
細胞外マトリックスタンパク質のアミノ酸配列RGDに特異的なヒト化抗体およびそれらの使用
本発明は、RGD配列を免疫特異的に認識するヒト化抗体を提供する。これらの抗体のいくつかはRGDタンパク質の生物学的機能を阻害し、それによって癌、例えば癌細胞の増殖および転移、および炎症疾患、例えば慢性関節リウマチ、骨関節炎、肝炎、子宮内膜症、気管支喘息、線維症、糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などを含めたRGDタンパク質と関係がある様々な障害または疾患に対する治療効果を示す。
【発明の詳細な説明】
【技術分野】
【0001】
本出願を通じて、いくつかの参照文献を本明細書で参照する。これらの参照文献の開示は、参照によってそれらの全容を本出願中にここで組み込む。
【0002】
本発明は、細胞外マトリックスタンパク質のアミノ酸配列RGD(Arg−Gly−Asp)を免疫特異的に認識するヒト化抗体、ならびに癌、炎症疾患、自己免疫疾患、感染疾患、骨疾患などを含めた様々な疾患または障害用のそれらの治療および診断用途に関する。
【背景技術】
【0003】
細胞接着は多細胞生物の生命を維持する際に重要な役割を果たす。多細胞生物の細胞接着は、細胞−細胞外マトリックス(本明細書では以後「ECM」と略す)接着および細胞間接着に分類される。細胞−ECM接着はインテグリンによって仲介され、かつ細胞間接着はカドヘリン、クラウジンおよびネクチンによって仲介されることが解明されている。
【0004】
インテグリンなどの膜貫通型接着タンパク質は細胞−ECM接着を構成する。インテグリンがα鎖とβ鎖のヘテロ二量体を形成することが報告されている。少なくとも18種(types)の鎖、8種のβ鎖および24種のαβヘテロ二量体がこれまでに同定され確認されている。それぞれの種類のインテグリンが、特異的リガンドを認識することが知られている。インテグリンを含めた膜貫通型接着タンパク質は、細胞接着以外に、ECMから細胞への細胞内シグナル伝達、ならびに増殖、移動性および分化の制御と関係がある(F.G Giancotti, et al., Science, 285, 1028-1032, 1999)。
【0005】
多くのタンパク質は、コラーゲン(I−XIX型など)、非コラーゲン性糖タンパク質(オステオポンチン(OPN)、ビトロネクチン、フィブロネクチン、フォンヴィレブランド因子、ラミニン、テナシン、フィブリノゲン、トロンボスポンジンなど)、エラスチンおよびプロテオグリカンに分類されるECMタンパク質として知られている。これらのECMタンパク質は対応するインテグリンと結合し、細胞内シグナル伝達経路を活性化して、細胞骨格系、移動性、増殖、分化などを制御する。ECMタンパク質結合インテグリンは、ECMタンパク質の種類(type)に応じて特異的シグナルを送ることにより、これらのシグナル活性化経路を制御する。RGD(アルギニン−グリシン−アスパラギン酸)配列は多くのECMタンパク質の細胞接着領域において一般に観察され、インテグリンと結合することによって様々な機能を示すようである。ECMタンパク質のRGD配列は薬剤標的であり得ると予想されており、いくつかの低分子化合物および人工ペプチドが提供されている。
【0006】
いくつかの型のインテグリン、α3β1インテグリン、α5β1インテグリン、α8β1インテグリン、ανβ1インテグリン、ανβ3インテグリン、ανβ5インテグリン、ανβ6インテグリン、ανβ8インテグリンなどは、RGD配列と結合することが知られている。α5β1インテグリンとその特異的リガンドフィブロネクチンの間の相互作用は、インテグリン仲介シグナル伝達の機構を調べるために明らかにされており、α5β1インテグリンは細胞接着および細胞移動だけでなく、細胞分化および細胞死亡率も制御することが報告される(S.M.Frisch et al., Curr.Opin.Cell Biol., 9, 701-706, 1997)。α5β1インテグリンは腫瘍細胞において高度に発現され、癌の悪性化と関係があることも示されている。それぞれのインテグリン仲介シグナルはECMタンパク質の結合に応じて異なる。例えば、増殖因子による刺激はフィブロネクチン結合内皮細胞の増殖を活性化するが、ラミニン−1結合内皮細胞の増殖は阻害する。さらに、ラミニン−10/11からα3β1インテグリンに伝わるシグナルはフィブロネクチンからα5β1インテグリンに伝わるシグナルと異なり、癌細胞の移動性を有意に高め(J.Gu et al., J.Biol.Chem., 276, 27090-27097, 2001)、血清飢餓によるアポトーシスを有意に回避する(J.Gu et al., J.Biol.Chem., 277, 19922-19928、2002)。RGD配列結合αvインテグリンの高発現は破骨細胞および新生血管において観察されており、RGD配列およびαvインテグリンの阻害は骨粗しょう症および癌に関する治療薬剤の標的として予想されている。α5β1インテグリンは腫瘍細胞において高度に発現され、癌の悪性化と関係があることが示されている。これらの発見から、抗α5β1インテグリン抗体(ボロシマブ)、抗α4インテグリン抗体(ナタリズマブ)、抗αvβ3インテグリン抗体(ビタキシン)が、インテグリンとECMタンパク質の間の相互作用を阻害するアンタゴニスト抗インテグリン抗体薬剤として開発されている。
【0007】
一方、コラーゲン、オステオポンチン(OPN)、ビトロネクチン、フィブロネクチン、フォンヴィレブランド因子、ラミニン、テナシン、フィブリノゲンおよびトロンボスポンジンなどのいくつかのECMタンパク質は、RGD配列を含むことが知られている。さらに、数種のウイルスおよび数種の細菌は、細胞と接着するためにRGD配列を有することが知られている。OPNは、骨中に多量に含有されるカルシウムとの結合性を有する酸性糖タンパク質である。OPNは、細胞接着、細胞移動、腫瘍形成、免疫応答および補体仲介細胞溶解において重要な役割を果たすことが報告される。OPNノックアウトマウスおよび抗OPN中和抗体の結果は、OPNが肝炎、慢性関節リウマチなどの自己免疫疾患、および癌の転移と関係があることを示す。したがって、ECMタンパク質と細胞の結合の阻害剤は、骨粗しょう症または癌の治療に使用することができることが予想される。したがって、インテグリンを標的化する前述のアンタゴニスト薬剤以外に、インテグリンの結合パートナーであるECMタンパク質を標的化するアンタゴニスト薬剤が開発されている。
【発明の概要】
【発明が解決しようとする課題】
【0008】
インテグリンとのRGD配列仲介相互作用を阻害する低分子、OPNに対する抗体、およびインテグリンに対する抗体などの薬剤は開発されているが、RGD配列を特異的に認識する抗体に関する報告は存在しない。RGD配列はECMタンパク質中の保存配列の1つなので、RGD配列を特異的に認識する抗体はヒトと治療モデル動物の両方において影響を有すると予想され、したがって治療剤の開発に非常に重要な活性成分として考えられる。したがって、RGD配列を特異的に認識する抗体が必要とされている。以前、本発明者らは、RGD配列を免疫特異的に認識しハイブリドーマクローン33E10および35B6(それぞれ寄託受託番号FERMBP−10440およびFERMBP−10441)によって産生されるマウスモノクローナル抗体を単離した。本明細書では、ハイブリドーマクローンの名称は、クローンによって産生されるモノクローナル抗体の名称として交互に使用する。全てのこれらのマウス抗RGD抗体はIgG1アイソタイプであった。これらのモノクローナル抗体は、オステオポンチンなどのECMタンパク質のRGD配列との結合によって、ECMと細胞の間でRGD配列によって仲介される結合に干渉することを観察した。したがって、これらの抗RGD抗体は、例えば癌、例えば癌細胞の増殖または転移、および炎症疾患、例えば慢性関節リウマチ、骨関節炎、感染疾患、肝炎、気管支喘息、線維症、糖尿病、動脈硬化症、多発性硬化症、肉芽腫、および炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患、骨粗しょう症などの、RGD配列関連疾患に対して治療または診断効果を示すと予想される。
【0009】
しかしながら、これらのモノクローナル抗体はマウス起源なので、ヒトにおけるそれらの免疫原性が原因で考えられる悪影響が、ヒトにおける診断または治療用途へのそれらの直接施用を妨げている。免疫原性を低減するために、本発明者らは、前記ヒト化抗体が由来する原型マウス抗RGD抗体によって示される活性に相当する、生物活性を有するヒト化抗体を調製した。
【課題を解決するための手段】
【0010】
したがって、本発明は、RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合断片を提供し、前記抗体は部分的に非ヒト起源に由来し部分的にヒト起源に由来する抗原結合領域を含む。本発明の一態様では、本発明のヒト化抗体またはその抗原結合断片は、33E10および35B6モノクローナル抗体などの非ヒト供給源(ドナー)に由来する相補性決定領域(本明細書では以後「CDR」と略す)、およびヒト供給源に由来するフレームワーク領域(本明細書では以後「FR」と略す)(アクセプター)を含む。前記ヒト化抗体またはその抗原結合断片は、RGD配列とそのリガンドの間の結合を阻害することができる。
【0011】
本発明の一態様では、RGD配列を免疫特異的に認識する前記ヒト化抗体またはその抗原結合断片は、(i)ヒトH鎖の可変領域(本明細書では以後「V領域」と略す)に由来する少なくとも1つのH鎖FR(本明細書では以後「FRH」と略す)、およびRGD配列を免疫特異的に認識する非ヒト抗体のCDRHの少なくとも1つに由来する少なくとも1つのH鎖CDR(本明細書では以後「CDRH」と略す)を含む重鎖(本明細書では以後「H鎖」と略す)、または(ii)ヒトL鎖のV領域に由来する少なくとも1つのL鎖FR(本明細書では以後「FRL」と略す)、およびRGD配列を免疫特異的に認識する非ヒト抗体のCDRLの少なくとも1つに由来する少なくとも1つのL鎖CDR(本明細書では以後「CDRL」と略す)を含む軽鎖(本明細書では以後「L鎖」と略す)、または前述の(i)および(ii)の両方を含む。
【0012】
本発明の好ましい態様では、本発明のヒト化抗体またはその抗原結合断片は、(i)VHを含むH鎖であって、前記VHが配列番号92、94または96のアミノ酸配列を含むH鎖、または(ii)VLを含むL鎖であって、前記VLが配列番号98または100のアミノ酸配列を含むL鎖、または(iii)前述の(i)および(ii)の両方を含む。本発明の別の態様では、本発明の前記ヒト化抗体またはその抗原結合断片は、配列番号91、92または93のヌクレオチド配列によってコードされるアミノ酸配列を含み、前記VLが配列番号97または99のヌクレオチド配列によってコードされるアミノ酸配列を含む。
【0013】
本発明の好ましい態様では、前記VHが配列番号91、92または93のヌクレオチド配列によってコードされるアミノ酸配列を含み、前記VLが配列番号97または99のヌクレオチド配列によってコードされるアミノ酸配列を含む。
【0014】
本発明の別の態様では、前記VHが配列番号94のアミノ酸配列を含み、前記VLが配列番号100のアミノ酸配列を含む。
【0015】
本発明の別の態様では、前記H鎖が配列番号25または27のアミノ酸配列を含み、前記L鎖が配列番号29または31のアミノ酸配列を含む。
【0016】
本発明のさらなる態様では、前記H鎖が配列番号25のアミノ酸配列を含み、前記L鎖が配列番号31のアミノ酸配列を含む。
【0017】
本発明は、RGD配列を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合断片のH鎖またはL鎖、または両方をコードするヌクレオチド配列を含むベクター、例えば発現ベクターをさらに提供する。このようなベクターでは、本発明のヌクレオチド配列は、1つまたは複数の制御エレメントに作動可能に連結することができる。本発明のヌクレオチド配列は、CDRが由来する非ヒトドナー抗体に固有のシグナルペプチド、または異種起源のシグナルペプチドをコードするヌクレオチド配列を含むことができる。
【0018】
さらに、本発明は、本発明の核酸分子を含むベクターを含む、本発明の核酸分子を含む宿主細胞を提供する。本発明の一態様では、本発明は、本発明のヒト化H鎖をコードする第一の核酸分子および本発明のヒト化L鎖をコードする第二の核酸分子を含む単離宿主細胞を提供し、前記第一および第二の核酸分子は、生物学的に機能性がある本発明のヒト化抗体またはその抗原結合断片が発現されるような形式で、それぞれ制御エレメントに作動可能に連結する。
【0019】
本発明は、本発明のヒト化抗体またはその抗原結合断片を調製するための方法であって、ヒト化抗体またはその抗原結合断片が発現されるような条件下で本発明の宿主細胞を培養すること、および生成したヒト化抗体を回収することを含む方法をさらに提供する。
【0020】
本発明は、本発明のヒト化抗体またはその抗原結合断片の少なくとも1つを含む組成物をさらに提供する。さらに本発明は、本発明のヒト化抗体またはその抗原結合断片の少なくとも1つ、および薬学的に許容される担体を含む、RGDタンパク質と関係がある障害または疾患を予防または治療するための医薬組成物を提供する。前記組成物のいずれかは、障害または疾患を相加的または相乗的に改善することができる別の活性化合物をさらに含むことができる。前記活性化合物には、抗炎症性化合物、化学療法化合物などがあるが、これらだけには限られない。前記活性化合物は、低分子化合物および抗体またはその抗原結合断片、ヒトα4インテグリン特異的抗体またはヒトα9インテグリン特異的抗体なども含む。
【0021】
別の態様では、本発明は、RGDタンパク質と関係または関与がある障害または疾患を予防または治療するための方法を提供し、前記方法は本発明のヒト化抗体またはその抗原結合断片の少なくとも1つを予防または治療有効量、その必要性がある対象に投与することを含む。このような用途のため、本発明のヒト化抗体またはその抗原結合断片を、ヒト化抗体またはその抗原結合断片の生物学的効果を高める治療成分と結合させることが可能である。このような治療成分の例には、別の抗体、細胞増殖抑制または細胞致死性である細胞毒素、放射性元素、および/または抗炎症剤、抗生物質を含めた他の治療剤などがある。
【0022】
さらに別の態様では、本発明は、RGDタンパク質と関係または関与がある対象中の障害または疾患を診断するための方法を提供し、前記方法は診断有効量の本発明のヒト化抗体またはその抗原結合断片を、検査する対象に投与することを含む。このような診断用途のため、本発明のヒト化抗体を、放射性元素などの検出可能マーカーで標識することができる。
【0023】
3.1.定義
本明細書で使用する用語「抗体」は、所望の抗原またはRGD配列などの所望の配列と免疫特異的に結合することができる抗体分子を指し、これは抗体分子全体を包含し、抗体の抗原結合断片を含んだ抗体の断片を包含することができる。
【0024】
本明細書で使用する用語「抗原結合断片」は、標的ポリペプチド、タンパク質または配列、特にRGD配列と免疫特異的に結合する能力を保持する抗体の任意の断片を指し、これは単鎖抗体、Fab断片、F(ab’)2断片、ジスルフィド結合Fvs、および標的ポリペプチド、タンパク質または配列と特異的に結合するVLおよび/またはVHまたはCDRのいずれかを含有する断片を含む。したがって、ヒト化抗体のこのような抗原結合断片は、部分または完全長ヒト定常領域を含むかまたは含まない可能性がある。前に記載した抗体断片を得るための様々な方法は当技術分野でよく知られている。
【0025】
本明細書で使用する用語「免疫特異的に認識する」は、標的ポリペプチド、タンパク質または配列、特にヒトRGD配列と特異的に結合する抗体またはその抗原結合断片の能力を指す。このような抗体が、他のポリペプチドまたはタンパク質と非特異的に結合することはない。しかしながら、標的ポリペプチドまたはタンパク質(例えば、RGDタンパク質)と免疫特異的に結合する抗体またはその抗原結合断片は、他の抗原と交差反応する可能性がある。例えば、ヒトRGDタンパク質を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合断片は、マウスRGDタンパク質と交差反応する可能性がある。RGDタンパク質を免疫特異的に認識する抗体またはその抗原結合断片は、他の抗原と交差反応しないことが好ましい。
【0026】
本明細書で使用する用語「ヒト供給源由来」または「非ヒト供給源由来」は、そのアミノ酸配列がそれぞれヒト抗体または非ヒト抗体の対応する部分に由来する抗体部分を指す。
【0027】
本明細書で使用する用語「アクセプター配列」は、通常非ヒト抗体であるドナー抗体由来のCDRに関するアクセプターとして働くヒト抗体VHまたはVL由来のFRの、ヌクレオチド配列またはアミノ酸配列を指す。
【0028】
本発明を例示する目的で、図1〜27は現在好ましい形態を反映する。しかしながら、本発明が図1〜27中に示す通りの形に限られないことは理解される。
【図面の簡単な説明】
【0029】
【図1】推定アミノ酸配列と共に示すマウス33E10VHcDNAのヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VHのN末端アミノ酸残基(E)には二重下線を引く。Kabat et al.(Sequences of Proteins of Immunological Interests, Fifth edition, NIH公開番号91−3242、米国厚生省、1991)の定義に従うCDR配列には下線を引く。
【図2】推定アミノ酸配列と共に示すマウス33E10VLcDNAのヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。
【図3】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接する設計33E10VH遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VHのN末端アミノ酸残基(E)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図4】推定アミノ酸配列と共に示すNheIおよびEcoRI部位(下線)と隣接する設計33E10VL遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図5】キメラおよびヒト化33E10抗体用の発現ベクター(集合的に「発現ベクター」)の概略構造を示す図である。頂上部のSalI部位から時計回りに進み、プラスミドは、抗体重鎖遺伝子の転写を開始するためのヒトサイトメガロウイルス(CMV)主要即時初期プロモーターおよびエンハンサー(CMVプロモーター)で始まる重鎖転写単位を含有する。CMVプロモーターに、VHエクソン、CH1を含むヒトγ−1重鎖定常領域を含有するゲノム配列、ヒンジ、介在イントロンを有するCH2およびCH3エクソン、およびCH3エクソンの後にポリアデニル化部位が続く。重鎖遺伝子配列の後、軽鎖転写単位が、CMVプロモーターで始まり、VLエクソン、およびそれに先行するイントロンの一部分を有するヒトkappa鎖定常領域エクソン(CL)を含有するゲノム配列、およびCLエクソン後のポリアデニル化部位が続く。次いで軽鎖遺伝子に、SV40初期プロモーター(SV40プロモーター)、大腸菌(E.coli)キサンチングアニンホスホリボシルトランスフェラーゼ遺伝子(gpt)、およびSV40ポリアデニル化部位(SV40ポリ(A)部位)を含有するセグメントが続く。最後に、プラスミドは、細菌複製起点(pUCori)およびβ−ラクタマーゼ遺伝子(βラクタマーゼ)を含むプラスミドpUC19の一部分を含有する。関連制限酵素部位の位置をこの図中に示す。
【図6】33E10VH、ヒト化33E10(Hu33E10)VHおよびヒトアクセプターU03400(GenBank受託番号)のアミノ酸配列のアラインメントを示す図である。アミノ酸残基は一文字コードで示す。配列の上の数字はKabat et al.(1991)に従う位置を示す。Kabat et al.(Sequences of Proteins of Immunological Interests, Fifth edition, NIH公開番号91−3242、米国厚生省、1991)によって定義されたCDR配列には下線を引く。二重下線を引いた残基はCDRと接触したと予想し、マウス残基はヒト化型でこれらの位置に保持された。U03400中のCDR残基はこの図中では省略する。
【図7】33E10VL、ヒト化33E10(Hu33E10)VLおよびヒトアクセプターX72452(GenBank受託番号)のアミノ酸配列のアラインメントを示す図である。アミノ酸残基は一文字コードで示す。配列の上の数字はKabat et al.(1991)に従う位置を示す。Kabat et al.(1991)によって定義したCDR配列には下線を引く。X72452中のCDR残基はこの図中では省略する。
【図8】Hu33E10VH1遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図9】Hu33E10VL1遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図10】Hu33E10VH1遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印はそれぞれのオリゴヌクレオチドの位置および方向(5’から3’)を示す。アミノ酸残基は一文字コードで示す。
【図11】Hu33E10VL1遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印はそれぞれのオリゴヌクレオチドの位置および方向(5’から3’)を示す。アミノ酸残基は一文字コードで示す。
【図12】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接するHu33E10VH1遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VHのN末端アミノ酸残基(E)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図13】推定アミノ酸配列と共に示すNheIおよびEcoRI部位(下線)と隣接するHu33E10VL1遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図14】推定アミノ酸配列と共に示すNheIおよびEcoRI部位(下線)と隣接するHu33E10VL6遺伝子のヌクレオチド配列を示す図である。シグナルペプチドコード領域中のスプライシングドナー部位を排除するためのサイレント突然変異には二重下線を引く。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図15A−B】hOPN5−BSAとキメラおよびヒト化33E10IgGl/κ抗体の結合のELISA分析を示す図である。精製Ch33E10、Hu33E10−VHl/VL6、Hu33E10−VH5/VL6およびHu33E10−VH7/VL6(図15A)ならびにCh33E10およびHu33E10−VH7.8/VL6(図15B)を、100μg/mlで始めて連続2倍希釈で、hOPN5−BSAとの結合に関して様々な濃度において試験した。実験は二連で実施した。
【図16】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接するHu33E10VH5遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。二重下線を引いたアミノ酸残基はVH1との違いを示す。
【図17】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接するHu33E10VH7遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。二重下線を引いたアミノ酸残基はVH1との違いを示す。
【図18】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接するHu33E10VH7.8遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。二重下線を引いたアミノ酸残基はVH1との違いを示す。
【図19】33E10VL、Hu33E10VLv2およびヒトアクセプターM29467(GenBank受託番号)のアミノ酸配列のアラインメントを示す図である。アミノ酸残基は一文字コードで示す。配列の上の数字はKabat et al.(1991)に従う位置を示す。Kabat et al.(1991)によって定義したCDR配列には下線を引く。二重下線を引いた残基はCDRと接触したと予想し、マウス残基はヒト化型でこれらの位置に保持された。M29467中のCDR残基はこの図中では省略する。
【図20】推定アミノ酸配列と共に示すNheIおよびEcoRI部位(下線)と隣接するHu33E10VLv2遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図21A−B】hOPN5−BSAとCh33E10およびHu33E10−VH7/VLv2IgGl/κ抗体の結合のELISA分析を示す図である。精製Ch33E10およびHu33E10−VH7/VLv2抗体を、200μg/ml(図21A)または100μg/ml(図21B)で始めて連続2倍希釈で、hOPN5−BSAとの結合に関して様々な濃度において試験した。二連での2つの独立した実験の結果を示す(図21Aおよび21B)。
【図22】Hu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチドの配列を示す図である。
【図23】推定アミノ酸配列と共に示すNS0−Hu33E10−4およびNS0−Hu33E10−6において発現されるVH7およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。終結コドンは「・」によって示す。
【図24】推定アミノ酸配列と共に示すNS0−Hu33E10−5において発現されるVH7.8およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。終結コドンは「・」によって示す。
【図25】推定アミノ酸配列と共に示すNS0−Hu33E10−4およびNS0−Hu33E10−5において発現されるVL6およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。終結コドンは「・」によって示す。
【図26】推定アミノ酸配列と共に示すNS0−Hu33E10−6において発現されるVLv2およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。終結コドンは「・」によって示す。
【図27】ヒト化抗OPN抗体が、hOPN5−BSAとMDA−MB−435Sの接着を阻害したことを示す図である。MDA−MB−435S(4×104細胞/ウェル)を、様々な濃度の抗体、ヒトIgG、Ch33E10、EJ3−1−1(Hu33E10−VH7.8/VL6)、R3−1(Hu33E10−VH7/VL6)、SP2−1(Hu33E10−VH7/VLv2)、およびm33E10の存在下において、hOPN5−BSA(2mg/ml)でプレコーティングした96ウェルプレートに接着させた。データは三連の実験の平均として表す。
【発明を実施するための形態】
【0030】
5.1 RGD配列に対する抗体の調製
RGD配列を免疫特異的に認識する抗体は、当技術分野で知られている任意の適切な方法によって作製することができる。
【0031】
RGDタンパク質またはペプチドは本発明の細胞接着「RGD」配列(本明細書では以後「RGDペプチド」と略す)を含み、または(1)RGDタンパク質を発現するヒトECMに由来、またはこれらのECMが存在する全組織に由来してよく、(2)細菌、酵母、動物細胞などの細胞系などへのトランスフェクションによるRGDタンパク質またはRGDペプチドをコードするDNA(好ましくはcDNA)の発現によって得られる組換えタンパク質またはペプチドであってよく、または(3)合成タンパク質またはペプチドであってよい。
【0032】
本発明で抗原として使用するRGDペプチドは、免疫処置によってRGD配列に対する抗体を産生することができる。RGDペプチドは、マウスECMタンパク質の細胞接着配列であるRGDペプチドアミノ酸配列CVDVPNGRGDSLAYGLR(配列番号71)を含む。RGDタンパク質またはRGDペプチドは、OPN、ビトロネクチン、フィブロネクチン、フォンヴィレブランド因子、コラーゲン、ラミニン、テナシン、フィブリノゲン、トロンボスポンジン、およびこれらの断片を含むRGDなどを含む。アミノ酸の置換、欠失、修飾および付加などの人為的または天然変異を、タンパク質またはペプチドがRGD−配列を含む限り、前記タンパク質または前記ペプチドに施すことができる。変異体タンパク質またはペプチドは、複数のアミノ酸、好ましくは1〜10個のアミノ酸、およびより好ましくは1〜数個(例えば、1〜5個)のアミノ酸が置換、欠失、修飾、付加または挿入されたアミノ酸配列を含むことができる。
【0033】
本明細書では、RGDペプチドは、少なくとも約5個のアミノ酸、好ましくは約5〜50個のアミノ酸、およびより好ましくは約10〜20個のアミノ酸を含む。本発明における抗原としてのRGDタンパク質またはRGDペプチドは、化学合成法、細胞培養法、遺伝子組換え法およびその適切な変形などの、当技術分野でよく知られている方法を使用することによって産生することができる。例えば、RGDペプチドは、適切にプロテアーゼでECMタンパク質を切断することによって得ることができる。RGDタンパク質またはRGDペプチドは、マウス、ラット、ウサギ、ブタ、ウシ、サルおよびヒトなどの哺乳動物に由来してよい。当技術分野でよく知られている任意の方法を、抗RGD抗体の調製に使用することができるRGDタンパク質またはRGDペプチドの調製に使用することができる。
【0034】
変異体ポリペプチドを生成するための方法の例には、合成オリゴヌクレオチド部位特異的突然変異誘発法(ギャップデュプレックス法)、硝酸塩または亜硝酸塩を用いた処理によるアトランダムな点突然変異の導入を含む点突然変異誘発法、Bal31酵素、または他の酵素を用いた欠失変異体の調製を含む方法、カセット突然変異誘発法、リンカースキャニング法、ミスインコーポレーション法、ミスマッチプライマー法、DNAセグメント合成法などがある。
【0035】
RGDペプチドは、チログロビン、キーホールリンペットヘモシアニン(KLH)、ウシ血清アルブミン(BSA)、オボアルブミン(OVA)またはウシグロブリン、好ましくはチログロビンなどの、他の生体高分子と結合させることが可能である。生体高分子にRGDペプチドを結合させるための方法は、活性エステル基およびマレイミド基を有する結合試薬などのカップリング試薬を使用することによって(活性エステル基はタンパク質またはペプチドのアミノ基と結合し、マレイミド基はタンパク質またはペプチドのチオール基と結合する、S.yoshirake et al., Eur.J.Biochem., 101, 395-399, 1979)、混成無水物法(B.F.Erlanger et al., J.Biol. Chem., 234, 1090-1094, 1954)を使用することによって、または活性エステル法(A.E.Karu et al., J.Agric.Food Chem., 42, 301-309, 1994)を使用することによって実施することができる。生体高分子にRGDペプチドを結合させるための方法は、カップリング試薬を使用することによって実施することが好ましい。
【0036】
抗原として、RGDタンパク質またはRGDペプチドを過剰発現する細胞自体を使用することもできる。RGDタンパク質またはRGDペプチドを過剰発現する細胞は、当技術分野でよく知られている組換えDNA技術によって調製することができる。
【0037】
前に記載したように調製した適切な抗原を使用して、RGD配列に特異的な抗体は、当技術分野でよく知られている様々な方法によって調製することができる。RGD配列に対するポリクローナル抗体は、当技術分野でよく知られている様々な手順によって産生することができる。例えば、対象の抗原を、ウサギ、マウス、ラットなどだけには限られないが、これらを含めた様々な宿主動物に投与して、抗原に特異的なポリクローナル抗体を含有する抗血清の産生を誘導することができる。フロイント(完全および不完全)アジュバント、水酸化アルミニウムなどのミネラルゲル、リゾレシチン、プルロニックポリマー、ポリアニオン、ペプチド、油エマルジョン、キーホールリンペットヘモシアニン、ジニトロフェノールなどの表面活性物質、およびBCG(カルメットゲラン桿菌(Bacille Calmette-Guerin)およびコリネバクテリウムパルバム(Corynebacterium parvum)などのヒトにおそらく有用なアジュバントだけには限られないが、これらを含めた様々なアジュバントを使用して、宿主の種類に応じて免疫応答を増大することができる。このようなアジュバントも当技術分野でよく知られている。
【0038】
モノクローナル抗体は、ハイブリドーマ、組換えおよびファージディスプレイ技術、またはこれらの組合せの使用を含めた、当技術分野で知られている広く様々な技術を使用することによって調製することができる。例えば、モノクローナル抗体は、当技術分野で知られており例えば(いずれもその全容を参照によって本明細書に組み込む)Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed.1988); Hammerling, et al., Monoclonal Antibodies and T-Cell Hybridomas, pp.563-681 (Elsevier, N.Y, 1981)中で教示された技法を含めた、ハイブリドーマ技法を使用することによって産生することができる。本明細書で使用する用語「モノクローナル抗体」は、ハイブリドーマ技術によって産生される抗体に限られない。用語「モノクローナル抗体」は単一クローンに由来する抗体を指し、それが産生される方法に限られず、任意の真核生物、原核生物またはファージクローンを含む。
【0039】
ハイブリドーマ技術を使用して特異的抗体を産生しスクリーニングするための方法は通常のことであり、当技術分野でよく知られている。非制限的な例では、対象の抗原またはこのような抗原を発現する細胞でマウスを免疫処置することができる。免疫応答を検出した後、例えば、抗原に特異的な抗体をマウス血清中で検出した後、マウスの脾臓を採取し脾臓細胞を単離した。次いで脾臓細胞は、よく知られている技法によって任意の適切なミエローマ細胞(例えば、P3U1、P3X63−Ag8、P3X63−Ag8−U1、P3NS1−Ag4、SP2/0−Ag14、P3X63−Ag8−653など)と融合させる。ハイブリドーマを選択し、限界希釈によってクローニングする。次いでハイブリドーマクローンを、当技術分野で知られている方法によって、抗原と結合することができる抗体を分泌する細胞に関してアッセイする。高レベルの抗体を一般に含有する腹水は、陽性ハイブリドーマクローンをマウスに腹腔内接種することによって生成することができる。
【0040】
特異的エピトープを認識する抗体断片は知られている技法によって作製することができる。例えば、FabおよびF(ab’)2断片は、パパイン(Fab断片産生用)またはペプシン(F(ab’)2断片産生用)などの酵素を使用して、免疫グロブリン分子のタンパク質分解切断によって産生することができる。F(ab’)2断片は完全L鎖、およびH鎖のV領域、CH1領域およびヒンジ領域を含有する。
【0041】
本発明の抗体またはその抗原結合断片は、抗体の合成に関して当技術分野で知られている任意の方法によって、特に化学合成によって、または好ましくは組換え発現技法によって産生することができる。
【0042】
抗体をコードするヌクレオチド配列は、当業者が入手可能な任意の情報から得ることができる(すなわちGenbank、文献から、または通常のクローニングおよび配列分析によって)。特定の抗体またはそのエピトープ結合断片をコードする核酸を含有するクローンが入手可能ではないが、抗体分子またはそのエピトープ結合断片の配列が知られている場合、免疫グロブリンをコードする核酸を、配列の5’末端とハイブリダイズ可能な合成プライマーを使用したPCR増幅によって、または特定の遺伝子配列に特異的なオリゴヌクレオチドプローブを使用したクローニングによって、化学合成して、または適切な供給源(例えば、抗体cDNAライブラリー、またはそこから作製したcDNAライブラリー、または任意の組織、もしくは抗体発現用に選択したハイブリドーマ細胞などの抗体を発現する細胞から単離した核酸、好ましくはポリA+RNA)から得て、例えば、抗体をコードするcDNAライブラリーからcDNAクローンを同定することができる。PCRにより作製した増幅核酸遺伝子は、当技術分野でよく知られている任意の方法を使用して、次いで複製可能クローニングベクターにクローニングすることができる。
【0043】
5.2 組換え抗体の調製
抗体のヌクレオチド配列は、ヌクレオチド配列の操作に関して当技術分野でよく知られている方法、例えば組換えDNA技法、部位特異的突然変異誘発法、PCRなどを使用することによって操作することができる(例えば、いずれもその全容を参照によって本明細書に組み込む、Sambrook et al., 上記、およびAusubel et al., eds., 1998, Current Protocols in Molecular Biology, John Wiley & Sons, NY中に記載された技法を参照)。抗体にはエピトープ結合ドメイン領域または任意の部分にアミノ酸の置換、欠失および/または挿入などの突然変異を導入して、生物活性を向上または低減することができる。
【0044】
抗体をコードするヌクレオチド配列を含有する発現ベクターは、抗体またはその抗原結合断片の組換え発現に使用することができる。抗体またはその抗原結合断片の産生用の、抗体分子、抗体またはその一部分のH鎖および/またはL鎖をコードするヌクレオチド配列を含むベクターは、前のセクションで論じたように当技術分野でよく知られている技法を使用して組換えDNA技術によって産生することができる。当業者によく知られている方法を使用して、抗体またはその抗原結合断片、コード配列、ならびに適切な転写および翻訳制御シグナルを含有する発現ベクターを構築することができる。これらの方法は、例えばin vitro組換えDNA技法、合成技法、およびin vivo遺伝子組換えを含む。VH、VL、VHとVLの両方、VHおよび/またはVLの抗原結合断片、または抗体の1つまたは複数のCDRをコードするヌクレオチド配列は、発現用のこのようなベクターにクローニングすることができる。このような配列は、原型抗体に固有または異種であり得るシグナルペプチドをコードするポリヌクレオチドと融合することができる。次いでこのように調製した発現ベクターを、抗体の発現に適した宿主細胞に導入することができる。したがって本発明は、RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合断片をコードするポリヌクレオチドを含有する宿主細胞を含む。
【0045】
宿主細胞は本発明の2つの発現ベクターでコトランスフェクトすることができ、この場合第一のベクターはH鎖由来ポリペプチドをコードし第二のベクターはL鎖由来ポリペプチドをコードする。2つのベクターは、H鎖およびL鎖ポリペプチドの等しい発現を可能にする同一の選択可能マーカー、または両プラスミドの維持を確実にするための異なる選択可能マーカーを含有することができる。あるいは、H鎖およびL鎖ポリペプチドの両方をコードし発現することができる、1つのベクターを使用することができる。H鎖およびL鎖のコード配列はcDNAまたはゲノムDNAを含むことができる。
【0046】
