
細胞内環境において向上した安定性を示す免疫グロブリンフレームワークおよびそれを同定する方法
【課題】非常に安定な可溶性単一鎖Fv抗体フラグメントを作成するためのフレームワークとして使用することができる組成物の提供。
【解決手段】フレームワークは、細胞内性能について選択されるため、安定性および可溶性が抗体フラグメントの性能に関して限定因子であるところで(たとえば、細胞の還元環境で)適用するためのscFv抗体フラグメントまたはscFv抗体ライブラリーを作成するために理想的に適合している。また、かかるフレームワークは、向上した可溶性および安定性を示す、高度に保存された残基およびコンセンサス配列を同定するために使用することができる。
【解決手段】フレームワークは、細胞内性能について選択されるため、安定性および可溶性が抗体フラグメントの性能に関して限定因子であるところで(たとえば、細胞の還元環境で)適用するためのscFv抗体フラグメントまたはscFv抗体ライブラリーを作成するために理想的に適合している。また、かかるフレームワークは、向上した可溶性および安定性を示す、高度に保存された残基およびコンセンサス配列を同定するために使用することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、タンパク質化学、分子生物学、および免疫学に関する。
【背景技術】
【0002】
抗体は、高い特異性と親和性をもって、ほとんどすべての分子を認識し、ターゲットと
することができる。この特徴は、これら天然のタンパク質を診断および治療用の強力なツ
ールに変化させるために利用されている。組換えDNA技術の進歩により、広範な種々の
非リンパ系細胞における抗体遺伝子の操作、クローニング、および発現が容易になってい
る(Skerra,1988; Martineau, 1998; Verma, 1998)。多くの様々な抗体フラグメントが
、種々の適用に最も適合するように構築されている。親の免疫グロブリン全体の完全な抗
原結合能力を保持する最小要素(entity)は、単一鎖Fvフラグメント(scFv)である(Bi
rd, 1988)。この抗体フラグメントは、単一遺伝子からのタンパク質の発現を可能にする
フレキシブルなペプチドリンカーにより連結された重鎖と軽鎖の可変領域を含む。
【0003】
抗体フラグメントは、免疫グロブリン分子全体と比較して幾つかの重要な利点を有する
。抗体フラグメントの方がサイズが小さいため、種々の発現宿主細胞(たとえばE.coli細
胞)においてその発現は容易であり、その産生量は増大する(Pluckthun, 1996)。更に
、抗体フラグメントは、インビボでの適用において腫瘍の侵入を改善することができ(Yo
kota, 1992)、治療アプローチのために種々のエフェクター分子と共有結合させることが
できる。
【0004】
形質細胞により分泌される天然に存在する抗体は、細胞外の酸化環境において機能する
ように発達する。天然に存在する抗体は、機能的な折りたたみ構造を得るために、別個の
ドメイン内にジスルフィド架橋を形成することが一般に必要であり、これは、免疫グロブ
リンの折りたたみの安定性にとって重要である。完全長の抗体とは対照的に、scFvまたは
Fab抗体フラグメントは、原則として、任意の細胞内の還元(reducing)環境において機
能的に発現し、任意のコンパートメントに誘導されて細胞内タンパク質をターゲットとし
、これにより特定の生物学的効果を誘発することができる(Biocca, 1991)。実際、イン
トラボディ(intrabodies)と称される幾つかの細胞内単一鎖抗体フラグメントが、様々
な生物学的システムにおいて細胞内ターゲットタンパク質の機能を調整するためにうまく
適用されている。これにより、ウイルス感染に対する抵抗性が植物バイオテクノロジーで
実証され(Tavladoraki,1993; Benvenuto, 1995)、HIVタンパク質に対するイントラボ
ディの結合が示され(Rondon, 1997)、癌遺伝子産物に対する結合が記載されている(Bi
occa, 1993;Cochet, 1998; Lener, 2000)。更に、細胞内抗体は、ヒトゲノムの配列決
定により今日同定されている多大な数の遺伝子の機能を特徴付ける際の有益なツールとな
る見込みがある(Richardson,1995; Marasco, 1997)。たとえば、細胞内抗体は、機能
的ゲノミクスアプローチで使用して、新たに同定されたタンパク質の活性を妨害したり調
整したりし、これによりタンパク質の機能の理解に貢献することができる。最後に、イン
トラボディは、たとえば遺伝子治療セッティングにおいて、潜在的な診断および治療の適
用を有している。
【0005】
このような大きな期待にもかかわらず、機能的なイントラボディの作成は、その不安定
性および不溶性または凝集傾向によりまだ限られている。細胞質の還元環境は、保存され
た鎖内ジスルフィド架橋の形成を妨害し、これにより高パーセンテージの抗体フラグメン
トを不安定なものにし、その結果、細胞内で機能的でない(Biocca, 1995; Proba, 1997
)。したがって、抗体フラグメントの安定性および溶解性は、インビボにおけるタンパク
質機能の潜在的モジュレーターとしてのイントラボディの適用にとって大きな障害である
。これまでのところ、細胞内環境において抗体フラグメントを機能的にする配列の要件に
関して予測することはできない。
【0006】
したがって、広範囲の様々な細胞タイプでうまく機能し、様々な結合特異性のためのフ
レームワークとして使用することができる抗体フラグメントに対する需要が存在する。か
かるフレームワークは、細胞内スクリーニングのためのライブラリーを構築するために使
用することができ、あるいは現存の抗体の結合部分のためのアクセプターとして機能させ
ることができる。
【0007】
非常に安定な可変ドメインフレームワークに基くかかる抗体フラグメントまたは抗体全
体は、細胞内適用のためにユニークに適合していることに加えて、多くの細胞外適用およ
びインビトロ適用において他の抗体よりも別個の利点を更に有する。かかるフレームワー
クを酸化環境で産生すると、ジスルフィド架橋を形成することができるため、更にその安
定性は高まり、凝集やプロテアーゼ分解に対する高い抵抗性を付与することができる。イ
ンビボでの半減期(および凝集や血清プロテアーゼによる分解に対する抵抗性)は、親和
性および特異性に加えて、治療または診断適用における抗体の成功にとって唯一最も重要
な因子である(Willuda,1999)。抗体フラグメントの半減期は、ポリエチレングリコー
ル(PEG)などのポリマー分子の共有結合により更に増大させることができる(Weir,200
2)。このタイプの安定な分子は、Fcの機能性を望まないときに特に(しかしそのときに
限定されず)、抗体の使用において有意な進歩を示す。
【0008】
抗体フラグメントライブラリーの実用的な重要性が、この分野における研究に刺激を与
えている。Winter(EP 0368684) は、抗体可変領域遺伝子の最初のクローニングと発現を
提供した。これら遺伝子からスタートして、Winterは、相補性決定領域(CDRs)とフレー
ムワーク領域の両方に高い多様性を有する大きな抗体ライブラリーを作成した。しかし、
Winterは、ライブラリー構築のための様々なフレームワークの有効性を開示していない。
【0009】
一方、Pluckthun(EP 0859841) は、フレームワークを限られた数の合成コンセンサス
配列に限定することにより、ライブラリーのデザインを改良することを試みた。合理的に
設計された多くの突然変異の導入を含むタンパク質操作の成果により、単離された可変免
疫グロブリンドメインの安定性を改良するための適切な手段として、各コンセンサス配列
に対する突然変異が既に示唆されている(Ohage 1999; Ohage 1999およびUS 5,854,027,
参照により本明細書に取り込む)。
【0010】
Pluckthun(EP 0859841) は、これらコンセンサス配列に基いて結合親和性を更に最適
化するための方法を開示する。また、Pluckthunの特許は、抗体に関する知識の継続的な
増大を認識しているため、ライブラリー設計においてかかる将来的発見を含めることを目
的とする。しかし、合成コンセンサスフレームワークの考えられる更なる改良は示唆され
ていない。
【0011】
Winter,Pluckthunおよびその他 (たとえば、Soderlind, WO 0175091) の教示により、
選択および酸化条件下における選択されたscFvsの適用に対するCDRsの高い多様性に焦点
を絞って、大きな抗体ライブラリーを作成することが試みられた。しかし、これらライブ
ラリーのすべては、細胞内適用のために最適化されてないため、選択および還元環境にお
ける適用、または発現抗体フラグメントの安定性と溶解性に対する特別な要件を設定する
他の状況に有効でない。
【0012】
抗体フラグメントが還元環境(たとえば、原核および真核細胞の細胞質)でうまく機能
するために必要な性質は明らかでない。したがって、細胞内抗体または「イントラボディ
」の適用は、安定性および溶解性の性質に影響を及ぼし得る、還元状況下での予測できな
いそれらの行動により現在制限されている(Biocca, 1995; Worn, 2000)。イントラボデ
ィの細胞内スクリーニングに関する現在の特許出願(EP1040201, EP1166121およびW00200
729)および出版物(Visintin,1999)は、スクリーニング技術に焦点を当てているが、
真核細胞(とりわけ酵母)で機能的であり、かつこの状況でライブラリー構築に有効であ
る特定の抗体配列を開示していない。
【0013】
VisintinおよびTseは、それぞれ独立して、いわゆる細胞内コンセンサス配列(ICS)の
単離について記載している(Visintin, 2002; Tse, 2002)。この配列は、酵母における
抗原−抗体−相互作用スクリーニングから単離された多くの配列から得られた。しかし、
細胞内スクリーニングへのインプットは、先のファージ−ディスプレイ選択のために大き
く偏っていた。よって、インプット配列の一つを除いて全ては、Visintin et alのケース
でVH 3サブグループに属していた。公表されたコンセンサス配列ICSは、Knappik(2000)
およびEP0859841に記載されるヒトVH3サブグループのコンセンサス配列と完全に一致す
る。また、ICSの62アミノ酸のうち60は、安定性の高い可変ドメインの構築のための基礎
としてSteipeにより提案された一般的なヒトVH-ドメインコンセンサス配列と一致する(
米国特許第6,262,238号、参照により本明細書に取り込む)。次に、これらの研究は、以
前の配列の収集(すなわちKabat, 1991)、および可変ドメインサブグループおよび構造
決定因子の規定(Tomlinson,1992; Williams, 1996; Chothia, 1989 および Chothia, 1
987)を基にした。しかし、イントラボディ選択へのインプットは、非常に大きく偏って
いたので(すなわち、Visintin et alのケースにおいてVHドメインの一つを除いて全てが
VH3であったので)、細胞内スクリーニングからのVH3配列の単離は、特に驚くべきことで
はない。これらのインプットライブラリーの大きな偏りのため、偏りのない調査により提
供され、かつヒトレパートリーに存在する有効なイントラボディフレームワークを同定す
るために必要とされる、ヒト可変ドメインレパートリーの完全な評価を、Tse et al.およ
びVisintinet al.の研究は提供していない。
【0014】
我々は、抗原結合特異性とは無関係に、酵母において安定かつ可溶性のイントラボディ
の選択を可能にするシステムを以前に記載している(Auf der Maur (2001), W00148017)
。このアプローチにより、scFvライブラリーの効率的なスクリーニング、および酵母細胞
の還元環境において安定かつ可溶性である特定のフレームワークの単離が可能となる。実
際にフレームワーク配列を単離し、そのパターンを使用して、第一の工程においてどの配
列タイプが還元環境において最も安定であるか予測し、第二の工程において分析、組換え
、更なるインビボおよびインビトロ実験により最適な配列を同定することが、目的として
残っている。
【発明の概要】
【課題を解決するための手段】
【0015】
本発明は、抗体作成の分野においてミッシングリンクを満たすものである。本発明は、
安定性および溶解性に関して優れた特徴を有する抗体可変ドメインフレームワーク配列を
提供する。これらは、多くの関連の適用にとって、たとえば診断、治療またはリサーチに
おいて重大な特徴である。これらフレームワークは、現存の結合特異性を移植するため、
または高い安定性および溶解性を有する抗体ライブラリーを作成するために使用すること
ができる。
【0016】
ScFvライブラリーは、酵母細胞の還元環境において安定かつ可溶性であるフレームワー
クを単離するために使用した。その後、単離されたフレームワークの性能を、ヒト細胞株
およびインビトロ実験において特徴づけた。記載されるフレームワークは、現存の結合特
異性のためのアクセプターバックボーンとして直接使用したり、または還元環境もしくは
他の挑戦的環境で使用するための一以上の超可変ループのランダム化によりCDRライブラ
リーを構築するために直接使用したりすることができる。単離された可変ドメイン配列は
、アラインメントにより更に分析し、好ましい配列ファミリーを同定した。好ましい可変
ドメイン配列ファミリーから、免疫グロブリンの折りたたみを混乱させるフレームワーク
残基を含有する配列を除外する構造解析に基いて、最適な配列を選択した。その後、同定
された可変ドメイン配列の候補を、あらゆる可能なバリエーションで再結合し、酵母、哺
乳類細胞およびインビトロでその性能を分析することにより、軽鎖と重鎖の可変ドメイン
の最適な組み合わせを選択した。
【0017】
これら最適化されたscFvsおよびそれを構成する可変ドメインフレームワーク、並びに
他の抗体フラグメントまたはそれに由来する抗体全体は、たとえば、現存の結合特異性の
ためのアクセプターバックボーンとして、または還元環境もしくは他の挑戦的環境で使用
するための一以上の超可変ループのランダム化によりCDRライブラリーを構築するために
理想的である。細胞内適用に適した抗体は、その名が示すとおり安定かつ可溶性である。
そのため、その使用は、細胞内環境の外での適用にも好都合である。
【0018】
本発明は、抗体可変ドメインのフレームワークおよび単一鎖Fv抗体(ScFv)フラグメン
トを含む組成物であって、種々の抗体フラグメントまたは抗体全体に組み込むことができ
る組成物を提供する。最も安定かつ可溶性であるために細胞内適用に最もよく適合した抗
体可変ドメインフラグメントのクラスが提供される。また、抗体可変ドメインとscFv抗体
フラグメントの特定のフレームワーク配列であって、細胞内アッセイにおいて最も高い性
能を示すフレームワーク配列が提供される。また本発明は、抗体可変ドメインの特定のフ
レームワーク配列およびscFvフラグメントの軽鎖と重鎖の可変ドメインの合成組み合わせ
であって、たとえば、細胞内適用にとって最適であり、安定性と溶解性に関してインビト
ロで最適な性能を示すものを提供する。
【0019】
本発明は、一般構造:
NH2-VL-リンカー-VH-COOHまたは
NH2-VH-リンカー-VL-COOH
を有する単一鎖フレームワーク試薬を提供する。
【0020】
本発明の別の態様において、単一鎖フレームワークは、第二のタンパク質部分に融合し
、一般構造:
NH2-VL-リンカー-VH-第二のタンパク質-COOHまたは
NH2-第二のタンパク質-VL-リンカー-VH-COOH
の融合構築物を産生してもよい。
【0021】
これら融合構築物においてVHおよびVL領域の方向性(orientation)は逆向きであって
もよい。
【0022】
本発明の別の態様において、可変ドメインは、Fabフラグメントに組み込まれていても
よく、これは更に第二のタンパク質部分に融合し、一般構造:
NH2-VH-CH-第二のタンパク質-COOHおよびNH2-VL-CL-COOH
の融合構築物を産生してもよい。
【0023】
第二のタンパク質は、重鎖または軽鎖のいずれかのN−末端またはC−末端のいずれか
に融合してもよい。
【0024】
好ましい態様において、単一鎖またはFabフレームワーク融合構築物の第二のタンパク
質は、直接的または転写活性を介して、細胞内アッセイの読出し部(a read-out)を提供
するタンパク質である。
【0025】
本発明の別の目的は、たとえば、還元環境もしくは他の挑戦的環境において機能的な抗
体を得るために、現存の抗体から超可変ループを移植するのに適している、抗体可変ドメ
インのフレームワーククラスおよび可変ドメインとscFvsの配列を提供することである。
【0026】
本発明の別の目的は、たとえば、かかるフレームワークの一以上の超可変ループのラン
ダム化を介して、還元環境または他の挑戦的環境で使用するためのライブラリーを作成す
るのに適している、抗体可変ドメインのフレームワーククラスおよび可変ドメインとscFv
sの配列を提供することである。
【0027】
本発明の別の目的は、保存された残基およびコンセンサス配列の同定において、開示さ
れる配列を使用することである。
【0028】
開示されるフレームワークの使用により得られる抗体または抗体フラグメントは、ター
ゲットの検証、並びにヒト、動物および植物の疾患の治療、予防および診断における試薬
として使用することができる。抗体は、タンパク質またはかかるタンパク質をコードする
DNAの形態で使用することができ、細胞内適用に限定されるものではない。
【図面の簡単な説明】
【0029】
【図1】図1は、lacZ発現の活性化によりアッセイされる、酵母における典型的な「クオリティーコントロール」スクリーニングの結果を示す(たとえば例1参照)。選択されたポジティブクローン(黒)は、幾つかの様々なスクリーニングで同定され、ポジティブクローンの対応の配列は、表1および2に見出すことができる。選択された配列を、ポジティブコントロールである非常に安定なラムダ−グラフト(ダークグレー)と比較する。
【図2】図2は、非常に安定なラムダ−グラフト(ダークグレー)と比較してルシフェラーゼ発現の活性化によりアッセイされる、酵母における典型的な「クオリティーコントロール」スクリーニングから単離したフレームワークのヒト細胞株Helaにおける性能を示す。ポジティブコントロールGal4-VPl6(白)は、このシステムにおいて最大限可能なレベルの転写活性化を示す。ルシフェラーゼ活性は、トランスフェクション効率について修正されている。
【図3】図3は、lacZ発現の活性化により酵母でアッセイされる、優れたフレームワークの組み合わせのインビボでの性能を示す。フレームワーク配列(黒)を、ポジティブコントロール(非常に安定なラムダ−グラフト(ダークグレー))と比較する。フレームワークのナンバリングは、表5に記載されるとおりである。
【図4】図4は、ルシフェラーゼ発現の活性化によりヒト細胞株HeLaでアッセイされ、非常に安定なラムダ−グラフト(ダークグレー)と比較して図示される、優れたフレームワークの組み合わせのインビボでの性能を示す。ポジティブコントロール、Gal4-VP16(白)は、このシステムにおいて最大限可能なレベルの転写活性化を示す。ルシフェラーゼ活性は、トランスフェクション効率について修正されている。
【図5】図5は、酵母株S.cerevisiae JPY9の細胞質で産生される可溶性タンパク質の量によりアッセイされる、優れたフレームワークの組み合わせのインビボでの性能を示す。
【図6】図6は、E.coliのペリプラズムにおける、選択されたフレームワークの組み合わせの発現行動を示す。矢印は、scFvフレームワークに対応するバンドの位置を示す。
【図7】図7は、3つのヒト細胞株(Hela(黒)、Saos-2(ダークグレー)、HEK 293(白))においてルシフェラーゼ発現の活性化によりアッセイされ、非常に安定なラムダ−グラフトと比較して図示される、選択された優れたフレームワークの組み合わせのインビボでの性能を示す。ポジティブコントロール、Gal4-VP16(白)は、このシステムにおいて最大限可能なレベルの転写活性化を示す。ルシフェラーゼ活性は、トランスフェクション効率について修正されている。
【図8】図8は、PBSバッファー中に示されるインキュベーション前およびインキュベーション後に存在するモノマータンパク質の量により定量される、選択されたフレームワークの組み合わせの37℃での凝集に対する抵抗性を示す。パネルAはフレームワーク2.4および5.2を表し、パネルBはフレームワーク4.4、6.4および7.3を表す。
【図9】図9は、長期インキュベーション前および長期インキュベーション後に存在する可溶性完全長タンパク質の量により定量される、選択されたフレームワークの組み合わせの37℃でのヒト血清におけるプロテアーゼ分解凝集に対する抵抗性を示す。
【図10】図10は、LacZ発現の活性化により酵母相互作用アッセイでアッセイされる、Fabコンテクスト(context)における新規フレームワーク7.3上の選択された2つのバインダーのインビボでの性能を示す。Fab鎖の発現は、ars/cenまたは2ミクロンベクター上の二方向性ガラクトース誘導プロモーターに由来する。Fabベクターからの発現により、抗体軽鎖およびVH-CH1-Gal4-AD融合タンパク質が産生される。バインダーは、ヒトPolo様キナーゼ1(hPLK1)に対して向けられる。ターゲットに対する結合を、無関係の抗原に対する非特異的結合および非ランダム化フレームワーク7.3の結合と比較する。参考のために含めた対応のscFvは、アクチンプロモーター(2ミクロン)から発現することに留意されたい。
