホスファターゼの活性は、ホスファターゼに結合し、その活性を刺激する分子に、T細胞を接触させることによって活性化することができる。たとえば、CD45に対する抗体を用いて、CD45をその表面に発現するT細胞におけるチロシンホスファターゼの活性を刺激することができる。したがって、ひとつの具体例においては、T細胞内においてプロテインチロシンリン酸化を阻害する作用剤は、抗CD45抗体またはCD45の活性を刺激する能力をもつそのフラグメントである。抗体フラグメントの例として、FabおよびF(ab')2フラグメントが挙げられる。抗体またはそのフラグメントは、多量体化または固定化などの刺激性形体で提供することができる。
【0020】
共刺激に関連する他の細胞内シグナルは、D−3ホスホイノシチド産生の阻害とともに阻害され、その結果としてT細胞応答を阻害することができる。たとえば、CD28の連結は、ホスホリパーゼCの活性の増加[ナン,J.らの(1993)Biochem.J.293:835〜842を参照)、および細胞内カルシウム濃度の増加[レドベター,J.A.らの(1990)Blood 75:1531〜1539および実施例を参照]に関連することがわかっている。したがって、PI3K活性を阻害する第1作用剤およびホスホリパーゼC活性を阻害および/または細胞内カルシウム濃度の増加を阻害する第2作用剤に、T細胞を接触させる。実施例に示すように、チロシンキナーゼインヒビターであるハービマイシンAもまたT細胞におけるCD28誘発性カルシウム代謝を阻害する。
【0021】
さらに、実施例に示すように、CD28を介する刺激の結果として、ハービマイシン耐性のT細胞によるIL−2の産生が起こり、これは、関係するシグナル変換経路に、プロテインセリンキナーゼなどのさらなるプロテインキナーゼが含まれることを示唆する。
【0022】
いくつかの研究によって、プロテインセリンキナーゼが、CD28に関連するシグナル変換経路に含まれることが示されている[シーゲル,J.N.らの(1993)J.Immunol.151:416〜4127およびパイ,S.V.らの(1994)J.Immunol.24:2364]。かくして、本発明の特定の具体例においては、PI3K活性を阻害する作用剤およびセリンキナーゼを阻害する作用剤に、T細胞を接触させることによって、T細胞応答が阻害される。別の具体例においては、このようなT細胞をさらに、プロテインチロシンキナーゼを阻害する作用剤に接触させる。さらに別の具体例においては、PI3K、プロテインチロシンキナーゼ、プロテインセリンキナーゼ、ホスホリパーゼC活性を阻害する作用剤の種々の組合せ、または細胞内カルシウムの増加を阻害する作用剤に、T細胞を接触させることによってT細胞応答を阻害する。
T細胞応答は、本発明方法にしたがってインビトロまたはインビボのいずれにおいても阻害することができる。たとえば、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤を、T細胞応答を阻害するのに十分な用量および期間において被験体に投与することができる。別法として、被験体からT細胞を採取し、インビトロにてT細胞を作用剤に接触させ、被験体に再投与することができる。「被験体」とは、ヒト、サル、イヌ、ネコ、ウサギ、ラット、マウスおよびそれらのトランスジェニック種などの哺乳動物などの免疫応答を起こす動物を意味する。T細胞応答を阻害しうる被験体には、免疫応答を下降調節することが望ましい被験体が含まれる。下降調節とは、リンホカイン産生およびT細胞増殖などのT細胞応答の部分的および完全阻害を意味する。たとえば、自己免疫疾患または免疫応答異常に関連する他の疾病の被験体、あるいは骨髄移植または他の臓器移植などの移植を受ける者に対し、本発明方法を適用することができる。
【0023】
本発明のひとつの具体例においては、T細胞内において共刺激性シグナルを阻害することによって抗原特異的T細胞不応答を誘発する。したがって、本発明の別の態様は、抗原に対するT細胞不応答を誘発する方法である。本明細書で用いる「T細胞不応答」とは、抗原(または抗原性部分)に対して、さらされた時にT細胞によるT細胞応答(増殖、リンホカイン分泌またはエフェクター機能の誘発など)が減少するかまたは応答しなくなり、T細胞が不応答になることを意味する。「T細胞不応答」および「T細胞アネルギー」は、本明細書においては互換的に用いられる。抗原に対する不応答は、T細胞における抗原特異的第1活性化シグナル(TCR/CD3複合体を介する活性化など)が、共刺激性シグナルが無い状態で引金を引かれることによって誘発することができる。本発明方法においては、共刺激に関連する細胞内シグナルを妨害する作用剤に、T細胞を接触させることによって、T細胞における共刺激性シグナルを遮断する。特に、共刺激性シグナルを阻害するためのT細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤の存在下に、T細胞における第1の活性化シグナルの引金をひくのに適した形体の抗原に、抗原特異的T細胞(すなわち、抗原を認識するTCRを発現しているT細胞)を接触させることによって、抗原に対するT細胞不応答を誘発することができる。該抗原は、たとえば、自己免疫反応を刺激する抗原または移植された細胞の拒絶反応を刺激するアロ抗原である。おそらく、D−3ホスホイノシチドの産生を阻害する作用剤が、ウォルトマンニンまたはクエルセチンなどのT細胞におけるPI3Kまたはその誘導体もしくは類縁体(LY294002など)の活性を阻害する。共刺激に関連する他の細胞内シグナルを阻害する追加の作用剤(前述)もまた、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤とともに用いることができる。たとえば、PI3KならびにハービマイシンAなどのプロテインチロシンキナーゼに、T細胞を接触させることができる。
【0024】
T細胞不応答を誘発するために、T細胞において第1の活性化シグナルの引金をひくのに適した形体の抗原に、抗原特異的T細胞を接触させるが、このことは、T細胞においてTCR/CD3複合体を介するシグナルが引金を引かれるように抗原をT細胞に対して付与することを意味する。たとえば、MHC分子に関連して細胞を付与する抗原によって、T細胞に対して抗原を付与することができる。B細胞、マクロファージ、モノサイト、樹状細胞、ランゲルハンス細胞またはT細胞に対して抗原を付与するその他の細胞などのシンジェネティックな抗原付与細胞を、抗原付与細胞がT細胞に対して抗原を付与するような抗原の存在下に、T細胞とともにインキュベートすることができる。別法として、アロ抗原に対するアネルギーを誘発するために、T細胞に対するアロ抗原を付与するアロジェネティックな細胞とともにT細胞をインキュベートすることができる。
【0025】
インビボにおいて抗原に対するT細胞不応答を誘発するために、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤を、抗原に対するT細胞不応答を誘発するのに十分な用量と期間で、被験体に投与する。作用剤投与に続いて、抗原(たとえば内因的に細胞によって発現される自己抗原)を抗原特異的T細胞に内因的に接触させる。別法として、抗原に対する不応答を、インビトロにて誘発することができる。この場合、T細胞を被験体から採取し、インビトロにて抗原ならびに抗原不応答を誘発する作用剤に接触させ、次いで被験体にT細胞を再投与する。たとえば、移植片を移植を受ける者に移植する前に、移植を受ける者から採取したT細胞を、T細胞におけるD−3ホスホイノシチド産生を阻害する作用剤(たとえばウォルトマンニン、クエルセチン、LY294002)および/または共刺激に関連する他の細胞内シグネルの1種あるいはそれ以上を阻害する1種あるいはそれ以上の作用剤とともに移植片ドナーからのアロジェニックな細胞に接触させて、アロ抗原特異的T細胞不応答を誘発することができる。次いで、ドナーの抗原に対して不応答になった移植を受ける者のT細胞を、該移植を受ける者に再投与する。別法として、骨髄移植の場合、移植される骨髄(いずれかの残留T細胞を含む)を、インビトロにて、D−3ホスホイノシチドの産生および/または他の共刺激に関連する細胞内シグナルを阻害する作用剤とともに骨髄移植を受ける者のアロジェネティックな細胞に接触させて、移植を受ける者のアロ抗原に対するドナーT細胞における不応答を誘発する。この前処理によって移植片と移植を受けた者の間で生じる疾患を阻害することができる。
【0026】
該T細胞不応答誘発方法は、臓器移植および骨髄移植などの移植、および自己免疫疾患および異常免疫応答に関連する他の疾病などの免疫応答を下降調節することが望ましい状況において治療的に適用することができる。自己免疫疾患または不適当あるいは異常免疫応答に関連する他の疾病の例として、リウマチ性関節炎、若年性リウマチ性関節炎、乾せん性関節炎、アレルギー、接触性皮膚炎、乾せん、癩病性逆反応、癩病性結節性紅斑、自己免疫性ぶどう膜炎、多発性硬化症、アレルギー性脳脊髄炎、全身性紅斑性狼瘡、急性壊死性出血性脳障害、突発性両側性進行性感覚神経聴力損失、再生不良性貧血、真正赤血球性貧血、突発性血小板血症、多発性軟骨炎、強皮症、ウェジナー肉芽腫症、慢性活動性肝炎、重症筋無力症、スチーブン−ジョンソン症候群、突発性スプルー、偏平苔せん、クローン病、グレイブス眼病、サルコイドーシス、原発性胆汁性肝硬変、原発性若年性糖尿病、シェーグレン症候群関連乾き眼、後ぶどう膜炎および間質性肺繊維症が挙げられる。
【0027】
本発明の他の態様は、共刺激性シグナルをT細胞に与えることによってT細胞応答を刺激することに関する。第1活性化シグナルにつづく共刺激シグナルの伝達によってT細胞応答を発生させることができる。本発明方法においては、共刺激性シグナルは、T細胞においてD−3ホスホイノシチドの産生を刺激する作用剤に、第1活性化シグナルを受けたT細胞を接触させることにより付与される。T細胞の「応答」とは、T細胞(IL−2など)による少なくとも1種のリンホカインの細胞および/またはT細胞の増殖を意味する。第1活性化シグナルは、抗CD3抗体またはMHC/抗原複合体などのTCR/CD3複合体を介してT細胞を刺激すること、またはホルボールエステル(PMA)などのこの刺激を模倣する作用剤を用いることによってT細胞に伝達される。本明細書中で用いる語句「作用剤」とは、T細胞表面受容体と共刺激分子の間に相互作用を要求することなくT細胞において共刺激シグナルを刺激する化学薬品および他の薬剤化合物を意味する。たとえば、該作用剤は、細胞内的に作用して、共刺激関連シグナルを刺激する。ひとつの具体例においては、該作用剤は非プロテイン性化合物である。本発明方法に用いる該作用剤は、天然の受容体:リガンド共刺激メカニズムをバイパスすることを意図しており、語句「作用剤」には、B7−1および/またはB7−2などの天然のリガンドCD28/CTLA4を発現している細胞を含まない。
【0028】
おそらく、T細胞におけるD−3ホスホイノシチドの産生は、PI3KのアクチベーターにT細胞を接触させることによって刺激される。PI3Kの活性化は、たとえば、後述の方法によって同定することができる。共刺激に関連する1種またはそれ以上の他の細胞内シグナルを刺激する追加の作用剤を、D−3ホスホイノシチド産生のアクチベーターとともに用いることができる。たとえば、PI3Kを刺激する第1作用剤およびT細胞内におけるプロテインチロシンリン酸化を刺激する第2作用剤の両方に、T細胞を接触させる。T細胞におけるプロテインチロシンリン酸化は、過バナジン酸塩などのプロテインチロシンキナーゼのアクチベーターに、T細胞を接触させることによって刺激することができる[オシア,J.J.らの(1992)Proc.Natl.Acad.Sci.USA89:10306〜10310;およびシーリスト,J.P.の(1993)J.Biol.Chem.268:5886〜5893を参照]。たとえば、PI3Kを刺激する第1作用剤、およびプロテインセリンキナーゼのアクチベーターにT細胞を接触させるなどのT細胞内におけるプロテインセリンリン酸化を刺激する第2作用剤にT細胞を接触させてもよい。別の具体例においては、プロテインチロシンリン酸化を刺激する第3の作用剤にさらにT細胞を接触させる。別法として、PI3Kのアクチベーター、およびT細胞における正味のプロテインチロシンリン酸化を増加するためのCD45などの細胞プロテインチロシンホスファターゼの活性および/または細胞プロテインセリンホスファターゼの活性を阻害する作用剤の両方にT細胞を接触させる。本発明には、ホスホリパーゼC活性の刺激および/または細胞内カルシウム濃度の増加などのT細胞の共刺激に関連する他の細胞内シグナルを刺激することも含まれる。
【0029】
本発明の他の具体例は、抗原特異的T細胞による抗原に対する特異的応答を刺激する方法を提供する。T細胞応答を刺激するために、抗原ならびにT細胞においてD−3ホスホイノシチドの産生を刺激する作用剤に、抗原特異的T細胞を接触させ、それによってT細胞における共刺激シグナルの引金をひく。おそらく、T細胞においてD−3ホスホイノシチドの産生を刺激する作用剤はPI3Kのアクチベーターである。MHC分子に関連の、T細胞における第1の活性化シグナルを刺激するのに適した形体の抗原に、T細胞を接触させる(TCR/CD3複合体を介するなど)。抗原付与細胞(B細胞、マクロファージ、モノサイト、樹状細胞、ランゲルハンス細胞またはT細胞に対して抗原を付与するその他の細胞など)を、抗原(可溶性抗原など)の存在下にT細胞とインキュベートすることができる。別法として、対象となる抗原を発現している細胞をT細胞とインキュベートすることができる。たとえば、腫瘍関連抗原を発現している腫瘍細胞を、T細胞ならびに細胞内共刺激シグナルを誘発する作用剤とともにインキュベートして腫瘍特異的応答を誘発することができる。別法として、ウイルスなどの病原性抗原を付与する病原体に感染した細胞を、作用剤の存在下にT細胞とインキュベートすることができる。T細胞においてD−3ホスホイノシチドの産生を刺激することに加えて、たとえば過バナジン酸塩またはセリンキナーゼのアクチベーターなどのプロテインチロシンキナーゼのアクチベーターなどのCD28連結に関連する1種またはそれ以上の追加の細胞内シグナルを刺激する1種またはそれ以上の作用剤に、T細胞を接触させることができる。
【0030】
PI3KのアクチベーターなどのT細胞においてCD28関連細胞内シグナルを刺激する作用剤を、インビボにて被検体に投与することができる。またあるいは別法として、被検体から採取したT細胞をインビトロにて刺激し、被検体に再投与してもよい。T細胞を刺激する方法は、免疫応答を上昇調節することが望ましい状況(たとえば応答の誘発または存在する応答の強化)において治療上有用である。たとえば、該方法を用いて腫瘍関連抗原に対するT細胞応答を強化することができる。被検体からの腫瘍細胞は、典型的に腫瘍関連抗原を発現するが、T細胞における共刺激シグナルを刺激することはできない(たとえば、それらは共刺激分子の発現が欠如しているからである)。したがって、被検体からのT細胞ならびにT細胞においてD−3ホスホイノシチドを刺激する作用剤に、腫瘍細胞を接触させてT細胞における共刺激の引金をひくことができる。別法として、本明細書中に記載したようにT細胞を刺激して、病原性作用剤(ウイルス(ヒト免疫不全ウイルスなど)、バクテリア、寄生虫および真菌など)に対する応答性を誘発または強化することができる。さらに、このような病原性作用剤に対するワクチンの効能を増強することもできる。たとえば、T細胞においてD−3ホスホイノシチドの産生を刺激する作用剤を、病原性作用剤に感染した被検体に投与することができ、またはワクチンとともに投与して、ワクチンの抗原に対するT細胞の応答性を強化することができる。別法として、病原性作用剤からの抗原を発現する抗原付与細胞ならびに共刺激に関連する細胞内シグナルを刺激する作用剤(PI3Kのアクチベーター)とともに、T細胞をインビトロで培養することができる。
【0031】
