
細胞外マトリックスタンパク質のアミノ酸配列RGDに特異的なヒト化抗体およびその使用
本発明は、RGD配列を免疫特異的に認識するヒト化抗体を提供する。こうした抗体の一部は、RGDタンパク質の生体機能を阻害することにより、癌、例えば、癌細胞の増殖および転移、ならびに炎症性疾患、例えば、関節リウマチ、骨関節炎、肝炎、子宮内膜症、気管支喘息、線維症、糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などを含めた、RGDタンパク質に関連する種々の障害または疾患に対して治療効果を示す。
【発明の詳細な説明】
【技術分野】
【0001】
1.発明の分野
本発明は、細胞外マトリックスタンパク質のアミノ酸配列RGD(Arg−Gly−Asp)を免疫特異的に認識するヒト化抗体、ならびに癌、炎症性疾患、自己免疫疾患、感染症、骨疾患などを含めた種々の疾患または障害に対するその治療および診断的使用に関する。
【0002】
本仮出願を通じて、いくつかの刊行物(特許および特許出願を含む)を参考文献として本明細書に引用する。これらの刊行物の開示はその全体が、本発明が属する分野の技術をより十分に説明するために、参照により本仮出願に組み込まれる。
【背景技術】
【0003】
2.発明の背景
細胞接着は、多細胞生物の生命維持に重要な役割を果たしている。多細胞生物の細胞接着は、細胞−細胞外マトリックス(以下「ECM」と略記する)間接着と細胞間接着に分類される。細胞−ECM間接着はインテグリンによって媒介され、細胞間接着はカドヘリン、クローディンおよびネクチンによって媒介されることが解明されている。
【0004】
インテグリンなどの膜貫通型接着タンパク質が細胞−ECM間接着を構成している。インテグリンは、α鎖およびβ鎖からなるヘテロ二量体を形成する。少なくとも18種類のα鎖、8種類のβ鎖および24種類のαβヘテロ二量体がこれまで同定され、確認されている。インテグリンのそれぞれの種類が、特異的なリガンドを認識する。インテグリンを含めた膜貫通型接着タンパク質は、細胞接着に加えて、ECMから細胞中への細胞内シグナル伝達ならびに増殖、移動および分化の調節に関連する(F.G.Giancottiら、Science、285、1028〜1032、1999)。
【0005】
ECMタンパク質として多数のタンパク質が知られており、これらはコラーゲン群(I型〜XIX型など)、非コラーゲン性糖タンパク質群(オステオポンチン(OPN)、ビトロネクチン、フィブロネクチン、フォンウィルブランド因子、ラミニン、テネイシン、フィブリノゲン、トロンボスポンジンなど)、エラスチン群およびプロテオグリカン群に分類される。これらのECMタンパク質は対応するインテグリンに結合し、細胞内シグナル伝達経路を活性化して細胞骨格形成、移動、増殖、分化などを調節する。ECMタンパク質が結合したインテグリンは、ECMタンパク質の種類によって特異的なシグナルを伝達することにより、これらのシグナル活性化経路を調節する。多数のECMタンパク質の細胞接着領域にRGD配列が共通に認められ、インテグリンと結合することによって様々な機能を発揮する。このECMタンパク質のRGD配列は薬物の可能な標的と考えられており、いくつもの小分子化合物や人工ペプチドが提供されている。
【0006】
α3β1インテグリン、α5β1インテグリン、α8β1インテグリン、αvβ1インテグリン、αvβ3インテグリン、αvβ5インテグリン、αvβ6インテグリン、αvβ8インテグリンなど、いくつかの種類のインテグリンがRGD配列に結合することが知られている。α5β1インテグリンとその特異的なリガンドであるフィブロネクチンの間の相互作用から、インテグリンを介したシグナル伝達の機序についての研究が盛んに行われている。このような研究から、α5β1インテグリンは細胞接着および細胞移動だけでなく、細胞分化および細胞死も調節することが示されている(S.M.Frischら、Curr.Opin.Cell Biol.9、701〜706、1997)。α5β1インテグリンは腫瘍細胞で高度に発現され、癌の悪性化に関連することも示されている。インテグリンを介したシグナルはそれぞれ、結合しているECMタンパク質によって異なり、例えば、増殖因子による刺激はフィブロネクチンが結合した内皮細胞の増殖を活性化するが、ラミニン−1が結合した内皮細胞の増殖を阻害する。また、ラミニン−10/11からα3β1インテグリンに伝達されるシグナルは、フィブロネクチンからα5β1インテグリンに伝達されるシグナルと異なり、癌性細胞の移動を著しく増強し(J.Guら、J.Biol.Chem.、276、27090〜27097、2001)、血液飢餓(blood starvation)によるアポトーシスを大きく回避する(J.Guら、J.Biol.Chem.、277、19922〜19928、2002)。RGD配列が結合したαvインテグリンの高発現は、破骨細胞および新生血管において認められており、RGD配列およびαvインテグリンの阻害が、骨粗鬆症および癌の治療薬の標的であると考えられている。α5β1インテグリンは腫瘍細胞で高度に発現され、癌の悪性化に関連することが示されている。これらの知見から、インテグリンとECMタンパク質の相互作用を阻害する抗インテグリンアンタゴニスト抗体薬として、抗α5β1インテグリン抗体(ボロシマブ(Volocimab))、抗α4インテグリン抗体(ナタリズマブ(Natalizumab))および抗αvβ3インテグリン抗体(ビタキシン(Vitaxin))が開発されている。
【0007】
一方、コラーゲン、オステオポンチン(OPN)、ビトロネクチン、フィブロネクチン、フォンウィルブランド因子、ラミニン、テネイシン、フィブリノゲン、トロンボスポンジンなど、いくつかのECMタンパク質はRGD配列を含むことが知られている。また、一部のウイルスおよび一部の細菌も、細胞に付着するためのRGD配列を有することが知られている。OPNは、骨に豊富に含有されるカルシウムに対する結合特性を有する酸性糖タンパク質である。OPNは、細胞接着、細胞遊走、腫瘍形成、免疫応答および補体媒介性細胞溶解に重要な役割を果たすことが報告されている。OPNノックアウトマウスおよび抗OPN中和抗体の結果から、OPNは肝炎、自己免疫疾患(関節リウマチなど)および癌の転移に関連することが示される。細胞に対するECMタンパク質の結合の阻害剤を骨粗鬆症または癌の治療に使用できることが知られている。したがって、インテグリンを標的とする上記の拮抗薬に加えて、インテグリンの結合パートナーであるECMタンパク質を標的とする拮抗薬が開発されている。
【発明の概要】
【発明が解決しようとする課題】
【0008】
3.発明の概要
RGD配列を介したインテグリンとの相互作用を阻害する小分子、OPNに対する抗体、インテグリンに対する抗体などの薬物は報告されているが、RGD配列を特異的に認識する抗体に関する報告はまだない。RGD配列はECMタンパク質の保存配列の1つであるので、RGD配列を特異的に認識する抗体は、ヒトと治療モデル動物の両方で効果を有する可能性があり、したがって治療薬の開発に非常に有用な有効成分とみなすこともできる。したがって、このようなRGD配列を特異的に認識する抗体が必要とされている。
【0009】
本発明者らは以前に、RGD配列を免疫特異的に認識し、ハイブリドーマクローン33E10および35B6(それぞれ寄託アクセッション番号FERM BP−10440およびFERM BP−10441)によって産生されるマウスモノクローナル抗体を単離した。本明細書では、ハイブリドーマクローンの名称は、そのクローンによって産生されるモノクローナル抗体の名称として互換的に使用する。これらのマウス抗RGD抗体はすべて、IgG1アイソタイプであった。こうしたモノクローナル抗体は、オステオポンチンなどのECMタンパク質のRGD配列に結合することにより、ECMと細胞の間のRGD配列を介した結合に干渉することが認められた。したがって、こうした抗RGD抗体は、癌、例えば、癌細胞の増殖または転移、および炎症性疾患、例えば、関節リウマチ、骨関節炎、感染症、肝炎、気管支喘息、線維症、真性糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患、骨粗鬆症などのRGD配列に関連する疾患に対して治療効果または診断効果を発揮する可能性がある。
【0010】
しかし、これらのモノクローナル抗体はマウス起源であるので、ヒトにおけるその免疫原性が原因で生じる可能な有害作用が、ヒトにおける診断または治療的使用にこの抗体を直接利用することの妨げとなった。その免疫原性を低下させるために、本発明者らは、前記ヒト化抗体が由来する元のマウス抗RGD抗体によって発揮される生物活性に相当する生物活性を有するヒト化抗体を調製した。
【課題を解決するための手段】
【0011】
したがって、本発明は、RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントであって、非ヒト起源に部分的に由来し、ヒト起源に部分的に由来する抗原結合性領域を含む抗体またはその抗原結合性フラグメントを提供する。一部の実施形態において、本発明のヒト化抗体またはその抗原結合性フラグメントは、33E10モノクローナル抗体、35B6モノクローナル抗体などの非ヒト供給源(ドナー)に由来する相補性決定領域(以下「CDR」と略記する)およびヒト供給源(アクセプター)に由来するフレームワーク領域(以下「FR」と略記する)を含む。前記ヒト化抗体またはその抗原結合性フラグメントは、RGD配列とそのリガンドの間の結合を阻害することができる。
【0012】
特定の実施形態において、RGD配列を免疫特異的に認識する前記ヒト化抗体またはその抗原結合性フラグメントは、(i)ヒト重鎖(以下「H鎖」と略記する)の可変領域(以下「V領域」と略記する)に由来する少なくとも1つのH鎖FR(以下「FRH」と略記する)と、RGD配列を免疫特異的に認識する非ヒト抗体のH鎖CDR(以下「CDRH」と略記する)のうちの少なくとも1つに由来する少なくとも1つのCDRHとを含むH鎖;または(ii)ヒト軽鎖(以下「L鎖」と略記する)のV領域に由来する少なくとも1つのL鎖FR(以下「FRL」と略記する)と、RGD配列を免疫特異的に認識する非ヒト抗体のL鎖CDR(以下「CDRL」と略記する)のうちの少なくとも1つに由来する少なくとも1つのCDRLとを含むL鎖;または上記の(i)と(ii)の両方を含む。一実施形態において、本発明のヒト化抗体のCDRHのうちの少なくとも1つおよび/またはCDRLのうちの少なくとも1つは、寄託アクセッション番号FERM BP−10440およびFERM BP−10441からなる群から選択されるハイブリドーマによって産生されたモノクローナル抗体に由来するものとすることができる。好ましい実施形態において、本発明のヒト化抗体またはその抗原結合性フラグメントは、(i)ヒトFRHに由来する少なくとも1つのFRH、ならびに配列番号1、2および3のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRH;または(ii)ヒトFRLに由来する少なくとも1つのFRL、ならびに配列番号4、5および6のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRL;または(iii)上記の(i)と(ii)の両方を含む。一部の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRH1に配列番号1、CDRH2に配列番号2、およびCDRH3に配列番号3を含むこともできる。一部の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRL1に配列番号4、CDRL2に配列番号5、およびCDRL3に配列番号6を含むこともできる。好ましくは、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRH1に配列番号1、CDRH2に配列番号2、CDRH3に配列番号3、CDRL1に配列番号4、CDRL2に配列番号5、およびCDRL3に配列番号6を含む。
【0013】
一部の特定の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、GenBankアクセッション番号X65891(配列番号13)によってコードされるヒトH鎖のV領域に由来するFRH、またはGenBankアクセッション番号X72441(配列番号18)によってコードされるヒトκ−L鎖のV領域に由来するFRLを含む。一部の実施形態において、本発明のヒト化抗体のFRHは、配列番号14、15、16および17のアミノ酸配列(それぞれX65891のFRH1、FRH2、FRH3およびFRH4のアミノ酸配列)からなる群から選択される少なくとも1つのアミノ酸配列を含む。一部の実施形態において、本発明のヒト化抗体のFRLは、配列番号19、20、21および22のアミノ酸配列(それぞれX72441のFRL1、FRL2、FRL3およびFRL4のアミノ酸配列)からなる群から選択される少なくとも1つのアミノ酸配列を含む。最も好ましい実施形態の1つにおいて、本発明のヒト化抗体またはその抗原結合性フラグメントは、(i)配列番号24のアミノ酸配列を含むH鎖のV領域(以下「VH」と略記する);または(ii)配列番号26のアミノ酸配列を含むL鎖のV領域(以下「VL」と略記する);または(iii)上記の(i)と(ii)の両方を含む。
【0014】
他の実施形態において、本発明のヒト化抗体またはその抗原結合性フラグメントは、(i)ヒトFRHに由来する少なくとも1つのFRH、ならびに配列番号7、8および9のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRH;または(ii)ヒトFRLに由来する少なくとも1つのFRL、ならびに配列番号10、11および12のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRL;または(iii)上記の(i)と(ii)の両方を含む。一部の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRH1に配列番号7、CDRH2に配列番号8、およびCDRH3に配列番号9を含むこともできる。一部の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRL1に配列番号10、CDRL2に配列番号11、およびCDRL3に配列番号12を含む。好ましくは、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRH1に配列番号7、CDRH2に配列番号8、CDRH3に配列番号9、CDRL1に配列番号10、CDRL2に配列番号11、およびCDRL3に配列番号12を含む。
【0015】
一部の特定の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、GenBankアクセッション番号X65891(配列番号13)によってコードされるヒトH鎖のV領域に由来するFRH、またはGenBankアクセッション番号X72441(配列番号18)によってコードされるヒトκ−L鎖のV領域に由来するFRLを含む。一部の実施形態において、本発明のヒト化抗体FRHは、配列番号14、15、16および17のアミノ酸配列(それぞれX65891のFRH1、FRH2、FRH3およびFRH4のアミノ酸配列)からなる群から選択される少なくとも1つのアミノ酸配列を含む。一部の実施形態において、本発明のヒト化抗体のFRLは、配列番号19、20、21および22のアミノ酸配列(それぞれX72441のFRL1、FRL2、FRL3およびFRL4のアミノ酸配列)からなる群から選択される少なくとも1つのアミノ酸配列を含む。最も好ましい実施形態の1つにおいて、本発明のヒト化抗体またはその抗原結合性フラグメントは、(i)配列番号28のアミノ酸配列を含むVH;または(ii)配列番号30のアミノ酸配列を含むVL;または(iii)上記の(i)と(ii)の両方を含む。
【0016】
本発明はさらに、RGD配列を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合性フラグメントをコードするヌクレオチド配列を含む単離された核酸分子を提供する。詳細には、本発明は、配列番号1、2、3、7、8および9からなる群から選択される少なくとも1つのアミノ酸配列を含むヒト化H鎖、または配列番号4、5、6、10、11および12からなる群から選択される少なくとも1つのアミノ酸配列を含むヒト化L鎖、または前記ヒト化H鎖と前記ヒト化L鎖の両方をコードするヌクレオチド配列を含む単離された核酸分子を提供する。好ましい特定の実施形態において、このような単離された核酸分子は、VHをコードする配列番号23のヌクレオチド配列、または配列番号24のアミノ酸配列をコードするヌクレオチド配列を含む。一部の好ましい特定の実施形態において、このような単離された核酸分子は、VLをコードする配列番号25のヌクレオチド配列、または配列番号26のアミノ酸配列をコードするヌクレオチド配列を含む。好ましくは、本発明の単離された核酸分子は、配列番号23と配列番号25の両方のヌクレオチド配列を含む。好ましい特定の実施形態において、本発明の単離された核酸分子は、配列番号32および34のアミノ酸配列などのドナー起源のシグナルペプチド、または異種起源のシグナルペプチドをコードするヌクレオチド配列をさらに含む。
【0017】
他の好ましい特定の実施形態において、このような単離された核酸分子は、VHをコードする配列番号27のヌクレオチド配列、または配列番号28のアミノ酸配列をコードするヌクレオチド配列を含む。一部の好ましい特定の実施形態において、このような単離された核酸分子は、VLをコードする配列番号29のヌクレオチド配列、または配列番号30のアミノ酸配列をコードするヌクレオチド配列を含む。好ましくは、本発明の単離された核酸分子は、配列番号27と配列番号29の両方のヌクレオチド配列を含む。好ましい特定の実施形態において、本発明の単離された核酸分子は、配列番号36および38のアミノ酸配列などのドナー起源のシグナルペプチド、または異種起源のシグナルペプチドをコードするヌクレオチド配列をさらに含む。
【0018】
本発明はさらに、RGD配列を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合性フラグメントのH鎖またはL鎖またはその両方をコードするヌクレオチド配列を含むベクター、例えば、発現ベクターを提供する。このようなベクターにおいて、本発明のヌクレオチド配列を1つまたは複数の調節エレメントと作動可能に連結することもできる。本発明のヌクレオチド配列は、CDRが由来する非ヒトドナー抗体の天然のシグナルペプチド、または異種起源のシグナルペプチドをコードするヌクレオチド配列を含むこともできる。
【0019】
さらに、本発明は、本発明の核酸分子を含むベクターを含めた、本発明の核酸分子を含む宿主細胞を提供する。一実施形態において、本発明は、本発明のヒト化H鎖をコードする第1の核酸分子および本発明のヒト化L鎖をコードする第2の核酸分子を含む単離された宿主細胞を提供し、前記第1および第2の核酸分子はそれぞれ、本発明の生物学的機能を有するヒト化抗体またはその抗原結合性フラグメントが発現されるように調節エレメントと作動可能に連結している。
【0020】
したがって、本発明はさらに、本発明のヒト化抗体またはその抗原結合性フラグメントを調製するための方法であって、本発明の宿主細胞をヒト化抗体またはその抗原結合性フラグメントが発現されるような条件下で培養すること、および産生されたヒト化抗体を収集することを含む方法を提供する。
【0021】
本発明はさらに、本発明のヒト化抗体またはその抗原結合性フラグメントのうちの少なくとも1つを含む組成物を提供する。さらに、本発明は、RGD−タンパク質に関連する障害または疾患を予防または治療するための医薬組成物であって、本発明のヒト化抗体またはその抗原結合性フラグメントのうちの少なくとも1つおよび薬学的に許容される担体を含む医薬組成物を提供する。前記組成物はいずれも、障害または疾患を相加的または相乗的に寛解させることができる別の活性化合物をさらに含むことができる。前記活性化合物としては、それだけには限定されないが、抗炎症性化合物、化学療法化合物などが挙げられる。前記活性化合物としてさらに、ヒトα4インテグリン特異的抗体またはヒトα9インテグリン特異的抗体などの、小分子化合物および抗体またはその抗原結合性フラグメントが挙げられる。
【0022】
別の態様において、本発明は、RGD−タンパク質に関連する、またはそれが関与する障害または疾患を予防または治療するための方法であって、予防または治療有効量の本発明のヒト化抗体またはその抗原結合性フラグメントのうちの少なくとも1つを、それを必要とする対象に投与することを含む方法を提供する。このような使用のために、本発明のヒト化抗体またはその抗原結合性フラグメントは、ヒト化抗体またはその抗原結合性フラグメントの生物学的効果を増強する治療用成分とコンジュゲートすることもできる。このような治療用成分の例としては、別の抗体、細胞増殖抑制性もしくは細胞破壊性を示す細胞毒、放射性元素および/または抗炎症薬、抗生物質などを含めた他の治療薬が挙げられる。
【0023】
さらに別の態様において、本発明は、対象においてRGD−タンパク質に関連する、またはそれが関与する障害または疾患を診断するための方法であって、診断有効量の本発明のヒト化抗体またはその抗原結合性フラグメントを被検対象に投与することを含む方法を提供する。このような診断的使用のために、本発明のヒト化抗体を放射性元素などの検出可能なマーカーで標識することもできる。
【0024】
3.1.定義
本明細書では、「抗体」という用語は、所望の抗原または所望の配列(RGD配列など)に免疫特異的に結合することができる抗体分子を指し、抗体分子全体または抗原結合性フラグメントを含めたそのフラグメントを包含する。
【0025】
本明細書で使用する「抗原結合性フラグメント」という用語は、VLおよび/もしくはVHまたは標的のポリペプチド、タンパク質もしくは配列に特異的に結合するCDRのいずれかを含有する、単鎖抗体、Fabフラグメント、F(ab’)2フラグメント、ジスルフィド結合したFvsおよびフラグメントを含めた、標的のポリペプチド、タンパク質または配列、特にRGD配列に免疫特異的に結合する能力を保持する抗体の任意のフラグメントを指す。したがって、ヒト化抗体のこのような抗原結合性フラグメントは、部分または全長のヒト定常領域を含んでも含まなくてもよい。当技術分野では上記の抗体フラグメントを得るための様々な方法がよく知られている。
【0026】
本明細書で使用する「免疫特異的に認識する」という用語は、抗体またはその抗原結合性フラグメントが標的のポリペプチド、タンパク質または配列、特にヒトRGD配列に特異的に結合できることを指す。このような抗体は、他のポリペプチドまたはタンパク質に非特異的に結合しない。しかし、標的のポリペプチドまたはタンパク質(例えば、RGD−タンパク質)に免疫特異的に結合する抗体またはその抗原結合性フラグメントは、他の抗原と交差反応することもできる。例えば、ヒトRGD−タンパク質を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合性フラグメントは、マウスRGD−タンパク質と交差反応することもできる。好ましくは、RGD−タンパク質を免疫特異的に認識する抗体またはその抗原結合性フラグメントは、他の抗原と交差反応しない。
【0027】
本明細書で使用する「ヒト供給源に由来する」または「非ヒト供給源に由来する」という用語は、そのアミノ酸配列がそれぞれヒト抗体または非ヒト抗体の対応する部分に由来する抗体部分を指す。
【0028】
本明細書で使用する「アクセプター配列」という用語は、通常非ヒト抗体であるドナー抗体由来のCDRのアクセプターとなるヒト抗体VHまたはVL由来のFRのヌクレオチド配列またはアミノ酸配列を指す。
【図面の簡単な説明】
【0029】
【図1】マウスOPNの部分ペプチドを使用したモノクローナル抗体4P11、11M6、25H15および35B6のエピトープ分析の結果を示す図である。
【図2】マウスOPNおよびヒトOPNの部分ペプチドを使用したモノクローナル抗体29R5、30C7、33E10および38I8のエピトープ分析の結果を示す図である。
【図3】RGD配列を含むマウスOPNの部分ペプチドを使用したモノクローナル抗体33E10および35B6のエピトープ分析の結果を示す図である。
【図4】マウスOPNの部分ペプチドを使用したモノクローナル抗体33E10および35B6のエピトープ分析の結果を示す図である。
【図5】マウスOPNの部分ペプチド(CGDSLAYGLR;配列番号79)を使用したモノクローナル抗体33E10および35B6のエピトープ分析の結果を示す図である。
【図6】抗RGDモノクローナル抗体のCDRH分析の結果を示す図である。この図で、33E10の99位のアミノ酸(F)および35B6の98位のアミノ酸(F)は、KでもRでもよい。
【図7】抗RGDモノクローナル抗体のCDRL分析の結果を示す図である。
【図8】抗RGD抗体とRGD配列を含む種々のECMタンパク質との結合親和性の結果を示す図である。
【図9】癌性細胞(NIH3T3細胞)に対するmOPNのN半分(N half)の結合の抗RGD抗体による阻害の結果を示す図である。
【図10A−10C】癌性細胞(NIH3T3細胞)に対する種々のECMタンパク質の結合の抗RGD抗体による阻害の結果を示す図である。
【図11】抗RGD抗体の肝炎抑制効果の結果を示す図である。
【図12A−12B】実験的転移モデルにおける抗RGD抗体の肺転移阻害効果の結果を示す図である。図12Aは転移細胞の数を示し、図12Bは体重の変化を示す。
【図13A−13C】自然発症転移モデルにおける抗RGD抗体の肺転移阻害効果の結果を示す図である。図13Aは癌のサイズを示し、図13Bは転移細胞の数を示し、図13Cは体重の変化を示す。
【図14】関節リウマチモデルにおける抗RGD抗体の治療効果の試験の結果を示す図である。
【図15】マウス33E10 VH cDNAのヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(E)は二重下線付きである。Kabatら(Sequences of Proteins of Immunological Interests、第5版、NIH公開番号91−3242、U.S.Department of Health and Human Services、1991)の定義に従うCDR配列は下線付きである。
【図16】マウス33E10 VL cDNAのヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。
【図17】SpeI部位およびHindIII部位(下線付き)が両端に隣接する、設計された33E10 VH遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(E)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図18】NheI部位およびEcoRI部位(下線付き)が両端に隣接する、設計された33E10 VL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図19】pCh33E10およびpHu33E10(総称して発現ベクターとする)の概略構造を示す図である。頂点のSalI部位から時計回りに進んで、プラスミドは、抗体重鎖遺伝子の転写を開始するためのヒトサイトメガロウイルス(CMV)主要最初期プロモーターおよびエンハンサー(CMVプロモーター)から始まる重鎖転写単位を含有する。CMVプロモーターにVHエキソンが続き、CH1、ヒンジ、CH2およびCH3エキソンを介在性イントロンと共に含むヒトγ−1重鎖定常領域を含有するゲノム配列、さらにはCH3エキソンの後にポリアデニル化部位がある。重鎖遺伝子配列の後に、軽鎖転写単位がCMVプロモーターから始まり、続いてVLエキソン、およびヒトκ鎖定常領域エキソン(CL)をそれに先行するイントロンの部分と共に含有するゲノム配列、さらにはCLエキソンに続くポリアデニル化部位がある。次いで、軽鎖遺伝子に続いて、SV40初期プロモーター(SV40プロモーター)、大腸菌キサンチングアニンホスホリボシルトランスフェラーゼ遺伝子(gpt)、およびSV40ポリアデニル化部位(SV40ポリ(A)部位)を含有するセグメントがある。最後に、プラスミドは、細菌の複製起点(pUC ori)およびβラクタマーゼ遺伝子(βラクタマーゼ)を含むプラスミドpUC19の一部を含有する。
【図20】33E10 VH、ヒト化33E10(Hu33E10)VHおよびヒトアクセプターU03400(GenBankアクセッション番号)のアミノ酸配列のアライメントを示す図である。アミノ酸残基は1文字表記で示す。配列上の数字はKabatら(1991)に従う位置を示す。Kabatら(Sequences of Proteins of Immunological Interests、第5版、NIH公開番号91−3242、U.S.Department of Health and Human Services、1991)によって定義されるCDR配列は下線付きである。二重下線付きの残基はCDRと接触すると予測され、マウス残基はこれらの位置においてヒト化された形で保持された。U03400中のCDR残基は図では省略する。
【図21】33E10 VL、ヒト化33E10(Hu33E10)VLおよびヒトアクセプターX72452(GenBankアクセッション番号)のアミノ酸配列のアライメントを示す図である。アミノ酸残基は1文字表記で示す。配列上の数字はKabatら(1991)に従う位置を示す。Kabatら(1991)によって定義されるCDR配列は下線付きである。X72452中のCDR残基は図では省略する。
【図22】Hu33E10 VH遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図23】Hu33E10 VL遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図24】Hu33E10 VH遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印は、各オリゴヌクレオチドの位置および方向(5’から3’)を表す。アミノ酸残基は1文字表記で示す。
【図25】Hu33E10 VL遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印は、各オリゴヌクレオチドの位置および方向(5’から3’)を表す。アミノ酸残基は1文字表記で示す。
【図26】SpeI部位およびHindIII部位(下線付き)が両端に隣接する、Hu33E10 VH遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(E)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図27】NheI部位およびEcoRI部位(下線付き)が両端に隣接する、Hu33E10 VL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図28】マウス35B6 VH cDNAのヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(Q)は二重下線付きである。Kabatら(Sequences of Proteins of Immunological Interests、第5版、NIH公開番号91−3242、U.S.Department of Health and Human Services、1991)の定義に従うCDR配列は下線付きである。
【図29】マウス35B6 VL cDNAのヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。
【図30】SpeI部位およびHindIII部位(下線付き)が両端に隣接する、設計された35B6 VH遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(Q)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図31】NheI部位およびEcoRI部位(下線付き)が両端に隣接する、設計された35B6 VL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図32】pCh35B6およびpHu35B6(総称して発現ベクターとする)の概略構造を示す図である。頂点のSalI部位から時計回りに進んで、プラスミドは、抗体重鎖遺伝子の転写を開始するためのヒトサイトメガロウイルス(CMV)主要最初期プロモーターおよびエンハンサー(CMVプロモーター)から始まる重鎖転写単位を含有する。CMVプロモーターにVHエキソンが続き、CH1、ヒンジ、CH2およびCH3エキソンを介在性イントロンと共に含むヒトγ−1重鎖定常領域を含有するゲノム配列、さらにはCH3エキソンの後にポリアデニル化部位がある。重鎖遺伝子配列の後に、軽鎖転写単位がCMVプロモーターから始まり、続いてVLエキソン、およびヒトκ鎖定常領域エキソン(CL)をそれに先行するイントロンの部分と共に含有するゲノム配列、さらにはCLエキソンに続くポリアデニル化部位がある。次いで、軽鎖遺伝子に続いて、SV40初期プロモーター(SV40プロモーター)、大腸菌(E.coli)キサンチングアニンホスホリボシルトランスフェラーゼ遺伝子(gpt)、およびSV40ポリアデニル化部位(SV40ポリ(A)部位)を含有するセグメントがある。最後に、プラスミドは、細菌の複製起点(pUC ori)およびβラクタマーゼ遺伝子(βラクタマーゼ)を含むプラスミドpUC19の一部を含有する。
【図33】35B6 VH、ヒト化35B6(Hu35B6)VHおよびヒトアクセプターZ47230(GenBankアクセッション番号)のアミノ酸配列のアライメントを示す図である。アミノ酸残基は1文字表記で示す。配列上の数字はKabatら(1991)に従う位置を示す。Kabatら(1991)によって定義されるCDR配列は下線付きである。二重下線付きの残基はCDRと接触すると予測され、マウス残基はこれらの位置においてヒト化された形で保持された。Z47230中のCDR残基は図では省略する。
【図34】35B6 VL、ヒト化35B6(Hu35B6)VLおよびヒトアクセプターX72479(GenBankアクセッション番号)のアミノ酸配列のアライメントを示す図である。アミノ酸残基は1文字表記で示す。配列上の数字はKabatら(1991)に従う位置を示す。Kabatら(1991)によって定義されるCDR配列は下線付きである。二重下線付きの残基はCDRと接触すると予測され、マウス残基はこれらの位置においてヒト化された形で保持された。X72479中のCDR残基は図では省略する。
【図35】Hu35B6 VH遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図36】Hu35B6 VL遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図37】Hu35B6 VH遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印は、各オリゴヌクレオチドの位置および方向(5’から3’)を表す。アミノ酸残基は1文字表記で示す。
【図38】Hu35B6 VL遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印は、各オリゴヌクレオチドの位置および方向(5’から3’)を表す。アミノ酸残基は1文字表記で示す。
【図39】SpeI部位およびHindIII部位(下線付き)が両端に隣接する、Hu35B6 VH遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(Q)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図40】NheI部位およびEcoRI部位(下線付き)が両端に隣接する、Hu35B6 VL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図41A−41B】キメラおよびヒト化35B6抗体とhOPN−BSAの結合をELISAによって分析した。各抗体を2.5μg/ml(図41A)または1.0μg/ml(図41B)の出発濃度および連続2倍希釈で試験した。実験は3連で実施した。各抗体濃度での平均吸光度値と標準偏差を図41A〜41Bに示す。
【発明を実施するための形態】
【0030】
5.発明の詳細な説明
5.1.RGD配列に対する抗体の調製
RGD配列を免疫特異的に認識する抗体は、当技術分野で知られている任意の適当な方法によって生成することができる。
【0031】
本発明における細胞接着性「RGD」配列を含むRGD−タンパク質またはRGD−ペプチド(以下「RGD−ペプチド」と略記する)は、(1)RGD−タンパク質を発現するヒトECMまたはこれらのECMが存在するすべての組織に由来するもの、(2)細菌、酵母、動物細胞などの細胞系などにトランスフェクトすることによる、RGD−タンパク質またはRGD−ペプチドをコードするDNA(好ましくはcDNA)の発現によって得られる組換えタンパク質またはペプチド、または(3)合成タンパク質またはペプチドであってもよい。
【0032】
本発明において抗原として使用されるRGD−ペプチドは、免疫化によるRGD配列に対する抗体の産生が可能であり得る。RGD−ペプチドは、マウスECMタンパク質の細胞接着性配列であるRGD−ペプチドアミノ酸配列CVDVPNGRGDSLAYGLR(配列番号71)を含む。RGD−タンパク質またはRGD−ペプチドとしては、OPN、ビトロネクチン、フィブロネクチン、フォンウィルブランド因子、コラーゲン、ラミニン、テネイシン、フィブリノゲン、トロンボスポンジンおよびRGDを含むその断片などが挙げられる。前記タンパク質または前記ペプチドがRGD配列を含む限り、タンパク質またはペプチドにアミノ酸の置換、欠失、改変、付加などの人工または天然の変異を適用することができる。変異タンパク質またはペプチドは、複数のアミノ酸、好ましくは1から10個のアミノ酸、より好ましくは1から数個(例えば、1から5個)のアミノ酸が置換、欠失、改変、付加または挿入されたアミノ酸配列を含むこともできる。
【0033】
本明細書では、RGD−ペプチドは、少なくとも約5個のアミノ酸、好ましくは約5から50個のアミノ酸、より好ましくは約10から20個のアミノ酸を含む。本発明における抗原としてのRGD−タンパク質またはRGD−ペプチドは、化学合成法、細胞培養法、遺伝子組換え法、その適切な改変などの当技術分野でよく知られている方法を使用することによって産生することができる。例えば、RGD−ペプチドは、ECMタンパク質をタンパク質分解酵素で適切に切断することによって得ることができる。RGD−タンパク質またはRGD−ペプチドは、マウス、ラット、ウサギ、ブタ、ウシ、サル、ヒトなどの哺乳動物に由来するものであってもよい。当技術分野でよく知られている任意の方法を使用して、抗RGD抗体の調製に使用することができるRGD−タンパク質またはRGD−ペプチドを調製することができる。
【0034】
変異ポリペプチドを生成するための方法の例としては、合成オリゴヌクレオチド部位特異的突然変異誘発(gapped duplex法)、亜硝酸または亜硫酸の処理によって点突然変異をランダムに導入することを含む点突然変異誘発法、Bal31酵素または他の酵素で欠失変異体を調製することを含む方法、カセット変異導入法、リンカースキャニング法(linker scanning method)、ミスインコーポレーション法(miss incorporation method)、ミスマッチプライマー法(mismatch primer method)、DNAセグメント合成法などが挙げられる。
【0035】