【図11】図11は、酵母株JPY9の細胞質で産生される可溶性タンパク質の量によりアッセイされる、Fabコンテクスト(context)におけるscFvフレームワークのインビボでの性能を示す。Gal4-AD-scFv融合体(アクチン/2ミクロン)の発現を、対応のFab構築物の発現、およびFabとしての親のフレームワーク7.3と比較する(そのいずれもが二つの異なるベクター(Gal-誘導、ars/cenおよび2ミクロン)に由来する)。Fabベクターからの発現により、抗体軽鎖およびVH-CH1-Gal4-AD融合タンパク質が産生され、これはこのブロットで検出される。
【発明を実施するための形態】
【0030】
特に規定しない限り、本明細書で使用される技術的および科学的用語はすべて、本発明
が属する分野の当業者により一般に理解されるのと同じ意味を有する。本明細書に記載さ
れるのと同様もしくは等価な方法および材料は、本発明を実施または試験する際に使用す
ることができるが、適切な方法および材料は以下に記載される。本明細書に記述される出
版物、特許出願、特許、および他の参考文献はすべて、その全体を参照により本明細書の
開示内容の一部とする。矛盾する場合、定義を含む本明細書がコントロールするでしょう
。更に、材料、方法、および実施例は、単に説明のためのものであって、限定するための
ものではない。
【0031】
本明細書で使用される「同一性(identity)」は、二つのポリペプチド、分子または二
つの核酸の間の配列の類似性をいう。二つの比較配列の両方における位置が、同じ塩基ま
たはアミノ酸モノマーサブユニットにより占められる場合(たとえば、二つのDNA分子の
それぞれにおける位置が、アデニンにより占められるか、あるいは、二つのポリペプチド
のそれぞれにおける位置がリシンにより占められる場合)、各々の分子はその位置におい
て相同である。二つの配列の間の「パーセンテージ同一性」は、二つの配列により共有さ
れる一致する位置の数を、比較した位置の数で割って100を乗じた関数である。たとえば
、二つの配列の10の位置のうち6つが一致している場合、二つの配列は60%の同一性を有
する。一例として、DNA配列CTGACTとCAGGTTは、50%の相同性を共有する(トータル6つの
位置のうち3つが一致する)。一般に、二つの配列を最大の相同性が得られるように整列
して比較を行う。かかる整列は、たとえば、Needleman et al., J. Mol Biol. 48: 443-4
53 (1970) の方法を用いて提供することができ、Alignprogram (DNAstar, Inc.) などの
コンピュータープログラムにより便利に実施される。
【0032】
「類似(similar)」配列は、整列したときに、同一および類似のアミノ酸残基を共有
するものであり、ここで類似の残基とは、整列させたレファレンス配列における対応のア
ミノ酸残基の、保存的置換または「許容される点突然変異」のことである。ここで、レフ
ァレンス配列における残基の「保存的置換」とは、対応のレファレンス残基に対して物理
的または機能的に類似する残基、たとえば、同様のサイズ、形状、電荷、共有結合もしく
は水素結合の形成能力などの化学的特性等を有する残基による置換のことである。よって
、「保存的置換修飾」配列は、一以上の保存的置換または許容される点突然変異が存在す
るという点において、レファレンス配列または野生型の配列とは異なるものである。二つ
の配列の間の「パーセンテージポジティブ」は、二つの配列により共有される一致残基ま
たは保存的置換を有する位置の数を、比較した位置の数で割って100を乗じた関数である
。たとえば、二つの配列の10の位置のうち6つが一致し、10の位置のうち2つが保存的置換
を有する場合、二つの配列は80%のポジティブ相同性を有する。
【0033】
「VHドメイン」は、免疫グロブリン分子の重鎖の可変部分をいう。
【0034】
「VLドメイン」は、免疫グロブリン分子の軽鎖の可変部分をいう。
【0035】
VHまたはVL「サブタイプ」は、Knappik(2000) に規定されるとおり、各コンセンサス
配列により規定されるサブタイプをいう。「サブファミリー」または「サブクラス」の用
語は、「サブタイプ」の同義語として使用される。本明細書で使用される「サブタイプ」
の用語は、当該サブタイプを代表する各コンセンサス配列に対して高度な一致性および類
似性を共有する配列をいう。ある可変ドメイン配列が、ある「サブタイプ」に属するか否
かは、各ドメインの公知のあらゆるヒト生殖細胞系セグメントまたは規定されるコンセン
サス配列の何れかとともに、その配列を整列し、その後最大ホモロジーを同定することに
より決定される。相同性を決定し、BLOSUM (Henikoff 1992) などのサーチマトリクスを
用いることにより配列をグループ分けする方法は、当業者に周知である。
【0036】
本明細書で使用される「アミノ酸コンセンサス配列」は、少なくとも二つ、好ましくは
より多くのアミノ酸配列のマトリクスを用いて作成することができ、整列の際にギャップ
を許容して、各位置において最も頻度の高いアミノ酸残基を決定することができる、アミ
ノ酸配列をいう。コンセンサス配列は、各位置において最も頻度が高く表示されるアミノ
酸を含む配列である。二以上のアミノ酸が単一の位置において等しく表示される場合には
、コンセンサス配列は、当該アミノ酸の両方またはすべてを含む。
【0037】
タンパク質のアミノ酸配列は、種々のレベルで分析することができる。たとえば、保存
性または変動性は、単一残基のレベル、複数残基のレベル、ギャップを有する複数残基の
レベルなどで示すことができる。残基は、同一の残基の保存性を示すことができ、あるい
はクラスレベルで残基を保存することができる。アミノ酸クラスの例は、極性があるが非
電荷のR基(セリン、トレオニン、アスパラギン、およびグルタミン);正に荷電したR
基(リシン、アルギニンおよびヒスチジン);負に荷電したR基(グルタミン酸およびア
スパラギン酸);疎水性R基(アラニン、イソロイシン、ロイシン、メチオニン、フェニ
ルアラニン、トリプトファン、バリンおよびチロシン);並びに特別なアミノ酸(システ
イン、グリシンおよびプロリン)を含む。他のクラスは、当業者に公知であり、構造決定
または他のデータを用いて代替可能性を評価して規定されてもよい。この意味において、
置換可能なアミノ酸とは、置換されて当該位置の機能的保存性を維持することができるア
ミノ酸をいう。
【0038】
本明細書で使用される「ポリヌクレオチドコンセンサス配列」は、少なくとも二つ、好
ましくはより多くの整列した核酸配列のマトリクスを用いて作成することができ、整列の
際にギャップを許容して、各位置において最も頻度の高いヌクレオチドを決定することが
できる、ヌクレオチド配列をいう。コンセンサス配列は、各位置において最も頻度が高く
表示されるヌクレオチドを含む配列である。二以上のヌクレオチドが単一の位置において
等しく表示される場合には、コンセンサス配列は、当該ヌクレオチドの両方またはすべて
を含む。
【0039】
本明細書で使用される「構造的サブエレメント」は、分子の規定の構造的または機能的
部分に対応するタンパク質またはポリペプチド内のアミノ酸残基の一区切り(stretch)
をいう。これらは、ループ(すなわち、抗体のCDRループ)であってもよいし、タンパク
質またはポリペプチド内の任意の他の二次構造または機能的構造(すなわち、ドメイン、
α−へリックス、β−シート、抗体のフレームワーク領域など)であってもよい。構造的
サブエレメントは、類似もしくは相同ポリペプチドの公知の構造を用いて、または整列し
たアミノ酸配列の上述のマトリクスを用いて、同定することができる。ここで、各位置に
おける変動性は、構造的サブエレメントに属するアミノ酸残基の一区切り(たとえば、抗
体の超可変領域)を決定するための基盤である。
【0040】
本明細書で使用される「サブ配列」は、少なくとも一の構造的サブエレメントをコード
する遺伝的モジュールをいう。これは、構造的サブエレメントと必ずしも同一ではない。
【0041】
本明細書で使用される「抗体CDR」は、Kabat et al. (1991) により規定されるとおり
、抗原結合ループから成る抗体の相補性決定領域をいう。抗体Fvフラグメントの二つの可
変ドメインはそれぞれ、たとえば、3つのCDRsを含有する。
【0042】
本明細書で使用される「抗体」は、「免疫グロブリン」の同義語である。本発明におい
て抗体は、免疫グロブリン全体であってもよいし、免疫グロブリンの少なくとも一の可変
ドメイン、たとえば単一の可変ドメイン、Fv(Skerra, 1988)、scFv(Bird, 1988; Hust
on,1988)、Fab、(Fab’)2を含むフラグメント、または当業者に周知の他のフラグメント
であってもよい。
【0043】
本明細書で使用される「抗体フレームワーク」は、可変ドメインの抗体結合ループのた
めの足場として機能する、可変ドメイン(VLまたはVHのいずれか)の一部をいう(Kabat
et al., 1991)。
【0044】
合理的に操作されたscFvフラグメントは、scFvフラグメントの熱力学的安定性とそのイ
ンビボでの性能との間に明らかな関係が実証されている(Worn, 2000; Auf der Maur, 20
01)。近年開発された「クオリティーコントロール(QualityControl)」と呼ばれるシ
ステム(Aufder Maur, 2001)を用いて、細胞内適用に適した特定の抗体可変ドメインフ
レームワークの配列が単離され(表1および2)、特徴づけられ(図1および2)、更に
改良された(図3〜9および表3)。我々の以前の実験で観察されるとおり、細胞内アッ
セイで選択された性能のよいフレームワークは、37℃での凝集およびプロテアーゼ分解に
対する抵抗性により実証されるようにインビトロで高い安定性を示す(図8および9)。
更に、フレームワークのサブファミリーに依存して、一般的なベースで、細胞内適用のた
めのフレームワークの選択を可能にするパターンが現れた(表4)。細胞内適用に有効な
特定の抗体可変ドメイン配列は、一般的なパターンとともに本明細書に開示される。一方
、これにより、これら配列を移植実験におけるフレームワークドナーとして使用し、ルー
プドナーの結合特異性を保持する機能的イントラボディを得ることが可能になる。加えて
、開示される配列をフレームワークとして用いて、抗体ライブラリーを構築することがで
きる。かかるライブラリーは、還元状況下(たとえば原核および真核細胞における還元状
況下)での細胞内選択システムに適している。加えて、開示される配列は、たとえば、保
存された配列または残基またはモチーフを同定するために使用してもよい。構造的サブエ
レメント(たとえば抗体の結合ループのサブエレメント)の移植(たとえばJung, 1997)
、並びに抗体またはそのフラグメントのライブラリーの作成(たとえばVaughan, 1996; K
nappik, 2000)は、詳細に記載されており、当業者に周知である。
【0045】
細胞内適用は、抗体フラグメントを非常に望ましくない状況(すなわち、高い温度、還
元環境)に晒すため、本発明に開示される配列は、当該配列を最も不利な状況に対して抵
抗性にする特徴を獲得した。したがって、「平均的」配列と比較すると、開示される配列
は、凝集およびプロテアーゼ分解に対する抵抗性により実証されるように、顕著な安定性
を有する(図8および9)。優れた発現量とともに、これらの特徴により、開示される抗
体フレームワーク配列は、細胞内の使用だけでなく、とりわけ長い半減期、頑強性、およ
び産生の容易性が大変重要である治療および診断のあらゆる適用にも独自に適している。
【0046】
本発明により、還元環境または他の挑戦的環境における適用に有効な抗体の可変部分を
少なくとも含むポリペプチド配列の設計が可能になる。第一の態様において、本発明は、
細胞内適用に適した抗体フレームワーク配列のコレクションを提供する(表1および2)
。第一の工程において、様々な配列のライブラリーを、酵母においてクオリティーコント
ロールシステムを用いて、結合親和性とは無関係にスクリーニングする。単離された配列
は、酵母および哺乳類細胞において細胞内性能について評価することができる(図1およ
び2)。
【0047】
本発明の一つの態様において、単離された配列のコレクションを、アラインメントによ
り分析し、細胞内適用に適した抗体可変ドメインサブクラスおよびコンセンサス配列を同
定する。
【0048】
本発明の更に好ましい態様において、上述の抗体フレームワーク配列のコレクションを
、互いにアラインメントにより更に分析し、サブファミリーにグループ分けする。一つの
サブタイプに属するフレームワークすべてを、酵母および哺乳類細胞において細胞内性能
に関して比較し(例として図1および2)、各サブタイプのコンセンサスに対する、アミ
ノ酸配列のネガティブ、ニュートラル、またはポジティブ交換の出現に関して比較する。
当業者であれば、免疫グロブリンドメインにおいて変化した特定の残基の構造環境に基い
て、ポジティブ、ニュートラル、およびネガティブ変化を識別することができる。その後
、最も優れた細胞内性能を示し、かつ各サブタイプのコンセンサスと比較してネガティブ
交換がない、可変抗体ドメインのフレームワーク配列を選択する。好ましくは、ポジティ
ブと考えられるアミノ酸交換を更に含有する配列を選択する。
【0049】
更に好ましい態様において、最も高い安定性と溶解性を備えた組み合わせを同定するた
めに、その後、重鎖および軽鎖の選択された抗体可変ドメインを、あらゆる可能な組み合
わせでscFvフラグメントに再結合する。この目的のために、新規な再結合scFvフラグメン
トを、酵母(図3)および哺乳類細胞株(図4および7)において細胞内相互作用アッセ
イで還元条件下のその性能について評価し、並びに酵母において可溶性細胞内発現(図5
)について評価する。E.coliにおけるペリプラズム発現量(図6)、増大した温度での凝
集に対する抵抗性(図8)、並びに37℃、ヒト血清中における長期インキュベーションに
よる凝集およびプロテアーゼ分解に対する抵抗性(図9)を分析することにより、有望な
組み合わせを、酸化条件下のその行動について更に評価する。これらデータは、細胞内ま
たは酸化条件下の何れかの任意の特定の適用に最適なscFvフレームワークを同定するため
に使用する。
【0050】
本明細書に開示される選択され最適化されたフレームワーク配列は、細胞内適用におい
てだけでなく、scFvの増大した安定性および/または溶解性から利益を得ることができる
あらゆる適用において、有意な利点を有する。その例として、診断適用に必要とされる高
濃度での長期保存、および(たとえば治療適用で必要とされる)37℃、血清中での長期間
の機能的半減期が挙げられる。
【0051】
本発明の一つの側面に従えば、一般構造:
NH2-VL-リンカー-VH-COOH;または
NH2-VH-リンカー-VL-COOH
(ここでVHフレームワークは、サブタイプ1a、1bまたは3である)を有する単一鎖フレー
ムワークを含むイントラボディのフレームワークが提供される。
【0052】
別の態様において、VHおよびVL領域の方向性(orientation)は、上述の単一鎖フレー
ムワークにおいて逆向きである。
【0053】
本発明の一つの側面に従えば、一般構造:
NH2-VL-リンカー-VH-COOH;または
NH2-VH-リンカー-VL-COOH
(ここでVHフレームワークは、サブタイプ1a、1bまたは3であり、VLフレームワークは、
サブタイプλ1、λ3またはκ1である)を有する単一鎖フレームワークを含むイントラボ
ディのフレームワークが提供される。
【0054】
別の態様において、本発明は、第二のタンパク質部分に融合し、一般構造:
NH2-VL-リンカー-VH-第二のタンパク質-COOH;または
NH2-第二のタンパク質-VL-リンカー-VH-COOH
(ここでVHフレームワークは、サブタイプ1a、1bまたは3であり、VLフレームワークは、
サブタイプλ1、λ3またはκ1である)の融合構築物を産生する、単一鎖フレームワーク
を提供する。
【0055】
別の態様において、これら融合構築物におけるVHおよびVL領域の方向性(orientation
)は、逆向きであってもよい。
【0056】
別の態様において、可変ドメインは、Fabフラグメントに組み込まれていてもよく、こ
れは更に第二のタンパク質部分に融合し、一般構造:
NH2-VH-CH-第二のタンパク質-COOHおよびNH2-VL-CL-COOH
の融合構築物を産生してもよい。
【0057】
第二のタンパク質は、重鎖または軽鎖のいずれかのN−末端またはC−末端のいずれか
に融合してもよい。
【0058】
本明細書に開示されるとおり、細胞内適用において、サブタイプ3、またサブタイプ1a
および1bのVHフレームワークが非常に強く優先される。軽鎖可変ドメイン(VL)に関して
は、カッパ1タイプのフレームワークが数的に明らかに優先され、またラムダ1および3も
豊富である。したがって、これらフレームワークのサブタイプ、すなわちカッパ1、ラム
ダ1または3 VLドメインと組み合わせたVH 1a、1bおよび3が、細胞内の使用およびscFv
の折りたたみ特性を必要とする他の適用に最も適合する。したがって、還元環境において
機能的でない分子の量を低減するために、細胞内スクリーニングシステムのためのライブ
ラリーは、これらフレームワークサブタイプの混合物から優先的に構築されるべきである
。
【0059】
好ましい態様において、本発明の抗体フラグメントのVHドメインは、サブタイプ1a、1b
または3である。
【0060】
好ましい態様において、本発明の抗体フラグメントのVLドメインは、サブタイプカッパ
1、ラムダ1または3である。
【0061】
好ましい態様において、フレームワークとして使用される抗体フラグメントは、表5に
記載される、1.1,2.1, 3.1, 4.1, 5.1, 1.2, 2.2, 3.2, 4.2, 5.2, 1.3, 2.3, 3.3, 4.3
, 5.3, 7.3,1.4, 2.4, 3.4, 4.4, 5.4, および6.4から成る群より選択される。
【0062】
本発明の一つの態様において、少なくとも二つ、好ましくはより多くのフレームワーク
が同定され、その後分析される。タンパク質配列のデータベースは、タンパク質配列を互
いに整列させて構築してもよい。このとき、アラインメントを使用して、たとえば、その
配列と(その情報が入手可能であれば)構造的配置の両方において高い類似度を示すフレ
ームワーク配列の残基、サブエレメント、サブ配列またはサブグループを規定することが
できる。
【0063】
サブエレメントの長さは、好ましくは、1アミノ酸(たとえば、酵素の活性部位または
構造決定残基における一残基)から150アミノ酸(たとえば、タンパク質ドメイン全体)
までの範囲であるが、これに限定されない。最も好ましくは、その長さは3から25アミノ
酸までの範囲であり、たとえば抗体のCDRループに最も一般に見出される。
【0064】
別の態様において、当該分析から推測されるコンセンサス核酸配列が合成される。これ
は、当業者に周知の幾つかの方法のいずれかにより、たとえばトータル遺伝子合成または
PCRベースのアプローチにより、達成することができる。
【0065】
別の態様において、核酸配列は、ベクターにクローニングされる。ベクターは、配列決
定ベクター、発現ベクターまたはディスプレイ(ファージディスプレイ)ベクターとする
ことができ、これらすべてが当業者に周知である。ベクターは、一つの核酸配列、または
二以上の核酸配列を含むことができ、異なるオペロンまたは同一のオペロンのいずれでも
よい。後者の場合、別々にクローニングされてもよいし、隣接配列であってもよい。
【0066】
一つの態様において、ポリペプチドは、特定の種に特徴的なアミノ酸パターンを有する
。これは、たとえば、たった一つの種の相同タンパク質のコレクションから、最も好まし
くはヒトタンパク質のコレクションから、コンセンサス配列を導き出すことにより達成す
ることができる。
【0067】
本発明の更なる態様は、上述のポリペプチド、並びに追加の部分の両方をコードするDN
A配列を提供することによる融合タンパク質に関する。
【0068】
更なる態様において、本発明は、核酸配列、該核酸配列を含有するベクター、該ベクタ
ーを含有する宿主細胞、および本明細書に記載される方法に従って得ることができるポリ
ペプチドを提供する。
【0069】
更なる態様において、本発明は、本発明の核酸配列の末端に制限部位を合成するか、ま
たはその他の方法で配置し、本発明の核酸配列を適切なベクターにクローニング可能にす
ることを提供する。
【0070】
更に好ましい態様において、本発明は、ポリペプチドをコードする核酸配列と適合する
ベクターシステムを提供する。該ベクターは、核酸配列をベクターにクローニングするの
に必須の制限部位を除いて、たとえばベクターシステム内においてユニークであり、ポリ
ペプチドをコードする核酸配列に組み込まれた制限部位に対して本質的にユニークである
制限部位を含む。
【0071】
別の態様において、本発明は、核酸配列の一以上のリスト、組換えベクター、ポリペプ
チド、および上記方法に従ったベクター、およびたとえばポリペプチドを産生するための
適切な宿主細胞を含むキットを提供する。
【0072】
本発明の上記態様のすべては、当業者に公知の分子生物学の標準的技術を用いて実施す
ることができる。
【0073】
別の態様において、核酸配列は、本発明のポリペプチドをコードすることができる任意
の配列である。
【0074】
別の態様において、本発明の核酸は、遺伝子治療において使用される。
【0075】
別の態様において、単一鎖フレームワークは、配列1.1, 2.1, 3.1, 4.1, 5.1, 1.2, 2.