さらに本発明のT細胞応答を刺激する方法は、CD28および/または共刺激に関連する他の細胞表面分子(CTLA4など)を介するシグナル変換が不良である患者に適用することができる。たとえば、突発性血小板減少症を患っている患者が、おそらく先天性欠陥であろうが、CD28介在性シグナル変換不良を呈することが報告されている[ペレッツ−ブラス,M.らの(1991)Clin.Exp.Immunol.85:424〜428]。CD28シグナリング能力不全の患者においては、正常CD28連結によって生成する細胞内シグナルを刺激する1種またはそれ以上の作用剤を、患者のT細胞に接触させることによって、該不良をバイパスし、CD28依存性T細胞活性化を回復することが可能である。たとえば、CD28連結によるD−3ホスホイノシチドの産生が低下または欠如していることによる不良をもつ患者を、T細胞においてD−3ホスホイノシチドの産生を刺激する作用剤に患者からのT細胞を接触させることによって、治療することができる。
【0032】
本発明のさらに別の態様は、PI3Kのインヒビターおよびアクチベーターを同定するためのスクリーニングアッセイであり、それらを用いてT細胞応答をそれぞれ阻害または刺激することができる。PI3Kは調節および触媒サブユニットからなるヘテロ二量体である。基質としてPtdIns(4,5)P2を優先的に用い、ウォルトマンニンによって阻害しうる2つの形体の酵素が記載されている[オツ,M.らの(1991)Cell65:91〜104;ヒュー,P.らの(1993)Mol.Cell.Biol.13:7677〜7688;およびハイルス,I.D.らの(1992)Cell70:419を参照]。基質としてPtdInsを優先的に用い、ウォルトマンニンによって阻害されない別の形体の酵素もまた記載されている[スティーブンス,L.らの(1994)Curr.Biol.4:203〜214を参照]。PI3Kの特異的インヒビターまたはアクチベーターの同定は、望まない逆の副作用を回避するための共刺激シグナルに必要とされるPI3Kの好適な細胞内形体に対して特異的でなければならないことが認識されよう。したがって、共刺激に必要とされるPI3Kの形体(たとえば、ウォルトマンニンによっても阻害しうる形体)を特異的に阻害または活性化する作用剤が好ましい。
【0033】
ひとつの具体例において、本発明のスクリーニングアッセイは、T細胞において少なくとも1種のD−3ホスホイノシチド(PtdIns(3,4,5)P3が好ましい)の産生をそれぞれ阻害または刺激する、PI3Kのインヒビターまたはアクチベーター能力に基づいている。PI3Kのインヒビターを同定するために、被検基質の存在あるいは不在下に、共刺激分子を結合する細胞表面受容体を介してT細胞を刺激する(すなわち、共刺激シグナルを受容したT細胞)。好ましくは、CD28を発現するT細胞をアッセイに用いる。別法として、CTLA4を発現するT細胞を用いてもよい。CD28またはCTLA4に対するリガンドに、T細胞を接触させることによって、T細胞における共刺激シグナルを刺激することができる。該リガンドは、T細胞表面受容体に対する抗体よりもむしろ膜結合B7−1またはB7−2などの生理学的リガンドであることが好ましい。天然にB7−1および/またはB7−2を発現する細胞を用いることができるが、共刺激分子を発現するようにトランスフェクトされた細胞(CHO細胞など)を用いるのが、より好ましい。PI3Kのインヒビターの存在下に、共刺激分子(CD28など)を結合する表面受容体を介するT細胞を刺激する結果として、該インヒビターの不在下における刺激と比較して、T細胞におけるD−3ホスホイノシチドの産生が低減化される。T細胞におけるD−3ホスホイノシチドの産生は、当業者には公知の適当な方法のいずれかによって測定することができる。たとえば、高圧液体クロマトグラフィーによって、T細胞におけるD−3ホスホイノシチドの産生を測定することができる(実施例に記載)。別法として、薄層クロマトグラフィーによってD−3ホスホイノシチドの産生を評価することができる[オカダ,T.らの(1994)J.Biol.Chem.269:3563〜3567]。細胞内産生を評価しえたD−3ホスホイノシチドとして、PtdIns(3)P、PtdIns(3,4)P2およびPtdIns(3,4,5)P3が挙げられる。被検基質の存在下に、T細胞におけるPtdIns(3,4,5)P3の産生を検出するのが好ましい。
【0034】
PI3Kのアクチベーターを同定するために、共刺激分子を結合する細胞表面受容体を発現するT細胞を、被検基質と接触させる。T細胞において少なくとも1種のD−3ホスホイノシチド(PtdIns(3,4,5)P3が好ましい)の産生を刺激する能力に基づいてPI3Kのアクチベーターを同定する。したがって、PI3Kアクチベーターの存在下において、T細胞におけるD−3ホスホイノシチドの量は、基質不在下におけるT細胞中のD−3ホスホイノシチドの量と比較して増加する。T細胞におけるD−3ホスホイノシチド[PtdIns(3)P、PtdIns(3,4)P2およびPtdIns(3,4,5)P3]の産生は、高圧液体クロマトグラフィーまたは薄層クロマトグラフィーなどの標準的方法によって評価することができる。
【0035】
本発明スクリーニングアッセイの別の具体例においては、細胞から単離されるPI3Kの活性を直接阻害または刺激するための基質の能力を評価し、次いで、PI3Kのインヒビターまたはアクチベーターとして同定された基質を、T細胞に接触させて、T細胞応答を阻害または刺激する基質の能力を決定する。たとえば、単離されたPI3Kを、放射標識されたホスフェートドナーおよび被検物質の存在下に、基質(PtdIns(4,5)P2など)とともにインキュベートする。PI3Kのキナーゼ活性のインヒビターは、インヒビター不在下におけるリン酸化と比較して、基質のリン酸化を低下させるであろう。一方、アクチベーターの不在下におけるリン酸化と比較して、アクチベーターは基質のリン酸化を増加するであろう。次いで、このように同定されたインヒビターまたはアクチベーターを、T細胞に接触させて、インヒビターまたはアクチベーターのT細胞応答(IL−2の産生など)の阻害または刺激能力をそれぞれ決定する。
【0036】
さらに本発明は、実施例に記載するように、ジャーカット細胞および末梢血液T細胞におけるCD28に関連する約67kDaのホスホチロシンプロテインに関する。67Kと称するこの新規なタンパク質は、itk/emtプロテインチロシンキナーゼ、lckおよびPI3Kとは異なると考えられ、CD28に関連するシグナル変換経路に含まれるようである。本発明の特定の具体例において、該67Kが含まれるCD28シグナル変換経路は、たとえば、そのリン酸化の遮断などによって67Kの活性化を遮断することによって阻害される。別法として、該67Kに関連するシグナル変換経路は、67KとCD28の相互作用を阻害することによって遮断される。本発明の別の具体例においては、67Kに関連するシグナル変換経路は、67Kを活性化することによるかまたは67KとCD28の相互作用を刺激することによって刺激される。67Kを含むシグナル変換経路の活性化または阻害を、上述の具体例のいずれかによって行うことができ、それによってT細胞応答が遮断されるかまたは別法においてはT細胞応答が刺激される。
【0037】
他の具体例
T細胞以外にも、CD28をその表面に発現する他のタイプの細胞が文献に記載されている。このような細胞のタイプには、プラズマ細胞[コズボール,D.らの(1987)J.Immunol.138:4128〜4132]および骨髄誘導マスト細胞が含まれる。CD28を介する他のCD28+細胞の刺激によっても、該細胞においてD−3ホスホイノシチドの産生および特異的細胞応答の発生が誘導される。本発明方法による、これらの細胞におけるD−3ホスホイノシチドの阻害および活性化もまた、他のCD28+細胞型による応答を阻害または刺激するのに有用である。
【図面の簡単な説明】
【0038】
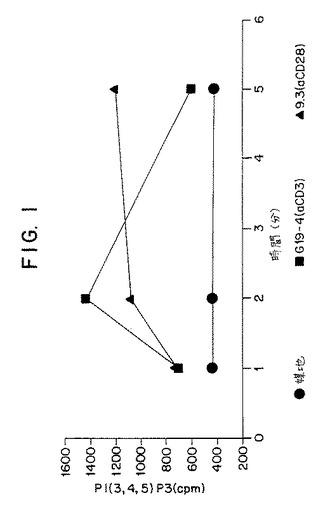
【図1】第1図は、CD28+T細胞(ジャーカット細胞)における、該細胞を培地、抗CD3抗体または抗CD28抗体で刺激した後のホスファチジルイノシトール(3,4,5)−トリリン酸塩の産生を示すグラフであり、CD3またはCD28を介した刺激におけるホスファチジルイノシトール3−キナーゼの活性化の独特の動力学を表している。
【図2】第2図は、CD28+T細胞(ジャーカット細胞)における、該細胞をB7−1またはB7−2を発現するようにトランスフェクトされたCHO細胞で刺激した後のホスファチジルイノシトール(3,4,5)−トリリン酸塩の産生を示すグラフであり、B7−1またはB7−2による刺激におけるホスファチジルイノシトール3−キナーゼの活性化の独特の動力学を表している。
【図3】第3図は、CD28+T細胞(ジャーカット細胞)における、該細胞をB7−2を発現するようにトランスフェクトされたCHO細胞で刺激した後のホスファチジルイノシトール(3,4,5)−トリリン酸塩の産生に関する種々の濃度のウォルトマンニン(0〜100mM)の効果を示すグラフである。
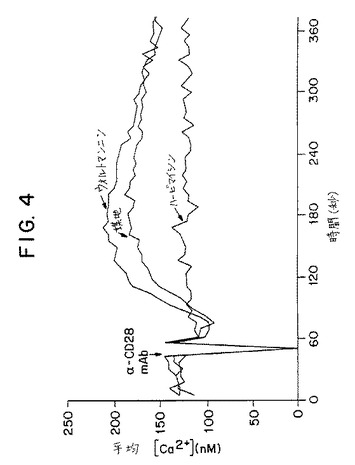
【図4】第4図は、CD28と抗CD28抗体の連結によって誘発されたT細胞におけるカルシウム代謝速度に関するウォルトマンニンまたはハービマイシンの効果を示すグラフである。
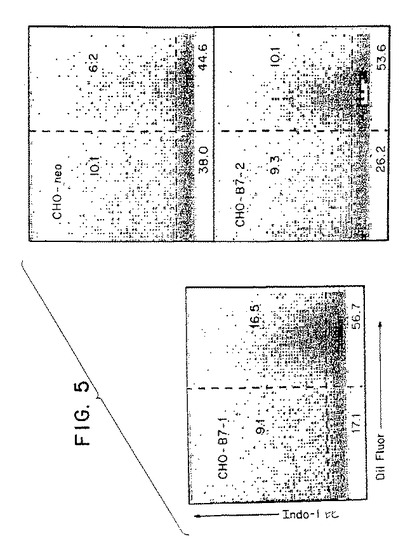
【図5】第5図は、ジャーカット細胞をCHO−neo、CHO−B7−1またはCHO−B7−2細胞のいずれかとともにインキュベートする細胞−コンジュゲートアッセイから得た一連のフローサイトメトリーのプロフィールである。カルシウム代謝速度をY軸に、細胞コンジュゲーションをX軸に示す。
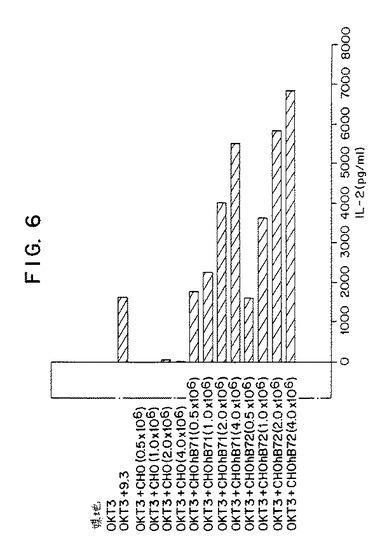
【図6】第6図は、精製ヒト末梢血液T細胞によるIL−2の産生に関する、抗CD3抗体と組み合わせた場合の膜結合B7−1およびB7−2の効果を示すグラフであり、B7−1またはB7−2のいずれかの共刺激によるIL−2の産生における用量依存性の増加を表している。
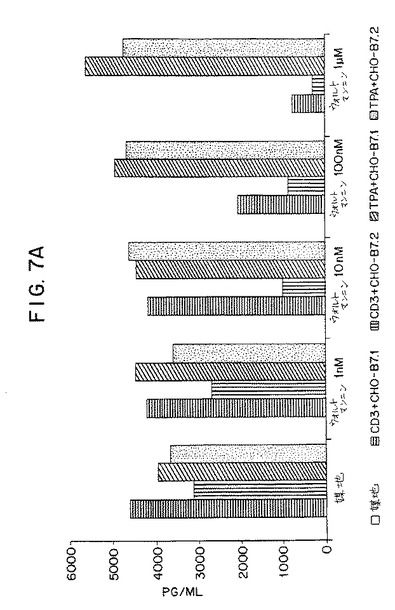
【図7A】第7A図は、培地、固定化抗CD3+CHO−B7−1、固定化抗CD3+CHO−B7−2、PMA+CHO−B7−1またはPMA+CHO−B7−2で休止期ヒトT細胞を刺激した24時間後の、該細胞によるIL−2の産生に関する、ウォルトマンニン処理(1nM〜1mM)の効果を示すグラフである。
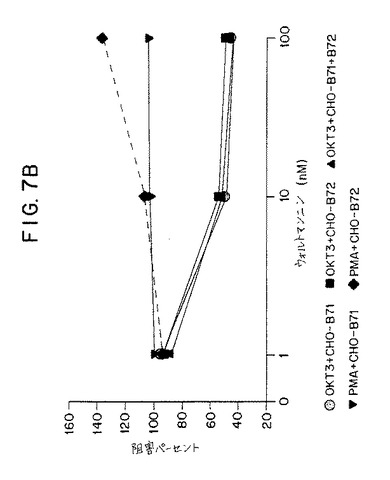
【図7B】第7B図は、抗CD3抗体(OKT3)ならびにB7−1、B7−2またはB7−1とB7−2の両方を発現するCHO細胞で24時間刺激されたヒトT細胞によるIL−産生、またはPMAならびにB7−1またはB7−2を発現するCHOで刺激されたT細胞によるIL−産生に関する、ウォルトマンニン(1〜100nM)による阻害パーセントを示すグラフである。
【図8】第8図は、抗CD28mAb(9.3)、CHO、CHO−B7−1またはCHO−B7−2で刺激されたジャーカット細胞によるホスファチジルイノシトール(3,4,5)−トリリン酸塩の産生に関する、ハービマイシン処理の効果を示すグラフである。
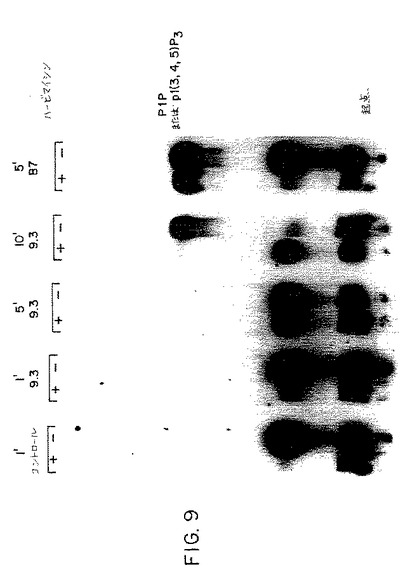
【図9】第9図は、ハービマイシンの存在(+)または不在(−)下で、抗CD28mAb(9.3)またはCHO−B7−1(B7)細胞で刺激した後の異なる時点におけるジャーカット細胞によって産生されたホスファチジルイノシトール(3,4,5)−トリリン酸塩の量を示す薄層クロマトグラフィーである。
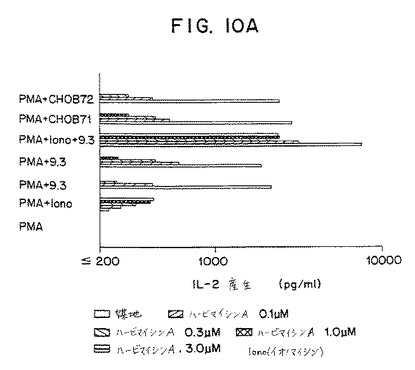
【図10A】第10A図は、ハービマイシンAの存在または不在下で、PMA単独またはPMAとCHO−B7−1(PMA+CHOB71)、PMAとCHO−B7−2(PMA+CHOB7−2)、PMAと9.3mAb溶液(PMA+9.3)、PMAとビーズ上の9.3mAb(PMA+9.3ビーズ)、PMAとイオノマイシン(PMA+イオノマイシン)あるいはPMAとイオノマイシンと可溶性9.3mAb(PMA+イオノ+9.3)で刺激された末梢血液T細胞によるIL−2分泌を示すグラフである。
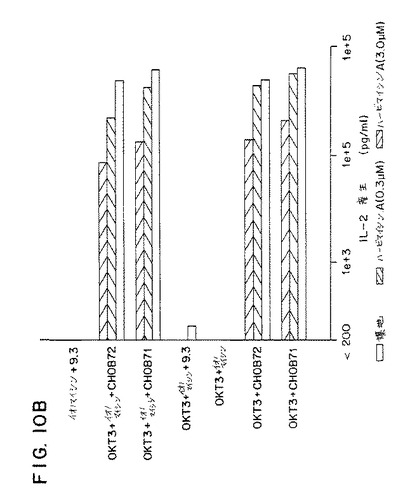
【図10B】第10B図は、イオノマイシンの存在(+イオノ)または不在下で、抗CD3(OKT3)単独またはOKT3とCHO−B7−1(OKT3+CHOB71)、OKT3とCHO−B7−2(OKT3+CHOB7−2)またはOKT3と9.3mAb(OKT3+9.3)で刺激された末梢血液T細胞によるIL−2分泌を示すグラフである。
【図11】第11図は、およそ6kDaのホスホチロシンプロテインが抗CD28抗体で共免疫沈降されることを示すイムノブロットのオートラジオグラフィーである。
【発明を実施するための形態】
【0039】
次の実施例によって本発明をさらに詳しく説明するが、本発明はこれらに限定されるものではない。本明細書に引用されたすべての参考文献、特許および特許出願公報はすべて、本発明の引例文献である。
【実施例1】
【0040】
CD28の連結はD−3ホスホイノシチドの産生を刺激する
この実施例では、CD28またはCD3を介するジャーカット細胞の刺激によるCD28+ジャーカット細胞におけるD−3ホスホイノシチドの産生を試験した。次のようにしてジャーカット細胞をキャリヤーフリーの32Pオルトホスフェート(32Pi)で標識した:ジャーカット細胞をホスフェートフリーの培地(DMEM/RPMI)で3回洗浄し、37℃で10分間、洗浄の間に10分間インキュベートした。細胞を20mMのHEPES,pH7.4および5%の透析したコウシ血清(生理的食塩水で一夜透析)を含む10mlのホスフェートフリーの培地に再懸濁した。細胞(1mCi/10ml細胞)にキャリヤーフリーの(32Pi)を加え、該細胞を37℃で4〜6時間インキュベートした。標識後、細胞をホスフェートフリーの培地で3回洗浄し、20mMHEPESを含むRPMI1640に再懸濁した。
【0041】
種々の実験において、32Pi標識ジャーカット細胞(0.15ml;2×107細胞)のアリコートを培地単独、抗CD3抗体(G19〜4)、抗CD28抗体(9.3)、非トランスフェクトCHO細胞またはCD28を発現するようにトランスフェクトされたCHO細胞(ヒトB7−1:CHO−B7−1またはヒトB7−2:CHO−B7−2のいずれか)で刺激した[Dr.G.フリーマンおよびL.ナドラーから入手;CHO細胞をヒトB7−1をコードするcDNA(該配列は、フリーマン,D.J.らの(1989)J.Immunol.143:2714〜2722に開示されている)またはヒトB7−2をコードするcDNA配列(該配列は、フリーマン,D.J.らの(1993)Science262:909〜911に開示されている)含有組換え発現ベクターによって標準的技術を用いてトランスフェクトした]。CHO細胞を刺激するために、ジャーカット細胞を107CHO細胞とともにインキュベートし、マイクロ遠心管による5秒間の低速度遠心分離によって細胞の接触を達成した。刺激に続いて種々の時間間隔(約1〜25分間の範囲)において、細胞を溶解し、ホスホリピッドを抽出し、脱アシル化し、アニオン交換HPLCによって分離した[基本的に、ウォード,S.G.らの(1992)J.Immunol.22:45の記載に準じ、下記の変更を加えた]。
【0042】
