
細胞性免疫原性が向上した抗ウイルスワクチン
本発明は、ウイルス感染症を治療又は予防するための組成物、方法、及びキットを提供する。本明細書に記載される多価(例えば、二価)ワクチンは、ワクチン接種を受けた対象において細胞性免疫応答の多様性又は幅及び深さを増加させることのできる、計算により最適化されたウイルスポリペプチドを組み込む。
【発明の詳細な説明】
【技術分野】
【0001】
連邦政府助成研究に関する記載
本研究は、NIH交付番号第U19−AI066305号及び同第U19−AI078526号から一部資金提供を受けている。政府は本発明に対して特定の権利を有する。
【0002】
発明の分野
本発明は、ウイルス感染症を治療又は予防するための組成物、方法、及びキットを提供する。本明細書に記載される多価(例えば、二価)ワクチンは、ワクチン接種を受けた対象における細胞性免疫応答の多様性又は幅及び深さを増加させることのできる、計算により最適化されたウイルスポリペプチドを組み込む。
【背景技術】
【0003】
ウイルスに対する細胞性免疫応答を誘発するワクチンは、ウイルス感染症を効果的に治療又は予防するため広域のウイルス多様性を反映しなければならない。例えば、強力で多様なHIV−1特異的T細胞応答の惹起は、有効なHIV−1ワクチンには極めて重要であると思われる。細胞傷害性Tリンパ球(CTL)応答はヒトにおける疾患進行の遅延と相関し、非ヒト霊長類ワクチン接種モデルにおけるCTL応答の重要性は十分に確立されている。大きいばらつきのあるエンベロープ(Env)がHIVに対する中和抗体の一次標的であり、ワクチン抗原もまた、それらの抗体応答を誘発するように作ることが必要となり得るが、T細胞ワクチン成分は、より保存されているタンパク質を標的化し、交差反応する可能性がより高い応答を引き起こすことができる。しかし、最も保存されているHIV−1タンパク質でさえ、ばらつきが問題となり得るのに十分な多様性を有する。コンセンサス及び祖先HIV−1配列などの人工的な中心配列のワクチン手法は、本質的に株間の「違いを分ける」もので、天然株ワクチンと比較して交差反応性が亢進した応答を刺激することができる。コンセンサス抗原は、あらゆる循環株の唯一最良の「平均」である合成抗原配列に相当する。こうした抗原は特異的細胞性免疫応答を誘発することができるが、その応答の幅及び強度は、これまでのワクチン戦略と比べて実質的に向上していない。ウイルス感染症を治療又は予防する次世代ワクチンの開発は、ワクチン接種が奏功する結果をもたらすことができるように、細胞性免疫の幅の増加を引き出すものでなければならない。かかるワクチンは、特にHIV−1の治療又は予防に差し迫って必要とされている。
【発明の概要】
【課題を解決するための手段】
【0004】
第1の態様において、本発明は、少なくとも2個の別個の最適化ウイルスポリペプチド(例えば、2、3、4、5個、又はそれ以上の別個の最適化ウイルスポリペプチド)を含む、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンを特徴とし、ここで最適化ウイルスポリペプチドは、同じウイルス遺伝子産物に対応する。一実施形態において、ウイルス感染症は、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる。他の実施形態において、レトロウイルスはヒト免疫不全ウイルス1型(HIV−1)であり、及びウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuを含む。さらなる実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuウイルス遺伝子産物のうちの1つに対応する最適化ウイルスポリペプチドを2個以下含む。別の実施形態において、ワクチンは、Gag及びNefに対応する最適化ウイルスポリペプチドを含まない。さらに別の実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuから選択される第1のウイルス遺伝子産物についての少なくとも2個の別個の最適化ウイルスポリペプチド(例えば、2、3、4、5個、又はそれ以上の別個の最適化ウイルスポリペプチド)と、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuから選択される第1のウイルス遺伝子産物と異なる第2のウイルス遺伝子産物についての1つ又は複数の別個の最適化ウイルスポリペプチド(例えば、2、3、4、5個、又はそれ以上の別個の最適化ウイルスポリペプチド)とを含む。
【0005】
第2の態様において、本発明は、ヒトなどの哺乳動物におけるヒト免疫不全ウイルス1型(HIV−1)感染症を治療し、又はそのリスクを低減するワクチンであって、配列番号1〜29に示される配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する最適化ウイルスポリペプチドを含むワクチンを特徴とする。一実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示される配列のいずれか1つとアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する。別の実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示される配列のいずれか1つのアミノ酸配列を有する。さらなる実施形態において、ワクチンは、a)〜k)群の任意の1つ又は複数から選択される少なくとも2個の最適化ウイルスポリペプチドを含む:a)配列番号1及び2;b)配列番号3、4、及び5;c)配列番号6及び7;d)配列番号8〜12;e)配列番号13、14、及び15;f)配列番号16、17、及び18;g)配列番号19及び20;h)配列番号21、22、及び23;i)配列番号24及び25;j)配列番号26及び27;並びにk)配列番号21〜22。別の実施形態において、ワクチンは、上記のa)〜k)群のいずれか1つから選択される最適化ウイルスポリペプチドと、同じ、又は異なるa)〜k)群からの1つ又は複数の異なる最適化ウイルスポリペプチドとの対を含むことができる。他の実施形態において、ワクチンは、a)〜k)群の1つ又は複数からの少なくとも3個又は4個又はそれ以上の最適化ウイルスポリペプチドを含むことができる。
【0006】
第3の態様において、本発明は、別個の最適化ウイルスポリペプチドの少なくとも2つの対を含む、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンを特徴とし、ここで最適化ウイルスポリペプチドの各対は同じウイルス遺伝子産物に対応し、及びここでワクチンに組み込まれた2個以下の最適化ウイルスポリペプチドは、同じウイルス遺伝子産物に対応する。一実施形態において、ワクチンは、別個の最適化ウイルスポリペプチドの少なくとも3つの対を含む。別の実施形態において、ワクチンは、別個の最適化ウイルスポリペプチドの少なくとも4つの対を含む。一実施形態において、ウイルス感染症は、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる。他の実施形態において、レトロウイルスはヒト免疫不全ウイルス1型(HIV−1)であり、及びウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuを含む。さらなる実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuウイルス遺伝子産物のうちの1つに対応する最適化ウイルスポリペプチドを2個以下含む。別の実施形態において、ワクチンは、Gag及びNefに対応する最適化ウイルスポリペプチドを含まない。さらなる実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuウイルス遺伝子産物のうちの任意の3つに対応する別個の最適化ウイルスポリペプチドの少なくとも3つの対を含む。別の実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuウイルス遺伝子産物のうちの任意の4つに対応する別個の最適化ウイルスポリペプチドの少なくとも4つの対を含む。
【0007】
本発明の最初の3つの態様のうちの任意の一実施形態において、ワクチンは、ウイルス遺伝子産物に対する細胞性免疫応答を誘発する。別の実施形態において、ワクチンは、HIV−1に対する細胞性免疫応答を誘発する。さらなる実施形態において、少なくとも1つの別個の最適化ウイルスポリペプチドのヌクレオチド配列は、核酸又はベクターによりコードされる。一実施形態において、ベクターは、アデノウイルス血清型26(Ad26)、アデノウイルス血清型34(Ad34)、アデノウイルス血清型35(Ad35)、アデノウイルス血清型48(Ad48)、又はアデノウイルス血清型5 HVR48(Ad5HVR48)などの組換えアデノウイルスである。さらなる実施形態において、ワクチンは、薬学的に許容可能な担体、賦形剤、又は希釈剤と組み合わせたものである。
【0008】
第4の態様において、本発明は、配列番号1〜29に示されるアミノ酸配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する最適化ウイルスポリペプチドのヌクレオチド配列を含む核酸を特徴とする。一実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示されるアミノ酸配列のいずれか1つと配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する。別の実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示されるアミノ酸配列のいずれか1つを有する。さらなる実施形態において、核酸はベクターを含む。一実施形態において、ベクターは、アデノウイルス血清型26(Ad26)、アデノウイルス血清型34(Ad34)、アデノウイルス血清型35(Ad35)、アデノウイルス血清型48(Ad48)、又はアデノウイルス血清型5 HVR48(Ad5HVR48)などの組換えアデノウイルスである。
【0009】
第5の態様において、本発明は、配列番号1〜29に示されるアミノ酸配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する最適化ウイルスポリペプチドを特徴とする。一実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示されるアミノ酸配列のいずれか1つと配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する。別の実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示されるアミノ酸配列のいずれか1つを有する。
【0010】
第6の態様において、本発明は、本発明のワクチン又は核酸を投与することにより、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減する方法を特徴とする。一実施形態において、ウイルス感染症は、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる。さらなる実施形態において、レトロウイルスはヒト免疫不全ウイルス1型(HIV−1)であり、及びウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuを含む。一実施形態において、ワクチン又は核酸は、ウイルス遺伝子産物に対する細胞性免疫応答を誘発する。
【0011】
第7の態様において、本発明は、本発明のワクチンを合成することによる、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンの製造方法を特徴とする。
【0012】
第8の態様において、本発明は、本発明の核酸を細胞と接触させ、及び最適化ウイルスポリペプチドを単離することによる、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンの製造方法を特徴とする。
【0013】
本発明の第7又は第8の態様の一実施形態において、最適化ウイルスポリペプチドは、哺乳動物に投与されると細胞性免疫応答を誘発する。細胞性免疫応答は、ウイルス遺伝子産物に対するものであってよい。別の実施形態において、ウイルス感染症は、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる。さらなる実施形態において、レトロウイルスはヒト免疫不全ウイルス1型(HIV−1)であり、及びウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuを含む。
【0014】
第9の態様において、本発明は、キットであって、本発明のワクチンと、薬学的に許容可能な担体、賦形剤、又は希釈剤と、キットの使用説明書とを含むキットを特徴とする。一実施形態において、キットはアジュバントも含む。
【0015】
最後の態様において、本発明は、キットであって、本発明の核酸と、薬学的に許容可能な担体、賦形剤、又は希釈剤と、キットの使用説明書とを含むキットを特徴とする。一実施形態において、キットはアジュバントも含む。
【0016】
本発明の全ての態様のある実施形態では、最適化ウイルスポリペプチドは、ヒトでの発現用に最適化されている核酸配列によりコードされる(例えば、配列番号5、10、11、12、15、18、及び23のうちの任意の1つ)。
【0017】
定義
「最適化ウイルスポリペプチド」又は「計算により最適化されたウイルスポリペプチド」とは、天然に存在するウイルスペプチド、ポリペプチド、又はタンパク質ではない免疫原性ポリペプチドを意味する。最適化ウイルスポリペプチド配列は、最初は、哺乳動物(例えば、ヒト)の免疫後に(例えば、本発明のワクチンに組み込まれたときに)生じる抗ウイルス免疫応答(例えば、細胞性又は体液性免疫応答)の幅、強度、深さ、又は寿命を増加させるように、1つ又は複数の天然に存在するウイルス遺伝子産物(例えば、ペプチド、ポリペプチド、及びタンパク質)のアミノ酸配列を修飾することによって生成される。従って、最適化ウイルスポリペプチドは「親」ウイルス遺伝子配列に対応し得る;或いは、最適化ウイルスポリペプチドは特定の「親」ウイルス遺伝子配列に対応しなくともよく、ウイルスの様々な株又は疑似種由来の類似配列に対応してもよい。最適化ウイルスポリペプチドに含めることのできるウイルス遺伝子配列に対する修飾としては、アミノ酸付加、置換、及び欠失が挙げられる。本発明の一実施形態において、最適化ウイルスポリペプチドは、2個以上の天然に存在するウイルス遺伝子産物(例えば、天然又は臨床ウイルス分離株)が複合した、又は合体したアミノ酸配列であり、ここでは潜在的な各エピトープ(例えば、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30又はそれ以上のアミノ酸長の各連続又は重複するアミノ酸配列)が分析及び修飾され、得られる最適化ウイルスポリペプチドの免疫原性が向上している。異なるウイルス遺伝子産物に対応する最適化ウイルスポリペプチドはまた、融合することによって本発明のワクチンへの組み込みを促進することができる。最適化ウイルスポリペプチドの生成方法は、例えば、参照により本明細書に援用されるFisherら「Polyvalent Vaccine for Optimal Coverage of Potential T−Cell Epitopes in Global HIV−1 Variants」、Nat.Med.13(1):100−106頁(2007年)及び国際公開第2007/024941号パンフレットに記載される。最適化ウイルスポリペプチド配列が生成されると、標準的な技術(例えば、参照により本明細書に援用される国際公開第2006/040330号パンフレット及び国際公開第2007/104792号パンフレットに開示されるアデノウイルスベクターなどの組換えウイルスベクター)により対応するポリペプチドを産生し、又は投与することができる。
【0018】
「薬学的に許容可能な担体」とは、それと共に投与される化合物の治療特性を維持しながら、治療を受ける哺乳動物にとって生理学的に許容可能である担体を意味する。例示的な薬学的に許容可能な担体の一つは生理食塩水である。他の生理学的に許容可能な担体及びそれらの製剤は当業者に公知であり、例えば、参照により本明細書に援用される「Remington’s Pharmaceutical Sciences」(第18版、A.Gennaro編、1990年、Mack Publishing Company、Easton、PA)に記載されている。
【0019】
「ベクター」とは、下流の遺伝子又はコード領域に作動可能に連結されたプロモーターを含むDNAコンストラクト(例えば、ポリペプチド又はポリペプチド断片をコードするcDNA又はゲノムDNA断片)を意味する。ベクターをレシピエント細胞(例えば、原核又は真核細胞、例えば、細菌、酵母、昆虫細胞、又は哺乳動物細胞、発現ベクター内のプロモーターに依存する)又は生体(例えばヒトを含む)に導入すると、細胞は、ベクターによってコードされるmRNAを発現することができ、次にそれが翻訳されて、コードされた本発明の最適化ウイルスポリペプチドとなる。インビトロ転写/翻訳用のベクターもまた当該技術分野において公知であり、本明細書においてさらに記載される。ベクターは、例えば、バクテリオファージ、アデノウイルス、レトロウイルス、ポックスウイルス、又はヘルペスウイルスに由来する遺伝的に改変されたプラスミド、ウイルス、又は人工染色体であってもよい。
【0020】
「ウイルス遺伝子産物」とは、任意の天然に存在するウイルスペプチド、ポリペプチド、又はタンパク質、又はその断片を意味する。本発明の一実施形態において、ウイルス遺伝子産物はヒト免疫不全ウイルス1型(HIV−1)に由来する。HIV−1ウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuポリペプチドを含む。
【図面の簡単な説明】
【0021】
【図1】計算により最適化されたHIV−1 Gag、Pol、及びEnvウイルスポリペプチドの、アカゲザルにおける全潜在的T細胞エピトープ(PTE)ペプチドに対する幅の拡大を示す図である。最適化ウイルスポリペプチド(青色)で免疫した動物が、最大数のリコールペプチドプールと反応した。
【図2】計算により修飾されたHIV−1 Gag、Pol、及びEnvウイルスポリペプチドがエピトープ特異的細胞性免疫応答の幅を拡大することを示す図である。
【図3】本発明の計算により修飾されたウイルスポリペプチドから得られたHIV−1ウイルス遺伝子産物Gag、Pol、及びEnvで免疫した後のアカゲザル、並びにコンセンサスHIV−1抗原又はHIV−1クレードC分離株抗原で免疫した動物で検出された細胞性免疫応答の幅を示す。最適化ウイルスポリペプチド(青色)で免疫した動物が、最大数のリコールペプチドプールと反応した。動物は非近交系であるため、プールは動物ごとに異なる。Gag、Pol、及びEnvは、各々が多くの細胞性免疫応答を誘発し、共通の反応性パターンを有し得る。
【図4】ウイルスポリペプチド(Pol(図4A)、Gag(図4B)、及びEnv(図4C))ごとの、種々の試験ワクチン(二価モザイク(Mos2)、Mコンセンサス(Mcon)、及び最適化クレードC(OptC))の間で共有される潜在的エピトープを示すグラフである。図4A〜図4Cは、種々のワクチン候補による現在のHIVデータベース完全長ゲノムセット及びPTEペプチドの相対的有効範囲を示す。
【図5】二価モザイク(Mos2)ワクチンに対するPTEペプチド応答数(ここでは重複にかかわらず各応答が独立したイベントと見なされる)が、M群コンセンサス(Mcon)ワクチン、及び最適な有効範囲のM群集合(OptC)ワクチン抗原をもたらすように選択された天然ウイルス株ワクチン(最適化クレードC(C天然(最適))に対する応答数と比べて多いことを示すグラフである。図5は、タンパク質、CD8+ T細胞、及びCD4+ T細胞ごとに動物当たりのPTEペプチド応答数を示す。統計的に、Mos2>Mcon〜OptC(Mconは、OptCより応答が多い傾向を示す)。Mconと比較したMos2のウィルコクソンp値:p値=0.001058。
【図6】T細胞応答を引き起こすPTEペプチド数を示す図である。CD8+ T細胞において応答を引き起こす二価モザイク(Mos2)ワクチンのPTEペプチドの中位数は16(値域;12〜29)であり、一方、CD8+ T細胞において応答を引き起こすMconペプチドの中位数は6(値域:0〜7)に過ぎず、及びOptCペプチドの中位数は3ペプチド(値域:0〜3)に過ぎない。CD4+ T細胞において応答を引き起こす二価モザイク(Mos2)ワクチンのPTEペプチドの中位数は4(値域;2〜6)であり、一方、CD4+ T細胞において応答を引き起こすMconペプチドの中位数は1(値域:0〜2)に過ぎず、及びOptCペプチドの中位数は0.5ペプチド(値域:0〜2)に過ぎない。従って、応答の傾向はMos2>Mcon>OptCである。
【図7】被験動物の各々に由来するT細胞によって認識される全てのCD8+ T細胞Gag PTEペプチドのマッピングを要約する概略図である(以下の実施例3を参照)。動物番号、ペプチドプール及びペプチド番号を、各反応性ペプチドの境界に表示する。記号は群を示す:*、Mos2;¥、ConM;±、OptC。ここでは例としてGagが挙げられる。動物が非近交系であっても、CD8応答はクラスタ化する傾向がある。モザイクは、単価ワクチンより有利である可能性がある。モザイクは、より多くの共通の変異体と反応する応答を刺激する確率がより高い。モザイクはまた、カクテル中に存在する種々の型に対する複数の応答を刺激する。従って、モザイクは共通のエスケープ経路を遮断する可能性を有する。本発明者らの研究では、モザイクワクチンは、より多くの重複ペプチドを認識するT細胞応答を刺激する傾向があった。反応性ペプチドが局在する多くのホットスポットがある。PTEペプチドは、ワクチンの評価に使用されるペプチド試薬におけるHIV−1 M群の潜在的エピトープ(又は9アミノ酸の連続するストレッチについての9−mer)の有効範囲を最大化するように設計される。必然的に、PTEペプチドには多数の重複が存在するが、そのアルゴリズムに起因して、重複は通常、いくらかのばらつきを伴う重複である。図7は配列番号42を開示する。
【図8】被験動物の各々に由来するT細胞により認識されるCD4+ T細胞Gag PTEペプチドのマッピングを要約する概略図である。図8は配列番号43を開示する。
【図9】ConMワクチン又は最適天然ワクチンに対するPTE応答の典型的なパターンを示す図であり、応答を誘発するペプチドをワクチンの関連領域と整列させている。これらのワクチンに対する反応を実現するには、ワクチンと標的PTEペプチドとの間の途切れのない同一のストレッチによる良好なマッチが必要である。図9は、掲載順にそれぞれ配列番号44〜57を開示する。
【図10】モザイクワクチンが、見かけ上の抗原競合なしに、且つ広範な局所的応答で、複数の変異重複ペプチドを認識する多くの応答を生じたことを示す図である。特に、4個の変異PTEペプチドが認識された。さらに、重複の領域では双方のモザイク型が、それら2つの組み合わせと同様に認識された。最後に、新規の型(S)が認識された。図10は、掲載順にそれぞれ配列番号58〜63を開示する。
【図11】モザイクワクチン接種を受けた動物(361−07)におけるCD8+ PTEペプチド応答の典型的なパターンを示す図である。22個のPTEペプチドを試験し、8個のCD8応答領域が同定された;5個の領域が、モザイクのいずれか1つのアミノ酸にマッチする変異ペプチドを含んだ。5個のCD4応答領域が同定された。このように、モザイクに対するT細胞応答は、所与の領域においてより多くの変異ペプチドが分かる。これは、特にCD8 T細胞応答に当てはまるように思われた。これは、エピトープの変異体を認識する複数のT細胞クローンをもたらし、それらが適応されるエスケープ経路を遮断し得る結果であり得る。より多くの応答があるのみならず、それらはより深く、且つより多くの変異体を網羅する。図11は、掲載順にそれぞれ配列番号64〜101としてCD8応答を開示する。CD4応答は、掲載順にそれぞれ配列番号102〜117として開示される。
【図12】ワクチンにより誘発されたT細胞によって標的化される領域にわたる重複変異PTEペプチドの数を示すグラフである。
【図13】2モザイク抗原ワクチンが、Mcon及びOptCワクチンと比べて、1つ又は複数の重複PTEペプチドを含む領域に対するより多くのT細胞応答をもたらすことを示すグラフである。図13は図5と同様であり、サルが右から左に同じ順序で示されるが、単一のペプチドではなく1つ又は複数の重複PTEペプチドを含む領域に対する応答数を反映して尺度を変えている。
【図14】二価モザイク(Mos2)、Mcon、及びOptCワクチンを投与した後の動物におけるT細胞応答数を示す図である。CD8+ T細胞において二価モザイク(Mos2)ワクチンが引き起こす応答の中位数は8であり、一方、Mcon及びOptCワクチンにより引き起こされるCD8+ T細胞応答の中位数は、それぞれ3ペプチド(値域:0〜6)及び1.5ペプチド(値域:0〜5)に過ぎない。CD4+ T細胞において二価モザイク(Mos2)ワクチンが引き起こす応答の中位数は3(値域;2〜5)であり、一方、Mcon及びOptCワクチンにより引き起こされるCD4+ T細胞応答の中位数は、それぞれ1(値域:0〜2)及び0.5(値域:0〜2)に過ぎない。従って、応答の傾向はMos2>Mcon>OptCである。
【図15】モザイクワクチンが、Cクレード天然ワクチンと比べて、Cクレード天然タンパク質と交差反応する応答をより多く誘発できることを示すグラフである:GAGプールペプチドが5個のタンパク質に相当する。M群コンセンサス又は最適な有効範囲のCクレード天然タンパク質によるワクチン接種を受けた動物は、これらのタンパク質に由来するペプチドに対する0〜2つの応答を有したが、一方、モザイクワクチン接種を受けた動物は、1〜5つのペプチドプールに対して応答することができた。モザイクワクチンは、M con又は最適Cのいずれと比べても、試験したタンパク質の各々に対するより多くの応答を誘発する。モザイクワクチンにより誘発されたT細胞応答はまた、実際のGagタンパク質にわたるプールペプチドセットをより多く認識した。10〜12サブプール=10×15merペプチド(5×20merペプチドである96ZM Gagを除く)。
【図16】モザイク設計が、時間の経過に伴うウイルスポリペプチドの変化に対してロバストであることを示すグラフである(例えば、Gag M)。
【図17】9−mer最適化を使用した有効範囲が、準(例えば、8〜12mer)最適化長さと比べてロバストであることを示すグラフである(Gagを図示)。
【図18】変異体数が増加すると有効範囲が大きくなるが、あくまでも収穫逓減を有することを示すグラフである(Gagを図示)。
【図19】PTEペプチドに対するエピトープ特異的Tリンパ球応答の幅及び大きさを示すグラフである。図19Aは、モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原を発現するrAd26ベクターの単回免疫後の個々のPTEペプチドに対するエピトープ特異的CD4+(上側)及びCD8+(下側)Tリンパ球応答の数を示すグラフである。個々のサルをx軸上に示す。各色の異なる陰影は、異なる抗原(Gag、Pol、Env)に対する応答を反映している。図19Bは、CD4+(上側)及びCD8+(下側)Tリンパ球応答領域の数を示すグラフである。
【図20A】HIV−1 Gag(図20A)(配列番号118)、Pol(図20B)(配列番号119)、及びEnv(図20C)(配列番号120)タンパク質配列にマッピングした免疫後4週目のPTEペプチドに対するCD8+ Tリンパ球応答を示す概略図を示す。色は、モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原の投与を受けたサルを示す。各エピトープについて、サル番号、抗原(G、Gag;P、Pol;E、Env)、サブプール番号、及び個々のPTEペプチド番号が示される。
【図20B−1】図20Aの続きである。
【図20B−2】図20B−1の続きである。
【図20C】図20B−2の続きである。
【図21A】HIV−1 Gag(図21A)(配列番号121)、Pol(図21B)(配列番号122)、及びEnv(図21C)(配列番号123)タンパク質配列にマッピングした免疫後4週目のPTEペプチドに対するCD4+ Tリンパ球応答を示す概略図を示す。色は、モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原の投与を受けたサルを示す。各エピトープについて、サル番号、抗原(G、Gag;P、Pol;E、Env)、サブプール番号、及び個々のPTEペプチド番号が示される。
【図21B】図21Aの続きである。
【図21C】図21Bの続きである。
【図22−1】モザイク、Mコンセンサス、クレードB+クレードC、又は最適天然クレードC HIV−1 Gag、Pol、及びEnv抗原を発現するrAd26ベクターによる免疫後4週目の全てのサルにおけるワクチン配列の反応性PTEペプチドとのアラインメントを示す概略図である。各サルについて、ワクチン配列を一番上に示し、抗原(G、Gag;P、Pol;E、Env)及びPTEペプチド番号により表される反応性PTEペプチドを、ワクチン配列の下に示す。最小重複領域は太字で示す。2つのモザイク又は2つのクレードB+クレードC抗原の間の配列多型は青色で示す。ワクチン配列と反応性PTEペプチドとの間の違いは赤色で示す。図22は、掲載順にそれぞれ配列番号124〜640を開示する。反応性ペプチドの間の重複が生じるときのその重複に基づき免疫応答エピトープを含む可能性があるペプチド内の最小領域は、ワクチンにおいて太字とする。重複ペプチドがない場合、エピトープがペプチドのどこにでもあり得ると仮定し、従って領域全体を太字とする。ペプチド内で種々の境界を有するエピトープを標的とする種々のT細胞応答、又は変異体が存在するときにばらつきを許容することのできるより雑多なクローン性T細胞応答の間を区別することはできない;いずれのシナリオも、ワクチン免疫応答において有利となり得る。標的化される領域の数は、データを考慮するのに必要な最小T細胞応答数に対応する。ワクチンとペプチドとがマッチしないアミノ酸は赤色で記す;アミノ酸がエピトープを含んでいる可能性のある領域内に収まる場合、それらは太字赤色とする。複数のペプチドが重複するときの重複領域外でのアミノ酸の違いは赤色で標示し、しかし太字にはしない。ワクチンが常に一番上である。各タンパク質の文字(GagはGであり、PolはPであり、エンベロープはEである)及びペプチド番号を用いて各反応性PTEペプチドを表示する。タンパク質及びHXB2番号が各ペプチドに続く。モザイク及びクレードB+Cワクチンについては2個の抗原があり、各々が双方ともアラインメントに含まれる;ワクチンにおけるアミノ酸の違いは青色で指示し、及び反応性ペプチドが第2のモザイクに変異アミノ酸を含む場合、それも青色とする。2つのワクチン抗原が異なる位置の各々において反応性ペプチドをさらに太字で標示し、2つの変異が含まれているその位置がワクチン免疫応答に影響を与え、より大きい幅及び深さをもたらした可能性があることを示す。例えば、要約される最初のワクチンはクレードB+Cワクチンであり、動物287−95が、それについての応答が掲載される最初の動物である。PTEペプチドに対してCD8応答が3つあり、CD4に対して1つあった。CD8ペプチドのうちの2つ、E26及びE282は実質的な重複を示し、従って双方が同じCTL応答を標的とし得る;このように、2つのCD8応答領域のみと、1つのCD4応答領域とがあることも記載する。各応答領域について領域当たりの重複ペプチドの数を書き出すことで(例えば、CD8:1 2 CD4:1)、応答の深さを評価する;2つは赤色とし、その重複の領域は反応性ペプチドにばらつきがあることを示す。ワクチンが異なる場合、2番目の反応性領域におけるD/Eのように、それを青色で標示する。重複の領域のみは太字である。E282におけるHはいずれのワクチンにも認められなかったため、赤色で標示する;これは重複の領域内にあるため、太字としている。各反応性ペプチドは、そのタンパク質及び対応するHXB2番号を有し、右側に記載する。
【図22−2】図22−1の続きである。
【図22−3】図22−2の続きである。
【図22−4】図22−3の続きである。
【図22−5】図22−4の続きである。
【図22−6】図22−5の続きである。
【図22−7】図22−6の続きである。
【図22−8】図22−7の続きである。
【図22−9】図22−8の続きである。
【図22−10】図22−9の続きである。
【図22−11】図22−10の続きである。
【図22−12】図22−11の続きである。
【図22−13】図22−12の続きである。
【図22−14】図22−13の続きである。
【図22−15】図22−14の続きである。
【図22−16】図22−15の続きである。
【図22−17】図22−16の続きである。
【図22−18】図22−17の続きである。
【図23】全てのGag、Pol、及びEnv特異的CD8+(図23A及び図23B)及びCD4+(図23C)Tリンパ球応答の大きさを、最低値から最高値まで並べて示すグラフである。
【図24A−B】PTEペプチドに対するエピトープ特異的Tリンパ球応答の深さを示す。図24Aは、最適天然クレードC抗原の投与を受けたサル366におけるマッピングしたTリンパ球応答の例を示す概略図である。図24Bは、二価モザイク抗原の投与を受けたサル361におけるマッピングしたTリンパ球応答の例を示す概略図である。図24A及び図24Bでは、ワクチン配列を一番上に示し(OptC;Mos1、Mos2)、抗原(G、Gag;P、Pol;E、Env)及びPTEペプチド番号により表される反応性PTEペプチドを、ワクチン配列の下に示す。最小重複領域は太字で示す。2つのモザイク抗原間の配列多型は青色で示す。ワクチン配列と反応性PTEペプチドとの違いは赤色で示す。応答領域ごとにまとめた全ての陽性ペプチドの完全アラインメントは、図22に示す。図24Cは、モザイク、Mコンセンサス、クレードB+クレードC、又は最適天然クレードC抗原を発現するrAd26ベクターで免疫した後のCD4+(上側)及びCD8+(下側)Tリンパ球応答の深さを示すグラフである。個々のサルをx軸上に示す。各エピトープ領域について、1つの応答変異体(薄い陰影)又は1つより多い応答変異体(濃い陰影)を示す。図24Aは、掲載順にそれぞれ配列番号641〜650を開示する。図24Bは、掲載順にそれぞれ配列番号651〜685を開示する。
【図24C】図24A−Bの続きである。
【図25】クレードA、B、及びC由来のHIV−1 Gagペプチドに対するエピトープ特異的Tリンパ球応答の幅を示すグラフである。細胞性免疫応答の幅を、以下のHIV−1 Gag株由来の重複ペプチドのサブプールを利用して評価した:クレードC DU422、クレードC ZM651、コンセンサスC、コンセンサスA、及びコンセンサスB。モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原を発現するrAd26ベクターの単回免疫後の陽性サブプールの数を示す。個々のサルをx軸上に示す。
【図26A−B】追加免疫後の細胞性及び体液性免疫応答を示すグラフである。各サルについてプライミング後4週目(各パネルの左側)及び追加免疫後44週目(各パネルの右側)の個々のTリンパ球応答の大きさ(図26A)及び幅(図26B)を示す。サルは、0週目にrAd26ベクターでプライミングし、40週目に、モザイク、Mコンセンサス、又は最適天然クレードC HIV−1 Gag、Pol、及びEnv抗原を発現するrAd5HVR48ベクターで追加免疫した。個々のサルをx軸上に示す。図26Aでは、赤色はCD8+ Tリンパ球応答を表し、青色はCD4+ Tリンパ球応答を表し、線は双方の時間点で観察された応答を表し、及び点は一方の時間点でのみ観察された応答を表す。図26Bでは、各色の異なる陰影が、異なる抗原(Gag、Pol、Env)に対する応答を反映している。図26Cは、0週目、4週目、及び44週目におけるEnv特異的ELISAエンドポイント力価を示すグラフである。図26Dは、44週目における第1層クレードA(DJ263.8)、クレードB(SF162.LS)、及びクレードC(MW965.26)ウイルスに対する中和抗体(NAb)力価を示すグラフである。陰性対照としてのマウス白血病ウイルスに対するNAb力価は、いずれの試料についても20未満であった。
【図26C−D】図26A−Bの続きである。
【図27】様々なワクチン抗原ごとのPTEペプチドの理論的な有効範囲を示すグラフである。モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原により網羅される9アミノ酸PTEペプチドのパーセンテージを示す。
【発明を実施するための形態】
【0022】
本発明は、天然に存在するウイルス遺伝子産物から計算により得られる最適化ウイルスポリペプチドを特徴とする。本発明の最適化ウイルスポリペプチドは、本発明の1つ若しくは複数の最適化ウイルスポリペプチド又は本発明の1つ若しくは複数の最適化ウイルスポリペプチドを組み込むワクチン(例えば、ベクター)で対象(例えば、ヒト)を免疫した後の、ウイルス特異的免疫(例えば、T細胞に基づく免疫応答などの細胞性免疫)の幅及び深さの増加を可能とする。本発明は、ウイルス感染症に感染した、又は感染するリスクがある対象(例えば、ヒト)に投与することのできるワクチンを提供する。本発明のワクチンは、代表される各対応するウイルス遺伝子産物についての少なくとも2個の別個の最適化ウイルスポリペプチドを組み込む。少なくとも2個の別個の最適化ウイルスポリペプチドを組み込むことにより、ワクチンにおける免疫原性エピトープの有効範囲及び代表性を高めることが可能となり、本発明者らが発見したこのことにより、対象のワクチン接種後にウイルス特異的免疫応答の総数の増加がもたらされる。本発明はまた、ワクチン、ベクター、及び最適化ウイルスポリペプチドを対象(例えば、ヒト)に投与する方法、及びそれらを製造する方法も提供する。本明細書に記載される組成物、方法、及びキットは、少なくとも2個の別個の最適化ウイルスポリペプチドを提供することによってウイルス特異的細胞性免疫応答の多様性、幅、及び/又は深さを実質的に増加させることができる。
【0023】
本発明の最適化ウイルスポリペプチド
本発明は、自然に循環しているウイルス遺伝子産物に対応し、且つそれから得られる計算により最適化されたウイルスポリペプチドを組み込む多価(例えば、二価)ワクチンを提供する。多価モザイクタンパク質は、天然配列からインシリコ組換えにより構築され、所与の結合価に対する潜在的なT細胞エピトープ(PTE)の最大の有効範囲を提供するように最適化される。モザイク抗原は、天然抗原の発現及びプロセシングを維持するように設計される完全長タンパク質である。
【0024】
本発明者らは、単一のウイルス遺伝子産物に対応し、且つそれから得られる2つの別個の最適化ウイルスポリペプチド(すなわち、二価ワクチン)で免疫すると、同じウイルス遺伝子産物から得られる天然に存在するポリペプチド(例えば、臨床分離株に基づく配列)、又は同じウイルス遺伝子産物から得られるかかる天然に存在するポリペプチドのコンセンサス配列を組み込む従来の単価又は多価ワクチンより実質的に多い数の細胞性免疫応答(例えば、T細胞応答)が誘発されることを発見した。従って、計算により最適化されたウイルスポリペプチドであって、その配列が循環ウイルス配列の希少でなく短いストレッチの最大有効範囲を提供する最適化ウイルスポリペプチドを組み込むワクチンは、免疫応答の幅及び深さを増加させることができる。
【0025】