本発明の他の態様では、当技術分野で知られている様々なファージディスプレイ法を使用して抗体を作製することもできる。ファージディスプレイ法では、機能的抗体ドメインを、それらをコードするポリヌクレオチド配列を保持するファージ粒子の表面上に提示する。本発明の特定の態様では、このようなファージを利用して、レパートリーまたはコンビナトリアル抗体ライブラリー(例えば、ヒトまたはマウス)から発現されるFabおよびFvまたはジスルフィド結合安定化Fvなどの抗原結合ドメインを提示することができる。対象の抗原と結合する抗原結合ドメインを発現するファージは、抗原を用いて、例えば標識抗原または固体表面もしくはビーズに結合もしくは捕捉した抗原を使用して、選択または同定することができる。これらの方法中で使用するファージは、典型的にはfdおよびM13を含めた繊維状ファージである。抗原結合ドメインは、ファージ遺伝子IIIまたは遺伝子VIIIタンパク質のいずれかとの組換え融合タンパク質として発現される。本発明の免疫グロブリン、またはその断片を作製するために使用することができるファージディスプレイ法の例には、その各々を参照によってそれらの全容を本明細書に組み込む、Brinkman et al., J.Immunol.Methods, 182: 41-50, 1995; Ames et al., J.Immunol.Methods, 184: 177-186, 1995; Kettleborough et al., Eur.J.Immunol., 24: 952-958, 1994; Persic et al., Gene, 187: 9-18, 1997; Burton et al., Advances in Immunology, 57: 191-280, 1994、PCT出願No.PCT/GB91/01134、PCT出願WO90/02809、WO91/10737、WO92/01047、WO92/18619、WO93/11236、WO95/15982、WO95/20401、ならびに米国特許第5,698,426号、同第5,223,409号、同第5,403,484号、同第5,580,717号、同第5,427,908号、同第5,750,753号、同第5,821,047号、同第5,571,698号、同第5,427,908号、同第5,516,637号、同第5,780,225号、同第5,658,727号、同第5,733,743および同第5,969,108号中に開示された方法がある。
【0047】
前述の参照文献中に記載されたように、ファージ選択後、例えば以下で詳細に記載するように、ファージ由来の領域をコードする抗体を単離し、これらを使用してヒト抗体、または任意の他の望ましい断片を含めた完全抗体を作製し、哺乳動物細胞、昆虫細胞、宿主細胞、酵母、および細菌を含めた任意の望ましい宿主中で発現させることが可能である。例えば、(その各々を参照によってそれらの全容を本明細書に組み込む)PCT公開WO92/22324、Mullinax et al., BioTechniques, 12(6): 864-869, 1992およびSawai et al., AJRI, 34: 26-34, 1995およびBetter et al., Science, 240: 1041-1043, 1988中に開示された方法などの当技術分野で知られている方法を使用して、Fab、Fab’およびF(ab)2断片を組換えによって産生するための技法を利用することもできる。単鎖Fvおよび抗体を産生するために使用することができる技法の例には、米国特許第4,946,778号および同第5,258,498号、Huston et al., Methods in Enzymology, 203: 46-88, 1991; Shu et al., PNAS, 90: 7995-7999, 1993およびSkerra et al., Science, 240: 1038-1040, 1988中に記載された技法がある。
【0048】
本発明の抗体分子を前に記載した任意の方法によって産生した後、次いでそれを、免疫グロブリン分子の精製に関して当技術分野で知られている任意の方法によって、例えばクロマトグラフィー(例えば、イオン交換、親和性、特にプロテインAまたはプロテインG精製後の特異的抗原に対する親和性によって、およびサイズ排除カラムクロマトグラフィー)、遠心分離、差次的溶解度によって、またはタンパク質を精製するための、任意の他の標準的技法によって精製することができる。さらに、本発明の抗体またはその断片を、本明細書に記載する異種ポリペプチド配列または当技術分野で知られている他の配列と融合させて、精製を容易にすることができる。
【0049】
ヒトにおける抗体のin vivo用途およびin vitro検出アッセイを含めた、いくつかの用途に関して、キメラ、ヒト化、またはヒト抗体を使用することが好ましい可能性がある。キメラ抗体およびヒト化抗体は以下のセクション5.3中で詳細に論じる。
【0050】
他の化合物または異種ポリペプチドと融合または結合した抗体は、in vitroイムノアッセイにおいて、精製法(例えば、親和性クロマトグラフィー)、およびin vivo治療または診断用途において使用することができる。例えば、参照によってそれらの全容を本明細書に組み込む、PCT公開番号WO93/21232、EP439,095、Naramura et al., Immunol.Lett., 39: 91-99, 1994、米国特許第5,474,981号、Gillies et al., PNAS, 89: 1428-1432, 1992、およびFell et al., J.Immunol., 146: 2446-2452, 1991を参照。例えば、知られている方法または市販のキット(例えば、ビオチン標識、FITC標識、APC標識)を使用して様々な方法で、抗体を標識することができる。別の例として、in vivoでの抗体の生物学的効果を高める治療成分に、抗体を結合させることが可能である。このような治療成分の例には、別の抗体、細胞増殖抑制または細胞致死性である細胞毒素、放射性元素、および/または抗炎症剤、抗生物質を含めた他の治療剤などがある。本発明では、ヒト化抗RGD抗体を別の抗体と結合させて二重特異性抗体を形成することが可能である。別の例として、本発明のヒト化抗体を、in vivo診断用途に、放射性元素などの検出可能マーカーで標識することができる。
【0051】
5.3 キメラおよびヒト化抗体
キメラ抗体は、抗体の異なる部分が異なる動物種に由来する分子、マウスモノクローナル抗体に由来するV領域およびヒト免疫グロブリン由来の定常領域を有する抗体などである。キメラ抗体を産生するための方法は当技術分野で知られている。例えば、参照によってそれらの全容を本明細書に組み込む、Morrison, Science, 229: 1202, 1985; Oi et al., BioTechniques, 4: 214 1986; Gillies et al., J. Immunol.Methods, 125: 191-202, 1989、米国特許第5,807,715号、同第4,816,567号、および同第4,816,397号を参照。
【0052】
ヒト化抗体は、望ましい抗原と結合し、非ヒト種由来の1つまたは複数のCDRおよびヒト免疫グロブリン分子由来の1つまたは複数のFRを含有するV領域を含む分子である。非ヒト抗体をヒト化するための典型的な方法は、いずれもその全容を参照によって本明細書に組み込む、Queen et al., 1989, Proc.Natl.Acad. Sci.USA86: 10029-10033および米国特許第5,585,089号および同第5,693,762号による参照文献、Riechmann et al., Nature, 332: 323, 1988による参照文献、およびTsurushita et al., Methods 36: 69-83, 2005による参照文献などの、様々な参照文献中に記載されている。例えば、Tsurushita et al.(2005、上記、本明細書では以後「Tsurushita」)による参照文献は、Queen et al.(1989、上記)により本来開発された抗体−ヒト化法に基づく、マウスモノクローナル抗体をヒト化するための実用的で有益なプロトコールを与える。Tsurushita中に開示された一般的プロトコールは以下で簡潔に要約する。
【0053】
5.3.1 ヒト化抗体を調製するための一般的プロトコール
マウスV遺伝子のクローニングおよび配列決定
様々な方法が、標的マウスモノクローナル抗体のVHおよびVLをコードするcDNAをクローニングするのに利用可能である。例えば、SMART RACE cDNA増幅キット(BD Biosciences、CA)またはGeneRacerキット(Invitrogen、CA)を使用した5’RACE(cDNA末端の迅速増幅)法が一般に使用されている。5’RACE用の遺伝子特異的プライマーは、それがVHおよびVLのすぐ下流に結合することができるように、標的モノクローナル抗体のH鎖およびL鎖のアイソタイプに基づいて調製することができる。したがって、5’RACEプライマーは、γ1、γ2a、γ2bまたはγ3などのマウスにおけるそれぞれの亜型に特異的であるように設計することができる。あるいは、全ての亜型に共通なプライマーを、亜型間のコンセンサスまたは非常に相同的な領域に基づいて設計することができる。Tsurushita中では、以下の5’RACEプライマーが例として開示される:
(i)5’−GCCAGTGGATAGACTGATGG−(配列番号82)(マウスγ1、γ2a、γ2bおよびγ3H鎖のクローニング用)
(ii)5’−GATGGATACAGTTGGTGCAGC−(配列番号83)(マウスkL鎖のクローニング用)。
【0054】
PCR増幅したV領域遺伝子断片は、例えばZero Blunt TOPO PCRクローニングキット(Invitrogen)を使用してプラスミドベクターに直接クローニングすることができ、それらのDNA配列を決定することができる。得られた配列は、例えばモデル241プロテインシークエンサー(Hewlett−Packard、CA)を使用して、例えばそれらのコードアミノ酸配列と、N末端アミノ酸配列決定により決定した標的モノクローナル抗体の配列を比較することによって確認しなければならない。典型的には、例えばエドマン分解による、標的抗体のN末端における少なくとも15〜20アミノ酸残基の決定は、クローニングDNA配列の真正を確認するのに十分である。Tsurushitaは、マウスにおける2つの最も一般的なN末端アミノ酸の1つであるグルタミンがN末端アミノ酸であるとき、それがピログルタミンに変換されてN末端における配列決定を阻害する可能性があることを警告する。その場合、N末端を脱保護して配列を得ることが必要である。
【0055】
V領域の三次元モデリング
VHおよびVLの配列に基づいて、CDRの立体配座構造を維持するのにおそらく重要である標的抗体のフレームワーク残基を、例えばR.Levy et al., 1989, Biochemistry28: 7168-7175によって記載された方法によって、およびB.Zilber et al., 1990, Biochemistry29: 10032-10041によって記載された方法によって最初に同定する。典型的には、VHおよびVLのそれぞれは14個の構造上有意なセグメントに分けられ、これらは免疫グロブリンスーパーファミリーのドメイン構造を含むβ鎖およびループ様構造である。標的抗体由来の各セグメントのアミノ酸配列を、PDBデータベースにおける知られている構造の対応する抗体セグメントと合わせる(H.M.Berman et al., 2000, Nucleic Acids Res.28: 235-342参照)。多重配列アラインメントによって、標的セグメントのそれぞれと最高の配列相同性を有する対応するセグメントを選択し、V領域の三次元モデルを構築する。構造を最適化するために、このモデルを多サイクルの共役勾配法によるエネルギー最小化に施す(例えば、ENCADを使用して、またはPress et al., 1990, "Numerical Recipes, Cambridge University Press, Cambridgeによって記載されたように、Weiner et al., 1981, J.Comp.Chem.2: 287-303によるAMBER、BioMolecularModellingで利用可能な3D-JIG-SAW、またはCancer Research UKによって運営される「BMM」ウェブサイト、またはSwiss Institute of Bioinformatics, Genevaによって運営されるExPASy Proteomics Serverウェブサイトで利用可能なSWISS-MODEL)。
【0056】
ヒトフレームワークの選択
V領域の構造のモデル化と並行して、それぞれマウスVHおよびVLのcDNAクローニングから推定したアミノ酸配列を、データベース、例えばKabatデータベース(Johnson et al., 2000, Nucleic Acids Res.28: 214-218を参照)、GenBankなどにおけるヒトV領域の配列と比較する。マウスの配列と少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、または少なくとも95%同一の全体配列同一率を有するヒトFRは、例えばSmith−Watermanのアルゴリズム(Gusfield, 1997, "Algorithms on Strings, Trees, and Sequences", Cambridge University Press, Cambridgeによる)、またはBLAST(Karlin et al., 1990, Proc.Natl.Acad.Sci.USA87: 2264-2268による)などを使用して検索することができる。これらのヒトの配列は、cDNAベースおよびタンパク質由来配列に基づいてよいが、しかしながら、生殖細胞系の使用が好ましいことが多い。cDNAベース、タンパク質由来配列の体細胞超突然変異と関係がある潜在的免疫原性を排除する際に、それが有用である可能性があるからである。代替では、Queen et al.(1989、上記)中に記載されたように、コンセンサスフレームワーク配列の使用によって、cDNAベースまたはタンパク質由来配列から得たフレームワーク中のこのような超突然変異残基を同定し除去することもできる。生殖細胞系VHセグメントをアクセプターフレームワークとして使用する場合、15および16ではなく第14染色体上にコードされるVHセグメントを使用しなければならない。第14染色体上のセグメントのみが機能的VHを生成するからである。
【0057】
ヒト化V領域の設計
Queen et al.(1989、上記)によれば、CDRの約4〜6Å以内のフレームワークアミノ酸を同定することが必要である。これらの残基は、正確なCDR構造を支えるおそらく重要なフレームワーク残基であると考えられるからである。このようなプロセスは、原子座標からの原子間距離を計算する国立科学財団(NSF)によって運営されるMolecular Visualization Freewareウェブサイトで利用可能なRASMOLなどのコンピュータプログラムを使用して、またはコンピュータモデルの手検測によって実施することができる。重要なフレームワーク位置におけるアミノ酸がマウスドナー配列とヒトアクセプター配列の間で異なる場合、マウスドナーのそれらを通常ヒト残基に置換する。しかしながら、このような残基がCDR構造を支持するのに最小の貢献度を有する場合、対応するヒト残基を典型的に使用する。さらに、選択したヒトアクセプターが、V領域配列の約10〜20%未満に生じる「異常な」アミノ酸を含有する場合、それらは親和性成熟中の体細胞超突然変異の結果である可能性があり、ドナー残基と置換してヒトにおける潜在的免疫原性を回避しなければならない。
【0058】
さらに、潜在的N−結合型グリコシル化シグナルの残基などの他の因子を注意深く考慮して、ヒト化V領域を設計する必要がある(詳細に関してはTsurushitaを参照)。
【0059】
ヒト化抗体は、治療用途に必要とされるかまたはそのために排除されるエフェクター機能に応じて、ヒト抗体、またはその変異体のヒトκもしくはλL鎖、および/またはγ1、γ2、γ3、γ4、μ、α1、α2、δ、もしくはεH鎖由来のヒト定常領域またはその一部分を含有することができる。例えば、突然変異を含有する定常領域のFc部分を本発明のキメラもしくはヒト化抗体のV領域と融合して、Fc受容体と抗体の結合を低減する、および/または補体を固定するその能力を低減することができる(例えば、Winter et al., GB2,209,757B、Morrison et al., WO89/07142、Morgan et al., WO94/29351を参照)。抗体分子のこのような操作は、セクション5.2中に記載したように組換えDNA技術によって実施することができる。
【0060】
生成するキメラまたはヒト化抗体は、非ヒトドナー抗体と同じ特異性、および非ヒトドナー抗体の親和性と同等またはその少なくとも約1/3、少なくとも約1/2、もしくは少なくとも約2/3の親和性を有することが好ましい。別の態様では、生成するキメラまたはヒト化抗体は、少なくとも約1×107M−1、好ましくは少なくとも約1×108M−1、および最も好ましくは少なくとも約1×109M−1の親和性定数を有する。
【0061】
前に記載した一般的プロトコール以外に、例えば、いずれもその全容を参照によって本明細書に組み込む、CDR移植(EP239,400;PCT公開WO91/09967、米国特許第5,225,539号、同第5,530,101号および同第5,585,089号)、ベニアリング(veneering)またはリサーフィシング(resurfacing)(EP592,106;EP519,596、Padlan, Molecular Immunology, 28(4/5): 489-498, 1991; Studnicka et al., Protein Engineering, 7(6): 805-814, 1994; Roguska et al., Proc Natl. Acad.Sci.USA, 91: 969-973, 1994)、および鎖シャッフリング(米国特許第5,565,332号)を含めた、当技術分野で知られている様々な技法を使用して、抗体をヒト化することができる。
【0062】
5.3.2.薬剤としてヒト化抗体を調製するための追加的考慮事項
薬剤としてヒト化抗体を与えるため、効率良く一貫した生成系をしたがって調製する必要がある。例えば、ヒト化抗体に適した発現ベクターはH鎖およびL鎖配列を挿入することによって調製し、発現ベクターでトランスフェクトした高生産性細胞系は、作業用細胞バンク(WCB)の安定した半永久的供給源として働く、マスター細胞バンク(MCB)の種細胞として得ることができる。次いで、ヒト化抗体は、WCB由来の作業用細胞を培養し培養培地を回収することによって調製することができる。
【0063】
適切な制御遺伝子を有する様々な発現ベクターを、このような生成細胞系の調製に使用することができる。宿主細胞として、哺乳動物タンパク質の発現に一般に使用される細胞をヒト化抗体の発現に使用することができる。このような宿主細胞の例には、チャイニーズハムスター卵巣(CHO)細胞、SP2/0−Agl4.19細胞、NSO細胞などがあるが、これらだけには限られない。ヒト化抗体の生産性は、発現ベクターと宿主細胞の最良の組合せを選択することによって最大にすることができる。さらに、宿主細胞によるヒト化抗体の発現を最適化することができるように、培養培地の組成を調べて、様々な無血清培養培地およびサプリメントから適切な培地を選択しなければならない。
【0064】
有効性および最終収率に基づいて、宿主細胞によって産生されるヒト化抗体を、親和性クロマトグラフィー、イオン交換クロマトグラフィー、疎水性相互作用クロマトグラフィーなどを含めた、当技術分野でよく知られている様々な方法を使用して培養上清から精製することができる。
【0065】
5.4 医薬組成物および治療用途
本発明は、RGD配列を免疫特異的に認識する前に記載したヒト化抗体またはその抗原結合断片を含む、医薬組成物を提供する。活性成分として本発明のヒト化抗体を含む医薬組成物は、癌、例えば癌細胞の増殖または転移、および炎症疾患、例えば慢性関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などだけには限られないが、これらを含めたRGDタンパク質と関係がある障害または疾患を予防および/または治療するための作用物質として使用することができる。
【0066】
本発明のヒト化抗体を含む医薬組成物を使用して、臓器移植後の慢性拒絶反応、および例えば全身性自己免疫疾患、エリテマトーデス、ブドウ膜炎、ベーチェット病、多発性筋炎、増殖性糸球体腎炎、サルコイドーシスなどの自己免疫疾患を治療することもできる。
【0067】
本発明のヒト化抗体を含む、前に記載した障害もしくは疾患を予防または治療するための予防および/または治療剤は低い毒性を有し、経口的または非経口的に、適切な溶媒中に混合することにより液体調製物として直接、または適切な剤形の医薬組成物としてヒトに投与することができる。
【0068】
前に記載した投与に使用する医薬組成物は、前述の抗体またはその塩、および薬学的に許容される担体、希釈剤または賦形剤を含有する。このような組成物は、経口または非経口投与に適した剤形で与えられる。
【0069】
用量は、投与する対象の年齢および大きさ、標的疾患、状態、投与の経路などに応じて変わる可能性がある。例えば成人患者において慢性関節リウマチを予防および/または治療するために抗体を使用するとき、通常約0.01〜約20mg/体重1kg、好ましくは約0.1〜約10mg/体重1kg、およびより好ましくは約0.1〜約5mg/体重1kgの一回用量で、1日当たりおよそ1〜5回、好ましくは1日当たりおよそ1〜3回、本発明の抗体を静脈内投与することが有益である。他の非経口投与および経口投与では、前に与えた用量に対応する用量で抗体を投与することができる。状態が特に重度であるとき、状態に応じて用量を増大することができる。
【0070】
様々な送達系、例えばリポソーム、マイクロ粒子、マイクロカプセルへの封入、変異ウイルスを発現することができる組換え細胞、受容体仲介エンドサイトーシスが知られており、これらを使用して本発明の医薬組成物を投与することができる(例えば、Wu and Wu, 1987, J.Biol.Chem.262: 4429 4432を参照)。導入の方法には、皮内、筋肉内、腹腔内、静脈内、皮下、鼻腔内、硬膜外、および経口投与があるが、これらだけには限られない。化合物は任意の好都合な経路によって、例えば注入またはボーラス注射によって、上皮または皮膚粘膜内側(例えば、経口粘膜、直腸および腸粘膜など)を介した吸収によって投与することができ、他の生物活性剤と一緒に投与することができる。投与は全身性または局所であってよい。例えば、吸入器または噴霧器、および製剤とエアロゾル剤を使用することによって、肺投与を利用することもできる。
【0071】
本発明の一態様では、治療の必要がある領域へ局所的に、本発明の医薬組成物を投与することが望ましい可能性がある。これは、例えばかつ非制限的に、外科手術中の局所注入、例えば外科手術後に創傷包帯を併用した局所施用によって、注射によって、カテーテルによって、座薬によって、鼻腔スプレーによって、インプラントによって(前記インプラントは多孔性、非多孔性、またはシアラスティック膜などの膜、またはファイバーを含めたゼラチン質材料である)実施することができる。本発明の一態様では、投与は部位(または感染前部位)感染組織への直接注射による投与であってよい。
【0072】
本発明の別の態様では、医薬組成物を小胞、特にリポソームで送達することができる(Langer, 1990, Science249: 1527-1533; Treat et al., Liposomes in the Therapy of Infectious Disease and Cancer, Lopez Berestein and Fidler(eds.), Liss, New York, pp.353-365(1989); Lopez-Berestein、既出、pp.317-327を参照、既出を一般に参照)。
【0073】
本発明のさらに別の態様では、医薬組成物を徐放システムで送達することができる。本発明の一態様では、ポンプを使用することができる(Langer、上記、Sefton, 1987, CRC Crit.Ref.Biomed.Eng. 14: 201; Buchwald et al., 1980, Surgery88: 507; およびSaudek et al., 1989, N.Engl.J.Med.321: 574を参照)。本発明の別の態様では、ポリマー物質を使用することができる(Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Florida(1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York (1984); Ranger and Peppas, J.Macromol.Sci.Rev.Macromol.Chem. 23: 61(1983)を参照、Levy et al., 1985, Science228: 190; During et al., 1989, Ann.Neurol.25: 351; Howard et al., 1989, J.Neurosurg. 71: 105も参照)。本発明のさらに別の態様では、徐放システムを組成物の標的の近くに置くことができ、したがってごくわずかな全身用量を必要とする(例えば、Goodson, Medical Applications of Controlled Release、上記、vol.2, pp.115-138(1984)を参照)。他の徐放システムはLanger(Science249: 1527-1533(1990))による総説中で論じられている。
【0074】
経口投与用の組成物の例には、固体または液体剤形、具体的には、錠剤(ドラジェおよびフィルムコーティング錠を含む)、ピル、顆粒、粉末調製物、カプセル(軟質カプセルを含む)、シロップ、乳濁液、縣濁液などがある。このような組成物は公に知られている方法によって製造され、製剤の分野で従来使用されている小胞、希釈剤または賦形剤を含有する。錠剤用の小胞または賦形剤の例は、ラクトース、スターチ、スクロース、ステアリン酸マグネシウムなどである。
【0075】
注射用調製物は、静脈内、皮下、皮内および筋肉内注射、点滴など用の剤形を含むことができる。これらの注射用調製物は、公に知られている方法によって調製することができる。注射用調製物は、例えば注射に従来使用される滅菌水性媒体または油状媒体中に、前に記載した抗体またはその塩を溶解、縣濁または乳化することによって調製することができる。注射用水性媒体として、例えば、アルコール(例えば、エタノール)、多価アルコール(例えば、プロピレングリコール、ポリエチレングリコール)などの適切な可溶化剤、非イオン性界面活性剤[例えば、ポリソルベート80、HCO−50(水素化ヒマシ油のポリオキシエチレン(50mol)付加生成物)]などと組合せて使用することができる、生理食塩水、グルコースおよび他の補助剤などを含有する等張液が存在する。油状媒体として、例えば、安息香酸ベンジル、ベンジルアルコールなどの可溶化剤などと組合せて使用することができる、ゴマ油、ダイズ油などが利用される。このように調製した注射用調製物は、適切なアンプル中に充填することが好ましい。直腸投与に使用する座薬は、前述の抗体またはその塩と従来の座薬基剤を混合することによって調製することができる。
【0076】
前に記載した経口または非経口用途の医薬組成物は、活性成分の投与に適合させるのに適した単位用量の剤形に調製することが有利である。このような単位用量の剤形には、例えば、錠剤、ピル、カプセル、注射(アンプル)、座薬などがある。含有される前述の抗体の量は、特に注射の形で単位用量の剤形当たり一般に約5〜500mgであり、前述の抗体は他の剤形に関して約5〜100mgおよび約10〜250mg含有されることが好ましい。
【0077】
製剤化が前に記載した抗体と何らかの悪い相互作用を引き起こさない限り、前に記載したそれぞれの組成物は他の活性成分をさらに含有することができる。
【0078】
本発明はさらに、活性成分としてRGD配列結合機能性分子(例えば、インテグリンなど)を含む細胞および/または組織再構築用の阻害剤および/または促進剤、およびRGDタンパク質発現細胞および/または組織(例えば、腫瘍細胞、好中球、平滑筋など)とRGDタンパク質結合機能性分子を接触させることを含む、細胞および/または組織の再構築を阻害および/または促進するための方法に関する。このような治療剤における活性成分の投与、薬剤調製などに関する用量、方法は、本発明のヒト化抗体を含む医薬品の前述の記載を参照することによって適切に決定することができる。
【0079】
前に記載したように、本発明は、RGDタンパク質と関係があるかまたはそれらに関与する障害もしくは疾患を予防または治療するための方法であって、少なくとも1つの本発明のヒト化抗体を有効量、その必要性がある対象に投与することを含む方法をさらに提供する。
【0080】
5.5 診断用途
本発明のヒト化抗体を含む医薬組成物は、癌、例えば癌細胞の増殖または転移、および炎症疾患、例えば慢性関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、糖尿病、癌転移、動脈硬化症、多発性硬化症、肉芽腫などの診断薬として、または臓器移植後の慢性拒絶反応、全身性自己免疫疾患、エリテマトーデス、ブドウ膜炎、ベーチェット病、多発性筋炎、増殖性糸球体腎炎、サルコイドーシスなどの自己免疫疾患の診断薬として使用することができる。本発明のヒト化抗体はRGD配列を特異的に認識することができ、したがってこれらを使用して、試験流体中のRGDタンパク質の定量化、特にサンドイッチイムノアッセイ、競合アッセイ、イムノメトリー、ネフェロメトリーなど、免疫染色などによる定量化を可能にすることができる。これらの免疫法を本発明のアッセイ法に適用する際に、何らかの特定条件、手順などを言及することは必要とされない。当技術分野で通常の技術的考慮事項を従来の条件および手順に加えることによって、アッセイシステムを構築することで十分である。これらの一般的な技術的意義の詳細に関しては、総説、教本などを参照することができる。
【0081】
前に記載したように、本発明の抗体を使用することによって、高感度でRGDタンパク質を定量化することができる。本発明のヒト化抗体は、in vivoでRGDタンパク質を定量化するための方法を適用することによって、RGDタンパク質と関係がある様々な疾患を診断するのに特に有用である。例えば、RGDタンパク質の発現レベルの増大または低下を検出する場合、人が現在RGDタンパク質と関係がある疾患、例えば癌または炎症疾患に罹患している可能性が非常に高い、または人が将来これらの疾患に罹患する可能性が非常に高いと診断することができる。したがって、本発明は、対象における、RGDタンパク質と関係があるかまたはそれらに関与する障害もしくは疾患を診断するための方法であって、本発明のヒト化抗体の少なくとも1つまたは両方を有効量、その必要性がある対象に投与することを含む方法も提供する。このようなin vivo診断に必要とされる用量は診断用途に必要とされる用量未満であってよく、通常の手順に従い当業者によって決定することができる。
【0082】
本発明のヒト化抗体は、試験サンプル(例えば、体液などの試験流体、組織など)中に存在するRGDタンパク質を特異的に検出するために使用することもできる。ヒト化抗体は、RGDタンパク質の精製用の抗体カラムの調製、精製時にそれぞれの分画中に含有されるRGDタンパク質の検出、または試験する細胞中のRGDタンパク質の挙動の分析に使用することもできる。
【0083】
本明細書の配列表中の配列識別番号は以下の配列を示す。
【0084】
[配列番号1]
これはマウス33E10およびヒト33E10と共通のCDRH1のアミノ酸配列を示す。
【0085】
[配列番号2]
これはマウス33E10およびヒト33E10と共通のCDRH2のアミノ酸配列を示す。
【0086】
[配列番号3]
これはマウス33E10およびヒト33E10と共通のCDRH3のアミノ酸配列を示す。
【0087】
[配列番号4]
これはマウス33E10およびヒト33E10と共通のCDRL1のアミノ酸配列を示す。
【0088】
[配列番号5]
これはマウス33E10およびヒト33E10と共通のCDRL2のアミノ酸配列を示す。
【0089】
[配列番号6]
これはマウス33E10およびヒト33E10と共通のCDRL3のアミノ酸配列を示す。
【0090】
[配列番号7]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VH5遺伝子のヌクレオチド配列を示す。
【0091】
[配列番号8]
これはシグナルペプチドを含むHu33E10VH5遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0092】
[配列番号9]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VH7遺伝子のヌクレオチド配列を示す。
【0093】
[配列番号10]
これはシグナルペプチドを含むHu33E10VH7遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0094】
[配列番号11]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VH7.8遺伝子のヌクレオチド配列を示す。
【0095】
[配列番号12]
これはシグナルペプチドを含むHu33E10VH7.8遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0096】
[配列番号13]
これはNheIおよびEcoRI部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VL6遺伝子のヌクレオチド配列を示す。
【0097】
[配列番号14]
これはシグナルペプチドを含むHu33E10VL6遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0098】
[配列番号15]
これはNheIおよびEcoRI部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VLv2遺伝子のヌクレオチド配列を示す。
【0099】
[配列番号16]
これはシグナルペプチドを含むHu33E10VLv2遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0100】
[配列番号17]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[CMV2]を示す。
【0101】
[配列番号18]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JΝΤ026]を示す。
【0102】
[配列番号19]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT080]を示す。
【0103】
[配列番号20]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT082]を示す。
【0104】
[配列番号21]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT084]を示す。
【0105】
[配列番号22]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT097]を示す。
【0106】
[配列番号23]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT098]を示す。
【0107】
[配列番号24]
これはNS0−Hu33E10−4およびNS0−Hu33E10−6において発現されるVH7およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列を示す。
【0108】
[配列番号25]
これはNS0−Hu33E10−4およびNS0−Hu33E10−6において発現されるVH7およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0109】
[配列番号26]
これはNS0−Hu33E10−5において発現されるVH7.8およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列を示す。
【0110】
[配列番号27]
これはNS0−Hu33E10−5において発現されるVH7.8およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0111】
[配列番号28]
これはNS0−Hu33E10−4およびNS0−Hu33E10−5において発現されるVL6およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列を示す。
【0112】
[配列番号29]
これはNS0−Hu33E10−4およびNS0−Hu33E10−5において発現されるVL6およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0113】
[配列番号30]
これはNS0−Hu33E10−6において発現されるVLv2およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列を示す。
【0114】
[配列番号31]
これはNS0−Hu33E10−6において発現されるVLv2およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0115】
[配列番号32]
これは33E10VLのアミノ酸配列を示す。
【0116】
[配列番号33]
これはHu33E10VLv2のアミノ酸配列を示す。
【0117】
[配列番号34]
これはマウス33E10VHcDNAのヌクレオチド配列を示す。
【0118】
[配列番号35]
これはマウス33E10VHcDNAのヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0119】
[配列番号36]
これはマウス33E10VLcDNAのヌクレオチド配列を示す。
【0120】
[配列番号37]
これはマウス33E10VLcDNAのヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0121】
[配列番号38]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含む設計33E10VH遺伝子のヌクレオチド配列を示す。
【0122】
[配列番号39]
これはシグナルペプチドを含む設計33E10VH遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0123】
[配列番号40]
これはNheIおよびEcoRI部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含む設計33E10VL遺伝子のヌクレオチド配列を示す。
【0124】
[配列番号41]
これはシグナルペプチドを含む設計33E10VL遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0125】
[配列番号42]
これは33E10VHのアミノ酸配列を示す。
【0126】
[配列番号43]
これはヒト化33E10(Hu33E10)VH1のアミノ酸配列を示す。
【0127】
[配列番号44]
これは33E10VLのアミノ酸配列を示す。
【0128】
[配列番号45]
これはヒト化33E10(Hu33E10)VL1のアミノ酸配列を示す。
【0129】