RGD−ペプチドは、サイログロブリン、キーホールリンペットヘモシアニン(KLH)、ウシ血清アルブミン(BSA)、オボアルブミン(OVA)またはウシグロブリンなど他の生体巨大分子、好ましくはサイログロブリンに結合させることができる。RGD−ペプチドを生体巨大分子に結合させる方法は、活性エステル基およびマレイミド基を有する結合試薬などのカップリング試薬(活性エステル基はタンパク質またはペプチドのアミノ基に結合し、マレイミド基はタンパク質またはペプチドのチオール基に結合する。S.Yoshirakeら、Eur.J.Biochem.、101、395〜399、1979)、混合酸無水物法(B.F.Erlangerら、J.Biol.Chem.、234、1090〜1094、1954)、または活性エステル法(A.E.Karuら、J.Agric.Food Chem.、42、301〜309、1994)を使用することによって実現することができる。RGD−ペプチドを生体巨大分子に結合させるための方法は、カップリング試薬を使用することによって達成することが好ましい。
【0036】
抗原として、RGD−タンパク質またはRGD−ペプチドを過剰発現する細胞それ自体を使用することもできる。RGD−タンパク質またはRGD−ペプチドを過剰発現する細胞は、当技術分野でよく知られている組換えDNA技術によって調製することができる。
【0037】
上記のとおり調製した適切な抗原を使用して、当技術分野でよく知られている様々な方法によってRGD配列に特異的な抗体を調製することができる。RGD配列に対するポリクローナル抗体は、当技術分野でよく知られている様々な手順によって産生することができる。例えば、対象の抗原は、それだけには限定されないが、ウサギ、マウス、ラットなどを含めた、種々の宿主動物に投与して、抗原に特異的なポリクローナル抗体を含有する抗血清の産生を誘発することができる。様々なアジュバントを使用して宿主種に応じて免疫応答を増大させることができ、そのアジュバントとしては、それだけには限定されないが、フロイント(完全および不完全)アジュバント、水酸化アルミニウムなどの無機ゲル(mineral gel)、リゾレシチンなどの界面活性物質、プルロニック(pluronic)ポリオール、ポリアニオン、ペプチド、オイルエマルジョン、キーホールリンペットヘモシアニン、ジニトロフェノールおよびBCG(カルメット・ゲラン桿菌)、コリネバクテリウム・パルバム(Corynebacterium parvum)などのヒトに潜在的に有用なアジュバントが挙げられる。このようなアジュバントも当技術分野でよく知られている。
【0038】
モノクローナル抗体は、ハイブリドーマ技術、組換え技術およびファージディスプレイ技術、またはその組合せの使用を含めた、当技術分野で知られている多種多様な技法を使用することによって調製することができる。例えば、モノクローナル抗体は、当技術分野で知られており、例えば、Harlowら、Antibodies:A Laboratory Manual、(Cold Spring Harbor Laboratory Press、第2版1988);Hammerlingら、in:Monoclonal Antibodies and T−Cell Hybridomas、563〜681ページ(Elsevier,N.Y.、1981)(両文献ともその全体が参照により組み込まれる)に教示されているものを含めて、ハイブリドーマ技法を使用することによって産生することができる。本明細書で使用する「モノクローナル抗体」という用語は、ハイブリドーマ技術によって産生される抗体に限定されない。「モノクローナル抗体」という用語は、単一のクローンに由来する抗体を指し、真核生物、原核生物またはファージの任意のクローンを含むが、それが産生される方法に限定されない。
【0039】
ハイブリドーマ技術を使用して特異的抗体を産生およびスクリーニングするための方法は、常用されており当技術分野でよく知られている。非限定的な例において、マウスは、対象の抗原またはこのような抗原を発現する細胞で免疫することができる。免疫応答が検出される、例えば、マウス血清中で抗原に特異的な抗体が検出されると、マウス脾臓が採取され、脾細胞が単離される。次いで、脾細胞は、よく知られている技法によって任意の適当な骨髄腫細胞(例えば、P3U1、P3X63−Ag8、P3X63−Ag8−U1、P3NS1−Ag4、SP2/0−Ag14、P3X63−Ag8−653など)と融合される。ハイブリドーマは、限界希釈によって選択され、クローニングされる。次いで、ハイブリドーマクローンは、当技術分野で知られている方法により、抗原に結合することができる抗体を分泌する細胞についてアッセイされる。高レベルの抗体を通常含有する腹水は、マウスの腹腔内に陽性のハイブリドーマクローンを接種することによって生成することができる。
【0040】
特異的エピトープを認識する抗体フラグメントは、既知の技法によって生成することもできる。例えば、FabおよびF(ab’)2フラグメントは、パパイン(Fabフラグメントを産生するため)またはペプシン(F(ab’)2フラグメントを産生するため)などの酵素を使用した、免疫グロブリン分子のタンパク質分解切断によって産生することもできる。F(ab’)2フラグメントは、完全L鎖、ならびにH鎖のV領域、CH1領域およびヒンジ領域を含有する。
【0041】
本発明の抗体またはその抗原結合性フラグメントは、当技術分野で知られている、抗体を合成するための任意の方法によって、特に、化学合成によって、または好ましくは組換え発現技法によって産生することもできる。
【0042】
抗体をコードするヌクレオチド配列は、当業者に利用可能な任意の情報から(すなわち、GenBank、文献から、または常用のクローニングおよび配列解析によって)得ることもできる。特定の抗体またはそのエピトープ結合性フラグメントをコードする核酸を含有するクローンが利用可能でないが、抗体分子またはそのエピトープ結合性フラグメントの配列が知られている場合、免疫グロブリンをコードする核酸は、化学的に合成するか、または配列の末端および5’末端にハイブリダイズ可能な合成プライマーを使用したPCR増幅によって、もしくは例えば、抗体をコードするcDNAライブラリー由来cDNAクローンを同定するための、特定の遺伝子配列に特異的なオリゴヌクレオチドプローブを使用したクローニングによって、適当な供給源(例えば、抗体cDNAライブラリー、または抗体を発現するように選択されたハイブリドーマ細胞など、抗体を発現する任意の組織もしくは細胞から作製されるcDNAライブラリー、またはそれから単離された核酸、好ましくはポリA+RNA)から得ることができる。次いで、PCRによって生成された増幅核酸は、当技術分野でよく知られている任意の方法を使用して複製可能なクローニングベクター中にクローニングすることもできる。
【0043】
5.2.組換え抗体の調製
抗体のヌクレオチド配列は、当技術分野でよく知られている、ヌクレオチド配列を操作するための方法、例えば、組換えDNA技術、部位特異的変異誘発、PCRなど(例えば、両文献ともその全体が参照により本明細書に組み込まれる、Sambrookら、前掲;およびAusubelら編、1998、Current Protocols in Molecular Biology、John Wiley & Sons、NYに記載の技法を参照されたい)を使用することによって操作することもできる。抗体のエピトープ結合ドメイン領域に、または生物活性を増強もしくは低減する任意の部分にアミノ酸の置換、欠失および/または挿入などの突然変異を導入することもできる。
【0044】
抗体をコードするヌクレオチド配列を含有する発現ベクターを、抗体またはその抗原結合性フラグメントの組換え発現に使用することができる。抗体またはその抗原結合性フラグメントを産生するための、抗体分子、抗体のH鎖および/もしくはL鎖、またはその一部分をコードするヌクレオチド配列を含むベクターは、以前のセクションで検討したとおり、当技術分野でよく知られている技法を使用して、組換えDNA技術によって産生することができる。当業者によく知られている方法を使用して、抗体またはその抗原結合性フラグメントをコードする配列ならびに適切な転写および翻訳調節シグナルを含有する発現ベクターを構築することができる。これらの方法としては、例えば、in vitro組換えDNA技術、合成的技法およびin vivo遺伝子組換えが挙げられる。抗体のVH、VL、VHとVLの両方、VHおよび/またはVLの抗原結合性フラグメント、または1つもしくは複数のCDRをコードするヌクレオチド配列は、このような発現用ベクター中にクローニングすることができる。このような配列は、元の抗体の天然のものであっても異種のものであってもよいシグナルペプチドをコードするポリヌクレオチドと融合することもできる。次いで、このようにして調製された発現ベクターは、抗体を発現させるための適切な宿主細胞中に導入することができる。したがって、本発明は、RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントをコードするポリヌクレオチドを含有する宿主細胞を含む。
【0045】
宿主細胞には、第1のベクターがH鎖由来のポリペプチドをコードし、第2のベクターがL鎖由来のポリペプチドをコードする本発明の2つの発現ベクターを同時トランスフェクトすることもできる。2つのベクターは、H鎖およびL鎖ポリペプチドの等しい発現を可能にする同一の選択マーカー、または両方のプラスミドの維持を確実にするための異なる選択マーカーを含有することもできる。あるいは、H鎖およびL鎖ポリペプチドの両方をコードし、発現することができる単一のベクターを使用することもできる。H鎖およびL鎖のコード配列は、cDNAまたはゲノムDNAを含むこともできる。
【0046】
別の実施形態において、抗体は、当技術分野で知られている種々のファージディスプレイ法を使用して作製することもできる。ファージディスプレイ法では、機能的抗体ドメインが、それをコードするポリヌクレオチド配列を保有するファージ粒子の表面上に提示される。特定の実施形態において、このようなファージを利用して、レパートリーまたはコンビナトリアル抗体ライブラリー(例えば、ヒトまたはマウス)から発現させた、FabおよびFvまたはジスルフィド結合安定化Fvなどの抗原結合性ドメインを提示することができる。対象の抗原に結合する抗原結合性ドメインを発現するファージは、抗原を用いて、例えば、標識された抗原、または固体表面もしくはビーズに結合もしくは捕捉した抗原を使用して選択または同定することができる。これらの方法に使用するファージは通常、fdおよびM13を含めた繊維状ファージである。抗原結合性ドメインは、ファージ遺伝子IIIまたは遺伝子VIIIタンパク質のいずれかと組換えによって融合したタンパク質として発現される。本発明の免疫グロブリンまたはそのフラグメントを作製するために使用することができるファージディスプレイ法の例としては、Brinkmanら、J.Immunol.Methods、182:41〜50、1995;Amesら、J.Immunol.Methods、184:177〜186、1995;Kettleboroughら、Eur.J.Immunol.、24:952〜958、1994;Persicら、Gene、187:9〜18、1997;Burtonら、Advances in Immunology、57:191〜280、1994;PCT出願番号PCT/GB91/01134;PCT公報WO90/02809;WO91/10737;WO92/01047;WO92/18619;WO93/11236;WO95/15982;WO95/20401;ならびに米国特許第5,698,426号;同第5,223,409号;同第5,403,484号;同第5,580,717号;同第5,427,908号;同第5,750,753号;同第5,821,047号;同第5,571,698号;同第5,427,908号;同第5,516,637号;同第5,780,225号;同第5,658,727号;同第5,733,743号および同第5,969,108号に開示されたものが挙げられ、それぞれその全体が参照により本明細書に組み込まれる。
【0047】
上記の参考文献に記載のとおり、ファージ選択後、ファージ由来の抗体コード領域を単離および使用して、例えば、以下に詳細に記載のとおり、ヒト抗体または任意の他の所望のフラグメントを含めた全抗体を作製し、哺乳動物細胞、昆虫細胞、植物細胞、酵母および細菌を含めた任意の所望の宿主中で発現させることができる。例えば、Fab、Fab’およびF(ab’)2フラグメントを組換えによって産生するための技法は、PCT公報WO92/22324;Mullinaxら、BioTechniques、12(6):864〜869、1992;およびSawaiら、AJRI、34:26〜34、1995;およびBetterら、Science、240:1041〜1043、1988(それぞれその全体が参照により組み込まれる)に開示されているものなど、当技術分野で知られている方法を使用して採用することもできる。単鎖Fvsおよび抗体を産生するために使用することができる技法の例としては、米国特許第4,946,778号および同第5,258,498号;Hustonら、Methods in Enzymology、203:46〜88、1991;Shuら、PNAS、90:7995〜7999、1993;およびSkerraら、Science、240:1038〜1040、1988に記載のものが挙げられる。
【0048】
本発明の抗体分子は、上記の任意の方法によって産生させた後、当技術分野で知られている、免疫グロブリン分子を精製するための任意の方法によって、例えば、クロマトグラフィー(例えば、イオン交換、親和性、特に、プロテインAまたはプロテインG精製後の特異的な抗原に対する親和性によって、およびサイジングカラムクロマトグラフィー(sizing column chromatography))、遠心分離、溶解度差(differential solubility)、またはタンパク質を精製するための任意の他の標準的な技法によって精製することができる。さらに、本発明の抗体またはそのフラグメントは、本明細書に記載のものか、そうでなければ当技術分野で知られている異種ポリペプチド配列と融合させて精製を容易にすることもできる。
【0049】
ヒトにおける抗体のin vivo使用およびin vitro検出アッセイを含めた一部の使用のために、キメラ抗体、ヒト化抗体またはヒト抗体を使用することが好ましいこともある。キメラ抗体およびヒト化抗体は、以下のセクション5.3で詳細に検討する。
【0050】
他の化合物または異種ポリペプチドと融合またはコンジュゲートした抗体は、in vitroイムノアッセイ、精製法(例えば、アフィニティークロマトグラフィー)、ならびにin vivoでの治療または診断的使用で使用することができる。例えば、その全体が参照により本明細書に組み込まれる、PCT公開番号WO93/21232;EP439,095;Naramuraら、Immunol.Lett.、39:91〜99、1994;米国特許第5,474,981号;Gilliesら、PNAS、89:1428〜1432、1992;およびFellら、J.Immunol.、146:2446〜2452、1991を参照されたい。例えば、抗体は、既知の方法または市販のキット(例えば、ビオチン標識、FITC標識、APC標識)を使用して様々な形で標識することができる。別の例として、抗体は、抗体の生物学的効果をin vivoで増強する治療用成分とコンジュゲートすることもできる。このような治療用成分の例としては、別の抗体、細胞増殖抑制性もしくは細胞破壊性を示す細胞毒、放射性元素および/または抗炎症薬を含めた他の治療薬、抗生物質などが挙げられる。本発明において、ヒト化抗RGD抗体は、別の抗体とコンジュゲートして二重特異性抗体を形成することもできる。別の例として、本発明のヒト化抗体は、in vivoでの診断的使用のために放射性元素などの検出可能なマーカーで標識することもできる。
【0051】
5.3.キメラ抗体およびヒト化抗体
キメラ抗体は、マウスモノクローナル抗体に由来するV領域およびヒト免疫グロブリンに由来する定常領域を有する抗体など、抗体の異なる部分が異なる動物種に由来する分子である。キメラ抗体を産生するための方法は当技術分野で知られている。例えば、その全体が参照により本明細書に組み込まれる、Morrison、Science、229:1202、1985;Oiら、BioTechniques、4:214 1986;Gilliesら、J.Immunol.Methods、125:191〜202、1989;米国特許第5,807,715号;同第4,816,567号;および同第4,816,397号を参照されたい。
【0052】
ヒト化抗体は、所望の抗原に結合し、非ヒト種に由来する1つまたは複数のCDRおよびヒト免疫グロブリン分子に由来する1つまたは複数のFRを含有するV領域を含む分子である。非ヒト抗体をヒト化するための典型的な方法は、Queenら、1989、Proc.Natl.Acad.Sci.USA 86:10029〜10033および米国特許第5,585,089号および同第5,693,762号;Riechmannら、Nature、332:323、1988;およびTsurushitaら、Methods 36:69〜83、2005の文献など、様々な参考文献に記載されており、すべてその全体が参照により本明細書に組み込まれる)。例えば、Tsurushitaら(2005、前掲;以下「Tsurushita」とする)の参考文献では、元々Queenら(1989、前掲)によって開発された抗体ヒト化方法に基づいてマウスモノクローナル抗体をヒト化するための実用的および指示的プロトコールが示されている。Tsurushitaに開示されている一般的なプロトコールは以下に簡潔に要約する。
【0053】
5.3.1.ヒト化抗体を調製するための一般的なプロトコール
マウスV遺伝子のクローニングおよび配列決定
標的のマウスモノクローナル抗体のVHおよびVLをコードするcDNAのクローニングに様々な方法が利用可能である。例えば、SMART RACE cDNA Amplificationキット(BD Biosciences、CA)またはGeneRacerキット(Invitrogen、CA)を使用した5’RACE(rapid amplification of cDNA ends)法が一般に使用されている。5’RACE用の遺伝子特異的プライマーは、VHおよびVLのすぐ下流に結合するように、標的のモノクローナル抗体のH鎖およびL鎖のアイソタイプに基づいて調製することができる。したがって、5’RACEプライマーは、γ1、γ2a、γ2bまたはγ3など、マウスのそれぞれのサブタイプに特異的であるように設計することもできる。あるいは、すべてのサブタイプ用の一般的なプライマーは、サブタイプの中で共通のまたは高度に相同な領域に基づいて設計することもできる。Tsurushitaでは、以下の5’RACEプライマーを例として開示する。
(i)5’−GCCAGTGGATAGACTGATGG−(配列番号82)(マウスγ1、γ2a、γ2bおよびγ3 H鎖のクローニング用)
(ii)5’−GATGGATACAGTTGGTGCAGC−(配列番号83)(マウスκL鎖のクローニング用)。
【0054】
PCR増幅されたV領域遺伝子フラグメントを、例えば、Zero Blunt TOPO PCR Cloning Kit(Invitrogen)を使用してプラスミドベクター中に直接的にクローニングし、そのDNA配列を決定することができる。得られた配列は、例えば、そのコードアミノ酸配列を、例えば、Model 241 Protein Sequencer(Hewlett−Packard、CA)を使用したN末端アミノ酸配列決定によって決定された標的のモノクローナル抗体の配列と比較することによって確認すべきである。通常、例えば、エドマン分解による標的抗体のN末端の少なくとも15〜20アミノ酸残基の決定は、クローン化DNA配列の確実性を確認するのに十分である。マウスにおいて2つの最も多くみられるN末端アミノ酸の1つであるグルタミンがN末端アミノ酸である場合、それはピログルタミンに変換される可能性があり、N末端における配列決定を阻止すると、Tsurushitaは注意を促している。その場合は、配列を得るためにN末端での阻止を解除することが必要である。
【0055】
V領域の三次元モデリング
まず、VHおよびVLの配列に基づいて、CDRの立体構造の維持において潜在的に重要である標的抗体のフレームワーク残基を、例えば、R.Levyら、1989、Biochemistry28:7168〜7175;およびB.Zilberら、1990、Biochemistry29:10032〜10041に記載されている方法によって同定する。通常、VHおよびVLはそれぞれ、構造的に意味ある14のセグメントに分けられ、それは、免疫グロブリンスーパーファミリーのドメイン構造を構成するβ鎖およびループ様構造である。標的抗体由来のセグメントのそれぞれのアミノ酸配列は、PDBデータベース(H.M.Bermanら、2000、Nucleic Acids Res.28:235〜342参照)における既知の構造を有する抗体の対応するセグメントとアラインメントする。複数の配列アライメントにより、標的セグメントのそれぞれの最高の配列相同性を有する対応するセグメントを選択し、V領域の三次元モデルを構築する。構造を最適化するために、モデルを複数サイクルの共役勾配エネルギー最小化(conjugate gradient energy minimization)に供する(例えば、ENCAD、またはPressら、1990、「Numerical Recipes、Cambridge University Press、Cambridgeに記載;Weinerら、1981、J.Comp.Chem.2:287〜303によるAMBER;BioMolecularModellingで利用可能な3D−JIG−SAW、またはCancer Research UKによって運営される「BMM」ウェブサイト;またはSwiss Institute of Bioinformatics、Genevaによって運営されるExPASy Proteomics Serverウェブサイトで利用可能なSWISS−MODELを使用)。
【0056】
ヒトフレームワークの選択
V領域の構造のモデリングと並行して、マウスVHおよびVLそれぞれのcDNAクローニングから推定されたアミノ酸配列をデータベース、例えば、Kabatデータベース(Johnsonら、2000、Johnsonら、2000、Nucleic Acids Res.28:214〜218を参照されたい)、GenBankなどにおいてヒトV領域配列と比較する。マウス配列と少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、または少なくとも95%の同一性という全体にわたる配列同一性を有するヒトFRは、例えば、Smith−Watermanアルゴリズム(Gusfield、1997、「Algorithms on Strings, Trees, and Sequences」掲載、Cambridge University Press、Cambridgeによる)、またはBLAST(Karlinら、1990、Proc.Natl.Acad.Sci.USA87:2264〜2268による)などを使用して検索ことができる。これらのヒト配列はcDNAベースの配列およびタンパク質由来の配列に基づくものであってもよいが、生殖系列の使用がしばしば好ましく、それは、この生殖系列は、cDNAベースの配列、タンパク質由来の配列の体細胞超変異に関連する潜在的な免疫原性の排除に有用である可能性があるからである。別の方法では、Queenら(1989、前掲)に記載のとおり、コンセンサスフレームワーク配列を使用することにより、cDNAベースの配列またはタンパク質由来の配列から得られたフレームワーク中のこのような超変異残基を同定および除去することもできる。生殖系列VHセグメントをアクセプターフレームワークとして使用する場合、第14染色体上にコードされるVHセグメントだけが機能的VHを産生するので、第15染色体および第16染色体ではなく第14染色体上のものを使用すべきである。
【0057】
ヒト化V領域の設計
Queenら(1989、前掲)によれば、CDRの約4〜6Å以内にあるフレームワークアミノ酸を同定することが必要であり、それは、これらの残基が正しいCDR構造を支持する潜在的な重要なフレームワーク残基であると考えられるからである。このようなプロセスは、National Science Foundation(NSF)によって支援されるMolecular Visualization Freewareウェブサイトで利用可能な、原子座標から原子間距離を計算するRASMOLなどのコンピュータプログラムを使用して、またはコンピュータモデルのマニュアル検査によって達成することができる。重要なフレームワーク位置のアミノ酸がマウスドナー配列とヒトアクセプター配列の間で異なる場合、通常ヒト残基がマウスドナーの残基によって置き換えられる。しかし、このような残基が、CDR構造の支持への寄与がわずかしかない場合、対応するヒト残基を通常使用する。また、選択されたヒトアクセプターがV領域配列の約10〜20%未満に存在する「非定型的な(atypical)」アミノ酸を含有する場合、これらのアミノ酸は親和性成熟の間の体細胞超変異の結果である可能性があり、ヒトにおける潜在的な免疫原性を回避するためにドナー残基で置き換えるべきである。
【0058】
さらに、潜在的なN結合型グリコシル化シグナルの残基などの他の因子を、ヒト化V領域を設計するために慎重に考慮する必要がある(詳細についてはTsurushitaを参照されたい)。
【0059】
ヒト化抗体は、治療的使用に必要とされる、または排除されるべきエフェクター機能に依存して、ヒト抗体またはその変異体のヒトκもしくはλL鎖、および/またはγ1、γ2、γ3、γ4、μ、α1、α2、δもしくはεH鎖由来のヒト定常領域またはその一部分を含有することができる。例えば、抗体とFc受容体の結合が低減し、かつ/または補体を固定する抗体の能力が低減するように、突然変異を含有する定常領域Fc部分を本発明のキメラまたはヒト化抗体のV領域と融合することもできる(例えば、Winterら、GB2,209,757B;Morrisonら、WO89/07142、Morganら、WO94/29351を参照されたい)。抗体分子のこのような操作は、セクション5.2に記載のとおり組換えDNA技術によって実施することができる。
【0060】
好ましくは、得られたキメラまたはヒト化抗体は、非ヒトドナー抗体と同一の特異性、および非ヒトドナー抗体の親和性に類似する、またはそれの少なくとも約1/3、少なくとも約1/2もしくは少なくとも約2/3の親和性を有する。別の態様において、得られたキメラまたはヒト化抗体は、少なくとも約1×107M−1、好ましくは少なくとも約1×108M−1、最も好ましくは少なくとも約1×109M−1の親和性定数を有する。
【0061】
上記の一般的なプロトコールに加えて、抗体は、例えば、CDRグラフティング(EP239,400;PCT公報WO91/09967;米国特許第5,225,539号;同第5,530,101号および同第5,585,089号)、ベニアリング(veneering)またはリサーフェシング(resurfacing)(EP592,106;EP519,596;Padlan、Molecular Immunology、28(4/5):489〜498、1991;Studnickaら、Protein Engineering、7(6):805〜814、1994;Roguskaら、Proc Natl.Acad.Sci.USA、91:969〜973、1994)、およびチェインシャッフリング(chain shuffling)(米国特許第5,565,332号)を含めた当技術分野で知られている種々の技法を使用してヒト化することができ、すべてその全体が参照により本明細書に組み込まれる。
【0062】
5.3.2.ヒト化抗体を医薬品として調製するための追加の考慮事項
ヒト化抗体を医薬品として提供するために、したがって、効率的で一貫した産生系を調製する必要がある。例えば、H鎖およびL鎖配列を挿入することによってヒト化抗体に適切な発現ベクターを調製し、発現ベクターをトランスフェクトした高生産性細胞株をマスターセルバンク(master cell bank)(MCB)の種細胞として得ることができ、それはワーキングセルバンク(working cell bank)(WCB)の安定で半永久的な供給源となる。次いで、WCB由来のワーキングセル(working cell)を培養し、培地を収集することによってヒト化抗体を調製することができる。
【0063】
適切な制御遺伝子を有する種々の発現ベクターをこのような産生細胞株の調製に使用することができる。宿主細胞として、哺乳動物タンパク質を発現させるのに一般に使用されるものをヒト化抗体の発現に使用することができる。このような宿主細胞の例としては、それだけには限定されないが、チャイニーズハムスター卵巣(CHO)細胞、SP2/0−Ag14.19細胞、NSO細胞などが挙げられる。ヒト化抗体の生産性は、発現ベクターと宿主細胞の最良の組合せを選択することによって最大化することができる。さらに、宿主細胞によるヒト化抗体の発現が最適化されるように、適した培地を選択するために種々の無血清培地および補充物質から培地の組成を検討すべきである。
【0064】
効率および最終収率に基づいて、宿主細胞によって産生されたヒト化抗体は、アフィニティークロマトグラフィー、イオン交換クロマトグラフィー、疎水的相互作用クロマトグラフィーなどを含めた当技術分野でよく知られている様々な方法を使用して、培養上清から精製することができる。
【0065】
5.4.医薬組成物および治療的使用
本発明は、RGD配列を免疫特異的に認識する上記のヒト化抗体またはその抗原結合性フラグメントを含む医薬組成物を提供する。本発明のヒト化抗体を有効成分として含む医薬組成物は、それだけには限定されないが、癌、例えば、癌細胞の増殖または転移、および炎症性疾患、例えば、関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、真性糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などを含めた、RGDタンパク質に関連する障害または疾患の予防薬および/または治療薬として使用することができる。
【0066】
本発明のヒト化抗体を含む医薬組成物を使用して、臓器移植後の慢性拒否反応、および全身性自己免疫疾患、エリテマトーデス、ブドウ膜炎、ベーチェット病、多発性筋炎、増殖性糸球体腎炎、サルコイドーシスなどの自己免疫疾患を治療することもできる。
【0067】
本発明のヒト化抗体を含む、上記の障害または疾患を予防または治療するための予防薬および/または治療薬は、低い毒性を有し、適当な溶媒中に混合することによる液体製剤として、または適切な剤形の医薬組成物として直接的にヒトに経口または非経口投与することができる。
【0068】
上記の投与に使用される医薬組成物は、前記の抗体またはその塩および薬学的に許容される担体、希釈剤または添加剤を含有する。このような組成物は、経口または非経口投与に適した剤形で提供される。
【0069】
用量は、投与される対象の年齢およびサイズ、標的の疾患、条件、投与経路などによって様々であってよい。抗体を、例えば、成人患者の関節リウマチの予防および/または治療に使用する場合、本発明の抗体を静脈内に通常、約0.01〜約20mg/kg体重、好ましくは約0.1〜約10mg/kg体重、より好ましくは約0.1〜約5mg/kg体重の単回用量で、1日に約1〜5回、好ましくは1日に約1〜3回投与することが有利である。他の非経口投与および経口投与においても、抗体を上記で示される用量に相当する用量で投与することができる。状態が特に重い場合、用量を状態に従って増加させることもできる。
【0070】
様々なデリバリーシステムが知られており、本発明の医薬組成物を投与するために使用することができ、例えば、リポソーム、微粒子、マイクロカプセル中への封入、突然変異ウイルスを発現することができる組換え細胞、受容体を介したエンドサイトーシスがある(例えば、WuおよびWu、1987、J.Biol.Chem.262:4429 4432を参照されたい)。導入方法としては、それだけには限定されないが、皮内、筋肉内、腹腔内、静脈内、皮下、鼻腔内、硬膜外、および経口の経路が挙げられる。これらの化合物は、任意の好都合な経路によって、例えば、注入またはボーラス注射によって、上皮層または皮膚粘膜層(例えば、口腔粘膜、直腸および腸管粘膜など)を通した吸収によって投与することができ、他の生物活性を有する作用物質と一緒に投与することもできる。投与は全身でも局所でもよい。例えば、吸入器またはネブライザーおよびエアロゾル化剤との配合の使用によって肺への投与を採用することもできる。
【0071】
特定の実施形態において、本発明の医薬組成物を、治療を必要とする領域に局所的に投与することが望ましいことがあり、これは、例えば、限定的なものではないが、手術中の局所注入、例えば、手術後の創傷包帯と組み合わせた外用塗布により、注射により、カテーテルを用いて、坐剤を用いて、鼻内噴霧を用いて、またはシアラスティック(sialastic)膜などの膜、または繊維を含めた、多孔性、非多孔性またはゼラチン状の材料のインプラントを用いて実現することができる。一実施形態において、感染組織の部位(または以前の部位)への直接注射によって投与することができる。
【0072】
別の実施形態において、医薬組成物は、小胞、特にリポソームとして送達することができる(Langer、1990、Science249:1527〜1533;Treatら、Liposomes in the Therapy of Infectious Disease and Cancer掲載、Lopez BeresteinおよびFidler(編)、Liss、New York、353〜365ページ(1989);Lopez−Berestein、同書、317〜327ページを参照されたい;同書を全体として参照されたい)。
【0073】
さらに別の実施形態では、医薬組成物を制御放出システムにおいて送達することができる。一実施形態において、ポンプを使用することもできる(Langer、前掲;Sefton、1987、CRC Crit.Ref.Biomed.Eng.14:201;Buchwaldら、1980、Surgery88:507;およびSaudekら、1989、N.Engl.J.Med.321:574を参照されたい)。別の実施形態において、ポリマー材料を使用することができる(Medical Applications of Controlled Release、LangerおよびWise(編)、CRC Pres.、Boca Raton、Florida(1974);Controlled Drug Bioavailability、Drug Product Design and Performance、SmolenおよびBall(編)、Wiley、New York(1984);RangerおよびPeppas、J.Macromol.Sci.Rev.Macromol.Chem.23:61(1983)を参照されたい;Levyら、1985、Science228:190;Duringら、1989、Ann.Neurol.25:351;Howardら、1989、J.Neurosurg.71:105も参照されたい)。さらに別の実施形態において、制御放出システムを組成物の標的の近くに配置することができ、したがって全身用量の一部しか必要とされない(例えば、Goodson、in Medical Applications of Controlled Release、前掲、2巻、115〜138ページ(1984)を参照されたい)。他の制御放出システムはLanger(Science249:1527〜1533(1990))による総説において検討されている。
【0074】
経口投与用の組成物の例としては、固形または液体の剤形、詳細には、錠剤(糖衣錠およびフィルムコーティング錠を含む)、丸剤、顆粒剤、粉末状の製剤、カプセル剤(軟カプセル剤を含む)、シロップ剤、乳剤、懸濁剤などが挙げられる。このような組成物は、公知の方法によって製造され、医薬製剤の分野で従来使用されているビヒクル、希釈剤または添加剤を含有する。錠剤のビヒクルまたは添加剤の例は、乳糖、デンプン、ショ糖、ステアリン酸マグネシウムなどである。
【0075】
注射用製剤としては、静脈内、皮下、皮内および筋肉内の注射、点滴注入などの剤形が挙げられる。これらの注射用製剤は、公知の方法によって調製することができる。注射用製剤は、例えば、上記の抗体またはその塩を、従来より注射剤に使用されている無菌水媒体または油性媒体に溶解、懸濁または乳化することによって調製することができる。注射剤の水媒体としては、例えば、生理的食塩水、ブドウ糖および他の助剤(auxiliary agent)などを含有する等張液などがあり、アルコール(例えば、エタノール)、ポリアルコール(例えば、プロピレングリコール、ポリエチレングリコール)、非イオン性界面活性剤[例えば、ポリソルベート80、HCO−50(硬化ヒマシ油のポリオキシエチレン(50mol)付加物)]などの適切な溶解補助剤(solubilizing agent)と組み合わせて使用することもできる。油性媒体としては、例えば、ゴマ油、ダイズ油などが採用され、安息香酸ベンジル、ベンジルアルコールなどの溶解補助剤と組み合わせて使用することもできる。このようにして調製される注射剤は、好ましくは、適切なアンプルに充填される。直腸投与に使用される坐剤は、前記の抗体またはその塩を坐剤用の常用の塩基とブレンドすることによって調製することができる。
【0076】
有利には、上記の経口または非経口用の医薬組成物は、有効成分の用量に適合するのに適した単位用量の剤形に調製される。このような単位用量の剤形としては、例えば、錠剤、丸剤、カプセル剤、注射剤(アンプル)、坐剤などが挙げられる。前記の抗体の含有量は一般に、単位用量の剤形当たり約5〜500mgであり、特に、注射剤の剤形では前記の抗体は約5〜100mg含有され、他の剤形では約10〜250mg含有されることが好ましい。
【0077】
上記の組成物はそれぞれ、配合により上記の抗体と任意の有害な相互作用を引き起こさない限り、他の活性成分をさらに含有することもできる。
【0078】
本発明はまた、RGD配列結合機能分子(例えば、インテグリンなど)を有効成分として含む、細胞および/または組織リモデリングの阻害剤および/または促進剤;ならびにRGDタンパク質を発現する細胞および/または組織(例えば、腫瘍細胞、好中球、平滑筋など)をRGDタンパク質結合機能分子と接触させることを含む、細胞および/または組織リモデリングを阻害および/または促進するための方法に関する。このような治療薬における有効成分の用量、投与方法、医薬製剤などは、本発明のヒト化抗体を含む医薬に関する前述の記載を参照することによって適切に決定することができる。
【0079】
上記のとおり、本発明はさらに、RGDタンパク質に関連する、またはそれが関与する障害または疾患を予防または治療するための方法であって、有効量の本発明のヒト化抗体のうちの少なくとも1つを、それを必要とする対象に投与することを含む方法を提供する。
【0080】
5.5.診断的使用
本発明のヒト化抗体を含む医薬組成物は、癌(例えば、癌細胞の増殖または転移)および/または炎症性疾患(例えば、関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、真性糖尿病、動脈硬化症、多発性硬化症、肉芽腫など)の診断用薬として、または臓器移植後の慢性拒否反応、全身性自己免疫疾患、エリテマトーデス、ブドウ膜炎、ベーチェット病、多発性筋炎、増殖性糸球体腎炎、サルコイドーシスなどの自己免疫疾患の診断用薬として使用することができる。本発明のヒト化抗体は、RGD配列を特異的に認識することができ、したがって試験体液中のRGDタンパク質を定量するために、特に、サンドイッチイムノアッセイ、競合的アッセイ、イムノメトリー(immunometry)、比濁分析(nephrometry)など、免疫染色などによる定量に使用することができる。これらの免疫学的方法を本発明のアッセイ法に適用する際は、いかなる特定の条件、手順なども説明する必要はない。当技術分野における通常の技術的考慮事項を従来の条件および手順に付加することによりアッセイ系を構築することが十分である。これらの一般的な技術的手段の詳細については、総説、テキストなどを参照することができる.