遺伝的アルゴリズムを使用して、入力として提供される天然に存在するウイルス遺伝子産物配列の任意のセットの断片の「モザイク」ブレンドとして最適化ウイルスポリペプチドのセットが作成される。この遺伝的アルゴリズム戦略は、入力データセットとして一般的なウイルス集団からの整列していないタンパク質配列を使用し、従って「アラインメントに依存しない」という長所を有する。これにより、自然界に見られるウイルスタンパク質に類似しているが、天然には存在しない人工的な最適化ウイルスポリペプチドが作り出される。遺伝的アルゴリズムは、目的とする標的の又は所望の免疫応答に応じて、種々の長さのウイルスポリペプチドを最適化するように調整することができる。ほとんどのT細胞エピトープは9アミノ酸長であるため、本発明の最適化ウイルスポリペプチドの設計に利用した遺伝的アルゴリズムは、所与のウイルス遺伝子産物(例えば、HIV−1 Gag)の各々の連続した9−merアミノ酸配列の最適化に基づいた。この手法に従えば、自然界に存在しない、又は極めて希少である9−mer(例えば)を除外することができる−これは、コンセンサス配列に基づくワクチン戦略と比べたとき、コンセンサス配列は、自然界にはまれにしか、又は全く存在しないいくつかの9−mer(例えば)を含み得るため、改良点である。遺伝的アルゴリズムに用いられる適応度の定義は、最も「適応する」多価カクテルが、集団中の全ての9merのなかで最良の有効範囲(最高の完全マッチ率)を与え、且つ集団に存在しない、又はまれである9merがないという制約を受ける入力ウイルス配列の組み合わせであることである。本発明の最適化ウイルスポリペプチドの生成に用いられる遺伝的アルゴリズムについては、参照により本明細書に援用される国際公開第2007/024941号パンフレットにさらに記載されている。
【0026】
一実施形態において、本発明は、単一の最適化されたHIV−1ポリペプチド(例えば、配列番号1〜29に示されるポリペプチド)を組み込む多価(例えば、二価)HIV−1ワクチンを提供する。別の実施形態において、本発明は、2個以上の最適化されたHIV−1ポリペプチドを組み込む多価ワクチンを特徴とする。いずれの場合も、最適化HIV−1ポリペプチドは、HIV−1メイン(M)群として知られる全体的循環における全てのHIV−1変異体に基づく。本発明者らは、遺伝子当たり2つの変異体(例えば、各々がGag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuについての2つのポリペプチド配列)のみを利用するM群モザイク遺伝子に基づき細胞性免疫の幅及び深さを増大させる最適化HIV−1ポリペプチドのセット(配列番号1〜29)を生成した。本発明者らは、アカゲザルにおいて、多価(例えば、二価)HIV−1 M群ワクチンにこれらの最適化HIV−1ポリペプチドを使用すると、2つの他の有力なワクチン抗原戦略(Mコンセンサス抗原及び最適天然クレードC抗原)と比較したとき有意に大きい幅及び深さのHIV−1特異的細胞性免疫応答が誘発されるという新規の、且つ意外な結果を得た。
【0027】
本発明は、異なるウイルス遺伝子産物に対応する最適化ウイルスポリペプチドの融合を提供する。上記の遺伝的アルゴリズムを用いて、本発明のワクチンに使用される融合ポリペプチドを生成することができる。例えば、Gag/Nef(配列番号19〜20)と、Gag/Pol(配列番号21〜27)と、Gag/Pol/Nef(配列番号28〜29)との最適化HIV−1ポリペプチド融合体を、HIV−1に感染した、又は感染するリスクがある対象(例えば、ヒト)に投与するため本発明のベクターに組み込むことができる。本発明のワクチンは(ポリペプチド形態であろうと、又は核酸形態であろうと)、例えば、最適クレードC配列(配列番号30〜36)又はコンセンサス配列(配列番号37〜39)などの非「モザイク」ポリペプチド(又はそれぞれ、それをコードする配列)の1つ又は複数も含むことができる。
【0028】
本発明に開示される最適化ウイルスポリペプチドは、Merrifield、J.Amer.Chem.Soc.85:2149頁(1963年)によって記載されるものなどの化学合成技法により従来どおり調製することができる(また、例えば、Stemmerら、164 Gene 49(1995年)も参照のこと)。例えば、ワクチンは、固相ペプチド合成(SPPS)を用いて容易に調製することができる。自動化された固相合成を、Applied Biosystems ABI 433Aペプチドシンセサイザーなどの、数多くの周知されている市販の自動シンセサイザーのいずれか一つを使用して行うことができる。或いは、本発明の最適化ウイルスポリペプチドは、最適化ウイルスポリペプチドの細胞内発現を可能にする核酸又はベクター(例えば、アデノウイルスなどのウイルスベクター)を細胞又は生体にトランスフェクト又は形質導入することにより組換え産生することができる。本発明の最適化ウイルスポリペプチドのヌクレオチド配列をコードする核酸及びベクターは、本明細書に記載されるものを含めた公知の組換えDNA技法により合成することができる。
【0029】
本発明のワクチン
本発明はまた、ウイルス(例えば、HIV−1)に感染した、又は感染するリスクがある患者に投与することのできるワクチンも特徴とする。本発明のワクチンは、本明細書において考察されるとおりの、本発明の最適化ウイルスポリペプチドの少なくとも1つを含む。本発明のワクチンは、本発明の2個以上の最適化ウイルスポリペプチドのヌクレオチド配列(例えば、組換えの(例えば、サブユニット)又は生体全体の(例えば、ウイルス全体)ウイルスベクターの免疫原性成分)をコードする核酸であってもよい。核酸は、本発明の2個以上の最適化ウイルスポリペプチドのヌクレオチド配列を組み込むベクター(例えば、アデノウイルスなどのウイルスベクター)を含む。本発明の最適化ウイルスポリペプチド、並びに最適化ウイルスポリペプチドを組み込むワクチン、核酸、及びベクターは、細胞若しくは生体において組換え発現させることができ、又はウイルスに感染した、若しくは感染するリスクがある対象(例えば、ヒト)に直接投与することができる。
【0030】
本発明のベクター
本発明はまた、本発明の1つ又は複数の最適化ウイルスポリペプチドのヌクレオチド配列(例えば、DNA又はRNA)をコードするベクターも特徴とする。ベクターは、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするヌクレオチド配列を含む担体(例えば、リポソーム)、プラスミド、コスミド、酵母人工染色体、又はウイルスであってもよい。ベクターは、いくつかの供給源由来のさらなる核酸配列を含むことができる。
【0031】
本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするベクターは、当該技術分野において公知の任意の組換え分子生物学的技法を用いて作成することができる。ベクターは、標的細胞又は生体のトランスフェクション又は形質導入後、染色体外にあってもよく、又は宿主細胞の染色体に組み込まれてもよい。ベクターの核酸成分は、標的細胞当たりのコピー数が単一又は複数であってよく、及び直鎖状、環状、又はコンカテマー状であってよい。
【0032】
本発明のベクターはまた、単一の核酸転写産物からの複数のペプチド鎖又はポリペプチド鎖の発現を可能にする内部リボソーム侵入部位(IRES)配列を含むこともできる。例えば、本発明のベクターは、本発明の1つ又は複数の最適化ウイルスポリペプチド並びに別のポリペプチド(例えば、緑色蛍光タンパク質(GFP)などの検出可能標識)をコードすることができる。
【0033】
本発明のベクターは、本発明の最適化ウイルスポリペプチドの発現を促進する遺伝子発現エレメントをさらに含む。本発明の最適化ウイルスポリペプチドをコードするベクターの発現に有用な遺伝子発現エレメントとしては、限定はされないが、(a)ウイルス転写プロモーター並びにそのエンハンサーエレメント、例えば、SV40初期プロモーター、ラウス肉腫ウイルスLTR、及びモロニーマウス白血病ウイルスLTRなどの調節配列;(b)SV40後期領域に由来するものなどのスプライス領域及びポリアデニル化部位;及び(c)SV40などにおけるポリアデニル化部位が挙げられる。また、プラスミド複製起点、抗生物質耐性又は選択遺伝子、多重クローニング部位(例えば、制限酵素切断位置)、及び他のウイルス遺伝子配列(例えば、HIV末端反復配列(LTR)などの、ウイルスの構造、機能、又は調節エレメントをコードする配列)も含む。
【0034】
本発明のベクターはまた、例えば、配列番号11、14〜18、及び23のいずれか一つなどの、ヒトにおける発現用に最適化されている本発明の最適化ウイルスポリペプチドを含むこともできる。
【0035】
本発明のベクターはまた、以下の酵素切断部位:XbaI−EcoRI−Kozak−Start...Stop−BamHI−NheI;及び以下の配列:TCTAGA GAATTC GCCACC[ATG遺伝子TAA TGA]GGATCC GCTAGCを有する多重クローニング部位(MCS)を含むように改変することもできる。このMCSを有するベクターは、内部的なXbaI、EcoRI、BamHI、NheI部位を有しないとともに、6個以上のC又はGのストレッチを有しない最適化ウイルスポリペプチドと共に用いることができる。
【0036】
生体内投与
本発明は、本発明の1つ又は複数のワクチン(例えば、本発明の2個以上の最適化ウイルスポリペプチドをコードするベクター)を対象(例えば、ヒト)に生体内投与して本発明の2個以上の最適化ウイルスポリペプチドの発現を促進する方法を特徴とする。ワクチンを対象に投与すると、ウイルス免疫原に対する防御的又は治療的免疫応答(例えば、細胞性又は体液性免疫応答)を誘発することのできる本発明の1つ又は複数の最適化ウイルスポリペプチドが発現する。
【0037】
いくつかのタイプのベクターを用いて、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするヌクレオチド配列を対象(例えば、ヒト)に直接送達することができる。本発明のベクターとしては、ウイルス、ネイキッドDNA、オリゴヌクレオチド、カチオン性脂質(例えば、リポソーム)、カチオン性ポリマー(例えば、ポリソーム)、ビロソーム、及びデンドリマーが挙げられる。本発明は、細胞(例えば、血球細胞)のエキソビボでのトランスフェクション又は形質導入を提供し、続いてそれらの細胞をドナー対象に投与し戻すことで、免疫原特性を有する本発明の最適化ウイルスポリペプチドの発現が可能となる。本発明の方法により単離し、エキソビボでトランスフェクト又は形質導入することのできる細胞としては、限定はされないが、血球細胞、皮膚細胞、線維芽細胞、内皮細胞、骨格筋細胞、肝実質細胞、前立腺上皮細胞、及び血管内皮細胞が挙げられる。幹細胞もまた、本発明のベクターの形質導入又はトランスフェクションに適した細胞である。本発明の方法によれば、骨髄前駆細胞及び造血幹細胞(HSC)を含む全能性、多能性、多分化能、又は単能性幹細胞を単離して、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするベクターをトランスフェクト又は形質導入し、対象に投与することができる。
【0038】
本発明の最適化ウイルスベクターを発現させるために用いられるトランスフェクション又は形質導入の方法は、トランスフェクト又は形質導入細胞における、及びそれに続いて細胞の投与を受ける対象におけるタンパク質発現の強度及び寿命に強い影響を有する。本発明は、本質的に一時的なベクター(例えば、アデノウイルスベクター)又は長命のベクター(例えば、レトロウイルスベクター)を提供する。タンパク質発現の調節に用いることのできる調節配列(例えば、プロモーター及びエンハンサー)が、当該技術分野において公知である。トランスフェクト又は形質導入される細胞のタイプもまた、タンパク質発現の強度及び寿命と強い関連を有する。例えば、代謝回転速度が高い細胞型は、タンパク質発現の期間がより短いと予想され得る。
【0039】
エキソビボトランスフェクション及び形質導入
本発明はまた、細胞(例えば、リンパ球などの血球細胞)をエキソビボでトランスフェクト及び形質導入し、続いてそれらの細胞を対象(例えば、ヒト)に投与する方法も特徴とする。一実施形態において、細胞は治療を受ける対象の自己由来である。細胞は、本発明の1つ又は複数の最適化ウイルスポリペプチドのヌクレオチド配列をコードする1つ又は複数のベクターをエキソビボでトランスフェクト又は形質導入することにより、治療対象における最適化ウイルスポリペプチドの一時的又は永久的な発現が可能なものとすることができる。このような修飾細胞を対象に投与すると、ウイルス免疫原に対する防御的又は治療的免疫応答(例えば、細胞性又は体液性免疫応答)を誘発することのできる本発明の1つ又は複数の最適化ウイルスベクターが発現する。
【0040】
いくつかのタイプのベクターを用いて、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするヌクレオチド配列を細胞(例えば、リンパ球などの血球細胞)に送達することができる。本発明のベクターとしては、ウイルス、ネイキッドDNA、オリゴヌクレオチド、カチオン性脂質(例えば、リポソーム)、カチオン性ポリマー(例えば、ポリソーム)、ビロソーム、及びデンドリマーが挙げられる。本発明は、細胞(例えば、血球細胞)のエキソビボでのトランスフェクション又は形質導入を提供し、続いてそれらの細胞をドナー対象に投与し戻すことで、免疫原特性を有する本発明の最適化ウイルスポリペプチドの発現が可能となる。本発明の方法により単離し、エキソビボでトランスフェクト又は形質導入することのできる細胞としては、限定はされないが、血球細胞、皮膚細胞、線維芽細胞、内皮細胞、骨格筋細胞、肝実質細胞、前立腺上皮細胞、及び血管内皮細胞が挙げられる。幹細胞もまた、本発明のベクターの形質導入又はトランスフェクションに適した細胞である。本発明の方法によれば、骨髄前駆細胞及び造血幹細胞(HSC)を含む全能性、多能性、多分化能、又は単能性幹細胞を単離して、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするベクターをトランスフェクト又は形質導入し、対象に投与することができる。
【0041】
本発明の最適化ウイルスベクターを発現させるために用いられるトランスフェクション又は形質導入の方法は、トランスフェクト又は形質導入細胞における、及びそれに続いて細胞の投与を受ける対象におけるタンパク質発現の強度及び寿命に強い影響を有する。本発明は、本質的に一時的なベクター(例えば、アデノウイルスベクター)又は長命のベクター(例えば、レトロウイルスベクター)を提供する。タンパク質発現の調節に用いることのできる調節配列(例えば、プロモーター及びエンハンサー)が、当該技術分野において公知である。トランスフェクト又は形質導入される細胞のタイプもまた、タンパク質発現の強度及び寿命と強い関連を有する。例えば、代謝回転速度が高い細胞型は、タンパク質発現の期間がより短いと予想され得る。
【0042】
ウイルスベクター
本発明の1つ又は複数の最適化ウイルスポリペプチドのヌクレオチド配列をコードするウイルスベクターは、本発明のワクチンとして使用することができる。例えば、本発明の1つ又は複数の最適化ウイルスポリペプチドのヌクレオチド配列は、対象の形質導入(例えば、生体内投与)又は対象から単離された細胞の形質導入(例えば、エキソビボ形質導入と、それに続く対象に細胞を戻す投与)に好適な天然の、又は修飾された(例えば、弱毒化された)ウイルスゲノムのヌクレオチド配列に組換え的に挿入することができる。ウイルスにさらなる修飾を加えることにより、感染力又はトロピズムを高め(例えば、シュードタイピング)、複製能を低下させ、若しくは除去し、又はウイルス成分の免疫原性(例えば、免疫原性ワクチン剤に関連しないあらゆる成分)を低下させることができる。本発明のベクターは形質導入細胞により発現し、細胞外空間に分泌されるか、又は発現細胞に(例えば、細胞内分子として、又は細胞表面に提示されて)留まることができる。キメラ又はシュードタイプのウイルスベクターを細胞の形質導入に使用して、本発明の1つ又は複数の最適化ウイルスポリペプチドの発現を可能にすることもできる。例示的ベクターを以下に記載する。
【0043】
アデノウイルス
組換えアデノウイルスは、本発明の1つ又は複数の最適化ウイルスポリペプチドを発現させるベクターとしての使用に対し、いくつかの重要な利点を提供する。このウイルスは高力価に調製することができ、非複製細胞に感染することができ、及び標的細胞集団との接触後、エキソビボでの標的細胞の高効率の形質導入をもたらすことができる。さらに、アデノウイルスは、そのDNAを宿主ゲノムに組み込むことがない。従って、それを発現ベクターとして使用すると、自発的増殖性疾患が惹起されるリスクが低減する。動物モデルでは、アデノウイルスベクターは、概して高レベル発現を約1週間にわたり媒介することが分かっている。導入遺伝子発現(本発明の最適化ウイルスポリペプチドをコードする核酸の発現)の持続期間は、細胞又は組織特異的プロモーターを使用することにより延ばすことができる。アデノウイルスベクターそれ自体の分子改変における他の改善点として、より持続的な導入遺伝子発現及び炎症低下がもたらされている。これは、さらなる初期アデノウイルス遺伝子に特異的突然変異を内包するいわゆる「第二世代」ベクター、及びCre−Lox戦略を利用して事実上全てのウイルス遺伝子を欠失させる「ガットレス」ベクターに見られる(Engelhardtら、Proc.Natl.Acad.Sci.USA 91:6196頁(1994年)及びKochanekら、Proc.Natl.Acad.Sci.USA 93:5731頁(1996年)、各々参照により本明細書に援用される)。
【0044】
国際公開第2006/040330号パンフレット及び国際公開第2007/104792号パンフレット(各々参照により本明細書に援用される)に開示されるまれな血清型及びキメラアデノウイルスベクターは、本発明のベクターとして特に有用である。例えば、組換えアデノウイルスrAd26、rAd34、rAd35、rAd48、及びrAd5HVR48は、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードすることができる。本発明の最適化ウイルスポリペプチドをコードする1つ又は複数の組換えウイルスベクターを対象に投与して、ウイルス感染症を治療又は予防することができる。
【0045】
アデノ随伴ウイルス(AAV)
非病原性パルボウイルスに由来するアデノ随伴ウイルス(rAAV)もまた、そのベクターは抗ベクター細胞性免疫応答をほぼ誘起せず、及びほとんどの実験系において数ヶ月持続する導入遺伝子発現をもたらすため、本発明の最適化ウイルスポリペプチドの発現に使用することができる。
【0046】
レトロウイルス
レトロウイルスは、本発明の最適化ウイルスポリペプチドの発現に有用である。アデノウイルスとは異なり、レトロウイルスゲノムはRNAベースである。レトロウイルスは、細胞に感染すると、そのRNAをいくつかの酵素と共に細胞に導入する。レトロウイルス由来のウイルスRNA分子は、逆転写と呼ばれる過程を介してプロウイルスと呼ばれる二本鎖DNAコピーを産生する。細胞核内に輸送された後、プロウイルスDNAは宿主細胞の染色体に組み込まれ、形質導入細胞及びその細胞から派生し得る任意の子孫細胞のゲノムを永久的に変化させる。遺伝子を細胞又は生体に永久的に導入する能力は、遺伝子治療に使用されるレトロウイルスを特徴付ける特性である。レトロウイルスとしては、ウイルス感染及びプロウイルスの組み込みを促進するいくつかのアクセサリータンパク質を含むヒト免疫不全ウイルス(HIV)を含むウイルスファミリーであるレンチウイルスが挙げられる。現在、「第三世代」レンチウイルスベクターは、完全な複製無能力、広いトロピズム、及び哺乳動物細胞に対する高い遺伝子導入能を特徴とする(例えば、Mangeat及びTrono、Human Gene Therapy 16(8):913頁(2005年)及びWiznerowicz及びTrono、Trends Biotechnol.23(1):42頁(2005年)(各々参照により本明細書に援用される)を参照)。
【0047】
他のウイルスベクター
アデノウイルス及びレトロウイルスベクターに加えて、細胞(例えば、リンパ球などの血球細胞)又は対象(例えば、ヒト)における本発明の最適化ウイルスポリペプチドの発現に使用することのできる他のウイルスベクター及び技法が、当該技術分野において公知である。こうしたウイルスとしては、ポックスウイルス(例えば、ワクシニアウイルス及び修飾ワクシニアウイルスのアンカラすなわち(MVA);例えば、米国特許第4,603,112号明細書及び同第5,762,938号明細書(各々参照によって本明細書に援用される)を参照)、ヘルペスウイルス、トガウイルス(例えば、ベネズエラウマ脳炎ウイルス;例えば、米国特許第5,643,576号明細書(参照によって本明細書に援用される)を参照)、ピコルナウイルス(例えば、ポリオウイルス;例えば、米国特許第5,639,649号明細書(参照によって本明細書に援用される)を参照)、バキュロウイルス、及びWattanapitayakul及びBauerにより記載される他のもの(Biomed.Pharmacother.54:487頁(2000年)(参照によって本明細書に援用される))が挙げられる。
【0048】
その他の発現ベクター:ネイキッドDNA及びオリゴヌクレオチド
本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするネイキッドDNA又はオリゴヌクレオチドもまた、細胞(例えば、リンパ球などの血球細胞)又は対象(例えば、ヒト)におけるそうしたポリペプチドの発現に使用することができる。例えば、Cohen、Science 259:1691−1692頁(1993年);Fynanら、Proc.Natl.Acad.Sci.USA、90:11478頁(1993年);及びWolffら、BioTechniques 11:474485頁(1991年)(各々参照により本明細書に援用される)を参照のこと。これは、非ウイルス性のトランスフェクションの最も単純な方法である。電気穿孔、並びに高圧ガス及び担体粒子(例えば、金)を使用してDNAコーティングされた金粒子を細胞に撃ち込む「遺伝子銃」の使用などの、ネイキッドDNAを送達する効率的な方法が存在する。
【0049】
リポプレックス及びポリプレックス
本発明の最適化ウイルスポリペプチドをコードする核酸の細胞又は対象への送達を向上させるため、リポプレックス(例えば、リポソーム)及びポリプレックスを使用して、トランスフェクション過程における望ましくない分解からベクターDNAを保護することができる。プラスミドDNAは、ミセル又はリポソームのように組織化された構造の脂質で被覆することができる。組織化された構造がDNAと複合体を形成すると、リポプレックスと称される。3タイプの脂質、すなわちアニオン性(負に帯電している)、中性、又はカチオン性(正に帯電している)がある。カチオン性脂質を利用するリポプレックスは、遺伝子導入に有用であることが分かっている。カチオン性脂質は正電荷であるため、負に帯電したDNAと自然に複合体を形成する。また、その電荷の結果として、カチオン性脂質は細胞膜と相互作用し、リポプレックスのエンドサイトーシスが起こり、細胞質中にDNAが放出される。カチオン性脂質はまた、細胞によるDNAの分解も保護する。
【0050】
ポリマーのDNAとの複合体はポリプレックスと称される。ほとんどのポリプレックスはカチオン性ポリマーからなり、その生成はイオン相互作用により調節される。ポリプレックスとリポプレックスとの働き方の一つの大きい違いは、ポリプレックスはそのDNA負荷を細胞質中に放出することができず、従ってそれを目的として不活化アデノウイルスなどのエンドソーム溶解剤のコトランスフェクションを(エンドサイトーシスの間に生じたエンドソームを溶解させるため)行わなければならない。しかしながら、必ずしもこれが常に該当するわけではない;ポリエチレンイミンなどのポリマーは、キトサン及びトリメチルキトサンがそうであるように、その独自のエンドソーム破壊方法を有する。
【0051】
本発明の最適化ウイルスポリペプチドをコードする核酸と組み合わせてリポプレックス、又はポリプレックスを形成するために使用することのできる例示的なカチオン性脂質及びポリマーとしては、限定はされないが、ポリエチレンイミン、リポフェクチン、リポフェクタミン、ポリリジン、キトサン、トリメチルキトサン、及びアルギン酸塩が挙げられる。
【0052】
ハイブリッド方法
遺伝子導入のいくつかのハイブリッド方法は、2つ以上の技法を組み合わせる。例えばビロソームは、リポプレックス(例えば、リポソーム)を不活化ウイルスと組み合わせる。この手法は、単独でのウイルスによる方法又はリポソームによる方法のいずれと比べても、呼吸上皮細胞においてより効率的な遺伝子導入をもたらすことが示されている。他の方法は、他のウイルスベクターをカチオン性脂質又はハイブリッド形成ウイルスと混合することを含む。これらの方法の各々を使用して、細胞(例えば、リンパ球などの血球細胞)又は対象(例えば、ヒト)への本発明の最適化ウイルスポリペプチドをコードする核酸の導入を促進することができる。
【0053】
デンドリマー
また、デンドリマーを使用して、本発明の最適化ウイルスポリペプチドをコードする核酸を細胞(例えば、リンパ球などの血球細胞)又は対象(例えば、ヒト)に導入してもよい。デンドリマーは、球形状の高度に枝分かれした巨大分子である。粒子の表面は、多くの方法で機能化することができ、得られるコンストラクトの特性の多くが、その表面によって決定される。特に、カチオン性デンドリマー(すなわち正の表面電荷を有するもの)を作成することが可能である。DNA又はRNAなどの遺伝物質が存在するとき、電荷の相補性により核酸のカチオン性デンドリマーとの一時的な結合がもたらされる。その目標に達すると、次にデンドリマー−核酸複合体は、エンドサイトーシスを介して細胞に取り込まれる。
【0054】
生体内投与
本発明はまた、対象(例えば、ヒト)を本発明のワクチンで免疫するためのインビボ方法も特徴とする。一実施形態において、本発明の1つ又は複数のワクチンを対象に直接投与して、ウイルス(例えば、HIV−1)に対する防御的又は治療的免疫応答(例えば、細胞性又は体液性免疫応答)を誘発することができる。或いは、上記のとおりの、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするベクターを対象に直接投与することにより、ウイルス感染症を予防又は治療することができる。1つ又は複数の細胞を生体内に効率的にトランスフェクト又は形質導入するベクター(例えば、ウイルスベクター)は、治療を受ける対象において広域で耐久性があり、且つ強力な免疫応答を誘発することができる。発現ベクターの核酸成分が宿主細胞(例えば、リンパ球などの血球細胞)に導入されると、宿主細胞は本発明のワクチンを産生して提示又は分泌し、次にそれが抗原提示細胞(APC)、T細胞、及びB細胞などの免疫系の成分を活性化する働きをし、その結果、免疫が構築される。
【0055】
医薬組成物
本発明は、1つ又は複数の薬学的に許容可能な賦形剤、希釈剤、緩衝剤、又は他の許容可能な担体と組み合わせた、本発明のワクチン、ベクター、及び最適化ウイルスポリペプチドを特徴とする。ワクチン、ベクター、又は最適化ウイルスポリペプチドの製剤化は、有効量の最適化ウイルスポリペプチド免疫原の発現を利用し、又は可能にし得る。すなわち、そこには、後のウイルス(例えば、HIV−1)に対する曝露からの防御を対象に提供し、又は既存のウイルス感染症を治療するための、特定の十分な免疫応答を治療対象(例えば、ヒト)に生じさせ得る量の抗原が含まれ得る。例えば、本発明のワクチンの製剤化により、広域の且つ特異的な細胞性免疫応答を対象に生じさせ得る量の抗原を発現させることが可能となり得る。本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドで治療される対象はまた、対象に防御的又は治療的利益を付与することのできる抗ウイルス抗体(例えば、中和抗体)を産生することもできる。本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドは、単独で、又は当該技術分野において公知の任意の薬学的に許容可能な担体、塩又は補助剤と組み合わせて、対象に直接投与することができる。
【0056】
薬学的に許容可能な塩は、製薬業界で一般に用いられている非毒性の酸付加塩又は金属錯体を含み得る。酸付加塩の例としては、酢酸、乳酸、パモン酸、マレイン酸、クエン酸、リンゴ酸、アスコルビン酸、コハク酸、安息香酸、パルミチン酸、スベリン酸、サリチル酸、酒石酸、メタンスルホン酸、トルエンスルホン酸、又はトリフルオロ酢酸などの有機酸;タンニン酸、カルボキシメチルセルロースなどのポリマー酸;及び塩酸、臭化水素酸、硫酸、リン酸などの無機酸が挙げられる。金属錯体としては、亜鉛、鉄などが挙げられる。薬学的に許容可能な担体の例示的な一つは、生理食塩水である。他の生理学的に許容可能な担体及びその製剤は当業者に公知であり、例えば、「Remington’s Pharmaceutical Sciences」、(第18版)、A.Gennaro編、1990年、Mack Publishing Company、Easton、PAに記載されている。
【0057】
予防上又は治療上有効量の本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの医薬製剤は、経口的に、非経口的に(例えば、筋肉内、腹腔内、静脈内、又は皮下注射、吸入、皮内、点眼、又は植え込み)、経鼻的に、経膣的に、経直腸的に、舌下に、又は局所的に、投与経路に適合した薬学的に許容可能な担体と混合して投与することができる。製剤中における本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの濃度は、約0.1〜100wt.%まで様々であり得る。
【0058】
本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドを含有する組成物の非経口投与製剤は、滅菌水性又は非水性溶液、懸濁液、又はエマルジョンを含む。好適な媒体の例としては、プロピレングリコール、ポリエチレングリコール、植物油、ゼラチン、水素化ナフタレン、及び注射用の有機エステル、例えばオレイン酸エチルが挙げられる。かかる製剤はまた、防腐剤、湿潤剤、乳化剤、及び分散剤などの補助剤を含有してもよい。生体適合性、生分解性のラクチドポリマー、ラクチド/グリコリドコポリマー、又はポリオキシエチレン−ポリオキシプロピレンコポリマーを使用して化合物の放出を制御してもよい。本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドを含有する組成物に潜在的に有用な他の非経口送達系としては、エチレン−酢酸ビニルコポリマー粒子、浸透圧ポンプ、植込み型注入システム、及びリポソームが挙げられる。
【0059】
液体製剤は、例えば、細菌保持フィルタを介したろ過によるか、組成物に滅菌剤を組み込むことによるか、又は組成物を照射若しくは加熱することにより、滅菌することができる。或いはまた、液体製剤は、使用直前に滅菌水又は他の何らかの無菌注射用媒質中に溶解することのできる無菌固形組成物の形態で製造されてもよい。
【0060】
直腸投与又は経膣投与用の本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドを含有する組成物は、好ましくは、作用物質に加えてカカオ脂又は坐薬ワックスなどの賦形剤を含有し得る坐薬である。経鼻投与又は舌下投与用の組成物もまた、当該技術分野において公知の標準的な賦形剤と共に調製される。吸入用製剤は、賦形剤、例えばラクトースを含有してもよく、又は点鼻液若しくは鼻内スプレーの形態で、又はゲルとして投与するため、例えばポリオキシエチレン−9−ラウリルエーテル、グリココール酸塩及びデオキシコール酸塩を含有する水溶液であってもよく、又は油性溶液であってもよい。
【0061】
本発明の組成物中における活性成分の量は様々であり得る。当業者は、投与されるペプチド、投与時間、投与経路、製剤の性質、排泄速度、対象の病態の性質、並びに患者の年齢、体重、健康状態、及び性別を含めた様々な要因に依存して、正確な個々の投薬量はいくらか調整され得ることを理解するであろう。加えて、ワクチン、ベクター、又は最適化ウイルスポリペプチドによって治療される病態の重症度もまた、投薬量レベルに影響を有し得る。一般に、0.1μg/kg体重〜100mg/kg体重の投薬量レベルが、単回用量として、又は複数回用量に分割して毎日投与される。好ましくは、一般的な投薬量範囲は、1日当たり250μg/kg体重〜5.0mg/kg体重である。様々な投与経路の種々の効率にかんがみて、必要な投薬量は幅広く異なることが予想される。例えば、経口投与は、概して静脈注射による投与より高い投薬量レベルが要求されると予想され得る。このような種々の投薬量レベルは、当該技術分野において公知の最適化のための標準的な実験ルーチンを用いて調整することができる。一般に、正確な予防上又は治療上の有効投薬量は、担当臨床医が上記に特定した要因を考慮して決定することができる。
【0062】
患者に投与される各用量中に存在する本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの量は、患者の年齢、体重、性別、全般的な健康状態などの考慮事項に関連して選択される。患者において深刻な有害副作用なしに免疫応答(例えば、細胞性免疫応答)を誘起する、又は外因性効果を生じさせるために必要なワクチン、ベクター、又は最適化ウイルスポリペプチドの量は、用いられる医薬組成物及びアジュバントの任意の存在に応じて異なる。場合により、初回用量に続き、所望の場合には追加免疫を繰り返すことができる。本方法は、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの長期投与を含むことができる。治療的用途又は予防的用途のため、年一回の追加免疫又は他の間隔での追加免疫など、免疫ワクチン、ベクター、又は最適化ウイルスポリペプチドの反復投薬量が望ましいこともある。投与される投薬量は、当然ながら、特定のワクチン、ベクター、又は最適化ウイルスポリペプチドの薬力学的特性、並びにその投与様式及び経路;被投与者の年齢、健康状態、及び体重;症状の性質及び程度、併用治療の種類、治療頻度、及び所望の効果などの公知の要因に応じて異なり得る。本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドは、針刺しによる急性感染又は母体感染のリスクがある対象の長期治療において投与することができる。かかる「急性」感染についての投薬頻度は、約6週間の期間にわたる静脈内又は筋肉内による1日1回投薬から週1回又は2回の範囲であってもよい。ワクチン、ベクター、又は最適化ウイルスポリペプチドはまた、ウイルス(例えば、HIV−1)による感染患者、又は感染が進行している患者の長期治療においても用いることができる。感染患者では、長期投与の頻度は、静脈内又は筋肉内で1日1回投薬から週1回又は2回の範囲であってもよく、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチド中に存在する免疫原の半減期に依存し得る。
【0063】
アジュバント
ウイルスに対するワクチンを必要とする哺乳動物(例えば、ヒト)のワクチン接種に使用される本発明のワクチンは、ワクチンの免疫原性を高める1つ又は複数の薬学的に許容可能なアジュバントと同時に、又は連続して投与することができる。ヒトへの使用が承認されているアジュバントとしては、アルミニウム塩(ミョウバン)が挙げられる。こうしたアジュバントは、B型肝炎、ジフテリア、ポリオ、狂犬病、及びインフルエンザを含むいくつかのワクチンに有用となっている。他の有用なアジュバントとしては、完全フロイントアジュバント(CFA)、不完全フロイントアジュバント(IFA)、ムラミルジペプチド(MDP)、MDPの合成類似体、N−アセチルムラミル−L−アラニル−D−イソグルタミル−L−アラニン−2−[1,2−ジパルミトイル−s−グリ−セロ−3−(ヒドロキシホスホリルオキシ)]エチルアミド(MTP−PE)並びに代謝可能な油及び乳化剤であって、実質的に全てが直径1ミクロン未満の油滴を有する水中油エマルジョンの形態で存在する油及び乳化剤を含有する組成物が挙げられる。
【0064】
キット
本発明は、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドと、薬学的に許容可能な担体とを含む医薬組成物を、ウイルス感染症の予防又は治療に治療上有効な量で含むキットを提供する。キットは、臨床従事者(例えば、医師又は看護師)がそこに含まれる組成物を投与することができるように説明書を含む。
【0065】
好ましくは、キットは、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの有効量を含有する1つ又は複数の単回用量医薬組成物の複数のパッケージを含む。場合により、1つ又は複数の医薬組成物の投与に必要な器具又は装置がキットに含まれてもよい。例えば、本発明のキットは、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの有効量を含む1つ又は複数のプレフィルドシリンジを提供してもよい。さらに、キットはまた、ウイルスに感染した、又は感染するリスクがある患者が、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドを含有する1つ又は複数の医薬組成物を使用するための説明書又は投与スケジュール表などのさらなる要素を含んでもよい。
【0066】
当業者には、本発明の趣旨及び範囲から逸脱することなく本発明の組成物、方法、及びキットに様々な修正及び変更を加え得ることは明らかであろう。従って、本発明は、本発明の修正及び変更を、それらが添付の特許請求の範囲及びその均等物の範囲内に含まれることを条件として包含することが意図される。
【実施例】
【0067】
以下の例により本発明を説明し、これらの例は、何ら本発明を限定することを意図するものではない。
【0068】
〔実施例1〕
上記で考察される遺伝的アルゴリズムを使用してモザイク抗原Gag、Pol、Nef、及びEnv配列(配列番号1〜8)を作成した。次にそれらの配列をワクチン開発に実用的なものとするため、Env(配列番号9〜11)において切断/融合活性を除去し、Pol(配列番号12〜14)において触媒活性を除去し、Nef(配列番号16〜18)においてミリスチル化部位を除去し、及びGagNef、GagPol、又はGagPolNef(配列番号19〜29)を含む融合コンストラクトを作成することにより修飾した。コンパレータ最適天然クレードC遺伝子もまた示される(配列番号30〜36)。
【0069】
〔実施例2〕
Mコンセンサス(第1群)、二価Mモザイク(第2群)、又は最適天然クレードC(第3群)配列由来のGag、Pol、及びEnv遺伝子を発現する3×1010個のvp rAd26ベクターで20匹のアカゲザルを免疫した。Mコンセンサス配列は、世界中の循環ウイルスの唯一最良の「平均」を表す合成配列に相当する。二価Mモザイク配列は上記に記載される。最適天然クレードC配列は、特性が最も「コンセンサス様」である実際のクレードC HIV−1ウイルス由来の天然に存在する配列である。全潜在的T細胞エピトープ(PTE)ペプチドセットからの応答ペプチド数を調べることにより、細胞性免疫幅を評価した。PTEペプチドは、全HIV−1配列の85%超に相当し、NIHから自由に入手することができる。
【0070】
結果は、新規二価Mモザイク配列が、これらの他の2つの有力な抗原コンセプトより著しく優れていたことを示す。表1に示されるとおり、二価Mモザイク抗原は、Mコンセンサス抗原及び最適天然クレードC抗原と比較して、Gag特異的、Env特異的、Pol特異的、及び合計Tリンパ球の応答幅の有意な増加を誘発した。(平均値は、各サル群における平均エピトープ数を表す;SEMは平均値の標準誤差を表す)。
【表1】