750mlのCHCl3/メタノール/水(32.6%/65.3%/2.1%v/v/v)を添加してインキュベーションを終了した。細胞の反応を停止するとすぐに、次の抽出中を通じてサンプルを氷上で保持した。725mlのCHCl3(10mgのフロックリピッドを含有;たとえばシグマ,カタログNo.B1502から入手)および172mlの2.4MHCl、5mMの硫酸テトラブチルアンモニウムを各サンプルに加えることによって相分離を行った。サンプルを渦巻き撹拌し、1000rpmで5分間遠心分離することによって相を分離した。下の相を除去し、1/2容の1M HCl、25mM Na2EDTA,pH7.5、0.5mM硫酸テトラブチルアンモニウムを管に加えた。サンプルを再遠心分離して相を分離し、底の相を除去し、清浄な管に入れ、サンプルを減圧乾燥した。サンプルに1mlのメチルアミン試薬(40%水/メタノール/n−ブタノール(4:4:1v/v/v)溶液)を加えて渦巻き撹拌し、53℃で40分間インキュベートして脱アシル化した。サンプルを氷上に置き、次いで減圧乾燥した。サンプルを0.5mlの滅菌蒸留水に再懸濁し、渦巻き撹拌して混合した。サンプルを0.7mlのn−ブタノール(40〜60%石油エーテル/ギ酸エチル(20:4:1v/v/v))で2回抽した。底の水相を減圧乾燥し、HPLC分析を行うまで−70℃で貯蔵した。緩衝液A(水)/B[1.25M(NH4)2HPO4](H3PO4にて25℃でpH3.8に調節)に基づく溶離勾配およびPartisphereSAXカラム(ワットマン社から購入)を用いてHPLCを行った。脱アシル化したホスホリピッドサンプルを0.1mlの蒸留水に再懸濁し、該カラムに注入した。溶出液をCanberra Packard A−500 Flo−Oneオンラインベータ線検出装置に導入し、3部のFlo−ScintIVシンチレーションカクテルと混合し、結果をFlo−Oneデータプログラム(ラジオマチック,USA)によって分析した。溶出したピークを[3H]PtdIns、[3H]PtdIns(4)Pおよび[3H]PtdIns(4,5)P2(アマーシャム・インターナショナル社から購入)から調製した標準液の保持時間と比較した。ウォード,S.G.らの(1992)J.Biol.Chem.267:23862の記載にしたがって、単離したホスファチジルイノシトール3−キナーゼをPtdIns(シグマ社から購入)とともにインキュベートすることによって標準の[32P]PtdIns(3)P、[32P]PtdIns(3,4)P2および[32P]PtdIns(3,4,5)P3を調製した。
【0043】
抗CD3または抗CD28抗体で1〜5分間刺激した後のジャーカット細胞におけるPtdIns(3,4,5)P3の産生を第1図に示す。その結果から、CD3またはCD28のいずれかを介する刺激はPtdIns(3,4,5)P3の産生を誘発するけれども、2つの経路の誘発動力学は別々であることがわかる。CD3による刺激によって早期(すなわち2分以内)にPI3Kの活性化(PtdIns(3,4,5)P3の産生によって評価される)が増加する。逆に、CD28刺激によって誘発されるPI3Kの活性化はCD3刺激による活性化よりも遅く(すなわち5分あるいはそれよりも遅くまで最大にはならない)、CD3刺激による活性化よりも長く持続する。このような結果から、CD3またはCD28連結のいずれかによって仲介されるPI3K活性化には別のメカニズムが含まれることが示される。
【0044】
CHO−B7−1またはCHO−B7−2細胞で0〜20分間刺激した後のジャーカット細胞におけるPtdIns(3,4,5)P3の産生を第2図に示す。その結果から、B7−1またはB7−2のいずれかによる刺激は有効な活性化(PtdIns(3,4,5)P3の産生によって評価される)を誘発することが示される。誘発の動力学は2つのCD28リガンドとはわずかに異なっている:B7−1とB7−2のプラトーは同じレベルであるが、B7−1はB7−2よりも早期に活性化を刺激する。B7−1またはB7−2のいずれかによるPtdIns(3,4,5)P3産生の刺激は非常に強く、持続的である(すなわち20分以上継続する)。
【0045】
この実施例は、CD28の天然リガンドであるB7−1およびB7−2または抗CD28抗体のいずれかによるCD28を介するT細胞の刺激が、T細胞内においてD−3ホスホイノシチドの産生を誘発することを証明し、CD28連結に基づくPI3Kの活性化が示される。さらに、この実施例は、ジャーカット細胞がCTLA4−ではなくCD28+であり、CTLA4を発現するようには誘発されえないので[リンステン,T.の(1993)J.Immunol.151:3489〜3499に示される]、CD28が通例B7−1とB7−2を生理的リガンドとして分けもつことを証明する。したがって、CTLA4は明らかにB7誘発シグナル変換において要求されず、B7−1とB7−2の両方がCD28に対する生理的リガンドである。
【実施例2】
【0046】
ホスファチジルイノシトール3−キナーゼインヒビターはCD28連結によって誘発されるD−3ホスホイノシチドの産生を阻害する
この実施例では、CD28が仲介するジャーカット細胞におけるD−3ホスホイノシチドの産生に関するホスファチジルイノシトール3−キナーゼのインヒビターとしての効果を試験した。実施例1に記載したようにジャーカット細胞をオルトホスフェートで標識し、B7−2を発現するようにトランスフェクトさせたCHO細胞で刺激した。さらに、刺激を行っている期間中、種々の濃度(0〜100mM)の真菌代謝物ウォルトマンニン(ホスファチジルイノシトール3−キナーゼのインヒビターである)の存在下で細胞をインキュベートした。ウォルトマンニンはシグマ・ケミカル・コーポレイションから購入し、10mMのDMSO溶液として−40℃で貯蔵した。それを培養中の細胞に添加する直前に培地で希釈した。ウォルトマンニンの存在あるいは不在下において刺激を行った後、実施例1に記載したようにHPLCによって細胞内に産生されたPtdIns(3,4,5)P3の量を測定した。結果を第3図に示す。第3図はCHO−B7−2で刺激する際にウォルトマンニン処理されたジャーカット細胞において検出されたPtdIns(3,4,5)P3の量を、CHO−B7−2で刺激された非処理ジャーカット細胞において検出されたPtdIns(3,4,5)P3の量に対するパーセントとしてプロットしてグラフ化したものである。その結果から、ジャーカット細胞を濃度増加ウォルトマンニンで処理することにより、B7−2によりCD28が連結した細胞において産生されるD−3ホスホイノシチドの量が減少することが示される。したがって、この実施例は、T細胞内におけるホスファチジルイノシトール3−キナーゼの活性を阻害することによって、CD28を介するT細胞の刺激の結果としての細胞内的D−3ホスホイノシチドの産生を阻害しうることを証明している。
【実施例3】
【0047】
CD28仲介カルシウム代謝における薬物的インヒビターの効果
この実施例では、CD28抗体によって誘発されるジャーカット細胞におけるカルシウム代謝速度に関する薬物的インヒビターの効果を試験した。研究した薬物的インヒビターはPI3Kの活性を阻害するウォルトマンニンおよびプロテインチロシンキナーゼの活性を阻害するハービマイシンAである。培地単独あるいはウォルトマンニンもしくはハービマイシンAの存在下のいずれかにおいて抗CD28抗体でジャーカット細胞を刺激し、細胞中の平均カルシウム濃度(nM)を刺激後数分にわたって測定した。第4図に示すように、ハービマイシンAはCD28抗体誘発カルシウム代謝を阻害する能力があった。逆に、ウォルトマンニンはCD28抗体誘発カルシウム流出を阻害することができなかった。これらの結果は、T細胞におけるウォルトマンニンの効果が、カルシウム代謝を妨害することによって仲介されないことを示している。さらにそのうえ、B7−1またはB7−2によって誘発されるIL−2の産生として測定されたところの“ウォルトマンニンが補刺激を阻害すること”を示している後記実施例5の結果、このデータは、CD28が誘発するカルシウム濃度上昇が、特異的にT細胞不応答(すなわちアネルギー)または共刺激を誘発する化合物の評価という目的に対して、おそらく読み取りの誤りによるものであろうということを示している。
【実施例4】
【0048】
天然のCD28リガンドによるカルシウム代謝の誘発
この実施例では、CHO−B7−1またはCHO−B7−2のジャーカット細胞への接着ならびにCHO−B7−1およびCHO−B7−2による刺激に応じたジャーカット細胞におけるカルシウム可動化を試験した。アベ,R.らの(1992)J.Exp.Med.176:459〜468に記載されているフローサイトメトリー細胞カルシウム−コンジュゲートアッセイを用いた。該細胞コンジュゲートアッセイでは、カルシウム感受性蛍光プローブindo−1(青色および緑色のシグナルを発する)でT細胞を着色する。コントロールプラスミド(CHO−neo)でトランスフェクトされたCHO細胞またはB7−1あるいはB7−2を発現するようにトランスフェクトされたCHO細胞をトレーサー着色剤DilC22(3)(赤色のシグナルを発する:モレキュラー・プローブ社から購入)で着色する。ジャーカット細胞−CHO複合体をフローサイトメトリーによって分析する。T細胞とCHO細胞からなる複合体は赤色のシグナルを補足することによって測定でき、カルシウム濃度は青色または緑色のシグナルを補足することによってT細胞内で測定できる。第5図に示すように、一連の2つのパラメーターの点プロットとして結果を表示する。カルシウム(indo−1比)をY軸に、細胞複合体(赤色トレーサー)をX軸に示す。右上のカドラントにおける細胞は、CHO細胞に複合したカルシウム濃度の高いジャーカット細胞を表す。右下のカドラントの細胞は、CHO細胞に複合したカルシウム濃度が正常であるジャーカット細胞を表す。左の上下のカドラントにおける細胞は、それぞれ高濃度または低濃度のカルシウムを含む非複合ジャーカット細胞を表す。
データは、B7−1およびB7−2の両方がジャーカット細胞への接着を仲介することができることを示している。しかし、両方のリガンドのカルシウム可動化の増加を引き起こす能力は乏しい。したがって、B7−1およびB7−2は、カルシウム可動化よりもPI3K活性化を誘発することにおける能力が高い(実施例1参照)。逆に、抗CD28抗体は、PI3K活性化(実施例1参照)およびカルシウム代謝(実施例3参照)の両方を刺激する能力がある。したがって、刺激を天然のリガンド(B7−1またはB7−2など)を用いて行うか抗体を用いて行うかによって、CD28の連結を介して発生する細胞内シグナルに違いがあると考えられる。ナンズ,J.らの(1993)Int.Immunol.5:311〜315に、CD28抗体が、T細胞のシグナル変換および活性化(IL−2の産生など)という生化学的局面において多重かつ異種の効果をもつことが既に記載されている。これらの観察から、CD28に関するこれまでの結果からの外挿に基づいてD−3代謝物の産生におけるCD28の天然のリガンド(すなわち膜結合B7−1およびB7−2)生化学的効果を予測することは不可能であることがさらに示される。
【実施例5】
【0049】
ホスファチジルイノシトール3−キナーゼインヒビターはCD28の連結によって誘発されるインターロイキン2の産生を阻害することができる
この実施例では、T細胞によるインターロイキン2のCD28依存性産生におけるホスファチジルイノシトール3−キナーゼのインヒビターとしての効果を試験した。最初の一連の実験では、ウォルトマンニンの不在下でのIL−2の産生における、CD28を介するT細胞刺激の効果(CD3を介する刺激に関連して)を評価した。高純度ヒト末梢血液T細胞を、固定化抗CD3抗体(OKT3)単独、または抗CD28抗体(9.3)あるいは非トランスフェクトまたはB7−1またはB7−2を発現するようにトランスフェクトされたマイトマイシンC処理CHO細胞とともに24時間インキュベートした。24時間培養後、ELISAにて標準的技術を用いてIL−2産生について培養上清をアッセイした。第6図に示すように、培地単独で、OKT3単独でまたはOKT3と非トランスフェクトCHO細胞とともにインキュベートされた細胞はIL−2を産生しなかった。逆に、OKT3およびB7−1またはB7−2を発現するようにトランスフェクトされたCHO細胞とともに培養された細胞は、用量依存的にIL−2の産生を刺激された。OKT3と9.3抗体との培養もまたIL−2の産生を刺激した。これらの結果から、たとえばB7−1またはB7−2刺激によるCD28の連結がリンホカイン産生に対する共刺激シグナルを提供することができることが確認される。
【0050】
次の一連の実験では、休止期ヒトT細胞をそれぞれ:1)固定化OKT3+CHO−B7−1,2、2)固定化OKT3+CHO−B7−2,3、3)固定化OKT3+CHO−B7−1+B7−2、4)PMA+CHO−B7−1、または5)PMA+B7−2で刺激した。培地単独中、またはウォルトマンニンを1nM〜1mMの濃度で含有する培地中でT細胞の刺激を行った。培養の24時間後、ELISAによってIL−2の産生について上清をアッセイした。結果を第7A図および第7B図に示す。該結果は、ウォルトマンニンが、CD3刺激に関連したB7−1またはB7−2のいずれかによって刺激されるIL−2の産生を阻害することができることを示す。IL−2のウォルトマンニン仲介阻害は用量依存性であった。B7−2仲介刺激の阻害のID50は約10nMであった。B7−1仲介刺激の阻害のID50は10〜100nMであった。これらのウォルトマンニンの用量は細胞に対して一般的に毒性ではなく、非特異的メカニズムによるIL−2の産生を阻害しない。PMAおよび膜結合B7−1またはB7−2によって刺激されるによって刺激されるIL−2の産生が、1mMという高濃度のウォルトマンニンによって阻害されないという事実が証拠である。この実施例は、TCR/CD3複合体およびCD28を介する刺激に応じたIL−2の産生によって評価されるように、T細胞内のホスファチジルイノシトール3−キナーゼの活性を阻害する作用剤でT細胞を処理することによってT細胞の活性化を阻害しうることを証明する。
【実施例6】
【0051】
抗CD28またはCD28の天然のリガンドで刺激したジャーカット細胞のPI3K活性化へのハービマイシンの異なる効果
PI3キナーゼ活性化の意味を評価するため、ジャーカットT細胞(20×106細胞)を抗CD28mAb9.3(10μg)またはB7−1またはB7−2(10×106細胞)を発現しているCHO細胞で刺激した。該細胞を実施例1で記載したように32Pオルトホスフェートで標識し、ついでビヒクル希釈薬または3μMのハービマイシンAを含む培地で前もってインキュベートした。5分後、該細胞は溶解し、PI3キナーゼを実施例1に記載したようにHPLCによって、PtdIns(3,4,5)P3の測定により評価した。
該結果を図8に示す。9.3抗体で刺激したジャーカット細胞によるPtdIns(3,4,5)P3の産生をハービマイシンの前処置によって妨げた。予期せず、PtdIns(3,4,5)P3の産生によって測定されたようにPI3キナーゼ活性を誘発したB7−1およびB7−2はハービマイシンの前処置に耐性を示した。これらの結果は、CD28によってPI3キナーゼの活性に関するハービマイシンの独立した経路が存在することを示し、さらにハービマイシンは様々なsrcファミリーキナーゼを抑制することを示しているのでCD28はsrcファミリープロテインチロシンキナーゼから独立してPI3キナーゼを活性化することができることを示す。
【0052】
この両分をさらに調査するため、20×106ジャーカット細胞を9.3mAb(10μg)または10×106CHO−B7−1細胞で刺激した。該細胞をハービマイシンA 3μMまたはビヒクル内で前もってインキュベートした。該CD28 mAbにさらにヤギ抗マウスIgGを結合させ、該細胞を5〜10分後溶解した。PI3キナーゼ活性化はイン・ビボリピッドキナーゼアッセイを使用して薄層クロマトグラフィーによって評価した(ウォード(Ward,S.G.)ら(1995)Eur.J.Immunol.25:526−532に記載のように)。
該結果を図9に示す。抗CD28 9.3mAbおよびCHO−B7細胞の両者は、コントロールレーンと10分後の9.3mAbレーンおよび5分後のB7レーンとの比較から明らかなようにPI3キナーゼ活性を誘発する。再び、PIの反対のキナーゼの産物は9.3mAb刺激後ハービマイシンによって抑制され、B7刺激後ハービマイシンに耐性を示した。
従って、ハービマイシンは抗CD28で刺激したジャーカット細胞でPI3Kのハービマイシンの活性化を遮断するが、CHO−B7−1またはCHO−B7−2細胞で刺激したジャーカット細胞においては遮断せず、CD28によるPI3キナーゼの活性化のハービマイシンの独立した経路が存在することを示した。
【実施例7】
【0053】
T細胞によるIL−2の分泌へのへルビマイシンの異なる効果
さらに、CD28受容体ライゲーションに関するシグナル変換経路を決定するため、CD28+T細胞(抹消血T細胞のネガティブセレクションによって調製した)を種々の濃度のハービマイシン(0.1〜3μM)にて一晩培養し、ついでPMAを3ng/mlだけまたは、1μg/mlの溶液内のmAb 9.3、mAb9.3コートしたビーズ、0.08μg/mlのイオノマイシン、CHO−B7−1またはCHO−B7−2細胞で刺激した。上清を24時間後に回収し、ELISAによってIL−2含有を分析した。
