
細胞培養物をモニターする方法
本発明は、細胞培養の生理学的状態をモニターする方法に関する。いくつかのパラメーター、例えば、細胞生存度、増殖、代謝プロフィール、および生産性が、細胞培養物の代謝フィンガープリントまたはメタボロミクスプロフィールを確立するためにモニターされうる。

【発明の詳細な説明】
【技術分野】
【0001】
本出願は、2009年3月10出願の米国仮出願第61/158,954号からの利益を主張し、その内容はその全体を引用により本明細書に含める。
【0002】
発明の分野
本発明は細胞培養(cell cultivation)の生理学的状態(physiological state)をモニターする方法に関する。いくつかのパラメーター、例えば、細胞生存度、増殖(growth)、代謝プロフィール(metabolic profile)、および生産性が、細胞培養物(cell culture)の代謝フィンガープリントまたはメタボロミクスプロフィール(metabolomic profile)を確立するためにモニターされうる。
【背景技術】
【0003】
発明の背景
哺乳類細胞培養物は患者における有効性を確実にするために複雑な翻訳後修飾が必要である治療用タンパク質の生産のために広く用いられてきている。今日まで、大規模流加回分(fed-batch)培養がいまだに治療用タンパク質生産の支配的形式である(Chu、et al.、Curr. Opin. Biotechnol. 12:180-187、2001)。高密度かん流培養 (Konstantinov、et al.、Adv. Biochem. Eng. Biotechnol. 101:75-98、2006; Konstantinov、et al.、Biotechnol. Prog. 12:100-109、1996; Trampler、et al.、Biotechnology 12:281-284,1994)は典型的には不安定な分子の場合に用いられ、そのためには上昇したバイオリアクター温度での最小の滞留時間が望ましい(例えば、図 1参照)。a)投与の要求(dosing demand)および価格圧力、b)規制当局の厳しいタンパク質品質要求、およびc)工業プロセス発達のための積極的な時系列を鑑みると、治療用タンパク質生産のための最新のプロセス開発プログラムの第一目的は、高い生産収率および一貫した生産物品質を特徴とするバイオリアクタープロセスの迅速な開発である。さらに、これらのプロセスの高い製造コストのために、細胞培養ロバスト性(robustness)のための正確かつ高感度の制御の同定および使用が望ましい。これらの制御はタンパク質生産性および/または培養の最後の前の最終品質における問題の早期警戒を提供しうる。現在、バイオリアクターモニタリングおよびプロセス改善の両方は、主に細胞増殖、代謝活性、およびタンパク質生産性データに基づいている。有用である一方で、この細胞特異的な比率(rate)に基づくアプローチの限界が認識されてきており、アプローチ、例えば、順リアルタイムの代謝フラックス分析が、細胞の生理学的状態のより強固な特徴決定のために示唆されてきている(Goudar、et al.、Adv. Biochem. Eng. Biotechnol. 101:99-118、2006; Konstantinov、Biotechnol. Bioeng. 52:271-289、1996)。
【発明の概要】
【発明が解決しようとする課題】
【0004】
したがってモニタリングプロセスの一貫性およびロバスト性のための測定の現在の常套のセットをより改善する、哺乳類細胞培養物の生理学的状態の包括的特徴決定を可能とする方法の開発と応用のための明らかな必要性が存在する。さらに、様々な細胞株および実験条件を用いる実験を可能とする連続プロセス改善プログラムの関係において用いられるため、これら方法は大きな生理学的データセットを作り出すことが出来、それはタンパク質生産および製造プロセスの全体の理解を促すであろう。かかる開発は、タンパク質生産性についてのもののみならず、タンパク質品質についての正確および高感度のマーカーの同定を導きうる。これらのマーカーは次いで培養の最後の前の最終タンパク質品質の予測のための製造プロセスにおいて用いられ得、一方、それらはまた、最終結果におけるバッチ間の(batch to batch)一貫性を確実にするためにタンパク質生産および製造プロセスを改善することにおいても役立ちうる。最後に、これらの生理学的特徴決定方法は、費用効率の高いプラットフォームに基づきうる。
【課題を解決するための手段】
【0005】
哺乳類細胞培養物の生理学的状態の特徴決定に取り組むために、ガスクロマトグラフィー-質量分析(GC-MS) メタボロミクスを、実験室規模および製造規模の両方で高細胞密度かん流リアクター(perfusion reactor)において培養された新生児ハムスター腎臓 (BHK) 細胞を分析するために利用した。メタボロミクスプロファイリング(metabolomic profiling)は、細胞齢、バイオリアクター規模および細胞源に基づく細胞培養物の識別を可能とし、一方、識別性の(differentiating)代謝産物の同定は、培養物のインビボ生理学的状態に関する重要な情報を提供した。したがって、メタボロミクスは細胞培養工学における価値ある分子分析ツールである。
【図面の簡単な説明】
【0006】
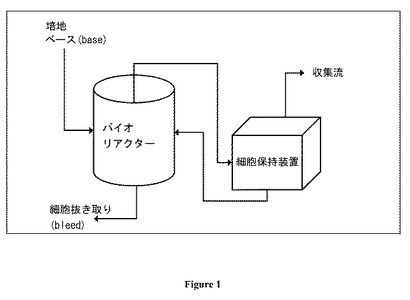
【図1】図 1:細胞培養物かん流システムの模式図。
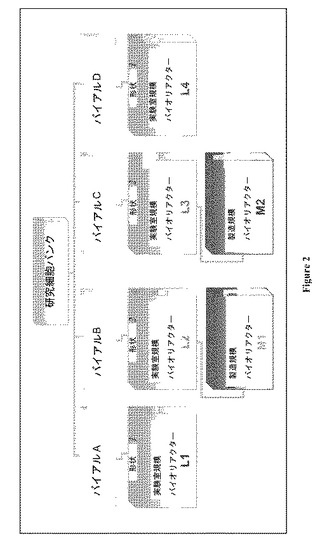
【図2】図 2:発酵プロセスの概観。バイアルおよびリアクターを連結する線は、実験室規模および製造規模のバイオリアクターについての接種源を示す。
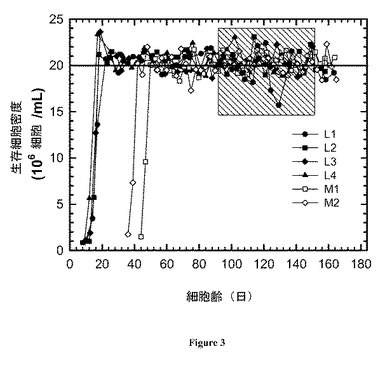
【図3】図 3:作動(operation)の全過程にわたるリアクターについての生存細胞密度の時間プロフィール。
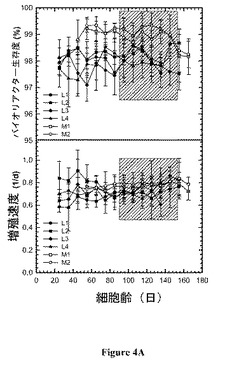
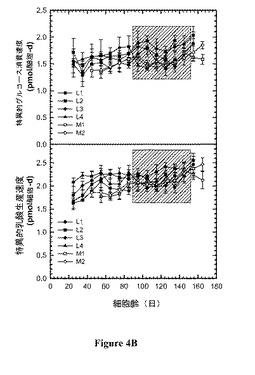
【図4】図 4:リアクターについての、 a)バイオリアクター生存度(viability)、b) 細胞増殖、c)特異的グルコース消費速度およびd) 特異的乳酸(lactate) 生産速度の時間プロフィール。リアクターについての培養の過程を定常状態への到達から開始して10-日間隔に分割し、各間隔についての平均値およびそれに伴う標準偏差を間隔の真ん中の時点にて表す。
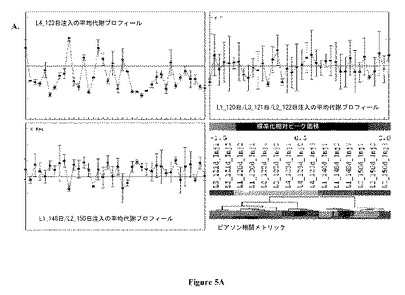
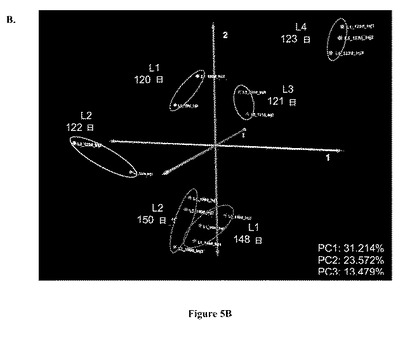
【図5】図 5:実験室-規模バイオリアクターのGC-MS 極性代謝プロフィールの、5A. 階層的クラスタリング(Hierarchical clustering) (HCL)および5B. 主成分分析 (PCA)。両方の分析は、式 (1)に規定するように、プロフィールにおける各代謝産物についての標準化相対ピーク面積に基づいていた。ピアソンの相関はHCLについて距離メトリックであった。図 5Aにおいて、3つの同定されたサブクラスターのそれぞれに含まれるプロフィールの重心(centroid)グラフも示される。PC1、PC2、およびPC3は、それぞれ、主成分1、2、および3によって持たれる元の実験空間におけるデータセットからの% 変動をさす。
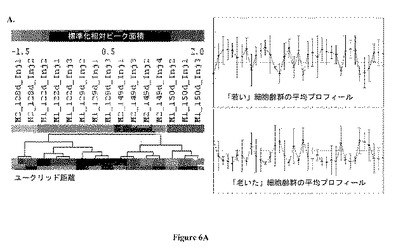
【図6】図 6:製造規模バイオリアクターのGC-MS 極性代謝プロフィールの、6A. 階層的クラスタリング (HCL)および6B. 主成分分析 (PCA)。両方の分析は、式 (1)に規定するように、プロフィールにおける各代謝産物についての標準化相対ピーク面積に基づいていた。ユークリッドはHCLについて距離メトリックであった。図 6Aにおいて、2つの同定されたサブクラスターのそれぞれに含まれるプロフィールの重心(centroid)グラフも示される。PC1、PC2、およびPC3は、それぞれ、主成分1、2、および3によって持たれる元の実験空間におけるデータセットからの% 変動をさす。
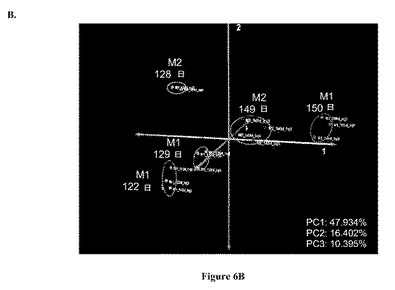
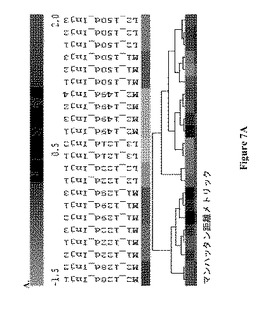
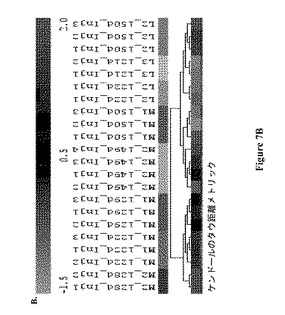
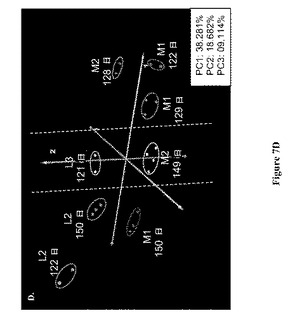
【図7】図 7: 製造規模バイオリアクターM1およびM2および実験室-規模バイオリアクターL2およびL3のGC-MS 極性代謝プロフィールの、7A.マンハッタン距離メトリックによる階層的クラスタリング (HCL)、7B.ケンドールのタウ(Kendal-Tau) 距離メトリックによる階層的クラスタリング、7C.スピアマンの相関(Spearmn Correlation)距離メトリックによる階層的クラスタリング、および 7D.主成分分析 (PCA)。すべての分析は、本文中式 (1)にて規定されるように、プロフィールにおける各代謝産物についての標準化相対ピーク面積に基づいていた。すべての示された記号は図5および6の説明において説明されているように用いられる。
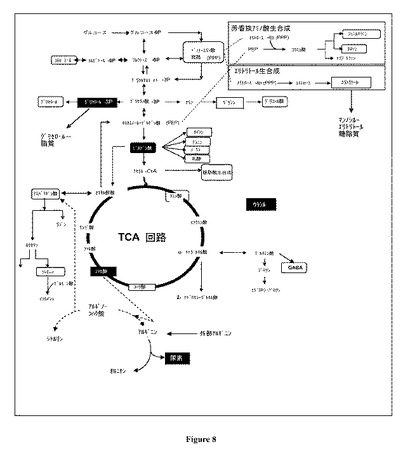
【図8】図 8:代謝経路網の関係においてリアクター M1 サンプルについて122日と比較して129日サンプルにおいて有意に上昇しているとしてその濃度が同定された代謝産物(黒い(bold)箱)。有意な代謝産物はδ = 1.64 および 0% FDR (メジアン)についてマイクロアレイの有意性分析(Significant Analysis for Microarrays (SAM))を用いて同定した。正規化および選別の後に分析に含めたが、SAMによっては有意であると同定されなかった代謝産物は白い(blank)箱にて示す。
【発明を実施するための形態】
【0007】
発明の説明
本発明は、記載された特定の方法、プロトコール、細胞株、動物種または属、コンストラクト、および試薬に限定されず、したがって変動しうることを理解されたい。本明細書において用いる専門用語は具体的な態様を説明するのみの目的のものであり、添付の特許請求の範囲によってのみ限定される本発明の範囲を制限する意図のものではないことも理解されるべきである。
【0008】
本明細書および添付の特許請求の範囲において用いる場合、単数形の「ある」、「1つの」および「その」は、特に断りのないかぎり複数の言及も含むことに注意しなければならない。したがって、例えば、「代謝産物」についての言及は1以上の代謝産物 (例えば、1、2、5、10、50、100またはそれを超える)についての言及であり、当業者に公知のその均等物なども含む。
【0009】
特に断りのないかぎり、本明細書において用いるすべての技術および科学用語は、本発明が属する技術分野の当業者に一般に理解されているものと同じ意味を有する。本明細書に記載のものと類似または同等のあらゆる方法、装置、および材料が本発明の実施または試験において用いることが出来るが、好ましい方法、装置および材料をここで記載する。
【0010】
本明細書において言及するすべての刊行物および特許は、例えば、このたび記載する本発明との関係において使用されうる刊行物に記載のコンストラクトおよび方法を記載および開示する目的のために引用により本明細書に含まれる。先におよび本明細書中に論じられる刊行物は本出願の出願日の前のそれらの開示のためにのみ提供される。ここに記載されるものはいずれも本発明者らが以前の発明によってかかる開示に先行するものでないと認めるものと解釈してはならない。
【0011】
細胞培養工学は、細胞培養物の性能に関する重要な疑問を解決することを試みて今日までトランスクリプトミクス(transcriptomic)、プロテオミクス、および代謝フラックス分析を用いてきた。しかし、哺乳類細胞培養物の生理学的状態の包括的特徴決定を可能とし、プロセスの一貫性およびロバスト性をモニターするための現在常套的な測定値のセットを超えて改善する、方法の開発および適用に対する明白な要求が存在する。