【0071】
〔実施例3〕
実施例2に記載されるMコンセンサス(第1群;n=7)、二価Mモザイク(第2群;n=7)、又は最適天然クレードC(第3群;n=6)配列由来のGag、Pol、及びEnv遺伝子を発現する3×1010個のvp rAd26ベクターでマカクザルを筋肉内免疫した。全潜在的T細胞エピトープ(PTE)ペプチドセットからの応答ペプチド数を調べることにより、細胞性免疫幅を評価した。
【0072】
読み出す情報として、プールPTEペプチドに対するCD4/CD8 IFNγ Elispot応答(大きさ)を評価した。15merのPTEペプチドを用いてエピトープを完全にマッピングし、陽性の数(陽性は、106個のPBMC当たり55個のスポット形成細胞(SFC)及びバックグラウンドの4倍として定義した)を評価した。完全なタンパク質のセットと応答を比較するため、5個のGagタンパク質にわたる重複ペプチドのプールセットも試験した。
【0073】
結果は、二価Mモザイク配列が、他の2つの有力な抗原コンセプト(Mcon及びOptC)より著しく優れていたことを示す。
【0074】
〔実施例4〕
モデリングを使用して、T細胞応答がモザイクワクチンの結果として増加するという本発明者らの観察を検証した。反応性ペプチド数をワクチン、ポリペプチド、及びT細胞型の関数として予測し、次に交互作用のステップワイズ除去を行ったポアソン回帰モデルをフィットした。モザイクワクチンは陽性PTE応答数の高度に有意な亢進をもたらしたが、それは、あらゆるポリタンパク質及びT細胞型にわたって多かれ少なかれ一様にもたらされたことが認められた。従って、動物が投与を受けるT細胞、ポリペプチド、及びワクチンのタイプに個別に依存する寄与分を合わせることにより、動物において陽性効果を有するペプチド数を予測し得る。
【0075】
これらのモデルはまた、動物間のばらつきを考慮するため変量効果も含んだ。これは、モデルの予測力を適切に割り当てることによりp値の信頼性を高めるために設計される予防措置である。
【0076】
以下の効果が認められた:
a)CD8応答はCD4応答と比べてはるかに多く、4.37倍である(p<2×10−16);
b)gp160ではgag又はpolと比べて応答が少なく、0.54倍であり(p=0.000830)、及びgagとpolとの間に(polはGagの2倍長く、従って応答する確率がより高いため、配列長さにより正規化した場合であっても)有意差はない;及び
c)モザイクワクチンはMconと比べて有意により多くの陽性応答をもたらし(3.6倍、p=6.26×10−11)、一方、OptCがもたらす応答はより少なく、但しMcon−OptCの差は有意ではない。
【0077】
〔実施例5〕
ワクチンにより誘発され、PTEペプチドにより検出される最小応答数だけを考え、従って変異に関わらず8アミノ酸以上重複する全てのペプチドを1回のみ数える場合にも、モザイクワクチンはなお、別個の領域に対するより多い応答数をもたらす。
【0078】
CD8について、各重複ペプチドセットを1回のみ数える:
統計概要:
Mos2>Mcon〜OptC(MconはOptCより応答が多い傾向を示す)
Mconと比較したMos2についてのウィルコクソンp値:p値=0.0009992
最適Cと比較したMconについてのウィルコクソンp値:p値=0.2351
【0079】
【0080】
CD4について、各重複ペプチドセットを1回のみ数える(CD4には重複はほとんどないため、これは最初のカウントとほぼ同じである)。
【0081】
統計概要:
Mos2>>Mcon〜OptC(MconはOptCより応答が多い傾向を示す)
Mconと比較したMos2についてのウィルコクソンp値:p値=0.00198
最適Cと比較したMconについてのウィルコクソンp値:0.099
【0082】
【0083】
〔実施例6〕各重複ペプチドセットを1回のみ数えるポアソン回帰
重複PTEペプチドを使用して以下を決定したが、これは、各陽性PTE応答を別のものとして数えた上記の実施例4で考察した結果と広く一致している:
a)CD8応答はCD4応答と比べてはるかに多く、約2.8倍である(p≒1×10−7;
b)モザイクワクチンはMconと比べて有意により多くの陽性応答をもたらし(2.84倍、p≒4.3×10−7)、一方OptCがもたらす応答はより少なく、但しMcon−OptCの差は有意ではない;及び
c)Gagに対する応答よりPolに対する応答が多く、及びgp160に対する応答よりGagに対する応答が多いが、有意であったのはPol−gp160の差のみで、約2倍であった(p<0.001)。
【0084】
〔実施例7〕
以下の表は、2モザイク(Mos2)又はMconのワクチン接種を受けた7動物、及び最適天然Cクレード(OptC)のワクチン接種を受けた6動物における3つのワクチンに対するGag、Pol、及びEnv応答に対する応答合計の集計表である:
【0085】
OptCワクチンは、全てのサルにわたり、タンパク質当たりのCD8+ T細胞応答より僅かに少ない平均応答をもたらした。Mconワクチンは、タンパク質当たり約1つの応答を示した。Mos2に関してのみタンパク質の違いが認められ、ここでEnvは、典型的にはGag又はPolのいずれと比べても応答が少ない。
【0086】
Mos2ワクチン中のタンパク質の各々は多くの応答を誘発し、全般的な応答に寄与した。不活化polに対する修飾、並びにEnvにおける切断及び融合ドメインの欠失後のコンセンサスタンパク質の相対的な長さは、以下のとおりであった:Envの671アミノ酸、Polの851、Gagの498(1.35:1.7:1)。
【0087】
要約
幅:2モザイクワクチンは、Mコンセンサス又は単一最適天然株よりはるかに多くのエピトープ領域の認識能を有するT細胞応答を誘発する。
【0088】
深さ:認識されたPTEペプチドの多様性は、2モザイクの双方の型が変異ペプチドに対する異なるT細胞応答を誘発し、交差反応の可能性が増加していることを示唆する。
【0089】
〔実施例8〕
本発明のモザイクHIV−1ワクチンは、アカゲザルにおける細胞性免疫応答の幅及び深さを拡大する。HIV−1 M群配列のPTE有効範囲を最適化したモザイクHIV−1 Gag、Pol、及びEnv抗原を作成した。HIV−1 M群配列は、ロスアラモスHIV−1配列データベース内の全ての主要なHIV−1クレード及び組換え系統を含む。二価モザイク戦略を利用して、理論的な有効範囲と実用性との競合問題について均衡をとった。二価モザイクHIV−1 Gag、Pol、及びEnv抗原は、アカゲザルにおいてコンセンサス配列及び天然配列のHIV−1抗原を使用して観察される免疫応答と比べて、アカゲザルにおけるエピトープ特異的CD8+及びCD4+ Tリンパ球応答の幅及び大きさ(深さ)を実質的に拡大した。
【0090】
27匹の非近交系アカゲザルを、以下の抗原を発現する組換えアデノウイルス血清型26(rAd26)ベクターの単回注射で免疫した:(i)二価モザイク(N=7)、(ii)Mコンセンサス(N=7)、(iii)クレードBとクレードC(N=7)とを組み合わせた二価、又は(iv)最適天然クレードC(N=6)HIV−1 Gag、Pol、及びEnv抗原。これらの抗原を発現するrAd26ベクターの3×1010個のウイルス粒子の全用量を各動物に1回筋肉内投与した。最適クレードC抗原は、ロスアラモスHIV−1配列データベースにおけるクレードC配列の最大PTE有効範囲を提供するように選択した天然株配列であった(以下の「材料及び方法」で考察する)。ワクチンにより誘発されたHIV−1特異的Tリンパ球応答の幅及び大きさ(深さ)を、HIV−1 M群配列の少なくとも15%に存在する全てのPTEを含むペプチドのプール及びサブプールを利用して、免疫後4週目にIFN−γ ELISPOTアッセイにより評価した。全ての個々のペプチド応答を解析し、細胞枯渇IFN−γ ELISPOTアッセイを実施して、反応性ペプチドがCD8+又はCD4+ Tリンパ球エピトープに相当するかどうかを判断した。
【0091】
モザイク抗原により誘発されたPTEペプチドに対するGag特異的、Pol特異的、及びEnv特異的細胞性免疫応答の総数は、コンセンサス又は天然配列抗原により誘起された応答数と比べて3.8倍高かった(図19A;P=1×10−11、モザイクを次に高い群のコンセンサス抗原との比較、ポアソン回帰モデルに基づく)。CD4+ Tリンパ球応答と比べてCD8+は4.4倍多く(P<10−11)、Gag又はPolに対する応答と比べてEnvに対する応答は少なかった(P<0.0007)。CD8+ Tリンパ球応答の中位数はモザイクワクチンについて最も高く、それにコンセンサス、組み合わせのB+C、及び天然クレードCワクチンが続いた(それぞれ、各群における動物当たりの応答の中央値は16、5、3、及び2)。全体ではCD4+ Tリンパ球応答はより少なかったが、CD4+ Tリンパ球応答に関しても、最高数がモザイクワクチンに対するもので、それにコンセンサス、組み合わせのB+C、及び天然クレードCワクチンが続く、同じ相対的パターンが現れた(それぞれ、各群における動物当たりの応答の中央値は4、1、1、及び0.5)。コンセンサス、組み合わせのB+C、及び天然クレードCワクチンにより誘発されたCD8+及びCD4+ Tリンパ球応答の数は、統計的には区別できなかった。
【0092】
PTEペプチドは、天然に存在するHIV−1配列多型を反映する複数の重複配列を含み、従ってPTEペプチド応答は、特定のエピトープの認識(幅)と当該エピトープの変異体の交差認識(深さ)との双方を包含する。サル当たりの反応性エピトープ領域の数を評価することにより幅の保存分析を実施し、ここでは8個以上のアミノ酸が重複する全ての反応性PTEペプチドを1イベントとして数えた。この保存分析においてもなお、モザイク抗原が、コンセンサス抗原又は天然配列抗原と比較したとき3.1倍多いGag、Pol、及びEnv反応性エピトープ領域数を誘発したことが観察された(図19B;P=1.6×10−7、ポアソン回帰)。エピトープ領域は、高いエピトープ密度の領域によって明らかなとおり、動物にわたっていくらかのクラスタ化を示した(図20A〜図20C及び図21A〜図21C)。応答領域ごとに整理した全ての陽性ペプチドの完全アラインメントを図22に示す。
【0093】
これらのデータは、モザイク抗原が、Mコンセンサス及び天然クレードC抗原と比較したとき、細胞性免疫応答の幅を実質的に増加させたことを示す。二価モザイク抗原はまた、クレードB及びクレードC抗原の二価の組み合わせより優れていることも分かり(図19A及び図19B)、幅の亢進がモザイク配列設計に起因したもので、単純にタンパク質当たりに2つの異なる抗原配列を使用したことを反映したものではないことが示された。モザイク抗原により誘起された幅の増加により応答の潜在能力が損なわれたかどうかを判断するため、全ての個々のCD8+及びCD4+ Tリンパ球応答の大きさを評価した。これらの応答の大きさはあらゆる群間で同程度であることが分かった(図23;それぞれP=0.58及びP=0.99、コルモゴロフ−スミルノフの両側検定)。このように、モザイク抗原は、個々のエピトープ特異的応答の大きさを損なうことなく細胞性免疫幅を拡大し、抗原競合及び免疫優性の制約が、この試験のモザイク抗原の免疫原性を制限しなかったことが示された。
【0094】
次に、様々なワクチンレジメンにより誘発された細胞性免疫応答の深さを特徴付けた。特定のエピトープ領域について同時に誘発された変異PTEペプチドの数として深さを定義した。複数の共通のエピトープ変異体に対する応答が誘起されると、感染ウイルス配列の免疫有効範囲は大きくなり、生体内で共通のエスケープ経路が遮断され、又は大きいフィットネスコストを被る三次エスケープ経路にウイルスが押し込まれ得る。コンセンサス及び天然配列抗原は、天然クレードC抗原の投与を受けたサル366における応答により例示されるとおりの、ワクチン配列と反応性PTEペプチドとの間の高度の配列同一性により特徴付けられる応答を誘発した(図24A;また図22も参照)。対照的に、モザイク抗原は、特定のエピトープ領域における複数の反応性PTEペプチドにより特徴付けられる応答を誘発した。これらのペプチドは共通の変異体を表し、多くの場合に、サル361における応答により例示されるとおり、モザイクワクチン配列に含まれる多型を反映した(図24B;また図22も参照)。これらの動物における全てのエピトープ特異的応答についての要約から、コンセンサス又は天然配列抗原と比較したとき、モザイク抗原は、2個以上の標的化された変異体を有するペプチドに対する細胞性免疫応答の頻度を増加させたことが実証される(図24C;P=0.001、モザイクを次に高い群のコンセンサス抗原と比較するウィルコクソン順位和検定)。
【0095】
PTEペプチドを利用する分析を補足するため、5個の異なるGag配列:クレードC DU422、クレードC ZM651、コンセンサスC、コンセンサスA、及びコンセンサスBを包含する従来の重複ペプチドによるワクチン接種を受けたサルにおける細胞性免疫応答の幅も評価した。細胞性免疫幅は、各Gag配列にわたる10個の重複ペプチドのサブプールに対する反応性を評価することにより決定した。モザイク抗原は、コンセンサス又は天然配列抗原と比較したとき、試験した全てのGag配列に対してTリンパ球応答の増幅を誘発した(図25;P=1×10−7、二項回帰)。このように、モザイク抗原は、PTEペプチドに対するだけでなく、クレードA、B、及びC由来の実際のGagペプチドに対する細胞性免疫幅も増大させた。モザイク抗原は、クレードC Gagペプチドに対する応答の誘起について最適天然クレードC抗原より優れていることまでもが分かった。さらに、モザイク抗原は複数のクレード由来のGagペプチドに対する同程度の応答を誘発したが、一方、天然クレードC抗原は、クレードA及びクレードB Gagペプチドに対する応答の低下を示した(図25)。
【0096】
これらの観察の耐久性を評価するため、モザイク、コンセンサス、及び最適天然クレードC抗原の投与を受けたサルを、初回の免疫で利用された配列と一致するHIV−1 Gag、Pol、及びEnv抗原を発現する異種ベクターrAd5HVR48の3×1010個のウイルス粒子の合計用量で40週目に追加免疫した。10個のPTEペプチドのサブプールに対する反応性を4週目(プライミング後)及び44週目(追加免疫後)に評価することにより、細胞性免疫幅を決定した。プライミング免疫後に観察されたCD8+及びCD4+ Tリンパ球応答の大部分が、追加免疫後に拡大し(図26A、赤色及び青色の線)、及び数多くの新規の応答も検出された(図26A、赤色及び青色の点)。44週目、個々の細胞性免疫応答の大きさは群間で同程度であることが分かった(図26A)。しかしながら、モザイク抗原により誘発されたサブプール応答の数は(動物当たりの応答の中央値27)、追加免疫後、コンセンサス抗原(動物当たりの応答の中央値11)又は最適天然クレードC抗原(動物当たりの応答の中央値10)により誘起されたサブプール応答の数と比べて実質的に高いままであった(図26B)。追加免疫の前も、及び後も、モザイクワクチンにより誘発された動物当たりの応答は、コンセンサス又は天然クレードCワクチンによるものより多かった(P<0.001、全てのペアワイズ比較についてのウィルコクソン順位和検定)。
【0097】
また、追加免疫後、ELISA(図26C)及びルシフェラーゼベースのシュードウイルス中和アッセイ(図26D)によりEnv特異的体液性免疫応答も測定した。全ての群がクレードC gp140に対する同程度のELISA力価を示し、第1層クレードCウイルスMW965.26に対する同程度の中和抗体(NAb)応答を示した。モザイク抗原は、コンセンサス又は天然クレードC抗原と比較したとき、第1層クレードBウイルスSF162.LSに対する僅かに高いNab応答を誘発したが(P=0.02、ウィルコクソン順位和検定)、しかしながらどの群においても、第2層ウイルスに対するNAb応答は検出されなかった。
【0098】
本発明者らのデータは、モザイクHIV−1 Gag、Pol、及びEnv抗原が、理論的予想と良好に一致して、アカゲザルにおいてコンセンサス又は天然配列抗原と比較したときにエピトープ特異的細胞性免疫応答の幅及び深さの双方を増大させたことを実証する(図27)。この試験におけるモザイク抗原に関する顕著な結果は、rAd26ベクターが、CD8+ Tリンパ球応答の誘発に特に効率的であるという事実、並びにモザイク抗原がCD8+ Tリンパ球幅の増大に特に有効であるように思われるという事実を反映したものであり得る(図19A及び図19B)。また、モザイク抗原に関してCD4+ Tリンパ球幅の亢進も認められたが、しかしながらそれらの応答数は実質的により少なかった。
【0099】
Gag特異的細胞性免疫応答の幅は、アカゲザルにおけるSIV制御及びヒトにおけるHIV−1制御にとって重要であることが示されている。さらに、第2b相STEP試験において、天然クレードB Gag、Pol、及びNef抗原を発現するrAd5ベースのHIV−1ワクチン候補は、限られた幅のHIV−1特異的細胞性免疫応答しか誘発せず、ワクチン有益性は認められなかった。STEP試験のワクチン被接種体は、Gagに対するわずか1のエピトープ特異的応答の中央値を含め、わずか2〜3のエピトープ特異的Tリンパ球応答の中央値を生じ、この極めて狭い幅の細胞性免疫応答が、感染ウイルスの多様性の不十分な免疫有効範囲をもたらしたように思われる。CD8+ Tリンパ球からのウイルスエスケープもまた、急性HIV−1感染において急速に起こることが報告されており、従って共通のエピトープ変異体に対するワクチンにより誘発された細胞性免疫応答もまた重要であることが分かり得る。まとめると、以上の試験は、細胞性免疫の幅及び深さを増大させるHIV−1ワクチン戦略を開発する必要性を強調するものである。
【0100】
本発明者らはこの試験においてモザイクHIV−1抗原を評価したため、SIV攻撃誘発に対するこうしたワクチンレジメンの防御効力を評価することはできなかった。しかしながら、本発明者らは、rAdベクターにより誘発されたSIV特異的細胞性免疫応答の幅が、アカゲザルにおいてSIV攻撃誘発に対する防御効力と相関していることを以前報告している(Liuら、Nature 457:87頁、2009年)。本発明者らはまた、変異エピトープに対する細胞性免疫応答が、アカゲザルにおいてインビボでSIV突然変異進化を阻止することができることも示しており(Barouchら、Nat.Immunol.6:247頁、2005年)、細胞性免疫の深さの拡大の生物学的関連性が示唆される。非ヒト霊長類におけるSIV攻撃誘発に対するモザイクワクチンの防御効力のモデリングには固有の限界があり、これは、SIVとHIV−1 M群配列とでは観察される多様性が実質的に異なり、影響を受ける基礎となる生物学が異なるためである。例えば、スーティーマンガベイなどの天然宿主におけるCD8+ Tリンパ球選択圧は、ヒトにおけるものと比べて実質的に低いものと思われる。従って、候補HIV−1ワクチンとしてのモザイク抗原のさらなる評価には、臨床治験が役立ち得る。
【0101】
要約すれば、本発明者らは、アカゲザルにおいて二価モザイクHIV−1 Gag、Pol、及びEnv抗原が細胞性免疫幅及び深さを実質的に拡大したことを実証する。全体的なウイルス多様性及び細胞性免疫応答からのウイルスのエスケープは、T細胞ベースのHIV−1ワクチンの開発における決定的な難関に相当するため、こうした知見は、HIV−1ワクチン開発に対する重要な含意を有する。モザイク抗原の二価カクテルはまた、臨床開発が実際的及び潜在的に実現可能でもある。モザイク抗原の結合価が増加すると、有効範囲がさらに向上し得る。最後に、このモザイク抗原戦略は一般化可能であり、HIV−1に加えて他の遺伝的に多様な病原体に利用される可能性がある。
【0102】
材料及び方法
抗原設計及びベクター産生。本質的に、ロスアラモスHIV−1配列データベースにおけるHIV−1 M群配列の最適な有効範囲を提供するように二価モザイクGag、Pol、及びEnv抗原を作成した。ロスアラモスHIV−1配列データベースにおける最適PTE有効範囲のクレードC配列を提供する配列となるように最適天然クレードC抗原を選択した(C.IN.−.70177 Gag、C.ZA.04.04ZASK208B1 Pol、C.SN.90.90SE_364 Env)。準コンセンサス又はコンセンサス配列となるようにクレードB抗原を選択し(B.CAM−1 Gag、B.IIIB Pol、B.Con Env)、それを用いて二価クレードB+Cワクチン手法に対する最適クレードC抗原を補足した。Pol抗原はRT及びINを含んでPRを含まず、及び記載されるとおり触媒活性を除去する点突然変異を含んだ(Priddyら、Clinical infectious diseases 46:1769頁、2008年)。Env gp140抗原は、切断及び融合活性を除去する点突然変異を含んだ。ワクチン配列を図27に示す。組換え複製無能アデノウイルス血清型26(rAd26)及びそれらの抗原を発現するヘキソン−キメラrAd5HVR48ベクターをPER.55K細胞で成長させて、本質的に記載されるとおり二重CsCl勾配沈降により精製した(Abbinkら、J.Virol.81:4654頁、2007年、及びRobertsら、Nature 441:239頁、2006年)。
【0103】
動物及び免疫化。MHCクラスI対立遺伝子Mamu−A*01を発現しなかった27匹の非近交系アカゲザルをNew England Primate Research Center(NEPRC)、Southborough、MAで飼育した。免疫化は、モザイク、Mコンセンサス、クレードB+クレードC、又は最適天然クレードC HIV−1 Gag、Pol、及びEnv抗原を発現する3×1010個のウイルス粒子rAd26又はrAd5HVR48ベクターを、1mlの筋肉内注射として0週目及び40週目に双方の大腿四頭筋に送達することを含んだ。全ての動物試験は、本発明者らの研究機関内の動物の管理及び使用に関する委員会(Institutional Animal Care and Use Committees:IACUC)により承認された。
【0104】
IFN−γ ELISPOTアッセイ。ワクチン接種を受けたサルにおけるHIV−1特異的細胞性免疫応答を、本質的に記載されるとおりインターフェロン−γ(IFN−γ)ELISPOTアッセイにより評価した(Robertsら、Nature 441:239頁、2006年、及びLiuら、Nature 457:87頁、2009年)。HIV−1 M群配列の少なくとも15%に存在するHIV−1 Gag、Pol、及びEnvの潜在的T細胞エピトープ(PTE)ペプチド、並びにクレードC DU422、クレードC ZM651、コンセンサスC、コンセンサスA、及びコンセンサスB株由来のHIV−1 Gagペプチドを、NIH AIDS Research and Reference Reagent Programから入手した。96ウェルマルチスクリーンプレート(Millipore)を、エンドトキシンフリーのダルベッコPBS(D−PBS)中の10μg/ml抗ヒトIFN−γ(BD Biosciences)の100μl/ウェルで一晩コートした。次にプレートを0.25%Tween−20含有D−PBS(D−PBS/Tween)で3回洗浄し、37℃で5%FBS含有D−PBSにより2時間ブロックし、D−PBS/Tweenで3回洗浄し、10%FBS含有RPMI 1640でリンスしてTween−20を取り除き、100μlの反応量で3通りに各2μg/mlのペプチド及び2×105PBMCと共にインキュベートした。37℃で18時間インキュベートした後、プレートをPBS/Tweenで9回及び蒸留水で1回洗浄した。次にプレートを、2μg/mlのビオチン化抗ヒトIFN−γ(BD Biosciences)と共に室温で2時間インキュベートし、PBS/Tweenで6回洗浄し、1:500希釈のストレプトアビジン−アルカリ性ホスファターゼ(Southern Biotechnology Associates)と共に2時間インキュベートした。PBS/Tweenで5回及びPBSで1回洗浄した後、プレートをニトロブルーテトラゾリウム/5−ブロモ−4−クロロ−3−インドリル−リン酸色原体(Pierce)で展開し、水道水で洗浄することにより停止させ、風乾し、ELISPOTリーダー(Cellular Technology Ltd)を使用して読み取った。106個のPBMC当たりのスポット形成細胞(SFC)を計算した。媒質バックグラウンドは、典型的には106PBMC当たり15SFC未満であった。陽性応答は、106個のPBMC当たり55個より多いSFC、且つバックグラウンドの4倍超として定義した。
【0105】
エピトープマッピング。NIH AIDS Research and Reference Reagent Programから入手したGag、Pol、及びEnv PTEペプチドを利用して、包括的なCD8+及びCD4+ Tリンパ球エピトープマッピングを実施した。最初に完全ペプチドプールで、並びに10個のPTEペプチドを含むサブプールで免疫した後4週目に、IFN−γ ELISPOTアッセイを行った。陽性応答を有する全てのペプチドサブプールをデコンボリューション処理し、個々の15アミノ酸PTEペプチドでエピトープが確認された。次に細胞枯渇IFN−γ ELISPOTアッセイを実施して、反応性ペプチドがCD8+又はCD4+ Tリンパ球エピトープに相当するかどうかを判断した。追加免疫の4週間後44週目にPTEサブプールを利用した部分的エピトープマッピングもまた実施した。ボーダーラインの応答は全て再試験し、確認された場合にのみ陽性と見なした。10個の重複Gagペプチドを含むサブプールを利用した部分的エピトープマッピングもまた実施し、様々なクレード由来のHIV−1 Gagの幅を評価した。
【0106】
体液性免疫アッセイ。本質的に記載されるとおり、HIV−1クレードC Env gp140及びルシフェラーゼベースのシュードウイルス中和アッセイを利用する直接ELISAにより、Env特異的体液性免疫応答について調べた(Montefiori、「Evaluating neutralizing antibodies against HIV, SIV and SHIV in luciferase reporter gene assays」、Current Protocols in Immunology、Coligan、Kruisbeek、Margulies、Shevach、Strober、及びCoico編(John Wiley & Sons、2004年、1〜15頁)。
【0107】
統計分析。全ての統計分析はパッケージR(Team、Foundation for Statisical Computing、Vienna、オーストリア、2009年)を使用して行った。細胞性免疫応答の幅を分析してPTEペプチドをマッピングするため(図19A)、ワクチン群、抗原(Gag、Pol、Env)、及びリンパ球部分母集団(CD4、CD8)の関数として反応性ペプチドの数を予想するポアソン回帰モデルをフィットさせた。本発明者らのモデルでは、動物間のばらつきに対応するため変量効果を含め、パッケージRのlme4ライブラリ(Pinheiro、Springer、New York(2000年))によりフィットさせた。データはモデルを良好にフィットし(分散パラメータ1.0)、3個の説明因子間に有意な交互作用はなかった。例えば、モザイク抗原の投与を受けたサルによって認識されたPTEペプチド数が、コンセンサス又は天然配列抗原の投与を受けた場合と比較したとき3.8倍亢進したことは(図19A)、Gag、Pol、及びEnv由来のPTEに等しく当てはまり、CD8+並びにCD4+ Tリンパ球による応答について維持された。反応性エピトープ領域数の分析(図19B)にもまた変量効果を伴うポアソン回帰モデルを含め、これもまた有意な交互作用なしに良好にフィットした(分散パラメータ0.87)。コルモゴロフ−スミルノフの両側検定を利用してCD8+及びCD4+ Tリンパ球応答の大きさの比較(図23)を実施した。サル当たりの応答の幅及び深さを種々のワクチン間で比較するノンパラメトリック検定もまた実施した(図19A及び図24C)。本発明者らは、4つのワクチン群間に違いがあるかどうかを判断するため最初にクラスカル・ワリス検定を用いた。いずれの場合も、これは高度に有意であったとともに、次に本発明者らはウィルコクソン順位和検定を使用して4つのワクチン群間の全てのペアワイズ比較を評価した。これらの比較の各々において、モザイクワクチンが誘発したサル当たりの応答は、他の3つのワクチンと比べて有意に多かった。様々なクレード由来のHIV−1 Gagに対する応答の幅を分析するため(図25)、データを二項回帰モデルにフィットさせた。これらのモデルでは説明変数としてワクチン群を使用し、動物間及び株間のばらつきを考慮するため変量効果を含めた。データは僅かに過小分散であったが、モザイクワクチンの投与を受けた動物はなお有意により多い応答数を誘発した。ロスアラモスHIV−1配列データベースで利用可能なツールを使用してPTE有効範囲の評価を実施した。
【0108】
配列別添資料
I.二価MモザイクENV GP160、GAG、POL、NEF配列
モザイクENV1 GP160(アミノ酸配列)
配列番号1
【0109】
モザイクENV2 GP160(アミノ酸配列)
配列番号2
【0110】
モザイクGAG1(アミノ酸配列)
配列番号3
【0111】
モザイクGAG2(アミノ酸配列)
配列番号4
【0112】
モザイクPOL1(アミノ酸配列)
配列番号5
【0113】
モザイクPOL2(アミノ酸配列)
配列番号6
【0114】
モザイクNEF1(アミノ酸配列)
配列番号7
【0115】
モザイクNEF2(アミノ酸配列)
配列番号8
【0116】
II.二価MモザイクENV GP140配列(切断/融合欠損)
モザイクENV1 GP140(アミノ酸配列)
配列番号9
【0117】
【0118】
モザイクENV2 GP140(アミノ酸配列)
配列番号10
【0119】
MOS3 ENV GP140(アミノ酸配列)
678アミノ酸
配列番号11
【0120】
III.二価MモザイクPOL配列(広範囲に不活化、PR欠失、9個のAの不活化突然変異による触媒活性の除去)
モザイクPOL1(アミノ酸配列)
配列番号12
【0121】
モザイクPOL2(アミノ酸配列)
配列番号13
【0122】
MOS3 POL V3(アミノ酸配列)
851アミノ酸
配列番号14
【0123】
IV.二価MモザイクGAG配列
MOS3 GAG(アミノ酸配列)
508アミノ酸
配列番号15
【0124】
V.二価MモザイクNEF配列(2位のGをAにすることによるミリスチル化部位の欠失)
MOS1 NEF
(206アミノ酸)
配列番号16
【0125】
MOS2 NEF
(206アミノ酸)−2位のGをAにすることによるミリスチル化部位の欠失
配列番号17
【0126】
MOS3 NEF
(208アミノ酸)
配列番号18
【0127】
VI.二価MモザイクGAGNEF融合配列
モザイクGAGNEF1(アミノ酸配列)
配列番号19
【0128】
モザイクGAGNEF2(アミノ酸配列)
配列番号20
【0129】
【0130】
VII.二価MモザイクGAGPOL融合配列(バージョン3;POLは広範囲に不活化、PR欠失、9個のAの不活化突然変異による触媒活性の除去)
モザイクGAGPOL1 V3(アミノ酸配列)
配列番号21
【0131】
モザイクGAGPOL2 V3(アミノ酸配列)
配列番号22
【0132】
【0133】
MOS3 GAG−POL V3(アミノ酸配列)
1359アミノ酸−完全GAGと修飾POLとのGAG−POL融合
配列番号23
【0134】
【0135】
VIII.二価MモザイクGAGPOL融合配列(バージョン4;POLは最小限の不活化、完全なPR−RT−IN)
モザイクGAGPOL1 V4(アミノ酸配列)
配列番号24
【0136】
モザイクGAGPOL2 V4(アミノ酸配列)
配列番号25
【0137】
【0138】
IX.二価MモザイクGAGPOL融合配列(バージョン5;POLは最小限の不活化、PR欠失)
モザイクGAGPOL1 V5(アミノ酸配列)
配列番号26
【0139】
【0140】
モザイクGAGPOL2 V5(アミノ酸配列)
配列番号27
【0141】
X.二価MモザイクGAGPOLNEF融合配列(POLは広範囲に不活化、PR欠失)
モザイクGagPolNef1(アミノ酸配列)
配列番号28
【0142】
【0143】
モザイクGAGPOLNEF2(アミノ酸配列)
配列番号29
【0144】
【0145】
XI.最適クレードC ENV GP160、GAG、POL、NEF配列
最適クレードC ENV GP160(SN90.90.SE364)(アミノ酸配列)
配列番号30
【0146】
最適クレードC GAG(IN.70177)(アミノ酸配列)
配列番号31
【0147】
最適クレードC POL(ZA.04.04ZASK208B1)(アミノ酸配列)
配列番号32
【0148】
【0149】
最適クレードC NEF(ZA00.1170MB)(アミノ酸配列)
配列番号33
【0150】
XII.最適クレードC ENV GP140配列(切断/融合欠損)
最適クレードC ENV GP140(SN90.90.SE364)(アミノ酸配列)
配列番号34
【0151】
XIII.最適クレードC POL 配列(広範囲に不活化、PR欠失)
最適クレードC POL(ZA.04.04ZASK208B1)(アミノ酸配列)
配列番号35
【0152】
XIV.最適クレードC GAGNEF融合配列
最適クレードC GAGNEF(IN.70177−ZA00.1170MB)(アミノ酸配列)
配列番号36
【0153】
XV.コンセンサス配列
MコンセンサスENV
配列番号37
【0154】
MコンセンサスGAG
配列番号38
【0155】
MコンセンサスPOL
配列番号39
【0156】
【0157】
他の実施形態
本発明はその特定の実施形態に関連して記載されているが、さらなる変更が可能であり、及び本願が、概して本発明の原理に従い、且つ本発明が関係する技術分野のなかの公知の又は通常の実施の範囲内に含まれるとともに本明細書で以上に示された本質的な特徴に該当し得る本開示からのかかる逸脱を含む本発明の任意の変形例、使用、又は適応を包含することを意図していることは理解されるであろう。
【0158】
本明細書において言及される全ての刊行物及び特許出願は、あたかも各々の独立した刊行物又は特許出願が、全体として参照により援用されるように具体的且つ個別的に示されたのと同じ程度まで、参照により本明細書に援用される。
【技術分野】
【0001】
連邦政府助成研究に関する記載
本研究は、NIH交付番号第U19−AI066305号及び同第U19−AI078526号から一部資金提供を受けている。政府は本発明に対して特定の権利を有する。
【0002】
発明の分野
本発明は、ウイルス感染症を治療又は予防するための組成物、方法、及びキットを提供する。本明細書に記載される多価(例えば、二価)ワクチンは、ワクチン接種を受けた対象における細胞性免疫応答の多様性又は幅及び深さを増加させることのできる、計算により最適化されたウイルスポリペプチドを組み込む。
【背景技術】
【0003】
ウイルスに対する細胞性免疫応答を誘発するワクチンは、ウイルス感染症を効果的に治療又は予防するため広域のウイルス多様性を反映しなければならない。例えば、強力で多様なHIV−1特異的T細胞応答の惹起は、有効なHIV−1ワクチンには極めて重要であると思われる。細胞傷害性Tリンパ球(CTL)応答はヒトにおける疾患進行の遅延と相関し、非ヒト霊長類ワクチン接種モデルにおけるCTL応答の重要性は十分に確立されている。大きいばらつきのあるエンベロープ(Env)がHIVに対する中和抗体の一次標的であり、ワクチン抗原もまた、それらの抗体応答を誘発するように作ることが必要となり得るが、T細胞ワクチン成分は、より保存されているタンパク質を標的化し、交差反応する可能性がより高い応答を引き起こすことができる。しかし、最も保存されているHIV−1タンパク質でさえ、ばらつきが問題となり得るのに十分な多様性を有する。コンセンサス及び祖先HIV−1配列などの人工的な中心配列のワクチン手法は、本質的に株間の「違いを分ける」もので、天然株ワクチンと比較して交差反応性が亢進した応答を刺激することができる。コンセンサス抗原は、あらゆる循環株の唯一最良の「平均」である合成抗原配列に相当する。こうした抗原は特異的細胞性免疫応答を誘発することができるが、その応答の幅及び強度は、これまでのワクチン戦略と比べて実質的に向上していない。ウイルス感染症を治療又は予防する次世代ワクチンの開発は、ワクチン接種が奏功する結果をもたらすことができるように、細胞性免疫の幅の増加を引き出すものでなければならない。かかるワクチンは、特にHIV−1の治療又は予防に差し迫って必要とされている。
【発明の概要】
【課題を解決するための手段】
【0004】
第1の態様において、本発明は、少なくとも2個の別個の最適化ウイルスポリペプチド(例えば、2、3、4、5個、又はそれ以上の別個の最適化ウイルスポリペプチド)を含む、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンを特徴とし、ここで最適化ウイルスポリペプチドは、同じウイルス遺伝子産物に対応する。一実施形態において、ウイルス感染症は、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる。他の実施形態において、レトロウイルスはヒト免疫不全ウイルス1型(HIV−1)であり、及びウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuを含む。さらなる実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuウイルス遺伝子産物のうちの1つに対応する最適化ウイルスポリペプチドを2個以下含む。別の実施形態において、ワクチンは、Gag及びNefに対応する最適化ウイルスポリペプチドを含まない。さらに別の実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuから選択される第1のウイルス遺伝子産物についての少なくとも2個の別個の最適化ウイルスポリペプチド(例えば、2、3、4、5個、又はそれ以上の別個の最適化ウイルスポリペプチド)と、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuから選択される第1のウイルス遺伝子産物と異なる第2のウイルス遺伝子産物についての1つ又は複数の別個の最適化ウイルスポリペプチド(例えば、2、3、4、5個、又はそれ以上の別個の最適化ウイルスポリペプチド)とを含む。
【0005】
第2の態様において、本発明は、ヒトなどの哺乳動物におけるヒト免疫不全ウイルス1型(HIV−1)感染症を治療し、又はそのリスクを低減するワクチンであって、配列番号1〜29に示される配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する最適化ウイルスポリペプチドを含むワクチンを特徴とする。一実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示される配列のいずれか1つとアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する。別の実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示される配列のいずれか1つのアミノ酸配列を有する。さらなる実施形態において、ワクチンは、a)〜k)群の任意の1つ又は複数から選択される少なくとも2個の最適化ウイルスポリペプチドを含む:a)配列番号1及び2;b)配列番号3、4、及び5;c)配列番号6及び7;d)配列番号8〜12;e)配列番号13、14、及び15;f)配列番号16、17、及び18;g)配列番号19及び20;h)配列番号21、22、及び23;i)配列番号24及び25;j)配列番号26及び27;並びにk)配列番号21〜22。別の実施形態において、ワクチンは、上記のa)〜k)群のいずれか1つから選択される最適化ウイルスポリペプチドと、同じ、又は異なるa)〜k)群からの1つ又は複数の異なる最適化ウイルスポリペプチドとの対を含むことができる。他の実施形態において、ワクチンは、a)〜k)群の1つ又は複数からの少なくとも3個又は4個又はそれ以上の最適化ウイルスポリペプチドを含むことができる。
【0006】
第3の態様において、本発明は、別個の最適化ウイルスポリペプチドの少なくとも2つの対を含む、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンを特徴とし、ここで最適化ウイルスポリペプチドの各対は同じウイルス遺伝子産物に対応し、及びここでワクチンに組み込まれた2個以下の最適化ウイルスポリペプチドは、同じウイルス遺伝子産物に対応する。一実施形態において、ワクチンは、別個の最適化ウイルスポリペプチドの少なくとも3つの対を含む。別の実施形態において、ワクチンは、別個の最適化ウイルスポリペプチドの少なくとも4つの対を含む。一実施形態において、ウイルス感染症は、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる。他の実施形態において、レトロウイルスはヒト免疫不全ウイルス1型(HIV−1)であり、及びウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuを含む。さらなる実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuウイルス遺伝子産物のうちの1つに対応する最適化ウイルスポリペプチドを2個以下含む。別の実施形態において、ワクチンは、Gag及びNefに対応する最適化ウイルスポリペプチドを含まない。さらなる実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuウイルス遺伝子産物のうちの任意の3つに対応する別個の最適化ウイルスポリペプチドの少なくとも3つの対を含む。別の実施形態において、ワクチンは、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuウイルス遺伝子産物のうちの任意の4つに対応する別個の最適化ウイルスポリペプチドの少なくとも4つの対を含む。
【0007】
本発明の最初の3つの態様のうちの任意の一実施形態において、ワクチンは、ウイルス遺伝子産物に対する細胞性免疫応答を誘発する。別の実施形態において、ワクチンは、HIV−1に対する細胞性免疫応答を誘発する。さらなる実施形態において、少なくとも1つの別個の最適化ウイルスポリペプチドのヌクレオチド配列は、核酸又はベクターによりコードされる。一実施形態において、ベクターは、アデノウイルス血清型26(Ad26)、アデノウイルス血清型34(Ad34)、アデノウイルス血清型35(Ad35)、アデノウイルス血清型48(Ad48)、又はアデノウイルス血清型5 HVR48(Ad5HVR48)などの組換えアデノウイルスである。さらなる実施形態において、ワクチンは、薬学的に許容可能な担体、賦形剤、又は希釈剤と組み合わせたものである。
【0008】