【0081】
上記のとおり、本発明の抗体を使用することにより、RGDタンパク質を高感度で定量することもできる。本発明のヒト化抗体は、RGDタンパク質をin vivoで定量するための方法を適用することにより、RGDタンパク質に関連する種々の疾患を診断するのに特に有用である。例えば、RGDタンパク質の発現レベルの増大または低下が検出された場合、RGDタンパク質に関連する疾患、例えば、癌または炎症性疾患に罹患している可能性が高い、または将来これらの疾患に罹患する可能性が高いと診断することができる。したがって、本発明はまた、対象においてRGDタンパク質に関連する、またはそれが関与する障害または疾患を診断するための方法であって、有効量の、本発明のヒト化抗体のうちの少なくとも1つまたは両方を、それを必要とする対象に投与することを含む方法を提供する。このようなin vivo診断に必要な投与量は、治療的使用に必要な投与量未満であってもよく、常用の手順に従って当業者によって決定することができる。
【0082】
本発明のヒト化抗体を使用して、体液、組織などの試験体液に存在するRGDタンパク質を特異的に検出することもできる。ヒト化抗体は、RGDタンパク質を精製するための抗体カラムの調製、精製時のそれぞれの画分に含有されるRGDタンパク質の検出、または試験対象の細胞中のRGDタンパク質の挙動の分析に使用することもできる。
【実施例】
【0083】
6.実施例
以下の実施例では、RGD配列を免疫特異的に認識するモノクローナル抗体の調製、モノクローナル抗体のV領域の配列決定、抗体のエピトープマッピングおよび他の特徴付け、ならびにこのような抗体のキメラ化およびヒト化、ならびに得られたキメラ抗体およびヒト化抗体の特徴付けを例示する。これらの実施例は、本発明の範囲を限定するものと解釈すべきではない。
【0084】
6.1.RGD配列に対するマウス抗体の調製
RGD配列に対するマウスモノクローナル抗体をサブトラクティブ免疫化法(subtractive immunization method)(Williams C.V.ら、1992、Biotechniques12:842〜847による)に従って調製した。RGD配列を含むアミノ酸配列CVDVPNGRGDSLAYGLR(配列番号71)およびECMタンパク質の細胞接着性配列であるアミノ酸配列SLAYGLR(配列番号72)の合成ペプチドとして抗原を調製した。抗原ペプチドをEMCS(Dojin)を介してサイログロブリンに結合させ、これを抗原としてアジュバントと共にマウスに免疫した。ハイブリドーマを当技術分野でよく知られている方法(例えば、Harlowら、Antibodies:A Laboratory Manual、(Cold Spring Harbor Laboratory Press、第2版1988);Hammerlingら、Monoclonal Antibodies and T−Cell Hybridomas掲載、563〜681ページ(Elsevier,N.Y.、1981)を参照されたい)によって調製した。4回免疫した後、脾細胞を収集し、骨髄腫細胞X63−Ag8−653と融合した。次いで、HAT培地を使用し、ELISA(抗原ペプチド固相)を用いて培養上清をスクリーニングすることにより、RGD配列に免疫特異的に反応性を示すモノクローナル抗体を産生するハイブリドーマクローンを選択した。RGD配列を免疫特異的に認識するモノクローナル抗体を産生するハイブリドーマクローンを単離すると、8つのハイブリドーマクローン4P11、11M6、25H15、29R5、30C17、33E10、35B6および38I8を樹立した。チオールセファロースビーズ(Amasham Bioscience)を使用することによって調製された抗原ペプチドカラムを使用することにより、抗体をハイブリドーマの上清から精製した。
【0085】
6.2.抗RGD配列モノクローナル抗体のエピトープ分析
アミノ酸配列CLPVKTDSGSSEEKLY(mOPN1)(配列番号73)、CVDVPNGRGDSLAYGLR(mOPN5)(配列番号71)、CVDVPNGRGDS(配列番号74)、CPNGRGD(配列番号75)、CGRGDSLAYGLR(配列番号76)、CGDSLAYG(配列番号77)、CGDSLAUGLR(配列番号78)およびCSLAYGLR(配列番号72)からなる、マウスOPNに由来する部分ペプチドを含むペプチド、アミノ酸配列CVDTYDGRGDSVVYGLRS(配列番号79)およびCSVVYGLR(配列番号80)からなる、ヒトOPNに由来する部分ペプチドを含むペプチド、ならびにアミノ酸配列CGRGDS(配列番号81)の、ヒトOPNおよびマウスOPNの共通のペプチド配列を含むペプチドを、EMCS(同仁化学研究所)を介してBSA(Sigma corporation)に結合させ、ELISAに使用した。
【0086】
96ウェルプレートにペプチド(10μg/ml)またはタンパク質(5μg/ml)を37℃で1時間培養することによって固定化し、0.1%BSA/PBS/0.05%NaN3溶液でブロッキングし、次いで様々な濃度の抗体と37℃で1時間反応させた。次に、プレートを二次抗体であるHRP標識抗マウスIgG抗体(Jackson ImmunoResearch Laboratories,Inc.)と37℃で30分間反応させ、OPDを色素原として添加し、1N H2SO4を添加して反応を停止させ、次いで490nmでの吸光度を検出した。
【0087】
図1、図2、図3、図4および図5に示すとおり、モノクローナル抗体4P11、11M6、25H5、35B6および33E10は、mOPN5およびhOPN5に結合し、RGDタンパク質のマウスおよびヒト部分ペプチドを認識する。モノクローナル抗体33E10は、GRGDS(配列番号81)、VDVPNGRGDS(配列番号74)およびPNGRGD(配列番号75)を認識したが、OPNのRGDに続く配列を含むSLAYGLR(配列番号72)またはSVVYGLR(配列番号80)を認識しなかった。モノクローナル抗体33E10は、GRGDS(配列番号81)、VDVPNGRGDS(配列番号74)およびPNGRGD(配列番号75)に共通に含まれるRGD配列を認識し、ヒトペプチドとマウスペプチドの両方に結合することができる。モノクローナル抗体35B6は、GRGDSLAYGLR(配列番号76)、GDSLAYG(配列番号77)およびGDSLAYGLR(配列番号78)を認識したが、GRGDS(配列番号81)、VDVPNGRGDS(配列番号74)またはPNGRGD(配列番号75)を認識しなかった。モノクローナル抗体35B6は、GDを含む、RGDに続く配列を認識する。モノクローナル抗体29R5、30C17および38I8は、GRGDS(配列番号81)、SLAYGLR(配列番号72)およびSVVYGLR(配列番号80)にはわずかな反応性しか示さなかったが、mOPN5にだけ反応し、それにより、これらのモノクローナル抗体はマウスOPNのVDVPNGRGDSLAYGLR(配列番号71)を認識することが示される。
【0088】
6.3.抗RGD抗体のCDR分析
モノクローナル抗体33E10および35B6のCDRのアミノ酸配列を以下の手順によって決定した。RNeasy Miniキット(Qiagen)を使用して対応するハイブリドーマからRNAを抽出し、First−strand cDNA synthesisキットを使用してcDNAを調製した。抗体のH鎖cDNAを、Heavy primer amplificationキット(Amasham Bioscience)を使用したPCRによって増幅し、pCRII−TOPOベクター(Invitrogen)中にクローニングし、次いでcDNA配列およびアミノ酸配列を決定した。ABG:Directory of 3D structures of antibodies(http://www.ibt.unam.mx/vir/structure/structures.html)によってCDRを決定した。V鎖およびL鎖のCDRは以下のとおりである(図6および図7にも示す)。
(H鎖)
[CDRH1]
33E10:GFTFTDYYMI(配列番号1)
35B6:GYTFTNYWMH(配列番号7)
[CDRH2]
33E10:WLGFIRNKANGYTTEYSASVKG(配列番号2)
35B6:WIGNINPRNGDSNYNEKFRS(配列番号8)
[CDRH3]
33E10:GAY(配列番号3)
35B6:GYFDV(配列番号9)
(L鎖)
[CDRL1]
33E10:RSSQSIVHSNGNTYLE(配列番号4)
35B6:KASQDINSYLS(配列番号10)
[CDRL2]
33E10:RVSNRFS(配列番号5)
35B6:RANRLVD(配列番号11)
[CDRL3]
33E10:GSFVPW(配列番号6)
35B6:YDEFPF(配列番号12)
【0089】
本実施例では、CDRをABGによって決定した。しかし、他のプログラムを使用してCDRを決定することもでき、それによりある程度異なる配列が得られることもあることは当技術分野でよく知られている。
【0090】
6.4.RGD配列を有するECMタンパク質に対する結合能
ヒトOPN(hOPN)遺伝子またはマウスOPN(mOPN)遺伝子をそれぞれ導入したCHO−K1細胞の培養上清から、抗OPN抗体カラムを使用することにより、hOPNまたはmOPNをそれぞれ精製した。ヒトビトロネクチン(以下「VN」と略記する)はAGC TECHNO GLASS Co.,Ltdから得た。ヒトフィブロネクチン(以下「FN」と略記する)、ヒトトロンボスポンジンおよびマウスラミニンはSigma Corporationから得た。
【0091】
上記で得たhOPN、mOPN、FN、VNまたはラミニンを固定化した96ウェルプレートを使用したELISAにより、ECMタンパク質に対するモノクローナル抗体33E10および35B6の結合能を検出した。96ウェルプレートにペプチド(10μg/ml)またはタンパク質(5μg/ml)を37℃で1時間培養することによって固定化し、0.1%BSA/PBS/0.05%NaN3溶液でブロッキングし、次いで様々な濃度の抗体と37℃で1時間反応させた。次に、プレートを二次抗体のHRP標識抗マウスIgG抗体(Jackson ImmunoResearch Laboratories,Inc.)と37℃で30分間反応させ、OPDを色素原として添加し、1N H2SO4を添加して反応を停止させ、次いで490nmでの吸光度を検出した。
【0092】
図8にその結果を示した。モノクローナル抗体33E10は、試験したすべてのECMタンパク質と交差反応性を示したが、ラミニンとは低い反応性を示した。モノクローナル抗体35B6は、hOPNおよびmOPNと反応性を示したが、ラミニンとは反応性を示さなかった。
【0093】
6.5.細胞接着阻害活性
細胞接着にはRGDペプチドとそのリガンド、すなわちインテグリンなどとの結合が関与することが知られているので、単離された抗RGD抗体をその細胞接着阻害活性について検討した。ヒトOPN(hOPN)遺伝子またはマウスOPN(mOPN)遺伝子をそれぞれ導入したCHO−K1細胞の培養上清から、抗OPN抗体カラムを使用することにより、hOPNまたはmOPNをそれぞれ精製した。大腸菌から単離することによる、トロンビン切断型mOPNのN末端部分とのグルタチオンSトランスフェラーゼ(GST)融合タンパク質として、mOPNのN半分を精製した。ヒトFNおよびヒトVNはSigma Corporationから得た。
【0094】
50μlのタンパク質を96ウェルプレートの各ウェルに添加し、37℃で1時間インキュベートし、プレート上に固定化した。プレートをブロッキング溶液(0.5%BSA/PBS)でブロッキングし、PBSで1回洗浄した後、NIH3T3細胞を0.25%BSA−最小必須培地(MEM)に懸濁させ、単離されたモノクローナル抗体を1.0×105細胞/mlの最終濃度で混合し、プレートに200μl/ウェルで添加し、5%CO2下、37℃で1時間インキュベートした。非接着細胞をPBSですすぎ、接着細胞を固定し、0.5%Crystal Violet(和光純薬工業、大阪、日本による)/20%メタノールで染色した。染色細胞を室温で30分間静置させた。プレートを蒸留水で洗浄し、それに20%酢酸溶液を添加して溶解させた。590nmでのODを測定することによって接着活性を定量した。
【0095】
mOPNのN半分を固定化した96ウェルプレートに、NIH3T3細胞とモノクローナル抗体33E10または35B6の混合物を添加し、NIH3T3細胞とmOPNのN半分との結合に及ぼす抗体の効果を検討した。mOPNのN半分、FNまたはVNをそれぞれ固定化した96ウェルプレートに、NIH3T3細胞とモノクローナル抗体33E10の混合物を添加し、NIH3T3細胞と各タンパク質との結合に及ぼす抗体の効果を検討した。
【0096】
図9および図10A〜10Cに示すとおり、NIH3T3細胞はmOPNのN半分に接着し、この接着を抗RGD抗体によって阻害した。モノクローナル抗体33E10は、モノクローナル抗体35B6と比較して強い阻害活性を示した。NIH3T3細胞は、検討したすべてのECMタンパク質に接着した。モノクローナル抗体33E10は、mOPNのN半分との細胞接着を阻害したが、FNまたはVNとの細胞接着を阻害しなかった。したがって、モノクローナル抗体33E10はOPNと細胞の間の接着を特異的に阻害することが示される。
【0097】
6.6.抗RGD抗体の治療効果
抗RGD抗体の治療効果をマウス系において検討した。抗RGDモノクローナル抗体(4P11、11M6、29R5、30C7、38I8、33E10および35B6)を抗RGD抗体についての記載(セクション6.1、前掲を参照されたい)と実質的に同様に調製した。
【0098】
6.6.1.肝炎に対する治療効果
WO02/081522は、OPN機能を阻害することによって肝炎を治療できることを開示している。したがって、マウス肝炎モデルにおいてマウス抗RGD抗体(4P11、11M6、29R5、30C7、38I8、33E10および35B6)を使用して抗RGD抗体の治療効果を試験した。マウス(マウス5匹/群)の血中ASTおよびALTレベルを、200μgのコンカナバリンA(ConA)(Vector)の静脈内投与から12時間後、GPT/ALT−PIIIおよびGOT/AST−PIII(富士フイルム)を使用して測定した。ConA注射の3時間前に抗体を200μg投与した。対照抗体としてマウスIgGを使用した。
【0099】
図11に示すとおり、モノクローナル抗体25H15はいかなる治療効果も示さなかったが、モノクローナル抗体4P11、11M6、29R5、30C17および38I8は治療効果を示した。モノクローナル抗体33E10または35B6を投与したマウスは、ASTおよびALTレベルの増大を示さなかった。したがって、この結果から、モノクローナル抗体33E10または35B6によって肝炎を治療する可能性があることが明らかになった。
【0100】
6.6.2.マウス癌細胞株の転移に及ぼす抗RGD抗体の効果
肺転移のマウス実験モデルおよび自然発症モデルにおいて転移に及ぼす抗RGD抗体の効果を試験した。
【0101】
実験モデルでは、400μg/マウスのモノクローナル抗体と混合したマウスメラノーマ細胞株B16−Luc細胞(1×105細胞/マウス)をC57BL/6マウスの尾静脈に注射し、注射から14日後、肺転移数をカウントした。対照として同じ種類の抗体(mIgG1)を使用した。
【0102】
自然発症モデルでは、マウスメラノーマ細胞株B16−BL6細胞(4×105細胞/マウス)をC57BL/6マウスの左足蹠に皮下注射した。注射から19日後、原発性腫瘍を外科的に切除し、切除から14日後(B16−BL6細胞の注射から33日後)、屠殺したマウス由来の肺における腫瘍コロニーの数をカウントした。200μg/マウスのモノクローナル抗体を腫瘍細胞の注射から3、5、7、9、11、13、15および17日後の8回、腹腔内投与した。原発性腫瘍のサイズは切除までに測定した。外科切除から14日後、肺における腫瘍コロニーの数をカウントした。対照として同じ種類の抗体(mIgG1)を使用した。
【0103】
図12A〜12Bにマウス実験モデルの結果を示す。モノクローナル抗体33E10または35B6を投与したマウスにおける肺転移の平均数は対照と比較して少なかった。モノクローナル抗体35B6は肺転移を有意に阻害した。
【0104】
図13A〜13Cは、マウス自然発症モデルにおける以下のフォーマット:原発性腫瘍サイズの経日変化、肺転移コロニーの数、および体重変化の結果を示す。モノクローナル抗体33E10または35B6のいずれかを投与したマウスにおける原発性腫瘍のサイズは、対照マウスと比較して小さかった。したがって、その結果から、モノクローナル抗体33E10または35B6によって腫瘍増殖を阻害する可能性があることが示される。対照群における5匹のマウスのうち2匹が非常に多数の肺転移コロニーを示したので、35B6に関しては統計学的有意差が示されなかった。しかし、モノクローナル抗体33E10または35B6を投与したマウスにおける肺転移の平均数は対照抗体と比較して少なかった。したがって、モノクローナル抗体33E10または35B6は癌転移を阻害することが示される。
【0105】
6.6.3.マウス関節リウマチモデルにおける抗RGD抗体の治療効果
II型コラーゲン特異的モノクローナル抗体(IBL、日本)のカクテルを供給元のプロトコールに従って使用して、マウスに関節リウマチを誘発した。すなわち、マウス(Balb/c)にII型コラーゲン特異的モノクローナル抗体のカクテルを注射し、注射の3日後、LPSを注射して関節リウマチを発症させた。抗RGDモノクローナル抗体または正常ハムスターIgG(NHG)をコラーゲン抗体注射の前日からコラーゲン抗体注射の6日後まで全8回200μg/マウス/日で腹腔内に投与した。マウスをコラーゲン抗体注射の日から毎日観察し、各足を関節の紅斑および腫脹に基づいて0から4までに類別することによって関節炎のレベルをスコア付けした(0=紅斑または腫脹なし;1=つま先などの1つの小さい関節に紅斑または腫脹あり;2=小さい関節の2つ以上に紅斑または腫脹あり、または手首または足首などのより大きい関節に紅斑または腫脹あり;3=足全体に紅斑または腫脹あり;4=足全体に完全な紅斑または腫脹あり;マウス(4足)1匹の最大スコア16)。
【0106】
その結果を図14に示す。対照NHGを注射したマウスは高スコアを示し、関節リウマチを発症したが、抗RGD抗体33E10または35B6をそれぞれ注射したマウスは低スコアを示し、関節リウマチの発症は完全に遮断された。したがって、この結果に照らして、抗RGD抗体は関節リウマチに対して予防的および治療的効果を有することが示された。
【0107】
6.6.4.マウス子宮内膜症モデルにおける抗RGD抗体の治療効果
子宮内膜症の症状は、嚢胞形成、周囲の間質における炎症、子宮内膜腺上皮の異所的な増殖による平滑筋化生、神経新生および血管形成を示す。免疫組織化学的方法により、オステオポンチン(OPN)がヒト子宮内膜症およびラット子宮内膜症モデルにおいて高度に発現されることが報告された。したがって、抗RGD抗体(33E10)による動物モデル対する治療効果を子宮内膜症に対する新たな療法として調べた。
【0108】
C57BL/6J雌性マウス(9週齢)を使用した。子宮内膜症モデルとして18匹のマウスを準備した。右の子宮を切除し、子宮の2mm×2mm角の2つ小片を腹部に自己移植した。左の子宮は参照用に処置しなかった。マウス抗RGD抗体(500μg/頭部、ip)を9匹のマウス(処置群)に週2回4週間(8回投与の全投与量:4000μg/頭部)施した。対照群(マウス9匹)には抗体を投与しなかった。投与後、形成された嚢胞の数を確認し、組織学的検査によって病的変化を観察した。
【0109】
その結果を表1に示した。2つの群に体重の差はなかった。処置群の出現した子宮内膜症の数は対照群よりも著しく少なかった。形成された嚢胞の重量は明らかに減少し、処置群は対照群の平均して約1/3であった。免疫組織化学的検査により、子宮内膜腺上皮におけるOPN発現が抗体処置によって抑制されることを確認することができた。処置群の間質における平滑筋の厚さは、対照群の1/2に減少した。
【0110】
マウス子宮内膜症モデルは抗RGD抗体の投与による治療効果を示した。
【0111】
【表1】

【0112】
6.7.非ヒト抗体のヒト化
6.7.1.マウス33E10可変領域遺伝子のクローニングおよび配列決定
7.5%CO2インキュベーターにおいて10%ウシ胎児血清(FBS;HyClone、Logan、UT)を含有するTIL培地I(免疫生物研究所、群馬、日本)中、37℃でマウス33E10ハイブリドーマ細胞を増殖させた。TRIzol試薬(Invitrogen、Carlsbad、CA)を供給元のプロトコールに従って使用して、約3×106ハイブリドーマ細胞から全RNAを抽出した。GeneRacer Kit(Invitrogen)を供給元のプロトコールに従って使用して、オリゴdTプライマーによるcDNAを合成した。35B6重鎖および軽鎖の可変領域cDNAは、ポリメラーゼ連鎖反応(PCR)により、マウスγ−1およびκ鎖定常領域にそれぞれアニーリングする3’プライマー、ならびにGeneRacer Kitにおいて提供されるGeneRacer5’プライマー(5’−CGACTGGAGCACGAGGACACTGA−3’)(配列番号84)を使用して、Phusion DNAポリメラーゼ(New England Biolabs、Beverly、MA)を用いて増幅した。VHのPCR増幅については、3’プライマーは配列5’−GCCAGTGGATAGACAGATGG−3’(配列番号85)を有する。VLのPCR増幅については、3’プライマーは配列5’−GATGGATACAGTTGGTGCAGC−3’(配列番号86)を有する。増幅されたVHおよびVL cDNAをpCR4Blunt−TOPOベクター(Invitrogen)中にクローニングして配列を決定した。可変領域のDNA配列決定をTocore(Menlo Park、CA)で実施した。いくつかの重鎖および軽鎖クローンの配列を決定し、典型的なマウス重鎖および軽鎖可変領域に相同である固有の配列を同定した。33E10 VHおよびVLのコンセンサスcDNA配列を推定アミノ酸配列と一緒にそれぞれ図15および16に示す。
【0113】
6.7.2.キメラ33E10 IgG1/κ抗体の構築
33E10 VHをコードする遺伝子は、PCRにより、鋳型として33E10 VH cDNA、5’プライマーとして5’−GGGACTAGTACCACCATGAAGTTGTGGCTGAACTGGATT−3’(SpeI部位は下線付き)(配列番号87)および3’プライマーとして5’−GGGAAGCTTGAAGTTAGGACTCACCTGCAGAGACAGTGACCAGAGTCCC−3’(HindIII部位は下線付き)(配列番号88)を使用して、スプライスドナーシグナルおよび適切なフランキング制限酵素部位を含むエキソンとして作製した(図17)。同様に、33E10 VLをコードする遺伝子は、PCRにより、鋳型として33E10 VL cDNA、5’プライマーとして5’−GGGGCTAGCACCACCATGAAGTTGCCTGTTAGGCTGTTG−3’(NheI部位は下線付き)(配列番号89)および3’プライマーとして5’−GGGGAATTCTTTGGATTCTACTTACGTTTGATTTCCAGCTTGGTGCCTCC−3’(EcoRI部位は下線付き)(配列番号90)を使用して、スプライスドナーシグナルおよび適切なフランキング制限酵素部位を含むエキソンとして作製した(図18)。33E10 VHおよびVLエキソンのスプライスドナーシグナルは、それぞれマウス生殖系列JH3およびJκ1配列に由来するものであった。QIAquick Gel Extraction Kit(Qiagen、Valencia、CA)を使用してPCR増幅断片をゲル精製し、SpeIおよびHindIII(VHについて)またはNheIおよびEcoRI(VLについて)で消化し、ヒトγ−1およびκ定常領域を担う哺乳動物発現ベクター中にクローニングしてキメラ33E10 IgG1/κ抗体を産生した。得られた発現ベクターpCh33E10の概略構造を図19に示す。
【0114】
6.7.3.ヒト化33E10 VHおよびVL遺伝子の作製
33E10可変領域のヒト化をQueenら(Proc.Natl.Acad.Sci.USA86:10029〜10033、1989)に概説されているとおりに実施した。まず、コンピュータプログラムを利用して33E10可変領域の分子モデルを構築した。次に、ヒト可変領域配列に対する相同性検索に基づいて、33E10 VHと高い相同性を有するU03400(GenBankアクセッション番号)のヒトアミノ酸配列を、ヒト化33E10 VHのフレームワークを提供するアクセプターとして選択した。同様に、X72452(GenBankアクセッション番号)のヒトアミノ酸配列を33E10 VLのヒト化用のアクセプターとして選択した。
【0115】
コンピュータモデルがCDRとの有効な接触を提案したフレームワーク位置で、ヒトフレームワークアミノ酸をマウス33E10可変領域のアミノ酸で置換した。30位および48位でこれを実施して、ヒト化33E10(Hu33E10)VHを作製した(図20)。軽鎖については、置き換えることを必要とせずに、ヒト化33E10(Hu33E10)VLを作製した(図21)。33E10、設計したHu33E10およびヒトアクセプターアミノ酸配列のアライメントをVHについては図20、VLについては図21に示す。
【0116】
Hu33E10 VHおよびVLのぞれぞれをコードする遺伝子を、シグナルペプチド、スプライスドナーシグナルおよび適切な制限酵素部位を含むエキソンとして設計して、続いて哺乳動物発現ベクター中にクローニングした。Hu33E10 VHおよびVLエキソンのスプライスドナーシグナルは、それぞれヒト生殖系列JH4およびJκ1配列に由来するものであった。マウス33E10 VL遺伝子のシグナルペプチド配列は、SIG−Predシグナルペプチド予測ソフトウェア(http://bmbpcu36.leeds.ac.uk/prot_analysis/Signal.html)により、正確な切断に部分最適であることが示された。したがって、SIG−Predソフトウェアによって効率的かつ正確に切断すると予測された、マウスモノクローナル抗体35B6のVL遺伝子のシグナルペプチド(ジーンテクノサイエンス)をHu33E10 VLエキソンにおいて使用した。Hu33E10 VHエキソンにおけるシグナルペプチド配列は、対応するマウス33E10 VH配列に由来するものであった。SIG−Predソフトウェアにより、Hu33E10 VH遺伝子のシグナルペプチドが効率的かつ正確に切断されることが示された。
【0117】
Hu33E10 VHおよびVL遺伝子は、Heら(J.Immunol.160:1029〜1035、1998)に概説されているとおり、Phusion DNAポリメラーゼを使用して、いくつかの重複している合成オリゴヌクレオチドプライマー(配列番号91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122および123)の伸長およびPCR増幅によって構築した。Hu33E10 VHおよびVL遺伝子の構築に使用されたオリゴヌクレオチドをそれぞれ図22および図23に記載する。Hu33E10 VHおよびVL遺伝子におけるオリゴヌクレオチドの位置をそれぞれ図24および25を示す。QIAquick Gel Extraction Kit(Qiagen)を使用してPCR増幅断片をゲル精製し、pCR4Blunt−TOPOベクター中にクローニングして配列を決定した。SpeIおよびHindIII(VHについて)またはNheIおよびEcoRI(VLについて)で消化した後、Hu33E10 VHおよびVL遺伝子を哺乳動物発現ベクター中の対応する部位にサブクローニングしてヒトIgG1/κの形で産生した。得られた発現ベクターpHu33E10の概略構造を図19に示す。得られたHu33E10 VHおよびVL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒にそれぞれ図26(配列番号52)および図27(配列番号54)に示す。
【0118】
6.7.4.キメラおよびヒト化33E10 IgG1/κの一過性発現
キメラおよびヒト化33E10 IgG1/κ抗体は、Durocherら(Nucl.Acids Res.30:e9、2002)に従ってポリエチレンイミンを使用してpCh35B6およびpHu35B6プラスミドDNAをそれぞれHEK293細胞にトランスフェクトすることによって一過性に発現させる。一過性にトランスフェクトされたHEK293細胞を、7.5%CO2インキュベーターにおいて10%FBSを含有するDMEM中、37℃で2日間維持する。培養上清におけるCh33E10およびHu33E10 IgG1/κ抗体のそれぞれの発現レベルをサンドイッチELISAによって測定する。ELISAプレートをPBS中2,000倍希釈したヤギ抗ヒトIgG Fcγ鎖特異的ポリクローナル抗体(SouthernBiotech、Birmingham、AL)を100μl/ウェルで加えて4℃で一晩コーティングし、Wash Buffer(0.05%Tween20を含有するPBS)で洗浄し、Blocking Buffer(2%Skim Milkおよび0.05%Tween20を含有するPBS)を300μl/ウェルで加えて室温で1時間ブロッキングする。Wash Bufferで洗浄した後、ELISA Buffer(1%Skim Milkおよび0.025%Tween20を含有するPBS)中で適切に希釈した試料を100μl/ウェルでELISAプレートにアプライする。ヒト骨髄腫血清(SouthernBiotech)から精製したヒトIgG1/κ抗体を標準物質として使用する。ELISAプレートを室温で2時間インキュベートし、Wash Bufferで洗浄した後、2,000倍希釈したHRPコンジュゲートヤギ抗ヒトκ鎖ポリクローナル抗体(SouthernBiotech)を100μl/ウェルで使用して、結合した抗体を検出する。室温で1時間インキュベートし、Wash Bufferで洗浄した後、ABTS基質(bioWORLD、Dublin、OH)を100μl/ウェルで添加することによって発色を実施する。2%シュウ酸を100μl/ウェルで添加することによって発色を停止させる。405nmでの吸光度を読み取る。
【0119】
6.7.5.ヒト化35B6の特徴付け
ELISAにより、ヒト化35B6 IgG1/κの親和性をキメラ33E10 IgG1/κと比較する。抗原として、ウシ血清アルブミンとコンジュゲートした合成オリゴペプチド(Cys−Val−Asp−Thr−Tyr−Asp−Gly−Arg−Gly−Asp−Ser−Val−Val−Tyr−Gly−Leu−Arg−Ser)(hOPN5−BSA)を使用する。典型的な実験において、ELISAプレートをPBS中1μg/ml hOPN−BSAを100μl/ウェルで加えて4℃で一晩コーティングし、Wash Bufferで洗浄し、Blocking Bufferを300μl/ウェルで加えて室温で1時間ブロッキングする。Wash Bufferで洗浄した後、ELISA Buffer中で適切に希釈した試料を100μl/ウェルでELISAプレートにアプライする。ELISAプレートを4℃で一晩インキュベートし、Wash Bufferで洗浄した後、2,000倍希釈したHRPコンジュゲートヤギ抗ヒトγ鎖ポリクローナル抗体(SouthernBiotech)を100μl/ウェルで使用して、結合した抗体を検出する。室温で1時間インキュベートし、Wash Bufferで洗浄した後、ABTS基質を100μl/ウェルで添加することによって発色を実施し、2%シュウ酸を100μl/ウェルで添加することによって停止させる。405nmでの吸光度を読み取る。