【0012】
メタボロミクスのハイスループット分子分析プラットフォームはこの要求を満たしうる。遊離低分子代謝産物プールの(相対的) 濃度の同時定量をいうメタボロミクスは、生物学的系の代謝フィンガープリントをモニターすることを可能とする (Fiehn、et al.、Nat. Biotechnol. 18:1157-1168、2000; Roessner、et al.、Plant J. 23:131-142、2000)。細胞機能全体との関係における代謝の役割を考慮すると、完全かつ正確な代謝プロフィール地図の定量がなぜ細胞培養工学研究において非常に重要であり得るかということは容易に理解できる。代謝産物濃度と代謝フラックスとは線形の関係にはないが、代謝プロファイリングはハイスループットであり、したがって一過性の代謝状態をモニターするために容易に用いることが出来る。さらに、代謝フラックス分析 (MFA)の場合におけるように、調べられた代謝経路網の構造および調節の知識は必要ではない。さらに、MFAは典型的には定常状態または偽-定常状態条件にのみ適用される一方、メタボロミクスは、その他の2つの主なオミクス(omic)プラットフォームであるトランスクリプトミクスおよびプロテオミクスのように一過性生理学的条件下で用いられ得る。また、トランスクリプトミクスおよびプロテオミクスとは異なり、メタボロミクスは特別な分析装置を必要としない。メタボロミクス(metabolomic)方法は、古典的な分析化学技術、例えば主に、核磁気共鳴 (NMR) 分光法および質量分析(MS) (例えば、ガスクロマトグラフィー-質量分析および液体クロマトグラフィー-質量分析)に基づくものであり、最もコストのかからないオミクス(omics)アプローチである(Kanani、et al.、J. Chromatogr B. Analyt. Technol. Biomed. Life Sci. 871:191-201、2008)。特に細胞培養系についてメタボロミクスがその他のオミクス技術よりも特に優れている点は、細胞内代謝状態および細胞外培地の組成の両方をモニターすることへのその適用可能性である。これは代謝経路網活性のよりよい理解を提供する。最後に、代謝は生物学的系の間で良好に保存されているので、比較メタボロミクス研究はより容易であり、サンプル間の凝った正規化を必要としない。
【0013】
分子フィンガープリンティングによってインビボでの代謝生理をモニターすることにより、従来の測定によっては直接的に観察することができなかったサンプル間の微妙な相違の判定が可能となる。細胞生理学的状態の増強された(enhanced)特徴決定は、現在用いられているプロセス制御方法を改善することが出来る代謝フィンガープリンティングによって可能となる。さらに、多変量統計解析により、その濃度変化が細胞培養物の生理学における相違を特徴づける代謝産物の同定が可能となる。この情報は一貫性のない(inconsistent)性能についての早期警戒マーカーを同定することを助けることが出来、最終的にはプロセス最適化を導く細胞生理学の理解もさらに助けることが出来る。
【0014】
メタボロミクスフィンガープリンティングは細胞培養工学における非常に有用な分子分析ツールでありうる。個別にまたは細胞機能のその他のレベルを評価するその他のハイスループット分子分析技術と組み合わせて、発酵プロセスの最適化および発酵の一連の活動の状況および品質の一貫性をモニターするために用いられている現在利用可能な測定値のセットの改善に向けての手がかりを提供しうる。
【0015】
本発明は、高密度かん流培養における哺乳類細胞生理をモニターするための細胞培養物のGC-MS メタボロミクスの使用に関する。GC-MS メタボロミクスは実験室および製造規模の両方について異なる培養段階での細胞の生理学的状態を分析するために用いることが出来る。
【実施例】
【0016】
本発明がよりよく理解され得るために、以下の実施例を示す。これらの実施例は例示の目的のみのためのものであり、いかなるようにも本発明の範囲を限定するものとして解釈されるべきではない。本明細書において言及するすべての刊行物はそれら全体を引用により本明細書に含める。
【0017】
実施例 1. 細胞かん流培養
BHK 細胞を主な炭素源としてグルコースおよびグルタミンを用いてかん流様式 (図 1)にて培養した。新生児ハムスター 腎臓 (BHK) 細胞の4つの独立のバイアル(図 2におけるA-D)をそれぞれ、4つの実験室-規模 15L かん流システム、L1-L4に接種するのに用いた。L1およびL4バイオリアクターは典型的な円筒状容器であり、一方、L2およびL3は底の小さい(small-bottom)リアクターであった。L2およびL3を、それぞれ2つの製造規模バイオリアクターであるM1およびM2に接種するのに用いた。すべての実験室規模および製造規模かん流システムはすべてのモニターした変数について同じ運転条件および設定値を有していた。バイオリアクター稼働の持続時間は113 - 155日の範囲であり、その結果、合計は826バイオリアクター日となる。各リアクターの作動の最後の40日の間に、サンプルを、表1に示すように様々な時点で収集した。細胞齢はバイアル解凍の日から測定し、同じリアクターにおける培養時間は各リアクターの開始日およびサンプル収集日の間の時間差として評価する。これらサンプルからの細胞ペレットをGC-MS メタボロミクスおよび多変量統計解析を用いて分析した。
【0018】
表1
【表1】
【0019】
バイオリアクター温度は35.5℃に維持し、47 RPM (製造規模バイオリアクターについては15 RPM)にて撹拌した。溶存酸素 (DO) 濃度は膜通気により50% 空気飽和(air saturation)に維持し、pHは6% Na2CO3の自動添加により6.8に維持した。バイオリアクターに出発細胞密度が〜1 x 106 細胞/mLとなるよう接種し、細胞を定常状態濃度の20 x 106 細胞/mLとなるまで蓄積させた。この標的定常状態細胞密度をバイオリアクターからの自動細胞抜き取り(bleed)により維持した。
【0020】
サンプルをCEDEX システム (Innovatis、Bielefeld、Germany)を用いる細胞密度および生存度分析のために各バイオリアクターから毎日収集した。これらのサンプルを次いで遠心分離にかけ(Beckman Coulter、Fullerton、CA)、上清を栄養素および代謝産物濃度について分析した。グルコース、乳酸、グルタミン、およびグルタミン酸濃度はYSI モデル 2700 分析器(Yellow Sprints Instruments、Yellow Springs、OH)を用いて決定し、一方、アンモニアはEktachem DT60 分析器 (Eastman Kodak、Rochester、NY)により測定した。pHおよびDOは格納式電極(Metler-Toledo Inc.、Columbus、OH)を用いてオンラインで測定し、その測定精度はRapidlab 248 血液ガス分析器 (Bayer HealthCare、Tarrytown、NY)におけるオフライン分析により確証した。同じ計器を用いて溶存 CO2 濃度を測定した。細胞密度のオンライン測定は格納式光学密度プローブ (Aquasant Messtechnik、Bubendorf、Switzerland)により行い、CEDEX システムからの細胞密度測定により較正した。
【0021】
実施例 2:メタボロミクスプロファイリング
バイオリアクターからのサンプルを氷上に引き取り(drawn)、あらかじめ冷却しておいたローターを用いて遠心分離した。遠心分離の後、上清を棄て、細胞ペレットを冷たいPBS バッファーで洗浄した。細胞ペレットを次いで70℃の水浴に15 分間入れておいた。ペレットを次いで減圧下で70℃で24 時間乾燥させた。サンプルからの乾燥細胞ペレットをメタボロミクス分析に用いた。
【0022】
乾燥細胞ペレットの極性(polar)代謝産物抽出物をメタノール/水抽出を用い(Kanani、et al.、2008; Roessner、et al.、2000)、リビトール (0.1mg/g乾燥細胞重量)および[U-13C]-グルコース (0.2 mg/g 乾燥細胞重量)を内部標準として得た。乾燥極性抽出物を、150μLのピリジン中のメトキシアミン塩酸塩溶液 (20 mg/mL)との90 分間の反応、次いで、300 μLのN-メチル-トリメチルシリル-トリフルオロアセタミド (MSTFA)との少なくとも 6 時間の室温での反応(Kanani、et al.、2008; Kanani、et al.、Metab. Eng.、9:39-51、2007)を介してそれらの(MeOx)TMS-誘導体へと誘導体化した。メタボロミクスプロフィール(metabolomic profile)を、Saturn 2200T ガスクロマトグラフ - (イオントラップ) 質量分析計 (Varian Inc.、CA)を用いて得た。ピーク同定および定量は、(Kanani、et al.、2007)に記載されているようにして行った。未加工の(raw)メタボロミクスデータセットは 91のピークから構成され、そのそれぞれは獲得されたメタボロミクスプロフィールの少なくとも1つにおいて検出され、化学的分類が公知の化合物に対応する(例えば、Kanani、et al.、2008; Kanani、et al.、2007参照)。
【0023】
すべての検出されたピークの相対的面積(RPA)を、内部標準であるリビトール (マーカーイオン: 217)による正規化(normalization)から見積もった。データ妥当性検証、正規化、および補正方法を、アミン基含有代謝産物からの複数の誘導体の形成に主に起因する誘導体化バイアスを説明するために適用した(Kanani、et al.、2008; Kanani、et al.、2007)。GC-MS 運転条件は、[U-13C]-グルコースの2つのピークの比に基づくサンプルの獲得された(acquired)メタボロミクスプロフィールの取得の際に確証した。第二に、95日のL4 サンプルのアミノ酸誘導体のプロフィールに基づいて評価された重み係数を用いて(表1参照)、同じアミン基含有代謝産物に対応する誘導体ピーク面積を併せて1つの累積 (有効) ピーク面積とした。イソロイシン、β-アラニン、およびグルタミン酸をさらなる分析から除去した。というのはそれらの利用可能な測定値が、すべての正の重み係数が見積もられることを可能としなかったからである。特定の誘導体化範囲において1つのみの誘導体が観察されたが1より多い誘導体が知られているアミノ酸は、分析の次の工程に含めた; 非常にしばしばそれらは、注入間の変動の高い係数のためにその後の工程において除去された。さらに、(a)公知のケトン基含有代謝産物の2つのMeOx ピークのうち最も小さいもの、(b)未知のアミン基含有代謝産物に対応するピーク、(c)誘導体化アーティファクトであると同定されたかまたは有意なキャリーオーバーを有するピーク、および(d) 一貫しては検出されなかった代謝産物ピークは分析から除去した。38の代謝産物を含むこれらの最終代謝産物 RPA プロフィールを、80%カットオフを用いるTM4 MeV (V4.0) データ分析ソフトウェア (Saeed、et al.、Biotechniques 34:374-378、2003)において用いた。あらゆる不明のRPAをTM4 MeV に備えられているためk近傍法を用いて帰属させた (Troyanskaya、et al.、Bioinformatics 17:520-525、2001)。
【0024】
獲得されたメタボロミクスプロフィールに対して適用された分析は代謝産物相対ピーク面積の標準値に基づくものとした(下記式 1参照)。これらの分析における標準化相対ピーク面積の使用は同じ代謝プロフィールにおける代謝産物の間の相対ピーク面積レベルの桁における大きな違いによって必要とされる。メタボロミクスプロフィール jにおける代謝産物 Mの標準化相対ピーク面積、RPAMjは以下に等しい:
【数1】
【0025】