第4の態様において、本発明は、配列番号1〜29に示されるアミノ酸配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する最適化ウイルスポリペプチドのヌクレオチド配列を含む核酸を特徴とする。一実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示されるアミノ酸配列のいずれか1つと配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する。別の実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示されるアミノ酸配列のいずれか1つを有する。さらなる実施形態において、核酸はベクターを含む。一実施形態において、ベクターは、アデノウイルス血清型26(Ad26)、アデノウイルス血清型34(Ad34)、アデノウイルス血清型35(Ad35)、アデノウイルス血清型48(Ad48)、又はアデノウイルス血清型5 HVR48(Ad5HVR48)などの組換えアデノウイルスである。
【0009】
第5の態様において、本発明は、配列番号1〜29に示されるアミノ酸配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する最適化ウイルスポリペプチドを特徴とする。一実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示されるアミノ酸配列のいずれか1つと配列同一性を有する少なくとも7個の連続するアミノ酸(例えば、少なくとも8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、50、100、150、175、200、250、300、350、400、450、500又はそれ以上の連続するアミノ酸長)を有する。別の実施形態において、最適化ウイルスポリペプチドは、配列番号1〜29に示されるアミノ酸配列のいずれか1つを有する。
【0010】
第6の態様において、本発明は、本発明のワクチン又は核酸を投与することにより、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減する方法を特徴とする。一実施形態において、ウイルス感染症は、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる。さらなる実施形態において、レトロウイルスはヒト免疫不全ウイルス1型(HIV−1)であり、及びウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuを含む。一実施形態において、ワクチン又は核酸は、ウイルス遺伝子産物に対する細胞性免疫応答を誘発する。
【0011】
第7の態様において、本発明は、本発明のワクチンを合成することによる、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンの製造方法を特徴とする。
【0012】
第8の態様において、本発明は、本発明の核酸を細胞と接触させ、及び最適化ウイルスポリペプチドを単離することによる、ヒトなどの哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンの製造方法を特徴とする。
【0013】
本発明の第7又は第8の態様の一実施形態において、最適化ウイルスポリペプチドは、哺乳動物に投与されると細胞性免疫応答を誘発する。細胞性免疫応答は、ウイルス遺伝子産物に対するものであってよい。別の実施形態において、ウイルス感染症は、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる。さらなる実施形態において、レトロウイルスはヒト免疫不全ウイルス1型(HIV−1)であり、及びウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、又はVpuを含む。
【0014】
第9の態様において、本発明は、キットであって、本発明のワクチンと、薬学的に許容可能な担体、賦形剤、又は希釈剤と、キットの使用説明書とを含むキットを特徴とする。一実施形態において、キットはアジュバントも含む。
【0015】
最後の態様において、本発明は、キットであって、本発明の核酸と、薬学的に許容可能な担体、賦形剤、又は希釈剤と、キットの使用説明書とを含むキットを特徴とする。一実施形態において、キットはアジュバントも含む。
【0016】
本発明の全ての態様のある実施形態では、最適化ウイルスポリペプチドは、ヒトでの発現用に最適化されている核酸配列によりコードされる(例えば、配列番号5、10、11、12、15、18、及び23のうちの任意の1つ)。
【0017】
定義
「最適化ウイルスポリペプチド」又は「計算により最適化されたウイルスポリペプチド」とは、天然に存在するウイルスペプチド、ポリペプチド、又はタンパク質ではない免疫原性ポリペプチドを意味する。最適化ウイルスポリペプチド配列は、最初は、哺乳動物(例えば、ヒト)の免疫後に(例えば、本発明のワクチンに組み込まれたときに)生じる抗ウイルス免疫応答(例えば、細胞性又は体液性免疫応答)の幅、強度、深さ、又は寿命を増加させるように、1つ又は複数の天然に存在するウイルス遺伝子産物(例えば、ペプチド、ポリペプチド、及びタンパク質)のアミノ酸配列を修飾することによって生成される。従って、最適化ウイルスポリペプチドは「親」ウイルス遺伝子配列に対応し得る;或いは、最適化ウイルスポリペプチドは特定の「親」ウイルス遺伝子配列に対応しなくともよく、ウイルスの様々な株又は疑似種由来の類似配列に対応してもよい。最適化ウイルスポリペプチドに含めることのできるウイルス遺伝子配列に対する修飾としては、アミノ酸付加、置換、及び欠失が挙げられる。本発明の一実施形態において、最適化ウイルスポリペプチドは、2個以上の天然に存在するウイルス遺伝子産物(例えば、天然又は臨床ウイルス分離株)が複合した、又は合体したアミノ酸配列であり、ここでは潜在的な各エピトープ(例えば、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30又はそれ以上のアミノ酸長の各連続又は重複するアミノ酸配列)が分析及び修飾され、得られる最適化ウイルスポリペプチドの免疫原性が向上している。異なるウイルス遺伝子産物に対応する最適化ウイルスポリペプチドはまた、融合することによって本発明のワクチンへの組み込みを促進することができる。最適化ウイルスポリペプチドの生成方法は、例えば、参照により本明細書に援用されるFisherら「Polyvalent Vaccine for Optimal Coverage of Potential T−Cell Epitopes in Global HIV−1 Variants」、Nat.Med.13(1):100−106頁(2007年)及び国際公開第2007/024941号パンフレットに記載される。最適化ウイルスポリペプチド配列が生成されると、標準的な技術(例えば、参照により本明細書に援用される国際公開第2006/040330号パンフレット及び国際公開第2007/104792号パンフレットに開示されるアデノウイルスベクターなどの組換えウイルスベクター)により対応するポリペプチドを産生し、又は投与することができる。
【0018】
「薬学的に許容可能な担体」とは、それと共に投与される化合物の治療特性を維持しながら、治療を受ける哺乳動物にとって生理学的に許容可能である担体を意味する。例示的な薬学的に許容可能な担体の一つは生理食塩水である。他の生理学的に許容可能な担体及びそれらの製剤は当業者に公知であり、例えば、参照により本明細書に援用される「Remington’s Pharmaceutical Sciences」(第18版、A.Gennaro編、1990年、Mack Publishing Company、Easton、PA)に記載されている。
【0019】
「ベクター」とは、下流の遺伝子又はコード領域に作動可能に連結されたプロモーターを含むDNAコンストラクト(例えば、ポリペプチド又はポリペプチド断片をコードするcDNA又はゲノムDNA断片)を意味する。ベクターをレシピエント細胞(例えば、原核又は真核細胞、例えば、細菌、酵母、昆虫細胞、又は哺乳動物細胞、発現ベクター内のプロモーターに依存する)又は生体(例えばヒトを含む)に導入すると、細胞は、ベクターによってコードされるmRNAを発現することができ、次にそれが翻訳されて、コードされた本発明の最適化ウイルスポリペプチドとなる。インビトロ転写/翻訳用のベクターもまた当該技術分野において公知であり、本明細書においてさらに記載される。ベクターは、例えば、バクテリオファージ、アデノウイルス、レトロウイルス、ポックスウイルス、又はヘルペスウイルスに由来する遺伝的に改変されたプラスミド、ウイルス、又は人工染色体であってもよい。
【0020】
「ウイルス遺伝子産物」とは、任意の天然に存在するウイルスペプチド、ポリペプチド、又はタンパク質、又はその断片を意味する。本発明の一実施形態において、ウイルス遺伝子産物はヒト免疫不全ウイルス1型(HIV−1)に由来する。HIV−1ウイルス遺伝子産物は、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuポリペプチドを含む。
【図面の簡単な説明】
【0021】
【図1】計算により最適化されたHIV−1 Gag、Pol、及びEnvウイルスポリペプチドの、アカゲザルにおける全潜在的T細胞エピトープ(PTE)ペプチドに対する幅の拡大を示す図である。最適化ウイルスポリペプチド(青色)で免疫した動物が、最大数のリコールペプチドプールと反応した。
【図2】計算により修飾されたHIV−1 Gag、Pol、及びEnvウイルスポリペプチドがエピトープ特異的細胞性免疫応答の幅を拡大することを示す図である。
【図3】本発明の計算により修飾されたウイルスポリペプチドから得られたHIV−1ウイルス遺伝子産物Gag、Pol、及びEnvで免疫した後のアカゲザル、並びにコンセンサスHIV−1抗原又はHIV−1クレードC分離株抗原で免疫した動物で検出された細胞性免疫応答の幅を示す。最適化ウイルスポリペプチド(青色)で免疫した動物が、最大数のリコールペプチドプールと反応した。動物は非近交系であるため、プールは動物ごとに異なる。Gag、Pol、及びEnvは、各々が多くの細胞性免疫応答を誘発し、共通の反応性パターンを有し得る。
【図4】ウイルスポリペプチド(Pol(図4A)、Gag(図4B)、及びEnv(図4C))ごとの、種々の試験ワクチン(二価モザイク(Mos2)、Mコンセンサス(Mcon)、及び最適化クレードC(OptC))の間で共有される潜在的エピトープを示すグラフである。図4A〜図4Cは、種々のワクチン候補による現在のHIVデータベース完全長ゲノムセット及びPTEペプチドの相対的有効範囲を示す。
【図5】二価モザイク(Mos2)ワクチンに対するPTEペプチド応答数(ここでは重複にかかわらず各応答が独立したイベントと見なされる)が、M群コンセンサス(Mcon)ワクチン、及び最適な有効範囲のM群集合(OptC)ワクチン抗原をもたらすように選択された天然ウイルス株ワクチン(最適化クレードC(C天然(最適))に対する応答数と比べて多いことを示すグラフである。図5は、タンパク質、CD8+ T細胞、及びCD4+ T細胞ごとに動物当たりのPTEペプチド応答数を示す。統計的に、Mos2>Mcon〜OptC(Mconは、OptCより応答が多い傾向を示す)。Mconと比較したMos2のウィルコクソンp値:p値=0.001058。
【図6】T細胞応答を引き起こすPTEペプチド数を示す図である。CD8+ T細胞において応答を引き起こす二価モザイク(Mos2)ワクチンのPTEペプチドの中位数は16(値域;12〜29)であり、一方、CD8+ T細胞において応答を引き起こすMconペプチドの中位数は6(値域:0〜7)に過ぎず、及びOptCペプチドの中位数は3ペプチド(値域:0〜3)に過ぎない。CD4+ T細胞において応答を引き起こす二価モザイク(Mos2)ワクチンのPTEペプチドの中位数は4(値域;2〜6)であり、一方、CD4+ T細胞において応答を引き起こすMconペプチドの中位数は1(値域:0〜2)に過ぎず、及びOptCペプチドの中位数は0.5ペプチド(値域:0〜2)に過ぎない。従って、応答の傾向はMos2>Mcon>OptCである。
【図7】被験動物の各々に由来するT細胞によって認識される全てのCD8+ T細胞Gag PTEペプチドのマッピングを要約する概略図である(以下の実施例3を参照)。動物番号、ペプチドプール及びペプチド番号を、各反応性ペプチドの境界に表示する。記号は群を示す:*、Mos2;¥、ConM;±、OptC。ここでは例としてGagが挙げられる。動物が非近交系であっても、CD8応答はクラスタ化する傾向がある。モザイクは、単価ワクチンより有利である可能性がある。モザイクは、より多くの共通の変異体と反応する応答を刺激する確率がより高い。モザイクはまた、カクテル中に存在する種々の型に対する複数の応答を刺激する。従って、モザイクは共通のエスケープ経路を遮断する可能性を有する。本発明者らの研究では、モザイクワクチンは、より多くの重複ペプチドを認識するT細胞応答を刺激する傾向があった。反応性ペプチドが局在する多くのホットスポットがある。PTEペプチドは、ワクチンの評価に使用されるペプチド試薬におけるHIV−1 M群の潜在的エピトープ(又は9アミノ酸の連続するストレッチについての9−mer)の有効範囲を最大化するように設計される。必然的に、PTEペプチドには多数の重複が存在するが、そのアルゴリズムに起因して、重複は通常、いくらかのばらつきを伴う重複である。図7は配列番号42を開示する。
【図8】被験動物の各々に由来するT細胞により認識されるCD4+ T細胞Gag PTEペプチドのマッピングを要約する概略図である。図8は配列番号43を開示する。
【図9】ConMワクチン又は最適天然ワクチンに対するPTE応答の典型的なパターンを示す図であり、応答を誘発するペプチドをワクチンの関連領域と整列させている。これらのワクチンに対する反応を実現するには、ワクチンと標的PTEペプチドとの間の途切れのない同一のストレッチによる良好なマッチが必要である。図9は、掲載順にそれぞれ配列番号44〜57を開示する。
【図10】モザイクワクチンが、見かけ上の抗原競合なしに、且つ広範な局所的応答で、複数の変異重複ペプチドを認識する多くの応答を生じたことを示す図である。特に、4個の変異PTEペプチドが認識された。さらに、重複の領域では双方のモザイク型が、それら2つの組み合わせと同様に認識された。最後に、新規の型(S)が認識された。図10は、掲載順にそれぞれ配列番号58〜63を開示する。
【図11】モザイクワクチン接種を受けた動物(361−07)におけるCD8+ PTEペプチド応答の典型的なパターンを示す図である。22個のPTEペプチドを試験し、8個のCD8応答領域が同定された;5個の領域が、モザイクのいずれか1つのアミノ酸にマッチする変異ペプチドを含んだ。5個のCD4応答領域が同定された。このように、モザイクに対するT細胞応答は、所与の領域においてより多くの変異ペプチドが分かる。これは、特にCD8 T細胞応答に当てはまるように思われた。これは、エピトープの変異体を認識する複数のT細胞クローンをもたらし、それらが適応されるエスケープ経路を遮断し得る結果であり得る。より多くの応答があるのみならず、それらはより深く、且つより多くの変異体を網羅する。図11は、掲載順にそれぞれ配列番号64〜101としてCD8応答を開示する。CD4応答は、掲載順にそれぞれ配列番号102〜117として開示される。
【図12】ワクチンにより誘発されたT細胞によって標的化される領域にわたる重複変異PTEペプチドの数を示すグラフである。
【図13】2モザイク抗原ワクチンが、Mcon及びOptCワクチンと比べて、1つ又は複数の重複PTEペプチドを含む領域に対するより多くのT細胞応答をもたらすことを示すグラフである。図13は図5と同様であり、サルが右から左に同じ順序で示されるが、単一のペプチドではなく1つ又は複数の重複PTEペプチドを含む領域に対する応答数を反映して尺度を変えている。
【図14】二価モザイク(Mos2)、Mcon、及びOptCワクチンを投与した後の動物におけるT細胞応答数を示す図である。CD8+ T細胞において二価モザイク(Mos2)ワクチンが引き起こす応答の中位数は8であり、一方、Mcon及びOptCワクチンにより引き起こされるCD8+ T細胞応答の中位数は、それぞれ3ペプチド(値域:0〜6)及び1.5ペプチド(値域:0〜5)に過ぎない。CD4+ T細胞において二価モザイク(Mos2)ワクチンが引き起こす応答の中位数は3(値域;2〜5)であり、一方、Mcon及びOptCワクチンにより引き起こされるCD4+ T細胞応答の中位数は、それぞれ1(値域:0〜2)及び0.5(値域:0〜2)に過ぎない。従って、応答の傾向はMos2>Mcon>OptCである。
【図15】モザイクワクチンが、Cクレード天然ワクチンと比べて、Cクレード天然タンパク質と交差反応する応答をより多く誘発できることを示すグラフである:GAGプールペプチドが5個のタンパク質に相当する。M群コンセンサス又は最適な有効範囲のCクレード天然タンパク質によるワクチン接種を受けた動物は、これらのタンパク質に由来するペプチドに対する0〜2つの応答を有したが、一方、モザイクワクチン接種を受けた動物は、1〜5つのペプチドプールに対して応答することができた。モザイクワクチンは、M con又は最適Cのいずれと比べても、試験したタンパク質の各々に対するより多くの応答を誘発する。モザイクワクチンにより誘発されたT細胞応答はまた、実際のGagタンパク質にわたるプールペプチドセットをより多く認識した。10〜12サブプール=10×15merペプチド(5×20merペプチドである96ZM Gagを除く)。
【図16】モザイク設計が、時間の経過に伴うウイルスポリペプチドの変化に対してロバストであることを示すグラフである(例えば、Gag M)。
【図17】9−mer最適化を使用した有効範囲が、準(例えば、8〜12mer)最適化長さと比べてロバストであることを示すグラフである(Gagを図示)。
【図18】変異体数が増加すると有効範囲が大きくなるが、あくまでも収穫逓減を有することを示すグラフである(Gagを図示)。
【図19】PTEペプチドに対するエピトープ特異的Tリンパ球応答の幅及び大きさを示すグラフである。図19Aは、モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原を発現するrAd26ベクターの単回免疫後の個々のPTEペプチドに対するエピトープ特異的CD4+(上側)及びCD8+(下側)Tリンパ球応答の数を示すグラフである。個々のサルをx軸上に示す。各色の異なる陰影は、異なる抗原(Gag、Pol、Env)に対する応答を反映している。図19Bは、CD4+(上側)及びCD8+(下側)Tリンパ球応答領域の数を示すグラフである。
【図20A】HIV−1 Gag(図20A)(配列番号118)、Pol(図20B)(配列番号119)、及びEnv(図20C)(配列番号120)タンパク質配列にマッピングした免疫後4週目のPTEペプチドに対するCD8+ Tリンパ球応答を示す概略図を示す。色は、モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原の投与を受けたサルを示す。各エピトープについて、サル番号、抗原(G、Gag;P、Pol;E、Env)、サブプール番号、及び個々のPTEペプチド番号が示される。
【図20B−1】図20Aの続きである。
【図20B−2】図20B−1の続きである。
【図20C】図20B−2の続きである。
【図21A】HIV−1 Gag(図21A)(配列番号121)、Pol(図21B)(配列番号122)、及びEnv(図21C)(配列番号123)タンパク質配列にマッピングした免疫後4週目のPTEペプチドに対するCD4+ Tリンパ球応答を示す概略図を示す。色は、モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原の投与を受けたサルを示す。各エピトープについて、サル番号、抗原(G、Gag;P、Pol;E、Env)、サブプール番号、及び個々のPTEペプチド番号が示される。
【図21B】図21Aの続きである。
【図21C】図21Bの続きである。
【図22−1】モザイク、Mコンセンサス、クレードB+クレードC、又は最適天然クレードC HIV−1 Gag、Pol、及びEnv抗原を発現するrAd26ベクターによる免疫後4週目の全てのサルにおけるワクチン配列の反応性PTEペプチドとのアラインメントを示す概略図である。各サルについて、ワクチン配列を一番上に示し、抗原(G、Gag;P、Pol;E、Env)及びPTEペプチド番号により表される反応性PTEペプチドを、ワクチン配列の下に示す。最小重複領域は太字で示す。2つのモザイク又は2つのクレードB+クレードC抗原の間の配列多型は青色で示す。ワクチン配列と反応性PTEペプチドとの間の違いは赤色で示す。図22は、掲載順にそれぞれ配列番号124〜640を開示する。反応性ペプチドの間の重複が生じるときのその重複に基づき免疫応答エピトープを含む可能性があるペプチド内の最小領域は、ワクチンにおいて太字とする。重複ペプチドがない場合、エピトープがペプチドのどこにでもあり得ると仮定し、従って領域全体を太字とする。ペプチド内で種々の境界を有するエピトープを標的とする種々のT細胞応答、又は変異体が存在するときにばらつきを許容することのできるより雑多なクローン性T細胞応答の間を区別することはできない;いずれのシナリオも、ワクチン免疫応答において有利となり得る。標的化される領域の数は、データを考慮するのに必要な最小T細胞応答数に対応する。ワクチンとペプチドとがマッチしないアミノ酸は赤色で記す;アミノ酸がエピトープを含んでいる可能性のある領域内に収まる場合、それらは太字赤色とする。複数のペプチドが重複するときの重複領域外でのアミノ酸の違いは赤色で標示し、しかし太字にはしない。ワクチンが常に一番上である。各タンパク質の文字(GagはGであり、PolはPであり、エンベロープはEである)及びペプチド番号を用いて各反応性PTEペプチドを表示する。タンパク質及びHXB2番号が各ペプチドに続く。モザイク及びクレードB+Cワクチンについては2個の抗原があり、各々が双方ともアラインメントに含まれる;ワクチンにおけるアミノ酸の違いは青色で指示し、及び反応性ペプチドが第2のモザイクに変異アミノ酸を含む場合、それも青色とする。2つのワクチン抗原が異なる位置の各々において反応性ペプチドをさらに太字で標示し、2つの変異が含まれているその位置がワクチン免疫応答に影響を与え、より大きい幅及び深さをもたらした可能性があることを示す。例えば、要約される最初のワクチンはクレードB+Cワクチンであり、動物287−95が、それについての応答が掲載される最初の動物である。PTEペプチドに対してCD8応答が3つあり、CD4に対して1つあった。CD8ペプチドのうちの2つ、E26及びE282は実質的な重複を示し、従って双方が同じCTL応答を標的とし得る;このように、2つのCD8応答領域のみと、1つのCD4応答領域とがあることも記載する。各応答領域について領域当たりの重複ペプチドの数を書き出すことで(例えば、CD8:1 2 CD4:1)、応答の深さを評価する;2つは赤色とし、その重複の領域は反応性ペプチドにばらつきがあることを示す。ワクチンが異なる場合、2番目の反応性領域におけるD/Eのように、それを青色で標示する。重複の領域のみは太字である。E282におけるHはいずれのワクチンにも認められなかったため、赤色で標示する;これは重複の領域内にあるため、太字としている。各反応性ペプチドは、そのタンパク質及び対応するHXB2番号を有し、右側に記載する。
【図22−2】図22−1の続きである。
【図22−3】図22−2の続きである。
【図22−4】図22−3の続きである。
【図22−5】図22−4の続きである。
【図22−6】図22−5の続きである。
【図22−7】図22−6の続きである。
【図22−8】図22−7の続きである。
【図22−9】図22−8の続きである。
【図22−10】図22−9の続きである。
【図22−11】図22−10の続きである。
【図22−12】図22−11の続きである。
【図22−13】図22−12の続きである。
【図22−14】図22−13の続きである。
【図22−15】図22−14の続きである。
【図22−16】図22−15の続きである。
【図22−17】図22−16の続きである。
【図22−18】図22−17の続きである。
【図23】全てのGag、Pol、及びEnv特異的CD8+(図23A及び図23B)及びCD4+(図23C)Tリンパ球応答の大きさを、最低値から最高値まで並べて示すグラフである。
【図24A−B】PTEペプチドに対するエピトープ特異的Tリンパ球応答の深さを示す。図24Aは、最適天然クレードC抗原の投与を受けたサル366におけるマッピングしたTリンパ球応答の例を示す概略図である。図24Bは、二価モザイク抗原の投与を受けたサル361におけるマッピングしたTリンパ球応答の例を示す概略図である。図24A及び図24Bでは、ワクチン配列を一番上に示し(OptC;Mos1、Mos2)、抗原(G、Gag;P、Pol;E、Env)及びPTEペプチド番号により表される反応性PTEペプチドを、ワクチン配列の下に示す。最小重複領域は太字で示す。2つのモザイク抗原間の配列多型は青色で示す。ワクチン配列と反応性PTEペプチドとの違いは赤色で示す。応答領域ごとにまとめた全ての陽性ペプチドの完全アラインメントは、図22に示す。図24Cは、モザイク、Mコンセンサス、クレードB+クレードC、又は最適天然クレードC抗原を発現するrAd26ベクターで免疫した後のCD4+(上側)及びCD8+(下側)Tリンパ球応答の深さを示すグラフである。個々のサルをx軸上に示す。各エピトープ領域について、1つの応答変異体(薄い陰影)又は1つより多い応答変異体(濃い陰影)を示す。図24Aは、掲載順にそれぞれ配列番号641〜650を開示する。図24Bは、掲載順にそれぞれ配列番号651〜685を開示する。
【図24C】図24A−Bの続きである。
【図25】クレードA、B、及びC由来のHIV−1 Gagペプチドに対するエピトープ特異的Tリンパ球応答の幅を示すグラフである。細胞性免疫応答の幅を、以下のHIV−1 Gag株由来の重複ペプチドのサブプールを利用して評価した:クレードC DU422、クレードC ZM651、コンセンサスC、コンセンサスA、及びコンセンサスB。モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原を発現するrAd26ベクターの単回免疫後の陽性サブプールの数を示す。個々のサルをx軸上に示す。
【図26A−B】追加免疫後の細胞性及び体液性免疫応答を示すグラフである。各サルについてプライミング後4週目(各パネルの左側)及び追加免疫後44週目(各パネルの右側)の個々のTリンパ球応答の大きさ(図26A)及び幅(図26B)を示す。サルは、0週目にrAd26ベクターでプライミングし、40週目に、モザイク、Mコンセンサス、又は最適天然クレードC HIV−1 Gag、Pol、及びEnv抗原を発現するrAd5HVR48ベクターで追加免疫した。個々のサルをx軸上に示す。図26Aでは、赤色はCD8+ Tリンパ球応答を表し、青色はCD4+ Tリンパ球応答を表し、線は双方の時間点で観察された応答を表し、及び点は一方の時間点でのみ観察された応答を表す。図26Bでは、各色の異なる陰影が、異なる抗原(Gag、Pol、Env)に対する応答を反映している。図26Cは、0週目、4週目、及び44週目におけるEnv特異的ELISAエンドポイント力価を示すグラフである。図26Dは、44週目における第1層クレードA(DJ263.8)、クレードB(SF162.LS)、及びクレードC(MW965.26)ウイルスに対する中和抗体(NAb)力価を示すグラフである。陰性対照としてのマウス白血病ウイルスに対するNAb力価は、いずれの試料についても20未満であった。
【図26C−D】図26A−Bの続きである。
【図27】様々なワクチン抗原ごとのPTEペプチドの理論的な有効範囲を示すグラフである。モザイク(青色)、Mコンセンサス(緑色)、クレードB+クレードC(紫色)、又は最適天然クレードC(赤色)HIV−1 Gag、Pol、及びEnv抗原により網羅される9アミノ酸PTEペプチドのパーセンテージを示す。
【発明を実施するための形態】
【0022】
本発明は、天然に存在するウイルス遺伝子産物から計算により得られる最適化ウイルスポリペプチドを特徴とする。本発明の最適化ウイルスポリペプチドは、本発明の1つ若しくは複数の最適化ウイルスポリペプチド又は本発明の1つ若しくは複数の最適化ウイルスポリペプチドを組み込むワクチン(例えば、ベクター)で対象(例えば、ヒト)を免疫した後の、ウイルス特異的免疫(例えば、T細胞に基づく免疫応答などの細胞性免疫)の幅及び深さの増加を可能とする。本発明は、ウイルス感染症に感染した、又は感染するリスクがある対象(例えば、ヒト)に投与することのできるワクチンを提供する。本発明のワクチンは、代表される各対応するウイルス遺伝子産物についての少なくとも2個の別個の最適化ウイルスポリペプチドを組み込む。少なくとも2個の別個の最適化ウイルスポリペプチドを組み込むことにより、ワクチンにおける免疫原性エピトープの有効範囲及び代表性を高めることが可能となり、本発明者らが発見したこのことにより、対象のワクチン接種後にウイルス特異的免疫応答の総数の増加がもたらされる。本発明はまた、ワクチン、ベクター、及び最適化ウイルスポリペプチドを対象(例えば、ヒト)に投与する方法、及びそれらを製造する方法も提供する。本明細書に記載される組成物、方法、及びキットは、少なくとも2個の別個の最適化ウイルスポリペプチドを提供することによってウイルス特異的細胞性免疫応答の多様性、幅、及び/又は深さを実質的に増加させることができる。
【0023】
本発明の最適化ウイルスポリペプチド
本発明は、自然に循環しているウイルス遺伝子産物に対応し、且つそれから得られる計算により最適化されたウイルスポリペプチドを組み込む多価(例えば、二価)ワクチンを提供する。多価モザイクタンパク質は、天然配列からインシリコ組換えにより構築され、所与の結合価に対する潜在的なT細胞エピトープ(PTE)の最大の有効範囲を提供するように最適化される。モザイク抗原は、天然抗原の発現及びプロセシングを維持するように設計される完全長タンパク質である。
【0024】
本発明者らは、単一のウイルス遺伝子産物に対応し、且つそれから得られる2つの別個の最適化ウイルスポリペプチド(すなわち、二価ワクチン)で免疫すると、同じウイルス遺伝子産物から得られる天然に存在するポリペプチド(例えば、臨床分離株に基づく配列)、又は同じウイルス遺伝子産物から得られるかかる天然に存在するポリペプチドのコンセンサス配列を組み込む従来の単価又は多価ワクチンより実質的に多い数の細胞性免疫応答(例えば、T細胞応答)が誘発されることを発見した。従って、計算により最適化されたウイルスポリペプチドであって、その配列が循環ウイルス配列の希少でなく短いストレッチの最大有効範囲を提供する最適化ウイルスポリペプチドを組み込むワクチンは、免疫応答の幅及び深さを増加させることができる。
【0025】
遺伝的アルゴリズムを使用して、入力として提供される天然に存在するウイルス遺伝子産物配列の任意のセットの断片の「モザイク」ブレンドとして最適化ウイルスポリペプチドのセットが作成される。この遺伝的アルゴリズム戦略は、入力データセットとして一般的なウイルス集団からの整列していないタンパク質配列を使用し、従って「アラインメントに依存しない」という長所を有する。これにより、自然界に見られるウイルスタンパク質に類似しているが、天然には存在しない人工的な最適化ウイルスポリペプチドが作り出される。遺伝的アルゴリズムは、目的とする標的の又は所望の免疫応答に応じて、種々の長さのウイルスポリペプチドを最適化するように調整することができる。ほとんどのT細胞エピトープは9アミノ酸長であるため、本発明の最適化ウイルスポリペプチドの設計に利用した遺伝的アルゴリズムは、所与のウイルス遺伝子産物(例えば、HIV−1 Gag)の各々の連続した9−merアミノ酸配列の最適化に基づいた。この手法に従えば、自然界に存在しない、又は極めて希少である9−mer(例えば)を除外することができる−これは、コンセンサス配列に基づくワクチン戦略と比べたとき、コンセンサス配列は、自然界にはまれにしか、又は全く存在しないいくつかの9−mer(例えば)を含み得るため、改良点である。遺伝的アルゴリズムに用いられる適応度の定義は、最も「適応する」多価カクテルが、集団中の全ての9merのなかで最良の有効範囲(最高の完全マッチ率)を与え、且つ集団に存在しない、又はまれである9merがないという制約を受ける入力ウイルス配列の組み合わせであることである。本発明の最適化ウイルスポリペプチドの生成に用いられる遺伝的アルゴリズムについては、参照により本明細書に援用される国際公開第2007/024941号パンフレットにさらに記載されている。
【0026】
一実施形態において、本発明は、単一の最適化されたHIV−1ポリペプチド(例えば、配列番号1〜29に示されるポリペプチド)を組み込む多価(例えば、二価)HIV−1ワクチンを提供する。別の実施形態において、本発明は、2個以上の最適化されたHIV−1ポリペプチドを組み込む多価ワクチンを特徴とする。いずれの場合も、最適化HIV−1ポリペプチドは、HIV−1メイン(M)群として知られる全体的循環における全てのHIV−1変異体に基づく。本発明者らは、遺伝子当たり2つの変異体(例えば、各々がGag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuについての2つのポリペプチド配列)のみを利用するM群モザイク遺伝子に基づき細胞性免疫の幅及び深さを増大させる最適化HIV−1ポリペプチドのセット(配列番号1〜29)を生成した。本発明者らは、アカゲザルにおいて、多価(例えば、二価)HIV−1 M群ワクチンにこれらの最適化HIV−1ポリペプチドを使用すると、2つの他の有力なワクチン抗原戦略(Mコンセンサス抗原及び最適天然クレードC抗原)と比較したとき有意に大きい幅及び深さのHIV−1特異的細胞性免疫応答が誘発されるという新規の、且つ意外な結果を得た。
【0027】
本発明は、異なるウイルス遺伝子産物に対応する最適化ウイルスポリペプチドの融合を提供する。上記の遺伝的アルゴリズムを用いて、本発明のワクチンに使用される融合ポリペプチドを生成することができる。例えば、Gag/Nef(配列番号19〜20)と、Gag/Pol(配列番号21〜27)と、Gag/Pol/Nef(配列番号28〜29)との最適化HIV−1ポリペプチド融合体を、HIV−1に感染した、又は感染するリスクがある対象(例えば、ヒト)に投与するため本発明のベクターに組み込むことができる。本発明のワクチンは(ポリペプチド形態であろうと、又は核酸形態であろうと)、例えば、最適クレードC配列(配列番号30〜36)又はコンセンサス配列(配列番号37〜39)などの非「モザイク」ポリペプチド(又はそれぞれ、それをコードする配列)の1つ又は複数も含むことができる。
【0028】
本発明に開示される最適化ウイルスポリペプチドは、Merrifield、J.Amer.Chem.Soc.85:2149頁(1963年)によって記載されるものなどの化学合成技法により従来どおり調製することができる(また、例えば、Stemmerら、164 Gene 49(1995年)も参照のこと)。例えば、ワクチンは、固相ペプチド合成(SPPS)を用いて容易に調製することができる。自動化された固相合成を、Applied Biosystems ABI 433Aペプチドシンセサイザーなどの、数多くの周知されている市販の自動シンセサイザーのいずれか一つを使用して行うことができる。或いは、本発明の最適化ウイルスポリペプチドは、最適化ウイルスポリペプチドの細胞内発現を可能にする核酸又はベクター(例えば、アデノウイルスなどのウイルスベクター)を細胞又は生体にトランスフェクト又は形質導入することにより組換え産生することができる。本発明の最適化ウイルスポリペプチドのヌクレオチド配列をコードする核酸及びベクターは、本明細書に記載されるものを含めた公知の組換えDNA技法により合成することができる。
【0029】
本発明のワクチン
本発明はまた、ウイルス(例えば、HIV−1)に感染した、又は感染するリスクがある患者に投与することのできるワクチンも特徴とする。本発明のワクチンは、本明細書において考察されるとおりの、本発明の最適化ウイルスポリペプチドの少なくとも1つを含む。本発明のワクチンは、本発明の2個以上の最適化ウイルスポリペプチドのヌクレオチド配列(例えば、組換えの(例えば、サブユニット)又は生体全体の(例えば、ウイルス全体)ウイルスベクターの免疫原性成分)をコードする核酸であってもよい。核酸は、本発明の2個以上の最適化ウイルスポリペプチドのヌクレオチド配列を組み込むベクター(例えば、アデノウイルスなどのウイルスベクター)を含む。本発明の最適化ウイルスポリペプチド、並びに最適化ウイルスポリペプチドを組み込むワクチン、核酸、及びベクターは、細胞若しくは生体において組換え発現させることができ、又はウイルスに感染した、若しくは感染するリスクがある対象(例えば、ヒト)に直接投与することができる。
【0030】
本発明のベクター
本発明はまた、本発明の1つ又は複数の最適化ウイルスポリペプチドのヌクレオチド配列(例えば、DNA又はRNA)をコードするベクターも特徴とする。ベクターは、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするヌクレオチド配列を含む担体(例えば、リポソーム)、プラスミド、コスミド、酵母人工染色体、又はウイルスであってもよい。ベクターは、いくつかの供給源由来のさらなる核酸配列を含むことができる。
【0031】
本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするベクターは、当該技術分野において公知の任意の組換え分子生物学的技法を用いて作成することができる。ベクターは、標的細胞又は生体のトランスフェクション又は形質導入後、染色体外にあってもよく、又は宿主細胞の染色体に組み込まれてもよい。ベクターの核酸成分は、標的細胞当たりのコピー数が単一又は複数であってよく、及び直鎖状、環状、又はコンカテマー状であってよい。
【0032】
本発明のベクターはまた、単一の核酸転写産物からの複数のペプチド鎖又はポリペプチド鎖の発現を可能にする内部リボソーム侵入部位(IRES)配列を含むこともできる。例えば、本発明のベクターは、本発明の1つ又は複数の最適化ウイルスポリペプチド並びに別のポリペプチド(例えば、緑色蛍光タンパク質(GFP)などの検出可能標識)をコードすることができる。
【0033】
本発明のベクターは、本発明の最適化ウイルスポリペプチドの発現を促進する遺伝子発現エレメントをさらに含む。本発明の最適化ウイルスポリペプチドをコードするベクターの発現に有用な遺伝子発現エレメントとしては、限定はされないが、(a)ウイルス転写プロモーター並びにそのエンハンサーエレメント、例えば、SV40初期プロモーター、ラウス肉腫ウイルスLTR、及びモロニーマウス白血病ウイルスLTRなどの調節配列;(b)SV40後期領域に由来するものなどのスプライス領域及びポリアデニル化部位;及び(c)SV40などにおけるポリアデニル化部位が挙げられる。また、プラスミド複製起点、抗生物質耐性又は選択遺伝子、多重クローニング部位(例えば、制限酵素切断位置)、及び他のウイルス遺伝子配列(例えば、HIV末端反復配列(LTR)などの、ウイルスの構造、機能、又は調節エレメントをコードする配列)も含む。
【0034】
本発明のベクターはまた、例えば、配列番号11、14〜18、及び23のいずれか一つなどの、ヒトにおける発現用に最適化されている本発明の最適化ウイルスポリペプチドを含むこともできる。
【0035】
本発明のベクターはまた、以下の酵素切断部位:XbaI−EcoRI−Kozak−Start...Stop−BamHI−NheI;及び以下の配列:TCTAGA GAATTC GCCACC[ATG遺伝子TAA TGA]GGATCC GCTAGCを有する多重クローニング部位(MCS)を含むように改変することもできる。このMCSを有するベクターは、内部的なXbaI、EcoRI、BamHI、NheI部位を有しないとともに、6個以上のC又はGのストレッチを有しない最適化ウイルスポリペプチドと共に用いることができる。
【0036】
生体内投与
本発明は、本発明の1つ又は複数のワクチン(例えば、本発明の2個以上の最適化ウイルスポリペプチドをコードするベクター)を対象(例えば、ヒト)に生体内投与して本発明の2個以上の最適化ウイルスポリペプチドの発現を促進する方法を特徴とする。ワクチンを対象に投与すると、ウイルス免疫原に対する防御的又は治療的免疫応答(例えば、細胞性又は体液性免疫応答)を誘発することのできる本発明の1つ又は複数の最適化ウイルスポリペプチドが発現する。
【0037】
いくつかのタイプのベクターを用いて、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするヌクレオチド配列を対象(例えば、ヒト)に直接送達することができる。本発明のベクターとしては、ウイルス、ネイキッドDNA、オリゴヌクレオチド、カチオン性脂質(例えば、リポソーム)、カチオン性ポリマー(例えば、ポリソーム)、ビロソーム、及びデンドリマーが挙げられる。本発明は、細胞(例えば、血球細胞)のエキソビボでのトランスフェクション又は形質導入を提供し、続いてそれらの細胞をドナー対象に投与し戻すことで、免疫原特性を有する本発明の最適化ウイルスポリペプチドの発現が可能となる。本発明の方法により単離し、エキソビボでトランスフェクト又は形質導入することのできる細胞としては、限定はされないが、血球細胞、皮膚細胞、線維芽細胞、内皮細胞、骨格筋細胞、肝実質細胞、前立腺上皮細胞、及び血管内皮細胞が挙げられる。幹細胞もまた、本発明のベクターの形質導入又はトランスフェクションに適した細胞である。本発明の方法によれば、骨髄前駆細胞及び造血幹細胞(HSC)を含む全能性、多能性、多分化能、又は単能性幹細胞を単離して、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするベクターをトランスフェクト又は形質導入し、対象に投与することができる。
【0038】
本発明の最適化ウイルスベクターを発現させるために用いられるトランスフェクション又は形質導入の方法は、トランスフェクト又は形質導入細胞における、及びそれに続いて細胞の投与を受ける対象におけるタンパク質発現の強度及び寿命に強い影響を有する。本発明は、本質的に一時的なベクター(例えば、アデノウイルスベクター)又は長命のベクター(例えば、レトロウイルスベクター)を提供する。タンパク質発現の調節に用いることのできる調節配列(例えば、プロモーター及びエンハンサー)が、当該技術分野において公知である。トランスフェクト又は形質導入される細胞のタイプもまた、タンパク質発現の強度及び寿命と強い関連を有する。例えば、代謝回転速度が高い細胞型は、タンパク質発現の期間がより短いと予想され得る。
【0039】
エキソビボトランスフェクション及び形質導入