該結果を図10パネルAに示す。該結果はPMAプラス9.3mAbが誘発したIL−2分泌が約0.2μMのID50を有するハービマイシンに感受性を有することを示す。同様に、PMAプラスCHO−B7−1細胞によって誘発したIL−2分泌はまたハービマイシンにも感受性があった。対照的に、IL−2分泌は細胞が3μMより大きいID50を有するPMAプラスイオノマイシンで刺激した場合、完全に耐性があった。ハービマイシンによる抑制のメカニズムをさらに試験するため、細胞をイオノマイシンの存在下または不在下でも刺激した。PMAプラスイオノマシインプラス9.3mAbで刺激した細胞におけるIL−2分泌はハービマイシン、3μMのハービマイシンAにさえ完全耐性となる。
【0054】
さらにハービマイシン感受性および耐性を有するIL−2分泌の結果を試験するため、T細胞を、CHO−B7−1もしくはCHO−B7−2細胞またはイオノマイシンの存在下もしくは不在下にて9.3mAbを有するビーズに不動化したCD3抗体OKT3で刺激した。上清を24時間後に回収し、ELISAによってIL−2の量を決定した。
該結果を図10パネルBに示す。PMAおよびB7−1またはB7−2でのT細胞の刺激による結果のハービマイシンに対して感受性を有すると分かったIL−2分泌に反して、PMAが抗CD3で置換された場合、IL−2分泌はハービマイシンに完全に耐性を有すると分かった。これらの結果は、CD28はハービマイシン耐性IL−2分泌をシグナルの状況に依存して開始することができることを示す。T細胞がカルシウムシグナル(すなわち、PMAプラス抗CD28)を開始しないような方法で活性化された場合、IL−2分泌はハービマイシンに感受性を有する。T細胞がカルシウムシグナル(すなわち、PMAプラスイオノマイシンプラスB7または抗CD28)の存在下で活性化された場合、該シグナルはハービマシンに対して耐性を示した。これはCD28はハービマイシン感受性のチロシンキナーゼによって提供される不可欠なシグナルを必要としないかまたは代わって、CD28によって開始される不可欠なチロシンキナーゼカスケードの必要性を無視することができるかもしれないことを示している。
【実施例8】
【0055】
CD28に関連した67kDaのチロシンホスホプロテインの同定
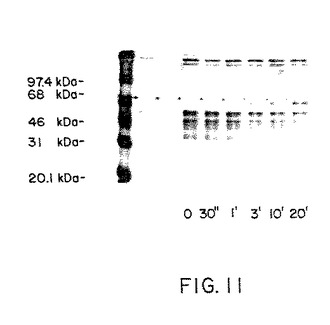
現在までに、CD28と物理的な関連を示す唯一報告されたプロテインはPI3キナーゼ(トルイット(Truitt,K.E.)ら(1994)J.Exp.Med.179:1071−1076)およびsrcファミリーキナーゼlck(オーガスト(August,A.)およびデュポン(Dupont,B.)(1994)Biochem.Biophys.Res.Commun.199:1466−1473)である。CD28媒介シグナル変換をさらに研究するため、CD28と物理的に関連のあるタンパク質に関するジャーカット細胞およびT細胞において研究を開始した。細胞を何度も9.3mAbで刺激し、NP−40清浄剤で溶解し、ブドウ球菌タンパク質Aで免疫沈降した。該免疫沈降物を洗浄し、抽出しついで還元条件のもとでSDS−PAGEによって分離した。トランスファーの後、西洋ワサビペルオキシダーゼとともに抗ホスホチロシンAb4G10を使用してイムノブロットを行った。チロシンリン酸化タンパク質をECLを使用して化学発光によって同定した。
該結果を図11に示すが、これはイムノブロットのオートラジオグラフィーを示している。〜67kDaの相対的な移動を示すバンドをCD28免疫沈降物内に同定した。ホスホプロテインに対応するバンドはCD28刺激後3分以内にジャーカット細胞に即座に現れた。同様のまたは同一のタンパク質をT細胞幼若細胞において同定した。このタンパク質はitk/emtプロテインチロシンキナーゼではないようであった。というのはitkに対する抗体はCD28免疫沈降物内にitkを明らかにしないからである。さらにその上、このCD28関連タンパク質の分子量がlckまたはPI3と一致していない。
従って、該結果は約67kDaの新規なホスホチロシンタンパク質がCD28を通じて刺激したジャーカット細胞およびT細胞幼若細胞内におけるCD28と関連を有することを示す。
【0056】
均等物
当業者は、本明細書に記載の発明の特定の態様に均等な多くのものについて認識するか、常套的な実験を使用するだけで確認することができるであろう。そのような均等物は以下の請求の範囲に包含するよう意図されている。
【技術分野】
【0001】
本発明は、細胞内シグナル変換操作によるT細胞応答の調節に関する。
【背景技術】
【0002】
抗原特異的T細胞応答は、T細胞上の細胞表面受容体と抗原付与細胞(APCs)上のリガンドとの間の多重の相互作用によって誘発される。第1の相互作用は、T細胞上のT細胞受容体(TCR)/CD3複合体と抗原付与細胞上の主要組織適合遺伝子複合体(MHC)分子/抗原ペプチド複合体のとの間の作用である。この相互作用は、T細胞における第1の、抗原特異的、活性化シグナルの引金となる。第1の活性化シグナルに加えて、T細胞応答を誘発するには、第2の、共刺激性(costimulatory)シグナルが必要である。適当な共刺激がないと、TCRシグナリングはT細胞にアネルギー状態を誘発する。アネルギーT細胞に対し、引き続いて抗原を適当に付与すると、特有の応答を起こさなくなる[シュワルツ,R.H.らの(1990)Science248:1349を参照]。
【0003】
共刺激シグナルは、T細胞内でCD28などのT細胞表面受容体を通って発信される。たとえば、T細胞のサブ最適ポリクローナル刺激[たとえば、抗CD3抗体またはホルボールエステル(これらは第1の活性化シグナルを提供しうる)による刺激]は、CD28と抗CD28抗体の架橋によって有効化されることが説明されている[リンスリー,P.S.らの(1991)J.Exp.Med.173:721;ギミ,C.D.らの(1991)Proc.Natl.Acad.Sci.U.S.A.88:6575]。さらに、CD28の刺激は、T細胞クローンにおけるアネルギー状態の誘発を妨げることができる[ハーデイング,F.A.の(1992)Nature356:607〜609]。CD28に対する天然のリガンドはAPCs上で同定されている。CD28リガンドには、B7−1(CD80)およびB7−2(CD70などのプロテインのB7ファミリーのメンバーが含まれる[フリードマン,A.S.らの(1987)J.Immunol.137:3260〜3267;フリーマン,G.J.らの(1989)J.Immunol.143:2714〜2722;フリーマン,G.J.らの(1991)J.Exp.Med.174:625〜631;フリーマン,G.J.らの(1993)Science262:909〜911;アズマ,M.らの(1993)Nature366:76〜79;フリーマン,G.J.らの(1993)J.Exp.Med.178:2185〜2192]。CD28に加えて、B7ファミリーのプロテインも、CTLA4と呼ばれるCD28に関連するT細胞上の他の表面受容体(これは、またT細胞共刺激においても役割を演じる)に結合することがわかっている[リンスリー,P.S.らの(1991)J.Exp.Med.174:561〜569;フリーマン,G.J.らの(1993)Science262:909〜911]。
【0004】
CD28/CTLA4とB7ファミリープロテインの受容体:リガンド相互関係の解明および共刺激におけるこの相互作用の役割の解明によって、共刺激分子を結合するというT細胞表面受容体の細胞外相互作用処置などの治療用途へのアプローチが導かれた。たとえば、B7−1とB7−2の両方に結合し、それらのCD28/CTLA4との相互作用を遮断するCTLA4Ig融合プロテインは、同種および異種移植における拒絶反応の阻害のために用いられている[ターカ,L.A.らの(1992)Proc.Natl.Acad.Sci.U.S.A.89:11102〜11105;レンスコー,D.J.らの(1992)Science257:789〜792などを参照]。同様に、B7−1および/またはB7−2と反応する抗体が、インビトロにおけるT細胞の増殖およびIL−2の産生の阻害およびインビトロにおける抗原に対する第1免疫応答の阻害に用いられている[ハスコック,K.S.らの(1993)Science262:905〜907;アズマ,M.らの(1993)Nature366:76〜79;パワーズ,G.D.らの(1994)Cell.Immunol.153:298〜311;チェン,C.らの(1994)J.Immunol.152:2105〜2114]。同時に、これらの研究では、B7−1およびB7−2などの共刺激性分子を結合するT細胞表面受容体が免疫応答を操作するための所望標的であることが示されている。
【0005】
CD28/CTLA4とそれらのリガンド間の細胞外相互作用は、ある程度詳細に解明されているけれども、これらの分子の連結に続いてT細胞内で起きる細胞内のイベントについては少ししかわかっていない。T細胞の共刺激経路はシクロスポリンAの阻害効果に対して耐性があるので、T細胞の共刺激は、TCRを経るシグナリングとは異なる細胞内シグナル変換経路をもつと考えられている[ジューン,C.H.らの(1990)Immunology Today11:211〜216を参照]。プロテインチロシンのリン酸化は、T細胞内でCD28との連結により起こることがわかっており、プロテインチロシンキナーゼインヒビターであるハービマイシンAがCD28誘発IL−2産生を阻害しうることが説明されている[ヴァンデンバーグ,P.らの(1992)J.Exp.Med.175:951〜960;ルー,Y.らの(1992)J.Immunol.149:24〜29]。
【0006】
CD28受容体は、抗原受容体に関係するシグナルならびに抗原受容体とは無関係のシグナルの引金をひくことができることが研究によってわかっている。さらに、CD28のオリゴマー形成の度合は、CD28媒介シグナル変換において重要な決定要素である[リンスリー,P.S.およびレドベター,J.A.の(1993)Annu.Rev.Immunol.11:191およびレドベター,J.A.らの(1990)Blood75:1531]。抗CD28および抗CD3モノクローナル抗体を用いる研究から、カルシウムシグナリング、ホスホリパーゼC(PLC)活性化、チロシンキナーゼ活性の増加およびp21ras活性化のすべてが、受容体結合後の早期に起こることがわかっている[ジューン,C.H.らの(1994)Immunology Today15:321;およびラッド,C.E.らの(1994)IImmunol.Today15:225]。プロテインチロシンキナーゼp72itkおよびp56lck[オーガスト,A.およびデュポン,B.の(1994)Biochem.Biophys.Res.Commun.199:1466およびオーガスト,A.らの(1994)Proc.Natl.Acad.Sci.U.S.A.91:9347]は、CD28とT細胞抗原受容体の両方によって活性化される、あるいはそれらと物理的に会合すると報告されている。CD28媒介カルシウム、p21rasおよびp70s6キナーゼシグナルは休止期のT細胞よりも幼若化T細胞において顕著であり、抗原受容体シグナルと比較して一時的に遅い[レドベター,J.A.らの(1987)Proc.Natl.Acad.Sci.U.S.A.84:1384;ナンズ,J.S.らの(1993)Biochem.J.293:835;シーゲル,J.N.らの(1993)J.Immunol.151:4116;およびパイ,S.V.らの(1994)J.Immunol.24:2364]。さらに、CD8+T細胞およびCD4+T細胞はその両方がCD28シグナルに応答するけれども、抗CD28誘発カルシウムシグナルはCD4+T細胞に対して第1に制限される[アベ,R.P.らの(1995)J.Immunol.印刷中]。これらの研究は共に、CD28シグナル変換が数個のシグナル変換カスケードに連結し、シグナルの性質がT細胞発達中に規定されることを示している。
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明は、細胞内シグナル変換操作によるT細胞応答の調節に関する。さらに詳しくは本発明は、T細胞の共刺激において起こる細胞内シグナリングイベントを操作することに関する。
【課題を解決するための手段】
【0008】
本発明は、共刺激性分子を結合するT細胞の表面受容体の連結から得られる1種あるいはそれ以上の細胞内シグナルを阻害または刺激することによるT細胞応答の調節方法を含む。現在、CD28受容体刺激によって、T細胞内にD3−ホスホイノシチドが産生されることが発見されている。さらに、T細胞内のホスファチジルイノシトール3−キナーゼの活性が、リンホカイン産生および細胞増殖などのT細胞応答を阻害しうることが発見されている。これらの発見は、共刺激性シグナル変換経路におけるD3−ホスホイノシチドの機能的役割を示し、T細胞応答を調節するための細胞内標的としてのホスファチジルイノシトール3−キナーゼを提供する。したがって、D3−ホスホイノシチドを含む細胞内シグナリングイベントは、共刺激シグナルの阻害およびそれによるT細胞不応答の誘発、あるいは共刺激シグナルの発信およびそれによるT細胞応答の産生のいずれかによって調節される。さらに、T細胞応答を阻害または刺激するのに用いうる、ホスファチジルイノシトール3−キナーゼのインヒビターまたはアクチベーターを同定するための新規なスクリーニングアッセイも本発明の範囲に含まれる。
【0009】
さらに本発明は、共刺激性分子を結合する表面受容体を発現するというT細胞による応答の阻害方法に関する。このような方法には、T細胞中のD3−ホスホイノシチドの産生を阻害する作用剤にT細胞を接触させることが包含される。ひとつの具体例においては、該作用剤は、真菌代謝物であるウォルトマンニンまたはバイオフラボノイドであるクエルセチンあるいはそれらの類縁体など(たとえばLY294002)のホスファチジルイノシトール3−キナーゼのインヒビターである。本発明方法の別の具体例においては、プロテインチロシンリン酸化などの共刺激に関連する異なる細胞内シグナルを阻害する作用剤の少なくとも1種にT細胞を接触させる。たとえば、ホスファチジルイノシトール3−キナーゼおよびプロテインチロシンキナーゼのインヒビターにT細胞を接触させることができる。好ましいプロテインチロシンキナーゼのインヒビターはハービマイシンAである。別法として、チロシンホスファターゼまたはチロシンホスファターゼのアクチベーターによってT細胞内でのプロテインチロシンリン酸化を阻害することができる。この具体例においては、ホスファチジルイノシトール3−キナーゼおよび細胞チロシンホスファターゼに結合して活性化させる抗体などの分子(たとえばCD45またはHcph)にT細胞を接触させることができる。本発明方法の別の好ましい具体例においては、プロテインセリンキナーゼのインヒビターまたはセリンホスファターゼのアクチベーターにT細胞を接触させる。
【0010】
さらに本発明は、T細胞内の共刺激に関連する細胞内シグナルを妨害しながらT細胞における第1の、抗原特異的シグナルの引金をひくことによってT細胞内に抗原に対する不応答を誘発する方法を提供する。共刺激性シグナル変換の妨害の結果として、抗原の存在下でT細胞が適当な共刺激性シグナルを受け取ることができず、抗原特異的不応答がT細胞内に誘発される。T細胞の不応答を誘発するために、T細胞における第1の活性化シグナルの刺激に適した形体の抗原およびT細胞内におけるD3−ホスホイノシチドの産生を阻害する作用剤に抗原特異的T細胞を接触させる。たとえば、APCによって供与される抗原およびウォルトマンニンまたはクエルセチンあるいはそれらの誘導体または類縁体など(たとえばLY294002)のホスファチジルイノシトール3−キナーゼのインヒビターにT細胞を接触させることができる。さらに、プロテインチロシンリン酸化および/またはプロテインセリンリン酸化などの共刺激に関連する他の細胞内シグナルをT細胞内で阻害することができる。
【0011】