階層的クラスタリング(Hierarchical clustering) (HCL)を用いてサンプルをそれらのメタボロミクスプロフィールに基づいてクラスタリング(cluster)した。HCLにおいて、代謝プロフィールは階層木においてクラスタリングされる。かかる木の最も低いレベルにおいて、各代謝プロフィールは別々のクラスターであるとみなされるが、すべてのサンプルはもっとも高いレベルにおいて1つのクラスターにグルーピングされる。もっとも低いレベルから出発して、入手可能なクラスターの各対についての相関係数がアルゴリズムの各ラウンドにて特定の距離メトリックに基づいて見積もられる。最も高い相関係数を有するクラスターはアルゴリズムの次のラウンドのために1つのクラスターへとグルーピングされる (Quackenbush、Nat. Genet. 2:418-427、2001; Eisen、et. al.、Proc. Natl. Acad. Sci. USA 95:14863-14868、1998)。獲得された階層木は、研究される生物学的問題の観点から解釈されなければならない。例えば、図 5Aにおいて、HCLは、1ヶ月離れた細胞齢に対応する代謝プロフィールの2つのクラスターを同定する。より低い細胞齢クラスターのなかで、リアクター L4の代謝プロフィールはその他の3つの実験室-規模リアクターの代謝プロフィールから離れている。
【0026】
主成分分析 (PCA)を用いて様々な細胞培養物サンプルがそれらのメタボロミクスプロフィールに基づいて識別可能かどうかを可視化した。PCAはデータマトリックスの特異値分解を伴い、それはデータセットにおける最大の分散(variance)をとらえる新しい座標系の上への元のデータセットの直交線形変換をいう(Strang、Introduction to Linear Algebra、Wellesley-Cambridge Press、Wellesley、Massachusetts、1993)。この変換は元の実験空間の回転および/または伸縮を伴う可能性がある。主成分 1は元のデータセットにおける最高の分散の方向に対応し、主成分 2は第二に高い分散の方向に対応し、以下同様である。ハイスループット生物学的データ分析において、PCAは簡略化の目的で座標化するために用いられており、それゆえ元のデータセットにおける分散の大部分は3-D 空間内に可視化される (Raychaudhuri、et. al.、Pac. Symp. Biocomput. 2000:455,466、2000)。少数の主成分がしばしばデータにおける構造のほとんどを説明するために十分である (Scholkopf、et al.、Learning with Kernels - Support Vector Machines、Regularization、Optimization and Beyond、The MIT Press、Cambridge、Massachusettes、2002)。PCAグラフを読む場合、個別に示された主成分のすべておよび個々によって示された空間内にとらえられている元のデータセットにおける分散のパーセンテージを考慮することが重要である。各主成分の単位(重み)はそれによって表される元のデータセットにおける分散のパーセンテージと等しい。したがって、例えば、主成分 1上で距離がxである2つのデータポイントは主成分 2上で同じ距離にある2つのデータポイントよりもそれらの生理学的状態において大きい差を表す。
【0027】
1つのセットの細胞培養物サンプルにおいて別のセットと比較してその濃度が有意により高かったかまたは低かった代謝産物を、それぞれ特定の比較の正にまたは負に有意な代謝産物と称した。1つの比較における有意な代謝産物を対応のないマイクロアレイの有意性分析(Significance Analysis of Microarrays)(SAM) アプローチ (Tusher、et al.、Proc. Natl. Acad. Sci. USA 98:5116-5121、2001)を用いて同定した。
【0028】
SAM (Tusher、et al.、2001; Larsson、et al.、BMC Bioinformatics 6:129、2005; Wu、Bioinformatics 21:1565-1571、2005)は、異なる生理学的条件を表す2つの測定セットの間で有意に異なる分子の量の同定のための置換に基づく(permutation-based)(ノンパラメトリック) 仮説検証方法である。SAMはDNA マイクロアレイに基づく転写プロファイリングデータの分析のために注文設計されてきており、その他のオミクス(omic)データセットの分析のために同様に用いられてきている(例えば、Dutta、et. al.、Biotechnol. Bioeng. 102:264-279、2009参照)。メタボロミクス分析の場合、SAMは2つのサンプルの間のその濃度における相違が、ランダムな変動のみに起因して予測されうる相違よりも大きい代謝産物を同定する:
【数2】
式中、diは観察された相違であり、deiは期待された相違であり、δは有意性閾値である。パラメトリック仮説検証方法とは異なり、置換に基づく(permutation-based)(ノンパラメトリック)方法ではデータが特定の分布に従う必要はない。それらはまた偽発見率 (FDR)の見積を提供し、偽発見率は、濃度において示差的に(differentially)変化しているとして同定された所与の代謝産物が偽陽性である確率である。さらに、SAM はFDRおよび多数の有意な(significant)代謝産物の閾値の変化に対する感度が決定され得るようにδの調整を可能とする。
【0029】
TM4 MeV v4.0 (Saeed et al.、2003)に実装されているアルゴリズムを用いた。2つの細胞培養物サンプルの間の代謝プロフィールにおける相違の全体論的視野を、適当に色分けされた代謝経路網において有意であると同定された代謝産物を位置づけすることによって得た。代謝経路網再構築はKEGG (KEGG Database、2008)およびEXPASY (EXPASY Database、2008)データベースからの情報に基づいた。
【0030】
プロセスの一貫性を確実にするためにかん流システムにおいて注意深く制御される必要がある基本変数の1つは生存細胞密度である。かん流システムが定常状態条件に到達した後は、培養の過程にわたってそれは典型的にはターゲット設定値に維持される。定常状態細胞密度はターゲット設定値である20 x 106 細胞/mL付近に維持された (表2、図 3)。図 3はそれらの作動の過程にわたってのリアクターについての生存細胞密度の時間プロフィールを示し、一方、定常状態平均は表2に示される。細胞増殖-、代謝活性-、および生産性-関連変数についての定常状態平均が示される。変動の係数は括弧内に示される。FVCD、sGCR、およびsLPRはそれぞれ、バイオリアクター生存細胞密度、特異的グルコース消費速度、および特異的乳酸生産速度である。すべてのリアクターの平均特異的生産性はL1 リアクターについての平均値との比較にて示される。
【0031】
細胞密度測定には〜8.5%の誤差が伴うことが示され、平均値の周囲の観察された変動(即ち、4.4% - 6.3%の範囲の変動の係数(CoV’s))は、良好な細胞密度制御を反映して非常に小さいことが示唆される。
【0032】
表2
【表2】
【0033】
図 4 は、この研究におけるリアクターについての、a)バイオリアクター生存度、b) 細胞増殖、c) 特異的グルコース消費速度、および、d) 特異的乳酸生産速度の時間プロフィールを提供する。それぞれのリアクターについての培養の過程は、定常状態に達した時点から開始して10-日間隔に分けた。それぞれの間隔にわたる平均値および伴う標準偏差は間隔の中央の時点にて表される。20 - 30日の細胞齢からのデータの平均をとり、25日の細胞齢に対応するものとして示した。有意な時間関連変動は、生存度および増殖速度についてはみられなかった。わずかに上昇する傾向が特異的グルコース消費および乳酸生産速度の時間プロフィールについてみられた。プロセスをモニターするための細胞培養工学において用いられた測定値のセットに基づいて、リアクターのそれぞれにおいて細胞齢に起因する生理の明らかな相違はない。リアクター間での一貫性はまた、平均特異的タンパク質生産性データ(表2)においても観察され、ここでリアクター L1の生産性は任意に1.0に設定され、比較のために用いた。0.92 - 1.04の範囲の特異的生産性は明らかにバイオリアクター間の一貫性を示す。さらに、バイオリアクターからの初期、中期および後期ステージからの材料を精製し、典型的なタンパク質品質特性について試験した。生成物品質データは仕様の範囲内(within specification)であり、バイオリアクターの間および培養の経過にわたっての一貫性を示唆している。
【0034】
実験室-規模バイオリアクターの分析からのデータを図5Aおよび5Bに示す。明らかな細胞齢に基づく差別化が主成分 2 (図 5B)上においてみられ、ここで、120-123 日サンプルの代謝プロフィールはグラフの上側部分にみられるが、148-150 日サンプルのものは下側部分にある。異なる実験室-規模リアクターから獲得された同じ細胞齢の培養物の代謝プロフィールは主成分 1の上で主に明らかに差別化されている。主成分 1は元のデータセットにおける相違のもっとも大きな部分を有するため、この差別化は、メタボロミクスプロフィールは、同じ細胞バンクにおけるバイアルとバイアルの間の変動および/または異なるバイオリアクターにおける細胞培養物増殖を識別する力がありうることを示す。これは、現在のモニターするためのツールボックスに基づいては観察可能ではない、生理におけるわずかな相違を同定する生体分子フィンガープリンティングの能力を支持する。さらに図 5Bの調査は、L3およびL4 サンプル(グラフの右側)の、両方の調べた細胞齢のL1およびL2 サンプル(グラフの左側)からの分離を示す。
【0035】
図 5Aに示す階層的クラスタリング (HCL) 分析結果は、PCAグラフにおける代謝プロフィールの間の可視化された相違を確認する。代謝プロフィールをそれらの形状に基づいて階層化するピアソンの相関距離メトリックを用いて、得られた階層的クラスタリング木は、2つの異なる細胞齢の代謝プロフィールに対応する2つの主な枝を含む。2つの代謝プロフィールの間のピアソンの相関係数であるrは、2つのプロフィールの共分散をそれらの標準偏差の積で割った値に等しい (Box et. al.、1978)。それは-1と1との間の値をとりうる。共分散は2つのプロフィールの間の線形依存性の尺度である。したがって、ピアソンの相関は、2つのプロフィールの形状の間の類似性を明らかにすることが期待される (Quackenbush、2001)。
【0036】
より低い細胞齢の枝(木の左側)のなかで、2つの別々のサブクラスターが同定された; 1つは123 日L4 サンプルの3注入(injection)からの代謝プロフィールを含み、別の木の代謝プロフィールではリアクターのサンプルが別の亜枝にクラスタリングしている。L4 代謝プロフィールデータポイントは、PCAグラフから示されるように実験空間における残りからさらに離れていた (図 5B)。図 5A に示すL4 注入の平均代謝プロフィール(この重心(centroid)プロフィールはTM4 MeVソフトウェアから作成した; Saeed、et. al.、2003)は、同様の細胞齢のリアクターの残りの平均代謝プロフィールとは異なっている。
【0037】
ユークリッド距離を用いるHCL 分析は、両方の細胞齢のL3およびL4 プロフィールのL1およびL2 プロフィールからの差別化を示した。ユークリッド距離メトリックは、ユークリッド空間における2つの地点の間の古典的な(最短の)距離であり、ピタゴラスの定理によって規定される。M-次元空間において、地点、x = (x1、x2、…、xM)および y = (y1、y2,…、yM)の間のユークリッド距離は以下のように規定される:
【数3】
したがって、2-次元空間において、基点からの同じユークリッド距離の点によって囲まれる領域は特定の距離と等しい半径を持った円である。
【0038】