本発明はまた、細胞(例えば、リンパ球などの血球細胞)をエキソビボでトランスフェクト及び形質導入し、続いてそれらの細胞を対象(例えば、ヒト)に投与する方法も特徴とする。一実施形態において、細胞は治療を受ける対象の自己由来である。細胞は、本発明の1つ又は複数の最適化ウイルスポリペプチドのヌクレオチド配列をコードする1つ又は複数のベクターをエキソビボでトランスフェクト又は形質導入することにより、治療対象における最適化ウイルスポリペプチドの一時的又は永久的な発現が可能なものとすることができる。このような修飾細胞を対象に投与すると、ウイルス免疫原に対する防御的又は治療的免疫応答(例えば、細胞性又は体液性免疫応答)を誘発することのできる本発明の1つ又は複数の最適化ウイルスベクターが発現する。
【0040】
いくつかのタイプのベクターを用いて、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするヌクレオチド配列を細胞(例えば、リンパ球などの血球細胞)に送達することができる。本発明のベクターとしては、ウイルス、ネイキッドDNA、オリゴヌクレオチド、カチオン性脂質(例えば、リポソーム)、カチオン性ポリマー(例えば、ポリソーム)、ビロソーム、及びデンドリマーが挙げられる。本発明は、細胞(例えば、血球細胞)のエキソビボでのトランスフェクション又は形質導入を提供し、続いてそれらの細胞をドナー対象に投与し戻すことで、免疫原特性を有する本発明の最適化ウイルスポリペプチドの発現が可能となる。本発明の方法により単離し、エキソビボでトランスフェクト又は形質導入することのできる細胞としては、限定はされないが、血球細胞、皮膚細胞、線維芽細胞、内皮細胞、骨格筋細胞、肝実質細胞、前立腺上皮細胞、及び血管内皮細胞が挙げられる。幹細胞もまた、本発明のベクターの形質導入又はトランスフェクションに適した細胞である。本発明の方法によれば、骨髄前駆細胞及び造血幹細胞(HSC)を含む全能性、多能性、多分化能、又は単能性幹細胞を単離して、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするベクターをトランスフェクト又は形質導入し、対象に投与することができる。
【0041】
本発明の最適化ウイルスベクターを発現させるために用いられるトランスフェクション又は形質導入の方法は、トランスフェクト又は形質導入細胞における、及びそれに続いて細胞の投与を受ける対象におけるタンパク質発現の強度及び寿命に強い影響を有する。本発明は、本質的に一時的なベクター(例えば、アデノウイルスベクター)又は長命のベクター(例えば、レトロウイルスベクター)を提供する。タンパク質発現の調節に用いることのできる調節配列(例えば、プロモーター及びエンハンサー)が、当該技術分野において公知である。トランスフェクト又は形質導入される細胞のタイプもまた、タンパク質発現の強度及び寿命と強い関連を有する。例えば、代謝回転速度が高い細胞型は、タンパク質発現の期間がより短いと予想され得る。
【0042】
ウイルスベクター
本発明の1つ又は複数の最適化ウイルスポリペプチドのヌクレオチド配列をコードするウイルスベクターは、本発明のワクチンとして使用することができる。例えば、本発明の1つ又は複数の最適化ウイルスポリペプチドのヌクレオチド配列は、対象の形質導入(例えば、生体内投与)又は対象から単離された細胞の形質導入(例えば、エキソビボ形質導入と、それに続く対象に細胞を戻す投与)に好適な天然の、又は修飾された(例えば、弱毒化された)ウイルスゲノムのヌクレオチド配列に組換え的に挿入することができる。ウイルスにさらなる修飾を加えることにより、感染力又はトロピズムを高め(例えば、シュードタイピング)、複製能を低下させ、若しくは除去し、又はウイルス成分の免疫原性(例えば、免疫原性ワクチン剤に関連しないあらゆる成分)を低下させることができる。本発明のベクターは形質導入細胞により発現し、細胞外空間に分泌されるか、又は発現細胞に(例えば、細胞内分子として、又は細胞表面に提示されて)留まることができる。キメラ又はシュードタイプのウイルスベクターを細胞の形質導入に使用して、本発明の1つ又は複数の最適化ウイルスポリペプチドの発現を可能にすることもできる。例示的ベクターを以下に記載する。
【0043】
アデノウイルス
組換えアデノウイルスは、本発明の1つ又は複数の最適化ウイルスポリペプチドを発現させるベクターとしての使用に対し、いくつかの重要な利点を提供する。このウイルスは高力価に調製することができ、非複製細胞に感染することができ、及び標的細胞集団との接触後、エキソビボでの標的細胞の高効率の形質導入をもたらすことができる。さらに、アデノウイルスは、そのDNAを宿主ゲノムに組み込むことがない。従って、それを発現ベクターとして使用すると、自発的増殖性疾患が惹起されるリスクが低減する。動物モデルでは、アデノウイルスベクターは、概して高レベル発現を約1週間にわたり媒介することが分かっている。導入遺伝子発現(本発明の最適化ウイルスポリペプチドをコードする核酸の発現)の持続期間は、細胞又は組織特異的プロモーターを使用することにより延ばすことができる。アデノウイルスベクターそれ自体の分子改変における他の改善点として、より持続的な導入遺伝子発現及び炎症低下がもたらされている。これは、さらなる初期アデノウイルス遺伝子に特異的突然変異を内包するいわゆる「第二世代」ベクター、及びCre−Lox戦略を利用して事実上全てのウイルス遺伝子を欠失させる「ガットレス」ベクターに見られる(Engelhardtら、Proc.Natl.Acad.Sci.USA 91:6196頁(1994年)及びKochanekら、Proc.Natl.Acad.Sci.USA 93:5731頁(1996年)、各々参照により本明細書に援用される)。
【0044】
国際公開第2006/040330号パンフレット及び国際公開第2007/104792号パンフレット(各々参照により本明細書に援用される)に開示されるまれな血清型及びキメラアデノウイルスベクターは、本発明のベクターとして特に有用である。例えば、組換えアデノウイルスrAd26、rAd34、rAd35、rAd48、及びrAd5HVR48は、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードすることができる。本発明の最適化ウイルスポリペプチドをコードする1つ又は複数の組換えウイルスベクターを対象に投与して、ウイルス感染症を治療又は予防することができる。
【0045】
アデノ随伴ウイルス(AAV)
非病原性パルボウイルスに由来するアデノ随伴ウイルス(rAAV)もまた、そのベクターは抗ベクター細胞性免疫応答をほぼ誘起せず、及びほとんどの実験系において数ヶ月持続する導入遺伝子発現をもたらすため、本発明の最適化ウイルスポリペプチドの発現に使用することができる。
【0046】
レトロウイルス
レトロウイルスは、本発明の最適化ウイルスポリペプチドの発現に有用である。アデノウイルスとは異なり、レトロウイルスゲノムはRNAベースである。レトロウイルスは、細胞に感染すると、そのRNAをいくつかの酵素と共に細胞に導入する。レトロウイルス由来のウイルスRNA分子は、逆転写と呼ばれる過程を介してプロウイルスと呼ばれる二本鎖DNAコピーを産生する。細胞核内に輸送された後、プロウイルスDNAは宿主細胞の染色体に組み込まれ、形質導入細胞及びその細胞から派生し得る任意の子孫細胞のゲノムを永久的に変化させる。遺伝子を細胞又は生体に永久的に導入する能力は、遺伝子治療に使用されるレトロウイルスを特徴付ける特性である。レトロウイルスとしては、ウイルス感染及びプロウイルスの組み込みを促進するいくつかのアクセサリータンパク質を含むヒト免疫不全ウイルス(HIV)を含むウイルスファミリーであるレンチウイルスが挙げられる。現在、「第三世代」レンチウイルスベクターは、完全な複製無能力、広いトロピズム、及び哺乳動物細胞に対する高い遺伝子導入能を特徴とする(例えば、Mangeat及びTrono、Human Gene Therapy 16(8):913頁(2005年)及びWiznerowicz及びTrono、Trends Biotechnol.23(1):42頁(2005年)(各々参照により本明細書に援用される)を参照)。
【0047】
他のウイルスベクター
アデノウイルス及びレトロウイルスベクターに加えて、細胞(例えば、リンパ球などの血球細胞)又は対象(例えば、ヒト)における本発明の最適化ウイルスポリペプチドの発現に使用することのできる他のウイルスベクター及び技法が、当該技術分野において公知である。こうしたウイルスとしては、ポックスウイルス(例えば、ワクシニアウイルス及び修飾ワクシニアウイルスのアンカラすなわち(MVA);例えば、米国特許第4,603,112号明細書及び同第5,762,938号明細書(各々参照によって本明細書に援用される)を参照)、ヘルペスウイルス、トガウイルス(例えば、ベネズエラウマ脳炎ウイルス;例えば、米国特許第5,643,576号明細書(参照によって本明細書に援用される)を参照)、ピコルナウイルス(例えば、ポリオウイルス;例えば、米国特許第5,639,649号明細書(参照によって本明細書に援用される)を参照)、バキュロウイルス、及びWattanapitayakul及びBauerにより記載される他のもの(Biomed.Pharmacother.54:487頁(2000年)(参照によって本明細書に援用される))が挙げられる。
【0048】
その他の発現ベクター:ネイキッドDNA及びオリゴヌクレオチド
本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするネイキッドDNA又はオリゴヌクレオチドもまた、細胞(例えば、リンパ球などの血球細胞)又は対象(例えば、ヒト)におけるそうしたポリペプチドの発現に使用することができる。例えば、Cohen、Science 259:1691−1692頁(1993年);Fynanら、Proc.Natl.Acad.Sci.USA、90:11478頁(1993年);及びWolffら、BioTechniques 11:474485頁(1991年)(各々参照により本明細書に援用される)を参照のこと。これは、非ウイルス性のトランスフェクションの最も単純な方法である。電気穿孔、並びに高圧ガス及び担体粒子(例えば、金)を使用してDNAコーティングされた金粒子を細胞に撃ち込む「遺伝子銃」の使用などの、ネイキッドDNAを送達する効率的な方法が存在する。
【0049】
リポプレックス及びポリプレックス
本発明の最適化ウイルスポリペプチドをコードする核酸の細胞又は対象への送達を向上させるため、リポプレックス(例えば、リポソーム)及びポリプレックスを使用して、トランスフェクション過程における望ましくない分解からベクターDNAを保護することができる。プラスミドDNAは、ミセル又はリポソームのように組織化された構造の脂質で被覆することができる。組織化された構造がDNAと複合体を形成すると、リポプレックスと称される。3タイプの脂質、すなわちアニオン性(負に帯電している)、中性、又はカチオン性(正に帯電している)がある。カチオン性脂質を利用するリポプレックスは、遺伝子導入に有用であることが分かっている。カチオン性脂質は正電荷であるため、負に帯電したDNAと自然に複合体を形成する。また、その電荷の結果として、カチオン性脂質は細胞膜と相互作用し、リポプレックスのエンドサイトーシスが起こり、細胞質中にDNAが放出される。カチオン性脂質はまた、細胞によるDNAの分解も保護する。
【0050】
ポリマーのDNAとの複合体はポリプレックスと称される。ほとんどのポリプレックスはカチオン性ポリマーからなり、その生成はイオン相互作用により調節される。ポリプレックスとリポプレックスとの働き方の一つの大きい違いは、ポリプレックスはそのDNA負荷を細胞質中に放出することができず、従ってそれを目的として不活化アデノウイルスなどのエンドソーム溶解剤のコトランスフェクションを(エンドサイトーシスの間に生じたエンドソームを溶解させるため)行わなければならない。しかしながら、必ずしもこれが常に該当するわけではない;ポリエチレンイミンなどのポリマーは、キトサン及びトリメチルキトサンがそうであるように、その独自のエンドソーム破壊方法を有する。
【0051】
本発明の最適化ウイルスポリペプチドをコードする核酸と組み合わせてリポプレックス、又はポリプレックスを形成するために使用することのできる例示的なカチオン性脂質及びポリマーとしては、限定はされないが、ポリエチレンイミン、リポフェクチン、リポフェクタミン、ポリリジン、キトサン、トリメチルキトサン、及びアルギン酸塩が挙げられる。
【0052】
ハイブリッド方法
遺伝子導入のいくつかのハイブリッド方法は、2つ以上の技法を組み合わせる。例えばビロソームは、リポプレックス(例えば、リポソーム)を不活化ウイルスと組み合わせる。この手法は、単独でのウイルスによる方法又はリポソームによる方法のいずれと比べても、呼吸上皮細胞においてより効率的な遺伝子導入をもたらすことが示されている。他の方法は、他のウイルスベクターをカチオン性脂質又はハイブリッド形成ウイルスと混合することを含む。これらの方法の各々を使用して、細胞(例えば、リンパ球などの血球細胞)又は対象(例えば、ヒト)への本発明の最適化ウイルスポリペプチドをコードする核酸の導入を促進することができる。
【0053】
デンドリマー
また、デンドリマーを使用して、本発明の最適化ウイルスポリペプチドをコードする核酸を細胞(例えば、リンパ球などの血球細胞)又は対象(例えば、ヒト)に導入してもよい。デンドリマーは、球形状の高度に枝分かれした巨大分子である。粒子の表面は、多くの方法で機能化することができ、得られるコンストラクトの特性の多くが、その表面によって決定される。特に、カチオン性デンドリマー(すなわち正の表面電荷を有するもの)を作成することが可能である。DNA又はRNAなどの遺伝物質が存在するとき、電荷の相補性により核酸のカチオン性デンドリマーとの一時的な結合がもたらされる。その目標に達すると、次にデンドリマー−核酸複合体は、エンドサイトーシスを介して細胞に取り込まれる。
【0054】
生体内投与
本発明はまた、対象(例えば、ヒト)を本発明のワクチンで免疫するためのインビボ方法も特徴とする。一実施形態において、本発明の1つ又は複数のワクチンを対象に直接投与して、ウイルス(例えば、HIV−1)に対する防御的又は治療的免疫応答(例えば、細胞性又は体液性免疫応答)を誘発することができる。或いは、上記のとおりの、本発明の1つ又は複数の最適化ウイルスポリペプチドをコードするベクターを対象に直接投与することにより、ウイルス感染症を予防又は治療することができる。1つ又は複数の細胞を生体内に効率的にトランスフェクト又は形質導入するベクター(例えば、ウイルスベクター)は、治療を受ける対象において広域で耐久性があり、且つ強力な免疫応答を誘発することができる。発現ベクターの核酸成分が宿主細胞(例えば、リンパ球などの血球細胞)に導入されると、宿主細胞は本発明のワクチンを産生して提示又は分泌し、次にそれが抗原提示細胞(APC)、T細胞、及びB細胞などの免疫系の成分を活性化する働きをし、その結果、免疫が構築される。
【0055】
医薬組成物
本発明は、1つ又は複数の薬学的に許容可能な賦形剤、希釈剤、緩衝剤、又は他の許容可能な担体と組み合わせた、本発明のワクチン、ベクター、及び最適化ウイルスポリペプチドを特徴とする。ワクチン、ベクター、又は最適化ウイルスポリペプチドの製剤化は、有効量の最適化ウイルスポリペプチド免疫原の発現を利用し、又は可能にし得る。すなわち、そこには、後のウイルス(例えば、HIV−1)に対する曝露からの防御を対象に提供し、又は既存のウイルス感染症を治療するための、特定の十分な免疫応答を治療対象(例えば、ヒト)に生じさせ得る量の抗原が含まれ得る。例えば、本発明のワクチンの製剤化により、広域の且つ特異的な細胞性免疫応答を対象に生じさせ得る量の抗原を発現させることが可能となり得る。本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドで治療される対象はまた、対象に防御的又は治療的利益を付与することのできる抗ウイルス抗体(例えば、中和抗体)を産生することもできる。本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドは、単独で、又は当該技術分野において公知の任意の薬学的に許容可能な担体、塩又は補助剤と組み合わせて、対象に直接投与することができる。
【0056】
薬学的に許容可能な塩は、製薬業界で一般に用いられている非毒性の酸付加塩又は金属錯体を含み得る。酸付加塩の例としては、酢酸、乳酸、パモン酸、マレイン酸、クエン酸、リンゴ酸、アスコルビン酸、コハク酸、安息香酸、パルミチン酸、スベリン酸、サリチル酸、酒石酸、メタンスルホン酸、トルエンスルホン酸、又はトリフルオロ酢酸などの有機酸;タンニン酸、カルボキシメチルセルロースなどのポリマー酸;及び塩酸、臭化水素酸、硫酸、リン酸などの無機酸が挙げられる。金属錯体としては、亜鉛、鉄などが挙げられる。薬学的に許容可能な担体の例示的な一つは、生理食塩水である。他の生理学的に許容可能な担体及びその製剤は当業者に公知であり、例えば、「Remington’s Pharmaceutical Sciences」、(第18版)、A.Gennaro編、1990年、Mack Publishing Company、Easton、PAに記載されている。
【0057】
予防上又は治療上有効量の本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの医薬製剤は、経口的に、非経口的に(例えば、筋肉内、腹腔内、静脈内、又は皮下注射、吸入、皮内、点眼、又は植え込み)、経鼻的に、経膣的に、経直腸的に、舌下に、又は局所的に、投与経路に適合した薬学的に許容可能な担体と混合して投与することができる。製剤中における本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの濃度は、約0.1〜100wt.%まで様々であり得る。
【0058】
本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドを含有する組成物の非経口投与製剤は、滅菌水性又は非水性溶液、懸濁液、又はエマルジョンを含む。好適な媒体の例としては、プロピレングリコール、ポリエチレングリコール、植物油、ゼラチン、水素化ナフタレン、及び注射用の有機エステル、例えばオレイン酸エチルが挙げられる。かかる製剤はまた、防腐剤、湿潤剤、乳化剤、及び分散剤などの補助剤を含有してもよい。生体適合性、生分解性のラクチドポリマー、ラクチド/グリコリドコポリマー、又はポリオキシエチレン−ポリオキシプロピレンコポリマーを使用して化合物の放出を制御してもよい。本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドを含有する組成物に潜在的に有用な他の非経口送達系としては、エチレン−酢酸ビニルコポリマー粒子、浸透圧ポンプ、植込み型注入システム、及びリポソームが挙げられる。
【0059】
液体製剤は、例えば、細菌保持フィルタを介したろ過によるか、組成物に滅菌剤を組み込むことによるか、又は組成物を照射若しくは加熱することにより、滅菌することができる。或いはまた、液体製剤は、使用直前に滅菌水又は他の何らかの無菌注射用媒質中に溶解することのできる無菌固形組成物の形態で製造されてもよい。
【0060】
直腸投与又は経膣投与用の本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドを含有する組成物は、好ましくは、作用物質に加えてカカオ脂又は坐薬ワックスなどの賦形剤を含有し得る坐薬である。経鼻投与又は舌下投与用の組成物もまた、当該技術分野において公知の標準的な賦形剤と共に調製される。吸入用製剤は、賦形剤、例えばラクトースを含有してもよく、又は点鼻液若しくは鼻内スプレーの形態で、又はゲルとして投与するため、例えばポリオキシエチレン−9−ラウリルエーテル、グリココール酸塩及びデオキシコール酸塩を含有する水溶液であってもよく、又は油性溶液であってもよい。
【0061】
本発明の組成物中における活性成分の量は様々であり得る。当業者は、投与されるペプチド、投与時間、投与経路、製剤の性質、排泄速度、対象の病態の性質、並びに患者の年齢、体重、健康状態、及び性別を含めた様々な要因に依存して、正確な個々の投薬量はいくらか調整され得ることを理解するであろう。加えて、ワクチン、ベクター、又は最適化ウイルスポリペプチドによって治療される病態の重症度もまた、投薬量レベルに影響を有し得る。一般に、0.1μg/kg体重〜100mg/kg体重の投薬量レベルが、単回用量として、又は複数回用量に分割して毎日投与される。好ましくは、一般的な投薬量範囲は、1日当たり250μg/kg体重〜5.0mg/kg体重である。様々な投与経路の種々の効率にかんがみて、必要な投薬量は幅広く異なることが予想される。例えば、経口投与は、概して静脈注射による投与より高い投薬量レベルが要求されると予想され得る。このような種々の投薬量レベルは、当該技術分野において公知の最適化のための標準的な実験ルーチンを用いて調整することができる。一般に、正確な予防上又は治療上の有効投薬量は、担当臨床医が上記に特定した要因を考慮して決定することができる。
【0062】
患者に投与される各用量中に存在する本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの量は、患者の年齢、体重、性別、全般的な健康状態などの考慮事項に関連して選択される。患者において深刻な有害副作用なしに免疫応答(例えば、細胞性免疫応答)を誘起する、又は外因性効果を生じさせるために必要なワクチン、ベクター、又は最適化ウイルスポリペプチドの量は、用いられる医薬組成物及びアジュバントの任意の存在に応じて異なる。場合により、初回用量に続き、所望の場合には追加免疫を繰り返すことができる。本方法は、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの長期投与を含むことができる。治療的用途又は予防的用途のため、年一回の追加免疫又は他の間隔での追加免疫など、免疫ワクチン、ベクター、又は最適化ウイルスポリペプチドの反復投薬量が望ましいこともある。投与される投薬量は、当然ながら、特定のワクチン、ベクター、又は最適化ウイルスポリペプチドの薬力学的特性、並びにその投与様式及び経路;被投与者の年齢、健康状態、及び体重;症状の性質及び程度、併用治療の種類、治療頻度、及び所望の効果などの公知の要因に応じて異なり得る。本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドは、針刺しによる急性感染又は母体感染のリスクがある対象の長期治療において投与することができる。かかる「急性」感染についての投薬頻度は、約6週間の期間にわたる静脈内又は筋肉内による1日1回投薬から週1回又は2回の範囲であってもよい。ワクチン、ベクター、又は最適化ウイルスポリペプチドはまた、ウイルス(例えば、HIV−1)による感染患者、又は感染が進行している患者の長期治療においても用いることができる。感染患者では、長期投与の頻度は、静脈内又は筋肉内で1日1回投薬から週1回又は2回の範囲であってもよく、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチド中に存在する免疫原の半減期に依存し得る。
【0063】
アジュバント
ウイルスに対するワクチンを必要とする哺乳動物(例えば、ヒト)のワクチン接種に使用される本発明のワクチンは、ワクチンの免疫原性を高める1つ又は複数の薬学的に許容可能なアジュバントと同時に、又は連続して投与することができる。ヒトへの使用が承認されているアジュバントとしては、アルミニウム塩(ミョウバン)が挙げられる。こうしたアジュバントは、B型肝炎、ジフテリア、ポリオ、狂犬病、及びインフルエンザを含むいくつかのワクチンに有用となっている。他の有用なアジュバントとしては、完全フロイントアジュバント(CFA)、不完全フロイントアジュバント(IFA)、ムラミルジペプチド(MDP)、MDPの合成類似体、N−アセチルムラミル−L−アラニル−D−イソグルタミル−L−アラニン−2−[1,2−ジパルミトイル−s−グリ−セロ−3−(ヒドロキシホスホリルオキシ)]エチルアミド(MTP−PE)並びに代謝可能な油及び乳化剤であって、実質的に全てが直径1ミクロン未満の油滴を有する水中油エマルジョンの形態で存在する油及び乳化剤を含有する組成物が挙げられる。
【0064】
キット
本発明は、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドと、薬学的に許容可能な担体とを含む医薬組成物を、ウイルス感染症の予防又は治療に治療上有効な量で含むキットを提供する。キットは、臨床従事者(例えば、医師又は看護師)がそこに含まれる組成物を投与することができるように説明書を含む。
【0065】
好ましくは、キットは、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの有効量を含有する1つ又は複数の単回用量医薬組成物の複数のパッケージを含む。場合により、1つ又は複数の医薬組成物の投与に必要な器具又は装置がキットに含まれてもよい。例えば、本発明のキットは、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドの有効量を含む1つ又は複数のプレフィルドシリンジを提供してもよい。さらに、キットはまた、ウイルスに感染した、又は感染するリスクがある患者が、本発明のワクチン、ベクター、又は最適化ウイルスポリペプチドを含有する1つ又は複数の医薬組成物を使用するための説明書又は投与スケジュール表などのさらなる要素を含んでもよい。
【0066】
当業者には、本発明の趣旨及び範囲から逸脱することなく本発明の組成物、方法、及びキットに様々な修正及び変更を加え得ることは明らかであろう。従って、本発明は、本発明の修正及び変更を、それらが添付の特許請求の範囲及びその均等物の範囲内に含まれることを条件として包含することが意図される。
【実施例】
【0067】
以下の例により本発明を説明し、これらの例は、何ら本発明を限定することを意図するものではない。
【0068】
〔実施例1〕
上記で考察される遺伝的アルゴリズムを使用してモザイク抗原Gag、Pol、Nef、及びEnv配列(配列番号1〜8)を作成した。次にそれらの配列をワクチン開発に実用的なものとするため、Env(配列番号9〜11)において切断/融合活性を除去し、Pol(配列番号12〜14)において触媒活性を除去し、Nef(配列番号16〜18)においてミリスチル化部位を除去し、及びGagNef、GagPol、又はGagPolNef(配列番号19〜29)を含む融合コンストラクトを作成することにより修飾した。コンパレータ最適天然クレードC遺伝子もまた示される(配列番号30〜36)。
【0069】
〔実施例2〕
Mコンセンサス(第1群)、二価Mモザイク(第2群)、又は最適天然クレードC(第3群)配列由来のGag、Pol、及びEnv遺伝子を発現する3×1010個のvp rAd26ベクターで20匹のアカゲザルを免疫した。Mコンセンサス配列は、世界中の循環ウイルスの唯一最良の「平均」を表す合成配列に相当する。二価Mモザイク配列は上記に記載される。最適天然クレードC配列は、特性が最も「コンセンサス様」である実際のクレードC HIV−1ウイルス由来の天然に存在する配列である。全潜在的T細胞エピトープ(PTE)ペプチドセットからの応答ペプチド数を調べることにより、細胞性免疫幅を評価した。PTEペプチドは、全HIV−1配列の85%超に相当し、NIHから自由に入手することができる。
【0070】
結果は、新規二価Mモザイク配列が、これらの他の2つの有力な抗原コンセプトより著しく優れていたことを示す。表1に示されるとおり、二価Mモザイク抗原は、Mコンセンサス抗原及び最適天然クレードC抗原と比較して、Gag特異的、Env特異的、Pol特異的、及び合計Tリンパ球の応答幅の有意な増加を誘発した。(平均値は、各サル群における平均エピトープ数を表す;SEMは平均値の標準誤差を表す)。
【表1】
【0071】
〔実施例3〕
実施例2に記載されるMコンセンサス(第1群;n=7)、二価Mモザイク(第2群;n=7)、又は最適天然クレードC(第3群;n=6)配列由来のGag、Pol、及びEnv遺伝子を発現する3×1010個のvp rAd26ベクターでマカクザルを筋肉内免疫した。全潜在的T細胞エピトープ(PTE)ペプチドセットからの応答ペプチド数を調べることにより、細胞性免疫幅を評価した。
【0072】
読み出す情報として、プールPTEペプチドに対するCD4/CD8 IFNγ Elispot応答(大きさ)を評価した。15merのPTEペプチドを用いてエピトープを完全にマッピングし、陽性の数(陽性は、106個のPBMC当たり55個のスポット形成細胞(SFC)及びバックグラウンドの4倍として定義した)を評価した。完全なタンパク質のセットと応答を比較するため、5個のGagタンパク質にわたる重複ペプチドのプールセットも試験した。
【0073】
結果は、二価Mモザイク配列が、他の2つの有力な抗原コンセプト(Mcon及びOptC)より著しく優れていたことを示す。
【0074】
〔実施例4〕
モデリングを使用して、T細胞応答がモザイクワクチンの結果として増加するという本発明者らの観察を検証した。反応性ペプチド数をワクチン、ポリペプチド、及びT細胞型の関数として予測し、次に交互作用のステップワイズ除去を行ったポアソン回帰モデルをフィットした。モザイクワクチンは陽性PTE応答数の高度に有意な亢進をもたらしたが、それは、あらゆるポリタンパク質及びT細胞型にわたって多かれ少なかれ一様にもたらされたことが認められた。従って、動物が投与を受けるT細胞、ポリペプチド、及びワクチンのタイプに個別に依存する寄与分を合わせることにより、動物において陽性効果を有するペプチド数を予測し得る。
【0075】
これらのモデルはまた、動物間のばらつきを考慮するため変量効果も含んだ。これは、モデルの予測力を適切に割り当てることによりp値の信頼性を高めるために設計される予防措置である。
【0076】
以下の効果が認められた:
a)CD8応答はCD4応答と比べてはるかに多く、4.37倍である(p<2×10−16);
b)gp160ではgag又はpolと比べて応答が少なく、0.54倍であり(p=0.000830)、及びgagとpolとの間に(polはGagの2倍長く、従って応答する確率がより高いため、配列長さにより正規化した場合であっても)有意差はない;及び
c)モザイクワクチンはMconと比べて有意により多くの陽性応答をもたらし(3.6倍、p=6.26×10−11)、一方、OptCがもたらす応答はより少なく、但しMcon−OptCの差は有意ではない。
【0077】
〔実施例5〕
ワクチンにより誘発され、PTEペプチドにより検出される最小応答数だけを考え、従って変異に関わらず8アミノ酸以上重複する全てのペプチドを1回のみ数える場合にも、モザイクワクチンはなお、別個の領域に対するより多い応答数をもたらす。
【0078】
CD8について、各重複ペプチドセットを1回のみ数える:
統計概要:
Mos2>Mcon〜OptC(MconはOptCより応答が多い傾向を示す)
Mconと比較したMos2についてのウィルコクソンp値:p値=0.0009992
最適Cと比較したMconについてのウィルコクソンp値:p値=0.2351
【0079】
【0080】
CD4について、各重複ペプチドセットを1回のみ数える(CD4には重複はほとんどないため、これは最初のカウントとほぼ同じである)。
【0081】
統計概要:
Mos2>>Mcon〜OptC(MconはOptCより応答が多い傾向を示す)
Mconと比較したMos2についてのウィルコクソンp値:p値=0.00198
最適Cと比較したMconについてのウィルコクソンp値:0.099
【0082】
【0083】
〔実施例6〕各重複ペプチドセットを1回のみ数えるポアソン回帰
重複PTEペプチドを使用して以下を決定したが、これは、各陽性PTE応答を別のものとして数えた上記の実施例4で考察した結果と広く一致している:
a)CD8応答はCD4応答と比べてはるかに多く、約2.8倍である(p≒1×10−7;
b)モザイクワクチンはMconと比べて有意により多くの陽性応答をもたらし(2.84倍、p≒4.3×10−7)、一方OptCがもたらす応答はより少なく、但しMcon−OptCの差は有意ではない;及び
c)Gagに対する応答よりPolに対する応答が多く、及びgp160に対する応答よりGagに対する応答が多いが、有意であったのはPol−gp160の差のみで、約2倍であった(p<0.001)。
【0084】
〔実施例7〕
以下の表は、2モザイク(Mos2)又はMconのワクチン接種を受けた7動物、及び最適天然Cクレード(OptC)のワクチン接種を受けた6動物における3つのワクチンに対するGag、Pol、及びEnv応答に対する応答合計の集計表である:
【0085】
OptCワクチンは、全てのサルにわたり、タンパク質当たりのCD8+ T細胞応答より僅かに少ない平均応答をもたらした。Mconワクチンは、タンパク質当たり約1つの応答を示した。Mos2に関してのみタンパク質の違いが認められ、ここでEnvは、典型的にはGag又はPolのいずれと比べても応答が少ない。
【0086】
Mos2ワクチン中のタンパク質の各々は多くの応答を誘発し、全般的な応答に寄与した。不活化polに対する修飾、並びにEnvにおける切断及び融合ドメインの欠失後のコンセンサスタンパク質の相対的な長さは、以下のとおりであった:Envの671アミノ酸、Polの851、Gagの498(1.35:1.7:1)。
【0087】
要約
幅:2モザイクワクチンは、Mコンセンサス又は単一最適天然株よりはるかに多くのエピトープ領域の認識能を有するT細胞応答を誘発する。
【0088】
深さ:認識されたPTEペプチドの多様性は、2モザイクの双方の型が変異ペプチドに対する異なるT細胞応答を誘発し、交差反応の可能性が増加していることを示唆する。
【0089】
〔実施例8〕
本発明のモザイクHIV−1ワクチンは、アカゲザルにおける細胞性免疫応答の幅及び深さを拡大する。HIV−1 M群配列のPTE有効範囲を最適化したモザイクHIV−1 Gag、Pol、及びEnv抗原を作成した。HIV−1 M群配列は、ロスアラモスHIV−1配列データベース内の全ての主要なHIV−1クレード及び組換え系統を含む。二価モザイク戦略を利用して、理論的な有効範囲と実用性との競合問題について均衡をとった。二価モザイクHIV−1 Gag、Pol、及びEnv抗原は、アカゲザルにおいてコンセンサス配列及び天然配列のHIV−1抗原を使用して観察される免疫応答と比べて、アカゲザルにおけるエピトープ特異的CD8+及びCD4+ Tリンパ球応答の幅及び大きさ(深さ)を実質的に拡大した。
【0090】
27匹の非近交系アカゲザルを、以下の抗原を発現する組換えアデノウイルス血清型26(rAd26)ベクターの単回注射で免疫した:(i)二価モザイク(N=7)、(ii)Mコンセンサス(N=7)、(iii)クレードBとクレードC(N=7)とを組み合わせた二価、又は(iv)最適天然クレードC(N=6)HIV−1 Gag、Pol、及びEnv抗原。これらの抗原を発現するrAd26ベクターの3×1010個のウイルス粒子の全用量を各動物に1回筋肉内投与した。最適クレードC抗原は、ロスアラモスHIV−1配列データベースにおけるクレードC配列の最大PTE有効範囲を提供するように選択した天然株配列であった(以下の「材料及び方法」で考察する)。ワクチンにより誘発されたHIV−1特異的Tリンパ球応答の幅及び大きさ(深さ)を、HIV−1 M群配列の少なくとも15%に存在する全てのPTEを含むペプチドのプール及びサブプールを利用して、免疫後4週目にIFN−γ ELISPOTアッセイにより評価した。全ての個々のペプチド応答を解析し、細胞枯渇IFN−γ ELISPOTアッセイを実施して、反応性ペプチドがCD8+又はCD4+ Tリンパ球エピトープに相当するかどうかを判断した。
【0091】
モザイク抗原により誘発されたPTEペプチドに対するGag特異的、Pol特異的、及びEnv特異的細胞性免疫応答の総数は、コンセンサス又は天然配列抗原により誘起された応答数と比べて3.8倍高かった(図19A;P=1×10−11、モザイクを次に高い群のコンセンサス抗原との比較、ポアソン回帰モデルに基づく)。CD4+ Tリンパ球応答と比べてCD8+は4.4倍多く(P<10−11)、Gag又はPolに対する応答と比べてEnvに対する応答は少なかった(P<0.0007)。CD8+ Tリンパ球応答の中位数はモザイクワクチンについて最も高く、それにコンセンサス、組み合わせのB+C、及び天然クレードCワクチンが続いた(それぞれ、各群における動物当たりの応答の中央値は16、5、3、及び2)。全体ではCD4+ Tリンパ球応答はより少なかったが、CD4+ Tリンパ球応答に関しても、最高数がモザイクワクチンに対するもので、それにコンセンサス、組み合わせのB+C、及び天然クレードCワクチンが続く、同じ相対的パターンが現れた(それぞれ、各群における動物当たりの応答の中央値は4、1、1、及び0.5)。コンセンサス、組み合わせのB+C、及び天然クレードCワクチンにより誘発されたCD8+及びCD4+ Tリンパ球応答の数は、統計的には区別できなかった。
【0092】
PTEペプチドは、天然に存在するHIV−1配列多型を反映する複数の重複配列を含み、従ってPTEペプチド応答は、特定のエピトープの認識(幅)と当該エピトープの変異体の交差認識(深さ)との双方を包含する。サル当たりの反応性エピトープ領域の数を評価することにより幅の保存分析を実施し、ここでは8個以上のアミノ酸が重複する全ての反応性PTEペプチドを1イベントとして数えた。この保存分析においてもなお、モザイク抗原が、コンセンサス抗原又は天然配列抗原と比較したとき3.1倍多いGag、Pol、及びEnv反応性エピトープ領域数を誘発したことが観察された(図19B;P=1.6×10−7、ポアソン回帰)。エピトープ領域は、高いエピトープ密度の領域によって明らかなとおり、動物にわたっていくらかのクラスタ化を示した(図20A〜図20C及び図21A〜図21C)。応答領域ごとに整理した全ての陽性ペプチドの完全アラインメントを図22に示す。
【0093】
これらのデータは、モザイク抗原が、Mコンセンサス及び天然クレードC抗原と比較したとき、細胞性免疫応答の幅を実質的に増加させたことを示す。二価モザイク抗原はまた、クレードB及びクレードC抗原の二価の組み合わせより優れていることも分かり(図19A及び図19B)、幅の亢進がモザイク配列設計に起因したもので、単純にタンパク質当たりに2つの異なる抗原配列を使用したことを反映したものではないことが示された。モザイク抗原により誘起された幅の増加により応答の潜在能力が損なわれたかどうかを判断するため、全ての個々のCD8+及びCD4+ Tリンパ球応答の大きさを評価した。これらの応答の大きさはあらゆる群間で同程度であることが分かった(図23;それぞれP=0.58及びP=0.99、コルモゴロフ−スミルノフの両側検定)。このように、モザイク抗原は、個々のエピトープ特異的応答の大きさを損なうことなく細胞性免疫幅を拡大し、抗原競合及び免疫優性の制約が、この試験のモザイク抗原の免疫原性を制限しなかったことが示された。
【0094】
次に、様々なワクチンレジメンにより誘発された細胞性免疫応答の深さを特徴付けた。特定のエピトープ領域について同時に誘発された変異PTEペプチドの数として深さを定義した。複数の共通のエピトープ変異体に対する応答が誘起されると、感染ウイルス配列の免疫有効範囲は大きくなり、生体内で共通のエスケープ経路が遮断され、又は大きいフィットネスコストを被る三次エスケープ経路にウイルスが押し込まれ得る。コンセンサス及び天然配列抗原は、天然クレードC抗原の投与を受けたサル366における応答により例示されるとおりの、ワクチン配列と反応性PTEペプチドとの間の高度の配列同一性により特徴付けられる応答を誘発した(図24A;また図22も参照)。対照的に、モザイク抗原は、特定のエピトープ領域における複数の反応性PTEペプチドにより特徴付けられる応答を誘発した。これらのペプチドは共通の変異体を表し、多くの場合に、サル361における応答により例示されるとおり、モザイクワクチン配列に含まれる多型を反映した(図24B;また図22も参照)。これらの動物における全てのエピトープ特異的応答についての要約から、コンセンサス又は天然配列抗原と比較したとき、モザイク抗原は、2個以上の標的化された変異体を有するペプチドに対する細胞性免疫応答の頻度を増加させたことが実証される(図24C;P=0.001、モザイクを次に高い群のコンセンサス抗原と比較するウィルコクソン順位和検定)。
【0095】
PTEペプチドを利用する分析を補足するため、5個の異なるGag配列:クレードC DU422、クレードC ZM651、コンセンサスC、コンセンサスA、及びコンセンサスBを包含する従来の重複ペプチドによるワクチン接種を受けたサルにおける細胞性免疫応答の幅も評価した。細胞性免疫幅は、各Gag配列にわたる10個の重複ペプチドのサブプールに対する反応性を評価することにより決定した。モザイク抗原は、コンセンサス又は天然配列抗原と比較したとき、試験した全てのGag配列に対してTリンパ球応答の増幅を誘発した(図25;P=1×10−7、二項回帰)。このように、モザイク抗原は、PTEペプチドに対するだけでなく、クレードA、B、及びC由来の実際のGagペプチドに対する細胞性免疫幅も増大させた。モザイク抗原は、クレードC Gagペプチドに対する応答の誘起について最適天然クレードC抗原より優れていることまでもが分かった。さらに、モザイク抗原は複数のクレード由来のGagペプチドに対する同程度の応答を誘発したが、一方、天然クレードC抗原は、クレードA及びクレードB Gagペプチドに対する応答の低下を示した(図25)。
【0096】
これらの観察の耐久性を評価するため、モザイク、コンセンサス、及び最適天然クレードC抗原の投与を受けたサルを、初回の免疫で利用された配列と一致するHIV−1 Gag、Pol、及びEnv抗原を発現する異種ベクターrAd5HVR48の3×1010個のウイルス粒子の合計用量で40週目に追加免疫した。10個のPTEペプチドのサブプールに対する反応性を4週目(プライミング後)及び44週目(追加免疫後)に評価することにより、細胞性免疫幅を決定した。プライミング免疫後に観察されたCD8+及びCD4+ Tリンパ球応答の大部分が、追加免疫後に拡大し(図26A、赤色及び青色の線)、及び数多くの新規の応答も検出された(図26A、赤色及び青色の点)。44週目、個々の細胞性免疫応答の大きさは群間で同程度であることが分かった(図26A)。しかしながら、モザイク抗原により誘発されたサブプール応答の数は(動物当たりの応答の中央値27)、追加免疫後、コンセンサス抗原(動物当たりの応答の中央値11)又は最適天然クレードC抗原(動物当たりの応答の中央値10)により誘起されたサブプール応答の数と比べて実質的に高いままであった(図26B)。追加免疫の前も、及び後も、モザイクワクチンにより誘発された動物当たりの応答は、コンセンサス又は天然クレードCワクチンによるものより多かった(P<0.001、全てのペアワイズ比較についてのウィルコクソン順位和検定)。
【0097】