[配列番号46]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ220]を示す。
【0130】
[配列番号47]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ206]を示す。
【0131】
[配列番号48]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ207]を示す。
【0132】
[配列番号49]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ208]を示す。
【0133】
[配列番号50]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ209]を示す。
【0134】
[配列番号51]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ210]を示す。
【0135】
[配列番号52]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ211]を示す。
【0136】
[配列番号53]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ212]を示す。
【0137】
[配列番号54]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ213]を示す。
【0138】
[配列番号55]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ214]を示す。
【0139】
[配列番号56]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ215]を示す。
【0140】
[配列番号57]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ216]を示す。
【0141】
[配列番号58]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ217]を示す。
【0142】
[配列番号59]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ218]を示す。
【0143】
[配列番号60]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ219]を示す。
【0144】
[配列番号61]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ221]を示す。
【0145】
[配列番号62]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ116]を示す。
【0146】
[配列番号63]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ193]を示す。
【0147】
[配列番号64]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ194]を示す。
【0148】
[配列番号65]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ195]を示す。
【0149】
[配列番号66]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ196]を示す。
【0150】
[配列番号67]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ197]を示す。
【0151】
[配列番号68]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ198]を示す。
【0152】
[配列番号69]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ199]を示す。
【0153】
[配列番号70]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ200]を示す。
【0154】
[配列番号71]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ201]を示す。
【0155】
[配列番号72]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ202]を示す。
【0156】
[配列番号73]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ203]を示す。
【0157】
[配列番号74]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ204]を示す。
【0158】
[配列番号75]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ205]を示す。
【0159】
[配列番号76]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ101]を示す。
【0160】
[配列番号77]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ117]を示す。
【0161】
[配列番号78]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VH1遺伝子のヌクレオチド配列を示す。
【0162】
[配列番号79]
これはシグナルペプチドを含むHu33E10VH1遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0163】
[配列番号80]
これはNheIおよびEcoRI部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VL1遺伝子のヌクレオチド配列を示す。
【0164】
[配列番号81]
これはシグナルペプチドを含むHu33E10VL1遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0165】
[配列番号82]
これは5’RACEプライマーのヌクレオチド配列を示す。
【0166】
[配列番号83]
これは5’RACEプライマーのヌクレオチド配列を示す。
【0167】
[配列番号84]
これはGeneRacer5’プライマーのヌクレオチド配列を示す。
【0168】
[配列番号85]
これは33E10VH3’プライマーのヌクレオチド配列を示す。
【0169】
[配列番号86]
これは33E10VL3’プライマーのヌクレオチド配列を示す。
【0170】
[配列番号87]
これは33E10VH5’プライマーのヌクレオチド配列を示す。
【0171】
[配列番号88]
これは33E10VH3’プライマーのヌクレオチド配列を示す。
【0172】
[配列番号89]
これは33E10VL5’プライマーのヌクレオチド配列を示す。
【0173】
[配列番号90]
これは33E10VL3’プライマーのヌクレオチド配列を示す。
【0174】
[配列番号91]
これはHu33E10VH5遺伝子のヌクレオチド配列を示す。
【0175】
[配列番号92]
これはHu33E10VH5遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0176】
[配列番号93]
これはHu33E10VH7遺伝子のヌクレオチド配列を示す。
【0177】
[配列番号94]
これはHu33E10VH7遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0178】
[配列番号95]
これはHu33E10VH7.8遺伝子のヌクレオチド配列を示す。
【0179】
[配列番号96]
これはHu33E10VH7.8のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0180】
[配列番号97]
これはHu33E10VL6遺伝子のヌクレオチド配列を示す。
【0181】
[配列番号98]
これはHu33E10VL6遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0182】
[配列番号99]
これはHu33E10VLv2遺伝子のヌクレオチド配列を示す。
【0183】
[配列番号100]
これはHu33E10VLv2遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【実施例】
【0184】
以下の実施例は、RGD配列を免疫特異的に認識するモノクローナル抗体の調製、モノクローナル抗体のV領域の配列決定、これらの抗体のエピトープマッピングおよび他の特徴付け、およびこのような抗体のキメラ化およびヒト化、ならびに生成したキメラおよびヒト化抗体の特徴付けを例示する。これらの実施例は、本発明の範囲を制限するものとして解釈するべきではない。
【0185】
6.1.マウス33E10可変領域遺伝子のクローニングおよび配列決定
マウス33E10ハイブリドーマ細胞を、7.5%CO2インキュベーター中で37℃において、10%ウシ胎児血清(FBS;HyClone、Logan、UT)を含有するTIL培地I(Immuno−Biological Laboratories、群馬、日本)中で増殖させた。全RNA(total RNA)は、供給者のプロトコールに従いTRIzol試薬(Invitrogen、Carlsbad、CA)を使用して約3×106個のハイブリドーマ細胞から抽出した。オリゴdTプライマー処理cDNAは、供給者のプロトコールに従いGeneRacerキット(Invitrogen)を使用して合成した。33E10重鎖および軽鎖に関する可変領域cDNAは、マウスγ−1およびkappa鎖定常領域とそれぞれアニーリングする3’プライマー、およびGeneRacerキット中に供給されたGeneRacer5’プライマー(5’−CGACTGGAGCACGAGGACACTGA−3’(配列番号84))を使用して、Phusion DNAポリメラーゼ(New England Biolabs、Beverly、MA)を用いてポリメラーゼ連鎖反応(PCR)によって増幅した。重鎖可変領域(VH)のPCR増幅用に、3’プライマーは配列5’−GCCAGTGGATAGACAGATGG−3’(配列番号85)を有する。軽鎖可変領域(VL)のPCR増幅用に、3’プライマーは配列5’−GATGGATACAGTTGGTGCAGC−3’(配列番号86)を有する。増幅したVHおよびVLcDNAは配列決定用にpCR4Blunt−TOPOベクター(Invitrogen)にクローニングした。可変領域のDNA配列決定はTocore(Menlo Park、CA)で実施した。数個の重鎖および軽鎖クローンを配列決定し、典型的なマウス重鎖および軽鎖可変領域と相同的な独自の配列を同定した。33E10VHおよびVLの推定アミノ酸配列と共にコンセンサスcDNA配列をそれぞれ図1および2中に示す。
【0186】
6.2.キメラ33E10IgG1/κ抗体の構築
33E10VHをコードする遺伝子を、鋳型として33E10VHcDNA、5’プライマーとして5’−GGGACTAGTACCACCATGAAGTTGTGGCTGAACTGGATT−3’(配列番号87)(SpeI部位に下線を引く)、および3’プライマーとして5’−GGGAAGCTTGAAGTTAGGACTCACCTGCAGAGACAGTGACCAGAGTCCC−3’(配列番号88)(HindIII部位に下線を引く)(図3)を使用してPCRによって、スプライスドナーシグナルおよび適切な隣接制限酵素部位を含むエクソンとして作製した。同様に、33E10VLをコードする遺伝子を、鋳型として33E10VLcDNA、5’プライマーとして5’−GGGGCTAGCACCACCATGAAGTTGCCTGTTAGGCTGTTG−3’(配列番号89)(NheI部位に下線を引く)、および3’プライマーとして5’−GGGGAATTCTTTGGATTCTACTTACGTTTGATTTCCAGCTTGGTGCCTCC−3’(配列番号90)(EcoRI部位に下線を引く)(図4)を使用してPCRによって、スプライスドナーシグナルおよび適切な隣接制限酵素部位を含むエクソンとして作製した。33E10VHおよびVLエクソンのスプライスドナーシグナルは、それぞれマウス生殖細胞JH3およびJK1配列に由来した(Kabat et al., Sequences of Proteins of Immunological Interests, Fifth edition, NIH公開番号91−3242、米国厚生省、1991)。PCR増幅断片はQIAquickゲル抽出キット(Qiagen、Valencia、CA)を使用してゲル精製し、(VHに関して)SpeIおよびHindIIIまたは(VLに関して)NheIおよびEcoRIで消化し、キメラ33E10IgGl/κ抗体の産生用にヒトγ−1およびkappa定常領域を保持する哺乳動物発現ベクターにクローニングした。生成した発現ベクター、pCh33E10の概略構造は図5中に示す。
【0187】
6.3.ヒト化33E10VHおよびVL遺伝子の作製
33E10可変領域のヒト化を、Queen et al.(Proc.Natl.Acad.Sci.USA86: 10029-10033、1989)によって概説されたように実施した。最初に、33E10可変領域の三次元分子モデルをコンピュータプログラムの助力によって構築した。次に、ヒト可変領域配列に対する相同性検索に基づいて、33E10VHと高い相同性を有するU03400(GenBank受託番号)のヒトVHアミノ酸配列を、ヒト化33E10VHのフレームワークを得るためのアクセプターとして選択した。同様に、X72452(GenBank受託番号)のヒトVLアミノ酸配列を33E10VLのヒト化用のアクセプターとして選択した。
【0188】
三次元モデルが相補性決定領域(CDR)との有意な接触を示したフレームワーク位置において、マウス33E10可変領域由来のアミノ酸残基をヒトフレームワーク残基に置換した。これは位置30および48で実施してヒト化33E10(Hu33E10)VH1を作製した(図6)。軽鎖に関しては、Hu33E10VL1を作製するのに置換を必要としなかった(図7)。33E10、設計Hu33E10およびヒトアクセプターアミノ酸配列のアラインメントは、VHに関しては図6中およびVLに関しては図7中に示す。
【0189】
Hu33E10VH1およびVL1のそれぞれをコードする遺伝子を、哺乳動物発現ベクターへの後のクローニング用に、シグナルペプチド、スプライスドナーシグナル、および適切な制限酵素部位を含むエクソンとして設計した。Hu33E10VH1およびVL1エクソンのスプライスドナーシグナルは、それぞれヒト生殖細胞JH3およびJk1配列に由来した(Kabat et al., 1991)。マウス33E10VL1遺伝子のシグナルペプチド配列は、SIG−Predシグナルペプチド予想ソフトウェア(http: //bmbpcu36.leeds.ac.uk/prot_analysis/Signal.html)によって正確な切断の最適下限であったことを示した。したがって、SIG−Predソフトウェアによって有効かつ正確に切断されると予想されたマウスモノクローナル抗体35B6(Gene Techno Science)のVL遺伝子のシグナルペプチドを、Hu33E10VL1エクソンにおいて使用した。Hu33E10VH1エクソン中のシグナルペプチド配列は対応するマウス33E10VH配列に由来した。SIG−Predソフトウェアは、Hu33E10VH1遺伝子のシグナルペプチドが有効かつ正確に切断されることを示した。
【0190】
Hu33E10VH1およびVL1遺伝子を、He et al.(J.Immunol.160: 1029-1035、1998)によって概説されたPhusion DNAポリメラーゼを使用して、数個の重複合成オリゴヌクレオチドプライマーの延長およびPCR増幅によって構築した。Hu33E10VH1およびVL1遺伝子の構築に使用したオリゴヌクレオチドは、それぞれ図8および9中に列挙する。Hu33E10VH1およびVL1遺伝子におけるオリゴヌクレオチドの位置は、それぞれ図10および11中に示す。PCR増幅断片はQIAquickゲル抽出キット(Qiagen)を使用してゲル精製し、配列決定用にpCR4Blunt−TOPOベクターにクローニングした。(VHに関して)SpeIおよびHindIIIまたは(VLに関して)NheIおよびEcoRIでの消化後、Hu33E10VH1およびVL1遺伝子を、ヒトIgG1/κ型の産生用に哺乳動物発現ベクター中の対応する部位にサブクローニングした。生成した発現ベクター、pHu33E10−1の概略構造は図5中に示す。推定アミノ酸配列と共に得られたHu33E10VH1およびVL1遺伝子のヌクレオチド配列は、それぞれ図12および13中に示す。
【0191】
6.4.キメラおよびヒト化33E10抗体の発現
キメラおよびヒト化33E10IgG1/κ抗体のそれぞれを安定的に産生する細胞系を得るために、発現ベクターpCh33E10およびpHu33E10−1をそれぞれ、マウスミエローマ細胞系NS0(European Collection of Animal Cell Cultures、Salisbury、Wiltshire、UK)の染色体に導入した。7.5%CO2インキュベーター中で37℃において、NS0細胞を10%FBSを含有するDME培地中で増殖させた。NS0細胞への安定したトランスフェクションは、Bebbington et al.(Bio/Technology10: 169-175、1992)中に記載されたエレクトロポレーションによって実施した。トランスフェクション前に、それぞれの発現ベクターはFspIを使用して線状化した。約107個の細胞を10μgの線状化プラスミドでトランスフェクトし、10%FBSを含有するDME培地中に縣濁し、数枚の96ウェルプレートに平板培養した。24〜48時間後、選択培地(10%FBSを含有するDME培地、HT培地サプリメント(Sigma、St.Louis、MO)、0.25mg/mlキサンチンおよび1μg/mlミコフェノール酸)を施した。選択開始後約10日で、培養上清を抗体産生に関してアッセイした。
【0192】
培養上清中のpCh33E10およびpHu33E10−1から発現した組換え抗体(それぞれCh33E10およびHu33E10−VH1/VL1)の発現レベルをサンドウィッチELISAによって測定した。典型的な実験では、PBSに溶かした100μl/ウェルの1/2,000希釈ヤギ抗ヒトIgGFcγ鎖特異的ポリクローナル抗体(SouthernBiotech、Birmingham、AL)で4℃において一晩、ELISAプレートをコーティングし、洗浄バッファー(0.05%のTween20を含有するPBS)で洗浄し、300μl/ウェルのブロッキングバッファー(2%のスキムミルクおよび0.05%のTween20を含有するPBS)で室温において1時間ブロッキングした。洗浄バッファーで洗浄した後、ELISAバッファー(1%のスキムミルクおよび0.025%のTween20を含有するPBS)に適切に希釈した100μl/ウェルのサンプルをELISAプレートに施した。ヒトミエローマ血清(SouthernBiotech)から精製したヒトIgG1/κ抗体または精製Ch33E10を標準として使用した。室温で2時間ELISAプレートをインキュベートし洗浄バッファーで洗浄した後、100μl/ウェルの1/2,000希釈HRP結合ヤギ抗ヒトkappa鎖ポリクローナル抗体(SouthernBiotech)を使用して、結合した抗体を検出した。室温で1時間のインキュベーションおよび洗浄バッファーでの洗浄後、発色現像は100μl/ウェルのABTS基質を加えることによって実施し、100μl/ウェルの2%シュウ酸で停止させた。吸光度は405nmで読み取った。
【0193】
高レベルのCh33E10(NSO−Ch33E102C11;L4−2C11とも呼ばれる)を産生するNS0安定トランスフェクタントの1つを、ハイブリドーマSFM(Invitrogen)を使用して無血清培地中での増殖に適合させ、約106/mlの密度までローラーボトル中で増殖させ、6mg/mlのUltrafiltrated Soy Hydrolyzate(Irvine Scientific カタログ番号96857)を供給し、細胞生存率が50%未満になるまでさらに増殖させた。遠心分離および濾過後、培養上清をプロテイン−Aセファロースカラム(GE Healthcare、Piscataway、NJ)に充填した。抗体を0.1Mのグリシン−HCl(pH3.0)で溶出する前に、カラムをPBSで洗浄した。1Mのトリス−HCl(pH8)での中和後、溶出抗体のバッファーを透析によってPBSに交換した。抗体濃度は280nmにおける吸光度を測定することによって決定した(1mg/ml=1.4OD)。
【0194】
Ch33E10を発現するNS0安定トランスフェクタントは容易に得たが、pHu33E10−1で安定的にトランスフェクトしたNS0トランスフェクタントはいずれも、Hu33E10−VH1/VL1をあまり産生しないことが分かった。重鎖および軽鎖mRNAの分析によって、VL1遺伝子のシグナルペプチドコード領域中の偶然生成したスプライスドナー部位のためVLコード領域を欠く、異常な軽鎖mRNAの存在が明らかになった。この問題を解決するため、シグナルペプチドコード領域中のスプライスドナー部位を、オーバーラップエクステンションPCR法を使用して部位特異的突然変異誘発によって排除した(Higuchi, R., PCR Technology. Principles and Applications for DNA Amplification.Erlich, H.A., ed.pp.61-70, Stockton Press, New York, 1989)。生成した変異Hu33E10VL遺伝子、VL6の配列は図14中に示す。VL領域のアミノ酸配列はVL1とVL6遺伝子の間で互いに同一である。VL6遺伝子は発現ベクターpHu33E10−1中のVL1に交換してpHu33E10−2を作製した。pHu33E10−2で安定的にトランスフェクトし高レベルの組換え抗体(Hu33E10−VH1/VL6)を産生したNS0細胞は、前に記載した方法によって得た。pHu33E10−2で安定的にトランスフェクトしたNS0細胞では、異常な軽鎖mRNAは観察しなかった。高レベルのHu33E10−VH1/VL6を産生するNS0安定トランスフェクタントの1つ(NS0−Hu33E10−2#22)を適合させ、ハイブリドーマSFM中で増殖させ、培養上清は前に記載したようにHu33E10−VH1/VL6の精製に使用した。
【0195】
6.5.Hu33E10−VHl/VL6の特徴付け
hOPN5−BSAとCh33E10およびHu33E10−VHl/VL6IgG1/κ抗体の結合をELISAによって調べた。抗原として、ウシ血清アルブミンと結合した合成オリゴペプチド(Cys−Val−Asp−Thr−Tyr−Asp−Gly−Arg−Gly−Asp−Ser−Val−Val−Tyr−Gly−Leu−Arg−Ser;Gene Techno Scienceにより提供された)(hOPN5−BSA)を使用した。典型的な実験では、0.2MのNa2C03バッファー(pH9.4)に溶かした100μl/ウェルの1μg/mlのhOPN5−BSAで4℃において一晩、ELISAプレートをコーティングし、洗浄バッファーで洗浄し、300μl/ウェルのブロッキングバッファーで室温において1時間ブロッキングした。洗浄バッファーで洗浄した後、ELISAバッファーに適切に希釈したサンプルを100μl/ウェルでELISAプレートに施した。4℃において一晩ELISAプレートをインキュベートし洗浄バッファーで洗浄した後、100μl/ウェルの1/2,000希釈HRP結合ヤギ抗ヒトγ鎖ポリクローナル抗体(SouthernBiotech)を使用して、結合した抗体を検出した。室温で1時間のインキュベーションおよび洗浄バッファーでの洗浄後、発色現像は100μl/ウェルのABTS基質を加えることによって実施し、100μl/ウェルの2%シュウ酸で停止させた。吸光度は405nmで読み取った。
【0196】
図15A中に示すように、Hu33E10−VH1/VL6は、Ch33E10よりhOPN5−BSAとの結合に関して約100倍弱かった。
【0197】
6.6.Hu33E10VH5、VH7およびVH7.8の作製および特徴付け
どのヒト化VH1およびVL6遺伝子がHu33E10−VHl/VL6の抗原結合親和性の消失の原因であるかを確認するために、1つがマウスVHおよびヒト化VL6(MoHu33E10)からなり、および他方がヒト化VH1およびマウスVL(HuMo33E10)からなる、2つの組換えIgG1/κ抗体を作製した。MoHu33E10およびHuMo33E10抗体の分析によって、Hu33E10−VH1/VL6の親和性の消失は大部分はVH1遺伝子に原因があり、部分的にはVL1遺伝子に原因があったことが分かった(ここでデータ示す)。
【0198】
VH1遺伝子の予備突然変異分析に基づくと、位置73〜77におけるアミノ酸は、抗原結合親和性を維持するのに重要である懸念があった(データ示さず)。オーバーラップエクステンションPCR法(Higuchi, 1989)を使用し部位特異的突然変異誘発によって構築したVH1遺伝子の3つの変異体(VH5、VH7およびVH7.8)は、さらなる分析用に選択した。VH1と比較して、位置77におけるIleはVH5中のSer(図6および16)、VH7中では位置76におけるSerおよび位置77におけるIleはそれぞれAsnおよびSer(図6および17)、およびVH7.8中では位置74におけるSer、位置76におけるSerおよび位置77におけるIleはそれぞれAla、AsnおよびSerに置換した(図6および18)。
【0199】
SpeIおよびHindIIIで消化したVH5、VH7およびVH7.8遺伝子のそれぞれをpHu33E10−2においてVH1に置換して、それぞれ発現ベクターpHu33E10−3、pHu33E10−4およびpHu33E10−5を構築した。pHu33E10−3、pHu33E10−4およびpHu33E10−5のそれぞれで安定的にトランスフェクトしたNS0細胞はセクション4中に記載したように生成した。Hu33E10−VH5/VL6を産生するNSO−Hu33E10−3#65、Hu33E10−VH7/VL6を産生するNS0−Hu33E10−45D3(R3−15D3とも呼ばれる)、およびHu33E10−VH7.8/VL6を産生するNS0−Hu33E10−51E5(EJ3−31E5とも呼ばれる)をハイブリドーマSFM中での増殖に適合させ、ローラーボトル中で増殖させた。Hu33E10−VH5/VL6、Hu33E10−VH7/VL6およびHu33E10−VH7.8/VL6抗体は、セクション4中に記載したようにプロテインAカラムを使用して対応する培養上清から精製した。
【0200】
hOPN5−BSAとHu33E10−VH5/VL6、Hu33E10−VH7/VL6およびHu33E10−VH7.8/VL6抗体の結合は、セクション5中に記載したようにELISAによってCh33E10の結合と比較した。hOPN5−BSAとHu33E10−VH5/VL6の親和性はHu33E10−VH1/VL6と比較したとき若干向上したが、それはCh33E10の親和性よりさらに一層弱かった(図15A)。hOPN5−BSAとのHu33E10−VH7/VL6の結合親和性は、Ch33E10より3〜4倍弱かった(図15A)。Hu33E10−VH7.8/VL6の親和性は、この実験中Ch33E10の親和性より約3倍低かった(図15B)。反復実験中、Hu33E10−VH7/VL6とHu33E10−VH7.8/VL6は、hOPN5−BSAとの結合に関して互いに同様に挙動した。
【0201】
6.7.Hu33E10VLv2の作製および特徴付け
VL1遺伝子の予備突然変異分析によって、Hu33E10−VH1/VL6の結合親和性の消失に貢献するいかなる特定のフレームワークアミノ酸残基も確認することはできなかったが(データ示さず)、VL1遺伝子は部分的には親和性消失の原因であることが疑われた。VL1遺伝子をさらに特徴付ける代わりに、異なるヒトフレームワークに基づく新たなヒト化33E10VL(VLv2)を設計した(図19)。VLv2用に、M29467のヒトアミノ酸配列(GenBank受託番号)をアクセプターとして選択した。CDRとの有意な相互作用が予想された位置22および37では、マウス33E10VLのアミノ酸残基はヒト化型で保持された。NheIおよびEcoRI部位と隣接するHu33E10VLv2遺伝子(図20)はGenScript USA Inc.(Piscataway、NJ)で合成された。VL6遺伝子とVLv2遺伝子は、同じシグナルペプチド配列およびスプライスドナーシグナルを共有する(図14および20)。
【0202】
NheIおよびEcoRIで消化したHu33E10VLv2遺伝子をpHu33E10−4においてVL1に置換して、新たな発現ベクターpHu33E10−6を構築した。Hu33E10−VH7/VLv2IgG1/κを産生するためにpHu33E10−6で安定的にトランスフェクトしたNS0細胞は、セクション4中に記載したように生成した。トランスフェクタントの1つ、NS0−Hu33E10−6I−15(SP2−1I−15とも呼ばれる)を適合させ、ハイブリドーマSFM中で増殖させ、Hu33E10−VH7/VLv2抗体は、セクション4中に記載した手順を使用して培養上清から精製した。
【0203】
hOPN5−BSAとHu33E10−VH7/VLv2の結合は、セクション5中に記載したようにELISAによって分析した。2つの独立した実験の結果を図21中に示す。両方の実験において、Hu33E10−VH7/VLv2の結合親和性はCh33E10の結合親和性の2倍以内であった。
【0204】
6.8.NS0安定トランスフェクタントの特徴付け
PCRマイコプラズマ検出セット(Takara Bio USA、Madison、WT)を用いた試験によって、NS0−Ch33E102C11、NS0−Hu33E10−45D3、NS0−Hu33E10−51E5およびNS0−Hu33E10−6−I−15はいずれも、マイコプラズマの存在に関して陰性であったことを示した。
【0205】
それぞれNS0−Hu33E10−45D3、NS0−Hu33E10−51E5およびNS0−Hu33E10−6−I−15において産生されたHu33E10−VH7/VL6、Hu33E10−VH7.8/VL6およびHu33E10−VH7/VLv2IgGl/κ抗体の重鎖および軽鎖の真正を、cDNA配列決定によって確認した。全RNAはTRIzol試薬(Invitrogen)を使用してこれらの細胞から抽出し、オリゴdTプライマー処理cDNAは、供給者のプロトコールに従いRT−PCR用のSuperScriptIII第一鎖合成システム(Invitrogen)を使用して合成した。γ−1重鎖のコード領域は、プライマーとしてCMV2およびJNT098(図22)ならびにPhusion DNAポリメラーゼを使用しPCRによって増幅した。PCR断片はゲル精製し、プライマーとしてCMV2、JNT082、JNT097およびJNT098を用いて配列決定に施した(図22)。同様に、kappa軽鎖のコード領域はCMV2およびJNT026を使用して増幅し(図22)、ゲル精製したDNA断片はプライマーとしてCMV2、JNT026、JNT080およびJNT084を用いて配列決定に施した(図22)。重鎖および軽鎖コード領域に関して得た配列は、NS0−Hu33E10−45D3、NS0−Hu33E10−51E5およびNS0−Hu33E10−6−I−15安定トランスフェクタントのそれぞれに関する発現ベクター中の対応する配列と完全に一致した。Hu33E10VH7およびVH7.8からなるγ−1重鎖の全コード領域に関する配列は、それぞれ図23および24中に示す。Hu33E10VL6およびVLv2からなるkappa軽鎖の全コード領域に関する配列は図25および26中に示す。
【0206】
さらに、NS0−Ch33E102C11において産生されたCh33E10IgG1/κ抗体のVHおよびVL領域の真正を、cDNA配列決定によって確認した。全RNAの抽出およびcDNA合成は前に記載したように実施した。VHコード領域は、プライマーとしてCMV2およびJNT082ならびにPhusionポリメラーゼを使用しPCRによって増幅した。PCR断片はゲル精製し、プライマーとしてCMV2およびJNT082を用いて配列決定に施した。同様に、VLコード領域はCMV2およびJNT026を使用して増幅し、ゲル精製したDNA断片はプライマーとしてCMV2およびJNT026を用いて配列決定に施した。得られたVHおよびVL領域のヌクレオチド配列は、pCh33E10発現ベクター中の対応する配列と完全に一致した(それぞれ図3および4)。
【0207】
6.9.結論
hOPN5−BSAに対するHu33E10−VH7/VLv2IgG1/κの親和性は、Ch33E10IgG1/κの親和性の3倍以内であったと決定した。
【0208】
6.10.細胞接着阻害アッセイ
図27は、ヒト化抗OPN抗体が、hOPN5−BSAとMDA−MB−435Sの接着を阻害したことを示す。MDA−MB−435Sヒト乳癌細胞系はアメリカンタイプカルチャーコレクション(ATCC)から得て、10%ウシ胎児血清を補充したDMEM中で培養した。96ウェルプレートは37℃において1時間hOPN5−BSA(2μg/ml)でコーティングし、次に室温において1時間0.5%BSAで処理した。ヒト免疫グロブリンG1kappa、Ch33E10、Hu33E10−VH7.8/VL6(EJ3−1−1)、Hu33E10−VH7/VL6(E3−1)、Hu33E10−VH7/VLv2(SP−2)、およびマウス33E10抗体を0.25%BSA、DMEM中に溶かし、100μl/mlで始めて様々な濃度で処理し、37℃において20分間連続2倍希釈した。2×104個のMDA−MB−435S細胞をそれぞれのウェルに加えた。37℃で1時間のインキュベーション後、培地を除去し、(37℃まで温めた)温かいPBSで軽く二回洗浄した。接着細胞は固定し、20%メタノールに溶かした0.5%クリスタルバイオレットで染色し、20%酢酸で溶かした。吸光度は590nmで測定した。抗体を加えずに、阻害率はこの吸光度で標準化した。
【0209】
7.寄託
マウス抗RGDモノクローナル抗体を産生する33E10および35B6として本明細書に示すハイブリドーマは、微生物の寄託に関するブダペスト条約に従い、2005年10月27日に茨城県つくば市東1−1−1中央第6(郵便番号:305−8566)に位置する産業技術総合研究所の特許生物寄託センターに寄託され、それぞれ受託番号FERMBP−10440およびFERMBP−10441を与えられ、これらはいずれもその全容を参照によって本明細書に組み込む。
【産業上の利用可能性】
【0210】
本発明のヒト化モノクローナル抗体は、RGDタンパク質の機能を阻害して、癌、例えば癌細胞の増殖または転移、および炎症疾患、例えば慢性関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、糖尿病、癌転移、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などに対する治療効果を示す。本発明の抗RGD抗体と抗インテグリン抗体の両方を含む医薬組成物は、癌および炎症性疾患に対してより改善された治療効果を発揮する。
【0211】
具体的な実施形態を本明細書で例示および記載してきたが、様々な他のおよび/または均等な履行を、本発明の範囲から逸脱せずに、記載し示した具体的な実施形態に置き換えることができることは、当業者によって理解されよう。本出願は、本明細書で論じる具体的な実施形態の任意の適合または変形を含むと考えられる。したがって、本発明は特許請求の範囲およびその均等物によってのみ制限されると考えられる。
【技術分野】
【0001】
本出願を通じて、いくつかの参照文献を本明細書で参照する。これらの参照文献の開示は、参照によってそれらの全容を本出願中にここで組み込む。
【0002】
本発明は、細胞外マトリックスタンパク質のアミノ酸配列RGD(Arg−Gly−Asp)を免疫特異的に認識するヒト化抗体、ならびに癌、炎症疾患、自己免疫疾患、感染疾患、骨疾患などを含めた様々な疾患または障害用のそれらの治療および診断用途に関する。
【背景技術】
【0003】
細胞接着は多細胞生物の生命を維持する際に重要な役割を果たす。多細胞生物の細胞接着は、細胞−細胞外マトリックス(本明細書では以後「ECM」と略す)接着および細胞間接着に分類される。細胞−ECM接着はインテグリンによって仲介され、かつ細胞間接着はカドヘリン、クラウジンおよびネクチンによって仲介されることが解明されている。
【0004】
インテグリンなどの膜貫通型接着タンパク質は細胞−ECM接着を構成する。インテグリンがα鎖とβ鎖のヘテロ二量体を形成することが報告されている。少なくとも18種(types)の鎖、8種のβ鎖および24種のαβヘテロ二量体がこれまでに同定され確認されている。それぞれの種類のインテグリンが、特異的リガンドを認識することが知られている。インテグリンを含めた膜貫通型接着タンパク質は、細胞接着以外に、ECMから細胞への細胞内シグナル伝達、ならびに増殖、移動性および分化の制御と関係がある(F.G Giancotti, et al., Science, 285, 1028-1032, 1999)。
【0005】
多くのタンパク質は、コラーゲン(I−XIX型など)、非コラーゲン性糖タンパク質(オステオポンチン(OPN)、ビトロネクチン、フィブロネクチン、フォンヴィレブランド因子、ラミニン、テナシン、フィブリノゲン、トロンボスポンジンなど)、エラスチンおよびプロテオグリカンに分類されるECMタンパク質として知られている。これらのECMタンパク質は対応するインテグリンと結合し、細胞内シグナル伝達経路を活性化して、細胞骨格系、移動性、増殖、分化などを制御する。ECMタンパク質結合インテグリンは、ECMタンパク質の種類(type)に応じて特異的シグナルを送ることにより、これらのシグナル活性化経路を制御する。RGD(アルギニン−グリシン−アスパラギン酸)配列は多くのECMタンパク質の細胞接着領域において一般に観察され、インテグリンと結合することによって様々な機能を示すようである。ECMタンパク質のRGD配列は薬剤標的であり得ると予想されており、いくつかの低分子化合物および人工ペプチドが提供されている。
【0006】
いくつかの型のインテグリン、α3β1インテグリン、α5β1インテグリン、α8β1インテグリン、ανβ1インテグリン、ανβ3インテグリン、ανβ5インテグリン、ανβ6インテグリン、ανβ8インテグリンなどは、RGD配列と結合することが知られている。α5β1インテグリンとその特異的リガンドフィブロネクチンの間の相互作用は、インテグリン仲介シグナル伝達の機構を調べるために明らかにされており、α5β1インテグリンは細胞接着および細胞移動だけでなく、細胞分化および細胞死亡率も制御することが報告される(S.M.Frisch et al., Curr.Opin.Cell Biol., 9, 701-706, 1997)。α5β1インテグリンは腫瘍細胞において高度に発現され、癌の悪性化と関係があることも示されている。それぞれのインテグリン仲介シグナルはECMタンパク質の結合に応じて異なる。例えば、増殖因子による刺激はフィブロネクチン結合内皮細胞の増殖を活性化するが、ラミニン−1結合内皮細胞の増殖は阻害する。さらに、ラミニン−10/11からα3β1インテグリンに伝わるシグナルはフィブロネクチンからα5β1インテグリンに伝わるシグナルと異なり、癌細胞の移動性を有意に高め(J.Gu et al., J.Biol.Chem., 276, 27090-27097, 2001)、血清飢餓によるアポトーシスを有意に回避する(J.Gu et al., J.Biol.Chem., 277, 19922-19928、2002)。RGD配列結合αvインテグリンの高発現は破骨細胞および新生血管において観察されており、RGD配列およびαvインテグリンの阻害は骨粗しょう症および癌に関する治療薬剤の標的として予想されている。α5β1インテグリンは腫瘍細胞において高度に発現され、癌の悪性化と関係があることが示されている。これらの発見から、抗α5β1インテグリン抗体(ボロシマブ)、抗α4インテグリン抗体(ナタリズマブ)、抗αvβ3インテグリン抗体(ビタキシン)が、インテグリンとECMタンパク質の間の相互作用を阻害するアンタゴニスト抗インテグリン抗体薬剤として開発されている。