PCAおよびHCL 分析の両方が、バイアルとバイアルの間の変動性および細胞齢がリアクター幾何学(標準的円筒状、対、底の小さい立体配置)よりも高い影響を細胞培養物サンプルの測定される極性代謝プロフィールに対して有することを示した。入手可能な実験室-規模培養サンプルにおいて変動するこれら3つのすべてのパラメーターのなかで、リアクター幾何学の代謝生理に対する効果は、サンプルのクラスタリングにおいて直接的に明らかではない。
【0039】
図6Aおよび6Bは、製造規模リアクターサンプルの代謝プロフィールについての、それぞれHCLおよびPCA 分析の結果を示す。サンプルの細胞齢に基づく分離が主成分 1においてみられる(図 6B)。この分離は HCL 分析 (図 6A)によっても明らかであり、ここで、階層的クラスタリング木の二つの主な枝は2つの細胞齢に対応する。図 6Aに示す2つの細胞齢の平均代謝プロフィールもまた、この差別化を支持する。
【0040】
122-129日および149-150 日サンプル群のなかで、HCLおよびPCAはともにM1およびM2 サンプルの間の明らかな差別化を示した。149-150 日サンプルのM1およびM2 代謝プロフィールの間の相違は122-129 日齢の群の対応するものよりも大きかった。最初の場合において、サンプルは主成分 1上で分離しており (48% 分散)、第二のセットのサンプルは主成分 2上で分離していた (16.5%分散)。
【0041】
製造規模バイオリアクターの38の極性代謝産物プロフィールを次いで、それらから接種された実験室-規模バイオリアクターのプロフィールと組み合わせて分析した。図7A-DはこのセットのサンプルについてのHCLおよびPCA 分析の結果を示す。いずれの距離メトリックに基づいたPCAおよび HCL 分析も、製造規模バイオリアクター、M1および M2の122-129日齢サンプルの代謝プロフィールを、すべてのその他のサンプルから別れたクラスターであると同定した。HCL 分析のいずれにおいても(図 7A-C)、これらサンプルは木の2つの主な枝の1つを形成しており、それらはまた、PCA グラフの右側に分離して示される(図 7D)。
【0042】
図 7における残りの5つのサンプルは、細胞齢、リアクタータイプ、および細胞源に基づいて分類され得た。2つのサンプル (L2およびL3)は121 - 122日の範囲の細胞齢を有しており、一方L2、M1、およびM2は149 - 150 日サンプルであった。3つのサンプルは実験室-規模バイオリアクターからのものであり、一方2つは製造規模システムからのものであった。サンプル対 L2-M1およびL3-M2は同じ細胞源からのものであった(それぞれ、バイアルBおよびC)。図 7Dにおいて、これらのサンプルは、リアクターサイズおよび細胞源に基づいて明らかに差別化されている。リアクターサイズに基づく差別化は主成分 2 上にあり、実験室-規模リアクターは正の側にあり、製造規模リアクターは負の側にあった。細胞源に基づく差別化は主成分 1上にあり、L2およびM1 サンプルはL3およびM2 サンプルよりも左側にクラスタリングされていた。
【0043】
細胞源に基づくクラスタリングも、ユークリッドまたはマンハッタン距離メトリックが用いられた場合、HCL 分析において明らかである。M-次元空間において、地点x = (x1、x2、…、xM)およびy = (y1、y2,…、yM)の間のマンハッタン距離は、座標軸上への(x,y) 線分の投影の長さの和として規定される:
【数4】
2-次元空間において、起点から同じマンハッタン距離の点によって囲まれる領域は、辺が座標軸に対して45°の方向である正方形である。マンハッタン距離は異常値に対してユークリッド距離よりも感度が低い(Filzmoser、et. al.、Comput. Stat. Data Anal. 52:1694-1711、2008)。マンハッタンとユークリッド距離とは共に入手可能なデータベクトル(この場合は、代謝プロフィール)の間の絶対差を測る。クラスタリング分析において用いた場合、両方のメトリックは、代謝プロフィールのピーク面積レベルの間の類似度を明らかにすると期待される。
【0044】
図 7Aにおいて、階層木の右側の枝は3つの枝に分かれており、1つはM2およびL3 サンプルをクラスタリングしており、もう1つは150日L2およびM1 サンプルを、残りの1つは122日L2 サンプルをクラスタリングしており、PCAグラフと類似している(図 7D)。HCL 分析におけるリアクターサイズに関する代謝プロフィールのクラスタリングは、ケンドールのタウ(Kendall’s Tau)距離メトリックを用いるとみられた。このメトリックは代謝プロフィールのランキングベクトルを指す。代謝プロフィールのランキングベクトルにおいて、各相対ピーク面積が、代謝プロフィールにおけるすべての相対ピーク面積のなかのそのランキングを示す整数に取って代わられる。この場合、2つのランキングベクトルの間の距離は2つのベクトルにおいて2つの整数が逆の順にある回数に基づいて測定される。もしこの数が0である場合、2つのランキングベクトルは同一である。
【0045】
図 7Bにおいて、階層木の右側の枝は、実験室-規模および製造規模サンプルに対応する2つの亜枝に分かれる。最後に、スピアマン相関距離メトリックを用いるHCL 分析は細胞齢に基づくサンプルクラスタリングを示した。この相関は、代謝プロフィールのランキングベクトルに対するピアソンの相関距離メトリックの適用を指す。図 7Cにおいて、階層木の右側の枝は2つの亜枝に分かれ、1つは121-122日L2およびL3 サンプルを含み、もう1つは149-150日L2、M1およびM2 サンプルを含む。
【0046】
マイクロアレイの有意性分析(SAM)を用いて、122 日サンプルと比較して129 日 M1 サンプルにおいてその濃度が有意に上昇 (正に有意)または低下(負に有意)した代謝産物を同定した。これらのサンプルは1週間しか離れていないにもかかわらず、それらはHCL およびPCA 分析から差別化できた。SAM (1.64のデルタ値および0% 偽発見率 (FDR)について)は、5つの正に有意なそして0の負に有意な代謝産物を同定し、それらは有意性の高さの順に、フマル酸(fumarate)、グリセロール-3-リン酸、尿素、ウラシル、およびピルビン酸(pyruvate)であった。4つの代謝産物を代謝経路網との関連において図 8に示す。高濃度のフマル酸および尿素は、122 日サンプルと比較して129 日における尿素サイクルのより高い活性を示し得、したがって窒素同化のより高い活性を示し得る。さらに、グリセロール-3-リン酸の濃度上昇はグリセロ脂質の生産上昇を示しうる。ウリジンの前駆体であって、ウリジンリン酸(UMP、UDPおよびUTP)の主要な構成成分であるウラシルは、炭水化物代謝、タンパク質グリコシル化および糖脂質形成に重要な役割を果たす。このタイプの情報はバイオリアクター作動の最適化において有用であり得ることが明白である。
【0047】
細胞生理学的状態のより高い分解能の特徴決定が代謝産物プロファイリングによって可能となった。メタボロミクスプロフィールは、目的のパラメーター、例えば、細胞齢についての1以上の識別力のある代謝産物の同定に利用可能であった。例えば、メタボロミクスプロフィールは目的のパラメーターについての1、2、5、10、50、100またはそれを超える識別力のある代謝産物の同定に利用することが可能であった。これらの識別力のある代謝産物は従来測定されてきた細胞培養物生理学的変数と組み合わせてバイオリアクター培養の最適化に用いることが出来、早期警戒またはプロセス混乱(upset)として役立ちうる。メタボロミクスは細胞培養工学において高感度のハイスループット分子分析ツールとして利用することが出来る。
【0048】
本発明のその他の態様は、本明細書の考慮または本明細書に開示する本発明の実施から当業者に明らかであろう。本明細書および実施例は例示的にのみ考慮されるべきであって、本発明の真の範囲と精神は、以下の請求の範囲によって示されることが意図される。
【技術分野】
【0001】
本出願は、2009年3月10出願の米国仮出願第61/158,954号からの利益を主張し、その内容はその全体を引用により本明細書に含める。
【0002】
発明の分野
本発明は細胞培養(cell cultivation)の生理学的状態(physiological state)をモニターする方法に関する。いくつかのパラメーター、例えば、細胞生存度、増殖(growth)、代謝プロフィール(metabolic profile)、および生産性が、細胞培養物(cell culture)の代謝フィンガープリントまたはメタボロミクスプロフィール(metabolomic profile)を確立するためにモニターされうる。
【背景技術】
【0003】
発明の背景
哺乳類細胞培養物は患者における有効性を確実にするために複雑な翻訳後修飾が必要である治療用タンパク質の生産のために広く用いられてきている。今日まで、大規模流加回分(fed-batch)培養がいまだに治療用タンパク質生産の支配的形式である(Chu、et al.、Curr. Opin. Biotechnol. 12:180-187、2001)。高密度かん流培養 (Konstantinov、et al.、Adv. Biochem. Eng. Biotechnol. 101:75-98、2006; Konstantinov、et al.、Biotechnol. Prog. 12:100-109、1996; Trampler、et al.、Biotechnology 12:281-284,1994)は典型的には不安定な分子の場合に用いられ、そのためには上昇したバイオリアクター温度での最小の滞留時間が望ましい(例えば、図 1参照)。a)投与の要求(dosing demand)および価格圧力、b)規制当局の厳しいタンパク質品質要求、およびc)工業プロセス発達のための積極的な時系列を鑑みると、治療用タンパク質生産のための最新のプロセス開発プログラムの第一目的は、高い生産収率および一貫した生産物品質を特徴とするバイオリアクタープロセスの迅速な開発である。さらに、これらのプロセスの高い製造コストのために、細胞培養ロバスト性(robustness)のための正確かつ高感度の制御の同定および使用が望ましい。これらの制御はタンパク質生産性および/または培養の最後の前の最終品質における問題の早期警戒を提供しうる。現在、バイオリアクターモニタリングおよびプロセス改善の両方は、主に細胞増殖、代謝活性、およびタンパク質生産性データに基づいている。有用である一方で、この細胞特異的な比率(rate)に基づくアプローチの限界が認識されてきており、アプローチ、例えば、順リアルタイムの代謝フラックス分析が、細胞の生理学的状態のより強固な特徴決定のために示唆されてきている(Goudar、et al.、Adv. Biochem. Eng. Biotechnol. 101:99-118、2006; Konstantinov、Biotechnol. Bioeng. 52:271-289、1996)。
【発明の概要】
【発明が解決しようとする課題】
【0004】
したがってモニタリングプロセスの一貫性およびロバスト性のための測定の現在の常套のセットをより改善する、哺乳類細胞培養物の生理学的状態の包括的特徴決定を可能とする方法の開発と応用のための明らかな必要性が存在する。さらに、様々な細胞株および実験条件を用いる実験を可能とする連続プロセス改善プログラムの関係において用いられるため、これら方法は大きな生理学的データセットを作り出すことが出来、それはタンパク質生産および製造プロセスの全体の理解を促すであろう。かかる開発は、タンパク質生産性についてのもののみならず、タンパク質品質についての正確および高感度のマーカーの同定を導きうる。これらのマーカーは次いで培養の最後の前の最終タンパク質品質の予測のための製造プロセスにおいて用いられ得、一方、それらはまた、最終結果におけるバッチ間の(batch to batch)一貫性を確実にするためにタンパク質生産および製造プロセスを改善することにおいても役立ちうる。最後に、これらの生理学的特徴決定方法は、費用効率の高いプラットフォームに基づきうる。
【課題を解決するための手段】
【0005】