また、追加免疫後、ELISA(図26C)及びルシフェラーゼベースのシュードウイルス中和アッセイ(図26D)によりEnv特異的体液性免疫応答も測定した。全ての群がクレードC gp140に対する同程度のELISA力価を示し、第1層クレードCウイルスMW965.26に対する同程度の中和抗体(NAb)応答を示した。モザイク抗原は、コンセンサス又は天然クレードC抗原と比較したとき、第1層クレードBウイルスSF162.LSに対する僅かに高いNab応答を誘発したが(P=0.02、ウィルコクソン順位和検定)、しかしながらどの群においても、第2層ウイルスに対するNAb応答は検出されなかった。
【0098】
本発明者らのデータは、モザイクHIV−1 Gag、Pol、及びEnv抗原が、理論的予想と良好に一致して、アカゲザルにおいてコンセンサス又は天然配列抗原と比較したときにエピトープ特異的細胞性免疫応答の幅及び深さの双方を増大させたことを実証する(図27)。この試験におけるモザイク抗原に関する顕著な結果は、rAd26ベクターが、CD8+ Tリンパ球応答の誘発に特に効率的であるという事実、並びにモザイク抗原がCD8+ Tリンパ球幅の増大に特に有効であるように思われるという事実を反映したものであり得る(図19A及び図19B)。また、モザイク抗原に関してCD4+ Tリンパ球幅の亢進も認められたが、しかしながらそれらの応答数は実質的により少なかった。
【0099】
Gag特異的細胞性免疫応答の幅は、アカゲザルにおけるSIV制御及びヒトにおけるHIV−1制御にとって重要であることが示されている。さらに、第2b相STEP試験において、天然クレードB Gag、Pol、及びNef抗原を発現するrAd5ベースのHIV−1ワクチン候補は、限られた幅のHIV−1特異的細胞性免疫応答しか誘発せず、ワクチン有益性は認められなかった。STEP試験のワクチン被接種体は、Gagに対するわずか1のエピトープ特異的応答の中央値を含め、わずか2〜3のエピトープ特異的Tリンパ球応答の中央値を生じ、この極めて狭い幅の細胞性免疫応答が、感染ウイルスの多様性の不十分な免疫有効範囲をもたらしたように思われる。CD8+ Tリンパ球からのウイルスエスケープもまた、急性HIV−1感染において急速に起こることが報告されており、従って共通のエピトープ変異体に対するワクチンにより誘発された細胞性免疫応答もまた重要であることが分かり得る。まとめると、以上の試験は、細胞性免疫の幅及び深さを増大させるHIV−1ワクチン戦略を開発する必要性を強調するものである。
【0100】
本発明者らはこの試験においてモザイクHIV−1抗原を評価したため、SIV攻撃誘発に対するこうしたワクチンレジメンの防御効力を評価することはできなかった。しかしながら、本発明者らは、rAdベクターにより誘発されたSIV特異的細胞性免疫応答の幅が、アカゲザルにおいてSIV攻撃誘発に対する防御効力と相関していることを以前報告している(Liuら、Nature 457:87頁、2009年)。本発明者らはまた、変異エピトープに対する細胞性免疫応答が、アカゲザルにおいてインビボでSIV突然変異進化を阻止することができることも示しており(Barouchら、Nat.Immunol.6:247頁、2005年)、細胞性免疫の深さの拡大の生物学的関連性が示唆される。非ヒト霊長類におけるSIV攻撃誘発に対するモザイクワクチンの防御効力のモデリングには固有の限界があり、これは、SIVとHIV−1 M群配列とでは観察される多様性が実質的に異なり、影響を受ける基礎となる生物学が異なるためである。例えば、スーティーマンガベイなどの天然宿主におけるCD8+ Tリンパ球選択圧は、ヒトにおけるものと比べて実質的に低いものと思われる。従って、候補HIV−1ワクチンとしてのモザイク抗原のさらなる評価には、臨床治験が役立ち得る。
【0101】
要約すれば、本発明者らは、アカゲザルにおいて二価モザイクHIV−1 Gag、Pol、及びEnv抗原が細胞性免疫幅及び深さを実質的に拡大したことを実証する。全体的なウイルス多様性及び細胞性免疫応答からのウイルスのエスケープは、T細胞ベースのHIV−1ワクチンの開発における決定的な難関に相当するため、こうした知見は、HIV−1ワクチン開発に対する重要な含意を有する。モザイク抗原の二価カクテルはまた、臨床開発が実際的及び潜在的に実現可能でもある。モザイク抗原の結合価が増加すると、有効範囲がさらに向上し得る。最後に、このモザイク抗原戦略は一般化可能であり、HIV−1に加えて他の遺伝的に多様な病原体に利用される可能性がある。
【0102】
材料及び方法
抗原設計及びベクター産生。本質的に、ロスアラモスHIV−1配列データベースにおけるHIV−1 M群配列の最適な有効範囲を提供するように二価モザイクGag、Pol、及びEnv抗原を作成した。ロスアラモスHIV−1配列データベースにおける最適PTE有効範囲のクレードC配列を提供する配列となるように最適天然クレードC抗原を選択した(C.IN.−.70177 Gag、C.ZA.04.04ZASK208B1 Pol、C.SN.90.90SE_364 Env)。準コンセンサス又はコンセンサス配列となるようにクレードB抗原を選択し(B.CAM−1 Gag、B.IIIB Pol、B.Con Env)、それを用いて二価クレードB+Cワクチン手法に対する最適クレードC抗原を補足した。Pol抗原はRT及びINを含んでPRを含まず、及び記載されるとおり触媒活性を除去する点突然変異を含んだ(Priddyら、Clinical infectious diseases 46:1769頁、2008年)。Env gp140抗原は、切断及び融合活性を除去する点突然変異を含んだ。ワクチン配列を図27に示す。組換え複製無能アデノウイルス血清型26(rAd26)及びそれらの抗原を発現するヘキソン−キメラrAd5HVR48ベクターをPER.55K細胞で成長させて、本質的に記載されるとおり二重CsCl勾配沈降により精製した(Abbinkら、J.Virol.81:4654頁、2007年、及びRobertsら、Nature 441:239頁、2006年)。
【0103】
動物及び免疫化。MHCクラスI対立遺伝子Mamu−A*01を発現しなかった27匹の非近交系アカゲザルをNew England Primate Research Center(NEPRC)、Southborough、MAで飼育した。免疫化は、モザイク、Mコンセンサス、クレードB+クレードC、又は最適天然クレードC HIV−1 Gag、Pol、及びEnv抗原を発現する3×1010個のウイルス粒子rAd26又はrAd5HVR48ベクターを、1mlの筋肉内注射として0週目及び40週目に双方の大腿四頭筋に送達することを含んだ。全ての動物試験は、本発明者らの研究機関内の動物の管理及び使用に関する委員会(Institutional Animal Care and Use Committees:IACUC)により承認された。
【0104】
IFN−γ ELISPOTアッセイ。ワクチン接種を受けたサルにおけるHIV−1特異的細胞性免疫応答を、本質的に記載されるとおりインターフェロン−γ(IFN−γ)ELISPOTアッセイにより評価した(Robertsら、Nature 441:239頁、2006年、及びLiuら、Nature 457:87頁、2009年)。HIV−1 M群配列の少なくとも15%に存在するHIV−1 Gag、Pol、及びEnvの潜在的T細胞エピトープ(PTE)ペプチド、並びにクレードC DU422、クレードC ZM651、コンセンサスC、コンセンサスA、及びコンセンサスB株由来のHIV−1 Gagペプチドを、NIH AIDS Research and Reference Reagent Programから入手した。96ウェルマルチスクリーンプレート(Millipore)を、エンドトキシンフリーのダルベッコPBS(D−PBS)中の10μg/ml抗ヒトIFN−γ(BD Biosciences)の100μl/ウェルで一晩コートした。次にプレートを0.25%Tween−20含有D−PBS(D−PBS/Tween)で3回洗浄し、37℃で5%FBS含有D−PBSにより2時間ブロックし、D−PBS/Tweenで3回洗浄し、10%FBS含有RPMI 1640でリンスしてTween−20を取り除き、100μlの反応量で3通りに各2μg/mlのペプチド及び2×105PBMCと共にインキュベートした。37℃で18時間インキュベートした後、プレートをPBS/Tweenで9回及び蒸留水で1回洗浄した。次にプレートを、2μg/mlのビオチン化抗ヒトIFN−γ(BD Biosciences)と共に室温で2時間インキュベートし、PBS/Tweenで6回洗浄し、1:500希釈のストレプトアビジン−アルカリ性ホスファターゼ(Southern Biotechnology Associates)と共に2時間インキュベートした。PBS/Tweenで5回及びPBSで1回洗浄した後、プレートをニトロブルーテトラゾリウム/5−ブロモ−4−クロロ−3−インドリル−リン酸色原体(Pierce)で展開し、水道水で洗浄することにより停止させ、風乾し、ELISPOTリーダー(Cellular Technology Ltd)を使用して読み取った。106個のPBMC当たりのスポット形成細胞(SFC)を計算した。媒質バックグラウンドは、典型的には106PBMC当たり15SFC未満であった。陽性応答は、106個のPBMC当たり55個より多いSFC、且つバックグラウンドの4倍超として定義した。
【0105】
エピトープマッピング。NIH AIDS Research and Reference Reagent Programから入手したGag、Pol、及びEnv PTEペプチドを利用して、包括的なCD8+及びCD4+ Tリンパ球エピトープマッピングを実施した。最初に完全ペプチドプールで、並びに10個のPTEペプチドを含むサブプールで免疫した後4週目に、IFN−γ ELISPOTアッセイを行った。陽性応答を有する全てのペプチドサブプールをデコンボリューション処理し、個々の15アミノ酸PTEペプチドでエピトープが確認された。次に細胞枯渇IFN−γ ELISPOTアッセイを実施して、反応性ペプチドがCD8+又はCD4+ Tリンパ球エピトープに相当するかどうかを判断した。追加免疫の4週間後44週目にPTEサブプールを利用した部分的エピトープマッピングもまた実施した。ボーダーラインの応答は全て再試験し、確認された場合にのみ陽性と見なした。10個の重複Gagペプチドを含むサブプールを利用した部分的エピトープマッピングもまた実施し、様々なクレード由来のHIV−1 Gagの幅を評価した。
【0106】
体液性免疫アッセイ。本質的に記載されるとおり、HIV−1クレードC Env gp140及びルシフェラーゼベースのシュードウイルス中和アッセイを利用する直接ELISAにより、Env特異的体液性免疫応答について調べた(Montefiori、「Evaluating neutralizing antibodies against HIV, SIV and SHIV in luciferase reporter gene assays」、Current Protocols in Immunology、Coligan、Kruisbeek、Margulies、Shevach、Strober、及びCoico編(John Wiley & Sons、2004年、1〜15頁)。
【0107】
統計分析。全ての統計分析はパッケージR(Team、Foundation for Statisical Computing、Vienna、オーストリア、2009年)を使用して行った。細胞性免疫応答の幅を分析してPTEペプチドをマッピングするため(図19A)、ワクチン群、抗原(Gag、Pol、Env)、及びリンパ球部分母集団(CD4、CD8)の関数として反応性ペプチドの数を予想するポアソン回帰モデルをフィットさせた。本発明者らのモデルでは、動物間のばらつきに対応するため変量効果を含め、パッケージRのlme4ライブラリ(Pinheiro、Springer、New York(2000年))によりフィットさせた。データはモデルを良好にフィットし(分散パラメータ1.0)、3個の説明因子間に有意な交互作用はなかった。例えば、モザイク抗原の投与を受けたサルによって認識されたPTEペプチド数が、コンセンサス又は天然配列抗原の投与を受けた場合と比較したとき3.8倍亢進したことは(図19A)、Gag、Pol、及びEnv由来のPTEに等しく当てはまり、CD8+並びにCD4+ Tリンパ球による応答について維持された。反応性エピトープ領域数の分析(図19B)にもまた変量効果を伴うポアソン回帰モデルを含め、これもまた有意な交互作用なしに良好にフィットした(分散パラメータ0.87)。コルモゴロフ−スミルノフの両側検定を利用してCD8+及びCD4+ Tリンパ球応答の大きさの比較(図23)を実施した。サル当たりの応答の幅及び深さを種々のワクチン間で比較するノンパラメトリック検定もまた実施した(図19A及び図24C)。本発明者らは、4つのワクチン群間に違いがあるかどうかを判断するため最初にクラスカル・ワリス検定を用いた。いずれの場合も、これは高度に有意であったとともに、次に本発明者らはウィルコクソン順位和検定を使用して4つのワクチン群間の全てのペアワイズ比較を評価した。これらの比較の各々において、モザイクワクチンが誘発したサル当たりの応答は、他の3つのワクチンと比べて有意に多かった。様々なクレード由来のHIV−1 Gagに対する応答の幅を分析するため(図25)、データを二項回帰モデルにフィットさせた。これらのモデルでは説明変数としてワクチン群を使用し、動物間及び株間のばらつきを考慮するため変量効果を含めた。データは僅かに過小分散であったが、モザイクワクチンの投与を受けた動物はなお有意により多い応答数を誘発した。ロスアラモスHIV−1配列データベースで利用可能なツールを使用してPTE有効範囲の評価を実施した。
【0108】
配列別添資料
I.二価MモザイクENV GP160、GAG、POL、NEF配列
モザイクENV1 GP160(アミノ酸配列)
配列番号1
【0109】
モザイクENV2 GP160(アミノ酸配列)
配列番号2
【0110】
モザイクGAG1(アミノ酸配列)
配列番号3
【0111】
モザイクGAG2(アミノ酸配列)
配列番号4
【0112】
モザイクPOL1(アミノ酸配列)
配列番号5
【0113】
モザイクPOL2(アミノ酸配列)
配列番号6
【0114】
モザイクNEF1(アミノ酸配列)
配列番号7
【0115】
モザイクNEF2(アミノ酸配列)
配列番号8
【0116】
II.二価MモザイクENV GP140配列(切断/融合欠損)
モザイクENV1 GP140(アミノ酸配列)
配列番号9
【0117】
【0118】
モザイクENV2 GP140(アミノ酸配列)
配列番号10
【0119】
MOS3 ENV GP140(アミノ酸配列)
678アミノ酸
配列番号11
【0120】
III.二価MモザイクPOL配列(広範囲に不活化、PR欠失、9個のAの不活化突然変異による触媒活性の除去)
モザイクPOL1(アミノ酸配列)
配列番号12
【0121】
モザイクPOL2(アミノ酸配列)
配列番号13
【0122】
MOS3 POL V3(アミノ酸配列)
851アミノ酸
配列番号14
【0123】
IV.二価MモザイクGAG配列
MOS3 GAG(アミノ酸配列)
508アミノ酸
配列番号15
【0124】
V.二価MモザイクNEF配列(2位のGをAにすることによるミリスチル化部位の欠失)
MOS1 NEF
(206アミノ酸)
配列番号16
【0125】
MOS2 NEF
(206アミノ酸)−2位のGをAにすることによるミリスチル化部位の欠失
配列番号17
【0126】
MOS3 NEF
(208アミノ酸)
配列番号18
【0127】
VI.二価MモザイクGAGNEF融合配列
モザイクGAGNEF1(アミノ酸配列)
配列番号19
【0128】
モザイクGAGNEF2(アミノ酸配列)
配列番号20
【0129】
【0130】
VII.二価MモザイクGAGPOL融合配列(バージョン3;POLは広範囲に不活化、PR欠失、9個のAの不活化突然変異による触媒活性の除去)
モザイクGAGPOL1 V3(アミノ酸配列)
配列番号21
【0131】
モザイクGAGPOL2 V3(アミノ酸配列)
配列番号22
【0132】
【0133】
MOS3 GAG−POL V3(アミノ酸配列)
1359アミノ酸−完全GAGと修飾POLとのGAG−POL融合
配列番号23
【0134】
【0135】
VIII.二価MモザイクGAGPOL融合配列(バージョン4;POLは最小限の不活化、完全なPR−RT−IN)
モザイクGAGPOL1 V4(アミノ酸配列)
配列番号24
【0136】
モザイクGAGPOL2 V4(アミノ酸配列)
配列番号25
【0137】
【0138】
IX.二価MモザイクGAGPOL融合配列(バージョン5;POLは最小限の不活化、PR欠失)
モザイクGAGPOL1 V5(アミノ酸配列)
配列番号26
【0139】
【0140】
モザイクGAGPOL2 V5(アミノ酸配列)
配列番号27
【0141】
X.二価MモザイクGAGPOLNEF融合配列(POLは広範囲に不活化、PR欠失)
モザイクGagPolNef1(アミノ酸配列)
配列番号28
【0142】
【0143】
モザイクGAGPOLNEF2(アミノ酸配列)
配列番号29
【0144】
【0145】
XI.最適クレードC ENV GP160、GAG、POL、NEF配列
最適クレードC ENV GP160(SN90.90.SE364)(アミノ酸配列)
配列番号30
【0146】
最適クレードC GAG(IN.70177)(アミノ酸配列)
配列番号31
【0147】
最適クレードC POL(ZA.04.04ZASK208B1)(アミノ酸配列)
配列番号32
【0148】
【0149】
最適クレードC NEF(ZA00.1170MB)(アミノ酸配列)
配列番号33
【0150】
XII.最適クレードC ENV GP140配列(切断/融合欠損)
最適クレードC ENV GP140(SN90.90.SE364)(アミノ酸配列)
配列番号34
【0151】
XIII.最適クレードC POL 配列(広範囲に不活化、PR欠失)
最適クレードC POL(ZA.04.04ZASK208B1)(アミノ酸配列)
配列番号35
【0152】
XIV.最適クレードC GAGNEF融合配列
最適クレードC GAGNEF(IN.70177−ZA00.1170MB)(アミノ酸配列)
配列番号36
【0153】
XV.コンセンサス配列
MコンセンサスENV
配列番号37
【0154】
MコンセンサスGAG
配列番号38
【0155】
MコンセンサスPOL
配列番号39
【0156】
【0157】
他の実施形態
本発明はその特定の実施形態に関連して記載されているが、さらなる変更が可能であり、及び本願が、概して本発明の原理に従い、且つ本発明が関係する技術分野のなかの公知の又は通常の実施の範囲内に含まれるとともに本明細書で以上に示された本質的な特徴に該当し得る本開示からのかかる逸脱を含む本発明の任意の変形例、使用、又は適応を包含することを意図していることは理解されるであろう。
【0158】
本明細書において言及される全ての刊行物及び特許出願は、あたかも各々の独立した刊行物又は特許出願が、全体として参照により援用されるように具体的且つ個別的に示されたのと同じ程度まで、参照により本明細書に援用される。
【特許請求の範囲】
【請求項1】
少なくとも2個の別個の最適化ウイルスポリペプチドを含む、哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンであって、前記最適化ウイルスポリペプチドが同じウイルス遺伝子産物に対応する、前記ワクチン。
【請求項2】
前記哺乳動物がヒトである、請求項1に記載のワクチン。
【請求項3】
前記ワクチンが、前記ウイルス遺伝子産物に対する細胞性免疫応答を誘発する、請求項1に記載のワクチン。
【請求項4】
前記ウイルス感染症が、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる、請求項1に記載のワクチン。
【請求項5】
前記レトロウイルスがヒト免疫不全ウイルス1型(HIV−1)である、請求項4に記載のワクチン。
【請求項6】
前記ウイルス遺伝子産物が、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される、請求項5に記載のワクチン。
【請求項7】
前記ワクチンが、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される前記ウイルス遺伝子産物のうちの1つに対応する最適化ウイルスポリペプチドを2個以下含む、請求項6に記載のワクチン。
【請求項8】
前記ワクチンが、Gag及びNefに対応する最適化ウイルスポリペプチドを含まない、請求項5又は6に記載のワクチン。
【請求項9】
配列番号1〜29に示される配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸を含む最適化ウイルスポリペプチドを含む、哺乳動物におけるヒト免疫不全ウイルス1型(HIV−1)感染症を治療し、又はそのリスクを低減するワクチン。
【請求項10】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示される配列のいずれか1つとのアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸を含む、請求項9に記載のワクチン。
【請求項11】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示される配列のいずれか1つのアミノ酸配列を含む、請求項9に記載のワクチン。
【請求項12】
前記哺乳動物がヒトである、請求項9に記載のワクチン。
【請求項13】
前記ワクチンが、HIV−1に対する細胞性免疫応答を誘発する、請求項9に記載のワクチン。
【請求項14】
前記ワクチンが、少なくとも2つの前記最適化ウイルスポリペプチドを含む、請求項9に記載のワクチン。
【請求項15】
前記少なくとも2つの前記最適化ウイルスポリペプチドが、
a) 配列番号1及び2;
b) 配列番号3及び4;
c) 配列番号5及び6;
d) 配列番号7及び8;
e) 配列番号9及び10;
f) 配列番号10及び11;
g) 配列番号9及び11;
h) 配列番号12及び13;
i) 配列番号13及び14;
j) 配列番号12及び14;
k) 配列番号3及び15;
l) 配列番号4及び15;
m) 配列番号16及び17;
n) 配列番号17及び18;
o) 配列番号16及び18;
p) 配列番号19及び20;
q) 配列番号21及び22;
r) 配列番号22及び23;
s) 配列番号21及び23;
t) 配列番号24及び25;
u) 配列番号26及び27;又は
v) 配列番号28及び29、
を含む、請求項14に記載のワクチン。
【請求項16】
前記ワクチンが、少なくとも3つの前記最適化ウイルスポリペプチドを含む、請求項9に記載のワクチン。
【請求項17】
前記ワクチンが、少なくとも4つの前記最適化ウイルスポリペプチドを含む、請求項9に記載のワクチン。
【請求項18】
別個の最適化ウイルスポリペプチドの少なくとも2つの対を含む、哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンであって、前記最適化ウイルスポリペプチドの各対が同じウイルス遺伝子産物に対応し、及び前記ワクチンに組み込まれる2個以下の最適化ウイルスポリペプチドが同じウイルス遺伝子産物に対応する、前記ワクチン。
【請求項19】
前記哺乳動物がヒトである、請求項18に記載のワクチン。
【請求項20】
前記ワクチンが、前記ウイルス遺伝子産物に対する細胞性免疫応答を誘発する、請求項18に記載のワクチン。
【請求項21】
別個の最適化ウイルスポリペプチドの少なくとも3つの対を含む、請求項18に記載のワクチン。
【請求項22】
別個の最適化ウイルスポリペプチドの少なくとも4つの対を含む、請求項18に記載のワクチン。
【請求項23】
前記ウイルス感染症が、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる、請求項18に記載のワクチン。
【請求項24】
前記レトロウイルスがヒト免疫不全ウイルス1型(HIV−1)である、請求項23に記載のワクチン。
【請求項25】
前記別個の最適化ウイルスポリペプチドの少なくとも2つの対が、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される前記ウイルス遺伝子産物のうちの任意の2つに対応する、請求項24に記載のワクチン。
【請求項26】
前記少なくとも2つのウイルス遺伝子産物が、Gag及びNefではない、請求項25に記載のワクチン。
【請求項27】
Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される前記ウイルス遺伝子産物のうちの任意の3つに対応する別個の最適化ウイルスポリペプチドの少なくとも3つの対を含む、請求項25に記載のワクチン。
【請求項28】
Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される前記ウイルス遺伝子産物のうちの任意の4つに対応する別個の最適化ウイルスポリペプチドの少なくとも4つの対を含む、請求項18に記載のワクチン。
【請求項29】
前記ワクチンが、HIV−1に対する細胞性免疫応答を誘発する、請求項24に記載のワクチン。
【請求項30】
少なくとも1つの別個の最適化ウイルスポリペプチドのヌクレオチド配列が核酸又はベクターによりコードされる、請求項1、9又は18に記載のワクチン。
【請求項31】
前記ベクターが組換えアデノウイルスである、請求項30に記載のワクチン。
【請求項32】
前記組換えアデノウイルスが、アデノウイルス血清型26(Ad26)、アデノウイルス血清型34(Ad34)、アデノウイルス血清型35(Ad35)、アデノウイルス血清型48(Ad48)、又はアデノウイルス血清型5 HVR48(Ad5HVR48)である、請求項31に記載のワクチン。
【請求項33】
薬学的に許容可能な担体、賦形剤、又は希釈剤とさらに組み合わされた、請求項1、9又は18に記載のワクチン。
【請求項34】
配列番号1〜29に示されるアミノ酸配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸を含む最適化ウイルスポリペプチドのヌクレオチド配列を含む核酸。
【請求項35】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示されるアミノ酸配列のいずれか1つと配列同一性を有する少なくとも7個の連続するアミノ酸を含む、請求項34に記載の核酸。
【請求項36】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示されるアミノ酸配列のいずれか1つを含む、請求項34に記載の核酸。
【請求項37】
ベクターをさらに含む、請求項34に記載の核酸。
【請求項38】
前記ベクターが組換えアデノウイルスである、請求項37に記載の核酸。
【請求項39】
前記組換えアデノウイルスが、アデノウイルス血清型26(Ad26)、アデノウイルス血清型34(Ad34)、アデノウイルス血清型35(Ad35)、アデノウイルス血清型48(Ad48)、又はアデノウイルス血清型5 HVR48(Ad5HVR48)である、請求項38に記載の核酸。
【請求項40】
配列番号1〜29に示されるアミノ酸配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸を含む最適化ウイルスポリペプチド。
【請求項41】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示されるアミノ酸配列のいずれか1つと配列同一性を有する少なくとも7個の連続するアミノ酸を含む、請求項40に記載の最適化ウイルスポリペプチド。
【請求項42】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示されるアミノ酸配列のいずれか1つを含む、請求項40に記載の最適化ウイルスポリペプチド。
【請求項43】
哺乳動物に対し、請求項1、9若しくは18に記載のワクチン又は請求項37に記載の核酸を投与することを含む、前記哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減する方法。
【請求項44】
前記哺乳動物がヒトである、請求項43に記載の方法。
【請求項45】
前記ワクチン又は核酸が、前記ウイルス遺伝子産物に対する細胞性免疫応答を誘発する、請求項43に記載の方法。
【請求項46】
前記ウイルス感染症が、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる、請求項43に記載の方法。
【請求項47】
前記レトロウイルスがヒト免疫不全ウイルス1型(HIV−1)である、請求項46に記載の方法。
【請求項48】
前記ウイルス遺伝子産物が、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される、請求項43に記載の方法。
【請求項49】
哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンの製造方法であって、請求項1、9又は18に記載のワクチンを合成することを含む、前記方法。
【請求項50】
哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンの製造方法であって、
a)請求項37に記載の核酸を細胞と接触させるステップと、
b)前記最適化ウイルスポリペプチドを単離するステップと、
を含む、前記方法。
【請求項51】
前記最適化ウイルスポリペプチドが、哺乳動物に投与されると細胞性免疫応答を誘発する、請求項49又は50に記載の方法。
【請求項52】
前記哺乳動物がヒトである、請求項49又は50に記載の方法。
【請求項53】
前記ワクチンが、前記ウイルス遺伝子産物に対する細胞性免疫応答を誘発する、請求項49又は50に記載の方法。
【請求項54】
前記ウイルス感染症が、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる、請求項49又は50に記載の方法。
【請求項55】
前記レトロウイルスがヒト免疫不全ウイルス1型(HIV−1)である、請求項54に記載の方法。
【請求項56】
前記ウイルス遺伝子産物が、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される、請求項55に記載の方法。
【請求項57】
a)請求項1、9又は18に記載のワクチンと、
b)薬学的に許容可能な担体、賦形剤、又は希釈剤と、
c)キットの使用説明書と、
を含むキット。
【請求項58】
アジュバントをさらに含む、請求項57に記載のキット。
【請求項59】
a)請求項37に記載の核酸と、
b)薬学的に許容可能な担体、賦形剤、又は希釈剤と、
c)キットの使用説明書と、
を含むキット。
【請求項60】
アジュバントをさらに含む、請求項59に記載のキット。
【請求項1】
少なくとも2個の別個の最適化ウイルスポリペプチドを含む、哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンであって、前記最適化ウイルスポリペプチドが同じウイルス遺伝子産物に対応する、前記ワクチン。
【請求項2】
前記哺乳動物がヒトである、請求項1に記載のワクチン。
【請求項3】
前記ワクチンが、前記ウイルス遺伝子産物に対する細胞性免疫応答を誘発する、請求項1に記載のワクチン。
【請求項4】
前記ウイルス感染症が、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる、請求項1に記載のワクチン。
【請求項5】
前記レトロウイルスがヒト免疫不全ウイルス1型(HIV−1)である、請求項4に記載のワクチン。
【請求項6】
前記ウイルス遺伝子産物が、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される、請求項5に記載のワクチン。
【請求項7】
前記ワクチンが、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される前記ウイルス遺伝子産物のうちの1つに対応する最適化ウイルスポリペプチドを2個以下含む、請求項6に記載のワクチン。
【請求項8】
前記ワクチンが、Gag及びNefに対応する最適化ウイルスポリペプチドを含まない、請求項5又は6に記載のワクチン。
【請求項9】
配列番号1〜29に示される配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸を含む最適化ウイルスポリペプチドを含む、哺乳動物におけるヒト免疫不全ウイルス1型(HIV−1)感染症を治療し、又はそのリスクを低減するワクチン。
【請求項10】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示される配列のいずれか1つとのアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸を含む、請求項9に記載のワクチン。
【請求項11】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示される配列のいずれか1つのアミノ酸配列を含む、請求項9に記載のワクチン。
【請求項12】
前記哺乳動物がヒトである、請求項9に記載のワクチン。
【請求項13】
前記ワクチンが、HIV−1に対する細胞性免疫応答を誘発する、請求項9に記載のワクチン。
【請求項14】
前記ワクチンが、少なくとも2つの前記最適化ウイルスポリペプチドを含む、請求項9に記載のワクチン。
【請求項15】
前記少なくとも2つの前記最適化ウイルスポリペプチドが、
a) 配列番号1及び2;
b) 配列番号3及び4;
c) 配列番号5及び6;
d) 配列番号7及び8;
e) 配列番号9及び10;
f) 配列番号10及び11;
g) 配列番号9及び11;
h) 配列番号12及び13;
i) 配列番号13及び14;
j) 配列番号12及び14;
k) 配列番号3及び15;
l) 配列番号4及び15;
m) 配列番号16及び17;
n) 配列番号17及び18;
o) 配列番号16及び18;
p) 配列番号19及び20;
q) 配列番号21及び22;
r) 配列番号22及び23;
s) 配列番号21及び23;
t) 配列番号24及び25;
u) 配列番号26及び27;又は
v) 配列番号28及び29、
を含む、請求項14に記載のワクチン。
【請求項16】
前記ワクチンが、少なくとも3つの前記最適化ウイルスポリペプチドを含む、請求項9に記載のワクチン。
【請求項17】
前記ワクチンが、少なくとも4つの前記最適化ウイルスポリペプチドを含む、請求項9に記載のワクチン。
【請求項18】
別個の最適化ウイルスポリペプチドの少なくとも2つの対を含む、哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンであって、前記最適化ウイルスポリペプチドの各対が同じウイルス遺伝子産物に対応し、及び前記ワクチンに組み込まれる2個以下の最適化ウイルスポリペプチドが同じウイルス遺伝子産物に対応する、前記ワクチン。
【請求項19】
前記哺乳動物がヒトである、請求項18に記載のワクチン。
【請求項20】
前記ワクチンが、前記ウイルス遺伝子産物に対する細胞性免疫応答を誘発する、請求項18に記載のワクチン。
【請求項21】
別個の最適化ウイルスポリペプチドの少なくとも3つの対を含む、請求項18に記載のワクチン。
【請求項22】
別個の最適化ウイルスポリペプチドの少なくとも4つの対を含む、請求項18に記載のワクチン。
【請求項23】
前記ウイルス感染症が、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる、請求項18に記載のワクチン。
【請求項24】
前記レトロウイルスがヒト免疫不全ウイルス1型(HIV−1)である、請求項23に記載のワクチン。
【請求項25】
前記別個の最適化ウイルスポリペプチドの少なくとも2つの対が、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される前記ウイルス遺伝子産物のうちの任意の2つに対応する、請求項24に記載のワクチン。
【請求項26】
前記少なくとも2つのウイルス遺伝子産物が、Gag及びNefではない、請求項25に記載のワクチン。
【請求項27】
Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される前記ウイルス遺伝子産物のうちの任意の3つに対応する別個の最適化ウイルスポリペプチドの少なくとも3つの対を含む、請求項25に記載のワクチン。
【請求項28】
Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される前記ウイルス遺伝子産物のうちの任意の4つに対応する別個の最適化ウイルスポリペプチドの少なくとも4つの対を含む、請求項18に記載のワクチン。
【請求項29】
前記ワクチンが、HIV−1に対する細胞性免疫応答を誘発する、請求項24に記載のワクチン。
【請求項30】
少なくとも1つの別個の最適化ウイルスポリペプチドのヌクレオチド配列が核酸又はベクターによりコードされる、請求項1、9又は18に記載のワクチン。
【請求項31】
前記ベクターが組換えアデノウイルスである、請求項30に記載のワクチン。
【請求項32】
前記組換えアデノウイルスが、アデノウイルス血清型26(Ad26)、アデノウイルス血清型34(Ad34)、アデノウイルス血清型35(Ad35)、アデノウイルス血清型48(Ad48)、又はアデノウイルス血清型5 HVR48(Ad5HVR48)である、請求項31に記載のワクチン。
【請求項33】
薬学的に許容可能な担体、賦形剤、又は希釈剤とさらに組み合わされた、請求項1、9又は18に記載のワクチン。
【請求項34】
配列番号1〜29に示されるアミノ酸配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸を含む最適化ウイルスポリペプチドのヌクレオチド配列を含む核酸。
【請求項35】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示されるアミノ酸配列のいずれか1つと配列同一性を有する少なくとも7個の連続するアミノ酸を含む、請求項34に記載の核酸。
【請求項36】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示されるアミノ酸配列のいずれか1つを含む、請求項34に記載の核酸。
【請求項37】
ベクターをさらに含む、請求項34に記載の核酸。
【請求項38】
前記ベクターが組換えアデノウイルスである、請求項37に記載の核酸。
【請求項39】
前記組換えアデノウイルスが、アデノウイルス血清型26(Ad26)、アデノウイルス血清型34(Ad34)、アデノウイルス血清型35(Ad35)、アデノウイルス血清型48(Ad48)、又はアデノウイルス血清型5 HVR48(Ad5HVR48)である、請求項38に記載の核酸。
【請求項40】
配列番号1〜29に示されるアミノ酸配列のいずれか1つと少なくとも85%のアミノ酸配列同一性を有する少なくとも7個の連続するアミノ酸を含む最適化ウイルスポリペプチド。
【請求項41】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示されるアミノ酸配列のいずれか1つと配列同一性を有する少なくとも7個の連続するアミノ酸を含む、請求項40に記載の最適化ウイルスポリペプチド。
【請求項42】
前記最適化ウイルスポリペプチドが、配列番号1〜29に示されるアミノ酸配列のいずれか1つを含む、請求項40に記載の最適化ウイルスポリペプチド。
【請求項43】
哺乳動物に対し、請求項1、9若しくは18に記載のワクチン又は請求項37に記載の核酸を投与することを含む、前記哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減する方法。
【請求項44】
前記哺乳動物がヒトである、請求項43に記載の方法。
【請求項45】
前記ワクチン又は核酸が、前記ウイルス遺伝子産物に対する細胞性免疫応答を誘発する、請求項43に記載の方法。
【請求項46】
前記ウイルス感染症が、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる、請求項43に記載の方法。
【請求項47】
前記レトロウイルスがヒト免疫不全ウイルス1型(HIV−1)である、請求項46に記載の方法。
【請求項48】
前記ウイルス遺伝子産物が、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される、請求項43に記載の方法。
【請求項49】
哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンの製造方法であって、請求項1、9又は18に記載のワクチンを合成することを含む、前記方法。
【請求項50】
哺乳動物におけるウイルス感染症を治療し、又はそのリスクを低減するワクチンの製造方法であって、
a)請求項37に記載の核酸を細胞と接触させるステップと、
b)前記最適化ウイルスポリペプチドを単離するステップと、
を含む、前記方法。
【請求項51】
前記最適化ウイルスポリペプチドが、哺乳動物に投与されると細胞性免疫応答を誘発する、請求項49又は50に記載の方法。
【請求項52】
前記哺乳動物がヒトである、請求項49又は50に記載の方法。
【請求項53】
前記ワクチンが、前記ウイルス遺伝子産物に対する細胞性免疫応答を誘発する、請求項49又は50に記載の方法。
【請求項54】
前記ウイルス感染症が、レトロウイルス、レオウイルス、ピコルナウイルス、トガウイルス、オルトミクソウイルス、パラミクソウイルス、カリシウイルス、アレナウイルス、フラビウイルス、フィロウイルス、ブニヤウイルス、コロナウイルス、アストロウイルス、アデノウイルス、パピローマウイルス、パルボウイルス、ヘルペスウイルス、ヘパドナウイルス、ポックスウイルス、又はポリオーマウイルスにより引き起こされる、請求項49又は50に記載の方法。
【請求項55】
前記レトロウイルスがヒト免疫不全ウイルス1型(HIV−1)である、請求項54に記載の方法。
【請求項56】
前記ウイルス遺伝子産物が、Gag、Pol、Env、Nef、Tat、Rev、Vif、Vpr、及びVpuからなる群から選択される、請求項55に記載の方法。
【請求項57】
a)請求項1、9又は18に記載のワクチンと、
b)薬学的に許容可能な担体、賦形剤、又は希釈剤と、
c)キットの使用説明書と、
を含むキット。
【請求項58】
アジュバントをさらに含む、請求項57に記載のキット。
【請求項59】
a)請求項37に記載の核酸と、
b)薬学的に許容可能な担体、賦形剤、又は希釈剤と、
c)キットの使用説明書と、
を含むキット。
【請求項60】
アジュバントをさらに含む、請求項59に記載のキット。
【図1】