2, 3.2, 4.2, 5.2,1.3, 2.3, 3.3, 4.3, 5.3, 7.3, 1.4, 2.4, 3.4, 4.4, 5.4, 6.4(表
5)の何れか一つの変異体であり、本明細書で使用される「変異体」は90%以上の同一性
を示し、増大した安定性を維持する配列をいう。
【0076】
別の態様において、単一鎖フレームワークは、配列1.1, 2.1, 3.1, 4.1, 5.1, 1.2, 2.
2, 3.2, 4.2,5.2, 1.3, 2.3, 3.3, 4.3, 5.3, 7.3, 1.4, 2.4, 3.4, 4.4, 5.4, 6.4(表
5)の何れか一つの誘導体であり、本明細書で使用される「誘導体」は、分子の機能およ
び安定性に重要なそれらのアミノ酸のみを維持する配列をいう。例3に記載されるフレー
ムワークにおけるニュートラルもしくはポジティブな単離された交換は、本発明の抗体フ
レームワークに関連のある変化とは考えられない。
【0077】
本発明の好ましい態様において、単一鎖フレームワークは、細胞内アッセイの読出し部
(aread-out)を提供する第二のタンパク質に融合される。読出し部は、直接的であり得
、たとえば、検出可能なタンパク質(たとえば、蛍光により観察可能である、GFP(緑色
蛍光タンパク質)、enhanced青色蛍光タンパク質、enhanced黄色蛍光タンパク質、enhanc
edシアン蛍光タンパク質)、または様々な検出方法による他の融合パートナーとの融合体
の形態であり得る。あるいは、読出し部は、レポーター遺伝子の転写活性化により達成す
ることができ、ここでscFv融合タンパク質の融合パートナーは、転写アクチベーター、た
とえばGal4活性化ドメイン、またはDNA結合タンパク質、たとえばlexA−もしくはGal4 D
NA−結合ドメインの何れかであり、これは、酵素(β−ガラクトシダーゼ、ルシフェラー
ゼ、α−ガラクトシダーゼ、β−グルクロニダーゼ、クロラムフェニコールアセチルトラ
ンスフェラーゼなど)のレポーター遺伝子の転写を活性化し、これが、順に読出し部(a
read out)を提供する。読出し部を提供する融合タンパク質は、当業者に周知である。
【0078】
本発明の別の態様は、本明細書に記載されるフレームワークを含む抗体である。
【0079】
本発明の別の態様は、本発明の抗体の使用である。
【0080】
本発明の更に好ましい態様は、還元環境または他の挑戦的環境において機能的である抗
体を得るために、現存の抗体から超可変ループを移植するための、抗体可変ドメインの記
載されるフレームワーククラス、並びに可変ドメインおよびscFvsの配列の使用である。
【0081】
本発明の更に好ましい別の態様は、たとえばフレームワークの一以上の超可変ループの
ランダム化による、還元環境または他の挑戦的環境における適用のためのライブラリーを
作成するための、抗体可変ドメインの記載されるフレームワーククラス、並びに可変ドメ
インおよびscFvsの配列の使用である。
【0082】
当業者に明らかであるとおり、本明細書に記載される本発明の分子は、診断および治療
適用、ターゲットの検証、および遺伝子治療において使用することができる。
【0083】
本発明は、以下の実施例により詳説され得るが、以下の実施例は、本発明の範囲を限定
するためのものではない。
(参考文献)
【0084】

本発明は、以下の非限定的な実施例において更に詳説される。
【0085】
例1
酵母において「クオリティーコントロール」システムを用いてヒトライブラリーをスク
リーニングすることによるイントラボディフレームワークの選択
安定なフレームワークのため「クオリティーコントロール」システムを用いたスクリー
ニングを、Aufder Maur (WO0148017、Auf der Maur 2001、それぞれ参照により本明細書
に組み込む) に詳細に記載されるとおり基本的に実施した。
【0086】
酵母でスクリーニングするためのscFv融合構築物の発現用プラスミドは、pESBA-Actか
ら得た(Worn, 2000)。それは、S.cerevisiaeにおける形質転換選択のための酵母TRP1遺
伝子、および高いコピー数を保証するための2ミクロン複製起点を含有する。更に、強力
な発現のための構成的アクチンプロモーターおよびGAL11転写終結配列を、マルチプルク
ローニングサイトにより隔てられて有する。細菌システムで扱うために、更に、細菌の複
製起点およびamp抵抗性遺伝子を有する。
【0087】
Gal4活性化ドメイン(ADアミノ酸768-881)は、pGAD424(Clontech)を鋳型として用い
て、Gal4-ADに対するSV40 T-抗原核局在シグナルN-末端を含むプライマーでPCRにより初
めに増幅した。Gal11Pのアミノ酸263-352をコードするDNAフラグメントを、PCRにより増
幅し、SV40-NLS-Gal4-AD-構築物に対するN-末端にフレームで(inframe)クローニング
した。他に記載されるとおりヒト脾臓細胞cDNAから増幅したヒトscFvライブラリー(Wel
schhof, 1995;Krebber, 1997; de Haard, 1999)を、StiI-部位を介してこの融合構築物
に対するN-末端に、VL-リンカー-VH(ここでリンカーは配列(GGGS)4を有する)の向き
で、フレームで(inframe)クローニングした。かかる発現により、一般構造scFv-Gal11
P-SV40 NLS-Gal4ADの融合タンパク質が産生される。
【0088】
スクリーニングは、異なる向きのLacZおよびHIS3レポーター遺伝子を、his3Δ200遺伝
子座へ、Gal1-GAL10調節配列由来の天然UASGの制御下に組み込むことによりJPY9株(Esch
er, 2000)から誘導した酵母株、S.cerevisiae YDE172 (MATα ura3-52 leu2Δ1 trp1d6
3 his3Δ200 lys2Δ385 gal4Δ 11) (Auf der Maur, 2001) で行った。レポーターシス
テムの転写活性化は、Gal11P部分とGal4-DNA結合ドメイン(DBD、アミノ酸1-100)との特
異的相互作用に続いて、scFv-融合構築物のGal4-AD部分により媒介される。Gal4-DBDは、
第二のプラスミドpMP83からの発現により提供される。それは、S.cerevisiaeにおける形
質転換選択のための酵母LEU2遺伝子およびARS CEN複製起点を含有する。更に、強力な発
現のための構成的アクチンプロモーターおよびGAL11転写終結配列を有する。細菌システ
ムで扱うために、更に、細菌の複製起点およびamp抵抗性遺伝子を有する。
【0089】
スクリーニングのために、酵母株S. cerevisiae YDE172を、pESBA-Act2ベクター上の
融合構築物としてのscFv-ライブラリーを用いて、一方Gal4-DBDを提供するpMP83ベクター
を用いて同時形質転換した(co-transform)。標準的な酢酸リチウム形質転換プロトコー
ルを使用した(Agatep,1998)。形質転換の後、細胞を、80 mM 3-アミノトリアゾールを
含有するドロップアウトプレート(-Trp/-Leu/-His)に播いた(plate)。30℃で3日間イ
ンキュベートした後コロニーを採取し、80 mM 3-アミノトリアゾールを含有するドロップ
アウトプレート(-Trp/-Leu/-His)に再び播いた(streak)。再度増殖した細胞を、基質
X-Galを含有するプレート上でのフィルターアッセイで、青色の発色によりLacZ発現につ
いて試験した。ポジティブクローンを、更なる解析のために採取し、該解析は、酵母から
のscFv-担持プラスミドの単離、E.coli DH5αへの形質転換、E.coliの単一コロニーか
らのプラスミドの単離、および後述のアッセイのために新たに調製した酵母株S. cerevis
iae YDE172への再形質転換を伴う。すべての方法は、当業者に周知の標準的手法に従っ
て行われた。
【0090】
更に、改変されたスクリーニング手法を使用し、scFvを、DNA結合ドメイン(LexAアミ
ノ酸1-202)と活性化ドメイン(Gal4、アミノ酸768-881)の両方に直接融合し、以下の構
造の融合構築物を産生した:scFv-LexA-NLS-Gal4AD。酵母におけるスクリーニングのため
のscFv融合構築物の発現用プラスミドは、pESBA-Act2から誘導した。それは、S.cerevisi
aeにおける形質転換選択のための酵母TRP1遺伝子、および高いコピー数を保証するための
2ミクロン複製起点を含有する。更に、(強力な発現のための)構成的アクチンプロモー
ターおよびGAL11転写終結配列を、マルチプルクローニングサイトにより隔てられて有す
る。細菌システムで扱うために、更に、細菌の複製起点およびamp抵抗性遺伝子を有する
。
【0091】
スクリーニングは、異なる向きのLacZおよびHIS3レポーター遺伝子を、his3Δ200遺伝
子座へ、6つのLex-A結合部位を有する二方向性プロモーターの制御下に組み込むことによ
り(インテグレイティングレポータープラスミド pDE200, Escher 2000)、そしてLEU2レ
ポーター遺伝子を、Leu2Δ1遺伝子座へ、(EGY48由来の)8つのLexA結合部位を有するプ
ロモーターの制御下に組み込むことにより、JPY5株から誘導した酵母株、S. cerevisiae
ImmunaLHB (MATαura3-52 leu2Δ1 trp1d63 his3Δ200 lys2Δ 385) で行った。レポー
ターシステムの転写活性化は、scFv融合構築物のGal4-AD部分により媒介される。スクリ
ーニングは、ドロップアウト培地(-Trp/-Leu/-His)および40 mMまでの3-アミノトリゾ
ール濃度を用いて基本的に上述のとおり行った。
【0092】
例2
インビボ性能の評価
a)酵母において
酵母において選択フレームワークの性能を定量分析するため(図1および3)、S.cere
visiae株ImmunaLHBを、標準的な酢酸リチウム形質転換プロトコールに従うことにより、
pESBA-Act2ベクター上のLexA-Gal4-AD-融合構築物としての単離scFvsを用いて形質転換し
た(Agatep,1998)。形質転換の後、細胞をドロップアウトプレート(-Trp)に播いた(
plate)。ドロップアウト培地(-Trp)中の2mL一晩培養物を、幾つかのコロニーを含有
する培養物(streaks)から2回接種し、30℃で生育させた。培養物を、600nm(OD600)
における光学濃度が0.7になるように、1mLドロップアウト培地(-Trp)で希釈した。そ
の後、30℃で2時間生育させた。アッセイのために、100μLの細胞培養物を採取し、900μ
Lバッファー、45μLクロロホルムおよび30μL 0.1%SDSと混合し、ボルテクスにかけ、
室温で5分間インキュベートした。0.2mL ONPG(4 mg/mL)の添加により発色を開始し、
0.5 mL Na2CO3(1M)で停止させた。アッセイ培養物のOD600、並びに使用した発色のイ
ンキュベーション時間および培養体積を考慮することにより活性を計算した。
【0093】
ポジティブコントロール(以前に記載された非常に安定なラムダ−グラフト(Worn, 20
00; Auf derMaur, 2001))と少なくとも同等、またはそれより優れたクローンを、配列
決定し、フレームワークサブタイプを同定した(Tomlinson, (1992), Cox,(1994) and Wi
lliams, (1996)に従ったフレームワークサブタイプの定義)。配列決定により、フレーム
ワークサブタイプの顕著に好ましいものが明らかにされた。重鎖可変ドメイン(VH)に関
して、フレームワークサブタイプ2および6は見られず、4はポジティブクローンのなかで
著しく減少していた。酵母細胞内アッセイにおいて、単離された配列の性能を修正すると
、サブタイプ3のVHフレームワークだけでなく、1aおよび1bも、細胞内適用において非常
に強力に好ましい。軽鎖可変ドメイン(VL)に関しては、カッパ1、ラムダ1、およびラム
ダ3サブタイプのフレームワークが明らかに好ましい。(表4)。
【0094】
したがって、これらフレームワークのサブタイプ、すなわちカッパ1、ラムダ1およびラ
ムダ3 VLドメインと組み合わせたVH1a、1bおよび3が、細胞内使用およびscFvの折りたた
み特性に関して厳格な要件を有する他の適用に最も適合している。細胞内スクリーニング
システムのためのライブラリーは、たとえば、これらフレームワークのサブタイプのみの
混合物から優先的に構築し、還元環境において機能的でない分子の量を低減すべきである
。
【0095】
b)哺乳類細胞において
ヒト細胞において、選択したフレームワークの性能を定量的に分析するためにHela細胞
株を使用した(図2、4および7)。ルシフェラーゼレポーター遺伝子は、天然Gal4 UA
Sの制御下にルシフェラーゼを含有する、同時トランスフェクトされるpGL3(Promega)レ
ポータープラスミドから提供された。一過性トランスフェクションのために使用される哺
乳類発現ベクターは、CMVプロモーターの制御下に、VP16-ADに対してC末端で融合したGa
l4(1-147)を含有する。単離されたscFvsを、Gal4(1-147)-VP16-融合物に対してC末端
にフレームでクローニングし、発現によりGal4(1-147)-VP16-scFv-融合タンパク質を産生
した。細胞を、2.5% FCSおよび2mM l-グルタミンを添加したDMEM中で培養した。一過
性トランスフェクションは、Polyfect-protocol (Qiagen) に従って、60 mm組織培養プレ
ートで、scFv構築物を含有する0.01-0.1μgのベクター、0.5μgのCMVプロモーター誘導Ga
l4(1-147)-VP16-scFv発現プラスミド、およびトランスフェクション効率のためのレファ
レンスとして0.5μgのLacZ発現ベクターを用いて行った。細胞を、トランスフェクション
から24〜48時間後に集め、1000μLバッファー中に再懸濁し、3回の凍結−融解サイクルに
より溶解させた。細胞溶解物を、遠心分離し、ルシフェラーゼアッセイ溶液(Promega)
を用いてルシフェラーゼ活性について、および標準的プロトコールに従ってLacZ活性につ
いて、上清をアッセイした。得られたルシフェラーゼ活性は、LacZ活性を用いて修正し、
トランスフェクション効率におけるばらつきを説明した。
【0096】
例3
マルチプルアラインメントおよび配列比較の分析
細胞内適用に適したフレームワーク配列の一般的パターンを解明するため、すべてのポ
ジティブクローン(すなわち、クオリティーコントロールシステムにおいて選択条件下で
生育したクローン)を単離し、scFvsをコードする部分を配列決定した。その後、scFv配
列を、軽鎖および重鎖の構成要素に分け、Honegger (2001) による免疫グロブリンドメイ
ンの構造調整ナンバリングスキームに従って、各ドメイン(表1および2)のアラインメ
ントを可能にした。
【0097】
得られたデータを評価するため、非選択ライブラリーを表すアラインメントを作成した
(表3)。非選択配列を得るために、ライブラリーを、scFv遺伝子を発現しないE. coli
細胞に形質転換し、プラスミド単離およびscFv配列決定のために、クローンをランダムに
採取した。ライブラリーは、予想されるとおりヒト抗体レパートリーを包含し、ヒトで一
般に見出される発現パターンにより予想される以外、特定のサブグループに対する偏りは
みられない。
【0098】
VHおよびVL配列を、サブグループに従ってグループ分けした。サブグループ特異的コン
センサス配列への変化が強調された。当業者であれば、特定の交換残基の構造的環境に基
づいて、ポジティブ、ニュートラルおよびネガティブ変化を識別することができる(たと
えば、Honegger,2001)。特定グループのアミノ酸に属する残基の同一グループの残基へ
の交換は、ニュートラルな交換として一般に認められる。タンパク質の疎水性コアに位置
する疎水性アミノ酸のグループに属する残基の、極性を有するが非電荷または正もしくは
負電荷のアミノ酸のグループの一のアミノ酸への交換は、不満足な水素ドナー/アクセプ
ター部位が疎水性コアの堅固なパッキングを乱すため非常に好ましくない。したがって、
かかる変化は、ネガティブと考えられる。免疫グロブリンドメインの表面における極性を
有するが非電荷の残基のグループに属する残基の、正もしくは負電荷の残基のグループの
アミノ酸への交換は、タンパク質の溶解度を増大させるため非常に好ましい。したがって
、かかる変化は、ポジティブと認められるが、極性から疎水性残基への交換は、タンパク
質の溶解度を減少させるため非常に好ましくないため、ネガティブと認められる。保存さ
れたポジティブなファイアングル(phi-angle)を有する位置において、任意のアミノ酸
のグリシンへの交換は、ポジティブと認められるが、グリシンの任意のアミノ酸への交換
は、ネガティブと認められ、これは、グリシンが、ポジティブなファイアングルを形成す
ることができる唯一のアミノ酸であるためである。正もしくは負電荷の残基のグループの
アミノ酸から非電荷のアミノ酸への交換による、45-53、45-100、77-100および108-137位
置の間の保存された塩橋の損失は、熱力学的安定性の低下につながるため、ネガティブと
考えられる。
【0099】
最後に、我々は、クオリティーコントロールの間に優先的に選択され(すなわち、コン
センサス配列から最小のネガティブと最大のポジティブ交換を示して、該サブグループに
当てはまり)、それぞれが酵母において高いインビボ性能を示す、7つのVLドメインおよ
び4つのVHドメインを選択した。その配列は、表5に要約され、2つのVκ1(k I 27 (1. x
) および k III 25(2. x))、2つのVκ3 (k IV 103 (3. x) および k IV135 (5. x))、1
つのVλ1 (kIV 107 (4. x))、2つのVλ3 (a33 (7. x) および a43 (6. x))、1つのVHlb
(a33 (x. 3))、および3つのVH3(a fw10 (x. 2), a43 (x. 4) および a44 (x. 1)) を含
む。これらVLおよびVHドメインは、シャッフルされ、scFvフォーマットにおいて22の新規
な組み合わせ(1.1, 2.1, 3.1, 4.1, 5.1, 1.2, 2.2, 3.2, 4.2, 5.2, 1.3, 2.3, 3.3, 4
.3, 5.3, 7.3,1.4, 2.4, 3.4, 4.4, 5.4, 6.4) を提供した。
【0100】
例4
シャッフルしたドメインのインビボ性能の評価
a)酵母および哺乳類細胞における細胞内アッセイでの性能
22の組み合わせを、例2に記載されるとおり、酵母および哺乳類細胞においてインビボ
性能について試験した(図3および4)。
【0101】
b)酵母における還元条件下での可溶性タンパク質の発現
還元条件下での発現による可溶性タンパク質の産生量を比較するため、選択されたフレ
ームワークを、酵母S.cerevisiaeの細胞質において、Gal4 ADとの融合体として発現させ
た。pESBA-Act2ベクター上の融合構築物は、一般構造Gal4 AD-scFvを有していた。その
融合構築物を、上述のとおり酵母S.cerevisiae株JPY9に形質転換し、-Trp, ドロップアウ
トプレートに播いた。
【0102】
ドロップアウト培地(-Trp)における5 mL一晩培養物を、幾つかのコロニーを含有する
培養物(streaks)から接種し、30℃で生育させた。培養物を、600nm(OD600)における
光学濃度が0.5になるように、50mLドロップアウト培地(-Trp)で希釈した。その後、30
℃で5時間生育させた。ネガティブ細胞抽出物については、OD600を3に規格化した2.5mL