哺乳類細胞培養物の生理学的状態の特徴決定に取り組むために、ガスクロマトグラフィー-質量分析(GC-MS) メタボロミクスを、実験室規模および製造規模の両方で高細胞密度かん流リアクター(perfusion reactor)において培養された新生児ハムスター腎臓 (BHK) 細胞を分析するために利用した。メタボロミクスプロファイリング(metabolomic profiling)は、細胞齢、バイオリアクター規模および細胞源に基づく細胞培養物の識別を可能とし、一方、識別性の(differentiating)代謝産物の同定は、培養物のインビボ生理学的状態に関する重要な情報を提供した。したがって、メタボロミクスは細胞培養工学における価値ある分子分析ツールである。
【図面の簡単な説明】
【0006】
【図1】図 1:細胞培養物かん流システムの模式図。
【図2】図 2:発酵プロセスの概観。バイアルおよびリアクターを連結する線は、実験室規模および製造規模のバイオリアクターについての接種源を示す。
【図3】図 3:作動(operation)の全過程にわたるリアクターについての生存細胞密度の時間プロフィール。
【図4】図 4:リアクターについての、 a)バイオリアクター生存度(viability)、b) 細胞増殖、c)特異的グルコース消費速度およびd) 特異的乳酸(lactate) 生産速度の時間プロフィール。リアクターについての培養の過程を定常状態への到達から開始して10-日間隔に分割し、各間隔についての平均値およびそれに伴う標準偏差を間隔の真ん中の時点にて表す。
【図5】図 5:実験室-規模バイオリアクターのGC-MS 極性代謝プロフィールの、5A. 階層的クラスタリング(Hierarchical clustering) (HCL)および5B. 主成分分析 (PCA)。両方の分析は、式 (1)に規定するように、プロフィールにおける各代謝産物についての標準化相対ピーク面積に基づいていた。ピアソンの相関はHCLについて距離メトリックであった。図 5Aにおいて、3つの同定されたサブクラスターのそれぞれに含まれるプロフィールの重心(centroid)グラフも示される。PC1、PC2、およびPC3は、それぞれ、主成分1、2、および3によって持たれる元の実験空間におけるデータセットからの% 変動をさす。
【図6】図 6:製造規模バイオリアクターのGC-MS 極性代謝プロフィールの、6A. 階層的クラスタリング (HCL)および6B. 主成分分析 (PCA)。両方の分析は、式 (1)に規定するように、プロフィールにおける各代謝産物についての標準化相対ピーク面積に基づいていた。ユークリッドはHCLについて距離メトリックであった。図 6Aにおいて、2つの同定されたサブクラスターのそれぞれに含まれるプロフィールの重心(centroid)グラフも示される。PC1、PC2、およびPC3は、それぞれ、主成分1、2、および3によって持たれる元の実験空間におけるデータセットからの% 変動をさす。
【図7】図 7: 製造規模バイオリアクターM1およびM2および実験室-規模バイオリアクターL2およびL3のGC-MS 極性代謝プロフィールの、7A.マンハッタン距離メトリックによる階層的クラスタリング (HCL)、7B.ケンドールのタウ(Kendal-Tau) 距離メトリックによる階層的クラスタリング、7C.スピアマンの相関(Spearmn Correlation)距離メトリックによる階層的クラスタリング、および 7D.主成分分析 (PCA)。すべての分析は、本文中式 (1)にて規定されるように、プロフィールにおける各代謝産物についての標準化相対ピーク面積に基づいていた。すべての示された記号は図5および6の説明において説明されているように用いられる。
【図8】図 8:代謝経路網の関係においてリアクター M1 サンプルについて122日と比較して129日サンプルにおいて有意に上昇しているとしてその濃度が同定された代謝産物(黒い(bold)箱)。有意な代謝産物はδ = 1.64 および 0% FDR (メジアン)についてマイクロアレイの有意性分析(Significant Analysis for Microarrays (SAM))を用いて同定した。正規化および選別の後に分析に含めたが、SAMによっては有意であると同定されなかった代謝産物は白い(blank)箱にて示す。
【発明を実施するための形態】
【0007】
発明の説明
本発明は、記載された特定の方法、プロトコール、細胞株、動物種または属、コンストラクト、および試薬に限定されず、したがって変動しうることを理解されたい。本明細書において用いる専門用語は具体的な態様を説明するのみの目的のものであり、添付の特許請求の範囲によってのみ限定される本発明の範囲を制限する意図のものではないことも理解されるべきである。
【0008】
本明細書および添付の特許請求の範囲において用いる場合、単数形の「ある」、「1つの」および「その」は、特に断りのないかぎり複数の言及も含むことに注意しなければならない。したがって、例えば、「代謝産物」についての言及は1以上の代謝産物 (例えば、1、2、5、10、50、100またはそれを超える)についての言及であり、当業者に公知のその均等物なども含む。
【0009】
特に断りのないかぎり、本明細書において用いるすべての技術および科学用語は、本発明が属する技術分野の当業者に一般に理解されているものと同じ意味を有する。本明細書に記載のものと類似または同等のあらゆる方法、装置、および材料が本発明の実施または試験において用いることが出来るが、好ましい方法、装置および材料をここで記載する。
【0010】
本明細書において言及するすべての刊行物および特許は、例えば、このたび記載する本発明との関係において使用されうる刊行物に記載のコンストラクトおよび方法を記載および開示する目的のために引用により本明細書に含まれる。先におよび本明細書中に論じられる刊行物は本出願の出願日の前のそれらの開示のためにのみ提供される。ここに記載されるものはいずれも本発明者らが以前の発明によってかかる開示に先行するものでないと認めるものと解釈してはならない。
【0011】
細胞培養工学は、細胞培養物の性能に関する重要な疑問を解決することを試みて今日までトランスクリプトミクス(transcriptomic)、プロテオミクス、および代謝フラックス分析を用いてきた。しかし、哺乳類細胞培養物の生理学的状態の包括的特徴決定を可能とし、プロセスの一貫性およびロバスト性をモニターするための現在常套的な測定値のセットを超えて改善する、方法の開発および適用に対する明白な要求が存在する。
【0012】
メタボロミクスのハイスループット分子分析プラットフォームはこの要求を満たしうる。遊離低分子代謝産物プールの(相対的) 濃度の同時定量をいうメタボロミクスは、生物学的系の代謝フィンガープリントをモニターすることを可能とする (Fiehn、et al.、Nat. Biotechnol. 18:1157-1168、2000; Roessner、et al.、Plant J. 23:131-142、2000)。細胞機能全体との関係における代謝の役割を考慮すると、完全かつ正確な代謝プロフィール地図の定量がなぜ細胞培養工学研究において非常に重要であり得るかということは容易に理解できる。代謝産物濃度と代謝フラックスとは線形の関係にはないが、代謝プロファイリングはハイスループットであり、したがって一過性の代謝状態をモニターするために容易に用いることが出来る。さらに、代謝フラックス分析 (MFA)の場合におけるように、調べられた代謝経路網の構造および調節の知識は必要ではない。さらに、MFAは典型的には定常状態または偽-定常状態条件にのみ適用される一方、メタボロミクスは、その他の2つの主なオミクス(omic)プラットフォームであるトランスクリプトミクスおよびプロテオミクスのように一過性生理学的条件下で用いられ得る。また、トランスクリプトミクスおよびプロテオミクスとは異なり、メタボロミクスは特別な分析装置を必要としない。メタボロミクス(metabolomic)方法は、古典的な分析化学技術、例えば主に、核磁気共鳴 (NMR) 分光法および質量分析(MS) (例えば、ガスクロマトグラフィー-質量分析および液体クロマトグラフィー-質量分析)に基づくものであり、最もコストのかからないオミクス(omics)アプローチである(Kanani、et al.、J. Chromatogr B. Analyt. Technol. Biomed. Life Sci. 871:191-201、2008)。特に細胞培養系についてメタボロミクスがその他のオミクス技術よりも特に優れている点は、細胞内代謝状態および細胞外培地の組成の両方をモニターすることへのその適用可能性である。これは代謝経路網活性のよりよい理解を提供する。最後に、代謝は生物学的系の間で良好に保存されているので、比較メタボロミクス研究はより容易であり、サンプル間の凝った正規化を必要としない。
【0013】
分子フィンガープリンティングによってインビボでの代謝生理をモニターすることにより、従来の測定によっては直接的に観察することができなかったサンプル間の微妙な相違の判定が可能となる。細胞生理学的状態の増強された(enhanced)特徴決定は、現在用いられているプロセス制御方法を改善することが出来る代謝フィンガープリンティングによって可能となる。さらに、多変量統計解析により、その濃度変化が細胞培養物の生理学における相違を特徴づける代謝産物の同定が可能となる。この情報は一貫性のない(inconsistent)性能についての早期警戒マーカーを同定することを助けることが出来、最終的にはプロセス最適化を導く細胞生理学の理解もさらに助けることが出来る。
【0014】
メタボロミクスフィンガープリンティングは細胞培養工学における非常に有用な分子分析ツールでありうる。個別にまたは細胞機能のその他のレベルを評価するその他のハイスループット分子分析技術と組み合わせて、発酵プロセスの最適化および発酵の一連の活動の状況および品質の一貫性をモニターするために用いられている現在利用可能な測定値のセットの改善に向けての手がかりを提供しうる。
【0015】
本発明は、高密度かん流培養における哺乳類細胞生理をモニターするための細胞培養物のGC-MS メタボロミクスの使用に関する。GC-MS メタボロミクスは実験室および製造規模の両方について異なる培養段階での細胞の生理学的状態を分析するために用いることが出来る。
【実施例】
【0016】
本発明がよりよく理解され得るために、以下の実施例を示す。これらの実施例は例示の目的のみのためのものであり、いかなるようにも本発明の範囲を限定するものとして解釈されるべきではない。本明細書において言及するすべての刊行物はそれら全体を引用により本明細書に含める。
【0017】
実施例 1. 細胞かん流培養
BHK 細胞を主な炭素源としてグルコースおよびグルタミンを用いてかん流様式 (図 1)にて培養した。新生児ハムスター 腎臓 (BHK) 細胞の4つの独立のバイアル(図 2におけるA-D)をそれぞれ、4つの実験室-規模 15L かん流システム、L1-L4に接種するのに用いた。L1およびL4バイオリアクターは典型的な円筒状容器であり、一方、L2およびL3は底の小さい(small-bottom)リアクターであった。L2およびL3を、それぞれ2つの製造規模バイオリアクターであるM1およびM2に接種するのに用いた。すべての実験室規模および製造規模かん流システムはすべてのモニターした変数について同じ運転条件および設定値を有していた。バイオリアクター稼働の持続時間は113 - 155日の範囲であり、その結果、合計は826バイオリアクター日となる。各リアクターの作動の最後の40日の間に、サンプルを、表1に示すように様々な時点で収集した。細胞齢はバイアル解凍の日から測定し、同じリアクターにおける培養時間は各リアクターの開始日およびサンプル収集日の間の時間差として評価する。これらサンプルからの細胞ペレットをGC-MS メタボロミクスおよび多変量統計解析を用いて分析した。
【0018】
表1
【表1】
【0019】
バイオリアクター温度は35.5℃に維持し、47 RPM (製造規模バイオリアクターについては15 RPM)にて撹拌した。溶存酸素 (DO) 濃度は膜通気により50% 空気飽和(air saturation)に維持し、pHは6% Na2CO3の自動添加により6.8に維持した。バイオリアクターに出発細胞密度が〜1 x 106 細胞/mLとなるよう接種し、細胞を定常状態濃度の20 x 106 細胞/mLとなるまで蓄積させた。この標的定常状態細胞密度をバイオリアクターからの自動細胞抜き取り(bleed)により維持した。