T細胞応答の阻害方法およびT細胞不応答の誘発方法は、たとえば、移植を受ける被験体(臓器移植または骨髄移植など)または自己免疫疾患あるいは異常免疫応答に関連する他の疾病に罹患している被験体などの免疫応答を下降方向に調節することが望ましい状況において有用である。共刺激に関連するシグナル変換を阻害する作用剤(たとえばイノシトールリン酸3−キナーゼのインヒビターなど)を被験体に投与することができ、また別法として被験体からT細胞を採取し、本明細書に記載したようにインビトロで処理し、それを被験体に投与することもできる。
【0012】
さらに本発明は、第1の活性化シグナルを受け、共刺激分子を結合する表面受容体を発現したT細胞によって応答を刺激する方法に関する。このような方法には、ホスファチジルイノシトール3−キナーゼなどのT細胞中のD−3ホスホイノシチドの産生を刺激する作用剤にT細胞を接触させることが包含される。別の具体例においては、D3−ホスホイノシチドの産生を刺激する作用剤およびプロテインチロシンリン酸化などの共刺激に関連する異なる細胞内シグナルを刺激する作用剤の少なくとも1種にT細胞を接触させる。たとえば、ホスファチジルイノシトール3−キナーゼおよび過バナジウム酸塩などのプロテインチロシンキナーゼのアクチベーターにT細胞を接触させることができる。本発明の別の具体例においては、さらに、プロテインセリンキナーゼを活性化する作用剤にT細胞を接触させる。別法として、CD45またはHcphなどの細胞ホスファターゼのインヒビターをPI3Kアクチベーターとともに用いることができる。さらなる本発明の他の具体例においては、抗原特異的T細胞を抗原およびT細胞内でD−3ホスホイノシチドの産生を刺激する作用剤に接触させることによって抗原特異的T細胞応答を刺激し、それによってT細胞内における第1の活性化シグナルおよび共刺激性シグナルの両方を刺激する。本発明の別の具体例においては、1種またはそれ以上のプロテインチロシンおよび/またはプロテインセリンキナーゼを活性化する作用剤に抗原特異的T細胞を接触させることによって、抗原特異的T細胞応答をさらに刺激する。
【0013】
T細胞応答を刺激する方法は、免疫応答を上昇方向に調節することが望ましい状況において有用である。たとえば、腫瘍ができている被験体における腫瘍に対する応答、あるいは被験体の病原体(たとえばバクテリア、HIVなどのウイルス、真菌、寄生虫など)に対する応答を刺激することができる。さらに、本発明方法は、ワクチン接種の効能を増強するのに用いることができる。共刺激に関連する細胞内シグナルを刺激する作用剤(イノシトールホスファターゼ3−キナーゼのアクチベーターなど)を被験体に投与するかまたは別法としてインビトロにてT細胞を刺激し、次いでそれを被験体に投与することができる。
【0014】
本発明はさらに、ホスファチジルイノシトール3−キナーゼのインヒビターまたはアクチベーターを同定するためのスクリーニングアッセイに関する。ひとつの具体例においては、共刺激性分子を結合する細胞表面受容体を発現するT細胞(CD28など)が用いられる。インヒビターを同定するには、T細胞における受容体に関連する細胞内シグナル変換経路を、被検作用剤の存在下に刺激し、T細胞におけるD−3ホスホイノシチドの少なくとも1種の産生を阻害する能力に基づいてインヒビターを同定する。アクチベーターを同定するには、被検作用剤にT細胞を接触させ、T細胞におけるD−3ホスホイノシチドの少なくとも1種の産生を刺激する能力に基づいてアクチベーターを同定する。
本発明はさらに、CD28を介して刺激されたT細胞におけるCD28に関連するおよそ67kDaの新規なチロシン−リン酸化プロテインに関する。該プロテインの活性化あるいは該プロテインのCD28との相互作用を遮断する方法、または別法として、該67Kプロテインの活性化あるいは該プロテインのCD28との相互作用を刺激する方法もまた本発明に包含される。
【0015】
本発明は、T細胞上の表面受容体の共刺激分子への結合などの共刺激によってT細胞において産生される細胞内シグナルを調節することによるT細胞応答の調節方法に関するものである。さらに詳しくは本発明は、共刺激性シグナルを阻害または刺激し、それによってT細胞応答を阻害または刺激するためのT細胞におけるD−3ホスホイノイシチド産生の調節に関する。共刺激に関連するシグナル変換の妨害による共刺激性シグナルの阻害を利用してさらに、T細胞不応答を誘発することができる。本発明は、CD28表面受容体を介するT細胞の刺激によってT細胞にD−3ホスホイノシチドが産生され、CD28が連結したT細胞においてホスファチジルイノシトール3−キナーゼ(本明細書においてPI3Kとも称する)のインヒビターがD−3ホスホイノシチドの産生を阻害するという発見に少なくとも部分的には基づいている。さらに本発明は、T細胞におけるPI3K活性を阻害すると、サイトカイン産生および細胞増殖などのT細胞応答が阻害されるという発見に少なくとも部分的には基づいている。
【0016】
したがって、さらに本発明は、シグナル変換に関連する細胞内シグナル変換を妨害することによるT細胞の応答を阻害する方法に関する。ひとつの具体例においては、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤に、共刺激性分子を結合する細胞表面受容体を発現しているT細胞を接触させることによって細胞内シグナルを阻害する。語句「共刺激性分子に結合する細胞表面受容体を発現しているT細胞」とは、CD28および/またはCRLA4、あるいはB7−1、B7−2または他のB7ファミリーのメンバーなどの共刺激性分子を結合する能力をもつ他の受容体を発現しているT細胞を意味する。
【0017】
T細胞による「応答」とは、T細胞における第1の活性化シグナルおよび共刺激性シグナルの引金をひくことによって生じるT細胞応答を意味し、リンホカインの産生(IL−2産生など)およびT細胞の増殖が含まれる。T細胞応答の阻害には、応答の完全遮断(無応答など)または応答の大きさの程度の減少(応答の部分的阻害など)も含まれる。
「D−3ホスホイノシチド」とは、イノシトール環のD−3位でリン酸化されるホスファチジルイノシトールの誘導体を意味し、ホスファチジルイノシトール(3)−モノリン酸塩(PtdIns(3)P)、ホスファチジルイノシトール(3,4)−ビスリン酸塩(PtdIns(3,4)P2)およびホスファチジルイノシトール(3,4,5)−トリリン酸塩(PtdIns(3,4,5)P3)などの化合物が含まれる。
D−3ホスホイノシチドはホスファチジルイノシトール3−キナーゼ(PI3K)の活性によって細胞内的に産生される。PI3Kは、そのSH2ドメインを介してチロシル−リン酸化プロテインを結合する85kDaのサブユニットおよび110kDaの触媒サブユニットからなるヘテロ二量体である。PI3Kは最初、ホスファチジルイノシトール、PtdIns(4)PおよびPtdIns(4,5)P2のイノシトール環のD−3位をリン酸化する脂質キナーゼとして同定された。最近の2つの研究では、PI3Kが、事実、脂質およびセリンキナーゼ活性の両方を有する2重特異的キナーゼであることが説明されている[ジャンド,R.らの(1991)EMBO J.13:522およびカーペンター,C.L.らの(1993)Mol.Crll Biol.13:1657]。
【0018】
したがって、ひとつの具体例においては、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤は、PI3Kの活性を阻害する作用剤である。T細胞におけるPI3K活性を阻害する好ましい作用剤は、真菌代謝物であるウォルトマンニンまたはその誘導体あるいは類縁体である。ウォルトマンニンはT・ウォルトマンニイ(T.wortmannii)(協和発酵株式会社)またはP.フミクロスム(P.fumiculsum)(シグマ)から誘導されるインヒビターである。ウォルトマンニン誘導体または類縁体には、PI3KおよびT細胞応答を阻害する能力をもつウォルトマンニンに構造的に関連する化合物が含まれる。ウォルトマンニン誘導体よび類縁体の例は、ワイシンガー,D.らの(1974)Experientia30:135〜136;クロッス,A.らの(1981)J.Med.Chem.24:1465〜1471;およびバッジョリーニ,M.らの(1987)Exp.Cell Res.169:408〜418に開示されている。使用しうるその他のPI3K活性のインヒビターは、バイオフラボノイドであるクエルセチンまたはその誘導体あるいは類縁体である。クエルセチン誘導体または類縁体には、PI3KおよびT細胞応答を阻害する能力をもつクエルセチンに構造的に関連する化合物が含まれる。クエルセチン誘導体および類縁体の例は、ブラホス,C.J.らの(1994)J.Biol.Chem.269:5241〜5284に開示されている。PI3K活性を阻害する好ましいクエルセチン誘導体は、LY29400[2−(4−モルホリニル)−8−フェニル−4H−1−ベンゾピラン−4−オン](リリー、インデイアナポリス、IN)(上記ブラホスらに記載)。別法として、後記方法で同定される他のPI3Kインヒビターを用いることもできる。
【0019】
本発明の別の態様としては、共刺激に関連する2種またはそれ以上の細胞内シグナリングイベントの妨害によるT細胞応答の阻害が含まれる。たとえば、CD28の刺激によってT細胞におけるプロテインチロシンリン酸化が起こることが示されている[ヴァンデンバーグ,P.らの(1992)J.Exp.Med.175:951〜960;ルー,Y.らの(1992)J.Immunol.149:24〜29などを参照]。したがって、ひとつの具体例においては、T細胞応答は、T細胞における少なくとも1種のD−3ホスホイノシチドの産生を阻害する第1作用剤およびT細胞におけるチロシンリン酸化を阻害する第2作用剤にT細胞を接触させることによって阻害される。たとえば、T細胞をPI3K活性を阻害する作用剤およびプロテインチロシンキナーゼ活性を阻害する作用剤の両方に接触させることができる。好ましいプロテインチロシンキナーゼインヒビターは、srcプロテインチロシンキナーゼを阻害するものである。ひとつの具体例においては、該srcプロテインチロシンキナーゼインヒビターは、ハービマイシンAまたはその誘導体もしくは類縁体である。ハービマイシンAの誘導体もしくは類縁体には、構造的にハービマイシンAに関連し、プロテインチロシンキナーゼを阻害する能力をもつ化合物が含まれる。別の具体例においては、プロテインチロシンリン酸化を阻害する作用剤は、プロテインチロシンホスファターゼまたはプロテインチロシンホスファターゼのアクチベーターである。T細胞におけるチロシンホスファターゼ活性の増加によって、プロテインチロシンリン酸化の正味量が減少する。プロテインチロシンホスファターゼは、T細胞内においてはCD45またはHcphなどの細胞プロテインチロシンホスファターゼとなりうる。T細胞上の細胞表面チロシンホスファターゼの活性は、ホスファターゼに結合し、その活性を刺激する分子に、T細胞を接触させることによって活性化することができる。たとえば、CD45に対する抗体を用いて、CD45をその表面に発現するT細胞におけるチロシンホスファターゼの活性を刺激することができる。したがって、ひとつの具体例においては、T細胞内においてプロテインチロシンリン酸化を阻害する作用剤は、抗CD45抗体またはCD45の活性を刺激する能力をもつそのフラグメントである。抗体フラグメントの例として、FabおよびF(ab')2フラグメントが挙げられる。抗体またはそのフラグメントは、多量体化または固定化などの刺激性形体で提供することができる。
【0020】
共刺激に関連する他の細胞内シグナルは、D−3ホスホイノシチド産生の阻害とともに阻害され、その結果としてT細胞応答を阻害することができる。たとえば、CD28の連結は、ホスホリパーゼCの活性の増加[ナン,J.らの(1993)Biochem.J.293:835〜842を参照)、および細胞内カルシウム濃度の増加[レドベター,J.A.らの(1990)Blood 75:1531〜1539および実施例を参照]に関連することがわかっている。したがって、PI3K活性を阻害する第1作用剤およびホスホリパーゼC活性を阻害および/または細胞内カルシウム濃度の増加を阻害する第2作用剤に、T細胞を接触させる。実施例に示すように、チロシンキナーゼインヒビターであるハービマイシンAもまたT細胞におけるCD28誘発性カルシウム代謝を阻害する。
【0021】
さらに、実施例に示すように、CD28を介する刺激の結果として、ハービマイシン耐性のT細胞によるIL−2の産生が起こり、これは、関係するシグナル変換経路に、プロテインセリンキナーゼなどのさらなるプロテインキナーゼが含まれることを示唆する。
【0022】
いくつかの研究によって、プロテインセリンキナーゼが、CD28に関連するシグナル変換経路に含まれることが示されている[シーゲル,J.N.らの(1993)J.Immunol.151:416〜4127およびパイ,S.V.らの(1994)J.Immunol.24:2364]。かくして、本発明の特定の具体例においては、PI3K活性を阻害する作用剤およびセリンキナーゼを阻害する作用剤に、T細胞を接触させることによって、T細胞応答が阻害される。別の具体例においては、このようなT細胞をさらに、プロテインチロシンキナーゼを阻害する作用剤に接触させる。さらに別の具体例においては、PI3K、プロテインチロシンキナーゼ、プロテインセリンキナーゼ、ホスホリパーゼC活性を阻害する作用剤の種々の組合せ、または細胞内カルシウムの増加を阻害する作用剤に、T細胞を接触させることによってT細胞応答を阻害する。
T細胞応答は、本発明方法にしたがってインビトロまたはインビボのいずれにおいても阻害することができる。たとえば、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤を、T細胞応答を阻害するのに十分な用量および期間において被験体に投与することができる。別法として、被験体からT細胞を採取し、インビトロにてT細胞を作用剤に接触させ、被験体に再投与することができる。「被験体」とは、ヒト、サル、イヌ、ネコ、ウサギ、ラット、マウスおよびそれらのトランスジェニック種などの哺乳動物などの免疫応答を起こす動物を意味する。T細胞応答を阻害しうる被験体には、免疫応答を下降調節することが望ましい被験体が含まれる。下降調節とは、リンホカイン産生およびT細胞増殖などのT細胞応答の部分的および完全阻害を意味する。たとえば、自己免疫疾患または免疫応答異常に関連する他の疾病の被験体、あるいは骨髄移植または他の臓器移植などの移植を受ける者に対し、本発明方法を適用することができる。
【0023】
本発明のひとつの具体例においては、T細胞内において共刺激性シグナルを阻害することによって抗原特異的T細胞不応答を誘発する。したがって、本発明の別の態様は、抗原に対するT細胞不応答を誘発する方法である。本明細書で用いる「T細胞不応答」とは、抗原(または抗原性部分)に対して、さらされた時にT細胞によるT細胞応答(増殖、リンホカイン分泌またはエフェクター機能の誘発など)が減少するかまたは応答しなくなり、T細胞が不応答になることを意味する。「T細胞不応答」および「T細胞アネルギー」は、本明細書においては互換的に用いられる。抗原に対する不応答は、T細胞における抗原特異的第1活性化シグナル(TCR/CD3複合体を介する活性化など)が、共刺激性シグナルが無い状態で引金を引かれることによって誘発することができる。本発明方法においては、共刺激に関連する細胞内シグナルを妨害する作用剤に、T細胞を接触させることによって、T細胞における共刺激性シグナルを遮断する。特に、共刺激性シグナルを阻害するためのT細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤の存在下に、T細胞における第1の活性化シグナルの引金をひくのに適した形体の抗原に、抗原特異的T細胞(すなわち、抗原を認識するTCRを発現しているT細胞)を接触させることによって、抗原に対するT細胞不応答を誘発することができる。該抗原は、たとえば、自己免疫反応を刺激する抗原または移植された細胞の拒絶反応を刺激するアロ抗原である。おそらく、D−3ホスホイノシチドの産生を阻害する作用剤が、ウォルトマンニンまたはクエルセチンなどのT細胞におけるPI3Kまたはその誘導体もしくは類縁体(LY294002など)の活性を阻害する。共刺激に関連する他の細胞内シグナルを阻害する追加の作用剤(前述)もまた、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤とともに用いることができる。たとえば、PI3KならびにハービマイシンAなどのプロテインチロシンキナーゼに、T細胞を接触させることができる。