【0120】
6.7.6.マウス35B6可変領域遺伝子のクローニングおよび配列決定
7.5%CO2インキュベーターにおいて10%ウシ胎児血清(FBS;HyClone、Logan、UT)を含有するTIL培地I(免疫生物研究所、群馬、日本)中、37℃でマウス35B6ハイブリドーマ細胞を増殖させた。TRIzol試薬(Invitrogen、Carlsbad、CA)を供給元のプロトコールに従って使用して、約3×106ハイブリドーマ細胞から全RNAを抽出した。GeneRacer Kit(Invitrogen)を供給元のプロトコールに従って使用して、オリゴdTプライマーによるcDNAを合成した。35B6重鎖および軽鎖の可変領域cDNAは、ポリメラーゼ連鎖反応(PCR)により、マウスγ−1およびκ鎖定常領域にそれぞれアニーリングする3’プライマー、ならびにGeneRacer Kitにおいて提供されるGeneRacer5’プライマー(5’−CGACTGGAGCACGAGGACACTGA−3’)(配列番号84)を使用して、Phusion DNAポリメラーゼ(New England Biolabs、Beverly、MA)を用いて増幅した。VHのPCR増幅については、3’プライマーは配列5’−GCCAGTGGATAGACAGATGG−3’(配列番号124)を有する。VLのPCR増幅については、3’プライマーは配列5’−GATGGATACAGTTGGTGCAGC−3’(配列番号125)を有する。増幅されたVHおよびVL cDNAをpCR4Blunt−TOPOベクター(Invitrogen)中にクローニングして配列を決定した。可変領域のDNA配列決定をTocore(Menlo Park、CA)で実施した。いくつかの重鎖および軽鎖クローンの配列を決定し、典型的なマウス重鎖および軽鎖可変領域に相同である固有の配列を同定した。35B6 VHおよびVLのコンセンサスcDNA配列を推定アミノ酸配列と一緒にそれぞれ図28および29に示す。
【0121】
6.7.7.キメラ35B6 IgG1/κ抗体の構築
35B6 VHをコードする遺伝子は、PCRにより、鋳型として35B6 VH cDNA、5’プライマーとして5’−GGGACTAGTACCACCATGGGATGGAGCTGTATCCTC−3’(SpeI部位は下線付き)(配列番号126)および3’プライマーとして5’−GGGAAGCTTAAAAAAAGCCAGCTTACCTGAGGAGACGGTGACCGTGGTCCC−3’(HindIII部位は下線付き)(配列番号127)を使用して、スプライスドナーシグナルおよび適切なフランキング制限酵素部位を含むエキソンとして作製した(図30)。同様に、35B6 VLをコードする遺伝子は、PCRにより、鋳型として35B6 VL cDNA、5’プライマーとして5’−GGGGCTAGCACCACCATGAGGACCCCTGCTCAGTTTCTT−3’(NheI部位は下線付き)(配列番号128)および3’プライマーとして5’−GGGGAATTCGCAAAAGTCTACTTACGTTTTATTTCCAACTTTGTCCCCGA−3’(EcoRI部位は下線付き)(配列番号129)を使用して、スプライスドナーシグナルおよび適切なフランキング制限酵素部位を含むエキソンとして作製した(図31)。35B6 VHおよびVLエキソンのスプライスドナーシグナルは、それぞれマウス生殖系列JH1およびJκ4配列に由来するものであった。QIAquick Gel Extraction Kit(Qiagen、Valencia、CA)を使用してPCR増幅断片をゲル精製し、SpeIおよびHindIII(VHについて)またはNheIおよびEcoRI(VLについて)で消化し、ヒトγ−1およびκ定常領域を担う哺乳動物発現ベクター中にクローニングしてキメラ35B6 IgG1/κ抗体を産生した。得られた発現ベクターpCh35B6の概略構造を図32に示す。
【0122】
6.7.8.ヒト化35B6 VHおよびVL遺伝子の作製
35B6可変領域のヒト化をQueenら(Proc.Natl.Acad.Sci.USA86:10029〜10033、1989)に概説されているとおりに実施した。まず、コンピュータプログラムを利用して35B6可変領域の分子モデルを構築した。次に、ヒト可変領域配列に対する相同性検索に基づいて、35B6 VHと高い相同性を有するZ47230(GenBankアクセッション番号)のヒトアミノ酸配列を、ヒト化35B6 VHのフレームワークを提供するアクセプターとして選択した。同様に、X72479(GenBankアクセッション番号)のヒトアミノ酸配列を35B6 VLのヒト化用のアクセプターとして選択した。
【0123】
コンピュータモデルがCDRとの有効な接触を提案したフレームワーク位置で、ヒトフレームワークアミノ酸をマウス35B6可変領域のアミノ酸で置換した。48位、66位、67位、68位、69位および71位でこれを実施して、ヒト化35B6(Hu35B6)VHを作製した(図33)。軽鎖については、46位および69位で置き換えて、ヒト化35B6(Hu35B6)VLを作製した(図34)。35B6、設計したHu35B6およびヒトアクセプターアミノ酸配列のアライメントをVHについては図33、VLについては図34に示す。
【0124】
Hu35B6 VHおよびVLのぞれぞれをコードする遺伝子を、シグナルペプチド、スプライスドナーシグナルおよび適切な制限酵素部位を含むエキソンとして設計して、続いて哺乳動物発現ベクター中にクローニングした。Hu35B6 VHおよびVLエキソンのスプライスドナーシグナルは、それぞれヒト生殖系列JH6およびJκ1配列に由来するものであった。マウス35B6 VH遺伝子のシグナルペプチド配列は、SIG−Predシグナルペプチド予測ソフトウェア(http://bmbpcu36.leeds.ac.uk/prot_analysis/Signal.html)により、正確な切断に部分最適であることが示された。したがって、SIG−Predソフトウェアによって効率的かつ正確に切断すると予測された、マウスモノクローナル抗体35E10のVH遺伝子のシグナルペプチド(ジーンテクノサイエンス)をHu35B6 VLエキソンにおいて使用した。ヒト化Hu35B6 VLエキソンにおけるシグナルペプチド配列は、対応するマウス35B6 VL配列に由来するものであった。SIG−Predソフトウェアにより、Hu35B6 VL遺伝子のシグナルペプチドが効率的かつ正確に切断されることが示された。
【0125】
Hu35B6 VHおよびVL遺伝子は、Heら(J.Immunol.160:1029〜1035、1998)に概説されているとおり、Phusion DNAポリメラーゼを使用して、いくつかの重複している合成オリゴヌクレオチドプライマー(配列番号91、92、130、131、132、133、134、135、136、137、138、139、140、141、142、143、107、108、144、145、146、147、148、149、150、151、152、153、154および155)の伸長およびPCR増幅によって構築した。Hu35B6 VHおよびVL遺伝子の構築に使用されたオリゴヌクレオチドをそれぞれ図35および図36に記載する。Hu35B6 VHおよびVL遺伝子におけるオリゴヌクレオチドの位置をそれぞれ図37および38を示す。QIAquick Gel Extraction Kit(Qiagen、Valencia、CA)を使用してPCR増幅断片をゲル精製し、pCR4Blunt−TOPOベクター中にクローニングして配列を決定した。SpeIおよびHindIII(VHについて)またはNheIおよびEcoRI(VLについて)で消化した後、Hu35B6 VHおよびVL遺伝子を哺乳動物発現ベクター中の対応する部位にサブクローニングしてヒトIgG1/κの形で産生した。得られた発現ベクターpHu35B6の概略構造を図32に示す。得られたHu35B6 VHおよびVL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒にそれぞれ図39(配列番号68)および図40(配列番号70)に示す。
【0126】
6.7.9.キメラおよびヒト化35B6 IgG1/κの一過性発現
キメラおよびヒト化35B6 IgG1/κ抗体は、Durocherら(Nucl.Acids Res.30:e9、2002)に従ってポリエチレンイミンを使用してpCh35B6およびpHu35B6プラスミドDNAをそれぞれHEK293細胞にトランスフェクトすることによって一過性に発現させた。一過性にトランスフェクトされたHEK293細胞を、7.5%CO2インキュベーターにおいて10%FBSを含有するDMEM中、37℃で2日間維持した。培養上清におけるCh35B6およびHu35B6 IgG1/κ抗体のそれぞれの発現レベルをサンドイッチELISAによって測定した。ELISAプレートをPBS中2,000倍希釈したヤギ抗ヒトIgG Fcγ鎖特異的ポリクローナル抗体(SouthernBiotech、Birmingham、AL)を100μl/ウェルで加えて4℃で一晩コーティングし、Wash Buffer(0.05%Tween20を含有するPBS)で洗浄し、Blocking Buffer(2%Skim Milkおよび0.05%Tween20を含有するPBS)を300μl/ウェルで加えて室温で1時間ブロッキングした。Wash Bufferで洗浄した後、ELISA Buffer(1%Skim Milkおよび0.025%Tween20を含有するPBS)中で適切に希釈した試料を100μl/ウェルでELISAプレートにアプライした。ヒト骨髄腫血清(SouthernBiotech)から精製したヒトIgG1/κ抗体を標準物質として使用した。ELISAプレートを室温で2時間インキュベートし、Wash Bufferで洗浄した後、2,000倍希釈したHRPコンジュゲートヤギ抗ヒトκ鎖ポリクローナル抗体(SouthernBiotech)を100μl/ウェルで使用して、結合した抗体を検出した。室温で1時間インキュベートし、Wash Bufferで洗浄した後、ABTS基質(bioWORLD、Dublin、OH)を100μl/ウェルで添加することによって発色を実施した。2%シュウ酸を100μl/ウェルで添加することによって発色を停止させた。405nmでの吸光度を読み取った。
【0127】
6.7.10.ヒト化35B6の特徴付け
ELISAにより、ヒト化35B6 IgG1/κの親和性をキメラ35B6 IgG1/κと比較した。抗原として、ウシ血清アルブミンとコンジュゲートした合成オリゴペプチ(Cys−Val−Asp−Thr−Tyr−Asp−Gly−Arg−Gly−Asp−Ser−Val−Val−Tyr−Gly−Leu−Arg−Ser)(配列番号79)(hOPN5−BSA)を使用した。典型的な実験において、ELISAプレートをPBS中1μg/ml hOPN−BSAを100μl/ウェルで加えて4℃で一晩コーティングし、Wash Bufferで洗浄し、Blocking Bufferを300μl/ウェルで加えて室温で1時間ブロッキングした。Wash Bufferで洗浄した後、ELISA Buffer中で適切に希釈した試料を100μl/ウェルでELISAプレートにアプライした。ELISAプレートを4℃で一晩インキュベートし、Wash Bufferで洗浄した後、2,000倍希釈したHRPコンジュゲートヤギ抗ヒトγ鎖ポリクローナル抗体(SouthernBiotech)を100μl/ウェルで使用して、結合した抗体を検出した。室温で1時間インキュベートし、Wash Bufferで洗浄した後、ABTS基質を100μl/ウェルで添加することによって発色を実施し、2%シュウ酸を100μl/ウェルで添加することによって停止させた。405nmでの吸光度を読み取った。図41A、41Bに示すとおり、ヒト化35B6 IgG1/κとhOPN5−BSAの結合は、キメラ35B6 IgG1/κに類似し(図41A)、または識別不可能であった(図41B)。この結果から、マウス35B6抗体のヒト化に成功したことが示される。
【0128】
7.寄託
本明細書で33E10および35B6と称した、マウス抗RGDモノクローナル抗体を産生するハイブリドーマは、微生物の寄託に関するブダペスト条約に従って、2005年10月27日、茨城県つくば市東1−1−1つくばセンター中央第6(郵便番号:305−8566)にある独立行政法人産業技術総合研究所特許生物寄託センターに寄託し、それぞれアクセッション番号FERM BP−10440およびFERM BP−10441を受けた。すべてその全体が参照により本明細書に組み込まれる。
【産業上の利用可能性】
【0129】
8.産業上の利用可能性
本発明のヒト化モノクローナル抗体は、RGDタンパク質の機能を阻害して、癌、例えば、癌細胞の増殖または転移、および炎症性疾患、例えば、関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、真性糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などに対して治療効果を発揮する。本発明の抗RGD抗体および抗インテグリン抗体の両方を含む医薬組成物は、癌および炎症性疾患に対してより改善された治療効果を及ぼす。
【0130】
9.配列の表
本明細書を通じて参照される配列を以下に要約する。
【0131】
【表2】
【技術分野】
【0001】
1.発明の分野
本発明は、細胞外マトリックスタンパク質のアミノ酸配列RGD(Arg−Gly−Asp)を免疫特異的に認識するヒト化抗体、ならびに癌、炎症性疾患、自己免疫疾患、感染症、骨疾患などを含めた種々の疾患または障害に対するその治療および診断的使用に関する。
【0002】
本仮出願を通じて、いくつかの刊行物(特許および特許出願を含む)を参考文献として本明細書に引用する。これらの刊行物の開示はその全体が、本発明が属する分野の技術をより十分に説明するために、参照により本仮出願に組み込まれる。
【背景技術】
【0003】
2.発明の背景
細胞接着は、多細胞生物の生命維持に重要な役割を果たしている。多細胞生物の細胞接着は、細胞−細胞外マトリックス(以下「ECM」と略記する)間接着と細胞間接着に分類される。細胞−ECM間接着はインテグリンによって媒介され、細胞間接着はカドヘリン、クローディンおよびネクチンによって媒介されることが解明されている。
【0004】
インテグリンなどの膜貫通型接着タンパク質が細胞−ECM間接着を構成している。インテグリンは、α鎖およびβ鎖からなるヘテロ二量体を形成する。少なくとも18種類のα鎖、8種類のβ鎖および24種類のαβヘテロ二量体がこれまで同定され、確認されている。インテグリンのそれぞれの種類が、特異的なリガンドを認識する。インテグリンを含めた膜貫通型接着タンパク質は、細胞接着に加えて、ECMから細胞中への細胞内シグナル伝達ならびに増殖、移動および分化の調節に関連する(F.G.Giancottiら、Science、285、1028〜1032、1999)。
【0005】
ECMタンパク質として多数のタンパク質が知られており、これらはコラーゲン群(I型〜XIX型など)、非コラーゲン性糖タンパク質群(オステオポンチン(OPN)、ビトロネクチン、フィブロネクチン、フォンウィルブランド因子、ラミニン、テネイシン、フィブリノゲン、トロンボスポンジンなど)、エラスチン群およびプロテオグリカン群に分類される。これらのECMタンパク質は対応するインテグリンに結合し、細胞内シグナル伝達経路を活性化して細胞骨格形成、移動、増殖、分化などを調節する。ECMタンパク質が結合したインテグリンは、ECMタンパク質の種類によって特異的なシグナルを伝達することにより、これらのシグナル活性化経路を調節する。多数のECMタンパク質の細胞接着領域にRGD配列が共通に認められ、インテグリンと結合することによって様々な機能を発揮する。このECMタンパク質のRGD配列は薬物の可能な標的と考えられており、いくつもの小分子化合物や人工ペプチドが提供されている。
【0006】
α3β1インテグリン、α5β1インテグリン、α8β1インテグリン、αvβ1インテグリン、αvβ3インテグリン、αvβ5インテグリン、αvβ6インテグリン、αvβ8インテグリンなど、いくつかの種類のインテグリンがRGD配列に結合することが知られている。α5β1インテグリンとその特異的なリガンドであるフィブロネクチンの間の相互作用から、インテグリンを介したシグナル伝達の機序についての研究が盛んに行われている。このような研究から、α5β1インテグリンは細胞接着および細胞移動だけでなく、細胞分化および細胞死も調節することが示されている(S.M.Frischら、Curr.Opin.Cell Biol.9、701〜706、1997)。α5β1インテグリンは腫瘍細胞で高度に発現され、癌の悪性化に関連することも示されている。インテグリンを介したシグナルはそれぞれ、結合しているECMタンパク質によって異なり、例えば、増殖因子による刺激はフィブロネクチンが結合した内皮細胞の増殖を活性化するが、ラミニン−1が結合した内皮細胞の増殖を阻害する。また、ラミニン−10/11からα3β1インテグリンに伝達されるシグナルは、フィブロネクチンからα5β1インテグリンに伝達されるシグナルと異なり、癌性細胞の移動を著しく増強し(J.Guら、J.Biol.Chem.、276、27090〜27097、2001)、血液飢餓(blood starvation)によるアポトーシスを大きく回避する(J.Guら、J.Biol.Chem.、277、19922〜19928、2002)。RGD配列が結合したαvインテグリンの高発現は、破骨細胞および新生血管において認められており、RGD配列およびαvインテグリンの阻害が、骨粗鬆症および癌の治療薬の標的であると考えられている。α5β1インテグリンは腫瘍細胞で高度に発現され、癌の悪性化に関連することが示されている。これらの知見から、インテグリンとECMタンパク質の相互作用を阻害する抗インテグリンアンタゴニスト抗体薬として、抗α5β1インテグリン抗体(ボロシマブ(Volocimab))、抗α4インテグリン抗体(ナタリズマブ(Natalizumab))および抗αvβ3インテグリン抗体(ビタキシン(Vitaxin))が開発されている。
【0007】
一方、コラーゲン、オステオポンチン(OPN)、ビトロネクチン、フィブロネクチン、フォンウィルブランド因子、ラミニン、テネイシン、フィブリノゲン、トロンボスポンジンなど、いくつかのECMタンパク質はRGD配列を含むことが知られている。また、一部のウイルスおよび一部の細菌も、細胞に付着するためのRGD配列を有することが知られている。OPNは、骨に豊富に含有されるカルシウムに対する結合特性を有する酸性糖タンパク質である。OPNは、細胞接着、細胞遊走、腫瘍形成、免疫応答および補体媒介性細胞溶解に重要な役割を果たすことが報告されている。OPNノックアウトマウスおよび抗OPN中和抗体の結果から、OPNは肝炎、自己免疫疾患(関節リウマチなど)および癌の転移に関連することが示される。細胞に対するECMタンパク質の結合の阻害剤を骨粗鬆症または癌の治療に使用できることが知られている。したがって、インテグリンを標的とする上記の拮抗薬に加えて、インテグリンの結合パートナーであるECMタンパク質を標的とする拮抗薬が開発されている。
【発明の概要】
【発明が解決しようとする課題】
【0008】
3.発明の概要
RGD配列を介したインテグリンとの相互作用を阻害する小分子、OPNに対する抗体、インテグリンに対する抗体などの薬物は報告されているが、RGD配列を特異的に認識する抗体に関する報告はまだない。RGD配列はECMタンパク質の保存配列の1つであるので、RGD配列を特異的に認識する抗体は、ヒトと治療モデル動物の両方で効果を有する可能性があり、したがって治療薬の開発に非常に有用な有効成分とみなすこともできる。したがって、このようなRGD配列を特異的に認識する抗体が必要とされている。
【0009】
本発明者らは以前に、RGD配列を免疫特異的に認識し、ハイブリドーマクローン33E10および35B6(それぞれ寄託アクセッション番号FERM BP−10440およびFERM BP−10441)によって産生されるマウスモノクローナル抗体を単離した。本明細書では、ハイブリドーマクローンの名称は、そのクローンによって産生されるモノクローナル抗体の名称として互換的に使用する。これらのマウス抗RGD抗体はすべて、IgG1アイソタイプであった。こうしたモノクローナル抗体は、オステオポンチンなどのECMタンパク質のRGD配列に結合することにより、ECMと細胞の間のRGD配列を介した結合に干渉することが認められた。したがって、こうした抗RGD抗体は、癌、例えば、癌細胞の増殖または転移、および炎症性疾患、例えば、関節リウマチ、骨関節炎、感染症、肝炎、気管支喘息、線維症、真性糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患、骨粗鬆症などのRGD配列に関連する疾患に対して治療効果または診断効果を発揮する可能性がある。
【0010】
しかし、これらのモノクローナル抗体はマウス起源であるので、ヒトにおけるその免疫原性が原因で生じる可能な有害作用が、ヒトにおける診断または治療的使用にこの抗体を直接利用することの妨げとなった。その免疫原性を低下させるために、本発明者らは、前記ヒト化抗体が由来する元のマウス抗RGD抗体によって発揮される生物活性に相当する生物活性を有するヒト化抗体を調製した。
【課題を解決するための手段】
【0011】
したがって、本発明は、RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントであって、非ヒト起源に部分的に由来し、ヒト起源に部分的に由来する抗原結合性領域を含む抗体またはその抗原結合性フラグメントを提供する。一部の実施形態において、本発明のヒト化抗体またはその抗原結合性フラグメントは、33E10モノクローナル抗体、35B6モノクローナル抗体などの非ヒト供給源(ドナー)に由来する相補性決定領域(以下「CDR」と略記する)およびヒト供給源(アクセプター)に由来するフレームワーク領域(以下「FR」と略記する)を含む。前記ヒト化抗体またはその抗原結合性フラグメントは、RGD配列とそのリガンドの間の結合を阻害することができる。
【0012】
特定の実施形態において、RGD配列を免疫特異的に認識する前記ヒト化抗体またはその抗原結合性フラグメントは、(i)ヒト重鎖(以下「H鎖」と略記する)の可変領域(以下「V領域」と略記する)に由来する少なくとも1つのH鎖FR(以下「FRH」と略記する)と、RGD配列を免疫特異的に認識する非ヒト抗体のH鎖CDR(以下「CDRH」と略記する)のうちの少なくとも1つに由来する少なくとも1つのCDRHとを含むH鎖;または(ii)ヒト軽鎖(以下「L鎖」と略記する)のV領域に由来する少なくとも1つのL鎖FR(以下「FRL」と略記する)と、RGD配列を免疫特異的に認識する非ヒト抗体のL鎖CDR(以下「CDRL」と略記する)のうちの少なくとも1つに由来する少なくとも1つのCDRLとを含むL鎖;または上記の(i)と(ii)の両方を含む。一実施形態において、本発明のヒト化抗体のCDRHのうちの少なくとも1つおよび/またはCDRLのうちの少なくとも1つは、寄託アクセッション番号FERM BP−10440およびFERM BP−10441からなる群から選択されるハイブリドーマによって産生されたモノクローナル抗体に由来するものとすることができる。好ましい実施形態において、本発明のヒト化抗体またはその抗原結合性フラグメントは、(i)ヒトFRHに由来する少なくとも1つのFRH、ならびに配列番号1、2および3のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRH;または(ii)ヒトFRLに由来する少なくとも1つのFRL、ならびに配列番号4、5および6のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRL;または(iii)上記の(i)と(ii)の両方を含む。一部の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRH1に配列番号1、CDRH2に配列番号2、およびCDRH3に配列番号3を含むこともできる。一部の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRL1に配列番号4、CDRL2に配列番号5、およびCDRL3に配列番号6を含むこともできる。好ましくは、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRH1に配列番号1、CDRH2に配列番号2、CDRH3に配列番号3、CDRL1に配列番号4、CDRL2に配列番号5、およびCDRL3に配列番号6を含む。
【0013】
一部の特定の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、GenBankアクセッション番号X65891(配列番号13)によってコードされるヒトH鎖のV領域に由来するFRH、またはGenBankアクセッション番号X72441(配列番号18)によってコードされるヒトκ−L鎖のV領域に由来するFRLを含む。一部の実施形態において、本発明のヒト化抗体のFRHは、配列番号14、15、16および17のアミノ酸配列(それぞれX65891のFRH1、FRH2、FRH3およびFRH4のアミノ酸配列)からなる群から選択される少なくとも1つのアミノ酸配列を含む。一部の実施形態において、本発明のヒト化抗体のFRLは、配列番号19、20、21および22のアミノ酸配列(それぞれX72441のFRL1、FRL2、FRL3およびFRL4のアミノ酸配列)からなる群から選択される少なくとも1つのアミノ酸配列を含む。最も好ましい実施形態の1つにおいて、本発明のヒト化抗体またはその抗原結合性フラグメントは、(i)配列番号24のアミノ酸配列を含むH鎖のV領域(以下「VH」と略記する);または(ii)配列番号26のアミノ酸配列を含むL鎖のV領域(以下「VL」と略記する);または(iii)上記の(i)と(ii)の両方を含む。
【0014】
他の実施形態において、本発明のヒト化抗体またはその抗原結合性フラグメントは、(i)ヒトFRHに由来する少なくとも1つのFRH、ならびに配列番号7、8および9のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRH;または(ii)ヒトFRLに由来する少なくとも1つのFRL、ならびに配列番号10、11および12のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRL;または(iii)上記の(i)と(ii)の両方を含む。一部の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRH1に配列番号7、CDRH2に配列番号8、およびCDRH3に配列番号9を含むこともできる。一部の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRL1に配列番号10、CDRL2に配列番号11、およびCDRL3に配列番号12を含む。好ましくは、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、CDRH1に配列番号7、CDRH2に配列番号8、CDRH3に配列番号9、CDRL1に配列番号10、CDRL2に配列番号11、およびCDRL3に配列番号12を含む。
【0015】
一部の特定の実施形態において、本発明の前記ヒト化抗体またはその抗原結合性フラグメントは、GenBankアクセッション番号X65891(配列番号13)によってコードされるヒトH鎖のV領域に由来するFRH、またはGenBankアクセッション番号X72441(配列番号18)によってコードされるヒトκ−L鎖のV領域に由来するFRLを含む。一部の実施形態において、本発明のヒト化抗体FRHは、配列番号14、15、16および17のアミノ酸配列(それぞれX65891のFRH1、FRH2、FRH3およびFRH4のアミノ酸配列)からなる群から選択される少なくとも1つのアミノ酸配列を含む。一部の実施形態において、本発明のヒト化抗体のFRLは、配列番号19、20、21および22のアミノ酸配列(それぞれX72441のFRL1、FRL2、FRL3およびFRL4のアミノ酸配列)からなる群から選択される少なくとも1つのアミノ酸配列を含む。最も好ましい実施形態の1つにおいて、本発明のヒト化抗体またはその抗原結合性フラグメントは、(i)配列番号28のアミノ酸配列を含むVH;または(ii)配列番号30のアミノ酸配列を含むVL;または(iii)上記の(i)と(ii)の両方を含む。
【0016】
本発明はさらに、RGD配列を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合性フラグメントをコードするヌクレオチド配列を含む単離された核酸分子を提供する。詳細には、本発明は、配列番号1、2、3、7、8および9からなる群から選択される少なくとも1つのアミノ酸配列を含むヒト化H鎖、または配列番号4、5、6、10、11および12からなる群から選択される少なくとも1つのアミノ酸配列を含むヒト化L鎖、または前記ヒト化H鎖と前記ヒト化L鎖の両方をコードするヌクレオチド配列を含む単離された核酸分子を提供する。好ましい特定の実施形態において、このような単離された核酸分子は、VHをコードする配列番号23のヌクレオチド配列、または配列番号24のアミノ酸配列をコードするヌクレオチド配列を含む。一部の好ましい特定の実施形態において、このような単離された核酸分子は、VLをコードする配列番号25のヌクレオチド配列、または配列番号26のアミノ酸配列をコードするヌクレオチド配列を含む。好ましくは、本発明の単離された核酸分子は、配列番号23と配列番号25の両方のヌクレオチド配列を含む。好ましい特定の実施形態において、本発明の単離された核酸分子は、配列番号32および34のアミノ酸配列などのドナー起源のシグナルペプチド、または異種起源のシグナルペプチドをコードするヌクレオチド配列をさらに含む。
【0017】
他の好ましい特定の実施形態において、このような単離された核酸分子は、VHをコードする配列番号27のヌクレオチド配列、または配列番号28のアミノ酸配列をコードするヌクレオチド配列を含む。一部の好ましい特定の実施形態において、このような単離された核酸分子は、VLをコードする配列番号29のヌクレオチド配列、または配列番号30のアミノ酸配列をコードするヌクレオチド配列を含む。好ましくは、本発明の単離された核酸分子は、配列番号27と配列番号29の両方のヌクレオチド配列を含む。好ましい特定の実施形態において、本発明の単離された核酸分子は、配列番号36および38のアミノ酸配列などのドナー起源のシグナルペプチド、または異種起源のシグナルペプチドをコードするヌクレオチド配列をさらに含む。
【0018】