【0020】
サンプルをCEDEX システム (Innovatis、Bielefeld、Germany)を用いる細胞密度および生存度分析のために各バイオリアクターから毎日収集した。これらのサンプルを次いで遠心分離にかけ(Beckman Coulter、Fullerton、CA)、上清を栄養素および代謝産物濃度について分析した。グルコース、乳酸、グルタミン、およびグルタミン酸濃度はYSI モデル 2700 分析器(Yellow Sprints Instruments、Yellow Springs、OH)を用いて決定し、一方、アンモニアはEktachem DT60 分析器 (Eastman Kodak、Rochester、NY)により測定した。pHおよびDOは格納式電極(Metler-Toledo Inc.、Columbus、OH)を用いてオンラインで測定し、その測定精度はRapidlab 248 血液ガス分析器 (Bayer HealthCare、Tarrytown、NY)におけるオフライン分析により確証した。同じ計器を用いて溶存 CO2 濃度を測定した。細胞密度のオンライン測定は格納式光学密度プローブ (Aquasant Messtechnik、Bubendorf、Switzerland)により行い、CEDEX システムからの細胞密度測定により較正した。
【0021】
実施例 2:メタボロミクスプロファイリング
バイオリアクターからのサンプルを氷上に引き取り(drawn)、あらかじめ冷却しておいたローターを用いて遠心分離した。遠心分離の後、上清を棄て、細胞ペレットを冷たいPBS バッファーで洗浄した。細胞ペレットを次いで70℃の水浴に15 分間入れておいた。ペレットを次いで減圧下で70℃で24 時間乾燥させた。サンプルからの乾燥細胞ペレットをメタボロミクス分析に用いた。
【0022】
乾燥細胞ペレットの極性(polar)代謝産物抽出物をメタノール/水抽出を用い(Kanani、et al.、2008; Roessner、et al.、2000)、リビトール (0.1mg/g乾燥細胞重量)および[U-13C]-グルコース (0.2 mg/g 乾燥細胞重量)を内部標準として得た。乾燥極性抽出物を、150μLのピリジン中のメトキシアミン塩酸塩溶液 (20 mg/mL)との90 分間の反応、次いで、300 μLのN-メチル-トリメチルシリル-トリフルオロアセタミド (MSTFA)との少なくとも 6 時間の室温での反応(Kanani、et al.、2008; Kanani、et al.、Metab. Eng.、9:39-51、2007)を介してそれらの(MeOx)TMS-誘導体へと誘導体化した。メタボロミクスプロフィール(metabolomic profile)を、Saturn 2200T ガスクロマトグラフ - (イオントラップ) 質量分析計 (Varian Inc.、CA)を用いて得た。ピーク同定および定量は、(Kanani、et al.、2007)に記載されているようにして行った。未加工の(raw)メタボロミクスデータセットは 91のピークから構成され、そのそれぞれは獲得されたメタボロミクスプロフィールの少なくとも1つにおいて検出され、化学的分類が公知の化合物に対応する(例えば、Kanani、et al.、2008; Kanani、et al.、2007参照)。
【0023】
すべての検出されたピークの相対的面積(RPA)を、内部標準であるリビトール (マーカーイオン: 217)による正規化(normalization)から見積もった。データ妥当性検証、正規化、および補正方法を、アミン基含有代謝産物からの複数の誘導体の形成に主に起因する誘導体化バイアスを説明するために適用した(Kanani、et al.、2008; Kanani、et al.、2007)。GC-MS 運転条件は、[U-13C]-グルコースの2つのピークの比に基づくサンプルの獲得された(acquired)メタボロミクスプロフィールの取得の際に確証した。第二に、95日のL4 サンプルのアミノ酸誘導体のプロフィールに基づいて評価された重み係数を用いて(表1参照)、同じアミン基含有代謝産物に対応する誘導体ピーク面積を併せて1つの累積 (有効) ピーク面積とした。イソロイシン、β-アラニン、およびグルタミン酸をさらなる分析から除去した。というのはそれらの利用可能な測定値が、すべての正の重み係数が見積もられることを可能としなかったからである。特定の誘導体化範囲において1つのみの誘導体が観察されたが1より多い誘導体が知られているアミノ酸は、分析の次の工程に含めた; 非常にしばしばそれらは、注入間の変動の高い係数のためにその後の工程において除去された。さらに、(a)公知のケトン基含有代謝産物の2つのMeOx ピークのうち最も小さいもの、(b)未知のアミン基含有代謝産物に対応するピーク、(c)誘導体化アーティファクトであると同定されたかまたは有意なキャリーオーバーを有するピーク、および(d) 一貫しては検出されなかった代謝産物ピークは分析から除去した。38の代謝産物を含むこれらの最終代謝産物 RPA プロフィールを、80%カットオフを用いるTM4 MeV (V4.0) データ分析ソフトウェア (Saeed、et al.、Biotechniques 34:374-378、2003)において用いた。あらゆる不明のRPAをTM4 MeV に備えられているためk近傍法を用いて帰属させた (Troyanskaya、et al.、Bioinformatics 17:520-525、2001)。
【0024】
獲得されたメタボロミクスプロフィールに対して適用された分析は代謝産物相対ピーク面積の標準値に基づくものとした(下記式 1参照)。これらの分析における標準化相対ピーク面積の使用は同じ代謝プロフィールにおける代謝産物の間の相対ピーク面積レベルの桁における大きな違いによって必要とされる。メタボロミクスプロフィール jにおける代謝産物 Mの標準化相対ピーク面積、RPAMjは以下に等しい:
【数1】
【0025】
階層的クラスタリング(Hierarchical clustering) (HCL)を用いてサンプルをそれらのメタボロミクスプロフィールに基づいてクラスタリング(cluster)した。HCLにおいて、代謝プロフィールは階層木においてクラスタリングされる。かかる木の最も低いレベルにおいて、各代謝プロフィールは別々のクラスターであるとみなされるが、すべてのサンプルはもっとも高いレベルにおいて1つのクラスターにグルーピングされる。もっとも低いレベルから出発して、入手可能なクラスターの各対についての相関係数がアルゴリズムの各ラウンドにて特定の距離メトリックに基づいて見積もられる。最も高い相関係数を有するクラスターはアルゴリズムの次のラウンドのために1つのクラスターへとグルーピングされる (Quackenbush、Nat. Genet. 2:418-427、2001; Eisen、et. al.、Proc. Natl. Acad. Sci. USA 95:14863-14868、1998)。獲得された階層木は、研究される生物学的問題の観点から解釈されなければならない。例えば、図 5Aにおいて、HCLは、1ヶ月離れた細胞齢に対応する代謝プロフィールの2つのクラスターを同定する。より低い細胞齢クラスターのなかで、リアクター L4の代謝プロフィールはその他の3つの実験室-規模リアクターの代謝プロフィールから離れている。
【0026】
主成分分析 (PCA)を用いて様々な細胞培養物サンプルがそれらのメタボロミクスプロフィールに基づいて識別可能かどうかを可視化した。PCAはデータマトリックスの特異値分解を伴い、それはデータセットにおける最大の分散(variance)をとらえる新しい座標系の上への元のデータセットの直交線形変換をいう(Strang、Introduction to Linear Algebra、Wellesley-Cambridge Press、Wellesley、Massachusetts、1993)。この変換は元の実験空間の回転および/または伸縮を伴う可能性がある。主成分 1は元のデータセットにおける最高の分散の方向に対応し、主成分 2は第二に高い分散の方向に対応し、以下同様である。ハイスループット生物学的データ分析において、PCAは簡略化の目的で座標化するために用いられており、それゆえ元のデータセットにおける分散の大部分は3-D 空間内に可視化される (Raychaudhuri、et. al.、Pac. Symp. Biocomput. 2000:455,466、2000)。少数の主成分がしばしばデータにおける構造のほとんどを説明するために十分である (Scholkopf、et al.、Learning with Kernels - Support Vector Machines、Regularization、Optimization and Beyond、The MIT Press、Cambridge、Massachusettes、2002)。PCAグラフを読む場合、個別に示された主成分のすべておよび個々によって示された空間内にとらえられている元のデータセットにおける分散のパーセンテージを考慮することが重要である。各主成分の単位(重み)はそれによって表される元のデータセットにおける分散のパーセンテージと等しい。したがって、例えば、主成分 1上で距離がxである2つのデータポイントは主成分 2上で同じ距離にある2つのデータポイントよりもそれらの生理学的状態において大きい差を表す。
【0027】
1つのセットの細胞培養物サンプルにおいて別のセットと比較してその濃度が有意により高かったかまたは低かった代謝産物を、それぞれ特定の比較の正にまたは負に有意な代謝産物と称した。1つの比較における有意な代謝産物を対応のないマイクロアレイの有意性分析(Significance Analysis of Microarrays)(SAM) アプローチ (Tusher、et al.、Proc. Natl. Acad. Sci. USA 98:5116-5121、2001)を用いて同定した。
【0028】
SAM (Tusher、et al.、2001; Larsson、et al.、BMC Bioinformatics 6:129、2005; Wu、Bioinformatics 21:1565-1571、2005)は、異なる生理学的条件を表す2つの測定セットの間で有意に異なる分子の量の同定のための置換に基づく(permutation-based)(ノンパラメトリック) 仮説検証方法である。SAMはDNA マイクロアレイに基づく転写プロファイリングデータの分析のために注文設計されてきており、その他のオミクス(omic)データセットの分析のために同様に用いられてきている(例えば、Dutta、et. al.、Biotechnol. Bioeng. 102:264-279、2009参照)。メタボロミクス分析の場合、SAMは2つのサンプルの間のその濃度における相違が、ランダムな変動のみに起因して予測されうる相違よりも大きい代謝産物を同定する:
【数2】
式中、diは観察された相違であり、deiは期待された相違であり、δは有意性閾値である。パラメトリック仮説検証方法とは異なり、置換に基づく(permutation-based)(ノンパラメトリック)方法ではデータが特定の分布に従う必要はない。それらはまた偽発見率 (FDR)の見積を提供し、偽発見率は、濃度において示差的に(differentially)変化しているとして同定された所与の代謝産物が偽陽性である確率である。さらに、SAM はFDRおよび多数の有意な(significant)代謝産物の閾値の変化に対する感度が決定され得るようにδの調整を可能とする。
【0029】
TM4 MeV v4.0 (Saeed et al.、2003)に実装されているアルゴリズムを用いた。2つの細胞培養物サンプルの間の代謝プロフィールにおける相違の全体論的視野を、適当に色分けされた代謝経路網において有意であると同定された代謝産物を位置づけすることによって得た。代謝経路網再構築はKEGG (KEGG Database、2008)およびEXPASY (EXPASY Database、2008)データベースからの情報に基づいた。