細胞培養物を、遠心分離により収集し、液体窒素で凍結し、その後、プロテアーゼ阻害剤
(PMSF)を含有する75μL Y-PER(Pierce)に再懸濁した。再懸濁した細胞の沈殿物を、
短時間ボルテクスにかけ、20℃で20分(わずかに振盪して)インキュベートした。不溶性
の凝集物を、4℃で10分間、エッペンドルフ遠心分離機において最大速度で沈殿させた。
上清をローディング色素と混合し、5分間100℃に加熱し、12% SDS-PAGEで分離した。可
溶性Gal4 AD-scFv融合構築物を、一次抗体として抗Gal4ADモノクローナルマウス抗体(S
anta CruzBiotechnology)、二次抗体として抗マウスペルオキシダーゼコンジュゲート
(Sigma)を用いて、化学発光基質(Pierce)を用いて、Gal4部分の検出を介したウェス
タンブロッティングにより視覚化した(図5)。SDS-PAGEおよびウェスタンブロッティン
グの手法は、当業者に周知である。
【0103】
c)E.coliのペリプラズムにおける発現行動
E.coliにおけるペリプラズム発現行動を評価するため(図6)、単離されたscFvs−フ
レームワークを、cam抵抗性遺伝子(catR)およびlacIリプレッサー遺伝子(Krebber,19
97)を担持する細菌ベクターに、N-末端のpelBリーダー配列およびC-末端のhis-tagを備
えて、lacプロモーター/オペレーターの制御下にクローニングした。コンピーテントE.co
li JM83を、これらプラスミドを用いて形質転換した。振盪フラスコ中で、35mg/Lクロ
ラムフェニコールを含有する50 mL dYT培地に、一晩培養物を1:40で接種し、30℃でイン
キュベートした。細胞を、1 mM IPTGを用いてOD600 0.8に誘導し、遠心分離による導入
の3時間後に収集した。沈殿を、50mM Tris, pH 7.5、500 mM NaClに再懸濁し、OD600を1
0に規格化した。各scFvフラグメントのサンプルを、直接(全抽出物)もしくは遠心分離
に続く音波処理の後(可溶性フラクション)に、SDS-PAGEにより分析した。可溶性タンパ
ク質の量は、クマシー染色されたゲルから評価した。
【0104】
例5
細胞外使用に関して優れた特性を有する5つの組み合わせの詳細な評価
酵母および哺乳類の細胞内アッセイの両方において優れた性能を示し、酵母およびE.co
liにおいて発現の間に可溶性タンパク質を産生し、クオリティー・コントロールの間に優
先的に選択されたサブグループに当てはまる、5つの組み合わせを例として選択した(2.4
, 4.4, 5.2, 6.4および7.3、詳細については表5参照)。我々は、これら組み合わせをよ
り詳細に分析し、還元条件および酸化条件下におけるそれらの使用を更に評価した。
【0105】
a)様々な哺乳類細胞における細胞内アッセイでの性能
ヒト細胞における5つの組み合わせの性能の定量分析は、例2で実施したとおり、Hela
細胞、ヒト骨肉腫細胞株Saos-2およびヒト胚性腎細胞株HEK293を用いて行った(図7)。
【0106】
b)インビトロにおける性能
発現および精製
インビトロ性能を評価するため、5つの優れた組み合わせを、E.coliのペリプラズムで
発現させた(図6)。振盪フラスコ中の35 mg/Lクロラムフェニコールを含有する0.1 L
dYT-培地に、一晩培養物を1:40で接種し、30℃でインキュベートした。細胞を、1mM IPT
Gを用いてOD550 1.5に誘導し、遠心分離による導入の2時間後に収集した。scFvsを精製
するため、細胞の沈殿物を再懸濁し、音波処理により溶解させた。20 krpm、4℃で30分間
、SS34で遠心分離した後、上清を、Ni-MC-アフィニティーカラム(Hi-TrapTM Chelating
HP, 1ml,Amersham Pharmacia) にpH 7.5で適用し、Amersham PharmaciaからのAkta Basi
c systemを用いて、200mMイミダゾールで溶出した。scFvフラグメントの純度は、SDS-PA
GEにより測定すると98%より高かった(データ示さず)。精製されたタンパク質の濃度は
、280 nmにおける計算減衰係数を用いて決定した。可溶性精製タンパク質の収率は、OD60
0 10の1 L培養体積に規格化し、8から55mgまで変動した。
【0107】
凝集に対する抵抗性
凝集に対する抵抗性は、インビトロにおける熱力学的安定性(Worn, 1999)およびマウ
スの異種移植腫瘍モデルにおける腫瘍局在の効率(Willuda, 1999)と関係することが示
された。安定性、凝集に対する抵抗性、およびアンフォールディング(unfolding)の可
逆性を試験するために、50 mM Tris, pH 7.5, 100 mM NaCl中、6μMの濃度の精製タンパ
ク質の200μLサンプルを、4℃で4日間または37℃で4日間または4℃で3日間維持し、その
後、100℃で15分または60分インキュベーションし、室温までゆっくり冷まし、4℃で一晩
インキュベートした。その後、各サンプルのオリゴマー状態を、50 mM Tris, pH 7.5, 10
0 mM NaClを用いて平衡化したゲルろ過カラムで分析し、モノマー物質に対して凝集した
量を評価した(図8)。タンパク質を、Akta Basicシステム(Amersham Pharmacia)で、
流速1 mL/分、100μLの体積で、Superdex-75カラム(AmershamPharmacia)上に注入した
。
【0108】
プロテアーゼ分解に対する抵抗性
治療適用にとって重要なパラメーターである、プロテアーゼ分解に対する単離フレーム
ワークの安定性を決定するため、我々はヒト血清中で37℃において精製フレームワークを
インキュベートした(図9)。
【0109】
50μMの濃度の精製his-タグscFvタンパク質(上記参照)を、ヒト血清に10倍希釈し、
最終濃度を90%血清中5μMとした。その後、サンプルを、37℃で3日もしくは1日インキュ
ベートするか、またはローディングのために直接採取した。ローディングの前に、不溶性
の凝集物質を、4℃で10分間、エッペンドルフ遠心分離機において最大速度で沈殿させた
。ゲルにロードされる血清の量を減らすため上清をローディング色素で6倍に希釈し、5分
間100℃に加熱し、12%SDS-PAGEで分離した。可溶性his-タグscFvフラグメントを、一次
抗体として抗hisモノクローナルマウス抗体(Qiagen)、二次抗体として抗マウスペルオ
キシダーゼコンジュゲート(Sigma)を用いて、化学発光基質(Pierce)を用いて、his-
タグの検出を介したウェスタンブロッティングにより視覚化した。SDS-PAGEおよびウェス
タンブロッティングの手法は、当業者に周知である。
【0110】
例6
酵母における相互作用スクリーニングシステムでの、フレームワーク7.3上のランダム
化CDRライブラリーのスクリーニングを介した抗原バインダーの選抜
抗原バインダーの相互作用システムを用いたスクリーニングは、以前に詳細に記載され
るとおりに本質的に実施した(Auf der Maur, 2002)。
【0111】
酵母でスクリーニングするためのscFv融合構築物の発現用プラスミドは、pESBA-Act2か
ら誘導した。それは、酵母TRP1栄養性マーカーおよび2ミクロン複製起点を含有する。更
に、強力な発現のための構成的アクチンプロモーターおよびGAL11転写終結配列を、マル
チプルクローニングサイトにより隔てられて有する。細菌システムで扱うために、更に、
細菌の複製起点およびamp抵抗性遺伝子を有する。
【0112】
Gal4活性化ドメイン(ADアミノ酸768-881)は、pGAD424(Clontech)を鋳型として用い
て、Gal4-ADに対するSV40 T-抗原核局在シグナルN-末端を含むプライマーでPCRにより初
めに増幅した。scFvライブラリーは、VHのCDR3内の7アミノ酸をランダム化するプライマ
ーを用いてscFv-フレームワーク7.3のPCR増幅により得た。得られたPCR産物は、フレーム
ワーク7.3にクローンニングされ、Gal4-ADに対するC-末端融合体として、VL-リンカー-VH
の向きでベクターに存在する。かかる発現により、一般構造Gal4-AD-scFvの融合タンパク
質が産生される。
【0113】
スクリーニングは、酵母株、S. cerevisiae Immuna LHB (MATα ura3-52 leu2Δ1 trp1
d63 his3Δ200lys2Δ 385) で行った。その酵母株は、異なる向きのLacZおよびHIS3レポ
ーター遺伝子を、his3Δ200遺伝子座へ、6つのLexA結合部位を有する二方向性プロモータ
ーの制御下に組み込むことにより(インテグレイティングレポータープラスミド pDE200,
Escher2000)、そしてLEU2レポーター遺伝子を、leu2Δ1遺伝子座へ、(EGY48由来の)
8つのLexA結合部位を有するプロモーターの制御下に組み込むことにより、JPY5株から誘
導した。
【0114】
レポーターシステムの転写活性化は、そのscFv部分とbait−融合タンパク質の抗原部分
との特異的相互作用に続いて、scFv融合構築物のGal4-AD部分により媒介される。bait−
融合タンパク質は、DNA結合LexAタンパク質にC末端で融合したヒトpolo様キナーゼ1(hpl
k1-KD)のキナーゼドメインから成る。キナーゼドメイン(アミノ酸2-332)は、上流プラ
イマー5'-tgctctagaagtgctgcagtgactgcag-3' (配列番号12) および下流プライマー 5'
-ggttgtcgacttacaggctgctgggagcaatcg-3'(配列番号13) を用いて、hPlk1 cDNAからPC
R増幅した。得られたPCR産物は、baitベクターに、XbaIおよびSalIを介してLexAのC末端
にクローニングした。baitベクターは、URA3栄養性マーカーおよびArs Cen複製起点を含
有する。bait−融合タンパク質の発現は、構成的に活性なアクチンプロモーターにより誘
導される。転写は、GAL11終結配列により終結される。また、baitベクターは、細菌シス
テムでの繁殖のために、細菌の複製起点およびamp抵抗性遺伝子を有する。
【0115】
スクリーニングのために、酵母株S. cerevisiae Immuna LHBを、標準的な酢酸リチウ
ム形質転換プロトコール(Agatep, 1998)に従って、pESBA-Act2ベクター上のGal4-ADと
の融合物としてのscFv-ライブラリーを用いて、およびLexA-hPLK1-KD融合物を提供するba
itベクターを用いて同時形質転換した(co-transform)。形質転換の後、細胞を、ドロッ
プアウトプレート(-Trp/-Leu/-Ura)に播いた(plate)。30℃で3〜5日間インキュベー
トした後コロニーを採取し、ドロップアウトプレート(-Trp/-Leu/-Ura)に再び播いた(
streak)。再度増殖した細胞を、基質X-Galを含有するプレート上でのフィルターアッセ
イで、青色の発色によりLacZ発現について試験した。ポジティブクローンを、更なる解析
のために採取し、該解析は、酵母からのscFv-担持プラスミドの単離、E.coli DH5αへの
形質転換、E.coliの単一コロニーからのプラスミドの単離、および後述のアッセイのた
めに新たに調製した酵母株S. cerevisiae Immuna LHBへの再形質転換を伴う。すべての
方法は、当業者に周知の標準的手法に従って行われた。
【0116】
例7
新規scFvフレームワークから誘導したFab構築物のインビボ性能の評価
様々な抗体フォーマットにおいて安定な可変ドメインフレームワークを使用する有利な
効果を評価するため、酵母相互作用スクリーニングで使用するためのFab発現ベクターを
構築した。
【0117】
a)酵母における細胞内スクリーニングのためのFab構築物
2種類の発現ベクターを構築し、異なる発現レベルを可能にした。ベクターは、yEplac
112(2ミクロン)またはyCplac22(ars/cen)バックボーンのいずれかに基づくものであ
る(Gietz,1998)。いずれのベクターも、細菌システムで扱うために、酵母TRP1栄養性
マーカー、誘導性二方向性Gal1/Gal10プロモーター、細菌の複製起点およびamp抵抗性遺
伝子を含有する。一方向において、フレームワーク7.3のVHドメインを、C末端システイ
ンを含むIgG1のCH1-ドメインに対してN末端にクローニングし、その後に、リンカー、お
よびSV40 T-抗原を含むGal4活性化ドメイン(ADアミノ酸768-881)が続く。一方、フレ
ームワーク7.3のVLドメインを、C末端システインを含むCL(ラムダ)ドメインに対して
N末端にクローニングした。ターミネーターは、重鎖の側ではGal11ターミネーターであ
り、軽鎖の側ではサイクリン1ターミネーターである。
【0118】
b)酵母における細胞内アッセイでの性能
酵母においてscFvおよびFabフォーマットにおける抗原バインダーの性能を定量的に分
析するため(図1および3)、S. cerevisiae株Immuna LHBを、標準的な酢酸リチウム形
質転換プロトコールに従うことにより、pESBA-Act2ベクター上のGal4-AD-融合構築物とし
ての単離scFvsと、LexA-hPLK1-KD融合物を含有するbaitベクターとを用いて同時形質転換
した(Agatep,1998)。形質転換の後、細胞をドロップアウトプレート(-Trp、-Ura、Gl
c)に播いた。ドロップアウト培地(-Trp、-Ura、Glc)中の2mL一晩培養物を、幾つかの
コロニーを含有する培養物(streaks)から2回接種し、30℃で生育させた。培養物を、60
0 nm(OD600)における光学濃度が0.7になるように、1mLドロップアウト培地(-Trp、-U
ra、Gal)で希釈した。それを30℃で5時間生育させた。アッセイは上述のとおり行った。
【0119】
c)酵母における還元条件下での可溶性タンパク質の発現
還元条件下での発現による可溶性タンパク質の産生量を比較するため、scFvとFab構築
物を、hPLK1-KD-baitベクターと一緒に、上述のとおり、酵母S.cerevisiaeの細胞質にお
いて発現させた。それを、上述のとおり酵母株YDE173に形質転換し、グルコースを含有す
る-Trp、-Ura、ドロップアウトプレートに播いた。
【0120】
ドロップアウト培地(-Trp、-Ura、Glc)における5 mL一晩培養物を、幾つかのコロニ
ーを含有する培養物(streaks)から接種し、30℃で生育させた。培養物を、600 nm(OD6
00)における光学濃度が0.5になるように、YPAGで希釈した。その後、30℃で7.5時間生育
させた。ネガティブ細胞抽出物については、OD600を3に規格化した2.5 mL細胞培養物を、
遠心分離により収集し、液体窒素で凍結し、その後、75μL Y-PER(Pierce)に再懸濁し
た。再懸濁した細胞の沈殿物を、短時間ボルテクスにかけ、20℃で20分わずかに振盪させ
てインキュベートした。その後、不溶性の凝集物を、4℃で10分間、エッペンドルフ遠心
分離機において最大速度で沈殿させた。上清をローディング色素と混合し、5分間100℃に
加熱し、12%SDS-PAGEで分離した。可溶性Gal4-AD-scFv融合物およびGal4-ADに融合した
Fabの重鎖部分を、一次抗体として抗Gal4-ADモノクローナルマウス抗体(SantaCruz Bio
technology)、二次抗体として抗マウスペルオキシダーゼコンジュゲート(Sigma)を用
いて、化学発光基質(Pierce)を用いて、Gal4部分の検出を介したウェスタンブロッティ
ングにより視覚化した(図11)。SDS-PAGEおよびウェスタンブロッティングの手法は、
当業者に周知である。
【0121】
表1は、酵母における種々の「クオリティーコントロール」スクリーニングから選択さ
れた全てのVHドメインフレームワーク配列のアラインメントを示す。
【表1】
【0122】
表2は、酵母における種々の「クオリティーコントロール」スクリーニングから選択さ
れた全てのVLドメインフレームワーク配列のアラインメントを示す。
【表2】
【0123】
表3は、ライブラリーからランダムに選ばれた配列のアラインメントを示す。
【表3】
【0124】
表4は、「クオリティーコントロール」システムを用いて単離された配列におけるVH-
およびVL-ドメインのサブグループ頻度の統計解析を示す。定量的な酵母アッセイでポジ
ティブであることがその後に見出された配列のみを考慮した。選択された配列を、限られ
た数のランダム配列(表3)から決定される非選択ライブラリーと比較する。
【表4】
【0125】
表5は、更なる組換えおよびscFvsの最良の組み合わせの評価のために使用した配列、
並びにそれらの各略語(abb.)、供給源およびサブファミリーを示す。
【表5−1】
【表5−2】
【技術分野】
【0001】
本発明は、タンパク質化学、分子生物学、および免疫学に関する。
【背景技術】
【0002】
抗体は、高い特異性と親和性をもって、ほとんどすべての分子を認識し、ターゲットと
することができる。この特徴は、これら天然のタンパク質を診断および治療用の強力なツ
ールに変化させるために利用されている。組換えDNA技術の進歩により、広範な種々の
非リンパ系細胞における抗体遺伝子の操作、クローニング、および発現が容易になってい
る(Skerra,1988; Martineau, 1998; Verma, 1998)。多くの様々な抗体フラグメントが
、種々の適用に最も適合するように構築されている。親の免疫グロブリン全体の完全な抗
原結合能力を保持する最小要素(entity)は、単一鎖Fvフラグメント(scFv)である(Bi
rd, 1988)。この抗体フラグメントは、単一遺伝子からのタンパク質の発現を可能にする
フレキシブルなペプチドリンカーにより連結された重鎖と軽鎖の可変領域を含む。
【0003】
抗体フラグメントは、免疫グロブリン分子全体と比較して幾つかの重要な利点を有する
。抗体フラグメントの方がサイズが小さいため、種々の発現宿主細胞(たとえばE.coli細
胞)においてその発現は容易であり、その産生量は増大する(Pluckthun, 1996)。更に
、抗体フラグメントは、インビボでの適用において腫瘍の侵入を改善することができ(Yo
kota, 1992)、治療アプローチのために種々のエフェクター分子と共有結合させることが
できる。
【0004】
形質細胞により分泌される天然に存在する抗体は、細胞外の酸化環境において機能する
ように発達する。天然に存在する抗体は、機能的な折りたたみ構造を得るために、別個の
ドメイン内にジスルフィド架橋を形成することが一般に必要であり、これは、免疫グロブ
リンの折りたたみの安定性にとって重要である。完全長の抗体とは対照的に、scFvまたは
Fab抗体フラグメントは、原則として、任意の細胞内の還元(reducing)環境において機
能的に発現し、任意のコンパートメントに誘導されて細胞内タンパク質をターゲットとし
、これにより特定の生物学的効果を誘発することができる(Biocca, 1991)。実際、イン
トラボディ(intrabodies)と称される幾つかの細胞内単一鎖抗体フラグメントが、様々
な生物学的システムにおいて細胞内ターゲットタンパク質の機能を調整するためにうまく
適用されている。これにより、ウイルス感染に対する抵抗性が植物バイオテクノロジーで
実証され(Tavladoraki,1993; Benvenuto, 1995)、HIVタンパク質に対するイントラボ
ディの結合が示され(Rondon, 1997)、癌遺伝子産物に対する結合が記載されている(Bi
occa, 1993;Cochet, 1998; Lener, 2000)。更に、細胞内抗体は、ヒトゲノムの配列決
定により今日同定されている多大な数の遺伝子の機能を特徴付ける際の有益なツールとな