【0024】
T細胞不応答を誘発するために、T細胞において第1の活性化シグナルの引金をひくのに適した形体の抗原に、抗原特異的T細胞を接触させるが、このことは、T細胞においてTCR/CD3複合体を介するシグナルが引金を引かれるように抗原をT細胞に対して付与することを意味する。たとえば、MHC分子に関連して細胞を付与する抗原によって、T細胞に対して抗原を付与することができる。B細胞、マクロファージ、モノサイト、樹状細胞、ランゲルハンス細胞またはT細胞に対して抗原を付与するその他の細胞などのシンジェネティックな抗原付与細胞を、抗原付与細胞がT細胞に対して抗原を付与するような抗原の存在下に、T細胞とともにインキュベートすることができる。別法として、アロ抗原に対するアネルギーを誘発するために、T細胞に対するアロ抗原を付与するアロジェネティックな細胞とともにT細胞をインキュベートすることができる。
【0025】
インビボにおいて抗原に対するT細胞不応答を誘発するために、T細胞におけるD−3ホスホイノシチドの産生を阻害する作用剤を、抗原に対するT細胞不応答を誘発するのに十分な用量と期間で、被験体に投与する。作用剤投与に続いて、抗原(たとえば内因的に細胞によって発現される自己抗原)を抗原特異的T細胞に内因的に接触させる。別法として、抗原に対する不応答を、インビトロにて誘発することができる。この場合、T細胞を被験体から採取し、インビトロにて抗原ならびに抗原不応答を誘発する作用剤に接触させ、次いで被験体にT細胞を再投与する。たとえば、移植片を移植を受ける者に移植する前に、移植を受ける者から採取したT細胞を、T細胞におけるD−3ホスホイノシチド産生を阻害する作用剤(たとえばウォルトマンニン、クエルセチン、LY294002)および/または共刺激に関連する他の細胞内シグネルの1種あるいはそれ以上を阻害する1種あるいはそれ以上の作用剤とともに移植片ドナーからのアロジェニックな細胞に接触させて、アロ抗原特異的T細胞不応答を誘発することができる。次いで、ドナーの抗原に対して不応答になった移植を受ける者のT細胞を、該移植を受ける者に再投与する。別法として、骨髄移植の場合、移植される骨髄(いずれかの残留T細胞を含む)を、インビトロにて、D−3ホスホイノシチドの産生および/または他の共刺激に関連する細胞内シグナルを阻害する作用剤とともに骨髄移植を受ける者のアロジェネティックな細胞に接触させて、移植を受ける者のアロ抗原に対するドナーT細胞における不応答を誘発する。この前処理によって移植片と移植を受けた者の間で生じる疾患を阻害することができる。
【0026】
該T細胞不応答誘発方法は、臓器移植および骨髄移植などの移植、および自己免疫疾患および異常免疫応答に関連する他の疾病などの免疫応答を下降調節することが望ましい状況において治療的に適用することができる。自己免疫疾患または不適当あるいは異常免疫応答に関連する他の疾病の例として、リウマチ性関節炎、若年性リウマチ性関節炎、乾せん性関節炎、アレルギー、接触性皮膚炎、乾せん、癩病性逆反応、癩病性結節性紅斑、自己免疫性ぶどう膜炎、多発性硬化症、アレルギー性脳脊髄炎、全身性紅斑性狼瘡、急性壊死性出血性脳障害、突発性両側性進行性感覚神経聴力損失、再生不良性貧血、真正赤血球性貧血、突発性血小板血症、多発性軟骨炎、強皮症、ウェジナー肉芽腫症、慢性活動性肝炎、重症筋無力症、スチーブン−ジョンソン症候群、突発性スプルー、偏平苔せん、クローン病、グレイブス眼病、サルコイドーシス、原発性胆汁性肝硬変、原発性若年性糖尿病、シェーグレン症候群関連乾き眼、後ぶどう膜炎および間質性肺繊維症が挙げられる。
【0027】
本発明の他の態様は、共刺激性シグナルをT細胞に与えることによってT細胞応答を刺激することに関する。第1活性化シグナルにつづく共刺激シグナルの伝達によってT細胞応答を発生させることができる。本発明方法においては、共刺激性シグナルは、T細胞においてD−3ホスホイノシチドの産生を刺激する作用剤に、第1活性化シグナルを受けたT細胞を接触させることにより付与される。T細胞の「応答」とは、T細胞(IL−2など)による少なくとも1種のリンホカインの細胞および/またはT細胞の増殖を意味する。第1活性化シグナルは、抗CD3抗体またはMHC/抗原複合体などのTCR/CD3複合体を介してT細胞を刺激すること、またはホルボールエステル(PMA)などのこの刺激を模倣する作用剤を用いることによってT細胞に伝達される。本明細書中で用いる語句「作用剤」とは、T細胞表面受容体と共刺激分子の間に相互作用を要求することなくT細胞において共刺激シグナルを刺激する化学薬品および他の薬剤化合物を意味する。たとえば、該作用剤は、細胞内的に作用して、共刺激関連シグナルを刺激する。ひとつの具体例においては、該作用剤は非プロテイン性化合物である。本発明方法に用いる該作用剤は、天然の受容体:リガンド共刺激メカニズムをバイパスすることを意図しており、語句「作用剤」には、B7−1および/またはB7−2などの天然のリガンドCD28/CTLA4を発現している細胞を含まない。
【0028】
おそらく、T細胞におけるD−3ホスホイノシチドの産生は、PI3KのアクチベーターにT細胞を接触させることによって刺激される。PI3Kの活性化は、たとえば、後述の方法によって同定することができる。共刺激に関連する1種またはそれ以上の他の細胞内シグナルを刺激する追加の作用剤を、D−3ホスホイノシチド産生のアクチベーターとともに用いることができる。たとえば、PI3Kを刺激する第1作用剤およびT細胞内におけるプロテインチロシンリン酸化を刺激する第2作用剤の両方に、T細胞を接触させる。T細胞におけるプロテインチロシンリン酸化は、過バナジン酸塩などのプロテインチロシンキナーゼのアクチベーターに、T細胞を接触させることによって刺激することができる[オシア,J.J.らの(1992)Proc.Natl.Acad.Sci.USA89:10306〜10310;およびシーリスト,J.P.の(1993)J.Biol.Chem.268:5886〜5893を参照]。たとえば、PI3Kを刺激する第1作用剤、およびプロテインセリンキナーゼのアクチベーターにT細胞を接触させるなどのT細胞内におけるプロテインセリンリン酸化を刺激する第2作用剤にT細胞を接触させてもよい。別の具体例においては、プロテインチロシンリン酸化を刺激する第3の作用剤にさらにT細胞を接触させる。別法として、PI3Kのアクチベーター、およびT細胞における正味のプロテインチロシンリン酸化を増加するためのCD45などの細胞プロテインチロシンホスファターゼの活性および/または細胞プロテインセリンホスファターゼの活性を阻害する作用剤の両方にT細胞を接触させる。本発明には、ホスホリパーゼC活性の刺激および/または細胞内カルシウム濃度の増加などのT細胞の共刺激に関連する他の細胞内シグナルを刺激することも含まれる。
【0029】
本発明の他の具体例は、抗原特異的T細胞による抗原に対する特異的応答を刺激する方法を提供する。T細胞応答を刺激するために、抗原ならびにT細胞においてD−3ホスホイノシチドの産生を刺激する作用剤に、抗原特異的T細胞を接触させ、それによってT細胞における共刺激シグナルの引金をひく。おそらく、T細胞においてD−3ホスホイノシチドの産生を刺激する作用剤はPI3Kのアクチベーターである。MHC分子に関連の、T細胞における第1の活性化シグナルを刺激するのに適した形体の抗原に、T細胞を接触させる(TCR/CD3複合体を介するなど)。抗原付与細胞(B細胞、マクロファージ、モノサイト、樹状細胞、ランゲルハンス細胞またはT細胞に対して抗原を付与するその他の細胞など)を、抗原(可溶性抗原など)の存在下にT細胞とインキュベートすることができる。別法として、対象となる抗原を発現している細胞をT細胞とインキュベートすることができる。たとえば、腫瘍関連抗原を発現している腫瘍細胞を、T細胞ならびに細胞内共刺激シグナルを誘発する作用剤とともにインキュベートして腫瘍特異的応答を誘発することができる。別法として、ウイルスなどの病原性抗原を付与する病原体に感染した細胞を、作用剤の存在下にT細胞とインキュベートすることができる。T細胞においてD−3ホスホイノシチドの産生を刺激することに加えて、たとえば過バナジン酸塩またはセリンキナーゼのアクチベーターなどのプロテインチロシンキナーゼのアクチベーターなどのCD28連結に関連する1種またはそれ以上の追加の細胞内シグナルを刺激する1種またはそれ以上の作用剤に、T細胞を接触させることができる。
【0030】
PI3KのアクチベーターなどのT細胞においてCD28関連細胞内シグナルを刺激する作用剤を、インビボにて被検体に投与することができる。またあるいは別法として、被検体から採取したT細胞をインビトロにて刺激し、被検体に再投与してもよい。T細胞を刺激する方法は、免疫応答を上昇調節することが望ましい状況(たとえば応答の誘発または存在する応答の強化)において治療上有用である。たとえば、該方法を用いて腫瘍関連抗原に対するT細胞応答を強化することができる。被検体からの腫瘍細胞は、典型的に腫瘍関連抗原を発現するが、T細胞における共刺激シグナルを刺激することはできない(たとえば、それらは共刺激分子の発現が欠如しているからである)。したがって、被検体からのT細胞ならびにT細胞においてD−3ホスホイノシチドを刺激する作用剤に、腫瘍細胞を接触させてT細胞における共刺激の引金をひくことができる。別法として、本明細書中に記載したようにT細胞を刺激して、病原性作用剤(ウイルス(ヒト免疫不全ウイルスなど)、バクテリア、寄生虫および真菌など)に対する応答性を誘発または強化することができる。さらに、このような病原性作用剤に対するワクチンの効能を増強することもできる。たとえば、T細胞においてD−3ホスホイノシチドの産生を刺激する作用剤を、病原性作用剤に感染した被検体に投与することができ、またはワクチンとともに投与して、ワクチンの抗原に対するT細胞の応答性を強化することができる。別法として、病原性作用剤からの抗原を発現する抗原付与細胞ならびに共刺激に関連する細胞内シグナルを刺激する作用剤(PI3Kのアクチベーター)とともに、T細胞をインビトロで培養することができる。
【0031】
さらに本発明のT細胞応答を刺激する方法は、CD28および/または共刺激に関連する他の細胞表面分子(CTLA4など)を介するシグナル変換が不良である患者に適用することができる。たとえば、突発性血小板減少症を患っている患者が、おそらく先天性欠陥であろうが、CD28介在性シグナル変換不良を呈することが報告されている[ペレッツ−ブラス,M.らの(1991)Clin.Exp.Immunol.85:424〜428]。CD28シグナリング能力不全の患者においては、正常CD28連結によって生成する細胞内シグナルを刺激する1種またはそれ以上の作用剤を、患者のT細胞に接触させることによって、該不良をバイパスし、CD28依存性T細胞活性化を回復することが可能である。たとえば、CD28連結によるD−3ホスホイノシチドの産生が低下または欠如していることによる不良をもつ患者を、T細胞においてD−3ホスホイノシチドの産生を刺激する作用剤に患者からのT細胞を接触させることによって、治療することができる。
【0032】
本発明のさらに別の態様は、PI3Kのインヒビターおよびアクチベーターを同定するためのスクリーニングアッセイであり、それらを用いてT細胞応答をそれぞれ阻害または刺激することができる。PI3Kは調節および触媒サブユニットからなるヘテロ二量体である。基質としてPtdIns(4,5)P2を優先的に用い、ウォルトマンニンによって阻害しうる2つの形体の酵素が記載されている[オツ,M.らの(1991)Cell65:91〜104;ヒュー,P.らの(1993)Mol.Cell.Biol.13:7677〜7688;およびハイルス,I.D.らの(1992)Cell70:419を参照]。基質としてPtdInsを優先的に用い、ウォルトマンニンによって阻害されない別の形体の酵素もまた記載されている[スティーブンス,L.らの(1994)Curr.Biol.4:203〜214を参照]。PI3Kの特異的インヒビターまたはアクチベーターの同定は、望まない逆の副作用を回避するための共刺激シグナルに必要とされるPI3Kの好適な細胞内形体に対して特異的でなければならないことが認識されよう。したがって、共刺激に必要とされるPI3Kの形体(たとえば、ウォルトマンニンによっても阻害しうる形体)を特異的に阻害または活性化する作用剤が好ましい。
【0033】
ひとつの具体例において、本発明のスクリーニングアッセイは、T細胞において少なくとも1種のD−3ホスホイノシチド(PtdIns(3,4,5)P3が好ましい)の産生をそれぞれ阻害または刺激する、PI3Kのインヒビターまたはアクチベーター能力に基づいている。PI3Kのインヒビターを同定するために、被検基質の存在あるいは不在下に、共刺激分子を結合する細胞表面受容体を介してT細胞を刺激する(すなわち、共刺激シグナルを受容したT細胞)。好ましくは、CD28を発現するT細胞をアッセイに用いる。別法として、CTLA4を発現するT細胞を用いてもよい。CD28またはCTLA4に対するリガンドに、T細胞を接触させることによって、T細胞における共刺激シグナルを刺激することができる。該リガンドは、T細胞表面受容体に対する抗体よりもむしろ膜結合B7−1またはB7−2などの生理学的リガンドであることが好ましい。天然にB7−1および/またはB7−2を発現する細胞を用いることができるが、共刺激分子を発現するようにトランスフェクトされた細胞(CHO細胞など)を用いるのが、より好ましい。PI3Kのインヒビターの存在下に、共刺激分子(CD28など)を結合する表面受容体を介するT細胞を刺激する結果として、該インヒビターの不在下における刺激と比較して、T細胞におけるD−3ホスホイノシチドの産生が低減化される。T細胞におけるD−3ホスホイノシチドの産生は、当業者には公知の適当な方法のいずれかによって測定することができる。たとえば、高圧液体クロマトグラフィーによって、T細胞におけるD−3ホスホイノシチドの産生を測定することができる(実施例に記載)。別法として、薄層クロマトグラフィーによってD−3ホスホイノシチドの産生を評価することができる[オカダ,T.らの(1994)J.Biol.Chem.269:3563〜3567]。細胞内産生を評価しえたD−3ホスホイノシチドとして、PtdIns(3)P、PtdIns(3,4)P2およびPtdIns(3,4,5)P3が挙げられる。被検基質の存在下に、T細胞におけるPtdIns(3,4,5)P3の産生を検出するのが好ましい。
【0034】
PI3Kのアクチベーターを同定するために、共刺激分子を結合する細胞表面受容体を発現するT細胞を、被検基質と接触させる。T細胞において少なくとも1種のD−3ホスホイノシチド(PtdIns(3,4,5)P3が好ましい)の産生を刺激する能力に基づいてPI3Kのアクチベーターを同定する。したがって、PI3Kアクチベーターの存在下において、T細胞におけるD−3ホスホイノシチドの量は、基質不在下におけるT細胞中のD−3ホスホイノシチドの量と比較して増加する。T細胞におけるD−3ホスホイノシチド[PtdIns(3)P、PtdIns(3,4)P2およびPtdIns(3,4,5)P3]の産生は、高圧液体クロマトグラフィーまたは薄層クロマトグラフィーなどの標準的方法によって評価することができる。
【0035】
本発明スクリーニングアッセイの別の具体例においては、細胞から単離されるPI3Kの活性を直接阻害または刺激するための基質の能力を評価し、次いで、PI3Kのインヒビターまたはアクチベーターとして同定された基質を、T細胞に接触させて、T細胞応答を阻害または刺激する基質の能力を決定する。たとえば、単離されたPI3Kを、放射標識されたホスフェートドナーおよび被検物質の存在下に、基質(PtdIns(4,5)P2など)とともにインキュベートする。PI3Kのキナーゼ活性のインヒビターは、インヒビター不在下におけるリン酸化と比較して、基質のリン酸化を低下させるであろう。一方、アクチベーターの不在下におけるリン酸化と比較して、アクチベーターは基質のリン酸化を増加するであろう。次いで、このように同定されたインヒビターまたはアクチベーターを、T細胞に接触させて、インヒビターまたはアクチベーターのT細胞応答(IL−2の産生など)の阻害または刺激能力をそれぞれ決定する。