【0030】
プロセスの一貫性を確実にするためにかん流システムにおいて注意深く制御される必要がある基本変数の1つは生存細胞密度である。かん流システムが定常状態条件に到達した後は、培養の過程にわたってそれは典型的にはターゲット設定値に維持される。定常状態細胞密度はターゲット設定値である20 x 106 細胞/mL付近に維持された (表2、図 3)。図 3はそれらの作動の過程にわたってのリアクターについての生存細胞密度の時間プロフィールを示し、一方、定常状態平均は表2に示される。細胞増殖-、代謝活性-、および生産性-関連変数についての定常状態平均が示される。変動の係数は括弧内に示される。FVCD、sGCR、およびsLPRはそれぞれ、バイオリアクター生存細胞密度、特異的グルコース消費速度、および特異的乳酸生産速度である。すべてのリアクターの平均特異的生産性はL1 リアクターについての平均値との比較にて示される。
【0031】
細胞密度測定には〜8.5%の誤差が伴うことが示され、平均値の周囲の観察された変動(即ち、4.4% - 6.3%の範囲の変動の係数(CoV’s))は、良好な細胞密度制御を反映して非常に小さいことが示唆される。
【0032】
表2
【表2】
【0033】
図 4 は、この研究におけるリアクターについての、a)バイオリアクター生存度、b) 細胞増殖、c) 特異的グルコース消費速度、および、d) 特異的乳酸生産速度の時間プロフィールを提供する。それぞれのリアクターについての培養の過程は、定常状態に達した時点から開始して10-日間隔に分けた。それぞれの間隔にわたる平均値および伴う標準偏差は間隔の中央の時点にて表される。20 - 30日の細胞齢からのデータの平均をとり、25日の細胞齢に対応するものとして示した。有意な時間関連変動は、生存度および増殖速度についてはみられなかった。わずかに上昇する傾向が特異的グルコース消費および乳酸生産速度の時間プロフィールについてみられた。プロセスをモニターするための細胞培養工学において用いられた測定値のセットに基づいて、リアクターのそれぞれにおいて細胞齢に起因する生理の明らかな相違はない。リアクター間での一貫性はまた、平均特異的タンパク質生産性データ(表2)においても観察され、ここでリアクター L1の生産性は任意に1.0に設定され、比較のために用いた。0.92 - 1.04の範囲の特異的生産性は明らかにバイオリアクター間の一貫性を示す。さらに、バイオリアクターからの初期、中期および後期ステージからの材料を精製し、典型的なタンパク質品質特性について試験した。生成物品質データは仕様の範囲内(within specification)であり、バイオリアクターの間および培養の経過にわたっての一貫性を示唆している。
【0034】
実験室-規模バイオリアクターの分析からのデータを図5Aおよび5Bに示す。明らかな細胞齢に基づく差別化が主成分 2 (図 5B)上においてみられ、ここで、120-123 日サンプルの代謝プロフィールはグラフの上側部分にみられるが、148-150 日サンプルのものは下側部分にある。異なる実験室-規模リアクターから獲得された同じ細胞齢の培養物の代謝プロフィールは主成分 1の上で主に明らかに差別化されている。主成分 1は元のデータセットにおける相違のもっとも大きな部分を有するため、この差別化は、メタボロミクスプロフィールは、同じ細胞バンクにおけるバイアルとバイアルの間の変動および/または異なるバイオリアクターにおける細胞培養物増殖を識別する力がありうることを示す。これは、現在のモニターするためのツールボックスに基づいては観察可能ではない、生理におけるわずかな相違を同定する生体分子フィンガープリンティングの能力を支持する。さらに図 5Bの調査は、L3およびL4 サンプル(グラフの右側)の、両方の調べた細胞齢のL1およびL2 サンプル(グラフの左側)からの分離を示す。
【0035】
図 5Aに示す階層的クラスタリング (HCL) 分析結果は、PCAグラフにおける代謝プロフィールの間の可視化された相違を確認する。代謝プロフィールをそれらの形状に基づいて階層化するピアソンの相関距離メトリックを用いて、得られた階層的クラスタリング木は、2つの異なる細胞齢の代謝プロフィールに対応する2つの主な枝を含む。2つの代謝プロフィールの間のピアソンの相関係数であるrは、2つのプロフィールの共分散をそれらの標準偏差の積で割った値に等しい (Box et. al.、1978)。それは-1と1との間の値をとりうる。共分散は2つのプロフィールの間の線形依存性の尺度である。したがって、ピアソンの相関は、2つのプロフィールの形状の間の類似性を明らかにすることが期待される (Quackenbush、2001)。
【0036】
より低い細胞齢の枝(木の左側)のなかで、2つの別々のサブクラスターが同定された; 1つは123 日L4 サンプルの3注入(injection)からの代謝プロフィールを含み、別の木の代謝プロフィールではリアクターのサンプルが別の亜枝にクラスタリングしている。L4 代謝プロフィールデータポイントは、PCAグラフから示されるように実験空間における残りからさらに離れていた (図 5B)。図 5A に示すL4 注入の平均代謝プロフィール(この重心(centroid)プロフィールはTM4 MeVソフトウェアから作成した; Saeed、et. al.、2003)は、同様の細胞齢のリアクターの残りの平均代謝プロフィールとは異なっている。
【0037】
ユークリッド距離を用いるHCL 分析は、両方の細胞齢のL3およびL4 プロフィールのL1およびL2 プロフィールからの差別化を示した。ユークリッド距離メトリックは、ユークリッド空間における2つの地点の間の古典的な(最短の)距離であり、ピタゴラスの定理によって規定される。M-次元空間において、地点、x = (x1、x2、…、xM)および y = (y1、y2,…、yM)の間のユークリッド距離は以下のように規定される:
【数3】
したがって、2-次元空間において、基点からの同じユークリッド距離の点によって囲まれる領域は特定の距離と等しい半径を持った円である。
【0038】
PCAおよびHCL 分析の両方が、バイアルとバイアルの間の変動性および細胞齢がリアクター幾何学(標準的円筒状、対、底の小さい立体配置)よりも高い影響を細胞培養物サンプルの測定される極性代謝プロフィールに対して有することを示した。入手可能な実験室-規模培養サンプルにおいて変動するこれら3つのすべてのパラメーターのなかで、リアクター幾何学の代謝生理に対する効果は、サンプルのクラスタリングにおいて直接的に明らかではない。
【0039】
図6Aおよび6Bは、製造規模リアクターサンプルの代謝プロフィールについての、それぞれHCLおよびPCA 分析の結果を示す。サンプルの細胞齢に基づく分離が主成分 1においてみられる(図 6B)。この分離は HCL 分析 (図 6A)によっても明らかであり、ここで、階層的クラスタリング木の二つの主な枝は2つの細胞齢に対応する。図 6Aに示す2つの細胞齢の平均代謝プロフィールもまた、この差別化を支持する。
【0040】
122-129日および149-150 日サンプル群のなかで、HCLおよびPCAはともにM1およびM2 サンプルの間の明らかな差別化を示した。149-150 日サンプルのM1およびM2 代謝プロフィールの間の相違は122-129 日齢の群の対応するものよりも大きかった。最初の場合において、サンプルは主成分 1上で分離しており (48% 分散)、第二のセットのサンプルは主成分 2上で分離していた (16.5%分散)。
【0041】
製造規模バイオリアクターの38の極性代謝産物プロフィールを次いで、それらから接種された実験室-規模バイオリアクターのプロフィールと組み合わせて分析した。図7A-DはこのセットのサンプルについてのHCLおよびPCA 分析の結果を示す。いずれの距離メトリックに基づいたPCAおよび HCL 分析も、製造規模バイオリアクター、M1および M2の122-129日齢サンプルの代謝プロフィールを、すべてのその他のサンプルから別れたクラスターであると同定した。HCL 分析のいずれにおいても(図 7A-C)、これらサンプルは木の2つの主な枝の1つを形成しており、それらはまた、PCA グラフの右側に分離して示される(図 7D)。
【0042】
図 7における残りの5つのサンプルは、細胞齢、リアクタータイプ、および細胞源に基づいて分類され得た。2つのサンプル (L2およびL3)は121 - 122日の範囲の細胞齢を有しており、一方L2、M1、およびM2は149 - 150 日サンプルであった。3つのサンプルは実験室-規模バイオリアクターからのものであり、一方2つは製造規模システムからのものであった。サンプル対 L2-M1およびL3-M2は同じ細胞源からのものであった(それぞれ、バイアルBおよびC)。図 7Dにおいて、これらのサンプルは、リアクターサイズおよび細胞源に基づいて明らかに差別化されている。リアクターサイズに基づく差別化は主成分 2 上にあり、実験室-規模リアクターは正の側にあり、製造規模リアクターは負の側にあった。細胞源に基づく差別化は主成分 1上にあり、L2およびM1 サンプルはL3およびM2 サンプルよりも左側にクラスタリングされていた。
【0043】
細胞源に基づくクラスタリングも、ユークリッドまたはマンハッタン距離メトリックが用いられた場合、HCL 分析において明らかである。M-次元空間において、地点x = (x1、x2、…、xM)およびy = (y1、y2,…、yM)の間のマンハッタン距離は、座標軸上への(x,y) 線分の投影の長さの和として規定される:
【数4】
2-次元空間において、起点から同じマンハッタン距離の点によって囲まれる領域は、辺が座標軸に対して45°の方向である正方形である。マンハッタン距離は異常値に対してユークリッド距離よりも感度が低い(Filzmoser、et. al.、Comput. Stat. Data Anal. 52:1694-1711、2008)。マンハッタンとユークリッド距離とは共に入手可能なデータベクトル(この場合は、代謝プロフィール)の間の絶対差を測る。クラスタリング分析において用いた場合、両方のメトリックは、代謝プロフィールのピーク面積レベルの間の類似度を明らかにすると期待される。
【0044】
図 7Aにおいて、階層木の右側の枝は3つの枝に分かれており、1つはM2およびL3 サンプルをクラスタリングしており、もう1つは150日L2およびM1 サンプルを、残りの1つは122日L2 サンプルをクラスタリングしており、PCAグラフと類似している(図 7D)。HCL 分析におけるリアクターサイズに関する代謝プロフィールのクラスタリングは、ケンドールのタウ(Kendall’s Tau)距離メトリックを用いるとみられた。このメトリックは代謝プロフィールのランキングベクトルを指す。代謝プロフィールのランキングベクトルにおいて、各相対ピーク面積が、代謝プロフィールにおけるすべての相対ピーク面積のなかのそのランキングを示す整数に取って代わられる。この場合、2つのランキングベクトルの間の距離は2つのベクトルにおいて2つの整数が逆の順にある回数に基づいて測定される。もしこの数が0である場合、2つのランキングベクトルは同一である。
【0045】
図 7Bにおいて、階層木の右側の枝は、実験室-規模および製造規模サンプルに対応する2つの亜枝に分かれる。最後に、スピアマン相関距離メトリックを用いるHCL 分析は細胞齢に基づくサンプルクラスタリングを示した。この相関は、代謝プロフィールのランキングベクトルに対するピアソンの相関距離メトリックの適用を指す。図 7Cにおいて、階層木の右側の枝は2つの亜枝に分かれ、1つは121-122日L2およびL3 サンプルを含み、もう1つは149-150日L2、M1およびM2 サンプルを含む。
【0046】