本発明はさらに、RGD配列を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合性フラグメントのH鎖またはL鎖またはその両方をコードするヌクレオチド配列を含むベクター、例えば、発現ベクターを提供する。このようなベクターにおいて、本発明のヌクレオチド配列を1つまたは複数の調節エレメントと作動可能に連結することもできる。本発明のヌクレオチド配列は、CDRが由来する非ヒトドナー抗体の天然のシグナルペプチド、または異種起源のシグナルペプチドをコードするヌクレオチド配列を含むこともできる。
【0019】
さらに、本発明は、本発明の核酸分子を含むベクターを含めた、本発明の核酸分子を含む宿主細胞を提供する。一実施形態において、本発明は、本発明のヒト化H鎖をコードする第1の核酸分子および本発明のヒト化L鎖をコードする第2の核酸分子を含む単離された宿主細胞を提供し、前記第1および第2の核酸分子はそれぞれ、本発明の生物学的機能を有するヒト化抗体またはその抗原結合性フラグメントが発現されるように調節エレメントと作動可能に連結している。
【0020】
したがって、本発明はさらに、本発明のヒト化抗体またはその抗原結合性フラグメントを調製するための方法であって、本発明の宿主細胞をヒト化抗体またはその抗原結合性フラグメントが発現されるような条件下で培養すること、および産生されたヒト化抗体を収集することを含む方法を提供する。
【0021】
本発明はさらに、本発明のヒト化抗体またはその抗原結合性フラグメントのうちの少なくとも1つを含む組成物を提供する。さらに、本発明は、RGD−タンパク質に関連する障害または疾患を予防または治療するための医薬組成物であって、本発明のヒト化抗体またはその抗原結合性フラグメントのうちの少なくとも1つおよび薬学的に許容される担体を含む医薬組成物を提供する。前記組成物はいずれも、障害または疾患を相加的または相乗的に寛解させることができる別の活性化合物をさらに含むことができる。前記活性化合物としては、それだけには限定されないが、抗炎症性化合物、化学療法化合物などが挙げられる。前記活性化合物としてさらに、ヒトα4インテグリン特異的抗体またはヒトα9インテグリン特異的抗体などの、小分子化合物および抗体またはその抗原結合性フラグメントが挙げられる。
【0022】
別の態様において、本発明は、RGD−タンパク質に関連する、またはそれが関与する障害または疾患を予防または治療するための方法であって、予防または治療有効量の本発明のヒト化抗体またはその抗原結合性フラグメントのうちの少なくとも1つを、それを必要とする対象に投与することを含む方法を提供する。このような使用のために、本発明のヒト化抗体またはその抗原結合性フラグメントは、ヒト化抗体またはその抗原結合性フラグメントの生物学的効果を増強する治療用成分とコンジュゲートすることもできる。このような治療用成分の例としては、別の抗体、細胞増殖抑制性もしくは細胞破壊性を示す細胞毒、放射性元素および/または抗炎症薬、抗生物質などを含めた他の治療薬が挙げられる。
【0023】
さらに別の態様において、本発明は、対象においてRGD−タンパク質に関連する、またはそれが関与する障害または疾患を診断するための方法であって、診断有効量の本発明のヒト化抗体またはその抗原結合性フラグメントを被検対象に投与することを含む方法を提供する。このような診断的使用のために、本発明のヒト化抗体を放射性元素などの検出可能なマーカーで標識することもできる。
【0024】
3.1.定義
本明細書では、「抗体」という用語は、所望の抗原または所望の配列(RGD配列など)に免疫特異的に結合することができる抗体分子を指し、抗体分子全体または抗原結合性フラグメントを含めたそのフラグメントを包含する。
【0025】
本明細書で使用する「抗原結合性フラグメント」という用語は、VLおよび/もしくはVHまたは標的のポリペプチド、タンパク質もしくは配列に特異的に結合するCDRのいずれかを含有する、単鎖抗体、Fabフラグメント、F(ab’)2フラグメント、ジスルフィド結合したFvsおよびフラグメントを含めた、標的のポリペプチド、タンパク質または配列、特にRGD配列に免疫特異的に結合する能力を保持する抗体の任意のフラグメントを指す。したがって、ヒト化抗体のこのような抗原結合性フラグメントは、部分または全長のヒト定常領域を含んでも含まなくてもよい。当技術分野では上記の抗体フラグメントを得るための様々な方法がよく知られている。
【0026】
本明細書で使用する「免疫特異的に認識する」という用語は、抗体またはその抗原結合性フラグメントが標的のポリペプチド、タンパク質または配列、特にヒトRGD配列に特異的に結合できることを指す。このような抗体は、他のポリペプチドまたはタンパク質に非特異的に結合しない。しかし、標的のポリペプチドまたはタンパク質(例えば、RGD−タンパク質)に免疫特異的に結合する抗体またはその抗原結合性フラグメントは、他の抗原と交差反応することもできる。例えば、ヒトRGD−タンパク質を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合性フラグメントは、マウスRGD−タンパク質と交差反応することもできる。好ましくは、RGD−タンパク質を免疫特異的に認識する抗体またはその抗原結合性フラグメントは、他の抗原と交差反応しない。
【0027】
本明細書で使用する「ヒト供給源に由来する」または「非ヒト供給源に由来する」という用語は、そのアミノ酸配列がそれぞれヒト抗体または非ヒト抗体の対応する部分に由来する抗体部分を指す。
【0028】
本明細書で使用する「アクセプター配列」という用語は、通常非ヒト抗体であるドナー抗体由来のCDRのアクセプターとなるヒト抗体VHまたはVL由来のFRのヌクレオチド配列またはアミノ酸配列を指す。
【図面の簡単な説明】
【0029】
【図1】マウスOPNの部分ペプチドを使用したモノクローナル抗体4P11、11M6、25H15および35B6のエピトープ分析の結果を示す図である。
【図2】マウスOPNおよびヒトOPNの部分ペプチドを使用したモノクローナル抗体29R5、30C7、33E10および38I8のエピトープ分析の結果を示す図である。
【図3】RGD配列を含むマウスOPNの部分ペプチドを使用したモノクローナル抗体33E10および35B6のエピトープ分析の結果を示す図である。
【図4】マウスOPNの部分ペプチドを使用したモノクローナル抗体33E10および35B6のエピトープ分析の結果を示す図である。
【図5】マウスOPNの部分ペプチド(CGDSLAYGLR;配列番号79)を使用したモノクローナル抗体33E10および35B6のエピトープ分析の結果を示す図である。
【図6】抗RGDモノクローナル抗体のCDRH分析の結果を示す図である。この図で、33E10の99位のアミノ酸(F)および35B6の98位のアミノ酸(F)は、KでもRでもよい。
【図7】抗RGDモノクローナル抗体のCDRL分析の結果を示す図である。
【図8】抗RGD抗体とRGD配列を含む種々のECMタンパク質との結合親和性の結果を示す図である。
【図9】癌性細胞(NIH3T3細胞)に対するmOPNのN半分(N half)の結合の抗RGD抗体による阻害の結果を示す図である。
【図10A−10C】癌性細胞(NIH3T3細胞)に対する種々のECMタンパク質の結合の抗RGD抗体による阻害の結果を示す図である。
【図11】抗RGD抗体の肝炎抑制効果の結果を示す図である。
【図12A−12B】実験的転移モデルにおける抗RGD抗体の肺転移阻害効果の結果を示す図である。図12Aは転移細胞の数を示し、図12Bは体重の変化を示す。
【図13A−13C】自然発症転移モデルにおける抗RGD抗体の肺転移阻害効果の結果を示す図である。図13Aは癌のサイズを示し、図13Bは転移細胞の数を示し、図13Cは体重の変化を示す。
【図14】関節リウマチモデルにおける抗RGD抗体の治療効果の試験の結果を示す図である。
【図15】マウス33E10 VH cDNAのヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(E)は二重下線付きである。Kabatら(Sequences of Proteins of Immunological Interests、第5版、NIH公開番号91−3242、U.S.Department of Health and Human Services、1991)の定義に従うCDR配列は下線付きである。
【図16】マウス33E10 VL cDNAのヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。
【図17】SpeI部位およびHindIII部位(下線付き)が両端に隣接する、設計された33E10 VH遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(E)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図18】NheI部位およびEcoRI部位(下線付き)が両端に隣接する、設計された33E10 VL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図19】pCh33E10およびpHu33E10(総称して発現ベクターとする)の概略構造を示す図である。頂点のSalI部位から時計回りに進んで、プラスミドは、抗体重鎖遺伝子の転写を開始するためのヒトサイトメガロウイルス(CMV)主要最初期プロモーターおよびエンハンサー(CMVプロモーター)から始まる重鎖転写単位を含有する。CMVプロモーターにVHエキソンが続き、CH1、ヒンジ、CH2およびCH3エキソンを介在性イントロンと共に含むヒトγ−1重鎖定常領域を含有するゲノム配列、さらにはCH3エキソンの後にポリアデニル化部位がある。重鎖遺伝子配列の後に、軽鎖転写単位がCMVプロモーターから始まり、続いてVLエキソン、およびヒトκ鎖定常領域エキソン(CL)をそれに先行するイントロンの部分と共に含有するゲノム配列、さらにはCLエキソンに続くポリアデニル化部位がある。次いで、軽鎖遺伝子に続いて、SV40初期プロモーター(SV40プロモーター)、大腸菌キサンチングアニンホスホリボシルトランスフェラーゼ遺伝子(gpt)、およびSV40ポリアデニル化部位(SV40ポリ(A)部位)を含有するセグメントがある。最後に、プラスミドは、細菌の複製起点(pUC ori)およびβラクタマーゼ遺伝子(βラクタマーゼ)を含むプラスミドpUC19の一部を含有する。
【図20】33E10 VH、ヒト化33E10(Hu33E10)VHおよびヒトアクセプターU03400(GenBankアクセッション番号)のアミノ酸配列のアライメントを示す図である。アミノ酸残基は1文字表記で示す。配列上の数字はKabatら(1991)に従う位置を示す。Kabatら(Sequences of Proteins of Immunological Interests、第5版、NIH公開番号91−3242、U.S.Department of Health and Human Services、1991)によって定義されるCDR配列は下線付きである。二重下線付きの残基はCDRと接触すると予測され、マウス残基はこれらの位置においてヒト化された形で保持された。U03400中のCDR残基は図では省略する。
【図21】33E10 VL、ヒト化33E10(Hu33E10)VLおよびヒトアクセプターX72452(GenBankアクセッション番号)のアミノ酸配列のアライメントを示す図である。アミノ酸残基は1文字表記で示す。配列上の数字はKabatら(1991)に従う位置を示す。Kabatら(1991)によって定義されるCDR配列は下線付きである。X72452中のCDR残基は図では省略する。
【図22】Hu33E10 VH遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図23】Hu33E10 VL遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図24】Hu33E10 VH遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印は、各オリゴヌクレオチドの位置および方向(5’から3’)を表す。アミノ酸残基は1文字表記で示す。
【図25】Hu33E10 VL遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印は、各オリゴヌクレオチドの位置および方向(5’から3’)を表す。アミノ酸残基は1文字表記で示す。
【図26】SpeI部位およびHindIII部位(下線付き)が両端に隣接する、Hu33E10 VH遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(E)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図27】NheI部位およびEcoRI部位(下線付き)が両端に隣接する、Hu33E10 VL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図28】マウス35B6 VH cDNAのヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(Q)は二重下線付きである。Kabatら(Sequences of Proteins of Immunological Interests、第5版、NIH公開番号91−3242、U.S.Department of Health and Human Services、1991)の定義に従うCDR配列は下線付きである。
【図29】マウス35B6 VL cDNAのヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。
【図30】SpeI部位およびHindIII部位(下線付き)が両端に隣接する、設計された35B6 VH遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(Q)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図31】NheI部位およびEcoRI部位(下線付き)が両端に隣接する、設計された35B6 VL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図32】pCh35B6およびpHu35B6(総称して発現ベクターとする)の概略構造を示す図である。頂点のSalI部位から時計回りに進んで、プラスミドは、抗体重鎖遺伝子の転写を開始するためのヒトサイトメガロウイルス(CMV)主要最初期プロモーターおよびエンハンサー(CMVプロモーター)から始まる重鎖転写単位を含有する。CMVプロモーターにVHエキソンが続き、CH1、ヒンジ、CH2およびCH3エキソンを介在性イントロンと共に含むヒトγ−1重鎖定常領域を含有するゲノム配列、さらにはCH3エキソンの後にポリアデニル化部位がある。重鎖遺伝子配列の後に、軽鎖転写単位がCMVプロモーターから始まり、続いてVLエキソン、およびヒトκ鎖定常領域エキソン(CL)をそれに先行するイントロンの部分と共に含有するゲノム配列、さらにはCLエキソンに続くポリアデニル化部位がある。次いで、軽鎖遺伝子に続いて、SV40初期プロモーター(SV40プロモーター)、大腸菌(E.coli)キサンチングアニンホスホリボシルトランスフェラーゼ遺伝子(gpt)、およびSV40ポリアデニル化部位(SV40ポリ(A)部位)を含有するセグメントがある。最後に、プラスミドは、細菌の複製起点(pUC ori)およびβラクタマーゼ遺伝子(βラクタマーゼ)を含むプラスミドpUC19の一部を含有する。
【図33】35B6 VH、ヒト化35B6(Hu35B6)VHおよびヒトアクセプターZ47230(GenBankアクセッション番号)のアミノ酸配列のアライメントを示す図である。アミノ酸残基は1文字表記で示す。配列上の数字はKabatら(1991)に従う位置を示す。Kabatら(1991)によって定義されるCDR配列は下線付きである。二重下線付きの残基はCDRと接触すると予測され、マウス残基はこれらの位置においてヒト化された形で保持された。Z47230中のCDR残基は図では省略する。
【図34】35B6 VL、ヒト化35B6(Hu35B6)VLおよびヒトアクセプターX72479(GenBankアクセッション番号)のアミノ酸配列のアライメントを示す図である。アミノ酸残基は1文字表記で示す。配列上の数字はKabatら(1991)に従う位置を示す。Kabatら(1991)によって定義されるCDR配列は下線付きである。二重下線付きの残基はCDRと接触すると予測され、マウス残基はこれらの位置においてヒト化された形で保持された。X72479中のCDR残基は図では省略する。
【図35】Hu35B6 VH遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図36】Hu35B6 VL遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図37】Hu35B6 VH遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印は、各オリゴヌクレオチドの位置および方向(5’から3’)を表す。アミノ酸残基は1文字表記で示す。
【図38】Hu35B6 VL遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印は、各オリゴヌクレオチドの位置および方向(5’から3’)を表す。アミノ酸残基は1文字表記で示す。
【図39】SpeI部位およびHindIII部位(下線付き)が両端に隣接する、Hu35B6 VH遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VHのN末端アミノ酸残基(Q)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図40】NheI部位およびEcoRI部位(下線付き)が両端に隣接する、Hu35B6 VL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒に示す図である。アミノ酸残基は1文字表記で示す。シグナルペプチド配列はイタリック体である。成熟VLのN末端アミノ酸残基(D)は二重下線付きである。Kabatら(1991)の定義に従うCDR配列は下線付きである。イントロン配列はイタリック体である。
【図41A−41B】キメラおよびヒト化35B6抗体とhOPN−BSAの結合をELISAによって分析した。各抗体を2.5μg/ml(図41A)または1.0μg/ml(図41B)の出発濃度および連続2倍希釈で試験した。実験は3連で実施した。各抗体濃度での平均吸光度値と標準偏差を図41A〜41Bに示す。
【発明を実施するための形態】
【0030】
5.発明の詳細な説明
5.1.RGD配列に対する抗体の調製
RGD配列を免疫特異的に認識する抗体は、当技術分野で知られている任意の適当な方法によって生成することができる。
【0031】
本発明における細胞接着性「RGD」配列を含むRGD−タンパク質またはRGD−ペプチド(以下「RGD−ペプチド」と略記する)は、(1)RGD−タンパク質を発現するヒトECMまたはこれらのECMが存在するすべての組織に由来するもの、(2)細菌、酵母、動物細胞などの細胞系などにトランスフェクトすることによる、RGD−タンパク質またはRGD−ペプチドをコードするDNA(好ましくはcDNA)の発現によって得られる組換えタンパク質またはペプチド、または(3)合成タンパク質またはペプチドであってもよい。
【0032】
本発明において抗原として使用されるRGD−ペプチドは、免疫化によるRGD配列に対する抗体の産生が可能であり得る。RGD−ペプチドは、マウスECMタンパク質の細胞接着性配列であるRGD−ペプチドアミノ酸配列CVDVPNGRGDSLAYGLR(配列番号71)を含む。RGD−タンパク質またはRGD−ペプチドとしては、OPN、ビトロネクチン、フィブロネクチン、フォンウィルブランド因子、コラーゲン、ラミニン、テネイシン、フィブリノゲン、トロンボスポンジンおよびRGDを含むその断片などが挙げられる。前記タンパク質または前記ペプチドがRGD配列を含む限り、タンパク質またはペプチドにアミノ酸の置換、欠失、改変、付加などの人工または天然の変異を適用することができる。変異タンパク質またはペプチドは、複数のアミノ酸、好ましくは1から10個のアミノ酸、より好ましくは1から数個(例えば、1から5個)のアミノ酸が置換、欠失、改変、付加または挿入されたアミノ酸配列を含むこともできる。
【0033】
本明細書では、RGD−ペプチドは、少なくとも約5個のアミノ酸、好ましくは約5から50個のアミノ酸、より好ましくは約10から20個のアミノ酸を含む。本発明における抗原としてのRGD−タンパク質またはRGD−ペプチドは、化学合成法、細胞培養法、遺伝子組換え法、その適切な改変などの当技術分野でよく知られている方法を使用することによって産生することができる。例えば、RGD−ペプチドは、ECMタンパク質をタンパク質分解酵素で適切に切断することによって得ることができる。RGD−タンパク質またはRGD−ペプチドは、マウス、ラット、ウサギ、ブタ、ウシ、サル、ヒトなどの哺乳動物に由来するものであってもよい。当技術分野でよく知られている任意の方法を使用して、抗RGD抗体の調製に使用することができるRGD−タンパク質またはRGD−ペプチドを調製することができる。
【0034】
変異ポリペプチドを生成するための方法の例としては、合成オリゴヌクレオチド部位特異的突然変異誘発(gapped duplex法)、亜硝酸または亜硫酸の処理によって点突然変異をランダムに導入することを含む点突然変異誘発法、Bal31酵素または他の酵素で欠失変異体を調製することを含む方法、カセット変異導入法、リンカースキャニング法(linker scanning method)、ミスインコーポレーション法(miss incorporation method)、ミスマッチプライマー法(mismatch primer method)、DNAセグメント合成法などが挙げられる。
【0035】
RGD−ペプチドは、サイログロブリン、キーホールリンペットヘモシアニン(KLH)、ウシ血清アルブミン(BSA)、オボアルブミン(OVA)またはウシグロブリンなど他の生体巨大分子、好ましくはサイログロブリンに結合させることができる。RGD−ペプチドを生体巨大分子に結合させる方法は、活性エステル基およびマレイミド基を有する結合試薬などのカップリング試薬(活性エステル基はタンパク質またはペプチドのアミノ基に結合し、マレイミド基はタンパク質またはペプチドのチオール基に結合する。S.Yoshirakeら、Eur.J.Biochem.、101、395〜399、1979)、混合酸無水物法(B.F.Erlangerら、J.Biol.Chem.、234、1090〜1094、1954)、または活性エステル法(A.E.Karuら、J.Agric.Food Chem.、42、301〜309、1994)を使用することによって実現することができる。RGD−ペプチドを生体巨大分子に結合させるための方法は、カップリング試薬を使用することによって達成することが好ましい。
【0036】
抗原として、RGD−タンパク質またはRGD−ペプチドを過剰発現する細胞それ自体を使用することもできる。RGD−タンパク質またはRGD−ペプチドを過剰発現する細胞は、当技術分野でよく知られている組換えDNA技術によって調製することができる。
【0037】
上記のとおり調製した適切な抗原を使用して、当技術分野でよく知られている様々な方法によってRGD配列に特異的な抗体を調製することができる。RGD配列に対するポリクローナル抗体は、当技術分野でよく知られている様々な手順によって産生することができる。例えば、対象の抗原は、それだけには限定されないが、ウサギ、マウス、ラットなどを含めた、種々の宿主動物に投与して、抗原に特異的なポリクローナル抗体を含有する抗血清の産生を誘発することができる。様々なアジュバントを使用して宿主種に応じて免疫応答を増大させることができ、そのアジュバントとしては、それだけには限定されないが、フロイント(完全および不完全)アジュバント、水酸化アルミニウムなどの無機ゲル(mineral gel)、リゾレシチンなどの界面活性物質、プルロニック(pluronic)ポリオール、ポリアニオン、ペプチド、オイルエマルジョン、キーホールリンペットヘモシアニン、ジニトロフェノールおよびBCG(カルメット・ゲラン桿菌)、コリネバクテリウム・パルバム(Corynebacterium parvum)などのヒトに潜在的に有用なアジュバントが挙げられる。このようなアジュバントも当技術分野でよく知られている。
【0038】
モノクローナル抗体は、ハイブリドーマ技術、組換え技術およびファージディスプレイ技術、またはその組合せの使用を含めた、当技術分野で知られている多種多様な技法を使用することによって調製することができる。例えば、モノクローナル抗体は、当技術分野で知られており、例えば、Harlowら、Antibodies:A Laboratory Manual、(Cold Spring Harbor Laboratory Press、第2版1988);Hammerlingら、in:Monoclonal Antibodies and T−Cell Hybridomas、563〜681ページ(Elsevier,N.Y.、1981)(両文献ともその全体が参照により組み込まれる)に教示されているものを含めて、ハイブリドーマ技法を使用することによって産生することができる。本明細書で使用する「モノクローナル抗体」という用語は、ハイブリドーマ技術によって産生される抗体に限定されない。「モノクローナル抗体」という用語は、単一のクローンに由来する抗体を指し、真核生物、原核生物またはファージの任意のクローンを含むが、それが産生される方法に限定されない。
【0039】
ハイブリドーマ技術を使用して特異的抗体を産生およびスクリーニングするための方法は、常用されており当技術分野でよく知られている。非限定的な例において、マウスは、対象の抗原またはこのような抗原を発現する細胞で免疫することができる。免疫応答が検出される、例えば、マウス血清中で抗原に特異的な抗体が検出されると、マウス脾臓が採取され、脾細胞が単離される。次いで、脾細胞は、よく知られている技法によって任意の適当な骨髄腫細胞(例えば、P3U1、P3X63−Ag8、P3X63−Ag8−U1、P3NS1−Ag4、SP2/0−Ag14、P3X63−Ag8−653など)と融合される。ハイブリドーマは、限界希釈によって選択され、クローニングされる。次いで、ハイブリドーマクローンは、当技術分野で知られている方法により、抗原に結合することができる抗体を分泌する細胞についてアッセイされる。高レベルの抗体を通常含有する腹水は、マウスの腹腔内に陽性のハイブリドーマクローンを接種することによって生成することができる。
【0040】
特異的エピトープを認識する抗体フラグメントは、既知の技法によって生成することもできる。例えば、FabおよびF(ab’)2フラグメントは、パパイン(Fabフラグメントを産生するため)またはペプシン(F(ab’)2フラグメントを産生するため)などの酵素を使用した、免疫グロブリン分子のタンパク質分解切断によって産生することもできる。F(ab’)2フラグメントは、完全L鎖、ならびにH鎖のV領域、CH1領域およびヒンジ領域を含有する。
【0041】
本発明の抗体またはその抗原結合性フラグメントは、当技術分野で知られている、抗体を合成するための任意の方法によって、特に、化学合成によって、または好ましくは組換え発現技法によって産生することもできる。
【0042】
抗体をコードするヌクレオチド配列は、当業者に利用可能な任意の情報から(すなわち、GenBank、文献から、または常用のクローニングおよび配列解析によって)得ることもできる。特定の抗体またはそのエピトープ結合性フラグメントをコードする核酸を含有するクローンが利用可能でないが、抗体分子またはそのエピトープ結合性フラグメントの配列が知られている場合、免疫グロブリンをコードする核酸は、化学的に合成するか、または配列の末端および5’末端にハイブリダイズ可能な合成プライマーを使用したPCR増幅によって、もしくは例えば、抗体をコードするcDNAライブラリー由来cDNAクローンを同定するための、特定の遺伝子配列に特異的なオリゴヌクレオチドプローブを使用したクローニングによって、適当な供給源(例えば、抗体cDNAライブラリー、または抗体を発現するように選択されたハイブリドーマ細胞など、抗体を発現する任意の組織もしくは細胞から作製されるcDNAライブラリー、またはそれから単離された核酸、好ましくはポリA+RNA)から得ることができる。次いで、PCRによって生成された増幅核酸は、当技術分野でよく知られている任意の方法を使用して複製可能なクローニングベクター中にクローニングすることもできる。
【0043】
5.2.組換え抗体の調製
抗体のヌクレオチド配列は、当技術分野でよく知られている、ヌクレオチド配列を操作するための方法、例えば、組換えDNA技術、部位特異的変異誘発、PCRなど(例えば、両文献ともその全体が参照により本明細書に組み込まれる、Sambrookら、前掲;およびAusubelら編、1998、Current Protocols in Molecular Biology、John Wiley & Sons、NYに記載の技法を参照されたい)を使用することによって操作することもできる。抗体のエピトープ結合ドメイン領域に、または生物活性を増強もしくは低減する任意の部分にアミノ酸の置換、欠失および/または挿入などの突然変異を導入することもできる。
【0044】
抗体をコードするヌクレオチド配列を含有する発現ベクターを、抗体またはその抗原結合性フラグメントの組換え発現に使用することができる。抗体またはその抗原結合性フラグメントを産生するための、抗体分子、抗体のH鎖および/もしくはL鎖、またはその一部分をコードするヌクレオチド配列を含むベクターは、以前のセクションで検討したとおり、当技術分野でよく知られている技法を使用して、組換えDNA技術によって産生することができる。当業者によく知られている方法を使用して、抗体またはその抗原結合性フラグメントをコードする配列ならびに適切な転写および翻訳調節シグナルを含有する発現ベクターを構築することができる。これらの方法としては、例えば、in vitro組換えDNA技術、合成的技法およびin vivo遺伝子組換えが挙げられる。抗体のVH、VL、VHとVLの両方、VHおよび/またはVLの抗原結合性フラグメント、または1つもしくは複数のCDRをコードするヌクレオチド配列は、このような発現用ベクター中にクローニングすることができる。このような配列は、元の抗体の天然のものであっても異種のものであってもよいシグナルペプチドをコードするポリヌクレオチドと融合することもできる。次いで、このようにして調製された発現ベクターは、抗体を発現させるための適切な宿主細胞中に導入することができる。したがって、本発明は、RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントをコードするポリヌクレオチドを含有する宿主細胞を含む。
【0045】