【0007】
一方、コラーゲン、オステオポンチン(OPN)、ビトロネクチン、フィブロネクチン、フォンヴィレブランド因子、ラミニン、テナシン、フィブリノゲンおよびトロンボスポンジンなどのいくつかのECMタンパク質は、RGD配列を含むことが知られている。さらに、数種のウイルスおよび数種の細菌は、細胞と接着するためにRGD配列を有することが知られている。OPNは、骨中に多量に含有されるカルシウムとの結合性を有する酸性糖タンパク質である。OPNは、細胞接着、細胞移動、腫瘍形成、免疫応答および補体仲介細胞溶解において重要な役割を果たすことが報告される。OPNノックアウトマウスおよび抗OPN中和抗体の結果は、OPNが肝炎、慢性関節リウマチなどの自己免疫疾患、および癌の転移と関係があることを示す。したがって、ECMタンパク質と細胞の結合の阻害剤は、骨粗しょう症または癌の治療に使用することができることが予想される。したがって、インテグリンを標的化する前述のアンタゴニスト薬剤以外に、インテグリンの結合パートナーであるECMタンパク質を標的化するアンタゴニスト薬剤が開発されている。
【発明の概要】
【発明が解決しようとする課題】
【0008】
インテグリンとのRGD配列仲介相互作用を阻害する低分子、OPNに対する抗体、およびインテグリンに対する抗体などの薬剤は開発されているが、RGD配列を特異的に認識する抗体に関する報告は存在しない。RGD配列はECMタンパク質中の保存配列の1つなので、RGD配列を特異的に認識する抗体はヒトと治療モデル動物の両方において影響を有すると予想され、したがって治療剤の開発に非常に重要な活性成分として考えられる。したがって、RGD配列を特異的に認識する抗体が必要とされている。以前、本発明者らは、RGD配列を免疫特異的に認識しハイブリドーマクローン33E10および35B6(それぞれ寄託受託番号FERMBP−10440およびFERMBP−10441)によって産生されるマウスモノクローナル抗体を単離した。本明細書では、ハイブリドーマクローンの名称は、クローンによって産生されるモノクローナル抗体の名称として交互に使用する。全てのこれらのマウス抗RGD抗体はIgG1アイソタイプであった。これらのモノクローナル抗体は、オステオポンチンなどのECMタンパク質のRGD配列との結合によって、ECMと細胞の間でRGD配列によって仲介される結合に干渉することを観察した。したがって、これらの抗RGD抗体は、例えば癌、例えば癌細胞の増殖または転移、および炎症疾患、例えば慢性関節リウマチ、骨関節炎、感染疾患、肝炎、気管支喘息、線維症、糖尿病、動脈硬化症、多発性硬化症、肉芽腫、および炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患、骨粗しょう症などの、RGD配列関連疾患に対して治療または診断効果を示すと予想される。
【0009】
しかしながら、これらのモノクローナル抗体はマウス起源なので、ヒトにおけるそれらの免疫原性が原因で考えられる悪影響が、ヒトにおける診断または治療用途へのそれらの直接施用を妨げている。免疫原性を低減するために、本発明者らは、前記ヒト化抗体が由来する原型マウス抗RGD抗体によって示される活性に相当する、生物活性を有するヒト化抗体を調製した。
【課題を解決するための手段】
【0010】
したがって、本発明は、RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合断片を提供し、前記抗体は部分的に非ヒト起源に由来し部分的にヒト起源に由来する抗原結合領域を含む。本発明の一態様では、本発明のヒト化抗体またはその抗原結合断片は、33E10および35B6モノクローナル抗体などの非ヒト供給源(ドナー)に由来する相補性決定領域(本明細書では以後「CDR」と略す)、およびヒト供給源に由来するフレームワーク領域(本明細書では以後「FR」と略す)(アクセプター)を含む。前記ヒト化抗体またはその抗原結合断片は、RGD配列とそのリガンドの間の結合を阻害することができる。
【0011】
本発明の一態様では、RGD配列を免疫特異的に認識する前記ヒト化抗体またはその抗原結合断片は、(i)ヒトH鎖の可変領域(本明細書では以後「V領域」と略す)に由来する少なくとも1つのH鎖FR(本明細書では以後「FRH」と略す)、およびRGD配列を免疫特異的に認識する非ヒト抗体のCDRHの少なくとも1つに由来する少なくとも1つのH鎖CDR(本明細書では以後「CDRH」と略す)を含む重鎖(本明細書では以後「H鎖」と略す)、または(ii)ヒトL鎖のV領域に由来する少なくとも1つのL鎖FR(本明細書では以後「FRL」と略す)、およびRGD配列を免疫特異的に認識する非ヒト抗体のCDRLの少なくとも1つに由来する少なくとも1つのL鎖CDR(本明細書では以後「CDRL」と略す)を含む軽鎖(本明細書では以後「L鎖」と略す)、または前述の(i)および(ii)の両方を含む。
【0012】
本発明の好ましい態様では、本発明のヒト化抗体またはその抗原結合断片は、(i)VHを含むH鎖であって、前記VHが配列番号92、94または96のアミノ酸配列を含むH鎖、または(ii)VLを含むL鎖であって、前記VLが配列番号98または100のアミノ酸配列を含むL鎖、または(iii)前述の(i)および(ii)の両方を含む。本発明の別の態様では、本発明の前記ヒト化抗体またはその抗原結合断片は、配列番号91、92または93のヌクレオチド配列によってコードされるアミノ酸配列を含み、前記VLが配列番号97または99のヌクレオチド配列によってコードされるアミノ酸配列を含む。
【0013】
本発明の好ましい態様では、前記VHが配列番号91、92または93のヌクレオチド配列によってコードされるアミノ酸配列を含み、前記VLが配列番号97または99のヌクレオチド配列によってコードされるアミノ酸配列を含む。
【0014】
本発明の別の態様では、前記VHが配列番号94のアミノ酸配列を含み、前記VLが配列番号100のアミノ酸配列を含む。
【0015】
本発明の別の態様では、前記H鎖が配列番号25または27のアミノ酸配列を含み、前記L鎖が配列番号29または31のアミノ酸配列を含む。
【0016】
本発明のさらなる態様では、前記H鎖が配列番号25のアミノ酸配列を含み、前記L鎖が配列番号31のアミノ酸配列を含む。
【0017】
本発明は、RGD配列を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合断片のH鎖またはL鎖、または両方をコードするヌクレオチド配列を含むベクター、例えば発現ベクターをさらに提供する。このようなベクターでは、本発明のヌクレオチド配列は、1つまたは複数の制御エレメントに作動可能に連結することができる。本発明のヌクレオチド配列は、CDRが由来する非ヒトドナー抗体に固有のシグナルペプチド、または異種起源のシグナルペプチドをコードするヌクレオチド配列を含むことができる。
【0018】
さらに、本発明は、本発明の核酸分子を含むベクターを含む、本発明の核酸分子を含む宿主細胞を提供する。本発明の一態様では、本発明は、本発明のヒト化H鎖をコードする第一の核酸分子および本発明のヒト化L鎖をコードする第二の核酸分子を含む単離宿主細胞を提供し、前記第一および第二の核酸分子は、生物学的に機能性がある本発明のヒト化抗体またはその抗原結合断片が発現されるような形式で、それぞれ制御エレメントに作動可能に連結する。
【0019】
本発明は、本発明のヒト化抗体またはその抗原結合断片を調製するための方法であって、ヒト化抗体またはその抗原結合断片が発現されるような条件下で本発明の宿主細胞を培養すること、および生成したヒト化抗体を回収することを含む方法をさらに提供する。
【0020】
本発明は、本発明のヒト化抗体またはその抗原結合断片の少なくとも1つを含む組成物をさらに提供する。さらに本発明は、本発明のヒト化抗体またはその抗原結合断片の少なくとも1つ、および薬学的に許容される担体を含む、RGDタンパク質と関係がある障害または疾患を予防または治療するための医薬組成物を提供する。前記組成物のいずれかは、障害または疾患を相加的または相乗的に改善することができる別の活性化合物をさらに含むことができる。前記活性化合物には、抗炎症性化合物、化学療法化合物などがあるが、これらだけには限られない。前記活性化合物は、低分子化合物および抗体またはその抗原結合断片、ヒトα4インテグリン特異的抗体またはヒトα9インテグリン特異的抗体なども含む。
【0021】
別の態様では、本発明は、RGDタンパク質と関係または関与がある障害または疾患を予防または治療するための方法を提供し、前記方法は本発明のヒト化抗体またはその抗原結合断片の少なくとも1つを予防または治療有効量、その必要性がある対象に投与することを含む。このような用途のため、本発明のヒト化抗体またはその抗原結合断片を、ヒト化抗体またはその抗原結合断片の生物学的効果を高める治療成分と結合させることが可能である。このような治療成分の例には、別の抗体、細胞増殖抑制または細胞致死性である細胞毒素、放射性元素、および/または抗炎症剤、抗生物質を含めた他の治療剤などがある。
【0022】
さらに別の態様では、本発明は、RGDタンパク質と関係または関与がある対象中の障害または疾患を診断するための方法を提供し、前記方法は診断有効量の本発明のヒト化抗体またはその抗原結合断片を、検査する対象に投与することを含む。このような診断用途のため、本発明のヒト化抗体を、放射性元素などの検出可能マーカーで標識することができる。
【0023】
3.1.定義
本明細書で使用する用語「抗体」は、所望の抗原またはRGD配列などの所望の配列と免疫特異的に結合することができる抗体分子を指し、これは抗体分子全体を包含し、抗体の抗原結合断片を含んだ抗体の断片を包含することができる。
【0024】
本明細書で使用する用語「抗原結合断片」は、標的ポリペプチド、タンパク質または配列、特にRGD配列と免疫特異的に結合する能力を保持する抗体の任意の断片を指し、これは単鎖抗体、Fab断片、F(ab’)2断片、ジスルフィド結合Fvs、および標的ポリペプチド、タンパク質または配列と特異的に結合するVLおよび/またはVHまたはCDRのいずれかを含有する断片を含む。したがって、ヒト化抗体のこのような抗原結合断片は、部分または完全長ヒト定常領域を含むかまたは含まない可能性がある。前に記載した抗体断片を得るための様々な方法は当技術分野でよく知られている。
【0025】
本明細書で使用する用語「免疫特異的に認識する」は、標的ポリペプチド、タンパク質または配列、特にヒトRGD配列と特異的に結合する抗体またはその抗原結合断片の能力を指す。このような抗体が、他のポリペプチドまたはタンパク質と非特異的に結合することはない。しかしながら、標的ポリペプチドまたはタンパク質(例えば、RGDタンパク質)と免疫特異的に結合する抗体またはその抗原結合断片は、他の抗原と交差反応する可能性がある。例えば、ヒトRGDタンパク質を免疫特異的に認識する本発明のヒト化抗体またはその抗原結合断片は、マウスRGDタンパク質と交差反応する可能性がある。RGDタンパク質を免疫特異的に認識する抗体またはその抗原結合断片は、他の抗原と交差反応しないことが好ましい。
【0026】
本明細書で使用する用語「ヒト供給源由来」または「非ヒト供給源由来」は、そのアミノ酸配列がそれぞれヒト抗体または非ヒト抗体の対応する部分に由来する抗体部分を指す。
【0027】
本明細書で使用する用語「アクセプター配列」は、通常非ヒト抗体であるドナー抗体由来のCDRに関するアクセプターとして働くヒト抗体VHまたはVL由来のFRの、ヌクレオチド配列またはアミノ酸配列を指す。
【0028】
本発明を例示する目的で、図1〜27は現在好ましい形態を反映する。しかしながら、本発明が図1〜27中に示す通りの形に限られないことは理解される。
【図面の簡単な説明】
【0029】
【図1】推定アミノ酸配列と共に示すマウス33E10VHcDNAのヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VHのN末端アミノ酸残基(E)には二重下線を引く。Kabat et al.(Sequences of Proteins of Immunological Interests, Fifth edition, NIH公開番号91−3242、米国厚生省、1991)の定義に従うCDR配列には下線を引く。
【図2】推定アミノ酸配列と共に示すマウス33E10VLcDNAのヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。
【図3】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接する設計33E10VH遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VHのN末端アミノ酸残基(E)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図4】推定アミノ酸配列と共に示すNheIおよびEcoRI部位(下線)と隣接する設計33E10VL遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図5】キメラおよびヒト化33E10抗体用の発現ベクター(集合的に「発現ベクター」)の概略構造を示す図である。頂上部のSalI部位から時計回りに進み、プラスミドは、抗体重鎖遺伝子の転写を開始するためのヒトサイトメガロウイルス(CMV)主要即時初期プロモーターおよびエンハンサー(CMVプロモーター)で始まる重鎖転写単位を含有する。CMVプロモーターに、VHエクソン、CH1を含むヒトγ−1重鎖定常領域を含有するゲノム配列、ヒンジ、介在イントロンを有するCH2およびCH3エクソン、およびCH3エクソンの後にポリアデニル化部位が続く。重鎖遺伝子配列の後、軽鎖転写単位が、CMVプロモーターで始まり、VLエクソン、およびそれに先行するイントロンの一部分を有するヒトkappa鎖定常領域エクソン(CL)を含有するゲノム配列、およびCLエクソン後のポリアデニル化部位が続く。次いで軽鎖遺伝子に、SV40初期プロモーター(SV40プロモーター)、大腸菌(E.coli)キサンチングアニンホスホリボシルトランスフェラーゼ遺伝子(gpt)、およびSV40ポリアデニル化部位(SV40ポリ(A)部位)を含有するセグメントが続く。最後に、プラスミドは、細菌複製起点(pUCori)およびβ−ラクタマーゼ遺伝子(βラクタマーゼ)を含むプラスミドpUC19の一部分を含有する。関連制限酵素部位の位置をこの図中に示す。
【図6】33E10VH、ヒト化33E10(Hu33E10)VHおよびヒトアクセプターU03400(GenBank受託番号)のアミノ酸配列のアラインメントを示す図である。アミノ酸残基は一文字コードで示す。配列の上の数字はKabat et al.(1991)に従う位置を示す。Kabat et al.(Sequences of Proteins of Immunological Interests, Fifth edition, NIH公開番号91−3242、米国厚生省、1991)によって定義されたCDR配列には下線を引く。二重下線を引いた残基はCDRと接触したと予想し、マウス残基はヒト化型でこれらの位置に保持された。U03400中のCDR残基はこの図中では省略する。
【図7】33E10VL、ヒト化33E10(Hu33E10)VLおよびヒトアクセプターX72452(GenBank受託番号)のアミノ酸配列のアラインメントを示す図である。アミノ酸残基は一文字コードで示す。配列の上の数字はKabat et al.(1991)に従う位置を示す。Kabat et al.(1991)によって定義したCDR配列には下線を引く。X72452中のCDR残基はこの図中では省略する。
【図8】Hu33E10VH1遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図9】Hu33E10VL1遺伝子の構築に使用したオリゴヌクレオチドを示す図である。
【図10】Hu33E10VH1遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印はそれぞれのオリゴヌクレオチドの位置および方向(5’から3’)を示す。アミノ酸残基は一文字コードで示す。
【図11】Hu33E10VL1遺伝子の構築に使用したオリゴヌクレオチドを示す図である。矢印はそれぞれのオリゴヌクレオチドの位置および方向(5’から3’)を示す。アミノ酸残基は一文字コードで示す。
【図12】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接するHu33E10VH1遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VHのN末端アミノ酸残基(E)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図13】推定アミノ酸配列と共に示すNheIおよびEcoRI部位(下線)と隣接するHu33E10VL1遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図14】推定アミノ酸配列と共に示すNheIおよびEcoRI部位(下線)と隣接するHu33E10VL6遺伝子のヌクレオチド配列を示す図である。シグナルペプチドコード領域中のスプライシングドナー部位を排除するためのサイレント突然変異には二重下線を引く。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図15A−B】hOPN5−BSAとキメラおよびヒト化33E10IgGl/κ抗体の結合のELISA分析を示す図である。精製Ch33E10、Hu33E10−VHl/VL6、Hu33E10−VH5/VL6およびHu33E10−VH7/VL6(図15A)ならびにCh33E10およびHu33E10−VH7.8/VL6(図15B)を、100μg/mlで始めて連続2倍希釈で、hOPN5−BSAとの結合に関して様々な濃度において試験した。実験は二連で実施した。
【図16】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接するHu33E10VH5遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。二重下線を引いたアミノ酸残基はVH1との違いを示す。
【図17】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接するHu33E10VH7遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。二重下線を引いたアミノ酸残基はVH1との違いを示す。
【図18】推定アミノ酸配列と共に示すSpeIおよびHindIII部位(下線)と隣接するHu33E10VH7.8遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。二重下線を引いたアミノ酸残基はVH1との違いを示す。
【図19】33E10VL、Hu33E10VLv2およびヒトアクセプターM29467(GenBank受託番号)のアミノ酸配列のアラインメントを示す図である。アミノ酸残基は一文字コードで示す。配列の上の数字はKabat et al.(1991)に従う位置を示す。Kabat et al.(1991)によって定義したCDR配列には下線を引く。二重下線を引いた残基はCDRと接触したと予想し、マウス残基はヒト化型でこれらの位置に保持された。M29467中のCDR残基はこの図中では省略する。
【図20】推定アミノ酸配列と共に示すNheIおよびEcoRI部位(下線)と隣接するHu33E10VLv2遺伝子のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。シグナルペプチド配列は斜字体である。成熟VLのN末端アミノ酸残基(D)には二重下線を引く。Kabat et al.(1991)の定義に従うCDR配列には下線を引く。イントロン配列は斜字体である。
【図21A−B】hOPN5−BSAとCh33E10およびHu33E10−VH7/VLv2IgGl/κ抗体の結合のELISA分析を示す図である。精製Ch33E10およびHu33E10−VH7/VLv2抗体を、200μg/ml(図21A)または100μg/ml(図21B)で始めて連続2倍希釈で、hOPN5−BSAとの結合に関して様々な濃度において試験した。二連での2つの独立した実験の結果を示す(図21Aおよび21B)。
【図22】Hu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチドの配列を示す図である。
【図23】推定アミノ酸配列と共に示すNS0−Hu33E10−4およびNS0−Hu33E10−6において発現されるVH7およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。終結コドンは「・」によって示す。
【図24】推定アミノ酸配列と共に示すNS0−Hu33E10−5において発現されるVH7.8およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。終結コドンは「・」によって示す。
【図25】推定アミノ酸配列と共に示すNS0−Hu33E10−4およびNS0−Hu33E10−5において発現されるVL6およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。終結コドンは「・」によって示す。
【図26】推定アミノ酸配列と共に示すNS0−Hu33E10−6において発現されるVLv2およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列を示す図である。アミノ酸残基は一文字コードで示す。終結コドンは「・」によって示す。
【図27】ヒト化抗OPN抗体が、hOPN5−BSAとMDA−MB−435Sの接着を阻害したことを示す図である。MDA−MB−435S(4×104細胞/ウェル)を、様々な濃度の抗体、ヒトIgG、Ch33E10、EJ3−1−1(Hu33E10−VH7.8/VL6)、R3−1(Hu33E10−VH7/VL6)、SP2−1(Hu33E10−VH7/VLv2)、およびm33E10の存在下において、hOPN5−BSA(2mg/ml)でプレコーティングした96ウェルプレートに接着させた。データは三連の実験の平均として表す。
【発明を実施するための形態】
【0030】
5.1 RGD配列に対する抗体の調製
RGD配列を免疫特異的に認識する抗体は、当技術分野で知られている任意の適切な方法によって作製することができる。
【0031】
RGDタンパク質またはペプチドは本発明の細胞接着「RGD」配列(本明細書では以後「RGDペプチド」と略す)を含み、または(1)RGDタンパク質を発現するヒトECMに由来、またはこれらのECMが存在する全組織に由来してよく、(2)細菌、酵母、動物細胞などの細胞系などへのトランスフェクションによるRGDタンパク質またはRGDペプチドをコードするDNA(好ましくはcDNA)の発現によって得られる組換えタンパク質またはペプチドであってよく、または(3)合成タンパク質またはペプチドであってよい。
【0032】
本発明で抗原として使用するRGDペプチドは、免疫処置によってRGD配列に対する抗体を産生することができる。RGDペプチドは、マウスECMタンパク質の細胞接着配列であるRGDペプチドアミノ酸配列CVDVPNGRGDSLAYGLR(配列番号71)を含む。RGDタンパク質またはRGDペプチドは、OPN、ビトロネクチン、フィブロネクチン、フォンヴィレブランド因子、コラーゲン、ラミニン、テナシン、フィブリノゲン、トロンボスポンジン、およびこれらの断片を含むRGDなどを含む。アミノ酸の置換、欠失、修飾および付加などの人為的または天然変異を、タンパク質またはペプチドがRGD−配列を含む限り、前記タンパク質または前記ペプチドに施すことができる。変異体タンパク質またはペプチドは、複数のアミノ酸、好ましくは1〜10個のアミノ酸、およびより好ましくは1〜数個(例えば、1〜5個)のアミノ酸が置換、欠失、修飾、付加または挿入されたアミノ酸配列を含むことができる。
【0033】
本明細書では、RGDペプチドは、少なくとも約5個のアミノ酸、好ましくは約5〜50個のアミノ酸、およびより好ましくは約10〜20個のアミノ酸を含む。本発明における抗原としてのRGDタンパク質またはRGDペプチドは、化学合成法、細胞培養法、遺伝子組換え法およびその適切な変形などの、当技術分野でよく知られている方法を使用することによって産生することができる。例えば、RGDペプチドは、適切にプロテアーゼでECMタンパク質を切断することによって得ることができる。RGDタンパク質またはRGDペプチドは、マウス、ラット、ウサギ、ブタ、ウシ、サルおよびヒトなどの哺乳動物に由来してよい。当技術分野でよく知られている任意の方法を、抗RGD抗体の調製に使用することができるRGDタンパク質またはRGDペプチドの調製に使用することができる。
【0034】
変異体ポリペプチドを生成するための方法の例には、合成オリゴヌクレオチド部位特異的突然変異誘発法(ギャップデュプレックス法)、硝酸塩または亜硝酸塩を用いた処理によるアトランダムな点突然変異の導入を含む点突然変異誘発法、Bal31酵素、または他の酵素を用いた欠失変異体の調製を含む方法、カセット突然変異誘発法、リンカースキャニング法、ミスインコーポレーション法、ミスマッチプライマー法、DNAセグメント合成法などがある。
【0035】
RGDペプチドは、チログロビン、キーホールリンペットヘモシアニン(KLH)、ウシ血清アルブミン(BSA)、オボアルブミン(OVA)またはウシグロブリン、好ましくはチログロビンなどの、他の生体高分子と結合させることが可能である。生体高分子にRGDペプチドを結合させるための方法は、活性エステル基およびマレイミド基を有する結合試薬などのカップリング試薬を使用することによって(活性エステル基はタンパク質またはペプチドのアミノ基と結合し、マレイミド基はタンパク質またはペプチドのチオール基と結合する、S.yoshirake et al., Eur.J.Biochem., 101, 395-399, 1979)、混成無水物法(B.F.Erlanger et al., J.Biol. Chem., 234, 1090-1094, 1954)を使用することによって、または活性エステル法(A.E.Karu et al., J.Agric.Food Chem., 42, 301-309, 1994)を使用することによって実施することができる。生体高分子にRGDペプチドを結合させるための方法は、カップリング試薬を使用することによって実施することが好ましい。
【0036】
抗原として、RGDタンパク質またはRGDペプチドを過剰発現する細胞自体を使用することもできる。RGDタンパク質またはRGDペプチドを過剰発現する細胞は、当技術分野でよく知られている組換えDNA技術によって調製することができる。
【0037】
前に記載したように調製した適切な抗原を使用して、RGD配列に特異的な抗体は、当技術分野でよく知られている様々な方法によって調製することができる。RGD配列に対するポリクローナル抗体は、当技術分野でよく知られている様々な手順によって産生することができる。例えば、対象の抗原を、ウサギ、マウス、ラットなどだけには限られないが、これらを含めた様々な宿主動物に投与して、抗原に特異的なポリクローナル抗体を含有する抗血清の産生を誘導することができる。フロイント(完全および不完全)アジュバント、水酸化アルミニウムなどのミネラルゲル、リゾレシチン、プルロニックポリマー、ポリアニオン、ペプチド、油エマルジョン、キーホールリンペットヘモシアニン、ジニトロフェノールなどの表面活性物質、およびBCG(カルメットゲラン桿菌(Bacille Calmette-Guerin)およびコリネバクテリウムパルバム(Corynebacterium parvum)などのヒトにおそらく有用なアジュバントだけには限られないが、これらを含めた様々なアジュバントを使用して、宿主の種類に応じて免疫応答を増大することができる。このようなアジュバントも当技術分野でよく知られている。
【0038】
モノクローナル抗体は、ハイブリドーマ、組換えおよびファージディスプレイ技術、またはこれらの組合せの使用を含めた、当技術分野で知られている広く様々な技術を使用することによって調製することができる。例えば、モノクローナル抗体は、当技術分野で知られており例えば(いずれもその全容を参照によって本明細書に組み込む)Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed.1988); Hammerling, et al., Monoclonal Antibodies and T-Cell Hybridomas, pp.563-681 (Elsevier, N.Y, 1981)中で教示された技法を含めた、ハイブリドーマ技法を使用することによって産生することができる。本明細書で使用する用語「モノクローナル抗体」は、ハイブリドーマ技術によって産生される抗体に限られない。用語「モノクローナル抗体」は単一クローンに由来する抗体を指し、それが産生される方法に限られず、任意の真核生物、原核生物またはファージクローンを含む。
【0039】
ハイブリドーマ技術を使用して特異的抗体を産生しスクリーニングするための方法は通常のことであり、当技術分野でよく知られている。非制限的な例では、対象の抗原またはこのような抗原を発現する細胞でマウスを免疫処置することができる。免疫応答を検出した後、例えば、抗原に特異的な抗体をマウス血清中で検出した後、マウスの脾臓を採取し脾臓細胞を単離した。次いで脾臓細胞は、よく知られている技法によって任意の適切なミエローマ細胞(例えば、P3U1、P3X63−Ag8、P3X63−Ag8−U1、P3NS1−Ag4、SP2/0−Ag14、P3X63−Ag8−653など)と融合させる。ハイブリドーマを選択し、限界希釈によってクローニングする。次いでハイブリドーマクローンを、当技術分野で知られている方法によって、抗原と結合することができる抗体を分泌する細胞に関してアッセイする。高レベルの抗体を一般に含有する腹水は、陽性ハイブリドーマクローンをマウスに腹腔内接種することによって生成することができる。
【0040】
特異的エピトープを認識する抗体断片は知られている技法によって作製することができる。例えば、FabおよびF(ab’)2断片は、パパイン(Fab断片産生用)またはペプシン(F(ab’)2断片産生用)などの酵素を使用して、免疫グロブリン分子のタンパク質分解切断によって産生することができる。F(ab’)2断片は完全L鎖、およびH鎖のV領域、CH1領域およびヒンジ領域を含有する。
【0041】
本発明の抗体またはその抗原結合断片は、抗体の合成に関して当技術分野で知られている任意の方法によって、特に化学合成によって、または好ましくは組換え発現技法によって産生することができる。
【0042】
抗体をコードするヌクレオチド配列は、当業者が入手可能な任意の情報から得ることができる(すなわちGenbank、文献から、または通常のクローニングおよび配列分析によって)。特定の抗体またはそのエピトープ結合断片をコードする核酸を含有するクローンが入手可能ではないが、抗体分子またはそのエピトープ結合断片の配列が知られている場合、免疫グロブリンをコードする核酸を、配列の5’末端とハイブリダイズ可能な合成プライマーを使用したPCR増幅によって、または特定の遺伝子配列に特異的なオリゴヌクレオチドプローブを使用したクローニングによって、化学合成して、または適切な供給源(例えば、抗体cDNAライブラリー、またはそこから作製したcDNAライブラリー、または任意の組織、もしくは抗体発現用に選択したハイブリドーマ細胞などの抗体を発現する細胞から単離した核酸、好ましくはポリA+RNA)から得て、例えば、抗体をコードするcDNAライブラリーからcDNAクローンを同定することができる。PCRにより作製した増幅核酸遺伝子は、当技術分野でよく知られている任意の方法を使用して、次いで複製可能クローニングベクターにクローニングすることができる。
【0043】
5.2 組換え抗体の調製
抗体のヌクレオチド配列は、ヌクレオチド配列の操作に関して当技術分野でよく知られている方法、例えば組換えDNA技法、部位特異的突然変異誘発法、PCRなどを使用することによって操作することができる(例えば、いずれもその全容を参照によって本明細書に組み込む、Sambrook et al., 上記、およびAusubel et al., eds., 1998, Current Protocols in Molecular Biology, John Wiley & Sons, NY中に記載された技法を参照)。抗体にはエピトープ結合ドメイン領域または任意の部分にアミノ酸の置換、欠失および/または挿入などの突然変異を導入して、生物活性を向上または低減することができる。
【0044】
抗体をコードするヌクレオチド配列を含有する発現ベクターは、抗体またはその抗原結合断片の組換え発現に使用することができる。抗体またはその抗原結合断片の産生用の、抗体分子、抗体またはその一部分のH鎖および/またはL鎖をコードするヌクレオチド配列を含むベクターは、前のセクションで論じたように当技術分野でよく知られている技法を使用して組換えDNA技術によって産生することができる。当業者によく知られている方法を使用して、抗体またはその抗原結合断片、コード配列、ならびに適切な転写および翻訳制御シグナルを含有する発現ベクターを構築することができる。これらの方法は、例えばin vitro組換えDNA技法、合成技法、およびin vivo遺伝子組換えを含む。VH、VL、VHとVLの両方、VHおよび/またはVLの抗原結合断片、または抗体の1つまたは複数のCDRをコードするヌクレオチド配列は、発現用のこのようなベクターにクローニングすることができる。このような配列は、原型抗体に固有または異種であり得るシグナルペプチドをコードするポリヌクレオチドと融合することができる。次いでこのように調製した発現ベクターを、抗体の発現に適した宿主細胞に導入することができる。したがって本発明は、RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合断片をコードするポリヌクレオチドを含有する宿主細胞を含む。
【0045】
宿主細胞は本発明の2つの発現ベクターでコトランスフェクトすることができ、この場合第一のベクターはH鎖由来ポリペプチドをコードし第二のベクターはL鎖由来ポリペプチドをコードする。2つのベクターは、H鎖およびL鎖ポリペプチドの等しい発現を可能にする同一の選択可能マーカー、または両プラスミドの維持を確実にするための異なる選択可能マーカーを含有することができる。あるいは、H鎖およびL鎖ポリペプチドの両方をコードし発現することができる、1つのベクターを使用することができる。H鎖およびL鎖のコード配列はcDNAまたはゲノムDNAを含むことができる。
【0046】
本発明の他の態様では、当技術分野で知られている様々なファージディスプレイ法を使用して抗体を作製することもできる。ファージディスプレイ法では、機能的抗体ドメインを、それらをコードするポリヌクレオチド配列を保持するファージ粒子の表面上に提示する。本発明の特定の態様では、このようなファージを利用して、レパートリーまたはコンビナトリアル抗体ライブラリー(例えば、ヒトまたはマウス)から発現されるFabおよびFvまたはジスルフィド結合安定化Fvなどの抗原結合ドメインを提示することができる。対象の抗原と結合する抗原結合ドメインを発現するファージは、抗原を用いて、例えば標識抗原または固体表面もしくはビーズに結合もしくは捕捉した抗原を使用して、選択または同定することができる。これらの方法中で使用するファージは、典型的にはfdおよびM13を含めた繊維状ファージである。抗原結合ドメインは、ファージ遺伝子IIIまたは遺伝子VIIIタンパク質のいずれかとの組換え融合タンパク質として発現される。本発明の免疫グロブリン、またはその断片を作製するために使用することができるファージディスプレイ法の例には、その各々を参照によってそれらの全容を本明細書に組み込む、Brinkman et al., J.Immunol.Methods, 182: 41-50, 1995; Ames et al., J.Immunol.Methods, 184: 177-186, 1995; Kettleborough et al., Eur.J.Immunol., 24: 952-958, 1994; Persic et al., Gene, 187: 9-18, 1997; Burton et al., Advances in Immunology, 57: 191-280, 1994、PCT出願No.PCT/GB91/01134、PCT出願WO90/02809、WO91/10737、WO92/01047、WO92/18619、WO93/11236、WO95/15982、WO95/20401、ならびに米国特許第5,698,426号、同第5,223,409号、同第5,403,484号、同第5,580,717号、同第5,427,908号、同第5,750,753号、同第5,821,047号、同第5,571,698号、同第5,427,908号、同第5,516,637号、同第5,780,225号、同第5,658,727号、同第5,733,743および同第5,969,108号中に開示された方法がある。
【0047】
前述の参照文献中に記載されたように、ファージ選択後、例えば以下で詳細に記載するように、ファージ由来の領域をコードする抗体を単離し、これらを使用してヒト抗体、または任意の他の望ましい断片を含めた完全抗体を作製し、哺乳動物細胞、昆虫細胞、宿主細胞、酵母、および細菌を含めた任意の望ましい宿主中で発現させることが可能である。例えば、(その各々を参照によってそれらの全容を本明細書に組み込む)PCT公開WO92/22324、Mullinax et al., BioTechniques, 12(6): 864-869, 1992およびSawai et al., AJRI, 34: 26-34, 1995およびBetter et al., Science, 240: 1041-1043, 1988中に開示された方法などの当技術分野で知られている方法を使用して、Fab、Fab’およびF(ab)2断片を組換えによって産生するための技法を利用することもできる。単鎖Fvおよび抗体を産生するために使用することができる技法の例には、米国特許第4,946,778号および同第5,258,498号、Huston et al., Methods in Enzymology, 203: 46-88, 1991; Shu et al., PNAS, 90: 7995-7999, 1993およびSkerra et al., Science, 240: 1038-1040, 1988中に記載された技法がある。
【0048】
本発明の抗体分子を前に記載した任意の方法によって産生した後、次いでそれを、免疫グロブリン分子の精製に関して当技術分野で知られている任意の方法によって、例えばクロマトグラフィー(例えば、イオン交換、親和性、特にプロテインAまたはプロテインG精製後の特異的抗原に対する親和性によって、およびサイズ排除カラムクロマトグラフィー)、遠心分離、差次的溶解度によって、またはタンパク質を精製するための、任意の他の標準的技法によって精製することができる。さらに、本発明の抗体またはその断片を、本明細書に記載する異種ポリペプチド配列または当技術分野で知られている他の配列と融合させて、精製を容易にすることができる。
【0049】