【0036】
さらに本発明は、実施例に記載するように、ジャーカット細胞および末梢血液T細胞におけるCD28に関連する約67kDaのホスホチロシンプロテインに関する。67Kと称するこの新規なタンパク質は、itk/emtプロテインチロシンキナーゼ、lckおよびPI3Kとは異なると考えられ、CD28に関連するシグナル変換経路に含まれるようである。本発明の特定の具体例において、該67Kが含まれるCD28シグナル変換経路は、たとえば、そのリン酸化の遮断などによって67Kの活性化を遮断することによって阻害される。別法として、該67Kに関連するシグナル変換経路は、67KとCD28の相互作用を阻害することによって遮断される。本発明の別の具体例においては、67Kに関連するシグナル変換経路は、67Kを活性化することによるかまたは67KとCD28の相互作用を刺激することによって刺激される。67Kを含むシグナル変換経路の活性化または阻害を、上述の具体例のいずれかによって行うことができ、それによってT細胞応答が遮断されるかまたは別法においてはT細胞応答が刺激される。
【0037】
他の具体例
T細胞以外にも、CD28をその表面に発現する他のタイプの細胞が文献に記載されている。このような細胞のタイプには、プラズマ細胞[コズボール,D.らの(1987)J.Immunol.138:4128〜4132]および骨髄誘導マスト細胞が含まれる。CD28を介する他のCD28+細胞の刺激によっても、該細胞においてD−3ホスホイノシチドの産生および特異的細胞応答の発生が誘導される。本発明方法による、これらの細胞におけるD−3ホスホイノシチドの阻害および活性化もまた、他のCD28+細胞型による応答を阻害または刺激するのに有用である。
【図面の簡単な説明】
【0038】
【図1】第1図は、CD28+T細胞(ジャーカット細胞)における、該細胞を培地、抗CD3抗体または抗CD28抗体で刺激した後のホスファチジルイノシトール(3,4,5)−トリリン酸塩の産生を示すグラフであり、CD3またはCD28を介した刺激におけるホスファチジルイノシトール3−キナーゼの活性化の独特の動力学を表している。
【図2】第2図は、CD28+T細胞(ジャーカット細胞)における、該細胞をB7−1またはB7−2を発現するようにトランスフェクトされたCHO細胞で刺激した後のホスファチジルイノシトール(3,4,5)−トリリン酸塩の産生を示すグラフであり、B7−1またはB7−2による刺激におけるホスファチジルイノシトール3−キナーゼの活性化の独特の動力学を表している。
【図3】第3図は、CD28+T細胞(ジャーカット細胞)における、該細胞をB7−2を発現するようにトランスフェクトされたCHO細胞で刺激した後のホスファチジルイノシトール(3,4,5)−トリリン酸塩の産生に関する種々の濃度のウォルトマンニン(0〜100mM)の効果を示すグラフである。
【図4】第4図は、CD28と抗CD28抗体の連結によって誘発されたT細胞におけるカルシウム代謝速度に関するウォルトマンニンまたはハービマイシンの効果を示すグラフである。
【図5】第5図は、ジャーカット細胞をCHO−neo、CHO−B7−1またはCHO−B7−2細胞のいずれかとともにインキュベートする細胞−コンジュゲートアッセイから得た一連のフローサイトメトリーのプロフィールである。カルシウム代謝速度をY軸に、細胞コンジュゲーションをX軸に示す。
【図6】第6図は、精製ヒト末梢血液T細胞によるIL−2の産生に関する、抗CD3抗体と組み合わせた場合の膜結合B7−1およびB7−2の効果を示すグラフであり、B7−1またはB7−2のいずれかの共刺激によるIL−2の産生における用量依存性の増加を表している。
【図7A】第7A図は、培地、固定化抗CD3+CHO−B7−1、固定化抗CD3+CHO−B7−2、PMA+CHO−B7−1またはPMA+CHO−B7−2で休止期ヒトT細胞を刺激した24時間後の、該細胞によるIL−2の産生に関する、ウォルトマンニン処理(1nM〜1mM)の効果を示すグラフである。
【図7B】第7B図は、抗CD3抗体(OKT3)ならびにB7−1、B7−2またはB7−1とB7−2の両方を発現するCHO細胞で24時間刺激されたヒトT細胞によるIL−産生、またはPMAならびにB7−1またはB7−2を発現するCHOで刺激されたT細胞によるIL−産生に関する、ウォルトマンニン(1〜100nM)による阻害パーセントを示すグラフである。
【図8】第8図は、抗CD28mAb(9.3)、CHO、CHO−B7−1またはCHO−B7−2で刺激されたジャーカット細胞によるホスファチジルイノシトール(3,4,5)−トリリン酸塩の産生に関する、ハービマイシン処理の効果を示すグラフである。
【図9】第9図は、ハービマイシンの存在(+)または不在(−)下で、抗CD28mAb(9.3)またはCHO−B7−1(B7)細胞で刺激した後の異なる時点におけるジャーカット細胞によって産生されたホスファチジルイノシトール(3,4,5)−トリリン酸塩の量を示す薄層クロマトグラフィーである。
【図10A】第10A図は、ハービマイシンAの存在または不在下で、PMA単独またはPMAとCHO−B7−1(PMA+CHOB71)、PMAとCHO−B7−2(PMA+CHOB7−2)、PMAと9.3mAb溶液(PMA+9.3)、PMAとビーズ上の9.3mAb(PMA+9.3ビーズ)、PMAとイオノマイシン(PMA+イオノマイシン)あるいはPMAとイオノマイシンと可溶性9.3mAb(PMA+イオノ+9.3)で刺激された末梢血液T細胞によるIL−2分泌を示すグラフである。
【図10B】第10B図は、イオノマイシンの存在(+イオノ)または不在下で、抗CD3(OKT3)単独またはOKT3とCHO−B7−1(OKT3+CHOB71)、OKT3とCHO−B7−2(OKT3+CHOB7−2)またはOKT3と9.3mAb(OKT3+9.3)で刺激された末梢血液T細胞によるIL−2分泌を示すグラフである。
【図11】第11図は、およそ6kDaのホスホチロシンプロテインが抗CD28抗体で共免疫沈降されることを示すイムノブロットのオートラジオグラフィーである。
【発明を実施するための形態】
【0039】
次の実施例によって本発明をさらに詳しく説明するが、本発明はこれらに限定されるものではない。本明細書に引用されたすべての参考文献、特許および特許出願公報はすべて、本発明の引例文献である。
【実施例1】
【0040】
CD28の連結はD−3ホスホイノシチドの産生を刺激する
この実施例では、CD28またはCD3を介するジャーカット細胞の刺激によるCD28+ジャーカット細胞におけるD−3ホスホイノシチドの産生を試験した。次のようにしてジャーカット細胞をキャリヤーフリーの32Pオルトホスフェート(32Pi)で標識した:ジャーカット細胞をホスフェートフリーの培地(DMEM/RPMI)で3回洗浄し、37℃で10分間、洗浄の間に10分間インキュベートした。細胞を20mMのHEPES,pH7.4および5%の透析したコウシ血清(生理的食塩水で一夜透析)を含む10mlのホスフェートフリーの培地に再懸濁した。細胞(1mCi/10ml細胞)にキャリヤーフリーの(32Pi)を加え、該細胞を37℃で4〜6時間インキュベートした。標識後、細胞をホスフェートフリーの培地で3回洗浄し、20mMHEPESを含むRPMI1640に再懸濁した。
【0041】
種々の実験において、32Pi標識ジャーカット細胞(0.15ml;2×107細胞)のアリコートを培地単独、抗CD3抗体(G19〜4)、抗CD28抗体(9.3)、非トランスフェクトCHO細胞またはCD28を発現するようにトランスフェクトされたCHO細胞(ヒトB7−1:CHO−B7−1またはヒトB7−2:CHO−B7−2のいずれか)で刺激した[Dr.G.フリーマンおよびL.ナドラーから入手;CHO細胞をヒトB7−1をコードするcDNA(該配列は、フリーマン,D.J.らの(1989)J.Immunol.143:2714〜2722に開示されている)またはヒトB7−2をコードするcDNA配列(該配列は、フリーマン,D.J.らの(1993)Science262:909〜911に開示されている)含有組換え発現ベクターによって標準的技術を用いてトランスフェクトした]。CHO細胞を刺激するために、ジャーカット細胞を107CHO細胞とともにインキュベートし、マイクロ遠心管による5秒間の低速度遠心分離によって細胞の接触を達成した。刺激に続いて種々の時間間隔(約1〜25分間の範囲)において、細胞を溶解し、ホスホリピッドを抽出し、脱アシル化し、アニオン交換HPLCによって分離した[基本的に、ウォード,S.G.らの(1992)J.Immunol.22:45の記載に準じ、下記の変更を加えた]。
【0042】
750mlのCHCl3/メタノール/水(32.6%/65.3%/2.1%v/v/v)を添加してインキュベーションを終了した。細胞の反応を停止するとすぐに、次の抽出中を通じてサンプルを氷上で保持した。725mlのCHCl3(10mgのフロックリピッドを含有;たとえばシグマ,カタログNo.B1502から入手)および172mlの2.4MHCl、5mMの硫酸テトラブチルアンモニウムを各サンプルに加えることによって相分離を行った。サンプルを渦巻き撹拌し、1000rpmで5分間遠心分離することによって相を分離した。下の相を除去し、1/2容の1M HCl、25mM Na2EDTA,pH7.5、0.5mM硫酸テトラブチルアンモニウムを管に加えた。サンプルを再遠心分離して相を分離し、底の相を除去し、清浄な管に入れ、サンプルを減圧乾燥した。サンプルに1mlのメチルアミン試薬(40%水/メタノール/n−ブタノール(4:4:1v/v/v)溶液)を加えて渦巻き撹拌し、53℃で40分間インキュベートして脱アシル化した。サンプルを氷上に置き、次いで減圧乾燥した。サンプルを0.5mlの滅菌蒸留水に再懸濁し、渦巻き撹拌して混合した。サンプルを0.7mlのn−ブタノール(40〜60%石油エーテル/ギ酸エチル(20:4:1v/v/v))で2回抽した。底の水相を減圧乾燥し、HPLC分析を行うまで−70℃で貯蔵した。緩衝液A(水)/B[1.25M(NH4)2HPO4](H3PO4にて25℃でpH3.8に調節)に基づく溶離勾配およびPartisphereSAXカラム(ワットマン社から購入)を用いてHPLCを行った。脱アシル化したホスホリピッドサンプルを0.1mlの蒸留水に再懸濁し、該カラムに注入した。溶出液をCanberra Packard A−500 Flo−Oneオンラインベータ線検出装置に導入し、3部のFlo−ScintIVシンチレーションカクテルと混合し、結果をFlo−Oneデータプログラム(ラジオマチック,USA)によって分析した。溶出したピークを[3H]PtdIns、[3H]PtdIns(4)Pおよび[3H]PtdIns(4,5)P2(アマーシャム・インターナショナル社から購入)から調製した標準液の保持時間と比較した。ウォード,S.G.らの(1992)J.Biol.Chem.267:23862の記載にしたがって、単離したホスファチジルイノシトール3−キナーゼをPtdIns(シグマ社から購入)とともにインキュベートすることによって標準の[32P]PtdIns(3)P、[32P]PtdIns(3,4)P2および[32P]PtdIns(3,4,5)P3を調製した。
【0043】
抗CD3または抗CD28抗体で1〜5分間刺激した後のジャーカット細胞におけるPtdIns(3,4,5)P3の産生を第1図に示す。その結果から、CD3またはCD28のいずれかを介する刺激はPtdIns(3,4,5)P3の産生を誘発するけれども、2つの経路の誘発動力学は別々であることがわかる。CD3による刺激によって早期(すなわち2分以内)にPI3Kの活性化(PtdIns(3,4,5)P3の産生によって評価される)が増加する。逆に、CD28刺激によって誘発されるPI3Kの活性化はCD3刺激による活性化よりも遅く(すなわち5分あるいはそれよりも遅くまで最大にはならない)、CD3刺激による活性化よりも長く持続する。このような結果から、CD3またはCD28連結のいずれかによって仲介されるPI3K活性化には別のメカニズムが含まれることが示される。
【0044】
CHO−B7−1またはCHO−B7−2細胞で0〜20分間刺激した後のジャーカット細胞におけるPtdIns(3,4,5)P3の産生を第2図に示す。その結果から、B7−1またはB7−2のいずれかによる刺激は有効な活性化(PtdIns(3,4,5)P3の産生によって評価される)を誘発することが示される。誘発の動力学は2つのCD28リガンドとはわずかに異なっている:B7−1とB7−2のプラトーは同じレベルであるが、B7−1はB7−2よりも早期に活性化を刺激する。B7−1またはB7−2のいずれかによるPtdIns(3,4,5)P3産生の刺激は非常に強く、持続的である(すなわち20分以上継続する)。
【0045】
この実施例は、CD28の天然リガンドであるB7−1およびB7−2または抗CD28抗体のいずれかによるCD28を介するT細胞の刺激が、T細胞内においてD−3ホスホイノシチドの産生を誘発することを証明し、CD28連結に基づくPI3Kの活性化が示される。さらに、この実施例は、ジャーカット細胞がCTLA4−ではなくCD28+であり、CTLA4を発現するようには誘発されえないので[リンステン,T.の(1993)J.Immunol.151:3489〜3499に示される]、CD28が通例B7−1とB7−2を生理的リガンドとして分けもつことを証明する。したがって、CTLA4は明らかにB7誘発シグナル変換において要求されず、B7−1とB7−2の両方がCD28に対する生理的リガンドである。
【実施例2】
【0046】
ホスファチジルイノシトール3−キナーゼインヒビターはCD28連結によって誘発されるD−3ホスホイノシチドの産生を阻害する
この実施例では、CD28が仲介するジャーカット細胞におけるD−3ホスホイノシチドの産生に関するホスファチジルイノシトール3−キナーゼのインヒビターとしての効果を試験した。実施例1に記載したようにジャーカット細胞をオルトホスフェートで標識し、B7−2を発現するようにトランスフェクトさせたCHO細胞で刺激した。さらに、刺激を行っている期間中、種々の濃度(0〜100mM)の真菌代謝物ウォルトマンニン(ホスファチジルイノシトール3−キナーゼのインヒビターである)の存在下で細胞をインキュベートした。ウォルトマンニンはシグマ・ケミカル・コーポレイションから購入し、10mMのDMSO溶液として−40℃で貯蔵した。それを培養中の細胞に添加する直前に培地で希釈した。ウォルトマンニンの存在あるいは不在下において刺激を行った後、実施例1に記載したようにHPLCによって細胞内に産生されたPtdIns(3,4,5)P3の量を測定した。結果を第3図に示す。第3図はCHO−B7−2で刺激する際にウォルトマンニン処理されたジャーカット細胞において検出されたPtdIns(3,4,5)P3の量を、CHO−B7−2で刺激された非処理ジャーカット細胞において検出されたPtdIns(3,4,5)P3の量に対するパーセントとしてプロットしてグラフ化したものである。その結果から、ジャーカット細胞を濃度増加ウォルトマンニンで処理することにより、B7−2によりCD28が連結した細胞において産生されるD−3ホスホイノシチドの量が減少することが示される。したがって、この実施例は、T細胞内におけるホスファチジルイノシトール3−キナーゼの活性を阻害することによって、CD28を介するT細胞の刺激の結果としての細胞内的D−3ホスホイノシチドの産生を阻害しうることを証明している。
【実施例3】
【0047】
CD28仲介カルシウム代謝における薬物的インヒビターの効果
この実施例では、CD28抗体によって誘発されるジャーカット細胞におけるカルシウム代謝速度に関する薬物的インヒビターの効果を試験した。研究した薬物的インヒビターはPI3Kの活性を阻害するウォルトマンニンおよびプロテインチロシンキナーゼの活性を阻害するハービマイシンAである。培地単独あるいはウォルトマンニンもしくはハービマイシンAの存在下のいずれかにおいて抗CD28抗体でジャーカット細胞を刺激し、細胞中の平均カルシウム濃度(nM)を刺激後数分にわたって測定した。第4図に示すように、ハービマイシンAはCD28抗体誘発カルシウム代謝を阻害する能力があった。逆に、ウォルトマンニンはCD28抗体誘発カルシウム流出を阻害することができなかった。これらの結果は、T細胞におけるウォルトマンニンの効果が、カルシウム代謝を妨害することによって仲介されないことを示している。さらにそのうえ、B7−1またはB7−2によって誘発されるIL−2の産生として測定されたところの“ウォルトマンニンが補刺激を阻害すること”を示している後記実施例5の結果、このデータは、CD28が誘発するカルシウム濃度上昇が、特異的にT細胞不応答(すなわちアネルギー)または共刺激を誘発する化合物の評価という目的に対して、おそらく読み取りの誤りによるものであろうということを示している。