宿主細胞には、第1のベクターがH鎖由来のポリペプチドをコードし、第2のベクターがL鎖由来のポリペプチドをコードする本発明の2つの発現ベクターを同時トランスフェクトすることもできる。2つのベクターは、H鎖およびL鎖ポリペプチドの等しい発現を可能にする同一の選択マーカー、または両方のプラスミドの維持を確実にするための異なる選択マーカーを含有することもできる。あるいは、H鎖およびL鎖ポリペプチドの両方をコードし、発現することができる単一のベクターを使用することもできる。H鎖およびL鎖のコード配列は、cDNAまたはゲノムDNAを含むこともできる。
【0046】
別の実施形態において、抗体は、当技術分野で知られている種々のファージディスプレイ法を使用して作製することもできる。ファージディスプレイ法では、機能的抗体ドメインが、それをコードするポリヌクレオチド配列を保有するファージ粒子の表面上に提示される。特定の実施形態において、このようなファージを利用して、レパートリーまたはコンビナトリアル抗体ライブラリー(例えば、ヒトまたはマウス)から発現させた、FabおよびFvまたはジスルフィド結合安定化Fvなどの抗原結合性ドメインを提示することができる。対象の抗原に結合する抗原結合性ドメインを発現するファージは、抗原を用いて、例えば、標識された抗原、または固体表面もしくはビーズに結合もしくは捕捉した抗原を使用して選択または同定することができる。これらの方法に使用するファージは通常、fdおよびM13を含めた繊維状ファージである。抗原結合性ドメインは、ファージ遺伝子IIIまたは遺伝子VIIIタンパク質のいずれかと組換えによって融合したタンパク質として発現される。本発明の免疫グロブリンまたはそのフラグメントを作製するために使用することができるファージディスプレイ法の例としては、Brinkmanら、J.Immunol.Methods、182:41〜50、1995;Amesら、J.Immunol.Methods、184:177〜186、1995;Kettleboroughら、Eur.J.Immunol.、24:952〜958、1994;Persicら、Gene、187:9〜18、1997;Burtonら、Advances in Immunology、57:191〜280、1994;PCT出願番号PCT/GB91/01134;PCT公報WO90/02809;WO91/10737;WO92/01047;WO92/18619;WO93/11236;WO95/15982;WO95/20401;ならびに米国特許第5,698,426号;同第5,223,409号;同第5,403,484号;同第5,580,717号;同第5,427,908号;同第5,750,753号;同第5,821,047号;同第5,571,698号;同第5,427,908号;同第5,516,637号;同第5,780,225号;同第5,658,727号;同第5,733,743号および同第5,969,108号に開示されたものが挙げられ、それぞれその全体が参照により本明細書に組み込まれる。
【0047】
上記の参考文献に記載のとおり、ファージ選択後、ファージ由来の抗体コード領域を単離および使用して、例えば、以下に詳細に記載のとおり、ヒト抗体または任意の他の所望のフラグメントを含めた全抗体を作製し、哺乳動物細胞、昆虫細胞、植物細胞、酵母および細菌を含めた任意の所望の宿主中で発現させることができる。例えば、Fab、Fab’およびF(ab’)2フラグメントを組換えによって産生するための技法は、PCT公報WO92/22324;Mullinaxら、BioTechniques、12(6):864〜869、1992;およびSawaiら、AJRI、34:26〜34、1995;およびBetterら、Science、240:1041〜1043、1988(それぞれその全体が参照により組み込まれる)に開示されているものなど、当技術分野で知られている方法を使用して採用することもできる。単鎖Fvsおよび抗体を産生するために使用することができる技法の例としては、米国特許第4,946,778号および同第5,258,498号;Hustonら、Methods in Enzymology、203:46〜88、1991;Shuら、PNAS、90:7995〜7999、1993;およびSkerraら、Science、240:1038〜1040、1988に記載のものが挙げられる。
【0048】
本発明の抗体分子は、上記の任意の方法によって産生させた後、当技術分野で知られている、免疫グロブリン分子を精製するための任意の方法によって、例えば、クロマトグラフィー(例えば、イオン交換、親和性、特に、プロテインAまたはプロテインG精製後の特異的な抗原に対する親和性によって、およびサイジングカラムクロマトグラフィー(sizing column chromatography))、遠心分離、溶解度差(differential solubility)、またはタンパク質を精製するための任意の他の標準的な技法によって精製することができる。さらに、本発明の抗体またはそのフラグメントは、本明細書に記載のものか、そうでなければ当技術分野で知られている異種ポリペプチド配列と融合させて精製を容易にすることもできる。
【0049】
ヒトにおける抗体のin vivo使用およびin vitro検出アッセイを含めた一部の使用のために、キメラ抗体、ヒト化抗体またはヒト抗体を使用することが好ましいこともある。キメラ抗体およびヒト化抗体は、以下のセクション5.3で詳細に検討する。
【0050】
他の化合物または異種ポリペプチドと融合またはコンジュゲートした抗体は、in vitroイムノアッセイ、精製法(例えば、アフィニティークロマトグラフィー)、ならびにin vivoでの治療または診断的使用で使用することができる。例えば、その全体が参照により本明細書に組み込まれる、PCT公開番号WO93/21232;EP439,095;Naramuraら、Immunol.Lett.、39:91〜99、1994;米国特許第5,474,981号;Gilliesら、PNAS、89:1428〜1432、1992;およびFellら、J.Immunol.、146:2446〜2452、1991を参照されたい。例えば、抗体は、既知の方法または市販のキット(例えば、ビオチン標識、FITC標識、APC標識)を使用して様々な形で標識することができる。別の例として、抗体は、抗体の生物学的効果をin vivoで増強する治療用成分とコンジュゲートすることもできる。このような治療用成分の例としては、別の抗体、細胞増殖抑制性もしくは細胞破壊性を示す細胞毒、放射性元素および/または抗炎症薬を含めた他の治療薬、抗生物質などが挙げられる。本発明において、ヒト化抗RGD抗体は、別の抗体とコンジュゲートして二重特異性抗体を形成することもできる。別の例として、本発明のヒト化抗体は、in vivoでの診断的使用のために放射性元素などの検出可能なマーカーで標識することもできる。
【0051】
5.3.キメラ抗体およびヒト化抗体
キメラ抗体は、マウスモノクローナル抗体に由来するV領域およびヒト免疫グロブリンに由来する定常領域を有する抗体など、抗体の異なる部分が異なる動物種に由来する分子である。キメラ抗体を産生するための方法は当技術分野で知られている。例えば、その全体が参照により本明細書に組み込まれる、Morrison、Science、229:1202、1985;Oiら、BioTechniques、4:214 1986;Gilliesら、J.Immunol.Methods、125:191〜202、1989;米国特許第5,807,715号;同第4,816,567号;および同第4,816,397号を参照されたい。
【0052】
ヒト化抗体は、所望の抗原に結合し、非ヒト種に由来する1つまたは複数のCDRおよびヒト免疫グロブリン分子に由来する1つまたは複数のFRを含有するV領域を含む分子である。非ヒト抗体をヒト化するための典型的な方法は、Queenら、1989、Proc.Natl.Acad.Sci.USA 86:10029〜10033および米国特許第5,585,089号および同第5,693,762号;Riechmannら、Nature、332:323、1988;およびTsurushitaら、Methods 36:69〜83、2005の文献など、様々な参考文献に記載されており、すべてその全体が参照により本明細書に組み込まれる)。例えば、Tsurushitaら(2005、前掲;以下「Tsurushita」とする)の参考文献では、元々Queenら(1989、前掲)によって開発された抗体ヒト化方法に基づいてマウスモノクローナル抗体をヒト化するための実用的および指示的プロトコールが示されている。Tsurushitaに開示されている一般的なプロトコールは以下に簡潔に要約する。
【0053】
5.3.1.ヒト化抗体を調製するための一般的なプロトコール
マウスV遺伝子のクローニングおよび配列決定
標的のマウスモノクローナル抗体のVHおよびVLをコードするcDNAのクローニングに様々な方法が利用可能である。例えば、SMART RACE cDNA Amplificationキット(BD Biosciences、CA)またはGeneRacerキット(Invitrogen、CA)を使用した5’RACE(rapid amplification of cDNA ends)法が一般に使用されている。5’RACE用の遺伝子特異的プライマーは、VHおよびVLのすぐ下流に結合するように、標的のモノクローナル抗体のH鎖およびL鎖のアイソタイプに基づいて調製することができる。したがって、5’RACEプライマーは、γ1、γ2a、γ2bまたはγ3など、マウスのそれぞれのサブタイプに特異的であるように設計することもできる。あるいは、すべてのサブタイプ用の一般的なプライマーは、サブタイプの中で共通のまたは高度に相同な領域に基づいて設計することもできる。Tsurushitaでは、以下の5’RACEプライマーを例として開示する。
(i)5’−GCCAGTGGATAGACTGATGG−(配列番号82)(マウスγ1、γ2a、γ2bおよびγ3 H鎖のクローニング用)
(ii)5’−GATGGATACAGTTGGTGCAGC−(配列番号83)(マウスκL鎖のクローニング用)。
【0054】
PCR増幅されたV領域遺伝子フラグメントを、例えば、Zero Blunt TOPO PCR Cloning Kit(Invitrogen)を使用してプラスミドベクター中に直接的にクローニングし、そのDNA配列を決定することができる。得られた配列は、例えば、そのコードアミノ酸配列を、例えば、Model 241 Protein Sequencer(Hewlett−Packard、CA)を使用したN末端アミノ酸配列決定によって決定された標的のモノクローナル抗体の配列と比較することによって確認すべきである。通常、例えば、エドマン分解による標的抗体のN末端の少なくとも15〜20アミノ酸残基の決定は、クローン化DNA配列の確実性を確認するのに十分である。マウスにおいて2つの最も多くみられるN末端アミノ酸の1つであるグルタミンがN末端アミノ酸である場合、それはピログルタミンに変換される可能性があり、N末端における配列決定を阻止すると、Tsurushitaは注意を促している。その場合は、配列を得るためにN末端での阻止を解除することが必要である。
【0055】
V領域の三次元モデリング
まず、VHおよびVLの配列に基づいて、CDRの立体構造の維持において潜在的に重要である標的抗体のフレームワーク残基を、例えば、R.Levyら、1989、Biochemistry28:7168〜7175;およびB.Zilberら、1990、Biochemistry29:10032〜10041に記載されている方法によって同定する。通常、VHおよびVLはそれぞれ、構造的に意味ある14のセグメントに分けられ、それは、免疫グロブリンスーパーファミリーのドメイン構造を構成するβ鎖およびループ様構造である。標的抗体由来のセグメントのそれぞれのアミノ酸配列は、PDBデータベース(H.M.Bermanら、2000、Nucleic Acids Res.28:235〜342参照)における既知の構造を有する抗体の対応するセグメントとアラインメントする。複数の配列アライメントにより、標的セグメントのそれぞれの最高の配列相同性を有する対応するセグメントを選択し、V領域の三次元モデルを構築する。構造を最適化するために、モデルを複数サイクルの共役勾配エネルギー最小化(conjugate gradient energy minimization)に供する(例えば、ENCAD、またはPressら、1990、「Numerical Recipes、Cambridge University Press、Cambridgeに記載;Weinerら、1981、J.Comp.Chem.2:287〜303によるAMBER;BioMolecularModellingで利用可能な3D−JIG−SAW、またはCancer Research UKによって運営される「BMM」ウェブサイト;またはSwiss Institute of Bioinformatics、Genevaによって運営されるExPASy Proteomics Serverウェブサイトで利用可能なSWISS−MODELを使用)。
【0056】
ヒトフレームワークの選択
V領域の構造のモデリングと並行して、マウスVHおよびVLそれぞれのcDNAクローニングから推定されたアミノ酸配列をデータベース、例えば、Kabatデータベース(Johnsonら、2000、Johnsonら、2000、Nucleic Acids Res.28:214〜218を参照されたい)、GenBankなどにおいてヒトV領域配列と比較する。マウス配列と少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、または少なくとも95%の同一性という全体にわたる配列同一性を有するヒトFRは、例えば、Smith−Watermanアルゴリズム(Gusfield、1997、「Algorithms on Strings, Trees, and Sequences」掲載、Cambridge University Press、Cambridgeによる)、またはBLAST(Karlinら、1990、Proc.Natl.Acad.Sci.USA87:2264〜2268による)などを使用して検索ことができる。これらのヒト配列はcDNAベースの配列およびタンパク質由来の配列に基づくものであってもよいが、生殖系列の使用がしばしば好ましく、それは、この生殖系列は、cDNAベースの配列、タンパク質由来の配列の体細胞超変異に関連する潜在的な免疫原性の排除に有用である可能性があるからである。別の方法では、Queenら(1989、前掲)に記載のとおり、コンセンサスフレームワーク配列を使用することにより、cDNAベースの配列またはタンパク質由来の配列から得られたフレームワーク中のこのような超変異残基を同定および除去することもできる。生殖系列VHセグメントをアクセプターフレームワークとして使用する場合、第14染色体上にコードされるVHセグメントだけが機能的VHを産生するので、第15染色体および第16染色体ではなく第14染色体上のものを使用すべきである。
【0057】
ヒト化V領域の設計
Queenら(1989、前掲)によれば、CDRの約4〜6Å以内にあるフレームワークアミノ酸を同定することが必要であり、それは、これらの残基が正しいCDR構造を支持する潜在的な重要なフレームワーク残基であると考えられるからである。このようなプロセスは、National Science Foundation(NSF)によって支援されるMolecular Visualization Freewareウェブサイトで利用可能な、原子座標から原子間距離を計算するRASMOLなどのコンピュータプログラムを使用して、またはコンピュータモデルのマニュアル検査によって達成することができる。重要なフレームワーク位置のアミノ酸がマウスドナー配列とヒトアクセプター配列の間で異なる場合、通常ヒト残基がマウスドナーの残基によって置き換えられる。しかし、このような残基が、CDR構造の支持への寄与がわずかしかない場合、対応するヒト残基を通常使用する。また、選択されたヒトアクセプターがV領域配列の約10〜20%未満に存在する「非定型的な(atypical)」アミノ酸を含有する場合、これらのアミノ酸は親和性成熟の間の体細胞超変異の結果である可能性があり、ヒトにおける潜在的な免疫原性を回避するためにドナー残基で置き換えるべきである。
【0058】
さらに、潜在的なN結合型グリコシル化シグナルの残基などの他の因子を、ヒト化V領域を設計するために慎重に考慮する必要がある(詳細についてはTsurushitaを参照されたい)。
【0059】
ヒト化抗体は、治療的使用に必要とされる、または排除されるべきエフェクター機能に依存して、ヒト抗体またはその変異体のヒトκもしくはλL鎖、および/またはγ1、γ2、γ3、γ4、μ、α1、α2、δもしくはεH鎖由来のヒト定常領域またはその一部分を含有することができる。例えば、抗体とFc受容体の結合が低減し、かつ/または補体を固定する抗体の能力が低減するように、突然変異を含有する定常領域Fc部分を本発明のキメラまたはヒト化抗体のV領域と融合することもできる(例えば、Winterら、GB2,209,757B;Morrisonら、WO89/07142、Morganら、WO94/29351を参照されたい)。抗体分子のこのような操作は、セクション5.2に記載のとおり組換えDNA技術によって実施することができる。
【0060】
好ましくは、得られたキメラまたはヒト化抗体は、非ヒトドナー抗体と同一の特異性、および非ヒトドナー抗体の親和性に類似する、またはそれの少なくとも約1/3、少なくとも約1/2もしくは少なくとも約2/3の親和性を有する。別の態様において、得られたキメラまたはヒト化抗体は、少なくとも約1×107M−1、好ましくは少なくとも約1×108M−1、最も好ましくは少なくとも約1×109M−1の親和性定数を有する。
【0061】
上記の一般的なプロトコールに加えて、抗体は、例えば、CDRグラフティング(EP239,400;PCT公報WO91/09967;米国特許第5,225,539号;同第5,530,101号および同第5,585,089号)、ベニアリング(veneering)またはリサーフェシング(resurfacing)(EP592,106;EP519,596;Padlan、Molecular Immunology、28(4/5):489〜498、1991;Studnickaら、Protein Engineering、7(6):805〜814、1994;Roguskaら、Proc Natl.Acad.Sci.USA、91:969〜973、1994)、およびチェインシャッフリング(chain shuffling)(米国特許第5,565,332号)を含めた当技術分野で知られている種々の技法を使用してヒト化することができ、すべてその全体が参照により本明細書に組み込まれる。
【0062】
5.3.2.ヒト化抗体を医薬品として調製するための追加の考慮事項
ヒト化抗体を医薬品として提供するために、したがって、効率的で一貫した産生系を調製する必要がある。例えば、H鎖およびL鎖配列を挿入することによってヒト化抗体に適切な発現ベクターを調製し、発現ベクターをトランスフェクトした高生産性細胞株をマスターセルバンク(master cell bank)(MCB)の種細胞として得ることができ、それはワーキングセルバンク(working cell bank)(WCB)の安定で半永久的な供給源となる。次いで、WCB由来のワーキングセル(working cell)を培養し、培地を収集することによってヒト化抗体を調製することができる。
【0063】
適切な制御遺伝子を有する種々の発現ベクターをこのような産生細胞株の調製に使用することができる。宿主細胞として、哺乳動物タンパク質を発現させるのに一般に使用されるものをヒト化抗体の発現に使用することができる。このような宿主細胞の例としては、それだけには限定されないが、チャイニーズハムスター卵巣(CHO)細胞、SP2/0−Ag14.19細胞、NSO細胞などが挙げられる。ヒト化抗体の生産性は、発現ベクターと宿主細胞の最良の組合せを選択することによって最大化することができる。さらに、宿主細胞によるヒト化抗体の発現が最適化されるように、適した培地を選択するために種々の無血清培地および補充物質から培地の組成を検討すべきである。
【0064】
効率および最終収率に基づいて、宿主細胞によって産生されたヒト化抗体は、アフィニティークロマトグラフィー、イオン交換クロマトグラフィー、疎水的相互作用クロマトグラフィーなどを含めた当技術分野でよく知られている様々な方法を使用して、培養上清から精製することができる。
【0065】
5.4.医薬組成物および治療的使用
本発明は、RGD配列を免疫特異的に認識する上記のヒト化抗体またはその抗原結合性フラグメントを含む医薬組成物を提供する。本発明のヒト化抗体を有効成分として含む医薬組成物は、それだけには限定されないが、癌、例えば、癌細胞の増殖または転移、および炎症性疾患、例えば、関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、真性糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などを含めた、RGDタンパク質に関連する障害または疾患の予防薬および/または治療薬として使用することができる。
【0066】
本発明のヒト化抗体を含む医薬組成物を使用して、臓器移植後の慢性拒否反応、および全身性自己免疫疾患、エリテマトーデス、ブドウ膜炎、ベーチェット病、多発性筋炎、増殖性糸球体腎炎、サルコイドーシスなどの自己免疫疾患を治療することもできる。
【0067】
本発明のヒト化抗体を含む、上記の障害または疾患を予防または治療するための予防薬および/または治療薬は、低い毒性を有し、適当な溶媒中に混合することによる液体製剤として、または適切な剤形の医薬組成物として直接的にヒトに経口または非経口投与することができる。
【0068】
上記の投与に使用される医薬組成物は、前記の抗体またはその塩および薬学的に許容される担体、希釈剤または添加剤を含有する。このような組成物は、経口または非経口投与に適した剤形で提供される。
【0069】
用量は、投与される対象の年齢およびサイズ、標的の疾患、条件、投与経路などによって様々であってよい。抗体を、例えば、成人患者の関節リウマチの予防および/または治療に使用する場合、本発明の抗体を静脈内に通常、約0.01〜約20mg/kg体重、好ましくは約0.1〜約10mg/kg体重、より好ましくは約0.1〜約5mg/kg体重の単回用量で、1日に約1〜5回、好ましくは1日に約1〜3回投与することが有利である。他の非経口投与および経口投与においても、抗体を上記で示される用量に相当する用量で投与することができる。状態が特に重い場合、用量を状態に従って増加させることもできる。
【0070】
様々なデリバリーシステムが知られており、本発明の医薬組成物を投与するために使用することができ、例えば、リポソーム、微粒子、マイクロカプセル中への封入、突然変異ウイルスを発現することができる組換え細胞、受容体を介したエンドサイトーシスがある(例えば、WuおよびWu、1987、J.Biol.Chem.262:4429 4432を参照されたい)。導入方法としては、それだけには限定されないが、皮内、筋肉内、腹腔内、静脈内、皮下、鼻腔内、硬膜外、および経口の経路が挙げられる。これらの化合物は、任意の好都合な経路によって、例えば、注入またはボーラス注射によって、上皮層または皮膚粘膜層(例えば、口腔粘膜、直腸および腸管粘膜など)を通した吸収によって投与することができ、他の生物活性を有する作用物質と一緒に投与することもできる。投与は全身でも局所でもよい。例えば、吸入器またはネブライザーおよびエアロゾル化剤との配合の使用によって肺への投与を採用することもできる。
【0071】
特定の実施形態において、本発明の医薬組成物を、治療を必要とする領域に局所的に投与することが望ましいことがあり、これは、例えば、限定的なものではないが、手術中の局所注入、例えば、手術後の創傷包帯と組み合わせた外用塗布により、注射により、カテーテルを用いて、坐剤を用いて、鼻内噴霧を用いて、またはシアラスティック(sialastic)膜などの膜、または繊維を含めた、多孔性、非多孔性またはゼラチン状の材料のインプラントを用いて実現することができる。一実施形態において、感染組織の部位(または以前の部位)への直接注射によって投与することができる。
【0072】
別の実施形態において、医薬組成物は、小胞、特にリポソームとして送達することができる(Langer、1990、Science249:1527〜1533;Treatら、Liposomes in the Therapy of Infectious Disease and Cancer掲載、Lopez BeresteinおよびFidler(編)、Liss、New York、353〜365ページ(1989);Lopez−Berestein、同書、317〜327ページを参照されたい;同書を全体として参照されたい)。
【0073】
さらに別の実施形態では、医薬組成物を制御放出システムにおいて送達することができる。一実施形態において、ポンプを使用することもできる(Langer、前掲;Sefton、1987、CRC Crit.Ref.Biomed.Eng.14:201;Buchwaldら、1980、Surgery88:507;およびSaudekら、1989、N.Engl.J.Med.321:574を参照されたい)。別の実施形態において、ポリマー材料を使用することができる(Medical Applications of Controlled Release、LangerおよびWise(編)、CRC Pres.、Boca Raton、Florida(1974);Controlled Drug Bioavailability、Drug Product Design and Performance、SmolenおよびBall(編)、Wiley、New York(1984);RangerおよびPeppas、J.Macromol.Sci.Rev.Macromol.Chem.23:61(1983)を参照されたい;Levyら、1985、Science228:190;Duringら、1989、Ann.Neurol.25:351;Howardら、1989、J.Neurosurg.71:105も参照されたい)。さらに別の実施形態において、制御放出システムを組成物の標的の近くに配置することができ、したがって全身用量の一部しか必要とされない(例えば、Goodson、in Medical Applications of Controlled Release、前掲、2巻、115〜138ページ(1984)を参照されたい)。他の制御放出システムはLanger(Science249:1527〜1533(1990))による総説において検討されている。
【0074】
経口投与用の組成物の例としては、固形または液体の剤形、詳細には、錠剤(糖衣錠およびフィルムコーティング錠を含む)、丸剤、顆粒剤、粉末状の製剤、カプセル剤(軟カプセル剤を含む)、シロップ剤、乳剤、懸濁剤などが挙げられる。このような組成物は、公知の方法によって製造され、医薬製剤の分野で従来使用されているビヒクル、希釈剤または添加剤を含有する。錠剤のビヒクルまたは添加剤の例は、乳糖、デンプン、ショ糖、ステアリン酸マグネシウムなどである。
【0075】
注射用製剤としては、静脈内、皮下、皮内および筋肉内の注射、点滴注入などの剤形が挙げられる。これらの注射用製剤は、公知の方法によって調製することができる。注射用製剤は、例えば、上記の抗体またはその塩を、従来より注射剤に使用されている無菌水媒体または油性媒体に溶解、懸濁または乳化することによって調製することができる。注射剤の水媒体としては、例えば、生理的食塩水、ブドウ糖および他の助剤(auxiliary agent)などを含有する等張液などがあり、アルコール(例えば、エタノール)、ポリアルコール(例えば、プロピレングリコール、ポリエチレングリコール)、非イオン性界面活性剤[例えば、ポリソルベート80、HCO−50(硬化ヒマシ油のポリオキシエチレン(50mol)付加物)]などの適切な溶解補助剤(solubilizing agent)と組み合わせて使用することもできる。油性媒体としては、例えば、ゴマ油、ダイズ油などが採用され、安息香酸ベンジル、ベンジルアルコールなどの溶解補助剤と組み合わせて使用することもできる。このようにして調製される注射剤は、好ましくは、適切なアンプルに充填される。直腸投与に使用される坐剤は、前記の抗体またはその塩を坐剤用の常用の塩基とブレンドすることによって調製することができる。
【0076】
有利には、上記の経口または非経口用の医薬組成物は、有効成分の用量に適合するのに適した単位用量の剤形に調製される。このような単位用量の剤形としては、例えば、錠剤、丸剤、カプセル剤、注射剤(アンプル)、坐剤などが挙げられる。前記の抗体の含有量は一般に、単位用量の剤形当たり約5〜500mgであり、特に、注射剤の剤形では前記の抗体は約5〜100mg含有され、他の剤形では約10〜250mg含有されることが好ましい。
【0077】
上記の組成物はそれぞれ、配合により上記の抗体と任意の有害な相互作用を引き起こさない限り、他の活性成分をさらに含有することもできる。
【0078】
本発明はまた、RGD配列結合機能分子(例えば、インテグリンなど)を有効成分として含む、細胞および/または組織リモデリングの阻害剤および/または促進剤;ならびにRGDタンパク質を発現する細胞および/または組織(例えば、腫瘍細胞、好中球、平滑筋など)をRGDタンパク質結合機能分子と接触させることを含む、細胞および/または組織リモデリングを阻害および/または促進するための方法に関する。このような治療薬における有効成分の用量、投与方法、医薬製剤などは、本発明のヒト化抗体を含む医薬に関する前述の記載を参照することによって適切に決定することができる。
【0079】
上記のとおり、本発明はさらに、RGDタンパク質に関連する、またはそれが関与する障害または疾患を予防または治療するための方法であって、有効量の本発明のヒト化抗体のうちの少なくとも1つを、それを必要とする対象に投与することを含む方法を提供する。
【0080】
5.5.診断的使用
本発明のヒト化抗体を含む医薬組成物は、癌(例えば、癌細胞の増殖または転移)および/または炎症性疾患(例えば、関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、真性糖尿病、動脈硬化症、多発性硬化症、肉芽腫など)の診断用薬として、または臓器移植後の慢性拒否反応、全身性自己免疫疾患、エリテマトーデス、ブドウ膜炎、ベーチェット病、多発性筋炎、増殖性糸球体腎炎、サルコイドーシスなどの自己免疫疾患の診断用薬として使用することができる。本発明のヒト化抗体は、RGD配列を特異的に認識することができ、したがって試験体液中のRGDタンパク質を定量するために、特に、サンドイッチイムノアッセイ、競合的アッセイ、イムノメトリー(immunometry)、比濁分析(nephrometry)など、免疫染色などによる定量に使用することができる。これらの免疫学的方法を本発明のアッセイ法に適用する際は、いかなる特定の条件、手順なども説明する必要はない。当技術分野における通常の技術的考慮事項を従来の条件および手順に付加することによりアッセイ系を構築することが十分である。これらの一般的な技術的手段の詳細については、総説、テキストなどを参照することができる.