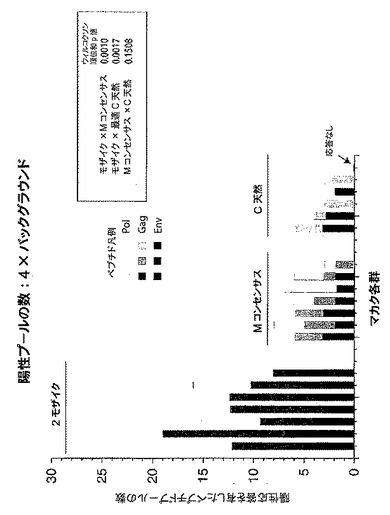
【図2】
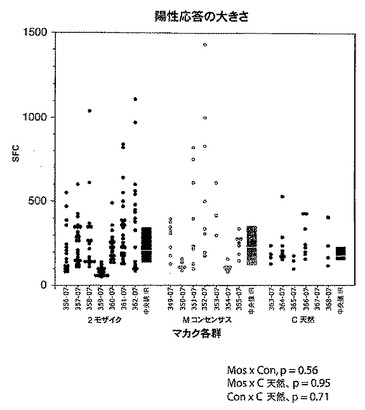
【図3】
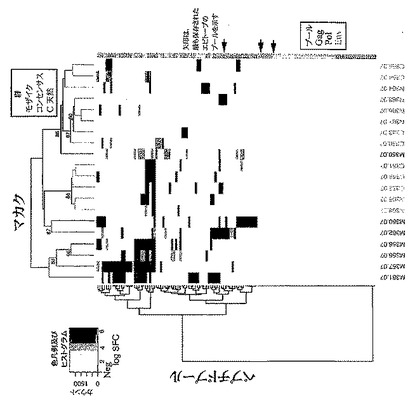
【図4】
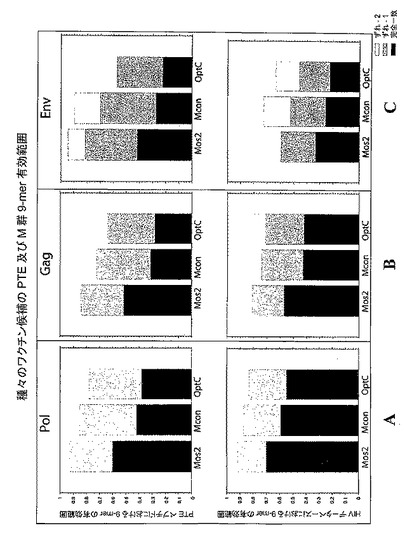
【図5】
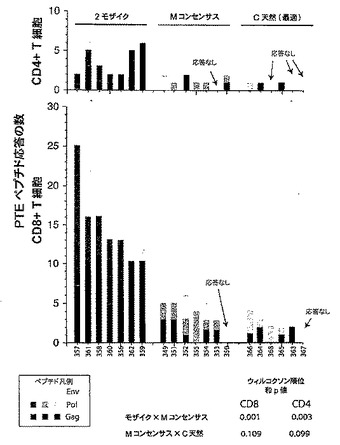
【図6】