ヒトにおける抗体のin vivo用途およびin vitro検出アッセイを含めた、いくつかの用途に関して、キメラ、ヒト化、またはヒト抗体を使用することが好ましい可能性がある。キメラ抗体およびヒト化抗体は以下のセクション5.3中で詳細に論じる。
【0050】
他の化合物または異種ポリペプチドと融合または結合した抗体は、in vitroイムノアッセイにおいて、精製法(例えば、親和性クロマトグラフィー)、およびin vivo治療または診断用途において使用することができる。例えば、参照によってそれらの全容を本明細書に組み込む、PCT公開番号WO93/21232、EP439,095、Naramura et al., Immunol.Lett., 39: 91-99, 1994、米国特許第5,474,981号、Gillies et al., PNAS, 89: 1428-1432, 1992、およびFell et al., J.Immunol., 146: 2446-2452, 1991を参照。例えば、知られている方法または市販のキット(例えば、ビオチン標識、FITC標識、APC標識)を使用して様々な方法で、抗体を標識することができる。別の例として、in vivoでの抗体の生物学的効果を高める治療成分に、抗体を結合させることが可能である。このような治療成分の例には、別の抗体、細胞増殖抑制または細胞致死性である細胞毒素、放射性元素、および/または抗炎症剤、抗生物質を含めた他の治療剤などがある。本発明では、ヒト化抗RGD抗体を別の抗体と結合させて二重特異性抗体を形成することが可能である。別の例として、本発明のヒト化抗体を、in vivo診断用途に、放射性元素などの検出可能マーカーで標識することができる。
【0051】
5.3 キメラおよびヒト化抗体
キメラ抗体は、抗体の異なる部分が異なる動物種に由来する分子、マウスモノクローナル抗体に由来するV領域およびヒト免疫グロブリン由来の定常領域を有する抗体などである。キメラ抗体を産生するための方法は当技術分野で知られている。例えば、参照によってそれらの全容を本明細書に組み込む、Morrison, Science, 229: 1202, 1985; Oi et al., BioTechniques, 4: 214 1986; Gillies et al., J. Immunol.Methods, 125: 191-202, 1989、米国特許第5,807,715号、同第4,816,567号、および同第4,816,397号を参照。
【0052】
ヒト化抗体は、望ましい抗原と結合し、非ヒト種由来の1つまたは複数のCDRおよびヒト免疫グロブリン分子由来の1つまたは複数のFRを含有するV領域を含む分子である。非ヒト抗体をヒト化するための典型的な方法は、いずれもその全容を参照によって本明細書に組み込む、Queen et al., 1989, Proc.Natl.Acad. Sci.USA86: 10029-10033および米国特許第5,585,089号および同第5,693,762号による参照文献、Riechmann et al., Nature, 332: 323, 1988による参照文献、およびTsurushita et al., Methods 36: 69-83, 2005による参照文献などの、様々な参照文献中に記載されている。例えば、Tsurushita et al.(2005、上記、本明細書では以後「Tsurushita」)による参照文献は、Queen et al.(1989、上記)により本来開発された抗体−ヒト化法に基づく、マウスモノクローナル抗体をヒト化するための実用的で有益なプロトコールを与える。Tsurushita中に開示された一般的プロトコールは以下で簡潔に要約する。
【0053】
5.3.1 ヒト化抗体を調製するための一般的プロトコール
マウスV遺伝子のクローニングおよび配列決定
様々な方法が、標的マウスモノクローナル抗体のVHおよびVLをコードするcDNAをクローニングするのに利用可能である。例えば、SMART RACE cDNA増幅キット(BD Biosciences、CA)またはGeneRacerキット(Invitrogen、CA)を使用した5’RACE(cDNA末端の迅速増幅)法が一般に使用されている。5’RACE用の遺伝子特異的プライマーは、それがVHおよびVLのすぐ下流に結合することができるように、標的モノクローナル抗体のH鎖およびL鎖のアイソタイプに基づいて調製することができる。したがって、5’RACEプライマーは、γ1、γ2a、γ2bまたはγ3などのマウスにおけるそれぞれの亜型に特異的であるように設計することができる。あるいは、全ての亜型に共通なプライマーを、亜型間のコンセンサスまたは非常に相同的な領域に基づいて設計することができる。Tsurushita中では、以下の5’RACEプライマーが例として開示される:
(i)5’−GCCAGTGGATAGACTGATGG−(配列番号82)(マウスγ1、γ2a、γ2bおよびγ3H鎖のクローニング用)
(ii)5’−GATGGATACAGTTGGTGCAGC−(配列番号83)(マウスkL鎖のクローニング用)。
【0054】
PCR増幅したV領域遺伝子断片は、例えばZero Blunt TOPO PCRクローニングキット(Invitrogen)を使用してプラスミドベクターに直接クローニングすることができ、それらのDNA配列を決定することができる。得られた配列は、例えばモデル241プロテインシークエンサー(Hewlett−Packard、CA)を使用して、例えばそれらのコードアミノ酸配列と、N末端アミノ酸配列決定により決定した標的モノクローナル抗体の配列を比較することによって確認しなければならない。典型的には、例えばエドマン分解による、標的抗体のN末端における少なくとも15〜20アミノ酸残基の決定は、クローニングDNA配列の真正を確認するのに十分である。Tsurushitaは、マウスにおける2つの最も一般的なN末端アミノ酸の1つであるグルタミンがN末端アミノ酸であるとき、それがピログルタミンに変換されてN末端における配列決定を阻害する可能性があることを警告する。その場合、N末端を脱保護して配列を得ることが必要である。
【0055】
V領域の三次元モデリング
VHおよびVLの配列に基づいて、CDRの立体配座構造を維持するのにおそらく重要である標的抗体のフレームワーク残基を、例えばR.Levy et al., 1989, Biochemistry28: 7168-7175によって記載された方法によって、およびB.Zilber et al., 1990, Biochemistry29: 10032-10041によって記載された方法によって最初に同定する。典型的には、VHおよびVLのそれぞれは14個の構造上有意なセグメントに分けられ、これらは免疫グロブリンスーパーファミリーのドメイン構造を含むβ鎖およびループ様構造である。標的抗体由来の各セグメントのアミノ酸配列を、PDBデータベースにおける知られている構造の対応する抗体セグメントと合わせる(H.M.Berman et al., 2000, Nucleic Acids Res.28: 235-342参照)。多重配列アラインメントによって、標的セグメントのそれぞれと最高の配列相同性を有する対応するセグメントを選択し、V領域の三次元モデルを構築する。構造を最適化するために、このモデルを多サイクルの共役勾配法によるエネルギー最小化に施す(例えば、ENCADを使用して、またはPress et al., 1990, "Numerical Recipes, Cambridge University Press, Cambridgeによって記載されたように、Weiner et al., 1981, J.Comp.Chem.2: 287-303によるAMBER、BioMolecularModellingで利用可能な3D-JIG-SAW、またはCancer Research UKによって運営される「BMM」ウェブサイト、またはSwiss Institute of Bioinformatics, Genevaによって運営されるExPASy Proteomics Serverウェブサイトで利用可能なSWISS-MODEL)。
【0056】
ヒトフレームワークの選択
V領域の構造のモデル化と並行して、それぞれマウスVHおよびVLのcDNAクローニングから推定したアミノ酸配列を、データベース、例えばKabatデータベース(Johnson et al., 2000, Nucleic Acids Res.28: 214-218を参照)、GenBankなどにおけるヒトV領域の配列と比較する。マウスの配列と少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%、または少なくとも95%同一の全体配列同一率を有するヒトFRは、例えばSmith−Watermanのアルゴリズム(Gusfield, 1997, "Algorithms on Strings, Trees, and Sequences", Cambridge University Press, Cambridgeによる)、またはBLAST(Karlin et al., 1990, Proc.Natl.Acad.Sci.USA87: 2264-2268による)などを使用して検索することができる。これらのヒトの配列は、cDNAベースおよびタンパク質由来配列に基づいてよいが、しかしながら、生殖細胞系の使用が好ましいことが多い。cDNAベース、タンパク質由来配列の体細胞超突然変異と関係がある潜在的免疫原性を排除する際に、それが有用である可能性があるからである。代替では、Queen et al.(1989、上記)中に記載されたように、コンセンサスフレームワーク配列の使用によって、cDNAベースまたはタンパク質由来配列から得たフレームワーク中のこのような超突然変異残基を同定し除去することもできる。生殖細胞系VHセグメントをアクセプターフレームワークとして使用する場合、15および16ではなく第14染色体上にコードされるVHセグメントを使用しなければならない。第14染色体上のセグメントのみが機能的VHを生成するからである。
【0057】
ヒト化V領域の設計
Queen et al.(1989、上記)によれば、CDRの約4〜6Å以内のフレームワークアミノ酸を同定することが必要である。これらの残基は、正確なCDR構造を支えるおそらく重要なフレームワーク残基であると考えられるからである。このようなプロセスは、原子座標からの原子間距離を計算する国立科学財団(NSF)によって運営されるMolecular Visualization Freewareウェブサイトで利用可能なRASMOLなどのコンピュータプログラムを使用して、またはコンピュータモデルの手検測によって実施することができる。重要なフレームワーク位置におけるアミノ酸がマウスドナー配列とヒトアクセプター配列の間で異なる場合、マウスドナーのそれらを通常ヒト残基に置換する。しかしながら、このような残基がCDR構造を支持するのに最小の貢献度を有する場合、対応するヒト残基を典型的に使用する。さらに、選択したヒトアクセプターが、V領域配列の約10〜20%未満に生じる「異常な」アミノ酸を含有する場合、それらは親和性成熟中の体細胞超突然変異の結果である可能性があり、ドナー残基と置換してヒトにおける潜在的免疫原性を回避しなければならない。
【0058】
さらに、潜在的N−結合型グリコシル化シグナルの残基などの他の因子を注意深く考慮して、ヒト化V領域を設計する必要がある(詳細に関してはTsurushitaを参照)。
【0059】
ヒト化抗体は、治療用途に必要とされるかまたはそのために排除されるエフェクター機能に応じて、ヒト抗体、またはその変異体のヒトκもしくはλL鎖、および/またはγ1、γ2、γ3、γ4、μ、α1、α2、δ、もしくはεH鎖由来のヒト定常領域またはその一部分を含有することができる。例えば、突然変異を含有する定常領域のFc部分を本発明のキメラもしくはヒト化抗体のV領域と融合して、Fc受容体と抗体の結合を低減する、および/または補体を固定するその能力を低減することができる(例えば、Winter et al., GB2,209,757B、Morrison et al., WO89/07142、Morgan et al., WO94/29351を参照)。抗体分子のこのような操作は、セクション5.2中に記載したように組換えDNA技術によって実施することができる。
【0060】
生成するキメラまたはヒト化抗体は、非ヒトドナー抗体と同じ特異性、および非ヒトドナー抗体の親和性と同等またはその少なくとも約1/3、少なくとも約1/2、もしくは少なくとも約2/3の親和性を有することが好ましい。別の態様では、生成するキメラまたはヒト化抗体は、少なくとも約1×107M−1、好ましくは少なくとも約1×108M−1、および最も好ましくは少なくとも約1×109M−1の親和性定数を有する。
【0061】
前に記載した一般的プロトコール以外に、例えば、いずれもその全容を参照によって本明細書に組み込む、CDR移植(EP239,400;PCT公開WO91/09967、米国特許第5,225,539号、同第5,530,101号および同第5,585,089号)、ベニアリング(veneering)またはリサーフィシング(resurfacing)(EP592,106;EP519,596、Padlan, Molecular Immunology, 28(4/5): 489-498, 1991; Studnicka et al., Protein Engineering, 7(6): 805-814, 1994; Roguska et al., Proc Natl. Acad.Sci.USA, 91: 969-973, 1994)、および鎖シャッフリング(米国特許第5,565,332号)を含めた、当技術分野で知られている様々な技法を使用して、抗体をヒト化することができる。
【0062】
5.3.2.薬剤としてヒト化抗体を調製するための追加的考慮事項
薬剤としてヒト化抗体を与えるため、効率良く一貫した生成系をしたがって調製する必要がある。例えば、ヒト化抗体に適した発現ベクターはH鎖およびL鎖配列を挿入することによって調製し、発現ベクターでトランスフェクトした高生産性細胞系は、作業用細胞バンク(WCB)の安定した半永久的供給源として働く、マスター細胞バンク(MCB)の種細胞として得ることができる。次いで、ヒト化抗体は、WCB由来の作業用細胞を培養し培養培地を回収することによって調製することができる。
【0063】
適切な制御遺伝子を有する様々な発現ベクターを、このような生成細胞系の調製に使用することができる。宿主細胞として、哺乳動物タンパク質の発現に一般に使用される細胞をヒト化抗体の発現に使用することができる。このような宿主細胞の例には、チャイニーズハムスター卵巣(CHO)細胞、SP2/0−Agl4.19細胞、NSO細胞などがあるが、これらだけには限られない。ヒト化抗体の生産性は、発現ベクターと宿主細胞の最良の組合せを選択することによって最大にすることができる。さらに、宿主細胞によるヒト化抗体の発現を最適化することができるように、培養培地の組成を調べて、様々な無血清培養培地およびサプリメントから適切な培地を選択しなければならない。
【0064】
有効性および最終収率に基づいて、宿主細胞によって産生されるヒト化抗体を、親和性クロマトグラフィー、イオン交換クロマトグラフィー、疎水性相互作用クロマトグラフィーなどを含めた、当技術分野でよく知られている様々な方法を使用して培養上清から精製することができる。
【0065】
5.4 医薬組成物および治療用途
本発明は、RGD配列を免疫特異的に認識する前に記載したヒト化抗体またはその抗原結合断片を含む、医薬組成物を提供する。活性成分として本発明のヒト化抗体を含む医薬組成物は、癌、例えば癌細胞の増殖または転移、および炎症疾患、例えば慢性関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、糖尿病、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などだけには限られないが、これらを含めたRGDタンパク質と関係がある障害または疾患を予防および/または治療するための作用物質として使用することができる。
【0066】
本発明のヒト化抗体を含む医薬組成物を使用して、臓器移植後の慢性拒絶反応、および例えば全身性自己免疫疾患、エリテマトーデス、ブドウ膜炎、ベーチェット病、多発性筋炎、増殖性糸球体腎炎、サルコイドーシスなどの自己免疫疾患を治療することもできる。
【0067】
本発明のヒト化抗体を含む、前に記載した障害もしくは疾患を予防または治療するための予防および/または治療剤は低い毒性を有し、経口的または非経口的に、適切な溶媒中に混合することにより液体調製物として直接、または適切な剤形の医薬組成物としてヒトに投与することができる。
【0068】
前に記載した投与に使用する医薬組成物は、前述の抗体またはその塩、および薬学的に許容される担体、希釈剤または賦形剤を含有する。このような組成物は、経口または非経口投与に適した剤形で与えられる。
【0069】
用量は、投与する対象の年齢および大きさ、標的疾患、状態、投与の経路などに応じて変わる可能性がある。例えば成人患者において慢性関節リウマチを予防および/または治療するために抗体を使用するとき、通常約0.01〜約20mg/体重1kg、好ましくは約0.1〜約10mg/体重1kg、およびより好ましくは約0.1〜約5mg/体重1kgの一回用量で、1日当たりおよそ1〜5回、好ましくは1日当たりおよそ1〜3回、本発明の抗体を静脈内投与することが有益である。他の非経口投与および経口投与では、前に与えた用量に対応する用量で抗体を投与することができる。状態が特に重度であるとき、状態に応じて用量を増大することができる。
【0070】
様々な送達系、例えばリポソーム、マイクロ粒子、マイクロカプセルへの封入、変異ウイルスを発現することができる組換え細胞、受容体仲介エンドサイトーシスが知られており、これらを使用して本発明の医薬組成物を投与することができる(例えば、Wu and Wu, 1987, J.Biol.Chem.262: 4429 4432を参照)。導入の方法には、皮内、筋肉内、腹腔内、静脈内、皮下、鼻腔内、硬膜外、および経口投与があるが、これらだけには限られない。化合物は任意の好都合な経路によって、例えば注入またはボーラス注射によって、上皮または皮膚粘膜内側(例えば、経口粘膜、直腸および腸粘膜など)を介した吸収によって投与することができ、他の生物活性剤と一緒に投与することができる。投与は全身性または局所であってよい。例えば、吸入器または噴霧器、および製剤とエアロゾル剤を使用することによって、肺投与を利用することもできる。
【0071】
本発明の一態様では、治療の必要がある領域へ局所的に、本発明の医薬組成物を投与することが望ましい可能性がある。これは、例えばかつ非制限的に、外科手術中の局所注入、例えば外科手術後に創傷包帯を併用した局所施用によって、注射によって、カテーテルによって、座薬によって、鼻腔スプレーによって、インプラントによって(前記インプラントは多孔性、非多孔性、またはシアラスティック膜などの膜、またはファイバーを含めたゼラチン質材料である)実施することができる。本発明の一態様では、投与は部位(または感染前部位)感染組織への直接注射による投与であってよい。
【0072】
本発明の別の態様では、医薬組成物を小胞、特にリポソームで送達することができる(Langer, 1990, Science249: 1527-1533; Treat et al., Liposomes in the Therapy of Infectious Disease and Cancer, Lopez Berestein and Fidler(eds.), Liss, New York, pp.353-365(1989); Lopez-Berestein、既出、pp.317-327を参照、既出を一般に参照)。
【0073】
本発明のさらに別の態様では、医薬組成物を徐放システムで送達することができる。本発明の一態様では、ポンプを使用することができる(Langer、上記、Sefton, 1987, CRC Crit.Ref.Biomed.Eng. 14: 201; Buchwald et al., 1980, Surgery88: 507; およびSaudek et al., 1989, N.Engl.J.Med.321: 574を参照)。本発明の別の態様では、ポリマー物質を使用することができる(Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Florida(1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York (1984); Ranger and Peppas, J.Macromol.Sci.Rev.Macromol.Chem. 23: 61(1983)を参照、Levy et al., 1985, Science228: 190; During et al., 1989, Ann.Neurol.25: 351; Howard et al., 1989, J.Neurosurg. 71: 105も参照)。本発明のさらに別の態様では、徐放システムを組成物の標的の近くに置くことができ、したがってごくわずかな全身用量を必要とする(例えば、Goodson, Medical Applications of Controlled Release、上記、vol.2, pp.115-138(1984)を参照)。他の徐放システムはLanger(Science249: 1527-1533(1990))による総説中で論じられている。
【0074】
経口投与用の組成物の例には、固体または液体剤形、具体的には、錠剤(ドラジェおよびフィルムコーティング錠を含む)、ピル、顆粒、粉末調製物、カプセル(軟質カプセルを含む)、シロップ、乳濁液、縣濁液などがある。このような組成物は公に知られている方法によって製造され、製剤の分野で従来使用されている小胞、希釈剤または賦形剤を含有する。錠剤用の小胞または賦形剤の例は、ラクトース、スターチ、スクロース、ステアリン酸マグネシウムなどである。
【0075】
注射用調製物は、静脈内、皮下、皮内および筋肉内注射、点滴など用の剤形を含むことができる。これらの注射用調製物は、公に知られている方法によって調製することができる。注射用調製物は、例えば注射に従来使用される滅菌水性媒体または油状媒体中に、前に記載した抗体またはその塩を溶解、縣濁または乳化することによって調製することができる。注射用水性媒体として、例えば、アルコール(例えば、エタノール)、多価アルコール(例えば、プロピレングリコール、ポリエチレングリコール)などの適切な可溶化剤、非イオン性界面活性剤[例えば、ポリソルベート80、HCO−50(水素化ヒマシ油のポリオキシエチレン(50mol)付加生成物)]などと組合せて使用することができる、生理食塩水、グルコースおよび他の補助剤などを含有する等張液が存在する。油状媒体として、例えば、安息香酸ベンジル、ベンジルアルコールなどの可溶化剤などと組合せて使用することができる、ゴマ油、ダイズ油などが利用される。このように調製した注射用調製物は、適切なアンプル中に充填することが好ましい。直腸投与に使用する座薬は、前述の抗体またはその塩と従来の座薬基剤を混合することによって調製することができる。
【0076】
前に記載した経口または非経口用途の医薬組成物は、活性成分の投与に適合させるのに適した単位用量の剤形に調製することが有利である。このような単位用量の剤形には、例えば、錠剤、ピル、カプセル、注射(アンプル)、座薬などがある。含有される前述の抗体の量は、特に注射の形で単位用量の剤形当たり一般に約5〜500mgであり、前述の抗体は他の剤形に関して約5〜100mgおよび約10〜250mg含有されることが好ましい。
【0077】
製剤化が前に記載した抗体と何らかの悪い相互作用を引き起こさない限り、前に記載したそれぞれの組成物は他の活性成分をさらに含有することができる。
【0078】
本発明はさらに、活性成分としてRGD配列結合機能性分子(例えば、インテグリンなど)を含む細胞および/または組織再構築用の阻害剤および/または促進剤、およびRGDタンパク質発現細胞および/または組織(例えば、腫瘍細胞、好中球、平滑筋など)とRGDタンパク質結合機能性分子を接触させることを含む、細胞および/または組織の再構築を阻害および/または促進するための方法に関する。このような治療剤における活性成分の投与、薬剤調製などに関する用量、方法は、本発明のヒト化抗体を含む医薬品の前述の記載を参照することによって適切に決定することができる。
【0079】
前に記載したように、本発明は、RGDタンパク質と関係があるかまたはそれらに関与する障害もしくは疾患を予防または治療するための方法であって、少なくとも1つの本発明のヒト化抗体を有効量、その必要性がある対象に投与することを含む方法をさらに提供する。
【0080】
5.5 診断用途
本発明のヒト化抗体を含む医薬組成物は、癌、例えば癌細胞の増殖または転移、および炎症疾患、例えば慢性関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、糖尿病、癌転移、動脈硬化症、多発性硬化症、肉芽腫などの診断薬として、または臓器移植後の慢性拒絶反応、全身性自己免疫疾患、エリテマトーデス、ブドウ膜炎、ベーチェット病、多発性筋炎、増殖性糸球体腎炎、サルコイドーシスなどの自己免疫疾患の診断薬として使用することができる。本発明のヒト化抗体はRGD配列を特異的に認識することができ、したがってこれらを使用して、試験流体中のRGDタンパク質の定量化、特にサンドイッチイムノアッセイ、競合アッセイ、イムノメトリー、ネフェロメトリーなど、免疫染色などによる定量化を可能にすることができる。これらの免疫法を本発明のアッセイ法に適用する際に、何らかの特定条件、手順などを言及することは必要とされない。当技術分野で通常の技術的考慮事項を従来の条件および手順に加えることによって、アッセイシステムを構築することで十分である。これらの一般的な技術的意義の詳細に関しては、総説、教本などを参照することができる。
【0081】
前に記載したように、本発明の抗体を使用することによって、高感度でRGDタンパク質を定量化することができる。本発明のヒト化抗体は、in vivoでRGDタンパク質を定量化するための方法を適用することによって、RGDタンパク質と関係がある様々な疾患を診断するのに特に有用である。例えば、RGDタンパク質の発現レベルの増大または低下を検出する場合、人が現在RGDタンパク質と関係がある疾患、例えば癌または炎症疾患に罹患している可能性が非常に高い、または人が将来これらの疾患に罹患する可能性が非常に高いと診断することができる。したがって、本発明は、対象における、RGDタンパク質と関係があるかまたはそれらに関与する障害もしくは疾患を診断するための方法であって、本発明のヒト化抗体の少なくとも1つまたは両方を有効量、その必要性がある対象に投与することを含む方法も提供する。このようなin vivo診断に必要とされる用量は診断用途に必要とされる用量未満であってよく、通常の手順に従い当業者によって決定することができる。
【0082】
本発明のヒト化抗体は、試験サンプル(例えば、体液などの試験流体、組織など)中に存在するRGDタンパク質を特異的に検出するために使用することもできる。ヒト化抗体は、RGDタンパク質の精製用の抗体カラムの調製、精製時にそれぞれの分画中に含有されるRGDタンパク質の検出、または試験する細胞中のRGDタンパク質の挙動の分析に使用することもできる。
【0083】
本明細書の配列表中の配列識別番号は以下の配列を示す。
【0084】
[配列番号1]
これはマウス33E10およびヒト33E10と共通のCDRH1のアミノ酸配列を示す。
【0085】
[配列番号2]
これはマウス33E10およびヒト33E10と共通のCDRH2のアミノ酸配列を示す。
【0086】
[配列番号3]
これはマウス33E10およびヒト33E10と共通のCDRH3のアミノ酸配列を示す。
【0087】
[配列番号4]
これはマウス33E10およびヒト33E10と共通のCDRL1のアミノ酸配列を示す。
【0088】
[配列番号5]
これはマウス33E10およびヒト33E10と共通のCDRL2のアミノ酸配列を示す。
【0089】
[配列番号6]
これはマウス33E10およびヒト33E10と共通のCDRL3のアミノ酸配列を示す。
【0090】
[配列番号7]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VH5遺伝子のヌクレオチド配列を示す。
【0091】
[配列番号8]
これはシグナルペプチドを含むHu33E10VH5遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0092】
[配列番号9]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VH7遺伝子のヌクレオチド配列を示す。
【0093】
[配列番号10]
これはシグナルペプチドを含むHu33E10VH7遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0094】
[配列番号11]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VH7.8遺伝子のヌクレオチド配列を示す。
【0095】
[配列番号12]
これはシグナルペプチドを含むHu33E10VH7.8遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0096】
[配列番号13]
これはNheIおよびEcoRI部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VL6遺伝子のヌクレオチド配列を示す。
【0097】
[配列番号14]
これはシグナルペプチドを含むHu33E10VL6遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0098】
[配列番号15]
これはNheIおよびEcoRI部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VLv2遺伝子のヌクレオチド配列を示す。
【0099】
[配列番号16]
これはシグナルペプチドを含むHu33E10VLv2遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0100】
[配列番号17]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[CMV2]を示す。
【0101】
[配列番号18]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JΝΤ026]を示す。
【0102】
[配列番号19]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT080]を示す。
【0103】
[配列番号20]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT082]を示す。
【0104】
[配列番号21]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT084]を示す。
【0105】
[配列番号22]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT097]を示す。
【0106】
[配列番号23]
これはHu33E10重鎖および軽鎖cDNAのPCR増幅および配列決定に使用したオリゴヌクレオチド[JNT098]を示す。
【0107】
[配列番号24]
これはNS0−Hu33E10−4およびNS0−Hu33E10−6において発現されるVH7およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列を示す。
【0108】
[配列番号25]
これはNS0−Hu33E10−4およびNS0−Hu33E10−6において発現されるVH7およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0109】
[配列番号26]
これはNS0−Hu33E10−5において発現されるVH7.8およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列を示す。
【0110】
[配列番号27]
これはNS0−Hu33E10−5において発現されるVH7.8およびγ−1重鎖定常領域に関するコード領域のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0111】
[配列番号28]
これはNS0−Hu33E10−4およびNS0−Hu33E10−5において発現されるVL6およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列を示す。
【0112】
[配列番号29]
これはNS0−Hu33E10−4およびNS0−Hu33E10−5において発現されるVL6およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0113】
[配列番号30]
これはNS0−Hu33E10−6において発現されるVLv2およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列を示す。
【0114】
[配列番号31]
これはNS0−Hu33E10−6において発現されるVLv2およびkappa軽鎖定常領域に関するコード領域のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0115】
[配列番号32]
これは33E10VLのアミノ酸配列を示す。
【0116】
[配列番号33]
これはHu33E10VLv2のアミノ酸配列を示す。
【0117】
[配列番号34]
これはマウス33E10VHcDNAのヌクレオチド配列を示す。
【0118】
[配列番号35]
これはマウス33E10VHcDNAのヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0119】
[配列番号36]
これはマウス33E10VLcDNAのヌクレオチド配列を示す。
【0120】
[配列番号37]
これはマウス33E10VLcDNAのヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0121】
[配列番号38]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含む設計33E10VH遺伝子のヌクレオチド配列を示す。
【0122】
[配列番号39]
これはシグナルペプチドを含む設計33E10VH遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0123】
[配列番号40]
これはNheIおよびEcoRI部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含む設計33E10VL遺伝子のヌクレオチド配列を示す。
【0124】
[配列番号41]
これはシグナルペプチドを含む設計33E10VL遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0125】
[配列番号42]
これは33E10VHのアミノ酸配列を示す。
【0126】
[配列番号43]
これはヒト化33E10(Hu33E10)VH1のアミノ酸配列を示す。
【0127】
[配列番号44]
これは33E10VLのアミノ酸配列を示す。
【0128】
[配列番号45]
これはヒト化33E10(Hu33E10)VL1のアミノ酸配列を示す。
【0129】
[配列番号46]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ220]を示す。
【0130】
[配列番号47]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ206]を示す。
【0131】
[配列番号48]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ207]を示す。
【0132】
[配列番号49]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ208]を示す。
【0133】
[配列番号50]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ209]を示す。
【0134】
[配列番号51]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ210]を示す。
【0135】
[配列番号52]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ211]を示す。
【0136】
[配列番号53]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ212]を示す。
【0137】
[配列番号54]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ213]を示す。
【0138】
[配列番号55]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ214]を示す。
【0139】
[配列番号56]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ215]を示す。
【0140】
[配列番号57]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ216]を示す。
【0141】
[配列番号58]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ217]を示す。
【0142】
[配列番号59]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ218]を示す。
【0143】
[配列番号60]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ219]を示す。
【0144】
[配列番号61]
これはHu33E10VH1遺伝子の構築に使用したオリゴヌクレオチド[JNJ221]を示す。
【0145】
[配列番号62]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ116]を示す。
【0146】
[配列番号63]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ193]を示す。
【0147】
[配列番号64]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ194]を示す。
【0148】
[配列番号65]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ195]を示す。
【0149】
[配列番号66]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ196]を示す。
【0150】
[配列番号67]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ197]を示す。
【0151】
[配列番号68]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ198]を示す。
【0152】
[配列番号69]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ199]を示す。
【0153】
[配列番号70]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ200]を示す。
【0154】
[配列番号71]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ201]を示す。
【0155】