マイクロアレイの有意性分析(SAM)を用いて、122 日サンプルと比較して129 日 M1 サンプルにおいてその濃度が有意に上昇 (正に有意)または低下(負に有意)した代謝産物を同定した。これらのサンプルは1週間しか離れていないにもかかわらず、それらはHCL およびPCA 分析から差別化できた。SAM (1.64のデルタ値および0% 偽発見率 (FDR)について)は、5つの正に有意なそして0の負に有意な代謝産物を同定し、それらは有意性の高さの順に、フマル酸(fumarate)、グリセロール-3-リン酸、尿素、ウラシル、およびピルビン酸(pyruvate)であった。4つの代謝産物を代謝経路網との関連において図 8に示す。高濃度のフマル酸および尿素は、122 日サンプルと比較して129 日における尿素サイクルのより高い活性を示し得、したがって窒素同化のより高い活性を示し得る。さらに、グリセロール-3-リン酸の濃度上昇はグリセロ脂質の生産上昇を示しうる。ウリジンの前駆体であって、ウリジンリン酸(UMP、UDPおよびUTP)の主要な構成成分であるウラシルは、炭水化物代謝、タンパク質グリコシル化および糖脂質形成に重要な役割を果たす。このタイプの情報はバイオリアクター作動の最適化において有用であり得ることが明白である。
【0047】
細胞生理学的状態のより高い分解能の特徴決定が代謝産物プロファイリングによって可能となった。メタボロミクスプロフィールは、目的のパラメーター、例えば、細胞齢についての1以上の識別力のある代謝産物の同定に利用可能であった。例えば、メタボロミクスプロフィールは目的のパラメーターについての1、2、5、10、50、100またはそれを超える識別力のある代謝産物の同定に利用することが可能であった。これらの識別力のある代謝産物は従来測定されてきた細胞培養物生理学的変数と組み合わせてバイオリアクター培養の最適化に用いることが出来、早期警戒またはプロセス混乱(upset)として役立ちうる。メタボロミクスは細胞培養工学において高感度のハイスループット分子分析ツールとして利用することが出来る。
【0048】
本発明のその他の態様は、本明細書の考慮または本明細書に開示する本発明の実施から当業者に明らかであろう。本明細書および実施例は例示的にのみ考慮されるべきであって、本発明の真の範囲と精神は、以下の請求の範囲によって示されることが意図される。
【特許請求の範囲】
【請求項1】
以下の工程を含む、細胞培養物の生理学的状態をモニターする方法:
(a)第1のバイオリアクターから採取した第1の培養サンプルにおける1以上の代謝産物のレベルを決定する工程;
(b)第2のバイオリアクターから採取した少なくとも第2の培養サンプルにおける1以上の代謝産物のレベルを決定する工程;および、
(c)第1の培養サンプルにおける1以上の代謝産物のレベルと第2の培養サンプルにおける代謝産物のレベルとを比較する工程;
ここで、第1の培養サンプルにおける1以上の代謝産物のレベルの、第2の培養サンプルにおける1以上の代謝産物のレベルと比較しての変化は、細胞培養物の生理学的状態の指標である。
【請求項2】
生理学的状態が、細胞増殖、代謝プロフィール、および細胞齢から選択される、請求項1の方法。
【請求項3】
細胞培養物が哺乳類細胞培養物である、請求項1の方法。
【請求項4】
哺乳類細胞培養物が新生児ハムスター腎臓細胞培養物である請求項3の方法。
【請求項5】
バイオリアクターが、実験室規模のバイオリアクター、製造規模のバイオリアクター、かん流バイオリアクター、および流加回分バイオリアクターから選択される、請求項1の方法。
【請求項6】
代謝産物のレベルが、質量分析およびNMRによって決定される、請求項1の方法。
【請求項7】
代謝産物のレベルが、ガスクロマトグラフィー-質量分析および液体クロマトグラフィー-質量分析によって決定される、請求項1の方法。
【請求項8】
代謝産物のレベルの比較が、多変量統計解析を用いて決定される、請求項1の方法。
【請求項9】
代謝産物のレベルの比較が、階層的クラスタリングおよび主成分分析から選択される方法を用いて決定される、請求項1の方法。
【請求項10】
代謝産物が、アミン基含有代謝産物およびケトン基含有代謝産物から選択される、請求項1の方法。
【請求項11】
代謝産物が、グルコース、乳酸、グルタミン、グルタミン酸、アンモニア、フマル酸、グリセロール-3-リン酸、尿素、ウラシル、およびピルビン酸から選択される、請求項1の方法。
【請求項12】
方法がハイスループット方法である請求項1の方法。
【請求項13】
代謝産物が細胞内または細胞外のものである請求項1の方法。
【請求項14】
以下の工程を含む、細胞培養物の生理学的状態をモニターする方法:
(a)バイオリアクターから採取した培養サンプルにおける1以上の代謝産物のレベルを決定する工程;および、
(b)培養サンプルにおける1以上の代謝産物のレベルを標準培養サンプルにおける代謝産物のレベルと比較する工程;
ここで、培養サンプルにおける1以上の代謝産物のレベルの標準培養サンプルにおける1以上の代謝産物のレベルと比較しての変化は、細胞培養物の生理学的状態の指標である。
【請求項15】
生理学的状態が、細胞増殖、代謝プロフィール、および細胞齢から選択される、請求項14の方法。
【請求項16】
細胞培養物が哺乳類細胞培養物である請求項14の方法。
【請求項17】
哺乳類細胞培養物が新生児ハムスター腎臓細胞培養物である請求項16の方法。
【請求項18】
バイオリアクターが、実験室規模のバイオリアクター、製造規模のバイオリアクター、かん流バイオリアクター、および流加回分バイオリアクターから選択される、請求項14の方法。
【請求項19】
代謝産物のレベルが、質量分析およびNMRにより決定される、請求項14の方法。
【請求項20】
代謝産物のレベルが、ガスクロマトグラフィー-質量分析および液体クロマトグラフィー-質量分析により決定される、請求項14の方法。
【請求項21】
代謝産物のレベルの比較が、多変量統計解析を用いて決定される、請求項14の方法。
【請求項22】
代謝産物のレベルの比較が、階層的クラスタリングおよび主成分分析から選択される方法を用いて決定される、請求項14の方法。
【請求項23】
代謝産物が、アミン基含有代謝産物およびケトン基含有代謝産物から選択される、請求項14の方法。
【請求項24】
代謝産物が、グルコース、乳酸、グルタミン、グルタミン酸、アンモニア、フマル酸、グリセロール-3-リン酸、尿素、ウラシル、およびピルビン酸から選択される、請求項14の方法。
【請求項25】
方法がハイスループット方法である請求項14の方法。
【請求項26】
代謝産物が細胞内または細胞外のものである請求項14の方法。
【請求項27】
請求項1または請求項14の方法により作成されたメタボロミクスプロフィール。
【請求項1】
以下の工程を含む、細胞培養物の生理学的状態をモニターする方法:
(a)第1のバイオリアクターから採取した第1の培養サンプルにおける1以上の代謝産物のレベルを決定する工程;
(b)第2のバイオリアクターから採取した少なくとも第2の培養サンプルにおける1以上の代謝産物のレベルを決定する工程;および、
(c)第1の培養サンプルにおける1以上の代謝産物のレベルと第2の培養サンプルにおける代謝産物のレベルとを比較する工程;
ここで、第1の培養サンプルにおける1以上の代謝産物のレベルの、第2の培養サンプルにおける1以上の代謝産物のレベルと比較しての変化は、細胞培養物の生理学的状態の指標である。
【請求項2】
生理学的状態が、細胞増殖、代謝プロフィール、および細胞齢から選択される、請求項1の方法。
【請求項3】
細胞培養物が哺乳類細胞培養物である、請求項1の方法。
【請求項4】
哺乳類細胞培養物が新生児ハムスター腎臓細胞培養物である請求項3の方法。
【請求項5】
バイオリアクターが、実験室規模のバイオリアクター、製造規模のバイオリアクター、かん流バイオリアクター、および流加回分バイオリアクターから選択される、請求項1の方法。
【請求項6】
代謝産物のレベルが、質量分析およびNMRによって決定される、請求項1の方法。
【請求項7】
代謝産物のレベルが、ガスクロマトグラフィー-質量分析および液体クロマトグラフィー-質量分析によって決定される、請求項1の方法。
【請求項8】
代謝産物のレベルの比較が、多変量統計解析を用いて決定される、請求項1の方法。
【請求項9】
代謝産物のレベルの比較が、階層的クラスタリングおよび主成分分析から選択される方法を用いて決定される、請求項1の方法。
【請求項10】
代謝産物が、アミン基含有代謝産物およびケトン基含有代謝産物から選択される、請求項1の方法。
【請求項11】
代謝産物が、グルコース、乳酸、グルタミン、グルタミン酸、アンモニア、フマル酸、グリセロール-3-リン酸、尿素、ウラシル、およびピルビン酸から選択される、請求項1の方法。
【請求項12】
方法がハイスループット方法である請求項1の方法。
【請求項13】
代謝産物が細胞内または細胞外のものである請求項1の方法。
【請求項14】
以下の工程を含む、細胞培養物の生理学的状態をモニターする方法:
(a)バイオリアクターから採取した培養サンプルにおける1以上の代謝産物のレベルを決定する工程;および、
(b)培養サンプルにおける1以上の代謝産物のレベルを標準培養サンプルにおける代謝産物のレベルと比較する工程;
ここで、培養サンプルにおける1以上の代謝産物のレベルの標準培養サンプルにおける1以上の代謝産物のレベルと比較しての変化は、細胞培養物の生理学的状態の指標である。
【請求項15】
生理学的状態が、細胞増殖、代謝プロフィール、および細胞齢から選択される、請求項14の方法。
【請求項16】
細胞培養物が哺乳類細胞培養物である請求項14の方法。
【請求項17】
哺乳類細胞培養物が新生児ハムスター腎臓細胞培養物である請求項16の方法。
【請求項18】
バイオリアクターが、実験室規模のバイオリアクター、製造規模のバイオリアクター、かん流バイオリアクター、および流加回分バイオリアクターから選択される、請求項14の方法。
【請求項19】
代謝産物のレベルが、質量分析およびNMRにより決定される、請求項14の方法。
【請求項20】
代謝産物のレベルが、ガスクロマトグラフィー-質量分析および液体クロマトグラフィー-質量分析により決定される、請求項14の方法。
【請求項21】
代謝産物のレベルの比較が、多変量統計解析を用いて決定される、請求項14の方法。
【請求項22】
代謝産物のレベルの比較が、階層的クラスタリングおよび主成分分析から選択される方法を用いて決定される、請求項14の方法。
【請求項23】
代謝産物が、アミン基含有代謝産物およびケトン基含有代謝産物から選択される、請求項14の方法。
【請求項24】
代謝産物が、グルコース、乳酸、グルタミン、グルタミン酸、アンモニア、フマル酸、グリセロール-3-リン酸、尿素、ウラシル、およびピルビン酸から選択される、請求項14の方法。
【請求項25】
方法がハイスループット方法である請求項14の方法。
【請求項26】
代謝産物が細胞内または細胞外のものである請求項14の方法。
【請求項27】
請求項1または請求項14の方法により作成されたメタボロミクスプロフィール。
【図1】

【図2】
【図3】
【図4−A】
【図4−B】
【図5−A】
【図5−B】
【図6−A】
【図6−B】
【図7−A】
【図7−B】
【図7−C】
【図7−D】
【図8】
【図2】
【図3】
【図4−A】
【図4−B】
【図5−A】
【図5−B】
【図6−A】
【図6−B】
【図7−A】
【図7−B】
【図7−C】
【図7−D】
【図8】
【公表番号】特表2012−520078(P2012−520078A)
【公表日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願番号】特願2011−554158(P2011−554158)
【出願日】平成22年3月10日(2010.3.10)
【国際出願番号】PCT/US2010/026843
【国際公開番号】WO2010/104963
【国際公開日】平成22年9月16日(2010.9.16)
【出願人】(503106111)バイエル・ヘルスケア・エルエルシー (154)
【Fターム(参考)】
【公表日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願日】平成22年3月10日(2010.3.10)
【国際出願番号】PCT/US2010/026843
【国際公開番号】WO2010/104963
【国際公開日】平成22年9月16日(2010.9.16)
【出願人】(503106111)バイエル・ヘルスケア・エルエルシー (154)
【Fターム(参考)】
[ Back to top ]