【実施例4】
【0048】
天然のCD28リガンドによるカルシウム代謝の誘発
この実施例では、CHO−B7−1またはCHO−B7−2のジャーカット細胞への接着ならびにCHO−B7−1およびCHO−B7−2による刺激に応じたジャーカット細胞におけるカルシウム可動化を試験した。アベ,R.らの(1992)J.Exp.Med.176:459〜468に記載されているフローサイトメトリー細胞カルシウム−コンジュゲートアッセイを用いた。該細胞コンジュゲートアッセイでは、カルシウム感受性蛍光プローブindo−1(青色および緑色のシグナルを発する)でT細胞を着色する。コントロールプラスミド(CHO−neo)でトランスフェクトされたCHO細胞またはB7−1あるいはB7−2を発現するようにトランスフェクトされたCHO細胞をトレーサー着色剤DilC22(3)(赤色のシグナルを発する:モレキュラー・プローブ社から購入)で着色する。ジャーカット細胞−CHO複合体をフローサイトメトリーによって分析する。T細胞とCHO細胞からなる複合体は赤色のシグナルを補足することによって測定でき、カルシウム濃度は青色または緑色のシグナルを補足することによってT細胞内で測定できる。第5図に示すように、一連の2つのパラメーターの点プロットとして結果を表示する。カルシウム(indo−1比)をY軸に、細胞複合体(赤色トレーサー)をX軸に示す。右上のカドラントにおける細胞は、CHO細胞に複合したカルシウム濃度の高いジャーカット細胞を表す。右下のカドラントの細胞は、CHO細胞に複合したカルシウム濃度が正常であるジャーカット細胞を表す。左の上下のカドラントにおける細胞は、それぞれ高濃度または低濃度のカルシウムを含む非複合ジャーカット細胞を表す。
データは、B7−1およびB7−2の両方がジャーカット細胞への接着を仲介することができることを示している。しかし、両方のリガンドのカルシウム可動化の増加を引き起こす能力は乏しい。したがって、B7−1およびB7−2は、カルシウム可動化よりもPI3K活性化を誘発することにおける能力が高い(実施例1参照)。逆に、抗CD28抗体は、PI3K活性化(実施例1参照)およびカルシウム代謝(実施例3参照)の両方を刺激する能力がある。したがって、刺激を天然のリガンド(B7−1またはB7−2など)を用いて行うか抗体を用いて行うかによって、CD28の連結を介して発生する細胞内シグナルに違いがあると考えられる。ナンズ,J.らの(1993)Int.Immunol.5:311〜315に、CD28抗体が、T細胞のシグナル変換および活性化(IL−2の産生など)という生化学的局面において多重かつ異種の効果をもつことが既に記載されている。これらの観察から、CD28に関するこれまでの結果からの外挿に基づいてD−3代謝物の産生におけるCD28の天然のリガンド(すなわち膜結合B7−1およびB7−2)生化学的効果を予測することは不可能であることがさらに示される。
【実施例5】
【0049】
ホスファチジルイノシトール3−キナーゼインヒビターはCD28の連結によって誘発されるインターロイキン2の産生を阻害することができる
この実施例では、T細胞によるインターロイキン2のCD28依存性産生におけるホスファチジルイノシトール3−キナーゼのインヒビターとしての効果を試験した。最初の一連の実験では、ウォルトマンニンの不在下でのIL−2の産生における、CD28を介するT細胞刺激の効果(CD3を介する刺激に関連して)を評価した。高純度ヒト末梢血液T細胞を、固定化抗CD3抗体(OKT3)単独、または抗CD28抗体(9.3)あるいは非トランスフェクトまたはB7−1またはB7−2を発現するようにトランスフェクトされたマイトマイシンC処理CHO細胞とともに24時間インキュベートした。24時間培養後、ELISAにて標準的技術を用いてIL−2産生について培養上清をアッセイした。第6図に示すように、培地単独で、OKT3単独でまたはOKT3と非トランスフェクトCHO細胞とともにインキュベートされた細胞はIL−2を産生しなかった。逆に、OKT3およびB7−1またはB7−2を発現するようにトランスフェクトされたCHO細胞とともに培養された細胞は、用量依存的にIL−2の産生を刺激された。OKT3と9.3抗体との培養もまたIL−2の産生を刺激した。これらの結果から、たとえばB7−1またはB7−2刺激によるCD28の連結がリンホカイン産生に対する共刺激シグナルを提供することができることが確認される。
【0050】
次の一連の実験では、休止期ヒトT細胞をそれぞれ:1)固定化OKT3+CHO−B7−1,2、2)固定化OKT3+CHO−B7−2,3、3)固定化OKT3+CHO−B7−1+B7−2、4)PMA+CHO−B7−1、または5)PMA+B7−2で刺激した。培地単独中、またはウォルトマンニンを1nM〜1mMの濃度で含有する培地中でT細胞の刺激を行った。培養の24時間後、ELISAによってIL−2の産生について上清をアッセイした。結果を第7A図および第7B図に示す。該結果は、ウォルトマンニンが、CD3刺激に関連したB7−1またはB7−2のいずれかによって刺激されるIL−2の産生を阻害することができることを示す。IL−2のウォルトマンニン仲介阻害は用量依存性であった。B7−2仲介刺激の阻害のID50は約10nMであった。B7−1仲介刺激の阻害のID50は10〜100nMであった。これらのウォルトマンニンの用量は細胞に対して一般的に毒性ではなく、非特異的メカニズムによるIL−2の産生を阻害しない。PMAおよび膜結合B7−1またはB7−2によって刺激されるによって刺激されるIL−2の産生が、1mMという高濃度のウォルトマンニンによって阻害されないという事実が証拠である。この実施例は、TCR/CD3複合体およびCD28を介する刺激に応じたIL−2の産生によって評価されるように、T細胞内のホスファチジルイノシトール3−キナーゼの活性を阻害する作用剤でT細胞を処理することによってT細胞の活性化を阻害しうることを証明する。
【実施例6】
【0051】
抗CD28またはCD28の天然のリガンドで刺激したジャーカット細胞のPI3K活性化へのハービマイシンの異なる効果
PI3キナーゼ活性化の意味を評価するため、ジャーカットT細胞(20×106細胞)を抗CD28mAb9.3(10μg)またはB7−1またはB7−2(10×106細胞)を発現しているCHO細胞で刺激した。該細胞を実施例1で記載したように32Pオルトホスフェートで標識し、ついでビヒクル希釈薬または3μMのハービマイシンAを含む培地で前もってインキュベートした。5分後、該細胞は溶解し、PI3キナーゼを実施例1に記載したようにHPLCによって、PtdIns(3,4,5)P3の測定により評価した。
該結果を図8に示す。9.3抗体で刺激したジャーカット細胞によるPtdIns(3,4,5)P3の産生をハービマイシンの前処置によって妨げた。予期せず、PtdIns(3,4,5)P3の産生によって測定されたようにPI3キナーゼ活性を誘発したB7−1およびB7−2はハービマイシンの前処置に耐性を示した。これらの結果は、CD28によってPI3キナーゼの活性に関するハービマイシンの独立した経路が存在することを示し、さらにハービマイシンは様々なsrcファミリーキナーゼを抑制することを示しているのでCD28はsrcファミリープロテインチロシンキナーゼから独立してPI3キナーゼを活性化することができることを示す。
【0052】
この両分をさらに調査するため、20×106ジャーカット細胞を9.3mAb(10μg)または10×106CHO−B7−1細胞で刺激した。該細胞をハービマイシンA 3μMまたはビヒクル内で前もってインキュベートした。該CD28 mAbにさらにヤギ抗マウスIgGを結合させ、該細胞を5〜10分後溶解した。PI3キナーゼ活性化はイン・ビボリピッドキナーゼアッセイを使用して薄層クロマトグラフィーによって評価した(ウォード(Ward,S.G.)ら(1995)Eur.J.Immunol.25:526−532に記載のように)。
該結果を図9に示す。抗CD28 9.3mAbおよびCHO−B7細胞の両者は、コントロールレーンと10分後の9.3mAbレーンおよび5分後のB7レーンとの比較から明らかなようにPI3キナーゼ活性を誘発する。再び、PIの反対のキナーゼの産物は9.3mAb刺激後ハービマイシンによって抑制され、B7刺激後ハービマイシンに耐性を示した。
従って、ハービマイシンは抗CD28で刺激したジャーカット細胞でPI3Kのハービマイシンの活性化を遮断するが、CHO−B7−1またはCHO−B7−2細胞で刺激したジャーカット細胞においては遮断せず、CD28によるPI3キナーゼの活性化のハービマイシンの独立した経路が存在することを示した。
【実施例7】
【0053】
T細胞によるIL−2の分泌へのへルビマイシンの異なる効果
さらに、CD28受容体ライゲーションに関するシグナル変換経路を決定するため、CD28+T細胞(抹消血T細胞のネガティブセレクションによって調製した)を種々の濃度のハービマイシン(0.1〜3μM)にて一晩培養し、ついでPMAを3ng/mlだけまたは、1μg/mlの溶液内のmAb 9.3、mAb9.3コートしたビーズ、0.08μg/mlのイオノマイシン、CHO−B7−1またはCHO−B7−2細胞で刺激した。上清を24時間後に回収し、ELISAによってIL−2含有を分析した。
該結果を図10パネルAに示す。該結果はPMAプラス9.3mAbが誘発したIL−2分泌が約0.2μMのID50を有するハービマイシンに感受性を有することを示す。同様に、PMAプラスCHO−B7−1細胞によって誘発したIL−2分泌はまたハービマイシンにも感受性があった。対照的に、IL−2分泌は細胞が3μMより大きいID50を有するPMAプラスイオノマイシンで刺激した場合、完全に耐性があった。ハービマイシンによる抑制のメカニズムをさらに試験するため、細胞をイオノマイシンの存在下または不在下でも刺激した。PMAプラスイオノマシインプラス9.3mAbで刺激した細胞におけるIL−2分泌はハービマイシン、3μMのハービマイシンAにさえ完全耐性となる。
【0054】
さらにハービマイシン感受性および耐性を有するIL−2分泌の結果を試験するため、T細胞を、CHO−B7−1もしくはCHO−B7−2細胞またはイオノマイシンの存在下もしくは不在下にて9.3mAbを有するビーズに不動化したCD3抗体OKT3で刺激した。上清を24時間後に回収し、ELISAによってIL−2の量を決定した。
該結果を図10パネルBに示す。PMAおよびB7−1またはB7−2でのT細胞の刺激による結果のハービマイシンに対して感受性を有すると分かったIL−2分泌に反して、PMAが抗CD3で置換された場合、IL−2分泌はハービマイシンに完全に耐性を有すると分かった。これらの結果は、CD28はハービマイシン耐性IL−2分泌をシグナルの状況に依存して開始することができることを示す。T細胞がカルシウムシグナル(すなわち、PMAプラス抗CD28)を開始しないような方法で活性化された場合、IL−2分泌はハービマイシンに感受性を有する。T細胞がカルシウムシグナル(すなわち、PMAプラスイオノマイシンプラスB7または抗CD28)の存在下で活性化された場合、該シグナルはハービマシンに対して耐性を示した。これはCD28はハービマイシン感受性のチロシンキナーゼによって提供される不可欠なシグナルを必要としないかまたは代わって、CD28によって開始される不可欠なチロシンキナーゼカスケードの必要性を無視することができるかもしれないことを示している。
【実施例8】
【0055】
CD28に関連した67kDaのチロシンホスホプロテインの同定
現在までに、CD28と物理的な関連を示す唯一報告されたプロテインはPI3キナーゼ(トルイット(Truitt,K.E.)ら(1994)J.Exp.Med.179:1071−1076)およびsrcファミリーキナーゼlck(オーガスト(August,A.)およびデュポン(Dupont,B.)(1994)Biochem.Biophys.Res.Commun.199:1466−1473)である。CD28媒介シグナル変換をさらに研究するため、CD28と物理的に関連のあるタンパク質に関するジャーカット細胞およびT細胞において研究を開始した。細胞を何度も9.3mAbで刺激し、NP−40清浄剤で溶解し、ブドウ球菌タンパク質Aで免疫沈降した。該免疫沈降物を洗浄し、抽出しついで還元条件のもとでSDS−PAGEによって分離した。トランスファーの後、西洋ワサビペルオキシダーゼとともに抗ホスホチロシンAb4G10を使用してイムノブロットを行った。チロシンリン酸化タンパク質をECLを使用して化学発光によって同定した。
該結果を図11に示すが、これはイムノブロットのオートラジオグラフィーを示している。〜67kDaの相対的な移動を示すバンドをCD28免疫沈降物内に同定した。ホスホプロテインに対応するバンドはCD28刺激後3分以内にジャーカット細胞に即座に現れた。同様のまたは同一のタンパク質をT細胞幼若細胞において同定した。このタンパク質はitk/emtプロテインチロシンキナーゼではないようであった。というのはitkに対する抗体はCD28免疫沈降物内にitkを明らかにしないからである。さらにその上、このCD28関連タンパク質の分子量がlckまたはPI3と一致していない。
従って、該結果は約67kDaの新規なホスホチロシンタンパク質がCD28を通じて刺激したジャーカット細胞およびT細胞幼若細胞内におけるCD28と関連を有することを示す。
【0056】
均等物
当業者は、本明細書に記載の発明の特定の態様に均等な多くのものについて認識するか、常套的な実験を使用するだけで確認することができるであろう。そのような均等物は以下の請求の範囲に包含するよう意図されている。
【特許請求の範囲】
【請求項1】
T細胞による応答を阻害するための医薬組成物の製造のための、共刺激分子に結合するCD28細胞表面受容体を発現しているT細胞においてD−3ホスホイノシチドの産生を細胞内で阻害するように働く作用剤の使用であって、該応答が、T細胞によるIL−2産生および/またはT細胞の増殖を含む使用。
【請求項1】
T細胞による応答を阻害するための医薬組成物の製造のための、共刺激分子に結合するCD28細胞表面受容体を発現しているT細胞においてD−3ホスホイノシチドの産生を細胞内で阻害するように働く作用剤の使用であって、該応答が、T細胞によるIL−2産生および/またはT細胞の増殖を含む使用。
【図1】


【図2】

【図3】

【図4】
【図5】
【図6】
【図7A】
【図7B】
【図8】

【図9】
【図10A】
【図10B】
【図11】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【図8】
【図9】
【図10A】
【図10B】
【図11】
【公開番号】特開2010−260868(P2010−260868A)
【公開日】平成22年11月18日(2010.11.18)
【国際特許分類】
【出願番号】特願2010−141945(P2010−141945)
【出願日】平成22年6月22日(2010.6.22)
【分割の表示】特願2006−315500(P2006−315500)の分割
【原出願日】平成7年5月1日(1995.5.1)
【出願人】(505477408)アメリカ合衆国 (6)
【氏名又は名称原語表記】THE UNITED STATES OF AMERICA
【Fターム(参考)】
【公開日】平成22年11月18日(2010.11.18)
【国際特許分類】
【出願日】平成22年6月22日(2010.6.22)
【分割の表示】特願2006−315500(P2006−315500)の分割
【原出願日】平成7年5月1日(1995.5.1)
【出願人】(505477408)アメリカ合衆国 (6)
【氏名又は名称原語表記】THE UNITED STATES OF AMERICA
【Fターム(参考)】
[ Back to top ]