[配列番号72]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ202]を示す。
【0156】
[配列番号73]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ203]を示す。
【0157】
[配列番号74]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ204]を示す。
【0158】
[配列番号75]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ205]を示す。
【0159】
[配列番号76]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ101]を示す。
【0160】
[配列番号77]
これはHu33E10VL1遺伝子の構築に使用したオリゴヌクレオチド[JNJ117]を示す。
【0161】
[配列番号78]
これはSpeIおよびHindIII部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VH1遺伝子のヌクレオチド配列を示す。
【0162】
[配列番号79]
これはシグナルペプチドを含むHu33E10VH1遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0163】
[配列番号80]
これはNheIおよびEcoRI部位と隣接するシグナルペプチドおよびイントロン配列をコードする配列を含むHu33E10VL1遺伝子のヌクレオチド配列を示す。
【0164】
[配列番号81]
これはシグナルペプチドを含むHu33E10VL1遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0165】
[配列番号82]
これは5’RACEプライマーのヌクレオチド配列を示す。
【0166】
[配列番号83]
これは5’RACEプライマーのヌクレオチド配列を示す。
【0167】
[配列番号84]
これはGeneRacer5’プライマーのヌクレオチド配列を示す。
【0168】
[配列番号85]
これは33E10VH3’プライマーのヌクレオチド配列を示す。
【0169】
[配列番号86]
これは33E10VL3’プライマーのヌクレオチド配列を示す。
【0170】
[配列番号87]
これは33E10VH5’プライマーのヌクレオチド配列を示す。
【0171】
[配列番号88]
これは33E10VH3’プライマーのヌクレオチド配列を示す。
【0172】
[配列番号89]
これは33E10VL5’プライマーのヌクレオチド配列を示す。
【0173】
[配列番号90]
これは33E10VL3’プライマーのヌクレオチド配列を示す。
【0174】
[配列番号91]
これはHu33E10VH5遺伝子のヌクレオチド配列を示す。
【0175】
[配列番号92]
これはHu33E10VH5遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0176】
[配列番号93]
これはHu33E10VH7遺伝子のヌクレオチド配列を示す。
【0177】
[配列番号94]
これはHu33E10VH7遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0178】
[配列番号95]
これはHu33E10VH7.8遺伝子のヌクレオチド配列を示す。
【0179】
[配列番号96]
これはHu33E10VH7.8のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0180】
[配列番号97]
これはHu33E10VL6遺伝子のヌクレオチド配列を示す。
【0181】
[配列番号98]
これはHu33E10VL6遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【0182】
[配列番号99]
これはHu33E10VLv2遺伝子のヌクレオチド配列を示す。
【0183】
[配列番号100]
これはHu33E10VLv2遺伝子のヌクレオチド配列によってコードされるアミノ酸配列を示す。
【実施例】
【0184】
以下の実施例は、RGD配列を免疫特異的に認識するモノクローナル抗体の調製、モノクローナル抗体のV領域の配列決定、これらの抗体のエピトープマッピングおよび他の特徴付け、およびこのような抗体のキメラ化およびヒト化、ならびに生成したキメラおよびヒト化抗体の特徴付けを例示する。これらの実施例は、本発明の範囲を制限するものとして解釈するべきではない。
【0185】
6.1.マウス33E10可変領域遺伝子のクローニングおよび配列決定
マウス33E10ハイブリドーマ細胞を、7.5%CO2インキュベーター中で37℃において、10%ウシ胎児血清(FBS;HyClone、Logan、UT)を含有するTIL培地I(Immuno−Biological Laboratories、群馬、日本)中で増殖させた。全RNA(total RNA)は、供給者のプロトコールに従いTRIzol試薬(Invitrogen、Carlsbad、CA)を使用して約3×106個のハイブリドーマ細胞から抽出した。オリゴdTプライマー処理cDNAは、供給者のプロトコールに従いGeneRacerキット(Invitrogen)を使用して合成した。33E10重鎖および軽鎖に関する可変領域cDNAは、マウスγ−1およびkappa鎖定常領域とそれぞれアニーリングする3’プライマー、およびGeneRacerキット中に供給されたGeneRacer5’プライマー(5’−CGACTGGAGCACGAGGACACTGA−3’(配列番号84))を使用して、Phusion DNAポリメラーゼ(New England Biolabs、Beverly、MA)を用いてポリメラーゼ連鎖反応(PCR)によって増幅した。重鎖可変領域(VH)のPCR増幅用に、3’プライマーは配列5’−GCCAGTGGATAGACAGATGG−3’(配列番号85)を有する。軽鎖可変領域(VL)のPCR増幅用に、3’プライマーは配列5’−GATGGATACAGTTGGTGCAGC−3’(配列番号86)を有する。増幅したVHおよびVLcDNAは配列決定用にpCR4Blunt−TOPOベクター(Invitrogen)にクローニングした。可変領域のDNA配列決定はTocore(Menlo Park、CA)で実施した。数個の重鎖および軽鎖クローンを配列決定し、典型的なマウス重鎖および軽鎖可変領域と相同的な独自の配列を同定した。33E10VHおよびVLの推定アミノ酸配列と共にコンセンサスcDNA配列をそれぞれ図1および2中に示す。
【0186】
6.2.キメラ33E10IgG1/κ抗体の構築
33E10VHをコードする遺伝子を、鋳型として33E10VHcDNA、5’プライマーとして5’−GGGACTAGTACCACCATGAAGTTGTGGCTGAACTGGATT−3’(配列番号87)(SpeI部位に下線を引く)、および3’プライマーとして5’−GGGAAGCTTGAAGTTAGGACTCACCTGCAGAGACAGTGACCAGAGTCCC−3’(配列番号88)(HindIII部位に下線を引く)(図3)を使用してPCRによって、スプライスドナーシグナルおよび適切な隣接制限酵素部位を含むエクソンとして作製した。同様に、33E10VLをコードする遺伝子を、鋳型として33E10VLcDNA、5’プライマーとして5’−GGGGCTAGCACCACCATGAAGTTGCCTGTTAGGCTGTTG−3’(配列番号89)(NheI部位に下線を引く)、および3’プライマーとして5’−GGGGAATTCTTTGGATTCTACTTACGTTTGATTTCCAGCTTGGTGCCTCC−3’(配列番号90)(EcoRI部位に下線を引く)(図4)を使用してPCRによって、スプライスドナーシグナルおよび適切な隣接制限酵素部位を含むエクソンとして作製した。33E10VHおよびVLエクソンのスプライスドナーシグナルは、それぞれマウス生殖細胞JH3およびJK1配列に由来した(Kabat et al., Sequences of Proteins of Immunological Interests, Fifth edition, NIH公開番号91−3242、米国厚生省、1991)。PCR増幅断片はQIAquickゲル抽出キット(Qiagen、Valencia、CA)を使用してゲル精製し、(VHに関して)SpeIおよびHindIIIまたは(VLに関して)NheIおよびEcoRIで消化し、キメラ33E10IgGl/κ抗体の産生用にヒトγ−1およびkappa定常領域を保持する哺乳動物発現ベクターにクローニングした。生成した発現ベクター、pCh33E10の概略構造は図5中に示す。
【0187】
6.3.ヒト化33E10VHおよびVL遺伝子の作製
33E10可変領域のヒト化を、Queen et al.(Proc.Natl.Acad.Sci.USA86: 10029-10033、1989)によって概説されたように実施した。最初に、33E10可変領域の三次元分子モデルをコンピュータプログラムの助力によって構築した。次に、ヒト可変領域配列に対する相同性検索に基づいて、33E10VHと高い相同性を有するU03400(GenBank受託番号)のヒトVHアミノ酸配列を、ヒト化33E10VHのフレームワークを得るためのアクセプターとして選択した。同様に、X72452(GenBank受託番号)のヒトVLアミノ酸配列を33E10VLのヒト化用のアクセプターとして選択した。
【0188】
三次元モデルが相補性決定領域(CDR)との有意な接触を示したフレームワーク位置において、マウス33E10可変領域由来のアミノ酸残基をヒトフレームワーク残基に置換した。これは位置30および48で実施してヒト化33E10(Hu33E10)VH1を作製した(図6)。軽鎖に関しては、Hu33E10VL1を作製するのに置換を必要としなかった(図7)。33E10、設計Hu33E10およびヒトアクセプターアミノ酸配列のアラインメントは、VHに関しては図6中およびVLに関しては図7中に示す。
【0189】
Hu33E10VH1およびVL1のそれぞれをコードする遺伝子を、哺乳動物発現ベクターへの後のクローニング用に、シグナルペプチド、スプライスドナーシグナル、および適切な制限酵素部位を含むエクソンとして設計した。Hu33E10VH1およびVL1エクソンのスプライスドナーシグナルは、それぞれヒト生殖細胞JH3およびJk1配列に由来した(Kabat et al., 1991)。マウス33E10VL1遺伝子のシグナルペプチド配列は、SIG−Predシグナルペプチド予想ソフトウェア(http: //bmbpcu36.leeds.ac.uk/prot_analysis/Signal.html)によって正確な切断の最適下限であったことを示した。したがって、SIG−Predソフトウェアによって有効かつ正確に切断されると予想されたマウスモノクローナル抗体35B6(Gene Techno Science)のVL遺伝子のシグナルペプチドを、Hu33E10VL1エクソンにおいて使用した。Hu33E10VH1エクソン中のシグナルペプチド配列は対応するマウス33E10VH配列に由来した。SIG−Predソフトウェアは、Hu33E10VH1遺伝子のシグナルペプチドが有効かつ正確に切断されることを示した。
【0190】
Hu33E10VH1およびVL1遺伝子を、He et al.(J.Immunol.160: 1029-1035、1998)によって概説されたPhusion DNAポリメラーゼを使用して、数個の重複合成オリゴヌクレオチドプライマーの延長およびPCR増幅によって構築した。Hu33E10VH1およびVL1遺伝子の構築に使用したオリゴヌクレオチドは、それぞれ図8および9中に列挙する。Hu33E10VH1およびVL1遺伝子におけるオリゴヌクレオチドの位置は、それぞれ図10および11中に示す。PCR増幅断片はQIAquickゲル抽出キット(Qiagen)を使用してゲル精製し、配列決定用にpCR4Blunt−TOPOベクターにクローニングした。(VHに関して)SpeIおよびHindIIIまたは(VLに関して)NheIおよびEcoRIでの消化後、Hu33E10VH1およびVL1遺伝子を、ヒトIgG1/κ型の産生用に哺乳動物発現ベクター中の対応する部位にサブクローニングした。生成した発現ベクター、pHu33E10−1の概略構造は図5中に示す。推定アミノ酸配列と共に得られたHu33E10VH1およびVL1遺伝子のヌクレオチド配列は、それぞれ図12および13中に示す。
【0191】
6.4.キメラおよびヒト化33E10抗体の発現
キメラおよびヒト化33E10IgG1/κ抗体のそれぞれを安定的に産生する細胞系を得るために、発現ベクターpCh33E10およびpHu33E10−1をそれぞれ、マウスミエローマ細胞系NS0(European Collection of Animal Cell Cultures、Salisbury、Wiltshire、UK)の染色体に導入した。7.5%CO2インキュベーター中で37℃において、NS0細胞を10%FBSを含有するDME培地中で増殖させた。NS0細胞への安定したトランスフェクションは、Bebbington et al.(Bio/Technology10: 169-175、1992)中に記載されたエレクトロポレーションによって実施した。トランスフェクション前に、それぞれの発現ベクターはFspIを使用して線状化した。約107個の細胞を10μgの線状化プラスミドでトランスフェクトし、10%FBSを含有するDME培地中に縣濁し、数枚の96ウェルプレートに平板培養した。24〜48時間後、選択培地(10%FBSを含有するDME培地、HT培地サプリメント(Sigma、St.Louis、MO)、0.25mg/mlキサンチンおよび1μg/mlミコフェノール酸)を施した。選択開始後約10日で、培養上清を抗体産生に関してアッセイした。
【0192】
培養上清中のpCh33E10およびpHu33E10−1から発現した組換え抗体(それぞれCh33E10およびHu33E10−VH1/VL1)の発現レベルをサンドウィッチELISAによって測定した。典型的な実験では、PBSに溶かした100μl/ウェルの1/2,000希釈ヤギ抗ヒトIgGFcγ鎖特異的ポリクローナル抗体(SouthernBiotech、Birmingham、AL)で4℃において一晩、ELISAプレートをコーティングし、洗浄バッファー(0.05%のTween20を含有するPBS)で洗浄し、300μl/ウェルのブロッキングバッファー(2%のスキムミルクおよび0.05%のTween20を含有するPBS)で室温において1時間ブロッキングした。洗浄バッファーで洗浄した後、ELISAバッファー(1%のスキムミルクおよび0.025%のTween20を含有するPBS)に適切に希釈した100μl/ウェルのサンプルをELISAプレートに施した。ヒトミエローマ血清(SouthernBiotech)から精製したヒトIgG1/κ抗体または精製Ch33E10を標準として使用した。室温で2時間ELISAプレートをインキュベートし洗浄バッファーで洗浄した後、100μl/ウェルの1/2,000希釈HRP結合ヤギ抗ヒトkappa鎖ポリクローナル抗体(SouthernBiotech)を使用して、結合した抗体を検出した。室温で1時間のインキュベーションおよび洗浄バッファーでの洗浄後、発色現像は100μl/ウェルのABTS基質を加えることによって実施し、100μl/ウェルの2%シュウ酸で停止させた。吸光度は405nmで読み取った。
【0193】
高レベルのCh33E10(NSO−Ch33E102C11;L4−2C11とも呼ばれる)を産生するNS0安定トランスフェクタントの1つを、ハイブリドーマSFM(Invitrogen)を使用して無血清培地中での増殖に適合させ、約106/mlの密度までローラーボトル中で増殖させ、6mg/mlのUltrafiltrated Soy Hydrolyzate(Irvine Scientific カタログ番号96857)を供給し、細胞生存率が50%未満になるまでさらに増殖させた。遠心分離および濾過後、培養上清をプロテイン−Aセファロースカラム(GE Healthcare、Piscataway、NJ)に充填した。抗体を0.1Mのグリシン−HCl(pH3.0)で溶出する前に、カラムをPBSで洗浄した。1Mのトリス−HCl(pH8)での中和後、溶出抗体のバッファーを透析によってPBSに交換した。抗体濃度は280nmにおける吸光度を測定することによって決定した(1mg/ml=1.4OD)。
【0194】
Ch33E10を発現するNS0安定トランスフェクタントは容易に得たが、pHu33E10−1で安定的にトランスフェクトしたNS0トランスフェクタントはいずれも、Hu33E10−VH1/VL1をあまり産生しないことが分かった。重鎖および軽鎖mRNAの分析によって、VL1遺伝子のシグナルペプチドコード領域中の偶然生成したスプライスドナー部位のためVLコード領域を欠く、異常な軽鎖mRNAの存在が明らかになった。この問題を解決するため、シグナルペプチドコード領域中のスプライスドナー部位を、オーバーラップエクステンションPCR法を使用して部位特異的突然変異誘発によって排除した(Higuchi, R., PCR Technology. Principles and Applications for DNA Amplification.Erlich, H.A., ed.pp.61-70, Stockton Press, New York, 1989)。生成した変異Hu33E10VL遺伝子、VL6の配列は図14中に示す。VL領域のアミノ酸配列はVL1とVL6遺伝子の間で互いに同一である。VL6遺伝子は発現ベクターpHu33E10−1中のVL1に交換してpHu33E10−2を作製した。pHu33E10−2で安定的にトランスフェクトし高レベルの組換え抗体(Hu33E10−VH1/VL6)を産生したNS0細胞は、前に記載した方法によって得た。pHu33E10−2で安定的にトランスフェクトしたNS0細胞では、異常な軽鎖mRNAは観察しなかった。高レベルのHu33E10−VH1/VL6を産生するNS0安定トランスフェクタントの1つ(NS0−Hu33E10−2#22)を適合させ、ハイブリドーマSFM中で増殖させ、培養上清は前に記載したようにHu33E10−VH1/VL6の精製に使用した。
【0195】
6.5.Hu33E10−VHl/VL6の特徴付け
hOPN5−BSAとCh33E10およびHu33E10−VHl/VL6IgG1/κ抗体の結合をELISAによって調べた。抗原として、ウシ血清アルブミンと結合した合成オリゴペプチド(Cys−Val−Asp−Thr−Tyr−Asp−Gly−Arg−Gly−Asp−Ser−Val−Val−Tyr−Gly−Leu−Arg−Ser;Gene Techno Scienceにより提供された)(hOPN5−BSA)を使用した。典型的な実験では、0.2MのNa2C03バッファー(pH9.4)に溶かした100μl/ウェルの1μg/mlのhOPN5−BSAで4℃において一晩、ELISAプレートをコーティングし、洗浄バッファーで洗浄し、300μl/ウェルのブロッキングバッファーで室温において1時間ブロッキングした。洗浄バッファーで洗浄した後、ELISAバッファーに適切に希釈したサンプルを100μl/ウェルでELISAプレートに施した。4℃において一晩ELISAプレートをインキュベートし洗浄バッファーで洗浄した後、100μl/ウェルの1/2,000希釈HRP結合ヤギ抗ヒトγ鎖ポリクローナル抗体(SouthernBiotech)を使用して、結合した抗体を検出した。室温で1時間のインキュベーションおよび洗浄バッファーでの洗浄後、発色現像は100μl/ウェルのABTS基質を加えることによって実施し、100μl/ウェルの2%シュウ酸で停止させた。吸光度は405nmで読み取った。
【0196】
図15A中に示すように、Hu33E10−VH1/VL6は、Ch33E10よりhOPN5−BSAとの結合に関して約100倍弱かった。
【0197】
6.6.Hu33E10VH5、VH7およびVH7.8の作製および特徴付け
どのヒト化VH1およびVL6遺伝子がHu33E10−VHl/VL6の抗原結合親和性の消失の原因であるかを確認するために、1つがマウスVHおよびヒト化VL6(MoHu33E10)からなり、および他方がヒト化VH1およびマウスVL(HuMo33E10)からなる、2つの組換えIgG1/κ抗体を作製した。MoHu33E10およびHuMo33E10抗体の分析によって、Hu33E10−VH1/VL6の親和性の消失は大部分はVH1遺伝子に原因があり、部分的にはVL1遺伝子に原因があったことが分かった(ここでデータ示す)。
【0198】
VH1遺伝子の予備突然変異分析に基づくと、位置73〜77におけるアミノ酸は、抗原結合親和性を維持するのに重要である懸念があった(データ示さず)。オーバーラップエクステンションPCR法(Higuchi, 1989)を使用し部位特異的突然変異誘発によって構築したVH1遺伝子の3つの変異体(VH5、VH7およびVH7.8)は、さらなる分析用に選択した。VH1と比較して、位置77におけるIleはVH5中のSer(図6および16)、VH7中では位置76におけるSerおよび位置77におけるIleはそれぞれAsnおよびSer(図6および17)、およびVH7.8中では位置74におけるSer、位置76におけるSerおよび位置77におけるIleはそれぞれAla、AsnおよびSerに置換した(図6および18)。
【0199】
SpeIおよびHindIIIで消化したVH5、VH7およびVH7.8遺伝子のそれぞれをpHu33E10−2においてVH1に置換して、それぞれ発現ベクターpHu33E10−3、pHu33E10−4およびpHu33E10−5を構築した。pHu33E10−3、pHu33E10−4およびpHu33E10−5のそれぞれで安定的にトランスフェクトしたNS0細胞はセクション4中に記載したように生成した。Hu33E10−VH5/VL6を産生するNSO−Hu33E10−3#65、Hu33E10−VH7/VL6を産生するNS0−Hu33E10−45D3(R3−15D3とも呼ばれる)、およびHu33E10−VH7.8/VL6を産生するNS0−Hu33E10−51E5(EJ3−31E5とも呼ばれる)をハイブリドーマSFM中での増殖に適合させ、ローラーボトル中で増殖させた。Hu33E10−VH5/VL6、Hu33E10−VH7/VL6およびHu33E10−VH7.8/VL6抗体は、セクション4中に記載したようにプロテインAカラムを使用して対応する培養上清から精製した。
【0200】
hOPN5−BSAとHu33E10−VH5/VL6、Hu33E10−VH7/VL6およびHu33E10−VH7.8/VL6抗体の結合は、セクション5中に記載したようにELISAによってCh33E10の結合と比較した。hOPN5−BSAとHu33E10−VH5/VL6の親和性はHu33E10−VH1/VL6と比較したとき若干向上したが、それはCh33E10の親和性よりさらに一層弱かった(図15A)。hOPN5−BSAとのHu33E10−VH7/VL6の結合親和性は、Ch33E10より3〜4倍弱かった(図15A)。Hu33E10−VH7.8/VL6の親和性は、この実験中Ch33E10の親和性より約3倍低かった(図15B)。反復実験中、Hu33E10−VH7/VL6とHu33E10−VH7.8/VL6は、hOPN5−BSAとの結合に関して互いに同様に挙動した。
【0201】
6.7.Hu33E10VLv2の作製および特徴付け
VL1遺伝子の予備突然変異分析によって、Hu33E10−VH1/VL6の結合親和性の消失に貢献するいかなる特定のフレームワークアミノ酸残基も確認することはできなかったが(データ示さず)、VL1遺伝子は部分的には親和性消失の原因であることが疑われた。VL1遺伝子をさらに特徴付ける代わりに、異なるヒトフレームワークに基づく新たなヒト化33E10VL(VLv2)を設計した(図19)。VLv2用に、M29467のヒトアミノ酸配列(GenBank受託番号)をアクセプターとして選択した。CDRとの有意な相互作用が予想された位置22および37では、マウス33E10VLのアミノ酸残基はヒト化型で保持された。NheIおよびEcoRI部位と隣接するHu33E10VLv2遺伝子(図20)はGenScript USA Inc.(Piscataway、NJ)で合成された。VL6遺伝子とVLv2遺伝子は、同じシグナルペプチド配列およびスプライスドナーシグナルを共有する(図14および20)。
【0202】
NheIおよびEcoRIで消化したHu33E10VLv2遺伝子をpHu33E10−4においてVL1に置換して、新たな発現ベクターpHu33E10−6を構築した。Hu33E10−VH7/VLv2IgG1/κを産生するためにpHu33E10−6で安定的にトランスフェクトしたNS0細胞は、セクション4中に記載したように生成した。トランスフェクタントの1つ、NS0−Hu33E10−6I−15(SP2−1I−15とも呼ばれる)を適合させ、ハイブリドーマSFM中で増殖させ、Hu33E10−VH7/VLv2抗体は、セクション4中に記載した手順を使用して培養上清から精製した。
【0203】
hOPN5−BSAとHu33E10−VH7/VLv2の結合は、セクション5中に記載したようにELISAによって分析した。2つの独立した実験の結果を図21中に示す。両方の実験において、Hu33E10−VH7/VLv2の結合親和性はCh33E10の結合親和性の2倍以内であった。
【0204】
6.8.NS0安定トランスフェクタントの特徴付け
PCRマイコプラズマ検出セット(Takara Bio USA、Madison、WT)を用いた試験によって、NS0−Ch33E102C11、NS0−Hu33E10−45D3、NS0−Hu33E10−51E5およびNS0−Hu33E10−6−I−15はいずれも、マイコプラズマの存在に関して陰性であったことを示した。
【0205】
それぞれNS0−Hu33E10−45D3、NS0−Hu33E10−51E5およびNS0−Hu33E10−6−I−15において産生されたHu33E10−VH7/VL6、Hu33E10−VH7.8/VL6およびHu33E10−VH7/VLv2IgGl/κ抗体の重鎖および軽鎖の真正を、cDNA配列決定によって確認した。全RNAはTRIzol試薬(Invitrogen)を使用してこれらの細胞から抽出し、オリゴdTプライマー処理cDNAは、供給者のプロトコールに従いRT−PCR用のSuperScriptIII第一鎖合成システム(Invitrogen)を使用して合成した。γ−1重鎖のコード領域は、プライマーとしてCMV2およびJNT098(図22)ならびにPhusion DNAポリメラーゼを使用しPCRによって増幅した。PCR断片はゲル精製し、プライマーとしてCMV2、JNT082、JNT097およびJNT098を用いて配列決定に施した(図22)。同様に、kappa軽鎖のコード領域はCMV2およびJNT026を使用して増幅し(図22)、ゲル精製したDNA断片はプライマーとしてCMV2、JNT026、JNT080およびJNT084を用いて配列決定に施した(図22)。重鎖および軽鎖コード領域に関して得た配列は、NS0−Hu33E10−45D3、NS0−Hu33E10−51E5およびNS0−Hu33E10−6−I−15安定トランスフェクタントのそれぞれに関する発現ベクター中の対応する配列と完全に一致した。Hu33E10VH7およびVH7.8からなるγ−1重鎖の全コード領域に関する配列は、それぞれ図23および24中に示す。Hu33E10VL6およびVLv2からなるkappa軽鎖の全コード領域に関する配列は図25および26中に示す。
【0206】
さらに、NS0−Ch33E102C11において産生されたCh33E10IgG1/κ抗体のVHおよびVL領域の真正を、cDNA配列決定によって確認した。全RNAの抽出およびcDNA合成は前に記載したように実施した。VHコード領域は、プライマーとしてCMV2およびJNT082ならびにPhusionポリメラーゼを使用しPCRによって増幅した。PCR断片はゲル精製し、プライマーとしてCMV2およびJNT082を用いて配列決定に施した。同様に、VLコード領域はCMV2およびJNT026を使用して増幅し、ゲル精製したDNA断片はプライマーとしてCMV2およびJNT026を用いて配列決定に施した。得られたVHおよびVL領域のヌクレオチド配列は、pCh33E10発現ベクター中の対応する配列と完全に一致した(それぞれ図3および4)。
【0207】
6.9.結論
hOPN5−BSAに対するHu33E10−VH7/VLv2IgG1/κの親和性は、Ch33E10IgG1/κの親和性の3倍以内であったと決定した。
【0208】
6.10.細胞接着阻害アッセイ
図27は、ヒト化抗OPN抗体が、hOPN5−BSAとMDA−MB−435Sの接着を阻害したことを示す。MDA−MB−435Sヒト乳癌細胞系はアメリカンタイプカルチャーコレクション(ATCC)から得て、10%ウシ胎児血清を補充したDMEM中で培養した。96ウェルプレートは37℃において1時間hOPN5−BSA(2μg/ml)でコーティングし、次に室温において1時間0.5%BSAで処理した。ヒト免疫グロブリンG1kappa、Ch33E10、Hu33E10−VH7.8/VL6(EJ3−1−1)、Hu33E10−VH7/VL6(E3−1)、Hu33E10−VH7/VLv2(SP−2)、およびマウス33E10抗体を0.25%BSA、DMEM中に溶かし、100μl/mlで始めて様々な濃度で処理し、37℃において20分間連続2倍希釈した。2×104個のMDA−MB−435S細胞をそれぞれのウェルに加えた。37℃で1時間のインキュベーション後、培地を除去し、(37℃まで温めた)温かいPBSで軽く二回洗浄した。接着細胞は固定し、20%メタノールに溶かした0.5%クリスタルバイオレットで染色し、20%酢酸で溶かした。吸光度は590nmで測定した。抗体を加えずに、阻害率はこの吸光度で標準化した。
【0209】
7.寄託
マウス抗RGDモノクローナル抗体を産生する33E10および35B6として本明細書に示すハイブリドーマは、微生物の寄託に関するブダペスト条約に従い、2005年10月27日に茨城県つくば市東1−1−1中央第6(郵便番号:305−8566)に位置する産業技術総合研究所の特許生物寄託センターに寄託され、それぞれ受託番号FERMBP−10440およびFERMBP−10441を与えられ、これらはいずれもその全容を参照によって本明細書に組み込む。
【産業上の利用可能性】
【0210】
本発明のヒト化モノクローナル抗体は、RGDタンパク質の機能を阻害して、癌、例えば癌細胞の増殖または転移、および炎症疾患、例えば慢性関節リウマチ、骨関節炎、肝炎、気管支喘息、線維症、糖尿病、癌転移、動脈硬化症、多発性硬化症、肉芽腫、炎症性腸疾患(潰瘍性大腸炎およびクローン病)、自己免疫疾患などに対する治療効果を示す。本発明の抗RGD抗体と抗インテグリン抗体の両方を含む医薬組成物は、癌および炎症性疾患に対してより改善された治療効果を発揮する。
【0211】
具体的な実施形態を本明細書で例示および記載してきたが、様々な他のおよび/または均等な履行を、本発明の範囲から逸脱せずに、記載し示した具体的な実施形態に置き換えることができることは、当業者によって理解されよう。本出願は、本明細書で論じる具体的な実施形態の任意の適合または変形を含むと考えられる。したがって、本発明は特許請求の範囲およびその均等物によってのみ制限されると考えられる。
【特許請求の範囲】
【請求項1】
RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合断片であって、
(i)VHを含むH鎖であって、前記VHが配列番号92、94または96のアミノ酸配列を含むH鎖、または
(ii)VLを含むL鎖であって、前記VLが配列番号98または100のアミノ酸配列を含むL鎖、または
(iii)前述の(i)および(ii)の両方を含むヒト化抗体またはその抗原結合断片。
【請求項2】
前記VHが配列番号91、93または95のヌクレオチド配列によってコードされるアミノ酸配列を含み、前記VLが配列番号97または99のヌクレオチド配列によってコードされるアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合断片。
【請求項3】
前記VHが配列番号94のアミノ酸配列を含み、前記VLが配列番号100のアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合断片。
【請求項4】
前記H鎖が配列番号25または27のアミノ酸配列を含み、前記L鎖が配列番号29または31のアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合断片。
【請求項5】
前記H鎖が配列番号25のアミノ酸配列を含み、前記L鎖が配列番号31のアミノ酸配列を含む、請求項1または2に記載のヒト化抗体またはその抗原結合断片。
【請求項6】
配列番号92、94または96のアミノ酸配列をコードするヌクレオチド配列を含む単離核酸分子。
【請求項7】
前記ヌクレオチド配列が配列番号91、93または95のヌクレオチド配列を有する、請求項6に記載の単離核酸分子。
【請求項8】
配列番号98または100のアミノ酸配列をコードするヌクレオチド配列を含む単離核酸分子。
【請求項9】
前記ヌクレオチド配列が配列番号97または99のヌクレオチド配列を有する、請求項8に記載の単離核酸分子。
【請求項10】
配列番号25または27のアミノ酸配列をコードするヌクレオチド配列を含む単離核酸分子。
【請求項11】
前記ヌクレオチド配列が配列番号24または26のヌクレオチド配列を有する、請求項10に記載の単離核酸分子。
【請求項12】
配列番号29または31のアミノ酸配列をコードするヌクレオチド配列を含む単離核酸分子。
【請求項13】
前記ヌクレオチド配列が配列番号28または30のヌクレオチド配列を有する、請求項10に記載の単離核酸分子。
【請求項14】
前記核酸分子が1つまたは複数の制御エレメントに作動可能に連結した、請求項6または8または両方に記載の核酸分子を含むベクター。
【請求項15】
前記核酸分子が1つまたは複数の制御エレメントに作動可能に連結した、請求項10または12または両方に記載の核酸分子を含むベクター。
【請求項16】
請求項14または15に記載のベクターを含む単離宿主細胞。
【請求項17】
ヒト化抗体またはその抗原結合断片を調製するための方法であって、ヒト化抗体またはその抗原結合断片が発現されるような条件下で請求項16に記載の宿主細胞を培養すること、および発現したヒト化抗体を回収することを含む方法。
【請求項18】
細胞接着を阻害するための組成物であって、請求項1から5のいずれか一項に記載のヒト化抗体または抗原結合断片を含む組成物。
【請求項19】
細胞との細胞外マトリックスタンパク質の結合に干渉するための組成物であって、請求項1から5のいずれか一項に記載のヒト化抗体またはその抗原結合断片を含む組成物。
【請求項20】
前記組成物がRGDタンパク質と関係がある障害または疾患の治療または診断に使用される、請求項18または19に記載の組成物。
【請求項21】
RGDタンパク質と関係がある障害または疾患を予防または治療するための方法であって、有効量の請求項1から5のいずれか一項に記載のヒト化抗体またはその抗原結合断片を、その必要性がある対象に投与することを含む方法。
【請求項22】
試験サンプル中のRGDタンパク質を検出するための方法であって、試験サンプル中のRGDタンパク質との、請求項1から5のいずれか一項に記載のヒト化抗体またはその抗原結合断片の結合をアッセイすることを含む方法。
【請求項23】
RGDタンパク質と関係がある障害または疾患をin vivoで診断するための方法であって、有効量の請求項1から5のいずれか一項に記載のヒト化抗体またはその抗原結合断片を、検査する対象に投与することを含む方法。
【請求項1】
RGD配列を免疫特異的に認識するヒト化抗体またはその抗原結合断片であって、
(i)VHを含むH鎖であって、前記VHが配列番号92、94または96のアミノ酸配列を含むH鎖、または
(ii)VLを含むL鎖であって、前記VLが配列番号98または100のアミノ酸配列を含むL鎖、または
(iii)前述の(i)および(ii)の両方を含むヒト化抗体またはその抗原結合断片。
【請求項2】
前記VHが配列番号91、93または95のヌクレオチド配列によってコードされるアミノ酸配列を含み、前記VLが配列番号97または99のヌクレオチド配列によってコードされるアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合断片。
【請求項3】
前記VHが配列番号94のアミノ酸配列を含み、前記VLが配列番号100のアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合断片。
【請求項4】
前記H鎖が配列番号25または27のアミノ酸配列を含み、前記L鎖が配列番号29または31のアミノ酸配列を含む、請求項1に記載のヒト化抗体またはその抗原結合断片。
【請求項5】
前記H鎖が配列番号25のアミノ酸配列を含み、前記L鎖が配列番号31のアミノ酸配列を含む、請求項1または2に記載のヒト化抗体またはその抗原結合断片。
【請求項6】
配列番号92、94または96のアミノ酸配列をコードするヌクレオチド配列を含む単離核酸分子。
【請求項7】
前記ヌクレオチド配列が配列番号91、93または95のヌクレオチド配列を有する、請求項6に記載の単離核酸分子。
【請求項8】
配列番号98または100のアミノ酸配列をコードするヌクレオチド配列を含む単離核酸分子。
【請求項9】
前記ヌクレオチド配列が配列番号97または99のヌクレオチド配列を有する、請求項8に記載の単離核酸分子。
【請求項10】
配列番号25または27のアミノ酸配列をコードするヌクレオチド配列を含む単離核酸分子。
【請求項11】
前記ヌクレオチド配列が配列番号24または26のヌクレオチド配列を有する、請求項10に記載の単離核酸分子。
【請求項12】
配列番号29または31のアミノ酸配列をコードするヌクレオチド配列を含む単離核酸分子。
【請求項13】
前記ヌクレオチド配列が配列番号28または30のヌクレオチド配列を有する、請求項10に記載の単離核酸分子。
【請求項14】
前記核酸分子が1つまたは複数の制御エレメントに作動可能に連結した、請求項6または8または両方に記載の核酸分子を含むベクター。
【請求項15】
前記核酸分子が1つまたは複数の制御エレメントに作動可能に連結した、請求項10または12または両方に記載の核酸分子を含むベクター。
【請求項16】
請求項14または15に記載のベクターを含む単離宿主細胞。
【請求項17】
ヒト化抗体またはその抗原結合断片を調製するための方法であって、ヒト化抗体またはその抗原結合断片が発現されるような条件下で請求項16に記載の宿主細胞を培養すること、および発現したヒト化抗体を回収することを含む方法。
【請求項18】
細胞接着を阻害するための組成物であって、請求項1から5のいずれか一項に記載のヒト化抗体または抗原結合断片を含む組成物。
【請求項19】
細胞との細胞外マトリックスタンパク質の結合に干渉するための組成物であって、請求項1から5のいずれか一項に記載のヒト化抗体またはその抗原結合断片を含む組成物。
【請求項20】
前記組成物がRGDタンパク質と関係がある障害または疾患の治療または診断に使用される、請求項18または19に記載の組成物。
【請求項21】
RGDタンパク質と関係がある障害または疾患を予防または治療するための方法であって、有効量の請求項1から5のいずれか一項に記載のヒト化抗体またはその抗原結合断片を、その必要性がある対象に投与することを含む方法。
【請求項22】
試験サンプル中のRGDタンパク質を検出するための方法であって、試験サンプル中のRGDタンパク質との、請求項1から5のいずれか一項に記載のヒト化抗体またはその抗原結合断片の結合をアッセイすることを含む方法。
【請求項23】
RGDタンパク質と関係がある障害または疾患をin vivoで診断するための方法であって、有効量の請求項1から5のいずれか一項に記載のヒト化抗体またはその抗原結合断片を、検査する対象に投与することを含む方法。
【図1】