【図7】

【図8】

【図9】
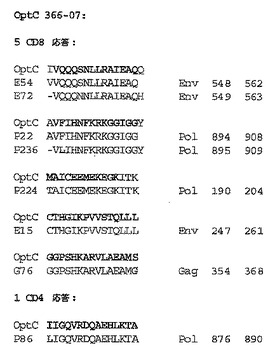
【図10】

【図11】

【図12】
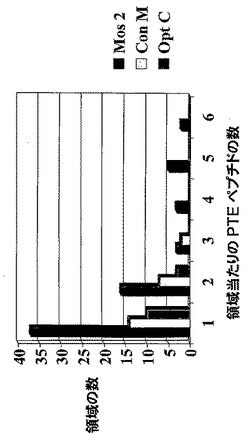
【図13】
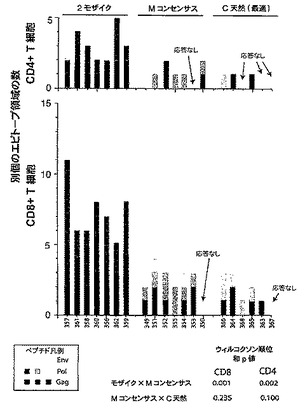
【図14】
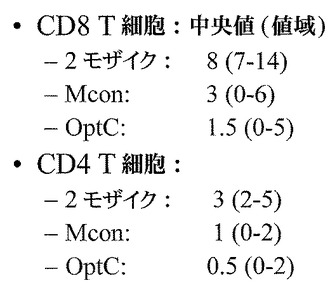
【図15】
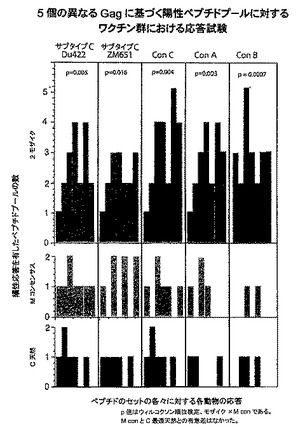
【図16】
【図17】
【図18】
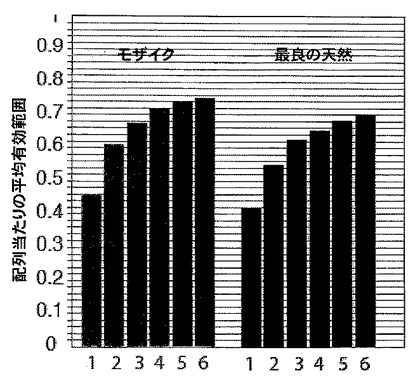
【図19】
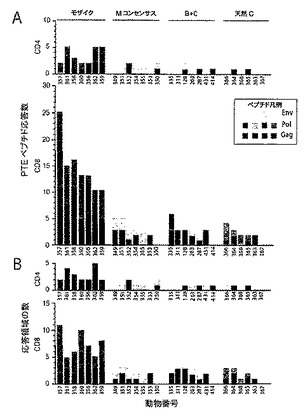
【図20A】

【図20B−1】

【図20B−2】
【図20C】

【図21A】

【図21B】

【図21C】
【図22−1】

【図22−2】

【図22−3】

【図22−4】

【図22−5】
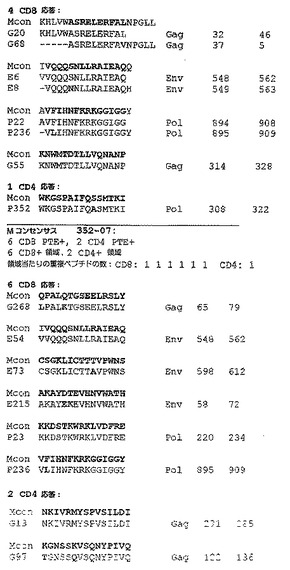
【図22−6】

【図22−7】

【図22−8】

【図22−9】

【図22−10】

【図22−11】
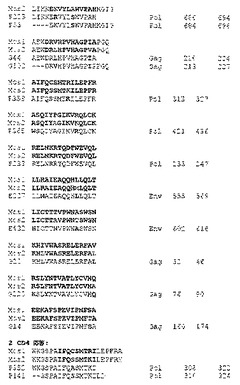
【図22−12】

【図22−13】

【図22−14】

【図22−15】

【図22−16】

【図22−17】

【図22−18】

【図23】
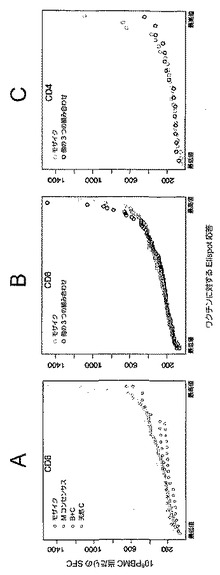
【図24A−B】
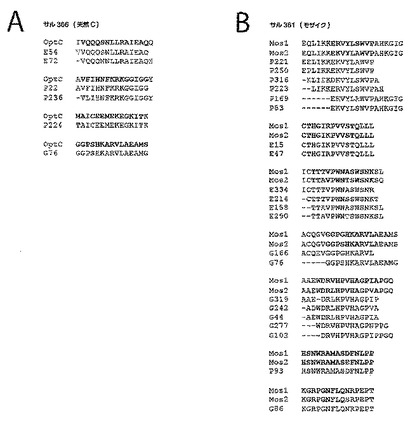
【図24C】
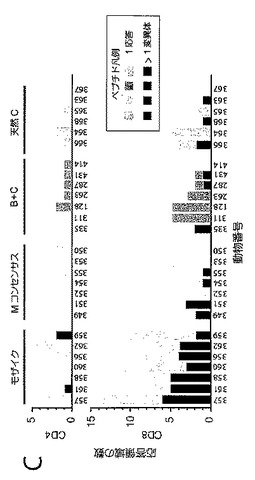
【図25】
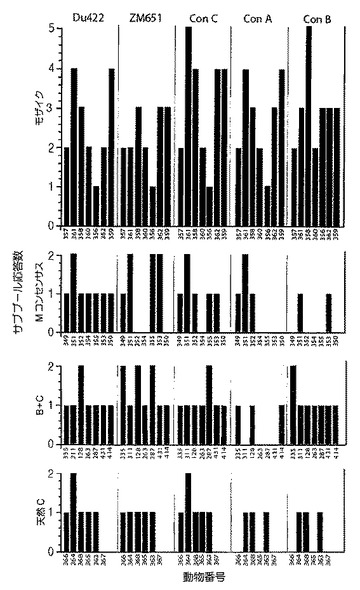
【図26A−B】
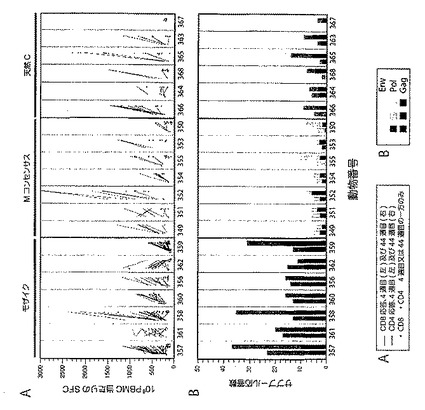
【図26C−D】
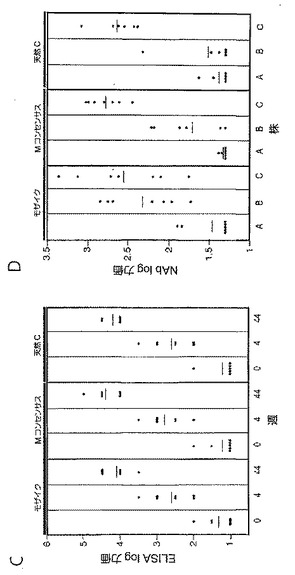
【図27】
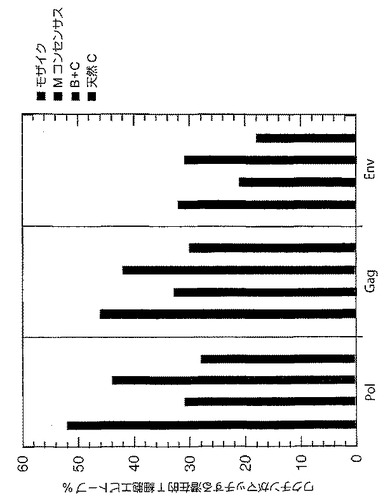
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20A】
【図20B−1】
【図20B−2】
【図20C】
【図21A】
【図21B】
【図21C】
【図22−1】
【図22−2】
【図22−3】
【図22−4】
【図22−5】
【図22−6】
【図22−7】
【図22−8】
【図22−9】
【図22−10】
【図22−11】
【図22−12】
【図22−13】
【図22−14】
【図22−15】
【図22−16】
【図22−17】
【図22−18】
【図23】
【図24A−B】
【図24C】
【図25】
【図26A−B】
【図26C−D】
【図27】
【公表番号】特表2012−509340(P2012−509340A)
【公表日】平成24年4月19日(2012.4.19)
【国際特許分類】
【出願番号】特願2011−537586(P2011−537586)
【出願日】平成21年11月18日(2009.11.18)
【国際出願番号】PCT/US2009/064999
【国際公開番号】WO2010/059732
【国際公開日】平成22年5月27日(2010.5.27)
【出願人】(303036197)ベス イスラエル デアコネス メディカル センター インコーポレイテッド (4)
【出願人】(511121447)ロス アラモス ナショナル セキュリティー,エルエルシー (1)
【Fターム(参考)】
【公表日】平成24年4月19日(2012.4.19)
【国際特許分類】
【出願日】平成21年11月18日(2009.11.18)
【国際出願番号】PCT/US2009/064999
【国際公開番号】WO2010/059732
【国際公開日】平成22年5月27日(2010.5.27)
【出願人】(303036197)ベス イスラエル デアコネス メディカル センター インコーポレイテッド (4)
【出願人】(511121447)ロス アラモス ナショナル セキュリティー,エルエルシー (1)
【Fターム(参考)】
[ Back to top ]