【0081】
上記のとおり、本発明の抗体を使用することにより、RGDタンパク質を高感度で定量することもできる。本発明のヒト化抗体は、RGDタンパク質をin vivoで定量するための方法を適用することにより、RGDタンパク質に関連する種々の疾患を診断するのに特に有用である。例えば、RGDタンパク質の発現レベルの増大または低下が検出された場合、RGDタンパク質に関連する疾患、例えば、癌または炎症性疾患に罹患している可能性が高い、または将来これらの疾患に罹患する可能性が高いと診断することができる。したがって、本発明はまた、対象においてRGDタンパク質に関連する、またはそれが関与する障害または疾患を診断するための方法であって、有効量の、本発明のヒト化抗体のうちの少なくとも1つまたは両方を、それを必要とする対象に投与することを含む方法を提供する。このようなin vivo診断に必要な投与量は、治療的使用に必要な投与量未満であってもよく、常用の手順に従って当業者によって決定することができる。
【0082】
本発明のヒト化抗体を使用して、体液、組織などの試験体液に存在するRGDタンパク質を特異的に検出することもできる。ヒト化抗体は、RGDタンパク質を精製するための抗体カラムの調製、精製時のそれぞれの画分に含有されるRGDタンパク質の検出、または試験対象の細胞中のRGDタンパク質の挙動の分析に使用することもできる。
【実施例】
【0083】
6.実施例
以下の実施例では、RGD配列を免疫特異的に認識するモノクローナル抗体の調製、モノクローナル抗体のV領域の配列決定、抗体のエピトープマッピングおよび他の特徴付け、ならびにこのような抗体のキメラ化およびヒト化、ならびに得られたキメラ抗体およびヒト化抗体の特徴付けを例示する。これらの実施例は、本発明の範囲を限定するものと解釈すべきではない。
【0084】
6.1.RGD配列に対するマウス抗体の調製
RGD配列に対するマウスモノクローナル抗体をサブトラクティブ免疫化法(subtractive immunization method)(Williams C.V.ら、1992、Biotechniques12:842〜847による)に従って調製した。RGD配列を含むアミノ酸配列CVDVPNGRGDSLAYGLR(配列番号71)およびECMタンパク質の細胞接着性配列であるアミノ酸配列SLAYGLR(配列番号72)の合成ペプチドとして抗原を調製した。抗原ペプチドをEMCS(Dojin)を介してサイログロブリンに結合させ、これを抗原としてアジュバントと共にマウスに免疫した。ハイブリドーマを当技術分野でよく知られている方法(例えば、Harlowら、Antibodies:A Laboratory Manual、(Cold Spring Harbor Laboratory Press、第2版1988);Hammerlingら、Monoclonal Antibodies and T−Cell Hybridomas掲載、563〜681ページ(Elsevier,N.Y.、1981)を参照されたい)によって調製した。4回免疫した後、脾細胞を収集し、骨髄腫細胞X63−Ag8−653と融合した。次いで、HAT培地を使用し、ELISA(抗原ペプチド固相)を用いて培養上清をスクリーニングすることにより、RGD配列に免疫特異的に反応性を示すモノクローナル抗体を産生するハイブリドーマクローンを選択した。RGD配列を免疫特異的に認識するモノクローナル抗体を産生するハイブリドーマクローンを単離すると、8つのハイブリドーマクローン4P11、11M6、25H15、29R5、30C17、33E10、35B6および38I8を樹立した。チオールセファロースビーズ(Amasham Bioscience)を使用することによって調製された抗原ペプチドカラムを使用することにより、抗体をハイブリドーマの上清から精製した。
【0085】
6.2.抗RGD配列モノクローナル抗体のエピトープ分析
アミノ酸配列CLPVKTDSGSSEEKLY(mOPN1)(配列番号73)、CVDVPNGRGDSLAYGLR(mOPN5)(配列番号71)、CVDVPNGRGDS(配列番号74)、CPNGRGD(配列番号75)、CGRGDSLAYGLR(配列番号76)、CGDSLAYG(配列番号77)、CGDSLAUGLR(配列番号78)およびCSLAYGLR(配列番号72)からなる、マウスOPNに由来する部分ペプチドを含むペプチド、アミノ酸配列CVDTYDGRGDSVVYGLRS(配列番号79)およびCSVVYGLR(配列番号80)からなる、ヒトOPNに由来する部分ペプチドを含むペプチド、ならびにアミノ酸配列CGRGDS(配列番号81)の、ヒトOPNおよびマウスOPNの共通のペプチド配列を含むペプチドを、EMCS(同仁化学研究所)を介してBSA(Sigma corporation)に結合させ、ELISAに使用した。
【0086】
96ウェルプレートにペプチド(10μg/ml)またはタンパク質(5μg/ml)を37℃で1時間培養することによって固定化し、0.1%BSA/PBS/0.05%NaN3溶液でブロッキングし、次いで様々な濃度の抗体と37℃で1時間反応させた。次に、プレートを二次抗体であるHRP標識抗マウスIgG抗体(Jackson ImmunoResearch Laboratories,Inc.)と37℃で30分間反応させ、OPDを色素原として添加し、1N H2SO4を添加して反応を停止させ、次いで490nmでの吸光度を検出した。
【0087】
図1、図2、図3、図4および図5に示すとおり、モノクローナル抗体4P11、11M6、25H5、35B6および33E10は、mOPN5およびhOPN5に結合し、RGDタンパク質のマウスおよびヒト部分ペプチドを認識する。モノクローナル抗体33E10は、GRGDS(配列番号81)、VDVPNGRGDS(配列番号74)およびPNGRGD(配列番号75)を認識したが、OPNのRGDに続く配列を含むSLAYGLR(配列番号72)またはSVVYGLR(配列番号80)を認識しなかった。モノクローナル抗体33E10は、GRGDS(配列番号81)、VDVPNGRGDS(配列番号74)およびPNGRGD(配列番号75)に共通に含まれるRGD配列を認識し、ヒトペプチドとマウスペプチドの両方に結合することができる。モノクローナル抗体35B6は、GRGDSLAYGLR(配列番号76)、GDSLAYG(配列番号77)およびGDSLAYGLR(配列番号78)を認識したが、GRGDS(配列番号81)、VDVPNGRGDS(配列番号74)またはPNGRGD(配列番号75)を認識しなかった。モノクローナル抗体35B6は、GDを含む、RGDに続く配列を認識する。モノクローナル抗体29R5、30C17および38I8は、GRGDS(配列番号81)、SLAYGLR(配列番号72)およびSVVYGLR(配列番号80)にはわずかな反応性しか示さなかったが、mOPN5にだけ反応し、それにより、これらのモノクローナル抗体はマウスOPNのVDVPNGRGDSLAYGLR(配列番号71)を認識することが示される。
【0088】
6.3.抗RGD抗体のCDR分析
モノクローナル抗体33E10および35B6のCDRのアミノ酸配列を以下の手順によって決定した。RNeasy Miniキット(Qiagen)を使用して対応するハイブリドーマからRNAを抽出し、First−strand cDNA synthesisキットを使用してcDNAを調製した。抗体のH鎖cDNAを、Heavy primer amplificationキット(Amasham Bioscience)を使用したPCRによって増幅し、pCRII−TOPOベクター(Invitrogen)中にクローニングし、次いでcDNA配列およびアミノ酸配列を決定した。ABG:Directory of 3D structures of antibodies(http://www.ibt.unam.mx/vir/structure/structures.html)によってCDRを決定した。V鎖およびL鎖のCDRは以下のとおりである(図6および図7にも示す)。
(H鎖)
[CDRH1]
33E10:GFTFTDYYMI(配列番号1)
35B6:GYTFTNYWMH(配列番号7)
[CDRH2]
33E10:WLGFIRNKANGYTTEYSASVKG(配列番号2)
35B6:WIGNINPRNGDSNYNEKFRS(配列番号8)
[CDRH3]
33E10:GAY(配列番号3)
35B6:GYFDV(配列番号9)
(L鎖)
[CDRL1]
33E10:RSSQSIVHSNGNTYLE(配列番号4)
35B6:KASQDINSYLS(配列番号10)
[CDRL2]
33E10:RVSNRFS(配列番号5)
35B6:RANRLVD(配列番号11)
[CDRL3]
33E10:GSFVPW(配列番号6)
35B6:YDEFPF(配列番号12)
【0089】
本実施例では、CDRをABGによって決定した。しかし、他のプログラムを使用してCDRを決定することもでき、それによりある程度異なる配列が得られることもあることは当技術分野でよく知られている。
【0090】
6.4.RGD配列を有するECMタンパク質に対する結合能
ヒトOPN(hOPN)遺伝子またはマウスOPN(mOPN)遺伝子をそれぞれ導入したCHO−K1細胞の培養上清から、抗OPN抗体カラムを使用することにより、hOPNまたはmOPNをそれぞれ精製した。ヒトビトロネクチン(以下「VN」と略記する)はAGC TECHNO GLASS Co.,Ltdから得た。ヒトフィブロネクチン(以下「FN」と略記する)、ヒトトロンボスポンジンおよびマウスラミニンはSigma Corporationから得た。
【0091】
上記で得たhOPN、mOPN、FN、VNまたはラミニンを固定化した96ウェルプレートを使用したELISAにより、ECMタンパク質に対するモノクローナル抗体33E10および35B6の結合能を検出した。96ウェルプレートにペプチド(10μg/ml)またはタンパク質(5μg/ml)を37℃で1時間培養することによって固定化し、0.1%BSA/PBS/0.05%NaN3溶液でブロッキングし、次いで様々な濃度の抗体と37℃で1時間反応させた。次に、プレートを二次抗体のHRP標識抗マウスIgG抗体(Jackson ImmunoResearch Laboratories,Inc.)と37℃で30分間反応させ、OPDを色素原として添加し、1N H2SO4を添加して反応を停止させ、次いで490nmでの吸光度を検出した。
【0092】
図8にその結果を示した。モノクローナル抗体33E10は、試験したすべてのECMタンパク質と交差反応性を示したが、ラミニンとは低い反応性を示した。モノクローナル抗体35B6は、hOPNおよびmOPNと反応性を示したが、ラミニンとは反応性を示さなかった。
【0093】
6.5.細胞接着阻害活性
細胞接着にはRGDペプチドとそのリガンド、すなわちインテグリンなどとの結合が関与することが知られているので、単離された抗RGD抗体をその細胞接着阻害活性について検討した。ヒトOPN(hOPN)遺伝子またはマウスOPN(mOPN)遺伝子をそれぞれ導入したCHO−K1細胞の培養上清から、抗OPN抗体カラムを使用することにより、hOPNまたはmOPNをそれぞれ精製した。大腸菌から単離することによる、トロンビン切断型mOPNのN末端部分とのグルタチオンSトランスフェラーゼ(GST)融合タンパク質として、mOPNのN半分を精製した。ヒトFNおよびヒトVNはSigma Corporationから得た。
【0094】
50μlのタンパク質を96ウェルプレートの各ウェルに添加し、37℃で1時間インキュベートし、プレート上に固定化した。プレートをブロッキング溶液(0.5%BSA/PBS)でブロッキングし、PBSで1回洗浄した後、NIH3T3細胞を0.25%BSA−最小必須培地(MEM)に懸濁させ、単離されたモノクローナル抗体を1.0×105細胞/mlの最終濃度で混合し、プレートに200μl/ウェルで添加し、5%CO2下、37℃で1時間インキュベートした。非接着細胞をPBSですすぎ、接着細胞を固定し、0.5%Crystal Violet(和光純薬工業、大阪、日本による)/20%メタノールで染色した。染色細胞を室温で30分間静置させた。プレートを蒸留水で洗浄し、それに20%酢酸溶液を添加して溶解させた。590nmでのODを測定することによって接着活性を定量した。
【0095】
mOPNのN半分を固定化した96ウェルプレートに、NIH3T3細胞とモノクローナル抗体33E10または35B6の混合物を添加し、NIH3T3細胞とmOPNのN半分との結合に及ぼす抗体の効果を検討した。mOPNのN半分、FNまたはVNをそれぞれ固定化した96ウェルプレートに、NIH3T3細胞とモノクローナル抗体33E10の混合物を添加し、NIH3T3細胞と各タンパク質との結合に及ぼす抗体の効果を検討した。
【0096】
図9および図10A〜10Cに示すとおり、NIH3T3細胞はmOPNのN半分に接着し、この接着を抗RGD抗体によって阻害した。モノクローナル抗体33E10は、モノクローナル抗体35B6と比較して強い阻害活性を示した。NIH3T3細胞は、検討したすべてのECMタンパク質に接着した。モノクローナル抗体33E10は、mOPNのN半分との細胞接着を阻害したが、FNまたはVNとの細胞接着を阻害しなかった。したがって、モノクローナル抗体33E10はOPNと細胞の間の接着を特異的に阻害することが示される。
【0097】
6.6.抗RGD抗体の治療効果
抗RGD抗体の治療効果をマウス系において検討した。抗RGDモノクローナル抗体(4P11、11M6、29R5、30C7、38I8、33E10および35B6)を抗RGD抗体についての記載(セクション6.1、前掲を参照されたい)と実質的に同様に調製した。
【0098】
6.6.1.肝炎に対する治療効果
WO02/081522は、OPN機能を阻害することによって肝炎を治療できることを開示している。したがって、マウス肝炎モデルにおいてマウス抗RGD抗体(4P11、11M6、29R5、30C7、38I8、33E10および35B6)を使用して抗RGD抗体の治療効果を試験した。マウス(マウス5匹/群)の血中ASTおよびALTレベルを、200μgのコンカナバリンA(ConA)(Vector)の静脈内投与から12時間後、GPT/ALT−PIIIおよびGOT/AST−PIII(富士フイルム)を使用して測定した。ConA注射の3時間前に抗体を200μg投与した。対照抗体としてマウスIgGを使用した。
【0099】
図11に示すとおり、モノクローナル抗体25H15はいかなる治療効果も示さなかったが、モノクローナル抗体4P11、11M6、29R5、30C17および38I8は治療効果を示した。モノクローナル抗体33E10または35B6を投与したマウスは、ASTおよびALTレベルの増大を示さなかった。したがって、この結果から、モノクローナル抗体33E10または35B6によって肝炎を治療する可能性があることが明らかになった。
【0100】
6.6.2.マウス癌細胞株の転移に及ぼす抗RGD抗体の効果
肺転移のマウス実験モデルおよび自然発症モデルにおいて転移に及ぼす抗RGD抗体の効果を試験した。
【0101】
実験モデルでは、400μg/マウスのモノクローナル抗体と混合したマウスメラノーマ細胞株B16−Luc細胞(1×105細胞/マウス)をC57BL/6マウスの尾静脈に注射し、注射から14日後、肺転移数をカウントした。対照として同じ種類の抗体(mIgG1)を使用した。
【0102】
自然発症モデルでは、マウスメラノーマ細胞株B16−BL6細胞(4×105細胞/マウス)をC57BL/6マウスの左足蹠に皮下注射した。注射から19日後、原発性腫瘍を外科的に切除し、切除から14日後(B16−BL6細胞の注射から33日後)、屠殺したマウス由来の肺における腫瘍コロニーの数をカウントした。200μg/マウスのモノクローナル抗体を腫瘍細胞の注射から3、5、7、9、11、13、15および17日後の8回、腹腔内投与した。原発性腫瘍のサイズは切除までに測定した。外科切除から14日後、肺における腫瘍コロニーの数をカウントした。対照として同じ種類の抗体(mIgG1)を使用した。
【0103】
図12A〜12Bにマウス実験モデルの結果を示す。モノクローナル抗体33E10または35B6を投与したマウスにおける肺転移の平均数は対照と比較して少なかった。モノクローナル抗体35B6は肺転移を有意に阻害した。
【0104】
図13A〜13Cは、マウス自然発症モデルにおける以下のフォーマット:原発性腫瘍サイズの経日変化、肺転移コロニーの数、および体重変化の結果を示す。モノクローナル抗体33E10または35B6のいずれかを投与したマウスにおける原発性腫瘍のサイズは、対照マウスと比較して小さかった。したがって、その結果から、モノクローナル抗体33E10または35B6によって腫瘍増殖を阻害する可能性があることが示される。対照群における5匹のマウスのうち2匹が非常に多数の肺転移コロニーを示したので、35B6に関しては統計学的有意差が示されなかった。しかし、モノクローナル抗体33E10または35B6を投与したマウスにおける肺転移の平均数は対照抗体と比較して少なかった。したがって、モノクローナル抗体33E10または35B6は癌転移を阻害することが示される。
【0105】
6.6.3.マウス関節リウマチモデルにおける抗RGD抗体の治療効果
II型コラーゲン特異的モノクローナル抗体(IBL、日本)のカクテルを供給元のプロトコールに従って使用して、マウスに関節リウマチを誘発した。すなわち、マウス(Balb/c)にII型コラーゲン特異的モノクローナル抗体のカクテルを注射し、注射の3日後、LPSを注射して関節リウマチを発症させた。抗RGDモノクローナル抗体または正常ハムスターIgG(NHG)をコラーゲン抗体注射の前日からコラーゲン抗体注射の6日後まで全8回200μg/マウス/日で腹腔内に投与した。マウスをコラーゲン抗体注射の日から毎日観察し、各足を関節の紅斑および腫脹に基づいて0から4までに類別することによって関節炎のレベルをスコア付けした(0=紅斑または腫脹なし;1=つま先などの1つの小さい関節に紅斑または腫脹あり;2=小さい関節の2つ以上に紅斑または腫脹あり、または手首または足首などのより大きい関節に紅斑または腫脹あり;3=足全体に紅斑または腫脹あり;4=足全体に完全な紅斑または腫脹あり;マウス(4足)1匹の最大スコア16)。
【0106】
その結果を図14に示す。対照NHGを注射したマウスは高スコアを示し、関節リウマチを発症したが、抗RGD抗体33E10または35B6をそれぞれ注射したマウスは低スコアを示し、関節リウマチの発症は完全に遮断された。したがって、この結果に照らして、抗RGD抗体は関節リウマチに対して予防的および治療的効果を有することが示された。
【0107】
6.6.4.マウス子宮内膜症モデルにおける抗RGD抗体の治療効果
子宮内膜症の症状は、嚢胞形成、周囲の間質における炎症、子宮内膜腺上皮の異所的な増殖による平滑筋化生、神経新生および血管形成を示す。免疫組織化学的方法により、オステオポンチン(OPN)がヒト子宮内膜症およびラット子宮内膜症モデルにおいて高度に発現されることが報告された。したがって、抗RGD抗体(33E10)による動物モデル対する治療効果を子宮内膜症に対する新たな療法として調べた。
【0108】
C57BL/6J雌性マウス(9週齢)を使用した。子宮内膜症モデルとして18匹のマウスを準備した。右の子宮を切除し、子宮の2mm×2mm角の2つ小片を腹部に自己移植した。左の子宮は参照用に処置しなかった。マウス抗RGD抗体(500μg/頭部、ip)を9匹のマウス(処置群)に週2回4週間(8回投与の全投与量:4000μg/頭部)施した。対照群(マウス9匹)には抗体を投与しなかった。投与後、形成された嚢胞の数を確認し、組織学的検査によって病的変化を観察した。
【0109】
その結果を表1に示した。2つの群に体重の差はなかった。処置群の出現した子宮内膜症の数は対照群よりも著しく少なかった。形成された嚢胞の重量は明らかに減少し、処置群は対照群の平均して約1/3であった。免疫組織化学的検査により、子宮内膜腺上皮におけるOPN発現が抗体処置によって抑制されることを確認することができた。処置群の間質における平滑筋の厚さは、対照群の1/2に減少した。
【0110】
マウス子宮内膜症モデルは抗RGD抗体の投与による治療効果を示した。
【0111】
【表1】
【0112】
6.7.非ヒト抗体のヒト化
6.7.1.マウス33E10可変領域遺伝子のクローニングおよび配列決定
7.5%CO2インキュベーターにおいて10%ウシ胎児血清(FBS;HyClone、Logan、UT)を含有するTIL培地I(免疫生物研究所、群馬、日本)中、37℃でマウス33E10ハイブリドーマ細胞を増殖させた。TRIzol試薬(Invitrogen、Carlsbad、CA)を供給元のプロトコールに従って使用して、約3×106ハイブリドーマ細胞から全RNAを抽出した。GeneRacer Kit(Invitrogen)を供給元のプロトコールに従って使用して、オリゴdTプライマーによるcDNAを合成した。35B6重鎖および軽鎖の可変領域cDNAは、ポリメラーゼ連鎖反応(PCR)により、マウスγ−1およびκ鎖定常領域にそれぞれアニーリングする3’プライマー、ならびにGeneRacer Kitにおいて提供されるGeneRacer5’プライマー(5’−CGACTGGAGCACGAGGACACTGA−3’)(配列番号84)を使用して、Phusion DNAポリメラーゼ(New England Biolabs、Beverly、MA)を用いて増幅した。VHのPCR増幅については、3’プライマーは配列5’−GCCAGTGGATAGACAGATGG−3’(配列番号85)を有する。VLのPCR増幅については、3’プライマーは配列5’−GATGGATACAGTTGGTGCAGC−3’(配列番号86)を有する。増幅されたVHおよびVL cDNAをpCR4Blunt−TOPOベクター(Invitrogen)中にクローニングして配列を決定した。可変領域のDNA配列決定をTocore(Menlo Park、CA)で実施した。いくつかの重鎖および軽鎖クローンの配列を決定し、典型的なマウス重鎖および軽鎖可変領域に相同である固有の配列を同定した。33E10 VHおよびVLのコンセンサスcDNA配列を推定アミノ酸配列と一緒にそれぞれ図15および16に示す。
【0113】
6.7.2.キメラ33E10 IgG1/κ抗体の構築
33E10 VHをコードする遺伝子は、PCRにより、鋳型として33E10 VH cDNA、5’プライマーとして5’−GGGACTAGTACCACCATGAAGTTGTGGCTGAACTGGATT−3’(SpeI部位は下線付き)(配列番号87)および3’プライマーとして5’−GGGAAGCTTGAAGTTAGGACTCACCTGCAGAGACAGTGACCAGAGTCCC−3’(HindIII部位は下線付き)(配列番号88)を使用して、スプライスドナーシグナルおよび適切なフランキング制限酵素部位を含むエキソンとして作製した(図17)。同様に、33E10 VLをコードする遺伝子は、PCRにより、鋳型として33E10 VL cDNA、5’プライマーとして5’−GGGGCTAGCACCACCATGAAGTTGCCTGTTAGGCTGTTG−3’(NheI部位は下線付き)(配列番号89)および3’プライマーとして5’−GGGGAATTCTTTGGATTCTACTTACGTTTGATTTCCAGCTTGGTGCCTCC−3’(EcoRI部位は下線付き)(配列番号90)を使用して、スプライスドナーシグナルおよび適切なフランキング制限酵素部位を含むエキソンとして作製した(図18)。33E10 VHおよびVLエキソンのスプライスドナーシグナルは、それぞれマウス生殖系列JH3およびJκ1配列に由来するものであった。QIAquick Gel Extraction Kit(Qiagen、Valencia、CA)を使用してPCR増幅断片をゲル精製し、SpeIおよびHindIII(VHについて)またはNheIおよびEcoRI(VLについて)で消化し、ヒトγ−1およびκ定常領域を担う哺乳動物発現ベクター中にクローニングしてキメラ33E10 IgG1/κ抗体を産生した。得られた発現ベクターpCh33E10の概略構造を図19に示す。
【0114】
6.7.3.ヒト化33E10 VHおよびVL遺伝子の作製
33E10可変領域のヒト化をQueenら(Proc.Natl.Acad.Sci.USA86:10029〜10033、1989)に概説されているとおりに実施した。まず、コンピュータプログラムを利用して33E10可変領域の分子モデルを構築した。次に、ヒト可変領域配列に対する相同性検索に基づいて、33E10 VHと高い相同性を有するU03400(GenBankアクセッション番号)のヒトアミノ酸配列を、ヒト化33E10 VHのフレームワークを提供するアクセプターとして選択した。同様に、X72452(GenBankアクセッション番号)のヒトアミノ酸配列を33E10 VLのヒト化用のアクセプターとして選択した。
【0115】
コンピュータモデルがCDRとの有効な接触を提案したフレームワーク位置で、ヒトフレームワークアミノ酸をマウス33E10可変領域のアミノ酸で置換した。30位および48位でこれを実施して、ヒト化33E10(Hu33E10)VHを作製した(図20)。軽鎖については、置き換えることを必要とせずに、ヒト化33E10(Hu33E10)VLを作製した(図21)。33E10、設計したHu33E10およびヒトアクセプターアミノ酸配列のアライメントをVHについては図20、VLについては図21に示す。
【0116】
Hu33E10 VHおよびVLのぞれぞれをコードする遺伝子を、シグナルペプチド、スプライスドナーシグナルおよび適切な制限酵素部位を含むエキソンとして設計して、続いて哺乳動物発現ベクター中にクローニングした。Hu33E10 VHおよびVLエキソンのスプライスドナーシグナルは、それぞれヒト生殖系列JH4およびJκ1配列に由来するものであった。マウス33E10 VL遺伝子のシグナルペプチド配列は、SIG−Predシグナルペプチド予測ソフトウェア(http://bmbpcu36.leeds.ac.uk/prot_analysis/Signal.html)により、正確な切断に部分最適であることが示された。したがって、SIG−Predソフトウェアによって効率的かつ正確に切断すると予測された、マウスモノクローナル抗体35B6のVL遺伝子のシグナルペプチド(ジーンテクノサイエンス)をHu33E10 VLエキソンにおいて使用した。Hu33E10 VHエキソンにおけるシグナルペプチド配列は、対応するマウス33E10 VH配列に由来するものであった。SIG−Predソフトウェアにより、Hu33E10 VH遺伝子のシグナルペプチドが効率的かつ正確に切断されることが示された。
【0117】
Hu33E10 VHおよびVL遺伝子は、Heら(J.Immunol.160:1029〜1035、1998)に概説されているとおり、Phusion DNAポリメラーゼを使用して、いくつかの重複している合成オリゴヌクレオチドプライマー(配列番号91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122および123)の伸長およびPCR増幅によって構築した。Hu33E10 VHおよびVL遺伝子の構築に使用されたオリゴヌクレオチドをそれぞれ図22および図23に記載する。Hu33E10 VHおよびVL遺伝子におけるオリゴヌクレオチドの位置をそれぞれ図24および25を示す。QIAquick Gel Extraction Kit(Qiagen)を使用してPCR増幅断片をゲル精製し、pCR4Blunt−TOPOベクター中にクローニングして配列を決定した。SpeIおよびHindIII(VHについて)またはNheIおよびEcoRI(VLについて)で消化した後、Hu33E10 VHおよびVL遺伝子を哺乳動物発現ベクター中の対応する部位にサブクローニングしてヒトIgG1/κの形で産生した。得られた発現ベクターpHu33E10の概略構造を図19に示す。得られたHu33E10 VHおよびVL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒にそれぞれ図26(配列番号52)および図27(配列番号54)に示す。
【0118】
6.7.4.キメラおよびヒト化33E10 IgG1/κの一過性発現
キメラおよびヒト化33E10 IgG1/κ抗体は、Durocherら(Nucl.Acids Res.30:e9、2002)に従ってポリエチレンイミンを使用してpCh35B6およびpHu35B6プラスミドDNAをそれぞれHEK293細胞にトランスフェクトすることによって一過性に発現させる。一過性にトランスフェクトされたHEK293細胞を、7.5%CO2インキュベーターにおいて10%FBSを含有するDMEM中、37℃で2日間維持する。培養上清におけるCh33E10およびHu33E10 IgG1/κ抗体のそれぞれの発現レベルをサンドイッチELISAによって測定する。ELISAプレートをPBS中2,000倍希釈したヤギ抗ヒトIgG Fcγ鎖特異的ポリクローナル抗体(SouthernBiotech、Birmingham、AL)を100μl/ウェルで加えて4℃で一晩コーティングし、Wash Buffer(0.05%Tween20を含有するPBS)で洗浄し、Blocking Buffer(2%Skim Milkおよび0.05%Tween20を含有するPBS)を300μl/ウェルで加えて室温で1時間ブロッキングする。Wash Bufferで洗浄した後、ELISA Buffer(1%Skim Milkおよび0.025%Tween20を含有するPBS)中で適切に希釈した試料を100μl/ウェルでELISAプレートにアプライする。ヒト骨髄腫血清(SouthernBiotech)から精製したヒトIgG1/κ抗体を標準物質として使用する。ELISAプレートを室温で2時間インキュベートし、Wash Bufferで洗浄した後、2,000倍希釈したHRPコンジュゲートヤギ抗ヒトκ鎖ポリクローナル抗体(SouthernBiotech)を100μl/ウェルで使用して、結合した抗体を検出する。室温で1時間インキュベートし、Wash Bufferで洗浄した後、ABTS基質(bioWORLD、Dublin、OH)を100μl/ウェルで添加することによって発色を実施する。2%シュウ酸を100μl/ウェルで添加することによって発色を停止させる。405nmでの吸光度を読み取る。
【0119】
6.7.5.ヒト化35B6の特徴付け
ELISAにより、ヒト化35B6 IgG1/κの親和性をキメラ33E10 IgG1/κと比較する。抗原として、ウシ血清アルブミンとコンジュゲートした合成オリゴペプチド(Cys−Val−Asp−Thr−Tyr−Asp−Gly−Arg−Gly−Asp−Ser−Val−Val−Tyr−Gly−Leu−Arg−Ser)(hOPN5−BSA)を使用する。典型的な実験において、ELISAプレートをPBS中1μg/ml hOPN−BSAを100μl/ウェルで加えて4℃で一晩コーティングし、Wash Bufferで洗浄し、Blocking Bufferを300μl/ウェルで加えて室温で1時間ブロッキングする。Wash Bufferで洗浄した後、ELISA Buffer中で適切に希釈した試料を100μl/ウェルでELISAプレートにアプライする。ELISAプレートを4℃で一晩インキュベートし、Wash Bufferで洗浄した後、2,000倍希釈したHRPコンジュゲートヤギ抗ヒトγ鎖ポリクローナル抗体(SouthernBiotech)を100μl/ウェルで使用して、結合した抗体を検出する。室温で1時間インキュベートし、Wash Bufferで洗浄した後、ABTS基質を100μl/ウェルで添加することによって発色を実施し、2%シュウ酸を100μl/ウェルで添加することによって停止させる。405nmでの吸光度を読み取る。
【0120】
6.7.6.マウス35B6可変領域遺伝子のクローニングおよび配列決定