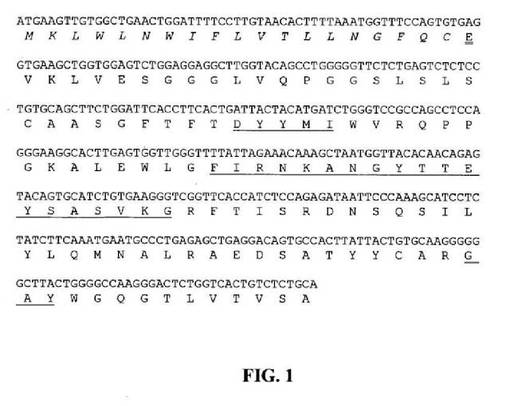
【図2】
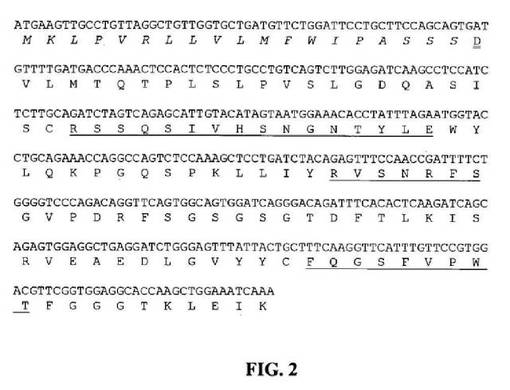
【図3】