る見込みがある(Richardson,1995; Marasco, 1997)。たとえば、細胞内抗体は、機能
的ゲノミクスアプローチで使用して、新たに同定されたタンパク質の活性を妨害したり調
整したりし、これによりタンパク質の機能の理解に貢献することができる。最後に、イン
トラボディは、たとえば遺伝子治療セッティングにおいて、潜在的な診断および治療の適
用を有している。
【0005】
このような大きな期待にもかかわらず、機能的なイントラボディの作成は、その不安定
性および不溶性または凝集傾向によりまだ限られている。細胞質の還元環境は、保存され
た鎖内ジスルフィド架橋の形成を妨害し、これにより高パーセンテージの抗体フラグメン
トを不安定なものにし、その結果、細胞内で機能的でない(Biocca, 1995; Proba, 1997
)。したがって、抗体フラグメントの安定性および溶解性は、インビボにおけるタンパク
質機能の潜在的モジュレーターとしてのイントラボディの適用にとって大きな障害である
。これまでのところ、細胞内環境において抗体フラグメントを機能的にする配列の要件に
関して予測することはできない。
【0006】
したがって、広範囲の様々な細胞タイプでうまく機能し、様々な結合特異性のためのフ
レームワークとして使用することができる抗体フラグメントに対する需要が存在する。か
かるフレームワークは、細胞内スクリーニングのためのライブラリーを構築するために使
用することができ、あるいは現存の抗体の結合部分のためのアクセプターとして機能させ
ることができる。
【0007】
非常に安定な可変ドメインフレームワークに基くかかる抗体フラグメントまたは抗体全
体は、細胞内適用のためにユニークに適合していることに加えて、多くの細胞外適用およ
びインビトロ適用において他の抗体よりも別個の利点を更に有する。かかるフレームワー
クを酸化環境で産生すると、ジスルフィド架橋を形成することができるため、更にその安
定性は高まり、凝集やプロテアーゼ分解に対する高い抵抗性を付与することができる。イ
ンビボでの半減期(および凝集や血清プロテアーゼによる分解に対する抵抗性)は、親和
性および特異性に加えて、治療または診断適用における抗体の成功にとって唯一最も重要
な因子である(Willuda,1999)。抗体フラグメントの半減期は、ポリエチレングリコー
ル(PEG)などのポリマー分子の共有結合により更に増大させることができる(Weir,200
2)。このタイプの安定な分子は、Fcの機能性を望まないときに特に(しかしそのときに
限定されず)、抗体の使用において有意な進歩を示す。
【0008】
抗体フラグメントライブラリーの実用的な重要性が、この分野における研究に刺激を与
えている。Winter(EP 0368684) は、抗体可変領域遺伝子の最初のクローニングと発現を
提供した。これら遺伝子からスタートして、Winterは、相補性決定領域(CDRs)とフレー
ムワーク領域の両方に高い多様性を有する大きな抗体ライブラリーを作成した。しかし、
Winterは、ライブラリー構築のための様々なフレームワークの有効性を開示していない。
【0009】
一方、Pluckthun(EP 0859841) は、フレームワークを限られた数の合成コンセンサス
配列に限定することにより、ライブラリーのデザインを改良することを試みた。合理的に
設計された多くの突然変異の導入を含むタンパク質操作の成果により、単離された可変免
疫グロブリンドメインの安定性を改良するための適切な手段として、各コンセンサス配列
に対する突然変異が既に示唆されている(Ohage 1999; Ohage 1999およびUS 5,854,027,
参照により本明細書に取り込む)。
【0010】
Pluckthun(EP 0859841) は、これらコンセンサス配列に基いて結合親和性を更に最適
化するための方法を開示する。また、Pluckthunの特許は、抗体に関する知識の継続的な
増大を認識しているため、ライブラリー設計においてかかる将来的発見を含めることを目
的とする。しかし、合成コンセンサスフレームワークの考えられる更なる改良は示唆され
ていない。
【0011】
Winter,Pluckthunおよびその他 (たとえば、Soderlind, WO 0175091) の教示により、
選択および酸化条件下における選択されたscFvsの適用に対するCDRsの高い多様性に焦点
を絞って、大きな抗体ライブラリーを作成することが試みられた。しかし、これらライブ
ラリーのすべては、細胞内適用のために最適化されてないため、選択および還元環境にお
ける適用、または発現抗体フラグメントの安定性と溶解性に対する特別な要件を設定する
他の状況に有効でない。
【0012】
抗体フラグメントが還元環境(たとえば、原核および真核細胞の細胞質)でうまく機能
するために必要な性質は明らかでない。したがって、細胞内抗体または「イントラボディ
」の適用は、安定性および溶解性の性質に影響を及ぼし得る、還元状況下での予測できな
いそれらの行動により現在制限されている(Biocca, 1995; Worn, 2000)。イントラボデ
ィの細胞内スクリーニングに関する現在の特許出願(EP1040201, EP1166121およびW00200
729)および出版物(Visintin,1999)は、スクリーニング技術に焦点を当てているが、
真核細胞(とりわけ酵母)で機能的であり、かつこの状況でライブラリー構築に有効であ
る特定の抗体配列を開示していない。
【0013】
VisintinおよびTseは、それぞれ独立して、いわゆる細胞内コンセンサス配列(ICS)の
単離について記載している(Visintin, 2002; Tse, 2002)。この配列は、酵母における
抗原−抗体−相互作用スクリーニングから単離された多くの配列から得られた。しかし、
細胞内スクリーニングへのインプットは、先のファージ−ディスプレイ選択のために大き
く偏っていた。よって、インプット配列の一つを除いて全ては、Visintin et alのケース
でVH 3サブグループに属していた。公表されたコンセンサス配列ICSは、Knappik(2000)
およびEP0859841に記載されるヒトVH3サブグループのコンセンサス配列と完全に一致す
る。また、ICSの62アミノ酸のうち60は、安定性の高い可変ドメインの構築のための基礎
としてSteipeにより提案された一般的なヒトVH-ドメインコンセンサス配列と一致する(
米国特許第6,262,238号、参照により本明細書に取り込む)。次に、これらの研究は、以
前の配列の収集(すなわちKabat, 1991)、および可変ドメインサブグループおよび構造
決定因子の規定(Tomlinson,1992; Williams, 1996; Chothia, 1989 および Chothia, 1
987)を基にした。しかし、イントラボディ選択へのインプットは、非常に大きく偏って
いたので(すなわち、Visintin et alのケースにおいてVHドメインの一つを除いて全てが
VH3であったので)、細胞内スクリーニングからのVH3配列の単離は、特に驚くべきことで
はない。これらのインプットライブラリーの大きな偏りのため、偏りのない調査により提
供され、かつヒトレパートリーに存在する有効なイントラボディフレームワークを同定す
るために必要とされる、ヒト可変ドメインレパートリーの完全な評価を、Tse et al.およ
びVisintinet al.の研究は提供していない。
【0014】
我々は、抗原結合特異性とは無関係に、酵母において安定かつ可溶性のイントラボディ
の選択を可能にするシステムを以前に記載している(Auf der Maur (2001), W00148017)
。このアプローチにより、scFvライブラリーの効率的なスクリーニング、および酵母細胞
の還元環境において安定かつ可溶性である特定のフレームワークの単離が可能となる。実
際にフレームワーク配列を単離し、そのパターンを使用して、第一の工程においてどの配
列タイプが還元環境において最も安定であるか予測し、第二の工程において分析、組換え
、更なるインビボおよびインビトロ実験により最適な配列を同定することが、目的として
残っている。
【発明の概要】
【課題を解決するための手段】
【0015】
本発明は、抗体作成の分野においてミッシングリンクを満たすものである。本発明は、
安定性および溶解性に関して優れた特徴を有する抗体可変ドメインフレームワーク配列を
提供する。これらは、多くの関連の適用にとって、たとえば診断、治療またはリサーチに
おいて重大な特徴である。これらフレームワークは、現存の結合特異性を移植するため、
または高い安定性および溶解性を有する抗体ライブラリーを作成するために使用すること
ができる。
【0016】
ScFvライブラリーは、酵母細胞の還元環境において安定かつ可溶性であるフレームワー
クを単離するために使用した。その後、単離されたフレームワークの性能を、ヒト細胞株
およびインビトロ実験において特徴づけた。記載されるフレームワークは、現存の結合特
異性のためのアクセプターバックボーンとして直接使用したり、または還元環境もしくは
他の挑戦的環境で使用するための一以上の超可変ループのランダム化によりCDRライブラ
リーを構築するために直接使用したりすることができる。単離された可変ドメイン配列は
、アラインメントにより更に分析し、好ましい配列ファミリーを同定した。好ましい可変
ドメイン配列ファミリーから、免疫グロブリンの折りたたみを混乱させるフレームワーク
残基を含有する配列を除外する構造解析に基いて、最適な配列を選択した。その後、同定
された可変ドメイン配列の候補を、あらゆる可能なバリエーションで再結合し、酵母、哺
乳類細胞およびインビトロでその性能を分析することにより、軽鎖と重鎖の可変ドメイン
の最適な組み合わせを選択した。
【0017】
これら最適化されたscFvsおよびそれを構成する可変ドメインフレームワーク、並びに
他の抗体フラグメントまたはそれに由来する抗体全体は、たとえば、現存の結合特異性の
ためのアクセプターバックボーンとして、または還元環境もしくは他の挑戦的環境で使用
するための一以上の超可変ループのランダム化によりCDRライブラリーを構築するために
理想的である。細胞内適用に適した抗体は、その名が示すとおり安定かつ可溶性である。
そのため、その使用は、細胞内環境の外での適用にも好都合である。
【0018】
本発明は、抗体可変ドメインのフレームワークおよび単一鎖Fv抗体(ScFv)フラグメン
トを含む組成物であって、種々の抗体フラグメントまたは抗体全体に組み込むことができ
る組成物を提供する。最も安定かつ可溶性であるために細胞内適用に最もよく適合した抗
体可変ドメインフラグメントのクラスが提供される。また、抗体可変ドメインとscFv抗体
フラグメントの特定のフレームワーク配列であって、細胞内アッセイにおいて最も高い性
能を示すフレームワーク配列が提供される。また本発明は、抗体可変ドメインの特定のフ
レームワーク配列およびscFvフラグメントの軽鎖と重鎖の可変ドメインの合成組み合わせ
であって、たとえば、細胞内適用にとって最適であり、安定性と溶解性に関してインビト
ロで最適な性能を示すものを提供する。
【0019】
本発明は、一般構造:
NH2-VL-リンカー-VH-COOHまたは
NH2-VH-リンカー-VL-COOH
を有する単一鎖フレームワーク試薬を提供する。
【0020】
本発明の別の態様において、単一鎖フレームワークは、第二のタンパク質部分に融合し
、一般構造:
NH2-VL-リンカー-VH-第二のタンパク質-COOHまたは
NH2-第二のタンパク質-VL-リンカー-VH-COOH
の融合構築物を産生してもよい。
【0021】
これら融合構築物においてVHおよびVL領域の方向性(orientation)は逆向きであって
もよい。
【0022】
本発明の別の態様において、可変ドメインは、Fabフラグメントに組み込まれていても
よく、これは更に第二のタンパク質部分に融合し、一般構造:
NH2-VH-CH-第二のタンパク質-COOHおよびNH2-VL-CL-COOH
の融合構築物を産生してもよい。
【0023】
第二のタンパク質は、重鎖または軽鎖のいずれかのN−末端またはC−末端のいずれか
に融合してもよい。
【0024】
好ましい態様において、単一鎖またはFabフレームワーク融合構築物の第二のタンパク
質は、直接的または転写活性を介して、細胞内アッセイの読出し部(a read-out)を提供
するタンパク質である。
【0025】
本発明の別の目的は、たとえば、還元環境もしくは他の挑戦的環境において機能的な抗
体を得るために、現存の抗体から超可変ループを移植するのに適している、抗体可変ドメ
インのフレームワーククラスおよび可変ドメインとscFvsの配列を提供することである。
【0026】
本発明の別の目的は、たとえば、かかるフレームワークの一以上の超可変ループのラン
ダム化を介して、還元環境または他の挑戦的環境で使用するためのライブラリーを作成す
るのに適している、抗体可変ドメインのフレームワーククラスおよび可変ドメインとscFv
sの配列を提供することである。
【0027】
本発明の別の目的は、保存された残基およびコンセンサス配列の同定において、開示さ
れる配列を使用することである。
【0028】
開示されるフレームワークの使用により得られる抗体または抗体フラグメントは、ター
ゲットの検証、並びにヒト、動物および植物の疾患の治療、予防および診断における試薬
として使用することができる。抗体は、タンパク質またはかかるタンパク質をコードする
DNAの形態で使用することができ、細胞内適用に限定されるものではない。
【図面の簡単な説明】
【0029】
【図1】図1は、lacZ発現の活性化によりアッセイされる、酵母における典型的な「クオリティーコントロール」スクリーニングの結果を示す(たとえば例1参照)。選択されたポジティブクローン(黒)は、幾つかの様々なスクリーニングで同定され、ポジティブクローンの対応の配列は、表1および2に見出すことができる。選択された配列を、ポジティブコントロールである非常に安定なラムダ−グラフト(ダークグレー)と比較する。
【図2】図2は、非常に安定なラムダ−グラフト(ダークグレー)と比較してルシフェラーゼ発現の活性化によりアッセイされる、酵母における典型的な「クオリティーコントロール」スクリーニングから単離したフレームワークのヒト細胞株Helaにおける性能を示す。ポジティブコントロールGal4-VPl6(白)は、このシステムにおいて最大限可能なレベルの転写活性化を示す。ルシフェラーゼ活性は、トランスフェクション効率について修正されている。
【図3】図3は、lacZ発現の活性化により酵母でアッセイされる、優れたフレームワークの組み合わせのインビボでの性能を示す。フレームワーク配列(黒)を、ポジティブコントロール(非常に安定なラムダ−グラフト(ダークグレー))と比較する。フレームワークのナンバリングは、表5に記載されるとおりである。
【図4】図4は、ルシフェラーゼ発現の活性化によりヒト細胞株HeLaでアッセイされ、非常に安定なラムダ−グラフト(ダークグレー)と比較して図示される、優れたフレームワークの組み合わせのインビボでの性能を示す。ポジティブコントロール、Gal4-VP16(白)は、このシステムにおいて最大限可能なレベルの転写活性化を示す。ルシフェラーゼ活性は、トランスフェクション効率について修正されている。
【図5】図5は、酵母株S.cerevisiae JPY9の細胞質で産生される可溶性タンパク質の量によりアッセイされる、優れたフレームワークの組み合わせのインビボでの性能を示す。
【図6】図6は、E.coliのペリプラズムにおける、選択されたフレームワークの組み合わせの発現行動を示す。矢印は、scFvフレームワークに対応するバンドの位置を示す。
【図7】図7は、3つのヒト細胞株(Hela(黒)、Saos-2(ダークグレー)、HEK 293(白))においてルシフェラーゼ発現の活性化によりアッセイされ、非常に安定なラムダ−グラフトと比較して図示される、選択された優れたフレームワークの組み合わせのインビボでの性能を示す。ポジティブコントロール、Gal4-VP16(白)は、このシステムにおいて最大限可能なレベルの転写活性化を示す。ルシフェラーゼ活性は、トランスフェクション効率について修正されている。
【図8】図8は、PBSバッファー中に示されるインキュベーション前およびインキュベーション後に存在するモノマータンパク質の量により定量される、選択されたフレームワークの組み合わせの37℃での凝集に対する抵抗性を示す。パネルAはフレームワーク2.4および5.2を表し、パネルBはフレームワーク4.4、6.4および7.3を表す。
【図9】図9は、長期インキュベーション前および長期インキュベーション後に存在する可溶性完全長タンパク質の量により定量される、選択されたフレームワークの組み合わせの37℃でのヒト血清におけるプロテアーゼ分解凝集に対する抵抗性を示す。
【図10】図10は、LacZ発現の活性化により酵母相互作用アッセイでアッセイされる、Fabコンテクスト(context)における新規フレームワーク7.3上の選択された2つのバインダーのインビボでの性能を示す。Fab鎖の発現は、ars/cenまたは2ミクロンベクター上の二方向性ガラクトース誘導プロモーターに由来する。Fabベクターからの発現により、抗体軽鎖およびVH-CH1-Gal4-AD融合タンパク質が産生される。バインダーは、ヒトPolo様キナーゼ1(hPLK1)に対して向けられる。ターゲットに対する結合を、無関係の抗原に対する非特異的結合および非ランダム化フレームワーク7.3の結合と比較する。参考のために含めた対応のscFvは、アクチンプロモーター(2ミクロン)から発現することに留意されたい。
【図11】図11は、酵母株JPY9の細胞質で産生される可溶性タンパク質の量によりアッセイされる、Fabコンテクスト(context)におけるscFvフレームワークのインビボでの性能を示す。Gal4-AD-scFv融合体(アクチン/2ミクロン)の発現を、対応のFab構築物の発現、およびFabとしての親のフレームワーク7.3と比較する(そのいずれもが二つの異なるベクター(Gal-誘導、ars/cenおよび2ミクロン)に由来する)。Fabベクターからの発現により、抗体軽鎖およびVH-CH1-Gal4-AD融合タンパク質が産生され、これはこのブロットで検出される。
【発明を実施するための形態】
【0030】
特に規定しない限り、本明細書で使用される技術的および科学的用語はすべて、本発明
が属する分野の当業者により一般に理解されるのと同じ意味を有する。本明細書に記載さ
れるのと同様もしくは等価な方法および材料は、本発明を実施または試験する際に使用す
ることができるが、適切な方法および材料は以下に記載される。本明細書に記述される出
版物、特許出願、特許、および他の参考文献はすべて、その全体を参照により本明細書の
開示内容の一部とする。矛盾する場合、定義を含む本明細書がコントロールするでしょう
。更に、材料、方法、および実施例は、単に説明のためのものであって、限定するための
ものではない。
【0031】
本明細書で使用される「同一性(identity)」は、二つのポリペプチド、分子または二
つの核酸の間の配列の類似性をいう。二つの比較配列の両方における位置が、同じ塩基ま
たはアミノ酸モノマーサブユニットにより占められる場合(たとえば、二つのDNA分子の
それぞれにおける位置が、アデニンにより占められるか、あるいは、二つのポリペプチド
のそれぞれにおける位置がリシンにより占められる場合)、各々の分子はその位置におい
て相同である。二つの配列の間の「パーセンテージ同一性」は、二つの配列により共有さ
れる一致する位置の数を、比較した位置の数で割って100を乗じた関数である。たとえば
、二つの配列の10の位置のうち6つが一致している場合、二つの配列は60%の同一性を有
する。一例として、DNA配列CTGACTとCAGGTTは、50%の相同性を共有する(トータル6つの
位置のうち3つが一致する)。一般に、二つの配列を最大の相同性が得られるように整列