7.5%CO2インキュベーターにおいて10%ウシ胎児血清(FBS;HyClone、Logan、UT)を含有するTIL培地I(免疫生物研究所、群馬、日本)中、37℃でマウス35B6ハイブリドーマ細胞を増殖させた。TRIzol試薬(Invitrogen、Carlsbad、CA)を供給元のプロトコールに従って使用して、約3×106ハイブリドーマ細胞から全RNAを抽出した。GeneRacer Kit(Invitrogen)を供給元のプロトコールに従って使用して、オリゴdTプライマーによるcDNAを合成した。35B6重鎖および軽鎖の可変領域cDNAは、ポリメラーゼ連鎖反応(PCR)により、マウスγ−1およびκ鎖定常領域にそれぞれアニーリングする3’プライマー、ならびにGeneRacer Kitにおいて提供されるGeneRacer5’プライマー(5’−CGACTGGAGCACGAGGACACTGA−3’)(配列番号84)を使用して、Phusion DNAポリメラーゼ(New England Biolabs、Beverly、MA)を用いて増幅した。VHのPCR増幅については、3’プライマーは配列5’−GCCAGTGGATAGACAGATGG−3’(配列番号124)を有する。VLのPCR増幅については、3’プライマーは配列5’−GATGGATACAGTTGGTGCAGC−3’(配列番号125)を有する。増幅されたVHおよびVL cDNAをpCR4Blunt−TOPOベクター(Invitrogen)中にクローニングして配列を決定した。可変領域のDNA配列決定をTocore(Menlo Park、CA)で実施した。いくつかの重鎖および軽鎖クローンの配列を決定し、典型的なマウス重鎖および軽鎖可変領域に相同である固有の配列を同定した。35B6 VHおよびVLのコンセンサスcDNA配列を推定アミノ酸配列と一緒にそれぞれ図28および29に示す。
【0121】
6.7.7.キメラ35B6 IgG1/κ抗体の構築
35B6 VHをコードする遺伝子は、PCRにより、鋳型として35B6 VH cDNA、5’プライマーとして5’−GGGACTAGTACCACCATGGGATGGAGCTGTATCCTC−3’(SpeI部位は下線付き)(配列番号126)および3’プライマーとして5’−GGGAAGCTTAAAAAAAGCCAGCTTACCTGAGGAGACGGTGACCGTGGTCCC−3’(HindIII部位は下線付き)(配列番号127)を使用して、スプライスドナーシグナルおよび適切なフランキング制限酵素部位を含むエキソンとして作製した(図30)。同様に、35B6 VLをコードする遺伝子は、PCRにより、鋳型として35B6 VL cDNA、5’プライマーとして5’−GGGGCTAGCACCACCATGAGGACCCCTGCTCAGTTTCTT−3’(NheI部位は下線付き)(配列番号128)および3’プライマーとして5’−GGGGAATTCGCAAAAGTCTACTTACGTTTTATTTCCAACTTTGTCCCCGA−3’(EcoRI部位は下線付き)(配列番号129)を使用して、スプライスドナーシグナルおよび適切なフランキング制限酵素部位を含むエキソンとして作製した(図31)。35B6 VHおよびVLエキソンのスプライスドナーシグナルは、それぞれマウス生殖系列JH1およびJκ4配列に由来するものであった。QIAquick Gel Extraction Kit(Qiagen、Valencia、CA)を使用してPCR増幅断片をゲル精製し、SpeIおよびHindIII(VHについて)またはNheIおよびEcoRI(VLについて)で消化し、ヒトγ−1およびκ定常領域を担う哺乳動物発現ベクター中にクローニングしてキメラ35B6 IgG1/κ抗体を産生した。得られた発現ベクターpCh35B6の概略構造を図32に示す。
【0122】
6.7.8.ヒト化35B6 VHおよびVL遺伝子の作製
35B6可変領域のヒト化をQueenら(Proc.Natl.Acad.Sci.USA86:10029〜10033、1989)に概説されているとおりに実施した。まず、コンピュータプログラムを利用して35B6可変領域の分子モデルを構築した。次に、ヒト可変領域配列に対する相同性検索に基づいて、35B6 VHと高い相同性を有するZ47230(GenBankアクセッション番号)のヒトアミノ酸配列を、ヒト化35B6 VHのフレームワークを提供するアクセプターとして選択した。同様に、X72479(GenBankアクセッション番号)のヒトアミノ酸配列を35B6 VLのヒト化用のアクセプターとして選択した。
【0123】
コンピュータモデルがCDRとの有効な接触を提案したフレームワーク位置で、ヒトフレームワークアミノ酸をマウス35B6可変領域のアミノ酸で置換した。48位、66位、67位、68位、69位および71位でこれを実施して、ヒト化35B6(Hu35B6)VHを作製した(図33)。軽鎖については、46位および69位で置き換えて、ヒト化35B6(Hu35B6)VLを作製した(図34)。35B6、設計したHu35B6およびヒトアクセプターアミノ酸配列のアライメントをVHについては図33、VLについては図34に示す。
【0124】
Hu35B6 VHおよびVLのぞれぞれをコードする遺伝子を、シグナルペプチド、スプライスドナーシグナルおよび適切な制限酵素部位を含むエキソンとして設計して、続いて哺乳動物発現ベクター中にクローニングした。Hu35B6 VHおよびVLエキソンのスプライスドナーシグナルは、それぞれヒト生殖系列JH6およびJκ1配列に由来するものであった。マウス35B6 VH遺伝子のシグナルペプチド配列は、SIG−Predシグナルペプチド予測ソフトウェア(http://bmbpcu36.leeds.ac.uk/prot_analysis/Signal.html)により、正確な切断に部分最適であることが示された。したがって、SIG−Predソフトウェアによって効率的かつ正確に切断すると予測された、マウスモノクローナル抗体35E10のVH遺伝子のシグナルペプチド(ジーンテクノサイエンス)をHu35B6 VLエキソンにおいて使用した。ヒト化Hu35B6 VLエキソンにおけるシグナルペプチド配列は、対応するマウス35B6 VL配列に由来するものであった。SIG−Predソフトウェアにより、Hu35B6 VL遺伝子のシグナルペプチドが効率的かつ正確に切断されることが示された。
【0125】
Hu35B6 VHおよびVL遺伝子は、Heら(J.Immunol.160:1029〜1035、1998)に概説されているとおり、Phusion DNAポリメラーゼを使用して、いくつかの重複している合成オリゴヌクレオチドプライマー(配列番号91、92、130、131、132、133、134、135、136、137、138、139、140、141、142、143、107、108、144、145、146、147、148、149、150、151、152、153、154および155)の伸長およびPCR増幅によって構築した。Hu35B6 VHおよびVL遺伝子の構築に使用されたオリゴヌクレオチドをそれぞれ図35および図36に記載する。Hu35B6 VHおよびVL遺伝子におけるオリゴヌクレオチドの位置をそれぞれ図37および38を示す。QIAquick Gel Extraction Kit(Qiagen、Valencia、CA)を使用してPCR増幅断片をゲル精製し、pCR4Blunt−TOPOベクター中にクローニングして配列を決定した。SpeIおよびHindIII(VHについて)またはNheIおよびEcoRI(VLについて)で消化した後、Hu35B6 VHおよびVL遺伝子を哺乳動物発現ベクター中の対応する部位にサブクローニングしてヒトIgG1/κの形で産生した。得られた発現ベクターpHu35B6の概略構造を図32に示す。得られたHu35B6 VHおよびVL遺伝子のヌクレオチド配列を推定アミノ酸配列と一緒にそれぞれ図39(配列番号68)および図40(配列番号70)に示す。
【0126】
6.7.9.キメラおよびヒト化35B6 IgG1/κの一過性発現
キメラおよびヒト化35B6 IgG1/κ抗体は、Durocherら(Nucl.Acids Res.30:e9、2002)に従ってポリエチレンイミンを使用してpCh35B6およびpHu35B6プラスミドDNAをそれぞれHEK293細胞にトランスフェクトすることによって一過性に発現させた。一過性にトランスフェクトされたHEK293細胞を、7.5%CO2インキュベーターにおいて10%FBSを含有するDMEM中、37℃で2日間維持した。培養上清におけるCh35B6およびHu35B6 IgG1/κ抗体のそれぞれの発現レベルをサンドイッチELISAによって測定した。ELISAプレートをPBS中2,000倍希釈したヤギ抗ヒトIgG Fcγ鎖特異的ポリクローナル抗体(SouthernBiotech、Birmingham、AL)を100μl/ウェルで加えて4℃で一晩コーティングし、Wash Buffer(0.05%Tween20を含有するPBS)で洗浄し、Blocking Buffer(2%Skim Milkおよび0.05%Tween20を含有するPBS)を300μl/ウェルで加えて室温で1時間ブロッキングした。Wash Bufferで洗浄した後、ELISA Buffer(1%Skim Milkおよび0.025%Tween20を含有するPBS)中で適切に希釈した試料を100μl/ウェルでELISAプレートにアプライした。ヒト骨髄腫血清(SouthernBiotech)から精製したヒトIgG1/κ抗体を標準物質として使用した。ELISAプレートを室温で2時間インキュベートし、Wash Bufferで洗浄した後、2,000倍希釈したHRPコンジュゲートヤギ抗ヒトκ鎖ポリクローナル抗体(SouthernBiotech)を100μl/ウェルで使用して、結合した抗体を検出した。室温で1時間インキュベートし、Wash Bufferで洗浄した後、ABTS基質(bioWORLD、Dublin、OH)を100μl/ウェルで添加することによって発色を実施した。2%シュウ酸を100μl/ウェルで添加することによって発色を停止させた。405nmでの吸光度を読み取った。
【0127】
6.7.10.ヒト化35B6の特徴付け
ELISAにより、ヒト化35B6 IgG1/κの親和性をキメラ35B6 IgG1/κと比較した。抗原として、ウシ血清アルブミンとコンジュゲートした合成オリゴペプチ(Cys−Val−Asp−Thr−Tyr−Asp−Gly−Arg−Gly−Asp−Ser−Val−Val−Tyr−Gly−Leu−Arg−Ser)(配列番号79)(hOPN5−BSA)を使用した。典型的な実験において、ELISAプレートをPBS中1μg/ml hOPN−BSAを100μl/ウェルで加えて4℃で一晩コーティングし、Wash Bufferで洗浄し、Blocking Bufferを300μl/ウェルで加えて室温で1時間ブロッキングした。Wash Bufferで洗浄した後、ELISA Buffer中で適切に希釈した試料を100μl/ウェルでELISAプレートにアプライした。ELISAプレートを4℃で一晩インキュベートし、Wash Bufferで洗浄した後、2,000倍希釈したHRPコンジュゲートヤギ抗ヒトγ鎖ポリクローナル抗体(SouthernBiotech)を100μl/ウェルで使用して、結合した抗体を検出した。室温で1時間インキュベートし、Wash Bufferで洗浄した後、ABTS基質を100μl/ウェルで添加することによって発色を実施し、2%シュウ酸を100μl/ウェルで添加することによって停止させた。405nmでの吸光度を読み取った。図41A、41Bに示すとおり、ヒト化35B6 IgG1/κとhOPN5−BSAの結合は、キメラ35B6 IgG1/κに類似し(図41A)、または識別不可能であった(図41B)。この結果から、マウス35B6抗体のヒト化に成功したことが示される。
【0128】
7.寄託
本明細書で33E10および35B6と称した、マウス抗RGDモノクローナル抗体を産生するハイブリドーマは、微生物の寄託に関するブダペスト条約に従って、2005年10月27日、茨城県つくば市東1−1−1つくばセンター中央第6(郵便番号:305−8566)にある独立行政法人産業技術総合研究所特許生物寄託センターに寄託し、それぞれアクセッション番号FERM BP−10440およびFERM BP−10441を受けた。すべてその全体が参照により本明細書に組み込まれる。
【産業上の利用可能性】
【0129】
8.産業上の利用可能性
本発明のヒト化モノクローナル抗体は、RGDタンパク質の機能を阻害して、癌、例えば、癌細胞の増殖または転移、および炎症性疾患、例えば、関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、真性糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などに対して治療効果を発揮する。本発明の抗RGD抗体および抗インテグリン抗体の両方を含む医薬組成物は、癌および炎症性疾患に対してより改善された治療効果を及ぼす。
【0130】
9.配列の表
本明細書を通じて参照される配列を以下に要約する。
【0131】
【表2】
【特許請求の範囲】
【請求項1】
RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントであって、
(i)ヒト抗体のVHに由来する少なくとも1つのFRHと、RGD配列を免疫特異的に認識する非ヒト抗体のCDRHのうちの少なくとも1つに由来する少なくとも1つのCDRHとを含むH鎖;または
(ii)ヒト抗体のVLに由来する少なくとも1つのFRLと、RGD配列を免疫特異的に認識する非ヒト抗体のCDRLのうちの少なくとも1つに由来する少なくとも1つのCDRLとを含むL鎖;または
(iii)上記の(i)と(ii)の両方
を含むヒト化抗体またはその抗原結合性フラグメント。
【請求項2】
前記非ヒト抗体が、寄託アクセッション番号FERM BP−10440およびFERM BP−10441からなる群から選択されるハイブリドーマによって産生されたマウスモノクローナル抗体である、請求項1に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項3】
前記ヒト化抗体の少なくとも1つのCDRHが、配列番号1、2および3のアミノ酸配列からなる群から選択されるアミノ酸配列を含み、前記ヒト化抗体の少なくとも1つのCDRLが、配列番号4、5および6のアミノ酸配列からなる群から選択されるアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項4】
前記ヒト化抗体の少なくとも1つのCDRHが、配列番号7、8および9のアミノ酸配列からなる群から選択されるアミノ酸配列を含み、前記ヒト化抗体の少なくとも1つのCDRLが、配列番号10、11および12のアミノ酸配列からなる群から選択されるアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項5】
ヒト抗体の前記VHが、GenBankアクセッション番号X65891(配列番号13)のヌクレオチド配列によってコードされるアミノ酸配列に由来するアミノ酸配列を含み、ヒト抗体の前記VLが、GenBankアクセッション番号X72441(配列番号18)のヌクレオチド配列によってコードされるアミノ酸配列に由来するアミノ酸配列を含む、請求項2から4のいずれか一項に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項6】
前記H鎖が配列番号24のアミノ酸配列を含み、前記L鎖が配列番号26のアミノ酸配列を含む、請求項4に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項7】
前記H鎖が配列番号28のアミノ酸配列を含み、前記L鎖が配列番号30のアミノ酸配列を含む、請求項4に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項8】
配列番号24のアミノ酸配列をコードするヌクレオチド配列を含む単離された核酸分子。
【請求項9】
前記ヌクレオチド配列が配列番号23のヌクレオチド配列を有する、請求項8に記載の核酸分子。
【請求項10】
シグナルペプチドをコードするヌクレオチド配列をさらに含む、請求項8に記載の核酸分子。
【請求項11】
前記シグナルペプチドが配列番号32のアミノ酸配列を含む、請求項10に記載の核酸分子。
【請求項12】
配列番号26のアミノ酸配列をコードするヌクレオチド配列を含む単離された核酸分子。
【請求項13】
前記ヌクレオチド配列が配列番号25のヌクレオチド配列を有する、請求項12に記載の核酸分子。
【請求項14】
シグナルペプチドをコードするヌクレオチド配列をさらに含む、請求項12に記載の核酸分子。
【請求項15】
前記シグナルペプチドが配列番号34のアミノ酸配列を含む、請求項14に記載の核酸分子。
【請求項16】
配列番号28のアミノ酸配列をコードするヌクレオチド配列を含む単離された核酸分子。
【請求項17】
前記ヌクレオチド配列が配列番号27のヌクレオチド配列を有する、請求項16に記載の核酸分子。
【請求項18】
シグナルペプチドをコードするヌクレオチド配列をさらに含む、請求項16に記載の核酸分子。
【請求項19】
前記シグナルペプチドが配列番号36のアミノ酸配列を含む、請求項18に記載の核酸分子。
【請求項20】
配列番号30のアミノ酸配列をコードするヌクレオチド配列を含む単離された核酸分子。
【請求項21】
前記ヌクレオチド配列が配列番号29のヌクレオチド配列を有する、請求項20に記載の核酸分子。
【請求項22】
シグナルペプチドをコードするヌクレオチド配列をさらに含む、請求項20に記載の核酸分子。
【請求項23】
前記シグナルペプチドが配列番号38のアミノ酸配列を含む、請求項18に記載の核酸分子。
【請求項24】
1つまたは複数の調節エレメントに作動可能に連結している、請求項8もしくは23に記載の核酸分子またはその両方を含むベクター。
【請求項25】
請求項24に記載のベクターを含む単離された宿主細胞。
【請求項26】
ヒト化抗体またはその抗原結合性フラグメントを調製するための方法であって、請求項25に記載の宿主細胞をヒト化抗体またはその抗原結合性フラグメントが発現される条件下で培養すること、および発現されたヒト化抗体を収集することを含む方法。
【請求項27】
請求項1、2、6または7のいずれか一項に記載のヒト化抗体またはその抗原結合性フラグメントおよび薬学的に許容される担体を含む医薬組成物。
【請求項28】
RGDタンパク質に関連する障害または疾患を予防または治療するための方法であって、有効量の請求項6または7に記載のヒト化抗体またはその抗原結合性フラグメントを、それを必要とする対象に投与することを含む方法。
【請求項29】
RGDタンパク質に関連する障害または疾患をin vivoで診断するための方法であって、有効量の請求項1または2に記載のヒト化抗体またはその抗原結合性フラグメントを被検対象に投与することを含む方法。
【請求項30】
RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントであって、
(i)ヒト抗体のVHに由来する少なくとも1つのFRHと、配列番号1、2および3のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRHとを含むH鎖、または
(ii)ヒト抗体のVLに由来する少なくとも1つのFRLと、配列番号4、5および6のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRLとを含むL鎖、または
(iii)上記の(i)と(ii)の両方
を含むヒト化抗体またはその抗原結合性フラグメント。
【請求項31】
RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントであって、
(i)ヒト抗体のVHに由来する少なくとも1つのFRHと、配列番号7、8および9のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRHとを含むH鎖、または
(ii)ヒト抗体のVLに由来する少なくとも1つのFRLと、配列番号10、11および12のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRLとを含むL鎖、または
(iii)上記の(i)と(ii)の両方
を含むヒト化抗体またはその抗原結合性フラグメント。
【請求項1】
RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントであって、
(i)ヒト抗体のVHに由来する少なくとも1つのFRHと、RGD配列を免疫特異的に認識する非ヒト抗体のCDRHのうちの少なくとも1つに由来する少なくとも1つのCDRHとを含むH鎖;または
(ii)ヒト抗体のVLに由来する少なくとも1つのFRLと、RGD配列を免疫特異的に認識する非ヒト抗体のCDRLのうちの少なくとも1つに由来する少なくとも1つのCDRLとを含むL鎖;または
(iii)上記の(i)と(ii)の両方
を含むヒト化抗体またはその抗原結合性フラグメント。
【請求項2】
前記非ヒト抗体が、寄託アクセッション番号FERM BP−10440およびFERM BP−10441からなる群から選択されるハイブリドーマによって産生されたマウスモノクローナル抗体である、請求項1に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項3】
前記ヒト化抗体の少なくとも1つのCDRHが、配列番号1、2および3のアミノ酸配列からなる群から選択されるアミノ酸配列を含み、前記ヒト化抗体の少なくとも1つのCDRLが、配列番号4、5および6のアミノ酸配列からなる群から選択されるアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項4】
前記ヒト化抗体の少なくとも1つのCDRHが、配列番号7、8および9のアミノ酸配列からなる群から選択されるアミノ酸配列を含み、前記ヒト化抗体の少なくとも1つのCDRLが、配列番号10、11および12のアミノ酸配列からなる群から選択されるアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項5】
ヒト抗体の前記VHが、GenBankアクセッション番号X65891(配列番号13)のヌクレオチド配列によってコードされるアミノ酸配列に由来するアミノ酸配列を含み、ヒト抗体の前記VLが、GenBankアクセッション番号X72441(配列番号18)のヌクレオチド配列によってコードされるアミノ酸配列に由来するアミノ酸配列を含む、請求項2から4のいずれか一項に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項6】
前記H鎖が配列番号24のアミノ酸配列を含み、前記L鎖が配列番号26のアミノ酸配列を含む、請求項4に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項7】
前記H鎖が配列番号28のアミノ酸配列を含み、前記L鎖が配列番号30のアミノ酸配列を含む、請求項4に記載のヒト化抗体またはその抗原結合性フラグメント。
【請求項8】
配列番号24のアミノ酸配列をコードするヌクレオチド配列を含む単離された核酸分子。
【請求項9】
前記ヌクレオチド配列が配列番号23のヌクレオチド配列を有する、請求項8に記載の核酸分子。
【請求項10】
シグナルペプチドをコードするヌクレオチド配列をさらに含む、請求項8に記載の核酸分子。
【請求項11】
前記シグナルペプチドが配列番号32のアミノ酸配列を含む、請求項10に記載の核酸分子。
【請求項12】
配列番号26のアミノ酸配列をコードするヌクレオチド配列を含む単離された核酸分子。
【請求項13】
前記ヌクレオチド配列が配列番号25のヌクレオチド配列を有する、請求項12に記載の核酸分子。
【請求項14】
シグナルペプチドをコードするヌクレオチド配列をさらに含む、請求項12に記載の核酸分子。
【請求項15】
前記シグナルペプチドが配列番号34のアミノ酸配列を含む、請求項14に記載の核酸分子。
【請求項16】
配列番号28のアミノ酸配列をコードするヌクレオチド配列を含む単離された核酸分子。
【請求項17】
前記ヌクレオチド配列が配列番号27のヌクレオチド配列を有する、請求項16に記載の核酸分子。
【請求項18】
シグナルペプチドをコードするヌクレオチド配列をさらに含む、請求項16に記載の核酸分子。
【請求項19】
前記シグナルペプチドが配列番号36のアミノ酸配列を含む、請求項18に記載の核酸分子。
【請求項20】
配列番号30のアミノ酸配列をコードするヌクレオチド配列を含む単離された核酸分子。
【請求項21】
前記ヌクレオチド配列が配列番号29のヌクレオチド配列を有する、請求項20に記載の核酸分子。
【請求項22】
シグナルペプチドをコードするヌクレオチド配列をさらに含む、請求項20に記載の核酸分子。
【請求項23】
前記シグナルペプチドが配列番号38のアミノ酸配列を含む、請求項18に記載の核酸分子。
【請求項24】
1つまたは複数の調節エレメントに作動可能に連結している、請求項8もしくは23に記載の核酸分子またはその両方を含むベクター。
【請求項25】
請求項24に記載のベクターを含む単離された宿主細胞。
【請求項26】
ヒト化抗体またはその抗原結合性フラグメントを調製するための方法であって、請求項25に記載の宿主細胞をヒト化抗体またはその抗原結合性フラグメントが発現される条件下で培養すること、および発現されたヒト化抗体を収集することを含む方法。
【請求項27】
請求項1、2、6または7のいずれか一項に記載のヒト化抗体またはその抗原結合性フラグメントおよび薬学的に許容される担体を含む医薬組成物。
【請求項28】
RGDタンパク質に関連する障害または疾患を予防または治療するための方法であって、有効量の請求項6または7に記載のヒト化抗体またはその抗原結合性フラグメントを、それを必要とする対象に投与することを含む方法。
【請求項29】
RGDタンパク質に関連する障害または疾患をin vivoで診断するための方法であって、有効量の請求項1または2に記載のヒト化抗体またはその抗原結合性フラグメントを被検対象に投与することを含む方法。
【請求項30】
RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントであって、
(i)ヒト抗体のVHに由来する少なくとも1つのFRHと、配列番号1、2および3のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRHとを含むH鎖、または
(ii)ヒト抗体のVLに由来する少なくとも1つのFRLと、配列番号4、5および6のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRLとを含むL鎖、または
(iii)上記の(i)と(ii)の両方
を含むヒト化抗体またはその抗原結合性フラグメント。
【請求項31】
RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合性フラグメントであって、
(i)ヒト抗体のVHに由来する少なくとも1つのFRHと、配列番号7、8および9のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRHとを含むH鎖、または
(ii)ヒト抗体のVLに由来する少なくとも1つのFRLと、配列番号10、11および12のアミノ酸配列からなる群から選択されるアミノ酸配列を含む少なくとも1つのCDRLとを含むL鎖、または
(iii)上記の(i)と(ii)の両方
を含むヒト化抗体またはその抗原結合性フラグメント。
【図1】

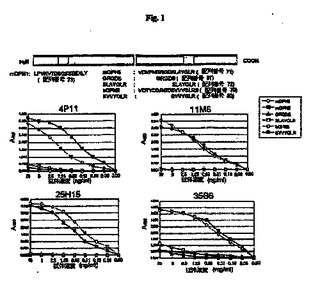
【図2】
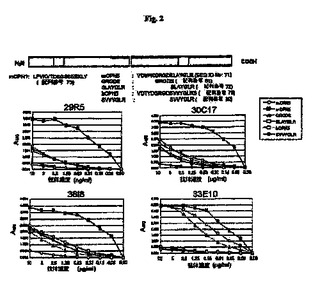
【図3】
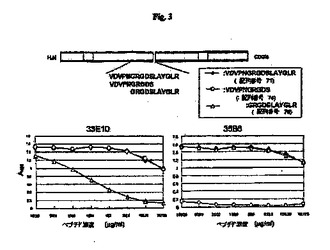
【図4】
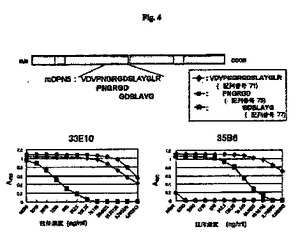
【図5】
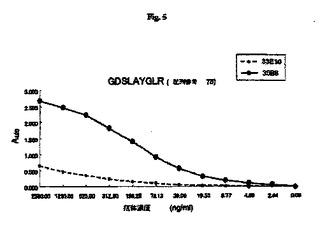
【図6】
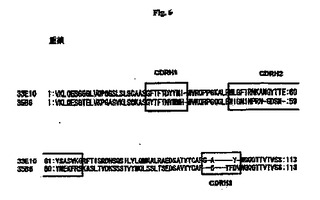
【図7】
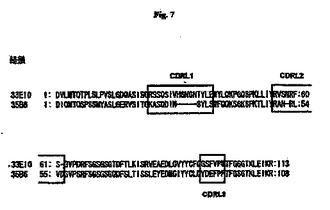
【図8】
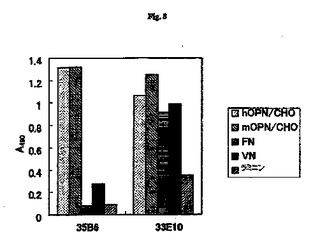
【図9】
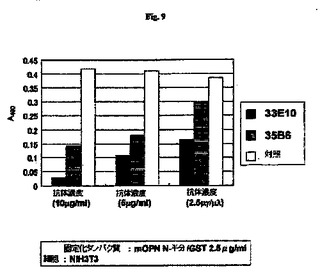
【図10】
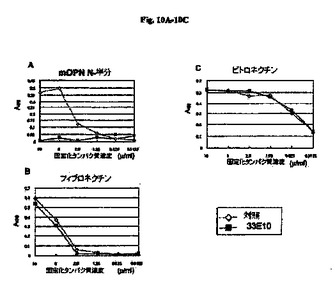
【図11】
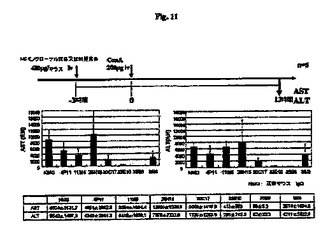
【図12】
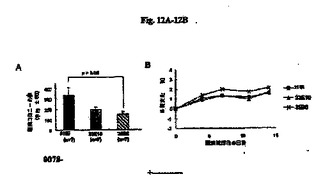
【図13】
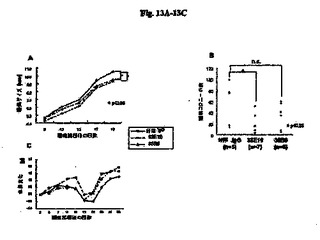
【図14】
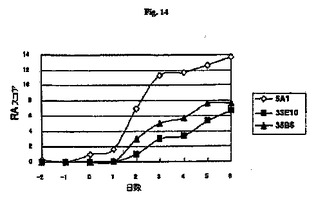
【図15】
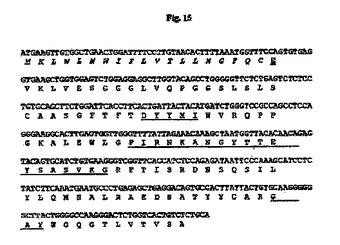
【図16】
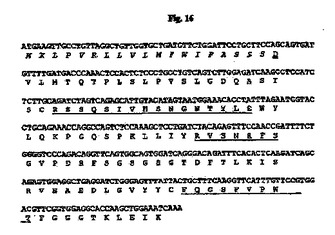
【図17】
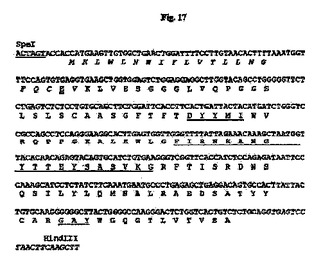
【図18】
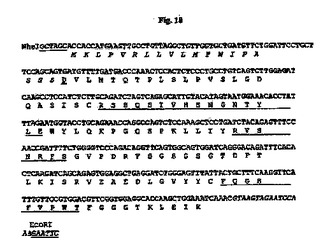
【図19】
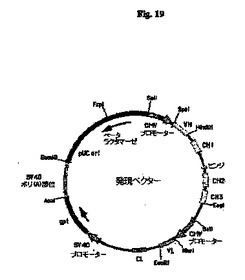
【図20】
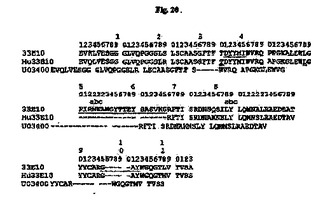
【図21】

【図22】

【図23】

【図24】

【図25】

【図26】
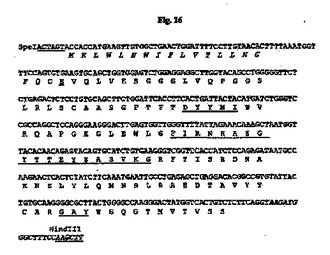
【図27】

【図28】
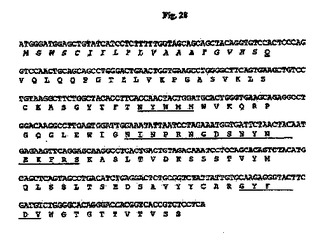
【図29】
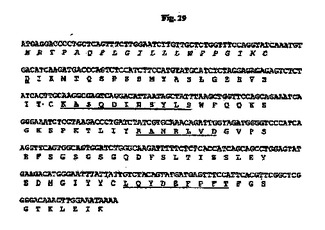
【図30】
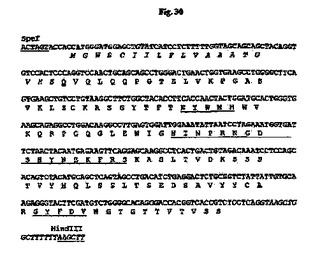
【図31】
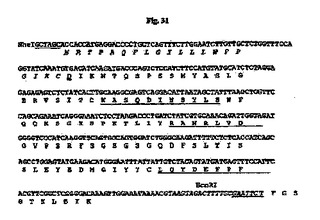
【図32】
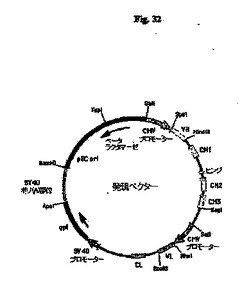
【図33】
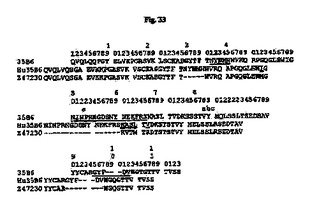
【図34】
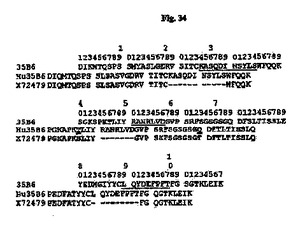
【図35】

【図36】

【図37】

【図38】

【図39】

【図40】
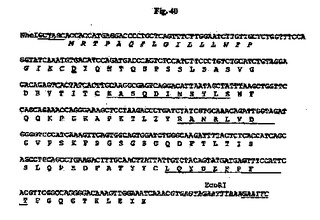
【図41】
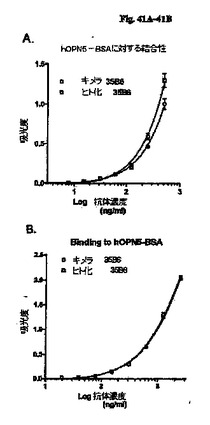
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【公表番号】特表2011−519547(P2011−519547A)
【公表日】平成23年7月14日(2011.7.14)
【国際特許分類】
【出願番号】特願2010−542452(P2010−542452)
【出願日】平成21年4月24日(2009.4.24)
【国際出願番号】PCT/JP2009/058604
【国際公開番号】WO2009/131256
【国際公開日】平成21年10月29日(2009.10.29)
【出願人】(501416243)株式会社ジーンテクノサイエンス (9)
【Fターム(参考)】
【公表日】平成23年7月14日(2011.7.14)
【国際特許分類】
【出願日】平成21年4月24日(2009.4.24)
【国際出願番号】PCT/JP2009/058604
【国際公開番号】WO2009/131256
【国際公開日】平成21年10月29日(2009.10.29)
【出願人】(501416243)株式会社ジーンテクノサイエンス (9)
【Fターム(参考)】
[ Back to top ]