【図4】
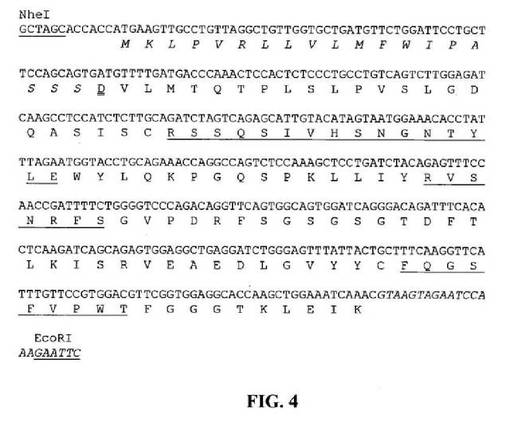
【図5】

【図6】

【図7】

【図8】

【図9】

【図10】

【図11】

【図12】

【図13】
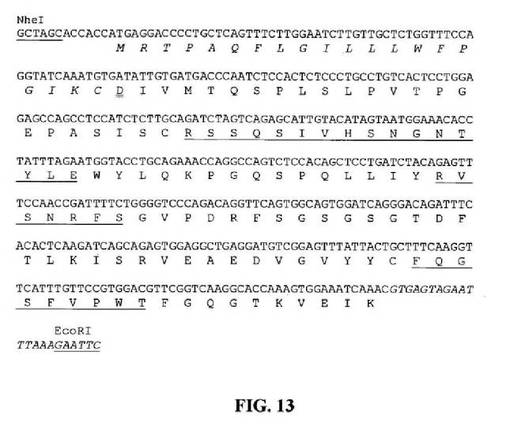
【図14】
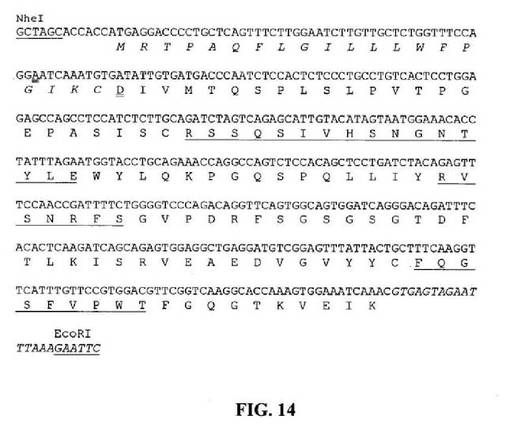
【図15A−B】
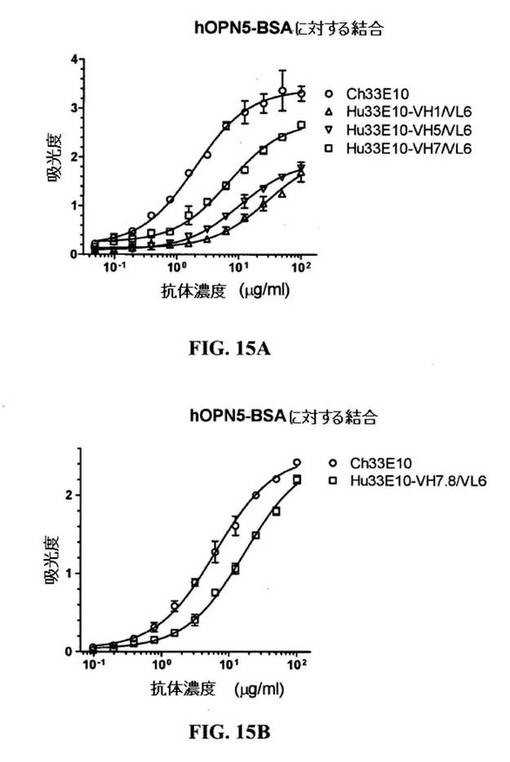
【図16】
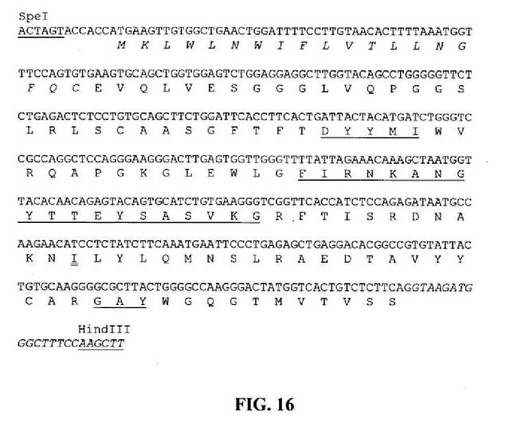
【図17】

【図18】
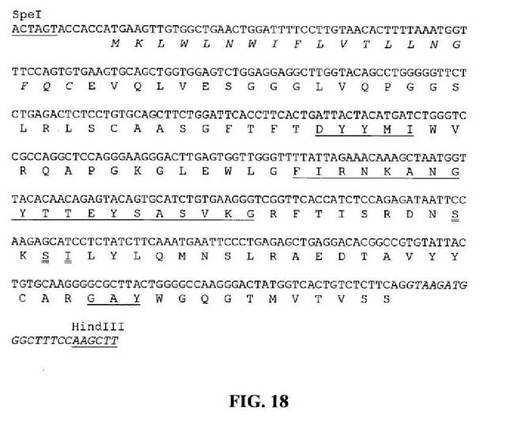
【図19】

【図20】

【図21A−B】
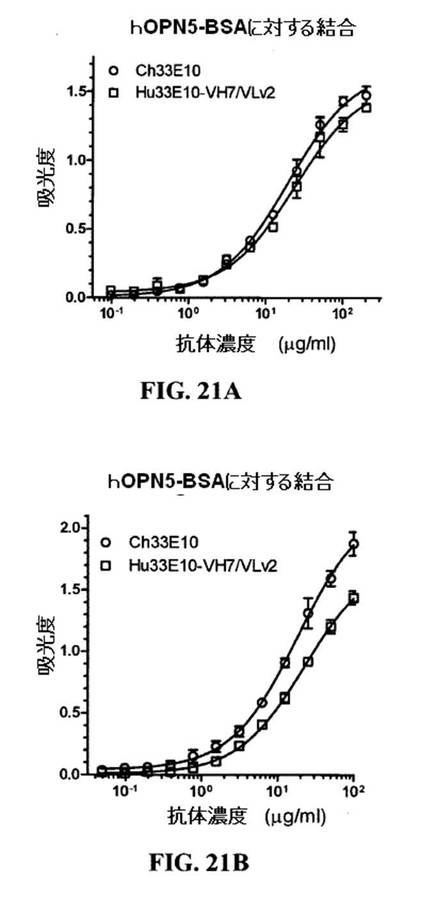
【図22】

【図23】

【図24】

【図25】

【図26】

【図27】
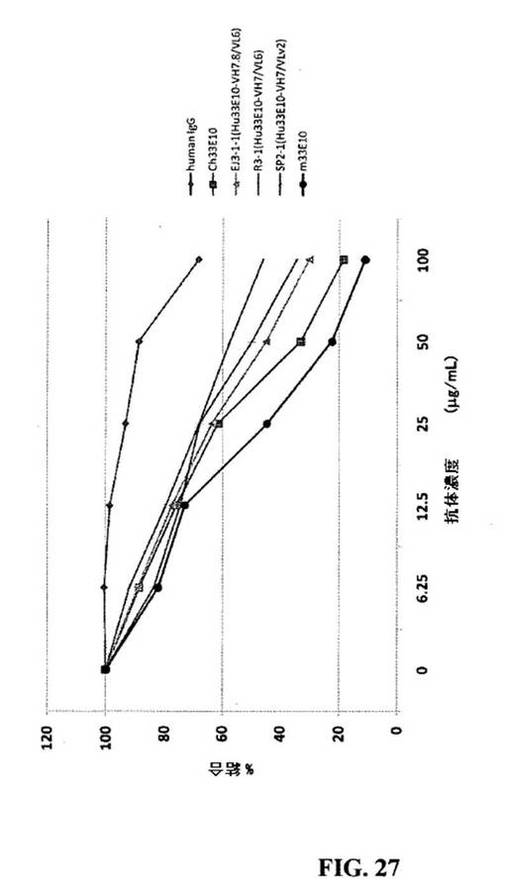
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15A−B】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21A−B】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【公表番号】特表2013−505702(P2013−505702A)
【公表日】平成25年2月21日(2013.2.21)
【国際特許分類】
【出願番号】特願2012−514276(P2012−514276)
【出願日】平成22年9月22日(2010.9.22)
【国際出願番号】PCT/JP2010/067017
【国際公開番号】WO2011/037271
【国際公開日】平成23年3月31日(2011.3.31)
【出願人】(501416243)株式会社ジーンテクノサイエンス (9)
【Fターム(参考)】
【公表日】平成25年2月21日(2013.2.21)
【国際特許分類】
【出願日】平成22年9月22日(2010.9.22)
【国際出願番号】PCT/JP2010/067017
【国際公開番号】WO2011/037271
【国際公開日】平成23年3月31日(2011.3.31)
【出願人】(501416243)株式会社ジーンテクノサイエンス (9)
【Fターム(参考)】
[ Back to top ]