して比較を行う。かかる整列は、たとえば、Needleman et al., J. Mol Biol. 48: 443-4
53 (1970) の方法を用いて提供することができ、Alignprogram (DNAstar, Inc.) などの
コンピュータープログラムにより便利に実施される。
【0032】
「類似(similar)」配列は、整列したときに、同一および類似のアミノ酸残基を共有
するものであり、ここで類似の残基とは、整列させたレファレンス配列における対応のア
ミノ酸残基の、保存的置換または「許容される点突然変異」のことである。ここで、レフ
ァレンス配列における残基の「保存的置換」とは、対応のレファレンス残基に対して物理
的または機能的に類似する残基、たとえば、同様のサイズ、形状、電荷、共有結合もしく
は水素結合の形成能力などの化学的特性等を有する残基による置換のことである。よって
、「保存的置換修飾」配列は、一以上の保存的置換または許容される点突然変異が存在す
るという点において、レファレンス配列または野生型の配列とは異なるものである。二つ
の配列の間の「パーセンテージポジティブ」は、二つの配列により共有される一致残基ま
たは保存的置換を有する位置の数を、比較した位置の数で割って100を乗じた関数である
。たとえば、二つの配列の10の位置のうち6つが一致し、10の位置のうち2つが保存的置換
を有する場合、二つの配列は80%のポジティブ相同性を有する。
【0033】
「VHドメイン」は、免疫グロブリン分子の重鎖の可変部分をいう。
【0034】
「VLドメイン」は、免疫グロブリン分子の軽鎖の可変部分をいう。
【0035】
VHまたはVL「サブタイプ」は、Knappik(2000) に規定されるとおり、各コンセンサス
配列により規定されるサブタイプをいう。「サブファミリー」または「サブクラス」の用
語は、「サブタイプ」の同義語として使用される。本明細書で使用される「サブタイプ」
の用語は、当該サブタイプを代表する各コンセンサス配列に対して高度な一致性および類
似性を共有する配列をいう。ある可変ドメイン配列が、ある「サブタイプ」に属するか否
かは、各ドメインの公知のあらゆるヒト生殖細胞系セグメントまたは規定されるコンセン
サス配列の何れかとともに、その配列を整列し、その後最大ホモロジーを同定することに
より決定される。相同性を決定し、BLOSUM (Henikoff 1992) などのサーチマトリクスを
用いることにより配列をグループ分けする方法は、当業者に周知である。
【0036】
本明細書で使用される「アミノ酸コンセンサス配列」は、少なくとも二つ、好ましくは
より多くのアミノ酸配列のマトリクスを用いて作成することができ、整列の際にギャップ
を許容して、各位置において最も頻度の高いアミノ酸残基を決定することができる、アミ
ノ酸配列をいう。コンセンサス配列は、各位置において最も頻度が高く表示されるアミノ
酸を含む配列である。二以上のアミノ酸が単一の位置において等しく表示される場合には
、コンセンサス配列は、当該アミノ酸の両方またはすべてを含む。
【0037】
タンパク質のアミノ酸配列は、種々のレベルで分析することができる。たとえば、保存
性または変動性は、単一残基のレベル、複数残基のレベル、ギャップを有する複数残基の
レベルなどで示すことができる。残基は、同一の残基の保存性を示すことができ、あるい
はクラスレベルで残基を保存することができる。アミノ酸クラスの例は、極性があるが非
電荷のR基(セリン、トレオニン、アスパラギン、およびグルタミン);正に荷電したR
基(リシン、アルギニンおよびヒスチジン);負に荷電したR基(グルタミン酸およびア
スパラギン酸);疎水性R基(アラニン、イソロイシン、ロイシン、メチオニン、フェニ
ルアラニン、トリプトファン、バリンおよびチロシン);並びに特別なアミノ酸(システ
イン、グリシンおよびプロリン)を含む。他のクラスは、当業者に公知であり、構造決定
または他のデータを用いて代替可能性を評価して規定されてもよい。この意味において、
置換可能なアミノ酸とは、置換されて当該位置の機能的保存性を維持することができるア
ミノ酸をいう。
【0038】
本明細書で使用される「ポリヌクレオチドコンセンサス配列」は、少なくとも二つ、好
ましくはより多くの整列した核酸配列のマトリクスを用いて作成することができ、整列の
際にギャップを許容して、各位置において最も頻度の高いヌクレオチドを決定することが
できる、ヌクレオチド配列をいう。コンセンサス配列は、各位置において最も頻度が高く
表示されるヌクレオチドを含む配列である。二以上のヌクレオチドが単一の位置において
等しく表示される場合には、コンセンサス配列は、当該ヌクレオチドの両方またはすべて
を含む。
【0039】
本明細書で使用される「構造的サブエレメント」は、分子の規定の構造的または機能的
部分に対応するタンパク質またはポリペプチド内のアミノ酸残基の一区切り(stretch)
をいう。これらは、ループ(すなわち、抗体のCDRループ)であってもよいし、タンパク
質またはポリペプチド内の任意の他の二次構造または機能的構造(すなわち、ドメイン、
α−へリックス、β−シート、抗体のフレームワーク領域など)であってもよい。構造的
サブエレメントは、類似もしくは相同ポリペプチドの公知の構造を用いて、または整列し
たアミノ酸配列の上述のマトリクスを用いて、同定することができる。ここで、各位置に
おける変動性は、構造的サブエレメントに属するアミノ酸残基の一区切り(たとえば、抗
体の超可変領域)を決定するための基盤である。
【0040】
本明細書で使用される「サブ配列」は、少なくとも一の構造的サブエレメントをコード
する遺伝的モジュールをいう。これは、構造的サブエレメントと必ずしも同一ではない。
【0041】
本明細書で使用される「抗体CDR」は、Kabat et al. (1991) により規定されるとおり
、抗原結合ループから成る抗体の相補性決定領域をいう。抗体Fvフラグメントの二つの可
変ドメインはそれぞれ、たとえば、3つのCDRsを含有する。
【0042】
本明細書で使用される「抗体」は、「免疫グロブリン」の同義語である。本発明におい
て抗体は、免疫グロブリン全体であってもよいし、免疫グロブリンの少なくとも一の可変
ドメイン、たとえば単一の可変ドメイン、Fv(Skerra, 1988)、scFv(Bird, 1988; Hust
on,1988)、Fab、(Fab’)2を含むフラグメント、または当業者に周知の他のフラグメント
であってもよい。
【0043】
本明細書で使用される「抗体フレームワーク」は、可変ドメインの抗体結合ループのた
めの足場として機能する、可変ドメイン(VLまたはVHのいずれか)の一部をいう(Kabat
et al., 1991)。
【0044】
合理的に操作されたscFvフラグメントは、scFvフラグメントの熱力学的安定性とそのイ
ンビボでの性能との間に明らかな関係が実証されている(Worn, 2000; Auf der Maur, 20
01)。近年開発された「クオリティーコントロール(QualityControl)」と呼ばれるシ
ステム(Aufder Maur, 2001)を用いて、細胞内適用に適した特定の抗体可変ドメインフ
レームワークの配列が単離され(表1および2)、特徴づけられ(図1および2)、更に
改良された(図3〜9および表3)。我々の以前の実験で観察されるとおり、細胞内アッ
セイで選択された性能のよいフレームワークは、37℃での凝集およびプロテアーゼ分解に
対する抵抗性により実証されるようにインビトロで高い安定性を示す(図8および9)。
更に、フレームワークのサブファミリーに依存して、一般的なベースで、細胞内適用のた
めのフレームワークの選択を可能にするパターンが現れた(表4)。細胞内適用に有効な
特定の抗体可変ドメイン配列は、一般的なパターンとともに本明細書に開示される。一方
、これにより、これら配列を移植実験におけるフレームワークドナーとして使用し、ルー
プドナーの結合特異性を保持する機能的イントラボディを得ることが可能になる。加えて
、開示される配列をフレームワークとして用いて、抗体ライブラリーを構築することがで
きる。かかるライブラリーは、還元状況下(たとえば原核および真核細胞における還元状
況下)での細胞内選択システムに適している。加えて、開示される配列は、たとえば、保
存された配列または残基またはモチーフを同定するために使用してもよい。構造的サブエ
レメント(たとえば抗体の結合ループのサブエレメント)の移植(たとえばJung, 1997)
、並びに抗体またはそのフラグメントのライブラリーの作成(たとえばVaughan, 1996; K
nappik, 2000)は、詳細に記載されており、当業者に周知である。
【0045】
細胞内適用は、抗体フラグメントを非常に望ましくない状況(すなわち、高い温度、還
元環境)に晒すため、本発明に開示される配列は、当該配列を最も不利な状況に対して抵
抗性にする特徴を獲得した。したがって、「平均的」配列と比較すると、開示される配列
は、凝集およびプロテアーゼ分解に対する抵抗性により実証されるように、顕著な安定性
を有する(図8および9)。優れた発現量とともに、これらの特徴により、開示される抗
体フレームワーク配列は、細胞内の使用だけでなく、とりわけ長い半減期、頑強性、およ
び産生の容易性が大変重要である治療および診断のあらゆる適用にも独自に適している。
【0046】
本発明により、還元環境または他の挑戦的環境における適用に有効な抗体の可変部分を
少なくとも含むポリペプチド配列の設計が可能になる。第一の態様において、本発明は、
細胞内適用に適した抗体フレームワーク配列のコレクションを提供する(表1および2)
。第一の工程において、様々な配列のライブラリーを、酵母においてクオリティーコント
ロールシステムを用いて、結合親和性とは無関係にスクリーニングする。単離された配列
は、酵母および哺乳類細胞において細胞内性能について評価することができる(図1およ
び2)。
【0047】
本発明の一つの態様において、単離された配列のコレクションを、アラインメントによ
り分析し、細胞内適用に適した抗体可変ドメインサブクラスおよびコンセンサス配列を同
定する。
【0048】
本発明の更に好ましい態様において、上述の抗体フレームワーク配列のコレクションを
、互いにアラインメントにより更に分析し、サブファミリーにグループ分けする。一つの
サブタイプに属するフレームワークすべてを、酵母および哺乳類細胞において細胞内性能
に関して比較し(例として図1および2)、各サブタイプのコンセンサスに対する、アミ
ノ酸配列のネガティブ、ニュートラル、またはポジティブ交換の出現に関して比較する。
当業者であれば、免疫グロブリンドメインにおいて変化した特定の残基の構造環境に基い
て、ポジティブ、ニュートラル、およびネガティブ変化を識別することができる。その後
、最も優れた細胞内性能を示し、かつ各サブタイプのコンセンサスと比較してネガティブ
交換がない、可変抗体ドメインのフレームワーク配列を選択する。好ましくは、ポジティ
ブと考えられるアミノ酸交換を更に含有する配列を選択する。
【0049】
更に好ましい態様において、最も高い安定性と溶解性を備えた組み合わせを同定するた
めに、その後、重鎖および軽鎖の選択された抗体可変ドメインを、あらゆる可能な組み合
わせでscFvフラグメントに再結合する。この目的のために、新規な再結合scFvフラグメン
トを、酵母(図3)および哺乳類細胞株(図4および7)において細胞内相互作用アッセ
イで還元条件下のその性能について評価し、並びに酵母において可溶性細胞内発現(図5
)について評価する。E.coliにおけるペリプラズム発現量(図6)、増大した温度での凝
集に対する抵抗性(図8)、並びに37℃、ヒト血清中における長期インキュベーションに
よる凝集およびプロテアーゼ分解に対する抵抗性(図9)を分析することにより、有望な
組み合わせを、酸化条件下のその行動について更に評価する。これらデータは、細胞内ま
たは酸化条件下の何れかの任意の特定の適用に最適なscFvフレームワークを同定するため
に使用する。
【0050】
本明細書に開示される選択され最適化されたフレームワーク配列は、細胞内適用におい
てだけでなく、scFvの増大した安定性および/または溶解性から利益を得ることができる
あらゆる適用において、有意な利点を有する。その例として、診断適用に必要とされる高
濃度での長期保存、および(たとえば治療適用で必要とされる)37℃、血清中での長期間
の機能的半減期が挙げられる。
【0051】
本発明の一つの側面に従えば、一般構造:
NH2-VL-リンカー-VH-COOH;または
NH2-VH-リンカー-VL-COOH
(ここでVHフレームワークは、サブタイプ1a、1bまたは3である)を有する単一鎖フレー
ムワークを含むイントラボディのフレームワークが提供される。
【0052】
別の態様において、VHおよびVL領域の方向性(orientation)は、上述の単一鎖フレー
ムワークにおいて逆向きである。
【0053】
本発明の一つの側面に従えば、一般構造:
NH2-VL-リンカー-VH-COOH;または
NH2-VH-リンカー-VL-COOH
(ここでVHフレームワークは、サブタイプ1a、1bまたは3であり、VLフレームワークは、
サブタイプλ1、λ3またはκ1である)を有する単一鎖フレームワークを含むイントラボ
ディのフレームワークが提供される。
【0054】
別の態様において、本発明は、第二のタンパク質部分に融合し、一般構造:
NH2-VL-リンカー-VH-第二のタンパク質-COOH;または
NH2-第二のタンパク質-VL-リンカー-VH-COOH
(ここでVHフレームワークは、サブタイプ1a、1bまたは3であり、VLフレームワークは、
サブタイプλ1、λ3またはκ1である)の融合構築物を産生する、単一鎖フレームワーク
を提供する。
【0055】
別の態様において、これら融合構築物におけるVHおよびVL領域の方向性(orientation
)は、逆向きであってもよい。
【0056】
別の態様において、可変ドメインは、Fabフラグメントに組み込まれていてもよく、こ
れは更に第二のタンパク質部分に融合し、一般構造:
NH2-VH-CH-第二のタンパク質-COOHおよびNH2-VL-CL-COOH
の融合構築物を産生してもよい。
【0057】
第二のタンパク質は、重鎖または軽鎖のいずれかのN−末端またはC−末端のいずれか
に融合してもよい。
【0058】
本明細書に開示されるとおり、細胞内適用において、サブタイプ3、またサブタイプ1a
および1bのVHフレームワークが非常に強く優先される。軽鎖可変ドメイン(VL)に関して
は、カッパ1タイプのフレームワークが数的に明らかに優先され、またラムダ1および3も
豊富である。したがって、これらフレームワークのサブタイプ、すなわちカッパ1、ラム
ダ1または3 VLドメインと組み合わせたVH 1a、1bおよび3が、細胞内の使用およびscFv
の折りたたみ特性を必要とする他の適用に最も適合する。したがって、還元環境において
機能的でない分子の量を低減するために、細胞内スクリーニングシステムのためのライブ
ラリーは、これらフレームワークサブタイプの混合物から優先的に構築されるべきである
。
【0059】
好ましい態様において、本発明の抗体フラグメントのVHドメインは、サブタイプ1a、1b
または3である。
【0060】
好ましい態様において、本発明の抗体フラグメントのVLドメインは、サブタイプカッパ
1、ラムダ1または3である。
【0061】
好ましい態様において、フレームワークとして使用される抗体フラグメントは、表5に
記載される、1.1,2.1, 3.1, 4.1, 5.1, 1.2, 2.2, 3.2, 4.2, 5.2, 1.3, 2.3, 3.3, 4.3
, 5.3, 7.3,1.4, 2.4, 3.4, 4.4, 5.4, および6.4から成る群より選択される。
【0062】
本発明の一つの態様において、少なくとも二つ、好ましくはより多くのフレームワーク
が同定され、その後分析される。タンパク質配列のデータベースは、タンパク質配列を互
いに整列させて構築してもよい。このとき、アラインメントを使用して、たとえば、その
配列と(その情報が入手可能であれば)構造的配置の両方において高い類似度を示すフレ
ームワーク配列の残基、サブエレメント、サブ配列またはサブグループを規定することが
できる。
【0063】
サブエレメントの長さは、好ましくは、1アミノ酸(たとえば、酵素の活性部位または
構造決定残基における一残基)から150アミノ酸(たとえば、タンパク質ドメイン全体)
までの範囲であるが、これに限定されない。最も好ましくは、その長さは3から25アミノ
酸までの範囲であり、たとえば抗体のCDRループに最も一般に見出される。
【0064】
別の態様において、当該分析から推測されるコンセンサス核酸配列が合成される。これ
は、当業者に周知の幾つかの方法のいずれかにより、たとえばトータル遺伝子合成または
PCRベースのアプローチにより、達成することができる。
【0065】
別の態様において、核酸配列は、ベクターにクローニングされる。ベクターは、配列決
定ベクター、発現ベクターまたはディスプレイ(ファージディスプレイ)ベクターとする
ことができ、これらすべてが当業者に周知である。ベクターは、一つの核酸配列、または
二以上の核酸配列を含むことができ、異なるオペロンまたは同一のオペロンのいずれでも
よい。後者の場合、別々にクローニングされてもよいし、隣接配列であってもよい。
【0066】
一つの態様において、ポリペプチドは、特定の種に特徴的なアミノ酸パターンを有する
。これは、たとえば、たった一つの種の相同タンパク質のコレクションから、最も好まし
くはヒトタンパク質のコレクションから、コンセンサス配列を導き出すことにより達成す
ることができる。
【0067】
本発明の更なる態様は、上述のポリペプチド、並びに追加の部分の両方をコードするDN
A配列を提供することによる融合タンパク質に関する。
【0068】
更なる態様において、本発明は、核酸配列、該核酸配列を含有するベクター、該ベクタ
ーを含有する宿主細胞、および本明細書に記載される方法に従って得ることができるポリ
ペプチドを提供する。
【0069】
更なる態様において、本発明は、本発明の核酸配列の末端に制限部位を合成するか、ま
たはその他の方法で配置し、本発明の核酸配列を適切なベクターにクローニング可能にす
ることを提供する。
【0070】
更に好ましい態様において、本発明は、ポリペプチドをコードする核酸配列と適合する
ベクターシステムを提供する。該ベクターは、核酸配列をベクターにクローニングするの
に必須の制限部位を除いて、たとえばベクターシステム内においてユニークであり、ポリ
ペプチドをコードする核酸配列に組み込まれた制限部位に対して本質的にユニークである
制限部位を含む。
【0071】
別の態様において、本発明は、核酸配列の一以上のリスト、組換えベクター、ポリペプ
チド、および上記方法に従ったベクター、およびたとえばポリペプチドを産生するための
適切な宿主細胞を含むキットを提供する。
【0072】
本発明の上記態様のすべては、当業者に公知の分子生物学の標準的技術を用いて実施す
ることができる。
【0073】
別の態様において、核酸配列は、本発明のポリペプチドをコードすることができる任意
の配列である。
【0074】
別の態様において、本発明の核酸は、遺伝子治療において使用される。
【0075】
別の態様において、単一鎖フレームワークは、配列1.1, 2.1, 3.1, 4.1, 5.1, 1.2, 2.
2, 3.2, 4.2, 5.2,1.3, 2.3, 3.3, 4.3, 5.3, 7.3, 1.4, 2.4, 3.4, 4.4, 5.4, 6.4(表
5)の何れか一つの変異体であり、本明細書で使用される「変異体」は90%以上の同一性
を示し、増大した安定性を維持する配列をいう。
【0076】
別の態様において、単一鎖フレームワークは、配列1.1, 2.1, 3.1, 4.1, 5.1, 1.2, 2.
2, 3.2, 4.2,5.2, 1.3, 2.3, 3.3, 4.3, 5.3, 7.3, 1.4, 2.4, 3.4, 4.4, 5.4, 6.4(表
5)の何れか一つの誘導体であり、本明細書で使用される「誘導体」は、分子の機能およ
び安定性に重要なそれらのアミノ酸のみを維持する配列をいう。例3に記載されるフレー
ムワークにおけるニュートラルもしくはポジティブな単離された交換は、本発明の抗体フ
レームワークに関連のある変化とは考えられない。
【0077】
本発明の好ましい態様において、単一鎖フレームワークは、細胞内アッセイの読出し部
(aread-out)を提供する第二のタンパク質に融合される。読出し部は、直接的であり得
、たとえば、検出可能なタンパク質(たとえば、蛍光により観察可能である、GFP(緑色
蛍光タンパク質)、enhanced青色蛍光タンパク質、enhanced黄色蛍光タンパク質、enhanc
edシアン蛍光タンパク質)、または様々な検出方法による他の融合パートナーとの融合体
の形態であり得る。あるいは、読出し部は、レポーター遺伝子の転写活性化により達成す
ることができ、ここでscFv融合タンパク質の融合パートナーは、転写アクチベーター、た
とえばGal4活性化ドメイン、またはDNA結合タンパク質、たとえばlexA−もしくはGal4 D
NA−結合ドメインの何れかであり、これは、酵素(β−ガラクトシダーゼ、ルシフェラー
ゼ、α−ガラクトシダーゼ、β−グルクロニダーゼ、クロラムフェニコールアセチルトラ
ンスフェラーゼなど)のレポーター遺伝子の転写を活性化し、これが、順に読出し部(a
read out)を提供する。読出し部を提供する融合タンパク質は、当業者に周知である。
【0078】
本発明の別の態様は、本明細書に記載されるフレームワークを含む抗体である。
【0079】
本発明の別の態様は、本発明の抗体の使用である。
【0080】
本発明の更に好ましい態様は、還元環境または他の挑戦的環境において機能的である抗
体を得るために、現存の抗体から超可変ループを移植するための、抗体可変ドメインの記
載されるフレームワーククラス、並びに可変ドメインおよびscFvsの配列の使用である。
【0081】
本発明の更に好ましい別の態様は、たとえばフレームワークの一以上の超可変ループの
ランダム化による、還元環境または他の挑戦的環境における適用のためのライブラリーを
作成するための、抗体可変ドメインの記載されるフレームワーククラス、並びに可変ドメ
インおよびscFvsの配列の使用である。
【0082】
当業者に明らかであるとおり、本明細書に記載される本発明の分子は、診断および治療
適用、ターゲットの検証、および遺伝子治療において使用することができる。
【0083】
本発明は、以下の実施例により詳説され得るが、以下の実施例は、本発明の範囲を限定
するためのものではない。
(参考文献)
【0084】
本発明は、以下の非限定的な実施例において更に詳説される。
【0085】
例1
酵母において「クオリティーコントロール」システムを用いてヒトライブラリーをスク
リーニングすることによるイントラボディフレームワークの選択
安定なフレームワークのため「クオリティーコントロール」システムを用いたスクリー
ニングを、Aufder Maur (WO0148017、Auf der Maur 2001、それぞれ参照により本明細書
に組み込む) に詳細に記載されるとおり基本的に実施した。
【0086】
酵母でスクリーニングするためのscFv融合構築物の発現用プラスミドは、pESBA-Actか
ら得た(Worn, 2000)。それは、S.cerevisiaeにおける形質転換選択のための酵母TRP1遺
伝子、および高いコピー数を保証するための2ミクロン複製起点を含有する。更に、強力
な発現のための構成的アクチンプロモーターおよびGAL11転写終結配列を、マルチプルク
ローニングサイトにより隔てられて有する。細菌システムで扱うために、更に、細菌の複
製起点およびamp抵抗性遺伝子を有する。
【0087】
Gal4活性化ドメイン(ADアミノ酸768-881)は、pGAD424(Clontech)を鋳型として用い
て、Gal4-ADに対するSV40 T-抗原核局在シグナルN-末端を含むプライマーでPCRにより初
めに増幅した。Gal11Pのアミノ酸263-352をコードするDNAフラグメントを、PCRにより増
幅し、SV40-NLS-Gal4-AD-構築物に対するN-末端にフレームで(inframe)クローニング
した。他に記載されるとおりヒト脾臓細胞cDNAから増幅したヒトscFvライブラリー(Wel
schhof, 1995;Krebber, 1997; de Haard, 1999)を、StiI-部位を介してこの融合構築物
に対するN-末端に、VL-リンカー-VH(ここでリンカーは配列(GGGS)4を有する)の向き
で、フレームで(inframe)クローニングした。かかる発現により、一般構造scFv-Gal11
P-SV40 NLS-Gal4ADの融合タンパク質が産生される。
【0088】
スクリーニングは、異なる向きのLacZおよびHIS3レポーター遺伝子を、his3Δ200遺伝
子座へ、Gal1-GAL10調節配列由来の天然UASGの制御下に組み込むことによりJPY9株(Esch
er, 2000)から誘導した酵母株、S.cerevisiae YDE172 (MATα ura3-52 leu2Δ1 trp1d6
3 his3Δ200 lys2Δ385 gal4Δ 11) (Auf der Maur, 2001) で行った。レポーターシス
テムの転写活性化は、Gal11P部分とGal4-DNA結合ドメイン(DBD、アミノ酸1-100)との特
異的相互作用に続いて、scFv-融合構築物のGal4-AD部分により媒介される。Gal4-DBDは、
第二のプラスミドpMP83からの発現により提供される。それは、S.cerevisiaeにおける形
質転換選択のための酵母LEU2遺伝子およびARS CEN複製起点を含有する。更に、強力な発
現のための構成的アクチンプロモーターおよびGAL11転写終結配列を有する。細菌システ
ムで扱うために、更に、細菌の複製起点およびamp抵抗性遺伝子を有する。
【0089】
スクリーニングのために、酵母株S. cerevisiae YDE172を、pESBA-Act2ベクター上の
融合構築物としてのscFv-ライブラリーを用いて、一方Gal4-DBDを提供するpMP83ベクター
を用いて同時形質転換した(co-transform)。標準的な酢酸リチウム形質転換プロトコー
ルを使用した(Agatep,1998)。形質転換の後、細胞を、80 mM 3-アミノトリアゾールを
含有するドロップアウトプレート(-Trp/-Leu/-His)に播いた(plate)。30℃で3日間イ
ンキュベートした後コロニーを採取し、80 mM 3-アミノトリアゾールを含有するドロップ
アウトプレート(-Trp/-Leu/-His)に再び播いた(streak)。再度増殖した細胞を、基質
X-Galを含有するプレート上でのフィルターアッセイで、青色の発色によりLacZ発現につ
いて試験した。ポジティブクローンを、更なる解析のために採取し、該解析は、酵母から
のscFv-担持プラスミドの単離、E.coli DH5αへの形質転換、E.coliの単一コロニーか
らのプラスミドの単離、および後述のアッセイのために新たに調製した酵母株S. cerevis
iae YDE172への再形質転換を伴う。すべての方法は、当業者に周知の標準的手法に従っ
て行われた。
【0090】
更に、改変されたスクリーニング手法を使用し、scFvを、DNA結合ドメイン(LexAアミ
ノ酸1-202)と活性化ドメイン(Gal4、アミノ酸768-881)の両方に直接融合し、以下の構
造の融合構築物を産生した:scFv-LexA-NLS-Gal4AD。酵母におけるスクリーニングのため
のscFv融合構築物の発現用プラスミドは、pESBA-Act2から誘導した。それは、S.cerevisi
aeにおける形質転換選択のための酵母TRP1遺伝子、および高いコピー数を保証するための
2ミクロン複製起点を含有する。更に、(強力な発現のための)構成的アクチンプロモー
ターおよびGAL11転写終結配列を、マルチプルクローニングサイトにより隔てられて有す
る。細菌システムで扱うために、更に、細菌の複製起点およびamp抵抗性遺伝子を有する
。
【0091】
スクリーニングは、異なる向きのLacZおよびHIS3レポーター遺伝子を、his3Δ200遺伝
子座へ、6つのLex-A結合部位を有する二方向性プロモーターの制御下に組み込むことによ
り(インテグレイティングレポータープラスミド pDE200, Escher 2000)、そしてLEU2レ
ポーター遺伝子を、Leu2Δ1遺伝子座へ、(EGY48由来の)8つのLexA結合部位を有するプ
ロモーターの制御下に組み込むことにより、JPY5株から誘導した酵母株、S. cerevisiae
ImmunaLHB (MATαura3-52 leu2Δ1 trp1d63 his3Δ200 lys2Δ 385) で行った。レポー
ターシステムの転写活性化は、scFv融合構築物のGal4-AD部分により媒介される。スクリ
ーニングは、ドロップアウト培地(-Trp/-Leu/-His)および40 mMまでの3-アミノトリゾ
ール濃度を用いて基本的に上述のとおり行った。
【0092】
例2
インビボ性能の評価
a)酵母において
酵母において選択フレームワークの性能を定量分析するため(図1および3)、S.cere
visiae株ImmunaLHBを、標準的な酢酸リチウム形質転換プロトコールに従うことにより、
pESBA-Act2ベクター上のLexA-Gal4-AD-融合構築物としての単離scFvsを用いて形質転換し
た(Agatep,1998)。形質転換の後、細胞をドロップアウトプレート(-Trp)に播いた(
plate)。ドロップアウト培地(-Trp)中の2mL一晩培養物を、幾つかのコロニーを含有
する培養物(streaks)から2回接種し、30℃で生育させた。培養物を、600nm(OD600)
における光学濃度が0.7になるように、1mLドロップアウト培地(-Trp)で希釈した。そ
の後、30℃で2時間生育させた。アッセイのために、100μLの細胞培養物を採取し、900μ
Lバッファー、45μLクロロホルムおよび30μL 0.1%SDSと混合し、ボルテクスにかけ、
室温で5分間インキュベートした。0.2mL ONPG(4 mg/mL)の添加により発色を開始し、
0.5 mL Na2CO3(1M)で停止させた。アッセイ培養物のOD600、並びに使用した発色のイ
ンキュベーション時間および培養体積を考慮することにより活性を計算した。
【0093】
ポジティブコントロール(以前に記載された非常に安定なラムダ−グラフト(Worn, 20
00; Auf derMaur, 2001))と少なくとも同等、またはそれより優れたクローンを、配列
決定し、フレームワークサブタイプを同定した(Tomlinson, (1992), Cox,(1994) and Wi
lliams, (1996)に従ったフレームワークサブタイプの定義)。配列決定により、フレーム
ワークサブタイプの顕著に好ましいものが明らかにされた。重鎖可変ドメイン(VH)に関
して、フレームワークサブタイプ2および6は見られず、4はポジティブクローンのなかで
著しく減少していた。酵母細胞内アッセイにおいて、単離された配列の性能を修正すると
、サブタイプ3のVHフレームワークだけでなく、1aおよび1bも、細胞内適用において非常
に強力に好ましい。軽鎖可変ドメイン(VL)に関しては、カッパ1、ラムダ1、およびラム
ダ3サブタイプのフレームワークが明らかに好ましい。(表4)。
【0094】
したがって、これらフレームワークのサブタイプ、すなわちカッパ1、ラムダ1およびラ
ムダ3 VLドメインと組み合わせたVH1a、1bおよび3が、細胞内使用およびscFvの折りたた
み特性に関して厳格な要件を有する他の適用に最も適合している。細胞内スクリーニング
システムのためのライブラリーは、たとえば、これらフレームワークのサブタイプのみの
混合物から優先的に構築し、還元環境において機能的でない分子の量を低減すべきである
。
【0095】
b)哺乳類細胞において
ヒト細胞において、選択したフレームワークの性能を定量的に分析するためにHela細胞
株を使用した(図2、4および7)。ルシフェラーゼレポーター遺伝子は、天然Gal4 UA
Sの制御下にルシフェラーゼを含有する、同時トランスフェクトされるpGL3(Promega)レ
ポータープラスミドから提供された。一過性トランスフェクションのために使用される哺
乳類発現ベクターは、CMVプロモーターの制御下に、VP16-ADに対してC末端で融合したGa
l4(1-147)を含有する。単離されたscFvsを、Gal4(1-147)-VP16-融合物に対してC末端
にフレームでクローニングし、発現によりGal4(1-147)-VP16-scFv-融合タンパク質を産生
した。細胞を、2.5% FCSおよび2mM l-グルタミンを添加したDMEM中で培養した。一過
性トランスフェクションは、Polyfect-protocol (Qiagen) に従って、60 mm組織培養プレ
ートで、scFv構築物を含有する0.01-0.1μgのベクター、0.5μgのCMVプロモーター誘導Ga
l4(1-147)-VP16-scFv発現プラスミド、およびトランスフェクション効率のためのレファ
レンスとして0.5μgのLacZ発現ベクターを用いて行った。細胞を、トランスフェクション
から24〜48時間後に集め、1000μLバッファー中に再懸濁し、3回の凍結−融解サイクルに
より溶解させた。細胞溶解物を、遠心分離し、ルシフェラーゼアッセイ溶液(Promega)
を用いてルシフェラーゼ活性について、および標準的プロトコールに従ってLacZ活性につ
いて、上清をアッセイした。得られたルシフェラーゼ活性は、LacZ活性を用いて修正し、
トランスフェクション効率におけるばらつきを説明した。
【0096】
例3
マルチプルアラインメントおよび配列比較の分析
細胞内適用に適したフレームワーク配列の一般的パターンを解明するため、すべてのポ
ジティブクローン(すなわち、クオリティーコントロールシステムにおいて選択条件下で
生育したクローン)を単離し、scFvsをコードする部分を配列決定した。その後、scFv配
列を、軽鎖および重鎖の構成要素に分け、Honegger (2001) による免疫グロブリンドメイ
ンの構造調整ナンバリングスキームに従って、各ドメイン(表1および2)のアラインメ
ントを可能にした。
【0097】
得られたデータを評価するため、非選択ライブラリーを表すアラインメントを作成した
(表3)。非選択配列を得るために、ライブラリーを、scFv遺伝子を発現しないE. coli
細胞に形質転換し、プラスミド単離およびscFv配列決定のために、クローンをランダムに
採取した。ライブラリーは、予想されるとおりヒト抗体レパートリーを包含し、ヒトで一
般に見出される発現パターンにより予想される以外、特定のサブグループに対する偏りは
みられない。
【0098】
VHおよびVL配列を、サブグループに従ってグループ分けした。サブグループ特異的コン
センサス配列への変化が強調された。当業者であれば、特定の交換残基の構造的環境に基
づいて、ポジティブ、ニュートラルおよびネガティブ変化を識別することができる(たと
えば、Honegger,2001)。特定グループのアミノ酸に属する残基の同一グループの残基へ
の交換は、ニュートラルな交換として一般に認められる。タンパク質の疎水性コアに位置
する疎水性アミノ酸のグループに属する残基の、極性を有するが非電荷または正もしくは
負電荷のアミノ酸のグループの一のアミノ酸への交換は、不満足な水素ドナー/アクセプ
ター部位が疎水性コアの堅固なパッキングを乱すため非常に好ましくない。したがって、
かかる変化は、ネガティブと考えられる。免疫グロブリンドメインの表面における極性を
有するが非電荷の残基のグループに属する残基の、正もしくは負電荷の残基のグループの
アミノ酸への交換は、タンパク質の溶解度を増大させるため非常に好ましい。したがって
、かかる変化は、ポジティブと認められるが、極性から疎水性残基への交換は、タンパク
質の溶解度を減少させるため非常に好ましくないため、ネガティブと認められる。保存さ
れたポジティブなファイアングル(phi-angle)を有する位置において、任意のアミノ酸
のグリシンへの交換は、ポジティブと認められるが、グリシンの任意のアミノ酸への交換
は、ネガティブと認められ、これは、グリシンが、ポジティブなファイアングルを形成す
ることができる唯一のアミノ酸であるためである。正もしくは負電荷の残基のグループの
アミノ酸から非電荷のアミノ酸への交換による、45-53、45-100、77-100および108-137位
置の間の保存された塩橋の損失は、熱力学的安定性の低下につながるため、ネガティブと
考えられる。
【0099】
最後に、我々は、クオリティーコントロールの間に優先的に選択され(すなわち、コン
センサス配列から最小のネガティブと最大のポジティブ交換を示して、該サブグループに
当てはまり)、それぞれが酵母において高いインビボ性能を示す、7つのVLドメインおよ
び4つのVHドメインを選択した。その配列は、表5に要約され、2つのVκ1(k I 27 (1. x
) および k III 25(2. x))、2つのVκ3 (k IV 103 (3. x) および k IV135 (5. x))、1
つのVλ1 (kIV 107 (4. x))、2つのVλ3 (a33 (7. x) および a43 (6. x))、1つのVHlb
(a33 (x. 3))、および3つのVH3(a fw10 (x. 2), a43 (x. 4) および a44 (x. 1)) を含
む。これらVLおよびVHドメインは、シャッフルされ、scFvフォーマットにおいて22の新規
な組み合わせ(1.1, 2.1, 3.1, 4.1, 5.1, 1.2, 2.2, 3.2, 4.2, 5.2, 1.3, 2.3, 3.3, 4
.3, 5.3, 7.3,1.4, 2.4, 3.4, 4.4, 5.4, 6.4) を提供した。
【0100】
例4
シャッフルしたドメインのインビボ性能の評価
a)酵母および哺乳類細胞における細胞内アッセイでの性能
22の組み合わせを、例2に記載されるとおり、酵母および哺乳類細胞においてインビボ
性能について試験した(図3および4)。
【0101】
b)酵母における還元条件下での可溶性タンパク質の発現
還元条件下での発現による可溶性タンパク質の産生量を比較するため、選択されたフレ
ームワークを、酵母S.cerevisiaeの細胞質において、Gal4 ADとの融合体として発現させ
た。pESBA-Act2ベクター上の融合構築物は、一般構造Gal4 AD-scFvを有していた。その
融合構築物を、上述のとおり酵母S.cerevisiae株JPY9に形質転換し、-Trp, ドロップアウ
トプレートに播いた。
【0102】
ドロップアウト培地(-Trp)における5 mL一晩培養物を、幾つかのコロニーを含有する
培養物(streaks)から接種し、30℃で生育させた。培養物を、600nm(OD600)における
光学濃度が0.5になるように、50mLドロップアウト培地(-Trp)で希釈した。その後、30
℃で5時間生育させた。ネガティブ細胞抽出物については、OD600を3に規格化した2.5mL
細胞培養物を、遠心分離により収集し、液体窒素で凍結し、その後、プロテアーゼ阻害剤
(PMSF)を含有する75μL Y-PER(Pierce)に再懸濁した。再懸濁した細胞の沈殿物を、
短時間ボルテクスにかけ、20℃で20分(わずかに振盪して)インキュベートした。不溶性
の凝集物を、4℃で10分間、エッペンドルフ遠心分離機において最大速度で沈殿させた。
上清をローディング色素と混合し、5分間100℃に加熱し、12% SDS-PAGEで分離した。可
溶性Gal4 AD-scFv融合構築物を、一次抗体として抗Gal4ADモノクローナルマウス抗体(S
anta CruzBiotechnology)、二次抗体として抗マウスペルオキシダーゼコンジュゲート
(Sigma)を用いて、化学発光基質(Pierce)を用いて、Gal4部分の検出を介したウェス
タンブロッティングにより視覚化した(図5)。SDS-PAGEおよびウェスタンブロッティン
グの手法は、当業者に周知である。
【0103】
c)E.coliのペリプラズムにおける発現行動
E.coliにおけるペリプラズム発現行動を評価するため(図6)、単離されたscFvs−フ
レームワークを、cam抵抗性遺伝子(catR)およびlacIリプレッサー遺伝子(Krebber,19
97)を担持する細菌ベクターに、N-末端のpelBリーダー配列およびC-末端のhis-tagを備
えて、lacプロモーター/オペレーターの制御下にクローニングした。コンピーテントE.co
li JM83を、これらプラスミドを用いて形質転換した。振盪フラスコ中で、35mg/Lクロ
ラムフェニコールを含有する50 mL dYT培地に、一晩培養物を1:40で接種し、30℃でイン
キュベートした。細胞を、1 mM IPTGを用いてOD600 0.8に誘導し、遠心分離による導入
の3時間後に収集した。沈殿を、50mM Tris, pH 7.5、500 mM NaClに再懸濁し、OD600を1
0に規格化した。各scFvフラグメントのサンプルを、直接(全抽出物)もしくは遠心分離
に続く音波処理の後(可溶性フラクション)に、SDS-PAGEにより分析した。可溶性タンパ
ク質の量は、クマシー染色されたゲルから評価した。
【0104】
例5
細胞外使用に関して優れた特性を有する5つの組み合わせの詳細な評価
酵母および哺乳類の細胞内アッセイの両方において優れた性能を示し、酵母およびE.co
liにおいて発現の間に可溶性タンパク質を産生し、クオリティー・コントロールの間に優
先的に選択されたサブグループに当てはまる、5つの組み合わせを例として選択した(2.4
, 4.4, 5.2, 6.4および7.3、詳細については表5参照)。我々は、これら組み合わせをよ
り詳細に分析し、還元条件および酸化条件下におけるそれらの使用を更に評価した。
【0105】
a)様々な哺乳類細胞における細胞内アッセイでの性能
ヒト細胞における5つの組み合わせの性能の定量分析は、例2で実施したとおり、Hela
細胞、ヒト骨肉腫細胞株Saos-2およびヒト胚性腎細胞株HEK293を用いて行った(図7)。
【0106】
b)インビトロにおける性能
発現および精製
インビトロ性能を評価するため、5つの優れた組み合わせを、E.coliのペリプラズムで
発現させた(図6)。振盪フラスコ中の35 mg/Lクロラムフェニコールを含有する0.1 L
dYT-培地に、一晩培養物を1:40で接種し、30℃でインキュベートした。細胞を、1mM IPT
Gを用いてOD550 1.5に誘導し、遠心分離による導入の2時間後に収集した。scFvsを精製
するため、細胞の沈殿物を再懸濁し、音波処理により溶解させた。20 krpm、4℃で30分間
、SS34で遠心分離した後、上清を、Ni-MC-アフィニティーカラム(Hi-TrapTM Chelating
HP, 1ml,Amersham Pharmacia) にpH 7.5で適用し、Amersham PharmaciaからのAkta Basi
c systemを用いて、200mMイミダゾールで溶出した。scFvフラグメントの純度は、SDS-PA
GEにより測定すると98%より高かった(データ示さず)。精製されたタンパク質の濃度は
、280 nmにおける計算減衰係数を用いて決定した。可溶性精製タンパク質の収率は、OD60
0 10の1 L培養体積に規格化し、8から55mgまで変動した。
【0107】
凝集に対する抵抗性
凝集に対する抵抗性は、インビトロにおける熱力学的安定性(Worn, 1999)およびマウ
スの異種移植腫瘍モデルにおける腫瘍局在の効率(Willuda, 1999)と関係することが示
された。安定性、凝集に対する抵抗性、およびアンフォールディング(unfolding)の可
逆性を試験するために、50 mM Tris, pH 7.5, 100 mM NaCl中、6μMの濃度の精製タンパ
ク質の200μLサンプルを、4℃で4日間または37℃で4日間または4℃で3日間維持し、その
後、100℃で15分または60分インキュベーションし、室温までゆっくり冷まし、4℃で一晩
インキュベートした。その後、各サンプルのオリゴマー状態を、50 mM Tris, pH 7.5, 10
0 mM NaClを用いて平衡化したゲルろ過カラムで分析し、モノマー物質に対して凝集した
量を評価した(図8)。タンパク質を、Akta Basicシステム(Amersham Pharmacia)で、
流速1 mL/分、100μLの体積で、Superdex-75カラム(AmershamPharmacia)上に注入した
。
【0108】
プロテアーゼ分解に対する抵抗性
治療適用にとって重要なパラメーターである、プロテアーゼ分解に対する単離フレーム
ワークの安定性を決定するため、我々はヒト血清中で37℃において精製フレームワークを
インキュベートした(図9)。
【0109】
50μMの濃度の精製his-タグscFvタンパク質(上記参照)を、ヒト血清に10倍希釈し、
最終濃度を90%血清中5μMとした。その後、サンプルを、37℃で3日もしくは1日インキュ
ベートするか、またはローディングのために直接採取した。ローディングの前に、不溶性
の凝集物質を、4℃で10分間、エッペンドルフ遠心分離機において最大速度で沈殿させた
。ゲルにロードされる血清の量を減らすため上清をローディング色素で6倍に希釈し、5分
間100℃に加熱し、12%SDS-PAGEで分離した。可溶性his-タグscFvフラグメントを、一次
抗体として抗hisモノクローナルマウス抗体(Qiagen)、二次抗体として抗マウスペルオ
キシダーゼコンジュゲート(Sigma)を用いて、化学発光基質(Pierce)を用いて、his-
タグの検出を介したウェスタンブロッティングにより視覚化した。SDS-PAGEおよびウェス
タンブロッティングの手法は、当業者に周知である。
【0110】
例6
酵母における相互作用スクリーニングシステムでの、フレームワーク7.3上のランダム
化CDRライブラリーのスクリーニングを介した抗原バインダーの選抜
抗原バインダーの相互作用システムを用いたスクリーニングは、以前に詳細に記載され
るとおりに本質的に実施した(Auf der Maur, 2002)。
【0111】
酵母でスクリーニングするためのscFv融合構築物の発現用プラスミドは、pESBA-Act2か
ら誘導した。それは、酵母TRP1栄養性マーカーおよび2ミクロン複製起点を含有する。更
に、強力な発現のための構成的アクチンプロモーターおよびGAL11転写終結配列を、マル
チプルクローニングサイトにより隔てられて有する。細菌システムで扱うために、更に、
細菌の複製起点およびamp抵抗性遺伝子を有する。
【0112】
Gal4活性化ドメイン(ADアミノ酸768-881)は、pGAD424(Clontech)を鋳型として用い
て、Gal4-ADに対するSV40 T-抗原核局在シグナルN-末端を含むプライマーでPCRにより初
めに増幅した。scFvライブラリーは、VHのCDR3内の7アミノ酸をランダム化するプライマ
ーを用いてscFv-フレームワーク7.3のPCR増幅により得た。得られたPCR産物は、フレーム
ワーク7.3にクローンニングされ、Gal4-ADに対するC-末端融合体として、VL-リンカー-VH
の向きでベクターに存在する。かかる発現により、一般構造Gal4-AD-scFvの融合タンパク
質が産生される。
【0113】
スクリーニングは、酵母株、S. cerevisiae Immuna LHB (MATα ura3-52 leu2Δ1 trp1
d63 his3Δ200lys2Δ 385) で行った。その酵母株は、異なる向きのLacZおよびHIS3レポ
ーター遺伝子を、his3Δ200遺伝子座へ、6つのLexA結合部位を有する二方向性プロモータ
ーの制御下に組み込むことにより(インテグレイティングレポータープラスミド pDE200,
Escher2000)、そしてLEU2レポーター遺伝子を、leu2Δ1遺伝子座へ、(EGY48由来の)
8つのLexA結合部位を有するプロモーターの制御下に組み込むことにより、JPY5株から誘
導した。
【0114】
レポーターシステムの転写活性化は、そのscFv部分とbait−融合タンパク質の抗原部分
との特異的相互作用に続いて、scFv融合構築物のGal4-AD部分により媒介される。bait−
融合タンパク質は、DNA結合LexAタンパク質にC末端で融合したヒトpolo様キナーゼ1(hpl
k1-KD)のキナーゼドメインから成る。キナーゼドメイン(アミノ酸2-332)は、上流プラ
イマー5'-tgctctagaagtgctgcagtgactgcag-3' (配列番号12) および下流プライマー 5'
-ggttgtcgacttacaggctgctgggagcaatcg-3'(配列番号13) を用いて、hPlk1 cDNAからPC
R増幅した。得られたPCR産物は、baitベクターに、XbaIおよびSalIを介してLexAのC末端
にクローニングした。baitベクターは、URA3栄養性マーカーおよびArs Cen複製起点を含
有する。bait−融合タンパク質の発現は、構成的に活性なアクチンプロモーターにより誘
導される。転写は、GAL11終結配列により終結される。また、baitベクターは、細菌シス
テムでの繁殖のために、細菌の複製起点およびamp抵抗性遺伝子を有する。
【0115】
スクリーニングのために、酵母株S. cerevisiae Immuna LHBを、標準的な酢酸リチウ
ム形質転換プロトコール(Agatep, 1998)に従って、pESBA-Act2ベクター上のGal4-ADと
の融合物としてのscFv-ライブラリーを用いて、およびLexA-hPLK1-KD融合物を提供するba
itベクターを用いて同時形質転換した(co-transform)。形質転換の後、細胞を、ドロッ
プアウトプレート(-Trp/-Leu/-Ura)に播いた(plate)。30℃で3〜5日間インキュベー
トした後コロニーを採取し、ドロップアウトプレート(-Trp/-Leu/-Ura)に再び播いた(
streak)。再度増殖した細胞を、基質X-Galを含有するプレート上でのフィルターアッセ
イで、青色の発色によりLacZ発現について試験した。ポジティブクローンを、更なる解析
のために採取し、該解析は、酵母からのscFv-担持プラスミドの単離、E.coli DH5αへの
形質転換、E.coliの単一コロニーからのプラスミドの単離、および後述のアッセイのた
めに新たに調製した酵母株S. cerevisiae Immuna LHBへの再形質転換を伴う。すべての
方法は、当業者に周知の標準的手法に従って行われた。
【0116】
例7
新規scFvフレームワークから誘導したFab構築物のインビボ性能の評価
様々な抗体フォーマットにおいて安定な可変ドメインフレームワークを使用する有利な
効果を評価するため、酵母相互作用スクリーニングで使用するためのFab発現ベクターを
構築した。
【0117】
a)酵母における細胞内スクリーニングのためのFab構築物
2種類の発現ベクターを構築し、異なる発現レベルを可能にした。ベクターは、yEplac
112(2ミクロン)またはyCplac22(ars/cen)バックボーンのいずれかに基づくものであ
る(Gietz,1998)。いずれのベクターも、細菌システムで扱うために、酵母TRP1栄養性
マーカー、誘導性二方向性Gal1/Gal10プロモーター、細菌の複製起点およびamp抵抗性遺
伝子を含有する。一方向において、フレームワーク7.3のVHドメインを、C末端システイ
ンを含むIgG1のCH1-ドメインに対してN末端にクローニングし、その後に、リンカー、お
よびSV40 T-抗原を含むGal4活性化ドメイン(ADアミノ酸768-881)が続く。一方、フレ
ームワーク7.3のVLドメインを、C末端システインを含むCL(ラムダ)ドメインに対して
N末端にクローニングした。ターミネーターは、重鎖の側ではGal11ターミネーターであ
り、軽鎖の側ではサイクリン1ターミネーターである。
【0118】
b)酵母における細胞内アッセイでの性能
酵母においてscFvおよびFabフォーマットにおける抗原バインダーの性能を定量的に分
析するため(図1および3)、S. cerevisiae株Immuna LHBを、標準的な酢酸リチウム形
質転換プロトコールに従うことにより、pESBA-Act2ベクター上のGal4-AD-融合構築物とし
ての単離scFvsと、LexA-hPLK1-KD融合物を含有するbaitベクターとを用いて同時形質転換
した(Agatep,1998)。形質転換の後、細胞をドロップアウトプレート(-Trp、-Ura、Gl
c)に播いた。ドロップアウト培地(-Trp、-Ura、Glc)中の2mL一晩培養物を、幾つかの
コロニーを含有する培養物(streaks)から2回接種し、30℃で生育させた。培養物を、60
0 nm(OD600)における光学濃度が0.7になるように、1mLドロップアウト培地(-Trp、-U
ra、Gal)で希釈した。それを30℃で5時間生育させた。アッセイは上述のとおり行った。
【0119】
c)酵母における還元条件下での可溶性タンパク質の発現
還元条件下での発現による可溶性タンパク質の産生量を比較するため、scFvとFab構築
物を、hPLK1-KD-baitベクターと一緒に、上述のとおり、酵母S.cerevisiaeの細胞質にお
いて発現させた。それを、上述のとおり酵母株YDE173に形質転換し、グルコースを含有す
る-Trp、-Ura、ドロップアウトプレートに播いた。
【0120】
ドロップアウト培地(-Trp、-Ura、Glc)における5 mL一晩培養物を、幾つかのコロニ
ーを含有する培養物(streaks)から接種し、30℃で生育させた。培養物を、600 nm(OD6
00)における光学濃度が0.5になるように、YPAGで希釈した。その後、30℃で7.5時間生育
させた。ネガティブ細胞抽出物については、OD600を3に規格化した2.5 mL細胞培養物を、
遠心分離により収集し、液体窒素で凍結し、その後、75μL Y-PER(Pierce)に再懸濁し
た。再懸濁した細胞の沈殿物を、短時間ボルテクスにかけ、20℃で20分わずかに振盪させ
てインキュベートした。その後、不溶性の凝集物を、4℃で10分間、エッペンドルフ遠心
分離機において最大速度で沈殿させた。上清をローディング色素と混合し、5分間100℃に
加熱し、12%SDS-PAGEで分離した。可溶性Gal4-AD-scFv融合物およびGal4-ADに融合した
Fabの重鎖部分を、一次抗体として抗Gal4-ADモノクローナルマウス抗体(SantaCruz Bio
technology)、二次抗体として抗マウスペルオキシダーゼコンジュゲート(Sigma)を用
いて、化学発光基質(Pierce)を用いて、Gal4部分の検出を介したウェスタンブロッティ
ングにより視覚化した(図11)。SDS-PAGEおよびウェスタンブロッティングの手法は、
当業者に周知である。
【0121】
表1は、酵母における種々の「クオリティーコントロール」スクリーニングから選択さ
れた全てのVHドメインフレームワーク配列のアラインメントを示す。
【表1】
【0122】
表2は、酵母における種々の「クオリティーコントロール」スクリーニングから選択さ
れた全てのVLドメインフレームワーク配列のアラインメントを示す。
【表2】
【0123】
表3は、ライブラリーからランダムに選ばれた配列のアラインメントを示す。
【表3】
【0124】
表4は、「クオリティーコントロール」システムを用いて単離された配列におけるVH-
およびVL-ドメインのサブグループ頻度の統計解析を示す。定量的な酵母アッセイでポジ
ティブであることがその後に見出された配列のみを考慮した。選択された配列を、限られ
た数のランダム配列(表3)から決定される非選択ライブラリーと比較する。
【表4】
【0125】
表5は、更なる組換えおよびscFvsの最良の組み合わせの評価のために使用した配列、
並びにそれらの各略語(abb.)、供給源およびサブファミリーを示す。
【表5−1】
【表5−2】
【特許請求の範囲】
【請求項1】
本願明細書に記載された発明。
【請求項1】
本願明細書に記載された発明。
【図1】

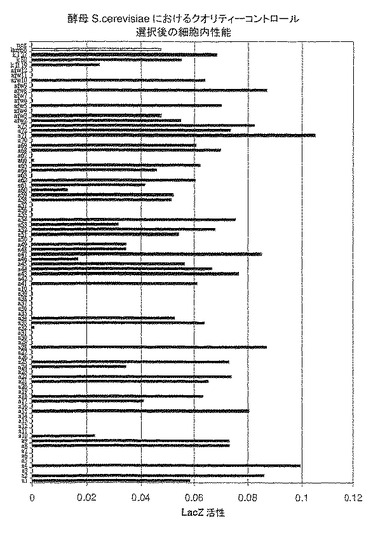
【図2】
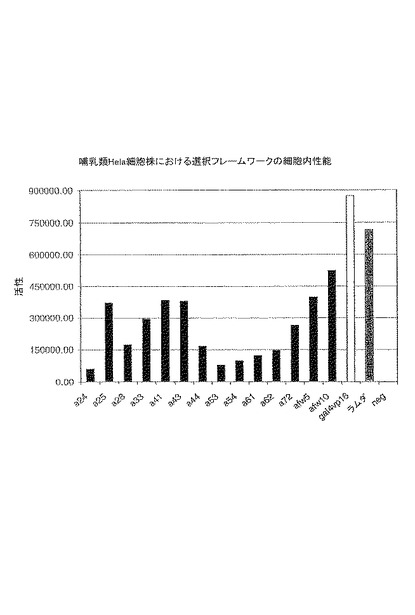
【図3】
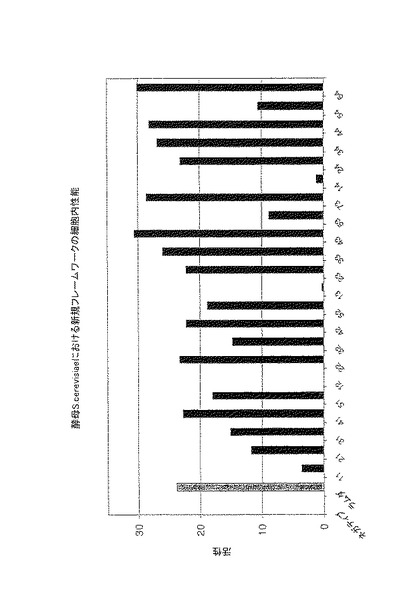
【図4】
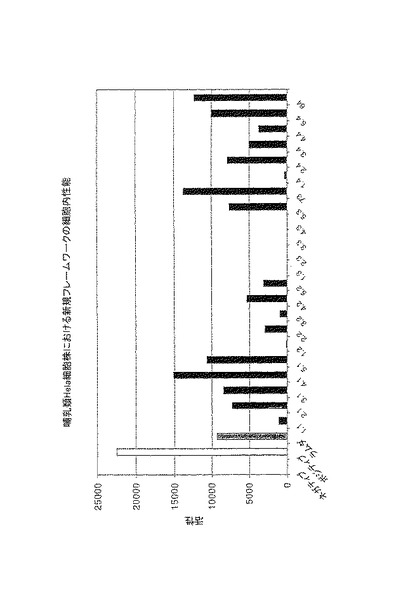
【図5】

【図6】
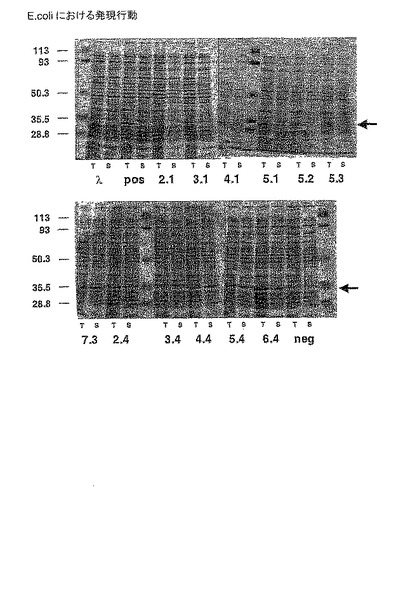
【図7】
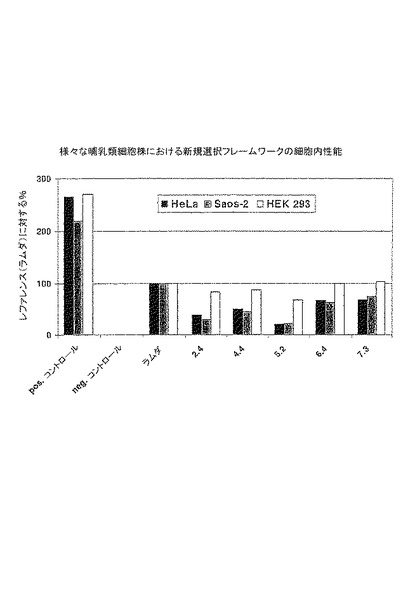
【図8】
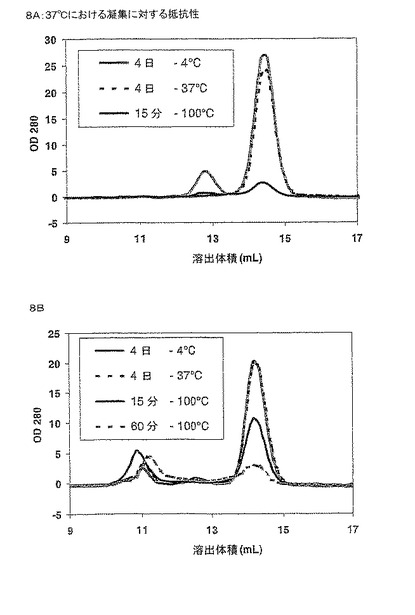
【図9】

【図10】
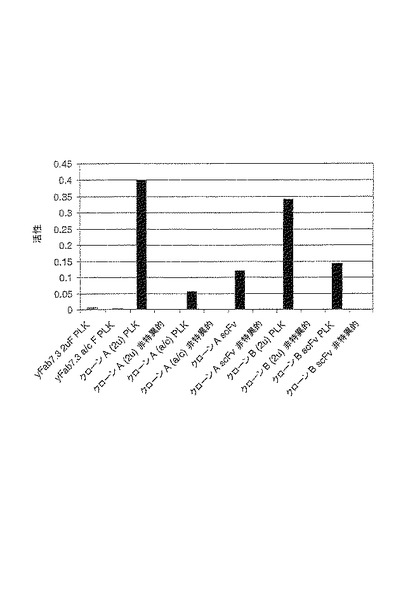
【図11】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2012−228272(P2012−228272A)
【公開日】平成24年11月22日(2012.11.22)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−187518(P2012−187518)
【出願日】平成24年8月28日(2012.8.28)
【分割の表示】特願2010−60713(P2010−60713)の分割
【原出願日】平成15年5月21日(2003.5.21)
【出願人】(502233344)エスバテック、アン アルコン バイオメディカル リサーチ ユニット、エルエルシー (19)
【氏名又は名称原語表記】ESBATech, an Alcon Biomedical Research Unit, LLC
【Fターム(参考)】
【公開日】平成24年11月22日(2012.11.22)
【国際特許分類】
【出願番号】特願2012−187518(P2012−187518)
【出願日】平成24年8月28日(2012.8.28)
【分割の表示】特願2010−60713(P2010−60713)の分割
【原出願日】平成15年5月21日(2003.5.21)
【出願人】(502233344)エスバテック、アン アルコン バイオメディカル リサーチ ユニット、エルエルシー (19)
【氏名又は名称原語表記】ESBATech, an Alcon Biomedical Research Unit, LLC
【Fターム(参考)】
[ Back to top ]